
植物におけるヘテロ‐オリゴマータンパク質の産生
植物体、植物組織、または植物細胞において、少なくとも第1および第2タンパク質サブユニットを含有する、ヘテロ‐オリゴマータンパク質を産生する方法であって、
(i)植物体、植物組織または植物細胞に、少なくとも第1および第2タンパク質サブユニットをコードする、プラス‐センス1本鎖RNAウイルスベクターを提供すること;または
(ii)植物体、植物組織または植物細胞に、第1および第2プラス‐センス1本鎖RNAウイルスベクターを提供するが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、少なくとも第1ウイルスベクターおよび第2ウイルスベクターは非競合ウイルスベクターであり、それにより、植物細胞に少なくとも第1および第2タンパク質サブユニットを発現させることを含む、方法。
(i)植物体、植物組織または植物細胞に、少なくとも第1および第2タンパク質サブユニットをコードする、プラス‐センス1本鎖RNAウイルスベクターを提供すること;または
(ii)植物体、植物組織または植物細胞に、第1および第2プラス‐センス1本鎖RNAウイルスベクターを提供するが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、少なくとも第1ウイルスベクターおよび第2ウイルスベクターは非競合ウイルスベクターであり、それにより、植物細胞に少なくとも第1および第2タンパク質サブユニットを発現させることを含む、方法。
【発明の詳細な説明】
【発明の開示】
【0001】
発明が属する技術分野
本発明は、プラス‐センス1本鎖RNAウイルスベクターを使用した、植物体、植物部分または植物細胞培養物におけるヘテロ‐オリゴマータンパク質の産生に関する。本発明に記載の方法およびベクターは、全長抗体、または別のタンパク質もしくはそれらのフラグメントとの融合物を含むそのヘテロ‐オリゴマー合成誘導体のような、高い収率の機能的ヘテロ‐オリゴマー組換えタンパク質を植物細胞に提供する。
【0002】
発明の背景
植物を基礎にした分子農業は、好ましくは、潜在的に低い生産費用、および工業的生産方法から生み出される動物‐由来タンパク質を除去するためのバイオ医薬品産業による試みのゆえに、ヒトおよび動物の健康分野で使用される予定の組換えタンパク質の産生にとっての魅力的な好機である。なぜなら、これらの産物は牛海綿状脳症(BSE)またはクロイツフェルト‐ヤコブ病(CJD、vCJD)のようなヒト病原体により汚染されうるからである。しかし、植物細胞における高収率のヘテロオリゴマータンパク質の産生は、以下の理由から、強力な構成的または組織‐特異的プロモーターの使用に基づいた標準発現系の協力により解決することができない難問である:第1に、そのような組換えタンパク質の大多数は植物の成長および発達に対して有害な作用を有し、それゆえ収率を著しく低下させる;第2に組織‐特異的(たとえば、種子‐特異的)プロモーターの使用は、食用になる作物植物(たとえば、イネ、トウモロコシ、コムギ)のゲノムへの医薬タンパク質をコードする遺伝子の安定した組み込みを必要とし、それは露地栽培の場合、トランスジーン流動制御に関する問題を引き起こす可能性がある。また、これらの系は産物の低い収率のため、閉鎖環境(温室)で使用する場合、商業的に実行可能ではない。
【0003】
植物ウイルスを基礎にした一過性の発現系(総説としては以下を参照されたい:Porta & Lomonossoff,1996,Mol.Biotechnol.,5,209−221;Yusibov et al.,1999,Curr.Top.Microbiol.Immunol.,240,81−94;Gleba et al.,2004,Curr.Opin.Plant Biol.,7,182−188)は、その技術が成長と産生ステージを分離させるため、植物葉組織における高発現レベルを提供することが可能であり、ある程度までは組換えタンパク質の細胞毒性、および植物発達に対するそれらの有害作用の難問に対処することができる。最も確立され、商業的に実行可能な系は、プラス‐センス1本鎖RNAウイルスベクター、好ましくはタバコモザイクウイルス(TMV)‐由来ベクター(Kumagai et al.,1994,Proc.Natl.Acad.Sci.USA,90,427−430;Mallory et al.,2002,Nature Biotechnol.20,622−625;US5316931;US5589367;US5866785;US5977438;WO02088369;WO02097080;WO0229068;US5491076)に基づく。しかし、これらの系は簡単で、比較的低分子のタンパク質の産生に使用が限定されるという深刻な制限を欠点として持つ。ウイルスベクターが1kbより大きな異種配列を持っている場合、このことはある程度は、それらの不安定性および野生型への高頻度での逆戻りによって引き起こされる。また、技術の深刻な制約は、最も有益な群の組換えタンパク質を代表する治療用モノクローナル抗体およびそれらの誘導体のような、複雑なヘテロオリゴマータンパク質の発現が可能なウイルスベクター系が無いことである。
【0004】
植物ウイルスベクターを使用した、植物における全長モノクローナル抗体の発現を扱った刊行物は1つしかない(Verch et al.,1998,J.Immunol.Meth.,220,69−75)。この論文は、全身葉部におけるモノクローナル抗体の重鎖および軽鎖発現のための2種のTMVを基礎にした全身ウイルスベクターを使用して、N.ベンサミアーナ植物体の同時感染時に、in vitroで合成された上記ベクターの転写物により、異なるベクターから異なる鎖が発現されることを説明する。しかし、上記の系における組換えタンパク質の収率は非常に低いため、集合したモノクローナル抗体の存在は、ウェスタンブロット法およびELISAのような感度の高い試験によって確認しなければならない。組換え抗体は収率が非常に低いため、この系は実際の適用に適切ではなく、商業的価値を持たない。感染された植物体の同じ植物組織中には、2種以上のTMVを基礎にしたウイルスベクターは通常存在しない(実施例1を参照されたい)ため、検出された抗原結合活性は、別々の細胞または組織に発現された抗体の重鎖および軽鎖からの分離手順中にin vitroで集合した抗体によるものである可能性がある。機能的抗体は変性し、弱めた抗体成分からin vitroで集められうることが以前に示された(Petersen & Dorrington,1974,J.Biol.Chem.,17,5633−5641;Maeda et al.,1996,Protein Engineering,9,95−100)。しかし、そのようなアセンブリーに都合のよい条件が存在しない場合、そのようなアセンブリーの効率は非常に低い。
【0005】
したがって、菌類または昆虫細胞発現系のような別の大規模発現系と市場で競い合うために十分な収率と効率の、植物における組換えヘテロオリゴマータンパク質の大規模発現系は存在しない。そのような植物発現はできる限り以下の基準を十分に満たさなければならない:
(i)できるだけ多くの植物組織および上記組織の中のできるだけ多くの細胞における、関心のあるヘテロオリゴマータンパク質の発現を含む、高収率;
(ii)植物の成長に対する組換えタンパク質発現の有害な作用を妨げるための、関心のあるタンパク質または産物の発現は所望する植物の発達段階で開始できるように、一過性(またはスイッチ可能)でなければならない;
(iii)植物細胞におけるヘテロオリゴマータンパク質の異なるサブユニットをコードするポリタンパク質の最適比を提供し、そうして上記サブユニットからの組換えタンパク質アセンブリーのレベルにおいて組換えタンパク質の高収率を維持すること。
【0006】
したがって、本発明の目的は植物体、植物部分、または植物細胞培養物における効率的なヘテロオリゴマータンパク質産生の方法を提供することである。別の目的は、ヘテロオリゴマータンパク質を発現することができる高収率植物発現系の提供である。本発明の別の目的は、同じ植物細胞中に1種より多い関心のあるポリペプチドを同時発現させる効率的な方法を提供することである。さらに、植物体、植物部分、または植物細胞培養物に抗体を発現させるための迅速で高収率の方法を提供することが本発明の目的である。
【0007】
発明の一般的説明
本発明は、植物体、植物組織、または植物細胞において、少なくとも第1および第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法を提供し、
(i)植物体、植物組織または植物細胞に、第1および第2プラス‐センス1本鎖RNAウイルスベクターを提供するが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、少なくとも第1ウイルスベクターおよび第2ウイルスベクターは非競合ウイルスベクターであり;または
(ii)植物体、植物組織または植物細胞に、少なくとも第1および第2タンパク質サブユニットをコードする、プラス‐センス1本鎖RNAウイルスベクターを提供すること
により、少なくとも第1および第2タンパク質サブユニットを植物組織に発現させることを含む。
【0008】
一態様では、第1ウイルスベクターおよび第2ウイルスベクターは、それらが異なるウイルスベクターであるという点で、非競合的である。別の態様では、第1ウイルスベクターおよび第2ウイルスベクターは、それらが同じRNAウイルスに由来しないという点で、非競合的である。別の態様では、第1ウイルスベクターおよび第2ウイルスベクターは、それらが第1および第2タンパク質サブユニットをコードする配列部分以外の配列部分において異なるという点で、非競合的である。別の態様では、第1ウイルスベクターおよび第2ウイルスベクターは、それらが異なるウイルス種に属するRNAウイルスに由来するという点で、非競合的である。別の態様では、第1ウイルスベクターおよび第2ウイルスベクターは、それらが異なるウイルス属に属するRNAウイルスに由来するという点で、非競合的である。
【0009】
本発明の別の態様は、抗体の重鎖および軽鎖のような、少なくとも第1および第2タンパク質サブユニットを含有する抗体を植物体、植物組織または植物細胞において産生する方法であって、
(i)第1プラス‐センス1本鎖RNAウイルスベクターのDNA前駆体および第2プラス‐センス1本鎖RNAウイルスベクターのDNA前駆体を、アグロバクテリウム媒介トランスフェクションにより植物体、植物組織、または植物細胞に提供するが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、少なくとも第1ウイルスベクターまたは第2ウイルスベクターは、植物体における第1または第2ウイルスベクターの全身移行に必要な機能的タンパク質をコードするオープンリーディングフレームを欠如し;または
(ii)少なくとも第1および第2タンパク質サブユニットをコードするプラス‐センス1本鎖RNAウイルスベクターのDNA前駆体を、アグロバクテリウム媒介トランスフェクションにより植物体、植物組織、または植物細胞に提供するが、ウイルスベクターは全身移行に必要な機能的タンパク質をコードするORFを欠如する
ことにより、少なくとも第1および第2タンパク質サブユニットを植物細胞に発現させることを含む。
【0010】
本発明はさらに、植物体、植物組織、または植物細胞において、少なくとも第1および第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法を提供し、方法は1種以上のプラス‐センス1本鎖RNAウイルスベクター(複数のベクター)由来の少なくとも第1および第2タンパク質サブユニットを植物細胞に発現させることを含む。
【0011】
本発明の別の態様は、特許請求の範囲および以下の説明に記載される。
本発明の発明者らは驚くべくことに、植物ウイルスベクターを使用して植物に高収率でヘテロオリゴマータンパク質を産生する方法を確認している。植物におけるヘテロオリゴマータンパク質の効率的な産生には、同じ植物細胞におけるヘテロオリゴマータンパク質の異なるタンパク質サブユニットの高収率の産生を必要とする。このようにして、小胞体(ER)の天然のタンパク質集合能力のような、上記細胞のそのような能力を使用して、上記の少なくとも2種のタンパク質サブユニットを発現している細胞にヘテロオリゴマータンパク質を効率的に集めることができる。したがって上記のヘテロオリゴマータンパク質の非効率的なin vitroでのアセンブリーは必要でない。本発明はステップ(i)により、またはステップ(ii)により、またはステップ(i)および(ii)の組み合わせにより同じ細胞に2種以上のタンパク質の効率的な同時発現を初めて成し遂げている。
【0012】
第1タンパク質サブユニットは第1異種(核酸)配列によってRNAウイルスベクターにコードされる。第2タンパク質サブユニットは第2異種(核酸)配列によってウイルスベクターにコードされる。これらの異種配列はしたがってウイルスベクター(複数のウイルスベクター)のRNA配列であり、典型的にはタンパク質サブユニットを発現するために必要な調節配列を含有するか、またはコードする。そのような調節配列の例としては、サブゲノムプロモーター、IRESエレメント、および3′‐非翻訳配列が挙げられる。本明細書では、ある配列がウイルスベクター(複数のウイルスベクター)が由来するウイルスに天然に存在しない場合、その配列は異種配列である。
【0013】
本発明に従って産生可能なヘテロオリゴマータンパク質は、少なくとも第1および第2サブユニットを有し、第1および第2サブユニットは異なるポリペプチド配列を有する。したがって、第1および第2サブユニットは典型的には異なる異種核酸配列から発現されなければならない。オリゴマータンパク質のサブユニットは会合して4次構造のオリゴマータンパク質を形成する。サブユニットのアセンブリーは典型的にはサブユニット間の非共有結合を包含する。さらに、共有結合、たとえばジスルフィド結合がタンパク質サブユニット間で形成されてもよい。
【0014】
一態様では、ヘテロオリゴマータンパク質はヘテロ‐二量体タンパク質、すなわち2種の異なるタンパク質サブユニットを有するタンパク質である。別の態様では、ヘテロオリゴマータンパク質は2種より多い異なるサブユニット、たとえば3種または4種の異なるサブユニット(それぞれヘテロトリマーまたはヘテロテトラマータンパク質)を有していてもよい。別の態様では、のヘテロオリゴマータンパク質は2種の異なるサブユニットを有していてもよく、サブユニットの中の1方または両方がヘテロオリゴマータンパク質中に、1回より多く存在してもよい。ヘテロオリゴマータンパク質のサブユニット構成の例としては、AaBb、AaBbCc、AaBbCcDdが挙げられ、ここでAは第1タンパク質サブユニットを表し、Bは第2タンパク質サブユニットを表し、そしてCおよびDは別のタンパク質サブユニットを表す。それぞれの大文字A、B、CおよびDは他のタンパク質サブユニットとは異なるタンパク質サブユニットを表し、そして小文字は少なくとも1の整数を表し、ヘテロオリゴマータンパク質中の個々のタンパク質サブユニットのコピー数を示す。A2B2のサブユニット構成を有するIgG抗体が例として挙げられ、ここでAは第1タンパク質サブユニット(たとえば重鎖)を表し、そしてBは第2タンパク質サブユニット(たとえば軽鎖)を表す。好ましくは、本発明に従って産生されたヘテロオリゴマータンパク質は2種または3種の異なるタンパク質サブユニットを有し、より好ましくは、それは2種の異なるタンパク質サブユニットを有する。
【0015】
本発明の方法では、少なくとも第1および第2タンパク質サブユニットは植物体、植物組織、または植物細胞の細胞中に、ステップ(i)または/およびステップ(ii)によって発現される。ステップ(i)および(ii)のそれぞれは、同じ細胞中に少なくとも第1および第2タンパク質サブユニットを発現させ、その結果ヘテロオリゴマータンパク質を細胞中に効率的に産生することができる。ステップ(i)および(ii)は、とりわけ3種、4種、またはそれより多い異なるタンパク質サブユニットを有するヘテロオリゴマータンパク質産生のために並行して行うことができる(実施例7を参照されたい)。
【0016】
本発明のプラス‐センス1本鎖RNAウイルスベクター(複数のウイルスベクター)はまた、本明細書では単に「ウイルスベクター」として表される。典型的には、プラス‐センス1本鎖RNAウイルスベクターはプラス‐センス1本鎖植物RNAウイルスに由来する。
【0017】
ステップ(ii)では、植物体、植物組織または植物細胞は、少なくとも第1および第2タンパク質サブユニットをコードするプラス‐センス1本鎖RNAウイルスベクターを提供される。少なくとも第1および第2タンパク質サブユニットをコードするウイルスベクターは、インサートとして第1タンパク質サブユニットをコードする第1異種配列を含有し、その発現は第1サブゲノムプロモーターの制御下にあってもよい。さらに、ウイルスベクターはインサートとして、第2タンパク質サブユニットをコードする第2異種配列を含有し、その発現は第2サブゲノムプロモーターの制御下にあってもよい。第1および第2タンパク質サブユニットの両方がサブゲノムプロモーターの制御下で発現される場合、これらのサブゲノムプロモーターは、好ましくは、植物細胞に望ましくない組換え事象を導くウイルスベクター中の自己相同性を回避するために配列が異なる。そのような異なるサブゲノムプロモーターは異なる株または種の植物ウイルスから採取されてもよく、たとえば1種のサブゲノムプロモーターはタバコモザイクウイルス(TMV)U1のコートタンパク質(CP)サブゲノムプロモーターであってもよく(またはそれに由来してもよく)、そして別のサブゲノムプロモーターは、TMV U5のCPサブゲノムプロモーターであってもよく(またはそれに由来してもよく)、またはアブラナ科植物‐感染トバモウイルス(cr‐TMV)に由来してもよい。
【0018】
第1または第2サブゲノムプロモーターのかわりに、第1または第2タンパク質サブユニットの翻訳がIRES(内部リボソーム侵入部位:internal ribosome entry site)エレメントの制御下にあってもよい。第1および第2タンパク質サブユニット両方の翻訳がIRESエレメントの制御下にあってもよいが、少なくともタンパク質サブユニットの1種はサブゲノムプロモーターを使用して発現される。本発明で使用されるIRESエレメントはcr‐TMVのような植物ウイルスまたは他の植物ウイルスから採取されてもよい(Proc Natl Acad Sci USA 2002,99,5301−6;Virology 1999,263,139−54;WO03020927;WO0229068)。
【0019】
ステップ(ii)のウイルスベクターは、好ましくは植物体または植物組織において全身移行をする能力がない。このことは、たとえばウイルス粒子アセンブリーの機能的起点を削除することにより成し遂げることができる。トバモウイルスではたとえば、ウイルス粒子アセンブリーの起点はMP ORFに位置する。したがって、ウイルス粒子アセンブリーの起点はウイルスベクターからMP ORFを完全に、または部分的に欠失することによって削除することができる。ウイルスベクターはしたがって、好ましくは機能的移行タンパク質(MP)ORFを持たない。より好ましくは、ウイルスベクターは、ウイルスベクターの全身移行に必要な機能的タンパク質を持たない。この態様では、ウイルスベクターは機能的コートタンパク質ORFを持たなくてもよく、そして最も好ましくは、ウイルスベクターは機能的移行タンパク質ORFおよび機能的コートタンパク質ORFの両方を持たない。ウイルスベクターからMP ORFおよび/またはCP ORFを除去することは、ウイルスベクター安定性を低下させずに、少なくとも第1および第2タンパク質サブユニットをコードするためのより多くの余地を提供する。この態様では、ウイルスベクターは好ましくは、多くの細胞の感染を成し遂げるために、植物体または植物組織の多くの細胞に提供される。このことはアグロバクテリウムを使用して、T−DNAのようなRNAウイルスベクターのDNA前駆体を提供することにより最も都合よく成し遂げられてもよい(以下を参照されたい)。
【0020】
ステップ(ii)のウイルスベクターが由来するウイルスはいずれかのプラス‐センス1本鎖植物RNAウイルス、たとえば「詳細な説明」の章で挙げられたものであってもよい。好ましいウイルス群はトバモウイルス、ポテクスウイルス、およびポティウイルスである。最も好ましいウイルスはTMVおよびPVXである。ウイルスベクターは少なくともそれが由来するウイルス由来の、ウイルスベクターの複製に必要なタンパク質をコードするORFを含有することになる。ウイルスベクターは典型的にはさらに、ウイルス複製のための調節エレメント、および少なくとも1種のサブゲノムプロモーターを含有する。
【0021】
ステップ(i)では、植物体、植物組織または植物細胞は、第1および第2プラス‐センス1本鎖RNAウイルスベクターを備える。第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードする。
【0022】
ステップ(i)は2種の異なるタンパク質サブユニットを有するヘテロオリゴマータンパク質の産生を可能にする。ステップ(i)はその上、2種より多い異なるサブユニット、たとえば3種または4種の異なるタンパク質サブユニットを有するヘテロオリゴマータンパク質の産生を可能にする。この場合、植物体、植物組織または植物細胞は、それぞれが上記の異なるタンパク質サブユニットの1種をコードする、第1、第2および第3ウイルスベクターを(そして場合により別のウイルスベクターを)備えてよい。3種または4種の異なるタンパク質サブユニットをステップ(i)で発現しなければならない場合、個々の3種または4種のウイルスベクターは好ましくは、すべてお互いに非競合ウイルスベクターである。3種のウイルスベクターの場合、第1ウイルスベクターはトバモウイルスに由来してもよく、第2ウイルスベクターはポティウイルスに由来してもよく、そして第3ウイルスベクターはポテクスウイルスに由来してもよい。
【0023】
2種のウイルスベクターは、同じ植物組織中でそれらがコードするタンパク質サブユニットを効率的に発現(同時発現)できる場合、非競合的である。同時発現は少なくとも2種の異なるウイルスベクターがコードする異種配列の実質的な量を発現する前は、それらは複製中にお互いに競合しないことを必要とする。上記の少なくとも2種のウイルスベクターのRNAレベルの配列の違いが大きいほど、それらは同じ植物細胞においていっそう非競合的である。第1ウイルスベクターおよび第2ウイルスベクターは異なるウイルスベクターであるため、第1ウイルスベクターおよび第2ウイルスベクターは非競合ウイルスベクターである。
【0024】
非競合的である場合、ある一般的な態様では、第1および第2ウイルスベクター(上記の2種の非競合ウイルスベクター)は、少なくともタンパク質サブユニットをコードする上記の異種配列以外の配列部分において異なる。好ましくは、上記の異種配列以外の配列部分におけるそのような違いは植物RNAウイルスに由来する配列部分であり、そのような配列部分は、ウイルス機能、たとえば植物細胞におけるウイルス複製(たとえばレプリカーゼをコードする配列)、ウイルスタンパク質の翻訳(たとえばサブゲノムプロモーター)またはウイルスの細胞間もしくは長距離移行に関与する配列部分である。
【0025】
非競合的である場合、別の一般的な態様では、第1および第2ウイルスベクター(上記の2種の非競合ウイルスベクター)は異なる植物ウイルスに由来する。この態様の一例では、上記の少なくとも2種の非競合ウイルスベクターは同じウイルス株のウイルスに由来しない。別の例では、上記の少なくとも2種の非競合ウイルスベクターは同じウイルス種のウイルスに由来しない。付加的な例では、上記の少なくとも2種の非競合ウイルスベクターは同じウイルス属のウイルスに由来しない。したがって、第1ウイルスベクターおよび第2ウイルスベクターは好ましくは異なる株のウイルス、より好ましくは異なる種のウイルス、最も好ましくは異なる属のウイルスに由来する。
【0026】
第1ウイルスベクターはポテクスウイルス属に属するウイルスに由来してもよく、そして第2ウイルスベクターはポティウイルス属に属するウイルスに由来してもよい。とりわけ、第1ウイルスベクターはジャガイモウイルスXに由来してもよく、そして第2ウイルスベクターはジャガイモウイルスYに由来してもよい。
【0027】
ポティウイルスベクターの場合、タンパク質サブユニットはウイルスポリタンパク質との融合物として発現してもよく、それによって本発明のタンパク質サブユニットはポティウイルスプロテアーゼ認識部位によりポリタンパク質から分離することができる。
【0028】
別の態様では、第1ウイルスベクターはポテクスウイルス属に属するウイルスに由来してもよく、そして第2ウイルスベクターはトバモウイルス属に属するウイルスに由来してもよい。とりわけ、第1ウイルスベクターはジャガイモウイルスXに由来してもよく、そして第2ウイルスベクターはタバコモザイクウイルスに由来してもよい。
【0029】
「ウイルスに由来している」とは、ウイルスの遺伝エレメントまたは配列部分をそのウイルスに由来するウイルスベクターが含有することを意味する。一態様では、ウイルスベクター(複数のウイルスベクター)はRNAウイルスから採取したレプリカーゼORF(オープンリーディングフレーム)を含有する。別の態様では、ウイルスベクターはRNAウイルス由来の移行タンパク質ORF、そして場合によりさらにレプリカーゼORFを含有する。RNAウイルスから採取したウイルス遺伝エレメントは、所望する場合、たとえばウイルスベクターのクローニングに必要な制限部位を導入するために変異してもよい。
【0030】
第1ウイルスベクターおよび第2ウイルスベクターが共にタバコモザイクウイルスTMV30Bに基づいている態様は、本発明の方法のステップ(i)から排除される。なぜなら、この場合、第1ウイルスベクターおよび第2ウイルスベクターは異なるウイルスベクターではなく、同じウイルスベクターであるからである(参照:Verch et al.,J.Immunological Methods 220(1998)69−75)。
【0031】
第1および第2ウイルスベクターは、好ましくは多くても90%、より好ましくは多くても80%、さらにより好ましくは多くても70%、そして最も好ましくは多くても60%のRNAレベルの配列相同性を有する。さらにとりわけ、第1ウイルスベクターの任意の100塩基の配列部分が好ましくは、第2ウイルスベクターの任意の100塩基の配列部分に多くても90%、好ましくは多くても80%、より好ましくは多くても70%、そして最も好ましくは多くても60%の配列相同性を有する。
【0032】
一態様では、第1ウイルスベクターのレプリカーゼORF(またはレプリカーゼが1種以上のORFによってコードされる場合、複数のORF)と第2ウイルスベクターのレプリカーゼORF(またはレプリカーゼが1種以上のORFによってコードされる場合、複数のORF)は、多くても90%、より好ましくは多くても80%、さらにより好ましくは多くても70%、そして最も好ましくは多くても60%の相同性を有する。別の態様では、第1ウイルスベクターのレプリカーゼORFと第2ウイルスベクターのレプリカーゼORFは多くても85%、より好ましくは多くても75%、さらにより好ましくは多くても65%、そして最も好ましくは多くても55%の同一性を有する。
【0033】
