
植物の種子休眠性を支配する遺伝子およびその利用
【課題】 植物における種子休眠性の程度を支配する新規な遺伝子を同定すること、並びに、同定された遺伝子を利用して、植物の種子休眠性の程度を効率的に判定する方法および種子休眠性が改変された植物を効率的に作出する方法を提供すること
【解決手段】 ポジショナルクローニングの手法により、オオムギの種子休眠性を支配するSD2遺伝子、およびこれに対応するコムギ遺伝子であるPhs1遺伝子を同定することに成功した。さらに、これら遺伝子において、その種子休眠性と相関する一塩基多型(SNP)を特定するとともに、弱種子休眠性型の遺伝子が優性であることを解明することにも成功した。同定した遺伝子を利用して、効率的に、植物の種子休眠性の程度を判定することや種子休眠性が改変された植物を作出することが可能であることを見出した。
【解決手段】 ポジショナルクローニングの手法により、オオムギの種子休眠性を支配するSD2遺伝子、およびこれに対応するコムギ遺伝子であるPhs1遺伝子を同定することに成功した。さらに、これら遺伝子において、その種子休眠性と相関する一塩基多型(SNP)を特定するとともに、弱種子休眠性型の遺伝子が優性であることを解明することにも成功した。同定した遺伝子を利用して、効率的に、植物の種子休眠性の程度を判定することや種子休眠性が改変された植物を作出することが可能であることを見出した。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、植物の種子休眠性を支配する遺伝子、並びに、それを利用した植物の種子休眠性の程度の判定および休眠性が改変された植物の作出に関する。
【背景技術】
【0002】
これまでに、オオムギやコムギなどの麦類の種子休眠性に関わる量的形質座(QTL)に関する報告は多数あり(非特許文献1〜2)、コムギでは、これまでに少なくとも30以上の種子休眠QTLが知られている。
【0003】
今までにオオムギのSD2領域である5H染色体長椀末端に種子休眠QTLが検出された報告は、Ullrichらの「Steptoe」と「Morex」の交配集団を使った例(非特許文献3)、武田らの「TR306」と「Harrington」の交配集団を使った例(非特許文献4)、Liらの「Chebec」と「Harrington」の交配集団を使った例(非特許文献5)、Pradaらの「Triumph」と「Morex」の交配集団を使った例(非特許文献6)、堀らの「はるな2条」と「OUH602」の交配集団を使った例(非特許文献7)などがあるが、いずれも種子休眠QTLの検出を報告しているだけで、原因遺伝子の単離は行われていない。
【0004】
SD2遺伝子の単離を目指した精密マッピングは、Ullrichのグループによって行われが(非特許文献8)、表現型の評価が難しく、精度の低いマッピングに終わっている。こうしたデータをもとに、イネとのシンテニー関係を利用した解析も行われ、SD2座の原因遺伝子が植物ホルモンであるジベレリンの生合成の律速段階を調整する酵素GA20-oxidaseである可能性が高いという報告もなされているが(非特許文献9)、推測に過ぎない。また、オオムギの大規模EST解析より、約3000マーカーが座乗する連鎖地図が作られ、SD2座と連鎖するマーカーが報告されているが(特許文献1)、SD2遺伝子本体を同定したわけではなく、既存のSD2のQTL解析のマーカーと同じくSD2座近傍のマーカーを同定したにすぎない。
【0005】
一方、コムギにおいては、効果の大きい主要なQTLが、複数の解析集団に共通して4A染色体の長腕(4AL)に検出されている(非特許文献10〜12)。これら4ALに検出されたQTLは、近傍のマーカー情報から互いに同一の遺伝子座である可能性が高いことが示唆された(非特許文献11)。その後、QTLが4ALに検出された報告例は続くものの、高精度マッピングやマップベース法による遺伝子単離へと発展した事例はない。これはコムギがA、B、Dゲノムからなる異質6倍体であり、イネの40倍にもおよぶゲノムサイズを有していることが要因であると考えられる。近年、イネとのシンテニーを利用することによりコムギでも特定の領域にDNAマーカーを設定できるようになり、高精度マッピングやマップベース法による遺伝子単離の成功例が報告されるようになった。コムギの種子休眠性については、戻し交雑集団と安定した登熟環境で種子休眠性を評価することにより、主働遺伝子(Phs1)を2.6cMの域内へマッピングすることに成功した(非特許文献13)。イネとのシンテニーを利用することにより、Phs1はさらに詳細にマッピングされた(非特許文献14)。しかしながら、ミクロなレベルでは、イネとコムギ間ではしばしば保存性が認められない。また、種子休眠性と密接に関連している植物ホルモンのアブシジン酸(ABA)のシグナリングに関わる遺伝子のホモログが、コムギにおいてマッピングされているが、4ALに相当する領域にはマッピングされなかった(非特許文献15)。以上のことから、既報の情報を利用してコムギPhs1の原因遺伝子を想定することは困難であり、マップベース法によりコムギPhs1遺伝子を同定するためには、コムギゲノムのさらなる絞込みとゲノム配列の解読および解析が必要である。
【0006】
このように、これまでオオムギにおけるSD2座やコムギにおけるPhs1座の報告は、QTLマッピングによる報告のみで、その困難性から、これら遺伝子の単離同定には至っていない。オオムギとコムギ以外の麦類についても、種子休眠QTLの原因遺伝子の単離同定が報告された例はない。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特許公開2005−229849号公報
【非特許文献】
【0008】
【非特許文献1】Hori et al. (2007) Theor Appl Genet 115:869-876
【非特許文献2】Flintham et al. (2002) Euphytica 126:39-45
【非特許文献3】Ullrich et al. (1992) Pre-harvest sprouting in cereals, p136-145.
【非特許文献4】Takeda (1995) 7th international symposium on pre-harvest sprouting in cereals, p205-212
【非特許文献5】Li et al. (2003) Australian Journal of Agricultural Research, vol.54, p1303-1313.
【非特許文献6】Prada et al. (2004) Theor Appl Genet, vol.109, p62-70.
【非特許文献7】Hori et al. (2007) Theor Appl Genet, vol.115, p869-876
【非特許文献8】Gao et al.(2003) Theor Appl Genet 107:552-559
【非特許文献9】Li et al. (2004) Funct Integr Genomics 4:84-93
【非特許文献10】Kato et al. (2001) Theor. Appl. Genet. 102:980-985.
【非特許文献11】Mares et al. (2005) Theor. Appl. Genet. 111:1357-1364.
【非特許文献12】Torada et al. (2005) Euphytica 143:251-255.
【非特許文献13】Torada et al. (2008) Genome. 51(6):426-32.
【非特許文献14】乕田ら 2010 大規模集団を用いたコムギ種子休眠性遺伝子座Phs1の精密マッピング 育種学研究 別冊2号 p.36
【非特許文献15】Nakamura et al. (2006) Theor. Appl. Genet. 114:1129-1139.
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、このような状況に鑑みてなされたものであり、その目的は、植物における種子休眠性の程度を支配する新規な遺伝子を同定することにある。また、本発明は、同定された遺伝子を利用して、植物の種子休眠性の程度を効率的に判定する方法を提供することを目的とする。さらなる本発明の目的は、同定された遺伝子を利用して、種子休眠性が改変された植物を効率的に作出する方法を提供することにある。
【課題を解決するための手段】
【0010】
本発明者らは、上記課題を解決すべく鋭意検討を行った結果、オオムギの種子休眠性を支配するSD2遺伝子、およびこれに対応するコムギ遺伝子であるPhs1遺伝子を同定することに成功した(以下、両遺伝子を合わせて「SD2/Phs1遺伝子」と称する)。さらに、本発明者らは、SD2/Phs1遺伝子において、その種子休眠性と相関する一塩基多型(SNP)を特定するとともに、弱種子休眠性型の遺伝子が優性であることを解明することにも成功した。そして、本発明者らは、同定したSD2/Phs1遺伝子を利用して、効率的に、植物の種子休眠性の程度を判定することや種子休眠性が改変された植物を作出することが可能であることを見出し、本発明を完成するに至った。
【0011】
本発明は、より詳しくは、下記を提供するものである。
【0012】
[1] 植物の種子休眠性の形質を弱める活性を有するタンパク質をコードする、下記(a)〜(d)のいずれかに記載のDNA。
(a)配列番号:2または4に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:1または3に記載の塩基配列のコード領域を含むDNA
(c)配列番号:2または4に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号:1または3に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNA
[2] 植物の種子休眠性の形質を強める活性を有するタンパク質をコードする、下記(a)〜(d)のいずれかに記載のDNA。
(a)配列番号:6または8に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:5または7に記載の塩基配列のコード領域を含むDNA
(c)配列番号:6または8に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号:5または7に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNA
[3] 植物の種子休眠性の形質を強める活性を有する、下記(a)〜(c)のいずれかに記載のDNA。
(a)[1]に記載のDNAの転写産物と相補的な二重鎖RNAをコードするDNA
(b)[1]に記載のDNAの転写産物と相補的なアンチセンスRNAをコードするDNA
(c)[1]に記載のDNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA
[4] [1]〜[3]のいずれかに記載のDNAを含むベクター。
【0013】
[5] [1]〜[3]のいずれかに記載のDNAが導入された植物細胞。
【0014】
[6] [5]に記載の細胞を含む植物植物体。
【0015】
[7] [6]に記載の植物体の子孫またはクローンである、植物植物体。
【0016】
[8] [6]または[7]に記載の植物植物体の繁殖材料。
【0017】
[9] 植物に[1]に記載のDNAを導入する工程を含む、種子休眠性の形質が弱められた植物の作出方法。
【0018】
[10] 植物における[1]に記載のDNAの発現または機能を抑制することを特徴とする、種子休眠性の形質が強められた植物の作出方法。
【0019】
[11] 植物に[2]または[3]に記載のDNAを導入する工程を含む、種子休眠性の形質が強められた植物の作出方法。
【0020】
[12] [1]に記載のDNA、または該DNAが挿入されたベクターを含む、植物の種子休眠性の形質を弱めるための薬剤。
【0021】
[13] [2]もしくは[3]に記載のDNA、または該DNAが挿入されたベクターを含む、植物の種子休眠性の形質を強めるための薬剤。
【0022】
[14] 植物における種子休眠性の程度を判定する方法であって、被検植物における[1]もしくは[2]に記載のDNAまたはその発現制御領域の塩基配列を解析し、対照の塩基配列と比較することを特徴とする方法。
【0023】
[15] 弱い種子休眠性の形質の植物を育種する方法であって、
(a)弱い種子休眠性の形質の植物品種と任意の植物品種とを交配させる工程、
(b)工程(a)における交配により得られた個体における種子休眠性の程度を、[14]に記載の方法により判定する工程、および
(c)弱い種子休眠性を有すると判定された品種を選抜する工程、を含む方法。
【0024】
[16] 強い種子休眠性の形質の植物を育種する方法であって、
(a)強い種子休眠性の形質の植物品種と任意の植物品種とを交配させる工程、
(b)工程(a)における交配により得られた個体における種子休眠性の程度を、[14]に記載の方法により判定する工程、および
(c)強い種子休眠性を有すると判定された品種を選抜する工程、を含む方法。
【0025】
なお、本発明において「種子休眠性」とは、種子の発芽に適した条件下においても発芽しない性質を意味する。また、本発明において「弱い種子休眠性の形質」とは、種子が成熟した後に休眠性の維持される期間が短く、発芽に適した検定条件下において高い発芽率を示すことを意味する。また、本発明において「強い種子休眠性の形質」とは、種子が成熟した後に休眠性の維持される期間が長く、発芽に適した検定条件下において低い発芽率を示すことを意味する。
【発明の効果】
【0026】
本発明によって、植物の種子休眠性を支配する新規な遺伝子SD2/Phs1が同定され、該遺伝子の染色体上の位置および構造が解明された。これによりSD2/Phs1遺伝子を標的とした植物の種子休眠性の程度の判定方法、および該判定方法を利用した種子休眠性が改変された植物の育種方法が提供された。さらに、SD2/Phs1遺伝子を利用した、休眠性が改変された植物の作出方法が提供された。本発明による植物の種子休眠性の程度の判定や休眠性が改変された植物の作出や育種は、種子休眠性の程度を支配するSD2/Phs1遺伝子に着目しているため、この遺伝子と連鎖するマーカーを利用した従来法に比して、精度が高い。このため従来法よりも、特異的かつ効率的に、植物の種子休眠性の程度の判定や休眠性が改変された植物の作出や育種を行うことが可能である。
【図面の簡単な説明】
【0027】
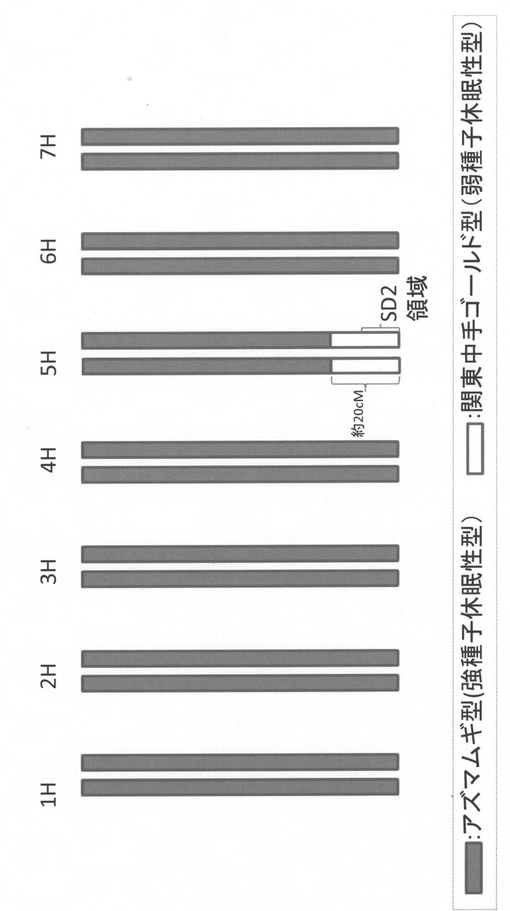
【図1】NIL#34の遺伝子型を示す図である。
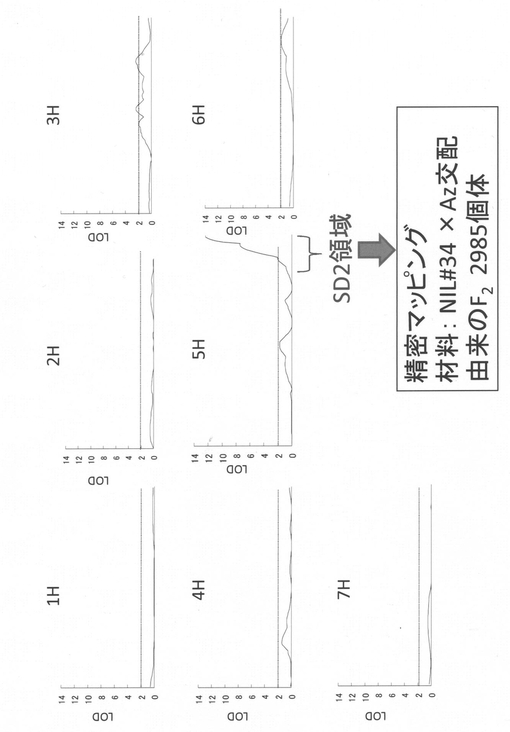
【図2】アズマムギ(Az)と関東中生ゴールド(KNG)の組換え近交系(RIL)を用いた種子休眠QTL解析法を示す図である。
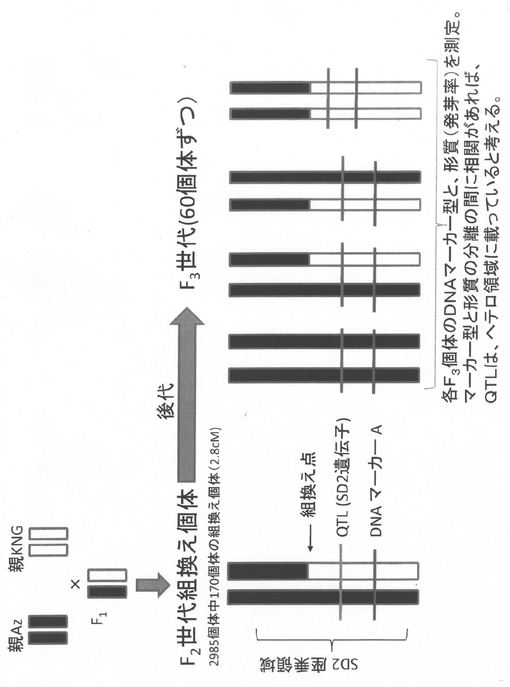
【図3】SD2遺伝子が座乗する領域を絞り込むための、アソシエーション解析法を示す図である。
【図4】アソシエーション解析結果とジェノタイピング結果のまとめの図である。
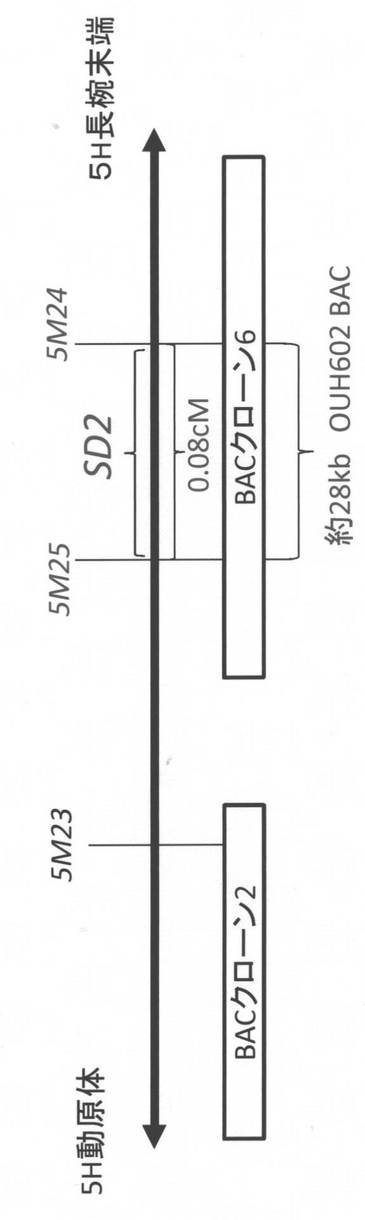
【図5】SD2遺伝子の精密マッピングとBACクローンとの対応関係を示す図である。
【図6】RiceGAASにより、SD2遺伝子が座乗する領域のゲノム配列中に存在する遺伝子を予測した結果を示す図である。
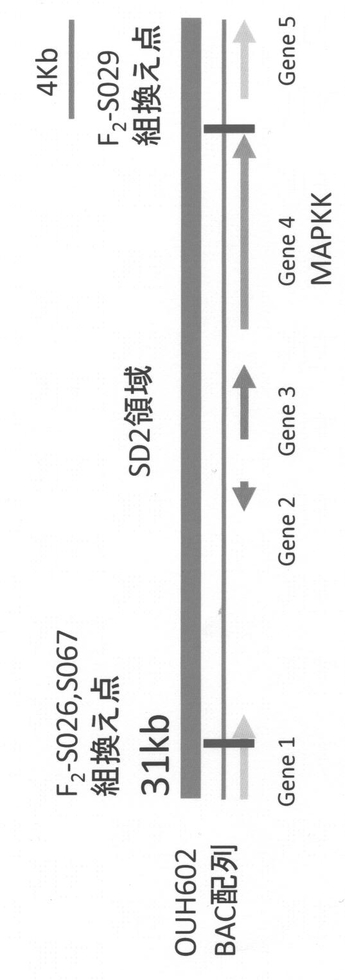
【図7】各オオムギ系統におけるGene4遺伝子のアミノ酸配列の比較を示す図である。Harunanijoは、はるな2条を、KNGは、関東中生ゴールドを、Azは、アズマムギを示す。三角は、植物間(イネ、トウモロコシ、ナズナ、ポプラ、タバコ、コケ、マツナ属Suaeda salsa、トウゴマ、ブドウ等)で高度に保存されているアミノ酸残基、丸は、植物間で相同性が高くないアミノ酸残基を示す。Harunanijoのアミノ酸配列の上に示したラインは、MAPKKドメインを示す。
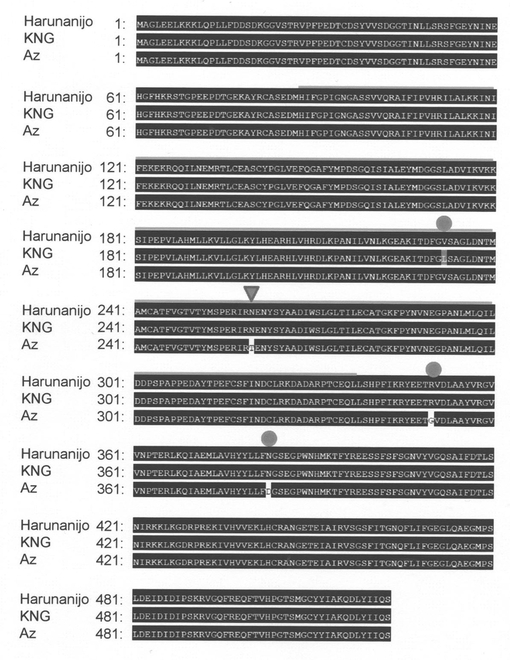
