
植物における生物学的に安全な一過性のタンパク質発現
植物又は植物の葉において目的配列から目的タンパク質を発現させることによって目的タンパク質を産生する方法であって:a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下で植物又は植物の葉に浸潤させることによって植物又は植物の葉にトランスフェクションし、レプリコンをコードする配列は、植物ウイルスに由来する、レプリコンのレプリコン機能に必要な配列、及びレプリコンから発現されるべき目的配列を含有し、b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、ことを含み、アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されている、前記方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、アグロバクテリウムによって標的植物に送達される植物ウイルスベクターに基づいた一過性発現を用いた、植物における生物学的に安全な組換えタンパク質の産生方法に関する。また、本発明は、標的宿主植物又は植物の葉における目的配列の発現方法に関する。本発明は、植物全体又は植物の部分において目的タンパク質の大規模産生を高収率で提供する、ウイルスベクターのアグロバクテリウム介在送達の使用について記載する。本発明の方法は、宿主植物へのアグロバクテリウム介在送達による一過性発現を制限する一方で、望ましくない生物への形質転換を制御する。更に、本発明は、植物又は植物の葉において目的配列を発現させるための生物学的に安全なアグロバクテリウムに関する。本発明は、生物学的安全性が上昇した、植物における高収率の工業的タンパク質産生を提供する。
【背景技術】
【0002】
発明の背景
現在、植物バイオテクノロジーは、植物において外来遺伝子を送達及び発現するための2つのアプローチ:安定な形質転換と一過性発現に依拠している。後者は、アグロ浸潤又はウイルス感染に基づいて構築することができる(概説として:Fischerら、1999、Biotechnol.Appl.Biochem.、30、113−116を参照されたい)。一過性発現は、構成的な、例えば35Sプロモーターの制御下で目的遺伝子の発現を駆動する標準の発現カセットを植物組織にアグロ浸潤させることによって(Vaqueroら、1999、Proc.Natl.Acad.Sci.USA.、96、11128−11133)、又は大規模では、植物にウイルスベクターをトランスフェクションすることによって達成することができる。通常、アグロ浸潤は高収率を提供しないが、p19又はHcProのような転写後遺伝子サイレンシング(PTGS)サプレッサーとの組み合わせにより、タンパク質発現レベルを50倍まで上昇させることができる(Voinnetら、2003、Plant J.、33、549−556)。依然として、ウイルス発現システムによって達成できる生物学的限界を遙かに下回っている(Marillonnetら、2004、Proc.Natl.Acad.Sci.USA、101、6852−6857)。植物組織において目的組換えタンパク質を一過性発現させるためのアグロ浸潤の使用に基づいたバイオリアクターについての記載は米国特許第6,740,526号に提供されている。しかしながら、この特許は、発現されるべきタンパク質の収率をいかにして改善するかについての記載がなく、当該技術分野において公知の他の方法に勝るものではない(Vaqueroら、1999、Proc.Natl.Acad.Sci.USA.、96、11128−11133)。一過性発現のより強力なアプローチは、ウイルスベクターの使用である。レポーター遺伝子(GFP又はDsRed)のTMVに基づいた発現は、システムの生物学的な収率限界に達することができ、新鮮な葉のバイオマス1gあたり数mgの組換えタンパク質を産生することが示された(Marillonnetら、2004、Proc.Natl.Acad.Sci.USA、101、6852−6857)。そのようなシステムの相対収率は、TSPの80%まで達することができ、下流処理を著しく容易にし、費用を低下させる。そのような高い相対収率は、ウイルスによって誘導される宿主タンパク質生合成の遮断により可能である。
【0003】
ウイルスベクターは、アグロ浸潤に用いる慣用のベクターよりも高い発現レベルを提供でき(概説として:Porta&Lomonossoff、1996、Mol.Biotechnol.、5、209−221;Yusibovら、1999、Curr.Top.Microbiol.Immunol.、240、81−94を参照されたい)、機能ゲノミクス研究の強力なツールである(Dalmayら、2000、Plant Cell、12、369−379;Ratcliffら、2001、Plant J.、25、237−245;Escobarら、2003、Plant Cell、15、1507−1523)ことは明らかである。当該技術分野における数多くの出版物及び特許がDNA及びRNAのウイルスベクターに基づいたシステムについて記載している(Kumagaiら、1994、Proc.Natl.Acad.Sci.USA、90、427−430;Malloryら、2002、Nature Biotechnol.20、622−625;Morら、2003、Biotechnol.Bioeng.、81、430−437;US5316931号;US5589367号;US5866785号;US5491076号;US5977438号;US5981236号;WO02/088369号;WO02/097080号;WO98/54342号)。既存のウイルスベクターシステムは、通常、最高性能の点から見ると狭い宿主範囲に限定され、最も好適な宿主におけるそのようなベクターの発現レベルでさえシステムの生物学的な上限を遙かに下回っている。ウイルスに基づいたシステムの重要な問題は、植物細胞へのウイルスレプリコンの送達方法である。大規模産生(例えば農場又は温室において多くの植物で同時に産生すること)に最も広く適用される送達法は、RNAウイルスベクターの感染性コピーの使用である(Kumagaiら、1995、Proc.Natl.Acad.Sci.USA、92、1679−1683)。組換えRNAウイルスベクターは複製サイクルのあいだに異種挿入物を喪失する傾向が比較的高いため、この方法はin vitroでDNA鋳型の転写を必要とし、したがって非効率的で高価である。
【0004】
一過性経路は、非常に速いが、ウイルスの低感染性、植物の大部分にトランスフェクションすることができないこと、及び遺伝子サイズの限界のために非常に限定されている。植物細胞へ感染性ウイルスベクターを送達するためのアグロ浸潤の使用(Liu&Lomonossof、2002、J.Virol.Methods、105、343−348)、又はアグロバクテリウムによって送達されるウイルスベクター成分からのウイルスベクターの組み立て(Marillonnetら、2004、Proc.Natl.Acad.Sci.USA、101、6852−6857)について記載する出版物がある。しかしながら、これらの出版物は、アグロ浸潤された植物組織のそれぞれの細胞におけるウイルスレプリコンの効率的な同調形成の問題に取り組んでおらず、更に、ベクターの伝播は細胞間移行及び浸透移行に関するウイルスの能力に依存している。この移行は比較的長期間を必要とし、通常、ウイルスベクターは感染後およそ10〜14日で最高の可能なベクター収率を提供する。これは、植物の細胞内プロセスを妨げる組換えタンパク質、特に制限酵素、プロテアーゼ、非特異的ヌクレアーゼ、多くの医薬タンパク質のような非常に細胞傷害性のタンパク質の産生には受け入れられない。逆に、アグロ浸潤によって送達される標準ベクターは、アグロ送達後3〜4日で最高の可能な発現レベルに達するが、そのようなベクターによって提供される収率は容認し難いほど低い。したがって、事実上アグロ浸潤された植物組織の全ての細胞において目的遺伝子の発現を提供するにもかかわらず、目的タンパク質を発現するための構成的プロモーターによって駆動される効率の低い標準の転写ベクターによって引き起こされる問題に直面する。反対に、アグロ送達されたウイルスベクターは高発現レベルを提供可能であるが、複製をほとんど開始せず、そのため大部分の細胞で発現を提供せず、ほんのわずかな画分のみである(アグロ浸潤された全組織の1%未満)。その結果、そのようなウイルスに基づいたベクターは発現を提供するのに著しく(3〜4倍)長期間を必要とするが、システムの生産性は、特に細胞障害性タンパク質の場合に、理論上の生産性よりも依然として低い。これは、アグロ浸潤された組織において同調発現を提供しないため、特に細胞障害性遺伝子を発現させる場合に、収率に影響を及ぼし、一過性発現システムにウイルスベクターを使用する上で重大な欠点である。また、感染した植物宿主はその組織の大部分においてウイルスベクターを含有しないため、産生プロセスから組織を除外する。更に、感染植物に対してウイルスベクターの最高の可能な伝播(及び発現レベル)を達成するのに要する時間は、標準のアグロ浸潤プロトコールと比較して3〜4倍長い。その上、一過性発現のための記載のシステムはいずれも、遺伝子操作された生物の使用を含めた工業的規模のタンパク質産生において重要な要素である、システムの生物学的安全性を高める問題に取り組んでいなかった。
【0005】
特許化技術を含め、当該技術分野には数多くの出版物があるにもかかわらず、主に2つの主要な理由により、依然として、商業的な高収率産生のために十分な効率と収率をもって作動するウイルスに基づいた大規模産生システムは存在しない:第1に、一過性の植物ウイルスに基づいた発現システムは、通常特定の宿主に制限され、環境因子の影響を受けやすいため大規模培養に好適であり得ない。その上、これらのシステムは、通常植物宿主のある部分に制限されるため、大部分の植物バイオマスを産生プロセスから除外し、その結果、単位植物バイオマスあたりの組換え産物の相対収率を、トランスジェニック植物において慣用の転写プロモーターによって達成可能なレベルに匹敵するレベルまで最小にする;第2に、各細胞に安定に組み込まれたウイルスレプリコン前駆体を有するトランスジェニック植物宿主を作製することによってウイルスに基づいた産生システムを拡大する試みは、特に、そのような位置におけるレプリコンの標準よりも低い働き、レプリコンから発現されるべき目的遺伝子の「漏出性」、そしてベクターの効率的スイッチシステムの欠失により、解決策を提供していない。RNAレプリコン形成の誘因としてPTGSサイレンシングのサプレッサーを用いることにより、PVXに基づいたベクターにおいて多少の進展が達成されたが(Malloryら、2002、Nature Biotechnol.、20、622−625)、このシステムは、ウイルスベクターの複製を誘発するスイッチ(PTGSサプレッサー)の効率的制御を提供する解決策がないため、依然として実用的価値に遠く及ばない。しかしながら、このシステムは、この種のシステムでこれまで知られた中で最高である、合計可溶性タンパク質(TSP)の3%に達するGUS遺伝子の発現レベルを提供したが、依然として強力なプロモーターの制御下の慣用のトランス遺伝子発現システムよりも良好ではない。植物3分節RNAウイルス、ブロムモザイクウイルス(BMV)に基づいた他の誘導性システム(Moriら、2001、Plant J.、27、79−86)は、目的タンパク質の収率が非常に低かった(3〜4μg/g新鮮重)。これは標準の転写プロモーターによって提供される収率と匹敵する。
【0006】
植物発現システムを用いてこれまでに達成された発現レベルの低さが、これらのシステムが細菌細胞、真菌細胞、又は昆虫細胞の発現システムのような他の発現システムとほとんど競合しない主な理由である。低発現レベルは、植物材料の巨大なバックグラウンドにおけるタンパク質の単離精製のための下流費用を非常に高くする。したがって、単位植物バイオマスあたりの目的タンパク質又は目的産物の収率が増加するにつれて、下流処理費用が急激に減少する。
【0007】
現在、細菌細胞、真菌細胞、又は昆虫細胞の発現システムのような他の大規模発現システムを用いた市場で競合するほど収率及び効率が十分に高い植物の大規模な一過性発現システムはない。そのような植物の発現は以下の基準をできるだけ良好に満たす必要があるであろう:
(i)できるだけ多くの植物組織及び組織の多くの細胞における目的タンパク質の発現を含め、高収率であること;
(ii)植物細胞の生存に対するタンパク質発現の有害な効果を防止するには、目的タンパク質又は目的産物の発現は、処置された植物又は植物組織の全ての植物細胞において同時に開始する必要がある。
【0008】
典型的には、目的タンパク質又は目的産物は、産物又はタンパク質を産生する各細胞にある時点まで蓄積する。しかしながら、蓄積のあいだ、分解過程が進むことが多く、目的タンパク質若しくは目的産物の収率又は品質が低下する傾向にある。したがって、発現スイッチが入った後、目的産物又は目的タンパク質を収穫すべき最適な時点がある。全体的な工程を効率的で収益性のあるものにするには、この最適な時点は、植物の全ての組織又は細胞において、そして選択したロットの全ての植物において同時に到達すべきである;
(iii)システムは、向上したバイオセイフティー性を組み込み、アグロ浸潤に用いるアグロバクテリウムは、少なくとも1つの以下の特徴を有するであろう:開かれた環境における生存率が低いか又はゼロであり、非標的生物又は非標的植物に対する感染性が低いか又はゼロである。
【0009】
したがって、本発明の目的は、大規模応用に拡大可能であり、発現すべきタンパク質の収率が高く、そして同時に、非標的生物を外来DNAでトランスフェクション又は形質転換する可能性が低い、生物学的に安全な、植物システムにおいてタンパク質を発現させる方法を提供することである。
【発明の開示】
【0010】
発明の一般的な説明
この目的は、標的植物又は標的植物の葉における目的配列からの目的タンパク質の一過性発現による目的タンパク質の産生方法によって達成され、この方法は:
レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉に浸潤させることによって植物又は植物の葉にトランスフェクションすることを含み、
レプリコンをコードする配列は、
(i)植物ウイルスに由来する、レプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、アグロバクテリウム株は、補完因子の非存在下では植物又は植物の葉を含めた生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されている。
【0011】
更に、本発明は、
(i)上で定義した通りのアグロバクテリウム株、及び
(ii)上で定義したアグロバクテリウム株の欠陥を補完可能な補完因子をコードする植物又はその種子、
を含むキット部品を提供する。
【0012】
更に、本発明は、
(i)上で定義した通りのアグロバクテリウム株、及び
(ii)上で定義したアグロバクテリウム株の欠陥を補完可能な補完因子をコードする第2のアグロバクテリウム株、
を含む、キット部品を提供する。
【0013】
更に、本発明は:
(i)補完因子の非存在下では植物又は植物の葉の細胞へのT−DNAの導入に必要な細菌の機能を不良にする第1の遺伝子改変を有し、
(ii)他の細菌へのプラスミド伝達の接合能が不良であり、そして
(iii)アグロバクテリウムの増殖に必要な主要代謝産物に対して栄養要求性である、
ことを特徴とする、本発明の方法のためのアグロバクテリウム属の細菌を提供する。
【0014】
自然の一般的原理によれば、1つの特性の改良は他の特性に負に影響する傾向があり、逆も同様であるため、システムの2以上の対立する特性を同時に向上させることは困難である。植物に基づいたタンパク質発現システムでは、発現されるべきタンパク質の高発現レベルを達成することと、高い生物学的安全性を達成することを同時に達成することは、高発現効率はシステムの選択性を低下させ、よって生物学的安全性が低い傾向にあるため困難である。例えば、高率発現システムを達成するために植物の感染に用いるアグロバクテリウム株を非常に感染性にする場合、アグロバクテリウム株は、非標的植物に形質移入する可能性が高く、組換えT−DNAが環境中に伝播する傾向があるが、これは回避しなければならない。一方、アグロバクテリウム株の感染性を低くすることによって生物学的安全性を改善する場合、標的植物における形質移入及び発現効率は劣るであろう。したがって、生物学的安全性と発現効率はそのように対立する特性である。
【0015】
植物に基づいたタンパク質発現システムへの要求は、システムを大規模応用、即ち多くの植物又は多くの植物の葉における同時発現に適合させる必要がある場合、より複雑になってさえいる。一方、大規模応用における生物学的安全性は、トランスジェニック生物が開放環境に拡張する可能性がより高まるため、より重要である。一方、高タンパク質発現効率及び高収率は、経済的に競合し得る方法を達成するには、大規模応において必須である。
【0016】
本発明は、目的タンパク質を産生するための、大規模で非常に効率的且つ生物学的に安全な方法を作製するための手段を初めて提供する。
本発明の方法では、目的タンパク質は、植物又は植物の葉において目的配列から一過性に発現される。一過性発現は、植物又は植物の葉の一過性トランスフェクションによって達成される。「一過性トランスフェクション」という用語は、異種DNA配列の導入が、植物染色体へ異種DNA配列を安定に組み込むためにトランスフェクションされた細胞を選択することなく行われることを意味する。目的配列は目的タンパク質をコードする。目的配列は、外来DNAを植物細胞へ導入可能なアグロバクテリウム株を用いて植物又は植物の葉へ導入される。目的配列は、レプリコンが由来する植物ウイルスに対して異種である。即ち、本発明の方法は、植物又は植物の葉を野生型植物ウイルスで形質転換する場合を含まない。DNA配列は異種目的配列を含有するため異種である。目的配列は、アグロバクテリウム株に対して固有の配列でないことが好ましい。
【0017】
植物又は植物の葉をアグロバクテリウム株で一過性にトランスフェクションする。植物又は植物の葉は、植物の多くの細胞及び多くの組織が1つの作業工程でトランスフェクションされるようにトランスフェクションされる必要がある。これは、アグロ浸潤によって最高に達成することができるが、植物又は植物の葉の主要部分のトランスフェクションを可能にする他の方法も適用することができる。したがって、植物又は植物の葉にアグロバクテリウム株を浸潤させることによって植物又は植物の葉に一過性にトランスフェクションすることが好ましい。浸潤は、システムの効率に顕著に寄与するため、本発明の重要な要素である。効率が高い程、植物又は植物の細胞のより多くの細胞が目的タンパク質を発現し、さまざまな組織及び細胞において発現開始が良好に同調する。浸潤は、植物又は植物の葉の多くの細胞を1つの作業工程で形質転換することを可能にし、それにより、目的タンパク質の発現がさまざまな植物組織及び植物のさまざまな部分において進む時点が実質的に同調する。更に、浸潤は、かなりの割合の植物又は植物の葉の細胞が形質転換される可能性を実質的に増加させる。したがって、レプリコンの遠隔距離移行の可能性は、全体的な効率を更に増加させることができるが、トランスフェクション及び発現は、植物又は植物の葉におけるレプリコンの遠隔距離移行に依存しない。
【0018】
本発明の方法は、植物への大規模な応用のため、即ち多くの植物で平行して実施するためにデザインされる。本発明の方法では、好ましくは少なくとも5、より好ましくは少なくとも10、より好ましくは少なくとも30、更により好ましくは少なくとも100、最も好ましくは少なくとも1000の植物にアグロバクテリウム株を平行してトランスフェクションする。
【0019】
本発明の方法を植物全体で実施する場合、トランスフェクション(好ましくは浸潤)は植物のいくつかの部分で行う。例えば、大部分の葉とトランスフェクションするか浸潤させる。全ての葉をトランスフェクションするか浸潤させることが好ましい。より好ましくは、植物の全ての葉及び茎をトランスフェクションするか浸潤させる。目的タンパク質を単離するためにその後収穫される全ての組織をトランスフェクションするか浸潤させるべきである。根における発現及び収穫はシステムの全体的な効率を更に増加させることができるが、通常、根を浸潤させる必要はない。アグロバクテリウム株の懸濁液中に植物を逆さにしてサッと浸し、真空を適用し、その後真空を急速に開放することによって植物全体又は少なくともその土の上の部分を浸潤させることができる。この手順は、規模を拡大することができ、多くの植物に順々に又は同時に適用することができる。
【0020】
本発明の方法を植物の葉で行う場合、葉の1以上の領域で浸潤を行う。好ましくは、葉に噴霧するか、アグロバクテリウム株の懸濁液にサッと浸し、好ましくは、その後アグロバクテリウム細胞を葉の組織に押し付けるか又は吸収させる。本発明にしたがい使用する植物の葉は、植物に付いていることが好ましい。本発明の方法は植物組織培養物で行わないことが好ましい。
【0021】
浸潤又はアグロ浸潤はアグロバクテリウム懸濁液を用いたトランスフェクション法として定義することができ、アグロバクテリウムを植物組織(細胞間隙)に押し付けるために圧力差を用いる。
【0022】
あるいは、ウイルス粒子を送達するために行ったようにして(Pogueら、2002、Annu.Rev.Phytopathol.、40、45−74)、アグロバクテリウムと研磨剤との混合物を植物へ展開するための高圧噴霧装置を用いてアグロバクテリウムを植物組織へ送達することができる。
【0023】
アグロバクテリウム株は、補完因子の非存在下ではT−DNAの生物への形質移入を不良にする第1の遺伝子改変を提供される。これは、生物学的安全性を向上させるための本発明の必須要素である。この生物は、異種DNA配列によるトランスフェクションが望ましくない生物である(「非標的生物」)。非標的生物は、とりわけ、植物及び細菌又は真菌のような微生物である。補完因子の非存在下では、本発明の植物種は非標的生物でもない。補完因子の非存在下では、アグロバクテリウム株は、そのような非標的生物に感染することも形質導入することも不良であり、好ましくは不可能である。
【0024】
アグロバクテリウム株、植物(又は植物の葉)、及び補完因子は、アグロバクテリウム株による植物の感染時、目的配列のトランスフェクション及び/又は発現を可能にするために選択され、一方で非標的生物のトランスフェクション及び/又は発現は起こりそうにない。非標的生物のトランスフェクションに関するアグロバクテリウム株欠陥、好ましくは不能は、アグロバクテリウム株の以後第1の遺伝子改変を称する遺伝子改変によって達成される。
【0025】
第1の遺伝子改変は、植物又は植物の葉の細胞へのT−DNAの導入に必要なアグロバクテリウム株の機能を不良にすることが好ましい。これは、標的生物のトランスフェクションに必要な機能をコードするアグロバクテリウム遺伝子を欠失又は分裂させることによって達成することができる。そのような機能は、とりわけ、Ti−プラスミド上、例えば植物又は植物の葉の細胞へ導入すべきT−DNAを有するTi−プラスミド上にコードされる。そのような遺伝子の例は、アグロバクテリウム遺伝子VirE2、GALLS、及びVirFである。
【0026】
他の態様では、第1の遺伝子改変は、アグロバクテリウム株を栄養要求性にし、それにより、その増殖を外部から加えられた代謝産物依存性にする。外部から加えられた代謝産物の非存在下では、アグロバクテリウム株は、生物にT−DNAをトランスフェクションすることができない。
【0027】
更なる態様では、第1の遺伝子改変として、生物のトランスフェクションに必要なアグロバクテリウム株の機能を、化学的に調節された異種プロモーターの制御下に置く。そのような遺伝子の例は、アグロバクテリウム遺伝子VirE2、GALLS、及びVirFであり、例えば、lacプロモーターの制御下に置くことができる。そのような態様では、補完因子は化学的に調節されたプロモーターを調節可能な小分子化合物である。lacプロモーターの場合、補完因子はIPTG又は乳糖であり得る。したがって、植物又は植物の葉に適用されるアグロバクテリウム懸濁液は、植物の感染、及び好ましくは植物の細胞へのT−DNAの導入を可能にする、IPTG又は乳糖のような小分子化合物を含有することができる。
【0028】
他の態様では、第1の遺伝子改変は、アグロバクテリウム株を条件的致死性にし、補完因子は、アグロバクテリウム株の生存に必要な、アグロバクテリウム株の主要代謝産物である。
【0029】
上記態様は組み合わせることができる。例えば、アグロバクテリウム株が、植物又は植物の葉の細胞へのT−DNAの導入に必要な機能が不良である場合、システムの生物学的安全性を高めるために、更に栄養要求性及び/又は条件的致死性であることができる。
【0030】
補完因子は、第1の遺伝子改変によって不良にされた機能を提供する。補完因子の存在下では、不良のアグロバクテリウム株による植物又は植物の葉の細胞へのT−DNAのトランスフェクションが回復する。補完因子は小分子化合物であることができ、又は第1の遺伝子改変に依存して、タンパク質又は該タンパク質をコードする核酸のようなポリマー性化合物であることもできる。アグロバクテリウム株が栄養要求性である場合、補完因子はアグロバクテリウム株の増殖及び感染性を可能にする。アグロバクテリウム株が、第1の遺伝子改変として不良のVirE2、GALLS、及びVirF遺伝子を有する場合、補完因子はそれぞれVirE2、GALLS、又はVirFであるか、それらをコードする。アグロバクテリウム株が化学的に調節されたプロモーターの制御下にある場合、補完因子は、化学的に調節されたプロモーターを調節可能な、好ましくは誘導可能な小分子化合物であることができる。
【0031】
補完因子は、その目的を遂行できるように提供される必要がある。補完因子が小分子化合物である場合、植物又は植物の葉の浸潤に用いるアグロバクテリウム株の水性懸濁液中に存在し得る。補完因子がタンパク質である場合も、この水性懸濁液に加えることができる。しかしながら、タンパク質である補完因子(例えばVirE2、GALLS、又はVirF)はDNAにコードされ、浸潤時、植物又は植物の葉に存在するように提供されることが好ましい。1つの態様では、植物又は植物の葉に、補完因子又はそれをコードするDNAを植物又は植物の葉に導入可能な第2のアグロバクテリウム株を浸潤させる。第2のアグロバクテリウム株は、第1の遺伝子改変を有さず、本発明の異種DNA配列も有さないアグロバクテリウム株であることが好ましい。本発明のアグロバクテリウム株及び第2のアグロバクテリウム株は、植物又は植物の葉を浸潤させる際、混合物として用いることができる。少量のそのような混合物が(好ましくは含まれている)本発明の方法を実施する環境を逃れるならば、そのようなアグロバクテリウム株の混合物は、2つの株が互いに近接する可能性を迅速に低下させるように迅速に希釈される。したがって、非標的生物は2つのアグロバクテリウム株に同時に触れず、よって、人工核酸も外来核酸も(例えば異種DNA配列)そのような非標的生物を形質転換しないであろう。
【0032】
他のより好ましい態様では、補完因子は植物又は植物の葉で安定に又は一過性に発現する。補完因子の安定な発現が好ましい。この態様では、植物は、補完因子をコードする遺伝子改変(以下及び特許請求の範囲において「第2の遺伝子改変」と称する)を提供される。アグロバクテリウム株は第2の遺伝子改変を有する植物を排他的に形質転換するため、高度な生物学的安全性はこのようにして達成することができる。アグロバクテリウム株が望ましくない生物又は望ましくない植物を形質転換する可能性は、これらの生物は第2の遺伝子改変を欠失しているため、非常に低い。
【0033】
これまでに記載した方法の生物学的安全性をさまざまな方法で更に向上させることができる。レプリコンをコードする配列部分は、植物又は植物の葉においてレプリコンの伝播を可能にする機能が不良のレプリコンをコードすることができる。当該機能は、レプリコンの遠隔距離移行又は細胞間移行のための機能、例えばそれぞれウイルスコートタンパク質又はウイルス移行タンパク質であり得る。レプリコンは少なくとも遠隔距離移行の可能性を欠失していることが好ましく、それにより、他の植物へのレプリコンの伝播を回避することができる。遠隔距離移行の可能性の欠失は、原理的にはタンパク質発現効率に不都合であり得る。しかしながら、本発明のシステムでは、本発明の他の要素によって、例えば、植物又は植物の葉の主要部分を浸潤させることによって、及び/又は以下に記載するRNAレプリコン形成の改良方法によって、この不都合を補完することができる。
【0034】
生物学的安全性を更に改良するための他のアプローチは、腫瘍抑制活性タンパク質Osa(REF)を発現する植物のトランスフェクションにアグロバクテリウム株を用いること、及びvirE2遺伝子を提供するための第2のアグロバクテリウム株を用いることである。更に、アグロバクテリウム株は、非標的生物へのT−DNA伝達を低下させるために、遺伝子改変された菌体数感知システム又は病原性誘発調節システムを有することができる。
【0035】
本発明の方法の生物学的安全性は、アグロバクテリウム株が、プラスミドDNAの他の細菌への接合性伝達を不良にする更なる遺伝子改変を有するように改良することもできる。接合性伝達は、以下のアグロバクテリウム遺伝子:oriT、traG、及びtraFのうちの少なくとも1つの機能を無効にすることによって達成することができる。
【0036】
異種DNA配列の配列部分はDNAレプリコンをコードしてもRNAレプリコンをコードしてもよい。RNAレプリコンが好ましく、以下の記載はほとんどRNAレプリコンをコードする配列部分に関係する。
【0037】
レプリコンをコードする配列部分は、レプリコン形成に必要な配列と、レプリコンから発現されるべき目的配列とを含有する。発現されるべき目的配列は、典型的には目的タンパク質をコードし、レプリコンから、又はRNAレプリコンのサブゲノムRNAから目的タンパク質を翻訳するための調節配列を含有することができる。
【0038】
レプリコンがRNAレプリコンである場合、レプリコン(i)のRNAレプリコン機能に必要な配列は、とりわけ前者が後者のDNAコピー(例えばcDNA)である場合におけるRNAウイルスの配列に相当する。DNAレプリコンの場合、レプリコン機能に関する配列は、細胞核で複製するための機能を有するDNAレプリコンを提供する;DNAレプリコンは、異種DNA配列又はその部分を鋳型として用いる複製によって異種DNA配列から形成することができる。RNAレプリコンの場合、レプリコン形成に関する配列は、細胞質で複製するための機能を有するRNAレプリコンを提供する。RNAレプリコン機能に関する配列は、典型的には、RNA依存性RNAポリメラーゼ(RdRp、レプリカーゼ)のような複製に関与する1以上のタンパク質をコードする。レプリコン機能に関する配列は、更に、サブゲノムプロモーター、転写エンハンサー、又は翻訳エンハンサーのようなレプリカーゼの発現に必要な機能であるか又はそれをコードすることができる。植物におけるRNAウイルスの細胞間伝播又は浸透伝播に関与する移行タンパク質又はコートタンパク質のようなタンパク質は、異種DNA配列中に存在し得るが、レプリコン機能を有さない。いずれにしても、レプリコンをコードする配列部分は野生型植物ウイルスをコードしない。レプリコン機能に関する配列は、植物RNAウイルスはレプリコン機能のための容易に接近可能な供給源であるため、植物RNAウイルスの配列に由来することが好ましい。RNAレプリコンの場合、「由来する」とは、レプリコン機能に関する配列が、RNAウイルスの対応する配列のDNAコピーであり、DNAコピーは細胞核に含有される異種DNA配列に組み込まれることを意味する。「由来する」とは、レプリコン機能に関する配列は、RNAウイルスの対応するRNA配列の正確なDNAコピーである必要はないが、以下に記載するように、機能保存的な相違を発揮できるという意味を含む。相違は機能保存的であるため、レプリコン機能に関する配列は、RNAウイルスに関してそうであるのと同様に、レプリコン機能を実施可能なタンパク質をコードすることが好ましい。
【0039】
レプリコンがタバコモザイクウイルスのようなトバモウイルスに基づく態様では、レプリコンはRdRpを含有するか又はコードする。このレプリコンは移行タンパク質を更にコードすることができる。このレプリコンは、コートタンパク質(CP)をコードせず、又は含有しないことが好ましい。
【0040】
更に好ましい態様では、本発明の方法の生物学的安全性は、高度に希釈したアグロバクテリウム株の懸濁液を用いることによって更に改善される。この態様は、アグロバクテリウム株の細胞が環境に広がる可能性を減少させるだけでなく、植物にとって病原体であるアグロバクテリウム株で感染させる際、植物又は植物の葉の露出及びストレスを減少させることによって本発明の方法の効率を改善させる。本発明者らは、驚くべきことに、ある限度内で、本発明の工程(a)で用いるアグロバクテリウム懸濁液の濃度の低下とともに方法の効率が増加することを発見した。とりわけ、植物の細胞で産生されたレプリコンの細胞間移行能は、これらアグロバクテリウム懸濁液濃度の低下とともに増加する。この予想外の現象の理由は未だ特定されていない。この現象はアグロバクテリウム感染に対する植物の反応によるものであり、低アグロバクテリウム濃度ではこの反応は起こらない(又はより少ない程度で起こる)と推測される。
【0041】
この好ましい態様では、600nmのOD(光学密度)が1.0のアグロバクテリウム株の細胞懸濁液を少なくとも25倍、好ましくは少なくとも100倍、より好ましくは少なくとも250倍、最も好ましくは少なくとも1000倍に希釈することによって得ることが可能なアグロバクテリウム細胞濃度のアグロバクテリウム株の細胞懸濁液で植物又は植物の葉を浸潤させる。そのような希釈は、600nmの算出OD値がそれぞれ高くても0.04、好ましくは高くても0.01、より好ましくは高くても0.004、最も好ましくは高くても0.001のアグロバクテリウム懸濁液をもたらす。
【0042】
次に、レプリコンがRNAレプリコンである場合、発現効率を顕著に増加させる重要な態様が記載される。この態様では、レプリコン機能に関する配列は、植物RNAウイルスの配列の選択された位置で、植物RNAウイルスの配列との機能保存的な相違を発揮し、その相違は、相違を発揮しないRNAレプリコンと比較して、レプリコン形成頻度を上昇させる。その相違は、全体的なプロセスのスイッチがいったん入ると、植物細胞におけるレプリコン形成頻度上昇の原因となる(以下を参照されたい)。レプリコン形成頻度上昇と相違とのあいだの因果関係を示す関連性は、相違を有するレプリコン機能に関する配列と相違を有さないレプリコン機能に関する配列とのあいだのレプリコン形成頻度を比較することによって実験的に試験することができる。そのような実験的比較は、実施例に記載するように、例えば目的配列を発現するプロトプラストを計数することによって行うことができる。容易に検出可能な緑色蛍光タンパク質(GFP)のようなレポータータンパク質をコードする目的配列をこの目的に用いることが好ましい。更に以下に記載するように、細胞間伝播不可能なRNAレプリコンとの実験的比較を行うことも好ましい。
【0043】
機能保存的な相違は、植物RNAウイルスの配列の選択された位置でレプリコン機能に関する配列に導入される。選択された位置は、核で転写されたRNAレプリコンが機能性レプリコンとして細胞質に出現する可能性の低さの原因である植物RNAウイルスのレプリコン機能に関する配列内に位置する。そのような選択された位置は、高A/T(U)含量、即ち高A含量及び/又は高T含量(RNAレベルでは高U含量)を有するか、又は潜在スプライシング部位、即ち核のスプライシング装置によってスプライシング部位として認識される配列部分を有することが好ましい。選択された位置は、以下に例示するように、RNAウイルスのRNAプロファイルを解析することによってRNAレプリコンが基づくRNAウイルス中に同定することができる。更に、選択された位置は、本発明による相違を発揮しないRNAレプリコンをコードする異種DNAでトランスフェクションした後、植物細胞で形成されるRNAを解析することによって実験的に同定することができる。この実験的解析は、RT−PCRによって、好ましくはRT−PCR産物の配列決定とともに行うことができる。このようにして、RNAレプリコンを破壊するスプライシングイベントを示す望ましくないスプライシング産物を同定することができる。更に、望ましくないスプライシングの正確な部位を同定し、そして機能保存的な相違を選択された位置に導入することによって是正することができる。
【0044】
機能保存的な相違は、RNAレプリコン形成頻度に対する選択された位置の有害効果を抑制することによってRNAレプリコン形成頻度を上昇させる。機能保存的な相違は、RNAレプリコンをコードする配列のレプリコン機能に関する配列において高A/T含量を低下させることによって、RNAレプリコンにおいて高A/U含量の低下を含むことができる。高A/U含量は、相違が機能保存的である限り、少なくとも部分的な欠失又はG/C塩基による少なくとも部分的な置換(例えば遺伝コードの縮重を用いて)によって低下させることができる。更に、植物RNAウイルス由来配列のA/Uに富んだ領域に隣接する潜在スプライシング部位は除去することができる。そのような機能保存的な相違は、1つの、好ましくはいくつかの選択された位置に導入することができる。
【0045】
好ましい機能保存的な相違は、植物RNAウイルスの配列に由来する配列のA/Uに富んだ位置の近傍又はその中への1以上のイントロンの挿入、最も好ましくは核イントロンの挿入、又は核イントロンを形成可能な1以上の配列の挿入を含む。驚くべきことに、A/Uに富んだ位置又はその近傍へのイントロンの導入は、RNAレプリコン形成頻度の上昇をもたらすことが発見された。いくつかのイントロンを導入することができ、さまざまな数の挿入イントロンに関し、本明細書に例示している。1より多いイントロンの効果は累積的である。更に、イントロンの挿入は、他の選択された位置で他の機能保存的な相違と組み合わせることができる。
【0046】
図11は、発現されるべき目的配列内ではあるが、核イントロンを形成可能な配列の導入例を示す。図11の例では、異種DNA配列の一部のリコンビナーゼによって触媒される反転によって2つの半イントロンからイントロンが形成される。この原理もRNAレプリコンのレプリコン機能に関する配列に応用することができる。同一細胞内で2つの異なるRNAレプリコンが形成される態様では、2つの異なるレプリコン間の組換えは、異なるレプリコン上に存在する2つの半イントロンから1つのイントロンを形成することができる。更に、RNAレプリコンは、いずれもレプリコンではない2つのレプリコン前駆体間の組換えによって形成することができる。この場合も、異なるレプリコン前駆体に由来する2つの半イントロンから1つのイントロンを組み立てることができる。
【0047】
RNAレプリコンをコードする配列部分を有する異種DNA配列は、転写がRNAレプリコン形成の必要条件であるため、転写プロモーターに機能可能に連結されているか又は連結可能である。アグロ浸潤させた全ての組織において目的配列の一過性発現を達成するには、転写プロモーターは構成的プロモーターであることが好ましい。構成的プロモーターの例は当該技術分野において公知である。
【0048】
本発明の1つの態様では、RNAレプリコンをコードする配列は、RNAレプリコンを共にコードする1以上のセグメントを有する。即ち、RNAレプリコンは1つの連続DNAによってコードされていない。実際、RNAレプリコンは2以上のセグメントによって不連続にコードされ、それにより、セグメントは同一T−DNA上に好ましくは互いに隣接して存在することができる。次に、RNAレプリコン形成は、例えば組換えによるセグメントの再配列を必要とすることができる。組換えのためのリコンビナーゼは、追加のアグロバクテリウム株によって、又は遺伝子操作された植物宿主によって提供することができ、よって一過性発現を植物宿主にとどめることができる。例として、レプリコン機能に関する配列の一部(例えばレプリカーゼをコードする配列の一部)が、そのような配列の他の部分に対して反転方向で異種DNA配列中に存在することができる。反転部分は組換え部位に隣接することができる。次に、レプリコン機能は提供できないため(例えば転写物は機能性レプリカーゼをコードしないため)、異種DNAの転写物はレプリコンではないであろう。組換え部位を認識する部位特異的リコンビナーゼを提供することによってセグメントの1つを反転させ、その結果レプリコンが連続的にコードされる。この態様では、リコンビナーゼを提供することにより、レプリコン形成と目的配列の発現に切り替えるためのスイッチとして機能し(更に以下を参照されたい)、高度な生物学的安全性に寄与することができる。
【0049】
あるいは、セグメントの1つはアグロバクテリウムのT−DNA上に存在することができ、他の1つは植物染色体に組み込むことができる。次に、RNAレプリコンの形成は、RNAレプリコンを組み立てるために、両方のセグメントの転写と、両方の転写物のトランススプライシングを必要とするであろう。この態様は、PCT/EP03/02986号に詳細に記載されているように、子孫植物又は細胞においてRNAレプリコンを共にコードするセグメントを迅速に分離するために用いることができ、システムの生物学的安全性に寄与することができる。
【0050】
本発明の方法は、本発明の異種DNA配列による植物又は植物の葉の一過性トランスフェクションを含む(工程(a))。「一過性トランスフェクション」という用語は、植物染色体に安定に組み込まれた異種DNAを有する形質転換されたトランスジェニック細胞を選択することなく異種DNAを導入することを意味する。一過性トランスフェクションは、通常、異種DNA配列によってコードされる遺伝子の一過性複製及び/又は一過性発現を提供する。本発明のトランスフェクションは、目的配列の発現を引き起こす。通常、発現は、植物又は植物の葉にアグロ浸潤にて異種DNA配列を提供後、自然発生的に起こる。必要に応じて、発現を引き起こすか又はスイッチを入れるさまざまな方法を用いることができる。本発明の方法のスイッチを入れるためにリコンビナーゼを用いる場合、リコンビナーゼは植物又は植物の葉に一過性に提供することができ、その提供が目的タンパク質の発現スイッチとして作用することができる。そのようなリコンビナーゼは植物細胞に安定にコードされ、構成的又はストレス誘導性プロモーターの制御下でリコンビナーゼを発現できることが好ましい。プロモーターの誘導によりリコンビナーゼ発現を誘導することにより、目的配列の発現を引き起こすことができる。
【0051】
本発明の方法の工程(b)は、工程(a)でトランスフェクションした、好ましくは浸潤させた植物又は植物の葉から目的タンパク質を単離することを含む。目的タンパク質の単離は、発現した目的タンパク質を含有する植物又は植物の葉の均質化を含むことが好ましい。タンパク質を植物から単離するためのさまざまな方法が、本明細書中に援用されるPCT/EP02/09605号、及びそこで引用されている文献に記載されている。
【0052】
本発明は、原理的には、感染性のDNA又はRNAウイルスが存在し、一過性発現を提供するアグロ浸潤が有効であるあらゆる植物に適用することができる。好ましい植物はベンサミアナタバコ及びタバコのようなタバコ種である;タバコ種以外の好ましい植物種は、ペチュニア、アブラナ、カラシナ、クレス、ルッコラ、マスタード、イチゴ、ホウレンソウ、アカザ、レタス、ヒマワリ、及びキュウリである。
