
植物および真菌において2−アセチル−1−ピロリン合成を増強する核酸
【課題】芳香族化合物2−アセチル−1−ピロリン(2AP)は、あらゆる香米の主要な強い香気成分であり、正確な生合成経路はまだ解明されていない2−アセチル−1−ピロリン合成に関与する遺伝子の同定、さらに植物および真菌において2−アセチル−1−ピロリンレベルを高めて芳香を増強するための方法を提供する。
【解決手段】2−アセチル−1−ピロリンが、天然に存在する非芳香性種で合成されるレベルよりも高いレベルで合成される非天然植物。さらに、より高いレベルの2−アセチル−1−ピロリンを合成する植物および真菌を選択するための分子マーカーとして使用することができる核酸。
【解決手段】2−アセチル−1−ピロリンが、天然に存在する非芳香性種で合成されるレベルよりも高いレベルで合成される非天然植物。さらに、より高いレベルの2−アセチル−1−ピロリンを合成する植物および真菌を選択するための分子マーカーとして使用することができる核酸。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一般に、植物分子遺伝学に関する。とりわけ、本発明は、高いレベルの2−アセチル−1−ピロリンを有する非天然植物および真菌、かかる植物および真菌を作製する方法、ならびに2−アセチル−1−ピロリン合成に関与する核酸に関する。
【背景技術】
【0002】
穀物の芳香は、高品質の米の最も魅力的な特徴であり、アジア市場で需要が増加しているだけでなく、欧州、そして世界中で広く認識されている。調理した米の芳香は、炭水化物、アルコール、アルデヒド、ケトン、酸、エステル、フェノール、ピリジン、ピラジンおよびそれ以外の化合物のような百を超える揮発性化合物からなっている(Yajima et al., 1978; Maga, 1984; Takashi et al., 1980; Paule and Power, 1989)。あらゆる香米、小麦パンおよびライ麦パンの皮の主要な強い香気成分として「ポップコーン様」の芳香族化合物、2−アセチル−1−ピロリン(2AP)が発見された(Buttery et al., 1982, 1983)。2−アセチル−1−ピロリンは、主として、数多くの香米品種に特有の芳香に関与している(Tanchotikul and Hsieh, 1991)。驚くべきことに、この米の芳香は、タコノキの葉(Buttery et al., 1983)、グルテン麦粉(Vallaris Glabra Ktze.)(Wongpornchai et al., 2003)、含水雑穀(wet millet)(Seitz et al., 1993)、ポップコーン(Schieberle, 1991)、バチルス・セレウス(Bacillus cereus)(Romanczyk et al., 1995)および真菌(Nagsuk et al., 2004)からも単離され、同定された。2−アセチル−1−ピロリンは、芳香性種のイネの、根を除く各部位(茎、葉、穀粒)に存在している(Lorieux et al., 1996)。芳香性種の穀粒にはこの芳香があるが、総ての穀粒にあるとは限らない。
【0003】
芳香族化合物、2−アセチル−1−ピロリンは、アミノ酸プロリンに類似したピロリン環を有している(図1)。アミノ酸プロリンを、2−アセチル−1−ピロリンを合成する前駆体として関連付ける最初の証拠が、細胞とカルス培養での試験によって見つけられた(Suprasanna et al., 1998; Suprasanna et al., 2002)。同位元素標識を用いた試験によって、タイ・ホーム・マリ(Thai Hom Mali)(THM)米において(Yoshihashi et al., 2002)、そしておそらく他の香米においても、穀粒2−アセチル−1−ピロリンの前駆体がアミノ酸プロリンである可能性が最も高いことが示され、その結果の裏付けが得られた。しかしながら、2−アセチル−1−ピロリンの正確な生合成経路はまだ解明されていない。従って、2−アセチル−1−ピロリン合成に関与する遺伝子を同定し、植物および真菌において2−アセチル−1−ピロリンレベルを高めて芳香を増強するための方法を提供する必要がある。
【発明の概要】
【0004】
本発明は、化合物2−アセチル−1−ピロリンが、対照植物および真菌で産生されるレベルよりも高いレベルで産生される非天然植物および真菌を提供することによりこれらの必要性に満たすものである。対照植物および真菌とは、化合物2−アセチル−1−ピロリンレベルが低い類似遺伝子型または関連遺伝子型の植物および真菌をさす。このような対照植物が非天然植物であってもよい。本発明はさらに、このような非天然植物および真菌、ならびに該植物から生じた種子についてスクリーニングし、作出する方法を提供する。
【0005】
化合物2−アセチル−1−ピロリンの正確な生合成経路は分かっていないが、その化合物はプロリンの誘導体である。プロリンは2−アセチル−1−ピロリンまたはグルタミン酸のいずれかに変換され得るため、グルタミン酸合成経路を阻害することによってプロリン(または中間体)の2−アセチル−1−ピロリン合成への利用可能性が高まる(図4A)という仮定が立てられる。Os2AP(オリザ・サティバ(oryza sativa)2−アセチル−1−ピロリン)と呼ばれる、米の芳香を制御するタンパク質をコードする遺伝子が、プロリンのグルタミン酸への変換において重要な役割を果たすと考えられているアルデヒトデヒドロゲナーゼファミリーのメンバーとして同定された。試験した総ての香米種では、この遺伝子に8個のヌクレオチドの欠失が存在する。この欠失は、ナンセンス変異を介したそのmRNAの分解を引き起こす早期停止コドンを与え、その結果、機能喪失表現型をもたらす。RNA干渉(RNAi)研究では、Os2AP遺伝子の転写破壊により、植物において高いレベルの2−アセチル−1−ピロリンが生じ、それとともに芳香が強まることが示された。
【0006】
本発明は、2−アセチル−1−ピロリン(2AP)遺伝子の発現を抑制し、2AP遺伝子のmRNAレベルを低減し、および/または2APタンパク質の活性を低下させることによって、高いレベルの2−アセチル−1−ピロリンが産生される非天然植物および真菌を提供する。2APタンパク質のレベルは、対照植物に対して、25パーセント、50パーセントまたは100パーセント低減する可能性がある。2AP遺伝子の発現の抑制または2AP遺伝子のmRNAの低減は、a)2AP遺伝子もしくはその断片をアンチセンス方向で発現させ、b)その遺伝子の一部をRNA干渉構築物にクローニングし、トランスジェニック植物においてこの構築物を発現させ、またはc)種々の方法(Targeting Induced Local Lesions IN Genomes(TILLING)法およびtDNA挿入突然変異誘発法を含む)によって突然変異誘発した後、PCRまたは他の方法により芳香性変異体についてスクリーニングすることにより、実現し得る。
【0007】
本発明はさらに、対照非トランスジェニックイネ植物が有するレベルよりも高いレベルの化合物2−アセチル−1−ピロリンを有するトランスジェニックイネ植物であって、該トランスジェニック植物における2AP遺伝子によりコードされるmRNAまたはタンパク質レベルを、該対照非トランスジェニック植物における2AP遺伝子によりコードされるmRNAまたはタンパク質レベルよりも、低減することによって、該トランスジェニック植物において該化合物のレベルが高められる、トランスジェニックイネ植物を提供する。一形式においては、mRNAおよびタンパク質レベルがRNA干渉によりまたはアンチセンスにより低減されている。本発明はさらに、本発明のイネから生じたトランスジェニックイネの種子に向けられる。
【0008】
後記の実施例は、イネにて実施した試験について記載しているが、本発明は、限定されるものではないが、コムギ、オオムギ、ライムギ、ココナッツ、ソルガムおよびオートムギを含む他の植物および真菌に関する。
【0009】
本発明はさらに、2AP遺伝子をコードする単離された核酸であって、少なくとも1回の0.1×SSCおよび0.1%SDSでの60〜65℃にて30分間の洗浄を含むハイブリダイゼーション条件下で、配列番号1、配列番号2、配列番号4もしくは配列番号5で示される核酸配列またはその相補配列とハイブリダイズする核酸、配列番号1、配列番号2、配列番号4もしくは配列番号5で示される配列と少なくとも70%同一、80%同一、90%同一、95%以上同一である核酸、および配列番号3または配列番号6で示されるアミノ酸配列と少なくとも70%、少なくとも80%、少なくとも90%、95%、または95%を超える割合で同一であるポリペプチドをコードする核酸を含む、単離された核酸を提供する。
【0010】
米中の2−アセチル−1−ピロリン量は、作況や土壌型によって変わり得る。非芳香性品種であるニホンバレ(Nipponbare)では、2−アセチル−1−ピロリンレベルが0〜0.1ppm(100万分の1)の範囲である。これに対して、香米品種であるタイ・ホーム・マリでは、2−アセチル−1−ピロリン量が1〜2.5ppmの範囲である。実施例2では、米ニホンバレ中の2−アセチル−1−ピロリンレベルを2.5ppmまで高めた、イネOs2AP遺伝子に対するRNA干渉試験について詳述している。よって、本発明は、非芳香性植物の芳香レベルを芳香族化合物レベルまで増強するための方法を提供する。
【0011】
本発明はさらに、プロモーターに作動可能なように連結され得る本発明の核酸を含有している組換え構築物および発現ベクターを提供する。1つの有用なプロモーターがカリフラワーモザイクウイルス(CaMV)プロモーターであり、このプロモーターは、ほとんどの植物組織において高レベルの発現を与える。
【0012】
本発明はさらに、本発明の核酸、構築物および発現ベクターを含有する宿主細胞を提供する。
【0013】
本発明はまた、植物および核酸を、2APタンパク質の発現または活性の低減を引き起こし、2AP化合物の産生が増加した結果として必然的に生じる芳香の増強をもたらす2AP遺伝子における突然変異についてスクリーニングするための方法も提供する。1つの特異的突然変異が、イネ2AP遺伝子の芳香性表現型と関連する8個のヌクレオチドの欠失である。この突然変異および他の突然変異についてのスクリーニングは、PCR、配列決定、ハイブリダイゼーションまたはマイクロアレイ試験のような種々の方法によって行うことができる。別の方法としては、2APタンパク質レベルの低減または2APタンパク質の構造もしくは活性の変化を探る方法がある。これについては、例えば、活性2APタンパク質と結合する抗体を使用することによって行えるし、2APタンパク質活性についてのアッセイによっても行える。
【0014】
本明細書に記載の配列は、核酸ハイブリダイゼーション試験におけるプローブまたはプライマーとして使用され得る。配列番号1、配列番号2、配列番号4もしくは配列番号5で示されるヌクレオチド配列の14ヌクレオチド長の連続したDNAセグメントと同じ配列を有するか、または配列番号1、配列番号2、配列番号4もしくは配列番号5で示されるヌクレオチド配列の14ヌクレオチド長の連続したDNAセグメントと相補的である少なくとも14ヌクレオチド長の連続した配列を含む核酸セグメントを使用することができる。
【0015】
より長い連続した同一または相補配列、例えば、約20、30、40、50、100、200、500、1,000、2,000、5,000、10,000ヌクレオチドのもの他(中間にあるあらゆる長さから全長配列を含むその長さまでを含む)もまた、特定の実施形態において有用である。
【0016】
このような核酸プローブの、2AP遺伝子配列と特異的にハイブリダイズする能力によって、それらが特定サンプルにおいて相補配列の存在を検出するのに有用であり得る。
【0017】
しかしながら、突然変異種プライマー、すなわち、他の遺伝的構造の調製用のプライマーの作製に用いる配列情報の使用をはじめとする他の使用が想定される。
【図面の説明】
【0018】
図1は、カオ・ド・マリ(Khao Dawk Mali)(KDML105)、タイジャスミン米(芳香性)およびコシヒカリ(Koshihikari)、非香米から、GC−MSにより得られた2−アセチル−1−ピロリンの化学構造を示す。
【0019】
図2は、穀物の芳香について分離する、単一F6植物由来の186のF9植物を用いたイネにおける芳香遺伝子の詳細なマッピングと穀物の芳香に関する遺伝子を含む物理的地図の構築を示す。
【0020】
図2の第1の部分は、単一F6植物由来の8のF11植物の図式遺伝子型を示す。第2の部分は、単一BACにおいて重要な領域を27kbに絞るための、単一F6植物由来の1116のF12植物を用いたウルトラスケールマッピングを示す。第3の部分は、3つのオープンリーディングフレームを示すKDML105のゲノム配列の注釈を示す。第4の部分は、KDML105とJHNの異種交配による177のF6植物を用いて、穀物の芳香および2−アセチル−1−ピロリン含量に大きな影響を及ぼす未知タンパク質遺伝子のエクソン7内に3つの二重組換え体が確認されることを示す。この未知タンパク質遺伝子は、Os2AP、オリザ・サティバ2−アセチル−1−ピロリンと名づけられた。「アロマーカー」は、穀物の芳香に特有の8塩基対の欠失と3つのSNP(一塩基変異多型)を定義するPCRベースのマーカーである。
【0021】
図3Aは、芳香性および非芳香性同質遺伝子系統の受粉の10、15および20日後に単離された全RNAからの、RT−PCRを用いた7つの候補遺伝子の発現を示す。この図において、Os2APは、ベタインアルデヒドデヒドロゲナーゼと見なされる。
【0022】
図3Bは、芳香性および非芳香性同質遺伝子系統の受粉の15日後に単離された全RNAからのOs2AP転写産物の葉、茎および根における示差的発現を示している。
【0023】
図3Cは、THM(KDML105)、芳香性同質遺伝子系統117、非芳香性同質遺伝子系統10およびそれらのF2(ISL117×ISL10)の穀粒における2−アセチル−1−ピロリンレベルの解析を示す。F1では、2−アセチル−1−ピロリンレベルの解析を葉で実施した。
【0024】
図3Dは、上のパネルは、Os2AP RNAi構築物を含有するトランスジェニックニホンバレにおける染色体4のOs2APホモログとアクチンの発現を示す。下のパネルは、芳香性および非芳香性同質遺伝子系統のイネにおいて示差的発現を示す、染色体4ではなく染色体8のOs2AP遺伝子(本明細書ではBADHと呼ぶ)を示す。
【0025】
図4Aは、プロリンおよびグルタミン酸間の代謝経路を示す。L−プロリンは、酵素P5CシンターゼおよびP5Cレダクターゼを用いてグルタミン酸から合成され、グルタミン酸は、プロリンデヒドロゲナーゼ(ProDH)およびP5Cデヒドロゲナーゼ(P5CDH)を用いてプロリンから合成される。提示された代謝シフトは、Os2APのナンセンス突然変異が介在するものであった。
【0026】
図4Bは、Microbiology Department of the University of Massachusetts, Amherst, USAのウェブサイトを通して利用されるプログラム、RASMOL(新しいバージョンがProtein Explorerである)を利用して作製されたOs2AP酵素の推定タンパク質構造を示す。
【0027】
図5A−1は、KDML105(芳香性)およびニホンバレ(非芳香性)由来のOs2AP遺伝子のゲノム配列比較を示す。図5A−2は、図5A−1の続きを示す。
【0028】
図5Bは、ニホンバレ、非芳香性系統と比べて、芳香性系統、タイ・ホーム・マリ(THM)における8塩基対の欠失を示すDNAアラインメントおよび2系統間のアミノ酸配列比較を示す。アラインメントしているヌクレオチド配列は、配列番号5のヌクレオチド701〜765と配列番号2のヌクレオチド701〜757である。
【0029】
図6は、単一F6植物由来の8のF11植物の図式遺伝子型と米粒子の2−アセチル−1−ピロリンレベルの解析を示す。
【0030】
図7Aは、RNA干渉構築物と形質転換用ベクターを示す。下のパネルは、構築物の発現確認を示す。本図面においてBADHとは、Os2APをさす。
【0031】
図7Bは、RNAi構築物、GFPおよび内在性Os2APの発現を示す。下のパネルは、2−アセチル−1−ピロリンレベルと芳香を示す。BADH−RNAi2は、Os2AP遺伝子に対するRNAi構築物である。
【0032】
図8Aは、「アロマーカー」プライマーセットを用いた、芳香の増強と関係しているOs2AP変異体についてのF6後代のスクリーニング結果を示す。
【0033】
図8Bは、芳香性Os2AP対立遺伝子についての種々の米品種のスクリーニング結果を示す。
【0034】
図9Aは、種々の系統のイネ由来のOs2APオーソログのDNA配列を示す。アラインメントしているヌクレオチド配列は、配列番号5のヌクレオチド701〜765と配列番号2のヌクレオチド701〜757である。
【0035】
図9Bは、22のBADH(Os2AP)オーソログの近隣結合樹を用いて構築した系統樹を示す。
【配列表の一部の配列の簡単な説明】
【0036】
配列番号1は、香米系統、タイ・ホーム・マリ由来のOs2AP遺伝子のゲノムヌクレオチド配列である。
配列番号1
【化1−1】

【化1−2】
【化1−3】
【0037】
配列番号2は、タイ・ホーム・マリOs2APのタンパク質をコードするヌクレオチド配列である。
配列番号2
【化2】
【0038】
配列番号3は、タイ・ホーム・マリOs2APタンパク質のアミノ酸配列である。
配列番号3
【化3】
【0039】
配列番号4は、非香米系統、ニホンバレ由来のOs2AP遺伝子のゲノムヌクレオチド配列である。
配列番号4
【化4−1】
【化4−2】
【0040】
配列番号5は、ニホンバレOs2APのタンパク質をコードするヌクレオチド配列である。
配列番号5
【化5】
【0041】
配列番号6は、ニホンバレOs2APタンパク質のアミノ酸配列である。
配列番号6
【化6】
【0042】
配列番号7〜88は、Os2APヌクレオチド配列の一部、GFPまたはアクチンを増幅するのに使用できるプライマーである(表1)。
【0043】
【化7】
【0044】
配列番号96は、一般アルデヒドデヒドロゲナーゼに高度に保存されているデカペプチドである。
Val-Thr-Leu-Glu-Leu-Gly-Gly-Lys-Ser-Pro
【発明の詳細な説明】
【0045】
本発明は、2−アセチル−1−ピロリン(2AP)遺伝子に関する。イネ、コムギ、トウモロコシ、オートムギ、タコノキの葉、芳香ココナッツおよび一部の細菌および真菌に見られる主要な芳香族化合物である、2−アセチル−1−ピロリンのレベルを高めるために、2AP遺伝子の核酸配列が用いられる。
【0046】
本発明の少なくとも1つの実施形態を詳細に説明する前に、本発明がその適用において、以下の説明において記載されている構造の詳細および要素の配列に限定されないことを理解すべきである。本発明は、他の実施形態が可能であり、または種々の方法での実践もしくは実施が可能である。さらに、本明細書において使用する表現および専門用語は、説明するためのものであって、限定を意図したものではないことも理解すべきである。
【0047】
この開示内容を通じて、種々の刊行物、特許および公開特許明細書が引用されるている。これらの刊行物、特許および公開特許明細書の開示内容は、本発明が関連する技術の状況をさらに十分説明するために、引用することにより本明細書の開示内容の一部とされる。本発明の実施では、特に断りのない限り、当技術分野の技術の範囲内である植物育種学、免疫学、分子生物学、微生物学、細胞生物学および組換えDNAの従来の技術を使用する。例えば、Sambrook, Fritsch and Maniatis, MOLECULAR CLONING: A LABORATORY MANUAL, 2nd edition (1989); CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F. M. Ausubel, et al eds., (1987); Plant Breeding: Principles and Prospects (Plant Breeding, Vol 1) M. D. Hayward, N. O. Bosemark, I. Romagosa; Chapman & Hall, (1993.); Coligan, Dunn, Ploegh, Speicher and Wingfeld, eds. (1995) CURRENT PROTOCOLS IN PROTEIN SCIENCE (John Wiley & Sons, Inc.); the series METHODS IN ENZYMOLOGY (Academic Press, Inc.): PCR 2: A PRACTICAL APPROACH (M. J. MacPherson, B. D. Hames and G. R. Taylor eds. (1995), Harlow and Lane, eds. (1988) ANTIBODIES, A LABORATORY MANUAL, and ANIMAL CELL CULTURE R. I. Freshney, ed. (1987)参照。
【0048】
特に断りのない限り、慣例的用法に従って専門用語を使用する。分子生物学において共通する用語の定義は、Lewin, Genes V, Oxford University Press, 1994 (SBN 0-19-854287-9)発行; Kendrew et al. (eds.), The Encyclopedia of Molecular Biology, Blackwell Science Ltd., 1994 (SBN 0-632-02182-9)発行;およびRobert A. Meyers (ed.), Molecular Biology and Biotechnology, a Comprehensive Desk Reference, VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8)発行; Ausubel et al. (1987) Current Protocols in Molecular Biology, Green Publishing; Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, New Yorkで見られる。植物生物学において共通する用語の定義は、Esau, Plant Anatomy, John Wiley & Sons (1977) (ISBN 0-471-24520-8)発行;およびSolomon et al, Biology, Saunders College Publishing (1993)発行、で見られる。
定義
【0049】
本発明の種々の実施形態の検討を容易にするため、以下の定義を提供する:
【0050】
2−アセチル−1−ピロリン(2AP)ポリヌクレオチド配列:本発明の遺伝子としては、本明細書において開示される全長配列だけでなく、本明細書において具体的に示される配列の特有の活性を保持するこのような配列の断片も挙げられる。
【0051】
イネ2AP遺伝子配列は、本明細書において開示されている。このイネ2AP遺伝子配列を用いたいくつかの手段を通じて、別の植物由来の2APポリヌクレオチド配列を容易に同定し、得ることができることは当業者には明らかである。特定の遺伝子またはその一部は、培養物寄託機関から得られ得、または合成により、例えば、遺伝子マシンを用いて構築され得る。
【0052】
このような遺伝子の変化は、点突然変異を作製するための標準的な技術を用いて容易に構築し得る。また、これらの遺伝子の断片も、標準的な手順に従って市販のエキソヌクレアーゼまたはエンドヌクレアーゼを用いて作製することができる。例えば、Bal31のような酵素または部位特異的突然変異誘発を利用して、これらの遺伝子の末端からヌクレオチドを意図的に切り出すことができる。また、種々の他の制限酵素を用いて、活性断片についてコードする遺伝子を得ることができる。種々の酵素を利用して、これらの2APポリヌクレオチド配列の活性断片を直接得てもよい。
【0053】
等価な2APポリヌクレオチド配列および/またはこれらの等価な2APポリヌクレオチド配列をコードする遺伝子も、本明細書において記載される教示を用いて、系統および/またはDNAライブラリーから単離され得る。例えば、本明細書において開示される2APタンパク質に対する抗体を使用して、タンパク質混合物から他の2APタンパク質を同定し、単離することができる。具体的に言えば、最も不変で、かつ、他のタンパク質と最も異なっている2APタンパク質の一部に対して抗体を作製してもよい。こうして作製した抗体を使用して、特に、特有の活性を有する等価な2APタンパク質を免疫沈降、酵素免疫測定法(ELISA)またはウエスタンブロッティングにより同定することができる。
【0054】
本発明の遺伝子を同定するためのさらなる方法は、オリゴヌクレオチドプローブの使用による方法である。これらのプローブは、検出可能な標識を有するヌクレオチド配列である。当技術分野では周知であるように、プローブ分子と核酸サンプルが2分子間で強い結合を形成することによってハイブリダイズするならば、プローブとサンプルが本質的に同じであると当然判断することができる。プローブの検出可能な標識は、ハイブリダイゼーションが起こったかどうかを既知の方法で判断するための手段を提供する。このようなプローブ解析は、本発明の遺伝子の迅速な同定法を提供する。典型的なプローブを表1に示している。
【0055】
本発明のプローブとして使用するヌクレオチドセグメントは、DNA合成装置を使用し、標準的な手順を用いて合成することができる。ヌクレオチドセグメントのプローブとしての使用においては、特定のプローブを、放射性および非放射性標識をはじめとする当業者には公知のあらゆる好適な標識で標識する。一般的な放射性標識としては、32P、125I、35Sなどが挙げられる。DNアーゼとDNAポリメラーゼを用いて、一般的なニックトランスレーション反応により、DNAサンプルと相補的であるヌクレオチド配列から放射性同位元素で標識したプローブを構築することができる。次いで、プローブおよびサンプルをハイブリダイゼーションバッファー溶液に入れて合わせ、アニーリングが起こるまで好適な温度に保つ。その後、メンブレンを洗浄し、異物を除いてサンプルを残し、結合したプローブ分子を、通常、オートラジオグラフィーおよび/または液体シンチレーション計測により検出し、定量する。
【0056】
非放射性標識としては、例えば、ビオチンもしくはチロキシンのようなリガンド、さらに、ヒドロラーゼもしくはペルオキシダーゼのような酵素、あるいはルシフェリンのような種々の化学発光体またはフルオレセインおよびその誘導体のような蛍光化合物も挙げられる。また、分離を容易にするために、プローブを、両末端において異なるタイプの標識で標識してもよく、例えば、一末端で、以上で記載した同位元素標識を、そして、もう一方の末端でビオチン標識を使用することによって標識してもよい。
【0057】
二重形成と安定性は、ハイブリッドの2鎖間の実質的な相補性に依存しており、以上で記載したように、ある程度のミスマッチは許容される。それゆえ、本発明のプローブは、上記配列の突然変異(単一突然変異と多重突然変異の両方)、欠失、挿入およびそれらの組合せ(前記突然変異、挿入および欠失によって目的の標的ポリヌクレオチドとの安定したハイブリッドが形成される)を含む。突然変異、挿入および欠失は、数多くの方法により、現在当業者に公知の方法により、そして、将来、広まるであろう他の方法により特定のポリヌクレオチド配列において作製することができる。
【0058】
表に記載したプローブの可能性ある変化は、ある程度、遺伝子コードの冗長性による。遺伝子コードの冗長性から、すなわち、タンパク質の作製に使用されるほとんどのアミノ酸に2つ以上のコーディングヌクレオチドトリプレット(コドン)が使用される。そのため、異なるヌクレオチド配列が特定のアミノ酸をコードする可能性がある。従って、2APタンパク質およびペプチドのアミノ酸配列を、タンパク質またはペプチドの同じアミノ酸配列をコードする等価なヌクレオチド配列により調製することができる。よって、本発明は、このような等価なヌクレオチド配列を含む。また、逆または相補配列も本発明の1つの態様であり、当業者ならば容易に使用することができる。さらに、構造および機能が同定されたタンパク質群が、アミノ酸配列を変えることにより、このような変化がタンパク質二次構造を改変しないという条件において、構築され得ることが示されている(Kaiser and Kezdy, 1984)。よって、本発明は、本明細書で示されるアミノ酸配列の、タンパク質二次構造が変化していないか、または構造が変化を受けているならば、生物活性が実質的に保持されている突然変異型を含む。さらに、本発明はまた、本発明の遺伝子をコードする2APポリヌクレオチド配列の総てまたは一部を受け入れる生物の突然変異体を含む。このような突然変異体は、当業者には周知の技術によって作製され得る。例えば、紫外線照射を利用して、宿主生物の突然変異体を作製することができるし、tDNA挿入突然変異誘発法を利用してもよいし、またはTILLING(Targeted Induced Local Lesion of Genome)法を利用してもよい。同様に、このような突然変異体としては、当技術分野で周知の手順によって作製することができる胞子無形成宿主細胞も含んでよい。
