
植物における改善された標的に向けたDNA挿入
【課題】外来性DNA断片の、植物ゲノム中のあらかじめ選ばれた挿入部位への特異的導入のための方法を提供する。
【解決手段】植物への標的指向DNA挿入を改善するための、レア切断を起こす「二重鎖切断」誘導酵素を用いた方法および手段。さらに改善されたI−SceIをコードするヌクレオチド配列。
【解決手段】植物への標的指向DNA挿入を改善するための、レア切断を起こす「二重鎖切断」誘導酵素を用いた方法および手段。さらに改善されたI−SceIをコードするヌクレオチド配列。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、分子の植物生物学の分野、より具体的には植物ゲノムエンジニアリングの分野に関する。外来性DNA断片の、植物ゲノム中のあらかじめ選ばれた挿入部位への特異的導入のための方法を提供する。特定の部位に挿入された外来性DNAを含む植物を、現在利用可能な標的に向けたDNA挿入法で可能なよりも、より高い頻度でおよびより高い精度で、今や得ることができる。さらに、結果として得られる大部分の植物においては、外来性DNAはあらかじめ選択された挿入部位のみに挿入されており、植物ゲノムの他の位置に無秩序に外来性DNAが挿入されてはいない。本発明の方法はこのように、先行技術の方法に対して定量的および定性的の両面での進歩である。さらに、キメラ遺伝子、プラスミド、ベクター、および本発明の方法中で用いられる他の手段も提供する。
【背景技術】
【0002】
前世紀80年代初めのAgrobacteriumを介する形質転換技術による遺伝子組換え植物の第一世代により、対象の外来性DNAまたは導入遺伝子を植物のゲノムへ導入する他の方法、例えばPEGを介するプロトプラストへのDNA取込、マイクロプロジェクタイル撃ち込み、シリコンホイスカーを介する形質転換その他、の開発に拍車がかけられた。
【0003】
しかし、すべての植物の形質転換法では共通して、植物ゲノムに組み入れられる導入遺伝子は、無作為な方式でまた予測できないコピー数だけ組み込まれる。しばしば、導入遺伝子は、導入遺伝子全体が、またはそれの部分が、リピートの形で組込まれる場合がある。そのような複雑な組込みパターンは、導入遺伝子の発現レベルに、例えば転写後遺伝子サイレンシング機構により転写されたRNAを破壊して、または導入されたDNAのメチル化を引き起こして、導入遺伝子の転写活性を下方制御することにより、影響を及ぼす可能性がある。さらに、組込み部位自体が、導入遺伝子の発現レベルに影響を及ぼす場合がある。これらの因子の組合せが、導入遺伝子または対象の外来性DNAの発現レベルの広汎な変化を、種々の遺伝子組換え植物細胞および植物系統中にもたらす。さらに、対象の外来性DNAの組込みが、組込みが生じるゲノム領域に破壊的な効果を有する可能性があり、その標的領域の正常な機能に影響を及ぼすかまたは妨害する場合があって、そのためにしばしば望ましくない副作用が生ずる。
【0004】
したがって、植物への特定の外来性DNA導入の影響を研究する場合は常に、有効な結果を得るために多数の遺伝子組換え植物系統を生成し分析することを必要とする。同様に、遺伝子組換え植物に所望の既知の表現型を与えるために、植物に対象の特定のDNAが導入される遺伝子組換え農作物の生成においては、多数の独立に形成された遺伝子組換え植物系統、即ちいわゆるイベントが形成されて、導入遺伝子が最適に発現しておりまた遺伝子組換え植物の表現型全体に亘って副作用が最小であるかまたは無い植物系統の選択が可能となる。特にこの分野で、望ましくない遺伝子組換えイベントの排除に必要な圃場試験の繰返しに伴う厄介な制御のための必要条件および高いコストを考慮すれば、この試行錯誤プロセスをより指向性のあるプロセスと置き換えることができれば有利であろう。更に、標的指向のDNA挿入が可能であることが、いわゆる導入遺伝子スタッキングのプロセスでさらに有益であることが明らかであろう。
【0005】
植物中への導入遺伝子組込みを制御する必要性が早い時期から認識されており、またいくつかの方法がこの必要を満たす努力の中で発展されてきた(総説としてKumar and Fladung, 2001, Trends in Plant Science, 6, ppl55-159 を参照のこと)。これらの方法はほとんど、原核生物および下等真核生物に成功裡に適用されてきた戦略である相同組換えに基づいた導入遺伝子組込みに依存している(例えば、EP0317509または対応したPaszkowski et al., 1988, EMBO J., 7,pp 4021-4026による文献を参照のこと)。しかし、植物については、導入遺伝子組込みの主な機構は、組換えをするDNA鎖間の相同性がほとんどない非正統的組換に基づいている。この領域における主な挑戦課題は、したがって、非正統的組換を介して導入される外来性DNAのはるかに効率的な組込みによって隠されている、レア相同組換えイベントの検出である。
【0006】
この問題を解決する1つの方法は、WO94/17176に例示されているような、非正統的組換によって生じた組込みイベント以外を選別することによる。
【0007】
この問題を解決する別の方法は、I−SceIなどのレア切断を起こすエンドヌクレアーゼによる二本鎖DNA切断の誘導による、標的座位の活性化および/またはドナーDNAの修復による。この技術はAgrobacteriumを用いて植物細胞に修復DNAを配送した場合に、相同組換え頻度を少なくとも二桁増加させることが示された(Puchta et 1996, Proc. Natl. Acad. U.S.A., 93, pp5055-5060; Chilton and Que, Plant Physiol., 2003)。
【0008】
WO96/14408が、酵素I−SceIをコードする単離されたDNAについて記述している。このDNA配列を、クローニングにより、発現ベクター、形質転換細胞系統、および遺伝子組換え動物、に組み入れることができる。このベクターは、遺伝子の遺伝子地図作製および部位特異的挿入に役立つ。
【0009】
WO00/46386が、I−Scel二重鎖切断を通じて遺伝子または細胞の他の染色体DNAを修飾し、修復し、減少させ、不活性化する方法について記述している。さらに、それが必要とされる個体中の遺伝病の治療または予防の方法を開示する。さらに、キメラの制限エンドヌクレアーゼを開示する。
【0010】
しかし、真核細胞のゲノム、特に植物細胞ゲノム中への外来性DNAの標的指向挿入の頻度を改善する必要性がいまだに残存している。これらの問題および他の問題が、以下に本発明の様々の詳細な実施態様ならびに請求項に記述するように、解決される。
【0011】
発明の開示
実施態様の1つにおいて本発明は、対象の外来性DNAであって、あらかじめ選択された部位に隣接するDNA領域と少なくとも80%の配列同一性を有するDNA領域と隣接している可能性のあるDNAを、トウモロコシ細胞のような植物細胞のゲノムの、I−SceI部位などのあらかじめ選択された部位へ導入する方法であって
(a)二本鎖DNA切断を細胞のゲノム中のあらかじめ選ばれた部位に、例えばI−SceIをコードする遺伝子を導入することによって引き起こす工程;
(b)対象の外来性DNAを植物細胞へ導入する工程;
を含み、外来性DNAが、対象の外来性DNAでコートされたマイクロプロジェクタイル撃ち込みにより遂行される直接DNA移入により送達されることを特徴とする方法を提供する。
I−SceIをコードする遺伝子は、配列番号1のアミノ酸配列をコードするヌクレオチド配列を含むことができ、前記ヌクレオチド配列は、約50%〜約60%のGC含量を有し、以下を条件とする。
i) ヌクレオチド配列が、
【表1】

からなる群より選択されるヌクレオチド配列を含まず;
ii) ヌクレオチドが、CCAAT、ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まず;
iii) ヌクレオチド配列が、ATTTA、AAGGT、AGGTA、GGTAまたはGCAGGからなる群より選択される配列を含まず;
iv) ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;
v) ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るATストレッチを含まず;および
vi) ヌクレオチド配列がコドンTTA、CTA、ATA、GTA、TCG、CCG、ACGおよびGCGを含まない。そのようなI−SceIをコードする遺伝子の一例は、配列番号4のヌクレオチド配列を含む。
植物細胞を工程(a)に先立って植物フェノール化合物中でインキュベーションしてもよい。
【0012】
別の実施態様において、本発明は、対象の外来性DNAを植物細胞ゲノムのあらかじめ選択された部位へ導入するための、以下の工程を含む方法に関する:
(a)二本鎖DNA切断を細胞のゲノム中のあらかじめ選択された部位に引き起こす工程;
(b)対象の外来性DNAを植物細胞中へ導入する工程:
を含み、二本鎖DNA切断が、ヌクレオチド配列によってコードされるレア切断エンドヌクレアーゼによって導入され、前記ヌクレオチド配列が、約50%〜約60%のGC含量を有し、
i)ヌクレオチド配列が、
【表2】
からなる群より選択されるヌクレオチド配列を含まず;
ii)ヌクレオチドが、CCAAT、ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まず;
iii)ヌクレオチド配列が、ATTTA、AAGGT、AGGTA、GGTAまたはGCAGGからなる群より選択されるヌクレオチド配列を含まず;
iv)ヌクレオチド配列が、GまたはCからなる群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;
v)ヌクレオチド配列が、AまたはTからなる群より選択される連続する5個のヌクレオチドから成るATストレッチを含まず;および
vi)ヌクレオチド配列が、コドンTTA、CTA、ATA、GTA、TCG、CCG、ACGおよびGCGを含まない、
ことを条件とすることを特徴とする。
【0013】
さらに別の実施態様において、本発明は、対象の外来性DNAを植物細胞のゲノムのあらかじめ選ばれた部位へ導入するための方法であって、
(a)二本鎖DNA切断を細胞のゲノム中のあらかじめ選ばれた部位に引き起こす工程;
(b)対象の外来性DNAを植物細胞へ導入する工程;
を含み、工程(a)に先立って植物細胞を、アセトシリンゴン(3,5−ジメトキシ−4−ヒドロキシアセトフェノン)、α−ヒドロキシ−アセトシリンゴン、シナピン酸(3,5ジメトキシ−4−ヒドロキシケイ皮酸)、シリング酸(4−ヒドロキシ−3,5ジメトキシ安息香酸)、フェルラ酸(4−ヒドロキシ−3−メトキシケイ皮酸)、カテコール(1,2−ジヒドロキシベンゼン)、p−ヒドロキシ安息香酸(4−ヒドロキシ安息香酸)、β−レソルシル酸(2,4ジヒドロキシ安息香酸)、プロトカテク酸(3,4−ジヒドロキシ安息香酸)、ピロ没食子酸(2,3,4−トリヒドロキシ安息香酸)、没食子酸(3,4,5−トリヒドロキシ安息香酸)、およびバニリン(3−メトキシ−4−ヒドロキシベンズアルデヒド)、の群より選択される植物フェノール化合物中でインキュベーションすることを特徴とする方法に関する。
【0014】
本発明は、さらに配列番号1のアミノ酸配列をコードするヌクレオチド配列を含む単離されたDNA断片であって、そのヌクレオチド配列が約50%〜約60%のGC含量を有し、以下を条件とする、DNA断片を提供する:
i)ヌクレオチド配列が、
【表3】
からなる群より選択されるヌクレオチド配列を含まず;
ii)ヌクレオチドが、CCAAT、ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まず;
iii)ヌクレオチド配列が、ATTTA,AAGGT,AGGTA、GGTAまたはGCAGGからなる群より選択される配列を含まず;
iv)ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;
v)ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るATストレッチを含まず;および
vi)前記ヌクレオチド配列の、ロイシン(Leu)、イソロイシン(Ile)、バリン(Val)、セリン(Ser)、プロリン(Pro)、トレオニン(Thr)、アラニン(Ala)をコードするコドンが、前記コドンの位置2および3にTAまたはGCという二連コードを含まない。
【0015】
本発明は、さらに配列番号4のヌクレオチド配列を含む単離されたDNA配列、および植物で発現可能なプロモーターに作動可能に連結された本発明による単離されたDNA断片を含むキメラ遺伝子、ならびに植物ゲノム中のI−SceI認識部位に外来性DNAを挿入するための、そのようなキメラ遺伝子の使用、を提供する。
【0016】
本発明のさらに別の実施態様において、対象の外来性DNAを、植物細胞のゲノムのあらかじめ選ばれた部位へ導入するための方法であって、
a)二本鎖DNA切断を、細胞のゲノム中のあらかじめ選ばれた部位に、レア切断エンドヌクレアーゼにより引き起こす工程;
b)対象の外来性DNAを植物細胞中へ導入する工程、
を含み、前記エンドヌクレアーゼが核移行シグナルを含むことを特徴とする方法を提供する。
【図面の簡単な説明】
【0017】
【図1】相同組換えを介する標的指向DNA挿入のための分析の中で用いられる、標的座位(A)および修復DNA(B)の模式的表示。組換え後の標的座位も表示されている(C)。DSB部位:二本鎖DNA切断部位;3’g7:A.tumefaciens 遺伝子7の転写終了およびポリアデニル化シグナル;neo:植物で発現可能なネオマイシンホスホトランスフェラーゼ;35S:CaMV35S転写のプロモーター;5’bar:ホスフィノトリシンアセチルトランスフェラーゼのアミノ末端部分をコードするDNA領域;3’nos:A. tumefaciensノパリン合成酵素遺伝子の転写終了およびポリアデニル化シグナル;Pnos:A. tumefaciensノパリン合成酵素遺伝子のプロモーター;3’ocs:A. tumefaciens オクトピン合成酵素遺伝子の3’転写終了およびポリアデニル化シグナル。
【0018】
発明の詳細な説明
本発明は次の発見に基づく:
a) 植物細胞中への、直接のDNA移入、特にマイクロプロジェクタイル撃ち込みにより挿入される外来性DNAの導入が、標的指向挿入イベントの頻度を予想外に増加させた。すべての得られた挿入イベントが標的指向DNA挿入イベントであって、二本鎖DNA切断が引き起こされた部位で生じた。さらに、すべてのこれらの標的指向挿入イベントは、二本鎖DNA切断に隣接する与えられた配列相同性の間の正確な組換えイベントであるように見えた。これらのイベントの約半分のみが、二本鎖DNA切断が引き起こされた部位とは異なる部位に、さらに追加の外来性DNA挿入を持っていた。
b)あらかじめ選択された一組の規則によって設計されたI−SceIなどの、レア切断エンドヌクレアーゼ用の合成コード領域を含むキメラ遺伝子によってコードされたI−SceIなどの、レア切断をする二重鎖切断誘導エンドヌクレアーゼの一時的発現による二本鎖DNA切断の誘導が、結果として生じる標的指向DNA挿入イベントの質(すなわち完全に標的指向DNA挿入イベントである頻度を)を驚くほど高めた。更に、エンドヌクレアーゼは核移行シグナルを備えていた。
c)アセトシリンゴンなどの植物フェノール化合物中で標的細胞をプレインキュベーションすることにより、さらに、植物細胞のゲノムに誘導された二本鎖DNA切断における標的指向挿入の頻度が増加した。
【0019】
上記の発見のいずれも、単独でまたは組合せて、相同組換えに基づいた標的指向挿入イベントが得られる頻度、ならびに回収されるイベントの質を改善する。
【0020】
このようにして、ある局面では本発明は、対象の外来性DNAを植物細胞のゲノムのあらかじめ選ばれた部位へ導入する方法であって
(a)二本鎖DNA切断を細胞のゲノム中のあらかじめ選ばれた部位に引き起こす工程;
(b)対象の外来性DNAを植物細胞中へ導入する工程;
を含み、外来性DNAが直接DNA移入により送達されることを特徴とする方法に関する。
【0021】
本明細書に用いられる「直接DNA移入」とは、植物細胞へDNAを導入することができる自然のAgrobacterium種の使用を含まない、植物細胞へのDNA導入の任意の方法である。これには、DNAのエレクトロポレーションによるプロトプラスト中への導入、DNAのエレクトロポレーションによるインタクトな植物細胞中への、または部分的に分解された組織または植物細胞への導入、DNAのPEGその他などの薬剤の作用によるプロトプラストへの導入、および特にDNAをコートしたマイクロプロジェクタイル撃ち込みなどの、従来技術において周知の方法が含まれる。DNAの直接移入による植物細胞への導入は、Agrobacteriumが媒介するDNA導入とは、少なくとも二本鎖DNAが植物細胞に入るという点で、入るDNAはいかなるタンパク質によってもコートされていないという点で、また植物細胞に入るDNAの量が相当に、より大きい可能性があるという点で異なる。さらに、二本鎖DNA切断誘導エンドヌクレアーゼをコードする導入されたキメラ遺伝子などの、直接移入法によって導入されたDNAは、転写により適していて、二本鎖DNA切断を誘導するタイミングがより良くなる可能性がある。本発明を特定の作用機序に制限するつもりはないが、効率的な相同性組換えに基づいた修復DNAまたは外来性DNAの植物細胞のゲノムへの挿入は、これらのパラメーターのうちのいずれかの組合せによるものと考えられる。
【0022】
都合よく、二本鎖DNA切断は、あらかじめ選ばれた部位で、レア切断を起こす二重鎖DNA切断導入酵素をコードする植物で発現可能な遺伝子の導入の後の一時的発現により引き起こすことができる。この文書の他の場所で述べるように、I−SceIを、I−SceI認識部位に外来性DNAを導入する目的で用いてもよい。しかし、他の二重鎖切断導入酵素もまた、外来性DNAをそれぞれの認識部位に挿入するために用いることができることは、当業者には直ちに明らかであろう。レア切断を起こすDSB誘導酵素およびそれぞれの認識部位のリストがWO 03/004659(17〜20ページ)(参照により本明細書に組み入れられる)の表Iに、提供されている。更に、選択した原則として任意の標的ヌクレオチド配列を認識する特別仕立てのレア切断を起こすエンドヌクレアーゼを設計するための方法を利用することができる。そのような方法については、例えばWO 03/080809、WO94/18313またはW095/09233に、およびIsalan et al., 2001, Nature Biotechnology 19,656-660; Liu et al. Proc. Natl. Acad. Sci. USA 94, 5525-5530に記載されている。
【0023】
したがって、本明細書で用いられる「あらかじめ選択された部位」とは、その座位に外来性DNAを挿入することが望まれる、植物の核ゲノム中の特定のヌクレオチド配列を表す。当業者は完全に、選択された標的ヌクレオチド配列を認識する二本鎖DNA切断誘導(「DSBI」)酵素を選ぶことができるか、またはそのようなDSBIエンドヌクレアーゼをエンジニアリングすることができるであろう。あるいは、任意の従来の形質転換法を用いて、またはゲノムにDSBIエンドヌクレアーゼ認識部位を有する植物系統を用いた従来の品種改良によって、植物ゲノムへDSBIエンドヌクレアーゼ認識部位を導入してもよく、また任意の所望の外来性DNAを後で、以前に導入されたあらかじめ選択された標的部位に導入してもよい。
【0024】
二重鎖切断誘導酵素をコードするDNA領域に作動可能に連結された植物発現可能プロモーター領域を含む植物発現可能なキメラ遺伝子を一過性に導入することにより、都合よく二本鎖DNA切断を引き起こすことができる。二重鎖切断誘導酵素をコードするDNA領域は合成DNA領域、例えば、コドンを本出願に別記するI−SceIコード領域の設計案に従って選択する合成DNA領域など、でもよい、しかしそれに限られない。
【0025】
二重鎖切断誘導酵素は、核移行シグナル(NLS)[Raikhel, Plant Physiol. 100: 1627-1632 (1992) およびその中の参照文献]、例えばSV40ラージT抗原のNLS[Kalderon et al., Cell 39: 499-509 (1984)]など、を含んでもよい(しかし含む必要はない)。核移行シグナルはタンパク質の任意の場所に位置してよいが、タンパク質のN末端に置くのが好都合である。核移行シグナルが、二重鎖切断誘導酵素のアミノ酸の1つ以上と置き換わってもよい。
【0026】
本明細書で用いる「対象の外来性DNA」とは、あらかじめ選択された部位への導入が望まれる任意のDNA断片を表す。厳密に必要とするわけではないが、対象の外来性DNAは、あらかじめ選択された部位に隣接するDNA領域に対して相同性を有する少なくとも1つのヌクレオチド配列領域と隣接してもよい。対象の外来性DNAは、あらかじめ選択された部位に隣接する両方のDNA領域に相同性を有するDNA領域により部位の両側を挟まれてもよい。したがって、植物細胞へ導入される修復DNA分子は、それぞれあらかじめ選択された部位の上流にまたは下流のDNA領域に対して相同性を有する1つまたは2つのフランキング配列によって隣接された外来性DNAを含んでもよい。これによって、外来性DNAの挿入をよりよく制御することが可能になる。確かに、相同組換えによる組込みが、外来性DNA断片を植物の核ゲノムへヌクレオチドレベルまで正確に接続することを可能にすることになる。
【0027】
隣接するヌクレオチド配列は、種々な長さでよいが、長さで少なくとも約10個のヌクレオチドでなければならい。しかしながら、フランキング領域は実際に可能な程度まで(例えば、完全なバクテリア人工染色体(BACs)のように約100〜150kbまで)長くてもよい。好ましくは、フランキング領域は約50bp〜約2000bpになるであろう。さらに、対象の外来性DNAに隣接する領域は、あらかじめ選択された部位に隣接するDNA領域と同一である必要はない。そして、あらかじめ選択された部位に隣接するDNA領域に、約80%〜約100%の配列同一性を、好ましくは約95%〜約100%の配列同一性を有していればよい。フランキング領域が長いほど、相同性の要求は、より厳格でない。さらに配列同一性は、外来性DNAの正確な挿入の位置の近くにおいて実際に可能な限度まで高いことが好ましい。
【0028】
さらに、対象の外来性DNAに隣接する領域は、あらかじめ選択された部位に直接隣接する領域に対して相同性を有する必要はなく、あらかじめ選択された部位からさらに遠い核ゲノムのDNA領域へ相同性があればよい。外来性DNAの挿入によって次に、あらかじめ選択された挿入部位と相同DNA領域の間の標的DNAが除去される結果になるであろう。言いかえれば、相同性領域間に位置した標的DNAは、対象の外来性DNAに置換されることになる。
【0029】
本発明の目的のためには、パーセンテージで表した2つの関連するヌクレオチドまたはアミノ酸配列の「配列同一性」とは、2つの最も良くアラインメントされた配列の、同一の残基を有する位置の数(×100)を比較した位置数で割ったものを指す。ギャップ、即ち残基が1つの配列中にはあり他方にはないアラインメント中の位置は、同一でない残基の位置と見なす。2つの配列のアラインメントは、ニードルマンとブンシュのアルゴリズム(Needleman and Wunsch 1970)によって、行なう。コンピューター支援配列アラインメントを Wisconsin Package 第10.1版(Genetics Computer Group, Madison, Wisconsin, USA)の一部であるGAPなどの標準ソフトウェアプログラムを用い、ギャップ生成ペナルティーを50、およびギャップ伸展ペナルティーを3とするデフォルトの得点マトリックスを用いて、便利に行なうことができる。
【0030】
別の局面では、本発明が、修飾されたI−SceIをコードするDNA断片、およびそれの、対象の外来性DNAを効率的に植物細胞ゲノムのあらかじめ選択された部位へ導入するための使用、に関する。この場合、修飾されたI−SceIをコードするDNA断片は、次の基準を満たすように設計されたヌクレオチド配列を有する:
a) ヌクレオチド配列が、配列番号1で与えられるアミノ酸配列を有するI−SceIエンドヌクレアーゼなどの、機能的なI−SceIエンドヌクレアーゼをコードする;
b)ヌクレオチド配列が、約50%〜約60%のGC含量を有する;
c)ヌクレオチド配列が、
【表4】
からなる群より選択されるヌクレオチド配列を含まない;
d)ヌクレオチドが、CCAAT、ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まない;
e)ヌクレオチド配列が、ATTTA、AAGGT、AGGTA、GGTAまたはGCAGGからなる群より選択されるヌクレオチド配列を含まない;
f)ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まない;
g)ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るGCストレッチを含まない;および
h)ヌクレオチド配列が、位置2および3にTAまたはCGという二連コードを含むLeu、Ile、Val、Ser、Pro、Thr、Alaをコードするコドン、を含まない(即ち、ヌクレオチド配列がコドンTTA、CTA、ATA、GTA、TCG、CCG、ACGおよびGCGを含まない)。
【0031】
I−SceIは、Saccharomyces cerevisea のミトコンドリア中のイントロン移動の原因となっている部位特異的なエンドヌクレアーゼである。酵素は、21S rRNA遺伝子のオプショナルイントロンScLSU.1によってコードされ、イントロン挿入部位で二本鎖DNA切断を開始し、3’OH張出しを有する4bpの互い違いの切断を生成する。I−SceIエンドヌクレアーゼの認識部位は、18bpの非対称配列上に伸展している(Colleaux et al. 1988 Proc. Natl. Acad. Sci. USA 85:: 6022-6026)。I−SceIのアミノ酸配列およびミトコンドリアのI−SceI遺伝子のユニバーサルコード等価物が、例えばWO 96/14408によって提供されている。
【0032】
WO96/14408は、I−SceIタンパク質の次の変異体がまだ機能的であることを開示している:
●位置1〜10を欠失させることができる。
●位置36:Gly(G)を許容する。
●位置40:Met(M)またはVal(V)を許容する。
●位置41:Ser(S)またはAsn(N)を許容する。
●位置43:Ala(A)を許容する。
●位置46:Val(V)またはN(Asn)を許容する。
●位置91:Ala(A)を許容する。
●位置123および156:Leu(L)を許容する。
●位置223:Ala(A)およびSer(S)を許容する。
また、そのような変異体I−SceI酵素をコードする合成ヌクレオチド配列を本発明に従って設計し用いることができる。
【0033】
