
植物の構成的遺伝子発現を増強する調節核酸分子
本発明は植物分子生物学の分野であって、核酸発現増強性核酸(NEENA)を前記プロモーターと機能的に連結することおよび/または植物中に導入することを特徴とする、高発現性プロモーターを生産するおよび核酸の発現が増強された植物を生産する方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は植物分子生物学の分野にあり、高発現性構成的プロモーターを産生するおよび核酸の構成的発現が増強された植物を産生する方法であって、核酸発現増強性核酸(NEENA)を前記プロモーターと機能的に連結するおよび/または植物中に導入することを特徴とする前記方法を提供する。
【背景技術】
【0002】
植物におけるトランスジーンの発現は様々な外的および内的因子の影響を強く受け、様々なかつ予想できないレベルのトランスジーン発現をもたらす。しばしば多数の形質転換体を産生させて分析し、所望の発現強度をもつ系統を同定しなければならない。所望の発現をもつ系統の形質転換とスクリーニングは高費用かつ労働集約的であるので、1以上のトランスジーンを植物で高発現させる必要性がある。特に問題になるのは、トランスジェニック植物において特定の効果を達成するためにいくつかの遺伝子を協調的に発現させる必要のある場合であって、それぞれおよび全ての遺伝子が強く発現された植物を同定しなければならないからである。
【0003】
例えば、トランスジーンの発現は、個々の形質転換事象における構築物設計およびT-DNA挿入遺伝子座の位置効果に応じて有意に変わりうる。強いプロモーターはこれらの課題を部分的に克服することができる。しかし、所望の特異性をもつ強い発現を示す好適なプロモーターの利用可能性は限られることが多い。所望の発現特異性をもつ十分なプロモーターの利用可能性を確実にするために、追加のプロモーターの同定と特徴付けがこのギャップを埋める助けになりうる。しかし、それぞれの特異性と強度をもつプロモーターの自然での利用可能性とプロモーター候補の特徴付けに費やす時間とが、好適な新しいプロモーターを同定する妨げとなる。
【0004】
これらの課題を克服するために、多様な遺伝エレメントおよび/またはモチーフが遺伝子発現にポジティブな影響を与えることが示されている。これらのなかにあって、いくつかのイントロンは遺伝子発現を改善する強い可能性をもつ遺伝エレメントであることが認識されている。その機構はほとんど判ってないが、いくつかのイントロンは、転写活性の増強、mRNA成熟の改善、核mRNAエキスポートの増強および/または翻訳開始の改善により、成熟mRNAの定常状態量にポジティブな影響を与えることが示されている(例えば、Huang and Gorman, 1990;Le Hir et al., 2003;Nott et al., 2004)。選択されたイントロンだけが発現を増加することが示されたので、スプライシングなどはおそらく観察された効果を説明することができない。
【0005】
イントロンがプロモーターに機能的に連結するときに観察される遺伝子発現の増加は、遺伝子発現のイントロン媒介性増強(IME)と呼ばれ、様々な単子葉類(例えば、Callis al., 1987;Vasil al., 1989;Bruce al., 1990;Lu al., 2008)および双子葉類植物(例えば、Chung al., 2006;Kim al., 2006;Rose al., 2008)において示されている。この点については、翻訳出発部位(ATG)との相対的なイントロンの位置が遺伝子発現のイントロン媒介性増強にとって重要であることが示されている(Rose al., 2004)。
【0006】
遺伝子発現を増強する能力の次に、少数のイントロンはまた、植物における生来のヌクレオチド環境における組織特異性に影響を与えることが示されている。
【0007】
レポーター遺伝子発現は2つまでのイントロンを含有するゲノム領域の存在に依存することが見出された(Sieburth al., 1997;Wang al., 2004)。5’UTRイントロンはまた、プロモーターエレメントの適当な機能性にとって重要であり、これはおそらくイントロンに存在する組織特異的遺伝子制御エレメントに因ることが報じられた(Fuet al., 1995a;Fuet al., 1995b;Vitaleet al., 2003;Kimet al., 2006)。しかし、これらの研究はまた、イントロンの異種プロモーターとの組み合わせが遺伝子発現の強度と異種特異性に強い負の影響を与えうることも示している(Vitaleet al., 2003;Kimet al., 2006、WO2006/003186、WO2007/098042)。
【0008】
例えば、強い構成的カリフラワーモザイクウイルスCaMV35Sプロモーターは、ゴマD2 5’UTRイントロンとの組み合わせを通して負の影響を受ける(Kimet al., 2006)。これらの観察とは対照的に、いくつかの文献は、それぞれのプロモーターの組織特異性に影響を与えることなくIMEによる核酸発現の増強を示している(Schuenmannet al., 2004)。
【発明の概要】
【課題を解決するための手段】
【0009】
本出願においては、構成的プロモーターと機能的に連結されるとそれらの特異性に影響を与えることなく前記プロモーターの発現を増強する、さらなる核酸分子を記載する。これらの核酸分子を本出願では「核酸発現増強性核酸」(NEENA)と記載する。イントロンはそれぞれのプレmRNAからスプライシングで除去される内因性特徴を有する。それとは対照的に、本出願で提示する核酸は、機能的に連結されたプロモーターから誘導される発現を増強するために、必ずしもmRNA中に含まれなくてもよいし、または、もしmRNA中に存在すれば、必ずしもmRNAからスプライシングで除去されなくてもよい。
【図面の簡単な説明】
【0010】
【図1】一過的に形質転換したシロイヌナズナ葉プロトプラストにおける、p-AtNit1プロモーターの制御下の、NEENAを含有しない(LJK132)および構成的に発現される遺伝子由来の推定NEENA分子を表すNEENAを含有する(LJK91〜LJK133)構築物のルシフェラーゼレポーター遺伝子発現分析である。規準化はウミシイタケ・ルシフェラーゼの共形質転換と分析により行い、発現値をNEENAを含まない対照構築物(LJK132=1)と比較して示した。発現値をNEENAを含まない対照構築物(LJK134=1)と比較して示した。
【図2】p-AtNit1プロモーターの制御下の、NEENAを含有しない(LJK138)または構成的に発現される遺伝子由来のNEENA分子を表すNEENAを含有する(LJK139〜LJK144)レポーター遺伝子構築物を保持する独立のトランスジェニックアブラナ植物系統の相対光単位(RLU)として、各サンプルのタンパク質含量(平均値、分析した20件の独立したトランスジェニック植物の組織)に対して規準化後に示したルシフェラーゼレポーター遺伝子活性の棒グラフである。A)葉組織、B)花、C)長角果。
【図3】p-AtNit1プロモーターの制御下の、NEENAを含まない(LJK138)または構成的に発現した遺伝子由来のNEENA分子を表すNEENAを含有する(LJK139〜LJK144)レポーター遺伝子構築物を保持する独立のトランスジェニックダイズ植物系統の、相対光単位(RLU)として、各サンプルのタンパク質含量(平均値、分析した10件の独立したトランスジェニック植物の組織)に対して規準化後に示した、ルシフェラーゼレポーター遺伝子活性の棒グラフである。A)葉組織、B)花、C)種子。
【図4】p-ZmUbiプロモーターの制御下の、NEENAを含まない(LJK309)または構成的に発現した遺伝子由来のNEENA分子を表すNEENAを含有する(LJK326〜LJK327)レポーター遺伝子構築物を保持する独立のトランスジェニックトウモロコシ植物系統の、相対光単位(RLU)(対数スケール)として、各サンプルのタンパク質含量(平均値、分析した15件の独立したトランスジェニック植物の組織)に対して規準化後に示した、ルシフェラーゼレポーター遺伝子活性の棒グラフである。A)葉組織、B)穀粒。
【図5】構成的な p-PRO239プロモーターの制御下の、NEENAを含まない(CD30963)または構成的に発現した遺伝子由来のNEENA分子を表すNEENAを含有する(CD30964)レポーター遺伝子構築物を保持する独立のトランスジェニックイネ植物系統の、相対光単位(RLU)として、各サンプルのタンパク質含量(平均値、分析した10件の独立したトランスジェニック植物の組織)に対して規準化後に示した、ルシフェラーゼレポーター遺伝子活性の棒グラフである。A)葉組織、B)種子。
【発明を実施するための形態】
【0011】
本発明の第1の実施形態は、プロモーターに1以上の核酸発現増強性核酸(NEENA)分子を機能的に連結するステップを含む高発現性構成的プロモーターを産生する方法であって、前記NEENA分子が
i) 配列番号1〜19、好ましくは配列番号1〜9のいずれかで定義した配列を有する核酸分子、または
ii) 配列番号1〜19、好ましくは配列番号1〜9で定義した配列のいずれかと80%以上の同一性をもつ、好ましくは、前記同一性が85%以上である、より好ましくは同一性が90%以上である、さらにより好ましくは、同一性が95%以上、96%以上、97%以上、98%以上または99%以上である、最も好ましい実施形態では、配列番号1〜19、好ましくは配列番号1〜9で定義した配列のいずれかとの同一性が100%である配列を有する核酸分子、または
iii) i)もしくはii)の核酸分子の100以上の連続する塩基、好ましくは150以上の連続する塩基、より好ましくは200以上の連続する塩基、さらにより好ましくは250以上の連続する塩基の断片であって、例えば、配列番号1〜19、好ましくは配列番号1〜9で定義した配列のいずれかの配列を有する対応する核酸分子の65%以上、好ましくは70%以上、より好ましくは75%以上、さらにより好ましくは80%以上、85%以上または90%以上の発現増強活性を有する、最も好ましい実施形態では、95%以上の発現増強活性を有する前記断片、または
iv) 先にi)〜iii)のもとで記載した核酸分子のいずれかの相補配列または逆相補配列である核酸分子、または
v) PCRにより、表2に示した配列番号20〜57、好ましくは配列番号20/21;26/27;30/31;38/39;42/43;44/45;46/47;50;51および56/57に記載のオリゴヌクレオチドプライマーを用いて取得可能な核酸分子、または
vi) 7%ドデシル硫酸ナトリウム(SDS)、0.5M NaPO4、1mM EDTA中の50℃におけるハイブリダイゼーションと、2 X SSC、0.1%SDS中の50℃または65℃、好ましくは65℃における洗浄と等価の条件のもとで、配列番号1〜19、好ましくは配列番号1〜9に記載の転写増強性ヌクレオチド配列またはそれらの相補配列の少なくとも50、好ましくは少なくとも100、より好ましくは少なくとも150、さらにより好ましくは少なくとも200、最も好ましくは少なくとも250個の連続するヌクレオチドを含む核酸分子とハイブリダイズする、100ヌクレオチド以上、150ヌクレオチド以上、200ヌクレオチド以上または250以上のヌクレオチドの核酸分子を含むものである前記方法を含む。
【0012】
好ましくは、前記核酸分子は、7%ドデシル硫酸ナトリウム(SDS)、0.5M NaPO4、1mM EDTA中の50℃におけるハイブリダイゼーションと、1 X SSC、0.1%SDS中の50℃または65℃、好ましくは65℃における洗浄と等価の条件のもとで、配列番号1〜19、好ましくは配列番号1〜9またはそれらの相補配列に記載の転写増強性ヌクレオチド配列の少なくとも50、好ましくは少なくとも100、より好ましくは少なくとも150、さらにより好ましくは少なくとも200、最も好ましくは少なくとも250個の連続するヌクレオチドを含む核酸分子とハイブリダイズし、より好ましくは、前記核酸分子は、7%ドデシル硫酸ナトリウム(SDS)、0.5M NaPO4、1mM EDTA中の50℃におけるハイブリダイゼーションと、0.1 X SSC、0.1%SDS中の50℃または65℃、好ましくは65℃における洗浄と等価の条件のもとで、配列番号1〜19、好ましくは配列番号1〜9に定義した配列またはそれらの相補配列のいずれかに記載の転写増強性ヌクレオチド配列の少なくとも50、好ましくは少なくとも100、より好ましくは少なくとも150、さらにより好ましくは少なくとも200、最も好ましくは少なくとも250個の連続するヌクレオチドを含む核酸分子とハイブリダイズする。
【0013】
一実施形態において、1以上のNEENAは、それが機能的に連結されたプロモーターに対して異種である。
【0014】
v)項にて記載した、表2に示した配列番号 20〜57、好ましくは 配列番号 20/21;26/27;30/31;38/39;42/43;44/45;46/47;50/51および56/57で定義したオリゴヌクレオチドを用いてPCRにより取得可能な核酸分子は、例えば、シロイヌナズナ植物由来のゲノムDNAから以下の実施例1に記載した条件を用いて取得可能である。
【0015】
当業者は、それぞれのNEENA分子を得るための温度プロファイル、サイクル数および/またはバッファー組成物または濃度のバリエーションに気付くであろう。それぞれのNEENA分子を得るそれぞれのPCR反応で用いるオリゴヌクレオチドの具体的な組み合わせを表2に記載した。
【0016】
当業者は一方向プロモーターを二方向プロモーターにする方法、およびプロモーターの相補配列または逆相補配列を用いて元来の配列に特異的な同じプロモーター特異性を有するプロモーターを作製する方法に気付くであろう。かかる方法は、例えば、構成的ならびに誘導プロモーターについて、Xie et al. (2001) “Bidirectionalization of polar promoters in plants” nature biotechnology 19 pages 677 - 679に記載されている。この著者らは、任意の所与のプロモーターの5'プライム末端に最小のプロモーターを加えるだけで、両方向に同じプロモーター特異性で発現を制御するプロモーターを得るのに十分であると記載している。従って、上記のNEENAと機能的に連結された高発現プロモーターは、相補配列または逆相補配列に機能性であり、それ故に、NEENAも相補配列または逆相補配列に機能性である。
【0017】
本明細書に記載した構成的プロモーターは、実質的に全植物組織に、実質的に植物もしくはその部分の全生存期間を通して発現されるプロモーターを意味する。実質的に全ての植物組織に発現されるプロモーターはまた、主な植物組織の少なくとも2つ、例えば、葉、幹および/または根に発現されるプロモーターを包含しうるし、そしていくつかまたは全てのマイナーな組織または細胞、例えば、表皮、気孔、毛状突起、花、種子または分裂組織で発現され得るかまたは発現され得ないプロモーターを包含する。好ましい実施形態において、本明細書で意味する構成的プロモーターは葉および幹などの緑色組織で少なくとも発現される。
【0018】
実質的に植物もしくはその部分の全生存期間を通して発現されるプロモーターはまた、若いおよび発達した組織において発現されるが、植物の生存期間中の特定の時点または特定の条件、例えば、発芽および/または老化の下で、または生物もしくは非生物ストレスの条件下、例えば真菌または細菌の感染、乾燥、熱または寒冷のもとで発現を欠くことがありうるプロモーターを包含する。好ましい実施形態において、実質的に植物の全生存期間に発現される構成的プロモーターは、少なくとも全ての拡大組織において老化の開始まで発現される。
【0019】
原則として、NEENAはいずれのプロモーターとも、例えば、組織特異的、誘導性、発生特異的または構成的プロモーターとも機能的に連結されうる。それぞれのNEENAは、少なくとも1つのNEENAが機能的に連結されたそれぞれのプロモーターの制御下の異種核酸の発現の増強に導きうる。構成的プロモーター以外のプロモーター、例えば組織特異的プロモーターの発現の増強は、これらのプロモーターを特異性にするであろう。前記核酸の転写物がNEENAがない場合に検出されなかったさらなる組織または発生段階において、それぞれのプロモーターの制御下の核酸の発現は検出可能になるであろう。それ故に、組織もしくは発生特異的なまたはいずれかの他のプロモーターを、上記NEENA分子の少なくとも1つを前記プロモーターと機能的に連結することにより構成的プロモーターにすることができる。それ故に本発明の他の実施形態は、それぞれのプロモーターをi)〜vi)で上に記載した配列を含むNEENA分子と連結することにより、植物中で機能性である任意の所与のプロモーターの特異性を構成的プロモーターにする方法を提供する。
【0020】
好ましくは、1以上のNEENAを任意の構成的プロモーターと機能的に連結し、前記プロモーターの制御下の核酸分子の発現を増強しうる。本発明の任意の方法で使用する構成的プロモーターは、植物、例えば単子葉または双子葉植物から、細菌および/またはウイルスから誘導してもよく、または合成プロモーターであってもよい。使用する構成的プロモーターは、例えば、P. crispum(WO 2003102198)由来のPcUbi-プロモーター、トウモロコシ由来のZmUbi-プロモーター、ニトリラーゼ1をコードするシロイヌナズナ遺伝子At3g44310由来のAtNit-プロモーター、フィグオート(figwort)モザイクウイルス由来の34S-プロモーター、タバコモザイクウイルス由来の35S-プロモーター、アグロバクテリア由来のnosおよびocs-プロモーター、フェレドキシンNADHレダクターゼをコードするシロイヌナズナ遺伝子At5g66190由来のScBV-プロモーター(US 5 994 123)、SUPER-プロモーター(Lee et al., 2007、Plant. Phys.)、 AtFNR-プロモーター、エンドウマメ由来のptxAプロモーター(WO2005085450)、リン酸トリオース輸送体をコードするシロイヌナズナ遺伝子At5g46110由来のAtTPT-プロモーター、シロイヌナズナ遺伝子At4g14880およびAt4g14890由来の二方向AtOASTL-プロモーター、グリセルアルデヒド-3-リン酸デヒドロゲナーゼをコードするシロイヌナズナ遺伝子At1g13440由来のPRO0194プロモーター、フルクトース-ビス-リン酸アルドラーゼをコードするシロイヌナズナ遺伝子At3g52930由来のPRO0162プロモーター、イネ由来のAHAS-プロモーター(WO2008124495)またはCaffeoylCoA-MTプロモーターおよびOsCP12(WO2006084868)である。
【0021】
NEENAと機能的に連結された本発明の高発現性構成的プロモーターは、例えば、藻類、シダ、裸子植物または被子植物、例えば単子葉または双子葉植物を含む任意の植物で使用することができる。好ましい実施形態においては、NEENAと機能的に連結された本発明の前記プロモーターを、単子葉または双子葉類植物、好ましくは作物植物、例えばトウモロコシ、ダイズ、アブラナ、ワタ、ジャガイモ、サトウダイコン、イネ、コムギ、モロコシ、オオムギ、バナナ、サトウキビ、ススキなどに使用することができる。本発明の好ましい実施形態においては、NEENAと機能的に連結された前記プロモーターを単子葉の作物植物、例えば、トウモロコシ、イネ、コムギ、モロコシ、バナナ、ススキ、サトウキビまたはオオムギに使用することができる。とりわけ好ましい実施形態においては、NEENAと機能的に連結されたプロモーターを、双子葉類の作物植物、例えば、ダイズ、アブラナ、ワタ、サトウダイコンまたはジャガイモに使用することができる。
【0022】
本出願に使用する高発現性プロモーターは、例えば、植物またはその部分においてプロモーターの構成的発現の増強を生じるNEENAと機能的に連結されたプロモーターであって、NEENAと機能的に連結されたそれぞれのプロモーターの制御下の核酸分子から誘導されるRNAの蓄積またはRNAの合成速度が、本発明のNEENAを欠く同じプロモーターにより生じる発現より高い、好ましくは有意に高い前記プロモーターを意味する。好ましくは、植物中のそれぞれの核酸のRNAの量および/またはRNA合成の速度および/またはRNA安定性は、本発明のNEENAと機能的に連結されてない同じ構成的プロモーターを含む同じ条件下で育てられた同じ齢の対照植物と比較して、50%以上、例えば100%以上、好ましくは200%以上、より好ましくは5倍以上、さらにより好ましくは10倍以上、最も好ましくは20倍以上、例えば50倍増加する。
【0023】
本明細書で使用する用語「有意に高い」は、統計的有意を意味し、当業者は、t-検定などの統計的検定をそれぞれのデータセットに応用することにより決定するやり方を理解している。
【0024】
プロモーターが与える発現を検出する方法は当技術分野で公知である。例えば、プロモーターをマーカー遺伝子、例えばGUS、GFPまたはルシフェラーゼと機能的に連結し、それぞれのマーカー遺伝子がコードするそれぞれのタンパク質の活性を植物またはその部分で確認することができる。代表例として、ルシフェラーゼを検出する方法を以下に詳しく記載する。他の方法としては、例えば、プロモーターにより制御される核酸分子のRNAの定常状態レベルまたは合成速度を、当技術分野で公知の方法、例えば、ノーザンブロット分析、qPCR、ラン-オンアッセイもしくは当技術分野で記載の他の方法により測定する方法がある。
【0025】
当業者は2以上の核酸分子を機能的に連結する様々な方法を知っている。かかる方法は制限/ライゲーション、リガーゼ非依存性クローニング、リコンビニアリング、組換えまたは合成を包含しうる。他の方法を使用して2以上の核酸分子を機能的に連結してもよい。
【0026】
本発明のさらなる実施形態は、それぞれの対照植物またはその部分と比較して、1以上の核酸分子の構成的発現が増強された植物またはその部分を産生する方法であって、植物またはその部分中に、i)〜vi)のもとで先に定義した核酸分子を含む1以上のNEENAを導入するステップ、ならびに、前記1以上のNEENAをプロモーター、好ましくは構成的プロモーターと、および前記プロモーター、好ましくは構成的プロモーターの制御下にある核酸分子(ここで、NEENAはこの核酸分子に対して異種である)と機能的に連結するステップを含むものである前記方法である。
【0027】
NEENAは、NEENAが機能的に連結された前記プロモーターの制御下にある核酸分子に対して異種であってもよく、またはNEENAはプロモーターおよび前記プロモーターの制御下の核酸分子の両方に対して異種であってもよい。
【0028】
核酸分子またはDNAに関する用語「異種」は、自然では作動しうる形で連結されていないかまたは自然では異なる位置で作動しうる形で連結されている第2の核酸分子と、作動しうる形で連結されるかまたは作動しうる形で連結されるように遺伝子操作された核酸分子を意味する。例えば、本発明のNEENAはその自然環境においてその生来のプロモーターと機能的に連結されているが、本発明において前記NEENAは同じ生物由来、異なる生物由来であってもよく、またはSUPER-プロモーターなどの合成プロモーターであってもよい他のプロモーターと連結される。また、本発明のNEENAはその生来のプロモーターと連結されているが、前記プロモーターの制御下の核酸分子はその生来のNEENAを含むプロモーターに対して異種であるということを意味する。さらに理解すべきことは、本発明のNEENAと機能的に連結された前記プロモーターの制御下のプロモーターおよび/または核酸分子は、その配列が例えば、挿入、欠失などの突然変異により遺伝子操作されると、前記プロモーターの制御下のプロモーターおよび/または核酸分子の自然配列が改変され、それ故に本発明のNEENAに対して異種になったので、前記NEENAに対して異種である。理解すべきことはまた、NEENAをその生来のプロモーターと機能的に連結し、ここで前記プロモーターに対するNEENAの位置を変えてプロモーターがかかる遺伝子操作後にさらに高い発現を示す場合も、NEENAは機能的に連結された核酸に対して異種である。
【0029】
本明細書で意味する核酸分子の増強された構成的発現を示す植物は、それぞれの核酸分子と機能的に連結されたそれぞれのNEENA無しに同じ条件下で育てた対照植物と比較して、より高い、好ましくは、統計的に有意に高い核酸分子の構成的発現を有する植物を意味する。かかる対照植物は、本発明の植物と同じ遺伝子を制御する同じプロモーターを含み、そのプロモーターが本発明のNEENAと連結されてない野生型植物またはトランスジェニック植物であってもよい。
【0030】
本明細書で使用する植物の産生は、安定して形質転換する、例えば、組換えDNA構築物を植物またはその部分中に、アグロバクテリウム媒介性形質転換、プロトプラスト形質転換、粒子ボンバードメントその他などの方法によって導入し、そして場合によっては引き続いてトランスジェニック植物を再生する方法を含むものである。これはまた、植物またはその部分をウイルス感染またはアグロバクテリウム浸潤などにより一過的に形質転換する方法も含む。当業者は植物またはその部分を、安定しておよび/または一過的に形質転換するさらなる方法を知っている。繁殖方法またはプロトプラスト融合などの手法も本発明の植物の生産に使うことができて、本発明に包含される。
【0031】
本発明の方法は、任意の植物、例えば裸子植物または被子植物、好ましくは被子植物、例えば双子葉または単子葉植物、好ましくは双子葉植物に応用することができる。好ましい単子葉植物は、例えばトウモロコシ、コムギ、イネ、オオムギ、モロコシ、バナナ、サトウキビ、ススキおよびブラキポディウムであり、とりわけ好ましい単子葉植物はトウモロコシ、コムギおよびイネである。好ましい双子葉植物は、例えばダイズ、アブラナ種子、アブラナ、亜麻仁、ワタ、ジャガイモ、サトウダイコン、マンジュギクおよびシロイヌナズナ、とりわけ好ましい双子葉植物はダイズ、アブラナ種子、アブラナおよびジャガイモである。
【0032】
本発明の一実施形態において、以上定義した方法は次のステップ:
a)先にi)〜vi)に定義した核酸分子を含む1以上のNEENAを植物またはその部分中に導入するステップ、および
b)前記1以上の NEENAを前記植物またはその部分のゲノム中に組込むステップであって、ここで前記1以上のNEENAは、前記1以上のNEENAに対して異種である内因性の、好ましくは、構成的に発現された核酸と機能的に連結されているステップ、および、任意に、
c)前記形質転換した細胞由来の前記1以上のNEENAを含む植物またはその部分を再生するステップ
を含むものである。
【0033】
NEENAは、NEENAが機能的に連結された前記プロモーターの制御下にある核酸分子に対して異種であってもよく、または、プロモーターおよび前記プロモーターの制御下の核酸分子の両方に対して異種であってもよい。
【0034】
1以上のNEENA分子を、植物またはその部分中に、粒子ボンバードメント、プロトプラストエレクトロポレーション、ウイルス感染、アグロバクテリウム媒介性形質転換または当技術分野で公知の任意の他の手法を用いて導入してもよい。NEENA分子を、例えば、プラスミドまたはウイルスDNAまたはウイルスRNA中に導入して組込んでもよい。NEENA分子をまた、BAC、YACまたは人工染色体に含ませた後、植物または植物の部分中に導入してもよい。これをまた、NEENA配列を含む直鎖の核酸分子として導入してもよく、その場合、核酸分子上のNEENA配列に隣接してさらなる配列が存在してもよい。NEENA配列に隣接するこれらの配列は約20bp、例えば20bp〜数100bp、例えば100bp以上であってもよく、そして例えば、相同組換えによりゲノム中への組込みを促進することができる。ゲノム組込みのための任意の他の方法を用いてもよく、標的化組込み手法、例えば相同性組換えまたは無作為組込み手法、例えば非正統的組換えであってもよい。
【0035】
NEENA分子が機能的に連結された、内因性の好ましくは構成的に発現される核酸は、任意の核酸、好ましくは任意の構成的に発現される核酸分子であってもよい。核酸分子は、タンパク質をコードする核酸分子または非コーディング分子、例えば、アンチセンスRNA、rRNA、tRNA、miRNA、ta-siRNA、siRNA、dsRNA、snRNA、snoRNAまたは当技術分野で公知の任意の他の非コーディングRNAであってもよい。
【0036】
当業者は、本発明の方法を、好ましくは、例えばマイクロアレイチップハイブリダイゼーション、qPCR、ノーザンブロット分析、次世代配列決定などにより適用することができる構成的に発現された核酸分子を同定する方法を知っている。
【0037】
本発明の方法を実施するさらなる手順は、
a)プロモーター、好ましくは先に定義した構成的プロモーターと機能的に連結され、かつ、1以上の核酸分子(この核酸分子は前記1以上のNEENAに対して異種でありかつ前記プロモーター、好ましくは構成的プロモーターの制御下にある)と機能的に連結された、1以上のNEENA(このNEENAは先にi)〜vi)に定義した核酸分子を含む)を含むものである発現構築物を提供するステップ、および
b)前記1以上のNEENAを含む発現構築物を、前記植物またはその部分のゲノム中に組込むステップ、および、任意に
c)前記形質転換した植物またはその部分から前記1以上の 発現 構築物を含む植物またはその部分を再生するステップ
であってもよい。
【0038】
NEENAは、このNEENAが機能的に連結されたプロモーターの制御下にある核酸分子に対して異種であってもよく、または前記プロモーターと前記プロモーターの制御下の核酸分子との両方に対して異種であってもよい。
【0039】
発現構築物は任意の当技術分野で公知の方法によってそれぞれの植物のゲノム中に組込むことができる。組込みは粒子ボンバードメントまたはアグロバクテリウム媒介性形質転換などの方法を用いて無作為であってもよい。好ましい実施形態において、組込みは、例えば相同性組換えによる標的化組込み経由であってもよい。後者の方法は、NEENAと機能的に連結された高発現プロモーターを含む発現構築物をお気に入りのゲノム領域中に組込むことを可能にする。好都合なゲノム領域は、例えば、高度に発現される遺伝子を含むことが公知のゲノム領域、例えば種子中であり、従って、転写活性を示さないゲノム領域と比較して前記発現構築物由来の発現を増加することができる。
【0040】
他の好ましい実施形態においては、前記1以上のNEENAをプロモーター、好ましくは前記異種核酸分子の転写出発部位に近い構成的プロモーターと機能的に連結する。
【0041】
本明細書で意味する「転写出発部位に近い」は、前記異種核酸分子の転写出発部位から2500bp以下、好ましくは2000bp以下、より好ましくは1500bp以下、さらにより好ましくは1000bp以下および最も好ましくは500bp以下離れたプロモーター、好ましくは構成的プロモーターと、1以上のNEENAを機能的に連結することを含む。理解すべきこととして、NEENAはそれぞれのプロモーターの転写出発部位のそれぞれの距離の上流または下流に組込むことができる。従って、1以上のNEENAは必ず、1以上のNEENAが機能的に連結された好ましくは構成的プロモーターの制御下のそれぞれの異種核酸の転写物中に必ずしも含まれなくてもよい。好ましくは、1以上の NEENAはそれぞれのプロモーター、好ましくは 構成的プロモーターの転写出発部位の下流に組込まれる。転写出発部位の下流の組込み部位は5'UTR5、3'UTR、エキソンまたはイントロンにあってもよく、またはイントロンまたは部分的にもしくは完全に、好ましくは構成的プロモーターの制御下の異種核酸の5'UTRまたは3'UTRを置き換えてもよい。好ましくは、1以上のNEENAは5'UTRまたはイントロンに組込まれるか、またはNEENAはイントロンまたは一部分もしくは完全な5'UTRと置き換えられ、最も好ましくは、それぞれの異種核酸の5'UTRに組込まれる。
【0042】
本発明のさらなる実施形態は先にi)〜vi)に定義した核酸分子を含む1以上のNEENAを含む組換え発現構築物を含むものである。
【0043】
組換え発現構築物はさらに、1以上のNEENAが機能的に連結された1以上のプロモーター、好ましくは構成的プロモーターならびに任意に1以上の発現される核酸分子(後者は前記1以上のNEENAに対して異種である)を含んでもよい。
【0044】
NEENAは、このNEENAが機能的に連結されたプロモーターの制御下にある核酸分子に対して異種であってもよく、または前記プロモーターと前記プロモーターの制御下の核酸分子との両方に対して異種であってもよい。
【0045】
発現構築物は、NEENAと機能的に連結されたプロモーター、好ましくは構成的プロモーターとそれぞれのNEENAに対して異種の発現される核酸分子との1以上、例えば2以上、例えば5以上、例えば10以上の組み合わせを含んでもよい。発現構築物はまた、それぞれのプロモーターに対して同族または異種の発現される核酸分子と機能的に連結された、NEENAを含まないさらなるプロモーターを含んでもよい。
【0046】
先に定義した1以上の組換え発現構築物を含む組換え発現ベクターは本発明の他の実施形態である。本発明に使用しうる複数の発現ベクターは当業者に知られている。例えばNEENAと機能的に連結されたプロモーターおよび、任意に、ターミネーターなどの他のエレメントを含む発現構築物を含むものであるベクターを植物のゲノム中に導入し、形質転換した細胞からトランスジェニック植物を回収する方法もまた、当技術分野で公知である。植物またはその部分を形質転換するために用いる方法に応じて、全ベクターを前記植物またはその部分のゲノム中に組込んでもよく、または、ベクターのある特定成分、例えばT-DNAなどをゲノム中に組込んでもよい。
【0047】
先にi)〜vi)に定義した1以上の異種NEENAを含むトランスジェニック植物またはその部分も本発明に包含される。NEENAは、もしそれが合成されたか、他生物由来であるか、または同じ生物であるがその自然のゲノム配置が対照植物、例えば野生型植物と比較して手を加えられていれば、その植物にとって異種であると理解すべきである。理解すべきこととして、手を加えられたゲノム配置とは、NEENAが他の染色体上にまたは同じ染色体であるが、その自然の、例えば野生型植物のゲノム配置から10kb以上、例えば10kb、好ましくは5kb以上、例えば5kb、より好ましくは1000bp以上、例えば1000bp、さらにより好ましくは500bp以上、例えば500bp、とりわけ好ましくは100bp以上、例えば100bp、最も好ましくは10bp以上、例えば10bpだけ異なった位置に配置されていることを意味する。
【0048】
先に定義した組換え発現ベクターまたは先に定義した組換え発現構築物を含むトランスジェニック細胞またはトランスジェニック植物またはその部分は本発明のさらなる実施形態である。トランスジェニック細胞、トランスジェニック植物またはその部分は、細菌、真菌、酵母または植物、昆虫または哺乳動物細胞または植物からなる群より選択することができる。好ましくはトランスジェニック細胞は、細菌、真菌、酵母または植物細胞である。好ましい細菌は、腸内細菌、例えば大腸菌およびアグロバクテリア属の細菌、例えばAgrobacterium tumefaciensおよびAgrobacterium rhizogenesである。好ましい植物は、単子葉または双子葉植物、例えば単子葉または双子葉作物植物、例えばトウモロコシ、ダイズ、アブラナ、ワタ、ジャガイモ、サトウダイコン、イネ、コムギ、モロコシ、オオムギ、ススキ、バナナ、サトウキビその他である。好ましい作物植物はトウモロコシ、イネ、コムギ、ダイズ、アブラナ、ワタまたはジャガイモである。とりわけ好ましい双子葉類の作物植物はダイズ、アブラナ、ワタまたはジャガイモである。
【0049】
とりわけ好ましい単子葉類の作物植物はトウモロコシ、コムギおよびイネである。
【0050】
前記異種の先にi)〜vi)に定義したNEENAまたは先に定義した前記組換え発現構築物または前記組換えベクターを含む、先に定義したトランスジェニック細胞または植物またはその部分から誘導されるトランスジェニック細胞培養、トランスジェニック種子、部分または繁殖材料は本発明の他の実施形態である。
【0051】
本明細書で意味するトランスジェニック部分または繁殖材料は、それぞれのNEENA、発現構築物または組換えベクターを含む切り穂、接ぎ穂、取り木、枝または若枝などの植物の繁殖および/または再生に有用である全ての組織および器官、例えば葉、幹および果実を含むものである。
【0052】
