
植物の成長の促進
【課題】植物の成長を促進する方法を提供する。
【解決手段】植物の成長を促進するために過敏反応誘発ポリペプチドまたはタンパク質を植物または植物種子に、または種子から成長した植物に施用することの変法として、トランスジェニック植物または植物種子が使用され得る。トランスジェニック植物を用いるとき、これには過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物を提供すること、および成長を促進するDNA分子が有効に用いられる条件下で植物を生育させること含む。他には、過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物種子が提供され、土壌に植えることができる。そして植物は、成長を促進するDNA分子の使用に有効な条件下、植えた種子から生育する。
【解決手段】植物の成長を促進するために過敏反応誘発ポリペプチドまたはタンパク質を植物または植物種子に、または種子から成長した植物に施用することの変法として、トランスジェニック植物または植物種子が使用され得る。トランスジェニック植物を用いるとき、これには過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物を提供すること、および成長を促進するDNA分子が有効に用いられる条件下で植物を生育させること含む。他には、過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物種子が提供され、土壌に植えることができる。そして植物は、成長を促進するDNA分子の使用に有効な条件下、植えた種子から生育する。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、1997年1月27日付け米国仮特許出願番号60/036,048の利益を請求する。
本発明は、USDA NRI Competitive Research Grant番号91-37303-6430のもと、米国政府より支持されている。
【0002】
本発明の分野
本発明は植物の成長の促進に関する。
【背景技術】
【0003】
発明の背景
有機肥料の施用による植物成長の改良は既知であり、数世紀にわたり実施されてきた(H. Marschner, “Mineral Nutrition of Higher Plants,” Academic Press: New York pg.674 (1986))。栽培者および農業者が成長促進の方法による効率向上のために、土壌または成長する作物に施用し得る、簡単な製品を生産するための複合的無機肥料生産システムを近世の人間は発達させてきた。植物の大きさ、色、成熟、および収穫は肥料製品の施用によって全て改善され得る。無機肥料には、通常、硝酸アンモニウムといった化学的な施用を含む。有機肥料には、動物の厩肥および堆肥にした芝の有機堆積物、他の多くの源を含み得る。
【0004】
近年、研究者たちは、生物学的産物の使用により植物成長を改良することを探求してきた。Beauveria bassianaおよびTrichoderma harizamumのような昆虫および疾病を制御する薬剤により、昆虫および疾病問題が制御されることが記されており、それゆえ間接的に植物成長および効率を改善する(Fravel et al., “Formulation of Microorganisms to Control Plant Diseases,” Formulation of Microbial Biopesticides, Beneficial Microorganisms, and Nematodes, H. D. Burges, ed. Chapman and Hall: London(1996))。
【0005】
微生物の施用または微生物生産物による方法によって直接植物成長の促進が示唆されている。新しい土地に植えるとき、根粒細菌はマメ科作物の種子に加えられる(Weaver et al., “Rhizobium,” Methods of Soil Analysis, Part 2, Chemical and Microbiological Properties, 2nd ed., American Society of Agronomy: Madison (1982))。これらの細菌は植物の根粒着生効率を改善し得、それゆえ窒素固定と呼ばれる過程、遊離窒素原子を利用できる形に変換する植物の能力を向上する。非マメ科作物は、大抵そのような処置による利益はない。Rhizobiumといった、加えた細菌は直接根毛に寄生し、そして保護および栄養を受けつつ、植物に利益提供することにより相利共生の関係を始める。
【0006】
菌根菌類はまた、多くの植物、特に栄養が枯渇した土壌での球果植物の意図的生育に必要な微生物であると認められている。植物ホルモンの生物学的合成(Frankenberger et al., “Biosynthesis of Indole-3-Acetic Acid by the Pine Ectomycorrhizal Fungas Pisolithus tinctorius,” Appl. Environ. Microbiol. 53:2908-13(1987))、ミネラル吸収の増加(Harley et al., “The Uptake of Phosphate by Excised Mycorrhizal Roots of Beech,” New Phytologist 49:388-97 (1950)およびHarley et al., “The Uptake of Phosphate by Excised Mycorrhizal Roots of Beech. IV. The Effect of Oxygen Concentration Upon Host and Fungus,” New Phytologist 52:124-32 (1953)および水(A. B. Hatch, “The Physical Basis of Mycotrophy in Pinus,” Black Rock Forest Bull. No. 6, 168pp. (1937))を含む機構が必要とされてきた。菌根菌類は根粒着生細菌程の使用頻度には到達していない、それは、菌根菌類が菌根菌の株毎に変動し、一定しない結果を与え、生物としての研究も困難を伴うからである。
【0007】
植物成長促進リゾバクテリア(PGPR)は、近年、植物の成長および生育を向上すると認められている。仮説の機構は、直接的な影響(例えば栄養吸収の増加)から間接的な機構(例えば病原体排除)にまでわたっている。PGPRの施用による成長促進は、通常、生存細菌の根組織への接種、および細菌生産性ホルモン効果、シデロホアを通じて、または抗生物質による疾病の防止、または競合による、成長改善の達成について言及されている。上記全ての場合において、その結果は、根のコロニー化、時折、種子コーティングの施用を通じて有効となる。PGPR株はノトバイオティック条件下、根の伸張を促進する直接的成長促進体となり得るという提唱をする情報には限界がある(Anderson et al., “Responses of Bean to Root Colonization With Pseudomonas putida in a Hydroponic System,” Phytopathology 75:992-95 (1985), Lifshitz et al., “Growth Promotion of Canola (rapeseed) Seedlings by a Strain of Pseudomonas putida Under Gnotobiotic Conditions,” Can. J. Microbiol. 33:390-95 (1987), Young et al., “PGPR: Is There Relationship Between Plant Growth Regulators and the Stimulation of Plant Growth or Biological Activity?,”Promoting Rhizobacteria: Progress and Prospects, Second International Workshop on Plant Growth-promoting Rhizobacteria, pp. 182-86 (1991), Loper et al., “Influence of Bacterial Sources of Indole-3-Acetic Acid on Root Elongation of Sugar Beet,” Phytopathology 76:386-89 (1986), および Mueller et al., “Hormonal Interactions in the Rhizosphere of Maize (Zea mays L.) and Their Effect on Plant Development,” Z. Pflanzenernaehrung Bodenkunde 152:247-54 (1989))。しかしながら、植物成長レギュレーターは、これらの効果を介する機構として提唱されている。多くの細菌はインビトロで様々な植物成長レギュレーター(Atzorn et al., “Production of Gibberellins and Indole-3-Acetic Acid by Rhizobium phaseoli in Relation to Nodulation of Phaseolus vulgaris Roots,” Planta 175:532-38 (1988) and M. E. Brown, “Plant Growth Substances Produced by MicroOrganism of Solid and Rhizosphere,” J. Appl. Bact. 35:443-51 (1972))、または抗生物質(Gardner et al., “Growth Promotion and Inhibition by Antibiotic-Producing Fluorescent Pseudomonads on Citrus Roots,” Plant Soil 77:103-13 (1984))を産生する。シデロホア産物は、幾つかのPGPR株に提唱されている他の機構である(Ahl et al., “Iron Bound-Siderophores, Cyanic Acid, and Antibiotics Involved in Suppression of Thievaliopsis basicola by a Pseudomonas fluorescens Strain,” J. Phytopathol. 116:121-34 (1986), Kloepper et al., “Enhanced Plant Growth by Siderophores Produced by Plant Growth-Promoting Rhizobacteria,” Nature 286:885-86 (1980), and Kloepper et al ., “Pseudomonas siderophores: A Mechanism Explaining Disease-Suppressive Soils,” Curr. Microbiol. 4:317-20 (1980))。根表面のコロニー化、すなわち表面における病原体細菌との直接的競合は作用の他の機構である(Kloepper et al., “Relationship of in vitro Antibiosis of Plant Growth-Promoting Rhizobacteria to Plant Growth and the Displacement of Root Microflora,” Phytopathology 71:1020-24 (1981), Weller, et al., “Increased Growth of Wheat by Seed Treatments With Fluorescent Pseudomonads, and Implications of Pythium Control,” Can. J. Microbiol. 8:328-34 (1986), and Suslow et al., “Rhizobacteria of Sugar Beets: Effects of Seed Application and Root Colonization on Yield,” Phytopathology 72:199-206 (1982))。キャノーラ(菜種)の研究は、収穫、幼苗発生(seedling emergence)および成長力、早生植物成長(early-season plant growth)(葉の数および主なランナー(runner)の長さ)、ならびに葉の面積を含む(Kloepper et al., “Plant Growth-Promoting Rhizobacteria on Canola (rapeseed),” Plant Disease 72:42-46 (1988))、PGPR増加植物成長パラメーターを示す。ジャガイモの研究で、Pseudomonas株をジャガイモの種子に施用すると、収穫が増えることが示されている(Burr et al., “Increased Potato Yields by Treatment of Seed Pieces With Specific Strains of Pseudomonas Fluorescens and P. putida,” Phytopathology 68:1377-83 (1978), Kloepper et al., “Effect of Seed Piece Inoculation With Plant Growth-Promoting Rhizobacteria on Populations of Erwinia carotovora on Potato Roots and in Daughter Tubers,” Phytopathology 73:217-19 (1983), Geels et al., “Reduction of Yield Depressions in High Frequency Potato Cropping Soil After Seed Tuber Treatments With Antagonistic Fluorescent Pseudomonas spp.,” Phytopathol. Z. 108:207-38 (1983), Howie et al., “Rhizobacteria: Influence of Cultivar and Soil Type on Plant Growth and Yield of Potato,” Soil Biol. Biochem. 15:127-32 (1983), and Vrany et al., “Growth and Yield of Potato Plants Inoculated With Rhizosphere Bacteria,” Folia Microbiol. 29:248-53 (1984))。収穫の増加は、恐らく抗生物質による塊茎における病原性細菌の排除に対するPGRPの競合効果のためであるように見える(Kloepper et al., “Effect of Seed Piece Inoculation With Plant Growth-Promoting Rhizobacteria on Populations of Erwinia carotovora on Potato Roots and in Daughter Tubers,” Phytopathology 73:217-19 (1983), Kloepper et al., “Effects of Rhizosphere Colonization by Plant Growth-Promoting Rhizobacteria on Potato Plant Development and Yield,” Phytopathology 70:1078-82 (1980), Kloepper et al., “Emergence-Promoting Rhizobacteria: Description and Implications for Agriculture,” pp. 155-164, Iron, Siderophores, and Plant Disease, T. R. Swinburne, ed. Plenum, New York (1986), and Kloepper et al., “Relationship of in vitro Antibiosis of Plant Growth-Promoting Rhizobacteria to Plant Growth and the Displacement of Root Microflora,” Phytopathology 71:1020-24 (1981))。幾つかの研究では、植物の毛状体はPGPRを用いて改良された(Tipping et al., “Development of Emergence-Promoting Rhizobacteria for Supersweet Corn,” Phytopathology 76:938-41 (1990) (abstract) and Kloepper et al., “Emergence-Promoting Rhizobacteria: Description and Implications for Agriculture,” pp. 155-164, Iron, Siderophores, and Plant Disease, T. R. Swinburne, ed. Plenum, New York (1986))。他の多くの研究は、植物の病原体の生態的防除のために、rhizobacteriaの処置によって植物の健康を改善することを示している(B. Schippers, “Biological Control of Pathogens With Rhizobacteria,” Phil. Trans. R. Soc. Lond. B. 318:283-93 (1988), Schroth et al., “Disease-Suppressive Soil and Root-Colonizing Bacteria,” Science 216:1376-81 (1982), Stutz et al., “Naturally Occurring Fluorescent Pseudomonads Involved in Suppression of Black Root Rot of Tobacco,” Phytopathology 76:181-85 (1986), and D. M. Weller, “Biological Control of Soilborne Plant Pathogens in the Rhizosphere With Bacteria,” Annu. Rev. Phytopathol. 26:379-407 (1988))。
【0008】
植物の病原体誘発性免疫(pathogen-induced immunization)により、成長が促進することが見出された。タバコ木質部への外からのPeronospora tabacinaの感染は、発育阻害を緩和するばかりでなく、成長および生育を促進する。温室および畑の両方の実験において、免疫化したタバコは、約40%高さが高くなり、乾燥重量が40%増加し、生体重が30%増加し、対照植物よりも葉が4-6枚多くなった(Tuzun, S., et al., “The Effect of Stem Injection with Peronospora tabacina and Metalaxyl Treatment on Growth of Tobacco and Protection Against Blue Mould in the Field,” Phytopathology, 74:804 (1984))。これらの植物は対照植物よりも約2-3週間早く花が咲いた(Tuzun, S., et al., “Movement of a Factor in Tobacco Infected with Peronospora tabacina Adam which Systemically Protects Against Blue Mould,” Physiological Plant Pathology, 26:321-30 (1985))。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は従来の植物の成長の促進方法を超える改良を示している。
【課題を解決するための手段】
【0010】
本発明の要旨
本発明は植物の成長を促進する方法に関する。この方法には、植物または植物種子に、植物または植物種子から成育した植物の成長の促進する条件下で、非感染型で、過敏反応誘発ポリペプチドまたはタンパク質を施用することを含む。
【0011】
植物の成長を促進するために過敏反応誘発ポリペプチドまたはタンパク質を植物または植物種子に、または種子から成長した植物に施用することの変法として、トランスジェニック植物または植物種子が使用され得る。トランスジェニック植物を用いるとき、これには過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物を提供すること、および成長を促進するDNA分子が有効に用いられる条件下で植物を生育させること含む。他には、過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物種子が提供され、土壌に植えることができる。そして植物は、成長を促進するDNA分子の使用に有効な条件下、植えた種子から生育する。
【0012】
本発明は植物の成長を高めまたは促進する何れの型にも有効となることを目的とする。これは、種子から植物の生長が始まる時のように早期にまたは植物の寿命の後期に行い得る。例えば、本発明の植物の成長には、収穫増加、生産された種子の量の増加、発芽する種子の割合の増加、植物の大きさの増進、より多くのバイオマス、より多くの果実およびより大きな果実、より早い果実の着色、ならびにより早い果実および植物の成熟を包含する。結果として、本発明は栽培者にとって意義ある経済的利益を提供する。例えば、早い発芽および早い成熟によって、作物は、成長不適季節であるためにその成長が妨げられる土地でも成長させ得る。種子の発芽割合の増加の結果、作物の植生の改良やより有効な種子の使用がもたらされる。増加した収穫、大きさの増進、およびバイオマス生産の促進により、与えられた土地の区画からより大きな収入を生み出すことになる。そして本発明は、農業効率において重要な利点を構成していることは明らかである。
【図面の簡単な説明】
【0013】
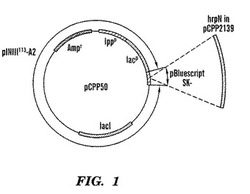
【図1】プラスミドベクターpCPP2139の地図であり、Erwinia amylovora過敏反応誘発物遺伝子を含んでいる。
【図2】プラスミドベクターpCPP50の地図であり、Erwinia amylovora過敏反応誘発物遺伝子は含んではいないが、それ以外図1に示したプラスミドベクターpCPP2139と同じである。Masui, et al., Bio/Technology 2:81-85 (1984)(それは、ここで引用することによりこの明細書の一部に加える)参照。
【発明を実施するための形態】
【0014】
本発明の詳細な説明
本発明は植物の成長を促進する方法に関する。この方法には、植物または植物種子の全てまたは一部に、植物または植物種子から成育した植物の成長の促進に有効な条件下、非感染型で、過敏反応誘発ポリペプチドまたはタンパク質を施用することを含む。他に、植物は種子を作成するためにこの方法で処理され得、その種子を植えたとき、子孫植物の成長は促進される。
【0015】