ステップ(i)では、第1ウイルスベクターは好ましくは、発現が第1サブゲノムプロモーターの制御下にあってもよい第1タンパク質サブユニットをコードする第1異種配列を含有する。第2ウイルスベクターは、発現が第2サブゲノムプロモーターの制御下にあってもよい第2タンパク質サブユニットをコードする第2異種配列を含有する。サブゲノムプロモーターの1種または両方はIRESエレメントによって置換されてもよい。先に記載のステップ(ii)の場合と同様に、第1および第2タンパク質サブユニットがサブゲノムプロモーターの制御下で発現される場合、植物細胞に望ましくない組換え事象を引き起こすことになる第1および第2(およびいずれか別のウイルスベクター)ウイルスベクター間の相同性を回避するために、これらのサブゲノムプロモーターは好ましくは配列が異なる。そのような異なるサブゲノムプロモーターは異なる株または種の植物ウイルスから採取されてもよく、たとえば1種のサブゲノムプロモーターはタバコモザイクウイルス(TMV)U1のコートタンパク質(CP)サブゲノムプロモーターであってもよく、そして別のサブゲノムプロモーターはTMV U5またはアブラナ科植物‐感染トバモウイルス(cr‐TMV)のCPサブゲノムプロモーターであってもよい。
【0034】
細胞間、もしくは長距離移行に必要な植物ウイルスのORFは本発明のウイルスベクター構築時に削除されてもよいため、上記の非競合ウイルスベクターの関連性はウイルスベクターのレプリカーゼORFを比較することにより確認してもよい。
【0035】
第1および第2タンパク質サブユニットをコードする第1および第2異種配列は、それぞれウイルスベクターに付加的な配列として添加されてもよい。異種配列は好ましくは高レベルの発現が成し遂げられるように添加される。この目的のために、異種配列はウイルスの3′末端に添加される。なぜなら3′ORFはしばしば多くのウイルスで最も高レベルで発現されるORFであるからである。好ましくは、しかし、異種配列は、ウイルスに生来ある配列、たとえばトバモウイルスのような多くのウイルスにおいてCP ORFであるウイルスの天然の3′ORFと入れ替わる。したがって、第1および/または第2ウイルスベクターは好ましくはウイルスベクターの全身移行のためのORFを欠如する。ウイルスベクターはさらに、トバモウイルスのMP ORFのような細胞間移行のためのORFを欠いてもよい。
【0036】
本発明では、ステップ(i)およびステップ(ii)は、とりわけヘテロオリゴマータンパク質の3種以上の異なるサブユニットを発現するために組み合わせてもよい。4種の異なるタンパク質サブユニットを有するヘテロオリゴマータンパク質が産生されることになる場合、2種のタンパク質サブユニットがステップ(i)に従って発現されてもよく、そして2種の付加的なタンパク質サブユニットがステップ(ii)に従って、植物体または植物組織の細胞中に産生されてもよい。しかし、好ましくは2種のタンパク質サブユニットが第1ウイルスベクターから発現されてもよく、そして2種のタンパク質サブユニットが第1ウイルスベクターに非競合的な第2ウイルスベクターから発現されてもよい。3種の異なるタンパク質サブユニットを有するヘテロオリゴマータンパク質が産生されることになる場合、上記の3種のタンパク質サブユニットは、ステップ(i)に従って、ステップ(ii)に関する記載と同様に第1ウイルスベクターから2種のタンパク質を発現し、そして非競合ウイルスベクターから第3のタンパク質サブユニットを発現することにより、発現されてもよい。
【0037】
本発明のウイルスベクターは、典型的にはDNAレベルで工学的に操作される。ウイルスベクターがRNAウイルスベクターのように植物体の細胞または植物組織の細胞に提供される場合、上記DNAは、たとえばT7ポリメラーゼのようなバクテリオファージポリメラーゼを適切なプロモーターと一緒に使用することにより、RNAウイルスベクターにin vitroで転写されてもよい。2種の異なるウイルスベクターは好ましくは、植物体の細胞が両方のウイルスベクターを備えることを確実にするために、混合物として植物体に提供される。好ましくは、しかし、植物体または植物組織をRNAウイルスベクターのDNA前駆体により形質転換することにより、本発明のウイルスベクターが植物体の細胞または植物組織の細胞に提供される。DNA前駆体は、植物体の細胞中で、DNA前駆体の転写によってウイルスベクターを形成することに対して活性な転写プロモーターを有する。最も好ましくは、DNA前駆体はアグロバクテリウムTiプラスミド中のT−DNAである。2種以上のウイルスベクターはその後、植物体をそれぞれの株が特定のウイルスベクターをコードするT−DNAを含有する2種以上のアグロバクテリウム株の混合物(たとえば、懸濁液)で処理することにより植物体または植物組織に提供されてもよい。そのようなアグロバクテリウム懸濁液で植物体の実質的な部分を処置することが、天然の植物ウイルスの全身移行機能および/または細胞間移行機能の代わりとなってもよい。
【0038】
アグロバクテリウム法を用いたウイルスベクターのDNA前駆体による植物体、植物組織または植物細胞の一過性トランスフェクションは、本発明において最も好ましい。しかし、ウイルスベクターのDNA前駆体が植物染色体DNAに安定に組み込まれてもよい。染色体DNAからのウイルスベクター(複数のウイルスベクター)の放出は誘導型プロモーターによって制御されてもよい。
【0039】
ウイルスベクターがDNA前駆体から植物体に提供される場合、それらが転写される細胞核から、ウイルスベクターが複製する細胞質へのウイルスベクターの移動効率を改善するための処置がとられることが好ましい。このことは、WO2005/71090として公開された国際特許出願PCT/EP05/000492(これを参照として本明細書に援用する)に詳細に記載されるように、DNA前駆体、とりわけウイルスベクターのレプリカーゼORFにイントロンを包含することにより成し遂げられてもよい。
【0040】
本発明の方法は、植物ウイルス発現系が存在するか、または将来作り出されることになるいずれかの植物に適用してもよい。植物は単子葉植物または双子葉植物であってもよい。双子葉植物の中では、ナス科、アブラナ科、アカザ科、およびマメ科が好ましい。ナス科の中では、N.タバカムまたはN.ベンサミアーナのようなニコチアナ属が好ましい。別の好ましい植物はメディカゴサティバおよびフダンソウのようなフダンソウ種である。
【0041】
本発明の方法は植物系においてヘテロオリゴマータンパク質を産生するために使用される。好ましいヘテロオリゴマータンパク質は以下のクラスのイムノグロブリン:イムノグロブリンG、イムノグロブリンA、イムノグロブリンM、イムノグロブリンD、およびイムノグロブリンEのようなイムノグロブリンである。これらのイムノグロブリンは少なくとも抗原結合ドメインの一部を含有してもよい。本発明に従って産生されるこれらのイムノグロブリンが少なくとも2種の異なるタンパク質サブユニットを含有するならば、必要に応じて、それらのイムノグロブリンは、生来の動物イムノグロブリンに関連して改変されてもよい。イムノグロブリンはイムノグロブリン重鎖と一緒に保護タンパク質を含有してもよく、ここで保護タンパク質はポリイムノグロブリン受容体の一部を含有する。別の好ましいヘテロオリゴマータンパク質はインスリンである。
【0042】
本発明のヘテロオリゴマータンパク質は、必要に応じて生来のタンパク質に関連して多くの異なる方法で改変されてもよい。ヘテロオリゴマータンパク質では、生来のヘテロオリゴマータンパク質の1種以上のタンパク質サブユニットの分泌シグナルを形成する生来のリーダー配列が植物‐特異的シグナルペプチドに置換されてもよい。上記の植物‐特異的シグナルペプチドはタバコカルレティキュリンおよび/またはイネアルファ‐アミラーゼに由来してもよい。ヘテロオリゴマータンパク質の少なくとも1種または少なくとも2種以上のサブユニットが植物細胞においてサブユニット由来のヘテロオリゴマータンパク質のアセンブリーを改善するための小胞体保持シグナルKDELを含有していてもよい。さらに、タンパク質サブユニットをコードする異種配列が変異してヘテロオリゴマータンパク質からグリコシル化部位を部分的に、または完全に除去してもよい。その上、発現されることになるヘテロオリゴマータンパク質のグリコシル化パターンが、たとえば1種以上のグリコシルトランスフェラーゼのような、植物グリコシル化装置の構成要素を工学的に操作することによって変えられてもよい。
【0043】
本発明のヘテロオリゴマータンパク質は一般に公知の手順に従って発現後に植物体、植物組織または植物細胞から分離されてもよい。上記のヘテロオリゴマータンパク質はその後実質的に均質に精製されてもよく、そのような状態はクーマシー染色したSDS‐PAGE上のヘテロオリゴマータンパク質に起因するバンドが、慣用のゲルリーダーにより測定したレーンの染色の少なくとも70%、好ましくは少なくとも80%、そして最も好ましくは少なくとも90%を占めると定義されてよい。
【0044】
発明の詳細な説明
本発明に記載のウイルスベクターは、上記のヘテロオリゴマータンパク質を形成するために必要なすべてのタンパク質サブユニットを単一のベクターがコードするウイルスベクター、または上記の少なくとも2種のウイルスベクターのそれぞれが、上記のヘテロオリゴマータンパク質を形成するために必要な異なるタンパク質サブユニットをコードする、少なくとも2種の異なる非競合ウイルスベクターのいずれかである。上記の非競合ウイルスベクターのそれぞれは、組換えヘテロオリゴマータンパク質の1種より多いサブユニットをコードする1種より多い異種核酸配列を発現することができる。上記のRNAウイルスベクターは植物細胞に一過性に送達されてもよく、あるいはDNA前駆体(複数の前駆体)として植物染色体DNAに安定に組み込まれてもよい。
【0045】
本発明は植物細胞における高収率のヘテロオリゴマータンパク質産生のための方法を提供する。この方法は、たとえば発現されることになる異種配列のサイズ制限、上記ベクターの高い不安定性および同じ植物細胞に異なる核酸配列を同時発現できないこと、のような既存のウイルスベクターを基礎にした発現系の制限を克服する。さらに、系からのウイルスコートタンパク質の除去が感染性ウイルス粒子の形成および野生型ウイルスへの逆戻りを妨げるため、上記方法はよりすぐれた生物学的安全性を提供する。本発明を実施することにより、関心のあるヘテロオリゴマータンパク質の高収率発現系の設計が実質的に任意の植物RNAウイルス‐由来レプリコンにとって可能であり、レプリコンは、関心のあるヘテロオリゴマータンパク質の異なるサブユニットをコードする、少なくとも2種の関心のある異種配列を発現可能にするためのレプリコンの改変によって、関心のある異種配列の発現に適切になる。あるいは、同じ植物細胞においてウイルスベクターと一緒に同時複製することができる別の非競合ウイルスベクターを見いだすことも可能である。
【0046】
異なる分類学的群に属するプラス‐センス1本鎖RNAウイルス(本明細書では、簡潔に「RNAウイルス」とも表す)は、本発明のプラス‐センス1本鎖RNAウイルスベクター(本明細書では、簡潔に「ウイルスベクター」とも表す)を構築するためにふさわしい。本明細書では、ウイルスベクターは植物細胞において複製する:すなわち鋳型としてRNAウイルスベクターを使用して、RNA‐依存的RNA重合により別のRNAベクター分子を形成することができるRNAベクターである。好ましくは、本発明のウイルスベクターはRNAウイルス機能を有する、少なくとも1種のウイルス配列エレメント、たとえば、レプリカーゼ、サブゲノムプロモーター、ウイルス粒子アセンブリーの起点、コートタンパク質ORF、または移行タンパク質ORFを含有する。さらに、ウイルスベクターはRNAウイルスIRESエレメントを有していてもよい。
【0047】
ウイルスベクターは、たとえばウイルスに制限部位を導入することにより、それが由来するウイルスから構築してもよく、制限部位は本発明の異種配列を導入するためにふさわしいクローニング部位を表す。当業者は、RNAウイルスまたはベクターに対する核酸の工学的操作は一般に、それぞれ上記のRNAウイルスまたはベクターのDNAコピーを使用して、DNAレベルで行われることを理解することになる。したがって、RNAウイルスベクターのDNAコピーは、RNAウイルスのDNAコピーに制限部位を導入することにより、それが由来するRNAウイルスのDNAコピーから構築することができ、制限部位はDNAレベルで本発明の異種配列のDNAコピーを導入するためにふさわしいクローニング部位を表す、と述べることがより正確である。これらの事柄は当業者には明白であり、一般に本明細書では強調されない。
【0048】
RNAウイルスのDNAコピーがクローニングにふさわしい制限部位(複数の制限部位)を天然に有する場合、ウイルス自体がウイルスベクターであってもよい。本明細書では、「ウイルスベクター」という用語は、異種配列がベクターに存在しない事例か、または本発明のタンパク質サブユニットをコードする配列がベクター中に全く存在しない事例を表す。異種配列または本発明のタンパク質サブユニットをコードする配列がウイルスベクターに挿入される事例は、そのようなインサートの存在を明白に特定することにより確認される。
【0049】
本明細書では、異種配列または本発明のタンパク質サブユニットをコードする配列が導入される以前は2種のウイルスベクターの配列が異なる場合、そのようなウイルスベクターは異なると表す。異種配列または本発明のタンパク質サブユニットをコードする配列が導入される前は2種のウイルスベクターの配列が同じ配列を有する場合、それらは同じであると表す。したがって、2種のウイルスベクターが異なるか、または同じであるかどうかを確認する場合、異種配列または本発明のタンパク質サブユニットをコードする配列は考慮にいれない。
【0050】
本明細書では、「レプリコン」または「ウイルスレプリコン」という用語は「ウイルスベクター」と同じ意味を有する。
本発明のウイルスベクターを工学的に操作するために使用可能なRNAウイルスのリストを以下に提示する。(イタリック体ではない)引用符に囲まれた分類名は、この分類がICTVにより国際的に認可された名称を持たないことを示す。種の名称(通称)は通常の活字体で示す。属または科への正式な帰属のないウイルスが示される。
【0051】
RNAウイルス:
ssRNAウイルス:
科:ブロモウイルス科、
属:アルファモウイルス、基準種:アルファルファモザイクウイルス、
属:イラルウイルス、基準種:タバコ条斑ウイルス、
属:ブロモウイルス、基準種:ブロモモザイクウイルス、
属:ククモウイルス、基準種:キュウリモザイクウイルス、
科:クロステロウイルス科、
属:クロステロウイルス、基準種:ビート萎黄ウイルス、
属:クリニウイルス、基準種:レタス伝染性黄斑ウイルス、
科:コモウイルス科、
属:コモウイルス、基準種:カウピーモザイクウイルス
属:ファバウイルス、基準種:ソラマメウィルトウイルス1,
属:ネポウイルス、基準種:タバコ輪点ウイルス;科:ポティウイルス科、
属:ポティウイルス、基準種:ジャガイモウイルスY、プラムポックスウイルス;タバコエッチウイルス;クローバー葉脈黄化ウイルス;タバコベインモットリングウイルス;
属:リモウイルス、基準種:ライグラスモザイクウイルス
属:ビモウイルス、基準種:オオムギ縞萎縮モザイクウイルス;
科:セキウイルス科、
属:セキウイルス、基準種:パースニップイエローフレックウイルス、
属:ワイカウイルス、基準種:ライスツングロスフェリカルウイルス;
科:トムブスウイルス
属:カルモウイルス、基準種:カーネーション斑紋ウイルス、
属:ディアンソウイルス、基準種:カーネーションリングスポットウイルス、
属:マクロモウイルス、基準種:メイズクロロティックモットルウイルス、
属:トムブスウイルス、基準種:トマトブッシースタントウイルス、
帰属されていないssRNAウイルスの属、
属:カピロウイルス、基準種:リンゴステムグルービングウイルス;
属:カルラウイルス、基準種:カーネーション潜在ウイルス;
属:エナモウイルス、基準種:ピーエネーションモザイクウイルス、
属:フロウイルス、基準種:ムギ類萎縮ウイルス、
属:ホルディウイルス、基準種:ムギ斑葉モザイクウイルス、
属:イダエオウイルス、基準種:ラズベリーブッシードワルフウイルス;
属:ルテオウイルス、基準種:オオムギ黄萎ウイルス、
属:マラフィウイルス、基準種:マイズラヤドフィノウイルス;
属:ポテクスウイルス、基準種:ジャガイモウイルスX;
属:ソベモウイルス、基準種:インゲンマメ南部モザイクウイルス、
属:テヌイウイルス、基準種:イネ縞葉枯ウイルス、
属:トバモウイルス、基準種:タバコモザイクウイルス、
属:トブラウイルス、基準種:タバコ茎えそウイルス、
属:トリコウイルス、基準種:リンゴクロロティックリーフスポットウイルス;
属:ティモウイルス、基準種:ターニップイエローモザイクウイルス;
属:ウムブラウイルス、基準種:キャロットモットルウイルス;
ネガティブssRNAウイルス:目:モノネガウイルス目、科:ラブドウイルス科、属:シトラブドウイルス、基準種:レタスネクロティックイエローズウイルス、
属:ヌクレオラブドロウイルス、基準種:ポテトイエロードワーフウイルス。
【0052】
RNAウイルスベクターは、関心のある遺伝子の発現を提供する異種RNAの極めて多数のコピーを植物細胞に提供することができる。しかし、そのようなベクターは、異種核酸配列のサイズがある範囲、通常1kbを越えて増大すると、極めて不安定になることが公知である。そのような制約のために、そのようなベクター系の適用は、今まで比較的単純な小〜中程度のサイズのタンパク質に限定されてきた。高分子または複雑なヘテロオリゴマータンパク質のいずれかを発現させる試みは首尾よい結果を導いてこなかった。我々は、驚くべきことに、以前には不可能であった複雑なヘテロオリゴマータンパク質の高収率発現のためにウイルスベクターを首尾よく採用できることを見いだしている。全長モノクローナル抗体を発現させる初期の試み(Verch et al.,1998,J.Immunol.Meth.,220,69−75)は、同じ細胞における関心のあるタンパク質の同時発現のために使用されるウイルスベクターの不適合性のために、満足すべき結果を提供しなかった。実施例1で、我々は、単一細胞レベルで、同じ植物RNAウイルス(TMV)由来のウイルスレプリコンから2種の異なる遺伝子(GFPとDsRed)の効率的な同時発現が不可能であることを証明している。図1(A‐右パネル)では、レポーター遺伝子‐DsRedおよびGFPの両方の発現を示すプロトプラストを検出できなかった。右下部パネルの一部のプロトプラストにおけるDsRedの弱い発現パターンは、強いGFP発現(右上部パネル)と一致し、そしてDsRed検出に使用されたフィルターの漏出による偽‐陽性の結果である。この漏出は高濃度のGFPを含有するプロトプラストの見かけの弱いDsRed蛍光を導く。したがって、同一であるか、または広範な相同性領域を共有するRNAレプリコンが異なる組換えタンパク質をコードする異なる異種核酸配列をたとえ持っていても、それらは1つの植物細胞に共存できない。この現象の理由は、今のところ解っていない。可能性のある説明として、1種のウイルスレプリコンのコピー数の急激な増加が、別のウイルスレプリコンを速やかに撃退することが考えられる。選択された細胞において2番目に複製を開始するレプリコンは、第1のレプリコンに追いつくことができず、したがって第1レプリコンを確認する事象は主に統計的特徴を有する。
【0053】
この問題に取り組むための我々の研究では、2種の異なるサブゲノムプロモーターの制御下で、異なるタンパク質をコードする2種の異なる異種核酸配列が上記のレプリコン中に存在するように、ウイルスレプリコンを工学的に操作してきた。そのようなRNAレプリコンをコードするT−DNA領域の一般的スキームは図2に示す。そのようなベクターの設計および最適化のために、我々は、便宜上、我々の初期の特許出願に記載のプロベクター法を使用してきた(WO02088369;Marillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857も参照されたい)。この方法は、部位特異的組換えにより、事前に作製されたモジュールに由来する最終ベクターのin plantaでの集合を可能にし、そうしてベクター設計およびベクター最適化を著しくスピードアップさせる。コンストラクトの設計は実施例2に記載し、図式表示は図3に示す。
【0054】
我々は驚くべきことに、最終レプリコンにおけるインサートの大きなサイズ、および2種の強いサブゲノムプロモーターの存在によるベクターの複雑な構造にもかかわらず、得られたRNAレプリコンは、in plantaでの高い安定性および同じ植物細胞における2種の異なる組換えタンパク質(GFPとDsRed)の同時発現を提供する能力を示すことを見いだしている。図4に示すように、同時発現頻度は感染した範囲ではすべての細胞のほとんど100%にまで到達可能である。そのようなコンストラクト中におけるレポーター遺伝子(GFPとDsRed)と、たとえばIgGの軽鎖および重鎖の置換、その上のインフィルトレーションされたN.ベンサミアーナ葉部における発現は驚くべき結果を生み出した。実施例3で記載するように、ウェスタンブロット解析は見事なほどの高濃度の集合したモノクローナル抗体(レーン6および7;図6)を明らかにした。我々の系によって提供される集合したモノクローナル抗体の収率は、現在の当該技術分野の系の状態によって生み出された収率とは比較にならないほど高い(図6、レーン4)。知る限りでは、これは、植物ウイルスベクターを基にした発現系を使用することによる、抗体のような複雑なヘテロオリゴマータンパク質発現の初めての証拠である。初期のすべての刊行物はモノクローナル抗体の簡単な人工誘導体の発現、たとえばTMV−ウイルスベクターを使用した(McCormick et al.,Proc Natl Acad Sci USA,96,703−708;McCormick et al.,2003,J.Immunol.Methods,278,95−104)およびPVXを基礎にした(Smolenska et al.,1998,FEBS Lett.441,379−382;Franconi et al.,1999,Immnotechnology,4,189−201;Hendy et al.,1999,J.Immunol.Methods,231,137−146;Roggero et al.,2001,Protein Expr.Purif.,22,70−74)1本鎖抗体(scFv)の発現に限定されていた。
【0055】
本発明では、好ましくは全身ウイルスベクターを使用しない。全身ウイルスベクターのかわりに、好ましくは植物系へのアグロバクテリウム媒介ウイルスベクター前駆体送達により、ウイルスベクターの能力を全身移行するように置換する。このことはウイルスコートタンパク質を、発現されることになる異種配列で置換することを可能にする。この方法は、ウイルスベクターの異種配列への適応性増大の可能性に貢献する。同時に、この方法はこの系の生産性を弱めることになる自発的な異種配列の欠失のために、ウイルスベクターが野生型ウイルスに変換される見込みを排除する。
【0056】
我々のウイルス‐レプリコンを基礎にした系は、先行技術の系より著しく改善された収率のモノクローナル抗体を産生するが、我々はウイルスレプリコンのサイズを減少させることによりそれ以上の収率増加の可能性を検討した。分泌抗体の2つの鎖を発現するウイルスレプリコンのサイズを減少させる可能性が限定されていることを考慮して、2種の異なるウイルスレプリコンからの抗体の重鎖および軽鎖の発現を試みてきた。同じウイルスを基礎にしたレプリコンは同じ植物細胞において2種の異なる異種配列を発現できない(上記参照、実施例1)ことを考慮して、非相同植物ウイルスであるタバコモザイクウイルス(TMV)とジャガイモウイルスX(PVX)に由来するウイルスベクターに、DsRedとGFP遺伝子を別々にクローニングすることにより、同じ植物細胞に2種の異なる遺伝子を同時発現させる可能性を試験した(実施例4を参照されたい)。ウイルスベクターのcDNAを含有するT−DNA領域の図式表示は図7および8(PVXのみ)に示す。実施例4では、便宜上、部位特異的組換えによりベクターモジュールからin plantaで集合させたTMVを基礎にしたウイルスベクターを使用した。驚くべくことに、共にインフィルトレーションされた植物葉領域から分離したプロトプラストはDsRedおよびGFPレポーター遺伝子の実質的に100%の同時発現頻度を示すことを我々は見いだしている(図9)。
【0057】
最近、異なって標識されたウイルスの空間的分離に関する研究がDietrichとMaiss(2003,J.Gen.Virology,84,2871−2876)によって提示された。この研究では、レポーターとして蛍光タンパク質が発現した。ヘテロオリゴマータンパク質の産生には言及しなかった。さらに、分離したプロトプラストレベルでの同時発現は検討しなかった。レポーター遺伝子産物は原形質連絡を介した拡散により隣接する細胞に拡散できるため、選択されたウイルス対がレポーター遺伝子を、同じ細胞で、または隣接する細胞で発現したかどうかについての情報は全く得られなかった。初期の刊行物は植物の感染時の野生型ウイルス間の相乗作用について述べているが、植物細胞におけるヘテロオリゴマータンパク質の発現については述べていない。(Rochow & Ross,1955,Virology,1,10−27;Goodman & Ross,1974,Virology,58,16−24;Goodman & Ross,1974,Virology,59,314−318;Goodman & Ross,1974,Virology,58,263−271)。同時感染した植物におけるウイルスの相乗的相互作用に関するこれらの初期の研究(Vance et al.,1995,Virlogy,206,583−590;Pruss et al.,1997,Plant Cell,9,859−868)のその後の発展は転写後遺伝子サイレンシング(PTGS)のサプレッサーの発見を導いた。組換えタンパク質発現系のさらに進んだ発展は、促進された組換えタンパク質産生のためのそのようなPTGSサプレッサーの研究に向けられた。相乗的ウイルスを基礎にしたウイルス発現系を使用した先行技術にはタンパク質発現に対する手がかりが全く存在しない。
【0058】
本発明は、非競合ウイルスベクターが植物細胞または植物体におけるヘテロオリゴマータンパク質産生の効率的な手段を表すことを証明する。2種以上のウイルスベクターが同じ植物細胞において複製可能であり、そして上記の複製および別の細胞からのトランスフェクション中にお互いに効力を低下させない場合、上記のウイルスベクターはお互いに非競合的である。効力低下がないとは、少なくとも10%、好ましくは50%、より好ましくは90%、そして最も好ましくは100%のトランスフェクションされた植物細胞が共にトランスフェクションされること、すなわち上記の2種以上の異なるウイルスベクターを含有することを意味する。好ましくは、上記のウイルスベクターは異種核酸配列を複製し、発現することが可能である。より好ましくは、上記の異種核酸配列が異なるタンパク質をコードする。よりいっそう好ましくは、上記の異なるタンパク質がヘテロオリゴマータンパク質のサブユニットである。ウイルスベクターが競合的であるか、または非競合的であるかを確認するために、植物細胞の同時トランスフェクションの頻度を測定することができる。このことは以下のプロトコルに従って行うことができる:2種のウイルスベクターは2種の異なるレポーター遺伝子、たとえばDsRedとGFPで標識される。上記の、異なって標識されたベクターは植物葉組織に(たとえばアグロ‐インフィルトレーション法により)同時送達され、3〜6日後、感染した領域から分離したプロトプラストを計数して、1種のレポーター遺伝子だけを発現しているプロトプラスト数に比較した、両方のレポーター遺伝子を同時発現しているプロトプラストの割合を決定することができる。そのような測定値を例示する実験は実施例2および4に記載する(図1Aおよび9も参照されたい)。通常、非競合ウイルスベクターは相乗的であるウイルスに由来することができ、たとえば、同じ植物宿主に首尾よく同時トランスフェクションすることができる。そのような相乗的RNAウイルス対の例としては以下のものが挙げられる:
ジャガイモウイルスX(PVX)/タバコモザイクウイルス(TMV);
PVX/タバコベインモットリングウイルス(TVMV);
PVX/タバコエッチウイルス(TEV);
PVX/クローバー葉脈黄化ウイルス(CIYVV);
PVX/プラムポックスウイルス(PPV);
PVX/ジャガイモウイルスY(PVY)。