本発明のさらなる実施形態は、先にi)〜vi)に定義したNEENAまたは先に定義した組換え構築物または組換えベクターの、植物またはその部分における発現の増強のための使用である。
【0053】
従って、本出願は、1以上のNEENAと機能的に連結された1以上のプロモーター、好ましくは、種子特異的および/または種子選好的プロモーターを含むものである、種子特異的および/または種子選好的な遺伝子発現を増強する核酸分子を提供する。さらに、かかる遺伝子発現を増強する核酸分子、ならびにかかる遺伝子発現を増強する核酸分子を含む発現構築物、発現ベクター、トランスジェニック植物またはその部分およびトランスジェニック細胞を提供する。
【0054】
先に定義したトランスジェニック細胞または植物またはその部分から誘導されるトランスジェニック細胞培養、トランスジェニック種子、部分または繁殖材料の、食糧、動物飼料、種子、医薬品またはファインケミカルを生産するための使用も本発明に包含される。
【0055】
定義
略語は次の通りである:NEENA、核酸発現増強性核酸;GFP、緑色蛍光タンパク質;GUS、β-グルクロニダーゼ;BAP、6-ベンジルアミノプリン;2,4-D、2,4-ジクロロフェノキシ酢酸;MS、MurashigeおよびSkoog培地;NAA、1-ナフタレン酢酸;MES、2-N-(モルホリノ)エタンスルホン酸;IAA、インドール酢酸;Kan、カナマイシン硫酸塩;GA3、ジベレリン酸;Timentin(登録商標)、チカルシリン二ナトリウム/カリウムクラブラネート;microl、マイクロリットル。
【0056】
理解すべきこととして、本発明は特別な手法またはプロトコルに限定されるものでない。理解すべきこととしてまた、本明細書で使用する用語法は特別な実施形態を記載することだけを目的とし、添付した請求の範囲が限定しうる本発明の範囲を限定することを意図しない。注意しなければならないこととして、本明細書および添付した請求の範囲における使用で、単数形の冠詞「a」「an」および「the」は、文脈が明確に断らない限り、複数の言及を含むものである。従って、例えば、「a」ベクターは1以上のベクターへの言及であり、当業者に公知のその等価物などを含むものである。本明細書で使用する用語「約」は、およそ、大雑把に、大体、またはその範囲のを意味する。用語「約」を数字の範囲と共に用いる場合、この用語は記載した数値の上限および下限の境界を拡げることにより範囲を改変する。一般に、本明細書では用語「約」を用いて、記載した値の上限および下限の数値を20%、好ましくは10%上方もしくは下方(高いもしくは低い)の分散をもつ値に改変する。本明細書で使用する用語「または」は特別なリストの任意の1つのメンバーを意味し、そのリストのメンバーの任意の組み合わせも含むものである。用語「含むものである(compriseとその変化形)」「含む(includeとその変化形)」は、本明細書および以下の請求の範囲で使用する場合、1以上の記載した特徴、整数、成分、またはステップの存在を説明することを意図するが、これらの用語は他の特徴、整数、成分、またはステップ、またはその群の存在もしくは付加を排除するものではない。明確にするために述べると、本明細書に使用するある特定の用語は次の通り定義されかつ使用される。
【0057】
アンチパラレル:ここで「アンチパラレル」は、5'-3'方向に走る1つのヌクレオチド配列と3'-5'方向に走る他のヌクレオチド配列との、リン酸ジエステル結合をもつ相補性塩基残基の間の水素結合を介して対合した2つのヌクレオチド配列を意味する。
【0058】
アンチセンス:用語「アンチセンス」は、転写または機能のためのその通常の配向に対して反転していて、そのために、宿主細胞内で発現される標的遺伝子mRNA分子と相補的であるRNA転写物を発現する(例えば、これは標的遺伝子mRNA分子または1本鎖ゲノムDNAとワットソン-クリック塩基対合を介してハイブリダイズすることができる)か、または標的DNA分子、例えば、宿主細胞に存在するゲノムDNAと相補的であるヌクレオチド配列を意味する。
【0059】
コード領域:本発明で使用する用語「コード領域」は、構造遺伝子を言及して用いる場合、mRNA分子の翻訳の結果として新生ポリペプチドに見出されるアミノ酸をコードするヌクレオチド配列を意味する。コード領域は、真核生物においては、5'側にヌクレオチドトリプレット「ATG」(イニシエーターのメチオニンをコードする)および3'側に停止コドンを表す3つのトリプレット(すなわち、TAA、TAG、TGA)の1つが結合されている。イントロンを含有するだけでなく、遺伝子のゲノム形態はまた、RNA転写物に存在する配列の5'と3'末端の両方に位置する配列も含みうる。これらの配列は、「隣接する(flanking)」配列または領域と呼ばれる(これらの隣接する領域はmRNA転写物に存在する非翻訳配列に対して5'もしくは3'に位置する)。5'に隣接する領域は、調節配列、例えば、遺伝子の転写を制御または影響するプロモーターおよびエンハンサーを含有しうる。3'に隣接する領域は、転写の終結、転写後の切断およびポリアデニル化を指令する配列を含有しうる。
【0060】
相補性:「相補的」または「相補性」は、アンチパラレルなヌクレオチド配列が相補的塩基残基間で水素結合を形成すると(塩基対合規則により)お互いに対合することができるアンチパラレルなヌクレオチド配列を含むものである、2つのヌクレオチド配列を意味する。例えば、配列 5'-AGT-3'は配列 5'-ACT-3'と相補的である。相補性は「部分的」であってもまたは「全体」であってもよい。「部分的」相補性は、1以上の核酸塩基が塩基対合規則によってマッチしない場合である。「全体」または「完全」相補性は、それぞれおよび全ての核酸塩基が他の塩基と、塩基対合規則のもとでマッチする場合である。核酸分子鎖間の相補性の程度は、核酸分子鎖間のハイブリダイゼーションの効率と強度に有意な効果を有する。本明細書で使用する核酸配列の「相補配列」は、その核酸分子が核酸配列の核酸分子に対して全体相補性を示すヌクレオチド配列を意味する。
【0061】
2本鎖RNA:「2本鎖 RNA」分子または「dsRNA」分子は、ヌクレオチド配列のセンスRNA断片とヌクレオチド配列のアンチセンスRNA断片を含むものであって、両方のRNA断片がお互いに相補的なヌクレオチド配列を含み、それにより、センスとアンチセンスRNA断片が対合して2本鎖RNA分子を形成することができる。
【0062】
内因性:「内因性」ヌクレオチド配列は、未翻訳の植物細胞のゲノム中に存在するヌクレオチド配列を意味する。
【0063】
発現の増強:植物細胞中の核酸分子の発現の「増強」または「増加」を本明細書では同じ意味で使用し、本発明の方法の適用後の植物、植物の部分または植物細胞における核酸分子の発現のレベルは、本発明の方法の適用前の植物、植物の部分または植物細胞におけるその発現のレベルよりまたは本発明の組換え核酸分子を欠く参照植物と比較して高いことを意味する。例えば、参照植物はそれぞれのNEENAを欠くだけである同じ構築物を含むものである。本明細書で使用する用語「増強」または「増加」は同義であり、ここでは発現すべき核酸分子のより高い、好ましくは有意により高い発現を意味する。本明細書で使用する、作用物質、例えばタンパク質、mRNAまたはRNAのレベルの「増強」または「増加」は、本発明の組換え核酸分子を欠く、例えば、本発明のNEENA分子、組換え構築物または組換えベクターを欠く、実質的に同一条件のもとで育てた実質的に同一の植物、植物の部分、または植物細胞と比較して、そのレベルが増加したことを意味する。本明細書で使用する、標的遺伝子により発現された作用物質、例えばpreRNA、mRNA、rRNA、tRNA、snoRNA、snRNAのおよび/またはそれがコードするタンパク質産物のレベルの「増強」または「増加」は、そのレベルが本発明の組換え核酸分子を欠く細胞または生物と比較して、50%以上、例えば100%以上、好ましくは200%以上、より好ましくは5倍以上、さらにより好ましくは10倍以上、最も好ましくは20倍以上例えば50倍増加することを意味する。その増強または増加は、当業者になじみのある方法により測定することができる。従って、核酸またはタンパク質の量の増強または増加はタンパク質の免疫学的検出により測定することができる。さらに、タンパク質アッセイ、蛍光、ノーザンハイブリダイゼーション、ヌクレアーゼ保護アッセイ、逆転写(定量RT-PCR)、ELISA(酵素結合免疫吸着アッセイ)、ウェスタンブロット、ラジオイムノアッセイ(RIA)または他のイムノアッセイおよび蛍光活性化細胞分析(FACS)などの技法を使って植物または植物細胞中の特定のタンパク質またはRNAを測定することができる。誘発されるタンパク質産物の型に応じて、生物または細胞の表現型に与えるその活性または効果を測定してもよい。タンパク質の量を測定する方法は当業者に公知である。記載しうる例には次のものがある:マイクロ-ビウレット法(Goa J (1953) Scand J Clin Lab Invest 5:218-222)、フォーリン-チオカルト法(Lowry OH ら (1951) J Biol Chem 193:265-275) または CBB G-250の吸収を測定する方法(Bradford MM (1976) Analyt Biochem 72:248-254)。タンパク質の活性を定量化する一例として、ルシフェラーゼ活性の検出を以下の実施例に記載した。
【0064】
発現:「発現」は細胞における、遺伝子産物の生合成、好ましくは、ヌクレオチド配列、例えば、内因性遺伝子または異種遺伝子の転写および翻訳を意味する。例えば、構造遺伝子の事例では、発現は構造遺伝子のmRNA中への転写および、場合によっては、引き続いてmRNAの1以上のポリペプチドへの翻訳に関わる。他の場合では、発現はRNA分子を保持するDNAの転写だけに関わりうる。
【0065】
発現構築物:本明細書で使用する「発現構築物」は、適当な植物の部分または植物細胞中の特別なヌクレオチド配列の発現を指令することができるDNA配列であって、目的のヌクレオチド配列と作動しうる形で連結された(場合によっては、終結シグナルと作動しうる形で連結された)、その中へ導入されるであろう前記植物の部分または植物細胞中で機能的なプロモーターを含む前記DNA配列を意味する。もし翻訳が必要であれば、発現構築物は典型的には、ヌクレオチド配列の適当な翻訳に必要な配列も含む。コード領域は目的のタンパク質をコードしうるがまた、目的の機能性RNA、例えばRNAa、siRNA、snoRNA、snRNA、マイクロRNA、ta-siRNAまたは任意の他の非コード調節性RNAをセンスまたはアンチセンス方向にコードしてもよい。目的のヌクレオチド配列を含む発現構築物はキメラ(1以上のその成分が1以上のその他の成分に対して異種であることを意味する)であってもよい。発現構築物はまた、天然であるが、異種発現にとって有用な組換え体の形態で得たものであってもよい。しかし典型的には、発現構築物は宿主に対して異種であり、すなわち、発現構築物の特別なDNA配列は宿主細胞で天然に生じるものでなく、宿主細胞中に宿主細胞の祖先中に形質転換事象によって導入されてなければならない。発現構築物中のヌクレオチド配列の発現は、構成的プロモーターまたは、宿主細胞をいくつかの特別な外界の刺激に曝したときにだけ転写を開始する誘導プロモーターの制御のもとで生じる。植物の事例では、プロモーターも特別な組織または器官または発生段階に特異的でありうる。
【0066】
外来:用語「外来」は、細胞のゲノム中に実験操作により導入されたいずれかの核酸分子(例えば、遺伝子配列)を意味し、導入された配列がいくつかの改変(例えば、点突然変異、選択マーカー遺伝子の存在など)を含有する限り、天然の配列とは異なる配列がその細胞中に見出される。
【0067】
機能的連結:用語「機能的連鎖」または「機能的連結」は、調節エレメント(例えば、プロモーター)の発現すべき核酸配列との、および、適宜、さらなる調節エレメント(例えば、ターミネーターまたはNEENAなど)との配列の配置であって、それぞれの調節エレメントがその意図する機能を完遂して前記核酸配列の発現を可能にし、改変し、促進しまたはさもなくば影響を与え得る前記配列の配置を意味すると理解される。同義語として、表現「作動しうる連鎖」または「作動しうる連結」を使用してもよい。発現は、核酸配列のセンスまたはアンチセンスRNAと関係した配置に依存してもたらされうる。この目的にとって、化学的意味の直接連結は必ずしも必要でない。例えば、エンハンサー配列などの遺伝子制御配列も、さらに離れた位置から、 または他のDNA分子からでも、標的配列に対するその機能を果たしうる。好ましい配置は、発現すべき核酸配列が組換えによってプロモーターとして作用する配列の後方に位置し、2つの配列がお互いに共有結合で連結される配置である。プロモーター配列と組換えで発現すべき核酸配列の間の距離は、好ましくは200塩基対未満、とりわけ好ましくは100塩基対未満、非常にとりわけ好ましくは50塩基対未満である。好ましい実施形態においては、転写開始が本発明のキメラRNAの所望の開始と同一であるように、転写すべき核酸配列をプロモーターの後ろに配置する。機能的連鎖、および発現構築物は、記載された通例の組換えおよびクローニング技法を用いて作製することができる(例えば、Maniatis T, Fritsch EF and Sambrook J (1989) Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor (NY);Silhavy et al. (1984) Expeiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor (NY);Ausubel et al., (1987) Current Protocols in Molecular Biology, Greene Publishing Assoc. and Wiley Interscience;Gelvin et al., (Eds) (1990) Plant Molecular Biology Manual; Kluwer Academic Publisher, Dordrecht, The Netherlands)。しかし、例えば、制限酵素に対する特異的切断部位をもつリンカーとして、またはシグナルペプチドとして作用するさらなる配列も2つの配列の間に配置することができる。配列の挿入はまた、融合タンパク質の発現をもたらしうる。好ましくは、調節領域、例えばプロモーターおよび発現すべき核酸配列の連鎖から成る発現構築物をベクターに組込まれた形で存在させてもよく、例えば、形質転換により植物ゲノム中に挿入することができる。
【0068】
遺伝子:用語「遺伝子」は、遺伝子産物(例えば、ポリペプチドまたは機能性RNA)の発現をいくつかの方式で調節することができる適当な調節配列と作動しうる形で接続された領域を意味する。遺伝子は、コード領域(オープンリードフレーム、ORF)に先行する(上流の)および後続する(下流の)DNAの非翻訳調節領域(例えば、プロモーター、エンハンサー、リプレッサーなど)ならびに、適用可能であれば、個々のコード領域(すなわち、液損)の間に介在配列(すなわち、イントロン)を含む。本明細書で使用する用語「構造遺伝子」は、mRNAに転写され、次いで特定のポリペプチドの特徴であるアミノ酸の配列に翻訳されるDNA配列を意味することを意図している。
【0069】
ゲノムとゲノムDNA:用語「ゲノム」または「ゲノムDNA」は、宿主生物の遺伝性の遺伝子情報を意味する。前記ゲノムDNAは核DNA(染色体DNAとも呼ばれる)を含むがまた、色素体(例えば、葉緑体)および他の細胞小器官(例えば、ミトコンドリア)のDNAも含む。好ましくは、用語「ゲノム」または「ゲノムDNA」は核の染色体DNAを意味する。
【0070】
異種:核酸分子またはDNAに関する用語「異種」は、自然では作動しうる形で連結されてないまたは自然では異なる位置で作動しうる形で連結された第2の核酸分子と、作動しうる形で連結された、または作動しうる形で連結されるように遺伝子操作された核酸分子を意味する。核酸分子およびそれと連結された1以上の調節核酸分子(プロモーターまたは転写終結シグナルなど)を含む異種発現構築物は、例えば、実験遺伝子操作に由来する構築物であって、a)前記核酸分子、またはb)前記調節核酸分子、またはc)両方(すなわち(a)と(b))がその自然(生来)の遺伝環境に位置しないか、または、実験遺伝子操作により改変されている(改変の例は1以上のヌクレオチド残基の置換、付加、欠失、反転または挿入である)前記構築物である。
【0071】
自然の遺伝環境は、起源の生物における自然の染色体遺伝子座、またはゲノムライブラリー中の存在を意味する。ゲノムライブラリーの場合、核酸分子の配列の自然の遺伝環境は好ましくは、少なくとも部分において保持される。前記環境は核酸配列に少なくとも1つの側において隣接し、そして少なくとも50bp、好ましくは少なくとも500bp、とりわけ好ましくは少なくとも1,000bp、非常にとりわけ好ましくは少なくとも5,000bpの長さの配列を有する。天然の発現構築物(例えば、プロモーターの対応する遺伝子との天然の組み合わせ)は、非自然、合成の「人工的」方法、例えば変異誘発により改変されると、トランスジェニック発現構築物になる。かかる方法は記載されている(US 5,565,350;WO 00/15815)。例えば、この分子の生来のプロモーターでないプロモーターと作動しうる形で連結されたタンパク質をコードする核酸分子は、プロモーターについて異種であるとみなされる。好ましくは、それを導入した細胞に対して内因性でないかまたは自然で随伴しないが、他の細胞から得られたかまたは合成された異種DNAは内因性でない。異種DNAはまた、内因性DNA配列のいくつかの改変、非天然、多コピーを含有する内因性DNA配列、それに物理的に連結された他のDNA配列を自然で随伴しないDNA配列を含む。一般に、必ずではないが、異種DNAはそれが発現される細胞により通常産生されないRNAまたはタンパク質をコードする。
【0072】
高発現性構成的プロモーター:本明細書で使用する「高発現性構成的プロモーター」は、植物またはその部分における構成的発現を引き起こすプロモーターであって、それぞれのプロモーターの制御下の核酸分子由来のRNAの蓄積または合成速度またはRNAの安定性が、本発明のNEENAを欠くプロモーターにより引き起こされる発現より高い、好ましくは、有意に高い前記プロモーターを意味する。好ましくは、RNAの量および/またはRNA合成の速度および/またはRNAの安定性が、本発明のNEENAを欠く構成的プロモーターと比較して、50%以上,例えば100%以上,好ましくは200%以上,より好ましくは5倍以上,さらにより好ましくは10倍以上,最も好ましくは20倍以上例えば50倍増加する。
【0073】
ハイブリダイゼーション:本明細書で使用する用語「ハイブリダイゼーション」は「核酸分子の1つの鎖が塩基対合を介して相補鎖に参加する任意のプロセス」を含む(J. Coombs (1994) Dictionary of Biotechnology, Stockton Press, New York)。ハイブリダイゼーションおよびハイブリダイゼーションの強度(すなわち、核酸分子間の連合の強度)は、核酸分子間の相補性の程度、関わる条件のストリンジェンシー、形成されるハイブリッドのTm、および核酸分子内のG:C比などの因子の影響を受ける。本明細書で使用する用語「Tm」は「融点」の意味で使用される。融点は2本鎖の核酸分子が半分に解離して1本鎖になる温度である。核酸分子のTmを計算する式は当技術分野において周知である。標準の参考文献に示されるように、核酸分子が1M NaClの水溶液中に含まれる場合、Tm値の単純な計算は式:Tm=81.5+0.41(% G+C)により計算することができる[例えば、Anderson and Young, Quantitative Filter Hybridization, in Nucleic Acid Hybridization (1985)を参照]。他の参考文献は、構造ならびに配列の特徴を考慮に入れてTmを計算するさらに複雑な計算を含む。厳密な条件は、当業者に公知であり、Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6に記載されている。
【0074】
「同一性」:「同一性」は、2以上の核酸またはアミノ酸分子の比較について使用する場合、前記分子がある特定の程度の配列類似性を共有し、配列が部分的に同一であることを意味する。
【0075】
2つのアミノ酸配列または2つの核酸分子のパーセント同一性(本明細書では相同性を互換的に使用する)を決定するために、配列の1つを他の配列の下に書いて最適な比較を行う(例えば、ギャップを1つのタンパク質の配列中に挿入し、他のタンパク質または他の核酸と最適なアラインメントを作り出すことができる)。
【0076】
対応するアミノ酸位置またはヌクレオチド位置のアミノ酸残基または核酸分子を次いで比較する。もし1つの配列の1つの位置が他配列の対応する位置と同じアミノ酸残基または同じ核酸分子により占められていれば、その分子はこの位置において相同性である(すなわち、本明細書の文脈で使用するアミノ酸または核酸「相同性」はアミノ酸または核酸「同一性」に対応する)。2つの配列間のパーセント同一性は、配列が共有する同一の位置の数の関数(すなわち、%同一性=同一の位置の数/位置の全数)である。用語「相同性」および「同一性」は従って、同義語と考えるべきである。
【0077】
2以上のアミノ酸または2以上のヌクレオチド配列のパーセント同一性 を確認するために、いくつかのコンピューターソフトウエアプログラムが開発されている。2以上配列の同一性は、例えば、ソフトウエアfastaを用いて計算することができ、これは現在バージョンfasta 3が使用されている(W. R. Pearson and D. J. Lipman, PNAS 85, 2444(1988);W. R. Pearson, Methods in Enzymology 183, 63 (1990);W. R. Pearson and D. J. Lipman, PNAS 85, 2444 (1988);W. R. Pearson, Enzymology 183, 63 (1990))。異なる配列の同一性を計算する他の有用なプログラムは標準blastプログラムであって、これはBiomax pedantソフトウエアに含まれている(Biomax、Munich、Federal Republic of Germany)。このソフトウエアは残念ながら、 blastが主題およびクエリの完全な配列を常に含まないので、最適性の劣る結果をもたらすことがある。それにも関わらず、このプログラムは非常に効率的であるので、膨大な数の配列を比較するために用いることができる。以下の設定は、配列のかかる比較のための典型的なものである。
【0078】
-p プログラム名 [文字列]; -d データベース [文字列]; デフォルト = nr; -i クエリーファイル[File In]; デフォルト = stdin; -e 期待値 (E) [実数]; デフォルト = 10.0; -m アラインメントビューオプション: 0 = ペアワイズ; 1 = 同一性を示すクエリー-アンカー; 2 = 同一性を示さないクエリー-アンカー; 3 = フラットクエリー-アンカー、同一性を示す; 4 = フラットクエリー-アンカー、同一性を示さない; 5 = フラットクエリー-アンカー、同一性を示さない、かつ平滑末端; 6 = フラットクエリー-アンカー、同一性を示さない、かつ平滑末端; 7 = XML Blast出力; 8 = 表形式; 9 コメントラインを含む表形式[整数]; デフォルト = 0; -o BLASTリポート出力ファイル[File Out] オプション; デフォルト = stdout; -F フィルタークエリー配列(blastnを用いるDUST、他を用いるSEG) [文字列]; デフォルト = T; -G ギャップを開くためのコスト(0はデフォルト行動を引き起こす) [整数]; デフォルト = 0; -E ギャップを拡張するためのコスト(0はデフォルト行動を引き起こす) [整数]; デフォルト = 0; -X ギャップ付加されたアラインメントに関するX下落値(ビット) (0はデフォルト行動を引き起こす); blastn 30、megablast 20、tblastx 0、他の全部15 [整数]; デフォルト = 0; -I デフラインGIを示す[T/F]; デフォルト = F; -q ヌクレオチド不一致に関するペナルティ(blastnのみ) [整数]; デフォルト = -3; -r ヌクレオチド一致に関する報酬(blastnのみ) [整数]; デフォルト = 1; -v (V)に関する1行記述を示すデータベース配列の数 [整数]; デフォルト = 500; -b (B)に関するアラインメントを示すデータベース配列の数 [整数]; デフォルト = 250; -f 拡張ヒットに関する閾値、デフォルト 0の場合; blastp 11、blastn 0、blastx 12、tblastn 13; tblastx 13、megablast 0 [整数]; デフォルト = 0; -g ギャップ付加アラインメントを実行(tblastxでは利用不可) [T/F]; デフォルト = T; -Q 使用するクエリー遺伝子コード[整数]; デフォルト = 1; -D DB遺伝子コード(tblast[nx]についてのみ) [整数]; デフォルト = 1; -a 使用するプロセッサーの数[整数]; デフォルト = 1; -O SeqAlignファイル[File Out] オプション; -J クエリーデフラインを信じる[T/F]; デフォルト = F; -M マトリックス[文字列]; デフォルト = BLOSUM62; -W ワードサイズ、デフォルト 0の場合 (blastn 11、megablast 28、他の全部 3) [整数]; デフォルト = 0; -z データベースの有効長さ(実際のサイズについては0を使用する) [実数]; デフォルト = 0; -K 保持する領域から得られる最良のヒットの数(デフォルトにより除去、使用する場合、100の値が推奨される) [整数]; デフォルト = 0; -P 複数ヒットについては0、単一ヒットについては1[整数]; デフォルト = 0; -Y 検索スペースの有効長さ(実際のサイズについては0を使用する) [実数]; デフォルト = 0; -S データベースに対して検索するためのクエリー・ストランド(blast[nx]、およびtblastxについて); 3は両方、1は上、2は下である[整数]; デフォルト = 3; -T HTML出力を作製 [T/F]; デフォルト = F; -l GIの一覧に対するデータベースの限定的検索 [文字列] オプション; -U FASTA配列のフィルタリングには小文字を使用する [T/F] オプション; デフォルト = F; -y ビットで示される非ギャップ付加拡張のためのX下落値(0.0はデフォルト行動を引き起こす); blastn 20、megablast 10、他の全部 7 [実数]; デフォルト = 0.0; -Z ビットで示される最終的なギャップ付加アラインメントに関するX下落値(0.0はデフォルト行動を引き起こす); blastn/megablast 50、tblastx 0、他の全部 25 [整数]; デフォルト = 0; -R PSI-TBLASTN チェックポイントファイル[File In] オプション; -n MegaBlast検索 [T/F]; デフォルト = F; -L クエリー配列上の位置[文字列] オプション; -A 複数ヒットウィンドウサイズ、デフォルト 0の場合(blastn/megablast 0、他の全部 40 [整数]; デフォルト = 0; -w フレームシフトペナルティ(blastxに関するOOFアルゴリズム) [整数]; デフォルト = 0; -t HSPsを連結するためにtblastnにおいて許容される最大のイントロンの長さ (0は連結不可) [整数]; デフォルト = 0。
【0079】
NeedlemanおよびWunsch、またはSmithおよびWatermanのアルゴリズムを用いることにより、高品質の結果が達成される。従って、前記アルゴリズムに基づくプログラムが好ましい。有利には、配列の比較を、プログラムPileUp(J. Mol. Evolution., 25, 351 (1987)、Higgins et al., CABIOS 5, 151 (1989))または、好ましくは、プログラム「Gap」および「Needle」(両方とも、NeedlemanおよびWunsch(J. Mol. Biol. 48; 443-453 (1970)のアルゴリズムに基づく)ならびに「BestFit」(SmithおよびWaterman(Adv. Appl. Math. 2; 482-489 (1981))のアルゴリズムに基づく)を用いて行うことができる。「Gap」および「Needle」はGCGソフトウェアパッケージ [Genetics Computer Group, 575 Science Drive, Madison, Wisconsin, USA 53711 (1991); Altschul ら(1997) Nucleic Acids Res. 25:3389 (1997)]の一部であり、「Needle」はThe European Molecular Biology Open Software Suite (EMBOSS)(Trends in Genetics 16 (6), 276 (2000))の一部である。従って、好ましくは、配列同一性のパーセントを決定する計算を、配列の全範囲にわたりプログラム「Gap」または「Needle」を用いて行う。核酸配列の比較のための以下の標準的な調整を「Needle」について用いた: マトリックス: EDNAFULL, ギャップ_ペナルティ: 10.0、 拡張_ペナルティ: 0.5。核酸配列の比較のための以下の標準的な調整を「Gap」について用いた:ギャップウエイト:50、長さウエイト:3、平均マッチ:10.000、平均ミスマッチ:0.000。
【0080】
例えば、核酸レベルで配列番号1の配列と80%同一性を有するといわれる配列は、配列番号1により表される配列と上記プログラム「Needle」により上記パラメーターセットを用いて比較すると80%同一性を有する配列を意味することがわかる。
【0081】
好ましくは、同一性はクエリー配列、例えば配列番号1の完全長さに基づいて計算する。
【0082】
イントロン:イントロンは、その遺伝子が産生するタンパク質の部分をコードせず、細胞核からエキスポートされる前に、遺伝子から転写されるmRNAからスプライシングで取除かれる遺伝子内のDNAのセクション(介在配列)を意味する。イントロン配列はイントロンの核酸配列を意味する。従って、イントロンは、コード配列(エキソン)と共に転写されるが、成熟mRNAの形成中に取り除かれるDNA配列の領域である。イントロンは実コード領域内にまたはプレmRNA(未スプライシングmRNA)の5'もしくは3'非翻訳リーダー内に位置しうる。一次転写物中のイントロンは切除され、コード配列は同時にかつ正確にライゲートされて成熟RNAを形成する。イントロンとエキソンの接合部がスプライス部位を形成する。イントロンの配列はGUで始まりかつAGで終わる。さらに、植物においては、2例のAU-AC イントロンが記載されている;シロイヌナズナ由来のRecA様タンパク質遺伝子の第14番目イントロンおよびG5遺伝子の第7イントロンはAT-ACイントロンである。イントロンを含有するプレmRNAは、(他の配列に加えて)正確にスプライスされるイントロンにとって必須である3つの短い配列を有する。これらの配列は5'スプライス位置、3'スプライス部位、および分岐点である。mRNAスプライシングは一次mRNA転写物中に存在する介在配列(イントロン)の除去およびエキソン配列の接続またはライゲーションである。これはまた、介在配列(イントロン)の除去と共に同じmRNA上の2つのエキソンを接続するcis-スプライシングとして公知である。イントロンの機能性エレメントは、スプライセソーム(例えば、イントロンの末端のスプライシングコンセンサス配列)の特異的タンパク質成分により認識されかつ結合される配列を含んでいる。機能性エレメントのスプライセオソームとの相互作用によって、成熟前mRNAからのイントロン配列の除去とエキソン配列の再接続がもたらされる。イントロンは3つの短い配列を有し、これらは、イントロンが正確にスプライスされるために(十分ではないが)必須である。これらの配列は、5'スプライス部位、3'スプライス部位および分岐点である。分岐点配列は植物におけるスプライシングおよびスプライス部位選択において重要である。分岐点配列は通常、3'スプライス部位の10〜60ヌクレオチド上流に位置する。
【0083】
同質遺伝子生物:異種DNA配列の存在または非存在によって異なりうることを除いて、遺伝的に同一である生物(例えば、植物)。
【0084】
単離された:本明細書で使用する用語「単離された」は、材料が人手により取り出されていて、その元来の、生来の環境から離れて存在し、それ故に自然の産物でないことを意味する。単離された材料または分子(DNA分子または酵素など)は精製された形態で存在しうるしまたは例えば、トランスジェニック宿主細胞などの非生来の環境で存在しうる。例えば、生存植物中に存在する天然のポリヌクレオチドまたはポリペプチドは単離されてないが、自然の系で共存する材料のいくつかのまたは全てから分離された同じポリヌクレオチドまたはポリペプチドは単離されている。かかるポリヌクレオチドはベクターの部分でありうるしおよび/またはかかるポリヌクレオチドまたはポリペプチドは組成物の部分でありうるのであって、かかるベクターまたは組成物が元来の環境の部分でないという点では単離されているであろう。好ましくは、核酸分子に関係して、「単離された核酸配列」のように使用される場合の「単離された」は、同定され、自然源で元来随伴している少なくとも1つの夾雑核酸分子から分離された核酸配列を意味する。単離された核酸分子は、自然で見出されるのとは異なる形態または設定で存在する核酸分子である。対照的に、単離されてない核酸分子は、それらが自然で存在する状態で見出されたDNAおよびRNAなどの核酸分子である。例えば、所与のDNA配列(例えば、遺伝子)は宿主細胞染色体において、隣接する遺伝子の近位で見出され;RNA配列、例えば、特定のタンパク質をコードする特定のmRNA配列は細胞において、多数のタンパク質をコードする数多くの他のmRNAとの混合物として見出される。しかし、例えば配列番号1を含む単離された核酸配列は、例として挙げれば、配列番号1を通常含有する細胞中のかかる核酸配列であって、その核酸配列が自然の細胞とは異なる染色体もしくは染色体外の位置にあるか、または、さもなくば、自然に見出されるのとは異なる核酸配列が隣接する前記核酸配列を含むものである。単離された核酸配列は1本鎖または2本鎖の形態で存在しうる。単離された核酸配列をタンパク質を発現するために利用する場合、その核酸配列は最小限、少なくともセンスまたはコード鎖の部分を含有してもよい(すなわち、核酸配列は1本鎖であってもよい)。あるいは、センスとアンチセンス鎖の両方を含有してもよい(すなわち、核酸配列は2本鎖であってもよい)。
【0085】
最小プロモーター:不活性であるかまたは非常に弱いプロモーター活性を有し、上流活性化が存在しないプロモーターエレメント、特にTATAエレメント。好適な転写因子が存在すると、最小プロモーターは転写を認める機能を果たす。
【0086】
NEENA:「核酸発現増強性核酸」を参照されたい。
【0087】
非コード:用語「非コード」は、発現されるタンパク質の部分もしくは全てをコードしない核酸分子の配列を意味する。非コード配列は、限定されるものでないが、イントロン、エンハンサー、プロモーター領域、3'非翻訳領域、および5'非翻訳領域を含む。
【0088】
核酸発現増強性核酸(NEENA):用語「核酸発現増強性核酸」は、NEENAが機能的に連結されたプロモーターの制御下の核酸の発現を増強する内因性の特性を有する特定の配列の配列および/または核酸分子を意味する。プロモーター配列と異なり、NEENAの類は発現を駆動することはできない。NEENAと機能的に連結された核酸分子の発現を増強する機能を遂行するために、NEENA自体がプロモーターと機能的に連結されていなければならない。当技術分野で公知のエンハンサー配列とは対照的に、NEENAはin cisで作用するが、in transでは作用しないし、発現される核酸の転写出発部位に近接して位置しなければならない。
【0089】