植物の成長を促進するために過敏反応誘発ポリペプチドまたはタンパク質を植物または植物種子に、または種子から成長した植物に施用することの変法として、トランスジェニック植物または植物種子が使用され得る。トランスジェニック植物を用いるとき、これには過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物を提供すること、および成長を促進するDNA分子が有効に用いられる条件下で植物を生育させること含む。他には、過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物種子が提供され、土壌に植えることができる。そして植物は、成長を促進するDNA分子の使用に有効な条件下、植えた種子から生育する。
【0016】
本発明で使用する過敏反応誘発ポリペプチドまたはタンパク質は、多くの種類の真菌性および細菌性病原体より誘導される過敏反応誘発ポリペプチドまたはタンパク質に相当し得る。そのようなポリペプチドまたはタンパク質は、誘発物と接触した植物組織において局所的な壊死を誘発し得る。
【0017】
ポリペプチドまたはタンパク質誘発物の適当な細菌性源の例には、Erwinia、Pseudomonas、およびXanthomonas種(例えば以下の細菌:Erwinia amylovora、Erwinia chrysanthemi、Erwinia stewartii、Erwinia carotovora、Pseudomonas syringae、Pseudomonas solanacearum、Xanthomonas campestris、およびそれらの混合)を含む。
【0018】
過敏反応誘発タンパク質またはポリペプチドの適当な真菌性源の例は、Phytophthoraである。Phytophthoraの適当な種には、Phytophthora pythium、Phytophthora cryptogea、Phytophthora cinnamomi、Phytophthora capsici、Phytophthora megasperma、およびPhytophthora citrophthoraが含まれる。
【0019】
過敏反応誘発ポリペプチドまたはタンパク質が植物または植物種子に施用される本発明の実施態様は、多くの方法で実施され得、1)単離した誘発ポリペプチドまたはタンパク質の施用;2)疾病の原因とはならない、過敏反応誘発ポリペプチドまたはタンパク質をコードしている遺伝子で形質転換された細菌の施用;および3)幾つかの植物種では疾病の原因となる(しかしそれらは施用される植物ではない)、天然に過敏反応誘発ポリペプチドまたはタンパク質をコードしている遺伝子を含む細菌の施用を含む。加えて、本発明の種子は、本発明に従って過敏反応誘発ポリペプチドまたはタンパク質で処理された植物から回収し得る。
【0020】
本発明のある実施態様では、過敏反応誘発ポリペプチドまたはタンパク質は、その相当する生物から単離され得、植物または植物種子に施用され得る。そういった単離方法は既知であり、Arlat, M., F. Van Gijsegem, J. C. Huet, J.C. Pemollet, and C. A. Boucher, “PopA1, a Protein which Induces a Hypersensitive-like Response in Specific Petunia Genotypes is Secreted via the Hrp Pathway of Pseudomonas solanacearum,” EMBO J. 13:543-553 (1994); He, S. Y., H. C. Huang, and A. Collmer, “Pseudomonas syringae pv. syringae HarpinPss:a Protein that is Secreted via the Hrp Pathway and Elicits the Hypersensitive Response in Plants,” Cell 73:1255-1266 (1993);およびWei, Z.-M., R. J. Laby, C. H. Zumoff, D. W. Bauer, S.-Y. He, A. Collmer, and S. V. Beer, “Harpin Elicitor of the Hypersensitive Response Produced by the Plant Pathogen Erwinia amylovora,” Science 257:85-88 (1992)(それらは、ここで引用することによりこの明細書の一部に加える)に記載されている。また仮米国特許出願番号08/200,024および08/062,024(それらは、ここで引用することによりこの明細書の一部に加える)参照。好ましくは、しかしながら、本発明の単離した過敏反応誘発ポリペプチドまたはタンパク質は、下記のように組換えで作成され、精製される。
【0021】
本発明の他の実施態様では、過敏反応誘発ポリペプチドまたはタンパク質は、過敏反応誘発ポリペプチドまたはタンパク質をコードする遺伝子を含む細菌を施用することにより種子または植物種子に施用し得る。そういった細菌は、誘発物が植物または植物種子の細胞に接触できるようにポリペプチドまたはタンパク質を分泌し、排出できなければならない。これらの実施態様では、過敏反応誘発ポリペプチドまたはタンパク質は植物または種子において細菌によって、または該細菌が植物または植物種子に導入される直前に産生される。
【0022】
本発明の細菌施用方法の1つの実施態様では、細菌は疾病の原因とはならず、過敏反応誘発ポリペプチドまたはタンパク質をコードする遺伝子で形質転換されている(例えば組換え)。例えば、植物に過敏反応を誘発しないE. coliは、過敏反応誘発ポリペプチドまたはタンパク質をコードしている遺伝子で形質転換され得、そして植物に施用され得る。E. coli以外の細菌種はまた、本発明のこの実施態様において使用し得る。
【0023】
本発明の細菌施用方法の他の実施態様では、細菌は疾病の原因となり、天然に過敏反応誘発ポリペプチドまたはタンパク質をコードしている遺伝子を含んでいる。そのような細菌の例は上記した通りである。しかしながら、この実施態様では、細菌が運ぶ疾病にかかりにくい植物またはそれらの種子に、これらの細菌を施用する。例えば、Erwinia amylovoraはリンゴまたは洋ナシでは疾病の原因となるが、トマトではならない。しかしながら、そういった細菌は、トマトにおいて過敏反応を誘発する。それゆえ、本発明のこの実施態様に従い、Erwinia amylovoraはトマトまたはその種子に施用され、その種において疾病の原因とはならずに成長を促進し得る。
【0024】
Erwinia chrysanthemi由来の過敏反応誘発ポリペプチドまたはタンパク質は、配列番号1に相当するアミノ酸配列を有し、その配列を以下に示す。
【化1】

【0025】
この過敏反応誘発ポリペプチドまたはタンパク質は分子量34kDaであり、熱に安定であり、グリシン含有は16%を超え、そして事実上システインが含まれていない。Erwinia chrysanthemi過敏反応誘発ポリペプチドまたはタンパク質は、配列番号2に相当するヌクレオチド配列を有するDNA分子によりコードされており、その配列を以下に示す。
【化2】
【化3】
【化4】
【0026】
Erwinia amylovoraから誘導された過敏反応誘発ポリペプチドまたはタンパク質は、配列番号3に相当するアミノ酸配列を有し、以下にその配列を示す。
【化5】
【化6】
【0027】
この過敏反応誘発ポリペプチドまたはタンパク質は、分子量39kDaであり、約pI4.3であり、そして100℃で少なくとも10分間加熱しても安定である。この過敏反応誘発ポリペプチドまたはタンパク質には事実上システインが含まれていない。Erwinia amylovoraから誘導された過敏反応誘発ポリペプチドまたはタンパク質は、さらにWei, Z.-M., R. J. Laby, C. H. Zumoff, D. W. Bauer, S.-Y. He, A. Collmer, and S. V. Beer, “Harpin, Elicitor of the Hypersensitive Response Produced by the Plant Pathogen Erwinia amylovora,” Science 257:85-88 (1992)(それは、ここで引用することによりこの明細書の一部に加える)に十分に記載されている。このポリペプチドまたはタンパク質をコードしているDNA分子は、配列番号4に相当するヌクレオチド配列を有し、それを以下に示す。
【化7】
【0028】
Pseudomonas syringaeから誘導された過敏反応誘発ポリペプチドまたはタンパク質は、配列番号5に相当するアミノ酸配列を有し、その配列を以下に示す。
【化8】
【0029】
この過敏反応誘発ポリペプチドまたはタンパク質は、分子量34-35kDaである。それはグリシンが豊富であり(約13.5%)、システインおよびチロシンを含まない。Pseudomonas syringaeから誘導される過敏反応誘発ポリペプチドまたはタンパク質についての更なる情報は、He, S. Y., H. C. Huang, and A. Collmer, “Pseudomonas syringae pv. syringae HarpinPss: a Protein that is Secreted via the Hrp Pathway and Elicits the Hypersensitive Response in Plants,” Cell 73:1255-1266 (1993)(それは、ここで引用することによりこの明細書の一部に加える)に見られる。過敏反応誘発物をコードするDNA分子は、配列番号6に相当するヌクレオチド配列を有し、それらを以下に示す。
【化9】
【0030】
Pseudomonas solanacearumから誘導された過敏反応誘発ポリペプチドまたはタンパク質は、配列番号7に相当するアミノ酸配列を有し、それを以下に示す。
【化10】
【0031】
それは配列番号8に相当するヌクレオチド配列を有するDNA分子によりコードされており、それを以下に示す。
【化11】
【0032】
Pseudomonas solanacearumから誘導された過敏反応誘発ポリペプチドまたはタンパク質に関する更なる情報は、Arlat, M., F. Van Gijsegem, J. C. Huet, J. C. Pemollet, and C. A. Boucher, “PopA1, a Protein which Induces a Hypersensitive-like Response in Specific Petunia Genotypes, is Secreted via the Hrp Pathway of Pseudomonas solanacearum,” EMBO J. 13:543-533 (1994)(それは、ここで引用することによりこの明細書の一部に加える)に示されている。
【0033】
Xanthomonas campestris pv. glycines由来の過敏反応誘発ポリペプチドまたはタンパク質は、配列番号9に相当するアミノ酸配列を有し、それを以下に示す。
【化12】
【0034】
この配列は、Xanthomonas campestris pv. glycinesの過敏反応誘発ポリペプチドまたはタンパク質のみからなる26残基のアミノ末端配列である。それは、他のXanthomonas campestris pathovarsにおいて決定されているふさ状サブユニットタンパク質(fimbrial subunit protein)に対合する(match)。
【0035】
Xanthomonas campestris pv. pelargonii由来の過敏反応誘発ポリペプチドまたはタンパク質は、熱に安定であり、プロテアーゼ感受性であり、分子量20kDaである。配列番号10に相当するアミノ酸配列を含み、それを以下に示す。
【化13】
【0036】
Erwinia carotovoraの過敏反応誘発ポリペプチドまたはタンパク質は、Cui et al., “The RsmA Mutants of Erwinia carotovora subsp. carotovora Strain Ecc71 Overexpress hrp NEcc and Elicit a Hypersensitive Reaction-like Response in Tobacco Leaves,” MPMI, 9(7):565-73 (1996)(それは、ここで引用することによりこの明細書の一部に加える)に記載されている。過敏反応誘発ポリペプチドまたはタンパク質は、Ahmad et al., “Harpin is Not Necessary for the Pathogenicity of Erwinia stewartii on Maize,” 8th Int'l. Cong. Molec. Plant-Microbe Interact., July 14-19, 1996 およびAhmad, et al., “Harpin is Not Necessary for the Pathogenicity of Erwinia stewartii on Maize,” Ann. Mtg. Am. Phytopath. Soc., July 27-31, 1996 (それらは、ここで引用することによりこの明細書の一部に加える)によって示されている。
【0037】
Phytophthora parasitica、Phytophthora cryptogea、Phytophthora cinnamoni、Phytophthora capsici、Phytophthora megasperma、およびPhytophthora citrophthora由来の過敏反応誘発ポリペプチドまたはタンパク質は、Kaman, et al., “Extracellular Protein Elicitors from Phytophthora: Most Specificity and Induction of Resistance to Bacterial and Fungal Phytopathogens,” Molec. Plant-Microbe Interact., 6(1):15-25 (1993), Ricci et al., “Structure and Activity of Proteins from Pathogenic Fungi Phytophthora Eliciting Necrosis and Acquired Resistance in Tobacco,” Eur. J. Biochem., 183:555-63 (1989)、Ricci et al., “Differential Production of Parasiticein, and Elicitor of Necrosis and Resistance in Tobacco, by Isolates of Phytophthora parasitica,” Plant Path. 41:298-307 (1992)、Baillreul et al., “A New Elicitor of the Hypersensitive Response in Tobacco: A Fungal Glycoprotein Elicits Cell Death, Expression of Defence Genes, Production of Salicylic Acid, and Induction of Systemic Acquired Resistance,” Plant J., 8(4):551-60 (1995)、およびBonnet et al., “Acquired Resistance Triggered by Elicitors in Tobacco and Other Plants,” Eur. J. Plant Path., 102:181-92 (1996)(それらは、ここで引用することによりこの明細書の一部に加える)に記載されている。
【0038】
上記誘発物は典型的なものである。他の誘発物は、誘発物をコードする遺伝子が発現する条件下、過敏反応を誘発する増殖真菌および細菌により同定され得る。培養上清の細胞フリー調製(Cell-free preparation)は、適当な植物組織への浸透にそれを用いることにより、誘発物活性(すなわち、局所的なネクローシス)の試験に用い得る。
【0039】
上記過敏反応誘発ポリペプチドまたはタンパク質のフラグメントおよび他の病原体の誘発物の全フラグメントを、本発明の方法で使用することもできる。
【0040】
適当なフラグメントは、幾つかの手段で作成され得る。最初に、既知誘発タンパク質をコードする遺伝子のサブクローンを、遺伝子フラグメントをサブクローニングする通常の分子遺伝学的手法によって作成する。そしてそのサブクローンが、インビトロまたはインビボの細菌細胞において発現すると、下記方法の誘発活性の試験を行い得る、より小さなタンパク質またはペプチドが得られる。
【0041】
他には、誘発タンパク質のフラグメントは、キモトリプシンまたはStaphylococcusプロテイナーゼA、またはトリプシンのようなタンパク質分解酵素で全長の誘発タンパク質を消化することにより作成され得る。異なるタンパク質分解酵素は、誘発タンパク質のアミノ酸配列に基き、異なる部位で誘発タンパク質を切断するのに適当である。タンパク質分解によって得られるフラグメントの幾つかは、活性な耐性誘発物でもある。
【0042】
タンパク質の1次構造の情報に基く、他のアプローチにおいて、誘発タンパク質遺伝子のフラグメントは、タンパク質の特定部位を選択した特定のプライマーの組み合わせと共に、PCR技術を用いて合成され得る。そして、これらは、トランケート(truncate)したペプチドまたはタンパク質の増殖および発現のために適当なベクターにクローニングされる。
【0043】
化学合成はまた、適当なフラグメントを作成するために使用され得る。そういった合成は、誘発物を産生するために既知のアミノ酸配列を用い実施される。他に、全長の誘発物を高温高圧にさらすとフラグメントが生ずる。そして、これらのフラグメントは、通常の方法により分離され得る(例えばクロマトグラフィー、SDS-PAGE)。
【0044】
有用なフラグメントの例は、Pseudomonas solanacearumの過敏反応誘発ポリペプチドまたはタンパク質のpopA1フラグメントである。Arlat, M., F. Van Gijsegem, J. C. Huet, J. C. Pemollet, and C. A. Boucher, “PopA1, a Protein Which Induces a Hypersensitive-like Response in Specific Petunia Genotypes is Secreted via the Hrp Pathway of Pseudomonas solanacearum,” EMBO J. 13:543-53 (1994)(それは、ここで引用することによりこの明細書の一部に加える)参照。Erwinia amylovoraに関して、適当なフラグメントは、例えば、配列番号3のアミノ酸1〜98の間に存在し、またはそれを含むポリペプチドと、配列番号3のアミノ酸137〜204の間に存在し、またはそれを含むポリペプチドの何れかであるか、または両方である。
【0045】
変異体はまた(あるいは別に)、ポリペプチドの性質、2次構造およびヒドロパシー性に最小限の影響しか与えずに、例えばアミノ酸の欠損または付加により修飾され得る。例えば、ポリペプチドは、タンパク質のN-末端でシグナル配列(またはリーダー配列)と連結し、共翻訳的(co-translationally)にまたは後翻訳的(post-translationally)にそのタンパク質の移動を指図し得る。ポリペプチドはまた、ポリペプチドの合成、精製または同定を容易にするためにリンカーまたは他の配列に連結し得る。
【0046】
本発明のタンパク質またはポリペプチドは、好ましくは通常技術によって精製された形(好ましくは少なくとも約60%、更に好ましくは80%の純度)で産生される。典型的に、本発明のタンパク質またはポリペプチドは産生されるが、組換え宿主細胞の増殖培地には分泌されない。他には、本発明のタンパク質またはポリペプチドは増殖培地に分泌する。分泌しないタンパク質の場合、タンパク質を単離するために、組換えプラスミドを有する宿主細胞(例えばE.coli)を増殖させ、超音波、加熱、化学的処置により溶解し、ホモジネートを遠心分離し、細菌の破壊物を除去する。そして上清を加熱処理し、過敏反応誘発タンパク質を遠心分離により分離する。本発明のポリペプチドまたはタンパク質を含む上清画分を、タンパク質分離のために適当な大きさのデキストランまたはポリアクリルアミドカラムでゲル濾過する。必要ならば、タンパク質画分は更にイオン交換またはHPLCにより精製し得る。
【0047】
過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子は、通常の組換えDNA技術を用いて細胞内に組込まれ得る。通常、これには、DNA分子が異質である(即ち、通常は存在しない)発現システムにDNA分子を組み入れることが含まれる。異種性DNA分子を適当なセンス方向および正しいリーディンフレームで発現システムまたはベクターに組込む。ベクターには挿入したタンパク質コーディング配列の転写および翻訳に必要なエレメントが含まれている。
【0048】
CohenおよびBoyerの米国特許番号4,237,224(それは、ここで引用することによりこの明細書の一部に加える)には、制限酵素の切断およびDNAリガーゼでの連結を用いた組換えプラスミドの形の発現システムの産生について記載されている。そして、これら組換えプラスミドは、形質転換法により組込まれ、原核生物および組織培養で増殖した真核細胞を含む単細胞培養中で複製される。
【0049】
組換え遺伝子はまた、ワクシニアウイルスのようなウイルスに導入され得る。組換えウイルスは、ウイルスに感染した細胞にプラスミドをトランスフェクトすることにより生じ得る。
【0050】
適当なベクターには、ラムダベクターシステムgt11、gt WES.tB、Charon 4のようなウイルスベクターおよびpBR322、pBR325、pACYC177、pACYC1084、pUC8、pUC9、pUC18、pUC19、pLG339、pR290、pKC37、pKC101、SV40、pBluescript II SK +/- またはKS +/-(Stratagene、La Jolla, Califの“Stratagene Cloning System” Catalog (1993)参照(それらは、ここで引用することによりこの明細書の一部に加える))、pQE、pIH821、pGEX、pETシリーズ(F. W. Studier et al., “Use of T7 RNA Polymerase to Direct Expression of Cloned genes,” Gene Expression Technology vol. 185 (1990)(それらは、ここで引用することによりこの明細書の一部に加える)参照)、およびそれらの誘導体を含むが、これらに限定されない。組換え分子は、形質転換、特にトランスダクション、接合、動員、またはエレクトロポレーションで細胞に導入され得る。DNA配列は、Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Springs Laboratory, Cold Springs Harbor, New York (1989)(それらは、ここで引用することによりこの明細書の一部に加える)に記載されている通りに、当分野の標準的クローニング法を用いてベクターにクローニングされる。
【0051】
様々な宿主ベクターシステムが使用され、タンパク質をコードする配列を発現し得る。第1に、ベクターシステムは、使用する宿主細胞に和合性がなければならない。宿主ベクターシステムには、バクテリオファージDNA、プラスミドDNAまたはコスミドDNAで形質転換した細菌;酵母ベクターを有する酵母のような微生物;ウイルス感染した哺乳類細胞システム(例えばワクシニアウイルス、アデノウイルスなど);ウイルス感染した昆虫細胞システム(例えばバキュロウイルスなど);および細菌に感染した植物細胞が含まれるが、これらに限らない。これらベクターの発現エレメントは、それらの威力および特異性が異なる。使用する宿主ベクターシステムに依存して、多くの適当な転写および翻訳エレメントのうち何れかを用い得る。
