
植物の生長を促進する変異型Myb関連遺伝子及びそれを用いた植物生長促進方法
【課題】変異型Myb転写制御因子の導入によって副作用を生じさせることなく目的の形質のみを獲得する方法を提供する。
【解決手段】特定塩基配列をコード領域内に含み、その産物が植物の生長に関与するMyb関連遺伝子を標的とするmiRNAがその遺伝子の発現抑制できなくなるように、前記塩基配列内にサイレント変異を導入した変異型Myb関連遺伝子。
【解決手段】特定塩基配列をコード領域内に含み、その産物が植物の生長に関与するMyb関連遺伝子を標的とするmiRNAがその遺伝子の発現抑制できなくなるように、前記塩基配列内にサイレント変異を導入した変異型Myb関連遺伝子。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、植物の生長を促進することのできる変異型Myb関連遺伝子及びそれを用いた植物生長促進方法並びに植物の生長を抑制することのできる前記Myb関連遺伝子を標的とするmiRNA及びそれを用いた植物生長抑制方法に関する。
【背景技術】
【0002】
Myb遺伝子は、酵母から動物及び植物に至るまで生物種を越えて広く存在する転写制御因子遺伝子である。本遺伝子は、当初、癌遺伝子c−Mybとしてトリ骨髄芽球症ウイルス(AMV)及びE26ウイルスから独立に単離された(非特許文献1)。しかし、その後の研究により、転写因子をコードすること(非特許文献2)、多数の関連遺伝子からなるファミリーを形成していること、脊椎動物のみならず、酵母、線虫、昆虫、植物に至るまで種を超えて広く存在することが明らかとなった(非特許文献3)。
【0003】
Myb遺伝子ファミリーに属する遺伝子は、いずれもMybドメインと呼ばれる約50塩基のへリックス‐ターン‐へリックス構造からなるDNA結合ドメインをコードすることを特徴としている。脊椎動物のMyb遺伝子産物は、a-Myb、b-Myb及びc-Mybの3つのメンバーが知られている。いずれの産物も3つのMybドメイン(R1、R2、R3)を有し、主に細胞増殖と分化に関わる転写制御因子として機能している(非特許文献4)。一方、植物には多数のMyb遺伝子が存在し、構造上の特徴から大きく3タイプに分類されている。すなわち、哺乳動物Myb遺伝子と同様に3つのMybドメインを有し、細胞周期の制御を介して細胞増殖を制御するR1R2R3タイプ(非特許文献5)、色素合成系、形態形成及び環境応答等の様々な生命現象を制御するR2R3タイプ(非特許文献6)、そして光による制御や形態形成及び病障害に拘るR1様又はR3様ドメインを1つもつタイプ(非特許文献7、8、9)である。
【0004】
ところで、植物では今世紀に入り多数の遺伝子情報が蓄積され、光合成、一次代謝及び二次代謝をはじめとする基本的生理機能に関与する遺伝子の転写制御に関わる因子とその機能の知見が急速に増大した。そして、次なる段階として植物に新たな機能を獲得させるために、遺伝子発現を人為的に制御するための様々な分子遺伝学的手法が開発されている。このような方法の一つとして、転写制御因子遺伝子のプロモータ領域を置換・改変したり、対象とする植物に変異型転写制御因子遺伝子を導入することによって所望の機能をその植物に付与する方法が知られている。しかし、このような方法は、たとえその目的を達成できたとしても、望ましくない又は有害な他の形質を副作用的に同時に誘導してしまう等の予想外の弊害を伴うという問題があった。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Roussel M.,1979, Nature,218:452-455
【非特許文献2】Biedenkapp H., et al., 1988, Nature, 335:835-837
【非特許文献3】岩淵雅樹, 篠崎一雄、植物ゲノム機能のダイナミズム: 転写因子による発現制御、シュプリンガー・ジャパン株式会社, 2001
【非特許文献4】Weston K., 1999,.Curr. Opin. Genet. Dev. 8, 76-81.
【非特許文献5】Ito M., 2005, J. Plant Res., 118(1):61-69
【非特許文献6】Jin H. and Martin C., 1999, Plant Mol. Biol., 41(5):577-588
【非特許文献7】Feldbrugge et al., 1997, Plant J. 11, 1079-1093.
【非特許文献8】Kirik V. and Baumlein H., 1996, Gene 183, 109-113.
【非特許文献9】Wang Z.Y., et al., 1997, Plant Cell 9, 491-507.
【発明の概要】
【発明が解決しようとする課題】
【0006】
変異型転写制御因子遺伝子の導入によって、副作用を生じさせることなく目的の形質のみを対象とする植物に獲得させる方法を開発することを課題とする。
【課題を解決するための手段】
【0007】
一般に転写制御因子は、様々な遺伝子の発現を統括的に制御する場合が多く、そのような転写制御因子への変異の導入は、結局のところ、その因子によって制御される他の多くの遺伝子の発現に少なからず影響を及ぼすことになる。それ故、変異型転写制御因子遺伝子の導入によって目的の形質のみを付与させることは、通常容易なことではない。
【0008】
しかし、近年、遺伝子の発現を翻訳段階で制御するsmall RNA(低分子RNA)、例えば、miRNA(micro RNA)やsiRNA (short interference RNA)等の存在及びRNAサイレンシング機構が明らかとなった(Lee R.C., et al., 1993, Cell, 75: 843-854、Baulcombe D, 2002, Science, 297:2002-2003)。このような機構の発見により、遺伝子発現は、転写調節によってのみ制御されているのではなく、転写段階と翻訳段階によって独立に制御され、それぞれの協調の結果として、もたらされることが判明した(Jackson R. J. and Standart N., Sci. STKE 2007,(367) re1)。
【0009】
そこで、本発明者らは、当該転写後制御機構を利用して前記課題を解決するための新規方法を開発すべく鋭意研究を重ねた。その結果、配列番号1で示される塩基配列をコード領域内に含み、かつ植物の生長に関与するMyb関連タンパク質をコードする遺伝子の発現を制御することのできるmiRNAを見出した。本発明は、当該知見に基づいて完成されたものであり、すなわち以下を提供する。
(1)植物細胞内への導入によって該植物の生長を促進できる変異型Myb関連遺伝子であって、配列番号1で示される塩基配列をコード領域内に含むMyb関連遺伝子において、前記塩基配列に結合するmiRNAを介した発現抑制が不能となるように前記塩基配列内にサイレント変異を導入した前記変異型Myb関連遺伝子。
(2)配列番号1で示される塩基配列が配列番号2で特定される、(1)に記載の変異型Myb関連遺伝子。
(3)前記サイレント変異の少なくとも一つが配列番号1における13番目のTをCに置換した変異である、前記(1)又は(2)に記載の変異型Myb関連遺伝子。
(4)少なくとも5個のサイレント変異を含む前記(1)〜(3)のいずれかに記載の変異型Myb関連遺伝子。
(5)前記Myb関連遺伝子が配列番号3で示される塩基配列又は該塩基酸配列と95%以上の同一性を有する塩基配列からなる、前記(1)〜(4)のいずれかに記載の変異型Myb関連遺伝子。
(6)前記(1)〜(5)のいずれかに記載の変異型Myb関連遺伝子を含む発現ベクター。
(7)前記(6)に記載の発現ベクターを宿主に導入した形質転換体又はその後代。
(8)宿主が微生物又は植物である、前記(7)に記載の形質転換体又はその後代。
(9)前記植物が、ユーカリプタス属(Eucalyptus;本願では以降、種名を除き、一般的略称である「ユーカリ」を使用する)植物又はタバコ属(Nicotiana)植物である、前記(8)に記載の形質転換体又はその後代。
(10)前記(6)に記載の発現ベクターを植物に導入して、該植物の生長を促進する方法。
(11)植物細胞内への導入によって該植物の生長を抑制できるmiRNAであって、配列番号7で示される塩基配列を含み、かつ(1)〜(5)のいずれかに記載のMyb関連遺伝子の転写産物を標的とするmiRNA。
(12)配列番号7で示される塩基配列が配列番号8で特定される、(11)に記載のmiRNA。
(13)前記(11)又は(12)に記載のmiRNAをコードするDNAを含む発現ベクター。
(14)前記(11)又は(12)に記載のmiRNAあるいは前記(13)に記載の発現ベクターを宿主に導入した形質転換体又はその後代。
(15)宿主が微生物又は植物である、(14)に記載の形質転換体又はその後代。
(16)前記植物が、ユーカリ属植物又はタバコ属植物である、(15)に記載の形質転換体又はその後代。
(17)前記(13)に記載の発現ベクターを植物に導入して、該植物の生長を抑制する方法。
【発明の効果】
【0010】
本発明の変異型Myb関連遺伝子及び本発明の植物の生長を促進する方法によれば、導入した植物に対して、他の副作用を発生させることなく、目的とする生長促進という形質のみを付与することができる。
【0011】
本発明のmiRNA及び本発明の植物の生長を抑制する方法によれば、内在性の植物生長抑制機構を特異的に増強することで、導入した植物の生長を抑制することができる。
【図面の簡単な説明】
【0012】
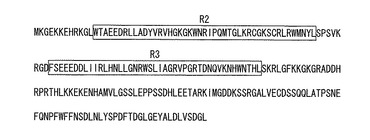
【図1】EcMybタンパク質のアミノ酸配列とR2-及びR3-Mybドメイン。
【図2】EcMyb関連遺伝子(EcMyb gene)の塩基配列及びEcMybタンパク質(EcMyb prt)のアミノ酸配列と前記遺伝子単離に使用したプライマーの位置、並びに配列番号2に相当する領域(Myb-miRNA結合部位)を示した図。
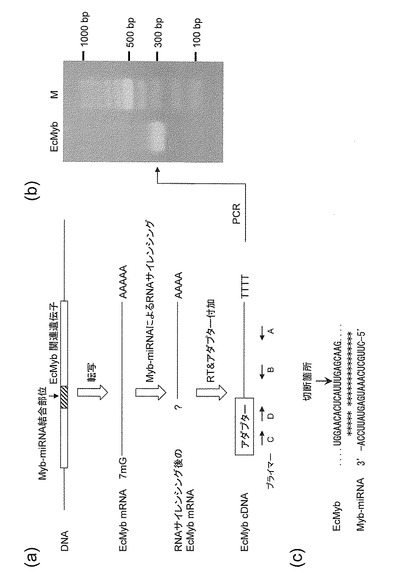
【図3】(a)Myb-miRNAによる切断部位検証の模式図、(b)Myb-miRNA結合部位とMyb-miRNAによる切断箇所、(c)Myb-miRNAによる切断を受けたEcMyb関連遺伝子転写産物を示すアガロースゲル電気泳動図。
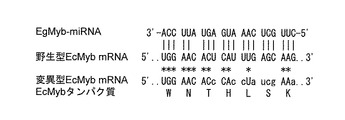
【図4】配列番号2で示されるMyb-miRNA結合部位に導入した9箇所のサイレント変異。このうち1箇所(6番目のC)は、EcMybにおける内在性のサイレント変異。
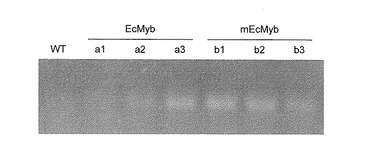
【図5】形質転換タバコにおけるEcMyb又はmEcMybの発現。
【図6】形質転換タバコの生長性(1)。
【図7】形質転換タバコの生長性(2)。
【発明を実施するための形態】
【0013】
1.変異型Myb関連遺伝子
本発明の一の態様は、植物細胞内への導入によって該植物の生長を促進できる変異型Myb関連遺伝子である。
【0014】
1−1.Myb関連遺伝子
本明細書の「Myb関連遺伝子」とは、植物のMyb遺伝子ファミリーにおいて配列番号1で示される塩基配列をコード領域内に含み、かつ植物の生長を促進する機能を有するR2R3タイプの野生型Myb関連遺伝子、その天然変異体又はそれらの断片をいう。
【0015】
「野生型Myb関連遺伝子」とは、自然界に存在するMyb関連タンパク質をコードし、同種集団内の対立遺伝子において最も多く存在する塩基配列を有するMyb関連遺伝子をいう。本遺伝子がコードするMyb関連タンパク質は、そのタンパク質本来の機能を有する。例えば、転写制御因子としての機能である。
【0016】
「R2R3タイプ」のMyb関連遺伝子とは、前述のように、その構造中に2つのMybドメイン(R2及びR3)をコードするMyb関連遺伝子である。例えば、毛根形成を負に制御するWEREWOLF(WER)(Lee et al., 1999, Cell, 99: 473-483)、トライコーム形成に関与するGLABRA1(GL1)(Oppenheimer et al., 1991, Cell 67(3): 483-493)及び種皮におけるタンニンの蓄積に機能するTRANSPARENT TESTA 2 (tt2)(Nesi et al., 2001, Plant Cell, 13(9), 2099-2114)等がこのタイプに属する。
【0017】
「その天然変異体」とは、自然界に存在する野生型Myb関連遺伝子の変異体である。例えば、野生型Myb関連遺伝子の塩基配列において1個又は数個のヌクレオチドが欠失、置換又は付加されたもの、前記塩基配列とそれぞれ95%以上、好ましくは98%以上、より好ましくは99%以上の同一性を有するもの、又は野生型Myb関連遺伝子の部分塩基配列に相補的な塩基配列からなる核酸断片とストリンジェントな条件下でハイブリダイズするものをいう。いずれも、野生型Mybタンパク質と同様に、植物の成長を促進する活性を有する。ここで前記「同一性」とは、2つの塩基配列にギャップを導入して又は導入しないでアラインメントさせたときに、一方の塩基配列の全塩基数に対する他方の塩基配列の同一塩基数の割合(%)をいう。「数個のヌクレオチド」とは、2〜10個、例えば、2〜7個、2〜5個、2〜4個又は2〜3個のヌクレオチドをいう。また、「ストリンジェントな条件」とは、非特異的なハイブリッドが形成されない条件を意味する。通常は低ストリンジェント〜高ストリンジェントな条件が挙げられるが、高ストリンジェントな条件が好ましい。低ストリンジェントな条件とは、ハイブリダイゼーション後の洗浄において、例えば42℃、5×SSC、0.1% SDSで洗浄する条件であり、好ましくは50℃、5×SSC及び0.1% SDSで洗浄する条件である。高ストリンジェントな条件とは、ハイブリダイゼーション後の洗浄において、例えば65℃、0.1×SSC及び0.1% SDSで洗浄する条件である。天然変異体の具体例としては、SNP(一塩基多型)等の多型に基づく変異体、スプライス変異体、遺伝暗号の縮重に基づく変異体等が挙げられる。
【0018】
「それらの断片」とは、前記野生型Myb関連遺伝子又はその天然変異体のDNA断片であって、配列番号1で示される塩基配列をコード領域内に含み、かつその断片がコードするMyb関連タンパク質の一部が野生型Myb関連遺伝子のコードするMyb関連タンパク質と同程度の生物学的機能、すなわち転写制御因子としての活性を保持しているものをいう。前記機能を保持している限り、DNA断片の塩基長は特に限定しない。
【0019】
「配列番号1で示される塩基配列」は、前記Myb関連遺伝子を標的とするmiRNA(本明細書では、以降便宜的に「Myb-miRNA」と呼ぶ)の結合部位(Myb-miRNA結合部位)に相当する。すなわち、本発明に記載のMyb関連遺伝子は、このMyb-miRNA結合部位へのmiRNAの結合を介して発現抑制を受ける。配列番号1において、R、Y及びNで示される塩基は、それぞれ、A(アデニン)又はG(グアニン)、C(シトシン)又はT(チミン)及びA、T、G又はCのいずれであってもよいことを示す。なお、Myb-miRNAについては、「5.Myb-miRNA」の項で詳述するため、ここではその説明を省略する。
【0020】
「コード領域内」とは、Myb関連タンパク質をコードする塩基配列の領域内を意味する。したがって、「配列番号1で示される塩基配列をコード領域内に含む」とは、配列番号1で示されるMyb-miRNA結合部位をコード領域内に含んでいることを意味する。
【0021】
「植物の生長」とは、茎(幹を含む)及び根の伸長並びに茎径(樹径を含む)の増大をいう。具体的には植物の成長点及び/又は維管束形成層における細胞の増殖、分化及び組織形成を意味する。「植物の生長を促進できる機能を有する」とは、前記植物の成長点及び/又は維管束形成層における細胞の増殖促進(細胞壁形成の制御を含む)に直接的に又は間接的に関与する機能を有することをいう。細胞の増殖促進に直接的に関与する機能とは、例えば、細胞周期制御に関与する因子のように、その機能が細胞の増殖に直接作用することをいう。細胞の増殖促進に間接的に関与する機能とは、例えば、細胞増殖に直接作用する遺伝子等の発現を制御することによって、間接的に細胞の増殖に寄与することをいう。
【0022】
配列番号1で示される塩基配列をコード領域内に含むR2R3タイプMyb関連遺伝子、すなわち、本発明の変異型Myb関連遺伝子の基礎となるMyb関連遺伝子の具体例としては、本発明者らによって今回新たに単離された配列番号3で示される塩基配列からなるユーカリプタス・カマルドレンシス(Eucalyptus camaldulensis)の野生型Myb関連遺伝子(本明細書では、以降便宜的に「EcMyb遺伝子」とする)又はその天然変異体が挙げられる。EcMyb遺伝子は、配列番号1に相当する部位として、配列番号2で示される塩基配列を、310〜331番目(開始コドンのAを1番目とする。以下、本明細書において同様とする。)の領域に有し、配列番号9で示される全長221アミノ酸からなるMyb関連タンパク質をコードする。また、Myb関連遺伝子は、配列番号3で示されるEcMyb遺伝子の塩基配列と95%以上、好ましくは98%以上、より好ましくは99%以上の同一性を有する塩基配列で示される、又はEcMyb遺伝子の部分塩基配列に相補的な塩基配列からなるプローブとストリンジェントな条件下でハイブリダイズする、他の植物種のMyb関連遺伝子、好ましくはEcMybオルソログ、であってもよい。例えば、配列番号4で示され、EcMyb遺伝子の塩基配列と97%の同一性を有するユーカリプタス・ガンニー(E. gunii)のMyb関連遺伝子(本明細書では、以降便宜的に「EgMyb遺伝子」とする)、GenBank XM_002302608.1として登録され、配列番号5で示されるポプラ(Populus trichocarpa)由来のMyb関連遺伝子(本明細書では、以降便宜的に「PtMyb遺伝子」とする)も本発明のMyb関連遺伝子に該当する。
