
植物における脂肪酸代謝の改変
【課題】新規のポリマーを作製するのために植物における脂肪酸の生合成および酸化を改変するための方法および系を提供する。
【解決手段】2つの酵素が必須である:D特異的エノイル−CoAヒドラターゼのようなヒドラターゼ(例えば、Aeromonas caviaeから得られるヒドラターゼ)およびβ−酸化酵素系。いくつかの植物は、その植物がこのヒドラターゼを発現するように操作される場合、ポリマー合成を改変するのに十分なβ−酸化酵素系を有する。実施例は、トランスジェニック植物におけるこれらの酵素の発現によるポリマーの生成を実証する。実施例はまた、脂肪酸生合成における改変を使用して植物の表現型を変化させ得、これにより、種子生産を減少させるか無くす、そして緑色植物の生物体量を増加する、ならびにポリヒドロキシアルカノエートを生成することを実証する。
【解決手段】2つの酵素が必須である:D特異的エノイル−CoAヒドラターゼのようなヒドラターゼ(例えば、Aeromonas caviaeから得られるヒドラターゼ)およびβ−酸化酵素系。いくつかの植物は、その植物がこのヒドラターゼを発現するように操作される場合、ポリマー合成を改変するのに十分なβ−酸化酵素系を有する。実施例は、トランスジェニック植物におけるこれらの酵素の発現によるポリマーの生成を実証する。実施例はまた、脂肪酸生合成における改変を使用して植物の表現型を変化させ得、これにより、種子生産を減少させるか無くす、そして緑色植物の生物体量を増加する、ならびにポリヒドロキシアルカノエートを生成することを実証する。
【発明の詳細な説明】
【背景技術】
【0001】
本発明は、一般的に、ポリヒドロキシアルカノエート物質の産生のためのトランスジェニック植物系、トリグリセリドおよび脂肪酸の改変、ならびに植物での種子生産を変更するための方法の分野に関する。
【0002】
作物栽培学的な作物についての安定なトランスジェニック植物を産生するための方法が、ここ最近15年にわたって開発されてきた。作物は、投入量および産出量の両方の形質を改良するために遺伝的に改変された。前者の形質において、特定の農薬に対する耐性が作物中で操作され、そして特定の天然の殺虫剤(例えば、Bacillus thuringenesisの毒素)が、植物中で直接発現された。ハイブリッド植物を産生するための、雄性不稔系の開発における有意な進歩もまた存在する。産出量の形質に関して、作物は、生産物、一般的には、植物の種子、穀粒または繊維の価値を増すために改変される。重要な代謝標的としては、デンプン、脂肪酸および油の生合成経路の改変が挙げられる。
【0003】
植物作物における微生物のポリヒドロキシアルカノエート(PHA)バイオポリマーの産生においてかなりの商業的な興味が存在する。例えば、PeoplesおよびSinskeyに対する米国特許第5,245,023号および同第5,250,430号;Brightらに対する米国特許第5,502,273号;PeoplesおよびSinskeyに対する米国特許第5,534,432号;Johnに対する米国特許第5,602,321号;Somervilleらに対する米国特許第5,610,041号;PCT WO91/00917;PCT WO92/19747;PCT WO93/02187;PCT WO93/02194;PCT WO94/12014;Poirierら、Science 256:520−23(1992);van der LeijおよびWitholt、Can.J.Microbiol.41(補遺):222−38(1995);NawrathおよびPoirier、The International Symposium on Bacterial Polyhydroxyalkanoate(Egginkら編)Davos Switzerland(1996年8月18〜23日);ならびにWilliamsおよびPeoples、CHEMTECH 26:38−44(1996)を参照のこと。PHAは、天然の、熱可塑性ポリエステルであり、そして非常に多くの適用(消費者包装、使い捨てダイアパーの裏地およびごみ袋、食物、ならびに医療製品を含む)に使用するための伝統的なポリマー技術により加工され得る。
【0004】
実験的な植物系であるArabidopsis thalianaの葉緑体でのポリヒドロキシ酪酸の産生における初期研究により、PHBとして、葉の乾燥重量で14%までの蓄積を生じた(Nawrathら、1993)。しかし、Arabidopsisは、作物栽培学的価値を有さない。さらに、作物栽培学的作物において経済的にPHAを産生するために、種子におけるPHAの産生が所望され、その結果、種子の収穫および加工のための現在の基幹施設が利用され得る。植物の種子からのPHAの回収(PCT WO97/15681)および最終用途の適用(WilliamsおよびPeoples、CHEMTECH 26:38−44(1996))の選択肢は、ポリマーの組成によって有意に影響を受ける。それゆえ、きちんと規定された組成を有するPHAポリマーを産生するトランスジェニック植物系を開発することは有利である。
【0005】
それらの基質特異性に基づいたPHA生合成酵素の慎重な選択は、トランスジェニック系において規定された組成のPHAポリマーの産生を可能にする(米国特許第5,229,279号;同第5,245,023号;同第5,250,430号;同第5,480,794号;同第5,512,669号;同第5,534,432号;同第5,661,026号;および同第5,663,063号)。
【0006】
細菌において、各PHA基は、特異的な経路により産生される。短いペンダント基PHAの場合では、以下の3つの酵素が関与する:β−ケトチオラーゼ、アセトアセチル−CoAレダクターゼ、およびPHAシンターゼ。例えば、ホモポリマーPHBは、アセチル補酵素Aの2つの分子の縮合により産生され、アセトアセチル補酵素Aを生じる。次いで、後者は、レダクターゼによりキラル中間体R−3−ヒドロキシブチリル補酵素Aに還元され、続いて、PHAシンターゼ酵素により重合される。PHAシンターゼは、4−ヒドロキシ酸ユニットおよび5−ヒドロキシ酸ユニットを両方とも含むC3〜C5ヒドロキシ酸モノマーの重合化を可能にする比較的広範な基質特異性を特に有する。この生合成経路は、多くの細菌(例えば、Alcaligenes eutrophus、A.latus、Azotobacter vinlandiiおよびZoogloea ramigera)において見出される。長いペンダント基PHAは、例えば、多くの異なるPseudomonas細菌により産生される。これらの生合成は、ヒドロキシアシル補酵素Aモノマー単位への経路として脂肪酸のβ−酸化および脂肪酸合成を含む。次いで、後者は、より大きなC6〜C14モノマー単位に有利な基質特異性を有するPHAシンターゼにより変換される(PeoplesおよびSinskey、1990)。
【0007】
脂肪酸での細胞増殖から通常産生されるPHB−コ−HXコポリマーの場合、これらの経路の組み合わせは、異なるモノマー単位の形成を担い得る。実際に、Aeromonas caviae(これは、コポリマーであるPHB−コ−3−ヒドロキシヘキサノエートを産生する)におけるPHAシンターゼ遺伝子をコードするDNA遺伝子座の分析を使用して、D−β−ヒドロキシブチリル−CoAおよびD−β−ヒドロキシヘキサノイル−CoA単位の産生を担うD特異的エノイル−CoAヒドラターゼをコードする遺伝子を同定した(FukuiおよびDoi、J.Bacteriol.179:4821−30(1997);Fukuiら、J.Bacteriol.180:667−73(1998))。このようなヒドラターゼ遺伝子および酵素の他の供給源としては、Alcaligenes、PseudomonasおよびRhodospirillumが挙げられる。
【0008】
A.eutrophusにおいて短いペンダント基PHAを産生する酵素である、PHAシンターゼ、アセトアセチル−CoAレダクターゼ、およびβ−ケトチオラーゼは、phbC−phbA−phbB遺伝子を含むオペロンによってコードされる(Peoplesら、1987;PeoplesおよびSinskey、1989)。Pseudomonas生物において、長いペンダント基PHAの産生を担うPHAシンターゼは、pha遺伝子座上に、特にphaA遺伝子およびphaC遺伝子によりコードされることが見出されている(米国特許第5,245,023号および同第5,250,430号;Huismanら、J.Biol.Chem.266:2191−98(1991))。これらの初期研究により、ある範囲のPHA生合成遺伝子が、単離および特徴づけされるか、またはゲノム配列決定プロジェクトから同定された。公知のPHA生合成遺伝子の例は、以下の参考文献に開示される:Aeronomas caviae(FukuiおよびDoi、1997、J.Bacteriol.179:4821−30);Alcaligenes eutrophus(米国特許第5,245,023号;同第5,250,430号;同第5,512,669号;および同第5,661,026号;PeoplesおよびSinsky、J.Biol.Chem.264:15298−03(1989));Acinetobacter(Schembriら、FEMS Microbiol.Lett.118:145−52(1994);Chromatium vinosum(LiebergesellおよびSteinbuchel、Eur.J.Biochem.209:135−50(1992)));Methylobacterium extorquens(ValentinおよびSteinbuchel、Appl.Microbiol.Biotechnol.39:309−17(1993));Nocardia corallina(GENBANK 受託番号AF019964;Hallら、1998、Can.J.Microbiol.44:687−69);Paracoccus denitrificans(Uedaら、J.Bacteriol.178:774−79(1996);Yabutaniら、FEMS Microbiol.Lett.133:85−90(1995));Pseudomonas acidophila(Umedaら、1998、Applied Biochemistry and Biotechnology、70−72:341−52);Pseudomonas sp.61−3(Matsusakiら、1998、J.Bacteriol.180:6459−67);Nocardia corallina;Pseudomonas aeruginosa(TimmおよびSteinbuchel、Eur.J.Biochem.209:15−30(1992));P.oleovorans(米国特許第5,245,023号および同第5,250,430号;Huismanら、J.Biol.Chem.266(4):2191−98(1991);Rhizobium etli(Cevallosら、J.Bacteriol.178:1646−54(1996));R meliloti(Tomboliniら、Microbiology 141:2553−59(1995));Rhodococcus ruber(Pieper−FurstおよびSteinbuchel、FEMS Microbiol.Lett.75:73−79(1992));Rhodospirillum rubrum(Hustedeら、FEMS Microbiol.Lett.93:285−90(1992));Rhodobacter sphaeroides(Hustedeら、FEMS Microbiol.Rev.9:217−30(1992));Biotechnol.Lett.15:709−14(1993);Synechocystis sp.(DNA Res.3:109−36(1996));Thiocapsiae violacea(Appl.Microbiol.Biotechnol.38:493−501(1993))およびZoogloea ramigera(Peoplesら、J.Biol.Chem.262:97−102(1987));PeoplesおよびSinskey、Molecular Microbiology3:349−57(1989))。これらの遺伝子または公開されたDNA配列の利用可能性は、PHA産生のためのある範囲の選択肢を提供するはずである。
【0009】
1〜99%HHモノマーを含むPHB−コ−HHコポリマーを産生するための適切なPHAシンターゼは、Rhodococcus ruber、Rhodospirillum rubrum、Thiocapsiae violacea、およびAeromonas caviaeのPHAシンターゼ遺伝子によりコードされる。R−3−ヒドロキシブチレートに加えて、6〜12の炭素原子の3−ヒドロキシ酸の取り込みのために有用な(すなわち、化学的に合成されたコポリマー(PCT WO95/20614、PCT WO95/20615およびPCT WO95/20621に記載される)に等価な生物学的ポリマーを産生するための)PHAシンターゼは、多くのPseudomonasおよびほかの細菌において同定されている(SteinbuechelおよびWiese、Appl.Microbiol.Biotechnol.37:691−97(1992);Valentinら、Appl.Microbiol.Biotechnol.36:507−14(1992);Valentinら、Appl.Microbiol.Biotechnol.40:710−16(1994);Leeら、Appl.Microbiol.Biotechnol.42:901−09(1995);Katoら、Appl.Microbiol.Biotechnol.45:363−70(1996);Abeら、Int.J.Biol.Macromol.16:115−19(1994);Valentinら、Appl.Microbiol.Biotechnol.46:261−67(1996))。そして、それらは、米国特許第5,245,023号および同第5,250,430号に記載されるように容易に単離され得る。P.oleovorans由来のPHAシンターゼ(米国特許第5,245,023号および同第5,250,430号;Huismanら、J.Biol.Chem.266(4):2191−98(1991))は、長いペンダント基PHAの産生に適切である。β−ケトチオラーゼをコードする植物遺伝子もまた、同定されている(VollackおよびBach、Plant Physiol.111:1097−107(1996))。
【先行技術文献】
【特許文献】
【0010】
【特許文献1】米国特許第5,245,023号
【特許文献2】米国特許第5,250,430号
【特許文献3】米国特許第5,502,273号
【特許文献4】米国特許第5,534,432号
【特許文献5】米国特許第5,602,321号
【特許文献6】米国特許第5,610,041号
【特許文献7】国際公開第91/00917号
【特許文献8】国際公開第92/19747号
【特許文献9】国際公開第93/02187号
【特許文献10】国際公開第93/02194号
【特許文献11】国際公開第94/12014号
【特許文献12】国際公開第97/15681号
【特許文献13】米国特許第5,229,279号
【特許文献14】米国特許第5,245,023号
【特許文献15】米国特許第5,480,794号
【特許文献16】米国特許第5,512,669号
【特許文献17】米国特許第5,661,026号
【特許文献18】米国特許第5,663,063号
【特許文献19】米国特許第5,245,023号
【非特許文献】
【0011】
【非特許文献1】Poirierら、Science 256:520−23(1992)
【非特許文献2】van der LeijおよびWitholt、Can.J.Microbiol.41(補遺):222−38(1995)
【非特許文献3】NawrathおよびPoirier、The International Symposium on Bacterial Polyhydroxyalkanoate(Egginkら編)Davos Switzerland(1996年8月18〜23日)
【非特許文献4】WilliamsおよびPeoples、CHEMTECH 26:38−44(1996)
【非特許文献5】FukuiおよびDoi、J.Bacteriol.179:4821−30(1997)
【非特許文献6】Fukuiら、J.Bacteriol.180:667−73(1998)
【非特許文献7】Huismanら、J.Biol.Chem.266:2191−98(1991)
【非特許文献8】FukuiおよびDoi、1997、J.Bacteriol.179:4821−30
【非特許文献9】PeoplesおよびSinsky、J.Biol.Chem.264:15298−03(1989)
【非特許文献10】Schembriら、FEMS Microbiol.Lett.118:145−52(1994)
【非特許文献11】LiebergesellおよびSteinbuchel、Eur.J.Biochem.209:135−50(1992)
【非特許文献12】ValentinおよびSteinbuchel、Appl.Microbiol.Biotechnol.39:309−17(1993)
【非特許文献13】Hallら、1998、Can.J.Microbiol.44:687−69
【非特許文献14】Uedaら、J.Bacteriol.178:774−79(1996)
【非特許文献15】Yabutaniら、FEMS Microbiol.Lett.133:85−90(1995)
【非特許文献16】Umedaら、1998、Applied Biochemistry and Biotechnology、70−72:341−52)
【非特許文献17】Matsusakiら、1998、J.Bacteriol.180:6459−67
【非特許文献18】TimmおよびSteinbuchel、Eur.J.Biochem.209:15−30(1992)
【非特許文献19】Huismanら、J.Biol.Chem.266(4):2191−98(1991)
【非特許文献20】Cevallosら、J.Bacteriol.178:1646−54(1996)
【非特許文献21】Tomboliniら、Microbiology 141:2553−59(1995)
【非特許文献22】Pieper−FurstおよびSteinbuchel、FEMS Microbiol.Lett.75:73−79(1992)
【非特許文献23】Hustedeら、FEMS Microbiol.Lett.93:285−90(1992)
【非特許文献24】Hustedeら、FEMS Microbiol.Rev.9:217−30(1992)
【非特許文献25】Hustedeら、Biotechnol.Lett.15:709−14(1993)
【非特許文献26】Hustedeら、Synechocystis sp.(DNA Res.3:109−36(1996)
【非特許文献27】Appl.Microbiol.Biotechnol.38:493−501(1993)
【非特許文献28】Peoplesら、J.Biol.Chem.262:97−102(1987)
【非特許文献29】PeoplesおよびSinskey、Molecular Microbiology3:349−57(1989)
【非特許文献30】SteinbuechelおよびWiese、Appl.Microbiol.Biotechnol.37:691−97(1992)
【非特許文献31】Valentinら、Appl.Microbiol.Biotechnol.36:507−14(1992)
【非特許文献32】Valentinら、Appl.Microbiol.Biotechnol.40:710−16(1994)
【非特許文献33】Leeら、Appl.Microbiol.Biotechnol.42:901−09(1995)
【非特許文献34】Katoら、Appl.Microbiol.Biotechnol.45:363−70(1996)
【非特許文献35】Abeら、Int.J.Biol.Macromol.16:115−19(1994)
【非特許文献36】Valentinら、Appl.Microbiol.Biotechnol.46:261−67(1996)
【非特許文献37】Huismanら、J.Biol.Chem.266(4):2191−98(1991)
【非特許文献38】VollackおよびBach、Plant Physiol.111:1097−107(1996)
【発明の概要】
【発明が解決しようとする課題】
【0012】
合成および基質の選択によりモノマー組成を改変する能力にもかかわらず、脂肪酸代謝に関する生合成のようなポリマー生合成の他の特徴を改変することが望ましい。
【課題を解決するための手段】
【0013】
従って、本発明の目的は、植物の細胞代謝を操作するための脂肪酸酸化酵素系を導入するための方法およびDNA構築物を提供することである。
【0014】
本発明の別の目的は、植物において、好ましくは、その脂肪種子においてPHAの産生を増強するための方法を提供することである。
【0015】
(発明の要旨)
新規なポリマーを作製するための、植物において脂肪酸生合成および酸化を改変するための方法および系が、記載される。2つの酵素が、必須である:D特異的エノイル−CoAヒドラターゼのようなヒドラターゼ(例えば、Aeromonas caviaeから得られたヒドラターゼ)およびβ−酸化酵素系。いくつかの植物は、植物が、ヒドラターゼを発現するように操作される場合、ポリマー合成を改変するのに十分なβ−酸化酵素系を有する。
【0016】
実施例は、トランスジェニック植物において、これらの酵素の発現によりポリマーの産生を実証する。実施例はまた、脂肪酸生合成における改変を使用して植物の表現型を変化させ得、これにより種子産生を減少または排除する、および緑色植物生物体量を増加させる、ならびにPHAを産生することを実証する。
【図面の簡単な説明】
【0017】
【図1】図1は、ポリヒドロキシアルカノエートモノマーを産生するための、脂肪酸β−酸化経路の模式図である。
【図2】図2は、プラスミド構築物pSBS2024およびpSBS2025を示す模式図である。
【図3A】図3Aは、プラスミド構築物pCGmf124およびpCGmf125を示す模式図である。
【図3B】図3Bは、プラスミド構築物pCGmf124およびpCGmf125を示す模式図である。
【図4A】図4Aは、プラスミド構築物pmf1249およびpmf1254を示す模式図である。
【図4B】図4Bは、プラスミド構築物pmf1249およびpmf1254を示す模式図である。
【図5A】図5Aは、プラスミド構築物pCGmf224およびpCGmf225を示す模式図である。
【図5B】図5Bは、プラスミド構築物pCGmf224およびpCGmf225を示す模式図である。
【図6A】図6Aは、プラスミド構築物pCGmf1P2SおよびpCGmf2P1Sを示す模式図である。
【図6B】図6Bは、プラスミド構築物pCGmf1P2SおよびpCGmf2P1Sを示す模式図である。
【発明を実施するための形態】
【0018】
(発明の詳細な説明)
発生中の脂肪種子または緑色組織の細胞質またはプラスチドに脂肪酸酸化酵素系を導入することによって植物の細胞性代謝を操作するための方法およびDNA構築物が提供される。脂肪酸酸化系は、代表的には、広範囲のβ−ケトアシル−CoA基質を利用するβ−ケトチオラーゼ酵素活性を含むいくつかの酵素活性を含む。
【0019】
驚くべきことに、豆のファゼオリン(phaseolin)プロモーターからの少なくとも1つのこれらの導入遺伝子の発現が、雄性不稔をもたらすことが見出された。興味深いことに、これらの植物は、種子をつけず、その代わりに正常よりも高いレベルの生物体量(例えば、葉、茎、柄)を生成した。それゆえ、本明細書中に記載される方法および構築物をまた用いて、例えば、ハイブリッド生成のために雄性不稔植物を作製し得るか、またはまぐさ(例えば、アルファルファまたはタバコ)の生物体量の生成を増加させ得る。これらの方法およびDNA構築物を用いて作製された植物は、ポリヒドロキシアルカノエートバイオポリマーを生成するため、または新規な油組成物を生成するために有用である。
【0020】
本明細書中に記載される方法は、さらなる導入遺伝子、特に、脂肪酸酸化またはポリイヒドロキシアルカノエート生合成に関与するさらなる酵素をコードする導入遺伝子のその後の取り込みを含む。ポリヒドロキシアルカノエート生合成については、この方法は、本明細書中に記載の方法およびDNA構築物を用いて、または伝統的な植物育種法によって生成されたトランスジェニック植物の続く形質転換による、酵素(例えば、NADHおよび/またはNADPHアセトアセチル−補酵素Aレダクターゼ、PHBシンターゼ、PHAシンターゼ、アセトアセチル−CoAチオラーゼ、ヒドロキシアシル−CoAエピメラーゼ、δ3−シス−δ2−トランスエノイル−CoAイソメラーゼ、アシル−CoAデヒドロゲナーゼ、アシル−CoAオキシダーゼ、およびエノイル−CoAヒドラターゼ)をコードする導入遺伝子の取り込みを含む。
【0021】
(I.植物発現系)