【0059】
イネ由来のものを含む2APポリヌクレオチド配列を、本発明のPCRプライマーを用いて得てもよい。これらのプライマーを表1に示しており、これらのプライマーは配列番号7〜配列番号79で示される配列と対応している。これらのプライマーの組合せを使用して、Os2AP遺伝子の異なる領域を増幅することができる。この増幅に有効なPCR条件は、以下のとおりである:10ngの鋳型DNA、0.1mM dNTP、0.5Mの各プライマー、0.5単位のTaqポリメラーゼ、2.0mM MgCl2および1×好熱性ポリメラーゼバッファー(Promega)を含有する10μL反応混合物。この混合物により、以下の時間および温度:94℃30秒間(変性)、60℃30秒間(アニーリング)および72℃2分間(伸長)を用いて30サイクルのPCRを行う必要がある。
【表1−1】
【表1−2】
【表1−3】
【0060】
オリザ・サティバ2−アセチル−1−ピロリン(Os2AP)ホモログ:本願にて記載したものと類似性を示す配列を、パブリックドメイン配列アラインメントアルゴリズムと配列データベースを検索するための配列類似性検索ツールを使用するコンピューターを利用した方法により同定することができる(パブリックドメインデータベースとしては、Genbank、EMBL、Swiss−Prot、PIRなどが挙げられる)。
【0061】
類似性検索を読み出し、解析しようとする標的配列(すなわち、質問配列)と比較する配列をアラインする。比較する配列の局所領域間の最適なアラインメントは、ローカルアラインメントとして知られている。配列比較アルゴリズムは、各アラインメントに総合スコアを与えるスコアリングマトリックスを使用している。
【0062】
ポリヌクレオチドおよびポリペプチド配列をアラインし、他のポリヌクレオチドおよびポリペプチド配列と比較した、指定領域における同一残基率を公的に入手可能なコンピューターアルゴリズムを利用して決定する。同一性パーセントスコアは、比較している配列のオーバーラップ領域の長さに依存する。
【0063】
2核酸配列間または2アミノ酸配列間の類似性は、配列同一性で表される(または、タンパク質の場合、配列類似性でも表される)。配列同一性は、同一性パーセントとして測定されることが多い;割合が高いほど、2配列の類似性が高いということである。本明細書において記載するように、2APタンパク質をコードする核酸分子のホモログおよび変異体を、本発明に使用してもよい。これらの核酸分子のホモログおよび変異体は、標準的な方法を使用してアラインした場合に、比較的高い配列同一性を有しているものである。このようなホモログおよび変異体は、高ストリンジェンシー条件下で互いにハイブリダイズする。
【0064】
比較用配列のアラインメント方法は、当技術分野ではよく知られている。種々のプログラムおよびアラインメントアルゴリズムについては、配列アラインメント方法と相同性計算の詳細にわたる検討を示しているSmith and Waterman (1981); Needleman and Wunsch (1970); Pearson and Lipman (1988); Higgins and Sharp (1988); Higgins and Sharp (1989); Corpet et al. (1988); Huang et al. (1992);およびPearson et al. (1994). Altschul et al.(1994)で記載されている。
【0065】
NCBIベーシックローカルアラインメント検索ツール(BLAST)(Altschul et al., 1990)は、National Center for Biotechnology Information(NCBI, Bethesda, MD)を含むいくつかの供給元からインターネット上で入手可能であり、配列解析プログラムblastp、blastn、blastx、tblastnおよびtblastxとともに用いられる。このツールはNCBIウェブサイトでアクセス可能である。このプログラムを利用した配列同一性決定方法についての解説は、NCBIウェブサイトで入手可能である。
【0066】
開示されたタンパク質配列のホモログは、一般に、開示された配列のアミノ酸配列との全長アラインメントにおいて、NCBI Blast2.0を使用し、ギャップ入りblastpをデフォルトパラメーターに設定して計算した場合に少なくとも40%配列同一性を有するという特徴を有するものである。調整パラメーターは、好ましくは、以下の値に設定する:オーバーラップスパン1、オーバーラップフラクション=0.125、ワード閾値(T)=11。HSP SおよびHSP S2パラメーターは動的値であり、それぞれの配列の構成と目的の配列が検索されるデータベースの構成に応じて、そのプログラム自体によって確立される。しかしながら、それらの値は、感度を高めるように調整される可能性がある。この方法を用いて評価した場合、参照配列との類似性がさらに高いタンパク質は、同一性パーセントの増加、例えば、少なくとも約50%、少なくとも約60%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%または少なくとも約95%配列の同一性を示す。
【0067】
開示された核酸配列のホモログは、一般に、開示された配列のアミノ酸配列との全長アラインメントにおいて、NCBI Blast2.0を使用し、ギャップ入りblastnをデフォルトパラメーターに設定して計算した場合に少なくとも40%配列同一性を有するという特徴を有するものである。さらに、このような配列は、高ストリンジェンシー条件下で相同配列とハイブリダイズする。好ましい方法は、WU−BLAST−2のBLASTNモジュールを使用し(Altschul et al., 1996)、デフォルトパラメーターに設定し、オーバーラップスパンおよびオーバーラップフラクションをそれぞれ1および0.125に設定する。この方法を用いて評価した場合、参照配列との類似性がさらに高い核酸配列は、同一性パーセントの増加、例えば、少なくとも約50%、少なくとも約60%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%または少なくとも約95%配列の同一性を示す。
【0068】
アラインメントは、アラインする配列へのギャップの挿入を含み得る。さらに、配列番号3もしくは配列番号6で示されるタンパク質より含有するアミノ酸が多い配列または配列番号3もしくは配列番号6で示されるタンパク質より含有するアミノ酸が少ない配列では、1つの実施形態において、配列同一性パーセントがアミノ酸総数に対する同一アミノ酸の数に基づいて決定されると理解される。よって、例えば、以下で述べる図面で示されたものより短い配列の配列同一性は、1つの実施形態において、長い方の配列のアミノ酸数を用いて決定される。同一性パーセントの計算において、相対重量は、挿入、欠失、置換他のような配列変化の多様な現れには関係していない。
【0069】
1つの実施形態においては、同一である場合だけにプラスの評点をつけ(+1)、ギャップを含むあらゆる形態の配列変化には「0」の値をつける。この場合、配列類似性計算に関して本明細書において記載する加重尺度またはパラメーターの必要性が除去される。配列同一性パーセントは、例えば、マッチしている同一残基数をアラインした領域の「短い方の」配列の残基総数で除し、100を乗じることにより計算することができる。「長い方の」配列は、アラインした領域に最も多くの現実の残基を有するものである。
【0070】
同じゲノム(パラログ)または異なるゲノム(オーソログ)において、タンパク質をそれらの配列関連性に従って他のタンパク質と区別することができる。オーソログ遺伝子は、共通の先祖遺伝子からの種形成によって生じた遺伝子である。これらの遺伝子は、通常、それらが導くものと同じ機能を保持している。パラログ遺伝子は、ゲノム内で複製される遺伝子である。これらの遺伝子は、初代のものに関係しているが、新しい特異性または改変された機能を獲得している。系統発生解析方法は、バイオインフォマティクスにおける通常の技量を有する人であれば周知である。
【0071】
当業者には明らかなように、本発明の配列は、配列決定エラーを含み得る。すなわち、不正確なアミノ酸配列、ヌクレオチド、フレームシフト、未知ヌクレオチドまたは他のタイプの配列決定エラーが配列中に存在する可能性がある。しかしながら、正確な配列が、核酸に関する本明細書における相同性およびストリンジェンシーの定義、ならびにタンパク質またはポリペプチドに関して記載されているタンパク質相同性の範囲に入る。
【0072】
2−アセチル−1−ピロリン(2AP)ポリペプチド:本明細書において「2APポリペプチド」とは、2APオーソログのアミノ酸配列を実質的に有する遺伝子産物を意味する用語である。2APポリペプチドは、1つには、その発現を低減し、そのmRNAレベルを低減し、またはタンパク質量もしくは活性を低減することによって、植物における化合物2−アセチル−1−ピロリンのレベルが高められるということを特徴とする。2APポリペプチドはまた、1つには、配列番号3または配列番号6で示されるアミノ酸配列と少なくとも約30%、40%、50%、60%、70%、80%、90%または95%アミノ酸同一性を有するアミノ酸配列を有していることも特徴とする。
【0073】
実質的に同一:「実質的に同一」とは、参照アミノ酸配列(例えば、配列番号3もしくは配列番号6で示されるアミノ酸配列)または核酸配列(例えば、配列番号1、配列番号2、配列番号4もしくは配列番号5で示される核酸配列)と少なくとも30%、好ましくは、50%、より好ましくは、80%、最も好ましくは、90%、あるいは95%相同性を示すポリペプチドまたは核酸を意味する。ポリペプチドの場合、比較配列の長さは、一般に、少なくとも16アミノ酸、好ましくは、少なくとも20アミノ酸、より好ましくは、少なくとも25アミノ酸、最も好ましくは、35アミノ酸以上である。核酸の場合、比較配列の長さは、一般に、少なくとも50ヌクレオチド、好ましくは、少なくとも60ヌクレオチド、より好ましくは、少なくとも75ヌクレオチド、最も好ましくは、110ヌクレオチド以上である。
【0074】
配列同一性は、一般に、配列解析ソフトウェア(例えば、Genetics Computer Group, University of Wisconsin Biotechnology Center, 1710 University Avenue, Madison, Wis. 53705の配列解析ソフトウェアパッケージ、BLASTまたはPILEUP/PRETTYBOXプログラム)を用いて測定する。例えば、このようなソフトウェアは、標準的なパラメーターに設定すると、種々の置換、欠失および/または他の改変に相同性度を与えることによって、同一または類似配列をマッチさせる。保存的置換としては、一般に、以下の群:グリシン、アラニン;バリン、イソロイシン、ロイシン;アスパラギン酸、グルタミン酸、アスパラギン、グルタミン;セリン、トレオニン;リシン、アルギニン;およびフェニルアラニン、チロシン内での置換が挙げられる。
【0075】
高いレベル:高いレベルとは、本明細書において、対応する天然植物における化合物2−アセチル−1−ピロリンの平均レベルと比べた際の非天然植物における化合物2−アセチル−1−ピロリンの平均レベルの上昇を意味する。当業者ならば、植物における化合物2−アセチル−1−ピロリンのレベルが数多くの変数に応じて植物によって異なるならば、非天然植物と対応する天然植物における化合物2−アセチル−1−ピロリンの平均レベルの比較においては、同様に制御された条件下で成長させた各タイプの植物の合理的な大きさのサンプル集団を比較するべきであることが分かるであろう。好ましくは、各集団からのいくつかの植物において化合物2−アセチル−1−ピロリンのレベルを測定し、それらを平均して、非天然植物が高いレベルのものを含有しているかどうかを判定する。本発明の非天然植物においての高いレベルとは、対応する天然植物におけるそのレベルよりも平均して、少なくとも約20%高い、40%高い、60%高い、80%高い、100%高い、150%高い、200%高い、250%高い、300%高い、400%高いまたは500%高いレベルである。
【0076】
プロモーター:構造遺伝子に発現制御エレメントを提供し、かつ、RNAポリメラーゼが特異的に結合し、それによりその遺伝子のRNA合成(転写)を開始するDNA配列またはDNA配列群にある認識部位。
【0077】
これらのプロモーターを用いた植物発現構築物の例が、Fraley et al., 米国特許第5,352,605号で見られる。トランスジェニック植物のほとんど組織では、CaMV 35Sプロモーターは強いプロモーターである(例えば、Odell et al., Nature 313: 810, 1985参照)。CaMVプロモーターは、単子葉植物においても極めて活性が高い(例えば、Dekeyser et al., Plant Cell 2: 591, 1990; Terada and Shimamoto, Mol. Gen. Genet. 220: 389, 1990参照)。さらに、このプロモーターの活性は、CaMV 35Sプロモーターの重複によってさらに高めることができる(すなわち、2〜10倍の間)(例えば、Kay et al., Science 236: 1299, 1987; Ow et al., Proc. Natl. Acad. Sci., U. S. A. 84: 4870, 1987;およびFang et al., Plant Cell 1: 141, 1989, and McPherson and Kay, 米国特許第5,378,142号参照)。
【0078】
他の有用な植物プロモーターとしては、限定されるものではないが、ノパリンシンターゼ(NOS)プロモーター(An et al., Plant Physiol. 88: 547, 1988およびRodgers and Fraley, 米国特許第5,034,322号)、オクトピンシンターゼプロモーター(Fromm et al., Plant Cell 1: 977, 1989)、ゴマノハグサモザイクウイルス(FMV)プロモーター(Rogers, 米国特許第5,378,619号)およびイネアクチンプロモーター(Wu and McElroy, WO91/09948)が挙げられる。
【0079】
典型的な単子葉植物プロモーターとしては、限定されるものではないが、ツユクサ黄色斑紋ウイルスプロモーター、サトウキビバドナウイルスプロモーター、イネツングロ桿菌状ウイルスプロモーター、トウモロコシ条斑ウイルスエレメントおよびコムギ萎縮ウイルスプロモーターが挙げられる。
【0080】
構築物:特に断りのない限り、「構築物」とは、本発明の1以上の単離ポリヌクレオチド配列を含んでなる組換え遺伝子分子をさす用語である。
【0081】
宿主生物における導入遺伝子の発現に使用される遺伝子構築物は、オープンリーディングフレームに作動可能なように連結された遺伝子プロモーター配列と、所望により、オープンリーディングフレームの3’下流に遺伝子終止配列を含む。オープンリーディングフレームは、遺伝子配列の使用目的に応じてセンス方向またはアンチセンス方向のいずれかに置かれている。構築物は、選択マーカー遺伝子と遺伝子発現用の他の調節エレメントも含んでいてもよい。
【0082】
ベクター:宿主細胞において複製可能であり、かつ/または付着したセグメントの複製が起こるように別のDNAセグメントが作動可能なように連結され得るDNA分子。プラスミド、ファージミド、コスミド、ファージ、ウイルス、YACおよびBACは総て典型的なベクターである。
【0083】
「ベクター」とは、ポリヌクレオチド配列を宿主細胞へと導入して、形質転換された宿主細胞を作製するために使用される核酸分子をさす用語である。「ベクター」は、上述の遺伝子構築物の他に、遺伝物質、例えば、1種以上の宿主細胞においてその複製を可能にする、複製起点のような1以上の核酸配列、選択マーカー遺伝子および当技術分野で公知の他の遺伝要素(例えば、遺伝物質を宿主細胞のゲノムに組み込むための配列など)を含んでいてよい。
【0084】
形質転換された:形質転換された細胞は、分子生物学技術によって核酸分子が導入されている細胞である。本明細書において、形質転換とは、核酸分子がこのような細胞、植物または動物細胞に導入され得る、ウイルスベクターによるトランスフェクション、アグロバクテリウム(Agrobacterium)による形質転換、プラスミドベクターの使用、ならびにエレクトロポレーション、リポフェクションおよび粒子銃加速による裸のDNAの導入をはじめとする総ての技術を包含する用語であり、一過性だけでなく安定な形質転換体も含む。
【0085】
植物細胞のDNA形質転換方法としては、アグロバクテリウムを介した植物形質転換、プロトプラスト形質転換、花粉への遺伝子導入、生殖器官への注入、未熟胚への注入および粒子衝撃が挙げられる。これらの方法は、それぞれ、異なる長所と短所を有している。そのため、特定の植物系統に遺伝子を導入する特定の一方法は、必ずしも別の植物系統に対して最も効果的な方法ではないが、それぞれの方法がどの植物系統に対して有効であるかは十分に分かっている。
【0086】
形質転換DNAセグメントを細胞に導入するための方法は数多くあるが、総てのものがDNAの植物細胞への送達に適しているわけではない。好適な方法としては、事実上、DNAが細胞に導入され得る、アグロバクテリウム感染、DNAの直接送達(例えば、プロトプラストのPEGを介した形質転換(Omirulleh et al., 1993)による、乾燥/阻害を介したDNA取り込みによる、エレクトロポレーションによる、カーボランダム繊維での攪拌による、DNAコーティング粒子の加速によるなど)によるなどの総ての方法が挙げられるであろう。特定の実施形態においては、加速法が好ましく、その方法としては、例えば、マイクロプロジェクタイル衝撃などが挙げられる。
【0087】
細胞へのDNA導入技術は、当業者には周知の技術である。遺伝子を細胞へと送達するための一般的な4通りの方法が記載されている:(1)化学的方法(Graham and van der Eb, 1973; Zatloukal et al., 1992);(2)マイクロインジェクション(Capecchi, 1 980)、エレクトロポレーション(Wong and Neumann, 1982; Fromm et al., 1985)および遺伝子銃(Johnston and Tang, 1994; Fynan et al., 1993)のような物理的方法;(3)ウイルスベクター法(Clapp, 1993; Lu et al., 1993; Eglitis and Anderson, 1988a; 1988b);ならびに(4)受容体を介する機構(Curiel et al., 1991; 1992; Wagner et al., 1992)。
【0088】
エレクトロポレーション:短時間の高電圧電気パルスの適用により種々の動物および植物細胞の原形質膜にナノメートルの大きさの穴を開ける。これらの穴を通じてか、またはその穴の閉鎖に伴って起こる膜成分の再分布の結果として、DNAが直接細胞質に取り入れられる。エレクトロポレーションは極めて効率が良く、クローニングした遺伝子の一過性発現にも、目的の遺伝子のコピーが組み込まれた細胞系統の確立にも使用することができる。エレクトロポレーションでは、リン酸カルシウムを介したトランスフェクションやプロトプラスト融合とは対照的に、外来DNAの1コピーまたは多くても数コピーが組み込まれた細胞系統が生じることが多い。
【0089】
エレクトロポレーションを用いたDNA導入は、当業者には周知のことである。この方法では、ペクチン分解酵素のような特定の細胞壁分解酵素を使用して、目的の受容細胞のエレクトロポレーションによる形質転換を未処理の細胞よりも起こりやすくする。あるいは、受容細胞の機械的損傷による形質転換を起こりやすくする。エレクトロポレーションによる形質転換を起こすために、細胞の懸濁培養物のようなもろい組織または胚発生カルスのいずれかを使用してもよいし、あるいは、未熟胚または他の組織化組織を直接形質転換してもよい。制御された方法で、選択した細胞の細胞壁をペクチン分解酵素(ペクトリアーゼ)または機械的損傷に曝すことによってそれらを部分的に分解する。このような分解を受けた細胞はこの段階で実施し得るエレクトロポレーションによるDNA移入を受け入れる。新たに組み込まれたDNAの性質に応じた好適な選択またはスクリーニングプロトコールによって、その後、形質転換された細胞を同定することができる。
【0090】
マイクロプロジェクタイル衝撃:形質転換用DNAセグメントを植物細胞へと送達するのにさらに有利な方法がマイクロプロジェクタイル衝撃である。この方法では、粒子を核酸でコーティングし、それを推進力によって細胞へと送達する。典型的な粒子としては、タングステン、金、白金などで構成されているものが挙げられる。
【0091】
マイクロプロジェクタイル衝撃(Bombardment)の利点は、安定形質転換単子葉植物を再生可能な方法で得る有効な手段であることの他、プロトプラストの単離(Cristou et al., 1988)もアグロバクテリウム感染に対する感受性も必要としないことである。加速法によりDNAをトウモロコシ細胞へと送達するための方法の実施形態がBiolistics Particle Delivery Systemである。このシステムを使用して、DNAまたは細胞でコーティングした粒子を、ステンレス鋼またはNytex社製スクリーンのようなスクリーンを通して、懸濁培養したトウモロコシ細胞で被覆したフィルター表面に推進することができる。粒子が大きな凝集体として受容細胞に送達されないようにこのスクリーンによって粒子を分散するものと考えられる。発射装置と衝撃しようとする細胞との間にあるスクリーンが発射物の凝集体の大きさを小さくし、大きすぎる発射物による受容細胞へのダメージを減少させることにより、形質転換の頻度を高めることへとつながる。
【0092】
衝撃では、懸濁細胞をフィルターまたは固体培養培地に集中させることが好ましい。あるいは、未熟胚または他の標的細胞を固体培養培地に配置してもよい。衝撃しようとする細胞をマクロプロジェクタイル停止プレートの下方、好適な距離に置く。要すれば、1以上のスクリーンも加速装置と衝撃しようとする細胞との間に配置する。本明細書で示される技術の使用を通じて、マーカー遺伝子を一過性発現している細胞の最大1000またはそれ以上の増殖巣を得ることができる。衝撃の48時間後に外来遺伝子産物を発現する増殖巣中の細胞数は、ほとんどの場合、1〜10および平均1〜3の範囲である。
【0093】
衝撃形質転換では、最大数の安定した形質転換体を得るために、衝撃前培養の条件および衝撃パラメーターを最適化することができる。衝撃に関する物理的パラメーターおよび生物学的パラメーターはいずれも、この技術において重要である。物理的因子としては、DNA/マイクロプロジェクタイル沈降の操作に関与するものまたはマクロプロジェクタイルもしくはマイクロプロジェクタイルのいずれかの飛行および速度に影響を及ぼすものがある。生物学的因子としては、衝撃前および衝撃直後の細胞の操作に関わる総ての工程、衝撃に伴う損傷の軽減のために役立つ標的細胞の浸透圧調整、さらに線状化DNAまたは完全なスーパーコイル状プラスミドのような形質転換用DNAの性質が挙げられる。未熟胚の形質転換を成功させるためには、衝撃前操作が特に重要であると考えられる。
【0094】
よって、条件を十分に最適化するためには小規模研究において衝撃パラメーターの一部を調整することが望ましいと考えられる。特に、ギャップ距離、飛行距離、組織距離およびヘリウム圧のような物理的パラメーターを調整することが望ましい。受容細胞の生理的状態に影響を及ぼし、その結果として、形質転換および組込み効率に影響を及ぼし得る条件を改変することにより、損傷低減係数(TRF)を最小限に抑えることもできる。例えば、最適な形質転換を行うために、受容細胞の浸透圧状態、組織の水和および継代培養段階または細胞周期を調整することができる。他の通常の調整の実施については、本開示内容に照らして、当業者には分かるであろう。
【0095】
アグロバクテリウムを介した導入:アグロバクテリウムを介した導入は、DNAをあらゆる植物組織に導入することができ、プロトプラストからの完全な植物を再分化させる必要性がなくなることから、応用の範囲が広い、遺伝子を植物細胞へと導入するためのシステムである。DNAを植物細胞に導入するためにアグロバクテリウムを介して植物に組み込むベクターを使用することは、当技術分野では周知のことである。例えば、記載されている方法参照(Fraley et al., 1985; Rogers et al., 1987)。さらに、Ti−DNAの組込みは、比較的正確なプロセスであり、再配列はほとんど起こらない。導入しようとするDNAの領域は、境界配列によって規定され、記載されている(Spielmann et al., 1986; Jorgensen et al., 1987)ように、介在するDNAが、通常、植物ゲノムに挿入される。
【0096】
最新のアグロバクテリウム形質転換ベクターは、大腸菌(E. coll)ならびにアグロバクテリウムでの複製が可能であり、記載されている(Klee et al., 1985)ように便宜的な操作が可能である。さらに、アグロバクテリウムを介した遺伝子導入用ベクターにおける最近の技術的進歩により、ベクターにおける遺伝子および制限部位の配列が改善され、種々のポリペプチドコード遺伝子を発現し得るベクターの構築が容易になった。記載されているベクター(Rogers et al., 1987)は、挿入されたポリペプチドコードグ遺伝子を直接発現させるために、プロモーターにフランキングする便宜的なマルチリンカー領域とポリアデニル化部位を有しており、このベクターは本目的に適している。さらに、形質転換には武装Ti遺伝子および非武装Ti遺伝子を含有するアグロバクテリウムを使用することができる。アグロバクテリウムを介した形質転換が有効である植物系統では、遺伝子導入が容易で明確なことからこの方法が一般的に好まれて用いられる。
【0097】
葉のディスクおよび子葉や胚軸のような他の組織のアグロバクテリウムを介した形質転換は、アグロバクテリウムが自然に感染する植物に限定されるようである。アグロバクテリウムを介した形質転換は、双子葉植物において最も効果的である。記載されている(Bytebier et al., 1987)ように、アグロバクテリウムベクターを用いてアスパラガスにおいてトランスジェニック植物が作製されているが、ほとんどの単子葉植物は、アグロバクテリウムの自然宿主ではないように思われる。そのため、イネ、トウモロコシおよびコムギのような商業的に重要な穀物は、通常、別の方法を用いて形質転換しなければならない。
【0098】
アグロバクテリウム形質転換法を用いて作製されたトランスジェニック植物は、一般に、一染色体上に単一遺伝子を有する。このようなトランスジェニック植物は、追加した遺伝子のヘテロ接合体であるといえる。しかしながら、「ヘテロ接合体」という言葉の使用には、通常、染色体対の第2の染色体の同じ遺伝子座に補足遺伝子が存在することが包含され、さらに、1個の追加された遺伝子を含有する植物にはこのような遺伝子がないため、追加された外来遺伝子が有糸分裂および減数分裂中に独立に分離することからも、このような植物に対するより正確な名前は、独立した分離体であると考えられる。
【0099】
追加された構造遺伝子のホモ接合体であるトランスジェニック植物;すなわち、2個の追加された遺伝子、染色体対の各染色体の同じ遺伝子座に1個の遺伝子を含有するトランスジェニック植物がより好ましい。ホモ接合体トランスジェニック植物は、単一追加遺伝子を有する独立した分離体であるトランスジェニック植物を雌雄交配(自殖)させ、生じた種子の一部を発芽させ、結果として得られた植物を、対照(天然非トランスジェニック)または独立した分離体であるトランスジェニック植物と比べて高い2−アセチル−1−ピロリンレベルについて解析することにより得ることができる。
【0100】
これらシステムの異なる植物系統への適用は、プロトプラストからその特定の植物系統を再分化させる能力に応じる。プロトプラストから穀物を再分化させる方法の例は記載されている(Fujimura et al., 1985; Toriyama et al., 1986; Yamada et al., 1986; Abdullah et al., 1986)。
