
植物のデオキシハイプシンシンターゼをコードするDNA、植物の真核開始因子5A、トランスジェニック植物、および植物におけるセネッセンスおよびプログラムされた細胞死をコントロールする方法
【課題】植物のデオキシハイプシンシンターゼをコードするDNA、トランスジェニック植物、および植物におけるセネッセンスおよびプログラムされた細胞死をコントロールする方法を提供することを目的とする。
【解決手段】セネッセンス誘導デオキシハイプシンシンターゼをコードする遺伝子または遺伝子フラグメントを植物ゲノムの中にアンチセンスの向きに組込むことによって、植物における、セネッセンスを含むプログラムされた細胞死の発現の調節は達成される。セネッセンス誘導デオキシハイプシンシンターゼをコードする植物遺伝子を同定し、そして単独のヌクレオチド配列を使用して、トランスジェニック植物におけるセネッセンスを変更する。
【解決手段】セネッセンス誘導デオキシハイプシンシンターゼをコードする遺伝子または遺伝子フラグメントを植物ゲノムの中にアンチセンスの向きに組込むことによって、植物における、セネッセンスを含むプログラムされた細胞死の発現の調節は達成される。セネッセンス誘導デオキシハイプシンシンターゼをコードする植物遺伝子を同定し、そして単独のヌクレオチド配列を使用して、トランスジェニック植物におけるセネッセンスを変更する。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、セネッセンス誘導発現を示す植物ポリペプチドをコードするポリヌクレオチドに関する。本発明は、また、アンチセンスの向きでポリヌクレオチドを含有するトランスジェニック植物、および植物における、セネッセンスを包含する、プログラムされた細胞死をコントロールする方法に関する。さらに詳しくは、本発明は、その発現がセネッセンスを包含するプログラムされた細胞死の開始により誘導される、セネッセンス誘導植物デオキシハイプシンシンターゼ遺伝子およびセネッセンス誘導elF−5A遺伝子、および植物におけるプログラムされた細胞死およびセネッセンスをコントロールするために、デオキシハイプシンシンターゼ遺伝子およびelF−5A遺伝子を、単独または組合わせで、使用することに関する。
【背景技術】
【0002】
先行技術の説明
セネッセンスは植物の寿命における生物学的発生の末期である。セネッセンスは死の前兆であり、全植物、器官、花および果実、組織および個々の細胞を包含する生物学的体制化の種々のレベルにおいて起こる。
【0003】
セネッセンスの開始は、内部および外部の両方の異なる因子により誘導されることがある。セネッセンスは、植物または植物組織、例えば、果実、花および葉の寿命において、複雑な、高度に調節された発生段階である。セネッセンスは、細胞膜および高分子の同調的破壊、および代謝物質の植物の他の部分への引き続く移動化を生ずる。
【0004】
正常の植物発生の間に起こるプログラムされたセネッセンスに加えて、細胞および組織の死および続いて起こる代謝物質の再移動化は、外部の、環境的因子に対する同調的応答として起こる。また壊死またはアポトーシスと呼ばれる、セネッセンスの早期の開始を誘導する外部の因子は、環境的ストレス、例えば、温度、乾燥、劣った光または栄養の供給、ならびに病原体の攻撃を包含する。環境的ストレスに暴露された植物組織は、また、ストレスエチレンとして普通に知られているエチレンを産生する(Buchanan−Wollaston、V.、1997、J.Exp.Botany、48:181−199;Wright、M.、1974、Plant、120:63−69)。エチレンはある植物においてセネッセンスを引き起こすことが知られている。
【0005】
セネッセンスは受動的プロセスではなく、むしろ、特異的遺伝子の共役発現を包含する、活性的に調節されたプロセスである。セネッセンスの間に、全RNAのレベルは減少し、多数の遺伝子の発現はスイッチが切られる(Bate他、1991、J.Exper.Botany、42:801−11;Hensel他、1993、The Plant Cell、5:553−64)。しかしながら、セネッセンスプロセスが核遺伝子の新規な転写に依存するという証拠が増加している。例えば、セネッセンスはmRNAおよびタンパク質合成および除核のインヒビターによりブロックされる。in vitro翻訳実験のためにセネッシング葉および緑色の葉からのmRNAを使用する分子の研究において、セネッシング葉において葉タンパク質産物の変化したパターンが示された(Thomas他、1992、J.Plant Physiol.、139:403−12)。
【0006】
ディファレンシャルスクリーニングおよび差引きハイブリダイゼーション技術を使用して、単子葉植物および双子葉植物、例えば、アラアビドプシス(Arabidopsis)、トウモロコシ、キュウリ、アスパラガス、トマト、イネおよびジャガイモを包含する、ある範囲の異なる植物から、セネッセンス誘導遺伝子を表す、多数のcDNAクローンが同定されてきている。セネッセンスの間に特異的に発現される遺伝子の同定は、セネッセンスの新たな転写を進行させるための要件の客観的な証拠である。
【0007】
セネッセンスの間に起こる事象は高度に同調して、壊死および死が起こる前の細胞成分の最大の使用を可能とする。特異的シグナルの感知および遺伝子発現のカスケードの誘導を包含する複雑な相互作用が起こって、このプロセスを調節するに違いない。セネッセンス関係タンパク質をコードする遺伝子の発現は、多分、普通のアクチベータータンパク質を介して調節され、引き続いて、アクチベータータンパク質はホルモンのシグナルにより直接的または間接的に活性化される。このプロセスの初期のシグナリングまたは引き続く同調に関係するメカニズムについてほとんど知られていない。
【0008】
同調的遺伝子発現は、開始因子を包含する、転写および翻訳に関係する因子を必要とする。翻訳開始因子の遺伝子は、植物を包含する、種々の生物において単離され、特性決定されてきている。真核生物の翻訳開始因子5A(elF−5A)はほぼ17kDaのサイズの必須タンパク質因子であり、これは真核細胞のタンパク質系合成の開始に関係する。それはelF−5Aの中にのみ存在することが知られているハイプシン[N−(4−アミノ−2−ヒドロキシブチル)リシン]、すなわち、ユニークな修飾されたアミノ酸、の存在により特徴づけられる。ポリアミン、スペリミジン、からelF−5A中の特定のリシン残基の側鎖アミノ基へのブチルアミノ基の転移およびヒドロキシル化を介して翻訳後にハイプシンは形成される。
【0009】
elF−5Aの活性化は、スペリミジンのブチルアミン残基のelF−5Aのリシンへの転移、ハイプシンの形成およびelF−5Aの活性化を包含する。真核生物において、デオキソハイプシンシンターゼ(DHS)はelF−5Aにおけるハイプシンの翻訳後の合成を仲介する。対応するDHS遺伝子は植物において同定されてきていないが、植物のelF−5Aはハイプシンを含有することが知られている。ハイプシンの修飾は、メチオニル−プロマイシンアッセイを使用するin vitro elF−5A活性のために必須であることが示された。
【0010】
ハイプシンはelF−5Aの中にユニークに存在し、すべての真核生物、いくつかの始原細菌(これらは真核生物に関係するように思われる)の中に見出されるが、真性細菌の中に見出されない。そのうえ、elF−5Aのアミノ酸配列は、特にハイプシン残基を取り囲む領域において、高度に保存され、elF−5Aおよびその活性化タンパク質、デオキシハイプシンシンターゼは真核細胞の生理学において基本的に重要な工程を実行することを示唆する(Joe他、JBC、270:22386−22392)。
【0011】
elF−5Aは、ヒト、アルファルファ、スライムのカビ、ニューロスポラ・クラッサ(Neurospora crassa)、タバコおよび酵母からクローニングされた。ウサギ網状赤血球ライゼイトのリボソームからの単離、およびメチオニン−プロマイシン合成を刺激するin vitro活性に基づいて、elF−5Aは本来一般的翻訳開始因子として同定された。しかしながら、elF−5Aは包括的タンパク質合成の翻訳開始因子ではなく、むしろmRNA集団の特定のサブセットの翻訳を促進する働きをすることが、より最近のデータにより示された。
【0012】
例えば、elF−5Aの1または2以上のイソ型は細胞増殖に関係するmRNAのサブセットの翻訳の仲介において必須の役割を演ずるということを示す強い証拠が、動物細胞および酵母を使用する実験から得られた。酵母において2つのイソ型が存在し、そして両方の遺伝子がサイレントである場合、細胞は分裂することができない(Park他、Biol. Signals、6:115−123、1997)。同様に、elF−5Aを活性化する酵母デオキシハイプシンシンターゼの発現をサイレンスすると、細胞分裂はブロックされる。事実、過増殖状態の治療において重要性を有すると思われるデオキシハイプシンシンターゼのインヒビターが開発された(Wolff他、JBC、272:15865−15871、1997)。
【0013】
他の研究において、elF−5Aの他のイソ型がHIV−1の複製におけるRev機能またはHTLV V複製におけるRex機能のために必須であるということが示された(Park他、Biol.Signals、6:115−123、1997)。また、タバコの中に少なくとも2つの発現されたelF−5A遺伝子が存在する。それらの両方は検査したすべての組織において発現されるが、各遺伝子は明確な発現パターンを有し、多分特定の転写物の翻訳を調節することを、遺伝子特異的プローブは示す(Chamot他、Nucl.Acids Res.、20:625−669、1992)。
【0014】
デオキシハイプシンシンターゼは、ラット精巣、HeLa細胞、ニューロスポラ・クラッサ(Neurospora crassa)および酵母から精製された。デオキシハイプシンシンターゼのアミノ酸配列は高度に保存され、そして異なる種からの酵素は同様な、触媒的性質を共有し、異種elF−5A前駆体と交差種反応性を示す(Park他、Biol.Signals、6:115−123、1997)。
【0015】
植物ポリアミンは、花の誘導、胚形成、病原体耐性、細胞の成長、分化および分裂を包含する、広範な種類の生理学的作用に関係づけられた(Evans他、1989、Ann.Rev.Plant Physiol.Plant Mol.Biol.、40:235−269;およびGalston他、1990、Plant Physiol.、94:406−10)。elF−5Aはポリアミンがその作用を発揮するとき通過する中間物であることが示唆された(Chamot他、1992、Nucl.Acids Res.、20(4):665−69)。
【0016】
タバコ(Nicotiana)からのelF−5Aのイソ型をコードする2つの遺伝子が同定された(NelF−5A1およびNelF−5A2)(Chamot他、1992、Nucl.Acids Res.、20(4):665−69)。これらの遺伝子は非常に類似することが示された。一方の遺伝子はmRNAレベルで構成的に発現されるが、他方の発現パターンは光合成活性の存在または非存在と相関する。遺伝子構造およびゲノムサザンマッピングに基づいて、タバコの中にNelF−5Aの多重遺伝子ファミリーが存在することが示唆された。セネッセンス/壊死特異的mRNA転写物のサブセットの翻訳を調節するelF−5Aのイソ型が存在すると思われる。
【0017】
現在、内部または外部の、例えば、環境的ストレスの、因子により引き起こされるプログラムされた細胞死(セネッセンスを包含する)の開始をコントロールする、広く適用可能な方法は存在しない。したがって、すべてのタイプの植物に適用可能でありかつセネッセンスに導く事象のカスケードにおいて最も早い段階において有効である、セネッセンスをモジュレートする技術を開発することが重要である。
【発明の概要】
【課題を解決するための手段】
【0018】
発明の要約
本発明は、トマトのセネッセンス誘導デオキシハイプシンシンターゼ(DHS)をコードする全長のcDNAクローン、ならびにアラアビドプシス(Arabidopsis)の葉およびカーネーションの花弁からの全長のセネッセンス誘導DHS cDNAクローンの発見およびクローニングに基づく。ヌクレオチド配列および対応するアミノ酸配列が本明細書において開示される。
【0019】
本発明は、また、一部分、トマト、アラアビドプシス(Arabidopsis)およびカーネーションからのセネッセンス誘導elF−5A遺伝子をコードする全長のcDNAクローンの発見およびクローニングに基づく。elF−5A cDNAの各々のヌクレオチド配列および対応するアミノ酸配列が本明細書において開示される。
【0020】
本発明は、年齢に関係するセネッセンスまたは環境的ストレス誘導セネッセンスである、セネッセンスの開始をコントロールするために、植物を遺伝的に修飾する方法を提供する。本発明のセネッセンス誘導DHSヌクレオチド配列、それらのフラグメント、またはこのようなフラグメントの組合わせを植物細胞の中に逆向きに導入して、内因的セネッセンス誘導DHS遺伝子の発現を阻害し、これにより内因的セネッセンス誘導DHSタンパク質のレベルを減少し、かつelF−5Aの活性化および引き続くセネッセンスを仲介する遺伝子の発現を減少および/または防止する。
【0021】
本発明の他の面において、本発明のセネッセンス誘導elF−5Aヌクレオチド配列、それらのフラグメント、またはこのようなフラグメントの組合わせを植物細胞の中に逆向きに導入して、内因的セネッセンス誘導elF−5A遺伝子の発現を阻害し、これにより内因的セネッセンス誘導elF−5Aタンパク質のレベルを減少し、かつelF−5Aの活性化および引き続くセネッセンスを仲介する遺伝子の発現を減少および/または防止する。あるいは、DHS配列およびelF−5A配列の両方を一緒に使用して、内因的DHSおよびelF−5Aタンパク質のレベルを減少させる。
【0022】
なお他の面において、本発明は、本発明のセネッセンス誘導elF−5Aヌクレオチド配列と本発明のセネッセンス誘導DHSヌクレオチド配列との組合わせを植物細胞の中に逆向きに導入して、内因的セネッセンス誘導elF−5A遺伝子およびセネッセンス誘導DHS遺伝子の発現を阻害し、これにより内因的セネッセンス誘導DHSタンパク質のレベルを減少し、かつelF−5Aの活性化および引き続くセネッセンスを仲介する遺伝子の発現を減少および/または防止することによって、年齢に関係するセネッセンスまたは環境的ストレス誘導セネッセンスである、セネッセンスの開始をコントロールするために、植物を遺伝的に修飾する方法に関する。
【0023】
本発明の方法を使用して、トランスジェニック植物を発生させ、成長、発生、および自然のセネッセンスまたは早期に誘導されたセネッセンスについてモニターする。セネッセンス誘導DHS、セネッセンス誘導elF−5Aまたは両方のレベルが減少するために延長した寿命または棚持ち(例えば、花の寿命の延長、果実または野菜の腐敗の減少、バイオマスの増強、種子収量の増加、種子老化の減少および/または葉の黄色化の減少)を示す植物または植物の分離した部分(例えば、挿木、花、野菜、果実、種子または葉)が、葉の黄色化の減少、花弁の離層の減少、輸送および貯蔵中の果実および野菜の腐敗の減少を包含する改良された性質を有する、所望の産物として選択される。これらのよりすぐれた植物を増殖させる。同様に、環境的ストレスに対する抵抗性が増加した、例えば、低温(冷却)、乾燥、感染、およびその他に対する感受性が減少した、および/または病原体に対する抵抗性が増加した植物がよりすぐれた産物として選択される。
【0024】
1つの面において、本発明は、セネッセンス誘導DHSをコードする単離されたDNA分子に関し、ここでDNA分子は配列番号1とハイブリダイゼーションするか、あるいは配列番号1とハイブリダイゼーションする単離されたDNA分子の機能的誘導体とハイブリダイゼーションする。本発明のこの面の1つの態様において、単離されたDNA分子は配列番号1のヌクレオチド配列、すなわち、配列番号1に対して100%の相補性(配列同一性)を有する。
【0025】
本発明は、また、セネッセンス誘導DHSをコードする単離されたDNA分子に関し、ここでDNA分子は配列番号9とハイブリダイゼーションするか、あるいは配列番号9とハイブリダイゼーションする単離されたDNA分子の機能的誘導体とハイブリダイゼーションする。本発明のこの面の1つの態様において、単離されたDNA分子は配列番号9のヌクレオチド配列、すなわち、配列番号9に対して100%の相補性(配列同一性)を有する。
【0026】
本発明は、また、セネッセンス誘導elF−5Aをコードする単離されたDNA分子に関し、ここでDNA分子は配列番号11、配列番号13または配列番号15とハイブリダイゼーションするか、あるいは配列番号11、配列番号13または配列番号15とハイブリダイゼーションする単離されたDNA分子の機能的誘導体とハイブリダイゼーションする。本発明のこの面の1つの態様において、単離されたDNA分子は配列番号11、配列番号13または配列番号15のヌクレオチド配列、すなわち、配列番号11、配列番号13または配列番号15に対して100%の相補性(配列同一性)を有する。
【0027】
本発明の他の態様において、本明細書において記載するDNA分子によりコードされる単離されたタンパク質、またはその機能的誘導体が提供される。好ましいタンパク質は配列番号2のアミノ酸配列を有するか、あるいはその機能的誘導体である。他の好ましいタンパク質は配列番号10のアミノ酸配列を有するか、あるいはその機能的誘導体である。本発明の他の好ましいタンパク質は、配列番号12、配列番号14または配列番号16のアミノ酸配列を有する。
【0028】
また、前述のDNA分子のRNA転写物の対応する部分に対して相補的であるRNA分子をコードするアンチセンスオリゴヌクレオチドまたはポリヌクレオチドが提供され、ここで内因的セネッセンス誘導DHSの発現が変更されるように、オリゴヌクレオチドまたはポリヌクレオチドはRNA転写物とハイブリダイゼーションする。本発明のこの面の他の態様において、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドは、内因的セネッセンス誘導elF−5Aの発現が変更されるように、前述のDNA分子のRNA転写物の対応する部分に対してハイブリダイゼーションするRNA分子である。アンチセンスオリゴヌクレオチドまたはポリヌクレオチドは全長であるか、あるいは好ましくは約6〜約100ヌクレオチドを有する。
【0029】
アンチセンスオリゴヌクレオチドまたはポリヌクレオチドはセネッセンス誘導DHSをコードするDNA分子の1つ鎖の対応する部分に対して実質的に相補的であることができ、ここでセネッセンス誘導DHSをコードするDNA分子は配列番号1、配列番号5、配列番号9、またはそれらの組合わせとハイブリダイゼーションするか、あるいはセネッセンス誘導DHSをコードするDNA分子によりコードされるRNA配列の少なくとも対応する部分に対して実質的に相補的である。
【0030】
本発明の1つの態様において、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドはヌクレオチド配列番号1、配列番号5、配列番号9の1つ鎖の対応する部分またはそれらの組合わせに対して実質的に相補的であるか、または配列番号1、配列番号5、配列番号9から転写されたRNA転写物またはそれらの組合わせに対して実質的に相補的である。他の態様において、アンチセンスオリゴヌクレオチドはセネッセンス誘導DHSをコードするDNA分子の1つ鎖の5’非コーディング部分または3’部分の対応する部分に対して実質的に相補的であり、ここでDNA分子は配列番号1、配列番号5、配列番号9またはそれらの組合わせとハイブリダイゼーションする。
【0031】
あるいは、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドはセネッセンス誘導elF−5AをコードするDNA分子の1つ鎖の対応する部分に対して実質的に相補的であることができ、ここでセネッセンス誘導elF−5AをコードするDNA分子は配列番号11、配列番号13、配列番号15、またはそれらの任意の組合わせとハイブリダイゼーションするか、あるいは配列番号11、配列番号13または配列番号15から転写されたRNA配列の少なくとも対応する部分に対して実質的に相補的である。
【0032】
本発明の1つの態様において、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドはヌクレオチド配列の配列番号11、配列番号13、配列番号15またはそれらの組合わせの1つ鎖の対応する部分に対して実質的に相補的であるか、あるいはコードされるRNA転写物はセネッセンス誘導elF−5AをコードするDNA分子によりコードされる調節配列の対応する部分に対して実質的に相補的である。他の態様において、アンチセンスオリゴヌクレオチドはセネッセンス誘導elF−5AをコードするDNA分子の1つ鎖の5’非コーディング領域または3’領域の対応する部分に対して実質的に相補的であり、ここでDNA分子は配列番号11、配列番号13、配列番号15またはそれらの組合わせとハイブリダイゼーションする。
【0033】
さらに、本発明は、下記の配列を含んでなる植物細胞の形質転換用ベクターに関する:
(a)(1)セネッセンス誘導DHSをコードするDNA分子の1つの鎖の対応する部分、ここでセネッセンス誘導DHSをコードするDNA分子は配列番号1、配列番号5または配列番号9とハイブリダイゼーションする、または(2)セネッセンス誘導DHSをコードするDNA分子によりコードされるRNA配列の対応する部分、に対して実質的に相補的であるアンチセンスオリゴヌクレオチドまたはポリヌクレオチド;および
(b)それがその中に形質転換されている植物細胞においてアンチセンスオリゴヌクレオチドまたはポリヌクレオチドが発現されるように、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドに作用可能に連鎖された調節配列。
【0034】
さらに、本発明は、下記の配列を含んでなる植物細胞の形質転換用ベクターに関する:
(a)(1)セネッセンス誘導elF−5AをコードするDNA分子の1つの鎖の対応する部分、ここでセネッセンス誘導elF−5AをコードするDNA分子は配列番号11、配列番号13、配列番号15とハイブリダイゼーションする、または(2)セネッセンス誘導elF−5AをコードするDNA分子によりコードされるRNA配列の対応する部分、に対して実質的に相補的であるアンチセンスオリゴヌクレオチドまたはポリヌクレオチド;および
(b)それがその中に形質転換されている植物細胞においてアンチセンスオリゴヌクレオチドまたはポリヌクレオチドが発現されるように、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドに作用可能に連鎖された調節配列。
【0035】
調節配列は形質転換された細胞中で機能的であるプロモーターを含み、このプロモーターは誘導可能または構成的であることができる。必要に応じて、調節配列はポリアデニル化シグナルを含む。
【0036】
本発明は、また、前述のベクターまたはベクターの組合わせで形質転換された植物細胞、このような細胞から発生した小植物または成熟植物、またはこのような小植物または成熟植物の植物部分を提供する。
【0037】
本発明は、下記の工程を含んでなる、非修飾植物に比較して、セネッセンス誘導DHS、セネッセンス誘導elF−5Aまたは両方のレベルが減少した植物を産生する方法に関する:
(1)植物を前述のベクターまたはベクターの組合わせで形質転換し;
(2)植物を少なくとも小植物段階に成長させ;
(3)形質転換された植物または小植物をセネッセンス誘導DHS活性および/またはelF−5A活性の変更および/またはセネッセンスの変更および/または環境的ストレス誘導セネッセンスおよび/または病原体誘導セネッセンスおよび/またはエチレン誘導セネッセンスの変更についてアッセイし;そして
(4)非形質転換植物に比較して、セネッセンス誘導DHS活性が変更されたおよび/またはelF−5A活性が減少したおよび/またはセネッセンスが変更したおよび/または環境的ストレス誘導セネッセンスが変更されたおよび/または病原体誘導セネッセンスおよび/またはエチレン誘導セネッセンスが変更した植物を選択し、そして成長させる。
【0038】
前述したように産生された植物、または子孫、ハイブリッド、クローンまたは植物部分は、好ましくは、セネッセンス誘導DHSの発現の減少、セネッセンス誘導elF−5A活性の減少、または両方およびセネッセンスの遅延および/または環境的ストレス誘導セネッセンスおよび/または病原体誘導セネッセンスおよび/またはエチレン誘導セネッセンスの遅延を示す。
【0039】
さらに、本発明は、下記の工程を含んでなる植物における内因的セネッセンス誘導DHSの発現を阻害する方法に関する:
(1)植物のゲノムの中にベクターを組込み、前記ベクターは、
(A)(i)内因的セネッセンス誘導DHSをコードするDNA分子の1つの鎖の対応する部分、ここで内因的セネッセンス誘導DHSをコードするDNA分子は配列番号1、配列番号5および/または配列番号9とハイブリダイゼーションする、または(ii)内因的セネッセンス誘導DHS遺伝子によりコードされるRNA配列の少なくとも一部分、に対して相補的であるアンチセンスオリゴヌクレオチドまたはポリヌクレオチド、および
(B)アンチセンスオリゴヌクレオチドまたはポリヌクレオチドが発現されるように、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドに作用可能に連鎖された調節配列、
を含んでなり;そして
(2)前記植物を成長させる、それにより前記アンチセンスオリゴヌクレオチドまたはポリヌクレオチドは転写され、前記内因的RNA配列に結合し、それにより前記セネッセンス誘導DHS遺伝子の発現は阻害される。
【0040】
本発明は、さらに、下記の工程を含んでなる植物における内因的セネッセンス誘導elF−5Aの発現を阻害する方法に関する:
(1)植物のゲノムの中にベクターを組込み、前記ベクターは、
(A)(i)内因的セネッセンス誘導elF−5AをコードするDNA分子の1つの鎖の対応する部分、ここで内因的セネッセンス誘導elF−5AをコードするDNA分子は配列番号11、配列番号15、配列番号17またはそれらの組合わせとハイブリダイゼーションする、または(ii)内因的セネッセンス誘導elF−5A遺伝子によりコードされるRNA配列の少なくとも一部分、に対して相補的であるアンチセンスオリゴヌクレオチドまたはポリヌクレオチド、および
【0041】
(B)アンチセンスオリゴヌクレオチドまたはポリヌクレオチドが発現されるように、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドに作用可能に連鎖された調節配列、
を含んでなり;そして
(2)前記植物を成長させる、それにより前記アンチセンスオリゴヌクレオチドまたはポリヌクレオチドは転写され、前記内因的RNA配列に結合し、それにより前記セネッセンス誘導elF−5A遺伝子の発現は阻害される。
【図面の簡単な説明】
【0042】
【図1】第1図は、セネッセンス誘導トマト葉DHS cDNA配列(配列番号1)およびトマト葉cDNAライブラリーから得られた誘導アミノ酸配列(配列番号2)のヌクレオチド配列を描写する。
【図2A】第2A図は、アラアビドプシス(Arabidopsis)遺伝子バンクにおいてトマトDHS配列を非同定ゲノム配列と整列させることによって得られたアラアビドプシス(Arabidopsis)DHS遺伝子のヌクレオチド配列を描写する(http://genome−www.stanford.edu/Arabidopsis/)(配列番号5)。アミノ酸配列間のギャップは予測されたイントロンである。
【図2B】第2B図は、誘導アラアビドプシス(Arabidopsis)DHSアミノ酸配列(配列番号6)を描写する。
【図2C】第2C図は、PCRにより得られた600塩基対のセネッセンス誘導アラアビドプシス(Arabidopsis)DHS cDNAのヌクレオチド配列を描写する。
【図2D】第2D図は、セネッセンス誘導アラアビドプシス(Arabidopsis)DHS cDNAフラグメントの誘導アミノ酸配列を描写する。
【図3】第3図は、誘導全長トマト葉セネッセンス誘導DHSアミノ酸配列(配列番号2)および誘導全長アラアビドプシス(Arabidopsis)セネッセンス誘導DHSアミノ酸配列と、ヒト、酵母、菌類、および始原細菌(Archaeobacteria)のDHSタンパク質の配列との整列である。配列の3つまたは4つの間で同一のアミノ酸はボックスで囲まれている。
【図4】第4図は、トマトDHS cDNAの制限地図である。
【図5】第5図は、トマト葉から単離され、32P−dCTP標識化全長トマトセネッセンス誘導DHS cDNAでプロービングしたゲノムDNAのサザンブロットである。
【0043】
【図6】第6図は、発生の異なる段階においてトマト花から単離されたRNAのノザンブロットである。第6A図は、全RNAの臭化エチジウム染色ゲルである。各レーンは10μgのRNAを含有する。第6B図は、32P−dCTP標識化全長トマトセネッセンス誘導DHS cDNAでプロービングしたノザンブロットのオートラジオグラフである。
【図7】第7図は、32P−dCTP標識化全長トマトセネッセンス誘導DHS cDNAでプロービングした成熟の種々の段階においてトマト果実から単離されたRNAのノザンブロットである。各レーンは10μgのRNAを含有する。
【図8】第8図は、2Mのソルビトールで6時間処理して乾燥ストレスさせたトマト葉から単離されたRNAのノザンブロットである。このブロットを32P−dCTP標識化全長トマトセネッセンス誘導DHS cDNAでプロービングした。
【図9】第9図は、冷却温度に暴露したトマト葉から単離されたRNAのノザンブロットである。第9A図は、全RNAの臭化エチジウム染色ゲルである。第9B図は、32P−dCTP標識化全長トマトセネッセンス誘導DHS cDNAでプロービングしたノザンブロットのオートラジオグラフである。第9C図は、葉透析質の導電率として測定した、対応する漏出データを示す。
【図10】第10図は、ポリAテイルおよび5’末端非コーディング領域を含まない、カーネーション全長(1384塩基対)cDNAクローンのヌクレオチド配列(配列番号9)である。誘導アミノ酸配列はヌクレオチド配列の下に示されている(373アミノ酸)(配列番号10)。
【0044】
【図11】第11図は、32P−dCTP標識化全長アラアビドプシス(Arabidopsis)セネッセンス誘導DHS cDNAでプロービングしたセネッセンシングアラアビドプシス(Arabidopsis)葉からの全RNAのノザンブロットである。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【図12】第12図は、種々の段階におけるカーネーション花から単離された全RNAのノザンブロットである。このブロットを32P−dCTP標識化全長カーネーションセネッセンス誘導DHS cDNAでプロービングした。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【図13】第13図は、トマト果実セネッセンス誘導elF−5A遺伝子のヌクレオチド(上部)(配列番号11)および誘導アミノ酸(下部)(配列番号12)配列である。
【図14】第14図は、カーネーションセネッセンス誘導elF−5A遺伝子のヌクレオチド(上部)(配列番号13)および誘導アミノ酸(下部)(配列番号14)配列である。
【図15】第15図は、アラアビドプシス(Arabidopsis)セネッセンス誘導elF−5A遺伝子のヌクレオチド(上部)(配列番号15)および誘導アミノ酸(下部)(配列番号16)配列である。
【0045】
【図16】第16図は、種々の発生段階におけるアラアビドプシス(Arabidopsis)植物から単離された全RNAのノザンブロットである。このブロットを32P−dCTP標識化全長アラアビドプシス(Arabidopsis)セネッセンス誘導DHS cDNAおよび全長セネッセンス誘導elF−5Aでプロービングした。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【図17】第17図は、発生のブレーカー(braker)(BK)、レッド−ファーム(red−firm)(RF)およびレッド−ソフト(red−soft)(RS)段階におけるトマト果実から単離された全RNAのノザンブロットである。このブロットを32P−dCTP標識化全長セネッセンス誘導DHS cDNAおよび全長セネッセンス誘導elF−5Aでプロービングした。DHSおよびelF−5Aは果実成熟と一致してレッド−ソフト果実において平行にアップレギュレートされる。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【図18】第18図は、ソルビトールで処理して乾燥ストレスを誘導したトマトの葉から単離された全RNAのノザンブロットである。Cは対照であり;Sはソルビトール処理である。このブロットを32P−dCTP標識化全長セネッセンス誘導DHS cDNAおよび全長セネッセンス誘導elF−5Aでプロービングした。elF−5AおよびDHSの両方は乾燥ストレスに応答してアップレギュレートされる。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【図19】第19図は、トマト植物の花芽および開いたセネッセンシング花から単離された全RNAのノザンブロットである。このブロットを32P−dCTP標識化全長セネッセンス誘導DHS cDNAおよび全長セネッセンス誘導elF−5Aでプロービングした。elF−5AおよびDHSの両方は開いた/セネッセンシング花においてアップレギュレートされる。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【図20】第20図は、冷却損傷したトマト葉から単離された全RNAのノザンブロットである。このブロットを32P−dCTP標識化全長セネッセンス誘導DHS cDNAおよび全長セネッセンス誘導elF−5Aでプロービングした。elF−5AおよびDHSの両方は再加温の間における冷却損傷植物の発生とともにアップレギュレートされる。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【0046】