【0052】
異なる遺伝学的シグナルおよびプロセッシング事象(processing event)により多くの遺伝子発現レベルが制御されている(例えばDNA転写およびメッセンジャーRNA(mRNA)翻訳)。
【0053】
DNAの転写は、RNAポリメラーゼの結合を指示し、mRNA合成を促進するDNA配列であるプロモーターの存在に依存する。真核生物のプロモーターのDNA配列は、原核生物のプロモーターとは異なる。更に、真核生物のプロモーターおよび付随する遺伝学的シグナルは、原核生物のシステム中では認識されず、または機能せず、更に原核生物のプロモーターは、真核性細胞の中では認識されず、機能しない。
【0054】
同様に、原核生物内でのmRNAの翻訳は、真核生物とは異なる適当な原核生物のシグナルの存在に依存する。原核生物におけるmRNAの翻訳を効率よくするには、mRNAにあるシャイン・ダルガルノ(“SD”)配列と呼ばれるリボゾーム結合部位が必要である。この配列は、mRNAの短いヌクレオチド配列であり、タンパク質のアミノ末端のメチオニンをコードする開始コドン(通常はAUG)の上流に位置する。SD配列は、16S rRNA(リボゾームRNA)の3'端と相補的であり、おそらく、rRNAと2本鎖となることによりmRNAとリボゾームとの結合を促進し、リボゾームを正確に位置させる。遺伝子発現を最大とする検討には、Roberts and Lauer, Methods in Enzymology, 68:473(1979)(それは、ここで引用することによりこの明細書の一部に加える)を参照のこと。
【0055】
プロモーターにより、“強さ”(即ち、転写を促進する能力)が変化する。クローニングした遺伝子を発現するという目的に対し、高レベルの転写、そして遺伝子の発現を得るためには、強力なプロモーターを用いることが望ましい。用いる宿主細胞システムに依存して、多くの適当なプロモーターのうちの何れかを使用し得る。例えば、E.coliでクローニングするとき、バクテリオファージ、またはプラスミド、T7ファージプロモーター、lacプロモーター、trpプロモーター、recAプロモーター、リボゾームRNAプロモーター、コリファージラムダおよび他のPRおよびPLプロモーター(lacUV5、ompF、bla、lppおよびその他が含まれるがこれらに限らない)といったプロモーターなどが、近接したDNAセグメントの転写を高レベルにするために使用し得る。加えて、組換えDNAまたは他のDNA合成技術により作成される、ハイブリッドtrp-lacUV5(tac)プロモーターまたは他のE.coliプロモーターが使用され得、挿入遺伝子を転写する。
【0056】
細菌性宿主細胞株および発現ベクターは、特別に誘導しない限りプロモーターの作動を抑制するように選択する。ある操作では、特定の誘発物の添加は、挿入遺伝子を効率よく転写するために必要となる。例えば、lacオペロンはラクトースまたはIPTG(イソプロピルチオ-β-D-ガラクトシド)の添加により誘発される。trp、pro等といった、様々な他のオペロンは他の異なる制御下に置かれる。
【0057】
特定の開始シグナルはまた、原核細胞中において効率よく転写および翻訳するために必要である。これらの転写および翻訳開始シグナルは、“強さ”が変化し得、遺伝子に特異なメッセンジャーRNAおよび合成されたタンパク質、それぞれの量により測定され得る。プロモーターを含むDNA発現ベクターにはまた、様々な“強力な”転写および/または翻訳開始シグナルの何れかの組み合わせが含まれている。例えば、E.coliにおいて翻訳の効率をよくするには、開始コドン(ATG)の約7-9塩基5'側にSD配列が必要であり、リボゾーム結合部位を供与することが必要である。それゆえ宿主細胞のリボゾームにより使用され得る何れかのSD-ATG組み合わせが用いられ得る。そういった組み合わせには、cro遺伝子もしくはコリファージラムダのN遺伝子のSD-ATG組合せ、またはE.coliトリプトファンE、D、C、BもしくはA遺伝子のSD-ATG組合せが含まれるが、これらに限らない。加えて、組換えDNAまたは合成ヌクレオチドの組み込みを含む他の技術により製造された何れかのSD-ATG組合わせが使用され得る。
【0058】
過敏反応誘発ポリペプチドまたはタンパク質をコードする単離DNA分子を1度発現システムにクローニングすれば、宿主細胞に容易に組込まれる。そのような組込みは、上記様々な型の形質転換で実施され得、それはベクター/宿主細胞システムに依存する。適当な宿主細胞には、細菌、ウイルス、酵母、哺乳類細胞、昆虫、植物等が含まれるが、これらに限らない。
【0059】
本発明の方法は、広く多様な植物またはその種子の成長を促進する処理に使用し得る。適当な植物には双子葉植物および単子葉植物が含まれる。更に特に有用な作物には、コメ、小麦、大麦、ライ麦、ワタ、ヒマワリ、ラッカセイ、トウモロコシ、ジャガイモ、サツマイモ、マメ、エンドウ、チコリー、レタス、エンダイブ、キャベツ、カリフラワー、ブロッコリー、カブ、ハツカダイコン、ホウレンソウ、タマネギ、ニンニク、ナス、トウガラシ、セロリ、ニンジン、カボチャ、ペポカボチャ、ズッキーニ、キュウリ、リンゴ、洋ナシ、メロン、イチゴ、ブドウ、ラズベリー、パイナップル、ダイズ、タバコ、トマト、サトウモロコシ、およびサトウキビが含まれ得る。適当な観賞用植物には、バラ、セントポーリア、ペチュニア、ペラルゴニューム、ポインセチア、キク、カーネーションおよびヒャクニチソウが含まれる。
【0060】
過敏反応誘発ポリペプチドまたはタンパク質の施用を含む本発明の方法は、植物の全てまたは一部(葉、茎、根等を含む)を処理するときに様々な方法で実施され得る。これは、植物内への過敏反応誘発ポリペプチドまたはタンパク質の浸透を含む(しかし必要ではない)。適当な施用方法には、局所的施用(例えば高または低圧噴霧(pressure spraying))、注入、散布、および誘発物の施用とほぼ同時の葉の表皮剥脱が含まれる。本発明の施用実施態様に従い、植物種子を処理するとき、過敏反応誘発ポリペプチドまたはタンパク質は、局所的施用(例えば高または低圧噴霧)、塗布、浸透、散布または注入により施用され得る。他の適当な施用方法は、過敏反応誘発ポリペプチドまたはタンパク質と植物細胞または植物種子を効果的に接触させることのできる当業者によって構想され得る。本発明の過敏反応誘発物で1度処理すると、種子は天然または農業用土壌に植えることができ、通常の方法を用いて栽培し植物を生産する。本発明に従って処理した種子から植物が生育し、その後植物は成長を促進するために1度またはそれ以上の施用により処理され得る。そういった成長した植物は、次に、成長を促進し得る植物を作成し得る種子または珠芽(例えば切枝(cutting))の作成に有用となり得る。
【0061】
本発明の過敏反応誘発ポリペプチドまたはタンパク質は、単独で、または他の物質との混合物で植物または植物種子に施用し得る。他に、過敏反応誘発ポリペプチドまたはタンパク質は、異なったときに施用される他の物質と共に、別々に植物に施用し得る。
【0062】
本発明の施用実施態様に従う、植物または植物種子の処理に適当な組成物は、担体中の過敏反応誘発ポリペプチドまたはタンパク質を含む。適当な担体は、水、水溶液、スラリーまたは乾燥粉末を含む。この実施態様において、組成物には、0.5nMを超える過敏反応誘発ポリペプチドまたはタンパク質が含まれる。
【0063】
必ずしも必要ではないけれども、この組成物には、肥料、殺虫剤、殺菌剤、殺線虫剤、除草剤、およびそれらの混合物を含む付加添加物が含まれる。適当な肥料には、(NH4)2NO3が含まれている。適当な殺虫剤の例にはマラチオンがある。有用な殺菌剤にはキャプタンが含まれる。
【0064】
他の適当な添加物は、緩衝剤、湿性剤、塗布剤、および剥離剤(abrading agent)を含む。これら物質は本発明の方法を容易にするのに使用し得る。加えて、過敏反応誘発ポリペプチドまたはタンパク質は、土およびポリサッカライドを含む他の慣用の種子用製剤または処理剤と共に、植物種子に施用し得る。
【0065】
トランスジェニック植物およびトランスジェニック種子の使用を含む本発明の他の実施態様において、過敏反応誘発ポリペプチドまたはタンパク質が植物または種子に局所的方法で施用される必要はない。代わりに、過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物は、微粒子銃またはAgrobacteriumを介する形質転換による方法といった当分野に既知の方法で作成される。適当な過敏反応誘発ポリペプチドまたはタンパク質およびそれらをコードするDNAの核酸配列の例は、上記に開示している。この型のトランスジェニック植物を1度作成すると、植物自身は、植物の成長を促進する過敏反応誘発物をコードする遺伝子を伴い、通常の方法によって生育され得る。他に、トランスジェニック種子はトランスジェニック植物から回収される。そしてこれら種子は土壌に植えられ、通常の方法を用い生育され、トランスジェニック植物となる。トランスジェニック植物は、成長の促進に効果的な条件下、植えられたトランスジェニック種子から生育する。理論によって縛られることは望まないが、そういった成長促進は、RNAを媒介するか、または誘発ポリペプチドまたはタンパク質の発現の結果として生じ得る。
【0066】
トランスジェニック植物および植物種子を本発明に従い使用したときでも、過敏反応誘発ポリペプチドまたはタンパク質を施用する植物および種子の処理に使用するのと同じ物質でさらにそれらは処理され得る。過敏反応誘発物を含む、これら他の物質は、高または低圧噴霧、注入、塗布、散布および浸透を含む上記方法によりトランスジェニック植物および植物種子に施用され得る。同様に、トランスジェニック植物種子から植物が生育すると、植物の成長を促進する過敏反応誘発物の一つまたはそれ以上の施用により処理され得る。そういった植物はまた、通常の植物処理剤(例えば殺虫剤、肥料等)で処理され得る。本発明のトランスジェニック植物は、成長の促進が可能な植物を作成する種子または珠芽(例えば切枝)の作成に有用である。
【実施例】
【0067】
実施例
実施例1−トマト種子のErwinia amylovora過敏反応誘発物処理による発芽率への作用
Marglobeトマト種の種子をErwinia amylovora過敏反応誘発物溶液(“ハルピン”)40mlに浸した。ハルピンは、プラスミドpCPP2139(図1参照)を含むE. coli株DH5を生育させ、超音波処理により細胞を融解し、沸騰水に5分間入れることにより熱処理し、続いて遠心して細胞残骸を除去し、タンパク質および他の熱不安定成分を除去して製造した。得られた沈殿(“CFEP”)を連続希釈した。これらの希釈(1:40、1:80、1:160、1:320、1:640)は、ウェスタンブロット分析を基本にしてそれぞれ20、10、5、2.5および1.25μgm/mlのハルピンを含んだ。種子をビーカー中のハルピンまたは緩衝液に日0に24時間、28℃の成育チャンバーで浸した。浸漬後、日1に種子を人工土壌の発芽用鉢に蒔いた。この方法を処理当たり100個の種子で行った。
【0068】
処理:
1.ハルピン中の種子(1:40)(20μgm/ml)。
2.ハルピン中の種子(1:80)(10μgm/ml)。
3.ハルピン中の種子(1:160)(5μgm/ml)。
4.ハルピン中の種子(1:320)(2.5μgm/ml)。
5.ハルピン中の種子(1:640)(1.25μgm/ml)。
6.緩衝液中の種子(5mM KPO4、pH6.8)。
【0069】
【表1】
【0070】
表1に示されるように、トマト種子のErwinia amylovora過敏反応誘発物での処理は、発芽に必要な時間を減らし、発芽の割合を非常に増加させた。
【0071】
実施例2−トマト種子のErwinia amylovora過敏反応誘発物処理によるトマト植物の高さへの作用
Marglobeトマト種の種子をビーカー中のハルピン(1:15、1:30、1:60および1:120)または緩衝液に、日0に24時間、28℃の成育チャンバーで浸した。浸漬後、日1に種子を人工土壌の発芽用鉢に蒔いた。
処理当たり10本の同一な見かけの植物を無作為に選択して測定した。苗木を土壌表面から植物の頂部まで定規で計った。
【0072】
処理:
1.ハルピン中(1:15)(52μgm/ml)。
2.ハルピン中(1:30)(26μgm/ml)。
3.ハルピン中(1:60)(13μgm/ml)。
4.ハルピン中(1:120)(6.5μgm/ml)。
5.緩衝液中(5mM KPO4、pH6.8)。
【0073】
【表2】
【0074】
【表3】
【0075】
【表4】
【0076】
表2−5に示されるように、トマト種子のErwinia amylovora過敏反応誘発物での処理は、植物の生育を促進させた。1:30希釈は最大の効果を有した---苗木の高さで16%増加。
【0077】
実施例3−トマト植物のErwinia amylovora過敏反応誘発物処理によるトマト植物の高さへの作用
Marglobeトマト植物が4週齢の時、それらに13μgm/ml(1:60)または8.7μgm/ml(1:90)のハルピンまたは緩衝液(5mM KPO4)を、6ml/植物で28℃の成育チャンバーで噴霧した。トマト植物の高さはハルピン噴霧後2週間(6週齢トマト植物)および噴霧2週間と5日後に測定した。処理当たり10本の同一な見かけの植物を無作為に選択して測定した。苗木を土壌表面から植物の頂部まで定規で計った。
【0078】
処理:
1.ハルピン(1:60)(13μgm/ml)。
2.ハルピン(1:90)(8.7μgm/ml)。
3.緩衝液(5mM KPO4、pH6.8)。
【0079】
【表5】
【0080】
表6に示されるように、トマト苗木へのErwinia amylovora過敏反応誘発物噴霧は、トマト植物の生育を促進させることができる。緩衝液処理コントロールと比較して、試験した二つの用量の過敏反応誘発物に関して同様の促進が示された。
【0081】
実施例4−トマト種子のErwinia amylovora過敏反応誘発物処理によるトマト植物の高さへの作用
Marglobeトマトの種子を、ビーカー内のErwinia amylovora過敏反応誘発物(“ハルピン”)(1:40、1:80、1:160、1:320および1:640)または緩衝液に、日0に24時間、28℃の成育チャンバーで浸した。種子をハルピンまたは緩衝液に浸漬後、日1に種子を人工土壌の発芽用鉢に蒔いた。処理当たり10本の同一な見かけの植物を無作為に選択して測定した。苗木を土壌表面から植物の頂部まで定規で計った。
【0082】
処理:
1.ハルピン(1:40)(20μgm/ml)。
2.ハルピン(1:80)(10μgm/ml)。
3.ハルピン(1:160)(5μgm/ml)。
4.ハルピン(1:320)(2.5μgm/ml)。
5.ハルピン(1:640)(1.25μgm/ml)。
6.緩衝液(5mM KPO4、pH6.8)。
【0083】
【表6】
【0084】
【表7】
【0085】
表7−10に示されるように、トマト種子のErwinia amylovora過敏反応誘発物での処理は、トマト植物の生育を促進できる。1:160希釈(5μg/mlハルピン)は最大の作用を有する--苗木の高さは緩衝液処理植物より20%以上増加した。
【0086】
実施例5−トマト種子のErwinia amylovora過敏反応誘発物処理による種子発芽率への作用
Marglobeトマト種の種子を、ビーカー中のErwinia amylovora過敏反応誘発物(“ハルピン”)溶液(1:50または1:100のE. coli DH5 (pCPP2139)由来のCFEP希釈、各々8μgm/mlおよび4μgm/mlの過敏反応誘発物を含む)および緩衝液に、日0に24時間、28℃の成育チャンバーで浸した。浸漬後、日1に種子を人工土壌の発芽用鉢に蒔いた。この方法を鉢当たり20個の種子で、処理当たり4鉢で行った。
【0087】
処理:
1.ハルピン(8μgm/ml)。
2.ハルピン(8μgm/ml)。
3.ハルピン(8μgm/ml)。
4.ハルピン(8μgm/ml)。
5.ハルピン(4μgm/ml)。
6.ハルピン(4μgm/ml)。
7.ハルピン(4μgm/ml)。
8.ハルピン(4μgm/ml)。
9.緩衝液(5mM KPO4、pH6.8)。
10.緩衝液(5mM KPO4、pH6.8)。
11.緩衝液(5mM KPO4、pH6.8)。
12.緩衝液(5mM KPO4、pH6.8)。
【0088】
【表8】
【0089】
表11に示されるように、トマト種子のErwinia amylovora過敏反応誘発物での処理は、トマト種子の発芽率およびレベルを増加できる。より高い用量の使用が、緩衝液よりも本実験の終了時にはより有効であるように見える。
【0090】
実施例6−過敏反応誘発物をコードする構築物であるpCPP2139またはプラスミドベクターpCPP50を含むE. coliから調製したタンパク質でのトマト種子処理の植物生育への作用
Marglobeトマトの種子を、Erwinia amylovora過敏反応誘発物(“ハルピン”)(E. coli DH5α (pCPP2139)(図1))またはコントロールとしてBSAタンパク質を添加したベクター製剤(DH5α(pCPP2139)(図2))に浸した。コントロールベクター製剤は、ml当たり、33.6μlのBSA(10mg/ml)を含み、ハルピンに由来するpCPP2139に含まれるタンパク質の量とほぼ同じである。1:50(8.0μg/ml)、1:100(4.0μg/ml)および1:200(2.0μg/ml)の希釈をビーカーで日1に調製し、種子を24時間、28℃で制御環境チャンバーで浸した。浸漬後、種子を、日2に人工土壌の発芽用鉢に蒔いた。処理当たり10本の同一な見かけの植物を移植後3回無作為に選択し、測定した。苗木を土壌表面から植物の頂部まで定規で計った。
【0091】
処理:
1.ハルピン 1:50(8.0μg/ml)。
2.ハルピン 1:100(4.0μg/ml)。
3.ハルピン 1:200(2.0μg/ml)。
4.ベクター+BSA 1:50(0ハルピン)。
5.ベクター+BSA 1:100(0ハルピン)。
6.ベクター+BSA 1:200(0ハルピン)。
【0092】
【表9】
【0093】
【表10】
【0094】
表12−15に示されるように、Erwinia amylovora過敏反応誘発物をコードする遺伝子を含むE. coliでの処理は、トマト植物の成長を促進できる。1:100希釈(4.0μg/ml)は最大の作用を有するが、それより高いかまたは低い濃度は作用が小さい。ハルピン4.0μg/mlでの処理による平均の苗木の高さは、同量の非ハルピンタンパク質を含むベクターコントロール製剤に対して約20%促進された。E. coli株DH5α(pCPP2139)中にhrpN遺伝子のベクターを保有する株E. coli DH5α(pCPP50)からの融解細胞製剤の成分は、ハルピン含有製剤と同じ成長促進作用を有せず、BSAタンパク質を、大量のハルピンタンパク質を含むDH5α(pCPP2139)製剤と同程度に添加したときでさえそうであった。
【0095】
実施例7−過敏反応誘発物をコードする構築物であるpCPP2139またはプラスミドベクターpCPP50を含むE. coliから調製したタンパク質でのトマト種子処理の植物生育への作用
Marglobeトマト種子を、1:25、1:50および1:100の希釈のErwinia amylovora過敏反応誘発物溶液(“ハルピン”)(ハルピンコードプラスミドpCPP2139ベクターに由来)およびpCPPベクター含有溶液に、ビーカー中で日1に24時間、28℃で生育チャンバーで浸した。浸漬後、種子を日2に人工土壌の発芽用鉢に蒔いた。処理当たり10本の同一な見かけの植物を移植後3回無作為に選択し、測定した。苗木を土壌表面から植物の頂部まで定規で計った。
【0096】
処理:
1.ハルピン16μgm/ml。
2.ハルピン8μgm/ml。
3.ハルピン4μgm/ml。
4.ベクター16μgm/ml。
5.ベクター8μgm/ml。
6.ベクター4μgm/ml。
【0097】
【表11】
【0098】
【表12】
【0099】
表16−18に見られるように、Erwinia amylovora過敏反応誘発物での処理は、トマト直物の成長を促進できる。1:50希釈(8μg/ml過敏反応誘発物)は最大の作用を有し、苗木の高さはコントロールの約20%上昇する。
【0100】
実施例8−ジャガイモの生育における無細胞Erwinia amylovora過敏反応誘発物の作用
3週齢ジャガイモ植物の種Norchipを、塊茎片から別々の容器で生育させた。各植物の葉に、1:50、1:100および1:200に希釈したErwinia amylovora過敏反応誘発物(“ハルピン”)またはE. coliおよびベクターcPP550のタンパク質を含むコントロール溶液(“ベクター”)を噴霧した。日20に、処理当たり12本の同一な見かけの植物を無作為に選択した。各処理由来の一本の植物を生育チャンバー中16℃で保持し、他方各処理由来の2本の植物を温室床箱で18−25℃で保持した。処理25日後、全植物の芽(茎)を個々に測定した。
【0101】
処理:
1.ハルピン 1:50 16μgm/ml。
2.ハルピン 1:100 8μgm/ml。
3.ハルピン 1:200 4μgm/ml。
4.ベクター 1:50 0ハルピン。
5.ベクター 1:100 0ハルピン。
6.ベクター 1:200 0ハルピン。
【0102】
【表13】
【0103】
表19および20に示されるように、Erwinia amylovora過敏反応誘発物でのジャガイモ植物の処理は、芽(茎)生育を促進する。従って、全体的な成長は、茎の数および平均の長さから判断して、温室および生育チャンバー生育植物の両方でハルピン処理植物で大きかった。中間の量のハルピン(8μgm/ml)で処理したジャガイモ植物は、その茎生育において、そのより高いまたはより低い用量よりも促進されたように見えた。中間の量のハルピンでの処理は、両方の生育条件下でより促進された成長をもたらした。
【0104】
実施例9−Erwinia amylovoraハルピンを含む無細胞誘発製剤のトマトへの噴霧作用
Marglobeトマト植物に、移植後8日に、ハルピン製剤(E. coli DH5α(pCPP2139))またはコントロールとしてBSAタンパク質を添加したベクター製剤(E. coli DH5α(pCPP50))を噴霧した。コントロールベクター製剤は、ml当たり、BSA(10mg/ml)を含み、ハルピンに起因するpCPP2139製剤に含まれるのと同じタンパク質の濃度を提供する。1:50(8.0μg/ml)、1:100(4.0μg/ml)および1:200(2.0μg/ml)の希釈を調製し、植物に、電動噴霧装置で流れ出すまで噴霧した。処理当たり15本の同一な見かけの植物を無作為選択し、処理に付した。植物を、処理前および後は28℃に制御環境チャンバーに保持した。
【0105】
全体の高さを処理後数回、土壌表面から植物の頂部まで定規で計った。トマト植物の頂部を土壌の表面近辺から茎を切断した直後に秤量した。
【0106】
処理:(希釈およびハルピン含量)
1.ハルピン 1:50(8.0μg/ml)
2.ハルピン 1:100(4.0μg/ml)
3.ハルピン 1:200(2.0μg/ml)
4.ベクター+BSA 1:50(0ハルピン)
5.ベクター+BSA 1:100(0ハルピン)
6.ベクター+BSA 1:200(0ハルピン)
【0107】
【表14】
【0108】
【表15】
【0109】
【表16】
【0110】
トマト苗木へのハルピンの1回噴霧は、一般に、BSAタンパク質を添加したコントロール(ベクター)製剤よりも大きな続く生育をもたらした。ハルピン処理植物での促進された生育は、植物高および生重量測定の両方で見られた。試験した3つの濃度で、低いほう二つは、最高の用量(8.0μg/ml)よりもより大きな植物生育を(いずれかの測定を基本にして)もたらした。低いほうの二つ(2および4μg/ml)の濃度で処理した植物の生育は僅かに異なった。E. coli株DH5α(pCPP2139)中のhrpN遺伝子のベクターを保持する株E. coli DH5α(pCPP50)由来の融解細胞製剤の成分は、大量のハルピンタンパク質を含むDH5α(pCPP2139)製剤と同程度に、BSAタンパク質を添加した場合でさえ、ハルピン含有製剤と同じ生育促進作用は有しなかった。従って、本実験はハルピンが植物生育を担うことを証明する。