【0023】
本発明者らは、Myb-miRNA結合部位をコード領域内に含むMyb関連遺伝子が翻訳段階でMyb-miRNAを介したRNAサイレンシング機構によってその発現を抑制されることを見出した。なお、miRNAについては、後述の「5.Myb-miRNA」の項を参照されたい。
【0024】
1−2.変異型Myb関連遺伝子
「変異型Myb関連遺伝子」とは、前記Myb関連遺伝子において配列番号1で示される塩基配列内、すなわち、Myb-miRNA結合部位内にサイレント変異を導入した遺伝子である。
【0025】
本発明におけるMyb-miRNAを介した前記Myb関連遺伝子の発現抑制は、後述する実施例2の結果から、Myb-miRNAがMyb関連遺伝子の転写産物上のMyb-miRNA結合部位と結合した後、Myb-miRNAと複合体を形成するRISC(RNA-induced silencing complex)/miRNPの作用によって配列番号1で示す塩基配列の13番目と14番目のT間(ただし、実際の標的分子はRNAであるため、13番目と14番目のU(ウリジン)間となる)が切断されることによって達成されると考えられる。なお、miRNAを介した遺伝子発現抑制機構の詳細については、後述の「5.Myb-miRNA」の項を参照されたい。本発明は、この機構に基づき、Myb-miRNAによる発現抑制が不能となる一方で、Myb関連遺伝子がコードするタンパク質のアミノ酸配列は変わらない変異型Myb関連遺伝子を提供する。
【0026】
「サイレント変異」とは、遺伝子の塩基配列には変異を生じるが、その遺伝子がコードするタンパク質のアミノ酸配列には変異を生じない変異、すなわち、縮重(縮退)コドン内における塩基の変異をいう。本発明においてMyb-miRNA結合部位は、WNTHLSK(配列番号6)からなるアミノ酸をコードする。したがって、本発明は、このアミノ酸配列を維持する全てのサイレント変異を包含する。例えば、配列番号1が配列番号2の塩基配列で特定される場合であれば、C6T(6番目のCをTに置換の意)、T9A/G/C(9番目のTをA、G又はCに置換の意)、T13-G15A/T/C(13番目がTのときにG15A/T/Cの意)若しくはT13C-G15A(13番目のTをCに置換したときにG15Aの意)、A16-C18T若しくはA16T-G17C-C18A/T/G、及び/又はG21Aの変異を導入することができる。
【0027】
「Myb-miRNAによる発現抑制が不能となる」とは、変異型Myb関連遺伝子がサイレント変異の導入によってMyb-miRNA耐性能力を獲得し、Myb-miRNAを介したRNAサイレンシングによる発現抑制を受けなくなる状態をいう。例えば、Myb-miRNAが変異型Myb関連遺伝子の転写産物を標的分子として認識できなくなること又は認識できても変異型Myb関連遺伝子の転写産物を切断できなくなることが挙げられる。
【0028】
「変異型Myb関連遺伝子の転写産物を標的分子として認識できなくなる」とは、Myb-miRNAが変異型Myb関連遺伝子の標的転写産物上のMyb-miRNA結合部位に結合できなくなることをいう。この場合、本発明の変異型Myb関連遺伝子を導入する植物において、その植物に内在するMyb-miRNAと前記変異型Myb関連遺伝子の転写産物上のMyb-miRNA結合部位との間で安定した塩基対合が形成されないようにサイレント変異を導入すればよい。例えば、配列番号1で示される塩基配列に少なくとも5個、最大で9個のサイレント変異を導入することが好ましい。Myb-miRNA結合部が配列番号2で示される塩基配列で特定されるEcMyb遺伝子の場合であれば、上述したサイレント変異を5個以上有していればよい(図4)。
【0029】
「認識できても変異型Myb関連遺伝子の転写産物を切断できなくなる」とは、Myb-miRNAが変異型Myb関連遺伝子の転写産物上のMyb-miRNA結合部位には結合できるが、上述した機序によって当該部位が切断されない状態をいう。例えば、Myb-miRNA結合部位において、生物種間で高度に保存された塩基にサイレント変異を導入する場合、例えば、配列番号1で示されるMyb-miRNA結合部位においてMyb-miRNAによって切断される位置に隣接する13番目のTをCに置換するサイレント変異(T13C)を導入した場合、が挙げられる。
【0030】
本発明の変異型Myb関連遺伝子を目的とする植物体中に導入することによって、内在のMyb-miRNAを介した発現抑制を特異的に回避し、かつ野生型Myb関連遺伝子と同じアミノ酸配列からなるMyb関連タンパク質を安定的にその植物に付与することができる。その結果、その植物の生長を促進することが可能となる。
【0031】
1−3.変異型Myb関連遺伝子の調製
本発明の変異型Myb関連遺伝子は、当該分野で公知の方法を用いて調製することができる。
一例を挙げると、まず、Myb関連遺伝子のcDNAを単離するために、目的とする植物体からmRNAを抽出する。抽出後、精製したcDNAをライブラリー化する。その後、PCR法(例えば、Inverse-PCR法、アンカーPCR法・TAIL-PCR法)、又はハイブリダイゼーション法を実施することにより、前記cDNAライブラリーから目的のMyb関連遺伝子を単離することができる。前記PCR法に使用するプライマー及びハイブリダイゼーション法に使用するプローブは、例えば、本明細書に記載の配列番号3(EcMyb遺伝子)、配列番号4(EgMyb遺伝子)、若しくは配列番号6(PtMyb遺伝子)、又はインターネットを介して一般的に利用可能なデータベース、例えば、NCBI(http://www.ncbi.nlm.nih.gov/)、理研(http://www.psc.riken.jp/database/index.html)、かずさDNA研究所DNA配列解析情報データベース(http://www.kazusa.or.jp/j/resources/database.html)等から入手できる既知の細胞壁転写制御因子の遺伝子情報から得た塩基配列に基づいて調製すればよい。特に、Myb-miRNA結合部位は、植物種間で比較的高度に保存されているため、当該部位をプローブ又は一方のプライマーとして利用すれば、異なる種のcDNAライブラリーからも目的のMyb関連遺伝子を単離することが可能となる。cDNAライブラリーの作製及び目的の遺伝子を単離する具体的な方法は、当該分野で公知である。例えば、Sambrook, J. et. al., (1989) Molecular Cloning: a Laboratory Manual Second Ed., Cold SpringHarbor Laboratory Press, Cold Spring Harbor, New Yorkを参照すればよい。
【0032】
次に、得られたMyb関連遺伝子において、配列番号1で示される塩基配列領域に必要なサイレント変異を導入する。この変異導入方法も当該分野で公知の方法に基づいて行うことができる。例えば、Inverse PCRを用いた方法、市販の突然変異誘発キット(例えば、Stratagene社のQuickChange Site-Directed Mutagenesis Kit、Promega社のAltered Sites II in vitro Mutagenesis System等)を利用して導入する方法を利用できる。また、いくつかのライフサイエンス系メーカー(例えば、タカラバイオ社)では、遺伝子等に所望の変異を導入する受託サービスを行っているのでそれを利用してもよい。
【0033】
2.変異型Myb関連遺伝子を含む発現ベクター
本発明の他の態様は、前記変異型Myb関連遺伝子を含む発現ベクター(以下、「変異型Myb関連遺伝子発現ベクター」とする)に関する。
【0034】
2−1.変異型Myb関連遺伝子発現ベクター
本発明において、「変異型Myb関連遺伝子発現ベクター」とは、前記変異型Myb関連遺伝子を適当な発現ベクター内に導入したものである。本ベクターは、適切な宿主細胞内に導入することで、適当な条件の下、挿入した変異型Myb関連遺伝子を発現させることが可能である。
【0035】
本発明の発現ベクターにおける骨格部分、すなわち、本発明の変異型Myb関連遺伝子以外の主要な構成部分は、特に限定はしないが、プラスミド又はウイルスが好ましい。プラスミドであれば、例えば、pBI系、pPZP系、pSMA系、pUC系、pBR系、pBluescript系(stratagene社)、pTriEXTM系(TaKaRa社)、又はpBI系若しくはpRI系のバイナリーベクター等を、ウイルスであれば、カリフラワーモザイクウイルス(CaMV)、インゲンマメゴールデンモザイクウイルス(BGMV)、タバコモザイクウイルス(TMV)等を利用することができる。これらは、導入する宿主に応じて適宜選択すればよい。導入する宿主としては、例えば、大腸菌、酵母、植物細胞(植物体、植物器官及び植物組織を含む)、昆虫細胞又は動物細胞が挙げられる。これらは、目的に応じて当該分野で公知のものを適宜選択すればよい。
【0036】
本発明の変異型Myb関連遺伝子発現ベクターの骨格部分には、変異型Myb関連遺伝子の他に、例えば、プロモータ、エンハンサ、若しくはターミネータ等の調節領域、又は選抜マーカー遺伝子等の他の標識的領域を連結することができる。それぞれの種類は、特に限定されない。導入する宿主(例えば、細菌、酵母、植物細胞又は昆虫細胞)に応じて又は宿主内での目的(例えば、クローニング又は遺伝子発現)に応じて当該分野で公知のものを適宜選択すればよい。それぞれについて、以下に例示して説明する。
【0037】
大腸菌中で作動可能なプロモータとしては、例えば、lac、trp若しくはtacプロモータ又はファージ・ラムダ由来のPR若しくはPLプロモータ等が挙げられる。酵母で作動可能なプロモータとしては、酵母解糖系遺伝子由来のプロモータ、アルコールデヒドロゲナーゼ遺伝子プロモータ、TPI1プロモータ、ADH2-4cプロモータ等が挙げられる。植物細胞で作動可能なプロモータとしては、カリフラワーモザイクウイルス(CaMV)の35Sプロモータ、ノパリン合成酵素遺伝子のプロモータ(Pnos)、トウモロコシ由来ユビキチンプロモータ、イネ由来のアクチンプロモータ、タバコ由来PRタンパク質プロモータ等が挙げられる。昆虫細胞で作動可能なプロモータとしては、ポリヘドリンプロモータ、P10プロモータ、オートグラファ・カリホルニカ・ポリヘドロシス塩基性タンパクプロモータ、バキュロウイルス即時型初期遺伝子1プロモータ、バキュロウイルス39K遅延型初期遺伝子プロモータ等が挙げられる。形質転換体の特定部位において局所的に変異型Myb関連遺伝子を発現させたい場合には、部位特異的プロモータを用いればよい。例えば、根の生長を促進させたい場合には、特開2007−77677号に記載の根特異的発現を誘導するプロモータを用いることができる。
【0038】
エンハンサとしては、CaMV 35Sプロモータ内の上流側の配列を含むエンハンサ領域及びCMVエンハンサ等が挙げられる。
【0039】
ターミネータとしては、ノパリン合成酵素(NOS)遺伝子のターミネータ、オクトピン合成酵素(OCS)遺伝子のターミネータ、CaMV 35Sターミネータ、大腸菌リポポリプロテインlppの3’ターミネータ、trpオペロンターミネータ、amyBターミネータ、ADH1遺伝子のターミネータ等が挙げられる。
【0040】
選抜マーカー遺伝子としては、薬剤耐性遺伝子(例えば、テトラサイクリン耐性遺伝子、アンピシリン耐性遺伝子、カナマイシン耐性遺伝子、ハイグロマイシン耐性遺伝子、スペクチノマイシン耐性遺伝子、クロラムフェニコール耐性遺伝子、又はネオマイシン耐性遺伝子)、蛍光又は発光レポーター遺伝子(例えば、ルシフェラーゼ、β-ガラクトシダーゼ、β-グルクロニターゼ(GUS)、又はグリーンフルオレッセンスプロテイン(GFP))、ネオマイシンホスホトランスフェラーゼII(NPT II)、ジヒドロ葉酸還元酵素等の酵素遺伝子が挙げられる。
【0041】
2−2.変異型Myb関連遺伝子発現ベクターの調製
前記変異型Myb関連遺伝子を発現ベクターの所定の部位に挿入する方法は、当該分野で公知の方法、例えば、Sambrook, J. et. al., (1989) Molecular Cloning: a Laboratory Manual Second Ed., Cold SpringHarbor Laboratory Press, Cold Spring Harbor, New Yorkに記載の方法に従って行えばよい。通常は、調製された変異型Myb関連遺伝子を適当な制限酵素で切断し、好適な発現ベクターの対応する制限酵素部位、マルチクローニングサイト、又はTaq DNAポリメラーゼ等による3'-A突出末端を有するPCR産物であれば5'-T突出末端に挿入して連結する方法等が採用される。その他、市販のシステム又はキットを用いる場合であれば、それらに特異的な方法によって調製することもできる。例えば、Gatewayシステム(Invitrogen社)が利用できる。
【0042】
本態様の変異型Myb関連遺伝子発現ベクターによれば、本ベクターをそれに適応する宿主に導入することで、次で説明する変異型Myb関連遺伝子を発現する形質転換体を得ることができる。
【0043】
3.形質転換体又はその後代
本発明の他の態様は、前記変異型Myb関連遺伝子を含む発現ベクターを宿主に導入した形質転換体又はその後代に関する。
【0044】
3−1.形質転換体
本明細書において「形質転換体」とは、前記変異型Myb関連遺伝子を含む発現ベクターの導入によって形質転換された宿主をいう。使用する宿主は、導入した発現ベクターがコードする本発明の変異型Myb関連遺伝子を発現できれば特に限定しない。一般的には、発現ベクターによって発現可能な宿主はある程度限定されるため、それに従えばよい。宿主の具体例を挙げると、細菌(例えば、大腸菌(Escherichia coli等)及び枯草菌(Bacillus subtilis))及び酵母(例えば、出芽酵母(Saccharomyces cerevisiae)、分裂酵母(Schizosaccharomyces pombe)又はメタノール資化性酵母(Pichia pastoris))のような微生物、真菌(例えば、コウジカビ(Aspergillus)及びアカパンカビ(Neurospora))、植物(植物体、その器官、組織、分化した細胞若しくは未分化状態の植物細胞(カルス)を含む)又は昆虫細胞(例えば、sf9又はsf21)が挙げられる。植物を宿主とする場合、コケ植物、シダ植物及び種子植物等を含み、その種類は問わない。
【0045】
なお、本発明の形質転換体は、同一の遺伝情報を有するクローン体を包含する。例えば、宿主が大腸菌や酵母等の無性生殖を行う単細胞微生物であれば、形質転換体第1世代から分裂又は出芽等によって新たに生じた細胞は、本発明の形質転換体に含まれるものとする。また、形質転換される宿主が植物であれば、形質転換体第1世代から採取した植物体の一部を挿し木、接木若しくは取り木したもの又は形質転換体第1世代から無性生殖で得られる栄養繁殖器官(例えば、根茎、塊根、球茎、ランナー等)より新たに生じた新たな栄養体も本発明の形質転換体に含まれる。
【0046】
3−2.形質転換体の後代
本明細書において「形質転換体の後代」とは、本発明の変異型Myb関連遺伝子を導入した形質転換体第1世代の有性生殖を介した子孫であって、本発明の変異型Myb関連遺伝子を発現可能な状態で保持しているものを意味する。例えば、形質転換体が植物の場合には、その形質転換体の実生が該当する。本発明の変異型Myb関連遺伝子を発現可能な状態で継代する限りにおいて後代の世代は問わない。
【0047】
3−3.形質転換方法
本発明の変異型Myb関連遺伝子発現ベクターを宿主に導入する方法は、当該分野で公知の方法を使用することができる。
【0048】
宿主が細菌であれば、ヒートショック法、カルシウムイオンを用いる方法(例えば、リン酸カルシウム法)、エレクトロポレーション法等を用いればよい。これらの技術は、いずれも当該分野で公知であり、例えば、Sambrook, J. et. al., (1989) Molecular Cloning: A Laboratory Manual Second Ed., ColdSpring HarborLaboratory Press, Cold Spring Harbor, New Yorkをはじめとする様々な文献に記載されている。
【0049】