【図21】第21図は、3.1週齢のアラアビドプシス(Arabidopsis)野生型(左)およびアンチセンスの向きでセネッセンスDHS遺伝子(第36図に示す配列)の3’末端を発現し、トランスジェニック植物において葉サイズの増加を示すトランスジェニック植物の写真である。
【図22】第22図は、4.6週齢のアラアビドプシス(Arabidopsis)野生型(左)およびアンチセンスの向きでセネッセンスDHS遺伝子(第36図に示す配列)の3’末端を発現し、トランスジェニック植物において葉サイズの増加を示すトランスジェニック植物の写真である。
【図23】第23図は、5.6週齢のアラアビドプシス(Arabidopsis)野生型(左)およびアンチセンスの向きでセネッセンスDHS遺伝子(第36図に示す配列)の3’末端を発現し、トランスジェニック植物において葉サイズの増加を示すトランスジェニック植物の写真である。
【図24】第24図は、6.1週齢のアラアビドプシス(Arabidopsis)野生型(左)およびアンチセンスの向きでセネッセンスDHS遺伝子(第36図に示す配列)の3’末端を発現し、トランスジェニック植物において葉サイズの増加を示すトランスジェニック植物の写真である。
【図25】第25図は、アンチセンスの向きでセネッセンス誘導DHS遺伝子を発現する3つのT1トランスジェニックアラアビドプシス(Arabidopsis)植物系統からの種子収量の増加を示すグラフである。種子収量は種子の体積として表されている。n=30についてのSEを野生型植物について示す。
【0047】
【図26】第26図は、アンチセンスの向きでセネッセンスDHS遺伝子(第36図に示す配列)の3’末端を発現するトランスジェニックトマト植物(左)およびトランスジェニック植物において葉サイズの増加および植物サイズの増加を示す野生型植物(右)の写真である。写真は小植物を土壌に移した後18日に撮った。
【図27】第27図は、アンチセンスの向きでセネッセンスDHS遺伝子(第36図に示す配列)の3’末端を発現するトランスジェニックトマト植物(左)およびトランスジェニック植物において葉サイズの増加および植物サイズの増加を示す野生型植物(右)の写真である。写真は小植物を土壌に移した後32日に撮った。
【図28】第28図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【図29】第29図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【図30】第30図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【0048】
【図31】第31図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【図32】第32図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【図33】第33図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【図34】第34図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【図35】第35図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【0049】
【図36】第36図は、植物を形質転換するためにアンチセンスの向きで使用したアラアビドプシス(Arabidopsis)セネッセンス誘導DHS遺伝子の3’末端のヌクレオチド(上部)(配列番号30)および誘導アミノ酸(下部)配列である。
【図37】第37図は、植物を形質転換するためにアンチセンスの向きで使用したトマトセネッセンス誘導DHS遺伝子の3’末端のヌクレオチド(上部)(配列番号31)および誘導アミノ酸(下部)配列である。
【図38】第38図は、全長アラアビドプシス(Arabidopsis)遺伝子を単離するために使用した600塩基対のアラアビドプシス(Arabidopsis)セネッセンス誘導DHSプローブのヌクレオチド(上部)(配列番号26)および誘導アミノ酸(下部)配列である。
【図39】第39図は、全長カーネーション遺伝子を単離するために使用した483塩基対のカーネーションセネッセンス誘導DHSプローブのヌクレオチド(上部)(配列番号27)および誘導アミノ酸(下部)配列である。
【発明を実施するための形態】
【0050】
発明の詳細な説明
植物細胞における1または2以上のセネッセンス誘導DHS遺伝子、1または2以上のセネッセンス誘導elF−5Aまたは両方の発現を変更する方法および組成物が提供される。植物におけるセネッセンス誘導DHSおよびセネッセンス誘導elF−5Aの、単独または組合わせにおける、発現の変更はセネッセンスの開始を遅延し、環境的ストレスおよび病原体に対する抵抗性を改良し、こうして植物の棚持ちおよび/または成長期間を延長する。
【0051】
セネッセンス誘導発現を示すトマトDHS遺伝子をコードする全長cDNA配列は、鋳型として冷却損傷したトマト葉から単離されたRNAを使用する逆転写酵素仲介ポリメラーゼ連鎖反応(RT−PCR)により単離し、RT−PCR生成物を使用して冷却損傷した、ソルビトール処理トマト葉cDNAライブラリーをスクリーニングする。単離されたトマト葉cDNA配列ならびに全長トマト葉cDNAの選択された領域に対応するポリヌクレオチドプローブを使用して、環境的ストレスに暴露された(冷却された)トマト葉、(脱水された)ソルビトール処理されたトマト葉、成熟するトマト果実およびセネッセンシングトマト花の中のDHS遺伝子をコードするmRNAの存在を決定する。
【0052】
アラアビドプシス(Arabidopsis)DHSゲノムクローンから設計したプライマーを使用することによって、鋳型としてセネッセンシングアラアビドプシス(Arabidopsis)葉cDNAライブラリーを使用してポリメラーゼ連鎖反応(PCR)生成物を発生させる。アラアビドプシス(Arabidopsis)ヌクレオチド配列は、セネッセンス誘導トマトDHSの対応する配列と73%のヌクレオチド配列同一性および81%のアミノ酸配列同一性を有する。
【0053】
RT−PCRを使用することによって、本発明のセネッセンス誘導トマトDHS遺伝子を単離した。トマトDHS遺伝子を単離するために使用する上流のプライマーは24ヌクレオチドのプライマーである:5’AG TCT AGA AGG TGC TCG TCC TGA T3’(配列番号3);下流のプライマーは34ヌクレオチドを含有する:5’G ACT GCA GTC GAC ATC GAT(T)153’(配列番号4)。100pmolの下流のプライマーを使用して、標準RT−PCRを使用してcDNAの第1鎖を単離した。次いで、上流のプライマーおよび下流のプライマーの両方を使用するRT−PCRにおいて、鋳型としてとして第1鎖を使用した。
【0054】
アガロースゲル上でRT−PCR生成物を分離すると、1.5kb〜600bpのサイズ範囲の3つの明確なバンドの存在が明らかにされた。それぞれ、上流のプライマーおよび下流のプライマーの中に存在するXbaIおよびSalIクローニング部位を使用して、3つのフラグメントをプラスミドベクター、pBluescript(Stratagene Cloning Systems、カリフォルニア州ラジョラ)の中にサブクローニングし、配列決定した。フラグメントの配列を遺伝子バンクのデータベースの中に存在する配列と比較し、整列させた。結果は1.5kbおよび1kbのフラグメントがトマトDHS配列であることを示した。また、600bpのフラグメントはヒト、酵母およびニューロスポラ(Neurospora)DHS配列と整列した。
【0055】
2Mのソルビトールで6時間処理して脱水を誘導したトマト葉から得られたRNAから作ったトマト(cv.Match F1ハイブリッド)cDNAライブラリーをスクリーニングするために、600bpのRT−PCRフラグメントを使用した。λZap(Stratagene Cloning Systems、カリフォルニア州ラジョラ)cDNAライブラリーキットを使用して、cDNAライブラリーを構築した。セネッセンス誘導DHS遺伝子に対応する3つの同一の陽性全長cDNAクローンが得られ、これらを配列決定した。セネッセンス誘導DHS cDNAクローンのヌクレオチド配列を配列番号1に示す。このcDNAクローンは、42.1kDaの計算した分子量を有する381アミノ酸のポリペプチド(配列番号2)をコードする。
【0056】
発生するおよびストレスに暴露されたトマト花、果実および葉の中の遺伝子の発現パターンに基づいて、それはセネッセンスに関係づけられる。
【0057】
トマトDHS cDNA配列をアラビドプシス・タリアナ(Arabidopsis thaliana)ゲノムバンクの中の未同定ゲノム配列と整列させた(http://genome−www.stanford.edu/Arabidopsis)。結果は未同定アラアビドプシス(Arabidopsis)ゲノム配列(AB107060)との整列を示した。この整列の情報を使用して、アラアビドプシス(Arabidopsis)配列中のオープンリーディングフレームを同定し、それから予測されたアミノ酸配列を発生させた。整列したアラアビドプシス(Arabidopsis)DHS遺伝子の生ずるヌクレオチド配列およびアミノ酸配列を、それぞれ、配列番号5(第2A図)および配列番号6と表示する。
【0058】
同定されたアラアビドプシス(Arabidopsis)DHS配列の短い領域に基づく2つのプライマーを発生させた:プライマー1、5’GGTGGTGTTGAGGAAGATC3’(配列番号7);およびプライマー2、5’GGTGCACGCCCTGATGAAGC3’(配列番号8)。標準PCRにおける2つのプライマーのための鋳型として、アラアビドプシス(Arabidopsis)セネッシング葉cDNAライブラリーを使用した。600bpのPCR生成物を単離し、配列決定し、これらの生成物はゲノムDHS配列の対応するフラグメントのそれと同一の配列を有することが示された。
【0059】
また、全長セネッセンス誘導トマトDHS cDNAを使用して、全長セネッセンス誘導アラアビドプシス(Arabidopsis)およびカーネーションDHS cDNAクローンを単離した。プローブとして全長トマトDHS cDNAクローンを使用して、それぞれ、セネッシングアラアビドプシス(Arabidopsis)葉cDNAライブラリーおよびセネッセンシングカーネーション花弁cDNAライブラリーをスクリーニングすることによって、アラアビドプシス(Arabidopsis)およびカーネーションDHS cDNAクローンを単離した。次いでcDNAライブラリーから得られたcDNAクローンを配列決定した。
【0060】
この方法において単離されたアラアビドプシス(Arabidopsis)全長cDNAクローンのヌクレオチド配列は、トマトcDNA配列との整列によりエンコーディングアラアビドプシス(Arabidopsis)DHSとして同定されたアラアビドプシス(Arabidopsis)ゲノム配列のコーディング領域と同一の配列を有する(第2A図、配列番号5)。全長カーネーション花弁セネッセンス誘導DHSクローンのヌクレオチド配列および誘導アミノ酸配列を第10図に示す(それぞれ、配列番号9および配列番号10)。
【0061】
トマト、カーネーションおよびアラアビドプシス(Arabidopsis)からのDHSをコードする、本発明のcDNA配列を同様な方法においてプローブとして使用して、他の植物からDHS遺伝子を単離し、次いでこれらはトランスジェニック植物における配列を変更するために使用することができる。
【0062】
セネッセンス誘導DHS遺伝子は、DHS遺伝子ファミリーのメンバーであるように思われる。ハイブリッド植物から抽出したゲノムDNAを使用して、トマト葉DNAのゲノムサザンブロット分析を実施した。DHS遺伝子のコーディング領域内の単一部位を認識するか、あるいはDHS遺伝子のオープンリーディングフレーム内のいずれの部位をも認識しない、種々の制限酵素を使用してDNAを切断した。トマトDHSの制限地図を第4図に示す。
【0063】
制限酵素消化したトマト葉ゲノムDNAを32P−dCTP標識化全長トマトDHS cDNAでプロービングした。高いストリンジェンシイの条件下のハイブリダイゼーションは、全長cDNAプローブが各制限酵素消化DNA試料についての3つの制限フラグメントに対してハイブリダイゼーションすることを示した。DHSのオープンリーディングフレーム(第4図)内に制限部位を有する、XbaIおよびEcoRIでトマト葉ゲノムDNAを消化したとき、2より多い制限フラグメントがサザンブロットにおいて検出可能であった(第5図)ことは特に注目すべきである。cv. Match F1、ハイブリッド変種、およびホモ接合性系統、UCT5、からのゲノムDNAは、制限消化の同一パターンを生じた。これらの結果が示唆するように、トマト植物の中に2またはそれ以上のイソ型が存在する。第3図に示すように、DHS遺伝子は種を通じて高度に保存されるので、任意の種内のイソ型の間で有意な量の保存が存在する。
【0064】
全長トマトcDNAでプロービングしたトマト花全RNAのノザンブロットは、セネッセンス誘導DHS遺伝子の発現がトマト花において有意に誘導されるが、発現は芽においてわずかに検出可能であることを示す(第6図)。トマト果実の種々の発生段階の間におけるDHS発現のノザンブロット分析は、DHS遺伝子がブレーカーおよびピンク色果実において低いレベルで発現され、これに対して赤色(成熟)トマト果実におけるDHS発現が有意に増強されることを証明する(第7図)。
【0065】
ノザンブロット分析は、また、セネッセンス誘導DHS遺伝子が環境的ストレス条件、例えば、脱水(第8図)および冷却(第9図)により誘導されることを証明する。脱水を誘導するように2Mのソルビトールで処理したトマト葉は、非処理葉に比較して脱水された葉におけるDHS発現の誘導を証明する(第8図)。冷却温度に暴露し、周囲温度に戻した植物は、冷却損傷症状(例えば、漏出)の発生と一致するセネッセンス誘導DHS遺伝子の発現の誘導を示す(第9図)。トマト植物および種々の植物組織、例えば、葉、果実および花における遺伝子発現の全体のパターンは、本発明のDHS遺伝子がこれらの植物および植物組織におけるセネッセンスの開始に関係することを証明する。
【0066】
DHS遺伝子発現の誘導による同様な結果は、アラアビドプシス(Arabidopsis)における葉セネッセンスおよびカーネーションの花弁セネッセンスの開始とともに観測される。種々の年齢の植物から単離されたアラアビドプシス(Arabidopsis)葉の全RNAのノザンブロット分析は、セネッセンス誘導DHS遺伝子の発現が若い(5週齢の)植物において明らかでないが、約6週において出現し始めることを示す。DHS遺伝子の発現は7週までに有意に誘導される。ノザンブロット分析は、植物が加齢するにつれてアラアビドプシス(Arabidopsis)DHS遺伝子が有意に増強されることを示す(第11図)。
【0067】
ノザンブロット分析は、また、DHS遺伝子が顕花植物、例えば、カーネーションにおいて同様に調節されることを示す(第12図)。種々の年齢のカーネーションの花弁から単離された全RNAのノザンブロット分析において、年齢誘導セネッセンスの症状、例えば、セネッセンスの最初の形態学的発現である、花弁のインローリング(inrolling)を有する花からの花弁において、カーネーションDHSの発現は有意に誘導されるが、堅い芽の花において発現は非常に低いことが示された。ちょうど開花し始めているカーネーションの花からの花弁は、堅い芽の段階における花よりも有意に多いDHS発現を有し、そして完全に開いた花からの花弁もまたDHS発現の増強を示す。
【0068】
こうして、植物組織におけるセネッセンス誘導DHS遺伝子の発現を実質的に表すか、あるいは変更することによって、劣化および腐敗を遅延し、腐敗しやすい果実、花、および野菜の棚持ちを増加することができ、そして植物およびそれらの組織をよりストレス抵抗性としかつ病原体抵抗性とすることができることが期待される。これは次のようにして達成することができる。好ましくは構成的プロモーター、例えば、CaMV 35Sプロモーターを使用するか、あるいは組織特異的またはセネッセンス/ストレス誘導プロモーターを使用して、DHS cDNAまたはそのオリゴヌクレオチドフラグメントが果実、花、葉および野菜においてアンチセンス立体配置において発現されるトランスジェニック植物を産生する。
【0069】
また、植物におけるセネッセンス関係形態学的変化の誘導に関係する、他の遺伝子、elF−5A、が単離され、ここにおいて配列決定され、そしてDHSと同様に、好ましくは、植物の中にアンチセンスの向きで導入することによって、植物におけるセネッセンスおよびセネッセンス関係プロセスを変更するためにそれを使用することができる。成熟するトマト果実、セネッシングアラアビドプシス(Arabidopsis)葉およびセネッシングカーネーション花cDNAライブラリーの各々から、全長セネッセンス誘導elF−5A cDNAを単離した。
【0070】
全長クローンの各々のヌクレオチド配列および誘導アミノ酸配列を第13図(トマトセネッセンス誘導elF−5A)、第14図(カーネーションセネッセンス誘導elF−5A)および第15図(アラアビドプシス(Arabidopsis)セネッセンス誘導elF−5A)に示す。また、これらのcDNAの各々のヌクレオチド配列を配列番号11(トマト)(第13図)、配列番号13(カーネーション)(第14図)および配列番号15(Arabidopsis)(第15図)として示す。遺伝子の各々の誘導アミノ酸配列を、それぞれ、配列番号12(第13図)、配列番号14(第14図)および配列番号16(第15図)として示す。
【0071】
本明細書に記載するDHS遺伝子配列を使用して場合におけるように、本発明のelF−5A配列を使用して他の植物からelF−5A遺伝子を単離することができる。単離されたelF−5A配列を使用して、植物におけるセネッセンスおよびセネッセンス関係プロセスを変更することができる。少なくとも約70%の交差種の配列類似性に基づいて、既知の方法を使用して、植物からのelF−5A配列の単離を達成することができる。
【0072】
elF−5AおよびDHSの平行誘導は植物においてセネッセンスの間に起こる。自然およびストレス誘導セネッセンスの両方の開始において、elF−5AはDHSと平行にアップレギュレートされることをノザンブロット分析は証明する(第16図〜第20図)。例えば、種々の段階におけるアラアビドプシス(Arabidopsis)植物の葉から単離された全RNAのノザンブロット分析において、葉セネッセンスが植物において明らかであるとき、elF−5Aの発現は誘導され、セネッセンスが進行するにつれて、発現は有意に増強されたことが証明される。結実植物、例えばトマトにおいて、elF−5AおよびDHSは果実の軟化および腐敗の開始と一致して赤色柔軟果実においてアップレギュレートされる(第17図)。
【0073】
また、ノザンブロット分析において、elF−5AおよびDHSは、環境的ストレス、例えば、乾燥(第18図)および冷却損傷(第20図)に応答して植物中で平行にアップレギュレートすることが証明される。同様に、顕花植物において、elF−5AおよびDHSは開いた花において平行にアップレギュレートされ、そして両方の遺伝子の発現は顕花の後の段階を通して増強され続ける。
【0074】
クローニングしたセネッセンス誘導DHS遺伝子、それらの1または2以上のフラグメント、あるいはクローニングしたセネッセンス誘導elF−5A遺伝子、それらの1または2以上のフラグメント、あるいはelF−5AとDHS配列との組合わせは、構成的プロモーター、例えば、イチジクイボモザイクウイルス35Sプロモーター、カリフラワーモザイクウイルスプロモーターCaMV35S、二重35SプロモーターまたはMASプロモーターの制御下に逆向き(アンチセンス)に導入されるとき、植物を遺伝的に修飾し、修飾された植物におけるセネッセンスを変更することができる。
【0075】
トマト、アラアビドプシス(Arabidopsis)またはカーネーションのセネッセンス誘導DHS遺伝子またはセネッセンス誘導elF−5A遺伝子と十分な配列同一性を共有する他の植物から選択されたアンチセンス配列を使用して、同様に遺伝的修飾を達成することができる。遺伝的修飾の1つの結果は、mRNAをコードする内因的翻訳可能セネッセンス誘導DHS、mRNAをコードするelF−5Aまたは両方の量の減少である。結局、植物細胞において産生されたセネッセンス誘導DHSおよび/またはセネッセンス誘導elF−5Aの量は減少され、これにより活性化されたelF−5Aの量は減少し、これは引き続いてセネッセンス誘導タンパク質、例えば、セネッセンス誘導リパーゼ、セネッセンス誘導プロテアーゼおよびセネッセンス誘導ヌクレアーゼの翻訳を減少させる。セネッセンスの開始のために新たなタンパク質合成が必要であるので、こうしてセネッセンスは阻害または遅延される。
【0076】
例えば、二重35Sプロモーターの調節下にアラアビドプシス(Arabidopsis)セネッセンス誘導DHS遺伝子(配列番号26)(第38図)の全長または3’領域をアンチセンスの向きに発現するベクターで形質転換されたアラアビドプシス(Arabidopsis)植物は、第21図〜第24図に示すように対照植物に比較して、成長のすべての段階を通じてバイオマスの増加、例えば、より大きい葉サイズおよび全体のより大きい植物成長、および葉セネッセンスの遅延を示す。
【0077】
トランスジェニックアラアビドプシス(Arabidopsis)植物においてアラアビドプシス(Arabidopsis)セネッセンス誘導DHS遺伝子の完全なまたは3’コーディング領域をアンチセンスの向きで発現することによって発生したセネッセンス誘導DHS遺伝子の発現の減少の効果は、また、トランスジェニック植物において種子収量の増加として見られる。アラアビドプシス(Arabidopsis)セネッセンス誘導DHS遺伝子のアンチセンス3’非コーディング領域を発現するアラアビドプシス(Arabidopsis)植物系統は、野生型植物よりも6倍までの種子を生産する(第25図)。
【0078】
アンチセンスの向きでかつ二重35Sプロモーターの調節下にトマトセネッセンス誘導DHS遺伝子(配列番号27)の3’末端で形質転換されたトマト植物を使用して、同様な結果が得られる。アンチセンスの向きで遺伝子の3’末端で形質転換された植物は、対照(非形質転換)植物に比較して葉サイズの増加および植物サイズの増加を示す(第26図および第27図)。
【0079】
全長トマトセネッセンス誘導DHSでアンチセンスの向きに形質転換されたトマト植物は、野生型植物に比較して遅延した軟化および腐敗を示す果実を産生する(第28図〜第35図)。こうして、本発明の方法および配列を使用して、果実の軟化および腐敗を遅延し、ならびに植物バイオマスおよび種子収量を増加し、一般に、植物においてセネッセンスを遅延することができる。
【0080】
本発明の単離されたヌクレオチド配列を使用して、他の植物または生物から実質的に相補的なDHSおよび/またはelF−5Aヌクレオチド配列を単離することができる。引き続いて、ここに示す単離されたヌクレオチド配列の使用とともに示した方法と同一の方法において、これらの配列を使用して、植物を形質転換し、これにより形質転換された植物のセネッセンスを変更することができる。
【0081】
DHS、elF−5A、それらのフラグメントまたはそれらの組合わせで植物を形質転換して得られる遺伝的修飾は、植物におけるセネッセンス誘導DHS、elF−5Aまたは両方のレベルを永久的に変化させることができ、そして自殖または他の生殖スキームにより子孫植物において増殖させることができる。遺伝的に変更された植物を使用して、変更が世代から世代に安定に伝達される植物の新しい変種または系統を産生する。本発明は、広い範囲の異なる植物においてセネッセンスの安定な遺伝的修飾を達成するために使用できる、適当なDNA配列を最初に提供する。
【0082】
セネッセンス誘導DHS遺伝子およびelF−5A遺伝子を同定し、単離するために、一般に、この分野においてよく知られている慣用法を使用して、プラスミドDNAの調製、制限酵素消化、DNAのアガロースゲル電気泳動、タンパク質のポリアクリルアミドゲル電気泳動、PCR、RT−PCR、サザンブロット、ノザンブロット、DNA結合および細菌の形質転換を実施した。例えば、下記の文献を参照のこと:Sambrook J. 他、Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor Press、Cold Spring Harbor、NY、1989。核酸のハイブリダイゼーション技術はSambrook(前掲)に開示されている。
【0083】
本明細書において使用するとき、用語「植物」は全植物、植物部分、植物細胞または植物細胞のグループを意味する。本発明の方法において使用できる植物の型は、例えば、下記のものを包含するが、これらに限定されない:エチレン感受性およびエチレン不感受性植物;結実植物、例えば、アプリコット、リンゴ、オレンジ、バナナ、グレープフルーツ、セイヨウナシ、トマト、イチゴ、アボカド、およびその他;野菜、例えば、ニンジン、エンドウ、レタス、キャベツ、カブ、ジャガイモ、ブロッコリ、アスパラガス、およびその他;花、例えば、カーネーション、バラ、キク、およびその他;耕種学的作物植物および森林種、例えば、トウモロコシ、イネ、大豆、アルファルファおよびその他;および一般に、本発明のDNA分子を吸収しかつ発現する任意の植物。それは種々の倍数レベル、例えば、一倍体、二倍体、四倍体および多倍数体の植物を包含することができる。植物は単子葉植物または双子葉植物であることができる。
【0084】
トランスジェニック植物は、ここにおいてある方法において遺伝的に修飾された植物として定義され、下記の植物を包含するが、これらに限定されない:異種または同種セネッセンス誘導DHS DNAまたは修飾されたDNA、あるいは異種セネッセンス誘導DHS DNAまたは同種セネッセンス誘導DHS DNAのある部分をそのゲノムの中に組込んで有する植物。あるいは、本発明のトランスジェニック植物は、異種または同種セネッセンス誘導elF−5A DNAまたは修飾されたDNA、あるいは異種セネッセンス誘導elF−5A DNAまたは同種セネッセンス誘導elF−5A DNAのある部分をそのゲノムの中に組込んで有することができる。
【0085】
本発明のトランスジェニック植物は、異種または同種セネッセンス誘導DHSおよびelF−5A DNAまたは修飾されたDNA、あるいは異種セネッセンス誘導DHSおよびelF−5A DNAまたは同種セネッセンス誘導DHSおよびelF−5A DNAのある部分、あるいは異種および同種のDHSおよびelF−5A配列の組合わせをそのゲノムの中に組込んで有することができる。変更された遺伝物質は、タンパク質をコードすることができ、調節配列または対照配列を含んでなることができるか、あるいはアンチセンス配列であるかまたはそれを含むか、あるいは内因的セネッセンス誘導DHSまたはelF−5AまたはmRNA配列またはその一部分に対してアンチセンスであるか、あるいはアンチセンスmRNA配列をコードすることができる。「トランスジーン」または「トランスジェニック配列」は、トランスジェニック植物の中に組込まれた外来遺伝子または部分的配列として定義される。
【0086】
用語「ハイブリダイゼーション」は、本明細書において使用するとき、一般に、プローブ配列およびターゲット配列の特質に依存して当業者にとって容易に明らかなように、適当なストリンジェンシイの条件における核酸のハイブリダイゼーションを意味するために使用される。ハイブリダイゼーションおよび洗浄の条件はこの分野においてよく知られており、そしてインキュベーション時間、温度および/または溶液のイオン強度を変化させることによって所望のストリンジェンシイに依存して条件を調節することは容易に達成される。例えば、下記の文献を参照のこと:Sambrook J. 他、Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor Press、Cold Spring Harbor、New York、1989。
【0087】
ハイブリダイゼーションする配列の長さ、特にプローブ配列の長さ、核酸の相対的G−C含量および許容すべき不一致の量により、条件を選択する。より低い程度の相補性を有する鎖間の部分的ハイブリダイゼーションを望むとき、低いストリンジェンシイの条件が好ましい。完全なまたはほぼ完全な相補性を望むとき、高いストリンジェンシイの条件が好ましい。典型的な高いストリンジェンシイの条件について、ハイブリダイゼーション溶液は6×SSC、0.01MのEDTA、1×デンハルト溶液および0.5%のSDSを含有する。ハイブリダイゼーションは約68℃においてクローニングされたDNAについて約3〜4時間実施し、そして全真核DNAについて約12〜約16時間実施する。より低いストリンジェンシイのために、ハイブリダイゼーション温度を二本鎖の溶融温度(TM)より約42℃低く低下させる。TMはG−C含量および二本鎖長さならびに溶液のイオン強度の関数であることが知られている。
【0088】
本明細書において使用するとき、用語「実質的配列同一性」または「実質的相同性」は、ヌクレオチド配列またはアミノ酸配列が他のヌクレオチド配列またはアミノ酸配列と実質的な構造的または機能的同等性を表すことを示すために使用される。実質的配列同一性または実質的相同性を有する配列間の構造的または機能的差は最小であろう;すなわち、それらは所望の用途において示すように配列が機能する能力に影響を与えないであろう。差は、例えば、異なる種間のコドン使用頻度における固有変動のためであることがある。
【0089】
2またはそれ以上の異なる配列間に有意な量の配列のオーバーラップまたは類似性が存在する場合、あるいは配列の長さまたは構造が異なる場合でさえ異なる配列が同様な物理的特性を示す場合、構造的差は最小であると考えられる。このような特性は、例えば、定められた条件下にハイブリダイゼーションする能力、またはタンパク質の場合において、免疫学的交差反応性、同様な酵素活性、およびその他を包含する。熟練した実施者は、既知の方法により、これらの特性の各々を容易に決定することができる。
【0090】
さらに、2つのヌクレオチド配列がそれらの間で少なくとも約70%、より好ましくは80%、最も好ましくは約90%の配列類似性を有する場合、それらの配列は「実質的に相補性」である。2つのアミノ酸配列がポリペプチドの活性部分間の少なくとも50%、好ましくは70%の類似性を有する場合、それらは実質的に相同性である。
【0091】
本明細書において使用するとき、DNAまたはRNA分子の「対応する部分に対してハイブリダイゼーションする」という句は、ハイブリダイゼーションする分子、例えば、オリゴヌクレオチド、ポリヌクレオチド、または任意のヌクレオチド配列が(センスまたはアンチセンスの向きで)ほぼ同一サイズの他の核酸分子中の配列を認識し、それに対してハイブリダイゼーションし、そして適当な条件下にハイブリダイゼーションを実行するために十分な類似性をその配列に対して有することを意味する。
【0092】
例えば、トマトDHSの3’コーディングまたは非コーディング領域からの100ヌクレオチド長さのアンチセンス分子は、2つの配列間に約70%またはそれより高い配列類似性が存在するかぎり、カーネーションDHS遺伝子または任意の他の植物DHS遺伝子のそれぞれ3’コーディングまたは非コーディング領域内のヌクレオチド配列のほぼ100ヌクレオチド部分を認識し、それに対してハイブリダイゼーションする。「対応する部分」がそれに対してハイブリダイゼーションする分子よりも小さいか、あるいは大きくあるように、例えば、20〜30%小さいか、あるいは大きく、好ましくは約12〜15%小さいか、あるいは大きくあるように、「対応する部分」のサイズがハイブリダイゼーションにおいて多少の不一致を可能とすることを理解すべきである。
【0093】
核酸(またはポリヌクレオチドまたはオリゴヌクレオチド)の「機能的誘導体」という用語は、ここにおいて、セネッセンス誘導DHSまたはセネッセンス誘導elF−5Aをコードする遺伝子またはヌクレオチド配列のフラグメント、変異型、相同体またはアナローグを意味するために使用する。機能的誘導体は、本発明による実用性を可能とするセネッセンス誘導DHSまたはelF−5AをコードするDNAの少なくとも一部分を保持することができる。
【0094】
このような機能は、低いストリンジェンシイの条件下に、天然のトマト、アラアビドプシス(Arabidopsis)またはカーネーションのセネッセンス誘導DHSまたはelF−5A、あるいはセネッセンス誘導DHSまたはelF−5Aをコードする他の植物からの実質的に相同性のDNAと、あるいはそれらのmRNA転写物と、または、アンチセンスの向きで、ハイブリダイゼーションして、植物セネッセンス誘導DHSまたはelF−5A mRNAの転写および/または翻訳、およびその他を阻害する能力を包含することができる。
【0095】