【図8】Leader(強種子休眠性型)/春よ恋(弱種子休眠性型)BC3F2個体の種子休眠性とPhs1の遺伝子型を示す図である。
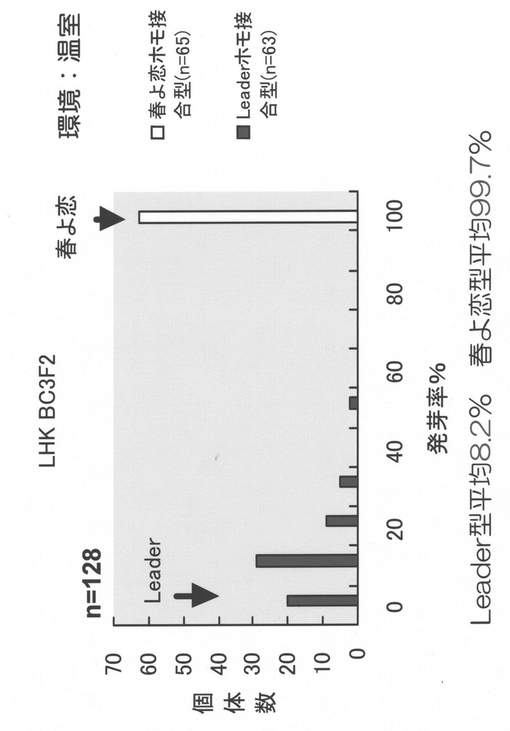
【図9】コムギ種子休眠性遺伝子座Phs1近傍の高精度連鎖地図とBACコンティグを示す図である。
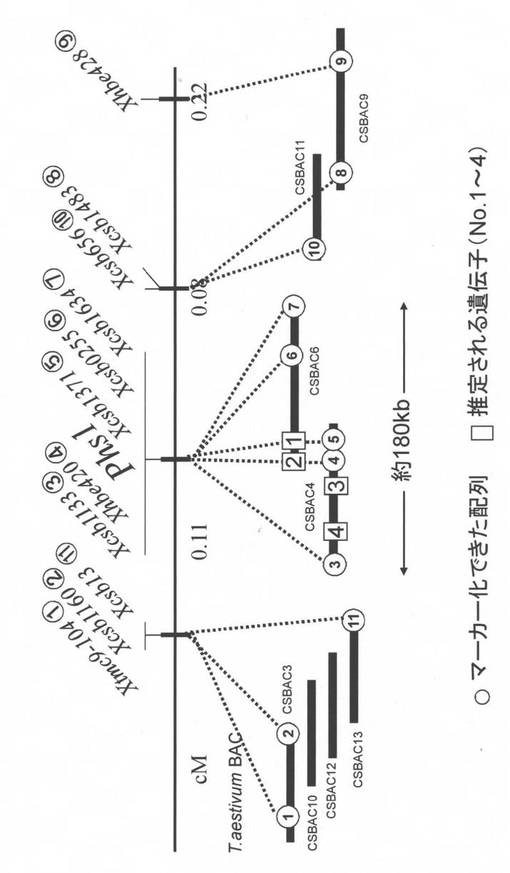
【図10】コムギPhs1候補領域を、イネとオオムギの対応領域と比較した地図である。
【図11】Phs1(TaMEK-A1)遺伝子の構造の模式図である。
【図12】休眠性の異なる14品種・系統における4遺伝子(No.1〜4)近傍配列の変異を示す図である。
【図13】Phs1(TaMEK-A1)タンパク質の構造の模式図である。
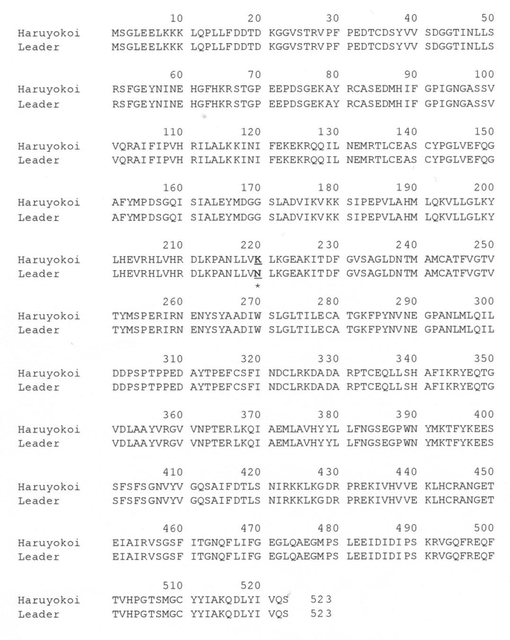
【図14】春よ恋とLeader間のPhs1(TaMEK-A1)遺伝子のアミノ酸配列の比較を示す図である。下線太字は、春よ恋とLeader間で異なるアミノ酸残基を示す。
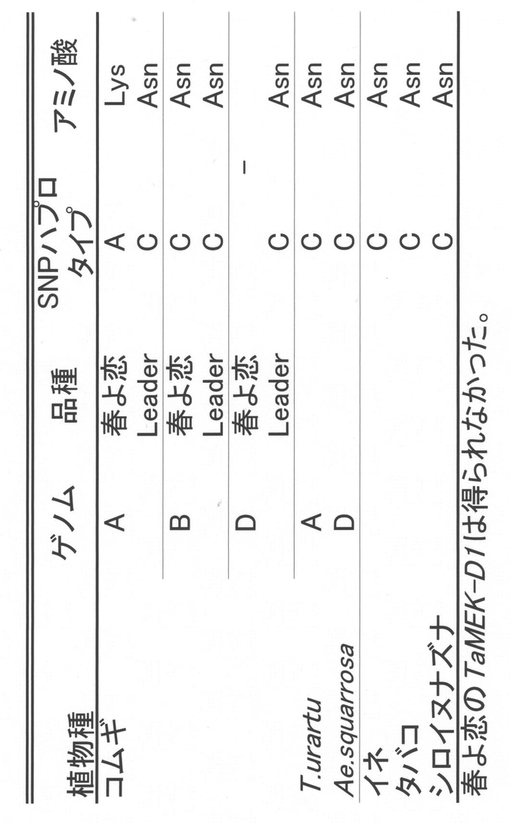
【図15】Phs1(TaMEK-A1)同祖遺伝子およびホモログにおけるSNPのハプロタイプを示す図である。
【図16】RT-PCRによるPhs1(TaMEK-A1)の発現解析結果を示す写真である。
【発明を実施するための形態】
【0028】
<弱種子休眠性型DNA、強種子休眠性型DNA>
本発明は、植物の種子休眠性の形質を弱める活性を有するタンパク質をコードするDNA(以下、「弱種子休眠性型DNA」と称する)を提供する。本発明者らにより同定された、オオムギの弱種子休眠性品種である関東中手ゴールド由来のSD2cDNAの塩基配列を配列番号:1に、該DNAがコードするタンパク質のアミノ酸配列を配列番号:2に示す。また、コムギの弱種子休眠性品種である春よ恋由来のPhs1cDNAの塩基配列を配列番号:3に、該DNAがコードするタンパク質のアミノ酸配列を配列番号:4に示す。
【0029】
本発明の弱種子休眠性型DNAの1つの態様は、配列番号:2に記載のアミノ酸配列からなるタンパク質をコードするDNA(典型的には、配列番号:1に記載の塩基配列のコード領域を含むDNA)であり、他の1つの態様は、配列番号:4に記載のアミノ酸配列からなるタンパク質をコードするDNA(典型的には、配列番号:2に記載の塩基配列のコード領域を含むDNA)である。
【0030】
本実施例において示されたように、オオムギにおいて、関東中手ゴールド由来のSD2タンパク質のアミノ酸配列(配列番号:2)とはるな二条由来のSD2タンパク質のアミノ酸配列とを比較すると1か所のアミノ酸が相違しているが(図7)、ともに弱種子休眠性型である。現在の技術水準においては、当業者であれば、特定の弱種子休眠性植物品種(例えば、オオムギの関東中手ゴールド、コムギの春よ恋)における弱種子休眠性型DNAの塩基配列情報が得られた場合、その塩基配列を改変し、そのコードするアミノ酸配列は異なるが、同じく弱種子休眠性型であるDNAを取得することが可能である。また、自然界においても、塩基配列の変異によりコードするタンパク質のアミノ酸配列が変異することは起こり得ることである。従って、本発明は、オオムギの関東中手ゴールドあるいはコムギの春よ恋におけるSD2/Phs1タンパク質のアミノ酸配列(配列番号:2または4)において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなり、植物の種子休眠性の形質を弱める活性を有するタンパク質をコードするDNAをも含むものである。ここで「複数」とは、改変後のSD2/Phs1タンパク質が植物の種子休眠性の形質を弱める活性を維持する範囲における、アミノ酸の改変数であり、通常、50アミノ酸以内、好ましくは30アミノ酸以内、さらに好ましくは10アミノ酸以内(例えば、5アミノ酸以内、3アミノ酸以内、2アミノ酸)である。
【0031】
さらに、現在の技術水準においては、当業者であれば、特定の弱種子休眠性植物品種(例えば、オオムギの関東中手ゴールド、コムギの春よ恋)から弱種子休眠性型DNAが得られた場合、その弱種子休眠性型DNAの塩基配列情報を利用して、他の植物品種(例えば、イネ、トウモロコシ、ダイズ、ポプラ、ブドウ、シロイヌナズナ、タバコなど)から、同じく弱種子休眠性型である相同遺伝子をコードするDNAを取得することが可能である。従って、本発明は、オオムギの関東中手ゴールドおよびコムギの春よ恋におけるSD2/Phs1DNA(配列番号:1または3)とストリンジェントな条件でハイブリダイズするDNAであって、植物の種子休眠性の形質を弱める活性を有するタンパク質をコードするDNAをも含むものである。
【0032】
こうして得られた変異DNAや相同DNAが、植物の種子休眠性の形質を弱める活性を有するタンパク質をコードするか否かは、例えば、遺伝子組み換え技術または交配により、これらDNAを導入した強種子休眠性型の品種から種子を採取し、本実施例に記載の発芽試験を行い、その発芽率が高まるか否かを検定することにより、判定することができる。発芽率が高くなれば、植物の種子休眠性の形質を弱める活性を有すると評価される。
【0033】
本発明は、また、植物の種子休眠性の形質を強める活性を有するタンパク質をコードするDNA(以下、「強種子休眠性型DNA」と称する)を提供する。本発明者らにより同定された、オオムギの強種子休眠性品種であるアズマムギ由来のPhs1cDNAの塩基配列を配列番号:5に、該DNAがコードするタンパク質のアミノ酸配列を配列番号:6に示す。また、また、コムギの強種子休眠性品種であるLeader由来のPhs1cDNAの塩基配列を配列番号:7に、該DNAがコードするタンパク質のアミノ酸配列を配列番号:8に示す。
【0034】
本発明の強種子休眠性型DNAの1つの態様は、配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA(典型的には、配列番号:5に記載の塩基配列のコード領域を含むDNA)であり、他の1つの態様は、配列番号:8に記載のアミノ酸配列からなるタンパク質をコードするDNA(典型的には、配列番号:7に記載の塩基配列のコード領域を含むDNA)である。
【0035】
本実施例において示されたように、オオムギのアズマムギ由来のSD2タンパク質のアミノ酸配列(配列番号:6)は、弱種子休眠性品種である関東中手ゴールド由来のSD2タンパク質のアミノ酸配列(配列番号:2)と比較すると260位においてアミノ酸の置換(N→T)が生じている(図7)。また、コムギのLeader由来のPhs1タンパク質のアミノ酸配列(配列番号:8)は、弱種子休眠性品種である春よ恋由来のPhs1タンパク質のアミノ酸配列(配列番号:4)と比較すると220位においてアミノ酸の置換(K→N)が生じている(図13、図14)。このため、本来のSD2/Phs1タンパク質の機能が抑制され、これにより個体に強種子休眠性の形質を付与していると考えられる。現在の技術水準においては、当業者であれば、特定の弱種子休眠性植物品種(例えば、オオムギのアズマムギ、コムギのLeader)のSD2/Phs1DNAの塩基配列において、そのコードするタンパク質の強種子休眠性の形質が維持されるような改変を行うことが可能である。また、自然界においても、塩基配列の変異によりコードするタンパク質のアミノ酸配列が変異することは起こり得ることである。従って、本発明は、オオムギのアズマムギあるいはコムギのLeaderにおけるSD2/Phs1タンパク質のアミノ酸配列(配列番号:6または8)において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質からなり、植物の種子休眠性の形質を強める活性を有するタンパク質をコードするDNAをも含むものである。ここで「複数」とは、改変後のSD2/Phs1タンパク質が植物の種子休眠性の形質を強める活性を有する範囲における、アミノ酸の改変数である。植物におけるSD2/Phs1タンパク質が本来の機能を発揮しなければ、強い種子休眠性の形質になると考えられるため、当該アミノ酸改変数は、本質的に制限はない。改変は、例えば、100アミノ酸以内(50アミノ酸以内、30アミノ酸以内、10アミノ酸以内、5アミノ酸以内、3アミノ酸以内、2アミノ酸)である。
【0036】
さらに、現在の技術水準においては、当業者であれば、特定の植物品種からSD2/Phs1DNAが得られた場合、そのDNAの塩基配列情報を利用して、他の植物品種(例えば、イネ、トウモロコシ、ダイズ、ポプラ、ブドウ、シロイヌナズナ、タバコなど)から、強種子休眠性型である相同遺伝子をコードするDNAを取得することができる。従って、本発明は、オオムギのアズマムギあるいはコムギのLeaderにおけるSD2/Phs1DNA(配列番号:5または7)とストリンジェントな条件でハイブリダイズするDNAであって、植物の種子休眠性の形質を強める活性を有するタンパク質をコードするDNAが含まれる。
【0037】
こうして得られた変異DNAや相同DNAが、植物の種子休眠性の形質を強める活性を有するタンパク質をコードするか否かは、例えば、当該DNAで、弱種子休眠性品種のSD2/Phs1遺伝子を組換え、当該DNAをホモで保持する植物を作出し、該植物から種子を採取し、本実施例に記載の発芽試験を行い、その発芽率が低くなるか否かを検定することにより、判定することができる。発芽率が低くなれば、植物の種子休眠性の形質を強める活性を有すると評価される。
【0038】
本発明の弱種子休眠性型DNAは、その導入により、植物の種子休眠性の形質を弱めることが可能であるという意味において、植物の種子休眠性の形質を弱めるための薬剤であり、一方、本発明の強種子休眠性型DNAは、その導入により、植物の種子休眠性の形質を強めることが可能であるという意味において、植物の種子休眠性を強めるための薬剤である。
【0039】
なお、上記した変異DNAを作製するための、DNAへの人為的な変異の導入は、例えば、部位特異的変異誘発(site-directed mutagenesis)法(Kramer, W. & Fritz, HJ., Methods Enzymol, 154:350-367, 1987)により行うことができる。
【0040】
また、上記した相同遺伝子を単離するための方法としては、例えば、ハイブリダイゼーション技術(Southern, E. M., Journal of Molecular Biology, 98:503, 1975)やポリメラーゼ連鎖反応(PCR)技術(Saiki, R. K., et al. Science, 230:1350-1354, 1985、Saiki, R. K. et al. Science, 239:487-491, 1988)が挙げられる。相同遺伝子をコードするDNAを単離するためには、通常ストリンジェントな条件下でハイブリダイゼーション反応を行なう。ストリンジェントなハイブリダイゼーション条件としては、6M尿素、0.4%SDS、0.5xSSCの条件またはこれと同等のストリンジェンシーのハイブリダイゼーション条件を例示できる。よりストリンジェンシーの高い条件、例えば、6M尿素、0.4%SDS、0.1xSSCの条件を用いれば、より相同性の高いDNAの単離を期待することができる。単離されたDNAは、核酸レベルあるいはアミノ酸配列レベルにおいて、少なくとも50%以上、さらに好ましくは70%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。配列の相同性は、BLASTN(核酸レベル)やBLASTX(アミノ酸レベル)のプログラム(Altschul et al. J. Mol. Biol., 215:403-410, 1990)を利用して決定することができる。該プログラムは、KarlinおよびAltschulによるアルゴリズムBLAST(Proc. Natl. Acad. Sci. USA, 87:2264-2268, 1990、Proc. Natl. Acad. Sci. USA, 90:5873-5877, 1993)に基づいている。BLASTNによって塩基配列を解析する場合には、パラメーターは例えばscore=100、wordlength=12とする。また、BLASTXによってアミノ酸配列を解析する場合には、パラメーターは例えばscore=50、wordlength=3とする。また、Gapped BLASTプログラムを用いて、アミノ酸配列を解析する場合は、Altschulら(Nucleic Acids Res. 25:3389-3402, 1997)に記載されているように行うことができる。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である。
【0041】
本発明のSD2/Phs1タンパク質をコードするDNAとしては、その形態に特に制限はなく、cDNAの他、ゲノムDNA、および化学合成DNAが含まれる。ゲノムDNAおよびcDNAの調製は、当業者にとって常套手段を利用して行うことが可能である。ゲノムDNAは、例えば、植物からゲノムDNAを抽出し、ゲノミックライブラリー(ベクターとしては、プラスミド、ファージ、コスミド、BAC、PACなどが利用できる)を作成し、これを展開して、SD2/Phs1遺伝子(例えば、配列番号:1,3,5または7のいずれかに記載のDNA)の塩基配列を基に調製したプローブを用いてコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより調製することが可能である。また、SD2/Phs1遺伝子に特異的なプライマーを作成し、これを利用したPCRを行うことによって調製することも可能である。また、cDNAは、例えば、植物から抽出したmRNAを基にcDNAを合成し、これをλZAP等のベクターに挿入してcDNAライブラリーを作成し、これを展開して、上記と同様にコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより、また、PCRを行うことにより調製することが可能である。
【0042】
<植物の弱種子休眠性型SD2/Phs1遺伝子の発現を抑制するために用いるDNA>
また、本発明は、植物の弱種子休眠性型SD2/Phs1遺伝子の発現を抑制するために用いるDNAを提供する。これらのDNAの導入により、植物の種子休眠性の形質を強めることが可能である。この意味において、植物の弱種子休眠性型SD2/Phs1遺伝子の発現を抑制するために用いるDNAは、植物の種子休眠性の形質を強めるための薬剤である。ここで「SD2/Phs1遺伝子の発現の抑制」には、遺伝子の転写の抑制およびタンパク質への翻訳の抑制の双方が含まれる。また、「発現の抑制」には、発現の完全な停止のみならず発現の減少も含まれる。
【0043】
植物の弱種子休眠性型SD2/Phs1遺伝子の発現を抑制するために用いるDNAの一つの態様は、上記した本発明の弱種子休眠性型DNAの転写産物と相補的なdsRNA(二重鎖RNA)をコードするDNAである。標的遺伝子配列と同一もしくは類似した配列を有するdsRNAを細胞内に導入することにより、導入した外来遺伝子および標的内因性遺伝子の発現がいずれも抑制される、RNAi(RNA干渉、RNA interference)と呼ばれる現象を引き起こすことができる。細胞に約40〜数百塩基対のdsRNAが導入されると、ヘリカーゼドメインを持つダイサー(Dicer)と呼ばれるRNaseIII様のヌクレアーゼが、ATP存在下で、dsRNAを3'末端から約21〜23塩基対ずつ切り出し、siRNA(short interference RNA)が生じる。このsiRNAに、特異的なタンパク質が結合して、ヌクレアーゼ複合体(RISC:RNA-induced silencing complex)が形成される。この複合体はsiRNAと同じ配列を認識して結合し、RNaseIII様の酵素活性によってsiRNAの中央部で標的遺伝子の転写産物(mRNA)を切断する。また、この経路とは別にsiRNAのアンチセンス鎖がmRNAに結合してRNA依存性RNAポリメラーゼ(RsRP)のプライマーとして作用し、dsRNAが合成される。このdsRNAが再びダイサーの基質となって、新たなsiRNAを生じて作用を増幅する経路も考えられている。
【0044】