核酸およびヌクレオチド:用語「核酸」および「ヌクレオチド」は天然または合成または人工の核酸またはヌクレオチドを意味する。用語「核酸」および「ヌクレオチド」はデオキシリボヌクレオチドまたはリボヌクレオチドまたは任意のヌクレオチド類似体およびポリマーまたはそれらのハイブリッドを、1本鎖もしくは2本鎖のセンスもしくはアンチセンス型で含む。特に断らない限り、特別な核酸配列はまた、暗黙のうちに保存して改変されたそれらの変異体(例えば、縮重コドン置換)および相補配列、ならびに明確に示した配列を包含する。用語「核酸」は、「遺伝子」、「cDNA」、「mRNA」、「オリゴヌクレオチド」、「ポリヌクレオチド」と本明細書では互換的に使用される。ヌクレオチド類似体は、塩基、糖および/またはリン酸塩の化学構造に改変を有するヌクレオチドを含み、前記改変には、限定されるものでないが、5-位置ピリミジン改変、8-位置プリン改変、シトシン環外アミン類における改変、5-ブロモ-ウラシルの置換など;および限定されるものでないが、2'-位置糖改変(2'-OHがH、OR、R、ハロ、SH、SR、NH2、NHR、NR2、またはCNから選択される群により置換された糖改変を含む)されたリボヌクレオチドが含まれる。ショートヘアピンRNA(shRNA)はまた、非自然エレメント、例えば、非自然塩基、例えばイオノシンおよびキサンチン、非自然糖類、例えば2'-メトキシリボース、または非自然リン酸ジエステル連鎖、例えばホスホン酸メチル、ホスホロチオアートおよびペプチドを含みうる。
【0090】
核酸配列:表現「核酸配列」は、5'から3'末端へ読まれたデオキシリボヌクレオチドまたはリボヌクレオチド塩基の1本鎖または2本鎖のポリマーを意味する。核酸配列は染色体のDNA、自己複製プラスミド、DNAまたはRNAの感染性ポリマーおよび主に構造的役割を果たすDNAまたはRNAを含む。「核酸配列」はまた、ヌクレオチドを表す略語、文字、記号または言語の継続的列挙を意味する。一実施形態において、核酸は長さが通常100ヌクレオチド未満の比較的短い核酸である「プローブ」であってもよい。核酸プローブは長さが約50ヌクレオチド〜長さが約10ヌクレオチドであることが多い。核酸の「標的領域」は目的のものであると同定された核酸の一部分である。核酸の「コード領域」は、適当な調節配列の制御下に置かれると、配列特異的な方式で転写されかつ翻訳され、特別なポリペプチドまたはタンパク質を産生する核酸の部分である。コード領域はかかるポリペプチドまたはタンパク質をコードすると言われる。
【0091】
オリゴヌクレオチド:用語「オリゴヌクレオチド」はリボ核酸(RNA)またはデオキシリボ核酸(DNA)またはそれらの擬似体のオリゴマーまたはポリマー、ならびに同様に機能する非天然部分を有するオリゴヌクレオチドを意味する。かかる改変されたまたは置換されたオリゴヌクレオチドは、所望の特性、例えば、細胞取込みの増強、核酸標的に対する親和性の増強およびヌクレアーゼの存在のもとでの安定性の故に、しばしば、生来の型よりも好ましい。オリゴヌクレオチドは、好ましくは、連鎖(例えば、リン酸ジエステル)または代わる連鎖によりお互いに共役結合された2以上のヌクレオモノマーを含む。
【0092】
オーバーハング:「オーバーハング」は、2本鎖のオリゴヌクレオチド分子の5'-または3'-ヒドロキシル末端の比較的短い1本鎖ヌクレオチド配列である(「伸長」「突出末端」または「粘着性末端」とも呼ばれる)。
【0093】
植物:植物は一般に、光合成の能力があるいずれかの真核生物の単または多細胞生物またはその細胞、組織、器官、部分または繁殖材料(例えば、種子または果実)を意味すると理解される。本発明の目的に含まれるのは、高等および下等植物の全ての属および種である。一年生、多年生、単子葉および双子葉植物が好ましい。この用語は、成熟植物、種子、シュートおよび実生およびそれらの誘導された部分、繁殖材料(種子または小胞子など)、植物器官、組織、プロトプラスト、カルスおよび他の培養、例えば細胞培養、および機能または構造単位を与える植物細胞分類のいずれか他の型を含む。成熟植物は実生の段階後の任意の所望の発生段階の植物を意味する。実生は、初期の発生段階の若い未成熟植物を意味する。一年生、多年生、単子葉および双子葉植物はトランスジェニック植物を作製するための好ましい宿主生物である。遺伝子の発現はさらに全ての観賞植物、有用または観賞用植物樹木、花、切り花、低木または芝生において有利である。例として挙げられる植物は、限定されるものでないが、被子植物、コケ植物、例えば、苔類(ゼニゴケ)および蘚類(藻類);
シダ植物、例えば、シダ類、トクサ類およびヒカゲノカズラ類;裸子植物、例えば、針葉樹類、ソテツ類、イチョウ類およびグネツム類;藻類、例えば、緑藻綱、褐藻網、紅藻網、藍藻網、黄緑藻綱、珪藻綱(珪藻)、およびミドリムシ綱である。好ましいのは、食品または飼料に用いられる植物であり、例えば、マメ科、例えば、エンドウマメ、アルファルファおよびダイズ;イネ科、例えば、イネ、トウモロコシ、コムギ、オオムギ、モロコシ、ミレット、ライムギ、ライコムギ、またはオート麦;セリ科、とりわけ、Daucus属、特にとりわけcarota種(ニンジン)およびApium属、特にとりわけGraveolensdulce種(セロリ)その他の多くの種;ナス科、とりわけLycopersicon属、特にとりわけesculentum種(トマト)およびナス属、特にとりわけtuberosum種(ジャガイモ)およびmelongena種(ナス)、その他の多くの種(タバコなど);およびCapsicum属、特にとりわけannuum種(コショウ)その他の多くの種;マメ科、とりわけGlycine属、特にとりわけmax種(ダイズ)、アルファルファ、エンドウマメ、ムラサキウマゴヤシ、マメまたはピーナッツおよびその他の多くの種; および十字花科(アブラナ科)、とりわけBrassica属、特にとりわけnapus種(ナタネ)、campestris種(ビート)、oleracea cv Tastie(キャベツ)、oleracea cv SnowballY(カリフラワー)およびoleracea cv Emperor(ブロッコリー);およびArabidopsis属、特にとりわけthaliana種(シロイヌナズナ)およびその他の多くの種;Compositae(キク)科、とりわけLactuca属、特にとりわけsativa種(レタス)およびその他の多くの種;Asteraceae(キク)科、例えば、ヒマワリ、マリーゴールド、レタスまたはCalendula種(ワタゲハナグルマ)およびその他の多くの種;ならびにウリ科、例えば、メロン、カボチャ/カボチャまたはズッキーニ、および亜麻仁である。さらなる好ましい植物は、ワタ、サトウキビ、アサ、亜麻、チリー、および様々な樹木、ナッツおよびワイン種である。
【0094】
ポリペプチド: 用語「ポリペプチド」、「ペプチド」、「オリゴペプチド」、「ポリペプチド」、「遺伝子産物」、「発現産物」および「タンパク質」は本明細書では互換的に使用されていて、連続するアミノ酸残基のポリマーまたはオリゴマーを意味する。
【0095】
プレタンパク質:通常、葉緑体などの細胞小器官を標的化し、まだ、その輸送ペプチドを含んでいるタンパク質。
【0096】
一次転写物:本明細書で使用する用語「一次転写物」は遺伝子の未成熟RNA転写物を意味する。「一次転写物」は例えば、まだ、イントロンを含有し、および/またはなおpolyAテールまたはcap構造を含まず、および/またはトリミングまたはエディッティングなどの転写物としての正しい機能に必要な他の改変が行われてない。
【0097】
プロモーター:「プロモーター」または「プロモーター配列」は等価であり、本明細書で使用する場合、目的のヌクレオチド配列に連結されると目的のヌクレオチド配列のRNA中への転写を制御することができるDNA配列を意味する。かかるプロモーターは、例えば、次の公共データベースに見出すことができる:http://www.grassius.org/grasspromdb.html、http://mendel.cs.rhul.ac.uk/mendel.php?topic=plantprom、http://ppdb.gene.nagoya-u.ac.jp/cgi-bin/index.cgi。そこに掲げられたプロモーターは本発明の方法について呼び出すことができ、これらは参照により本明細書に組み込まれる。プロモーターは、目的のヌクレオチド配列(mRNA中へのその転写を制御する)の転写出発部位の5'(すなわち、上流)の近位に位置し、そして転写開始のためのRNAポリメラーゼおよび他の転写因子による特異的結合のための部位を提供する。前記プロモーターは、例えば、転写出発部位に対して、例えば少なくとも10kb、例えば5kbまたは2kb近位にある。これはまた、転写出発部位に対して、少なくとも1500bp近位、好ましくは少なくとも1000bp、より好ましくは少なくとも500bp、さらにより好ましくは少なくとも400bp、少なくとも300bp、少なくとも200bpまたは少なくとも100bpにあってもよい。さらなる好ましい実施形態において、プロモーターは転写出発部位に対して、少なくとも50bp近位、例えば、少なくとも25bpであってもよい。プロモーターはエキソンおよび/またはイントロン領域または5'非翻訳領域を含まない。プロモーターはそれぞれの植物に対して異種または相同性であってもよい。ポリヌクレオチド配列は、もし外来種に由来するか、またはもし同じ種由来であってその元来の形態から改変されていれば、生物または第2のポリヌクレオチド配列に対して異種である。例えば、異種コード配列と作動しうる形で連結されたプロモーターという場合、そのコード配列は、そのプロモーターが由来する種と異なる種由来のコード配列、または、もし同じ種由来であれば、そのプロモーターに自然では随伴しないコード配列(例えば、遺伝子操作で作られたコード配列または異なるエコタイプもしくは変異体からの対立遺伝子)を意味する。好適なプロモーターは、発現が起こるべき宿主細胞の遺伝子からまたは宿主細胞に対する病原体(例えば、植物または植物ウイルスのような植物病原体)から誘導することができる。植物特異的プロモーターは植物中の発現を調節するのに好適なプロモーターである。これは植物からであってもよいが、植物病原体からであってもよくまたは人手により設計された合成プロモーターであってもよい。もしプロモーターが誘導プロモーターであれば、誘導剤に応答して転写速度が増加する。また、プロモーターを組織特異的または組織に好ましい方式で調節して、特定の組織型、例えば、葉、根または分裂組織の関係するコード領域を転写するのだけにまたは優性に活性があるようにすることができる。プロモーターに適用する用語「組織特異的」は、色々な型の組織(例えば、根)において同じ目的のヌクレオチド配列が発現しないのと比較して、特定の型の組織(例えば、花弁)において目的のヌクレオチド配列の選択的発現を指令することができるプロモーターを意味する。プロモーターの組織特異性は、例えば、レポーター遺伝子をレポーター構築物を作製するプロモーター配列と作動しうる形で連結し、そのレポーター遺伝子が植物のゲノム中に導入して得られるトランスジェニック植物の全ての組織中に組み込まれるようにし、そしてトランスジェニック植物の色々な組織中のレポーター遺伝子の発現(例えば、レポーター遺伝子がコードするmRNA、タンパク質、またはタンパク質の活性)を検出することにより評価することができる。1以上の組織におけるレポーター遺伝子の発現のレベルが他の組織におけるレポーター遺伝子の発現のレベルより高いことを検出すれば、そのプロモーターが高い発現のレベルを検出した組織にとって特異的であることが示される。プロモーターに適用する用語「細胞型特異的」は、同じ組織内の色々な型の細胞において同じ目的のヌクレオチド配列が発現しないのと比較して、特定の型の細胞において目的のヌクレオチド配列の選択的発現を指令することができるプロモーターを意味する。プロモーターに適用する用語「細胞型特異的」はまた、1つの組織内のある領域のヌクレオチド配列の選択的発現を促進することができるプロモーターを意味する。プロモーターの細胞型特異性は、当技術分野で周知の方法、例えば、GUS活性染色、GFPタンパク質または免疫組織化学的染色を用いて評価することができる。プロモーターまたはプロモーターに由来する発現に適用する用語「構成的」は、植物組織および細胞の大部分において、植物または植物の部分の実質的に全生存期間を通して、刺激(例えば、熱ショック、化学品、光など)の非存在のもとで、プロモーターが作動しうる形で連結された核酸分子の転写を指令することができることを意味する。典型的には、構成的プロモーターは実質的にいずれの細胞およびいずれの組織においても、トランスジーンの発現を指令することができる。
【0098】
プロモーター特異性:プロモーターを言及する場合、用語「特異性」はそれぞれのプロモーターが与える発現のパターンを意味する。特異性は、そのプロモーターが、それぞれのプロモーターの制御下で核酸分子の発現を与える植物またはその部分の組織および/または発生状態を記載する。プロモーターの特異性はまた、そのもとでプロモーターが活性化または下方調節されうる環境条件、例えば、寒冷、乾燥、創傷または感染などの生物学的または環境ストレスによる誘導または抑圧を含みうる。
【0099】
精製された:本明細書で使用する用語「精製された」は、その自然環境から取り出され、単離されるかまたは分離された分子、核酸もしくはアミノ酸配列を意味する。「実質的に精製された」分子は、それが自然で随伴する他成分の少なくとも60%を含有しない、好ましくは少なくとも75%を含有しない、そしてより好ましくは少なくとも90%を含有しない。精製された核酸配列は単離された核酸配列であってもよい。
【0100】
組換え体:核酸分子に関する用語「組換え体」は組換えDNA技法により産生された核酸分子を意味する。好ましくは、「組換え核酸分子」は、天然の核酸分子とは少なくとも1つの核酸が異なる非天然の核酸分子である。「組換え核酸分子」はまた、好ましくは作動しうる形で連結された自然ではその順で存在しない核酸分子の配列を含む「組換え構築物」を含みうる。前記組換え核酸分子を産生する好ましい方法は、クローニング技法、定方向または非部位特異的な突然変異誘発、合成または組換え技法を含みうる。
【0101】
センス:用語「センス」は標的配列、例えば、タンパク質転写因子と結合する、および所与の遺伝子の発現に関わる配列と相補性または同一である配列を有する核酸分子を意味すると理解される。好ましい実施形態によれば、核酸分子は目的の遺伝子および前記目的の遺伝子の発現を可能にするエレメントを含む。
【0102】
有意な増加または減少:測定技法に固有の誤差限界より大きい、例えば酵素活性または遺伝子発現の増加または減少、好ましくは、対照酵素の活性または対照細胞の発現の約2倍以上の増加または減少、より好ましくは、約5倍以上の増加または減少、そして最も好ましくは、約10倍以上の増加または減少を意味する。
【0103】
小核酸分子:「小核酸分子」は、核酸またはその誘導体から成る分子、例えばRNAまたはDNAであると理解される。これらは2本鎖または1本鎖であってもよく、そして、約15〜約30bp、例えば15〜30bp、より好ましくは約19〜約26bp、例えば19〜26bp、さらにより好ましくは約20〜約25bp例えば20〜25bpである。とりわけ 好ましい 実施形態において、オリゴヌクレオチドは約21〜約24bp、例えば21〜24bpである。最も好ましくは実施形態において、小核酸分子は約21および約24bp、例えば21および24bpである。
【0104】
実質的に相補的な:その最も広い意味で、用語「実質的に相補的な」は、参照または標的ヌクレオチド配列と比較してヌクレオチド配列について本明細書で使用する場合、前記参照または標的ヌクレオチド配列の実質的に相補的なヌクレオチド配列と正確に相補的な配列の間のパーセント同一性は少なくとも60%、より望ましくは少なくとも70%、より望ましくは少なくとも80%または85%、好ましくは少なくとも90%、より好ましくは少なくとも93%、さらにより好ましくは少なくとも95%または96%、なおさらにより好ましくは少なくとも97%または98%、またさらにより好ましくは少なくとも99%または最も好ましくは100%(最後の場合、この文脈では用語「同一」と等しい)であるヌクレオチド配列を意味する。(以下で特に断らなければ)好ましくは、同一性を核酸配列の少なくとも19ヌクレオチド、好ましくは少なくとも50ヌクレオチド、より好ましくは全長にわたり前記参照配列に対して評価する。配列比較は、デフォールトGAP分析を用いて、GAPのSEQWEBアプリケーション(University of Wisconsin GCG)により、NeedlemanおよびWunsch(Needleman and Wunsch (1970) J Mol. Biol. 48: 443-453;先に定義した通り)のアルゴリズムに基づいて実施する。参照ヌクレオチド配列に「実質的に相補的な」ヌクレオチド配列は参照ヌクレオチド配列と低ストリンジェンシー条件、好ましくは中ストリンジェンシー条件、最も好ましくは高ストリンジェンシー条件(先に定義した通り)のもとでハイブリダイズする。
【0105】
トランスジーン:本明細書で使用する用語「トランスジーン」は、実験遺伝子操作により細胞のゲノム中に導入される任意の核酸配列を意味する。トランスジーンは「内因性DNA配列」または「異種DNA配列」(すなわち「外来のDNA」)であってもよい。用語「内因性DNA配列」は、天然の配列に関係して何らかの改変(例えば、点突然変異、選択マーカー遺伝子の存在など)のない限り、導入される細胞において自然で見出されるヌクレオチド配列を意味する。
【0106】
トランスジェニック:生物に言及する場合、用語「トランスジェニック」は、形質転換された、好ましくは目的のDNA配列と作動しうる形で連結された好適なプロモーターを含む組換えDNA分子によって、好ましくは安定して形質転換されたことを意味する。
【0107】
ベクター:用語「ベクター」は、連結されている他の核酸分子を輸送することができる核酸分子を意味する。ベクターの1つの型はゲノムに組み込まれたベクター、すなわち、宿主細胞の染色体DNA中に組み込むことができる「組み込みベクター」である。ベクターの他の型はエピソームベクター、すなわち、染色体外複製の能力がある核酸分子である。作動しうる形で連結された遺伝子の発現を指令する能力のあるベクターを本明細書では「発現ベクター」と呼ぶ。本明細書においては、文脈からそうでないことが明らかでない限り、「プラスミド」と「ベクター」を互換的に使用する。本明細書に記載のRNAを産生するように設計された発現ベクターは、in vivoまたはin vitroでRNAポリメラーゼ(ミトコンドリアのRNAポリメラーゼ、RNA pol I、RNA pol II、および RNA pol IIIを含む)により認識される配列を含有しうる。これらのベクターを用いて本発明に従い、細胞において所望のRNA分子を転写することができる。植物形質転換ベクターは植物形質転換のプロセスに好適なベクターと理解されたい。
【0108】
野生型:生物、ポリペプチド、または 核酸配列に関する用語「野生型」、「自然の」または「自然の起源」は、前記生物が天然であるかまたは、少なくとも、改変、突然変異誘発またはさもなくば人手による遺伝子操作が行われていない天然の生物において入手できるものを意味する。
【実施例】
【0109】
化学品および共通の方法
特に断らない限り、本発明の目的で行ったクローニング手順(制限消化、アガロースゲル電気泳動、核酸の精製、核酸のライゲーション、細菌の形質転換、選択および培養を含む)は記載の通り実施した(Sambrooket al., 1989)。組換えDNAの配列解析は、レーザー蛍光 DNAシーケンサー(Applied Biosystems, Foster City, CA, USA)により、Sangerの技法(Sangeret al., 1977)を用いて実施した。特に断らない限り、化学品および試薬はSigma Aldrich(Sigma Aldrich, St. Louis, USA)から、 Promega(Madison, WI, USA)、Duchefa(Haarlem, The Netherlands)またはInvitrogen(Carlsbad, CA, USA)から入手した。制限エンドヌクレアーゼはNew England Biolabs(Ipswich, MA, USA)またはRoche Diagnostics GmbH(Penzberg, Germany)から入手した。オリゴヌクレオチドはEurofins MWG Operon(Ebersberg, Germany)が合成した。
【0110】
実施例1
構成的発現をする遺伝子からの核酸発現増強性核酸(NEENA)の同定
1.1 シロイヌナズナ(A. thaliana)遺伝子からのNEENA分子の同定
公的に利用可能なゲノムDNA配列(例えば、http://www.ncbi.nlm.nih.gov/genoms/PLANTS/PlantList.html)および転写物発現データ(例えば、http://www.weigelworld.org/resources/microarray/AtGenExpress/)を用いて、シロイヌナズナ転写物由来の18個の潜在NEENA候補の1セットを、高度に発現する構成的遺伝子から選択し、詳細な分析を行った。さらに、パセリ由来の推定NEENA分子もこの分析に加えた。候補名は次の通りである。
【0111】
表1: 構成的NEENA候補(NEENAc)
【表1】

【0112】
1.2 NEENA候補の単離
ゲノムDNAを、シロイヌナズナ緑色組織からQiagen DNeasy植物ミニキット(Qiagen, Hilden, Germany)を用いて抽出した。配列番号1をもつ推定NEENA分子に対しては、ベクター構築物1bxPcUbi4-2GUS(WO 2003102198)のDNAを用いた。推定NEENA分子を含有するゲノムDNA断片を、通常のポリメラーゼ連鎖反応(PCR)により単離した。ポリメラーゼ連鎖反応は19セットのプライマーを含むものであった(表2)。プライマーは多数のNEENA候補をもつシロイヌナズナのゲノム 配列に基づいて設計した。ベクター構築物1bxPcUbi4-2GUS(WO 2003102198)のヌクレオチド配列を、配列番号1(表2)をもつNEENA候補を増幅するプライマー(配列番号56および57)の設計に用いた。ポリメラーゼ連鎖反応は、Phusion高信頼性DNAポリメラーゼ(Cat No F-540L, New England Biolabs, Ipswich, MA, USA)に概説されたプロトコルに従った。単離したDNAを、次のプライマーを用いるPCR増幅のテンプレートDNAとして使用した。
【0113】
表 2:プライマー配列
【表2】
【0114】
PCR中の増幅は次の組成物(50 microl)を用いて行った:
3.00 microl シロイヌナズナゲノムDNA(50 ng/microl ゲノムDNA、5 ng/microl ベクター構築物)
10.00 microl 5x Phusion HF バッファー
4.00 microl dNTP(2.5 mM)
2.50 microl for プライマー(10 microM)
2.50 microl rev プライマー(10 microM)
0.50 microl Phusion HF DNA ポリメラーゼ(2U/microl)
次のパラメーターによるPCRに、タッチダウン手法を使った:98.0℃にて30秒(1サイクル)、98.0℃にて30秒、56.0℃にて30秒、および72.0℃にて60秒(4サイクル)、4回の追加サイクル、それぞれ54.0℃、51.0℃および49.0℃のアニーリング温度、引き続いて20サイクル98.0℃にて30秒、46.0℃にて30秒および72.0℃にて60秒(4サイクル)ならびに72.0℃にて5分。増幅産物を2%(w/v)アガロースゲルに供給し、80Vで分離した。PCR産物をゲルから切り取り、Qiagenゲル抽出キット(Qiagen, Hilden, Germany)で精製した。KpnI(10U/microl)およびNcoI(10U/microl)またはEcoRV(10U/microl)制限エンドヌクレアーゼによるDNA制限消化後、消化産物を再びQiagenゲル抽出キット(Qiagen, Hilden, Germany)で精製した。
【0115】
1.3 ベクター構築
1.3.1潜在NEENA分子をもつベクター構築物の作製
マルチサイトGatewayシステム(Invitrogen, Carlsbad, CA, USA)を用いて、プロモーター::NEENA::レポーター遺伝子カセットを植物形質転換用の2成分系構築物中にアセンブルした。シロイヌナズナp-AtNit1(At3g44310、GenBankX86454;WO03008596、接頭辞p-でプロモーターを表示)プロモーターをレポーター遺伝子構築物に用い、分析すべき推定NEENA分子の発現増強効果を定量的に測定するレポータータンパク質としてホタルルシフェラーゼ(Promega、Madison、WI、USA)を使った。
【0116】
製造業者マニュアル(Invitrogen, Carlsbad, CA, USA)に従って、p-AtNit1プロモーターを保持するpENTR/Aベクターを、pDONR/Aベクターとp-AtNit1増幅産物の間の部位特異的組換え(BP-反応)を介し、プライマーp-AtNit1-forおよびp-AtNit1-rev(表3)を用いて、部位特異的組換え部位を両端にもつゲノムDNA(上記参照)上にクローニングした。ポジティブpENTR/Aクローンを配列分析してp-AtNit1 プロモーターの正しさを保証した。
【0117】
表3:プライマー配列(p-AtNit1)
【表3】
【0118】
ホタルルシフェラーゼをコードする配列(Promega, Madison, WI, USA)と続いてt-nosノパリン合成酵素転写ターミネーター(Genbank V00087)を含有するENTR/Bベクターを作製した。NEENA候補PCR断片(上記参照)を別々に、ホタルルシフェラーゼをコードする配列の上流に、KpnIおよびNcoIまたはEcoRV制限酵素を用いてクローニングした。得られるpENTR/Bベクターを、接頭辞p-を有するプロモーター、接頭辞c-を有するコード配列、および接頭辞t-を有するターミネーターと共に表4に総括した。
【0119】
表4:全てのpENTR/Bベクター プラスおよびマイナスNEENA候補
【表4】
【0120】
pENTR/Cベクターを、KpnIおよびHindIII制限部位を介するマルチクローニングサイト(配列番号60)の導入により構築した。製造業者(Invitrogen, Carlsbad, CA, USA)のマルチサイトGatewayマニュアルに従い、部位特異的組換え(LR-反応)を実施することにより、作製したpENTR/A、pENTR/B および pENTR/CをpSUN デスティネーションベクター(pSUN誘導体)と組み合わせた。この反応によって、p-AtNit1プロモーター、ホタルルシフェラーゼをコードするc-LUCおよびt-nosターミネーター、ならびに、ホタルルシフェラーゼコード配列の直ぐ上流に配列番号1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18および19をコードする配列を保持する19個のベクターをもつ1つの二成分ベクターを得た(表5)。これと配列番号1との組み合わせを例示する(配列番号61)。全てのベクター(表5)は、ヌクレオチド配列配列番号2〜19が変わることを除くと、同一である。得られる植物形質転換ベクターを表5に総括した。
【0121】
表5:シロイヌナズナ形質転換用の植物発現ベクター
【表5】
【0122】
得られるベクターを引き続いて使用し、シロイヌナズナの葉のプロトプラストを一過的に形質転換した。
【0123】
1.3.2 ウミシイタケルシフェラーゼ対照構築物
ウミシイタケルシフェラーゼcDNAを、DNAテンプレートとしてPromega(Madison, WI, USA)由来のプラスミドpRL-nullの10ng、ならびにプライマーR-LUC_forおよびR-LUC_rev(表 6)を用いて、先に記載のPCRパラメーターにより増幅した。
【0124】
表6:プライマー配列(c-RLUC)
【表6】
【0125】
KpnI(10U/microl)およびSacI(10U/microl)制限エンドヌクレアーゼによるDNA制限消化の後、消化した産物を再びQiagenゲル抽出キット(Qiagen, Hilden, Germany)を用いて精製した。
【0126】
その断片を、ノパリン合成酵素構成的プロモーターp-nos(GenbankV00087)、引き続いてt-nos nopalin 合成酵素転写ターミネーター(Genbank V00087)を含有するENTR/Bベクター中に、KpnIおよびSacI制限部位を介してクローニングし、pENTR/Bクローンを得て、それを配列分析してウミシイタケルシフェラーゼを含有する発現カセットの正しさを保証した。
【0127】
実施例2
シロイヌナズナの一過的に形質転換された葉プロトプラストにおける、遺伝子発現を増強するNEENA候補分子のスクリーニング
本実施例は選択されたNEENA分子だけが遺伝子発現を増強する能力があることを説明する。
【0128】
2.1シロイヌナズナ葉プロトプラストの単離と一過的形質転換
シロイヌナズナ葉プロトプラストの単離と一過的形質転換を、確立されたプロトコル(Damm and Willmitzer, 1988; Damm et al., 1989)に従って修正した。4週齢のシロイヌナズナ植物の葉を、剃刀の刃を用いて小片にカットし、1.5%セルラーゼR10(Duchefa, Haarlem, The Netherlands)、0.3%マセロザイムR10(Duchefa, Haarlem, The Netherlands)、400mMマンニトール、20mM KCl、20mM MES、10mM CaCl2、pH5.7の溶液中に移し、一晩、室温にてインキュベートした。一過的シロイヌナズナ葉プロトプラスト形質転換は可変性であるので、ウミシイタケルシフェラーゼ(Dual-Luciferase(登録商標) Reporter Assay System, Promega, Madison, WI, USA)を用いて、上記構築物のホタルルシフェラーゼ発現能力を規準化した。NEENAを含まない(LJK132)および各NEENAを含有するベクター構築物(LJK66-LJK114)の一過的形質転換を6 microgのプラスミドDNAを用いて3重で実施し、これを25 microgのウミシイタケルシフェラーゼを含有する構築物と混合した後に、PEG(ポリエチレングリコール)と1x104プロトプラストを用いて形質転換した。
【0129】
2.2二重ルシフェラーゼレポーター遺伝子アッセイ
形質転換したシロイヌナズナプロトプラストを遠心分離により100gにて採集し、上清を除去した後、液体窒素にて凍結した。形質転換した細胞中のホタルおよびウミシイタケルシフェラーゼ活性を検出するアッセイを、製造業者(Promega、Madison、WI、USA)の二重ルシフェラーゼレポーターアッセイマニュアルに従って実施した。発光測定はMicroLumat Plus LB96V(Berthold Technologies、Bad Wildbad、Germany)で実施し、ルシフェラーゼ基質を加えた後に記録した。両方のルシフェラーゼ記録の計器読みはホタルルシフェラーゼとウミシイタケルシフェラーゼの間の比を作製することにより規準化した。3実験から得たデータを、各構築物について平均し、これらの平均発現値に基づいて、倍変化値を計算して、それぞれの推定NEENAを欠くレポーター遺伝子構築物を超える、推定NEENAの存在の影響を評価した。p-AtNit1プロモーターだけでNEENAを含まないレポーター遺伝子構築物と比較して、19個の試験したNEENA候補を含む構築物は、ホタルルシフェラーゼ活性でネガティブならびに0.1-倍〜18.1-倍の範囲のポジティブな効果を示した(図1)。全体で、 配列番号1、NO2、NO3、NO4、NO5、NO6、NO7、NO8 および NO9をもつ配列を含む 9個の推定 NEENA 分子は、NEENAを含まないプロモーターだけのレポーター遺伝子の構築物と比較して、ルシフェラーゼレポーター遺伝子活性に基づいて遺伝子発現の2倍以上の増加を与え(図1)、従って機能的NEENA分子であった。いくつかの試験したNEENA候補分子は遺伝子発現の増強に対して限界的なまたはむしろネガティブな効果を有するので、全ての推定NEENA分子が共通の刺激的効果を媒介するわけではなく、むしろ、選択されたNEENA配列が遺伝子発現の有意な増強を与える(配列番号1〜9)のである。
【0130】
実施例3
アブラナ植物において遺伝子発現を増強するNEENA分子の試験
本実施例はNEENA分子を用いて、NEENAを含まないプロモーターだけの手法と比較して、種全体にわたり、試験した全ての組織において遺伝子発現を増強できることを説明する。
【0131】
プレスクリーニングで遺伝子発現の最強の増強を媒介するNEENA分子(実施例2の配列番号1、2、3、4および5を参照)を選択して、トランスジェニックアブラナ植物における遺伝子発現に与える増強効果を測定した。
【0132】
3.1セイヨウアブラナ(Brassica napus)植物を形質転換するためのベクター構築
アブラナ植物を形質転換するために、遺伝子発現制御分子(配列番号1〜5)有りまたは無しのレポーター遺伝子カセットを、pENTR/Cベクター内にトランスジェニック植物系統を検出するための選択マーカーを運ぶ遺伝子発現カセットと組み合わせた。先に記載(上記1.4を参照)の部位特異的組換え(IR-反応)を実施することにより、選択マーカーカセットを運ぶpENTR/A、pENTR/BおよびpENTR/Cを、pSUNデスティネーションベクターと、製造業者(Invitrogen, Carlsbad, CA, USA)のマルチサイトGatewayマニュアルに従い、組み合わせた。反応は、p-AtNit1プロモーター、ホタルルシフェラーゼをコードする配列c-LUC、t-nosターミネーターおよび選択マーカーカセット、ならびに、ホタルルシフェラーゼをコードする配列の直ぐ上流に配列番号1、2、3、4、および5を保持する5つのベクターをもつ1つの二成分ベクターをもたらした(表7)。これに対する配列番号1との組み合わせを例示として与える(配列番号64)。配列番号2〜5が変わることを除くと、このヌクレオチド配列は全てのベクターで同一である。得られる植物形質転換ベクターを表7に総括する。
【0133】
表7:セイヨウアブラナ形質転換用の植物発現ベクター
【表7】
【0134】
3.2 トランスジェニックアブラナ植物の作製(Moloney et al., 1992, Plant Cell Reports, 8: 238-242による修正プロトコル)
トランスジェニックアブラナ植物を作製するための調製において、二成分ベクターをAgrobacterium tumefaciens C58C1:pGV2260中に形質転換した(Deblaere et al., 1985, Nucl. Acids. Res. 13: 4777-4788)。それぞれの二成分構築物を保持するアグロバクテリアの一晩培養の1:50希釈物を、3%ショ糖(3MS-培地)を補充したMurashige-Skoog培地(Murashige および Skoog、1962、Physiol. Plant 15、473)中で増殖した。アブラナ植物を形質転換するために、無菌植物の葉柄または低子葉を、1:50アグロバクテリウム溶液と5〜10分間インキュベートし、引き続いて3日間、暗所で25℃にて0.8%バクト-寒天を補充した3MS培地で共インキュベートした。3日後、その外植体を、 500 mg/l Claforan(セフォタキシム-ナトリウム)、100 nMイマゼタピル、20 microMベンジルアミノプリン(BAP)および1.6g/lグルコースを含有するMS培地に移し、16時間明/8時間暗を繰り返す光環境に1週間置いた。成長するシュートを、2 % ショ糖、250 mg/lクラフォランおよび0.8%バクト-寒天を含有するMS-培地に移した。3週後、成長ホルモン2-インドール酪酸を培地に加えて根形成を促進した。シュートを土壌に移し、根発生後、2週間成長室で成長させ、そして成熟まで温室条件で成長させた。
【0135】
3.3 植物分析