好ましい実施態様では、脂肪酸酸化導入遺伝子は、種子特異的プロモーターから発現され、そしてタンパク質は、発生中の脂肪種子の細胞質において発現される。別の好ましい実施態様では、脂肪酸酸化導入遺伝子は、種子特異的プロモーターから発現され、そして発現されたタンパク質は、プラスチド標的化シグナルを用いてプラスチドに指向される。別の好ましい実施態様では、脂肪酸酸化導入遺伝子は、プラスチド染色体から直接発現され、ここで、これらは相同組換えによって取り込まれる。脂肪酸酸化導入遺伝子はまた、構成性プロモーターから、植物組織全体を通して発現され得る。圃場において作物への農薬または他の活性成分の適用後に活性化され得るプロモーターを用いることにより、これらの導入遺伝子の発現を制御し得ることもまた有用である。本明細書中に記載される方法により含まれるこれらの遺伝子の発現のさらなる制御としては、植物染色体の特異的染色体部位への導入遺伝子の標的化挿入についての組換え酵素技術の使用、またはこの導入遺伝子の発現の調節が挙げられる。
【0022】
本明細書中に記載される方法としては、以下を含むゲノムを有する植物種子が挙げられる:(a)第1のDNA配列および3’非翻訳領域に作動可能に連結されたプロモーター(この第1のDNA配列は、脂肪酸酸化ポリペプチドをコードする)、そして必要に応じて(b)第2のDNA配列および3’非翻訳領域に作動可能に連結されたプロモーター(この第2のDNA配列は、脂肪酸酸化ポリペプチドをコードする)。2つの導入遺伝子の発現は、細胞質、またはペルオキシソームもしくはグリオキシソーム以外のプラスチドにおいて少なくともβ−ケトチオラーゼ活性、デヒドロゲナーゼ活性、およびヒドラターゼ活性を有する機能的脂肪酸β酸化系を植物に提供する。第1のDNA配列および/または第2のDNA配列は、細菌、酵母、真菌、藻類、植物、または動物から単離され得る。少なくとも1つのDNA配列が、少なくとも2つ、そして好ましくは3つの酵素活性を有するポリペプチドをコードすることが好ましい。
【0023】
(形質転換ベクター)
本明細書中に記載される方法において有用なDNA構築物としては、植物へ導入遺伝子を導入し得る形質転換ベクターが挙げられる。いくつかの植物形質転換ベクターの選択肢が利用可能であり、「Gene Transfer to Plants」(Potrykusら編)Springer−Verlag Berlin Heidelberg New York(1995);「Transgenic Plants:A Production System for Industrial and Pharmaceutical Proteins」(Owenら編)John Wiley & Sons,Ltd.England(1996);および「Methods in Plant Molecular Biology:A Laboratory Course Manual」(Maligaら編)Cold Spring Laboratory Press,New York(1995)に記載されるベクターが含まれる。植物形質転換ベクターは、一般に、5’調節配列および3’調節配列(プロモーター、転写終結シグナルおよび/またはポリアデニル化シグナルを含む)の転写制御下の目的の1以上のコード配列、ならびに選択マーカー遺伝子もしくはスクリーニング可能なマーカー遺伝子を含む。5’調節配列についての通常の必要条件は、プロモーター、転写終結シグナルおよび/またはポリアデニル化シグナルを含む。単一の転写物からの2以上のポリペプチドの発現のために、さらなるRNAプロセシングシグナルおよびリボザイム配列が構築物中に操作され得る(米国特許第5,519,164号)。このアプローチは、複数の導入遺伝子を単一の遺伝子座に局在化するという利点を有する。これは、続いての植物育種の試みにおいて有利である。さらなるアプローチは、相同組換えによって植物プラスチド染色体を特異的に形質転換するためのベクターを使用すること(米国特許第5,545,818号)であり、この場合、プラスチドゲノムの原核生物の性質を利用し、そして多数の導入遺伝子をオペロンとして挿入することが可能である。
【0024】
(プロモーター)
多数の植物プロモーターが公知であり、そして目的の遺伝子の構成的発現、または環境によってもしくは発生によって調節される発現のいずれかをもたらす。植物プロモーターを選択して、異なる植物組織または細胞小器官における導入遺伝子の発現を制御し得る。これらのための方法は全て、当業者に公知である(GasserおよびFraley,Science 244:1293−99(1989))。導入遺伝子の5’末端は、導入遺伝子とインフレームで連結された、プラスチドまたは他の細胞内小器官標的化ペプチドをコードする配列を含むように操作され得る。適切な構成性植物プロモーターとしては、カリフラワーモザイクウイルス35Sプロモーター(CaMV)および増強されたCaMVプロモーター(Odellら,Nature,313:810(1985))、アクチンプロモーター(McElroyら,Plant Cell 2:163−71(1990))、AdhIプロモーター(Frommら,Bio/Technology 8:833−39(1990);Kyozukaら、Mol.Gen.Genet.228:40−48(1991))、ユビキチンプロモーター、ゴマノハグサモザイクウイルスプロモーター、マンノピン(mannopine)シンターゼプロモーター、ノパリンシンターゼプロモーター、およびオクトピンシンターゼプロモーターが挙げられる。有用な調節可能なプロモーター系としては、ホウレンソウ硝酸誘導性プロモーター、熱ショックプロモーター、リブロースビスリン酸カルボキシラーゼ(ribulose biphosphate carboxylase)プロモーターの小サブユニット、および化学誘導性プロモーター(Hersheyらに対する米国特許第5,364,780号)が挙げられる。
【0025】
本明細書中に記載される方法の好ましい実施態様では、導入遺伝子は、発生中の種子においてのみ発現される。この目的のために適切なプロモーターとしては、ナピン遺伝子プロモーター(米国特許第5,420,034号および同第5,608,152号)、アセチル−CoAカルボキシラーゼプロモーター(米国特許第5,420,034号および同第5,608,152号)、2Sアルブミンプロモーター、種子貯蔵タンパク質プロモーター、ファゼオリンプロモーター(Slightomら,Proc.Natl.Acad.Sci.USA 80:1897−1901(1983))、オレオシン(oleosin)プロモーター(Plantら,Plant Mol.Biol.25:193−205(1994);Rowleyら,Biochim.Biophys.Acta.1345:1−4(1997);米国特許第5,650,554号;およびPCT WO 93/20216)、ゼインプロモーター、グルテリンプロモーター、デンプンシンターゼプロモーター、およびデンプン分枝酵素プロモーターが挙げられる。
【0026】
これらのベクターを用いる適切な作物栽培植物宿主の形質転換は、種々の方法および植物組織を用いて達成され得る。本明細書中に開示される方法において有用な代表的な植物としては、napus、rappa、sp.carinataおよびjunceaを含むBrassica科;トウモロコシ;ダイズ;ワタの実;ヒマワリ;ヤシ;ココヤシ;ベニバナ;ラッカセイ;Sinapis albaを含むカラシ;およびアマが挙げられる。生物体量(例えば、サイレージトウモロコシ、アルファルファ、またはタバコ)として収穫される作物もまた、本明細書中に開示される方法について有用である。これらのベクターを用いる形質転換のための代表的な組織としては、プロトプラスト、細胞、カルス組織、リーフディスク、花粉、および分裂組織が挙げられる。代表的な形質転換手順としては、Agrobacterium媒介形質転換、バイオリスチック(biolistic)、マイクロインジェクション、エレクトロポレーション、ポリエチレングリコール媒介プロトプラスト形質転換、リポソーム媒介形質転換、およびケイ素繊維媒介形質転換(米国特許第5,464,765号;「Gene Transfer to Plants」(Potrykusら編)Springer−Verlag Berlin Heidelberg New York(1995);「Transgenic Plants:A Production System for Industrial and Pharmaceutical Proteins」(Owenら編)John Wiley & Sons Ltd.England(1996);および「Methods in Plant Molecular Biology:A Laboratory Course Manual」(Maligaら編)Cold Spring Laboratory Press,New York(1995))が挙げられる。
【0027】
(II.トランスジェニック植物を作製し、そしてスクリーニングするための方法)
本明細書中に記載される構築物を用いてトランスジェニック植物を作製するために、以下の手順を用いて、形質転換後に導入遺伝子を発現する、形質転換された植物を入手し得る:選択培地上で形質転換された植物細胞を選択する;形質転換された植物細胞を再生して、分化した植物を生成する;所望のポリペプチドが所望の組織および細胞の位置において得られるようなレベルで導入遺伝子を発現する、形質転換された植物を選択する。
【0028】
記載された方法を実施するために有用な特定の作物について、形質転換手順が、例えば、以下に記載されるように、確立されている:「Gene Transfer to Plants」(Potrykusら編)Springer−Verlag Berlin Heidelberg New York(1995);「Transgenic Plants:A Production System for Industrial and Pharmaceutical Proteins」(Owenら編)John Wiley & Sons,Ltd.England(1996);および「Methods in Plant Molecular Biology:A Laboratory Course Manual」(Maligaら編)Cold Spring Laboratory Press,New York(1995)。
【0029】
Brassica napusは、例えば、米国特許第5,188,958号および同第5,463,174号に記載される通りに形質転換され得る。他のBrassica(例えば、rappa、carinata、およびjuncea)ならびにSinapis albaは、Moloneyら,Plant Cell Reports 8:238−42(1989)に記載される通りに形質転換され得る。ダイズは、多数の報告された手順(米国特許第5,015,580号;同第5,015,944号;同第5,024,944号;同第5,322,783号;同第5,416,011号;および同第5,169,770号)によって形質転換され得る。いくつかの形質転換手順(花粉の形質転換(米国特許第5,629,183号)、ケイ素繊維媒介形質転換(米国特許第5,464,765号)、プロトプラストのエレクトロポレーション(米国特許第5,231,019号;同第5,472,869号;および同第5,384,253号)、遺伝子銃(米国特許第5,538,877号および同第5,538,880号)、ならびにAgrobacterium媒介形質転換(EP 0 604 662 A1;PCT WO 94/00977)を含む)が、トランスジェニックトウモロコシ植物の生成について報告されている。Agrobacterium媒介手順は、特に好ましい。なぜなら、導入遺伝子構築物の単一の組み込み事象が、この手順を用いて、より容易に得られ、そしてこのことは、その後の植物育種を大いに容易にするからである。ワタは、粒子ボンバードメント(米国特許第5,004,863号および同第5,159,135号)によって形質転換され得る。ヒマワリは、粒子ボンバードメントとAgrobacterium感染との組合せを用いて形質転換され得る(EP 0 486 233 A2;米国特許第5,030,572号)。アマは、粒子ボンバードメントまたはAgrobacterium媒介形質転換のいずれかによって形質転換され得る。組換え酵素技術としては、cre−lox系、FLP/FRT系、およびGin系が挙げられる。これらの技術を利用する方法は、例えば、Hodgesらに対する米国特許第5,527,695号;DaleおよびOw,Proc.Natl.Acad.Sci.USA 88:10558−62(1991);Medberryら,Nucleic Acids Res.23:485−90(1995)に記載される。
【0030】
(選択マーカー遺伝子)
本明細書中に記載される方法を実施する際に有用な選択マーカー遺伝子としては、ネオマイシンホスホトランスフェラーゼ遺伝子nptII(米国特許第5,034,322号および同第5,530,196号)、ハイグロマイシン耐性遺伝子(米国特許第5,668,298号)、ホスフィノトリシンに対する耐性をコードするbar遺伝子(米国特許第5,276,268号)が挙げられる。EP 0 530 129 A1は、増殖培地に添加された不活性な化合物を活性化する酵素をコードする導入遺伝子を発現することにより、形質転換された植物が、非形質転換系統から成長して抜け出すのを可能にするポジティブ選択系を記載する。本明細書中の方法において有用なスクリーニング可能マーカー遺伝子としては、β−グルクロニダーゼ遺伝子(Jeffersonら,EMBO J.6:3901−07(1987);米国特許第5,268,463号)およびネイティブなグリーン蛍光タンパク質遺伝子または改変されたグリーン蛍光タンパク質遺伝子(Cubittら,Trends Biochem Sci.20:448−55(1995);Pangら,Plant Physiol.112:893−900(1996))が挙げられる。これらのマーカーのうちのいくつかは、形質(例えば、除草剤耐性)を目的の植物に導入し、それによって、投入側にさらなる作物栽培的価値を提供するという付加的な利点を有する。
【0031】
本明細書中に記載される方法の好ましい実施態様では、1より多くの遺伝子産物が植物中で発現される。この発現は、以下を含む、多数の異なる方法を介して達成され得る:(1)単一の形質転換事象においてコードDNAを導入する方法であって、ここで全ての必要なDNAが単一のベクター上に存在する、方法;(2)同時形質転換事象においてコードDNAを導入する方法であって、ここで全ての必要なDNAが別個のベクター上に存在するが、同時に植物細胞に導入される、方法;(3)独立した形質転換事象によって連続的に植物細胞にコードDNAを導入する方法(すなわち、さらなるDNA構築物を用いての1以上のコードDNAを発現するトランスジェニック植物細胞の形質転換);ならびに(4)別個の形質転換事象による、必要とされるDNA構築物の各々の形質転換によって、個々のタンパク質を発現するトランスジェニック植物を得、そして伝統的な植物育種法を用いて全ての経路を単一の植物に組み込む方法。
【0032】
(III.β酸化酵素経路)
植物の細胞質ゾルにおけるPHAの産生は、R−3−ヒドロキシアシルCoAチオエステルをPHAシンターゼについての基質として産生し得る酵素の細胞質ゾル局在化を必要とする。真核生物および原核生物の両方が、脂肪アシルCoAチオエステルをアセチルCoAへと変換する一連の酵素からなる脂肪酸分解についてのβ酸化経路を有する。これらの経路は、中間体3−ヒドロキシアシルCoAを介して進行するが、この中間体の立体配置は生物間で異なる。例えば、細菌のβ酸化経路および高等真核生物のペルオキシソーム経路は、S−3−ヒドロキシアシルCoAを介して脂肪酸をアセチルCoAへと分解する(Schultz,「Oxidation of Fatty Acids」、Biochemistry of Lipids,Lipoproteins and Membranes(Vanceら編)101〜06頁(Elsevier,Amsterdam 1991))。Escherichia coliでは、β酸化多機能酵素複合体によってコードされるエピメラーゼ活性は、S−3−ヒドロキシアシルCoAをR−3−ヒドロキシアシルCoAへと変換し得る。酵母は、中間体R−3−ヒドロキシアシルCoAを介して進行するペルオキシソームに局在した脂肪酸分解経路を保有し(Hiltunenら J.Biol.Chem.267:6646−53(1992);Filppulaら J.Biol.Chem.270:27453−57(1995))、その結果、PHAを生成するためにエピメラーゼ活性は必要とされない。
【0033】
植物は、他の高等真核生物と同様に、ペルオキシソーム中に細胞下に局在した脂肪酸分解のためのβ酸化経路を有する(Gerhardt,「Catabolism of Fatty Acids[α and β Oxidation]」、Lipid Metabolism in Plants(Moore,Jr.編)527−65頁(CRC Press,Boca Raton,Florida 1993))。それゆえ、植物の細胞質ゾルにおけるPHAの生成は、正確な鎖長のR−3−ヒドロキシアシルCoAチオエステルへの脂肪酸の変換のために、β酸化経路の細胞質ゾルでの発現、ならびにR−3−ヒドロキシアシルCoAをポリマーへと重合させるために適切なPHAシンターゼの細胞質ゾル発現を必要とする。
【0034】
脂肪酸は、植物のプラスチドにおいて飽和アシル−ACPチオエステルとして合成される(Hartwood,「Plant Lipid Metabolism」、Plant Biochemistry(Deyら編)237−72頁(Academic Press,San Diego 1997))。プラスチドから細胞質ゾルへの輸送の前に、大多数の脂肪酸は、Δ9デサチュラーゼを介して脱飽和される。大部分の脂肪種子作物における新たに合成された脂肪酸のプールは、主に、オレイン酸(シス9−オクタデセン酸)、ステアリン酸(オクタデカン酸)、およびパルミチン酸(ヘキサデカン酸)からなる。しかし、いくつかの植物(例えば、ココヤシおよびパーム核)は、より短い鎖の脂肪酸(C8〜14)を合成する。この脂肪酸は、ACPからチオエステラーゼを介して放出され、続いてプラスチド膜に局在するアシルCoAシンテターゼを介してアシルCoAチオエステルへと変換される(Andrewsら,「Fatty acid and lipid biosynthesis and degradation」Plant Physiology,Biochemistry,and Molecular Biology(Dennisら編)345−46頁(Longman Scientific & Technical,Essex,England 1990);Harwood,「Plant Lipid Metabolism」Plant Biochemistry(Deyら編)246頁(Academic Press,San Diego 1997))。
【0035】
脂肪酸分解経路を介する、新たに合成されたアシルCoAチオエステルのプールの細胞質ゾル変換、およびPHAシンターゼについてのR−3−ヒドロキシアシル−CoA基質へのこれらの一連の反応由来の中間体の変換は、図1に概説した酵素反応を介して達成され得る。PHAシンターゼ基質はC4〜C16 R−3−ヒドロキシアシルCoAである。飽和脂肪アシルCoAについては、脂肪酸分解経路を用いてのR−3−ヒドロキシアシルCoAチオエステルへの変換は、以下の一連の反応を必要とする:トランス−2−エノイル−CoAへのアシルCoAチオエステルの変換(反応1)、R−3−ヒドロキシ(hyddroxy)アシルCoAへのトランス−2−エノイル−CoAの水和(反応2a、例えば、酵母系は、この経路を通して作動し、そしてAeromonas caviae D特異的ヒドラターゼはC4〜C7 R−3ヒドロキシアシル−CoAを生じる)、S−3−ヒドロキシアシルCoAへのトランス−2−エノイル−CoAの水和(反応2b)、およびR−3−ヒドロキシアシルCoAへのS−3−ヒドロキシアシルCoAのエピマー化(反応5、例えば、キュウリ四機能タンパク質、細菌系)。3−ヒドロキシアシルCoAは、PHAを形成するPHAシンターゼによって重合されない場合、これは、残りのβ酸化経路を通して以下の通りに進行し得る:β−ケトアシルCoAを形成する、3−ヒドロキシアシルCoAの酸化(反応3)、続いてアセチルCoAおよび2炭素単位分短い飽和アシルCoAチオエステルを生じるCoA存在下でのチオ開裂(反応4)。反応4において生成されるアシルCoAチオエステルは、反応1のβ酸化経路に自由に再度侵入し、そして生成されたアセチルCoAは、β−ケトチオラーゼ(反応7)およびNADHアセトアセチル−CoAレダクターゼまたはNADPHアセトアセチル−CoAレダクターゼ(反応6)の作用によってR−3−ヒドロキシアシルCoAへと変換され得る。この後者の経路は、R−3−ヒドロキシブチリル−CoA、R−3−ヒドロキシバレリル−CoA、およびR−3−ヒドロキシヘキサノイル−CoAを生成するために有用である。この一連の酵素反応によって生成される4〜16炭素原子のR−3−ヒドロキシ酸は、導入遺伝子(または2つのサブユニットシンターゼ酵素の場合は複数の導入遺伝子)から発現されるPHAシンターゼによってPHAポリマーへと重合され得る。
【0036】
Δ9不飽和脂肪アシルCoAについては、記載される種々の一連の反応が必要とされる。図1に詳述されるように、3サイクルのβ酸化は、6個の炭素単位を除去して、3位にシス二重結合を有する不飽和アシルCoAチオエステルを生じる。Δ3−シス−Δ2−トランス−エノイルCoAイソメラーゼによって触媒される、2位でのトランス二重結合への3位でのシス二重結合の変換は、図1に概説した一連のβ酸化反応が進行するのを可能にする。この酵素活性は、微生物のβ酸化複合体および植物の四機能タンパク質に存在するが、酵母fox1には存在しない。
【0037】
アシルCoAチオエステルはまた、分解されてβ−ケトアシルCoAになり、そしてNADH依存性レダクターゼまたはNACPH依存性レダクターゼを介してR−3−ヒドロキシアシルCoAへと変換され得る(反応6)。
【0038】
S特異的ヒドラターゼ活性、S特異的デヒドロゲナーゼ活性、β−ケトチオラーゼ活性、エピメラーゼ活性、およびΔ3−シス−Δ2−トランス−エノイルCoAイソメラーゼ活性をコードする多機能性酵素は、Escherichia coli(Sprattら,J.Bacteriol.158:535−42(1984))およびPseudomonas fragi(Immureら,J.Biochem.107:184−89(1990))のような細菌において見出されている。多機能性酵素複合体は、触媒的に活性なタンパク質がヘテロテトラマーを形成するような、2コピーの各々2つのサブユニットからなる。ヒドラターゼ活性、デヒドロゲナーゼ活性、エピメラーゼ活性、およびΔ3−シス−Δ2−トランス−エノイルCoAイソメラーゼ活性は、一方のサブユニット上に存在し、一方、チオラーゼは別のサブユニットに存在する。生物(例えば、E.coli(Sprattら,J.Bacteriol.158:535−42(1984));DiRusso,J.Bacteriol.172:6459−68(1990))およびP.fragi(Satoら,J.Biochem.111:8−15(1992))由来の酵素をコードする遺伝子は、単離および配列決定されており、そして本明細書中に記載される方法を実施するために適切である。さらに、E.coli酵素系は、個々の酵素活性に重要なアミノ酸残基を同定するために部位特異的変異誘発分析に供されている(HeおよびYang,Biochemistry 35:9625−30(1996);Yangら,Biochemistry 34:6641−47(1995);HeおよびYang,Biochemistry 36:11044−49(1997);Heら、Biochemistry 36:261−68(1997);YangおよびElzinga,J.Biol.Chem.268:6588−92(1993))。これらの変異遺伝子はまた、本明細書中に記載される方法のいくつかの実施態様において用いられ得る。
【0039】
