
植物のバイオマスを増やし、鉄濃度を高め、病原体に対する耐性を向上させるための組成物および方法
植物のバイオマスをより多く生産する方法、植物の乾燥耐性を向上させる方法、植物のリグニン濃度を低下させる方法、植物の鉄濃度を上昇させる方法、または植物の真菌感染を阻害する方法はそれぞれ、未処理植物と比較して、バイオマスを増加させる、乾燥耐性を向上させる、リグニン濃度を低下させる、鉄濃度を上昇させる、または植物の真菌感染を阻害するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む。バチルス・スブチリス(Bacillus subtilis)FB17を含む農業用担体および種子コーティングを提供する。バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物のバイオマスは、バイオ燃料に変換してもよいし、あるいは、食用作物としてまたは他の用途で使用してもよい。

【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
本出願は、あらゆる目的のためその全体を本明細書に援用する2010年3月1日に出願された米国特許出願第61/309,134号、2010年11月16日に出願された米国特許出願第61/414,108号、および2010年11月22日に出願された米国特許出願第61/416,039号に対する優先権を主張する。
【0002】
政府支援に関する記載
開示された本発明に至る研究は、University of DelawareのGeorgetown,DEのUSDA Experimental Field Stationからの技術支援と共に、国立科学財団(National Science Foundation:NSF)Grant No.0923806およびNSF Grant No.IOS−0814477により一部資金提供を受けた。したがって、米国政府は、本発明に一定の権利を有する場合がある。
【0003】
本発明は一般にバイオマスの増加、乾燥耐性の向上、リグニン含有量の減少、種子発芽の促進、鉄濃度の上昇、および病原体に対する耐性の向上など植物生育の様々な特徴を向上させる植物生育促進根圏細菌の使用に関する。特に、本発明の実施形態は、バチルス・スブチリス(Bacillus subtilis)FB17の植物への投与に関する。その結果得られる植物は、バイオ燃料、食物の生産または他の目的に使用することができる。
【背景技術】
【0004】
食料安全保障は世界中で常に最優先課題であり、作物生産の環境影響に対する懸念の増大に伴い、環境を保護しつつ、生産性を高める新規な方法の開発および使用が必要とされている 植物生物学者は、増加する世界各地の人々が必要不可欠な原料を確実に入手できることを目指して作物植物を十分に生産するための戦略を開発し実行している。しかしながら、バイオ燃料および再生可能技術がますます重要な優先課題となってきているため、バイオ燃料および再生可能技術の開発がこの課題を大きくしている。したがって、田畑の多様な条件で作物収穫量を増大させる改良されたアプローチに対する必要性が増している。
【0005】
作物植物のバイオマスおよび収穫可能性を増加させるにあたっては、世界中に多くの様々な地理的環境および気候があり、異なる種類の課題が提示されている。乾燥は、作物生産を世界的に制約する重要な要因である。生育期の長期乾燥または短期乾燥は、作物生産を大きく制限したり、あるいはゼロにしたりする可能性さえある。世界の主要な作付け地域でも、世界的な気象パターンの変化が、乾燥の頻度および強さに影響を与えている。
【0006】
また、養分利用率(nutrient availability)も作物生産を制約する。養分による土壌の改善(soil augmentation)は費用がかかり、エネルギー集約的であるうえ、養分を十分な量で利用できるときでも、作物植物は養分取り込みが非効率的である場合がある。必須養分の取り込みが不十分であると、収穫量が低下し、食用作物の栄養的価値が低下する。たとえば、米(単子葉植物オリザ・サティバ(Oryza sativa)またはオリザ・グラベリマ(Oryza glaberrima)の種子)は、世界の人口の3分の2以上にとって最も重要な主食であり、消費されるカロリーのかなりの割合を供給している。米は世界人口の大半にとって主要な主食であり、世界的にヒトの最も一般的な微量栄養素欠乏症(micronutrient deficiencies)の1つが鉄欠乏症であることから、より高レベルの鉄を含む米を生産すれば、世界中で微量栄養素栄養失調の抑制に大きな影響を与える可能性がある。
【0007】
また、病原体ストレスも生産性を制約する。植物は病原体の攻撃から身を守るためエネルギーを投資しなければならず、このエネルギーの流用の結果、収穫量が減少する。また、植物は病害の進行を抑えるためその組成を変えるものであり、これらの変化が、多くの場合、作物の加工を難しくしている。さらに、一部の作物病原体は、遺伝的多様性により効率的に抑えることも、化学的駆除もできず、作物生産に世界的に大きな影響を与える。
【0008】
イネいもち病(rice blast)(マグナポルテ・グリセア(Magnaporthe grisea)またはマグナポルテ・オリザエ(Magnaporthe oryzae))は、米を侵す深刻な病害を引き起こす植物病原性真菌である。イネいもち病は経済的に大きな作物被害を毎年引き起こし、作物収穫量の40%を占めると推定される。イネいもち病は、成長期になるたびに世界中で数百万の人々の食料となるのに十分な米をだめにする。米は世界の大半で重要な主食であるため、イネいもち病の影響は、ヒトの健康および環境に広範に及ぶ。米不足はヒトの飢餓に直接つながる。イネいもち病はさらに作物の減少にもつながり、減少した収穫量を補う別の資源を使用することが必要になる。栄養が増加したより多くの量の食物が世界の人々に入手できるように、さらにバイオ燃料の生産など他の重要な利益のために、多様な生育条件で、乾燥ストレスに対する耐性、病原体の影響に対する耐性、養分利用率、および最終的な作物収穫量など植物生育の様々な特徴を向上させる戦略が引き続き強く求められている。
【発明の概要】
【課題を解決するための手段】
【0009】
本発明の実施形態は、植物のバイオマスをより多く生産する方法であって、未処理植物と比較して植物のバイオマスをより多く生産するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは植物の種子の周囲の土壌に投与することを含む方法を提供する。
【0010】
別の実施形態は、植物の乾燥耐性を向上させる方法であって、未処理植物と比較して植物の乾燥耐性を向上させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは植物の種子の周囲の土壌に投与することを含む方法を提供する。
【0011】
別の実施形態は、植物のリグニン濃度を低下させる方法であって、未処理植物と比較して植物のリグニン濃度を低下させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは植物の種子の周囲の土壌に投与することを含む方法を提供する。
【0012】
別の実施形態は、植物の種子発芽を促進する方法であって、未処理植物と比較して植物の種子発芽を促進するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは植物の種子の周囲の土壌に投与することを含む方法を提供する。
【0013】
別の実施形態は、植物の、特に米植物の鉄濃度を上昇させる方法であって、未処理植物と比較して植物の鉄濃度を上昇させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。
【0014】
別の実施形態は、植物病原真菌(plant fungal pathogen)の増殖と、病原真菌、特にイネいもち病による植物、特に米植物の感染とを阻害する方法であって、病原真菌による植物の感染を阻害するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。
【0015】
追加の実施形態は、バチルス・スブチリス(Bacillus subtilis)FB17を含む農業用担体および種子コーティングを提供する。バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物のバイオマスはバイオ燃料に変換することができ、生産された作物は、ヒトもしくは動物の食糧(foodstock)、または他の目的に安全に使用することができる。
【図面の簡単な説明】
【0016】
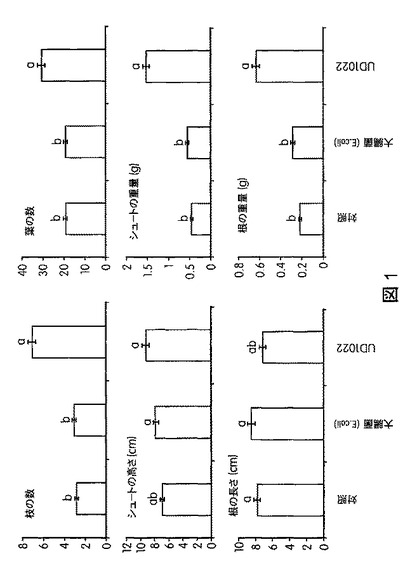
【図1】B.スブチリス(B.subtilis)FB17で処理したセイヨウヤマカモジ(Brachypodium distachyon)(Bd2−1)植物を対照と比較した、形態計測分析(枝の数、葉の数、シュートの高さ、シュートの重量、根の長さ、根の重量)を示す。これにより、B.スブチリス(B.subtilis)FB17の接種が植物の形態を高めることが示される。
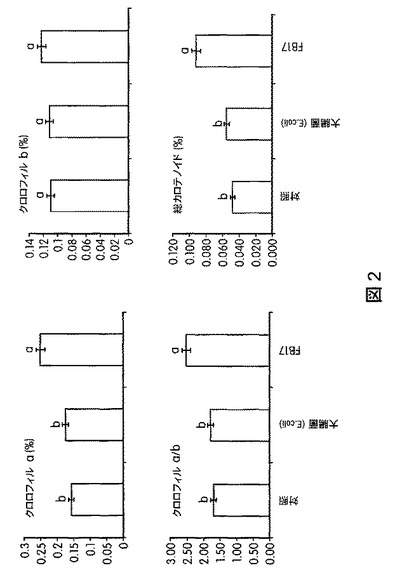
【図2】B.スブチリス(B.subtilis)FB17で処理したセイヨウヤマカモジ(Brachypodium distachyon)植物を総クロロフィルおよびカロテノイドにより測定し、対照と比較した生化学的解析を示す。これにより、B.スブチリス(B.subtilis)FB17の接種が、植物の光エネルギーを集める能力にプラスの影響を与えることが示される。
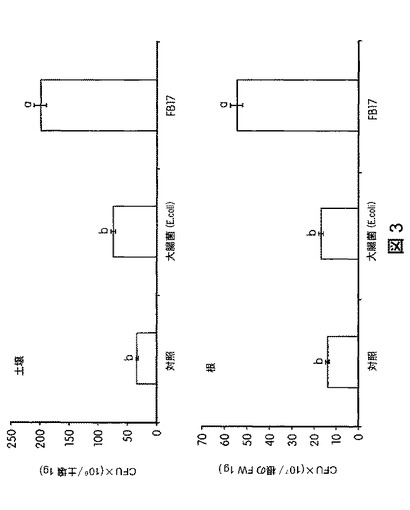
【図3】土壌およびセイヨウヤマカモジ(Brachypodium distachyon)の根から回収されたB.スブチリス(B.subtilis)FB17と対照との量を示す。これにより、B.スブチリス(B.subtilis)FB17を接種した植物の根との関係が安定していることが明らかにされる。
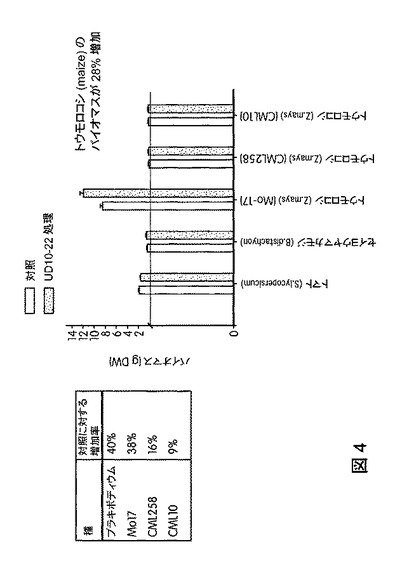
【図4】B.スブチリス(B.subtilis)FB17で処理した様々な植物種のバイオマスの総増加率を示す。トウモロコシ(Z.mays)(MO17)では、空中のバイオマスおよび根のバイオマスの両方で有意な増加(約28%)が観察された。
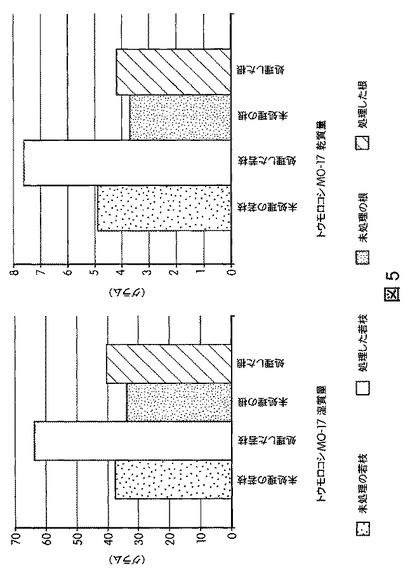
【図5】B.スブチリス(B.subtilis)FB17で種子処理したトウモロコシ(Z.mays)Mo−17植物の根およびシュートのバイオマスの増加を示す定量データである。
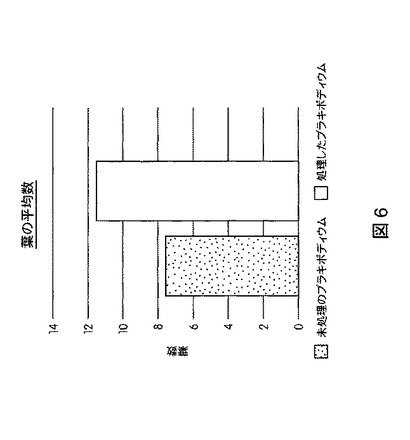
【図6】B.スブチリス(B.subtilis)FB17で種子処理したバイオエネルギー作物セイヨウヤマカモジ(Brachypodium distachyon)(遺伝子型Bd2−1)植物の葉数の増加を示す定量データである。
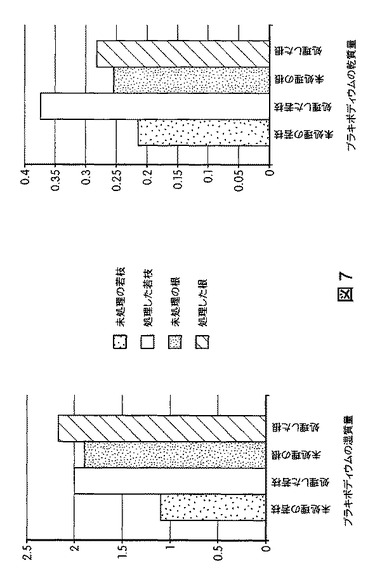
【図7】B.スブチリス(B.subtilis)FB17で種子処理したバイオエネルギー作物セイヨウヤマカモジ(Brachypodium distachyon)(遺伝子型Bd2−1)植物の根およびシュートのバイオマスの増加を示す定量データである。
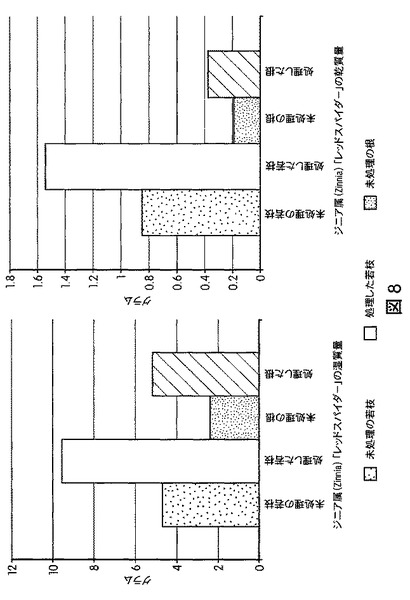
【図8】B.スブチリス(B.subtilis)FB17で種子処理したジニア・エスピー(Zinnia sp.)「レッドスパイダー」の根およびシュートのバイオマスの増加を示す定量データである。
【図9】B.スブチリス(B.subtilis)FB17で種子処理したジニア・エスピー(Zinnia sp.)「レッドスパイダー」の根およびシュートのバイオマスの増加を示す定量データである。
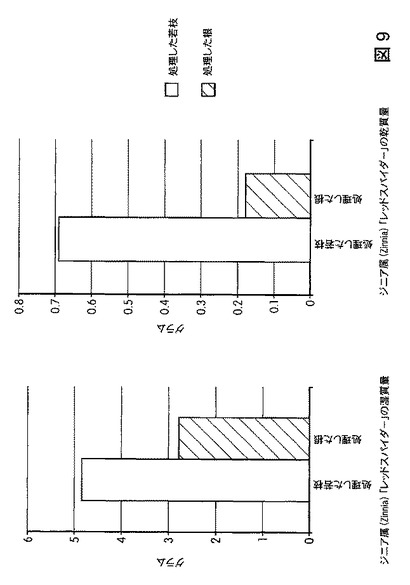
【図10】B.スブチリス(B.subtilis)FB17で処理した植物の総クロロフィル含有量を示す。FB17処理後、総クロロフィル含有量の有意な増加が、トマト(14%)、トウモロコシ(Z.mays)CML10(72%)およびCML258(87%)で観察された。
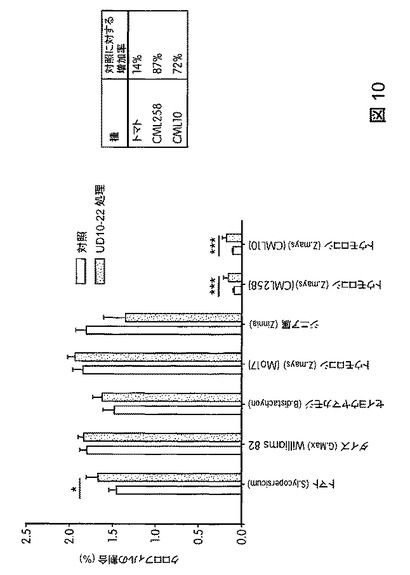
【図11】B.スブチリス(B.subtilis)FB17で処理した植物の総カロテノイド含有量を示す。FB17処理後、総カロテノイド含有量の有意な増加がダイズ(31%)およびトウモロコシ(Z.mays)MO17(82%)で観察された。
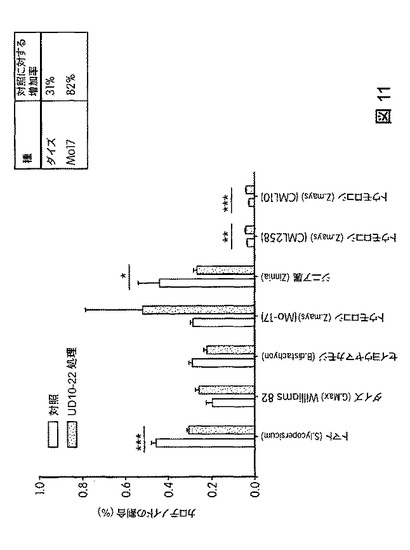
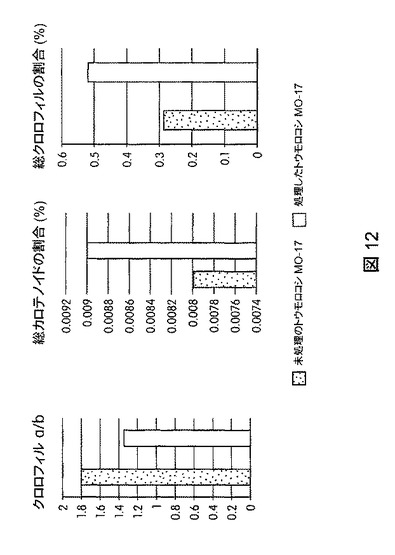
【図12】B.スブチリス(B.subtilis)FB17で種子処理したMo−17植物の光合成効率の上昇を示す定量データである。
【図13】B.スブチリス(B.subtilis)FB17で種子処理したバイオエネルギー作物セイヨウヤマカモジ(Brachypodium distachyon)(遺伝子型Bd2−1)植物の光合成効率の上昇を示す定量データである。
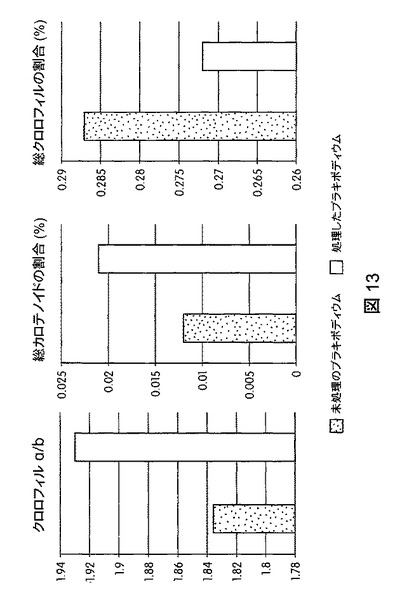
【図14】B.スブチリス(B.subtilis)FB17で種子処理したジニア・エスピー(Zinnia sp.)「レッドスパイダー」の光合成効率の上昇を示す定量データである。
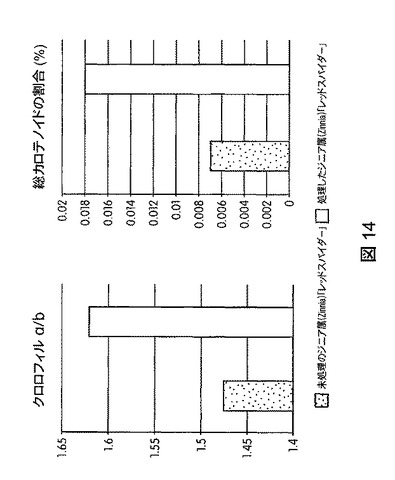
【図15】B.スブチリス(B.subtilis)FB17で種子処理した外来トウモロコシCML10およびCML258の光合成効率の上昇を示す定量データである。
【図16】B.スブチリス(B.subtilis)FB17で処理した種子の発芽率の上昇を示す。FB17処理後、総発芽含有率(plant−pathogenic fungus)の有意な増加がトマト(6%)、トウモロコシ(Z.mays)MO17(2.1%)およびCML258(14%)で観察された。特に外来種トウモロコシCML258の発芽率が劇的に上昇した。
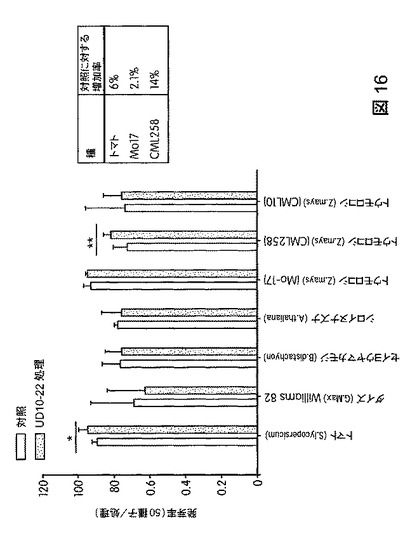
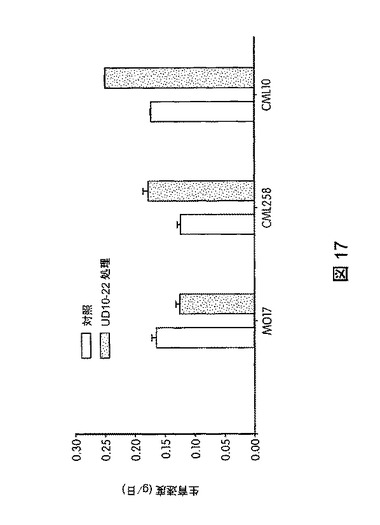
【図17】B.スブチリス(B.subtilis)FB17で処理したトウモロコシ(Zea mays)の生育速度を示す。
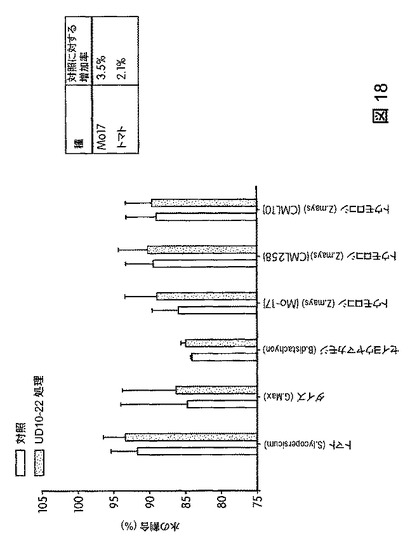
【図18】B.スブチリス(B.subtilis)FB17で処理した植物の保水能力を示す。FB17処理後、総保水能力および保水率の有意な増加がトマト(2.1%)およびトウモロコシ(Z.mays)MO17(3.5%)で観察された。
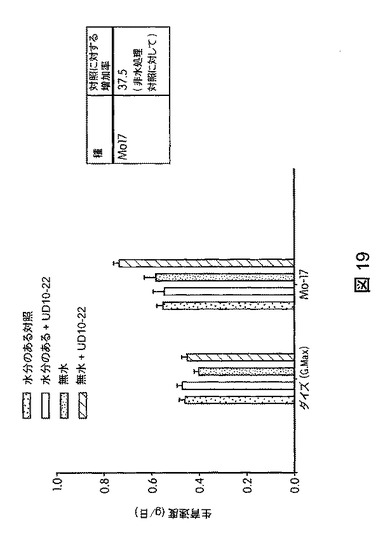
【図19】B.スブチリス(B.subtilis)FB17で処理した植物の乾燥耐性を示す。FB17処理後、MO17では乾燥ストレス(無水)非接種処理対照に対して、乾燥処理(無水)下での生育速度の有意な増加、すなわち、37.5%の増加が観察された
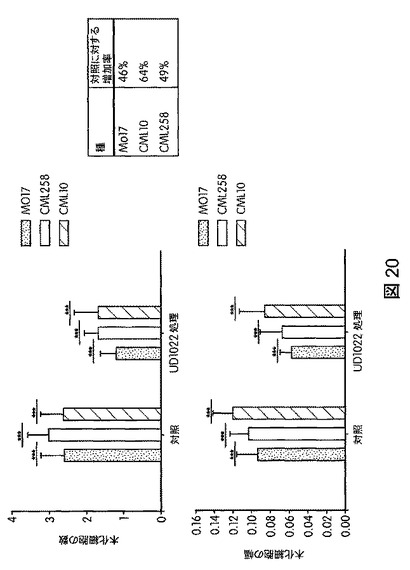
【図20】B.スブチリス(B.subtilis)FB17の種子処理によりトウモロコシのリグニン含有量が減少する。FB17処理後、非ストレス条件で総リグニン含有量の有意な減少がトウモロコシ(Z.mays)(MO17=46%;CML10=64%およびCML58=49%)で観察された。
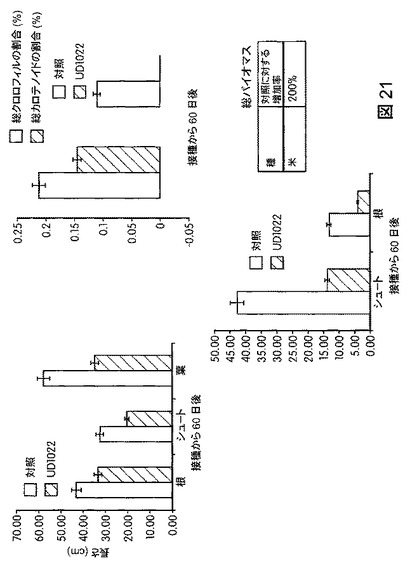
【図21】B.スブチリス(B.subtilis)FB17で処理したオリザ・サティバ(Oryza sativa)(日本晴(Nipponbare))の空中および根のバイオマスの増加を示す。FB17処理後、総バイオマスの有意な増加がO.サティバ(O.sativa)(米;品種日本晴(Nipponbare))で観察された(200%超)。
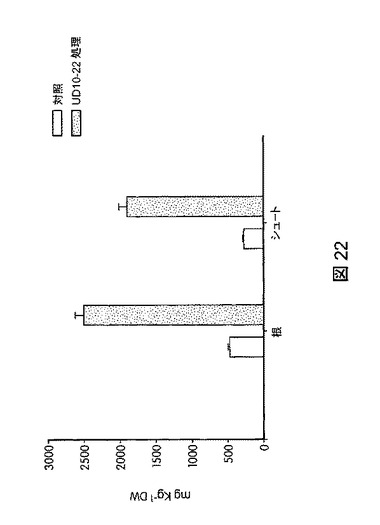