宿主が植物細胞であり、前記発現ベクターがプラスミドベクターである場合には、形質転換方法としてプロトプラスト法、パーティクルガン法又はアグロバクテリウム(Agrobacterium)法等を利用することができる。プロトプラスト法とは、セルラーゼ等の酵素的処理によって細胞壁を除去した植物細胞(プロトプラスト)を用いて、目的の遺伝子を植物細胞中に導入する方法であって、遺伝子導入の方法により、さらに、エレクトロポレーション法、マイクロインジェクション法又はポリエチレングリコール法等に細分することができる。エレクトロポレーション法は、プロトプラストと目的遺伝子の混合液に電気パルスを与えてプロトプラスト内に遺伝子を導入する方法である。また、マイクロインジェクション法は、微針を用いて顕微鏡下でプロトプラスト中に目的の遺伝子を直接導入する方法である。そして、ポリエチレングリコール法は、ポリエチレングリコールを作用させてプロトプラストに目的の遺伝子を導入する方法である。一方、パーティクルガン法とは、金又はタングステン等の微粒子に目的の遺伝子を付着させて、それを高圧ガスにより植物組織細胞内に打ち込み、目的の遺伝子を細胞内に導入する方法であって、宿主細胞のゲノムDNA中に目的の遺伝子が取り込まれた形質転換体を得ることができる。アグロバクテリウム法とは、形質転換因子としてアグロバクテリウム属の菌(例えば、アグロバクテリウム・ツメファシエンス(A. tumefaciens)、アグロバクテリウム・リゾゲネス(A. rhizogenes)等)及びそれに由来するTiプラスミドを用いる植物細胞の形質転換方法であって、目的の遺伝子を宿主植物細胞のゲノムDNA中に導入することができる。いずれの方法も当該分野においては、公知の方法であり、詳細については植物代謝工学ハンドブック(2002年、NTS社)又は新版モデル植物の実験プロトコール:遺伝学的手法からゲノム解析まで(2001年秀潤社)等を参照すればよい。
【0050】
また、宿主が植物細胞で、かつ前記発現ベクターがウイルスベクター(例えば、前述のCaMV、BGMV、TMV等)の場合には、そのウイルスベクターを植物細胞に感染させることによって、変異型Myb関連遺伝子を導入することができる。例えば、まず、植物ウイルスゲノムを大腸菌由来のベクター等のクローニングベクターに挿入して組換え体を調製する。次に、組換え体のウイルスゲノム中に変異型Myb関連遺伝子を挿入し、クローニングする。続いて、制限酵素によって組換え体から植物ウイルスゲノム領域を切り出し、得られたウイルスゲノムを目的の植物細胞に感染させる。以上により、目的の遺伝子を植物細胞内に導入できる。このようなウイルスベクターを用いた遺伝子導入方法の詳細については、Hohnらの方法(Molecular Biology of Plant Tumors(Academic Press、New York)1982、pp549)、米国特許第4,407,956号明細書等を参考にすればよい。
【0051】
宿主が植物細胞の場合、植物の種類については特に限定しない。好ましくは、配列番号7で示されるMyb-miRNA及びその標的遺伝子である配列番号1で示されるMyb-miRNA結合部位をコード領域内に有するMyb遺伝子を内在する植物である。例えば、ユーカリ属植物又はタバコ属植物由来の植物細胞が挙げられる。ユーカリ属植物としては、例えば、ユーカリプタス・グランディス (E. grandis)、ユーカリプタス・ユーロフィラ(E. urophylla)、ユーカリプタス・カマルドレンシス(E. camaludulensis)、ユーカリプタス・グロブラス(E. globulus)、ユーカリプタス・ニテンス(E. nitens)、ユーカリプタス・ガンニー(E. gunni)、ユーカリプタス・ラジアータ(E.radiata)、ユーカリプタス・アンプリフォリア(E. amplifolia)、ユーカリプタス・アーチェリ(E. archeri)、ユーカリプタス・ブリジェシアーナ(E. bridgesiana)、ユーカリプタス・バクステリ(E. baxteri)、ユーカリプタス・ビコスタータ(E. bicostata)、ユーカリプタス・ブラッケリー(E. blakelyi)、ユーカリプタス・ボツリオイデス(E. botryoides)等が挙げられる。タバコ属植物としては、例えば、ニコチアナ・タバカム(N. tabacum)、ニコチアナ・ルスチカ(N. rustica)等が挙げられる。この他、シロイヌナズナ(A. thaliana)、ポプラ属(Populus)植物、例えば、ポプルス・トリコカルパ(P. trichocarpa)、ポプルス・アルバ(P. alba)、ポプルス・ニグラ(P. nigra)、ポプルス・カンジカンス(P. candicans)、ポプルス・トレムラ(P.tremula)、ポプルス・シーボルディ(P. sieboldii)等が挙げられる。
【0052】
3−4.後代取得法
本発明の形質転換体から後代を得る方法は、その形質転換体の宿主である生物種において後代を得るために用いられる通常の方法で行えばよい。例えば、形質転換体の宿主が大腸菌や酵母であれば、適当な公知培地で培養することによって得ることができる。例えば、Sambrook, J. et. al., (1989) Molecular Cloning: A Laboratory Manual Second Ed., ColdSpring HarborLaboratory Press, Cold Spring Harbor, New Yorkを参照すればよい。また、形質転換体の宿主が植物であれば、一般的には土耕栽培(播種)により、実生を得ることは容易である。本発明の形質転換植物体から種子を得る方法の一例として、形質転換植物体を適当な培地において発根させ、その発根体を水分含有の土を入れたポットに移植する。適当な栽培条件下で生育させ、当該分野で通常の方法を用いて種子を形成させる。得られた種子を、例えば、水分含有の適当な土に播種し、適切な栽培条件下で生育させることにより本発明の形質転換体の後代を得ることができる。
【0053】
4.植物生長促進方法
本発明の他の態様は、前記変異型Myb関連遺伝子発現ベクターを植物体に導入して、該植物の生長を促進する方法である。
【0054】
4−1.対象植物
本発明の原理は、本発明の変異型Myb関連遺伝子を目的とする植物に導入して、その植物細胞内で発現させることによって、内在性のMyb-miRNAによる発現抑制制御を受けることなく、野生型アミノ酸配列を有する変異型Myb関連遺伝子タンパク質を付与することに基づく。したがって、本方法の対象植物は、本発明に記載の配列番号1で示す塩基配列を含み、植物の成長を促進するMyb関連遺伝子及び本発明のMyb-miRNAをゲノム中に一以上有する植物種であれば、特に限定しない。例えば、前記形質転換体の「3−2.形質転換方法」の項で説明したユーカリ属各種又はタバコ属各種又はポプラ属各種が挙げられる。
【0055】
4−2.方法
本発明は、前記「2.変異型Myb関連遺伝子を含む発現ベクター」の項で説明した変異型Myb関連遺伝子発現ベクターを目的とする植物に導入し、前記「3.形質転換体」の項で説明した形質転換植物を得ることによって達成される。変異型Myb関連遺伝子発現ベクターを目的とする植物に導入する方法については、前記「3−2.形質転換方法」に記載の方法に準じて行えばよい。このとき、形質転換体が植物であるため、使用する変異型Myb関連遺伝子発現ベクターは、植物細胞内で発現可能なベクターを用いるようにする。例えば、プラスミドベクターであれば、pBI系又はpRI系のバイナリーベクター等、またウイルスであれば、カリフラワーモザイクウイルス(CaMV)、インゲンマメモザイクウイルス(BGMV)、タバコモザイクウイルス(TMV)等である。また、これらのベクターを導入する宿主は、当然ながら目的とする植物となる。
【0056】
変異型Myb関連遺伝子発現ベクターで目的とする植物を形質転換する方法は、使用する発現ベクターの種類によって異なる。発現ベクターがプラスミドベクターである場合には、前述したプロトプラスト法、パーティクルガン法、又はアグロバクテリウム法等を用いることができる。これらの方法は、いずれも当該分野においては、公知の方法であり、詳細については前記例示の文献を参照すればよい。また、発現ベクターがウイルスベクターである場合にHohnらの方法(Molecular Biology of Plant Tumors(Academic Press、New York)1982、pp549)、米国特許第4,407,956号等を参考にすることができる。
【0057】
前記形質転換方法は、通常、カルス又は組織片を用いた植物組織培養を介するインビトロ法が用いられる。この場合、形質転換した植物細胞から植物体を再生する必要がある。本方法についても当該分野では公知であり、上述の植物代謝工学ハンドブック(2002年、NTS社)又は新版モデル植物の実験プロトコル:遺伝学的手法からゲノム解析まで(2001年秀潤社)等を参照することができる。また、目的とする植物がユーカリ属植物である場合には、形質転換したユーカリ属の植物細胞から植物体を再生する方法は、土肥らの方法(特開平11-127025号公報)を参照することができる。あるいは、前記形質転換方法は、カルスや細胞培養のステップを経ることなく、目的の植物個体の細胞に直接、本発明のMyb関連遺伝子を導入する形質転換法、いわゆるin planta法を用いてもよい。
【0058】
5.Myb-miRNA
本発明の別の態様は、植物細胞内への導入によって該植物の生長を抑制できるMyb-miRNAである。
本明細書で「Myb-miRNA」とは、配列番号7で示される21塩基からなる塩基配列を含み、本明細書に記載のMyb関連遺伝子の転写産物を標的とするmiRNAをいう。
【0059】
「miRNA」(マイクロRNA:micro RNA)とは、生物種間で進化的に保存された19〜25塩基からなる内在性ノン・コーディングRNA(non-cording RNA:タンパク質をコードしないRNA)であって、それに相補的な塩基配列を有する標的遺伝子の発現を転写後翻訳前の段階で制御するsmall RNAである。miRNAは、通常、数百〜数千塩基からなるpri-miRNA(primary miRNA)前駆体としてゲノムから転写され、核内でDrosha及びPasha等によって60〜100塩基程度のヘアピン型pre-miRNA(precursor-miRNA)へと変換される。続いて、細胞質に輸送され、Dicerによってヘアピンのループ部分が切断されて、19〜25塩基の二本鎖miRNA(miRNA duplex)となる。その後、標的遺伝子のmRNAに相補的な配列を有する側鎖が成熟一本鎖miRNAとしてRISC/miRNPに取り込まれ、この成熟miRNAの塩基配列との結合を介して標的mRNAが認識され、RISC及びmiRNPによって切断及び翻訳阻害されることで標的遺伝子の発現が抑制される(David P. Bartel, Cell, Vol. 116, 281-297, January 23, 2004,)。
【0060】
「配列番号7で示される塩基配列」部分は、成熟一本鎖Myb-miRNA部分に相当する。一本鎖Myb-miRNAは、自己と相補的な塩基配列である配列番号1で示されるMyb-miRNA結合部位(完全相補ではなく、1又は2つの不対合を有していてもよい)を認識することで、それを含むMyb関連遺伝子の発現を抑制的に制御している(実施例2参照)。配列番号7において、R、Y及びNで示される塩基は、それぞれ、A又はG、C又はU及びA、U、G又はCのいずれであってもよいことを示す。具体的な例としては、配列番号8で示される塩基配列を含むユーカリプタス・グランディス(E. grandis)由来のMyb-miRNA(以下、便宜的に「EgMyb-miRNA」とする)が挙げられる。なお、miRNAによる遺伝子の発現抑制機構については、当該分野で公知である。例えば、Eulalio et al., 2008, Cell, 132:9-14を参照されたい。
【0061】
6.Myb-miRNAをコードするDNAを含む発現ベクター
本発明の他の態様は、前記Myb-miRNAをコードするDNAを含む発現ベクター(以下、「Myb-miRNA発現ベクター」とする)を導入した形質転換体である。
【0062】
6−1.Myb-miRNAをコードするDNA
本明細書で「Myb-miRNAをコードするDNA」とは、転写後に本発明のMyb-miRNAを機能可能な状態でコードするDNAをいう。「機能可能な状態」とは、本発明のMyb-miRNAが転写された後、Myb-miRNAとしての機能を発揮し得る状態をいう。一般に、成熟一本鎖miRNA部分に相当する配列番号7で示される塩基配列のみをコードするDNAでは、本発明のMyb-miRNAとしての機能は発揮し得ない。転写後に二本鎖RNAが形成されないためである。したがって、本発明のMyb-miRNAを転写後に機能可能な状態とするためには、DNAが配列番号7で示す塩基配列を一方の鎖に有する二本鎖RNAが形成されるようにコードしていなければならない。
【0063】
転写後に本発明のMyb-miRNAを機能可能な状態でコードするDNAとしては、例えば、Myb-shRNA(short hairpin RNA)をコードするDNAが挙げられる。「shRNA」とは、標的とする遺伝子の一部(例えば、本発明のMyb-miRNA結合部位)に対応する塩基配列とそのアンチセンス鎖(例えば、本発明のMyb-miRNA)が短いスペーサ配列で連結された一本鎖RNAである。shRNAは、一分子内でセンス領域とアンチセンス領域が互いに塩基対合し、同時に前記スペーサ配列がループ構造をとることによって、分子全体としてヘアピン型のステム−ループ構造を有する。このようなMyb-shRNAをコードするDNAの設計は、例えば、配列番号7で示される塩基配列をコードするDNA断片、それに相補的な塩基配列を有するDNA断片及びこれらの一方のDNA断片の3’末端と他方のDNA断片の5’末端とを連結する適当な塩基配列からなるスペーサ配列から構成される。スペーサ配列は、通常3〜24塩基、好ましくは、4〜15塩基あればよい。shRNAをコードするDNAの調製は、多くのライフサイエンス系メーカー(例えば、タカラバイオ社、インビトロジェン社等)が受託製造サービスを行っており、それらを利用することもできる。この他、Myb-miRNAをコードするDNAは、Myb-miRNAの前駆体であるpri-Myb-miRNA若しくはpri-Myb-miRNAをコードするDNAであってもよい。
【0064】
他の例として、配列番号7で示す塩基配列を有するヌクレオチドとそれに相補する塩基配列を有するヌクレオチドをそれぞれコードするDNAを独立に発現可能なようにしたものであってもよい。それぞれのヌクレオチドは、シス(例えば、同一発現ベクター内)又はトランス(例えば、異なる発現ベクター内)のいずれで発現されても構わないが、両ヌクレオチドの発現が同程度になるように、例えば、同一のプロモータを使用することが好ましい。この場合、それぞれのDNAを目的の細胞内に導入して発現させることによって、細胞内で2本鎖RNAが形成され、結果として、本発明のMyb-miRNAを機能可能な状態にすることができる。
【0065】
6−2.Myb-miRNA発現ベクター
本発明において、「Myb-miRNA発現ベクター」とは、本発明のMyb-miRNAをコードするDNAを適当な発現ベクター内に導入したものである。適切な宿主細胞に導入することで、適当な条件の下、挿入したMyb-miRNAを発現させることが可能である。発現ベクターの種類は特に限定されない。導入する宿主、例えば、大腸菌、酵母、植物細胞(植物体、植物器官及び植物組織を含む)、昆虫細胞又は動物細胞によって適宜選択すればよい。
【0066】
本発明の発現ベクターにおける骨格部分、すなわち本発明のMyb-miRNA以外の主要な構成部分(プロモータ、エンハンサ、若しくはターミネータ等の調節領域、又は選抜マーカー遺伝子等を含む)については、上記「2−1.変異型Myb関連遺伝子発現ベクター」の項で説明した発現ベクターを同様に利用することができる。したがって、説明の重複する部分については省略をする。ただし、本発明の発現ベクターを後述する「植物の生長を抑制する方法」で使用する場合、用いるプロモータは植物細胞内でMyb-miRNAを過剰に発現できるプロモータであることが好ましい。効率的に生長を抑制させるためである。例えば、細胞分裂が旺盛な組織、あるいは細胞伸長に掛かる木部肥厚組織等において特異的に発現することが可能なプロモータ等が好ましい。
【0067】
6−2.Myb-miRNA発現ベクターの調製
前記Myb-miRNAをコードするDNAを発現ベクターの所定の部分に挿入し、Myb-miRNA発現ベクターを調製する方法は、当該分野で公知の方法、例えば、Sambrook, J. et. al., (1989) Molecular Cloning: a Laboratory Manual Second Ed., Cold SpringHarbor Laboratory Press, Cold Spring Harbor, New Yorkに記載の方法を用いることができる。
【0068】
7.形質転換体
本発明の他の態様は、前記Myb-miRNA発現ベクターを導入した形質転換体に関する。
【0069】
7−1.形質転換体
本明細書において「形質転換体」とは、前記Myb-miRNA発現ベクターによって形質転換された宿主及びそのクローン体をいう。使用する宿主は、使用したベクターに適する宿主であって、本発明のMyb-miRNAを発現できるものであれば特に限定されない。例えば、上記「3−1.形質転換体」の項で記載した宿主が挙げられる。好ましくは、微生物又は植物細胞(ユーカリ属植物又はタバコ属植物を含む)である。
【0070】
7−2.形質転換体の後代
本明細書において「形質転換体の後代」とは、本発明のMyb-miRNA発現ベクターを導入した形質転換体第1世代の有性生殖を介した子孫であって、本発明のMyb-miRNAをコードするDNAを発現可能な状態で保持するものを意味する。例えば、形質転換体が種子植物の場合には、その形質転換体の実生が該当する。本発明のMyb-miRNAをコードするDNAを発現可能な状態で継代する限りにおいて、後代の世代は問わない。
【0071】