ラットのような哺乳動物は、それらのペルオキシソーム中に、ヒドラターゼ、デヒドロゲナーゼ、およびΔ3−シス−Δ2−トランス−エノイルCoAイソメラーゼ活性を含む、3機能性のβ−酸化酵素を有する。3機能性の酵素は、ラットの肝臓から単離され、そして78kDaの分子量を有する単量体であることが見出された(Palosaariら、J.Biol.Chem.265:2446〜49(1990))。細菌系と異なり、チオラーゼ活性は、多酵素タンパク質の一部ではない(Schultz、「Oxidation of Fatty Acids」Biochemistry of Lipids、Lipoproteins and Membranes(Vanceら、編)95頁、Elsevier,Amsterdam(1991))。ラットにおけるエピマー化は、2つの異なるヒドラターゼの活性の組み合わせによって生じ、その1つは、R−3ヒドロキシアシルCoAをトランス−2−エノイルCoAに転換し、そして別の1つは、トランス−2−エノイルCoAをS−3−ヒドロキシアシルCoAに転換する(Smelandら、Biochemical and Biophysical Research Communications 160:988〜92(1989))。哺乳動物はまた、中間体S−3−ヒドロキシアシルCoAを介して脂肪酸をアセチルCoAに分解する、β−酸化経路をそれらのミトコンドリアに有する(Schultz、「Oxidation of Fatty Acids」Biochemistry of Lipids,Lipoproteins and Membranes(Vanceら、編)96頁(Elsevier,Amsterdam(1991)))。ミトコンドリアβ−酸化活性をコードする遺伝子は、いくつかの動物から単離されており、これらには、ラットミトコンドリア長鎖アシルCoAヒドラターゼ/3−ヒドロキシアシルCoAデヒドロゲナーゼ(GENBANK登録番号D16478)およびラットミトコンドリアチオラーゼ(GENBANK登録番号D13921およびD00511)が含まれる。
【0040】
酵母は、多機能酵素であるFox2を有し、これは、S−3−ヒドロキシアシルCoAの代わりにR−3−ヒドロキシアシルCoA中間体を介して進行するという点で、細菌および高等真核生物のβ−酸化複合体とは異なる(Hiltunenら、J.Biol.Chem.267:6646〜53(1992))。Fox2は、R−特異的ヒドラターゼ酵素活性およびR−特異的デヒドロゲナーゼ酵素活性を有する。この酵素は、Δ9−シス−ヒドロキシアシルCoAを分解してR−3−ヒドロキシアシルCoAを形成するのに必要な、Δ3−シス−Δ2−トランス−エノイルCoAイソメラーゼ活性を有さない。fox2をコードする酵母由来のこの遺伝子は単離され、そして配列決定されており、そして900アミノ酸タンパク質をコードする。この構造遺伝子のDNA配列およびコードされたポリペプチドのアミノ酸配列は、配列番号1および配列番号2に示される。
【0041】
植物は、酵母Fox2に類似するがまた、Δ3−シス−Δ2−トランス−エノイルCoAイソメラーゼ活性もコードする、4機能タンパク質を有する(Mullerら、J.Biol.Chem.269:20475〜81(1994))。このcDNAのDNA配列およびコードされるポリペプチドのアミノ酸配列は、配列番号3および配列番号4に示される。
【0042】
(IV.脂肪種子作物の細胞質への酵素の標的化)
植物の細胞質中のPHA産生を操作することは、植物の細胞質へのβ−酸化の発現を指示する工程を必要とする。faoABのような細菌系においては標的化シグナルは存在しない。真菌、酵母、植物、および哺乳動物においては、β−酸化は、細胞下オルガネラにおいて起こる。代表的には、その遺伝子は、核の染色体において発現され、そして細胞質において合成されるポリペプチドは、特定のアミノ酸配列の存在によってこれらのオルガネラに指向される。真核生物供給源から単離された遺伝子(例えば、酵母、真菌、植物、および哺乳動物のような真核生物供給源由来の脂肪酸酸化酵素)を使用して、本明細書に記載される方法を実行するために、細胞下標的化シグナルの除去または改変が、この酵素を細胞質ゾルに指向させるために必要である。タンパク質を小胞体に指向させるためのシグナルを付加することが有用であり得る。このプロセスにおいて有用なペプチドは、当該分野で周知である。一般的なアプローチは、小胞体に標的化するペプチド配列に特定化したDNA配列を挿入してキメラ遺伝子を形成することによって、導入遺伝子を改変することである。
【0043】
真核生物アシルCoAデヒドロゲナーゼおよび他のミトコンドリアタンパク質は、通常20〜60アミノ酸長であるタンパク質のN−末端のリーダーペプチドを介してミトコンドリアに標的化される(Horwich,Current Opinion in Cell Biology,2:625〜33(1990))。ミトコンドリア移入リーダーペプチドについての明白なコンセンサス配列の欠如にもかかわらず、リーダー配列において鍵となる残基の変異誘発は、ミトコンドリアタンパク質の移入を妨害することが実証された。例えば、Saccharomyces cerevisiae F1−ATPaseの移入は、そのリーダー配列の変異誘発によって妨害され、これは、細胞質における改変された前駆体タンパク質の蓄積を生じた(Bedwellら、Mol.Cell.Biol.9:1014〜25(1989))。
【0044】
3つの真核生物ペルオキシソーム標的化シグナルが報告されている(Gouldら、J.Cell.Biol.108:1657〜64(1989);Bricknerら、J.Plant Physiol.,113:1213〜21(1997))。トリペプチド標的化シグナルS/A/C−K/H/R−Lは、多くのペルオキシソームタンパク質のC−末端に生じる(Gouldら、J.Cell Biol.108:1657〜64(1989))。この配列の変異誘発は、ペルオキシソームへのタンパク質の移入を妨害することが示されてきた。いくつかのペルオキシソームタンパク質は、そのタンパク質のC−末端にこのトリペプチドを含まない。これらのタンパク質では、タンパク質配列中の内部の位置のトリペプチドを介してか(Gouldら、J.Cell.Biol.108:1657〜64(1989))、または未知の、無関係の配列を介して(Bricknerら、J.Plant Physiol.113:1213〜21(1997))、標的化が生じることが示唆されている。Candida tropicalis由来のアシルCoAオキシダーゼのフラグメントを用いる、インビトロでのペルオキシダーゼ標的化実験の結果は、後者の理論を支持するようであり、そしてポリペプチドの内部アミノ酸配列中に2つの別個の標的化シグナルが存在することを示唆する(Smallら、The EMBO Journal 7:1167〜73(1988))。前述の研究において、標的化シグナルは、長さにおいて118アミノ酸の2つの領域に局在し、そしていずれの領域も標的化シグナルS/A/C−K/H/R−Lを含むことが見出されなかった。少数のペルオキシソームのタンパク質は、ペルオキシソームに移入するためのアミノ末端リーダー配列を含むようである(Bricknerら、J.Plant Physiol.113:1213〜21(1997))。これらの標的化シグナルは、部位特異的変異誘発によって欠失させ得るか、または変更され得る。
【0045】
(V.トランスジェニック植物の培養および収穫)
トランスジェニック植物は、標準的な培養技術を使用して生育され得る。植物または植物部分はまた、標準的な装置および方法を使用して回収され得る。PHAは、植物または植物部分から、公知の技術(例えば、PCT WO97/15681に記載されるような、伝統的な播種プロセス技術と組み合わせた溶媒抽出)を使用して回収され得るか、または、例えば、動物の食物として、直接的に使用され得る。ここでは、植物生物体量からのPHAを抽出することは不要である。
【0046】
種子を産生しなかったいくつかの系統は、はるかにより高いレベルの生物体量を産生した。従って、この表現型は、牧草、飼料、または他の生物体量作物について、1エーカーあたりに産生される緑の生物体量の量を増加させる手段として有用であり得る。末端の使用は、動物の餌のための、または電気的な動力を生成するためのエネルギー作物としての、飼料作物の最もコスト的に効果的な産生、を含む。他の使用は、工業製品の引き続く回収(例えば、抽出によるPHA)のために、作物(例えば、アルファルファまたはタバコ)の生物体量レベルを増加させる工程を含む。
【0047】
本明細書中に記載される組成物およびその調製方法およびその使用方法は、以下の非限定的な実施例によってさらに記載される。
【実施例】
【0048】
(実施例1:Pseudomononas putida faoAB遺伝子およびFao酵素の単離および特徴付け)
PCR、DNA配列決定、E.coli形質転換、およびプラスミド精製を含む。すべてのDNA操作を、例えば、Sambrookら、Molecular Cloning:A Laboratory Manual(Cold Spring Harbor Laboratory Press、New York(1989))に記載されるような標準的な方法を用いて行った。Pseudomonas putida由来のfaoABをコードする遺伝子を、Pseudomonas fragi由来のfaoB(Satoら、J.Biochem.111:8−15(1992))に相同性を有するプライマー1および2を使用するPCR(ポリメラーゼ連鎖反応)によってP.putidaゲノムDNAから生成されたプローブを用いて単離した。
プライマー1:5’−gat ggg ccg ctc caa ggg tgg−3’(配列番号5)
プライマー2:5’−caa ccc gaa ggt gcc gcc att−3’(配列番号6)。
【0049】
1.1kb DNAフラグメントを、PCR反応から精製し、そしてλZAP発現系(Stratagene)を使用して、pBKCMVプラスミドに構築されたP.putidaゲノムライブラリーをスクリーニングするためにプローブとして使用した。プラスミドpMFX1を、ポジティブクローンから選択し、そしてfaoAB遺伝子を含むインサートのDNA配列および隣接配列を決定した。この配列を配列番号7に示す。以下のように、faoABを含むフラグメントを、ネイティブなP.putidaリボソーム結合部位を用いてインタクトに、発現ベクターpTRCNにサブクローニングして、プラスミドpMFX3を形成した。プラスミドpMFX1を、BsrG1を用いて消化した。得られる突出末端をクレノウで平滑化した。HindIIIを用いる消化は、FaoABをコードする、3.39kbの平滑末端/HindIIIフラグメントを産生した。発現ベクターpTRCNを、SmaI/HindIIIを用いて消化し、そしてfaoABフラグメントを用いて連結し、7.57kbのプラスミドpMFX3を形成した。
【0050】
FaoAB多酵素複合体の酵素を、以下のようにアッセイした。ヒドラターゼ活性を、アッセイがCoAの存在下で行われた以外は、以前に記載されたように(Filppulaら、J.Biol.Chem.270:27453〜57(1995))、カップリング酵素L−β−ヒドロキシアシルCoAデヒドロゲナーゼを使用して、NADからNADHへの転換をモニタリングすることによってアッセイした。カップリング酵素の激しい生成物阻害が、CoAの非存在下で観察された。このアッセイは、(最終容量1mLに)60μM クロトニル(crotonyl)CoA、50μM Tris−Cl、pH9、50μg/mlのウシ血清アルブミン、50mM KCl、1mM NAD、7μg/ml ブタ心臓由来L−特異的β−ヒドロキシアシルCoA デヒドロゲナーゼ、および0.25mM CoAを含んだ。このアッセイは、アッセイ混合液中へのFaoABの添加で開始された。コントロールアッセイを、生成されるヒドラターゼ産物、S−ヒドロキシブチリルCoAの非存在下でNADの消費速度を決定するために、基質なしで行った。活性の1ユニットは、1分間あたりのNADの1μモルの消費として定義される(ε340=6220M−1cm−1)。
【0051】
ヒドロキシアシルCoAデヒドロゲナーゼを、基質としてアセトアセチルCoAを用いて、340nmにおけるNADHからNADへの転換をモニタリングすることにより(Binstockら、Methods in Enzymology、71:403(1981))、逆方向でアッセイした。このアッセイは、(最終容量1mL中に)0.1M KH2PO4、PH7、0.2mg/mL ウシ血清アルブミン、0.1mM NADH、および33μM アセトアセチルCoAを含んだ。このアッセイを、アッセイ混合液へのFaoABの添加で開始した。必要な場合、酵素サンプルを、1mg/mLウシ血清アルブミンを含む、0.1M KH2PO4、pH7中に希釈した。コントロールアッセイを、ヒドロキシアシルCoAデヒドロゲナーゼ以外の酵素による粗サンプル中のNADHの消費速度を検出するために、基質アセトアセチルCoAなしで実行した。活性の1ユニットは、1分間あたりのNADHの1μモルの消費として定義される(ε340=6220M−1cm−1)。
【0052】
ヒドロキシアシルCoAデヒドロゲナーゼを、基質としてクロトニルCoAを用いて、340nmにおけるNADからNADHへの転換をモニタリングすることにより(Binstockら、Methods in Enzymology、71:403(1981))、正方向でアッセイした。このアッセイ混合液は、(最終容量1mL中に)0.1M KH2PO4、PH8、0.3mg/mL ウシ血清アルブミン、2mM β−メルカプトエタノール、0.25mM CoA、30μM クロトニルCoA、およびFaoABのアリコートを含んだ。反応を、S−ヒドロキシブチリルCoAのインサイチュでの形成を可能にするために数分間プレインキュベーションした。次いで、このアッセイを、NAD(0.45mM)の添加によって開始した。コントロールアッセイを、ヒドロキシアシルCoAデヒドロゲナーゼ以外の酵素によるNADの消費速度を検出するために、基質なしで実行した。活性の1ユニットは、1分間あたりのNADの1μモルの消費として定義される(ε340=6220M−1cm−1)。
【0053】
チオラーゼ活性を、いくつかの改変を伴って、以前に記載されたように(Palmerら、J.Biol.Chem.266:1〜7(1991))、基質アセトアセチルCoAの消費による304nmにおける吸収の減少をモニタリングすることによって決定した。アッセイは、(最終容量1mL中に)62.4mM Tris−Cl、pH 8.1、4.8mM MgCl2、62.5μM CoA、および62.5μMアセトアセチルCoAを含んだ。このアッセイは、アッセイ混合液へのFaoABの添加によって開始した。酵素を有さないコントロールサンプルを、酵素の非存在下でpH8.1の基質分解速度を検出するために、各々のアッセイについて実行した。活性の1ユニットは、1分間あたりの基質アセトアセチルCoAの1μモルの消費として定義される(ε340=16900M−1cm−1)。
【0054】
エピメラーゼ活性を、R−3−ヒドロキシアシルCoAチオエステルを、D,L−3−ヒドロキシアシルCoA混合物の代わりに使用する以外は、以前に記載されたように(Binstockら、Methods in Enzymology、71:403(1981))アッセイした。このアッセイは、(最終容量1mL中に)30μM R−3−ヒドロキシアシルCoA、150mM KH2PO4(pH8)、0.3mg/mL BSA、0.5mM NAD、0.1mM CoA、および7μg/mL ブタ心臓由来L−特異的β−ヒドロキシアシルCoAデヒドロゲナーゼを含んだ。このアッセイは、FaoABの添加によって開始した。
【0055】
DH5α/pMFX3におけるFaoABの発現のために、培養物を2×TY培地中で30℃で増殖させた。2×TY培地は、(1リットルあたり)16gトリプトン、10g酵母、および5gNaClを含む。スターター培養を、一晩増殖させ、そして新鮮な培地(小規模増殖では250mLエルレンマイヤーフラスコ中に100mL;大規模増殖には、2.8Lフラスコ中に1.5L)に接種(1%接種材料)するために使用した。600nmにおける吸光度が0.4〜0.6の範囲に達した場合、細胞を0.4mM IPTGを用いて誘導した。収穫に先立ち、細胞をさらに4時間培養した。細胞を超音波処理によって溶菌し、そして遠心分離によって不溶性物質を可溶性タンパク質から取り除いた。アシルCoAデヒドロゲナーゼ活性を、FaoAサブユニット(配列番号31)の活性を保証するために逆方向でモニタリングし、そしてチオラーゼ活性をFaoサブユニットの活性を決定するためにアッセイした。DH5α/pMFX3におけるFaoABは、それぞれ、4.3U/mgおよび0.99U/mgの値のデヒドロゲナーゼ活性およびチオラーゼ活性を含んだ。これは、コントロール株DH5α/pTRCNにおいてデヒドロゲナーゼおよびチオラーゼについて観察された、それぞれ0.0074U/mgおよび0.0033U/mgよりも顕著に高かった。
【0056】
FaoABを、Pseudomonas fragi由来のFaoABの精製について以前に記載された改変された手順(Imamuraら、J.Biochem.107:184〜89(1990))を用いて、DH5α/pMFX3から精製した。チオラーゼ活性(正方向においてアッセイされた)およびデヒドロゲナーゼ活性(逆方向においてアッセイされた)を、精製の全体を通じてモニタリングした。3リットルのDH5α/pMFX3細胞(2.8Lのエーレンマイヤーフラスコ中2×1.5Lアリコート)を、酵素活性分析のために調製された細胞について以前に記載された細胞増殖手順を用いて、2×TY培地中で増殖させた。細胞(15.8g)を、32mLの10mM KH2PO4、pH7中に再懸濁し、そして超音波処理によって溶菌した。可溶性タンパク質を、遠心分離(18,000RPM、30分間、4℃)によって不溶性の細胞の破片から除去した。可溶性の抽出物を、50%アセトン中で作製し、そして沈澱したタンパク質を、遠心分離によって単離し、そして10mM KH2PO4、pH7中に再溶解した。このサンプルを、(NH4)2SO4を用いて33%飽和に調整し、そして可溶性および不溶性のタンパク質を、遠心分離によって分離した。得られた上清を、(NH4)2SO4で56%飽和に調整し、そして不溶性ペレットを遠心分離によって単離し、そして10mM KH2PO4、pH7中に溶解した。このサンプルを、50℃で30分間加熱し、そして可溶性タンパク質を遠心分離によって単離し、そして6000〜8000分子量カットオフメンブレン中で、10mM KH2PO4、pH7において透析した(2×3L;20時間)。このサンプルを、あらかじめ10mM KH2PO4、pH7で平衡化したToyo Jozo DEAE FPLCカラム(3cm×14cm)上にロードした。このタンパク質を、直線勾配(100mLずつ;10 KH2PO4、pH7中、0〜500mM NaCl)を用いて、流速3mL/分で溶出した。FaoABは、300と325mM NaClとの間に溶出した。このサンプルを、事前に10mM KH2PO4、pH7で平衡化したマクロ−プレップ ハイドロキシアパタイト18/30(Biorad)FPLCカラム(2cm×15cm)にロードする前に、50,000分子量カットオフメンブレン中で、10mM KH2PO4、pH7において透析した(1×2L;15時間)。タンパク質を、3mL/分の流速で、直線勾配(250mLずつ;10〜500mM KH2PO4、pH7)を用いて溶出した。FaoABは、70と130mMのKH2PO4との間に溶出した。活性を含む画分を、MILLIPORETM100,000分子量カットオフ濃縮器を用いて9mLまで濃縮した。緩衝液を、20%ショ糖を含む10mM KH2PO4、pH7を用いて3回交換し、そして−70℃で凍結した。ハイドロキシアパタイト精製画分の酵素活性を、基質の範囲を用いてアッセイした。結果を、以下の表1に示す。
【0057】
【表1】

【0058】
(実施例2:FaoABおよびFaoABポリペプチドに対する抗体の産生)
実施例1に記載されるようなFaoABタンパク質の精製後、サンプルをSDS−PAGEによって分離した。FaoA(配列番号31)およびFaoB(配列番号26)に対応するタンパク質バンドを切り出し、そしてフロイント完全アジュバントを用いてニュージーランド白ウサギを免疫した。追加免疫を、フロイント不完全アジュバントを用いて3週間間隔で行った。抗体を、血清からプロテインAカラム(Pharmacia)のアフィニティークロマトグラフィーによって回収し、ウェスタンブロッティング手順によって抗原に対して試験した。Brassica種子のコントロール抽出物を使用して、植物タンパク質に対する交差反応について試験した。交差反応は検出されなかった。
【0059】
(実施例3:トランスジェニック脂肪種子におけるPseudomonas putido faoAB遺伝子の発現のためのプラスミドの構築)
(pSBS2024の構築)
マメのファゼオリン(phaseolin)プロモーター(配列番号10;Slightomら、1983)の5’−および3’−末端(下線を付した)それぞれに隣接する配列に相同な、 オリゴヌクレオチドプライマーGVR471
【0060】
【化1】
およびGVR472
【0061】
【化2】
を、それぞれ、GVR471およびGVR472の5’−末端にKpnI(斜字体、配列番号8のヌクレオチド1〜7)およびSwaI(斜字体、配列番号9のヌクレオチド1〜9)を付加するように設計した。これらの制限部位を、クローニングを容易にするために組み込んだ。これらのプライマーを使用して、1.4kbファゼオリンプロモーターを増幅した。このプロモーターを、平滑末端連結によってpUC19のSmaI部位にクローニングした。命名されたプラスミド、pCPPI(図2を参照のこと)を、SalIおよびSwaIを用いて切断し、そしてSalI/SwaIファゼオリンターミネーター(配列番号27)に連結した。ポリアデニル化シグナルを含むマメのファゼオリンターミネーター配列を、以下のPCRプライマーを使用して増幅した:GVR396:5’−GATTTAAATGCAAGCTTAAATAAGTATGAACTAAAATGC−3’(配列番号22)
およびGVR3975’−CGGTACCTTAGTTGGTAGGGTGCTA−3’(配列番号23)
そして1.2kbフラグメント(配列番号27)をpCCP1のSalI−SalI部位にクローニングして、pSBS2024を得た(図2)。クローニングのための独特のHindIII部位を含む、得られるプラスミドを、pSBS2024と命名した(図2)。
【0062】
(pSBS2024の構築)
ダイズオレオシンプロモーターフラグメント(配列番号11;Rowleyら、1997)を、DNA配列に隣接するプライマーを用いて簡易化した。
【0063】
プライマーJA408
【0064】
【化3】
は、5’末端(下線を付した)に相補的な配列を含む。
プライマーnp1
【0065】
【化4】
は、プロモーターフラグメントの3’末端(下線を付した)に相同な配列を含む。制限部位XbaI(斜字体)およびSwaI(斜字体)を、クローニングを容易にするために、JA408およびnpIの5’末端にそれぞれ組み込んだ。プライマーを、975bpプロモーターフラグメントを増幅するために使用し、次いでpUC19のSmaI部位にクローニングした(図2を参照のこと)。得られるプラスミド、pCSPIを、SalIおよびSwaIを用いて切断し、そしてダイズターミネーター(配列番号28)に連結した。ダイズオレオシンターミネーターを、以下のプライマーを使用して、PCRによって増幅した:JA410:5’−AAGCTTACGTGATGAGTATTAATGTGTTGTTATG−3’(配列番号29)
およびJA411:5’−TCTAGACAATTCATCAAATACAAATCACATTGCC−3’(配列番号30)
そして、225bpフラグメントをpCSPIのSalI−SwaI部位にクローニングして、プラスミドpSBS2025(図6)を得た。命名されたプラスミド、pSBS2025は、クローニングのための独特のHindIII部位を有する(図2)。
【0066】
(プロモーターコード配列融合体の構築)
以下の2つのオリゴヌクレオチドプライマーを合成した:プラスミドpmfx3のmfl(faoA、配列番号24)の5’隣接配列のヌクレオチド553〜557に相同なnp2
【0067】
【化5】
および3’末端に隣接するヌクレオチド2700〜2683に相補的なnp3
【0068】
【化6】
クローニングを容易にするために、HindIII(イタリック体)部位を、プライマーnp2およびnp3の5’末端に導入した。さらに、植物の翻訳開始コドンを包囲するより有利な配列を得るために、3bpのAAA配列(太字)を組み込んだ。プライマーnp2およびnp3を使用して、フラグメントを増幅し、そしてpUC19のSmaI部位にクローニングした。生じたプラスミドをpCmfI(図3Aおよび3B)と称した。プラスミドpBmf2を、同様のプロセスで構築した(図5Aおよび5B)。クローニングのために、mf2(faoB)遺伝子(配列番号25)の5’および3’末端で、HindIII(イタリック体)を生じるために、合成プライマーの第二のセットを設計した。
【0069】
プラスミドpmfx3のmf2(faoB、配列番号25)の5’(ヌクレオチド2732〜2752bp)に相補的なプライマーnp4
【0070】
【化7】
および3’(ヌクレオチド3907〜3886bp)配列に相同なプライマーnp5
【0071】
【化8】
をPCR反応に使用して、1.17kbのDNAフラグメントを増幅した。生じたPCR産物をpBluescriptのEcoRVにクローニングした。このプラスミドを、pBmf2と称した。
【0072】