【0053】
好適な植物/DNA又はRNAウイルスのペアは、以下に示すDNA及びRNAウイルスのリストに由来することができる。特に、本発明によるRNAレプリコン形成が非常に高率であるため、本発明を実施する際、植物ウイルスの植物種特異性はほとんど目立たない。同様に、本発明は、あらゆるRNAウイルスに基づいたRNAレプリコンにより用いることができる。RNAウイルスは、通常、宿主植物の細胞核の外部で展開し、レプリコンを細胞核内で産生する場合、そのようなウイルスに基づいたレプリコンを効果のないものにする選択された位置を有するであろう。本発明は、異なる植物RNAウイルスのあいだで改良レベルがさまざまであり得るが、全てのRNAウイルスに適用することができる。本発明が基づくことができる最も好ましい植物RNAウイルスは、トバモウイルス、特にタバコモザイクウイルス、及びジャガイモXウイルスなどのポテックスウイルスである。タバコモザイクウイルスの場合、通常、発現されるべき目的配列によって置換されるコートタンパク質であろう。移行タンパク質は、除去するか、又は発現されるべき目的配列によって置換することもできる。しかしながら、好ましくは、タバコモザイクウイルスに由来するRNAレプリコは、移行タンパク質をコードし、コートタンパク質が目的配列によって置換されている必要がある。
【0054】
植物とRNAレプリコンが由来するウイルスとの好ましい組み合わせは:タバコ種とトバモウイルス、タバコ種とタバコモザイクウイルスなどである。
DNAウイルスの中で、最も好ましいウイルスベクターはジェミニウイルスで構築することができる。ビーンゴールデンモザイクウイルス(BGMV)に基づくウイルス発現ベクターについての記載は、実施例15に提供されている。
【0055】
本発明の主要な応用は、目的タンパク質の植物又は植物の葉における産生である。本発明の方法を植物で実施する場合、タバコ種のようにヒト又は動物の食物連鎖に入らない植物が好ましい。アグロ浸潤後の植物全体又は植物の部分は、閉鎖環境、例えば温室又は特別に設計されたチャンバーに、所望の発現レベルを提供するのに必要なインキュベーション時間、閉じ込められるであろう。
【0056】
本発明の効率は、植物発現システムの新たな局面を達成するようなものである。本発明により達成可能な発現レベルは、下流処理費用(目的タンパク質の分離・精製を含む)が本発明の方法を他の大規模発現システムと競合させるのに十分なほど低いものである。安定に形質転換された植物を用いた先行技術の発現システムでは、細胞の小さな画分でレプリコンを産生するため、ウイルスに基づいたベクターを用いた場合であっても発現レベルは低い。植物中に伝播するレプリコンは、特に遠隔距離に対する伝播が遅いため、この問題を是正することができない。したがって、植物において発現は均一に進行せず、植物のある部分では目的タンパク質の分解が既に起こるが、他ではタンパク質発現が開始してもいない。更に、先行技術のトランスフェクション又はアグロ浸潤に基づいた一過性発現システムは、1つの産生システムで高発現レベルも高い生物学的安全性も提供しない。本発明は、コンピテント植物宿主のアグロ介在送達を介して植物全体でも分離した植物の部分(例えば葉)でも均一に発現を誘引させる。レプリコンを産生しない細胞の小さな画分を、隣接細胞由来のレプリコンによって速やかに感染させることができる。本発明は、大規模で使用できる初めての高収率植物発現システムを提供する。本発明は、一過性発現システムを特異的に遺伝子操作されたコンピテント植物宿主に限定することによって高い生物学的安全性も提供し、それにより非コンピテント(非標的)生物において発現する確立を低下させ、システムの生物学的安全性を増加させる。
【発明を実施するための最良の形態】
【0057】
発明の好ましい態様
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
RNAレプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下で植物又は植物の葉に浸潤させることによって植物又は植物の葉にトランスフェクションすることを含み、
レプリコンをコードする配列は、
(−) 植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(−) 発現されるべき目的配列、
を含有し、
アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されている。
【0058】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
好ましくはRNAレプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下で植物又は植物の葉に浸潤させることによって、植物又は植物の葉をトランスフェクションすることを含み、
レプリコンをコードする配列は、
(−) 植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(−) 発現されるべき目的配列、
を含有し、
アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変を提供されており、そして
植物又は植物の葉は、補完因子をコードし、植物又は植物の葉においてそれを発現する第2の遺伝子改変を有する。
【0059】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
600nmの算出光学密度が高くても0.04、好ましくは高くても0.01、より好ましくは高くても0.004、最も好ましくは高くても0.001に相当する細胞濃度を有する、RNAレプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株の細胞懸濁液を補完因子の存在下で植物又は植物の葉に浸潤させることによって植物又は植物の葉にトランスフェクションすることを含み、
レプリコンをコードする配列は、
(−) 植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(−) 発現されるべき目的配列、
を含有し、
アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されており、そして
レプリコン機能に必要な配列は、選択された位置で、植物RNAウイルスとの機能保存的な相違を発揮し、その相違は、相違を発揮しないRNAレプリコンと比較して、レプリコン形成頻度を上昇させる。
【0060】
更に好ましい態様を特許請求の範囲に定義する。
発明の詳細な説明
本発明者らは、土壌細菌であり、有効な媒介生物であるアグロバクテリウムを用いてRNAウイルスベクターをDNA前駆体として送達するための、高度に活性な合成鋳型を開発した。本明細書において、この改良「アグロ感染」法を用いて、植物の実質的に全ての成熟した葉において一過性遺伝子増幅及び高レベル発現を同時に開始することができ、そしてそのようなトランスフェクション経路を工業的規模で安価に実施することができるを示す。産生を遺伝子操作された「コンピテント」植物宿主に限定し、産生方法の一部ではない他の植物又は微生物へのアグロバクテリウムの感染能を制限することができるような方法で標的植物宿主生物、アグロバクテリウム、及び/又は植物ウイルスを遺伝子操作することによって、生物学的安全性の問題にも取り組む。この技術は、3つの生物学的システムの利点を組み合わせる:ウイルスの速度と発現レベル/収率、アグロバクテリウムのトランスフェクション効率と浸透送達、及び植物の翻訳後能力と低産生コスト。提案した方法は、目的産物をコードする異種核酸による植物の安定な遺伝子改変を必要としない工業的産生経路を可能にし、より安全で制御され、近年の産業基盤に適合している。
【0061】
本発明は、ウイルスレプリコン又はその前駆体のアグロバクテリウム介在送達を用いた目的タンパク質の高収率大規模産生のための一過性発現システムについて記載する。レプリコンは目的配列を発現可能であり、システムは組み込まれた生物学的安全性を有し、システムの3つの主要成分のうちの少なくとも2つ(遺伝子操作された植物宿主、アグロバクテリウム、レプリコン)を相互依存にする。
【0062】
驚くべきことに、システムの主要成分のうちの1つから他の1つへ機能を部分的に委譲することによって生ずるそのような相互依存を、システムの効率に悪影響を及ぼさずに方法全体をよりよく制御し、生物学的安全性を増大させるようになし得ることを発見した。本発明の基本原理を図1(A)に示す。ウイルスレプリコン前駆体のアグロ浸潤に基づいた大規模タンパク質産生方法は、目的タンパク質の発現に必要な機能を実施する際にアグロ浸潤に用いるアグロバクテリウム及び/又はレプリコンを補完するトランスジェニック宿主植物にて実施することができる。RNAレプリコンに関するそのような補完の1つの例を実施例4に記載する。この実施例では、植物全体を機能性移行タンパク質(MP)を欠失したRNAレプリコンをコードする異種DNA配列(図5)により一過性に形質転換した。このレプリコンは細胞間移行できなず、その結果、ウイルスMPを有する発現カセット(pICH10745、図5)を有さない植物宿主では高収率を提供することができない。
【0063】
ウイルスレプリコンの活性化若しくは操作、又は一過性トランスフェクションに必要な機能を植物又は他のアグロバクテリウム株によってトランスで提供する場合、核から細胞質へのRNA放出効率を増加させるための本明細書に記載の基準を用いたとき、発現効率に悪影響は全く見られない。植物イントロンをウイルスRNAレプリコンをコードするある領域へ組み込むことにより、又はレプリコン機能に関する配列内の潜在イントロンの除去若しくは置換により、宿主植物におけるレプリコンの効率を劇的に相加させることができる(少なくとも102倍)。そのような効率の増加は少なくとも1つの容易に測定可能なパラメータに反映された:ベクターの複製を示す細胞の相対的割合、即ち、レプリコン形成頻度の上昇。RNAレプリコン形成開始のそのような最適化は、実質的に植物全体において目的配列の発現スイッチを同時に入れる能力をもたらし、非改変ベクターよりも短時間で目的タンパク質の収率が劇的に増加する。本発明の実施例6〜14は、核から細胞質へのRNAウイルスベクターの放出効率を増加させるためのベクターの改良アプローチについて記載する。ウイルスベクターの改良に関する詳細な説明は、近年、Marillonnet及び同僚らによって公表された(2005、Nature Biotechnol.、23、718−723)。この改良は、RNAレプリコン、植物宿主、及びアグロバクテリウムのあいだで機能を効率的にやりとりさせるため、本発明の重要な態様である。興味深いことに、TMVに基づいたそのような改良ベクターは、ペチュニア、アブラナ、カラシナ、並びにクレス、ルッコラ、及びマスタードのようなタバコ種以外の植物種において効率的に作用する。最も発現する非ナス科種は、イチゴ、ホウレンソウ、アカザである。レタス(特に苗木)及びヒマワリのようなキク科並びにウリ科のキュウリは、さまざまな植物科由来のいくつかの植物種を、好ましくは更に方法を最適化した後で成功裏に用いることができることを示している。
【0064】
組換えDNAのコード領域へのイントロンの組み込みによる核のトランス遺伝子発現の増加に関する出版物(Mascarenhasら、1990、Plant Mol.Biol.、15、913−920;Bourdonら、2001、EMBO Reports、2、394−398;Rose,AB.、2002、RNA、8、1444−1453;US5,955,330号)にもかかわらず、先行技術には、ウイルスRNAレプリコン前駆体へのイントロンの組み込みが、ウイルスレプリコンの形成頻度及びレプリコンからの目的配列の発現レベルに良い効果を与えるであろうことを示す暗示は全くない。この効果は、核のmRNA転写とウイルスRNAの複製とは異なる細胞内区画で起こることを考慮すると驚くべきことである。RNAレプリコンのcDNAコピーを核内におく場合でさえ、ウイルスレプリコン前駆体の最初のコピーのみが核において産生され、次いで細胞質において核内とは異なる条件下で増幅される。先行技術では、野生型ウイルスcDNAを用いたクローニングのあいだ、大腸菌におけるウイルス遺伝子の「漏出性」発現の細胞障害効果を妨げるために、イントロンの使用が記載されている(Johansen,I.E.、1996、Proc.Natl.Acad.Sci.USA、93、12400−12405;Yangら、1998、Arch.Virol.、143、2443−2451;Lopez−Moya&Garcia、2000、Virus Res.、68、99−107)。イントロンinclusionが、ウイルスcDNAクローンからのレプリコン形成頻度を上昇させることができることを示す暗示は全くない。
【0065】
本発明は、DNA前駆体の転写によって派生し、目的配列の発現のためにデザインされている、RNAウイルス由来レプリコンの形成頻度を根本的に増加させる方法を提供する。この方法は、発現されるべき異種配列のサイズの限界やベクターの高度な不安定性のような、既存のウイルスベクターに基づいた発現システムの限界を克服する。更に、この方法は、より良い生物学的安全性を提供し、トランス遺伝子発現に対する漏出検査制御のデザインを可能にする(非誘導状態でゼロ発現レベル)。そのようなデザインは、RNAウイルス由来レプリコンをデザインするための戦略に統合された部分であるためである。RNAウイルス由来レプリコンの高頻度形成を提供することによって、本明細書に記載のアプローチは、植物全体、又はRNAレプリコンをコードする異種DNA配列を含有する葉のような植物の一部において目的配列の発現を速やかに開始させる。本発明を実施することにより、植物又は植物の葉のアグロ浸潤を介した異種目的配列の発現のためにデザインされた、植物RNAウイルスに由来する実質的に全てのレプリコンの性能を、レプリコン形成頻度の劇的な上昇によって顕著に向上させることができる。
【0066】
さまざまな分類群に属するDNA及びRNAウイルスが、本発明によるRNAレプリコンの構築に好適である。本発明を適用できるDNA及びRNAウイルスのリストを以下に提示する。引用符付きの分類群名(イタリック体ではない)は、この分類群にはICTVで国際的に承認された名称がないことを示している。種名(俗名)を標準体で示す。属又は科について正式に帰属されていないウイルスが示されている:
DNAウイルス:
環状2本鎖DNAウイルス:科:カリモウイルス科、属:バドナウイルス属、基準種:ツユクサ黄斑ウイルス、属:カリモウイルス属、基準種:カリフラワーモザイクウイルス、属:“SbCMV様ウイルス”、基準種:ダイズ退緑斑紋ウイルス、属:“CsVMV様ウイルス”、基準種:キャッサバ葉脈モザイクウイルス、属:“RTBV様ウイルス”、基準種:イネtungro bacilliformウイルス、属:“ペチュニア葉脈透化様ウイルス”、基準種:ペチュニア葉脈透化ウイルス;
環状1本鎖DNAウイルス:科:ジェミニウイルス科、属:マストレウイルス属(サブグループI ジェミニウイルス)、基準種:トウモロコシ条斑ウイルス、属:クルトウイルス属(サブグループII ジェミニウイルス)、基準種:ビートカーリートップウイルス、属:ベゴモウイルス属(サブグループIII ジェミニウイルス)、基準種:ビーンゴールデンモザイクウイルス;
RNAウイルス:
1本鎖RNAウイルス:科:ブロモウイルス科、属:アルファモウイルス属、基準種:アルファルファモザイクウイルス、属:イラルウイルス属、基準種:タバコ条斑ウイルス、属:ブロモウイルス属、基準種:ブロムモザイクウイルス、属:ククモウイルス属、基準種:キュウリモザイクウイルス;
科:クロステロウイルス科、属:クロステロウイルス属、基準種:ビート黄斑ウイルス、属:クリニウイルス属、基準種:レタス伝染性黄斑ウイルス、科:コモウイルス科、属:コモウイルス属、基準種:ササゲモザイクウイルス、属:ファバウイルス属、基準種:ソラマメ壊疽ウイルス1、属:ネポウイルス属、基準種:タバコ輪点ウイルス;
科:ポティウイルス科、属:ポティウイルス属、基準種:ジャガイモYウイルス、属:ライモウイルス属、基準種:ライグサモザイクウイルス、属:バイモウイルス属、基準種:オオムギ縞萎縮ウイルス;
科:セキウイルス科、属:セキウイルス属、基準種:パースニップ黄斑ウイルス、属:ワイカウイルス属、基準種:イネtungro sphericalウイルス、科:トンブスウイルス科、属:カルモウイルス属、基準種:カーネーション斑紋ウイルス、属:ダイアンソウイルス属、基準種:カーネーション輪点ウイルス、属:マクロモウイルス属、基準種:トウモロコシ黄色斑紋ウイルス、属:ネクロウイルス属、基準種:タバコ壊死ウイルス、属:トンブスウイルス属、基準種:トマトbushy矮化ウイルス、属が未分類の1本鎖RNAウイルス、属:カピロウイルス属、基準種:リンゴステムグルービングウイルス;
属:カーライルウイルス属、基準種:カーネーション潜在ウイルス、属:エナモウイルス属、基準種:エンドウひだ葉モザイクウイルス、
属:フロウイルス属、基準種:土壌伝染性コムギモザイクウイルス、属:ホルデイウイルス属、基準種:オオムギ斑葉モザイクウイルス、属:イダエオウイルス属、基準種:ラズベリーbushy萎縮ウイルス;
属:ルテオウイルス属、基準種:オオムギ黄萎ウイルス、属:マラフィウイルス属、基準種:トウモロコシrayado finoウイルス、属:ポテックスウイルス属、基準種:ジャガイモXウイルス;属:ソベモウイルス属、基準種:Southern bean モザイクウイルス、属:テヌイウイルス属、基準種:イネ縞葉枯ウイルス、
属:トバモウイルス属、基準種:タバコモザイクウイルス、
属:トブラウイルス属、基準種:タバコ茎壊疽ウイルス、
属:トリコウイルス属、基準種:リンゴchlorotic leaf spotウイルス、属:ティモウイルス属、基準種:カブ黄斑モザイクウイルス、属:アンブラウイルス属、基準種:ニンジン斑紋ウイルス;マイナスssRNAウイルス:目:モノネガウイルス目、科:ラブドウイルス科、属:サイトラブドウイルス属、基準種:レタス壊死性黄斑病ウイルス、属:ヌクレオラブドウイルス属、基準種:ジャガイモ黄萎ウイルス;
マイナス1本鎖RNAウイルス:科:ブニヤウイルス科、属:トスポウイルス属、基準種:トマト黄化壊疽ウイルス;
2本鎖RNAウイルス:科:パーティティウイルス科、属:アルファクリプトウイルス属、基準種:シロツメクサ潜在ウイルス1、属:ベータクリプトウイルス属、基準種:シロツメクサ潜在ウイルス2、科:レオウイルス科、属:フィジーウイルス属、基準種:フィジー病ウイルス、属:ファイトレオウイルス属、基準種:創傷腫瘍ウイルス、属:オリザウイルス属、基準種:イネragged stuntウイルス;
未分類ウイルス:
ゲノム:1本鎖RNA、種:ニンニクウイルスA、B、C、D、種:ブドウ葉脈fleckウイルス、種:トウモロコシwhite lineモザイクウイルス、種:オリーブ潜在ウイルス2、種:ウルミアメロンウイルス、種:ペラルゴニウムzonate spotウイルス。
【0067】
本発明の一般的原理を図1(A)に示す。本発明の実施に多くのさまざまなアプローチを用いることができる。これらのアプローチは一過性発現を提供不可能なアグロバクテリウム変異体の使用に基づくことができ、変異体はトランスジェニック宿主植物又は第2のアグロバクテリウム株によってトランスに補完することができる。
【0068】
システムの生物学的安全性を高めるために、本発明では感染しやすい植物へのアグロバクテリウムの感染を妨げ、それにより、一過性発現方法に対する制御を高める。植物宿主へのアグロバクテリウムのT−DNA伝達メカニズムに関する包括的な研究があり、これは本発明に成功裏に適用できる。
【0069】
1つの可能なアプローチは、ウイルスレプリコンをコードする異種DNA配列を有するアグロバクテリウム株を遺伝子操作された植物宿主に明確に限定することである。プラスミドpSAのosa遺伝子は、T−DNA輸送に必要なvirE2タンパク質を抑制することによって一過性発現を抑制し、更に植物の核DNAへの組み込みを抑制できる腫瘍抑制活性タンパク質をコードする(Lee,L−Y.ら、1999、J.Bacteriol.、181、186−196)。しかしながら、この抑制は、osa遺伝子を発現するアグロバクテリウムを野生型virE2を有するがosa遺伝子のないアグロバクテリウムと混合することによって、例えば変異体表現型をトランスで補完することによって逆転することができる。virE2を発現するトランスジェニック植物を作製することによって同様の結果を達成することができる。T−DNAは、その5’端に共有結合的に結合したVirD2とともに1本鎖中間体として植物細胞へ輸送されるが、virE2を欠失するアグロバクテリウム変異体は発癌性ではないため、virE2はこの方法の重要な成分である。そのような変異体の発癌性は、virE2を発現するトランスジェニック植物に接種することによって回復させることができる(Citovsky,V.ら、1992、Science、256、1802−1805)。更に、完全にアセトシリンゴン依存性のアグロバクテリウム変異体がある。本発明の実施例5では、本発明の異種DNA配列を有するアグロバクテリウム株の能力がvirE2タンパク質をトランスで補完するための遺伝子操作された植物宿又は野生型アグロバクテリウムに依存する一過性発現方法について記載する。また、アグロバクテリウム・リゾジェネスGALLSタンパク質は、VirE2の機能を補完することができる(Hodgesら、2004、J.Bacteriol.、186、3065−3077)。そのようなシステムは、異種DNAの伝播制御を顕著に改良し、システムの生物学的安全性を高める。この実施例に記載のように、実際に、virE2を欠失するアグロバクテリウムによる野生型ベンサミアナタバコ植物のアグロ浸潤後、目的タンパク質(GFP)の発現は全く検出されなかった。しかしながら、virE2を発現する補完された宿主植物をアグロ浸潤したとき、又はこの第1の遺伝子改変を有するアグロバクテリウム株との混合物中で野生型アグロバクテリウムをアグロ浸潤したとき、GFPは効率的に発現した。実際に、virE2を欠失するアグロバクテリウムによるT−DNA送達効率は、野生型アグロバクテリウムやトランスで補完された変異体アグロバクテリウムと比較して、約10,000倍低下した(実施例5、図17を参照されたい)。
【0070】
生物学的安全性を高めたシステムを設計するためのアプローチは、virE2機能のトランス補完に限定されない。virF遺伝子もvirE2と同様に、virF欠失アグロバクテリウムを補完することによって一過性発現に適した植物宿主を作製するために用いることができる(Regensburg−Tuink,AJ.&Hooykaas,PJ.、1993、Nature、363、69−71)。興味深いことに、osaタンパク質は、virE2タンパク質とvirFタンパク質の分泌をともに妨げる(Chen,L.ら、2000、Proc.Natl.Acad.Sci.USA、97、7545−7550)。あるいは、アグロバクテリウム・ツメファシエンスのIncP型又はIncN型接合伝達システムを異なる細菌、大腸菌株へ導入することができ、この菌株は、安全化アグロバクテリウム・ツメファシエンス株の存在下でT−DNA伝達を提供することができる(Pappas,KM.&Winans,SC.、2003、Appl.Envir.Microbiol.、69、6731−6739)。後者は、コンピテントアクセプター宿主及び当該コンピテント宿主に限定された1種又は複数の細菌ドナー株(例えばドナー株とヘルパー株)を有する、生物学的安全性を高めたシステムを設計するための多くのさまざまな可能性があることを示している。このアプローチは、他の生物への望ましくないT−DNA伝達の可能性を顕著に制限する。
【0071】
植物細胞へのアグロバクテリウムのT−DNA伝達能に加えて、アグロバクテリウムは、多くの他の細菌と同様、他の細菌との接合を介して遺伝物質(通常染色体外プラスミドDNA)を伝達することもできる。工業的アグロバクテリウム株から他のアグロバクテリウム株(例えば野生型アグロバクテリウム)へのそのようなプラスミドDNA(例えばバイナリーベクター)の伝達能は、環境へのトランス遺伝子放出に対する制御を危険にさらすであろう。多くの場合、そのような接合伝達は、多くのバイナリーベクターは接合伝達に必要な構造要素(例えば伝達起点を表すoriT)を含有せず、したがって、大腸菌から目的アグロバクテリウム株へのバイナリープラスミドの接合伝達に基づいた三親性接合を介さず、直接形質転換によってのみアグロバクテリウム株に導入することができるため、トランス遺伝子を有するT−DNA領域を有するバイナリーベクターを安全化Tiプラスミドと組換える場合にのみ可能である。さまざまなバイナリーベクターに関する概説として、Hellens,R.、Mullineaux,Ph.&Klee,H.(2000、Trend Plant Sci.、5、446−451)を参照されたい。しかしながら、バイナリーベクターがoriTを含有しない場合でさえ、依然として、バイナリーベクターと居住安全化Tiプラスミドとのあいだで組換わる可能性がある。そのような組換えは、トランス遺伝子を有するT−DNA領域を含有するプラスミドの形成をもたらし得、それにより、プラスミドは異なる細菌間で接合伝達可能であり得る。いくつかの遺伝子がアグロバクテリウム細胞間の接合伝達に関与している(Cookら、1997、J.Bacteriol.、179、1291−1297;Beckら、1989、J.Bacteriol.、171、5281−5289)。しかしながら、最も必須の領域(oriT、traG、traF)は全て居住Tiプラスミド上に局在している(Oger&Farrand、2002、J.Bacteriol.、184、1121−1131;Farrand、Hwang&Cook、1996、J.Bacteriol、178、4233−4247;Li&Farrand、2000、J.Bacteriol.、182、179−188)。相同組換えによるこの領域の除去は、居住Tiプラスミドとバイナリーベクターとの相同組換えが起こる場合でさえ、細菌間接合伝達を低下させるであろう。接合伝達に関与する領域を欠失するアグロバクテリウム株のデザインは、上記参考文献及び標準の分子生物学的技術を用いた本発明の実施例16に提供された記載にしたがい、容易に行うことができる。実施例16は、接合性プラスミド伝達に関与する遺伝子の欠失が、アグロバクテリウムから植物細胞へのT−DNA伝達効率に影響を及ぼさないことを示している(図20も参照されたい)。
【0072】
本発明による発現システムの発現効率を高めるための更なるアプローチは、アグロバクテリウム感染を限定する宿主細胞因子とアグロバクテリウム因子の使用である。多くのそのような因子が、アグロバクテリウムが介在する安定な植物トランスフェクションに関する概説に記載されている(Gelvin,SB.、2003、Trends Biotechnol.、21、95−98;Gelvin,SB.、2003、Microbiol.Mol.Biol.Rev.、67、16−37)。例えば、トランスジェニックタバコ植物においてシロイヌナズナ遺伝子VIP1を過剰発現させることにより、アグロバクテリウムによる一過性の及び安定な遺伝子トランスフェクションに対してより顕著に感受性にすることを見出した(Tzfira,T.、Vaidya,M.&Citovsky,V.、2002、Proc.Natl.Acad.Sci.USA、99、10435−10440)。植物のトランスフェクション効率を向上させる要因は、アグロバクテリウム病原性遺伝子の構成的発現であり得る。通常、そのような遺伝子は、環境因子又はvir遺伝子の発現を誘引するカスケードの制御下にあり、フェノール化合物、糖、pH変化によってスイッチを入れることができる。これらの因子、VirG のVirAタンパク質介在リン酸化を引き起こし、次いで他のvir遺伝子のプロモーターを活性化することができる。VirG内の単一アミノ酸置換によって起こる突然変異は、VirAとは独立して他のvir遺伝子の構成的発現を引き起こすことができる(Hansen,G.、Das,A.&Chilton,MD.、1994、Proc.Natl.Acad.Sci.USA、91、7603−7607)。この文献では、アグロバクテリウムの変異体VirG株を用いたGUSレポーター遺伝子による一過性発現実験で、タバコ及びトウモロコシの組織においてGUSを発現する焦点数の顕著な増加を検出した。タバコにおいて一過性発現効率を促進するために複数コピーの野生型VirGを用いたとき、同様の効果を観察したが、トウモロコシにおいては実際のところ効果がなかった。著者らは、そのようなVirG変異体株は、扱いにくい植物種の形質転換に有用であり得ることを示唆している。これは、それらが、通常、形質転換が困難な植物種のためのウイルスベクターに基づいた一過性発現システムの開発にも有用であることを意味している。植物細胞へのT−DNA伝達を刺激する、フェノール化合物(Melchersら、1989、Mol.Microbiol.、3、969−977)、アセトシリンゴン、若しくはオパインなどのさまざまな化学因子とともに、又はそれらとは独立した、上記のようなVirG変異体遺伝子の使用は、一過性発現効率を高めるための有用なアプローチである(Berthelotら、1998、Phytochemistry、49、1537−1548)。
【0073】
本発明の一過性発現システム及び方法の生物学的安全性を顕著に高めることができる他の要因は、栄養要求性アグロバクテリウム株の使用である。そのような栄養要求性株は開かれた環境で生存する確率が低いためである。X線処理による栄養要求性アグロバクテリウム株の作製は、Dirks&Peeters(US6323396号)に記載されている。この特許にはいくつかのそのような株(メチオニン若しくはシステイン要求株、又はヒスチジン及びアデニン要求株)が記載されている。生長にメチオニンを必要とする変異体の1つは、ホモシステイン・メチルトランスフェラーゼの機能が欠損している。データベースで完全なアグロバクテリウムゲノム配列を利用可能であることを考慮すると、X線処理によって得られるあまり予測できない結果にかわり、十分に特徴付けられた(配列決定された)アグロバクテリウム株のそのような変異体を、相同組換えによる遺伝子置換を用いて作製することは直接的である。例えば、アグロバクテリウム・ツメファシエンスC58ゲノム配列(GeneBank受託番号NC00305)は、5−メチルテトラヒドロプテロイル−トリグルタミン酸−ホモシステイン・メチルトランスフェラーゼをコードする遺伝子(遺伝子ID 1135697)を含有する。利用可能な配列情報を用いて変異体型遺伝子を作製し、アグロバクテリウムへ更に送達し、目的の栄養要求性変異体 (例えば、ホモシステイン・メチルトランスフェラーゼ変異体型遺伝子の場合、メチオニン代謝欠損変異体) を選択するために、隣接配列とともに、対抗選択可能マーカー遺伝子sacBRを含有する自殺ベクターへクローニングすることができる(Berger&Christie、1993、J.Bacteriology、175、1723−1734)。記載のアプローチ及びさまざまな栄養要求性アグロバクテリウム株に関する公的に利用可能な情報を用いて多くの他の変異体を作製することができる。Collens及び同僚ら(Biotechnol.Prog.20(2004)、890−896)は、挿入変異によって栄養要求性アグロバクテリウム変異体を作製した。そのような変異体株は、アデニン、ロイシン、システイン、又はチアミンの非存在下では増殖できない。これれらの変異体のいくつかは、T−DNA送達効率が低下しているが、他(cys−32)は、野生型アグロバクテリウムよりも高くさえある植物細胞へのT−DNA送達能を有する。影響を受けたことが予想されるアグロバクテリウム遺伝子はおそらくleuB、thiD並びにcysE及びcysKである。当然ながら、栄養要求性株の選択は、植物細胞への効率的T−DNA伝達のためのその適合性に基づくであろう。また、より高い生物学的安全性レベル(野生型への復帰変異頻度が低い)のために、2以上の異なる栄養素供給にたいする栄養要求性株(例えば2以上の変異遺伝子を有する)を作製することができる。栄養要求性アグロバクテリウム株(DleuB及びDpyrE)の作製は、実施例17に記載されている。
【0074】
要求される機能(例えばT−DNA伝達)のみを実施し、及び/又はトランス補完因子(例えばvirE2タンパク質、代謝化合物の添加)の存在下でのみ生存できるアグロバクテリウムの変異体株に加えて、細菌細胞の能動生物学的封じ込め(ABC)システムに基づいたアプローチを用いることができる。そのようなシステムの例は、Ronchel&Ramosによって記載されている(2001、Appl.Environ.Microbiol.、67、2649−2656)。この文献において、著者らは、汚染物質の非存在下で細菌集団の生存又は死を細胞死の誘導を介して自由自在に制御するためにデザインしたABCシステムについて記載している。このシステムは、大腸菌gef遺伝子によってコードされるポリン誘導タンパク質のような死滅遺伝子の使用に基づいている。この遺伝子は、lacI誘導性プロモーターの制御下に置かれる。このシステムにおけるLacI発現は、環境シグナルに依存するプロモーターの制御下にある。ABCシステムをデザインするための他の死滅遺伝子を使用する類似スキームは、数多くの文献に記載されている(例えば、Knudsenら(1995)Appl.Environ.Microbiol.61、985−991;Ronchelら(1998)Appl.Environ.Microbiol.64、4904−4911;Torresら(2003)Microbiology 149、3595−3601)。ストレプトアビジンに基づいた封じ込めシステムは、Szafranski及び同僚ら(1997、Proc.Natl.Acad.Sci.USA 94、1059−1063)、並びにKaplan及び同僚ら(1999、Biomol.Eng.、16、135−140)によって記載されている。
【0075】
あるいは、細菌細胞において機能性の誘導性システムを用いて、特定機能に関与する遺伝子の発現を制御することができる(例えばT−DNA伝達又はアミノ酸などの代謝産物の生合成)。実際に、変異体株(例えば栄養要求性変異体又はvirE2遺伝子機能欠損変異体)を作製するために細菌細胞から遺伝子を削除するかわりに、遺伝子を調節可能プロモーターの制御下に置くことができる。そのような調節可能プロモーターからの目的遺伝子の発現は、この場合テトラサイクリン又はIPTGのような小分子であり得る補完因子の適用を介して制御することができる。例えば十分に記述されたtet及びlacリプレッサーに基づいた細菌の調節可能システムのさまざまな改変を用いて、アグロバクテリウム細胞において目的遺伝子の発現を制御することができる。
【0076】
アグロバクテリウムに加えて、本発明は、植物細胞へのT−DNA伝達のために遺伝子操作された他の微生物に拡大適用することができる。例えば、近年、アグロバクテリウム属以外の共生細菌種を改変して、多くの多様な植物への遺伝子伝達を介在することができることが示された(Broothaertsら、(2005)Nature、433、629−633)。これらの細菌は、安全化Ti−プラスミド及びバイナリーベクターを獲得することによって、遺伝子伝達に適任となっている。
【0077】
植物RNAウイルス(例外はウイロイド−植物細胞核で増幅する小さな非コードRNA−である。概説として、Diener,T.O.、1999、Arch.Virol.Suppl.、15、203−220;Flores,R.、2001、CR Acad.Sci.III、324、943−952を参照されたい)は細胞核では発生せず、細胞質で発生することが知られている。したがって、RNAウイルスの配列は、プレmRNA、rRNA及びtRNAの前駆体を含めたプロセシングされたRNAの、細胞質内の輸送を含めたプロセシング工程に関与するモチーフの存在により、核のRNAプロセシングに抵抗するように適合されていない。5’端キャッピング、スプライシング、3’端生成、ポリアデニル化、分解、塩基及び糖による修飾、並びにエディティング(プラスチド及びミトコンドリア内)などのプロセシングイべントが集中的に研究されている。しかしながら、そのようなイベントの多くの要素は依然として不明確のままである。核におけるプレmRNAの最も劇的な変化は、介在RNA配列(イントロン)が初期転写産物から除去され、同時にエクソンが連結されるプロセスである、プレmRNAのスプライシングのあいだに起こる。スプライシングは、ウリジン(uridilate)に富んだ核の小さなリボ核タンパク質粒子を含む複合体構造であるスプライソームによって介在される。スプライソームは2つの連続工程でスプライシング反応を行う:第1工程−上流エクソン/イントロン接合部の5’スプライス部位を切断してラリアート形成し、第2工程−イントロン/下流エクソン接合部の3’スプライス部位を切断して上流と下流のエクソンを連結する(概説として:Kramer,A.、1996、Annu.Rew.Biochem.、65、367−409;Simpson,GG.&Filipowicz,W.1996、Plant.Mol.Biol.、32、1−41を参照されたい)。イントロン配列に隣接する5’及び3’スプライシング部位のジヌクレオチド(5’/GU;AG/3’)は高等植物で高度に保存されており、単一G置換により関与する部位のスプライシング活性を放棄するであろう。驚くべきことに、植物と動物とのあいだでスプライシング部位が高度に保存されているにもかかわらず、植物において異種イントロンは通常スプライシングされないか、又は不正確にスプライシングされる(van Santen,VL.ら、1987、Gene、56、253−265;Wiebauer,K.、Herrero,J.J.、Filipowicz,W.1988、Mol.Cel.Biol.、8、2042−2051)。植物ウイルスRNAは、核のRNAプロセシング装置に抵抗するための進化的圧力下になかったことを考慮すると、これらのRNAは、いったん核環境に置かれると、スプライシングを含めたそのようなプロセシングの対象となる可能性が非常に高い。この状況は、核遺伝子によってコードされるRNA転写産物とは全く異なる。後者の転写産物は、一連のRNA修飾が核で起こるにもかかわらず、機能性を保つように進化的に適合しているためである。しかしながら、そのような修飾は、ウイルスRNAレプリコン形成に劇的な結果を招くことができる。異種遺伝子のための発現ベクターを作製するために植物ウイルス遺伝子を再操作すると、ウイルス起源のRNA配列と相互作用する配列を更に追加するため、RNAウイルスに基づいたレプリコンの不安定性が更に増し、複製できない欠損RNAを産生するであろう。本発明は、発現ベクターをDNA前駆体として植物又は植物細胞に導入し、一過性発現又は植物染色体DNAへの安定な組み込みを提供する場合に機能性RNAレプリコンの形成頻度を顕著に上昇させる改変を発現ベクターに施すことによって、こうした問題に取り組んでいる。ウイルス由来配列の改変は、RNAウイルスに基づいたレプリコンの効率を高めるための最善策であり得る。改変は、直接的(例えばRNAウイルス由来配列内)でも間接的(例えば非ウイルス起源の配列内)でもよいが、間接的改変は、ウイルス起源の配列に安定効果をもたらし得る。本発明では、RNAレプリコン形成効率の上昇に不可欠であるため、RNAウイルス由来配列内の改変に主に焦点をあてる。
【0078】
驚くべきことに、潜在的に問題のある領域が存在する証拠を発見するための第1の試みは成功した。そして、更に驚くべきことに、予想外にも桁違いの改善を発見したことにより、実験的に確認した。発現ベクターpICH8543(実施例1、図2A)のRNAウイルス由来配列を、潜在イントロンとRNAスプライシング部位の存在に関し、NetgeneIIサーバープログラム(http://www.cbs.dtu.dk/services/NetGene2/)を用いて解析することにより、核RNAプロセシング装置によってスプライシングされ得るイントロン様領域の存在が示された(図6の丸で囲んだ領域を参照されたい)。植物ウイルスRNA配列内で潜在的に問題のある領域(選択された位置)を同定するために使用できるプログラムは、さまざまな生物のためのエクソン/イントロン予測プログラム(http://genes.mit.edu/GENSCAN.html)又はスプライシングシグナル予測プログラム(http://125.itba.mi.cnr.it/〜webgene/wwwspliceview.html)のように、他にも多数存在する。
【0079】
既存のプログラムは全て理想的ではなく、誤りがちであることを考慮すると、潜在的に問題のある領域は実験的に決定することもできる。これは、RT−PCRで(Frohman,MA.、1989、Methods Enzymol.、218、340−356)、又はさまざまな転写産物の濃度を正確に定量するのに好適な、リアルタイムPCRと呼ばれる進歩したバージョン(Gibsonら、1996、Genome Res.、6、995−1001)で、核環境で試験されているDNAベクター由来の転写産物を解析することによって行うことができ、好ましくはPCR増幅産物の配列決定を伴う。本発明の機能保存的な相違は、例えばA/Uに富んだ領域(イントロン様)からG/Cに富んだ領域(エクソン様)への置換を伴うサイレント変異を導入することにより、例えばイントロン様配列をエクソン様配列配列で置換することによって、RNAプロファイルを劇的に変化させる(図3の丸で囲んだ領域を参照されたい)。植物のイントロンはエクソンと異なり、通常A/Uに富んでいる(Csank,C.ら、1990、Nucl.Acid Res.、18、5133−5141;Goodall&Filipowicz、1989、Cell、58、473−483)が、例外がある。例えば単子葉植物では、G/Cに富んだイントロンが発見されている(Goodall&Filipowicz、1989、Cell、58、473−483;Goodall&Filipowicz、1991、EMBO J.、10、2635−2644)。本発明のこの態様を実施するには、A/Uに富んだ領域には、A/U含量が少なくとも55%以上の、少なくとも長さ20ヌクレオチドの配列である長い区間だけでなく、純粋にA/Uを含有する配列の並びの6〜19ヌクレオチドの、より短い区間(「アイランド」)も含まれる。本発明におけるA/U内容は、UよりもAに富んだ配列、又はAのみに富んだ配列、そして逆にこの定義によってカバーされる全ての配列を意味する。実施例6では、A/Uに富んだ領域の改変が、GFPを発現する細胞数を少なくとも10倍増加させることを立証する。これは、図10において、pICH15466(改変ベクター、図2A)でアグロ浸潤させた領域と、pICH14833(対照ベクター、図2A)でアグロ浸潤させた領域とを比較することによって立証される。移行タンパク質(MP)の除去は、初期感染部位から隣接細胞への細胞間移行は起こらないため、機能性RNAレプリコンを有する初代培養細胞の正確な計数を可能にする。実施例7では、多くの潜在スプライス部位(図6B)を含有し、移行タンパク質(MP)のサブゲノムプロモーターをカバーする他のUに富んだイントロン様領域の改変を行った(図8、丸で囲んである)。この改変は、ウイルスベクターpICH15900からのレプリコン形成頻度上昇に劇的な効果を及ぼす。プロトプラスト計数実験(実施例7)で立証したように、試験した2つのタバコ種−ベンサミアナタバコ及びタバコに関し、非改変ベクターpICH14833と比較して、およそ100倍増加した(図10A、Bの対応の浸潤領域を参照されたい)。一般に、本発明に記載のアプローチを用いることにより、RNAレプリコン形成頻度をおよそ300倍上昇させ、即ち、機能性レプリコンを有する細胞の割合を約0.2%(対照ベクター)から50%(改変ベクター)以上に増加させることができた。これは限界ではなく、100%の頻度に達することも非常に現実的であると考えられる。