遺伝子またはDNA配列の「フラグメント」は、分子のサブセット、例えば、より短いポリヌクレオチドまたはオリゴヌクレオチドを意味する。「変異型」は、全遺伝子またはそのフラグメントに実質的に類似する分子、例えば、1または2以上の置換したヌクレオチドを有するが、特定の遺伝子とハイブリダイゼーションする能力または自然DNAとハイブリダイゼーションするmRNA転写物をコードする能力を維持するヌクレオチド置換変異型を意味する。「相同体」は、異なる植物の属または種からのフラグメントまたは変異型配列を意味する。「アナローグ」は、全分子、その変異型またはフラグメントに実質的に類似するか、あるいは機能的にそれらに関係する、非天然分子を意味する。
【0096】
遺伝子、例えば、セネッセンス誘導DHS遺伝子またはセネッセンス誘導elF−5A遺伝子の「変更された発現」または「修飾された発現」とは、遺伝子の正常の発現、例えば、非修飾結実植物、顕花植物または他の植物における起こる発現がある方法において変化される、任意のプロセスまたは結果を意味する。本明細書において意図するとき、遺伝子発現の変更は、セネッセンス誘導DHS遺伝子またはセネッセンス誘導elF−5A遺伝子または両方の発現の完全なまたは部分的減少であるが、また、発現のタイミングの変化、あるいはセネッセンス誘導DHS遺伝子またはセネッセンス誘導elF−5A遺伝子または両方の発現が非修飾植物または栽培品種において最も自然に起こりやすい発現と異なる他の状態を包含することができる。好ましい変更は、非修飾植物における産生に比較して植物によるセネッセンス誘導DHS産生またはセネッセンス誘導elF−5A産生または両方を減少させる変更である。
【0097】
本発明に従い遺伝的に変更された植物を生産するとき、所望の特性、一般にセネッセンス誘導DHSの発現または産生の減少またはセネッセンス誘導elF−5Aの発現の減少または両方により個々の小植物または植物を選択することが好ましい。セネッセンス誘導DHSおよびセネッセンス誘導elF−5Aの発現は、例えば、トランスジェニック植物におけるセネッセンスの遅延または減少の観測により決定することができる。また、既知の方法を使用して対照(正常、非トランスジェニック)植物に比較してトランスジェニック植物におけるDHSおよび/またはelF−5Aの活性を定量することが可能である。
【0098】
新しく挿入された遺伝子またはDNA配列を発現させ、それがコードするタンパク質を産生するか、あるいは転写すべきアンチセンスDNAの場合において、アンチセンスRNA分子を産生するために、適切な調節因子を遺伝子またはDNA配列に関して適切な位置にかつ向きで存在させるべきである。調節領域はプロモーター、5’非翻訳リーダー配列および3’ポリアデニル化配列ならびにエンハンサーおよび他の調節配列を包含することができる。
【0099】
遺伝子のセンスまたはアンチセンス転写物を発生させるためにセネッセンス誘導DHS遺伝子と組み合わせて有効であるプロモーター調節因子は、一般に任意のプロモーター、特に構成的プロモーター、例えば、イチジクイボモザイクウイルス35Sプロモーター、カリフラワーモザイクウイルスプロモーター、CaMV35Sプロモーター、またはMASプロモーター、または組織特異的またはセネッセンス誘導プロモーター、例えば、カーネーション花弁GST1プロモーターまたはアラアビドプシス(Arabidopsis)SAG12プロモーターを包含する。
【0100】
(例えば、下記の文献を参照のこと:J.C.Palaqui他、Plant Physiol. 112:1447−1456(1996);Morton他、Molecular Breeding、1:123−132(1995);Fobert他、Plant Journal、6:567−577(1994);およびGan他、Plant Physiol.、113:313(1997)、引用することによって本明細書の一部とされる)。好ましくは、本発明において使用するプロモーターは構成的プロモーター、最も好ましくは二重35Sプロモーターを使用する。
【0101】
本発明のために有効であるプロモーターからの発現レベルは、慣用の発現系を使用して、例えば、プロモーター/リポーター遺伝子が導入されたトランスジェニック植物の葉、花、果実または他の組織の抽出物中のリポーター遺伝子産物、例えば、タンパク質またはmRNAのレベルを測定することによって試験することができる。
【0102】
本発明は、トマトセネッセンス誘導DHS、カーネーションセネッセンス誘導DHS、アラアビドプシス(Arabidopsis)セネッセンス誘導DHSをコードする遺伝子に対して相補的であるか、あるいは低い〜高いストリンジェンシイの条件下にトマト、カーネーションまたはアラアビドプシス(Arabidopsis)のセネッセンス誘導DHS遺伝子とハイブリダイゼーションする、他の植物からの遺伝子または遺伝子フラグメントに対して相補的である、アンチセンスオリゴヌクレオチドおよびポリヌクレオチドを提供する。
【0103】
本発明は、また、トマトセネッセンス誘導elF−5A、カーネーションセネッセンス誘導elF−5A、アラアビドプシス(Arabidopsis)セネッセンス誘導elF−5Aをコードする遺伝子に対して相補的であるか、あるいは低い〜高いストリンジェンシイの条件下にトマト、カーネーションまたはアラアビドプシス(Arabidopsis)のセネッセンス誘導elF−5A遺伝子とハイブリダイゼーションする、他の植物からの遺伝子または遺伝子フラグメントに対して相補的である、アンチセンスオリゴヌクレオチドおよびポリヌクレオチドを提供する。
【0104】
このようなアンチセンスオリゴヌクレオチドは少なくとも約6ヌクレオチド長さであってハイブリダイゼーションの最小の特異性を提供し、そしてセネッセンス誘導遺伝子またはその一部分をコードするDNAまたはmRNAの1つの鎖に対して、あるいはセネッセンス誘導DHSまたはelF−5A遺伝子の発現の調節に関係するゲノムDNA中のフランキング配列に対して相補的であることができる。アンチセンスオリゴヌクレオチドは100ヌクレオチドまたはそれより大きい程度に大きく、そしてそれに対してアンチセンスである完全なコーディング配列までかつそれを越える長さに伸長することができる。アンチセンスオリゴヌクレオチドはDNAまたはRNAまたはそれらのキメラ混合物または誘導体または修飾されたバージョン、一本鎖または二本鎖であることができる。
【0105】
アンチセンスオリゴヌクレオチドの作用は、細胞におけるセネッセンス誘導DHSの発現、セネッセンス誘導elF−5Aの発現または両方の変更、主として阻害を生ずることがある。アンチセンスの一般的開示については、下記の文献を参照のこと:Albert他、Molecular Biology of the Cell、第2版、Garland Publishing,Inc.、New York、New York、1989(特にpp.195−196、引用することによって本明細書の一部とされる)。
【0106】
アンチセンスオリゴヌクレオチドは、セネッセンス誘導DHSまたはelF−5A遺伝子の任意の対応する部分に対して相補的であることができる。他の態様において、アンチセンスオリゴヌクレオチドは6〜100ヌクレオチド長さであり、例えば、セネッセンス誘導DHSまたはelF−5A配列の3’末端内の5’非コーディングまたは配列に対して相補的であることができる。主として5’非コーディング配列に対して相補的であるアンチセンスオリゴヌクレオチドは、転写因子をコードする遺伝子の発現の有効なインヒビターであることが知られている。Branch、M.A.、Molec.Cell.Biol.,13:4284−4290(1993)。
【0107】
好ましいアンチセンスオリゴヌクレオチドはセネッセンス誘導DHSまたはセネッセンス誘導elF−5AをコードするmRNAの一部分に対して実質的に相補的であり、mRNAの一部分はアンチセンスオリゴヌクレオチドとほぼ同一のサイズである。例えば、セネッセンス誘導DHSまたはelF−5Aをコードする全長cDNAクローンを植物の中にアンチセンスの向きで導入すると、セネッセンス誘導DHSおよび/またはelF−5A遺伝子の発現を首尾よく変更することが期待される。そのうえ、第21図〜第35図において証明されるように、セネッセンス誘導DHS遺伝子またはセネッセンス誘導elF−5A遺伝子または両方の特定の部分をターゲットとした、部分的配列の導入は等しく有効であることがある。
【0108】
本発明が必要とする相同性の最小量は、特定のターゲットRNAまたはDNAを認識しかつその翻訳または機能を阻害または減少するために十分な相補性を生ずると同時に他のRNAまたはDNA分子の機能および他の遺伝子の発現に影響を与えないために十分である量である。本発明のアンチセンスオリゴヌクレオチドはセネッセンス誘導DHS遺伝子またはセネッセンス誘導elF−5A遺伝子のRNA転写物の対応する部分に対して相補的である配列を含んでなるが、絶対的相補性は、好ましいが、不必要である。
【0109】
ハイブリダイゼーションする能力は、アンチセンスオリゴヌクレオチドの長さおよび相補性の程度に依存することがある。一般に、ハイブリダイゼーションする核酸が長いほど、それが含有し、なお安定な二本鎖を形成することができる、セネッセンス誘導DHSターゲット配列に対してより多くの塩基が一致しなくなる。当業者は標準手順を使用して、例えば、ハイブリッドの複合体の溶融温度を測定することによって、許容可能な不一致の程度を確認することができる。
【0110】
外因的に導入された核酸配列からの転写により、アンチセンスRNAオリゴヌクレオチドを細胞内で発生させることができる。適当な調節因子、例えば、プロモーターに作用可能に連鎖されたアンチセンスセネッセンス誘導DHS配列をコードするDNAが組込まれたベクター、例えば、プラスミドまたはウイルスを使用する形質転換、トランスフェクションまたは感染により、アンチセンス分子を細胞の中に送達することができる。細胞内で外因的DNA配列が発現され、セネッセンス誘導DHS遺伝子のアンチセンスRNAが産生される。
【0111】
ベクターは好ましくはプラスミドであることができるか、あるいは植物細胞または細菌細胞中で複製し、コードされる遺伝子を発現することがこの分野において知られているウイルスまたは他のベクターであることができる。ベクターは所望のアンチセンスセネッセンス誘導DHS RNAを産生するように転写されることができるように、染色体的に統合されるようになる。このようなプラスミドまたはウイルスベクターは、この分野において標準的である組換えDNA技術的方法により構築することができる。例えば、ベクターは原核宿主中で機能的である複製系および本発明によるアンチセンスオリゴヌクレオチドまたはポリヌクレオチドを含有するプラスミドベクターであることができる。
【0112】
あるいは、ベクターはアグロバクテリウム(Agrobacterium)において機能的である複製系および本発明によるアンチセンスオリゴヌクレオチドまたはポリヌクレオチドを含有するプラスミドであることができる。アグロバクテリウム(Agrobacterium)中で複製することができるプラスミドはこの分野においてよく知られている。下記の文献を参照のこと:Miki他、Procedures of Introducing Foreign DNA Into Plants、Methods in Plant Molecular Biology and Biotechnology、B. .GlickおよびJ.E.Thompson編、CRC Press(1993)、pp. 67−83。
【0113】
トマトDHS遺伝子を下記の方法においてプラスミドベクターの中にアンチセンスの向きでクローニングした。pUC18バックボーンから構築され、カリフラワーモザイクウイルス(CaMV)および引き続いて多重クローニング部位およびオクタパインシンターゼ終結配列を含有するpCDプラスミドを使用して、トマトDHS遺伝子をクローニングした。XhoIおよびSacI制限部位を使用してpCDプラスミドの中に全長トマトDHS遺伝子をサブクローニングすることによって、pCd−DHS(アンチセンス)プラスミドを構築した。
【0114】
好ましくは約6〜約100ヌクレオチド長さでありかつセネッセンス誘導DHSまたはセネッセンス誘導elF−5Aのターゲット配列に対して相補的である、オリゴヌクレオチドは組換えヌクレオチド技術により調製することができるか、あるいは、例えば、モノヌクレオチドまたはより短いオリゴヌクレオチドから合成することができる。自動化合成装置を本発明のオリゴヌクレオチドおよびポリヌクレオチドの化学的合成に適用可能である。
【0115】
本発明に従い組換えヌクレオチド分子を構築する手順は下記の文献に開示されている:Sambrook他、Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor Press、Cold Spring Harbor、N.Y.(1989)、これは引用することによって本明細書の一部とされる。セネッセンス誘導デオキシハイプシンシンターゼ配列に対して相補的であるアンチセンスRNAをコードするオリゴヌクレオチドは、この分野においてよく知られている手順を使用して調製することができる。このような手順は下記の文献に詳述されている:Maniatis、T.他、Molecular mechanisms in the Control of Gene expression、Nierilich他、編、Academic Press、N.Y.(1976)。
【0116】
本発明の別の態様において、内因的植物セネッセンス誘導DHS、セネッセンス誘導elF−5Aまたは両方の発現の阻害は、植物細胞の中に導入された外因的セネッセンス誘導DHSまたはelF−5A遺伝子または遺伝子フラグメントまたは両方の過剰発現による共抑制の結果である。本発明のこの態様において、アンチセンス分子について本明細書において記載した方法と同一の方法により、センスの向きでセネッセンス誘導DHS、セネッセンス誘導elF−5Aまたは両方をコードするベクターを細胞の中に導入する。好ましくは、セネッセンス誘導DHSまたはセネッセンス誘導elF−5Aを強い構成的プロモーター、例えば、イチジクイボモザイクウイルスプロモーターまたはCaMV35Sまたは二重35Sプロモーターに作用可能に連鎖させる。
【0117】
本発明の他の態様において、内因的植物セネッセンス誘導DHS、セネッセンス誘導elF−5Aまたは両方の発現の阻害はリボザイムを使用して実施される。リボザイムは配列特異的エンドリボヌクレアーゼ活性を示すRNA分子である。1例はハンマーヘッドリボザイムであり、これはターゲットRNA中のUH(ここでHはA、CまたはU残基である)認識部位において切断し、かつリボザイムの触媒ドメインを基質RNAのターゲット部位に向ける塩基対合領域を含有する。リボザイムは高度にターゲット特異的であり、そして多重遺伝子ファミリーの1メンバーを不活性化するように設計するか、あるいは関係するmRNAの保存された領域にターゲットすることができる(下記の文献を参照のこと:Merlo他、The Plant Cell、10:1603−1621、1998)。
【0118】
リボザイム分子は、適当な調節因子、例えば、プラスミドに作用可能に連鎖されたリボザイムが組込まれたベクター、例えば、プラスミドまたはウイルスを使用する形質転換、トランスフェクションまたは感染により、細胞に送達すことができる。このようなリボザイム構築物は塩基対合アームを含有し、これらのアームはリボザイム構築物をセネッセンス誘導DHS mRNA、またはセネッセンス誘導elF−5A mRNA内の切断部位に向け、DHSまたはelF−5A mRNAの切断およびセネッセンス誘導DHSおよび/またはelF−5A発現の阻害を生ずる。
【0119】
本発明に従い作られるトランスジェニック植物は、この分野において知られている植物形質転換の任意の方法を使用するDNA形質転換により調製することができる。植物形質転換法は下記の方法を包含する:植物、組織または細胞とアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)との同時培養または直接的感染(Miki他、Meth.in Plant Mol.Biol.and Biotechnology、(1993)、p.67−88);プロトプラストの中への直接的遺伝子転移またはプロトプラスト吸収(Paszkowsk他、EMBO J.、12:2717(1984);エレクトロポレーション(Fromm他、Nature、319:719(1986);粒子衝撃(Klein他、Bio Technology、6:559−563(1988);実生および植物の分裂組織の中への注入(De LaPena他、Nature、325:274−276(1987);培養した細胞および組織のプロトプラストの中への注入(Reich他、Bio Technology、4:1001−1004(1986))。
【0120】
一般に、完全な植物は形質転換プロセスから得られる。プロトプラスト、カルス、組織の部分または外植体、およびその他から植物を再生する。セネッセンス誘導DHS、セネッセンス誘導elF−5Aまたは両方の発現が変更された再生した植物から得られた植物部分、例えば、葉、花、果実、種子およびその他は、本明細書において使用する「植物」の定義の中に含まれる。再生植物の子孫、変異型および突然変異もまた「植物」の定義の中に含まれる。
【0121】
トマト、カーネーションまたはアラアビドプシス(Arabidopsis)セネッセンス誘導DHSタンパク質またはそれらの機能的誘導体、およびトマト、カーネーションまたはアラアビドプシス(Arabidopsis)セネッセンス誘導elF−5Aタンパク質またはそれらの機能的誘導体は、必要に応じて化学的合成法と組み合わせて、組換え技術により産生されることが好ましい。本発明の1つの態様において、セネッセンス誘導DHSは融合タンパク質として発現され、この融合タンパク質はマルトース結合性植物と融合されたセネッセンス誘導DHSから成ることが好ましい。
【0122】
本明細書において記載するセネッセンス誘導DHSまたはセネッセンス誘導elF−5Aタンパク質の「機能的誘導体」は、それぞれ、セネッセンス誘導DHSまたはセネッセンス誘導elF−5Aのフラグメント、変異型、アナローグ、または化学的誘導体であり、これらはセネッセンス誘導DHSまたはelF−5A活性またはそれぞれセネッセンス誘導DHSまたはセネッセンス誘導elF−5Aに対して特異的な抗体との交差反応性の少なくとも一部分を保持する。セネッセンス誘導DHSまたはセネッセンス誘導elF−5Aタンパク質のフラグメントは、この分子の任意のサブセットを意味する。変異型ペプチドは直接的化学的合成により、例えば、この分野においてよく知られている方法を使用して作ることができる。
【0123】
セネッセンス誘導DHSまたはセネッセンス誘導elF−5Aのアナローグは、全タンパク質またはそのフラグメントに実質的に類似する非天然タンパク質を意味する。セネッセンス誘導DHSまたはセネッセンス誘導elF−5Aの化学的誘導体は、通常ペプチドまたはペプチドフラグメントの一部分でない追加の化学的部分を含有する。選択した側鎖または末端の残基と反応することができる有機誘導体化因子とペプチドのターゲッテッドアミノ酸残基を反応させることによって、修飾をペプチドまたはそのフラグメントの中に導入することができる。
【0124】
本発明のヌクレオチド配列で形質転換された細胞(センス向きで)を培養し、細胞がタンパク質を合成するようにさせ、次いで、使用するクローニングプロトコルに依存して、遊離タンパク質または融合タンパク質として、培地または細胞抽出物からタンパク質を単離することによって、本発明によるセネッセンス誘導DHSまたはセネッセンス誘導elF−5Aタンパク質またはペプチドを産生することができる。あるいは、タンパク質は無細胞系において産生することができる。Ranu他、Meth. Enzymol.、60:459−484、(1979)。
【0125】
本発明を一般に記載したが、下記の実施例を参照することによって本発明はいっそう容易に理解されるであろう。これらの実施例は例示であり、本発明を限定することを意図しない。
【実施例】
【0126】
実施例1.メッセンジャーRNA(mRNA)の単離
種々の発生段階におけるトマトの花およびトマトの果実から、そして葉(未処理または冷却またはソルビトール処理後の)から全RNAを単離した。簡単に述べると、組織(5g)を液体窒素中で粉砕した。粉砕した粉末を30mlのグアニジニウム緩衝液(4Mのグアニジニウムイソチオシアネート、2.5mMのNaOAc pH8.5、0.8%のβ−メルカプトエタノール)と混合した。この混合物を4層のチーズクロスを通して濾過し、10,000×g、4℃において30分間遠心した。次いで上清を塩化セシウム密度勾配で26,000×gにおいて20時間遠心した。
【0127】
ペレット化したRNAを75%のエタノールですすぎ、600μlのDEPC処理水の中に再懸濁させ、−70℃において0.75mlの95%のエタノールおよび30μlの3MのNaOAcでRNAを沈降させた。10μgのRNAを1.2%の変性ホルムアルデヒドアガロースゲル上で分画し、ナイロン膜に移した。ランダムにプライムした32P−dCTP標識化全長DHS cDNA(配列番号1)を使用して、膜を42℃において一夜プロービングした。次いで0.1%のSDSを含有する1×SSCで室温において膜を15分間1回洗浄し、0.1%のSDSを含有する0.2×SSCで65℃において各15分間3回洗浄した。膜を−70℃においてX線フィルムに一夜露出した。
【0128】
プロメガ(Promega)から入手可能なポリA+トラクトmRNA単離システムを使用して、tRNAからポリA+mRNAを単離した。ストラタジーン(Stratagene)(カリフォルニア州ラジョラ)から入手可能なZAP ExpressR cDNA合成システムを使用するcDNA合成の鋳型として、ポリA+mRNAを使用した。
【0129】
トマト葉cDNAライブラリーのスクリーニング
2Mのソルビトールに6時間暴露したMatch F1ハイブリッドトマト葉から単離したmRNAを使用して作ったcDNAライブラリーを、ほぼ5×106PFU/mlに希釈した。32P標識化600bpのRT−PCRフラグメントを使用して、cDNAライブラリーをスクリーニングした。3つの陽性cDNAクローンを切除し、製造業者のインストラクション中の方法を使用してpBK−CMVR(Stratagene)ファージミドの中に再循環させた。全長cDNAをpBK−CMVベクターの中に挿入した。
【0130】
プラスミドDNAの単離、DNAのスクリーニング
Sambrook他(前掲)が記載するアルカリ溶解法を使用してプラスミドDNAを単離した。デオキシ配列決定法を使用して、全長の陽性cDNAクローンを配列決定した。Sanger他、Proc. Natl. Acad. Sci. USA、74:5463−5467。オープンリーディングフレームを収集し、BLAST検索(GenBank、マリイランド州ベセスダ)を使用して分析し、下記の方法を使用して5つの最も相同性のタンパク質とコード化遺伝子の誘導アミノ酸配列との整列を達成した:BCM Search Launcher:Multiple Sequence Alignments Pattern−Induced Multiple Alignments Methods(参照:F.Corpet、Nucl.Acids Res.、16:10881−10890(1987))。誘導アミノ酸配列の中に存在する機能的モチーフをMultiFinderにより同定した。
【0131】
トマトRNAのノザンブロットハイブリダイゼーション
種々の段階におけるトマトの花(広く開いたまたは乾燥している芽および花およびセネッシング花弁)、トマトの葉、およびトマトの果実種々の成熟段階における(ブレーカー、すなわち、10%より少ない赤色の緑色果実、ピンク色、すなわち、全果実はオレンジ色またはピンク色である、柔らかいか、あるいは堅い)から単離された全RNAの10μgを1%の変性ホルムアルデヒドアガロースゲル上で分離し、ナイロン膜上に固定化した。
【0132】
ランダムプライマーキット(Boehringer Mannheim)を使用して32P−dCTPで標識化した全長トマトcDNAを使用して、フィルター(7×107cpm)をプロービングした。フィルターを1×SSC、0.1%SDSで室温において1回洗浄し、0.2×SSC、0.1%SDSで65℃において3回洗浄した。フィルターを乾燥し、−70℃においてX線フィルムに一夜露出した。結果を第6図、第7図、第8図および第9図に示す。
【0133】
アラアビドプシス(Arabidopsis)RNAのノザンブロットハイブリダイゼーション
5週齢(レーン1)、6週齢(レーン2)および7週齢(レーン3)のアラアビドプシス(Arabidopsis)植物の葉からの全RNAを前述したように単離し、1%の変性ホルムアルデヒドアガロースゲル上で分離し、ナイロン膜上に固定化した。ランダムプライマーキット(Boehringer Mannheim)を使用して32P−dCTPで標識化した全長アラアビドプシス(Arabidopsis)セネッセンス誘導DHS cDNAを使用して、フィルター(7×107cpm)をプロービングした。フィルターを1×SSC、0.1%SDSで室温において1回洗浄し、0.2×SSC、0.1%SDSで65℃において3回洗浄した。フィルターを乾燥し、−70℃においてX線フィルムに一夜露出した。結果を第11図に示す。
【0134】
カーネーションRNAのノザンブロットハイブリダイゼーション
種々の花発生段階、すなわち、堅い芽の花(レーン1)、開花し始めた花(レーン2)、完全に開いた花(レーン3)、インローリング(inrolling)花弁(レーン4)のカーネーション植物の花弁からの全RNAを前述したように単離し、1%の変性ホルムアルデヒドアガロースゲル上で分離し、ナイロン膜上に固定化した。ランダムプライマーキット(Boehringer Mannheim)を使用して32P−dCTPで標識化した全長カーネーションセネッセンス誘導DHS cDNAを使用して、フィルター(7×107cpm)をプロービングした。フィルターを1×SSC、0.1%SDSで室温において1回洗浄し、0.2×SSC、0.1%SDSで65℃において3回洗浄した。フィルターを乾燥し、−70℃においてX線フィルムに一夜露出した。結果を第12図に示す。
【0135】
実施例2.トマトセネッセンス誘導DHS遺伝子のソルビトール誘導
トマトの葉を密閉したチャンバー内において2Mのソルビトールで6時間処理した。次のようにしてRNAをソルビトール処理した葉から抽出した。
【0136】
葉(5g)を液体窒素中で粉砕した。粉砕した粉末を30mlのグアニジニウム緩衝液(4Mのグアニジニウムイソチオシアネート、2.5mMのNaOAc pH8.5、0.8%のβ−メルカプトエタノール)と混合した。この混合物を4層のチーズクロスを通して濾過し、10,000×g、4℃において30分間遠心した。次いで上清を塩化セシウム密度勾配で26,000×gにおいて20時間遠心した。
【0137】
ペレット化したRNAを75%のエタノールですすぎ、600μlのDEPC処理水の中に再懸濁させ、−70℃において0.75mlの95%のエタノールおよび30μlの3MのNaOAcでRNAを沈降させた。10μgのRNAを1.2%の変性ホルムアルデヒドアガロースゲル上で分画し、ナイロン膜に移した。ランダムにプライムした32P−dCTP標識化全長DHS cDNA(配列番号1)を使用して、膜を42℃において一夜プロービングした。次いで0.1%のSDSを含有する1×SSCで室温において膜を15分間1回洗浄し、0.1%のSDSを含有する0.2×SSCで65℃において各15分間3回洗浄した。膜を−70℃においてX線フィルムに一夜露出した。
結果を第8図に示す。見ることができるように、DHSの転写は葉においてソルビトールにより誘導される。
【0138】
実施例3.セネッシング花中のトマトDHS遺伝子の誘導
トマト植物の堅い花の芽および開いた、セネッシング花を収穫し、実施例2におけるようにRNAを単離した。10μgのRNAを1.2%の変性ホルムアルデヒドアガロースゲル上で分画し、ナイロン膜に移した。ランダムにプライムした32P−dCTP標識化全長DHS cDNA(配列番号1)を使用して、膜を42℃において一夜プロービングした。次いで0.1%のSDSを含有する1×SSCで室温において膜を15分間1回洗浄し、0.1%のSDSを含有する0.2×SSCで65℃において各15分間3回洗浄した。膜を−70℃においてX線フィルムに一夜露出した。
【0139】
結果を第6図に示す。見ることができるように、DHSの転写はセネッシング花において誘導される。
【0140】
実施例4.成熟する果実におけるトマトDHS遺伝子の誘導
実施例2におけるように、ブレーカー、ピンク色および成熟した果実からRNAを単離した。10μgのRNAを1.2%の変性ホルムアルデヒドアガロースゲル上で分画し、ナイロン膜に移した。ランダムにプライムした32P−dCTP標識化全長DHS cDNA(配列番号1)を使用して、膜を42℃において一夜プロービングした。次いで0.1%のSDSを含有する1×SSCで室温において膜を15分間1回洗浄し、0.1%のSDSを含有する0.2×SSCで65℃において各15分間3回洗浄した。膜を−70℃においてX線フィルムに一夜露出した。
【0141】
結果を第7図に示す。見ることができるように、DHSの転写はセネッセンスの開始直前の成熟した、赤色の果実において最強であり、腐敗に導く。
【0142】
実施例5.冷却によるトマトセネッセンス誘導DHS遺伝子の誘導
鉢中のトマト植物(7〜8週齢)を6℃に成長チャンバー内で2日間、3日間または6日間暴露した。光サイクルを8時間の暗闇および16時間の光に設定した。植物を温室に戻すことによって、再加温した。再加温しない植物を成長チャンバーから取出した直後に収穫した。次のようにしてRNAを葉から抽出した。
【0143】
葉(5g)を液体窒素中で粉砕した。粉砕した粉末を30mlのグアニジニウム緩衝液(4Mのグアニジニウムイソチオシアネート、2.5mMのNaOAc pH8.5、0.8%のβ−メルカプトエタノール)と混合した。この混合物を4層のチーズクロスを通して濾過し、10,000×g、4℃において30分間遠心した。次いで上清を塩化セシウム密度勾配で26,000×gにおいて20時間遠心した。ペレット化したRNAを75%のエタノールですすぎ、600μlのDEPC処理水の中に再懸濁させ、−70℃において0.75mlの95%のエタノールおよび30μlの3MのNaOAcでRNAを沈降させた。
【0144】
10μgのRNAを1.2%の変性ホルムアルデヒドアガロースゲル上で分画し、ナイロン膜に移した。ランダムにプライムした32P−dCTP標識化全長DHS cDNA(配列番号1)を使用して、膜を42℃において一夜プロービングした。次いで0.1%のSDSを含有する1×SSCで室温において膜を15分間1回洗浄し、0.1%のSDSを含有する0.2×SSCで65℃において各15分間3回洗浄した。膜を−70℃においてX線フィルムに一夜露出した。
【0145】
結果を第9図に示す。見ることができるように、冷却温度に暴露し、引き続いて再加温することによって、DHSの転写は葉において誘導され、そして増強された転写は膜漏出として測定した冷却損傷と相関する。
【0146】
実施例6.未同定アラアビドプシス(Arabidopsis)ゲノム配列に基づくプライマーを使用するアラアビドプシス(Arabidopsis)PCR生成物の発生
アラアビドプシス(Arabidopsis)ゲノム配列から設計した1対のオリゴヌクレオチドプライマーを使用するPCRにより、アラアビドプシス(Arabidopsis)cDNA鋳型から部分的長さのセネッセンス誘導DHS配列を発生させた。5’プライマーは配列、5’−GGTGGTGTTGAGGAAGATC(配列番号7)を有する19マーである;3’プライマーは配列、GGTGCACGCCCTGATGAAGC−3’(配列番号8)を有する20マーである。Expand High Fidelity PCR System(Boehringer Mannheim)および鋳型としてアラアビドプシス(Arabidopsis)セネッシング葉cDNAライブラリーを使用するポリメラーゼ連鎖反応を次のようにして実施した。
【0147】
反応成分:
cENA 1μl(5×107pfu)
dNTP(各々10mM) 1μl
MgCl2(5mM)+10×緩衝液 1μl
プライマー1および2(各々100μM) 2μl
Expand High Fidelity DNAプライマー 1.75U
反応体積 50μl
【0148】
反応パラメーター:
94℃、3分間
94℃/1分、58℃/1分、72℃/2分、45サイクル
72℃、15分間。
【0149】
実施例7.ゲノムDNAの単離およびサザン分析
10gのトマト葉組織を液体窒素中で微細粉末に粉砕することによって、ゲノムDNAをトマト葉から抽出した。25mlの均質化緩衝液を含有する37.5mlの混合物[100mMのTris−HCl、pH8.0、100mMのEDTA、250mMのNaCl、1%のサルコシル、1%の2−メルカプトエタノール、10μg/mlのRNアーゼおよび12.5mlのフェノール]を60℃に前もって加温し、粉砕した組織に添加した。
【0150】
この混合物を15分間震盪した。追加の12.5mlのクロロホルム/イソアミルアルコール(24:1)を混合物に添加し、さらに15分間震盪した。この混合物を遠心し、水性相を25mlのフェノール/クロロホルム/イソアミルアルコール(25:24:1)およびクロロホルム/イソアミルアルコール(24:1)で再抽出した。室温において15mlのイソプロパノールを使用する沈澱により、核酸を回収した。沈澱を1mlの水の中に再懸濁させた。
【0151】
ゲノムDNAを次のようにして制限酵素消化した:10μgのゲノムDNA、40μlの10×反応緩衝液および100Uの制限酵素(XbaI、EcoRI、EcoRVまたはHindIII)を、400μlの全反応体積で5〜6時間反応させた。次いでこの混合物をフェノール抽出し、エタノール沈降させた。消化したDNAを0.8%のアガロースゲル上で15ボルトにおいてほぼ15時間アガロースゲル電気泳動に付した。ゲルを変性緩衝液[87.66gのNaClおよび20gのNaOH/リットル]の中におだやかに撹拌しながら30分間沈め、蒸留水ですすぎ、中性緩衝液[87.66gのNaClおよび60.55gのTris−HCl、pH7.5/リットル]の中におだやかに撹拌しながら30分間沈めた。DNAを毛管ブロッティングによりHybond−H+ナイロン膜に移した。