【0059】
ウイルスベクターの競合性または非競合性の機構的理由は既知ではない。可能性のある1つの説明としては、相乗的ウイルスベクターに由来するウイルスレプリコンが異なる細胞下コンパートメントにおいてウイルス複製複合体(VRC)を形成する(Kawakami et al.,2004,Proc.Natl.Acad.Sci.USA,101,6291−6296)ことが挙げられ、したがってそれらはVRCを形成するために必要な空間に対してお互いに競合しない。実験的に証明されているように、非競合ウイルスベクターは、核酸配列レベルが著しく異なる、異なるウイルス種に由来してもよい。上記の非競合ウイルスベクターは、同じ植物宿主に首尾よく同時感染できる相乗的植物ウイルスに由来してもよい。相乗的ウイルスは通常異なる属に属する。たとえば、PVXはポテクスウイルス属に属するが、それに対して相乗的なウイルス、たとえばTVMV、TEV、PPV、およびCIEVVはポティウイルス属に属する。PVXウイルスに相乗的(非競合的)な別のウイルス、たとえばTMV、TVCV、およびcrTMVはトバモウイルス属に属する。明らかに、異なる属の典型間のゲノム相同性はかなり低く、通常、RNAレベルで50%同一性より低い。
【0060】
異なるウイルスベクター(たとえば、PVXおよびTMVを基礎にしたもの)へのモノクローナル抗体重鎖および軽鎖のクローニング、続いて同時感染した植物組織における上記の鎖の付加的な同時発現は実施例5に記載する。ベクターの図式表示は図10に示す。本発明および別の発現系により産生されたモノクローナル抗体の収率の比較ELISA測定は、2種の非競合ウイルスベクターを基礎にした上記の系の最も好ましい成果を示した(図11)。ウイルスベクター(複数のウイルスベクター)の補助による植物細胞での組換えヘテロオリゴマータンパク質の効率的な発現は先行技術では公知でない。
【0061】
我々のデータに基づいた場合、ウイルスベクターのウイルスに由来する配列は、上記ウイルスがお互いに関連するとみなすために十分な相同性を示さないことになる。そのようなウイルスの例としては、TMVとPVXのウイルス対が挙げられ、そしてウイルスベクターはそれに基づく。これらのウイルスベクターはそのような相同性を全く示さない。受容可能なウイルス対を選択するための別の必要条件は、植物宿主に同時感染中の、元の野生型ウイルスの相乗性であってもよい。
【0062】
上で論じた実験は、植物細胞へのアグロバクテリウムに媒介されたDNA前駆体送達に基づいた一過性発現系により行われた。しかし、本発明の代わりの適用としては、植物核ヌクレオソームに安定に組み込まれた上記のRNAレプリコン(複数のレプリコン)のDNA前駆体によるトランスジェニック植物への適用が挙げられる。これは、植物ウイルスベクターを基礎にした系の多くの制約、たとえば、ウイルスベクターが耐えうる異種配列の最大サイズに対する制限を克服する。DNA前駆体はトランスジェニック植物のそれぞれの細胞に存在することになるため、RNAレプリコンの全身移行または細胞間移行(レプリコンスプレッディング)のための絶対的な要求性が存在しない。これは、高い効率の本発明のRNAレプリコン形成および細胞質への輸送によって補われる。しかし、RNAレプリコン形成がすべての細胞でいつも起こるわけではないので、ベクターの細胞間移行の能力は付加的な価値のものであってもよい。
【0063】
異なる方法を使用して、植物体、植物組織または植物細胞に異種DNAを提供してもよい。上記のベクターは、アグロバクテリウム(US5,591,616;US4,940,838;US5,464,763)または粒子もしくはマイクロプロジェクタイルボンバードメント(US05100792;EP00444882B1;EP00434616B1)によって運ばれるTi−プラスミドベクターによって、植物細胞を形質転換してよい。別の植物形質転換法、たとえばマイクロインジェクション(WO09209696;WO09400583A1;EP175966B1)、エレクトロポレーション(EP00564595B1;EP00290395B1;WO08706614A1)またはプロトプラストのPEG‐媒介形質転換なども使用することができる。ベクター送達方法の選択は、形質転換されることになる植物種に依存してもよい。たとえば、マイクロプロジェクタイルボンバードメントは一般に単子葉植物の形質転換に好ましいが、双子葉植物には、一般にアグロバクテリウム媒介形質転換がより好ましい結果を示す。
【0064】
本発明の実施例では、ニコチアナ細胞へのベクター(上記の異種DNA)の一過性アグロバクテリウム媒介送達を使用した。しかし、上記ベクターは、関心のある植物種の安定な、または一過性形質転換にふさわしい標準技術のいずれかに従って、植物体に安定に導入されてもよい。双子葉植物のための形質転換技術は当該技術分野で公知であり、アグロバクテリウムを基礎にした技術とアグロバクテリウムを必要としない技術が挙げられる。アグロバクテリウムを使用しない技術には、プロトプラストまたは細胞による直接的な外来遺伝物質の取り込みが挙げられる。これらの技術には、PEGまたはエレクトロポレーション媒介取り込み、粒子ボンバードメント‐媒介送達およびマイクロインジェクションが挙げられる。これらの技術の例は、Paszkowski et al.,EMBO J3,2717−2722(1984)、Potrykus et al.,Mol.Gen.Genet.199,169−177(1985)、Reich et al.,Biotechnology 4:1001−1004(1986)、およびKlein et al.,Nature 327,70−73(1987)に記載される。それぞれの場合、形質転換された細胞は標準技術を使用して、全植物体に再生される。
【0065】
アグロバクテリウム媒介形質転換は、その高い形質転換効率と多くの異なる種に対するその広範な有用性のために、双子葉植物の形質転換に好ましい技術である。通常アグロバクテリウムによって形質転換することができる多くの作物種には、タバコ、トマト、ヒマワリ、綿花、ナタネ、ジャガイモ、ダイズ、アルファルファおよびポプラが挙げられる(EP0317511(綿花)、EP0249432(トマト)、WO87/07299(ナタネ)、米国特許第4,795,855号(ポプラ))。
【0066】
本発明の実施例では、関心のある遺伝子(複数の遺伝子)の一過性発現にT−DNAのアグロバクテリウム媒介送達(Vaquero et al.,1999,Proc.Natl.Acad.Sci.USA,96,11128−11133)が使用された。この方法は、小規模〜中規模の組換えタンパク質産生系にとってだけでなく、大規模発現にとっても極めて有用な手段である。
【0067】
植物染色体DNAに安定に組み込まれたウイルスレプリコン前駆体の放出は、誘導型またはいずれか別の調節された(たとえば発達的に調節された)プロモーターを使用して行うことができる。誘導型プロモーターはそれらの誘導条件に従って、2つのカテゴリーに分類することができる:非生物的因子(温度、pH、化学物質)によって誘導されるものおよび生物因子、たとえば病原体または害虫攻撃によって誘導されるもの。第1のカテゴリーの例としては、熱‐誘導型(US05187287)および低温‐誘導型(US05847102)プロモーター、銅‐誘導系(Mett et al.,1993,Proc.Natl.Acad.Sci.,90,4567−4571)、ステロイド‐誘導系(Aoyama & Chua,1997,Plant J.,11,605−612;Mcmellis et al.,1998,Plant J.,14,247−257;US06063985)、エタノール‐誘導系(Caddick et al.,1997,Nature Biotech.,16,177−180;WO09321334)、およびテトラサイクリン誘導系(Weinmann et al.,1994,Plant J.,5,559−569)が挙げられる。植物の化学的誘導系の分野での最も最近の成果の1つは、グルココルチコイドデキサメタゾンによりスイッチを入れ、テトラサイクリンによりスイッチを切ることができるキメラ化プロモーターである(Bohner et al.,1999,Plant J.,19,87−95)。化学的誘導系に関する総説としては以下を参照されたい:Zuo & Chua,(2000,Current Opin.Biothechnol.,11,146−151)およびPadidam,M(2003,Curr.Opin.Plant Biol.,6,169−177)。誘導型プロモーターの別の例としては、植物における病原性‐関連(PR)遺伝子の発現を制御するプロモーターが挙げられる。これらのプロモーターは、病原菌攻撃に反応する植物シグナリング経路の重要な構成要素であるサリチル酸、またはPR遺伝子発現を引き起こすことが可能な別の化合物(ベンゾー1,2,3−チアジアゾールまたはイソニコチン酸)(US05942662)による植物の処理によって誘導することができる。
【0068】
本発明は実施例に記載のTMVおよびPVXを基礎にしたベクターに限定されないが、上記ウイルスレプリコンに由来する発現系の確立に役立つ、別の植物RNAウイルスに由来するレプリコンに適用可能である。同時感染された植物において最も詳しく研究された相乗作用はウイルスのPVX/PVY対に関して公知である(Rochow & Ross,1955,Virology,1,10−27;Goodman & Ross,1974,Virology,58,16−24)。おそらく、多くのそれらのウイルスは同じ植物細胞中で共に複製可能である。Dietrich & Maiss(2003,J.Gen.Virol.,84,2871−2876)は、異なって標識されたウイルス対、たとえばプラムポックスウイルス(PPV)とジャガイモウイルスX(PVX)、タバコベインモットリングウイルス(TVMV)とPVX、クローバー葉脈黄化ウイルス(CIYVV)とPVXは、植物組織中の同じ感染領域において異なるレポーター遺伝子を同時発現できることを示している。本発明に記載のストラテジーを使用して、同じ植物細胞中で同時複製可能な、実質的に任意の対の植物プラスセンス1本鎖RNAウイルス‐由来レプリコンに対する組換えヘテロオリゴマータンパク質発現系が開発可能である。たとえば、アルファモウイルス属のアルファルファモザイクウイルス(AMV)に基づいたウイルスベクターが本発明で使用可能である。
【0069】
複雑な(ヘテロオリゴマー)タンパク質、それらの(機能的または非機能的)フラグメントならびにそれらの人工誘導体および融合物をコードする関心のある遺伝子は、本発明を使用して、植物体または植物細胞に発現することができる。多くの商業的に価値のあるヘテロオリゴマータンパク質の群は本発明を使用して生産し、精製することができる。それらの群には、工業用および研究用タンパク質、ならびにヒトまたは動物の健康分野における適用のためのタンパク質が挙げられるが、それらに限定されない。しかし、免疫反応タンパク質、とりわけ異なるクラスのイムノグロブリン(IgG、IgM、IgAおよびIgD)から選択されたモノクローナル抗体ならびにそれらの合成誘導体、たとえば変異型および別のタンパク質またはその一部との異なる型の融合体が最も好ましい。
【0070】
実施例
以下の実施例は本発明を説明するために提示される。本発明の意図および範囲から逸脱せずに改変および変更を行ってもよい。
【0071】
実施例1
同じ植物RNAウイルスに由来する2種のTMVを基礎にしたRNAウイルスベクターからのGFPとDsRedの同時発現の欠如
葉組織は同じ植物RNAウイルスに由来するが、関心のある2種の異なる遺伝子を含有する、2種のTMVを基礎にしたRNAウイルスベクターに感染された(図1)。TMVを基礎にしたウイルスベクターにクローニングした可視マーカー遺伝子GFPおよびDsRedを使用して、この方法を試験した。第1コンストラクト、pICH17272(Marillonnet et al.,2005,Nat.Biotechnol.,23,718−723の図1を参照されたい)はGFPを含有し、そしてベクター中のRdRPコーディング配列が14個よりむしろ9個の植物イントロンを含有すること以外は、pICH18711(国際特許出願PCT/EP03/12530,WO2005049839として公開;Marillonnet et al.,2005,Nat. Biotechnol.,23,718−723を参照されたい)に類似する。pICH18722(Marillonnet et al.,2005,Nat.Biotechnol.,23,718−723)はTMVの中の2種の密接に関連した株、CrTMV(Dorokhov et al.,1994,FEBS Lett.350,5−8)およびTVCV(Lartey et al.,1994,Arch.Viol.138,287−298)の骨格に構築され、コートタンパク質配列が異種配列によって置換されている、植物プロモーターの制御下にあるウイルスゲノムを含有する。RdRPおよびMP中の11個のイントロンの存在は、ベクターの葉組織へのアグロバクテリウム媒介送達後のウイルス複製開始の効率を増し、したがって同じ細胞における2種のウイルスベクターの同時発現の見込みを増大させる。GFPコーディング配列がXho1−Not1制限部位を使用してDsRedのコーディング配列によって置換されていること以外は、pICH18505は、pICH17272に類似する(図1)。pICH18505およびpICH17272のT−DNA領域の詳細な制限地図は図1Cに示す。pICH17272およびpICH18505はアグロバクテリウム株GV3101を形質転換し、先に記載(Marillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857)のように、ニコチアナベンサミアーナ葉組織がインフィルトレーションされた。プロトプラストはインフィルトレーション7日後にインフィルトレーション部位から作製し、顕微鏡下、青色または赤色光下で観察した。プロトプラストが数日後にインフィルトレーション部位から作製された場合でも、GFPとDsRedの両方を発現したのは非常にわずかのプロトプラストだけであった。このことは細胞が第1のTMVを基礎にしたウイルスベクターにひとたび感染すれば、第2のTMVを基礎にしたウイルスベクターによる再感染は不可能になることを示唆する。
【0072】
実施例2
単一ウイルスレプリコンからのGFPおよびDsRedの同時発現
2種の別々のサブゲノムプロモーターの制御下、関心のある2種の遺伝子がRNAウイルスベクターから発現された(図2)。両方のサブゲノムプロモーターは同一であってもよいが、ウイルス複製中におけるコンストラクト中の反復配列間の欠失のリスクを回避するために、関連するTMV株由来のサブゲノムプロモーターを使用することが好ましく;この場合、第2のサブゲノムプロモーターおよび3′非翻訳領域はTMGMV株U5に由来する(Shivprasad et al.,1999,Virology,255,312−323;Marillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857)。また、クローニングの便宜上、GFPおよびDsRedは、完全にアセンブルされたベクターよりむしろ、ウイルスプロベクター(WO02/088369およびMarillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857に記載)にクローニングされた。2種のプロベクター、pICH17388およびpICH15933(図3)は、in plantaで、両方のプロベクターモジュール間の部位特異的組換えにより完全に機能的なTMVを基礎にしたウイルスベクターに変換された。pICH17388は、(pICH17272と同じように)ウイルス配列に11個のイントロンが存在すること以外はpICHNOP(Marillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857)に類似する。pICH15933は、DsRedのコーディング配列によってTMGMV U5のコーディング配列が置換されている以外は、pICHGFPSYS(Marillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857)に相当する。
【0073】
pICH15933はアグロバクテリウム株GV3101に形質転換され、pICH17388およびpICH10881(図3)と一緒にニコチアナベンサミアーナにインフィルトレーションした。インフィルトレーション5日後に、すべてのGFP‐発現部位はDsRedも発現し、同じ植物細胞中における2種の遺伝子の卓越した同時発現を示した(図4)。
【0074】
実施例3
単一のウイルスレプリコンから抗体の発現
pICH15933におけるGFPおよびDsRedのコーディング配列は、IgG抗体軽鎖および重鎖によってそれぞれ置換され、コンストラクトpICH20241(図5)を得た。対照として、重鎖および軽鎖はTMVを基礎にしたプロベクターにクローニングし、pICH1740のコーディング配列を置換し、コンストラクトpICH20421およびpICH20431(図5)を得た。pICH20241はpICH17388およびpICH10881と一緒にニコチアナベンサミアーナ葉部にインフィルトレーションした(図3)。
【0075】
インフィルトレーションされた葉部から抽出した全可溶性タンパク質のウェスタンブロット解析は、ホースラディッシュペルオキシダーゼ(HRP)(Sigma)により標識した、1:6000に希釈した抗‐ヒトIgGウサギ抗体により行った。解析の結果は図6Aに示す。単一のウイルスベクターの補助によって達成された抗体の発現レベル(レーン6&7、図6)は、PTGSサプレッサーP19の存在においてさえ、強力な構成プロモーターの補助により成し遂げられたものより著しく高い(レーン3〜4、図6)。
【0076】
実施例4
TMVおよびPVXを基礎にしたウイルスベクターによるGFPおよびDsRedの同時発現
2種の遺伝子の同時発現のための別のストラテジーは、同じ細胞に同時感染し、複製することが可能な、異なるウイルスに基づいて構築された別々のウイルスベクターを使用することである。例として、ジャガイモウイルスX(PVX)に基づいた発現ベクターはTMVと一緒に同じ細胞に共に存在することができる。2種のそのような非競合ウイルスベクターの図式表示は図7に示す。
【0077】
PVX(PV0014株)の接種材料は、感染した乾燥葉材料として、German Collection of Microorganisms and Cell Cultutes(DSMZ)から得て、ニコチアナベンサミアーナ植物体の接種に使用した。ウイルス症状を示した接種植物の全身葉部は全RNA調製に使用した。第1鎖cDNAはプライマーpvxpr2、pvxpr4およびpvxpr6を使用して作製した。3種のcDNAフラグメントはPfu−Taqポリメラーゼミックスを使用して、PCRによりPVXcDNAから増幅した:
フラグメント1はプライマー:
【0078】
【化1】

【0079】
により増幅した。
フラグメント1はBsaIにより消化した。
フラグメント2はプライマー:
【0080】
【化2】
【0081】
により増幅した。
フラグメント2はEsp3Iにより消化した。
フラグメント3はプライマー:
【0082】
【化3】
【0083】
により増幅した。
フラグメント3はBpiI AatIIにより消化した。
アラビドプシスアクチン2遺伝子プロモーターを含有するフラグメント4(An et al.,1996,Plant J.10,107−121)はアラビドプシスゲノムDNAからプライマー:
【0084】
【化4】
【0085】
により増幅した。
フラグメント4はMlu1およびBsa1により消化した。
アラビドプシスアクチン2プロモーターの代わりとして、さらにCaMV35プロモーターを使用して、PVXを基礎にしたウイルスベクターの転写を行った(図10D、pICH20788;pICH21380)。PVXを基礎にしたベクターを作製する一般的ストラテジーはBaulcombe and colleagues(1995,Plant Journal,7:1045−1053)に記載のように使用した。
【0086】
4種すべてのフラグメントは、Mlu1およびAatIIにより消化したバイナリーベクター中に一緒にクローニングした。得られたコンストラクトの20個のクローンはアグロバクテリウム株GV3101に形質転換し、それぞれのクローンはニコチアナベンサミアーナ植物体の1枚の葉にインフィルトレーションした。1週間後、ウイルス感染症状の表現型に対してニコチアナベンサミアーナをスクリーニングし、陽性のプラスミドクローンを保存した(pICHPVX)。
【0087】
クローニングベクターは、CPサブゲノムプロモーター下流の、関心のある遺伝子をクローニングするためのこの完全に機能的なcDNAから作製した。GFP遺伝子は例としてクローニングした。CPサブゲノム領域中のATGはAGG(Genbank受託番号M95516中の位置5651)に変異させた。GFP遺伝子はNhe1部位の3′(座標5662〜5667)にクローニングした。PVXの3′末端(座標5618〜6435)はGFP配列の下流にクローニングし、この配列に12〜24Aのストレッチが続いた。このコンストラクト(pICH0130,図8)はニコチアナベンサミアーナ葉部にアグロインフィルトレーションした。GFP蛍光はインフィルトレーション後2〜3日目に出現した。GFPの全身移行は数日後に出現した。
【0088】
pICH0130はベクターpICH17388、pICH10580およびpICH10881(図3)と一緒にニコチアナベンサミアーナ葉部に同時インフィルトレーションし、DsRed発現を提供した。同時インフィルトレーション後6日目にインフィルトレーションした領域からプロトプラストを作製し、青色光の下、顕微鏡下で観察した。ほとんどすべてのプロトプラストはGFPおよびDsRedを両方とも発現し、卓越した同時発現レベルを示した(図9)。
【0089】
実施例5
TMVおよびPVXを基礎にしたベクターによるモノクローナル抗体の発現
IgGの重鎖および軽鎖はPVXベクターにクローニングし、pIC0130中のGFPのコーディング配列を置換して、それぞれ重鎖または軽鎖のいずれかを含有する2種のコンストラクト、pICH21240およびpICH21370を生み出した(図10A)。また、抗体の軽鎖または重鎖のいずれかを含有するTMVプロベクター部分のクローンが構築された(pICH20431およびpICH20421,図10A)。2種の異なる鎖の発現型であるPVXおよびTMVを基礎にしたベクターを提供するアグロバクテリウムの混合物はN.ベンサミアーナ葉部にインフィルトレーションし、抗体の発現レベルは接種後10日目にELISAによって測定した。結果(図11)は、PVXおよびTMVを基礎にしたベクター由来の重鎖および軽鎖の同時発現の事例における、集合した機能的抗体の極めて高レベルを示す。これらのレベルは両方の鎖を発現しているTMVベクターから得られたものより著しく高かった(実施例3)。
【0090】
IgG1サブクラスに属するさらに別のヒト腫瘍‐特異的モノクローナル抗体、抗癌mabは以下のように、TMVおよびPVXを基礎にしたベクターにサブクローニングした:抗癌mab軽鎖コーディング配列を含有する、730NcoI−NotIフラグメントはNotl部位で平滑化し、NcoI−XbaIにより開き、Xbal部位で平滑化した、TMVを基礎にした3′モジュールクローニングベクターpICH11599(図10B)にリゲーションし、pICH21910を得た。同じストラテジーを使用して、抗癌mab重鎖コーディング配列を持つ1421bpのNcoI−NotIフラグメントをpICH11599(図10B)にサブクローニングし、pICH21920(図10B)コンストラクトを得た。GFP発現のためのPVXを基礎にした3′モジュールpICH21282(図10C)はTMVを基礎にした3′モジュールクローニングベクターpICH10990(図10C)の基部に形成された。それは、GFPコーディング配列のpICH10990(図10C)への挿入および結果として起こるTMV3′非翻訳領域のPVX3′非翻訳領域による置換を介したいくつかのクローニングステップにより成し遂げられた。同様に、TMVを基礎にした3′プロベクターモジュールpICH21920(図10B)は、TMV3′非翻訳領域をPVXの3′非翻訳領域で置換することにより、PVXを基礎にした3′プロベクターコンストラクトpICH22250(図10C)に変換された。それは、HindIII部位で平滑化された、pICH21282(図10C)の575bp HindIII−NdeIフラグメントと、Sall部位で平滑化されたpICH21920のSaII−NdeIフラグメントのリゲーションによって成し遂げられた。抗癌mab軽鎖のコーディング配列を含有する723bp NcoI−EcoRIフラグメントは、NcoI−EcoRIにより消化されたpICH22250(図10C)にサブクローニングし、pICH22240(図10C)を得た。
【0091】
5′プロベクターpICH21380(図10D)は以下のように作製した:コンストラクトpICH17388(図3B)を鋳型として使用し、プライマーpv5p5F(SEQ ID NO:9):
【0092】
【化5】
【0093】
とpv5p5R(SEQ ID NO:10)
【0094】
【化6】
【0095】
の補助によりフラグメントをPCR増幅し、Nhe1およびSac1制限酵素により消化し、pICH20788の大きなNhe1‐Sac1フラグメントとリゲーションし、PVXベクターの3′部位をイントロンおよびAttP組換え部位の5′末端と置換した。クローニングのスキームは図10Dに示す。部位特異的組換えによりin plantaでウイルスベクターを集合させるために、5′プロベクターと3′プロベクターを使用するストラテジーは、以前に記載されたもののとおりであった(Marillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857)。
【0096】
アグロインフィルトレーション手順
得られたすべてのコンストラクトは、標準エレクトロポレーションプロトコルを使用してアグロバクテリウム株GV3101に形質転換し、さらにニコチアナベンサミアーナ葉部のアグロインフィルトレーションに使用した。OD600が1.8〜2.5の範囲の、一晩培養したアグロバクテリウム培養物の等しい容量を混合し、6000gで3分間沈殿させた。ペレットは、10mM MES(pH5.5)および10mM MgSO4を含有する溶液中に再懸濁し、それぞれ個々の培養物の10-1希釈物を得た。針の無いシリンジを使用して、温室で6〜8週間育てたニコチアナベンサミアーナ植物体葉部にインフィルトレーションした。アグロインフィルトレーションのためのプロベクターを使用する場合、アグロバクテリウムミックスは通常5種の異なるアグロバクテリウム株を含み、それらのそれぞれが以下の構成要素の中の1種を提供する:レコンビナーゼ(インテグラーゼ)供与源(pICH10881);5′PVXプロベクター(pICH21380);1種のIgG鎖を提供する1種の3′PVXプロベクター(重鎖のためのpICH22250または軽鎖のためのpICH22240のいずれか、TMVプロベクターによってどの相補鎖が提供されるかに依存)、;5′TMVプロベクター(pICH17388);1種の3′TMVプロベクター(軽鎖をコードするpICH21910または重鎖をコードするpICH21920)。
【0097】
SDS-PAGEとウェスタンブロット