アミノ末端に位置する4つのアミノ酸が核移行シグナルによって置換されたI−SceIのアミノ酸配列(配列番号1)をコードするヌクレオチド配列は、したがって、R1〜R244で表わすことのできる244のトリヌクレオチドから成る。これらの各位置に対して、同一のアミノ酸をコードするトリヌクレオチドの1〜6の可能な選択枝を選ぶことができる。表1は、配列番号1のアミノ酸配列をコードするトリヌクレオチドに対して可能な選択枝を記載し、また上に述べた「禁制ヌクレオチド配列」が合成DNA配列へ取り込まれるのを回避することを可能にする、構造上の必要条件(条件付きの、または絶対的な)を提供する。さらに、UIPACコードによる隣接するトリヌクレオチドのヌクレオチド配列を提供する。
【0034】
本明細書で用いられるUIPACコードの記号は、それらの通常の意味を有している、即ち:N=AまたはCまたはGまたはT;R=AまたはG;Y=CまたはT;B=CまたはGまたはT(Aではない);V=AまたはCまたはG(Tではない);D=AまたはGまたはT(Cではない);H=AまたはCまたはT(Gではない);K=GまたはT;M=AまたはC;S=GまたはC;W=AまたはT。
【0035】
したがって本発明の実施態様の1つでは、配列番号2に記載されているヌクレオチド配列を含む単離された合成DNA断片を提供する。この場合、コドンを、与えられた選択枝の中から約50%〜約60%の、好ましくは約54%〜55%の全GC含量を有するヌクレオチド配列を得るように選ぶ。ただし以下を条件とする:位置28〜位置30のヌクレオチド配列がAAGでなく;もし位置34〜位置36のヌクレオチド配列がAATである場合は、位置37〜位置39のヌクレオチド配列がATTまたはATAでなく;もし位置34〜位置36のヌクレオチド配列がAACである場合は、位置37〜位置39のヌクレオチド配列がATTでなく、同時に位置40〜位置42のヌクレオチド配列がAAAであり;もし位置34〜位置36のヌクレオチド配列がAACである場合は、位置37〜位置39のヌクレオチド配列がATAでなく;もし位置37〜位置39のヌクレオチド配列がATTまたはATAである場合は、位置40〜位置42のヌクレオチド配列がAAAでなく;位置49〜位置51のヌクレオチド配列がCAAでなく;位置52〜位置54のヌクレオチド配列がGTAでなく;位置58〜位置63のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置67〜位置69のヌクレオチド配列がCCCである場合は、位置70〜位置72のヌクレオチド配列がAATでなく;もし位置76〜位置78のヌクレオチド配列がAAAである場合は、位置79〜位置81のヌクレオチド配列がTTGでなく、同時に位置82〜位置84のヌクレオチド配列がCTNであり;もし位置79〜位置81のヌクレオチド配列がTTAまたはCTAである場合は、位置82〜位置84のヌクレオチド配列がTTAでなく;位置88〜位置90のヌクレオチド配列がGAAではなく;もし位置91〜位置93のヌクレオチド配列がTATである場合は、位置94〜位置96のヌクレオチド配列がAAAでなく;もし位置97〜位置99のヌクレオチド配列がTCCまたはTCGまたはAGCである場合は、位置100〜位置102のヌクレオチド配列がCCAでなく、同時に位置103〜位置105のヌクレオチド配列がTTRであり;もし位置100〜位置102のヌクレオチド配列がCAAである場合は、位置103〜位置105のヌクレオチド配列がTTAでなく;もし位置109〜位置111のヌクレオチド配列がGAAである場合は、位置112〜位置114のヌクレオチド配列がTTAでなく;もし位置115〜位置117のヌクレオチド配列がAATである場合は、位置118〜位置120のヌクレオチド配列がATTまたはATAでなく;位置121〜123のヌクレオチド配列がGAGである場合、位置124〜位置126のヌクレオチド配列は;位置133〜135のヌクレオチド配列がGCAでなく;位置139〜位置141のヌクレオチド配列がATTでなく;もし位置142〜位置144のヌクレオチド配列がGGAである場合は、位置145〜位置147のヌクレオチド配列がTTAでなく;もし位置145〜位置147のヌクレオチド配列がTTAである場合は、位置148〜位置150のヌクレオチド配列がATAでなく、同時に位置151〜位置153のヌクレオチド配列がTTRであり;もし位置145〜位置147のヌクレオチド配列がCTAである場合は、位置148〜位置150のヌクレオチド配列がATAでなく、同時に位置151〜位置153のヌクレオチド配列がTTRであり;もし位置148〜位置150のヌクレオチド配列がATAである場合は、位置151〜位置153のヌクレオチド配列がCTAまたはTTGでなく;もし位置160〜位置162のヌクレオチド配列がGCAである場合は、位置163〜位置165のヌクレオチド配列がTACでなく;もし位置163〜位置165のヌクレオチド配列がTATである場合は、位置166〜位置168のヌクレオチド配列がATAでなく、同時に位置169〜位置171のヌクレオチド配列がAGRであり;位置172〜位置177のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がGCAGGを含まないようになる選択に従って選ばれ;位置178〜位置186のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がAGGTAを含まないようになる選択に従って選ばれ;もし位置193〜位置195のヌクレオチド配列がTATである場合は、位置196〜位置198のヌクレオチド配列がTGCでなく;位置202〜位置204のヌクレオチド配列がCAAでなく;位置217〜位置219のヌクレオチド配列がAATでなく;もし位置220〜位置222のヌクレオチド配列がAAAである場合は、位置223〜位置225のヌクレオチド配列がGCAでなく;もし位置223〜位置225のヌクレオチド配列がGCAである場合は、位置226〜位置228のヌクレオチド配列がTACでなく;もし位置253〜位置255のヌクレオチド配列がGACである場合は、位置256〜位置258のヌクレオチド配列がCAAでなく;もし位置277〜位置279のヌクレオチド配列がCATである場合は、位置280〜位置282のヌクレオチド配列がAAAでなく;位置298〜位置303のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置304〜位置306のヌクレオチド配列がGGCである場合は、位置307〜位置309のヌクレオチド配列がAATでなく;位置307〜位置312のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;位置334〜位置342のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置340〜位置342のヌクレオチド配列がAAGである場合は、位置343〜位置345のヌクレオチド配列がCATでなく;もし位置346〜位置348のヌクレオチド配列がCAAである場合は、位置349〜位置351のヌクレオチド配列がGCAでなく;位置349〜位置357のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;位置355〜位置357のヌクレオチド配列がAATでなく;
もし位置358〜位置360のヌクレオチド配列がAAAである場合は、位置361〜位置363のヌクレオチド配列がTTGでなく;もし位置364〜位置366のヌクレオチド配列がGCCである場合は、位置367〜位置369のヌクレオチド配列がAATでなく;位置367〜位置378のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置382〜位置384のヌクレオチド配列がAATである場合は、位置385〜位置387のヌクレオチド配列がAATでなく;位置385〜位置387のヌクレオチド配列がAATでなく;もし位置400〜位置402のヌクレオチド配列がCCCである場合は、位置403〜位置405のヌクレオチド配列がAATでなく;もし位置403〜位置405のヌクレオチド配列がAATである場合は、位置406〜位置408のヌクレオチド配列がAATでなく;位置406〜位置411のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;位置421〜位置426のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;位置430〜位置432のヌクレオチド配列がCCAでなく;もし位置436〜位置438のヌクレオチド配列がTCAである場合は、位置439〜位置441のヌクレオチド配列がTTGでなく;位置445〜位置447のヌクレオチド配列がTATでなく;位置481〜483のヌクレオチド配列がAATでなく;もし位置484〜位置486のヌクレオチド配列がAAAである場合は、位置487〜位置489のヌクレオチド配列がAATでなく、同時に位置490〜位置492のヌクレオチド配列がAGYであり;もし位置490〜位置492のヌクレオチド配列がTCAである場合は、位置493〜位置495のヌクレオチド配列がACCでなく、同時に位置496〜位置498のヌクレオチド配列がAAYであり;もし位置493〜位置495のヌクレオチド配列がACCである場合は、位置496〜位置498のヌクレオチド配列がAATでなく;位置496〜位置498のヌクレオチド配列がAATでなく;もし位置499〜位置501のヌクレオチド配列がAAAである場合は、位置502〜位置504のヌクレオチド配列がTCAまたはAGCでなく;もし位置508〜位置510のヌクレオチド配列がGTAである場合は、位置511〜位置513のヌクレオチド配列がTTAでなく;もし位置514〜位置516のヌクレオチド配列がAATである場合は、位置517〜位置519のヌクレオチド配列がACAでなく;もし位置517〜位置519のヌクレオチド配列がACCまたはACGである場合は、位置520〜位置522のヌクレオチド配列がCAAでなく、同時に位置523〜位置525のヌクレオチド配列がTCNであり;位置523〜位置531のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置544〜位置546のヌクレオチド配列がGAAである場合は、位置547〜位置549のヌクレオチド配列がTATでなく、同時に位置550〜位置552のヌクレオチド配列がTTRであり;位置547〜位置552のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置559〜位置561のヌクレオチド配列がGGAである場合は、位置562〜位置564のヌクレオチド配列がTTGでなく、同時に位置565〜位置567のヌクレオチド配列がCGNであり;もし位置565〜位置567のヌクレオチド配列がCGCである場合は、位置568〜位置570のヌクレオチド配列がAATでなく;位置568〜位置570のヌクレオチド配列がAATでなく;もし位置574〜位置576のヌクレオチド配列がTTCである場合は、位置577〜位置579のヌクレオチド配列がCAAでなく、同時に位置580〜位置582のヌクレオチド配列がTTRであり;もし位置577〜位置579のヌクレオチド配列がCAAである場合は、位置580〜位置582のヌクレオチド配列がTTAでなく;
もし位置583〜位置585のヌクレオチド配列がAATである場合は、位置586〜位置588のヌクレオチド配列がTGCでなく;位置595〜位置597のヌクレオチド配列がAAAでなく;もし位置598〜位置600のヌクレオチド配列がATTである場合は、位置601〜位置603のヌクレオチド配列がAATでなく;位置598〜位置600のヌクレオチド配列がATAでなく;位置601〜位置603のヌクレオチド配列がAATでなく;もし位置604〜位置606のヌクレオチド配列がAAAである場合は、位置607〜位置609のヌクレオチド配列がAATでなく;位置607〜位置609のヌクレオチド配列がAATでなく;位置613〜位置615のヌクレオチド配列がCCAでなく;もし位置613〜位置615のヌクレオチド配列がCCGである場合は、位置616〜位置618のヌクレオチド配列がATAでなく;もし位置616〜位置618のヌクレオチド配列がATAである場合は、位置619〜位置621のヌクレオチド配列がATAでなく;もし位置619〜位置621のヌクレオチド配列がATAである場合は、位置622〜位置624のヌクレオチド配列がTACでなく;位置619〜位置621のヌクレオチド配列がATTでなく;位置640〜位置645のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置643〜位置645のヌクレオチド配列がTTAである場合は、位置646〜位置648のヌクレオチド配列がATAでなく;もし位置643〜位置645のヌクレオチド配列がCTAである場合は、位置646〜位置648のヌクレオチド配列がATAでなく;位置655〜位置660のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置658〜位置660のヌクレオチド配列がTTAまたはCTAである場合は、位置661〜位置663のヌクレオチド配列がATTまたはATCでなく;位置661〜位置663のヌクレオチド配列がATAでなく;もし位置661〜位置663のヌクレオチド配列がATTである場合は、位置664〜位置666のヌクレオチド配列がAAAでなく;位置670〜位置675のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置691〜位置693のヌクレオチド配列がTATである場合は、位置694〜位置696のヌクレオチド配列がAAAでなく;もし位置694〜位置696のヌクレオチド配列がAAAである場合は、位置697〜位置699のヌクレオチド配列がTTGでなく;もし位置700〜位置702のヌクレオチド配列がCCCである場合は、位置703〜位置705のヌクレオチド配列がAATでなく;もし位置703〜位置705のヌクレオチド配列がAATである場合は、位置706〜位置708のヌクレオチド配列がACAまたはACTでなく;もし位置706〜位置708のヌクレオチド配列がACAである場合は、位置709〜位置711のヌクレオチド配列がATAでなく、同時に位置712〜位置714のヌクレオチド配列がAGYであり;ヌクレオチド配列がコドンTTA、CTA、ATA、GTA、TCG、CCG、ACG、およびGCGを含まず;前記ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;および前記ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るATストレッチを含まない。
【0036】
合成ヌクレオチド配列の好ましい群を表2に記載する。そのヌクレオチド配列は配列番号3に記載されているヌクレオチド配列を含むように与えられる単離した合成DNA断片に対応する。この場合、コドンは、約50%〜約60%の、好ましくは約54%〜55%の全体GC含量を有するヌクレオチド配列を得る、かつ以下の条件を満たす選択肢の中から選択される:もし位置121〜位置123のヌクレオチド配列がGAGである場合は、位置124〜126のヌクレオチド配列がCAAでなく;もし位置253〜位置255のヌクレオチド配列がGACである場合は、位置256〜258のヌクレオチド配列がCAAでなく;もし位置277〜位置279のヌクレオチド配列がCATである場合は、位置280〜282のヌクレオチド配列がAAAでなく;もし位置340〜位置342のヌクレオチド配列がAAGである場合は、位置343〜345のヌクレオチド配列がCATでなく;もし位置490〜位置492のヌクレオチド配列がTCAである場合は、位置493〜495のヌクレオチド配列がACCでなく;もし位置499〜位置501のヌクレオチド配列がAAAである場合は、位置502〜504のヌクレオチド配列がTCAまたはAGCでなく;位置517〜位置519のヌクレオチド配列がACCである場合は、位置520〜522のヌクレオチド配列がCAAでなく、同時に位置523〜525のヌクレオチド配列がTCNであり;もし位置661〜位置663のヌクレオチド配列がATTである場合は、位置664〜666のヌクレオチド配列がAAAでなく;位置7〜位置15のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置61〜位置69のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置130〜位置138のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置268〜位置279のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置322〜位置333のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置460〜位置468のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置13〜位置27のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置37〜位置48のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置184〜位置192のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置214〜位置219のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置277〜位置285のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置388〜位置396のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置466〜位置474のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置484〜位置489のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置571〜位置576のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置598〜位置603のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置604〜位置609のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置613〜位置621のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置646〜位置651のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置661〜位置666のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;および位置706〜位置714のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選択される。
【0037】
配列番号4のヌクレオチド配列は、もはや回避するべきヌクレオチド配列またはコドンのどれも含んでいないI−SceIエンドヌクレアーゼをコードする合成ヌクレオチド配列の例である。しかしながら、当業者が、選択されるヌクレオチドの1つ以上を(2〜20を)配列番号3のヌクレオチド配列の中で提供される選択肢のいずれかに置換(前の段落に記述した禁止組合せのいずれも除外して)することにより、I−SceIをコードする同様の配列を容易に得ることができ、かつ使用して同様の効果を得ることができることは明らかであろう。
【0038】
植物細胞中の発現のために、I−SceIをコードする合成DNA断片を、植物で発現できるキメラ遺伝子を得るために、植物発現可能なプロモーターに作動可能に連結させてもよい。
【0039】
当業者は、本発明のこの局面については、修復DNAおよび/または、DSBIエンドヌクレアーゼをコードするDNAを直接のDNA移入法によって植物細胞へ導入することは必要でなく、従来技術において利用可能なAgrobacteriumを媒介とする形質転換法によって、DNAを植物細胞へこのように導入できることを直ちに認識するであろう。
【0040】
さらに別の局面では本発明は、対象の外来性DNAを植物細胞のゲノムのあらかじめ選択された部位へ導入する方法であって、
(a)二重鎖切断を細胞のゲノム中のあらかじめ選ばれた部位に引き起こす工程;および
(b)対象の外来性DNAを植物細胞へ導入する工程;
を含み、工程(a)に先立って植物細胞を植物フェノール化合物中でインキュベーションすることを特徴とする方法に関する。
【0041】
本発明に好適な「植物フェノール化合物」あるいは、「植物フェノール類」とは、正の走化性応答を引き起こすことができる置換フェノール分子、特にTiプラスミドを含むAgrobacterium種中で、特にTiプラスミドを含むAgrobacterium tumefaciens中で、vir遺伝子発現の増加を引き起こすことのできる分子である。植物フェノール化合物への走化性応答を測定する方法については、Ashby et al. (1988 J. Bacteriol. 170: 4181-4187)に記述されており、またvir遺伝子発現の誘導を測定する方法も周知である(Stachel et al., 1985 Nature 318: 624-629; Bolton et al.1986 Science 232: 983-98)。好ましい植物フェノール化合物は植物細胞の創傷滲出液で見出されるものである。最もよく知られている植物フェノール化合物の1つがアセトシリンゴンであり、それは様々な植物の多数の傷ついた細胞または完全な細胞に、様々な濃度とはいえ、存在する。しかしながら、アセトシリンゴン(3,5−ジメトキシ−4−ヒドロキシアセトフェノン)が、vir遺伝子の発現を引き起こすことができるただ一つの植物フェノールではない。他の例は、α−ヒドロキシ−アセトシリンゴン、シナピン酸(3,5ジメトキシ−4−ヒドロキシケイ皮酸)、シリング酸(4−ヒドロキシ−3,5ジメトキシ安息香酸)、フェルラ酸(4−ヒドロキシ−3−メトキシケイ皮酸)、カテコール(1,2−ジヒドロキシベンゼン)、p−ヒドロキシ安息香酸酸(4−ヒドロキシ安息香酸酸)、β−レソルシル酸(2,4ジヒドロキシ安息香酸)、プロトカテク酸(3,4−ジヒドロキシ安息香酸)、ピロ没食子酸(2,3,4−トリヒドロキシ安息香酸)、没食子酸(3,4,5−トリヒドロキシ安息香酸)、およびバニリン(3−メトキシ−4−ヒドロキシベンズアルデヒド)である。本明細書で用いられ、言及された分子を植物フェノール化合物と呼ぶ。植物フェノール化合物を、植物栽培培地に単独でまたは他の植物フェノール化合物と組み合わせて加えることができる。本発明を特定の作用機序に制限する意図はないが、これらの植物フェノール類の細胞分裂に対する(したがってゲノム複製に対しても)明らかな促進効果は、外来性DNAの標的指向挿入を促進している可能性があると思われる。
【0042】
植物細胞を、植物フェノール化合物中でまたはその上で、約1日または2日間インキュベーションすることで十分であろうと予想されるが、好ましくは約1週間植物フェノール化合物中でインキュベーションする。植物細胞を細胞分裂を促進するために十分な時間だけインキュベーションするべきである。Guivarc'h et al., (1993, Protoplasma 174: 10-18)によれば、植物細胞のほんの10分間ほどのインキュベーションによって、既にそのような効力を得ることができる。
【0043】
上述の相同組換えに基づく標的指向DNA挿入のための改善された方法は、直接DNA移入法により、特にマイクロプロジェクタイル撃ち込みにより得られる遺伝子組換え植物細胞および植物の、質を改善するためにも適用できる。マイクロプロジェクタイル撃ち込みによるDNAの導入により、しばしば導入されるDNAの複雑な組込みパターン(完全なまたは部分的な対象外来性DNAの複数のコピーの組込み、繰り返し構造の生成)がもたらされることは、従来技術において周知である。それにもかかわらず、いくつかの植物の遺伝子型または変種は、マイクロプロジェクタイル撃ち込みを用いる形質転換の方が、例えば Agrobacterium tumefaciens を使用する形質転換よりも、適している可能性がある。マイクロプロジェクタイル撃ち込みを通じて得られる遺伝子組換え植物細胞または植物の、質を改善することができれば、即ち外来性DNA組込みのパターンがより単純になるような影響を受ければ、有利であろう。
【0044】
上に述べた発見、即ち核ゲノム中に誘導された二本鎖DNA切断の存在下でマイクロプロジェクタイル撃ち込みにより外来性DNAを導入し、その際、外来性DNAは二本鎖DNA切断に隣接する配列に対して相同性を有している場合は、しばしば(得られるイベントの約50%が)単純な組込みパターン(予測可能な方式による単一コピーの挿入があり、外来性DNAの追加の断片の挿入がない)となる結果をもたらす、という発見は、植物細胞の核ゲノム中の外来性DNAの挿入の複雑さを単純化する方法の基礎を与える。
【0045】
したがって、本発明はまた、次の工程を含むマイクロプロジェクタイル撃ち込みによって遺伝子組換え植物を生成する方法に関する:
(a)本明細書に別記する方法または従来技術において利用可能な方法に従って、二本鎖DNA切断を、植物細胞のゲノム中のあらかじめ選択された部位に引き起こす工程;および
(b)対象の外来性DNAをマイクロプロジェクタイル撃ち込みによって植物細胞へ導入する工程(ここで前記対象の外来性DNAは、植物ゲノム中のあらかじめ選択された部位に隣接するDNA領域へ少なくとも80%の配列同一性を有する2つのDNA領域に挟まれている)。
このように得られる遺伝子組換え植物の個体数の多くの部分が、植物細胞のゲノム中に外来性DNAの単純な組込みパターンを有し、より詳しくいえば、遺伝子組換え植物のかなりの部分は、1コピーの外来性DNA挿入のみを、植物ゲノム中のあらかじめ選択された部位に隣接する2つのDNA領域の間に正確に有することになる。この部分は、二本鎖DNA切断を引き起こすことなしに、および外来性DNAにあらかじめ選択された部位に隣接するゲノム領域との相同性を与えることなしに、単純なマイクロプロジェクタイル撃ち込みによって植物が得られる場合における単純な組込みパターンを有する遺伝子組換え植物の個体数よりも高い。
【0046】
本発明の好都合の実施態様では、標的となる植物細胞が、そのゲノム中に、レア切断を起こす二本鎖DNA切断誘起エンドヌクレアーゼの2つの認識部位が各側に1つずつ隣接するマーカー遺伝子を含む。このマーカーDNAは、対象の植物細胞のゲノムに、形質転換の任意の方法を用いて導入してもよいし、または既に別の植物系統または変異種(形質転換が容易な植物系統または変異種など)の植物細胞のゲノムへ導入されていて、それを古典的育種技術により対象の植物細胞へ導入してもよい。好ましくは、レア切断を起こす二重鎖切断誘導エンドヌクレアーゼの2つの認識部位を両側面に有するマーカー遺伝子を含む遺伝子組換え植物または植物細胞の集団は、マ−カー遺伝子の発現パターン(高い発現、時間的または空間的に調節された発現など)についてすでに分析されており、望みの発現パターンを有する植物系統が同定されている。