本発明のdsRNAをコードするDNAは、標的遺伝子の転写産物(mRNA)のいずれかの領域に対するアンチセンスRNAをコードしたアンチセンスDNAと、該mRNAのいずれかの領域のセンスRNAをコードしたセンスDNAを含み、該アンチセンスDNAおよび該センスDNAより、それぞれアンチセンスRNAおよびセンスRNAを発現させることができる。また、これらのアンチセンスRNAおよびセンスRNAよりdsRNAを作成することができる。
【0045】
本発明のdsRNAの発現システムをベクター等に保持させる場合の構成としては、同一のベクターからアンチセンスRNAおよびセンスRNAを発現させる場合と、異なるベクターからそれぞれアンチセンスRNAとセンスRNAを発現させる場合がある。同一のベクターからアンチセンスRNAおよびセンスRNAを発現させる構成としては、例えば、アンチセンスDNAおよびセンスDNAの上流にそれぞれpolIII系のような短いRNAを発現し得るプロモーターを連結させたアンチセンスRNA発現カセットとセンスRNA発現カセットをそれぞれ構築し、これらカセットを同方向にあるいは逆方向にベクターに挿入する構成である。
【0046】
また、異なる鎖上に対向するように、アンチセンスDNAとセンスDNAとを逆向きに配置した発現システムを構成することもできる。この構成では、アンチセンスRNAコード鎖とセンスRNAコード鎖とが対となった一つの二本鎖DNA(siRNAコードDNA)が備えられ、その両側にそれぞれの鎖からアンチセンスRNAとセンスRNAとを発現し得るようにプロモーターを対向して備える。この場合には、センスRNAとアンチセンスRNAの下流に余分な配列が付加されることを避けるために、それぞれの鎖(アンチセンスRNAコード鎖、センスRNAコード鎖)の3'末端にターミネーターをそれぞれ備えることが好ましい。このターミネーターは、A(アデニン)塩基を4つ以上連続させた配列などを用いることができる。また、このパリンドロームスタイルの発現システムでは、二つのプロモーターの種類は異なっていることが好ましい。
【0047】
また、異なるベクターからアンチセンスRNAおよびセンスRNAを発現させる構成としては、例えば、アンチセンスDNAおよびセンスDNAの上流にそれぞれpolIII系のような短いRNAを発現し得るプロモーターを連結させたアンチセンスRNA発現カセットとセンスRNA発現カセットとをそれぞれ構築し、これらカセットを異なるベクターに保持させる構成である。
【0048】
本発明に用いるdsRNAとしては、siRNAが好ましい。「siRNA」は、細胞内で毒性を示さない範囲の短鎖からなる二重鎖RNAを意味する。標的SD2/Phs1遺伝子の発現を抑制することができ、かつ、毒性を示さなければ、その鎖長に特に制限はない。dsRNAの鎖長は、例えば、15〜49塩基対であり、好適には15〜35塩基対でり、さらに好適には21〜30塩基対である。
【0049】
本発明のdsRNAをコードするDNAとしては、標的配列のインバーテッドリピートの間に適当な配列(イントロン配列が望ましい)を挿入し、ヘアピン構造を持つダブルストランドRNA(self-complementary 'hairpin' RNA(hpRNA))を作るようなコンストラクト(Smith, N.A., et al. Nature, 407:319, 2000、Wesley, S. V. et al. Plant J. 27:581, 2001、Piccin, A. et al. Nucleic Acids Res. 29:E55, 2001)を用いることもできる。
【0050】
本発明のdsRNAをコードするDNAは、標的SD2/Phs1遺伝子の塩基配列と完全に同一である必要はないが、少なくとも70%以上、好ましくは80%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。配列の同一性は上述した手法(BLASTプログラム)により決定できる。
【0051】
dsRNAにおけるRNA同士が対合した二重鎖RNAの部分は、完全に対合しているものに限らず、ミスマッチ(対応する塩基が相補的でない)、バルジ(一方の鎖に対応する塩基がない)などにより不対合部分が含まれていてもよい。本発明においては、dsRNAにおけるRNA同士が対合する二重鎖RNA領域中に、バルジおよびミスマッチの両方が含まれていてもよい。
【0052】
植物の弱種子休眠性型SD2/Phs1遺伝子の発現を抑制するために用いるDNAの他の態様は、上記した本発明の弱種子休眠性型DNAの転写産物と相補的なアンチセンスRNAをコードするDNA(アンチセンスDNA)である。アンチセンスDNAが標的遺伝子の発現を抑制する作用としては、三重鎖形成による転写開始阻害、RNAポリメラーゼによって局部的に開状ループ構造がつくられた部位とのハイブリッド形成による転写抑制、合成の進みつつあるRNAとのハイブリッド形成による転写阻害、イントロンとエキソンとの接合点でのハイブリッド形成によるスプライシング抑制、スプライソソーム形成部位とのハイブリッド形成によるスプライシング抑制、mRNAとのハイブリッド形成による核から細胞質への移行抑制、キャッピング部位やポリ(A)付加部位とのハイブリッド形成によるスプライシング抑制、翻訳開始因子結合部位とのハイブリッド形成による翻訳開始抑制、開始コドン近傍のリボソーム結合部位とのハイブリッド形成による翻訳抑制、mRNAの翻訳領域やポリソーム結合部位とのハイブリッド形成によるペプチド鎖の伸長阻止、および核酸とタンパク質との相互作用部位とのハイブリッド形成による遺伝子発現抑制などが挙げられる。これらは、転写、スプライシング、または翻訳の過程を阻害して、標的遺伝子の発現を抑制する(平島および井上「新生化学実験講座2 核酸IV 遺伝子の複製と発現」,日本生化学会編,東京化学同人, pp.319-347, 1993)。本発明で用いられるアンチセンスDNAは、上記のいずれの作用で標的SD2/Phs1遺伝子の発現を抑制してもよい。一つの態様としては、標的遺伝子のmRNAの5'端近傍の非翻訳領域に相補的なアンチセンス配列を設計すれば、遺伝子の翻訳阻害に効果的であろう。しかし、コード領域もしくは3'側の非翻訳領域に相補的な配列も使用し得る。このように、遺伝子の翻訳領域だけでなく非翻訳領域の配列のアンチセンス配列を含むDNAも、本発明で利用されるアンチセンスDNAに含まれる。使用されるアンチセンスDNAは、適当なプロモーターの下流に連結され、好ましくは3'側に転写終結シグナルを含む配列が連結される。
【0053】
アンチセンスDNAは、本発明の弱種子休眠性型DNA(例えば、配列番号:1または3に記載の塩基配列からなるDNA)の配列情報を基にホスホロチオネート法(Stein, Nucleic Acids Res., 16:3209-3221, 1988)などにより調製することが可能である。調製されたDNAは、後述する公知の方法で、植物へ導入できる。アンチセンスDNAの配列は、植物が持つ内因性の弱種子休眠性SD2/Phs1遺伝子の転写産物と相補的な配列であることが好ましいが、遺伝子の発現を有効に阻害できる限り、完全に相補的でなくてもよい。転写されたRNAは、標的とする遺伝子の転写産物に対して好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の相補性を有する。効果的に標的遺伝子の発現を阻害するには、アンチセンスDNAの長さは、少なくとも15塩基以上であり、好ましくは100塩基以上であり、さらに好ましくは500塩基以上である。通常、用いられるアンチセンスDNAの長さは5kbよりも短く、好ましくは2.5kbよりも短い。
【0054】
植物の弱種子休眠性型SD2/Phs1遺伝子の発現を抑制するために用いるDNAの他の態様は、本発明の弱種子休眠性型DNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNAである。リボザイムには、グループIイントロン型や、RNasePに含まれるM1RNAのように400ヌクレオチド以上の大きさのものもあるが、ハンマーヘッド型やヘアピン型と呼ばれる40ヌクレオチド程度の活性ドメインを有するものもある(小泉誠および大塚栄子、蛋白質核酸酵素, 35:2191, 1990)。
【0055】
例えば、ハンマーヘッド型リボザイムの自己切断ドメインは、G13U14C15のC15の3'側を切断するが、活性にはU14が9位のAと塩基対を形成することが重要とされ、15位の塩基はCの他にAまたはUでも切断されることが示されている(Koizumi et. al., FEBS Lett. 228:225, 1988)。リボザイムの基質結合部を標的部位近傍のRNA配列と相補的になるように設計すれば、標的RNA中のUC、UUまたはUAという配列を認識する制限酵素的なRNA切断リボザイムを作出することが可能である(Koizumi et. al., FEBS Lett. 239:285, 1988、小泉誠および大塚栄子,蛋白質核酸酵素,35:2191, 1990、Koizumi et. al., Nucleic. Acids. Res. 17:7059, 1989)。
【0056】
また、ヘアピン型リボザイムも、本発明の目的のために有用である。ヘアピン型リボザイムは、例えばタバコリングスポットウイルスのサテライトRNAのマイナス鎖に見出される(Buzayan, Nature 323:349, 1986)。このリボザイムも、標的特異的なRNA切断を起こすように設計できることが示されている(Kikuchi and Sasaki, Nucleic Acids Res. 19:6751, 1992、菊池洋,化学と生物 30:112, 1992)。標的を切断できるよう設計されたリボザイムは、植物細胞中で転写されるようにカリフラワーモザイクウイルスの35Sプロモーターなどのプロモーターおよび転写終結配列に連結される。このような構成単位をタンデムに並べ、標的遺伝子内の複数の部位を切断できるようにして、より効果を高めることもできる(Yuyama et al., Biochem. Biophys. Res. Commun. 186:1271, 1992)。このようなリボザイムを用いて標的となるSD2/Phs1遺伝子の転写産物を特異的に切断し、該遺伝子の発現を抑制することができる。
【0057】
<ベクター、形質転換植物細胞、形質転換植物植物体>
本発明は、また、上記本発明のDNA(弱種子休眠性型DNA、強種子休眠性型DNA、SD2/Phs1遺伝子の発現を抑制するためのDNA)を含むベクター、上記本発明のDNAまたはそれを含むベクターが導入された植物細胞、該細胞を含む植物植物体、該植物体の子孫またはクローンである植物植物体、および、これら植物植物体の繁殖材料を提供する。
【0058】
本発明のベクターとしては、植物細胞内で挿入遺伝子を発現させることが可能なものであれば特に制限はない。本発明のベクターは、本発明のDNAを恒常的または誘導的に発現させるためのプロモーターを含有しうる。恒常的に発現させるためのプロモーターとしては、例えば、カリフラワーモザイクウイルスの35Sプロモーター、イネのアクチンプロモーター、トウモロコシのユビキチンプロモーターなどが挙げられる。また、誘導的に発現させるためのプロモーターとしては、例えば、糸状菌・細菌・ウイルスの感染や侵入、低温、高温、乾燥、紫外線の照射、特定の化合物の散布などの外因によって発現することが知られているプロモーターなどが挙げられる。このようなプロモーターとしては、例えば、糸状菌・細菌・ウイルスの感染や侵入によって発現するイネキチナーゼ遺伝子のプロモーターやタバコのPRタンパク質遺伝子のプロモーター、低温によって誘導されるイネのlip19遺伝子のプロモーター、高温によって誘導されるイネのhsp80遺伝子とhsp72遺伝子のプロモーター、乾燥によって誘導されるシロイヌナズナのrab16遺伝子のプロモーター、紫外線の照射によって誘導されるパセリのカルコン合成酵素遺伝子のプロモーター、嫌気的条件で誘導されるトウモロコシのアルコールデヒドロゲナーゼ遺伝子のプロモーターなどが挙げられる。また、イネキチナーゼ遺伝子のプロモーターとタバコのPRタンパク質遺伝子のプロモーターはサリチル酸などの特定の化合物によって、rab16は植物ホルモンのアブシジン酸の散布によっても誘導される。
【0059】
本発明のベクターを導入する植物細胞の由来する植物としては特に制限はなく、例えば、オオムギ、コムギ、ライムギ、ライコムギ、エンバク、イネ、トウモロコシ、ダイズ、ポプラ、ブドウ、シロイヌナズナ、タバコなどが挙げられる。好ましくは、オオムギ、コムギ、ライムギ、ライコムギ、エンバク、イネ、トウモロコシなどのイネ科植物であり、より好ましくは、オオムギ、コムギ、ライムギ、ライコムギ、エンバクなどの麦類であり、最も好ましくはオオムギおよびコムギである。本発明の植物細胞には、培養細胞の他、植物体中の細胞も含まれる。また、種々の形態の植物細胞、例えば、懸濁培養細胞、プロトプラスト、葉の切片、カルス、未熟胚、花粉などが含まれる。植物細胞へのベクターの導入は、ポリエチレングリコール法、電気穿孔法(エレクトロポーレーション)、アグロバクテリウムを介する方法、パーティクルガン法など当業者に公知の種々の方法を用いることができる。
【0060】
形質転換植物細胞からの植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。例えば、オオムギに関する形質転換植物体を作出する手法としては、Tingayら(Tingay S. et al. Plant J. 11: 1369-1376, 1997)、Murrayら(Murray F et al. Plant Cell Report 22: 397-402, 2004)、およびTravallaら(Travalla S et al. Plant Cell Report 23: 780-789, 2005)に記載された方法を挙げることができる。また、イネにおいて、形質転換植物体を作出する手法については、ポリエチレングリコールによりプロトプラストへ遺伝子導入し、植物体を再生させる方法(Datta, S. K. In Gene Transfer To Plants(Potrykus I and Spangenberg Eds.) pp66-74, 1995)、電気パルスによりプロトプラストへ遺伝子導入し、植物体を再生させる方法(Toki et al. Plant Physiol. 100, 1503-1507, 1992)、パーティクルガン法により細胞へ遺伝子を直接導入し、植物体を再生させる方法(Christou et al. Bio/technology, 9: 957-962, 1991)およびアグロバクテリウムを介して遺伝子を導入し、植物体を再生させる方法(Hiei et al. Plant J. 6: 271-282, 1994)など、いくつかの技術が既に確立し、本願発明の技術分野において広く用いられている。また、シロイヌナズナであれば、Akamaら(Akama et al. Plant Cell Reports 12: 7-11, 1992)の方法が挙げられる。本発明においては、これらの方法を好適に用いることができる。
【0061】
一旦、ゲノム内に本発明の閉花性型DNAが導入された形質転換植物体が得られれば、該植物体から有性生殖または無性生殖により子孫を得ることが可能である。また、該植物体やその子孫あるいはクローンから繁殖材料(例えば、種子、果実、切穂、株、カルス、プロトプラスト等)を得て、それらを基に該植物体を量産することも可能である。本発明には、上記本発明のDNAが導入された植物細胞、該細胞を含む植物体、該植物体の子孫およびクローン、ならびに該植物体、その子孫およびクローンの繁殖材料が含まれる。
【0062】
<種子休眠性の形質が改変された植物の作出方法>
本発明は、また、種子休眠性の形質が改変された植物の作出方法を提供する。本発明の方法の1つの態様は、植物に本発明の弱種子休眠性型DNAを導入する工程を含む、種子休眠性の形質が弱められた植物の作出方法である。本発明において、植物の種子休眠性の形質を「弱める」とは、強種子休眠性の形質を有する品種の種子休眠性の形質を弱めることのみならず、既に、一定の弱種子休眠性の形質を有している品種の種子休眠性をさらに減弱させることをも含む意である。
【0063】