【0111】
実施例10−小果実の早期着色および早期成熟
野外試験を、ラズベリー栽培種Canbyの収率および成熟パラメーターにおける過敏反応誘発物(“ハルピン”)処理の作用を評価するために行った。定着した植物に、区画40フィート長×3フィート幅(1植物幅)に2.5mg/100平方フィートでハルピンでの処理、非処理(“チェック”)または産業標準化学物質Ronilanを推奨された割合(“Ronilan”)での処理を行った。処理は4回繰り返し、実験野外部位のrepによりアレンジした。処置は、5−10%開花期に開始し、続いて7−10日間隔で2回行った。最初の2回の収穫は疾病コントロールに使用し、果実収率データの評価は最後の2回の収穫から集めた。観察は、ハルピン処理果実が非処理果実より大きく、赤いことを示し、成熟が1−2週間早まったことを示した。最少10個の果実を含む房当たりの成熟果実の数をこの時点で測定し、表26に要約する。ハルピン処理した区画はチェックまたはRonilan処理よりも10個のベリーの房当たり、より多い成熟果実を有した。最後の2回の収穫の合わせた収量は、ハルピンおよびRonilan処理区画の収量が、非処理コントロールよりも増加したことを示した(表27)。
【0112】
【表17】
【0113】
【表18】
【0114】
実施例11−さや豆の成長促進
Bush Blue Lake種のさや豆を種々の方法で処理し、商品の鉢植え用混合物を入れたプラスチック鉢に25cm2で植え、生育パラメーターの評価のために開放温室に置いた。処理は、非処理豆種子(“チェック”)、希釈剤としての水で調製した1.5%メチルセルロースのスラリーで処理した種子(“M/C”)、1.5%メチルセルロースで種子を処理し、続いて過敏反応誘発物(“ハルピン”)を0.125mg/mlで葉に投与した種子(“M/C+H”)、および1.5%メチルセルロース+50種子当たり5.0μgハルピンで乾燥したハルピンスプレーで種子を処理し、続いてハルピンを0.125mg/mlで葉に投与した種子(“M/C-SD+H”)を含む。種子を日0に土壌に蒔き、鉢当たり3個植え、発芽して鉢当たり1本の植物に間引いた。処理を開放温室内で10回繰り返し、repにより無作為化した。豆莢を64日後に収穫し、売り物になる大きさ(>10cm×5cmの大きさ)の豆莢の生重量を収量として集めた。処理手段を分けるために使用したFisher's LSDでの種々の分析によりデータを分析した。
【0115】
【表19】
1 売り物になる収量は10cm×0.5cm以上の全ての豆莢を含んだ。同じ文字の意味は、Fisher's LSDでP=0.05の有意な差がなかった。
【0116】
表28に示されるように、種々の施用法でのErwinia amylovoraハルピンの施用は、売り物になる大きさのさや豆の収量の増加をもたらした。メチルセルロース単独での処理はまた豆の収量の増加をもたらしたが、種子(噴霧乾燥)および葉処理としてハルピンと組み合わせた場合、実質的に増加する。
【0117】
実施例12−キュウリへのHP-1000TMの葉施用によるキュウリの収量増加
キュウリ幼苗および移植苗を、HP-1000TM(EDEN Bioscience, Bothell, Washington)(Erwinia amylovora過敏反応誘発製剤)の15、30または60μg/ml活性成分(a.i.)の量の葉噴霧で処理した。最初の噴霧は、最初の本葉が完全に伸びたときに行った。2回目の施用は、最初の噴霧から10日後に行った。全噴霧はバックパック噴霧器を使用して行い、非処理コントロール(UTC)もまた試験に含んだ。2回目のHP-1000TMの施用3日後、各処理の10本の植物を野外区画に無作為に移植し、3回繰り返した。これにより処理当たり全30本の植物を生産した。移植7日後、3回目のHP-1000TM葉噴霧を行った。ひどい旱魃がかなりの水ストレスをもたらしたが、全6回の収穫が標準の業務上収穫パターンに従ってなされた。各処理から収穫した果実の全重量を表29に示す。結果は、15および30μg/mlの量でHP-1000TM処理した植物が、UTCよりも有意に多い実を生産した。HP-1000TMで処理した植物は、収穫が僅かに増えたことを示す。これらの結果は、HP-1000TM処理が非処理植物よりも旱魃負荷条件に明らかにより耐性であることを示した。
【0118】
【表20】
1 活性成分(a.i.)。2 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0119】
実施例13−HP-1000TM処理による綿の収量増加
綿を12×12フィートの同一野外区画に、無作為完全ブロック(RCB)野外試験方式で植え、4回繰り返した。植物をHP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)、HP-1000TM+Pix(登録商標)(Pix(登録商標)(BASF Corp., Mount Olive, N. J.)は綿植物の高さが小さく保たれるように施用する成長調節剤である)またはEarly Harvest(登録商標)(Griffen Corp., Valdosta, Ga.)(拮抗的成長促進剤)で処理した。非処理コントロール(UTC)もまた試験に含んだ。バックパック噴霧器を使用して、葉施用を、作物の3つの成長段階の全ての処理で行った;第一本葉、開花前および開花早期。全ての肥料および雑草制御製剤を、全処理の慣用の農作業方式に従って施用した。収穫10週間前の綿玉の数は、他の処理と比較してHP-1000TM処理植物が明らかに多かった。収穫により、HP-1000TM処理植物は、UTCと比較して有意に増加したリント収率を(43%)を示した(表30)。HP-1000TMをPix(登録商標)と組み合わせたとき、リント収量はUTCを20%超えた。Pix(登録商標)は大エーカー数の綿に施用するため、この結果はHP-1000TMがPix(登録商標)と十分タンク混合され得ることを示す。拮抗的成長促進剤であるEarly Harvest(登録商標)の施用は、UTCと比較して僅かに9%増加させた。
【0120】
【表21】
1 HP-1000TMの量は活性成分(a.i.)として;Early Harvest(登録商標)およびPix(登録商標)の量は製剤製品として。
【0121】
実施例14−HP-1000TM処理による中国ナスの収量増加
苗床生育中国ナス苗木に、1回、HP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)を15、30または60μg/ml(a.i.)で噴霧し、次いで、野外区画に移植し、各処理を3回繰り返した。移植2週間後、2回目のHP-1000TM施用を行った。3回目の最終HP-1000TM施用を2回目の約2週間後に行った。全噴霧をバックパック噴霧器を使用して行った;非処理コントロール(UTC)もまた試験に含んだ。季節の進行と共に、各処理から全8回の収穫を行った。これらの収穫物からのデータは、HP-1000TMでの処理が植物当たりより多い実の収量をもたらすことを示す。
【0122】
【表22】
【0123】
実施例15−HP-1000TM処理による米の収量増加
稲苗を野外区画に移植し、3回繰り返し、次いでバックパック噴霧器を使用して3つの異なる量のHP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)で処理した。非処理コントロール(UTC)もまた試験に含んだ。最初のHP-1000TM施用は移植1週間後、2回目は1回目の3週間後であった。3回目の最終噴霧は米穀粒が穂に入り出す直前に行った。収穫の結果は、HP-1000TMの施用が30および60μg/mlの両方で収率を有意にそれぞれ47%および56%増加させることを証明した(表32)。
【0124】
【表23】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0125】
実施例16−HP-1000TM処理によるダイズの収量増加
ダイズを無作為化野外区画に植え、各処理を3回繰り返した。バックパック噴霧器をHP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)の葉面噴霧施用に使用し、非処理コントロール(UTC)もまた試験に含んだ。3つの量のHP-1000TMを、植物が約8インチの高さになったとき、4枚の本葉の始めに施用した。HP-1000TMの2回目の噴霧は、1回目の施用の10日後に、3回目の噴霧は2回目の10日後に行った。1回目の噴霧10日後に測定した植物の高さは、HP-1000TMの噴霧が有意な成長促進をもたらすことを示した(表33)。加えて、60μg/mlのHP-1000TMで処理した植物は、他の処理より5日速く開花し始めた。3回目の噴霧の約10日後、植物当たりのダイズ莢の数を繰り返し当たり無作為に選択した10本の植物から計数した。これらの結果は、HP-1000TMでの処理に由来する成長促進は、有意に多い収量をもたらすことを示した(表34)。
【0126】
【表24】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0127】
【表25】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0128】
実施例17−HP-1000TM処理によるイチゴの収量増加
HP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)での二つの野外試験を、二つのイチゴ種であるCamarosaおよびSelvaで行った。各種に関して、無作為完全ブロック(RCB)設計を、商業用のイチゴを生産する畑で、区画(5.33×10フィート)/処理で確立し、4回繰り返した。各区画内で、イチゴ植物を2列配置で植えた。未処理コントロール(UTC)もまた試験に含んだ。施用を開始する前に、全ての植物を他の花およびベリー類から離した。40μg/mlの量のHP-1000TMの噴霧を6週毎にバックパック噴霧器を使用して行った。各噴霧施用前に、各処理由来の全成熟果実を収穫し、秤量し、商業的標準に従って等級付した。HP-1000TMのSelvaイチゴ植物への施用3週間以内に、成長促進が、可視的に大きな地上生物量およびより元気で、緑で健康的な見かけとして識別できた。6回の収穫(即ち、これらの植物の予定の寿命)の後、全ての収量データを合計し、分析した。Camarosa種において、HP-1000TM処理植物からの売り物になる果実の収量は、最後の4回の収穫を平均したとき、UTCより有意に(27%)多かった(表35)。処理間の有意な差は最初の2回の収穫ではこの種では明らかではなかった。Selva種はHP-1000TMでの処理により反応性であった;Selvaイチゴ植物は、6回の収穫を平均したとき、UTCと比較して統計学的に有意な64%多い売り物になる果実を生産した(表35)。
【0129】
【表26】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0130】
実施例18−HP-1000TM処理によるトマトの早期成熟および収量増加
生販売用トマト(Solar Set種)を、区画(2×30フィート)で生育させ、5回無作為完全ブロック(RCB)野外試験で、商品トマト生産畑で繰り返した。処理はHP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)、実験的競合産物(ActigardTM (Novartis, Greensboro, N. C.))および化学標準(Kocide(登録商標) (Griffen Corp., Valdosta, GA))+Maneb(登録商標) (DuPont Agricultural Products, Wilmington, D. E.))を疾病コントロールのために含んだ。HP-1000TMの最初の施用は、移植直後に苗木に直接注ぐ50ml浸液(30μg/ml a.i.)により行った。その後、11週毎に葉噴霧をバックパック噴霧器を使用して行った。全処理由来の最初の収穫は移植後約6週で行い、完全に赤く熟したトマトのみを各処理から収穫した。結果は、HP-1000TM処理植物が、最初の収穫に間に合うトマトの有意に多い量を生産したことを示した(表36)。HP-1000TM処理植物から収穫したトマトは、他の植物より10−14日進んでいると評価された。
【0131】
【表27】
1 Kocide(登録商標)およびManeb(登録商標)の量は製剤製品。2 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0132】
実施例19−非消毒土壌に植えたときの、HP-1000TM処理によるイチゴの早期開花および成長促進
イチゴ植物(“プラグ(plugs)”および“根付き(bare-root)”)の栽培種Commanderを区画(2×30フィート)に移植し、無作為完全ブロック野外試験で5回繰り返した。約60本の別々の植物を各繰り返しで植えた。この野外試験での処理は下記に示す:
【表28】
1 非消毒土壌はソラマメ属を前の2年栽培していた。
【0133】
移植は、晩秋に、寒い気候が植物の生育を遅らせ始めたときに行った。移植2週間後、HP-1000TMの最初の葉施用を、バックパック噴霧器で40μg/ml(a.i.)で行った。移植3週間後、予備的結果を集め、臭化メチルおよびUTCとHP-1000TMの比較を、各繰り返しの全イチゴ“プラグ”植物の花の数を計数することにより行った。開花が“根付き”植物でまだ起こっていないため、この処理での繰り返しの各植物を、葉の先端から3枚の葉の塊の真中の葉の茎までの長さを測定することにより評価した。結果(表37および38)は、HP-1000TMが早期に促進された花の生育および葉の大きさを、“プラグ”および“根付き”イチゴ植物でそれぞれ提供することを示した。
【0134】
【表29】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0135】
【表30】
【0136】
実施例20−HP-1000TM処理によるハラペーニョトウガラシの早期成長促進
ハラペーニョトウガラシ(Mittlya栽培種)移植植物をHP-1000(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)(30μg/ml a.i.)での1時間の根浸漬により処理し、続いて4回繰り返した。非処理コントロール(UTC)もまた含んだ。移植14日に開始して、処理植物は3回HP-1000TMを、14日間隔で、バックパック噴霧器により受けた。HP-1000TMの3回目の施用の1週間後(移植54日後)、植物の高さを繰り返し当たり無作為に選択した4本の植物で測定した。これらの測定の結果は、HP-1000TM処理植物がUTC植物よりも約26%背が高いことを示した(表39)。加えて、各植物の芽、花または実の数を計数した。これらの結果は、HP-1000TM処理植物が、UTC植物と比較して、61%多い花、実または芽を有することを示した(表40)。
【0137】
【表31】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0138】
【表32】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0139】
実施例21−HP-1000TM施用によるタバコの成長促進
タバコ苗木を無作為化野外区画に植え、3回繰り返した。移植後、HP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)を3つの量のいずれか一つで葉噴霧した:15、30または60μg/ml a.i.。60日後、2回目のHP-1000の葉施用を行った。2回目の施用2日後、植物の高さ、植物当たりの葉の数および葉の大きさ(面積)を処理当たり無作為に選択した10本について測定した。これらの測定の結果は、HP-1000TMでの処理が有意に植物生育を促進させることを示した(表41、42および43)。植物の高さは6−13%増加し、30および60μg/mlでHP-1000TM処理した植物は平均UTCより植物当たり1枚葉が多かった。しかしながら、最も有意には、15、30および60μg/ml のHP-1000TMでの処理は、葉面積の対応する増加をもたらした。植物当たり1枚の余分な葉および増加した平均葉サイズ(面積)のタバコ植物は、商業的に有意な反応を示す。
【0140】
【表33】
【表34】
【表35】
【0141】
実施例22−HP-1000TM施用による冬小麦の成長促進
冬小麦種子に乾燥HP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)を、種子100ポンド当たり製剤製品(3%a.i.)3オンス“散布”し、続いて慣用の播種装置を使用して11.7フィート×100フィート長の試験無作為試験区画に植えた。更なる処理は、種子100ポンドあたりHP-1000TM粉末(3%a.i.)1オンスの“散布”、20μg/ml活性成分の濃度のHP-1000TM溶液への4時間の種子浸漬、続く植える前の空気乾燥、標準化学(Dividend(登録商標))殺菌剤“散布”および非処理コントロール(UTC)を含んだ。蒔いて8日後、HP-1000TM処理種子は発芽し始め、一方UTCおよび化学標準処理種子は蒔いて約14日後まで発芽せず、通常の時間経過であった。蒔いて41日後、苗木を地面から抜き、評価した。3オンス/100ポンドでの“散布”としてHP-1000TM処理した種子を目で調べ、他の処理のいずれよりも約2倍大きいと判断した。
【0142】
野外試験に続き、温室実験を計画し、これらの結果の確認をした。処理は、100ポンドの種子当たり3オンスの量での乾燥HP-1000TM(10%a.i.)の散布、20mg/mlの濃度の溶液中のHP-1000TMへの移植前4時間の浸漬および非処理コントロール(UTC)を含んだ。各処理由来の小麦種子を鉢当たり25個の種子で蒔き、5つの鉢を各処理の繰り返しとして提供した。蒔いて15日後、各処理鉢当たり無作為に選択した10本の苗木を抜き、注意深く洗い、根の長さを測定した。個々の苗木の地上部分は処理の効果を示さなかっため、HP-1000TMでの処理の根の生育の増加はサンプルの選択に影響しなかった。HP-1000TM処理の根の生育の増加は、UTCより有意に大きかった(表49);しかしながら、種子散布処理は、僅かに良い結果をもたらした。
【0143】
【表36】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0144】
実施例23−HP-1000TM施用によるキュウリの成長促進
商品として生産されるキュウリの野外試験は、4回の処理、2種の濃度(20または40μg/ml)でのHP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)、疾病コントロールの化学表品(Bravo(登録商標) (Zeneca Ag Products, Wilmington, Del.) + Maneb(登録商標))および非処理コントロール(UTC)であった。各処理を、3×75フィート区画で、各処理当たり約2フィートの植物の空間で4回行った。HP-1000TMの葉噴霧は、最初の本葉のときに開始し、14日間隔で、最後の収穫まで合計6回施用で繰り返した。標準殺菌剤混合物を7日毎または状況的な根拠があったら直ぐに施用した。商品の収穫はHP-1000TMの最初の施用から約2ヶ月後(HP-1000TMの5回噴霧後)に開始し、最終収穫は最初の収穫の約14日後であった。
【0145】
最初の収穫の結果は、HP-1000TMでの処理が、個々のキュウリの平均重量ではなく、収穫した平均キュウリ収量をキュウリの総数を増加させることにより増加させたことを示した(表45−47)。同じ傾向が最終収穫(表48−49)でも見られた。HP-1000TM処理によりもたらされる収量増加がキュウリ平均サイズの明白な増加により達成されるものでないことは商業的に重要であった。
【0146】
【表37】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0147】
【表38】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0148】
【表39】
【0149】
【表40】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0150】
【表41】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0151】
【表42】
【0152】
実施例24−トマトにおけるPseudomonas syringae pv syringae由来のハルピンpss誘導成長促進
ハルピンpss(即ち、Pseudomonas syringae pv syringae由来の過敏反応誘発物)(引用して本明細書に包含させるHe, S. Y., et al., “Pseudomonas syringae pv syringae Harpinpss. A Protein that is Secreted via the Hrp Pathway and Elicits the Hypersensitive Response in Plants,” Cell 73:1255-66 (1993))がまた植物生育を促進するか試験をするために、トマト種子(Marglobe種)を人工土壌の8インチ鉢に蒔いた。蒔いて10日後、苗木を別々の鉢に移植した。実験を通して、肥料、水の潅水、温度および土壌湿度は植物間で均質に維持した。移植16日後、最初の植物の高さを測定し、ハルピンpssの最初の施用を行い、日0とした。2回目の施用は日15に行った。更なる成長データを日10および日30に集めた。最終データ採集は日30で、植物の高さおよび生重量を含んだ。
【0153】
実験中の施用に使用したハルピンpssは、ハルピンpss(即ち、hrpZ)をコードする遺伝子のプラスミドを含むE. coli DH5を発酵させて、製造した。細胞を回収し、5mM リン酸カリウム緩衝液に再懸濁させ、超音波処理により破壊した。超音波処理物質を5分沸騰させ、次いで10分、10,000rpmで遠心した。懸濁液を無細胞誘発製剤(CFEP)と見なした。20および50μg/mlハルピンpss溶液を同じ緩衝液から調製し、細胞懸濁に使用した。同じプラスミドを含むが、hrpZ遺伝子を含まない同じ株から調製した同じCFEPをコントロール処理の材料として使用した。
【0154】
湿潤剤であるPinene II (Drexel Chemical Co., Memphis, Tenn.)をハルピンpss溶液に0.1%の濃度で添加し、次いでハルピンpssをトマト植物に流れ落ちるまで噴霧した。
【0155】
表51はハルピンpss処理グループとコントロールグループの間に有意な差があったことを示す。ハルピンpss処理トマトは高さで10%以上増加した。データは、ハルピンpssが、トマトおよび他の多くの植物種に施用したとき、成長促進作用があるという点で、Erwinia amylovora由来の過敏反応誘発物と同様に作用するという請求の範囲を支持する。トマトの高さの有意な増加に加えて、ハルピンpss処理トマトはより大きな生物量、より大きな葉、早期開花および相対的なより健康な見かけを有した。
【0156】
【表43】
1 植物の高さは最も近い0.5cmで測定した。日0は最初の植物の高さを記録し、最初の施用を行った日を意味する。
2 平均は括弧内のSDと共に示す(全処理グループでn=20)
3 異なる文字(aおよびb)は平均間の有意差(P 0.05)を意味する。差はFisher LSDに続くANOVAにより評価した。
【0157】
本発明は説明の目的で詳細に記載しているが、このような細部は単にこの目的のためであって、以下の請求の範囲に定義の本発明の精神および範囲を逸脱することなく、当業者はその変化をなし得ることは理解される。
【技術分野】