両方のプラスミドを個々にHindIIIで切断し、そしてそれらの挿入体を、同じ制限酵素で予め直線化したプラスミドpSBS2024およびpSBS2025にクローニングした。その結果、以下のプラスミドを生成した:pmf124およびpmf125(図3Aおよび3B)ならびにpmf224およびpmf225(図5Aおよび5B)。これらは、ファゼオリンまたはダイズプロモーターのいずれかで融合されたFao遺伝子(mf1およびmf2)を含む。DNA配列分析により、pmf124、pmf125、pmf224、およびpmf225について正しいプロモーター−コード配列−終結配列融合体が確認された。
【0073】
(実施例4:プロモーター−コード配列融合体の植物形質転換ベクターへのアセンブリ)
プラスミドpmf124、pmf125、pmf224、およびpmf225を得た後に、プロモーター−コード配列融合体を、二成分ベクターpCGN1559(McBrideおよびSummerfelt、1990)(これは、NPTII遺伝子(抗生物質カナマイシンに対する耐性を与える)の発現を駆動するCaMV35Sプロモーターを含む)およびpSBS2004(これは、除草剤フォスフィノスリシンに対する耐性を与えるPPT遺伝子を駆動するパセリユビキチンプロモーターを含む)に別々にクローニングした。種々の選択マーカーを有するこの目的に適した二成分ベクターは、いくつかの供給源から入手され得る。
【0074】
ファゼオリン−mf21融合カセットを、XbaIで親プラスミドから放出し、そして同じ制限酵素で直線化したpCGN1559と連結した。得られたプラスミドをpCGmf124と命名した(図3Aおよび3B)。ダイズ−mf1融合体を含むプラスミドpCGmf125を、pmf125およびpCGN1559の両方を連結前にBamHIで切断したことを除き、同様の方法で構築した(図3Aおよび3B)。
【0075】
(pmf1249およびpmf1254の構築)
プラスミドpSBS2004を、ダイズ−mf1融合体を含むBamHIフラグメントと共に直線化した。このプラスミドをpmf1254と命名した(図4Aおよび4B)。同様に、XbaIファゼオリン−mf1融合フラグメントを、同じ制限酵素で直線化したpSBS2004と連結した。得られたプラスミドをpmf1249と命名した(図4Aおよび4B)。
【0076】
(pCGmf224およびpCGmf225の構築)
ファゼオリン−mf2およびダイズ−mf2融合体を、BamHIまたはXbaIのいずれかで切断することによってベクターから融合体を切り出すことにより構築し、そしていずれかの制限酵素で直線化しておいたpCGN1559にクローニングした(図5Aおよび5B)。
【0077】
(pCGmf1P2SおよびpCGmf2P1Sの構築)
プロモーター−コード配列融合体を含む2つの発現カセットを、以下のように同じ二成分ベクター上でアセンブルした。:ファゼオリン−mf1融合体を含むプラスミドpmf124をBamHIで切断し、そしてpCGN1559のBamHI部位にクローニングして、pCGmfB124を作製した。次いで、このプラスミドをXbaIで直線化し、そしてダイズ−mf2融合体を含むpmf225のXbaIフラグメントに連結した。最終プラスミドを、pCGmf1P2Sと命名した(図6Aおよび6B)。プラスミドpCGmF2P1Sを、同様の様式でアセンブルした。ファゼオリン−mf2融合体をBamHIで切断することによりpmf224から放出し、そしてpCGN1559のBamHI部位でクローニングした。得られたプラスミドpCGmfB224をXbaIで直線化し、そしてダイズ−mf1融合体を含むpmf125のXbaIフラグメントに連結した(図6Aおよび6B)。
【0078】
(実施例5:Brassicaの形質転換)
Brassica種子を、10%の市販の漂白剤(Javex,Colgate−Palmolive)中で30分間、穏やかに振盪して、表面滅菌した。その種子を滅菌蒸留水中で3回洗浄した。種子を、Murashige−Skoog(MS)塩およびビタミン、3%(w/v)スクロースおよび0.7%(w/v)フィトアガー(phytoagar)を含む発芽培地(pH5.8)中に、プレートあたり20個の密度で配置し、そして24℃で、かつ60〜80μEm−2s−1の光強度での16時間の明期/8時間の暗期の光周期で、4〜5日間維持した。
【0079】
各々の構築物、pCGmf124、pCGmf125、pCGmf224、pCGmf1P2S、およびpCGmf2P1Sを、Agrobacterium tumefaciansEHA101株(Hoodら、J.Bacteriol.168:1291−1301(1986))内にエレクトロポレーションにより導入した。子葉の葉柄の形質転換前に、各々の構築物を有するEHA101株の単一コロニーを、100mg/lのカナマイシンおよび100mg/lのゲンタマイシンを補充した5mlの最小培地中、28℃で48時間増殖させた。1mlの細菌懸濁液を微量遠心管中で1分間の遠心分離によりペレット化した。そのペレットを1mlの最小培地中に再懸濁した。
【0080】
形質転換のために、子葉を、その基部に約2mmの葉柄を含むように、4日齢(いくつかの場合においては、5日齢)の実生から切り取った。それらの葉柄の切断表面を有する個々の子葉を、希釈した細菌懸濁液に1秒間浸けて、そしてただちに同時培養培地(3%(w/v)スクロースおよび0.7%(w/v)フィトアガーを含み、そして20μMのベンジルアデニンを富化したMS培地)中に約2mmの深さに包理した。接種した子葉を、プレートあたり10個の密度で配置し、そして同じ生長条件下で48時間インキュベートした。同時培養後、次いで子葉を、3%(w/v)スクロース、20μMベンジルアデニン、0.7%(w/v)フィトアガー(pH5.8)、300mg/lのチメンチニン(timentinin)、および20mg/lの硫酸カナマイシンを補充したMS培地を含む再生培地に移した。
【0081】
2〜3週間後、得られた再生苗条を切断し、そしてMagentaジャー中の「苗条伸長」培地(3%スクロース、300mg/lのチメンチン(timentin)、0.7%(w/v)フィトアガー、300mg/lのチメンチニン、および20mg/lの硫酸カナマイシンを含むMS培地(pH5.8))上で維持した。伸長した苗条を、「発根」培地(MS培地、3%スクロース、2mg/lのインドール酪酸、0.7%フィトアガー、および500mg/lのカルベニシリンを含む)に移した。根が出現した後、小植物をポッティングミックス(Redi Earth,W.R.Grace and Co.)に移した。植物を、噴霧チャンバー(75%の相対湿度)中で同じ生長条件下で維持した。生長2〜3週間後、葉のサンプルを、ネオマイシンホスホトランスフェラーゼ(NPTII)アッセイ(Moloneyら、Plant Cell Reports 8:238−42(1989))のために採取した。
【0082】
FaoAおよびFaoBトランスジェニック系統由来の種子を、抗FaoA抗体および抗FaoB抗体を使用するウェスタンブロッティングによって、脂肪酸酸化ポリペプチドの発現について分析し得る。FaoBポリペプチド(配列番号26)は、FaoA遺伝子産物の非存在下で機能的でない;しかしFaoAB遺伝子産物は酵素活性を有する。
【0083】
FaoAおよびFaoB複合体を発現するトランスジェニック系統を、個々のポリペプチドを発現するFaoAトランスジェニック系統およびFaoBトランスジェニック系統を交雑して獲得し、そして種子を、記載されるようなウエスタンブロッティングおよび酵素アッセイにより分析する。
【0084】
(実施例6:B.napus cv.Westarの形質転換およびトランスジェニック系統の分析)
(形質転換)
使用するプロトコルは、Moloneyら(1989)に記載される手順から採用した。Brassica napus cv.Westarの種子を、10%の市販の漂白剤(Javex,Colgate−Palmolive Canada Inc.)中で30分間、穏やかに振盪して、表面滅菌した。その種子を滅菌蒸留水中で3回洗浄した。種子を、Murashige−Skoog(MS)塩およびビタミン、3%スクロースおよび0.7%フィトアガーを含む発芽培地(pH5.8)中に、プレートあたり20個の密度で配置し、そして24℃で、かつ60〜80μEm−2s−1の光強度での16時間の明期/8時間の暗期の光周期で、4〜5日間維持した。
【0085】
各々の構築物、pCGmf124、pCGmf125、pCGmf224、pCGmf225、pCGmf1P2S、およびpCGmf2P1Sを、Agrobacterium tumefaciensEHA101株(Hoodら、1986)内にエレクトロポレーションにより導入した。子葉の葉柄の形質転換前に、各々の構築物を有する株EHA101の単一コロニーを、100mg/lのカナマイシンおよび100mg/lのゲンタマイシンを補充した5mlの最小培地中、28℃で48時間増殖させた。1mlの細菌懸濁液を微量遠心管中で1分間の遠心分離によりペレット化した。そのペレットを1mlの最小培地で再懸濁した。
【0086】
形質転換のために、子葉を、その基部に約2mmの葉柄を含むように、4日齢(いくつかの場合においては、5日齢)の実生から切り取った。それらの葉柄の切断表面を有する個々の子葉を、希釈した細菌懸濁液に1秒間浸けて、そしてただちに同時培養培地(3%スクロースおよび0.7%フィトアガーを含み、そして20μlMのベンジルアデニンを富化したMS培地)中に約2mmの深さに包理した。接種した子葉を、プレートあたり10個の密度で配置し、そして同じ生長条件下で48時間インキュベートした。同時培養後、次いで子葉を、3%スクロース、20μMベンジルアデニン、0.7%フィトアガー(pH5.8)、300mg/lのチメンチニン、および20mg/lの硫酸カナマイシンで補充したMS培地を含む再生培地に移した。
【0087】
2〜3週間後、再生苗条を獲得し、切断し、そしてMagentaジャー中の「苗条伸長」培地(3%スクロース、300mg/lのチメンチン、0.7%フィトアガー、および20mg/lのカナマイシンを含むMS培地(pH5.8))上で維持した。次いで、伸長した苗条を、「発根」培地(3%スクロース、2mg/lのインドール酪酸、0.7%フィトアガー、および500mg/lのカルベニシリンを含むMS培地)に移した。根が出現した後、小植物をポッティングミックス(Redi Earth,W.R.Grace and Co.Canada Ltd.)に移した。植物を、噴霧チャンバー(75%のRH)中で同じ生長条件下で維持した。生長2〜3週間後、葉のサンプルを、ネオマイシンホスホトランスフェラーゼ(NPTII)アッセイ(Moloneyら、1989)のために採取した。結果を以下の表2に示す。データは形質転換されたことが確認された植物の数を示す。
【0088】
【表2】
【0089】
形質転換DNAの発生運命を16の無作為に選択したトランスジェニック系統について調査した。サザンDNAハイブリダイゼーション分析により、FaoAおよび/またはFaoBが、試験したトランスジェニック系統のゲノムに組込まれたことが示された。
【0090】
FaoA遺伝子が強力なマメファゼオリンプロモーターから発現される、約80%のpmf124のトランスジェニック植物は、雄性不稔であることが観察された。このプロモーターからのFaoA遺伝子の明らかな高レベルの発現は、FaoA遺伝子産物の機能的発現を生じ、これは種子および/または花粉の発達を障害する。この結果は、非常に予想外のことであった。なぜなら、その植物細胞は、細胞質ゾル中のβ−酸化経路の第一段階を行うことが可能であることが予測されなかったからである。しかし、この結果は、ハイブリッド産生のための雄性不稔のために、または種子の産生を妨げるために、植物におけるβ−酸化遺伝子の発現のためのさらなる適用を提供する。正常なトランスジェニック系統との並行した比較において、pmf124株は、おそらく種子発達の排除に起因して、さらに高いレベルの生物体量を生成することもまた留意された。従って、この表現型は、サイレージ、飼料、または他の生物体量作物についての1エーカーあたりに産生される緑色の生物体量の量を増加するための手段として有用であり得る。ここで、誘導性プロモーター系またはリコンビナーゼ技術の使用が定植のための種子を生成するために使用され得る。これら不稔植物の7つが、pmf225トランスジェニック系統由来の花粉で首尾良く他家受粉され、そして種子を生じた。
【0091】
ファゼオリンプロモーター−FaoB構築物を含むpmf224株からの種子由来のRNAについてのノーザン分析により、コントロールを除く試験した全てのサンプルにおいて、予期された1.2kbの転写物を示すシグナルが示された。弱いダイズオレオシン(oleolsin)プロモーター−FaoA構築物を含むpmf125株からの種子由来のRNAについてのノーザン分析により、2.1kbの予期された大きさの転写物が明らかにされた。FaoA遺伝子が比較的弱いダイズオレオシンプロモーターから発現される、pmf125植物の約80%の種子由来の300〜500μgのタンパク質についてのウエスタンブロッティングは、結論付けには到らなかったが、弱いシグナルが1つのトランスジェニック系統において検出された。
【0092】
(脂肪酸分析)
種子において強力なマメファゼオリンプロモーターからFaoA遺伝子を発現することの強力な代謝効果を示した予想外の結果が与えられたため、弱いダイズオレオシンプロモーターからFaoA遺伝子を発現するトランスジェニック系統からの種子の脂肪酸プロフィールを分析した。マメファゼオリンプロモーターからFaoA遺伝子のみを発現する種子またはFaoB遺伝子も発現する種子を試験した。分析は、Millarら、The Plant Cell 11:1889−902(1998)に記載されるように行った。種子の脂肪酸メチルエステル(FAMES)を、B.napusの10個の種子を15×45−mmねじ口ガラス管に配置し、そして0.75mLの1Nメタノール性(methanolic)HCl試薬(Supelco,PA)および10μLの1mg/mLの17:0メチルエステル(内部標準)中、80℃で一晩加熱することによって調製した。そのサンプルを冷却後、FAMESを、0.3mLのヘキサンおよび0.5mLの0.9%NaClで激しくボルテックスすることにより抽出した。そのサンプルを、相を分離するために静置させ、そして300μLの有機相を抽出し、そしてHewlett−Packardガスクロマトグラフィー上で分析した。
【0093】
脂肪酸プロフィール分析により、その脂質プロフィールにおけるさらなる成分または増強された成分の存在が、FaoA遺伝子(配列番号24)を発現する全てのトランスジェニック植物において示され、これらはコントロール植物には存在しなかった。この結果はまた、FaoA遺伝子が転写されて、そして翻訳されたこと、およびFaoAポリペプチド(配列番号27)が、触媒活性を有することを結論的に証明する。このピークはまた、SoyP−FaoA遺伝子、PhaP−FaoA−SoyP−FaoB遺伝子、SoyP−FaoA−PhaP−FaoB遺伝子を有するさらなる11のトランスジェニック植物およびSoyP−FaoBで他家受粉された不稔(PhaP−FaoA)植物において観察された。これらのデータは、FaoA遺伝子の機能的発現、およびたとえ非常に低レベルの発現でもその種子の脂質プロフィールを変化するのに十分であることを明らかに実証する。本明細書中で記載される方法を適応することにより、当業者は、Arabidopsis oleosinプロモーター、ナピン(napin)プロモーター、またはクルシフェリン(cruciferin)プロモーターのような他のプロモーターを使用して、ファゼオリンプロモーターとダイズオレオシンプロモーターとの間で得られるレベルの中間のレベルでこれらの遺伝子を発現し得、そして誘導性プロモーター系またはリコンビナーゼ技術を使用して、脂肪酸酸化の導入遺伝子が発現される時期を制御し得る。
【0094】
(実施例7:酵母のβ−酸化の多機能酵素複合体)
S.cerevisiaeは、細菌および高等真核生物において観察されるS−3−ヒドロキシアシルCoAよりもむしろR−ヒドロキシアシルCoAを経て進行するβ−酸化経路を含む。酵母由来のfox2遺伝子は、ヒドラターゼ(これはトランス−2−エノイル−CoAからR−3−ヒドロキシアシルCoAを生成する)およびデヒドロゲナーゼ(これは、R−3−ヒドロキシアシルCoAを利用してβ−ケトアシルCoAを生成する)をコードする。
【0095】
fox2遺伝子(配列番号1に示される配列)を、S.cerevisiaeゲノムDNAからPCRにより2つの小片で単離した。プライマーN−fox2bおよびN−bamfox2bを利用して、Fox2のN末端領域をコードする1.1kbのSmaI/BamHIフラグメントをPCRし、そしてプライマーC−fox2およびC−bamfox2を利用して、C末端Fox2領域をコードする1.6kbのBamHI/XbaIフラグメントをPCRした。全長fox2遺伝子を、ベクターpTRCNへのサブクローニングを経て再構築した。
【0096】
【化9】
【0097】
しかし、fox1遺伝子は、β−ケトチオラーゼ活性を保持せず、そしてこの活性は第二の導入遺伝子により供給されなければならない。このような遺伝子の代表的な供給源としては、藻類、細菌、酵母、植物および哺乳動物が挙げられる。細菌Alcaligenes eutrophusは、本明細書中に記載される方法における使用に適切な広い特異性のβ−ケトチオラーゼ遺伝子を有する。それは、Peoplesらに対する米国特許第5,661,026号に記載のように、ハイブリダイゼーションプローブとしてアセトアセチル−CoAチオラーゼ遺伝子を使用して容易に単離され得る。この酵素はまた精製されており(Haywoodら、FEMS Micro.Lett.52:91(1988))、そしてその精製酵素は、抗体の調製または遺伝子単離のための基礎としてのタンパク質配列情報を決定するために有用である。
【0098】
(実施例8:植物のβ−酸化遺伝子)
β−酸化の4官能性タンパク質をコードするcDNAのDNA配列(配列番号4に示される)は、Preisig−Mullerら、J.Biol.Chem.269:20475−81(1994)に記載されるように単離され得る。等価な遺伝子は、類似の手順を使用するか、またはゲノムライブラリー(これらの多くは、例えば、Clontech Laboratories Inc.,Palo Alto,California,USA.から市販されている)をスクリーニングすることにより、他の植物種(Arabidopsis、Brassica、ダイズ、ヒマワリ、およびトウモロコシを含む)から単離され得る。ペルオキシソーム標的配列P−R−Mは、このタンパク質のカルボキシ末端で同定された。植物の細胞質ゾル中での発現に適切な構築物は、この配列を欠失するよう設計されたプライマーを使用する、この遺伝子のPCR増幅により調製され得る。
【背景技術】
【0001】
本発明は、一般的に、ポリヒドロキシアルカノエート物質の産生のためのトランスジェニック植物系、トリグリセリドおよび脂肪酸の改変、ならびに植物での種子生産を変更するための方法の分野に関する。
【0002】
作物栽培学的な作物についての安定なトランスジェニック植物を産生するための方法が、ここ最近15年にわたって開発されてきた。作物は、投入量および産出量の両方の形質を改良するために遺伝的に改変された。前者の形質において、特定の農薬に対する耐性が作物中で操作され、そして特定の天然の殺虫剤(例えば、Bacillus thuringenesisの毒素)が、植物中で直接発現された。ハイブリッド植物を産生するための、雄性不稔系の開発における有意な進歩もまた存在する。産出量の形質に関して、作物は、生産物、一般的には、植物の種子、穀粒または繊維の価値を増すために改変される。重要な代謝標的としては、デンプン、脂肪酸および油の生合成経路の改変が挙げられる。
【0003】
植物作物における微生物のポリヒドロキシアルカノエート(PHA)バイオポリマーの産生においてかなりの商業的な興味が存在する。例えば、PeoplesおよびSinskeyに対する米国特許第5,245,023号および同第5,250,430号;Brightらに対する米国特許第5,502,273号;PeoplesおよびSinskeyに対する米国特許第5,534,432号;Johnに対する米国特許第5,602,321号;Somervilleらに対する米国特許第5,610,041号;PCT WO91/00917;PCT WO92/19747;PCT WO93/02187;PCT WO93/02194;PCT WO94/12014;Poirierら、Science 256:520−23(1992);van der LeijおよびWitholt、Can.J.Microbiol.41(補遺):222−38(1995);NawrathおよびPoirier、The International Symposium on Bacterial Polyhydroxyalkanoate(Egginkら編)Davos Switzerland(1996年8月18〜23日);ならびにWilliamsおよびPeoples、CHEMTECH 26:38−44(1996)を参照のこと。PHAは、天然の、熱可塑性ポリエステルであり、そして非常に多くの適用(消費者包装、使い捨てダイアパーの裏地およびごみ袋、食物、ならびに医療製品を含む)に使用するための伝統的なポリマー技術により加工され得る。
【0004】
実験的な植物系であるArabidopsis thalianaの葉緑体でのポリヒドロキシ酪酸の産生における初期研究により、PHBとして、葉の乾燥重量で14%までの蓄積を生じた(Nawrathら、1993)。しかし、Arabidopsisは、作物栽培学的価値を有さない。さらに、作物栽培学的作物において経済的にPHAを産生するために、種子におけるPHAの産生が所望され、その結果、種子の収穫および加工のための現在の基幹施設が利用され得る。植物の種子からのPHAの回収(PCT WO97/15681)および最終用途の適用(WilliamsおよびPeoples、CHEMTECH 26:38−44(1996))の選択肢は、ポリマーの組成によって有意に影響を受ける。それゆえ、きちんと規定された組成を有するPHAポリマーを産生するトランスジェニック植物系を開発することは有利である。
【0005】
それらの基質特異性に基づいたPHA生合成酵素の慎重な選択は、トランスジェニック系において規定された組成のPHAポリマーの産生を可能にする(米国特許第5,229,279号;同第5,245,023号;同第5,250,430号;同第5,480,794号;同第5,512,669号;同第5,534,432号;同第5,661,026号;および同第5,663,063号)。
【0006】
細菌において、各PHA基は、特異的な経路により産生される。短いペンダント基PHAの場合では、以下の3つの酵素が関与する:β−ケトチオラーゼ、アセトアセチル−CoAレダクターゼ、およびPHAシンターゼ。例えば、ホモポリマーPHBは、アセチル補酵素Aの2つの分子の縮合により産生され、アセトアセチル補酵素Aを生じる。次いで、後者は、レダクターゼによりキラル中間体R−3−ヒドロキシブチリル補酵素Aに還元され、続いて、PHAシンターゼ酵素により重合される。PHAシンターゼは、4−ヒドロキシ酸ユニットおよび5−ヒドロキシ酸ユニットを両方とも含むC3〜C5ヒドロキシ酸モノマーの重合化を可能にする比較的広範な基質特異性を特に有する。この生合成経路は、多くの細菌(例えば、Alcaligenes eutrophus、A.latus、Azotobacter vinlandiiおよびZoogloea ramigera)において見出される。長いペンダント基PHAは、例えば、多くの異なるPseudomonas細菌により産生される。これらの生合成は、ヒドロキシアシル補酵素Aモノマー単位への経路として脂肪酸のβ−酸化および脂肪酸合成を含む。次いで、後者は、より大きなC6〜C14モノマー単位に有利な基質特異性を有するPHAシンターゼにより変換される(PeoplesおよびSinskey、1990)。
【0007】
脂肪酸での細胞増殖から通常産生されるPHB−コ−HXコポリマーの場合、これらの経路の組み合わせは、異なるモノマー単位の形成を担い得る。実際に、Aeromonas caviae(これは、コポリマーであるPHB−コ−3−ヒドロキシヘキサノエートを産生する)におけるPHAシンターゼ遺伝子をコードするDNA遺伝子座の分析を使用して、D−β−ヒドロキシブチリル−CoAおよびD−β−ヒドロキシヘキサノイル−CoA単位の産生を担うD特異的エノイル−CoAヒドラターゼをコードする遺伝子を同定した(FukuiおよびDoi、J.Bacteriol.179:4821−30(1997);Fukuiら、J.Bacteriol.180:667−73(1998))。このようなヒドラターゼ遺伝子および酵素の他の供給源としては、Alcaligenes、PseudomonasおよびRhodospirillumが挙げられる。
【0008】