【0101】
プロトプラストからの再分化がうまくいかなかった植物系統を形質転換するために、DNAを完全な細胞または組織に導入する他の方法を利用してもよい。例えば、未熟胚または外植片からの穀物の再分化は、記載されている(Vasil, 1988)ように行われる。さらに、「粒子銃」または高速マイクロプロジェクタイル技術を利用してもよい(Vasil, 1992)。
【0102】
後者の技術を用いて、記載されているように、金属小粒子の表面に付けたDNAを細胞壁を通過して細胞質に運び込む(Klein et al., 1987; Klein et al., 1988; McCabe et al., 1988)。金属粒子は数層の細胞を貫通するため、組織外植片内での細胞の形質転換が可能となる。
【0103】
単離された:「単離された」生物学的成分(核酸もしくはタンパク質または細胞小器官など)は、その成分が自然に存在する細胞または生物体における他の生物学的成分、すなわち、他の染色体および染色体外DNAおよびRNA、タンパク質ならびに細胞小器官から実質的に分離されているか、または精製されている。「単離された」核酸およびタンパク質としては、標準的な精製方法により精製された核酸およびタンパク質が挙げられる。この用語は化学合成された核酸を含む核酸を包含し、また、in vitroでの組換え発現または宿主細胞での組換え発現により調製されたタンパク質および以下に定義する組換え核酸も包含する。一例として、コンティグのような大きなゲノムDNA断片の遺伝子は、平均コンティグに見られる余分のDNAが比較的多量にあるため、他の生物学的成分から、単離されたと考えられるほど十分には精製されない。以下に概説するように、「組換え核酸」および「組換えタンパク質」もまた上記のように「単離される」。
【0104】
組換え:「組換え核酸」とは、本明細書において、天然には存在していない配列を有する核酸、または配列の2つの別の分離セグメントを人為的に組み合わせることによって作製された配列を有する核酸を意味する。この人為的な組合せは、化学合成により、または、一般的には、核酸の人為的操作により、例えば、制限酵素、リガーゼ、リコンビナーゼおよび/またはポリメラーゼによる少なくとも1つの核酸の操作によるような遺伝子工学技術により行われることが多い。宿主細胞に導入されると、組換え核酸が宿主細胞によって複製される;しかしながら、細胞で一度複製された組換え核酸は、依然として本発明の目的での組換え核酸である。「組換えタンパク質」とは、本明細書において、組換え核酸を使用する方法によって作製されたタンパク質を意味する。以上で概説したように、「組換え核酸」および「組換えタンパク質」もまた上記のように「単離される」。コンティグのような大きな断片の遺伝子は、このような人為的な組合せがその遺伝子と関係ない場合には、「組換え核酸」ではないだろう。しかしながら、コンティグのある遺伝子の周囲または内部の配列がその遺伝子に関連する目的で(すなわち、その遺伝子がコンティグの末端近くにあるという理由だけでなく)操作されている場合には、核酸の組換え部分と問題の遺伝子とが相対的に近いことから、コンティグのそのような遺伝子は「組換え核酸」の構成要素となるであろう。
【0105】
非天然植物:本明細書において、植物に関して使用する「非天然」とは、植物における化合物2−アセチル−1−ピロリンのレベルを改変するために人間の介入による遺伝子操作を受けた植物を意味する語である。遺伝子操作を受けた天然植物とは、「対照」植物をさす。本発明において、対照植物がトランスジェニック植物であってもよい。例えば、対照植物が除草剤耐性トランスジェニック植物であってもよい。この例においては、非天然植物が除草剤耐性であり、対照除草剤耐性植物よりも高いレベルの化合物2−アセチル−1−ピロリンを有するであろう。本発明のトランスジェニック植物は、例えば、2AP遺伝子またはその断片をコードする外来核酸分子を含有する非天然植物であり、それゆえ、人間の介入による遺伝子操作を受けたものである。さらに、例えば、突然変異誘発物質、例えば、化学的突然変異誘発物質またはトランスポゾンまたはT−DNAのような「挿入突然変異誘発物質」に計画的に曝露させた結果として2AP遺伝子調節エレメントまたはコーディング配列に突然変異を有する植物も、人間の介入による遺伝子操作を受けたということから非天然植物と考えられる。これに対し、偶発または自然発生突然変異しか有していない植物は、本明細書にて定義する「非天然植物」ではない。非天然植物は、一般に、天然植物と比べて変化した核酸配列を有しているが、同時に、非天然植物に、その核酸配列を変えることなく、例えば、そのメチル化パターンを改変することにより、人間の介入による遺伝子操作を行い得ることは当業者には明白である。
【0106】
トランスジェニック植物:形質転換された植物細胞またはプロトプラストから得られた、その植物DNAが同じ系統のネイティブな非トランスジェニック植物にはもともと存在していない導入された外来DNA分子を有している植物またはその後代。用語「トランスジェニック植物」および「形質転換された植物」とは、当技術分野において、その植物のDNAが外来DNA分子を有している場合の植物を定義するための同義語として使用されることがある。しかしながら、形質転換された植物細胞またはプロトプラストから得られた再分化植物またはカルスをトランスジェニック植物と呼ぶことがより科学的に正しいと考えられ、その使用法は、本明細書に従うものとする。
I.2−アセチル−1−ピロリン(2AP)遺伝子
【0107】
A.オーソログ/ホモログの単離:イネにおけるOs2AP遺伝子の単離および特性決定を記載している以下の実施例節は、2AP遺伝子を単離するための典型的な一般アプローチである。このような2AP遺伝子は2APタンパク質をコードする。単離された遺伝子は、その後、植物における2AP遺伝子発現を低減するための組換えベクターの構築に使用することができる。
【0108】
一般に、以下に記載する組換えDNA技術における用語および実験室手順は十分に知られているものであり、当技術分野では一般に使用されている。クローニング、DNAおよびRNA単離、増幅ならびに精製には標準的な技術を用いる。一般に、DNAリガーゼ、DNAポリメラーゼ、制限エンドヌクレアーゼなどを必要とする酵素反応は、製造業者の仕様書に従って実施する。これらの技術および種々の他の技術は、一般に、Sambrook et al., Molecular Colning - A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, N. Y., (1989)に従って実施する。
【0109】
2AP遺伝子の単離は、数多くの技術によって行うことができる。例えば、本明細書において開示された配列に基づくオリゴヌクレオチドプローブを使用して、cDNAまたはゲノムDNAライブラリー内の所望の遺伝子を同定することができる。ゲノムライブラリーを構築するため、ゲノムDNAの大きなセグメントを、例えば、制限エンドヌクレアーゼを使用したランダム断片化により作製し、それをベクターDNAと連結して、好適なベクターにパッケージングし得るコンカテマーを作出する。cDNAライブラリーを作成するため、mRNAを葉のような所望の器官から単離し、mRNAから2AP遺伝子転写物を含むcDNAライブラリーを作成する。あるいは、2AP遺伝子またはホモログが発現される他の組織から抽出されたmRNAよりcDNAを調製してもよい。
【0110】
本明細書において開示されたイネOs2AP遺伝子のようなクローニングされる2AP遺伝子の配列に基づくプローブを使用して、cDNAまたはゲノムライブラリーを、その後、スクリーニングすることができる。同じまたは異なる植物種の相同遺伝子を単離するために、プローブを使用してゲノムDNAまたはcDNA配列とハイブリダイズさせてもよい。
【0111】
10〜14、15〜20、30、50ヌクレオチド、場合によっては、およそ100〜200ヌクレオチドの、配列番号1、配列番号2、配列番号4もしくは配列番号5で示されるDNA配列と同じか、または配列番号1、配列番号2、配列番号4もしくは配列番号5で示されるDNA配列と相補的である、連続したヌクレオチドストレッチで構成されている配列領域を有する核酸分子が、例えば、サザンおよびノーザンブロット法で用いるハイブリダイゼーションプローブとして、特に意図される。連続した相補的領域の長さが約10〜14から約100または200ヌクレオチドまでのように可変であるより小さな断片は、一般に、ハイブリダイゼーション実施形態において有用であるが、検出が望まれる相補配列の長さに応じて、より大きな連続した相補的ストレッチを使用してもよい。
【0112】
約14ヌクレオチド長のハイブリダイゼーションプローブを使用することで安定した選択できる二本鎖分子が形成される。しかしながら、ハイブリッドの安定性と選択性を高めることによって、得られる特異的なハイブリッド分子の品質と程度を向上させるためには、14塩基長を上回るストレッチに対する連続した相補配列を有する分子が一般に好ましい。一般的には、15〜20個の連続したヌクレオチド、要すれば、より長いヌクレオチドの遺伝子相補的なストレッチを有する核酸分子を設計することが好ましい。
【0113】
当然のことながら、例えば、機械的剪断または制限酵素消化によるなどの他の技術によっても断片を得ることができる。小さな核酸セグメントまたは断片は、例えば、自動オリゴヌクレオチド合成装置を使用して一般に実施されているように、化学的手段によって断片を直接合成することによって、容易に調製され得る。また、米国特許第4,683,195号および第4,683,202号(それぞれ引用することにより本明細書の一部とされる)のPCR.TM.法のような核酸複製技術を適用することにより断片を得てもよいし、選択した配列を組換え生産用組換えベクターに導入することにより断片を得てもよいし、分子生物学分野における通常の技術を有する人には一般に知られている他の組換えDNA技術により断片を得てもよい。
【0114】
よって、DNA断片の相補的ストレッチと選択的に二本鎖分子を形成させるために本発明の核酸配列を使用することができる。想定される用途に応じて、ハイブリダイゼーション条件を変え、標的配列に向けるプローブの選択性の程度を変えることが望ましい。高い選択性を必要とする用途では、一般に、比較的ストリンジェントな条件を使用して、ハイブリッドを形成させることが望ましい。例えば、約0.02M〜約0.15M NaCl、約50℃〜約70℃の温度にて提供されるような比較的低塩および/または高温条件を選択する。このような選択条件は、プローブと鋳型または標的鎖間のミスマッチがもしあるとしてもほとんど許容しないため、2APポリペプチドをコードするDNAセグメントまたはタンパク質をコードするDNAセグメントの単離に特に好適であろう。ハイブリダイゼーションを介したDNAセグメントの検出は、当業者には周知のことであるが、米国特許第4,965,188号および第5,176,995号(それぞれ引用することにより本明細書の一部とされる)の教示がハイブリダイゼーション解析の典型的な方法である。Maloy et al., 1994; Segal 1976; Prokop, 1991;およびKuby, 1994の本文中に記載されている内容のような教示が特に適切である。
【0115】
当然のことながら、例えば、対象鋳型とハイブリダイズする突然変異型プライマー鎖を使用して、突然変異体を調製することが望ましく、また、関連種、機能的同等物などから2APタンパク質をコードする配列を単離しようとする一部の用途では、ヘテロ二本鎖を形成させるために、一般に、より低いストリンジェントなハイブリダイゼーション条件が必要である。これらの状況においては、約0.15M〜約0.9M塩、約20℃〜約55℃の範囲の温度のような条件を使用することが望まれる。制御ハイブリダイゼーションでは、クロスハイブリダイズしている種をハイブリダイズししているシグナルとして容易に確認することができる。どんな場合でも、温度上昇と同様に、ハイブリッド二本鎖を不安定にする働きをする漸増量のホルムアミドの添加によってより高いストリンジェントな条件にすることができることは一般に知られている。このように、ハイブリダイゼーション条件は容易に操作し得るものであることから、一般には、所望する結果に応じて方法が選択できる。
【0116】
特定の実施形態においては、本発明の核酸配列をハイブリダイゼーションを判定するための標識のような好適な手段と組み合わせて使用することが有利である。多種多様な好適な表示手段が当技術分野で公知であり、これらの手段としては、検出可能なシグナルを与え得る、アビジン/ビオチンのような、蛍光、放射性、酵素リガンドまたは他のリガンドが挙げられる。好ましい実施形態において、放射性試薬または他の環境的に望ましくない試薬の代わりに蛍光標識またはウレアーゼ、アルカリ性ホスファターゼもしくはペルオキシダーゼのような酵素タグを使用することが望ましいと思われる。酵素タグの場合、熱量表示基質が知られており、それを使用して、人間の眼でまたは分光光度法で確認できる手段を提供し、相補的核酸を含有するサンプルとの特異的ハイブリダイゼーションを確認することができる。
【0117】
一般に、本明細書において記載するハイブリダイゼーションプローブは、溶液ハイブリダイゼーションにおける試薬としても、固相を使用する実施形態においても有用であると想定される。固相に関連する実施形態では、試験DNA(またはRNA)を、選択したマトリックスまたは表面に吸着させるか、そうでなければ付着させる。この固定された一本鎖核酸を、続いて、所望の条件下で、選択したプローブとの特異的ハイブリダイゼーションに付す。選択する条件は、必要な特定の判定基準に基づいた特定の状況によって変わる(例えば、G+C含量、標的核酸のタイプ、核酸の起源、ハイブリダイゼーションプローブの大きさ他に応じる)。非特異的に結合しているプローブ分子を除去するためにハイブリダイズした表面を洗浄した後、特異的ハイブリダイゼーションを検出する。場合によっては、標識を用いて定量する。
【0118】
また、増幅技術を用いて、核酸サンプルから目的の核酸を増幅することができる。例えば、ポリメラーゼ連鎖反応(PCR)技術を使用して、2AP遺伝子の配列を、mRNAから、cDNAから、ゲノムライブラリーまたはcDNAライブラリーから直接増幅することができる。PCRおよび他のin vitro増幅法は、例えば、発現させようとするタンパク質をコードする核酸配列をクローニングするのに、サンプル中の所望のmRNAの存在を検出する目的で、核酸配列を決定する目的で、または他の目的でプローブとして使用する核酸を作製するのにも有用であり得る。
【0119】
植物組織由来の2AP遺伝子配列を同定するのに好適なプライマーおよびプローブは、本明細書において提供する配列の比較により得られる。PCRの一般概要に関しては、引用することにより本明細書の一部とされるPCRプロトコール:A Guide to Methods and Applications. (Innis, M, Gelfand, D., Sninsky, J. and White, T., eds.), Academic Press, San Diego (1990)参照。
【0120】
技術文献に記載されている周知の技術によりポリヌクレオチドも合成し得る。例えば、Carruthers et al., Cold Spring Harbor Symp. Quant. Biol. 47: 411-418 (1982)およびAdams et al., J. Am. Chem. Soc. 105: 661 (1983)参照。さらに、相補鎖を合成し、好適な条件下でそれらの鎖を一緒にアニーリングすることによるか、または好適なプライマー配列とともにDNAポリメラーゼを用いて相補鎖を合成することにより二本鎖DNA断片が得られる。
【0121】
B.2AP遺伝子オーソログを単離する生物:トウモロコシ属(Zea)、カラスムギ属(Avena)、オオムギ属(Hordeum)、ライムギ属(Secale)、コムギ属(Triticum)およびモロコシ属(Sorghum)の種を含むいろいろな種類の植物および真菌を使用することができる。2−アセチル−1−ピロリンは、あらゆる香米、小麦およびライ麦パン(Buttery et al., 1982, 1983)、含水雑穀(Seitz et al., 1993)、ポップコーン(Schieberle et al., 1991)、バチルス・セレウス(Romanczyk et al., 1995)ならびにアスペルギルス・オリザエ(Aspergillus oryzae)、アスペルギルス・アワモリ(Aspergillus awamori)およびスポロボルス・ビルギニカス(Sporobolus virginicus)のような一部の真菌種(Nagsuk et al., 2004)の主要な強い香気化合物であることが分かっている。ゲノムレベルでの穀物間でのシンテニー、さらに、植物および微生物においてよく保存されたプロリンおよびポリアミン代謝経路を示すいくつかの報告がある(Kyoto Encyclopedia of Genes and Genomes, Kyoto University, Bioinformatics Center, Institute for Chemical Researchのホームページにてオンラインで見つけられる)。そのため、2AP遺伝子のオーソログは、穀物用植物間で同様に作用するかもしれず、本発明にて示す教示によって、イネと同様に他の植物においても化合物2−アセチル−1−ピロリンの蓄積が増強する可能性がある。
II.植物における2−アセチル−1−ピロリンレベルを上昇させるための2AP遺伝子発現または2APタンパク質レベルもしくは活性の低減
【0122】
A.突然変異誘発:EMS(メタンスルホン酸、エチルエステル)のような化学的突然変異誘発物質ならびにγ線および高速中性子照射によってDNAに突然変異が生じる可能性がある。突然変異のいくつかの例が、欠失、挿入およびミスセンス突然変異である。突然変異後、スクリーニングを行って、早期停止コドンを与えるか、そうでなければ機能しない2AP遺伝子を生じる欠失を確認することができる。配列決定によるか、または本発明によって提供される、2AP遺伝子またはタンパク質に特異的なプローブもしくはプライマーを使用することにより突然変異体のスクリーニングを行うことができる。2AP遺伝子における特異的突然変異は、TILLING(Targeted Induced Local Lesion of Genome)(Till et al., 2003)およびtDNA挿入によっても起こし得る。このような突然変異の結果として、2AP遺伝子発現の低下、2AP mRNAの安定性の低下または2APタンパク質の活性および/もしくは安定性の低下が起こり得る。本明細書において定義するような植物は、天然に存在するものではない。
【0123】
B.アンチセンス技術:アンチセンス技術は、2AP mRNAのような標的mRNAの翻訳を妨げる。そのようなものとして、アンチセンス技術は、一般に、標的タンパク質、本明細書では2APタンパク質のレベルを低減する。2AP遺伝子またはその断片は、植物にアンチセンス方向で導入することができる。断片は、18ヌクレオチドといった小さなものであってもよいし、3,000ヌクレオチドといった大きなものであってもよいし、あるいはそれより大きくてもよい。2AP遺伝子のcDNA断片は、ベクター(例えば、pCAMBIA1302)に天然遺伝子とは逆方向にクローニングすることができる。逆転写物は、天然の2AP遺伝子転写物を含むヘテロ二本鎖構造を形成しているが、その後、その遺伝子転写物は翻訳前に分解される。センスおよびアンチセンス配列は同一である必要はなく、標的遺伝子発現の抑制を実現するには部分的な相同性で十分である。従って、イネOs2AP由来の配列(配列番号1、配列番号2、配列番号4および配列番号5)を使用して、他の植物由来の2AP遺伝子オーソログの配列が分からなくとも、その植物における2AP遺伝子発現を抑制することができる。
【0124】
C.RNA干渉:RNA干渉(RNAi)は、標的mRNAの排除に関連するもう1つの技術である。RNAi用のベクターの構築は様々であるが、基本的にはRNAにおいてヘアピンループ構造の形成が起こる(Horiguchi 2004)。実施例2では、そのcDNAとは逆方向にエクソン6、7および8を含むOs2AP核酸配列の断片をベクターにクローニングして、mRNAにおいて逆ヘアピン構造をもたらした。これらのヘアピン構造はDicerと呼ばれるエンドヌクレアーゼによって小断片に切断される。このDicerの機能は、間違った転写産物が翻訳されないようにすることである(Hamilton and Baulcombe, 1999, Matzke et al., 2001)。mRNAの小断片は、一般に、約20〜21ヌクレオチドの大きさであり、siRNAまたは小干渉RNAと呼ばれている。より大きな断片もより小さな断片も使用することができる。これらのsiRNAは、相同遺伝子の発現をダウンレギュレートし得る。この場合もまた、相同性が完全である必要はないため、本発明において詳述するイネ由来の配列を使用して、他の植物のRNAi構築物を作製することができる。実施例2は、非香米系統におけるOs2AP遺伝子を標的としたRNAi試験によって、いかにして、香米に見られるレベルまでに高められた2−アセチル−1−ピロリンレベルおよび芳香が得られるのかを示している。
III.トランスジェニック植物の作製
【0125】
これまでの技術において単離された2AP遺伝子配列を使用するために、植物細胞の形質転換に好適な組換えDNAベクターを調製することができる。多種多様な高等植物種を形質転換する技術はよく知られたものであり、技術文献および科学文献に記載されている。例えば、Weising et al. Ann. Rev. Genet. 22: 421-477 (1988)参照。
【0126】
このようなDNA構築物を、種々の従来の技術により所望の植物宿主のゲノムに導入し得る。例えば、DNA構築物を、植物細胞プロトプラストまたは胚発生カルスのエレクトロポレーション、PEGポレーション、粒子衝撃およびマイクロインジェクションのような技術を用いて植物細胞のゲノムDNAに直接導入してもよいし、またはDNA構築物を、DNA粒子衝撃のような弾道学的方法を用いて植物組織に直接導入することもできる。あるいは、DNA構築物を好適なTDNAフランキング領域と組み合わせ、一般的なアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)宿主ベクターに導入してもよい。アグロバクテリウム・ツメファシエンス宿主の毒性機能が、細胞が細菌類に感染したときに、構築物と隣接するマーカーの植物細胞DNAへの挿入を導く。
【0127】
マイクロインジェクション技術は、当技術分野では公知のものであり、科学文献および特許文献で詳しく記載されている。ポリエチレングリコール沈降を用いたDNA構築物の導入は、Paszkowski et al. Embo J. 3: 2717-2722 (1984)で記載されている。エレクトロポレーション技術は、Fromm et al. Proc. Natl. Acad. Sci. USA 82: 5824 (1985)で記載されている。弾道学的形質転換技術は、Klein et al. Nature 327: 70-73 (1987)で記載されている。数多くのアプローチを用いて、ライムギ(de la Pena et al., Nature 325: 274-276 (1987))、トウモロコシ(Rhodes et al., Science 240: 204-207 (1988))およびイネ(エレクトロポレーションによるShimamoto et al., Nature 338: 274-276 (1989); 弾道学的技術によるLi et al. Plant Cell Rep. 12: 250-255 (1993))のような穀物種を形質転換することができる。
【0128】
アグロバクテリウム・ツメファシエンスを介した形質転換技術は、科学文献で詳しく記載されている。例えば、Horsch et al. Science 233: 496-498 (1984)およびFraley et al. Proc. Natl. Acad. Sci. USA 80: 4803 (1983)参照。アグロバクテリウムは、主として、双子葉植物において有用であるが、アグロバクテリウムによって形質転換することができる単子葉植物もある。例えば、イネのアグロバクテリウム形質転換は、Hiei et al., Plant J. 6: 271-282 (1994)により記載されている。
【0129】
上記形質転換技術のいずれかによって誘導された形質転換植物細胞を培養して、形質転換された遺伝子型を有し、その結果としてより高いレベルの2−アセチル−1−ピロリンと増強された芳香を有する完全な植物体を再分化させることができる。このような再分化技術は、一般に、2APヌクレオチド配列とともに導入された殺生物剤および/または除草剤マーカーに依存して、組織培養増殖培地における特定の植物ホルモンの操作によって行うものである。培養プロトプラストからの植物再分化は、Evans et al., Protoplasts Isolation and Culture, Handbook of Plant Cell Culture, pp. 124-176, MacMillilan Publishing Company, New York, 1983;およびBinding, Regeneration of Plants, Plant Protoplasts, pp. 21-73, CRC Press, Boca Raton, 1985で記載されている。植物カルス、外植片、器官またはその一部からも再分化が可能である。このような再分化技術は、一般に、Klee et al. Ann. Rev. of Plant Phys. 38: 467-486 (1987)で記載されている。
【0130】
熟練者ならば、発現カセットがトランスジェニック植物に安定して組み込まれ、機能しうることが確認された後、それが雌雄交配により他の植物に導入することができることが分かるであろう。交配しようとする種に応じ、数多くの標準的な繁殖方法のいずれを使用してもよい。
IV.2−アセチル−1−ピロリンレベルについての試験
【0131】
A.官能試験:乾燥種子、調理した米またはすりつぶした葉の揮発性芳香は、感覚によって識別することができる。(Dhulappanavar, 1976, Ghose et al., 1952, Kadam et al., 1938).このような感覚識別法としては、味覚試験が挙げられる。米育種家の間で一般に実施されているもう1つのものが、水中で葉組織を加熱した後、希釈KOH溶液を添加する方法である(Sood and Siddiq, 1978)。これらの試験は必ずしも一貫性と信頼性があるとが限らず、人的エラーが起こりやすく、偏りがちである。
【0132】
B.クロマトグラフィー:調理した米100グラムから揮発性化合物を定量するためのガスクロマトグラフィーを使用したより信頼性のある方法を開発した(Petrov et al., 1995)。最近、たった1グラムの米から1ppb(10億分の1)といった低い2−アセチル−1−ピロリンレベルを分析するために、ガスクロマトグラフィー/質量分析(GCMS)法が開発された(Mahatheeranont et al., 1995)。そのようなものとして、GCMSを使用して、天然および非天然植物における2−アセチル−1−ピロリンのレベルを調べることができる。図1参照。
【実施例】
【0133】
好ましい実施形態の説明
実施例1:米の芳香を制御する遺伝子のポジショナルクローニング
米の芳香特性は、これまでに、定性的芳香測定法と定量的芳香測定法の両方に基づいて染色体8にマッピングされている(Lorieux et al. 1996)。芳香遺伝子は、RG28から4.5cM離れた位置にマッピングされ(Ahn et al. 1992)、RG28とRG1の間、12cM内にマッピングされている(Lorieux et al. 1996)。最近、公開イネゲノム配列から得られた一塩基変異多型(SNP)マーカーRSP04が芳香遺伝子から2cM離れた位置にマッピングされた(Jin et al. 2003)。
【0134】
単一QTLは、染色体8のRFLPマーカーRG1およびRG28によってフランキングされる4.5cM領域に位置していた(Lorieux et al., 1996; Lanceras et al. 2000)。KDML105由来のEcoRI BAGライブラリーは、17の一倍体ゲノムを示す75,264クローンで構成されたものであった。25BAGクローンからなる開始BAGコンティグを、領域内のマッピングされている6個のマーカーを用いることにより選択し(Wanchana et al., 2005)、HindIIIフィンガープリンティングによりさらに絞り込んだ。2−アセチル−1−ピロリンの蓄積に関与する遺伝子を同定するため、重要な領域を以下の戦略により狭めた。