【0001】
本出願は、1997年1月27日付け米国仮特許出願番号60/036,048の利益を請求する。
本発明は、USDA NRI Competitive Research Grant番号91-37303-6430のもと、米国政府より支持されている。
【0002】
本発明の分野
本発明は植物の成長の促進に関する。
【背景技術】
【0003】
発明の背景
有機肥料の施用による植物成長の改良は既知であり、数世紀にわたり実施されてきた(H. Marschner, “Mineral Nutrition of Higher Plants,” Academic Press: New York pg.674 (1986))。栽培者および農業者が成長促進の方法による効率向上のために、土壌または成長する作物に施用し得る、簡単な製品を生産するための複合的無機肥料生産システムを近世の人間は発達させてきた。植物の大きさ、色、成熟、および収穫は肥料製品の施用によって全て改善され得る。無機肥料には、通常、硝酸アンモニウムといった化学的な施用を含む。有機肥料には、動物の厩肥および堆肥にした芝の有機堆積物、他の多くの源を含み得る。
【0004】
近年、研究者たちは、生物学的産物の使用により植物成長を改良することを探求してきた。Beauveria bassianaおよびTrichoderma harizamumのような昆虫および疾病を制御する薬剤により、昆虫および疾病問題が制御されることが記されており、それゆえ間接的に植物成長および効率を改善する(Fravel et al., “Formulation of Microorganisms to Control Plant Diseases,” Formulation of Microbial Biopesticides, Beneficial Microorganisms, and Nematodes, H. D. Burges, ed. Chapman and Hall: London(1996))。
【0005】
微生物の施用または微生物生産物による方法によって直接植物成長の促進が示唆されている。新しい土地に植えるとき、根粒細菌はマメ科作物の種子に加えられる(Weaver et al., “Rhizobium,” Methods of Soil Analysis, Part 2, Chemical and Microbiological Properties, 2nd ed., American Society of Agronomy: Madison (1982))。これらの細菌は植物の根粒着生効率を改善し得、それゆえ窒素固定と呼ばれる過程、遊離窒素原子を利用できる形に変換する植物の能力を向上する。非マメ科作物は、大抵そのような処置による利益はない。Rhizobiumといった、加えた細菌は直接根毛に寄生し、そして保護および栄養を受けつつ、植物に利益提供することにより相利共生の関係を始める。
【0006】
菌根菌類はまた、多くの植物、特に栄養が枯渇した土壌での球果植物の意図的生育に必要な微生物であると認められている。植物ホルモンの生物学的合成(Frankenberger et al., “Biosynthesis of Indole-3-Acetic Acid by the Pine Ectomycorrhizal Fungas Pisolithus tinctorius,” Appl. Environ. Microbiol. 53:2908-13(1987))、ミネラル吸収の増加(Harley et al., “The Uptake of Phosphate by Excised Mycorrhizal Roots of Beech,” New Phytologist 49:388-97 (1950)およびHarley et al., “The Uptake of Phosphate by Excised Mycorrhizal Roots of Beech. IV. The Effect of Oxygen Concentration Upon Host and Fungus,” New Phytologist 52:124-32 (1953)および水(A. B. Hatch, “The Physical Basis of Mycotrophy in Pinus,” Black Rock Forest Bull. No. 6, 168pp. (1937))を含む機構が必要とされてきた。菌根菌類は根粒着生細菌程の使用頻度には到達していない、それは、菌根菌類が菌根菌の株毎に変動し、一定しない結果を与え、生物としての研究も困難を伴うからである。
【0007】
植物成長促進リゾバクテリア(PGPR)は、近年、植物の成長および生育を向上すると認められている。仮説の機構は、直接的な影響(例えば栄養吸収の増加)から間接的な機構(例えば病原体排除)にまでわたっている。PGPRの施用による成長促進は、通常、生存細菌の根組織への接種、および細菌生産性ホルモン効果、シデロホアを通じて、または抗生物質による疾病の防止、または競合による、成長改善の達成について言及されている。上記全ての場合において、その結果は、根のコロニー化、時折、種子コーティングの施用を通じて有効となる。PGPR株はノトバイオティック条件下、根の伸張を促進する直接的成長促進体となり得るという提唱をする情報には限界がある(Anderson et al., “Responses of Bean to Root Colonization With Pseudomonas putida in a Hydroponic System,” Phytopathology 75:992-95 (1985), Lifshitz et al., “Growth Promotion of Canola (rapeseed) Seedlings by a Strain of Pseudomonas putida Under Gnotobiotic Conditions,” Can. J. Microbiol. 33:390-95 (1987), Young et al., “PGPR: Is There Relationship Between Plant Growth Regulators and the Stimulation of Plant Growth or Biological Activity?,”Promoting Rhizobacteria: Progress and Prospects, Second International Workshop on Plant Growth-promoting Rhizobacteria, pp. 182-86 (1991), Loper et al., “Influence of Bacterial Sources of Indole-3-Acetic Acid on Root Elongation of Sugar Beet,” Phytopathology 76:386-89 (1986), および Mueller et al., “Hormonal Interactions in the Rhizosphere of Maize (Zea mays L.) and Their Effect on Plant Development,” Z. Pflanzenernaehrung Bodenkunde 152:247-54 (1989))。しかしながら、植物成長レギュレーターは、これらの効果を介する機構として提唱されている。多くの細菌はインビトロで様々な植物成長レギュレーター(Atzorn et al., “Production of Gibberellins and Indole-3-Acetic Acid by Rhizobium phaseoli in Relation to Nodulation of Phaseolus vulgaris Roots,” Planta 175:532-38 (1988) and M. E. Brown, “Plant Growth Substances Produced by MicroOrganism of Solid and Rhizosphere,” J. Appl. Bact. 35:443-51 (1972))、または抗生物質(Gardner et al., “Growth Promotion and Inhibition by Antibiotic-Producing Fluorescent Pseudomonads on Citrus Roots,” Plant Soil 77:103-13 (1984))を産生する。シデロホア産物は、幾つかのPGPR株に提唱されている他の機構である(Ahl et al., “Iron Bound-Siderophores, Cyanic Acid, and Antibiotics Involved in Suppression of Thievaliopsis basicola by a Pseudomonas fluorescens Strain,” J. Phytopathol. 116:121-34 (1986), Kloepper et al., “Enhanced Plant Growth by Siderophores Produced by Plant Growth-Promoting Rhizobacteria,” Nature 286:885-86 (1980), and Kloepper et al ., “Pseudomonas siderophores: A Mechanism Explaining Disease-Suppressive Soils,” Curr. Microbiol. 4:317-20 (1980))。根表面のコロニー化、すなわち表面における病原体細菌との直接的競合は作用の他の機構である(Kloepper et al., “Relationship of in vitro Antibiosis of Plant Growth-Promoting Rhizobacteria to Plant Growth and the Displacement of Root Microflora,” Phytopathology 71:1020-24 (1981), Weller, et al., “Increased Growth of Wheat by Seed Treatments With Fluorescent Pseudomonads, and Implications of Pythium Control,” Can. J. Microbiol. 8:328-34 (1986), and Suslow et al., “Rhizobacteria of Sugar Beets: Effects of Seed Application and Root Colonization on Yield,” Phytopathology 72:199-206 (1982))。キャノーラ(菜種)の研究は、収穫、幼苗発生(seedling emergence)および成長力、早生植物成長(early-season plant growth)(葉の数および主なランナー(runner)の長さ)、ならびに葉の面積を含む(Kloepper et al., “Plant Growth-Promoting Rhizobacteria on Canola (rapeseed),” Plant Disease 72:42-46 (1988))、PGPR増加植物成長パラメーターを示す。ジャガイモの研究で、Pseudomonas株をジャガイモの種子に施用すると、収穫が増えることが示されている(Burr et al., “Increased Potato Yields by Treatment of Seed Pieces With Specific Strains of Pseudomonas Fluorescens and P. putida,” Phytopathology 68:1377-83 (1978), Kloepper et al., “Effect of Seed Piece Inoculation With Plant Growth-Promoting Rhizobacteria on Populations of Erwinia carotovora on Potato Roots and in Daughter Tubers,” Phytopathology 73:217-19 (1983), Geels et al., “Reduction of Yield Depressions in High Frequency Potato Cropping Soil After Seed Tuber Treatments With Antagonistic Fluorescent Pseudomonas spp.,” Phytopathol. Z. 108:207-38 (1983), Howie et al., “Rhizobacteria: Influence of Cultivar and Soil Type on Plant Growth and Yield of Potato,” Soil Biol. Biochem. 15:127-32 (1983), and Vrany et al., “Growth and Yield of Potato Plants Inoculated With Rhizosphere Bacteria,” Folia Microbiol. 29:248-53 (1984))。収穫の増加は、恐らく抗生物質による塊茎における病原性細菌の排除に対するPGRPの競合効果のためであるように見える(Kloepper et al., “Effect of Seed Piece Inoculation With Plant Growth-Promoting Rhizobacteria on Populations of Erwinia carotovora on Potato Roots and in Daughter Tubers,” Phytopathology 73:217-19 (1983), Kloepper et al., “Effects of Rhizosphere Colonization by Plant Growth-Promoting Rhizobacteria on Potato Plant Development and Yield,” Phytopathology 70:1078-82 (1980), Kloepper et al., “Emergence-Promoting Rhizobacteria: Description and Implications for Agriculture,” pp. 155-164, Iron, Siderophores, and Plant Disease, T. R. Swinburne, ed. Plenum, New York (1986), and Kloepper et al., “Relationship of in vitro Antibiosis of Plant Growth-Promoting Rhizobacteria to Plant Growth and the Displacement of Root Microflora,” Phytopathology 71:1020-24 (1981))。幾つかの研究では、植物の毛状体はPGPRを用いて改良された(Tipping et al., “Development of Emergence-Promoting Rhizobacteria for Supersweet Corn,” Phytopathology 76:938-41 (1990) (abstract) and Kloepper et al., “Emergence-Promoting Rhizobacteria: Description and Implications for Agriculture,” pp. 155-164, Iron, Siderophores, and Plant Disease, T. R. Swinburne, ed. Plenum, New York (1986))。他の多くの研究は、植物の病原体の生態的防除のために、rhizobacteriaの処置によって植物の健康を改善することを示している(B. Schippers, “Biological Control of Pathogens With Rhizobacteria,” Phil. Trans. R. Soc. Lond. B. 318:283-93 (1988), Schroth et al., “Disease-Suppressive Soil and Root-Colonizing Bacteria,” Science 216:1376-81 (1982), Stutz et al., “Naturally Occurring Fluorescent Pseudomonads Involved in Suppression of Black Root Rot of Tobacco,” Phytopathology 76:181-85 (1986), and D. M. Weller, “Biological Control of Soilborne Plant Pathogens in the Rhizosphere With Bacteria,” Annu. Rev. Phytopathol. 26:379-407 (1988))。
【0008】
植物の病原体誘発性免疫(pathogen-induced immunization)により、成長が促進することが見出された。タバコ木質部への外からのPeronospora tabacinaの感染は、発育阻害を緩和するばかりでなく、成長および生育を促進する。温室および畑の両方の実験において、免疫化したタバコは、約40%高さが高くなり、乾燥重量が40%増加し、生体重が30%増加し、対照植物よりも葉が4-6枚多くなった(Tuzun, S., et al., “The Effect of Stem Injection with Peronospora tabacina and Metalaxyl Treatment on Growth of Tobacco and Protection Against Blue Mould in the Field,” Phytopathology, 74:804 (1984))。これらの植物は対照植物よりも約2-3週間早く花が咲いた(Tuzun, S., et al., “Movement of a Factor in Tobacco Infected with Peronospora tabacina Adam which Systemically Protects Against Blue Mould,” Physiological Plant Pathology, 26:321-30 (1985))。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は従来の植物の成長の促進方法を超える改良を示している。
【課題を解決するための手段】
【0010】
本発明の要旨
本発明は植物の成長を促進する方法に関する。この方法には、植物または植物種子に、植物または植物種子から成育した植物の成長の促進する条件下で、非感染型で、過敏反応誘発ポリペプチドまたはタンパク質を施用することを含む。
【0011】
植物の成長を促進するために過敏反応誘発ポリペプチドまたはタンパク質を植物または植物種子に、または種子から成長した植物に施用することの変法として、トランスジェニック植物または植物種子が使用され得る。トランスジェニック植物を用いるとき、これには過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物を提供すること、および成長を促進するDNA分子が有効に用いられる条件下で植物を生育させること含む。他には、過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物種子が提供され、土壌に植えることができる。そして植物は、成長を促進するDNA分子の使用に有効な条件下、植えた種子から生育する。
【0012】
本発明は植物の成長を高めまたは促進する何れの型にも有効となることを目的とする。これは、種子から植物の生長が始まる時のように早期にまたは植物の寿命の後期に行い得る。例えば、本発明の植物の成長には、収穫増加、生産された種子の量の増加、発芽する種子の割合の増加、植物の大きさの増進、より多くのバイオマス、より多くの果実およびより大きな果実、より早い果実の着色、ならびにより早い果実および植物の成熟を包含する。結果として、本発明は栽培者にとって意義ある経済的利益を提供する。例えば、早い発芽および早い成熟によって、作物は、成長不適季節であるためにその成長が妨げられる土地でも成長させ得る。種子の発芽割合の増加の結果、作物の植生の改良やより有効な種子の使用がもたらされる。増加した収穫、大きさの増進、およびバイオマス生産の促進により、与えられた土地の区画からより大きな収入を生み出すことになる。そして本発明は、農業効率において重要な利点を構成していることは明らかである。
【図面の簡単な説明】
【0013】
【図1】プラスミドベクターpCPP2139の地図であり、Erwinia amylovora過敏反応誘発物遺伝子を含んでいる。
【図2】プラスミドベクターpCPP50の地図であり、Erwinia amylovora過敏反応誘発物遺伝子は含んではいないが、それ以外図1に示したプラスミドベクターpCPP2139と同じである。Masui, et al., Bio/Technology 2:81-85 (1984)(それは、ここで引用することによりこの明細書の一部に加える)参照。
【発明を実施するための形態】
【0014】
本発明の詳細な説明
本発明は植物の成長を促進する方法に関する。この方法には、植物または植物種子の全てまたは一部に、植物または植物種子から成育した植物の成長の促進に有効な条件下、非感染型で、過敏反応誘発ポリペプチドまたはタンパク質を施用することを含む。他に、植物は種子を作成するためにこの方法で処理され得、その種子を植えたとき、子孫植物の成長は促進される。
【0015】
植物の成長を促進するために過敏反応誘発ポリペプチドまたはタンパク質を植物または植物種子に、または種子から成長した植物に施用することの変法として、トランスジェニック植物または植物種子が使用され得る。トランスジェニック植物を用いるとき、これには過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物を提供すること、および成長を促進するDNA分子が有効に用いられる条件下で植物を生育させること含む。他には、過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物種子が提供され、土壌に植えることができる。そして植物は、成長を促進するDNA分子の使用に有効な条件下、植えた種子から生育する。
【0016】
本発明で使用する過敏反応誘発ポリペプチドまたはタンパク質は、多くの種類の真菌性および細菌性病原体より誘導される過敏反応誘発ポリペプチドまたはタンパク質に相当し得る。そのようなポリペプチドまたはタンパク質は、誘発物と接触した植物組織において局所的な壊死を誘発し得る。
【0017】
ポリペプチドまたはタンパク質誘発物の適当な細菌性源の例には、Erwinia、Pseudomonas、およびXanthomonas種(例えば以下の細菌:Erwinia amylovora、Erwinia chrysanthemi、Erwinia stewartii、Erwinia carotovora、Pseudomonas syringae、Pseudomonas solanacearum、Xanthomonas campestris、およびそれらの混合)を含む。
【0018】
過敏反応誘発タンパク質またはポリペプチドの適当な真菌性源の例は、Phytophthoraである。Phytophthoraの適当な種には、Phytophthora pythium、Phytophthora cryptogea、Phytophthora cinnamomi、Phytophthora capsici、Phytophthora megasperma、およびPhytophthora citrophthoraが含まれる。