アグロバクテリウムにインフィルトレーションされたN.ベンサミアーナ葉部の試料100mgは、300μlタンパク質抽出バッファー(0.1M Tris、150mM NaClおよび0.1%Tween20、pH8.0)を含む液体窒素中ですりつぶした。葉の粗抽出物は、Laemmliのバッファー系を使用して10%(非還元状態)または12%(還元状態)ポリアクリルアミドゲルに溶解し、その後クーマシーブリリアントブルー染色を行った。ウェスタンブロット解析のために、SDS-PAGE由来のタンパク質は、ブロッテイングバッファー(25mM Tris、192mM グリシン、20%メタノール、pH8.3)を使用して、電気泳動的にHybondP膜(Amersham Biosciences,UK)に移した。膜はTBST(50mM Tris、100mM NaCl、0.05% Tween20、pH7.4)中5%スキムミルクで遮断し、それぞれ2.5%スキムミルク/TBS 1:10000および1:5000で希釈した、ヤギ抗‐ヒトラムダ軽鎖HRP‐結合抗体(Sigma,UK)またはヤギ抗‐ヒトガンマ鎖特異的HRP‐結合抗体(Sigma、UK)により精査した。結合した抗体はECL Detection Reagent(Amersham Biosciences,UK)を使用して検出した。電気泳動による分離およびウェスタンブロット解析の結果は図11Bに示す。IgG標準物との比較は植物組織におけるモノクローナル抗体の収率が新鮮な葉生物量1グラムにつき0.2〜0.35mgに達することを示した。
【0098】
植物体由来組換えMabの単離
全可溶性タンパク質は100mM リン酸ナトリウム、150mM NaClおよび0.05% Tween20(pH8.0)を含有するバッファーにより、アグロ接種したニコチアナベンサミアーナ葉領域から抽出した。小規模な粗タンパク質抽出物からの抗癌Mabの分離は、取扱説明書に従って、Protein A Magnetic Beads(New England Biolabs,USA)によるワンステップアフィニティー精製を使用して行った。精製した植物体由来Mabのクーマシー染色ゲルは図11Bに示す(パネルB)
【0099】
実施例6
TMVおよびPVXを基礎にしたベクターによる卵胞刺激ホルモン(FSH)の発現
ウシFSHのアルファ(GenBank受託番号NM_173901のコーディング配列101〜463に相当するヌクレオチド)およびベータ(GenBank受託番号NM_174060のコーディング配列70〜459に相当するヌクレオチド)ポリペプチドをコードする遺伝子(cDNA)は制限部位(FSH‐アルファおよびベータサブユニットに対して5′Nco1−3′EcoR1)を隣に配置して合成し、TMVプロベクター(pICH20431、pICH11599の誘導体、図10B)およびPVXベクター(pICH21240)にクローニングし、IgGの重鎖および軽鎖のコーディング配列を置換して、それぞれサブユニットアルファおよびサブユニットベータをコードする配列を含有する2種のコンストラクト、pICH−FSHAおよびpICH−FSHBを得た(図12)。PVXおよびTMVを基礎にしたベクターから発現される2種の異なるサブユニットを提供するアグロバクテリウムの混合物を作製し、先に記載のようにN.ベンサミアーナ葉部にインフィルトレーションし、ヘテロダイマーFSHの発現レベルは、市販のFSHダイマー特異的(FSH117)抗体を使用して接種10日後にELISAにより測定し、Tropix化学ルミネセンスシステム(Tropix,Bedford,MA)により検出した。
【0100】
実施例7
TMVおよびPVXを基礎にしたベクターによるIgMの発現
pICH15933におけるGFPおよびDsRedのコーディング配列はそれぞれIgM軽鎖およびJ鎖をコードする配列によって置換し、コンストラクトpICH−MLCJを得た(図13)。IgM重鎖をコードする配列はPVXベクターにクローニングし(pICH21240)、IgG重鎖のコーディング配列と置換し、コンストラクトpICH−MHCを得た(図13)。PVXおよびTMVを基礎にしたベクターから発現される3種の異なるIgM鎖を提供するアグロバクテリウム混合物はN.ベンサミアーナ葉部にインフィルトレーションし、ヘテロオリゴマー複合体の発現レベルは、市販のIgM特異的抗体を使用して接種後10日目にELISAにより測定した。
【0101】
欧州特許出願第05001819.1号および米国特許出願第60/593,606号の内容は、その優先権を本特許出願によって主張し、本明細書にそのまま参照として援用する。
【図面の簡単な説明】
【0102】
【図1A】(A)インフィルトレーションされたN.ベンサミアーナ葉部における2種の異なるTMVを基礎にしたベクター由来のGFPおよびDsRedの発現分布パターンを示す。左の写真:上部左側の明るいスポットはpICH17272にインフィルトレーションされた領域のGFP蛍光である。下部の弱く明るいスポットはベクターpICH18505にインフィルトレーションされた領域の赤色蛍光である。左の写真の底部右側の明るいスポットは、pICH17272+pICH18505にインフィルトレーションされた領域である。右側の写真は、2種の異なるベクターに同時インフィルトレーションされた葉部領域から分離されたプロトプラストを示す(上部:GFP蛍光検出条件下におけるプロトプラスト;下部パネル:DsRed蛍光検出条件下における同じプロトプラスト)。
【図1B】(B)pICH17272およびpICH18505のT−DNA領域の図式表示である。P‐転写プロモーター;T‐転写停止領域;RdRP ウイルスRNA‐依存的RNAポリメラーゼ;MP‐ウイルス移行タンパク質;3′NTR‐ウイルス3′非翻訳領域;Cr‐sgp‐crTMV株のCPサブゲノムプロモーター領域;GOI‐関心のある遺伝子。
【図1C】(C)pICH17272およびpICH18505のT−DNA領域の制限地図の図式表示である。
【図2】異なるサブゲノムプロモーター由来の2種の異なるトランスジーン(GOI‐1およびGOI‐2)の同時発現用に設計されたウイルスベクターのT−DNA領域の図式表示を表す。P‐転写プロモーター;T‐転写停止領域;RdRP ウイルスRNA‐依存的RNAポリメラーゼ;MP‐ウイルス移行タンパク質;3′NTR‐ウイルス3′非翻訳領域;Cr−sgp‐crTMV株のCPサブゲノムプロモーター領域;3PK‐トリプルシュードノット(pseudo knot)領域;U5spg‐TMV‐U5株のCPサブゲノムプロモーター。GOI‐関心のある遺伝子。
【図3A】(A)コンストラクトpICH17388、pICH17123,pICH15933、pICH7410、pICH10580、pICH10881およびpICH10745のT−DNA領域の図式表示を表す。RS‐PhiC31インテグラーゼにより認識される組換え部位。灰色の縦棒はイントロンを表す。
【図3B】(B)pICH17388、pICH17123、pICH15933のT−DNA領域の制限地図を表す。
【図3C】(C)pICH7410、pICH10580、pICH10881およびpICH10745のT−DNA領域の制限地図を表す。
【図4】(A)はDNA前駆体pICH17388およびpICH15933をレコンビナーゼ供与源と一緒にアグロバクテリウム送達して6日後のN.ベンサミアーナ葉部領域の蛍光顕微鏡写真を示す。RS‐PhiC31インテグラーゼにより認識される組換え部位。
【図5】抗体の重鎖および軽鎖発現のための異なるベクター系のT−DNA領域の図式表示を表す。RS‐PhiC31インテグラーゼにより認識される組換え部位。
【図6】ウイルスプロベクター系を使用した、ニコチアナベンサミアーナ葉部におけるIgG発現のウェスタンブロットを示す。TSPの電気泳動による分離は非還元条件下、12%ゲル上で行った。発現したタンパク質の検出は、6x103倍希釈した、HRP(Sigma)と結合したウサギ由来の抗‐ヒトIgG画分により行った。レーン:1‐非感染葉組織;レーン2‐サイトソルに発現したIgG重鎖(pICH17388);レーン3‐IgG重鎖および軽鎖、35Sプロモーターコンストラクト(pIC0123+pICH19846)による同時発現;レーン4‐P19(pICH6692)を用いる以外は、レーン3と同様;レーン5‐35SプロモーターコンストラクトおよびP19(pICH5290+pICH6692)により発現したGFP;レーン6‐バイシストロンコンストラクトpICH19860により同時発現したIgG重鎖および軽鎖;レーン7‐バイシストロンコンストラクトpICH19860(MP欠乏5′プロ‐ベクターpICH17123、MP in trans pICH10745)により同時発現したIgG重鎖および軽鎖;レーン8‐サイトソルに同時発現したGFPおよびdsRED、MPはin transで提供される(pICH17123+pICH19919+pICHI10745)。
【図7】同じ植物細胞において関心のある異なるタンパク質サブユニットの同時発現用に設計された非‐競合ウイルスベクターをコードするT−DNA領域の図式表示を表す。P‐転写プロモーター;T‐転写停止領域;TMV RdRP タバコモザイクウイルスのRNA‐依存的RNAポリメラーゼ;PVX RdRP ジャガイモウイルスXのウイルスRNA‐依存的RNAポリメラーゼ;MP‐ウイルス移行タンパク質;3′NTR‐ウイルス3′非翻訳領域;Cr‐sgp‐crTMV株のCPサブゲノムプロモーター領域;GOI‐関心のあるタンパク質サブユニットをコードする関心のある遺伝子。
【図8】バイナリーベクターpIC0130のT−DNA領域の図式表示を表す。
【図9】TMV(pICH17388+pICHI10580)およびPVX(pIC0130)を基礎にしたベクターを使用した、植物細胞におけるGFPおよびdsREDの同時発現を示す:アグロバクテリウムを接種したN.ベンサミアーナ葉部から分離したプロトプラストにおけるGFPおよびdsREDの視覚化。
【図10A】(A)バイナリーベクターpICH17620、pICH20431、pICH21240、pICH20421、およびpICH21370のT−DNA領域の図式表示を表す。
【図10B】(B)バイナリーベクターpICH11599、pICH21910、およびpICH21920のT−DNA領域の図式表示である。
【図10C】(C)バイナリーベクターpICH21282、pICH10990、pICH22250、およびpICH22240のT−DNA領域の図式表示である。LvおよびLc‐軽鎖の可変および保存領域;HvおよびHc‐重鎖の可変および保存領域。
【図10D】(D)PVX‐由来プロベクターpICH21380のクローニングスキームの図式表示である。
【図11A】(a)PVXおよびcrTMVベクターを使用したIgG軽鎖および重鎖の同時発現レベル測定のためのELISA試験の結果を示す。左側の組織培養プレートのウェルの番号付けは、ヒストグラムの棒の番号付けに相当する。ヒストグラムは405nmにおけるウェルのODを示す。1、2−非感染植物組織;3、4、5−PVXにおけるカルレティキュリンSP‐重鎖(それぞれpICH21240‐1、5および14クローン);6、7、8‐PVXにおけるカルレティキュリンSP‐IgGの軽鎖(LC)(N‐末端欠失、それぞれpICH21370−18、19、31);9、10、11‐PVXにおけるカルレティキュリンSP‐IgG軽鎖(それぞれpICH21370‐40、44、45);12‐ブランクコントロール(植物タンパク質非適用);13、14、15‐crTMVにおけるカルレティキュリンSP‐IgG軽鎖(pICH17620+pICH10881+pICH20431)と同時発現したPVXにおけるカルレティキュリンSP‐IgG重鎖(HC)(それぞれpICH21240‐1、5および14クローン)。16、17、18‐crTMVにおけるカルレティキュリン‐HC(pICH17620+pICH10881+pICH20421)と同時発現したPVXにおけるカルレティキュリン‐IgGのLC(N‐末端欠失、それぞれpICH21370−18、19、31)。19、20、21‐crTMVを基礎にしたベクター中におけるカルレティキュリンSP‐重鎖(pICH17620+pICH10881+pICH20421)と同時発現したPVXにおけるカルレティキュリンSP‐IgGLC(それぞれpICH21370−40、44、45)。22‐PVXにより発現したGFP(pIC0130);23‐PVXにより発現したGFP(pICH20799);24‐crTMVだけ(pICH17620+pICH10881+pICH20431)から発現したカルレティキュリンSP‐IgG軽鎖;25‐crTMVを基礎にしたベクター(pICH17620+pICH10881+pICH20421)から発現したカルレティキュリンSP‐IgG重鎖;26‐PTGSサプレッサーP19の存在下、強力な35Sプロモーター制御下で同時発現したIgGの軽鎖および重鎖27‐crTMVを基礎にしたベクター(pICH17388+pICH10881=pICH20421)からのIgG軽鎖および重鎖同時発現;28‐crTMVを基礎にしたベクター(pICH17388+pICH10881=pICH20421)、in transで提供されたMP(pICH10745)からの軽鎖および重鎖の同時発現;*‐測定可能な値をかなり超えたOD405値。
【図11B】(b)N.ベンサミアーナ葉部に発現した抗癌抗体の電気泳動およびウェスタンブロット解析を示す。(A)TMVとPVXプロベクターが同時感染したニコチアナベンサミアーナ葉部における抗癌抗体の重鎖および軽鎖の蓄積。軽鎖はPVXにより発現し、重鎖はTMVにより発現する。タンパク質は還元条件下、12%ポリアクリルアミドゲル中で分離する。上部パネルはクーマシー染色を示す;中間パネルはHRP‐結合ヤギ抗‐ヒトIgG(ガンマ鎖特異的)抗体(Sigma)によるウェスタンブロットを示す。下部パネルはHRP‐結合ウサギ抗ヒトIgG(ラムダ鎖‐特異的)抗体(Sigma)によるウェスタンブロットを示す。Uninf、非感染組織;2〜11、接種後の日数;S、抗癌mab(モノクローナル抗体)標準品;LC、TMVプロベクターだけにより発現した軽鎖(6dpi);HC、TMVプロベクターだけにより発現した重鎖(6dpi)。(B)プロテインA磁気ビーズ(NEB)を使用した抗癌mabの精製。(A)クーマシー染色ゲル上、還元条件下、12%ポリアクリルアミドゲル中で移動する植物‐由来(レーン1)および標準(レーン2)mab。(C)HCを発現するTMVプロベクターおよびLCを発現するPVXプロベクターに同時感染したニコチアナベンサミアーナ葉部における集合した抗癌mabの蓄積。タンパク質は非還元条件下、10%ポリアクリルアミドゲル中で分離する。ウェスタンブロットはヤギ抗ヒトIgG(ガンマ鎖特異的)抗体により同定した。Uninf、非感染組織;2〜11、接種後の日数;S、抗癌mab(モノクローナル抗体)標準品;LC、TMVプロベクター(6dpi)だけにより発現した軽鎖;HC、TMVプロベクター(6dpi)だけにより発現した重鎖;H2L2:2つの重鎖および2つの軽鎖を含有するIgGヘテロテトラマー、H2L‐2つの重鎖および1つの軽鎖を含有するIgGヘテロトリマー、H2‐重鎖ホモダイマー、L2‐軽鎖ホモダイマー。
【図12】バイナリーベクターpICH17620、pICH−FSHAおよびpICH−FSHBのT−DNA領域の図式表示を表す。
【図13】バイナリーベクターpICH17388、pICH−MLCJおよびpICH−MHCのT−DNA領域の図式表示を表す。
【発明の開示】
【0001】
発明が属する技術分野
本発明は、プラス‐センス1本鎖RNAウイルスベクターを使用した、植物体、植物部分または植物細胞培養物におけるヘテロ‐オリゴマータンパク質の産生に関する。本発明に記載の方法およびベクターは、全長抗体、または別のタンパク質もしくはそれらのフラグメントとの融合物を含むそのヘテロ‐オリゴマー合成誘導体のような、高い収率の機能的ヘテロ‐オリゴマー組換えタンパク質を植物細胞に提供する。
【0002】
発明の背景
植物を基礎にした分子農業は、好ましくは、潜在的に低い生産費用、および工業的生産方法から生み出される動物‐由来タンパク質を除去するためのバイオ医薬品産業による試みのゆえに、ヒトおよび動物の健康分野で使用される予定の組換えタンパク質の産生にとっての魅力的な好機である。なぜなら、これらの産物は牛海綿状脳症(BSE)またはクロイツフェルト‐ヤコブ病(CJD、vCJD)のようなヒト病原体により汚染されうるからである。しかし、植物細胞における高収率のヘテロオリゴマータンパク質の産生は、以下の理由から、強力な構成的または組織‐特異的プロモーターの使用に基づいた標準発現系の協力により解決することができない難問である:第1に、そのような組換えタンパク質の大多数は植物の成長および発達に対して有害な作用を有し、それゆえ収率を著しく低下させる;第2に組織‐特異的(たとえば、種子‐特異的)プロモーターの使用は、食用になる作物植物(たとえば、イネ、トウモロコシ、コムギ)のゲノムへの医薬タンパク質をコードする遺伝子の安定した組み込みを必要とし、それは露地栽培の場合、トランスジーン流動制御に関する問題を引き起こす可能性がある。また、これらの系は産物の低い収率のため、閉鎖環境(温室)で使用する場合、商業的に実行可能ではない。
【0003】
植物ウイルスを基礎にした一過性の発現系(総説としては以下を参照されたい:Porta & Lomonossoff,1996,Mol.Biotechnol.,5,209−221;Yusibov et al.,1999,Curr.Top.Microbiol.Immunol.,240,81−94;Gleba et al.,2004,Curr.Opin.Plant Biol.,7,182−188)は、その技術が成長と産生ステージを分離させるため、植物葉組織における高発現レベルを提供することが可能であり、ある程度までは組換えタンパク質の細胞毒性、および植物発達に対するそれらの有害作用の難問に対処することができる。最も確立され、商業的に実行可能な系は、プラス‐センス1本鎖RNAウイルスベクター、好ましくはタバコモザイクウイルス(TMV)‐由来ベクター(Kumagai et al.,1994,Proc.Natl.Acad.Sci.USA,90,427−430;Mallory et al.,2002,Nature Biotechnol.20,622−625;US5316931;US5589367;US5866785;US5977438;WO02088369;WO02097080;WO0229068;US5491076)に基づく。しかし、これらの系は簡単で、比較的低分子のタンパク質の産生に使用が限定されるという深刻な制限を欠点として持つ。ウイルスベクターが1kbより大きな異種配列を持っている場合、このことはある程度は、それらの不安定性および野生型への高頻度での逆戻りによって引き起こされる。また、技術の深刻な制約は、最も有益な群の組換えタンパク質を代表する治療用モノクローナル抗体およびそれらの誘導体のような、複雑なヘテロオリゴマータンパク質の発現が可能なウイルスベクター系が無いことである。
【0004】
植物ウイルスベクターを使用した、植物における全長モノクローナル抗体の発現を扱った刊行物は1つしかない(Verch et al.,1998,J.Immunol.Meth.,220,69−75)。この論文は、全身葉部におけるモノクローナル抗体の重鎖および軽鎖発現のための2種のTMVを基礎にした全身ウイルスベクターを使用して、N.ベンサミアーナ植物体の同時感染時に、in vitroで合成された上記ベクターの転写物により、異なるベクターから異なる鎖が発現されることを説明する。しかし、上記の系における組換えタンパク質の収率は非常に低いため、集合したモノクローナル抗体の存在は、ウェスタンブロット法およびELISAのような感度の高い試験によって確認しなければならない。組換え抗体は収率が非常に低いため、この系は実際の適用に適切ではなく、商業的価値を持たない。感染された植物体の同じ植物組織中には、2種以上のTMVを基礎にしたウイルスベクターは通常存在しない(実施例1を参照されたい)ため、検出された抗原結合活性は、別々の細胞または組織に発現された抗体の重鎖および軽鎖からの分離手順中にin vitroで集合した抗体によるものである可能性がある。機能的抗体は変性し、弱めた抗体成分からin vitroで集められうることが以前に示された(Petersen & Dorrington,1974,J.Biol.Chem.,17,5633−5641;Maeda et al.,1996,Protein Engineering,9,95−100)。しかし、そのようなアセンブリーに都合のよい条件が存在しない場合、そのようなアセンブリーの効率は非常に低い。
【0005】
したがって、菌類または昆虫細胞発現系のような別の大規模発現系と市場で競い合うために十分な収率と効率の、植物における組換えヘテロオリゴマータンパク質の大規模発現系は存在しない。そのような植物発現はできる限り以下の基準を十分に満たさなければならない:
(i)できるだけ多くの植物組織および上記組織の中のできるだけ多くの細胞における、関心のあるヘテロオリゴマータンパク質の発現を含む、高収率;
(ii)植物の成長に対する組換えタンパク質発現の有害な作用を妨げるための、関心のあるタンパク質または産物の発現は所望する植物の発達段階で開始できるように、一過性(またはスイッチ可能)でなければならない;
(iii)植物細胞におけるヘテロオリゴマータンパク質の異なるサブユニットをコードするポリタンパク質の最適比を提供し、そうして上記サブユニットからの組換えタンパク質アセンブリーのレベルにおいて組換えタンパク質の高収率を維持すること。
【0006】
したがって、本発明の目的は植物体、植物部分、または植物細胞培養物における効率的なヘテロオリゴマータンパク質産生の方法を提供することである。別の目的は、ヘテロオリゴマータンパク質を発現することができる高収率植物発現系の提供である。本発明の別の目的は、同じ植物細胞中に1種より多い関心のあるポリペプチドを同時発現させる効率的な方法を提供することである。さらに、植物体、植物部分、または植物細胞培養物に抗体を発現させるための迅速で高収率の方法を提供することが本発明の目的である。
【0007】
発明の一般的説明
本発明は、植物体、植物組織、または植物細胞において、少なくとも第1および第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法を提供し、
(i)植物体、植物組織または植物細胞に、第1および第2プラス‐センス1本鎖RNAウイルスベクターを提供するが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、少なくとも第1ウイルスベクターおよび第2ウイルスベクターは非競合ウイルスベクターであり;または
(ii)植物体、植物組織または植物細胞に、少なくとも第1および第2タンパク質サブユニットをコードする、プラス‐センス1本鎖RNAウイルスベクターを提供すること
により、少なくとも第1および第2タンパク質サブユニットを植物組織に発現させることを含む。
【0008】
一態様では、第1ウイルスベクターおよび第2ウイルスベクターは、それらが異なるウイルスベクターであるという点で、非競合的である。別の態様では、第1ウイルスベクターおよび第2ウイルスベクターは、それらが同じRNAウイルスに由来しないという点で、非競合的である。別の態様では、第1ウイルスベクターおよび第2ウイルスベクターは、それらが第1および第2タンパク質サブユニットをコードする配列部分以外の配列部分において異なるという点で、非競合的である。別の態様では、第1ウイルスベクターおよび第2ウイルスベクターは、それらが異なるウイルス種に属するRNAウイルスに由来するという点で、非競合的である。別の態様では、第1ウイルスベクターおよび第2ウイルスベクターは、それらが異なるウイルス属に属するRNAウイルスに由来するという点で、非競合的である。
【0009】
本発明の別の態様は、抗体の重鎖および軽鎖のような、少なくとも第1および第2タンパク質サブユニットを含有する抗体を植物体、植物組織または植物細胞において産生する方法であって、
(i)第1プラス‐センス1本鎖RNAウイルスベクターのDNA前駆体および第2プラス‐センス1本鎖RNAウイルスベクターのDNA前駆体を、アグロバクテリウム媒介トランスフェクションにより植物体、植物組織、または植物細胞に提供するが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、少なくとも第1ウイルスベクターまたは第2ウイルスベクターは、植物体における第1または第2ウイルスベクターの全身移行に必要な機能的タンパク質をコードするオープンリーディングフレームを欠如し;または
(ii)少なくとも第1および第2タンパク質サブユニットをコードするプラス‐センス1本鎖RNAウイルスベクターのDNA前駆体を、アグロバクテリウム媒介トランスフェクションにより植物体、植物組織、または植物細胞に提供するが、ウイルスベクターは全身移行に必要な機能的タンパク質をコードするORFを欠如する
ことにより、少なくとも第1および第2タンパク質サブユニットを植物細胞に発現させることを含む。
【0010】
本発明はさらに、植物体、植物組織、または植物細胞において、少なくとも第1および第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法を提供し、方法は1種以上のプラス‐センス1本鎖RNAウイルスベクター(複数のベクター)由来の少なくとも第1および第2タンパク質サブユニットを植物細胞に発現させることを含む。
【0011】
本発明の別の態様は、特許請求の範囲および以下の説明に記載される。
本発明の発明者らは驚くべくことに、植物ウイルスベクターを使用して植物に高収率でヘテロオリゴマータンパク質を産生する方法を確認している。植物におけるヘテロオリゴマータンパク質の効率的な産生には、同じ植物細胞におけるヘテロオリゴマータンパク質の異なるタンパク質サブユニットの高収率の産生を必要とする。このようにして、小胞体(ER)の天然のタンパク質集合能力のような、上記細胞のそのような能力を使用して、上記の少なくとも2種のタンパク質サブユニットを発現している細胞にヘテロオリゴマータンパク質を効率的に集めることができる。したがって上記のヘテロオリゴマータンパク質の非効率的なin vitroでのアセンブリーは必要でない。本発明はステップ(i)により、またはステップ(ii)により、またはステップ(i)および(ii)の組み合わせにより同じ細胞に2種以上のタンパク質の効率的な同時発現を初めて成し遂げている。
【0012】
第1タンパク質サブユニットは第1異種(核酸)配列によってRNAウイルスベクターにコードされる。第2タンパク質サブユニットは第2異種(核酸)配列によってウイルスベクターにコードされる。これらの異種配列はしたがってウイルスベクター(複数のウイルスベクター)のRNA配列であり、典型的にはタンパク質サブユニットを発現するために必要な調節配列を含有するか、またはコードする。そのような調節配列の例としては、サブゲノムプロモーター、IRESエレメント、および3′‐非翻訳配列が挙げられる。本明細書では、ある配列がウイルスベクター(複数のウイルスベクター)が由来するウイルスに天然に存在しない場合、その配列は異種配列である。
【0013】
本発明に従って産生可能なヘテロオリゴマータンパク質は、少なくとも第1および第2サブユニットを有し、第1および第2サブユニットは異なるポリペプチド配列を有する。したがって、第1および第2サブユニットは典型的には異なる異種核酸配列から発現されなければならない。オリゴマータンパク質のサブユニットは会合して4次構造のオリゴマータンパク質を形成する。サブユニットのアセンブリーは典型的にはサブユニット間の非共有結合を包含する。さらに、共有結合、たとえばジスルフィド結合がタンパク質サブユニット間で形成されてもよい。
【0014】