【0135】
最初に、重要な領域内に新規組換え体を生み出すための起源として単一のF6植物を選択した(そのF7において穀物の芳香について分離した)。F7〜F1から、各世代に対して、穀物の芳香とマーカー遺伝子型に基づいてヘテロ接合体植物を選択した。F10世代では、374個体のF10植物をKDML105BAG末端配列から得られた18個の多型マーカーを用いてスクリーニングすることにより重要な領域が380kbに狭められた。274個体のF11植物をスクリーニングすることによりその領域を120kbにさらに絞った(図2)。図式遺伝子型決定および2−アセチル−1−ピロリン量に基づいて、8のF11植物を選択して、芳香性、非芳香性およびヘテロ接合体同質遺伝子系統を作製した(図2)。この段階で、3つのKDML BAGクローン、155L11、68L13および167M23をショットガン配列決定用に選択した。KDML由来の配列と対応するニホンバレゲノムコンティグのアラインメントにより、F12世代におけるさらなるスクリーニングに用いられる21個の挿入/欠失(「indel」)マーカーが同定された。1,116個体のF12植物を使用することにより、標的領域を58kb BAG,167M23内の27kbに狭めた(図2)。この交配では、これ以上の組換えは確認されなかった。米粒子において2−アセチル−1−ピロリン蓄積に違いがはっきりと示される数系統の同質遺伝子系統が作出された(図6)。
【0136】
配列解析では、BAC 167M23が19個の遺伝子を含有していることが分かった。しかしながら、10個の遺伝子だけが既知タンパク質またはコーディング配列との類似性を有するものであった。27kb領域内では、3個の候補遺伝子が同定された:3−メチルクロトニル−CoAカルボキシラーゼ(MCCアーゼ)、仮定的遺伝子および未知タンパク質。KDML105とJaoHom Nin(JHN)、非芳香性黒色米の交配による177のF6植物を用いて、穀物の芳香および2−アセチル−1−ピロリンレベルに影響を及ぼす未知タンパク質のエクソン7内に組換え部位が確認された。BAG 167M23とBAG 68L13両方のこれらの7個の候補遺伝子の発現を調べるため、2−アセチル−1−ピロリンが米粒子に蓄積する登熟期間中にRT−PCRを行った。芳香性および非芳香性同質遺伝子系統の受粉の10、15および20日後にイネの穂から全RNAを採取した。NBS/LRR遺伝子の発現は検出されなかった。ほとんどの候補遺伝子については、芳香性および非芳香性同質遺伝子系統間での発現差は示されなかった。Os2APの場合においてのみ、芳香性同質遺伝子系統で受粉の15日後に遺伝子発現が急激に低下した(図3A)。Os2AP遺伝子発現の低下は、葉、茎および根においても顕著であった(図3B)。そのため、発現研究とポジショナルクローニングの両方により、穀物の芳香の蓄積および2−アセチル−1−ピロリンのin vivo合成に関与するレギュレーターとしてOs2APを確認する。
【0137】
これまでに、研究者によって穀物の芳香を制御する1つの劣性核内遺伝子が報告されている(Berner and Hoff, 1986; Yanjuan et al., 1992; Ali et al., 1993)。このことは、本明細書において報告する研究結果と一致する。2−アセチル−1−ピロリンの蓄積を、芳香性および非芳香性同質遺伝子系統とF1植物で比較した。これらの結果からは、F1葉では2−アセチル−1−ピロリンが検出されなかったことが示された。F2世代で分離が起こったことから、2−アセチル−1−ピロリンレベルは、その供与体である親に見られるレベルの4分の1であった(図3C)。その結果、古典的遺伝学および分子遺伝学の両方から、エクソン7における突然変異が植物における2−アセチル−1−ピロリン蓄積を調節する分子機構であることが裏付けられた。
【0138】
Os2AP遺伝子の構造を、KDML105、芳香性同質遺伝子系統、非芳香性同質遺伝子系統およびニホンバレ間で比較した。5.8kbのOs2AP遺伝子は、15エクソンで構成されており、いくつかの同義的な突然変異がエクソン2に見られた(図5A)。KDML105および芳香性同質遺伝子系統では、2つの重要な突然変異事象がエクソン7で確認された。まず、2つの過渡的な突然変異が730位(AからTへ)と732位(TからAへ)に見られ、続いて、8塩基対の欠失,GATTAGGC,が734位から開始していた。8同質遺伝子系統間での2−アセチル−1−ピロリンレベルの解析により、8塩基対の欠失がコメ粒子における2−アセチル−1−ピロリンの蓄積と関係していることが示された(図6A)。この突然変異は、729位からフレームシフト翻訳開始させ、753位から開始する早期停止コドンを与えた(図5B)。ニホンバレでは、Os2AP全長cDNAは503個のアミノ酸に翻訳される。欠失により252個のアミノ酸の末端切断型ペプチドが作出された(図5B)。KDML105とJHNから誘導された177のF6植物では、8塩基対の欠失にフランキングする二重組換えによって、米粒子において2−アセチル−1−ピロリンを生産できないことが説明される。
【0139】
この早期停止コドンは、Os2APの発現に大きな影響を与え得る。早期翻訳停止コドンを含む大部分のmRNAは、ほとんどの場合翻訳されない。その結果として、それらは、遺伝子発現におけるエラーを減少させるという働きをする監視システムとしてナンセンス変異を介したmRNAの分解(NMD)を引き起こすことが多い(Pulak and Anderson, 1993)。この現象によって、香米においてOs2AP遺伝子発現のレベルが低いことが説明される。
【0140】
2APの直接の前駆体は、まだ確認されていない。しかしながら、Os2APのピロリン環の前駆体は、同位体希釈により決定されたようにアミノ酸プロリンである(Yoshihashi et al., 2002)。また、1−ピロリンおよびピロリン−2−カルボン酸(P2C)のようなピロリン含有代謝産物が直接の前駆体としても供給される可能性もあるかもしれない。穀物の芳香の強さを増強するための1つの方法が、より多くの前駆体または中間体が蓄積され、粒子成長期間に2APへの特異的な変換により多く利用できるようになるように、関連生合成経路に関与する酵素の発現を操作することである。例としては、酵素P5Cシンターゼ、P5Cレダクターゼの発現を増強すると同時にプロリンおよびP5Cデヒドロゲナーゼを抑制することがある。ポリアミン生合成経路におけるいくつかの酵素を増強して、米粒の成長時により多くの1−ピロリンを生じさせることも可能である。代謝工学アプローチに加え、米において利用できる胚乳特異的なプロモーターを用いることにより、米胚乳における酵素発現を増強してもよい。これらの技術によって、精米においてより芳香性の高い米系統を生み出すことができる。
【0141】
実施例2:Os2AP遺伝子を用いた植物形質転換:
Os2AP遺伝子をRNAi構築物に組み込み、それを用いてOs2AP遺伝子発現をダウンレギュレートし、イネにおける2−アセチル−1−ピロリンレベルを高めた。
【0142】
Os2AP−ihpRNA含有ベクターの構築
Os2AP遺伝子のセンス−アンチセンス断片の発現用にpCAMBIA1302ベクターを使用した。KDML105由来のゲノムDNAおよび全RNAそれぞれを鋳型として用い、PCRによりセンスおよびアンチセンス断片を作製した。エクソン6〜9に対応するゲノムDNA配列を含有するセンス断片を、NcoIおよびSpeI制限部位(下線)を有するプライマー(順方向:AATTCCATGGGGTTGGTCTTCCTTCAGGTG(配列番号80);逆方向:AATTACTAGTTTCCACCAAGTTCCAGTGAA(配列番号81))を用いて増幅した。エクソン6〜8に対応するcDNA配列を含有するアンチセンス断片を、NcoIおよびNheI制限部位を有するプライマー(順方向:AATTCCATGGGGTTGGTCTTCCTTCAGGTG(配列番号80);逆方向:AATTGCTAGCGGTCCAAAAGCAACCAAAGA(配列番号82)を用いて増幅した。PCR産物をまず、SpeIおよびNheIで消化し、その後、T4 DNAリガーゼを用いて連結した。次いで、連結した断片をNcoIで消化した。精製した連結断片をpCAMBIA1302ベクターにNcoIクローニング部位にてクローニングした(図7A)。
【0143】
イネ形質転換
非香米品種(オリザ・サティバ L.日本型品種ニホンバレ)の胚発生カルスを粒子衝撃形質転換の標的組織として使用した(Nimlek, 1999)。ハイグロマイシン耐性カルスを、プライマーOs2AP−エクソン7.1−欠失_FおよびR(U:5'-TGCTCCTTTGTCATCACACC-3'(配列番号54)およびL:5'-TTTCCACCAAGTTCCAGTGA-3'(配列番号55))とGFPのプライマー(U:5'-CTTGTTGAATTAGATGGTGATGTT-3'(配列番号83)およびL:5'GTTGTGGGAGTTGTAGTTGTATTC-3'(配列番号84))を用いてPCRによりスクリーニングした。
【0144】
サザンブロット解析
トランスジェニック植物(R0)および対照(ニホンバレ)植物の葉から全DNAを単離した。Os2AP−ihpRNA断片の検出用にゲノムDNA(10mg)をNcoIで消化した。陽性対照として、プラスミドOs2AP−RNAiから単離されたDNAをNcoIで消化した。0.8%アガロースゲルでの電気泳動後、DNAをHybond−N+ナイロンメンブレン(Southern 1975)に移した。プローブとのハイブリダイゼーションを製造業者(Amersham)の使用説明書に従って実施した。(α−32P)−dCTPを用いてランダムプライマー法により放射性プローブを調製した。プローブは、Os2AP遺伝子のコード領域で構成されていた(pOs2AP−RNAi,210ヌクレオチドのSpeI−NcoI断片)。
【0145】
RT−PCR
異なる成長段階における転写レベルと異なる組織における転写レベルを調べるため、ニホンバレおよびトランスジェニック植物の若い植物(10日)、成葉(30日)および根(14日)および開花期の穂から全RNAを抽出し、RT−PCR解析に使用した。染色体8のOs2AP遺伝子の芳香遺伝子座を、Os2AP−エクソン7.1プライマー(U:5'-TGCTCCTTTGTCATCACACC-3'(配列番号54)およびL:5'-TTTCCACCAAGTTCCAGTGA-3'(配列番号55))を用いてPCRにより増幅した。BADH(Os2AP−chr4;Genbank受託番号AB001348)とも呼ばれる染色体4のOs2AP遺伝子のホモログを、Os2APch4プライマー(U:5'-TAGCTTCACATCCCCATGTG-3'(配列番号85)およびL:5'-GCACCTTCACATCTTGCTGT-3'(配列番号86)を用いてPCRにより増幅した。対照として、イネアクチン遺伝子(Genbank受託番号:X16280)を、アクチンU:5'-ACATCGCCCTGGACTATGAC-3'(配列番号87)およびアクチンL:5'-TGCTGAGAGATGCCAAGA TG-3'(配列番号88)を用いて増幅した。
【0146】
リアルタイム定量的PCR
Taqman chemistryおよびABI PRISM 7700配列検出システム(Applied Biosystems, Foster City, CA, USA)を利用することにより、製造業者の使用説明書に従ってリアルタイム定量的PCRを実施した。Os2APのコピー数とイネアクチンのものの比として相対量を算出した。Os2AP遺伝子の芳香性対立遺伝子と非芳香性対立遺伝子を区別するようにTaqmanプローブを設計した。
【0147】
結果
4つのハイグロマイシン−GFP陽性小植物が再分化された。トランスジェニックイネにおけるGFPの発現は様々であった。T0RNAi2(10)、GFPの最高の発現が示される系統、の内在性Os2APの発現は最も低い(図7B)。2−アセチル−1−ピロリン含量の解析とKOH法を用いた官能試験には、十分な相関関係があった。T0RNAi2(10)は、最大量の2−アセチル−1−ピロリンを含有しており、芳香性葉が最も多かった。干渉配列の発現量が少ないT0RNAi5(20)とT0RNAi3(30)は、2−アセチル−1−ピロリンの蓄積が少なく、芳香性葉スコアが低かった(図7B)。T1RNAi2(10)の穀物の芳香の分離比分析によりこの結果を確認した。その結果、RNA干渉によるOs2AP遺伝子発現の抑制により、イネにおける2−アセチル−1−ピロリンの蓄積が増加する。このことにより、8塩基対の欠失によって起こる香米におけるNMDによるOs2AP遺伝子発現の自然抑制が香米の芳香の直接原因であることが示唆される。
【0148】
実施例3:イネのOs2AP/BADH遺伝子座の特性決定
Os2APは、これまでに染色体4で見つけられたBADH(ベタインアルデヒドデヒドロゲナーゼ)のホモログである(Nagamura et al., 1997)。本発明者らは、Genbank配列受託番号AB001348とAP004463の、それぞれ、染色体4のBADH遺伝子と染色体8のOs2AP遺伝子のゲノム配列を特徴付けた。イネGAASの遺伝子注釈結果に従って(イネゲノム自動注釈システム, Sakata et al., 2002)、BADHおよびOs2AP両方の構造遺伝子は、14イントロンによって分断された15エクソンを含んでなる。BADHおよびOs2APそれぞれのORFは、505および503アミノ酸残基のタンパク質をコードする。両タンパク質は、アミノ酸レベルでは77%同一であり、他の植物BADHタンパク質と88%〜99%同一性を共有している。さらに、これらの2つのタンパク質は、一般アルデヒドデヒドロゲナーゼに高度に保存されているデカペプチドVal-Thr-Leu-Glu-Leu-Gly-Gly-Lys-Ser-Pro(配列番号96)をコードするヌクレオチド配列を含有している(Weretilnyk et al. 1990)。タンパク質配列のPfam解析によれば、BADHもOs2APもアルデヒドデヒドロゲナーゼファミリーに属する。このファミリーに属するタンパク質は、特定植物種におけるグリシンベタインの合成に関して推定機能を有する。しかしながら、イネ、トウモロコシおよびコムギのようなグリシンベタインを蓄積しない穀物でのこれらのタンパク質の機能は分かっていない。本発明者らは、プロリンが2−アセチル−1−ピロリンの前駆体として報告されていることから、プロリン異化において機能するピロリン−5−カルボン酸デヒドロゲナーゼ(P5CDH)(Genbank受託番号P30038)もこのファミリーに含まれることに驚いた。Os2APの機能がベタインアルデヒド以外のアルデヒドの脱水素化を触媒することである可能性がある。
【0149】
Os2AP遺伝子発現および米の芳香:両Os2AP遺伝子が芳香生合成に関連しているかどうかを確認するため、芳香性および非芳香性同質遺伝子系統(それらのゲノム背景は芳香遺伝子領域を除いて同一であった)の発現レベルを解析した。結果として、本発明者らは、BADHは両同質遺伝子系統間での発現差は示さないが、一方、Os2APは芳香性同質遺伝子系統の総ての組織において遺伝子抑制を一貫して示すことが分かった(図3D)。本発明者らはさらに、同質遺伝子系統ならびにニホンバレおよびK105(芳香性系統)における両タンパク質の発現についても研究した。Os2AP遺伝子発現は、芳香性同質遺伝子系統およびKDMLL105においてのみダウンレギュレートされた。これに対し、非芳香性同質遺伝子系統およびニホンバレ米(非芳香性系統)においては、Os2AP遺伝子発現のダウンレギュレーションは検出されなかった。
【0150】
実施例4:マーカーを利用した選択
通常の異種交配により新規香米を開発するために、香米供与体とより生産性の高い受容体米品種と交配してもよい。その交配による後代は、数ある形質の中で特に穀物の芳香について分離する。この状況は育種家を混乱させ、より優れた香米についての通常の育種が成功しにくくなる。芳香遺伝子の発見は、従来の育種植物の新しいパラダイムとなる。何千もの植物を最高の精度でスクリーニングし得るように、特異的な分子マーカーを芳香遺伝子を検出するために開発することができる。育種家は、DNAマーカー法によって、Os2AP遺伝子の芳香性対立遺伝子を早い段階で高感度にて検出し得る。このことにより、育種家が後の段階で芳香性植物に好ましい形質を与えるためのより多くの機会が残される。イネにおいて確認される芳香の分子基盤は、おそらく他の穀物でも見つけられるため、他の穀物のDNAマーカーを開発する方法も開かれる。
【0151】
実施例5:Os2AP遺伝子の系統発生解析
プロリンおよびポリアミン生合成は、植物界において高度に保存された経路である。本明細書において提示する2−アセチル−1−ピロリンの生合成は、他の2−アセチル−1−ピロリン産生植物が利用する共通のテーマである。この点について例証するために、マルチプル配列アラインメントを利用して2AP遺伝子のアミノ酸配列を比較した。結果として得られた系統樹により、イネ、コムギおよびオオムギ由来の2AP遺伝子が共通の祖先を有し得ることが分かる(図9B)。それゆえ、穀物における2−アセチル−1−ピロリンの生合成が共通のテーマを有している可能性がある。この点について例証するためには、将来、他の穀物において2AP遺伝子オーソログを用いた試験を行う必要がある。
【0152】
Os2AP遺伝子がどこから来たのかを遡るのは興味深い。「アロマーカー」PCRプライマーを用いて95の在来品種を細かく調べることにより、本発明者らは8塩基対の欠失を有する対立遺伝子が芳香の増強と厳密に相関していることが分かった。また、本発明者らは、オリザ・ニバラ(Oryza nivara)およびO.ルフィポゴン(O. rufipogon)をはじめとする野生種間で8塩基対の欠失に関連するエクソン7を配列決定した(図9A)。本発明者らは、タイジャスミン(Thai Jasmine)、バスマティ(Basmati)およびアズセナ(Azucena)をはじめとする最も芳香性の高い品種がその欠失を有しており、それゆえ、古代にまで遡り得る共通の祖先を有することが分かった。本発明者らは、芳香性野生米においても芳香性対立遺伝子を確認した。よって、本発明者らが今日知っている香米は、人間が栽培するようになるずっと以前に、単一突然変異が起こって生み出されたのである。
【参照文献】
【0153】
以下の参照文献は、それらの全開示内容が引用することにより本明細書の一部とされる。
【図面の簡単な説明】
【0154】
【図1】カオ・ド・マリ(KDML105)、タイジャスミン米(芳香性)およびコシヒカリ、非香米から、GC−MSにより得られた2−アセチル−1−ピロリンの化学構造。
【図2】穀物の芳香について分離する、単一F6植物由来の186のF9植物を用いたイネにおける芳香遺伝子の詳細なマッピングと穀物の芳香に関する遺伝子を含む物理的地図の構築。第1の部分は、単一F6植物由来の8のF11植物の図式遺伝子型を示す。第2の部分は、単一BACにおいて重要な領域を27kbに絞るための、単一F6植物由来の1116のF12植物を用いたウルトラスケールマッピングを示す。第3の部分は、3つのオープンリーディングフレームを示すKDML105のゲノム配列の注釈を示す。第4の部分は、KDML105とJHNの交配による177のF6植物を用いて、穀物の芳香および2−アセチル−1−ピロリン含量に大きな影響を及ぼす未知タンパク質遺伝子のエクソン7内に3つの二重組換え体が確認されることを示す。この未知タンパク質遺伝子は、Os2APと名づけられた。「アロマーカー」は、穀物の芳香に特有の8塩基対の欠失と3つのSNP(一塩基変異多型)を定義するPCRベースのマーカーである。
【図3A】芳香性および非芳香性同質遺伝子系統の受粉の10、15および20日後に単離された全RNAからの、RT−PCRを用いた7つの候補遺伝子の発現。この図において、Os2APは、ベタインアルデヒドデヒドロゲナーゼと見なされる。
【図3B】芳香性および非芳香性同質遺伝子系統の受粉の15日後に単離された全RNAからのOs2AP転写産物の葉、茎および根における示差的発現。
【図3C】THM(KDML105)、芳香性同質遺伝子系統117、非芳香性同質遺伝子系統10およびそれらのF2(ISL117×ISL10)の穀粒における2−アセチル−1−ピロリンレベルの解析。F1では、2−アセチル−1−ピロリンレベルの解析を葉で実施した。
【図3D】上のパネル:Os2AP RNAi構築物を含有するトランスジェニックニホンバレにおける染色体4のOs2APホモログとアクチンの発現。下のパネル:染色体4ではなく染色体8のOs2AP遺伝子(本明細書ではBADHと呼ぶ)が、芳香性および非芳香性同質遺伝子系統のイネにおいて示差的発現を示している。
【図4A】プロリンおよびグルタミン酸間の代謝経路。L−プロリンは、酵素P5CシンターゼおよびP5Cレダクターゼを用いてグルタミン酸から合成され、グルタミン酸は、プロリンデヒドロゲナーゼ(ProDH)およびP5Cデヒドロゲナーゼ(P5CDH)を用いてプロリンから合成される。提示された代謝シフトは、Os2APのナンセンス突然変異が介在するものであった。
【図4B】Os2AP酵素の推定タンパク質構造を示す。
【図5A−1】KDML105(芳香性)およびニホンバレ(非芳香性)由来のOs2AP遺伝子のゲノム配列比較。
【図5A−2】図5Aの続き。
【図5B】ニホンバレ、非芳香性系統と比べて、芳香性系統、タイ・ホーム・マリ(THM)における8塩基対の欠失を示すDNAアラインメントおよび2系統間のアミノ酸配列比較。
【図6】単一F6植物由来の8のF11植物の図式遺伝子型と米粒子の2−アセチル−1−ピロリンレベルの解析。
【図7A】RNA干渉構築物と形質転換用ベクター。下のパネルは、構築物の発現確認を示す。本図面においてBADHとは、Os2APをさす。
【図7B】RNAi構築物、GFPおよび内在性Os2APの発現。下のパネルは、2−アセチル−1−ピロリンレベルと芳香を示す。BADH−RNAi2は、Os2AP遺伝子に対するRNAi構築物である。
【図8A】「アロマーカー」プライマーセットを用いた、芳香の増強と関係しているOs2AP変異体についてのF6後代のスクリーニング。
【図8B】芳香性Os2AP対立遺伝子についての種々の米品種のスクリーニング。
【図9A】種々の系統のイネ由来のOs2APオーソログのDNA配列を示す。
【図9B】22のBADH(Os2AP)オーソログの近隣結合樹を用いて構築した系統樹。
【技術分野】
【0001】
本発明は、一般に、植物分子遺伝学に関する。とりわけ、本発明は、高いレベルの2−アセチル−1−ピロリンを有する非天然植物および真菌、かかる植物および真菌を作製する方法、ならびに2−アセチル−1−ピロリン合成に関与する核酸に関する。
【背景技術】
【0002】
穀物の芳香は、高品質の米の最も魅力的な特徴であり、アジア市場で需要が増加しているだけでなく、欧州、そして世界中で広く認識されている。調理した米の芳香は、炭水化物、アルコール、アルデヒド、ケトン、酸、エステル、フェノール、ピリジン、ピラジンおよびそれ以外の化合物のような百を超える揮発性化合物からなっている(Yajima et al., 1978; Maga, 1984; Takashi et al., 1980; Paule and Power, 1989)。あらゆる香米、小麦パンおよびライ麦パンの皮の主要な強い香気成分として「ポップコーン様」の芳香族化合物、2−アセチル−1−ピロリン(2AP)が発見された(Buttery et al., 1982, 1983)。2−アセチル−1−ピロリンは、主として、数多くの香米品種に特有の芳香に関与している(Tanchotikul and Hsieh, 1991)。驚くべきことに、この米の芳香は、タコノキの葉(Buttery et al., 1983)、グルテン麦粉(Vallaris Glabra Ktze.)(Wongpornchai et al., 2003)、含水雑穀(wet millet)(Seitz et al., 1993)、ポップコーン(Schieberle, 1991)、バチルス・セレウス(Bacillus cereus)(Romanczyk et al., 1995)および真菌(Nagsuk et al., 2004)からも単離され、同定された。2−アセチル−1−ピロリンは、芳香性種のイネの、根を除く各部位(茎、葉、穀粒)に存在している(Lorieux et al., 1996)。芳香性種の穀粒にはこの芳香があるが、総ての穀粒にあるとは限らない。
【0003】
芳香族化合物、2−アセチル−1−ピロリンは、アミノ酸プロリンに類似したピロリン環を有している(図1)。アミノ酸プロリンを、2−アセチル−1−ピロリンを合成する前駆体として関連付ける最初の証拠が、細胞とカルス培養での試験によって見つけられた(Suprasanna et al., 1998; Suprasanna et al., 2002)。同位元素標識を用いた試験によって、タイ・ホーム・マリ(Thai Hom Mali)(THM)米において(Yoshihashi et al., 2002)、そしておそらく他の香米においても、穀粒2−アセチル−1−ピロリンの前駆体がアミノ酸プロリンである可能性が最も高いことが示され、その結果の裏付けが得られた。しかしながら、2−アセチル−1−ピロリンの正確な生合成経路はまだ解明されていない。従って、2−アセチル−1−ピロリン合成に関与する遺伝子を同定し、植物および真菌において2−アセチル−1−ピロリンレベルを高めて芳香を増強するための方法を提供する必要がある。
【発明の概要】
【0004】
本発明は、化合物2−アセチル−1−ピロリンが、対照植物および真菌で産生されるレベルよりも高いレベルで産生される非天然植物および真菌を提供することによりこれらの必要性に満たすものである。対照植物および真菌とは、化合物2−アセチル−1−ピロリンレベルが低い類似遺伝子型または関連遺伝子型の植物および真菌をさす。このような対照植物が非天然植物であってもよい。本発明はさらに、このような非天然植物および真菌、ならびに該植物から生じた種子についてスクリーニングし、作出する方法を提供する。
【0005】
化合物2−アセチル−1−ピロリンの正確な生合成経路は分かっていないが、その化合物はプロリンの誘導体である。プロリンは2−アセチル−1−ピロリンまたはグルタミン酸のいずれかに変換され得るため、グルタミン酸合成経路を阻害することによってプロリン(または中間体)の2−アセチル−1−ピロリン合成への利用可能性が高まる(図4A)という仮定が立てられる。Os2AP(オリザ・サティバ(oryza sativa)2−アセチル−1−ピロリン)と呼ばれる、米の芳香を制御するタンパク質をコードする遺伝子が、プロリンのグルタミン酸への変換において重要な役割を果たすと考えられているアルデヒトデヒドロゲナーゼファミリーのメンバーとして同定された。試験した総ての香米種では、この遺伝子に8個のヌクレオチドの欠失が存在する。この欠失は、ナンセンス変異を介したそのmRNAの分解を引き起こす早期停止コドンを与え、その結果、機能喪失表現型をもたらす。RNA干渉(RNAi)研究では、Os2AP遺伝子の転写破壊により、植物において高いレベルの2−アセチル−1−ピロリンが生じ、それとともに芳香が強まることが示された。
【0006】
本発明は、2−アセチル−1−ピロリン(2AP)遺伝子の発現を抑制し、2AP遺伝子のmRNAレベルを低減し、および/または2APタンパク質の活性を低下させることによって、高いレベルの2−アセチル−1−ピロリンが産生される非天然植物および真菌を提供する。2APタンパク質のレベルは、対照植物に対して、25パーセント、50パーセントまたは100パーセント低減する可能性がある。2AP遺伝子の発現の抑制または2AP遺伝子のmRNAの低減は、a)2AP遺伝子もしくはその断片をアンチセンス方向で発現させ、b)その遺伝子の一部をRNA干渉構築物にクローニングし、トランスジェニック植物においてこの構築物を発現させ、またはc)種々の方法(Targeting Induced Local Lesions IN Genomes(TILLING)法およびtDNA挿入突然変異誘発法を含む)によって突然変異誘発した後、PCRまたは他の方法により芳香性変異体についてスクリーニングすることにより、実現し得る。
【0007】
本発明はさらに、対照非トランスジェニックイネ植物が有するレベルよりも高いレベルの化合物2−アセチル−1−ピロリンを有するトランスジェニックイネ植物であって、該トランスジェニック植物における2AP遺伝子によりコードされるmRNAまたはタンパク質レベルを、該対照非トランスジェニック植物における2AP遺伝子によりコードされるmRNAまたはタンパク質レベルよりも、低減することによって、該トランスジェニック植物において該化合物のレベルが高められる、トランスジェニックイネ植物を提供する。一形式においては、mRNAおよびタンパク質レベルがRNA干渉によりまたはアンチセンスにより低減されている。本発明はさらに、本発明のイネから生じたトランスジェニックイネの種子に向けられる。