【0019】
過敏反応誘発ポリペプチドまたはタンパク質が植物または植物種子に施用される本発明の実施態様は、多くの方法で実施され得、1)単離した誘発ポリペプチドまたはタンパク質の施用;2)疾病の原因とはならない、過敏反応誘発ポリペプチドまたはタンパク質をコードしている遺伝子で形質転換された細菌の施用;および3)幾つかの植物種では疾病の原因となる(しかしそれらは施用される植物ではない)、天然に過敏反応誘発ポリペプチドまたはタンパク質をコードしている遺伝子を含む細菌の施用を含む。加えて、本発明の種子は、本発明に従って過敏反応誘発ポリペプチドまたはタンパク質で処理された植物から回収し得る。
【0020】
本発明のある実施態様では、過敏反応誘発ポリペプチドまたはタンパク質は、その相当する生物から単離され得、植物または植物種子に施用され得る。そういった単離方法は既知であり、Arlat, M., F. Van Gijsegem, J. C. Huet, J.C. Pemollet, and C. A. Boucher, “PopA1, a Protein which Induces a Hypersensitive-like Response in Specific Petunia Genotypes is Secreted via the Hrp Pathway of Pseudomonas solanacearum,” EMBO J. 13:543-553 (1994); He, S. Y., H. C. Huang, and A. Collmer, “Pseudomonas syringae pv. syringae HarpinPss:a Protein that is Secreted via the Hrp Pathway and Elicits the Hypersensitive Response in Plants,” Cell 73:1255-1266 (1993);およびWei, Z.-M., R. J. Laby, C. H. Zumoff, D. W. Bauer, S.-Y. He, A. Collmer, and S. V. Beer, “Harpin Elicitor of the Hypersensitive Response Produced by the Plant Pathogen Erwinia amylovora,” Science 257:85-88 (1992)(それらは、ここで引用することによりこの明細書の一部に加える)に記載されている。また仮米国特許出願番号08/200,024および08/062,024(それらは、ここで引用することによりこの明細書の一部に加える)参照。好ましくは、しかしながら、本発明の単離した過敏反応誘発ポリペプチドまたはタンパク質は、下記のように組換えで作成され、精製される。
【0021】
本発明の他の実施態様では、過敏反応誘発ポリペプチドまたはタンパク質は、過敏反応誘発ポリペプチドまたはタンパク質をコードする遺伝子を含む細菌を施用することにより種子または植物種子に施用し得る。そういった細菌は、誘発物が植物または植物種子の細胞に接触できるようにポリペプチドまたはタンパク質を分泌し、排出できなければならない。これらの実施態様では、過敏反応誘発ポリペプチドまたはタンパク質は植物または種子において細菌によって、または該細菌が植物または植物種子に導入される直前に産生される。
【0022】
本発明の細菌施用方法の1つの実施態様では、細菌は疾病の原因とはならず、過敏反応誘発ポリペプチドまたはタンパク質をコードする遺伝子で形質転換されている(例えば組換え)。例えば、植物に過敏反応を誘発しないE. coliは、過敏反応誘発ポリペプチドまたはタンパク質をコードしている遺伝子で形質転換され得、そして植物に施用され得る。E. coli以外の細菌種はまた、本発明のこの実施態様において使用し得る。
【0023】
本発明の細菌施用方法の他の実施態様では、細菌は疾病の原因となり、天然に過敏反応誘発ポリペプチドまたはタンパク質をコードしている遺伝子を含んでいる。そのような細菌の例は上記した通りである。しかしながら、この実施態様では、細菌が運ぶ疾病にかかりにくい植物またはそれらの種子に、これらの細菌を施用する。例えば、Erwinia amylovoraはリンゴまたは洋ナシでは疾病の原因となるが、トマトではならない。しかしながら、そういった細菌は、トマトにおいて過敏反応を誘発する。それゆえ、本発明のこの実施態様に従い、Erwinia amylovoraはトマトまたはその種子に施用され、その種において疾病の原因とはならずに成長を促進し得る。
【0024】
Erwinia chrysanthemi由来の過敏反応誘発ポリペプチドまたはタンパク質は、配列番号1に相当するアミノ酸配列を有し、その配列を以下に示す。
【化1】
【0025】
この過敏反応誘発ポリペプチドまたはタンパク質は分子量34kDaであり、熱に安定であり、グリシン含有は16%を超え、そして事実上システインが含まれていない。Erwinia chrysanthemi過敏反応誘発ポリペプチドまたはタンパク質は、配列番号2に相当するヌクレオチド配列を有するDNA分子によりコードされており、その配列を以下に示す。
【化2】
【化3】
【化4】
【0026】
Erwinia amylovoraから誘導された過敏反応誘発ポリペプチドまたはタンパク質は、配列番号3に相当するアミノ酸配列を有し、以下にその配列を示す。
【化5】
【化6】
【0027】
この過敏反応誘発ポリペプチドまたはタンパク質は、分子量39kDaであり、約pI4.3であり、そして100℃で少なくとも10分間加熱しても安定である。この過敏反応誘発ポリペプチドまたはタンパク質には事実上システインが含まれていない。Erwinia amylovoraから誘導された過敏反応誘発ポリペプチドまたはタンパク質は、さらにWei, Z.-M., R. J. Laby, C. H. Zumoff, D. W. Bauer, S.-Y. He, A. Collmer, and S. V. Beer, “Harpin, Elicitor of the Hypersensitive Response Produced by the Plant Pathogen Erwinia amylovora,” Science 257:85-88 (1992)(それは、ここで引用することによりこの明細書の一部に加える)に十分に記載されている。このポリペプチドまたはタンパク質をコードしているDNA分子は、配列番号4に相当するヌクレオチド配列を有し、それを以下に示す。
【化7】
【0028】
Pseudomonas syringaeから誘導された過敏反応誘発ポリペプチドまたはタンパク質は、配列番号5に相当するアミノ酸配列を有し、その配列を以下に示す。
【化8】
【0029】
この過敏反応誘発ポリペプチドまたはタンパク質は、分子量34-35kDaである。それはグリシンが豊富であり(約13.5%)、システインおよびチロシンを含まない。Pseudomonas syringaeから誘導される過敏反応誘発ポリペプチドまたはタンパク質についての更なる情報は、He, S. Y., H. C. Huang, and A. Collmer, “Pseudomonas syringae pv. syringae HarpinPss: a Protein that is Secreted via the Hrp Pathway and Elicits the Hypersensitive Response in Plants,” Cell 73:1255-1266 (1993)(それは、ここで引用することによりこの明細書の一部に加える)に見られる。過敏反応誘発物をコードするDNA分子は、配列番号6に相当するヌクレオチド配列を有し、それらを以下に示す。
【化9】
【0030】
Pseudomonas solanacearumから誘導された過敏反応誘発ポリペプチドまたはタンパク質は、配列番号7に相当するアミノ酸配列を有し、それを以下に示す。
【化10】
【0031】
それは配列番号8に相当するヌクレオチド配列を有するDNA分子によりコードされており、それを以下に示す。
【化11】
【0032】
Pseudomonas solanacearumから誘導された過敏反応誘発ポリペプチドまたはタンパク質に関する更なる情報は、Arlat, M., F. Van Gijsegem, J. C. Huet, J. C. Pemollet, and C. A. Boucher, “PopA1, a Protein which Induces a Hypersensitive-like Response in Specific Petunia Genotypes, is Secreted via the Hrp Pathway of Pseudomonas solanacearum,” EMBO J. 13:543-533 (1994)(それは、ここで引用することによりこの明細書の一部に加える)に示されている。
【0033】
Xanthomonas campestris pv. glycines由来の過敏反応誘発ポリペプチドまたはタンパク質は、配列番号9に相当するアミノ酸配列を有し、それを以下に示す。
【化12】
【0034】
この配列は、Xanthomonas campestris pv. glycinesの過敏反応誘発ポリペプチドまたはタンパク質のみからなる26残基のアミノ末端配列である。それは、他のXanthomonas campestris pathovarsにおいて決定されているふさ状サブユニットタンパク質(fimbrial subunit protein)に対合する(match)。
【0035】
Xanthomonas campestris pv. pelargonii由来の過敏反応誘発ポリペプチドまたはタンパク質は、熱に安定であり、プロテアーゼ感受性であり、分子量20kDaである。配列番号10に相当するアミノ酸配列を含み、それを以下に示す。
【化13】
【0036】
Erwinia carotovoraの過敏反応誘発ポリペプチドまたはタンパク質は、Cui et al., “The RsmA Mutants of Erwinia carotovora subsp. carotovora Strain Ecc71 Overexpress hrp NEcc and Elicit a Hypersensitive Reaction-like Response in Tobacco Leaves,” MPMI, 9(7):565-73 (1996)(それは、ここで引用することによりこの明細書の一部に加える)に記載されている。過敏反応誘発ポリペプチドまたはタンパク質は、Ahmad et al., “Harpin is Not Necessary for the Pathogenicity of Erwinia stewartii on Maize,” 8th Int'l. Cong. Molec. Plant-Microbe Interact., July 14-19, 1996 およびAhmad, et al., “Harpin is Not Necessary for the Pathogenicity of Erwinia stewartii on Maize,” Ann. Mtg. Am. Phytopath. Soc., July 27-31, 1996 (それらは、ここで引用することによりこの明細書の一部に加える)によって示されている。
【0037】
Phytophthora parasitica、Phytophthora cryptogea、Phytophthora cinnamoni、Phytophthora capsici、Phytophthora megasperma、およびPhytophthora citrophthora由来の過敏反応誘発ポリペプチドまたはタンパク質は、Kaman, et al., “Extracellular Protein Elicitors from Phytophthora: Most Specificity and Induction of Resistance to Bacterial and Fungal Phytopathogens,” Molec. Plant-Microbe Interact., 6(1):15-25 (1993), Ricci et al., “Structure and Activity of Proteins from Pathogenic Fungi Phytophthora Eliciting Necrosis and Acquired Resistance in Tobacco,” Eur. J. Biochem., 183:555-63 (1989)、Ricci et al., “Differential Production of Parasiticein, and Elicitor of Necrosis and Resistance in Tobacco, by Isolates of Phytophthora parasitica,” Plant Path. 41:298-307 (1992)、Baillreul et al., “A New Elicitor of the Hypersensitive Response in Tobacco: A Fungal Glycoprotein Elicits Cell Death, Expression of Defence Genes, Production of Salicylic Acid, and Induction of Systemic Acquired Resistance,” Plant J., 8(4):551-60 (1995)、およびBonnet et al., “Acquired Resistance Triggered by Elicitors in Tobacco and Other Plants,” Eur. J. Plant Path., 102:181-92 (1996)(それらは、ここで引用することによりこの明細書の一部に加える)に記載されている。
【0038】
上記誘発物は典型的なものである。他の誘発物は、誘発物をコードする遺伝子が発現する条件下、過敏反応を誘発する増殖真菌および細菌により同定され得る。培養上清の細胞フリー調製(Cell-free preparation)は、適当な植物組織への浸透にそれを用いることにより、誘発物活性(すなわち、局所的なネクローシス)の試験に用い得る。
【0039】
上記過敏反応誘発ポリペプチドまたはタンパク質のフラグメントおよび他の病原体の誘発物の全フラグメントを、本発明の方法で使用することもできる。
【0040】
適当なフラグメントは、幾つかの手段で作成され得る。最初に、既知誘発タンパク質をコードする遺伝子のサブクローンを、遺伝子フラグメントをサブクローニングする通常の分子遺伝学的手法によって作成する。そしてそのサブクローンが、インビトロまたはインビボの細菌細胞において発現すると、下記方法の誘発活性の試験を行い得る、より小さなタンパク質またはペプチドが得られる。
【0041】
他には、誘発タンパク質のフラグメントは、キモトリプシンまたはStaphylococcusプロテイナーゼA、またはトリプシンのようなタンパク質分解酵素で全長の誘発タンパク質を消化することにより作成され得る。異なるタンパク質分解酵素は、誘発タンパク質のアミノ酸配列に基き、異なる部位で誘発タンパク質を切断するのに適当である。タンパク質分解によって得られるフラグメントの幾つかは、活性な耐性誘発物でもある。
【0042】
タンパク質の1次構造の情報に基く、他のアプローチにおいて、誘発タンパク質遺伝子のフラグメントは、タンパク質の特定部位を選択した特定のプライマーの組み合わせと共に、PCR技術を用いて合成され得る。そして、これらは、トランケート(truncate)したペプチドまたはタンパク質の増殖および発現のために適当なベクターにクローニングされる。
【0043】
化学合成はまた、適当なフラグメントを作成するために使用され得る。そういった合成は、誘発物を産生するために既知のアミノ酸配列を用い実施される。他に、全長の誘発物を高温高圧にさらすとフラグメントが生ずる。そして、これらのフラグメントは、通常の方法により分離され得る(例えばクロマトグラフィー、SDS-PAGE)。
【0044】
有用なフラグメントの例は、Pseudomonas solanacearumの過敏反応誘発ポリペプチドまたはタンパク質のpopA1フラグメントである。Arlat, M., F. Van Gijsegem, J. C. Huet, J. C. Pemollet, and C. A. Boucher, “PopA1, a Protein Which Induces a Hypersensitive-like Response in Specific Petunia Genotypes is Secreted via the Hrp Pathway of Pseudomonas solanacearum,” EMBO J. 13:543-53 (1994)(それは、ここで引用することによりこの明細書の一部に加える)参照。Erwinia amylovoraに関して、適当なフラグメントは、例えば、配列番号3のアミノ酸1〜98の間に存在し、またはそれを含むポリペプチドと、配列番号3のアミノ酸137〜204の間に存在し、またはそれを含むポリペプチドの何れかであるか、または両方である。
【0045】
変異体はまた(あるいは別に)、ポリペプチドの性質、2次構造およびヒドロパシー性に最小限の影響しか与えずに、例えばアミノ酸の欠損または付加により修飾され得る。例えば、ポリペプチドは、タンパク質のN-末端でシグナル配列(またはリーダー配列)と連結し、共翻訳的(co-translationally)にまたは後翻訳的(post-translationally)にそのタンパク質の移動を指図し得る。ポリペプチドはまた、ポリペプチドの合成、精製または同定を容易にするためにリンカーまたは他の配列に連結し得る。
【0046】
本発明のタンパク質またはポリペプチドは、好ましくは通常技術によって精製された形(好ましくは少なくとも約60%、更に好ましくは80%の純度)で産生される。典型的に、本発明のタンパク質またはポリペプチドは産生されるが、組換え宿主細胞の増殖培地には分泌されない。他には、本発明のタンパク質またはポリペプチドは増殖培地に分泌する。分泌しないタンパク質の場合、タンパク質を単離するために、組換えプラスミドを有する宿主細胞(例えばE.coli)を増殖させ、超音波、加熱、化学的処置により溶解し、ホモジネートを遠心分離し、細菌の破壊物を除去する。そして上清を加熱処理し、過敏反応誘発タンパク質を遠心分離により分離する。本発明のポリペプチドまたはタンパク質を含む上清画分を、タンパク質分離のために適当な大きさのデキストランまたはポリアクリルアミドカラムでゲル濾過する。必要ならば、タンパク質画分は更にイオン交換またはHPLCにより精製し得る。
【0047】
過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子は、通常の組換えDNA技術を用いて細胞内に組込まれ得る。通常、これには、DNA分子が異質である(即ち、通常は存在しない)発現システムにDNA分子を組み入れることが含まれる。異種性DNA分子を適当なセンス方向および正しいリーディンフレームで発現システムまたはベクターに組込む。ベクターには挿入したタンパク質コーディング配列の転写および翻訳に必要なエレメントが含まれている。
【0048】
CohenおよびBoyerの米国特許番号4,237,224(それは、ここで引用することによりこの明細書の一部に加える)には、制限酵素の切断およびDNAリガーゼでの連結を用いた組換えプラスミドの形の発現システムの産生について記載されている。そして、これら組換えプラスミドは、形質転換法により組込まれ、原核生物および組織培養で増殖した真核細胞を含む単細胞培養中で複製される。
【0049】
組換え遺伝子はまた、ワクシニアウイルスのようなウイルスに導入され得る。組換えウイルスは、ウイルスに感染した細胞にプラスミドをトランスフェクトすることにより生じ得る。
【0050】
適当なベクターには、ラムダベクターシステムgt11、gt WES.tB、Charon 4のようなウイルスベクターおよびpBR322、pBR325、pACYC177、pACYC1084、pUC8、pUC9、pUC18、pUC19、pLG339、pR290、pKC37、pKC101、SV40、pBluescript II SK +/- またはKS +/-(Stratagene、La Jolla, Califの“Stratagene Cloning System” Catalog (1993)参照(それらは、ここで引用することによりこの明細書の一部に加える))、pQE、pIH821、pGEX、pETシリーズ(F. W. Studier et al., “Use of T7 RNA Polymerase to Direct Expression of Cloned genes,” Gene Expression Technology vol. 185 (1990)(それらは、ここで引用することによりこの明細書の一部に加える)参照)、およびそれらの誘導体を含むが、これらに限定されない。組換え分子は、形質転換、特にトランスダクション、接合、動員、またはエレクトロポレーションで細胞に導入され得る。DNA配列は、Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Springs Laboratory, Cold Springs Harbor, New York (1989)(それらは、ここで引用することによりこの明細書の一部に加える)に記載されている通りに、当分野の標準的クローニング法を用いてベクターにクローニングされる。
【0051】
様々な宿主ベクターシステムが使用され、タンパク質をコードする配列を発現し得る。第1に、ベクターシステムは、使用する宿主細胞に和合性がなければならない。宿主ベクターシステムには、バクテリオファージDNA、プラスミドDNAまたはコスミドDNAで形質転換した細菌;酵母ベクターを有する酵母のような微生物;ウイルス感染した哺乳類細胞システム(例えばワクシニアウイルス、アデノウイルスなど);ウイルス感染した昆虫細胞システム(例えばバキュロウイルスなど);および細菌に感染した植物細胞が含まれるが、これらに限らない。これらベクターの発現エレメントは、それらの威力および特異性が異なる。使用する宿主ベクターシステムに依存して、多くの適当な転写および翻訳エレメントのうち何れかを用い得る。
【0052】
異なる遺伝学的シグナルおよびプロセッシング事象(processing event)により多くの遺伝子発現レベルが制御されている(例えばDNA転写およびメッセンジャーRNA(mRNA)翻訳)。
【0053】
DNAの転写は、RNAポリメラーゼの結合を指示し、mRNA合成を促進するDNA配列であるプロモーターの存在に依存する。真核生物のプロモーターのDNA配列は、原核生物のプロモーターとは異なる。更に、真核生物のプロモーターおよび付随する遺伝学的シグナルは、原核生物のシステム中では認識されず、または機能せず、更に原核生物のプロモーターは、真核性細胞の中では認識されず、機能しない。
【0054】
同様に、原核生物内でのmRNAの翻訳は、真核生物とは異なる適当な原核生物のシグナルの存在に依存する。原核生物におけるmRNAの翻訳を効率よくするには、mRNAにあるシャイン・ダルガルノ(“SD”)配列と呼ばれるリボゾーム結合部位が必要である。この配列は、mRNAの短いヌクレオチド配列であり、タンパク質のアミノ末端のメチオニンをコードする開始コドン(通常はAUG)の上流に位置する。SD配列は、16S rRNA(リボゾームRNA)の3'端と相補的であり、おそらく、rRNAと2本鎖となることによりmRNAとリボゾームとの結合を促進し、リボゾームを正確に位置させる。遺伝子発現を最大とする検討には、Roberts and Lauer, Methods in Enzymology, 68:473(1979)(それは、ここで引用することによりこの明細書の一部に加える)を参照のこと。
【0055】