A.eutrophusにおいて短いペンダント基PHAを産生する酵素である、PHAシンターゼ、アセトアセチル−CoAレダクターゼ、およびβ−ケトチオラーゼは、phbC−phbA−phbB遺伝子を含むオペロンによってコードされる(Peoplesら、1987;PeoplesおよびSinskey、1989)。Pseudomonas生物において、長いペンダント基PHAの産生を担うPHAシンターゼは、pha遺伝子座上に、特にphaA遺伝子およびphaC遺伝子によりコードされることが見出されている(米国特許第5,245,023号および同第5,250,430号;Huismanら、J.Biol.Chem.266:2191−98(1991))。これらの初期研究により、ある範囲のPHA生合成遺伝子が、単離および特徴づけされるか、またはゲノム配列決定プロジェクトから同定された。公知のPHA生合成遺伝子の例は、以下の参考文献に開示される:Aeronomas caviae(FukuiおよびDoi、1997、J.Bacteriol.179:4821−30);Alcaligenes eutrophus(米国特許第5,245,023号;同第5,250,430号;同第5,512,669号;および同第5,661,026号;PeoplesおよびSinsky、J.Biol.Chem.264:15298−03(1989));Acinetobacter(Schembriら、FEMS Microbiol.Lett.118:145−52(1994);Chromatium vinosum(LiebergesellおよびSteinbuchel、Eur.J.Biochem.209:135−50(1992)));Methylobacterium extorquens(ValentinおよびSteinbuchel、Appl.Microbiol.Biotechnol.39:309−17(1993));Nocardia corallina(GENBANK 受託番号AF019964;Hallら、1998、Can.J.Microbiol.44:687−69);Paracoccus denitrificans(Uedaら、J.Bacteriol.178:774−79(1996);Yabutaniら、FEMS Microbiol.Lett.133:85−90(1995));Pseudomonas acidophila(Umedaら、1998、Applied Biochemistry and Biotechnology、70−72:341−52);Pseudomonas sp.61−3(Matsusakiら、1998、J.Bacteriol.180:6459−67);Nocardia corallina;Pseudomonas aeruginosa(TimmおよびSteinbuchel、Eur.J.Biochem.209:15−30(1992));P.oleovorans(米国特許第5,245,023号および同第5,250,430号;Huismanら、J.Biol.Chem.266(4):2191−98(1991);Rhizobium etli(Cevallosら、J.Bacteriol.178:1646−54(1996));R meliloti(Tomboliniら、Microbiology 141:2553−59(1995));Rhodococcus ruber(Pieper−FurstおよびSteinbuchel、FEMS Microbiol.Lett.75:73−79(1992));Rhodospirillum rubrum(Hustedeら、FEMS Microbiol.Lett.93:285−90(1992));Rhodobacter sphaeroides(Hustedeら、FEMS Microbiol.Rev.9:217−30(1992));Biotechnol.Lett.15:709−14(1993);Synechocystis sp.(DNA Res.3:109−36(1996));Thiocapsiae violacea(Appl.Microbiol.Biotechnol.38:493−501(1993))およびZoogloea ramigera(Peoplesら、J.Biol.Chem.262:97−102(1987));PeoplesおよびSinskey、Molecular Microbiology3:349−57(1989))。これらの遺伝子または公開されたDNA配列の利用可能性は、PHA産生のためのある範囲の選択肢を提供するはずである。
【0009】
1〜99%HHモノマーを含むPHB−コ−HHコポリマーを産生するための適切なPHAシンターゼは、Rhodococcus ruber、Rhodospirillum rubrum、Thiocapsiae violacea、およびAeromonas caviaeのPHAシンターゼ遺伝子によりコードされる。R−3−ヒドロキシブチレートに加えて、6〜12の炭素原子の3−ヒドロキシ酸の取り込みのために有用な(すなわち、化学的に合成されたコポリマー(PCT WO95/20614、PCT WO95/20615およびPCT WO95/20621に記載される)に等価な生物学的ポリマーを産生するための)PHAシンターゼは、多くのPseudomonasおよびほかの細菌において同定されている(SteinbuechelおよびWiese、Appl.Microbiol.Biotechnol.37:691−97(1992);Valentinら、Appl.Microbiol.Biotechnol.36:507−14(1992);Valentinら、Appl.Microbiol.Biotechnol.40:710−16(1994);Leeら、Appl.Microbiol.Biotechnol.42:901−09(1995);Katoら、Appl.Microbiol.Biotechnol.45:363−70(1996);Abeら、Int.J.Biol.Macromol.16:115−19(1994);Valentinら、Appl.Microbiol.Biotechnol.46:261−67(1996))。そして、それらは、米国特許第5,245,023号および同第5,250,430号に記載されるように容易に単離され得る。P.oleovorans由来のPHAシンターゼ(米国特許第5,245,023号および同第5,250,430号;Huismanら、J.Biol.Chem.266(4):2191−98(1991))は、長いペンダント基PHAの産生に適切である。β−ケトチオラーゼをコードする植物遺伝子もまた、同定されている(VollackおよびBach、Plant Physiol.111:1097−107(1996))。
【先行技術文献】
【特許文献】
【0010】
【特許文献1】米国特許第5,245,023号
【特許文献2】米国特許第5,250,430号
【特許文献3】米国特許第5,502,273号
【特許文献4】米国特許第5,534,432号
【特許文献5】米国特許第5,602,321号
【特許文献6】米国特許第5,610,041号
【特許文献7】国際公開第91/00917号
【特許文献8】国際公開第92/19747号
【特許文献9】国際公開第93/02187号
【特許文献10】国際公開第93/02194号
【特許文献11】国際公開第94/12014号
【特許文献12】国際公開第97/15681号
【特許文献13】米国特許第5,229,279号
【特許文献14】米国特許第5,245,023号
【特許文献15】米国特許第5,480,794号
【特許文献16】米国特許第5,512,669号
【特許文献17】米国特許第5,661,026号
【特許文献18】米国特許第5,663,063号
【特許文献19】米国特許第5,245,023号
【非特許文献】
【0011】
【非特許文献1】Poirierら、Science 256:520−23(1992)
【非特許文献2】van der LeijおよびWitholt、Can.J.Microbiol.41(補遺):222−38(1995)
【非特許文献3】NawrathおよびPoirier、The International Symposium on Bacterial Polyhydroxyalkanoate(Egginkら編)Davos Switzerland(1996年8月18〜23日)
【非特許文献4】WilliamsおよびPeoples、CHEMTECH 26:38−44(1996)
【非特許文献5】FukuiおよびDoi、J.Bacteriol.179:4821−30(1997)
【非特許文献6】Fukuiら、J.Bacteriol.180:667−73(1998)
【非特許文献7】Huismanら、J.Biol.Chem.266:2191−98(1991)
【非特許文献8】FukuiおよびDoi、1997、J.Bacteriol.179:4821−30
【非特許文献9】PeoplesおよびSinsky、J.Biol.Chem.264:15298−03(1989)
【非特許文献10】Schembriら、FEMS Microbiol.Lett.118:145−52(1994)
【非特許文献11】LiebergesellおよびSteinbuchel、Eur.J.Biochem.209:135−50(1992)
【非特許文献12】ValentinおよびSteinbuchel、Appl.Microbiol.Biotechnol.39:309−17(1993)
【非特許文献13】Hallら、1998、Can.J.Microbiol.44:687−69
【非特許文献14】Uedaら、J.Bacteriol.178:774−79(1996)
【非特許文献15】Yabutaniら、FEMS Microbiol.Lett.133:85−90(1995)
【非特許文献16】Umedaら、1998、Applied Biochemistry and Biotechnology、70−72:341−52)
【非特許文献17】Matsusakiら、1998、J.Bacteriol.180:6459−67
【非特許文献18】TimmおよびSteinbuchel、Eur.J.Biochem.209:15−30(1992)
【非特許文献19】Huismanら、J.Biol.Chem.266(4):2191−98(1991)
【非特許文献20】Cevallosら、J.Bacteriol.178:1646−54(1996)
【非特許文献21】Tomboliniら、Microbiology 141:2553−59(1995)
【非特許文献22】Pieper−FurstおよびSteinbuchel、FEMS Microbiol.Lett.75:73−79(1992)
【非特許文献23】Hustedeら、FEMS Microbiol.Lett.93:285−90(1992)
【非特許文献24】Hustedeら、FEMS Microbiol.Rev.9:217−30(1992)
【非特許文献25】Hustedeら、Biotechnol.Lett.15:709−14(1993)
【非特許文献26】Hustedeら、Synechocystis sp.(DNA Res.3:109−36(1996)
【非特許文献27】Appl.Microbiol.Biotechnol.38:493−501(1993)
【非特許文献28】Peoplesら、J.Biol.Chem.262:97−102(1987)
【非特許文献29】PeoplesおよびSinskey、Molecular Microbiology3:349−57(1989)
【非特許文献30】SteinbuechelおよびWiese、Appl.Microbiol.Biotechnol.37:691−97(1992)
【非特許文献31】Valentinら、Appl.Microbiol.Biotechnol.36:507−14(1992)
【非特許文献32】Valentinら、Appl.Microbiol.Biotechnol.40:710−16(1994)
【非特許文献33】Leeら、Appl.Microbiol.Biotechnol.42:901−09(1995)
【非特許文献34】Katoら、Appl.Microbiol.Biotechnol.45:363−70(1996)
【非特許文献35】Abeら、Int.J.Biol.Macromol.16:115−19(1994)
【非特許文献36】Valentinら、Appl.Microbiol.Biotechnol.46:261−67(1996)
【非特許文献37】Huismanら、J.Biol.Chem.266(4):2191−98(1991)
【非特許文献38】VollackおよびBach、Plant Physiol.111:1097−107(1996)
【発明の概要】
【発明が解決しようとする課題】
【0012】
合成および基質の選択によりモノマー組成を改変する能力にもかかわらず、脂肪酸代謝に関する生合成のようなポリマー生合成の他の特徴を改変することが望ましい。
【課題を解決するための手段】
【0013】
従って、本発明の目的は、植物の細胞代謝を操作するための脂肪酸酸化酵素系を導入するための方法およびDNA構築物を提供することである。
【0014】
本発明の別の目的は、植物において、好ましくは、その脂肪種子においてPHAの産生を増強するための方法を提供することである。
【0015】
(発明の要旨)
新規なポリマーを作製するための、植物において脂肪酸生合成および酸化を改変するための方法および系が、記載される。2つの酵素が、必須である:D特異的エノイル−CoAヒドラターゼのようなヒドラターゼ(例えば、Aeromonas caviaeから得られたヒドラターゼ)およびβ−酸化酵素系。いくつかの植物は、植物が、ヒドラターゼを発現するように操作される場合、ポリマー合成を改変するのに十分なβ−酸化酵素系を有する。
【0016】
実施例は、トランスジェニック植物において、これらの酵素の発現によりポリマーの産生を実証する。実施例はまた、脂肪酸生合成における改変を使用して植物の表現型を変化させ得、これにより種子産生を減少または排除する、および緑色植物生物体量を増加させる、ならびにPHAを産生することを実証する。
【図面の簡単な説明】
【0017】
【図1】図1は、ポリヒドロキシアルカノエートモノマーを産生するための、脂肪酸β−酸化経路の模式図である。
【図2】図2は、プラスミド構築物pSBS2024およびpSBS2025を示す模式図である。
【図3A】図3Aは、プラスミド構築物pCGmf124およびpCGmf125を示す模式図である。
【図3B】図3Bは、プラスミド構築物pCGmf124およびpCGmf125を示す模式図である。
【図4A】図4Aは、プラスミド構築物pmf1249およびpmf1254を示す模式図である。
【図4B】図4Bは、プラスミド構築物pmf1249およびpmf1254を示す模式図である。
【図5A】図5Aは、プラスミド構築物pCGmf224およびpCGmf225を示す模式図である。
【図5B】図5Bは、プラスミド構築物pCGmf224およびpCGmf225を示す模式図である。
【図6A】図6Aは、プラスミド構築物pCGmf1P2SおよびpCGmf2P1Sを示す模式図である。
【図6B】図6Bは、プラスミド構築物pCGmf1P2SおよびpCGmf2P1Sを示す模式図である。
【発明を実施するための形態】
【0018】
(発明の詳細な説明)
発生中の脂肪種子または緑色組織の細胞質またはプラスチドに脂肪酸酸化酵素系を導入することによって植物の細胞性代謝を操作するための方法およびDNA構築物が提供される。脂肪酸酸化系は、代表的には、広範囲のβ−ケトアシル−CoA基質を利用するβ−ケトチオラーゼ酵素活性を含むいくつかの酵素活性を含む。
【0019】
驚くべきことに、豆のファゼオリン(phaseolin)プロモーターからの少なくとも1つのこれらの導入遺伝子の発現が、雄性不稔をもたらすことが見出された。興味深いことに、これらの植物は、種子をつけず、その代わりに正常よりも高いレベルの生物体量(例えば、葉、茎、柄)を生成した。それゆえ、本明細書中に記載される方法および構築物をまた用いて、例えば、ハイブリッド生成のために雄性不稔植物を作製し得るか、またはまぐさ(例えば、アルファルファまたはタバコ)の生物体量の生成を増加させ得る。これらの方法およびDNA構築物を用いて作製された植物は、ポリヒドロキシアルカノエートバイオポリマーを生成するため、または新規な油組成物を生成するために有用である。
【0020】
本明細書中に記載される方法は、さらなる導入遺伝子、特に、脂肪酸酸化またはポリイヒドロキシアルカノエート生合成に関与するさらなる酵素をコードする導入遺伝子のその後の取り込みを含む。ポリヒドロキシアルカノエート生合成については、この方法は、本明細書中に記載の方法およびDNA構築物を用いて、または伝統的な植物育種法によって生成されたトランスジェニック植物の続く形質転換による、酵素(例えば、NADHおよび/またはNADPHアセトアセチル−補酵素Aレダクターゼ、PHBシンターゼ、PHAシンターゼ、アセトアセチル−CoAチオラーゼ、ヒドロキシアシル−CoAエピメラーゼ、δ3−シス−δ2−トランスエノイル−CoAイソメラーゼ、アシル−CoAデヒドロゲナーゼ、アシル−CoAオキシダーゼ、およびエノイル−CoAヒドラターゼ)をコードする導入遺伝子の取り込みを含む。
【0021】
(I.植物発現系)
好ましい実施態様では、脂肪酸酸化導入遺伝子は、種子特異的プロモーターから発現され、そしてタンパク質は、発生中の脂肪種子の細胞質において発現される。別の好ましい実施態様では、脂肪酸酸化導入遺伝子は、種子特異的プロモーターから発現され、そして発現されたタンパク質は、プラスチド標的化シグナルを用いてプラスチドに指向される。別の好ましい実施態様では、脂肪酸酸化導入遺伝子は、プラスチド染色体から直接発現され、ここで、これらは相同組換えによって取り込まれる。脂肪酸酸化導入遺伝子はまた、構成性プロモーターから、植物組織全体を通して発現され得る。圃場において作物への農薬または他の活性成分の適用後に活性化され得るプロモーターを用いることにより、これらの導入遺伝子の発現を制御し得ることもまた有用である。本明細書中に記載される方法により含まれるこれらの遺伝子の発現のさらなる制御としては、植物染色体の特異的染色体部位への導入遺伝子の標的化挿入についての組換え酵素技術の使用、またはこの導入遺伝子の発現の調節が挙げられる。
【0022】
本明細書中に記載される方法としては、以下を含むゲノムを有する植物種子が挙げられる:(a)第1のDNA配列および3’非翻訳領域に作動可能に連結されたプロモーター(この第1のDNA配列は、脂肪酸酸化ポリペプチドをコードする)、そして必要に応じて(b)第2のDNA配列および3’非翻訳領域に作動可能に連結されたプロモーター(この第2のDNA配列は、脂肪酸酸化ポリペプチドをコードする)。2つの導入遺伝子の発現は、細胞質、またはペルオキシソームもしくはグリオキシソーム以外のプラスチドにおいて少なくともβ−ケトチオラーゼ活性、デヒドロゲナーゼ活性、およびヒドラターゼ活性を有する機能的脂肪酸β酸化系を植物に提供する。第1のDNA配列および/または第2のDNA配列は、細菌、酵母、真菌、藻類、植物、または動物から単離され得る。少なくとも1つのDNA配列が、少なくとも2つ、そして好ましくは3つの酵素活性を有するポリペプチドをコードすることが好ましい。
【0023】
(形質転換ベクター)
本明細書中に記載される方法において有用なDNA構築物としては、植物へ導入遺伝子を導入し得る形質転換ベクターが挙げられる。いくつかの植物形質転換ベクターの選択肢が利用可能であり、「Gene Transfer to Plants」(Potrykusら編)Springer−Verlag Berlin Heidelberg New York(1995);「Transgenic Plants:A Production System for Industrial and Pharmaceutical Proteins」(Owenら編)John Wiley & Sons,Ltd.England(1996);および「Methods in Plant Molecular Biology:A Laboratory Course Manual」(Maligaら編)Cold Spring Laboratory Press,New York(1995)に記載されるベクターが含まれる。植物形質転換ベクターは、一般に、5’調節配列および3’調節配列(プロモーター、転写終結シグナルおよび/またはポリアデニル化シグナルを含む)の転写制御下の目的の1以上のコード配列、ならびに選択マーカー遺伝子もしくはスクリーニング可能なマーカー遺伝子を含む。5’調節配列についての通常の必要条件は、プロモーター、転写終結シグナルおよび/またはポリアデニル化シグナルを含む。単一の転写物からの2以上のポリペプチドの発現のために、さらなるRNAプロセシングシグナルおよびリボザイム配列が構築物中に操作され得る(米国特許第5,519,164号)。このアプローチは、複数の導入遺伝子を単一の遺伝子座に局在化するという利点を有する。これは、続いての植物育種の試みにおいて有利である。さらなるアプローチは、相同組換えによって植物プラスチド染色体を特異的に形質転換するためのベクターを使用すること(米国特許第5,545,818号)であり、この場合、プラスチドゲノムの原核生物の性質を利用し、そして多数の導入遺伝子をオペロンとして挿入することが可能である。
【0024】
(プロモーター)
多数の植物プロモーターが公知であり、そして目的の遺伝子の構成的発現、または環境によってもしくは発生によって調節される発現のいずれかをもたらす。植物プロモーターを選択して、異なる植物組織または細胞小器官における導入遺伝子の発現を制御し得る。これらのための方法は全て、当業者に公知である(GasserおよびFraley,Science 244:1293−99(1989))。導入遺伝子の5’末端は、導入遺伝子とインフレームで連結された、プラスチドまたは他の細胞内小器官標的化ペプチドをコードする配列を含むように操作され得る。適切な構成性植物プロモーターとしては、カリフラワーモザイクウイルス35Sプロモーター(CaMV)および増強されたCaMVプロモーター(Odellら,Nature,313:810(1985))、アクチンプロモーター(McElroyら,Plant Cell 2:163−71(1990))、AdhIプロモーター(Frommら,Bio/Technology 8:833−39(1990);Kyozukaら、Mol.Gen.Genet.228:40−48(1991))、ユビキチンプロモーター、ゴマノハグサモザイクウイルスプロモーター、マンノピン(mannopine)シンターゼプロモーター、ノパリンシンターゼプロモーター、およびオクトピンシンターゼプロモーターが挙げられる。有用な調節可能なプロモーター系としては、ホウレンソウ硝酸誘導性プロモーター、熱ショックプロモーター、リブロースビスリン酸カルボキシラーゼ(ribulose biphosphate carboxylase)プロモーターの小サブユニット、および化学誘導性プロモーター(Hersheyらに対する米国特許第5,364,780号)が挙げられる。
【0025】
本明細書中に記載される方法の好ましい実施態様では、導入遺伝子は、発生中の種子においてのみ発現される。この目的のために適切なプロモーターとしては、ナピン遺伝子プロモーター(米国特許第5,420,034号および同第5,608,152号)、アセチル−CoAカルボキシラーゼプロモーター(米国特許第5,420,034号および同第5,608,152号)、2Sアルブミンプロモーター、種子貯蔵タンパク質プロモーター、ファゼオリンプロモーター(Slightomら,Proc.Natl.Acad.Sci.USA 80:1897−1901(1983))、オレオシン(oleosin)プロモーター(Plantら,Plant Mol.Biol.25:193−205(1994);Rowleyら,Biochim.Biophys.Acta.1345:1−4(1997);米国特許第5,650,554号;およびPCT WO 93/20216)、ゼインプロモーター、グルテリンプロモーター、デンプンシンターゼプロモーター、およびデンプン分枝酵素プロモーターが挙げられる。
【0026】
これらのベクターを用いる適切な作物栽培植物宿主の形質転換は、種々の方法および植物組織を用いて達成され得る。本明細書中に開示される方法において有用な代表的な植物としては、napus、rappa、sp.carinataおよびjunceaを含むBrassica科;トウモロコシ;ダイズ;ワタの実;ヒマワリ;ヤシ;ココヤシ;ベニバナ;ラッカセイ;Sinapis albaを含むカラシ;およびアマが挙げられる。生物体量(例えば、サイレージトウモロコシ、アルファルファ、またはタバコ)として収穫される作物もまた、本明細書中に開示される方法について有用である。これらのベクターを用いる形質転換のための代表的な組織としては、プロトプラスト、細胞、カルス組織、リーフディスク、花粉、および分裂組織が挙げられる。代表的な形質転換手順としては、Agrobacterium媒介形質転換、バイオリスチック(biolistic)、マイクロインジェクション、エレクトロポレーション、ポリエチレングリコール媒介プロトプラスト形質転換、リポソーム媒介形質転換、およびケイ素繊維媒介形質転換(米国特許第5,464,765号;「Gene Transfer to Plants」(Potrykusら編)Springer−Verlag Berlin Heidelberg New York(1995);「Transgenic Plants:A Production System for Industrial and Pharmaceutical Proteins」(Owenら編)John Wiley & Sons Ltd.England(1996);および「Methods in Plant Molecular Biology:A Laboratory Course Manual」(Maligaら編)Cold Spring Laboratory Press,New York(1995))が挙げられる。