【0080】
そのように高率のレプリコン形成は、同一植物細胞内で2つの異なるRNAレプリコンから2以上の異なる遺伝子を発現させる、例えば植物RNAウイルスに基づいたベクターを用いて異なる遺伝子を共発現させる道を開く。同一細胞で同時に2以上のレプリコンの同調した開放を達成することは、「先着順」の原則はウイルスベクターでは特に真実であるため、そのような共発現に重要である。異なるウイルスベクターは通常伝播領域で重ならないため、又はそのような重なりはわずかであるため、浸透移行も細胞間移行も役立たない。単純な計算によって、同一植物細胞において2つのレプリコンから2つの目的配列の共発現を達成するための、本発明に記載の技術の重要性を立証する。機能性レプリコン形成頻度が全細胞の0.2%である非最適化ウイルスベクターの場合、2つの異なるRNAレプリコンから2つの遺伝子を共発現する細胞の割合は0.2×0.2=0.04%であるが、機能性RNAレプリコン形成頻度が上昇した構築体の場合(全細胞の50%即ち1/2)、上記細胞の割合は0.5×0.5=0.25、即ち25%であり、例えば約625倍高いであろう。最高性能ベクターを用いると(例えばpICH16191、図10C)、機能性レプリコンを有する細胞の割合はおよそ90%に達することもある(図10C、右上)。これは、2つの独立したレプリコンから2つの異なる目的配列を発現させるためにそのようなベクターを用いることにより、全細胞の80%で共発現が起こり得ることを意味している。技術を更に改善し、100%共発現を達成できる可能性は非常に高いと考えられる。
【0081】
RNAレプリコンから発現されるべき異種目的配列における機能保存的な相違は、特にレプリコン機能に関する配列における相違と組み合わせて、RNAレプリコン形成頻度を上昇させるために使用し得ることに留意することは重要である。例えば、レプリコンの形成及び/又はプロセシングに必要な改変を目的配列内に導入することができる。
【0082】
本発明の他の態様では、核イントロンをレプリコン機能に関する配列に挿入することにより、レプリコン形成頻度が改善された(実施例8)。ウイルスRNA依存性RNAポリメラーゼ(RdRP)のコード領域へのイントロンの組み込み(実施例8及び12)は、改変ベクター(図2A、BのpICH15034、pICH15025、pICH15499)からのレプリコン形成頻度を顕著に(少なくとも50倍)上昇させた(図10A、B)。シロイヌナズナ由来の6つの挿入イントロンを含有するベクターのRNAプロファイルを図9に示す。他の実施例(実施例11)では、MP配列へのイントロンの挿入により、レプリコン形成頻度を少なくとも100倍上昇させる。
【0083】
本発明を実施するために多くの核イントロンを用いることができる。そのようなイントロンの例としては、限定されるものではないが、イネtpi Act1遺伝子及びsalT遺伝子由来のイントロン(Rethmeierら、1997、Plant J.、12、895−899;Xuら、1994、Plant Physiol.、100、459−467;McElroyら、1990、Plant Cell、2、163−171);トウモロコシAdh1遺伝子、GapA1遺伝子、アクチン遺伝子、及びBz1遺伝子由来のイントロン(Callisら、1987、Genes Dev.、1、1183−11200;Donathら、1995、Plant Mol.Biol.、28、667−676;Maasら、1991、Plant Mol.Biol.、16、199−207;WE Cohn、K Moldave監修、『核酸研究と分子生物学の進歩』、第42巻、Academic Press、ニューヨーク、pp.229−257中のSinibaldi&Mettler、1992);ペチュニア ルビスコ遺伝子SSU301由来のイントロン(Deanら、1989、Plant Cell、1、201−208);シロイヌナズナA1 EF1α遺伝子、UBQ10遺伝子、UBQ3遺伝子、PAT1遺伝子由来のイントロン(Curieら、1993、Mol.Gen.Genet.228、428−436;Norrisら、1993、Plant Mol.Biol.、21、895−906;Rose&Last、1997、Plant J.、11、455−464);並びに他の多くのものが挙げられる。合成イントロンも本発明に用いることができる。最小の使用可能イントロン又はそれらの部分は、通常、内部のイントロン配列に隣接する、スプライスドナー部位及びアクセプター部位に限定され得る。好ましくはイントロンのサイズは少なくとも50ヌクレオチド、より好ましくは100〜200ヌクレオチドである必要があるが、実際にはイントロンのサイズに限定はない。しかしながら、構築体のサイズは遺伝子操作に好適なように維持される必要がある。イントロンの由来、その構造及びサイズは、ベクターの性質に応じて個々に選択することができる。選択したイントロン又は対応のイントロン部分の効率を試験するために一過性発現実験を用いることができる。
【0084】
上記改変の効果は累積的であり、例えばイントロンの挿入をMPサブゲノムプロモーターの改変と組み合わせると、レプリコン形成頻度をおよそ300倍に上昇させることができる(実施例9)。RNAレプリコン形成頻度を上昇させるためのイントロン挿入に好ましい領域は、本明細書において選択された領域という。そのような位置は「イントロン様」構造を含有することができる。これは、MP、実際にはMPサブゲノムプロモーターのように問題のある領域の極近傍にイントロンを挿入することによって確認される(実施例11)。レプリコン形成頻度に100倍の上昇が観察された。「エクソン様」領域へのイントロンの挿入は、イントロン様領域への挿入のような明白な効果を有さない(実施例10)。
【0085】
目的配列の一過性発現に適格な、即ち本発明の欠損アグロバクテリウム株の補完に適格な植物宿主の作製に必要な代理機能を有するトランスジェニック植物宿主を作製するためにさまざまな方法を用いることができる。アグロバクテリウムによって運ばれるTi−プラスミドにより(US5,591,616号;US4,940,838号;US5,464,763号)、又は粒子銃若しくは微粒子銃(US05100792号;EP00444882B1号;EP00434616B1号)により、植物細胞をベクターで形質転換することができる。マイクロインジェクション(WO09/209696号;WO09/400583A1号;EP175966B1号)、エレクトロポレーション(EP00564595B1号;EP00290395B1号;WO08/706614A1号)、又はプロトプラストのPEG介在形質転換などのような他の植物形質転換法も用いることができる。ベクター送達方法の選択は、形質転換される植物種に依存し得る。例えば、微粒子銃は通常単子葉植物の形質転換に好ましいが、双子葉植物には、アグロバクテリウム介在形質転換が一般に良好な結果を与える。
【0086】
以下の実施例では、タバコ細胞へのアグロバクテリウム介在ベクター(異種DNA配列)送達を用いた。しかしながら、ベクターは、目的植物種の安定な又は一過性の形質転換又はトランスフェクションに好適な標準技術にしたがい、植物に組み込むことができる。双子葉植物の形質転換技術は当該技術分野において周知であり、アグロバクテリウムに基づいた技術とアグロバクテリウムを必要としない技術が含まれる。非アグロバクテリウム技術には、プロトプラスト又は細胞による外来遺伝物質の直接取り込みが含まれる。これらの技術には、PEG又はエレクトロポレーションを介在する取り込み、粒子銃介在送達、及びマイクロインジェクションが含まれる。これらの技術の例は、Paszkowskiら、EMBO J.、3、2717−2722(1984);Potrykusら、Mol.Gen.Genet.、199、169−177(1985);Reichら、Biotechnology 4:1001−1004(1986);及びKleinら、Nature 327、70−73(1987)に記載されている。いずれの場合にも、形質転換細胞は標準技術を用いて完全な植物に再生される。
【0087】
アグロバクテリウム介在トランスフェクションは、そのトランスフェクション効率の高さと多くのさまざまな種を用いる広範な有用性のために、双子葉植物のトランスフェクションに好ましい技術である。アグロバクテリウムによって日常的に形質転換され得る多くの作物種には、タバコ、トマト、ヒマワリ、ワタ、アブラナ、ジャガイモ、ダイズ、アルファルファ、及びポプラが含まれる(EP 0 317 511号(ワタ)、EP 0 249 432号(トマト)、WO87/07299号(アブラナ属)、米国特許第4,795,855号(ポプラ))。
【0088】
本発明の例では、目的遺伝子を一過性発現させるためのT−DNAのアグロバクテリウム介在送達法であるアグロ接種を用いた(Vaqueroら、1999、Proc.Natl.Acad.Sci.USA、96、11128−11133)。アグロ接種は、小規模から中規模の組換えタンパク質産生システムだけでなく、ベクター最適化の要素としても非常に有用なツールであり、構築体のさまざまな変異体を用いて迅速な結果を得ることが可能である。
【0089】
アグロ浸潤を実施するには、先行技術に記載のように、通常、アグロバクテリウムの一晩培養物を準備する(Marillonnetら、2004、Proc.Natl.Acad.Sci.USA.、101、6853−6857)。通常、一晩培養物の光学密度(O.D.)は波長600nmで3〜3.5単位に達し、アグロ浸潤の前に3〜5倍に希釈され、5〜9×109コロニー形成単位を生ずる(Turpenら、1993、J.Virol.Methods、42、227−240)。発明者らは、102倍、103倍、及び104倍希釈も、特に本明細書に記載のような機能保存的な相違を有するレプリコン機能に関する配列との組み合わせにおいて、非常に効率的に作用することを発見した。驚くべきことに、浸潤させたタバコの葉におけるベクターはその性能を更に向上させ、形質転換アグロバクテリウムの希釈率を上げるとGFPの良好な収率を与えた。例えば、103倍希釈は102倍希釈よりも良好な結果を与えた。102倍希釈は10倍希釈よりも良好なGFP収率を生ずる。この現象に対する可能な説明は、おそらく高濃度の病原性細菌に対する植物の応答の結果としての、非常に濃縮したアグロバクテリウム懸濁液の、ウイルスベクターの機能、例えば細胞間移行に対する負の効果である。この現象は、アグロ浸潤を介する組換えタンパク質産生に必要なアグロバクテリウム量を、先行技術の方法と比較して、少なくとも1桁軽減させるため、大規模な工業的タンパク質発現方法に特別な価値がある。
【0090】
実施例13では、不活化ウイルスRNAに基づいたレプリコンのDNA前駆体について示す。レプリコンは本発明にしたがって最適化される。更に、レプリコンは、非コンピテント植物、例えばゲノムDNAに安定に組み込まれたリコンビナーゼを提供する発現カセットを含有しない植物へアグロ送達された場合に目的配列の発現を妨げる構造を含有する。機能性RNAレプリコンの形成と同様に、発現は、部位特異的組換えにより構築体の一部を反転させることによって誘引することができる。この反転は、2つのイントロンの形成と、機能性目的配列の集合をもたらすことができる。実施例13に記載のシステムは、ウイルスベクターの最適化だけでなく、工業的タンパク質又は医薬用タンパク質を発現させるための植物発現システムを用いる高度な生物学的安全性基準の達成方法を示している。
【0091】
リコンビナーゼの転写は、誘導性の又は他の調節された(例えば発生学的に調節された)プロモーターの制御下に置くことができる。誘導性プロモーターは、その誘導条件によって2つのカテゴリーに分けることができる:非生物的因子(温度、光、化学物質)によって誘導されるものと、生物的因子、例えば病原体又は害虫の攻撃によって誘導できるものである。第1のカテゴリーの例は、熱誘導性(US05187287号)及び低温誘導性(US05847102号)のプロモーター、銅誘導性システム(Mettら、1993、Proc.Natl.Acad.Sci.USA、90、4567−4571)、ステロイド誘導性システム(Aoyama&Chua、1997、Plant J.、11、605−612;McNellisら、1998、Plant J.、14、247−257;US06063985号)、エタノール誘導性システム(Caddickら、1997、Nature Biotech.、16、177−180;WO09/321334号)、並びにテトラサイクリン誘導性システム(Weinmannら、1994、Plant J.、5、559−569)である。植物の化学誘導性システムの分野における最新の発展の1つは、グルココルチコイド デキサメサゾンによってスイッチを入れることができ、テトラサイクリンによってスイッチを切ることができるキメラプロモーターである(Bohnerら、1999、Plant J.、19、87−95)。化学的誘導性システムの概説として:Zuo&Chua(2000、Current Opin.Biotechnol.、11、146−151)及びPadidam,M(2003、Curr.Opin.Plant Biol.、6、169−177)を参照されたい。誘導性プロモーターの他の例は、植物の病原性関連(PR)遺伝子の発現を制御するプロモーターである。これらのプロモーターは、植物を、病原体の攻撃に対する応答における植物シグナル伝達経路の重要成分であるサリチル酸で、又はPR遺伝子発現を誘引可能な他の化学化合物(ベンゾ−1,2,3−チアジアゾール又はイソニコチン酸)(US05942662号)で処理することによって誘導することができる。
【0092】
本発明は、実施例に記載のTMVに基づいたベクターに限定されず、他の植物RNAウイルスに基づいたレプリコンにも適用可能である。他の植物ウイルスRNA配列の解析(実施例14、図13、図14)により、TMVに関して記載したものと類似の選択された位置及び植物核遺伝子のプレmRNAの配列を示す(図12)。これは、本発明に記載のアプローチを用いて、問題のある領域を除去/置換することにより、及び/又は核イントロンを挿入することにより、実際に植物RNAウイルス由来レプリコンを解消することができる強力な証拠である。
【0093】
本発明を用いて植物又は植物細胞において発現させることができる目的タンパク質、それらの断片(機能性又は非機能性)、及びそれらの人工誘導体には、限定されるものではないが、以下が含まれる:デンプン修飾酵素(デンプン合成酵素、デンプンリン酸化酵素、脱分岐酵素、デンプン分岐酵素、デンプン分岐酵素II、顆粒結合型デンプン合成酵素)、ショ糖リン酸合成酵素、ショ糖ホスホリラーゼ、ポリガラクツロナーゼ、ポリフルクタンスクラーゼ、ADPグルコース ピロホスホリラーゼ、シクロデキストリン グリコシルトランスフェラーゼ、フルクトシル トランスフェラーゼ、グリコーゲン合成酵素、ペクチン エステラーゼ、アプロチニン、アビジン、細菌レバンスクラーゼ、大腸菌glgAタンパク質、MAPK4と相同分子種、窒素同化/代謝酵素、グルタミン合成酵素、植物オスモチン、2Sアルブミン、タウマチン、部位特異的リコンビナーゼ/インテグラーゼ(FLP、Cre、Rリコンビナーゼ、Int、SSVIインテグラーゼR、インテグラーゼφC31、又はそれらの活性断片若しくは変異体)、オイル修飾酵素(脂肪酸不飽和化酵素、エロンガーゼなどのような)、イソペンテニル トランスフェラーゼ、Sca M5(ダイズ カルモジュリン)、甲虫型毒素又は殺虫活性断片、ユビキチン結合酵素(E2)融合タンパク質、脂質、アミノ酸、糖、核酸、及び多糖を代謝する酵素、スーパーオキシドジスムターゼ、不活性プロ酵素型プロテアーゼ、植物タンパク質毒素、繊維産生植物における改質繊維、バチルス・スリンギエンシス由来の甲虫活性毒素(Bt2毒素、殺虫性結晶タンパク質(ICP)、CrylC毒素、δエンドトキシン、ポリオペプチド毒素、プロトキシンなど)、昆虫特異的毒素AaIT、セルロース分解酵素、Acidothermus celluloticus由来のE1セルラーゼ、リグニン修飾酵素、シンナモイル アルコール脱水素酵素、トレハロース−6−リン酸合成酵素、サイトカイニン代謝経路の酵素、HMG−CoA 還元酵素、大腸菌無機ピロホスファターゼ、種子貯蔵タンパク質、エルウィニア・ヘルビコラ リコピン合成酵素、ACCオキシダーゼ、pTOM36にコードされるタンパク質、フィターゼ、ケトヒドロラーゼ、アセトアセチルCoA還元酵素、PHB(ポリヒドロキシブタノエート)合成酵素、ポリヒドロキシルアルカノエート(PHA)の合成に関与する酵素、アシルキャリアータンパク質、ナピン、EA9、非高等植物フィトエン合成酵素、pTOM5にコードされるタンパク質、ETR(エチレン受容体)、色素体ピルビン酸リン酸ジキナーゼ、線虫誘導性膜貫通ポアタンパク質、植物細胞の光合成機能又はプラスチド機能を高める形質、スチルベン合成酵素、フェノールをヒドロキシル化可能な酵素、カテコール ジオキシゲナーゼ、カテコール 2,3−ジオキシゲナーゼ、クロロムコネート シクロイソメラーゼ、アントラニル酸合成酵素、アブラナ属AGL15タンパク質、フルクトース 1,6−ビホスファターゼ(FBPアーゼ)、AMV RNA3、PVYレプリカーゼ、PLRVレプリカーゼ、ポティウイルスコートタンパク質、CMVコートタンパク質、TMVコートタンパク質、ルテオウイルス レプリカーゼ、MDMVメッセンジャーRNA、変異体ジェミニウイルス レプリカーゼ、Umbellularia californica C12:0選択性アシル−ACPチオエステラーゼ、植物のC10又はC12:0選択性アシル−ACPチオエステラーゼ、C14:0選択性アシル−ACPチオエステラーゼ(luxD)、植物合成酵素因子A、植物合成酵素因子B、D6−不飽和化酵素、脂肪酸の生合成及び修飾、例えば植物細胞における脂肪酸のペルオキシソームβ酸化において酵素活性を有するタンパク質、アシル−CoAオキシダーゼ、3−ケトアシル−CoAチオラーゼ、リパーゼ、トウモロコシ アセチル−CoA−カルボキシラーゼ、など;5−エノールピルビルシキミ酸−3−リン酸合成酵素(EPSP)、ホスフィノスリシン アセチルトランスフェラーゼ(BAR、PAT)、CP4タンパク質、ACC脱アミノ酵素、翻訳後切断部位を有するタンパク質、スルホンアミド耐性を付与するDHPS遺伝子、細菌ニトリラーゼ、2,4−D モノオキシゲナーゼ、アセト乳酸合成酵素又はアセトヒドロキシ酸合成酵素(ALS、AHAS)、ポリガラクツロナーゼ、Taqポリメラーゼ、細菌ニトリラーゼ、制限酵素、メチラーゼ、DNA及びRNAリガーゼ、DNA及びRNAポリメラーゼ、逆転写酵素、ヌクレアーゼ(DNA分解酵素及びRNA分解酵素)、ホスファターゼ、トランスフェラーゼなどを含めた細菌又はファージ由来の他の多くの酵素。
【0094】
本発明は、工業的酵素(セルラーゼ、リパーゼ、プロテアーゼ、フィターゼなど)、及び線維性タンパク質(コラーゲン、クモの糸タンパク質など)を含めた、商業的に価値があり、医薬的に重要なタンパク質の分子農業及び精製のために用いることができる。ヒト又は動物の健常なタンパク質は、本発明で記載したアプローチを用いて発現させ、精製することができる。そのような目的タンパク質の例としては、とりわけ、免疫応答タンパク質(モノクローナル抗体、1本鎖抗体、T細胞受容体など)、病原性微生物由来のものを含めた抗原、コロニー刺激因子、レラキシン、ソマトトロピン(HGH)及びプロインスリンを含めたポリペプチドホルモン、サイトカインとその受容体、インターフェロン、成長因子及び凝固因子、酵素学的に活性なリソソーム酵素、線溶性ポリペプチド、血液凝固因子、トリプシン、トリプシノーゲン、a1−アンチトリプシン(AAT)、ヒト血清アルブミン、グルコセレブロシダーゼ、天然コレラ毒素B、並びに上記タンパク質の融合体、変異体型、及び合成誘導体のような機能保存的タンパク質、トロンビン、ヒト胃リパーゼ、顆粒球−マクロファージコロニー刺激因子(GM−CMF)、セルピン、ラクトフェリン、リゾチーム、オレオシン、プロトロンビン、α−ガラクトシダーゼが挙げられる。
【0095】
本特許出願で優先権を主張する欧州特許出願第04016011.1号の内容は、本明細書にその全体が援用される。
実施例
以下の実施例は、本発明を具体的に説明するために提示するものである。必要に応じて改変や変形を行うことができる。
【実施例1】
【0096】
TMVに基づいたRNAベクターの構築
アブラナ科植物感染性のトバモウイルス(cr−TMV;Dorokhovら、1994、FEBS Lett.350、5−8)及びカブ葉脈透化ウイルス(TVCV;Larteyら、1994、Arch.Virol.138.287−298)のクローン化cDNAは、ロシアのモスクワ大学のAtabekov教授から入手した。緑色蛍光タンパク質(GFP)遺伝子を含有するウイルスベクターは、いくつかのクローニング工程で作製した。得られた構築体pICH8543(図2A)は、順に:シロイヌナズナ アクチン2プロモーター由来の787bp断片(ACT2、参考文献Anら、1996、GenBank受託番号AB026654、bp57962〜58748)、TVCVの5’端(GenBank受託番号BRU03387、bp1〜5455)、cr−TMV断片(GenBank受託番号Z29370、bp5457〜5677、コートタンパク質CPの開始コドンを除去するためチミン5606がシトシンに変わっている)、配列「taa teg ata act cga g」、合成GFP(sGFP)遺伝子、cr−TMV 3’−非翻訳領域(3’−NTR;GenBank受託番号Z29370、bp6078〜6312)、最後にノパリン合成酵素(Nos)ターミネーターを含有する。断片全体を、CarbR pBIN19由来のバイナリーベクターであるpICBV10のT−DNAの左の境界(LB)と右の境界(RB)とのあいだクローニングした。pICH8543でアグロバクテリウム株GV3101を形質転換し、ベンサミアナタバコの葉に浸潤させた。3dpiで出現したGFP蛍光の焦点は、増殖してコンフルエントになった。驚くべきことに、たとえ浸潤領域の大部分の細胞がウイルスの複製と移行により最終的にGFPを発現しても、GFPを発現する多くの独立した焦点によって検出されたように、ウイルス複製を開始したのはわずかな画分の細胞のみであった。ベンサミアナタバコの葉に35Sプロモーター制御下のGFP遺伝子を浸潤させることにより、浸潤領域のほとんど全ての細胞においてGFPが発現するため(図示していない)、限定要因は植物細胞へのDNA送達でないことが明らかになった。
【0097】
この観察結果について確認するため、MPに変異を含有するウイルスベクター構築体を作製した。pICH14833と呼ばれるこの構築体は、pICH8543に類似するが、MP遺伝子における、MPに存在するEcoRI部位の上流の389bpの欠失により異なっている。この欠失を含めたNcoI−EcoRI断片の配列を配列番号1として付録に示す。ウイルス構築体全体(ACT2プロモーターからNosターミネーターまで)を、pBIN19由来KanR バイナリーベクターであるpICBV49のT−DNAの左の境界と右の境界とのあいだにクローニングした。MP内の欠失のために、この構築体から産生されるレプリコンは細胞間移行できないが、細胞内で自律複製可能である。MPを、例えばカリフラワーモザイクウイルス35Sプロモーターのような構成的プロモーターからトランスで提供すると、細胞間移行を回復させることができる。
【0098】
MP発現構築体を作製するために、クローン化TVCV cDNA(GenBank受託番号Z29370、bp4802〜5628)からPCRにてTVCV MP遺伝子を増幅させ、35Sプロモーターの制御下でバイナリーベクターにサブクローニングした。pICH10745と呼ばれる得られた構築体(図示していない)及びpICH14833でアグロバクテリウム株GV3101を形質転換し、ベンサミアナタバコの葉に浸潤させた。pICH14833単独で浸潤させることにより、浸潤領域内にわずかなGFP発現細胞が出現した。浸潤領域から作製されたプロトプラストを計数することにより、合計500個プロトプラストのうち、わずか1〜3個のプロトプラストがGFPを発現することを発見した(0.2〜0.6%)。pICH14833とpICH10745との共浸潤により、それぞれの初期GFP発現細胞から増殖した、GFPを発現する焦点が形成された。最終的に、細胞間移行により、浸潤領域のかなりの割合の細胞がGFPを発現した(図10A)。
【0099】
ロシアのモスクワ大学のAtabekov教授から入手した、上記アブラナ科植物感染性のトバモウイルス(cr−TMV;Dorokhovら、1994、FEBS Lett.350、5−8)及びカブ葉脈透化ウイルス(TVCV;Larteyら、1994、Arch.Virol.138、287−298)の同一クローン化cDNAを用いて、いくつかの更なるウイルスベクターを構築した。いくつかのクローニング工程で緑色蛍光タンパク質(GFP)遺伝子を含有するウイルスベクターを作製した。得られた構築体pICH16707(図2C)は、順に:シロイヌナズナ アクチン2プロモーター由来の787bp断片(ACT2、参考文献Anら、1996、GenBank受託番号AB026654、bp57962〜58748)、TVCVの5’端(GenBank受託番号BRU03387、bp1〜5455)、cr−TMVの断片(GenBank受託番号Z29370、bp5457〜5677、コートタンパク質CPの開始コドンを除去するためにチミン5606がシトシンに変わっている)、配列「taa tcg ata act cga g」、合成GFP(sGFP)遺伝子、cr−TMV 3’−非翻訳領域(3’−NTR;GenBank受託番号Z29370、bp6078〜6312)、最後にノパリン合成酵素(Nos)ターミネーターを含有する。完全な断片を、KanR pBIN19由来バイナリーベクターであるpICBV29のT−DNAの左の境界(LB)と右の境界(RB)とのあいだにクローニングした。pICH16707でアグロバクテリウム株GV3101を形質転換した。形質転換アグロバクテリウム株を飽和するまでLB培地中で一晩増殖させた。アグロバクテリウムを増殖させた200μlの一晩培養物を、マイクロ遠心機にて8000rpmで3分間沈殿させた。沈殿を、10mM MES(pH5.5)、10mM MgSO4を含有する1mlの溶液に再懸濁させ、ODをおよそ0.7とした。温室栽培ベンサミアナタバコ植物の葉に針の無いシリンジで浸潤させた。浸潤後3日目にGFP蛍光の焦点が出現した。蛍光の焦点は増殖して数日後にコンフルエントになった。
【0100】
OD0.3〜0.4のアグロバクテリウムを有する大量の浸潤溶液(3リットル)を調製した。ポットを用いてベンサミアナタバコ植物全体を逆さにして溶液にサッと浸し、乾燥器に入れ、十分な溶液が葉に浸透するまで1〜2分間真空を施した。次に植物を取り出し、温室に置いて再生させた。浸潤後3〜4日目に、シリンジを用いて手で浸潤させた領域のように、浸潤領域に蛍光の斑点が出現した。写真は、浸潤後10日目のそのような植物を示す(図3)。
【0101】
トバモウイルスなどのRNAウイルスは細胞質で複製し、決して核には入らない。したがって、核のプレmRNAプロセシング装置に曝露されない環境で展開する。その結果、人工ウイルス構築体から核において産生されるRNAレプリコン転写物は、RNAプロセシング装置によって認識され正しくプロセシングされることができない。その上、ウイルスベクター由来のRNAレプリコンは非常に大きい:TMVに基づいたレプリコンの場合およそ7,000ヌクレオチド。そのように大きなサイズの植物遺伝子は非常にわずかであり、そのような遺伝子の大部分は、プレmRNAのプロセシングを容易にし、核から輸送し、プロセシングされた転写産物の安定性を向上させるイントロンを含有する。したがって、正確なプロセシングと、正確にプロセシングされた転写産物の核から細胞質への輸送の効率を向上させるプレmRNAの改変は、ウイルス複製を開始する細胞数の増加をもたらすであろうという仮説を立てた。結局、核へのDNA送達 後、ウイルス複製をより効率的に開始できるRNAウイルスに基づいたベクターを作製するために使用できるアプローチは2つあることがわかる:(1)1つのアプローチは、無用のプロセシングイベント(潜在スプライス部位を用いた選択的スプライシングイベント、又は中途終止イベントなど)を誘導し得る配列特徴の除去である;(2)第2のアプローチは、正しくプロセシングされる転写産物量を増加させ、RNAの核から細胞質への輸送を改善させ、及び/又は転写物の安定性を向上させるイントロンの付加である。
【実施例2】
【0102】
TMVに基づいた改良RNAベクターの構築
目的配列が浸潤領域で発現する速度を向上させるために、91〜443ヌクレオチドのサイズの16のシロイヌナズナ イントロンをベクターの位置1〜16に付加した:(TVCV配列に対する位置、GenBank受託番号BRU03387):1、bp209;2、bp423;3、bp828;4、bp1169;5、bp1378;6、bp1622;7、bp1844;8、bp2228;9、bp2589;10、bp2944;11、bp3143;12、bp3381;13、bp3672;14、bp3850;15、bp4299;16、bp4497;17、bp5099;18、bp5287;19、bp5444。得られた構築体pICH18711(図2C)を、さまざまな段階の(播種後17〜35日目)いくつかのベンサミアナタバコ植物の真空浸潤に用いた。GFP蛍光は2日後に出現し、浸潤後6日目に非常に強かった。写真は、浸潤後4日目を示す(図4)。GFP蛍光は、非改良ベクターpICH16707よりも非常に速く(浸潤後4〜6日)葉領域全体を覆った。改良ウイルスベクターの詳細な説明は、近年公表された(Marillonnetら、2005、Nature Biotechnol.、23、718−723)。
【0103】
植物全体の代わりに、ベンサミアナタバコ植物の枝(茎及び葉)を真空浸潤させた。幹の基部を1杯の水の中に入れることによって幹を回復させた。GFPは3日後に葉に出現し、植物全体ではなく植物の部分を目的遺伝子の発現に用いることができることを示した。
【実施例3】
【0104】
アグロバクテリウムによって送達されたウイルスベクターの真空浸潤を用いた薬用タンパク質の発現
医薬的に関心のある2つのタンパク質を、GFPの代わりに上記改良ベクター中にクローニングした。第1のタンパク質ヒト成長ホルモン(hGH)(配列 Genbank受託番号NM_000515)のクローニングによりベクターpICH17991を得た(図2C)。第2のタンパク質ヒトインターフェロンα(配列 Genbank受託番号V00548)のクローニングによりベクターpICH19081を得た(図2C)。この構築体では、インターフェロンの最初の17アミノ酸(LLVALLVLSCKSSCSVG)が、シロイヌナズナ カルレティキュリン アポプラスト標的配列(matqrranpsslhlitvfsllvavvsaev)で置換されている。構築体をアグロバクテリウム株 GV3101に挿入し、植物全体の浸潤に用いた。両方のタンパク質に関し、高レベルのタンパク質発現が得られた。pICH17991に関し、たとえタンパク質の毒性が浸潤領域で細胞死をもたらしても、浸潤葉組織1gあたり1mgのhGHという高レベルの発現が得られた。
【実施例4】
【0105】
MPを欠失するベクターによる植物全体の浸潤
pICH17811におけるMPの初めのAvrII部位にフレームシフトを作製し(図2C)、構築体pICH18722を得た(図5)。このフレームシフトはMP機能を完全に排除しており、この構築体の浸潤により、個々の細胞のみにおいて複製と発現をもたらす。しかしながら、イントロンの存在のために、多数の細胞が依然としてGFPを発現する。
【0106】
TVCVのMPコード配列を、クローン化TVCV cDNA(GenBank受託番号Z29370、bp4802〜5628)からPCRにて増幅し、35Sプロモーターの制御下、バイナリーベクターにサブクローニングしてプラスミドpICH10745を得た。pICH18722とpICH10745との共浸潤により、少なくともMPが一過性に発現しているあいだは細胞間移行が完全に回復する。pICH10745にてベンサミアナタバコを安定に形質転換した。MPを発現する完全な形質転換体にpICH18722を浸潤させることにより、野生型植物の浸潤はウイルスベクターの細胞間移行をもたらさないが、野生型植物全体にpICH18711を浸潤させた場合と同様のGFP発現をもたらした(図16を参照されたい)。
【実施例5】
【0107】
目的遺伝子の一過性発現を提供するための変異体アグロバクテリウムの補完
a)VirE2を発現するトランスジェニック植物宿主によるトランス補完
VirE2遺伝子を、アグロバクテリウム・ツメファシエンスC58 T−DNA(Gene Bank受託番号AE009437、b.p6368〜8038)のDNA調製物からPCRで増幅し、TVCVのMP遺伝子を置換したベクターpICH10745にクローニングした。得られた構築体pICHVirE2(図15)を、選択剤として50mg/Lのカナマイシン(Km)を用いて、アグロバクテリウム・ツメファシエンス(GV3101)、及びベンサミアナタバコ植物のアグロバクテリウム介在葉ディスクトランスフェクションに用いた、機能性VirE2遺伝子欠失アグロバクテリウム・ツメファシエンス固定した(Horshら、1985、Science、227、1229−1231)。MPにフレームシフトを有する構築体pICH18722(図5)にてVirE2機能欠損アグロバクテリウム・ツメファシエンス株を形質転換した。この欠損は、VirE2遺伝子の欠失により引き起こし、野生型植物とpICH10745で形質転換した植物のアグロ浸潤実験に用いた。
【0108】
コンピテント及び野生型宿主を用いて、葉表面の同一領域あたりのGFP発現細胞数を計数することにより、又は(より正確には)対照及び改変ウイルスベクターを用いたウイルス複製の開始頻度を比較するために図10Cに示すように、コンピテント及び野生型宿主におけるGFP発現プロトプラストの割合を測定して比較することにより、一過性発現効率の比較を行った。一過性発現を提供するT−DNA送達のそのような相対頻度は、生物学的安全性を構築していないシステムとの比較において、システムの生物学的安全性レベルの測定として用いることができる。
【0109】
VirE2機能を欠損した他の多くのアグロバクテリウム株、例えばVirE2の挿入変異を有する株(Christieら、1988、J.Bacteriol.、170、2659−2667)又はvirE2機能を抑制するosa遺伝子を有するプラスミドpSAを有する株(Lee、L−Yら、1999、J.Bacteriol.、181、186−196)をこの実験に用いることができる。更に、Tiプラスミド(例えばGeneBank受託番号NC003065;NC001277)及び染色体DNA(例えばGeneBank受託番号NC003063;NC003063)を含めたアグロバクテリウムゲノム配列情報の利用可能性により、相同組換えによって機能性遺伝子をその変異体型に置換することで、あらゆる種類の変異体を作製することができる。さまざまに希釈した、VirE2含有及びVirE2欠損アグロバクテリウム株の一晩培養物を比較することによって、植物細胞へのT−DNA伝達効率に対するVirE2遺伝子の効果について試験した。アグロバクテリウム株はともにpICH18711ウイルスベクターをバイナリーベクターのT−DNA領域内に含有する(図2C)。そのような比較の結果を図17に示す。VirE2欠損アグロバクテリウム株によるT−DNA伝達効率が、機能性VirE2遺伝子含有アグロバクテリウムと比較して、少なくとも10,0000倍低下していることは写真から明らかである。コンピテント(pICHVIRE2で形質転換されている)及び野生型のベンサミアナタバコ植物の葉における一過性発現効率の比較を行い、pICHVirE2で形質転換したトランスジェニック植物からのVirE2遺伝子機能の非常に効率的なトランス補完を示した(図18)。異なる希釈系列(示していない)を用いて実験することにより、トランスジェニック宿主からのVirE2遺伝子機能のトランス補完効率が、機能性VirE2遺伝子を有するアグロバクテリウム株の使用に匹敵することを立証した。
【0110】
b)VirE2を発現する野生型アグロバクテリウム株との混合によるトランス補完
pICH18722を有するvirE2機能欠損株を、virE2機能をトランスで補完するアグロバクテリウム・ツメファシエンス株C58と混合することによって、virE2機能の補完を達成した以外は、先の実施例に記載の実験を反復した。新たに接種した両アグロバクテリウム株の培養物を光学密度OD600が2.5〜3.0になるまで一晩増殖させ、植物組織の浸潤のために等量で混合した。浸潤用最終混合物を50倍、100倍、1000倍、及び10000倍に希釈した。植物組織の浸潤をベンサミアナタバコ及びタバコの葉にて行った。pICH18711バイナリーを有するΔVirE2株(機能性MPを有する)を用いて同様の実験を行った。この実験の結果を図19に示す。pICH18711バイナリーを有する変異体株(T−DNA+、ΔVirE2)と野生型株(T−DNA−、VirE2)との共浸潤は、野生型株によるT−DNAの直接送達(T−DNA+、VirE2)よりも、T−DNA送達がまさに効率的であることは明らかである。
【実施例6】
【0111】
イントロン様配列の除去により、細胞質におけるウイルスRNAレプリコン形成頻度が上昇する
pICH4351由来のRNAレプリコンの配列をNetgeneIIサーバープログラム(http://www.cbs.dtu.dk/services/NetGene2/)を用いて解析したところ、選択的スプライシングイベントを誘導し得るいくつかのイントロン様配列特徴に気づいた。そのような特徴の1つは、RdRPの初めにある、0.6kbのウリジンに富んだ領域(GenBank受託番号BRU03387のヌクレオチド827〜1462に対応)である(図6A)。pICH14833において、オリジナル配列とは異なる、PCRで変異させた配列で、54ヌクレオチド置換によりこの領域を置換した(付録に配列番号2として示す配列、図7)。52ヌクレオチド置換は、Tに富んだ配列を、よりGCに富んだ配列に置換するために行った。RdRPのタンパク質配列を変えないように、全てのヌクレオチド置換をサイレントにした。この変異断片も、推定潜在スプライスドナー部位とアクセプター部位をそれぞれ除去するために導入された2つのヌクレオチド置換(829位と1459位;GenBank受託番号BRU03387に対する座標)を含有する。これらの変異の効果を試験するために、得られたクローンpICH15466(図2A)を、pICH10745(トランス移行タンパク質)と共に、又は伴わずに、ベンサミアナタバコの葉にアグロ浸潤させた。浸潤後8日目に、pICH15466を浸潤させた領域において、GFP発現細胞数の10倍増加を観察した(pICH14833と比較、図10)。これは、ウイルスレプリコンからイントロン様配列を除去することにより、無用の選択的スプライシングイベントを妨げ、より効率的にウイルス複製を開始することを示唆している。pICH15466とpICH10745との共浸潤は、非改変レプリコンと同様の速度で改変レプリコンの細胞間移行をもたらす。これは、RNA配列の改変がウイルスベクターの細胞間移行に影響を及ぼさなかったことを示している。
【実施例7】
【0112】
MPサブゲノムプロモーターにおけるイントロン様配列の除去
第2の潜在的に問題のある領域はMPサブゲノムプロモーターに対応する(図6B)。この領域は非常にTに富んでおり、イントロン配列に非常に近似している。その結果、多くの潜在スプライスドナー部位及びアクセプター部位が、イントロン予測プログラムによって隣接配列中に予測される。残念ながら、サブゲノムプロモーター機能に影響を及ぼすことなくこの領域を容易に改変することはできない。サブゲノムプロモーターを構わずに全領域を完全に変異させ、MPをトランスで提供し、予想されるMP発現の喪失を補償することにした。この構築体からMPは発現しないため、目的遺伝子の発現を駆動するために必要なCPサブゲノムプロモーターを含有する3’配列以外は、MP配列の大部分も削除した。したがって、pICH14833の383bp断片(GenBank受託番号BRU03387のbp4584〜5455)を297bp変異断片で置換した。得られた構築体pICH15900(図2A)を、pICH10745と共に、又は伴わずに、ベンサミアナタバコの葉にアグロ浸潤させた。興味深いことに、pICH14833を浸潤させた葉領域と比較して、複製開始細胞数の大幅な増加を検出した。浸潤葉領域から調製したGFP発現プロトプラストを計数することにより、この改変で、未改変pICH14833と比較して、ウイルス複製開始細胞数が80〜100倍増加すると概算している。pICH15900をpICH10745(p35S−MP発現カセット)を共浸潤させ、細胞間移行によるGFP蛍光の増加を検出した。しかしながら、この増加は、細胞間移行がなくても非常に多くの細胞が既にGFPを発現していたため、非常に限られていた。pICH15900含有アグロバクテリウム懸濁液の1000倍希釈を、pICH10745含有アグロバクテリウムの未希釈懸濁液と共浸潤させることにより、分離したGFP発現焦点を生じさせた。蛍光焦点は、pICH14833を用いて得た対照焦点と同様の明るさ及び同一サイズであった。これは、pICH15900の改変及びMPのトランス送達は、複製レベル、目的配列の発現、及び細胞間移行に関し、レプリコンの機能性を傷つけないことを示している。同一構築体(pICH14933及びpICH15900、pICH10745と共に、又は伴わずに)をタバコの葉に共浸潤させた。pICH15900の改変は、ベンサミアナタバコの場合と同様に複製開始細胞数を増加させた(pICH14833との比較)。
【実施例8】
【0113】
イントロン付加は細胞質における機能性RNAレプリコンの形成頻度を上昇させる
次に、レプリコンをコードする配列部分へのイントロン付加が複製開始頻度を上昇させるかどうかについて試験した。2つの構築体pICH15025及びpICH15034(図2A)を作製した。いずれも、3つの異なるシロイヌナズナ イントロンをRdRPの2つの異なる領域中に含有する。pICH15025を、RdRPの中央にイントロンを含有するようにデザインした。一方、pICH15034は、RdRPの3’端であるMPサブゲノムプロモーターの上流にイントロンを含有する。イントロンをシロイヌナズナ ゲノムDNAからPCRで増幅し、予定のイントロン/エクソン接合部が重複するプライマーを用いて、PCRでウイルス配列に組み込んだ。イントロン含有断片をAvaI/HindIII断片(付録の配列番号4)としてpICH14833にサブクローニングしてpICH15025を作製するか、又はPstI/Ncol断片(付録の配列番号5)としてサブクローニングしてpICH15034を作製した。
【0114】