【図22】バチルス・スブチリス(Bacillus subtilis)FB17で処理した米植物で観察された鉄濃度を未処理米植物と比較して示す。このデータは、FB17の接種の結果、収穫量が増加し、米粒の鉄濃度が上昇することを示す。
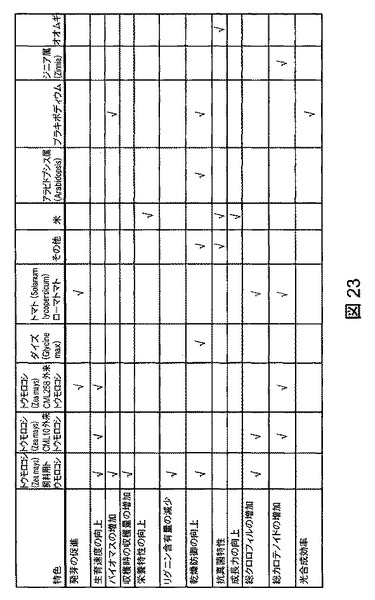
【図23】複数の植物種の様々な特色に対するB.スブチリス(B.subtilis)FB17の作用をまとめたものである。
【発明を実施するための形態】
【0017】
本出願人らは、植物に投与した際に驚くべき作用を示す植物生育促進根圏細菌(PGPR)、バチルス・スブチリス(Bacillus subtilis)FB17の株を発見した。B.スブチリス(B.subtilis)株FB17は、北アメリカのレッドビートの根から最初に単離された(本明細書に援用するFall et al.2004 System Appli.Microbiol.27:372−379を参照されたい)。この株は、表面のバイオフィルムおよび樹枝状の成長を形成する能力に基づきビートの根から単離された。
【0018】
特に、バチルス・スブチリス(Bacillus subtilis)FB17は、乾燥条件で様々な系統の植物のバイオマスを驚くほど向上させるほか、光合成効率上昇させ、生育速度を高めた。また、バチルス・スブチリス(Bacillus subtilis)FB17を植物に投与すると、植物のリグニン濃度が低下しており、これはバイオエネルギーの分野で重要な利点となり得る。リグニンは、植物バイオマスをバイオ燃料に変換する際の大きな障害の1つであるためである。米植物では、バチルス・スブチリス(Bacillus subtilis)FB17は、米の鉄濃度を驚くほど上昇させ、さらに世界中で米作物をだめにする病原真菌、イネいもち病の増殖を減弱させることも示されている。本発明は、様々な植物、特にトウモロコシ植物、ダイズ植物、および米植物などの作物植物のバイオマスを増加させる、乾燥耐性を向上させる、リグニン含有量を減少させる、種子発芽を促進する、鉄濃度を上昇させる、および病原体に対する耐性を向上させる方法を提供する。本発明はまた、バチルス・スブチリス(Bacillus subtilis)FB17を含む農業用担体および種子コーティングを提供する。
【0019】
本発明の実施形態は、植物のバイオマスをより多く生産する方法であって、未処理植物と比較して植物のバイオマスをより多く生産するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。本明細書で使用する場合、植物のバイオマスとは、植物の物質の全質量をいう。他に記載がない限り、バイオマスは、地上バイオマス(すなわち、以下に限定されるものではないが、茎、葉、および/または穀粒など空中バイオマス)および地下バイオマス(すなわち、根)の両方を含む。バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物のバイオマスは、公知の方法により測定することができる。一実施形態では、植物のバイオマスをグラム単位の植物の乾燥重量(DW)により測定する。
【0020】
バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物のバイオマスは、バチルス・スブチリス(Bacillus subtilis)FB17の植物への投与後、約7日〜約100日、約10日〜約75日、または約15日〜約35日の時点で測定してもよい。あるいは、バチルス・スブチリス(Bacillus subtilis)FB17を投与した作物植物のバイオマスは、植物を収穫してその穀粒または農産物を集める時点で、すなわち、トウモロコシ、ダイズまたはトマト植物などの成熟した作物植物を田畑から集める時点で測定してもよい。一例を挙げると、本発明の方法に従いバチルス・スブチリス(Bacillus subtilis)FB17を投与した作物植物は、グラム単位の乾燥重量で測定する場合、未処理植物より少なくとも約1%、約5%〜約200%、約5%〜約100%、約7.5%〜約75%、約15%〜約60%、または約30%〜約55%多い量で地上および地下の全バイオマスの量を多く生産する。一実施形態では、1つの方法は、バチルス・スブチリス(Bacillus subtilis)FB17の投与後、未処理植物より約5%〜約100%多い量で植物のバイオマスをより多く生産するのに効果的な量で、土壌に種子を播く前にバチルス・スブチリス(Bacillus subtilis)FB17を植物の種子に投与することを含む。たとえば、図4に図示したように、バチルス・スブチリス(Bacillus subtilis)FB17で処理したトウモロコシでは、処理から15日後、未処理トウモロコシと比較して空中および根のバイオマスが約28%増加することが観察された。
【0021】
本発明の別の実施形態は、植物の乾燥耐性を高める方法であって、未処理植物と比較して植物の乾燥耐性を高めるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。乾燥とは、土壌水分を枯渇させ、植物を害するだけの期間降雨がないか、または灌漑が行われないことをいう。植物の水分喪失が植物の根の吸水能力を上回る場合、および通常の植物のプロセスを阻害するのに十分に植物の含水量が減少する場合に乾燥ストレスが生じる。植物は、生育を停止し、光合成および水の使用を抑えるため他の植物のプロセスを抑制することにより水不足に対応する。本明細書で使用する場合、乾燥耐性とは、水の非存在下での植物の1日の生育速度、たとえば、未処理植物と比較してバチルス・スブチリス(Bacillus subtilis)FB17を接種した植物のバイオマスが増加した1日のグラム数をいう。たとえば、図19に図示したように、水の非存在下でバチルス・スブチリス(Bacillus subtilis)FB17で処理したトウモロコシ植物の種子は、処理から15日後、未処理植物と比較して1日の生育速度が約37.5%高まった。一実施形態では、1つの方法は、土壌に種子を播く前に、バチルス・スブチリス(Bacillus subtilis)FB17の投与後、未処理植物より少なくとも約10%高く植物の乾燥耐性を高めるのに効果的な量で、前記バチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の周囲の土壌、または植物の種子に投与することを含む。
【0022】
本発明の別の実施形態は、植物のリグニン濃度を低下させる方法であって、未処理植物と比較して植物のリグニン濃度を低下させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。リグニン濃度は、公知の方法により測定することができる。たとえば、図20に図示したように、バチルス・スブチリス(Bacillus subtilis)FB17で処理した植物は、未処理植物で観察された木化細胞の数が約46%〜約64%の減少を示した。リグニンは植物に不可欠な成分であり、植物細胞壁内、および植物細胞の間に見られる。リグニンは、植物バイオマスをバイオ燃料に変換する際の大きな障害の1つである。現在、もう1つの植物成分であるセルロースが、バイオ燃料の主な供給源である。セルロースはアルコールに発酵させやすいのに対し、リグニンは既存の発酵プロセスを用いて変換しないため、発酵性糖の抽出を難しくしている。したがって、リグニンの濃度が低い植物を生産することは有益である。本発明は、本発明の方法のいずれかによりバチルス・スブチリス(Bacillus subtilis)FB17を投与した植物のバイオマスのいずれか(たとえば、植物の全バイオマスまたは植物のバイオマスの任意の一部)をバイオ燃料に変換することにより生産されたバイオ燃料を提供する。バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物のバイオマスは、植物の糖成分の発酵によるなど任意の公知の方法によりバイオ燃料に変換することができる。
【0023】
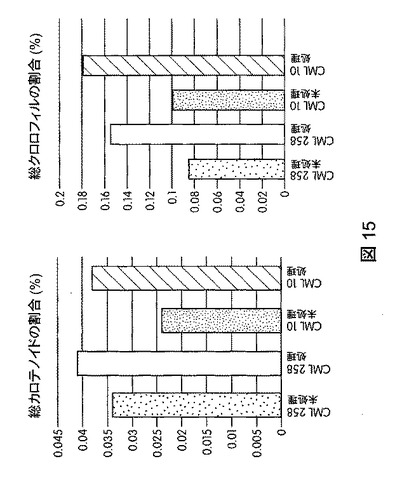
本発明の別の実施形態は、植物の種子発芽率を高める方法であって、未処理植物と比較して植物の種子発芽を促進するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは植物の種子の周囲の土壌に投与することを含む方法を提供する。たとえば、図16に図示したように、バチルス・スブチリス(Bacillus subtilis)FB17による投与後、トマト植物およびトウモロコシ植物で総発芽率の上昇が観察された。
【0024】
本発明の別の実施形態は、植物、特に米植物の鉄濃度を上昇させる方法であって、未処理植物と比較して植物の鉄濃度を上昇させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。鉄欠乏症は、ヒトの最も一般的な微量栄養素欠乏症の1つであり、かつ米は世界の人口の大部分にとって最も重要な主食であるため、本発明の方法により生産される米植物は、世界中で重要な栄養効果をもたらすことができる。バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物の鉄濃度は、誘導結合プラズマ発光分光分析法(ICP−AES)、誘導結合プラズマ質量分析法(ICP−MS)、または他の標準的な方法など公知の方法により測定することができる。一実施形態では、植物の乾燥重量1キログラム当たり鉄のミリグラム数により植物の鉄濃度を測定する。図22に図示したように、FB17で処理した米植物では、植物の乾燥重量1kg当たりの鉄のmgで測定すると、未処理植物と比較して鉄含有量が約81%増加することが観察された。
【0025】
本発明に使用するのに好適な米植物として、オリザ・サティバ(Oryza sativa)、オリザ・グラベリマ(Oryza glaberrima)、およびこれらのすべての亜種および品種が挙げられる。バチルス・スブチリス(Bacillus subtilis)FB17を投与した米植物の鉄濃度は、米を収穫してその穀粒または農産物を集める時点で、すなわち、成熟した米粒を田畑から集める時点で測定してもよい。あるいは、バチルス・スブチリス(Bacillus subtilis)FB17を投与した米植物の鉄濃度は、たとえば、バチルス・スブチリス(Bacillus subtilis)FB17を米植物に投与してから約1週間〜約5ヶ月、好ましくは約3ヶ月の間の時点で測定してもよい。本発明の方法に従いバチルス・スブチリス(Bacillus subtilis)FB17を投与した米植物は、たとえば、米植物の地上および地下バイオマスの乾燥重量1グラム当たりの鉄のグラム数で測定すると、鉄の量をより多く生産する。
【0026】
一例を挙げると、本発明の方法に従いバチルス・スブチリス(Bacillus subtilis)FB17を投与した米植物は、未処理植物より少なくとも約5%、約10%〜約200%、約25%〜約150%、約50%〜約100%、約70%〜約90%、約75%〜約85%、または約80%多い量で、米植物の地上および地下バイオマス乾燥重量当たりの鉄の量を多く生産する。たとえば、一実施形態では、1つの方法は、土壌に米の種子を播く前に、バチルス・スブチリス(Bacillus subtilis)FB17の投与後、未処理植物より少なくとも約25%高く米植物の鉄濃度を高くするのに効果的な量で前記バチルス・スブチリス(Bacillus subtilis)FB17を米の種子に投与することを含む。
【0027】
本発明の別の実施形態は、病原真菌による植物の感染を阻害する方法であって、未処理植物と比較して病原真菌による植物の感染を阻害するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。植物の例として、米の品種日本晴(Nipponbare)などの米植物およびオオムギ植物が挙げられる。特定の実施形態では、本発明は、病原真菌による米植物の感染症、特にイネいもち病を阻害する方法であって、未処理米植物と比較して病原真菌による米植物の感染症を阻害するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を米植物、米植物の種子、または米植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。本明細書で使用する場合、「イネいもち病」とは、植物病原性真菌(plant−pathogenic fungus)マグナポルテ・グリセア(Magnaporthe grisea)またはマグナポルテ・オリザエ(Magnaporthe oryzae)をいう。
【0028】
イネいもち病の症状として、植物のあらゆる部分、特に葉など植物の空中部、すなわち地上部に発生する病変または斑(たとえば、白色または灰色であることがある)がある。本明細書で使用する場合、「感染症を阻害する」とは、真菌感染症の軽減、たとえば、未処理植物と比較して米植物の空中部分の病変数の減少、または病変の一部または全部の大きさの縮小で判定した場合、米植物の真菌感染症を軽減することをいう。たとえば、特定の実施形態では、イネいもち病が原因の米植物の病変数を、未処理米植物と比較して約5%〜約100%、約10%〜約80%、約20%〜約60%または約25%〜約45%減少させるのに効果的な量で、バチルス・スブチリス(Bacillus subtilis)FB17を米植物、米植物の種子、または米植物もしくは種子の周囲の土壌に投与する。どのような理論にも拘泥するわけではないが、B.スブチリス(B.subtilis)FB17は、M.オリザエ(M.oryzae)の増殖を減弱させるまたは抑制する揮発性抗真菌化合物を生成すると考えられる。特定の実施形態では、イネいもち病の増殖および米植物の感染症を阻害するため、約1×107CFU/種子〜約1×109CFU/種子、一層好ましくは約1×108CFU/種子の量でバチルス・スブチリス(Bacillus subtilis)FB17を米の種子に投与し、その後種子を土壌に播く。
【0029】
本明細書で使用する場合、「未処理植物」とは、バチルス・スブチリス(Bacillus subtilis)FB17を投与していないこと以外は、本発明の方法に従いバチルス・スブチリス(Bacillus subtilis)FB17を投与した植物と同じ種で、実質的に同じ条件下(たとえば、同じ期間、同じ気候、同じ材料を用い同じ方法により栽培し、同じ方法により測定されたバイオマス、乾燥耐性、リグニン濃度、鉄濃度、真菌感染、および他の特徴を持つ)で生育させた植物をいう。本明細書で使用する場合、未処理植物と比較した、バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物の特徴、たとえばバイオマスの増加、乾燥耐性の向上、リグニン濃度の低下、鉄濃度の上昇または真菌感染症の軽減とは、それぞれ同じ時点で測定したバイオマスの増加、乾燥耐性の向上、リグニン濃度の低下、鉄濃度の上昇または真菌感染症の軽減をいう。
【0030】
本明細書に記載の方法のある種の実施形態では、バチルス・スブチリス(Bacillus subtilis)FB17の接種菌液約1ml/kg(すなわち、SmartSpec Bio Rad分光光度計を用いて測定した波長600nmの光学密度(OD)0.5の、LB培地で一晩増殖させたバチルス・スブチリス(Bacillus subtilis)FB17 1ml/kg)〜約50ml/kg、好ましくは約5ml/kg〜約25ml/kg、一層好ましくは約10ml/kg〜約15ml/kg、最も好ましくは約12.5ml/kgの量でバチルス・スブチリス(Bacillus subtilis)FB17を種子に投与する。代替の実施形態では、約1×106CFU/種子〜約1×109CFU/種子、一層好ましくは約1×107CFU/種子〜約1×108CFU/種子の量でバチルス・スブチリス(Bacillus subtilis)FB17を種子に投与する。
【0031】