本発明の方法の他の1つの態様は、植物において、本発明の弱種子休眠性型DNA(弱種子休眠性型SD2/Phs1遺伝子)の発現または機能を抑制する工程を含む、種子休眠性の形質が強められた植物の作出方法である。本発明において、植物の種子休眠性の形質を「強める」とは、弱種子休眠性の形質を有する品種の種子休眠性の形質を強めることのみならず、既に、一定の強種子休眠性の形質を有している品種の種子休眠性をさらに増大させることをも含む意である。
【0064】
植物における本発明の弱種子休眠性型DNAの発現または機能の抑制は、植物に、上記本発明の強種子休眠性型DNAを導入することにより実施することができる。植物における強種子休眠性の形質は、単一劣性遺伝子支配であるため、植物に強種子休眠性の形質の形質を付与するためには、通常、個体におけるSD2/Phs1対立遺伝子の双方を強種子休眠性型DNAにする必要がある。これにより個体中で強種子休眠性型DNAのみが発現し、植物の種子休眠性の形質を強めることができる。植物染色体への本発明の強種子休眠性型DNAの導入は、例えば、交配や相同組換えにより行うことができる。強種子休眠性型DNAを導入することに代えて、植物染色体上の弱種子休眠性型DNAに、特定のDNA配列を導入し、その機能を破壊してもよい。
【0065】
また、植物における本発明の弱種子休眠性型DNAの発現または機能の抑制は、植物に上記本発明のSD2/Phs1遺伝子の発現を抑制するためのDNA(例えば、dsRNAをコードするDNA、アンチセンスDNA、リボザイム活性を有するRNAをコードするDNAなど)を導入することにより実施することができる。これにより個体中の弱種子休眠性型DNAから、弱種子休眠性型の翻訳産物が生産されなくなるため、植物の種子休眠性の形質を強めることができる。
【0066】
植物における本発明の弱種子休眠性型DNAの発現または機能の抑制には、例えば、弱種子休眠性型DNAの発現を抑制する薬剤や弱種子休眠性型の翻訳産物に結合し、その機能を抑制する薬剤の利用も考えられる。
【0067】
<植物における種子休眠性の程度を判定する方法>
本発明は、また、植物の種子休眠性の程度を判定する方法を提供する。本発明の判定方法の一つの態様は、植物におけるSD2/Phs1遺伝子またはその発現制御領域の塩基配列を解析し、対照の塩基配列と比較することを特徴とする方法である。
【0068】
SD2/Phs1遺伝子またはその発現制御領域の塩基配列の解析に際しては、SD2/Phs1遺伝子またはその発現制御領域のDNAをPCRにより増幅した増幅産物を用いることができる。前記PCRを実施する場合において、用いられるプライマーは、SD2/Phs1遺伝子またはその発現制御領域を特異的に増幅できるものである限り制限はなく、SD2/Phs1遺伝子またはその発現制御領域の配列情報(例えば、配列番号:1,3,5または7)に基づいて適宜設計することができる。好適なプライマーとしては、配列番号:9〜12に記載のプライマーが挙げられる。設計したプライマーを適宜組み合わせて、SD2/Phs1遺伝子またはその発現制御領域の特定の塩基配列を増幅することができる。
【0069】
被検植物におけるSD2/Phs1遺伝子の塩基配列と比較する「対照の塩基配列」は、典型的には、弱種子休眠性型品種または強種子休眠性型品種におけるSD2/Phs1遺伝子の塩基配列である。決定したSD2/Phs1遺伝子の塩基配列と弱種子休眠性型品種における塩基配列(例えば、配列番号:1、3)または強種子休眠性型品種における塩基配列(例えば、配列番号:5、7)とを比較することにより、被検植物におけるSD2/Phs1遺伝子が、強種子休眠性型であるか弱種子休眠性型であるかを評価することができる。本実施例において示されたように、オオムギの強種子休眠性品種であるアズマムギ由来のSD2タンパク質のアミノ酸配列(配列番号:6)は、弱種子休眠性品種である関東中手ゴールド由来のSD2タンパク質のアミノ酸配列(配列番号:2)と比較すると260位においてアミノ酸の置換(N→T)が生じている(図7)。また、コムギの強種子休眠性品種であるLeader由来のPhs1タンパク質のアミノ酸配列(配列番号:8)は、弱種子休眠性品種である春よ恋由来のPhs1タンパク質のアミノ酸配列(配列番号:4)と比較すると220位においてアミノ酸の置換(K→N)が生じている(図13、図14)。従って、特に、オオムギにおける260位およびコムギにおける220位のアミノ酸は、このような評価の好ましい指標となる。
【0070】
また、SD2/Phs1遺伝子においては、弱種子休眠性型DNAが優性であるため、被検DNAがコードするタンパク質の機能を喪失させるような大きな変異が存在する場合(例えば、新たな終止コドンの出現やフレームシフトにより、コードするタンパク質の分子量やアミノ酸配列に大きな変化が生じる場合)には、強種子休眠性型である蓋然性が高いと判定される。
【0071】
また、発現制御領域の塩基配列において、SD2/Phs1遺伝子の発現を抑制するような変異が存在する場合、強種子休眠性型である蓋然性が高いと判定される。発現制御領域の塩基配列における変異がSD2/Phs1遺伝子の発現に影響を与えるか否かは、当該変異を有する発現制御領域の下流にレポーター遺伝子を発現可能に連結したベクターを構築し、当該ベクターを植物細胞に導入し、レポーター活性を検出することにより、判定することができる。
【0072】
被検植物におけるSD2/Phs1遺伝子またはその発現制御領域の塩基配列が、対照の塩基配列と相違するか否かは、上記した直接的な塩基配列の決定以外に、種々の方法により間接的に解析することができる。このような方法としては、例えば、制限酵素断片長多型(Restriction Fragment Length Polymorphism/RFLP)を利用したRFLP法やPCR-RFLP法、PCR-SSCP(single-strand conformation polymorphism、一本鎖高次構造多型)法、変性剤濃度勾配ゲル電気泳動法(denaturant gradient gel electrophoresis:DGGE)、アレル特異的オリゴヌクレオチド(Allele Specific Oligonucleotide/ASO)ハイブリダイゼーション法、リボヌクレアーゼAミスマッチ切断法が挙げられる。例えば、PCR-RFLP法において、配列番号:9と10に記載のプライマーを用いてコムギのPhs1遺伝子を増幅し、制限酵素SalIで処理した場合、春よ恋型のDNAのみが切断される。一方、配列番号:11と12プライマーに記載のプライマーを用いてコムギのPhs1遺伝子を増幅し、制限酵素DraIで処理した場合、Leader型のDNAのみが切断される。こうして制限酵素処理したPCR産物をアガロースゲルで検出することにより、簡便にPhs1遺伝子のタイプ判定することができる。
【0073】
なお、本発明の判定方法における、被検植物からのDNAの調製は、常法、例えば、CTAB法を用いて行うことができる。DNAの調製には、例えば、植物の種子、幼植物体、成長した植物体を用いることができる。また、塩基配列の決定は、常法、例えば、ジデオキシ法やマキサム-ギルバート法などにより行なうことができる。塩基配列の決定においては、市販のシークエンスキットおよびシークエンサーを利用することができる。
【0074】
<種子休眠性の形質が改変された植物を育種する方法>
本発明は、また、種子休眠性の形質が改変された植物を育種する方法を提供する。本発明の育種方法の1つの態様は、強種子休眠性の形質の植物を育種する方法であり、(a)強種子休眠性の形質の植物品種と任意の植物品種とを交配させる工程、(b)交配により得られた個体における種子休眠性の程度を、上記本発明の判定方法により判定する工程、および(c)強種子休眠性を有すると判定された品種を選抜する工程、を含む。
【0075】
強種子休眠性の形質の植物品種と交配させる「任意の植物品種」としては、例えば、弱種子休眠性の形質を有する品種、弱種子休眠性の形質を有する品種と強種子休眠性の形質を有する品種との交配により得られた個体が挙げられるが、これらに制限されない。
【0076】
本発明の育種方法の他の1つの態様は、弱種子休眠性の形質の植物を育種する方法であり、(a)弱種子休眠性の形質の植物品種と任意の植物品種とを交配させる工程、(b)交配により得られた個体における種子休眠性の程度を、上記本発明の判定方法により判定する工程、および(c)弱種子休眠性を有すると判定された品種を選抜する工程、を含む。
【0077】
弱種子休眠性の形質の植物品種と交配させる「任意の植物品種」としては、例えば、強種子休眠性の形質を有する品種、強種子休眠性の形質を有する品種と弱種子休眠性の形質を有する品種との交配により得られた個体が挙げられるが、これらに制限されない。
【0078】
本発明の育種方法を利用すれば、種子休眠性の形質が改変された植物を、種子や幼植物の段階で選抜することが可能となり、種子休眠性の形質が改変された品種の育成を短期間で行うことが可能となる。
【実施例】
【0079】
[実施例1] オオムギSD2遺伝子の同定
オオムギ材料は、圃場(独立行政法人 農業生物資源研究所)に植えて育成した。オオムギ品種および系統としては、次のものを用いた。(i)強種子休眠性型の6条品種「アズマムギ」(Az)、(ii)および弱種子休眠性型の2条品種「関東中生ゴールド」(KNG)、(iii)AzとKNGの交配由来の組換え近交系(RIL)99系統(Mano et al. (2001) Genome 44:284-292)、(iv)5H染色体長椀末端領域約20cMがKNGの染色体断片に置き換わっているAzの系統(NIL#34;図1)、(v)NIL#34とAzの交配由来のF2個体(2985個体)。
【0080】
発芽試験においては、生理的成熟期および完熟期の穂を採取し、30℃で1週間乾燥させた。その後、一穂から得られた種子を、No.2濾紙を2枚敷いたシャーレ(直径9cm高さ1.5cm)に撒き、2000分の1のベンレートを加えた4.5mlの蒸留水を加え、15℃一週間置いた。その後、各シャーレの発芽種子粒数および全種子粒数を数え、「発芽粒数/全粒数 × 100」の式で発芽率(%)を算出した。
【0081】
(1)量的形質座(QTL)解析
NIL#34とAzとを交配させて得た2985のF2個体(RIL99系統)について実施した発芽試験の結果およびManoらの文献(Mano et al. (2001) Genome 44:284-292)に記載の連鎖地図を用いてQTL解析を行った。QTLの検出は、MapQTL5解析ソフト(Biometris, Wageningen, The Netherlands, http://www.joinmap.nl)を用いて、シンプルインターバルマッピング法により行った。その結果、5H染色体長椀末端のSD2座に、作用力の大きな種子休眠QTLが検出された(図2)。
【0082】
(2)アソシエーション解析
SD2座近傍で組換えが起きているF2個体に由来する後代(F3個体)60個体を用いて、SD2座近傍の遺伝子型の同定と発芽率の測定を行い、遺伝子型と発芽率の相関を解析して、SD2座の遺伝子型と座乗している領域を同定した(図3)。新規マーカーの開発も行って、SD2座を絞り込んだ結果、最終的に、SD2座は、5M24と5M25のマーカーに挟まれた0.08cMの領域に座乗することが判明した(図4)。
【0083】
(3)オオムギBAC(バクテリア人工染色体)クローンの単離
5M24マーカーを用いて「OUH602」および「Morex」のゲノムBACライブラリー(Yu et al. (2000) Theor Appl Genet 101:1093-1099)をPCR法によりスクリーニングした。選抜されたBACクローンについてショットガンシークエンスにより配列を決定した。得られたゲノム配列は、Rice Genome Automated Annotation System(RiceGAAS) website(http://ricegaas.dna.affrc.go.jp/;Sakata et al. (2002) Nucleic Acids Res. 30:98-102)を用いて解析した。その結果、OUH602のゲノムBACライブラリーより、約70kbのゲノム配列を持つ「BACクローン6」が見出された。このBACクローン上には、5M25マーカーも座乗しており、SD2座が、5M24マーカーおよび5M25マーカーの間の約28kbのゲノム領域に座乗していることが明らかになった(図5)。
【0084】
(4)SD2遺伝子の同定
RiceGAAS(Rice Genome Automated Annotation System)を用いた分析により、絞り込まれた領域内およびその近傍に、5つの遺伝子が見出された(図6)。このうちGene1、Gene4、Gene5は、対応する完全長配列が見い出され、機能している遺伝子であることが確認できた。一方、Gene2とGene3は、完全長データベースに登録されていない遺伝子であった。Gene4とGene5は、完熟種子で発現していることが確認された。Gene3は、トランスポゾンによく見られるPMD(plant mobile domain)を保持していた。
【0085】
Gene4のアミノ酸配列をAzとKNGの間で比較すると、植物間で高度に保存されているアスパラギン(N)が、アズマムギではトレオニン(T)に置換されていることが明らかになった(図7)。
【0086】
また、Gene4が、Az型(強休眠性型)とKNG型(弱休眠性型)のヘテロとなっている個体につき、その種子の発芽率を測定し、いずれの型が優性であるかの検証を行った。その結果、KNG型(弱休眠性型)が優性であることが判明した。
【0087】
以上から、Mitogen-activated Kinase kinase(MAPKK)をコードするGene4が、SD2遺伝子であると考えられた。
【0088】
[実施例2] コムギPhs1(Pre-Harvest Sprouting 1)遺伝子の同定
カナダの穂発芽難品種(強種子休眠性型)の「Leader」を一回親とし、穂発芽中(弱種子休眠性型)品種「春よ恋」を反復親に配した戻し交雑によって、準同質遺伝子系統(NILs)を作製した。BC1F1を育苗箱で1個体ずつ養成し、緑葉からDNeasy Plant Mini Kit(キアゲン)を用いてDNAを抽出した。Phs1近傍に位置するPCRマーカーの遺伝子型を決定した。その際、マーカー遺伝子型がヘテロ接合型を示す個体を選抜し、幼苗をポットへ移植した。選抜個体を温室内で養成し、「春よ恋」との交配を繰り返した。この方法により2組合せについてBC4まで世代を進めた。選抜個体を自殖して得られたBC4F2個体の中からマーカーの遺伝子型により、Phs1近傍領域が、「Leader」ホモ接合型、「春よ恋」ホモ接合型にそれぞれ固定した個体、ならびにマーカー間で組換えを生じた個体を選抜し、種子休眠性試験に供試した。
【0089】
温室の温度は15〜25℃の範囲で変動させ、2月から6月の間に登熟した種子を休眠性の評価に用いた。日長感応性を示す材料については陽光ランプで補光し、出穂後は自然日長で登熟させた。
【0090】
種子休眠性の評価用種子の調製にあたっては、穂を成熟期に収集し、室温で7日間乾燥した。採取種子は傷がつかないように手で脱穀した後、試験まで-30℃で保存した。発芽試験においては、直径9cm、深さ2cmのシャーレ(テルモ)に濾紙(ワットマンNo.3)を一枚敷き、7mlの蒸留水を添加した。発芽試験に用いる種子は、胚が上向きになるように1粒ずつシャーレに置床した。発芽試験は15℃もしくは20℃のインキュベータ内で10日間行い、発芽を調査した。成熟乾燥後の種子休眠性検定から、コムギ4A染色体に座乗するQTLが主働遺伝子Phs1として2.6cMの域内にマッピングされている(非特許文献13)。Phs1近傍のDNAマーカーがLeader型ホモに固定している63個体、春よ恋型に固定している65個体を選抜し、種子休眠性を評価した結果、Phs1がLeader型と春よ恋型で明瞭に種子休眠性(発芽率)の差異が認められた(図8)。
【0091】
(1)Phs1遺伝子のマッピングによる絞込み
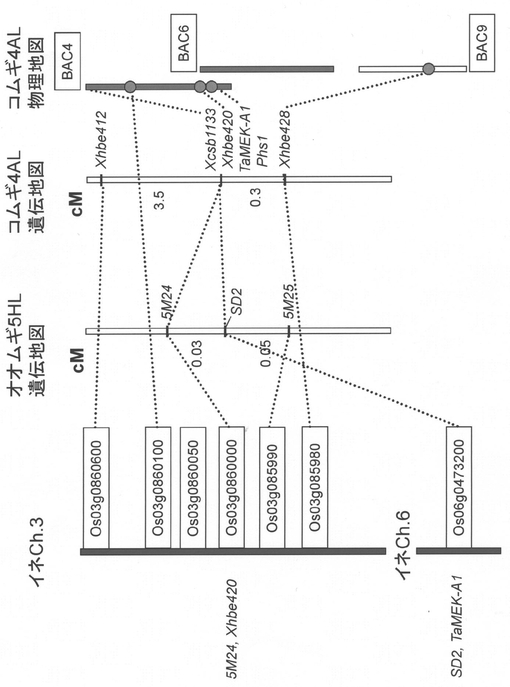
(i)大規模連鎖解析
6倍体パンコムギ(Triticum aestivum L.)品種Chinese Springから作成したBACライブラリー(Allouis et al. (2003) Cereal Research Communications. 31:331-338)からAゲノム特異的プライマーを用いてクローンを選抜し、末端配列および内部配列を解読して新たなマーカーおよびBACスクリーニング用プライマーを作成した。単離された代表的なBACクロ−ンの位置関係を図9に示した。