(a)本明細書に別記する方法または従来技術において利用可能な方法に従って、植物細胞のゲノム中のあらかじめ選ばれた部位に二本鎖DNA切断を引き起こす工程;および
(b)対象の外来性DNAをマイクロプロジェクタイル撃ち込みによって植物細胞へ導入する工程(ここで前記対象の外来性DNAは、植物ゲノム中のあらかじめ選ばれた部位に隣接するDNA領域と少なくとも80%の配列同一性を有する2つのDNA領域に挟まれている)、
を含むマイクロプロジェクタイル撃ち込みによる遺伝子組換え植物の産生が、マーカー遺伝子が対象の外来性DNAによって置換された遺伝子組換え植物細胞および植物をもたらすことになる。
【0047】
マーカー遺伝子は、選択可能なまたはスクリーニング可能な植物で発現可能なマーカー遺伝子でよく、それには従来のキメラのマーカー遺伝子が好適である。キメラのマーカー遺伝子は、プロモーター、好ましくはCaMV35Sプロモーターなどの植物で発現可能な構成的プロモーター、またはルビスコの小サブユニットをコードする遺伝子のプロモーターなどの光誘導可能なプロモーターの制御下にある、またはそのプロモーターの5’末端に動作可能なように連結され;またその3’末端で、適当な植物転写終了およびポリアデニル化シグナルに動作可能なように連結された、マーカーDNAを含むことができる。マーカーDNAは、植物の細胞中で発現された時、マーカーDNAを発現していない細胞から容易にそのような細胞を分離することを可能にするRNA、タンパク質またはポリペプチドをコードすることが好ましい。マーカーDNAの選択は決定的に重要ではなく、周知の方式で任意の適当なマーカーDNAを選択することができる。例えば、マーカーDNAは、A1遺伝子(Meyer et al., (1987), Nature 330:677)のように形質転換された植物細胞に識別可能な色彩を与えるタンパク質をコードすることができ、蛍光タンパク質[Chalfie et al., Science 263: 802-805 (1994); Crameri et al., Nature Biotechnology 14:315-319 (1996)]をコードすることができ、ホスフィノトリシンに対する耐性を与えるPATをコードするbar遺伝子(EP 0242246)のように、形質転換された植物細胞へ除草剤耐性を与えるタンパク質をコードすることができ、またはゲンタマイシンに対する耐性を与えるGATをコードするaac(6’)遺伝子(WO94/01560)のように、形質転換細胞に抗生物質耐性を与えるタンパク質をコードすることができる。そのような選択可能なマーカー遺伝子は一般に、細胞には普通有毒な抗生物質または他の化合物に対する細胞耐性を与えるタンパク質をコードする。植物では、選択可能なマーカー遺伝子がこのように、さらに有効成分としてグルタミン合成酵素阻害剤(例えば、ホスフィノトリシン)を含む除草剤などの、除草剤に対する耐性を与えるタンパク質をコードしてもよい。そのような遺伝子の例は、sfrまたはsfrv遺伝子(EP 242236; EP 242246; De Block et al., 1987 EMBO J. 6: 2513-2518)などの、ホスフィノトリシンアセチルトランスフェラーゼをコードする遺伝子である。
【0048】
導入された修復DNAはさらに、あらかじめ選ばれた部位での相同組換えによる組込みと、ゲノムの他の場所への組込みとを識別することを可能にするマーカー遺伝子を含んでもよい。そのようなマーカー遺伝子が従来技術において利用可能であり、それには、選択的な条件下でマーカー遺伝子の不存在を積極的に選択できるマーカー遺伝子(例えば、codA、5−フルオロシトシンに対する感受性を与える E. coli から得られるシトシンデアミナーゼ、Perera et al., 1993 Plant Mol. Biol. 23, 793; Stougaard (1993) Plant J: 755)が含まれる。修復DNAは、核ゲノム中へ修復DNAがランダムに組込まれると、マーカー遺伝子が存在する結果となり、一方で相同組換えによって修復DNAが組込まれると、マーカー遺伝子が存在しない結果となるような方式で、マーカー遺伝子を含む必要がある。
【0049】
マーカー遺伝子中またはその近くに位置する二重鎖切断を引き起こすただ1つのあらかじめ選択された部位を用いて、同じ結果を得ることができることは直ちに明らかであろう。その場合、相同性のフランキング領域を、マーカー遺伝子を不活性化するかまたはマーカー遺伝子を欠失させて、挿入される外来性DNAによって置き換えるような方式で選択することが好ましい。
【0050】
本発明の手段および方法は、特にトウモロコシに役立つが、他の植物に、特に小麦、オート麦、大麦、ライ麦、米、ターフグラス、ソルガム、アワ類またはサトウキビ植物を含む穀類植物に用いてもよく、同様の効果が得られることを認識することになろう。本発明の方法を、綿、タバコ、キャノーラ、アブラナ、ダイズ、蔬菜類、ジャガイモ、ウキクサ種、ハナタバコ種、アラビドプシス、アルファルファ、オオムギ、豆、トウモロコシ、綿、アマ、エンドウ、アブラナ、コメ、ライムギ、ベニバナ、モロコシ、ダイズ、ヒマワリ、タバコ、小麦、アスパラガス、ビート、ブロッコリ、キャベツ、ニンジン、カリフラワー、セロリ、キュウリ、ナス、レタス、タマネギ、アブラナ、コショウ、ジャガイモ、カボチャ、ハツカダイコン、ホウレンソウ、セイヨウカボチャ、トマト、ズッキーニ、アーモンド、リンゴ、アンズ、バナナ、クロイチゴ、ブルーベリー、カカオ、サクランボ、ココナッツ、クランベリー、ナツメヤシ、ブドウ、グレープフルーツ、グアバ、キウイ、レモン、ライム、マンゴー、メロン、ネクタリン、オレンジ、パパイヤ、パッションフルーツ、モモ、ピーナツ、セイヨウナシ、パイナップル、ピスタチオ、プラム、ラズベリー、イチゴ、タンジェリン、クルミ、およびスイカを含み、しかしこれらに限らない任意の植物にもまた、適用することができる。
【0051】
本発明の方法に従ってあらかじめ選択された部位に挿入された外来性DNA分子を含む植物細胞および植物を提供することもまた、本発明の目的である。標的指向DNA挿入イベントを含む、従来の育種方法によって産生される植物の配偶子、種子、胚、接合子または体細胞である子孫またはハイブリッドもまた、本発明の範囲内に含まれる。
【0052】
本明細書に記述した方法によって得られた植物をさらに従来の育種技術によって他の植物と交配して、本発明により得られた標的指向DNA挿入イベントを含む子孫植物を得ることができる。
【0053】
次の制限されない実施例は、修飾されたI−SceIをコードするキメラ遺伝子の設計、および、その植物ゲノムのあらかじめ選択された部位に外来性DNAを挿入するための使用について記述する。
【0054】
もし実施例において異なるように記述しない場合は、組換DNA技術をすべて、Sambrook et al., (1989) Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press, NY、および Ausubel et al., (1994) Current Protocols in Molecular Biology, Current Protocols, USA の第1および第2巻に記載されている標準プロトコルにより実施する。植物の分子的研究の標準材料および方法は、R.D.D. CroyによるBIOS Scientific Publications Ltd (UK) および Blackwell Scientific Publications, UK の共同出版の Plant Molecular Biology Labfax (1993)に記述されている。標準的分子生物学技術のための他の参考文献には、Sambrook and Russell (2001) Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Laboratory Press, NY、Brown (1998) Molecular Biology LabFax, Second Edition, Academic Press (UK) のの第Iおよび第II巻が含まれる。ポリメラーゼ連鎖反応の標準材料および方法は、Dieffenbach and Dveksler (1995) PCR Primer: A Laboratory Manual, Cold Spring Harbor Laboratory Press, および McPherson et al., (2000) PCR-Basics: From Background to Bench, First Edition, Springer Verlag, Germany の中に見出すことができる。
【0055】
明細書および実施例の全体にわたって、次の配列を参照する:
【0056】
配列番号1:I−SceIタンパク質に連結された核移行シグナルを含み、4個のアミノ末端のアミノ酸を欠くキメラI−SceIのアミノ酸配列。
配列番号2:I−SceIコード領域のヌクレオチド配列(UIPACコード)。
配列番号3:合成I−SceIコード領域のヌクレオチド配列(UIPACコード)。
配列番号4:合成I−SceIコード領域のヌクレオチド配列。
配列番号5:pTTAM78のT−DNA(標的座位)のヌクレオチド配列。
配列番号6:pTTA82のT−DNA(修復DNA)のヌクレオチド配列。
配列番号7:pCV78のヌクレオチド配列。
【0057】
【表5】
【0058】
【表6】
【実施例1】
【0059】
I−SceIをコードする、植物で発現可能なキメラ遺伝子の設計、合成および解析
4個のアミノ末端アミノ酸が核移行シグナルで置換されたI−SceIのコード領域を、次のプロセスを用いて最適化した:
1.I−SceIタンパク質のアミノ酸配列を変更せずに、Synergy Geneoptimizer(登録商標)を用いて、トウモロコシに対して最も好ましいコドン使用にコドンを変換するプロセス;
2.特異的な制限部位を作成または除去するために配列を調節して、合成I−SceIコード領域をユニバーサルコードのI−SceI遺伝子で交換するプロセス;
3.前躯体mRNAスプライシングに影響する可能性のあるRNAの二次構造形成を回避するために、6bpより長いGCストレッチおよび4bpより長いATストレッチをすべて除去するプロセス;
4.コドンの中の2と3の位置のCGおよびTAという二連コードを回避するプロセス;
5.他の調節要素、例えば未成熟のポリアデニル化シグナルの候補
【表7】
隠れたイントロンスプライシング部位(AAGGTAAGTおよびTGCAGG)、ATTTA五量体、およびCCAATボックス配列(CCAAT、ATTGG、CGAAT、およびATTCG)を回避するプロセス;
6.適合されたコード領域が上に述べた基準をすべて満たすかどうかを再確認するプロセス。
【0060】
そのようなヌクレオチド配列の可能な一例を、配列番号4に示す。配列番号4のヌクレオチド配列を有する合成DNA断片を合成し、CaMV35SプロモーターおよびCaMV35S3’終了およびポリアデニル化シグナルに作動可能に連結させた(プラスミドpCV78;配列番号7を生成した)。
【0061】
合成I−SceIコード領域をバクテリアの発現ベクター中に(アミロースビーズ上でのタンパク質濃縮を可能にする融合タンパク質として)クローニングした。半精製されたI−SceIタンパク質の、I−SceI認識部位を含むプラスミドをインビトロで切断する能力を確認した。
【実施例2】
【0062】
I−SceI部位が先行しプロモーターを欠くbar遺伝子を含むトウモロコシ細胞系統の単離
二本鎖DNA切断が誘導する相同性を介する組換えの分析を発展させるために、核ゲノム中に単一コピーで組み込まれた、I−SceI認識部位が先行しプロモーターを欠くbar遺伝子を含むトウモロコシ細胞の懸濁液を分離した。I−SceIエンドヌクレアーゼをコードする植物で発現可能なキメラ遺伝子の送達、およびbar遺伝子の5’末端に作動可能に連結されたCaMV35Sプロモーターを含む修復DNAの共同送達、による二本鎖DNA切断誘導によって、35Sプロモーターが相同性を介する標的指向DNA挿入により挿入され、その結果ホスフィノトリシン(PPT)に対する耐性を与える機能的なbar遺伝子が得られる。分析を模式的に図1に表わす。
【0063】
標的座位を、従来のクローニング技術によって作動可能に次のDNA領域を連結することにより構成した:
a)ノパリン合成酵素遺伝子からの3’末端終止シグナルおよびポリアデニル化シグナル、
b)プロモーターを欠くbarをコードするDNA領域、
c)I−SceI認識部位を含むDNA領域、
d)A. tumefaciens 遺伝子7からの3’末端終止およびポリアデニル化シグナル(3’g7)、
e)ノパリン合成酵素プロモーター、ネオマイシンホスホトランスフェラーゼ遺伝子、および3’ocsシグナルを含む植物で発現可能なネオマイシン耐性遺伝子。
このDNA領域を、T−DNAベクターのT−DNA境界間に挿入した。T−DNAベクターをpTTAM78と名付けた(T−DNAのヌクレオチド配列については配列番号5を参照のこと)。
【0064】
T−DNAベクターを直接用いて、EP0 469 273に記述された方法によりHe89由来のトウモロコシ細胞懸濁液を用いてトウモロコシのプロトプラストを形質転換した。T−DNAベクターをまた Agrobacterium tumefaciens C58ClRif (pEHA101) へも導入して、得られたAgrobacteriumをHe89由来の細胞系統を形質転換するために用いた。T24(プロトプラストの形質転換により得られる)および、14−1および1−20系統(Agrobacteriumを介する形質転換により得られる)などの標的座位コンストラクトpTTAM78の単一コピーを含む多数の標的系統を同定した。
【0065】
N6M細胞懸濁培地中のこれらの標的系統から細胞懸濁液を確立し、振盪機(120 rpm)で光を当て25℃で増殖させた。懸濁液を毎週継代培養した。
【実施例3】
【0066】
相同性に基づく標的指向挿入
修復DNA、pTTA82は、T−DNA境界間に作動可能に連結された下記のDNA領域を含むT−DNAベクターである:
a)bar遺伝子のアミノ末端部分のみをコードするDNA領域;
b)部分的なI−SceI認識部位(認識部位の5’末端に位置する13個のヌクレオチド)を含むDNA領域;
c)CaMV35Sプロモーター領域;
d)部分的なI−SceI認識部位(認識部位の3’末端に位置する9個のヌクレオチド)を含むDNA領域;
e)A. tumefaciens 遺伝子7由来の3’末端終止およびポリアデニル化シグナル(3’g7);
f)キメラの植物発現可能なネオマイシン耐性遺伝子;
g)CaMV35Sプロモーターの制御下の欠落を有するI−SceIエンドヌクレアーゼをコードする遺伝子。
【0067】
pTTA82のT−DNAヌクレオチド配列を配列番号6に示す。
【0068】
この修復DNAを、ろ紙上に薄層として塗布した懸濁液由来の細胞中へ粒子を撃ち込むことによって、pCV78(実施例1を参照のこと)と共同送達した。ろ紙をMahql VII基質上にプレートした。
【0069】
DNAを細胞へ、PDS−1000/He Biolistics装置を用いて撃ち込んだ。マイクロキャリアの調製およびその上へのDNAのコーティングは、ほとんどSanford et al., 1992 記載の通りに行った。粒子撃ち込みパラメーターは次の通りであった:標的距離9cm、撃ち込み圧力1350psi、ギャップ距離1/4”、およびマイクロキャリア飛行距離11cm。撃ち込み直後に組織を、選択的でないMhilVII基質上に移した。粒子撃ち込みによる成功したDNA送達の対照として、3つの標的系統にも、CaMV35Sプロモーター制御下のキメラbar遺伝子を含むプラスミドDNA(pRVA52)でコートされたマイクロキャリアによって撃ち込みを行った。
【0070】
撃ち込み4日後に、フィルタを、25mg/L PPTを補ったMhlVII基質上に、または50mg/L PPTを補ったAhx1.5VIIino1000基質上に移した。
【0071】
14日後に、フィルタを、標的系統T24および14−1用の10mg/L のPPTを有する新鮮なMhlVII培地、および標的系統1−20用の25mg/LのPPTを有するMhlVII基質の上に移した。
【0072】
2週間後に、標的指向挿入である可能性のあるイベントを、PPTに対するそれらの耐性に基づいて評価した。これらのPPT耐性イベントは、Liberly Link Corn Leaf/Seedテスト(Strategic Diagnostics Inc.)においても陽性であった。
【0073】
撃ち込み38日後のPPT耐性カルスの数:
【表8】
【0074】
PPT耐性イベントを、さらに10mg/LのPPTを含むMhlVII基質上で継代培養し、またカルス材料を分子分析に用いた。20の独立のTSI候補を、サザン解析によって、35Sプロモーターならびにノパリン合成酵素遺伝子由来の3’末端終止およびポリアデニル化シグナルをプローブとして用いて解析した。予期された断片のサイズに基づいて、イベントはすべて完全な標的指向配列挿入イベントであるように見えた。その上、標的指向配列挿入イベントの約半分についてのさらに詳しい解析は、修復DNAまたはI−SceIをコードするDNAのいずれについても追加の非標的指向組込みを示さなかった。
【0075】
標的指向挿入イベントのうちの8つから増幅されたDNAの配列分析が、これらのイベントが確かに完全な相同組換えに基づいたTSIイベントであることを実証した。
【0076】
これらのデータに基づくと、相同組換えに基づいたDNA挿入対「正常な」非正統的組換の比率は、1−20に対する約30%から、14−1に対する17%、および24に対する約1%まで変化する。
【0077】
Puchta et al., 1996 (上掲)に記述された、I−SceI誘導二本鎖DNA切断存在下でエレクトロポレーションによってタバコプロトプラストに送達されたベクターと同様のベクターを用いた場合は、相同組換えに基づいたDNA挿入対正常な挿入の比率は、約15%であった。しかし、33の特徴決定したイベントのうちの1だけが、相同組換えが二重鎖切断の両側で完全な相同性仲介標的指向配列挿入イベントであった。
【0078】
実施例2から得たベクター、しかし核移行シグナルを含む「ユニバーサルコードI−SceIコンストラクト」を備えたベクターを用いると、相同組換えに基づいたDNA挿入対正常な挿入の比率が、様々な標的系統について、エレクトロポレーションまたは Agrobacteriumを介するDNA送達を用いた両方の場合で、0.032%〜16%の間で変化した。完全な標的指向挿入イベントの相対度数は、種々の標的系統間で異なり、エレクトロポレーションを介するDNA送達に対して8〜70%の間で、また Agrobacteriumを介するDNA送達に対して73〜90%の間で変化した。
【実施例4】
【0079】
アセトシリンゴンによるプレインキュベーションが、標的指向挿入イベントの回収頻度を改善する。
実施例3に記述した撃ち込みの1週間前に、細胞懸濁液を、200μM アセトシリンゴンを補ったN6M培地またはLSIDhy1.5培地で希釈した。他の点では、実施例3に記述した方法を用いた。次の表に要約された結果からわかるように、形質転換される細胞のアセトシリンゴンとのプレインキュベーションは、標的指向PPT耐性挿入イベントの回収に有益な効果を有した。
【0080】
【表9】
【実施例5】
【0081】
Agrobacteriumを媒介とした修復DNAの送達による、DSBを介するトウモロコシへの標的指向配列挿入
修復DNAがAgrobacteriumに媒介される形質転換により送達される、トウモロコシ中へのDSBを介する標的指向配列挿入を解析するために、欠陥I−SceIを実施例1の合成I−SceIコード遺伝子と置換したT−DNAベクターをpTTA82(実施例3を参照のこと)と同様に構成した。T−DNAベクターは、さらにT−DNA境界の外側に、Agrobacterium tumefaciens のvirGおよびvirC(pTCV83)、または、virG、virC、およびvirB(pTCV87)の1コピ−を含んでいた。これらのT−DNAベクターを、ヘルパーTiプラスミドpAL4404を含むLBA4404に挿入し、それによってAgrobacterium 菌株A4995およびA4996がそれぞれ生み出された。
【0082】
実施例2の標的細胞系統の浮遊培養、ならびに実施例2に記述したと同様の方法で得られた他の標的細胞系統を、Agrobacterium 菌株と共栽培し、その後多数のプレートに平板培養した。プレートする数を、細胞懸濁液の密度により決定した。形質転換効率用の対照として、細胞懸濁液を、並行して行った実験で、ヘルパーTiプラスミドpAL4404およびCaMV35Sベクターの制御下のキメラのホスフィノトリシン耐性遺伝子(bar遺伝子)を有するT−DNAベクターを含むAgrobacterium strain LBA4404 と共培養した。T−DNAベクターは、さらにT−DNA境界の外に、Agrobacterium tumeficans のvirG、virC、およびvirB遺伝子の1コピーを含んでいた。4つの異なる独立した実験の結果を下の表に要約する:
【0083】
【表10】
【0084】
【表11】
【0085】
【表12】
【0086】
【表13】
【0087】
このように、Agrobacteriumに媒介された修復DNA送達は明白に実現可能であるが、標的指向配列挿入(TSI)イベントの頻度が粒子撃ち込みを媒介とした修復DNA送達と比較して、より低いことが明らかである。23の推定TSIイベントで行なわれたサザン解析により、20のTSIのイベントが完全であることが、断片のサイズに基づいて示された。しかしながら、実施例3におけるマイクロプロジェクタイル撃ち込みによって得られたイベントと対照的に、20のイベントのうちの6のみが、修復DNAの追加の挿入断片を含んでいなかった。9つのイベントが、修復DNAの1〜3の追加の挿入を含んでいた。また5つのイベントが、修復DNAの多くの追加の挿入を含んでいた。
【0088】
粒子撃ち込みを介する修復DNAの送達はまた、Agrobacteriumによる修復DNAの送達に比較して、DSBを介するTSIイベントのよりよい質をもたらす。これは、粒子撃ち込みを介する「正常な形質転換DNA」の送達(Agrobacteriumを介する形質転換と比較した場合に、形質転換体のより劣った質(複雑な組込みパターン)によって特徴づけられる)とは対照的である。
【0089】
これは、粒子撃ち込みまたは他の直接DNA送達法によって得られる形質転換体の質を、DSBを介した配列の挿入によって改善することができることを表している。この結果は次の実験によっても確認される:修復DNA中に標的座位と相同性を有するフランキング配列がない状態で、DSBを介する35Sプロモーターの標的指向配列挿入を行ったときに、我々は、修復DNAのエレクトロポレーションを介した送達に際して、TSIイベントの少数だけが付加的な35Sプロモーターの非標的指向挿入を含むこと(16の解析されたTSIイベントのうちの2つのTSIイベントが付加的なランダムな35Sプロモーター挿入を示す)を観察した。対照的に、35Sプロモーターのランダム挿入が、Agrobacteriumを介する35Sプロモーターの送達によって得られたTSIイベントでは相当より高かった(22の解析されたTSIイベントのうち17が付加されたランダムな35Sプロモーターの挿入を示した)。
【実施例6】
【0090】
培地の組成
Mahq1VII:100mg/L カゼイン加水分解物、6mM L−プロリン、0.5g/L 2−(N−モルホリノ)エタンスルホン酸(MES)、0.2M マニトール、0.2M ソルビトール、2% 蔗糖、lmg/L 2,4−ジクロロフェノキシ酢酸(2,4−D)、を補ったN6培地 (Chu et al., 1975)、pH5.8に調節し、2.5g/L Gelrite(登録商標)で凝固させた。
【0091】
Mhi1VII:0.5g/L 2−(N−モルホリノ)エタンスルホン酸(MES)、0.2M マニトール、2% 蔗糖、lmg/L 2,4−ジクロロフェノキシ酢酸(2,4−D)、を補ったN6培地 (Chu et al., 1975)、pH5.8に調節し、2.5g/L Gelrite(登録商標)で凝固させた。
【0092】
Mh1VII:0.2M マニトールを含まない以外は、Mhi1VII基質に同じ。
【0093】
Ahxl.5VIIino1000:1000mg/L ミオ−イノシトール、0.1mg/L チアミン−HCl、0.5mg/L ニコチン酸、0.5mg/L ピリドキシン−HCl、0.5g/L MES、30g/L 蔗糖、10g/L グルコース、1.5mg/L 2,4−D、を補ったMS塩類、pH5.8に調節し、2.5g/L Gelrite(登録商標)で凝固させた。
【0094】
LSIDhyl.5:0.5mg/L ニコチン酸、0.5mg/L ピリドキシン−HCl、1mg/L チアミン−HCl、100mg/L ミオ−イノシトール、6mM プロリン、0.5g/L MES、20g/L 蔗糖、10g/L グルコース、1.5mg/L 2,4−D、を補ったMS塩類、pH5.2に調節した。
【0095】
N6M:マクロ要素:2830mg/L KNO3、433mg/L(NH4)2SO4;166mg/L CaCl2.2H2O;250mg/L MgSO4.7H2O;400mg/L KH2PO4;37.3mg/L Na2EDTA;27.3mg/L FeSO4.7H2O、 MSミクロ要素:500mg/L バクトトリプトン、0.5g/L MES、lmg/L チアミン−HCl、0.5mg/L ニコチン酸、0.5mg/L ピリドキシン−HCl、2mg/L グリシン、100mg/L ミオ−イノシトール、3%蔗糖、0.5mg/L 2,4−D、pH5.8に調節した。
【0096】
表1は、合成I−SceIコード領域に対する可能なトリヌクレオチド(コドン)の選択を表わす(配列番号2中のヌクレオチド配列も参照のこと)。
表2は、合成I−SceIコード領域に対する好ましい可能なトリヌクレオチドの選択を表わす(配列番号3中のヌクレオチド配列も参照のこと)。
【技術分野】
【0001】
本発明は、分子の植物生物学の分野、より具体的には植物ゲノムエンジニアリングの分野に関する。外来性DNA断片の、植物ゲノム中のあらかじめ選ばれた挿入部位への特異的導入のための方法を提供する。特定の部位に挿入された外来性DNAを含む植物を、現在利用可能な標的に向けたDNA挿入法で可能なよりも、より高い頻度でおよびより高い精度で、今や得ることができる。さらに、結果として得られる大部分の植物においては、外来性DNAはあらかじめ選択された挿入部位のみに挿入されており、植物ゲノムの他の位置に無秩序に外来性DNAが挿入されてはいない。本発明の方法はこのように、先行技術の方法に対して定量的および定性的の両面での進歩である。さらに、キメラ遺伝子、プラスミド、ベクター、および本発明の方法中で用いられる他の手段も提供する。
【背景技術】
【0002】
前世紀80年代初めのAgrobacteriumを介する形質転換技術による遺伝子組換え植物の第一世代により、対象の外来性DNAまたは導入遺伝子を植物のゲノムへ導入する他の方法、例えばPEGを介するプロトプラストへのDNA取込、マイクロプロジェクタイル撃ち込み、シリコンホイスカーを介する形質転換その他、の開発に拍車がかけられた。