本発明の方法は、バイオマスの増加、乾燥耐性の向上、リグニン含有量の減少、種子発芽の促進、鉄濃度の上昇、および病原体に対する耐性の向上を目的として多くの種類の植物(およびその種子または周囲の土壌)を処理するために使用してもよい。植物として、単子葉植物または双子葉植物が挙げられる。植物としては特に、トウモロコシ、ダイズ、トマト、米またはオオムギなどの作物が挙げられる。本発明の方法により処理してもよい植物の別の例としては、シロイヌナズナ(Arabidopsis thaliana)およびジニア属(Zinnia)のほか、バイオエネルギー作物植物、すなわち、セイヨウヤマカモジ(Brachypodium distachyon)などバイオエネルギー源として現在使用されている、または使用される可能性がある植物(たとえば、バイオ燃料の生産に有用な植物)が挙げられる。
【0032】
本発明によれば、バチルス・スブチリス(Bacillus subtilis)FB17は、根、種子または葉の接種によるなど、植物の全部または一部を処理する任意の公知の方法により植物に投与してもよい。たとえば、バチルス・スブチリス(Bacillus subtilis)FB17は、土壌に種子を播く前に葉および茎など植物の空中部分、植物の根、植物の種子に投与しても、または植物もしくは植物の種子の周囲の土壌に投与してもよい。投与の方法には、ドレンチング(drenching)、噴霧、コーティング、注射または当業者に公知の他の方法がある。本明細書で使用する場合、バチルス・スブチリス(Bacillus subtilis)FB17を投与するとは、1回投与、反復投与(すなわち、バチルス・スブチリス(Bacillus subtilis)FB17を1回より多く投与する)、または連続投与のいずれかをいう。バチルス・スブチリス(Bacillus subtilis)FB17は、植物の生活環の任意の時点(たとえば、発芽前または後)で投与してもよい。たとえば、バチルス・スブチリス(Bacillus subtilis)FB17は、土壌に種子を播く前、および発芽の前に植物の種子に投与してもよい。あるいは、バチルス・スブチリス(Bacillus subtilis)FB17は、発芽が生じた後に植物、植物の種子、または植物の周囲の土壌に投与してもよい。種子をバチルス・スブチリス(Bacillus subtilis)FB17で処理したら、土壌に播き、植物を生育させる従来の方法を用いて栽培すればよい。
【0033】
本発明の実施形態によれば、バチルス・スブチリス(Bacillus subtilis)FB17は、植物、植物の種子または土壌に単独投与しても、または他の材料との混合物として投与してもよい。たとえば、バチルス・スブチリス(Bacillus subtilis)FB17は、追加の添加剤または材料をまったく含まない増殖培地中のバチルス・スブチリス(Bacillus subtilis)FB17から本質的になる組成物として投与してもよい。あるいは、バチルス・スブチリス(Bacillus subtilis)FB17は、増殖培地中のバチルス・スブチリス(Bacillus subtilis)FB17、水などの担体、水溶液または粉末を含む組成物として投与してもよい。増殖培地、担体、水溶液または粉末は、殺虫剤または殺菌剤など追加の添加剤を含んでもよい。あるいは、バチルス・スブチリス(Bacillus subtilis)FB17は、異なる時間に適用される他の添加剤または材料と一緒に個別に投与してもよい。ある種の実施形態では、バチルス・スブチリス(Bacillus subtilis)FB17を、約1ml/kg(すなわち、SmartSpec Bio Rad分光光度計を用いて測定した波長600nmの光学密度(OD)0.5 の、LB培地で一晩増殖させたバチルス・スブチリス(Bacillus subtilis)FB17 1ml/kg)〜約50ml/kg、好ましくは約5ml/kg〜約25ml/kg、一層好ましくは約10ml/kg〜約15ml/kg、最も好ましくは約12.5ml/kgの量で、バチルス・スブチリス(Bacillus subtilis)FB17を含む組成物として投与する。代替の実施形態では、バチルス・スブチリス(Bacillus subtilis)FB17を約1×106CFU/種子〜約1×109CFU/種子、一層好ましくは約1×107CFU/種子〜約1×108CFU/種子の量で含む組成物としてバチルス・スブチリス(Bacillus subtilis)FB17を投与する。
【0034】
本発明はさらに、播く前に植物(たとえば、根)、植物の周囲の土壌、または種子に適用できる、バチルス・スブチリス(Bacillus subtilis)FB17を含む農業用担体と、植物種子に適用できる、バチルス・スブチリス(Bacillus subtilis)FB17を含む種子コーティングとを提供する。本発明はまた、種子の全部または一部がバチルス・スブチリス(Bacillus subtilis)FB17を含むコーティングまたは薄膜を有するように、バチルス・スブチリス(Bacillus subtilis)FB17でコートした植物の種子、好ましくは作物植物の種子(たとえば、トウモロコシ植物、ダイズ植物、米植物、トマト植物、またはセイヨウヤマカモジ(Brachypodium distachyon)などのバイオエネルギー作物植物の種子)を提供する。農業用担体は、バチルス・スブチリス(Bacillus subtilis)FB17接種菌液約1ml/kg(すなわち、SmartSpec Bio Rad分光光度計を用いて測定した波長600nmの光学密度(OD)0.5の、LB培地で一晩増殖させたバチルス・スブチリス(Bacillus subtilis)FB17 1ml/kg)〜約50ml/kg、約5ml/kg〜約25ml/kg、約10ml/kg〜約15ml/kg、または約12.5ml/kgの量でバチルス・スブチリス(Bacillus subtilis)FB17を含んでもよい。種子コーティングは、約1×106CFU/種子〜約1×108CFU/種子、一層好ましくは 約1×107CFU/種子の量でバチルス・スブチリス(Bacillus subtilis)FB17を含んでもよい。農業用担体および種子コーティングは各々、追加の添加剤または材料をまったく含まない増殖培地中のバチルス・スブチリス(Bacillus subtilis)FB17から本質的になっていてもよい。あるいは、農業用担体および種子コーティングは各々、水などの増殖培地、水溶液または粉末中にバチルス・スブチリス(Bacillus subtilis)FB17を含んでもよい。増殖培地、水溶液または粉末は、殺虫剤または殺菌剤など追加の添加剤を含んでもよい。
【0035】
本発明には、基本的用途と応用的用途との両方がある。本明細書に記載の方法を使用すれば、広い意味で(たとえば、バイオ燃料に使用される代替植物種で、あるいは、作物植物の収穫可能性に影響を与えるため)バイオマスを増加させ、乾燥耐性を向上させることができる。これらの方法は、遺伝子導入アプローチと比較して遺伝子の同定、遺伝子導入系の構築および特徴付けの時間を必要とせずに任意の植物に直ちに適用でき、導入遺伝子の使用に関する規制上および社会上の問題を伴わない。本明細書に記載の方法は、伝統的な栽培法(化学肥料および水の利用)の使用と比較して資源消費、および栽培者の労働がより少なく、環境により好ましい。さらに、化学肥料の利用は、急速で弱々しい生育を誘導することにより作物病害感受性を高めることが知られている一方、本方法で生育させる植物は、病害感受性が高まることが示されていない。FB17は、種子の処理に使用される他の根圏細菌と比較して、有益な結果を得るのに低菌量接種菌液を必要とする。最後に、こうした方法は有機農法との適合性があるのに対し、上述の他の方法(たとえば、化学肥料の利用)は適合性がない。
【0036】
B.スブチリス(B.subtilis)株FB17の寄託物は、Delaware Biotechnology Institute,15 Innovation way,Room #145,Newark,DE 19711で2010年3月1日より前から入手することができる。また、B.スブチリス(B.subtilis)株FB17は、アメリカンタイプカルチャーコレクション(ATCC),10801 University Boulevard,Manassas,Va.20110−2209 USAに寄託される。このATCC寄託物の利用は、本出願の係属中、特許商標庁長官(Commissioner of Patents and Trademarks)および長官によりその権利を有すると決定された者に対して、請求に応じて可能となる。寄託物は、30年の期間、または直近の請求から5年間、または特許の権利行使可能期間のいずれか長い期間、公的寄託機関であるATCC寄託機関で保存され、その期間中に生育不能なった場合、再寄託される。寄託物またはその子孫は、本出願の副本が出願されている国の外国特許法の定めるところにより利用できる。
【0037】
さらに、本寄託物は、微生物の寄託物に関するブダペスト条約の規定に従い保存され、一般に提供される、すなわち、本寄託物は、寄託物のサンプルの分譲に関する直近の請求から少なくとも5年の期間、さらにどのような場合でも、寄託日から少なくとも30年の期間、または発行する可能性がある、培養物を開示する任意の特許の権利行使可能期間、生存可能で汚染されない状態を保つのに必要な十分な注意を払って保存される。
【0038】
以下の例は、本発明をより詳細に記載するために提供するものであり、本発明を説明することを意図しており、本発明を限定するものではない。下記の図の一部に記載された「UD10−22」とは、バチルス・スブチリス(Bacillus subtilis)FB17をいう。
【実施例】
【0039】
実施例1
セイヨウヤマカモジ(Brachypodium distachyon)植物およびトウモロコシ植物を発芽させ、21日間生育させた。5日に1回(3回)、ポットごとにOD0.5のB.スブチリス(B.subtilis)FB17 5mlを加えた。対照では、OD0.5の大腸菌(E.coli)OP50 5mlをポットごとに加えた。FB17およびOP50をLB培地で一晩生育させ、SmartSpec(Bio Rad)分光光度計を使用して波長(600nm)の光学密度(OD)を得た。最終処理から10日後、植物を解析した。本明細書のすべての実験に記載される対照とは、細菌で処理しなかったか、または大腸菌(E.coli)OP50で処理した植物をいう。
【0040】
B.スブチリス(B.subtilis)FB17、細菌の対照大腸菌(E.coli)、または模擬処理(mock treatment)で処理したセイヨウヤマカモジ(Brachypodium distachyon)(Bd2−1)植物およびトウモロコシ植物を、処理後30日間、標準的な条件(22〜25℃、湿度60%、明期16時間/暗期8時間の明暗周期)で4×4インチのポットにて生育させた。エネルギー作物セイヨウヤマカモジ(B.distachyon)の空中および根のバイオマスは、FB17処理により増加した。図1は、FB17で処理したセイヨウヤマカモジ(B.distachyon)のバイオマスが統計学的レベルで増加したことを示す。図2は、FB17で処理したセイヨウヤマカモジ(B.distachyon)で観察された光合成効率の向上を図示する。FB17で処理したセイヨウヤマカモジ(B.distachyon)は、対照より多くのクロロフィルと総カロエテノイドとを含んでおり、植物の健康状態が健全であることを示す。図3は、セイヨウヤマカモジ(B.distachyon)の根の周囲の土壌から回収されたFB17の量を示す。図から、FB17は、大腸菌(E.coli)と比較してセイヨウヤマカモジ(B.distachyon)の根に非常に強く関連しており、FB17の根圏細菌の本来の性質を示唆することが明らかにされる。
【0041】
また、トウモロコシ植物も、B.スブチリス(B.subtilis)FB17、細菌の対照大腸菌(E.coli)OP50、または模擬処理により処理してから30日間の生育後、空中および根のバイオマスの増加を示した。
【0042】
実施例2
シロイヌナズナ(Arabidopsis thaliana)の種子を発芽させ、21日間生育させた。5日に1回(3回)、OD0.5のB.スブチリス(B.subtilis)FB17 5mlをポットごとに加えた。対照では、OD0.5の大腸菌(E.coli)OP50 5mlを ポットごとに加えた。FB17およびOP50をLB培地で一晩生育させ、SmartSpec(Bio Rad)分光光度計を使用して波長(600nm)の光学密度(OD)を得た。最終処理から10日後、植物を25℃、湿度40%で4週間乾燥に供した(すなわち、水を加えなかった)。処理から30日後、未処理植物のstay green表現型の喪失をFB17で処理した植物と比較して乾燥を評価したところ、FB17がアラビドプシス属(Arabidopsis)の乾燥耐性を向上させることが示唆された。
【0043】
実施例3
B.スブチリス(B.subtilis)FB17の種子処理は、トウモロコシMo17、CML258、CML10、ジニア属(Zinnia)、およびセイヨウヤマカモジ(Brachypodium distachyon)のバイオマスの向上を促進する。
【0044】
トウモロコシ(Mo17、CML258、CML10)、ダイズ(Will−82)、トマト(Solanum lycopersicum)、ジニア属(Zinnia)、およびセイヨウヤマカモジ(Brachypodium distachyon)(エネルギー作物モデル)のバイオマスの向上に対するB.スブチリス(B.subtilis)FB17の作用を試験するため、植物種ごとに50の種子(n=50)を、B.スブチリス(B.subtilis)FB17(約1×107cfu/種子、またはSmartSpec Bio Rad分光光度計を用いて測定した波長600nmの光学密度(OD)0.5の、LB培地で一晩増殖させたバチルス・スブチリス(Bacillus subtilis)FB17 12.5ml/kg)で種子処理した。種子処理後、発芽およびバイオマスの研究のため、混合土壌を含むポット(4×4インチ)に種子を一つずつ播種した。興味深いことに、B.スブチリス(B.subtilis)FB17の種子処理は、試験した全作物種で根およびシュートの生育を促進した。測定値は、処理から15日後に得た。
【0045】
種子処理した植物が根のバイオマスの増加を促進し、その結果、根の長さが増加するよりもむしろ根系密度が高くなった。根系密度が高くなったのは側根および根毛が増加し、より多くの水および栄養素を取り込めるようになったことに起因する。
【0046】
トウモロコシ(Zea mays)var.CML258は、空中バイオマスが対照より約16%増加した。トウモロコシ(Zea mays)var.CML10は、空中バイオマスが対照より約9%増加した。トウモロコシ(Zea mays)var.Mo−17は、空中バイオマスが対照より約38%増加した。ブラキポディウム(Brachypodium)は、空中バイオマスが対照より約40%増加した。トウモロコシ(Z.mays)(MO17)の場合、空中および根の総バイオマスが対照より約28%という有意な増加が観察された。図4は、B.スブチリス(B.subtilis)FB17で処理した植物のバイオマスの総増加率を図示する。
【0047】
図5は、B.スブチリス(B.subtilis)FB17で種子処理したMo−17植物における根およびシュートのバイオマスの増加を示す定量データを図示する。図6は、B.スブチリス(B.subtilis)FB17で種子処理したバイオエネルギー作物セイヨウヤマカモジ(Brachypodium distachyon)(遺伝子型Bd2−1)植物における葉数の増加を示す定量データを図示する。図7は、B.スブチリス(B.subtilis)FB17で種子処理したバイオエネルギー作物セイヨウヤマカモジ(Brachypodium distachyon)(遺伝子型Bd2−1)植物における根およびシュートのバイオマスの増加を示す定量データを図示する。図8および図9は、B.スブチリス(B.subtilis)FB17で種子処理したジニア・エスピー(Zinnia sp.)「レッドスパイダー」における根およびシュートのバイオマスの増加を示す定量データを図示する。
【0048】
実施例4
B.スブチリス(B.subtilis)FB17の種子処理は、トウモロコシおよびトマトの光合成効率を促進する。
【0049】