組織サンプルを、葉、花および長角果から作製したトランスジェニック植物から採集し、-80℃にて冷凍庫で保存し、ルシフェラーゼレポーター遺伝子アッセイ(Ow et al., 1986後の修正プロトコル)にかけた。粉砕後、凍結組織サンプルを800microlのバッファーI(0.1M リン酸バッファー pH7.8、1mM DTT(Sigma Aldrich, St. Louis, MO, USA)、0.05% Tween 20(Sigma Aldrich, St. Louis, MO, USA))に再懸濁し、続いて10000gにて10分間、遠心分離し、75microlの水性の上清を96ウエルプレートに移した。25microlのバッファーII(80 mMグリシン-グリシル(Carl Roth, Karlsruhe, Germany)、40mM MgSO4(Duchefa, Haarlem, The Netherlands)、60mM ATP(Sigma Aldrich, St. Louis, MO, USA)、pH 7.8)および0.5 mMの最終濃度までのD-ルシフェリン(cat No: L-8220, BioSynth, Staad, Switzerland)を加えた後に、発光を1分間当たり相対発光量の単位、すなわちRLU(RLU/min)で与えるMicroLumat Plus LB96V(Berthold Technologies, Bad Wildbad, Germany)で記録した。
【0136】
サンプル間のルシフェラーゼ活性を規準化する目的で、水性上清中のタンパク質濃度を、ルシフェラーゼ活性と平行して測定した(Bradford, 1976, Anal. Biochem. 72, 248から適用した)。5 microlのバッファーI中の水性細胞抽出物を250 microlのBradford 試薬(Sigma Aldrich, St. Louis, MO, USA)と混合し、10分間室温にてインキュベートした。吸収をプレートリーダー(Thermo Electron Corporation, Multiskan Ascent 354)で595nmにて測定した。サンプル中の全タンパク質量を先に作製した標準濃度曲線を用いて計算した。RLU/minとmgタンパク質/mlサンプルの比から得た値を、同一の構築物を保持するトランスジェニック植物について平均化し、倍変化値を計算してNEENA分子存在のNEENAを含まないレポーター遺伝子構築物を超える影響を評価した。
【0137】
3.4 NEENA配列はアブラナ植物における遺伝子発現の強い増強を媒介する
アブラナ 植物における選択したNEENA分子(配列番号1、2、3、4および5)の遺伝子発現を増強する潜在能力を評価するために、同一の発生段階を有しかつ同一の成長条件のもとで育てた植物の葉、花、および植物の種子を保持する長角果を採集した。サンプルは、プロモーターだけのレポーター遺伝子構築物または NEENA(配列番号1、2、3、4および5)を含有するルシフェラーゼレポーター遺伝子構築物を保持する個々のトランスジェニックアブラナ植物から採取した。10個の種子を各トランスジェニック事象から採集し、処理し、そして先に記載した(実施例3.3)ルシフェラーゼ活性について分析した。
【0138】
構成的なp-AtNit1プロモーターだけでNEENAを含まないレポーター遺伝子構築物と比較して、5つの試験したNEENA分子は全て、葉組織の遺伝子発現に強い増強を媒介した(図2、a)。NEENAs(配列番号1、2、3、4および5)により媒介される遺伝子発現の比較しうる増強は、アブラナの花および種子を含む長角果において検出された(図2b、およびc)。
【0139】
実施例4
ダイズ植物における遺伝子発現の構成的増強の分析
本実施例はNEENA分子を用いると、NEENAを含まないプロモーターだけの手法と比較して、広範囲の植物種においてかつ色々な植物ファミリーの種境界を越えて、全組織の遺伝子発現を増強できることを説明する。
【0140】
プレスクリーニングにおいて遺伝子発現の最強の増強を媒介するNEENA配列分子(実施例2を参照、配列番号1、2、3、4 および5)を選択し、トランスジェニックダイズ植物における遺伝子発現レベルの増強を測定した。植物発現ベクターLJK138、LJK139、LJK141、LJK142、LJK143およびLJK144(実施例3.1を参照)を用いて安定なダイズ形質転換を行った。
【0141】
4.1トランスジェニックダイズ植物の作製(WO2005/121345;Olhoftet al., 2007による修正プロトコル)
ダイズ種子発芽、繁殖、土壌細菌A. rhizogenesおよび腋分裂組織外植体調製、および接種を、先に記載(WO2005/121345; Olhoft et al., 2007)の通り実施した(但し、構築物LJK138、LJK139、LJK141、LJK142、LJK143およびLJK144(実施例3.1を参照)は、選択のためにイミダゾリン除草剤に対する耐性を媒介するパセリのユビキチンプロモーターPcUbi4-2により駆動される変異AHAS遺伝子をそれぞれ含有することを除く)。
【0142】
4.2 NEENA 配列はダイズ植物における遺伝子発現の強い増強を媒介する
組織サンプルを葉、花および種子から作製したトランスジェニック植物から採集した。組織サンプルを処理し、上記の通り分析した(実施例3.3を参照)。
【0143】
構成的な p-AtNit1 プロモーターだけでNEENAを含まない レポーター遺伝子構築物と比較して、5個の試験したNEENA分子は全て葉における強力な増強を媒介した(図3a)。NEENA(配列番号1〜5)により媒介される遺伝子発現の比較しうる増強がダイズ花および長角果において検出された(図3、bおよびc)。
【0144】
実施例5
単子葉植物におけるNEENA活性の分析
本実施例は単子葉植物における配列番号 1、2、3、4 および5をもつNEENA配列の分析を記載する。
【0145】
5.1 ベクター 構築
単子葉植物において配列番号1、2、3、4および5をもつNEENA配列を分析するために、トウモロコシ由来のNEENAを含まない構成的な単子葉プロモーターp-Ubiで構成される発現カセットを保持するpUCに基づく発現ベクターを、β-グルクロニダーゼ(GUS)遺伝子のコード配列と続くノパリン合成酵素(NOS)転写のターミネーターと組み合わせる。ゲノムDNAはシロイヌナズナ緑色組織からQiagen DNeasy Plant Miniキット(Qiagen, Hilden, Germany)を用いて抽出する。NEENA分子を含有するゲノムDNA断片を通常のポリメラーゼ連鎖反応(PCR)により単離する。プライマーは複数のNEENA候補をもつシロイヌナズナゲノム配列に基づいて設計する。反応物は5セットのプライマー(表8)を含み、Phusion High Fidelity DNAポリメラーゼ(cat No F-540L, New England Biolabs, Ipswich, MA, USA)に概説されたプロトコルに従い、次のプライマーを用いる。
【0146】
表8:プライマー配列
【表8】
【0147】
PCR中の増幅および増幅産物の精製は先に詳説した通り行う(実施例1.2)。AscI(10 U/microl)制限エンドヌクレアーゼによるDNA制限消化の後、消化産物をQiagenゲル抽出キット(Qiagen, Hilden, Germany)で精製した。
【0148】
NEENA PCR断片(前記参照)を、AscI制限部位を用いてβ-グルクロニダーゼのコード配列の上流に分離してクローニングした。この反応は、p-Ubiプロモーター、β-グルクロニダーゼをコードする配列c-GUSおよびt-nosターミネーター、ならびに、β-グルクロニダーゼをコードする配列の直ぐ上流に配列番号1、NO2、NO3、NO4および NO5を保持する5つのベクターをもつ1つの二成分ベクターを生じた(表9)。これに対する配列番号1との組み合わせを例示として与える(配列番号75)。配列番号2〜5が変わるのを除くと、全てのベクターのヌクレオチド配列は同一である(表9)。得られるベクターは表9に総括したが、ここでプロモーター分子は接頭辞p-、コード配列は接頭辞c-、およびターミネーター分子は接頭辞t-を有する。
【0149】
表9:植物発現ベクター
【表9】
【0150】
得られるベクターを用いて、以下に概説した実験(実施例5.2)でNEENA分子を分析した。
【0151】
5.2 単子葉植物組織において遺伝子発現を増強するNEENA分子の分析
これらの実験は、単子葉植物組織または培養細胞のボンバードメント(実施例6.2.1)により、DNAの植物プロトプラストへのPEG媒介(または類似の技法)導入(実施例6.2.2)により、またはアグロバクテリウム媒介形質転換(実施例6.3.3)により実施した。これらの実験の標的組織は、植物組織(例えば、葉組織)、培養植物細胞(例えば、トウモロコシ、Black Mexican Sweetcorn(BMS)、またはアグロバクテリウムプロトコル用の植物胚芽であってもよい。
【0152】
5.2.1 微粒子ボンバードメントを用いる一過性アッセイ
プラスミド構築物をQiagenプラスミド キット(cat# 12143)を用いて単離する。DNAを、Sanford ら(1993)(Optimizing the biolistic プロセス for different 生物学的 applications. 方法 in Enzymology、217: 483-509)に記載のプロトコルに従って0.6microM 金粒子(Bio-Rad cat# 165 -2262)上に沈降させ、 PDS-1000/He系デバイス(Bio-Rad)を用いて標的組織(例えば、2週齢のトウモロコシ 葉、BMS培養細胞など)上に加速させる。全てのDNA沈降およびボンバードメントステップを無菌条件下で室温にて実施する。Black Mexican Sweetトウモロコシ(BMS)懸濁液培養細胞をBMS細胞培養液体培地[Murashige および Skoog(MS) 塩(4.3 g/L)、3%(w/v)スクロース、myo-イノシトール(100 mg/L)、3 mg/L 2,4-ジクロロフェノキシ酢酸(2,4-D)、カゼイン 加水分解酵素(1g/L)、チアミン(10mg/L)およびL-プロリン(1.15g/L)、pH5.8]中で増殖する。毎週10mLの定常細胞の培養を40mLの新しい培地に移し。250mLフラスコ中で110rpmで27℃にて運転する回転シェーカーで培養する。
【0153】
シリコン処理したエッペンドルフチューブ中の60mgの金粒子を、100%エタノールで再懸濁し、次いで30秒間遠心分離する。ペレットを100%エタノール中で1回、無菌水中で2回漱ぎ洗いし、それぞれの洗浄後に遠心分離する。ペレットを最終的には1mL無菌50%グリセロールに再懸濁する。金懸濁液を次いで50microlアリコートに分割し、4℃にて保存する。次の試薬を1つのアリコートに加える: 5 microLの1 microg/microL全 DNA、50 microLの2.5 M CaCl2、20 microLの0.1 M スペルミジン、フリーベース。DNA溶液を1分間ボルテックス攪拌し、-80℃にて3分間静置し、次いで10秒間遠心分離する。上清を取り除く。ペレットを軽く叩いて注意深く1 mL 100% エタノールに再懸濁し次いで10秒間遠心分離する。上清を除去し、ペレットを注意深く50microLの100%エタノールに再懸濁し、使用するまで(ボンバードメント前の30分〜4時間)-80℃に置く。もし金凝集物が溶液中で可視であれば、使用直前にチューブを1秒間水浴のソニケーターにおいて超音波処理する。
【0154】
ボンバードメントについては、2週齢のトウモロコシ葉をカットしておよび1cm長さの小片とし、向軸側を上にして浸透圧誘導培地M-N6-702[N6塩(3.96g/L)、3%(w/v)スクロース、1.5mg/Lの2,4-ジクロロフェノキシ酢酸(2,4-D)、カゼイン加水分解産物(100mg/L)、およびL-プロリン(2.9g/L)、MSビタミンストック溶液(1mL/L)、0.2Mマンニトール、0.2Mソルビトール、pH 5.8]上に置く。
【0155】
小片を1〜2時間インキュベートする。
【0156】
BMS培養細胞の場合、1週齢の懸濁液細胞を1000gにてBeckman/Coulter Avanti J25遠心分離機でペレット化し、上清は捨てる。細胞を、円形の無灰No 42 Whatmanフィルター上にスパチュラを用いて1/16インチ厚さの層として置く。植物材料を保持するろ紙を浸透圧誘導培地上に27℃にて暗所で1〜2時間置いた後にボンバードメントを行う。ボンバードメント直前にフィルターを培地から取り除いて無菌ろ紙のスタック上に置いてカルス表面を部分的に乾燥させる。
【0157】
各プレートを6microLの金-DNA溶液で2回、葉材料向けて1,800psiにて、およびBMS培養細胞向けて1,100psiにてショットする。植物材料の位置を保つために、無菌のワイヤーメッシュスクリーンをサンプルの頂上に敷く。ボンバードメント後に、サンプルを保持するフィルターを、マンにトールおよびソルビトールを欠くM-N6-702 培地上に移し、2日間、暗所で27℃にてインキュベートし、その後に一過性アッセイを行う。
【0158】
他の単子葉植物の微粒子ボンバードメントを介する一過性形質転換は、例えば、Wang et al., 1988 (Transient expression of foreign genes in rice, wheat and soybean cells following particle bombardment. Plant Molecular Biology, 11 (4), 433-439), Christou, 1997 (Rice transformation: bombardment. Plant Mol Biol. 35 (1-2))に記載の技法を用いて行う。
【0159】
上記(実施例5.1)の構築物で発現された遺伝子の発現レベルを、当技術分野のプロトコルを用いて、GUS 染色、発光/蛍光の定量化、RT-PCR および タンパク質存在量(特異的抗体の検出)により測定する。GUS染色は、植物材料をGUS溶液[100 mM NaHPO4、10 mM EDTA、0.05% Triton X100、0.025% X-Gluc 溶液(DMSOに溶解した5-ブロモ-4-クロロ -3-インドリル-β-D-グルクロン酸)、10%メタノール、pH 7.0]中で37℃にて16〜24時間インキュベートすることにより行う。植物組織を2回15分間真空浸潤して一様な染色を助ける。ルシフェラーゼ活性の分析は先に記載の通り実施する(実施例2および3.3)。
【0160】
これらのアッセイにおいてNEENA分子は全て、構成的なp-UbiプロモーターだけでNEENAを含まないレポーター遺伝子構築物と比較して、遺伝子発現の強い増強を媒介する。
【0161】
5.2.2 プロトプラストを用いる一過的アッセイ
プロトプラストの単離を、Sheen(1990)(Metabolic Repression of Transcription in Higher Plants. The Plant Cell 2 (10)、1027-1038)が開発した次のプロトコルにより実施する。トウモロコシ実生を暗所で25℃にて10日間置き、そして20時間照明後にプロトプラストを調製した。葉の中央部を0.5mmストリップ(長さで約6cm)にカットし、1%(w/v)セルロースRS、0.1%(w/v)マセロザイムR10(両方ともYakult Honsha、Nishinomiya、Japanから入手)、0.6Mマンニトール、10mM Mes(pH 5.7)、1mM CaCl2、1mM MgCl2、10mM β-メルカプトエタノール、および0.1% BSA(w/v)を含有する酵素溶液中で、3時間、23℃にてインキュベートし、次いで80rpmにて10分間静かに振とうしてプロトプラストを遊離させた。
【0162】
プロトプラストを100 x gにて2分間遠心分離により採集し、1回冷0.6Mマンニトール溶液で洗浄し、遠心分離し、そして冷0.6Mマンニトール(2 x 106/mL)に再懸濁した。
【0163】
合計100microLの無菌水中の合計50microgのプラスミドDNAを、0.5mLのトウモロコシプロトプラスト(1 x 106 細胞s/mL)の懸濁液中に加え、静かに混合する。0.5mLのPEG溶液(40% PEG 4,000、100 mM CaNO3、0.5 マンニトール)を加え、静かに振とうしながら70℃にて予熱し、次いで4.5 mL MM溶液(0.6 M マンニトール、15 mM MgCl2、および0.1% MES)を加える。この混合物を15分間、室温にてインキュベートする。プロトプラストを2回、600rpmにて5分間ペレット化することにより洗浄し、 1.0 mLのMMB溶液[0.6Mマンニトール、4mM Mes(pH 5.7)、および ブロムモザイクウイルス(BMV)塩(オプション)]に再懸濁し、そして暗所で25℃にて48時間インキュベートする。最後の洗浄ステップ後に、プロトプラストを3 mLのMMB培地に採集し、そして暗所で25℃にて48時間インキュベートする。
【0164】
他の 単子葉植物のプロトプラストの一過性形質転換を、例えば、Hodgeset al., 1991(Transformation and regeneration of rice protoplasts. Biotechnology in agriculture No. 6, Rice Biotechnology. International Rice Research Institute, ISBN: 0-85198-712-5)またはLeeet al., 1990(Transient gene expression in wheat (Triticum aestivum) protoplasts. Biotechnology in agriculture and forestry 13 - Wheat. Springer Verlag, ISBN-10: 3540518096)が記載した技法を用いて行う。
【0165】
先に記載の構築物(実施例5.1)中の発現された遺伝子の発現レベルをGUS染色、発光/蛍光の定量、RT-PCRまたはタンパク質存在量(特異的抗体による検出)により、当技術分野のプロトコルを用いて測定する。GUS染色は、植物材料をGUS溶液[100 mM NaHPO4、10 mM EDTA、0.05% Triton X100、0.025% X-Gluc 溶液(DMSOに溶解した5-ブロモ-4-クロロ -3-インドリル-β-D-グルクロン酸)、10%メタノール、pH 7.0]中で37℃にて16〜24時間インキュベートすることにより行う。ルシフェラーゼ活性の分析は先に記載の通り実施する(実施例2および3.3)。
【0166】
構成的なp-UbiプロモーターだけでNEENAを含まないレポーター遺伝子構築物と比較して、NEENA分子はこれらのアッセイにおける遺伝子発現の強い増強を媒介する。
【0167】
5.2.3 単子葉作物植物の形質転換および 再生
標準 形質転換 および 再生 技法を用いるアグロバクテリウム媒介性植物形質転換はまた、作物植物を形質転換する目的で行うことができる(Gelvin and Schilperoort, 1995, Plant Molecular Biology Manual, 2nd Edition, Dordrecht: Kluwer Academic Publ. ISBN 0-7923-2731-4; Glick and Thompson (1993) Methods in Plant Molecular Biology and Biotechnology, Boca Raton: CRC Press, ISBN 0-8493-5164-2)。トウモロコシまたは他の単子葉植物の形質転換は、例えば、米国特許第5,591,616号に記載の技法を用いて行うことができる。粒子ボンバードメント、ポリエチレングリコール媒介性DNA取込みまたはシリコンカーボネート繊維を介する技法を用いる植物の形質転換は、例えば、Freeling & Walbot (1993) ”The maize handbook” ISBN 3-540-97826-7に記載されている。
【0168】
先に記載の構築物(実施例5.1)中の発現された遺伝子の発現レベルをGUS染色、発光/蛍光の定量、RT-PCRまたはタンパク質存在量(特異的抗体による検出)により、当技術分野のプロトコルを用いて測定する。GUS染色は、植物材料をGUS溶液[100 mM NaHPO4、10 mM EDTA、0.05% Triton X100、0.025% X-Gluc 溶液(DMSOに溶解した5-ブロモ-4-クロロ -3-インドリル-β-D-グルクロン酸)、10%メタノール、pH 7.0]中で37℃にて16〜24時間インキュベートすることにより行う。植物組織を2回15分間真空浸潤して一様な染色を助ける。ルシフェラーゼ活性の分析は先に記載の通り実施する(実施例2および3.3)。
【0169】
構成的なp-UbiプロモーターだけでNEENAを含まないレポーター遺伝子構築物と比較して、NEENA分子は植物における遺伝子発現の強い増強を媒介する。
【0170】
実施例6
トウモロコシ植物におけるNEENA活性の定量分析
本実施例はトウモロコシ植物における配列番号1および2をもつNEENA配列の分析を記載する。
【0171】
6.1 ベクター 構築
単子葉植物において配列番号1および2をもつNEENA配列を定量的に分析するために、トウモロコシ由来のNEENAを含まない構成的な単子葉プロモーターp-Ubiで構成される発現カセットを保持するpUCに基づく発現ベクターを、ホタルルシフェラーゼ(LUC)遺伝子(Promega, Madison, WI, USA)と続くノパリン合成酵素(NOS)転写のターミネーターと組み合わせた。ゲノムDNAはシロイヌナズナ緑色組織からQiagen DNeasy Plant Miniキット(Qiagen, Hilden, Germany)を用いて抽出した。NEENA分子を含有するゲノムDNA断片を通常のポリメラーゼ連鎖反応(PCR)により単離した。プライマーは複数のNEENA候補をもつシロイヌナズナゲノム配列に基づいて設計した。反応物は2セットのプライマー(表10)を含み、Phusion High Fidelity DNAポリメラーゼ(cat No F-540L, New England Biolabs, Ipswich, MA, USA)に概説されたプロトコルに従い、次のプライマーを用いた。
【0172】
表10:プライマー配列
【表10】
【0173】
PCR中の増幅および増幅産物の精製は先に詳説した通り行った(実施例1.2)。MluI(10 U/microl) および AscI(10 U/microl)制限エンドヌクレアーゼによるDNA制限消化の後、消化産物をQiagenゲル抽出キット(Qiagen, Hilden, Germany)で精製した。
【0174】
NEENA PCR断片(前記参照)を、AscI制限部位を用いてホタル ルシフェラーゼのコード配列の上流に分離してクローニングした。この反応は、p-Ubiプロモーター、ホタル ルシフェラーゼをコードする配列c-LUCおよびt-nosターミネーター、ならびに、ホタル ルシフェラーゼをコードする配列の直ぐ上流に配列番号1および2を保持する2つのベクターをもつ1つの二成分ベクターを生じた(表11);これに対する配列番号1との組み合わせを例示として与える(配列番号80)。配列番号2が変わるのを除くと、ヌクレオチド配列は同一である(表11)。得られるベクターは表11に総括したが、ここでプロモーター分子は接頭辞p-、コード配列は接頭辞c-、およびターミネーター分子は接頭辞t-を有する。
【0175】
表11:植物発現ベクター
【表11】
【0176】
得られるベクターを用いて、以下に概説した実験(実施例6.2)でNEENA分子を分析した。
【0177】
6.2 トランスジェニックトウモロコシ植物の作製
トウモロコシ発芽、繁殖、土壌細菌A. tumefaciens調製および接種を、先に記載(WO2006136596、US20090249514)の通り実施したが、ただし構築物LJK309、LJK326およびLJK327(実施例6.1を参照)は、選択のためにイミダゾリン除草剤に対する耐性を媒介するトウモロコシのユビキチンプロモーターp-Ubiにより駆動される変異AHAS遺伝子をそれぞれ含有することを除く。
【0178】
6.3 NEENA配列はトウモロコシ植物における遺伝子発現の強い増強を媒介する
組織サンプルを、葉および穀粒から作製したトランスジェニック植物から採集した。組織サンプルを処理し、上記の通り分析した(実施例3.3を参照)。
【0179】
構成的な p-UbiプロモーターだけでNEENAを含まない レポーター遺伝子 構築物と比較して、2個の試験したNEENA分子は葉における強力な増強を媒介した(図4a)。NEENA(配列番号1〜2)により媒介される遺伝子発現の比較しうる増強がトウモロコシ穀粒において検出された(図4b)。
【0180】
実施例7
イネ植物におけるNEENA活性の定量分析
本実施例はイネ 植物における配列番号1をもつNEENA配列の分析を記載する。
【0181】
7.1 ベクター 構築
イネ 植物において配列番号1をもつNEENA配列を定量的に分析するために、pENTR/B ベクターLJK1およびLJK4(実施例1.3を参照)を、組換え部位の上流に構成的な PRO0239を保持するデスティネーション ベクターと、製造業者(Invitrogen、Carlsbad、CA、USA) Gatewayマニュアルに従って組み合わせた。反応物はPRO0239プロモーター、ホタルルシフェラーゼをコードする配列c-LUCおよびt-nosターミネーター、ならびに、ホタルルシフェラーゼをコードする配列の直ぐ上流に配列番号1を保持する1つのベクターをもつ1つの二成分ベクター(表12)をもたらした。
【0182】
得られるベクターを表12に総括したが、ここでプロモーター分子は接頭辞p-、コード配列は接頭辞c-、およびターミネーター分子は接頭辞t-を有する。
【0183】
表12:植物発現ベクター
【表12】
【0184】
得られるベクターを用いて、以下に概説した実験(実施例7.2)でNEENA分子を分析した。
【0185】
7.2トランスジェニックイネ植物の作製
それぞれの発現ベクターを含有するアグロバクテリアを用いてイネ(Oryza sativa)植物を形質転換した。イネ japonica 栽培品種Nipponbareの成熟乾燥種子の皮を剥いだ。1分間70%エタノール中で、次いで30分間0.2% HgCl2中でインキュベートして殺菌を行い、次いで6回15分間、無菌蒸留水で洗浄した。無菌種子を次いで 2,4-Dを含有する培地(カルス培地)上で発芽させた。暗所で4週間インキュベーション後、胚形成した、胚盤で誘導されたカルスを切除し、同じ培地上で繁殖させた。2週後、カルスを継代培養によって同じ培地上でさらに2週間増殖または繁殖させた。胚形成カルス片を新しい培地上で3日間、継代培養した後、共胚葉した(細胞分裂活性をブーストするために)。
【0186】
それぞれの発現ベクターを含有するアグロバクテリウム菌株LBA4404を共培養に用いた。アグロバクテリウムを適当な 抗生物質と共にAB培地上に接種し、3日間28℃にて培養した。細菌を次いで採集し、約1の密度(OD600)まで液体共培養培地に懸濁させた。懸濁液を次いでペトリ皿に移し、カルスを懸濁液中に15分間浸漬した。カルス組織を次いでろ紙上にブロットして乾かし、そして固化した共培養培地に移し、3日間暗所で25℃にてインキュベートした。共栽培したカルスを2,4-Dを含有する培地で4週間暗所で28℃にて選択剤の存在のもとで成長させた。この期間に、急速成長する抵抗性カルスアイランドが発生した。この材料を再生培地へ移し、明所でインキュベーションした後、胚形成ポテンシャルが放出されてシュートが次の4〜5週に発生する。シュートをカルスから切除し、2〜3週間オーキシンを含有する培地でインキュベートし、それからこれらを土壌に移した。硬くなったシュートを温室内で高湿度のもとでかつ短期間に成長させた。
【0187】
およそ35の独立したT0イネ形質転換体を1つの構築物のために作製した。
【0188】
一次形質転換体を組織培養チャンバーから温室へ移した。T-DNAインサートのコピー数を立証する定量PCR分析の後、選択剤に耐性を示す唯一のコピートランスジェニック植物を保ってT1種子を収穫した。種子を次いで移植後3〜5か月にわたって収穫した。この方法によって、単一の遺伝子座形質転換体を50%を超える比率で得た(Aldemita and Hodges1996, Chan et al. 1993, Hiei et al. 1994)。
【0189】
7.3 NEENA配列はイネ植物において遺伝子発現の強い増強を媒介する
組織サンプルを、葉および穀粒から作製したトランスジェニック植物から採集した。その組織サンプルを処理し、上記の通り分析した(実施例3.3参照)
構成的なp-PRO239プロモーターだけでNEENAを含まないレポーター遺伝子構築物と比較して、試験したNEENA分子(配列番号1)は葉の遺伝子発現の強い増強を媒介した(図5a)。NEENA(配列番号1)が媒介する遺伝子発現の強い増強はイネ種子において検出された(図5b)。
【技術分野】
【0001】
本発明は植物分子生物学の分野にあり、高発現性構成的プロモーターを産生するおよび核酸の構成的発現が増強された植物を産生する方法であって、核酸発現増強性核酸(NEENA)を前記プロモーターと機能的に連結するおよび/または植物中に導入することを特徴とする前記方法を提供する。
【背景技術】
【0002】
植物におけるトランスジーンの発現は様々な外的および内的因子の影響を強く受け、様々なかつ予想できないレベルのトランスジーン発現をもたらす。しばしば多数の形質転換体を産生させて分析し、所望の発現強度をもつ系統を同定しなければならない。所望の発現をもつ系統の形質転換とスクリーニングは高費用かつ労働集約的であるので、1以上のトランスジーンを植物で高発現させる必要性がある。特に問題になるのは、トランスジェニック植物において特定の効果を達成するためにいくつかの遺伝子を協調的に発現させる必要のある場合であって、それぞれおよび全ての遺伝子が強く発現された植物を同定しなければならないからである。
【0003】
例えば、トランスジーンの発現は、個々の形質転換事象における構築物設計およびT-DNA挿入遺伝子座の位置効果に応じて有意に変わりうる。強いプロモーターはこれらの課題を部分的に克服することができる。しかし、所望の特異性をもつ強い発現を示す好適なプロモーターの利用可能性は限られることが多い。所望の発現特異性をもつ十分なプロモーターの利用可能性を確実にするために、追加のプロモーターの同定と特徴付けがこのギャップを埋める助けになりうる。しかし、それぞれの特異性と強度をもつプロモーターの自然での利用可能性とプロモーター候補の特徴付けに費やす時間とが、好適な新しいプロモーターを同定する妨げとなる。
【0004】
これらの課題を克服するために、多様な遺伝エレメントおよび/またはモチーフが遺伝子発現にポジティブな影響を与えることが示されている。これらのなかにあって、いくつかのイントロンは遺伝子発現を改善する強い可能性をもつ遺伝エレメントであることが認識されている。その機構はほとんど判ってないが、いくつかのイントロンは、転写活性の増強、mRNA成熟の改善、核mRNAエキスポートの増強および/または翻訳開始の改善により、成熟mRNAの定常状態量にポジティブな影響を与えることが示されている(例えば、Huang and Gorman, 1990;Le Hir et al., 2003;Nott et al., 2004)。選択されたイントロンだけが発現を増加することが示されたので、スプライシングなどはおそらく観察された効果を説明することができない。
【0005】
イントロンがプロモーターに機能的に連結するときに観察される遺伝子発現の増加は、遺伝子発現のイントロン媒介性増強(IME)と呼ばれ、様々な単子葉類(例えば、Callis al., 1987;Vasil al., 1989;Bruce al., 1990;Lu al., 2008)および双子葉類植物(例えば、Chung al., 2006;Kim al., 2006;Rose al., 2008)において示されている。この点については、翻訳出発部位(ATG)との相対的なイントロンの位置が遺伝子発現のイントロン媒介性増強にとって重要であることが示されている(Rose al., 2004)。
【0006】
遺伝子発現を増強する能力の次に、少数のイントロンはまた、植物における生来のヌクレオチド環境における組織特異性に影響を与えることが示されている。
【0007】
レポーター遺伝子発現は2つまでのイントロンを含有するゲノム領域の存在に依存することが見出された(Sieburth al., 1997;Wang al., 2004)。5’UTRイントロンはまた、プロモーターエレメントの適当な機能性にとって重要であり、これはおそらくイントロンに存在する組織特異的遺伝子制御エレメントに因ることが報じられた(Fuet al., 1995a;Fuet al., 1995b;Vitaleet al., 2003;Kimet al., 2006)。しかし、これらの研究はまた、イントロンの異種プロモーターとの組み合わせが遺伝子発現の強度と異種特異性に強い負の影響を与えうることも示している(Vitaleet al., 2003;Kimet al., 2006、WO2006/003186、WO2007/098042)。
【0008】
例えば、強い構成的カリフラワーモザイクウイルスCaMV35Sプロモーターは、ゴマD2 5’UTRイントロンとの組み合わせを通して負の影響を受ける(Kimet al., 2006)。これらの観察とは対照的に、いくつかの文献は、それぞれのプロモーターの組織特異性に影響を与えることなくIMEによる核酸発現の増強を示している(Schuenmannet al., 2004)。
【発明の概要】
【課題を解決するための手段】
【0009】
本出願においては、構成的プロモーターと機能的に連結されるとそれらの特異性に影響を与えることなく前記プロモーターの発現を増強する、さらなる核酸分子を記載する。これらの核酸分子を本出願では「核酸発現増強性核酸」(NEENA)と記載する。イントロンはそれぞれのプレmRNAからスプライシングで除去される内因性特徴を有する。それとは対照的に、本出願で提示する核酸は、機能的に連結されたプロモーターから誘導される発現を増強するために、必ずしもmRNA中に含まれなくてもよいし、または、もしmRNA中に存在すれば、必ずしもmRNAからスプライシングで除去されなくてもよい。
【図面の簡単な説明】
【0010】
【図1】一過的に形質転換したシロイヌナズナ葉プロトプラストにおける、p-AtNit1プロモーターの制御下の、NEENAを含有しない(LJK132)および構成的に発現される遺伝子由来の推定NEENA分子を表すNEENAを含有する(LJK91〜LJK133)構築物のルシフェラーゼレポーター遺伝子発現分析である。規準化はウミシイタケ・ルシフェラーゼの共形質転換と分析により行い、発現値をNEENAを含まない対照構築物(LJK132=1)と比較して示した。発現値をNEENAを含まない対照構築物(LJK134=1)と比較して示した。
【図2】p-AtNit1プロモーターの制御下の、NEENAを含有しない(LJK138)または構成的に発現される遺伝子由来のNEENA分子を表すNEENAを含有する(LJK139〜LJK144)レポーター遺伝子構築物を保持する独立のトランスジェニックアブラナ植物系統の相対光単位(RLU)として、各サンプルのタンパク質含量(平均値、分析した20件の独立したトランスジェニック植物の組織)に対して規準化後に示したルシフェラーゼレポーター遺伝子活性の棒グラフである。A)葉組織、B)花、C)長角果。
【図3】p-AtNit1プロモーターの制御下の、NEENAを含まない(LJK138)または構成的に発現した遺伝子由来のNEENA分子を表すNEENAを含有する(LJK139〜LJK144)レポーター遺伝子構築物を保持する独立のトランスジェニックダイズ植物系統の、相対光単位(RLU)として、各サンプルのタンパク質含量(平均値、分析した10件の独立したトランスジェニック植物の組織)に対して規準化後に示した、ルシフェラーゼレポーター遺伝子活性の棒グラフである。A)葉組織、B)花、C)種子。
【図4】p-ZmUbiプロモーターの制御下の、NEENAを含まない(LJK309)または構成的に発現した遺伝子由来のNEENA分子を表すNEENAを含有する(LJK326〜LJK327)レポーター遺伝子構築物を保持する独立のトランスジェニックトウモロコシ植物系統の、相対光単位(RLU)(対数スケール)として、各サンプルのタンパク質含量(平均値、分析した15件の独立したトランスジェニック植物の組織)に対して規準化後に示した、ルシフェラーゼレポーター遺伝子活性の棒グラフである。A)葉組織、B)穀粒。