2つの構築体をベンサミアナタバコの葉にアグロ浸潤させ、pICH14833と比較した。2つの構築体はウイルス複製開始細胞数を顕著に増加させた(図10A)。この増加は、pICH14833に対して約50倍の改善であると概算した。また、2つの構築体をMP発現クローンとともに共浸潤させ、細胞間移行はイントロンのないクローンと同じであることがわかった。2つの構築体をタバコにおいても試験し、ベンサミアナタバコと同じ改善を観察した(図10B)。
【0115】
6つのイントロン全てを含有する第3のクローンpICH15499を作製した(図9、2B、10A、10B)。この構築体をベンサミアナタバコ及びタバコにおいて試験した。この構築体は、3つのイントロンを有する個々の構築体よりも効率的であったが、その改善は相加的ではなかった。
【実施例9】
【0116】
イントロン付加とイントロン様配列の除去は、細胞質における機能性RNAレプリコン形成頻度を上昇させる
1つの構築体におけるイントロン様特徴の除去と、追加イントロンの付加は、両タイプの改変がウイルス複製開始の向上に寄与できることを示した。pICH15499の6つのイントロンを、変異MPサブゲノムプロモーター領域を含有するpICH15900へサブクローニングした。得られたクローンpICH15860(図2B)をベンサミアナタバコの葉に浸潤させたところ、GFPを発現する全てのプロトプラストのおよそ50%〜90%の範囲で、それぞれの親クローンよりも顕著に良好に作用することを発見した(図10)。最高性能の構築体は、RdRP領域と改変MPサブゲノムプロモーター領域にイントロンを含有する(pICH16191、図10C)。未改変クローンとの比較で、これは80〜300倍の改善に相当する。また、この構築体を、MP発現構築体(pICH10745)と共に共浸潤させたところ、改変が細胞間移行も複製も傷つけないことを発見した。
【実施例10】
【0117】
全てのイントロン付加が細胞質における機能性RNAレプリコンの出現頻度を上昇させるわけではない
2つの異なるシロイヌナズナ イントロンをRdRPの初めに挿入し、クローンpICH15477を得た(図2B)(この領域の配列を配列番号7として付録に示す)。この領域の配列は、イントロンを付加する前に、既に非常に「エクソン様」(例えばGCに富んでおり、潜在スプライス部位がない)に見える。この構築体で、ウイルス複製の開始に全く改善は見られなかった。したがって、あらゆるイントロン付加がウイルスベクターを改善するわけではない。イントロン挿入又は変異生成のために選択した位置は重要なパラメータであるように見える。例えば、MPサブゲノムプロモーターのような問題のある構造の近傍領域に作製した全てのイントロン挿入又はヌクレオチド置換は大きな改善をもたらしたが、既に「エクソン様」である配列へのイントロン挿入は顕著にウイルス複製開始を改善しなかった。
【実施例11】
【0118】
MP配列へのイントロン挿入はウイルスレプリコン形成頻度を上昇させる
初めに、制限酵素AvrIIで消化し、充填し、再連結することによって、MPにフレームシフトを作製した。次にMPに2つのイントロンを挿入した。得られたクローンpICH16422(図2B)をベンサミアナタバコの葉に浸潤させた。機能性ウイルスレプリコン含有細胞数に約100倍の増加を検出した。
【実施例12】
【0119】
MP含有ベクターへのイントロン挿入は自律機能性クローンのウイルス複製開始頻度を上昇させる
pICH15499のKpnI/EcoRI断片pICH8543へサブクローニングした。得られたクローン16700(図2B)は、RdRPに6つのイントロンを有する完全なウイルスベクターを含有する。このクローンをベンサミアナタバコの葉に浸潤させたところ、効率的に複製を開始した。また、このクローンは、追加MPをトランスで提供ことを必要とせずに細胞間移行可能であって。
【実施例13】
【0120】
遺伝子操作された宿主植物におけるアグロ送達された不活性レプリコンの活性化
イントロン含有ウイルスベクター前駆体をトランスジェニック植物へアグロ浸潤させることも可能である。目的タンパク質の産生を運命付けられていない植物(例えば開放環境の植物)の偶発的感染を回避するために、ベクターの一部をアンチセンス方向に存在させた不活性クローンを作製することができる(図11)。反転断片の先端に組換え部位とイントロン配列とを組み込むことにより、適切なリコンビナーゼを用いることによってこの断片を正しい方向に「反転」させることが可能である。組換え部位はスプライシングによってレプリコンから完全に排除されるであろう。プロレプリコン中のイントロンは、組換えと転写の後、効率的に複製を開始させる。1つの具体例では、組換え部位が、目的遺伝子内であってプロレプリコンの下流に局在する。そのような立体配置は、組換え前の遺伝子発現を妨げる。RdRP中やプロモーターの上流のように、プロレプリコンの他の領域に組換え部位が局在する場合、他の立体配置を検討することができる。組換え部位のイントロン配列は、先に記載したように、組換え部位をレプリコンから完全に除去させる利点を有するだけでなく、ウイルス複製効率を上昇させる。リコンビナーゼ活性を提供するトランスジェニック植物宿主は、遺伝子操作して、不活性ウイルスベクター前駆体を含有するベクターと共にアグロ浸潤に用いることができる。ウイルスベクター前駆体は、遺伝子操作された植物宿主の細胞においてのみ活性化することができる。
【実施例14】
【0121】
植物ウイルスRNA配列は潜在的に不安定領域を含有する
選択された植物RNAウイルスのRNAプロファイルを、十分に特徴付けられた植物遺伝子(AtDMC1)のものと同様に、NetgeneIIサーバープログラム(http://www.cbs.dtu.dk/services/NetGene2/)を用いて解析した。AtDMC1について図12に示すRNAプロファイルは、先にcDNA配列とゲノムDNA配列との比較によって同定した14のイントロンの存在を明確に反映している(丸で囲んである)。2つの植物ウイルスのRNAプロファイルは、植物の核環境に置かれた場合、RNAの安定性に問題を引き起こし得る領域を有することは明らかである(図13、図14を参照されたい)。ブロモモザイクウイルス、異なるTMV株、及び他の多くのものなど、いくつかの他の代表的植物RNAウイルス(示していない)のRNAプロファイルを解析した。これらは全て、DNA前駆体として植物細胞へ送達された場合に植物RNAウイルスに基づいたレプリコン形成効率を落とし得る、潜在的に問題のある領域を有している。
【実施例15】
【0122】
ジェミニウイルスベクターによる植物全体の浸潤
ビーンゴールデンモザイクウイルス(BGMV)単離体DSMZ PV−0094に基づいた発現ベクターpICH4300、pICH5202/3、及びpICH5170(図15)を、WO02/077246号の実施例10及び11に記載したように作製した。構築体pICH5170を、実施例5に記載のようにベンサミアナタバコ植物及びタバコ植物の安定な形質転換に用いた。構築体pICH5202/3及びpICH5170を、VirE2機能欠損アグロバクテリウム・ツメファシエンス及びアグロバクテリウム・ツメファシエンスC58株に固定した。固定化ウイルスレプリコン(pICH5202/3)を有する株を、pICH5170で形質転換したタバコ植物全体のアグロ浸潤に用いた。トランスジェニック植物宿主は、いわゆる共通領域(CR)に局在するジェミニウイルスの複製起点を有するGFPを有するDNAレプリコンの複製を提供した。VirE2機能欠損アグロバクテリウム株の場合、非形質転換アグロバクテリウム・ツメファシエンスC58と共にアグロ浸潤を行った。あるいは、pICHVirE2で形質転換したベンサミアナタバコ植物を、pICH5170で形質転換させた植物と交配させた。両構築体由来のT−DNA領域を含有するF1子孫をPCRで選択し、アグロバクテリウム機能を有し、pICH5202/3を有するアグロ浸潤に用いた。
【0123】
固定化ウイルスレプリコン(pICH5170)を有する株をタバコ植物全体のアグロ浸潤に用いた。
上記実験の結果は、トランスジェニック宿主植物又はVirE2発現アグロバクテリウムによって機能を補完することなくVirE2機能欠損株を用いた場合、浸潤植物においてGFPを全く発現しないことを示した。しかしながら、VirE2機能のトランス補完を必要とする実験における発現レベルは、ベクターをアグロバクテリウム・ツメファシエンスC58に固定した場合と顕著な差を示さなかった。
【実施例16】
【0124】
細菌プラスミド接合性伝達欠損変異体の作製
tra遺伝子及びtrb遺伝子は、Tiプラスミドの接合性伝達に関与すると記載されている(Farrandら、1996、J.Bacteriol.、178、4233−4247;Liら、1999、J.Bacteriol.、181、5033−41)。アグロバクテリウム・ツメファシエンス株C58のTiプラスミドからこれらの遺伝子を削除するために、隣接DNA領域を含有するベクターを形質転換に用い、2つの連続的組換えイベントにより、欠失を導入した。欠失ベクターは、正の選択マーカーとしてカルベニシリン耐性を、負の選択マーカーとしてショ糖不耐性を引き起こすsacB遺伝子を含有するベクターpDNR−1r(クローンテック)に基づいている。
【0125】
tra遺伝子(traG、traD、traC、oriT、traA、traF、traB、traH、traM)を欠失させるために、およそ700bpの隣接領域をアグロバクテリウムC58 DNAから、oSM570プライマー(5’−gaggatccaacgtttaggagaaccag−3’)+oSM571プライマー(5’−ttggtctcacggtatacgcacactgaacatgcg−3’)、及びoSM572プライマー(5’−ttggtctcaaccggtttccgtttgtctc−3’)+oSM573プライマー(5’−acgtctagagatcgcgttccagaccaac−3’)を用いてPCRで増幅させた。PCR断片をBsaIで制限処理した後連結し、追加ヌクレオチドを導入することなくBst11071制限部位を作製した;外端にプライマーを用いてBamHI部位とXbaI部位を導入した。BamHIとXbaIで制限処理したベクターpDNR−1rに2つのPCR断片を連結した。得られたプラスミドpICF12721を、エレクトロポレーションによるアグロバクテリウム形質転換に用い、初めはカルベニシリンで、その後抗生物質はないが5%ショ糖の存在下で選択し、2重組換えイベントを有する、ベクター骨格を含有しない細胞を対抗選択した。
【0126】
trb遺伝子(trbB、trbC、trbD、trbE、trbJ)を欠失させるために、およそ700bpの隣接領域を、アグロバクテリウムC58 DNAから、oSM566プライマー(5’−ctgaattcaggcaaacgcaccgtgagatg−3’)+oSM567プライマー(5’−tcaccatgggtcacgcggcactcctg−3’)、及びoSM568プライマー(5’−tcaccatggcccaggcccggcgtgaac−3’)+oSM569プライマー(5’−acatctagatgccggcatcgaagatgttg−3’)を用いてPCRで増幅させた。PCR断片の連結により、追加のヌクレオチドを導入することなくNcoI制限部位を作製した;外端にEcoRI部位及びXbaI部位をプライマーによって導入した。2つのPCR断片を、EcoRII及びXbaIで制限処理したベクターpDNR−1rに連結した。得られたプラスミドpICF12711を上記のようにアグロバクテリウム形質転換に用いた。
【0127】
アグロバクテリウム間の接合伝達を測定するために、オパイン結合リプレッサーをコードするaccR遺伝子をカナマイシン耐性カセットで置換することによって、構成的伝達を示す選択可能Tiプラスミドを構築した。これは、やはりベクターpDNR−1rに基づき、アグロバクテリウムC58 DNAからoSM554プライマー(5’−gagctagctccgtccttcacctgggc−3’)+oSM555プライマー(5’−ttggtctcaccggccgatagccaaaaactgc−3’)、及びoSM556プライマー(5’−ttggtctcgccggccaaactccggtttgc−3’)+oSM557プライマー(5’−atgggcccttcgaacgcaattcctgttgc−3’)を用いてPCRで増幅させた2つの隣接領域、及びそれらのあいだのXmaIII部位にカナマイシン耐性(oSM584プライマー(5’−cctcggccgcgaacggcctcac−3’)及びoSM585プライマー(5’−ctacggccgctgacagctaaaacaattcatcc−3’)を用いてバイナリーベクターpICH18711からPCRで増幅させた)を含有するベクターpICF12741でアグロバクテリウム・ツメファシエンス株C58を形質転換することによって作製した。カナマイシン耐性Tiプラスミドを有する株を、染色体リファンピシン耐性を有するがTiプラスミドのないアクセプター株GV3101と共に、ニトロセルロースメンブラン上で28℃にて2時間、Piper&Farrand(1999、Appl Environ Microbiol.、65、2798−2801)に記載されたのと同様にインキュベーションした。リファンピシンとカナマイシンを添加したプレートでトランス接合個体を選択した。aceを欠失しているが無傷のtra遺伝子及びtrb遺伝子を有するTiプラスミドの場合、投入ドナーあたりの接合効率はおよそ3〜4×10−4トランス接合個体であった。tra及びtrb変異体の場合、接合効率は検出限界以下であった(<10−7)。
【0128】
ベンサミアナタバコ植物に、GFPをコードするウイルスベクターpICH18711を含有する細菌を浸潤させることにより、植物へのDNA伝達に対する変異の影響について試験した。細菌は、葉で単一蛍光スポットを観察できるように、さまざまな希釈率で用いた。植物へのDNA伝達効率は、ace、tra、又はtrb欠失のない細菌と匹敵するものであった。
【実施例17】
【0129】
栄養要求性変異体の作製
栄養要求性アグロバクテリウム株を作製するために、ロイシンの生合成に関与する3−イソプロピルリンゴ酸脱水素酵素をコードするleuB遺伝子とウリジンの生合成に関与するオロチン酸ホスホリボシルトランスフェラーゼをコードするpyrEを選択した。leuBのコード配列は、アグロバクテリウム染色体の逆鎖に局在する仮想タンパク質ATU2792の3’と重複するため、完全に欠失させることができない。したがって、染色体1のbp2792384−2793470を、上流配列の775bpと下流配列の834bpとを、leuBコード配列の5’端に天然に生ずるBspHI制限部位を介して接続することにより欠失させるために選択した。ベクターpDNR−1r(クローンテック)を用いて、アグロバクテリウムの標的遺伝子内に欠失を作製した。
【0130】
leuBと同様に、pyrE遺伝子の異化補完は可能ではない。ウラシルホスホリボシルトランスフェラーゼ遺伝子がアグロバクテリウムに存在するため、ウリジン添加は安価なウラシルで置き換え得る。更に、pyrE欠失は正の選択マーカーとして用いることができる。選択は、野生型アグロバクテリウムは毒性のウラシル類似体である5−フルオロオロチン酸に対して感受性であるが、pyrE変異体は当然耐性であるという事実を利用する。
【0131】
pyrEの開始コドンは、異なるデータベースにおいて異なって注記されている(bp394734−395534、受託番号NP_531105とbp394836−395534、受託番号Q8UI98)。しかしながら、他のpyrE配列に対する相同性はbp394836の前からは開始しない。それにもかかわらず、2つの欠損変異体(欠失は1番染色体の394760位又は394849位から開始し、いずれも395531位で終わる)をデザインした。欠失含有断片を、ベクターpDNR−1rのEcoRI−BamHI部位にクローニングし、ベクターpICF1263(ΔleuB bp2792384−2793470)、pICF1264(ΔpyrE bp394760−395531)、及びpICF1265(ΔpyrE bp394848−395531)を得た(図21を参照されたい)。
【0132】
アグロバクテリウム株GV3101を、プラスミドpICF1263−3(ΔleuB bp2792384−2793470)、pICF1264−1(ΔpyrE bp394760−395531)、及びpICF1265−10(ΔpyrE bp394848−395531)で形質転換した。カルベニシリン含有培地での選択2日目に少量の形質転換体(6〜10cfu/μg)を得た。LB+リファンピシン液及びLB+リファンピシン+サッカロース液中で選択したクローンを連続して増殖させることによって共組換え体を決定した。標的遺伝子に所望の欠失を含有するアグロバクテリウム変異体を、LB+リファンピシン+サッカロース培養物から単離したアグロバクテリウムDNAのPCR解析によって確認した。
【0133】
変異体株1263−3−3及び1263−3−10並びに野生型親株GV3101を、30mg/l及び150mg/l ロイシンの添加の有無において、LB富栄養培地及びM9最少培地中で増殖させた。変異体株と野生型株とのあいだでLB培地での増殖に差はなかったが、M9培地では野生型株のみ増殖できた。M9培地にロイシンを添加した場合、野生型株も変異体株も増殖できた。添加ありと添加なしの培地でのさまざまな変異体株の増殖を図22及び図23に示す。ロイシン濃度の増加とともに増殖率が増加した。アグロバクテリウム変異体1263−3−3をGFPプラスミドpICH18711で形質転換し、ベンサミアナタバコにおける感染効率について解析した。
【0134】
付録
配列番号1(pICH14833のNcoI−EcoRI断片):
【0135】
【化1】

配列番号2(pICH15466の部分):
【0136】
【化2】
配列番号3(pICH15900の部分):
【0137】
【化3】
配列番号4(pICH15025の部分):
【0138】
【化4】
配列番号5(pICH15034の部分):
【0139】
【化5】
配列番号6(pICH15477の断片):
【0140】
【化6】
【図面の簡単な説明】
【0141】
【図1A】図1Aは、生物学的安全性の上昇(A)に基づいた、本発明の一般的原理を表す。
【図1B】図1Bは、RNAウイルスに基づくレプリコン形成頻度の上昇(B)に基づいた、本発明の一般的原理を表す。
【図2A】図2Aは、未改変の及び本発明にしたがい改変したウイルス構築体を図式的に示す。Act2−シロイヌナズナ アクチン2遺伝子のプロモーター;RdRP−ウイルスRNA依存性RNAポリメラーゼ;MP−ウイルス移行タンパク質;NTR−ウイルス3’非翻訳領域;GFP−緑色蛍光タンパク質;インターフェロン−ヒトインターフェロンα 2b;hGH−ヒト成長ホルモン;nos−ノパリン合成酵素遺伝の転写終止領域。
【図2B】図2Bは、未改変の及び本発明にしたがい改変したウイルス構築体を図式的に示す。Act2−シロイヌナズナ アクチン2遺伝子のプロモーター;RdRP−ウイルスRNA依存性RNAポリメラーゼ;MP−ウイルス移行タンパク質;NTR−ウイルス3’非翻訳領域;GFP−緑色蛍光タンパク質;インターフェロン−ヒトインターフェロンα 2b;hGH−ヒト成長ホルモン;nos−ノパリン合成酵素遺伝の転写終止領域。
【図2C】図2Cは、未改変の及び本発明にしたがい改変したウイルス構築体を図式的に示す。Act2−シロイヌナズナ アクチン2遺伝子のプロモーター;RdRP−ウイルスRNA依存性RNAポリメラーゼ;MP−ウイルス移行タンパク質;NTR−ウイルス3’非翻訳領域;GFP−緑色蛍光タンパク質;インターフェロン−ヒトインターフェロンα 2b;hGH−ヒト成長ホルモン;nos−ノパリン合成酵素遺伝の転写終止領域。
【図3】図3は、ベクターpICH16707のアグロ浸潤後10日目のベンサミアナタバコ植物をUV光下にて示す。
【図4】図4は、ベクターpICH18711のアグロ浸潤後4日目の、17日(A)、22日(B)、28日(C)、及び35日(D)齢のベンサミアナタバコ植物をUV光下にて示す。
【図5】図5は、構築体pICH18722及びpICH10745のT−DNA領域を図式的に示す。Act2−シロイヌナズナ アクチン2遺伝子のプロモーター;RdRP−ウイルスRNA依存性RNAポリメラーゼ;MP−ウイルス移行タンパク質;NTR−ウイルス3’非翻訳領域;GFP−緑色蛍光タンパク質;nosT−ノパリン合成酵素遺伝子の転写終止領域;nosP−ノパリン合成酵素遺伝子の転写プロモーター;oscT−オクトピン合成酵素遺伝子の転写終止領域;NPTII−ネオマイシン ホスホトランスフェラーゼII遺伝子のコード配列;TVCV MP−カブ葉脈透化ウイルスの移行タンパク質。
【図6A】図6は、ベクターpICH8543の転写領域のイントロン予測プロファイルを示す。ヌクレオチド番号を横軸に示す。縦軸は、対応の配列/配列領域がコード配列(コード)であるか、ドナー部位(ドナー)若しくはアクセプター部位(アクセプター)として作用する確率を示す。丸で囲んだ部分は、機能保存的な相違を導入すべき選択された位置に相当する。
【図6B】図6は、ベクターpICH8543の転写領域のイントロン予測プロファイルを示す。ヌクレオチド番号を横軸に示す。縦軸は、対応の配列/配列領域がコード配列(コード)であるか、ドナー部位(ドナー)若しくはアクセプター部位(アクセプター)として作用する確率を示す。丸で囲んだ部分は、機能保存的な相違を導入すべき選択された位置に相当する。
【図7】図7は、ベクターpICH15466の転写領域(転写領域の前半の半分)のイントロン予測プロファイルを示す。丸で囲んだ領域は本発明に記載のように改変されている(図6Aとの比較)。
【図8】図8は、pICH15900の転写領域(転写領域の第2の半分)のイントロン予測プロファイルを示す。丸で囲んだ領域は本発明に記載のように改変されている(図6Bとの比較)。
【図9A】図9は、pICH15499の転写領域のイントロン予測プロファイルを示す。丸で囲んだ領域は、6つの挿入された植物核イントロンに相当する。
【図9B】図9は、pICH15499の転写領域のイントロン予測プロファイルを示す。丸で囲んだ領域は、6つの挿入された植物核イントロンに相当する。
【図10A】図10は、ベンサミアナタバコ及びタバコの葉におけるウイルス構築体のアグロ浸潤後のGFP発現を示す。各浸潤領域におけるベクター同定番号を示す。A−ベンサミアナタバコ、アグロ浸潤後8日目。
【図10B】図10は、ベンサミアナタバコ及びタバコの葉におけるウイルス構築体のアグロ浸潤後のGFP発現を示す。各浸潤領域におけるベクター同定番号を示す。B−タバコ、アグロ浸潤後8日目。
【図10C】図10は、ベンサミアナタバコ及びタバコの葉におけるウイルス構築体のアグロ浸潤後のGFP発現を示す。各浸潤領域におけるベクター同定番号を示す。C−アグロ浸潤後5日目に単離したベンサミアナタバコのプロトプラスト。右の写真の多くの光点は、非常に高頻度のレプリコン形成及びGFP発現を示す。
【図11】図11は、本発明にしたがいデザインされたRNAウイルスに基づいたレプリコン前駆体の略図であり、非誘導状態では目的配列(GFP、Gと示す)の発現レベルがゼロである。P−転写プロモーター;T−転写終止領域;SM−選択可能マーカー遺伝子;Ac2−シロイヌナズナ アクチン2遺伝子のプロモーター;RdRP−ウイルスRNA依存性RNAポリメラーゼ;MP−ウイルス移行タンパク質;NTR−ウイルス3’非翻訳領域。
【図12】図12は、直鎖(+鎖)を用いたシロイヌナズナ減数分裂特異的遺伝子AtDMC1(GenBank受託番号U76670)のイントロン予測プロファイルを示す。イントロンコード領域を丸で囲んである。
【図13A】図13Aは、ジャガイモXウイルス(PVX)ゲノム(GenBank受託番号AF172259)の直鎖(+鎖)内の潜在的に問題のある領域(丸で囲んである)の予測プロファイルを示す。
【図13B】図13Bは、ジャガイモXウイルス(PVX)ゲノム(GenBank受託番号AF172259)の直鎖(+鎖)内の潜在的に問題のある領域(丸で囲んである)の予測プロファイルを示す。
【図14A】図14は、アルファルファモザイクウイルスゲノムRNA1(GenBank受託番号K02703)の直鎖(+鎖)の潜在的に問題のある領域(丸で囲んである)の予測プロファイルを示す。
【図14B】図14は、アルファルファモザイクウイルスゲノムRNA2(GenBank受託番号K02702)の直鎖(+鎖)の潜在的に問題のある領域(丸で囲んである)の予測プロファイルを示す。
【図14C】図14は、アルファルファモザイクウイルスゲノムRNA3(GenBank受託番号L00163)の直鎖(+鎖)の潜在的に問題のある領域(丸で囲んである)の予測プロファイルを示す。
【図15】図15は、構築体pICHVirE2、pICH4300、pICH5202/3、及びpICH5170のT−DNA領域の略図を示す。
【図16】図16は、機能性MP(移行タンパク質)欠失ウイルスベクターの細胞間移行のトランスジェニック植物宿主(ベンサミアナタバコ)からのトランス補完の効果を示す。上部のラベルは植物を表し、下部のラベルはアグロ浸潤に用いたベクターを示す。左の写真は図示したように処理した植物の典型的な葉を示す。右の写真はトランスジェニック植物の全体を示す。WT:野生型植物;pICH10745トランスジェニック:TVCV移行タンパク質(MP)を発現するトランスジェニック植物;pICH18722:細胞間移行不良のウイルスベクターによる浸潤;pICH18711:細胞間移行可能なウイルスベクターによる浸潤。GFP発現を検出するために植物をUV光下にて示す。
【図17】図17は、バイナリーベクターpICH18711からのT−DNA伝達効率に対するVirE2遺伝子の効果を示す。VirE2:機能性VirE2遺伝子を含有するアグロバクテリウムを浸潤させたベンサミアナタバコの葉;ΔvirE2:VirE2遺伝子機能不良のアグロバクテリウムを浸潤させたベンサミアナタバコの葉;102〜107の数は浸潤前のアグロバクテリウムストックの希釈を示す(ΔVirE2株とVirE2株のストックは同一OD600)。矢印は同一のT−DNA伝達頻度を発揮する条件を結び付けており、ΔVirE2株は102倍希釈、VirE2株は107倍希釈に相当する。
【図18】図18は、野生型(WT)植物、及びpICH18711バイナリーベクターを含有するΔVirE2アグロバクテリウム株を浸潤させたトランスジェニック(pICHVirE2)植物を示す。写真は、10dpi(浸潤後日数)にUV光下にて撮影した。同一のアグロバクテリウム一晩培養物を、WTの浸潤には10倍に希釈し、トランスジェニック植物の浸潤には1000倍に希釈した。
【図19】図19は、野生型植物への変異体アグロバクテリウム(機能性virE2遺伝子を欠失)と野生型アグロバクテリウム(VirE2遺伝子機能を維持)との共浸潤によるVirE2遺伝子機能のトランス補完を示す。変異体及び野生型のアグロバクテリウムをさまざまな組み合わせで浸潤させたベンサミアナタバコの葉をUV光下にて示す。四角内のラベルは、アグロバクテリウムの個々の種類を表す。四角のあいだの+印は、葉の同一の場所に2種類のアグロバクテリウムを共浸潤させたことを示す。「T−DNA」の前の+印又は−印は、それぞれ、GFPを有するT−DNAの存在及び不存在を示す。機能性VirE2の不存在をΔで示す。
【図20】図20は、aceオペロンを欠失したアグロバクテリウムからのT−DNA伝達効率試験の結果を示す。TiプラスミドpMP90(対照)又はpTiC58Δacc(acc欠失)を有するアグロバクテリウム株GV3101を、GFPをコードするウイルスベクターpICH18711で形質転換し、一晩培養物のさまざまな希釈物を植物の浸潤に用いた。写真は接種後7日目にUV光下にて撮影した。
【図21】図21は、アグロバクテリウムのΔleuB変異体及びΔpyrE変異体を作製するための戦略を表す。
【図22】図22は、補完の有無における、さまざまな培地でのアグロバクテリウム株1264(ΔpyrE)の増殖速度を示す。ΔpyrE変異体株1264−1−2及び1264−1−8はウラシル含有培地においてのみ増殖する。縦軸上の数値はOD600を示す。
【図23】図23は、補完の有無における、さまざまな培地でのアグロバクテリウム株1263(ΔleuB)の増殖速度を示す。ΔleuB変異体株1263−3−3及び1263−3−10はロイシン依存性であるが、野生型親株GV3101はロイシン依存性ではない。縦軸上の数値はOD600を示す。
【技術分野】
【0001】
本発明は、アグロバクテリウムによって標的植物に送達される植物ウイルスベクターに基づいた一過性発現を用いた、植物における生物学的に安全な組換えタンパク質の産生方法に関する。また、本発明は、標的宿主植物又は植物の葉における目的配列の発現方法に関する。本発明は、植物全体又は植物の部分において目的タンパク質の大規模産生を高収率で提供する、ウイルスベクターのアグロバクテリウム介在送達の使用について記載する。本発明の方法は、宿主植物へのアグロバクテリウム介在送達による一過性発現を制限する一方で、望ましくない生物への形質転換を制御する。更に、本発明は、植物又は植物の葉において目的配列を発現させるための生物学的に安全なアグロバクテリウムに関する。本発明は、生物学的安全性が上昇した、植物における高収率の工業的タンパク質産生を提供する。
【背景技術】
【0002】
発明の背景
現在、植物バイオテクノロジーは、植物において外来遺伝子を送達及び発現するための2つのアプローチ:安定な形質転換と一過性発現に依拠している。後者は、アグロ浸潤又はウイルス感染に基づいて構築することができる(概説として:Fischerら、1999、Biotechnol.Appl.Biochem.、30、113−116を参照されたい)。一過性発現は、構成的な、例えば35Sプロモーターの制御下で目的遺伝子の発現を駆動する標準の発現カセットを植物組織にアグロ浸潤させることによって(Vaqueroら、1999、Proc.Natl.Acad.Sci.USA.、96、11128−11133)、又は大規模では、植物にウイルスベクターをトランスフェクションすることによって達成することができる。通常、アグロ浸潤は高収率を提供しないが、p19又はHcProのような転写後遺伝子サイレンシング(PTGS)サプレッサーとの組み合わせにより、タンパク質発現レベルを50倍まで上昇させることができる(Voinnetら、2003、Plant J.、33、549−556)。依然として、ウイルス発現システムによって達成できる生物学的限界を遙かに下回っている(Marillonnetら、2004、Proc.Natl.Acad.Sci.USA、101、6852−6857)。植物組織において目的組換えタンパク質を一過性発現させるためのアグロ浸潤の使用に基づいたバイオリアクターについての記載は米国特許第6,740,526号に提供されている。しかしながら、この特許は、発現されるべきタンパク質の収率をいかにして改善するかについての記載がなく、当該技術分野において公知の他の方法に勝るものではない(Vaqueroら、1999、Proc.Natl.Acad.Sci.USA.、96、11128−11133)。一過性発現のより強力なアプローチは、ウイルスベクターの使用である。レポーター遺伝子(GFP又はDsRed)のTMVに基づいた発現は、システムの生物学的な収率限界に達することができ、新鮮な葉のバイオマス1gあたり数mgの組換えタンパク質を産生することが示された(Marillonnetら、2004、Proc.Natl.Acad.Sci.USA、101、6852−6857)。そのようなシステムの相対収率は、TSPの80%まで達することができ、下流処理を著しく容易にし、費用を低下させる。そのような高い相対収率は、ウイルスによって誘導される宿主タンパク質生合成の遮断により可能である。
【0003】
ウイルスベクターは、アグロ浸潤に用いる慣用のベクターよりも高い発現レベルを提供でき(概説として:Porta&Lomonossoff、1996、Mol.Biotechnol.、5、209−221;Yusibovら、1999、Curr.Top.Microbiol.Immunol.、240、81−94を参照されたい)、機能ゲノミクス研究の強力なツールである(Dalmayら、2000、Plant Cell、12、369−379;Ratcliffら、2001、Plant J.、25、237−245;Escobarら、2003、Plant Cell、15、1507−1523)ことは明らかである。当該技術分野における数多くの出版物及び特許がDNA及びRNAのウイルスベクターに基づいたシステムについて記載している(Kumagaiら、1994、Proc.Natl.Acad.Sci.USA、90、427−430;Malloryら、2002、Nature Biotechnol.20、622−625;Morら、2003、Biotechnol.Bioeng.、81、430−437;US5316931号;US5589367号;US5866785号;US5491076号;US5977438号;US5981236号;WO02/088369号;WO02/097080号;WO98/54342号)。既存のウイルスベクターシステムは、通常、最高性能の点から見ると狭い宿主範囲に限定され、最も好適な宿主におけるそのようなベクターの発現レベルでさえシステムの生物学的な上限を遙かに下回っている。ウイルスに基づいたシステムの重要な問題は、植物細胞へのウイルスレプリコンの送達方法である。大規模産生(例えば農場又は温室において多くの植物で同時に産生すること)に最も広く適用される送達法は、RNAウイルスベクターの感染性コピーの使用である(Kumagaiら、1995、Proc.Natl.Acad.Sci.USA、92、1679−1683)。組換えRNAウイルスベクターは複製サイクルのあいだに異種挿入物を喪失する傾向が比較的高いため、この方法はin vitroでDNA鋳型の転写を必要とし、したがって非効率的で高価である。
【0004】
一過性経路は、非常に速いが、ウイルスの低感染性、植物の大部分にトランスフェクションすることができないこと、及び遺伝子サイズの限界のために非常に限定されている。植物細胞へ感染性ウイルスベクターを送達するためのアグロ浸潤の使用(Liu&Lomonossof、2002、J.Virol.Methods、105、343−348)、又はアグロバクテリウムによって送達されるウイルスベクター成分からのウイルスベクターの組み立て(Marillonnetら、2004、Proc.Natl.Acad.Sci.USA、101、6852−6857)について記載する出版物がある。しかしながら、これらの出版物は、アグロ浸潤された植物組織のそれぞれの細胞におけるウイルスレプリコンの効率的な同調形成の問題に取り組んでおらず、更に、ベクターの伝播は細胞間移行及び浸透移行に関するウイルスの能力に依存している。この移行は比較的長期間を必要とし、通常、ウイルスベクターは感染後およそ10〜14日で最高の可能なベクター収率を提供する。これは、植物の細胞内プロセスを妨げる組換えタンパク質、特に制限酵素、プロテアーゼ、非特異的ヌクレアーゼ、多くの医薬タンパク質のような非常に細胞傷害性のタンパク質の産生には受け入れられない。逆に、アグロ浸潤によって送達される標準ベクターは、アグロ送達後3〜4日で最高の可能な発現レベルに達するが、そのようなベクターによって提供される収率は容認し難いほど低い。したがって、事実上アグロ浸潤された植物組織の全ての細胞において目的遺伝子の発現を提供するにもかかわらず、目的タンパク質を発現するための構成的プロモーターによって駆動される効率の低い標準の転写ベクターによって引き起こされる問題に直面する。反対に、アグロ送達されたウイルスベクターは高発現レベルを提供可能であるが、複製をほとんど開始せず、そのため大部分の細胞で発現を提供せず、ほんのわずかな画分のみである(アグロ浸潤された全組織の1%未満)。その結果、そのようなウイルスに基づいたベクターは発現を提供するのに著しく(3〜4倍)長期間を必要とするが、システムの生産性は、特に細胞障害性タンパク質の場合に、理論上の生産性よりも依然として低い。これは、アグロ浸潤された組織において同調発現を提供しないため、特に細胞障害性遺伝子を発現させる場合に、収率に影響を及ぼし、一過性発現システムにウイルスベクターを使用する上で重大な欠点である。また、感染した植物宿主はその組織の大部分においてウイルスベクターを含有しないため、産生プロセスから組織を除外する。更に、感染植物に対してウイルスベクターの最高の可能な伝播(及び発現レベル)を達成するのに要する時間は、標準のアグロ浸潤プロトコールと比較して3〜4倍長い。その上、一過性発現のための記載のシステムはいずれも、遺伝子操作された生物の使用を含めた工業的規模のタンパク質産生において重要な要素である、システムの生物学的安全性を高める問題に取り組んでいなかった。
【0005】
特許化技術を含め、当該技術分野には数多くの出版物があるにもかかわらず、主に2つの主要な理由により、依然として、商業的な高収率産生のために十分な効率と収率をもって作動するウイルスに基づいた大規模産生システムは存在しない:第1に、一過性の植物ウイルスに基づいた発現システムは、通常特定の宿主に制限され、環境因子の影響を受けやすいため大規模培養に好適であり得ない。その上、これらのシステムは、通常植物宿主のある部分に制限されるため、大部分の植物バイオマスを産生プロセスから除外し、その結果、単位植物バイオマスあたりの組換え産物の相対収率を、トランスジェニック植物において慣用の転写プロモーターによって達成可能なレベルに匹敵するレベルまで最小にする;第2に、各細胞に安定に組み込まれたウイルスレプリコン前駆体を有するトランスジェニック植物宿主を作製することによってウイルスに基づいた産生システムを拡大する試みは、特に、そのような位置におけるレプリコンの標準よりも低い働き、レプリコンから発現されるべき目的遺伝子の「漏出性」、そしてベクターの効率的スイッチシステムの欠失により、解決策を提供していない。RNAレプリコン形成の誘因としてPTGSサイレンシングのサプレッサーを用いることにより、PVXに基づいたベクターにおいて多少の進展が達成されたが(Malloryら、2002、Nature Biotechnol.、20、622−625)、このシステムは、ウイルスベクターの複製を誘発するスイッチ(PTGSサプレッサー)の効率的制御を提供する解決策がないため、依然として実用的価値に遠く及ばない。しかしながら、このシステムは、この種のシステムでこれまで知られた中で最高である、合計可溶性タンパク質(TSP)の3%に達するGUS遺伝子の発現レベルを提供したが、依然として強力なプロモーターの制御下の慣用のトランス遺伝子発現システムよりも良好ではない。植物3分節RNAウイルス、ブロムモザイクウイルス(BMV)に基づいた他の誘導性システム(Moriら、2001、Plant J.、27、79−86)は、目的タンパク質の収率が非常に低かった(3〜4μg/g新鮮重)。これは標準の転写プロモーターによって提供される収率と匹敵する。
【0006】
植物発現システムを用いてこれまでに達成された発現レベルの低さが、これらのシステムが細菌細胞、真菌細胞、又は昆虫細胞の発現システムのような他の発現システムとほとんど競合しない主な理由である。低発現レベルは、植物材料の巨大なバックグラウンドにおけるタンパク質の単離精製のための下流費用を非常に高くする。したがって、単位植物バイオマスあたりの目的タンパク質又は目的産物の収率が増加するにつれて、下流処理費用が急激に減少する。
【0007】
現在、細菌細胞、真菌細胞、又は昆虫細胞の発現システムのような他の大規模発現システムを用いた市場で競合するほど収率及び効率が十分に高い植物の大規模な一過性発現システムはない。そのような植物の発現は以下の基準をできるだけ良好に満たす必要があるであろう:
(i)できるだけ多くの植物組織及び組織の多くの細胞における目的タンパク質の発現を含め、高収率であること;
(ii)植物細胞の生存に対するタンパク質発現の有害な効果を防止するには、目的タンパク質又は目的産物の発現は、処置された植物又は植物組織の全ての植物細胞において同時に開始する必要がある。
【0008】
典型的には、目的タンパク質又は目的産物は、産物又はタンパク質を産生する各細胞にある時点まで蓄積する。しかしながら、蓄積のあいだ、分解過程が進むことが多く、目的タンパク質若しくは目的産物の収率又は品質が低下する傾向にある。したがって、発現スイッチが入った後、目的産物又は目的タンパク質を収穫すべき最適な時点がある。全体的な工程を効率的で収益性のあるものにするには、この最適な時点は、植物の全ての組織又は細胞において、そして選択したロットの全ての植物において同時に到達すべきである;
(iii)システムは、向上したバイオセイフティー性を組み込み、アグロ浸潤に用いるアグロバクテリウムは、少なくとも1つの以下の特徴を有するであろう:開かれた環境における生存率が低いか又はゼロであり、非標的生物又は非標的植物に対する感染性が低いか又はゼロである。
【0009】
したがって、本発明の目的は、大規模応用に拡大可能であり、発現すべきタンパク質の収率が高く、そして同時に、非標的生物を外来DNAでトランスフェクション又は形質転換する可能性が低い、生物学的に安全な、植物システムにおいてタンパク質を発現させる方法を提供することである。
【発明の開示】
【0010】
発明の一般的な説明
この目的は、標的植物又は標的植物の葉における目的配列からの目的タンパク質の一過性発現による目的タンパク質の産生方法によって達成され、この方法は:
レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉に浸潤させることによって植物又は植物の葉にトランスフェクションすることを含み、
レプリコンをコードする配列は、
(i)植物ウイルスに由来する、レプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、アグロバクテリウム株は、補完因子の非存在下では植物又は植物の葉を含めた生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されている。
【0011】
更に、本発明は、
(i)上で定義した通りのアグロバクテリウム株、及び
(ii)上で定義したアグロバクテリウム株の欠陥を補完可能な補完因子をコードする植物又はその種子、
を含むキット部品を提供する。
【0012】
更に、本発明は、
(i)上で定義した通りのアグロバクテリウム株、及び
(ii)上で定義したアグロバクテリウム株の欠陥を補完可能な補完因子をコードする第2のアグロバクテリウム株、
を含む、キット部品を提供する。
【0013】
更に、本発明は:
(i)補完因子の非存在下では植物又は植物の葉の細胞へのT−DNAの導入に必要な細菌の機能を不良にする第1の遺伝子改変を有し、
(ii)他の細菌へのプラスミド伝達の接合能が不良であり、そして
(iii)アグロバクテリウムの増殖に必要な主要代謝産物に対して栄養要求性である、
ことを特徴とする、本発明の方法のためのアグロバクテリウム属の細菌を提供する。
【0014】
自然の一般的原理によれば、1つの特性の改良は他の特性に負に影響する傾向があり、逆も同様であるため、システムの2以上の対立する特性を同時に向上させることは困難である。植物に基づいたタンパク質発現システムでは、発現されるべきタンパク質の高発現レベルを達成することと、高い生物学的安全性を達成することを同時に達成することは、高発現効率はシステムの選択性を低下させ、よって生物学的安全性が低い傾向にあるため困難である。例えば、高率発現システムを達成するために植物の感染に用いるアグロバクテリウム株を非常に感染性にする場合、アグロバクテリウム株は、非標的植物に形質移入する可能性が高く、組換えT−DNAが環境中に伝播する傾向があるが、これは回避しなければならない。一方、アグロバクテリウム株の感染性を低くすることによって生物学的安全性を改善する場合、標的植物における形質移入及び発現効率は劣るであろう。したがって、生物学的安全性と発現効率はそのように対立する特性である。