【0092】
一方、Leader(強種子休眠性型)と春よ恋(弱種子休眠性型)間から作出したBC4F2集団(一部BC3F2個体含む)3,235個体を用いて、文献(非特許文献13)に記載の種子休眠性評価方法によりPhs1を0.19cMの域内に絞り込んだ(非特許文献14)。すなわち、マーカー間で組み換えを生じたF2個体からF3系統10〜20個体を展開し、春よ恋型およびLeader型に固定した個体を複数個体選抜した。選抜された個体の種子休眠性を評価し、分離が認められればF2のPhs1遺伝子型がヘテロ、春よ恋型(発芽率が高い)およびLeader型(発芽率が低い)に分離が認められなければ、F2のPhs1遺伝子型がそれぞれ、春よ恋ホモ、Leaderホモと判定しマッピングした。
【0093】
Phs1をカバーする連続的なBACコンティグの作成には至っていなかったが、CSBAC4からCSBAC6にわたる約180kbの領域がPhs1と共分離したことから、このBACクローンの塩基配列をショットガンシーケンス法により決定した。その結果、4種の遺伝子(No.1〜4)が想定された(図9)。遺伝子の推定にあたっては、RAP-DB(http://rapdb.dna.affrc.go.jp/)、NCBI(http://blast.ncbi.nlm.nih.gov/Blast.cgi)、GrainGenes (http://wheat.pw.usda.gov/GG2/index.shtml)のデータベースをサーベイした。
【0094】
(ii)オオムギ、イネとの比較ゲノム解析
イネのゲノム情報を通じて、オオムギゲノムとの比較解析を行った。オオムギSD2遺伝子近傍にマップされたマーカーとコムギPhs1近傍にマップされたマーカーのうち、イネの第3染色体に座乗するOs03g0860000(候補遺伝子No2に相当)、第6染色体に座乗するOs06g0473200(候補遺伝子No1に相当)が共通にマップされかつ同じ順序で並んでいた(図10)。このことから、コムギPhs1とオオムギSD2遺伝子は共通の遺伝子に制御されていることが強く示唆された。4種の候補遺伝子のうち、SD2遺伝子の有力な原因遺伝子と共通のMitogen-activated protein kinase kinase(MAPKK)遺伝子(候補遺伝子No1に相当)がPhs1の原因遺伝子と考えられ、コムギ「TaMEK-A1」と名づけた。なお、コムギ4ALは6倍体パンコムギへの進化の過程で逆位や5A染色体と7B染色体との部分的な相互転座が生じている(Devos et al. (1995) Theor Appl Genet 91:282-288)。Phs1近傍領域は5A末端領域が逆向きになっているため、オオムギ5HL染色体とはテロメア側とセントロメア側が逆になっている。
【0095】
(2)候補遺伝子TaMEK-A1の解析
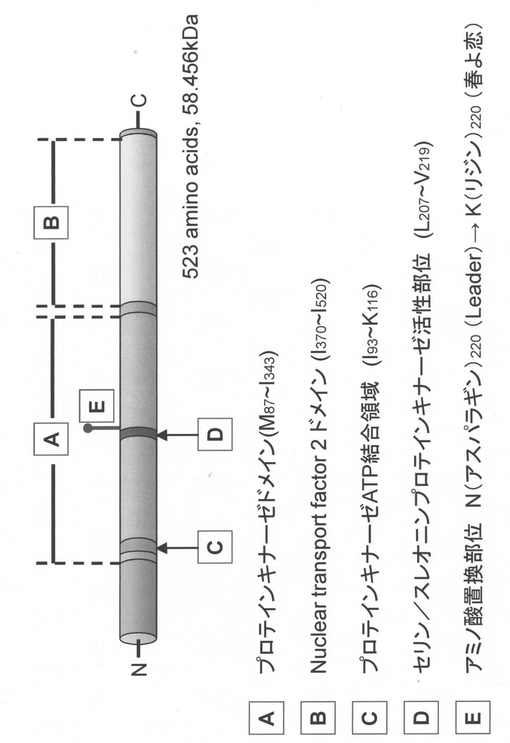
(i)候補遺伝子TaMEK-A1の塩基配列決定
CSBAC4およびCSBAC6の内部塩基配列情報をもとにして、Leaderと春よ恋のゲノムDNAを鋳型にしたPCR産物の塩基配列を決定した。TaMEK-A1は、10のイントロンと11のエキソンからなる。ORF長は1572bpと推定され、推定翻訳産物は523残基からなるアミノ酸をコードする(図11、13)。
【0096】
また、春よ恋の種子胚由来RNAからSuperScriptIII逆転写酵素(インビトロジェン)およびSMART RACE cDNA Amplification Kit(クローンテック)を用いて調製したcDNAから、TaMEK-A1に相当する完全長cDNAを得た。得られたcDNAを鋳型にして得られたTaMEK-A1遺伝子に相当するPCR産物の塩基配列を決定した。
【0097】
春よ恋およびLeaderの塩基配列の解読方法は以下の通りである。ゲノムDNAを鋳型にしてPCRを行い、目的領域に含まれる産物のみをクローニングして、シーケンスを決定した。クローニングにあたってはPCR産物をpGEM T-Easy Vector(プロメガ)にライゲーションし、大腸菌XL10-Goldに形質転換した(ストラタジーン)。コロニーPCRにより、組換え型クローンを選び、インサートDNAのシーケンスを決定した。また、染色体欠失系統を利用しても目的の産物を確認できなかった場合は、マッピング集団の両親のそれぞれについてPCR産物を得て、クローニングし、6〜8クローンを無作為に選んでシーケンスを決定した。Aゲノムの祖先種であるTriticum urartuのDNAを鋳型とするPCR産物も同時にシーケンスすることにより、Aゲノムシーケンスを同定した。得られたシーケンスのアライメントとグルーピングは、Sequencher ver. 4.1.2(Gene Code)を用いて行った。以下の(ii)、(iii)の実験も同様に行った。アミノ酸配列の解析は、BLAST-p, tblastn(http://blast.ncbi.nlm.nih.gov/Blast.cgi)を用いて行った。
【0098】
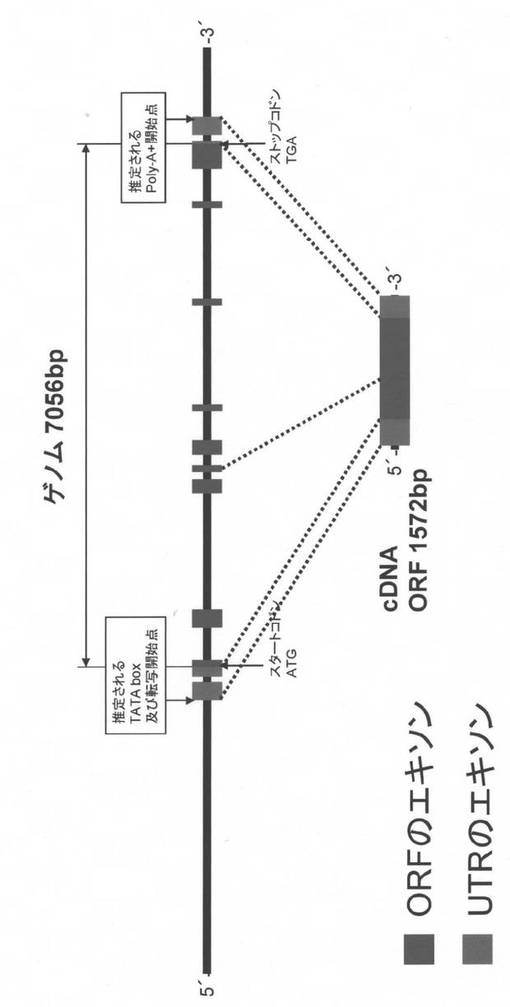
(ii)TAMEK-A1遺伝子の塩基配列と推定されるアミノ酸配列の品種間差
4A染色体のPhs1近傍にQTLが報告されている14品種・系統を用いて、TaMEK-A1およびNo2〜4遺伝子の想定されるエキソンおよび5′側の上流域について、春よ恋とLeader間に見出された塩基配列変異を調査した。その結果、TaMEK-A1のエキソン内に認められたSNPだけが唯一休眠性の品種間差とSNPの変異とがよく一致した(図12)。
【0099】
なお、弱種子休眠性品種のハルユタカ(日本)、Munstertaler(スイス)、Janz、Cascades、Cunningham、Cranbrook(以上オーストラリア)のTaMEK-A1のアミノ酸配列は、春よ恋(日本)と同一であった。また、強種子休眠性品種・系統のきたもえ、OS21-5(日本)、AC Domain(カナダ)、SW95-50213、AUS1408、AUS1490(以上オーストラリア)のTaMEK-A1のアミノ酸配列はLeader(カナダ)と同一であった。
【0100】
春よ恋およびLeaderのゲノムDNAを鋳型にして、5′上流域を含めると全長8,892bpのTaMEK-A1配列を決定した。2654番目の塩基に位置するエキソンに認められたSNPはTaMEK-A1遺伝子の推定アミノ酸配列内に予測されるセリン/スレオニン型タンパク質リン酸化酵素の活性部位に隣接するアミノ酸の置換を伴う変異であった。すなわちLeaderは塩基がCであるため、アミノ酸でアスパラギン(N)に翻訳されるのに対して春よ恋は塩基がAであるため、アミノ酸がリシン(K)に翻訳される(図13、図14)。タンパク質モチーフの予測はScanProsite(http://au.expasy.org/tools/scanprosite/)を利用した。
【0101】
(iii)TaMEK1のゲノム間および種間の比較
TaMEK-A1遺伝子のアミノ酸置換を伴う1塩基多型(SNP)だけが14品種の休眠性の差異とよく一致した。そこで、TaMEK1遺伝子のコムギ同祖遺伝子の単離を試みたところ、3種の異なる配列を同定した(TaMEK-B1、TAMEK-D1)。これらが同祖遺伝子の候補としてSNPのハプロタイプを判定した結果、B、Dゲノム(春よ恋のDゲノム遺伝子は未単離)のSNPはいずれもAゲノムのLeader(強種子休眠性型)と同じハプロタイプ(DNAでC、アミノ酸でアスパラギン)であった(図15)。また、Aゲノム祖先種T.urartu、Dゲノム祖先種のAe.squarrosaデータベースに登録されているホモログのハプロタイプはいずれもLeader型であった。従って、これまでのところLeader型のアリルが野生型で、Phs1の座乗するAゲノム遺伝子の春よ恋(弱種子休眠性型)アリルが変異型と考えられた。
【0102】
(iv)遺伝子発現
開花後40日目の種子胚からRNeasy Plant mini kit(キアゲン)を用いてRNAを抽出し、SuperScriptIII逆転写酵素(インビトロジェン)を用いてcDNAを合成した。cDNAを鋳型にしてTaMEK-A1遺伝子のゲノム特異的なプライマーを用いたRT-PCRにより遺伝子発現の解析を行った。その結果、春よ恋、LeaderともにTaMEK-A1遺伝子は種子胚で発現していることが明らかになった(図16)。
【0103】
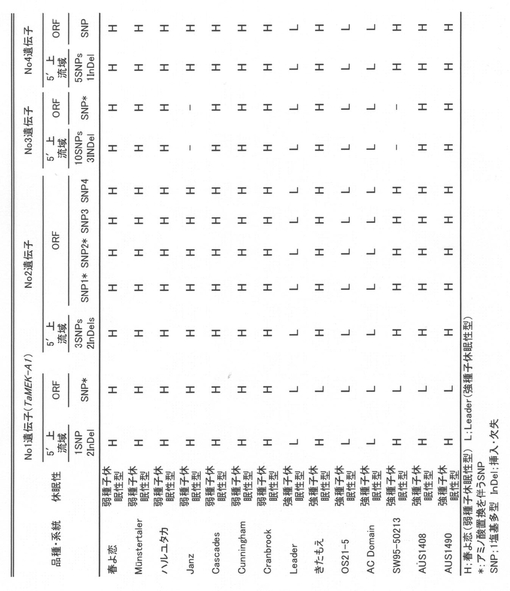
(v)TaMEK-A1の判定方法
TaMEK-A1のエキソン内に見出されたSNPをもとにdCAPS(derived cleaved amplified polymorphic sequence)マーカーを作成した。ゲノムDNAに対してプライマー配列A(フォワードTAAAGCCAGCAAATTTACTGGTCGA/配列番号:9、リバースGCTCATATTTAAGGAATATCAAAAGAAC/配列番号:10)を用いたPCRを行い(アニーリング温度55℃)、制限酵素SalIで処理した場合に春よ恋型の塩基Cタイプのみが切断される。一方、プライマー配列B(フォワードTCTAAAGCCAGCAAATTTACTGGGTAA/配列番号:11、リバースGCTCATATTTAAGGAATATCAAAAGAAC/配列番号:12)を用いたPCR産物(アニーリング温度55℃)を制限酵素DraIで処理するとLeader型のDNAのみが切断される。制限酵素処理したPCR産物をアガロースゲルで検出することができる。この2種を組み合わせることにより、Phs1遺伝子のタイプを簡易に判定できる。
【0104】
PCRの条件は以下の通りである。ゲノムDNAはDneasy Plant mini kit(キアゲン)およびMagAttract96 DNA Plant Core kit(キアゲン)を用いてDNAを抽出した。次に鋳型DNA、2.5mM MgCl2、1.5mM dNTP、1.5μM プライマー 1unitのTaqポリメラーゼを混合して反応液とした。PCRは94℃3分後、「94℃40秒55℃40秒72℃1分」を30サイクル繰り返し、最後に72℃7分伸長反応を行った。PCR反応液を制限酵素で処理後、アガロースゲルで電気泳動しSYBR green I(Cambrex Bio Science)で染色した後、多型を検出した。
【産業上の利用可能性】
【0105】
上記した通り、本発明により、植物の種子休眠性の程度を支配する新規な遺伝子SD2/Phs1が同定され、同定されたSD2/Phs1遺伝子を利用して、植物の種子休眠性の程度の判定や種子休眠性が改変された植物の作出・育種が可能となった。本発明における種子休眠性の程度の判定は、それを支配する遺伝子を標的としており、しかも、育成早期の個体(例えば、種子)を用いて実施することが可能である。このため、本発明の判定方法を利用すれば、種子休眠性が改変された品種を特異的かつ効率的に育種することが可能である。
【0106】
こうして作出・育種された弱種子休眠性の植物は、休眠が深くて発芽が著しく遅延する場合や乾燥地域で降水量が少ないために発芽が遅延する場合に、十分かつ早期に発芽個体を確保できる点で有用である。また、強種子休眠性の植物は、降雨の多い地域で穂発芽による種子の品質劣化を防げる点で有用である。従って、本発明は、特に農作物の収穫量の向上及び品質劣化による経済的損失の軽減に大きく貢献することができる。
【配列表フリーテキスト】
【0107】
配列番号9〜12
<223> 人工的に合成されたプライマーの配列
【技術分野】
【0001】
本発明は、植物の種子休眠性を支配する遺伝子、並びに、それを利用した植物の種子休眠性の程度の判定および休眠性が改変された植物の作出に関する。
【背景技術】
【0002】
これまでに、オオムギやコムギなどの麦類の種子休眠性に関わる量的形質座(QTL)に関する報告は多数あり(非特許文献1〜2)、コムギでは、これまでに少なくとも30以上の種子休眠QTLが知られている。
【0003】
今までにオオムギのSD2領域である5H染色体長椀末端に種子休眠QTLが検出された報告は、Ullrichらの「Steptoe」と「Morex」の交配集団を使った例(非特許文献3)、武田らの「TR306」と「Harrington」の交配集団を使った例(非特許文献4)、Liらの「Chebec」と「Harrington」の交配集団を使った例(非特許文献5)、Pradaらの「Triumph」と「Morex」の交配集団を使った例(非特許文献6)、堀らの「はるな2条」と「OUH602」の交配集団を使った例(非特許文献7)などがあるが、いずれも種子休眠QTLの検出を報告しているだけで、原因遺伝子の単離は行われていない。
【0004】
SD2遺伝子の単離を目指した精密マッピングは、Ullrichのグループによって行われが(非特許文献8)、表現型の評価が難しく、精度の低いマッピングに終わっている。こうしたデータをもとに、イネとのシンテニー関係を利用した解析も行われ、SD2座の原因遺伝子が植物ホルモンであるジベレリンの生合成の律速段階を調整する酵素GA20-oxidaseである可能性が高いという報告もなされているが(非特許文献9)、推測に過ぎない。また、オオムギの大規模EST解析より、約3000マーカーが座乗する連鎖地図が作られ、SD2座と連鎖するマーカーが報告されているが(特許文献1)、SD2遺伝子本体を同定したわけではなく、既存のSD2のQTL解析のマーカーと同じくSD2座近傍のマーカーを同定したにすぎない。
【0005】
一方、コムギにおいては、効果の大きい主要なQTLが、複数の解析集団に共通して4A染色体の長腕(4AL)に検出されている(非特許文献10〜12)。これら4ALに検出されたQTLは、近傍のマーカー情報から互いに同一の遺伝子座である可能性が高いことが示唆された(非特許文献11)。その後、QTLが4ALに検出された報告例は続くものの、高精度マッピングやマップベース法による遺伝子単離へと発展した事例はない。これはコムギがA、B、Dゲノムからなる異質6倍体であり、イネの40倍にもおよぶゲノムサイズを有していることが要因であると考えられる。近年、イネとのシンテニーを利用することによりコムギでも特定の領域にDNAマーカーを設定できるようになり、高精度マッピングやマップベース法による遺伝子単離の成功例が報告されるようになった。コムギの種子休眠性については、戻し交雑集団と安定した登熟環境で種子休眠性を評価することにより、主働遺伝子(Phs1)を2.6cMの域内へマッピングすることに成功した(非特許文献13)。イネとのシンテニーを利用することにより、Phs1はさらに詳細にマッピングされた(非特許文献14)。しかしながら、ミクロなレベルでは、イネとコムギ間ではしばしば保存性が認められない。また、種子休眠性と密接に関連している植物ホルモンのアブシジン酸(ABA)のシグナリングに関わる遺伝子のホモログが、コムギにおいてマッピングされているが、4ALに相当する領域にはマッピングされなかった(非特許文献15)。以上のことから、既報の情報を利用してコムギPhs1の原因遺伝子を想定することは困難であり、マップベース法によりコムギPhs1遺伝子を同定するためには、コムギゲノムのさらなる絞込みとゲノム配列の解読および解析が必要である。
【0006】
このように、これまでオオムギにおけるSD2座やコムギにおけるPhs1座の報告は、QTLマッピングによる報告のみで、その困難性から、これら遺伝子の単離同定には至っていない。オオムギとコムギ以外の麦類についても、種子休眠QTLの原因遺伝子の単離同定が報告された例はない。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特許公開2005−229849号公報
【非特許文献】
【0008】
【非特許文献1】Hori et al. (2007) Theor Appl Genet 115:869-876
【非特許文献2】Flintham et al. (2002) Euphytica 126:39-45
【非特許文献3】Ullrich et al. (1992) Pre-harvest sprouting in cereals, p136-145.