【0152】
1×106cpm/mlの32P−dCTP標識化DHS cDNAまたはDHS cDNAクローンの3’非コーディング領域を使用して42℃において、ハイブリダイゼーションを一夜実施した。50%のホルムアミド、6×SSC、5×デンハルト溶液、0.1%SDSおよび100mg/mlの変性サケ精子DNAを含有する緩衝液中でプレハイブリダイゼーションおよびハイブリダイゼーションを実施した。膜を2〜4時間プレハイブリダイゼーションした;ハイブリダイゼーションを一夜実施した。
【0153】
ハイブリダイゼーションが完結した後、膜を2×SSCおよび0.1%のSDS中ですすぎ、次いで2×SSCおよび0.1%のSDS中で15分間洗浄し、0.2×SSCおよび0.1%のSDS中で15分間洗浄した。次いで膜を−80℃においてX線フィルムに一夜露出した。結果を第5図に示す。
【0154】
実施例8.アラアビドプシス(Arabidopsis)からのセネッセンス誘導elF−5A遺伝子の単離
鋳型としてアラアビドプシス(Arabidopsis)セネッシング葉cDNAライブラリーを使用するPCRにより、アラアビドプシス(Arabidopsis)葉中で発現されたセネッセンス誘導elF−5A遺伝子の全長cDNAクローンを得た。最初に、ベクターT7プライマー<AATACGACTCACTATAG>(配列番号18)と対合したデジェネレイト上流のプライマー<AAARRYCGMCCYTGCAAGGT>(配列番号17)、およびベクターT3プライマー<ATTAACCCTCACTAAAG>(配列番号20)と対合したデジェネレイト下流のプライマー<TCYTTNCCYTCMKCTAAHCC>(配列番号19)を使用して、遺伝子の5’および3’末端に対応するPCR生成物を作った。
【0155】
PCR生成物を配列決定のためにpBluescriptの中にサブクローニングした。次いで、3’特異的プライマー<AGAAGAAGTATAAAAACCATC>(配列番号22)と対合した5’特異的プライマー<CTGTTACCAAAAAATCTGTACC>(配列番号21)を使用して全長cDNAを作り、配列決定のためにpBluescriptの中にサブクローニングした。
【0156】
実施例9.トマト果実からのセネッセンス誘導elF−5A遺伝子の単離
鋳型としてトマト果実cDNAライブラリーを使用するPCRにより、トマト果実中で発現されたセネッセンス誘導elF−5A遺伝子の全長cDNAクローンを得た。最初に、ベクターT7プライマー(配列番号18)と対合したデジェネレイト上流のプライマー(配列番号17)、およびベクターT3プライマー(配列番号20)と対合したデジェネレイト下流のプライマー(配列番号19)を使用して、遺伝子の5’および3’末端に対応するPCR生成物を作った。PCR生成物を配列決定のためにpBluescriptの中にサブクローニングした。次いで、ベクターT7プライマー(配列番号18)と対合した5’特異的プライマー<AAAGAATCCTAGAGAGAGAAAGG>(配列番号23)を使用して全長cDNAを作り、配列決定のためにpBluescriptの中にサブクローニングした。
【0157】
実施例10.カーネーションからのセネッセンス誘導elF−5A遺伝子の単離
鋳型としてカーネーション花cDNAライブラリーを使用するPCRにより、カーネーション花中で発現されたセネッセンス誘導elF−5A遺伝子の全長cDNAクローンを得た。最初に、ベクターT7プライマー(配列番号18)と対合したデジェネレイト上流のプライマー(配列番号17)、およびベクターT3プライマー(配列番号20)と対合したデジェネレイト下流のプライマー(配列番号19)を使用して、遺伝子の5’および3’末端に対応するPCR生成物を作った。
【0158】
PCR生成物を配列決定のためにpBluescriptの中にサブクローニングした。次いで、3’特異的プライマー<ACCAAAACCTGTGTTATAACTCC>(配列番号25)と対合した5’特異的プライマー<TTTTACATCAATCGAAAATACC>(配列番号24)を使用して全長cDNAを作り、配列決定のためにpBluescriptの中にサブクローニングした。
【0159】
実施例11.アラアビドプシス(Arabidopsis)からのセネッセンス誘導DHS遺伝子の単離
アラアビドプシス(Arabidopsis)セネッシング葉cDNAライブラリーをスクリーニングすることによって、アラアビドプシス(Arabidopsis)葉中で発現されたセネッセンス誘導DHS遺伝子の全長cDNAクローンを得た。スクリーニングに使用したプローブ(配列番号26)の配列を第38図に示す。鋳型としてセネッセンス葉cDNAライブラリーを使用しかつGenBankにおける未同定ゲノム配列(AB017060)から設計したプライマー(第38図に下線が引かれている領域として示す)を使用するPCRにより、このプローブを得た。PCR生成物を配列決定のためにpBluescriptの中にサブクローニングした。
【0160】
実施例12.カーネーションからのセネッセンス誘導DHS遺伝子の単離
カーネーションセネッシング花弁cDNAライブラリーをスクリーニングすることによって、カーネーション花弁中で発現されたセネッセンス誘導DHS遺伝子の全長cDNAクローンを得た。スクリーニングに使用したプローブ(配列番号27)の配列を第39図に示す。鋳型としてセネッセンス花弁cDNAライブラリーおよびデジェネレイトプライマー(上流:5’TTG ARG AAG ATY CAT MAA RTG CCT3’)(配列番号28);下流:5’CCA TCA AAY TCY TGK GCR GTG TT3’)(配列番号29))を使用するPCRにより、このプローブを得た。PCR生成物を配列決定のためにpBluescriptの中にサブクローニングした。
【0161】
実施例13.アンチセンスの向きにアラアビドプシス(Arabidopsis)DHSの全長または3’領域を使用するアラアビドプシス(Arabidopsis)の形質転換
二重35Sプロモーターの調節下に、全長セネッセンス誘導アラアビドプシス(Arabidopsis)DHS cDNA配列またはDHS遺伝子の3’末端(配列番号30)(第36図)(両方はアンチセンス立体配置で発現された)を含有するバイナリーベクター、pKYLX71、で、アグロバクテリア(Agrobacteria)を形質転換した。アラアビドプシス(Arabidopsis)植物を形質転換されたアグロバクテリア(Agrobacteria)で真空浸潤により形質転換し、生ずるT0植物からの形質転換された種子をアンピシリン上で選択した。
【0162】
第21図〜第24図は形質転換されたアラアビドプシス(Arabidopsis)植物の写真であり、形質転換された植物におけるアンチセンスの向きのDHS遺伝子またはその3’末端の発現がバイオマスを増加させ、例えば、葉を大きくしかつ植物のサイズを大きくすることを示す。第25図は、トランスジェニックアラアビドプシス(Arabidopsis)植物が種子収量を増加したことを図解する。
【0163】
実施例14.アンチセンスの向きにトマトDHSの全長または3’領域を使用するトマト植物の形質転換
二重35Sプロモーターの調節下に、全長セネッセンス誘導トマトDHS cDNA配列またはDHS遺伝子の3’末端(配列番号31)(第37図)(両方はアンチセンス立体配置で発現された)を含有するバイナリーベクター、pKYLX71、で、アグロバクテリア(Agrobacteria)を形質転換した。これらのアグロバクテリア(Agrobacteria)を使用してトマト葉の外植体を形成し、標準組織培養法によりカルスおよび小植物を発生させ、選択した。形質転換された植物を温室の条件下に成熟果実産生T1植物に成長させた。
【0164】
第26図〜第35図は、形質転換されたアラアビドプシス(Arabidopsis)植物において見られるように、形質転換された植物中のセネッセンス誘導トマトDHS遺伝子の発現の減少がバイオマスを増加させ、例えば、葉のサイズを大きくしかつ植物を大きくすること、ならびにトマト果実の軟化および腐敗を遅延することを示す写真である。
【技術分野】
【0001】
発明の分野
本発明は、セネッセンス誘導発現を示す植物ポリペプチドをコードするポリヌクレオチドに関する。本発明は、また、アンチセンスの向きでポリヌクレオチドを含有するトランスジェニック植物、および植物における、セネッセンスを包含する、プログラムされた細胞死をコントロールする方法に関する。さらに詳しくは、本発明は、その発現がセネッセンスを包含するプログラムされた細胞死の開始により誘導される、セネッセンス誘導植物デオキシハイプシンシンターゼ遺伝子およびセネッセンス誘導elF−5A遺伝子、および植物におけるプログラムされた細胞死およびセネッセンスをコントロールするために、デオキシハイプシンシンターゼ遺伝子およびelF−5A遺伝子を、単独または組合わせで、使用することに関する。
【背景技術】
【0002】
先行技術の説明
セネッセンスは植物の寿命における生物学的発生の末期である。セネッセンスは死の前兆であり、全植物、器官、花および果実、組織および個々の細胞を包含する生物学的体制化の種々のレベルにおいて起こる。
【0003】
セネッセンスの開始は、内部および外部の両方の異なる因子により誘導されることがある。セネッセンスは、植物または植物組織、例えば、果実、花および葉の寿命において、複雑な、高度に調節された発生段階である。セネッセンスは、細胞膜および高分子の同調的破壊、および代謝物質の植物の他の部分への引き続く移動化を生ずる。
【0004】
正常の植物発生の間に起こるプログラムされたセネッセンスに加えて、細胞および組織の死および続いて起こる代謝物質の再移動化は、外部の、環境的因子に対する同調的応答として起こる。また壊死またはアポトーシスと呼ばれる、セネッセンスの早期の開始を誘導する外部の因子は、環境的ストレス、例えば、温度、乾燥、劣った光または栄養の供給、ならびに病原体の攻撃を包含する。環境的ストレスに暴露された植物組織は、また、ストレスエチレンとして普通に知られているエチレンを産生する(Buchanan−Wollaston、V.、1997、J.Exp.Botany、48:181−199;Wright、M.、1974、Plant、120:63−69)。エチレンはある植物においてセネッセンスを引き起こすことが知られている。
【0005】
セネッセンスは受動的プロセスではなく、むしろ、特異的遺伝子の共役発現を包含する、活性的に調節されたプロセスである。セネッセンスの間に、全RNAのレベルは減少し、多数の遺伝子の発現はスイッチが切られる(Bate他、1991、J.Exper.Botany、42:801−11;Hensel他、1993、The Plant Cell、5:553−64)。しかしながら、セネッセンスプロセスが核遺伝子の新規な転写に依存するという証拠が増加している。例えば、セネッセンスはmRNAおよびタンパク質合成および除核のインヒビターによりブロックされる。in vitro翻訳実験のためにセネッシング葉および緑色の葉からのmRNAを使用する分子の研究において、セネッシング葉において葉タンパク質産物の変化したパターンが示された(Thomas他、1992、J.Plant Physiol.、139:403−12)。
【0006】
ディファレンシャルスクリーニングおよび差引きハイブリダイゼーション技術を使用して、単子葉植物および双子葉植物、例えば、アラアビドプシス(Arabidopsis)、トウモロコシ、キュウリ、アスパラガス、トマト、イネおよびジャガイモを包含する、ある範囲の異なる植物から、セネッセンス誘導遺伝子を表す、多数のcDNAクローンが同定されてきている。セネッセンスの間に特異的に発現される遺伝子の同定は、セネッセンスの新たな転写を進行させるための要件の客観的な証拠である。
【0007】
セネッセンスの間に起こる事象は高度に同調して、壊死および死が起こる前の細胞成分の最大の使用を可能とする。特異的シグナルの感知および遺伝子発現のカスケードの誘導を包含する複雑な相互作用が起こって、このプロセスを調節するに違いない。セネッセンス関係タンパク質をコードする遺伝子の発現は、多分、普通のアクチベータータンパク質を介して調節され、引き続いて、アクチベータータンパク質はホルモンのシグナルにより直接的または間接的に活性化される。このプロセスの初期のシグナリングまたは引き続く同調に関係するメカニズムについてほとんど知られていない。
【0008】
同調的遺伝子発現は、開始因子を包含する、転写および翻訳に関係する因子を必要とする。翻訳開始因子の遺伝子は、植物を包含する、種々の生物において単離され、特性決定されてきている。真核生物の翻訳開始因子5A(elF−5A)はほぼ17kDaのサイズの必須タンパク質因子であり、これは真核細胞のタンパク質系合成の開始に関係する。それはelF−5Aの中にのみ存在することが知られているハイプシン[N−(4−アミノ−2−ヒドロキシブチル)リシン]、すなわち、ユニークな修飾されたアミノ酸、の存在により特徴づけられる。ポリアミン、スペリミジン、からelF−5A中の特定のリシン残基の側鎖アミノ基へのブチルアミノ基の転移およびヒドロキシル化を介して翻訳後にハイプシンは形成される。
【0009】
elF−5Aの活性化は、スペリミジンのブチルアミン残基のelF−5Aのリシンへの転移、ハイプシンの形成およびelF−5Aの活性化を包含する。真核生物において、デオキソハイプシンシンターゼ(DHS)はelF−5Aにおけるハイプシンの翻訳後の合成を仲介する。対応するDHS遺伝子は植物において同定されてきていないが、植物のelF−5Aはハイプシンを含有することが知られている。ハイプシンの修飾は、メチオニル−プロマイシンアッセイを使用するin vitro elF−5A活性のために必須であることが示された。
【0010】
ハイプシンはelF−5Aの中にユニークに存在し、すべての真核生物、いくつかの始原細菌(これらは真核生物に関係するように思われる)の中に見出されるが、真性細菌の中に見出されない。そのうえ、elF−5Aのアミノ酸配列は、特にハイプシン残基を取り囲む領域において、高度に保存され、elF−5Aおよびその活性化タンパク質、デオキシハイプシンシンターゼは真核細胞の生理学において基本的に重要な工程を実行することを示唆する(Joe他、JBC、270:22386−22392)。
【0011】
elF−5Aは、ヒト、アルファルファ、スライムのカビ、ニューロスポラ・クラッサ(Neurospora crassa)、タバコおよび酵母からクローニングされた。ウサギ網状赤血球ライゼイトのリボソームからの単離、およびメチオニン−プロマイシン合成を刺激するin vitro活性に基づいて、elF−5Aは本来一般的翻訳開始因子として同定された。しかしながら、elF−5Aは包括的タンパク質合成の翻訳開始因子ではなく、むしろmRNA集団の特定のサブセットの翻訳を促進する働きをすることが、より最近のデータにより示された。
【0012】
例えば、elF−5Aの1または2以上のイソ型は細胞増殖に関係するmRNAのサブセットの翻訳の仲介において必須の役割を演ずるということを示す強い証拠が、動物細胞および酵母を使用する実験から得られた。酵母において2つのイソ型が存在し、そして両方の遺伝子がサイレントである場合、細胞は分裂することができない(Park他、Biol. Signals、6:115−123、1997)。同様に、elF−5Aを活性化する酵母デオキシハイプシンシンターゼの発現をサイレンスすると、細胞分裂はブロックされる。事実、過増殖状態の治療において重要性を有すると思われるデオキシハイプシンシンターゼのインヒビターが開発された(Wolff他、JBC、272:15865−15871、1997)。
【0013】
他の研究において、elF−5Aの他のイソ型がHIV−1の複製におけるRev機能またはHTLV V複製におけるRex機能のために必須であるということが示された(Park他、Biol.Signals、6:115−123、1997)。また、タバコの中に少なくとも2つの発現されたelF−5A遺伝子が存在する。それらの両方は検査したすべての組織において発現されるが、各遺伝子は明確な発現パターンを有し、多分特定の転写物の翻訳を調節することを、遺伝子特異的プローブは示す(Chamot他、Nucl.Acids Res.、20:625−669、1992)。
【0014】
デオキシハイプシンシンターゼは、ラット精巣、HeLa細胞、ニューロスポラ・クラッサ(Neurospora crassa)および酵母から精製された。デオキシハイプシンシンターゼのアミノ酸配列は高度に保存され、そして異なる種からの酵素は同様な、触媒的性質を共有し、異種elF−5A前駆体と交差種反応性を示す(Park他、Biol.Signals、6:115−123、1997)。
【0015】
植物ポリアミンは、花の誘導、胚形成、病原体耐性、細胞の成長、分化および分裂を包含する、広範な種類の生理学的作用に関係づけられた(Evans他、1989、Ann.Rev.Plant Physiol.Plant Mol.Biol.、40:235−269;およびGalston他、1990、Plant Physiol.、94:406−10)。elF−5Aはポリアミンがその作用を発揮するとき通過する中間物であることが示唆された(Chamot他、1992、Nucl.Acids Res.、20(4):665−69)。
【0016】
タバコ(Nicotiana)からのelF−5Aのイソ型をコードする2つの遺伝子が同定された(NelF−5A1およびNelF−5A2)(Chamot他、1992、Nucl.Acids Res.、20(4):665−69)。これらの遺伝子は非常に類似することが示された。一方の遺伝子はmRNAレベルで構成的に発現されるが、他方の発現パターンは光合成活性の存在または非存在と相関する。遺伝子構造およびゲノムサザンマッピングに基づいて、タバコの中にNelF−5Aの多重遺伝子ファミリーが存在することが示唆された。セネッセンス/壊死特異的mRNA転写物のサブセットの翻訳を調節するelF−5Aのイソ型が存在すると思われる。
【0017】
現在、内部または外部の、例えば、環境的ストレスの、因子により引き起こされるプログラムされた細胞死(セネッセンスを包含する)の開始をコントロールする、広く適用可能な方法は存在しない。したがって、すべてのタイプの植物に適用可能でありかつセネッセンスに導く事象のカスケードにおいて最も早い段階において有効である、セネッセンスをモジュレートする技術を開発することが重要である。
【発明の概要】
【課題を解決するための手段】
【0018】
発明の要約
本発明は、トマトのセネッセンス誘導デオキシハイプシンシンターゼ(DHS)をコードする全長のcDNAクローン、ならびにアラアビドプシス(Arabidopsis)の葉およびカーネーションの花弁からの全長のセネッセンス誘導DHS cDNAクローンの発見およびクローニングに基づく。ヌクレオチド配列および対応するアミノ酸配列が本明細書において開示される。
【0019】
本発明は、また、一部分、トマト、アラアビドプシス(Arabidopsis)およびカーネーションからのセネッセンス誘導elF−5A遺伝子をコードする全長のcDNAクローンの発見およびクローニングに基づく。elF−5A cDNAの各々のヌクレオチド配列および対応するアミノ酸配列が本明細書において開示される。
【0020】
本発明は、年齢に関係するセネッセンスまたは環境的ストレス誘導セネッセンスである、セネッセンスの開始をコントロールするために、植物を遺伝的に修飾する方法を提供する。本発明のセネッセンス誘導DHSヌクレオチド配列、それらのフラグメント、またはこのようなフラグメントの組合わせを植物細胞の中に逆向きに導入して、内因的セネッセンス誘導DHS遺伝子の発現を阻害し、これにより内因的セネッセンス誘導DHSタンパク質のレベルを減少し、かつelF−5Aの活性化および引き続くセネッセンスを仲介する遺伝子の発現を減少および/または防止する。
【0021】
本発明の他の面において、本発明のセネッセンス誘導elF−5Aヌクレオチド配列、それらのフラグメント、またはこのようなフラグメントの組合わせを植物細胞の中に逆向きに導入して、内因的セネッセンス誘導elF−5A遺伝子の発現を阻害し、これにより内因的セネッセンス誘導elF−5Aタンパク質のレベルを減少し、かつelF−5Aの活性化および引き続くセネッセンスを仲介する遺伝子の発現を減少および/または防止する。あるいは、DHS配列およびelF−5A配列の両方を一緒に使用して、内因的DHSおよびelF−5Aタンパク質のレベルを減少させる。
【0022】
なお他の面において、本発明は、本発明のセネッセンス誘導elF−5Aヌクレオチド配列と本発明のセネッセンス誘導DHSヌクレオチド配列との組合わせを植物細胞の中に逆向きに導入して、内因的セネッセンス誘導elF−5A遺伝子およびセネッセンス誘導DHS遺伝子の発現を阻害し、これにより内因的セネッセンス誘導DHSタンパク質のレベルを減少し、かつelF−5Aの活性化および引き続くセネッセンスを仲介する遺伝子の発現を減少および/または防止することによって、年齢に関係するセネッセンスまたは環境的ストレス誘導セネッセンスである、セネッセンスの開始をコントロールするために、植物を遺伝的に修飾する方法に関する。
【0023】
本発明の方法を使用して、トランスジェニック植物を発生させ、成長、発生、および自然のセネッセンスまたは早期に誘導されたセネッセンスについてモニターする。セネッセンス誘導DHS、セネッセンス誘導elF−5Aまたは両方のレベルが減少するために延長した寿命または棚持ち(例えば、花の寿命の延長、果実または野菜の腐敗の減少、バイオマスの増強、種子収量の増加、種子老化の減少および/または葉の黄色化の減少)を示す植物または植物の分離した部分(例えば、挿木、花、野菜、果実、種子または葉)が、葉の黄色化の減少、花弁の離層の減少、輸送および貯蔵中の果実および野菜の腐敗の減少を包含する改良された性質を有する、所望の産物として選択される。これらのよりすぐれた植物を増殖させる。同様に、環境的ストレスに対する抵抗性が増加した、例えば、低温(冷却)、乾燥、感染、およびその他に対する感受性が減少した、および/または病原体に対する抵抗性が増加した植物がよりすぐれた産物として選択される。
【0024】
1つの面において、本発明は、セネッセンス誘導DHSをコードする単離されたDNA分子に関し、ここでDNA分子は配列番号1とハイブリダイゼーションするか、あるいは配列番号1とハイブリダイゼーションする単離されたDNA分子の機能的誘導体とハイブリダイゼーションする。本発明のこの面の1つの態様において、単離されたDNA分子は配列番号1のヌクレオチド配列、すなわち、配列番号1に対して100%の相補性(配列同一性)を有する。
【0025】
本発明は、また、セネッセンス誘導DHSをコードする単離されたDNA分子に関し、ここでDNA分子は配列番号9とハイブリダイゼーションするか、あるいは配列番号9とハイブリダイゼーションする単離されたDNA分子の機能的誘導体とハイブリダイゼーションする。本発明のこの面の1つの態様において、単離されたDNA分子は配列番号9のヌクレオチド配列、すなわち、配列番号9に対して100%の相補性(配列同一性)を有する。
【0026】
本発明は、また、セネッセンス誘導elF−5Aをコードする単離されたDNA分子に関し、ここでDNA分子は配列番号11、配列番号13または配列番号15とハイブリダイゼーションするか、あるいは配列番号11、配列番号13または配列番号15とハイブリダイゼーションする単離されたDNA分子の機能的誘導体とハイブリダイゼーションする。本発明のこの面の1つの態様において、単離されたDNA分子は配列番号11、配列番号13または配列番号15のヌクレオチド配列、すなわち、配列番号11、配列番号13または配列番号15に対して100%の相補性(配列同一性)を有する。
【0027】
本発明の他の態様において、本明細書において記載するDNA分子によりコードされる単離されたタンパク質、またはその機能的誘導体が提供される。好ましいタンパク質は配列番号2のアミノ酸配列を有するか、あるいはその機能的誘導体である。他の好ましいタンパク質は配列番号10のアミノ酸配列を有するか、あるいはその機能的誘導体である。本発明の他の好ましいタンパク質は、配列番号12、配列番号14または配列番号16のアミノ酸配列を有する。
【0028】
また、前述のDNA分子のRNA転写物の対応する部分に対して相補的であるRNA分子をコードするアンチセンスオリゴヌクレオチドまたはポリヌクレオチドが提供され、ここで内因的セネッセンス誘導DHSの発現が変更されるように、オリゴヌクレオチドまたはポリヌクレオチドはRNA転写物とハイブリダイゼーションする。本発明のこの面の他の態様において、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドは、内因的セネッセンス誘導elF−5Aの発現が変更されるように、前述のDNA分子のRNA転写物の対応する部分に対してハイブリダイゼーションするRNA分子である。アンチセンスオリゴヌクレオチドまたはポリヌクレオチドは全長であるか、あるいは好ましくは約6〜約100ヌクレオチドを有する。
【0029】
アンチセンスオリゴヌクレオチドまたはポリヌクレオチドはセネッセンス誘導DHSをコードするDNA分子の1つ鎖の対応する部分に対して実質的に相補的であることができ、ここでセネッセンス誘導DHSをコードするDNA分子は配列番号1、配列番号5、配列番号9、またはそれらの組合わせとハイブリダイゼーションするか、あるいはセネッセンス誘導DHSをコードするDNA分子によりコードされるRNA配列の少なくとも対応する部分に対して実質的に相補的である。
【0030】
本発明の1つの態様において、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドはヌクレオチド配列番号1、配列番号5、配列番号9の1つ鎖の対応する部分またはそれらの組合わせに対して実質的に相補的であるか、または配列番号1、配列番号5、配列番号9から転写されたRNA転写物またはそれらの組合わせに対して実質的に相補的である。他の態様において、アンチセンスオリゴヌクレオチドはセネッセンス誘導DHSをコードするDNA分子の1つ鎖の5’非コーディング部分または3’部分の対応する部分に対して実質的に相補的であり、ここでDNA分子は配列番号1、配列番号5、配列番号9またはそれらの組合わせとハイブリダイゼーションする。
【0031】
あるいは、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドはセネッセンス誘導elF−5AをコードするDNA分子の1つ鎖の対応する部分に対して実質的に相補的であることができ、ここでセネッセンス誘導elF−5AをコードするDNA分子は配列番号11、配列番号13、配列番号15、またはそれらの任意の組合わせとハイブリダイゼーションするか、あるいは配列番号11、配列番号13または配列番号15から転写されたRNA配列の少なくとも対応する部分に対して実質的に相補的である。
【0032】
本発明の1つの態様において、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドはヌクレオチド配列の配列番号11、配列番号13、配列番号15またはそれらの組合わせの1つ鎖の対応する部分に対して実質的に相補的であるか、あるいはコードされるRNA転写物はセネッセンス誘導elF−5AをコードするDNA分子によりコードされる調節配列の対応する部分に対して実質的に相補的である。他の態様において、アンチセンスオリゴヌクレオチドはセネッセンス誘導elF−5AをコードするDNA分子の1つ鎖の5’非コーディング領域または3’領域の対応する部分に対して実質的に相補的であり、ここでDNA分子は配列番号11、配列番号13、配列番号15またはそれらの組合わせとハイブリダイゼーションする。
【0033】
さらに、本発明は、下記の配列を含んでなる植物細胞の形質転換用ベクターに関する:
(a)(1)セネッセンス誘導DHSをコードするDNA分子の1つの鎖の対応する部分、ここでセネッセンス誘導DHSをコードするDNA分子は配列番号1、配列番号5または配列番号9とハイブリダイゼーションする、または(2)セネッセンス誘導DHSをコードするDNA分子によりコードされるRNA配列の対応する部分、に対して実質的に相補的であるアンチセンスオリゴヌクレオチドまたはポリヌクレオチド;および
(b)それがその中に形質転換されている植物細胞においてアンチセンスオリゴヌクレオチドまたはポリヌクレオチドが発現されるように、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドに作用可能に連鎖された調節配列。
【0034】
さらに、本発明は、下記の配列を含んでなる植物細胞の形質転換用ベクターに関する:
(a)(1)セネッセンス誘導elF−5AをコードするDNA分子の1つの鎖の対応する部分、ここでセネッセンス誘導elF−5AをコードするDNA分子は配列番号11、配列番号13、配列番号15とハイブリダイゼーションする、または(2)セネッセンス誘導elF−5AをコードするDNA分子によりコードされるRNA配列の対応する部分、に対して実質的に相補的であるアンチセンスオリゴヌクレオチドまたはポリヌクレオチド;および
(b)それがその中に形質転換されている植物細胞においてアンチセンスオリゴヌクレオチドまたはポリヌクレオチドが発現されるように、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドに作用可能に連鎖された調節配列。
【0035】
調節配列は形質転換された細胞中で機能的であるプロモーターを含み、このプロモーターは誘導可能または構成的であることができる。必要に応じて、調節配列はポリアデニル化シグナルを含む。
【0036】
本発明は、また、前述のベクターまたはベクターの組合わせで形質転換された植物細胞、このような細胞から発生した小植物または成熟植物、またはこのような小植物または成熟植物の植物部分を提供する。
【0037】
本発明は、下記の工程を含んでなる、非修飾植物に比較して、セネッセンス誘導DHS、セネッセンス誘導elF−5Aまたは両方のレベルが減少した植物を産生する方法に関する:
(1)植物を前述のベクターまたはベクターの組合わせで形質転換し;
(2)植物を少なくとも小植物段階に成長させ;
(3)形質転換された植物または小植物をセネッセンス誘導DHS活性および/またはelF−5A活性の変更および/またはセネッセンスの変更および/または環境的ストレス誘導セネッセンスおよび/または病原体誘導セネッセンスおよび/またはエチレン誘導セネッセンスの変更についてアッセイし;そして
(4)非形質転換植物に比較して、セネッセンス誘導DHS活性が変更されたおよび/またはelF−5A活性が減少したおよび/またはセネッセンスが変更したおよび/または環境的ストレス誘導セネッセンスが変更されたおよび/または病原体誘導セネッセンスおよび/またはエチレン誘導セネッセンスが変更した植物を選択し、そして成長させる。
【0038】
前述したように産生された植物、または子孫、ハイブリッド、クローンまたは植物部分は、好ましくは、セネッセンス誘導DHSの発現の減少、セネッセンス誘導elF−5A活性の減少、または両方およびセネッセンスの遅延および/または環境的ストレス誘導セネッセンスおよび/または病原体誘導セネッセンスおよび/またはエチレン誘導セネッセンスの遅延を示す。
【0039】
さらに、本発明は、下記の工程を含んでなる植物における内因的セネッセンス誘導DHSの発現を阻害する方法に関する:
(1)植物のゲノムの中にベクターを組込み、前記ベクターは、
(A)(i)内因的セネッセンス誘導DHSをコードするDNA分子の1つの鎖の対応する部分、ここで内因的セネッセンス誘導DHSをコードするDNA分子は配列番号1、配列番号5および/または配列番号9とハイブリダイゼーションする、または(ii)内因的セネッセンス誘導DHS遺伝子によりコードされるRNA配列の少なくとも一部分、に対して相補的であるアンチセンスオリゴヌクレオチドまたはポリヌクレオチド、および
(B)アンチセンスオリゴヌクレオチドまたはポリヌクレオチドが発現されるように、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドに作用可能に連鎖された調節配列、
を含んでなり;そして
(2)前記植物を成長させる、それにより前記アンチセンスオリゴヌクレオチドまたはポリヌクレオチドは転写され、前記内因的RNA配列に結合し、それにより前記セネッセンス誘導DHS遺伝子の発現は阻害される。
【0040】
本発明は、さらに、下記の工程を含んでなる植物における内因的セネッセンス誘導elF−5Aの発現を阻害する方法に関する:
(1)植物のゲノムの中にベクターを組込み、前記ベクターは、
(A)(i)内因的セネッセンス誘導elF−5AをコードするDNA分子の1つの鎖の対応する部分、ここで内因的セネッセンス誘導elF−5AをコードするDNA分子は配列番号11、配列番号15、配列番号17またはそれらの組合わせとハイブリダイゼーションする、または(ii)内因的セネッセンス誘導elF−5A遺伝子によりコードされるRNA配列の少なくとも一部分、に対して相補的であるアンチセンスオリゴヌクレオチドまたはポリヌクレオチド、および
【0041】
(B)アンチセンスオリゴヌクレオチドまたはポリヌクレオチドが発現されるように、アンチセンスオリゴヌクレオチドまたはポリヌクレオチドに作用可能に連鎖された調節配列、
を含んでなり;そして
(2)前記植物を成長させる、それにより前記アンチセンスオリゴヌクレオチドまたはポリヌクレオチドは転写され、前記内因的RNA配列に結合し、それにより前記セネッセンス誘導elF−5A遺伝子の発現は阻害される。