7−3.形質転換方法
本発明のMyb-miRNA発現ベクターを宿主内に導入する方法は、前記「3−3.形質転換方法」の項に記載した方法に準じて行うことができる。
【0072】
7−4.後代取得法
本発明の形質転換体から後代を得る方法は、前記「3−4.後代取得法」の項に記載した方法に準じて行うことができる。
【0073】
8.植物の生長を抑制する方法
本発明の他の態様は、前記変異型Myb関連遺伝子又は前記発現ベクターを植物体に導入して、該植物の生長を促進する方法である。
【0074】
本発明は、前記「4.植物の生長を促進する方法」とは逆の原理に基づく。すなわち、本発明のMyb-miRNAをコードするDNAを目的とする植物に導入してその植物体内でMyb-miRNAを過剰発現させることによって、内在性の標的Myb関連遺伝子の発現を抑制させることに基づく。したがって、本方法の対象植物は、本発明に記載の配列番号1で示す塩基配列を含み、植物の成長を促進するMyb関連遺伝子をゲノム中に一以上有する植物種であれば、特に限定しない。例えば、前記形質転換体の「3−2.形質転換方法」の項で説明したユーカリ属各種又はタバコ属各種又はポプラ属各種等が挙げられる。
【0075】
本発明の方法の基本的操作は、前記「4.植物の生長を促進する方法」で説明した方法と同一であるため、前述の方法に準じて行えばよい。
【実施例】
【0076】
<実施例1>
ユーカリMyb関連遺伝子(EcMyb遺伝子)の取得
(1)総RNAの調製
王子製紙株式会社森林資源研究所内において成育中の5年生ユーカリプタス・カマルドレンシス(E. camaldulensis)の樹幹部、形成層組織を採取し、液体窒素中で粉砕した後、Concert Plant RNA Regent (インビトロジェン社)を用いて総RNAを抽出した。抽出手順については、添付のプロトコルに従った。その結果、1mgの総RNAを得た。
【0077】
(2)mRNAの精製
PolyA Tract mRNA Isolation System (プロメガ社)を用いて前記総RNAからmRNAの精製を行った。抽出手順については、添付のプロトコルに従った。その結果、1mgの総RNAから6μgのmRNAを得た。
【0078】
(3)cDNAライブラリーの作製
E. camaldulensisのcDNAライブラリーの作製は、SMART cDNA Library Construction Kit (タカラバイオ社)を用いて、添付のプロトコルに従って行った。具体的には、前記精製したmRNAのうち1μgにoligo dTプライマーをアニールさせた後、逆転写酵素によってcDNAを合成した。得られたcDNAを制限酵素Sfi Iで消化した後、カラムによるサイズ分画を行い、λTriplEx2ベクターと連結した。続いて、Gigapack Gold III (Stratagene社)を用いてλ-ファージパッケージング反応を行い、5×105の独立クローンからなるcDNAライブラリーを作製した。
【0079】
(4)EcMyb遺伝子の単離
前記独立クローンから約2万個のcDNAクローンを任意に抽出し、各クローンの塩基配列をCEQ8000遺伝子解析システム(ベックマン・コールター社製)により決定した。得られた塩基配列をBlast検索(http://blast.ncbi.nlm.nih.gov/Blast.cgi)で日本DNAデータバンク(DDBJ)の遺伝子データベースと照合した。その結果、シロイヌナズナ、イネ、ポプラ等において、分化・形態形成を制御していると考えられているWEREWOLF遺伝子(R2R3タイプMyb遺伝子)と高い相同性が認められるcDNAクローンを見出した。このcDNAクローンの配列を決定したところ、5’並びに3’非翻訳領域を含むアミノ酸コード領域全体を含んでいることが明らかとなった。この新たに単離されたE. camaldulensisのMyb関連遺伝子(EcMyb遺伝子:配列番号3)は、121アミノ酸からなるMyb関連タンパク質(EcMybタンパク質)をコードする(配列番号9)。EcMybタンパク質は、N末端側に2つのMybドメイン(R2、R3)を含んでおり、Myb66タンパク質と同様にR2R3タイプのMybファミリーに属する転写制御因子であることが判明した(図1)。
【0080】
<実施例2>
Myb遺伝子の発現抑制に関わるmiRNA(Myb-miRNA)の単離とその活性の検証
(1)ユーカリmiRNAの精製と解析
王子製紙(株)森林資源研究所内において成育中の5年生ユーカリプタス・グランディス(E. grandis)の花芽を採取し、液体窒素中で粉砕した後、Concert Plant RNA Regent (インビトロジェン社)を用いて総RNAを抽出した。この総RNAを15%ポリアクリルアミドゲルで常法により分離し、泳動マーカーのサイズから換算した約18〜26塩基長に相当するRNA領域のバンドをゲルから切り出し、FrashPAGE Fractinator(アンビオン社製)を用いて、ゲルからRNAを回収した。次に、MPSS法(タカラバイオ社, Bio View, 2005,No.50:17-19)により、約100万個の低分子RNAの塩基配列を決定した。解析の結果、これらの配列は、約13万種に分類できることが判明した。続いて、E. grandisのゲノム配列と相同性検索を行い、ゲノム配列に一致しない配列を除去し、さらに既知のRNA配列と相同性のある配列を除去した。その結果、約3万種の低分子RNAをmiRNAの候補として分離した。続いて、これらの配列の前後を含む約500bpの塩基配列情報をユーカリのゲノム配列より取得し、取得した領域内における各候補の2次構造予測を行い、miRNAに特徴的な構造、すなわち、ヘアピン構造を有するRNA分子を208個同定した。これらをユーカリmiRNA群とした。
【0081】
(2)EcMyb遺伝子の発現抑制に関わるmiRNAの単離
前記ユーカリmiRNA群から、実施例1で単離したEcMyb1遺伝子を標的分子とし得るmiRNAを探索した。その結果、配列番号8で示される塩基配列を含む一のユーカリmiRNA(EgMyb-miRNA)が得られた。
【0082】
(3)Myb-miRNAによるMyb関連遺伝子の発現抑制の検証
実施例1で単離したEcMyb遺伝子が、E.camaldulensisに内在すると予想されるEgMyb-miRNAオルソログEcMyb-miRNAによって発現制御を受けることを確認した。
【0083】
まず、E.camaldulensisの葉から総RNAを抽出し、オリゴdTプライマーを用いてcDNAを合成した。次に、合成したcDNAの5'末端にFirst Choice RLM-RACE Kit(Ambion社)を用いてアダプターを連結した(図3a)。基本的な方法については、添付のプロトコルに従った。続いて、アダプターを付加したcDNAを鋳型にして図1及び図2に示すEcMybに特異的なプライマー(アウタープライマーA及びインナープライマーB:それぞれ配列番号10及び11)と前記アダプターに特異的なプライマー(アウタープライマーC及びインナープライマーD:それぞれ配列番号 12及び13)を用いてPCR反応((変成反応(94℃、30秒)、アニーリング反応(55℃、30秒)、伸長反応(68℃、90秒)を1サイクルとして、35サイクル実施)を独立に10回(n=10)行った。続いて、得られたPCR産物を1.0%アガロースゲルで電気泳動して確認すると共に(図3b)、インビトロジェン社製のクローニングキット(TAクローニングキット)を用いて獲得した後、その塩基配列を決定した。その結果、EcMyb mRNAは、いずれの反応サンプルにおいて配列番号2に相当するMyb-miRNA結合部位の13番目と14番目のU間で切断されていることが明らかとなった(図3c)。この結果からEcMyb mRNAは、EcMyb-miRNAによる発現抑制を受けていることが立証された。
【0084】
<実施例3>
変異型EcMyb関連遺伝子発現ベクターの調製
(1)変異型EcMyb関連遺伝子
EcMyb mRNAに含まれる配列番号2で示される塩基配列からなる部位(Myb-miRNA結合部位)にMyb-miRNAが結合できないようにサイレント変異を導入した。ここで、EcMyb-miRNAは、未同定であることから、本実施例ではEcMyb-miRNAと相同性を有することが予測されるEgMyb-miRNAと配列番号2で示される塩基配列との間で塩基対合できないような9箇所のサイレント変異を導入した(図4)を作製した。実際の作製においては、タカラバイオ社へ配列番号14に示すDNA合成を委託した。
【0085】
(2)変異型EcMyb関連遺伝子発現ベクターの調製
得られた変異型EcMyb関連遺伝子mEcMyb(配列番号14)及び実施例1で調製した野生型のEcMyb遺伝子のそれぞれをGatewayTMエントリークローンpENTR/D/TOPOベクター(Invitrogen社)内にクローニングした後、LR反応によってバイナリーベクター(pGWB2;Nakagawaら,2007, J Biosci Bioeng, 34-41)に導入した。上記Gatewayシステム(Invitrogen社)を利用した一連の操作手順については、添付の説明書に従った。以上によって、目的とする発現ベクターpGWB2-mEcMyb及びpGWB2-EcMybを調製した。これらの発現ベクターをそれぞれアグロバクテリウムLBA4404株にバイオラッド社製ジーンパルサーを用いて、添付の実施方法に準拠して導入した。植物への形質転換までの間は、グリセロール保存の状態で維持した。
【0086】
<実施例4>
内在性Myb-miRNAを介した発現抑制の回避による植物生長促進
(1)形質転換タバコの作製
mEcMybの内在性Myb-miRNAによるRNAiの回避能力及びmEcMybを導入した植物の生長促進効果を確認するため、本実施例の(1)で調製したpGWB2-EcMyb及びpGWB2-mEcMybを保持したアグロバクテリウムを常法(参照:松本省吾・町田泰則 (1990) 植物形質転換法. 現代化学 6月:25〜29)に従ってタバコ葉(N. tabacum BY4)にそれぞれ感染させ、ハイグロマイシン耐性を指標とした選抜を行い、形質転換タバコを作製した。
【0087】
ここで、ユーカリに代えてタバコを用いた理由は、主にユーカリよりも生長速度が速いこと、イネ科以外の草本性有用植物の中では形質転換をはじめとする遺伝子操作技術が既に確立していること、及び木本性植物の形質転換が一般に草本性植物のそれと比較して困難なことによる。なお、タバコにおいてEcMybのオルソログとなる具体的な報告は無いものの、ゲノム解読が完了した又は進行中の他の植物種において配列比較を行った結果、EcMybオルソログ候補遺伝子は全ての植物に普遍的に存在すると類推された。したがって、タバコにおいてもEcMybのオルソログ遺伝子の存在が予測されたことから、本実施例で用いた。
【0088】
(2)mEcMyb遺伝子の発現確認
本態様の(1)で得られたpGWB2-EcMyb又はpGWB2-mEcMybをそれぞれ保持する形質転換タバコにおいてEcMyb又はmEcMybの発現を調べるため、各形質転換タバコの葉から総RNAを抽出し、常法に従ってRT-PCRを行った。PCRで用いたFw及びRvプライマーは、EcMybの158〜180番目のセンス鎖配列に相当する塩基配列(配列番号15)及びEcMybの385〜404番目のアンチセンス鎖配列に相当する塩基配列(配列番号16)をそれぞれ有する。また、Myb-miRNA結合部位は、両プライマー間(310〜331番目のアミノ酸に相当)に存在する。PCRサイクルは、変成反応(94℃、30秒)、アニーリング反応(55℃、30秒)、伸長反応(68℃、90秒)を1サイクルとして、35サイクル実施した。RT-PCRは、それぞれ独立に3回(n=3)行った。RT-PCR後、各サンプル2μlを1.0%のアガロースゲルで電気泳動した。
【0089】
結果を図5に示す。Wtは非形質転換の野生株タバコ(BY4)を、WtEcは野生型EcMyb形質転換体を、そしてmEcは変異型EcMyb形質転換体を示す。変異型EcMyb形質転換体中には、野生型EcMyb形質転換体と比較してMyb-miRNA結合部位が保持されたmRNAが多量に存在することが明らかとなった。これは、外来性のmEcMyb mRNAが、タバコの内在性Myb-miRNAによる切断作用、すなわちRNAiを受けることなく、タバコ細胞内において安定して機能し得ることを示している。一方、EcMyb mRNAは、発現ベクターからの過剰発現により内在性のNtMyb mRNAよりも多く検出されたが、mEcMyb mRNAの量と比較すると少なかった。これは、一部がタバコの内在性Myb-miRNAによって切断されたためと考えられる。以上の実験から、本発明の変異型Myb関連遺伝子は、その転写産物を標的とするMyb-miRNAが内在する植物において、そのMyb-miRNAによるRNAiを回避できることが明らかとなった。
【0090】
(3)形質転換タバコの生育調査
本態様の(1)で得られたpGWB2-mEcMybを導入した形質転換体と非形質転換体の野生株(BY4)をその後6ヶ月間通常方法にて育成し、それぞれの生長性を比較した。
【0091】
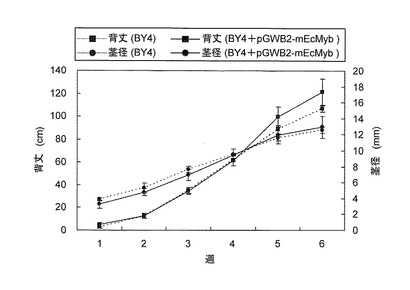
結果を図6及び図7に示す。図6における実線は形質転換体タバコ(BY4+pGWB2-mEcMyb)を、破線は野生型タバコ(BY4)を示し、四角プロットは背丈(cm)(左縦軸)を及び丸プロットは茎径(mm)(右縦軸)をそれぞれ示す。図7におけるaは野生型タバコ(BY4)を、b及びcは形質転換体タバコ(BY4+pGWB2-mEcMyb)をそれぞれ示す。
【0092】
pGWB2-mEcMyb遺伝子の導入により、非導入の野生株と比較して6ヶ月後には背丈及び茎径共に有意な生長差が見られた(図6、図7)。この結果から本発明の変異型Myb関連遺伝子を用いれば、植物が本来有する当該Myb関連遺伝子に対するRNAiを人為的に阻止できることが判明した。特に、本発明の変異型遺伝子を導入した植物では、その生長が通常よりも促進されること、また、本発明の変異型Myb関連遺伝子が植物種を超えて作用し得ることも明らかとなった。したがって、本発明の変異型Myb関連遺伝子によって事業植林用の樹木、例えば、ユーカリ属を形質転換すれば、木質バイオマスとしての活用がより有効に実施できる。
【技術分野】
【0001】
本発明は、植物の生長を促進することのできる変異型Myb関連遺伝子及びそれを用いた植物生長促進方法並びに植物の生長を抑制することのできる前記Myb関連遺伝子を標的とするmiRNA及びそれを用いた植物生長抑制方法に関する。
【背景技術】
【0002】
Myb遺伝子は、酵母から動物及び植物に至るまで生物種を越えて広く存在する転写制御因子遺伝子である。本遺伝子は、当初、癌遺伝子c−Mybとしてトリ骨髄芽球症ウイルス(AMV)及びE26ウイルスから独立に単離された(非特許文献1)。しかし、その後の研究により、転写因子をコードすること(非特許文献2)、多数の関連遺伝子からなるファミリーを形成していること、脊椎動物のみならず、酵母、線虫、昆虫、植物に至るまで種を超えて広く存在することが明らかとなった(非特許文献3)。
【0003】
Myb遺伝子ファミリーに属する遺伝子は、いずれもMybドメインと呼ばれる約50塩基のへリックス‐ターン‐へリックス構造からなるDNA結合ドメインをコードすることを特徴としている。脊椎動物のMyb遺伝子産物は、a-Myb、b-Myb及びc-Mybの3つのメンバーが知られている。いずれの産物も3つのMybドメイン(R1、R2、R3)を有し、主に細胞増殖と分化に関わる転写制御因子として機能している(非特許文献4)。一方、植物には多数のMyb遺伝子が存在し、構造上の特徴から大きく3タイプに分類されている。すなわち、哺乳動物Myb遺伝子と同様に3つのMybドメインを有し、細胞周期の制御を介して細胞増殖を制御するR1R2R3タイプ(非特許文献5)、色素合成系、形態形成及び環境応答等の様々な生命現象を制御するR2R3タイプ(非特許文献6)、そして光による制御や形態形成及び病障害に拘るR1様又はR3様ドメインを1つもつタイプ(非特許文献7、8、9)である。
【0004】
ところで、植物では今世紀に入り多数の遺伝子情報が蓄積され、光合成、一次代謝及び二次代謝をはじめとする基本的生理機能に関与する遺伝子の転写制御に関わる因子とその機能の知見が急速に増大した。そして、次なる段階として植物に新たな機能を獲得させるために、遺伝子発現を人為的に制御するための様々な分子遺伝学的手法が開発されている。このような方法の一つとして、転写制御因子遺伝子のプロモータ領域を置換・改変したり、対象とする植物に変異型転写制御因子遺伝子を導入することによって所望の機能をその植物に付与する方法が知られている。しかし、このような方法は、たとえその目的を達成できたとしても、望ましくない又は有害な他の形質を副作用的に同時に誘導してしまう等の予想外の弊害を伴うという問題があった。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Roussel M.,1979, Nature,218:452-455
【非特許文献2】Biedenkapp H., et al., 1988, Nature, 335:835-837
【非特許文献3】岩淵雅樹, 篠崎一雄、植物ゲノム機能のダイナミズム: 転写因子による発現制御、シュプリンガー・ジャパン株式会社, 2001
【非特許文献4】Weston K., 1999,.Curr. Opin. Genet. Dev. 8, 76-81.