【非特許文献4】Takeda (1995) 7th international symposium on pre-harvest sprouting in cereals, p205-212
【非特許文献5】Li et al. (2003) Australian Journal of Agricultural Research, vol.54, p1303-1313.
【非特許文献6】Prada et al. (2004) Theor Appl Genet, vol.109, p62-70.
【非特許文献7】Hori et al. (2007) Theor Appl Genet, vol.115, p869-876
【非特許文献8】Gao et al.(2003) Theor Appl Genet 107:552-559
【非特許文献9】Li et al. (2004) Funct Integr Genomics 4:84-93
【非特許文献10】Kato et al. (2001) Theor. Appl. Genet. 102:980-985.
【非特許文献11】Mares et al. (2005) Theor. Appl. Genet. 111:1357-1364.
【非特許文献12】Torada et al. (2005) Euphytica 143:251-255.
【非特許文献13】Torada et al. (2008) Genome. 51(6):426-32.
【非特許文献14】乕田ら 2010 大規模集団を用いたコムギ種子休眠性遺伝子座Phs1の精密マッピング 育種学研究 別冊2号 p.36
【非特許文献15】Nakamura et al. (2006) Theor. Appl. Genet. 114:1129-1139.
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、このような状況に鑑みてなされたものであり、その目的は、植物における種子休眠性の程度を支配する新規な遺伝子を同定することにある。また、本発明は、同定された遺伝子を利用して、植物の種子休眠性の程度を効率的に判定する方法を提供することを目的とする。さらなる本発明の目的は、同定された遺伝子を利用して、種子休眠性が改変された植物を効率的に作出する方法を提供することにある。
【課題を解決するための手段】
【0010】
本発明者らは、上記課題を解決すべく鋭意検討を行った結果、オオムギの種子休眠性を支配するSD2遺伝子、およびこれに対応するコムギ遺伝子であるPhs1遺伝子を同定することに成功した(以下、両遺伝子を合わせて「SD2/Phs1遺伝子」と称する)。さらに、本発明者らは、SD2/Phs1遺伝子において、その種子休眠性と相関する一塩基多型(SNP)を特定するとともに、弱種子休眠性型の遺伝子が優性であることを解明することにも成功した。そして、本発明者らは、同定したSD2/Phs1遺伝子を利用して、効率的に、植物の種子休眠性の程度を判定することや種子休眠性が改変された植物を作出することが可能であることを見出し、本発明を完成するに至った。
【0011】
本発明は、より詳しくは、下記を提供するものである。
【0012】
[1] 植物の種子休眠性の形質を弱める活性を有するタンパク質をコードする、下記(a)〜(d)のいずれかに記載のDNA。
(a)配列番号:2または4に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:1または3に記載の塩基配列のコード領域を含むDNA
(c)配列番号:2または4に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号:1または3に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNA
[2] 植物の種子休眠性の形質を強める活性を有するタンパク質をコードする、下記(a)〜(d)のいずれかに記載のDNA。
(a)配列番号:6または8に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:5または7に記載の塩基配列のコード領域を含むDNA
(c)配列番号:6または8に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号:5または7に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNA
[3] 植物の種子休眠性の形質を強める活性を有する、下記(a)〜(c)のいずれかに記載のDNA。
(a)[1]に記載のDNAの転写産物と相補的な二重鎖RNAをコードするDNA
(b)[1]に記載のDNAの転写産物と相補的なアンチセンスRNAをコードするDNA
(c)[1]に記載のDNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA
[4] [1]〜[3]のいずれかに記載のDNAを含むベクター。
【0013】
[5] [1]〜[3]のいずれかに記載のDNAが導入された植物細胞。
【0014】
[6] [5]に記載の細胞を含む植物植物体。
【0015】
[7] [6]に記載の植物体の子孫またはクローンである、植物植物体。
【0016】
[8] [6]または[7]に記載の植物植物体の繁殖材料。
【0017】
[9] 植物に[1]に記載のDNAを導入する工程を含む、種子休眠性の形質が弱められた植物の作出方法。
【0018】
[10] 植物における[1]に記載のDNAの発現または機能を抑制することを特徴とする、種子休眠性の形質が強められた植物の作出方法。
【0019】
[11] 植物に[2]または[3]に記載のDNAを導入する工程を含む、種子休眠性の形質が強められた植物の作出方法。
【0020】
[12] [1]に記載のDNA、または該DNAが挿入されたベクターを含む、植物の種子休眠性の形質を弱めるための薬剤。
【0021】
[13] [2]もしくは[3]に記載のDNA、または該DNAが挿入されたベクターを含む、植物の種子休眠性の形質を強めるための薬剤。
【0022】
[14] 植物における種子休眠性の程度を判定する方法であって、被検植物における[1]もしくは[2]に記載のDNAまたはその発現制御領域の塩基配列を解析し、対照の塩基配列と比較することを特徴とする方法。
【0023】
[15] 弱い種子休眠性の形質の植物を育種する方法であって、
(a)弱い種子休眠性の形質の植物品種と任意の植物品種とを交配させる工程、
(b)工程(a)における交配により得られた個体における種子休眠性の程度を、[14]に記載の方法により判定する工程、および
(c)弱い種子休眠性を有すると判定された品種を選抜する工程、を含む方法。
【0024】
[16] 強い種子休眠性の形質の植物を育種する方法であって、
(a)強い種子休眠性の形質の植物品種と任意の植物品種とを交配させる工程、
(b)工程(a)における交配により得られた個体における種子休眠性の程度を、[14]に記載の方法により判定する工程、および
(c)強い種子休眠性を有すると判定された品種を選抜する工程、を含む方法。
【0025】
なお、本発明において「種子休眠性」とは、種子の発芽に適した条件下においても発芽しない性質を意味する。また、本発明において「弱い種子休眠性の形質」とは、種子が成熟した後に休眠性の維持される期間が短く、発芽に適した検定条件下において高い発芽率を示すことを意味する。また、本発明において「強い種子休眠性の形質」とは、種子が成熟した後に休眠性の維持される期間が長く、発芽に適した検定条件下において低い発芽率を示すことを意味する。
【発明の効果】
【0026】
本発明によって、植物の種子休眠性を支配する新規な遺伝子SD2/Phs1が同定され、該遺伝子の染色体上の位置および構造が解明された。これによりSD2/Phs1遺伝子を標的とした植物の種子休眠性の程度の判定方法、および該判定方法を利用した種子休眠性が改変された植物の育種方法が提供された。さらに、SD2/Phs1遺伝子を利用した、休眠性が改変された植物の作出方法が提供された。本発明による植物の種子休眠性の程度の判定や休眠性が改変された植物の作出や育種は、種子休眠性の程度を支配するSD2/Phs1遺伝子に着目しているため、この遺伝子と連鎖するマーカーを利用した従来法に比して、精度が高い。このため従来法よりも、特異的かつ効率的に、植物の種子休眠性の程度の判定や休眠性が改変された植物の作出や育種を行うことが可能である。
【図面の簡単な説明】
【0027】
【図1】NIL#34の遺伝子型を示す図である。
【図2】アズマムギ(Az)と関東中生ゴールド(KNG)の組換え近交系(RIL)を用いた種子休眠QTL解析法を示す図である。
【図3】SD2遺伝子が座乗する領域を絞り込むための、アソシエーション解析法を示す図である。
【図4】アソシエーション解析結果とジェノタイピング結果のまとめの図である。
【図5】SD2遺伝子の精密マッピングとBACクローンとの対応関係を示す図である。
【図6】RiceGAASにより、SD2遺伝子が座乗する領域のゲノム配列中に存在する遺伝子を予測した結果を示す図である。
【図7】各オオムギ系統におけるGene4遺伝子のアミノ酸配列の比較を示す図である。Harunanijoは、はるな2条を、KNGは、関東中生ゴールドを、Azは、アズマムギを示す。三角は、植物間(イネ、トウモロコシ、ナズナ、ポプラ、タバコ、コケ、マツナ属Suaeda salsa、トウゴマ、ブドウ等)で高度に保存されているアミノ酸残基、丸は、植物間で相同性が高くないアミノ酸残基を示す。Harunanijoのアミノ酸配列の上に示したラインは、MAPKKドメインを示す。
【図8】Leader(強種子休眠性型)/春よ恋(弱種子休眠性型)BC3F2個体の種子休眠性とPhs1の遺伝子型を示す図である。
【図9】コムギ種子休眠性遺伝子座Phs1近傍の高精度連鎖地図とBACコンティグを示す図である。
【図10】コムギPhs1候補領域を、イネとオオムギの対応領域と比較した地図である。
【図11】Phs1(TaMEK-A1)遺伝子の構造の模式図である。
【図12】休眠性の異なる14品種・系統における4遺伝子(No.1〜4)近傍配列の変異を示す図である。
【図13】Phs1(TaMEK-A1)タンパク質の構造の模式図である。
【図14】春よ恋とLeader間のPhs1(TaMEK-A1)遺伝子のアミノ酸配列の比較を示す図である。下線太字は、春よ恋とLeader間で異なるアミノ酸残基を示す。
【図15】Phs1(TaMEK-A1)同祖遺伝子およびホモログにおけるSNPのハプロタイプを示す図である。
【図16】RT-PCRによるPhs1(TaMEK-A1)の発現解析結果を示す写真である。
【発明を実施するための形態】
【0028】
<弱種子休眠性型DNA、強種子休眠性型DNA>
本発明は、植物の種子休眠性の形質を弱める活性を有するタンパク質をコードするDNA(以下、「弱種子休眠性型DNA」と称する)を提供する。本発明者らにより同定された、オオムギの弱種子休眠性品種である関東中手ゴールド由来のSD2cDNAの塩基配列を配列番号:1に、該DNAがコードするタンパク質のアミノ酸配列を配列番号:2に示す。また、コムギの弱種子休眠性品種である春よ恋由来のPhs1cDNAの塩基配列を配列番号:3に、該DNAがコードするタンパク質のアミノ酸配列を配列番号:4に示す。
【0029】
本発明の弱種子休眠性型DNAの1つの態様は、配列番号:2に記載のアミノ酸配列からなるタンパク質をコードするDNA(典型的には、配列番号:1に記載の塩基配列のコード領域を含むDNA)であり、他の1つの態様は、配列番号:4に記載のアミノ酸配列からなるタンパク質をコードするDNA(典型的には、配列番号:2に記載の塩基配列のコード領域を含むDNA)である。
【0030】
本実施例において示されたように、オオムギにおいて、関東中手ゴールド由来のSD2タンパク質のアミノ酸配列(配列番号:2)とはるな二条由来のSD2タンパク質のアミノ酸配列とを比較すると1か所のアミノ酸が相違しているが(図7)、ともに弱種子休眠性型である。現在の技術水準においては、当業者であれば、特定の弱種子休眠性植物品種(例えば、オオムギの関東中手ゴールド、コムギの春よ恋)における弱種子休眠性型DNAの塩基配列情報が得られた場合、その塩基配列を改変し、そのコードするアミノ酸配列は異なるが、同じく弱種子休眠性型であるDNAを取得することが可能である。また、自然界においても、塩基配列の変異によりコードするタンパク質のアミノ酸配列が変異することは起こり得ることである。従って、本発明は、オオムギの関東中手ゴールドあるいはコムギの春よ恋におけるSD2/Phs1タンパク質のアミノ酸配列(配列番号:2または4)において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなり、植物の種子休眠性の形質を弱める活性を有するタンパク質をコードするDNAをも含むものである。ここで「複数」とは、改変後のSD2/Phs1タンパク質が植物の種子休眠性の形質を弱める活性を維持する範囲における、アミノ酸の改変数であり、通常、50アミノ酸以内、好ましくは30アミノ酸以内、さらに好ましくは10アミノ酸以内(例えば、5アミノ酸以内、3アミノ酸以内、2アミノ酸)である。
【0031】
さらに、現在の技術水準においては、当業者であれば、特定の弱種子休眠性植物品種(例えば、オオムギの関東中手ゴールド、コムギの春よ恋)から弱種子休眠性型DNAが得られた場合、その弱種子休眠性型DNAの塩基配列情報を利用して、他の植物品種(例えば、イネ、トウモロコシ、ダイズ、ポプラ、ブドウ、シロイヌナズナ、タバコなど)から、同じく弱種子休眠性型である相同遺伝子をコードするDNAを取得することが可能である。従って、本発明は、オオムギの関東中手ゴールドおよびコムギの春よ恋におけるSD2/Phs1DNA(配列番号:1または3)とストリンジェントな条件でハイブリダイズするDNAであって、植物の種子休眠性の形質を弱める活性を有するタンパク質をコードするDNAをも含むものである。
【0032】
こうして得られた変異DNAや相同DNAが、植物の種子休眠性の形質を弱める活性を有するタンパク質をコードするか否かは、例えば、遺伝子組み換え技術または交配により、これらDNAを導入した強種子休眠性型の品種から種子を採取し、本実施例に記載の発芽試験を行い、その発芽率が高まるか否かを検定することにより、判定することができる。発芽率が高くなれば、植物の種子休眠性の形質を弱める活性を有すると評価される。
【0033】
本発明は、また、植物の種子休眠性の形質を強める活性を有するタンパク質をコードするDNA(以下、「強種子休眠性型DNA」と称する)を提供する。本発明者らにより同定された、オオムギの強種子休眠性品種であるアズマムギ由来のPhs1cDNAの塩基配列を配列番号:5に、該DNAがコードするタンパク質のアミノ酸配列を配列番号:6に示す。また、また、コムギの強種子休眠性品種であるLeader由来のPhs1cDNAの塩基配列を配列番号:7に、該DNAがコードするタンパク質のアミノ酸配列を配列番号:8に示す。
【0034】
本発明の強種子休眠性型DNAの1つの態様は、配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA(典型的には、配列番号:5に記載の塩基配列のコード領域を含むDNA)であり、他の1つの態様は、配列番号:8に記載のアミノ酸配列からなるタンパク質をコードするDNA(典型的には、配列番号:7に記載の塩基配列のコード領域を含むDNA)である。
【0035】
本実施例において示されたように、オオムギのアズマムギ由来のSD2タンパク質のアミノ酸配列(配列番号:6)は、弱種子休眠性品種である関東中手ゴールド由来のSD2タンパク質のアミノ酸配列(配列番号:2)と比較すると260位においてアミノ酸の置換(N→T)が生じている(図7)。また、コムギのLeader由来のPhs1タンパク質のアミノ酸配列(配列番号:8)は、弱種子休眠性品種である春よ恋由来のPhs1タンパク質のアミノ酸配列(配列番号:4)と比較すると220位においてアミノ酸の置換(K→N)が生じている(図13、図14)。このため、本来のSD2/Phs1タンパク質の機能が抑制され、これにより個体に強種子休眠性の形質を付与していると考えられる。現在の技術水準においては、当業者であれば、特定の弱種子休眠性植物品種(例えば、オオムギのアズマムギ、コムギのLeader)のSD2/Phs1DNAの塩基配列において、そのコードするタンパク質の強種子休眠性の形質が維持されるような改変を行うことが可能である。また、自然界においても、塩基配列の変異によりコードするタンパク質のアミノ酸配列が変異することは起こり得ることである。従って、本発明は、オオムギのアズマムギあるいはコムギのLeaderにおけるSD2/Phs1タンパク質のアミノ酸配列(配列番号:6または8)において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質からなり、植物の種子休眠性の形質を強める活性を有するタンパク質をコードするDNAをも含むものである。ここで「複数」とは、改変後のSD2/Phs1タンパク質が植物の種子休眠性の形質を強める活性を有する範囲における、アミノ酸の改変数である。植物におけるSD2/Phs1タンパク質が本来の機能を発揮しなければ、強い種子休眠性の形質になると考えられるため、当該アミノ酸改変数は、本質的に制限はない。改変は、例えば、100アミノ酸以内(50アミノ酸以内、30アミノ酸以内、10アミノ酸以内、5アミノ酸以内、3アミノ酸以内、2アミノ酸)である。
【0036】
さらに、現在の技術水準においては、当業者であれば、特定の植物品種からSD2/Phs1DNAが得られた場合、そのDNAの塩基配列情報を利用して、他の植物品種(例えば、イネ、トウモロコシ、ダイズ、ポプラ、ブドウ、シロイヌナズナ、タバコなど)から、強種子休眠性型である相同遺伝子をコードするDNAを取得することができる。従って、本発明は、オオムギのアズマムギあるいはコムギのLeaderにおけるSD2/Phs1DNA(配列番号:5または7)とストリンジェントな条件でハイブリダイズするDNAであって、植物の種子休眠性の形質を強める活性を有するタンパク質をコードするDNAが含まれる。
【0037】
こうして得られた変異DNAや相同DNAが、植物の種子休眠性の形質を強める活性を有するタンパク質をコードするか否かは、例えば、当該DNAで、弱種子休眠性品種のSD2/Phs1遺伝子を組換え、当該DNAをホモで保持する植物を作出し、該植物から種子を採取し、本実施例に記載の発芽試験を行い、その発芽率が低くなるか否かを検定することにより、判定することができる。発芽率が低くなれば、植物の種子休眠性の形質を強める活性を有すると評価される。
【0038】
本発明の弱種子休眠性型DNAは、その導入により、植物の種子休眠性の形質を弱めることが可能であるという意味において、植物の種子休眠性の形質を弱めるための薬剤であり、一方、本発明の強種子休眠性型DNAは、その導入により、植物の種子休眠性の形質を強めることが可能であるという意味において、植物の種子休眠性を強めるための薬剤である。