【図面の簡単な説明】
【0042】
【図1】第1図は、セネッセンス誘導トマト葉DHS cDNA配列(配列番号1)およびトマト葉cDNAライブラリーから得られた誘導アミノ酸配列(配列番号2)のヌクレオチド配列を描写する。
【図2A】第2A図は、アラアビドプシス(Arabidopsis)遺伝子バンクにおいてトマトDHS配列を非同定ゲノム配列と整列させることによって得られたアラアビドプシス(Arabidopsis)DHS遺伝子のヌクレオチド配列を描写する(http://genome−www.stanford.edu/Arabidopsis/)(配列番号5)。アミノ酸配列間のギャップは予測されたイントロンである。
【図2B】第2B図は、誘導アラアビドプシス(Arabidopsis)DHSアミノ酸配列(配列番号6)を描写する。
【図2C】第2C図は、PCRにより得られた600塩基対のセネッセンス誘導アラアビドプシス(Arabidopsis)DHS cDNAのヌクレオチド配列を描写する。
【図2D】第2D図は、セネッセンス誘導アラアビドプシス(Arabidopsis)DHS cDNAフラグメントの誘導アミノ酸配列を描写する。
【図3】第3図は、誘導全長トマト葉セネッセンス誘導DHSアミノ酸配列(配列番号2)および誘導全長アラアビドプシス(Arabidopsis)セネッセンス誘導DHSアミノ酸配列と、ヒト、酵母、菌類、および始原細菌(Archaeobacteria)のDHSタンパク質の配列との整列である。配列の3つまたは4つの間で同一のアミノ酸はボックスで囲まれている。
【図4】第4図は、トマトDHS cDNAの制限地図である。
【図5】第5図は、トマト葉から単離され、32P−dCTP標識化全長トマトセネッセンス誘導DHS cDNAでプロービングしたゲノムDNAのサザンブロットである。
【0043】
【図6】第6図は、発生の異なる段階においてトマト花から単離されたRNAのノザンブロットである。第6A図は、全RNAの臭化エチジウム染色ゲルである。各レーンは10μgのRNAを含有する。第6B図は、32P−dCTP標識化全長トマトセネッセンス誘導DHS cDNAでプロービングしたノザンブロットのオートラジオグラフである。
【図7】第7図は、32P−dCTP標識化全長トマトセネッセンス誘導DHS cDNAでプロービングした成熟の種々の段階においてトマト果実から単離されたRNAのノザンブロットである。各レーンは10μgのRNAを含有する。
【図8】第8図は、2Mのソルビトールで6時間処理して乾燥ストレスさせたトマト葉から単離されたRNAのノザンブロットである。このブロットを32P−dCTP標識化全長トマトセネッセンス誘導DHS cDNAでプロービングした。
【図9】第9図は、冷却温度に暴露したトマト葉から単離されたRNAのノザンブロットである。第9A図は、全RNAの臭化エチジウム染色ゲルである。第9B図は、32P−dCTP標識化全長トマトセネッセンス誘導DHS cDNAでプロービングしたノザンブロットのオートラジオグラフである。第9C図は、葉透析質の導電率として測定した、対応する漏出データを示す。
【図10】第10図は、ポリAテイルおよび5’末端非コーディング領域を含まない、カーネーション全長(1384塩基対)cDNAクローンのヌクレオチド配列(配列番号9)である。誘導アミノ酸配列はヌクレオチド配列の下に示されている(373アミノ酸)(配列番号10)。
【0044】
【図11】第11図は、32P−dCTP標識化全長アラアビドプシス(Arabidopsis)セネッセンス誘導DHS cDNAでプロービングしたセネッセンシングアラアビドプシス(Arabidopsis)葉からの全RNAのノザンブロットである。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【図12】第12図は、種々の段階におけるカーネーション花から単離された全RNAのノザンブロットである。このブロットを32P−dCTP標識化全長カーネーションセネッセンス誘導DHS cDNAでプロービングした。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【図13】第13図は、トマト果実セネッセンス誘導elF−5A遺伝子のヌクレオチド(上部)(配列番号11)および誘導アミノ酸(下部)(配列番号12)配列である。
【図14】第14図は、カーネーションセネッセンス誘導elF−5A遺伝子のヌクレオチド(上部)(配列番号13)および誘導アミノ酸(下部)(配列番号14)配列である。
【図15】第15図は、アラアビドプシス(Arabidopsis)セネッセンス誘導elF−5A遺伝子のヌクレオチド(上部)(配列番号15)および誘導アミノ酸(下部)(配列番号16)配列である。
【0045】
【図16】第16図は、種々の発生段階におけるアラアビドプシス(Arabidopsis)植物から単離された全RNAのノザンブロットである。このブロットを32P−dCTP標識化全長アラアビドプシス(Arabidopsis)セネッセンス誘導DHS cDNAおよび全長セネッセンス誘導elF−5Aでプロービングした。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【図17】第17図は、発生のブレーカー(braker)(BK)、レッド−ファーム(red−firm)(RF)およびレッド−ソフト(red−soft)(RS)段階におけるトマト果実から単離された全RNAのノザンブロットである。このブロットを32P−dCTP標識化全長セネッセンス誘導DHS cDNAおよび全長セネッセンス誘導elF−5Aでプロービングした。DHSおよびelF−5Aは果実成熟と一致してレッド−ソフト果実において平行にアップレギュレートされる。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【図18】第18図は、ソルビトールで処理して乾燥ストレスを誘導したトマトの葉から単離された全RNAのノザンブロットである。Cは対照であり;Sはソルビトール処理である。このブロットを32P−dCTP標識化全長セネッセンス誘導DHS cDNAおよび全長セネッセンス誘導elF−5Aでプロービングした。elF−5AおよびDHSの両方は乾燥ストレスに応答してアップレギュレートされる。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【図19】第19図は、トマト植物の花芽および開いたセネッセンシング花から単離された全RNAのノザンブロットである。このブロットを32P−dCTP標識化全長セネッセンス誘導DHS cDNAおよび全長セネッセンス誘導elF−5Aでプロービングした。elF−5AおよびDHSの両方は開いた/セネッセンシング花においてアップレギュレートされる。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【図20】第20図は、冷却損傷したトマト葉から単離された全RNAのノザンブロットである。このブロットを32P−dCTP標識化全長セネッセンス誘導DHS cDNAおよび全長セネッセンス誘導elF−5Aでプロービングした。elF−5AおよびDHSの両方は再加温の間における冷却損傷植物の発生とともにアップレギュレートされる。オートラジオグラフは上部に存在し、エチジウム染色ゲルは下に存在する。
【0046】
【図21】第21図は、3.1週齢のアラアビドプシス(Arabidopsis)野生型(左)およびアンチセンスの向きでセネッセンスDHS遺伝子(第36図に示す配列)の3’末端を発現し、トランスジェニック植物において葉サイズの増加を示すトランスジェニック植物の写真である。
【図22】第22図は、4.6週齢のアラアビドプシス(Arabidopsis)野生型(左)およびアンチセンスの向きでセネッセンスDHS遺伝子(第36図に示す配列)の3’末端を発現し、トランスジェニック植物において葉サイズの増加を示すトランスジェニック植物の写真である。
【図23】第23図は、5.6週齢のアラアビドプシス(Arabidopsis)野生型(左)およびアンチセンスの向きでセネッセンスDHS遺伝子(第36図に示す配列)の3’末端を発現し、トランスジェニック植物において葉サイズの増加を示すトランスジェニック植物の写真である。
【図24】第24図は、6.1週齢のアラアビドプシス(Arabidopsis)野生型(左)およびアンチセンスの向きでセネッセンスDHS遺伝子(第36図に示す配列)の3’末端を発現し、トランスジェニック植物において葉サイズの増加を示すトランスジェニック植物の写真である。
【図25】第25図は、アンチセンスの向きでセネッセンス誘導DHS遺伝子を発現する3つのT1トランスジェニックアラアビドプシス(Arabidopsis)植物系統からの種子収量の増加を示すグラフである。種子収量は種子の体積として表されている。n=30についてのSEを野生型植物について示す。
【0047】
【図26】第26図は、アンチセンスの向きでセネッセンスDHS遺伝子(第36図に示す配列)の3’末端を発現するトランスジェニックトマト植物(左)およびトランスジェニック植物において葉サイズの増加および植物サイズの増加を示す野生型植物(右)の写真である。写真は小植物を土壌に移した後18日に撮った。
【図27】第27図は、アンチセンスの向きでセネッセンスDHS遺伝子(第36図に示す配列)の3’末端を発現するトランスジェニックトマト植物(左)およびトランスジェニック植物において葉サイズの増加および植物サイズの増加を示す野生型植物(右)の写真である。写真は小植物を土壌に移した後32日に撮った。
【図28】第28図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【図29】第29図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【図30】第30図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【0048】
【図31】第31図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【図32】第32図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【図33】第33図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【図34】第34図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【図35】第35図は、野生型からのトマト果実(上部パネル)およびアンチセンスの向きで全長セネッセンスDHS遺伝子を発現するトランスジェニック植物(下部パネル)の写真である。果実を発生のブレーカー段階において収穫し、成長チャンバー内で成熟させた。収集後の日数を各パネルの上左コーナーに示す。
【0049】
【図36】第36図は、植物を形質転換するためにアンチセンスの向きで使用したアラアビドプシス(Arabidopsis)セネッセンス誘導DHS遺伝子の3’末端のヌクレオチド(上部)(配列番号30)および誘導アミノ酸(下部)配列である。
【図37】第37図は、植物を形質転換するためにアンチセンスの向きで使用したトマトセネッセンス誘導DHS遺伝子の3’末端のヌクレオチド(上部)(配列番号31)および誘導アミノ酸(下部)配列である。
【図38】第38図は、全長アラアビドプシス(Arabidopsis)遺伝子を単離するために使用した600塩基対のアラアビドプシス(Arabidopsis)セネッセンス誘導DHSプローブのヌクレオチド(上部)(配列番号26)および誘導アミノ酸(下部)配列である。
【図39】第39図は、全長カーネーション遺伝子を単離するために使用した483塩基対のカーネーションセネッセンス誘導DHSプローブのヌクレオチド(上部)(配列番号27)および誘導アミノ酸(下部)配列である。
【発明を実施するための形態】
【0050】
発明の詳細な説明
植物細胞における1または2以上のセネッセンス誘導DHS遺伝子、1または2以上のセネッセンス誘導elF−5Aまたは両方の発現を変更する方法および組成物が提供される。植物におけるセネッセンス誘導DHSおよびセネッセンス誘導elF−5Aの、単独または組合わせにおける、発現の変更はセネッセンスの開始を遅延し、環境的ストレスおよび病原体に対する抵抗性を改良し、こうして植物の棚持ちおよび/または成長期間を延長する。
【0051】
セネッセンス誘導発現を示すトマトDHS遺伝子をコードする全長cDNA配列は、鋳型として冷却損傷したトマト葉から単離されたRNAを使用する逆転写酵素仲介ポリメラーゼ連鎖反応(RT−PCR)により単離し、RT−PCR生成物を使用して冷却損傷した、ソルビトール処理トマト葉cDNAライブラリーをスクリーニングする。単離されたトマト葉cDNA配列ならびに全長トマト葉cDNAの選択された領域に対応するポリヌクレオチドプローブを使用して、環境的ストレスに暴露された(冷却された)トマト葉、(脱水された)ソルビトール処理されたトマト葉、成熟するトマト果実およびセネッセンシングトマト花の中のDHS遺伝子をコードするmRNAの存在を決定する。
【0052】
アラアビドプシス(Arabidopsis)DHSゲノムクローンから設計したプライマーを使用することによって、鋳型としてセネッセンシングアラアビドプシス(Arabidopsis)葉cDNAライブラリーを使用してポリメラーゼ連鎖反応(PCR)生成物を発生させる。アラアビドプシス(Arabidopsis)ヌクレオチド配列は、セネッセンス誘導トマトDHSの対応する配列と73%のヌクレオチド配列同一性および81%のアミノ酸配列同一性を有する。
【0053】
RT−PCRを使用することによって、本発明のセネッセンス誘導トマトDHS遺伝子を単離した。トマトDHS遺伝子を単離するために使用する上流のプライマーは24ヌクレオチドのプライマーである:5’AG TCT AGA AGG TGC TCG TCC TGA T3’(配列番号3);下流のプライマーは34ヌクレオチドを含有する:5’G ACT GCA GTC GAC ATC GAT(T)153’(配列番号4)。100pmolの下流のプライマーを使用して、標準RT−PCRを使用してcDNAの第1鎖を単離した。次いで、上流のプライマーおよび下流のプライマーの両方を使用するRT−PCRにおいて、鋳型としてとして第1鎖を使用した。
【0054】
アガロースゲル上でRT−PCR生成物を分離すると、1.5kb〜600bpのサイズ範囲の3つの明確なバンドの存在が明らかにされた。それぞれ、上流のプライマーおよび下流のプライマーの中に存在するXbaIおよびSalIクローニング部位を使用して、3つのフラグメントをプラスミドベクター、pBluescript(Stratagene Cloning Systems、カリフォルニア州ラジョラ)の中にサブクローニングし、配列決定した。フラグメントの配列を遺伝子バンクのデータベースの中に存在する配列と比較し、整列させた。結果は1.5kbおよび1kbのフラグメントがトマトDHS配列であることを示した。また、600bpのフラグメントはヒト、酵母およびニューロスポラ(Neurospora)DHS配列と整列した。
【0055】
2Mのソルビトールで6時間処理して脱水を誘導したトマト葉から得られたRNAから作ったトマト(cv.Match F1ハイブリッド)cDNAライブラリーをスクリーニングするために、600bpのRT−PCRフラグメントを使用した。λZap(Stratagene Cloning Systems、カリフォルニア州ラジョラ)cDNAライブラリーキットを使用して、cDNAライブラリーを構築した。セネッセンス誘導DHS遺伝子に対応する3つの同一の陽性全長cDNAクローンが得られ、これらを配列決定した。セネッセンス誘導DHS cDNAクローンのヌクレオチド配列を配列番号1に示す。このcDNAクローンは、42.1kDaの計算した分子量を有する381アミノ酸のポリペプチド(配列番号2)をコードする。
【0056】
発生するおよびストレスに暴露されたトマト花、果実および葉の中の遺伝子の発現パターンに基づいて、それはセネッセンスに関係づけられる。
【0057】
トマトDHS cDNA配列をアラビドプシス・タリアナ(Arabidopsis thaliana)ゲノムバンクの中の未同定ゲノム配列と整列させた(http://genome−www.stanford.edu/Arabidopsis)。結果は未同定アラアビドプシス(Arabidopsis)ゲノム配列(AB107060)との整列を示した。この整列の情報を使用して、アラアビドプシス(Arabidopsis)配列中のオープンリーディングフレームを同定し、それから予測されたアミノ酸配列を発生させた。整列したアラアビドプシス(Arabidopsis)DHS遺伝子の生ずるヌクレオチド配列およびアミノ酸配列を、それぞれ、配列番号5(第2A図)および配列番号6と表示する。
【0058】
同定されたアラアビドプシス(Arabidopsis)DHS配列の短い領域に基づく2つのプライマーを発生させた:プライマー1、5’GGTGGTGTTGAGGAAGATC3’(配列番号7);およびプライマー2、5’GGTGCACGCCCTGATGAAGC3’(配列番号8)。標準PCRにおける2つのプライマーのための鋳型として、アラアビドプシス(Arabidopsis)セネッシング葉cDNAライブラリーを使用した。600bpのPCR生成物を単離し、配列決定し、これらの生成物はゲノムDHS配列の対応するフラグメントのそれと同一の配列を有することが示された。
【0059】
また、全長セネッセンス誘導トマトDHS cDNAを使用して、全長セネッセンス誘導アラアビドプシス(Arabidopsis)およびカーネーションDHS cDNAクローンを単離した。プローブとして全長トマトDHS cDNAクローンを使用して、それぞれ、セネッシングアラアビドプシス(Arabidopsis)葉cDNAライブラリーおよびセネッセンシングカーネーション花弁cDNAライブラリーをスクリーニングすることによって、アラアビドプシス(Arabidopsis)およびカーネーションDHS cDNAクローンを単離した。次いでcDNAライブラリーから得られたcDNAクローンを配列決定した。
【0060】
この方法において単離されたアラアビドプシス(Arabidopsis)全長cDNAクローンのヌクレオチド配列は、トマトcDNA配列との整列によりエンコーディングアラアビドプシス(Arabidopsis)DHSとして同定されたアラアビドプシス(Arabidopsis)ゲノム配列のコーディング領域と同一の配列を有する(第2A図、配列番号5)。全長カーネーション花弁セネッセンス誘導DHSクローンのヌクレオチド配列および誘導アミノ酸配列を第10図に示す(それぞれ、配列番号9および配列番号10)。
【0061】
トマト、カーネーションおよびアラアビドプシス(Arabidopsis)からのDHSをコードする、本発明のcDNA配列を同様な方法においてプローブとして使用して、他の植物からDHS遺伝子を単離し、次いでこれらはトランスジェニック植物における配列を変更するために使用することができる。
【0062】
セネッセンス誘導DHS遺伝子は、DHS遺伝子ファミリーのメンバーであるように思われる。ハイブリッド植物から抽出したゲノムDNAを使用して、トマト葉DNAのゲノムサザンブロット分析を実施した。DHS遺伝子のコーディング領域内の単一部位を認識するか、あるいはDHS遺伝子のオープンリーディングフレーム内のいずれの部位をも認識しない、種々の制限酵素を使用してDNAを切断した。トマトDHSの制限地図を第4図に示す。
【0063】
制限酵素消化したトマト葉ゲノムDNAを32P−dCTP標識化全長トマトDHS cDNAでプロービングした。高いストリンジェンシイの条件下のハイブリダイゼーションは、全長cDNAプローブが各制限酵素消化DNA試料についての3つの制限フラグメントに対してハイブリダイゼーションすることを示した。DHSのオープンリーディングフレーム(第4図)内に制限部位を有する、XbaIおよびEcoRIでトマト葉ゲノムDNAを消化したとき、2より多い制限フラグメントがサザンブロットにおいて検出可能であった(第5図)ことは特に注目すべきである。cv. Match F1、ハイブリッド変種、およびホモ接合性系統、UCT5、からのゲノムDNAは、制限消化の同一パターンを生じた。これらの結果が示唆するように、トマト植物の中に2またはそれ以上のイソ型が存在する。第3図に示すように、DHS遺伝子は種を通じて高度に保存されるので、任意の種内のイソ型の間で有意な量の保存が存在する。
【0064】
全長トマトcDNAでプロービングしたトマト花全RNAのノザンブロットは、セネッセンス誘導DHS遺伝子の発現がトマト花において有意に誘導されるが、発現は芽においてわずかに検出可能であることを示す(第6図)。トマト果実の種々の発生段階の間におけるDHS発現のノザンブロット分析は、DHS遺伝子がブレーカーおよびピンク色果実において低いレベルで発現され、これに対して赤色(成熟)トマト果実におけるDHS発現が有意に増強されることを証明する(第7図)。
【0065】
ノザンブロット分析は、また、セネッセンス誘導DHS遺伝子が環境的ストレス条件、例えば、脱水(第8図)および冷却(第9図)により誘導されることを証明する。脱水を誘導するように2Mのソルビトールで処理したトマト葉は、非処理葉に比較して脱水された葉におけるDHS発現の誘導を証明する(第8図)。冷却温度に暴露し、周囲温度に戻した植物は、冷却損傷症状(例えば、漏出)の発生と一致するセネッセンス誘導DHS遺伝子の発現の誘導を示す(第9図)。トマト植物および種々の植物組織、例えば、葉、果実および花における遺伝子発現の全体のパターンは、本発明のDHS遺伝子がこれらの植物および植物組織におけるセネッセンスの開始に関係することを証明する。
【0066】
DHS遺伝子発現の誘導による同様な結果は、アラアビドプシス(Arabidopsis)における葉セネッセンスおよびカーネーションの花弁セネッセンスの開始とともに観測される。種々の年齢の植物から単離されたアラアビドプシス(Arabidopsis)葉の全RNAのノザンブロット分析は、セネッセンス誘導DHS遺伝子の発現が若い(5週齢の)植物において明らかでないが、約6週において出現し始めることを示す。DHS遺伝子の発現は7週までに有意に誘導される。ノザンブロット分析は、植物が加齢するにつれてアラアビドプシス(Arabidopsis)DHS遺伝子が有意に増強されることを示す(第11図)。
【0067】
ノザンブロット分析は、また、DHS遺伝子が顕花植物、例えば、カーネーションにおいて同様に調節されることを示す(第12図)。種々の年齢のカーネーションの花弁から単離された全RNAのノザンブロット分析において、年齢誘導セネッセンスの症状、例えば、セネッセンスの最初の形態学的発現である、花弁のインローリング(inrolling)を有する花からの花弁において、カーネーションDHSの発現は有意に誘導されるが、堅い芽の花において発現は非常に低いことが示された。ちょうど開花し始めているカーネーションの花からの花弁は、堅い芽の段階における花よりも有意に多いDHS発現を有し、そして完全に開いた花からの花弁もまたDHS発現の増強を示す。
【0068】
こうして、植物組織におけるセネッセンス誘導DHS遺伝子の発現を実質的に表すか、あるいは変更することによって、劣化および腐敗を遅延し、腐敗しやすい果実、花、および野菜の棚持ちを増加することができ、そして植物およびそれらの組織をよりストレス抵抗性としかつ病原体抵抗性とすることができることが期待される。これは次のようにして達成することができる。好ましくは構成的プロモーター、例えば、CaMV 35Sプロモーターを使用するか、あるいは組織特異的またはセネッセンス/ストレス誘導プロモーターを使用して、DHS cDNAまたはそのオリゴヌクレオチドフラグメントが果実、花、葉および野菜においてアンチセンス立体配置において発現されるトランスジェニック植物を産生する。
【0069】
また、植物におけるセネッセンス関係形態学的変化の誘導に関係する、他の遺伝子、elF−5A、が単離され、ここにおいて配列決定され、そしてDHSと同様に、好ましくは、植物の中にアンチセンスの向きで導入することによって、植物におけるセネッセンスおよびセネッセンス関係プロセスを変更するためにそれを使用することができる。成熟するトマト果実、セネッシングアラアビドプシス(Arabidopsis)葉およびセネッシングカーネーション花cDNAライブラリーの各々から、全長セネッセンス誘導elF−5A cDNAを単離した。
【0070】
全長クローンの各々のヌクレオチド配列および誘導アミノ酸配列を第13図(トマトセネッセンス誘導elF−5A)、第14図(カーネーションセネッセンス誘導elF−5A)および第15図(アラアビドプシス(Arabidopsis)セネッセンス誘導elF−5A)に示す。また、これらのcDNAの各々のヌクレオチド配列を配列番号11(トマト)(第13図)、配列番号13(カーネーション)(第14図)および配列番号15(Arabidopsis)(第15図)として示す。遺伝子の各々の誘導アミノ酸配列を、それぞれ、配列番号12(第13図)、配列番号14(第14図)および配列番号16(第15図)として示す。
【0071】
本明細書に記載するDHS遺伝子配列を使用して場合におけるように、本発明のelF−5A配列を使用して他の植物からelF−5A遺伝子を単離することができる。単離されたelF−5A配列を使用して、植物におけるセネッセンスおよびセネッセンス関係プロセスを変更することができる。少なくとも約70%の交差種の配列類似性に基づいて、既知の方法を使用して、植物からのelF−5A配列の単離を達成することができる。
【0072】
elF−5AおよびDHSの平行誘導は植物においてセネッセンスの間に起こる。自然およびストレス誘導セネッセンスの両方の開始において、elF−5AはDHSと平行にアップレギュレートされることをノザンブロット分析は証明する(第16図〜第20図)。例えば、種々の段階におけるアラアビドプシス(Arabidopsis)植物の葉から単離された全RNAのノザンブロット分析において、葉セネッセンスが植物において明らかであるとき、elF−5Aの発現は誘導され、セネッセンスが進行するにつれて、発現は有意に増強されたことが証明される。結実植物、例えばトマトにおいて、elF−5AおよびDHSは果実の軟化および腐敗の開始と一致して赤色柔軟果実においてアップレギュレートされる(第17図)。
【0073】
また、ノザンブロット分析において、elF−5AおよびDHSは、環境的ストレス、例えば、乾燥(第18図)および冷却損傷(第20図)に応答して植物中で平行にアップレギュレートすることが証明される。同様に、顕花植物において、elF−5AおよびDHSは開いた花において平行にアップレギュレートされ、そして両方の遺伝子の発現は顕花の後の段階を通して増強され続ける。
【0074】
クローニングしたセネッセンス誘導DHS遺伝子、それらの1または2以上のフラグメント、あるいはクローニングしたセネッセンス誘導elF−5A遺伝子、それらの1または2以上のフラグメント、あるいはelF−5AとDHS配列との組合わせは、構成的プロモーター、例えば、イチジクイボモザイクウイルス35Sプロモーター、カリフラワーモザイクウイルスプロモーターCaMV35S、二重35SプロモーターまたはMASプロモーターの制御下に逆向き(アンチセンス)に導入されるとき、植物を遺伝的に修飾し、修飾された植物におけるセネッセンスを変更することができる。
【0075】
トマト、アラアビドプシス(Arabidopsis)またはカーネーションのセネッセンス誘導DHS遺伝子またはセネッセンス誘導elF−5A遺伝子と十分な配列同一性を共有する他の植物から選択されたアンチセンス配列を使用して、同様に遺伝的修飾を達成することができる。遺伝的修飾の1つの結果は、mRNAをコードする内因的翻訳可能セネッセンス誘導DHS、mRNAをコードするelF−5Aまたは両方の量の減少である。結局、植物細胞において産生されたセネッセンス誘導DHSおよび/またはセネッセンス誘導elF−5Aの量は減少され、これにより活性化されたelF−5Aの量は減少し、これは引き続いてセネッセンス誘導タンパク質、例えば、セネッセンス誘導リパーゼ、セネッセンス誘導プロテアーゼおよびセネッセンス誘導ヌクレアーゼの翻訳を減少させる。セネッセンスの開始のために新たなタンパク質合成が必要であるので、こうしてセネッセンスは阻害または遅延される。
【0076】
例えば、二重35Sプロモーターの調節下にアラアビドプシス(Arabidopsis)セネッセンス誘導DHS遺伝子(配列番号26)(第38図)の全長または3’領域をアンチセンスの向きに発現するベクターで形質転換されたアラアビドプシス(Arabidopsis)植物は、第21図〜第24図に示すように対照植物に比較して、成長のすべての段階を通じてバイオマスの増加、例えば、より大きい葉サイズおよび全体のより大きい植物成長、および葉セネッセンスの遅延を示す。
【0077】
トランスジェニックアラアビドプシス(Arabidopsis)植物においてアラアビドプシス(Arabidopsis)セネッセンス誘導DHS遺伝子の完全なまたは3’コーディング領域をアンチセンスの向きで発現することによって発生したセネッセンス誘導DHS遺伝子の発現の減少の効果は、また、トランスジェニック植物において種子収量の増加として見られる。アラアビドプシス(Arabidopsis)セネッセンス誘導DHS遺伝子のアンチセンス3’非コーディング領域を発現するアラアビドプシス(Arabidopsis)植物系統は、野生型植物よりも6倍までの種子を生産する(第25図)。
【0078】
アンチセンスの向きでかつ二重35Sプロモーターの調節下にトマトセネッセンス誘導DHS遺伝子(配列番号27)の3’末端で形質転換されたトマト植物を使用して、同様な結果が得られる。アンチセンスの向きで遺伝子の3’末端で形質転換された植物は、対照(非形質転換)植物に比較して葉サイズの増加および植物サイズの増加を示す(第26図および第27図)。
【0079】
全長トマトセネッセンス誘導DHSでアンチセンスの向きに形質転換されたトマト植物は、野生型植物に比較して遅延した軟化および腐敗を示す果実を産生する(第28図〜第35図)。こうして、本発明の方法および配列を使用して、果実の軟化および腐敗を遅延し、ならびに植物バイオマスおよび種子収量を増加し、一般に、植物においてセネッセンスを遅延することができる。
【0080】
本発明の単離されたヌクレオチド配列を使用して、他の植物または生物から実質的に相補的なDHSおよび/またはelF−5Aヌクレオチド配列を単離することができる。引き続いて、ここに示す単離されたヌクレオチド配列の使用とともに示した方法と同一の方法において、これらの配列を使用して、植物を形質転換し、これにより形質転換された植物のセネッセンスを変更することができる。
【0081】
DHS、elF−5A、それらのフラグメントまたはそれらの組合わせで植物を形質転換して得られる遺伝的修飾は、植物におけるセネッセンス誘導DHS、elF−5Aまたは両方のレベルを永久的に変化させることができ、そして自殖または他の生殖スキームにより子孫植物において増殖させることができる。遺伝的に変更された植物を使用して、変更が世代から世代に安定に伝達される植物の新しい変種または系統を産生する。本発明は、広い範囲の異なる植物においてセネッセンスの安定な遺伝的修飾を達成するために使用できる、適当なDNA配列を最初に提供する。
【0082】
セネッセンス誘導DHS遺伝子およびelF−5A遺伝子を同定し、単離するために、一般に、この分野においてよく知られている慣用法を使用して、プラスミドDNAの調製、制限酵素消化、DNAのアガロースゲル電気泳動、タンパク質のポリアクリルアミドゲル電気泳動、PCR、RT−PCR、サザンブロット、ノザンブロット、DNA結合および細菌の形質転換を実施した。例えば、下記の文献を参照のこと:Sambrook J. 他、Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor Press、Cold Spring Harbor、NY、1989。核酸のハイブリダイゼーション技術はSambrook(前掲)に開示されている。
【0083】
本明細書において使用するとき、用語「植物」は全植物、植物部分、植物細胞または植物細胞のグループを意味する。本発明の方法において使用できる植物の型は、例えば、下記のものを包含するが、これらに限定されない:エチレン感受性およびエチレン不感受性植物;結実植物、例えば、アプリコット、リンゴ、オレンジ、バナナ、グレープフルーツ、セイヨウナシ、トマト、イチゴ、アボカド、およびその他;野菜、例えば、ニンジン、エンドウ、レタス、キャベツ、カブ、ジャガイモ、ブロッコリ、アスパラガス、およびその他;花、例えば、カーネーション、バラ、キク、およびその他;耕種学的作物植物および森林種、例えば、トウモロコシ、イネ、大豆、アルファルファおよびその他;および一般に、本発明のDNA分子を吸収しかつ発現する任意の植物。それは種々の倍数レベル、例えば、一倍体、二倍体、四倍体および多倍数体の植物を包含することができる。植物は単子葉植物または双子葉植物であることができる。
【0084】
トランスジェニック植物は、ここにおいてある方法において遺伝的に修飾された植物として定義され、下記の植物を包含するが、これらに限定されない:異種または同種セネッセンス誘導DHS DNAまたは修飾されたDNA、あるいは異種セネッセンス誘導DHS DNAまたは同種セネッセンス誘導DHS DNAのある部分をそのゲノムの中に組込んで有する植物。あるいは、本発明のトランスジェニック植物は、異種または同種セネッセンス誘導elF−5A DNAまたは修飾されたDNA、あるいは異種セネッセンス誘導elF−5A DNAまたは同種セネッセンス誘導elF−5A DNAのある部分をそのゲノムの中に組込んで有することができる。