【0003】
しかし、すべての植物の形質転換法では共通して、植物ゲノムに組み入れられる導入遺伝子は、無作為な方式でまた予測できないコピー数だけ組み込まれる。しばしば、導入遺伝子は、導入遺伝子全体が、またはそれの部分が、リピートの形で組込まれる場合がある。そのような複雑な組込みパターンは、導入遺伝子の発現レベルに、例えば転写後遺伝子サイレンシング機構により転写されたRNAを破壊して、または導入されたDNAのメチル化を引き起こして、導入遺伝子の転写活性を下方制御することにより、影響を及ぼす可能性がある。さらに、組込み部位自体が、導入遺伝子の発現レベルに影響を及ぼす場合がある。これらの因子の組合せが、導入遺伝子または対象の外来性DNAの発現レベルの広汎な変化を、種々の遺伝子組換え植物細胞および植物系統中にもたらす。さらに、対象の外来性DNAの組込みが、組込みが生じるゲノム領域に破壊的な効果を有する可能性があり、その標的領域の正常な機能に影響を及ぼすかまたは妨害する場合があって、そのためにしばしば望ましくない副作用が生ずる。
【0004】
したがって、植物への特定の外来性DNA導入の影響を研究する場合は常に、有効な結果を得るために多数の遺伝子組換え植物系統を生成し分析することを必要とする。同様に、遺伝子組換え植物に所望の既知の表現型を与えるために、植物に対象の特定のDNAが導入される遺伝子組換え農作物の生成においては、多数の独立に形成された遺伝子組換え植物系統、即ちいわゆるイベントが形成されて、導入遺伝子が最適に発現しておりまた遺伝子組換え植物の表現型全体に亘って副作用が最小であるかまたは無い植物系統の選択が可能となる。特にこの分野で、望ましくない遺伝子組換えイベントの排除に必要な圃場試験の繰返しに伴う厄介な制御のための必要条件および高いコストを考慮すれば、この試行錯誤プロセスをより指向性のあるプロセスと置き換えることができれば有利であろう。更に、標的指向のDNA挿入が可能であることが、いわゆる導入遺伝子スタッキングのプロセスでさらに有益であることが明らかであろう。
【0005】
植物中への導入遺伝子組込みを制御する必要性が早い時期から認識されており、またいくつかの方法がこの必要を満たす努力の中で発展されてきた(総説としてKumar and Fladung, 2001, Trends in Plant Science, 6, ppl55-159 を参照のこと)。これらの方法はほとんど、原核生物および下等真核生物に成功裡に適用されてきた戦略である相同組換えに基づいた導入遺伝子組込みに依存している(例えば、EP0317509または対応したPaszkowski et al., 1988, EMBO J., 7,pp 4021-4026による文献を参照のこと)。しかし、植物については、導入遺伝子組込みの主な機構は、組換えをするDNA鎖間の相同性がほとんどない非正統的組換に基づいている。この領域における主な挑戦課題は、したがって、非正統的組換を介して導入される外来性DNAのはるかに効率的な組込みによって隠されている、レア相同組換えイベントの検出である。
【0006】
この問題を解決する1つの方法は、WO94/17176に例示されているような、非正統的組換によって生じた組込みイベント以外を選別することによる。
【0007】
この問題を解決する別の方法は、I−SceIなどのレア切断を起こすエンドヌクレアーゼによる二本鎖DNA切断の誘導による、標的座位の活性化および/またはドナーDNAの修復による。この技術はAgrobacteriumを用いて植物細胞に修復DNAを配送した場合に、相同組換え頻度を少なくとも二桁増加させることが示された(Puchta et 1996, Proc. Natl. Acad. U.S.A., 93, pp5055-5060; Chilton and Que, Plant Physiol., 2003)。
【0008】
WO96/14408が、酵素I−SceIをコードする単離されたDNAについて記述している。このDNA配列を、クローニングにより、発現ベクター、形質転換細胞系統、および遺伝子組換え動物、に組み入れることができる。このベクターは、遺伝子の遺伝子地図作製および部位特異的挿入に役立つ。
【0009】
WO00/46386が、I−Scel二重鎖切断を通じて遺伝子または細胞の他の染色体DNAを修飾し、修復し、減少させ、不活性化する方法について記述している。さらに、それが必要とされる個体中の遺伝病の治療または予防の方法を開示する。さらに、キメラの制限エンドヌクレアーゼを開示する。
【0010】
しかし、真核細胞のゲノム、特に植物細胞ゲノム中への外来性DNAの標的指向挿入の頻度を改善する必要性がいまだに残存している。これらの問題および他の問題が、以下に本発明の様々の詳細な実施態様ならびに請求項に記述するように、解決される。
【0011】
発明の開示
実施態様の1つにおいて本発明は、対象の外来性DNAであって、あらかじめ選択された部位に隣接するDNA領域と少なくとも80%の配列同一性を有するDNA領域と隣接している可能性のあるDNAを、トウモロコシ細胞のような植物細胞のゲノムの、I−SceI部位などのあらかじめ選択された部位へ導入する方法であって
(a)二本鎖DNA切断を細胞のゲノム中のあらかじめ選ばれた部位に、例えばI−SceIをコードする遺伝子を導入することによって引き起こす工程;
(b)対象の外来性DNAを植物細胞へ導入する工程;
を含み、外来性DNAが、対象の外来性DNAでコートされたマイクロプロジェクタイル撃ち込みにより遂行される直接DNA移入により送達されることを特徴とする方法を提供する。
I−SceIをコードする遺伝子は、配列番号1のアミノ酸配列をコードするヌクレオチド配列を含むことができ、前記ヌクレオチド配列は、約50%〜約60%のGC含量を有し、以下を条件とする。
i) ヌクレオチド配列が、
【表1】
からなる群より選択されるヌクレオチド配列を含まず;
ii) ヌクレオチドが、CCAAT、ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まず;
iii) ヌクレオチド配列が、ATTTA、AAGGT、AGGTA、GGTAまたはGCAGGからなる群より選択される配列を含まず;
iv) ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;
v) ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るATストレッチを含まず;および
vi) ヌクレオチド配列がコドンTTA、CTA、ATA、GTA、TCG、CCG、ACGおよびGCGを含まない。そのようなI−SceIをコードする遺伝子の一例は、配列番号4のヌクレオチド配列を含む。
植物細胞を工程(a)に先立って植物フェノール化合物中でインキュベーションしてもよい。
【0012】
別の実施態様において、本発明は、対象の外来性DNAを植物細胞ゲノムのあらかじめ選択された部位へ導入するための、以下の工程を含む方法に関する:
(a)二本鎖DNA切断を細胞のゲノム中のあらかじめ選択された部位に引き起こす工程;
(b)対象の外来性DNAを植物細胞中へ導入する工程:
を含み、二本鎖DNA切断が、ヌクレオチド配列によってコードされるレア切断エンドヌクレアーゼによって導入され、前記ヌクレオチド配列が、約50%〜約60%のGC含量を有し、
i)ヌクレオチド配列が、
【表2】
からなる群より選択されるヌクレオチド配列を含まず;
ii)ヌクレオチドが、CCAAT、ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まず;
iii)ヌクレオチド配列が、ATTTA、AAGGT、AGGTA、GGTAまたはGCAGGからなる群より選択されるヌクレオチド配列を含まず;
iv)ヌクレオチド配列が、GまたはCからなる群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;
v)ヌクレオチド配列が、AまたはTからなる群より選択される連続する5個のヌクレオチドから成るATストレッチを含まず;および
vi)ヌクレオチド配列が、コドンTTA、CTA、ATA、GTA、TCG、CCG、ACGおよびGCGを含まない、
ことを条件とすることを特徴とする。
【0013】
さらに別の実施態様において、本発明は、対象の外来性DNAを植物細胞のゲノムのあらかじめ選ばれた部位へ導入するための方法であって、
(a)二本鎖DNA切断を細胞のゲノム中のあらかじめ選ばれた部位に引き起こす工程;
(b)対象の外来性DNAを植物細胞へ導入する工程;
を含み、工程(a)に先立って植物細胞を、アセトシリンゴン(3,5−ジメトキシ−4−ヒドロキシアセトフェノン)、α−ヒドロキシ−アセトシリンゴン、シナピン酸(3,5ジメトキシ−4−ヒドロキシケイ皮酸)、シリング酸(4−ヒドロキシ−3,5ジメトキシ安息香酸)、フェルラ酸(4−ヒドロキシ−3−メトキシケイ皮酸)、カテコール(1,2−ジヒドロキシベンゼン)、p−ヒドロキシ安息香酸(4−ヒドロキシ安息香酸)、β−レソルシル酸(2,4ジヒドロキシ安息香酸)、プロトカテク酸(3,4−ジヒドロキシ安息香酸)、ピロ没食子酸(2,3,4−トリヒドロキシ安息香酸)、没食子酸(3,4,5−トリヒドロキシ安息香酸)、およびバニリン(3−メトキシ−4−ヒドロキシベンズアルデヒド)、の群より選択される植物フェノール化合物中でインキュベーションすることを特徴とする方法に関する。
【0014】
本発明は、さらに配列番号1のアミノ酸配列をコードするヌクレオチド配列を含む単離されたDNA断片であって、そのヌクレオチド配列が約50%〜約60%のGC含量を有し、以下を条件とする、DNA断片を提供する:
i)ヌクレオチド配列が、
【表3】
からなる群より選択されるヌクレオチド配列を含まず;
ii)ヌクレオチドが、CCAAT、ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まず;
iii)ヌクレオチド配列が、ATTTA,AAGGT,AGGTA、GGTAまたはGCAGGからなる群より選択される配列を含まず;
iv)ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;
v)ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るATストレッチを含まず;および
vi)前記ヌクレオチド配列の、ロイシン(Leu)、イソロイシン(Ile)、バリン(Val)、セリン(Ser)、プロリン(Pro)、トレオニン(Thr)、アラニン(Ala)をコードするコドンが、前記コドンの位置2および3にTAまたはGCという二連コードを含まない。
【0015】
本発明は、さらに配列番号4のヌクレオチド配列を含む単離されたDNA配列、および植物で発現可能なプロモーターに作動可能に連結された本発明による単離されたDNA断片を含むキメラ遺伝子、ならびに植物ゲノム中のI−SceI認識部位に外来性DNAを挿入するための、そのようなキメラ遺伝子の使用、を提供する。
【0016】
本発明のさらに別の実施態様において、対象の外来性DNAを、植物細胞のゲノムのあらかじめ選ばれた部位へ導入するための方法であって、
a)二本鎖DNA切断を、細胞のゲノム中のあらかじめ選ばれた部位に、レア切断エンドヌクレアーゼにより引き起こす工程;
b)対象の外来性DNAを植物細胞中へ導入する工程、
を含み、前記エンドヌクレアーゼが核移行シグナルを含むことを特徴とする方法を提供する。
【図面の簡単な説明】
【0017】
【図1】相同組換えを介する標的指向DNA挿入のための分析の中で用いられる、標的座位(A)および修復DNA(B)の模式的表示。組換え後の標的座位も表示されている(C)。DSB部位:二本鎖DNA切断部位;3’g7:A.tumefaciens 遺伝子7の転写終了およびポリアデニル化シグナル;neo:植物で発現可能なネオマイシンホスホトランスフェラーゼ;35S:CaMV35S転写のプロモーター;5’bar:ホスフィノトリシンアセチルトランスフェラーゼのアミノ末端部分をコードするDNA領域;3’nos:A. tumefaciensノパリン合成酵素遺伝子の転写終了およびポリアデニル化シグナル;Pnos:A. tumefaciensノパリン合成酵素遺伝子のプロモーター;3’ocs:A. tumefaciens オクトピン合成酵素遺伝子の3’転写終了およびポリアデニル化シグナル。
【0018】
発明の詳細な説明
本発明は次の発見に基づく:
a) 植物細胞中への、直接のDNA移入、特にマイクロプロジェクタイル撃ち込みにより挿入される外来性DNAの導入が、標的指向挿入イベントの頻度を予想外に増加させた。すべての得られた挿入イベントが標的指向DNA挿入イベントであって、二本鎖DNA切断が引き起こされた部位で生じた。さらに、すべてのこれらの標的指向挿入イベントは、二本鎖DNA切断に隣接する与えられた配列相同性の間の正確な組換えイベントであるように見えた。これらのイベントの約半分のみが、二本鎖DNA切断が引き起こされた部位とは異なる部位に、さらに追加の外来性DNA挿入を持っていた。
b)あらかじめ選択された一組の規則によって設計されたI−SceIなどの、レア切断エンドヌクレアーゼ用の合成コード領域を含むキメラ遺伝子によってコードされたI−SceIなどの、レア切断をする二重鎖切断誘導エンドヌクレアーゼの一時的発現による二本鎖DNA切断の誘導が、結果として生じる標的指向DNA挿入イベントの質(すなわち完全に標的指向DNA挿入イベントである頻度を)を驚くほど高めた。更に、エンドヌクレアーゼは核移行シグナルを備えていた。
c)アセトシリンゴンなどの植物フェノール化合物中で標的細胞をプレインキュベーションすることにより、さらに、植物細胞のゲノムに誘導された二本鎖DNA切断における標的指向挿入の頻度が増加した。
【0019】
上記の発見のいずれも、単独でまたは組合せて、相同組換えに基づいた標的指向挿入イベントが得られる頻度、ならびに回収されるイベントの質を改善する。
【0020】
このようにして、ある局面では本発明は、対象の外来性DNAを植物細胞のゲノムのあらかじめ選ばれた部位へ導入する方法であって
(a)二本鎖DNA切断を細胞のゲノム中のあらかじめ選ばれた部位に引き起こす工程;
(b)対象の外来性DNAを植物細胞中へ導入する工程;
を含み、外来性DNAが直接DNA移入により送達されることを特徴とする方法に関する。
【0021】
本明細書に用いられる「直接DNA移入」とは、植物細胞へDNAを導入することができる自然のAgrobacterium種の使用を含まない、植物細胞へのDNA導入の任意の方法である。これには、DNAのエレクトロポレーションによるプロトプラスト中への導入、DNAのエレクトロポレーションによるインタクトな植物細胞中への、または部分的に分解された組織または植物細胞への導入、DNAのPEGその他などの薬剤の作用によるプロトプラストへの導入、および特にDNAをコートしたマイクロプロジェクタイル撃ち込みなどの、従来技術において周知の方法が含まれる。DNAの直接移入による植物細胞への導入は、Agrobacteriumが媒介するDNA導入とは、少なくとも二本鎖DNAが植物細胞に入るという点で、入るDNAはいかなるタンパク質によってもコートされていないという点で、また植物細胞に入るDNAの量が相当に、より大きい可能性があるという点で異なる。さらに、二本鎖DNA切断誘導エンドヌクレアーゼをコードする導入されたキメラ遺伝子などの、直接移入法によって導入されたDNAは、転写により適していて、二本鎖DNA切断を誘導するタイミングがより良くなる可能性がある。本発明を特定の作用機序に制限するつもりはないが、効率的な相同性組換えに基づいた修復DNAまたは外来性DNAの植物細胞のゲノムへの挿入は、これらのパラメーターのうちのいずれかの組合せによるものと考えられる。
【0022】
都合よく、二本鎖DNA切断は、あらかじめ選ばれた部位で、レア切断を起こす二重鎖DNA切断導入酵素をコードする植物で発現可能な遺伝子の導入の後の一時的発現により引き起こすことができる。この文書の他の場所で述べるように、I−SceIを、I−SceI認識部位に外来性DNAを導入する目的で用いてもよい。しかし、他の二重鎖切断導入酵素もまた、外来性DNAをそれぞれの認識部位に挿入するために用いることができることは、当業者には直ちに明らかであろう。レア切断を起こすDSB誘導酵素およびそれぞれの認識部位のリストがWO 03/004659(17〜20ページ)(参照により本明細書に組み入れられる)の表Iに、提供されている。更に、選択した原則として任意の標的ヌクレオチド配列を認識する特別仕立てのレア切断を起こすエンドヌクレアーゼを設計するための方法を利用することができる。そのような方法については、例えばWO 03/080809、WO94/18313またはW095/09233に、およびIsalan et al., 2001, Nature Biotechnology 19,656-660; Liu et al. Proc. Natl. Acad. Sci. USA 94, 5525-5530に記載されている。
【0023】
したがって、本明細書で用いられる「あらかじめ選択された部位」とは、その座位に外来性DNAを挿入することが望まれる、植物の核ゲノム中の特定のヌクレオチド配列を表す。当業者は完全に、選択された標的ヌクレオチド配列を認識する二本鎖DNA切断誘導(「DSBI」)酵素を選ぶことができるか、またはそのようなDSBIエンドヌクレアーゼをエンジニアリングすることができるであろう。あるいは、任意の従来の形質転換法を用いて、またはゲノムにDSBIエンドヌクレアーゼ認識部位を有する植物系統を用いた従来の品種改良によって、植物ゲノムへDSBIエンドヌクレアーゼ認識部位を導入してもよく、また任意の所望の外来性DNAを後で、以前に導入されたあらかじめ選択された標的部位に導入してもよい。
【0024】
二重鎖切断誘導酵素をコードするDNA領域に作動可能に連結された植物発現可能プロモーター領域を含む植物発現可能なキメラ遺伝子を一過性に導入することにより、都合よく二本鎖DNA切断を引き起こすことができる。二重鎖切断誘導酵素をコードするDNA領域は合成DNA領域、例えば、コドンを本出願に別記するI−SceIコード領域の設計案に従って選択する合成DNA領域など、でもよい、しかしそれに限られない。
【0025】
二重鎖切断誘導酵素は、核移行シグナル(NLS)[Raikhel, Plant Physiol. 100: 1627-1632 (1992) およびその中の参照文献]、例えばSV40ラージT抗原のNLS[Kalderon et al., Cell 39: 499-509 (1984)]など、を含んでもよい(しかし含む必要はない)。核移行シグナルはタンパク質の任意の場所に位置してよいが、タンパク質のN末端に置くのが好都合である。核移行シグナルが、二重鎖切断誘導酵素のアミノ酸の1つ以上と置き換わってもよい。
【0026】
本明細書で用いる「対象の外来性DNA」とは、あらかじめ選択された部位への導入が望まれる任意のDNA断片を表す。厳密に必要とするわけではないが、対象の外来性DNAは、あらかじめ選択された部位に隣接するDNA領域に対して相同性を有する少なくとも1つのヌクレオチド配列領域と隣接してもよい。対象の外来性DNAは、あらかじめ選択された部位に隣接する両方のDNA領域に相同性を有するDNA領域により部位の両側を挟まれてもよい。したがって、植物細胞へ導入される修復DNA分子は、それぞれあらかじめ選択された部位の上流にまたは下流のDNA領域に対して相同性を有する1つまたは2つのフランキング配列によって隣接された外来性DNAを含んでもよい。これによって、外来性DNAの挿入をよりよく制御することが可能になる。確かに、相同組換えによる組込みが、外来性DNA断片を植物の核ゲノムへヌクレオチドレベルまで正確に接続することを可能にすることになる。
【0027】
隣接するヌクレオチド配列は、種々な長さでよいが、長さで少なくとも約10個のヌクレオチドでなければならい。しかしながら、フランキング領域は実際に可能な程度まで(例えば、完全なバクテリア人工染色体(BACs)のように約100〜150kbまで)長くてもよい。好ましくは、フランキング領域は約50bp〜約2000bpになるであろう。さらに、対象の外来性DNAに隣接する領域は、あらかじめ選択された部位に隣接するDNA領域と同一である必要はない。そして、あらかじめ選択された部位に隣接するDNA領域に、約80%〜約100%の配列同一性を、好ましくは約95%〜約100%の配列同一性を有していればよい。フランキング領域が長いほど、相同性の要求は、より厳格でない。さらに配列同一性は、外来性DNAの正確な挿入の位置の近くにおいて実際に可能な限度まで高いことが好ましい。
【0028】
さらに、対象の外来性DNAに隣接する領域は、あらかじめ選択された部位に直接隣接する領域に対して相同性を有する必要はなく、あらかじめ選択された部位からさらに遠い核ゲノムのDNA領域へ相同性があればよい。外来性DNAの挿入によって次に、あらかじめ選択された挿入部位と相同DNA領域の間の標的DNAが除去される結果になるであろう。言いかえれば、相同性領域間に位置した標的DNAは、対象の外来性DNAに置換されることになる。
【0029】
本発明の目的のためには、パーセンテージで表した2つの関連するヌクレオチドまたはアミノ酸配列の「配列同一性」とは、2つの最も良くアラインメントされた配列の、同一の残基を有する位置の数(×100)を比較した位置数で割ったものを指す。ギャップ、即ち残基が1つの配列中にはあり他方にはないアラインメント中の位置は、同一でない残基の位置と見なす。2つの配列のアラインメントは、ニードルマンとブンシュのアルゴリズム(Needleman and Wunsch 1970)によって、行なう。コンピューター支援配列アラインメントを Wisconsin Package 第10.1版(Genetics Computer Group, Madison, Wisconsin, USA)の一部であるGAPなどの標準ソフトウェアプログラムを用い、ギャップ生成ペナルティーを50、およびギャップ伸展ペナルティーを3とするデフォルトの得点マトリックスを用いて、便利に行なうことができる。
【0030】
別の局面では、本発明が、修飾されたI−SceIをコードするDNA断片、およびそれの、対象の外来性DNAを効率的に植物細胞ゲノムのあらかじめ選択された部位へ導入するための使用、に関する。この場合、修飾されたI−SceIをコードするDNA断片は、次の基準を満たすように設計されたヌクレオチド配列を有する:
a) ヌクレオチド配列が、配列番号1で与えられるアミノ酸配列を有するI−SceIエンドヌクレアーゼなどの、機能的なI−SceIエンドヌクレアーゼをコードする;
b)ヌクレオチド配列が、約50%〜約60%のGC含量を有する;
c)ヌクレオチド配列が、
【表4】
からなる群より選択されるヌクレオチド配列を含まない;
d)ヌクレオチドが、CCAAT、ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まない;
e)ヌクレオチド配列が、ATTTA、AAGGT、AGGTA、GGTAまたはGCAGGからなる群より選択されるヌクレオチド配列を含まない;
f)ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まない;
g)ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るGCストレッチを含まない;および
h)ヌクレオチド配列が、位置2および3にTAまたはCGという二連コードを含むLeu、Ile、Val、Ser、Pro、Thr、Alaをコードするコドン、を含まない(即ち、ヌクレオチド配列がコドンTTA、CTA、ATA、GTA、TCG、CCG、ACGおよびGCGを含まない)。
【0031】
I−SceIは、Saccharomyces cerevisea のミトコンドリア中のイントロン移動の原因となっている部位特異的なエンドヌクレアーゼである。酵素は、21S rRNA遺伝子のオプショナルイントロンScLSU.1によってコードされ、イントロン挿入部位で二本鎖DNA切断を開始し、3’OH張出しを有する4bpの互い違いの切断を生成する。I−SceIエンドヌクレアーゼの認識部位は、18bpの非対称配列上に伸展している(Colleaux et al. 1988 Proc. Natl. Acad. Sci. USA 85:: 6022-6026)。I−SceIのアミノ酸配列およびミトコンドリアのI−SceI遺伝子のユニバーサルコード等価物が、例えばWO 96/14408によって提供されている。
【0032】
WO96/14408は、I−SceIタンパク質の次の変異体がまだ機能的であることを開示している:
●位置1〜10を欠失させることができる。
●位置36:Gly(G)を許容する。
●位置40:Met(M)またはVal(V)を許容する。
●位置41:Ser(S)またはAsn(N)を許容する。
●位置43:Ala(A)を許容する。
●位置46:Val(V)またはN(Asn)を許容する。
●位置91:Ala(A)を許容する。
●位置123および156:Leu(L)を許容する。
●位置223:Ala(A)およびSer(S)を許容する。
また、そのような変異体I−SceI酵素をコードする合成ヌクレオチド配列を本発明に従って設計し用いることができる。
【0033】
アミノ末端に位置する4つのアミノ酸が核移行シグナルによって置換されたI−SceIのアミノ酸配列(配列番号1)をコードするヌクレオチド配列は、したがって、R1〜R244で表わすことのできる244のトリヌクレオチドから成る。これらの各位置に対して、同一のアミノ酸をコードするトリヌクレオチドの1〜6の可能な選択枝を選ぶことができる。表1は、配列番号1のアミノ酸配列をコードするトリヌクレオチドに対して可能な選択枝を記載し、また上に述べた「禁制ヌクレオチド配列」が合成DNA配列へ取り込まれるのを回避することを可能にする、構造上の必要条件(条件付きの、または絶対的な)を提供する。さらに、UIPACコードによる隣接するトリヌクレオチドのヌクレオチド配列を提供する。
【0034】
本明細書で用いられるUIPACコードの記号は、それらの通常の意味を有している、即ち:N=AまたはCまたはGまたはT;R=AまたはG;Y=CまたはT;B=CまたはGまたはT(Aではない);V=AまたはCまたはG(Tではない);D=AまたはGまたはT(Cではない);H=AまたはCまたはT(Gではない);K=GまたはT;M=AまたはC;S=GまたはC;W=AまたはT。
【0035】