一態様では、ヘテロオリゴマータンパク質はヘテロ‐二量体タンパク質、すなわち2種の異なるタンパク質サブユニットを有するタンパク質である。別の態様では、ヘテロオリゴマータンパク質は2種より多い異なるサブユニット、たとえば3種または4種の異なるサブユニット(それぞれヘテロトリマーまたはヘテロテトラマータンパク質)を有していてもよい。別の態様では、のヘテロオリゴマータンパク質は2種の異なるサブユニットを有していてもよく、サブユニットの中の1方または両方がヘテロオリゴマータンパク質中に、1回より多く存在してもよい。ヘテロオリゴマータンパク質のサブユニット構成の例としては、AaBb、AaBbCc、AaBbCcDdが挙げられ、ここでAは第1タンパク質サブユニットを表し、Bは第2タンパク質サブユニットを表し、そしてCおよびDは別のタンパク質サブユニットを表す。それぞれの大文字A、B、CおよびDは他のタンパク質サブユニットとは異なるタンパク質サブユニットを表し、そして小文字は少なくとも1の整数を表し、ヘテロオリゴマータンパク質中の個々のタンパク質サブユニットのコピー数を示す。A2B2のサブユニット構成を有するIgG抗体が例として挙げられ、ここでAは第1タンパク質サブユニット(たとえば重鎖)を表し、そしてBは第2タンパク質サブユニット(たとえば軽鎖)を表す。好ましくは、本発明に従って産生されたヘテロオリゴマータンパク質は2種または3種の異なるタンパク質サブユニットを有し、より好ましくは、それは2種の異なるタンパク質サブユニットを有する。
【0015】
本発明の方法では、少なくとも第1および第2タンパク質サブユニットは植物体、植物組織、または植物細胞の細胞中に、ステップ(i)または/およびステップ(ii)によって発現される。ステップ(i)および(ii)のそれぞれは、同じ細胞中に少なくとも第1および第2タンパク質サブユニットを発現させ、その結果ヘテロオリゴマータンパク質を細胞中に効率的に産生することができる。ステップ(i)および(ii)は、とりわけ3種、4種、またはそれより多い異なるタンパク質サブユニットを有するヘテロオリゴマータンパク質産生のために並行して行うことができる(実施例7を参照されたい)。
【0016】
本発明のプラス‐センス1本鎖RNAウイルスベクター(複数のウイルスベクター)はまた、本明細書では単に「ウイルスベクター」として表される。典型的には、プラス‐センス1本鎖RNAウイルスベクターはプラス‐センス1本鎖植物RNAウイルスに由来する。
【0017】
ステップ(ii)では、植物体、植物組織または植物細胞は、少なくとも第1および第2タンパク質サブユニットをコードするプラス‐センス1本鎖RNAウイルスベクターを提供される。少なくとも第1および第2タンパク質サブユニットをコードするウイルスベクターは、インサートとして第1タンパク質サブユニットをコードする第1異種配列を含有し、その発現は第1サブゲノムプロモーターの制御下にあってもよい。さらに、ウイルスベクターはインサートとして、第2タンパク質サブユニットをコードする第2異種配列を含有し、その発現は第2サブゲノムプロモーターの制御下にあってもよい。第1および第2タンパク質サブユニットの両方がサブゲノムプロモーターの制御下で発現される場合、これらのサブゲノムプロモーターは、好ましくは、植物細胞に望ましくない組換え事象を導くウイルスベクター中の自己相同性を回避するために配列が異なる。そのような異なるサブゲノムプロモーターは異なる株または種の植物ウイルスから採取されてもよく、たとえば1種のサブゲノムプロモーターはタバコモザイクウイルス(TMV)U1のコートタンパク質(CP)サブゲノムプロモーターであってもよく(またはそれに由来してもよく)、そして別のサブゲノムプロモーターは、TMV U5のCPサブゲノムプロモーターであってもよく(またはそれに由来してもよく)、またはアブラナ科植物‐感染トバモウイルス(cr‐TMV)に由来してもよい。
【0018】
第1または第2サブゲノムプロモーターのかわりに、第1または第2タンパク質サブユニットの翻訳がIRES(内部リボソーム侵入部位:internal ribosome entry site)エレメントの制御下にあってもよい。第1および第2タンパク質サブユニット両方の翻訳がIRESエレメントの制御下にあってもよいが、少なくともタンパク質サブユニットの1種はサブゲノムプロモーターを使用して発現される。本発明で使用されるIRESエレメントはcr‐TMVのような植物ウイルスまたは他の植物ウイルスから採取されてもよい(Proc Natl Acad Sci USA 2002,99,5301−6;Virology 1999,263,139−54;WO03020927;WO0229068)。
【0019】
ステップ(ii)のウイルスベクターは、好ましくは植物体または植物組織において全身移行をする能力がない。このことは、たとえばウイルス粒子アセンブリーの機能的起点を削除することにより成し遂げることができる。トバモウイルスではたとえば、ウイルス粒子アセンブリーの起点はMP ORFに位置する。したがって、ウイルス粒子アセンブリーの起点はウイルスベクターからMP ORFを完全に、または部分的に欠失することによって削除することができる。ウイルスベクターはしたがって、好ましくは機能的移行タンパク質(MP)ORFを持たない。より好ましくは、ウイルスベクターは、ウイルスベクターの全身移行に必要な機能的タンパク質を持たない。この態様では、ウイルスベクターは機能的コートタンパク質ORFを持たなくてもよく、そして最も好ましくは、ウイルスベクターは機能的移行タンパク質ORFおよび機能的コートタンパク質ORFの両方を持たない。ウイルスベクターからMP ORFおよび/またはCP ORFを除去することは、ウイルスベクター安定性を低下させずに、少なくとも第1および第2タンパク質サブユニットをコードするためのより多くの余地を提供する。この態様では、ウイルスベクターは好ましくは、多くの細胞の感染を成し遂げるために、植物体または植物組織の多くの細胞に提供される。このことはアグロバクテリウムを使用して、T−DNAのようなRNAウイルスベクターのDNA前駆体を提供することにより最も都合よく成し遂げられてもよい(以下を参照されたい)。
【0020】
ステップ(ii)のウイルスベクターが由来するウイルスはいずれかのプラス‐センス1本鎖植物RNAウイルス、たとえば「詳細な説明」の章で挙げられたものであってもよい。好ましいウイルス群はトバモウイルス、ポテクスウイルス、およびポティウイルスである。最も好ましいウイルスはTMVおよびPVXである。ウイルスベクターは少なくともそれが由来するウイルス由来の、ウイルスベクターの複製に必要なタンパク質をコードするORFを含有することになる。ウイルスベクターは典型的にはさらに、ウイルス複製のための調節エレメント、および少なくとも1種のサブゲノムプロモーターを含有する。
【0021】
ステップ(i)では、植物体、植物組織または植物細胞は、第1および第2プラス‐センス1本鎖RNAウイルスベクターを備える。第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードする。
【0022】
ステップ(i)は2種の異なるタンパク質サブユニットを有するヘテロオリゴマータンパク質の産生を可能にする。ステップ(i)はその上、2種より多い異なるサブユニット、たとえば3種または4種の異なるタンパク質サブユニットを有するヘテロオリゴマータンパク質の産生を可能にする。この場合、植物体、植物組織または植物細胞は、それぞれが上記の異なるタンパク質サブユニットの1種をコードする、第1、第2および第3ウイルスベクターを(そして場合により別のウイルスベクターを)備えてよい。3種または4種の異なるタンパク質サブユニットをステップ(i)で発現しなければならない場合、個々の3種または4種のウイルスベクターは好ましくは、すべてお互いに非競合ウイルスベクターである。3種のウイルスベクターの場合、第1ウイルスベクターはトバモウイルスに由来してもよく、第2ウイルスベクターはポティウイルスに由来してもよく、そして第3ウイルスベクターはポテクスウイルスに由来してもよい。
【0023】
2種のウイルスベクターは、同じ植物組織中でそれらがコードするタンパク質サブユニットを効率的に発現(同時発現)できる場合、非競合的である。同時発現は少なくとも2種の異なるウイルスベクターがコードする異種配列の実質的な量を発現する前は、それらは複製中にお互いに競合しないことを必要とする。上記の少なくとも2種のウイルスベクターのRNAレベルの配列の違いが大きいほど、それらは同じ植物細胞においていっそう非競合的である。第1ウイルスベクターおよび第2ウイルスベクターは異なるウイルスベクターであるため、第1ウイルスベクターおよび第2ウイルスベクターは非競合ウイルスベクターである。
【0024】
非競合的である場合、ある一般的な態様では、第1および第2ウイルスベクター(上記の2種の非競合ウイルスベクター)は、少なくともタンパク質サブユニットをコードする上記の異種配列以外の配列部分において異なる。好ましくは、上記の異種配列以外の配列部分におけるそのような違いは植物RNAウイルスに由来する配列部分であり、そのような配列部分は、ウイルス機能、たとえば植物細胞におけるウイルス複製(たとえばレプリカーゼをコードする配列)、ウイルスタンパク質の翻訳(たとえばサブゲノムプロモーター)またはウイルスの細胞間もしくは長距離移行に関与する配列部分である。
【0025】
非競合的である場合、別の一般的な態様では、第1および第2ウイルスベクター(上記の2種の非競合ウイルスベクター)は異なる植物ウイルスに由来する。この態様の一例では、上記の少なくとも2種の非競合ウイルスベクターは同じウイルス株のウイルスに由来しない。別の例では、上記の少なくとも2種の非競合ウイルスベクターは同じウイルス種のウイルスに由来しない。付加的な例では、上記の少なくとも2種の非競合ウイルスベクターは同じウイルス属のウイルスに由来しない。したがって、第1ウイルスベクターおよび第2ウイルスベクターは好ましくは異なる株のウイルス、より好ましくは異なる種のウイルス、最も好ましくは異なる属のウイルスに由来する。
【0026】
第1ウイルスベクターはポテクスウイルス属に属するウイルスに由来してもよく、そして第2ウイルスベクターはポティウイルス属に属するウイルスに由来してもよい。とりわけ、第1ウイルスベクターはジャガイモウイルスXに由来してもよく、そして第2ウイルスベクターはジャガイモウイルスYに由来してもよい。
【0027】
ポティウイルスベクターの場合、タンパク質サブユニットはウイルスポリタンパク質との融合物として発現してもよく、それによって本発明のタンパク質サブユニットはポティウイルスプロテアーゼ認識部位によりポリタンパク質から分離することができる。
【0028】
別の態様では、第1ウイルスベクターはポテクスウイルス属に属するウイルスに由来してもよく、そして第2ウイルスベクターはトバモウイルス属に属するウイルスに由来してもよい。とりわけ、第1ウイルスベクターはジャガイモウイルスXに由来してもよく、そして第2ウイルスベクターはタバコモザイクウイルスに由来してもよい。
【0029】
「ウイルスに由来している」とは、ウイルスの遺伝エレメントまたは配列部分をそのウイルスに由来するウイルスベクターが含有することを意味する。一態様では、ウイルスベクター(複数のウイルスベクター)はRNAウイルスから採取したレプリカーゼORF(オープンリーディングフレーム)を含有する。別の態様では、ウイルスベクターはRNAウイルス由来の移行タンパク質ORF、そして場合によりさらにレプリカーゼORFを含有する。RNAウイルスから採取したウイルス遺伝エレメントは、所望する場合、たとえばウイルスベクターのクローニングに必要な制限部位を導入するために変異してもよい。
【0030】
第1ウイルスベクターおよび第2ウイルスベクターが共にタバコモザイクウイルスTMV30Bに基づいている態様は、本発明の方法のステップ(i)から排除される。なぜなら、この場合、第1ウイルスベクターおよび第2ウイルスベクターは異なるウイルスベクターではなく、同じウイルスベクターであるからである(参照:Verch et al.,J.Immunological Methods 220(1998)69−75)。
【0031】
第1および第2ウイルスベクターは、好ましくは多くても90%、より好ましくは多くても80%、さらにより好ましくは多くても70%、そして最も好ましくは多くても60%のRNAレベルの配列相同性を有する。さらにとりわけ、第1ウイルスベクターの任意の100塩基の配列部分が好ましくは、第2ウイルスベクターの任意の100塩基の配列部分に多くても90%、好ましくは多くても80%、より好ましくは多くても70%、そして最も好ましくは多くても60%の配列相同性を有する。
【0032】
一態様では、第1ウイルスベクターのレプリカーゼORF(またはレプリカーゼが1種以上のORFによってコードされる場合、複数のORF)と第2ウイルスベクターのレプリカーゼORF(またはレプリカーゼが1種以上のORFによってコードされる場合、複数のORF)は、多くても90%、より好ましくは多くても80%、さらにより好ましくは多くても70%、そして最も好ましくは多くても60%の相同性を有する。別の態様では、第1ウイルスベクターのレプリカーゼORFと第2ウイルスベクターのレプリカーゼORFは多くても85%、より好ましくは多くても75%、さらにより好ましくは多くても65%、そして最も好ましくは多くても55%の同一性を有する。
【0033】
ステップ(i)では、第1ウイルスベクターは好ましくは、発現が第1サブゲノムプロモーターの制御下にあってもよい第1タンパク質サブユニットをコードする第1異種配列を含有する。第2ウイルスベクターは、発現が第2サブゲノムプロモーターの制御下にあってもよい第2タンパク質サブユニットをコードする第2異種配列を含有する。サブゲノムプロモーターの1種または両方はIRESエレメントによって置換されてもよい。先に記載のステップ(ii)の場合と同様に、第1および第2タンパク質サブユニットがサブゲノムプロモーターの制御下で発現される場合、植物細胞に望ましくない組換え事象を引き起こすことになる第1および第2(およびいずれか別のウイルスベクター)ウイルスベクター間の相同性を回避するために、これらのサブゲノムプロモーターは好ましくは配列が異なる。そのような異なるサブゲノムプロモーターは異なる株または種の植物ウイルスから採取されてもよく、たとえば1種のサブゲノムプロモーターはタバコモザイクウイルス(TMV)U1のコートタンパク質(CP)サブゲノムプロモーターであってもよく、そして別のサブゲノムプロモーターはTMV U5またはアブラナ科植物‐感染トバモウイルス(cr‐TMV)のCPサブゲノムプロモーターであってもよい。
【0034】
細胞間、もしくは長距離移行に必要な植物ウイルスのORFは本発明のウイルスベクター構築時に削除されてもよいため、上記の非競合ウイルスベクターの関連性はウイルスベクターのレプリカーゼORFを比較することにより確認してもよい。
【0035】
第1および第2タンパク質サブユニットをコードする第1および第2異種配列は、それぞれウイルスベクターに付加的な配列として添加されてもよい。異種配列は好ましくは高レベルの発現が成し遂げられるように添加される。この目的のために、異種配列はウイルスの3′末端に添加される。なぜなら3′ORFはしばしば多くのウイルスで最も高レベルで発現されるORFであるからである。好ましくは、しかし、異種配列は、ウイルスに生来ある配列、たとえばトバモウイルスのような多くのウイルスにおいてCP ORFであるウイルスの天然の3′ORFと入れ替わる。したがって、第1および/または第2ウイルスベクターは好ましくはウイルスベクターの全身移行のためのORFを欠如する。ウイルスベクターはさらに、トバモウイルスのMP ORFのような細胞間移行のためのORFを欠いてもよい。
【0036】
本発明では、ステップ(i)およびステップ(ii)は、とりわけヘテロオリゴマータンパク質の3種以上の異なるサブユニットを発現するために組み合わせてもよい。4種の異なるタンパク質サブユニットを有するヘテロオリゴマータンパク質が産生されることになる場合、2種のタンパク質サブユニットがステップ(i)に従って発現されてもよく、そして2種の付加的なタンパク質サブユニットがステップ(ii)に従って、植物体または植物組織の細胞中に産生されてもよい。しかし、好ましくは2種のタンパク質サブユニットが第1ウイルスベクターから発現されてもよく、そして2種のタンパク質サブユニットが第1ウイルスベクターに非競合的な第2ウイルスベクターから発現されてもよい。3種の異なるタンパク質サブユニットを有するヘテロオリゴマータンパク質が産生されることになる場合、上記の3種のタンパク質サブユニットは、ステップ(i)に従って、ステップ(ii)に関する記載と同様に第1ウイルスベクターから2種のタンパク質を発現し、そして非競合ウイルスベクターから第3のタンパク質サブユニットを発現することにより、発現されてもよい。
【0037】
本発明のウイルスベクターは、典型的にはDNAレベルで工学的に操作される。ウイルスベクターがRNAウイルスベクターのように植物体の細胞または植物組織の細胞に提供される場合、上記DNAは、たとえばT7ポリメラーゼのようなバクテリオファージポリメラーゼを適切なプロモーターと一緒に使用することにより、RNAウイルスベクターにin vitroで転写されてもよい。2種の異なるウイルスベクターは好ましくは、植物体の細胞が両方のウイルスベクターを備えることを確実にするために、混合物として植物体に提供される。好ましくは、しかし、植物体または植物組織をRNAウイルスベクターのDNA前駆体により形質転換することにより、本発明のウイルスベクターが植物体の細胞または植物組織の細胞に提供される。DNA前駆体は、植物体の細胞中で、DNA前駆体の転写によってウイルスベクターを形成することに対して活性な転写プロモーターを有する。最も好ましくは、DNA前駆体はアグロバクテリウムTiプラスミド中のT−DNAである。2種以上のウイルスベクターはその後、植物体をそれぞれの株が特定のウイルスベクターをコードするT−DNAを含有する2種以上のアグロバクテリウム株の混合物(たとえば、懸濁液)で処理することにより植物体または植物組織に提供されてもよい。そのようなアグロバクテリウム懸濁液で植物体の実質的な部分を処置することが、天然の植物ウイルスの全身移行機能および/または細胞間移行機能の代わりとなってもよい。
【0038】
アグロバクテリウム法を用いたウイルスベクターのDNA前駆体による植物体、植物組織または植物細胞の一過性トランスフェクションは、本発明において最も好ましい。しかし、ウイルスベクターのDNA前駆体が植物染色体DNAに安定に組み込まれてもよい。染色体DNAからのウイルスベクター(複数のウイルスベクター)の放出は誘導型プロモーターによって制御されてもよい。
【0039】
ウイルスベクターがDNA前駆体から植物体に提供される場合、それらが転写される細胞核から、ウイルスベクターが複製する細胞質へのウイルスベクターの移動効率を改善するための処置がとられることが好ましい。このことは、WO2005/71090として公開された国際特許出願PCT/EP05/000492(これを参照として本明細書に援用する)に詳細に記載されるように、DNA前駆体、とりわけウイルスベクターのレプリカーゼORFにイントロンを包含することにより成し遂げられてもよい。
【0040】
本発明の方法は、植物ウイルス発現系が存在するか、または将来作り出されることになるいずれかの植物に適用してもよい。植物は単子葉植物または双子葉植物であってもよい。双子葉植物の中では、ナス科、アブラナ科、アカザ科、およびマメ科が好ましい。ナス科の中では、N.タバカムまたはN.ベンサミアーナのようなニコチアナ属が好ましい。別の好ましい植物はメディカゴサティバおよびフダンソウのようなフダンソウ種である。
【0041】
本発明の方法は植物系においてヘテロオリゴマータンパク質を産生するために使用される。好ましいヘテロオリゴマータンパク質は以下のクラスのイムノグロブリン:イムノグロブリンG、イムノグロブリンA、イムノグロブリンM、イムノグロブリンD、およびイムノグロブリンEのようなイムノグロブリンである。これらのイムノグロブリンは少なくとも抗原結合ドメインの一部を含有してもよい。本発明に従って産生されるこれらのイムノグロブリンが少なくとも2種の異なるタンパク質サブユニットを含有するならば、必要に応じて、それらのイムノグロブリンは、生来の動物イムノグロブリンに関連して改変されてもよい。イムノグロブリンはイムノグロブリン重鎖と一緒に保護タンパク質を含有してもよく、ここで保護タンパク質はポリイムノグロブリン受容体の一部を含有する。別の好ましいヘテロオリゴマータンパク質はインスリンである。
【0042】
本発明のヘテロオリゴマータンパク質は、必要に応じて生来のタンパク質に関連して多くの異なる方法で改変されてもよい。ヘテロオリゴマータンパク質では、生来のヘテロオリゴマータンパク質の1種以上のタンパク質サブユニットの分泌シグナルを形成する生来のリーダー配列が植物‐特異的シグナルペプチドに置換されてもよい。上記の植物‐特異的シグナルペプチドはタバコカルレティキュリンおよび/またはイネアルファ‐アミラーゼに由来してもよい。ヘテロオリゴマータンパク質の少なくとも1種または少なくとも2種以上のサブユニットが植物細胞においてサブユニット由来のヘテロオリゴマータンパク質のアセンブリーを改善するための小胞体保持シグナルKDELを含有していてもよい。さらに、タンパク質サブユニットをコードする異種配列が変異してヘテロオリゴマータンパク質からグリコシル化部位を部分的に、または完全に除去してもよい。その上、発現されることになるヘテロオリゴマータンパク質のグリコシル化パターンが、たとえば1種以上のグリコシルトランスフェラーゼのような、植物グリコシル化装置の構成要素を工学的に操作することによって変えられてもよい。
【0043】
本発明のヘテロオリゴマータンパク質は一般に公知の手順に従って発現後に植物体、植物組織または植物細胞から分離されてもよい。上記のヘテロオリゴマータンパク質はその後実質的に均質に精製されてもよく、そのような状態はクーマシー染色したSDS‐PAGE上のヘテロオリゴマータンパク質に起因するバンドが、慣用のゲルリーダーにより測定したレーンの染色の少なくとも70%、好ましくは少なくとも80%、そして最も好ましくは少なくとも90%を占めると定義されてよい。
【0044】
発明の詳細な説明
本発明に記載のウイルスベクターは、上記のヘテロオリゴマータンパク質を形成するために必要なすべてのタンパク質サブユニットを単一のベクターがコードするウイルスベクター、または上記の少なくとも2種のウイルスベクターのそれぞれが、上記のヘテロオリゴマータンパク質を形成するために必要な異なるタンパク質サブユニットをコードする、少なくとも2種の異なる非競合ウイルスベクターのいずれかである。上記の非競合ウイルスベクターのそれぞれは、組換えヘテロオリゴマータンパク質の1種より多いサブユニットをコードする1種より多い異種核酸配列を発現することができる。上記のRNAウイルスベクターは植物細胞に一過性に送達されてもよく、あるいはDNA前駆体(複数の前駆体)として植物染色体DNAに安定に組み込まれてもよい。
【0045】
本発明は植物細胞における高収率のヘテロオリゴマータンパク質産生のための方法を提供する。この方法は、たとえば発現されることになる異種配列のサイズ制限、上記ベクターの高い不安定性および同じ植物細胞に異なる核酸配列を同時発現できないこと、のような既存のウイルスベクターを基礎にした発現系の制限を克服する。さらに、系からのウイルスコートタンパク質の除去が感染性ウイルス粒子の形成および野生型ウイルスへの逆戻りを妨げるため、上記方法はよりすぐれた生物学的安全性を提供する。本発明を実施することにより、関心のあるヘテロオリゴマータンパク質の高収率発現系の設計が実質的に任意の植物RNAウイルス‐由来レプリコンにとって可能であり、レプリコンは、関心のあるヘテロオリゴマータンパク質の異なるサブユニットをコードする、少なくとも2種の関心のある異種配列を発現可能にするためのレプリコンの改変によって、関心のある異種配列の発現に適切になる。あるいは、同じ植物細胞においてウイルスベクターと一緒に同時複製することができる別の非競合ウイルスベクターを見いだすことも可能である。
【0046】
異なる分類学的群に属するプラス‐センス1本鎖RNAウイルス(本明細書では、簡潔に「RNAウイルス」とも表す)は、本発明のプラス‐センス1本鎖RNAウイルスベクター(本明細書では、簡潔に「ウイルスベクター」とも表す)を構築するためにふさわしい。本明細書では、ウイルスベクターは植物細胞において複製する:すなわち鋳型としてRNAウイルスベクターを使用して、RNA‐依存的RNA重合により別のRNAベクター分子を形成することができるRNAベクターである。好ましくは、本発明のウイルスベクターはRNAウイルス機能を有する、少なくとも1種のウイルス配列エレメント、たとえば、レプリカーゼ、サブゲノムプロモーター、ウイルス粒子アセンブリーの起点、コートタンパク質ORF、または移行タンパク質ORFを含有する。さらに、ウイルスベクターはRNAウイルスIRESエレメントを有していてもよい。
【0047】
ウイルスベクターは、たとえばウイルスに制限部位を導入することにより、それが由来するウイルスから構築してもよく、制限部位は本発明の異種配列を導入するためにふさわしいクローニング部位を表す。当業者は、RNAウイルスまたはベクターに対する核酸の工学的操作は一般に、それぞれ上記のRNAウイルスまたはベクターのDNAコピーを使用して、DNAレベルで行われることを理解することになる。したがって、RNAウイルスベクターのDNAコピーは、RNAウイルスのDNAコピーに制限部位を導入することにより、それが由来するRNAウイルスのDNAコピーから構築することができ、制限部位はDNAレベルで本発明の異種配列のDNAコピーを導入するためにふさわしいクローニング部位を表す、と述べることがより正確である。これらの事柄は当業者には明白であり、一般に本明細書では強調されない。
【0048】
RNAウイルスのDNAコピーがクローニングにふさわしい制限部位(複数の制限部位)を天然に有する場合、ウイルス自体がウイルスベクターであってもよい。本明細書では、「ウイルスベクター」という用語は、異種配列がベクターに存在しない事例か、または本発明のタンパク質サブユニットをコードする配列がベクター中に全く存在しない事例を表す。異種配列または本発明のタンパク質サブユニットをコードする配列がウイルスベクターに挿入される事例は、そのようなインサートの存在を明白に特定することにより確認される。
【0049】
本明細書では、異種配列または本発明のタンパク質サブユニットをコードする配列が導入される以前は2種のウイルスベクターの配列が異なる場合、そのようなウイルスベクターは異なると表す。異種配列または本発明のタンパク質サブユニットをコードする配列が導入される前は2種のウイルスベクターの配列が同じ配列を有する場合、それらは同じであると表す。したがって、2種のウイルスベクターが異なるか、または同じであるかどうかを確認する場合、異種配列または本発明のタンパク質サブユニットをコードする配列は考慮にいれない。
【0050】
本明細書では、「レプリコン」または「ウイルスレプリコン」という用語は「ウイルスベクター」と同じ意味を有する。
本発明のウイルスベクターを工学的に操作するために使用可能なRNAウイルスのリストを以下に提示する。(イタリック体ではない)引用符に囲まれた分類名は、この分類がICTVにより国際的に認可された名称を持たないことを示す。種の名称(通称)は通常の活字体で示す。属または科への正式な帰属のないウイルスが示される。
【0051】
RNAウイルス:
ssRNAウイルス:
科:ブロモウイルス科、
属:アルファモウイルス、基準種:アルファルファモザイクウイルス、
属:イラルウイルス、基準種:タバコ条斑ウイルス、
属:ブロモウイルス、基準種:ブロモモザイクウイルス、
属:ククモウイルス、基準種:キュウリモザイクウイルス、
科:クロステロウイルス科、
属:クロステロウイルス、基準種:ビート萎黄ウイルス、
属:クリニウイルス、基準種:レタス伝染性黄斑ウイルス、
科:コモウイルス科、
属:コモウイルス、基準種:カウピーモザイクウイルス
属:ファバウイルス、基準種:ソラマメウィルトウイルス1,
属:ネポウイルス、基準種:タバコ輪点ウイルス;科:ポティウイルス科、
属:ポティウイルス、基準種:ジャガイモウイルスY、プラムポックスウイルス;タバコエッチウイルス;クローバー葉脈黄化ウイルス;タバコベインモットリングウイルス;