【0085】
本発明のトランスジェニック植物は、異種または同種セネッセンス誘導DHSおよびelF−5A DNAまたは修飾されたDNA、あるいは異種セネッセンス誘導DHSおよびelF−5A DNAまたは同種セネッセンス誘導DHSおよびelF−5A DNAのある部分、あるいは異種および同種のDHSおよびelF−5A配列の組合わせをそのゲノムの中に組込んで有することができる。変更された遺伝物質は、タンパク質をコードすることができ、調節配列または対照配列を含んでなることができるか、あるいはアンチセンス配列であるかまたはそれを含むか、あるいは内因的セネッセンス誘導DHSまたはelF−5AまたはmRNA配列またはその一部分に対してアンチセンスであるか、あるいはアンチセンスmRNA配列をコードすることができる。「トランスジーン」または「トランスジェニック配列」は、トランスジェニック植物の中に組込まれた外来遺伝子または部分的配列として定義される。
【0086】
用語「ハイブリダイゼーション」は、本明細書において使用するとき、一般に、プローブ配列およびターゲット配列の特質に依存して当業者にとって容易に明らかなように、適当なストリンジェンシイの条件における核酸のハイブリダイゼーションを意味するために使用される。ハイブリダイゼーションおよび洗浄の条件はこの分野においてよく知られており、そしてインキュベーション時間、温度および/または溶液のイオン強度を変化させることによって所望のストリンジェンシイに依存して条件を調節することは容易に達成される。例えば、下記の文献を参照のこと:Sambrook J. 他、Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor Press、Cold Spring Harbor、New York、1989。
【0087】
ハイブリダイゼーションする配列の長さ、特にプローブ配列の長さ、核酸の相対的G−C含量および許容すべき不一致の量により、条件を選択する。より低い程度の相補性を有する鎖間の部分的ハイブリダイゼーションを望むとき、低いストリンジェンシイの条件が好ましい。完全なまたはほぼ完全な相補性を望むとき、高いストリンジェンシイの条件が好ましい。典型的な高いストリンジェンシイの条件について、ハイブリダイゼーション溶液は6×SSC、0.01MのEDTA、1×デンハルト溶液および0.5%のSDSを含有する。ハイブリダイゼーションは約68℃においてクローニングされたDNAについて約3〜4時間実施し、そして全真核DNAについて約12〜約16時間実施する。より低いストリンジェンシイのために、ハイブリダイゼーション温度を二本鎖の溶融温度(TM)より約42℃低く低下させる。TMはG−C含量および二本鎖長さならびに溶液のイオン強度の関数であることが知られている。
【0088】
本明細書において使用するとき、用語「実質的配列同一性」または「実質的相同性」は、ヌクレオチド配列またはアミノ酸配列が他のヌクレオチド配列またはアミノ酸配列と実質的な構造的または機能的同等性を表すことを示すために使用される。実質的配列同一性または実質的相同性を有する配列間の構造的または機能的差は最小であろう;すなわち、それらは所望の用途において示すように配列が機能する能力に影響を与えないであろう。差は、例えば、異なる種間のコドン使用頻度における固有変動のためであることがある。
【0089】
2またはそれ以上の異なる配列間に有意な量の配列のオーバーラップまたは類似性が存在する場合、あるいは配列の長さまたは構造が異なる場合でさえ異なる配列が同様な物理的特性を示す場合、構造的差は最小であると考えられる。このような特性は、例えば、定められた条件下にハイブリダイゼーションする能力、またはタンパク質の場合において、免疫学的交差反応性、同様な酵素活性、およびその他を包含する。熟練した実施者は、既知の方法により、これらの特性の各々を容易に決定することができる。
【0090】
さらに、2つのヌクレオチド配列がそれらの間で少なくとも約70%、より好ましくは80%、最も好ましくは約90%の配列類似性を有する場合、それらの配列は「実質的に相補性」である。2つのアミノ酸配列がポリペプチドの活性部分間の少なくとも50%、好ましくは70%の類似性を有する場合、それらは実質的に相同性である。
【0091】
本明細書において使用するとき、DNAまたはRNA分子の「対応する部分に対してハイブリダイゼーションする」という句は、ハイブリダイゼーションする分子、例えば、オリゴヌクレオチド、ポリヌクレオチド、または任意のヌクレオチド配列が(センスまたはアンチセンスの向きで)ほぼ同一サイズの他の核酸分子中の配列を認識し、それに対してハイブリダイゼーションし、そして適当な条件下にハイブリダイゼーションを実行するために十分な類似性をその配列に対して有することを意味する。
【0092】
例えば、トマトDHSの3’コーディングまたは非コーディング領域からの100ヌクレオチド長さのアンチセンス分子は、2つの配列間に約70%またはそれより高い配列類似性が存在するかぎり、カーネーションDHS遺伝子または任意の他の植物DHS遺伝子のそれぞれ3’コーディングまたは非コーディング領域内のヌクレオチド配列のほぼ100ヌクレオチド部分を認識し、それに対してハイブリダイゼーションする。「対応する部分」がそれに対してハイブリダイゼーションする分子よりも小さいか、あるいは大きくあるように、例えば、20〜30%小さいか、あるいは大きく、好ましくは約12〜15%小さいか、あるいは大きくあるように、「対応する部分」のサイズがハイブリダイゼーションにおいて多少の不一致を可能とすることを理解すべきである。
【0093】
核酸(またはポリヌクレオチドまたはオリゴヌクレオチド)の「機能的誘導体」という用語は、ここにおいて、セネッセンス誘導DHSまたはセネッセンス誘導elF−5Aをコードする遺伝子またはヌクレオチド配列のフラグメント、変異型、相同体またはアナローグを意味するために使用する。機能的誘導体は、本発明による実用性を可能とするセネッセンス誘導DHSまたはelF−5AをコードするDNAの少なくとも一部分を保持することができる。
【0094】
このような機能は、低いストリンジェンシイの条件下に、天然のトマト、アラアビドプシス(Arabidopsis)またはカーネーションのセネッセンス誘導DHSまたはelF−5A、あるいはセネッセンス誘導DHSまたはelF−5Aをコードする他の植物からの実質的に相同性のDNAと、あるいはそれらのmRNA転写物と、または、アンチセンスの向きで、ハイブリダイゼーションして、植物セネッセンス誘導DHSまたはelF−5A mRNAの転写および/または翻訳、およびその他を阻害する能力を包含することができる。
【0095】
遺伝子またはDNA配列の「フラグメント」は、分子のサブセット、例えば、より短いポリヌクレオチドまたはオリゴヌクレオチドを意味する。「変異型」は、全遺伝子またはそのフラグメントに実質的に類似する分子、例えば、1または2以上の置換したヌクレオチドを有するが、特定の遺伝子とハイブリダイゼーションする能力または自然DNAとハイブリダイゼーションするmRNA転写物をコードする能力を維持するヌクレオチド置換変異型を意味する。「相同体」は、異なる植物の属または種からのフラグメントまたは変異型配列を意味する。「アナローグ」は、全分子、その変異型またはフラグメントに実質的に類似するか、あるいは機能的にそれらに関係する、非天然分子を意味する。
【0096】
遺伝子、例えば、セネッセンス誘導DHS遺伝子またはセネッセンス誘導elF−5A遺伝子の「変更された発現」または「修飾された発現」とは、遺伝子の正常の発現、例えば、非修飾結実植物、顕花植物または他の植物における起こる発現がある方法において変化される、任意のプロセスまたは結果を意味する。本明細書において意図するとき、遺伝子発現の変更は、セネッセンス誘導DHS遺伝子またはセネッセンス誘導elF−5A遺伝子または両方の発現の完全なまたは部分的減少であるが、また、発現のタイミングの変化、あるいはセネッセンス誘導DHS遺伝子またはセネッセンス誘導elF−5A遺伝子または両方の発現が非修飾植物または栽培品種において最も自然に起こりやすい発現と異なる他の状態を包含することができる。好ましい変更は、非修飾植物における産生に比較して植物によるセネッセンス誘導DHS産生またはセネッセンス誘導elF−5A産生または両方を減少させる変更である。
【0097】
本発明に従い遺伝的に変更された植物を生産するとき、所望の特性、一般にセネッセンス誘導DHSの発現または産生の減少またはセネッセンス誘導elF−5Aの発現の減少または両方により個々の小植物または植物を選択することが好ましい。セネッセンス誘導DHSおよびセネッセンス誘導elF−5Aの発現は、例えば、トランスジェニック植物におけるセネッセンスの遅延または減少の観測により決定することができる。また、既知の方法を使用して対照(正常、非トランスジェニック)植物に比較してトランスジェニック植物におけるDHSおよび/またはelF−5Aの活性を定量することが可能である。
【0098】
新しく挿入された遺伝子またはDNA配列を発現させ、それがコードするタンパク質を産生するか、あるいは転写すべきアンチセンスDNAの場合において、アンチセンスRNA分子を産生するために、適切な調節因子を遺伝子またはDNA配列に関して適切な位置にかつ向きで存在させるべきである。調節領域はプロモーター、5’非翻訳リーダー配列および3’ポリアデニル化配列ならびにエンハンサーおよび他の調節配列を包含することができる。
【0099】
遺伝子のセンスまたはアンチセンス転写物を発生させるためにセネッセンス誘導DHS遺伝子と組み合わせて有効であるプロモーター調節因子は、一般に任意のプロモーター、特に構成的プロモーター、例えば、イチジクイボモザイクウイルス35Sプロモーター、カリフラワーモザイクウイルスプロモーター、CaMV35Sプロモーター、またはMASプロモーター、または組織特異的またはセネッセンス誘導プロモーター、例えば、カーネーション花弁GST1プロモーターまたはアラアビドプシス(Arabidopsis)SAG12プロモーターを包含する。
【0100】
(例えば、下記の文献を参照のこと:J.C.Palaqui他、Plant Physiol. 112:1447−1456(1996);Morton他、Molecular Breeding、1:123−132(1995);Fobert他、Plant Journal、6:567−577(1994);およびGan他、Plant Physiol.、113:313(1997)、引用することによって本明細書の一部とされる)。好ましくは、本発明において使用するプロモーターは構成的プロモーター、最も好ましくは二重35Sプロモーターを使用する。
【0101】
本発明のために有効であるプロモーターからの発現レベルは、慣用の発現系を使用して、例えば、プロモーター/リポーター遺伝子が導入されたトランスジェニック植物の葉、花、果実または他の組織の抽出物中のリポーター遺伝子産物、例えば、タンパク質またはmRNAのレベルを測定することによって試験することができる。
【0102】
本発明は、トマトセネッセンス誘導DHS、カーネーションセネッセンス誘導DHS、アラアビドプシス(Arabidopsis)セネッセンス誘導DHSをコードする遺伝子に対して相補的であるか、あるいは低い〜高いストリンジェンシイの条件下にトマト、カーネーションまたはアラアビドプシス(Arabidopsis)のセネッセンス誘導DHS遺伝子とハイブリダイゼーションする、他の植物からの遺伝子または遺伝子フラグメントに対して相補的である、アンチセンスオリゴヌクレオチドおよびポリヌクレオチドを提供する。
【0103】
本発明は、また、トマトセネッセンス誘導elF−5A、カーネーションセネッセンス誘導elF−5A、アラアビドプシス(Arabidopsis)セネッセンス誘導elF−5Aをコードする遺伝子に対して相補的であるか、あるいは低い〜高いストリンジェンシイの条件下にトマト、カーネーションまたはアラアビドプシス(Arabidopsis)のセネッセンス誘導elF−5A遺伝子とハイブリダイゼーションする、他の植物からの遺伝子または遺伝子フラグメントに対して相補的である、アンチセンスオリゴヌクレオチドおよびポリヌクレオチドを提供する。
【0104】
このようなアンチセンスオリゴヌクレオチドは少なくとも約6ヌクレオチド長さであってハイブリダイゼーションの最小の特異性を提供し、そしてセネッセンス誘導遺伝子またはその一部分をコードするDNAまたはmRNAの1つの鎖に対して、あるいはセネッセンス誘導DHSまたはelF−5A遺伝子の発現の調節に関係するゲノムDNA中のフランキング配列に対して相補的であることができる。アンチセンスオリゴヌクレオチドは100ヌクレオチドまたはそれより大きい程度に大きく、そしてそれに対してアンチセンスである完全なコーディング配列までかつそれを越える長さに伸長することができる。アンチセンスオリゴヌクレオチドはDNAまたはRNAまたはそれらのキメラ混合物または誘導体または修飾されたバージョン、一本鎖または二本鎖であることができる。
【0105】
アンチセンスオリゴヌクレオチドの作用は、細胞におけるセネッセンス誘導DHSの発現、セネッセンス誘導elF−5Aの発現または両方の変更、主として阻害を生ずることがある。アンチセンスの一般的開示については、下記の文献を参照のこと:Albert他、Molecular Biology of the Cell、第2版、Garland Publishing,Inc.、New York、New York、1989(特にpp.195−196、引用することによって本明細書の一部とされる)。
【0106】
アンチセンスオリゴヌクレオチドは、セネッセンス誘導DHSまたはelF−5A遺伝子の任意の対応する部分に対して相補的であることができる。他の態様において、アンチセンスオリゴヌクレオチドは6〜100ヌクレオチド長さであり、例えば、セネッセンス誘導DHSまたはelF−5A配列の3’末端内の5’非コーディングまたは配列に対して相補的であることができる。主として5’非コーディング配列に対して相補的であるアンチセンスオリゴヌクレオチドは、転写因子をコードする遺伝子の発現の有効なインヒビターであることが知られている。Branch、M.A.、Molec.Cell.Biol.,13:4284−4290(1993)。
【0107】
好ましいアンチセンスオリゴヌクレオチドはセネッセンス誘導DHSまたはセネッセンス誘導elF−5AをコードするmRNAの一部分に対して実質的に相補的であり、mRNAの一部分はアンチセンスオリゴヌクレオチドとほぼ同一のサイズである。例えば、セネッセンス誘導DHSまたはelF−5Aをコードする全長cDNAクローンを植物の中にアンチセンスの向きで導入すると、セネッセンス誘導DHSおよび/またはelF−5A遺伝子の発現を首尾よく変更することが期待される。そのうえ、第21図〜第35図において証明されるように、セネッセンス誘導DHS遺伝子またはセネッセンス誘導elF−5A遺伝子または両方の特定の部分をターゲットとした、部分的配列の導入は等しく有効であることがある。
【0108】
本発明が必要とする相同性の最小量は、特定のターゲットRNAまたはDNAを認識しかつその翻訳または機能を阻害または減少するために十分な相補性を生ずると同時に他のRNAまたはDNA分子の機能および他の遺伝子の発現に影響を与えないために十分である量である。本発明のアンチセンスオリゴヌクレオチドはセネッセンス誘導DHS遺伝子またはセネッセンス誘導elF−5A遺伝子のRNA転写物の対応する部分に対して相補的である配列を含んでなるが、絶対的相補性は、好ましいが、不必要である。
【0109】
ハイブリダイゼーションする能力は、アンチセンスオリゴヌクレオチドの長さおよび相補性の程度に依存することがある。一般に、ハイブリダイゼーションする核酸が長いほど、それが含有し、なお安定な二本鎖を形成することができる、セネッセンス誘導DHSターゲット配列に対してより多くの塩基が一致しなくなる。当業者は標準手順を使用して、例えば、ハイブリッドの複合体の溶融温度を測定することによって、許容可能な不一致の程度を確認することができる。
【0110】
外因的に導入された核酸配列からの転写により、アンチセンスRNAオリゴヌクレオチドを細胞内で発生させることができる。適当な調節因子、例えば、プロモーターに作用可能に連鎖されたアンチセンスセネッセンス誘導DHS配列をコードするDNAが組込まれたベクター、例えば、プラスミドまたはウイルスを使用する形質転換、トランスフェクションまたは感染により、アンチセンス分子を細胞の中に送達することができる。細胞内で外因的DNA配列が発現され、セネッセンス誘導DHS遺伝子のアンチセンスRNAが産生される。
【0111】
ベクターは好ましくはプラスミドであることができるか、あるいは植物細胞または細菌細胞中で複製し、コードされる遺伝子を発現することがこの分野において知られているウイルスまたは他のベクターであることができる。ベクターは所望のアンチセンスセネッセンス誘導DHS RNAを産生するように転写されることができるように、染色体的に統合されるようになる。このようなプラスミドまたはウイルスベクターは、この分野において標準的である組換えDNA技術的方法により構築することができる。例えば、ベクターは原核宿主中で機能的である複製系および本発明によるアンチセンスオリゴヌクレオチドまたはポリヌクレオチドを含有するプラスミドベクターであることができる。
【0112】
あるいは、ベクターはアグロバクテリウム(Agrobacterium)において機能的である複製系および本発明によるアンチセンスオリゴヌクレオチドまたはポリヌクレオチドを含有するプラスミドであることができる。アグロバクテリウム(Agrobacterium)中で複製することができるプラスミドはこの分野においてよく知られている。下記の文献を参照のこと:Miki他、Procedures of Introducing Foreign DNA Into Plants、Methods in Plant Molecular Biology and Biotechnology、B. .GlickおよびJ.E.Thompson編、CRC Press(1993)、pp. 67−83。
【0113】
トマトDHS遺伝子を下記の方法においてプラスミドベクターの中にアンチセンスの向きでクローニングした。pUC18バックボーンから構築され、カリフラワーモザイクウイルス(CaMV)および引き続いて多重クローニング部位およびオクタパインシンターゼ終結配列を含有するpCDプラスミドを使用して、トマトDHS遺伝子をクローニングした。XhoIおよびSacI制限部位を使用してpCDプラスミドの中に全長トマトDHS遺伝子をサブクローニングすることによって、pCd−DHS(アンチセンス)プラスミドを構築した。
【0114】
好ましくは約6〜約100ヌクレオチド長さでありかつセネッセンス誘導DHSまたはセネッセンス誘導elF−5Aのターゲット配列に対して相補的である、オリゴヌクレオチドは組換えヌクレオチド技術により調製することができるか、あるいは、例えば、モノヌクレオチドまたはより短いオリゴヌクレオチドから合成することができる。自動化合成装置を本発明のオリゴヌクレオチドおよびポリヌクレオチドの化学的合成に適用可能である。
【0115】
本発明に従い組換えヌクレオチド分子を構築する手順は下記の文献に開示されている:Sambrook他、Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor Press、Cold Spring Harbor、N.Y.(1989)、これは引用することによって本明細書の一部とされる。セネッセンス誘導デオキシハイプシンシンターゼ配列に対して相補的であるアンチセンスRNAをコードするオリゴヌクレオチドは、この分野においてよく知られている手順を使用して調製することができる。このような手順は下記の文献に詳述されている:Maniatis、T.他、Molecular mechanisms in the Control of Gene expression、Nierilich他、編、Academic Press、N.Y.(1976)。
【0116】
本発明の別の態様において、内因的植物セネッセンス誘導DHS、セネッセンス誘導elF−5Aまたは両方の発現の阻害は、植物細胞の中に導入された外因的セネッセンス誘導DHSまたはelF−5A遺伝子または遺伝子フラグメントまたは両方の過剰発現による共抑制の結果である。本発明のこの態様において、アンチセンス分子について本明細書において記載した方法と同一の方法により、センスの向きでセネッセンス誘導DHS、セネッセンス誘導elF−5Aまたは両方をコードするベクターを細胞の中に導入する。好ましくは、セネッセンス誘導DHSまたはセネッセンス誘導elF−5Aを強い構成的プロモーター、例えば、イチジクイボモザイクウイルスプロモーターまたはCaMV35Sまたは二重35Sプロモーターに作用可能に連鎖させる。
【0117】
本発明の他の態様において、内因的植物セネッセンス誘導DHS、セネッセンス誘導elF−5Aまたは両方の発現の阻害はリボザイムを使用して実施される。リボザイムは配列特異的エンドリボヌクレアーゼ活性を示すRNA分子である。1例はハンマーヘッドリボザイムであり、これはターゲットRNA中のUH(ここでHはA、CまたはU残基である)認識部位において切断し、かつリボザイムの触媒ドメインを基質RNAのターゲット部位に向ける塩基対合領域を含有する。リボザイムは高度にターゲット特異的であり、そして多重遺伝子ファミリーの1メンバーを不活性化するように設計するか、あるいは関係するmRNAの保存された領域にターゲットすることができる(下記の文献を参照のこと:Merlo他、The Plant Cell、10:1603−1621、1998)。
【0118】
リボザイム分子は、適当な調節因子、例えば、プラスミドに作用可能に連鎖されたリボザイムが組込まれたベクター、例えば、プラスミドまたはウイルスを使用する形質転換、トランスフェクションまたは感染により、細胞に送達すことができる。このようなリボザイム構築物は塩基対合アームを含有し、これらのアームはリボザイム構築物をセネッセンス誘導DHS mRNA、またはセネッセンス誘導elF−5A mRNA内の切断部位に向け、DHSまたはelF−5A mRNAの切断およびセネッセンス誘導DHSおよび/またはelF−5A発現の阻害を生ずる。
【0119】
本発明に従い作られるトランスジェニック植物は、この分野において知られている植物形質転換の任意の方法を使用するDNA形質転換により調製することができる。植物形質転換法は下記の方法を包含する:植物、組織または細胞とアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)との同時培養または直接的感染(Miki他、Meth.in Plant Mol.Biol.and Biotechnology、(1993)、p.67−88);プロトプラストの中への直接的遺伝子転移またはプロトプラスト吸収(Paszkowsk他、EMBO J.、12:2717(1984);エレクトロポレーション(Fromm他、Nature、319:719(1986);粒子衝撃(Klein他、Bio Technology、6:559−563(1988);実生および植物の分裂組織の中への注入(De LaPena他、Nature、325:274−276(1987);培養した細胞および組織のプロトプラストの中への注入(Reich他、Bio Technology、4:1001−1004(1986))。
【0120】
一般に、完全な植物は形質転換プロセスから得られる。プロトプラスト、カルス、組織の部分または外植体、およびその他から植物を再生する。セネッセンス誘導DHS、セネッセンス誘導elF−5Aまたは両方の発現が変更された再生した植物から得られた植物部分、例えば、葉、花、果実、種子およびその他は、本明細書において使用する「植物」の定義の中に含まれる。再生植物の子孫、変異型および突然変異もまた「植物」の定義の中に含まれる。
【0121】
トマト、カーネーションまたはアラアビドプシス(Arabidopsis)セネッセンス誘導DHSタンパク質またはそれらの機能的誘導体、およびトマト、カーネーションまたはアラアビドプシス(Arabidopsis)セネッセンス誘導elF−5Aタンパク質またはそれらの機能的誘導体は、必要に応じて化学的合成法と組み合わせて、組換え技術により産生されることが好ましい。本発明の1つの態様において、セネッセンス誘導DHSは融合タンパク質として発現され、この融合タンパク質はマルトース結合性植物と融合されたセネッセンス誘導DHSから成ることが好ましい。
【0122】
本明細書において記載するセネッセンス誘導DHSまたはセネッセンス誘導elF−5Aタンパク質の「機能的誘導体」は、それぞれ、セネッセンス誘導DHSまたはセネッセンス誘導elF−5Aのフラグメント、変異型、アナローグ、または化学的誘導体であり、これらはセネッセンス誘導DHSまたはelF−5A活性またはそれぞれセネッセンス誘導DHSまたはセネッセンス誘導elF−5Aに対して特異的な抗体との交差反応性の少なくとも一部分を保持する。セネッセンス誘導DHSまたはセネッセンス誘導elF−5Aタンパク質のフラグメントは、この分子の任意のサブセットを意味する。変異型ペプチドは直接的化学的合成により、例えば、この分野においてよく知られている方法を使用して作ることができる。
【0123】
セネッセンス誘導DHSまたはセネッセンス誘導elF−5Aのアナローグは、全タンパク質またはそのフラグメントに実質的に類似する非天然タンパク質を意味する。セネッセンス誘導DHSまたはセネッセンス誘導elF−5Aの化学的誘導体は、通常ペプチドまたはペプチドフラグメントの一部分でない追加の化学的部分を含有する。選択した側鎖または末端の残基と反応することができる有機誘導体化因子とペプチドのターゲッテッドアミノ酸残基を反応させることによって、修飾をペプチドまたはそのフラグメントの中に導入することができる。
【0124】
本発明のヌクレオチド配列で形質転換された細胞(センス向きで)を培養し、細胞がタンパク質を合成するようにさせ、次いで、使用するクローニングプロトコルに依存して、遊離タンパク質または融合タンパク質として、培地または細胞抽出物からタンパク質を単離することによって、本発明によるセネッセンス誘導DHSまたはセネッセンス誘導elF−5Aタンパク質またはペプチドを産生することができる。あるいは、タンパク質は無細胞系において産生することができる。Ranu他、Meth. Enzymol.、60:459−484、(1979)。
【0125】
本発明を一般に記載したが、下記の実施例を参照することによって本発明はいっそう容易に理解されるであろう。これらの実施例は例示であり、本発明を限定することを意図しない。
【実施例】
【0126】
実施例1.メッセンジャーRNA(mRNA)の単離
種々の発生段階におけるトマトの花およびトマトの果実から、そして葉(未処理または冷却またはソルビトール処理後の)から全RNAを単離した。簡単に述べると、組織(5g)を液体窒素中で粉砕した。粉砕した粉末を30mlのグアニジニウム緩衝液(4Mのグアニジニウムイソチオシアネート、2.5mMのNaOAc pH8.5、0.8%のβ−メルカプトエタノール)と混合した。この混合物を4層のチーズクロスを通して濾過し、10,000×g、4℃において30分間遠心した。次いで上清を塩化セシウム密度勾配で26,000×gにおいて20時間遠心した。
【0127】
ペレット化したRNAを75%のエタノールですすぎ、600μlのDEPC処理水の中に再懸濁させ、−70℃において0.75mlの95%のエタノールおよび30μlの3MのNaOAcでRNAを沈降させた。10μgのRNAを1.2%の変性ホルムアルデヒドアガロースゲル上で分画し、ナイロン膜に移した。ランダムにプライムした32P−dCTP標識化全長DHS cDNA(配列番号1)を使用して、膜を42℃において一夜プロービングした。次いで0.1%のSDSを含有する1×SSCで室温において膜を15分間1回洗浄し、0.1%のSDSを含有する0.2×SSCで65℃において各15分間3回洗浄した。膜を−70℃においてX線フィルムに一夜露出した。
【0128】
プロメガ(Promega)から入手可能なポリA+トラクトmRNA単離システムを使用して、tRNAからポリA+mRNAを単離した。ストラタジーン(Stratagene)(カリフォルニア州ラジョラ)から入手可能なZAP ExpressR cDNA合成システムを使用するcDNA合成の鋳型として、ポリA+mRNAを使用した。
【0129】
トマト葉cDNAライブラリーのスクリーニング
2Mのソルビトールに6時間暴露したMatch F1ハイブリッドトマト葉から単離したmRNAを使用して作ったcDNAライブラリーを、ほぼ5×106PFU/mlに希釈した。32P標識化600bpのRT−PCRフラグメントを使用して、cDNAライブラリーをスクリーニングした。3つの陽性cDNAクローンを切除し、製造業者のインストラクション中の方法を使用してpBK−CMVR(Stratagene)ファージミドの中に再循環させた。全長cDNAをpBK−CMVベクターの中に挿入した。
【0130】
プラスミドDNAの単離、DNAのスクリーニング
Sambrook他(前掲)が記載するアルカリ溶解法を使用してプラスミドDNAを単離した。デオキシ配列決定法を使用して、全長の陽性cDNAクローンを配列決定した。Sanger他、Proc. Natl. Acad. Sci. USA、74:5463−5467。オープンリーディングフレームを収集し、BLAST検索(GenBank、マリイランド州ベセスダ)を使用して分析し、下記の方法を使用して5つの最も相同性のタンパク質とコード化遺伝子の誘導アミノ酸配列との整列を達成した:BCM Search Launcher:Multiple Sequence Alignments Pattern−Induced Multiple Alignments Methods(参照:F.Corpet、Nucl.Acids Res.、16:10881−10890(1987))。誘導アミノ酸配列の中に存在する機能的モチーフをMultiFinderにより同定した。
【0131】
トマトRNAのノザンブロットハイブリダイゼーション
種々の段階におけるトマトの花(広く開いたまたは乾燥している芽および花およびセネッシング花弁)、トマトの葉、およびトマトの果実種々の成熟段階における(ブレーカー、すなわち、10%より少ない赤色の緑色果実、ピンク色、すなわち、全果実はオレンジ色またはピンク色である、柔らかいか、あるいは堅い)から単離された全RNAの10μgを1%の変性ホルムアルデヒドアガロースゲル上で分離し、ナイロン膜上に固定化した。
【0132】
ランダムプライマーキット(Boehringer Mannheim)を使用して32P−dCTPで標識化した全長トマトcDNAを使用して、フィルター(7×107cpm)をプロービングした。フィルターを1×SSC、0.1%SDSで室温において1回洗浄し、0.2×SSC、0.1%SDSで65℃において3回洗浄した。フィルターを乾燥し、−70℃においてX線フィルムに一夜露出した。結果を第6図、第7図、第8図および第9図に示す。
【0133】
アラアビドプシス(Arabidopsis)RNAのノザンブロットハイブリダイゼーション
5週齢(レーン1)、6週齢(レーン2)および7週齢(レーン3)のアラアビドプシス(Arabidopsis)植物の葉からの全RNAを前述したように単離し、1%の変性ホルムアルデヒドアガロースゲル上で分離し、ナイロン膜上に固定化した。ランダムプライマーキット(Boehringer Mannheim)を使用して32P−dCTPで標識化した全長アラアビドプシス(Arabidopsis)セネッセンス誘導DHS cDNAを使用して、フィルター(7×107cpm)をプロービングした。フィルターを1×SSC、0.1%SDSで室温において1回洗浄し、0.2×SSC、0.1%SDSで65℃において3回洗浄した。フィルターを乾燥し、−70℃においてX線フィルムに一夜露出した。結果を第11図に示す。
【0134】
カーネーションRNAのノザンブロットハイブリダイゼーション
種々の花発生段階、すなわち、堅い芽の花(レーン1)、開花し始めた花(レーン2)、完全に開いた花(レーン3)、インローリング(inrolling)花弁(レーン4)のカーネーション植物の花弁からの全RNAを前述したように単離し、1%の変性ホルムアルデヒドアガロースゲル上で分離し、ナイロン膜上に固定化した。ランダムプライマーキット(Boehringer Mannheim)を使用して32P−dCTPで標識化した全長カーネーションセネッセンス誘導DHS cDNAを使用して、フィルター(7×107cpm)をプロービングした。フィルターを1×SSC、0.1%SDSで室温において1回洗浄し、0.2×SSC、0.1%SDSで65℃において3回洗浄した。フィルターを乾燥し、−70℃においてX線フィルムに一夜露出した。結果を第12図に示す。
【0135】
実施例2.トマトセネッセンス誘導DHS遺伝子のソルビトール誘導
トマトの葉を密閉したチャンバー内において2Mのソルビトールで6時間処理した。次のようにしてRNAをソルビトール処理した葉から抽出した。
【0136】
葉(5g)を液体窒素中で粉砕した。粉砕した粉末を30mlのグアニジニウム緩衝液(4Mのグアニジニウムイソチオシアネート、2.5mMのNaOAc pH8.5、0.8%のβ−メルカプトエタノール)と混合した。この混合物を4層のチーズクロスを通して濾過し、10,000×g、4℃において30分間遠心した。次いで上清を塩化セシウム密度勾配で26,000×gにおいて20時間遠心した。
【0137】