【図5】構成的な p-PRO239プロモーターの制御下の、NEENAを含まない(CD30963)または構成的に発現した遺伝子由来のNEENA分子を表すNEENAを含有する(CD30964)レポーター遺伝子構築物を保持する独立のトランスジェニックイネ植物系統の、相対光単位(RLU)として、各サンプルのタンパク質含量(平均値、分析した10件の独立したトランスジェニック植物の組織)に対して規準化後に示した、ルシフェラーゼレポーター遺伝子活性の棒グラフである。A)葉組織、B)種子。
【発明を実施するための形態】
【0011】
本発明の第1の実施形態は、プロモーターに1以上の核酸発現増強性核酸(NEENA)分子を機能的に連結するステップを含む高発現性構成的プロモーターを産生する方法であって、前記NEENA分子が
i) 配列番号1〜19、好ましくは配列番号1〜9のいずれかで定義した配列を有する核酸分子、または
ii) 配列番号1〜19、好ましくは配列番号1〜9で定義した配列のいずれかと80%以上の同一性をもつ、好ましくは、前記同一性が85%以上である、より好ましくは同一性が90%以上である、さらにより好ましくは、同一性が95%以上、96%以上、97%以上、98%以上または99%以上である、最も好ましい実施形態では、配列番号1〜19、好ましくは配列番号1〜9で定義した配列のいずれかとの同一性が100%である配列を有する核酸分子、または
iii) i)もしくはii)の核酸分子の100以上の連続する塩基、好ましくは150以上の連続する塩基、より好ましくは200以上の連続する塩基、さらにより好ましくは250以上の連続する塩基の断片であって、例えば、配列番号1〜19、好ましくは配列番号1〜9で定義した配列のいずれかの配列を有する対応する核酸分子の65%以上、好ましくは70%以上、より好ましくは75%以上、さらにより好ましくは80%以上、85%以上または90%以上の発現増強活性を有する、最も好ましい実施形態では、95%以上の発現増強活性を有する前記断片、または
iv) 先にi)〜iii)のもとで記載した核酸分子のいずれかの相補配列または逆相補配列である核酸分子、または
v) PCRにより、表2に示した配列番号20〜57、好ましくは配列番号20/21;26/27;30/31;38/39;42/43;44/45;46/47;50;51および56/57に記載のオリゴヌクレオチドプライマーを用いて取得可能な核酸分子、または
vi) 7%ドデシル硫酸ナトリウム(SDS)、0.5M NaPO4、1mM EDTA中の50℃におけるハイブリダイゼーションと、2 X SSC、0.1%SDS中の50℃または65℃、好ましくは65℃における洗浄と等価の条件のもとで、配列番号1〜19、好ましくは配列番号1〜9に記載の転写増強性ヌクレオチド配列またはそれらの相補配列の少なくとも50、好ましくは少なくとも100、より好ましくは少なくとも150、さらにより好ましくは少なくとも200、最も好ましくは少なくとも250個の連続するヌクレオチドを含む核酸分子とハイブリダイズする、100ヌクレオチド以上、150ヌクレオチド以上、200ヌクレオチド以上または250以上のヌクレオチドの核酸分子を含むものである前記方法を含む。
【0012】
好ましくは、前記核酸分子は、7%ドデシル硫酸ナトリウム(SDS)、0.5M NaPO4、1mM EDTA中の50℃におけるハイブリダイゼーションと、1 X SSC、0.1%SDS中の50℃または65℃、好ましくは65℃における洗浄と等価の条件のもとで、配列番号1〜19、好ましくは配列番号1〜9またはそれらの相補配列に記載の転写増強性ヌクレオチド配列の少なくとも50、好ましくは少なくとも100、より好ましくは少なくとも150、さらにより好ましくは少なくとも200、最も好ましくは少なくとも250個の連続するヌクレオチドを含む核酸分子とハイブリダイズし、より好ましくは、前記核酸分子は、7%ドデシル硫酸ナトリウム(SDS)、0.5M NaPO4、1mM EDTA中の50℃におけるハイブリダイゼーションと、0.1 X SSC、0.1%SDS中の50℃または65℃、好ましくは65℃における洗浄と等価の条件のもとで、配列番号1〜19、好ましくは配列番号1〜9に定義した配列またはそれらの相補配列のいずれかに記載の転写増強性ヌクレオチド配列の少なくとも50、好ましくは少なくとも100、より好ましくは少なくとも150、さらにより好ましくは少なくとも200、最も好ましくは少なくとも250個の連続するヌクレオチドを含む核酸分子とハイブリダイズする。
【0013】
一実施形態において、1以上のNEENAは、それが機能的に連結されたプロモーターに対して異種である。
【0014】
v)項にて記載した、表2に示した配列番号 20〜57、好ましくは 配列番号 20/21;26/27;30/31;38/39;42/43;44/45;46/47;50/51および56/57で定義したオリゴヌクレオチドを用いてPCRにより取得可能な核酸分子は、例えば、シロイヌナズナ植物由来のゲノムDNAから以下の実施例1に記載した条件を用いて取得可能である。
【0015】
当業者は、それぞれのNEENA分子を得るための温度プロファイル、サイクル数および/またはバッファー組成物または濃度のバリエーションに気付くであろう。それぞれのNEENA分子を得るそれぞれのPCR反応で用いるオリゴヌクレオチドの具体的な組み合わせを表2に記載した。
【0016】
当業者は一方向プロモーターを二方向プロモーターにする方法、およびプロモーターの相補配列または逆相補配列を用いて元来の配列に特異的な同じプロモーター特異性を有するプロモーターを作製する方法に気付くであろう。かかる方法は、例えば、構成的ならびに誘導プロモーターについて、Xie et al. (2001) “Bidirectionalization of polar promoters in plants” nature biotechnology 19 pages 677 - 679に記載されている。この著者らは、任意の所与のプロモーターの5'プライム末端に最小のプロモーターを加えるだけで、両方向に同じプロモーター特異性で発現を制御するプロモーターを得るのに十分であると記載している。従って、上記のNEENAと機能的に連結された高発現プロモーターは、相補配列または逆相補配列に機能性であり、それ故に、NEENAも相補配列または逆相補配列に機能性である。
【0017】
本明細書に記載した構成的プロモーターは、実質的に全植物組織に、実質的に植物もしくはその部分の全生存期間を通して発現されるプロモーターを意味する。実質的に全ての植物組織に発現されるプロモーターはまた、主な植物組織の少なくとも2つ、例えば、葉、幹および/または根に発現されるプロモーターを包含しうるし、そしていくつかまたは全てのマイナーな組織または細胞、例えば、表皮、気孔、毛状突起、花、種子または分裂組織で発現され得るかまたは発現され得ないプロモーターを包含する。好ましい実施形態において、本明細書で意味する構成的プロモーターは葉および幹などの緑色組織で少なくとも発現される。
【0018】
実質的に植物もしくはその部分の全生存期間を通して発現されるプロモーターはまた、若いおよび発達した組織において発現されるが、植物の生存期間中の特定の時点または特定の条件、例えば、発芽および/または老化の下で、または生物もしくは非生物ストレスの条件下、例えば真菌または細菌の感染、乾燥、熱または寒冷のもとで発現を欠くことがありうるプロモーターを包含する。好ましい実施形態において、実質的に植物の全生存期間に発現される構成的プロモーターは、少なくとも全ての拡大組織において老化の開始まで発現される。
【0019】
原則として、NEENAはいずれのプロモーターとも、例えば、組織特異的、誘導性、発生特異的または構成的プロモーターとも機能的に連結されうる。それぞれのNEENAは、少なくとも1つのNEENAが機能的に連結されたそれぞれのプロモーターの制御下の異種核酸の発現の増強に導きうる。構成的プロモーター以外のプロモーター、例えば組織特異的プロモーターの発現の増強は、これらのプロモーターを特異性にするであろう。前記核酸の転写物がNEENAがない場合に検出されなかったさらなる組織または発生段階において、それぞれのプロモーターの制御下の核酸の発現は検出可能になるであろう。それ故に、組織もしくは発生特異的なまたはいずれかの他のプロモーターを、上記NEENA分子の少なくとも1つを前記プロモーターと機能的に連結することにより構成的プロモーターにすることができる。それ故に本発明の他の実施形態は、それぞれのプロモーターをi)〜vi)で上に記載した配列を含むNEENA分子と連結することにより、植物中で機能性である任意の所与のプロモーターの特異性を構成的プロモーターにする方法を提供する。
【0020】
好ましくは、1以上のNEENAを任意の構成的プロモーターと機能的に連結し、前記プロモーターの制御下の核酸分子の発現を増強しうる。本発明の任意の方法で使用する構成的プロモーターは、植物、例えば単子葉または双子葉植物から、細菌および/またはウイルスから誘導してもよく、または合成プロモーターであってもよい。使用する構成的プロモーターは、例えば、P. crispum(WO 2003102198)由来のPcUbi-プロモーター、トウモロコシ由来のZmUbi-プロモーター、ニトリラーゼ1をコードするシロイヌナズナ遺伝子At3g44310由来のAtNit-プロモーター、フィグオート(figwort)モザイクウイルス由来の34S-プロモーター、タバコモザイクウイルス由来の35S-プロモーター、アグロバクテリア由来のnosおよびocs-プロモーター、フェレドキシンNADHレダクターゼをコードするシロイヌナズナ遺伝子At5g66190由来のScBV-プロモーター(US 5 994 123)、SUPER-プロモーター(Lee et al., 2007、Plant. Phys.)、 AtFNR-プロモーター、エンドウマメ由来のptxAプロモーター(WO2005085450)、リン酸トリオース輸送体をコードするシロイヌナズナ遺伝子At5g46110由来のAtTPT-プロモーター、シロイヌナズナ遺伝子At4g14880およびAt4g14890由来の二方向AtOASTL-プロモーター、グリセルアルデヒド-3-リン酸デヒドロゲナーゼをコードするシロイヌナズナ遺伝子At1g13440由来のPRO0194プロモーター、フルクトース-ビス-リン酸アルドラーゼをコードするシロイヌナズナ遺伝子At3g52930由来のPRO0162プロモーター、イネ由来のAHAS-プロモーター(WO2008124495)またはCaffeoylCoA-MTプロモーターおよびOsCP12(WO2006084868)である。
【0021】
NEENAと機能的に連結された本発明の高発現性構成的プロモーターは、例えば、藻類、シダ、裸子植物または被子植物、例えば単子葉または双子葉植物を含む任意の植物で使用することができる。好ましい実施形態においては、NEENAと機能的に連結された本発明の前記プロモーターを、単子葉または双子葉類植物、好ましくは作物植物、例えばトウモロコシ、ダイズ、アブラナ、ワタ、ジャガイモ、サトウダイコン、イネ、コムギ、モロコシ、オオムギ、バナナ、サトウキビ、ススキなどに使用することができる。本発明の好ましい実施形態においては、NEENAと機能的に連結された前記プロモーターを単子葉の作物植物、例えば、トウモロコシ、イネ、コムギ、モロコシ、バナナ、ススキ、サトウキビまたはオオムギに使用することができる。とりわけ好ましい実施形態においては、NEENAと機能的に連結されたプロモーターを、双子葉類の作物植物、例えば、ダイズ、アブラナ、ワタ、サトウダイコンまたはジャガイモに使用することができる。
【0022】
本出願に使用する高発現性プロモーターは、例えば、植物またはその部分においてプロモーターの構成的発現の増強を生じるNEENAと機能的に連結されたプロモーターであって、NEENAと機能的に連結されたそれぞれのプロモーターの制御下の核酸分子から誘導されるRNAの蓄積またはRNAの合成速度が、本発明のNEENAを欠く同じプロモーターにより生じる発現より高い、好ましくは有意に高い前記プロモーターを意味する。好ましくは、植物中のそれぞれの核酸のRNAの量および/またはRNA合成の速度および/またはRNA安定性は、本発明のNEENAと機能的に連結されてない同じ構成的プロモーターを含む同じ条件下で育てられた同じ齢の対照植物と比較して、50%以上、例えば100%以上、好ましくは200%以上、より好ましくは5倍以上、さらにより好ましくは10倍以上、最も好ましくは20倍以上、例えば50倍増加する。
【0023】
本明細書で使用する用語「有意に高い」は、統計的有意を意味し、当業者は、t-検定などの統計的検定をそれぞれのデータセットに応用することにより決定するやり方を理解している。
【0024】
プロモーターが与える発現を検出する方法は当技術分野で公知である。例えば、プロモーターをマーカー遺伝子、例えばGUS、GFPまたはルシフェラーゼと機能的に連結し、それぞれのマーカー遺伝子がコードするそれぞれのタンパク質の活性を植物またはその部分で確認することができる。代表例として、ルシフェラーゼを検出する方法を以下に詳しく記載する。他の方法としては、例えば、プロモーターにより制御される核酸分子のRNAの定常状態レベルまたは合成速度を、当技術分野で公知の方法、例えば、ノーザンブロット分析、qPCR、ラン-オンアッセイもしくは当技術分野で記載の他の方法により測定する方法がある。
【0025】
当業者は2以上の核酸分子を機能的に連結する様々な方法を知っている。かかる方法は制限/ライゲーション、リガーゼ非依存性クローニング、リコンビニアリング、組換えまたは合成を包含しうる。他の方法を使用して2以上の核酸分子を機能的に連結してもよい。
【0026】
本発明のさらなる実施形態は、それぞれの対照植物またはその部分と比較して、1以上の核酸分子の構成的発現が増強された植物またはその部分を産生する方法であって、植物またはその部分中に、i)〜vi)のもとで先に定義した核酸分子を含む1以上のNEENAを導入するステップ、ならびに、前記1以上のNEENAをプロモーター、好ましくは構成的プロモーターと、および前記プロモーター、好ましくは構成的プロモーターの制御下にある核酸分子(ここで、NEENAはこの核酸分子に対して異種である)と機能的に連結するステップを含むものである前記方法である。
【0027】
NEENAは、NEENAが機能的に連結された前記プロモーターの制御下にある核酸分子に対して異種であってもよく、またはNEENAはプロモーターおよび前記プロモーターの制御下の核酸分子の両方に対して異種であってもよい。
【0028】
核酸分子またはDNAに関する用語「異種」は、自然では作動しうる形で連結されていないかまたは自然では異なる位置で作動しうる形で連結されている第2の核酸分子と、作動しうる形で連結されるかまたは作動しうる形で連結されるように遺伝子操作された核酸分子を意味する。例えば、本発明のNEENAはその自然環境においてその生来のプロモーターと機能的に連結されているが、本発明において前記NEENAは同じ生物由来、異なる生物由来であってもよく、またはSUPER-プロモーターなどの合成プロモーターであってもよい他のプロモーターと連結される。また、本発明のNEENAはその生来のプロモーターと連結されているが、前記プロモーターの制御下の核酸分子はその生来のNEENAを含むプロモーターに対して異種であるということを意味する。さらに理解すべきことは、本発明のNEENAと機能的に連結された前記プロモーターの制御下のプロモーターおよび/または核酸分子は、その配列が例えば、挿入、欠失などの突然変異により遺伝子操作されると、前記プロモーターの制御下のプロモーターおよび/または核酸分子の自然配列が改変され、それ故に本発明のNEENAに対して異種になったので、前記NEENAに対して異種である。理解すべきことはまた、NEENAをその生来のプロモーターと機能的に連結し、ここで前記プロモーターに対するNEENAの位置を変えてプロモーターがかかる遺伝子操作後にさらに高い発現を示す場合も、NEENAは機能的に連結された核酸に対して異種である。
【0029】
本明細書で意味する核酸分子の増強された構成的発現を示す植物は、それぞれの核酸分子と機能的に連結されたそれぞれのNEENA無しに同じ条件下で育てた対照植物と比較して、より高い、好ましくは、統計的に有意に高い核酸分子の構成的発現を有する植物を意味する。かかる対照植物は、本発明の植物と同じ遺伝子を制御する同じプロモーターを含み、そのプロモーターが本発明のNEENAと連結されてない野生型植物またはトランスジェニック植物であってもよい。
【0030】
本明細書で使用する植物の産生は、安定して形質転換する、例えば、組換えDNA構築物を植物またはその部分中に、アグロバクテリウム媒介性形質転換、プロトプラスト形質転換、粒子ボンバードメントその他などの方法によって導入し、そして場合によっては引き続いてトランスジェニック植物を再生する方法を含むものである。これはまた、植物またはその部分をウイルス感染またはアグロバクテリウム浸潤などにより一過的に形質転換する方法も含む。当業者は植物またはその部分を、安定しておよび/または一過的に形質転換するさらなる方法を知っている。繁殖方法またはプロトプラスト融合などの手法も本発明の植物の生産に使うことができて、本発明に包含される。
【0031】
本発明の方法は、任意の植物、例えば裸子植物または被子植物、好ましくは被子植物、例えば双子葉または単子葉植物、好ましくは双子葉植物に応用することができる。好ましい単子葉植物は、例えばトウモロコシ、コムギ、イネ、オオムギ、モロコシ、バナナ、サトウキビ、ススキおよびブラキポディウムであり、とりわけ好ましい単子葉植物はトウモロコシ、コムギおよびイネである。好ましい双子葉植物は、例えばダイズ、アブラナ種子、アブラナ、亜麻仁、ワタ、ジャガイモ、サトウダイコン、マンジュギクおよびシロイヌナズナ、とりわけ好ましい双子葉植物はダイズ、アブラナ種子、アブラナおよびジャガイモである。
【0032】
本発明の一実施形態において、以上定義した方法は次のステップ:
a)先にi)〜vi)に定義した核酸分子を含む1以上のNEENAを植物またはその部分中に導入するステップ、および
b)前記1以上の NEENAを前記植物またはその部分のゲノム中に組込むステップであって、ここで前記1以上のNEENAは、前記1以上のNEENAに対して異種である内因性の、好ましくは、構成的に発現された核酸と機能的に連結されているステップ、および、任意に、
c)前記形質転換した細胞由来の前記1以上のNEENAを含む植物またはその部分を再生するステップ
を含むものである。
【0033】
NEENAは、NEENAが機能的に連結された前記プロモーターの制御下にある核酸分子に対して異種であってもよく、または、プロモーターおよび前記プロモーターの制御下の核酸分子の両方に対して異種であってもよい。
【0034】
1以上のNEENA分子を、植物またはその部分中に、粒子ボンバードメント、プロトプラストエレクトロポレーション、ウイルス感染、アグロバクテリウム媒介性形質転換または当技術分野で公知の任意の他の手法を用いて導入してもよい。NEENA分子を、例えば、プラスミドまたはウイルスDNAまたはウイルスRNA中に導入して組込んでもよい。NEENA分子をまた、BAC、YACまたは人工染色体に含ませた後、植物または植物の部分中に導入してもよい。これをまた、NEENA配列を含む直鎖の核酸分子として導入してもよく、その場合、核酸分子上のNEENA配列に隣接してさらなる配列が存在してもよい。NEENA配列に隣接するこれらの配列は約20bp、例えば20bp〜数100bp、例えば100bp以上であってもよく、そして例えば、相同組換えによりゲノム中への組込みを促進することができる。ゲノム組込みのための任意の他の方法を用いてもよく、標的化組込み手法、例えば相同性組換えまたは無作為組込み手法、例えば非正統的組換えであってもよい。
【0035】
NEENA分子が機能的に連結された、内因性の好ましくは構成的に発現される核酸は、任意の核酸、好ましくは任意の構成的に発現される核酸分子であってもよい。核酸分子は、タンパク質をコードする核酸分子または非コーディング分子、例えば、アンチセンスRNA、rRNA、tRNA、miRNA、ta-siRNA、siRNA、dsRNA、snRNA、snoRNAまたは当技術分野で公知の任意の他の非コーディングRNAであってもよい。
【0036】
当業者は、本発明の方法を、好ましくは、例えばマイクロアレイチップハイブリダイゼーション、qPCR、ノーザンブロット分析、次世代配列決定などにより適用することができる構成的に発現された核酸分子を同定する方法を知っている。
【0037】
本発明の方法を実施するさらなる手順は、
a)プロモーター、好ましくは先に定義した構成的プロモーターと機能的に連結され、かつ、1以上の核酸分子(この核酸分子は前記1以上のNEENAに対して異種でありかつ前記プロモーター、好ましくは構成的プロモーターの制御下にある)と機能的に連結された、1以上のNEENA(このNEENAは先にi)〜vi)に定義した核酸分子を含む)を含むものである発現構築物を提供するステップ、および
b)前記1以上のNEENAを含む発現構築物を、前記植物またはその部分のゲノム中に組込むステップ、および、任意に
c)前記形質転換した植物またはその部分から前記1以上の 発現 構築物を含む植物またはその部分を再生するステップ
であってもよい。
【0038】
NEENAは、このNEENAが機能的に連結されたプロモーターの制御下にある核酸分子に対して異種であってもよく、または前記プロモーターと前記プロモーターの制御下の核酸分子との両方に対して異種であってもよい。
【0039】
発現構築物は任意の当技術分野で公知の方法によってそれぞれの植物のゲノム中に組込むことができる。組込みは粒子ボンバードメントまたはアグロバクテリウム媒介性形質転換などの方法を用いて無作為であってもよい。好ましい実施形態において、組込みは、例えば相同性組換えによる標的化組込み経由であってもよい。後者の方法は、NEENAと機能的に連結された高発現プロモーターを含む発現構築物をお気に入りのゲノム領域中に組込むことを可能にする。好都合なゲノム領域は、例えば、高度に発現される遺伝子を含むことが公知のゲノム領域、例えば種子中であり、従って、転写活性を示さないゲノム領域と比較して前記発現構築物由来の発現を増加することができる。
【0040】
他の好ましい実施形態においては、前記1以上のNEENAをプロモーター、好ましくは前記異種核酸分子の転写出発部位に近い構成的プロモーターと機能的に連結する。
【0041】
本明細書で意味する「転写出発部位に近い」は、前記異種核酸分子の転写出発部位から2500bp以下、好ましくは2000bp以下、より好ましくは1500bp以下、さらにより好ましくは1000bp以下および最も好ましくは500bp以下離れたプロモーター、好ましくは構成的プロモーターと、1以上のNEENAを機能的に連結することを含む。理解すべきこととして、NEENAはそれぞれのプロモーターの転写出発部位のそれぞれの距離の上流または下流に組込むことができる。従って、1以上のNEENAは必ず、1以上のNEENAが機能的に連結された好ましくは構成的プロモーターの制御下のそれぞれの異種核酸の転写物中に必ずしも含まれなくてもよい。好ましくは、1以上の NEENAはそれぞれのプロモーター、好ましくは 構成的プロモーターの転写出発部位の下流に組込まれる。転写出発部位の下流の組込み部位は5'UTR5、3'UTR、エキソンまたはイントロンにあってもよく、またはイントロンまたは部分的にもしくは完全に、好ましくは構成的プロモーターの制御下の異種核酸の5'UTRまたは3'UTRを置き換えてもよい。好ましくは、1以上のNEENAは5'UTRまたはイントロンに組込まれるか、またはNEENAはイントロンまたは一部分もしくは完全な5'UTRと置き換えられ、最も好ましくは、それぞれの異種核酸の5'UTRに組込まれる。
【0042】
本発明のさらなる実施形態は先にi)〜vi)に定義した核酸分子を含む1以上のNEENAを含む組換え発現構築物を含むものである。
【0043】
組換え発現構築物はさらに、1以上のNEENAが機能的に連結された1以上のプロモーター、好ましくは構成的プロモーターならびに任意に1以上の発現される核酸分子(後者は前記1以上のNEENAに対して異種である)を含んでもよい。
【0044】
NEENAは、このNEENAが機能的に連結されたプロモーターの制御下にある核酸分子に対して異種であってもよく、または前記プロモーターと前記プロモーターの制御下の核酸分子との両方に対して異種であってもよい。
【0045】
発現構築物は、NEENAと機能的に連結されたプロモーター、好ましくは構成的プロモーターとそれぞれのNEENAに対して異種の発現される核酸分子との1以上、例えば2以上、例えば5以上、例えば10以上の組み合わせを含んでもよい。発現構築物はまた、それぞれのプロモーターに対して同族または異種の発現される核酸分子と機能的に連結された、NEENAを含まないさらなるプロモーターを含んでもよい。
【0046】
先に定義した1以上の組換え発現構築物を含む組換え発現ベクターは本発明の他の実施形態である。本発明に使用しうる複数の発現ベクターは当業者に知られている。例えばNEENAと機能的に連結されたプロモーターおよび、任意に、ターミネーターなどの他のエレメントを含む発現構築物を含むものであるベクターを植物のゲノム中に導入し、形質転換した細胞からトランスジェニック植物を回収する方法もまた、当技術分野で公知である。植物またはその部分を形質転換するために用いる方法に応じて、全ベクターを前記植物またはその部分のゲノム中に組込んでもよく、または、ベクターのある特定成分、例えばT-DNAなどをゲノム中に組込んでもよい。
【0047】
先にi)〜vi)に定義した1以上の異種NEENAを含むトランスジェニック植物またはその部分も本発明に包含される。NEENAは、もしそれが合成されたか、他生物由来であるか、または同じ生物であるがその自然のゲノム配置が対照植物、例えば野生型植物と比較して手を加えられていれば、その植物にとって異種であると理解すべきである。理解すべきこととして、手を加えられたゲノム配置とは、NEENAが他の染色体上にまたは同じ染色体であるが、その自然の、例えば野生型植物のゲノム配置から10kb以上、例えば10kb、好ましくは5kb以上、例えば5kb、より好ましくは1000bp以上、例えば1000bp、さらにより好ましくは500bp以上、例えば500bp、とりわけ好ましくは100bp以上、例えば100bp、最も好ましくは10bp以上、例えば10bpだけ異なった位置に配置されていることを意味する。
【0048】
先に定義した組換え発現ベクターまたは先に定義した組換え発現構築物を含むトランスジェニック細胞またはトランスジェニック植物またはその部分は本発明のさらなる実施形態である。トランスジェニック細胞、トランスジェニック植物またはその部分は、細菌、真菌、酵母または植物、昆虫または哺乳動物細胞または植物からなる群より選択することができる。好ましくはトランスジェニック細胞は、細菌、真菌、酵母または植物細胞である。好ましい細菌は、腸内細菌、例えば大腸菌およびアグロバクテリア属の細菌、例えばAgrobacterium tumefaciensおよびAgrobacterium rhizogenesである。好ましい植物は、単子葉または双子葉植物、例えば単子葉または双子葉作物植物、例えばトウモロコシ、ダイズ、アブラナ、ワタ、ジャガイモ、サトウダイコン、イネ、コムギ、モロコシ、オオムギ、ススキ、バナナ、サトウキビその他である。好ましい作物植物はトウモロコシ、イネ、コムギ、ダイズ、アブラナ、ワタまたはジャガイモである。とりわけ好ましい双子葉類の作物植物はダイズ、アブラナ、ワタまたはジャガイモである。
【0049】
とりわけ好ましい単子葉類の作物植物はトウモロコシ、コムギおよびイネである。
【0050】
前記異種の先にi)〜vi)に定義したNEENAまたは先に定義した前記組換え発現構築物または前記組換えベクターを含む、先に定義したトランスジェニック細胞または植物またはその部分から誘導されるトランスジェニック細胞培養、トランスジェニック種子、部分または繁殖材料は本発明の他の実施形態である。
【0051】
本明細書で意味するトランスジェニック部分または繁殖材料は、それぞれのNEENA、発現構築物または組換えベクターを含む切り穂、接ぎ穂、取り木、枝または若枝などの植物の繁殖および/または再生に有用である全ての組織および器官、例えば葉、幹および果実を含むものである。
【0052】
本発明のさらなる実施形態は、先にi)〜vi)に定義したNEENAまたは先に定義した組換え構築物または組換えベクターの、植物またはその部分における発現の増強のための使用である。
【0053】
従って、本出願は、1以上のNEENAと機能的に連結された1以上のプロモーター、好ましくは、種子特異的および/または種子選好的プロモーターを含むものである、種子特異的および/または種子選好的な遺伝子発現を増強する核酸分子を提供する。さらに、かかる遺伝子発現を増強する核酸分子、ならびにかかる遺伝子発現を増強する核酸分子を含む発現構築物、発現ベクター、トランスジェニック植物またはその部分およびトランスジェニック細胞を提供する。
【0054】
先に定義したトランスジェニック細胞または植物またはその部分から誘導されるトランスジェニック細胞培養、トランスジェニック種子、部分または繁殖材料の、食糧、動物飼料、種子、医薬品またはファインケミカルを生産するための使用も本発明に包含される。
【0055】
定義
略語は次の通りである:NEENA、核酸発現増強性核酸;GFP、緑色蛍光タンパク質;GUS、β-グルクロニダーゼ;BAP、6-ベンジルアミノプリン;2,4-D、2,4-ジクロロフェノキシ酢酸;MS、MurashigeおよびSkoog培地;NAA、1-ナフタレン酢酸;MES、2-N-(モルホリノ)エタンスルホン酸;IAA、インドール酢酸;Kan、カナマイシン硫酸塩;GA3、ジベレリン酸;Timentin(登録商標)、チカルシリン二ナトリウム/カリウムクラブラネート;microl、マイクロリットル。
【0056】
理解すべきこととして、本発明は特別な手法またはプロトコルに限定されるものでない。理解すべきこととしてまた、本明細書で使用する用語法は特別な実施形態を記載することだけを目的とし、添付した請求の範囲が限定しうる本発明の範囲を限定することを意図しない。注意しなければならないこととして、本明細書および添付した請求の範囲における使用で、単数形の冠詞「a」「an」および「the」は、文脈が明確に断らない限り、複数の言及を含むものである。従って、例えば、「a」ベクターは1以上のベクターへの言及であり、当業者に公知のその等価物などを含むものである。本明細書で使用する用語「約」は、およそ、大雑把に、大体、またはその範囲のを意味する。用語「約」を数字の範囲と共に用いる場合、この用語は記載した数値の上限および下限の境界を拡げることにより範囲を改変する。一般に、本明細書では用語「約」を用いて、記載した値の上限および下限の数値を20%、好ましくは10%上方もしくは下方(高いもしくは低い)の分散をもつ値に改変する。本明細書で使用する用語「または」は特別なリストの任意の1つのメンバーを意味し、そのリストのメンバーの任意の組み合わせも含むものである。用語「含むものである(compriseとその変化形)」「含む(includeとその変化形)」は、本明細書および以下の請求の範囲で使用する場合、1以上の記載した特徴、整数、成分、またはステップの存在を説明することを意図するが、これらの用語は他の特徴、整数、成分、またはステップ、またはその群の存在もしくは付加を排除するものではない。明確にするために述べると、本明細書に使用するある特定の用語は次の通り定義されかつ使用される。
【0057】
アンチパラレル:ここで「アンチパラレル」は、5'-3'方向に走る1つのヌクレオチド配列と3'-5'方向に走る他のヌクレオチド配列との、リン酸ジエステル結合をもつ相補性塩基残基の間の水素結合を介して対合した2つのヌクレオチド配列を意味する。
【0058】
アンチセンス:用語「アンチセンス」は、転写または機能のためのその通常の配向に対して反転していて、そのために、宿主細胞内で発現される標的遺伝子mRNA分子と相補的であるRNA転写物を発現する(例えば、これは標的遺伝子mRNA分子または1本鎖ゲノムDNAとワットソン-クリック塩基対合を介してハイブリダイズすることができる)か、または標的DNA分子、例えば、宿主細胞に存在するゲノムDNAと相補的であるヌクレオチド配列を意味する。
【0059】
コード領域:本発明で使用する用語「コード領域」は、構造遺伝子を言及して用いる場合、mRNA分子の翻訳の結果として新生ポリペプチドに見出されるアミノ酸をコードするヌクレオチド配列を意味する。コード領域は、真核生物においては、5'側にヌクレオチドトリプレット「ATG」(イニシエーターのメチオニンをコードする)および3'側に停止コドンを表す3つのトリプレット(すなわち、TAA、TAG、TGA)の1つが結合されている。イントロンを含有するだけでなく、遺伝子のゲノム形態はまた、RNA転写物に存在する配列の5'と3'末端の両方に位置する配列も含みうる。これらの配列は、「隣接する(flanking)」配列または領域と呼ばれる(これらの隣接する領域はmRNA転写物に存在する非翻訳配列に対して5'もしくは3'に位置する)。5'に隣接する領域は、調節配列、例えば、遺伝子の転写を制御または影響するプロモーターおよびエンハンサーを含有しうる。3'に隣接する領域は、転写の終結、転写後の切断およびポリアデニル化を指令する配列を含有しうる。
【0060】
相補性:「相補的」または「相補性」は、アンチパラレルなヌクレオチド配列が相補的塩基残基間で水素結合を形成すると(塩基対合規則により)お互いに対合することができるアンチパラレルなヌクレオチド配列を含むものである、2つのヌクレオチド配列を意味する。例えば、配列 5'-AGT-3'は配列 5'-ACT-3'と相補的である。相補性は「部分的」であってもまたは「全体」であってもよい。「部分的」相補性は、1以上の核酸塩基が塩基対合規則によってマッチしない場合である。「全体」または「完全」相補性は、それぞれおよび全ての核酸塩基が他の塩基と、塩基対合規則のもとでマッチする場合である。核酸分子鎖間の相補性の程度は、核酸分子鎖間のハイブリダイゼーションの効率と強度に有意な効果を有する。本明細書で使用する核酸配列の「相補配列」は、その核酸分子が核酸配列の核酸分子に対して全体相補性を示すヌクレオチド配列を意味する。
【0061】
2本鎖RNA:「2本鎖 RNA」分子または「dsRNA」分子は、ヌクレオチド配列のセンスRNA断片とヌクレオチド配列のアンチセンスRNA断片を含むものであって、両方のRNA断片がお互いに相補的なヌクレオチド配列を含み、それにより、センスとアンチセンスRNA断片が対合して2本鎖RNA分子を形成することができる。
【0062】
内因性:「内因性」ヌクレオチド配列は、未翻訳の植物細胞のゲノム中に存在するヌクレオチド配列を意味する。
【0063】
発現の増強:植物細胞中の核酸分子の発現の「増強」または「増加」を本明細書では同じ意味で使用し、本発明の方法の適用後の植物、植物の部分または植物細胞における核酸分子の発現のレベルは、本発明の方法の適用前の植物、植物の部分または植物細胞におけるその発現のレベルよりまたは本発明の組換え核酸分子を欠く参照植物と比較して高いことを意味する。例えば、参照植物はそれぞれのNEENAを欠くだけである同じ構築物を含むものである。本明細書で使用する用語「増強」または「増加」は同義であり、ここでは発現すべき核酸分子のより高い、好ましくは有意により高い発現を意味する。本明細書で使用する、作用物質、例えばタンパク質、mRNAまたはRNAのレベルの「増強」または「増加」は、本発明の組換え核酸分子を欠く、例えば、本発明のNEENA分子、組換え構築物または組換えベクターを欠く、実質的に同一条件のもとで育てた実質的に同一の植物、植物の部分、または植物細胞と比較して、そのレベルが増加したことを意味する。本明細書で使用する、標的遺伝子により発現された作用物質、例えばpreRNA、mRNA、rRNA、tRNA、snoRNA、snRNAのおよび/またはそれがコードするタンパク質産物のレベルの「増強」または「増加」は、そのレベルが本発明の組換え核酸分子を欠く細胞または生物と比較して、50%以上、例えば100%以上、好ましくは200%以上、より好ましくは5倍以上、さらにより好ましくは10倍以上、最も好ましくは20倍以上例えば50倍増加することを意味する。その増強または増加は、当業者になじみのある方法により測定することができる。従って、核酸またはタンパク質の量の増強または増加はタンパク質の免疫学的検出により測定することができる。さらに、タンパク質アッセイ、蛍光、ノーザンハイブリダイゼーション、ヌクレアーゼ保護アッセイ、逆転写(定量RT-PCR)、ELISA(酵素結合免疫吸着アッセイ)、ウェスタンブロット、ラジオイムノアッセイ(RIA)または他のイムノアッセイおよび蛍光活性化細胞分析(FACS)などの技法を使って植物または植物細胞中の特定のタンパク質またはRNAを測定することができる。誘発されるタンパク質産物の型に応じて、生物または細胞の表現型に与えるその活性または効果を測定してもよい。タンパク質の量を測定する方法は当業者に公知である。記載しうる例には次のものがある:マイクロ-ビウレット法(Goa J (1953) Scand J Clin Lab Invest 5:218-222)、フォーリン-チオカルト法(Lowry OH ら (1951) J Biol Chem 193:265-275) または CBB G-250の吸収を測定する方法(Bradford MM (1976) Analyt Biochem 72:248-254)。タンパク質の活性を定量化する一例として、ルシフェラーゼ活性の検出を以下の実施例に記載した。