属:リモウイルス、基準種:ライグラスモザイクウイルス
属:ビモウイルス、基準種:オオムギ縞萎縮モザイクウイルス;
科:セキウイルス科、
属:セキウイルス、基準種:パースニップイエローフレックウイルス、
属:ワイカウイルス、基準種:ライスツングロスフェリカルウイルス;
科:トムブスウイルス
属:カルモウイルス、基準種:カーネーション斑紋ウイルス、
属:ディアンソウイルス、基準種:カーネーションリングスポットウイルス、
属:マクロモウイルス、基準種:メイズクロロティックモットルウイルス、
属:トムブスウイルス、基準種:トマトブッシースタントウイルス、
帰属されていないssRNAウイルスの属、
属:カピロウイルス、基準種:リンゴステムグルービングウイルス;
属:カルラウイルス、基準種:カーネーション潜在ウイルス;
属:エナモウイルス、基準種:ピーエネーションモザイクウイルス、
属:フロウイルス、基準種:ムギ類萎縮ウイルス、
属:ホルディウイルス、基準種:ムギ斑葉モザイクウイルス、
属:イダエオウイルス、基準種:ラズベリーブッシードワルフウイルス;
属:ルテオウイルス、基準種:オオムギ黄萎ウイルス、
属:マラフィウイルス、基準種:マイズラヤドフィノウイルス;
属:ポテクスウイルス、基準種:ジャガイモウイルスX;
属:ソベモウイルス、基準種:インゲンマメ南部モザイクウイルス、
属:テヌイウイルス、基準種:イネ縞葉枯ウイルス、
属:トバモウイルス、基準種:タバコモザイクウイルス、
属:トブラウイルス、基準種:タバコ茎えそウイルス、
属:トリコウイルス、基準種:リンゴクロロティックリーフスポットウイルス;
属:ティモウイルス、基準種:ターニップイエローモザイクウイルス;
属:ウムブラウイルス、基準種:キャロットモットルウイルス;
ネガティブssRNAウイルス:目:モノネガウイルス目、科:ラブドウイルス科、属:シトラブドウイルス、基準種:レタスネクロティックイエローズウイルス、
属:ヌクレオラブドロウイルス、基準種:ポテトイエロードワーフウイルス。
【0052】
RNAウイルスベクターは、関心のある遺伝子の発現を提供する異種RNAの極めて多数のコピーを植物細胞に提供することができる。しかし、そのようなベクターは、異種核酸配列のサイズがある範囲、通常1kbを越えて増大すると、極めて不安定になることが公知である。そのような制約のために、そのようなベクター系の適用は、今まで比較的単純な小〜中程度のサイズのタンパク質に限定されてきた。高分子または複雑なヘテロオリゴマータンパク質のいずれかを発現させる試みは首尾よい結果を導いてこなかった。我々は、驚くべきことに、以前には不可能であった複雑なヘテロオリゴマータンパク質の高収率発現のためにウイルスベクターを首尾よく採用できることを見いだしている。全長モノクローナル抗体を発現させる初期の試み(Verch et al.,1998,J.Immunol.Meth.,220,69−75)は、同じ細胞における関心のあるタンパク質の同時発現のために使用されるウイルスベクターの不適合性のために、満足すべき結果を提供しなかった。実施例1で、我々は、単一細胞レベルで、同じ植物RNAウイルス(TMV)由来のウイルスレプリコンから2種の異なる遺伝子(GFPとDsRed)の効率的な同時発現が不可能であることを証明している。図1(A‐右パネル)では、レポーター遺伝子‐DsRedおよびGFPの両方の発現を示すプロトプラストを検出できなかった。右下部パネルの一部のプロトプラストにおけるDsRedの弱い発現パターンは、強いGFP発現(右上部パネル)と一致し、そしてDsRed検出に使用されたフィルターの漏出による偽‐陽性の結果である。この漏出は高濃度のGFPを含有するプロトプラストの見かけの弱いDsRed蛍光を導く。したがって、同一であるか、または広範な相同性領域を共有するRNAレプリコンが異なる組換えタンパク質をコードする異なる異種核酸配列をたとえ持っていても、それらは1つの植物細胞に共存できない。この現象の理由は、今のところ解っていない。可能性のある説明として、1種のウイルスレプリコンのコピー数の急激な増加が、別のウイルスレプリコンを速やかに撃退することが考えられる。選択された細胞において2番目に複製を開始するレプリコンは、第1のレプリコンに追いつくことができず、したがって第1レプリコンを確認する事象は主に統計的特徴を有する。
【0053】
この問題に取り組むための我々の研究では、2種の異なるサブゲノムプロモーターの制御下で、異なるタンパク質をコードする2種の異なる異種核酸配列が上記のレプリコン中に存在するように、ウイルスレプリコンを工学的に操作してきた。そのようなRNAレプリコンをコードするT−DNA領域の一般的スキームは図2に示す。そのようなベクターの設計および最適化のために、我々は、便宜上、我々の初期の特許出願に記載のプロベクター法を使用してきた(WO02088369;Marillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857も参照されたい)。この方法は、部位特異的組換えにより、事前に作製されたモジュールに由来する最終ベクターのin plantaでの集合を可能にし、そうしてベクター設計およびベクター最適化を著しくスピードアップさせる。コンストラクトの設計は実施例2に記載し、図式表示は図3に示す。
【0054】
我々は驚くべきことに、最終レプリコンにおけるインサートの大きなサイズ、および2種の強いサブゲノムプロモーターの存在によるベクターの複雑な構造にもかかわらず、得られたRNAレプリコンは、in plantaでの高い安定性および同じ植物細胞における2種の異なる組換えタンパク質(GFPとDsRed)の同時発現を提供する能力を示すことを見いだしている。図4に示すように、同時発現頻度は感染した範囲ではすべての細胞のほとんど100%にまで到達可能である。そのようなコンストラクト中におけるレポーター遺伝子(GFPとDsRed)と、たとえばIgGの軽鎖および重鎖の置換、その上のインフィルトレーションされたN.ベンサミアーナ葉部における発現は驚くべき結果を生み出した。実施例3で記載するように、ウェスタンブロット解析は見事なほどの高濃度の集合したモノクローナル抗体(レーン6および7;図6)を明らかにした。我々の系によって提供される集合したモノクローナル抗体の収率は、現在の当該技術分野の系の状態によって生み出された収率とは比較にならないほど高い(図6、レーン4)。知る限りでは、これは、植物ウイルスベクターを基にした発現系を使用することによる、抗体のような複雑なヘテロオリゴマータンパク質発現の初めての証拠である。初期のすべての刊行物はモノクローナル抗体の簡単な人工誘導体の発現、たとえばTMV−ウイルスベクターを使用した(McCormick et al.,Proc Natl Acad Sci USA,96,703−708;McCormick et al.,2003,J.Immunol.Methods,278,95−104)およびPVXを基礎にした(Smolenska et al.,1998,FEBS Lett.441,379−382;Franconi et al.,1999,Immnotechnology,4,189−201;Hendy et al.,1999,J.Immunol.Methods,231,137−146;Roggero et al.,2001,Protein Expr.Purif.,22,70−74)1本鎖抗体(scFv)の発現に限定されていた。
【0055】
本発明では、好ましくは全身ウイルスベクターを使用しない。全身ウイルスベクターのかわりに、好ましくは植物系へのアグロバクテリウム媒介ウイルスベクター前駆体送達により、ウイルスベクターの能力を全身移行するように置換する。このことはウイルスコートタンパク質を、発現されることになる異種配列で置換することを可能にする。この方法は、ウイルスベクターの異種配列への適応性増大の可能性に貢献する。同時に、この方法はこの系の生産性を弱めることになる自発的な異種配列の欠失のために、ウイルスベクターが野生型ウイルスに変換される見込みを排除する。
【0056】
我々のウイルス‐レプリコンを基礎にした系は、先行技術の系より著しく改善された収率のモノクローナル抗体を産生するが、我々はウイルスレプリコンのサイズを減少させることによりそれ以上の収率増加の可能性を検討した。分泌抗体の2つの鎖を発現するウイルスレプリコンのサイズを減少させる可能性が限定されていることを考慮して、2種の異なるウイルスレプリコンからの抗体の重鎖および軽鎖の発現を試みてきた。同じウイルスを基礎にしたレプリコンは同じ植物細胞において2種の異なる異種配列を発現できない(上記参照、実施例1)ことを考慮して、非相同植物ウイルスであるタバコモザイクウイルス(TMV)とジャガイモウイルスX(PVX)に由来するウイルスベクターに、DsRedとGFP遺伝子を別々にクローニングすることにより、同じ植物細胞に2種の異なる遺伝子を同時発現させる可能性を試験した(実施例4を参照されたい)。ウイルスベクターのcDNAを含有するT−DNA領域の図式表示は図7および8(PVXのみ)に示す。実施例4では、便宜上、部位特異的組換えによりベクターモジュールからin plantaで集合させたTMVを基礎にしたウイルスベクターを使用した。驚くべくことに、共にインフィルトレーションされた植物葉領域から分離したプロトプラストはDsRedおよびGFPレポーター遺伝子の実質的に100%の同時発現頻度を示すことを我々は見いだしている(図9)。
【0057】
最近、異なって標識されたウイルスの空間的分離に関する研究がDietrichとMaiss(2003,J.Gen.Virology,84,2871−2876)によって提示された。この研究では、レポーターとして蛍光タンパク質が発現した。ヘテロオリゴマータンパク質の産生には言及しなかった。さらに、分離したプロトプラストレベルでの同時発現は検討しなかった。レポーター遺伝子産物は原形質連絡を介した拡散により隣接する細胞に拡散できるため、選択されたウイルス対がレポーター遺伝子を、同じ細胞で、または隣接する細胞で発現したかどうかについての情報は全く得られなかった。初期の刊行物は植物の感染時の野生型ウイルス間の相乗作用について述べているが、植物細胞におけるヘテロオリゴマータンパク質の発現については述べていない。(Rochow & Ross,1955,Virology,1,10−27;Goodman & Ross,1974,Virology,58,16−24;Goodman & Ross,1974,Virology,59,314−318;Goodman & Ross,1974,Virology,58,263−271)。同時感染した植物におけるウイルスの相乗的相互作用に関するこれらの初期の研究(Vance et al.,1995,Virlogy,206,583−590;Pruss et al.,1997,Plant Cell,9,859−868)のその後の発展は転写後遺伝子サイレンシング(PTGS)のサプレッサーの発見を導いた。組換えタンパク質発現系のさらに進んだ発展は、促進された組換えタンパク質産生のためのそのようなPTGSサプレッサーの研究に向けられた。相乗的ウイルスを基礎にしたウイルス発現系を使用した先行技術にはタンパク質発現に対する手がかりが全く存在しない。
【0058】
本発明は、非競合ウイルスベクターが植物細胞または植物体におけるヘテロオリゴマータンパク質産生の効率的な手段を表すことを証明する。2種以上のウイルスベクターが同じ植物細胞において複製可能であり、そして上記の複製および別の細胞からのトランスフェクション中にお互いに効力を低下させない場合、上記のウイルスベクターはお互いに非競合的である。効力低下がないとは、少なくとも10%、好ましくは50%、より好ましくは90%、そして最も好ましくは100%のトランスフェクションされた植物細胞が共にトランスフェクションされること、すなわち上記の2種以上の異なるウイルスベクターを含有することを意味する。好ましくは、上記のウイルスベクターは異種核酸配列を複製し、発現することが可能である。より好ましくは、上記の異種核酸配列が異なるタンパク質をコードする。よりいっそう好ましくは、上記の異なるタンパク質がヘテロオリゴマータンパク質のサブユニットである。ウイルスベクターが競合的であるか、または非競合的であるかを確認するために、植物細胞の同時トランスフェクションの頻度を測定することができる。このことは以下のプロトコルに従って行うことができる:2種のウイルスベクターは2種の異なるレポーター遺伝子、たとえばDsRedとGFPで標識される。上記の、異なって標識されたベクターは植物葉組織に(たとえばアグロ‐インフィルトレーション法により)同時送達され、3〜6日後、感染した領域から分離したプロトプラストを計数して、1種のレポーター遺伝子だけを発現しているプロトプラスト数に比較した、両方のレポーター遺伝子を同時発現しているプロトプラストの割合を決定することができる。そのような測定値を例示する実験は実施例2および4に記載する(図1Aおよび9も参照されたい)。通常、非競合ウイルスベクターは相乗的であるウイルスに由来することができ、たとえば、同じ植物宿主に首尾よく同時トランスフェクションすることができる。そのような相乗的RNAウイルス対の例としては以下のものが挙げられる:
ジャガイモウイルスX(PVX)/タバコモザイクウイルス(TMV);
PVX/タバコベインモットリングウイルス(TVMV);
PVX/タバコエッチウイルス(TEV);
PVX/クローバー葉脈黄化ウイルス(CIYVV);
PVX/プラムポックスウイルス(PPV);
PVX/ジャガイモウイルスY(PVY)。
【0059】
ウイルスベクターの競合性または非競合性の機構的理由は既知ではない。可能性のある1つの説明としては、相乗的ウイルスベクターに由来するウイルスレプリコンが異なる細胞下コンパートメントにおいてウイルス複製複合体(VRC)を形成する(Kawakami et al.,2004,Proc.Natl.Acad.Sci.USA,101,6291−6296)ことが挙げられ、したがってそれらはVRCを形成するために必要な空間に対してお互いに競合しない。実験的に証明されているように、非競合ウイルスベクターは、核酸配列レベルが著しく異なる、異なるウイルス種に由来してもよい。上記の非競合ウイルスベクターは、同じ植物宿主に首尾よく同時感染できる相乗的植物ウイルスに由来してもよい。相乗的ウイルスは通常異なる属に属する。たとえば、PVXはポテクスウイルス属に属するが、それに対して相乗的なウイルス、たとえばTVMV、TEV、PPV、およびCIEVVはポティウイルス属に属する。PVXウイルスに相乗的(非競合的)な別のウイルス、たとえばTMV、TVCV、およびcrTMVはトバモウイルス属に属する。明らかに、異なる属の典型間のゲノム相同性はかなり低く、通常、RNAレベルで50%同一性より低い。
【0060】
異なるウイルスベクター(たとえば、PVXおよびTMVを基礎にしたもの)へのモノクローナル抗体重鎖および軽鎖のクローニング、続いて同時感染した植物組織における上記の鎖の付加的な同時発現は実施例5に記載する。ベクターの図式表示は図10に示す。本発明および別の発現系により産生されたモノクローナル抗体の収率の比較ELISA測定は、2種の非競合ウイルスベクターを基礎にした上記の系の最も好ましい成果を示した(図11)。ウイルスベクター(複数のウイルスベクター)の補助による植物細胞での組換えヘテロオリゴマータンパク質の効率的な発現は先行技術では公知でない。
【0061】
我々のデータに基づいた場合、ウイルスベクターのウイルスに由来する配列は、上記ウイルスがお互いに関連するとみなすために十分な相同性を示さないことになる。そのようなウイルスの例としては、TMVとPVXのウイルス対が挙げられ、そしてウイルスベクターはそれに基づく。これらのウイルスベクターはそのような相同性を全く示さない。受容可能なウイルス対を選択するための別の必要条件は、植物宿主に同時感染中の、元の野生型ウイルスの相乗性であってもよい。
【0062】
上で論じた実験は、植物細胞へのアグロバクテリウムに媒介されたDNA前駆体送達に基づいた一過性発現系により行われた。しかし、本発明の代わりの適用としては、植物核ヌクレオソームに安定に組み込まれた上記のRNAレプリコン(複数のレプリコン)のDNA前駆体によるトランスジェニック植物への適用が挙げられる。これは、植物ウイルスベクターを基礎にした系の多くの制約、たとえば、ウイルスベクターが耐えうる異種配列の最大サイズに対する制限を克服する。DNA前駆体はトランスジェニック植物のそれぞれの細胞に存在することになるため、RNAレプリコンの全身移行または細胞間移行(レプリコンスプレッディング)のための絶対的な要求性が存在しない。これは、高い効率の本発明のRNAレプリコン形成および細胞質への輸送によって補われる。しかし、RNAレプリコン形成がすべての細胞でいつも起こるわけではないので、ベクターの細胞間移行の能力は付加的な価値のものであってもよい。
【0063】
異なる方法を使用して、植物体、植物組織または植物細胞に異種DNAを提供してもよい。上記のベクターは、アグロバクテリウム(US5,591,616;US4,940,838;US5,464,763)または粒子もしくはマイクロプロジェクタイルボンバードメント(US05100792;EP00444882B1;EP00434616B1)によって運ばれるTi−プラスミドベクターによって、植物細胞を形質転換してよい。別の植物形質転換法、たとえばマイクロインジェクション(WO09209696;WO09400583A1;EP175966B1)、エレクトロポレーション(EP00564595B1;EP00290395B1;WO08706614A1)またはプロトプラストのPEG‐媒介形質転換なども使用することができる。ベクター送達方法の選択は、形質転換されることになる植物種に依存してもよい。たとえば、マイクロプロジェクタイルボンバードメントは一般に単子葉植物の形質転換に好ましいが、双子葉植物には、一般にアグロバクテリウム媒介形質転換がより好ましい結果を示す。
【0064】
本発明の実施例では、ニコチアナ細胞へのベクター(上記の異種DNA)の一過性アグロバクテリウム媒介送達を使用した。しかし、上記ベクターは、関心のある植物種の安定な、または一過性形質転換にふさわしい標準技術のいずれかに従って、植物体に安定に導入されてもよい。双子葉植物のための形質転換技術は当該技術分野で公知であり、アグロバクテリウムを基礎にした技術とアグロバクテリウムを必要としない技術が挙げられる。アグロバクテリウムを使用しない技術には、プロトプラストまたは細胞による直接的な外来遺伝物質の取り込みが挙げられる。これらの技術には、PEGまたはエレクトロポレーション媒介取り込み、粒子ボンバードメント‐媒介送達およびマイクロインジェクションが挙げられる。これらの技術の例は、Paszkowski et al.,EMBO J3,2717−2722(1984)、Potrykus et al.,Mol.Gen.Genet.199,169−177(1985)、Reich et al.,Biotechnology 4:1001−1004(1986)、およびKlein et al.,Nature 327,70−73(1987)に記載される。それぞれの場合、形質転換された細胞は標準技術を使用して、全植物体に再生される。
【0065】
アグロバクテリウム媒介形質転換は、その高い形質転換効率と多くの異なる種に対するその広範な有用性のために、双子葉植物の形質転換に好ましい技術である。通常アグロバクテリウムによって形質転換することができる多くの作物種には、タバコ、トマト、ヒマワリ、綿花、ナタネ、ジャガイモ、ダイズ、アルファルファおよびポプラが挙げられる(EP0317511(綿花)、EP0249432(トマト)、WO87/07299(ナタネ)、米国特許第4,795,855号(ポプラ))。
【0066】
本発明の実施例では、関心のある遺伝子(複数の遺伝子)の一過性発現にT−DNAのアグロバクテリウム媒介送達(Vaquero et al.,1999,Proc.Natl.Acad.Sci.USA,96,11128−11133)が使用された。この方法は、小規模〜中規模の組換えタンパク質産生系にとってだけでなく、大規模発現にとっても極めて有用な手段である。
【0067】
植物染色体DNAに安定に組み込まれたウイルスレプリコン前駆体の放出は、誘導型またはいずれか別の調節された(たとえば発達的に調節された)プロモーターを使用して行うことができる。誘導型プロモーターはそれらの誘導条件に従って、2つのカテゴリーに分類することができる:非生物的因子(温度、pH、化学物質)によって誘導されるものおよび生物因子、たとえば病原体または害虫攻撃によって誘導されるもの。第1のカテゴリーの例としては、熱‐誘導型(US05187287)および低温‐誘導型(US05847102)プロモーター、銅‐誘導系(Mett et al.,1993,Proc.Natl.Acad.Sci.,90,4567−4571)、ステロイド‐誘導系(Aoyama & Chua,1997,Plant J.,11,605−612;Mcmellis et al.,1998,Plant J.,14,247−257;US06063985)、エタノール‐誘導系(Caddick et al.,1997,Nature Biotech.,16,177−180;WO09321334)、およびテトラサイクリン誘導系(Weinmann et al.,1994,Plant J.,5,559−569)が挙げられる。植物の化学的誘導系の分野での最も最近の成果の1つは、グルココルチコイドデキサメタゾンによりスイッチを入れ、テトラサイクリンによりスイッチを切ることができるキメラ化プロモーターである(Bohner et al.,1999,Plant J.,19,87−95)。化学的誘導系に関する総説としては以下を参照されたい:Zuo & Chua,(2000,Current Opin.Biothechnol.,11,146−151)およびPadidam,M(2003,Curr.Opin.Plant Biol.,6,169−177)。誘導型プロモーターの別の例としては、植物における病原性‐関連(PR)遺伝子の発現を制御するプロモーターが挙げられる。これらのプロモーターは、病原菌攻撃に反応する植物シグナリング経路の重要な構成要素であるサリチル酸、またはPR遺伝子発現を引き起こすことが可能な別の化合物(ベンゾー1,2,3−チアジアゾールまたはイソニコチン酸)(US05942662)による植物の処理によって誘導することができる。
【0068】
本発明は実施例に記載のTMVおよびPVXを基礎にしたベクターに限定されないが、上記ウイルスレプリコンに由来する発現系の確立に役立つ、別の植物RNAウイルスに由来するレプリコンに適用可能である。同時感染された植物において最も詳しく研究された相乗作用はウイルスのPVX/PVY対に関して公知である(Rochow & Ross,1955,Virology,1,10−27;Goodman & Ross,1974,Virology,58,16−24)。おそらく、多くのそれらのウイルスは同じ植物細胞中で共に複製可能である。Dietrich & Maiss(2003,J.Gen.Virol.,84,2871−2876)は、異なって標識されたウイルス対、たとえばプラムポックスウイルス(PPV)とジャガイモウイルスX(PVX)、タバコベインモットリングウイルス(TVMV)とPVX、クローバー葉脈黄化ウイルス(CIYVV)とPVXは、植物組織中の同じ感染領域において異なるレポーター遺伝子を同時発現できることを示している。本発明に記載のストラテジーを使用して、同じ植物細胞中で同時複製可能な、実質的に任意の対の植物プラスセンス1本鎖RNAウイルス‐由来レプリコンに対する組換えヘテロオリゴマータンパク質発現系が開発可能である。たとえば、アルファモウイルス属のアルファルファモザイクウイルス(AMV)に基づいたウイルスベクターが本発明で使用可能である。
【0069】
複雑な(ヘテロオリゴマー)タンパク質、それらの(機能的または非機能的)フラグメントならびにそれらの人工誘導体および融合物をコードする関心のある遺伝子は、本発明を使用して、植物体または植物細胞に発現することができる。多くの商業的に価値のあるヘテロオリゴマータンパク質の群は本発明を使用して生産し、精製することができる。それらの群には、工業用および研究用タンパク質、ならびにヒトまたは動物の健康分野における適用のためのタンパク質が挙げられるが、それらに限定されない。しかし、免疫反応タンパク質、とりわけ異なるクラスのイムノグロブリン(IgG、IgM、IgAおよびIgD)から選択されたモノクローナル抗体ならびにそれらの合成誘導体、たとえば変異型および別のタンパク質またはその一部との異なる型の融合体が最も好ましい。
【0070】
実施例
以下の実施例は本発明を説明するために提示される。本発明の意図および範囲から逸脱せずに改変および変更を行ってもよい。
【0071】
実施例1
同じ植物RNAウイルスに由来する2種のTMVを基礎にしたRNAウイルスベクターからのGFPとDsRedの同時発現の欠如
葉組織は同じ植物RNAウイルスに由来するが、関心のある2種の異なる遺伝子を含有する、2種のTMVを基礎にしたRNAウイルスベクターに感染された(図1)。TMVを基礎にしたウイルスベクターにクローニングした可視マーカー遺伝子GFPおよびDsRedを使用して、この方法を試験した。第1コンストラクト、pICH17272(Marillonnet et al.,2005,Nat.Biotechnol.,23,718−723の図1を参照されたい)はGFPを含有し、そしてベクター中のRdRPコーディング配列が14個よりむしろ9個の植物イントロンを含有すること以外は、pICH18711(国際特許出願PCT/EP03/12530,WO2005049839として公開;Marillonnet et al.,2005,Nat. Biotechnol.,23,718−723を参照されたい)に類似する。pICH18722(Marillonnet et al.,2005,Nat.Biotechnol.,23,718−723)はTMVの中の2種の密接に関連した株、CrTMV(Dorokhov et al.,1994,FEBS Lett.350,5−8)およびTVCV(Lartey et al.,1994,Arch.Viol.138,287−298)の骨格に構築され、コートタンパク質配列が異種配列によって置換されている、植物プロモーターの制御下にあるウイルスゲノムを含有する。RdRPおよびMP中の11個のイントロンの存在は、ベクターの葉組織へのアグロバクテリウム媒介送達後のウイルス複製開始の効率を増し、したがって同じ細胞における2種のウイルスベクターの同時発現の見込みを増大させる。GFPコーディング配列がXho1−Not1制限部位を使用してDsRedのコーディング配列によって置換されていること以外は、pICH18505は、pICH17272に類似する(図1)。pICH18505およびpICH17272のT−DNA領域の詳細な制限地図は図1Cに示す。pICH17272およびpICH18505はアグロバクテリウム株GV3101を形質転換し、先に記載(Marillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857)のように、ニコチアナベンサミアーナ葉組織がインフィルトレーションされた。プロトプラストはインフィルトレーション7日後にインフィルトレーション部位から作製し、顕微鏡下、青色または赤色光下で観察した。プロトプラストが数日後にインフィルトレーション部位から作製された場合でも、GFPとDsRedの両方を発現したのは非常にわずかのプロトプラストだけであった。このことは細胞が第1のTMVを基礎にしたウイルスベクターにひとたび感染すれば、第2のTMVを基礎にしたウイルスベクターによる再感染は不可能になることを示唆する。
【0072】
実施例2
単一ウイルスレプリコンからのGFPおよびDsRedの同時発現