【非特許文献5】Ito M., 2005, J. Plant Res., 118(1):61-69
【非特許文献6】Jin H. and Martin C., 1999, Plant Mol. Biol., 41(5):577-588
【非特許文献7】Feldbrugge et al., 1997, Plant J. 11, 1079-1093.
【非特許文献8】Kirik V. and Baumlein H., 1996, Gene 183, 109-113.
【非特許文献9】Wang Z.Y., et al., 1997, Plant Cell 9, 491-507.
【発明の概要】
【発明が解決しようとする課題】
【0006】
変異型転写制御因子遺伝子の導入によって、副作用を生じさせることなく目的の形質のみを対象とする植物に獲得させる方法を開発することを課題とする。
【課題を解決するための手段】
【0007】
一般に転写制御因子は、様々な遺伝子の発現を統括的に制御する場合が多く、そのような転写制御因子への変異の導入は、結局のところ、その因子によって制御される他の多くの遺伝子の発現に少なからず影響を及ぼすことになる。それ故、変異型転写制御因子遺伝子の導入によって目的の形質のみを付与させることは、通常容易なことではない。
【0008】
しかし、近年、遺伝子の発現を翻訳段階で制御するsmall RNA(低分子RNA)、例えば、miRNA(micro RNA)やsiRNA (short interference RNA)等の存在及びRNAサイレンシング機構が明らかとなった(Lee R.C., et al., 1993, Cell, 75: 843-854、Baulcombe D, 2002, Science, 297:2002-2003)。このような機構の発見により、遺伝子発現は、転写調節によってのみ制御されているのではなく、転写段階と翻訳段階によって独立に制御され、それぞれの協調の結果として、もたらされることが判明した(Jackson R. J. and Standart N., Sci. STKE 2007,(367) re1)。
【0009】
そこで、本発明者らは、当該転写後制御機構を利用して前記課題を解決するための新規方法を開発すべく鋭意研究を重ねた。その結果、配列番号1で示される塩基配列をコード領域内に含み、かつ植物の生長に関与するMyb関連タンパク質をコードする遺伝子の発現を制御することのできるmiRNAを見出した。本発明は、当該知見に基づいて完成されたものであり、すなわち以下を提供する。
(1)植物細胞内への導入によって該植物の生長を促進できる変異型Myb関連遺伝子であって、配列番号1で示される塩基配列をコード領域内に含むMyb関連遺伝子において、前記塩基配列に結合するmiRNAを介した発現抑制が不能となるように前記塩基配列内にサイレント変異を導入した前記変異型Myb関連遺伝子。
(2)配列番号1で示される塩基配列が配列番号2で特定される、(1)に記載の変異型Myb関連遺伝子。
(3)前記サイレント変異の少なくとも一つが配列番号1における13番目のTをCに置換した変異である、前記(1)又は(2)に記載の変異型Myb関連遺伝子。
(4)少なくとも5個のサイレント変異を含む前記(1)〜(3)のいずれかに記載の変異型Myb関連遺伝子。
(5)前記Myb関連遺伝子が配列番号3で示される塩基配列又は該塩基酸配列と95%以上の同一性を有する塩基配列からなる、前記(1)〜(4)のいずれかに記載の変異型Myb関連遺伝子。
(6)前記(1)〜(5)のいずれかに記載の変異型Myb関連遺伝子を含む発現ベクター。
(7)前記(6)に記載の発現ベクターを宿主に導入した形質転換体又はその後代。
(8)宿主が微生物又は植物である、前記(7)に記載の形質転換体又はその後代。
(9)前記植物が、ユーカリプタス属(Eucalyptus;本願では以降、種名を除き、一般的略称である「ユーカリ」を使用する)植物又はタバコ属(Nicotiana)植物である、前記(8)に記載の形質転換体又はその後代。
(10)前記(6)に記載の発現ベクターを植物に導入して、該植物の生長を促進する方法。
(11)植物細胞内への導入によって該植物の生長を抑制できるmiRNAであって、配列番号7で示される塩基配列を含み、かつ(1)〜(5)のいずれかに記載のMyb関連遺伝子の転写産物を標的とするmiRNA。
(12)配列番号7で示される塩基配列が配列番号8で特定される、(11)に記載のmiRNA。
(13)前記(11)又は(12)に記載のmiRNAをコードするDNAを含む発現ベクター。
(14)前記(11)又は(12)に記載のmiRNAあるいは前記(13)に記載の発現ベクターを宿主に導入した形質転換体又はその後代。
(15)宿主が微生物又は植物である、(14)に記載の形質転換体又はその後代。
(16)前記植物が、ユーカリ属植物又はタバコ属植物である、(15)に記載の形質転換体又はその後代。
(17)前記(13)に記載の発現ベクターを植物に導入して、該植物の生長を抑制する方法。
【発明の効果】
【0010】
本発明の変異型Myb関連遺伝子及び本発明の植物の生長を促進する方法によれば、導入した植物に対して、他の副作用を発生させることなく、目的とする生長促進という形質のみを付与することができる。
【0011】
本発明のmiRNA及び本発明の植物の生長を抑制する方法によれば、内在性の植物生長抑制機構を特異的に増強することで、導入した植物の生長を抑制することができる。
【図面の簡単な説明】
【0012】
【図1】EcMybタンパク質のアミノ酸配列とR2-及びR3-Mybドメイン。
【図2】EcMyb関連遺伝子(EcMyb gene)の塩基配列及びEcMybタンパク質(EcMyb prt)のアミノ酸配列と前記遺伝子単離に使用したプライマーの位置、並びに配列番号2に相当する領域(Myb-miRNA結合部位)を示した図。
【図3】(a)Myb-miRNAによる切断部位検証の模式図、(b)Myb-miRNA結合部位とMyb-miRNAによる切断箇所、(c)Myb-miRNAによる切断を受けたEcMyb関連遺伝子転写産物を示すアガロースゲル電気泳動図。
【図4】配列番号2で示されるMyb-miRNA結合部位に導入した9箇所のサイレント変異。このうち1箇所(6番目のC)は、EcMybにおける内在性のサイレント変異。
【図5】形質転換タバコにおけるEcMyb又はmEcMybの発現。
【図6】形質転換タバコの生長性(1)。
【図7】形質転換タバコの生長性(2)。
【発明を実施するための形態】
【0013】
1.変異型Myb関連遺伝子
本発明の一の態様は、植物細胞内への導入によって該植物の生長を促進できる変異型Myb関連遺伝子である。
【0014】
1−1.Myb関連遺伝子
本明細書の「Myb関連遺伝子」とは、植物のMyb遺伝子ファミリーにおいて配列番号1で示される塩基配列をコード領域内に含み、かつ植物の生長を促進する機能を有するR2R3タイプの野生型Myb関連遺伝子、その天然変異体又はそれらの断片をいう。
【0015】
「野生型Myb関連遺伝子」とは、自然界に存在するMyb関連タンパク質をコードし、同種集団内の対立遺伝子において最も多く存在する塩基配列を有するMyb関連遺伝子をいう。本遺伝子がコードするMyb関連タンパク質は、そのタンパク質本来の機能を有する。例えば、転写制御因子としての機能である。
【0016】
「R2R3タイプ」のMyb関連遺伝子とは、前述のように、その構造中に2つのMybドメイン(R2及びR3)をコードするMyb関連遺伝子である。例えば、毛根形成を負に制御するWEREWOLF(WER)(Lee et al., 1999, Cell, 99: 473-483)、トライコーム形成に関与するGLABRA1(GL1)(Oppenheimer et al., 1991, Cell 67(3): 483-493)及び種皮におけるタンニンの蓄積に機能するTRANSPARENT TESTA 2 (tt2)(Nesi et al., 2001, Plant Cell, 13(9), 2099-2114)等がこのタイプに属する。
【0017】
「その天然変異体」とは、自然界に存在する野生型Myb関連遺伝子の変異体である。例えば、野生型Myb関連遺伝子の塩基配列において1個又は数個のヌクレオチドが欠失、置換又は付加されたもの、前記塩基配列とそれぞれ95%以上、好ましくは98%以上、より好ましくは99%以上の同一性を有するもの、又は野生型Myb関連遺伝子の部分塩基配列に相補的な塩基配列からなる核酸断片とストリンジェントな条件下でハイブリダイズするものをいう。いずれも、野生型Mybタンパク質と同様に、植物の成長を促進する活性を有する。ここで前記「同一性」とは、2つの塩基配列にギャップを導入して又は導入しないでアラインメントさせたときに、一方の塩基配列の全塩基数に対する他方の塩基配列の同一塩基数の割合(%)をいう。「数個のヌクレオチド」とは、2〜10個、例えば、2〜7個、2〜5個、2〜4個又は2〜3個のヌクレオチドをいう。また、「ストリンジェントな条件」とは、非特異的なハイブリッドが形成されない条件を意味する。通常は低ストリンジェント〜高ストリンジェントな条件が挙げられるが、高ストリンジェントな条件が好ましい。低ストリンジェントな条件とは、ハイブリダイゼーション後の洗浄において、例えば42℃、5×SSC、0.1% SDSで洗浄する条件であり、好ましくは50℃、5×SSC及び0.1% SDSで洗浄する条件である。高ストリンジェントな条件とは、ハイブリダイゼーション後の洗浄において、例えば65℃、0.1×SSC及び0.1% SDSで洗浄する条件である。天然変異体の具体例としては、SNP(一塩基多型)等の多型に基づく変異体、スプライス変異体、遺伝暗号の縮重に基づく変異体等が挙げられる。
【0018】
「それらの断片」とは、前記野生型Myb関連遺伝子又はその天然変異体のDNA断片であって、配列番号1で示される塩基配列をコード領域内に含み、かつその断片がコードするMyb関連タンパク質の一部が野生型Myb関連遺伝子のコードするMyb関連タンパク質と同程度の生物学的機能、すなわち転写制御因子としての活性を保持しているものをいう。前記機能を保持している限り、DNA断片の塩基長は特に限定しない。
【0019】
「配列番号1で示される塩基配列」は、前記Myb関連遺伝子を標的とするmiRNA(本明細書では、以降便宜的に「Myb-miRNA」と呼ぶ)の結合部位(Myb-miRNA結合部位)に相当する。すなわち、本発明に記載のMyb関連遺伝子は、このMyb-miRNA結合部位へのmiRNAの結合を介して発現抑制を受ける。配列番号1において、R、Y及びNで示される塩基は、それぞれ、A(アデニン)又はG(グアニン)、C(シトシン)又はT(チミン)及びA、T、G又はCのいずれであってもよいことを示す。なお、Myb-miRNAについては、「5.Myb-miRNA」の項で詳述するため、ここではその説明を省略する。
【0020】
「コード領域内」とは、Myb関連タンパク質をコードする塩基配列の領域内を意味する。したがって、「配列番号1で示される塩基配列をコード領域内に含む」とは、配列番号1で示されるMyb-miRNA結合部位をコード領域内に含んでいることを意味する。
【0021】
「植物の生長」とは、茎(幹を含む)及び根の伸長並びに茎径(樹径を含む)の増大をいう。具体的には植物の成長点及び/又は維管束形成層における細胞の増殖、分化及び組織形成を意味する。「植物の生長を促進できる機能を有する」とは、前記植物の成長点及び/又は維管束形成層における細胞の増殖促進(細胞壁形成の制御を含む)に直接的に又は間接的に関与する機能を有することをいう。細胞の増殖促進に直接的に関与する機能とは、例えば、細胞周期制御に関与する因子のように、その機能が細胞の増殖に直接作用することをいう。細胞の増殖促進に間接的に関与する機能とは、例えば、細胞増殖に直接作用する遺伝子等の発現を制御することによって、間接的に細胞の増殖に寄与することをいう。
【0022】
配列番号1で示される塩基配列をコード領域内に含むR2R3タイプMyb関連遺伝子、すなわち、本発明の変異型Myb関連遺伝子の基礎となるMyb関連遺伝子の具体例としては、本発明者らによって今回新たに単離された配列番号3で示される塩基配列からなるユーカリプタス・カマルドレンシス(Eucalyptus camaldulensis)の野生型Myb関連遺伝子(本明細書では、以降便宜的に「EcMyb遺伝子」とする)又はその天然変異体が挙げられる。EcMyb遺伝子は、配列番号1に相当する部位として、配列番号2で示される塩基配列を、310〜331番目(開始コドンのAを1番目とする。以下、本明細書において同様とする。)の領域に有し、配列番号9で示される全長221アミノ酸からなるMyb関連タンパク質をコードする。また、Myb関連遺伝子は、配列番号3で示されるEcMyb遺伝子の塩基配列と95%以上、好ましくは98%以上、より好ましくは99%以上の同一性を有する塩基配列で示される、又はEcMyb遺伝子の部分塩基配列に相補的な塩基配列からなるプローブとストリンジェントな条件下でハイブリダイズする、他の植物種のMyb関連遺伝子、好ましくはEcMybオルソログ、であってもよい。例えば、配列番号4で示され、EcMyb遺伝子の塩基配列と97%の同一性を有するユーカリプタス・ガンニー(E. gunii)のMyb関連遺伝子(本明細書では、以降便宜的に「EgMyb遺伝子」とする)、GenBank XM_002302608.1として登録され、配列番号5で示されるポプラ(Populus trichocarpa)由来のMyb関連遺伝子(本明細書では、以降便宜的に「PtMyb遺伝子」とする)も本発明のMyb関連遺伝子に該当する。
【0023】
本発明者らは、Myb-miRNA結合部位をコード領域内に含むMyb関連遺伝子が翻訳段階でMyb-miRNAを介したRNAサイレンシング機構によってその発現を抑制されることを見出した。なお、miRNAについては、後述の「5.Myb-miRNA」の項を参照されたい。
【0024】
1−2.変異型Myb関連遺伝子
「変異型Myb関連遺伝子」とは、前記Myb関連遺伝子において配列番号1で示される塩基配列内、すなわち、Myb-miRNA結合部位内にサイレント変異を導入した遺伝子である。
【0025】
本発明におけるMyb-miRNAを介した前記Myb関連遺伝子の発現抑制は、後述する実施例2の結果から、Myb-miRNAがMyb関連遺伝子の転写産物上のMyb-miRNA結合部位と結合した後、Myb-miRNAと複合体を形成するRISC(RNA-induced silencing complex)/miRNPの作用によって配列番号1で示す塩基配列の13番目と14番目のT間(ただし、実際の標的分子はRNAであるため、13番目と14番目のU(ウリジン)間となる)が切断されることによって達成されると考えられる。なお、miRNAを介した遺伝子発現抑制機構の詳細については、後述の「5.Myb-miRNA」の項を参照されたい。本発明は、この機構に基づき、Myb-miRNAによる発現抑制が不能となる一方で、Myb関連遺伝子がコードするタンパク質のアミノ酸配列は変わらない変異型Myb関連遺伝子を提供する。
【0026】
「サイレント変異」とは、遺伝子の塩基配列には変異を生じるが、その遺伝子がコードするタンパク質のアミノ酸配列には変異を生じない変異、すなわち、縮重(縮退)コドン内における塩基の変異をいう。本発明においてMyb-miRNA結合部位は、WNTHLSK(配列番号6)からなるアミノ酸をコードする。したがって、本発明は、このアミノ酸配列を維持する全てのサイレント変異を包含する。例えば、配列番号1が配列番号2の塩基配列で特定される場合であれば、C6T(6番目のCをTに置換の意)、T9A/G/C(9番目のTをA、G又はCに置換の意)、T13-G15A/T/C(13番目がTのときにG15A/T/Cの意)若しくはT13C-G15A(13番目のTをCに置換したときにG15Aの意)、A16-C18T若しくはA16T-G17C-C18A/T/G、及び/又はG21Aの変異を導入することができる。
【0027】
「Myb-miRNAによる発現抑制が不能となる」とは、変異型Myb関連遺伝子がサイレント変異の導入によってMyb-miRNA耐性能力を獲得し、Myb-miRNAを介したRNAサイレンシングによる発現抑制を受けなくなる状態をいう。例えば、Myb-miRNAが変異型Myb関連遺伝子の転写産物を標的分子として認識できなくなること又は認識できても変異型Myb関連遺伝子の転写産物を切断できなくなることが挙げられる。
【0028】
「変異型Myb関連遺伝子の転写産物を標的分子として認識できなくなる」とは、Myb-miRNAが変異型Myb関連遺伝子の標的転写産物上のMyb-miRNA結合部位に結合できなくなることをいう。この場合、本発明の変異型Myb関連遺伝子を導入する植物において、その植物に内在するMyb-miRNAと前記変異型Myb関連遺伝子の転写産物上のMyb-miRNA結合部位との間で安定した塩基対合が形成されないようにサイレント変異を導入すればよい。例えば、配列番号1で示される塩基配列に少なくとも5個、最大で9個のサイレント変異を導入することが好ましい。Myb-miRNA結合部が配列番号2で示される塩基配列で特定されるEcMyb遺伝子の場合であれば、上述したサイレント変異を5個以上有していればよい(図4)。
【0029】