【0039】
なお、上記した変異DNAを作製するための、DNAへの人為的な変異の導入は、例えば、部位特異的変異誘発(site-directed mutagenesis)法(Kramer, W. & Fritz, HJ., Methods Enzymol, 154:350-367, 1987)により行うことができる。
【0040】
また、上記した相同遺伝子を単離するための方法としては、例えば、ハイブリダイゼーション技術(Southern, E. M., Journal of Molecular Biology, 98:503, 1975)やポリメラーゼ連鎖反応(PCR)技術(Saiki, R. K., et al. Science, 230:1350-1354, 1985、Saiki, R. K. et al. Science, 239:487-491, 1988)が挙げられる。相同遺伝子をコードするDNAを単離するためには、通常ストリンジェントな条件下でハイブリダイゼーション反応を行なう。ストリンジェントなハイブリダイゼーション条件としては、6M尿素、0.4%SDS、0.5xSSCの条件またはこれと同等のストリンジェンシーのハイブリダイゼーション条件を例示できる。よりストリンジェンシーの高い条件、例えば、6M尿素、0.4%SDS、0.1xSSCの条件を用いれば、より相同性の高いDNAの単離を期待することができる。単離されたDNAは、核酸レベルあるいはアミノ酸配列レベルにおいて、少なくとも50%以上、さらに好ましくは70%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。配列の相同性は、BLASTN(核酸レベル)やBLASTX(アミノ酸レベル)のプログラム(Altschul et al. J. Mol. Biol., 215:403-410, 1990)を利用して決定することができる。該プログラムは、KarlinおよびAltschulによるアルゴリズムBLAST(Proc. Natl. Acad. Sci. USA, 87:2264-2268, 1990、Proc. Natl. Acad. Sci. USA, 90:5873-5877, 1993)に基づいている。BLASTNによって塩基配列を解析する場合には、パラメーターは例えばscore=100、wordlength=12とする。また、BLASTXによってアミノ酸配列を解析する場合には、パラメーターは例えばscore=50、wordlength=3とする。また、Gapped BLASTプログラムを用いて、アミノ酸配列を解析する場合は、Altschulら(Nucleic Acids Res. 25:3389-3402, 1997)に記載されているように行うことができる。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である。
【0041】
本発明のSD2/Phs1タンパク質をコードするDNAとしては、その形態に特に制限はなく、cDNAの他、ゲノムDNA、および化学合成DNAが含まれる。ゲノムDNAおよびcDNAの調製は、当業者にとって常套手段を利用して行うことが可能である。ゲノムDNAは、例えば、植物からゲノムDNAを抽出し、ゲノミックライブラリー(ベクターとしては、プラスミド、ファージ、コスミド、BAC、PACなどが利用できる)を作成し、これを展開して、SD2/Phs1遺伝子(例えば、配列番号:1,3,5または7のいずれかに記載のDNA)の塩基配列を基に調製したプローブを用いてコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより調製することが可能である。また、SD2/Phs1遺伝子に特異的なプライマーを作成し、これを利用したPCRを行うことによって調製することも可能である。また、cDNAは、例えば、植物から抽出したmRNAを基にcDNAを合成し、これをλZAP等のベクターに挿入してcDNAライブラリーを作成し、これを展開して、上記と同様にコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより、また、PCRを行うことにより調製することが可能である。
【0042】
<植物の弱種子休眠性型SD2/Phs1遺伝子の発現を抑制するために用いるDNA>
また、本発明は、植物の弱種子休眠性型SD2/Phs1遺伝子の発現を抑制するために用いるDNAを提供する。これらのDNAの導入により、植物の種子休眠性の形質を強めることが可能である。この意味において、植物の弱種子休眠性型SD2/Phs1遺伝子の発現を抑制するために用いるDNAは、植物の種子休眠性の形質を強めるための薬剤である。ここで「SD2/Phs1遺伝子の発現の抑制」には、遺伝子の転写の抑制およびタンパク質への翻訳の抑制の双方が含まれる。また、「発現の抑制」には、発現の完全な停止のみならず発現の減少も含まれる。
【0043】
植物の弱種子休眠性型SD2/Phs1遺伝子の発現を抑制するために用いるDNAの一つの態様は、上記した本発明の弱種子休眠性型DNAの転写産物と相補的なdsRNA(二重鎖RNA)をコードするDNAである。標的遺伝子配列と同一もしくは類似した配列を有するdsRNAを細胞内に導入することにより、導入した外来遺伝子および標的内因性遺伝子の発現がいずれも抑制される、RNAi(RNA干渉、RNA interference)と呼ばれる現象を引き起こすことができる。細胞に約40〜数百塩基対のdsRNAが導入されると、ヘリカーゼドメインを持つダイサー(Dicer)と呼ばれるRNaseIII様のヌクレアーゼが、ATP存在下で、dsRNAを3'末端から約21〜23塩基対ずつ切り出し、siRNA(short interference RNA)が生じる。このsiRNAに、特異的なタンパク質が結合して、ヌクレアーゼ複合体(RISC:RNA-induced silencing complex)が形成される。この複合体はsiRNAと同じ配列を認識して結合し、RNaseIII様の酵素活性によってsiRNAの中央部で標的遺伝子の転写産物(mRNA)を切断する。また、この経路とは別にsiRNAのアンチセンス鎖がmRNAに結合してRNA依存性RNAポリメラーゼ(RsRP)のプライマーとして作用し、dsRNAが合成される。このdsRNAが再びダイサーの基質となって、新たなsiRNAを生じて作用を増幅する経路も考えられている。
【0044】
本発明のdsRNAをコードするDNAは、標的遺伝子の転写産物(mRNA)のいずれかの領域に対するアンチセンスRNAをコードしたアンチセンスDNAと、該mRNAのいずれかの領域のセンスRNAをコードしたセンスDNAを含み、該アンチセンスDNAおよび該センスDNAより、それぞれアンチセンスRNAおよびセンスRNAを発現させることができる。また、これらのアンチセンスRNAおよびセンスRNAよりdsRNAを作成することができる。
【0045】
本発明のdsRNAの発現システムをベクター等に保持させる場合の構成としては、同一のベクターからアンチセンスRNAおよびセンスRNAを発現させる場合と、異なるベクターからそれぞれアンチセンスRNAとセンスRNAを発現させる場合がある。同一のベクターからアンチセンスRNAおよびセンスRNAを発現させる構成としては、例えば、アンチセンスDNAおよびセンスDNAの上流にそれぞれpolIII系のような短いRNAを発現し得るプロモーターを連結させたアンチセンスRNA発現カセットとセンスRNA発現カセットをそれぞれ構築し、これらカセットを同方向にあるいは逆方向にベクターに挿入する構成である。
【0046】
また、異なる鎖上に対向するように、アンチセンスDNAとセンスDNAとを逆向きに配置した発現システムを構成することもできる。この構成では、アンチセンスRNAコード鎖とセンスRNAコード鎖とが対となった一つの二本鎖DNA(siRNAコードDNA)が備えられ、その両側にそれぞれの鎖からアンチセンスRNAとセンスRNAとを発現し得るようにプロモーターを対向して備える。この場合には、センスRNAとアンチセンスRNAの下流に余分な配列が付加されることを避けるために、それぞれの鎖(アンチセンスRNAコード鎖、センスRNAコード鎖)の3'末端にターミネーターをそれぞれ備えることが好ましい。このターミネーターは、A(アデニン)塩基を4つ以上連続させた配列などを用いることができる。また、このパリンドロームスタイルの発現システムでは、二つのプロモーターの種類は異なっていることが好ましい。
【0047】
また、異なるベクターからアンチセンスRNAおよびセンスRNAを発現させる構成としては、例えば、アンチセンスDNAおよびセンスDNAの上流にそれぞれpolIII系のような短いRNAを発現し得るプロモーターを連結させたアンチセンスRNA発現カセットとセンスRNA発現カセットとをそれぞれ構築し、これらカセットを異なるベクターに保持させる構成である。
【0048】
本発明に用いるdsRNAとしては、siRNAが好ましい。「siRNA」は、細胞内で毒性を示さない範囲の短鎖からなる二重鎖RNAを意味する。標的SD2/Phs1遺伝子の発現を抑制することができ、かつ、毒性を示さなければ、その鎖長に特に制限はない。dsRNAの鎖長は、例えば、15〜49塩基対であり、好適には15〜35塩基対でり、さらに好適には21〜30塩基対である。
【0049】
本発明のdsRNAをコードするDNAとしては、標的配列のインバーテッドリピートの間に適当な配列(イントロン配列が望ましい)を挿入し、ヘアピン構造を持つダブルストランドRNA(self-complementary 'hairpin' RNA(hpRNA))を作るようなコンストラクト(Smith, N.A., et al. Nature, 407:319, 2000、Wesley, S. V. et al. Plant J. 27:581, 2001、Piccin, A. et al. Nucleic Acids Res. 29:E55, 2001)を用いることもできる。
【0050】
本発明のdsRNAをコードするDNAは、標的SD2/Phs1遺伝子の塩基配列と完全に同一である必要はないが、少なくとも70%以上、好ましくは80%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。配列の同一性は上述した手法(BLASTプログラム)により決定できる。
【0051】
dsRNAにおけるRNA同士が対合した二重鎖RNAの部分は、完全に対合しているものに限らず、ミスマッチ(対応する塩基が相補的でない)、バルジ(一方の鎖に対応する塩基がない)などにより不対合部分が含まれていてもよい。本発明においては、dsRNAにおけるRNA同士が対合する二重鎖RNA領域中に、バルジおよびミスマッチの両方が含まれていてもよい。
【0052】
植物の弱種子休眠性型SD2/Phs1遺伝子の発現を抑制するために用いるDNAの他の態様は、上記した本発明の弱種子休眠性型DNAの転写産物と相補的なアンチセンスRNAをコードするDNA(アンチセンスDNA)である。アンチセンスDNAが標的遺伝子の発現を抑制する作用としては、三重鎖形成による転写開始阻害、RNAポリメラーゼによって局部的に開状ループ構造がつくられた部位とのハイブリッド形成による転写抑制、合成の進みつつあるRNAとのハイブリッド形成による転写阻害、イントロンとエキソンとの接合点でのハイブリッド形成によるスプライシング抑制、スプライソソーム形成部位とのハイブリッド形成によるスプライシング抑制、mRNAとのハイブリッド形成による核から細胞質への移行抑制、キャッピング部位やポリ(A)付加部位とのハイブリッド形成によるスプライシング抑制、翻訳開始因子結合部位とのハイブリッド形成による翻訳開始抑制、開始コドン近傍のリボソーム結合部位とのハイブリッド形成による翻訳抑制、mRNAの翻訳領域やポリソーム結合部位とのハイブリッド形成によるペプチド鎖の伸長阻止、および核酸とタンパク質との相互作用部位とのハイブリッド形成による遺伝子発現抑制などが挙げられる。これらは、転写、スプライシング、または翻訳の過程を阻害して、標的遺伝子の発現を抑制する(平島および井上「新生化学実験講座2 核酸IV 遺伝子の複製と発現」,日本生化学会編,東京化学同人, pp.319-347, 1993)。本発明で用いられるアンチセンスDNAは、上記のいずれの作用で標的SD2/Phs1遺伝子の発現を抑制してもよい。一つの態様としては、標的遺伝子のmRNAの5'端近傍の非翻訳領域に相補的なアンチセンス配列を設計すれば、遺伝子の翻訳阻害に効果的であろう。しかし、コード領域もしくは3'側の非翻訳領域に相補的な配列も使用し得る。このように、遺伝子の翻訳領域だけでなく非翻訳領域の配列のアンチセンス配列を含むDNAも、本発明で利用されるアンチセンスDNAに含まれる。使用されるアンチセンスDNAは、適当なプロモーターの下流に連結され、好ましくは3'側に転写終結シグナルを含む配列が連結される。
【0053】
アンチセンスDNAは、本発明の弱種子休眠性型DNA(例えば、配列番号:1または3に記載の塩基配列からなるDNA)の配列情報を基にホスホロチオネート法(Stein, Nucleic Acids Res., 16:3209-3221, 1988)などにより調製することが可能である。調製されたDNAは、後述する公知の方法で、植物へ導入できる。アンチセンスDNAの配列は、植物が持つ内因性の弱種子休眠性SD2/Phs1遺伝子の転写産物と相補的な配列であることが好ましいが、遺伝子の発現を有効に阻害できる限り、完全に相補的でなくてもよい。転写されたRNAは、標的とする遺伝子の転写産物に対して好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の相補性を有する。効果的に標的遺伝子の発現を阻害するには、アンチセンスDNAの長さは、少なくとも15塩基以上であり、好ましくは100塩基以上であり、さらに好ましくは500塩基以上である。通常、用いられるアンチセンスDNAの長さは5kbよりも短く、好ましくは2.5kbよりも短い。
【0054】
植物の弱種子休眠性型SD2/Phs1遺伝子の発現を抑制するために用いるDNAの他の態様は、本発明の弱種子休眠性型DNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNAである。リボザイムには、グループIイントロン型や、RNasePに含まれるM1RNAのように400ヌクレオチド以上の大きさのものもあるが、ハンマーヘッド型やヘアピン型と呼ばれる40ヌクレオチド程度の活性ドメインを有するものもある(小泉誠および大塚栄子、蛋白質核酸酵素, 35:2191, 1990)。
【0055】
例えば、ハンマーヘッド型リボザイムの自己切断ドメインは、G13U14C15のC15の3'側を切断するが、活性にはU14が9位のAと塩基対を形成することが重要とされ、15位の塩基はCの他にAまたはUでも切断されることが示されている(Koizumi et. al., FEBS Lett. 228:225, 1988)。リボザイムの基質結合部を標的部位近傍のRNA配列と相補的になるように設計すれば、標的RNA中のUC、UUまたはUAという配列を認識する制限酵素的なRNA切断リボザイムを作出することが可能である(Koizumi et. al., FEBS Lett. 239:285, 1988、小泉誠および大塚栄子,蛋白質核酸酵素,35:2191, 1990、Koizumi et. al., Nucleic. Acids. Res. 17:7059, 1989)。
【0056】
また、ヘアピン型リボザイムも、本発明の目的のために有用である。ヘアピン型リボザイムは、例えばタバコリングスポットウイルスのサテライトRNAのマイナス鎖に見出される(Buzayan, Nature 323:349, 1986)。このリボザイムも、標的特異的なRNA切断を起こすように設計できることが示されている(Kikuchi and Sasaki, Nucleic Acids Res. 19:6751, 1992、菊池洋,化学と生物 30:112, 1992)。標的を切断できるよう設計されたリボザイムは、植物細胞中で転写されるようにカリフラワーモザイクウイルスの35Sプロモーターなどのプロモーターおよび転写終結配列に連結される。このような構成単位をタンデムに並べ、標的遺伝子内の複数の部位を切断できるようにして、より効果を高めることもできる(Yuyama et al., Biochem. Biophys. Res. Commun. 186:1271, 1992)。このようなリボザイムを用いて標的となるSD2/Phs1遺伝子の転写産物を特異的に切断し、該遺伝子の発現を抑制することができる。
【0057】
<ベクター、形質転換植物細胞、形質転換植物植物体>
本発明は、また、上記本発明のDNA(弱種子休眠性型DNA、強種子休眠性型DNA、SD2/Phs1遺伝子の発現を抑制するためのDNA)を含むベクター、上記本発明のDNAまたはそれを含むベクターが導入された植物細胞、該細胞を含む植物植物体、該植物体の子孫またはクローンである植物植物体、および、これら植物植物体の繁殖材料を提供する。
【0058】
本発明のベクターとしては、植物細胞内で挿入遺伝子を発現させることが可能なものであれば特に制限はない。本発明のベクターは、本発明のDNAを恒常的または誘導的に発現させるためのプロモーターを含有しうる。恒常的に発現させるためのプロモーターとしては、例えば、カリフラワーモザイクウイルスの35Sプロモーター、イネのアクチンプロモーター、トウモロコシのユビキチンプロモーターなどが挙げられる。また、誘導的に発現させるためのプロモーターとしては、例えば、糸状菌・細菌・ウイルスの感染や侵入、低温、高温、乾燥、紫外線の照射、特定の化合物の散布などの外因によって発現することが知られているプロモーターなどが挙げられる。このようなプロモーターとしては、例えば、糸状菌・細菌・ウイルスの感染や侵入によって発現するイネキチナーゼ遺伝子のプロモーターやタバコのPRタンパク質遺伝子のプロモーター、低温によって誘導されるイネのlip19遺伝子のプロモーター、高温によって誘導されるイネのhsp80遺伝子とhsp72遺伝子のプロモーター、乾燥によって誘導されるシロイヌナズナのrab16遺伝子のプロモーター、紫外線の照射によって誘導されるパセリのカルコン合成酵素遺伝子のプロモーター、嫌気的条件で誘導されるトウモロコシのアルコールデヒドロゲナーゼ遺伝子のプロモーターなどが挙げられる。また、イネキチナーゼ遺伝子のプロモーターとタバコのPRタンパク質遺伝子のプロモーターはサリチル酸などの特定の化合物によって、rab16は植物ホルモンのアブシジン酸の散布によっても誘導される。
【0059】