プロモーターにより、“強さ”(即ち、転写を促進する能力)が変化する。クローニングした遺伝子を発現するという目的に対し、高レベルの転写、そして遺伝子の発現を得るためには、強力なプロモーターを用いることが望ましい。用いる宿主細胞システムに依存して、多くの適当なプロモーターのうちの何れかを使用し得る。例えば、E.coliでクローニングするとき、バクテリオファージ、またはプラスミド、T7ファージプロモーター、lacプロモーター、trpプロモーター、recAプロモーター、リボゾームRNAプロモーター、コリファージラムダおよび他のPRおよびPLプロモーター(lacUV5、ompF、bla、lppおよびその他が含まれるがこれらに限らない)といったプロモーターなどが、近接したDNAセグメントの転写を高レベルにするために使用し得る。加えて、組換えDNAまたは他のDNA合成技術により作成される、ハイブリッドtrp-lacUV5(tac)プロモーターまたは他のE.coliプロモーターが使用され得、挿入遺伝子を転写する。
【0056】
細菌性宿主細胞株および発現ベクターは、特別に誘導しない限りプロモーターの作動を抑制するように選択する。ある操作では、特定の誘発物の添加は、挿入遺伝子を効率よく転写するために必要となる。例えば、lacオペロンはラクトースまたはIPTG(イソプロピルチオ-β-D-ガラクトシド)の添加により誘発される。trp、pro等といった、様々な他のオペロンは他の異なる制御下に置かれる。
【0057】
特定の開始シグナルはまた、原核細胞中において効率よく転写および翻訳するために必要である。これらの転写および翻訳開始シグナルは、“強さ”が変化し得、遺伝子に特異なメッセンジャーRNAおよび合成されたタンパク質、それぞれの量により測定され得る。プロモーターを含むDNA発現ベクターにはまた、様々な“強力な”転写および/または翻訳開始シグナルの何れかの組み合わせが含まれている。例えば、E.coliにおいて翻訳の効率をよくするには、開始コドン(ATG)の約7-9塩基5'側にSD配列が必要であり、リボゾーム結合部位を供与することが必要である。それゆえ宿主細胞のリボゾームにより使用され得る何れかのSD-ATG組み合わせが用いられ得る。そういった組み合わせには、cro遺伝子もしくはコリファージラムダのN遺伝子のSD-ATG組合せ、またはE.coliトリプトファンE、D、C、BもしくはA遺伝子のSD-ATG組合せが含まれるが、これらに限らない。加えて、組換えDNAまたは合成ヌクレオチドの組み込みを含む他の技術により製造された何れかのSD-ATG組合わせが使用され得る。
【0058】
過敏反応誘発ポリペプチドまたはタンパク質をコードする単離DNA分子を1度発現システムにクローニングすれば、宿主細胞に容易に組込まれる。そのような組込みは、上記様々な型の形質転換で実施され得、それはベクター/宿主細胞システムに依存する。適当な宿主細胞には、細菌、ウイルス、酵母、哺乳類細胞、昆虫、植物等が含まれるが、これらに限らない。
【0059】
本発明の方法は、広く多様な植物またはその種子の成長を促進する処理に使用し得る。適当な植物には双子葉植物および単子葉植物が含まれる。更に特に有用な作物には、コメ、小麦、大麦、ライ麦、ワタ、ヒマワリ、ラッカセイ、トウモロコシ、ジャガイモ、サツマイモ、マメ、エンドウ、チコリー、レタス、エンダイブ、キャベツ、カリフラワー、ブロッコリー、カブ、ハツカダイコン、ホウレンソウ、タマネギ、ニンニク、ナス、トウガラシ、セロリ、ニンジン、カボチャ、ペポカボチャ、ズッキーニ、キュウリ、リンゴ、洋ナシ、メロン、イチゴ、ブドウ、ラズベリー、パイナップル、ダイズ、タバコ、トマト、サトウモロコシ、およびサトウキビが含まれ得る。適当な観賞用植物には、バラ、セントポーリア、ペチュニア、ペラルゴニューム、ポインセチア、キク、カーネーションおよびヒャクニチソウが含まれる。
【0060】
過敏反応誘発ポリペプチドまたはタンパク質の施用を含む本発明の方法は、植物の全てまたは一部(葉、茎、根等を含む)を処理するときに様々な方法で実施され得る。これは、植物内への過敏反応誘発ポリペプチドまたはタンパク質の浸透を含む(しかし必要ではない)。適当な施用方法には、局所的施用(例えば高または低圧噴霧(pressure spraying))、注入、散布、および誘発物の施用とほぼ同時の葉の表皮剥脱が含まれる。本発明の施用実施態様に従い、植物種子を処理するとき、過敏反応誘発ポリペプチドまたはタンパク質は、局所的施用(例えば高または低圧噴霧)、塗布、浸透、散布または注入により施用され得る。他の適当な施用方法は、過敏反応誘発ポリペプチドまたはタンパク質と植物細胞または植物種子を効果的に接触させることのできる当業者によって構想され得る。本発明の過敏反応誘発物で1度処理すると、種子は天然または農業用土壌に植えることができ、通常の方法を用いて栽培し植物を生産する。本発明に従って処理した種子から植物が生育し、その後植物は成長を促進するために1度またはそれ以上の施用により処理され得る。そういった成長した植物は、次に、成長を促進し得る植物を作成し得る種子または珠芽(例えば切枝(cutting))の作成に有用となり得る。
【0061】
本発明の過敏反応誘発ポリペプチドまたはタンパク質は、単独で、または他の物質との混合物で植物または植物種子に施用し得る。他に、過敏反応誘発ポリペプチドまたはタンパク質は、異なったときに施用される他の物質と共に、別々に植物に施用し得る。
【0062】
本発明の施用実施態様に従う、植物または植物種子の処理に適当な組成物は、担体中の過敏反応誘発ポリペプチドまたはタンパク質を含む。適当な担体は、水、水溶液、スラリーまたは乾燥粉末を含む。この実施態様において、組成物には、0.5nMを超える過敏反応誘発ポリペプチドまたはタンパク質が含まれる。
【0063】
必ずしも必要ではないけれども、この組成物には、肥料、殺虫剤、殺菌剤、殺線虫剤、除草剤、およびそれらの混合物を含む付加添加物が含まれる。適当な肥料には、(NH4)2NO3が含まれている。適当な殺虫剤の例にはマラチオンがある。有用な殺菌剤にはキャプタンが含まれる。
【0064】
他の適当な添加物は、緩衝剤、湿性剤、塗布剤、および剥離剤(abrading agent)を含む。これら物質は本発明の方法を容易にするのに使用し得る。加えて、過敏反応誘発ポリペプチドまたはタンパク質は、土およびポリサッカライドを含む他の慣用の種子用製剤または処理剤と共に、植物種子に施用し得る。
【0065】
トランスジェニック植物およびトランスジェニック種子の使用を含む本発明の他の実施態様において、過敏反応誘発ポリペプチドまたはタンパク質が植物または種子に局所的方法で施用される必要はない。代わりに、過敏反応誘発ポリペプチドまたはタンパク質をコードするDNA分子で形質転換されたトランスジェニック植物は、微粒子銃またはAgrobacteriumを介する形質転換による方法といった当分野に既知の方法で作成される。適当な過敏反応誘発ポリペプチドまたはタンパク質およびそれらをコードするDNAの核酸配列の例は、上記に開示している。この型のトランスジェニック植物を1度作成すると、植物自身は、植物の成長を促進する過敏反応誘発物をコードする遺伝子を伴い、通常の方法によって生育され得る。他に、トランスジェニック種子はトランスジェニック植物から回収される。そしてこれら種子は土壌に植えられ、通常の方法を用い生育され、トランスジェニック植物となる。トランスジェニック植物は、成長の促進に効果的な条件下、植えられたトランスジェニック種子から生育する。理論によって縛られることは望まないが、そういった成長促進は、RNAを媒介するか、または誘発ポリペプチドまたはタンパク質の発現の結果として生じ得る。
【0066】
トランスジェニック植物および植物種子を本発明に従い使用したときでも、過敏反応誘発ポリペプチドまたはタンパク質を施用する植物および種子の処理に使用するのと同じ物質でさらにそれらは処理され得る。過敏反応誘発物を含む、これら他の物質は、高または低圧噴霧、注入、塗布、散布および浸透を含む上記方法によりトランスジェニック植物および植物種子に施用され得る。同様に、トランスジェニック植物種子から植物が生育すると、植物の成長を促進する過敏反応誘発物の一つまたはそれ以上の施用により処理され得る。そういった植物はまた、通常の植物処理剤(例えば殺虫剤、肥料等)で処理され得る。本発明のトランスジェニック植物は、成長の促進が可能な植物を作成する種子または珠芽(例えば切枝)の作成に有用である。
【実施例】
【0067】
実施例
実施例1−トマト種子のErwinia amylovora過敏反応誘発物処理による発芽率への作用
Marglobeトマト種の種子をErwinia amylovora過敏反応誘発物溶液(“ハルピン”)40mlに浸した。ハルピンは、プラスミドpCPP2139(図1参照)を含むE. coli株DH5を生育させ、超音波処理により細胞を融解し、沸騰水に5分間入れることにより熱処理し、続いて遠心して細胞残骸を除去し、タンパク質および他の熱不安定成分を除去して製造した。得られた沈殿(“CFEP”)を連続希釈した。これらの希釈(1:40、1:80、1:160、1:320、1:640)は、ウェスタンブロット分析を基本にしてそれぞれ20、10、5、2.5および1.25μgm/mlのハルピンを含んだ。種子をビーカー中のハルピンまたは緩衝液に日0に24時間、28℃の成育チャンバーで浸した。浸漬後、日1に種子を人工土壌の発芽用鉢に蒔いた。この方法を処理当たり100個の種子で行った。
【0068】
処理:
1.ハルピン中の種子(1:40)(20μgm/ml)。
2.ハルピン中の種子(1:80)(10μgm/ml)。
3.ハルピン中の種子(1:160)(5μgm/ml)。
4.ハルピン中の種子(1:320)(2.5μgm/ml)。
5.ハルピン中の種子(1:640)(1.25μgm/ml)。
6.緩衝液中の種子(5mM KPO4、pH6.8)。
【0069】
【表1】
【0070】
表1に示されるように、トマト種子のErwinia amylovora過敏反応誘発物での処理は、発芽に必要な時間を減らし、発芽の割合を非常に増加させた。
【0071】
実施例2−トマト種子のErwinia amylovora過敏反応誘発物処理によるトマト植物の高さへの作用
Marglobeトマト種の種子をビーカー中のハルピン(1:15、1:30、1:60および1:120)または緩衝液に、日0に24時間、28℃の成育チャンバーで浸した。浸漬後、日1に種子を人工土壌の発芽用鉢に蒔いた。
処理当たり10本の同一な見かけの植物を無作為に選択して測定した。苗木を土壌表面から植物の頂部まで定規で計った。
【0072】
処理:
1.ハルピン中(1:15)(52μgm/ml)。
2.ハルピン中(1:30)(26μgm/ml)。
3.ハルピン中(1:60)(13μgm/ml)。
4.ハルピン中(1:120)(6.5μgm/ml)。
5.緩衝液中(5mM KPO4、pH6.8)。
【0073】
【表2】
【0074】
【表3】
【0075】
【表4】
【0076】
表2−5に示されるように、トマト種子のErwinia amylovora過敏反応誘発物での処理は、植物の生育を促進させた。1:30希釈は最大の効果を有した---苗木の高さで16%増加。
【0077】
実施例3−トマト植物のErwinia amylovora過敏反応誘発物処理によるトマト植物の高さへの作用
Marglobeトマト植物が4週齢の時、それらに13μgm/ml(1:60)または8.7μgm/ml(1:90)のハルピンまたは緩衝液(5mM KPO4)を、6ml/植物で28℃の成育チャンバーで噴霧した。トマト植物の高さはハルピン噴霧後2週間(6週齢トマト植物)および噴霧2週間と5日後に測定した。処理当たり10本の同一な見かけの植物を無作為に選択して測定した。苗木を土壌表面から植物の頂部まで定規で計った。
【0078】
処理:
1.ハルピン(1:60)(13μgm/ml)。
2.ハルピン(1:90)(8.7μgm/ml)。
3.緩衝液(5mM KPO4、pH6.8)。
【0079】
【表5】
【0080】
表6に示されるように、トマト苗木へのErwinia amylovora過敏反応誘発物噴霧は、トマト植物の生育を促進させることができる。緩衝液処理コントロールと比較して、試験した二つの用量の過敏反応誘発物に関して同様の促進が示された。
【0081】
実施例4−トマト種子のErwinia amylovora過敏反応誘発物処理によるトマト植物の高さへの作用
Marglobeトマトの種子を、ビーカー内のErwinia amylovora過敏反応誘発物(“ハルピン”)(1:40、1:80、1:160、1:320および1:640)または緩衝液に、日0に24時間、28℃の成育チャンバーで浸した。種子をハルピンまたは緩衝液に浸漬後、日1に種子を人工土壌の発芽用鉢に蒔いた。処理当たり10本の同一な見かけの植物を無作為に選択して測定した。苗木を土壌表面から植物の頂部まで定規で計った。
【0082】
処理:
1.ハルピン(1:40)(20μgm/ml)。
2.ハルピン(1:80)(10μgm/ml)。
3.ハルピン(1:160)(5μgm/ml)。
4.ハルピン(1:320)(2.5μgm/ml)。
5.ハルピン(1:640)(1.25μgm/ml)。
6.緩衝液(5mM KPO4、pH6.8)。
【0083】
【表6】
【0084】
【表7】
【0085】
表7−10に示されるように、トマト種子のErwinia amylovora過敏反応誘発物での処理は、トマト植物の生育を促進できる。1:160希釈(5μg/mlハルピン)は最大の作用を有する--苗木の高さは緩衝液処理植物より20%以上増加した。
【0086】
実施例5−トマト種子のErwinia amylovora過敏反応誘発物処理による種子発芽率への作用
Marglobeトマト種の種子を、ビーカー中のErwinia amylovora過敏反応誘発物(“ハルピン”)溶液(1:50または1:100のE. coli DH5 (pCPP2139)由来のCFEP希釈、各々8μgm/mlおよび4μgm/mlの過敏反応誘発物を含む)および緩衝液に、日0に24時間、28℃の成育チャンバーで浸した。浸漬後、日1に種子を人工土壌の発芽用鉢に蒔いた。この方法を鉢当たり20個の種子で、処理当たり4鉢で行った。
【0087】
処理:
1.ハルピン(8μgm/ml)。
2.ハルピン(8μgm/ml)。
3.ハルピン(8μgm/ml)。
4.ハルピン(8μgm/ml)。
5.ハルピン(4μgm/ml)。
6.ハルピン(4μgm/ml)。
7.ハルピン(4μgm/ml)。
8.ハルピン(4μgm/ml)。
9.緩衝液(5mM KPO4、pH6.8)。
10.緩衝液(5mM KPO4、pH6.8)。
11.緩衝液(5mM KPO4、pH6.8)。
12.緩衝液(5mM KPO4、pH6.8)。
【0088】
【表8】
【0089】
表11に示されるように、トマト種子のErwinia amylovora過敏反応誘発物での処理は、トマト種子の発芽率およびレベルを増加できる。より高い用量の使用が、緩衝液よりも本実験の終了時にはより有効であるように見える。
【0090】
実施例6−過敏反応誘発物をコードする構築物であるpCPP2139またはプラスミドベクターpCPP50を含むE. coliから調製したタンパク質でのトマト種子処理の植物生育への作用
Marglobeトマトの種子を、Erwinia amylovora過敏反応誘発物(“ハルピン”)(E. coli DH5α (pCPP2139)(図1))またはコントロールとしてBSAタンパク質を添加したベクター製剤(DH5α(pCPP2139)(図2))に浸した。コントロールベクター製剤は、ml当たり、33.6μlのBSA(10mg/ml)を含み、ハルピンに由来するpCPP2139に含まれるタンパク質の量とほぼ同じである。1:50(8.0μg/ml)、1:100(4.0μg/ml)および1:200(2.0μg/ml)の希釈をビーカーで日1に調製し、種子を24時間、28℃で制御環境チャンバーで浸した。浸漬後、種子を、日2に人工土壌の発芽用鉢に蒔いた。処理当たり10本の同一な見かけの植物を移植後3回無作為に選択し、測定した。苗木を土壌表面から植物の頂部まで定規で計った。
【0091】
処理:
1.ハルピン 1:50(8.0μg/ml)。
2.ハルピン 1:100(4.0μg/ml)。
3.ハルピン 1:200(2.0μg/ml)。
4.ベクター+BSA 1:50(0ハルピン)。
5.ベクター+BSA 1:100(0ハルピン)。
6.ベクター+BSA 1:200(0ハルピン)。
【0092】
【表9】
【0093】
【表10】
【0094】
表12−15に示されるように、Erwinia amylovora過敏反応誘発物をコードする遺伝子を含むE. coliでの処理は、トマト植物の成長を促進できる。1:100希釈(4.0μg/ml)は最大の作用を有するが、それより高いかまたは低い濃度は作用が小さい。ハルピン4.0μg/mlでの処理による平均の苗木の高さは、同量の非ハルピンタンパク質を含むベクターコントロール製剤に対して約20%促進された。E. coli株DH5α(pCPP2139)中にhrpN遺伝子のベクターを保有する株E. coli DH5α(pCPP50)からの融解細胞製剤の成分は、ハルピン含有製剤と同じ成長促進作用を有せず、BSAタンパク質を、大量のハルピンタンパク質を含むDH5α(pCPP2139)製剤と同程度に添加したときでさえそうであった。
【0095】
実施例7−過敏反応誘発物をコードする構築物であるpCPP2139またはプラスミドベクターpCPP50を含むE. coliから調製したタンパク質でのトマト種子処理の植物生育への作用
Marglobeトマト種子を、1:25、1:50および1:100の希釈のErwinia amylovora過敏反応誘発物溶液(“ハルピン”)(ハルピンコードプラスミドpCPP2139ベクターに由来)およびpCPPベクター含有溶液に、ビーカー中で日1に24時間、28℃で生育チャンバーで浸した。浸漬後、種子を日2に人工土壌の発芽用鉢に蒔いた。処理当たり10本の同一な見かけの植物を移植後3回無作為に選択し、測定した。苗木を土壌表面から植物の頂部まで定規で計った。
【0096】
処理:
1.ハルピン16μgm/ml。
2.ハルピン8μgm/ml。
3.ハルピン4μgm/ml。
4.ベクター16μgm/ml。
5.ベクター8μgm/ml。
6.ベクター4μgm/ml。
【0097】
【表11】
【0098】
【表12】
【0099】
表16−18に見られるように、Erwinia amylovora過敏反応誘発物での処理は、トマト直物の成長を促進できる。1:50希釈(8μg/ml過敏反応誘発物)は最大の作用を有し、苗木の高さはコントロールの約20%上昇する。
【0100】
実施例8−ジャガイモの生育における無細胞Erwinia amylovora過敏反応誘発物の作用
3週齢ジャガイモ植物の種Norchipを、塊茎片から別々の容器で生育させた。各植物の葉に、1:50、1:100および1:200に希釈したErwinia amylovora過敏反応誘発物(“ハルピン”)またはE. coliおよびベクターcPP550のタンパク質を含むコントロール溶液(“ベクター”)を噴霧した。日20に、処理当たり12本の同一な見かけの植物を無作為に選択した。各処理由来の一本の植物を生育チャンバー中16℃で保持し、他方各処理由来の2本の植物を温室床箱で18−25℃で保持した。処理25日後、全植物の芽(茎)を個々に測定した。
【0101】
処理:
1.ハルピン 1:50 16μgm/ml。
2.ハルピン 1:100 8μgm/ml。
3.ハルピン 1:200 4μgm/ml。
4.ベクター 1:50 0ハルピン。
5.ベクター 1:100 0ハルピン。
6.ベクター 1:200 0ハルピン。
【0102】
【表13】
【0103】
表19および20に示されるように、Erwinia amylovora過敏反応誘発物でのジャガイモ植物の処理は、芽(茎)生育を促進する。従って、全体的な成長は、茎の数および平均の長さから判断して、温室および生育チャンバー生育植物の両方でハルピン処理植物で大きかった。中間の量のハルピン(8μgm/ml)で処理したジャガイモ植物は、その茎生育において、そのより高いまたはより低い用量よりも促進されたように見えた。中間の量のハルピンでの処理は、両方の生育条件下でより促進された成長をもたらした。
【0104】
実施例9−Erwinia amylovoraハルピンを含む無細胞誘発製剤のトマトへの噴霧作用
Marglobeトマト植物に、移植後8日に、ハルピン製剤(E. coli DH5α(pCPP2139))またはコントロールとしてBSAタンパク質を添加したベクター製剤(E. coli DH5α(pCPP50))を噴霧した。コントロールベクター製剤は、ml当たり、BSA(10mg/ml)を含み、ハルピンに起因するpCPP2139製剤に含まれるのと同じタンパク質の濃度を提供する。1:50(8.0μg/ml)、1:100(4.0μg/ml)および1:200(2.0μg/ml)の希釈を調製し、植物に、電動噴霧装置で流れ出すまで噴霧した。処理当たり15本の同一な見かけの植物を無作為選択し、処理に付した。植物を、処理前および後は28℃に制御環境チャンバーに保持した。
【0105】
全体の高さを処理後数回、土壌表面から植物の頂部まで定規で計った。トマト植物の頂部を土壌の表面近辺から茎を切断した直後に秤量した。
【0106】
処理:(希釈およびハルピン含量)
1.ハルピン 1:50(8.0μg/ml)
2.ハルピン 1:100(4.0μg/ml)
3.ハルピン 1:200(2.0μg/ml)
4.ベクター+BSA 1:50(0ハルピン)
5.ベクター+BSA 1:100(0ハルピン)
6.ベクター+BSA 1:200(0ハルピン)
【0107】
【表14】
【0108】
【表15】
【0109】
【表16】
【0110】
トマト苗木へのハルピンの1回噴霧は、一般に、BSAタンパク質を添加したコントロール(ベクター)製剤よりも大きな続く生育をもたらした。ハルピン処理植物での促進された生育は、植物高および生重量測定の両方で見られた。試験した3つの濃度で、低いほう二つは、最高の用量(8.0μg/ml)よりもより大きな植物生育を(いずれかの測定を基本にして)もたらした。低いほうの二つ(2および4μg/ml)の濃度で処理した植物の生育は僅かに異なった。E. coli株DH5α(pCPP2139)中のhrpN遺伝子のベクターを保持する株E. coli DH5α(pCPP50)由来の融解細胞製剤の成分は、大量のハルピンタンパク質を含むDH5α(pCPP2139)製剤と同程度に、BSAタンパク質を添加した場合でさえ、ハルピン含有製剤と同じ生育促進作用は有しなかった。従って、本実験はハルピンが植物生育を担うことを証明する。
【0111】
実施例10−小果実の早期着色および早期成熟
野外試験を、ラズベリー栽培種Canbyの収率および成熟パラメーターにおける過敏反応誘発物(“ハルピン”)処理の作用を評価するために行った。定着した植物に、区画40フィート長×3フィート幅(1植物幅)に2.5mg/100平方フィートでハルピンでの処理、非処理(“チェック”)または産業標準化学物質Ronilanを推奨された割合(“Ronilan”)での処理を行った。処理は4回繰り返し、実験野外部位のrepによりアレンジした。処置は、5−10%開花期に開始し、続いて7−10日間隔で2回行った。最初の2回の収穫は疾病コントロールに使用し、果実収率データの評価は最後の2回の収穫から集めた。観察は、ハルピン処理果実が非処理果実より大きく、赤いことを示し、成熟が1−2週間早まったことを示した。最少10個の果実を含む房当たりの成熟果実の数をこの時点で測定し、表26に要約する。ハルピン処理した区画はチェックまたはRonilan処理よりも10個のベリーの房当たり、より多い成熟果実を有した。最後の2回の収穫の合わせた収量は、ハルピンおよびRonilan処理区画の収量が、非処理コントロールよりも増加したことを示した(表27)。
【0112】
【表17】
【0113】
【表18】
【0114】
実施例11−さや豆の成長促進