【0015】
植物に基づいたタンパク質発現システムへの要求は、システムを大規模応用、即ち多くの植物又は多くの植物の葉における同時発現に適合させる必要がある場合、より複雑になってさえいる。一方、大規模応用における生物学的安全性は、トランスジェニック生物が開放環境に拡張する可能性がより高まるため、より重要である。一方、高タンパク質発現効率及び高収率は、経済的に競合し得る方法を達成するには、大規模応において必須である。
【0016】
本発明は、目的タンパク質を産生するための、大規模で非常に効率的且つ生物学的に安全な方法を作製するための手段を初めて提供する。
本発明の方法では、目的タンパク質は、植物又は植物の葉において目的配列から一過性に発現される。一過性発現は、植物又は植物の葉の一過性トランスフェクションによって達成される。「一過性トランスフェクション」という用語は、異種DNA配列の導入が、植物染色体へ異種DNA配列を安定に組み込むためにトランスフェクションされた細胞を選択することなく行われることを意味する。目的配列は目的タンパク質をコードする。目的配列は、外来DNAを植物細胞へ導入可能なアグロバクテリウム株を用いて植物又は植物の葉へ導入される。目的配列は、レプリコンが由来する植物ウイルスに対して異種である。即ち、本発明の方法は、植物又は植物の葉を野生型植物ウイルスで形質転換する場合を含まない。DNA配列は異種目的配列を含有するため異種である。目的配列は、アグロバクテリウム株に対して固有の配列でないことが好ましい。
【0017】
植物又は植物の葉をアグロバクテリウム株で一過性にトランスフェクションする。植物又は植物の葉は、植物の多くの細胞及び多くの組織が1つの作業工程でトランスフェクションされるようにトランスフェクションされる必要がある。これは、アグロ浸潤によって最高に達成することができるが、植物又は植物の葉の主要部分のトランスフェクションを可能にする他の方法も適用することができる。したがって、植物又は植物の葉にアグロバクテリウム株を浸潤させることによって植物又は植物の葉に一過性にトランスフェクションすることが好ましい。浸潤は、システムの効率に顕著に寄与するため、本発明の重要な要素である。効率が高い程、植物又は植物の細胞のより多くの細胞が目的タンパク質を発現し、さまざまな組織及び細胞において発現開始が良好に同調する。浸潤は、植物又は植物の葉の多くの細胞を1つの作業工程で形質転換することを可能にし、それにより、目的タンパク質の発現がさまざまな植物組織及び植物のさまざまな部分において進む時点が実質的に同調する。更に、浸潤は、かなりの割合の植物又は植物の葉の細胞が形質転換される可能性を実質的に増加させる。したがって、レプリコンの遠隔距離移行の可能性は、全体的な効率を更に増加させることができるが、トランスフェクション及び発現は、植物又は植物の葉におけるレプリコンの遠隔距離移行に依存しない。
【0018】
本発明の方法は、植物への大規模な応用のため、即ち多くの植物で平行して実施するためにデザインされる。本発明の方法では、好ましくは少なくとも5、より好ましくは少なくとも10、より好ましくは少なくとも30、更により好ましくは少なくとも100、最も好ましくは少なくとも1000の植物にアグロバクテリウム株を平行してトランスフェクションする。
【0019】
本発明の方法を植物全体で実施する場合、トランスフェクション(好ましくは浸潤)は植物のいくつかの部分で行う。例えば、大部分の葉とトランスフェクションするか浸潤させる。全ての葉をトランスフェクションするか浸潤させることが好ましい。より好ましくは、植物の全ての葉及び茎をトランスフェクションするか浸潤させる。目的タンパク質を単離するためにその後収穫される全ての組織をトランスフェクションするか浸潤させるべきである。根における発現及び収穫はシステムの全体的な効率を更に増加させることができるが、通常、根を浸潤させる必要はない。アグロバクテリウム株の懸濁液中に植物を逆さにしてサッと浸し、真空を適用し、その後真空を急速に開放することによって植物全体又は少なくともその土の上の部分を浸潤させることができる。この手順は、規模を拡大することができ、多くの植物に順々に又は同時に適用することができる。
【0020】
本発明の方法を植物の葉で行う場合、葉の1以上の領域で浸潤を行う。好ましくは、葉に噴霧するか、アグロバクテリウム株の懸濁液にサッと浸し、好ましくは、その後アグロバクテリウム細胞を葉の組織に押し付けるか又は吸収させる。本発明にしたがい使用する植物の葉は、植物に付いていることが好ましい。本発明の方法は植物組織培養物で行わないことが好ましい。
【0021】
浸潤又はアグロ浸潤はアグロバクテリウム懸濁液を用いたトランスフェクション法として定義することができ、アグロバクテリウムを植物組織(細胞間隙)に押し付けるために圧力差を用いる。
【0022】
あるいは、ウイルス粒子を送達するために行ったようにして(Pogueら、2002、Annu.Rev.Phytopathol.、40、45−74)、アグロバクテリウムと研磨剤との混合物を植物へ展開するための高圧噴霧装置を用いてアグロバクテリウムを植物組織へ送達することができる。
【0023】
アグロバクテリウム株は、補完因子の非存在下ではT−DNAの生物への形質移入を不良にする第1の遺伝子改変を提供される。これは、生物学的安全性を向上させるための本発明の必須要素である。この生物は、異種DNA配列によるトランスフェクションが望ましくない生物である(「非標的生物」)。非標的生物は、とりわけ、植物及び細菌又は真菌のような微生物である。補完因子の非存在下では、本発明の植物種は非標的生物でもない。補完因子の非存在下では、アグロバクテリウム株は、そのような非標的生物に感染することも形質導入することも不良であり、好ましくは不可能である。
【0024】
アグロバクテリウム株、植物(又は植物の葉)、及び補完因子は、アグロバクテリウム株による植物の感染時、目的配列のトランスフェクション及び/又は発現を可能にするために選択され、一方で非標的生物のトランスフェクション及び/又は発現は起こりそうにない。非標的生物のトランスフェクションに関するアグロバクテリウム株欠陥、好ましくは不能は、アグロバクテリウム株の以後第1の遺伝子改変を称する遺伝子改変によって達成される。
【0025】
第1の遺伝子改変は、植物又は植物の葉の細胞へのT−DNAの導入に必要なアグロバクテリウム株の機能を不良にすることが好ましい。これは、標的生物のトランスフェクションに必要な機能をコードするアグロバクテリウム遺伝子を欠失又は分裂させることによって達成することができる。そのような機能は、とりわけ、Ti−プラスミド上、例えば植物又は植物の葉の細胞へ導入すべきT−DNAを有するTi−プラスミド上にコードされる。そのような遺伝子の例は、アグロバクテリウム遺伝子VirE2、GALLS、及びVirFである。
【0026】
他の態様では、第1の遺伝子改変は、アグロバクテリウム株を栄養要求性にし、それにより、その増殖を外部から加えられた代謝産物依存性にする。外部から加えられた代謝産物の非存在下では、アグロバクテリウム株は、生物にT−DNAをトランスフェクションすることができない。
【0027】
更なる態様では、第1の遺伝子改変として、生物のトランスフェクションに必要なアグロバクテリウム株の機能を、化学的に調節された異種プロモーターの制御下に置く。そのような遺伝子の例は、アグロバクテリウム遺伝子VirE2、GALLS、及びVirFであり、例えば、lacプロモーターの制御下に置くことができる。そのような態様では、補完因子は化学的に調節されたプロモーターを調節可能な小分子化合物である。lacプロモーターの場合、補完因子はIPTG又は乳糖であり得る。したがって、植物又は植物の葉に適用されるアグロバクテリウム懸濁液は、植物の感染、及び好ましくは植物の細胞へのT−DNAの導入を可能にする、IPTG又は乳糖のような小分子化合物を含有することができる。
【0028】
他の態様では、第1の遺伝子改変は、アグロバクテリウム株を条件的致死性にし、補完因子は、アグロバクテリウム株の生存に必要な、アグロバクテリウム株の主要代謝産物である。
【0029】
上記態様は組み合わせることができる。例えば、アグロバクテリウム株が、植物又は植物の葉の細胞へのT−DNAの導入に必要な機能が不良である場合、システムの生物学的安全性を高めるために、更に栄養要求性及び/又は条件的致死性であることができる。
【0030】
補完因子は、第1の遺伝子改変によって不良にされた機能を提供する。補完因子の存在下では、不良のアグロバクテリウム株による植物又は植物の葉の細胞へのT−DNAのトランスフェクションが回復する。補完因子は小分子化合物であることができ、又は第1の遺伝子改変に依存して、タンパク質又は該タンパク質をコードする核酸のようなポリマー性化合物であることもできる。アグロバクテリウム株が栄養要求性である場合、補完因子はアグロバクテリウム株の増殖及び感染性を可能にする。アグロバクテリウム株が、第1の遺伝子改変として不良のVirE2、GALLS、及びVirF遺伝子を有する場合、補完因子はそれぞれVirE2、GALLS、又はVirFであるか、それらをコードする。アグロバクテリウム株が化学的に調節されたプロモーターの制御下にある場合、補完因子は、化学的に調節されたプロモーターを調節可能な、好ましくは誘導可能な小分子化合物であることができる。
【0031】
補完因子は、その目的を遂行できるように提供される必要がある。補完因子が小分子化合物である場合、植物又は植物の葉の浸潤に用いるアグロバクテリウム株の水性懸濁液中に存在し得る。補完因子がタンパク質である場合も、この水性懸濁液に加えることができる。しかしながら、タンパク質である補完因子(例えばVirE2、GALLS、又はVirF)はDNAにコードされ、浸潤時、植物又は植物の葉に存在するように提供されることが好ましい。1つの態様では、植物又は植物の葉に、補完因子又はそれをコードするDNAを植物又は植物の葉に導入可能な第2のアグロバクテリウム株を浸潤させる。第2のアグロバクテリウム株は、第1の遺伝子改変を有さず、本発明の異種DNA配列も有さないアグロバクテリウム株であることが好ましい。本発明のアグロバクテリウム株及び第2のアグロバクテリウム株は、植物又は植物の葉を浸潤させる際、混合物として用いることができる。少量のそのような混合物が(好ましくは含まれている)本発明の方法を実施する環境を逃れるならば、そのようなアグロバクテリウム株の混合物は、2つの株が互いに近接する可能性を迅速に低下させるように迅速に希釈される。したがって、非標的生物は2つのアグロバクテリウム株に同時に触れず、よって、人工核酸も外来核酸も(例えば異種DNA配列)そのような非標的生物を形質転換しないであろう。
【0032】
他のより好ましい態様では、補完因子は植物又は植物の葉で安定に又は一過性に発現する。補完因子の安定な発現が好ましい。この態様では、植物は、補完因子をコードする遺伝子改変(以下及び特許請求の範囲において「第2の遺伝子改変」と称する)を提供される。アグロバクテリウム株は第2の遺伝子改変を有する植物を排他的に形質転換するため、高度な生物学的安全性はこのようにして達成することができる。アグロバクテリウム株が望ましくない生物又は望ましくない植物を形質転換する可能性は、これらの生物は第2の遺伝子改変を欠失しているため、非常に低い。
【0033】
これまでに記載した方法の生物学的安全性をさまざまな方法で更に向上させることができる。レプリコンをコードする配列部分は、植物又は植物の葉においてレプリコンの伝播を可能にする機能が不良のレプリコンをコードすることができる。当該機能は、レプリコンの遠隔距離移行又は細胞間移行のための機能、例えばそれぞれウイルスコートタンパク質又はウイルス移行タンパク質であり得る。レプリコンは少なくとも遠隔距離移行の可能性を欠失していることが好ましく、それにより、他の植物へのレプリコンの伝播を回避することができる。遠隔距離移行の可能性の欠失は、原理的にはタンパク質発現効率に不都合であり得る。しかしながら、本発明のシステムでは、本発明の他の要素によって、例えば、植物又は植物の葉の主要部分を浸潤させることによって、及び/又は以下に記載するRNAレプリコン形成の改良方法によって、この不都合を補完することができる。
【0034】
生物学的安全性を更に改良するための他のアプローチは、腫瘍抑制活性タンパク質Osa(REF)を発現する植物のトランスフェクションにアグロバクテリウム株を用いること、及びvirE2遺伝子を提供するための第2のアグロバクテリウム株を用いることである。更に、アグロバクテリウム株は、非標的生物へのT−DNA伝達を低下させるために、遺伝子改変された菌体数感知システム又は病原性誘発調節システムを有することができる。
【0035】
本発明の方法の生物学的安全性は、アグロバクテリウム株が、プラスミドDNAの他の細菌への接合性伝達を不良にする更なる遺伝子改変を有するように改良することもできる。接合性伝達は、以下のアグロバクテリウム遺伝子:oriT、traG、及びtraFのうちの少なくとも1つの機能を無効にすることによって達成することができる。
【0036】
異種DNA配列の配列部分はDNAレプリコンをコードしてもRNAレプリコンをコードしてもよい。RNAレプリコンが好ましく、以下の記載はほとんどRNAレプリコンをコードする配列部分に関係する。
【0037】
レプリコンをコードする配列部分は、レプリコン形成に必要な配列と、レプリコンから発現されるべき目的配列とを含有する。発現されるべき目的配列は、典型的には目的タンパク質をコードし、レプリコンから、又はRNAレプリコンのサブゲノムRNAから目的タンパク質を翻訳するための調節配列を含有することができる。
【0038】
レプリコンがRNAレプリコンである場合、レプリコン(i)のRNAレプリコン機能に必要な配列は、とりわけ前者が後者のDNAコピー(例えばcDNA)である場合におけるRNAウイルスの配列に相当する。DNAレプリコンの場合、レプリコン機能に関する配列は、細胞核で複製するための機能を有するDNAレプリコンを提供する;DNAレプリコンは、異種DNA配列又はその部分を鋳型として用いる複製によって異種DNA配列から形成することができる。RNAレプリコンの場合、レプリコン形成に関する配列は、細胞質で複製するための機能を有するRNAレプリコンを提供する。RNAレプリコン機能に関する配列は、典型的には、RNA依存性RNAポリメラーゼ(RdRp、レプリカーゼ)のような複製に関与する1以上のタンパク質をコードする。レプリコン機能に関する配列は、更に、サブゲノムプロモーター、転写エンハンサー、又は翻訳エンハンサーのようなレプリカーゼの発現に必要な機能であるか又はそれをコードすることができる。植物におけるRNAウイルスの細胞間伝播又は浸透伝播に関与する移行タンパク質又はコートタンパク質のようなタンパク質は、異種DNA配列中に存在し得るが、レプリコン機能を有さない。いずれにしても、レプリコンをコードする配列部分は野生型植物ウイルスをコードしない。レプリコン機能に関する配列は、植物RNAウイルスはレプリコン機能のための容易に接近可能な供給源であるため、植物RNAウイルスの配列に由来することが好ましい。RNAレプリコンの場合、「由来する」とは、レプリコン機能に関する配列が、RNAウイルスの対応する配列のDNAコピーであり、DNAコピーは細胞核に含有される異種DNA配列に組み込まれることを意味する。「由来する」とは、レプリコン機能に関する配列は、RNAウイルスの対応するRNA配列の正確なDNAコピーである必要はないが、以下に記載するように、機能保存的な相違を発揮できるという意味を含む。相違は機能保存的であるため、レプリコン機能に関する配列は、RNAウイルスに関してそうであるのと同様に、レプリコン機能を実施可能なタンパク質をコードすることが好ましい。
【0039】
レプリコンがタバコモザイクウイルスのようなトバモウイルスに基づく態様では、レプリコンはRdRpを含有するか又はコードする。このレプリコンは移行タンパク質を更にコードすることができる。このレプリコンは、コートタンパク質(CP)をコードせず、又は含有しないことが好ましい。
【0040】
更に好ましい態様では、本発明の方法の生物学的安全性は、高度に希釈したアグロバクテリウム株の懸濁液を用いることによって更に改善される。この態様は、アグロバクテリウム株の細胞が環境に広がる可能性を減少させるだけでなく、植物にとって病原体であるアグロバクテリウム株で感染させる際、植物又は植物の葉の露出及びストレスを減少させることによって本発明の方法の効率を改善させる。本発明者らは、驚くべきことに、ある限度内で、本発明の工程(a)で用いるアグロバクテリウム懸濁液の濃度の低下とともに方法の効率が増加することを発見した。とりわけ、植物の細胞で産生されたレプリコンの細胞間移行能は、これらアグロバクテリウム懸濁液濃度の低下とともに増加する。この予想外の現象の理由は未だ特定されていない。この現象はアグロバクテリウム感染に対する植物の反応によるものであり、低アグロバクテリウム濃度ではこの反応は起こらない(又はより少ない程度で起こる)と推測される。
【0041】
この好ましい態様では、600nmのOD(光学密度)が1.0のアグロバクテリウム株の細胞懸濁液を少なくとも25倍、好ましくは少なくとも100倍、より好ましくは少なくとも250倍、最も好ましくは少なくとも1000倍に希釈することによって得ることが可能なアグロバクテリウム細胞濃度のアグロバクテリウム株の細胞懸濁液で植物又は植物の葉を浸潤させる。そのような希釈は、600nmの算出OD値がそれぞれ高くても0.04、好ましくは高くても0.01、より好ましくは高くても0.004、最も好ましくは高くても0.001のアグロバクテリウム懸濁液をもたらす。
【0042】
次に、レプリコンがRNAレプリコンである場合、発現効率を顕著に増加させる重要な態様が記載される。この態様では、レプリコン機能に関する配列は、植物RNAウイルスの配列の選択された位置で、植物RNAウイルスの配列との機能保存的な相違を発揮し、その相違は、相違を発揮しないRNAレプリコンと比較して、レプリコン形成頻度を上昇させる。その相違は、全体的なプロセスのスイッチがいったん入ると、植物細胞におけるレプリコン形成頻度上昇の原因となる(以下を参照されたい)。レプリコン形成頻度上昇と相違とのあいだの因果関係を示す関連性は、相違を有するレプリコン機能に関する配列と相違を有さないレプリコン機能に関する配列とのあいだのレプリコン形成頻度を比較することによって実験的に試験することができる。そのような実験的比較は、実施例に記載するように、例えば目的配列を発現するプロトプラストを計数することによって行うことができる。容易に検出可能な緑色蛍光タンパク質(GFP)のようなレポータータンパク質をコードする目的配列をこの目的に用いることが好ましい。更に以下に記載するように、細胞間伝播不可能なRNAレプリコンとの実験的比較を行うことも好ましい。
【0043】
機能保存的な相違は、植物RNAウイルスの配列の選択された位置でレプリコン機能に関する配列に導入される。選択された位置は、核で転写されたRNAレプリコンが機能性レプリコンとして細胞質に出現する可能性の低さの原因である植物RNAウイルスのレプリコン機能に関する配列内に位置する。そのような選択された位置は、高A/T(U)含量、即ち高A含量及び/又は高T含量(RNAレベルでは高U含量)を有するか、又は潜在スプライシング部位、即ち核のスプライシング装置によってスプライシング部位として認識される配列部分を有することが好ましい。選択された位置は、以下に例示するように、RNAウイルスのRNAプロファイルを解析することによってRNAレプリコンが基づくRNAウイルス中に同定することができる。更に、選択された位置は、本発明による相違を発揮しないRNAレプリコンをコードする異種DNAでトランスフェクションした後、植物細胞で形成されるRNAを解析することによって実験的に同定することができる。この実験的解析は、RT−PCRによって、好ましくはRT−PCR産物の配列決定とともに行うことができる。このようにして、RNAレプリコンを破壊するスプライシングイベントを示す望ましくないスプライシング産物を同定することができる。更に、望ましくないスプライシングの正確な部位を同定し、そして機能保存的な相違を選択された位置に導入することによって是正することができる。
【0044】
機能保存的な相違は、RNAレプリコン形成頻度に対する選択された位置の有害効果を抑制することによってRNAレプリコン形成頻度を上昇させる。機能保存的な相違は、RNAレプリコンをコードする配列のレプリコン機能に関する配列において高A/T含量を低下させることによって、RNAレプリコンにおいて高A/U含量の低下を含むことができる。高A/U含量は、相違が機能保存的である限り、少なくとも部分的な欠失又はG/C塩基による少なくとも部分的な置換(例えば遺伝コードの縮重を用いて)によって低下させることができる。更に、植物RNAウイルス由来配列のA/Uに富んだ領域に隣接する潜在スプライシング部位は除去することができる。そのような機能保存的な相違は、1つの、好ましくはいくつかの選択された位置に導入することができる。
【0045】
好ましい機能保存的な相違は、植物RNAウイルスの配列に由来する配列のA/Uに富んだ位置の近傍又はその中への1以上のイントロンの挿入、最も好ましくは核イントロンの挿入、又は核イントロンを形成可能な1以上の配列の挿入を含む。驚くべきことに、A/Uに富んだ位置又はその近傍へのイントロンの導入は、RNAレプリコン形成頻度の上昇をもたらすことが発見された。いくつかのイントロンを導入することができ、さまざまな数の挿入イントロンに関し、本明細書に例示している。1より多いイントロンの効果は累積的である。更に、イントロンの挿入は、他の選択された位置で他の機能保存的な相違と組み合わせることができる。
【0046】
図11は、発現されるべき目的配列内ではあるが、核イントロンを形成可能な配列の導入例を示す。図11の例では、異種DNA配列の一部のリコンビナーゼによって触媒される反転によって2つの半イントロンからイントロンが形成される。この原理もRNAレプリコンのレプリコン機能に関する配列に応用することができる。同一細胞内で2つの異なるRNAレプリコンが形成される態様では、2つの異なるレプリコン間の組換えは、異なるレプリコン上に存在する2つの半イントロンから1つのイントロンを形成することができる。更に、RNAレプリコンは、いずれもレプリコンではない2つのレプリコン前駆体間の組換えによって形成することができる。この場合も、異なるレプリコン前駆体に由来する2つの半イントロンから1つのイントロンを組み立てることができる。
【0047】
RNAレプリコンをコードする配列部分を有する異種DNA配列は、転写がRNAレプリコン形成の必要条件であるため、転写プロモーターに機能可能に連結されているか又は連結可能である。アグロ浸潤させた全ての組織において目的配列の一過性発現を達成するには、転写プロモーターは構成的プロモーターであることが好ましい。構成的プロモーターの例は当該技術分野において公知である。
【0048】
本発明の1つの態様では、RNAレプリコンをコードする配列は、RNAレプリコンを共にコードする1以上のセグメントを有する。即ち、RNAレプリコンは1つの連続DNAによってコードされていない。実際、RNAレプリコンは2以上のセグメントによって不連続にコードされ、それにより、セグメントは同一T−DNA上に好ましくは互いに隣接して存在することができる。次に、RNAレプリコン形成は、例えば組換えによるセグメントの再配列を必要とすることができる。組換えのためのリコンビナーゼは、追加のアグロバクテリウム株によって、又は遺伝子操作された植物宿主によって提供することができ、よって一過性発現を植物宿主にとどめることができる。例として、レプリコン機能に関する配列の一部(例えばレプリカーゼをコードする配列の一部)が、そのような配列の他の部分に対して反転方向で異種DNA配列中に存在することができる。反転部分は組換え部位に隣接することができる。次に、レプリコン機能は提供できないため(例えば転写物は機能性レプリカーゼをコードしないため)、異種DNAの転写物はレプリコンではないであろう。組換え部位を認識する部位特異的リコンビナーゼを提供することによってセグメントの1つを反転させ、その結果レプリコンが連続的にコードされる。この態様では、リコンビナーゼを提供することにより、レプリコン形成と目的配列の発現に切り替えるためのスイッチとして機能し(更に以下を参照されたい)、高度な生物学的安全性に寄与することができる。
【0049】
あるいは、セグメントの1つはアグロバクテリウムのT−DNA上に存在することができ、他の1つは植物染色体に組み込むことができる。次に、RNAレプリコンの形成は、RNAレプリコンを組み立てるために、両方のセグメントの転写と、両方の転写物のトランススプライシングを必要とするであろう。この態様は、PCT/EP03/02986号に詳細に記載されているように、子孫植物又は細胞においてRNAレプリコンを共にコードするセグメントを迅速に分離するために用いることができ、システムの生物学的安全性に寄与することができる。
【0050】
本発明の方法は、本発明の異種DNA配列による植物又は植物の葉の一過性トランスフェクションを含む(工程(a))。「一過性トランスフェクション」という用語は、植物染色体に安定に組み込まれた異種DNAを有する形質転換されたトランスジェニック細胞を選択することなく異種DNAを導入することを意味する。一過性トランスフェクションは、通常、異種DNA配列によってコードされる遺伝子の一過性複製及び/又は一過性発現を提供する。本発明のトランスフェクションは、目的配列の発現を引き起こす。通常、発現は、植物又は植物の葉にアグロ浸潤にて異種DNA配列を提供後、自然発生的に起こる。必要に応じて、発現を引き起こすか又はスイッチを入れるさまざまな方法を用いることができる。本発明の方法のスイッチを入れるためにリコンビナーゼを用いる場合、リコンビナーゼは植物又は植物の葉に一過性に提供することができ、その提供が目的タンパク質の発現スイッチとして作用することができる。そのようなリコンビナーゼは植物細胞に安定にコードされ、構成的又はストレス誘導性プロモーターの制御下でリコンビナーゼを発現できることが好ましい。プロモーターの誘導によりリコンビナーゼ発現を誘導することにより、目的配列の発現を引き起こすことができる。
【0051】
本発明の方法の工程(b)は、工程(a)でトランスフェクションした、好ましくは浸潤させた植物又は植物の葉から目的タンパク質を単離することを含む。目的タンパク質の単離は、発現した目的タンパク質を含有する植物又は植物の葉の均質化を含むことが好ましい。タンパク質を植物から単離するためのさまざまな方法が、本明細書中に援用されるPCT/EP02/09605号、及びそこで引用されている文献に記載されている。
【0052】
本発明は、原理的には、感染性のDNA又はRNAウイルスが存在し、一過性発現を提供するアグロ浸潤が有効であるあらゆる植物に適用することができる。好ましい植物はベンサミアナタバコ及びタバコのようなタバコ種である;タバコ種以外の好ましい植物種は、ペチュニア、アブラナ、カラシナ、クレス、ルッコラ、マスタード、イチゴ、ホウレンソウ、アカザ、レタス、ヒマワリ、及びキュウリである。
【0053】
好適な植物/DNA又はRNAウイルスのペアは、以下に示すDNA及びRNAウイルスのリストに由来することができる。特に、本発明によるRNAレプリコン形成が非常に高率であるため、本発明を実施する際、植物ウイルスの植物種特異性はほとんど目立たない。同様に、本発明は、あらゆるRNAウイルスに基づいたRNAレプリコンにより用いることができる。RNAウイルスは、通常、宿主植物の細胞核の外部で展開し、レプリコンを細胞核内で産生する場合、そのようなウイルスに基づいたレプリコンを効果のないものにする選択された位置を有するであろう。本発明は、異なる植物RNAウイルスのあいだで改良レベルがさまざまであり得るが、全てのRNAウイルスに適用することができる。本発明が基づくことができる最も好ましい植物RNAウイルスは、トバモウイルス、特にタバコモザイクウイルス、及びジャガイモXウイルスなどのポテックスウイルスである。タバコモザイクウイルスの場合、通常、発現されるべき目的配列によって置換されるコートタンパク質であろう。移行タンパク質は、除去するか、又は発現されるべき目的配列によって置換することもできる。しかしながら、好ましくは、タバコモザイクウイルスに由来するRNAレプリコは、移行タンパク質をコードし、コートタンパク質が目的配列によって置換されている必要がある。
【0054】
植物とRNAレプリコンが由来するウイルスとの好ましい組み合わせは:タバコ種とトバモウイルス、タバコ種とタバコモザイクウイルスなどである。
DNAウイルスの中で、最も好ましいウイルスベクターはジェミニウイルスで構築することができる。ビーンゴールデンモザイクウイルス(BGMV)に基づくウイルス発現ベクターについての記載は、実施例15に提供されている。
【0055】
本発明の主要な応用は、目的タンパク質の植物又は植物の葉における産生である。本発明の方法を植物で実施する場合、タバコ種のようにヒト又は動物の食物連鎖に入らない植物が好ましい。アグロ浸潤後の植物全体又は植物の部分は、閉鎖環境、例えば温室又は特別に設計されたチャンバーに、所望の発現レベルを提供するのに必要なインキュベーション時間、閉じ込められるであろう。
【0056】
本発明の効率は、植物発現システムの新たな局面を達成するようなものである。本発明により達成可能な発現レベルは、下流処理費用(目的タンパク質の分離・精製を含む)が本発明の方法を他の大規模発現システムと競合させるのに十分なほど低いものである。安定に形質転換された植物を用いた先行技術の発現システムでは、細胞の小さな画分でレプリコンを産生するため、ウイルスに基づいたベクターを用いた場合であっても発現レベルは低い。植物中に伝播するレプリコンは、特に遠隔距離に対する伝播が遅いため、この問題を是正することができない。したがって、植物において発現は均一に進行せず、植物のある部分では目的タンパク質の分解が既に起こるが、他ではタンパク質発現が開始してもいない。更に、先行技術のトランスフェクション又はアグロ浸潤に基づいた一過性発現システムは、1つの産生システムで高発現レベルも高い生物学的安全性も提供しない。本発明は、コンピテント植物宿主のアグロ介在送達を介して植物全体でも分離した植物の部分(例えば葉)でも均一に発現を誘引させる。レプリコンを産生しない細胞の小さな画分を、隣接細胞由来のレプリコンによって速やかに感染させることができる。本発明は、大規模で使用できる初めての高収率植物発現システムを提供する。本発明は、一過性発現システムを特異的に遺伝子操作されたコンピテント植物宿主に限定することによって高い生物学的安全性も提供し、それにより非コンピテント(非標的)生物において発現する確立を低下させ、システムの生物学的安全性を増加させる。
【発明を実施するための最良の形態】
【0057】
発明の好ましい態様
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
RNAレプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下で植物又は植物の葉に浸潤させることによって植物又は植物の葉にトランスフェクションすることを含み、
レプリコンをコードする配列は、
(−) 植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(−) 発現されるべき目的配列、
を含有し、
アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されている。
【0058】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
好ましくはRNAレプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下で植物又は植物の葉に浸潤させることによって、植物又は植物の葉をトランスフェクションすることを含み、
レプリコンをコードする配列は、
(−) 植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(−) 発現されるべき目的配列、
を含有し、
アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変を提供されており、そして
植物又は植物の葉は、補完因子をコードし、植物又は植物の葉においてそれを発現する第2の遺伝子改変を有する。
【0059】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
600nmの算出光学密度が高くても0.04、好ましくは高くても0.01、より好ましくは高くても0.004、最も好ましくは高くても0.001に相当する細胞濃度を有する、RNAレプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株の細胞懸濁液を補完因子の存在下で植物又は植物の葉に浸潤させることによって植物又は植物の葉にトランスフェクションすることを含み、
レプリコンをコードする配列は、
(−) 植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(−) 発現されるべき目的配列、
を含有し、
アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されており、そして
レプリコン機能に必要な配列は、選択された位置で、植物RNAウイルスとの機能保存的な相違を発揮し、その相違は、相違を発揮しないRNAレプリコンと比較して、レプリコン形成頻度を上昇させる。
【0060】
更に好ましい態様を特許請求の範囲に定義する。
発明の詳細な説明
本発明者らは、土壌細菌であり、有効な媒介生物であるアグロバクテリウムを用いてRNAウイルスベクターをDNA前駆体として送達するための、高度に活性な合成鋳型を開発した。本明細書において、この改良「アグロ感染」法を用いて、植物の実質的に全ての成熟した葉において一過性遺伝子増幅及び高レベル発現を同時に開始することができ、そしてそのようなトランスフェクション経路を工業的規模で安価に実施することができるを示す。産生を遺伝子操作された「コンピテント」植物宿主に限定し、産生方法の一部ではない他の植物又は微生物へのアグロバクテリウムの感染能を制限することができるような方法で標的植物宿主生物、アグロバクテリウム、及び/又は植物ウイルスを遺伝子操作することによって、生物学的安全性の問題にも取り組む。この技術は、3つの生物学的システムの利点を組み合わせる:ウイルスの速度と発現レベル/収率、アグロバクテリウムのトランスフェクション効率と浸透送達、及び植物の翻訳後能力と低産生コスト。提案した方法は、目的産物をコードする異種核酸による植物の安定な遺伝子改変を必要としない工業的産生経路を可能にし、より安全で制御され、近年の産業基盤に適合している。
【0061】
本発明は、ウイルスレプリコン又はその前駆体のアグロバクテリウム介在送達を用いた目的タンパク質の高収率大規模産生のための一過性発現システムについて記載する。レプリコンは目的配列を発現可能であり、システムは組み込まれた生物学的安全性を有し、システムの3つの主要成分のうちの少なくとも2つ(遺伝子操作された植物宿主、アグロバクテリウム、レプリコン)を相互依存にする。
【0062】
驚くべきことに、システムの主要成分のうちの1つから他の1つへ機能を部分的に委譲することによって生ずるそのような相互依存を、システムの効率に悪影響を及ぼさずに方法全体をよりよく制御し、生物学的安全性を増大させるようになし得ることを発見した。本発明の基本原理を図1(A)に示す。ウイルスレプリコン前駆体のアグロ浸潤に基づいた大規模タンパク質産生方法は、目的タンパク質の発現に必要な機能を実施する際にアグロ浸潤に用いるアグロバクテリウム及び/又はレプリコンを補完するトランスジェニック宿主植物にて実施することができる。RNAレプリコンに関するそのような補完の1つの例を実施例4に記載する。この実施例では、植物全体を機能性移行タンパク質(MP)を欠失したRNAレプリコンをコードする異種DNA配列(図5)により一過性に形質転換した。このレプリコンは細胞間移行できなず、その結果、ウイルスMPを有する発現カセット(pICH10745、図5)を有さない植物宿主では高収率を提供することができない。
【0063】
ウイルスレプリコンの活性化若しくは操作、又は一過性トランスフェクションに必要な機能を植物又は他のアグロバクテリウム株によってトランスで提供する場合、核から細胞質へのRNA放出効率を増加させるための本明細書に記載の基準を用いたとき、発現効率に悪影響は全く見られない。植物イントロンをウイルスRNAレプリコンをコードするある領域へ組み込むことにより、又はレプリコン機能に関する配列内の潜在イントロンの除去若しくは置換により、宿主植物におけるレプリコンの効率を劇的に相加させることができる(少なくとも102倍)。そのような効率の増加は少なくとも1つの容易に測定可能なパラメータに反映された:ベクターの複製を示す細胞の相対的割合、即ち、レプリコン形成頻度の上昇。RNAレプリコン形成開始のそのような最適化は、実質的に植物全体において目的配列の発現スイッチを同時に入れる能力をもたらし、非改変ベクターよりも短時間で目的タンパク質の収率が劇的に増加する。本発明の実施例6〜14は、核から細胞質へのRNAウイルスベクターの放出効率を増加させるためのベクターの改良アプローチについて記載する。ウイルスベクターの改良に関する詳細な説明は、近年、Marillonnet及び同僚らによって公表された(2005、Nature Biotechnol.、23、718−723)。この改良は、RNAレプリコン、植物宿主、及びアグロバクテリウムのあいだで機能を効率的にやりとりさせるため、本発明の重要な態様である。興味深いことに、TMVに基づいたそのような改良ベクターは、ペチュニア、アブラナ、カラシナ、並びにクレス、ルッコラ、及びマスタードのようなタバコ種以外の植物種において効率的に作用する。最も発現する非ナス科種は、イチゴ、ホウレンソウ、アカザである。レタス(特に苗木)及びヒマワリのようなキク科並びにウリ科のキュウリは、さまざまな植物科由来のいくつかの植物種を、好ましくは更に方法を最適化した後で成功裏に用いることができることを示している。
【0064】
組換えDNAのコード領域へのイントロンの組み込みによる核のトランス遺伝子発現の増加に関する出版物(Mascarenhasら、1990、Plant Mol.Biol.、15、913−920;Bourdonら、2001、EMBO Reports、2、394−398;Rose,AB.、2002、RNA、8、1444−1453;US5,955,330号)にもかかわらず、先行技術には、ウイルスRNAレプリコン前駆体へのイントロンの組み込みが、ウイルスレプリコンの形成頻度及びレプリコンからの目的配列の発現レベルに良い効果を与えるであろうことを示す暗示は全くない。この効果は、核のmRNA転写とウイルスRNAの複製とは異なる細胞内区画で起こることを考慮すると驚くべきことである。RNAレプリコンのcDNAコピーを核内におく場合でさえ、ウイルスレプリコン前駆体の最初のコピーのみが核において産生され、次いで細胞質において核内とは異なる条件下で増幅される。先行技術では、野生型ウイルスcDNAを用いたクローニングのあいだ、大腸菌におけるウイルス遺伝子の「漏出性」発現の細胞障害効果を妨げるために、イントロンの使用が記載されている(Johansen,I.E.、1996、Proc.Natl.Acad.Sci.USA、93、12400−12405;Yangら、1998、Arch.Virol.、143、2443−2451;Lopez−Moya&Garcia、2000、Virus Res.、68、99−107)。イントロンinclusionが、ウイルスcDNAクローンからのレプリコン形成頻度を上昇させることができることを示す暗示は全くない。
【0065】
本発明は、DNA前駆体の転写によって派生し、目的配列の発現のためにデザインされている、RNAウイルス由来レプリコンの形成頻度を根本的に増加させる方法を提供する。この方法は、発現されるべき異種配列のサイズの限界やベクターの高度な不安定性のような、既存のウイルスベクターに基づいた発現システムの限界を克服する。更に、この方法は、より良い生物学的安全性を提供し、トランス遺伝子発現に対する漏出検査制御のデザインを可能にする(非誘導状態でゼロ発現レベル)。そのようなデザインは、RNAウイルス由来レプリコンをデザインするための戦略に統合された部分であるためである。RNAウイルス由来レプリコンの高頻度形成を提供することによって、本明細書に記載のアプローチは、植物全体、又はRNAレプリコンをコードする異種DNA配列を含有する葉のような植物の一部において目的配列の発現を速やかに開始させる。本発明を実施することにより、植物又は植物の葉のアグロ浸潤を介した異種目的配列の発現のためにデザインされた、植物RNAウイルスに由来する実質的に全てのレプリコンの性能を、レプリコン形成頻度の劇的な上昇によって顕著に向上させることができる。
【0066】
さまざまな分類群に属するDNA及びRNAウイルスが、本発明によるRNAレプリコンの構築に好適である。本発明を適用できるDNA及びRNAウイルスのリストを以下に提示する。引用符付きの分類群名(イタリック体ではない)は、この分類群にはICTVで国際的に承認された名称がないことを示している。種名(俗名)を標準体で示す。属又は科について正式に帰属されていないウイルスが示されている:
DNAウイルス:
環状2本鎖DNAウイルス:科:カリモウイルス科、属:バドナウイルス属、基準種:ツユクサ黄斑ウイルス、属:カリモウイルス属、基準種:カリフラワーモザイクウイルス、属:“SbCMV様ウイルス”、基準種:ダイズ退緑斑紋ウイルス、属:“CsVMV様ウイルス”、基準種:キャッサバ葉脈モザイクウイルス、属:“RTBV様ウイルス”、基準種:イネtungro bacilliformウイルス、属:“ペチュニア葉脈透化様ウイルス”、基準種:ペチュニア葉脈透化ウイルス;
環状1本鎖DNAウイルス:科:ジェミニウイルス科、属:マストレウイルス属(サブグループI ジェミニウイルス)、基準種:トウモロコシ条斑ウイルス、属:クルトウイルス属(サブグループII ジェミニウイルス)、基準種:ビートカーリートップウイルス、属:ベゴモウイルス属(サブグループIII ジェミニウイルス)、基準種:ビーンゴールデンモザイクウイルス;
RNAウイルス:
1本鎖RNAウイルス:科:ブロモウイルス科、属:アルファモウイルス属、基準種:アルファルファモザイクウイルス、属:イラルウイルス属、基準種:タバコ条斑ウイルス、属:ブロモウイルス属、基準種:ブロムモザイクウイルス、属:ククモウイルス属、基準種:キュウリモザイクウイルス;
科:クロステロウイルス科、属:クロステロウイルス属、基準種:ビート黄斑ウイルス、属:クリニウイルス属、基準種:レタス伝染性黄斑ウイルス、科:コモウイルス科、属:コモウイルス属、基準種:ササゲモザイクウイルス、属:ファバウイルス属、基準種:ソラマメ壊疽ウイルス1、属:ネポウイルス属、基準種:タバコ輪点ウイルス;