「認識できても変異型Myb関連遺伝子の転写産物を切断できなくなる」とは、Myb-miRNAが変異型Myb関連遺伝子の転写産物上のMyb-miRNA結合部位には結合できるが、上述した機序によって当該部位が切断されない状態をいう。例えば、Myb-miRNA結合部位において、生物種間で高度に保存された塩基にサイレント変異を導入する場合、例えば、配列番号1で示されるMyb-miRNA結合部位においてMyb-miRNAによって切断される位置に隣接する13番目のTをCに置換するサイレント変異(T13C)を導入した場合、が挙げられる。
【0030】
本発明の変異型Myb関連遺伝子を目的とする植物体中に導入することによって、内在のMyb-miRNAを介した発現抑制を特異的に回避し、かつ野生型Myb関連遺伝子と同じアミノ酸配列からなるMyb関連タンパク質を安定的にその植物に付与することができる。その結果、その植物の生長を促進することが可能となる。
【0031】
1−3.変異型Myb関連遺伝子の調製
本発明の変異型Myb関連遺伝子は、当該分野で公知の方法を用いて調製することができる。
一例を挙げると、まず、Myb関連遺伝子のcDNAを単離するために、目的とする植物体からmRNAを抽出する。抽出後、精製したcDNAをライブラリー化する。その後、PCR法(例えば、Inverse-PCR法、アンカーPCR法・TAIL-PCR法)、又はハイブリダイゼーション法を実施することにより、前記cDNAライブラリーから目的のMyb関連遺伝子を単離することができる。前記PCR法に使用するプライマー及びハイブリダイゼーション法に使用するプローブは、例えば、本明細書に記載の配列番号3(EcMyb遺伝子)、配列番号4(EgMyb遺伝子)、若しくは配列番号6(PtMyb遺伝子)、又はインターネットを介して一般的に利用可能なデータベース、例えば、NCBI(http://www.ncbi.nlm.nih.gov/)、理研(http://www.psc.riken.jp/database/index.html)、かずさDNA研究所DNA配列解析情報データベース(http://www.kazusa.or.jp/j/resources/database.html)等から入手できる既知の細胞壁転写制御因子の遺伝子情報から得た塩基配列に基づいて調製すればよい。特に、Myb-miRNA結合部位は、植物種間で比較的高度に保存されているため、当該部位をプローブ又は一方のプライマーとして利用すれば、異なる種のcDNAライブラリーからも目的のMyb関連遺伝子を単離することが可能となる。cDNAライブラリーの作製及び目的の遺伝子を単離する具体的な方法は、当該分野で公知である。例えば、Sambrook, J. et. al., (1989) Molecular Cloning: a Laboratory Manual Second Ed., Cold SpringHarbor Laboratory Press, Cold Spring Harbor, New Yorkを参照すればよい。
【0032】
次に、得られたMyb関連遺伝子において、配列番号1で示される塩基配列領域に必要なサイレント変異を導入する。この変異導入方法も当該分野で公知の方法に基づいて行うことができる。例えば、Inverse PCRを用いた方法、市販の突然変異誘発キット(例えば、Stratagene社のQuickChange Site-Directed Mutagenesis Kit、Promega社のAltered Sites II in vitro Mutagenesis System等)を利用して導入する方法を利用できる。また、いくつかのライフサイエンス系メーカー(例えば、タカラバイオ社)では、遺伝子等に所望の変異を導入する受託サービスを行っているのでそれを利用してもよい。
【0033】
2.変異型Myb関連遺伝子を含む発現ベクター
本発明の他の態様は、前記変異型Myb関連遺伝子を含む発現ベクター(以下、「変異型Myb関連遺伝子発現ベクター」とする)に関する。
【0034】
2−1.変異型Myb関連遺伝子発現ベクター
本発明において、「変異型Myb関連遺伝子発現ベクター」とは、前記変異型Myb関連遺伝子を適当な発現ベクター内に導入したものである。本ベクターは、適切な宿主細胞内に導入することで、適当な条件の下、挿入した変異型Myb関連遺伝子を発現させることが可能である。
【0035】
本発明の発現ベクターにおける骨格部分、すなわち、本発明の変異型Myb関連遺伝子以外の主要な構成部分は、特に限定はしないが、プラスミド又はウイルスが好ましい。プラスミドであれば、例えば、pBI系、pPZP系、pSMA系、pUC系、pBR系、pBluescript系(stratagene社)、pTriEXTM系(TaKaRa社)、又はpBI系若しくはpRI系のバイナリーベクター等を、ウイルスであれば、カリフラワーモザイクウイルス(CaMV)、インゲンマメゴールデンモザイクウイルス(BGMV)、タバコモザイクウイルス(TMV)等を利用することができる。これらは、導入する宿主に応じて適宜選択すればよい。導入する宿主としては、例えば、大腸菌、酵母、植物細胞(植物体、植物器官及び植物組織を含む)、昆虫細胞又は動物細胞が挙げられる。これらは、目的に応じて当該分野で公知のものを適宜選択すればよい。
【0036】
本発明の変異型Myb関連遺伝子発現ベクターの骨格部分には、変異型Myb関連遺伝子の他に、例えば、プロモータ、エンハンサ、若しくはターミネータ等の調節領域、又は選抜マーカー遺伝子等の他の標識的領域を連結することができる。それぞれの種類は、特に限定されない。導入する宿主(例えば、細菌、酵母、植物細胞又は昆虫細胞)に応じて又は宿主内での目的(例えば、クローニング又は遺伝子発現)に応じて当該分野で公知のものを適宜選択すればよい。それぞれについて、以下に例示して説明する。
【0037】
大腸菌中で作動可能なプロモータとしては、例えば、lac、trp若しくはtacプロモータ又はファージ・ラムダ由来のPR若しくはPLプロモータ等が挙げられる。酵母で作動可能なプロモータとしては、酵母解糖系遺伝子由来のプロモータ、アルコールデヒドロゲナーゼ遺伝子プロモータ、TPI1プロモータ、ADH2-4cプロモータ等が挙げられる。植物細胞で作動可能なプロモータとしては、カリフラワーモザイクウイルス(CaMV)の35Sプロモータ、ノパリン合成酵素遺伝子のプロモータ(Pnos)、トウモロコシ由来ユビキチンプロモータ、イネ由来のアクチンプロモータ、タバコ由来PRタンパク質プロモータ等が挙げられる。昆虫細胞で作動可能なプロモータとしては、ポリヘドリンプロモータ、P10プロモータ、オートグラファ・カリホルニカ・ポリヘドロシス塩基性タンパクプロモータ、バキュロウイルス即時型初期遺伝子1プロモータ、バキュロウイルス39K遅延型初期遺伝子プロモータ等が挙げられる。形質転換体の特定部位において局所的に変異型Myb関連遺伝子を発現させたい場合には、部位特異的プロモータを用いればよい。例えば、根の生長を促進させたい場合には、特開2007−77677号に記載の根特異的発現を誘導するプロモータを用いることができる。
【0038】
エンハンサとしては、CaMV 35Sプロモータ内の上流側の配列を含むエンハンサ領域及びCMVエンハンサ等が挙げられる。
【0039】
ターミネータとしては、ノパリン合成酵素(NOS)遺伝子のターミネータ、オクトピン合成酵素(OCS)遺伝子のターミネータ、CaMV 35Sターミネータ、大腸菌リポポリプロテインlppの3’ターミネータ、trpオペロンターミネータ、amyBターミネータ、ADH1遺伝子のターミネータ等が挙げられる。
【0040】
選抜マーカー遺伝子としては、薬剤耐性遺伝子(例えば、テトラサイクリン耐性遺伝子、アンピシリン耐性遺伝子、カナマイシン耐性遺伝子、ハイグロマイシン耐性遺伝子、スペクチノマイシン耐性遺伝子、クロラムフェニコール耐性遺伝子、又はネオマイシン耐性遺伝子)、蛍光又は発光レポーター遺伝子(例えば、ルシフェラーゼ、β-ガラクトシダーゼ、β-グルクロニターゼ(GUS)、又はグリーンフルオレッセンスプロテイン(GFP))、ネオマイシンホスホトランスフェラーゼII(NPT II)、ジヒドロ葉酸還元酵素等の酵素遺伝子が挙げられる。
【0041】
2−2.変異型Myb関連遺伝子発現ベクターの調製
前記変異型Myb関連遺伝子を発現ベクターの所定の部位に挿入する方法は、当該分野で公知の方法、例えば、Sambrook, J. et. al., (1989) Molecular Cloning: a Laboratory Manual Second Ed., Cold SpringHarbor Laboratory Press, Cold Spring Harbor, New Yorkに記載の方法に従って行えばよい。通常は、調製された変異型Myb関連遺伝子を適当な制限酵素で切断し、好適な発現ベクターの対応する制限酵素部位、マルチクローニングサイト、又はTaq DNAポリメラーゼ等による3'-A突出末端を有するPCR産物であれば5'-T突出末端に挿入して連結する方法等が採用される。その他、市販のシステム又はキットを用いる場合であれば、それらに特異的な方法によって調製することもできる。例えば、Gatewayシステム(Invitrogen社)が利用できる。
【0042】
本態様の変異型Myb関連遺伝子発現ベクターによれば、本ベクターをそれに適応する宿主に導入することで、次で説明する変異型Myb関連遺伝子を発現する形質転換体を得ることができる。
【0043】
3.形質転換体又はその後代
本発明の他の態様は、前記変異型Myb関連遺伝子を含む発現ベクターを宿主に導入した形質転換体又はその後代に関する。
【0044】
3−1.形質転換体
本明細書において「形質転換体」とは、前記変異型Myb関連遺伝子を含む発現ベクターの導入によって形質転換された宿主をいう。使用する宿主は、導入した発現ベクターがコードする本発明の変異型Myb関連遺伝子を発現できれば特に限定しない。一般的には、発現ベクターによって発現可能な宿主はある程度限定されるため、それに従えばよい。宿主の具体例を挙げると、細菌(例えば、大腸菌(Escherichia coli等)及び枯草菌(Bacillus subtilis))及び酵母(例えば、出芽酵母(Saccharomyces cerevisiae)、分裂酵母(Schizosaccharomyces pombe)又はメタノール資化性酵母(Pichia pastoris))のような微生物、真菌(例えば、コウジカビ(Aspergillus)及びアカパンカビ(Neurospora))、植物(植物体、その器官、組織、分化した細胞若しくは未分化状態の植物細胞(カルス)を含む)又は昆虫細胞(例えば、sf9又はsf21)が挙げられる。植物を宿主とする場合、コケ植物、シダ植物及び種子植物等を含み、その種類は問わない。
【0045】
なお、本発明の形質転換体は、同一の遺伝情報を有するクローン体を包含する。例えば、宿主が大腸菌や酵母等の無性生殖を行う単細胞微生物であれば、形質転換体第1世代から分裂又は出芽等によって新たに生じた細胞は、本発明の形質転換体に含まれるものとする。また、形質転換される宿主が植物であれば、形質転換体第1世代から採取した植物体の一部を挿し木、接木若しくは取り木したもの又は形質転換体第1世代から無性生殖で得られる栄養繁殖器官(例えば、根茎、塊根、球茎、ランナー等)より新たに生じた新たな栄養体も本発明の形質転換体に含まれる。
【0046】
3−2.形質転換体の後代
本明細書において「形質転換体の後代」とは、本発明の変異型Myb関連遺伝子を導入した形質転換体第1世代の有性生殖を介した子孫であって、本発明の変異型Myb関連遺伝子を発現可能な状態で保持しているものを意味する。例えば、形質転換体が植物の場合には、その形質転換体の実生が該当する。本発明の変異型Myb関連遺伝子を発現可能な状態で継代する限りにおいて後代の世代は問わない。
【0047】
3−3.形質転換方法
本発明の変異型Myb関連遺伝子発現ベクターを宿主に導入する方法は、当該分野で公知の方法を使用することができる。
【0048】
宿主が細菌であれば、ヒートショック法、カルシウムイオンを用いる方法(例えば、リン酸カルシウム法)、エレクトロポレーション法等を用いればよい。これらの技術は、いずれも当該分野で公知であり、例えば、Sambrook, J. et. al., (1989) Molecular Cloning: A Laboratory Manual Second Ed., ColdSpring HarborLaboratory Press, Cold Spring Harbor, New Yorkをはじめとする様々な文献に記載されている。
【0049】
宿主が植物細胞であり、前記発現ベクターがプラスミドベクターである場合には、形質転換方法としてプロトプラスト法、パーティクルガン法又はアグロバクテリウム(Agrobacterium)法等を利用することができる。プロトプラスト法とは、セルラーゼ等の酵素的処理によって細胞壁を除去した植物細胞(プロトプラスト)を用いて、目的の遺伝子を植物細胞中に導入する方法であって、遺伝子導入の方法により、さらに、エレクトロポレーション法、マイクロインジェクション法又はポリエチレングリコール法等に細分することができる。エレクトロポレーション法は、プロトプラストと目的遺伝子の混合液に電気パルスを与えてプロトプラスト内に遺伝子を導入する方法である。また、マイクロインジェクション法は、微針を用いて顕微鏡下でプロトプラスト中に目的の遺伝子を直接導入する方法である。そして、ポリエチレングリコール法は、ポリエチレングリコールを作用させてプロトプラストに目的の遺伝子を導入する方法である。一方、パーティクルガン法とは、金又はタングステン等の微粒子に目的の遺伝子を付着させて、それを高圧ガスにより植物組織細胞内に打ち込み、目的の遺伝子を細胞内に導入する方法であって、宿主細胞のゲノムDNA中に目的の遺伝子が取り込まれた形質転換体を得ることができる。アグロバクテリウム法とは、形質転換因子としてアグロバクテリウム属の菌(例えば、アグロバクテリウム・ツメファシエンス(A. tumefaciens)、アグロバクテリウム・リゾゲネス(A. rhizogenes)等)及びそれに由来するTiプラスミドを用いる植物細胞の形質転換方法であって、目的の遺伝子を宿主植物細胞のゲノムDNA中に導入することができる。いずれの方法も当該分野においては、公知の方法であり、詳細については植物代謝工学ハンドブック(2002年、NTS社)又は新版モデル植物の実験プロトコール:遺伝学的手法からゲノム解析まで(2001年秀潤社)等を参照すればよい。
【0050】
また、宿主が植物細胞で、かつ前記発現ベクターがウイルスベクター(例えば、前述のCaMV、BGMV、TMV等)の場合には、そのウイルスベクターを植物細胞に感染させることによって、変異型Myb関連遺伝子を導入することができる。例えば、まず、植物ウイルスゲノムを大腸菌由来のベクター等のクローニングベクターに挿入して組換え体を調製する。次に、組換え体のウイルスゲノム中に変異型Myb関連遺伝子を挿入し、クローニングする。続いて、制限酵素によって組換え体から植物ウイルスゲノム領域を切り出し、得られたウイルスゲノムを目的の植物細胞に感染させる。以上により、目的の遺伝子を植物細胞内に導入できる。このようなウイルスベクターを用いた遺伝子導入方法の詳細については、Hohnらの方法(Molecular Biology of Plant Tumors(Academic Press、New York)1982、pp549)、米国特許第4,407,956号明細書等を参考にすればよい。
【0051】
宿主が植物細胞の場合、植物の種類については特に限定しない。好ましくは、配列番号7で示されるMyb-miRNA及びその標的遺伝子である配列番号1で示されるMyb-miRNA結合部位をコード領域内に有するMyb遺伝子を内在する植物である。例えば、ユーカリ属植物又はタバコ属植物由来の植物細胞が挙げられる。ユーカリ属植物としては、例えば、ユーカリプタス・グランディス (E. grandis)、ユーカリプタス・ユーロフィラ(E. urophylla)、ユーカリプタス・カマルドレンシス(E. camaludulensis)、ユーカリプタス・グロブラス(E. globulus)、ユーカリプタス・ニテンス(E. nitens)、ユーカリプタス・ガンニー(E. gunni)、ユーカリプタス・ラジアータ(E.radiata)、ユーカリプタス・アンプリフォリア(E. amplifolia)、ユーカリプタス・アーチェリ(E. archeri)、ユーカリプタス・ブリジェシアーナ(E. bridgesiana)、ユーカリプタス・バクステリ(E. baxteri)、ユーカリプタス・ビコスタータ(E. bicostata)、ユーカリプタス・ブラッケリー(E. blakelyi)、ユーカリプタス・ボツリオイデス(E. botryoides)等が挙げられる。タバコ属植物としては、例えば、ニコチアナ・タバカム(N. tabacum)、ニコチアナ・ルスチカ(N. rustica)等が挙げられる。この他、シロイヌナズナ(A. thaliana)、ポプラ属(Populus)植物、例えば、ポプルス・トリコカルパ(P. trichocarpa)、ポプルス・アルバ(P. alba)、ポプルス・ニグラ(P. nigra)、ポプルス・カンジカンス(P. candicans)、ポプルス・トレムラ(P.tremula)、ポプルス・シーボルディ(P. sieboldii)等が挙げられる。