【0064】
発現:「発現」は細胞における、遺伝子産物の生合成、好ましくは、ヌクレオチド配列、例えば、内因性遺伝子または異種遺伝子の転写および翻訳を意味する。例えば、構造遺伝子の事例では、発現は構造遺伝子のmRNA中への転写および、場合によっては、引き続いてmRNAの1以上のポリペプチドへの翻訳に関わる。他の場合では、発現はRNA分子を保持するDNAの転写だけに関わりうる。
【0065】
発現構築物:本明細書で使用する「発現構築物」は、適当な植物の部分または植物細胞中の特別なヌクレオチド配列の発現を指令することができるDNA配列であって、目的のヌクレオチド配列と作動しうる形で連結された(場合によっては、終結シグナルと作動しうる形で連結された)、その中へ導入されるであろう前記植物の部分または植物細胞中で機能的なプロモーターを含む前記DNA配列を意味する。もし翻訳が必要であれば、発現構築物は典型的には、ヌクレオチド配列の適当な翻訳に必要な配列も含む。コード領域は目的のタンパク質をコードしうるがまた、目的の機能性RNA、例えばRNAa、siRNA、snoRNA、snRNA、マイクロRNA、ta-siRNAまたは任意の他の非コード調節性RNAをセンスまたはアンチセンス方向にコードしてもよい。目的のヌクレオチド配列を含む発現構築物はキメラ(1以上のその成分が1以上のその他の成分に対して異種であることを意味する)であってもよい。発現構築物はまた、天然であるが、異種発現にとって有用な組換え体の形態で得たものであってもよい。しかし典型的には、発現構築物は宿主に対して異種であり、すなわち、発現構築物の特別なDNA配列は宿主細胞で天然に生じるものでなく、宿主細胞中に宿主細胞の祖先中に形質転換事象によって導入されてなければならない。発現構築物中のヌクレオチド配列の発現は、構成的プロモーターまたは、宿主細胞をいくつかの特別な外界の刺激に曝したときにだけ転写を開始する誘導プロモーターの制御のもとで生じる。植物の事例では、プロモーターも特別な組織または器官または発生段階に特異的でありうる。
【0066】
外来:用語「外来」は、細胞のゲノム中に実験操作により導入されたいずれかの核酸分子(例えば、遺伝子配列)を意味し、導入された配列がいくつかの改変(例えば、点突然変異、選択マーカー遺伝子の存在など)を含有する限り、天然の配列とは異なる配列がその細胞中に見出される。
【0067】
機能的連結:用語「機能的連鎖」または「機能的連結」は、調節エレメント(例えば、プロモーター)の発現すべき核酸配列との、および、適宜、さらなる調節エレメント(例えば、ターミネーターまたはNEENAなど)との配列の配置であって、それぞれの調節エレメントがその意図する機能を完遂して前記核酸配列の発現を可能にし、改変し、促進しまたはさもなくば影響を与え得る前記配列の配置を意味すると理解される。同義語として、表現「作動しうる連鎖」または「作動しうる連結」を使用してもよい。発現は、核酸配列のセンスまたはアンチセンスRNAと関係した配置に依存してもたらされうる。この目的にとって、化学的意味の直接連結は必ずしも必要でない。例えば、エンハンサー配列などの遺伝子制御配列も、さらに離れた位置から、 または他のDNA分子からでも、標的配列に対するその機能を果たしうる。好ましい配置は、発現すべき核酸配列が組換えによってプロモーターとして作用する配列の後方に位置し、2つの配列がお互いに共有結合で連結される配置である。プロモーター配列と組換えで発現すべき核酸配列の間の距離は、好ましくは200塩基対未満、とりわけ好ましくは100塩基対未満、非常にとりわけ好ましくは50塩基対未満である。好ましい実施形態においては、転写開始が本発明のキメラRNAの所望の開始と同一であるように、転写すべき核酸配列をプロモーターの後ろに配置する。機能的連鎖、および発現構築物は、記載された通例の組換えおよびクローニング技法を用いて作製することができる(例えば、Maniatis T, Fritsch EF and Sambrook J (1989) Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor (NY);Silhavy et al. (1984) Expeiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor (NY);Ausubel et al., (1987) Current Protocols in Molecular Biology, Greene Publishing Assoc. and Wiley Interscience;Gelvin et al., (Eds) (1990) Plant Molecular Biology Manual; Kluwer Academic Publisher, Dordrecht, The Netherlands)。しかし、例えば、制限酵素に対する特異的切断部位をもつリンカーとして、またはシグナルペプチドとして作用するさらなる配列も2つの配列の間に配置することができる。配列の挿入はまた、融合タンパク質の発現をもたらしうる。好ましくは、調節領域、例えばプロモーターおよび発現すべき核酸配列の連鎖から成る発現構築物をベクターに組込まれた形で存在させてもよく、例えば、形質転換により植物ゲノム中に挿入することができる。
【0068】
遺伝子:用語「遺伝子」は、遺伝子産物(例えば、ポリペプチドまたは機能性RNA)の発現をいくつかの方式で調節することができる適当な調節配列と作動しうる形で接続された領域を意味する。遺伝子は、コード領域(オープンリードフレーム、ORF)に先行する(上流の)および後続する(下流の)DNAの非翻訳調節領域(例えば、プロモーター、エンハンサー、リプレッサーなど)ならびに、適用可能であれば、個々のコード領域(すなわち、液損)の間に介在配列(すなわち、イントロン)を含む。本明細書で使用する用語「構造遺伝子」は、mRNAに転写され、次いで特定のポリペプチドの特徴であるアミノ酸の配列に翻訳されるDNA配列を意味することを意図している。
【0069】
ゲノムとゲノムDNA:用語「ゲノム」または「ゲノムDNA」は、宿主生物の遺伝性の遺伝子情報を意味する。前記ゲノムDNAは核DNA(染色体DNAとも呼ばれる)を含むがまた、色素体(例えば、葉緑体)および他の細胞小器官(例えば、ミトコンドリア)のDNAも含む。好ましくは、用語「ゲノム」または「ゲノムDNA」は核の染色体DNAを意味する。
【0070】
異種:核酸分子またはDNAに関する用語「異種」は、自然では作動しうる形で連結されてないまたは自然では異なる位置で作動しうる形で連結された第2の核酸分子と、作動しうる形で連結された、または作動しうる形で連結されるように遺伝子操作された核酸分子を意味する。核酸分子およびそれと連結された1以上の調節核酸分子(プロモーターまたは転写終結シグナルなど)を含む異種発現構築物は、例えば、実験遺伝子操作に由来する構築物であって、a)前記核酸分子、またはb)前記調節核酸分子、またはc)両方(すなわち(a)と(b))がその自然(生来)の遺伝環境に位置しないか、または、実験遺伝子操作により改変されている(改変の例は1以上のヌクレオチド残基の置換、付加、欠失、反転または挿入である)前記構築物である。
【0071】
自然の遺伝環境は、起源の生物における自然の染色体遺伝子座、またはゲノムライブラリー中の存在を意味する。ゲノムライブラリーの場合、核酸分子の配列の自然の遺伝環境は好ましくは、少なくとも部分において保持される。前記環境は核酸配列に少なくとも1つの側において隣接し、そして少なくとも50bp、好ましくは少なくとも500bp、とりわけ好ましくは少なくとも1,000bp、非常にとりわけ好ましくは少なくとも5,000bpの長さの配列を有する。天然の発現構築物(例えば、プロモーターの対応する遺伝子との天然の組み合わせ)は、非自然、合成の「人工的」方法、例えば変異誘発により改変されると、トランスジェニック発現構築物になる。かかる方法は記載されている(US 5,565,350;WO 00/15815)。例えば、この分子の生来のプロモーターでないプロモーターと作動しうる形で連結されたタンパク質をコードする核酸分子は、プロモーターについて異種であるとみなされる。好ましくは、それを導入した細胞に対して内因性でないかまたは自然で随伴しないが、他の細胞から得られたかまたは合成された異種DNAは内因性でない。異種DNAはまた、内因性DNA配列のいくつかの改変、非天然、多コピーを含有する内因性DNA配列、それに物理的に連結された他のDNA配列を自然で随伴しないDNA配列を含む。一般に、必ずではないが、異種DNAはそれが発現される細胞により通常産生されないRNAまたはタンパク質をコードする。
【0072】
高発現性構成的プロモーター:本明細書で使用する「高発現性構成的プロモーター」は、植物またはその部分における構成的発現を引き起こすプロモーターであって、それぞれのプロモーターの制御下の核酸分子由来のRNAの蓄積または合成速度またはRNAの安定性が、本発明のNEENAを欠くプロモーターにより引き起こされる発現より高い、好ましくは、有意に高い前記プロモーターを意味する。好ましくは、RNAの量および/またはRNA合成の速度および/またはRNAの安定性が、本発明のNEENAを欠く構成的プロモーターと比較して、50%以上,例えば100%以上,好ましくは200%以上,より好ましくは5倍以上,さらにより好ましくは10倍以上,最も好ましくは20倍以上例えば50倍増加する。
【0073】
ハイブリダイゼーション:本明細書で使用する用語「ハイブリダイゼーション」は「核酸分子の1つの鎖が塩基対合を介して相補鎖に参加する任意のプロセス」を含む(J. Coombs (1994) Dictionary of Biotechnology, Stockton Press, New York)。ハイブリダイゼーションおよびハイブリダイゼーションの強度(すなわち、核酸分子間の連合の強度)は、核酸分子間の相補性の程度、関わる条件のストリンジェンシー、形成されるハイブリッドのTm、および核酸分子内のG:C比などの因子の影響を受ける。本明細書で使用する用語「Tm」は「融点」の意味で使用される。融点は2本鎖の核酸分子が半分に解離して1本鎖になる温度である。核酸分子のTmを計算する式は当技術分野において周知である。標準の参考文献に示されるように、核酸分子が1M NaClの水溶液中に含まれる場合、Tm値の単純な計算は式:Tm=81.5+0.41(% G+C)により計算することができる[例えば、Anderson and Young, Quantitative Filter Hybridization, in Nucleic Acid Hybridization (1985)を参照]。他の参考文献は、構造ならびに配列の特徴を考慮に入れてTmを計算するさらに複雑な計算を含む。厳密な条件は、当業者に公知であり、Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6に記載されている。
【0074】
「同一性」:「同一性」は、2以上の核酸またはアミノ酸分子の比較について使用する場合、前記分子がある特定の程度の配列類似性を共有し、配列が部分的に同一であることを意味する。
【0075】
2つのアミノ酸配列または2つの核酸分子のパーセント同一性(本明細書では相同性を互換的に使用する)を決定するために、配列の1つを他の配列の下に書いて最適な比較を行う(例えば、ギャップを1つのタンパク質の配列中に挿入し、他のタンパク質または他の核酸と最適なアラインメントを作り出すことができる)。
【0076】
対応するアミノ酸位置またはヌクレオチド位置のアミノ酸残基または核酸分子を次いで比較する。もし1つの配列の1つの位置が他配列の対応する位置と同じアミノ酸残基または同じ核酸分子により占められていれば、その分子はこの位置において相同性である(すなわち、本明細書の文脈で使用するアミノ酸または核酸「相同性」はアミノ酸または核酸「同一性」に対応する)。2つの配列間のパーセント同一性は、配列が共有する同一の位置の数の関数(すなわち、%同一性=同一の位置の数/位置の全数)である。用語「相同性」および「同一性」は従って、同義語と考えるべきである。
【0077】
2以上のアミノ酸または2以上のヌクレオチド配列のパーセント同一性 を確認するために、いくつかのコンピューターソフトウエアプログラムが開発されている。2以上配列の同一性は、例えば、ソフトウエアfastaを用いて計算することができ、これは現在バージョンfasta 3が使用されている(W. R. Pearson and D. J. Lipman, PNAS 85, 2444(1988);W. R. Pearson, Methods in Enzymology 183, 63 (1990);W. R. Pearson and D. J. Lipman, PNAS 85, 2444 (1988);W. R. Pearson, Enzymology 183, 63 (1990))。異なる配列の同一性を計算する他の有用なプログラムは標準blastプログラムであって、これはBiomax pedantソフトウエアに含まれている(Biomax、Munich、Federal Republic of Germany)。このソフトウエアは残念ながら、 blastが主題およびクエリの完全な配列を常に含まないので、最適性の劣る結果をもたらすことがある。それにも関わらず、このプログラムは非常に効率的であるので、膨大な数の配列を比較するために用いることができる。以下の設定は、配列のかかる比較のための典型的なものである。
【0078】
-p プログラム名 [文字列]; -d データベース [文字列]; デフォルト = nr; -i クエリーファイル[File In]; デフォルト = stdin; -e 期待値 (E) [実数]; デフォルト = 10.0; -m アラインメントビューオプション: 0 = ペアワイズ; 1 = 同一性を示すクエリー-アンカー; 2 = 同一性を示さないクエリー-アンカー; 3 = フラットクエリー-アンカー、同一性を示す; 4 = フラットクエリー-アンカー、同一性を示さない; 5 = フラットクエリー-アンカー、同一性を示さない、かつ平滑末端; 6 = フラットクエリー-アンカー、同一性を示さない、かつ平滑末端; 7 = XML Blast出力; 8 = 表形式; 9 コメントラインを含む表形式[整数]; デフォルト = 0; -o BLASTリポート出力ファイル[File Out] オプション; デフォルト = stdout; -F フィルタークエリー配列(blastnを用いるDUST、他を用いるSEG) [文字列]; デフォルト = T; -G ギャップを開くためのコスト(0はデフォルト行動を引き起こす) [整数]; デフォルト = 0; -E ギャップを拡張するためのコスト(0はデフォルト行動を引き起こす) [整数]; デフォルト = 0; -X ギャップ付加されたアラインメントに関するX下落値(ビット) (0はデフォルト行動を引き起こす); blastn 30、megablast 20、tblastx 0、他の全部15 [整数]; デフォルト = 0; -I デフラインGIを示す[T/F]; デフォルト = F; -q ヌクレオチド不一致に関するペナルティ(blastnのみ) [整数]; デフォルト = -3; -r ヌクレオチド一致に関する報酬(blastnのみ) [整数]; デフォルト = 1; -v (V)に関する1行記述を示すデータベース配列の数 [整数]; デフォルト = 500; -b (B)に関するアラインメントを示すデータベース配列の数 [整数]; デフォルト = 250; -f 拡張ヒットに関する閾値、デフォルト 0の場合; blastp 11、blastn 0、blastx 12、tblastn 13; tblastx 13、megablast 0 [整数]; デフォルト = 0; -g ギャップ付加アラインメントを実行(tblastxでは利用不可) [T/F]; デフォルト = T; -Q 使用するクエリー遺伝子コード[整数]; デフォルト = 1; -D DB遺伝子コード(tblast[nx]についてのみ) [整数]; デフォルト = 1; -a 使用するプロセッサーの数[整数]; デフォルト = 1; -O SeqAlignファイル[File Out] オプション; -J クエリーデフラインを信じる[T/F]; デフォルト = F; -M マトリックス[文字列]; デフォルト = BLOSUM62; -W ワードサイズ、デフォルト 0の場合 (blastn 11、megablast 28、他の全部 3) [整数]; デフォルト = 0; -z データベースの有効長さ(実際のサイズについては0を使用する) [実数]; デフォルト = 0; -K 保持する領域から得られる最良のヒットの数(デフォルトにより除去、使用する場合、100の値が推奨される) [整数]; デフォルト = 0; -P 複数ヒットについては0、単一ヒットについては1[整数]; デフォルト = 0; -Y 検索スペースの有効長さ(実際のサイズについては0を使用する) [実数]; デフォルト = 0; -S データベースに対して検索するためのクエリー・ストランド(blast[nx]、およびtblastxについて); 3は両方、1は上、2は下である[整数]; デフォルト = 3; -T HTML出力を作製 [T/F]; デフォルト = F; -l GIの一覧に対するデータベースの限定的検索 [文字列] オプション; -U FASTA配列のフィルタリングには小文字を使用する [T/F] オプション; デフォルト = F; -y ビットで示される非ギャップ付加拡張のためのX下落値(0.0はデフォルト行動を引き起こす); blastn 20、megablast 10、他の全部 7 [実数]; デフォルト = 0.0; -Z ビットで示される最終的なギャップ付加アラインメントに関するX下落値(0.0はデフォルト行動を引き起こす); blastn/megablast 50、tblastx 0、他の全部 25 [整数]; デフォルト = 0; -R PSI-TBLASTN チェックポイントファイル[File In] オプション; -n MegaBlast検索 [T/F]; デフォルト = F; -L クエリー配列上の位置[文字列] オプション; -A 複数ヒットウィンドウサイズ、デフォルト 0の場合(blastn/megablast 0、他の全部 40 [整数]; デフォルト = 0; -w フレームシフトペナルティ(blastxに関するOOFアルゴリズム) [整数]; デフォルト = 0; -t HSPsを連結するためにtblastnにおいて許容される最大のイントロンの長さ (0は連結不可) [整数]; デフォルト = 0。
【0079】
NeedlemanおよびWunsch、またはSmithおよびWatermanのアルゴリズムを用いることにより、高品質の結果が達成される。従って、前記アルゴリズムに基づくプログラムが好ましい。有利には、配列の比較を、プログラムPileUp(J. Mol. Evolution., 25, 351 (1987)、Higgins et al., CABIOS 5, 151 (1989))または、好ましくは、プログラム「Gap」および「Needle」(両方とも、NeedlemanおよびWunsch(J. Mol. Biol. 48; 443-453 (1970)のアルゴリズムに基づく)ならびに「BestFit」(SmithおよびWaterman(Adv. Appl. Math. 2; 482-489 (1981))のアルゴリズムに基づく)を用いて行うことができる。「Gap」および「Needle」はGCGソフトウェアパッケージ [Genetics Computer Group, 575 Science Drive, Madison, Wisconsin, USA 53711 (1991); Altschul ら(1997) Nucleic Acids Res. 25:3389 (1997)]の一部であり、「Needle」はThe European Molecular Biology Open Software Suite (EMBOSS)(Trends in Genetics 16 (6), 276 (2000))の一部である。従って、好ましくは、配列同一性のパーセントを決定する計算を、配列の全範囲にわたりプログラム「Gap」または「Needle」を用いて行う。核酸配列の比較のための以下の標準的な調整を「Needle」について用いた: マトリックス: EDNAFULL, ギャップ_ペナルティ: 10.0、 拡張_ペナルティ: 0.5。核酸配列の比較のための以下の標準的な調整を「Gap」について用いた:ギャップウエイト:50、長さウエイト:3、平均マッチ:10.000、平均ミスマッチ:0.000。
【0080】
例えば、核酸レベルで配列番号1の配列と80%同一性を有するといわれる配列は、配列番号1により表される配列と上記プログラム「Needle」により上記パラメーターセットを用いて比較すると80%同一性を有する配列を意味することがわかる。
【0081】
好ましくは、同一性はクエリー配列、例えば配列番号1の完全長さに基づいて計算する。
【0082】
イントロン:イントロンは、その遺伝子が産生するタンパク質の部分をコードせず、細胞核からエキスポートされる前に、遺伝子から転写されるmRNAからスプライシングで取除かれる遺伝子内のDNAのセクション(介在配列)を意味する。イントロン配列はイントロンの核酸配列を意味する。従って、イントロンは、コード配列(エキソン)と共に転写されるが、成熟mRNAの形成中に取り除かれるDNA配列の領域である。イントロンは実コード領域内にまたはプレmRNA(未スプライシングmRNA)の5'もしくは3'非翻訳リーダー内に位置しうる。一次転写物中のイントロンは切除され、コード配列は同時にかつ正確にライゲートされて成熟RNAを形成する。イントロンとエキソンの接合部がスプライス部位を形成する。イントロンの配列はGUで始まりかつAGで終わる。さらに、植物においては、2例のAU-AC イントロンが記載されている;シロイヌナズナ由来のRecA様タンパク質遺伝子の第14番目イントロンおよびG5遺伝子の第7イントロンはAT-ACイントロンである。イントロンを含有するプレmRNAは、(他の配列に加えて)正確にスプライスされるイントロンにとって必須である3つの短い配列を有する。これらの配列は5'スプライス位置、3'スプライス部位、および分岐点である。mRNAスプライシングは一次mRNA転写物中に存在する介在配列(イントロン)の除去およびエキソン配列の接続またはライゲーションである。これはまた、介在配列(イントロン)の除去と共に同じmRNA上の2つのエキソンを接続するcis-スプライシングとして公知である。イントロンの機能性エレメントは、スプライセソーム(例えば、イントロンの末端のスプライシングコンセンサス配列)の特異的タンパク質成分により認識されかつ結合される配列を含んでいる。機能性エレメントのスプライセオソームとの相互作用によって、成熟前mRNAからのイントロン配列の除去とエキソン配列の再接続がもたらされる。イントロンは3つの短い配列を有し、これらは、イントロンが正確にスプライスされるために(十分ではないが)必須である。これらの配列は、5'スプライス部位、3'スプライス部位および分岐点である。分岐点配列は植物におけるスプライシングおよびスプライス部位選択において重要である。分岐点配列は通常、3'スプライス部位の10〜60ヌクレオチド上流に位置する。
【0083】
同質遺伝子生物:異種DNA配列の存在または非存在によって異なりうることを除いて、遺伝的に同一である生物(例えば、植物)。
【0084】
単離された:本明細書で使用する用語「単離された」は、材料が人手により取り出されていて、その元来の、生来の環境から離れて存在し、それ故に自然の産物でないことを意味する。単離された材料または分子(DNA分子または酵素など)は精製された形態で存在しうるしまたは例えば、トランスジェニック宿主細胞などの非生来の環境で存在しうる。例えば、生存植物中に存在する天然のポリヌクレオチドまたはポリペプチドは単離されてないが、自然の系で共存する材料のいくつかのまたは全てから分離された同じポリヌクレオチドまたはポリペプチドは単離されている。かかるポリヌクレオチドはベクターの部分でありうるしおよび/またはかかるポリヌクレオチドまたはポリペプチドは組成物の部分でありうるのであって、かかるベクターまたは組成物が元来の環境の部分でないという点では単離されているであろう。好ましくは、核酸分子に関係して、「単離された核酸配列」のように使用される場合の「単離された」は、同定され、自然源で元来随伴している少なくとも1つの夾雑核酸分子から分離された核酸配列を意味する。単離された核酸分子は、自然で見出されるのとは異なる形態または設定で存在する核酸分子である。対照的に、単離されてない核酸分子は、それらが自然で存在する状態で見出されたDNAおよびRNAなどの核酸分子である。例えば、所与のDNA配列(例えば、遺伝子)は宿主細胞染色体において、隣接する遺伝子の近位で見出され;RNA配列、例えば、特定のタンパク質をコードする特定のmRNA配列は細胞において、多数のタンパク質をコードする数多くの他のmRNAとの混合物として見出される。しかし、例えば配列番号1を含む単離された核酸配列は、例として挙げれば、配列番号1を通常含有する細胞中のかかる核酸配列であって、その核酸配列が自然の細胞とは異なる染色体もしくは染色体外の位置にあるか、または、さもなくば、自然に見出されるのとは異なる核酸配列が隣接する前記核酸配列を含むものである。単離された核酸配列は1本鎖または2本鎖の形態で存在しうる。単離された核酸配列をタンパク質を発現するために利用する場合、その核酸配列は最小限、少なくともセンスまたはコード鎖の部分を含有してもよい(すなわち、核酸配列は1本鎖であってもよい)。あるいは、センスとアンチセンス鎖の両方を含有してもよい(すなわち、核酸配列は2本鎖であってもよい)。
【0085】
最小プロモーター:不活性であるかまたは非常に弱いプロモーター活性を有し、上流活性化が存在しないプロモーターエレメント、特にTATAエレメント。好適な転写因子が存在すると、最小プロモーターは転写を認める機能を果たす。
【0086】
NEENA:「核酸発現増強性核酸」を参照されたい。
【0087】
非コード:用語「非コード」は、発現されるタンパク質の部分もしくは全てをコードしない核酸分子の配列を意味する。非コード配列は、限定されるものでないが、イントロン、エンハンサー、プロモーター領域、3'非翻訳領域、および5'非翻訳領域を含む。
【0088】
核酸発現増強性核酸(NEENA):用語「核酸発現増強性核酸」は、NEENAが機能的に連結されたプロモーターの制御下の核酸の発現を増強する内因性の特性を有する特定の配列の配列および/または核酸分子を意味する。プロモーター配列と異なり、NEENAの類は発現を駆動することはできない。NEENAと機能的に連結された核酸分子の発現を増強する機能を遂行するために、NEENA自体がプロモーターと機能的に連結されていなければならない。当技術分野で公知のエンハンサー配列とは対照的に、NEENAはin cisで作用するが、in transでは作用しないし、発現される核酸の転写出発部位に近接して位置しなければならない。
【0089】
核酸およびヌクレオチド:用語「核酸」および「ヌクレオチド」は天然または合成または人工の核酸またはヌクレオチドを意味する。用語「核酸」および「ヌクレオチド」はデオキシリボヌクレオチドまたはリボヌクレオチドまたは任意のヌクレオチド類似体およびポリマーまたはそれらのハイブリッドを、1本鎖もしくは2本鎖のセンスもしくはアンチセンス型で含む。特に断らない限り、特別な核酸配列はまた、暗黙のうちに保存して改変されたそれらの変異体(例えば、縮重コドン置換)および相補配列、ならびに明確に示した配列を包含する。用語「核酸」は、「遺伝子」、「cDNA」、「mRNA」、「オリゴヌクレオチド」、「ポリヌクレオチド」と本明細書では互換的に使用される。ヌクレオチド類似体は、塩基、糖および/またはリン酸塩の化学構造に改変を有するヌクレオチドを含み、前記改変には、限定されるものでないが、5-位置ピリミジン改変、8-位置プリン改変、シトシン環外アミン類における改変、5-ブロモ-ウラシルの置換など;および限定されるものでないが、2'-位置糖改変(2'-OHがH、OR、R、ハロ、SH、SR、NH2、NHR、NR2、またはCNから選択される群により置換された糖改変を含む)されたリボヌクレオチドが含まれる。ショートヘアピンRNA(shRNA)はまた、非自然エレメント、例えば、非自然塩基、例えばイオノシンおよびキサンチン、非自然糖類、例えば2'-メトキシリボース、または非自然リン酸ジエステル連鎖、例えばホスホン酸メチル、ホスホロチオアートおよびペプチドを含みうる。
【0090】
核酸配列:表現「核酸配列」は、5'から3'末端へ読まれたデオキシリボヌクレオチドまたはリボヌクレオチド塩基の1本鎖または2本鎖のポリマーを意味する。核酸配列は染色体のDNA、自己複製プラスミド、DNAまたはRNAの感染性ポリマーおよび主に構造的役割を果たすDNAまたはRNAを含む。「核酸配列」はまた、ヌクレオチドを表す略語、文字、記号または言語の継続的列挙を意味する。一実施形態において、核酸は長さが通常100ヌクレオチド未満の比較的短い核酸である「プローブ」であってもよい。核酸プローブは長さが約50ヌクレオチド〜長さが約10ヌクレオチドであることが多い。核酸の「標的領域」は目的のものであると同定された核酸の一部分である。核酸の「コード領域」は、適当な調節配列の制御下に置かれると、配列特異的な方式で転写されかつ翻訳され、特別なポリペプチドまたはタンパク質を産生する核酸の部分である。コード領域はかかるポリペプチドまたはタンパク質をコードすると言われる。
【0091】
オリゴヌクレオチド:用語「オリゴヌクレオチド」はリボ核酸(RNA)またはデオキシリボ核酸(DNA)またはそれらの擬似体のオリゴマーまたはポリマー、ならびに同様に機能する非天然部分を有するオリゴヌクレオチドを意味する。かかる改変されたまたは置換されたオリゴヌクレオチドは、所望の特性、例えば、細胞取込みの増強、核酸標的に対する親和性の増強およびヌクレアーゼの存在のもとでの安定性の故に、しばしば、生来の型よりも好ましい。オリゴヌクレオチドは、好ましくは、連鎖(例えば、リン酸ジエステル)または代わる連鎖によりお互いに共役結合された2以上のヌクレオモノマーを含む。
【0092】
オーバーハング:「オーバーハング」は、2本鎖のオリゴヌクレオチド分子の5'-または3'-ヒドロキシル末端の比較的短い1本鎖ヌクレオチド配列である(「伸長」「突出末端」または「粘着性末端」とも呼ばれる)。
【0093】
植物:植物は一般に、光合成の能力があるいずれかの真核生物の単または多細胞生物またはその細胞、組織、器官、部分または繁殖材料(例えば、種子または果実)を意味すると理解される。本発明の目的に含まれるのは、高等および下等植物の全ての属および種である。一年生、多年生、単子葉および双子葉植物が好ましい。この用語は、成熟植物、種子、シュートおよび実生およびそれらの誘導された部分、繁殖材料(種子または小胞子など)、植物器官、組織、プロトプラスト、カルスおよび他の培養、例えば細胞培養、および機能または構造単位を与える植物細胞分類のいずれか他の型を含む。成熟植物は実生の段階後の任意の所望の発生段階の植物を意味する。実生は、初期の発生段階の若い未成熟植物を意味する。一年生、多年生、単子葉および双子葉植物はトランスジェニック植物を作製するための好ましい宿主生物である。遺伝子の発現はさらに全ての観賞植物、有用または観賞用植物樹木、花、切り花、低木または芝生において有利である。例として挙げられる植物は、限定されるものでないが、被子植物、コケ植物、例えば、苔類(ゼニゴケ)および蘚類(藻類);
シダ植物、例えば、シダ類、トクサ類およびヒカゲノカズラ類;裸子植物、例えば、針葉樹類、ソテツ類、イチョウ類およびグネツム類;藻類、例えば、緑藻綱、褐藻網、紅藻網、藍藻網、黄緑藻綱、珪藻綱(珪藻)、およびミドリムシ綱である。好ましいのは、食品または飼料に用いられる植物であり、例えば、マメ科、例えば、エンドウマメ、アルファルファおよびダイズ;イネ科、例えば、イネ、トウモロコシ、コムギ、オオムギ、モロコシ、ミレット、ライムギ、ライコムギ、またはオート麦;セリ科、とりわけ、Daucus属、特にとりわけcarota種(ニンジン)およびApium属、特にとりわけGraveolensdulce種(セロリ)その他の多くの種;ナス科、とりわけLycopersicon属、特にとりわけesculentum種(トマト)およびナス属、特にとりわけtuberosum種(ジャガイモ)およびmelongena種(ナス)、その他の多くの種(タバコなど);およびCapsicum属、特にとりわけannuum種(コショウ)その他の多くの種;マメ科、とりわけGlycine属、特にとりわけmax種(ダイズ)、アルファルファ、エンドウマメ、ムラサキウマゴヤシ、マメまたはピーナッツおよびその他の多くの種; および十字花科(アブラナ科)、とりわけBrassica属、特にとりわけnapus種(ナタネ)、campestris種(ビート)、oleracea cv Tastie(キャベツ)、oleracea cv SnowballY(カリフラワー)およびoleracea cv Emperor(ブロッコリー);およびArabidopsis属、特にとりわけthaliana種(シロイヌナズナ)およびその他の多くの種;Compositae(キク)科、とりわけLactuca属、特にとりわけsativa種(レタス)およびその他の多くの種;Asteraceae(キク)科、例えば、ヒマワリ、マリーゴールド、レタスまたはCalendula種(ワタゲハナグルマ)およびその他の多くの種;ならびにウリ科、例えば、メロン、カボチャ/カボチャまたはズッキーニ、および亜麻仁である。さらなる好ましい植物は、ワタ、サトウキビ、アサ、亜麻、チリー、および様々な樹木、ナッツおよびワイン種である。
【0094】
ポリペプチド: 用語「ポリペプチド」、「ペプチド」、「オリゴペプチド」、「ポリペプチド」、「遺伝子産物」、「発現産物」および「タンパク質」は本明細書では互換的に使用されていて、連続するアミノ酸残基のポリマーまたはオリゴマーを意味する。
【0095】
プレタンパク質:通常、葉緑体などの細胞小器官を標的化し、まだ、その輸送ペプチドを含んでいるタンパク質。
【0096】
一次転写物:本明細書で使用する用語「一次転写物」は遺伝子の未成熟RNA転写物を意味する。「一次転写物」は例えば、まだ、イントロンを含有し、および/またはなおpolyAテールまたはcap構造を含まず、および/またはトリミングまたはエディッティングなどの転写物としての正しい機能に必要な他の改変が行われてない。
【0097】