したがって本発明の実施態様の1つでは、配列番号2に記載されているヌクレオチド配列を含む単離された合成DNA断片を提供する。この場合、コドンを、与えられた選択枝の中から約50%〜約60%の、好ましくは約54%〜55%の全GC含量を有するヌクレオチド配列を得るように選ぶ。ただし以下を条件とする:位置28〜位置30のヌクレオチド配列がAAGでなく;もし位置34〜位置36のヌクレオチド配列がAATである場合は、位置37〜位置39のヌクレオチド配列がATTまたはATAでなく;もし位置34〜位置36のヌクレオチド配列がAACである場合は、位置37〜位置39のヌクレオチド配列がATTでなく、同時に位置40〜位置42のヌクレオチド配列がAAAであり;もし位置34〜位置36のヌクレオチド配列がAACである場合は、位置37〜位置39のヌクレオチド配列がATAでなく;もし位置37〜位置39のヌクレオチド配列がATTまたはATAである場合は、位置40〜位置42のヌクレオチド配列がAAAでなく;位置49〜位置51のヌクレオチド配列がCAAでなく;位置52〜位置54のヌクレオチド配列がGTAでなく;位置58〜位置63のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置67〜位置69のヌクレオチド配列がCCCである場合は、位置70〜位置72のヌクレオチド配列がAATでなく;もし位置76〜位置78のヌクレオチド配列がAAAである場合は、位置79〜位置81のヌクレオチド配列がTTGでなく、同時に位置82〜位置84のヌクレオチド配列がCTNであり;もし位置79〜位置81のヌクレオチド配列がTTAまたはCTAである場合は、位置82〜位置84のヌクレオチド配列がTTAでなく;位置88〜位置90のヌクレオチド配列がGAAではなく;もし位置91〜位置93のヌクレオチド配列がTATである場合は、位置94〜位置96のヌクレオチド配列がAAAでなく;もし位置97〜位置99のヌクレオチド配列がTCCまたはTCGまたはAGCである場合は、位置100〜位置102のヌクレオチド配列がCCAでなく、同時に位置103〜位置105のヌクレオチド配列がTTRであり;もし位置100〜位置102のヌクレオチド配列がCAAである場合は、位置103〜位置105のヌクレオチド配列がTTAでなく;もし位置109〜位置111のヌクレオチド配列がGAAである場合は、位置112〜位置114のヌクレオチド配列がTTAでなく;もし位置115〜位置117のヌクレオチド配列がAATである場合は、位置118〜位置120のヌクレオチド配列がATTまたはATAでなく;位置121〜123のヌクレオチド配列がGAGである場合、位置124〜位置126のヌクレオチド配列は;位置133〜135のヌクレオチド配列がGCAでなく;位置139〜位置141のヌクレオチド配列がATTでなく;もし位置142〜位置144のヌクレオチド配列がGGAである場合は、位置145〜位置147のヌクレオチド配列がTTAでなく;もし位置145〜位置147のヌクレオチド配列がTTAである場合は、位置148〜位置150のヌクレオチド配列がATAでなく、同時に位置151〜位置153のヌクレオチド配列がTTRであり;もし位置145〜位置147のヌクレオチド配列がCTAである場合は、位置148〜位置150のヌクレオチド配列がATAでなく、同時に位置151〜位置153のヌクレオチド配列がTTRであり;もし位置148〜位置150のヌクレオチド配列がATAである場合は、位置151〜位置153のヌクレオチド配列がCTAまたはTTGでなく;もし位置160〜位置162のヌクレオチド配列がGCAである場合は、位置163〜位置165のヌクレオチド配列がTACでなく;もし位置163〜位置165のヌクレオチド配列がTATである場合は、位置166〜位置168のヌクレオチド配列がATAでなく、同時に位置169〜位置171のヌクレオチド配列がAGRであり;位置172〜位置177のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がGCAGGを含まないようになる選択に従って選ばれ;位置178〜位置186のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がAGGTAを含まないようになる選択に従って選ばれ;もし位置193〜位置195のヌクレオチド配列がTATである場合は、位置196〜位置198のヌクレオチド配列がTGCでなく;位置202〜位置204のヌクレオチド配列がCAAでなく;位置217〜位置219のヌクレオチド配列がAATでなく;もし位置220〜位置222のヌクレオチド配列がAAAである場合は、位置223〜位置225のヌクレオチド配列がGCAでなく;もし位置223〜位置225のヌクレオチド配列がGCAである場合は、位置226〜位置228のヌクレオチド配列がTACでなく;もし位置253〜位置255のヌクレオチド配列がGACである場合は、位置256〜位置258のヌクレオチド配列がCAAでなく;もし位置277〜位置279のヌクレオチド配列がCATである場合は、位置280〜位置282のヌクレオチド配列がAAAでなく;位置298〜位置303のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置304〜位置306のヌクレオチド配列がGGCである場合は、位置307〜位置309のヌクレオチド配列がAATでなく;位置307〜位置312のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;位置334〜位置342のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置340〜位置342のヌクレオチド配列がAAGである場合は、位置343〜位置345のヌクレオチド配列がCATでなく;もし位置346〜位置348のヌクレオチド配列がCAAである場合は、位置349〜位置351のヌクレオチド配列がGCAでなく;位置349〜位置357のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;位置355〜位置357のヌクレオチド配列がAATでなく;
もし位置358〜位置360のヌクレオチド配列がAAAである場合は、位置361〜位置363のヌクレオチド配列がTTGでなく;もし位置364〜位置366のヌクレオチド配列がGCCである場合は、位置367〜位置369のヌクレオチド配列がAATでなく;位置367〜位置378のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置382〜位置384のヌクレオチド配列がAATである場合は、位置385〜位置387のヌクレオチド配列がAATでなく;位置385〜位置387のヌクレオチド配列がAATでなく;もし位置400〜位置402のヌクレオチド配列がCCCである場合は、位置403〜位置405のヌクレオチド配列がAATでなく;もし位置403〜位置405のヌクレオチド配列がAATである場合は、位置406〜位置408のヌクレオチド配列がAATでなく;位置406〜位置411のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;位置421〜位置426のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;位置430〜位置432のヌクレオチド配列がCCAでなく;もし位置436〜位置438のヌクレオチド配列がTCAである場合は、位置439〜位置441のヌクレオチド配列がTTGでなく;位置445〜位置447のヌクレオチド配列がTATでなく;位置481〜483のヌクレオチド配列がAATでなく;もし位置484〜位置486のヌクレオチド配列がAAAである場合は、位置487〜位置489のヌクレオチド配列がAATでなく、同時に位置490〜位置492のヌクレオチド配列がAGYであり;もし位置490〜位置492のヌクレオチド配列がTCAである場合は、位置493〜位置495のヌクレオチド配列がACCでなく、同時に位置496〜位置498のヌクレオチド配列がAAYであり;もし位置493〜位置495のヌクレオチド配列がACCである場合は、位置496〜位置498のヌクレオチド配列がAATでなく;位置496〜位置498のヌクレオチド配列がAATでなく;もし位置499〜位置501のヌクレオチド配列がAAAである場合は、位置502〜位置504のヌクレオチド配列がTCAまたはAGCでなく;もし位置508〜位置510のヌクレオチド配列がGTAである場合は、位置511〜位置513のヌクレオチド配列がTTAでなく;もし位置514〜位置516のヌクレオチド配列がAATである場合は、位置517〜位置519のヌクレオチド配列がACAでなく;もし位置517〜位置519のヌクレオチド配列がACCまたはACGである場合は、位置520〜位置522のヌクレオチド配列がCAAでなく、同時に位置523〜位置525のヌクレオチド配列がTCNであり;位置523〜位置531のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置544〜位置546のヌクレオチド配列がGAAである場合は、位置547〜位置549のヌクレオチド配列がTATでなく、同時に位置550〜位置552のヌクレオチド配列がTTRであり;位置547〜位置552のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置559〜位置561のヌクレオチド配列がGGAである場合は、位置562〜位置564のヌクレオチド配列がTTGでなく、同時に位置565〜位置567のヌクレオチド配列がCGNであり;もし位置565〜位置567のヌクレオチド配列がCGCである場合は、位置568〜位置570のヌクレオチド配列がAATでなく;位置568〜位置570のヌクレオチド配列がAATでなく;もし位置574〜位置576のヌクレオチド配列がTTCである場合は、位置577〜位置579のヌクレオチド配列がCAAでなく、同時に位置580〜位置582のヌクレオチド配列がTTRであり;もし位置577〜位置579のヌクレオチド配列がCAAである場合は、位置580〜位置582のヌクレオチド配列がTTAでなく;
もし位置583〜位置585のヌクレオチド配列がAATである場合は、位置586〜位置588のヌクレオチド配列がTGCでなく;位置595〜位置597のヌクレオチド配列がAAAでなく;もし位置598〜位置600のヌクレオチド配列がATTである場合は、位置601〜位置603のヌクレオチド配列がAATでなく;位置598〜位置600のヌクレオチド配列がATAでなく;位置601〜位置603のヌクレオチド配列がAATでなく;もし位置604〜位置606のヌクレオチド配列がAAAである場合は、位置607〜位置609のヌクレオチド配列がAATでなく;位置607〜位置609のヌクレオチド配列がAATでなく;位置613〜位置615のヌクレオチド配列がCCAでなく;もし位置613〜位置615のヌクレオチド配列がCCGである場合は、位置616〜位置618のヌクレオチド配列がATAでなく;もし位置616〜位置618のヌクレオチド配列がATAである場合は、位置619〜位置621のヌクレオチド配列がATAでなく;もし位置619〜位置621のヌクレオチド配列がATAである場合は、位置622〜位置624のヌクレオチド配列がTACでなく;位置619〜位置621のヌクレオチド配列がATTでなく;位置640〜位置645のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置643〜位置645のヌクレオチド配列がTTAである場合は、位置646〜位置648のヌクレオチド配列がATAでなく;もし位置643〜位置645のヌクレオチド配列がCTAである場合は、位置646〜位置648のヌクレオチド配列がATAでなく;位置655〜位置660のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置658〜位置660のヌクレオチド配列がTTAまたはCTAである場合は、位置661〜位置663のヌクレオチド配列がATTまたはATCでなく;位置661〜位置663のヌクレオチド配列がATAでなく;もし位置661〜位置663のヌクレオチド配列がATTである場合は、位置664〜位置666のヌクレオチド配列がAAAでなく;位置670〜位置675のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;もし位置691〜位置693のヌクレオチド配列がTATである場合は、位置694〜位置696のヌクレオチド配列がAAAでなく;もし位置694〜位置696のヌクレオチド配列がAAAである場合は、位置697〜位置699のヌクレオチド配列がTTGでなく;もし位置700〜位置702のヌクレオチド配列がCCCである場合は、位置703〜位置705のヌクレオチド配列がAATでなく;もし位置703〜位置705のヌクレオチド配列がAATである場合は、位置706〜位置708のヌクレオチド配列がACAまたはACTでなく;もし位置706〜位置708のヌクレオチド配列がACAである場合は、位置709〜位置711のヌクレオチド配列がATAでなく、同時に位置712〜位置714のヌクレオチド配列がAGYであり;ヌクレオチド配列がコドンTTA、CTA、ATA、GTA、TCG、CCG、ACG、およびGCGを含まず;前記ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;および前記ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るATストレッチを含まない。
【0036】
合成ヌクレオチド配列の好ましい群を表2に記載する。そのヌクレオチド配列は配列番号3に記載されているヌクレオチド配列を含むように与えられる単離した合成DNA断片に対応する。この場合、コドンは、約50%〜約60%の、好ましくは約54%〜55%の全体GC含量を有するヌクレオチド配列を得る、かつ以下の条件を満たす選択肢の中から選択される:もし位置121〜位置123のヌクレオチド配列がGAGである場合は、位置124〜126のヌクレオチド配列がCAAでなく;もし位置253〜位置255のヌクレオチド配列がGACである場合は、位置256〜258のヌクレオチド配列がCAAでなく;もし位置277〜位置279のヌクレオチド配列がCATである場合は、位置280〜282のヌクレオチド配列がAAAでなく;もし位置340〜位置342のヌクレオチド配列がAAGである場合は、位置343〜345のヌクレオチド配列がCATでなく;もし位置490〜位置492のヌクレオチド配列がTCAである場合は、位置493〜495のヌクレオチド配列がACCでなく;もし位置499〜位置501のヌクレオチド配列がAAAである場合は、位置502〜504のヌクレオチド配列がTCAまたはAGCでなく;位置517〜位置519のヌクレオチド配列がACCである場合は、位置520〜522のヌクレオチド配列がCAAでなく、同時に位置523〜525のヌクレオチド配列がTCNであり;もし位置661〜位置663のヌクレオチド配列がATTである場合は、位置664〜666のヌクレオチド配列がAAAでなく;位置7〜位置15のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置61〜位置69のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置130〜位置138のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置268〜位置279のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置322〜位置333のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置460〜位置468のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置13〜位置27のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置37〜位置48のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置184〜位置192のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置214〜位置219のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置277〜位置285のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置388〜位置396のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置466〜位置474のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置484〜位置489のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置571〜位置576のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置598〜位置603のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置604〜位置609のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置613〜位置621のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置646〜位置651のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;位置661〜位置666のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;および位置706〜位置714のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選択される。
【0037】
配列番号4のヌクレオチド配列は、もはや回避するべきヌクレオチド配列またはコドンのどれも含んでいないI−SceIエンドヌクレアーゼをコードする合成ヌクレオチド配列の例である。しかしながら、当業者が、選択されるヌクレオチドの1つ以上を(2〜20を)配列番号3のヌクレオチド配列の中で提供される選択肢のいずれかに置換(前の段落に記述した禁止組合せのいずれも除外して)することにより、I−SceIをコードする同様の配列を容易に得ることができ、かつ使用して同様の効果を得ることができることは明らかであろう。
【0038】
植物細胞中の発現のために、I−SceIをコードする合成DNA断片を、植物で発現できるキメラ遺伝子を得るために、植物発現可能なプロモーターに作動可能に連結させてもよい。
【0039】
当業者は、本発明のこの局面については、修復DNAおよび/または、DSBIエンドヌクレアーゼをコードするDNAを直接のDNA移入法によって植物細胞へ導入することは必要でなく、従来技術において利用可能なAgrobacteriumを媒介とする形質転換法によって、DNAを植物細胞へこのように導入できることを直ちに認識するであろう。
【0040】
さらに別の局面では本発明は、対象の外来性DNAを植物細胞のゲノムのあらかじめ選択された部位へ導入する方法であって、
(a)二重鎖切断を細胞のゲノム中のあらかじめ選ばれた部位に引き起こす工程;および
(b)対象の外来性DNAを植物細胞へ導入する工程;
を含み、工程(a)に先立って植物細胞を植物フェノール化合物中でインキュベーションすることを特徴とする方法に関する。
【0041】
本発明に好適な「植物フェノール化合物」あるいは、「植物フェノール類」とは、正の走化性応答を引き起こすことができる置換フェノール分子、特にTiプラスミドを含むAgrobacterium種中で、特にTiプラスミドを含むAgrobacterium tumefaciens中で、vir遺伝子発現の増加を引き起こすことのできる分子である。植物フェノール化合物への走化性応答を測定する方法については、Ashby et al. (1988 J. Bacteriol. 170: 4181-4187)に記述されており、またvir遺伝子発現の誘導を測定する方法も周知である(Stachel et al., 1985 Nature 318: 624-629; Bolton et al.1986 Science 232: 983-98)。好ましい植物フェノール化合物は植物細胞の創傷滲出液で見出されるものである。最もよく知られている植物フェノール化合物の1つがアセトシリンゴンであり、それは様々な植物の多数の傷ついた細胞または完全な細胞に、様々な濃度とはいえ、存在する。しかしながら、アセトシリンゴン(3,5−ジメトキシ−4−ヒドロキシアセトフェノン)が、vir遺伝子の発現を引き起こすことができるただ一つの植物フェノールではない。他の例は、α−ヒドロキシ−アセトシリンゴン、シナピン酸(3,5ジメトキシ−4−ヒドロキシケイ皮酸)、シリング酸(4−ヒドロキシ−3,5ジメトキシ安息香酸)、フェルラ酸(4−ヒドロキシ−3−メトキシケイ皮酸)、カテコール(1,2−ジヒドロキシベンゼン)、p−ヒドロキシ安息香酸酸(4−ヒドロキシ安息香酸酸)、β−レソルシル酸(2,4ジヒドロキシ安息香酸)、プロトカテク酸(3,4−ジヒドロキシ安息香酸)、ピロ没食子酸(2,3,4−トリヒドロキシ安息香酸)、没食子酸(3,4,5−トリヒドロキシ安息香酸)、およびバニリン(3−メトキシ−4−ヒドロキシベンズアルデヒド)である。本明細書で用いられ、言及された分子を植物フェノール化合物と呼ぶ。植物フェノール化合物を、植物栽培培地に単独でまたは他の植物フェノール化合物と組み合わせて加えることができる。本発明を特定の作用機序に制限する意図はないが、これらの植物フェノール類の細胞分裂に対する(したがってゲノム複製に対しても)明らかな促進効果は、外来性DNAの標的指向挿入を促進している可能性があると思われる。
【0042】
植物細胞を、植物フェノール化合物中でまたはその上で、約1日または2日間インキュベーションすることで十分であろうと予想されるが、好ましくは約1週間植物フェノール化合物中でインキュベーションする。植物細胞を細胞分裂を促進するために十分な時間だけインキュベーションするべきである。Guivarc'h et al., (1993, Protoplasma 174: 10-18)によれば、植物細胞のほんの10分間ほどのインキュベーションによって、既にそのような効力を得ることができる。
【0043】
上述の相同組換えに基づく標的指向DNA挿入のための改善された方法は、直接DNA移入法により、特にマイクロプロジェクタイル撃ち込みにより得られる遺伝子組換え植物細胞および植物の、質を改善するためにも適用できる。マイクロプロジェクタイル撃ち込みによるDNAの導入により、しばしば導入されるDNAの複雑な組込みパターン(完全なまたは部分的な対象外来性DNAの複数のコピーの組込み、繰り返し構造の生成)がもたらされることは、従来技術において周知である。それにもかかわらず、いくつかの植物の遺伝子型または変種は、マイクロプロジェクタイル撃ち込みを用いる形質転換の方が、例えば Agrobacterium tumefaciens を使用する形質転換よりも、適している可能性がある。マイクロプロジェクタイル撃ち込みを通じて得られる遺伝子組換え植物細胞または植物の、質を改善することができれば、即ち外来性DNA組込みのパターンがより単純になるような影響を受ければ、有利であろう。
【0044】
上に述べた発見、即ち核ゲノム中に誘導された二本鎖DNA切断の存在下でマイクロプロジェクタイル撃ち込みにより外来性DNAを導入し、その際、外来性DNAは二本鎖DNA切断に隣接する配列に対して相同性を有している場合は、しばしば(得られるイベントの約50%が)単純な組込みパターン(予測可能な方式による単一コピーの挿入があり、外来性DNAの追加の断片の挿入がない)となる結果をもたらす、という発見は、植物細胞の核ゲノム中の外来性DNAの挿入の複雑さを単純化する方法の基礎を与える。
【0045】
したがって、本発明はまた、次の工程を含むマイクロプロジェクタイル撃ち込みによって遺伝子組換え植物を生成する方法に関する:
(a)本明細書に別記する方法または従来技術において利用可能な方法に従って、二本鎖DNA切断を、植物細胞のゲノム中のあらかじめ選択された部位に引き起こす工程;および
(b)対象の外来性DNAをマイクロプロジェクタイル撃ち込みによって植物細胞へ導入する工程(ここで前記対象の外来性DNAは、植物ゲノム中のあらかじめ選択された部位に隣接するDNA領域へ少なくとも80%の配列同一性を有する2つのDNA領域に挟まれている)。
このように得られる遺伝子組換え植物の個体数の多くの部分が、植物細胞のゲノム中に外来性DNAの単純な組込みパターンを有し、より詳しくいえば、遺伝子組換え植物のかなりの部分は、1コピーの外来性DNA挿入のみを、植物ゲノム中のあらかじめ選択された部位に隣接する2つのDNA領域の間に正確に有することになる。この部分は、二本鎖DNA切断を引き起こすことなしに、および外来性DNAにあらかじめ選択された部位に隣接するゲノム領域との相同性を与えることなしに、単純なマイクロプロジェクタイル撃ち込みによって植物が得られる場合における単純な組込みパターンを有する遺伝子組換え植物の個体数よりも高い。
【0046】
本発明の好都合の実施態様では、標的となる植物細胞が、そのゲノム中に、レア切断を起こす二本鎖DNA切断誘起エンドヌクレアーゼの2つの認識部位が各側に1つずつ隣接するマーカー遺伝子を含む。このマーカーDNAは、対象の植物細胞のゲノムに、形質転換の任意の方法を用いて導入してもよいし、または既に別の植物系統または変異種(形質転換が容易な植物系統または変異種など)の植物細胞のゲノムへ導入されていて、それを古典的育種技術により対象の植物細胞へ導入してもよい。好ましくは、レア切断を起こす二重鎖切断誘導エンドヌクレアーゼの2つの認識部位を両側面に有するマーカー遺伝子を含む遺伝子組換え植物または植物細胞の集団は、マ−カー遺伝子の発現パターン(高い発現、時間的または空間的に調節された発現など)についてすでに分析されており、望みの発現パターンを有する植物系統が同定されている。
(a)本明細書に別記する方法または従来技術において利用可能な方法に従って、植物細胞のゲノム中のあらかじめ選ばれた部位に二本鎖DNA切断を引き起こす工程;および
(b)対象の外来性DNAをマイクロプロジェクタイル撃ち込みによって植物細胞へ導入する工程(ここで前記対象の外来性DNAは、植物ゲノム中のあらかじめ選ばれた部位に隣接するDNA領域と少なくとも80%の配列同一性を有する2つのDNA領域に挟まれている)、
を含むマイクロプロジェクタイル撃ち込みによる遺伝子組換え植物の産生が、マーカー遺伝子が対象の外来性DNAによって置換された遺伝子組換え植物細胞および植物をもたらすことになる。
【0047】