ペレット化したRNAを75%のエタノールですすぎ、600μlのDEPC処理水の中に再懸濁させ、−70℃において0.75mlの95%のエタノールおよび30μlの3MのNaOAcでRNAを沈降させた。10μgのRNAを1.2%の変性ホルムアルデヒドアガロースゲル上で分画し、ナイロン膜に移した。ランダムにプライムした32P−dCTP標識化全長DHS cDNA(配列番号1)を使用して、膜を42℃において一夜プロービングした。次いで0.1%のSDSを含有する1×SSCで室温において膜を15分間1回洗浄し、0.1%のSDSを含有する0.2×SSCで65℃において各15分間3回洗浄した。膜を−70℃においてX線フィルムに一夜露出した。
結果を第8図に示す。見ることができるように、DHSの転写は葉においてソルビトールにより誘導される。
【0138】
実施例3.セネッシング花中のトマトDHS遺伝子の誘導
トマト植物の堅い花の芽および開いた、セネッシング花を収穫し、実施例2におけるようにRNAを単離した。10μgのRNAを1.2%の変性ホルムアルデヒドアガロースゲル上で分画し、ナイロン膜に移した。ランダムにプライムした32P−dCTP標識化全長DHS cDNA(配列番号1)を使用して、膜を42℃において一夜プロービングした。次いで0.1%のSDSを含有する1×SSCで室温において膜を15分間1回洗浄し、0.1%のSDSを含有する0.2×SSCで65℃において各15分間3回洗浄した。膜を−70℃においてX線フィルムに一夜露出した。
【0139】
結果を第6図に示す。見ることができるように、DHSの転写はセネッシング花において誘導される。
【0140】
実施例4.成熟する果実におけるトマトDHS遺伝子の誘導
実施例2におけるように、ブレーカー、ピンク色および成熟した果実からRNAを単離した。10μgのRNAを1.2%の変性ホルムアルデヒドアガロースゲル上で分画し、ナイロン膜に移した。ランダムにプライムした32P−dCTP標識化全長DHS cDNA(配列番号1)を使用して、膜を42℃において一夜プロービングした。次いで0.1%のSDSを含有する1×SSCで室温において膜を15分間1回洗浄し、0.1%のSDSを含有する0.2×SSCで65℃において各15分間3回洗浄した。膜を−70℃においてX線フィルムに一夜露出した。
【0141】
結果を第7図に示す。見ることができるように、DHSの転写はセネッセンスの開始直前の成熟した、赤色の果実において最強であり、腐敗に導く。
【0142】
実施例5.冷却によるトマトセネッセンス誘導DHS遺伝子の誘導
鉢中のトマト植物(7〜8週齢)を6℃に成長チャンバー内で2日間、3日間または6日間暴露した。光サイクルを8時間の暗闇および16時間の光に設定した。植物を温室に戻すことによって、再加温した。再加温しない植物を成長チャンバーから取出した直後に収穫した。次のようにしてRNAを葉から抽出した。
【0143】
葉(5g)を液体窒素中で粉砕した。粉砕した粉末を30mlのグアニジニウム緩衝液(4Mのグアニジニウムイソチオシアネート、2.5mMのNaOAc pH8.5、0.8%のβ−メルカプトエタノール)と混合した。この混合物を4層のチーズクロスを通して濾過し、10,000×g、4℃において30分間遠心した。次いで上清を塩化セシウム密度勾配で26,000×gにおいて20時間遠心した。ペレット化したRNAを75%のエタノールですすぎ、600μlのDEPC処理水の中に再懸濁させ、−70℃において0.75mlの95%のエタノールおよび30μlの3MのNaOAcでRNAを沈降させた。
【0144】
10μgのRNAを1.2%の変性ホルムアルデヒドアガロースゲル上で分画し、ナイロン膜に移した。ランダムにプライムした32P−dCTP標識化全長DHS cDNA(配列番号1)を使用して、膜を42℃において一夜プロービングした。次いで0.1%のSDSを含有する1×SSCで室温において膜を15分間1回洗浄し、0.1%のSDSを含有する0.2×SSCで65℃において各15分間3回洗浄した。膜を−70℃においてX線フィルムに一夜露出した。
【0145】
結果を第9図に示す。見ることができるように、冷却温度に暴露し、引き続いて再加温することによって、DHSの転写は葉において誘導され、そして増強された転写は膜漏出として測定した冷却損傷と相関する。
【0146】
実施例6.未同定アラアビドプシス(Arabidopsis)ゲノム配列に基づくプライマーを使用するアラアビドプシス(Arabidopsis)PCR生成物の発生
アラアビドプシス(Arabidopsis)ゲノム配列から設計した1対のオリゴヌクレオチドプライマーを使用するPCRにより、アラアビドプシス(Arabidopsis)cDNA鋳型から部分的長さのセネッセンス誘導DHS配列を発生させた。5’プライマーは配列、5’−GGTGGTGTTGAGGAAGATC(配列番号7)を有する19マーである;3’プライマーは配列、GGTGCACGCCCTGATGAAGC−3’(配列番号8)を有する20マーである。Expand High Fidelity PCR System(Boehringer Mannheim)および鋳型としてアラアビドプシス(Arabidopsis)セネッシング葉cDNAライブラリーを使用するポリメラーゼ連鎖反応を次のようにして実施した。
【0147】
反応成分:
cENA 1μl(5×107pfu)
dNTP(各々10mM) 1μl
MgCl2(5mM)+10×緩衝液 1μl
プライマー1および2(各々100μM) 2μl
Expand High Fidelity DNAプライマー 1.75U
反応体積 50μl
【0148】
反応パラメーター:
94℃、3分間
94℃/1分、58℃/1分、72℃/2分、45サイクル
72℃、15分間。
【0149】
実施例7.ゲノムDNAの単離およびサザン分析
10gのトマト葉組織を液体窒素中で微細粉末に粉砕することによって、ゲノムDNAをトマト葉から抽出した。25mlの均質化緩衝液を含有する37.5mlの混合物[100mMのTris−HCl、pH8.0、100mMのEDTA、250mMのNaCl、1%のサルコシル、1%の2−メルカプトエタノール、10μg/mlのRNアーゼおよび12.5mlのフェノール]を60℃に前もって加温し、粉砕した組織に添加した。
【0150】
この混合物を15分間震盪した。追加の12.5mlのクロロホルム/イソアミルアルコール(24:1)を混合物に添加し、さらに15分間震盪した。この混合物を遠心し、水性相を25mlのフェノール/クロロホルム/イソアミルアルコール(25:24:1)およびクロロホルム/イソアミルアルコール(24:1)で再抽出した。室温において15mlのイソプロパノールを使用する沈澱により、核酸を回収した。沈澱を1mlの水の中に再懸濁させた。
【0151】
ゲノムDNAを次のようにして制限酵素消化した:10μgのゲノムDNA、40μlの10×反応緩衝液および100Uの制限酵素(XbaI、EcoRI、EcoRVまたはHindIII)を、400μlの全反応体積で5〜6時間反応させた。次いでこの混合物をフェノール抽出し、エタノール沈降させた。消化したDNAを0.8%のアガロースゲル上で15ボルトにおいてほぼ15時間アガロースゲル電気泳動に付した。ゲルを変性緩衝液[87.66gのNaClおよび20gのNaOH/リットル]の中におだやかに撹拌しながら30分間沈め、蒸留水ですすぎ、中性緩衝液[87.66gのNaClおよび60.55gのTris−HCl、pH7.5/リットル]の中におだやかに撹拌しながら30分間沈めた。DNAを毛管ブロッティングによりHybond−H+ナイロン膜に移した。
【0152】
1×106cpm/mlの32P−dCTP標識化DHS cDNAまたはDHS cDNAクローンの3’非コーディング領域を使用して42℃において、ハイブリダイゼーションを一夜実施した。50%のホルムアミド、6×SSC、5×デンハルト溶液、0.1%SDSおよび100mg/mlの変性サケ精子DNAを含有する緩衝液中でプレハイブリダイゼーションおよびハイブリダイゼーションを実施した。膜を2〜4時間プレハイブリダイゼーションした;ハイブリダイゼーションを一夜実施した。
【0153】
ハイブリダイゼーションが完結した後、膜を2×SSCおよび0.1%のSDS中ですすぎ、次いで2×SSCおよび0.1%のSDS中で15分間洗浄し、0.2×SSCおよび0.1%のSDS中で15分間洗浄した。次いで膜を−80℃においてX線フィルムに一夜露出した。結果を第5図に示す。
【0154】
実施例8.アラアビドプシス(Arabidopsis)からのセネッセンス誘導elF−5A遺伝子の単離
鋳型としてアラアビドプシス(Arabidopsis)セネッシング葉cDNAライブラリーを使用するPCRにより、アラアビドプシス(Arabidopsis)葉中で発現されたセネッセンス誘導elF−5A遺伝子の全長cDNAクローンを得た。最初に、ベクターT7プライマー<AATACGACTCACTATAG>(配列番号18)と対合したデジェネレイト上流のプライマー<AAARRYCGMCCYTGCAAGGT>(配列番号17)、およびベクターT3プライマー<ATTAACCCTCACTAAAG>(配列番号20)と対合したデジェネレイト下流のプライマー<TCYTTNCCYTCMKCTAAHCC>(配列番号19)を使用して、遺伝子の5’および3’末端に対応するPCR生成物を作った。
【0155】
PCR生成物を配列決定のためにpBluescriptの中にサブクローニングした。次いで、3’特異的プライマー<AGAAGAAGTATAAAAACCATC>(配列番号22)と対合した5’特異的プライマー<CTGTTACCAAAAAATCTGTACC>(配列番号21)を使用して全長cDNAを作り、配列決定のためにpBluescriptの中にサブクローニングした。
【0156】
実施例9.トマト果実からのセネッセンス誘導elF−5A遺伝子の単離
鋳型としてトマト果実cDNAライブラリーを使用するPCRにより、トマト果実中で発現されたセネッセンス誘導elF−5A遺伝子の全長cDNAクローンを得た。最初に、ベクターT7プライマー(配列番号18)と対合したデジェネレイト上流のプライマー(配列番号17)、およびベクターT3プライマー(配列番号20)と対合したデジェネレイト下流のプライマー(配列番号19)を使用して、遺伝子の5’および3’末端に対応するPCR生成物を作った。PCR生成物を配列決定のためにpBluescriptの中にサブクローニングした。次いで、ベクターT7プライマー(配列番号18)と対合した5’特異的プライマー<AAAGAATCCTAGAGAGAGAAAGG>(配列番号23)を使用して全長cDNAを作り、配列決定のためにpBluescriptの中にサブクローニングした。
【0157】
実施例10.カーネーションからのセネッセンス誘導elF−5A遺伝子の単離
鋳型としてカーネーション花cDNAライブラリーを使用するPCRにより、カーネーション花中で発現されたセネッセンス誘導elF−5A遺伝子の全長cDNAクローンを得た。最初に、ベクターT7プライマー(配列番号18)と対合したデジェネレイト上流のプライマー(配列番号17)、およびベクターT3プライマー(配列番号20)と対合したデジェネレイト下流のプライマー(配列番号19)を使用して、遺伝子の5’および3’末端に対応するPCR生成物を作った。
【0158】
PCR生成物を配列決定のためにpBluescriptの中にサブクローニングした。次いで、3’特異的プライマー<ACCAAAACCTGTGTTATAACTCC>(配列番号25)と対合した5’特異的プライマー<TTTTACATCAATCGAAAATACC>(配列番号24)を使用して全長cDNAを作り、配列決定のためにpBluescriptの中にサブクローニングした。
【0159】
実施例11.アラアビドプシス(Arabidopsis)からのセネッセンス誘導DHS遺伝子の単離
アラアビドプシス(Arabidopsis)セネッシング葉cDNAライブラリーをスクリーニングすることによって、アラアビドプシス(Arabidopsis)葉中で発現されたセネッセンス誘導DHS遺伝子の全長cDNAクローンを得た。スクリーニングに使用したプローブ(配列番号26)の配列を第38図に示す。鋳型としてセネッセンス葉cDNAライブラリーを使用しかつGenBankにおける未同定ゲノム配列(AB017060)から設計したプライマー(第38図に下線が引かれている領域として示す)を使用するPCRにより、このプローブを得た。PCR生成物を配列決定のためにpBluescriptの中にサブクローニングした。
【0160】
実施例12.カーネーションからのセネッセンス誘導DHS遺伝子の単離
カーネーションセネッシング花弁cDNAライブラリーをスクリーニングすることによって、カーネーション花弁中で発現されたセネッセンス誘導DHS遺伝子の全長cDNAクローンを得た。スクリーニングに使用したプローブ(配列番号27)の配列を第39図に示す。鋳型としてセネッセンス花弁cDNAライブラリーおよびデジェネレイトプライマー(上流:5’TTG ARG AAG ATY CAT MAA RTG CCT3’)(配列番号28);下流:5’CCA TCA AAY TCY TGK GCR GTG TT3’)(配列番号29))を使用するPCRにより、このプローブを得た。PCR生成物を配列決定のためにpBluescriptの中にサブクローニングした。
【0161】
実施例13.アンチセンスの向きにアラアビドプシス(Arabidopsis)DHSの全長または3’領域を使用するアラアビドプシス(Arabidopsis)の形質転換
二重35Sプロモーターの調節下に、全長セネッセンス誘導アラアビドプシス(Arabidopsis)DHS cDNA配列またはDHS遺伝子の3’末端(配列番号30)(第36図)(両方はアンチセンス立体配置で発現された)を含有するバイナリーベクター、pKYLX71、で、アグロバクテリア(Agrobacteria)を形質転換した。アラアビドプシス(Arabidopsis)植物を形質転換されたアグロバクテリア(Agrobacteria)で真空浸潤により形質転換し、生ずるT0植物からの形質転換された種子をアンピシリン上で選択した。
【0162】
第21図〜第24図は形質転換されたアラアビドプシス(Arabidopsis)植物の写真であり、形質転換された植物におけるアンチセンスの向きのDHS遺伝子またはその3’末端の発現がバイオマスを増加させ、例えば、葉を大きくしかつ植物のサイズを大きくすることを示す。第25図は、トランスジェニックアラアビドプシス(Arabidopsis)植物が種子収量を増加したことを図解する。
【0163】
実施例14.アンチセンスの向きにトマトDHSの全長または3’領域を使用するトマト植物の形質転換
二重35Sプロモーターの調節下に、全長セネッセンス誘導トマトDHS cDNA配列またはDHS遺伝子の3’末端(配列番号31)(第37図)(両方はアンチセンス立体配置で発現された)を含有するバイナリーベクター、pKYLX71、で、アグロバクテリア(Agrobacteria)を形質転換した。これらのアグロバクテリア(Agrobacteria)を使用してトマト葉の外植体を形成し、標準組織培養法によりカルスおよび小植物を発生させ、選択した。形質転換された植物を温室の条件下に成熟果実産生T1植物に成長させた。
【0164】
第26図〜第35図は、形質転換されたアラアビドプシス(Arabidopsis)植物において見られるように、形質転換された植物中のセネッセンス誘導トマトDHS遺伝子の発現の減少がバイオマスを増加させ、例えば、葉のサイズを大きくしかつ植物を大きくすること、ならびにトマト果実の軟化および腐敗を遅延することを示す写真である。
【特許請求の範囲】
【請求項1】
配列番号1または配列番号9のヌクレオチド配列を含む、セネッセンス誘導デオキシハイプシンシンターゼをコードする単離されたDNA分子。
【請求項2】
前記DNA分子が配列番号1または配列番号9のヌクレオチド配列からなる、請求項1に記載の単離されたDNA分子。
【請求項3】
配列番号2または配列番号10を含むセネッセンス誘導デオキシハイプシンシンターゼをコードする単離されたDNA分子。
【請求項4】
配列番号1または配列番号9を含むヌクレオチド配列によってコードされたセネッセンス誘導デオキシハイプシンシンターゼ。
【請求項5】
配列番号2または配列番号10のアミノ酸配列を含む、請求項4に記載のセネッセンス誘導デオキシハイプシンシンターゼ。
【請求項6】
(a)配列番号1、配列番号5、または配列番号9を含むヌクレオチド配列によってコードされたセネッセンス誘導デオキシハイプシンシンターゼ;そして
(b)その中に形質転換されている植物細胞において前記デオキシハイプシンシンターゼが発現されるように、DNA分子に作用可能に連鎖された調節配列
をコードするDNA分子を含むベクター。
【請求項7】
下記の配列を含んでなる植物細胞の形質転換用ベクター:
(a)(1)配列番号1、配列番号5および/または配列番号9のヌクレオチド配列を含む、セネッセンス誘導デオキシハイプシンシンターゼをコードするDNA分子の1つの鎖、または(2)セネッセンス誘導デオキシハイプシンシンターゼをコードするDNA分子によりコードされるRNA配列の全部または部分に対して実質的に相補的であるアンチセンスヌクレオチド配列;および
(b)それがその中に形質転換されている植物細胞においてアンチセンスヌクレオチド配列が発現されるように、アンチセンスヌクレオチド配列に作用可能に連鎖された調節配列。
【請求項8】
調節配列がプロモーターおよび転写終結領域を含んでなる、請求項6または7に記載のベクター。
【請求項9】
調節配列が構成的プロモーターを含んでなる、請求項6または7に記載のベクター。
【請求項10】
調節配列が植物組織特異的プロモーターを含んでなる、請求項6または7に記載のベクター。
【請求項11】
調節配列がセネッセンス誘導植物プロモーターを含んでなる、請求項6または7に記載のベクター。
【請求項12】
調節配列がウイルスのプロモーターを含んでなる、請求項6または7に記載のベクター。
【請求項13】
調節配列が構成的プロモーターをさらに含んでなる、請求項11に記載のウイルス。
【請求項14】
(i)配列番号1、配列番号5または配列番号9のヌクレオチド配列を含む植物セネッセンス誘導デオキシハイプシンシンターゼ遺伝子のRNA転写物の全部または部分に対して相補的であるRNA分子をコードするアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項15】
オリゴヌクレオチドまたはポリヌクレオチドが約6〜約100ヌクレオチドを含んでなる、請求項14に記載のアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項16】
アンチセンスオリゴヌクレオチドまたはポリヌクレオチドがRNA転写物の5’非コーディング領域の対応する部分に対して相補的である、請求項14に記載のアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項17】
アンチセンスオリゴヌクレオチドまたはポリヌクレオチドがRNA転写物の3’末端の対応する部分に対して実質的に相補的である、請求項14に記載のアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項18】
アンチセンスオリゴヌクレオチドまたはポリヌクレオチドがアラアビドプシス(Arabidopsis)セネッセンス誘導DHS遺伝子の3’末端に対して実質的に相補的である、請求項14に記載のアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項19】
アンチセンスオリゴヌクレオチドまたはポリヌクレオチドが配列番号23に対して相補的である、請求項18に記載のアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項20】
アンチセンスオリゴヌクレオチドまたはポリヌクレオチドが配列番号30に対して相補的である、請求項18に記載のアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項21】
工程:
(1)植物のゲノムの中にベクターを組込み、前記ベクターは、
(A)(i)配列番号1、配列番号5、または配列番号9を含むヌクレオチド配列によってコードされた内因的セネッセンス誘導デオキシハイプシンシンターゼをコードするDNA分子、または(ii)内因的セネッセンス誘導デオキシハイプシンシンターゼ遺伝子によりコードされるRNA配列、の全部または部分に対して相補的なアンチセンスヌクレオチド配列、あるいは
(B)アンチセンスヌクレオチド配列が発現されるように、アンチセンスヌクレオチド配列に作用可能に連鎖された調節配列、
を含んでなり;そして
(2)前記植物を成長させる、それにより前記アンチセンスヌクレオチド配列は転写され、前記RNA配列に結合し、それによりセネッセンス誘導デオキシハイプシンシンターゼ遺伝子の発現は阻害される;
を含んでなる、植物における内因的セネッセンス誘導デオキシハイプシンシンターゼの発現を阻害する方法。
【請求項22】
アンチセンスヌクレオチド配列がそれに対して相補的であるDNAの部分またはRNAの部分が5’非コーディング配列または3’コーディング配列および/または非コーディング配列を含んでなる、請求項21に記載の方法。
【請求項23】
アンチセンスヌクレオチド配列が配列番号23に対して実質的に相補的である、請求項21に記載の方法。
【請求項24】
アンチセンスヌクレオチド配列が配列番号30に対して実質的に相補的である、請求項21に記載の方法。
【請求項25】
前記阻害が植物の変更されたセネッセンスを生ずる、請求項21に記載の方法。
【請求項26】
前記阻害が環境ストレス誘導および/または病原体誘導セネッセンスに対する前記植物の耐性を増加させる、請求項21に記載の方法。
【請求項27】
前記阻害が前記植物のバイオマスを増加させる、請求項21に記載の方法。
【請求項28】
前記阻害が前記植物における果実の軟化および腐敗を遅延させる、請求項21に記載の方法。
【請求項29】
前記阻害が前記植物の種子収量を増加させる、請求項21に記載の方法。
【請求項30】
調節配列が植物において活性である構成的プロモーターを含んでなる、請求項21に記載の方法。
【請求項31】
調節配列が植物において活性である組織特異的プロモーターを含んでなる、請求項21に記載の方法。
【請求項32】
調節配列が植物において活性であるセネッセンス誘導プロモーターを含んでなる、請求項21に記載の方法。
【請求項33】
前記植物が結実植物、顕花植物、野菜、栽培作物の植物および森林種から成る群より選択される、請求項21に記載の方法。
【請求項34】
植物がトマトである、請求項21に記載の方法。
【請求項35】
植物が顕花植物である、請求項21に記載の方法。
【請求項36】
工程:
(1)植物の少なくとも1つの細胞のゲノムび中にベクターを組込み、前記ベクターは、
(A)外因的セネッセンス誘導デオキシハイプシンシンターゼをコードする単離されたDNA分子、ここで前記DNA分子は配列番号1、配列番号5、または配列番号9のヌクレオチド配列を含む;および
(B)それによりコードされる外因的セネッセンス誘導デオキシハイプシンシンターゼが発現されるように、DNA分子に作用可能に連鎖された調節配列;を含んでなる;そして
(2)前記植物を成長させる、それにより前記DNA分子は過剰発現され、そして内因的セネッセンス誘導デオキシハイプシンシンターゼ遺伝子は外因的セネッセンス誘導デオキシハイプシンシンターゼにより阻害される;
を含んでなる、植物細胞における内因的セネッセンス誘導デオキシハイプシンシンターゼ遺伝子の発現を阻害する方法。
【請求項37】
調節配列が構成的プロモーターを含んでなる、請求項36に記載の方法。
【請求項38】
工程:
(1)植物のゲノムの中にベクターを組込み、前記ベクターは、
(A)(i)内因的セネッセンス誘導デオキシハイプシンシンターゼをコードするDNA分子、ここで前記内因的セネッセンス誘導デオキシハイプシンシンターゼをコードするDNAは配列番号1、配列番号5、または配列番号9のヌクレオチド配列を含む、または(ii)内因的セネッセンス誘導デオキシハイプシンシンターゼ遺伝子をコードするRNA配列、の全部または部分に対して相補的であるアンチセンスヌクレオチド配列、
(B)アンチセンスヌクレオチド配列が発現されるように、アンチセンスヌクレオチド配列に作用可能に連鎖された調節配列、を含んでなり;そして
(2)前記植物を成長させる、それにより前記アンチセンスヌクレオチド配列は転写され、前記RNA配列に結合し、そして前記デオキシハイプシンシンターゼの発現は阻害される;
を含んでなる、植物における年齢に関係するセネッセンスおよび/または環境的ストレスに関係するセネッセンスを変更する方法。
【請求項39】
請求項6もしくは7に記載のベクター、または前記ベクターの組合わせを含んでなるトランスジェニック植物細胞。
【請求項1】
配列番号1または配列番号9のヌクレオチド配列を含む、セネッセンス誘導デオキシハイプシンシンターゼをコードする単離されたDNA分子。
【請求項2】
前記DNA分子が配列番号1または配列番号9のヌクレオチド配列からなる、請求項1に記載の単離されたDNA分子。
【請求項3】
配列番号2または配列番号10を含むセネッセンス誘導デオキシハイプシンシンターゼをコードする単離されたDNA分子。
【請求項4】
配列番号1または配列番号9を含むヌクレオチド配列によってコードされたセネッセンス誘導デオキシハイプシンシンターゼ。
【請求項5】
配列番号2または配列番号10のアミノ酸配列を含む、請求項4に記載のセネッセンス誘導デオキシハイプシンシンターゼ。
【請求項6】
(a)配列番号1、配列番号5、または配列番号9を含むヌクレオチド配列によってコードされたセネッセンス誘導デオキシハイプシンシンターゼ;そして
(b)その中に形質転換されている植物細胞において前記デオキシハイプシンシンターゼが発現されるように、DNA分子に作用可能に連鎖された調節配列
をコードするDNA分子を含むベクター。
【請求項7】
下記の配列を含んでなる植物細胞の形質転換用ベクター:
(a)(1)配列番号1、配列番号5および/または配列番号9のヌクレオチド配列を含む、セネッセンス誘導デオキシハイプシンシンターゼをコードするDNA分子の1つの鎖、または(2)セネッセンス誘導デオキシハイプシンシンターゼをコードするDNA分子によりコードされるRNA配列の全部または部分に対して実質的に相補的であるアンチセンスヌクレオチド配列;および
(b)それがその中に形質転換されている植物細胞においてアンチセンスヌクレオチド配列が発現されるように、アンチセンスヌクレオチド配列に作用可能に連鎖された調節配列。
【請求項8】
調節配列がプロモーターおよび転写終結領域を含んでなる、請求項6または7に記載のベクター。
【請求項9】
調節配列が構成的プロモーターを含んでなる、請求項6または7に記載のベクター。
【請求項10】
調節配列が植物組織特異的プロモーターを含んでなる、請求項6または7に記載のベクター。
【請求項11】
調節配列がセネッセンス誘導植物プロモーターを含んでなる、請求項6または7に記載のベクター。
【請求項12】
調節配列がウイルスのプロモーターを含んでなる、請求項6または7に記載のベクター。
【請求項13】
調節配列が構成的プロモーターをさらに含んでなる、請求項11に記載のウイルス。
【請求項14】
(i)配列番号1、配列番号5または配列番号9のヌクレオチド配列を含む植物セネッセンス誘導デオキシハイプシンシンターゼ遺伝子のRNA転写物の全部または部分に対して相補的であるRNA分子をコードするアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項15】
オリゴヌクレオチドまたはポリヌクレオチドが約6〜約100ヌクレオチドを含んでなる、請求項14に記載のアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項16】
アンチセンスオリゴヌクレオチドまたはポリヌクレオチドがRNA転写物の5’非コーディング領域の対応する部分に対して相補的である、請求項14に記載のアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項17】
アンチセンスオリゴヌクレオチドまたはポリヌクレオチドがRNA転写物の3’末端の対応する部分に対して実質的に相補的である、請求項14に記載のアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項18】
アンチセンスオリゴヌクレオチドまたはポリヌクレオチドがアラアビドプシス(Arabidopsis)セネッセンス誘導DHS遺伝子の3’末端に対して実質的に相補的である、請求項14に記載のアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項19】
アンチセンスオリゴヌクレオチドまたはポリヌクレオチドが配列番号23に対して相補的である、請求項18に記載のアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項20】
アンチセンスオリゴヌクレオチドまたはポリヌクレオチドが配列番号30に対して相補的である、請求項18に記載のアンチセンスオリゴヌクレオチドまたはポリヌクレオチド。
【請求項21】
工程:
(1)植物のゲノムの中にベクターを組込み、前記ベクターは、
(A)(i)配列番号1、配列番号5、または配列番号9を含むヌクレオチド配列によってコードされた内因的セネッセンス誘導デオキシハイプシンシンターゼをコードするDNA分子、または(ii)内因的セネッセンス誘導デオキシハイプシンシンターゼ遺伝子によりコードされるRNA配列、の全部または部分に対して相補的なアンチセンスヌクレオチド配列、あるいは
(B)アンチセンスヌクレオチド配列が発現されるように、アンチセンスヌクレオチド配列に作用可能に連鎖された調節配列、
を含んでなり;そして
(2)前記植物を成長させる、それにより前記アンチセンスヌクレオチド配列は転写され、前記RNA配列に結合し、それによりセネッセンス誘導デオキシハイプシンシンターゼ遺伝子の発現は阻害される;
を含んでなる、植物における内因的セネッセンス誘導デオキシハイプシンシンターゼの発現を阻害する方法。
【請求項22】
アンチセンスヌクレオチド配列がそれに対して相補的であるDNAの部分またはRNAの部分が5’非コーディング配列または3’コーディング配列および/または非コーディング配列を含んでなる、請求項21に記載の方法。
【請求項23】
アンチセンスヌクレオチド配列が配列番号23に対して実質的に相補的である、請求項21に記載の方法。
【請求項24】
アンチセンスヌクレオチド配列が配列番号30に対して実質的に相補的である、請求項21に記載の方法。
【請求項25】
前記阻害が植物の変更されたセネッセンスを生ずる、請求項21に記載の方法。
【請求項26】
前記阻害が環境ストレス誘導および/または病原体誘導セネッセンスに対する前記植物の耐性を増加させる、請求項21に記載の方法。
【請求項27】
前記阻害が前記植物のバイオマスを増加させる、請求項21に記載の方法。
【請求項28】
前記阻害が前記植物における果実の軟化および腐敗を遅延させる、請求項21に記載の方法。
【請求項29】
前記阻害が前記植物の種子収量を増加させる、請求項21に記載の方法。
【請求項30】
調節配列が植物において活性である構成的プロモーターを含んでなる、請求項21に記載の方法。
【請求項31】
調節配列が植物において活性である組織特異的プロモーターを含んでなる、請求項21に記載の方法。
【請求項32】
調節配列が植物において活性であるセネッセンス誘導プロモーターを含んでなる、請求項21に記載の方法。
【請求項33】
前記植物が結実植物、顕花植物、野菜、栽培作物の植物および森林種から成る群より選択される、請求項21に記載の方法。
【請求項34】
植物がトマトである、請求項21に記載の方法。
【請求項35】
植物が顕花植物である、請求項21に記載の方法。
【請求項36】
工程:
(1)植物の少なくとも1つの細胞のゲノムび中にベクターを組込み、前記ベクターは、
(A)外因的セネッセンス誘導デオキシハイプシンシンターゼをコードする単離されたDNA分子、ここで前記DNA分子は配列番号1、配列番号5、または配列番号9のヌクレオチド配列を含む;および
(B)それによりコードされる外因的セネッセンス誘導デオキシハイプシンシンターゼが発現されるように、DNA分子に作用可能に連鎖された調節配列;を含んでなる;そして
(2)前記植物を成長させる、それにより前記DNA分子は過剰発現され、そして内因的セネッセンス誘導デオキシハイプシンシンターゼ遺伝子は外因的セネッセンス誘導デオキシハイプシンシンターゼにより阻害される;
を含んでなる、植物細胞における内因的セネッセンス誘導デオキシハイプシンシンターゼ遺伝子の発現を阻害する方法。
【請求項37】
調節配列が構成的プロモーターを含んでなる、請求項36に記載の方法。
【請求項38】
工程:
(1)植物のゲノムの中にベクターを組込み、前記ベクターは、
(A)(i)内因的セネッセンス誘導デオキシハイプシンシンターゼをコードするDNA分子、ここで前記内因的セネッセンス誘導デオキシハイプシンシンターゼをコードするDNAは配列番号1、配列番号5、または配列番号9のヌクレオチド配列を含む、または(ii)内因的セネッセンス誘導デオキシハイプシンシンターゼ遺伝子をコードするRNA配列、の全部または部分に対して相補的であるアンチセンスヌクレオチド配列、
(B)アンチセンスヌクレオチド配列が発現されるように、アンチセンスヌクレオチド配列に作用可能に連鎖された調節配列、を含んでなり;そして
(2)前記植物を成長させる、それにより前記アンチセンスヌクレオチド配列は転写され、前記RNA配列に結合し、そして前記デオキシハイプシンシンターゼの発現は阻害される;
を含んでなる、植物における年齢に関係するセネッセンスおよび/または環境的ストレスに関係するセネッセンスを変更する方法。
【請求項39】
請求項6もしくは7に記載のベクター、または前記ベクターの組合わせを含んでなるトランスジェニック植物細胞。
【図1】