【0008】
後記の実施例は、イネにて実施した試験について記載しているが、本発明は、限定されるものではないが、コムギ、オオムギ、ライムギ、ココナッツ、ソルガムおよびオートムギを含む他の植物および真菌に関する。
【0009】
本発明はさらに、2AP遺伝子をコードする単離された核酸であって、少なくとも1回の0.1×SSCおよび0.1%SDSでの60〜65℃にて30分間の洗浄を含むハイブリダイゼーション条件下で、配列番号1、配列番号2、配列番号4もしくは配列番号5で示される核酸配列またはその相補配列とハイブリダイズする核酸、配列番号1、配列番号2、配列番号4もしくは配列番号5で示される配列と少なくとも70%同一、80%同一、90%同一、95%以上同一である核酸、および配列番号3または配列番号6で示されるアミノ酸配列と少なくとも70%、少なくとも80%、少なくとも90%、95%、または95%を超える割合で同一であるポリペプチドをコードする核酸を含む、単離された核酸を提供する。
【0010】
米中の2−アセチル−1−ピロリン量は、作況や土壌型によって変わり得る。非芳香性品種であるニホンバレ(Nipponbare)では、2−アセチル−1−ピロリンレベルが0〜0.1ppm(100万分の1)の範囲である。これに対して、香米品種であるタイ・ホーム・マリでは、2−アセチル−1−ピロリン量が1〜2.5ppmの範囲である。実施例2では、米ニホンバレ中の2−アセチル−1−ピロリンレベルを2.5ppmまで高めた、イネOs2AP遺伝子に対するRNA干渉試験について詳述している。よって、本発明は、非芳香性植物の芳香レベルを芳香族化合物レベルまで増強するための方法を提供する。
【0011】
本発明はさらに、プロモーターに作動可能なように連結され得る本発明の核酸を含有している組換え構築物および発現ベクターを提供する。1つの有用なプロモーターがカリフラワーモザイクウイルス(CaMV)プロモーターであり、このプロモーターは、ほとんどの植物組織において高レベルの発現を与える。
【0012】
本発明はさらに、本発明の核酸、構築物および発現ベクターを含有する宿主細胞を提供する。
【0013】
本発明はまた、植物および核酸を、2APタンパク質の発現または活性の低減を引き起こし、2AP化合物の産生が増加した結果として必然的に生じる芳香の増強をもたらす2AP遺伝子における突然変異についてスクリーニングするための方法も提供する。1つの特異的突然変異が、イネ2AP遺伝子の芳香性表現型と関連する8個のヌクレオチドの欠失である。この突然変異および他の突然変異についてのスクリーニングは、PCR、配列決定、ハイブリダイゼーションまたはマイクロアレイ試験のような種々の方法によって行うことができる。別の方法としては、2APタンパク質レベルの低減または2APタンパク質の構造もしくは活性の変化を探る方法がある。これについては、例えば、活性2APタンパク質と結合する抗体を使用することによって行えるし、2APタンパク質活性についてのアッセイによっても行える。
【0014】
本明細書に記載の配列は、核酸ハイブリダイゼーション試験におけるプローブまたはプライマーとして使用され得る。配列番号1、配列番号2、配列番号4もしくは配列番号5で示されるヌクレオチド配列の14ヌクレオチド長の連続したDNAセグメントと同じ配列を有するか、または配列番号1、配列番号2、配列番号4もしくは配列番号5で示されるヌクレオチド配列の14ヌクレオチド長の連続したDNAセグメントと相補的である少なくとも14ヌクレオチド長の連続した配列を含む核酸セグメントを使用することができる。
【0015】
より長い連続した同一または相補配列、例えば、約20、30、40、50、100、200、500、1,000、2,000、5,000、10,000ヌクレオチドのもの他(中間にあるあらゆる長さから全長配列を含むその長さまでを含む)もまた、特定の実施形態において有用である。
【0016】
このような核酸プローブの、2AP遺伝子配列と特異的にハイブリダイズする能力によって、それらが特定サンプルにおいて相補配列の存在を検出するのに有用であり得る。
【0017】
しかしながら、突然変異種プライマー、すなわち、他の遺伝的構造の調製用のプライマーの作製に用いる配列情報の使用をはじめとする他の使用が想定される。
【図面の説明】
【0018】
図1は、カオ・ド・マリ(Khao Dawk Mali)(KDML105)、タイジャスミン米(芳香性)およびコシヒカリ(Koshihikari)、非香米から、GC−MSにより得られた2−アセチル−1−ピロリンの化学構造を示す。
【0019】
図2は、穀物の芳香について分離する、単一F6植物由来の186のF9植物を用いたイネにおける芳香遺伝子の詳細なマッピングと穀物の芳香に関する遺伝子を含む物理的地図の構築を示す。
【0020】
図2の第1の部分は、単一F6植物由来の8のF11植物の図式遺伝子型を示す。第2の部分は、単一BACにおいて重要な領域を27kbに絞るための、単一F6植物由来の1116のF12植物を用いたウルトラスケールマッピングを示す。第3の部分は、3つのオープンリーディングフレームを示すKDML105のゲノム配列の注釈を示す。第4の部分は、KDML105とJHNの異種交配による177のF6植物を用いて、穀物の芳香および2−アセチル−1−ピロリン含量に大きな影響を及ぼす未知タンパク質遺伝子のエクソン7内に3つの二重組換え体が確認されることを示す。この未知タンパク質遺伝子は、Os2AP、オリザ・サティバ2−アセチル−1−ピロリンと名づけられた。「アロマーカー」は、穀物の芳香に特有の8塩基対の欠失と3つのSNP(一塩基変異多型)を定義するPCRベースのマーカーである。
【0021】
図3Aは、芳香性および非芳香性同質遺伝子系統の受粉の10、15および20日後に単離された全RNAからの、RT−PCRを用いた7つの候補遺伝子の発現を示す。この図において、Os2APは、ベタインアルデヒドデヒドロゲナーゼと見なされる。
【0022】
図3Bは、芳香性および非芳香性同質遺伝子系統の受粉の15日後に単離された全RNAからのOs2AP転写産物の葉、茎および根における示差的発現を示している。
【0023】
図3Cは、THM(KDML105)、芳香性同質遺伝子系統117、非芳香性同質遺伝子系統10およびそれらのF2(ISL117×ISL10)の穀粒における2−アセチル−1−ピロリンレベルの解析を示す。F1では、2−アセチル−1−ピロリンレベルの解析を葉で実施した。
【0024】
図3Dは、上のパネルは、Os2AP RNAi構築物を含有するトランスジェニックニホンバレにおける染色体4のOs2APホモログとアクチンの発現を示す。下のパネルは、芳香性および非芳香性同質遺伝子系統のイネにおいて示差的発現を示す、染色体4ではなく染色体8のOs2AP遺伝子(本明細書ではBADHと呼ぶ)を示す。
【0025】
図4Aは、プロリンおよびグルタミン酸間の代謝経路を示す。L−プロリンは、酵素P5CシンターゼおよびP5Cレダクターゼを用いてグルタミン酸から合成され、グルタミン酸は、プロリンデヒドロゲナーゼ(ProDH)およびP5Cデヒドロゲナーゼ(P5CDH)を用いてプロリンから合成される。提示された代謝シフトは、Os2APのナンセンス突然変異が介在するものであった。
【0026】
図4Bは、Microbiology Department of the University of Massachusetts, Amherst, USAのウェブサイトを通して利用されるプログラム、RASMOL(新しいバージョンがProtein Explorerである)を利用して作製されたOs2AP酵素の推定タンパク質構造を示す。
【0027】
図5A−1は、KDML105(芳香性)およびニホンバレ(非芳香性)由来のOs2AP遺伝子のゲノム配列比較を示す。図5A−2は、図5A−1の続きを示す。
【0028】
図5Bは、ニホンバレ、非芳香性系統と比べて、芳香性系統、タイ・ホーム・マリ(THM)における8塩基対の欠失を示すDNAアラインメントおよび2系統間のアミノ酸配列比較を示す。アラインメントしているヌクレオチド配列は、配列番号5のヌクレオチド701〜765と配列番号2のヌクレオチド701〜757である。
【0029】
図6は、単一F6植物由来の8のF11植物の図式遺伝子型と米粒子の2−アセチル−1−ピロリンレベルの解析を示す。
【0030】
図7Aは、RNA干渉構築物と形質転換用ベクターを示す。下のパネルは、構築物の発現確認を示す。本図面においてBADHとは、Os2APをさす。
【0031】
図7Bは、RNAi構築物、GFPおよび内在性Os2APの発現を示す。下のパネルは、2−アセチル−1−ピロリンレベルと芳香を示す。BADH−RNAi2は、Os2AP遺伝子に対するRNAi構築物である。
【0032】
図8Aは、「アロマーカー」プライマーセットを用いた、芳香の増強と関係しているOs2AP変異体についてのF6後代のスクリーニング結果を示す。
【0033】
図8Bは、芳香性Os2AP対立遺伝子についての種々の米品種のスクリーニング結果を示す。
【0034】
図9Aは、種々の系統のイネ由来のOs2APオーソログのDNA配列を示す。アラインメントしているヌクレオチド配列は、配列番号5のヌクレオチド701〜765と配列番号2のヌクレオチド701〜757である。
【0035】
図9Bは、22のBADH(Os2AP)オーソログの近隣結合樹を用いて構築した系統樹を示す。
【配列表の一部の配列の簡単な説明】
【0036】
配列番号1は、香米系統、タイ・ホーム・マリ由来のOs2AP遺伝子のゲノムヌクレオチド配列である。
配列番号1
【化1−1】
【化1−2】
【化1−3】
【0037】
配列番号2は、タイ・ホーム・マリOs2APのタンパク質をコードするヌクレオチド配列である。
配列番号2
【化2】
【0038】
配列番号3は、タイ・ホーム・マリOs2APタンパク質のアミノ酸配列である。
配列番号3
【化3】
【0039】
配列番号4は、非香米系統、ニホンバレ由来のOs2AP遺伝子のゲノムヌクレオチド配列である。
配列番号4
【化4−1】
【化4−2】
【0040】
配列番号5は、ニホンバレOs2APのタンパク質をコードするヌクレオチド配列である。
配列番号5
【化5】
【0041】
配列番号6は、ニホンバレOs2APタンパク質のアミノ酸配列である。
配列番号6
【化6】
【0042】
配列番号7〜88は、Os2APヌクレオチド配列の一部、GFPまたはアクチンを増幅するのに使用できるプライマーである(表1)。
【0043】
【化7】
【0044】
配列番号96は、一般アルデヒドデヒドロゲナーゼに高度に保存されているデカペプチドである。
Val-Thr-Leu-Glu-Leu-Gly-Gly-Lys-Ser-Pro
【発明の詳細な説明】
【0045】
本発明は、2−アセチル−1−ピロリン(2AP)遺伝子に関する。イネ、コムギ、トウモロコシ、オートムギ、タコノキの葉、芳香ココナッツおよび一部の細菌および真菌に見られる主要な芳香族化合物である、2−アセチル−1−ピロリンのレベルを高めるために、2AP遺伝子の核酸配列が用いられる。
【0046】
本発明の少なくとも1つの実施形態を詳細に説明する前に、本発明がその適用において、以下の説明において記載されている構造の詳細および要素の配列に限定されないことを理解すべきである。本発明は、他の実施形態が可能であり、または種々の方法での実践もしくは実施が可能である。さらに、本明細書において使用する表現および専門用語は、説明するためのものであって、限定を意図したものではないことも理解すべきである。
【0047】
この開示内容を通じて、種々の刊行物、特許および公開特許明細書が引用されるている。これらの刊行物、特許および公開特許明細書の開示内容は、本発明が関連する技術の状況をさらに十分説明するために、引用することにより本明細書の開示内容の一部とされる。本発明の実施では、特に断りのない限り、当技術分野の技術の範囲内である植物育種学、免疫学、分子生物学、微生物学、細胞生物学および組換えDNAの従来の技術を使用する。例えば、Sambrook, Fritsch and Maniatis, MOLECULAR CLONING: A LABORATORY MANUAL, 2nd edition (1989); CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F. M. Ausubel, et al eds., (1987); Plant Breeding: Principles and Prospects (Plant Breeding, Vol 1) M. D. Hayward, N. O. Bosemark, I. Romagosa; Chapman & Hall, (1993.); Coligan, Dunn, Ploegh, Speicher and Wingfeld, eds. (1995) CURRENT PROTOCOLS IN PROTEIN SCIENCE (John Wiley & Sons, Inc.); the series METHODS IN ENZYMOLOGY (Academic Press, Inc.): PCR 2: A PRACTICAL APPROACH (M. J. MacPherson, B. D. Hames and G. R. Taylor eds. (1995), Harlow and Lane, eds. (1988) ANTIBODIES, A LABORATORY MANUAL, and ANIMAL CELL CULTURE R. I. Freshney, ed. (1987)参照。
【0048】
特に断りのない限り、慣例的用法に従って専門用語を使用する。分子生物学において共通する用語の定義は、Lewin, Genes V, Oxford University Press, 1994 (SBN 0-19-854287-9)発行; Kendrew et al. (eds.), The Encyclopedia of Molecular Biology, Blackwell Science Ltd., 1994 (SBN 0-632-02182-9)発行;およびRobert A. Meyers (ed.), Molecular Biology and Biotechnology, a Comprehensive Desk Reference, VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8)発行; Ausubel et al. (1987) Current Protocols in Molecular Biology, Green Publishing; Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, New Yorkで見られる。植物生物学において共通する用語の定義は、Esau, Plant Anatomy, John Wiley & Sons (1977) (ISBN 0-471-24520-8)発行;およびSolomon et al, Biology, Saunders College Publishing (1993)発行、で見られる。
定義
【0049】
本発明の種々の実施形態の検討を容易にするため、以下の定義を提供する:
【0050】
2−アセチル−1−ピロリン(2AP)ポリヌクレオチド配列:本発明の遺伝子としては、本明細書において開示される全長配列だけでなく、本明細書において具体的に示される配列の特有の活性を保持するこのような配列の断片も挙げられる。
【0051】
イネ2AP遺伝子配列は、本明細書において開示されている。このイネ2AP遺伝子配列を用いたいくつかの手段を通じて、別の植物由来の2APポリヌクレオチド配列を容易に同定し、得ることができることは当業者には明らかである。特定の遺伝子またはその一部は、培養物寄託機関から得られ得、または合成により、例えば、遺伝子マシンを用いて構築され得る。
【0052】
このような遺伝子の変化は、点突然変異を作製するための標準的な技術を用いて容易に構築し得る。また、これらの遺伝子の断片も、標準的な手順に従って市販のエキソヌクレアーゼまたはエンドヌクレアーゼを用いて作製することができる。例えば、Bal31のような酵素または部位特異的突然変異誘発を利用して、これらの遺伝子の末端からヌクレオチドを意図的に切り出すことができる。また、種々の他の制限酵素を用いて、活性断片についてコードする遺伝子を得ることができる。種々の酵素を利用して、これらの2APポリヌクレオチド配列の活性断片を直接得てもよい。
【0053】
等価な2APポリヌクレオチド配列および/またはこれらの等価な2APポリヌクレオチド配列をコードする遺伝子も、本明細書において記載される教示を用いて、系統および/またはDNAライブラリーから単離され得る。例えば、本明細書において開示される2APタンパク質に対する抗体を使用して、タンパク質混合物から他の2APタンパク質を同定し、単離することができる。具体的に言えば、最も不変で、かつ、他のタンパク質と最も異なっている2APタンパク質の一部に対して抗体を作製してもよい。こうして作製した抗体を使用して、特に、特有の活性を有する等価な2APタンパク質を免疫沈降、酵素免疫測定法(ELISA)またはウエスタンブロッティングにより同定することができる。
【0054】
本発明の遺伝子を同定するためのさらなる方法は、オリゴヌクレオチドプローブの使用による方法である。これらのプローブは、検出可能な標識を有するヌクレオチド配列である。当技術分野では周知であるように、プローブ分子と核酸サンプルが2分子間で強い結合を形成することによってハイブリダイズするならば、プローブとサンプルが本質的に同じであると当然判断することができる。プローブの検出可能な標識は、ハイブリダイゼーションが起こったかどうかを既知の方法で判断するための手段を提供する。このようなプローブ解析は、本発明の遺伝子の迅速な同定法を提供する。典型的なプローブを表1に示している。
【0055】
本発明のプローブとして使用するヌクレオチドセグメントは、DNA合成装置を使用し、標準的な手順を用いて合成することができる。ヌクレオチドセグメントのプローブとしての使用においては、特定のプローブを、放射性および非放射性標識をはじめとする当業者には公知のあらゆる好適な標識で標識する。一般的な放射性標識としては、32P、125I、35Sなどが挙げられる。DNアーゼとDNAポリメラーゼを用いて、一般的なニックトランスレーション反応により、DNAサンプルと相補的であるヌクレオチド配列から放射性同位元素で標識したプローブを構築することができる。次いで、プローブおよびサンプルをハイブリダイゼーションバッファー溶液に入れて合わせ、アニーリングが起こるまで好適な温度に保つ。その後、メンブレンを洗浄し、異物を除いてサンプルを残し、結合したプローブ分子を、通常、オートラジオグラフィーおよび/または液体シンチレーション計測により検出し、定量する。
【0056】
非放射性標識としては、例えば、ビオチンもしくはチロキシンのようなリガンド、さらに、ヒドロラーゼもしくはペルオキシダーゼのような酵素、あるいはルシフェリンのような種々の化学発光体またはフルオレセインおよびその誘導体のような蛍光化合物も挙げられる。また、分離を容易にするために、プローブを、両末端において異なるタイプの標識で標識してもよく、例えば、一末端で、以上で記載した同位元素標識を、そして、もう一方の末端でビオチン標識を使用することによって標識してもよい。
【0057】
二重形成と安定性は、ハイブリッドの2鎖間の実質的な相補性に依存しており、以上で記載したように、ある程度のミスマッチは許容される。それゆえ、本発明のプローブは、上記配列の突然変異(単一突然変異と多重突然変異の両方)、欠失、挿入およびそれらの組合せ(前記突然変異、挿入および欠失によって目的の標的ポリヌクレオチドとの安定したハイブリッドが形成される)を含む。突然変異、挿入および欠失は、数多くの方法により、現在当業者に公知の方法により、そして、将来、広まるであろう他の方法により特定のポリヌクレオチド配列において作製することができる。
【0058】
表に記載したプローブの可能性ある変化は、ある程度、遺伝子コードの冗長性による。遺伝子コードの冗長性から、すなわち、タンパク質の作製に使用されるほとんどのアミノ酸に2つ以上のコーディングヌクレオチドトリプレット(コドン)が使用される。そのため、異なるヌクレオチド配列が特定のアミノ酸をコードする可能性がある。従って、2APタンパク質およびペプチドのアミノ酸配列を、タンパク質またはペプチドの同じアミノ酸配列をコードする等価なヌクレオチド配列により調製することができる。よって、本発明は、このような等価なヌクレオチド配列を含む。また、逆または相補配列も本発明の1つの態様であり、当業者ならば容易に使用することができる。さらに、構造および機能が同定されたタンパク質群が、アミノ酸配列を変えることにより、このような変化がタンパク質二次構造を改変しないという条件において、構築され得ることが示されている(Kaiser and Kezdy, 1984)。よって、本発明は、本明細書で示されるアミノ酸配列の、タンパク質二次構造が変化していないか、または構造が変化を受けているならば、生物活性が実質的に保持されている突然変異型を含む。さらに、本発明はまた、本発明の遺伝子をコードする2APポリヌクレオチド配列の総てまたは一部を受け入れる生物の突然変異体を含む。このような突然変異体は、当業者には周知の技術によって作製され得る。例えば、紫外線照射を利用して、宿主生物の突然変異体を作製することができるし、tDNA挿入突然変異誘発法を利用してもよいし、またはTILLING(Targeted Induced Local Lesion of Genome)法を利用してもよい。同様に、このような突然変異体としては、当技術分野で周知の手順によって作製することができる胞子無形成宿主細胞も含んでよい。
【0059】
イネ由来のものを含む2APポリヌクレオチド配列を、本発明のPCRプライマーを用いて得てもよい。これらのプライマーを表1に示しており、これらのプライマーは配列番号7〜配列番号79で示される配列と対応している。これらのプライマーの組合せを使用して、Os2AP遺伝子の異なる領域を増幅することができる。この増幅に有効なPCR条件は、以下のとおりである:10ngの鋳型DNA、0.1mM dNTP、0.5Mの各プライマー、0.5単位のTaqポリメラーゼ、2.0mM MgCl2および1×好熱性ポリメラーゼバッファー(Promega)を含有する10μL反応混合物。この混合物により、以下の時間および温度:94℃30秒間(変性)、60℃30秒間(アニーリング)および72℃2分間(伸長)を用いて30サイクルのPCRを行う必要がある。
【表1−1】
【表1−2】
【表1−3】
【0060】
オリザ・サティバ2−アセチル−1−ピロリン(Os2AP)ホモログ:本願にて記載したものと類似性を示す配列を、パブリックドメイン配列アラインメントアルゴリズムと配列データベースを検索するための配列類似性検索ツールを使用するコンピューターを利用した方法により同定することができる(パブリックドメインデータベースとしては、Genbank、EMBL、Swiss−Prot、PIRなどが挙げられる)。
【0061】
類似性検索を読み出し、解析しようとする標的配列(すなわち、質問配列)と比較する配列をアラインする。比較する配列の局所領域間の最適なアラインメントは、ローカルアラインメントとして知られている。配列比較アルゴリズムは、各アラインメントに総合スコアを与えるスコアリングマトリックスを使用している。
【0062】
ポリヌクレオチドおよびポリペプチド配列をアラインし、他のポリヌクレオチドおよびポリペプチド配列と比較した、指定領域における同一残基率を公的に入手可能なコンピューターアルゴリズムを利用して決定する。同一性パーセントスコアは、比較している配列のオーバーラップ領域の長さに依存する。
【0063】
2核酸配列間または2アミノ酸配列間の類似性は、配列同一性で表される(または、タンパク質の場合、配列類似性でも表される)。配列同一性は、同一性パーセントとして測定されることが多い;割合が高いほど、2配列の類似性が高いということである。本明細書において記載するように、2APタンパク質をコードする核酸分子のホモログおよび変異体を、本発明に使用してもよい。これらの核酸分子のホモログおよび変異体は、標準的な方法を使用してアラインした場合に、比較的高い配列同一性を有しているものである。このようなホモログおよび変異体は、高ストリンジェンシー条件下で互いにハイブリダイズする。
【0064】
比較用配列のアラインメント方法は、当技術分野ではよく知られている。種々のプログラムおよびアラインメントアルゴリズムについては、配列アラインメント方法と相同性計算の詳細にわたる検討を示しているSmith and Waterman (1981); Needleman and Wunsch (1970); Pearson and Lipman (1988); Higgins and Sharp (1988); Higgins and Sharp (1989); Corpet et al. (1988); Huang et al. (1992);およびPearson et al. (1994). Altschul et al.(1994)で記載されている。
【0065】
NCBIベーシックローカルアラインメント検索ツール(BLAST)(Altschul et al., 1990)は、National Center for Biotechnology Information(NCBI, Bethesda, MD)を含むいくつかの供給元からインターネット上で入手可能であり、配列解析プログラムblastp、blastn、blastx、tblastnおよびtblastxとともに用いられる。このツールはNCBIウェブサイトでアクセス可能である。このプログラムを利用した配列同一性決定方法についての解説は、NCBIウェブサイトで入手可能である。
【0066】
開示されたタンパク質配列のホモログは、一般に、開示された配列のアミノ酸配列との全長アラインメントにおいて、NCBI Blast2.0を使用し、ギャップ入りblastpをデフォルトパラメーターに設定して計算した場合に少なくとも40%配列同一性を有するという特徴を有するものである。調整パラメーターは、好ましくは、以下の値に設定する:オーバーラップスパン1、オーバーラップフラクション=0.125、ワード閾値(T)=11。HSP SおよびHSP S2パラメーターは動的値であり、それぞれの配列の構成と目的の配列が検索されるデータベースの構成に応じて、そのプログラム自体によって確立される。しかしながら、それらの値は、感度を高めるように調整される可能性がある。この方法を用いて評価した場合、参照配列との類似性がさらに高いタンパク質は、同一性パーセントの増加、例えば、少なくとも約50%、少なくとも約60%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%または少なくとも約95%配列の同一性を示す。
【0067】
開示された核酸配列のホモログは、一般に、開示された配列のアミノ酸配列との全長アラインメントにおいて、NCBI Blast2.0を使用し、ギャップ入りblastnをデフォルトパラメーターに設定して計算した場合に少なくとも40%配列同一性を有するという特徴を有するものである。さらに、このような配列は、高ストリンジェンシー条件下で相同配列とハイブリダイズする。好ましい方法は、WU−BLAST−2のBLASTNモジュールを使用し(Altschul et al., 1996)、デフォルトパラメーターに設定し、オーバーラップスパンおよびオーバーラップフラクションをそれぞれ1および0.125に設定する。この方法を用いて評価した場合、参照配列との類似性がさらに高い核酸配列は、同一性パーセントの増加、例えば、少なくとも約50%、少なくとも約60%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%または少なくとも約95%配列の同一性を示す。
【0068】
アラインメントは、アラインする配列へのギャップの挿入を含み得る。さらに、配列番号3もしくは配列番号6で示されるタンパク質より含有するアミノ酸が多い配列または配列番号3もしくは配列番号6で示されるタンパク質より含有するアミノ酸が少ない配列では、1つの実施形態において、配列同一性パーセントがアミノ酸総数に対する同一アミノ酸の数に基づいて決定されると理解される。よって、例えば、以下で述べる図面で示されたものより短い配列の配列同一性は、1つの実施形態において、長い方の配列のアミノ酸数を用いて決定される。同一性パーセントの計算において、相対重量は、挿入、欠失、置換他のような配列変化の多様な現れには関係していない。