2種の別々のサブゲノムプロモーターの制御下、関心のある2種の遺伝子がRNAウイルスベクターから発現された(図2)。両方のサブゲノムプロモーターは同一であってもよいが、ウイルス複製中におけるコンストラクト中の反復配列間の欠失のリスクを回避するために、関連するTMV株由来のサブゲノムプロモーターを使用することが好ましく;この場合、第2のサブゲノムプロモーターおよび3′非翻訳領域はTMGMV株U5に由来する(Shivprasad et al.,1999,Virology,255,312−323;Marillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857)。また、クローニングの便宜上、GFPおよびDsRedは、完全にアセンブルされたベクターよりむしろ、ウイルスプロベクター(WO02/088369およびMarillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857に記載)にクローニングされた。2種のプロベクター、pICH17388およびpICH15933(図3)は、in plantaで、両方のプロベクターモジュール間の部位特異的組換えにより完全に機能的なTMVを基礎にしたウイルスベクターに変換された。pICH17388は、(pICH17272と同じように)ウイルス配列に11個のイントロンが存在すること以外はpICHNOP(Marillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857)に類似する。pICH15933は、DsRedのコーディング配列によってTMGMV U5のコーディング配列が置換されている以外は、pICHGFPSYS(Marillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857)に相当する。
【0073】
pICH15933はアグロバクテリウム株GV3101に形質転換され、pICH17388およびpICH10881(図3)と一緒にニコチアナベンサミアーナにインフィルトレーションした。インフィルトレーション5日後に、すべてのGFP‐発現部位はDsRedも発現し、同じ植物細胞中における2種の遺伝子の卓越した同時発現を示した(図4)。
【0074】
実施例3
単一のウイルスレプリコンから抗体の発現
pICH15933におけるGFPおよびDsRedのコーディング配列は、IgG抗体軽鎖および重鎖によってそれぞれ置換され、コンストラクトpICH20241(図5)を得た。対照として、重鎖および軽鎖はTMVを基礎にしたプロベクターにクローニングし、pICH1740のコーディング配列を置換し、コンストラクトpICH20421およびpICH20431(図5)を得た。pICH20241はpICH17388およびpICH10881と一緒にニコチアナベンサミアーナ葉部にインフィルトレーションした(図3)。
【0075】
インフィルトレーションされた葉部から抽出した全可溶性タンパク質のウェスタンブロット解析は、ホースラディッシュペルオキシダーゼ(HRP)(Sigma)により標識した、1:6000に希釈した抗‐ヒトIgGウサギ抗体により行った。解析の結果は図6Aに示す。単一のウイルスベクターの補助によって達成された抗体の発現レベル(レーン6&7、図6)は、PTGSサプレッサーP19の存在においてさえ、強力な構成プロモーターの補助により成し遂げられたものより著しく高い(レーン3〜4、図6)。
【0076】
実施例4
TMVおよびPVXを基礎にしたウイルスベクターによるGFPおよびDsRedの同時発現
2種の遺伝子の同時発現のための別のストラテジーは、同じ細胞に同時感染し、複製することが可能な、異なるウイルスに基づいて構築された別々のウイルスベクターを使用することである。例として、ジャガイモウイルスX(PVX)に基づいた発現ベクターはTMVと一緒に同じ細胞に共に存在することができる。2種のそのような非競合ウイルスベクターの図式表示は図7に示す。
【0077】
PVX(PV0014株)の接種材料は、感染した乾燥葉材料として、German Collection of Microorganisms and Cell Cultutes(DSMZ)から得て、ニコチアナベンサミアーナ植物体の接種に使用した。ウイルス症状を示した接種植物の全身葉部は全RNA調製に使用した。第1鎖cDNAはプライマーpvxpr2、pvxpr4およびpvxpr6を使用して作製した。3種のcDNAフラグメントはPfu−Taqポリメラーゼミックスを使用して、PCRによりPVXcDNAから増幅した:
フラグメント1はプライマー:
【0078】
【化1】
【0079】
により増幅した。
フラグメント1はBsaIにより消化した。
フラグメント2はプライマー:
【0080】
【化2】
【0081】
により増幅した。
フラグメント2はEsp3Iにより消化した。
フラグメント3はプライマー:
【0082】
【化3】
【0083】
により増幅した。
フラグメント3はBpiI AatIIにより消化した。
アラビドプシスアクチン2遺伝子プロモーターを含有するフラグメント4(An et al.,1996,Plant J.10,107−121)はアラビドプシスゲノムDNAからプライマー:
【0084】
【化4】
【0085】
により増幅した。
フラグメント4はMlu1およびBsa1により消化した。
アラビドプシスアクチン2プロモーターの代わりとして、さらにCaMV35プロモーターを使用して、PVXを基礎にしたウイルスベクターの転写を行った(図10D、pICH20788;pICH21380)。PVXを基礎にしたベクターを作製する一般的ストラテジーはBaulcombe and colleagues(1995,Plant Journal,7:1045−1053)に記載のように使用した。
【0086】
4種すべてのフラグメントは、Mlu1およびAatIIにより消化したバイナリーベクター中に一緒にクローニングした。得られたコンストラクトの20個のクローンはアグロバクテリウム株GV3101に形質転換し、それぞれのクローンはニコチアナベンサミアーナ植物体の1枚の葉にインフィルトレーションした。1週間後、ウイルス感染症状の表現型に対してニコチアナベンサミアーナをスクリーニングし、陽性のプラスミドクローンを保存した(pICHPVX)。
【0087】
クローニングベクターは、CPサブゲノムプロモーター下流の、関心のある遺伝子をクローニングするためのこの完全に機能的なcDNAから作製した。GFP遺伝子は例としてクローニングした。CPサブゲノム領域中のATGはAGG(Genbank受託番号M95516中の位置5651)に変異させた。GFP遺伝子はNhe1部位の3′(座標5662〜5667)にクローニングした。PVXの3′末端(座標5618〜6435)はGFP配列の下流にクローニングし、この配列に12〜24Aのストレッチが続いた。このコンストラクト(pICH0130,図8)はニコチアナベンサミアーナ葉部にアグロインフィルトレーションした。GFP蛍光はインフィルトレーション後2〜3日目に出現した。GFPの全身移行は数日後に出現した。
【0088】
pICH0130はベクターpICH17388、pICH10580およびpICH10881(図3)と一緒にニコチアナベンサミアーナ葉部に同時インフィルトレーションし、DsRed発現を提供した。同時インフィルトレーション後6日目にインフィルトレーションした領域からプロトプラストを作製し、青色光の下、顕微鏡下で観察した。ほとんどすべてのプロトプラストはGFPおよびDsRedを両方とも発現し、卓越した同時発現レベルを示した(図9)。
【0089】
実施例5
TMVおよびPVXを基礎にしたベクターによるモノクローナル抗体の発現
IgGの重鎖および軽鎖はPVXベクターにクローニングし、pIC0130中のGFPのコーディング配列を置換して、それぞれ重鎖または軽鎖のいずれかを含有する2種のコンストラクト、pICH21240およびpICH21370を生み出した(図10A)。また、抗体の軽鎖または重鎖のいずれかを含有するTMVプロベクター部分のクローンが構築された(pICH20431およびpICH20421,図10A)。2種の異なる鎖の発現型であるPVXおよびTMVを基礎にしたベクターを提供するアグロバクテリウムの混合物はN.ベンサミアーナ葉部にインフィルトレーションし、抗体の発現レベルは接種後10日目にELISAによって測定した。結果(図11)は、PVXおよびTMVを基礎にしたベクター由来の重鎖および軽鎖の同時発現の事例における、集合した機能的抗体の極めて高レベルを示す。これらのレベルは両方の鎖を発現しているTMVベクターから得られたものより著しく高かった(実施例3)。
【0090】
IgG1サブクラスに属するさらに別のヒト腫瘍‐特異的モノクローナル抗体、抗癌mabは以下のように、TMVおよびPVXを基礎にしたベクターにサブクローニングした:抗癌mab軽鎖コーディング配列を含有する、730NcoI−NotIフラグメントはNotl部位で平滑化し、NcoI−XbaIにより開き、Xbal部位で平滑化した、TMVを基礎にした3′モジュールクローニングベクターpICH11599(図10B)にリゲーションし、pICH21910を得た。同じストラテジーを使用して、抗癌mab重鎖コーディング配列を持つ1421bpのNcoI−NotIフラグメントをpICH11599(図10B)にサブクローニングし、pICH21920(図10B)コンストラクトを得た。GFP発現のためのPVXを基礎にした3′モジュールpICH21282(図10C)はTMVを基礎にした3′モジュールクローニングベクターpICH10990(図10C)の基部に形成された。それは、GFPコーディング配列のpICH10990(図10C)への挿入および結果として起こるTMV3′非翻訳領域のPVX3′非翻訳領域による置換を介したいくつかのクローニングステップにより成し遂げられた。同様に、TMVを基礎にした3′プロベクターモジュールpICH21920(図10B)は、TMV3′非翻訳領域をPVXの3′非翻訳領域で置換することにより、PVXを基礎にした3′プロベクターコンストラクトpICH22250(図10C)に変換された。それは、HindIII部位で平滑化された、pICH21282(図10C)の575bp HindIII−NdeIフラグメントと、Sall部位で平滑化されたpICH21920のSaII−NdeIフラグメントのリゲーションによって成し遂げられた。抗癌mab軽鎖のコーディング配列を含有する723bp NcoI−EcoRIフラグメントは、NcoI−EcoRIにより消化されたpICH22250(図10C)にサブクローニングし、pICH22240(図10C)を得た。
【0091】
5′プロベクターpICH21380(図10D)は以下のように作製した:コンストラクトpICH17388(図3B)を鋳型として使用し、プライマーpv5p5F(SEQ ID NO:9):
【0092】
【化5】
【0093】
とpv5p5R(SEQ ID NO:10)
【0094】
【化6】
【0095】
の補助によりフラグメントをPCR増幅し、Nhe1およびSac1制限酵素により消化し、pICH20788の大きなNhe1‐Sac1フラグメントとリゲーションし、PVXベクターの3′部位をイントロンおよびAttP組換え部位の5′末端と置換した。クローニングのスキームは図10Dに示す。部位特異的組換えによりin plantaでウイルスベクターを集合させるために、5′プロベクターと3′プロベクターを使用するストラテジーは、以前に記載されたもののとおりであった(Marillonnet et al.,2004,Proc.Natl.Acad.Sci.USA,101,6852−6857)。
【0096】
アグロインフィルトレーション手順
得られたすべてのコンストラクトは、標準エレクトロポレーションプロトコルを使用してアグロバクテリウム株GV3101に形質転換し、さらにニコチアナベンサミアーナ葉部のアグロインフィルトレーションに使用した。OD600が1.8〜2.5の範囲の、一晩培養したアグロバクテリウム培養物の等しい容量を混合し、6000gで3分間沈殿させた。ペレットは、10mM MES(pH5.5)および10mM MgSO4を含有する溶液中に再懸濁し、それぞれ個々の培養物の10-1希釈物を得た。針の無いシリンジを使用して、温室で6〜8週間育てたニコチアナベンサミアーナ植物体葉部にインフィルトレーションした。アグロインフィルトレーションのためのプロベクターを使用する場合、アグロバクテリウムミックスは通常5種の異なるアグロバクテリウム株を含み、それらのそれぞれが以下の構成要素の中の1種を提供する:レコンビナーゼ(インテグラーゼ)供与源(pICH10881);5′PVXプロベクター(pICH21380);1種のIgG鎖を提供する1種の3′PVXプロベクター(重鎖のためのpICH22250または軽鎖のためのpICH22240のいずれか、TMVプロベクターによってどの相補鎖が提供されるかに依存)、;5′TMVプロベクター(pICH17388);1種の3′TMVプロベクター(軽鎖をコードするpICH21910または重鎖をコードするpICH21920)。
【0097】
SDS-PAGEとウェスタンブロット
アグロバクテリウムにインフィルトレーションされたN.ベンサミアーナ葉部の試料100mgは、300μlタンパク質抽出バッファー(0.1M Tris、150mM NaClおよび0.1%Tween20、pH8.0)を含む液体窒素中ですりつぶした。葉の粗抽出物は、Laemmliのバッファー系を使用して10%(非還元状態)または12%(還元状態)ポリアクリルアミドゲルに溶解し、その後クーマシーブリリアントブルー染色を行った。ウェスタンブロット解析のために、SDS-PAGE由来のタンパク質は、ブロッテイングバッファー(25mM Tris、192mM グリシン、20%メタノール、pH8.3)を使用して、電気泳動的にHybondP膜(Amersham Biosciences,UK)に移した。膜はTBST(50mM Tris、100mM NaCl、0.05% Tween20、pH7.4)中5%スキムミルクで遮断し、それぞれ2.5%スキムミルク/TBS 1:10000および1:5000で希釈した、ヤギ抗‐ヒトラムダ軽鎖HRP‐結合抗体(Sigma,UK)またはヤギ抗‐ヒトガンマ鎖特異的HRP‐結合抗体(Sigma、UK)により精査した。結合した抗体はECL Detection Reagent(Amersham Biosciences,UK)を使用して検出した。電気泳動による分離およびウェスタンブロット解析の結果は図11Bに示す。IgG標準物との比較は植物組織におけるモノクローナル抗体の収率が新鮮な葉生物量1グラムにつき0.2〜0.35mgに達することを示した。
【0098】
植物体由来組換えMabの単離
全可溶性タンパク質は100mM リン酸ナトリウム、150mM NaClおよび0.05% Tween20(pH8.0)を含有するバッファーにより、アグロ接種したニコチアナベンサミアーナ葉領域から抽出した。小規模な粗タンパク質抽出物からの抗癌Mabの分離は、取扱説明書に従って、Protein A Magnetic Beads(New England Biolabs,USA)によるワンステップアフィニティー精製を使用して行った。精製した植物体由来Mabのクーマシー染色ゲルは図11Bに示す(パネルB)
【0099】
実施例6
TMVおよびPVXを基礎にしたベクターによる卵胞刺激ホルモン(FSH)の発現
ウシFSHのアルファ(GenBank受託番号NM_173901のコーディング配列101〜463に相当するヌクレオチド)およびベータ(GenBank受託番号NM_174060のコーディング配列70〜459に相当するヌクレオチド)ポリペプチドをコードする遺伝子(cDNA)は制限部位(FSH‐アルファおよびベータサブユニットに対して5′Nco1−3′EcoR1)を隣に配置して合成し、TMVプロベクター(pICH20431、pICH11599の誘導体、図10B)およびPVXベクター(pICH21240)にクローニングし、IgGの重鎖および軽鎖のコーディング配列を置換して、それぞれサブユニットアルファおよびサブユニットベータをコードする配列を含有する2種のコンストラクト、pICH−FSHAおよびpICH−FSHBを得た(図12)。PVXおよびTMVを基礎にしたベクターから発現される2種の異なるサブユニットを提供するアグロバクテリウムの混合物を作製し、先に記載のようにN.ベンサミアーナ葉部にインフィルトレーションし、ヘテロダイマーFSHの発現レベルは、市販のFSHダイマー特異的(FSH117)抗体を使用して接種10日後にELISAにより測定し、Tropix化学ルミネセンスシステム(Tropix,Bedford,MA)により検出した。
【0100】
実施例7
TMVおよびPVXを基礎にしたベクターによるIgMの発現
pICH15933におけるGFPおよびDsRedのコーディング配列はそれぞれIgM軽鎖およびJ鎖をコードする配列によって置換し、コンストラクトpICH−MLCJを得た(図13)。IgM重鎖をコードする配列はPVXベクターにクローニングし(pICH21240)、IgG重鎖のコーディング配列と置換し、コンストラクトpICH−MHCを得た(図13)。PVXおよびTMVを基礎にしたベクターから発現される3種の異なるIgM鎖を提供するアグロバクテリウム混合物はN.ベンサミアーナ葉部にインフィルトレーションし、ヘテロオリゴマー複合体の発現レベルは、市販のIgM特異的抗体を使用して接種後10日目にELISAにより測定した。
【0101】
欧州特許出願第05001819.1号および米国特許出願第60/593,606号の内容は、その優先権を本特許出願によって主張し、本明細書にそのまま参照として援用する。
【図面の簡単な説明】
【0102】
【図1A】(A)インフィルトレーションされたN.ベンサミアーナ葉部における2種の異なるTMVを基礎にしたベクター由来のGFPおよびDsRedの発現分布パターンを示す。左の写真:上部左側の明るいスポットはpICH17272にインフィルトレーションされた領域のGFP蛍光である。下部の弱く明るいスポットはベクターpICH18505にインフィルトレーションされた領域の赤色蛍光である。左の写真の底部右側の明るいスポットは、pICH17272+pICH18505にインフィルトレーションされた領域である。右側の写真は、2種の異なるベクターに同時インフィルトレーションされた葉部領域から分離されたプロトプラストを示す(上部:GFP蛍光検出条件下におけるプロトプラスト;下部パネル:DsRed蛍光検出条件下における同じプロトプラスト)。
【図1B】(B)pICH17272およびpICH18505のT−DNA領域の図式表示である。P‐転写プロモーター;T‐転写停止領域;RdRP ウイルスRNA‐依存的RNAポリメラーゼ;MP‐ウイルス移行タンパク質;3′NTR‐ウイルス3′非翻訳領域;Cr‐sgp‐crTMV株のCPサブゲノムプロモーター領域;GOI‐関心のある遺伝子。
【図1C】(C)pICH17272およびpICH18505のT−DNA領域の制限地図の図式表示である。
【図2】異なるサブゲノムプロモーター由来の2種の異なるトランスジーン(GOI‐1およびGOI‐2)の同時発現用に設計されたウイルスベクターのT−DNA領域の図式表示を表す。P‐転写プロモーター;T‐転写停止領域;RdRP ウイルスRNA‐依存的RNAポリメラーゼ;MP‐ウイルス移行タンパク質;3′NTR‐ウイルス3′非翻訳領域;Cr−sgp‐crTMV株のCPサブゲノムプロモーター領域;3PK‐トリプルシュードノット(pseudo knot)領域;U5spg‐TMV‐U5株のCPサブゲノムプロモーター。GOI‐関心のある遺伝子。
【図3A】(A)コンストラクトpICH17388、pICH17123,pICH15933、pICH7410、pICH10580、pICH10881およびpICH10745のT−DNA領域の図式表示を表す。RS‐PhiC31インテグラーゼにより認識される組換え部位。灰色の縦棒はイントロンを表す。
【図3B】(B)pICH17388、pICH17123、pICH15933のT−DNA領域の制限地図を表す。
【図3C】(C)pICH7410、pICH10580、pICH10881およびpICH10745のT−DNA領域の制限地図を表す。
【図4】(A)はDNA前駆体pICH17388およびpICH15933をレコンビナーゼ供与源と一緒にアグロバクテリウム送達して6日後のN.ベンサミアーナ葉部領域の蛍光顕微鏡写真を示す。RS‐PhiC31インテグラーゼにより認識される組換え部位。
【図5】抗体の重鎖および軽鎖発現のための異なるベクター系のT−DNA領域の図式表示を表す。RS‐PhiC31インテグラーゼにより認識される組換え部位。
【図6】ウイルスプロベクター系を使用した、ニコチアナベンサミアーナ葉部におけるIgG発現のウェスタンブロットを示す。TSPの電気泳動による分離は非還元条件下、12%ゲル上で行った。発現したタンパク質の検出は、6x103倍希釈した、HRP(Sigma)と結合したウサギ由来の抗‐ヒトIgG画分により行った。レーン:1‐非感染葉組織;レーン2‐サイトソルに発現したIgG重鎖(pICH17388);レーン3‐IgG重鎖および軽鎖、35Sプロモーターコンストラクト(pIC0123+pICH19846)による同時発現;レーン4‐P19(pICH6692)を用いる以外は、レーン3と同様;レーン5‐35SプロモーターコンストラクトおよびP19(pICH5290+pICH6692)により発現したGFP;レーン6‐バイシストロンコンストラクトpICH19860により同時発現したIgG重鎖および軽鎖;レーン7‐バイシストロンコンストラクトpICH19860(MP欠乏5′プロ‐ベクターpICH17123、MP in trans pICH10745)により同時発現したIgG重鎖および軽鎖;レーン8‐サイトソルに同時発現したGFPおよびdsRED、MPはin transで提供される(pICH17123+pICH19919+pICHI10745)。
【図7】同じ植物細胞において関心のある異なるタンパク質サブユニットの同時発現用に設計された非‐競合ウイルスベクターをコードするT−DNA領域の図式表示を表す。P‐転写プロモーター;T‐転写停止領域;TMV RdRP タバコモザイクウイルスのRNA‐依存的RNAポリメラーゼ;PVX RdRP ジャガイモウイルスXのウイルスRNA‐依存的RNAポリメラーゼ;MP‐ウイルス移行タンパク質;3′NTR‐ウイルス3′非翻訳領域;Cr‐sgp‐crTMV株のCPサブゲノムプロモーター領域;GOI‐関心のあるタンパク質サブユニットをコードする関心のある遺伝子。
【図8】バイナリーベクターpIC0130のT−DNA領域の図式表示を表す。
【図9】TMV(pICH17388+pICHI10580)およびPVX(pIC0130)を基礎にしたベクターを使用した、植物細胞におけるGFPおよびdsREDの同時発現を示す:アグロバクテリウムを接種したN.ベンサミアーナ葉部から分離したプロトプラストにおけるGFPおよびdsREDの視覚化。
【図10A】(A)バイナリーベクターpICH17620、pICH20431、pICH21240、pICH20421、およびpICH21370のT−DNA領域の図式表示を表す。
【図10B】(B)バイナリーベクターpICH11599、pICH21910、およびpICH21920のT−DNA領域の図式表示である。
【図10C】(C)バイナリーベクターpICH21282、pICH10990、pICH22250、およびpICH22240のT−DNA領域の図式表示である。LvおよびLc‐軽鎖の可変および保存領域;HvおよびHc‐重鎖の可変および保存領域。
【図10D】(D)PVX‐由来プロベクターpICH21380のクローニングスキームの図式表示である。
【図11A】(a)PVXおよびcrTMVベクターを使用したIgG軽鎖および重鎖の同時発現レベル測定のためのELISA試験の結果を示す。左側の組織培養プレートのウェルの番号付けは、ヒストグラムの棒の番号付けに相当する。ヒストグラムは405nmにおけるウェルのODを示す。1、2−非感染植物組織;3、4、5−PVXにおけるカルレティキュリンSP‐重鎖(それぞれpICH21240‐1、5および14クローン);6、7、8‐PVXにおけるカルレティキュリンSP‐IgGの軽鎖(LC)(N‐末端欠失、それぞれpICH21370−18、19、31);9、10、11‐PVXにおけるカルレティキュリンSP‐IgG軽鎖(それぞれpICH21370‐40、44、45);12‐ブランクコントロール(植物タンパク質非適用);13、14、15‐crTMVにおけるカルレティキュリンSP‐IgG軽鎖(pICH17620+pICH10881+pICH20431)と同時発現したPVXにおけるカルレティキュリンSP‐IgG重鎖(HC)(それぞれpICH21240‐1、5および14クローン)。16、17、18‐crTMVにおけるカルレティキュリン‐HC(pICH17620+pICH10881+pICH20421)と同時発現したPVXにおけるカルレティキュリン‐IgGのLC(N‐末端欠失、それぞれpICH21370−18、19、31)。19、20、21‐crTMVを基礎にしたベクター中におけるカルレティキュリンSP‐重鎖(pICH17620+pICH10881+pICH20421)と同時発現したPVXにおけるカルレティキュリンSP‐IgGLC(それぞれpICH21370−40、44、45)。22‐PVXにより発現したGFP(pIC0130);23‐PVXにより発現したGFP(pICH20799);24‐crTMVだけ(pICH17620+pICH10881+pICH20431)から発現したカルレティキュリンSP‐IgG軽鎖;25‐crTMVを基礎にしたベクター(pICH17620+pICH10881+pICH20421)から発現したカルレティキュリンSP‐IgG重鎖;26‐PTGSサプレッサーP19の存在下、強力な35Sプロモーター制御下で同時発現したIgGの軽鎖および重鎖27‐crTMVを基礎にしたベクター(pICH17388+pICH10881=pICH20421)からのIgG軽鎖および重鎖同時発現;28‐crTMVを基礎にしたベクター(pICH17388+pICH10881=pICH20421)、in transで提供されたMP(pICH10745)からの軽鎖および重鎖の同時発現;*‐測定可能な値をかなり超えたOD405値。
【図11B】(b)N.ベンサミアーナ葉部に発現した抗癌抗体の電気泳動およびウェスタンブロット解析を示す。(A)TMVとPVXプロベクターが同時感染したニコチアナベンサミアーナ葉部における抗癌抗体の重鎖および軽鎖の蓄積。軽鎖はPVXにより発現し、重鎖はTMVにより発現する。タンパク質は還元条件下、12%ポリアクリルアミドゲル中で分離する。上部パネルはクーマシー染色を示す;中間パネルはHRP‐結合ヤギ抗‐ヒトIgG(ガンマ鎖特異的)抗体(Sigma)によるウェスタンブロットを示す。下部パネルはHRP‐結合ウサギ抗ヒトIgG(ラムダ鎖‐特異的)抗体(Sigma)によるウェスタンブロットを示す。Uninf、非感染組織;2〜11、接種後の日数;S、抗癌mab(モノクローナル抗体)標準品;LC、TMVプロベクターだけにより発現した軽鎖(6dpi);HC、TMVプロベクターだけにより発現した重鎖(6dpi)。(B)プロテインA磁気ビーズ(NEB)を使用した抗癌mabの精製。(A)クーマシー染色ゲル上、還元条件下、12%ポリアクリルアミドゲル中で移動する植物‐由来(レーン1)および標準(レーン2)mab。(C)HCを発現するTMVプロベクターおよびLCを発現するPVXプロベクターに同時感染したニコチアナベンサミアーナ葉部における集合した抗癌mabの蓄積。タンパク質は非還元条件下、10%ポリアクリルアミドゲル中で分離する。ウェスタンブロットはヤギ抗ヒトIgG(ガンマ鎖特異的)抗体により同定した。Uninf、非感染組織;2〜11、接種後の日数;S、抗癌mab(モノクローナル抗体)標準品;LC、TMVプロベクター(6dpi)だけにより発現した軽鎖;HC、TMVプロベクター(6dpi)だけにより発現した重鎖;H2L2:2つの重鎖および2つの軽鎖を含有するIgGヘテロテトラマー、H2L‐2つの重鎖および1つの軽鎖を含有するIgGヘテロトリマー、H2‐重鎖ホモダイマー、L2‐軽鎖ホモダイマー。