マーカー遺伝子は、選択可能なまたはスクリーニング可能な植物で発現可能なマーカー遺伝子でよく、それには従来のキメラのマーカー遺伝子が好適である。キメラのマーカー遺伝子は、プロモーター、好ましくはCaMV35Sプロモーターなどの植物で発現可能な構成的プロモーター、またはルビスコの小サブユニットをコードする遺伝子のプロモーターなどの光誘導可能なプロモーターの制御下にある、またはそのプロモーターの5’末端に動作可能なように連結され;またその3’末端で、適当な植物転写終了およびポリアデニル化シグナルに動作可能なように連結された、マーカーDNAを含むことができる。マーカーDNAは、植物の細胞中で発現された時、マーカーDNAを発現していない細胞から容易にそのような細胞を分離することを可能にするRNA、タンパク質またはポリペプチドをコードすることが好ましい。マーカーDNAの選択は決定的に重要ではなく、周知の方式で任意の適当なマーカーDNAを選択することができる。例えば、マーカーDNAは、A1遺伝子(Meyer et al., (1987), Nature 330:677)のように形質転換された植物細胞に識別可能な色彩を与えるタンパク質をコードすることができ、蛍光タンパク質[Chalfie et al., Science 263: 802-805 (1994); Crameri et al., Nature Biotechnology 14:315-319 (1996)]をコードすることができ、ホスフィノトリシンに対する耐性を与えるPATをコードするbar遺伝子(EP 0242246)のように、形質転換された植物細胞へ除草剤耐性を与えるタンパク質をコードすることができ、またはゲンタマイシンに対する耐性を与えるGATをコードするaac(6’)遺伝子(WO94/01560)のように、形質転換細胞に抗生物質耐性を与えるタンパク質をコードすることができる。そのような選択可能なマーカー遺伝子は一般に、細胞には普通有毒な抗生物質または他の化合物に対する細胞耐性を与えるタンパク質をコードする。植物では、選択可能なマーカー遺伝子がこのように、さらに有効成分としてグルタミン合成酵素阻害剤(例えば、ホスフィノトリシン)を含む除草剤などの、除草剤に対する耐性を与えるタンパク質をコードしてもよい。そのような遺伝子の例は、sfrまたはsfrv遺伝子(EP 242236; EP 242246; De Block et al., 1987 EMBO J. 6: 2513-2518)などの、ホスフィノトリシンアセチルトランスフェラーゼをコードする遺伝子である。
【0048】
導入された修復DNAはさらに、あらかじめ選ばれた部位での相同組換えによる組込みと、ゲノムの他の場所への組込みとを識別することを可能にするマーカー遺伝子を含んでもよい。そのようなマーカー遺伝子が従来技術において利用可能であり、それには、選択的な条件下でマーカー遺伝子の不存在を積極的に選択できるマーカー遺伝子(例えば、codA、5−フルオロシトシンに対する感受性を与える E. coli から得られるシトシンデアミナーゼ、Perera et al., 1993 Plant Mol. Biol. 23, 793; Stougaard (1993) Plant J: 755)が含まれる。修復DNAは、核ゲノム中へ修復DNAがランダムに組込まれると、マーカー遺伝子が存在する結果となり、一方で相同組換えによって修復DNAが組込まれると、マーカー遺伝子が存在しない結果となるような方式で、マーカー遺伝子を含む必要がある。
【0049】
マーカー遺伝子中またはその近くに位置する二重鎖切断を引き起こすただ1つのあらかじめ選択された部位を用いて、同じ結果を得ることができることは直ちに明らかであろう。その場合、相同性のフランキング領域を、マーカー遺伝子を不活性化するかまたはマーカー遺伝子を欠失させて、挿入される外来性DNAによって置き換えるような方式で選択することが好ましい。
【0050】
本発明の手段および方法は、特にトウモロコシに役立つが、他の植物に、特に小麦、オート麦、大麦、ライ麦、米、ターフグラス、ソルガム、アワ類またはサトウキビ植物を含む穀類植物に用いてもよく、同様の効果が得られることを認識することになろう。本発明の方法を、綿、タバコ、キャノーラ、アブラナ、ダイズ、蔬菜類、ジャガイモ、ウキクサ種、ハナタバコ種、アラビドプシス、アルファルファ、オオムギ、豆、トウモロコシ、綿、アマ、エンドウ、アブラナ、コメ、ライムギ、ベニバナ、モロコシ、ダイズ、ヒマワリ、タバコ、小麦、アスパラガス、ビート、ブロッコリ、キャベツ、ニンジン、カリフラワー、セロリ、キュウリ、ナス、レタス、タマネギ、アブラナ、コショウ、ジャガイモ、カボチャ、ハツカダイコン、ホウレンソウ、セイヨウカボチャ、トマト、ズッキーニ、アーモンド、リンゴ、アンズ、バナナ、クロイチゴ、ブルーベリー、カカオ、サクランボ、ココナッツ、クランベリー、ナツメヤシ、ブドウ、グレープフルーツ、グアバ、キウイ、レモン、ライム、マンゴー、メロン、ネクタリン、オレンジ、パパイヤ、パッションフルーツ、モモ、ピーナツ、セイヨウナシ、パイナップル、ピスタチオ、プラム、ラズベリー、イチゴ、タンジェリン、クルミ、およびスイカを含み、しかしこれらに限らない任意の植物にもまた、適用することができる。
【0051】
本発明の方法に従ってあらかじめ選択された部位に挿入された外来性DNA分子を含む植物細胞および植物を提供することもまた、本発明の目的である。標的指向DNA挿入イベントを含む、従来の育種方法によって産生される植物の配偶子、種子、胚、接合子または体細胞である子孫またはハイブリッドもまた、本発明の範囲内に含まれる。
【0052】
本明細書に記述した方法によって得られた植物をさらに従来の育種技術によって他の植物と交配して、本発明により得られた標的指向DNA挿入イベントを含む子孫植物を得ることができる。
【0053】
次の制限されない実施例は、修飾されたI−SceIをコードするキメラ遺伝子の設計、および、その植物ゲノムのあらかじめ選択された部位に外来性DNAを挿入するための使用について記述する。
【0054】
もし実施例において異なるように記述しない場合は、組換DNA技術をすべて、Sambrook et al., (1989) Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press, NY、および Ausubel et al., (1994) Current Protocols in Molecular Biology, Current Protocols, USA の第1および第2巻に記載されている標準プロトコルにより実施する。植物の分子的研究の標準材料および方法は、R.D.D. CroyによるBIOS Scientific Publications Ltd (UK) および Blackwell Scientific Publications, UK の共同出版の Plant Molecular Biology Labfax (1993)に記述されている。標準的分子生物学技術のための他の参考文献には、Sambrook and Russell (2001) Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Laboratory Press, NY、Brown (1998) Molecular Biology LabFax, Second Edition, Academic Press (UK) のの第Iおよび第II巻が含まれる。ポリメラーゼ連鎖反応の標準材料および方法は、Dieffenbach and Dveksler (1995) PCR Primer: A Laboratory Manual, Cold Spring Harbor Laboratory Press, および McPherson et al., (2000) PCR-Basics: From Background to Bench, First Edition, Springer Verlag, Germany の中に見出すことができる。
【0055】
明細書および実施例の全体にわたって、次の配列を参照する:
【0056】
配列番号1:I−SceIタンパク質に連結された核移行シグナルを含み、4個のアミノ末端のアミノ酸を欠くキメラI−SceIのアミノ酸配列。
配列番号2:I−SceIコード領域のヌクレオチド配列(UIPACコード)。
配列番号3:合成I−SceIコード領域のヌクレオチド配列(UIPACコード)。
配列番号4:合成I−SceIコード領域のヌクレオチド配列。
配列番号5:pTTAM78のT−DNA(標的座位)のヌクレオチド配列。
配列番号6:pTTA82のT−DNA(修復DNA)のヌクレオチド配列。
配列番号7:pCV78のヌクレオチド配列。
【0057】
【表5】
【0058】
【表6】
【実施例1】
【0059】
I−SceIをコードする、植物で発現可能なキメラ遺伝子の設計、合成および解析
4個のアミノ末端アミノ酸が核移行シグナルで置換されたI−SceIのコード領域を、次のプロセスを用いて最適化した:
1.I−SceIタンパク質のアミノ酸配列を変更せずに、Synergy Geneoptimizer(登録商標)を用いて、トウモロコシに対して最も好ましいコドン使用にコドンを変換するプロセス;
2.特異的な制限部位を作成または除去するために配列を調節して、合成I−SceIコード領域をユニバーサルコードのI−SceI遺伝子で交換するプロセス;
3.前躯体mRNAスプライシングに影響する可能性のあるRNAの二次構造形成を回避するために、6bpより長いGCストレッチおよび4bpより長いATストレッチをすべて除去するプロセス;
4.コドンの中の2と3の位置のCGおよびTAという二連コードを回避するプロセス;
5.他の調節要素、例えば未成熟のポリアデニル化シグナルの候補
【表7】
隠れたイントロンスプライシング部位(AAGGTAAGTおよびTGCAGG)、ATTTA五量体、およびCCAATボックス配列(CCAAT、ATTGG、CGAAT、およびATTCG)を回避するプロセス;
6.適合されたコード領域が上に述べた基準をすべて満たすかどうかを再確認するプロセス。
【0060】
そのようなヌクレオチド配列の可能な一例を、配列番号4に示す。配列番号4のヌクレオチド配列を有する合成DNA断片を合成し、CaMV35SプロモーターおよびCaMV35S3’終了およびポリアデニル化シグナルに作動可能に連結させた(プラスミドpCV78;配列番号7を生成した)。
【0061】
合成I−SceIコード領域をバクテリアの発現ベクター中に(アミロースビーズ上でのタンパク質濃縮を可能にする融合タンパク質として)クローニングした。半精製されたI−SceIタンパク質の、I−SceI認識部位を含むプラスミドをインビトロで切断する能力を確認した。
【実施例2】
【0062】
I−SceI部位が先行しプロモーターを欠くbar遺伝子を含むトウモロコシ細胞系統の単離
二本鎖DNA切断が誘導する相同性を介する組換えの分析を発展させるために、核ゲノム中に単一コピーで組み込まれた、I−SceI認識部位が先行しプロモーターを欠くbar遺伝子を含むトウモロコシ細胞の懸濁液を分離した。I−SceIエンドヌクレアーゼをコードする植物で発現可能なキメラ遺伝子の送達、およびbar遺伝子の5’末端に作動可能に連結されたCaMV35Sプロモーターを含む修復DNAの共同送達、による二本鎖DNA切断誘導によって、35Sプロモーターが相同性を介する標的指向DNA挿入により挿入され、その結果ホスフィノトリシン(PPT)に対する耐性を与える機能的なbar遺伝子が得られる。分析を模式的に図1に表わす。
【0063】
標的座位を、従来のクローニング技術によって作動可能に次のDNA領域を連結することにより構成した:
a)ノパリン合成酵素遺伝子からの3’末端終止シグナルおよびポリアデニル化シグナル、
b)プロモーターを欠くbarをコードするDNA領域、
c)I−SceI認識部位を含むDNA領域、
d)A. tumefaciens 遺伝子7からの3’末端終止およびポリアデニル化シグナル(3’g7)、
e)ノパリン合成酵素プロモーター、ネオマイシンホスホトランスフェラーゼ遺伝子、および3’ocsシグナルを含む植物で発現可能なネオマイシン耐性遺伝子。
このDNA領域を、T−DNAベクターのT−DNA境界間に挿入した。T−DNAベクターをpTTAM78と名付けた(T−DNAのヌクレオチド配列については配列番号5を参照のこと)。
【0064】
T−DNAベクターを直接用いて、EP0 469 273に記述された方法によりHe89由来のトウモロコシ細胞懸濁液を用いてトウモロコシのプロトプラストを形質転換した。T−DNAベクターをまた Agrobacterium tumefaciens C58ClRif (pEHA101) へも導入して、得られたAgrobacteriumをHe89由来の細胞系統を形質転換するために用いた。T24(プロトプラストの形質転換により得られる)および、14−1および1−20系統(Agrobacteriumを介する形質転換により得られる)などの標的座位コンストラクトpTTAM78の単一コピーを含む多数の標的系統を同定した。
【0065】
N6M細胞懸濁培地中のこれらの標的系統から細胞懸濁液を確立し、振盪機(120 rpm)で光を当て25℃で増殖させた。懸濁液を毎週継代培養した。
【実施例3】
【0066】
相同性に基づく標的指向挿入
修復DNA、pTTA82は、T−DNA境界間に作動可能に連結された下記のDNA領域を含むT−DNAベクターである:
a)bar遺伝子のアミノ末端部分のみをコードするDNA領域;
b)部分的なI−SceI認識部位(認識部位の5’末端に位置する13個のヌクレオチド)を含むDNA領域;
c)CaMV35Sプロモーター領域;
d)部分的なI−SceI認識部位(認識部位の3’末端に位置する9個のヌクレオチド)を含むDNA領域;
e)A. tumefaciens 遺伝子7由来の3’末端終止およびポリアデニル化シグナル(3’g7);
f)キメラの植物発現可能なネオマイシン耐性遺伝子;
g)CaMV35Sプロモーターの制御下の欠落を有するI−SceIエンドヌクレアーゼをコードする遺伝子。
【0067】
pTTA82のT−DNAヌクレオチド配列を配列番号6に示す。
【0068】
この修復DNAを、ろ紙上に薄層として塗布した懸濁液由来の細胞中へ粒子を撃ち込むことによって、pCV78(実施例1を参照のこと)と共同送達した。ろ紙をMahql VII基質上にプレートした。
【0069】
DNAを細胞へ、PDS−1000/He Biolistics装置を用いて撃ち込んだ。マイクロキャリアの調製およびその上へのDNAのコーティングは、ほとんどSanford et al., 1992 記載の通りに行った。粒子撃ち込みパラメーターは次の通りであった:標的距離9cm、撃ち込み圧力1350psi、ギャップ距離1/4”、およびマイクロキャリア飛行距離11cm。撃ち込み直後に組織を、選択的でないMhilVII基質上に移した。粒子撃ち込みによる成功したDNA送達の対照として、3つの標的系統にも、CaMV35Sプロモーター制御下のキメラbar遺伝子を含むプラスミドDNA(pRVA52)でコートされたマイクロキャリアによって撃ち込みを行った。
【0070】
撃ち込み4日後に、フィルタを、25mg/L PPTを補ったMhlVII基質上に、または50mg/L PPTを補ったAhx1.5VIIino1000基質上に移した。
【0071】
14日後に、フィルタを、標的系統T24および14−1用の10mg/L のPPTを有する新鮮なMhlVII培地、および標的系統1−20用の25mg/LのPPTを有するMhlVII基質の上に移した。
【0072】
2週間後に、標的指向挿入である可能性のあるイベントを、PPTに対するそれらの耐性に基づいて評価した。これらのPPT耐性イベントは、Liberly Link Corn Leaf/Seedテスト(Strategic Diagnostics Inc.)においても陽性であった。
【0073】
撃ち込み38日後のPPT耐性カルスの数:
【表8】
【0074】
PPT耐性イベントを、さらに10mg/LのPPTを含むMhlVII基質上で継代培養し、またカルス材料を分子分析に用いた。20の独立のTSI候補を、サザン解析によって、35Sプロモーターならびにノパリン合成酵素遺伝子由来の3’末端終止およびポリアデニル化シグナルをプローブとして用いて解析した。予期された断片のサイズに基づいて、イベントはすべて完全な標的指向配列挿入イベントであるように見えた。その上、標的指向配列挿入イベントの約半分についてのさらに詳しい解析は、修復DNAまたはI−SceIをコードするDNAのいずれについても追加の非標的指向組込みを示さなかった。
【0075】
標的指向挿入イベントのうちの8つから増幅されたDNAの配列分析が、これらのイベントが確かに完全な相同組換えに基づいたTSIイベントであることを実証した。
【0076】
これらのデータに基づくと、相同組換えに基づいたDNA挿入対「正常な」非正統的組換の比率は、1−20に対する約30%から、14−1に対する17%、および24に対する約1%まで変化する。
【0077】
Puchta et al., 1996 (上掲)に記述された、I−SceI誘導二本鎖DNA切断存在下でエレクトロポレーションによってタバコプロトプラストに送達されたベクターと同様のベクターを用いた場合は、相同組換えに基づいたDNA挿入対正常な挿入の比率は、約15%であった。しかし、33の特徴決定したイベントのうちの1だけが、相同組換えが二重鎖切断の両側で完全な相同性仲介標的指向配列挿入イベントであった。
【0078】
実施例2から得たベクター、しかし核移行シグナルを含む「ユニバーサルコードI−SceIコンストラクト」を備えたベクターを用いると、相同組換えに基づいたDNA挿入対正常な挿入の比率が、様々な標的系統について、エレクトロポレーションまたは Agrobacteriumを介するDNA送達を用いた両方の場合で、0.032%〜16%の間で変化した。完全な標的指向挿入イベントの相対度数は、種々の標的系統間で異なり、エレクトロポレーションを介するDNA送達に対して8〜70%の間で、また Agrobacteriumを介するDNA送達に対して73〜90%の間で変化した。
【実施例4】
【0079】
アセトシリンゴンによるプレインキュベーションが、標的指向挿入イベントの回収頻度を改善する。
実施例3に記述した撃ち込みの1週間前に、細胞懸濁液を、200μM アセトシリンゴンを補ったN6M培地またはLSIDhy1.5培地で希釈した。他の点では、実施例3に記述した方法を用いた。次の表に要約された結果からわかるように、形質転換される細胞のアセトシリンゴンとのプレインキュベーションは、標的指向PPT耐性挿入イベントの回収に有益な効果を有した。
【0080】
【表9】
【実施例5】
【0081】
Agrobacteriumを媒介とした修復DNAの送達による、DSBを介するトウモロコシへの標的指向配列挿入
修復DNAがAgrobacteriumに媒介される形質転換により送達される、トウモロコシ中へのDSBを介する標的指向配列挿入を解析するために、欠陥I−SceIを実施例1の合成I−SceIコード遺伝子と置換したT−DNAベクターをpTTA82(実施例3を参照のこと)と同様に構成した。T−DNAベクターは、さらにT−DNA境界の外側に、Agrobacterium tumefaciens のvirGおよびvirC(pTCV83)、または、virG、virC、およびvirB(pTCV87)の1コピ−を含んでいた。これらのT−DNAベクターを、ヘルパーTiプラスミドpAL4404を含むLBA4404に挿入し、それによってAgrobacterium 菌株A4995およびA4996がそれぞれ生み出された。
【0082】
実施例2の標的細胞系統の浮遊培養、ならびに実施例2に記述したと同様の方法で得られた他の標的細胞系統を、Agrobacterium 菌株と共栽培し、その後多数のプレートに平板培養した。プレートする数を、細胞懸濁液の密度により決定した。形質転換効率用の対照として、細胞懸濁液を、並行して行った実験で、ヘルパーTiプラスミドpAL4404およびCaMV35Sベクターの制御下のキメラのホスフィノトリシン耐性遺伝子(bar遺伝子)を有するT−DNAベクターを含むAgrobacterium strain LBA4404 と共培養した。T−DNAベクターは、さらにT−DNA境界の外に、Agrobacterium tumeficans のvirG、virC、およびvirB遺伝子の1コピーを含んでいた。4つの異なる独立した実験の結果を下の表に要約する:
【0083】
【表10】
【0084】
【表11】
【0085】
【表12】
【0086】
【表13】
【0087】
このように、Agrobacteriumに媒介された修復DNA送達は明白に実現可能であるが、標的指向配列挿入(TSI)イベントの頻度が粒子撃ち込みを媒介とした修復DNA送達と比較して、より低いことが明らかである。23の推定TSIイベントで行なわれたサザン解析により、20のTSIのイベントが完全であることが、断片のサイズに基づいて示された。しかしながら、実施例3におけるマイクロプロジェクタイル撃ち込みによって得られたイベントと対照的に、20のイベントのうちの6のみが、修復DNAの追加の挿入断片を含んでいなかった。9つのイベントが、修復DNAの1〜3の追加の挿入を含んでいた。また5つのイベントが、修復DNAの多くの追加の挿入を含んでいた。
【0088】
粒子撃ち込みを介する修復DNAの送達はまた、Agrobacteriumによる修復DNAの送達に比較して、DSBを介するTSIイベントのよりよい質をもたらす。これは、粒子撃ち込みを介する「正常な形質転換DNA」の送達(Agrobacteriumを介する形質転換と比較した場合に、形質転換体のより劣った質(複雑な組込みパターン)によって特徴づけられる)とは対照的である。
【0089】
これは、粒子撃ち込みまたは他の直接DNA送達法によって得られる形質転換体の質を、DSBを介した配列の挿入によって改善することができることを表している。この結果は次の実験によっても確認される:修復DNA中に標的座位と相同性を有するフランキング配列がない状態で、DSBを介する35Sプロモーターの標的指向配列挿入を行ったときに、我々は、修復DNAのエレクトロポレーションを介した送達に際して、TSIイベントの少数だけが付加的な35Sプロモーターの非標的指向挿入を含むこと(16の解析されたTSIイベントのうちの2つのTSIイベントが付加的なランダムな35Sプロモーター挿入を示す)を観察した。対照的に、35Sプロモーターのランダム挿入が、Agrobacteriumを介する35Sプロモーターの送達によって得られたTSIイベントでは相当より高かった(22の解析されたTSIイベントのうち17が付加されたランダムな35Sプロモーターの挿入を示した)。
【実施例6】
【0090】
培地の組成
Mahq1VII:100mg/L カゼイン加水分解物、6mM L−プロリン、0.5g/L 2−(N−モルホリノ)エタンスルホン酸(MES)、0.2M マニトール、0.2M ソルビトール、2% 蔗糖、lmg/L 2,4−ジクロロフェノキシ酢酸(2,4−D)、を補ったN6培地 (Chu et al., 1975)、pH5.8に調節し、2.5g/L Gelrite(登録商標)で凝固させた。
【0091】
Mhi1VII:0.5g/L 2−(N−モルホリノ)エタンスルホン酸(MES)、0.2M マニトール、2% 蔗糖、lmg/L 2,4−ジクロロフェノキシ酢酸(2,4−D)、を補ったN6培地 (Chu et al., 1975)、pH5.8に調節し、2.5g/L Gelrite(登録商標)で凝固させた。
【0092】
Mh1VII:0.2M マニトールを含まない以外は、Mhi1VII基質に同じ。
【0093】
Ahxl.5VIIino1000:1000mg/L ミオ−イノシトール、0.1mg/L チアミン−HCl、0.5mg/L ニコチン酸、0.5mg/L ピリドキシン−HCl、0.5g/L MES、30g/L 蔗糖、10g/L グルコース、1.5mg/L 2,4−D、を補ったMS塩類、pH5.8に調節し、2.5g/L Gelrite(登録商標)で凝固させた。
【0094】
LSIDhyl.5:0.5mg/L ニコチン酸、0.5mg/L ピリドキシン−HCl、1mg/L チアミン−HCl、100mg/L ミオ−イノシトール、6mM プロリン、0.5g/L MES、20g/L 蔗糖、10g/L グルコース、1.5mg/L 2,4−D、を補ったMS塩類、pH5.2に調節した。
【0095】
N6M:マクロ要素:2830mg/L KNO3、433mg/L(NH4)2SO4;166mg/L CaCl2.2H2O;250mg/L MgSO4.7H2O;400mg/L KH2PO4;37.3mg/L Na2EDTA;27.3mg/L FeSO4.7H2O、 MSミクロ要素:500mg/L バクトトリプトン、0.5g/L MES、lmg/L チアミン−HCl、0.5mg/L ニコチン酸、0.5mg/L ピリドキシン−HCl、2mg/L グリシン、100mg/L ミオ−イノシトール、3%蔗糖、0.5mg/L 2,4−D、pH5.8に調節した。
【0096】
表1は、合成I−SceIコード領域に対する可能なトリヌクレオチド(コドン)の選択を表わす(配列番号2中のヌクレオチド配列も参照のこと)。
表2は、合成I−SceIコード領域に対する好ましい可能なトリヌクレオチドの選択を表わす(配列番号3中のヌクレオチド配列も参照のこと)。
【特許請求の範囲】
【請求項1】
対象の外来性DNAを、植物細胞ゲノムのあらかじめ選択された部位へ導入するための方法であって、
(a)二本鎖DNA切断を細胞のゲノム中のあらかじめ選ばれた部位に引き起こす工程;
(b)対象の外来性DNAを植物細胞中へ導入する工程;
を含み、外来性DNAを直接DNA移入によって送達することを特徴とする方法。
【請求項2】
前記直接DNA移入が、対象の外来性DNAでコートされたマイクロプロジェクタイルの撃ち込みにより遂行される、請求項1記載の方法。
【請求項3】
前記対象の外来性DNAは、あらかじめ選択された部位に隣接するDNA領域と少なくとも80%の配列同一性を有するDNA領域と隣接している、請求項1または2記載の方法。
【請求項4】
前記二本鎖DNA切断が、I−SceIをコードする遺伝子の導入により引き起される、請求項1〜3のいずれか1項記載の方法。
【請求項5】
前記I−SceIをコードする遺伝子が配列番号1のアミノ酸配列をコードするヌクレオチド配列を含む、請求項4記載の方法であって、前記ヌクレオチド配列は、約50%〜約60%のGC含量を有し、ただし、
vii)前記ヌクレオチド配列が、
【表14】
からなる群より選択されるヌクレオチド配列を含まず;
viii)前記ヌクレオチドが、CCAAT、ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まず;
ix)前記ヌクレオチド配列が、ATTTA,AAGGT,AGGTA、GGTAまたはGCAGGからなる群より選択される配列を含まず;
x)前記ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;
xi)前記ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るATストレッチを含まず;および
xii)前記ヌクレオチド配列がコドンTTA、CTA、ATA、GTA、TCG、CCG、ACGおよびGCGを含まない、
ことを条件とする方法。
【請求項6】
I−SceIをコードする遺伝子が配列番号4のヌクレオチド配列を含む、請求項5記載の方法。
【請求項7】
植物細胞がトウモロコシ細胞である、請求項1〜6のいずれかに記載の方法。
【請求項8】
トウモロコシ細胞が細胞懸濁液中に含まれる、請求項7記載の方法。
【請求項9】
前記植物細胞を、工程(a)に先立って植物フェノール化合物中でインキュベーションする、請求項1〜8のいずれかに記載の方法。
【請求項10】
前記植物フェノール化合物がアセトシリンゴンである、請求項9記載の方法。
【請求項11】
対象の外来性DNAを、植物細胞のゲノムのあらかじめ選択された部位へ導入する方法であって、
(a)二本鎖DNA切断を細胞のゲノム中のあらかじめ選択された部位に引き起こす工程;
(b)対象の外来性DNAを植物細胞中へ導入する工程;