【0069】
1つの実施形態においては、同一である場合だけにプラスの評点をつけ(+1)、ギャップを含むあらゆる形態の配列変化には「0」の値をつける。この場合、配列類似性計算に関して本明細書において記載する加重尺度またはパラメーターの必要性が除去される。配列同一性パーセントは、例えば、マッチしている同一残基数をアラインした領域の「短い方の」配列の残基総数で除し、100を乗じることにより計算することができる。「長い方の」配列は、アラインした領域に最も多くの現実の残基を有するものである。
【0070】
同じゲノム(パラログ)または異なるゲノム(オーソログ)において、タンパク質をそれらの配列関連性に従って他のタンパク質と区別することができる。オーソログ遺伝子は、共通の先祖遺伝子からの種形成によって生じた遺伝子である。これらの遺伝子は、通常、それらが導くものと同じ機能を保持している。パラログ遺伝子は、ゲノム内で複製される遺伝子である。これらの遺伝子は、初代のものに関係しているが、新しい特異性または改変された機能を獲得している。系統発生解析方法は、バイオインフォマティクスにおける通常の技量を有する人であれば周知である。
【0071】
当業者には明らかなように、本発明の配列は、配列決定エラーを含み得る。すなわち、不正確なアミノ酸配列、ヌクレオチド、フレームシフト、未知ヌクレオチドまたは他のタイプの配列決定エラーが配列中に存在する可能性がある。しかしながら、正確な配列が、核酸に関する本明細書における相同性およびストリンジェンシーの定義、ならびにタンパク質またはポリペプチドに関して記載されているタンパク質相同性の範囲に入る。
【0072】
2−アセチル−1−ピロリン(2AP)ポリペプチド:本明細書において「2APポリペプチド」とは、2APオーソログのアミノ酸配列を実質的に有する遺伝子産物を意味する用語である。2APポリペプチドは、1つには、その発現を低減し、そのmRNAレベルを低減し、またはタンパク質量もしくは活性を低減することによって、植物における化合物2−アセチル−1−ピロリンのレベルが高められるということを特徴とする。2APポリペプチドはまた、1つには、配列番号3または配列番号6で示されるアミノ酸配列と少なくとも約30%、40%、50%、60%、70%、80%、90%または95%アミノ酸同一性を有するアミノ酸配列を有していることも特徴とする。
【0073】
実質的に同一:「実質的に同一」とは、参照アミノ酸配列(例えば、配列番号3もしくは配列番号6で示されるアミノ酸配列)または核酸配列(例えば、配列番号1、配列番号2、配列番号4もしくは配列番号5で示される核酸配列)と少なくとも30%、好ましくは、50%、より好ましくは、80%、最も好ましくは、90%、あるいは95%相同性を示すポリペプチドまたは核酸を意味する。ポリペプチドの場合、比較配列の長さは、一般に、少なくとも16アミノ酸、好ましくは、少なくとも20アミノ酸、より好ましくは、少なくとも25アミノ酸、最も好ましくは、35アミノ酸以上である。核酸の場合、比較配列の長さは、一般に、少なくとも50ヌクレオチド、好ましくは、少なくとも60ヌクレオチド、より好ましくは、少なくとも75ヌクレオチド、最も好ましくは、110ヌクレオチド以上である。
【0074】
配列同一性は、一般に、配列解析ソフトウェア(例えば、Genetics Computer Group, University of Wisconsin Biotechnology Center, 1710 University Avenue, Madison, Wis. 53705の配列解析ソフトウェアパッケージ、BLASTまたはPILEUP/PRETTYBOXプログラム)を用いて測定する。例えば、このようなソフトウェアは、標準的なパラメーターに設定すると、種々の置換、欠失および/または他の改変に相同性度を与えることによって、同一または類似配列をマッチさせる。保存的置換としては、一般に、以下の群:グリシン、アラニン;バリン、イソロイシン、ロイシン;アスパラギン酸、グルタミン酸、アスパラギン、グルタミン;セリン、トレオニン;リシン、アルギニン;およびフェニルアラニン、チロシン内での置換が挙げられる。
【0075】
高いレベル:高いレベルとは、本明細書において、対応する天然植物における化合物2−アセチル−1−ピロリンの平均レベルと比べた際の非天然植物における化合物2−アセチル−1−ピロリンの平均レベルの上昇を意味する。当業者ならば、植物における化合物2−アセチル−1−ピロリンのレベルが数多くの変数に応じて植物によって異なるならば、非天然植物と対応する天然植物における化合物2−アセチル−1−ピロリンの平均レベルの比較においては、同様に制御された条件下で成長させた各タイプの植物の合理的な大きさのサンプル集団を比較するべきであることが分かるであろう。好ましくは、各集団からのいくつかの植物において化合物2−アセチル−1−ピロリンのレベルを測定し、それらを平均して、非天然植物が高いレベルのものを含有しているかどうかを判定する。本発明の非天然植物においての高いレベルとは、対応する天然植物におけるそのレベルよりも平均して、少なくとも約20%高い、40%高い、60%高い、80%高い、100%高い、150%高い、200%高い、250%高い、300%高い、400%高いまたは500%高いレベルである。
【0076】
プロモーター:構造遺伝子に発現制御エレメントを提供し、かつ、RNAポリメラーゼが特異的に結合し、それによりその遺伝子のRNA合成(転写)を開始するDNA配列またはDNA配列群にある認識部位。
【0077】
これらのプロモーターを用いた植物発現構築物の例が、Fraley et al., 米国特許第5,352,605号で見られる。トランスジェニック植物のほとんど組織では、CaMV 35Sプロモーターは強いプロモーターである(例えば、Odell et al., Nature 313: 810, 1985参照)。CaMVプロモーターは、単子葉植物においても極めて活性が高い(例えば、Dekeyser et al., Plant Cell 2: 591, 1990; Terada and Shimamoto, Mol. Gen. Genet. 220: 389, 1990参照)。さらに、このプロモーターの活性は、CaMV 35Sプロモーターの重複によってさらに高めることができる(すなわち、2〜10倍の間)(例えば、Kay et al., Science 236: 1299, 1987; Ow et al., Proc. Natl. Acad. Sci., U. S. A. 84: 4870, 1987;およびFang et al., Plant Cell 1: 141, 1989, and McPherson and Kay, 米国特許第5,378,142号参照)。
【0078】
他の有用な植物プロモーターとしては、限定されるものではないが、ノパリンシンターゼ(NOS)プロモーター(An et al., Plant Physiol. 88: 547, 1988およびRodgers and Fraley, 米国特許第5,034,322号)、オクトピンシンターゼプロモーター(Fromm et al., Plant Cell 1: 977, 1989)、ゴマノハグサモザイクウイルス(FMV)プロモーター(Rogers, 米国特許第5,378,619号)およびイネアクチンプロモーター(Wu and McElroy, WO91/09948)が挙げられる。
【0079】
典型的な単子葉植物プロモーターとしては、限定されるものではないが、ツユクサ黄色斑紋ウイルスプロモーター、サトウキビバドナウイルスプロモーター、イネツングロ桿菌状ウイルスプロモーター、トウモロコシ条斑ウイルスエレメントおよびコムギ萎縮ウイルスプロモーターが挙げられる。
【0080】
構築物:特に断りのない限り、「構築物」とは、本発明の1以上の単離ポリヌクレオチド配列を含んでなる組換え遺伝子分子をさす用語である。
【0081】
宿主生物における導入遺伝子の発現に使用される遺伝子構築物は、オープンリーディングフレームに作動可能なように連結された遺伝子プロモーター配列と、所望により、オープンリーディングフレームの3’下流に遺伝子終止配列を含む。オープンリーディングフレームは、遺伝子配列の使用目的に応じてセンス方向またはアンチセンス方向のいずれかに置かれている。構築物は、選択マーカー遺伝子と遺伝子発現用の他の調節エレメントも含んでいてもよい。
【0082】
ベクター:宿主細胞において複製可能であり、かつ/または付着したセグメントの複製が起こるように別のDNAセグメントが作動可能なように連結され得るDNA分子。プラスミド、ファージミド、コスミド、ファージ、ウイルス、YACおよびBACは総て典型的なベクターである。
【0083】
「ベクター」とは、ポリヌクレオチド配列を宿主細胞へと導入して、形質転換された宿主細胞を作製するために使用される核酸分子をさす用語である。「ベクター」は、上述の遺伝子構築物の他に、遺伝物質、例えば、1種以上の宿主細胞においてその複製を可能にする、複製起点のような1以上の核酸配列、選択マーカー遺伝子および当技術分野で公知の他の遺伝要素(例えば、遺伝物質を宿主細胞のゲノムに組み込むための配列など)を含んでいてよい。
【0084】
形質転換された:形質転換された細胞は、分子生物学技術によって核酸分子が導入されている細胞である。本明細書において、形質転換とは、核酸分子がこのような細胞、植物または動物細胞に導入され得る、ウイルスベクターによるトランスフェクション、アグロバクテリウム(Agrobacterium)による形質転換、プラスミドベクターの使用、ならびにエレクトロポレーション、リポフェクションおよび粒子銃加速による裸のDNAの導入をはじめとする総ての技術を包含する用語であり、一過性だけでなく安定な形質転換体も含む。
【0085】
植物細胞のDNA形質転換方法としては、アグロバクテリウムを介した植物形質転換、プロトプラスト形質転換、花粉への遺伝子導入、生殖器官への注入、未熟胚への注入および粒子衝撃が挙げられる。これらの方法は、それぞれ、異なる長所と短所を有している。そのため、特定の植物系統に遺伝子を導入する特定の一方法は、必ずしも別の植物系統に対して最も効果的な方法ではないが、それぞれの方法がどの植物系統に対して有効であるかは十分に分かっている。
【0086】
形質転換DNAセグメントを細胞に導入するための方法は数多くあるが、総てのものがDNAの植物細胞への送達に適しているわけではない。好適な方法としては、事実上、DNAが細胞に導入され得る、アグロバクテリウム感染、DNAの直接送達(例えば、プロトプラストのPEGを介した形質転換(Omirulleh et al., 1993)による、乾燥/阻害を介したDNA取り込みによる、エレクトロポレーションによる、カーボランダム繊維での攪拌による、DNAコーティング粒子の加速によるなど)によるなどの総ての方法が挙げられるであろう。特定の実施形態においては、加速法が好ましく、その方法としては、例えば、マイクロプロジェクタイル衝撃などが挙げられる。
【0087】
細胞へのDNA導入技術は、当業者には周知の技術である。遺伝子を細胞へと送達するための一般的な4通りの方法が記載されている:(1)化学的方法(Graham and van der Eb, 1973; Zatloukal et al., 1992);(2)マイクロインジェクション(Capecchi, 1 980)、エレクトロポレーション(Wong and Neumann, 1982; Fromm et al., 1985)および遺伝子銃(Johnston and Tang, 1994; Fynan et al., 1993)のような物理的方法;(3)ウイルスベクター法(Clapp, 1993; Lu et al., 1993; Eglitis and Anderson, 1988a; 1988b);ならびに(4)受容体を介する機構(Curiel et al., 1991; 1992; Wagner et al., 1992)。
【0088】
エレクトロポレーション:短時間の高電圧電気パルスの適用により種々の動物および植物細胞の原形質膜にナノメートルの大きさの穴を開ける。これらの穴を通じてか、またはその穴の閉鎖に伴って起こる膜成分の再分布の結果として、DNAが直接細胞質に取り入れられる。エレクトロポレーションは極めて効率が良く、クローニングした遺伝子の一過性発現にも、目的の遺伝子のコピーが組み込まれた細胞系統の確立にも使用することができる。エレクトロポレーションでは、リン酸カルシウムを介したトランスフェクションやプロトプラスト融合とは対照的に、外来DNAの1コピーまたは多くても数コピーが組み込まれた細胞系統が生じることが多い。
【0089】
エレクトロポレーションを用いたDNA導入は、当業者には周知のことである。この方法では、ペクチン分解酵素のような特定の細胞壁分解酵素を使用して、目的の受容細胞のエレクトロポレーションによる形質転換を未処理の細胞よりも起こりやすくする。あるいは、受容細胞の機械的損傷による形質転換を起こりやすくする。エレクトロポレーションによる形質転換を起こすために、細胞の懸濁培養物のようなもろい組織または胚発生カルスのいずれかを使用してもよいし、あるいは、未熟胚または他の組織化組織を直接形質転換してもよい。制御された方法で、選択した細胞の細胞壁をペクチン分解酵素(ペクトリアーゼ)または機械的損傷に曝すことによってそれらを部分的に分解する。このような分解を受けた細胞はこの段階で実施し得るエレクトロポレーションによるDNA移入を受け入れる。新たに組み込まれたDNAの性質に応じた好適な選択またはスクリーニングプロトコールによって、その後、形質転換された細胞を同定することができる。
【0090】
マイクロプロジェクタイル衝撃:形質転換用DNAセグメントを植物細胞へと送達するのにさらに有利な方法がマイクロプロジェクタイル衝撃である。この方法では、粒子を核酸でコーティングし、それを推進力によって細胞へと送達する。典型的な粒子としては、タングステン、金、白金などで構成されているものが挙げられる。
【0091】
マイクロプロジェクタイル衝撃(Bombardment)の利点は、安定形質転換単子葉植物を再生可能な方法で得る有効な手段であることの他、プロトプラストの単離(Cristou et al., 1988)もアグロバクテリウム感染に対する感受性も必要としないことである。加速法によりDNAをトウモロコシ細胞へと送達するための方法の実施形態がBiolistics Particle Delivery Systemである。このシステムを使用して、DNAまたは細胞でコーティングした粒子を、ステンレス鋼またはNytex社製スクリーンのようなスクリーンを通して、懸濁培養したトウモロコシ細胞で被覆したフィルター表面に推進することができる。粒子が大きな凝集体として受容細胞に送達されないようにこのスクリーンによって粒子を分散するものと考えられる。発射装置と衝撃しようとする細胞との間にあるスクリーンが発射物の凝集体の大きさを小さくし、大きすぎる発射物による受容細胞へのダメージを減少させることにより、形質転換の頻度を高めることへとつながる。
【0092】
衝撃では、懸濁細胞をフィルターまたは固体培養培地に集中させることが好ましい。あるいは、未熟胚または他の標的細胞を固体培養培地に配置してもよい。衝撃しようとする細胞をマクロプロジェクタイル停止プレートの下方、好適な距離に置く。要すれば、1以上のスクリーンも加速装置と衝撃しようとする細胞との間に配置する。本明細書で示される技術の使用を通じて、マーカー遺伝子を一過性発現している細胞の最大1000またはそれ以上の増殖巣を得ることができる。衝撃の48時間後に外来遺伝子産物を発現する増殖巣中の細胞数は、ほとんどの場合、1〜10および平均1〜3の範囲である。
【0093】
衝撃形質転換では、最大数の安定した形質転換体を得るために、衝撃前培養の条件および衝撃パラメーターを最適化することができる。衝撃に関する物理的パラメーターおよび生物学的パラメーターはいずれも、この技術において重要である。物理的因子としては、DNA/マイクロプロジェクタイル沈降の操作に関与するものまたはマクロプロジェクタイルもしくはマイクロプロジェクタイルのいずれかの飛行および速度に影響を及ぼすものがある。生物学的因子としては、衝撃前および衝撃直後の細胞の操作に関わる総ての工程、衝撃に伴う損傷の軽減のために役立つ標的細胞の浸透圧調整、さらに線状化DNAまたは完全なスーパーコイル状プラスミドのような形質転換用DNAの性質が挙げられる。未熟胚の形質転換を成功させるためには、衝撃前操作が特に重要であると考えられる。
【0094】
よって、条件を十分に最適化するためには小規模研究において衝撃パラメーターの一部を調整することが望ましいと考えられる。特に、ギャップ距離、飛行距離、組織距離およびヘリウム圧のような物理的パラメーターを調整することが望ましい。受容細胞の生理的状態に影響を及ぼし、その結果として、形質転換および組込み効率に影響を及ぼし得る条件を改変することにより、損傷低減係数(TRF)を最小限に抑えることもできる。例えば、最適な形質転換を行うために、受容細胞の浸透圧状態、組織の水和および継代培養段階または細胞周期を調整することができる。他の通常の調整の実施については、本開示内容に照らして、当業者には分かるであろう。
【0095】
アグロバクテリウムを介した導入:アグロバクテリウムを介した導入は、DNAをあらゆる植物組織に導入することができ、プロトプラストからの完全な植物を再分化させる必要性がなくなることから、応用の範囲が広い、遺伝子を植物細胞へと導入するためのシステムである。DNAを植物細胞に導入するためにアグロバクテリウムを介して植物に組み込むベクターを使用することは、当技術分野では周知のことである。例えば、記載されている方法参照(Fraley et al., 1985; Rogers et al., 1987)。さらに、Ti−DNAの組込みは、比較的正確なプロセスであり、再配列はほとんど起こらない。導入しようとするDNAの領域は、境界配列によって規定され、記載されている(Spielmann et al., 1986; Jorgensen et al., 1987)ように、介在するDNAが、通常、植物ゲノムに挿入される。
【0096】
最新のアグロバクテリウム形質転換ベクターは、大腸菌(E. coll)ならびにアグロバクテリウムでの複製が可能であり、記載されている(Klee et al., 1985)ように便宜的な操作が可能である。さらに、アグロバクテリウムを介した遺伝子導入用ベクターにおける最近の技術的進歩により、ベクターにおける遺伝子および制限部位の配列が改善され、種々のポリペプチドコード遺伝子を発現し得るベクターの構築が容易になった。記載されているベクター(Rogers et al., 1987)は、挿入されたポリペプチドコードグ遺伝子を直接発現させるために、プロモーターにフランキングする便宜的なマルチリンカー領域とポリアデニル化部位を有しており、このベクターは本目的に適している。さらに、形質転換には武装Ti遺伝子および非武装Ti遺伝子を含有するアグロバクテリウムを使用することができる。アグロバクテリウムを介した形質転換が有効である植物系統では、遺伝子導入が容易で明確なことからこの方法が一般的に好まれて用いられる。
【0097】
葉のディスクおよび子葉や胚軸のような他の組織のアグロバクテリウムを介した形質転換は、アグロバクテリウムが自然に感染する植物に限定されるようである。アグロバクテリウムを介した形質転換は、双子葉植物において最も効果的である。記載されている(Bytebier et al., 1987)ように、アグロバクテリウムベクターを用いてアスパラガスにおいてトランスジェニック植物が作製されているが、ほとんどの単子葉植物は、アグロバクテリウムの自然宿主ではないように思われる。そのため、イネ、トウモロコシおよびコムギのような商業的に重要な穀物は、通常、別の方法を用いて形質転換しなければならない。
【0098】
アグロバクテリウム形質転換法を用いて作製されたトランスジェニック植物は、一般に、一染色体上に単一遺伝子を有する。このようなトランスジェニック植物は、追加した遺伝子のヘテロ接合体であるといえる。しかしながら、「ヘテロ接合体」という言葉の使用には、通常、染色体対の第2の染色体の同じ遺伝子座に補足遺伝子が存在することが包含され、さらに、1個の追加された遺伝子を含有する植物にはこのような遺伝子がないため、追加された外来遺伝子が有糸分裂および減数分裂中に独立に分離することからも、このような植物に対するより正確な名前は、独立した分離体であると考えられる。
【0099】
追加された構造遺伝子のホモ接合体であるトランスジェニック植物;すなわち、2個の追加された遺伝子、染色体対の各染色体の同じ遺伝子座に1個の遺伝子を含有するトランスジェニック植物がより好ましい。ホモ接合体トランスジェニック植物は、単一追加遺伝子を有する独立した分離体であるトランスジェニック植物を雌雄交配(自殖)させ、生じた種子の一部を発芽させ、結果として得られた植物を、対照(天然非トランスジェニック)または独立した分離体であるトランスジェニック植物と比べて高い2−アセチル−1−ピロリンレベルについて解析することにより得ることができる。
【0100】
これらシステムの異なる植物系統への適用は、プロトプラストからその特定の植物系統を再分化させる能力に応じる。プロトプラストから穀物を再分化させる方法の例は記載されている(Fujimura et al., 1985; Toriyama et al., 1986; Yamada et al., 1986; Abdullah et al., 1986)。
【0101】
プロトプラストからの再分化がうまくいかなかった植物系統を形質転換するために、DNAを完全な細胞または組織に導入する他の方法を利用してもよい。例えば、未熟胚または外植片からの穀物の再分化は、記載されている(Vasil, 1988)ように行われる。さらに、「粒子銃」または高速マイクロプロジェクタイル技術を利用してもよい(Vasil, 1992)。
【0102】
後者の技術を用いて、記載されているように、金属小粒子の表面に付けたDNAを細胞壁を通過して細胞質に運び込む(Klein et al., 1987; Klein et al., 1988; McCabe et al., 1988)。金属粒子は数層の細胞を貫通するため、組織外植片内での細胞の形質転換が可能となる。
【0103】
単離された:「単離された」生物学的成分(核酸もしくはタンパク質または細胞小器官など)は、その成分が自然に存在する細胞または生物体における他の生物学的成分、すなわち、他の染色体および染色体外DNAおよびRNA、タンパク質ならびに細胞小器官から実質的に分離されているか、または精製されている。「単離された」核酸およびタンパク質としては、標準的な精製方法により精製された核酸およびタンパク質が挙げられる。この用語は化学合成された核酸を含む核酸を包含し、また、in vitroでの組換え発現または宿主細胞での組換え発現により調製されたタンパク質および以下に定義する組換え核酸も包含する。一例として、コンティグのような大きなゲノムDNA断片の遺伝子は、平均コンティグに見られる余分のDNAが比較的多量にあるため、他の生物学的成分から、単離されたと考えられるほど十分には精製されない。以下に概説するように、「組換え核酸」および「組換えタンパク質」もまた上記のように「単離される」。
【0104】
組換え:「組換え核酸」とは、本明細書において、天然には存在していない配列を有する核酸、または配列の2つの別の分離セグメントを人為的に組み合わせることによって作製された配列を有する核酸を意味する。この人為的な組合せは、化学合成により、または、一般的には、核酸の人為的操作により、例えば、制限酵素、リガーゼ、リコンビナーゼおよび/またはポリメラーゼによる少なくとも1つの核酸の操作によるような遺伝子工学技術により行われることが多い。宿主細胞に導入されると、組換え核酸が宿主細胞によって複製される;しかしながら、細胞で一度複製された組換え核酸は、依然として本発明の目的での組換え核酸である。「組換えタンパク質」とは、本明細書において、組換え核酸を使用する方法によって作製されたタンパク質を意味する。以上で概説したように、「組換え核酸」および「組換えタンパク質」もまた上記のように「単離される」。コンティグのような大きな断片の遺伝子は、このような人為的な組合せがその遺伝子と関係ない場合には、「組換え核酸」ではないだろう。しかしながら、コンティグのある遺伝子の周囲または内部の配列がその遺伝子に関連する目的で(すなわち、その遺伝子がコンティグの末端近くにあるという理由だけでなく)操作されている場合には、核酸の組換え部分と問題の遺伝子とが相対的に近いことから、コンティグのそのような遺伝子は「組換え核酸」の構成要素となるであろう。
【0105】
非天然植物:本明細書において、植物に関して使用する「非天然」とは、植物における化合物2−アセチル−1−ピロリンのレベルを改変するために人間の介入による遺伝子操作を受けた植物を意味する語である。遺伝子操作を受けた天然植物とは、「対照」植物をさす。本発明において、対照植物がトランスジェニック植物であってもよい。例えば、対照植物が除草剤耐性トランスジェニック植物であってもよい。この例においては、非天然植物が除草剤耐性であり、対照除草剤耐性植物よりも高いレベルの化合物2−アセチル−1−ピロリンを有するであろう。本発明のトランスジェニック植物は、例えば、2AP遺伝子またはその断片をコードする外来核酸分子を含有する非天然植物であり、それゆえ、人間の介入による遺伝子操作を受けたものである。さらに、例えば、突然変異誘発物質、例えば、化学的突然変異誘発物質またはトランスポゾンまたはT−DNAのような「挿入突然変異誘発物質」に計画的に曝露させた結果として2AP遺伝子調節エレメントまたはコーディング配列に突然変異を有する植物も、人間の介入による遺伝子操作を受けたということから非天然植物と考えられる。これに対し、偶発または自然発生突然変異しか有していない植物は、本明細書にて定義する「非天然植物」ではない。非天然植物は、一般に、天然植物と比べて変化した核酸配列を有しているが、同時に、非天然植物に、その核酸配列を変えることなく、例えば、そのメチル化パターンを改変することにより、人間の介入による遺伝子操作を行い得ることは当業者には明白である。
【0106】
トランスジェニック植物:形質転換された植物細胞またはプロトプラストから得られた、その植物DNAが同じ系統のネイティブな非トランスジェニック植物にはもともと存在していない導入された外来DNA分子を有している植物またはその後代。用語「トランスジェニック植物」および「形質転換された植物」とは、当技術分野において、その植物のDNAが外来DNA分子を有している場合の植物を定義するための同義語として使用されることがある。しかしながら、形質転換された植物細胞またはプロトプラストから得られた再分化植物またはカルスをトランスジェニック植物と呼ぶことがより科学的に正しいと考えられ、その使用法は、本明細書に従うものとする。
I.2−アセチル−1−ピロリン(2AP)遺伝子
【0107】
A.オーソログ/ホモログの単離:イネにおけるOs2AP遺伝子の単離および特性決定を記載している以下の実施例節は、2AP遺伝子を単離するための典型的な一般アプローチである。このような2AP遺伝子は2APタンパク質をコードする。単離された遺伝子は、その後、植物における2AP遺伝子発現を低減するための組換えベクターの構築に使用することができる。
【0108】
一般に、以下に記載する組換えDNA技術における用語および実験室手順は十分に知られているものであり、当技術分野では一般に使用されている。クローニング、DNAおよびRNA単離、増幅ならびに精製には標準的な技術を用いる。一般に、DNAリガーゼ、DNAポリメラーゼ、制限エンドヌクレアーゼなどを必要とする酵素反応は、製造業者の仕様書に従って実施する。これらの技術および種々の他の技術は、一般に、Sambrook et al., Molecular Colning - A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, N. Y., (1989)に従って実施する。
【0109】
2AP遺伝子の単離は、数多くの技術によって行うことができる。例えば、本明細書において開示された配列に基づくオリゴヌクレオチドプローブを使用して、cDNAまたはゲノムDNAライブラリー内の所望の遺伝子を同定することができる。ゲノムライブラリーを構築するため、ゲノムDNAの大きなセグメントを、例えば、制限エンドヌクレアーゼを使用したランダム断片化により作製し、それをベクターDNAと連結して、好適なベクターにパッケージングし得るコンカテマーを作出する。cDNAライブラリーを作成するため、mRNAを葉のような所望の器官から単離し、mRNAから2AP遺伝子転写物を含むcDNAライブラリーを作成する。あるいは、2AP遺伝子またはホモログが発現される他の組織から抽出されたmRNAよりcDNAを調製してもよい。