トウモロコシ(Mo17、CML258、CML10)、ダイズ(Will−82)、トマト(Solanum lycopersicum)、ジニア属(Zinnia)、およびブラキポディウム(Brachypodium)(エネルギー作物モデル)の光合成効率に対するB.スブチリス(B.subtilis)FB17の作用を試験するため、植物種ごとに50の種子(n=50)を、B.スブチリス(B.subtilis)FB17(12.5ml/kgまたは1e7cfu/種子)で種子処理した。処理から15〜32日後の葉を収穫し、総クロロフィル含有量について解析した。結果から、図10および図11に図示したように、B.スブチリス(B.subtilis)FB17を接種したトウモロコシ植物およびトマト植物(トマトおよび外来種のトウモロコシCML258およびCML10)は未処理サンプルと比較してクロロフィルおよびカロテノイド含有量の増加を示すことが明らかになった。
【0050】
総クロロフィル値の増加は、CML258およびCML10で見られたように成長力およびバイオマスの増加を促進する可能性がある。B.スブチリス(B.subtilis)FB17を種子接種したトマトの総クロロフィル含有量は、約14%増加した。より一層顕著なのは外来種のトウモロコシCML258およびCML10であり、それぞれ約87%および約72%増加した。
【0051】
総クロロフィル含有量は増加しているものの、そのことは、総カロテノイド含有量も増加した値に対応することを意味するものではない。トマトおよびジニア属(Zinnia)は、B.スブチリス(B.subtilis)FB17を接種して未処理種子と比較すると、総カロテノイドの割合が有意に低下した。トウモロコシCML258およびCML10は総カロテノイドの割合が有意に増加した一方、ダイズ、トウモロコシMo17、およびブラキポディウム(Brachypodium)は、処理した種子と未処理の種子との間で統計学的に有意な差を示さなかった。
【0052】
図12は、B.スブチリス(B.subtilis)FB17で種子処理したMo−17植物における光合成効率の上昇を示す定量データを図示する。図13は、B.スブチリス(B.subtilis)FB17で種子処理したバイオエネルギー作物セイヨウヤマカモジ(Brachypodium distachyon)(遺伝子型Bd2−1)植物における光合成効率の上昇を示す定量データを図示する。図14は、B.スブチリス(B.subtilis)FB17で種子処理したジニア・エスピー(Zinnia sp.)「レッドスパイダー」における光合成効率の上昇を示す定量データを図示する。図15は、B.スブチリス(B.subtilis)FB17で種子処理した外来トウモロコシCML10およびCML258における光合成効率の上昇を示す定量データを図示する。
【0053】
実施例5
B.スブチリス(B.subtilis)FB17の種子処理は、トウモロコシ植物およびトマト植物の発芽を促進する。
【0054】
トウモロコシ(Mo17、CML258、CML10)、ダイズ(Will−82)、トマト(Solanum lycopersicum)、ジニア属(Zinnia)、およびブラキポディウム(Brachypodium)(エネルギー作物モデル)の発芽増加率に対するB.スブチリス(B.subtilis)FB17の作用を試験するため、植物種ごとに50の種子(n=50)を、B.スブチリス(B.subtilis)FB17(12.5ml/kgまたは1e7cfu/種子)で種子処理した。最終的な発芽率を播種した日から8日後にスコア化した。結果から、B.スブチリス(B.subtilis)FB17処理は、図16に図示したようにトマトおよびトウモロコシの発芽反応を統計学的に有意に促進することが明らかになった。
【0055】
B.スブチリス(B.subtilis)FB17で処理したトマトの種子および外来種トウモロコシCML258はそれぞれ発芽率が5.9%および14%上昇した。
【0056】
B.スブチリス(B.subtilis)FB17の種子処理は、試験した全作物種について中立およびプラスの作用を有した。種子処理を適用した場合、発芽率に対するマイナスの反応は、統計学的に認められなかった。
【0057】
実施例6
トウモロコシ(Mo17、CML258、CML10)、ダイズ(Will−82)、トマト(Solanum lycopersicum)、ジニア属(Zinnia)、およびブラキポディウム(Brachypodium)(エネルギー作物モデル)におけるB.スブチリス(B.subtilis)FB17の作用を試験するため、植物種ごとに50の種子(n=50)を、B.スブチリス(B.subtilis)FB17(1e7cfu/種子またはSmartSpec Bio Rad分光光度計を用いて測定した波長600nmの光学密度(OD)0.5の、LB培地で一晩増殖させたバチルス・スブチリス(Bacillus subtilis)FB17 12.5ml/kg)で種子処理した。種子処理後、種子を一つずつポット(4×4インチ)播種した。測定は、処理から15日後に行った。図17は、B.スブチリス(B.subtilis)FB17で処理後のトウモロコシ(Zea mays)の生育速度を図示する。図18は、B.スブチリス(B.subtilis)FB17で処理した植物の保水能力を図示する。FB17処理後、総保水能力および保水率が、トマト(2.1%)およびトウモロコシ(Z.mays)MO17(3.5%)で有意に高まることが観察された。図19は、B.スブチリス(B.subtilis)FB17で処理した植物の乾燥耐性を図示する。MO17の生育速度がFB17処理後、乾燥処理下で有意に高まることが観察された(非水処理対照に対して37.5%増)。図20は、B.スブチリス(B.subtilis)FB17の種子処理がトウモロコシのリグニン含有量を減少させることを図示する。トウモロコシ(Z.mays)ではFB17処理後、総リグニン含有量が有意に減少することが観察された(MO17で約46%減少;CML10で約64%減少、およびCML58で約49%減少)。
【0058】
実施例7
図21は、B.スブチリス(B.subtilis)FB17で処理した米植物オリザ・サティバ(Oryza sativa)(日本晴(Nipponbare))の空中および根のバイオマスが接種から60日後に増加することを図示する。LBで一晩生育させたFB17の培養液を用いて108細胞/mlの接種菌液を作製した。FB17の補充には、水耕法で生育させ4週齢の米植物(品種日本晴(Nipponbare))を使用した。B.スブチリス(B.subtilis)FB17を投与した米植物は、未処理米植物と比較して約200%のバイオマスの増加を示した。
【0059】
実施例8
バチルス・スブチリス(Bacillus subtilis)FB17が米の根に定着するかどうかを評価するため、米植物(品種日本晴(Nipponbare))にバチルス・スブチリス(Bacillus subtilis)FB17を接種し、接種から96時間後に米植物の根を共焦点走査型レーザー顕微鏡により観察した。観察結果から、この有益な根圏細菌(バチルス・スブチリス(Bacillus subtilis)FB17)は、植物体にバイオフィルムを形成することが確認された。特に、データからは、バチルス・スブチリス(Bacillus subtilis)FB17が処理の96時間後に米の根に効率的に定着することが示唆されることから、米の根が有益な微生物の定着を支持することが示される。
【0060】
米植物の根圏細菌処理が気孔開口に何らかの変化を与えるかどうかを評価するため、出願人らは、根圏細菌で処理した米植物を解析した。結果から、バチルス・スブチリス(Bacillus subtilis)FB17で米を根圏細菌処理すると、処理した米植物(品種日本晴(Nipponbare))の気孔開口が大きく減少することが明らかになった。バチルス・スブチリス(Bacillus subtilis)FB17処理の場合、バチルス・スブチリス(Bacillus subtilis)FB17の添加から1週後に孔辺細胞が観察された。これらの結果から、B.スブチリス(B.subtilis)FB17(バチルス・スブチリス(Bacillus subtilis)FB17)は、シロイヌナズナ(A.thaliana)および米の両方で証明されたように、単子葉植物および双子葉植物の両方で一般的な気孔閉鎖反応を与えることが示唆される。
【0061】
バチルス・スブチリス(Bacillus subtilis)FB17がイネいもち病の増殖を減弱させるかどうかを評価するため、出願人らは、マグナポルテ・オリザエ(Magnaporthe oryzae)をバチルス・スブチリス(Bacillus subtilis)FB17培養液に曝露した。定性コンパートメントプレート(qualitative compartment plate)および定量データから、バチルス・スブチリス(Bacillus subtilis)FB17曝露真菌培養液での放射状の増殖の低下により示されたように、バチルス・スブチリス(Bacillus subtilis)FB17はM.オリザエ(M.oryzae)の増殖を減弱させることが示された。対照(TYおよびLB)との比較から、処理しない場合に病原体が増殖すると考えられる程度が明らかにされる。表1に示すように、バチルス・スブチリス(Bacillus subtilis)FB17はインビトロでM.オリザエ(M.oryzae)の増殖を約25%抑制した。これらの結果から、B.スブチリス(B.subtilis)FB17は、M.オリザエ(M.oryzae)の増殖を減弱させるか、または阻害することができる揮発性抗真菌化合物を生成することが示唆される。
【0062】
【表1】
バチルス・スブチリス(Bacillus subtilis)FB17は、米およびオオムギでマグナポルテ・オリザエ(Magnaporthe oryzae)に対する全身抵抗性を誘導した。M.オリザエ(M.oryzae)に曝露された米植物およびオオムギ植物はどちらも、表2に示すように対照と比較してFB17で処理した植物で米の葉およびオオムギの子葉の病変形成がそれぞれ抑制された(いもち病に典型的なダイヤモンド状の病変が葉に少なくとも1つ存在する場合に「感染した」と定義した)。
【0063】
【表2】
実施例9
バチルス・スブチリス(Bacillus subtilis)FB17が米の鉄強化を促進するかどうかを評価するため、本出願人らは、誘導結合プラズマ発光分光分析法(ICP−AES)を用いて、バチルス・スブチリス(Bacillus subtilis)FB17を補充した植物の米の葉、根、および穀粒における全鉄含有量を解析した。結果から、バチルス・スブチリス(Bacillus subtilis)FB17を米に補充すると、植物体に鉄が集まりやすくなる、すなわち、必須元素の鉄が、植物の生育および成長に利用される植物に積極的に取り込まれることが明らかになった。図22に図示したように、植物の乾燥重量1kg当たりの鉄のmg数で測定した場合、FB17で処理した米植物では未処理対照と比較して鉄含有量が81%増加することが観察された(図1に示したように「UD1022」とはバチルス・スブチリス(Bacillus subtilis)FB17をいう)。したがって、バチルス・スブチリス(Bacillus subtilis)FB17を植物、特に米植物に投与すると、食物中の鉄濃度の上昇により食物の栄養価を大きく高めることができる。
【0064】
図23は、上述の複数の植物種の様々な特色に対するB.スブチリス(B.subtilis)FB17の作用をまとめたものである。
【0065】
本発明について特定の実施形態と共に記載してきたが、特許請求の範囲に記載の本発明は、こうした特定の実施形態に必要以上に限定されるべきものではないことを理解すべきである。実際、本発明の記載された組成物および方法の様々な修正および変形は、当業者には明らかであり、添付の特許請求の範囲の範囲内にあることを意図している。
【技術分野】
【0001】
関連出願の相互参照
本出願は、あらゆる目的のためその全体を本明細書に援用する2010年3月1日に出願された米国特許出願第61/309,134号、2010年11月16日に出願された米国特許出願第61/414,108号、および2010年11月22日に出願された米国特許出願第61/416,039号に対する優先権を主張する。
【0002】
政府支援に関する記載
開示された本発明に至る研究は、University of DelawareのGeorgetown,DEのUSDA Experimental Field Stationからの技術支援と共に、国立科学財団(National Science Foundation:NSF)Grant No.0923806およびNSF Grant No.IOS−0814477により一部資金提供を受けた。したがって、米国政府は、本発明に一定の権利を有する場合がある。
【0003】
本発明は一般にバイオマスの増加、乾燥耐性の向上、リグニン含有量の減少、種子発芽の促進、鉄濃度の上昇、および病原体に対する耐性の向上など植物生育の様々な特徴を向上させる植物生育促進根圏細菌の使用に関する。特に、本発明の実施形態は、バチルス・スブチリス(Bacillus subtilis)FB17の植物への投与に関する。その結果得られる植物は、バイオ燃料、食物の生産または他の目的に使用することができる。
【背景技術】
【0004】
食料安全保障は世界中で常に最優先課題であり、作物生産の環境影響に対する懸念の増大に伴い、環境を保護しつつ、生産性を高める新規な方法の開発および使用が必要とされている 植物生物学者は、増加する世界各地の人々が必要不可欠な原料を確実に入手できることを目指して作物植物を十分に生産するための戦略を開発し実行している。しかしながら、バイオ燃料および再生可能技術がますます重要な優先課題となってきているため、バイオ燃料および再生可能技術の開発がこの課題を大きくしている。したがって、田畑の多様な条件で作物収穫量を増大させる改良されたアプローチに対する必要性が増している。
【0005】
作物植物のバイオマスおよび収穫可能性を増加させるにあたっては、世界中に多くの様々な地理的環境および気候があり、異なる種類の課題が提示されている。乾燥は、作物生産を世界的に制約する重要な要因である。生育期の長期乾燥または短期乾燥は、作物生産を大きく制限したり、あるいはゼロにしたりする可能性さえある。世界の主要な作付け地域でも、世界的な気象パターンの変化が、乾燥の頻度および強さに影響を与えている。
【0006】
また、養分利用率(nutrient availability)も作物生産を制約する。養分による土壌の改善(soil augmentation)は費用がかかり、エネルギー集約的であるうえ、養分を十分な量で利用できるときでも、作物植物は養分取り込みが非効率的である場合がある。必須養分の取り込みが不十分であると、収穫量が低下し、食用作物の栄養的価値が低下する。たとえば、米(単子葉植物オリザ・サティバ(Oryza sativa)またはオリザ・グラベリマ(Oryza glaberrima)の種子)は、世界の人口の3分の2以上にとって最も重要な主食であり、消費されるカロリーのかなりの割合を供給している。米は世界人口の大半にとって主要な主食であり、世界的にヒトの最も一般的な微量栄養素欠乏症(micronutrient deficiencies)の1つが鉄欠乏症であることから、より高レベルの鉄を含む米を生産すれば、世界中で微量栄養素栄養失調の抑制に大きな影響を与える可能性がある。
【0007】
また、病原体ストレスも生産性を制約する。植物は病原体の攻撃から身を守るためエネルギーを投資しなければならず、このエネルギーの流用の結果、収穫量が減少する。また、植物は病害の進行を抑えるためその組成を変えるものであり、これらの変化が、多くの場合、作物の加工を難しくしている。さらに、一部の作物病原体は、遺伝的多様性により効率的に抑えることも、化学的駆除もできず、作物生産に世界的に大きな影響を与える。
【0008】
イネいもち病(rice blast)(マグナポルテ・グリセア(Magnaporthe grisea)またはマグナポルテ・オリザエ(Magnaporthe oryzae))は、米を侵す深刻な病害を引き起こす植物病原性真菌である。イネいもち病は経済的に大きな作物被害を毎年引き起こし、作物収穫量の40%を占めると推定される。イネいもち病は、成長期になるたびに世界中で数百万の人々の食料となるのに十分な米をだめにする。米は世界の大半で重要な主食であるため、イネいもち病の影響は、ヒトの健康および環境に広範に及ぶ。米不足はヒトの飢餓に直接つながる。イネいもち病はさらに作物の減少にもつながり、減少した収穫量を補う別の資源を使用することが必要になる。栄養が増加したより多くの量の食物が世界の人々に入手できるように、さらにバイオ燃料の生産など他の重要な利益のために、多様な生育条件で、乾燥ストレスに対する耐性、病原体の影響に対する耐性、養分利用率、および最終的な作物収穫量など植物生育の様々な特徴を向上させる戦略が引き続き強く求められている。