プロモーター:「プロモーター」または「プロモーター配列」は等価であり、本明細書で使用する場合、目的のヌクレオチド配列に連結されると目的のヌクレオチド配列のRNA中への転写を制御することができるDNA配列を意味する。かかるプロモーターは、例えば、次の公共データベースに見出すことができる:http://www.grassius.org/grasspromdb.html、http://mendel.cs.rhul.ac.uk/mendel.php?topic=plantprom、http://ppdb.gene.nagoya-u.ac.jp/cgi-bin/index.cgi。そこに掲げられたプロモーターは本発明の方法について呼び出すことができ、これらは参照により本明細書に組み込まれる。プロモーターは、目的のヌクレオチド配列(mRNA中へのその転写を制御する)の転写出発部位の5'(すなわち、上流)の近位に位置し、そして転写開始のためのRNAポリメラーゼおよび他の転写因子による特異的結合のための部位を提供する。前記プロモーターは、例えば、転写出発部位に対して、例えば少なくとも10kb、例えば5kbまたは2kb近位にある。これはまた、転写出発部位に対して、少なくとも1500bp近位、好ましくは少なくとも1000bp、より好ましくは少なくとも500bp、さらにより好ましくは少なくとも400bp、少なくとも300bp、少なくとも200bpまたは少なくとも100bpにあってもよい。さらなる好ましい実施形態において、プロモーターは転写出発部位に対して、少なくとも50bp近位、例えば、少なくとも25bpであってもよい。プロモーターはエキソンおよび/またはイントロン領域または5'非翻訳領域を含まない。プロモーターはそれぞれの植物に対して異種または相同性であってもよい。ポリヌクレオチド配列は、もし外来種に由来するか、またはもし同じ種由来であってその元来の形態から改変されていれば、生物または第2のポリヌクレオチド配列に対して異種である。例えば、異種コード配列と作動しうる形で連結されたプロモーターという場合、そのコード配列は、そのプロモーターが由来する種と異なる種由来のコード配列、または、もし同じ種由来であれば、そのプロモーターに自然では随伴しないコード配列(例えば、遺伝子操作で作られたコード配列または異なるエコタイプもしくは変異体からの対立遺伝子)を意味する。好適なプロモーターは、発現が起こるべき宿主細胞の遺伝子からまたは宿主細胞に対する病原体(例えば、植物または植物ウイルスのような植物病原体)から誘導することができる。植物特異的プロモーターは植物中の発現を調節するのに好適なプロモーターである。これは植物からであってもよいが、植物病原体からであってもよくまたは人手により設計された合成プロモーターであってもよい。もしプロモーターが誘導プロモーターであれば、誘導剤に応答して転写速度が増加する。また、プロモーターを組織特異的または組織に好ましい方式で調節して、特定の組織型、例えば、葉、根または分裂組織の関係するコード領域を転写するのだけにまたは優性に活性があるようにすることができる。プロモーターに適用する用語「組織特異的」は、色々な型の組織(例えば、根)において同じ目的のヌクレオチド配列が発現しないのと比較して、特定の型の組織(例えば、花弁)において目的のヌクレオチド配列の選択的発現を指令することができるプロモーターを意味する。プロモーターの組織特異性は、例えば、レポーター遺伝子をレポーター構築物を作製するプロモーター配列と作動しうる形で連結し、そのレポーター遺伝子が植物のゲノム中に導入して得られるトランスジェニック植物の全ての組織中に組み込まれるようにし、そしてトランスジェニック植物の色々な組織中のレポーター遺伝子の発現(例えば、レポーター遺伝子がコードするmRNA、タンパク質、またはタンパク質の活性)を検出することにより評価することができる。1以上の組織におけるレポーター遺伝子の発現のレベルが他の組織におけるレポーター遺伝子の発現のレベルより高いことを検出すれば、そのプロモーターが高い発現のレベルを検出した組織にとって特異的であることが示される。プロモーターに適用する用語「細胞型特異的」は、同じ組織内の色々な型の細胞において同じ目的のヌクレオチド配列が発現しないのと比較して、特定の型の細胞において目的のヌクレオチド配列の選択的発現を指令することができるプロモーターを意味する。プロモーターに適用する用語「細胞型特異的」はまた、1つの組織内のある領域のヌクレオチド配列の選択的発現を促進することができるプロモーターを意味する。プロモーターの細胞型特異性は、当技術分野で周知の方法、例えば、GUS活性染色、GFPタンパク質または免疫組織化学的染色を用いて評価することができる。プロモーターまたはプロモーターに由来する発現に適用する用語「構成的」は、植物組織および細胞の大部分において、植物または植物の部分の実質的に全生存期間を通して、刺激(例えば、熱ショック、化学品、光など)の非存在のもとで、プロモーターが作動しうる形で連結された核酸分子の転写を指令することができることを意味する。典型的には、構成的プロモーターは実質的にいずれの細胞およびいずれの組織においても、トランスジーンの発現を指令することができる。
【0098】
プロモーター特異性:プロモーターを言及する場合、用語「特異性」はそれぞれのプロモーターが与える発現のパターンを意味する。特異性は、そのプロモーターが、それぞれのプロモーターの制御下で核酸分子の発現を与える植物またはその部分の組織および/または発生状態を記載する。プロモーターの特異性はまた、そのもとでプロモーターが活性化または下方調節されうる環境条件、例えば、寒冷、乾燥、創傷または感染などの生物学的または環境ストレスによる誘導または抑圧を含みうる。
【0099】
精製された:本明細書で使用する用語「精製された」は、その自然環境から取り出され、単離されるかまたは分離された分子、核酸もしくはアミノ酸配列を意味する。「実質的に精製された」分子は、それが自然で随伴する他成分の少なくとも60%を含有しない、好ましくは少なくとも75%を含有しない、そしてより好ましくは少なくとも90%を含有しない。精製された核酸配列は単離された核酸配列であってもよい。
【0100】
組換え体:核酸分子に関する用語「組換え体」は組換えDNA技法により産生された核酸分子を意味する。好ましくは、「組換え核酸分子」は、天然の核酸分子とは少なくとも1つの核酸が異なる非天然の核酸分子である。「組換え核酸分子」はまた、好ましくは作動しうる形で連結された自然ではその順で存在しない核酸分子の配列を含む「組換え構築物」を含みうる。前記組換え核酸分子を産生する好ましい方法は、クローニング技法、定方向または非部位特異的な突然変異誘発、合成または組換え技法を含みうる。
【0101】
センス:用語「センス」は標的配列、例えば、タンパク質転写因子と結合する、および所与の遺伝子の発現に関わる配列と相補性または同一である配列を有する核酸分子を意味すると理解される。好ましい実施形態によれば、核酸分子は目的の遺伝子および前記目的の遺伝子の発現を可能にするエレメントを含む。
【0102】
有意な増加または減少:測定技法に固有の誤差限界より大きい、例えば酵素活性または遺伝子発現の増加または減少、好ましくは、対照酵素の活性または対照細胞の発現の約2倍以上の増加または減少、より好ましくは、約5倍以上の増加または減少、そして最も好ましくは、約10倍以上の増加または減少を意味する。
【0103】
小核酸分子:「小核酸分子」は、核酸またはその誘導体から成る分子、例えばRNAまたはDNAであると理解される。これらは2本鎖または1本鎖であってもよく、そして、約15〜約30bp、例えば15〜30bp、より好ましくは約19〜約26bp、例えば19〜26bp、さらにより好ましくは約20〜約25bp例えば20〜25bpである。とりわけ 好ましい 実施形態において、オリゴヌクレオチドは約21〜約24bp、例えば21〜24bpである。最も好ましくは実施形態において、小核酸分子は約21および約24bp、例えば21および24bpである。
【0104】
実質的に相補的な:その最も広い意味で、用語「実質的に相補的な」は、参照または標的ヌクレオチド配列と比較してヌクレオチド配列について本明細書で使用する場合、前記参照または標的ヌクレオチド配列の実質的に相補的なヌクレオチド配列と正確に相補的な配列の間のパーセント同一性は少なくとも60%、より望ましくは少なくとも70%、より望ましくは少なくとも80%または85%、好ましくは少なくとも90%、より好ましくは少なくとも93%、さらにより好ましくは少なくとも95%または96%、なおさらにより好ましくは少なくとも97%または98%、またさらにより好ましくは少なくとも99%または最も好ましくは100%(最後の場合、この文脈では用語「同一」と等しい)であるヌクレオチド配列を意味する。(以下で特に断らなければ)好ましくは、同一性を核酸配列の少なくとも19ヌクレオチド、好ましくは少なくとも50ヌクレオチド、より好ましくは全長にわたり前記参照配列に対して評価する。配列比較は、デフォールトGAP分析を用いて、GAPのSEQWEBアプリケーション(University of Wisconsin GCG)により、NeedlemanおよびWunsch(Needleman and Wunsch (1970) J Mol. Biol. 48: 443-453;先に定義した通り)のアルゴリズムに基づいて実施する。参照ヌクレオチド配列に「実質的に相補的な」ヌクレオチド配列は参照ヌクレオチド配列と低ストリンジェンシー条件、好ましくは中ストリンジェンシー条件、最も好ましくは高ストリンジェンシー条件(先に定義した通り)のもとでハイブリダイズする。
【0105】
トランスジーン:本明細書で使用する用語「トランスジーン」は、実験遺伝子操作により細胞のゲノム中に導入される任意の核酸配列を意味する。トランスジーンは「内因性DNA配列」または「異種DNA配列」(すなわち「外来のDNA」)であってもよい。用語「内因性DNA配列」は、天然の配列に関係して何らかの改変(例えば、点突然変異、選択マーカー遺伝子の存在など)のない限り、導入される細胞において自然で見出されるヌクレオチド配列を意味する。
【0106】
トランスジェニック:生物に言及する場合、用語「トランスジェニック」は、形質転換された、好ましくは目的のDNA配列と作動しうる形で連結された好適なプロモーターを含む組換えDNA分子によって、好ましくは安定して形質転換されたことを意味する。
【0107】
ベクター:用語「ベクター」は、連結されている他の核酸分子を輸送することができる核酸分子を意味する。ベクターの1つの型はゲノムに組み込まれたベクター、すなわち、宿主細胞の染色体DNA中に組み込むことができる「組み込みベクター」である。ベクターの他の型はエピソームベクター、すなわち、染色体外複製の能力がある核酸分子である。作動しうる形で連結された遺伝子の発現を指令する能力のあるベクターを本明細書では「発現ベクター」と呼ぶ。本明細書においては、文脈からそうでないことが明らかでない限り、「プラスミド」と「ベクター」を互換的に使用する。本明細書に記載のRNAを産生するように設計された発現ベクターは、in vivoまたはin vitroでRNAポリメラーゼ(ミトコンドリアのRNAポリメラーゼ、RNA pol I、RNA pol II、および RNA pol IIIを含む)により認識される配列を含有しうる。これらのベクターを用いて本発明に従い、細胞において所望のRNA分子を転写することができる。植物形質転換ベクターは植物形質転換のプロセスに好適なベクターと理解されたい。
【0108】
野生型:生物、ポリペプチド、または 核酸配列に関する用語「野生型」、「自然の」または「自然の起源」は、前記生物が天然であるかまたは、少なくとも、改変、突然変異誘発またはさもなくば人手による遺伝子操作が行われていない天然の生物において入手できるものを意味する。
【実施例】
【0109】
化学品および共通の方法
特に断らない限り、本発明の目的で行ったクローニング手順(制限消化、アガロースゲル電気泳動、核酸の精製、核酸のライゲーション、細菌の形質転換、選択および培養を含む)は記載の通り実施した(Sambrooket al., 1989)。組換えDNAの配列解析は、レーザー蛍光 DNAシーケンサー(Applied Biosystems, Foster City, CA, USA)により、Sangerの技法(Sangeret al., 1977)を用いて実施した。特に断らない限り、化学品および試薬はSigma Aldrich(Sigma Aldrich, St. Louis, USA)から、 Promega(Madison, WI, USA)、Duchefa(Haarlem, The Netherlands)またはInvitrogen(Carlsbad, CA, USA)から入手した。制限エンドヌクレアーゼはNew England Biolabs(Ipswich, MA, USA)またはRoche Diagnostics GmbH(Penzberg, Germany)から入手した。オリゴヌクレオチドはEurofins MWG Operon(Ebersberg, Germany)が合成した。
【0110】
実施例1
構成的発現をする遺伝子からの核酸発現増強性核酸(NEENA)の同定
1.1 シロイヌナズナ(A. thaliana)遺伝子からのNEENA分子の同定
公的に利用可能なゲノムDNA配列(例えば、http://www.ncbi.nlm.nih.gov/genoms/PLANTS/PlantList.html)および転写物発現データ(例えば、http://www.weigelworld.org/resources/microarray/AtGenExpress/)を用いて、シロイヌナズナ転写物由来の18個の潜在NEENA候補の1セットを、高度に発現する構成的遺伝子から選択し、詳細な分析を行った。さらに、パセリ由来の推定NEENA分子もこの分析に加えた。候補名は次の通りである。
【0111】
表1: 構成的NEENA候補(NEENAc)
【表1】
【0112】
1.2 NEENA候補の単離
ゲノムDNAを、シロイヌナズナ緑色組織からQiagen DNeasy植物ミニキット(Qiagen, Hilden, Germany)を用いて抽出した。配列番号1をもつ推定NEENA分子に対しては、ベクター構築物1bxPcUbi4-2GUS(WO 2003102198)のDNAを用いた。推定NEENA分子を含有するゲノムDNA断片を、通常のポリメラーゼ連鎖反応(PCR)により単離した。ポリメラーゼ連鎖反応は19セットのプライマーを含むものであった(表2)。プライマーは多数のNEENA候補をもつシロイヌナズナのゲノム 配列に基づいて設計した。ベクター構築物1bxPcUbi4-2GUS(WO 2003102198)のヌクレオチド配列を、配列番号1(表2)をもつNEENA候補を増幅するプライマー(配列番号56および57)の設計に用いた。ポリメラーゼ連鎖反応は、Phusion高信頼性DNAポリメラーゼ(Cat No F-540L, New England Biolabs, Ipswich, MA, USA)に概説されたプロトコルに従った。単離したDNAを、次のプライマーを用いるPCR増幅のテンプレートDNAとして使用した。
【0113】
表 2:プライマー配列
【表2】
【0114】
PCR中の増幅は次の組成物(50 microl)を用いて行った:
3.00 microl シロイヌナズナゲノムDNA(50 ng/microl ゲノムDNA、5 ng/microl ベクター構築物)
10.00 microl 5x Phusion HF バッファー
4.00 microl dNTP(2.5 mM)
2.50 microl for プライマー(10 microM)
2.50 microl rev プライマー(10 microM)
0.50 microl Phusion HF DNA ポリメラーゼ(2U/microl)
次のパラメーターによるPCRに、タッチダウン手法を使った:98.0℃にて30秒(1サイクル)、98.0℃にて30秒、56.0℃にて30秒、および72.0℃にて60秒(4サイクル)、4回の追加サイクル、それぞれ54.0℃、51.0℃および49.0℃のアニーリング温度、引き続いて20サイクル98.0℃にて30秒、46.0℃にて30秒および72.0℃にて60秒(4サイクル)ならびに72.0℃にて5分。増幅産物を2%(w/v)アガロースゲルに供給し、80Vで分離した。PCR産物をゲルから切り取り、Qiagenゲル抽出キット(Qiagen, Hilden, Germany)で精製した。KpnI(10U/microl)およびNcoI(10U/microl)またはEcoRV(10U/microl)制限エンドヌクレアーゼによるDNA制限消化後、消化産物を再びQiagenゲル抽出キット(Qiagen, Hilden, Germany)で精製した。
【0115】
1.3 ベクター構築
1.3.1潜在NEENA分子をもつベクター構築物の作製
マルチサイトGatewayシステム(Invitrogen, Carlsbad, CA, USA)を用いて、プロモーター::NEENA::レポーター遺伝子カセットを植物形質転換用の2成分系構築物中にアセンブルした。シロイヌナズナp-AtNit1(At3g44310、GenBankX86454;WO03008596、接頭辞p-でプロモーターを表示)プロモーターをレポーター遺伝子構築物に用い、分析すべき推定NEENA分子の発現増強効果を定量的に測定するレポータータンパク質としてホタルルシフェラーゼ(Promega、Madison、WI、USA)を使った。
【0116】
製造業者マニュアル(Invitrogen, Carlsbad, CA, USA)に従って、p-AtNit1プロモーターを保持するpENTR/Aベクターを、pDONR/Aベクターとp-AtNit1増幅産物の間の部位特異的組換え(BP-反応)を介し、プライマーp-AtNit1-forおよびp-AtNit1-rev(表3)を用いて、部位特異的組換え部位を両端にもつゲノムDNA(上記参照)上にクローニングした。ポジティブpENTR/Aクローンを配列分析してp-AtNit1 プロモーターの正しさを保証した。
【0117】
表3:プライマー配列(p-AtNit1)
【表3】
【0118】
ホタルルシフェラーゼをコードする配列(Promega, Madison, WI, USA)と続いてt-nosノパリン合成酵素転写ターミネーター(Genbank V00087)を含有するENTR/Bベクターを作製した。NEENA候補PCR断片(上記参照)を別々に、ホタルルシフェラーゼをコードする配列の上流に、KpnIおよびNcoIまたはEcoRV制限酵素を用いてクローニングした。得られるpENTR/Bベクターを、接頭辞p-を有するプロモーター、接頭辞c-を有するコード配列、および接頭辞t-を有するターミネーターと共に表4に総括した。
【0119】
表4:全てのpENTR/Bベクター プラスおよびマイナスNEENA候補
【表4】
【0120】
pENTR/Cベクターを、KpnIおよびHindIII制限部位を介するマルチクローニングサイト(配列番号60)の導入により構築した。製造業者(Invitrogen, Carlsbad, CA, USA)のマルチサイトGatewayマニュアルに従い、部位特異的組換え(LR-反応)を実施することにより、作製したpENTR/A、pENTR/B および pENTR/CをpSUN デスティネーションベクター(pSUN誘導体)と組み合わせた。この反応によって、p-AtNit1プロモーター、ホタルルシフェラーゼをコードするc-LUCおよびt-nosターミネーター、ならびに、ホタルルシフェラーゼコード配列の直ぐ上流に配列番号1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18および19をコードする配列を保持する19個のベクターをもつ1つの二成分ベクターを得た(表5)。これと配列番号1との組み合わせを例示する(配列番号61)。全てのベクター(表5)は、ヌクレオチド配列配列番号2〜19が変わることを除くと、同一である。得られる植物形質転換ベクターを表5に総括した。
【0121】
表5:シロイヌナズナ形質転換用の植物発現ベクター
【表5】
【0122】
得られるベクターを引き続いて使用し、シロイヌナズナの葉のプロトプラストを一過的に形質転換した。
【0123】
1.3.2 ウミシイタケルシフェラーゼ対照構築物
ウミシイタケルシフェラーゼcDNAを、DNAテンプレートとしてPromega(Madison, WI, USA)由来のプラスミドpRL-nullの10ng、ならびにプライマーR-LUC_forおよびR-LUC_rev(表 6)を用いて、先に記載のPCRパラメーターにより増幅した。
【0124】
表6:プライマー配列(c-RLUC)
【表6】
【0125】
KpnI(10U/microl)およびSacI(10U/microl)制限エンドヌクレアーゼによるDNA制限消化の後、消化した産物を再びQiagenゲル抽出キット(Qiagen, Hilden, Germany)を用いて精製した。
【0126】
その断片を、ノパリン合成酵素構成的プロモーターp-nos(GenbankV00087)、引き続いてt-nos nopalin 合成酵素転写ターミネーター(Genbank V00087)を含有するENTR/Bベクター中に、KpnIおよびSacI制限部位を介してクローニングし、pENTR/Bクローンを得て、それを配列分析してウミシイタケルシフェラーゼを含有する発現カセットの正しさを保証した。
【0127】
実施例2
シロイヌナズナの一過的に形質転換された葉プロトプラストにおける、遺伝子発現を増強するNEENA候補分子のスクリーニング
本実施例は選択されたNEENA分子だけが遺伝子発現を増強する能力があることを説明する。
【0128】
2.1シロイヌナズナ葉プロトプラストの単離と一過的形質転換
シロイヌナズナ葉プロトプラストの単離と一過的形質転換を、確立されたプロトコル(Damm and Willmitzer, 1988; Damm et al., 1989)に従って修正した。4週齢のシロイヌナズナ植物の葉を、剃刀の刃を用いて小片にカットし、1.5%セルラーゼR10(Duchefa, Haarlem, The Netherlands)、0.3%マセロザイムR10(Duchefa, Haarlem, The Netherlands)、400mMマンニトール、20mM KCl、20mM MES、10mM CaCl2、pH5.7の溶液中に移し、一晩、室温にてインキュベートした。一過的シロイヌナズナ葉プロトプラスト形質転換は可変性であるので、ウミシイタケルシフェラーゼ(Dual-Luciferase(登録商標) Reporter Assay System, Promega, Madison, WI, USA)を用いて、上記構築物のホタルルシフェラーゼ発現能力を規準化した。NEENAを含まない(LJK132)および各NEENAを含有するベクター構築物(LJK66-LJK114)の一過的形質転換を6 microgのプラスミドDNAを用いて3重で実施し、これを25 microgのウミシイタケルシフェラーゼを含有する構築物と混合した後に、PEG(ポリエチレングリコール)と1x104プロトプラストを用いて形質転換した。
【0129】
2.2二重ルシフェラーゼレポーター遺伝子アッセイ
形質転換したシロイヌナズナプロトプラストを遠心分離により100gにて採集し、上清を除去した後、液体窒素にて凍結した。形質転換した細胞中のホタルおよびウミシイタケルシフェラーゼ活性を検出するアッセイを、製造業者(Promega、Madison、WI、USA)の二重ルシフェラーゼレポーターアッセイマニュアルに従って実施した。発光測定はMicroLumat Plus LB96V(Berthold Technologies、Bad Wildbad、Germany)で実施し、ルシフェラーゼ基質を加えた後に記録した。両方のルシフェラーゼ記録の計器読みはホタルルシフェラーゼとウミシイタケルシフェラーゼの間の比を作製することにより規準化した。3実験から得たデータを、各構築物について平均し、これらの平均発現値に基づいて、倍変化値を計算して、それぞれの推定NEENAを欠くレポーター遺伝子構築物を超える、推定NEENAの存在の影響を評価した。p-AtNit1プロモーターだけでNEENAを含まないレポーター遺伝子構築物と比較して、19個の試験したNEENA候補を含む構築物は、ホタルルシフェラーゼ活性でネガティブならびに0.1-倍〜18.1-倍の範囲のポジティブな効果を示した(図1)。全体で、 配列番号1、NO2、NO3、NO4、NO5、NO6、NO7、NO8 および NO9をもつ配列を含む 9個の推定 NEENA 分子は、NEENAを含まないプロモーターだけのレポーター遺伝子の構築物と比較して、ルシフェラーゼレポーター遺伝子活性に基づいて遺伝子発現の2倍以上の増加を与え(図1)、従って機能的NEENA分子であった。いくつかの試験したNEENA候補分子は遺伝子発現の増強に対して限界的なまたはむしろネガティブな効果を有するので、全ての推定NEENA分子が共通の刺激的効果を媒介するわけではなく、むしろ、選択されたNEENA配列が遺伝子発現の有意な増強を与える(配列番号1〜9)のである。
【0130】
実施例3
アブラナ植物において遺伝子発現を増強するNEENA分子の試験
本実施例はNEENA分子を用いて、NEENAを含まないプロモーターだけの手法と比較して、種全体にわたり、試験した全ての組織において遺伝子発現を増強できることを説明する。
【0131】
プレスクリーニングで遺伝子発現の最強の増強を媒介するNEENA分子(実施例2の配列番号1、2、3、4および5を参照)を選択して、トランスジェニックアブラナ植物における遺伝子発現に与える増強効果を測定した。
【0132】
3.1セイヨウアブラナ(Brassica napus)植物を形質転換するためのベクター構築
アブラナ植物を形質転換するために、遺伝子発現制御分子(配列番号1〜5)有りまたは無しのレポーター遺伝子カセットを、pENTR/Cベクター内にトランスジェニック植物系統を検出するための選択マーカーを運ぶ遺伝子発現カセットと組み合わせた。先に記載(上記1.4を参照)の部位特異的組換え(IR-反応)を実施することにより、選択マーカーカセットを運ぶpENTR/A、pENTR/BおよびpENTR/Cを、pSUNデスティネーションベクターと、製造業者(Invitrogen, Carlsbad, CA, USA)のマルチサイトGatewayマニュアルに従い、組み合わせた。反応は、p-AtNit1プロモーター、ホタルルシフェラーゼをコードする配列c-LUC、t-nosターミネーターおよび選択マーカーカセット、ならびに、ホタルルシフェラーゼをコードする配列の直ぐ上流に配列番号1、2、3、4、および5を保持する5つのベクターをもつ1つの二成分ベクターをもたらした(表7)。これに対する配列番号1との組み合わせを例示として与える(配列番号64)。配列番号2〜5が変わることを除くと、このヌクレオチド配列は全てのベクターで同一である。得られる植物形質転換ベクターを表7に総括する。
【0133】
表7:セイヨウアブラナ形質転換用の植物発現ベクター
【表7】
【0134】
3.2 トランスジェニックアブラナ植物の作製(Moloney et al., 1992, Plant Cell Reports, 8: 238-242による修正プロトコル)
トランスジェニックアブラナ植物を作製するための調製において、二成分ベクターをAgrobacterium tumefaciens C58C1:pGV2260中に形質転換した(Deblaere et al., 1985, Nucl. Acids. Res. 13: 4777-4788)。それぞれの二成分構築物を保持するアグロバクテリアの一晩培養の1:50希釈物を、3%ショ糖(3MS-培地)を補充したMurashige-Skoog培地(Murashige および Skoog、1962、Physiol. Plant 15、473)中で増殖した。アブラナ植物を形質転換するために、無菌植物の葉柄または低子葉を、1:50アグロバクテリウム溶液と5〜10分間インキュベートし、引き続いて3日間、暗所で25℃にて0.8%バクト-寒天を補充した3MS培地で共インキュベートした。3日後、その外植体を、 500 mg/l Claforan(セフォタキシム-ナトリウム)、100 nMイマゼタピル、20 microMベンジルアミノプリン(BAP)および1.6g/lグルコースを含有するMS培地に移し、16時間明/8時間暗を繰り返す光環境に1週間置いた。成長するシュートを、2 % ショ糖、250 mg/lクラフォランおよび0.8%バクト-寒天を含有するMS-培地に移した。3週後、成長ホルモン2-インドール酪酸を培地に加えて根形成を促進した。シュートを土壌に移し、根発生後、2週間成長室で成長させ、そして成熟まで温室条件で成長させた。
【0135】
3.3 植物分析
組織サンプルを、葉、花および長角果から作製したトランスジェニック植物から採集し、-80℃にて冷凍庫で保存し、ルシフェラーゼレポーター遺伝子アッセイ(Ow et al., 1986後の修正プロトコル)にかけた。粉砕後、凍結組織サンプルを800microlのバッファーI(0.1M リン酸バッファー pH7.8、1mM DTT(Sigma Aldrich, St. Louis, MO, USA)、0.05% Tween 20(Sigma Aldrich, St. Louis, MO, USA))に再懸濁し、続いて10000gにて10分間、遠心分離し、75microlの水性の上清を96ウエルプレートに移した。25microlのバッファーII(80 mMグリシン-グリシル(Carl Roth, Karlsruhe, Germany)、40mM MgSO4(Duchefa, Haarlem, The Netherlands)、60mM ATP(Sigma Aldrich, St. Louis, MO, USA)、pH 7.8)および0.5 mMの最終濃度までのD-ルシフェリン(cat No: L-8220, BioSynth, Staad, Switzerland)を加えた後に、発光を1分間当たり相対発光量の単位、すなわちRLU(RLU/min)で与えるMicroLumat Plus LB96V(Berthold Technologies, Bad Wildbad, Germany)で記録した。
【0136】
サンプル間のルシフェラーゼ活性を規準化する目的で、水性上清中のタンパク質濃度を、ルシフェラーゼ活性と平行して測定した(Bradford, 1976, Anal. Biochem. 72, 248から適用した)。5 microlのバッファーI中の水性細胞抽出物を250 microlのBradford 試薬(Sigma Aldrich, St. Louis, MO, USA)と混合し、10分間室温にてインキュベートした。吸収をプレートリーダー(Thermo Electron Corporation, Multiskan Ascent 354)で595nmにて測定した。サンプル中の全タンパク質量を先に作製した標準濃度曲線を用いて計算した。RLU/minとmgタンパク質/mlサンプルの比から得た値を、同一の構築物を保持するトランスジェニック植物について平均化し、倍変化値を計算してNEENA分子存在のNEENAを含まないレポーター遺伝子構築物を超える影響を評価した。
【0137】
3.4 NEENA配列はアブラナ植物における遺伝子発現の強い増強を媒介する
アブラナ 植物における選択したNEENA分子(配列番号1、2、3、4および5)の遺伝子発現を増強する潜在能力を評価するために、同一の発生段階を有しかつ同一の成長条件のもとで育てた植物の葉、花、および植物の種子を保持する長角果を採集した。サンプルは、プロモーターだけのレポーター遺伝子構築物または NEENA(配列番号1、2、3、4および5)を含有するルシフェラーゼレポーター遺伝子構築物を保持する個々のトランスジェニックアブラナ植物から採取した。10個の種子を各トランスジェニック事象から採集し、処理し、そして先に記載した(実施例3.3)ルシフェラーゼ活性について分析した。
【0138】
構成的なp-AtNit1プロモーターだけでNEENAを含まないレポーター遺伝子構築物と比較して、5つの試験したNEENA分子は全て、葉組織の遺伝子発現に強い増強を媒介した(図2、a)。NEENAs(配列番号1、2、3、4および5)により媒介される遺伝子発現の比較しうる増強は、アブラナの花および種子を含む長角果において検出された(図2b、およびc)。
【0139】
実施例4
ダイズ植物における遺伝子発現の構成的増強の分析
本実施例はNEENA分子を用いると、NEENAを含まないプロモーターだけの手法と比較して、広範囲の植物種においてかつ色々な植物ファミリーの種境界を越えて、全組織の遺伝子発現を増強できることを説明する。
【0140】
プレスクリーニングにおいて遺伝子発現の最強の増強を媒介するNEENA配列分子(実施例2を参照、配列番号1、2、3、4 および5)を選択し、トランスジェニックダイズ植物における遺伝子発現レベルの増強を測定した。植物発現ベクターLJK138、LJK139、LJK141、LJK142、LJK143およびLJK144(実施例3.1を参照)を用いて安定なダイズ形質転換を行った。
【0141】
4.1トランスジェニックダイズ植物の作製(WO2005/121345;Olhoftet al., 2007による修正プロトコル)
ダイズ種子発芽、繁殖、土壌細菌A. rhizogenesおよび腋分裂組織外植体調製、および接種を、先に記載(WO2005/121345; Olhoft et al., 2007)の通り実施した(但し、構築物LJK138、LJK139、LJK141、LJK142、LJK143およびLJK144(実施例3.1を参照)は、選択のためにイミダゾリン除草剤に対する耐性を媒介するパセリのユビキチンプロモーターPcUbi4-2により駆動される変異AHAS遺伝子をそれぞれ含有することを除く)。
【0142】
4.2 NEENA 配列はダイズ植物における遺伝子発現の強い増強を媒介する
組織サンプルを葉、花および種子から作製したトランスジェニック植物から採集した。組織サンプルを処理し、上記の通り分析した(実施例3.3を参照)。
【0143】
構成的な p-AtNit1 プロモーターだけでNEENAを含まない レポーター遺伝子構築物と比較して、5個の試験したNEENA分子は全て葉における強力な増強を媒介した(図3a)。NEENA(配列番号1〜5)により媒介される遺伝子発現の比較しうる増強がダイズ花および長角果において検出された(図3、bおよびc)。
【0144】
実施例5
単子葉植物におけるNEENA活性の分析
本実施例は単子葉植物における配列番号 1、2、3、4 および5をもつNEENA配列の分析を記載する。
【0145】
5.1 ベクター 構築
単子葉植物において配列番号1、2、3、4および5をもつNEENA配列を分析するために、トウモロコシ由来のNEENAを含まない構成的な単子葉プロモーターp-Ubiで構成される発現カセットを保持するpUCに基づく発現ベクターを、β-グルクロニダーゼ(GUS)遺伝子のコード配列と続くノパリン合成酵素(NOS)転写のターミネーターと組み合わせる。ゲノムDNAはシロイヌナズナ緑色組織からQiagen DNeasy Plant Miniキット(Qiagen, Hilden, Germany)を用いて抽出する。NEENA分子を含有するゲノムDNA断片を通常のポリメラーゼ連鎖反応(PCR)により単離する。プライマーは複数のNEENA候補をもつシロイヌナズナゲノム配列に基づいて設計する。反応物は5セットのプライマー(表8)を含み、Phusion High Fidelity DNAポリメラーゼ(cat No F-540L, New England Biolabs, Ipswich, MA, USA)に概説されたプロトコルに従い、次のプライマーを用いる。
【0146】
表8:プライマー配列
【表8】
【0147】
PCR中の増幅および増幅産物の精製は先に詳説した通り行う(実施例1.2)。AscI(10 U/microl)制限エンドヌクレアーゼによるDNA制限消化の後、消化産物をQiagenゲル抽出キット(Qiagen, Hilden, Germany)で精製した。
【0148】
NEENA PCR断片(前記参照)を、AscI制限部位を用いてβ-グルクロニダーゼのコード配列の上流に分離してクローニングした。この反応は、p-Ubiプロモーター、β-グルクロニダーゼをコードする配列c-GUSおよびt-nosターミネーター、ならびに、β-グルクロニダーゼをコードする配列の直ぐ上流に配列番号1、NO2、NO3、NO4および NO5を保持する5つのベクターをもつ1つの二成分ベクターを生じた(表9)。これに対する配列番号1との組み合わせを例示として与える(配列番号75)。配列番号2〜5が変わるのを除くと、全てのベクターのヌクレオチド配列は同一である(表9)。得られるベクターは表9に総括したが、ここでプロモーター分子は接頭辞p-、コード配列は接頭辞c-、およびターミネーター分子は接頭辞t-を有する。
【0149】
表9:植物発現ベクター
【表9】
【0150】
得られるベクターを用いて、以下に概説した実験(実施例5.2)でNEENA分子を分析した。
【0151】