科:ポティウイルス科、属:ポティウイルス属、基準種:ジャガイモYウイルス、属:ライモウイルス属、基準種:ライグサモザイクウイルス、属:バイモウイルス属、基準種:オオムギ縞萎縮ウイルス;
科:セキウイルス科、属:セキウイルス属、基準種:パースニップ黄斑ウイルス、属:ワイカウイルス属、基準種:イネtungro sphericalウイルス、科:トンブスウイルス科、属:カルモウイルス属、基準種:カーネーション斑紋ウイルス、属:ダイアンソウイルス属、基準種:カーネーション輪点ウイルス、属:マクロモウイルス属、基準種:トウモロコシ黄色斑紋ウイルス、属:ネクロウイルス属、基準種:タバコ壊死ウイルス、属:トンブスウイルス属、基準種:トマトbushy矮化ウイルス、属が未分類の1本鎖RNAウイルス、属:カピロウイルス属、基準種:リンゴステムグルービングウイルス;
属:カーライルウイルス属、基準種:カーネーション潜在ウイルス、属:エナモウイルス属、基準種:エンドウひだ葉モザイクウイルス、
属:フロウイルス属、基準種:土壌伝染性コムギモザイクウイルス、属:ホルデイウイルス属、基準種:オオムギ斑葉モザイクウイルス、属:イダエオウイルス属、基準種:ラズベリーbushy萎縮ウイルス;
属:ルテオウイルス属、基準種:オオムギ黄萎ウイルス、属:マラフィウイルス属、基準種:トウモロコシrayado finoウイルス、属:ポテックスウイルス属、基準種:ジャガイモXウイルス;属:ソベモウイルス属、基準種:Southern bean モザイクウイルス、属:テヌイウイルス属、基準種:イネ縞葉枯ウイルス、
属:トバモウイルス属、基準種:タバコモザイクウイルス、
属:トブラウイルス属、基準種:タバコ茎壊疽ウイルス、
属:トリコウイルス属、基準種:リンゴchlorotic leaf spotウイルス、属:ティモウイルス属、基準種:カブ黄斑モザイクウイルス、属:アンブラウイルス属、基準種:ニンジン斑紋ウイルス;マイナスssRNAウイルス:目:モノネガウイルス目、科:ラブドウイルス科、属:サイトラブドウイルス属、基準種:レタス壊死性黄斑病ウイルス、属:ヌクレオラブドウイルス属、基準種:ジャガイモ黄萎ウイルス;
マイナス1本鎖RNAウイルス:科:ブニヤウイルス科、属:トスポウイルス属、基準種:トマト黄化壊疽ウイルス;
2本鎖RNAウイルス:科:パーティティウイルス科、属:アルファクリプトウイルス属、基準種:シロツメクサ潜在ウイルス1、属:ベータクリプトウイルス属、基準種:シロツメクサ潜在ウイルス2、科:レオウイルス科、属:フィジーウイルス属、基準種:フィジー病ウイルス、属:ファイトレオウイルス属、基準種:創傷腫瘍ウイルス、属:オリザウイルス属、基準種:イネragged stuntウイルス;
未分類ウイルス:
ゲノム:1本鎖RNA、種:ニンニクウイルスA、B、C、D、種:ブドウ葉脈fleckウイルス、種:トウモロコシwhite lineモザイクウイルス、種:オリーブ潜在ウイルス2、種:ウルミアメロンウイルス、種:ペラルゴニウムzonate spotウイルス。
【0067】
本発明の一般的原理を図1(A)に示す。本発明の実施に多くのさまざまなアプローチを用いることができる。これらのアプローチは一過性発現を提供不可能なアグロバクテリウム変異体の使用に基づくことができ、変異体はトランスジェニック宿主植物又は第2のアグロバクテリウム株によってトランスに補完することができる。
【0068】
システムの生物学的安全性を高めるために、本発明では感染しやすい植物へのアグロバクテリウムの感染を妨げ、それにより、一過性発現方法に対する制御を高める。植物宿主へのアグロバクテリウムのT−DNA伝達メカニズムに関する包括的な研究があり、これは本発明に成功裏に適用できる。
【0069】
1つの可能なアプローチは、ウイルスレプリコンをコードする異種DNA配列を有するアグロバクテリウム株を遺伝子操作された植物宿主に明確に限定することである。プラスミドpSAのosa遺伝子は、T−DNA輸送に必要なvirE2タンパク質を抑制することによって一過性発現を抑制し、更に植物の核DNAへの組み込みを抑制できる腫瘍抑制活性タンパク質をコードする(Lee,L−Y.ら、1999、J.Bacteriol.、181、186−196)。しかしながら、この抑制は、osa遺伝子を発現するアグロバクテリウムを野生型virE2を有するがosa遺伝子のないアグロバクテリウムと混合することによって、例えば変異体表現型をトランスで補完することによって逆転することができる。virE2を発現するトランスジェニック植物を作製することによって同様の結果を達成することができる。T−DNAは、その5’端に共有結合的に結合したVirD2とともに1本鎖中間体として植物細胞へ輸送されるが、virE2を欠失するアグロバクテリウム変異体は発癌性ではないため、virE2はこの方法の重要な成分である。そのような変異体の発癌性は、virE2を発現するトランスジェニック植物に接種することによって回復させることができる(Citovsky,V.ら、1992、Science、256、1802−1805)。更に、完全にアセトシリンゴン依存性のアグロバクテリウム変異体がある。本発明の実施例5では、本発明の異種DNA配列を有するアグロバクテリウム株の能力がvirE2タンパク質をトランスで補完するための遺伝子操作された植物宿又は野生型アグロバクテリウムに依存する一過性発現方法について記載する。また、アグロバクテリウム・リゾジェネスGALLSタンパク質は、VirE2の機能を補完することができる(Hodgesら、2004、J.Bacteriol.、186、3065−3077)。そのようなシステムは、異種DNAの伝播制御を顕著に改良し、システムの生物学的安全性を高める。この実施例に記載のように、実際に、virE2を欠失するアグロバクテリウムによる野生型ベンサミアナタバコ植物のアグロ浸潤後、目的タンパク質(GFP)の発現は全く検出されなかった。しかしながら、virE2を発現する補完された宿主植物をアグロ浸潤したとき、又はこの第1の遺伝子改変を有するアグロバクテリウム株との混合物中で野生型アグロバクテリウムをアグロ浸潤したとき、GFPは効率的に発現した。実際に、virE2を欠失するアグロバクテリウムによるT−DNA送達効率は、野生型アグロバクテリウムやトランスで補完された変異体アグロバクテリウムと比較して、約10,000倍低下した(実施例5、図17を参照されたい)。
【0070】
生物学的安全性を高めたシステムを設計するためのアプローチは、virE2機能のトランス補完に限定されない。virF遺伝子もvirE2と同様に、virF欠失アグロバクテリウムを補完することによって一過性発現に適した植物宿主を作製するために用いることができる(Regensburg−Tuink,AJ.&Hooykaas,PJ.、1993、Nature、363、69−71)。興味深いことに、osaタンパク質は、virE2タンパク質とvirFタンパク質の分泌をともに妨げる(Chen,L.ら、2000、Proc.Natl.Acad.Sci.USA、97、7545−7550)。あるいは、アグロバクテリウム・ツメファシエンスのIncP型又はIncN型接合伝達システムを異なる細菌、大腸菌株へ導入することができ、この菌株は、安全化アグロバクテリウム・ツメファシエンス株の存在下でT−DNA伝達を提供することができる(Pappas,KM.&Winans,SC.、2003、Appl.Envir.Microbiol.、69、6731−6739)。後者は、コンピテントアクセプター宿主及び当該コンピテント宿主に限定された1種又は複数の細菌ドナー株(例えばドナー株とヘルパー株)を有する、生物学的安全性を高めたシステムを設計するための多くのさまざまな可能性があることを示している。このアプローチは、他の生物への望ましくないT−DNA伝達の可能性を顕著に制限する。
【0071】
植物細胞へのアグロバクテリウムのT−DNA伝達能に加えて、アグロバクテリウムは、多くの他の細菌と同様、他の細菌との接合を介して遺伝物質(通常染色体外プラスミドDNA)を伝達することもできる。工業的アグロバクテリウム株から他のアグロバクテリウム株(例えば野生型アグロバクテリウム)へのそのようなプラスミドDNA(例えばバイナリーベクター)の伝達能は、環境へのトランス遺伝子放出に対する制御を危険にさらすであろう。多くの場合、そのような接合伝達は、多くのバイナリーベクターは接合伝達に必要な構造要素(例えば伝達起点を表すoriT)を含有せず、したがって、大腸菌から目的アグロバクテリウム株へのバイナリープラスミドの接合伝達に基づいた三親性接合を介さず、直接形質転換によってのみアグロバクテリウム株に導入することができるため、トランス遺伝子を有するT−DNA領域を有するバイナリーベクターを安全化Tiプラスミドと組換える場合にのみ可能である。さまざまなバイナリーベクターに関する概説として、Hellens,R.、Mullineaux,Ph.&Klee,H.(2000、Trend Plant Sci.、5、446−451)を参照されたい。しかしながら、バイナリーベクターがoriTを含有しない場合でさえ、依然として、バイナリーベクターと居住安全化Tiプラスミドとのあいだで組換わる可能性がある。そのような組換えは、トランス遺伝子を有するT−DNA領域を含有するプラスミドの形成をもたらし得、それにより、プラスミドは異なる細菌間で接合伝達可能であり得る。いくつかの遺伝子がアグロバクテリウム細胞間の接合伝達に関与している(Cookら、1997、J.Bacteriol.、179、1291−1297;Beckら、1989、J.Bacteriol.、171、5281−5289)。しかしながら、最も必須の領域(oriT、traG、traF)は全て居住Tiプラスミド上に局在している(Oger&Farrand、2002、J.Bacteriol.、184、1121−1131;Farrand、Hwang&Cook、1996、J.Bacteriol、178、4233−4247;Li&Farrand、2000、J.Bacteriol.、182、179−188)。相同組換えによるこの領域の除去は、居住Tiプラスミドとバイナリーベクターとの相同組換えが起こる場合でさえ、細菌間接合伝達を低下させるであろう。接合伝達に関与する領域を欠失するアグロバクテリウム株のデザインは、上記参考文献及び標準の分子生物学的技術を用いた本発明の実施例16に提供された記載にしたがい、容易に行うことができる。実施例16は、接合性プラスミド伝達に関与する遺伝子の欠失が、アグロバクテリウムから植物細胞へのT−DNA伝達効率に影響を及ぼさないことを示している(図20も参照されたい)。
【0072】
本発明による発現システムの発現効率を高めるための更なるアプローチは、アグロバクテリウム感染を限定する宿主細胞因子とアグロバクテリウム因子の使用である。多くのそのような因子が、アグロバクテリウムが介在する安定な植物トランスフェクションに関する概説に記載されている(Gelvin,SB.、2003、Trends Biotechnol.、21、95−98;Gelvin,SB.、2003、Microbiol.Mol.Biol.Rev.、67、16−37)。例えば、トランスジェニックタバコ植物においてシロイヌナズナ遺伝子VIP1を過剰発現させることにより、アグロバクテリウムによる一過性の及び安定な遺伝子トランスフェクションに対してより顕著に感受性にすることを見出した(Tzfira,T.、Vaidya,M.&Citovsky,V.、2002、Proc.Natl.Acad.Sci.USA、99、10435−10440)。植物のトランスフェクション効率を向上させる要因は、アグロバクテリウム病原性遺伝子の構成的発現であり得る。通常、そのような遺伝子は、環境因子又はvir遺伝子の発現を誘引するカスケードの制御下にあり、フェノール化合物、糖、pH変化によってスイッチを入れることができる。これらの因子、VirG のVirAタンパク質介在リン酸化を引き起こし、次いで他のvir遺伝子のプロモーターを活性化することができる。VirG内の単一アミノ酸置換によって起こる突然変異は、VirAとは独立して他のvir遺伝子の構成的発現を引き起こすことができる(Hansen,G.、Das,A.&Chilton,MD.、1994、Proc.Natl.Acad.Sci.USA、91、7603−7607)。この文献では、アグロバクテリウムの変異体VirG株を用いたGUSレポーター遺伝子による一過性発現実験で、タバコ及びトウモロコシの組織においてGUSを発現する焦点数の顕著な増加を検出した。タバコにおいて一過性発現効率を促進するために複数コピーの野生型VirGを用いたとき、同様の効果を観察したが、トウモロコシにおいては実際のところ効果がなかった。著者らは、そのようなVirG変異体株は、扱いにくい植物種の形質転換に有用であり得ることを示唆している。これは、それらが、通常、形質転換が困難な植物種のためのウイルスベクターに基づいた一過性発現システムの開発にも有用であることを意味している。植物細胞へのT−DNA伝達を刺激する、フェノール化合物(Melchersら、1989、Mol.Microbiol.、3、969−977)、アセトシリンゴン、若しくはオパインなどのさまざまな化学因子とともに、又はそれらとは独立した、上記のようなVirG変異体遺伝子の使用は、一過性発現効率を高めるための有用なアプローチである(Berthelotら、1998、Phytochemistry、49、1537−1548)。
【0073】
本発明の一過性発現システム及び方法の生物学的安全性を顕著に高めることができる他の要因は、栄養要求性アグロバクテリウム株の使用である。そのような栄養要求性株は開かれた環境で生存する確率が低いためである。X線処理による栄養要求性アグロバクテリウム株の作製は、Dirks&Peeters(US6323396号)に記載されている。この特許にはいくつかのそのような株(メチオニン若しくはシステイン要求株、又はヒスチジン及びアデニン要求株)が記載されている。生長にメチオニンを必要とする変異体の1つは、ホモシステイン・メチルトランスフェラーゼの機能が欠損している。データベースで完全なアグロバクテリウムゲノム配列を利用可能であることを考慮すると、X線処理によって得られるあまり予測できない結果にかわり、十分に特徴付けられた(配列決定された)アグロバクテリウム株のそのような変異体を、相同組換えによる遺伝子置換を用いて作製することは直接的である。例えば、アグロバクテリウム・ツメファシエンスC58ゲノム配列(GeneBank受託番号NC00305)は、5−メチルテトラヒドロプテロイル−トリグルタミン酸−ホモシステイン・メチルトランスフェラーゼをコードする遺伝子(遺伝子ID 1135697)を含有する。利用可能な配列情報を用いて変異体型遺伝子を作製し、アグロバクテリウムへ更に送達し、目的の栄養要求性変異体 (例えば、ホモシステイン・メチルトランスフェラーゼ変異体型遺伝子の場合、メチオニン代謝欠損変異体) を選択するために、隣接配列とともに、対抗選択可能マーカー遺伝子sacBRを含有する自殺ベクターへクローニングすることができる(Berger&Christie、1993、J.Bacteriology、175、1723−1734)。記載のアプローチ及びさまざまな栄養要求性アグロバクテリウム株に関する公的に利用可能な情報を用いて多くの他の変異体を作製することができる。Collens及び同僚ら(Biotechnol.Prog.20(2004)、890−896)は、挿入変異によって栄養要求性アグロバクテリウム変異体を作製した。そのような変異体株は、アデニン、ロイシン、システイン、又はチアミンの非存在下では増殖できない。これれらの変異体のいくつかは、T−DNA送達効率が低下しているが、他(cys−32)は、野生型アグロバクテリウムよりも高くさえある植物細胞へのT−DNA送達能を有する。影響を受けたことが予想されるアグロバクテリウム遺伝子はおそらくleuB、thiD並びにcysE及びcysKである。当然ながら、栄養要求性株の選択は、植物細胞への効率的T−DNA伝達のためのその適合性に基づくであろう。また、より高い生物学的安全性レベル(野生型への復帰変異頻度が低い)のために、2以上の異なる栄養素供給にたいする栄養要求性株(例えば2以上の変異遺伝子を有する)を作製することができる。栄養要求性アグロバクテリウム株(DleuB及びDpyrE)の作製は、実施例17に記載されている。
【0074】
要求される機能(例えばT−DNA伝達)のみを実施し、及び/又はトランス補完因子(例えばvirE2タンパク質、代謝化合物の添加)の存在下でのみ生存できるアグロバクテリウムの変異体株に加えて、細菌細胞の能動生物学的封じ込め(ABC)システムに基づいたアプローチを用いることができる。そのようなシステムの例は、Ronchel&Ramosによって記載されている(2001、Appl.Environ.Microbiol.、67、2649−2656)。この文献において、著者らは、汚染物質の非存在下で細菌集団の生存又は死を細胞死の誘導を介して自由自在に制御するためにデザインしたABCシステムについて記載している。このシステムは、大腸菌gef遺伝子によってコードされるポリン誘導タンパク質のような死滅遺伝子の使用に基づいている。この遺伝子は、lacI誘導性プロモーターの制御下に置かれる。このシステムにおけるLacI発現は、環境シグナルに依存するプロモーターの制御下にある。ABCシステムをデザインするための他の死滅遺伝子を使用する類似スキームは、数多くの文献に記載されている(例えば、Knudsenら(1995)Appl.Environ.Microbiol.61、985−991;Ronchelら(1998)Appl.Environ.Microbiol.64、4904−4911;Torresら(2003)Microbiology 149、3595−3601)。ストレプトアビジンに基づいた封じ込めシステムは、Szafranski及び同僚ら(1997、Proc.Natl.Acad.Sci.USA 94、1059−1063)、並びにKaplan及び同僚ら(1999、Biomol.Eng.、16、135−140)によって記載されている。
【0075】
あるいは、細菌細胞において機能性の誘導性システムを用いて、特定機能に関与する遺伝子の発現を制御することができる(例えばT−DNA伝達又はアミノ酸などの代謝産物の生合成)。実際に、変異体株(例えば栄養要求性変異体又はvirE2遺伝子機能欠損変異体)を作製するために細菌細胞から遺伝子を削除するかわりに、遺伝子を調節可能プロモーターの制御下に置くことができる。そのような調節可能プロモーターからの目的遺伝子の発現は、この場合テトラサイクリン又はIPTGのような小分子であり得る補完因子の適用を介して制御することができる。例えば十分に記述されたtet及びlacリプレッサーに基づいた細菌の調節可能システムのさまざまな改変を用いて、アグロバクテリウム細胞において目的遺伝子の発現を制御することができる。
【0076】
アグロバクテリウムに加えて、本発明は、植物細胞へのT−DNA伝達のために遺伝子操作された他の微生物に拡大適用することができる。例えば、近年、アグロバクテリウム属以外の共生細菌種を改変して、多くの多様な植物への遺伝子伝達を介在することができることが示された(Broothaertsら、(2005)Nature、433、629−633)。これらの細菌は、安全化Ti−プラスミド及びバイナリーベクターを獲得することによって、遺伝子伝達に適任となっている。
【0077】
植物RNAウイルス(例外はウイロイド−植物細胞核で増幅する小さな非コードRNA−である。概説として、Diener,T.O.、1999、Arch.Virol.Suppl.、15、203−220;Flores,R.、2001、CR Acad.Sci.III、324、943−952を参照されたい)は細胞核では発生せず、細胞質で発生することが知られている。したがって、RNAウイルスの配列は、プレmRNA、rRNA及びtRNAの前駆体を含めたプロセシングされたRNAの、細胞質内の輸送を含めたプロセシング工程に関与するモチーフの存在により、核のRNAプロセシングに抵抗するように適合されていない。5’端キャッピング、スプライシング、3’端生成、ポリアデニル化、分解、塩基及び糖による修飾、並びにエディティング(プラスチド及びミトコンドリア内)などのプロセシングイべントが集中的に研究されている。しかしながら、そのようなイベントの多くの要素は依然として不明確のままである。核におけるプレmRNAの最も劇的な変化は、介在RNA配列(イントロン)が初期転写産物から除去され、同時にエクソンが連結されるプロセスである、プレmRNAのスプライシングのあいだに起こる。スプライシングは、ウリジン(uridilate)に富んだ核の小さなリボ核タンパク質粒子を含む複合体構造であるスプライソームによって介在される。スプライソームは2つの連続工程でスプライシング反応を行う:第1工程−上流エクソン/イントロン接合部の5’スプライス部位を切断してラリアート形成し、第2工程−イントロン/下流エクソン接合部の3’スプライス部位を切断して上流と下流のエクソンを連結する(概説として:Kramer,A.、1996、Annu.Rew.Biochem.、65、367−409;Simpson,GG.&Filipowicz,W.1996、Plant.Mol.Biol.、32、1−41を参照されたい)。イントロン配列に隣接する5’及び3’スプライシング部位のジヌクレオチド(5’/GU;AG/3’)は高等植物で高度に保存されており、単一G置換により関与する部位のスプライシング活性を放棄するであろう。驚くべきことに、植物と動物とのあいだでスプライシング部位が高度に保存されているにもかかわらず、植物において異種イントロンは通常スプライシングされないか、又は不正確にスプライシングされる(van Santen,VL.ら、1987、Gene、56、253−265;Wiebauer,K.、Herrero,J.J.、Filipowicz,W.1988、Mol.Cel.Biol.、8、2042−2051)。植物ウイルスRNAは、核のRNAプロセシング装置に抵抗するための進化的圧力下になかったことを考慮すると、これらのRNAは、いったん核環境に置かれると、スプライシングを含めたそのようなプロセシングの対象となる可能性が非常に高い。この状況は、核遺伝子によってコードされるRNA転写産物とは全く異なる。後者の転写産物は、一連のRNA修飾が核で起こるにもかかわらず、機能性を保つように進化的に適合しているためである。しかしながら、そのような修飾は、ウイルスRNAレプリコン形成に劇的な結果を招くことができる。異種遺伝子のための発現ベクターを作製するために植物ウイルス遺伝子を再操作すると、ウイルス起源のRNA配列と相互作用する配列を更に追加するため、RNAウイルスに基づいたレプリコンの不安定性が更に増し、複製できない欠損RNAを産生するであろう。本発明は、発現ベクターをDNA前駆体として植物又は植物細胞に導入し、一過性発現又は植物染色体DNAへの安定な組み込みを提供する場合に機能性RNAレプリコンの形成頻度を顕著に上昇させる改変を発現ベクターに施すことによって、こうした問題に取り組んでいる。ウイルス由来配列の改変は、RNAウイルスに基づいたレプリコンの効率を高めるための最善策であり得る。改変は、直接的(例えばRNAウイルス由来配列内)でも間接的(例えば非ウイルス起源の配列内)でもよいが、間接的改変は、ウイルス起源の配列に安定効果をもたらし得る。本発明では、RNAレプリコン形成効率の上昇に不可欠であるため、RNAウイルス由来配列内の改変に主に焦点をあてる。
【0078】
驚くべきことに、潜在的に問題のある領域が存在する証拠を発見するための第1の試みは成功した。そして、更に驚くべきことに、予想外にも桁違いの改善を発見したことにより、実験的に確認した。発現ベクターpICH8543(実施例1、図2A)のRNAウイルス由来配列を、潜在イントロンとRNAスプライシング部位の存在に関し、NetgeneIIサーバープログラム(http://www.cbs.dtu.dk/services/NetGene2/)を用いて解析することにより、核RNAプロセシング装置によってスプライシングされ得るイントロン様領域の存在が示された(図6の丸で囲んだ領域を参照されたい)。植物ウイルスRNA配列内で潜在的に問題のある領域(選択された位置)を同定するために使用できるプログラムは、さまざまな生物のためのエクソン/イントロン予測プログラム(http://genes.mit.edu/GENSCAN.html)又はスプライシングシグナル予測プログラム(http://125.itba.mi.cnr.it/〜webgene/wwwspliceview.html)のように、他にも多数存在する。
【0079】
既存のプログラムは全て理想的ではなく、誤りがちであることを考慮すると、潜在的に問題のある領域は実験的に決定することもできる。これは、RT−PCRで(Frohman,MA.、1989、Methods Enzymol.、218、340−356)、又はさまざまな転写産物の濃度を正確に定量するのに好適な、リアルタイムPCRと呼ばれる進歩したバージョン(Gibsonら、1996、Genome Res.、6、995−1001)で、核環境で試験されているDNAベクター由来の転写産物を解析することによって行うことができ、好ましくはPCR増幅産物の配列決定を伴う。本発明の機能保存的な相違は、例えばA/Uに富んだ領域(イントロン様)からG/Cに富んだ領域(エクソン様)への置換を伴うサイレント変異を導入することにより、例えばイントロン様配列をエクソン様配列配列で置換することによって、RNAプロファイルを劇的に変化させる(図3の丸で囲んだ領域を参照されたい)。植物のイントロンはエクソンと異なり、通常A/Uに富んでいる(Csank,C.ら、1990、Nucl.Acid Res.、18、5133−5141;Goodall&Filipowicz、1989、Cell、58、473−483)が、例外がある。例えば単子葉植物では、G/Cに富んだイントロンが発見されている(Goodall&Filipowicz、1989、Cell、58、473−483;Goodall&Filipowicz、1991、EMBO J.、10、2635−2644)。本発明のこの態様を実施するには、A/Uに富んだ領域には、A/U含量が少なくとも55%以上の、少なくとも長さ20ヌクレオチドの配列である長い区間だけでなく、純粋にA/Uを含有する配列の並びの6〜19ヌクレオチドの、より短い区間(「アイランド」)も含まれる。本発明におけるA/U内容は、UよりもAに富んだ配列、又はAのみに富んだ配列、そして逆にこの定義によってカバーされる全ての配列を意味する。実施例6では、A/Uに富んだ領域の改変が、GFPを発現する細胞数を少なくとも10倍増加させることを立証する。これは、図10において、pICH15466(改変ベクター、図2A)でアグロ浸潤させた領域と、pICH14833(対照ベクター、図2A)でアグロ浸潤させた領域とを比較することによって立証される。移行タンパク質(MP)の除去は、初期感染部位から隣接細胞への細胞間移行は起こらないため、機能性RNAレプリコンを有する初代培養細胞の正確な計数を可能にする。実施例7では、多くの潜在スプライス部位(図6B)を含有し、移行タンパク質(MP)のサブゲノムプロモーターをカバーする他のUに富んだイントロン様領域の改変を行った(図8、丸で囲んである)。この改変は、ウイルスベクターpICH15900からのレプリコン形成頻度上昇に劇的な効果を及ぼす。プロトプラスト計数実験(実施例7)で立証したように、試験した2つのタバコ種−ベンサミアナタバコ及びタバコに関し、非改変ベクターpICH14833と比較して、およそ100倍増加した(図10A、Bの対応の浸潤領域を参照されたい)。一般に、本発明に記載のアプローチを用いることにより、RNAレプリコン形成頻度をおよそ300倍上昇させ、即ち、機能性レプリコンを有する細胞の割合を約0.2%(対照ベクター)から50%(改変ベクター)以上に増加させることができた。これは限界ではなく、100%の頻度に達することも非常に現実的であると考えられる。
【0080】
そのように高率のレプリコン形成は、同一植物細胞内で2つの異なるRNAレプリコンから2以上の異なる遺伝子を発現させる、例えば植物RNAウイルスに基づいたベクターを用いて異なる遺伝子を共発現させる道を開く。同一細胞で同時に2以上のレプリコンの同調した開放を達成することは、「先着順」の原則はウイルスベクターでは特に真実であるため、そのような共発現に重要である。異なるウイルスベクターは通常伝播領域で重ならないため、又はそのような重なりはわずかであるため、浸透移行も細胞間移行も役立たない。単純な計算によって、同一植物細胞において2つのレプリコンから2つの目的配列の共発現を達成するための、本発明に記載の技術の重要性を立証する。機能性レプリコン形成頻度が全細胞の0.2%である非最適化ウイルスベクターの場合、2つの異なるRNAレプリコンから2つの遺伝子を共発現する細胞の割合は0.2×0.2=0.04%であるが、機能性RNAレプリコン形成頻度が上昇した構築体の場合(全細胞の50%即ち1/2)、上記細胞の割合は0.5×0.5=0.25、即ち25%であり、例えば約625倍高いであろう。最高性能ベクターを用いると(例えばpICH16191、図10C)、機能性レプリコンを有する細胞の割合はおよそ90%に達することもある(図10C、右上)。これは、2つの独立したレプリコンから2つの異なる目的配列を発現させるためにそのようなベクターを用いることにより、全細胞の80%で共発現が起こり得ることを意味している。技術を更に改善し、100%共発現を達成できる可能性は非常に高いと考えられる。
【0081】
RNAレプリコンから発現されるべき異種目的配列における機能保存的な相違は、特にレプリコン機能に関する配列における相違と組み合わせて、RNAレプリコン形成頻度を上昇させるために使用し得ることに留意することは重要である。例えば、レプリコンの形成及び/又はプロセシングに必要な改変を目的配列内に導入することができる。
【0082】
本発明の他の態様では、核イントロンをレプリコン機能に関する配列に挿入することにより、レプリコン形成頻度が改善された(実施例8)。ウイルスRNA依存性RNAポリメラーゼ(RdRP)のコード領域へのイントロンの組み込み(実施例8及び12)は、改変ベクター(図2A、BのpICH15034、pICH15025、pICH15499)からのレプリコン形成頻度を顕著に(少なくとも50倍)上昇させた(図10A、B)。シロイヌナズナ由来の6つの挿入イントロンを含有するベクターのRNAプロファイルを図9に示す。他の実施例(実施例11)では、MP配列へのイントロンの挿入により、レプリコン形成頻度を少なくとも100倍上昇させる。
【0083】
本発明を実施するために多くの核イントロンを用いることができる。そのようなイントロンの例としては、限定されるものではないが、イネtpi Act1遺伝子及びsalT遺伝子由来のイントロン(Rethmeierら、1997、Plant J.、12、895−899;Xuら、1994、Plant Physiol.、100、459−467;McElroyら、1990、Plant Cell、2、163−171);トウモロコシAdh1遺伝子、GapA1遺伝子、アクチン遺伝子、及びBz1遺伝子由来のイントロン(Callisら、1987、Genes Dev.、1、1183−11200;Donathら、1995、Plant Mol.Biol.、28、667−676;Maasら、1991、Plant Mol.Biol.、16、199−207;WE Cohn、K Moldave監修、『核酸研究と分子生物学の進歩』、第42巻、Academic Press、ニューヨーク、pp.229−257中のSinibaldi&Mettler、1992);ペチュニア ルビスコ遺伝子SSU301由来のイントロン(Deanら、1989、Plant Cell、1、201−208);シロイヌナズナA1 EF1α遺伝子、UBQ10遺伝子、UBQ3遺伝子、PAT1遺伝子由来のイントロン(Curieら、1993、Mol.Gen.Genet.228、428−436;Norrisら、1993、Plant Mol.Biol.、21、895−906;Rose&Last、1997、Plant J.、11、455−464);並びに他の多くのものが挙げられる。合成イントロンも本発明に用いることができる。最小の使用可能イントロン又はそれらの部分は、通常、内部のイントロン配列に隣接する、スプライスドナー部位及びアクセプター部位に限定され得る。好ましくはイントロンのサイズは少なくとも50ヌクレオチド、より好ましくは100〜200ヌクレオチドである必要があるが、実際にはイントロンのサイズに限定はない。しかしながら、構築体のサイズは遺伝子操作に好適なように維持される必要がある。イントロンの由来、その構造及びサイズは、ベクターの性質に応じて個々に選択することができる。選択したイントロン又は対応のイントロン部分の効率を試験するために一過性発現実験を用いることができる。
【0084】
上記改変の効果は累積的であり、例えばイントロンの挿入をMPサブゲノムプロモーターの改変と組み合わせると、レプリコン形成頻度をおよそ300倍に上昇させることができる(実施例9)。RNAレプリコン形成頻度を上昇させるためのイントロン挿入に好ましい領域は、本明細書において選択された領域という。そのような位置は「イントロン様」構造を含有することができる。これは、MP、実際にはMPサブゲノムプロモーターのように問題のある領域の極近傍にイントロンを挿入することによって確認される(実施例11)。レプリコン形成頻度に100倍の上昇が観察された。「エクソン様」領域へのイントロンの挿入は、イントロン様領域への挿入のような明白な効果を有さない(実施例10)。
【0085】
目的配列の一過性発現に適格な、即ち本発明の欠損アグロバクテリウム株の補完に適格な植物宿主の作製に必要な代理機能を有するトランスジェニック植物宿主を作製するためにさまざまな方法を用いることができる。アグロバクテリウムによって運ばれるTi−プラスミドにより(US5,591,616号;US4,940,838号;US5,464,763号)、又は粒子銃若しくは微粒子銃(US05100792号;EP00444882B1号;EP00434616B1号)により、植物細胞をベクターで形質転換することができる。マイクロインジェクション(WO09/209696号;WO09/400583A1号;EP175966B1号)、エレクトロポレーション(EP00564595B1号;EP00290395B1号;WO08/706614A1号)、又はプロトプラストのPEG介在形質転換などのような他の植物形質転換法も用いることができる。ベクター送達方法の選択は、形質転換される植物種に依存し得る。例えば、微粒子銃は通常単子葉植物の形質転換に好ましいが、双子葉植物には、アグロバクテリウム介在形質転換が一般に良好な結果を与える。
【0086】
以下の実施例では、タバコ細胞へのアグロバクテリウム介在ベクター(異種DNA配列)送達を用いた。しかしながら、ベクターは、目的植物種の安定な又は一過性の形質転換又はトランスフェクションに好適な標準技術にしたがい、植物に組み込むことができる。双子葉植物の形質転換技術は当該技術分野において周知であり、アグロバクテリウムに基づいた技術とアグロバクテリウムを必要としない技術が含まれる。非アグロバクテリウム技術には、プロトプラスト又は細胞による外来遺伝物質の直接取り込みが含まれる。これらの技術には、PEG又はエレクトロポレーションを介在する取り込み、粒子銃介在送達、及びマイクロインジェクションが含まれる。これらの技術の例は、Paszkowskiら、EMBO J.、3、2717−2722(1984);Potrykusら、Mol.Gen.Genet.、199、169−177(1985);Reichら、Biotechnology 4:1001−1004(1986);及びKleinら、Nature 327、70−73(1987)に記載されている。いずれの場合にも、形質転換細胞は標準技術を用いて完全な植物に再生される。
【0087】
アグロバクテリウム介在トランスフェクションは、そのトランスフェクション効率の高さと多くのさまざまな種を用いる広範な有用性のために、双子葉植物のトランスフェクションに好ましい技術である。アグロバクテリウムによって日常的に形質転換され得る多くの作物種には、タバコ、トマト、ヒマワリ、ワタ、アブラナ、ジャガイモ、ダイズ、アルファルファ、及びポプラが含まれる(EP 0 317 511号(ワタ)、EP 0 249 432号(トマト)、WO87/07299号(アブラナ属)、米国特許第4,795,855号(ポプラ))。
【0088】
本発明の例では、目的遺伝子を一過性発現させるためのT−DNAのアグロバクテリウム介在送達法であるアグロ接種を用いた(Vaqueroら、1999、Proc.Natl.Acad.Sci.USA、96、11128−11133)。アグロ接種は、小規模から中規模の組換えタンパク質産生システムだけでなく、ベクター最適化の要素としても非常に有用なツールであり、構築体のさまざまな変異体を用いて迅速な結果を得ることが可能である。
【0089】
アグロ浸潤を実施するには、先行技術に記載のように、通常、アグロバクテリウムの一晩培養物を準備する(Marillonnetら、2004、Proc.Natl.Acad.Sci.USA.、101、6853−6857)。通常、一晩培養物の光学密度(O.D.)は波長600nmで3〜3.5単位に達し、アグロ浸潤の前に3〜5倍に希釈され、5〜9×109コロニー形成単位を生ずる(Turpenら、1993、J.Virol.Methods、42、227−240)。発明者らは、102倍、103倍、及び104倍希釈も、特に本明細書に記載のような機能保存的な相違を有するレプリコン機能に関する配列との組み合わせにおいて、非常に効率的に作用することを発見した。驚くべきことに、浸潤させたタバコの葉におけるベクターはその性能を更に向上させ、形質転換アグロバクテリウムの希釈率を上げるとGFPの良好な収率を与えた。例えば、103倍希釈は102倍希釈よりも良好な結果を与えた。102倍希釈は10倍希釈よりも良好なGFP収率を生ずる。この現象に対する可能な説明は、おそらく高濃度の病原性細菌に対する植物の応答の結果としての、非常に濃縮したアグロバクテリウム懸濁液の、ウイルスベクターの機能、例えば細胞間移行に対する負の効果である。この現象は、アグロ浸潤を介する組換えタンパク質産生に必要なアグロバクテリウム量を、先行技術の方法と比較して、少なくとも1桁軽減させるため、大規模な工業的タンパク質発現方法に特別な価値がある。
【0090】
実施例13では、不活化ウイルスRNAに基づいたレプリコンのDNA前駆体について示す。レプリコンは本発明にしたがって最適化される。更に、レプリコンは、非コンピテント植物、例えばゲノムDNAに安定に組み込まれたリコンビナーゼを提供する発現カセットを含有しない植物へアグロ送達された場合に目的配列の発現を妨げる構造を含有する。機能性RNAレプリコンの形成と同様に、発現は、部位特異的組換えにより構築体の一部を反転させることによって誘引することができる。この反転は、2つのイントロンの形成と、機能性目的配列の集合をもたらすことができる。実施例13に記載のシステムは、ウイルスベクターの最適化だけでなく、工業的タンパク質又は医薬用タンパク質を発現させるための植物発現システムを用いる高度な生物学的安全性基準の達成方法を示している。
【0091】
リコンビナーゼの転写は、誘導性の又は他の調節された(例えば発生学的に調節された)プロモーターの制御下に置くことができる。誘導性プロモーターは、その誘導条件によって2つのカテゴリーに分けることができる:非生物的因子(温度、光、化学物質)によって誘導されるものと、生物的因子、例えば病原体又は害虫の攻撃によって誘導できるものである。第1のカテゴリーの例は、熱誘導性(US05187287号)及び低温誘導性(US05847102号)のプロモーター、銅誘導性システム(Mettら、1993、Proc.Natl.Acad.Sci.USA、90、4567−4571)、ステロイド誘導性システム(Aoyama&Chua、1997、Plant J.、11、605−612;McNellisら、1998、Plant J.、14、247−257;US06063985号)、エタノール誘導性システム(Caddickら、1997、Nature Biotech.、16、177−180;WO09/321334号)、並びにテトラサイクリン誘導性システム(Weinmannら、1994、Plant J.、5、559−569)である。植物の化学誘導性システムの分野における最新の発展の1つは、グルココルチコイド デキサメサゾンによってスイッチを入れることができ、テトラサイクリンによってスイッチを切ることができるキメラプロモーターである(Bohnerら、1999、Plant J.、19、87−95)。化学的誘導性システムの概説として:Zuo&Chua(2000、Current Opin.Biotechnol.、11、146−151)及びPadidam,M(2003、Curr.Opin.Plant Biol.、6、169−177)を参照されたい。誘導性プロモーターの他の例は、植物の病原性関連(PR)遺伝子の発現を制御するプロモーターである。これらのプロモーターは、植物を、病原体の攻撃に対する応答における植物シグナル伝達経路の重要成分であるサリチル酸で、又はPR遺伝子発現を誘引可能な他の化学化合物(ベンゾ−1,2,3−チアジアゾール又はイソニコチン酸)(US05942662号)で処理することによって誘導することができる。
【0092】
本発明は、実施例に記載のTMVに基づいたベクターに限定されず、他の植物RNAウイルスに基づいたレプリコンにも適用可能である。他の植物ウイルスRNA配列の解析(実施例14、図13、図14)により、TMVに関して記載したものと類似の選択された位置及び植物核遺伝子のプレmRNAの配列を示す(図12)。これは、本発明に記載のアプローチを用いて、問題のある領域を除去/置換することにより、及び/又は核イントロンを挿入することにより、実際に植物RNAウイルス由来レプリコンを解消することができる強力な証拠である。
【0093】