【図2A】

【図2B】

【図2C】

【図2D】

【図3】

【図4】
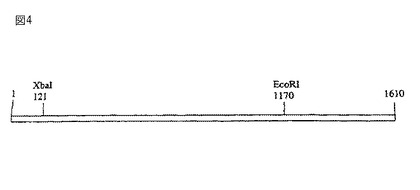
【図5】

【図6】
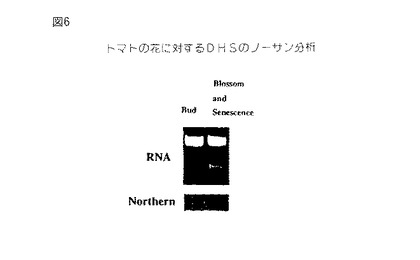
【図7】
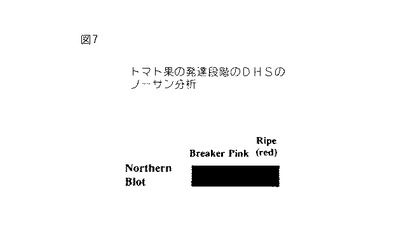
【図8】
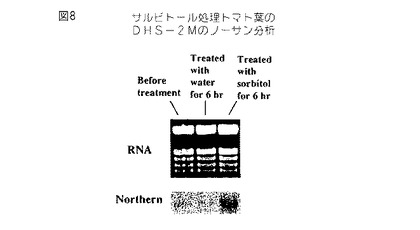
【図9】
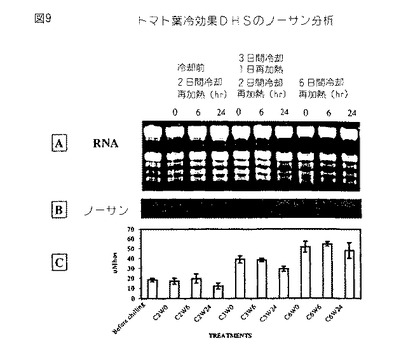
【図10】

【図11】
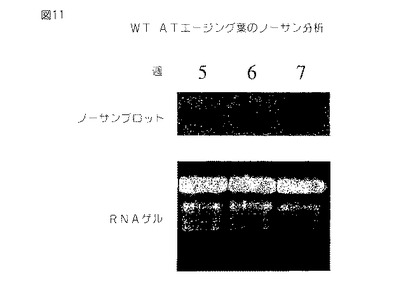
【図12】
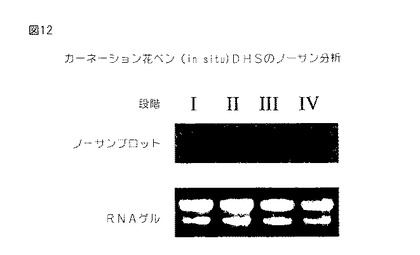
【図13】

【図14】

【図15】

【図16】
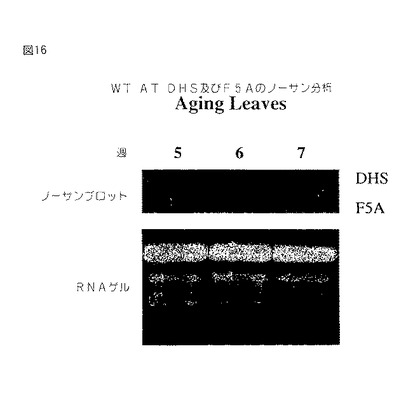
【図17】

【図18】

【図19】

【図20】

【図21】

【図22】

【図23】

【図24】

【図25】

【図26】
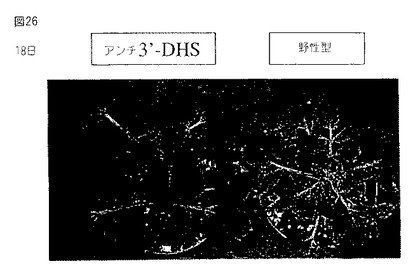
【図27】
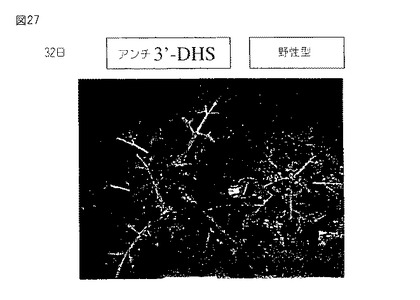
【図28】
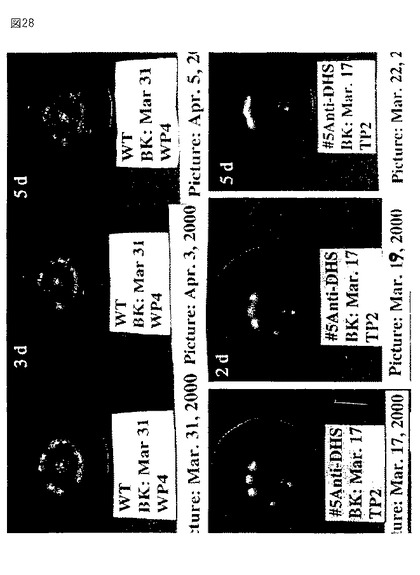
【図29】
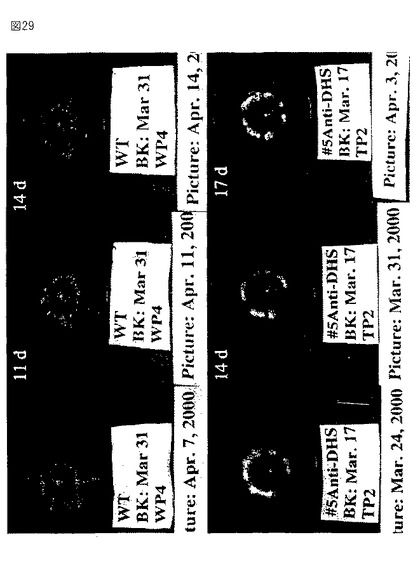
【図30】
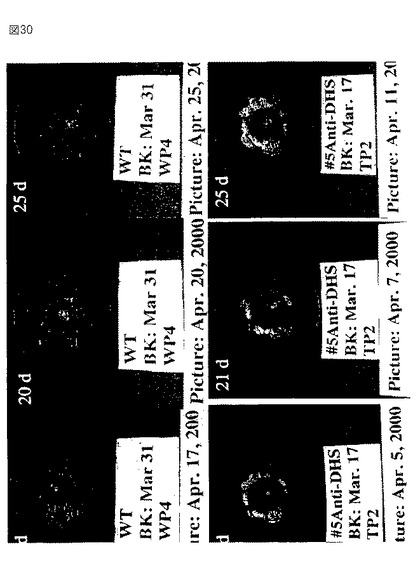
【図31】
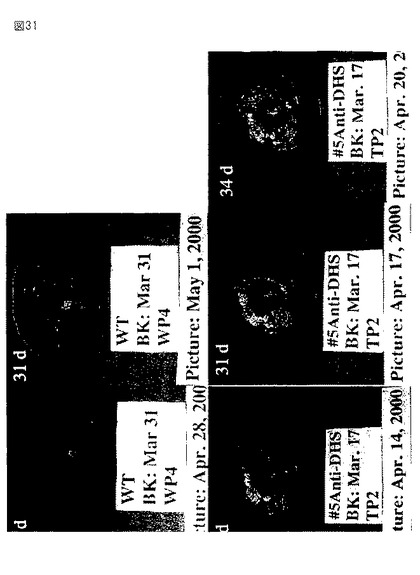
【図32】

【図33】

【図34】

【図35】

【図36】

【図37】

【図38】

【図39】

【図2A】
【図2B】
【図2C】
【図2D】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【公開番号】特開2012−187113(P2012−187113A)
【公開日】平成24年10月4日(2012.10.4)
【国際特許分類】
【出願番号】特願2012−127234(P2012−127234)
【出願日】平成24年6月4日(2012.6.4)
【分割の表示】特願2010−137640(P2010−137640)の分割
【原出願日】平成12年7月6日(2000.7.6)
【出願人】(502458132)セネスコ テクノロジーズ,インコーポレイティド (16)
【Fターム(参考)】
【公開日】平成24年10月4日(2012.10.4)
【国際特許分類】
【出願日】平成24年6月4日(2012.6.4)
【分割の表示】特願2010−137640(P2010−137640)の分割
【原出願日】平成12年7月6日(2000.7.6)
【出願人】(502458132)セネスコ テクノロジーズ,インコーポレイティド (16)
【Fターム(参考)】
[ Back to top ]