【0052】
3−4.後代取得法
本発明の形質転換体から後代を得る方法は、その形質転換体の宿主である生物種において後代を得るために用いられる通常の方法で行えばよい。例えば、形質転換体の宿主が大腸菌や酵母であれば、適当な公知培地で培養することによって得ることができる。例えば、Sambrook, J. et. al., (1989) Molecular Cloning: A Laboratory Manual Second Ed., ColdSpring HarborLaboratory Press, Cold Spring Harbor, New Yorkを参照すればよい。また、形質転換体の宿主が植物であれば、一般的には土耕栽培(播種)により、実生を得ることは容易である。本発明の形質転換植物体から種子を得る方法の一例として、形質転換植物体を適当な培地において発根させ、その発根体を水分含有の土を入れたポットに移植する。適当な栽培条件下で生育させ、当該分野で通常の方法を用いて種子を形成させる。得られた種子を、例えば、水分含有の適当な土に播種し、適切な栽培条件下で生育させることにより本発明の形質転換体の後代を得ることができる。
【0053】
4.植物生長促進方法
本発明の他の態様は、前記変異型Myb関連遺伝子発現ベクターを植物体に導入して、該植物の生長を促進する方法である。
【0054】
4−1.対象植物
本発明の原理は、本発明の変異型Myb関連遺伝子を目的とする植物に導入して、その植物細胞内で発現させることによって、内在性のMyb-miRNAによる発現抑制制御を受けることなく、野生型アミノ酸配列を有する変異型Myb関連遺伝子タンパク質を付与することに基づく。したがって、本方法の対象植物は、本発明に記載の配列番号1で示す塩基配列を含み、植物の成長を促進するMyb関連遺伝子及び本発明のMyb-miRNAをゲノム中に一以上有する植物種であれば、特に限定しない。例えば、前記形質転換体の「3−2.形質転換方法」の項で説明したユーカリ属各種又はタバコ属各種又はポプラ属各種が挙げられる。
【0055】
4−2.方法
本発明は、前記「2.変異型Myb関連遺伝子を含む発現ベクター」の項で説明した変異型Myb関連遺伝子発現ベクターを目的とする植物に導入し、前記「3.形質転換体」の項で説明した形質転換植物を得ることによって達成される。変異型Myb関連遺伝子発現ベクターを目的とする植物に導入する方法については、前記「3−2.形質転換方法」に記載の方法に準じて行えばよい。このとき、形質転換体が植物であるため、使用する変異型Myb関連遺伝子発現ベクターは、植物細胞内で発現可能なベクターを用いるようにする。例えば、プラスミドベクターであれば、pBI系又はpRI系のバイナリーベクター等、またウイルスであれば、カリフラワーモザイクウイルス(CaMV)、インゲンマメモザイクウイルス(BGMV)、タバコモザイクウイルス(TMV)等である。また、これらのベクターを導入する宿主は、当然ながら目的とする植物となる。
【0056】
変異型Myb関連遺伝子発現ベクターで目的とする植物を形質転換する方法は、使用する発現ベクターの種類によって異なる。発現ベクターがプラスミドベクターである場合には、前述したプロトプラスト法、パーティクルガン法、又はアグロバクテリウム法等を用いることができる。これらの方法は、いずれも当該分野においては、公知の方法であり、詳細については前記例示の文献を参照すればよい。また、発現ベクターがウイルスベクターである場合にHohnらの方法(Molecular Biology of Plant Tumors(Academic Press、New York)1982、pp549)、米国特許第4,407,956号等を参考にすることができる。
【0057】
前記形質転換方法は、通常、カルス又は組織片を用いた植物組織培養を介するインビトロ法が用いられる。この場合、形質転換した植物細胞から植物体を再生する必要がある。本方法についても当該分野では公知であり、上述の植物代謝工学ハンドブック(2002年、NTS社)又は新版モデル植物の実験プロトコル:遺伝学的手法からゲノム解析まで(2001年秀潤社)等を参照することができる。また、目的とする植物がユーカリ属植物である場合には、形質転換したユーカリ属の植物細胞から植物体を再生する方法は、土肥らの方法(特開平11-127025号公報)を参照することができる。あるいは、前記形質転換方法は、カルスや細胞培養のステップを経ることなく、目的の植物個体の細胞に直接、本発明のMyb関連遺伝子を導入する形質転換法、いわゆるin planta法を用いてもよい。
【0058】
5.Myb-miRNA
本発明の別の態様は、植物細胞内への導入によって該植物の生長を抑制できるMyb-miRNAである。
本明細書で「Myb-miRNA」とは、配列番号7で示される21塩基からなる塩基配列を含み、本明細書に記載のMyb関連遺伝子の転写産物を標的とするmiRNAをいう。
【0059】
「miRNA」(マイクロRNA:micro RNA)とは、生物種間で進化的に保存された19〜25塩基からなる内在性ノン・コーディングRNA(non-cording RNA:タンパク質をコードしないRNA)であって、それに相補的な塩基配列を有する標的遺伝子の発現を転写後翻訳前の段階で制御するsmall RNAである。miRNAは、通常、数百〜数千塩基からなるpri-miRNA(primary miRNA)前駆体としてゲノムから転写され、核内でDrosha及びPasha等によって60〜100塩基程度のヘアピン型pre-miRNA(precursor-miRNA)へと変換される。続いて、細胞質に輸送され、Dicerによってヘアピンのループ部分が切断されて、19〜25塩基の二本鎖miRNA(miRNA duplex)となる。その後、標的遺伝子のmRNAに相補的な配列を有する側鎖が成熟一本鎖miRNAとしてRISC/miRNPに取り込まれ、この成熟miRNAの塩基配列との結合を介して標的mRNAが認識され、RISC及びmiRNPによって切断及び翻訳阻害されることで標的遺伝子の発現が抑制される(David P. Bartel, Cell, Vol. 116, 281-297, January 23, 2004,)。
【0060】
「配列番号7で示される塩基配列」部分は、成熟一本鎖Myb-miRNA部分に相当する。一本鎖Myb-miRNAは、自己と相補的な塩基配列である配列番号1で示されるMyb-miRNA結合部位(完全相補ではなく、1又は2つの不対合を有していてもよい)を認識することで、それを含むMyb関連遺伝子の発現を抑制的に制御している(実施例2参照)。配列番号7において、R、Y及びNで示される塩基は、それぞれ、A又はG、C又はU及びA、U、G又はCのいずれであってもよいことを示す。具体的な例としては、配列番号8で示される塩基配列を含むユーカリプタス・グランディス(E. grandis)由来のMyb-miRNA(以下、便宜的に「EgMyb-miRNA」とする)が挙げられる。なお、miRNAによる遺伝子の発現抑制機構については、当該分野で公知である。例えば、Eulalio et al., 2008, Cell, 132:9-14を参照されたい。
【0061】
6.Myb-miRNAをコードするDNAを含む発現ベクター
本発明の他の態様は、前記Myb-miRNAをコードするDNAを含む発現ベクター(以下、「Myb-miRNA発現ベクター」とする)を導入した形質転換体である。
【0062】
6−1.Myb-miRNAをコードするDNA
本明細書で「Myb-miRNAをコードするDNA」とは、転写後に本発明のMyb-miRNAを機能可能な状態でコードするDNAをいう。「機能可能な状態」とは、本発明のMyb-miRNAが転写された後、Myb-miRNAとしての機能を発揮し得る状態をいう。一般に、成熟一本鎖miRNA部分に相当する配列番号7で示される塩基配列のみをコードするDNAでは、本発明のMyb-miRNAとしての機能は発揮し得ない。転写後に二本鎖RNAが形成されないためである。したがって、本発明のMyb-miRNAを転写後に機能可能な状態とするためには、DNAが配列番号7で示す塩基配列を一方の鎖に有する二本鎖RNAが形成されるようにコードしていなければならない。
【0063】
転写後に本発明のMyb-miRNAを機能可能な状態でコードするDNAとしては、例えば、Myb-shRNA(short hairpin RNA)をコードするDNAが挙げられる。「shRNA」とは、標的とする遺伝子の一部(例えば、本発明のMyb-miRNA結合部位)に対応する塩基配列とそのアンチセンス鎖(例えば、本発明のMyb-miRNA)が短いスペーサ配列で連結された一本鎖RNAである。shRNAは、一分子内でセンス領域とアンチセンス領域が互いに塩基対合し、同時に前記スペーサ配列がループ構造をとることによって、分子全体としてヘアピン型のステム−ループ構造を有する。このようなMyb-shRNAをコードするDNAの設計は、例えば、配列番号7で示される塩基配列をコードするDNA断片、それに相補的な塩基配列を有するDNA断片及びこれらの一方のDNA断片の3’末端と他方のDNA断片の5’末端とを連結する適当な塩基配列からなるスペーサ配列から構成される。スペーサ配列は、通常3〜24塩基、好ましくは、4〜15塩基あればよい。shRNAをコードするDNAの調製は、多くのライフサイエンス系メーカー(例えば、タカラバイオ社、インビトロジェン社等)が受託製造サービスを行っており、それらを利用することもできる。この他、Myb-miRNAをコードするDNAは、Myb-miRNAの前駆体であるpri-Myb-miRNA若しくはpri-Myb-miRNAをコードするDNAであってもよい。
【0064】
他の例として、配列番号7で示す塩基配列を有するヌクレオチドとそれに相補する塩基配列を有するヌクレオチドをそれぞれコードするDNAを独立に発現可能なようにしたものであってもよい。それぞれのヌクレオチドは、シス(例えば、同一発現ベクター内)又はトランス(例えば、異なる発現ベクター内)のいずれで発現されても構わないが、両ヌクレオチドの発現が同程度になるように、例えば、同一のプロモータを使用することが好ましい。この場合、それぞれのDNAを目的の細胞内に導入して発現させることによって、細胞内で2本鎖RNAが形成され、結果として、本発明のMyb-miRNAを機能可能な状態にすることができる。
【0065】
6−2.Myb-miRNA発現ベクター
本発明において、「Myb-miRNA発現ベクター」とは、本発明のMyb-miRNAをコードするDNAを適当な発現ベクター内に導入したものである。適切な宿主細胞に導入することで、適当な条件の下、挿入したMyb-miRNAを発現させることが可能である。発現ベクターの種類は特に限定されない。導入する宿主、例えば、大腸菌、酵母、植物細胞(植物体、植物器官及び植物組織を含む)、昆虫細胞又は動物細胞によって適宜選択すればよい。
【0066】
本発明の発現ベクターにおける骨格部分、すなわち本発明のMyb-miRNA以外の主要な構成部分(プロモータ、エンハンサ、若しくはターミネータ等の調節領域、又は選抜マーカー遺伝子等を含む)については、上記「2−1.変異型Myb関連遺伝子発現ベクター」の項で説明した発現ベクターを同様に利用することができる。したがって、説明の重複する部分については省略をする。ただし、本発明の発現ベクターを後述する「植物の生長を抑制する方法」で使用する場合、用いるプロモータは植物細胞内でMyb-miRNAを過剰に発現できるプロモータであることが好ましい。効率的に生長を抑制させるためである。例えば、細胞分裂が旺盛な組織、あるいは細胞伸長に掛かる木部肥厚組織等において特異的に発現することが可能なプロモータ等が好ましい。
【0067】
6−2.Myb-miRNA発現ベクターの調製
前記Myb-miRNAをコードするDNAを発現ベクターの所定の部分に挿入し、Myb-miRNA発現ベクターを調製する方法は、当該分野で公知の方法、例えば、Sambrook, J. et. al., (1989) Molecular Cloning: a Laboratory Manual Second Ed., Cold SpringHarbor Laboratory Press, Cold Spring Harbor, New Yorkに記載の方法を用いることができる。
【0068】
7.形質転換体
本発明の他の態様は、前記Myb-miRNA発現ベクターを導入した形質転換体に関する。
【0069】
7−1.形質転換体
本明細書において「形質転換体」とは、前記Myb-miRNA発現ベクターによって形質転換された宿主及びそのクローン体をいう。使用する宿主は、使用したベクターに適する宿主であって、本発明のMyb-miRNAを発現できるものであれば特に限定されない。例えば、上記「3−1.形質転換体」の項で記載した宿主が挙げられる。好ましくは、微生物又は植物細胞(ユーカリ属植物又はタバコ属植物を含む)である。
【0070】
7−2.形質転換体の後代
本明細書において「形質転換体の後代」とは、本発明のMyb-miRNA発現ベクターを導入した形質転換体第1世代の有性生殖を介した子孫であって、本発明のMyb-miRNAをコードするDNAを発現可能な状態で保持するものを意味する。例えば、形質転換体が種子植物の場合には、その形質転換体の実生が該当する。本発明のMyb-miRNAをコードするDNAを発現可能な状態で継代する限りにおいて、後代の世代は問わない。
【0071】
7−3.形質転換方法
本発明のMyb-miRNA発現ベクターを宿主内に導入する方法は、前記「3−3.形質転換方法」の項に記載した方法に準じて行うことができる。
【0072】
7−4.後代取得法
本発明の形質転換体から後代を得る方法は、前記「3−4.後代取得法」の項に記載した方法に準じて行うことができる。
【0073】
8.植物の生長を抑制する方法
本発明の他の態様は、前記変異型Myb関連遺伝子又は前記発現ベクターを植物体に導入して、該植物の生長を促進する方法である。
【0074】
本発明は、前記「4.植物の生長を促進する方法」とは逆の原理に基づく。すなわち、本発明のMyb-miRNAをコードするDNAを目的とする植物に導入してその植物体内でMyb-miRNAを過剰発現させることによって、内在性の標的Myb関連遺伝子の発現を抑制させることに基づく。したがって、本方法の対象植物は、本発明に記載の配列番号1で示す塩基配列を含み、植物の成長を促進するMyb関連遺伝子をゲノム中に一以上有する植物種であれば、特に限定しない。例えば、前記形質転換体の「3−2.形質転換方法」の項で説明したユーカリ属各種又はタバコ属各種又はポプラ属各種等が挙げられる。
【0075】
本発明の方法の基本的操作は、前記「4.植物の生長を促進する方法」で説明した方法と同一であるため、前述の方法に準じて行えばよい。
【実施例】
【0076】
<実施例1>
ユーカリMyb関連遺伝子(EcMyb遺伝子)の取得
(1)総RNAの調製
王子製紙株式会社森林資源研究所内において成育中の5年生ユーカリプタス・カマルドレンシス(E. camaldulensis)の樹幹部、形成層組織を採取し、液体窒素中で粉砕した後、Concert Plant RNA Regent (インビトロジェン社)を用いて総RNAを抽出した。抽出手順については、添付のプロトコルに従った。その結果、1mgの総RNAを得た。
【0077】
(2)mRNAの精製
PolyA Tract mRNA Isolation System (プロメガ社)を用いて前記総RNAからmRNAの精製を行った。抽出手順については、添付のプロトコルに従った。その結果、1mgの総RNAから6μgのmRNAを得た。
【0078】
(3)cDNAライブラリーの作製
E. camaldulensisのcDNAライブラリーの作製は、SMART cDNA Library Construction Kit (タカラバイオ社)を用いて、添付のプロトコルに従って行った。具体的には、前記精製したmRNAのうち1μgにoligo dTプライマーをアニールさせた後、逆転写酵素によってcDNAを合成した。得られたcDNAを制限酵素Sfi Iで消化した後、カラムによるサイズ分画を行い、λTriplEx2ベクターと連結した。続いて、Gigapack Gold III (Stratagene社)を用いてλ-ファージパッケージング反応を行い、5×105の独立クローンからなるcDNAライブラリーを作製した。
【0079】
(4)EcMyb遺伝子の単離
前記独立クローンから約2万個のcDNAクローンを任意に抽出し、各クローンの塩基配列をCEQ8000遺伝子解析システム(ベックマン・コールター社製)により決定した。得られた塩基配列をBlast検索(http://blast.ncbi.nlm.nih.gov/Blast.cgi)で日本DNAデータバンク(DDBJ)の遺伝子データベースと照合した。その結果、シロイヌナズナ、イネ、ポプラ等において、分化・形態形成を制御していると考えられているWEREWOLF遺伝子(R2R3タイプMyb遺伝子)と高い相同性が認められるcDNAクローンを見出した。このcDNAクローンの配列を決定したところ、5’並びに3’非翻訳領域を含むアミノ酸コード領域全体を含んでいることが明らかとなった。この新たに単離されたE. camaldulensisのMyb関連遺伝子(EcMyb遺伝子:配列番号3)は、121アミノ酸からなるMyb関連タンパク質(EcMybタンパク質)をコードする(配列番号9)。EcMybタンパク質は、N末端側に2つのMybドメイン(R2、R3)を含んでおり、Myb66タンパク質と同様にR2R3タイプのMybファミリーに属する転写制御因子であることが判明した(図1)。