【図12】バイナリーベクターpICH17620、pICH−FSHAおよびpICH−FSHBのT−DNA領域の図式表示を表す。
【図13】バイナリーベクターpICH17388、pICH−MLCJおよびpICH−MHCのT−DNA領域の図式表示を表す。
【特許請求の範囲】
【請求項1】
植物体、植物組織、または植物細胞において、少なくとも第1および第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法であって、
(i)植物体、植物組織または植物細胞に、第1および第2プラス‐センス1本鎖RNAウイルスベクターを提供するが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、第1ウイルスベクターおよび第2ウイルスベクターは異なるウイルスベクターであり;または
(ii)植物体、植物組織または植物細胞に、少なくとも第1および第2タンパク質サブユニットをコードするプラス‐センス1本鎖RNAウイルスベクターを提供する
ことにより、植物細胞に少なくとも第1および第2タンパク質サブユニットを発現させることを含む方法。
【請求項2】
植物体、植物組織または植物細胞由来のヘテロ‐オリゴマータンパク質を分離することを次に行う、請求項1に記載の方法。
【請求項3】
少なくとも第1および第2タンパク質サブユニットをコードするRNAウイルスベクターが、第1タンパク質サブユニットをコードする第1異種配列および第2タンパク質サブユニットをコードする第2異種配列を含有し、第1および/または第2タンパク質サブユニットの発現がサブゲノムプロモーターの制御下にある、請求項1または2に記載の方法。
【請求項4】
少なくとも第1および第2タンパク質サブユニットをコードするRNAウイルスベクターが、第1タンパク質サブユニットをコードする第1異種配列および第2タンパク質サブユニットをコードする第2異種配列を含有し、第1および/または第2タンパク質サブユニットの発現がIRESエレメントの制御下にある、請求項1または2に記載の方法。
【請求項5】
少なくとも第1および第2タンパク質サブユニットをコードするRNAウイルスベクターが、ウイルスベクターの全身移行のために機能するタンパク質をコードするORFを欠如する、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
第1ウイルスベクターが第1タンパク質サブユニットをコードする第1異種配列を含有し、第2ウイルスベクターが第2タンパク質サブユニットをコードする第2異種配列を含有し、第1および/または第2タンパク質サブユニットの発現がサブゲノムプロモーターの制御下にある、請求項1または2に記載の方法。
【請求項7】
第1ウイルスベクターが第1タンパク質サブユニットをコードする第1異種配列を含有し、第2ウイルスベクターが第2タンパク質サブユニットをコードする第2異種配列を含有し、第1および/または第2タンパク質サブユニットの発現がIRESエレメントの制御下にある、請求項1または2に記載の方法。
【請求項8】
少なくとも第1および第2タンパク質サブユニットをコードするウイルスベクター、第1ウイルスベクター、および/または第2ウイルスベクターが
機能的コートタンパク質ORF、
機能的移行タンパク質ORF、および/または
ウイルス粒子アセンブリーの機能的起点
を持たない、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
第1ウイルスベクターがポテクスウイルス属に属するウイルスに由来し、第2ウイルスベクターがポティウイルス属に属するウイルスに由来する、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
第1ウイルスベクターがジャガイモウイルスXに由来し、そして第2ウイルスベクターがジャガイモウイルスYに由来する、請求項9に記載の方法。
【請求項11】
第1ウイルスベクターがポテクスウイルス属に属するウイルスに由来し、そして第2ウイルスベクターがトバモウイルス属に属するウイルスに由来する、請求項1〜8のいずれか1項に記載の方法。
【請求項12】
第1ウイルスベクターがジャガイモウイルスXに由来し、そして第2ウイルスベクターがタバコモザイクウイルスに由来する、請求項11に記載の方法。
【請求項13】
第1および第2ウイルスベクターが多くても60%の配列相同性を有する、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
ヘテロオリゴマータンパク質が抗体である、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
ウイルスベクターが植物体、植物組織、または植物細胞に一過性に提供される、請求項1〜14のいずれか1項に記載の方法。
【請求項16】
ウイルスベクターがアグロバクテリウムトランスフェクションにより植物体、植物組織、または植物細胞に一過性に提供される、請求項1〜15のいずれか1項に記載の方法。
【請求項17】
RNAウイルスベクターがRNAウイルスベクターのDNA前駆体として植物染色体DNAに安定に組み込まれる、請求項1〜14のいずれか1項に記載の方法。
【請求項18】
DNA前駆体由来のRNAウイルスベクターの制御された放出が誘導型プロモーターによって提供される、請求項17に記載の方法。
【請求項19】
少なくとも第1または第2タンパク質サブユニットがER‐ターゲッティングシグナルとしての植物‐特異的シグナルペプチドを有する、請求項1〜18のいずれか1項に記載の方法。
【請求項20】
植物‐特異的シグナルペプチドがタバコカルレティキュリンおよび/イネアルファ‐アミラーゼに由来する、請求項19に記載の方法。
【請求項21】
抗体が少なくとも抗原結合ドメインの一部を含有するイムノグロブリンである、請求項14に記載の方法。
【請求項22】
抗体がポリイムノグロブリン受容体の一部を含有する保護タンパク質をイムノグロブリン重鎖と一緒に含有する、請求項14または21に記載の方法。
【請求項23】
抗体またはその誘導体がイムノグロブリンGクラス、イムノグロブリンAクラス、イムノグロブリンMクラス、イムノグロブリンDクラス、またはイムノグロブリンEクラスに属する、請求項14、21、または22のいずれか1項に記載の方法。
【請求項24】
ヘテロオリゴマータンパク質がインスリンである、請求項1〜12のいずれか1項に記載の方法。
【請求項25】
少なくとも1つまたは少なくとも2つ以上のヘテロオリゴマータンパク質のサブユニットが小胞体保留シグナルKDELを含有する、請求項1〜24のいずれか1項に記載の方法。
【請求項26】
異種核酸配列が変異して、ヘテロオリゴマータンパク質から部分的に、または完全にグリコシル化部位を除去している、請求項1〜25のいずれか1項に記載の方法。
【請求項27】
植物体、植物組織、または植物細胞が工学的に操作されて、ヘテロオリゴマータンパク質のグリコシル化パターンを変化させる、請求項1〜26のいずれか1項に記載の方法。
【請求項28】
植物体が単子葉または双子葉植物である、請求項1〜27のいずれか1項に記載の方法。
【請求項29】
双子葉植物がナス科に属する、請求項28に記載の方法。
【請求項30】
双子葉植物がニコチアナ種である、請求項28に記載の方法。
【請求項31】
双子葉植物がニコチアナタバカムまたはニコチアナベンサミアーナである、請求項28に記載の方法。
【請求項32】
双子葉植物がアブラナ科に属する、請求項28に記載の方法。
【請求項33】
双子葉植物がマメ科に属する、請求項28に記載の方法。
【請求項34】
双子葉植物がメディカゴサティバ(Medicago sativa)である、請求項28に記載の方法。
【請求項35】
双子葉植物がアカザ科に属する、請求項28に記載の方法。
【請求項36】
双子葉植物がフダンソウである、請求項28に記載の方法。
【請求項37】
植物体、植物組織、または植物細胞において少なくとも第1および第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法であって、1種以上のプラス‐センス1本鎖RNAウイルスベクター由来の少なくとも第1および第2タンパク質サブユニットを植物細胞に同時発現させることを含む、方法。
【請求項38】
少なくとも第1タンパク質サブユニットおよび第2タンパク質サブユニットを含有する抗体を植物体または植物組織において産生する方法であって、
(i)第1プラス‐センス1本鎖RNAウイルスベクターのDNA前駆体および第2プラス‐センス1本鎖RNAウイルスベクターのDNA前駆体を、アグロバクテリウム媒介トランスフェクションにより植物体、植物組織、または植物細胞に提供するが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、少なくとも第1ウイルスベクターまたは第2ウイルスベクターは、植物体における第1または第2ウイルスベクターの全身移行に必要な機能的タンパク質をコードするオープンリーディングフレームを欠如し;または
(ii)少なくとも第1および第2タンパク質サブユニットをコードするプラス‐センス1本鎖RNAウイルスベクターのDNA前駆体を、アグロバクテリウム媒介トランスフェクションにより植物体、植物組織、または植物細胞に提供するが、ウイルスベクターは全身移行に必要な機能的タンパク質をコードするORFを欠如する
ことにより、少なくとも第1および第2タンパク質サブユニットを植物細胞に発現させることを含む、方法
【請求項39】
植物体、植物組織、または植物細胞において、少なくとも第1タンパク質サブユニットおよび第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法であって、少なくとも第1および第2プラス‐センス1本鎖RNAウイルスベクターを植物体、植物組織、または植物細胞に提供することにより、植物細胞に少なくとも第1および第2タンパク質サブユニットを発現させることを含むが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、ここで第1ウイルスベクターおよび第2ウイルスベクターは異なるプラス‐センス1本鎖植物RNAウイルスに由来する、方法。
【請求項40】
植物体、植物組織、または植物細胞において、少なくとも第1タンパク質サブユニットおよび第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法であって、少なくとも第1および第2プラス‐センス1本鎖RNAウイルスベクターを植物体、植物組織、または植物細胞に提供することにより、植物細胞に少なくとも第1および第2タンパク質サブユニットを発現させることを含むが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、ここで、第1ウイルスベクターおよび第2ウイルスベクターは、第1または第2タンパク質サブユニットをコードする配列部分以外の配列部分が異なる、方法。
【請求項41】
植物体、植物組織、または植物細胞において、少なくとも第1タンパク質サブユニットおよび第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法であって、少なくとも第1および第2プラス‐センス1本鎖RNAウイルスベクターを植物体、植物組織、または植物細胞に提供することにより、植物細胞に少なくとも第1および第2タンパク質サブユニットを発現させることを含むが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、ここで、第1ウイルスベクターおよび第2ウイルスベクターは、異なる種に属するプラス‐センス1本鎖RNAウイルスに由来する、方法。
【請求項42】
植物体、植物組織、または植物細胞において、少なくとも第1および第2タンパク質サブユニットを含有する抗体を産生する方法であって、少なくとも第1および第2プラス‐センス1本鎖RNAウイルスベクターを植物体、植物組織、または植物細胞に提供することにより、植物細胞に少なくとも第1および第2タンパク質サブユニットを発現させることを含むが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、第1ウイルスベクターおよび第2ウイルスベクターは、異なるウイルスベクターである、方法。
【請求項1】
植物体、植物組織、または植物細胞において、少なくとも第1および第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法であって、
(i)植物体、植物組織または植物細胞に、第1および第2プラス‐センス1本鎖RNAウイルスベクターを提供するが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、第1ウイルスベクターおよび第2ウイルスベクターは異なるウイルスベクターであり;または
(ii)植物体、植物組織または植物細胞に、少なくとも第1および第2タンパク質サブユニットをコードするプラス‐センス1本鎖RNAウイルスベクターを提供する
ことにより、植物細胞に少なくとも第1および第2タンパク質サブユニットを発現させることを含む方法。
【請求項2】
植物体、植物組織または植物細胞由来のヘテロ‐オリゴマータンパク質を分離することを次に行う、請求項1に記載の方法。
【請求項3】
少なくとも第1および第2タンパク質サブユニットをコードするRNAウイルスベクターが、第1タンパク質サブユニットをコードする第1異種配列および第2タンパク質サブユニットをコードする第2異種配列を含有し、第1および/または第2タンパク質サブユニットの発現がサブゲノムプロモーターの制御下にある、請求項1または2に記載の方法。
【請求項4】
少なくとも第1および第2タンパク質サブユニットをコードするRNAウイルスベクターが、第1タンパク質サブユニットをコードする第1異種配列および第2タンパク質サブユニットをコードする第2異種配列を含有し、第1および/または第2タンパク質サブユニットの発現がIRESエレメントの制御下にある、請求項1または2に記載の方法。
【請求項5】
少なくとも第1および第2タンパク質サブユニットをコードするRNAウイルスベクターが、ウイルスベクターの全身移行のために機能するタンパク質をコードするORFを欠如する、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
第1ウイルスベクターが第1タンパク質サブユニットをコードする第1異種配列を含有し、第2ウイルスベクターが第2タンパク質サブユニットをコードする第2異種配列を含有し、第1および/または第2タンパク質サブユニットの発現がサブゲノムプロモーターの制御下にある、請求項1または2に記載の方法。
【請求項7】
第1ウイルスベクターが第1タンパク質サブユニットをコードする第1異種配列を含有し、第2ウイルスベクターが第2タンパク質サブユニットをコードする第2異種配列を含有し、第1および/または第2タンパク質サブユニットの発現がIRESエレメントの制御下にある、請求項1または2に記載の方法。
【請求項8】
少なくとも第1および第2タンパク質サブユニットをコードするウイルスベクター、第1ウイルスベクター、および/または第2ウイルスベクターが
機能的コートタンパク質ORF、
機能的移行タンパク質ORF、および/または
ウイルス粒子アセンブリーの機能的起点
を持たない、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
第1ウイルスベクターがポテクスウイルス属に属するウイルスに由来し、第2ウイルスベクターがポティウイルス属に属するウイルスに由来する、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
第1ウイルスベクターがジャガイモウイルスXに由来し、そして第2ウイルスベクターがジャガイモウイルスYに由来する、請求項9に記載の方法。
【請求項11】
第1ウイルスベクターがポテクスウイルス属に属するウイルスに由来し、そして第2ウイルスベクターがトバモウイルス属に属するウイルスに由来する、請求項1〜8のいずれか1項に記載の方法。
【請求項12】
第1ウイルスベクターがジャガイモウイルスXに由来し、そして第2ウイルスベクターがタバコモザイクウイルスに由来する、請求項11に記載の方法。
【請求項13】
第1および第2ウイルスベクターが多くても60%の配列相同性を有する、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
ヘテロオリゴマータンパク質が抗体である、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
ウイルスベクターが植物体、植物組織、または植物細胞に一過性に提供される、請求項1〜14のいずれか1項に記載の方法。
【請求項16】
ウイルスベクターがアグロバクテリウムトランスフェクションにより植物体、植物組織、または植物細胞に一過性に提供される、請求項1〜15のいずれか1項に記載の方法。
【請求項17】
RNAウイルスベクターがRNAウイルスベクターのDNA前駆体として植物染色体DNAに安定に組み込まれる、請求項1〜14のいずれか1項に記載の方法。
【請求項18】
DNA前駆体由来のRNAウイルスベクターの制御された放出が誘導型プロモーターによって提供される、請求項17に記載の方法。
【請求項19】
少なくとも第1または第2タンパク質サブユニットがER‐ターゲッティングシグナルとしての植物‐特異的シグナルペプチドを有する、請求項1〜18のいずれか1項に記載の方法。
【請求項20】
植物‐特異的シグナルペプチドがタバコカルレティキュリンおよび/イネアルファ‐アミラーゼに由来する、請求項19に記載の方法。
【請求項21】
抗体が少なくとも抗原結合ドメインの一部を含有するイムノグロブリンである、請求項14に記載の方法。
【請求項22】
抗体がポリイムノグロブリン受容体の一部を含有する保護タンパク質をイムノグロブリン重鎖と一緒に含有する、請求項14または21に記載の方法。
【請求項23】
抗体またはその誘導体がイムノグロブリンGクラス、イムノグロブリンAクラス、イムノグロブリンMクラス、イムノグロブリンDクラス、またはイムノグロブリンEクラスに属する、請求項14、21、または22のいずれか1項に記載の方法。
【請求項24】
ヘテロオリゴマータンパク質がインスリンである、請求項1〜12のいずれか1項に記載の方法。
【請求項25】
少なくとも1つまたは少なくとも2つ以上のヘテロオリゴマータンパク質のサブユニットが小胞体保留シグナルKDELを含有する、請求項1〜24のいずれか1項に記載の方法。
【請求項26】
異種核酸配列が変異して、ヘテロオリゴマータンパク質から部分的に、または完全にグリコシル化部位を除去している、請求項1〜25のいずれか1項に記載の方法。
【請求項27】
植物体、植物組織、または植物細胞が工学的に操作されて、ヘテロオリゴマータンパク質のグリコシル化パターンを変化させる、請求項1〜26のいずれか1項に記載の方法。
【請求項28】
植物体が単子葉または双子葉植物である、請求項1〜27のいずれか1項に記載の方法。
【請求項29】
双子葉植物がナス科に属する、請求項28に記載の方法。
【請求項30】
双子葉植物がニコチアナ種である、請求項28に記載の方法。
【請求項31】
双子葉植物がニコチアナタバカムまたはニコチアナベンサミアーナである、請求項28に記載の方法。
【請求項32】
双子葉植物がアブラナ科に属する、請求項28に記載の方法。
【請求項33】
双子葉植物がマメ科に属する、請求項28に記載の方法。
【請求項34】
双子葉植物がメディカゴサティバ(Medicago sativa)である、請求項28に記載の方法。
【請求項35】
双子葉植物がアカザ科に属する、請求項28に記載の方法。
【請求項36】
双子葉植物がフダンソウである、請求項28に記載の方法。
【請求項37】
植物体、植物組織、または植物細胞において少なくとも第1および第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法であって、1種以上のプラス‐センス1本鎖RNAウイルスベクター由来の少なくとも第1および第2タンパク質サブユニットを植物細胞に同時発現させることを含む、方法。
【請求項38】
少なくとも第1タンパク質サブユニットおよび第2タンパク質サブユニットを含有する抗体を植物体または植物組織において産生する方法であって、
(i)第1プラス‐センス1本鎖RNAウイルスベクターのDNA前駆体および第2プラス‐センス1本鎖RNAウイルスベクターのDNA前駆体を、アグロバクテリウム媒介トランスフェクションにより植物体、植物組織、または植物細胞に提供するが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、少なくとも第1ウイルスベクターまたは第2ウイルスベクターは、植物体における第1または第2ウイルスベクターの全身移行に必要な機能的タンパク質をコードするオープンリーディングフレームを欠如し;または
(ii)少なくとも第1および第2タンパク質サブユニットをコードするプラス‐センス1本鎖RNAウイルスベクターのDNA前駆体を、アグロバクテリウム媒介トランスフェクションにより植物体、植物組織、または植物細胞に提供するが、ウイルスベクターは全身移行に必要な機能的タンパク質をコードするORFを欠如する
ことにより、少なくとも第1および第2タンパク質サブユニットを植物細胞に発現させることを含む、方法
【請求項39】
植物体、植物組織、または植物細胞において、少なくとも第1タンパク質サブユニットおよび第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法であって、少なくとも第1および第2プラス‐センス1本鎖RNAウイルスベクターを植物体、植物組織、または植物細胞に提供することにより、植物細胞に少なくとも第1および第2タンパク質サブユニットを発現させることを含むが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、ここで第1ウイルスベクターおよび第2ウイルスベクターは異なるプラス‐センス1本鎖植物RNAウイルスに由来する、方法。
【請求項40】
植物体、植物組織、または植物細胞において、少なくとも第1タンパク質サブユニットおよび第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法であって、少なくとも第1および第2プラス‐センス1本鎖RNAウイルスベクターを植物体、植物組織、または植物細胞に提供することにより、植物細胞に少なくとも第1および第2タンパク質サブユニットを発現させることを含むが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、ここで、第1ウイルスベクターおよび第2ウイルスベクターは、第1または第2タンパク質サブユニットをコードする配列部分以外の配列部分が異なる、方法。
【請求項41】
植物体、植物組織、または植物細胞において、少なくとも第1タンパク質サブユニットおよび第2タンパク質サブユニットを含有するヘテロ‐オリゴマータンパク質を産生する方法であって、少なくとも第1および第2プラス‐センス1本鎖RNAウイルスベクターを植物体、植物組織、または植物細胞に提供することにより、植物細胞に少なくとも第1および第2タンパク質サブユニットを発現させることを含むが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、ここで、第1ウイルスベクターおよび第2ウイルスベクターは、異なる種に属するプラス‐センス1本鎖RNAウイルスに由来する、方法。
【請求項42】
植物体、植物組織、または植物細胞において、少なくとも第1および第2タンパク質サブユニットを含有する抗体を産生する方法であって、少なくとも第1および第2プラス‐センス1本鎖RNAウイルスベクターを植物体、植物組織、または植物細胞に提供することにより、植物細胞に少なくとも第1および第2タンパク質サブユニットを発現させることを含むが、第1ウイルスベクターは少なくとも第1タンパク質サブユニットをコードし、第2ウイルスベクターは少なくとも第2タンパク質サブユニットをコードし、第1ウイルスベクターおよび第2ウイルスベクターは、異なるウイルスベクターである、方法。
【図1A】

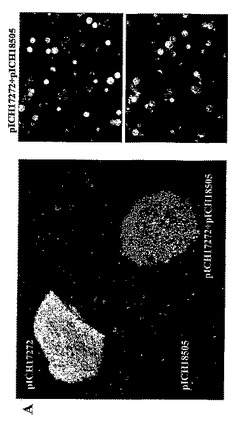
【図1B】
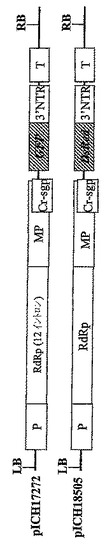
【図1C】
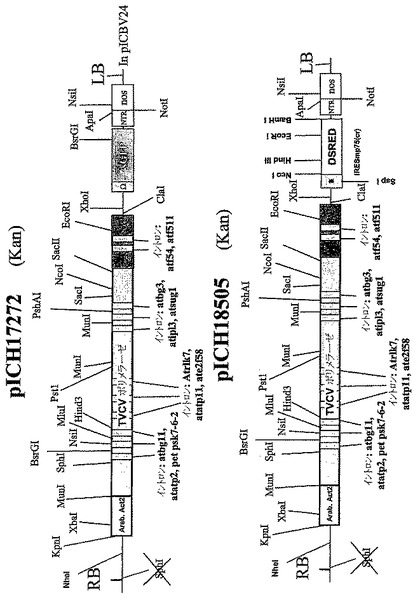
【図2】
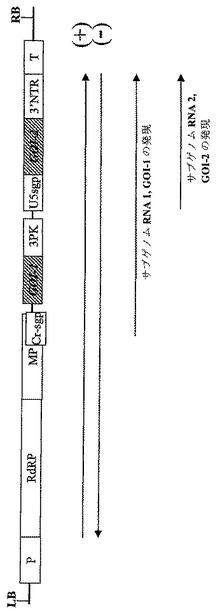
【図3A】
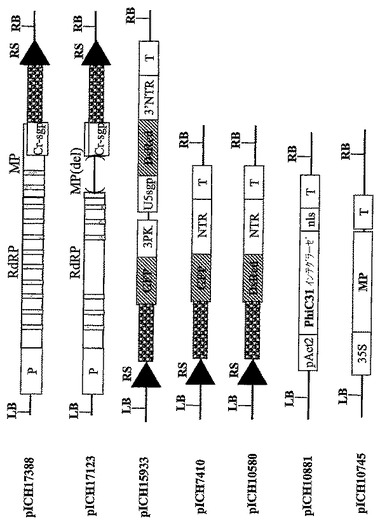
【図3B】
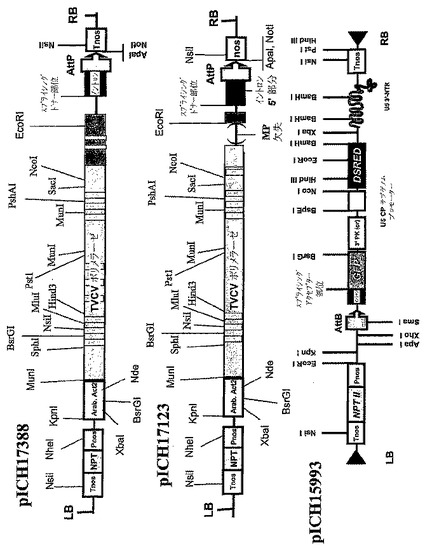
【図3C】
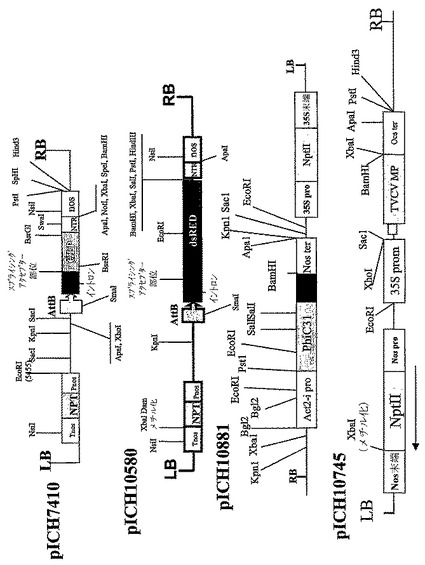
【図4】
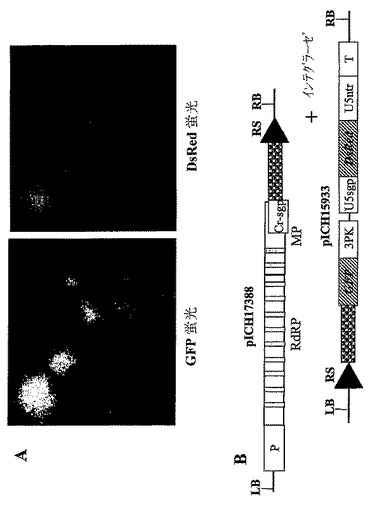
【図5】
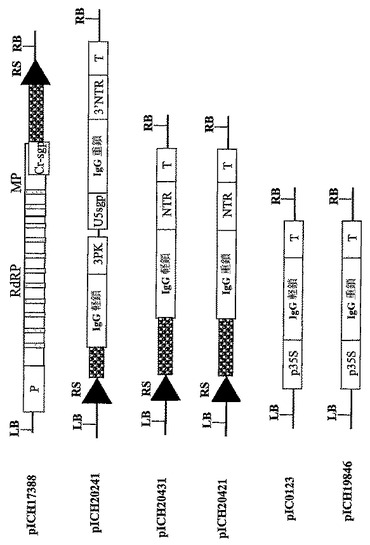
【図6】
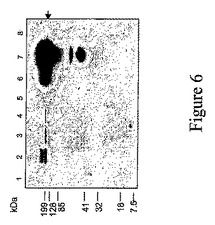
【図7】
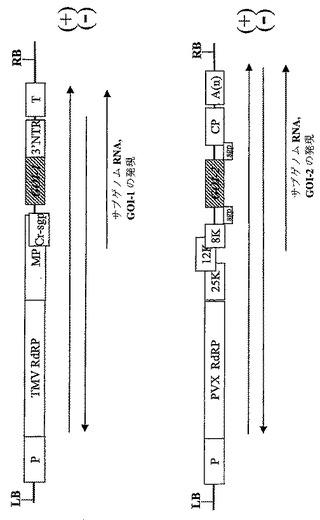
【図8】
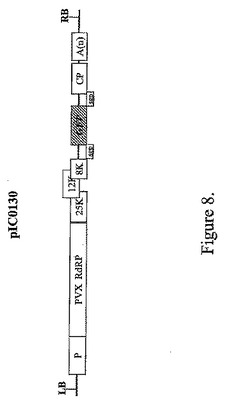
【図9】
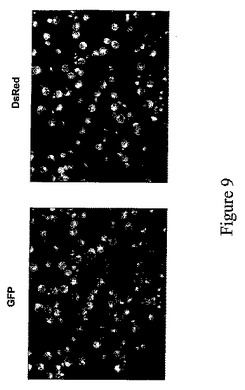
【図10A】
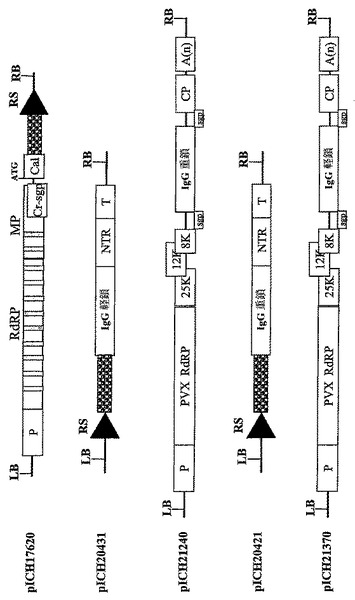
【図10B】
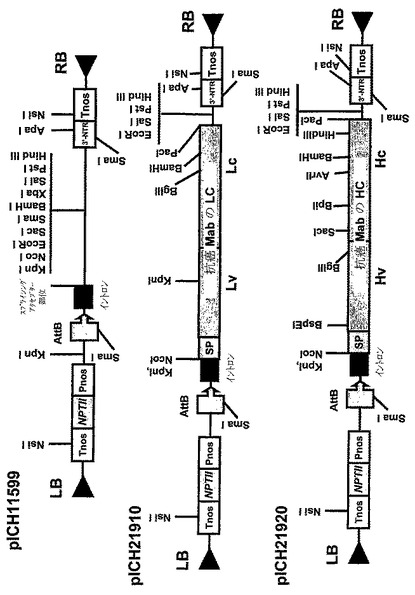
【図10C】
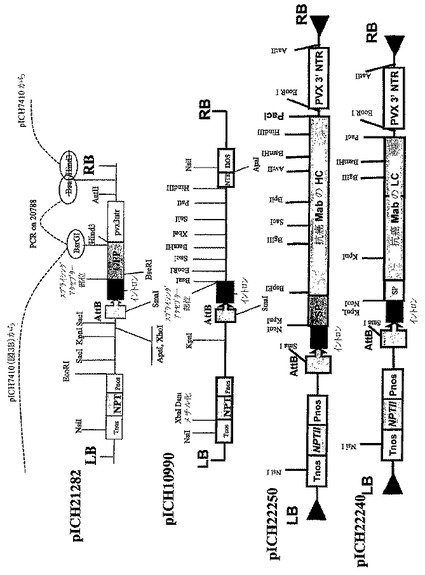
【図10D】
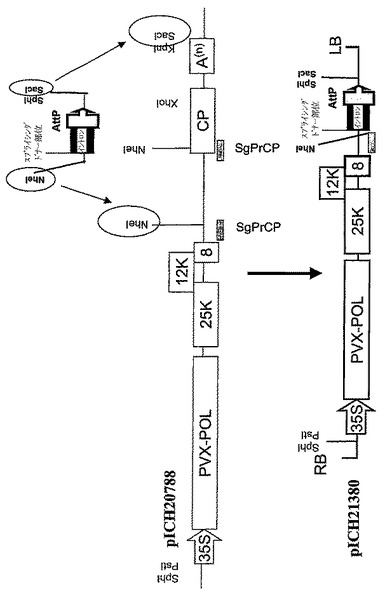
【図11A】
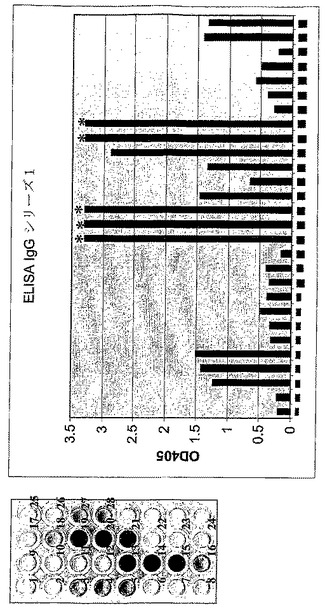
【図11B】
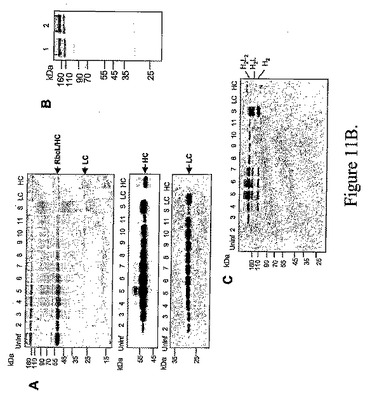
【図12】
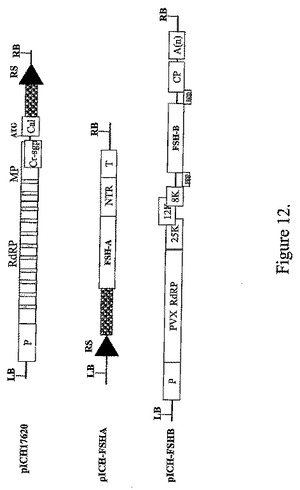
【図13】
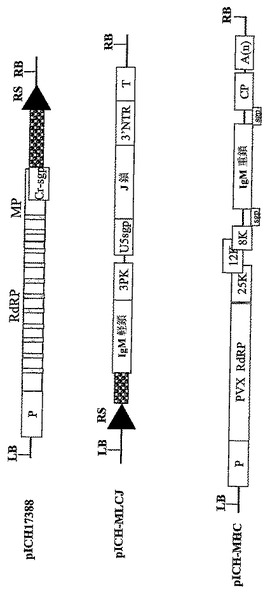
【図1B】
【図1C】
【図2】
【図3A】
【図3B】
【図3C】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10A】
【図10B】
【図10C】
【図10D】
【図11A】
【図11B】
【図12】
【図13】
【公表番号】特表2008−535473(P2008−535473A)
【公表日】平成20年9月4日(2008.9.4)
【国際特許分類】
【出願番号】特願2007−552582(P2007−552582)
【出願日】平成18年1月27日(2006.1.27)
【国際出願番号】PCT/EP2006/000721
【国際公開番号】WO2006/079546
【国際公開日】平成18年8月3日(2006.8.3)
【出願人】(507254997)アイコン・ジェネティクス・ゲーエムベーハー (5)
【Fターム(参考)】
【公表日】平成20年9月4日(2008.9.4)
【国際特許分類】
【出願日】平成18年1月27日(2006.1.27)
【国際出願番号】PCT/EP2006/000721
【国際公開番号】WO2006/079546
【国際公開日】平成18年8月3日(2006.8.3)
【出願人】(507254997)アイコン・ジェネティクス・ゲーエムベーハー (5)
【Fターム(参考)】
[ Back to top ]