【発明の概要】
【課題を解決するための手段】
【0009】
本発明の実施形態は、植物のバイオマスをより多く生産する方法であって、未処理植物と比較して植物のバイオマスをより多く生産するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは植物の種子の周囲の土壌に投与することを含む方法を提供する。
【0010】
別の実施形態は、植物の乾燥耐性を向上させる方法であって、未処理植物と比較して植物の乾燥耐性を向上させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは植物の種子の周囲の土壌に投与することを含む方法を提供する。
【0011】
別の実施形態は、植物のリグニン濃度を低下させる方法であって、未処理植物と比較して植物のリグニン濃度を低下させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは植物の種子の周囲の土壌に投与することを含む方法を提供する。
【0012】
別の実施形態は、植物の種子発芽を促進する方法であって、未処理植物と比較して植物の種子発芽を促進するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは植物の種子の周囲の土壌に投与することを含む方法を提供する。
【0013】
別の実施形態は、植物の、特に米植物の鉄濃度を上昇させる方法であって、未処理植物と比較して植物の鉄濃度を上昇させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。
【0014】
別の実施形態は、植物病原真菌(plant fungal pathogen)の増殖と、病原真菌、特にイネいもち病による植物、特に米植物の感染とを阻害する方法であって、病原真菌による植物の感染を阻害するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。
【0015】
追加の実施形態は、バチルス・スブチリス(Bacillus subtilis)FB17を含む農業用担体および種子コーティングを提供する。バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物のバイオマスはバイオ燃料に変換することができ、生産された作物は、ヒトもしくは動物の食糧(foodstock)、または他の目的に安全に使用することができる。
【図面の簡単な説明】
【0016】
【図1】B.スブチリス(B.subtilis)FB17で処理したセイヨウヤマカモジ(Brachypodium distachyon)(Bd2−1)植物を対照と比較した、形態計測分析(枝の数、葉の数、シュートの高さ、シュートの重量、根の長さ、根の重量)を示す。これにより、B.スブチリス(B.subtilis)FB17の接種が植物の形態を高めることが示される。
【図2】B.スブチリス(B.subtilis)FB17で処理したセイヨウヤマカモジ(Brachypodium distachyon)植物を総クロロフィルおよびカロテノイドにより測定し、対照と比較した生化学的解析を示す。これにより、B.スブチリス(B.subtilis)FB17の接種が、植物の光エネルギーを集める能力にプラスの影響を与えることが示される。
【図3】土壌およびセイヨウヤマカモジ(Brachypodium distachyon)の根から回収されたB.スブチリス(B.subtilis)FB17と対照との量を示す。これにより、B.スブチリス(B.subtilis)FB17を接種した植物の根との関係が安定していることが明らかにされる。
【図4】B.スブチリス(B.subtilis)FB17で処理した様々な植物種のバイオマスの総増加率を示す。トウモロコシ(Z.mays)(MO17)では、空中のバイオマスおよび根のバイオマスの両方で有意な増加(約28%)が観察された。
【図5】B.スブチリス(B.subtilis)FB17で種子処理したトウモロコシ(Z.mays)Mo−17植物の根およびシュートのバイオマスの増加を示す定量データである。
【図6】B.スブチリス(B.subtilis)FB17で種子処理したバイオエネルギー作物セイヨウヤマカモジ(Brachypodium distachyon)(遺伝子型Bd2−1)植物の葉数の増加を示す定量データである。
【図7】B.スブチリス(B.subtilis)FB17で種子処理したバイオエネルギー作物セイヨウヤマカモジ(Brachypodium distachyon)(遺伝子型Bd2−1)植物の根およびシュートのバイオマスの増加を示す定量データである。
【図8】B.スブチリス(B.subtilis)FB17で種子処理したジニア・エスピー(Zinnia sp.)「レッドスパイダー」の根およびシュートのバイオマスの増加を示す定量データである。
【図9】B.スブチリス(B.subtilis)FB17で種子処理したジニア・エスピー(Zinnia sp.)「レッドスパイダー」の根およびシュートのバイオマスの増加を示す定量データである。
【図10】B.スブチリス(B.subtilis)FB17で処理した植物の総クロロフィル含有量を示す。FB17処理後、総クロロフィル含有量の有意な増加が、トマト(14%)、トウモロコシ(Z.mays)CML10(72%)およびCML258(87%)で観察された。
【図11】B.スブチリス(B.subtilis)FB17で処理した植物の総カロテノイド含有量を示す。FB17処理後、総カロテノイド含有量の有意な増加がダイズ(31%)およびトウモロコシ(Z.mays)MO17(82%)で観察された。
【図12】B.スブチリス(B.subtilis)FB17で種子処理したMo−17植物の光合成効率の上昇を示す定量データである。
【図13】B.スブチリス(B.subtilis)FB17で種子処理したバイオエネルギー作物セイヨウヤマカモジ(Brachypodium distachyon)(遺伝子型Bd2−1)植物の光合成効率の上昇を示す定量データである。
【図14】B.スブチリス(B.subtilis)FB17で種子処理したジニア・エスピー(Zinnia sp.)「レッドスパイダー」の光合成効率の上昇を示す定量データである。
【図15】B.スブチリス(B.subtilis)FB17で種子処理した外来トウモロコシCML10およびCML258の光合成効率の上昇を示す定量データである。
【図16】B.スブチリス(B.subtilis)FB17で処理した種子の発芽率の上昇を示す。FB17処理後、総発芽含有率(plant−pathogenic fungus)の有意な増加がトマト(6%)、トウモロコシ(Z.mays)MO17(2.1%)およびCML258(14%)で観察された。特に外来種トウモロコシCML258の発芽率が劇的に上昇した。
【図17】B.スブチリス(B.subtilis)FB17で処理したトウモロコシ(Zea mays)の生育速度を示す。
【図18】B.スブチリス(B.subtilis)FB17で処理した植物の保水能力を示す。FB17処理後、総保水能力および保水率の有意な増加がトマト(2.1%)およびトウモロコシ(Z.mays)MO17(3.5%)で観察された。
【図19】B.スブチリス(B.subtilis)FB17で処理した植物の乾燥耐性を示す。FB17処理後、MO17では乾燥ストレス(無水)非接種処理対照に対して、乾燥処理(無水)下での生育速度の有意な増加、すなわち、37.5%の増加が観察された
【図20】B.スブチリス(B.subtilis)FB17の種子処理によりトウモロコシのリグニン含有量が減少する。FB17処理後、非ストレス条件で総リグニン含有量の有意な減少がトウモロコシ(Z.mays)(MO17=46%;CML10=64%およびCML58=49%)で観察された。
【図21】B.スブチリス(B.subtilis)FB17で処理したオリザ・サティバ(Oryza sativa)(日本晴(Nipponbare))の空中および根のバイオマスの増加を示す。FB17処理後、総バイオマスの有意な増加がO.サティバ(O.sativa)(米;品種日本晴(Nipponbare))で観察された(200%超)。
【図22】バチルス・スブチリス(Bacillus subtilis)FB17で処理した米植物で観察された鉄濃度を未処理米植物と比較して示す。このデータは、FB17の接種の結果、収穫量が増加し、米粒の鉄濃度が上昇することを示す。
【図23】複数の植物種の様々な特色に対するB.スブチリス(B.subtilis)FB17の作用をまとめたものである。
【発明を実施するための形態】
【0017】
本出願人らは、植物に投与した際に驚くべき作用を示す植物生育促進根圏細菌(PGPR)、バチルス・スブチリス(Bacillus subtilis)FB17の株を発見した。B.スブチリス(B.subtilis)株FB17は、北アメリカのレッドビートの根から最初に単離された(本明細書に援用するFall et al.2004 System Appli.Microbiol.27:372−379を参照されたい)。この株は、表面のバイオフィルムおよび樹枝状の成長を形成する能力に基づきビートの根から単離された。
【0018】
特に、バチルス・スブチリス(Bacillus subtilis)FB17は、乾燥条件で様々な系統の植物のバイオマスを驚くほど向上させるほか、光合成効率上昇させ、生育速度を高めた。また、バチルス・スブチリス(Bacillus subtilis)FB17を植物に投与すると、植物のリグニン濃度が低下しており、これはバイオエネルギーの分野で重要な利点となり得る。リグニンは、植物バイオマスをバイオ燃料に変換する際の大きな障害の1つであるためである。米植物では、バチルス・スブチリス(Bacillus subtilis)FB17は、米の鉄濃度を驚くほど上昇させ、さらに世界中で米作物をだめにする病原真菌、イネいもち病の増殖を減弱させることも示されている。本発明は、様々な植物、特にトウモロコシ植物、ダイズ植物、および米植物などの作物植物のバイオマスを増加させる、乾燥耐性を向上させる、リグニン含有量を減少させる、種子発芽を促進する、鉄濃度を上昇させる、および病原体に対する耐性を向上させる方法を提供する。本発明はまた、バチルス・スブチリス(Bacillus subtilis)FB17を含む農業用担体および種子コーティングを提供する。
【0019】
本発明の実施形態は、植物のバイオマスをより多く生産する方法であって、未処理植物と比較して植物のバイオマスをより多く生産するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。本明細書で使用する場合、植物のバイオマスとは、植物の物質の全質量をいう。他に記載がない限り、バイオマスは、地上バイオマス(すなわち、以下に限定されるものではないが、茎、葉、および/または穀粒など空中バイオマス)および地下バイオマス(すなわち、根)の両方を含む。バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物のバイオマスは、公知の方法により測定することができる。一実施形態では、植物のバイオマスをグラム単位の植物の乾燥重量(DW)により測定する。
【0020】
バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物のバイオマスは、バチルス・スブチリス(Bacillus subtilis)FB17の植物への投与後、約7日〜約100日、約10日〜約75日、または約15日〜約35日の時点で測定してもよい。あるいは、バチルス・スブチリス(Bacillus subtilis)FB17を投与した作物植物のバイオマスは、植物を収穫してその穀粒または農産物を集める時点で、すなわち、トウモロコシ、ダイズまたはトマト植物などの成熟した作物植物を田畑から集める時点で測定してもよい。一例を挙げると、本発明の方法に従いバチルス・スブチリス(Bacillus subtilis)FB17を投与した作物植物は、グラム単位の乾燥重量で測定する場合、未処理植物より少なくとも約1%、約5%〜約200%、約5%〜約100%、約7.5%〜約75%、約15%〜約60%、または約30%〜約55%多い量で地上および地下の全バイオマスの量を多く生産する。一実施形態では、1つの方法は、バチルス・スブチリス(Bacillus subtilis)FB17の投与後、未処理植物より約5%〜約100%多い量で植物のバイオマスをより多く生産するのに効果的な量で、土壌に種子を播く前にバチルス・スブチリス(Bacillus subtilis)FB17を植物の種子に投与することを含む。たとえば、図4に図示したように、バチルス・スブチリス(Bacillus subtilis)FB17で処理したトウモロコシでは、処理から15日後、未処理トウモロコシと比較して空中および根のバイオマスが約28%増加することが観察された。
【0021】
本発明の別の実施形態は、植物の乾燥耐性を高める方法であって、未処理植物と比較して植物の乾燥耐性を高めるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。乾燥とは、土壌水分を枯渇させ、植物を害するだけの期間降雨がないか、または灌漑が行われないことをいう。植物の水分喪失が植物の根の吸水能力を上回る場合、および通常の植物のプロセスを阻害するのに十分に植物の含水量が減少する場合に乾燥ストレスが生じる。植物は、生育を停止し、光合成および水の使用を抑えるため他の植物のプロセスを抑制することにより水不足に対応する。本明細書で使用する場合、乾燥耐性とは、水の非存在下での植物の1日の生育速度、たとえば、未処理植物と比較してバチルス・スブチリス(Bacillus subtilis)FB17を接種した植物のバイオマスが増加した1日のグラム数をいう。たとえば、図19に図示したように、水の非存在下でバチルス・スブチリス(Bacillus subtilis)FB17で処理したトウモロコシ植物の種子は、処理から15日後、未処理植物と比較して1日の生育速度が約37.5%高まった。一実施形態では、1つの方法は、土壌に種子を播く前に、バチルス・スブチリス(Bacillus subtilis)FB17の投与後、未処理植物より少なくとも約10%高く植物の乾燥耐性を高めるのに効果的な量で、前記バチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の周囲の土壌、または植物の種子に投与することを含む。
【0022】
本発明の別の実施形態は、植物のリグニン濃度を低下させる方法であって、未処理植物と比較して植物のリグニン濃度を低下させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。リグニン濃度は、公知の方法により測定することができる。たとえば、図20に図示したように、バチルス・スブチリス(Bacillus subtilis)FB17で処理した植物は、未処理植物で観察された木化細胞の数が約46%〜約64%の減少を示した。リグニンは植物に不可欠な成分であり、植物細胞壁内、および植物細胞の間に見られる。リグニンは、植物バイオマスをバイオ燃料に変換する際の大きな障害の1つである。現在、もう1つの植物成分であるセルロースが、バイオ燃料の主な供給源である。セルロースはアルコールに発酵させやすいのに対し、リグニンは既存の発酵プロセスを用いて変換しないため、発酵性糖の抽出を難しくしている。したがって、リグニンの濃度が低い植物を生産することは有益である。本発明は、本発明の方法のいずれかによりバチルス・スブチリス(Bacillus subtilis)FB17を投与した植物のバイオマスのいずれか(たとえば、植物の全バイオマスまたは植物のバイオマスの任意の一部)をバイオ燃料に変換することにより生産されたバイオ燃料を提供する。バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物のバイオマスは、植物の糖成分の発酵によるなど任意の公知の方法によりバイオ燃料に変換することができる。