【0110】
本明細書において開示されたイネOs2AP遺伝子のようなクローニングされる2AP遺伝子の配列に基づくプローブを使用して、cDNAまたはゲノムライブラリーを、その後、スクリーニングすることができる。同じまたは異なる植物種の相同遺伝子を単離するために、プローブを使用してゲノムDNAまたはcDNA配列とハイブリダイズさせてもよい。
【0111】
10〜14、15〜20、30、50ヌクレオチド、場合によっては、およそ100〜200ヌクレオチドの、配列番号1、配列番号2、配列番号4もしくは配列番号5で示されるDNA配列と同じか、または配列番号1、配列番号2、配列番号4もしくは配列番号5で示されるDNA配列と相補的である、連続したヌクレオチドストレッチで構成されている配列領域を有する核酸分子が、例えば、サザンおよびノーザンブロット法で用いるハイブリダイゼーションプローブとして、特に意図される。連続した相補的領域の長さが約10〜14から約100または200ヌクレオチドまでのように可変であるより小さな断片は、一般に、ハイブリダイゼーション実施形態において有用であるが、検出が望まれる相補配列の長さに応じて、より大きな連続した相補的ストレッチを使用してもよい。
【0112】
約14ヌクレオチド長のハイブリダイゼーションプローブを使用することで安定した選択できる二本鎖分子が形成される。しかしながら、ハイブリッドの安定性と選択性を高めることによって、得られる特異的なハイブリッド分子の品質と程度を向上させるためには、14塩基長を上回るストレッチに対する連続した相補配列を有する分子が一般に好ましい。一般的には、15〜20個の連続したヌクレオチド、要すれば、より長いヌクレオチドの遺伝子相補的なストレッチを有する核酸分子を設計することが好ましい。
【0113】
当然のことながら、例えば、機械的剪断または制限酵素消化によるなどの他の技術によっても断片を得ることができる。小さな核酸セグメントまたは断片は、例えば、自動オリゴヌクレオチド合成装置を使用して一般に実施されているように、化学的手段によって断片を直接合成することによって、容易に調製され得る。また、米国特許第4,683,195号および第4,683,202号(それぞれ引用することにより本明細書の一部とされる)のPCR.TM.法のような核酸複製技術を適用することにより断片を得てもよいし、選択した配列を組換え生産用組換えベクターに導入することにより断片を得てもよいし、分子生物学分野における通常の技術を有する人には一般に知られている他の組換えDNA技術により断片を得てもよい。
【0114】
よって、DNA断片の相補的ストレッチと選択的に二本鎖分子を形成させるために本発明の核酸配列を使用することができる。想定される用途に応じて、ハイブリダイゼーション条件を変え、標的配列に向けるプローブの選択性の程度を変えることが望ましい。高い選択性を必要とする用途では、一般に、比較的ストリンジェントな条件を使用して、ハイブリッドを形成させることが望ましい。例えば、約0.02M〜約0.15M NaCl、約50℃〜約70℃の温度にて提供されるような比較的低塩および/または高温条件を選択する。このような選択条件は、プローブと鋳型または標的鎖間のミスマッチがもしあるとしてもほとんど許容しないため、2APポリペプチドをコードするDNAセグメントまたはタンパク質をコードするDNAセグメントの単離に特に好適であろう。ハイブリダイゼーションを介したDNAセグメントの検出は、当業者には周知のことであるが、米国特許第4,965,188号および第5,176,995号(それぞれ引用することにより本明細書の一部とされる)の教示がハイブリダイゼーション解析の典型的な方法である。Maloy et al., 1994; Segal 1976; Prokop, 1991;およびKuby, 1994の本文中に記載されている内容のような教示が特に適切である。
【0115】
当然のことながら、例えば、対象鋳型とハイブリダイズする突然変異型プライマー鎖を使用して、突然変異体を調製することが望ましく、また、関連種、機能的同等物などから2APタンパク質をコードする配列を単離しようとする一部の用途では、ヘテロ二本鎖を形成させるために、一般に、より低いストリンジェントなハイブリダイゼーション条件が必要である。これらの状況においては、約0.15M〜約0.9M塩、約20℃〜約55℃の範囲の温度のような条件を使用することが望まれる。制御ハイブリダイゼーションでは、クロスハイブリダイズしている種をハイブリダイズししているシグナルとして容易に確認することができる。どんな場合でも、温度上昇と同様に、ハイブリッド二本鎖を不安定にする働きをする漸増量のホルムアミドの添加によってより高いストリンジェントな条件にすることができることは一般に知られている。このように、ハイブリダイゼーション条件は容易に操作し得るものであることから、一般には、所望する結果に応じて方法が選択できる。
【0116】
特定の実施形態においては、本発明の核酸配列をハイブリダイゼーションを判定するための標識のような好適な手段と組み合わせて使用することが有利である。多種多様な好適な表示手段が当技術分野で公知であり、これらの手段としては、検出可能なシグナルを与え得る、アビジン/ビオチンのような、蛍光、放射性、酵素リガンドまたは他のリガンドが挙げられる。好ましい実施形態において、放射性試薬または他の環境的に望ましくない試薬の代わりに蛍光標識またはウレアーゼ、アルカリ性ホスファターゼもしくはペルオキシダーゼのような酵素タグを使用することが望ましいと思われる。酵素タグの場合、熱量表示基質が知られており、それを使用して、人間の眼でまたは分光光度法で確認できる手段を提供し、相補的核酸を含有するサンプルとの特異的ハイブリダイゼーションを確認することができる。
【0117】
一般に、本明細書において記載するハイブリダイゼーションプローブは、溶液ハイブリダイゼーションにおける試薬としても、固相を使用する実施形態においても有用であると想定される。固相に関連する実施形態では、試験DNA(またはRNA)を、選択したマトリックスまたは表面に吸着させるか、そうでなければ付着させる。この固定された一本鎖核酸を、続いて、所望の条件下で、選択したプローブとの特異的ハイブリダイゼーションに付す。選択する条件は、必要な特定の判定基準に基づいた特定の状況によって変わる(例えば、G+C含量、標的核酸のタイプ、核酸の起源、ハイブリダイゼーションプローブの大きさ他に応じる)。非特異的に結合しているプローブ分子を除去するためにハイブリダイズした表面を洗浄した後、特異的ハイブリダイゼーションを検出する。場合によっては、標識を用いて定量する。
【0118】
また、増幅技術を用いて、核酸サンプルから目的の核酸を増幅することができる。例えば、ポリメラーゼ連鎖反応(PCR)技術を使用して、2AP遺伝子の配列を、mRNAから、cDNAから、ゲノムライブラリーまたはcDNAライブラリーから直接増幅することができる。PCRおよび他のin vitro増幅法は、例えば、発現させようとするタンパク質をコードする核酸配列をクローニングするのに、サンプル中の所望のmRNAの存在を検出する目的で、核酸配列を決定する目的で、または他の目的でプローブとして使用する核酸を作製するのにも有用であり得る。
【0119】
植物組織由来の2AP遺伝子配列を同定するのに好適なプライマーおよびプローブは、本明細書において提供する配列の比較により得られる。PCRの一般概要に関しては、引用することにより本明細書の一部とされるPCRプロトコール:A Guide to Methods and Applications. (Innis, M, Gelfand, D., Sninsky, J. and White, T., eds.), Academic Press, San Diego (1990)参照。
【0120】
技術文献に記載されている周知の技術によりポリヌクレオチドも合成し得る。例えば、Carruthers et al., Cold Spring Harbor Symp. Quant. Biol. 47: 411-418 (1982)およびAdams et al., J. Am. Chem. Soc. 105: 661 (1983)参照。さらに、相補鎖を合成し、好適な条件下でそれらの鎖を一緒にアニーリングすることによるか、または好適なプライマー配列とともにDNAポリメラーゼを用いて相補鎖を合成することにより二本鎖DNA断片が得られる。
【0121】
B.2AP遺伝子オーソログを単離する生物:トウモロコシ属(Zea)、カラスムギ属(Avena)、オオムギ属(Hordeum)、ライムギ属(Secale)、コムギ属(Triticum)およびモロコシ属(Sorghum)の種を含むいろいろな種類の植物および真菌を使用することができる。2−アセチル−1−ピロリンは、あらゆる香米、小麦およびライ麦パン(Buttery et al., 1982, 1983)、含水雑穀(Seitz et al., 1993)、ポップコーン(Schieberle et al., 1991)、バチルス・セレウス(Romanczyk et al., 1995)ならびにアスペルギルス・オリザエ(Aspergillus oryzae)、アスペルギルス・アワモリ(Aspergillus awamori)およびスポロボルス・ビルギニカス(Sporobolus virginicus)のような一部の真菌種(Nagsuk et al., 2004)の主要な強い香気化合物であることが分かっている。ゲノムレベルでの穀物間でのシンテニー、さらに、植物および微生物においてよく保存されたプロリンおよびポリアミン代謝経路を示すいくつかの報告がある(Kyoto Encyclopedia of Genes and Genomes, Kyoto University, Bioinformatics Center, Institute for Chemical Researchのホームページにてオンラインで見つけられる)。そのため、2AP遺伝子のオーソログは、穀物用植物間で同様に作用するかもしれず、本発明にて示す教示によって、イネと同様に他の植物においても化合物2−アセチル−1−ピロリンの蓄積が増強する可能性がある。
II.植物における2−アセチル−1−ピロリンレベルを上昇させるための2AP遺伝子発現または2APタンパク質レベルもしくは活性の低減
【0122】
A.突然変異誘発:EMS(メタンスルホン酸、エチルエステル)のような化学的突然変異誘発物質ならびにγ線および高速中性子照射によってDNAに突然変異が生じる可能性がある。突然変異のいくつかの例が、欠失、挿入およびミスセンス突然変異である。突然変異後、スクリーニングを行って、早期停止コドンを与えるか、そうでなければ機能しない2AP遺伝子を生じる欠失を確認することができる。配列決定によるか、または本発明によって提供される、2AP遺伝子またはタンパク質に特異的なプローブもしくはプライマーを使用することにより突然変異体のスクリーニングを行うことができる。2AP遺伝子における特異的突然変異は、TILLING(Targeted Induced Local Lesion of Genome)(Till et al., 2003)およびtDNA挿入によっても起こし得る。このような突然変異の結果として、2AP遺伝子発現の低下、2AP mRNAの安定性の低下または2APタンパク質の活性および/もしくは安定性の低下が起こり得る。本明細書において定義するような植物は、天然に存在するものではない。
【0123】
B.アンチセンス技術:アンチセンス技術は、2AP mRNAのような標的mRNAの翻訳を妨げる。そのようなものとして、アンチセンス技術は、一般に、標的タンパク質、本明細書では2APタンパク質のレベルを低減する。2AP遺伝子またはその断片は、植物にアンチセンス方向で導入することができる。断片は、18ヌクレオチドといった小さなものであってもよいし、3,000ヌクレオチドといった大きなものであってもよいし、あるいはそれより大きくてもよい。2AP遺伝子のcDNA断片は、ベクター(例えば、pCAMBIA1302)に天然遺伝子とは逆方向にクローニングすることができる。逆転写物は、天然の2AP遺伝子転写物を含むヘテロ二本鎖構造を形成しているが、その後、その遺伝子転写物は翻訳前に分解される。センスおよびアンチセンス配列は同一である必要はなく、標的遺伝子発現の抑制を実現するには部分的な相同性で十分である。従って、イネOs2AP由来の配列(配列番号1、配列番号2、配列番号4および配列番号5)を使用して、他の植物由来の2AP遺伝子オーソログの配列が分からなくとも、その植物における2AP遺伝子発現を抑制することができる。
【0124】
C.RNA干渉:RNA干渉(RNAi)は、標的mRNAの排除に関連するもう1つの技術である。RNAi用のベクターの構築は様々であるが、基本的にはRNAにおいてヘアピンループ構造の形成が起こる(Horiguchi 2004)。実施例2では、そのcDNAとは逆方向にエクソン6、7および8を含むOs2AP核酸配列の断片をベクターにクローニングして、mRNAにおいて逆ヘアピン構造をもたらした。これらのヘアピン構造はDicerと呼ばれるエンドヌクレアーゼによって小断片に切断される。このDicerの機能は、間違った転写産物が翻訳されないようにすることである(Hamilton and Baulcombe, 1999, Matzke et al., 2001)。mRNAの小断片は、一般に、約20〜21ヌクレオチドの大きさであり、siRNAまたは小干渉RNAと呼ばれている。より大きな断片もより小さな断片も使用することができる。これらのsiRNAは、相同遺伝子の発現をダウンレギュレートし得る。この場合もまた、相同性が完全である必要はないため、本発明において詳述するイネ由来の配列を使用して、他の植物のRNAi構築物を作製することができる。実施例2は、非香米系統におけるOs2AP遺伝子を標的としたRNAi試験によって、いかにして、香米に見られるレベルまでに高められた2−アセチル−1−ピロリンレベルおよび芳香が得られるのかを示している。
III.トランスジェニック植物の作製
【0125】
これまでの技術において単離された2AP遺伝子配列を使用するために、植物細胞の形質転換に好適な組換えDNAベクターを調製することができる。多種多様な高等植物種を形質転換する技術はよく知られたものであり、技術文献および科学文献に記載されている。例えば、Weising et al. Ann. Rev. Genet. 22: 421-477 (1988)参照。
【0126】
このようなDNA構築物を、種々の従来の技術により所望の植物宿主のゲノムに導入し得る。例えば、DNA構築物を、植物細胞プロトプラストまたは胚発生カルスのエレクトロポレーション、PEGポレーション、粒子衝撃およびマイクロインジェクションのような技術を用いて植物細胞のゲノムDNAに直接導入してもよいし、またはDNA構築物を、DNA粒子衝撃のような弾道学的方法を用いて植物組織に直接導入することもできる。あるいは、DNA構築物を好適なTDNAフランキング領域と組み合わせ、一般的なアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)宿主ベクターに導入してもよい。アグロバクテリウム・ツメファシエンス宿主の毒性機能が、細胞が細菌類に感染したときに、構築物と隣接するマーカーの植物細胞DNAへの挿入を導く。
【0127】
マイクロインジェクション技術は、当技術分野では公知のものであり、科学文献および特許文献で詳しく記載されている。ポリエチレングリコール沈降を用いたDNA構築物の導入は、Paszkowski et al. Embo J. 3: 2717-2722 (1984)で記載されている。エレクトロポレーション技術は、Fromm et al. Proc. Natl. Acad. Sci. USA 82: 5824 (1985)で記載されている。弾道学的形質転換技術は、Klein et al. Nature 327: 70-73 (1987)で記載されている。数多くのアプローチを用いて、ライムギ(de la Pena et al., Nature 325: 274-276 (1987))、トウモロコシ(Rhodes et al., Science 240: 204-207 (1988))およびイネ(エレクトロポレーションによるShimamoto et al., Nature 338: 274-276 (1989); 弾道学的技術によるLi et al. Plant Cell Rep. 12: 250-255 (1993))のような穀物種を形質転換することができる。
【0128】
アグロバクテリウム・ツメファシエンスを介した形質転換技術は、科学文献で詳しく記載されている。例えば、Horsch et al. Science 233: 496-498 (1984)およびFraley et al. Proc. Natl. Acad. Sci. USA 80: 4803 (1983)参照。アグロバクテリウムは、主として、双子葉植物において有用であるが、アグロバクテリウムによって形質転換することができる単子葉植物もある。例えば、イネのアグロバクテリウム形質転換は、Hiei et al., Plant J. 6: 271-282 (1994)により記載されている。
【0129】
上記形質転換技術のいずれかによって誘導された形質転換植物細胞を培養して、形質転換された遺伝子型を有し、その結果としてより高いレベルの2−アセチル−1−ピロリンと増強された芳香を有する完全な植物体を再分化させることができる。このような再分化技術は、一般に、2APヌクレオチド配列とともに導入された殺生物剤および/または除草剤マーカーに依存して、組織培養増殖培地における特定の植物ホルモンの操作によって行うものである。培養プロトプラストからの植物再分化は、Evans et al., Protoplasts Isolation and Culture, Handbook of Plant Cell Culture, pp. 124-176, MacMillilan Publishing Company, New York, 1983;およびBinding, Regeneration of Plants, Plant Protoplasts, pp. 21-73, CRC Press, Boca Raton, 1985で記載されている。植物カルス、外植片、器官またはその一部からも再分化が可能である。このような再分化技術は、一般に、Klee et al. Ann. Rev. of Plant Phys. 38: 467-486 (1987)で記載されている。
【0130】
熟練者ならば、発現カセットがトランスジェニック植物に安定して組み込まれ、機能しうることが確認された後、それが雌雄交配により他の植物に導入することができることが分かるであろう。交配しようとする種に応じ、数多くの標準的な繁殖方法のいずれを使用してもよい。
IV.2−アセチル−1−ピロリンレベルについての試験
【0131】
A.官能試験:乾燥種子、調理した米またはすりつぶした葉の揮発性芳香は、感覚によって識別することができる。(Dhulappanavar, 1976, Ghose et al., 1952, Kadam et al., 1938).このような感覚識別法としては、味覚試験が挙げられる。米育種家の間で一般に実施されているもう1つのものが、水中で葉組織を加熱した後、希釈KOH溶液を添加する方法である(Sood and Siddiq, 1978)。これらの試験は必ずしも一貫性と信頼性があるとが限らず、人的エラーが起こりやすく、偏りがちである。
【0132】
B.クロマトグラフィー:調理した米100グラムから揮発性化合物を定量するためのガスクロマトグラフィーを使用したより信頼性のある方法を開発した(Petrov et al., 1995)。最近、たった1グラムの米から1ppb(10億分の1)といった低い2−アセチル−1−ピロリンレベルを分析するために、ガスクロマトグラフィー/質量分析(GCMS)法が開発された(Mahatheeranont et al., 1995)。そのようなものとして、GCMSを使用して、天然および非天然植物における2−アセチル−1−ピロリンのレベルを調べることができる。図1参照。
【実施例】
【0133】
好ましい実施形態の説明
実施例1:米の芳香を制御する遺伝子のポジショナルクローニング
米の芳香特性は、これまでに、定性的芳香測定法と定量的芳香測定法の両方に基づいて染色体8にマッピングされている(Lorieux et al. 1996)。芳香遺伝子は、RG28から4.5cM離れた位置にマッピングされ(Ahn et al. 1992)、RG28とRG1の間、12cM内にマッピングされている(Lorieux et al. 1996)。最近、公開イネゲノム配列から得られた一塩基変異多型(SNP)マーカーRSP04が芳香遺伝子から2cM離れた位置にマッピングされた(Jin et al. 2003)。
【0134】
単一QTLは、染色体8のRFLPマーカーRG1およびRG28によってフランキングされる4.5cM領域に位置していた(Lorieux et al., 1996; Lanceras et al. 2000)。KDML105由来のEcoRI BAGライブラリーは、17の一倍体ゲノムを示す75,264クローンで構成されたものであった。25BAGクローンからなる開始BAGコンティグを、領域内のマッピングされている6個のマーカーを用いることにより選択し(Wanchana et al., 2005)、HindIIIフィンガープリンティングによりさらに絞り込んだ。2−アセチル−1−ピロリンの蓄積に関与する遺伝子を同定するため、重要な領域を以下の戦略により狭めた。
【0135】
最初に、重要な領域内に新規組換え体を生み出すための起源として単一のF6植物を選択した(そのF7において穀物の芳香について分離した)。F7〜F1から、各世代に対して、穀物の芳香とマーカー遺伝子型に基づいてヘテロ接合体植物を選択した。F10世代では、374個体のF10植物をKDML105BAG末端配列から得られた18個の多型マーカーを用いてスクリーニングすることにより重要な領域が380kbに狭められた。274個体のF11植物をスクリーニングすることによりその領域を120kbにさらに絞った(図2)。図式遺伝子型決定および2−アセチル−1−ピロリン量に基づいて、8のF11植物を選択して、芳香性、非芳香性およびヘテロ接合体同質遺伝子系統を作製した(図2)。この段階で、3つのKDML BAGクローン、155L11、68L13および167M23をショットガン配列決定用に選択した。KDML由来の配列と対応するニホンバレゲノムコンティグのアラインメントにより、F12世代におけるさらなるスクリーニングに用いられる21個の挿入/欠失(「indel」)マーカーが同定された。1,116個体のF12植物を使用することにより、標的領域を58kb BAG,167M23内の27kbに狭めた(図2)。この交配では、これ以上の組換えは確認されなかった。米粒子において2−アセチル−1−ピロリン蓄積に違いがはっきりと示される数系統の同質遺伝子系統が作出された(図6)。
【0136】
配列解析では、BAC 167M23が19個の遺伝子を含有していることが分かった。しかしながら、10個の遺伝子だけが既知タンパク質またはコーディング配列との類似性を有するものであった。27kb領域内では、3個の候補遺伝子が同定された:3−メチルクロトニル−CoAカルボキシラーゼ(MCCアーゼ)、仮定的遺伝子および未知タンパク質。KDML105とJaoHom Nin(JHN)、非芳香性黒色米の交配による177のF6植物を用いて、穀物の芳香および2−アセチル−1−ピロリンレベルに影響を及ぼす未知タンパク質のエクソン7内に組換え部位が確認された。BAG 167M23とBAG 68L13両方のこれらの7個の候補遺伝子の発現を調べるため、2−アセチル−1−ピロリンが米粒子に蓄積する登熟期間中にRT−PCRを行った。芳香性および非芳香性同質遺伝子系統の受粉の10、15および20日後にイネの穂から全RNAを採取した。NBS/LRR遺伝子の発現は検出されなかった。ほとんどの候補遺伝子については、芳香性および非芳香性同質遺伝子系統間での発現差は示されなかった。Os2APの場合においてのみ、芳香性同質遺伝子系統で受粉の15日後に遺伝子発現が急激に低下した(図3A)。Os2AP遺伝子発現の低下は、葉、茎および根においても顕著であった(図3B)。そのため、発現研究とポジショナルクローニングの両方により、穀物の芳香の蓄積および2−アセチル−1−ピロリンのin vivo合成に関与するレギュレーターとしてOs2APを確認する。
【0137】
これまでに、研究者によって穀物の芳香を制御する1つの劣性核内遺伝子が報告されている(Berner and Hoff, 1986; Yanjuan et al., 1992; Ali et al., 1993)。このことは、本明細書において報告する研究結果と一致する。2−アセチル−1−ピロリンの蓄積を、芳香性および非芳香性同質遺伝子系統とF1植物で比較した。これらの結果からは、F1葉では2−アセチル−1−ピロリンが検出されなかったことが示された。F2世代で分離が起こったことから、2−アセチル−1−ピロリンレベルは、その供与体である親に見られるレベルの4分の1であった(図3C)。その結果、古典的遺伝学および分子遺伝学の両方から、エクソン7における突然変異が植物における2−アセチル−1−ピロリン蓄積を調節する分子機構であることが裏付けられた。
【0138】
Os2AP遺伝子の構造を、KDML105、芳香性同質遺伝子系統、非芳香性同質遺伝子系統およびニホンバレ間で比較した。5.8kbのOs2AP遺伝子は、15エクソンで構成されており、いくつかの同義的な突然変異がエクソン2に見られた(図5A)。KDML105および芳香性同質遺伝子系統では、2つの重要な突然変異事象がエクソン7で確認された。まず、2つの過渡的な突然変異が730位(AからTへ)と732位(TからAへ)に見られ、続いて、8塩基対の欠失,GATTAGGC,が734位から開始していた。8同質遺伝子系統間での2−アセチル−1−ピロリンレベルの解析により、8塩基対の欠失がコメ粒子における2−アセチル−1−ピロリンの蓄積と関係していることが示された(図6A)。この突然変異は、729位からフレームシフト翻訳開始させ、753位から開始する早期停止コドンを与えた(図5B)。ニホンバレでは、Os2AP全長cDNAは503個のアミノ酸に翻訳される。欠失により252個のアミノ酸の末端切断型ペプチドが作出された(図5B)。KDML105とJHNから誘導された177のF6植物では、8塩基対の欠失にフランキングする二重組換えによって、米粒子において2−アセチル−1−ピロリンを生産できないことが説明される。
【0139】
この早期停止コドンは、Os2APの発現に大きな影響を与え得る。早期翻訳停止コドンを含む大部分のmRNAは、ほとんどの場合翻訳されない。その結果として、それらは、遺伝子発現におけるエラーを減少させるという働きをする監視システムとしてナンセンス変異を介したmRNAの分解(NMD)を引き起こすことが多い(Pulak and Anderson, 1993)。この現象によって、香米においてOs2AP遺伝子発現のレベルが低いことが説明される。
【0140】
2APの直接の前駆体は、まだ確認されていない。しかしながら、Os2APのピロリン環の前駆体は、同位体希釈により決定されたようにアミノ酸プロリンである(Yoshihashi et al., 2002)。また、1−ピロリンおよびピロリン−2−カルボン酸(P2C)のようなピロリン含有代謝産物が直接の前駆体としても供給される可能性もあるかもしれない。穀物の芳香の強さを増強するための1つの方法が、より多くの前駆体または中間体が蓄積され、粒子成長期間に2APへの特異的な変換により多く利用できるようになるように、関連生合成経路に関与する酵素の発現を操作することである。例としては、酵素P5Cシンターゼ、P5Cレダクターゼの発現を増強すると同時にプロリンおよびP5Cデヒドロゲナーゼを抑制することがある。ポリアミン生合成経路におけるいくつかの酵素を増強して、米粒の成長時により多くの1−ピロリンを生じさせることも可能である。代謝工学アプローチに加え、米において利用できる胚乳特異的なプロモーターを用いることにより、米胚乳における酵素発現を増強してもよい。これらの技術によって、精米においてより芳香性の高い米系統を生み出すことができる。
【0141】
実施例2:Os2AP遺伝子を用いた植物形質転換:
Os2AP遺伝子をRNAi構築物に組み込み、それを用いてOs2AP遺伝子発現をダウンレギュレートし、イネにおける2−アセチル−1−ピロリンレベルを高めた。
【0142】
Os2AP−ihpRNA含有ベクターの構築