【0080】
<実施例2>
Myb遺伝子の発現抑制に関わるmiRNA(Myb-miRNA)の単離とその活性の検証
(1)ユーカリmiRNAの精製と解析
王子製紙(株)森林資源研究所内において成育中の5年生ユーカリプタス・グランディス(E. grandis)の花芽を採取し、液体窒素中で粉砕した後、Concert Plant RNA Regent (インビトロジェン社)を用いて総RNAを抽出した。この総RNAを15%ポリアクリルアミドゲルで常法により分離し、泳動マーカーのサイズから換算した約18〜26塩基長に相当するRNA領域のバンドをゲルから切り出し、FrashPAGE Fractinator(アンビオン社製)を用いて、ゲルからRNAを回収した。次に、MPSS法(タカラバイオ社, Bio View, 2005,No.50:17-19)により、約100万個の低分子RNAの塩基配列を決定した。解析の結果、これらの配列は、約13万種に分類できることが判明した。続いて、E. grandisのゲノム配列と相同性検索を行い、ゲノム配列に一致しない配列を除去し、さらに既知のRNA配列と相同性のある配列を除去した。その結果、約3万種の低分子RNAをmiRNAの候補として分離した。続いて、これらの配列の前後を含む約500bpの塩基配列情報をユーカリのゲノム配列より取得し、取得した領域内における各候補の2次構造予測を行い、miRNAに特徴的な構造、すなわち、ヘアピン構造を有するRNA分子を208個同定した。これらをユーカリmiRNA群とした。
【0081】
(2)EcMyb遺伝子の発現抑制に関わるmiRNAの単離
前記ユーカリmiRNA群から、実施例1で単離したEcMyb1遺伝子を標的分子とし得るmiRNAを探索した。その結果、配列番号8で示される塩基配列を含む一のユーカリmiRNA(EgMyb-miRNA)が得られた。
【0082】
(3)Myb-miRNAによるMyb関連遺伝子の発現抑制の検証
実施例1で単離したEcMyb遺伝子が、E.camaldulensisに内在すると予想されるEgMyb-miRNAオルソログEcMyb-miRNAによって発現制御を受けることを確認した。
【0083】
まず、E.camaldulensisの葉から総RNAを抽出し、オリゴdTプライマーを用いてcDNAを合成した。次に、合成したcDNAの5'末端にFirst Choice RLM-RACE Kit(Ambion社)を用いてアダプターを連結した(図3a)。基本的な方法については、添付のプロトコルに従った。続いて、アダプターを付加したcDNAを鋳型にして図1及び図2に示すEcMybに特異的なプライマー(アウタープライマーA及びインナープライマーB:それぞれ配列番号10及び11)と前記アダプターに特異的なプライマー(アウタープライマーC及びインナープライマーD:それぞれ配列番号 12及び13)を用いてPCR反応((変成反応(94℃、30秒)、アニーリング反応(55℃、30秒)、伸長反応(68℃、90秒)を1サイクルとして、35サイクル実施)を独立に10回(n=10)行った。続いて、得られたPCR産物を1.0%アガロースゲルで電気泳動して確認すると共に(図3b)、インビトロジェン社製のクローニングキット(TAクローニングキット)を用いて獲得した後、その塩基配列を決定した。その結果、EcMyb mRNAは、いずれの反応サンプルにおいて配列番号2に相当するMyb-miRNA結合部位の13番目と14番目のU間で切断されていることが明らかとなった(図3c)。この結果からEcMyb mRNAは、EcMyb-miRNAによる発現抑制を受けていることが立証された。
【0084】
<実施例3>
変異型EcMyb関連遺伝子発現ベクターの調製
(1)変異型EcMyb関連遺伝子
EcMyb mRNAに含まれる配列番号2で示される塩基配列からなる部位(Myb-miRNA結合部位)にMyb-miRNAが結合できないようにサイレント変異を導入した。ここで、EcMyb-miRNAは、未同定であることから、本実施例ではEcMyb-miRNAと相同性を有することが予測されるEgMyb-miRNAと配列番号2で示される塩基配列との間で塩基対合できないような9箇所のサイレント変異を導入した(図4)を作製した。実際の作製においては、タカラバイオ社へ配列番号14に示すDNA合成を委託した。
【0085】
(2)変異型EcMyb関連遺伝子発現ベクターの調製
得られた変異型EcMyb関連遺伝子mEcMyb(配列番号14)及び実施例1で調製した野生型のEcMyb遺伝子のそれぞれをGatewayTMエントリークローンpENTR/D/TOPOベクター(Invitrogen社)内にクローニングした後、LR反応によってバイナリーベクター(pGWB2;Nakagawaら,2007, J Biosci Bioeng, 34-41)に導入した。上記Gatewayシステム(Invitrogen社)を利用した一連の操作手順については、添付の説明書に従った。以上によって、目的とする発現ベクターpGWB2-mEcMyb及びpGWB2-EcMybを調製した。これらの発現ベクターをそれぞれアグロバクテリウムLBA4404株にバイオラッド社製ジーンパルサーを用いて、添付の実施方法に準拠して導入した。植物への形質転換までの間は、グリセロール保存の状態で維持した。
【0086】
<実施例4>
内在性Myb-miRNAを介した発現抑制の回避による植物生長促進
(1)形質転換タバコの作製
mEcMybの内在性Myb-miRNAによるRNAiの回避能力及びmEcMybを導入した植物の生長促進効果を確認するため、本実施例の(1)で調製したpGWB2-EcMyb及びpGWB2-mEcMybを保持したアグロバクテリウムを常法(参照:松本省吾・町田泰則 (1990) 植物形質転換法. 現代化学 6月:25〜29)に従ってタバコ葉(N. tabacum BY4)にそれぞれ感染させ、ハイグロマイシン耐性を指標とした選抜を行い、形質転換タバコを作製した。
【0087】
ここで、ユーカリに代えてタバコを用いた理由は、主にユーカリよりも生長速度が速いこと、イネ科以外の草本性有用植物の中では形質転換をはじめとする遺伝子操作技術が既に確立していること、及び木本性植物の形質転換が一般に草本性植物のそれと比較して困難なことによる。なお、タバコにおいてEcMybのオルソログとなる具体的な報告は無いものの、ゲノム解読が完了した又は進行中の他の植物種において配列比較を行った結果、EcMybオルソログ候補遺伝子は全ての植物に普遍的に存在すると類推された。したがって、タバコにおいてもEcMybのオルソログ遺伝子の存在が予測されたことから、本実施例で用いた。
【0088】
(2)mEcMyb遺伝子の発現確認
本態様の(1)で得られたpGWB2-EcMyb又はpGWB2-mEcMybをそれぞれ保持する形質転換タバコにおいてEcMyb又はmEcMybの発現を調べるため、各形質転換タバコの葉から総RNAを抽出し、常法に従ってRT-PCRを行った。PCRで用いたFw及びRvプライマーは、EcMybの158〜180番目のセンス鎖配列に相当する塩基配列(配列番号15)及びEcMybの385〜404番目のアンチセンス鎖配列に相当する塩基配列(配列番号16)をそれぞれ有する。また、Myb-miRNA結合部位は、両プライマー間(310〜331番目のアミノ酸に相当)に存在する。PCRサイクルは、変成反応(94℃、30秒)、アニーリング反応(55℃、30秒)、伸長反応(68℃、90秒)を1サイクルとして、35サイクル実施した。RT-PCRは、それぞれ独立に3回(n=3)行った。RT-PCR後、各サンプル2μlを1.0%のアガロースゲルで電気泳動した。
【0089】
結果を図5に示す。Wtは非形質転換の野生株タバコ(BY4)を、WtEcは野生型EcMyb形質転換体を、そしてmEcは変異型EcMyb形質転換体を示す。変異型EcMyb形質転換体中には、野生型EcMyb形質転換体と比較してMyb-miRNA結合部位が保持されたmRNAが多量に存在することが明らかとなった。これは、外来性のmEcMyb mRNAが、タバコの内在性Myb-miRNAによる切断作用、すなわちRNAiを受けることなく、タバコ細胞内において安定して機能し得ることを示している。一方、EcMyb mRNAは、発現ベクターからの過剰発現により内在性のNtMyb mRNAよりも多く検出されたが、mEcMyb mRNAの量と比較すると少なかった。これは、一部がタバコの内在性Myb-miRNAによって切断されたためと考えられる。以上の実験から、本発明の変異型Myb関連遺伝子は、その転写産物を標的とするMyb-miRNAが内在する植物において、そのMyb-miRNAによるRNAiを回避できることが明らかとなった。
【0090】
(3)形質転換タバコの生育調査
本態様の(1)で得られたpGWB2-mEcMybを導入した形質転換体と非形質転換体の野生株(BY4)をその後6ヶ月間通常方法にて育成し、それぞれの生長性を比較した。
【0091】
結果を図6及び図7に示す。図6における実線は形質転換体タバコ(BY4+pGWB2-mEcMyb)を、破線は野生型タバコ(BY4)を示し、四角プロットは背丈(cm)(左縦軸)を及び丸プロットは茎径(mm)(右縦軸)をそれぞれ示す。図7におけるaは野生型タバコ(BY4)を、b及びcは形質転換体タバコ(BY4+pGWB2-mEcMyb)をそれぞれ示す。
【0092】
pGWB2-mEcMyb遺伝子の導入により、非導入の野生株と比較して6ヶ月後には背丈及び茎径共に有意な生長差が見られた(図6、図7)。この結果から本発明の変異型Myb関連遺伝子を用いれば、植物が本来有する当該Myb関連遺伝子に対するRNAiを人為的に阻止できることが判明した。特に、本発明の変異型遺伝子を導入した植物では、その生長が通常よりも促進されること、また、本発明の変異型Myb関連遺伝子が植物種を超えて作用し得ることも明らかとなった。したがって、本発明の変異型Myb関連遺伝子によって事業植林用の樹木、例えば、ユーカリ属を形質転換すれば、木質バイオマスとしての活用がより有効に実施できる。
【特許請求の範囲】
【請求項1】
植物細胞内への導入によって該植物の生長を促進できる変異型Myb関連遺伝子であって、
配列番号1で示される塩基配列をコード領域内に含むMyb関連遺伝子において、前記塩基配列に結合するmiRNAを介した発現抑制が不能となるように前記塩基配列内にサイレント変異を導入した前記変異型Myb関連遺伝子。
【請求項2】
配列番号1で示される塩基配列が配列番号2で特定される、請求項1に記載の変異型Myb関連遺伝子。
【請求項3】
サイレント変異の少なくとも一つが配列番号1における13番目のTをCに置換した変異である、請求項1又は2に記載の変異型Myb関連遺伝子。
【請求項4】
少なくとも5個のサイレント変異を含む、請求項1〜3のいずれか1項に記載の変異型Myb関連遺伝子。
【請求項5】
前記Myb関連遺伝子が配列番号3で示される塩基配列又は該塩基酸配列と95%以上の同一性を有する塩基配列からなる、請求項1〜4のいずれか1項に記載の変異型Myb関連遺伝子。
【請求項6】
請求項1〜5のいずれか1項に記載の変異型Myb関連遺伝子を含む発現ベクター。
【請求項7】
請求項6に記載の発現ベクターを宿主に導入した形質転換体又はその後代。
【請求項8】
宿主が微生物又は植物である、請求項7に記載の形質転換体又はその後代。
【請求項9】
前記植物がユーカリ属(Eucalyptus)植物又はタバコ属(Nicotiana)植物である、請求項8に記載の形質転換体又はその後代。
【請求項10】
請求項6に記載の発現ベクターを植物に導入して、該植物の生長を促進する方法。
【請求項11】
植物細胞内への導入によって該植物の生長を抑制できるmiRNAであって、
配列番号7で示される塩基配列を含み、かつ請求項1〜5のいずれか1項に記載のMyb関連遺伝子の転写産物を標的とするmiRNA。
【請求項12】
配列番号7で示される塩基配列が配列番号8で特定される、請求項11に記載のmiRNA。
【請求項13】
請求項11又は12に記載のmiRNAをコードするDNAを含む発現ベクター。
【請求項14】
請求項11又は12に記載のmiRNAあるいは請求項13に記載の発現ベクターを宿主に導入した形質転換体又はその後代。
【請求項15】
宿主が微生物又は植物である、請求項14に記載の形質転換体又はその後代。
【請求項16】
前記植物がユーカリ属植物又はタバコ属植物である、請求項15に記載の形質転換体又はその後代。
【請求項17】
請求項13に記載の発現ベクターを植物に導入して、該植物の生長を抑制する方法。
【請求項1】
植物細胞内への導入によって該植物の生長を促進できる変異型Myb関連遺伝子であって、
配列番号1で示される塩基配列をコード領域内に含むMyb関連遺伝子において、前記塩基配列に結合するmiRNAを介した発現抑制が不能となるように前記塩基配列内にサイレント変異を導入した前記変異型Myb関連遺伝子。
【請求項2】
配列番号1で示される塩基配列が配列番号2で特定される、請求項1に記載の変異型Myb関連遺伝子。
【請求項3】
サイレント変異の少なくとも一つが配列番号1における13番目のTをCに置換した変異である、請求項1又は2に記載の変異型Myb関連遺伝子。
【請求項4】
少なくとも5個のサイレント変異を含む、請求項1〜3のいずれか1項に記載の変異型Myb関連遺伝子。
【請求項5】
前記Myb関連遺伝子が配列番号3で示される塩基配列又は該塩基酸配列と95%以上の同一性を有する塩基配列からなる、請求項1〜4のいずれか1項に記載の変異型Myb関連遺伝子。
【請求項6】
請求項1〜5のいずれか1項に記載の変異型Myb関連遺伝子を含む発現ベクター。
【請求項7】
請求項6に記載の発現ベクターを宿主に導入した形質転換体又はその後代。
【請求項8】
宿主が微生物又は植物である、請求項7に記載の形質転換体又はその後代。
【請求項9】
前記植物がユーカリ属(Eucalyptus)植物又はタバコ属(Nicotiana)植物である、請求項8に記載の形質転換体又はその後代。
【請求項10】
請求項6に記載の発現ベクターを植物に導入して、該植物の生長を促進する方法。
【請求項11】
植物細胞内への導入によって該植物の生長を抑制できるmiRNAであって、
配列番号7で示される塩基配列を含み、かつ請求項1〜5のいずれか1項に記載のMyb関連遺伝子の転写産物を標的とするmiRNA。
【請求項12】
配列番号7で示される塩基配列が配列番号8で特定される、請求項11に記載のmiRNA。
【請求項13】
請求項11又は12に記載のmiRNAをコードするDNAを含む発現ベクター。
【請求項14】
請求項11又は12に記載のmiRNAあるいは請求項13に記載の発現ベクターを宿主に導入した形質転換体又はその後代。
【請求項15】
宿主が微生物又は植物である、請求項14に記載の形質転換体又はその後代。
【請求項16】
前記植物がユーカリ属植物又はタバコ属植物である、請求項15に記載の形質転換体又はその後代。
【請求項17】
請求項13に記載の発現ベクターを植物に導入して、該植物の生長を抑制する方法。
【図1】


【図2】

【図3】
【図4】
【図5】
【図6】
【図7】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2011−15645(P2011−15645A)
【公開日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願番号】特願2009−163138(P2009−163138)
【出願日】平成21年7月9日(2009.7.9)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成21年度、独立行政法人新エネルギー・産業技術総合開発機構、受託研究「環境安心イノベーションプログラム/植物機能を活用した高度モノづくり基盤技術開発/植物の物質生産プロセス制御基盤技術開発/循環型工業原料木質バイオマス統括的生産制御技術の研究開発」、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000122298)王子製紙株式会社 (2,055)
【Fターム(参考)】
【公開日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願日】平成21年7月9日(2009.7.9)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成21年度、独立行政法人新エネルギー・産業技術総合開発機構、受託研究「環境安心イノベーションプログラム/植物機能を活用した高度モノづくり基盤技術開発/植物の物質生産プロセス制御基盤技術開発/循環型工業原料木質バイオマス統括的生産制御技術の研究開発」、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000122298)王子製紙株式会社 (2,055)
【Fターム(参考)】
[ Back to top ]