【0023】
本発明の別の実施形態は、植物の種子発芽率を高める方法であって、未処理植物と比較して植物の種子発芽を促進するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは植物の種子の周囲の土壌に投与することを含む方法を提供する。たとえば、図16に図示したように、バチルス・スブチリス(Bacillus subtilis)FB17による投与後、トマト植物およびトウモロコシ植物で総発芽率の上昇が観察された。
【0024】
本発明の別の実施形態は、植物、特に米植物の鉄濃度を上昇させる方法であって、未処理植物と比較して植物の鉄濃度を上昇させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。鉄欠乏症は、ヒトの最も一般的な微量栄養素欠乏症の1つであり、かつ米は世界の人口の大部分にとって最も重要な主食であるため、本発明の方法により生産される米植物は、世界中で重要な栄養効果をもたらすことができる。バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物の鉄濃度は、誘導結合プラズマ発光分光分析法(ICP−AES)、誘導結合プラズマ質量分析法(ICP−MS)、または他の標準的な方法など公知の方法により測定することができる。一実施形態では、植物の乾燥重量1キログラム当たり鉄のミリグラム数により植物の鉄濃度を測定する。図22に図示したように、FB17で処理した米植物では、植物の乾燥重量1kg当たりの鉄のmgで測定すると、未処理植物と比較して鉄含有量が約81%増加することが観察された。
【0025】
本発明に使用するのに好適な米植物として、オリザ・サティバ(Oryza sativa)、オリザ・グラベリマ(Oryza glaberrima)、およびこれらのすべての亜種および品種が挙げられる。バチルス・スブチリス(Bacillus subtilis)FB17を投与した米植物の鉄濃度は、米を収穫してその穀粒または農産物を集める時点で、すなわち、成熟した米粒を田畑から集める時点で測定してもよい。あるいは、バチルス・スブチリス(Bacillus subtilis)FB17を投与した米植物の鉄濃度は、たとえば、バチルス・スブチリス(Bacillus subtilis)FB17を米植物に投与してから約1週間〜約5ヶ月、好ましくは約3ヶ月の間の時点で測定してもよい。本発明の方法に従いバチルス・スブチリス(Bacillus subtilis)FB17を投与した米植物は、たとえば、米植物の地上および地下バイオマスの乾燥重量1グラム当たりの鉄のグラム数で測定すると、鉄の量をより多く生産する。
【0026】
一例を挙げると、本発明の方法に従いバチルス・スブチリス(Bacillus subtilis)FB17を投与した米植物は、未処理植物より少なくとも約5%、約10%〜約200%、約25%〜約150%、約50%〜約100%、約70%〜約90%、約75%〜約85%、または約80%多い量で、米植物の地上および地下バイオマス乾燥重量当たりの鉄の量を多く生産する。たとえば、一実施形態では、1つの方法は、土壌に米の種子を播く前に、バチルス・スブチリス(Bacillus subtilis)FB17の投与後、未処理植物より少なくとも約25%高く米植物の鉄濃度を高くするのに効果的な量で前記バチルス・スブチリス(Bacillus subtilis)FB17を米の種子に投与することを含む。
【0027】
本発明の別の実施形態は、病原真菌による植物の感染を阻害する方法であって、未処理植物と比較して病原真菌による植物の感染を阻害するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を植物、植物の種子、または植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。植物の例として、米の品種日本晴(Nipponbare)などの米植物およびオオムギ植物が挙げられる。特定の実施形態では、本発明は、病原真菌による米植物の感染症、特にイネいもち病を阻害する方法であって、未処理米植物と比較して病原真菌による米植物の感染症を阻害するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を米植物、米植物の種子、または米植物もしくは種子の周囲の土壌に投与することを含む方法を提供する。本明細書で使用する場合、「イネいもち病」とは、植物病原性真菌(plant−pathogenic fungus)マグナポルテ・グリセア(Magnaporthe grisea)またはマグナポルテ・オリザエ(Magnaporthe oryzae)をいう。
【0028】
イネいもち病の症状として、植物のあらゆる部分、特に葉など植物の空中部、すなわち地上部に発生する病変または斑(たとえば、白色または灰色であることがある)がある。本明細書で使用する場合、「感染症を阻害する」とは、真菌感染症の軽減、たとえば、未処理植物と比較して米植物の空中部分の病変数の減少、または病変の一部または全部の大きさの縮小で判定した場合、米植物の真菌感染症を軽減することをいう。たとえば、特定の実施形態では、イネいもち病が原因の米植物の病変数を、未処理米植物と比較して約5%〜約100%、約10%〜約80%、約20%〜約60%または約25%〜約45%減少させるのに効果的な量で、バチルス・スブチリス(Bacillus subtilis)FB17を米植物、米植物の種子、または米植物もしくは種子の周囲の土壌に投与する。どのような理論にも拘泥するわけではないが、B.スブチリス(B.subtilis)FB17は、M.オリザエ(M.oryzae)の増殖を減弱させるまたは抑制する揮発性抗真菌化合物を生成すると考えられる。特定の実施形態では、イネいもち病の増殖および米植物の感染症を阻害するため、約1×107CFU/種子〜約1×109CFU/種子、一層好ましくは約1×108CFU/種子の量でバチルス・スブチリス(Bacillus subtilis)FB17を米の種子に投与し、その後種子を土壌に播く。
【0029】
本明細書で使用する場合、「未処理植物」とは、バチルス・スブチリス(Bacillus subtilis)FB17を投与していないこと以外は、本発明の方法に従いバチルス・スブチリス(Bacillus subtilis)FB17を投与した植物と同じ種で、実質的に同じ条件下(たとえば、同じ期間、同じ気候、同じ材料を用い同じ方法により栽培し、同じ方法により測定されたバイオマス、乾燥耐性、リグニン濃度、鉄濃度、真菌感染、および他の特徴を持つ)で生育させた植物をいう。本明細書で使用する場合、未処理植物と比較した、バチルス・スブチリス(Bacillus subtilis)FB17を投与した植物の特徴、たとえばバイオマスの増加、乾燥耐性の向上、リグニン濃度の低下、鉄濃度の上昇または真菌感染症の軽減とは、それぞれ同じ時点で測定したバイオマスの増加、乾燥耐性の向上、リグニン濃度の低下、鉄濃度の上昇または真菌感染症の軽減をいう。
【0030】
本明細書に記載の方法のある種の実施形態では、バチルス・スブチリス(Bacillus subtilis)FB17の接種菌液約1ml/kg(すなわち、SmartSpec Bio Rad分光光度計を用いて測定した波長600nmの光学密度(OD)0.5の、LB培地で一晩増殖させたバチルス・スブチリス(Bacillus subtilis)FB17 1ml/kg)〜約50ml/kg、好ましくは約5ml/kg〜約25ml/kg、一層好ましくは約10ml/kg〜約15ml/kg、最も好ましくは約12.5ml/kgの量でバチルス・スブチリス(Bacillus subtilis)FB17を種子に投与する。代替の実施形態では、約1×106CFU/種子〜約1×109CFU/種子、一層好ましくは約1×107CFU/種子〜約1×108CFU/種子の量でバチルス・スブチリス(Bacillus subtilis)FB17を種子に投与する。
【0031】
本発明の方法は、バイオマスの増加、乾燥耐性の向上、リグニン含有量の減少、種子発芽の促進、鉄濃度の上昇、および病原体に対する耐性の向上を目的として多くの種類の植物(およびその種子または周囲の土壌)を処理するために使用してもよい。植物として、単子葉植物または双子葉植物が挙げられる。植物としては特に、トウモロコシ、ダイズ、トマト、米またはオオムギなどの作物が挙げられる。本発明の方法により処理してもよい植物の別の例としては、シロイヌナズナ(Arabidopsis thaliana)およびジニア属(Zinnia)のほか、バイオエネルギー作物植物、すなわち、セイヨウヤマカモジ(Brachypodium distachyon)などバイオエネルギー源として現在使用されている、または使用される可能性がある植物(たとえば、バイオ燃料の生産に有用な植物)が挙げられる。
【0032】
本発明によれば、バチルス・スブチリス(Bacillus subtilis)FB17は、根、種子または葉の接種によるなど、植物の全部または一部を処理する任意の公知の方法により植物に投与してもよい。たとえば、バチルス・スブチリス(Bacillus subtilis)FB17は、土壌に種子を播く前に葉および茎など植物の空中部分、植物の根、植物の種子に投与しても、または植物もしくは植物の種子の周囲の土壌に投与してもよい。投与の方法には、ドレンチング(drenching)、噴霧、コーティング、注射または当業者に公知の他の方法がある。本明細書で使用する場合、バチルス・スブチリス(Bacillus subtilis)FB17を投与するとは、1回投与、反復投与(すなわち、バチルス・スブチリス(Bacillus subtilis)FB17を1回より多く投与する)、または連続投与のいずれかをいう。バチルス・スブチリス(Bacillus subtilis)FB17は、植物の生活環の任意の時点(たとえば、発芽前または後)で投与してもよい。たとえば、バチルス・スブチリス(Bacillus subtilis)FB17は、土壌に種子を播く前、および発芽の前に植物の種子に投与してもよい。あるいは、バチルス・スブチリス(Bacillus subtilis)FB17は、発芽が生じた後に植物、植物の種子、または植物の周囲の土壌に投与してもよい。種子をバチルス・スブチリス(Bacillus subtilis)FB17で処理したら、土壌に播き、植物を生育させる従来の方法を用いて栽培すればよい。
【0033】
本発明の実施形態によれば、バチルス・スブチリス(Bacillus subtilis)FB17は、植物、植物の種子または土壌に単独投与しても、または他の材料との混合物として投与してもよい。たとえば、バチルス・スブチリス(Bacillus subtilis)FB17は、追加の添加剤または材料をまったく含まない増殖培地中のバチルス・スブチリス(Bacillus subtilis)FB17から本質的になる組成物として投与してもよい。あるいは、バチルス・スブチリス(Bacillus subtilis)FB17は、増殖培地中のバチルス・スブチリス(Bacillus subtilis)FB17、水などの担体、水溶液または粉末を含む組成物として投与してもよい。増殖培地、担体、水溶液または粉末は、殺虫剤または殺菌剤など追加の添加剤を含んでもよい。あるいは、バチルス・スブチリス(Bacillus subtilis)FB17は、異なる時間に適用される他の添加剤または材料と一緒に個別に投与してもよい。ある種の実施形態では、バチルス・スブチリス(Bacillus subtilis)FB17を、約1ml/kg(すなわち、SmartSpec Bio Rad分光光度計を用いて測定した波長600nmの光学密度(OD)0.5 の、LB培地で一晩増殖させたバチルス・スブチリス(Bacillus subtilis)FB17 1ml/kg)〜約50ml/kg、好ましくは約5ml/kg〜約25ml/kg、一層好ましくは約10ml/kg〜約15ml/kg、最も好ましくは約12.5ml/kgの量で、バチルス・スブチリス(Bacillus subtilis)FB17を含む組成物として投与する。代替の実施形態では、バチルス・スブチリス(Bacillus subtilis)FB17を約1×106CFU/種子〜約1×109CFU/種子、一層好ましくは約1×107CFU/種子〜約1×108CFU/種子の量で含む組成物としてバチルス・スブチリス(Bacillus subtilis)FB17を投与する。
【0034】
本発明はさらに、播く前に植物(たとえば、根)、植物の周囲の土壌、または種子に適用できる、バチルス・スブチリス(Bacillus subtilis)FB17を含む農業用担体と、植物種子に適用できる、バチルス・スブチリス(Bacillus subtilis)FB17を含む種子コーティングとを提供する。本発明はまた、種子の全部または一部がバチルス・スブチリス(Bacillus subtilis)FB17を含むコーティングまたは薄膜を有するように、バチルス・スブチリス(Bacillus subtilis)FB17でコートした植物の種子、好ましくは作物植物の種子(たとえば、トウモロコシ植物、ダイズ植物、米植物、トマト植物、またはセイヨウヤマカモジ(Brachypodium distachyon)などのバイオエネルギー作物植物の種子)を提供する。農業用担体は、バチルス・スブチリス(Bacillus subtilis)FB17接種菌液約1ml/kg(すなわち、SmartSpec Bio Rad分光光度計を用いて測定した波長600nmの光学密度(OD)0.5の、LB培地で一晩増殖させたバチルス・スブチリス(Bacillus subtilis)FB17 1ml/kg)〜約50ml/kg、約5ml/kg〜約25ml/kg、約10ml/kg〜約15ml/kg、または約12.5ml/kgの量でバチルス・スブチリス(Bacillus subtilis)FB17を含んでもよい。種子コーティングは、約1×106CFU/種子〜約1×108CFU/種子、一層好ましくは 約1×107CFU/種子の量でバチルス・スブチリス(Bacillus subtilis)FB17を含んでもよい。農業用担体および種子コーティングは各々、追加の添加剤または材料をまったく含まない増殖培地中のバチルス・スブチリス(Bacillus subtilis)FB17から本質的になっていてもよい。あるいは、農業用担体および種子コーティングは各々、水などの増殖培地、水溶液または粉末中にバチルス・スブチリス(Bacillus subtilis)FB17を含んでもよい。増殖培地、水溶液または粉末は、殺虫剤または殺菌剤など追加の添加剤を含んでもよい。
【0035】
本発明には、基本的用途と応用的用途との両方がある。本明細書に記載の方法を使用すれば、広い意味で(たとえば、バイオ燃料に使用される代替植物種で、あるいは、作物植物の収穫可能性に影響を与えるため)バイオマスを増加させ、乾燥耐性を向上させることができる。これらの方法は、遺伝子導入アプローチと比較して遺伝子の同定、遺伝子導入系の構築および特徴付けの時間を必要とせずに任意の植物に直ちに適用でき、導入遺伝子の使用に関する規制上および社会上の問題を伴わない。本明細書に記載の方法は、伝統的な栽培法(化学肥料および水の利用)の使用と比較して資源消費、および栽培者の労働がより少なく、環境により好ましい。さらに、化学肥料の利用は、急速で弱々しい生育を誘導することにより作物病害感受性を高めることが知られている一方、本方法で生育させる植物は、病害感受性が高まることが示されていない。FB17は、種子の処理に使用される他の根圏細菌と比較して、有益な結果を得るのに低菌量接種菌液を必要とする。最後に、こうした方法は有機農法との適合性があるのに対し、上述の他の方法(たとえば、化学肥料の利用)は適合性がない。
【0036】