【0027】
(II.トランスジェニック植物を作製し、そしてスクリーニングするための方法)
本明細書中に記載される構築物を用いてトランスジェニック植物を作製するために、以下の手順を用いて、形質転換後に導入遺伝子を発現する、形質転換された植物を入手し得る:選択培地上で形質転換された植物細胞を選択する;形質転換された植物細胞を再生して、分化した植物を生成する;所望のポリペプチドが所望の組織および細胞の位置において得られるようなレベルで導入遺伝子を発現する、形質転換された植物を選択する。
【0028】
記載された方法を実施するために有用な特定の作物について、形質転換手順が、例えば、以下に記載されるように、確立されている:「Gene Transfer to Plants」(Potrykusら編)Springer−Verlag Berlin Heidelberg New York(1995);「Transgenic Plants:A Production System for Industrial and Pharmaceutical Proteins」(Owenら編)John Wiley & Sons,Ltd.England(1996);および「Methods in Plant Molecular Biology:A Laboratory Course Manual」(Maligaら編)Cold Spring Laboratory Press,New York(1995)。
【0029】
Brassica napusは、例えば、米国特許第5,188,958号および同第5,463,174号に記載される通りに形質転換され得る。他のBrassica(例えば、rappa、carinata、およびjuncea)ならびにSinapis albaは、Moloneyら,Plant Cell Reports 8:238−42(1989)に記載される通りに形質転換され得る。ダイズは、多数の報告された手順(米国特許第5,015,580号;同第5,015,944号;同第5,024,944号;同第5,322,783号;同第5,416,011号;および同第5,169,770号)によって形質転換され得る。いくつかの形質転換手順(花粉の形質転換(米国特許第5,629,183号)、ケイ素繊維媒介形質転換(米国特許第5,464,765号)、プロトプラストのエレクトロポレーション(米国特許第5,231,019号;同第5,472,869号;および同第5,384,253号)、遺伝子銃(米国特許第5,538,877号および同第5,538,880号)、ならびにAgrobacterium媒介形質転換(EP 0 604 662 A1;PCT WO 94/00977)を含む)が、トランスジェニックトウモロコシ植物の生成について報告されている。Agrobacterium媒介手順は、特に好ましい。なぜなら、導入遺伝子構築物の単一の組み込み事象が、この手順を用いて、より容易に得られ、そしてこのことは、その後の植物育種を大いに容易にするからである。ワタは、粒子ボンバードメント(米国特許第5,004,863号および同第5,159,135号)によって形質転換され得る。ヒマワリは、粒子ボンバードメントとAgrobacterium感染との組合せを用いて形質転換され得る(EP 0 486 233 A2;米国特許第5,030,572号)。アマは、粒子ボンバードメントまたはAgrobacterium媒介形質転換のいずれかによって形質転換され得る。組換え酵素技術としては、cre−lox系、FLP/FRT系、およびGin系が挙げられる。これらの技術を利用する方法は、例えば、Hodgesらに対する米国特許第5,527,695号;DaleおよびOw,Proc.Natl.Acad.Sci.USA 88:10558−62(1991);Medberryら,Nucleic Acids Res.23:485−90(1995)に記載される。
【0030】
(選択マーカー遺伝子)
本明細書中に記載される方法を実施する際に有用な選択マーカー遺伝子としては、ネオマイシンホスホトランスフェラーゼ遺伝子nptII(米国特許第5,034,322号および同第5,530,196号)、ハイグロマイシン耐性遺伝子(米国特許第5,668,298号)、ホスフィノトリシンに対する耐性をコードするbar遺伝子(米国特許第5,276,268号)が挙げられる。EP 0 530 129 A1は、増殖培地に添加された不活性な化合物を活性化する酵素をコードする導入遺伝子を発現することにより、形質転換された植物が、非形質転換系統から成長して抜け出すのを可能にするポジティブ選択系を記載する。本明細書中の方法において有用なスクリーニング可能マーカー遺伝子としては、β−グルクロニダーゼ遺伝子(Jeffersonら,EMBO J.6:3901−07(1987);米国特許第5,268,463号)およびネイティブなグリーン蛍光タンパク質遺伝子または改変されたグリーン蛍光タンパク質遺伝子(Cubittら,Trends Biochem Sci.20:448−55(1995);Pangら,Plant Physiol.112:893−900(1996))が挙げられる。これらのマーカーのうちのいくつかは、形質(例えば、除草剤耐性)を目的の植物に導入し、それによって、投入側にさらなる作物栽培的価値を提供するという付加的な利点を有する。
【0031】
本明細書中に記載される方法の好ましい実施態様では、1より多くの遺伝子産物が植物中で発現される。この発現は、以下を含む、多数の異なる方法を介して達成され得る:(1)単一の形質転換事象においてコードDNAを導入する方法であって、ここで全ての必要なDNAが単一のベクター上に存在する、方法;(2)同時形質転換事象においてコードDNAを導入する方法であって、ここで全ての必要なDNAが別個のベクター上に存在するが、同時に植物細胞に導入される、方法;(3)独立した形質転換事象によって連続的に植物細胞にコードDNAを導入する方法(すなわち、さらなるDNA構築物を用いての1以上のコードDNAを発現するトランスジェニック植物細胞の形質転換);ならびに(4)別個の形質転換事象による、必要とされるDNA構築物の各々の形質転換によって、個々のタンパク質を発現するトランスジェニック植物を得、そして伝統的な植物育種法を用いて全ての経路を単一の植物に組み込む方法。
【0032】
(III.β酸化酵素経路)
植物の細胞質ゾルにおけるPHAの産生は、R−3−ヒドロキシアシルCoAチオエステルをPHAシンターゼについての基質として産生し得る酵素の細胞質ゾル局在化を必要とする。真核生物および原核生物の両方が、脂肪アシルCoAチオエステルをアセチルCoAへと変換する一連の酵素からなる脂肪酸分解についてのβ酸化経路を有する。これらの経路は、中間体3−ヒドロキシアシルCoAを介して進行するが、この中間体の立体配置は生物間で異なる。例えば、細菌のβ酸化経路および高等真核生物のペルオキシソーム経路は、S−3−ヒドロキシアシルCoAを介して脂肪酸をアセチルCoAへと分解する(Schultz,「Oxidation of Fatty Acids」、Biochemistry of Lipids,Lipoproteins and Membranes(Vanceら編)101〜06頁(Elsevier,Amsterdam 1991))。Escherichia coliでは、β酸化多機能酵素複合体によってコードされるエピメラーゼ活性は、S−3−ヒドロキシアシルCoAをR−3−ヒドロキシアシルCoAへと変換し得る。酵母は、中間体R−3−ヒドロキシアシルCoAを介して進行するペルオキシソームに局在した脂肪酸分解経路を保有し(Hiltunenら J.Biol.Chem.267:6646−53(1992);Filppulaら J.Biol.Chem.270:27453−57(1995))、その結果、PHAを生成するためにエピメラーゼ活性は必要とされない。
【0033】
植物は、他の高等真核生物と同様に、ペルオキシソーム中に細胞下に局在した脂肪酸分解のためのβ酸化経路を有する(Gerhardt,「Catabolism of Fatty Acids[α and β Oxidation]」、Lipid Metabolism in Plants(Moore,Jr.編)527−65頁(CRC Press,Boca Raton,Florida 1993))。それゆえ、植物の細胞質ゾルにおけるPHAの生成は、正確な鎖長のR−3−ヒドロキシアシルCoAチオエステルへの脂肪酸の変換のために、β酸化経路の細胞質ゾルでの発現、ならびにR−3−ヒドロキシアシルCoAをポリマーへと重合させるために適切なPHAシンターゼの細胞質ゾル発現を必要とする。
【0034】
脂肪酸は、植物のプラスチドにおいて飽和アシル−ACPチオエステルとして合成される(Hartwood,「Plant Lipid Metabolism」、Plant Biochemistry(Deyら編)237−72頁(Academic Press,San Diego 1997))。プラスチドから細胞質ゾルへの輸送の前に、大多数の脂肪酸は、Δ9デサチュラーゼを介して脱飽和される。大部分の脂肪種子作物における新たに合成された脂肪酸のプールは、主に、オレイン酸(シス9−オクタデセン酸)、ステアリン酸(オクタデカン酸)、およびパルミチン酸(ヘキサデカン酸)からなる。しかし、いくつかの植物(例えば、ココヤシおよびパーム核)は、より短い鎖の脂肪酸(C8〜14)を合成する。この脂肪酸は、ACPからチオエステラーゼを介して放出され、続いてプラスチド膜に局在するアシルCoAシンテターゼを介してアシルCoAチオエステルへと変換される(Andrewsら,「Fatty acid and lipid biosynthesis and degradation」Plant Physiology,Biochemistry,and Molecular Biology(Dennisら編)345−46頁(Longman Scientific & Technical,Essex,England 1990);Harwood,「Plant Lipid Metabolism」Plant Biochemistry(Deyら編)246頁(Academic Press,San Diego 1997))。
【0035】
脂肪酸分解経路を介する、新たに合成されたアシルCoAチオエステルのプールの細胞質ゾル変換、およびPHAシンターゼについてのR−3−ヒドロキシアシル−CoA基質へのこれらの一連の反応由来の中間体の変換は、図1に概説した酵素反応を介して達成され得る。PHAシンターゼ基質はC4〜C16 R−3−ヒドロキシアシルCoAである。飽和脂肪アシルCoAについては、脂肪酸分解経路を用いてのR−3−ヒドロキシアシルCoAチオエステルへの変換は、以下の一連の反応を必要とする:トランス−2−エノイル−CoAへのアシルCoAチオエステルの変換(反応1)、R−3−ヒドロキシ(hyddroxy)アシルCoAへのトランス−2−エノイル−CoAの水和(反応2a、例えば、酵母系は、この経路を通して作動し、そしてAeromonas caviae D特異的ヒドラターゼはC4〜C7 R−3ヒドロキシアシル−CoAを生じる)、S−3−ヒドロキシアシルCoAへのトランス−2−エノイル−CoAの水和(反応2b)、およびR−3−ヒドロキシアシルCoAへのS−3−ヒドロキシアシルCoAのエピマー化(反応5、例えば、キュウリ四機能タンパク質、細菌系)。3−ヒドロキシアシルCoAは、PHAを形成するPHAシンターゼによって重合されない場合、これは、残りのβ酸化経路を通して以下の通りに進行し得る:β−ケトアシルCoAを形成する、3−ヒドロキシアシルCoAの酸化(反応3)、続いてアセチルCoAおよび2炭素単位分短い飽和アシルCoAチオエステルを生じるCoA存在下でのチオ開裂(反応4)。反応4において生成されるアシルCoAチオエステルは、反応1のβ酸化経路に自由に再度侵入し、そして生成されたアセチルCoAは、β−ケトチオラーゼ(反応7)およびNADHアセトアセチル−CoAレダクターゼまたはNADPHアセトアセチル−CoAレダクターゼ(反応6)の作用によってR−3−ヒドロキシアシルCoAへと変換され得る。この後者の経路は、R−3−ヒドロキシブチリル−CoA、R−3−ヒドロキシバレリル−CoA、およびR−3−ヒドロキシヘキサノイル−CoAを生成するために有用である。この一連の酵素反応によって生成される4〜16炭素原子のR−3−ヒドロキシ酸は、導入遺伝子(または2つのサブユニットシンターゼ酵素の場合は複数の導入遺伝子)から発現されるPHAシンターゼによってPHAポリマーへと重合され得る。
【0036】
Δ9不飽和脂肪アシルCoAについては、記載される種々の一連の反応が必要とされる。図1に詳述されるように、3サイクルのβ酸化は、6個の炭素単位を除去して、3位にシス二重結合を有する不飽和アシルCoAチオエステルを生じる。Δ3−シス−Δ2−トランス−エノイルCoAイソメラーゼによって触媒される、2位でのトランス二重結合への3位でのシス二重結合の変換は、図1に概説した一連のβ酸化反応が進行するのを可能にする。この酵素活性は、微生物のβ酸化複合体および植物の四機能タンパク質に存在するが、酵母fox1には存在しない。
【0037】
アシルCoAチオエステルはまた、分解されてβ−ケトアシルCoAになり、そしてNADH依存性レダクターゼまたはNACPH依存性レダクターゼを介してR−3−ヒドロキシアシルCoAへと変換され得る(反応6)。
【0038】
S特異的ヒドラターゼ活性、S特異的デヒドロゲナーゼ活性、β−ケトチオラーゼ活性、エピメラーゼ活性、およびΔ3−シス−Δ2−トランス−エノイルCoAイソメラーゼ活性をコードする多機能性酵素は、Escherichia coli(Sprattら,J.Bacteriol.158:535−42(1984))およびPseudomonas fragi(Immureら,J.Biochem.107:184−89(1990))のような細菌において見出されている。多機能性酵素複合体は、触媒的に活性なタンパク質がヘテロテトラマーを形成するような、2コピーの各々2つのサブユニットからなる。ヒドラターゼ活性、デヒドロゲナーゼ活性、エピメラーゼ活性、およびΔ3−シス−Δ2−トランス−エノイルCoAイソメラーゼ活性は、一方のサブユニット上に存在し、一方、チオラーゼは別のサブユニットに存在する。生物(例えば、E.coli(Sprattら,J.Bacteriol.158:535−42(1984));DiRusso,J.Bacteriol.172:6459−68(1990))およびP.fragi(Satoら,J.Biochem.111:8−15(1992))由来の酵素をコードする遺伝子は、単離および配列決定されており、そして本明細書中に記載される方法を実施するために適切である。さらに、E.coli酵素系は、個々の酵素活性に重要なアミノ酸残基を同定するために部位特異的変異誘発分析に供されている(HeおよびYang,Biochemistry 35:9625−30(1996);Yangら,Biochemistry 34:6641−47(1995);HeおよびYang,Biochemistry 36:11044−49(1997);Heら、Biochemistry 36:261−68(1997);YangおよびElzinga,J.Biol.Chem.268:6588−92(1993))。これらの変異遺伝子はまた、本明細書中に記載される方法のいくつかの実施態様において用いられ得る。
【0039】
ラットのような哺乳動物は、それらのペルオキシソーム中に、ヒドラターゼ、デヒドロゲナーゼ、およびΔ3−シス−Δ2−トランス−エノイルCoAイソメラーゼ活性を含む、3機能性のβ−酸化酵素を有する。3機能性の酵素は、ラットの肝臓から単離され、そして78kDaの分子量を有する単量体であることが見出された(Palosaariら、J.Biol.Chem.265:2446〜49(1990))。細菌系と異なり、チオラーゼ活性は、多酵素タンパク質の一部ではない(Schultz、「Oxidation of Fatty Acids」Biochemistry of Lipids、Lipoproteins and Membranes(Vanceら、編)95頁、Elsevier,Amsterdam(1991))。ラットにおけるエピマー化は、2つの異なるヒドラターゼの活性の組み合わせによって生じ、その1つは、R−3ヒドロキシアシルCoAをトランス−2−エノイルCoAに転換し、そして別の1つは、トランス−2−エノイルCoAをS−3−ヒドロキシアシルCoAに転換する(Smelandら、Biochemical and Biophysical Research Communications 160:988〜92(1989))。哺乳動物はまた、中間体S−3−ヒドロキシアシルCoAを介して脂肪酸をアセチルCoAに分解する、β−酸化経路をそれらのミトコンドリアに有する(Schultz、「Oxidation of Fatty Acids」Biochemistry of Lipids,Lipoproteins and Membranes(Vanceら、編)96頁(Elsevier,Amsterdam(1991)))。ミトコンドリアβ−酸化活性をコードする遺伝子は、いくつかの動物から単離されており、これらには、ラットミトコンドリア長鎖アシルCoAヒドラターゼ/3−ヒドロキシアシルCoAデヒドロゲナーゼ(GENBANK登録番号D16478)およびラットミトコンドリアチオラーゼ(GENBANK登録番号D13921およびD00511)が含まれる。
【0040】
酵母は、多機能酵素であるFox2を有し、これは、S−3−ヒドロキシアシルCoAの代わりにR−3−ヒドロキシアシルCoA中間体を介して進行するという点で、細菌および高等真核生物のβ−酸化複合体とは異なる(Hiltunenら、J.Biol.Chem.267:6646〜53(1992))。Fox2は、R−特異的ヒドラターゼ酵素活性およびR−特異的デヒドロゲナーゼ酵素活性を有する。この酵素は、Δ9−シス−ヒドロキシアシルCoAを分解してR−3−ヒドロキシアシルCoAを形成するのに必要な、Δ3−シス−Δ2−トランス−エノイルCoAイソメラーゼ活性を有さない。fox2をコードする酵母由来のこの遺伝子は単離され、そして配列決定されており、そして900アミノ酸タンパク質をコードする。この構造遺伝子のDNA配列およびコードされたポリペプチドのアミノ酸配列は、配列番号1および配列番号2に示される。
【0041】
植物は、酵母Fox2に類似するがまた、Δ3−シス−Δ2−トランス−エノイルCoAイソメラーゼ活性もコードする、4機能タンパク質を有する(Mullerら、J.Biol.Chem.269:20475〜81(1994))。このcDNAのDNA配列およびコードされるポリペプチドのアミノ酸配列は、配列番号3および配列番号4に示される。
【0042】
(IV.脂肪種子作物の細胞質への酵素の標的化)
植物の細胞質中のPHA産生を操作することは、植物の細胞質へのβ−酸化の発現を指示する工程を必要とする。faoABのような細菌系においては標的化シグナルは存在しない。真菌、酵母、植物、および哺乳動物においては、β−酸化は、細胞下オルガネラにおいて起こる。代表的には、その遺伝子は、核の染色体において発現され、そして細胞質において合成されるポリペプチドは、特定のアミノ酸配列の存在によってこれらのオルガネラに指向される。真核生物供給源から単離された遺伝子(例えば、酵母、真菌、植物、および哺乳動物のような真核生物供給源由来の脂肪酸酸化酵素)を使用して、本明細書に記載される方法を実行するために、細胞下標的化シグナルの除去または改変が、この酵素を細胞質ゾルに指向させるために必要である。タンパク質を小胞体に指向させるためのシグナルを付加することが有用であり得る。このプロセスにおいて有用なペプチドは、当該分野で周知である。一般的なアプローチは、小胞体に標的化するペプチド配列に特定化したDNA配列を挿入してキメラ遺伝子を形成することによって、導入遺伝子を改変することである。
【0043】
真核生物アシルCoAデヒドロゲナーゼおよび他のミトコンドリアタンパク質は、通常20〜60アミノ酸長であるタンパク質のN−末端のリーダーペプチドを介してミトコンドリアに標的化される(Horwich,Current Opinion in Cell Biology,2:625〜33(1990))。ミトコンドリア移入リーダーペプチドについての明白なコンセンサス配列の欠如にもかかわらず、リーダー配列において鍵となる残基の変異誘発は、ミトコンドリアタンパク質の移入を妨害することが実証された。例えば、Saccharomyces cerevisiae F1−ATPaseの移入は、そのリーダー配列の変異誘発によって妨害され、これは、細胞質における改変された前駆体タンパク質の蓄積を生じた(Bedwellら、Mol.Cell.Biol.9:1014〜25(1989))。
【0044】
3つの真核生物ペルオキシソーム標的化シグナルが報告されている(Gouldら、J.Cell.Biol.108:1657〜64(1989);Bricknerら、J.Plant Physiol.,113:1213〜21(1997))。トリペプチド標的化シグナルS/A/C−K/H/R−Lは、多くのペルオキシソームタンパク質のC−末端に生じる(Gouldら、J.Cell Biol.108:1657〜64(1989))。この配列の変異誘発は、ペルオキシソームへのタンパク質の移入を妨害することが示されてきた。いくつかのペルオキシソームタンパク質は、そのタンパク質のC−末端にこのトリペプチドを含まない。これらのタンパク質では、タンパク質配列中の内部の位置のトリペプチドを介してか(Gouldら、J.Cell.Biol.108:1657〜64(1989))、または未知の、無関係の配列を介して(Bricknerら、J.Plant Physiol.113:1213〜21(1997))、標的化が生じることが示唆されている。Candida tropicalis由来のアシルCoAオキシダーゼのフラグメントを用いる、インビトロでのペルオキシダーゼ標的化実験の結果は、後者の理論を支持するようであり、そしてポリペプチドの内部アミノ酸配列中に2つの別個の標的化シグナルが存在することを示唆する(Smallら、The EMBO Journal 7:1167〜73(1988))。前述の研究において、標的化シグナルは、長さにおいて118アミノ酸の2つの領域に局在し、そしていずれの領域も標的化シグナルS/A/C−K/H/R−Lを含むことが見出されなかった。少数のペルオキシソームのタンパク質は、ペルオキシソームに移入するためのアミノ末端リーダー配列を含むようである(Bricknerら、J.Plant Physiol.113:1213〜21(1997))。これらの標的化シグナルは、部位特異的変異誘発によって欠失させ得るか、または変更され得る。
【0045】
(V.トランスジェニック植物の培養および収穫)
トランスジェニック植物は、標準的な培養技術を使用して生育され得る。植物または植物部分はまた、標準的な装置および方法を使用して回収され得る。PHAは、植物または植物部分から、公知の技術(例えば、PCT WO97/15681に記載されるような、伝統的な播種プロセス技術と組み合わせた溶媒抽出)を使用して回収され得るか、または、例えば、動物の食物として、直接的に使用され得る。ここでは、植物生物体量からのPHAを抽出することは不要である。
【0046】
種子を産生しなかったいくつかの系統は、はるかにより高いレベルの生物体量を産生した。従って、この表現型は、牧草、飼料、または他の生物体量作物について、1エーカーあたりに産生される緑の生物体量の量を増加させる手段として有用であり得る。末端の使用は、動物の餌のための、または電気的な動力を生成するためのエネルギー作物としての、飼料作物の最もコスト的に効果的な産生、を含む。他の使用は、工業製品の引き続く回収(例えば、抽出によるPHA)のために、作物(例えば、アルファルファまたはタバコ)の生物体量レベルを増加させる工程を含む。
【0047】
本明細書中に記載される組成物およびその調製方法およびその使用方法は、以下の非限定的な実施例によってさらに記載される。
【実施例】
【0048】
(実施例1:Pseudomononas putida faoAB遺伝子およびFao酵素の単離および特徴付け)