Os2AP遺伝子のセンス−アンチセンス断片の発現用にpCAMBIA1302ベクターを使用した。KDML105由来のゲノムDNAおよび全RNAそれぞれを鋳型として用い、PCRによりセンスおよびアンチセンス断片を作製した。エクソン6〜9に対応するゲノムDNA配列を含有するセンス断片を、NcoIおよびSpeI制限部位(下線)を有するプライマー(順方向:AATTCCATGGGGTTGGTCTTCCTTCAGGTG(配列番号80);逆方向:AATTACTAGTTTCCACCAAGTTCCAGTGAA(配列番号81))を用いて増幅した。エクソン6〜8に対応するcDNA配列を含有するアンチセンス断片を、NcoIおよびNheI制限部位を有するプライマー(順方向:AATTCCATGGGGTTGGTCTTCCTTCAGGTG(配列番号80);逆方向:AATTGCTAGCGGTCCAAAAGCAACCAAAGA(配列番号82)を用いて増幅した。PCR産物をまず、SpeIおよびNheIで消化し、その後、T4 DNAリガーゼを用いて連結した。次いで、連結した断片をNcoIで消化した。精製した連結断片をpCAMBIA1302ベクターにNcoIクローニング部位にてクローニングした(図7A)。
【0143】
イネ形質転換
非香米品種(オリザ・サティバ L.日本型品種ニホンバレ)の胚発生カルスを粒子衝撃形質転換の標的組織として使用した(Nimlek, 1999)。ハイグロマイシン耐性カルスを、プライマーOs2AP−エクソン7.1−欠失_FおよびR(U:5'-TGCTCCTTTGTCATCACACC-3'(配列番号54)およびL:5'-TTTCCACCAAGTTCCAGTGA-3'(配列番号55))とGFPのプライマー(U:5'-CTTGTTGAATTAGATGGTGATGTT-3'(配列番号83)およびL:5'GTTGTGGGAGTTGTAGTTGTATTC-3'(配列番号84))を用いてPCRによりスクリーニングした。
【0144】
サザンブロット解析
トランスジェニック植物(R0)および対照(ニホンバレ)植物の葉から全DNAを単離した。Os2AP−ihpRNA断片の検出用にゲノムDNA(10mg)をNcoIで消化した。陽性対照として、プラスミドOs2AP−RNAiから単離されたDNAをNcoIで消化した。0.8%アガロースゲルでの電気泳動後、DNAをHybond−N+ナイロンメンブレン(Southern 1975)に移した。プローブとのハイブリダイゼーションを製造業者(Amersham)の使用説明書に従って実施した。(α−32P)−dCTPを用いてランダムプライマー法により放射性プローブを調製した。プローブは、Os2AP遺伝子のコード領域で構成されていた(pOs2AP−RNAi,210ヌクレオチドのSpeI−NcoI断片)。
【0145】
RT−PCR
異なる成長段階における転写レベルと異なる組織における転写レベルを調べるため、ニホンバレおよびトランスジェニック植物の若い植物(10日)、成葉(30日)および根(14日)および開花期の穂から全RNAを抽出し、RT−PCR解析に使用した。染色体8のOs2AP遺伝子の芳香遺伝子座を、Os2AP−エクソン7.1プライマー(U:5'-TGCTCCTTTGTCATCACACC-3'(配列番号54)およびL:5'-TTTCCACCAAGTTCCAGTGA-3'(配列番号55))を用いてPCRにより増幅した。BADH(Os2AP−chr4;Genbank受託番号AB001348)とも呼ばれる染色体4のOs2AP遺伝子のホモログを、Os2APch4プライマー(U:5'-TAGCTTCACATCCCCATGTG-3'(配列番号85)およびL:5'-GCACCTTCACATCTTGCTGT-3'(配列番号86)を用いてPCRにより増幅した。対照として、イネアクチン遺伝子(Genbank受託番号:X16280)を、アクチンU:5'-ACATCGCCCTGGACTATGAC-3'(配列番号87)およびアクチンL:5'-TGCTGAGAGATGCCAAGA TG-3'(配列番号88)を用いて増幅した。
【0146】
リアルタイム定量的PCR
Taqman chemistryおよびABI PRISM 7700配列検出システム(Applied Biosystems, Foster City, CA, USA)を利用することにより、製造業者の使用説明書に従ってリアルタイム定量的PCRを実施した。Os2APのコピー数とイネアクチンのものの比として相対量を算出した。Os2AP遺伝子の芳香性対立遺伝子と非芳香性対立遺伝子を区別するようにTaqmanプローブを設計した。
【0147】
結果
4つのハイグロマイシン−GFP陽性小植物が再分化された。トランスジェニックイネにおけるGFPの発現は様々であった。T0RNAi2(10)、GFPの最高の発現が示される系統、の内在性Os2APの発現は最も低い(図7B)。2−アセチル−1−ピロリン含量の解析とKOH法を用いた官能試験には、十分な相関関係があった。T0RNAi2(10)は、最大量の2−アセチル−1−ピロリンを含有しており、芳香性葉が最も多かった。干渉配列の発現量が少ないT0RNAi5(20)とT0RNAi3(30)は、2−アセチル−1−ピロリンの蓄積が少なく、芳香性葉スコアが低かった(図7B)。T1RNAi2(10)の穀物の芳香の分離比分析によりこの結果を確認した。その結果、RNA干渉によるOs2AP遺伝子発現の抑制により、イネにおける2−アセチル−1−ピロリンの蓄積が増加する。このことにより、8塩基対の欠失によって起こる香米におけるNMDによるOs2AP遺伝子発現の自然抑制が香米の芳香の直接原因であることが示唆される。
【0148】
実施例3:イネのOs2AP/BADH遺伝子座の特性決定
Os2APは、これまでに染色体4で見つけられたBADH(ベタインアルデヒドデヒドロゲナーゼ)のホモログである(Nagamura et al., 1997)。本発明者らは、Genbank配列受託番号AB001348とAP004463の、それぞれ、染色体4のBADH遺伝子と染色体8のOs2AP遺伝子のゲノム配列を特徴付けた。イネGAASの遺伝子注釈結果に従って(イネゲノム自動注釈システム, Sakata et al., 2002)、BADHおよびOs2AP両方の構造遺伝子は、14イントロンによって分断された15エクソンを含んでなる。BADHおよびOs2APそれぞれのORFは、505および503アミノ酸残基のタンパク質をコードする。両タンパク質は、アミノ酸レベルでは77%同一であり、他の植物BADHタンパク質と88%〜99%同一性を共有している。さらに、これらの2つのタンパク質は、一般アルデヒドデヒドロゲナーゼに高度に保存されているデカペプチドVal-Thr-Leu-Glu-Leu-Gly-Gly-Lys-Ser-Pro(配列番号96)をコードするヌクレオチド配列を含有している(Weretilnyk et al. 1990)。タンパク質配列のPfam解析によれば、BADHもOs2APもアルデヒドデヒドロゲナーゼファミリーに属する。このファミリーに属するタンパク質は、特定植物種におけるグリシンベタインの合成に関して推定機能を有する。しかしながら、イネ、トウモロコシおよびコムギのようなグリシンベタインを蓄積しない穀物でのこれらのタンパク質の機能は分かっていない。本発明者らは、プロリンが2−アセチル−1−ピロリンの前駆体として報告されていることから、プロリン異化において機能するピロリン−5−カルボン酸デヒドロゲナーゼ(P5CDH)(Genbank受託番号P30038)もこのファミリーに含まれることに驚いた。Os2APの機能がベタインアルデヒド以外のアルデヒドの脱水素化を触媒することである可能性がある。
【0149】
Os2AP遺伝子発現および米の芳香:両Os2AP遺伝子が芳香生合成に関連しているかどうかを確認するため、芳香性および非芳香性同質遺伝子系統(それらのゲノム背景は芳香遺伝子領域を除いて同一であった)の発現レベルを解析した。結果として、本発明者らは、BADHは両同質遺伝子系統間での発現差は示さないが、一方、Os2APは芳香性同質遺伝子系統の総ての組織において遺伝子抑制を一貫して示すことが分かった(図3D)。本発明者らはさらに、同質遺伝子系統ならびにニホンバレおよびK105(芳香性系統)における両タンパク質の発現についても研究した。Os2AP遺伝子発現は、芳香性同質遺伝子系統およびKDMLL105においてのみダウンレギュレートされた。これに対し、非芳香性同質遺伝子系統およびニホンバレ米(非芳香性系統)においては、Os2AP遺伝子発現のダウンレギュレーションは検出されなかった。
【0150】
実施例4:マーカーを利用した選択
通常の異種交配により新規香米を開発するために、香米供与体とより生産性の高い受容体米品種と交配してもよい。その交配による後代は、数ある形質の中で特に穀物の芳香について分離する。この状況は育種家を混乱させ、より優れた香米についての通常の育種が成功しにくくなる。芳香遺伝子の発見は、従来の育種植物の新しいパラダイムとなる。何千もの植物を最高の精度でスクリーニングし得るように、特異的な分子マーカーを芳香遺伝子を検出するために開発することができる。育種家は、DNAマーカー法によって、Os2AP遺伝子の芳香性対立遺伝子を早い段階で高感度にて検出し得る。このことにより、育種家が後の段階で芳香性植物に好ましい形質を与えるためのより多くの機会が残される。イネにおいて確認される芳香の分子基盤は、おそらく他の穀物でも見つけられるため、他の穀物のDNAマーカーを開発する方法も開かれる。
【0151】
実施例5:Os2AP遺伝子の系統発生解析
プロリンおよびポリアミン生合成は、植物界において高度に保存された経路である。本明細書において提示する2−アセチル−1−ピロリンの生合成は、他の2−アセチル−1−ピロリン産生植物が利用する共通のテーマである。この点について例証するために、マルチプル配列アラインメントを利用して2AP遺伝子のアミノ酸配列を比較した。結果として得られた系統樹により、イネ、コムギおよびオオムギ由来の2AP遺伝子が共通の祖先を有し得ることが分かる(図9B)。それゆえ、穀物における2−アセチル−1−ピロリンの生合成が共通のテーマを有している可能性がある。この点について例証するためには、将来、他の穀物において2AP遺伝子オーソログを用いた試験を行う必要がある。
【0152】
Os2AP遺伝子がどこから来たのかを遡るのは興味深い。「アロマーカー」PCRプライマーを用いて95の在来品種を細かく調べることにより、本発明者らは8塩基対の欠失を有する対立遺伝子が芳香の増強と厳密に相関していることが分かった。また、本発明者らは、オリザ・ニバラ(Oryza nivara)およびO.ルフィポゴン(O. rufipogon)をはじめとする野生種間で8塩基対の欠失に関連するエクソン7を配列決定した(図9A)。本発明者らは、タイジャスミン(Thai Jasmine)、バスマティ(Basmati)およびアズセナ(Azucena)をはじめとする最も芳香性の高い品種がその欠失を有しており、それゆえ、古代にまで遡り得る共通の祖先を有することが分かった。本発明者らは、芳香性野生米においても芳香性対立遺伝子を確認した。よって、本発明者らが今日知っている香米は、人間が栽培するようになるずっと以前に、単一突然変異が起こって生み出されたのである。
【参照文献】
【0153】
以下の参照文献は、それらの全開示内容が引用することにより本明細書の一部とされる。
【図面の簡単な説明】
【0154】
【図1】カオ・ド・マリ(KDML105)、タイジャスミン米(芳香性)およびコシヒカリ、非香米から、GC−MSにより得られた2−アセチル−1−ピロリンの化学構造。
【図2】穀物の芳香について分離する、単一F6植物由来の186のF9植物を用いたイネにおける芳香遺伝子の詳細なマッピングと穀物の芳香に関する遺伝子を含む物理的地図の構築。第1の部分は、単一F6植物由来の8のF11植物の図式遺伝子型を示す。第2の部分は、単一BACにおいて重要な領域を27kbに絞るための、単一F6植物由来の1116のF12植物を用いたウルトラスケールマッピングを示す。第3の部分は、3つのオープンリーディングフレームを示すKDML105のゲノム配列の注釈を示す。第4の部分は、KDML105とJHNの交配による177のF6植物を用いて、穀物の芳香および2−アセチル−1−ピロリン含量に大きな影響を及ぼす未知タンパク質遺伝子のエクソン7内に3つの二重組換え体が確認されることを示す。この未知タンパク質遺伝子は、Os2APと名づけられた。「アロマーカー」は、穀物の芳香に特有の8塩基対の欠失と3つのSNP(一塩基変異多型)を定義するPCRベースのマーカーである。
【図3A】芳香性および非芳香性同質遺伝子系統の受粉の10、15および20日後に単離された全RNAからの、RT−PCRを用いた7つの候補遺伝子の発現。この図において、Os2APは、ベタインアルデヒドデヒドロゲナーゼと見なされる。
【図3B】芳香性および非芳香性同質遺伝子系統の受粉の15日後に単離された全RNAからのOs2AP転写産物の葉、茎および根における示差的発現。
【図3C】THM(KDML105)、芳香性同質遺伝子系統117、非芳香性同質遺伝子系統10およびそれらのF2(ISL117×ISL10)の穀粒における2−アセチル−1−ピロリンレベルの解析。F1では、2−アセチル−1−ピロリンレベルの解析を葉で実施した。
【図3D】上のパネル:Os2AP RNAi構築物を含有するトランスジェニックニホンバレにおける染色体4のOs2APホモログとアクチンの発現。下のパネル:染色体4ではなく染色体8のOs2AP遺伝子(本明細書ではBADHと呼ぶ)が、芳香性および非芳香性同質遺伝子系統のイネにおいて示差的発現を示している。
【図4A】プロリンおよびグルタミン酸間の代謝経路。L−プロリンは、酵素P5CシンターゼおよびP5Cレダクターゼを用いてグルタミン酸から合成され、グルタミン酸は、プロリンデヒドロゲナーゼ(ProDH)およびP5Cデヒドロゲナーゼ(P5CDH)を用いてプロリンから合成される。提示された代謝シフトは、Os2APのナンセンス突然変異が介在するものであった。
【図4B】Os2AP酵素の推定タンパク質構造を示す。
【図5A−1】KDML105(芳香性)およびニホンバレ(非芳香性)由来のOs2AP遺伝子のゲノム配列比較。
【図5A−2】図5Aの続き。
【図5B】ニホンバレ、非芳香性系統と比べて、芳香性系統、タイ・ホーム・マリ(THM)における8塩基対の欠失を示すDNAアラインメントおよび2系統間のアミノ酸配列比較。
【図6】単一F6植物由来の8のF11植物の図式遺伝子型と米粒子の2−アセチル−1−ピロリンレベルの解析。
【図7A】RNA干渉構築物と形質転換用ベクター。下のパネルは、構築物の発現確認を示す。本図面においてBADHとは、Os2APをさす。
【図7B】RNAi構築物、GFPおよび内在性Os2APの発現。下のパネルは、2−アセチル−1−ピロリンレベルと芳香を示す。BADH−RNAi2は、Os2AP遺伝子に対するRNAi構築物である。
【図8A】「アロマーカー」プライマーセットを用いた、芳香の増強と関係しているOs2AP変異体についてのF6後代のスクリーニング。
【図8B】芳香性Os2AP対立遺伝子についての種々の米品種のスクリーニング。
【図9A】種々の系統のイネ由来のOs2APオーソログのDNA配列を示す。
【図9B】22のBADH(Os2AP)オーソログの近隣結合樹を用いて構築した系統樹。
【特許請求の範囲】
【請求項1】
対照非トランスジェニックイネ植物におけるレベルよりも高いレベルの化合物2−アセチル−1−ピロリンを含んでなるトランスジェニックイネ植物であって、該化合物のレベルが、該トランスジェニック植物において、該トランスジェニックイネ植物における2−アセチル−1−ピロリン遺伝子によりコードされるmRNAまたはタンパク質レベルを、該対照非トランスジェニックイネ植物における2−アセチル−1−ピロリン遺伝子によりコードされるmRNAまたはタンパク質レベルよりも低減することによって高められる、トランスジェニックイネ植物。
【請求項2】
該2−アセチル−1−ピロリン遺伝子のmRNAレベルが、該トランスジェニック植物において、2−アセチル−1−ピロリン遺伝子もしくはその一部のアンチセンス方向での発現により、またはRNA干渉により低減されている、請求項1に記載のトランスジェニックイネ植物。
【請求項3】
該2−アセチル−1−ピロリン遺伝子が配列番号5で示される配列を含んでなる、請求項1に記載のトランスジェニックイネ植物。
【請求項4】
請求項1に記載のトランスジェニック植物由来の種子。
【請求項5】
植物2−アセチル−1−ピロリンタンパク質をコードする単離された核酸であって、
該核酸が、少なくとも1回の0.1×SSCおよび0.1%SDSでの60〜65℃にて30分間の洗浄を含むハイブリダイゼーション条件下で、配列番号2で示される核酸配列またはその相補配列とハイブリダイズする、単離された核酸。
【請求項6】
配列番号2で示される核酸配列と70%より高い割合、80%より高い割合、90%より高い割合または95%より高い割合で同一である、請求項5に記載の単離された核酸。
【請求項7】
配列番号2で示される配列と実質的に同一である、請求項5に記載の単離された核酸。
【請求項8】
配列番号3で示されるアミノ酸配列と実質的に同一であるポリペプチドをコードする、請求項5に記載の単離された核酸。
【請求項9】
配列番号3で示されるアミノ酸配列と少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、または95%を超える割合で同一であるポリペプチドをコードする、請求項5に記載の単離された核酸。
【請求項10】
プロモーターに作動可能なように連結されている、請求項5に記載の単離された核酸。
【請求項11】
プロモーターが構成的プロモーター、発生的調節プロモーターまたは誘導プロモーターである、請求項10に記載の単離された核酸。
【請求項12】
プロモーターが、ツユクサ黄色斑紋ウイルスプロモーター、サトウキビバドナウイルスプロモーター、イネツングロ桿菌状ウイルスプロモーター、トウモロコシ条斑ウイルスエレメント、コムギ萎縮ウイルスプロモーター、カリフラワーモザイクウイルス(CaMV)プロモーター、ノパリンシンターゼ(NOS)プロモーター、オクトピンシンターゼプロモーター、ゴマノハグサモザイクウイルス(FMV)プロモーターおよびイネアクチンプロモーターから選択される、請求項10に記載の単離された核酸。
【請求項13】
請求項5に記載の単離された核酸を含んでなる、細胞。
【請求項14】
対照天然植物におけるレベルよりも高いレベルの化合物2−アセチル−1−ピロリンを含んでなる非天然植物であって、
化合物2−アセチル−1−ピロリンのレベルが、対照植物よりも、1以上の2−アセチル−1−ピロリン遺伝子の発現を低減し、該1以上の2−アセチル−1−ピロリン遺伝子によってもたらされるmRNAレベルを低減し、および/または1以上の2−アセチル−1−ピロリンタンパク質の活性もしくは量を低減することによって高められる、非天然植物。
【請求項15】
2−アセチル−1−ピロリン遺伝子の発現が、2−アセチル−1−ピロリン遺伝子もしくはその一部のアンチセンス方向での発現により、またはRNA干渉により低減される、請求項14に記載の非天然植物。
【請求項16】
2−アセチル−1−ピロリン遺伝子の発現が、突然変異誘発により低減される、請求項14に記載の非天然植物。
【請求項17】
植物がイネ、コムギ、オオムギ、ライムギ、ココナッツ、ソルガムおよびオートムギからなる群から選択される、請求項14に記載の非天然植物。
【請求項18】
請求項14に記載の非天然植物から生じた、種子。
【請求項19】
植物または植物の一部における2−アセチル−1−ピロリン遺伝子の芳香性対立遺伝子の存在を検出する方法であって、2−アセチル−1−ピロリン遺伝子が2−アセチル−1−ピロリンタンパク質の量、機能または活性を低減する突然変異を有しているかどうかを検出することを含む、方法。
【請求項20】
突然変異がOs2APの芳香性対立遺伝子に見られる8塩基対の欠失である、請求項19に記載の方法。
【請求項21】
突然変異を検出する方法がPCRおよび/またはマイクロアレイの使用を含む、請求項19に記載の方法。
【請求項22】
突然変異を検出する方法がハイブリダイゼーションまたは配列決定を含む、請求項19に記載の方法。
【請求項23】
突然変異を検出する方法が2−アセチル−1−ピロリンタンパク質レベル、構造または活性を検出することを含む、請求項22に記載の方法。
【請求項1】
対照非トランスジェニックイネ植物におけるレベルよりも高いレベルの化合物2−アセチル−1−ピロリンを含んでなるトランスジェニックイネ植物であって、該化合物のレベルが、該トランスジェニック植物において、該トランスジェニックイネ植物における2−アセチル−1−ピロリン遺伝子によりコードされるmRNAまたはタンパク質レベルを、該対照非トランスジェニックイネ植物における2−アセチル−1−ピロリン遺伝子によりコードされるmRNAまたはタンパク質レベルよりも低減することによって高められる、トランスジェニックイネ植物。
【請求項2】
該2−アセチル−1−ピロリン遺伝子のmRNAレベルが、該トランスジェニック植物において、2−アセチル−1−ピロリン遺伝子もしくはその一部のアンチセンス方向での発現により、またはRNA干渉により低減されている、請求項1に記載のトランスジェニックイネ植物。
【請求項3】
該2−アセチル−1−ピロリン遺伝子が配列番号5で示される配列を含んでなる、請求項1に記載のトランスジェニックイネ植物。
【請求項4】
請求項1に記載のトランスジェニック植物由来の種子。
【請求項5】
植物2−アセチル−1−ピロリンタンパク質をコードする単離された核酸であって、
該核酸が、少なくとも1回の0.1×SSCおよび0.1%SDSでの60〜65℃にて30分間の洗浄を含むハイブリダイゼーション条件下で、配列番号2で示される核酸配列またはその相補配列とハイブリダイズする、単離された核酸。
【請求項6】
配列番号2で示される核酸配列と70%より高い割合、80%より高い割合、90%より高い割合または95%より高い割合で同一である、請求項5に記載の単離された核酸。
【請求項7】
配列番号2で示される配列と実質的に同一である、請求項5に記載の単離された核酸。
【請求項8】
配列番号3で示されるアミノ酸配列と実質的に同一であるポリペプチドをコードする、請求項5に記載の単離された核酸。
【請求項9】
配列番号3で示されるアミノ酸配列と少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、または95%を超える割合で同一であるポリペプチドをコードする、請求項5に記載の単離された核酸。
【請求項10】
プロモーターに作動可能なように連結されている、請求項5に記載の単離された核酸。
【請求項11】
プロモーターが構成的プロモーター、発生的調節プロモーターまたは誘導プロモーターである、請求項10に記載の単離された核酸。
【請求項12】
プロモーターが、ツユクサ黄色斑紋ウイルスプロモーター、サトウキビバドナウイルスプロモーター、イネツングロ桿菌状ウイルスプロモーター、トウモロコシ条斑ウイルスエレメント、コムギ萎縮ウイルスプロモーター、カリフラワーモザイクウイルス(CaMV)プロモーター、ノパリンシンターゼ(NOS)プロモーター、オクトピンシンターゼプロモーター、ゴマノハグサモザイクウイルス(FMV)プロモーターおよびイネアクチンプロモーターから選択される、請求項10に記載の単離された核酸。
【請求項13】
請求項5に記載の単離された核酸を含んでなる、細胞。
【請求項14】
対照天然植物におけるレベルよりも高いレベルの化合物2−アセチル−1−ピロリンを含んでなる非天然植物であって、
化合物2−アセチル−1−ピロリンのレベルが、対照植物よりも、1以上の2−アセチル−1−ピロリン遺伝子の発現を低減し、該1以上の2−アセチル−1−ピロリン遺伝子によってもたらされるmRNAレベルを低減し、および/または1以上の2−アセチル−1−ピロリンタンパク質の活性もしくは量を低減することによって高められる、非天然植物。
【請求項15】
2−アセチル−1−ピロリン遺伝子の発現が、2−アセチル−1−ピロリン遺伝子もしくはその一部のアンチセンス方向での発現により、またはRNA干渉により低減される、請求項14に記載の非天然植物。
【請求項16】
2−アセチル−1−ピロリン遺伝子の発現が、突然変異誘発により低減される、請求項14に記載の非天然植物。
【請求項17】
植物がイネ、コムギ、オオムギ、ライムギ、ココナッツ、ソルガムおよびオートムギからなる群から選択される、請求項14に記載の非天然植物。
【請求項18】
請求項14に記載の非天然植物から生じた、種子。
【請求項19】
植物または植物の一部における2−アセチル−1−ピロリン遺伝子の芳香性対立遺伝子の存在を検出する方法であって、2−アセチル−1−ピロリン遺伝子が2−アセチル−1−ピロリンタンパク質の量、機能または活性を低減する突然変異を有しているかどうかを検出することを含む、方法。
【請求項20】
突然変異がOs2APの芳香性対立遺伝子に見られる8塩基対の欠失である、請求項19に記載の方法。
【請求項21】
突然変異を検出する方法がPCRおよび/またはマイクロアレイの使用を含む、請求項19に記載の方法。
【請求項22】
突然変異を検出する方法がハイブリダイゼーションまたは配列決定を含む、請求項19に記載の方法。
【請求項23】
突然変異を検出する方法が2−アセチル−1−ピロリンタンパク質レベル、構造または活性を検出することを含む、請求項22に記載の方法。
【図1】

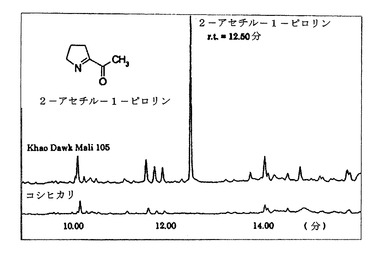
【図2】
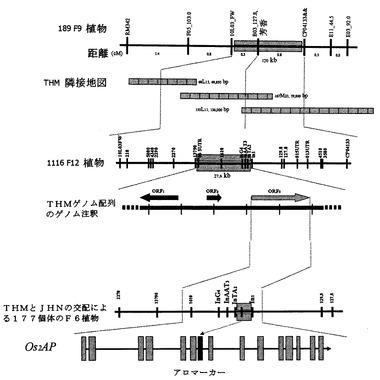
【図3C】
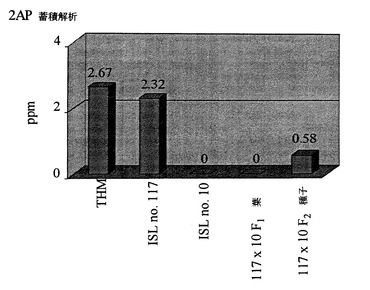
【図4A】
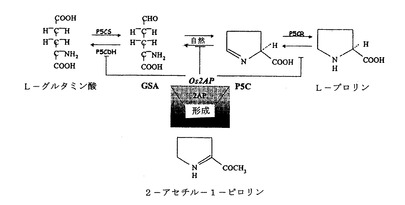
【図5A−1】

【図5A−2】

【図5B】
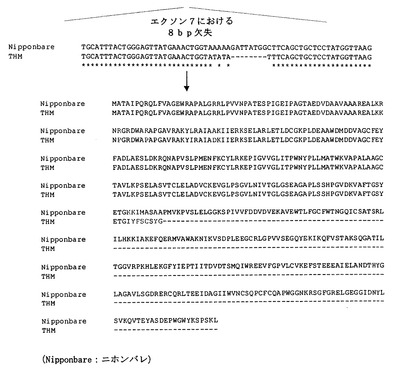
【図6】
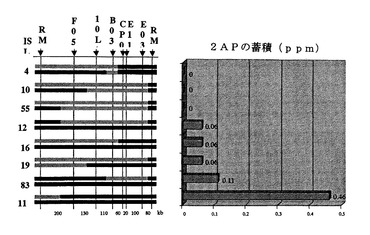
【図8B】

【図9A】

【図9B】
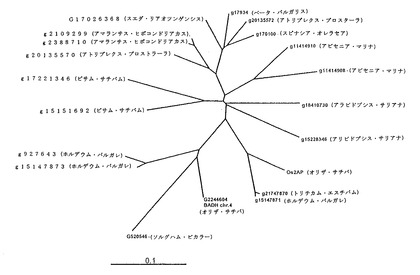
【図3A】
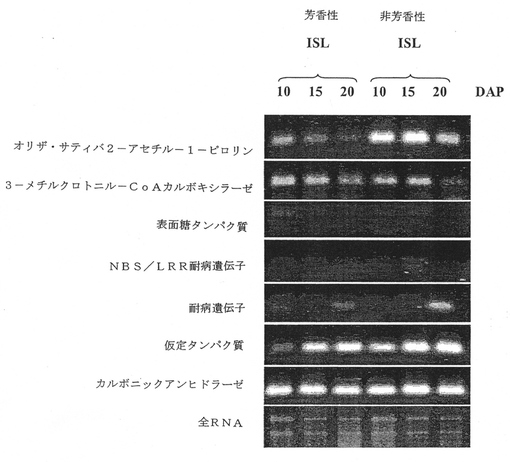
【図3B】
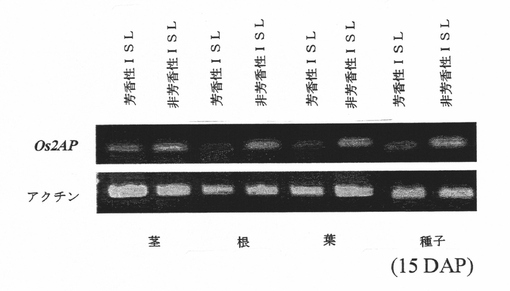
【図3D】
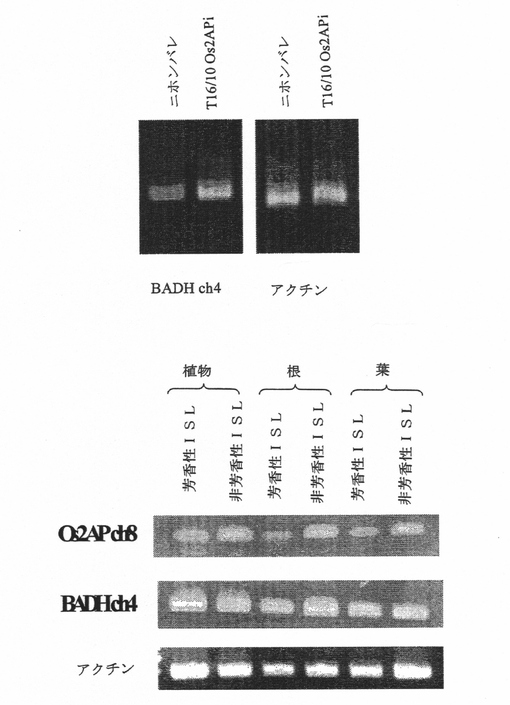
【図4B】
【図7A】
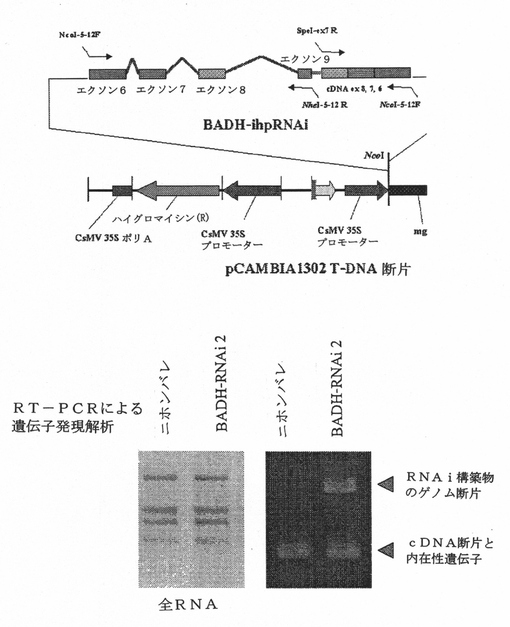
【図7B】
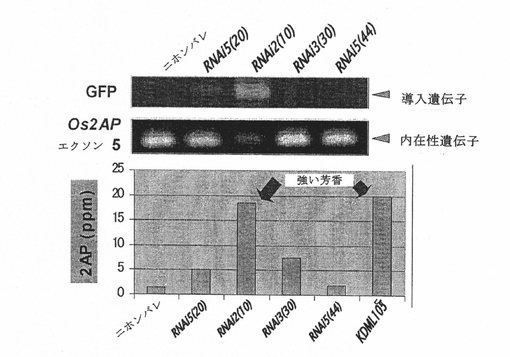
【図8A】
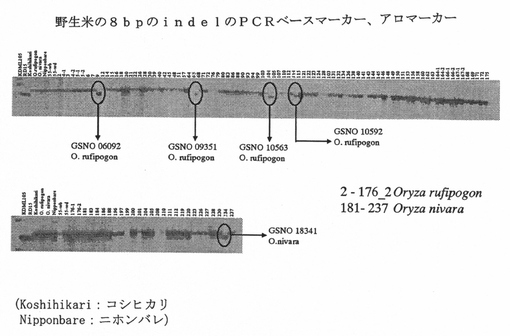
【図2】
【図3C】
【図4A】
【図5A−1】
【図5A−2】
【図5B】
【図6】
【図8B】
【図9A】
【図9B】
【図3A】
【図3B】
【図3D】
【図4B】
【図7A】
【図7B】
【図8A】
【公開番号】特開2011−4752(P2011−4752A)
【公開日】平成23年1月13日(2011.1.13)
【国際特許分類】
【外国語出願】
【出願番号】特願2010−174002(P2010−174002)
【出願日】平成22年8月2日(2010.8.2)
【分割の表示】特願2006−16908(P2006−16908)の分割
【原出願日】平成18年1月25日(2006.1.25)
【出願人】(506028144)ナショナル、サイエンス、アンド、テクノロジー、ディベロップメント、エイジェンシー (4)
【氏名又は名称原語表記】NATIONAL SCIENCE AND TECHNOLOGY DEVELOPMENT AGENCY
【出願人】(506028155)カセトサート、ユニバーシティー (2)
【氏名又は名称原語表記】KASETSART UNIVERSITY
【Fターム(参考)】
【公開日】平成23年1月13日(2011.1.13)
【国際特許分類】
【出願番号】特願2010−174002(P2010−174002)
【出願日】平成22年8月2日(2010.8.2)
【分割の表示】特願2006−16908(P2006−16908)の分割
【原出願日】平成18年1月25日(2006.1.25)
【出願人】(506028144)ナショナル、サイエンス、アンド、テクノロジー、ディベロップメント、エイジェンシー (4)
【氏名又は名称原語表記】NATIONAL SCIENCE AND TECHNOLOGY DEVELOPMENT AGENCY
【出願人】(506028155)カセトサート、ユニバーシティー (2)
【氏名又は名称原語表記】KASETSART UNIVERSITY
【Fターム(参考)】
[ Back to top ]