B.スブチリス(B.subtilis)株FB17の寄託物は、Delaware Biotechnology Institute,15 Innovation way,Room #145,Newark,DE 19711で2010年3月1日より前から入手することができる。また、B.スブチリス(B.subtilis)株FB17は、アメリカンタイプカルチャーコレクション(ATCC),10801 University Boulevard,Manassas,Va.20110−2209 USAに寄託される。このATCC寄託物の利用は、本出願の係属中、特許商標庁長官(Commissioner of Patents and Trademarks)および長官によりその権利を有すると決定された者に対して、請求に応じて可能となる。寄託物は、30年の期間、または直近の請求から5年間、または特許の権利行使可能期間のいずれか長い期間、公的寄託機関であるATCC寄託機関で保存され、その期間中に生育不能なった場合、再寄託される。寄託物またはその子孫は、本出願の副本が出願されている国の外国特許法の定めるところにより利用できる。
【0037】
さらに、本寄託物は、微生物の寄託物に関するブダペスト条約の規定に従い保存され、一般に提供される、すなわち、本寄託物は、寄託物のサンプルの分譲に関する直近の請求から少なくとも5年の期間、さらにどのような場合でも、寄託日から少なくとも30年の期間、または発行する可能性がある、培養物を開示する任意の特許の権利行使可能期間、生存可能で汚染されない状態を保つのに必要な十分な注意を払って保存される。
【0038】
以下の例は、本発明をより詳細に記載するために提供するものであり、本発明を説明することを意図しており、本発明を限定するものではない。下記の図の一部に記載された「UD10−22」とは、バチルス・スブチリス(Bacillus subtilis)FB17をいう。
【実施例】
【0039】
実施例1
セイヨウヤマカモジ(Brachypodium distachyon)植物およびトウモロコシ植物を発芽させ、21日間生育させた。5日に1回(3回)、ポットごとにOD0.5のB.スブチリス(B.subtilis)FB17 5mlを加えた。対照では、OD0.5の大腸菌(E.coli)OP50 5mlをポットごとに加えた。FB17およびOP50をLB培地で一晩生育させ、SmartSpec(Bio Rad)分光光度計を使用して波長(600nm)の光学密度(OD)を得た。最終処理から10日後、植物を解析した。本明細書のすべての実験に記載される対照とは、細菌で処理しなかったか、または大腸菌(E.coli)OP50で処理した植物をいう。
【0040】
B.スブチリス(B.subtilis)FB17、細菌の対照大腸菌(E.coli)、または模擬処理(mock treatment)で処理したセイヨウヤマカモジ(Brachypodium distachyon)(Bd2−1)植物およびトウモロコシ植物を、処理後30日間、標準的な条件(22〜25℃、湿度60%、明期16時間/暗期8時間の明暗周期)で4×4インチのポットにて生育させた。エネルギー作物セイヨウヤマカモジ(B.distachyon)の空中および根のバイオマスは、FB17処理により増加した。図1は、FB17で処理したセイヨウヤマカモジ(B.distachyon)のバイオマスが統計学的レベルで増加したことを示す。図2は、FB17で処理したセイヨウヤマカモジ(B.distachyon)で観察された光合成効率の向上を図示する。FB17で処理したセイヨウヤマカモジ(B.distachyon)は、対照より多くのクロロフィルと総カロエテノイドとを含んでおり、植物の健康状態が健全であることを示す。図3は、セイヨウヤマカモジ(B.distachyon)の根の周囲の土壌から回収されたFB17の量を示す。図から、FB17は、大腸菌(E.coli)と比較してセイヨウヤマカモジ(B.distachyon)の根に非常に強く関連しており、FB17の根圏細菌の本来の性質を示唆することが明らかにされる。
【0041】
また、トウモロコシ植物も、B.スブチリス(B.subtilis)FB17、細菌の対照大腸菌(E.coli)OP50、または模擬処理により処理してから30日間の生育後、空中および根のバイオマスの増加を示した。
【0042】
実施例2
シロイヌナズナ(Arabidopsis thaliana)の種子を発芽させ、21日間生育させた。5日に1回(3回)、OD0.5のB.スブチリス(B.subtilis)FB17 5mlをポットごとに加えた。対照では、OD0.5の大腸菌(E.coli)OP50 5mlを ポットごとに加えた。FB17およびOP50をLB培地で一晩生育させ、SmartSpec(Bio Rad)分光光度計を使用して波長(600nm)の光学密度(OD)を得た。最終処理から10日後、植物を25℃、湿度40%で4週間乾燥に供した(すなわち、水を加えなかった)。処理から30日後、未処理植物のstay green表現型の喪失をFB17で処理した植物と比較して乾燥を評価したところ、FB17がアラビドプシス属(Arabidopsis)の乾燥耐性を向上させることが示唆された。
【0043】
実施例3
B.スブチリス(B.subtilis)FB17の種子処理は、トウモロコシMo17、CML258、CML10、ジニア属(Zinnia)、およびセイヨウヤマカモジ(Brachypodium distachyon)のバイオマスの向上を促進する。
【0044】
トウモロコシ(Mo17、CML258、CML10)、ダイズ(Will−82)、トマト(Solanum lycopersicum)、ジニア属(Zinnia)、およびセイヨウヤマカモジ(Brachypodium distachyon)(エネルギー作物モデル)のバイオマスの向上に対するB.スブチリス(B.subtilis)FB17の作用を試験するため、植物種ごとに50の種子(n=50)を、B.スブチリス(B.subtilis)FB17(約1×107cfu/種子、またはSmartSpec Bio Rad分光光度計を用いて測定した波長600nmの光学密度(OD)0.5の、LB培地で一晩増殖させたバチルス・スブチリス(Bacillus subtilis)FB17 12.5ml/kg)で種子処理した。種子処理後、発芽およびバイオマスの研究のため、混合土壌を含むポット(4×4インチ)に種子を一つずつ播種した。興味深いことに、B.スブチリス(B.subtilis)FB17の種子処理は、試験した全作物種で根およびシュートの生育を促進した。測定値は、処理から15日後に得た。
【0045】
種子処理した植物が根のバイオマスの増加を促進し、その結果、根の長さが増加するよりもむしろ根系密度が高くなった。根系密度が高くなったのは側根および根毛が増加し、より多くの水および栄養素を取り込めるようになったことに起因する。
【0046】
トウモロコシ(Zea mays)var.CML258は、空中バイオマスが対照より約16%増加した。トウモロコシ(Zea mays)var.CML10は、空中バイオマスが対照より約9%増加した。トウモロコシ(Zea mays)var.Mo−17は、空中バイオマスが対照より約38%増加した。ブラキポディウム(Brachypodium)は、空中バイオマスが対照より約40%増加した。トウモロコシ(Z.mays)(MO17)の場合、空中および根の総バイオマスが対照より約28%という有意な増加が観察された。図4は、B.スブチリス(B.subtilis)FB17で処理した植物のバイオマスの総増加率を図示する。
【0047】
図5は、B.スブチリス(B.subtilis)FB17で種子処理したMo−17植物における根およびシュートのバイオマスの増加を示す定量データを図示する。図6は、B.スブチリス(B.subtilis)FB17で種子処理したバイオエネルギー作物セイヨウヤマカモジ(Brachypodium distachyon)(遺伝子型Bd2−1)植物における葉数の増加を示す定量データを図示する。図7は、B.スブチリス(B.subtilis)FB17で種子処理したバイオエネルギー作物セイヨウヤマカモジ(Brachypodium distachyon)(遺伝子型Bd2−1)植物における根およびシュートのバイオマスの増加を示す定量データを図示する。図8および図9は、B.スブチリス(B.subtilis)FB17で種子処理したジニア・エスピー(Zinnia sp.)「レッドスパイダー」における根およびシュートのバイオマスの増加を示す定量データを図示する。
【0048】
実施例4
B.スブチリス(B.subtilis)FB17の種子処理は、トウモロコシおよびトマトの光合成効率を促進する。
【0049】
トウモロコシ(Mo17、CML258、CML10)、ダイズ(Will−82)、トマト(Solanum lycopersicum)、ジニア属(Zinnia)、およびブラキポディウム(Brachypodium)(エネルギー作物モデル)の光合成効率に対するB.スブチリス(B.subtilis)FB17の作用を試験するため、植物種ごとに50の種子(n=50)を、B.スブチリス(B.subtilis)FB17(12.5ml/kgまたは1e7cfu/種子)で種子処理した。処理から15〜32日後の葉を収穫し、総クロロフィル含有量について解析した。結果から、図10および図11に図示したように、B.スブチリス(B.subtilis)FB17を接種したトウモロコシ植物およびトマト植物(トマトおよび外来種のトウモロコシCML258およびCML10)は未処理サンプルと比較してクロロフィルおよびカロテノイド含有量の増加を示すことが明らかになった。
【0050】
総クロロフィル値の増加は、CML258およびCML10で見られたように成長力およびバイオマスの増加を促進する可能性がある。B.スブチリス(B.subtilis)FB17を種子接種したトマトの総クロロフィル含有量は、約14%増加した。より一層顕著なのは外来種のトウモロコシCML258およびCML10であり、それぞれ約87%および約72%増加した。
【0051】
総クロロフィル含有量は増加しているものの、そのことは、総カロテノイド含有量も増加した値に対応することを意味するものではない。トマトおよびジニア属(Zinnia)は、B.スブチリス(B.subtilis)FB17を接種して未処理種子と比較すると、総カロテノイドの割合が有意に低下した。トウモロコシCML258およびCML10は総カロテノイドの割合が有意に増加した一方、ダイズ、トウモロコシMo17、およびブラキポディウム(Brachypodium)は、処理した種子と未処理の種子との間で統計学的に有意な差を示さなかった。
【0052】
図12は、B.スブチリス(B.subtilis)FB17で種子処理したMo−17植物における光合成効率の上昇を示す定量データを図示する。図13は、B.スブチリス(B.subtilis)FB17で種子処理したバイオエネルギー作物セイヨウヤマカモジ(Brachypodium distachyon)(遺伝子型Bd2−1)植物における光合成効率の上昇を示す定量データを図示する。図14は、B.スブチリス(B.subtilis)FB17で種子処理したジニア・エスピー(Zinnia sp.)「レッドスパイダー」における光合成効率の上昇を示す定量データを図示する。図15は、B.スブチリス(B.subtilis)FB17で種子処理した外来トウモロコシCML10およびCML258における光合成効率の上昇を示す定量データを図示する。
【0053】
実施例5
B.スブチリス(B.subtilis)FB17の種子処理は、トウモロコシ植物およびトマト植物の発芽を促進する。
【0054】
トウモロコシ(Mo17、CML258、CML10)、ダイズ(Will−82)、トマト(Solanum lycopersicum)、ジニア属(Zinnia)、およびブラキポディウム(Brachypodium)(エネルギー作物モデル)の発芽増加率に対するB.スブチリス(B.subtilis)FB17の作用を試験するため、植物種ごとに50の種子(n=50)を、B.スブチリス(B.subtilis)FB17(12.5ml/kgまたは1e7cfu/種子)で種子処理した。最終的な発芽率を播種した日から8日後にスコア化した。結果から、B.スブチリス(B.subtilis)FB17処理は、図16に図示したようにトマトおよびトウモロコシの発芽反応を統計学的に有意に促進することが明らかになった。
【0055】
B.スブチリス(B.subtilis)FB17で処理したトマトの種子および外来種トウモロコシCML258はそれぞれ発芽率が5.9%および14%上昇した。
【0056】
B.スブチリス(B.subtilis)FB17の種子処理は、試験した全作物種について中立およびプラスの作用を有した。種子処理を適用した場合、発芽率に対するマイナスの反応は、統計学的に認められなかった。
【0057】
実施例6
トウモロコシ(Mo17、CML258、CML10)、ダイズ(Will−82)、トマト(Solanum lycopersicum)、ジニア属(Zinnia)、およびブラキポディウム(Brachypodium)(エネルギー作物モデル)におけるB.スブチリス(B.subtilis)FB17の作用を試験するため、植物種ごとに50の種子(n=50)を、B.スブチリス(B.subtilis)FB17(1e7cfu/種子またはSmartSpec Bio Rad分光光度計を用いて測定した波長600nmの光学密度(OD)0.5の、LB培地で一晩増殖させたバチルス・スブチリス(Bacillus subtilis)FB17 12.5ml/kg)で種子処理した。種子処理後、種子を一つずつポット(4×4インチ)播種した。測定は、処理から15日後に行った。図17は、B.スブチリス(B.subtilis)FB17で処理後のトウモロコシ(Zea mays)の生育速度を図示する。図18は、B.スブチリス(B.subtilis)FB17で処理した植物の保水能力を図示する。FB17処理後、総保水能力および保水率が、トマト(2.1%)およびトウモロコシ(Z.mays)MO17(3.5%)で有意に高まることが観察された。図19は、B.スブチリス(B.subtilis)FB17で処理した植物の乾燥耐性を図示する。MO17の生育速度がFB17処理後、乾燥処理下で有意に高まることが観察された(非水処理対照に対して37.5%増)。図20は、B.スブチリス(B.subtilis)FB17の種子処理がトウモロコシのリグニン含有量を減少させることを図示する。トウモロコシ(Z.mays)ではFB17処理後、総リグニン含有量が有意に減少することが観察された(MO17で約46%減少;CML10で約64%減少、およびCML58で約49%減少)。
【0058】
実施例7
図21は、B.スブチリス(B.subtilis)FB17で処理した米植物オリザ・サティバ(Oryza sativa)(日本晴(Nipponbare))の空中および根のバイオマスが接種から60日後に増加することを図示する。LBで一晩生育させたFB17の培養液を用いて108細胞/mlの接種菌液を作製した。FB17の補充には、水耕法で生育させ4週齢の米植物(品種日本晴(Nipponbare))を使用した。B.スブチリス(B.subtilis)FB17を投与した米植物は、未処理米植物と比較して約200%のバイオマスの増加を示した。
【0059】
実施例8
バチルス・スブチリス(Bacillus subtilis)FB17が米の根に定着するかどうかを評価するため、米植物(品種日本晴(Nipponbare))にバチルス・スブチリス(Bacillus subtilis)FB17を接種し、接種から96時間後に米植物の根を共焦点走査型レーザー顕微鏡により観察した。観察結果から、この有益な根圏細菌(バチルス・スブチリス(Bacillus subtilis)FB17)は、植物体にバイオフィルムを形成することが確認された。特に、データからは、バチルス・スブチリス(Bacillus subtilis)FB17が処理の96時間後に米の根に効率的に定着することが示唆されることから、米の根が有益な微生物の定着を支持することが示される。
【0060】
米植物の根圏細菌処理が気孔開口に何らかの変化を与えるかどうかを評価するため、出願人らは、根圏細菌で処理した米植物を解析した。結果から、バチルス・スブチリス(Bacillus subtilis)FB17で米を根圏細菌処理すると、処理した米植物(品種日本晴(Nipponbare))の気孔開口が大きく減少することが明らかになった。バチルス・スブチリス(Bacillus subtilis)FB17処理の場合、バチルス・スブチリス(Bacillus subtilis)FB17の添加から1週後に孔辺細胞が観察された。これらの結果から、B.スブチリス(B.subtilis)FB17(バチルス・スブチリス(Bacillus subtilis)FB17)は、シロイヌナズナ(A.thaliana)および米の両方で証明されたように、単子葉植物および双子葉植物の両方で一般的な気孔閉鎖反応を与えることが示唆される。