5.2 単子葉植物組織において遺伝子発現を増強するNEENA分子の分析
これらの実験は、単子葉植物組織または培養細胞のボンバードメント(実施例6.2.1)により、DNAの植物プロトプラストへのPEG媒介(または類似の技法)導入(実施例6.2.2)により、またはアグロバクテリウム媒介形質転換(実施例6.3.3)により実施した。これらの実験の標的組織は、植物組織(例えば、葉組織)、培養植物細胞(例えば、トウモロコシ、Black Mexican Sweetcorn(BMS)、またはアグロバクテリウムプロトコル用の植物胚芽であってもよい。
【0152】
5.2.1 微粒子ボンバードメントを用いる一過性アッセイ
プラスミド構築物をQiagenプラスミド キット(cat# 12143)を用いて単離する。DNAを、Sanford ら(1993)(Optimizing the biolistic プロセス for different 生物学的 applications. 方法 in Enzymology、217: 483-509)に記載のプロトコルに従って0.6microM 金粒子(Bio-Rad cat# 165 -2262)上に沈降させ、 PDS-1000/He系デバイス(Bio-Rad)を用いて標的組織(例えば、2週齢のトウモロコシ 葉、BMS培養細胞など)上に加速させる。全てのDNA沈降およびボンバードメントステップを無菌条件下で室温にて実施する。Black Mexican Sweetトウモロコシ(BMS)懸濁液培養細胞をBMS細胞培養液体培地[Murashige および Skoog(MS) 塩(4.3 g/L)、3%(w/v)スクロース、myo-イノシトール(100 mg/L)、3 mg/L 2,4-ジクロロフェノキシ酢酸(2,4-D)、カゼイン 加水分解酵素(1g/L)、チアミン(10mg/L)およびL-プロリン(1.15g/L)、pH5.8]中で増殖する。毎週10mLの定常細胞の培養を40mLの新しい培地に移し。250mLフラスコ中で110rpmで27℃にて運転する回転シェーカーで培養する。
【0153】
シリコン処理したエッペンドルフチューブ中の60mgの金粒子を、100%エタノールで再懸濁し、次いで30秒間遠心分離する。ペレットを100%エタノール中で1回、無菌水中で2回漱ぎ洗いし、それぞれの洗浄後に遠心分離する。ペレットを最終的には1mL無菌50%グリセロールに再懸濁する。金懸濁液を次いで50microlアリコートに分割し、4℃にて保存する。次の試薬を1つのアリコートに加える: 5 microLの1 microg/microL全 DNA、50 microLの2.5 M CaCl2、20 microLの0.1 M スペルミジン、フリーベース。DNA溶液を1分間ボルテックス攪拌し、-80℃にて3分間静置し、次いで10秒間遠心分離する。上清を取り除く。ペレットを軽く叩いて注意深く1 mL 100% エタノールに再懸濁し次いで10秒間遠心分離する。上清を除去し、ペレットを注意深く50microLの100%エタノールに再懸濁し、使用するまで(ボンバードメント前の30分〜4時間)-80℃に置く。もし金凝集物が溶液中で可視であれば、使用直前にチューブを1秒間水浴のソニケーターにおいて超音波処理する。
【0154】
ボンバードメントについては、2週齢のトウモロコシ葉をカットしておよび1cm長さの小片とし、向軸側を上にして浸透圧誘導培地M-N6-702[N6塩(3.96g/L)、3%(w/v)スクロース、1.5mg/Lの2,4-ジクロロフェノキシ酢酸(2,4-D)、カゼイン加水分解産物(100mg/L)、およびL-プロリン(2.9g/L)、MSビタミンストック溶液(1mL/L)、0.2Mマンニトール、0.2Mソルビトール、pH 5.8]上に置く。
【0155】
小片を1〜2時間インキュベートする。
【0156】
BMS培養細胞の場合、1週齢の懸濁液細胞を1000gにてBeckman/Coulter Avanti J25遠心分離機でペレット化し、上清は捨てる。細胞を、円形の無灰No 42 Whatmanフィルター上にスパチュラを用いて1/16インチ厚さの層として置く。植物材料を保持するろ紙を浸透圧誘導培地上に27℃にて暗所で1〜2時間置いた後にボンバードメントを行う。ボンバードメント直前にフィルターを培地から取り除いて無菌ろ紙のスタック上に置いてカルス表面を部分的に乾燥させる。
【0157】
各プレートを6microLの金-DNA溶液で2回、葉材料向けて1,800psiにて、およびBMS培養細胞向けて1,100psiにてショットする。植物材料の位置を保つために、無菌のワイヤーメッシュスクリーンをサンプルの頂上に敷く。ボンバードメント後に、サンプルを保持するフィルターを、マンにトールおよびソルビトールを欠くM-N6-702 培地上に移し、2日間、暗所で27℃にてインキュベートし、その後に一過性アッセイを行う。
【0158】
他の単子葉植物の微粒子ボンバードメントを介する一過性形質転換は、例えば、Wang et al., 1988 (Transient expression of foreign genes in rice, wheat and soybean cells following particle bombardment. Plant Molecular Biology, 11 (4), 433-439), Christou, 1997 (Rice transformation: bombardment. Plant Mol Biol. 35 (1-2))に記載の技法を用いて行う。
【0159】
上記(実施例5.1)の構築物で発現された遺伝子の発現レベルを、当技術分野のプロトコルを用いて、GUS 染色、発光/蛍光の定量化、RT-PCR および タンパク質存在量(特異的抗体の検出)により測定する。GUS染色は、植物材料をGUS溶液[100 mM NaHPO4、10 mM EDTA、0.05% Triton X100、0.025% X-Gluc 溶液(DMSOに溶解した5-ブロモ-4-クロロ -3-インドリル-β-D-グルクロン酸)、10%メタノール、pH 7.0]中で37℃にて16〜24時間インキュベートすることにより行う。植物組織を2回15分間真空浸潤して一様な染色を助ける。ルシフェラーゼ活性の分析は先に記載の通り実施する(実施例2および3.3)。
【0160】
これらのアッセイにおいてNEENA分子は全て、構成的なp-UbiプロモーターだけでNEENAを含まないレポーター遺伝子構築物と比較して、遺伝子発現の強い増強を媒介する。
【0161】
5.2.2 プロトプラストを用いる一過的アッセイ
プロトプラストの単離を、Sheen(1990)(Metabolic Repression of Transcription in Higher Plants. The Plant Cell 2 (10)、1027-1038)が開発した次のプロトコルにより実施する。トウモロコシ実生を暗所で25℃にて10日間置き、そして20時間照明後にプロトプラストを調製した。葉の中央部を0.5mmストリップ(長さで約6cm)にカットし、1%(w/v)セルロースRS、0.1%(w/v)マセロザイムR10(両方ともYakult Honsha、Nishinomiya、Japanから入手)、0.6Mマンニトール、10mM Mes(pH 5.7)、1mM CaCl2、1mM MgCl2、10mM β-メルカプトエタノール、および0.1% BSA(w/v)を含有する酵素溶液中で、3時間、23℃にてインキュベートし、次いで80rpmにて10分間静かに振とうしてプロトプラストを遊離させた。
【0162】
プロトプラストを100 x gにて2分間遠心分離により採集し、1回冷0.6Mマンニトール溶液で洗浄し、遠心分離し、そして冷0.6Mマンニトール(2 x 106/mL)に再懸濁した。
【0163】
合計100microLの無菌水中の合計50microgのプラスミドDNAを、0.5mLのトウモロコシプロトプラスト(1 x 106 細胞s/mL)の懸濁液中に加え、静かに混合する。0.5mLのPEG溶液(40% PEG 4,000、100 mM CaNO3、0.5 マンニトール)を加え、静かに振とうしながら70℃にて予熱し、次いで4.5 mL MM溶液(0.6 M マンニトール、15 mM MgCl2、および0.1% MES)を加える。この混合物を15分間、室温にてインキュベートする。プロトプラストを2回、600rpmにて5分間ペレット化することにより洗浄し、 1.0 mLのMMB溶液[0.6Mマンニトール、4mM Mes(pH 5.7)、および ブロムモザイクウイルス(BMV)塩(オプション)]に再懸濁し、そして暗所で25℃にて48時間インキュベートする。最後の洗浄ステップ後に、プロトプラストを3 mLのMMB培地に採集し、そして暗所で25℃にて48時間インキュベートする。
【0164】
他の 単子葉植物のプロトプラストの一過性形質転換を、例えば、Hodgeset al., 1991(Transformation and regeneration of rice protoplasts. Biotechnology in agriculture No. 6, Rice Biotechnology. International Rice Research Institute, ISBN: 0-85198-712-5)またはLeeet al., 1990(Transient gene expression in wheat (Triticum aestivum) protoplasts. Biotechnology in agriculture and forestry 13 - Wheat. Springer Verlag, ISBN-10: 3540518096)が記載した技法を用いて行う。
【0165】
先に記載の構築物(実施例5.1)中の発現された遺伝子の発現レベルをGUS染色、発光/蛍光の定量、RT-PCRまたはタンパク質存在量(特異的抗体による検出)により、当技術分野のプロトコルを用いて測定する。GUS染色は、植物材料をGUS溶液[100 mM NaHPO4、10 mM EDTA、0.05% Triton X100、0.025% X-Gluc 溶液(DMSOに溶解した5-ブロモ-4-クロロ -3-インドリル-β-D-グルクロン酸)、10%メタノール、pH 7.0]中で37℃にて16〜24時間インキュベートすることにより行う。ルシフェラーゼ活性の分析は先に記載の通り実施する(実施例2および3.3)。
【0166】
構成的なp-UbiプロモーターだけでNEENAを含まないレポーター遺伝子構築物と比較して、NEENA分子はこれらのアッセイにおける遺伝子発現の強い増強を媒介する。
【0167】
5.2.3 単子葉作物植物の形質転換および 再生
標準 形質転換 および 再生 技法を用いるアグロバクテリウム媒介性植物形質転換はまた、作物植物を形質転換する目的で行うことができる(Gelvin and Schilperoort, 1995, Plant Molecular Biology Manual, 2nd Edition, Dordrecht: Kluwer Academic Publ. ISBN 0-7923-2731-4; Glick and Thompson (1993) Methods in Plant Molecular Biology and Biotechnology, Boca Raton: CRC Press, ISBN 0-8493-5164-2)。トウモロコシまたは他の単子葉植物の形質転換は、例えば、米国特許第5,591,616号に記載の技法を用いて行うことができる。粒子ボンバードメント、ポリエチレングリコール媒介性DNA取込みまたはシリコンカーボネート繊維を介する技法を用いる植物の形質転換は、例えば、Freeling & Walbot (1993) ”The maize handbook” ISBN 3-540-97826-7に記載されている。
【0168】
先に記載の構築物(実施例5.1)中の発現された遺伝子の発現レベルをGUS染色、発光/蛍光の定量、RT-PCRまたはタンパク質存在量(特異的抗体による検出)により、当技術分野のプロトコルを用いて測定する。GUS染色は、植物材料をGUS溶液[100 mM NaHPO4、10 mM EDTA、0.05% Triton X100、0.025% X-Gluc 溶液(DMSOに溶解した5-ブロモ-4-クロロ -3-インドリル-β-D-グルクロン酸)、10%メタノール、pH 7.0]中で37℃にて16〜24時間インキュベートすることにより行う。植物組織を2回15分間真空浸潤して一様な染色を助ける。ルシフェラーゼ活性の分析は先に記載の通り実施する(実施例2および3.3)。
【0169】
構成的なp-UbiプロモーターだけでNEENAを含まないレポーター遺伝子構築物と比較して、NEENA分子は植物における遺伝子発現の強い増強を媒介する。
【0170】
実施例6
トウモロコシ植物におけるNEENA活性の定量分析
本実施例はトウモロコシ植物における配列番号1および2をもつNEENA配列の分析を記載する。
【0171】
6.1 ベクター 構築
単子葉植物において配列番号1および2をもつNEENA配列を定量的に分析するために、トウモロコシ由来のNEENAを含まない構成的な単子葉プロモーターp-Ubiで構成される発現カセットを保持するpUCに基づく発現ベクターを、ホタルルシフェラーゼ(LUC)遺伝子(Promega, Madison, WI, USA)と続くノパリン合成酵素(NOS)転写のターミネーターと組み合わせた。ゲノムDNAはシロイヌナズナ緑色組織からQiagen DNeasy Plant Miniキット(Qiagen, Hilden, Germany)を用いて抽出した。NEENA分子を含有するゲノムDNA断片を通常のポリメラーゼ連鎖反応(PCR)により単離した。プライマーは複数のNEENA候補をもつシロイヌナズナゲノム配列に基づいて設計した。反応物は2セットのプライマー(表10)を含み、Phusion High Fidelity DNAポリメラーゼ(cat No F-540L, New England Biolabs, Ipswich, MA, USA)に概説されたプロトコルに従い、次のプライマーを用いた。
【0172】
表10:プライマー配列
【表10】
【0173】
PCR中の増幅および増幅産物の精製は先に詳説した通り行った(実施例1.2)。MluI(10 U/microl) および AscI(10 U/microl)制限エンドヌクレアーゼによるDNA制限消化の後、消化産物をQiagenゲル抽出キット(Qiagen, Hilden, Germany)で精製した。
【0174】
NEENA PCR断片(前記参照)を、AscI制限部位を用いてホタル ルシフェラーゼのコード配列の上流に分離してクローニングした。この反応は、p-Ubiプロモーター、ホタル ルシフェラーゼをコードする配列c-LUCおよびt-nosターミネーター、ならびに、ホタル ルシフェラーゼをコードする配列の直ぐ上流に配列番号1および2を保持する2つのベクターをもつ1つの二成分ベクターを生じた(表11);これに対する配列番号1との組み合わせを例示として与える(配列番号80)。配列番号2が変わるのを除くと、ヌクレオチド配列は同一である(表11)。得られるベクターは表11に総括したが、ここでプロモーター分子は接頭辞p-、コード配列は接頭辞c-、およびターミネーター分子は接頭辞t-を有する。
【0175】
表11:植物発現ベクター
【表11】
【0176】
得られるベクターを用いて、以下に概説した実験(実施例6.2)でNEENA分子を分析した。
【0177】
6.2 トランスジェニックトウモロコシ植物の作製
トウモロコシ発芽、繁殖、土壌細菌A. tumefaciens調製および接種を、先に記載(WO2006136596、US20090249514)の通り実施したが、ただし構築物LJK309、LJK326およびLJK327(実施例6.1を参照)は、選択のためにイミダゾリン除草剤に対する耐性を媒介するトウモロコシのユビキチンプロモーターp-Ubiにより駆動される変異AHAS遺伝子をそれぞれ含有することを除く。
【0178】
6.3 NEENA配列はトウモロコシ植物における遺伝子発現の強い増強を媒介する
組織サンプルを、葉および穀粒から作製したトランスジェニック植物から採集した。組織サンプルを処理し、上記の通り分析した(実施例3.3を参照)。
【0179】
構成的な p-UbiプロモーターだけでNEENAを含まない レポーター遺伝子 構築物と比較して、2個の試験したNEENA分子は葉における強力な増強を媒介した(図4a)。NEENA(配列番号1〜2)により媒介される遺伝子発現の比較しうる増強がトウモロコシ穀粒において検出された(図4b)。
【0180】
実施例7
イネ植物におけるNEENA活性の定量分析
本実施例はイネ 植物における配列番号1をもつNEENA配列の分析を記載する。
【0181】
7.1 ベクター 構築
イネ 植物において配列番号1をもつNEENA配列を定量的に分析するために、pENTR/B ベクターLJK1およびLJK4(実施例1.3を参照)を、組換え部位の上流に構成的な PRO0239を保持するデスティネーション ベクターと、製造業者(Invitrogen、Carlsbad、CA、USA) Gatewayマニュアルに従って組み合わせた。反応物はPRO0239プロモーター、ホタルルシフェラーゼをコードする配列c-LUCおよびt-nosターミネーター、ならびに、ホタルルシフェラーゼをコードする配列の直ぐ上流に配列番号1を保持する1つのベクターをもつ1つの二成分ベクター(表12)をもたらした。
【0182】
得られるベクターを表12に総括したが、ここでプロモーター分子は接頭辞p-、コード配列は接頭辞c-、およびターミネーター分子は接頭辞t-を有する。
【0183】
表12:植物発現ベクター
【表12】
【0184】
得られるベクターを用いて、以下に概説した実験(実施例7.2)でNEENA分子を分析した。
【0185】
7.2トランスジェニックイネ植物の作製
それぞれの発現ベクターを含有するアグロバクテリアを用いてイネ(Oryza sativa)植物を形質転換した。イネ japonica 栽培品種Nipponbareの成熟乾燥種子の皮を剥いだ。1分間70%エタノール中で、次いで30分間0.2% HgCl2中でインキュベートして殺菌を行い、次いで6回15分間、無菌蒸留水で洗浄した。無菌種子を次いで 2,4-Dを含有する培地(カルス培地)上で発芽させた。暗所で4週間インキュベーション後、胚形成した、胚盤で誘導されたカルスを切除し、同じ培地上で繁殖させた。2週後、カルスを継代培養によって同じ培地上でさらに2週間増殖または繁殖させた。胚形成カルス片を新しい培地上で3日間、継代培養した後、共胚葉した(細胞分裂活性をブーストするために)。
【0186】
それぞれの発現ベクターを含有するアグロバクテリウム菌株LBA4404を共培養に用いた。アグロバクテリウムを適当な 抗生物質と共にAB培地上に接種し、3日間28℃にて培養した。細菌を次いで採集し、約1の密度(OD600)まで液体共培養培地に懸濁させた。懸濁液を次いでペトリ皿に移し、カルスを懸濁液中に15分間浸漬した。カルス組織を次いでろ紙上にブロットして乾かし、そして固化した共培養培地に移し、3日間暗所で25℃にてインキュベートした。共栽培したカルスを2,4-Dを含有する培地で4週間暗所で28℃にて選択剤の存在のもとで成長させた。この期間に、急速成長する抵抗性カルスアイランドが発生した。この材料を再生培地へ移し、明所でインキュベーションした後、胚形成ポテンシャルが放出されてシュートが次の4〜5週に発生する。シュートをカルスから切除し、2〜3週間オーキシンを含有する培地でインキュベートし、それからこれらを土壌に移した。硬くなったシュートを温室内で高湿度のもとでかつ短期間に成長させた。
【0187】
およそ35の独立したT0イネ形質転換体を1つの構築物のために作製した。
【0188】
一次形質転換体を組織培養チャンバーから温室へ移した。T-DNAインサートのコピー数を立証する定量PCR分析の後、選択剤に耐性を示す唯一のコピートランスジェニック植物を保ってT1種子を収穫した。種子を次いで移植後3〜5か月にわたって収穫した。この方法によって、単一の遺伝子座形質転換体を50%を超える比率で得た(Aldemita and Hodges1996, Chan et al. 1993, Hiei et al. 1994)。
【0189】
7.3 NEENA配列はイネ植物において遺伝子発現の強い増強を媒介する
組織サンプルを、葉および穀粒から作製したトランスジェニック植物から採集した。その組織サンプルを処理し、上記の通り分析した(実施例3.3参照)
構成的なp-PRO239プロモーターだけでNEENAを含まないレポーター遺伝子構築物と比較して、試験したNEENA分子(配列番号1)は葉の遺伝子発現の強い増強を媒介した(図5a)。NEENA(配列番号1)が媒介する遺伝子発現の強い増強はイネ種子において検出された(図5b)。
【特許請求の範囲】
【請求項1】
プロモーターに、該プロモーターに対して異種の1以上の核酸発現増強性核酸(NEENA)分子を機能的に連結するステップを含む高発現性構成的植物プロモーターを産生する方法であって、
前記NEENA分子が
i)配列番号1〜19、好ましくは配列番号1〜9のいずれかで定義した配列を有する核酸分子、または
ii)配列番号1〜19、好ましくは配列番号1〜9のいずれかと少なくとも80%の同一性を有する配列を有する核酸分子、または
iii)配列番号1〜19、好ましくは配列番号1〜9のいずれかの配列を有する対応する核酸分子のような発現増強活性を有する、i)もしくはii)の核酸分子の100以上の連続する塩基の断片、または
iv) i)もしくはii)のもとで先に記載した核酸分子のいずれかの相補配列または逆相補配列である核酸分子、または
v)PCRにより、表2に示した配列番号20〜57、好ましくは配列番号20/21;26/27;30/31;38/39;42/43;44/45;46/47;50;51および56/57に記載のオリゴヌクレオチドプライマーを用いて取得可能な核酸分子、または
vi)7%ドデシル硫酸ナトリウム(SDS)、0.5M NaPO4、1mM EDTA中、50℃におけるハイブリダイゼーションおよび2 X SSC、0.1%SDS中、50℃における洗浄と等価の条件のもとで、配列番号1〜19、好ましくは配列番号1〜9に記載の転写増強性ヌクレオチド配列またはそれらの相補配列の少なくとも50個の連続するヌクレオチドを含む核酸分子とハイブリダイズする核酸分子、
を含む、上記方法。
【請求項2】
それぞれの対照植物またはその部分と比較して、1以上の核酸分子の構成的発現が増強された植物またはその部分を産生する方法であって、
a)植物またはその部分中に、請求項1のi)〜vi)のもとで定義した核酸分子を含む1以上のNEENAを導入するステップ、および
b) 該1以上のNEENAを、構成的プロモーターとおよび該構成的プロモーターの制御下にある核酸分子と機能的に連結し、ここで、該EENAは該核酸分子に対して異種である、ステップ、
を含む、上記方法
【請求項3】
a)植物またはその部分中に、請求項1のi)〜vi)のもとで定義した核酸分子を含む1以上のNEENAを導入するステップ、および
b)該1以上のNEENAを該植物またはその部分のゲノム中に組み込み、ここで該1以上のNEENAは、該1以上のNEENAに対して異種の内因性で構成的に発現される核酸と機能的に連結されている、ステップ、および、任意に
c)形質転換した細胞からの該1以上のNEENAを含む植物またはその部分を再生するステップ、
を含む、請求項1および2に記載の方法。
【請求項4】
a)1以上のNEENAを含む発現構築物であって、構成的プロモーターと1以上の核酸分子と機能的に連結された請求項1のi)〜vi)に定義された核酸分子を含み、該1以上の核酸分子は該1以上のNEENAに対して異種でありかつ該構成的プロモーターの制御下にある、上記発現構築物を提供するステップ、および
b)該1以上のNEENAを含む該発現構築物を、前記植物またはその部分のゲノム中に組み込むステップ、および、任意に
c)該1以上の発現構築物を含む植物またはその部分を、形質転換した植物またはその部分から再生するステップ、
を含む、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
植物が単子葉植物または双子葉植物である、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
植物が双子葉植物である、請求項5に記載の方法。
【請求項7】
植物が単子葉植物である、請求項5に記載の方法。
【請求項8】
前記1以上のNEENAが前記異種核酸分子の転写出発部位に近接した構成的プロモーターと機能的に連結されている、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
前記1以上のNEENAが前記異種核酸分子の転写出発部位から2500bp以下、好ましくは2000bp以下、より好ましくは1500bp以下、さらにより好ましくは1000bp以下そして最も好ましくは500bp以下だけ離れている構成的プロモーターと機能的に連結されている、請求項8に記載の方法。
【請求項10】
前記1以上のNEENAが、その発現が前記構成的プロモーターの制御下にある核酸分子の翻訳出発部位の上流の構成プロモーターと機能的に連結されている、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
前記1以上のNEENAが、その発現が前記構成的プロモーターの制御下にある核酸分子の5'UTR内の構成的プロモーターと機能的に連結されている、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
請求項1のi)〜vi)のいずれかに定義された核酸分子を含む1以上のNEENAを含む、組換え発現構築物。
【請求項13】
1以上の構成的プロモーターおよび1以上の発現される核酸分子と機能的に連結された、請求項1のi)〜vi)のいずれかに定義された核酸分子を含む1以上のNEENAを含み、該1以上の発現される核酸分子は前記1以上のNEENAに対して異種である、請求項12に記載の組換え発現構築物。
【請求項14】
請求項12〜13のいずれか1項に記載の1以上の組換え発現構築物を含む組換え発現ベクター。
【請求項15】
請求項1のi)〜vi)のいずれかに定義された1以上の異種NEENAを含む、トランスジェニック植物またはその部分。
【請求項16】
請求項14に記載の組換え発現ベクターまたは請求項12〜13のいずれか1項に記載の組換え発現構築物を含む、トランスジェニック細胞またはトランスジェニック植物もしくはその部分。
【請求項17】
細菌、真菌、酵母または植物からなる群より選択または誘導された、請求項16に記載のトランスジェニック細胞またはトランスジェニック植物もしくはその部分。
【請求項18】
前記植物またはその部分が双子葉植物である、請求項17に記載のトランスジェニック植物またはその部分。
【請求項19】
前記植物またはその部分が単子葉植物である、請求項17に記載のトランスジェニック植物またはその部分。
【請求項20】
請求項1のi)〜vi)に定義された前記異種NEENA、請求項12もしくは13に記載の組換え発現構築物または請求項14に記載の組換えベクターを含む、請求項15〜19に記載のトランスジェニック細胞または植物もしくはその部分から誘導されるトランスジェニック細胞培養、トランスジェニック種子、部分または繁殖材料。
【請求項21】
請求項1のi)〜vi)に定義されたNEENA、請求項12〜14のいずれか1項に定義された組換え発現構築物または組換えベクターの、植物またはその部分の発現を増強するための使用。
【請求項22】
請求項20に記載のトランスジェニック細胞または植物から誘導されたトランスジェニック細胞培養、トランスジェニック種子、トランスジェニック植物、部分または繁殖材料の、食糧、動物飼料、種子、医薬品またはファインケミカルを生産するための使用。
【請求項1】
プロモーターに、該プロモーターに対して異種の1以上の核酸発現増強性核酸(NEENA)分子を機能的に連結するステップを含む高発現性構成的植物プロモーターを産生する方法であって、
前記NEENA分子が
i)配列番号1〜19、好ましくは配列番号1〜9のいずれかで定義した配列を有する核酸分子、または
ii)配列番号1〜19、好ましくは配列番号1〜9のいずれかと少なくとも80%の同一性を有する配列を有する核酸分子、または
iii)配列番号1〜19、好ましくは配列番号1〜9のいずれかの配列を有する対応する核酸分子のような発現増強活性を有する、i)もしくはii)の核酸分子の100以上の連続する塩基の断片、または
iv) i)もしくはii)のもとで先に記載した核酸分子のいずれかの相補配列または逆相補配列である核酸分子、または
v)PCRにより、表2に示した配列番号20〜57、好ましくは配列番号20/21;26/27;30/31;38/39;42/43;44/45;46/47;50;51および56/57に記載のオリゴヌクレオチドプライマーを用いて取得可能な核酸分子、または
vi)7%ドデシル硫酸ナトリウム(SDS)、0.5M NaPO4、1mM EDTA中、50℃におけるハイブリダイゼーションおよび2 X SSC、0.1%SDS中、50℃における洗浄と等価の条件のもとで、配列番号1〜19、好ましくは配列番号1〜9に記載の転写増強性ヌクレオチド配列またはそれらの相補配列の少なくとも50個の連続するヌクレオチドを含む核酸分子とハイブリダイズする核酸分子、
を含む、上記方法。
【請求項2】
それぞれの対照植物またはその部分と比較して、1以上の核酸分子の構成的発現が増強された植物またはその部分を産生する方法であって、
a)植物またはその部分中に、請求項1のi)〜vi)のもとで定義した核酸分子を含む1以上のNEENAを導入するステップ、および
b) 該1以上のNEENAを、構成的プロモーターとおよび該構成的プロモーターの制御下にある核酸分子と機能的に連結し、ここで、該EENAは該核酸分子に対して異種である、ステップ、
を含む、上記方法
【請求項3】
a)植物またはその部分中に、請求項1のi)〜vi)のもとで定義した核酸分子を含む1以上のNEENAを導入するステップ、および
b)該1以上のNEENAを該植物またはその部分のゲノム中に組み込み、ここで該1以上のNEENAは、該1以上のNEENAに対して異種の内因性で構成的に発現される核酸と機能的に連結されている、ステップ、および、任意に
c)形質転換した細胞からの該1以上のNEENAを含む植物またはその部分を再生するステップ、
を含む、請求項1および2に記載の方法。
【請求項4】
a)1以上のNEENAを含む発現構築物であって、構成的プロモーターと1以上の核酸分子と機能的に連結された請求項1のi)〜vi)に定義された核酸分子を含み、該1以上の核酸分子は該1以上のNEENAに対して異種でありかつ該構成的プロモーターの制御下にある、上記発現構築物を提供するステップ、および
b)該1以上のNEENAを含む該発現構築物を、前記植物またはその部分のゲノム中に組み込むステップ、および、任意に
c)該1以上の発現構築物を含む植物またはその部分を、形質転換した植物またはその部分から再生するステップ、
を含む、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
植物が単子葉植物または双子葉植物である、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
植物が双子葉植物である、請求項5に記載の方法。
【請求項7】
植物が単子葉植物である、請求項5に記載の方法。
【請求項8】
前記1以上のNEENAが前記異種核酸分子の転写出発部位に近接した構成的プロモーターと機能的に連結されている、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
前記1以上のNEENAが前記異種核酸分子の転写出発部位から2500bp以下、好ましくは2000bp以下、より好ましくは1500bp以下、さらにより好ましくは1000bp以下そして最も好ましくは500bp以下だけ離れている構成的プロモーターと機能的に連結されている、請求項8に記載の方法。
【請求項10】
前記1以上のNEENAが、その発現が前記構成的プロモーターの制御下にある核酸分子の翻訳出発部位の上流の構成プロモーターと機能的に連結されている、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
前記1以上のNEENAが、その発現が前記構成的プロモーターの制御下にある核酸分子の5'UTR内の構成的プロモーターと機能的に連結されている、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
請求項1のi)〜vi)のいずれかに定義された核酸分子を含む1以上のNEENAを含む、組換え発現構築物。
【請求項13】
1以上の構成的プロモーターおよび1以上の発現される核酸分子と機能的に連結された、請求項1のi)〜vi)のいずれかに定義された核酸分子を含む1以上のNEENAを含み、該1以上の発現される核酸分子は前記1以上のNEENAに対して異種である、請求項12に記載の組換え発現構築物。
【請求項14】
請求項12〜13のいずれか1項に記載の1以上の組換え発現構築物を含む組換え発現ベクター。
【請求項15】
請求項1のi)〜vi)のいずれかに定義された1以上の異種NEENAを含む、トランスジェニック植物またはその部分。
【請求項16】
請求項14に記載の組換え発現ベクターまたは請求項12〜13のいずれか1項に記載の組換え発現構築物を含む、トランスジェニック細胞またはトランスジェニック植物もしくはその部分。
【請求項17】
細菌、真菌、酵母または植物からなる群より選択または誘導された、請求項16に記載のトランスジェニック細胞またはトランスジェニック植物もしくはその部分。
【請求項18】
前記植物またはその部分が双子葉植物である、請求項17に記載のトランスジェニック植物またはその部分。
【請求項19】
前記植物またはその部分が単子葉植物である、請求項17に記載のトランスジェニック植物またはその部分。
【請求項20】
請求項1のi)〜vi)に定義された前記異種NEENA、請求項12もしくは13に記載の組換え発現構築物または請求項14に記載の組換えベクターを含む、請求項15〜19に記載のトランスジェニック細胞または植物もしくはその部分から誘導されるトランスジェニック細胞培養、トランスジェニック種子、部分または繁殖材料。
【請求項21】
請求項1のi)〜vi)に定義されたNEENA、請求項12〜14のいずれか1項に定義された組換え発現構築物または組換えベクターの、植物またはその部分の発現を増強するための使用。
【請求項22】
請求項20に記載のトランスジェニック細胞または植物から誘導されたトランスジェニック細胞培養、トランスジェニック種子、トランスジェニック植物、部分または繁殖材料の、食糧、動物飼料、種子、医薬品またはファインケミカルを生産するための使用。
【図1】

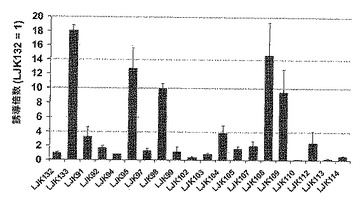
【図2】

【図3】

【図4】
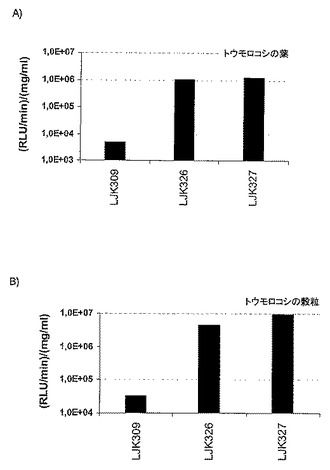
【図5】
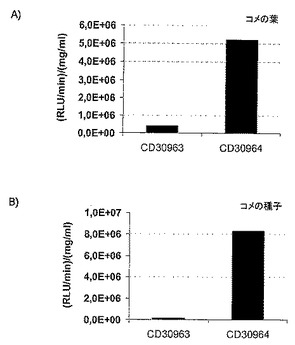
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2013−502907(P2013−502907A)
【公表日】平成25年1月31日(2013.1.31)
【国際特許分類】
【出願番号】特願2012−525978(P2012−525978)
【出願日】平成22年8月11日(2010.8.11)
【国際出願番号】PCT/EP2010/061659
【国際公開番号】WO2011/023537
【国際公開日】平成23年3月3日(2011.3.3)
【出願人】(512005634)ビーエーエスエフ プラント サイエンス カンパニー ゲーエムベーハー (8)
【Fターム(参考)】
【公表日】平成25年1月31日(2013.1.31)
【国際特許分類】
【出願日】平成22年8月11日(2010.8.11)
【国際出願番号】PCT/EP2010/061659
【国際公開番号】WO2011/023537
【国際公開日】平成23年3月3日(2011.3.3)
【出願人】(512005634)ビーエーエスエフ プラント サイエンス カンパニー ゲーエムベーハー (8)
【Fターム(参考)】
[ Back to top ]