を含み、二本鎖DNA切断が、ヌクレオチド配列によってコードされるレア切断エンドヌクレアーゼによって導入され、前記ヌクレオチド配列が約50%〜約60%のGC含量を有し、ただし、
i)前記ヌクレオチド配列が、
【表15】
からなる群より選択されるヌクレオチド配列を含まず;
ii)前記ヌクレオチドが、CCAAT,ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まず;
iii)前記ヌクレオチド配列が、ATTTA,AAGGT,AGGTA、GGTAまたはGCAGGからなる群より選択される配列を含まず;
iv)前記ヌクレオチド配列が、GまたはCからなる群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;
v)前記ヌクレオチド配列が、AまたはTからなる群より選択される連続する5個のヌクレオチドから成るATストレッチを含まず;および
vi)前記ヌクレオチド配列が、コドンTTA、CTA、ATA、GTA、TCG、CCG、ACGおよびGCGを含まない、
ことを条件とすることを特徴とする方法。
【請求項12】
ヌクレオチド配列が配列番号4のヌクレオチド配列を含む、請求項11記載の方法。
【請求項13】
対象の外来性DNAが、直接のDNA移入によって前記植物細胞中へ導入される、請求項11または12記載の方法。
【請求項14】
前記直接のDNA移入が、対象の外来性DNAでコートされたマイクロプロジェクタイルの撃ち込みにより遂行される、請求項11〜13のいずれか1項記載の方法。
【請求項15】
前記対象の外来性DNAが、あらかじめ選択された部位に隣接するDNA領域と少なくとも80%の配列同一性を有するDNA領域と隣接している、請求項11〜14のいずれか1項記載の方法。
【請求項16】
前記二本鎖DNA切断が、I−SceIをコードする遺伝子の導入により引き起される、請求項11〜15のいずれか1項記載の方法。
【請求項17】
植物細胞がトウモロコシ細胞である、請求項11〜16のいずれかに記載の方法。
【請求項18】
トウモロコシ細胞が細胞懸濁液中に含まれる、請求項17記載の方法。
【請求項19】
前記植物細胞を、工程(a)に先立って植物フェノール化合物中でインキュベーションする、請求項11〜18のいずれかに記載の方法。
【請求項20】
前記植物フェノール化合物がアセトシリンゴンである、請求項19記載の方法。
【請求項21】
対象の外来性DNAを植物細胞のゲノムのあらかじめ選択された部位へ導入するための方法であって、
(a)二本鎖DNA切断を、細胞のゲノム中のあらかじめ選ばれた部位に引き起こす工程;
(b)対象の外来性DNAを植物へ導入する工程;
を含み、工程(a)に先立って植物細胞を植物フェノール化合物中でインキュベーションすることを特徴とする方法。
【請求項22】
前記植物フェノール化合物が、アセトシリンゴン(3,5−ジメトキシ−4−ヒドロキシアセトフェノン)、α−ヒドロキシ−アセトシリンゴン、シナピン酸(3,5ジメトキシ−4−ヒドロキシケイ皮酸)、シリング酸(4−ヒドロキシ−3,5ジメトキシ安息香酸)、フェルラ酸(4−ヒドロキシ−3−メトキシケイ皮酸)、カテコール(1,2−ジヒドロキシベンゼン)、p−ヒドロキシ安息香酸(4−ヒドロキシ安息香酸)、β−レソルシル酸(2,4ジヒドロキシ安息香酸)、プロトカテク酸(3,4−ジヒドロキシ安息香酸)、ピロ没食子酸(2,3,4−トリヒドロキシ安息香酸)、没食子酸(3,4,5−トリヒドロキシ安息香酸)、およびバニリン(3−メトキシ−4−ヒドロキシベンズアルデヒド)、の群より選択される、請求項21に記載の方法、
【請求項23】
対象の外来性DNAを植物細胞のゲノムのあらかじめ選択された部位へ導入するための方法であって、
(a)二本鎖DNA切断を、細胞のゲノム中のあらかじめ選択された部位で、レア切断エンドヌクレアーゼによって引き起こす工程;
(b)対象の外来性DNAを植物へ導入する工程;
を含み、前記エンドヌクレアーゼが核移行シグナルを含むことを特徴とする方法。
【請求項24】
配列番号1のアミノ酸配列をコードするヌクレオチド配列を含む単離されたDNA断片であって、そのヌクレオチド配列が、約50%〜約60%のGC含量を有し、ただし、
i)前記ヌクレオチド配列が、
【表16】
からなる群より選択されるヌクレオチド配列を含まず;
ii)前記ヌクレオチドが、CCAAT、ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まず;
iii)前記ヌクレオチド配列が、ATTTA、AAGGT、AGGTA、GGTAまたはGCAGGからなる群より選択される配列を含まず;
iv)前記ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;
v)前記ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るATストレッチを含まず;および
vi)前記ヌクレオチド配列の、Leu、Ile、Val、Ser、Pro、Thr、Alaをコードするコドンが、前記コドンの位置2および3にTAまたはGCという二連コードを含まない;
ことを条件とする、DNA断片。
【請求項25】
配列番号2のヌクレオチド配列を含む単離されたDNA断片であって、前記ヌクレオチド配列のGC含量が、約50〜約60%であり、
i)前記ヌクレオチド配列の位置28〜位置30がAAGでなく;
ii)位置34〜位置36のヌクレオチド配列がAATである場合は、位置37〜位置39のヌクレオチド配列がATTまたはATAでなく;
iii)位置34〜位置36のヌクレオチド配列がAACである場合は、位置37〜位置39のヌクレオチド配列がATTでなく、同時に位置40〜位置42のヌクレオチド配列がAAAであり;
iv)位置34〜位置36のヌクレオチド配列がAACである場合は、位置37〜位置39のヌクレオチド配列がATAでなく;
v)位置37〜位置39のヌクレオチド配列がATTまたはATAである場合は、位置40〜位置42のヌクレオチド配列がAAAでなく;
vi)位置49〜位置51のヌクレオチド配列がCAAでなく;
vii)位置52〜位置54のヌクレオチド配列がGTAでなく;
viii)位置58〜位置63のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
ix)位置67〜位置69のヌクレオチド配列がCCCである場合は、位置70〜位置72のヌクレオチド配列がAATでなく;
x)位置76〜位置78のヌクレオチド配列がAAAである場合は、位置79〜位置81のヌクレオチド配列がTTGでなく、同時に位置82〜位置84のヌクレオチド配列がCTNであり;
xi)位置79〜位置81のヌクレオチド配列がTTAまたはCTAである場合は、位置82〜位置84のヌクレオチド配列がTTAでなく;
xii)位置88〜位置90のヌクレオチド配列がGAAではなく;
xiii)位置91〜位置93のヌクレオチド配列がTATである場合は、位置94〜位置96のヌクレオチド配列がAAAでなく;
xiv)位置97〜位置99のヌクレオチド配列がTCCまたはTCGまたはAGCである場合は、位置100〜位置102のヌクレオチド配列がCCAでなく、同時に位置103〜位置105のヌクレオチド配列がTTRであり;
xv)位置100〜位置102のヌクレオチド配列がCAAである場合は、位置103〜位置105のヌクレオチド配列がTTAでなく;
xvi)位置109〜位置111のヌクレオチド配列がGAAである場合は、位置112〜位置114のヌクレオチド配列がTTAでなく;
xvii)位置115〜117のヌクレオチド配列がAATである場合は、位置118〜位置120のヌクレオチド配列がATTまたはATAでなく;
xviii)位置121〜123のヌクレオチド配列がGAGである場合は、位置124〜位置126のヌクレオチド配列が;
xix)位置133〜135のヌクレオチド配列がGCAでなく;
xx)位置139〜位置141のヌクレオチド配列がATTでなく;
xxi)位置142〜位置144のヌクレオチド配列がGGAである場合は、位置145〜位置147のヌクレオチド配列がTTAでなく;
xxii)位置145〜位置147のヌクレオチド配列がTTAである場合は、位置148〜位置150のヌクレオチド配列がATAでなく、同時に位置151〜位置153のヌクレオチド配列がTTRであり;
xxiii)位置145〜位置147のヌクレオチド配列がCTAである場合は、位置148〜位置150のヌクレオチド配列がATAでなく、同時に位置151〜位置153のヌクレオチド配列がTTRであり;
xxiv)位置148〜位置150のヌクレオチド配列がATAである場合は、位置151〜位置153のヌクレオチド配列がCTAまたはTTGでなく;
xxv)位置160〜位置162のヌクレオチド配列がGCAである場合は、位置163〜位置165のヌクレオチド配列がTACでなく;
xxvi)位置163〜位置165のヌクレオチド配列がTATである場合は、位置166〜位置168のヌクレオチド配列がATAでなく、同時に位置169〜位置171のヌクレオチド配列がAGRであり;
xxvii)位置172〜位置177のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がGCAGGを含まないようになる選択に従って選ばれ;
xxviii)位置178〜位置186のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がAGGTAを含まないようになる選択に従って選ばれ;
xxix)位置193〜位置195のヌクレオチド配列がTATである場合は、位置196〜位置198のヌクレオチド配列がTGCでなく;
xxx)位置202〜位置204のヌクレオチド配列がCAAでなく;
xxxi)位置217〜位置219のヌクレオチド配列がAATでなく;
xxxii)位置220〜位置222のヌクレオチド配列がAAAである場合は、位置223〜位置225のヌクレオチド配列がGCAでなく;
xxxiii)位置223〜位置225のヌクレオチド配列がGCAである場合は、位置226〜位置228のヌクレオチド配列がTACでなく;
xxxiv)位置253〜位置255のヌクレオチド配列がGACである場合は、位置256〜位置258のヌクレオチド配列がCAAでなく;
xxxv)位置277〜位置279のヌクレオチド配列がCATである場合は、位置280〜位置282のヌクレオチド配列がAAAでなく;
xxxvi)位置298〜位置303のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
xxxvii)位置304〜位置306のヌクレオチド配列がGGCである場合は、位置307〜位置309のヌクレオチド配列がAATでなく;
xxxviii)位置307〜位置312のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
xxxix)位置334〜位置342のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
xl)位置340〜位置342のヌクレオチド配列がAAGである場合は、位置343〜位置345のヌクレオチド配列がCATでなく;
xli)位置346〜位置348のヌクレオチド配列がCAAである場合は、位置349〜位置351のヌクレオチド配列がGCAでなく;
xlii)位置349〜位置357のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
xliii)位置355〜位置357のヌクレオチド配列がAATでなく;
xliv)位置358〜位置360のヌクレオチド配列がAAAである場合は、位置361〜位置363のヌクレオチド配列がTTGでなく;
xlv)位置364〜位置366のヌクレオチド配列がGCCである場合は、位置367〜位置369のヌクレオチド配列がAATでなく;
xlvi)位置367〜位置378のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
xlvii)位置382〜位置384のヌクレオチド配列がAATである場合は、位置385〜位置387のヌクレオチド配列がAATでなく;
xlviii)位置385〜位置387のヌクレオチド配列がAATでなく;
xlix)位置400〜位置402のヌクレオチド配列がCCCである場合は、位置403〜位置405のヌクレオチド配列がAATでなく;
l) 位置403〜位置405のヌクレオチド配列がAATである場合は、位置406〜位置408のヌクレオチド配列がAATでなく;
li)位置406〜位置411のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
lii)位置421〜位置426のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
liii)位置430〜位置432のヌクレオチド配列がCCAでなく;
liv)位置436〜位置438のヌクレオチド配列がTCAである場合は、位置439〜位置441のヌクレオチド配列がTTGでなく;
lv)位置445〜位置447のヌクレオチド配列がTATでなく;
lvi)位置481〜位置483のヌクレオチド配列がAATでなく;
lvii)位置484〜位置486のヌクレオチド配列がAAAである場合は、位置487〜位置489のヌクレオチド配列がAATでなく、同時に位置490〜位置492のヌクレオチド配列がAGYであり;
lviii)位置490〜位置492のヌクレオチド配列がTCAである場合は、位置493〜位置495のヌクレオチド配列がACCでなく、同時に位置496〜位置498のヌクレオチド配列がAAYであり;
lix)位置493〜位置495のヌクレオチド配列がACCである場合は、位置496〜位置498のヌクレオチド配列がAATでなく;
lx)位置496〜位置498のヌクレオチド配列がAATでなく;
lxi)位置499〜位置501のヌクレオチド配列がAAAである場合は、位置502〜位置504のヌクレオチド配列がTCAまたはAGCでなく;
lxii)位置508〜位置510のヌクレオチド配列がGTAである場合は、位置511〜位置513のヌクレオチド配列がTTAでなく;
lxiii)位置514〜位置516のヌクレオチド配列がAATである場合は、位置517〜位置519のヌクレオチド配列がACAでなく;
lxiv)位置517〜位置519のヌクレオチド配列がACCまたはACGである場合は、位置520〜位置522のヌクレオチド配列がCAAでなく、同時に位置523〜位置525のヌクレオチド配列がTCNであり;
lxv)位置523〜位置531のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
lxvi)位置544〜位置546のヌクレオチド配列がGAAである場合は、位置547〜位置549のヌクレオチド配列がTATでなく、同時に位置550〜位置552のヌクレオチド配列がTTRであり;
lxvii)位置547〜位置552のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
lxviii)位置559〜位置561のヌクレオチド配列がGGAである場合は、位置562〜位置564のヌクレオチド配列がTTGでなく、同時に位置565〜位置567のヌクレオチド配列がCGNであり;
lxix)位置565〜位置567のヌクレオチド配列がCGCである場合は、位置568〜位置570のヌクレオチド配列がAATでなく;
lxx)位置568〜位置570のヌクレオチド配列がAATでなく;
lxxi)位置574〜位置576のヌクレオチド配列がTTCである場合は、位置577〜位置579のヌクレオチド配列がCAAでなく、同時に位置580〜位置582のヌクレオチド配列がTTRであり;
lxxii)位置577〜位置579のヌクレオチド配列がCAAである場合は、位置580〜位置582のヌクレオチド配列がTTAでなく;
lxxiii)位置583〜位置585のヌクレオチド配列がAATである場合は、位置586〜位置588のヌクレオチド配列がTGCでなく;
lxxiv)位置595〜位置597のヌクレオチド配列がAAAでなく;
lxxv)位置598〜位置600のヌクレオチド配列がATTである場合は、位置601〜位置603のヌクレオチド配列がAATでなく;
lxxvi)位置598〜位置600のヌクレオチド配列がATAでなく;
lxxvii)位置601〜位置603のヌクレオチド配列がAATでなく;
lxxviii)位置604〜位置606のヌクレオチド配列がAAAである場合は、位置607〜位置609のヌクレオチド配列がAATでなく;
lxxix)位置607〜位置609のヌクレオチド配列がAATでなく;
lxxx)位置613〜位置615のヌクレオチド配列がCCAでなく;
lxxxi)位置613〜位置615のヌクレオチド配列がCCGである場合は、位置616〜位置618のヌクレオチド配列がATAでなく;
lxxxii)位置616〜位置618のヌクレオチド配列がATAである場合は、位置619〜位置621のヌクレオチド配列がATAでなく;
lxxxiii)位置619〜位置621のヌクレオチド配列がATAである場合は、位置622〜位置624のヌクレオチド配列がTACでなく;
lxxxiv)位置619〜位置621のヌクレオチド配列がATTでなく;
lxxxv)位置640〜位置645のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
lxxxvi)位置643〜位置645のヌクレオチド配列がTTAである場合は、位置646〜位置648のヌクレオチド配列がATAでなく;
lxxxvii)位置643〜位置645のヌクレオチド配列がCTAである場合は、位置646〜位置648のヌクレオチド配列がATAでなく;
lxxxviii)位置655〜位置660のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
lxxxix)位置658〜位置660のヌクレオチド配列がTTAまたはCTAである場合は、位置661〜位置663のヌクレオチド配列がATTまたはATCでなく;
xc)位置661〜位置663のヌクレオチド配列がATAでなく;
xci)位置661〜位置663のヌクレオチド配列がATTである場合は、位置664〜位置666のヌクレオチド配列がAAAでなく;
xcii)位置670〜位置675のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
xciii)位置691〜位置693のヌクレオチド配列がTATである場合は、位置694〜位置696のヌクレオチド配列がAAAでなく;
xciv)位置694〜位置696のヌクレオチド配列がAAAである場合は、位置697〜位置699のヌクレオチド配列がTTGでなく;
xcv)位置700〜位置702のヌクレオチド配列がCCCである場合は、位置703〜位置705のヌクレオチド配列がAATでなく;
xcvi)位置703〜位置705のヌクレオチド配列がAATである場合は、位置706〜位置708のヌクレオチド配列がACAまたはACTでなく;
xcvii)位置706〜位置708のヌクレオチド配列がACAである場合は、位置709〜位置711のヌクレオチド配列がATAでなく、同時に位置712〜位置714のヌクレオチド配列がAGYであり;
xcviii)前記ヌクレオチド配列がコドンTTA、CTA、ATA、GTA、TCG、CCG、ACG、およびGCGを含まず;
xcix)前記ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;および
c) 前記ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るATストレッチを含まない、
ことを条件とする、DNA断片。
【請求項26】
配列番号3のヌクレオチド配列を含む単離されたDNA断片であって、前記ヌクレオチド配列のGC含量が約50〜約60%であり、
a)位置121〜位置123のヌクレオチド配列がGAGである場合は、位置124〜126のヌクレオチド配列がCAAでなく;
b)位置253〜位置255のヌクレオチド配列がGACである場合は、位置256〜258のヌクレオチド配列がCAAでなく;
c)位置277〜位置279のヌクレオチド配列がCATである場合は、位置280〜282のヌクレオチド配列がAAAでなく;
d)位置340〜位置342のヌクレオチド配列がAAGである場合は、位置343〜345のヌクレオチド配列がCATでなく;
e)位置490〜位置492のヌクレオチド配列がTCAである場合は、位置493〜495のヌクレオチド配列がACCでなく;
f)位置499〜位置501のヌクレオチド配列がAAAである場合は、位置502〜504のヌクレオチド配列がTCAまたはAGCでなく;
g)位置517〜位置519のヌクレオチド配列がACCである場合は、位置520〜522のヌクレオチド配列がCAAでなく、同時に位置523〜525のヌクレオチド配列がTCNであり;
h)位置661〜位置663のヌクレオチド配列がATTである場合は、位置664〜666のヌクレオチド配列がAAAでなく;
i)位置7〜位置15のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
j)位置61〜位置69のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
k)位置130〜位置138のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
l)位置268〜位置279のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのドストレッチを含まないようになる選択に従って選ばれ;
m)位置322〜位置333のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
n)位置460〜位置468のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
o)位置13〜位置27のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
p)位置37〜位置48のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
q)位置184〜位置192のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
r)位置214〜位置219のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
s)位置277〜位置285のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
t)位置388〜位置396のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
u)位置466〜位置474のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
v)位置484〜位置489のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
w)位置571〜位置576のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
x)位置598〜位置603のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
y)位置604〜位置609のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
z)位置613〜位置621のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
aa)位置646〜位置651のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
bb)位置661〜位置666のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;および
cc)位置706〜位置714のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれる、
ことを条件とする、DNA断片。
【請求項27】
1の位置のみが配列番号4のヌクレオチド配列と異なるヌクレオチド配列を含むことを特徴とする、請求項26記載の単離されたDNA配列。
【請求項28】
10の位置のみが配列番号4のヌクレオチド配列と異なるヌクレオチド配列を含むことを特徴とする、請求項26記載の単離されたDNA配列。
【請求項29】
配列番号4のヌクレオチド配列を含む、単離されたDNA配列。
【請求項30】
植物で発現可能なプロモーターに作動可能に連結された請求項24〜29のいずれか1項記載の単離されたDNA断片を含む、キメラ遺伝子。
【請求項31】
外来性DNAを、植物ゲノムのI−SceI認識部位に挿入するための、請求項30記載のキメラ遺伝子の使用。
【請求項1】
対象の外来性DNAを、植物細胞ゲノムのあらかじめ選択された部位へ導入するための方法であって、
(a)二本鎖DNA切断を細胞のゲノム中のあらかじめ選ばれた部位に引き起こす工程;
(b)対象の外来性DNAを植物細胞中へ導入する工程;
を含み、外来性DNAを直接DNA移入によって送達することを特徴とする方法。
【請求項2】
前記直接DNA移入が、対象の外来性DNAでコートされたマイクロプロジェクタイルの撃ち込みにより遂行される、請求項1記載の方法。
【請求項3】
前記対象の外来性DNAは、あらかじめ選択された部位に隣接するDNA領域と少なくとも80%の配列同一性を有するDNA領域と隣接している、請求項1または2記載の方法。
【請求項4】
前記二本鎖DNA切断が、I−SceIをコードする遺伝子の導入により引き起される、請求項1〜3のいずれか1項記載の方法。
【請求項5】
前記I−SceIをコードする遺伝子が配列番号1のアミノ酸配列をコードするヌクレオチド配列を含む、請求項4記載の方法であって、前記ヌクレオチド配列は、約50%〜約60%のGC含量を有し、ただし、
vii)前記ヌクレオチド配列が、
【表14】
からなる群より選択されるヌクレオチド配列を含まず;
viii)前記ヌクレオチドが、CCAAT、ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まず;
ix)前記ヌクレオチド配列が、ATTTA,AAGGT,AGGTA、GGTAまたはGCAGGからなる群より選択される配列を含まず;
x)前記ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;
xi)前記ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るATストレッチを含まず;および
xii)前記ヌクレオチド配列がコドンTTA、CTA、ATA、GTA、TCG、CCG、ACGおよびGCGを含まない、
ことを条件とする方法。
【請求項6】
I−SceIをコードする遺伝子が配列番号4のヌクレオチド配列を含む、請求項5記載の方法。
【請求項7】
植物細胞がトウモロコシ細胞である、請求項1〜6のいずれかに記載の方法。