本発明を用いて植物又は植物細胞において発現させることができる目的タンパク質、それらの断片(機能性又は非機能性)、及びそれらの人工誘導体には、限定されるものではないが、以下が含まれる:デンプン修飾酵素(デンプン合成酵素、デンプンリン酸化酵素、脱分岐酵素、デンプン分岐酵素、デンプン分岐酵素II、顆粒結合型デンプン合成酵素)、ショ糖リン酸合成酵素、ショ糖ホスホリラーゼ、ポリガラクツロナーゼ、ポリフルクタンスクラーゼ、ADPグルコース ピロホスホリラーゼ、シクロデキストリン グリコシルトランスフェラーゼ、フルクトシル トランスフェラーゼ、グリコーゲン合成酵素、ペクチン エステラーゼ、アプロチニン、アビジン、細菌レバンスクラーゼ、大腸菌glgAタンパク質、MAPK4と相同分子種、窒素同化/代謝酵素、グルタミン合成酵素、植物オスモチン、2Sアルブミン、タウマチン、部位特異的リコンビナーゼ/インテグラーゼ(FLP、Cre、Rリコンビナーゼ、Int、SSVIインテグラーゼR、インテグラーゼφC31、又はそれらの活性断片若しくは変異体)、オイル修飾酵素(脂肪酸不飽和化酵素、エロンガーゼなどのような)、イソペンテニル トランスフェラーゼ、Sca M5(ダイズ カルモジュリン)、甲虫型毒素又は殺虫活性断片、ユビキチン結合酵素(E2)融合タンパク質、脂質、アミノ酸、糖、核酸、及び多糖を代謝する酵素、スーパーオキシドジスムターゼ、不活性プロ酵素型プロテアーゼ、植物タンパク質毒素、繊維産生植物における改質繊維、バチルス・スリンギエンシス由来の甲虫活性毒素(Bt2毒素、殺虫性結晶タンパク質(ICP)、CrylC毒素、δエンドトキシン、ポリオペプチド毒素、プロトキシンなど)、昆虫特異的毒素AaIT、セルロース分解酵素、Acidothermus celluloticus由来のE1セルラーゼ、リグニン修飾酵素、シンナモイル アルコール脱水素酵素、トレハロース−6−リン酸合成酵素、サイトカイニン代謝経路の酵素、HMG−CoA 還元酵素、大腸菌無機ピロホスファターゼ、種子貯蔵タンパク質、エルウィニア・ヘルビコラ リコピン合成酵素、ACCオキシダーゼ、pTOM36にコードされるタンパク質、フィターゼ、ケトヒドロラーゼ、アセトアセチルCoA還元酵素、PHB(ポリヒドロキシブタノエート)合成酵素、ポリヒドロキシルアルカノエート(PHA)の合成に関与する酵素、アシルキャリアータンパク質、ナピン、EA9、非高等植物フィトエン合成酵素、pTOM5にコードされるタンパク質、ETR(エチレン受容体)、色素体ピルビン酸リン酸ジキナーゼ、線虫誘導性膜貫通ポアタンパク質、植物細胞の光合成機能又はプラスチド機能を高める形質、スチルベン合成酵素、フェノールをヒドロキシル化可能な酵素、カテコール ジオキシゲナーゼ、カテコール 2,3−ジオキシゲナーゼ、クロロムコネート シクロイソメラーゼ、アントラニル酸合成酵素、アブラナ属AGL15タンパク質、フルクトース 1,6−ビホスファターゼ(FBPアーゼ)、AMV RNA3、PVYレプリカーゼ、PLRVレプリカーゼ、ポティウイルスコートタンパク質、CMVコートタンパク質、TMVコートタンパク質、ルテオウイルス レプリカーゼ、MDMVメッセンジャーRNA、変異体ジェミニウイルス レプリカーゼ、Umbellularia californica C12:0選択性アシル−ACPチオエステラーゼ、植物のC10又はC12:0選択性アシル−ACPチオエステラーゼ、C14:0選択性アシル−ACPチオエステラーゼ(luxD)、植物合成酵素因子A、植物合成酵素因子B、D6−不飽和化酵素、脂肪酸の生合成及び修飾、例えば植物細胞における脂肪酸のペルオキシソームβ酸化において酵素活性を有するタンパク質、アシル−CoAオキシダーゼ、3−ケトアシル−CoAチオラーゼ、リパーゼ、トウモロコシ アセチル−CoA−カルボキシラーゼ、など;5−エノールピルビルシキミ酸−3−リン酸合成酵素(EPSP)、ホスフィノスリシン アセチルトランスフェラーゼ(BAR、PAT)、CP4タンパク質、ACC脱アミノ酵素、翻訳後切断部位を有するタンパク質、スルホンアミド耐性を付与するDHPS遺伝子、細菌ニトリラーゼ、2,4−D モノオキシゲナーゼ、アセト乳酸合成酵素又はアセトヒドロキシ酸合成酵素(ALS、AHAS)、ポリガラクツロナーゼ、Taqポリメラーゼ、細菌ニトリラーゼ、制限酵素、メチラーゼ、DNA及びRNAリガーゼ、DNA及びRNAポリメラーゼ、逆転写酵素、ヌクレアーゼ(DNA分解酵素及びRNA分解酵素)、ホスファターゼ、トランスフェラーゼなどを含めた細菌又はファージ由来の他の多くの酵素。
【0094】
本発明は、工業的酵素(セルラーゼ、リパーゼ、プロテアーゼ、フィターゼなど)、及び線維性タンパク質(コラーゲン、クモの糸タンパク質など)を含めた、商業的に価値があり、医薬的に重要なタンパク質の分子農業及び精製のために用いることができる。ヒト又は動物の健常なタンパク質は、本発明で記載したアプローチを用いて発現させ、精製することができる。そのような目的タンパク質の例としては、とりわけ、免疫応答タンパク質(モノクローナル抗体、1本鎖抗体、T細胞受容体など)、病原性微生物由来のものを含めた抗原、コロニー刺激因子、レラキシン、ソマトトロピン(HGH)及びプロインスリンを含めたポリペプチドホルモン、サイトカインとその受容体、インターフェロン、成長因子及び凝固因子、酵素学的に活性なリソソーム酵素、線溶性ポリペプチド、血液凝固因子、トリプシン、トリプシノーゲン、a1−アンチトリプシン(AAT)、ヒト血清アルブミン、グルコセレブロシダーゼ、天然コレラ毒素B、並びに上記タンパク質の融合体、変異体型、及び合成誘導体のような機能保存的タンパク質、トロンビン、ヒト胃リパーゼ、顆粒球−マクロファージコロニー刺激因子(GM−CMF)、セルピン、ラクトフェリン、リゾチーム、オレオシン、プロトロンビン、α−ガラクトシダーゼが挙げられる。
【0095】
本特許出願で優先権を主張する欧州特許出願第04016011.1号の内容は、本明細書にその全体が援用される。
実施例
以下の実施例は、本発明を具体的に説明するために提示するものである。必要に応じて改変や変形を行うことができる。
【実施例1】
【0096】
TMVに基づいたRNAベクターの構築
アブラナ科植物感染性のトバモウイルス(cr−TMV;Dorokhovら、1994、FEBS Lett.350、5−8)及びカブ葉脈透化ウイルス(TVCV;Larteyら、1994、Arch.Virol.138.287−298)のクローン化cDNAは、ロシアのモスクワ大学のAtabekov教授から入手した。緑色蛍光タンパク質(GFP)遺伝子を含有するウイルスベクターは、いくつかのクローニング工程で作製した。得られた構築体pICH8543(図2A)は、順に:シロイヌナズナ アクチン2プロモーター由来の787bp断片(ACT2、参考文献Anら、1996、GenBank受託番号AB026654、bp57962〜58748)、TVCVの5’端(GenBank受託番号BRU03387、bp1〜5455)、cr−TMV断片(GenBank受託番号Z29370、bp5457〜5677、コートタンパク質CPの開始コドンを除去するためチミン5606がシトシンに変わっている)、配列「taa teg ata act cga g」、合成GFP(sGFP)遺伝子、cr−TMV 3’−非翻訳領域(3’−NTR;GenBank受託番号Z29370、bp6078〜6312)、最後にノパリン合成酵素(Nos)ターミネーターを含有する。断片全体を、CarbR pBIN19由来のバイナリーベクターであるpICBV10のT−DNAの左の境界(LB)と右の境界(RB)とのあいだクローニングした。pICH8543でアグロバクテリウム株GV3101を形質転換し、ベンサミアナタバコの葉に浸潤させた。3dpiで出現したGFP蛍光の焦点は、増殖してコンフルエントになった。驚くべきことに、たとえ浸潤領域の大部分の細胞がウイルスの複製と移行により最終的にGFPを発現しても、GFPを発現する多くの独立した焦点によって検出されたように、ウイルス複製を開始したのはわずかな画分の細胞のみであった。ベンサミアナタバコの葉に35Sプロモーター制御下のGFP遺伝子を浸潤させることにより、浸潤領域のほとんど全ての細胞においてGFPが発現するため(図示していない)、限定要因は植物細胞へのDNA送達でないことが明らかになった。
【0097】
この観察結果について確認するため、MPに変異を含有するウイルスベクター構築体を作製した。pICH14833と呼ばれるこの構築体は、pICH8543に類似するが、MP遺伝子における、MPに存在するEcoRI部位の上流の389bpの欠失により異なっている。この欠失を含めたNcoI−EcoRI断片の配列を配列番号1として付録に示す。ウイルス構築体全体(ACT2プロモーターからNosターミネーターまで)を、pBIN19由来KanR バイナリーベクターであるpICBV49のT−DNAの左の境界と右の境界とのあいだにクローニングした。MP内の欠失のために、この構築体から産生されるレプリコンは細胞間移行できないが、細胞内で自律複製可能である。MPを、例えばカリフラワーモザイクウイルス35Sプロモーターのような構成的プロモーターからトランスで提供すると、細胞間移行を回復させることができる。
【0098】
MP発現構築体を作製するために、クローン化TVCV cDNA(GenBank受託番号Z29370、bp4802〜5628)からPCRにてTVCV MP遺伝子を増幅させ、35Sプロモーターの制御下でバイナリーベクターにサブクローニングした。pICH10745と呼ばれる得られた構築体(図示していない)及びpICH14833でアグロバクテリウム株GV3101を形質転換し、ベンサミアナタバコの葉に浸潤させた。pICH14833単独で浸潤させることにより、浸潤領域内にわずかなGFP発現細胞が出現した。浸潤領域から作製されたプロトプラストを計数することにより、合計500個プロトプラストのうち、わずか1〜3個のプロトプラストがGFPを発現することを発見した(0.2〜0.6%)。pICH14833とpICH10745との共浸潤により、それぞれの初期GFP発現細胞から増殖した、GFPを発現する焦点が形成された。最終的に、細胞間移行により、浸潤領域のかなりの割合の細胞がGFPを発現した(図10A)。
【0099】
ロシアのモスクワ大学のAtabekov教授から入手した、上記アブラナ科植物感染性のトバモウイルス(cr−TMV;Dorokhovら、1994、FEBS Lett.350、5−8)及びカブ葉脈透化ウイルス(TVCV;Larteyら、1994、Arch.Virol.138、287−298)の同一クローン化cDNAを用いて、いくつかの更なるウイルスベクターを構築した。いくつかのクローニング工程で緑色蛍光タンパク質(GFP)遺伝子を含有するウイルスベクターを作製した。得られた構築体pICH16707(図2C)は、順に:シロイヌナズナ アクチン2プロモーター由来の787bp断片(ACT2、参考文献Anら、1996、GenBank受託番号AB026654、bp57962〜58748)、TVCVの5’端(GenBank受託番号BRU03387、bp1〜5455)、cr−TMVの断片(GenBank受託番号Z29370、bp5457〜5677、コートタンパク質CPの開始コドンを除去するためにチミン5606がシトシンに変わっている)、配列「taa tcg ata act cga g」、合成GFP(sGFP)遺伝子、cr−TMV 3’−非翻訳領域(3’−NTR;GenBank受託番号Z29370、bp6078〜6312)、最後にノパリン合成酵素(Nos)ターミネーターを含有する。完全な断片を、KanR pBIN19由来バイナリーベクターであるpICBV29のT−DNAの左の境界(LB)と右の境界(RB)とのあいだにクローニングした。pICH16707でアグロバクテリウム株GV3101を形質転換した。形質転換アグロバクテリウム株を飽和するまでLB培地中で一晩増殖させた。アグロバクテリウムを増殖させた200μlの一晩培養物を、マイクロ遠心機にて8000rpmで3分間沈殿させた。沈殿を、10mM MES(pH5.5)、10mM MgSO4を含有する1mlの溶液に再懸濁させ、ODをおよそ0.7とした。温室栽培ベンサミアナタバコ植物の葉に針の無いシリンジで浸潤させた。浸潤後3日目にGFP蛍光の焦点が出現した。蛍光の焦点は増殖して数日後にコンフルエントになった。
【0100】
OD0.3〜0.4のアグロバクテリウムを有する大量の浸潤溶液(3リットル)を調製した。ポットを用いてベンサミアナタバコ植物全体を逆さにして溶液にサッと浸し、乾燥器に入れ、十分な溶液が葉に浸透するまで1〜2分間真空を施した。次に植物を取り出し、温室に置いて再生させた。浸潤後3〜4日目に、シリンジを用いて手で浸潤させた領域のように、浸潤領域に蛍光の斑点が出現した。写真は、浸潤後10日目のそのような植物を示す(図3)。
【0101】
トバモウイルスなどのRNAウイルスは細胞質で複製し、決して核には入らない。したがって、核のプレmRNAプロセシング装置に曝露されない環境で展開する。その結果、人工ウイルス構築体から核において産生されるRNAレプリコン転写物は、RNAプロセシング装置によって認識され正しくプロセシングされることができない。その上、ウイルスベクター由来のRNAレプリコンは非常に大きい:TMVに基づいたレプリコンの場合およそ7,000ヌクレオチド。そのように大きなサイズの植物遺伝子は非常にわずかであり、そのような遺伝子の大部分は、プレmRNAのプロセシングを容易にし、核から輸送し、プロセシングされた転写産物の安定性を向上させるイントロンを含有する。したがって、正確なプロセシングと、正確にプロセシングされた転写産物の核から細胞質への輸送の効率を向上させるプレmRNAの改変は、ウイルス複製を開始する細胞数の増加をもたらすであろうという仮説を立てた。結局、核へのDNA送達 後、ウイルス複製をより効率的に開始できるRNAウイルスに基づいたベクターを作製するために使用できるアプローチは2つあることがわかる:(1)1つのアプローチは、無用のプロセシングイベント(潜在スプライス部位を用いた選択的スプライシングイベント、又は中途終止イベントなど)を誘導し得る配列特徴の除去である;(2)第2のアプローチは、正しくプロセシングされる転写産物量を増加させ、RNAの核から細胞質への輸送を改善させ、及び/又は転写物の安定性を向上させるイントロンの付加である。
【実施例2】
【0102】
TMVに基づいた改良RNAベクターの構築
目的配列が浸潤領域で発現する速度を向上させるために、91〜443ヌクレオチドのサイズの16のシロイヌナズナ イントロンをベクターの位置1〜16に付加した:(TVCV配列に対する位置、GenBank受託番号BRU03387):1、bp209;2、bp423;3、bp828;4、bp1169;5、bp1378;6、bp1622;7、bp1844;8、bp2228;9、bp2589;10、bp2944;11、bp3143;12、bp3381;13、bp3672;14、bp3850;15、bp4299;16、bp4497;17、bp5099;18、bp5287;19、bp5444。得られた構築体pICH18711(図2C)を、さまざまな段階の(播種後17〜35日目)いくつかのベンサミアナタバコ植物の真空浸潤に用いた。GFP蛍光は2日後に出現し、浸潤後6日目に非常に強かった。写真は、浸潤後4日目を示す(図4)。GFP蛍光は、非改良ベクターpICH16707よりも非常に速く(浸潤後4〜6日)葉領域全体を覆った。改良ウイルスベクターの詳細な説明は、近年公表された(Marillonnetら、2005、Nature Biotechnol.、23、718−723)。
【0103】
植物全体の代わりに、ベンサミアナタバコ植物の枝(茎及び葉)を真空浸潤させた。幹の基部を1杯の水の中に入れることによって幹を回復させた。GFPは3日後に葉に出現し、植物全体ではなく植物の部分を目的遺伝子の発現に用いることができることを示した。
【実施例3】
【0104】
アグロバクテリウムによって送達されたウイルスベクターの真空浸潤を用いた薬用タンパク質の発現
医薬的に関心のある2つのタンパク質を、GFPの代わりに上記改良ベクター中にクローニングした。第1のタンパク質ヒト成長ホルモン(hGH)(配列 Genbank受託番号NM_000515)のクローニングによりベクターpICH17991を得た(図2C)。第2のタンパク質ヒトインターフェロンα(配列 Genbank受託番号V00548)のクローニングによりベクターpICH19081を得た(図2C)。この構築体では、インターフェロンの最初の17アミノ酸(LLVALLVLSCKSSCSVG)が、シロイヌナズナ カルレティキュリン アポプラスト標的配列(matqrranpsslhlitvfsllvavvsaev)で置換されている。構築体をアグロバクテリウム株 GV3101に挿入し、植物全体の浸潤に用いた。両方のタンパク質に関し、高レベルのタンパク質発現が得られた。pICH17991に関し、たとえタンパク質の毒性が浸潤領域で細胞死をもたらしても、浸潤葉組織1gあたり1mgのhGHという高レベルの発現が得られた。
【実施例4】
【0105】
MPを欠失するベクターによる植物全体の浸潤
pICH17811におけるMPの初めのAvrII部位にフレームシフトを作製し(図2C)、構築体pICH18722を得た(図5)。このフレームシフトはMP機能を完全に排除しており、この構築体の浸潤により、個々の細胞のみにおいて複製と発現をもたらす。しかしながら、イントロンの存在のために、多数の細胞が依然としてGFPを発現する。
【0106】
TVCVのMPコード配列を、クローン化TVCV cDNA(GenBank受託番号Z29370、bp4802〜5628)からPCRにて増幅し、35Sプロモーターの制御下、バイナリーベクターにサブクローニングしてプラスミドpICH10745を得た。pICH18722とpICH10745との共浸潤により、少なくともMPが一過性に発現しているあいだは細胞間移行が完全に回復する。pICH10745にてベンサミアナタバコを安定に形質転換した。MPを発現する完全な形質転換体にpICH18722を浸潤させることにより、野生型植物の浸潤はウイルスベクターの細胞間移行をもたらさないが、野生型植物全体にpICH18711を浸潤させた場合と同様のGFP発現をもたらした(図16を参照されたい)。
【実施例5】
【0107】
目的遺伝子の一過性発現を提供するための変異体アグロバクテリウムの補完
a)VirE2を発現するトランスジェニック植物宿主によるトランス補完
VirE2遺伝子を、アグロバクテリウム・ツメファシエンスC58 T−DNA(Gene Bank受託番号AE009437、b.p6368〜8038)のDNA調製物からPCRで増幅し、TVCVのMP遺伝子を置換したベクターpICH10745にクローニングした。得られた構築体pICHVirE2(図15)を、選択剤として50mg/Lのカナマイシン(Km)を用いて、アグロバクテリウム・ツメファシエンス(GV3101)、及びベンサミアナタバコ植物のアグロバクテリウム介在葉ディスクトランスフェクションに用いた、機能性VirE2遺伝子欠失アグロバクテリウム・ツメファシエンス固定した(Horshら、1985、Science、227、1229−1231)。MPにフレームシフトを有する構築体pICH18722(図5)にてVirE2機能欠損アグロバクテリウム・ツメファシエンス株を形質転換した。この欠損は、VirE2遺伝子の欠失により引き起こし、野生型植物とpICH10745で形質転換した植物のアグロ浸潤実験に用いた。
【0108】
コンピテント及び野生型宿主を用いて、葉表面の同一領域あたりのGFP発現細胞数を計数することにより、又は(より正確には)対照及び改変ウイルスベクターを用いたウイルス複製の開始頻度を比較するために図10Cに示すように、コンピテント及び野生型宿主におけるGFP発現プロトプラストの割合を測定して比較することにより、一過性発現効率の比較を行った。一過性発現を提供するT−DNA送達のそのような相対頻度は、生物学的安全性を構築していないシステムとの比較において、システムの生物学的安全性レベルの測定として用いることができる。
【0109】
VirE2機能を欠損した他の多くのアグロバクテリウム株、例えばVirE2の挿入変異を有する株(Christieら、1988、J.Bacteriol.、170、2659−2667)又はvirE2機能を抑制するosa遺伝子を有するプラスミドpSAを有する株(Lee、L−Yら、1999、J.Bacteriol.、181、186−196)をこの実験に用いることができる。更に、Tiプラスミド(例えばGeneBank受託番号NC003065;NC001277)及び染色体DNA(例えばGeneBank受託番号NC003063;NC003063)を含めたアグロバクテリウムゲノム配列情報の利用可能性により、相同組換えによって機能性遺伝子をその変異体型に置換することで、あらゆる種類の変異体を作製することができる。さまざまに希釈した、VirE2含有及びVirE2欠損アグロバクテリウム株の一晩培養物を比較することによって、植物細胞へのT−DNA伝達効率に対するVirE2遺伝子の効果について試験した。アグロバクテリウム株はともにpICH18711ウイルスベクターをバイナリーベクターのT−DNA領域内に含有する(図2C)。そのような比較の結果を図17に示す。VirE2欠損アグロバクテリウム株によるT−DNA伝達効率が、機能性VirE2遺伝子含有アグロバクテリウムと比較して、少なくとも10,0000倍低下していることは写真から明らかである。コンピテント(pICHVIRE2で形質転換されている)及び野生型のベンサミアナタバコ植物の葉における一過性発現効率の比較を行い、pICHVirE2で形質転換したトランスジェニック植物からのVirE2遺伝子機能の非常に効率的なトランス補完を示した(図18)。異なる希釈系列(示していない)を用いて実験することにより、トランスジェニック宿主からのVirE2遺伝子機能のトランス補完効率が、機能性VirE2遺伝子を有するアグロバクテリウム株の使用に匹敵することを立証した。
【0110】
b)VirE2を発現する野生型アグロバクテリウム株との混合によるトランス補完
pICH18722を有するvirE2機能欠損株を、virE2機能をトランスで補完するアグロバクテリウム・ツメファシエンス株C58と混合することによって、virE2機能の補完を達成した以外は、先の実施例に記載の実験を反復した。新たに接種した両アグロバクテリウム株の培養物を光学密度OD600が2.5〜3.0になるまで一晩増殖させ、植物組織の浸潤のために等量で混合した。浸潤用最終混合物を50倍、100倍、1000倍、及び10000倍に希釈した。植物組織の浸潤をベンサミアナタバコ及びタバコの葉にて行った。pICH18711バイナリーを有するΔVirE2株(機能性MPを有する)を用いて同様の実験を行った。この実験の結果を図19に示す。pICH18711バイナリーを有する変異体株(T−DNA+、ΔVirE2)と野生型株(T−DNA−、VirE2)との共浸潤は、野生型株によるT−DNAの直接送達(T−DNA+、VirE2)よりも、T−DNA送達がまさに効率的であることは明らかである。
【実施例6】
【0111】
イントロン様配列の除去により、細胞質におけるウイルスRNAレプリコン形成頻度が上昇する
pICH4351由来のRNAレプリコンの配列をNetgeneIIサーバープログラム(http://www.cbs.dtu.dk/services/NetGene2/)を用いて解析したところ、選択的スプライシングイベントを誘導し得るいくつかのイントロン様配列特徴に気づいた。そのような特徴の1つは、RdRPの初めにある、0.6kbのウリジンに富んだ領域(GenBank受託番号BRU03387のヌクレオチド827〜1462に対応)である(図6A)。pICH14833において、オリジナル配列とは異なる、PCRで変異させた配列で、54ヌクレオチド置換によりこの領域を置換した(付録に配列番号2として示す配列、図7)。52ヌクレオチド置換は、Tに富んだ配列を、よりGCに富んだ配列に置換するために行った。RdRPのタンパク質配列を変えないように、全てのヌクレオチド置換をサイレントにした。この変異断片も、推定潜在スプライスドナー部位とアクセプター部位をそれぞれ除去するために導入された2つのヌクレオチド置換(829位と1459位;GenBank受託番号BRU03387に対する座標)を含有する。これらの変異の効果を試験するために、得られたクローンpICH15466(図2A)を、pICH10745(トランス移行タンパク質)と共に、又は伴わずに、ベンサミアナタバコの葉にアグロ浸潤させた。浸潤後8日目に、pICH15466を浸潤させた領域において、GFP発現細胞数の10倍増加を観察した(pICH14833と比較、図10)。これは、ウイルスレプリコンからイントロン様配列を除去することにより、無用の選択的スプライシングイベントを妨げ、より効率的にウイルス複製を開始することを示唆している。pICH15466とpICH10745との共浸潤は、非改変レプリコンと同様の速度で改変レプリコンの細胞間移行をもたらす。これは、RNA配列の改変がウイルスベクターの細胞間移行に影響を及ぼさなかったことを示している。
【実施例7】
【0112】
MPサブゲノムプロモーターにおけるイントロン様配列の除去
第2の潜在的に問題のある領域はMPサブゲノムプロモーターに対応する(図6B)。この領域は非常にTに富んでおり、イントロン配列に非常に近似している。その結果、多くの潜在スプライスドナー部位及びアクセプター部位が、イントロン予測プログラムによって隣接配列中に予測される。残念ながら、サブゲノムプロモーター機能に影響を及ぼすことなくこの領域を容易に改変することはできない。サブゲノムプロモーターを構わずに全領域を完全に変異させ、MPをトランスで提供し、予想されるMP発現の喪失を補償することにした。この構築体からMPは発現しないため、目的遺伝子の発現を駆動するために必要なCPサブゲノムプロモーターを含有する3’配列以外は、MP配列の大部分も削除した。したがって、pICH14833の383bp断片(GenBank受託番号BRU03387のbp4584〜5455)を297bp変異断片で置換した。得られた構築体pICH15900(図2A)を、pICH10745と共に、又は伴わずに、ベンサミアナタバコの葉にアグロ浸潤させた。興味深いことに、pICH14833を浸潤させた葉領域と比較して、複製開始細胞数の大幅な増加を検出した。浸潤葉領域から調製したGFP発現プロトプラストを計数することにより、この改変で、未改変pICH14833と比較して、ウイルス複製開始細胞数が80〜100倍増加すると概算している。pICH15900をpICH10745(p35S−MP発現カセット)を共浸潤させ、細胞間移行によるGFP蛍光の増加を検出した。しかしながら、この増加は、細胞間移行がなくても非常に多くの細胞が既にGFPを発現していたため、非常に限られていた。pICH15900含有アグロバクテリウム懸濁液の1000倍希釈を、pICH10745含有アグロバクテリウムの未希釈懸濁液と共浸潤させることにより、分離したGFP発現焦点を生じさせた。蛍光焦点は、pICH14833を用いて得た対照焦点と同様の明るさ及び同一サイズであった。これは、pICH15900の改変及びMPのトランス送達は、複製レベル、目的配列の発現、及び細胞間移行に関し、レプリコンの機能性を傷つけないことを示している。同一構築体(pICH14933及びpICH15900、pICH10745と共に、又は伴わずに)をタバコの葉に共浸潤させた。pICH15900の改変は、ベンサミアナタバコの場合と同様に複製開始細胞数を増加させた(pICH14833との比較)。
【実施例8】
【0113】
イントロン付加は細胞質における機能性RNAレプリコンの形成頻度を上昇させる
次に、レプリコンをコードする配列部分へのイントロン付加が複製開始頻度を上昇させるかどうかについて試験した。2つの構築体pICH15025及びpICH15034(図2A)を作製した。いずれも、3つの異なるシロイヌナズナ イントロンをRdRPの2つの異なる領域中に含有する。pICH15025を、RdRPの中央にイントロンを含有するようにデザインした。一方、pICH15034は、RdRPの3’端であるMPサブゲノムプロモーターの上流にイントロンを含有する。イントロンをシロイヌナズナ ゲノムDNAからPCRで増幅し、予定のイントロン/エクソン接合部が重複するプライマーを用いて、PCRでウイルス配列に組み込んだ。イントロン含有断片をAvaI/HindIII断片(付録の配列番号4)としてpICH14833にサブクローニングしてpICH15025を作製するか、又はPstI/Ncol断片(付録の配列番号5)としてサブクローニングしてpICH15034を作製した。
【0114】
2つの構築体をベンサミアナタバコの葉にアグロ浸潤させ、pICH14833と比較した。2つの構築体はウイルス複製開始細胞数を顕著に増加させた(図10A)。この増加は、pICH14833に対して約50倍の改善であると概算した。また、2つの構築体をMP発現クローンとともに共浸潤させ、細胞間移行はイントロンのないクローンと同じであることがわかった。2つの構築体をタバコにおいても試験し、ベンサミアナタバコと同じ改善を観察した(図10B)。
【0115】
6つのイントロン全てを含有する第3のクローンpICH15499を作製した(図9、2B、10A、10B)。この構築体をベンサミアナタバコ及びタバコにおいて試験した。この構築体は、3つのイントロンを有する個々の構築体よりも効率的であったが、その改善は相加的ではなかった。
【実施例9】
【0116】
イントロン付加とイントロン様配列の除去は、細胞質における機能性RNAレプリコン形成頻度を上昇させる
1つの構築体におけるイントロン様特徴の除去と、追加イントロンの付加は、両タイプの改変がウイルス複製開始の向上に寄与できることを示した。pICH15499の6つのイントロンを、変異MPサブゲノムプロモーター領域を含有するpICH15900へサブクローニングした。得られたクローンpICH15860(図2B)をベンサミアナタバコの葉に浸潤させたところ、GFPを発現する全てのプロトプラストのおよそ50%〜90%の範囲で、それぞれの親クローンよりも顕著に良好に作用することを発見した(図10)。最高性能の構築体は、RdRP領域と改変MPサブゲノムプロモーター領域にイントロンを含有する(pICH16191、図10C)。未改変クローンとの比較で、これは80〜300倍の改善に相当する。また、この構築体を、MP発現構築体(pICH10745)と共に共浸潤させたところ、改変が細胞間移行も複製も傷つけないことを発見した。
【実施例10】
【0117】
全てのイントロン付加が細胞質における機能性RNAレプリコンの出現頻度を上昇させるわけではない
2つの異なるシロイヌナズナ イントロンをRdRPの初めに挿入し、クローンpICH15477を得た(図2B)(この領域の配列を配列番号7として付録に示す)。この領域の配列は、イントロンを付加する前に、既に非常に「エクソン様」(例えばGCに富んでおり、潜在スプライス部位がない)に見える。この構築体で、ウイルス複製の開始に全く改善は見られなかった。したがって、あらゆるイントロン付加がウイルスベクターを改善するわけではない。イントロン挿入又は変異生成のために選択した位置は重要なパラメータであるように見える。例えば、MPサブゲノムプロモーターのような問題のある構造の近傍領域に作製した全てのイントロン挿入又はヌクレオチド置換は大きな改善をもたらしたが、既に「エクソン様」である配列へのイントロン挿入は顕著にウイルス複製開始を改善しなかった。
【実施例11】
【0118】
MP配列へのイントロン挿入はウイルスレプリコン形成頻度を上昇させる
初めに、制限酵素AvrIIで消化し、充填し、再連結することによって、MPにフレームシフトを作製した。次にMPに2つのイントロンを挿入した。得られたクローンpICH16422(図2B)をベンサミアナタバコの葉に浸潤させた。機能性ウイルスレプリコン含有細胞数に約100倍の増加を検出した。
【実施例12】
【0119】
MP含有ベクターへのイントロン挿入は自律機能性クローンのウイルス複製開始頻度を上昇させる
pICH15499のKpnI/EcoRI断片pICH8543へサブクローニングした。得られたクローン16700(図2B)は、RdRPに6つのイントロンを有する完全なウイルスベクターを含有する。このクローンをベンサミアナタバコの葉に浸潤させたところ、効率的に複製を開始した。また、このクローンは、追加MPをトランスで提供ことを必要とせずに細胞間移行可能であって。
【実施例13】
【0120】
遺伝子操作された宿主植物におけるアグロ送達された不活性レプリコンの活性化
イントロン含有ウイルスベクター前駆体をトランスジェニック植物へアグロ浸潤させることも可能である。目的タンパク質の産生を運命付けられていない植物(例えば開放環境の植物)の偶発的感染を回避するために、ベクターの一部をアンチセンス方向に存在させた不活性クローンを作製することができる(図11)。反転断片の先端に組換え部位とイントロン配列とを組み込むことにより、適切なリコンビナーゼを用いることによってこの断片を正しい方向に「反転」させることが可能である。組換え部位はスプライシングによってレプリコンから完全に排除されるであろう。プロレプリコン中のイントロンは、組換えと転写の後、効率的に複製を開始させる。1つの具体例では、組換え部位が、目的遺伝子内であってプロレプリコンの下流に局在する。そのような立体配置は、組換え前の遺伝子発現を妨げる。RdRP中やプロモーターの上流のように、プロレプリコンの他の領域に組換え部位が局在する場合、他の立体配置を検討することができる。組換え部位のイントロン配列は、先に記載したように、組換え部位をレプリコンから完全に除去させる利点を有するだけでなく、ウイルス複製効率を上昇させる。リコンビナーゼ活性を提供するトランスジェニック植物宿主は、遺伝子操作して、不活性ウイルスベクター前駆体を含有するベクターと共にアグロ浸潤に用いることができる。ウイルスベクター前駆体は、遺伝子操作された植物宿主の細胞においてのみ活性化することができる。
【実施例14】
【0121】
植物ウイルスRNA配列は潜在的に不安定領域を含有する
選択された植物RNAウイルスのRNAプロファイルを、十分に特徴付けられた植物遺伝子(AtDMC1)のものと同様に、NetgeneIIサーバープログラム(http://www.cbs.dtu.dk/services/NetGene2/)を用いて解析した。AtDMC1について図12に示すRNAプロファイルは、先にcDNA配列とゲノムDNA配列との比較によって同定した14のイントロンの存在を明確に反映している(丸で囲んである)。2つの植物ウイルスのRNAプロファイルは、植物の核環境に置かれた場合、RNAの安定性に問題を引き起こし得る領域を有することは明らかである(図13、図14を参照されたい)。ブロモモザイクウイルス、異なるTMV株、及び他の多くのものなど、いくつかの他の代表的植物RNAウイルス(示していない)のRNAプロファイルを解析した。これらは全て、DNA前駆体として植物細胞へ送達された場合に植物RNAウイルスに基づいたレプリコン形成効率を落とし得る、潜在的に問題のある領域を有している。
【実施例15】
【0122】
ジェミニウイルスベクターによる植物全体の浸潤
ビーンゴールデンモザイクウイルス(BGMV)単離体DSMZ PV−0094に基づいた発現ベクターpICH4300、pICH5202/3、及びpICH5170(図15)を、WO02/077246号の実施例10及び11に記載したように作製した。構築体pICH5170を、実施例5に記載のようにベンサミアナタバコ植物及びタバコ植物の安定な形質転換に用いた。構築体pICH5202/3及びpICH5170を、VirE2機能欠損アグロバクテリウム・ツメファシエンス及びアグロバクテリウム・ツメファシエンスC58株に固定した。固定化ウイルスレプリコン(pICH5202/3)を有する株を、pICH5170で形質転換したタバコ植物全体のアグロ浸潤に用いた。トランスジェニック植物宿主は、いわゆる共通領域(CR)に局在するジェミニウイルスの複製起点を有するGFPを有するDNAレプリコンの複製を提供した。VirE2機能欠損アグロバクテリウム株の場合、非形質転換アグロバクテリウム・ツメファシエンスC58と共にアグロ浸潤を行った。あるいは、pICHVirE2で形質転換したベンサミアナタバコ植物を、pICH5170で形質転換させた植物と交配させた。両構築体由来のT−DNA領域を含有するF1子孫をPCRで選択し、アグロバクテリウム機能を有し、pICH5202/3を有するアグロ浸潤に用いた。
【0123】
固定化ウイルスレプリコン(pICH5170)を有する株をタバコ植物全体のアグロ浸潤に用いた。
上記実験の結果は、トランスジェニック宿主植物又はVirE2発現アグロバクテリウムによって機能を補完することなくVirE2機能欠損株を用いた場合、浸潤植物においてGFPを全く発現しないことを示した。しかしながら、VirE2機能のトランス補完を必要とする実験における発現レベルは、ベクターをアグロバクテリウム・ツメファシエンスC58に固定した場合と顕著な差を示さなかった。
【実施例16】
【0124】
細菌プラスミド接合性伝達欠損変異体の作製
tra遺伝子及びtrb遺伝子は、Tiプラスミドの接合性伝達に関与すると記載されている(Farrandら、1996、J.Bacteriol.、178、4233−4247;Liら、1999、J.Bacteriol.、181、5033−41)。アグロバクテリウム・ツメファシエンス株C58のTiプラスミドからこれらの遺伝子を削除するために、隣接DNA領域を含有するベクターを形質転換に用い、2つの連続的組換えイベントにより、欠失を導入した。欠失ベクターは、正の選択マーカーとしてカルベニシリン耐性を、負の選択マーカーとしてショ糖不耐性を引き起こすsacB遺伝子を含有するベクターpDNR−1r(クローンテック)に基づいている。
【0125】
tra遺伝子(traG、traD、traC、oriT、traA、traF、traB、traH、traM)を欠失させるために、およそ700bpの隣接領域をアグロバクテリウムC58 DNAから、oSM570プライマー(5’−gaggatccaacgtttaggagaaccag−3’)+oSM571プライマー(5’−ttggtctcacggtatacgcacactgaacatgcg−3’)、及びoSM572プライマー(5’−ttggtctcaaccggtttccgtttgtctc−3’)+oSM573プライマー(5’−acgtctagagatcgcgttccagaccaac−3’)を用いてPCRで増幅させた。PCR断片をBsaIで制限処理した後連結し、追加ヌクレオチドを導入することなくBst11071制限部位を作製した;外端にプライマーを用いてBamHI部位とXbaI部位を導入した。BamHIとXbaIで制限処理したベクターpDNR−1rに2つのPCR断片を連結した。得られたプラスミドpICF12721を、エレクトロポレーションによるアグロバクテリウム形質転換に用い、初めはカルベニシリンで、その後抗生物質はないが5%ショ糖の存在下で選択し、2重組換えイベントを有する、ベクター骨格を含有しない細胞を対抗選択した。
【0126】
trb遺伝子(trbB、trbC、trbD、trbE、trbJ)を欠失させるために、およそ700bpの隣接領域を、アグロバクテリウムC58 DNAから、oSM566プライマー(5’−ctgaattcaggcaaacgcaccgtgagatg−3’)+oSM567プライマー(5’−tcaccatgggtcacgcggcactcctg−3’)、及びoSM568プライマー(5’−tcaccatggcccaggcccggcgtgaac−3’)+oSM569プライマー(5’−acatctagatgccggcatcgaagatgttg−3’)を用いてPCRで増幅させた。PCR断片の連結により、追加のヌクレオチドを導入することなくNcoI制限部位を作製した;外端にEcoRI部位及びXbaI部位をプライマーによって導入した。2つのPCR断片を、EcoRII及びXbaIで制限処理したベクターpDNR−1rに連結した。得られたプラスミドpICF12711を上記のようにアグロバクテリウム形質転換に用いた。
【0127】
アグロバクテリウム間の接合伝達を測定するために、オパイン結合リプレッサーをコードするaccR遺伝子をカナマイシン耐性カセットで置換することによって、構成的伝達を示す選択可能Tiプラスミドを構築した。これは、やはりベクターpDNR−1rに基づき、アグロバクテリウムC58 DNAからoSM554プライマー(5’−gagctagctccgtccttcacctgggc−3’)+oSM555プライマー(5’−ttggtctcaccggccgatagccaaaaactgc−3’)、及びoSM556プライマー(5’−ttggtctcgccggccaaactccggtttgc−3’)+oSM557プライマー(5’−atgggcccttcgaacgcaattcctgttgc−3’)を用いてPCRで増幅させた2つの隣接領域、及びそれらのあいだのXmaIII部位にカナマイシン耐性(oSM584プライマー(5’−cctcggccgcgaacggcctcac−3’)及びoSM585プライマー(5’−ctacggccgctgacagctaaaacaattcatcc−3’)を用いてバイナリーベクターpICH18711からPCRで増幅させた)を含有するベクターpICF12741でアグロバクテリウム・ツメファシエンス株C58を形質転換することによって作製した。カナマイシン耐性Tiプラスミドを有する株を、染色体リファンピシン耐性を有するがTiプラスミドのないアクセプター株GV3101と共に、ニトロセルロースメンブラン上で28℃にて2時間、Piper&Farrand(1999、Appl Environ Microbiol.、65、2798−2801)に記載されたのと同様にインキュベーションした。リファンピシンとカナマイシンを添加したプレートでトランス接合個体を選択した。aceを欠失しているが無傷のtra遺伝子及びtrb遺伝子を有するTiプラスミドの場合、投入ドナーあたりの接合効率はおよそ3〜4×10−4トランス接合個体であった。tra及びtrb変異体の場合、接合効率は検出限界以下であった(<10−7)。
【0128】
ベンサミアナタバコ植物に、GFPをコードするウイルスベクターpICH18711を含有する細菌を浸潤させることにより、植物へのDNA伝達に対する変異の影響について試験した。細菌は、葉で単一蛍光スポットを観察できるように、さまざまな希釈率で用いた。植物へのDNA伝達効率は、ace、tra、又はtrb欠失のない細菌と匹敵するものであった。
【実施例17】
【0129】
栄養要求性変異体の作製
栄養要求性アグロバクテリウム株を作製するために、ロイシンの生合成に関与する3−イソプロピルリンゴ酸脱水素酵素をコードするleuB遺伝子とウリジンの生合成に関与するオロチン酸ホスホリボシルトランスフェラーゼをコードするpyrEを選択した。leuBのコード配列は、アグロバクテリウム染色体の逆鎖に局在する仮想タンパク質ATU2792の3’と重複するため、完全に欠失させることができない。したがって、染色体1のbp2792384−2793470を、上流配列の775bpと下流配列の834bpとを、leuBコード配列の5’端に天然に生ずるBspHI制限部位を介して接続することにより欠失させるために選択した。ベクターpDNR−1r(クローンテック)を用いて、アグロバクテリウムの標的遺伝子内に欠失を作製した。
【0130】
leuBと同様に、pyrE遺伝子の異化補完は可能ではない。ウラシルホスホリボシルトランスフェラーゼ遺伝子がアグロバクテリウムに存在するため、ウリジン添加は安価なウラシルで置き換え得る。更に、pyrE欠失は正の選択マーカーとして用いることができる。選択は、野生型アグロバクテリウムは毒性のウラシル類似体である5−フルオロオロチン酸に対して感受性であるが、pyrE変異体は当然耐性であるという事実を利用する。
【0131】
pyrEの開始コドンは、異なるデータベースにおいて異なって注記されている(bp394734−395534、受託番号NP_531105とbp394836−395534、受託番号Q8UI98)。しかしながら、他のpyrE配列に対する相同性はbp394836の前からは開始しない。それにもかかわらず、2つの欠損変異体(欠失は1番染色体の394760位又は394849位から開始し、いずれも395531位で終わる)をデザインした。欠失含有断片を、ベクターpDNR−1rのEcoRI−BamHI部位にクローニングし、ベクターpICF1263(ΔleuB bp2792384−2793470)、pICF1264(ΔpyrE bp394760−395531)、及びpICF1265(ΔpyrE bp394848−395531)を得た(図21を参照されたい)。
【0132】