【0061】
バチルス・スブチリス(Bacillus subtilis)FB17がイネいもち病の増殖を減弱させるかどうかを評価するため、出願人らは、マグナポルテ・オリザエ(Magnaporthe oryzae)をバチルス・スブチリス(Bacillus subtilis)FB17培養液に曝露した。定性コンパートメントプレート(qualitative compartment plate)および定量データから、バチルス・スブチリス(Bacillus subtilis)FB17曝露真菌培養液での放射状の増殖の低下により示されたように、バチルス・スブチリス(Bacillus subtilis)FB17はM.オリザエ(M.oryzae)の増殖を減弱させることが示された。対照(TYおよびLB)との比較から、処理しない場合に病原体が増殖すると考えられる程度が明らかにされる。表1に示すように、バチルス・スブチリス(Bacillus subtilis)FB17はインビトロでM.オリザエ(M.oryzae)の増殖を約25%抑制した。これらの結果から、B.スブチリス(B.subtilis)FB17は、M.オリザエ(M.oryzae)の増殖を減弱させるか、または阻害することができる揮発性抗真菌化合物を生成することが示唆される。
【0062】
【表1】
バチルス・スブチリス(Bacillus subtilis)FB17は、米およびオオムギでマグナポルテ・オリザエ(Magnaporthe oryzae)に対する全身抵抗性を誘導した。M.オリザエ(M.oryzae)に曝露された米植物およびオオムギ植物はどちらも、表2に示すように対照と比較してFB17で処理した植物で米の葉およびオオムギの子葉の病変形成がそれぞれ抑制された(いもち病に典型的なダイヤモンド状の病変が葉に少なくとも1つ存在する場合に「感染した」と定義した)。
【0063】
【表2】
実施例9
バチルス・スブチリス(Bacillus subtilis)FB17が米の鉄強化を促進するかどうかを評価するため、本出願人らは、誘導結合プラズマ発光分光分析法(ICP−AES)を用いて、バチルス・スブチリス(Bacillus subtilis)FB17を補充した植物の米の葉、根、および穀粒における全鉄含有量を解析した。結果から、バチルス・スブチリス(Bacillus subtilis)FB17を米に補充すると、植物体に鉄が集まりやすくなる、すなわち、必須元素の鉄が、植物の生育および成長に利用される植物に積極的に取り込まれることが明らかになった。図22に図示したように、植物の乾燥重量1kg当たりの鉄のmg数で測定した場合、FB17で処理した米植物では未処理対照と比較して鉄含有量が81%増加することが観察された(図1に示したように「UD1022」とはバチルス・スブチリス(Bacillus subtilis)FB17をいう)。したがって、バチルス・スブチリス(Bacillus subtilis)FB17を植物、特に米植物に投与すると、食物中の鉄濃度の上昇により食物の栄養価を大きく高めることができる。
【0064】
図23は、上述の複数の植物種の様々な特色に対するB.スブチリス(B.subtilis)FB17の作用をまとめたものである。
【0065】
本発明について特定の実施形態と共に記載してきたが、特許請求の範囲に記載の本発明は、こうした特定の実施形態に必要以上に限定されるべきものではないことを理解すべきである。実際、本発明の記載された組成物および方法の様々な修正および変形は、当業者には明らかであり、添付の特許請求の範囲の範囲内にあることを意図している。
【特許請求の範囲】
【請求項1】
植物のバイオマスをより多く生産する方法であって、未処理植物と比較して前記植物のバイオマスをより多く生産するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を前記植物、前記植物の種子、または前記植物もしくは前記種子の周囲の土壌に投与することを含む方法。
【請求項2】
前記バチルス・スブチリス(Bacillus subtilis)FB17を前記植物の前記種子に投与することを含む、請求項1に記載の方法。
【請求項3】
前記植物はトウモロコシ植物、ダイズ植物、米植物、およびトマト植物からなる群から選択される、請求項1に記載の方法。
【請求項4】
前記植物はバイオエネルギー作物植物である、請求項1に記載の方法。
【請求項5】
前記植物はセイヨウヤマカモジ(Brachypodium distachyon)である、請求項4に記載の方法。
【請求項6】
未処理植物と比較して前記植物のバイオマスを約5%〜約100%多く生産するのに効果的な量で前記バチルス・スブチリス(Bacillus subtilis)FB17を投与することを含む、請求項1に記載の方法。
【請求項7】
約1×106CFU/種子〜約1×108CFU/種子の量で前記バチルス・スブチリス(Bacillus subtilis)FB17を前記種子に投与することを含む、請求項2に記載の方法。
【請求項8】
植物の乾燥耐性を高める方法であって、未処理植物と比較して前記植物の乾燥耐性を高めるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を前記植物、前記植物の種子、または前記植物もしくは前記種子の周囲の土壌に投与することを含む方法。
【請求項9】
前記バチルス・スブチリス(Bacillus subtilis)FB17を前記植物の前記種子に投与することを含む、請求項8に記載の方法。
【請求項10】
前記植物はトウモロコシ植物、ダイズ植物、米植物、およびトマト植物からなる群から選択される、請求項8に記載の方法
【請求項11】
植物のリグニン濃度を低下させる方法であって、未処理植物と比較して前記植物のリグニン濃度を低下させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を前記植物、前記植物の種子、または前記植物もしくは前記種子の周囲の土壌に投与することを含む方法。
【請求項12】
前記バチルス・スブチリス(Bacillus subtilis)FB17を前記植物の前記種子に投与することを含む、請求項11に記載の方法。
【請求項13】
前記植物はトウモロコシ植物、ダイズ植物、米植物、およびトマト植物からなる群から選択される、請求項11に記載の方法。
【請求項14】
約1×106CFU/種子〜約1×108CFU/種子の量で前記バチルス・スブチリス(Bacillus subtilis)FB17を前記種子に投与することを含む、請求項12に記載の方法。
【請求項15】
バイオ燃料を生産する方法であって、バチルス・スブチリス(Bacillus subtilis)FB17が投与された請求項11に記載の植物のバイオマスを前記バイオ燃料に変換することを含む方法。
【請求項16】
請求項15に従い生産されたバイオ燃料。
【請求項17】
植物の鉄濃度を上昇させる方法であって、未処理植物と比較して前記植物の鉄濃度を上昇させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を前記植物、前記植物の種子、または前記植物もしくは前記種子の周囲の土壌に投与することを含む方法。
【請求項18】
前記植物は米植物である、請求項17に記載の方法。
【請求項19】
播く前に前記バチルス・スブチリス(Bacillus subtilis)FB17を前記米植物の前記種子に投与することを含む、請求項18に記載の方法。
【請求項20】
未処理米植物と比較して前記米植物の鉄濃度を少なくとも約25%上昇させるのに効果的な量で前記バチルス・スブチリス(Bacillus subtilis)FB17を投与することを含む、請求項18に記載の方法。
【請求項21】
約1×106CFU/種子〜約1×108CFU/種子の量で前記バチルス・スブチリス(Bacillus subtilis)FB17を前記種子に投与することを含む、請求項19に記載の方法。
【請求項22】
病原真菌による植物の感染を阻害する方法であって、未処理植物と比較して前記病原真菌による前記植物の感染を阻害する量でバチルス・スブチリス(Bacillus subtilis)FB17を前記植物、前記植物の種子、または前記植物もしくは前記種子の周囲の土壌に投与することを含む方法。
【請求項23】
前記植物は米植物であり、前記病原真菌はイネいもち病である、請求項22に記載の方法。
【請求項24】
播く前に前記バチルス・スブチリス(Bacillus subtilis)FB17を前記米植物の前記種子に投与することを含む、請求項23に記載の方法。
【請求項25】
約1×106CFU/種子〜約1×108CFU/種子の量で前記バチルス・スブチリス(Bacillus subtilis)FB17を前記米植物の前記種子に投与することを含む、請求項24に記載の方法。
【請求項26】
前記イネいもち病の症状は未処理植物と比較して約5%〜約100%軽減される、請求項23に記載の方法。
【請求項27】
バチルス・スブチリス(Bacillus subtilis)FB17を含む農業用担体。
【請求項28】
バチルス・スブチリス(Bacillus subtilis)FB17を含む植物種子コーティング。
【請求項29】
請求項28に記載のコーティングを含む植物の種子。
【請求項1】
植物のバイオマスをより多く生産する方法であって、未処理植物と比較して前記植物のバイオマスをより多く生産するのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を前記植物、前記植物の種子、または前記植物もしくは前記種子の周囲の土壌に投与することを含む方法。
【請求項2】
前記バチルス・スブチリス(Bacillus subtilis)FB17を前記植物の前記種子に投与することを含む、請求項1に記載の方法。
【請求項3】
前記植物はトウモロコシ植物、ダイズ植物、米植物、およびトマト植物からなる群から選択される、請求項1に記載の方法。
【請求項4】
前記植物はバイオエネルギー作物植物である、請求項1に記載の方法。
【請求項5】
前記植物はセイヨウヤマカモジ(Brachypodium distachyon)である、請求項4に記載の方法。
【請求項6】
未処理植物と比較して前記植物のバイオマスを約5%〜約100%多く生産するのに効果的な量で前記バチルス・スブチリス(Bacillus subtilis)FB17を投与することを含む、請求項1に記載の方法。
【請求項7】
約1×106CFU/種子〜約1×108CFU/種子の量で前記バチルス・スブチリス(Bacillus subtilis)FB17を前記種子に投与することを含む、請求項2に記載の方法。
【請求項8】
植物の乾燥耐性を高める方法であって、未処理植物と比較して前記植物の乾燥耐性を高めるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を前記植物、前記植物の種子、または前記植物もしくは前記種子の周囲の土壌に投与することを含む方法。
【請求項9】
前記バチルス・スブチリス(Bacillus subtilis)FB17を前記植物の前記種子に投与することを含む、請求項8に記載の方法。
【請求項10】
前記植物はトウモロコシ植物、ダイズ植物、米植物、およびトマト植物からなる群から選択される、請求項8に記載の方法
【請求項11】
植物のリグニン濃度を低下させる方法であって、未処理植物と比較して前記植物のリグニン濃度を低下させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を前記植物、前記植物の種子、または前記植物もしくは前記種子の周囲の土壌に投与することを含む方法。
【請求項12】
前記バチルス・スブチリス(Bacillus subtilis)FB17を前記植物の前記種子に投与することを含む、請求項11に記載の方法。
【請求項13】
前記植物はトウモロコシ植物、ダイズ植物、米植物、およびトマト植物からなる群から選択される、請求項11に記載の方法。
【請求項14】
約1×106CFU/種子〜約1×108CFU/種子の量で前記バチルス・スブチリス(Bacillus subtilis)FB17を前記種子に投与することを含む、請求項12に記載の方法。
【請求項15】
バイオ燃料を生産する方法であって、バチルス・スブチリス(Bacillus subtilis)FB17が投与された請求項11に記載の植物のバイオマスを前記バイオ燃料に変換することを含む方法。
【請求項16】
請求項15に従い生産されたバイオ燃料。
【請求項17】
植物の鉄濃度を上昇させる方法であって、未処理植物と比較して前記植物の鉄濃度を上昇させるのに効果的な量でバチルス・スブチリス(Bacillus subtilis)FB17を前記植物、前記植物の種子、または前記植物もしくは前記種子の周囲の土壌に投与することを含む方法。
【請求項18】
前記植物は米植物である、請求項17に記載の方法。
【請求項19】
播く前に前記バチルス・スブチリス(Bacillus subtilis)FB17を前記米植物の前記種子に投与することを含む、請求項18に記載の方法。
【請求項20】
未処理米植物と比較して前記米植物の鉄濃度を少なくとも約25%上昇させるのに効果的な量で前記バチルス・スブチリス(Bacillus subtilis)FB17を投与することを含む、請求項18に記載の方法。
【請求項21】
約1×106CFU/種子〜約1×108CFU/種子の量で前記バチルス・スブチリス(Bacillus subtilis)FB17を前記種子に投与することを含む、請求項19に記載の方法。
【請求項22】
病原真菌による植物の感染を阻害する方法であって、未処理植物と比較して前記病原真菌による前記植物の感染を阻害する量でバチルス・スブチリス(Bacillus subtilis)FB17を前記植物、前記植物の種子、または前記植物もしくは前記種子の周囲の土壌に投与することを含む方法。
【請求項23】
前記植物は米植物であり、前記病原真菌はイネいもち病である、請求項22に記載の方法。
【請求項24】
播く前に前記バチルス・スブチリス(Bacillus subtilis)FB17を前記米植物の前記種子に投与することを含む、請求項23に記載の方法。
【請求項25】
約1×106CFU/種子〜約1×108CFU/種子の量で前記バチルス・スブチリス(Bacillus subtilis)FB17を前記米植物の前記種子に投与することを含む、請求項24に記載の方法。
【請求項26】
前記イネいもち病の症状は未処理植物と比較して約5%〜約100%軽減される、請求項23に記載の方法。
【請求項27】
バチルス・スブチリス(Bacillus subtilis)FB17を含む農業用担体。
【請求項28】
バチルス・スブチリス(Bacillus subtilis)FB17を含む植物種子コーティング。
【請求項29】
請求項28に記載のコーティングを含む植物の種子。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【公表番号】特表2013−521298(P2013−521298A)
【公表日】平成25年6月10日(2013.6.10)
【国際特許分類】
【出願番号】特願2012−556172(P2012−556172)
【出願日】平成23年3月1日(2011.3.1)
【国際出願番号】PCT/US2011/026683
【国際公開番号】WO2011/109395
【国際公開日】平成23年9月9日(2011.9.9)
【出願人】(512226882)ユニバーシティー オブ デラウェア (1)
【Fターム(参考)】
【公表日】平成25年6月10日(2013.6.10)
【国際特許分類】
【出願日】平成23年3月1日(2011.3.1)
【国際出願番号】PCT/US2011/026683
【国際公開番号】WO2011/109395
【国際公開日】平成23年9月9日(2011.9.9)
【出願人】(512226882)ユニバーシティー オブ デラウェア (1)
【Fターム(参考)】
[ Back to top ]