【請求項8】
トウモロコシ細胞が細胞懸濁液中に含まれる、請求項7記載の方法。
【請求項9】
前記植物細胞を、工程(a)に先立って植物フェノール化合物中でインキュベーションする、請求項1〜8のいずれかに記載の方法。
【請求項10】
前記植物フェノール化合物がアセトシリンゴンである、請求項9記載の方法。
【請求項11】
対象の外来性DNAを、植物細胞のゲノムのあらかじめ選択された部位へ導入する方法であって、
(a)二本鎖DNA切断を細胞のゲノム中のあらかじめ選択された部位に引き起こす工程;
(b)対象の外来性DNAを植物細胞中へ導入する工程;
を含み、二本鎖DNA切断が、ヌクレオチド配列によってコードされるレア切断エンドヌクレアーゼによって導入され、前記ヌクレオチド配列が約50%〜約60%のGC含量を有し、ただし、
i)前記ヌクレオチド配列が、
【表15】
からなる群より選択されるヌクレオチド配列を含まず;
ii)前記ヌクレオチドが、CCAAT,ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まず;
iii)前記ヌクレオチド配列が、ATTTA,AAGGT,AGGTA、GGTAまたはGCAGGからなる群より選択される配列を含まず;
iv)前記ヌクレオチド配列が、GまたはCからなる群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;
v)前記ヌクレオチド配列が、AまたはTからなる群より選択される連続する5個のヌクレオチドから成るATストレッチを含まず;および
vi)前記ヌクレオチド配列が、コドンTTA、CTA、ATA、GTA、TCG、CCG、ACGおよびGCGを含まない、
ことを条件とすることを特徴とする方法。
【請求項12】
ヌクレオチド配列が配列番号4のヌクレオチド配列を含む、請求項11記載の方法。
【請求項13】
対象の外来性DNAが、直接のDNA移入によって前記植物細胞中へ導入される、請求項11または12記載の方法。
【請求項14】
前記直接のDNA移入が、対象の外来性DNAでコートされたマイクロプロジェクタイルの撃ち込みにより遂行される、請求項11〜13のいずれか1項記載の方法。
【請求項15】
前記対象の外来性DNAが、あらかじめ選択された部位に隣接するDNA領域と少なくとも80%の配列同一性を有するDNA領域と隣接している、請求項11〜14のいずれか1項記載の方法。
【請求項16】
前記二本鎖DNA切断が、I−SceIをコードする遺伝子の導入により引き起される、請求項11〜15のいずれか1項記載の方法。
【請求項17】
植物細胞がトウモロコシ細胞である、請求項11〜16のいずれかに記載の方法。
【請求項18】
トウモロコシ細胞が細胞懸濁液中に含まれる、請求項17記載の方法。
【請求項19】
前記植物細胞を、工程(a)に先立って植物フェノール化合物中でインキュベーションする、請求項11〜18のいずれかに記載の方法。
【請求項20】
前記植物フェノール化合物がアセトシリンゴンである、請求項19記載の方法。
【請求項21】
対象の外来性DNAを植物細胞のゲノムのあらかじめ選択された部位へ導入するための方法であって、
(a)二本鎖DNA切断を、細胞のゲノム中のあらかじめ選ばれた部位に引き起こす工程;
(b)対象の外来性DNAを植物へ導入する工程;
を含み、工程(a)に先立って植物細胞を植物フェノール化合物中でインキュベーションすることを特徴とする方法。
【請求項22】
前記植物フェノール化合物が、アセトシリンゴン(3,5−ジメトキシ−4−ヒドロキシアセトフェノン)、α−ヒドロキシ−アセトシリンゴン、シナピン酸(3,5ジメトキシ−4−ヒドロキシケイ皮酸)、シリング酸(4−ヒドロキシ−3,5ジメトキシ安息香酸)、フェルラ酸(4−ヒドロキシ−3−メトキシケイ皮酸)、カテコール(1,2−ジヒドロキシベンゼン)、p−ヒドロキシ安息香酸(4−ヒドロキシ安息香酸)、β−レソルシル酸(2,4ジヒドロキシ安息香酸)、プロトカテク酸(3,4−ジヒドロキシ安息香酸)、ピロ没食子酸(2,3,4−トリヒドロキシ安息香酸)、没食子酸(3,4,5−トリヒドロキシ安息香酸)、およびバニリン(3−メトキシ−4−ヒドロキシベンズアルデヒド)、の群より選択される、請求項21に記載の方法、
【請求項23】
対象の外来性DNAを植物細胞のゲノムのあらかじめ選択された部位へ導入するための方法であって、
(a)二本鎖DNA切断を、細胞のゲノム中のあらかじめ選択された部位で、レア切断エンドヌクレアーゼによって引き起こす工程;
(b)対象の外来性DNAを植物へ導入する工程;
を含み、前記エンドヌクレアーゼが核移行シグナルを含むことを特徴とする方法。
【請求項24】
配列番号1のアミノ酸配列をコードするヌクレオチド配列を含む単離されたDNA断片であって、そのヌクレオチド配列が、約50%〜約60%のGC含量を有し、ただし、
i)前記ヌクレオチド配列が、
【表16】
からなる群より選択されるヌクレオチド配列を含まず;
ii)前記ヌクレオチドが、CCAAT、ATTGG、GCAATおよびATTGCからなる群より選択されるヌクレオチド配列を含まず;
iii)前記ヌクレオチド配列が、ATTTA、AAGGT、AGGTA、GGTAまたはGCAGGからなる群より選択される配列を含まず;
iv)前記ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;
v)前記ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るATストレッチを含まず;および
vi)前記ヌクレオチド配列の、Leu、Ile、Val、Ser、Pro、Thr、Alaをコードするコドンが、前記コドンの位置2および3にTAまたはGCという二連コードを含まない;
ことを条件とする、DNA断片。
【請求項25】
配列番号2のヌクレオチド配列を含む単離されたDNA断片であって、前記ヌクレオチド配列のGC含量が、約50〜約60%であり、
i)前記ヌクレオチド配列の位置28〜位置30がAAGでなく;
ii)位置34〜位置36のヌクレオチド配列がAATである場合は、位置37〜位置39のヌクレオチド配列がATTまたはATAでなく;
iii)位置34〜位置36のヌクレオチド配列がAACである場合は、位置37〜位置39のヌクレオチド配列がATTでなく、同時に位置40〜位置42のヌクレオチド配列がAAAであり;
iv)位置34〜位置36のヌクレオチド配列がAACである場合は、位置37〜位置39のヌクレオチド配列がATAでなく;
v)位置37〜位置39のヌクレオチド配列がATTまたはATAである場合は、位置40〜位置42のヌクレオチド配列がAAAでなく;
vi)位置49〜位置51のヌクレオチド配列がCAAでなく;
vii)位置52〜位置54のヌクレオチド配列がGTAでなく;
viii)位置58〜位置63のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
ix)位置67〜位置69のヌクレオチド配列がCCCである場合は、位置70〜位置72のヌクレオチド配列がAATでなく;
x)位置76〜位置78のヌクレオチド配列がAAAである場合は、位置79〜位置81のヌクレオチド配列がTTGでなく、同時に位置82〜位置84のヌクレオチド配列がCTNであり;
xi)位置79〜位置81のヌクレオチド配列がTTAまたはCTAである場合は、位置82〜位置84のヌクレオチド配列がTTAでなく;
xii)位置88〜位置90のヌクレオチド配列がGAAではなく;
xiii)位置91〜位置93のヌクレオチド配列がTATである場合は、位置94〜位置96のヌクレオチド配列がAAAでなく;
xiv)位置97〜位置99のヌクレオチド配列がTCCまたはTCGまたはAGCである場合は、位置100〜位置102のヌクレオチド配列がCCAでなく、同時に位置103〜位置105のヌクレオチド配列がTTRであり;
xv)位置100〜位置102のヌクレオチド配列がCAAである場合は、位置103〜位置105のヌクレオチド配列がTTAでなく;
xvi)位置109〜位置111のヌクレオチド配列がGAAである場合は、位置112〜位置114のヌクレオチド配列がTTAでなく;
xvii)位置115〜117のヌクレオチド配列がAATである場合は、位置118〜位置120のヌクレオチド配列がATTまたはATAでなく;
xviii)位置121〜123のヌクレオチド配列がGAGである場合は、位置124〜位置126のヌクレオチド配列が;
xix)位置133〜135のヌクレオチド配列がGCAでなく;
xx)位置139〜位置141のヌクレオチド配列がATTでなく;
xxi)位置142〜位置144のヌクレオチド配列がGGAである場合は、位置145〜位置147のヌクレオチド配列がTTAでなく;
xxii)位置145〜位置147のヌクレオチド配列がTTAである場合は、位置148〜位置150のヌクレオチド配列がATAでなく、同時に位置151〜位置153のヌクレオチド配列がTTRであり;
xxiii)位置145〜位置147のヌクレオチド配列がCTAである場合は、位置148〜位置150のヌクレオチド配列がATAでなく、同時に位置151〜位置153のヌクレオチド配列がTTRであり;
xxiv)位置148〜位置150のヌクレオチド配列がATAである場合は、位置151〜位置153のヌクレオチド配列がCTAまたはTTGでなく;
xxv)位置160〜位置162のヌクレオチド配列がGCAである場合は、位置163〜位置165のヌクレオチド配列がTACでなく;
xxvi)位置163〜位置165のヌクレオチド配列がTATである場合は、位置166〜位置168のヌクレオチド配列がATAでなく、同時に位置169〜位置171のヌクレオチド配列がAGRであり;
xxvii)位置172〜位置177のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がGCAGGを含まないようになる選択に従って選ばれ;
xxviii)位置178〜位置186のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がAGGTAを含まないようになる選択に従って選ばれ;
xxix)位置193〜位置195のヌクレオチド配列がTATである場合は、位置196〜位置198のヌクレオチド配列がTGCでなく;
xxx)位置202〜位置204のヌクレオチド配列がCAAでなく;
xxxi)位置217〜位置219のヌクレオチド配列がAATでなく;
xxxii)位置220〜位置222のヌクレオチド配列がAAAである場合は、位置223〜位置225のヌクレオチド配列がGCAでなく;
xxxiii)位置223〜位置225のヌクレオチド配列がGCAである場合は、位置226〜位置228のヌクレオチド配列がTACでなく;
xxxiv)位置253〜位置255のヌクレオチド配列がGACである場合は、位置256〜位置258のヌクレオチド配列がCAAでなく;
xxxv)位置277〜位置279のヌクレオチド配列がCATである場合は、位置280〜位置282のヌクレオチド配列がAAAでなく;
xxxvi)位置298〜位置303のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
xxxvii)位置304〜位置306のヌクレオチド配列がGGCである場合は、位置307〜位置309のヌクレオチド配列がAATでなく;
xxxviii)位置307〜位置312のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
xxxix)位置334〜位置342のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
xl)位置340〜位置342のヌクレオチド配列がAAGである場合は、位置343〜位置345のヌクレオチド配列がCATでなく;
xli)位置346〜位置348のヌクレオチド配列がCAAである場合は、位置349〜位置351のヌクレオチド配列がGCAでなく;
xlii)位置349〜位置357のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
xliii)位置355〜位置357のヌクレオチド配列がAATでなく;
xliv)位置358〜位置360のヌクレオチド配列がAAAである場合は、位置361〜位置363のヌクレオチド配列がTTGでなく;
xlv)位置364〜位置366のヌクレオチド配列がGCCである場合は、位置367〜位置369のヌクレオチド配列がAATでなく;
xlvi)位置367〜位置378のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
xlvii)位置382〜位置384のヌクレオチド配列がAATである場合は、位置385〜位置387のヌクレオチド配列がAATでなく;
xlviii)位置385〜位置387のヌクレオチド配列がAATでなく;
xlix)位置400〜位置402のヌクレオチド配列がCCCである場合は、位置403〜位置405のヌクレオチド配列がAATでなく;
l) 位置403〜位置405のヌクレオチド配列がAATである場合は、位置406〜位置408のヌクレオチド配列がAATでなく;
li)位置406〜位置411のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
lii)位置421〜位置426のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
liii)位置430〜位置432のヌクレオチド配列がCCAでなく;
liv)位置436〜位置438のヌクレオチド配列がTCAである場合は、位置439〜位置441のヌクレオチド配列がTTGでなく;
lv)位置445〜位置447のヌクレオチド配列がTATでなく;
lvi)位置481〜位置483のヌクレオチド配列がAATでなく;
lvii)位置484〜位置486のヌクレオチド配列がAAAである場合は、位置487〜位置489のヌクレオチド配列がAATでなく、同時に位置490〜位置492のヌクレオチド配列がAGYであり;
lviii)位置490〜位置492のヌクレオチド配列がTCAである場合は、位置493〜位置495のヌクレオチド配列がACCでなく、同時に位置496〜位置498のヌクレオチド配列がAAYであり;
lix)位置493〜位置495のヌクレオチド配列がACCである場合は、位置496〜位置498のヌクレオチド配列がAATでなく;
lx)位置496〜位置498のヌクレオチド配列がAATでなく;
lxi)位置499〜位置501のヌクレオチド配列がAAAである場合は、位置502〜位置504のヌクレオチド配列がTCAまたはAGCでなく;
lxii)位置508〜位置510のヌクレオチド配列がGTAである場合は、位置511〜位置513のヌクレオチド配列がTTAでなく;
lxiii)位置514〜位置516のヌクレオチド配列がAATである場合は、位置517〜位置519のヌクレオチド配列がACAでなく;
lxiv)位置517〜位置519のヌクレオチド配列がACCまたはACGである場合は、位置520〜位置522のヌクレオチド配列がCAAでなく、同時に位置523〜位置525のヌクレオチド配列がTCNであり;
lxv)位置523〜位置531のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
lxvi)位置544〜位置546のヌクレオチド配列がGAAである場合は、位置547〜位置549のヌクレオチド配列がTATでなく、同時に位置550〜位置552のヌクレオチド配列がTTRであり;
lxvii)位置547〜位置552のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
lxviii)位置559〜位置561のヌクレオチド配列がGGAである場合は、位置562〜位置564のヌクレオチド配列がTTGでなく、同時に位置565〜位置567のヌクレオチド配列がCGNであり;
lxix)位置565〜位置567のヌクレオチド配列がCGCである場合は、位置568〜位置570のヌクレオチド配列がAATでなく;
lxx)位置568〜位置570のヌクレオチド配列がAATでなく;
lxxi)位置574〜位置576のヌクレオチド配列がTTCである場合は、位置577〜位置579のヌクレオチド配列がCAAでなく、同時に位置580〜位置582のヌクレオチド配列がTTRであり;
lxxii)位置577〜位置579のヌクレオチド配列がCAAである場合は、位置580〜位置582のヌクレオチド配列がTTAでなく;
lxxiii)位置583〜位置585のヌクレオチド配列がAATである場合は、位置586〜位置588のヌクレオチド配列がTGCでなく;
lxxiv)位置595〜位置597のヌクレオチド配列がAAAでなく;
lxxv)位置598〜位置600のヌクレオチド配列がATTである場合は、位置601〜位置603のヌクレオチド配列がAATでなく;
lxxvi)位置598〜位置600のヌクレオチド配列がATAでなく;
lxxvii)位置601〜位置603のヌクレオチド配列がAATでなく;
lxxviii)位置604〜位置606のヌクレオチド配列がAAAである場合は、位置607〜位置609のヌクレオチド配列がAATでなく;
lxxix)位置607〜位置609のヌクレオチド配列がAATでなく;
lxxx)位置613〜位置615のヌクレオチド配列がCCAでなく;
lxxxi)位置613〜位置615のヌクレオチド配列がCCGである場合は、位置616〜位置618のヌクレオチド配列がATAでなく;
lxxxii)位置616〜位置618のヌクレオチド配列がATAである場合は、位置619〜位置621のヌクレオチド配列がATAでなく;
lxxxiii)位置619〜位置621のヌクレオチド配列がATAである場合は、位置622〜位置624のヌクレオチド配列がTACでなく;
lxxxiv)位置619〜位置621のヌクレオチド配列がATTでなく;
lxxxv)位置640〜位置645のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
lxxxvi)位置643〜位置645のヌクレオチド配列がTTAである場合は、位置646〜位置648のヌクレオチド配列がATAでなく;
lxxxvii)位置643〜位置645のヌクレオチド配列がCTAである場合は、位置646〜位置648のヌクレオチド配列がATAでなく;
lxxxviii)位置655〜位置660のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
lxxxix)位置658〜位置660のヌクレオチド配列がTTAまたはCTAである場合は、位置661〜位置663のヌクレオチド配列がATTまたはATCでなく;
xc)位置661〜位置663のヌクレオチド配列がATAでなく;
xci)位置661〜位置663のヌクレオチド配列がATTである場合は、位置664〜位置666のヌクレオチド配列がAAAでなく;
xcii)位置670〜位置675のヌクレオチド配列のコドンが、その結果のヌクレオチド配列がATTTAを含まないようになる選択に従って選ばれ;
xciii)位置691〜位置693のヌクレオチド配列がTATである場合は、位置694〜位置696のヌクレオチド配列がAAAでなく;
xciv)位置694〜位置696のヌクレオチド配列がAAAである場合は、位置697〜位置699のヌクレオチド配列がTTGでなく;
xcv)位置700〜位置702のヌクレオチド配列がCCCである場合は、位置703〜位置705のヌクレオチド配列がAATでなく;
xcvi)位置703〜位置705のヌクレオチド配列がAATである場合は、位置706〜位置708のヌクレオチド配列がACAまたはACTでなく;
xcvii)位置706〜位置708のヌクレオチド配列がACAである場合は、位置709〜位置711のヌクレオチド配列がATAでなく、同時に位置712〜位置714のヌクレオチド配列がAGYであり;
xcviii)前記ヌクレオチド配列がコドンTTA、CTA、ATA、GTA、TCG、CCG、ACG、およびGCGを含まず;
xcix)前記ヌクレオチド配列が、GまたはCの群より選択される連続する7個のヌクレオチドから成るGCストレッチを含まず;および
c) 前記ヌクレオチド配列が、AまたはTの群より選択される連続する5個のヌクレオチドから成るATストレッチを含まない、
ことを条件とする、DNA断片。
【請求項26】
配列番号3のヌクレオチド配列を含む単離されたDNA断片であって、前記ヌクレオチド配列のGC含量が約50〜約60%であり、
a)位置121〜位置123のヌクレオチド配列がGAGである場合は、位置124〜126のヌクレオチド配列がCAAでなく;
b)位置253〜位置255のヌクレオチド配列がGACである場合は、位置256〜258のヌクレオチド配列がCAAでなく;
c)位置277〜位置279のヌクレオチド配列がCATである場合は、位置280〜282のヌクレオチド配列がAAAでなく;
d)位置340〜位置342のヌクレオチド配列がAAGである場合は、位置343〜345のヌクレオチド配列がCATでなく;
e)位置490〜位置492のヌクレオチド配列がTCAである場合は、位置493〜495のヌクレオチド配列がACCでなく;
f)位置499〜位置501のヌクレオチド配列がAAAである場合は、位置502〜504のヌクレオチド配列がTCAまたはAGCでなく;
g)位置517〜位置519のヌクレオチド配列がACCである場合は、位置520〜522のヌクレオチド配列がCAAでなく、同時に位置523〜525のヌクレオチド配列がTCNであり;
h)位置661〜位置663のヌクレオチド配列がATTである場合は、位置664〜666のヌクレオチド配列がAAAでなく;
i)位置7〜位置15のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
j)位置61〜位置69のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
k)位置130〜位置138のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
l)位置268〜位置279のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのドストレッチを含まないようになる選択に従って選ばれ;
m)位置322〜位置333のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
n)位置460〜位置468のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、GまたはCの群からの連続する7個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
o)位置13〜位置27のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
p)位置37〜位置48のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
q)位置184〜位置192のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
r)位置214〜位置219のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
s)位置277〜位置285のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
t)位置388〜位置396のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
u)位置466〜位置474のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
v)位置484〜位置489のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
w)位置571〜位置576のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
x)位置598〜位置603のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
y)位置604〜位置609のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
z)位置613〜位置621のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
aa)位置646〜位置651のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;
bb)位置661〜位置666のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれ;および
cc)位置706〜位置714のヌクレオチド配列のコドンが、その結果生じるヌクレオチド配列が、AまたはTの群からの連続する5個のヌクレオチドのストレッチを含まないようになる選択に従って選ばれる、
ことを条件とする、DNA断片。
【請求項27】
1の位置のみが配列番号4のヌクレオチド配列と異なるヌクレオチド配列を含むことを特徴とする、請求項26記載の単離されたDNA配列。
【請求項28】
10の位置のみが配列番号4のヌクレオチド配列と異なるヌクレオチド配列を含むことを特徴とする、請求項26記載の単離されたDNA配列。
【請求項29】
配列番号4のヌクレオチド配列を含む、単離されたDNA配列。
【請求項30】
植物で発現可能なプロモーターに作動可能に連結された請求項24〜29のいずれか1項記載の単離されたDNA断片を含む、キメラ遺伝子。
【請求項31】
外来性DNAを、植物ゲノムのI−SceI認識部位に挿入するための、請求項30記載のキメラ遺伝子の使用。
【図1】

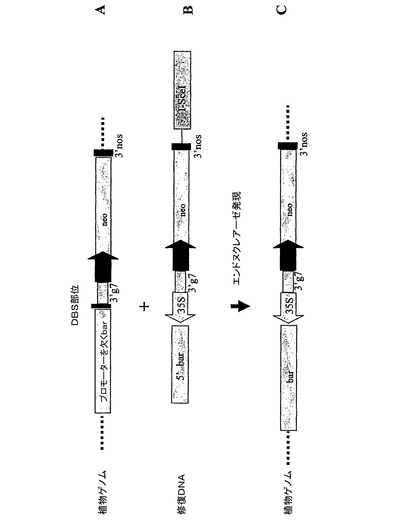
【公開番号】特開2011−177191(P2011−177191A)
【公開日】平成23年9月15日(2011.9.15)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−108596(P2011−108596)
【出願日】平成23年5月13日(2011.5.13)
【分割の表示】特願2006−540331(P2006−540331)の分割
【原出願日】平成16年11月17日(2004.11.17)
【出願人】(501018298)バイエル・バイオサイエンス・エヌ・ヴェー (12)
【氏名又は名称原語表記】Bayer BioScience N.V.
【Fターム(参考)】
【公開日】平成23年9月15日(2011.9.15)
【国際特許分類】
【出願番号】特願2011−108596(P2011−108596)
【出願日】平成23年5月13日(2011.5.13)
【分割の表示】特願2006−540331(P2006−540331)の分割
【原出願日】平成16年11月17日(2004.11.17)
【出願人】(501018298)バイエル・バイオサイエンス・エヌ・ヴェー (12)
【氏名又は名称原語表記】Bayer BioScience N.V.
【Fターム(参考)】
[ Back to top ]