アグロバクテリウム株GV3101を、プラスミドpICF1263−3(ΔleuB bp2792384−2793470)、pICF1264−1(ΔpyrE bp394760−395531)、及びpICF1265−10(ΔpyrE bp394848−395531)で形質転換した。カルベニシリン含有培地での選択2日目に少量の形質転換体(6〜10cfu/μg)を得た。LB+リファンピシン液及びLB+リファンピシン+サッカロース液中で選択したクローンを連続して増殖させることによって共組換え体を決定した。標的遺伝子に所望の欠失を含有するアグロバクテリウム変異体を、LB+リファンピシン+サッカロース培養物から単離したアグロバクテリウムDNAのPCR解析によって確認した。
【0133】
変異体株1263−3−3及び1263−3−10並びに野生型親株GV3101を、30mg/l及び150mg/l ロイシンの添加の有無において、LB富栄養培地及びM9最少培地中で増殖させた。変異体株と野生型株とのあいだでLB培地での増殖に差はなかったが、M9培地では野生型株のみ増殖できた。M9培地にロイシンを添加した場合、野生型株も変異体株も増殖できた。添加ありと添加なしの培地でのさまざまな変異体株の増殖を図22及び図23に示す。ロイシン濃度の増加とともに増殖率が増加した。アグロバクテリウム変異体1263−3−3をGFPプラスミドpICH18711で形質転換し、ベンサミアナタバコにおける感染効率について解析した。
【0134】
付録
配列番号1(pICH14833のNcoI−EcoRI断片):
【0135】
【化1】
配列番号2(pICH15466の部分):
【0136】
【化2】
配列番号3(pICH15900の部分):
【0137】
【化3】
配列番号4(pICH15025の部分):
【0138】
【化4】
配列番号5(pICH15034の部分):
【0139】
【化5】
配列番号6(pICH15477の断片):
【0140】
【化6】
【図面の簡単な説明】
【0141】
【図1A】図1Aは、生物学的安全性の上昇(A)に基づいた、本発明の一般的原理を表す。
【図1B】図1Bは、RNAウイルスに基づくレプリコン形成頻度の上昇(B)に基づいた、本発明の一般的原理を表す。
【図2A】図2Aは、未改変の及び本発明にしたがい改変したウイルス構築体を図式的に示す。Act2−シロイヌナズナ アクチン2遺伝子のプロモーター;RdRP−ウイルスRNA依存性RNAポリメラーゼ;MP−ウイルス移行タンパク質;NTR−ウイルス3’非翻訳領域;GFP−緑色蛍光タンパク質;インターフェロン−ヒトインターフェロンα 2b;hGH−ヒト成長ホルモン;nos−ノパリン合成酵素遺伝の転写終止領域。
【図2B】図2Bは、未改変の及び本発明にしたがい改変したウイルス構築体を図式的に示す。Act2−シロイヌナズナ アクチン2遺伝子のプロモーター;RdRP−ウイルスRNA依存性RNAポリメラーゼ;MP−ウイルス移行タンパク質;NTR−ウイルス3’非翻訳領域;GFP−緑色蛍光タンパク質;インターフェロン−ヒトインターフェロンα 2b;hGH−ヒト成長ホルモン;nos−ノパリン合成酵素遺伝の転写終止領域。
【図2C】図2Cは、未改変の及び本発明にしたがい改変したウイルス構築体を図式的に示す。Act2−シロイヌナズナ アクチン2遺伝子のプロモーター;RdRP−ウイルスRNA依存性RNAポリメラーゼ;MP−ウイルス移行タンパク質;NTR−ウイルス3’非翻訳領域;GFP−緑色蛍光タンパク質;インターフェロン−ヒトインターフェロンα 2b;hGH−ヒト成長ホルモン;nos−ノパリン合成酵素遺伝の転写終止領域。
【図3】図3は、ベクターpICH16707のアグロ浸潤後10日目のベンサミアナタバコ植物をUV光下にて示す。
【図4】図4は、ベクターpICH18711のアグロ浸潤後4日目の、17日(A)、22日(B)、28日(C)、及び35日(D)齢のベンサミアナタバコ植物をUV光下にて示す。
【図5】図5は、構築体pICH18722及びpICH10745のT−DNA領域を図式的に示す。Act2−シロイヌナズナ アクチン2遺伝子のプロモーター;RdRP−ウイルスRNA依存性RNAポリメラーゼ;MP−ウイルス移行タンパク質;NTR−ウイルス3’非翻訳領域;GFP−緑色蛍光タンパク質;nosT−ノパリン合成酵素遺伝子の転写終止領域;nosP−ノパリン合成酵素遺伝子の転写プロモーター;oscT−オクトピン合成酵素遺伝子の転写終止領域;NPTII−ネオマイシン ホスホトランスフェラーゼII遺伝子のコード配列;TVCV MP−カブ葉脈透化ウイルスの移行タンパク質。
【図6A】図6は、ベクターpICH8543の転写領域のイントロン予測プロファイルを示す。ヌクレオチド番号を横軸に示す。縦軸は、対応の配列/配列領域がコード配列(コード)であるか、ドナー部位(ドナー)若しくはアクセプター部位(アクセプター)として作用する確率を示す。丸で囲んだ部分は、機能保存的な相違を導入すべき選択された位置に相当する。
【図6B】図6は、ベクターpICH8543の転写領域のイントロン予測プロファイルを示す。ヌクレオチド番号を横軸に示す。縦軸は、対応の配列/配列領域がコード配列(コード)であるか、ドナー部位(ドナー)若しくはアクセプター部位(アクセプター)として作用する確率を示す。丸で囲んだ部分は、機能保存的な相違を導入すべき選択された位置に相当する。
【図7】図7は、ベクターpICH15466の転写領域(転写領域の前半の半分)のイントロン予測プロファイルを示す。丸で囲んだ領域は本発明に記載のように改変されている(図6Aとの比較)。
【図8】図8は、pICH15900の転写領域(転写領域の第2の半分)のイントロン予測プロファイルを示す。丸で囲んだ領域は本発明に記載のように改変されている(図6Bとの比較)。
【図9A】図9は、pICH15499の転写領域のイントロン予測プロファイルを示す。丸で囲んだ領域は、6つの挿入された植物核イントロンに相当する。
【図9B】図9は、pICH15499の転写領域のイントロン予測プロファイルを示す。丸で囲んだ領域は、6つの挿入された植物核イントロンに相当する。
【図10A】図10は、ベンサミアナタバコ及びタバコの葉におけるウイルス構築体のアグロ浸潤後のGFP発現を示す。各浸潤領域におけるベクター同定番号を示す。A−ベンサミアナタバコ、アグロ浸潤後8日目。
【図10B】図10は、ベンサミアナタバコ及びタバコの葉におけるウイルス構築体のアグロ浸潤後のGFP発現を示す。各浸潤領域におけるベクター同定番号を示す。B−タバコ、アグロ浸潤後8日目。
【図10C】図10は、ベンサミアナタバコ及びタバコの葉におけるウイルス構築体のアグロ浸潤後のGFP発現を示す。各浸潤領域におけるベクター同定番号を示す。C−アグロ浸潤後5日目に単離したベンサミアナタバコのプロトプラスト。右の写真の多くの光点は、非常に高頻度のレプリコン形成及びGFP発現を示す。
【図11】図11は、本発明にしたがいデザインされたRNAウイルスに基づいたレプリコン前駆体の略図であり、非誘導状態では目的配列(GFP、Gと示す)の発現レベルがゼロである。P−転写プロモーター;T−転写終止領域;SM−選択可能マーカー遺伝子;Ac2−シロイヌナズナ アクチン2遺伝子のプロモーター;RdRP−ウイルスRNA依存性RNAポリメラーゼ;MP−ウイルス移行タンパク質;NTR−ウイルス3’非翻訳領域。
【図12】図12は、直鎖(+鎖)を用いたシロイヌナズナ減数分裂特異的遺伝子AtDMC1(GenBank受託番号U76670)のイントロン予測プロファイルを示す。イントロンコード領域を丸で囲んである。
【図13A】図13Aは、ジャガイモXウイルス(PVX)ゲノム(GenBank受託番号AF172259)の直鎖(+鎖)内の潜在的に問題のある領域(丸で囲んである)の予測プロファイルを示す。
【図13B】図13Bは、ジャガイモXウイルス(PVX)ゲノム(GenBank受託番号AF172259)の直鎖(+鎖)内の潜在的に問題のある領域(丸で囲んである)の予測プロファイルを示す。
【図14A】図14は、アルファルファモザイクウイルスゲノムRNA1(GenBank受託番号K02703)の直鎖(+鎖)の潜在的に問題のある領域(丸で囲んである)の予測プロファイルを示す。
【図14B】図14は、アルファルファモザイクウイルスゲノムRNA2(GenBank受託番号K02702)の直鎖(+鎖)の潜在的に問題のある領域(丸で囲んである)の予測プロファイルを示す。
【図14C】図14は、アルファルファモザイクウイルスゲノムRNA3(GenBank受託番号L00163)の直鎖(+鎖)の潜在的に問題のある領域(丸で囲んである)の予測プロファイルを示す。
【図15】図15は、構築体pICHVirE2、pICH4300、pICH5202/3、及びpICH5170のT−DNA領域の略図を示す。
【図16】図16は、機能性MP(移行タンパク質)欠失ウイルスベクターの細胞間移行のトランスジェニック植物宿主(ベンサミアナタバコ)からのトランス補完の効果を示す。上部のラベルは植物を表し、下部のラベルはアグロ浸潤に用いたベクターを示す。左の写真は図示したように処理した植物の典型的な葉を示す。右の写真はトランスジェニック植物の全体を示す。WT:野生型植物;pICH10745トランスジェニック:TVCV移行タンパク質(MP)を発現するトランスジェニック植物;pICH18722:細胞間移行不良のウイルスベクターによる浸潤;pICH18711:細胞間移行可能なウイルスベクターによる浸潤。GFP発現を検出するために植物をUV光下にて示す。
【図17】図17は、バイナリーベクターpICH18711からのT−DNA伝達効率に対するVirE2遺伝子の効果を示す。VirE2:機能性VirE2遺伝子を含有するアグロバクテリウムを浸潤させたベンサミアナタバコの葉;ΔvirE2:VirE2遺伝子機能不良のアグロバクテリウムを浸潤させたベンサミアナタバコの葉;102〜107の数は浸潤前のアグロバクテリウムストックの希釈を示す(ΔVirE2株とVirE2株のストックは同一OD600)。矢印は同一のT−DNA伝達頻度を発揮する条件を結び付けており、ΔVirE2株は102倍希釈、VirE2株は107倍希釈に相当する。
【図18】図18は、野生型(WT)植物、及びpICH18711バイナリーベクターを含有するΔVirE2アグロバクテリウム株を浸潤させたトランスジェニック(pICHVirE2)植物を示す。写真は、10dpi(浸潤後日数)にUV光下にて撮影した。同一のアグロバクテリウム一晩培養物を、WTの浸潤には10倍に希釈し、トランスジェニック植物の浸潤には1000倍に希釈した。
【図19】図19は、野生型植物への変異体アグロバクテリウム(機能性virE2遺伝子を欠失)と野生型アグロバクテリウム(VirE2遺伝子機能を維持)との共浸潤によるVirE2遺伝子機能のトランス補完を示す。変異体及び野生型のアグロバクテリウムをさまざまな組み合わせで浸潤させたベンサミアナタバコの葉をUV光下にて示す。四角内のラベルは、アグロバクテリウムの個々の種類を表す。四角のあいだの+印は、葉の同一の場所に2種類のアグロバクテリウムを共浸潤させたことを示す。「T−DNA」の前の+印又は−印は、それぞれ、GFPを有するT−DNAの存在及び不存在を示す。機能性VirE2の不存在をΔで示す。
【図20】図20は、aceオペロンを欠失したアグロバクテリウムからのT−DNA伝達効率試験の結果を示す。TiプラスミドpMP90(対照)又はpTiC58Δacc(acc欠失)を有するアグロバクテリウム株GV3101を、GFPをコードするウイルスベクターpICH18711で形質転換し、一晩培養物のさまざまな希釈物を植物の浸潤に用いた。写真は接種後7日目にUV光下にて撮影した。
【図21】図21は、アグロバクテリウムのΔleuB変異体及びΔpyrE変異体を作製するための戦略を表す。
【図22】図22は、補完の有無における、さまざまな培地でのアグロバクテリウム株1264(ΔpyrE)の増殖速度を示す。ΔpyrE変異体株1264−1−2及び1264−1−8はウラシル含有培地においてのみ増殖する。縦軸上の数値はOD600を示す。
【図23】図23は、補完の有無における、さまざまな培地でのアグロバクテリウム株1263(ΔleuB)の増殖速度を示す。ΔleuB変異体株1263−3−3及び1263−3−10はロイシン依存性であるが、野生型親株GV3101はロイシン依存性ではない。縦軸上の数値はOD600を示す。
【特許請求の範囲】
【請求項1】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
(a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉にトランスフェクションし、レプリコンをコードする配列は、
(i)植物ウイルスに由来する、レプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、
(b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、
ことを含み、
アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されている、前記方法。
【請求項2】
第1の遺伝子改変が、植物又は植物の葉の細胞へのT−DNAの導入に必要なアグロバクテリウム株の機能を不良にする、請求項1に記載の方法。
【請求項3】
補完因子が、アグロバクテリウム株による植物又は植物の葉の細胞へのT−DNAのトランスフェクションに必要な機能を提供する、請求項1又は2に記載の方法。
【請求項4】
植物又は植物の葉の細胞へのT−DNAのトランスフェクションに必要な機能が、VirE2、GALLS、及びVirFから成る群より選択される遺伝子又はタンパク質によって提供される、請求項3に記載の方法。
【請求項5】
第1の遺伝子改変として、生物へのトランスフェクションに必要なアグロバクテリウム株の機能が、化学的に調節された異種プロモーターの制御下に置かれており;補完因子が、化学的に調節されたプロモーターを調節可能な小分子化合物である、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
補完因子が、第2のアグロバクテリウム株によって植物又は植物の葉に提供される、請求項1〜4のいずれか1項に記載の方法。
【請求項7】
植物に、補完因子であるか又は補完因子をコードする第2の遺伝子改変を提供する、請求項1〜4のいずれか1項に記載の方法。
【請求項8】
アグロバクテリウム株が栄養要求性であり、アグロバクテリウム株の栄養要求性増殖に必要な主要代謝産物の存在下で植物又は植物の葉に提供される、請求項2〜8のいずれか1項に記載の方法。
【請求項9】
第1の遺伝子改変がアグロバクテリウム株を栄養要求性にし、補完因子が、アグロバクテリウム株の栄養要求性増殖に必要なアグロバクテリウム株の主要代謝産物である、請求項1に記載の方法。
【請求項10】
アグロバクテリウム株が条件的致死性であり、アグロバクテリウム株の生存に必要な代謝産物の存在下で増殖し、及び/又は植物又は植物の葉に提供される、請求項2〜7のいずれか1項に記載の方法。
【請求項11】
第1の遺伝子改変がアグロバクテリウム株を条件的致死性にし、補完因子が、アグロバクテリウム株の生存に必要なアグロバクテリウム株の主要代謝産物である、請求項1に記載の方法。
【請求項12】
アグロバクテリウム株が、アグロバクテリウムから又はアグロバクテリウムへのプラスミドの接合性伝達を不良にする更なる遺伝子改変を有する、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
接合性伝達の不良が、以下の遺伝子:oriT、traG、及びtraFのうちの少なくとも1つの機能を欠失することによって起こる、請求項12に記載の方法。
【請求項14】
レプリコンをコードする配列部分が、植物又は植物の葉におけるレプリコンの伝播に必要な機能を欠損したレプリコンをコードする、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
配列部分が、レプリコンの遠隔距離移行又は細胞間移行に必要な機能を欠損したレプリコンをコードする、請求項14に記載の方法。
【請求項16】
レプリコンの伝播に必要な機能が、植物又は植物の葉において異種DNA配列以外の核酸配列から機能を発現させることによって植物又は植物の葉に提供される、請求項14又は15に記載の方法。
【請求項17】
アグロバクテリウム株が腫瘍抑制活性タンパク質Osaを発現する、請求項1〜16のいずれか1項に記載の方法。
【請求項18】
アグロバクテリウム株が、非標的生物へのT−DNA伝達を減少させるための、遺伝子改変された菌体数感知システム又は病原性誘導調節システムを有する、請求項1〜17のいずれか1項に記載の方法。
【請求項19】
補完因子が、植物又は植物の葉において2つのRNA配列のトランススプライシングによって提供され、2つのRNA配列のうちの多くても1つがアグロバクテリウム株にコードされる、請求項1〜18のいずれか1項に記載の方法。
【請求項20】
レプリコンがDNAレプリコンである、請求項1〜19のいずれか1項に記載の方法。
【請求項21】
レプリコンがRNAレプリコンであり、RNAレプリコンをコードする配列部分が転写プロモーターに機能可能に連結している、請求項1〜19のいずれか1項に記載の方法。
【請求項22】
レプリコン機能に必要な配列が、選択された位置で、植物RNAウイルスとの機能保存的な相違を発揮し、その相違は、相違を発揮しないRNAレプリコンと比較してレプリコン形成頻度を上昇させる、請求項21に記載の方法。
【請求項23】
機能保存的な相違が、A/Uに富んだ領域に隣接する潜在スプライシング部位の除去を含む、請求項22に記載の方法。
【請求項24】
機能保存的な相違が、レプリコン形成に必要な配列のA/Uに富んだ位置の近傍又はその中に1以上の核イントロン又は核イントロンを形成可能な1以上の配列の挿入を含む、請求項22又は23に記載の方法。
【請求項25】
レプリコンが、植物又は植物の葉において細胞間移行不良である、請求項1〜24のいずれか1項に記載の方法。
【請求項26】
植物ウイルスがトバモウイルスであり、好ましくはタバコモザイクウイルスである、請求項21〜25のいずれか1項に記載の方法。
【請求項27】
アグロバクテリウム株がアグロバクテリウム・ツメファシエンス若しくはアグロバクテリウム・リゾゲネス、又はアグロバクテリウム由来機能性T−DNA伝達発生装置を含有するように操作された他の微生物である、請求項1〜26のいずれか1項に記載の方法。
【請求項28】
遺伝子改変が細菌染色体及び/又はプラスミド染色体上になされる、請求項1〜27のいずれか1項に記載の方法。
【請求項29】
植物又は植物の葉を、工程(a)において、600nmの算出光学密度が高くても0.04、好ましくは高くても0.01、より好ましくは高くても0.004、最も好ましくは高くても0.001に相当するアグロバクテリウム株の細胞濃度を有するアグロバクテリウム株の懸濁液に浸潤させ、算出光学密度は、600nmのODが1.0のアグロバクテリウム懸濁液のそれぞれ少なくとも25倍希釈、好ましくは少なくとも100倍希釈、より好ましくは少なくとも250倍希釈、最も好ましくは少なくとも1000倍希釈によって規定される、請求項1〜28のいずれか1項に記載の方法。
【請求項30】
植物又は植物の葉が以下の種:ベンサミアナタバコ、タバコ、ペチュニア、アブラナ、カラシナ、クレス、ルッコラ、マスタード、イチゴ、ホウレンソウ、アカザ、レタス、ヒマワリ、及びキュウリのうちの1つに属する、請求項1〜29のいずれか1項に記載の方法。
【請求項31】
植物又は植物の葉を浸潤させることによってトランスフェクションを行う、請求項1〜30のいずれか1項に記載の方法。
【請求項32】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
(a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉にトランスフェクションし、RNAレプリコンをコードする配列は、
(i)植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、
(b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、
ことを含み、
アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されており、そして
レプリコン機能に必要な配列は、選択された位置で、植物RNAウイルスとの機能保存的な相違を発揮し、その相違は、相違を発揮しないRNAレプリコンと比較してレプリコン形成頻度を上昇させる、前記方法。
【請求項33】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
(a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉にトランスフェクションし、RNAレプリコンをコードする配列は、
(i)植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、
(b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、
ことを含み、
アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されており、そして
レプリコン機能に必要な配列は1以上の核イントロンを含有する、前記方法。
【請求項34】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
(a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉にトランスフェクションし、RNAレプリコンをコードする配列は、
(i)植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、
(b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、
ことを含み、
アグロバクテリウム株は、補完因子の非存在下では植物又は植物の葉の細胞へのT−DNAの導入に必要なアグロバクテリウム株の機能を不良にする第1の遺伝子改変が提供されており、そして
アグロバクテリウム株は、他の細菌へのプラスミドDNAの接合性伝達を不良にする更なる遺伝子改変を有する、前記方法。
【請求項35】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
(a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉にトランスフェクションし、RNAレプリコンをコードする配列は、
(i)植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、
(b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、
ことを含み、
アグロバクテリウム株は、補完因子の非存在下では植物又は植物の葉の細胞へのT−DNAの導入に必要なアグロバクテリウム株の機能を不良にする第1の遺伝子改変が提供されており、そして、
アグロバクテリウム株は、アグロバクテリウム株の栄養要求性増殖に必要な主要代謝産物に対して栄養要求性である、前記方法。
【請求項36】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
(a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉にトランスフェクションし、RNAレプリコンをコードする配列は、
(i)植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、
(b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、
ことを含み、
アグロバクテリウム株は、補完因子の非存在下では植物又は植物の葉の細胞へのT−DNAの導入に必要なアグロバクテリウム株の機能を不良にする第1の遺伝子改変が提供されており、
アグロバクテリウム株は、アグロバクテリウム株の栄養要求性増殖に必要な主要代謝産物に対して栄養要求性であり、そして
レプリコン機能に必要な配列は1以上の核イントロンを含有する、前記方法。
【請求項37】
(i)請求項1に定義した通りのアグロバクテリウム株、及び
(ii)請求項1に定義したアグロバクテリウム株の欠陥を補完可能な補完因子をコードする植物又はその種子、
を含む、キット部品。
【請求項38】
(i)請求項1に定義した通りのアグロバクテリウム株、及び
(ii)請求項1で定義したアグロバクテリウム株の欠陥を補完可能な補完因子をコードする第2のアグロバクテリウム株、
を含む、キット部品。
【請求項39】
(i)補完因子の非存在下では植物又は植物の葉の細胞へのT−DNAの導入に必要な細菌の機能を欠損した第1の遺伝子改変を有し、
(ii)他の細菌へのプラスミド伝達の接合能を欠損しており、そして
(iii)栄養要求性である、
ことを特徴とする、アグロバクテリウム属の細菌。
【請求項40】
アグロバクテリウムが、レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有し、レプリコンをコードする配列は、
(i) 植物ウイルスに由来する、植物のレプリコンのレプリコン機能に必要な配列、及び
(ii) 細菌を適用した植物において発現されるべき目的配列、
を含有する、請求項39に記載の細菌。
【請求項1】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
(a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉にトランスフェクションし、レプリコンをコードする配列は、
(i)植物ウイルスに由来する、レプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、
(b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、
ことを含み、
アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されている、前記方法。
【請求項2】
第1の遺伝子改変が、植物又は植物の葉の細胞へのT−DNAの導入に必要なアグロバクテリウム株の機能を不良にする、請求項1に記載の方法。
【請求項3】
補完因子が、アグロバクテリウム株による植物又は植物の葉の細胞へのT−DNAのトランスフェクションに必要な機能を提供する、請求項1又は2に記載の方法。
【請求項4】
植物又は植物の葉の細胞へのT−DNAのトランスフェクションに必要な機能が、VirE2、GALLS、及びVirFから成る群より選択される遺伝子又はタンパク質によって提供される、請求項3に記載の方法。
【請求項5】
第1の遺伝子改変として、生物へのトランスフェクションに必要なアグロバクテリウム株の機能が、化学的に調節された異種プロモーターの制御下に置かれており;補完因子が、化学的に調節されたプロモーターを調節可能な小分子化合物である、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
補完因子が、第2のアグロバクテリウム株によって植物又は植物の葉に提供される、請求項1〜4のいずれか1項に記載の方法。
【請求項7】
植物に、補完因子であるか又は補完因子をコードする第2の遺伝子改変を提供する、請求項1〜4のいずれか1項に記載の方法。
【請求項8】
アグロバクテリウム株が栄養要求性であり、アグロバクテリウム株の栄養要求性増殖に必要な主要代謝産物の存在下で植物又は植物の葉に提供される、請求項2〜8のいずれか1項に記載の方法。
【請求項9】
第1の遺伝子改変がアグロバクテリウム株を栄養要求性にし、補完因子が、アグロバクテリウム株の栄養要求性増殖に必要なアグロバクテリウム株の主要代謝産物である、請求項1に記載の方法。
【請求項10】
アグロバクテリウム株が条件的致死性であり、アグロバクテリウム株の生存に必要な代謝産物の存在下で増殖し、及び/又は植物又は植物の葉に提供される、請求項2〜7のいずれか1項に記載の方法。
【請求項11】
第1の遺伝子改変がアグロバクテリウム株を条件的致死性にし、補完因子が、アグロバクテリウム株の生存に必要なアグロバクテリウム株の主要代謝産物である、請求項1に記載の方法。
【請求項12】
アグロバクテリウム株が、アグロバクテリウムから又はアグロバクテリウムへのプラスミドの接合性伝達を不良にする更なる遺伝子改変を有する、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
接合性伝達の不良が、以下の遺伝子:oriT、traG、及びtraFのうちの少なくとも1つの機能を欠失することによって起こる、請求項12に記載の方法。
【請求項14】
レプリコンをコードする配列部分が、植物又は植物の葉におけるレプリコンの伝播に必要な機能を欠損したレプリコンをコードする、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
配列部分が、レプリコンの遠隔距離移行又は細胞間移行に必要な機能を欠損したレプリコンをコードする、請求項14に記載の方法。
【請求項16】
レプリコンの伝播に必要な機能が、植物又は植物の葉において異種DNA配列以外の核酸配列から機能を発現させることによって植物又は植物の葉に提供される、請求項14又は15に記載の方法。
【請求項17】
アグロバクテリウム株が腫瘍抑制活性タンパク質Osaを発現する、請求項1〜16のいずれか1項に記載の方法。
【請求項18】
アグロバクテリウム株が、非標的生物へのT−DNA伝達を減少させるための、遺伝子改変された菌体数感知システム又は病原性誘導調節システムを有する、請求項1〜17のいずれか1項に記載の方法。
【請求項19】
補完因子が、植物又は植物の葉において2つのRNA配列のトランススプライシングによって提供され、2つのRNA配列のうちの多くても1つがアグロバクテリウム株にコードされる、請求項1〜18のいずれか1項に記載の方法。
【請求項20】
レプリコンがDNAレプリコンである、請求項1〜19のいずれか1項に記載の方法。
【請求項21】
レプリコンがRNAレプリコンであり、RNAレプリコンをコードする配列部分が転写プロモーターに機能可能に連結している、請求項1〜19のいずれか1項に記載の方法。
【請求項22】
レプリコン機能に必要な配列が、選択された位置で、植物RNAウイルスとの機能保存的な相違を発揮し、その相違は、相違を発揮しないRNAレプリコンと比較してレプリコン形成頻度を上昇させる、請求項21に記載の方法。
【請求項23】
機能保存的な相違が、A/Uに富んだ領域に隣接する潜在スプライシング部位の除去を含む、請求項22に記載の方法。
【請求項24】
機能保存的な相違が、レプリコン形成に必要な配列のA/Uに富んだ位置の近傍又はその中に1以上の核イントロン又は核イントロンを形成可能な1以上の配列の挿入を含む、請求項22又は23に記載の方法。
【請求項25】
レプリコンが、植物又は植物の葉において細胞間移行不良である、請求項1〜24のいずれか1項に記載の方法。
【請求項26】
植物ウイルスがトバモウイルスであり、好ましくはタバコモザイクウイルスである、請求項21〜25のいずれか1項に記載の方法。
【請求項27】
アグロバクテリウム株がアグロバクテリウム・ツメファシエンス若しくはアグロバクテリウム・リゾゲネス、又はアグロバクテリウム由来機能性T−DNA伝達発生装置を含有するように操作された他の微生物である、請求項1〜26のいずれか1項に記載の方法。
【請求項28】
遺伝子改変が細菌染色体及び/又はプラスミド染色体上になされる、請求項1〜27のいずれか1項に記載の方法。
【請求項29】
植物又は植物の葉を、工程(a)において、600nmの算出光学密度が高くても0.04、好ましくは高くても0.01、より好ましくは高くても0.004、最も好ましくは高くても0.001に相当するアグロバクテリウム株の細胞濃度を有するアグロバクテリウム株の懸濁液に浸潤させ、算出光学密度は、600nmのODが1.0のアグロバクテリウム懸濁液のそれぞれ少なくとも25倍希釈、好ましくは少なくとも100倍希釈、より好ましくは少なくとも250倍希釈、最も好ましくは少なくとも1000倍希釈によって規定される、請求項1〜28のいずれか1項に記載の方法。
【請求項30】
植物又は植物の葉が以下の種:ベンサミアナタバコ、タバコ、ペチュニア、アブラナ、カラシナ、クレス、ルッコラ、マスタード、イチゴ、ホウレンソウ、アカザ、レタス、ヒマワリ、及びキュウリのうちの1つに属する、請求項1〜29のいずれか1項に記載の方法。
【請求項31】
植物又は植物の葉を浸潤させることによってトランスフェクションを行う、請求項1〜30のいずれか1項に記載の方法。
【請求項32】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
(a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉にトランスフェクションし、RNAレプリコンをコードする配列は、
(i)植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、
(b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、
ことを含み、
アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されており、そして
レプリコン機能に必要な配列は、選択された位置で、植物RNAウイルスとの機能保存的な相違を発揮し、その相違は、相違を発揮しないRNAレプリコンと比較してレプリコン形成頻度を上昇させる、前記方法。
【請求項33】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
(a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉にトランスフェクションし、RNAレプリコンをコードする配列は、
(i)植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、
(b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、
ことを含み、
アグロバクテリウム株は、補完因子の非存在下では生物へのT−DNAのトランスフェクションを不良にする第1の遺伝子改変が提供されており、そして
レプリコン機能に必要な配列は1以上の核イントロンを含有する、前記方法。
【請求項34】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
(a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉にトランスフェクションし、RNAレプリコンをコードする配列は、
(i)植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、
(b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、
ことを含み、
アグロバクテリウム株は、補完因子の非存在下では植物又は植物の葉の細胞へのT−DNAの導入に必要なアグロバクテリウム株の機能を不良にする第1の遺伝子改変が提供されており、そして
アグロバクテリウム株は、他の細菌へのプラスミドDNAの接合性伝達を不良にする更なる遺伝子改変を有する、前記方法。
【請求項35】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
(a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉にトランスフェクションし、RNAレプリコンをコードする配列は、
(i)植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、
(b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、
ことを含み、
アグロバクテリウム株は、補完因子の非存在下では植物又は植物の葉の細胞へのT−DNAの導入に必要なアグロバクテリウム株の機能を不良にする第1の遺伝子改変が提供されており、そして、
アグロバクテリウム株は、アグロバクテリウム株の栄養要求性増殖に必要な主要代謝産物に対して栄養要求性である、前記方法。
【請求項36】
目的配列を植物又は植物の葉において発現させることによって目的タンパク質を産生する方法であって:
(a)レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有するアグロバクテリウム株を補完因子の存在下又は非存在下で植物又は植物の葉にトランスフェクションし、RNAレプリコンをコードする配列は、
(i)植物ウイルスに由来する、RNAレプリコンのレプリコン機能に必要な配列、及び
(ii)発現されるべき目的配列、
を含有し、
(b)場合により、工程(a)で浸潤させた植物又は植物の葉から目的タンパク質を単離する、
ことを含み、
アグロバクテリウム株は、補完因子の非存在下では植物又は植物の葉の細胞へのT−DNAの導入に必要なアグロバクテリウム株の機能を不良にする第1の遺伝子改変が提供されており、
アグロバクテリウム株は、アグロバクテリウム株の栄養要求性増殖に必要な主要代謝産物に対して栄養要求性であり、そして
レプリコン機能に必要な配列は1以上の核イントロンを含有する、前記方法。
【請求項37】
(i)請求項1に定義した通りのアグロバクテリウム株、及び
(ii)請求項1に定義したアグロバクテリウム株の欠陥を補完可能な補完因子をコードする植物又はその種子、
を含む、キット部品。
【請求項38】
(i)請求項1に定義した通りのアグロバクテリウム株、及び
(ii)請求項1で定義したアグロバクテリウム株の欠陥を補完可能な補完因子をコードする第2のアグロバクテリウム株、
を含む、キット部品。
【請求項39】
(i)補完因子の非存在下では植物又は植物の葉の細胞へのT−DNAの導入に必要な細菌の機能を欠損した第1の遺伝子改変を有し、
(ii)他の細菌へのプラスミド伝達の接合能を欠損しており、そして
(iii)栄養要求性である、
ことを特徴とする、アグロバクテリウム属の細菌。
【請求項40】
アグロバクテリウムが、レプリコンをコードする配列部分を有する異種DNA配列をT−DNA中に含有し、レプリコンをコードする配列は、
(i) 植物ウイルスに由来する、植物のレプリコンのレプリコン機能に必要な配列、及び
(ii) 細菌を適用した植物において発現されるべき目的配列、
を含有する、請求項39に記載の細菌。
【図1A】

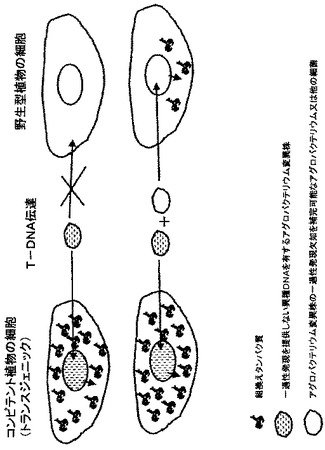
【図1B】
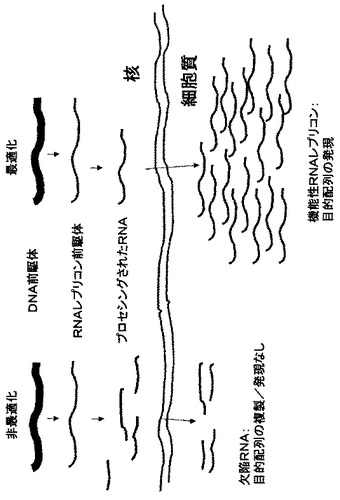
【図2A】
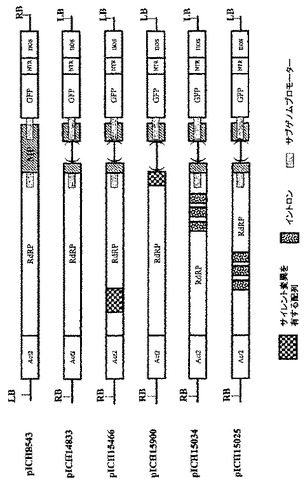
【図2B】
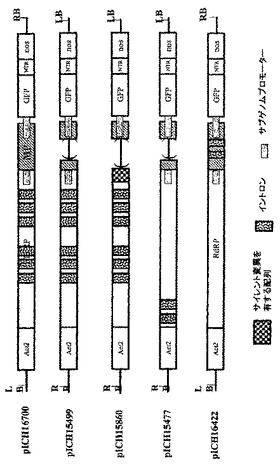
【図2C】
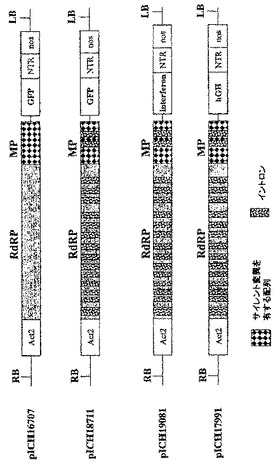
【図3】
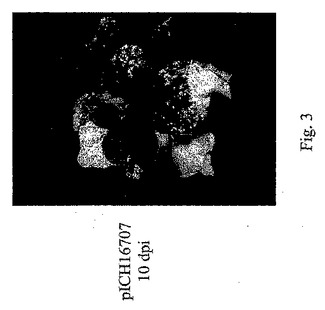
【図4】
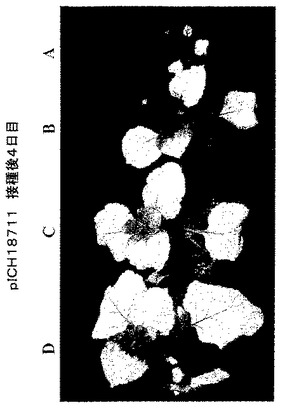
【図5】
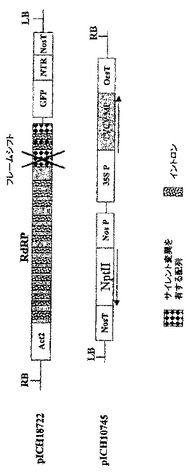
【図6A】
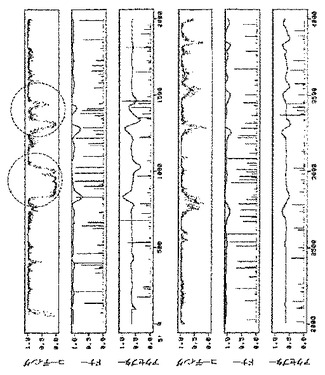
【図6B】
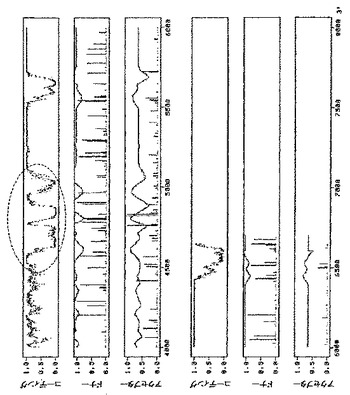
【図7】
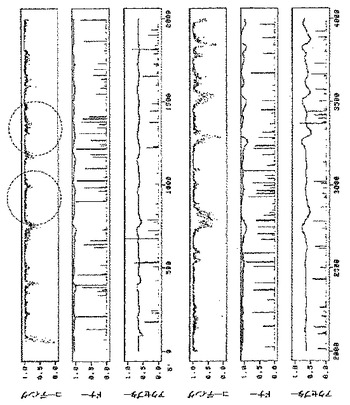
【図8】
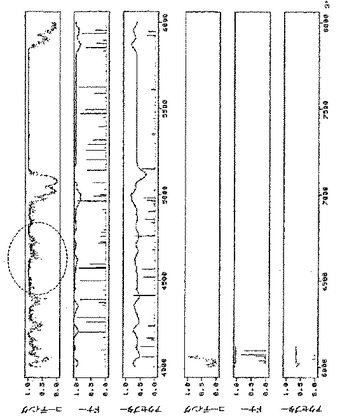
【図9A】
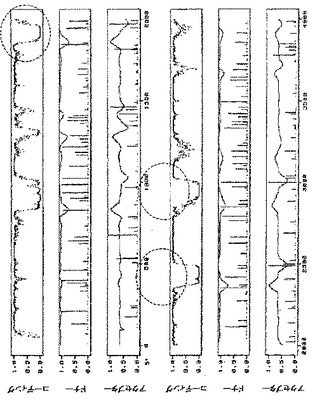
【図9B】
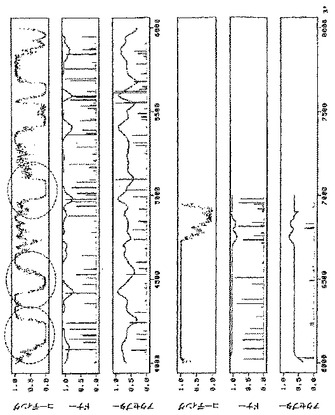
【図10A】
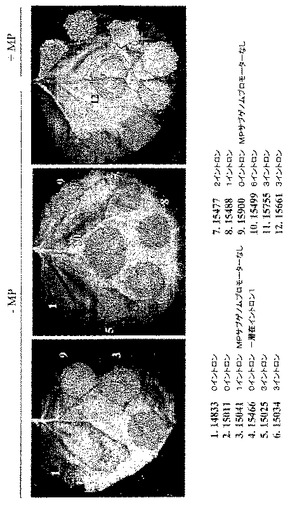
【図10B】

【図10C】
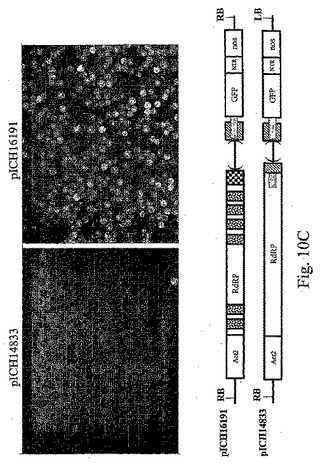
【図11】
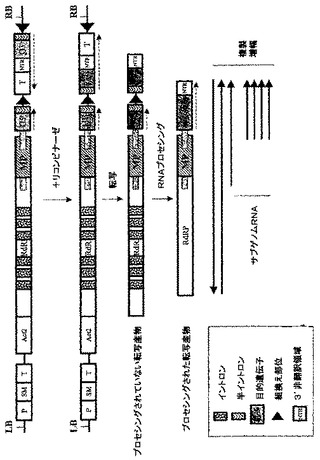
【図12】

【図13A】
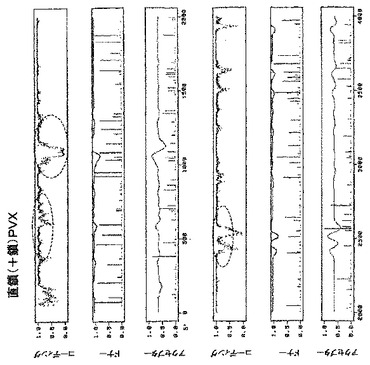
【図13B】

【図14A】
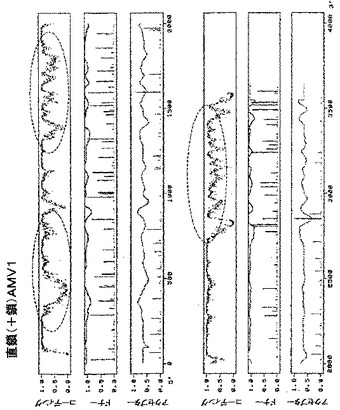
【図14B】
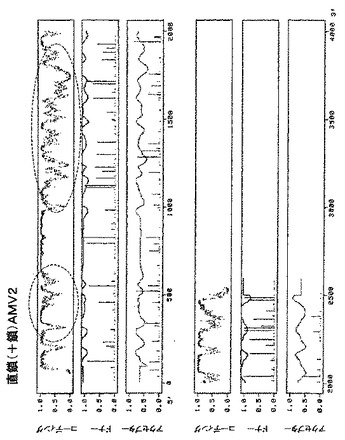
【図14C】
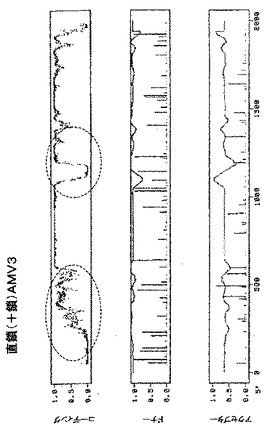
【図15】
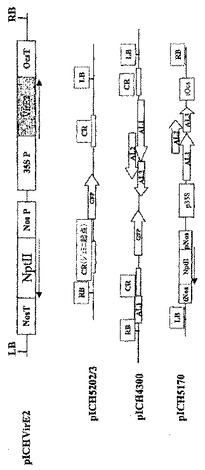
【図16】
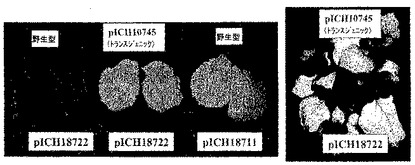
【図17】
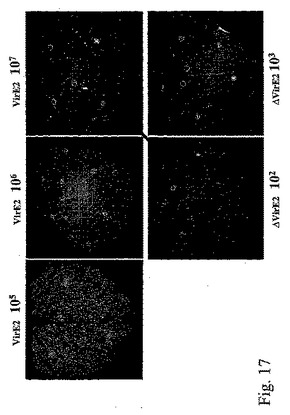
【図18】
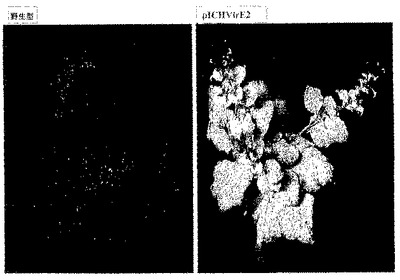
【図19】
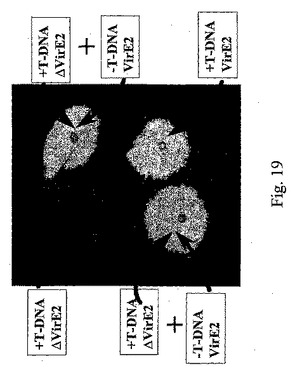
【図20】

【図21】
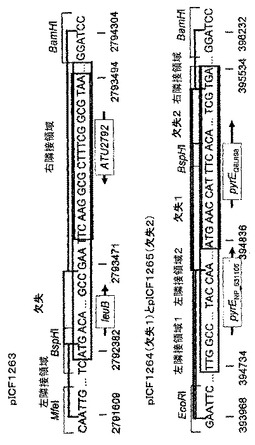
【図22】
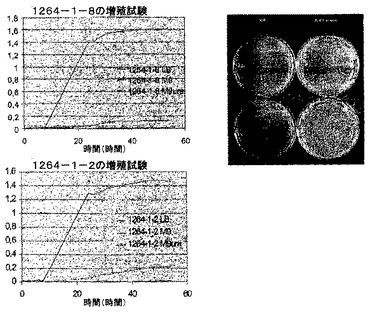
【図23】
【図1B】
【図2A】
【図2B】
【図2C】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図7】
【図8】
【図9A】
【図9B】
【図10A】
【図10B】
【図10C】
【図11】
【図12】
【図13A】
【図13B】
【図14A】
【図14B】
【図14C】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【公表番号】特表2008−505622(P2008−505622A)
【公表日】平成20年2月28日(2008.2.28)
【国際特許分類】
【出願番号】特願2007−519727(P2007−519727)
【出願日】平成17年7月7日(2005.7.7)
【国際出願番号】PCT/EP2005/007361
【国際公開番号】WO2006/003018
【国際公開日】平成18年1月12日(2006.1.12)
【出願人】(504402991)アイコン ジェネティクス アクチェンゲゼルシャフト (2)
【Fターム(参考)】
【公表日】平成20年2月28日(2008.2.28)
【国際特許分類】
【出願日】平成17年7月7日(2005.7.7)
【国際出願番号】PCT/EP2005/007361
【国際公開番号】WO2006/003018
【国際公開日】平成18年1月12日(2006.1.12)
【出願人】(504402991)アイコン ジェネティクス アクチェンゲゼルシャフト (2)
【Fターム(参考)】
[ Back to top ]