Bush Blue Lake種のさや豆を種々の方法で処理し、商品の鉢植え用混合物を入れたプラスチック鉢に25cm2で植え、生育パラメーターの評価のために開放温室に置いた。処理は、非処理豆種子(“チェック”)、希釈剤としての水で調製した1.5%メチルセルロースのスラリーで処理した種子(“M/C”)、1.5%メチルセルロースで種子を処理し、続いて過敏反応誘発物(“ハルピン”)を0.125mg/mlで葉に投与した種子(“M/C+H”)、および1.5%メチルセルロース+50種子当たり5.0μgハルピンで乾燥したハルピンスプレーで種子を処理し、続いてハルピンを0.125mg/mlで葉に投与した種子(“M/C-SD+H”)を含む。種子を日0に土壌に蒔き、鉢当たり3個植え、発芽して鉢当たり1本の植物に間引いた。処理を開放温室内で10回繰り返し、repにより無作為化した。豆莢を64日後に収穫し、売り物になる大きさ(>10cm×5cmの大きさ)の豆莢の生重量を収量として集めた。処理手段を分けるために使用したFisher's LSDでの種々の分析によりデータを分析した。
【0115】
【表19】
1 売り物になる収量は10cm×0.5cm以上の全ての豆莢を含んだ。同じ文字の意味は、Fisher's LSDでP=0.05の有意な差がなかった。
【0116】
表28に示されるように、種々の施用法でのErwinia amylovoraハルピンの施用は、売り物になる大きさのさや豆の収量の増加をもたらした。メチルセルロース単独での処理はまた豆の収量の増加をもたらしたが、種子(噴霧乾燥)および葉処理としてハルピンと組み合わせた場合、実質的に増加する。
【0117】
実施例12−キュウリへのHP-1000TMの葉施用によるキュウリの収量増加
キュウリ幼苗および移植苗を、HP-1000TM(EDEN Bioscience, Bothell, Washington)(Erwinia amylovora過敏反応誘発製剤)の15、30または60μg/ml活性成分(a.i.)の量の葉噴霧で処理した。最初の噴霧は、最初の本葉が完全に伸びたときに行った。2回目の施用は、最初の噴霧から10日後に行った。全噴霧はバックパック噴霧器を使用して行い、非処理コントロール(UTC)もまた試験に含んだ。2回目のHP-1000TMの施用3日後、各処理の10本の植物を野外区画に無作為に移植し、3回繰り返した。これにより処理当たり全30本の植物を生産した。移植7日後、3回目のHP-1000TM葉噴霧を行った。ひどい旱魃がかなりの水ストレスをもたらしたが、全6回の収穫が標準の業務上収穫パターンに従ってなされた。各処理から収穫した果実の全重量を表29に示す。結果は、15および30μg/mlの量でHP-1000TM処理した植物が、UTCよりも有意に多い実を生産した。HP-1000TMで処理した植物は、収穫が僅かに増えたことを示す。これらの結果は、HP-1000TM処理が非処理植物よりも旱魃負荷条件に明らかにより耐性であることを示した。
【0118】
【表20】
1 活性成分(a.i.)。2 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0119】
実施例13−HP-1000TM処理による綿の収量増加
綿を12×12フィートの同一野外区画に、無作為完全ブロック(RCB)野外試験方式で植え、4回繰り返した。植物をHP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)、HP-1000TM+Pix(登録商標)(Pix(登録商標)(BASF Corp., Mount Olive, N. J.)は綿植物の高さが小さく保たれるように施用する成長調節剤である)またはEarly Harvest(登録商標)(Griffen Corp., Valdosta, Ga.)(拮抗的成長促進剤)で処理した。非処理コントロール(UTC)もまた試験に含んだ。バックパック噴霧器を使用して、葉施用を、作物の3つの成長段階の全ての処理で行った;第一本葉、開花前および開花早期。全ての肥料および雑草制御製剤を、全処理の慣用の農作業方式に従って施用した。収穫10週間前の綿玉の数は、他の処理と比較してHP-1000TM処理植物が明らかに多かった。収穫により、HP-1000TM処理植物は、UTCと比較して有意に増加したリント収率を(43%)を示した(表30)。HP-1000TMをPix(登録商標)と組み合わせたとき、リント収量はUTCを20%超えた。Pix(登録商標)は大エーカー数の綿に施用するため、この結果はHP-1000TMがPix(登録商標)と十分タンク混合され得ることを示す。拮抗的成長促進剤であるEarly Harvest(登録商標)の施用は、UTCと比較して僅かに9%増加させた。
【0120】
【表21】
1 HP-1000TMの量は活性成分(a.i.)として;Early Harvest(登録商標)およびPix(登録商標)の量は製剤製品として。
【0121】
実施例14−HP-1000TM処理による中国ナスの収量増加
苗床生育中国ナス苗木に、1回、HP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)を15、30または60μg/ml(a.i.)で噴霧し、次いで、野外区画に移植し、各処理を3回繰り返した。移植2週間後、2回目のHP-1000TM施用を行った。3回目の最終HP-1000TM施用を2回目の約2週間後に行った。全噴霧をバックパック噴霧器を使用して行った;非処理コントロール(UTC)もまた試験に含んだ。季節の進行と共に、各処理から全8回の収穫を行った。これらの収穫物からのデータは、HP-1000TMでの処理が植物当たりより多い実の収量をもたらすことを示す。
【0122】
【表22】
【0123】
実施例15−HP-1000TM処理による米の収量増加
稲苗を野外区画に移植し、3回繰り返し、次いでバックパック噴霧器を使用して3つの異なる量のHP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)で処理した。非処理コントロール(UTC)もまた試験に含んだ。最初のHP-1000TM施用は移植1週間後、2回目は1回目の3週間後であった。3回目の最終噴霧は米穀粒が穂に入り出す直前に行った。収穫の結果は、HP-1000TMの施用が30および60μg/mlの両方で収率を有意にそれぞれ47%および56%増加させることを証明した(表32)。
【0124】
【表23】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0125】
実施例16−HP-1000TM処理によるダイズの収量増加
ダイズを無作為化野外区画に植え、各処理を3回繰り返した。バックパック噴霧器をHP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)の葉面噴霧施用に使用し、非処理コントロール(UTC)もまた試験に含んだ。3つの量のHP-1000TMを、植物が約8インチの高さになったとき、4枚の本葉の始めに施用した。HP-1000TMの2回目の噴霧は、1回目の施用の10日後に、3回目の噴霧は2回目の10日後に行った。1回目の噴霧10日後に測定した植物の高さは、HP-1000TMの噴霧が有意な成長促進をもたらすことを示した(表33)。加えて、60μg/mlのHP-1000TMで処理した植物は、他の処理より5日速く開花し始めた。3回目の噴霧の約10日後、植物当たりのダイズ莢の数を繰り返し当たり無作為に選択した10本の植物から計数した。これらの結果は、HP-1000TMでの処理に由来する成長促進は、有意に多い収量をもたらすことを示した(表34)。
【0126】
【表24】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0127】
【表25】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0128】
実施例17−HP-1000TM処理によるイチゴの収量増加
HP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)での二つの野外試験を、二つのイチゴ種であるCamarosaおよびSelvaで行った。各種に関して、無作為完全ブロック(RCB)設計を、商業用のイチゴを生産する畑で、区画(5.33×10フィート)/処理で確立し、4回繰り返した。各区画内で、イチゴ植物を2列配置で植えた。未処理コントロール(UTC)もまた試験に含んだ。施用を開始する前に、全ての植物を他の花およびベリー類から離した。40μg/mlの量のHP-1000TMの噴霧を6週毎にバックパック噴霧器を使用して行った。各噴霧施用前に、各処理由来の全成熟果実を収穫し、秤量し、商業的標準に従って等級付した。HP-1000TMのSelvaイチゴ植物への施用3週間以内に、成長促進が、可視的に大きな地上生物量およびより元気で、緑で健康的な見かけとして識別できた。6回の収穫(即ち、これらの植物の予定の寿命)の後、全ての収量データを合計し、分析した。Camarosa種において、HP-1000TM処理植物からの売り物になる果実の収量は、最後の4回の収穫を平均したとき、UTCより有意に(27%)多かった(表35)。処理間の有意な差は最初の2回の収穫ではこの種では明らかではなかった。Selva種はHP-1000TMでの処理により反応性であった;Selvaイチゴ植物は、6回の収穫を平均したとき、UTCと比較して統計学的に有意な64%多い売り物になる果実を生産した(表35)。
【0129】
【表26】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0130】
実施例18−HP-1000TM処理によるトマトの早期成熟および収量増加
生販売用トマト(Solar Set種)を、区画(2×30フィート)で生育させ、5回無作為完全ブロック(RCB)野外試験で、商品トマト生産畑で繰り返した。処理はHP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)、実験的競合産物(ActigardTM (Novartis, Greensboro, N. C.))および化学標準(Kocide(登録商標) (Griffen Corp., Valdosta, GA))+Maneb(登録商標) (DuPont Agricultural Products, Wilmington, D. E.))を疾病コントロールのために含んだ。HP-1000TMの最初の施用は、移植直後に苗木に直接注ぐ50ml浸液(30μg/ml a.i.)により行った。その後、11週毎に葉噴霧をバックパック噴霧器を使用して行った。全処理由来の最初の収穫は移植後約6週で行い、完全に赤く熟したトマトのみを各処理から収穫した。結果は、HP-1000TM処理植物が、最初の収穫に間に合うトマトの有意に多い量を生産したことを示した(表36)。HP-1000TM処理植物から収穫したトマトは、他の植物より10−14日進んでいると評価された。
【0131】
【表27】
1 Kocide(登録商標)およびManeb(登録商標)の量は製剤製品。2 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0132】
実施例19−非消毒土壌に植えたときの、HP-1000TM処理によるイチゴの早期開花および成長促進
イチゴ植物(“プラグ(plugs)”および“根付き(bare-root)”)の栽培種Commanderを区画(2×30フィート)に移植し、無作為完全ブロック野外試験で5回繰り返した。約60本の別々の植物を各繰り返しで植えた。この野外試験での処理は下記に示す:
【表28】
1 非消毒土壌はソラマメ属を前の2年栽培していた。
【0133】
移植は、晩秋に、寒い気候が植物の生育を遅らせ始めたときに行った。移植2週間後、HP-1000TMの最初の葉施用を、バックパック噴霧器で40μg/ml(a.i.)で行った。移植3週間後、予備的結果を集め、臭化メチルおよびUTCとHP-1000TMの比較を、各繰り返しの全イチゴ“プラグ”植物の花の数を計数することにより行った。開花が“根付き”植物でまだ起こっていないため、この処理での繰り返しの各植物を、葉の先端から3枚の葉の塊の真中の葉の茎までの長さを測定することにより評価した。結果(表37および38)は、HP-1000TMが早期に促進された花の生育および葉の大きさを、“プラグ”および“根付き”イチゴ植物でそれぞれ提供することを示した。
【0134】
【表29】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0135】
【表30】
【0136】
実施例20−HP-1000TM処理によるハラペーニョトウガラシの早期成長促進
ハラペーニョトウガラシ(Mittlya栽培種)移植植物をHP-1000(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)(30μg/ml a.i.)での1時間の根浸漬により処理し、続いて4回繰り返した。非処理コントロール(UTC)もまた含んだ。移植14日に開始して、処理植物は3回HP-1000TMを、14日間隔で、バックパック噴霧器により受けた。HP-1000TMの3回目の施用の1週間後(移植54日後)、植物の高さを繰り返し当たり無作為に選択した4本の植物で測定した。これらの測定の結果は、HP-1000TM処理植物がUTC植物よりも約26%背が高いことを示した(表39)。加えて、各植物の芽、花または実の数を計数した。これらの結果は、HP-1000TM処理植物が、UTC植物と比較して、61%多い花、実または芽を有することを示した(表40)。
【0137】
【表31】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0138】
【表32】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0139】
実施例21−HP-1000TM施用によるタバコの成長促進
タバコ苗木を無作為化野外区画に植え、3回繰り返した。移植後、HP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)を3つの量のいずれか一つで葉噴霧した:15、30または60μg/ml a.i.。60日後、2回目のHP-1000の葉施用を行った。2回目の施用2日後、植物の高さ、植物当たりの葉の数および葉の大きさ(面積)を処理当たり無作為に選択した10本について測定した。これらの測定の結果は、HP-1000TMでの処理が有意に植物生育を促進させることを示した(表41、42および43)。植物の高さは6−13%増加し、30および60μg/mlでHP-1000TM処理した植物は平均UTCより植物当たり1枚葉が多かった。しかしながら、最も有意には、15、30および60μg/ml のHP-1000TMでの処理は、葉面積の対応する増加をもたらした。植物当たり1枚の余分な葉および増加した平均葉サイズ(面積)のタバコ植物は、商業的に有意な反応を示す。
【0140】
【表33】
【表34】
【表35】
【0141】
実施例22−HP-1000TM施用による冬小麦の成長促進
冬小麦種子に乾燥HP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)を、種子100ポンド当たり製剤製品(3%a.i.)3オンス“散布”し、続いて慣用の播種装置を使用して11.7フィート×100フィート長の試験無作為試験区画に植えた。更なる処理は、種子100ポンドあたりHP-1000TM粉末(3%a.i.)1オンスの“散布”、20μg/ml活性成分の濃度のHP-1000TM溶液への4時間の種子浸漬、続く植える前の空気乾燥、標準化学(Dividend(登録商標))殺菌剤“散布”および非処理コントロール(UTC)を含んだ。蒔いて8日後、HP-1000TM処理種子は発芽し始め、一方UTCおよび化学標準処理種子は蒔いて約14日後まで発芽せず、通常の時間経過であった。蒔いて41日後、苗木を地面から抜き、評価した。3オンス/100ポンドでの“散布”としてHP-1000TM処理した種子を目で調べ、他の処理のいずれよりも約2倍大きいと判断した。
【0142】
野外試験に続き、温室実験を計画し、これらの結果の確認をした。処理は、100ポンドの種子当たり3オンスの量での乾燥HP-1000TM(10%a.i.)の散布、20mg/mlの濃度の溶液中のHP-1000TMへの移植前4時間の浸漬および非処理コントロール(UTC)を含んだ。各処理由来の小麦種子を鉢当たり25個の種子で蒔き、5つの鉢を各処理の繰り返しとして提供した。蒔いて15日後、各処理鉢当たり無作為に選択した10本の苗木を抜き、注意深く洗い、根の長さを測定した。個々の苗木の地上部分は処理の効果を示さなかっため、HP-1000TMでの処理の根の生育の増加はサンプルの選択に影響しなかった。HP-1000TM処理の根の生育の増加は、UTCより有意に大きかった(表49);しかしながら、種子散布処理は、僅かに良い結果をもたらした。
【0143】
【表36】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0144】
実施例23−HP-1000TM施用によるキュウリの成長促進
商品として生産されるキュウリの野外試験は、4回の処理、2種の濃度(20または40μg/ml)でのHP-1000TM(EDEN Bioscience)(Erwinia amylovora過敏反応誘発製剤)、疾病コントロールの化学表品(Bravo(登録商標) (Zeneca Ag Products, Wilmington, Del.) + Maneb(登録商標))および非処理コントロール(UTC)であった。各処理を、3×75フィート区画で、各処理当たり約2フィートの植物の空間で4回行った。HP-1000TMの葉噴霧は、最初の本葉のときに開始し、14日間隔で、最後の収穫まで合計6回施用で繰り返した。標準殺菌剤混合物を7日毎または状況的な根拠があったら直ぐに施用した。商品の収穫はHP-1000TMの最初の施用から約2ヶ月後(HP-1000TMの5回噴霧後)に開始し、最終収穫は最初の収穫の約14日後であった。
【0145】
最初の収穫の結果は、HP-1000TMでの処理が、個々のキュウリの平均重量ではなく、収穫した平均キュウリ収量をキュウリの総数を増加させることにより増加させたことを示した(表45−47)。同じ傾向が最終収穫(表48−49)でも見られた。HP-1000TM処理によりもたらされる収量増加がキュウリ平均サイズの明白な増加により達成されるものでないことは商業的に重要であった。
【0146】
【表37】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0147】
【表38】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0148】
【表39】
【0149】
【表40】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0150】
【表41】
1 異なる文字の意味は、Duncan's MRTに従った有意差、P=0.05である。
【0151】
【表42】
【0152】
実施例24−トマトにおけるPseudomonas syringae pv syringae由来のハルピンpss誘導成長促進
ハルピンpss(即ち、Pseudomonas syringae pv syringae由来の過敏反応誘発物)(引用して本明細書に包含させるHe, S. Y., et al., “Pseudomonas syringae pv syringae Harpinpss. A Protein that is Secreted via the Hrp Pathway and Elicits the Hypersensitive Response in Plants,” Cell 73:1255-66 (1993))がまた植物生育を促進するか試験をするために、トマト種子(Marglobe種)を人工土壌の8インチ鉢に蒔いた。蒔いて10日後、苗木を別々の鉢に移植した。実験を通して、肥料、水の潅水、温度および土壌湿度は植物間で均質に維持した。移植16日後、最初の植物の高さを測定し、ハルピンpssの最初の施用を行い、日0とした。2回目の施用は日15に行った。更なる成長データを日10および日30に集めた。最終データ採集は日30で、植物の高さおよび生重量を含んだ。
【0153】
実験中の施用に使用したハルピンpssは、ハルピンpss(即ち、hrpZ)をコードする遺伝子のプラスミドを含むE. coli DH5を発酵させて、製造した。細胞を回収し、5mM リン酸カリウム緩衝液に再懸濁させ、超音波処理により破壊した。超音波処理物質を5分沸騰させ、次いで10分、10,000rpmで遠心した。懸濁液を無細胞誘発製剤(CFEP)と見なした。20および50μg/mlハルピンpss溶液を同じ緩衝液から調製し、細胞懸濁に使用した。同じプラスミドを含むが、hrpZ遺伝子を含まない同じ株から調製した同じCFEPをコントロール処理の材料として使用した。
【0154】
湿潤剤であるPinene II (Drexel Chemical Co., Memphis, Tenn.)をハルピンpss溶液に0.1%の濃度で添加し、次いでハルピンpssをトマト植物に流れ落ちるまで噴霧した。
【0155】
表51はハルピンpss処理グループとコントロールグループの間に有意な差があったことを示す。ハルピンpss処理トマトは高さで10%以上増加した。データは、ハルピンpssが、トマトおよび他の多くの植物種に施用したとき、成長促進作用があるという点で、Erwinia amylovora由来の過敏反応誘発物と同様に作用するという請求の範囲を支持する。トマトの高さの有意な増加に加えて、ハルピンpss処理トマトはより大きな生物量、より大きな葉、早期開花および相対的なより健康な見かけを有した。
【0156】
【表43】
1 植物の高さは最も近い0.5cmで測定した。日0は最初の植物の高さを記録し、最初の施用を行った日を意味する。
2 平均は括弧内のSDと共に示す(全処理グループでn=20)
3 異なる文字(aおよびb)は平均間の有意差(P 0.05)を意味する。差はFisher LSDに続くANOVAにより評価した。
【0157】
本発明は説明の目的で詳細に記載しているが、このような細部は単にこの目的のためであって、以下の請求の範囲に定義の本発明の精神および範囲を逸脱することなく、当業者はその変化をなし得ることは理解される。
【特許請求の範囲】
【請求項1】
植物または植物種子に、植物または植物種子から成育した植物の成長の促進に有効な条件下で、非感染型で、過敏反応誘発ポリペプチドまたはタンパク質を適用することを含む、植物の成長を促進する方法。
【請求項1】
植物または植物種子に、植物または植物種子から成育した植物の成長の促進に有効な条件下で、非感染型で、過敏反応誘発ポリペプチドまたはタンパク質を適用することを含む、植物の成長を促進する方法。
【図1】

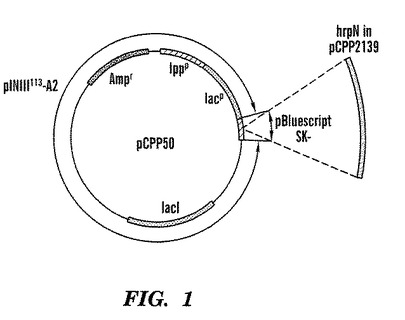
【図2】
【図2】
【公開番号】特開2011−92193(P2011−92193A)
【公開日】平成23年5月12日(2011.5.12)
【国際特許分類】
【出願番号】特願2010−260009(P2010−260009)
【出願日】平成22年11月22日(2010.11.22)
【分割の表示】特願2009−167530(P2009−167530)の分割
【原出願日】平成10年1月27日(1998.1.27)
【出願人】(592035453)コーネル・リサーチ・ファンデーション・インコーポレイテッド (39)
【氏名又は名称原語表記】CORNELL RESEARCH FOUNDATION, INCORPORATED
【Fターム(参考)】
【公開日】平成23年5月12日(2011.5.12)
【国際特許分類】
【出願日】平成22年11月22日(2010.11.22)
【分割の表示】特願2009−167530(P2009−167530)の分割
【原出願日】平成10年1月27日(1998.1.27)
【出願人】(592035453)コーネル・リサーチ・ファンデーション・インコーポレイテッド (39)
【氏名又は名称原語表記】CORNELL RESEARCH FOUNDATION, INCORPORATED
【Fターム(参考)】
[ Back to top ]