PCR、DNA配列決定、E.coli形質転換、およびプラスミド精製を含む。すべてのDNA操作を、例えば、Sambrookら、Molecular Cloning:A Laboratory Manual(Cold Spring Harbor Laboratory Press、New York(1989))に記載されるような標準的な方法を用いて行った。Pseudomonas putida由来のfaoABをコードする遺伝子を、Pseudomonas fragi由来のfaoB(Satoら、J.Biochem.111:8−15(1992))に相同性を有するプライマー1および2を使用するPCR(ポリメラーゼ連鎖反応)によってP.putidaゲノムDNAから生成されたプローブを用いて単離した。
プライマー1:5’−gat ggg ccg ctc caa ggg tgg−3’(配列番号5)
プライマー2:5’−caa ccc gaa ggt gcc gcc att−3’(配列番号6)。
【0049】
1.1kb DNAフラグメントを、PCR反応から精製し、そしてλZAP発現系(Stratagene)を使用して、pBKCMVプラスミドに構築されたP.putidaゲノムライブラリーをスクリーニングするためにプローブとして使用した。プラスミドpMFX1を、ポジティブクローンから選択し、そしてfaoAB遺伝子を含むインサートのDNA配列および隣接配列を決定した。この配列を配列番号7に示す。以下のように、faoABを含むフラグメントを、ネイティブなP.putidaリボソーム結合部位を用いてインタクトに、発現ベクターpTRCNにサブクローニングして、プラスミドpMFX3を形成した。プラスミドpMFX1を、BsrG1を用いて消化した。得られる突出末端をクレノウで平滑化した。HindIIIを用いる消化は、FaoABをコードする、3.39kbの平滑末端/HindIIIフラグメントを産生した。発現ベクターpTRCNを、SmaI/HindIIIを用いて消化し、そしてfaoABフラグメントを用いて連結し、7.57kbのプラスミドpMFX3を形成した。
【0050】
FaoAB多酵素複合体の酵素を、以下のようにアッセイした。ヒドラターゼ活性を、アッセイがCoAの存在下で行われた以外は、以前に記載されたように(Filppulaら、J.Biol.Chem.270:27453〜57(1995))、カップリング酵素L−β−ヒドロキシアシルCoAデヒドロゲナーゼを使用して、NADからNADHへの転換をモニタリングすることによってアッセイした。カップリング酵素の激しい生成物阻害が、CoAの非存在下で観察された。このアッセイは、(最終容量1mLに)60μM クロトニル(crotonyl)CoA、50μM Tris−Cl、pH9、50μg/mlのウシ血清アルブミン、50mM KCl、1mM NAD、7μg/ml ブタ心臓由来L−特異的β−ヒドロキシアシルCoA デヒドロゲナーゼ、および0.25mM CoAを含んだ。このアッセイは、アッセイ混合液中へのFaoABの添加で開始された。コントロールアッセイを、生成されるヒドラターゼ産物、S−ヒドロキシブチリルCoAの非存在下でNADの消費速度を決定するために、基質なしで行った。活性の1ユニットは、1分間あたりのNADの1μモルの消費として定義される(ε340=6220M−1cm−1)。
【0051】
ヒドロキシアシルCoAデヒドロゲナーゼを、基質としてアセトアセチルCoAを用いて、340nmにおけるNADHからNADへの転換をモニタリングすることにより(Binstockら、Methods in Enzymology、71:403(1981))、逆方向でアッセイした。このアッセイは、(最終容量1mL中に)0.1M KH2PO4、PH7、0.2mg/mL ウシ血清アルブミン、0.1mM NADH、および33μM アセトアセチルCoAを含んだ。このアッセイを、アッセイ混合液へのFaoABの添加で開始した。必要な場合、酵素サンプルを、1mg/mLウシ血清アルブミンを含む、0.1M KH2PO4、pH7中に希釈した。コントロールアッセイを、ヒドロキシアシルCoAデヒドロゲナーゼ以外の酵素による粗サンプル中のNADHの消費速度を検出するために、基質アセトアセチルCoAなしで実行した。活性の1ユニットは、1分間あたりのNADHの1μモルの消費として定義される(ε340=6220M−1cm−1)。
【0052】
ヒドロキシアシルCoAデヒドロゲナーゼを、基質としてクロトニルCoAを用いて、340nmにおけるNADからNADHへの転換をモニタリングすることにより(Binstockら、Methods in Enzymology、71:403(1981))、正方向でアッセイした。このアッセイ混合液は、(最終容量1mL中に)0.1M KH2PO4、PH8、0.3mg/mL ウシ血清アルブミン、2mM β−メルカプトエタノール、0.25mM CoA、30μM クロトニルCoA、およびFaoABのアリコートを含んだ。反応を、S−ヒドロキシブチリルCoAのインサイチュでの形成を可能にするために数分間プレインキュベーションした。次いで、このアッセイを、NAD(0.45mM)の添加によって開始した。コントロールアッセイを、ヒドロキシアシルCoAデヒドロゲナーゼ以外の酵素によるNADの消費速度を検出するために、基質なしで実行した。活性の1ユニットは、1分間あたりのNADの1μモルの消費として定義される(ε340=6220M−1cm−1)。
【0053】
チオラーゼ活性を、いくつかの改変を伴って、以前に記載されたように(Palmerら、J.Biol.Chem.266:1〜7(1991))、基質アセトアセチルCoAの消費による304nmにおける吸収の減少をモニタリングすることによって決定した。アッセイは、(最終容量1mL中に)62.4mM Tris−Cl、pH 8.1、4.8mM MgCl2、62.5μM CoA、および62.5μMアセトアセチルCoAを含んだ。このアッセイは、アッセイ混合液へのFaoABの添加によって開始した。酵素を有さないコントロールサンプルを、酵素の非存在下でpH8.1の基質分解速度を検出するために、各々のアッセイについて実行した。活性の1ユニットは、1分間あたりの基質アセトアセチルCoAの1μモルの消費として定義される(ε340=16900M−1cm−1)。
【0054】
エピメラーゼ活性を、R−3−ヒドロキシアシルCoAチオエステルを、D,L−3−ヒドロキシアシルCoA混合物の代わりに使用する以外は、以前に記載されたように(Binstockら、Methods in Enzymology、71:403(1981))アッセイした。このアッセイは、(最終容量1mL中に)30μM R−3−ヒドロキシアシルCoA、150mM KH2PO4(pH8)、0.3mg/mL BSA、0.5mM NAD、0.1mM CoA、および7μg/mL ブタ心臓由来L−特異的β−ヒドロキシアシルCoAデヒドロゲナーゼを含んだ。このアッセイは、FaoABの添加によって開始した。
【0055】
DH5α/pMFX3におけるFaoABの発現のために、培養物を2×TY培地中で30℃で増殖させた。2×TY培地は、(1リットルあたり)16gトリプトン、10g酵母、および5gNaClを含む。スターター培養を、一晩増殖させ、そして新鮮な培地(小規模増殖では250mLエルレンマイヤーフラスコ中に100mL;大規模増殖には、2.8Lフラスコ中に1.5L)に接種(1%接種材料)するために使用した。600nmにおける吸光度が0.4〜0.6の範囲に達した場合、細胞を0.4mM IPTGを用いて誘導した。収穫に先立ち、細胞をさらに4時間培養した。細胞を超音波処理によって溶菌し、そして遠心分離によって不溶性物質を可溶性タンパク質から取り除いた。アシルCoAデヒドロゲナーゼ活性を、FaoAサブユニット(配列番号31)の活性を保証するために逆方向でモニタリングし、そしてチオラーゼ活性をFaoサブユニットの活性を決定するためにアッセイした。DH5α/pMFX3におけるFaoABは、それぞれ、4.3U/mgおよび0.99U/mgの値のデヒドロゲナーゼ活性およびチオラーゼ活性を含んだ。これは、コントロール株DH5α/pTRCNにおいてデヒドロゲナーゼおよびチオラーゼについて観察された、それぞれ0.0074U/mgおよび0.0033U/mgよりも顕著に高かった。
【0056】
FaoABを、Pseudomonas fragi由来のFaoABの精製について以前に記載された改変された手順(Imamuraら、J.Biochem.107:184〜89(1990))を用いて、DH5α/pMFX3から精製した。チオラーゼ活性(正方向においてアッセイされた)およびデヒドロゲナーゼ活性(逆方向においてアッセイされた)を、精製の全体を通じてモニタリングした。3リットルのDH5α/pMFX3細胞(2.8Lのエーレンマイヤーフラスコ中2×1.5Lアリコート)を、酵素活性分析のために調製された細胞について以前に記載された細胞増殖手順を用いて、2×TY培地中で増殖させた。細胞(15.8g)を、32mLの10mM KH2PO4、pH7中に再懸濁し、そして超音波処理によって溶菌した。可溶性タンパク質を、遠心分離(18,000RPM、30分間、4℃)によって不溶性の細胞の破片から除去した。可溶性の抽出物を、50%アセトン中で作製し、そして沈澱したタンパク質を、遠心分離によって単離し、そして10mM KH2PO4、pH7中に再溶解した。このサンプルを、(NH4)2SO4を用いて33%飽和に調整し、そして可溶性および不溶性のタンパク質を、遠心分離によって分離した。得られた上清を、(NH4)2SO4で56%飽和に調整し、そして不溶性ペレットを遠心分離によって単離し、そして10mM KH2PO4、pH7中に溶解した。このサンプルを、50℃で30分間加熱し、そして可溶性タンパク質を遠心分離によって単離し、そして6000〜8000分子量カットオフメンブレン中で、10mM KH2PO4、pH7において透析した(2×3L;20時間)。このサンプルを、あらかじめ10mM KH2PO4、pH7で平衡化したToyo Jozo DEAE FPLCカラム(3cm×14cm)上にロードした。このタンパク質を、直線勾配(100mLずつ;10 KH2PO4、pH7中、0〜500mM NaCl)を用いて、流速3mL/分で溶出した。FaoABは、300と325mM NaClとの間に溶出した。このサンプルを、事前に10mM KH2PO4、pH7で平衡化したマクロ−プレップ ハイドロキシアパタイト18/30(Biorad)FPLCカラム(2cm×15cm)にロードする前に、50,000分子量カットオフメンブレン中で、10mM KH2PO4、pH7において透析した(1×2L;15時間)。タンパク質を、3mL/分の流速で、直線勾配(250mLずつ;10〜500mM KH2PO4、pH7)を用いて溶出した。FaoABは、70と130mMのKH2PO4との間に溶出した。活性を含む画分を、MILLIPORETM100,000分子量カットオフ濃縮器を用いて9mLまで濃縮した。緩衝液を、20%ショ糖を含む10mM KH2PO4、pH7を用いて3回交換し、そして−70℃で凍結した。ハイドロキシアパタイト精製画分の酵素活性を、基質の範囲を用いてアッセイした。結果を、以下の表1に示す。
【0057】
【表1】
【0058】
(実施例2:FaoABおよびFaoABポリペプチドに対する抗体の産生)
実施例1に記載されるようなFaoABタンパク質の精製後、サンプルをSDS−PAGEによって分離した。FaoA(配列番号31)およびFaoB(配列番号26)に対応するタンパク質バンドを切り出し、そしてフロイント完全アジュバントを用いてニュージーランド白ウサギを免疫した。追加免疫を、フロイント不完全アジュバントを用いて3週間間隔で行った。抗体を、血清からプロテインAカラム(Pharmacia)のアフィニティークロマトグラフィーによって回収し、ウェスタンブロッティング手順によって抗原に対して試験した。Brassica種子のコントロール抽出物を使用して、植物タンパク質に対する交差反応について試験した。交差反応は検出されなかった。
【0059】
(実施例3:トランスジェニック脂肪種子におけるPseudomonas putido faoAB遺伝子の発現のためのプラスミドの構築)
(pSBS2024の構築)
マメのファゼオリン(phaseolin)プロモーター(配列番号10;Slightomら、1983)の5’−および3’−末端(下線を付した)それぞれに隣接する配列に相同な、 オリゴヌクレオチドプライマーGVR471
【0060】
【化1】
およびGVR472
【0061】
【化2】
を、それぞれ、GVR471およびGVR472の5’−末端にKpnI(斜字体、配列番号8のヌクレオチド1〜7)およびSwaI(斜字体、配列番号9のヌクレオチド1〜9)を付加するように設計した。これらの制限部位を、クローニングを容易にするために組み込んだ。これらのプライマーを使用して、1.4kbファゼオリンプロモーターを増幅した。このプロモーターを、平滑末端連結によってpUC19のSmaI部位にクローニングした。命名されたプラスミド、pCPPI(図2を参照のこと)を、SalIおよびSwaIを用いて切断し、そしてSalI/SwaIファゼオリンターミネーター(配列番号27)に連結した。ポリアデニル化シグナルを含むマメのファゼオリンターミネーター配列を、以下のPCRプライマーを使用して増幅した:GVR396:5’−GATTTAAATGCAAGCTTAAATAAGTATGAACTAAAATGC−3’(配列番号22)
およびGVR3975’−CGGTACCTTAGTTGGTAGGGTGCTA−3’(配列番号23)
そして1.2kbフラグメント(配列番号27)をpCCP1のSalI−SalI部位にクローニングして、pSBS2024を得た(図2)。クローニングのための独特のHindIII部位を含む、得られるプラスミドを、pSBS2024と命名した(図2)。
【0062】
(pSBS2024の構築)
ダイズオレオシンプロモーターフラグメント(配列番号11;Rowleyら、1997)を、DNA配列に隣接するプライマーを用いて簡易化した。
【0063】
プライマーJA408
【0064】
【化3】
は、5’末端(下線を付した)に相補的な配列を含む。
プライマーnp1
【0065】
【化4】
は、プロモーターフラグメントの3’末端(下線を付した)に相同な配列を含む。制限部位XbaI(斜字体)およびSwaI(斜字体)を、クローニングを容易にするために、JA408およびnpIの5’末端にそれぞれ組み込んだ。プライマーを、975bpプロモーターフラグメントを増幅するために使用し、次いでpUC19のSmaI部位にクローニングした(図2を参照のこと)。得られるプラスミド、pCSPIを、SalIおよびSwaIを用いて切断し、そしてダイズターミネーター(配列番号28)に連結した。ダイズオレオシンターミネーターを、以下のプライマーを使用して、PCRによって増幅した:JA410:5’−AAGCTTACGTGATGAGTATTAATGTGTTGTTATG−3’(配列番号29)
およびJA411:5’−TCTAGACAATTCATCAAATACAAATCACATTGCC−3’(配列番号30)
そして、225bpフラグメントをpCSPIのSalI−SwaI部位にクローニングして、プラスミドpSBS2025(図6)を得た。命名されたプラスミド、pSBS2025は、クローニングのための独特のHindIII部位を有する(図2)。
【0066】
(プロモーターコード配列融合体の構築)
以下の2つのオリゴヌクレオチドプライマーを合成した:プラスミドpmfx3のmfl(faoA、配列番号24)の5’隣接配列のヌクレオチド553〜557に相同なnp2
【0067】
【化5】
および3’末端に隣接するヌクレオチド2700〜2683に相補的なnp3
【0068】
【化6】
クローニングを容易にするために、HindIII(イタリック体)部位を、プライマーnp2およびnp3の5’末端に導入した。さらに、植物の翻訳開始コドンを包囲するより有利な配列を得るために、3bpのAAA配列(太字)を組み込んだ。プライマーnp2およびnp3を使用して、フラグメントを増幅し、そしてpUC19のSmaI部位にクローニングした。生じたプラスミドをpCmfI(図3Aおよび3B)と称した。プラスミドpBmf2を、同様のプロセスで構築した(図5Aおよび5B)。クローニングのために、mf2(faoB)遺伝子(配列番号25)の5’および3’末端で、HindIII(イタリック体)を生じるために、合成プライマーの第二のセットを設計した。
【0069】
プラスミドpmfx3のmf2(faoB、配列番号25)の5’(ヌクレオチド2732〜2752bp)に相補的なプライマーnp4
【0070】
【化7】
および3’(ヌクレオチド3907〜3886bp)配列に相同なプライマーnp5
【0071】
【化8】
をPCR反応に使用して、1.17kbのDNAフラグメントを増幅した。生じたPCR産物をpBluescriptのEcoRVにクローニングした。このプラスミドを、pBmf2と称した。
【0072】
両方のプラスミドを個々にHindIIIで切断し、そしてそれらの挿入体を、同じ制限酵素で予め直線化したプラスミドpSBS2024およびpSBS2025にクローニングした。その結果、以下のプラスミドを生成した:pmf124およびpmf125(図3Aおよび3B)ならびにpmf224およびpmf225(図5Aおよび5B)。これらは、ファゼオリンまたはダイズプロモーターのいずれかで融合されたFao遺伝子(mf1およびmf2)を含む。DNA配列分析により、pmf124、pmf125、pmf224、およびpmf225について正しいプロモーター−コード配列−終結配列融合体が確認された。
【0073】
(実施例4:プロモーター−コード配列融合体の植物形質転換ベクターへのアセンブリ)
プラスミドpmf124、pmf125、pmf224、およびpmf225を得た後に、プロモーター−コード配列融合体を、二成分ベクターpCGN1559(McBrideおよびSummerfelt、1990)(これは、NPTII遺伝子(抗生物質カナマイシンに対する耐性を与える)の発現を駆動するCaMV35Sプロモーターを含む)およびpSBS2004(これは、除草剤フォスフィノスリシンに対する耐性を与えるPPT遺伝子を駆動するパセリユビキチンプロモーターを含む)に別々にクローニングした。種々の選択マーカーを有するこの目的に適した二成分ベクターは、いくつかの供給源から入手され得る。
【0074】
ファゼオリン−mf21融合カセットを、XbaIで親プラスミドから放出し、そして同じ制限酵素で直線化したpCGN1559と連結した。得られたプラスミドをpCGmf124と命名した(図3Aおよび3B)。ダイズ−mf1融合体を含むプラスミドpCGmf125を、pmf125およびpCGN1559の両方を連結前にBamHIで切断したことを除き、同様の方法で構築した(図3Aおよび3B)。
【0075】
(pmf1249およびpmf1254の構築)
プラスミドpSBS2004を、ダイズ−mf1融合体を含むBamHIフラグメントと共に直線化した。このプラスミドをpmf1254と命名した(図4Aおよび4B)。同様に、XbaIファゼオリン−mf1融合フラグメントを、同じ制限酵素で直線化したpSBS2004と連結した。得られたプラスミドをpmf1249と命名した(図4Aおよび4B)。
【0076】
(pCGmf224およびpCGmf225の構築)
ファゼオリン−mf2およびダイズ−mf2融合体を、BamHIまたはXbaIのいずれかで切断することによってベクターから融合体を切り出すことにより構築し、そしていずれかの制限酵素で直線化しておいたpCGN1559にクローニングした(図5Aおよび5B)。
【0077】
(pCGmf1P2SおよびpCGmf2P1Sの構築)
プロモーター−コード配列融合体を含む2つの発現カセットを、以下のように同じ二成分ベクター上でアセンブルした。:ファゼオリン−mf1融合体を含むプラスミドpmf124をBamHIで切断し、そしてpCGN1559のBamHI部位にクローニングして、pCGmfB124を作製した。次いで、このプラスミドをXbaIで直線化し、そしてダイズ−mf2融合体を含むpmf225のXbaIフラグメントに連結した。最終プラスミドを、pCGmf1P2Sと命名した(図6Aおよび6B)。プラスミドpCGmF2P1Sを、同様の様式でアセンブルした。ファゼオリン−mf2融合体をBamHIで切断することによりpmf224から放出し、そしてpCGN1559のBamHI部位でクローニングした。得られたプラスミドpCGmfB224をXbaIで直線化し、そしてダイズ−mf1融合体を含むpmf125のXbaIフラグメントに連結した(図6Aおよび6B)。
【0078】
(実施例5:Brassicaの形質転換)
Brassica種子を、10%の市販の漂白剤(Javex,Colgate−Palmolive)中で30分間、穏やかに振盪して、表面滅菌した。その種子を滅菌蒸留水中で3回洗浄した。種子を、Murashige−Skoog(MS)塩およびビタミン、3%(w/v)スクロースおよび0.7%(w/v)フィトアガー(phytoagar)を含む発芽培地(pH5.8)中に、プレートあたり20個の密度で配置し、そして24℃で、かつ60〜80μEm−2s−1の光強度での16時間の明期/8時間の暗期の光周期で、4〜5日間維持した。
【0079】
各々の構築物、pCGmf124、pCGmf125、pCGmf224、pCGmf1P2S、およびpCGmf2P1Sを、Agrobacterium tumefaciansEHA101株(Hoodら、J.Bacteriol.168:1291−1301(1986))内にエレクトロポレーションにより導入した。子葉の葉柄の形質転換前に、各々の構築物を有するEHA101株の単一コロニーを、100mg/lのカナマイシンおよび100mg/lのゲンタマイシンを補充した5mlの最小培地中、28℃で48時間増殖させた。1mlの細菌懸濁液を微量遠心管中で1分間の遠心分離によりペレット化した。そのペレットを1mlの最小培地中に再懸濁した。
【0080】
形質転換のために、子葉を、その基部に約2mmの葉柄を含むように、4日齢(いくつかの場合においては、5日齢)の実生から切り取った。それらの葉柄の切断表面を有する個々の子葉を、希釈した細菌懸濁液に1秒間浸けて、そしてただちに同時培養培地(3%(w/v)スクロースおよび0.7%(w/v)フィトアガーを含み、そして20μMのベンジルアデニンを富化したMS培地)中に約2mmの深さに包理した。接種した子葉を、プレートあたり10個の密度で配置し、そして同じ生長条件下で48時間インキュベートした。同時培養後、次いで子葉を、3%(w/v)スクロース、20μMベンジルアデニン、0.7%(w/v)フィトアガー(pH5.8)、300mg/lのチメンチニン(timentinin)、および20mg/lの硫酸カナマイシンを補充したMS培地を含む再生培地に移した。
【0081】
2〜3週間後、得られた再生苗条を切断し、そしてMagentaジャー中の「苗条伸長」培地(3%スクロース、300mg/lのチメンチン(timentin)、0.7%(w/v)フィトアガー、300mg/lのチメンチニン、および20mg/lの硫酸カナマイシンを含むMS培地(pH5.8))上で維持した。伸長した苗条を、「発根」培地(MS培地、3%スクロース、2mg/lのインドール酪酸、0.7%フィトアガー、および500mg/lのカルベニシリンを含む)に移した。根が出現した後、小植物をポッティングミックス(Redi Earth,W.R.Grace and Co.)に移した。植物を、噴霧チャンバー(75%の相対湿度)中で同じ生長条件下で維持した。生長2〜3週間後、葉のサンプルを、ネオマイシンホスホトランスフェラーゼ(NPTII)アッセイ(Moloneyら、Plant Cell Reports 8:238−42(1989))のために採取した。
【0082】
FaoAおよびFaoBトランスジェニック系統由来の種子を、抗FaoA抗体および抗FaoB抗体を使用するウェスタンブロッティングによって、脂肪酸酸化ポリペプチドの発現について分析し得る。FaoBポリペプチド(配列番号26)は、FaoA遺伝子産物の非存在下で機能的でない;しかしFaoAB遺伝子産物は酵素活性を有する。
【0083】
FaoAおよびFaoB複合体を発現するトランスジェニック系統を、個々のポリペプチドを発現するFaoAトランスジェニック系統およびFaoBトランスジェニック系統を交雑して獲得し、そして種子を、記載されるようなウエスタンブロッティングおよび酵素アッセイにより分析する。
【0084】
(実施例6:B.napus cv.Westarの形質転換およびトランスジェニック系統の分析)
(形質転換)
使用するプロトコルは、Moloneyら(1989)に記載される手順から採用した。Brassica napus cv.Westarの種子を、10%の市販の漂白剤(Javex,Colgate−Palmolive Canada Inc.)中で30分間、穏やかに振盪して、表面滅菌した。その種子を滅菌蒸留水中で3回洗浄した。種子を、Murashige−Skoog(MS)塩およびビタミン、3%スクロースおよび0.7%フィトアガーを含む発芽培地(pH5.8)中に、プレートあたり20個の密度で配置し、そして24℃で、かつ60〜80μEm−2s−1の光強度での16時間の明期/8時間の暗期の光周期で、4〜5日間維持した。
【0085】
各々の構築物、pCGmf124、pCGmf125、pCGmf224、pCGmf225、pCGmf1P2S、およびpCGmf2P1Sを、Agrobacterium tumefaciensEHA101株(Hoodら、1986)内にエレクトロポレーションにより導入した。子葉の葉柄の形質転換前に、各々の構築物を有する株EHA101の単一コロニーを、100mg/lのカナマイシンおよび100mg/lのゲンタマイシンを補充した5mlの最小培地中、28℃で48時間増殖させた。1mlの細菌懸濁液を微量遠心管中で1分間の遠心分離によりペレット化した。そのペレットを1mlの最小培地で再懸濁した。
【0086】
形質転換のために、子葉を、その基部に約2mmの葉柄を含むように、4日齢(いくつかの場合においては、5日齢)の実生から切り取った。それらの葉柄の切断表面を有する個々の子葉を、希釈した細菌懸濁液に1秒間浸けて、そしてただちに同時培養培地(3%スクロースおよび0.7%フィトアガーを含み、そして20μlMのベンジルアデニンを富化したMS培地)中に約2mmの深さに包理した。接種した子葉を、プレートあたり10個の密度で配置し、そして同じ生長条件下で48時間インキュベートした。同時培養後、次いで子葉を、3%スクロース、20μMベンジルアデニン、0.7%フィトアガー(pH5.8)、300mg/lのチメンチニン、および20mg/lの硫酸カナマイシンで補充したMS培地を含む再生培地に移した。
【0087】
2〜3週間後、再生苗条を獲得し、切断し、そしてMagentaジャー中の「苗条伸長」培地(3%スクロース、300mg/lのチメンチン、0.7%フィトアガー、および20mg/lのカナマイシンを含むMS培地(pH5.8))上で維持した。次いで、伸長した苗条を、「発根」培地(3%スクロース、2mg/lのインドール酪酸、0.7%フィトアガー、および500mg/lのカルベニシリンを含むMS培地)に移した。根が出現した後、小植物をポッティングミックス(Redi Earth,W.R.Grace and Co.Canada Ltd.)に移した。植物を、噴霧チャンバー(75%のRH)中で同じ生長条件下で維持した。生長2〜3週間後、葉のサンプルを、ネオマイシンホスホトランスフェラーゼ(NPTII)アッセイ(Moloneyら、1989)のために採取した。結果を以下の表2に示す。データは形質転換されたことが確認された植物の数を示す。
【0088】
【表2】
【0089】
形質転換DNAの発生運命を16の無作為に選択したトランスジェニック系統について調査した。サザンDNAハイブリダイゼーション分析により、FaoAおよび/またはFaoBが、試験したトランスジェニック系統のゲノムに組込まれたことが示された。
【0090】
FaoA遺伝子が強力なマメファゼオリンプロモーターから発現される、約80%のpmf124のトランスジェニック植物は、雄性不稔であることが観察された。このプロモーターからのFaoA遺伝子の明らかな高レベルの発現は、FaoA遺伝子産物の機能的発現を生じ、これは種子および/または花粉の発達を障害する。この結果は、非常に予想外のことであった。なぜなら、その植物細胞は、細胞質ゾル中のβ−酸化経路の第一段階を行うことが可能であることが予測されなかったからである。しかし、この結果は、ハイブリッド産生のための雄性不稔のために、または種子の産生を妨げるために、植物におけるβ−酸化遺伝子の発現のためのさらなる適用を提供する。正常なトランスジェニック系統との並行した比較において、pmf124株は、おそらく種子発達の排除に起因して、さらに高いレベルの生物体量を生成することもまた留意された。従って、この表現型は、サイレージ、飼料、または他の生物体量作物についての1エーカーあたりに産生される緑色の生物体量の量を増加するための手段として有用であり得る。ここで、誘導性プロモーター系またはリコンビナーゼ技術の使用が定植のための種子を生成するために使用され得る。これら不稔植物の7つが、pmf225トランスジェニック系統由来の花粉で首尾良く他家受粉され、そして種子を生じた。
【0091】
ファゼオリンプロモーター−FaoB構築物を含むpmf224株からの種子由来のRNAについてのノーザン分析により、コントロールを除く試験した全てのサンプルにおいて、予期された1.2kbの転写物を示すシグナルが示された。弱いダイズオレオシン(oleolsin)プロモーター−FaoA構築物を含むpmf125株からの種子由来のRNAについてのノーザン分析により、2.1kbの予期された大きさの転写物が明らかにされた。FaoA遺伝子が比較的弱いダイズオレオシンプロモーターから発現される、pmf125植物の約80%の種子由来の300〜500μgのタンパク質についてのウエスタンブロッティングは、結論付けには到らなかったが、弱いシグナルが1つのトランスジェニック系統において検出された。
【0092】
(脂肪酸分析)
種子において強力なマメファゼオリンプロモーターからFaoA遺伝子を発現することの強力な代謝効果を示した予想外の結果が与えられたため、弱いダイズオレオシンプロモーターからFaoA遺伝子を発現するトランスジェニック系統からの種子の脂肪酸プロフィールを分析した。マメファゼオリンプロモーターからFaoA遺伝子のみを発現する種子またはFaoB遺伝子も発現する種子を試験した。分析は、Millarら、The Plant Cell 11:1889−902(1998)に記載されるように行った。種子の脂肪酸メチルエステル(FAMES)を、B.napusの10個の種子を15×45−mmねじ口ガラス管に配置し、そして0.75mLの1Nメタノール性(methanolic)HCl試薬(Supelco,PA)および10μLの1mg/mLの17:0メチルエステル(内部標準)中、80℃で一晩加熱することによって調製した。そのサンプルを冷却後、FAMESを、0.3mLのヘキサンおよび0.5mLの0.9%NaClで激しくボルテックスすることにより抽出した。そのサンプルを、相を分離するために静置させ、そして300μLの有機相を抽出し、そしてHewlett−Packardガスクロマトグラフィー上で分析した。
【0093】
脂肪酸プロフィール分析により、その脂質プロフィールにおけるさらなる成分または増強された成分の存在が、FaoA遺伝子(配列番号24)を発現する全てのトランスジェニック植物において示され、これらはコントロール植物には存在しなかった。この結果はまた、FaoA遺伝子が転写されて、そして翻訳されたこと、およびFaoAポリペプチド(配列番号27)が、触媒活性を有することを結論的に証明する。このピークはまた、SoyP−FaoA遺伝子、PhaP−FaoA−SoyP−FaoB遺伝子、SoyP−FaoA−PhaP−FaoB遺伝子を有するさらなる11のトランスジェニック植物およびSoyP−FaoBで他家受粉された不稔(PhaP−FaoA)植物において観察された。これらのデータは、FaoA遺伝子の機能的発現、およびたとえ非常に低レベルの発現でもその種子の脂質プロフィールを変化するのに十分であることを明らかに実証する。本明細書中で記載される方法を適応することにより、当業者は、Arabidopsis oleosinプロモーター、ナピン(napin)プロモーター、またはクルシフェリン(cruciferin)プロモーターのような他のプロモーターを使用して、ファゼオリンプロモーターとダイズオレオシンプロモーターとの間で得られるレベルの中間のレベルでこれらの遺伝子を発現し得、そして誘導性プロモーター系またはリコンビナーゼ技術を使用して、脂肪酸酸化の導入遺伝子が発現される時期を制御し得る。
【0094】
(実施例7:酵母のβ−酸化の多機能酵素複合体)
S.cerevisiaeは、細菌および高等真核生物において観察されるS−3−ヒドロキシアシルCoAよりもむしろR−ヒドロキシアシルCoAを経て進行するβ−酸化経路を含む。酵母由来のfox2遺伝子は、ヒドラターゼ(これはトランス−2−エノイル−CoAからR−3−ヒドロキシアシルCoAを生成する)およびデヒドロゲナーゼ(これは、R−3−ヒドロキシアシルCoAを利用してβ−ケトアシルCoAを生成する)をコードする。
【0095】
fox2遺伝子(配列番号1に示される配列)を、S.cerevisiaeゲノムDNAからPCRにより2つの小片で単離した。プライマーN−fox2bおよびN−bamfox2bを利用して、Fox2のN末端領域をコードする1.1kbのSmaI/BamHIフラグメントをPCRし、そしてプライマーC−fox2およびC−bamfox2を利用して、C末端Fox2領域をコードする1.6kbのBamHI/XbaIフラグメントをPCRした。全長fox2遺伝子を、ベクターpTRCNへのサブクローニングを経て再構築した。
【0096】
【化9】
【0097】
しかし、fox1遺伝子は、β−ケトチオラーゼ活性を保持せず、そしてこの活性は第二の導入遺伝子により供給されなければならない。このような遺伝子の代表的な供給源としては、藻類、細菌、酵母、植物および哺乳動物が挙げられる。細菌Alcaligenes eutrophusは、本明細書中に記載される方法における使用に適切な広い特異性のβ−ケトチオラーゼ遺伝子を有する。それは、Peoplesらに対する米国特許第5,661,026号に記載のように、ハイブリダイゼーションプローブとしてアセトアセチル−CoAチオラーゼ遺伝子を使用して容易に単離され得る。この酵素はまた精製されており(Haywoodら、FEMS Micro.Lett.52:91(1988))、そしてその精製酵素は、抗体の調製または遺伝子単離のための基礎としてのタンパク質配列情報を決定するために有用である。
【0098】
(実施例8:植物のβ−酸化遺伝子)
β−酸化の4官能性タンパク質をコードするcDNAのDNA配列(配列番号4に示される)は、Preisig−Mullerら、J.Biol.Chem.269:20475−81(1994)に記載されるように単離され得る。等価な遺伝子は、類似の手順を使用するか、またはゲノムライブラリー(これらの多くは、例えば、Clontech Laboratories Inc.,Palo Alto,California,USA.から市販されている)をスクリーニングすることにより、他の植物種(Arabidopsis、Brassica、ダイズ、ヒマワリ、およびトウモロコシを含む)から単離され得る。ペルオキシソーム標的配列P−R−Mは、このタンパク質のカルボキシ末端で同定された。植物の細胞質ゾル中での発現に適切な構築物は、この配列を欠失するよう設計されたプライマーを使用する、この遺伝子のPCR増幅により調製され得る。
【特許請求の範囲】
【請求項1】
植物の代謝を操作するための方法であって、該植物の細胞質ゾル、ペルオキシソームまたはグリオキシソーム以外のプラスチド、あるいはミトコンドリアにおいて脂肪酸β−酸化酵素活性をコードする導入遺伝子を発現する工程を含む、方法。
【請求項2】
前記脂肪酸β−酸化酵素が、細菌、酵母、真菌、植物および哺乳動物からなる群から選択される遺伝子から発現される、請求項1に記載の方法。
【請求項3】
前記脂肪酸β−酸化酵素が、Escherichia、Pseudomonas、AlcaligenesおよびCoryneformからなる群から選択される細菌由来の遺伝子から発現される、請求項1に記載の方法。
【請求項4】
前記遺伝子が、Pseudomonas putida faoABである、請求項3に記載の方法。
【請求項5】
ポリヒドロキシアルカノエートシンターゼ、アセトアセチル−CoAレダクターゼ、β−ケトアシル−CoAチオラーゼ、およびエノイル−CoAヒドラターゼからなる群から選択される酵素をコードする、細菌、真菌、酵母、植物、または動物起源の遺伝子を発現する工程をさらに含み、ここで、これらの遺伝子によってコードされる該酵素が、前記植物の細胞質ゾル、ペルオキシソームまたはグリオキシソーム以外のプラスチド、あるいはミトコンドリアに対して指向される、請求項1に記載の方法。
【請求項6】
植物細胞の代謝を操作する方法において使用するためのDNA構築物であって、
(a)植物において機能的なプロモーター領域;
(b)少なくとも1つの脂肪酸β−酸化酵素活性をコードする構造的DNA配列;および
(c)植物において天然に発現される遺伝子の3’非翻訳領域であって、該非翻訳領域が、mRNAのポリアデニル化についてのシグナル配列をコードする、3’非翻訳領域、を同じ相で含む、DNA構築物。
【請求項7】
前記プロモーターが、種子特異的プロモーターである、請求項6に記載のDNA構築物。
【請求項8】
前記種子特異的プロモーターが、ナピンプロモーター、ファゼオリンプロモーター、オレオシンプロモーター、2Sアルブミンプロモーター、ゼインプロモーター、β−コングリシニンプロモーター、アシルキャリアタンパク質プロモーター、および脂肪酸デサチュラーゼプロモーターからなる群から選択される、請求項7に記載のDNA構築物。
【請求項9】
前記プロモーターが、構成的プロモーターである、請求項6に記載のDNA構築物。
【請求項10】
前記プロモーターが、CaMV 35Sプロモーター、増強されたCaMV 35Sプロモーターおよびユビキチンプロモーターからなる群から選択される、請求項6に記載のDNA構築物。
【請求項11】
トランスジェニック植物において、ポリヒドロキシアルカノエートの生物学的生成を増強するための方法であって、該植物の細胞質ゾル、ペルオキシソームまたはグリオキシソーム以外のプラスチド、あるいはミトコンドリアにおいて異種の脂肪酸β−酸化酵素をコードする遺伝子を発現する工程を含む、方法。
【請求項12】
前記トランスジェニック植物が、Brassica、トウモロコシ、ダイズ、綿実、ヒマワリ、ヤシ、ココヤシ、ベニバナ、ラッカセイ、カラシ、アマ、タバコ、およびアルファルファからなる群から選択される、請求項11に記載の方法。
【請求項13】
前記植物の細胞質ゾル、ペルオキシソームまたはグリオキシソーム以外のプラスチド、あるいはミトコンドリアにおいて脂肪酸β−酸化酵素をコードする異種の遺伝子を含む、トランスジェニック植物またはその一部。
【請求項14】
前記脂肪酸β−酸化酵素が、細菌、酵母、真菌、植物、および哺乳動物からなる群から選択される遺伝子から発現される、請求項13に記載の植物またはその一部。
【請求項15】
前記脂肪酸酸化酵素が、Escherichia、Pseudomonas、Alcaligenes、およびCoryneformからなる群から選択される細菌由来の遺伝子から発現される、請求項14に記載の植物またはその一部。
【請求項16】
前記遺伝子が、Pseudomonas putida faoABである、請求項15に記載の植物またはその一部。
【請求項17】
ポリヒドロキシアルカノエートシンターゼ、アセトアセチル−CoA レダクターゼ、β−ケトアシル−CoA チオラーゼ、およびエノイル−CoA ヒドラターゼからなる群から選択される酵素をコードする遺伝子をさらに含む、請求項13に記載の植物またはその一部。
【請求項18】
前記植物が、Brassica、トウモロコシ、ダイズ、綿実、ヒマワリ、ヤシ、ココヤシ、ベニバナ、ラッカセイ、カラシ、アマ、タバコ、およびアルファルファからなる群から選択される、請求項13に記載の植物またはその一部。
【請求項19】
(a)植物において機能的なプロモーター領域;
(b)少なくとも1つの脂肪酸β−酸化酵素活性をコードする構造的DNA配列;および
(c)植物において天然に発現される遺伝子の3’非翻訳領域であって、ここで該非翻訳領域が、mRNAのポリアデニル化についてのシグナル配列をコードする、3’非翻訳領域、を同じ相で含むDNA構築物を含む、請求項13に記載の植物またはその一部。
【請求項20】
前記プロモーターが種子特異的プロモーターである、請求項19に記載の植物またはその一部。
【請求項21】
前記種子特異的プロモーターが、ナピンプロモーター、ファゼオリンプロモーター、オレオシンプロモーター、2Sアルブミンプロモーター、ゼインプロモーター、β−コングリシニンプロモーター、アシルキャリアタンパク質プロモーター、および脂肪酸デサチュラーゼプロモーターからなる群から選択される、請求項20に記載の植物またはその一部。
【請求項22】
前記プロモーターが、構成的プロモーターである、請求項19に記載の植物またはその一部。
【請求項23】
前記プロモーターが、CaMV 35Sプロモーター、増強されたCaMV 35Sプロモーターおよびユビキチンプロモーターからなる群から選択される、請求項20に記載の植物またはその一部。
【請求項24】
植物における種子生産を、予防または抑制する方法であって、該植物の細胞質ゾル、またはペルオキシソーム、グリオキシソーム以外のプラスチド、またはミトコンドリアにおいて脂肪酸β−酸化酵素をコードする異種の遺伝子を発現する工程を含む、方法。
【請求項1】
植物の代謝を操作するための方法であって、該植物の細胞質ゾル、ペルオキシソームまたはグリオキシソーム以外のプラスチド、あるいはミトコンドリアにおいて脂肪酸β−酸化酵素活性をコードする導入遺伝子を発現する工程を含む、方法。
【請求項2】
前記脂肪酸β−酸化酵素が、細菌、酵母、真菌、植物および哺乳動物からなる群から選択される遺伝子から発現される、請求項1に記載の方法。
【請求項3】
前記脂肪酸β−酸化酵素が、Escherichia、Pseudomonas、AlcaligenesおよびCoryneformからなる群から選択される細菌由来の遺伝子から発現される、請求項1に記載の方法。
【請求項4】
前記遺伝子が、Pseudomonas putida faoABである、請求項3に記載の方法。
【請求項5】
ポリヒドロキシアルカノエートシンターゼ、アセトアセチル−CoAレダクターゼ、β−ケトアシル−CoAチオラーゼ、およびエノイル−CoAヒドラターゼからなる群から選択される酵素をコードする、細菌、真菌、酵母、植物、または動物起源の遺伝子を発現する工程をさらに含み、ここで、これらの遺伝子によってコードされる該酵素が、前記植物の細胞質ゾル、ペルオキシソームまたはグリオキシソーム以外のプラスチド、あるいはミトコンドリアに対して指向される、請求項1に記載の方法。
【請求項6】
植物細胞の代謝を操作する方法において使用するためのDNA構築物であって、
(a)植物において機能的なプロモーター領域;
(b)少なくとも1つの脂肪酸β−酸化酵素活性をコードする構造的DNA配列;および
(c)植物において天然に発現される遺伝子の3’非翻訳領域であって、該非翻訳領域が、mRNAのポリアデニル化についてのシグナル配列をコードする、3’非翻訳領域、を同じ相で含む、DNA構築物。
【請求項7】
前記プロモーターが、種子特異的プロモーターである、請求項6に記載のDNA構築物。
【請求項8】
前記種子特異的プロモーターが、ナピンプロモーター、ファゼオリンプロモーター、オレオシンプロモーター、2Sアルブミンプロモーター、ゼインプロモーター、β−コングリシニンプロモーター、アシルキャリアタンパク質プロモーター、および脂肪酸デサチュラーゼプロモーターからなる群から選択される、請求項7に記載のDNA構築物。
【請求項9】
前記プロモーターが、構成的プロモーターである、請求項6に記載のDNA構築物。
【請求項10】
前記プロモーターが、CaMV 35Sプロモーター、増強されたCaMV 35Sプロモーターおよびユビキチンプロモーターからなる群から選択される、請求項6に記載のDNA構築物。
【請求項11】
トランスジェニック植物において、ポリヒドロキシアルカノエートの生物学的生成を増強するための方法であって、該植物の細胞質ゾル、ペルオキシソームまたはグリオキシソーム以外のプラスチド、あるいはミトコンドリアにおいて異種の脂肪酸β−酸化酵素をコードする遺伝子を発現する工程を含む、方法。
【請求項12】
前記トランスジェニック植物が、Brassica、トウモロコシ、ダイズ、綿実、ヒマワリ、ヤシ、ココヤシ、ベニバナ、ラッカセイ、カラシ、アマ、タバコ、およびアルファルファからなる群から選択される、請求項11に記載の方法。
【請求項13】
前記植物の細胞質ゾル、ペルオキシソームまたはグリオキシソーム以外のプラスチド、あるいはミトコンドリアにおいて脂肪酸β−酸化酵素をコードする異種の遺伝子を含む、トランスジェニック植物またはその一部。
【請求項14】
前記脂肪酸β−酸化酵素が、細菌、酵母、真菌、植物、および哺乳動物からなる群から選択される遺伝子から発現される、請求項13に記載の植物またはその一部。
【請求項15】
前記脂肪酸酸化酵素が、Escherichia、Pseudomonas、Alcaligenes、およびCoryneformからなる群から選択される細菌由来の遺伝子から発現される、請求項14に記載の植物またはその一部。
【請求項16】
前記遺伝子が、Pseudomonas putida faoABである、請求項15に記載の植物またはその一部。
【請求項17】
ポリヒドロキシアルカノエートシンターゼ、アセトアセチル−CoA レダクターゼ、β−ケトアシル−CoA チオラーゼ、およびエノイル−CoA ヒドラターゼからなる群から選択される酵素をコードする遺伝子をさらに含む、請求項13に記載の植物またはその一部。
【請求項18】
前記植物が、Brassica、トウモロコシ、ダイズ、綿実、ヒマワリ、ヤシ、ココヤシ、ベニバナ、ラッカセイ、カラシ、アマ、タバコ、およびアルファルファからなる群から選択される、請求項13に記載の植物またはその一部。
【請求項19】
(a)植物において機能的なプロモーター領域;
(b)少なくとも1つの脂肪酸β−酸化酵素活性をコードする構造的DNA配列;および
(c)植物において天然に発現される遺伝子の3’非翻訳領域であって、ここで該非翻訳領域が、mRNAのポリアデニル化についてのシグナル配列をコードする、3’非翻訳領域、を同じ相で含むDNA構築物を含む、請求項13に記載の植物またはその一部。
【請求項20】
前記プロモーターが種子特異的プロモーターである、請求項19に記載の植物またはその一部。
【請求項21】
前記種子特異的プロモーターが、ナピンプロモーター、ファゼオリンプロモーター、オレオシンプロモーター、2Sアルブミンプロモーター、ゼインプロモーター、β−コングリシニンプロモーター、アシルキャリアタンパク質プロモーター、および脂肪酸デサチュラーゼプロモーターからなる群から選択される、請求項20に記載の植物またはその一部。
【請求項22】
前記プロモーターが、構成的プロモーターである、請求項19に記載の植物またはその一部。
【請求項23】
前記プロモーターが、CaMV 35Sプロモーター、増強されたCaMV 35Sプロモーターおよびユビキチンプロモーターからなる群から選択される、請求項20に記載の植物またはその一部。
【請求項24】
植物における種子生産を、予防または抑制する方法であって、該植物の細胞質ゾル、またはペルオキシソーム、グリオキシソーム以外のプラスチド、またはミトコンドリアにおいて脂肪酸β−酸化酵素をコードする異種の遺伝子を発現する工程を含む、方法。
【図1】

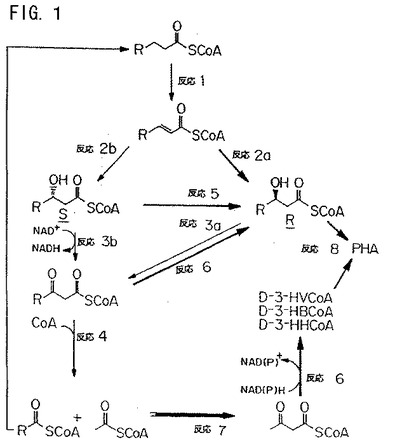
【図2】
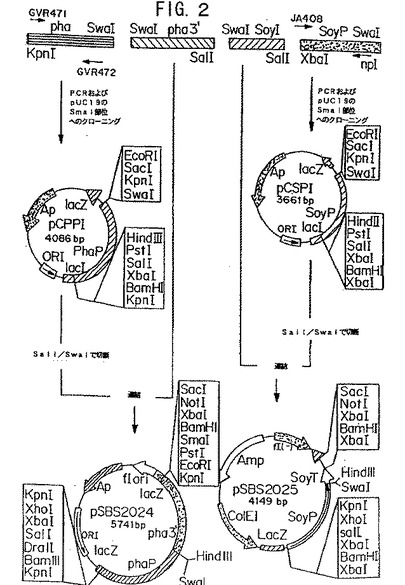
【図3A】
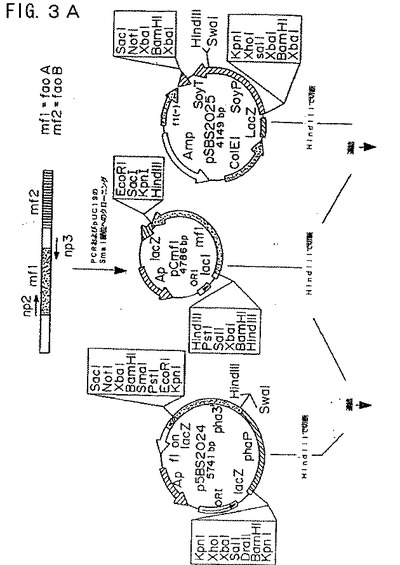
【図3B】
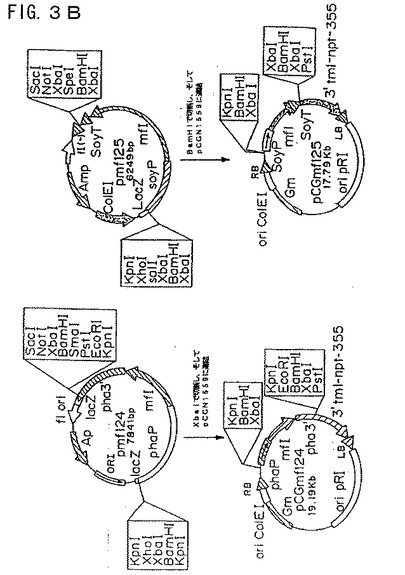
【図4A】
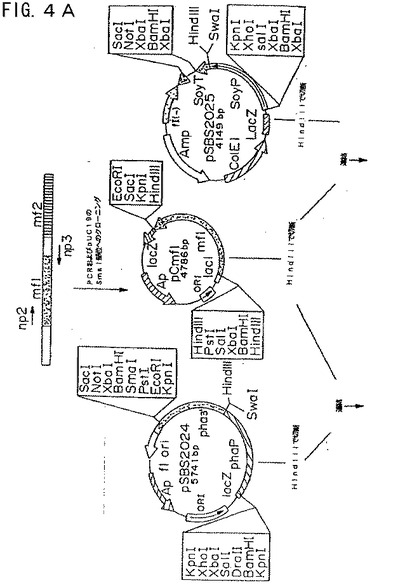
【図4B】
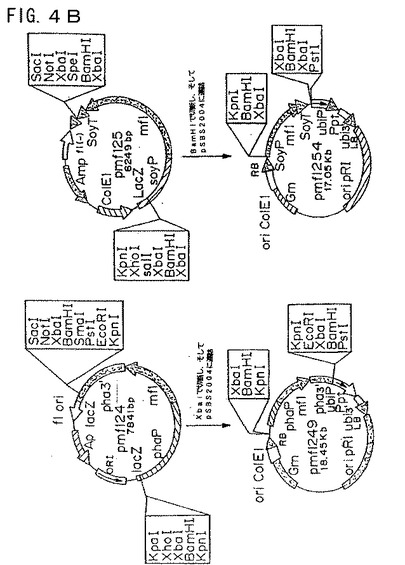
【図5A】
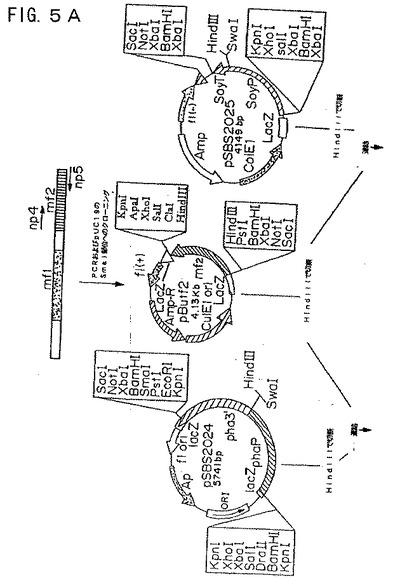
【図5B】
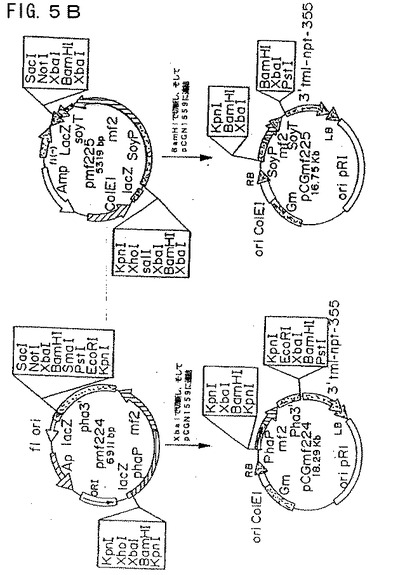
【図6A】
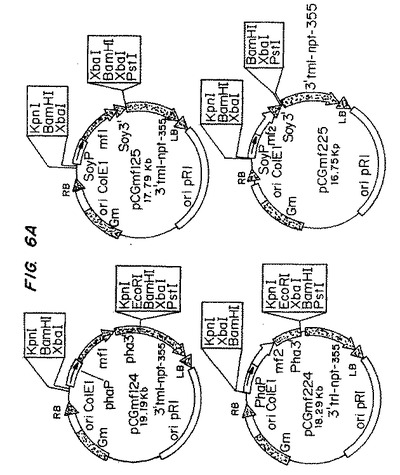
【図6B】
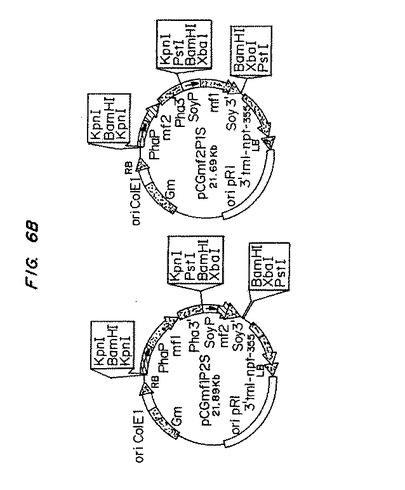
【図2】
【図3A】
【図3B】
【図4A】
【図4B】
【図5A】
【図5B】
【図6A】
【図6B】
【公開番号】特開2009−291204(P2009−291204A)
【公開日】平成21年12月17日(2009.12.17)
【国際特許分類】
【出願番号】特願2009−181255(P2009−181255)
【出願日】平成21年8月4日(2009.8.4)
【分割の表示】特願2000−534653(P2000−534653)の分割
【原出願日】平成11年3月5日(1999.3.5)
【出願人】(398055233)メタボリックス,インコーポレイテッド (18)
【Fターム(参考)】
【公開日】平成21年12月17日(2009.12.17)
【国際特許分類】
【出願日】平成21年8月4日(2009.8.4)
【分割の表示】特願2000−534653(P2000−534653)の分割
【原出願日】平成11年3月5日(1999.3.5)
【出願人】(398055233)メタボリックス,インコーポレイテッド (18)
【Fターム(参考)】
[ Back to top ]