本発明のベクターを導入する植物細胞の由来する植物としては特に制限はなく、例えば、オオムギ、コムギ、ライムギ、ライコムギ、エンバク、イネ、トウモロコシ、ダイズ、ポプラ、ブドウ、シロイヌナズナ、タバコなどが挙げられる。好ましくは、オオムギ、コムギ、ライムギ、ライコムギ、エンバク、イネ、トウモロコシなどのイネ科植物であり、より好ましくは、オオムギ、コムギ、ライムギ、ライコムギ、エンバクなどの麦類であり、最も好ましくはオオムギおよびコムギである。本発明の植物細胞には、培養細胞の他、植物体中の細胞も含まれる。また、種々の形態の植物細胞、例えば、懸濁培養細胞、プロトプラスト、葉の切片、カルス、未熟胚、花粉などが含まれる。植物細胞へのベクターの導入は、ポリエチレングリコール法、電気穿孔法(エレクトロポーレーション)、アグロバクテリウムを介する方法、パーティクルガン法など当業者に公知の種々の方法を用いることができる。
【0060】
形質転換植物細胞からの植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。例えば、オオムギに関する形質転換植物体を作出する手法としては、Tingayら(Tingay S. et al. Plant J. 11: 1369-1376, 1997)、Murrayら(Murray F et al. Plant Cell Report 22: 397-402, 2004)、およびTravallaら(Travalla S et al. Plant Cell Report 23: 780-789, 2005)に記載された方法を挙げることができる。また、イネにおいて、形質転換植物体を作出する手法については、ポリエチレングリコールによりプロトプラストへ遺伝子導入し、植物体を再生させる方法(Datta, S. K. In Gene Transfer To Plants(Potrykus I and Spangenberg Eds.) pp66-74, 1995)、電気パルスによりプロトプラストへ遺伝子導入し、植物体を再生させる方法(Toki et al. Plant Physiol. 100, 1503-1507, 1992)、パーティクルガン法により細胞へ遺伝子を直接導入し、植物体を再生させる方法(Christou et al. Bio/technology, 9: 957-962, 1991)およびアグロバクテリウムを介して遺伝子を導入し、植物体を再生させる方法(Hiei et al. Plant J. 6: 271-282, 1994)など、いくつかの技術が既に確立し、本願発明の技術分野において広く用いられている。また、シロイヌナズナであれば、Akamaら(Akama et al. Plant Cell Reports 12: 7-11, 1992)の方法が挙げられる。本発明においては、これらの方法を好適に用いることができる。
【0061】
一旦、ゲノム内に本発明の閉花性型DNAが導入された形質転換植物体が得られれば、該植物体から有性生殖または無性生殖により子孫を得ることが可能である。また、該植物体やその子孫あるいはクローンから繁殖材料(例えば、種子、果実、切穂、株、カルス、プロトプラスト等)を得て、それらを基に該植物体を量産することも可能である。本発明には、上記本発明のDNAが導入された植物細胞、該細胞を含む植物体、該植物体の子孫およびクローン、ならびに該植物体、その子孫およびクローンの繁殖材料が含まれる。
【0062】
<種子休眠性の形質が改変された植物の作出方法>
本発明は、また、種子休眠性の形質が改変された植物の作出方法を提供する。本発明の方法の1つの態様は、植物に本発明の弱種子休眠性型DNAを導入する工程を含む、種子休眠性の形質が弱められた植物の作出方法である。本発明において、植物の種子休眠性の形質を「弱める」とは、強種子休眠性の形質を有する品種の種子休眠性の形質を弱めることのみならず、既に、一定の弱種子休眠性の形質を有している品種の種子休眠性をさらに減弱させることをも含む意である。
【0063】
本発明の方法の他の1つの態様は、植物において、本発明の弱種子休眠性型DNA(弱種子休眠性型SD2/Phs1遺伝子)の発現または機能を抑制する工程を含む、種子休眠性の形質が強められた植物の作出方法である。本発明において、植物の種子休眠性の形質を「強める」とは、弱種子休眠性の形質を有する品種の種子休眠性の形質を強めることのみならず、既に、一定の強種子休眠性の形質を有している品種の種子休眠性をさらに増大させることをも含む意である。
【0064】
植物における本発明の弱種子休眠性型DNAの発現または機能の抑制は、植物に、上記本発明の強種子休眠性型DNAを導入することにより実施することができる。植物における強種子休眠性の形質は、単一劣性遺伝子支配であるため、植物に強種子休眠性の形質の形質を付与するためには、通常、個体におけるSD2/Phs1対立遺伝子の双方を強種子休眠性型DNAにする必要がある。これにより個体中で強種子休眠性型DNAのみが発現し、植物の種子休眠性の形質を強めることができる。植物染色体への本発明の強種子休眠性型DNAの導入は、例えば、交配や相同組換えにより行うことができる。強種子休眠性型DNAを導入することに代えて、植物染色体上の弱種子休眠性型DNAに、特定のDNA配列を導入し、その機能を破壊してもよい。
【0065】
また、植物における本発明の弱種子休眠性型DNAの発現または機能の抑制は、植物に上記本発明のSD2/Phs1遺伝子の発現を抑制するためのDNA(例えば、dsRNAをコードするDNA、アンチセンスDNA、リボザイム活性を有するRNAをコードするDNAなど)を導入することにより実施することができる。これにより個体中の弱種子休眠性型DNAから、弱種子休眠性型の翻訳産物が生産されなくなるため、植物の種子休眠性の形質を強めることができる。
【0066】
植物における本発明の弱種子休眠性型DNAの発現または機能の抑制には、例えば、弱種子休眠性型DNAの発現を抑制する薬剤や弱種子休眠性型の翻訳産物に結合し、その機能を抑制する薬剤の利用も考えられる。
【0067】
<植物における種子休眠性の程度を判定する方法>
本発明は、また、植物の種子休眠性の程度を判定する方法を提供する。本発明の判定方法の一つの態様は、植物におけるSD2/Phs1遺伝子またはその発現制御領域の塩基配列を解析し、対照の塩基配列と比較することを特徴とする方法である。
【0068】
SD2/Phs1遺伝子またはその発現制御領域の塩基配列の解析に際しては、SD2/Phs1遺伝子またはその発現制御領域のDNAをPCRにより増幅した増幅産物を用いることができる。前記PCRを実施する場合において、用いられるプライマーは、SD2/Phs1遺伝子またはその発現制御領域を特異的に増幅できるものである限り制限はなく、SD2/Phs1遺伝子またはその発現制御領域の配列情報(例えば、配列番号:1,3,5または7)に基づいて適宜設計することができる。好適なプライマーとしては、配列番号:9〜12に記載のプライマーが挙げられる。設計したプライマーを適宜組み合わせて、SD2/Phs1遺伝子またはその発現制御領域の特定の塩基配列を増幅することができる。
【0069】
被検植物におけるSD2/Phs1遺伝子の塩基配列と比較する「対照の塩基配列」は、典型的には、弱種子休眠性型品種または強種子休眠性型品種におけるSD2/Phs1遺伝子の塩基配列である。決定したSD2/Phs1遺伝子の塩基配列と弱種子休眠性型品種における塩基配列(例えば、配列番号:1、3)または強種子休眠性型品種における塩基配列(例えば、配列番号:5、7)とを比較することにより、被検植物におけるSD2/Phs1遺伝子が、強種子休眠性型であるか弱種子休眠性型であるかを評価することができる。本実施例において示されたように、オオムギの強種子休眠性品種であるアズマムギ由来のSD2タンパク質のアミノ酸配列(配列番号:6)は、弱種子休眠性品種である関東中手ゴールド由来のSD2タンパク質のアミノ酸配列(配列番号:2)と比較すると260位においてアミノ酸の置換(N→T)が生じている(図7)。また、コムギの強種子休眠性品種であるLeader由来のPhs1タンパク質のアミノ酸配列(配列番号:8)は、弱種子休眠性品種である春よ恋由来のPhs1タンパク質のアミノ酸配列(配列番号:4)と比較すると220位においてアミノ酸の置換(K→N)が生じている(図13、図14)。従って、特に、オオムギにおける260位およびコムギにおける220位のアミノ酸は、このような評価の好ましい指標となる。
【0070】
また、SD2/Phs1遺伝子においては、弱種子休眠性型DNAが優性であるため、被検DNAがコードするタンパク質の機能を喪失させるような大きな変異が存在する場合(例えば、新たな終止コドンの出現やフレームシフトにより、コードするタンパク質の分子量やアミノ酸配列に大きな変化が生じる場合)には、強種子休眠性型である蓋然性が高いと判定される。
【0071】
また、発現制御領域の塩基配列において、SD2/Phs1遺伝子の発現を抑制するような変異が存在する場合、強種子休眠性型である蓋然性が高いと判定される。発現制御領域の塩基配列における変異がSD2/Phs1遺伝子の発現に影響を与えるか否かは、当該変異を有する発現制御領域の下流にレポーター遺伝子を発現可能に連結したベクターを構築し、当該ベクターを植物細胞に導入し、レポーター活性を検出することにより、判定することができる。
【0072】
被検植物におけるSD2/Phs1遺伝子またはその発現制御領域の塩基配列が、対照の塩基配列と相違するか否かは、上記した直接的な塩基配列の決定以外に、種々の方法により間接的に解析することができる。このような方法としては、例えば、制限酵素断片長多型(Restriction Fragment Length Polymorphism/RFLP)を利用したRFLP法やPCR-RFLP法、PCR-SSCP(single-strand conformation polymorphism、一本鎖高次構造多型)法、変性剤濃度勾配ゲル電気泳動法(denaturant gradient gel electrophoresis:DGGE)、アレル特異的オリゴヌクレオチド(Allele Specific Oligonucleotide/ASO)ハイブリダイゼーション法、リボヌクレアーゼAミスマッチ切断法が挙げられる。例えば、PCR-RFLP法において、配列番号:9と10に記載のプライマーを用いてコムギのPhs1遺伝子を増幅し、制限酵素SalIで処理した場合、春よ恋型のDNAのみが切断される。一方、配列番号:11と12プライマーに記載のプライマーを用いてコムギのPhs1遺伝子を増幅し、制限酵素DraIで処理した場合、Leader型のDNAのみが切断される。こうして制限酵素処理したPCR産物をアガロースゲルで検出することにより、簡便にPhs1遺伝子のタイプ判定することができる。
【0073】
なお、本発明の判定方法における、被検植物からのDNAの調製は、常法、例えば、CTAB法を用いて行うことができる。DNAの調製には、例えば、植物の種子、幼植物体、成長した植物体を用いることができる。また、塩基配列の決定は、常法、例えば、ジデオキシ法やマキサム-ギルバート法などにより行なうことができる。塩基配列の決定においては、市販のシークエンスキットおよびシークエンサーを利用することができる。
【0074】
<種子休眠性の形質が改変された植物を育種する方法>
本発明は、また、種子休眠性の形質が改変された植物を育種する方法を提供する。本発明の育種方法の1つの態様は、強種子休眠性の形質の植物を育種する方法であり、(a)強種子休眠性の形質の植物品種と任意の植物品種とを交配させる工程、(b)交配により得られた個体における種子休眠性の程度を、上記本発明の判定方法により判定する工程、および(c)強種子休眠性を有すると判定された品種を選抜する工程、を含む。
【0075】
強種子休眠性の形質の植物品種と交配させる「任意の植物品種」としては、例えば、弱種子休眠性の形質を有する品種、弱種子休眠性の形質を有する品種と強種子休眠性の形質を有する品種との交配により得られた個体が挙げられるが、これらに制限されない。
【0076】
本発明の育種方法の他の1つの態様は、弱種子休眠性の形質の植物を育種する方法であり、(a)弱種子休眠性の形質の植物品種と任意の植物品種とを交配させる工程、(b)交配により得られた個体における種子休眠性の程度を、上記本発明の判定方法により判定する工程、および(c)弱種子休眠性を有すると判定された品種を選抜する工程、を含む。
【0077】
弱種子休眠性の形質の植物品種と交配させる「任意の植物品種」としては、例えば、強種子休眠性の形質を有する品種、強種子休眠性の形質を有する品種と弱種子休眠性の形質を有する品種との交配により得られた個体が挙げられるが、これらに制限されない。
【0078】
本発明の育種方法を利用すれば、種子休眠性の形質が改変された植物を、種子や幼植物の段階で選抜することが可能となり、種子休眠性の形質が改変された品種の育成を短期間で行うことが可能となる。
【実施例】
【0079】
[実施例1] オオムギSD2遺伝子の同定
オオムギ材料は、圃場(独立行政法人 農業生物資源研究所)に植えて育成した。オオムギ品種および系統としては、次のものを用いた。(i)強種子休眠性型の6条品種「アズマムギ」(Az)、(ii)および弱種子休眠性型の2条品種「関東中生ゴールド」(KNG)、(iii)AzとKNGの交配由来の組換え近交系(RIL)99系統(Mano et al. (2001) Genome 44:284-292)、(iv)5H染色体長椀末端領域約20cMがKNGの染色体断片に置き換わっているAzの系統(NIL#34;図1)、(v)NIL#34とAzの交配由来のF2個体(2985個体)。
【0080】
発芽試験においては、生理的成熟期および完熟期の穂を採取し、30℃で1週間乾燥させた。その後、一穂から得られた種子を、No.2濾紙を2枚敷いたシャーレ(直径9cm高さ1.5cm)に撒き、2000分の1のベンレートを加えた4.5mlの蒸留水を加え、15℃一週間置いた。その後、各シャーレの発芽種子粒数および全種子粒数を数え、「発芽粒数/全粒数 × 100」の式で発芽率(%)を算出した。
【0081】
(1)量的形質座(QTL)解析
NIL#34とAzとを交配させて得た2985のF2個体(RIL99系統)について実施した発芽試験の結果およびManoらの文献(Mano et al. (2001) Genome 44:284-292)に記載の連鎖地図を用いてQTL解析を行った。QTLの検出は、MapQTL5解析ソフト(Biometris, Wageningen, The Netherlands, http://www.joinmap.nl)を用いて、シンプルインターバルマッピング法により行った。その結果、5H染色体長椀末端のSD2座に、作用力の大きな種子休眠QTLが検出された(図2)。
【0082】
(2)アソシエーション解析
SD2座近傍で組換えが起きているF2個体に由来する後代(F3個体)60個体を用いて、SD2座近傍の遺伝子型の同定と発芽率の測定を行い、遺伝子型と発芽率の相関を解析して、SD2座の遺伝子型と座乗している領域を同定した(図3)。新規マーカーの開発も行って、SD2座を絞り込んだ結果、最終的に、SD2座は、5M24と5M25のマーカーに挟まれた0.08cMの領域に座乗することが判明した(図4)。
【0083】
(3)オオムギBAC(バクテリア人工染色体)クローンの単離
5M24マーカーを用いて「OUH602」および「Morex」のゲノムBACライブラリー(Yu et al. (2000) Theor Appl Genet 101:1093-1099)をPCR法によりスクリーニングした。選抜されたBACクローンについてショットガンシークエンスにより配列を決定した。得られたゲノム配列は、Rice Genome Automated Annotation System(RiceGAAS) website(http://ricegaas.dna.affrc.go.jp/;Sakata et al. (2002) Nucleic Acids Res. 30:98-102)を用いて解析した。その結果、OUH602のゲノムBACライブラリーより、約70kbのゲノム配列を持つ「BACクローン6」が見出された。このBACクローン上には、5M25マーカーも座乗しており、SD2座が、5M24マーカーおよび5M25マーカーの間の約28kbのゲノム領域に座乗していることが明らかになった(図5)。
【0084】
(4)SD2遺伝子の同定
RiceGAAS(Rice Genome Automated Annotation System)を用いた分析により、絞り込まれた領域内およびその近傍に、5つの遺伝子が見出された(図6)。このうちGene1、Gene4、Gene5は、対応する完全長配列が見い出され、機能している遺伝子であることが確認できた。一方、Gene2とGene3は、完全長データベースに登録されていない遺伝子であった。Gene4とGene5は、完熟種子で発現していることが確認された。Gene3は、トランスポゾンによく見られるPMD(plant mobile domain)を保持していた。
【0085】
Gene4のアミノ酸配列をAzとKNGの間で比較すると、植物間で高度に保存されているアスパラギン(N)が、アズマムギではトレオニン(T)に置換されていることが明らかになった(図7)。
【0086】
また、Gene4が、Az型(強休眠性型)とKNG型(弱休眠性型)のヘテロとなっている個体につき、その種子の発芽率を測定し、いずれの型が優性であるかの検証を行った。その結果、KNG型(弱休眠性型)が優性であることが判明した。
【0087】
以上から、Mitogen-activated Kinase kinase(MAPKK)をコードするGene4が、SD2遺伝子であると考えられた。
【0088】
[実施例2] コムギPhs1(Pre-Harvest Sprouting 1)遺伝子の同定
カナダの穂発芽難品種(強種子休眠性型)の「Leader」を一回親とし、穂発芽中(弱種子休眠性型)品種「春よ恋」を反復親に配した戻し交雑によって、準同質遺伝子系統(NILs)を作製した。BC1F1を育苗箱で1個体ずつ養成し、緑葉からDNeasy Plant Mini Kit(キアゲン)を用いてDNAを抽出した。Phs1近傍に位置するPCRマーカーの遺伝子型を決定した。その際、マーカー遺伝子型がヘテロ接合型を示す個体を選抜し、幼苗をポットへ移植した。選抜個体を温室内で養成し、「春よ恋」との交配を繰り返した。この方法により2組合せについてBC4まで世代を進めた。選抜個体を自殖して得られたBC4F2個体の中からマーカーの遺伝子型により、Phs1近傍領域が、「Leader」ホモ接合型、「春よ恋」ホモ接合型にそれぞれ固定した個体、ならびにマーカー間で組換えを生じた個体を選抜し、種子休眠性試験に供試した。
【0089】
温室の温度は15〜25℃の範囲で変動させ、2月から6月の間に登熟した種子を休眠性の評価に用いた。日長感応性を示す材料については陽光ランプで補光し、出穂後は自然日長で登熟させた。
【0090】
種子休眠性の評価用種子の調製にあたっては、穂を成熟期に収集し、室温で7日間乾燥した。採取種子は傷がつかないように手で脱穀した後、試験まで-30℃で保存した。発芽試験においては、直径9cm、深さ2cmのシャーレ(テルモ)に濾紙(ワットマンNo.3)を一枚敷き、7mlの蒸留水を添加した。発芽試験に用いる種子は、胚が上向きになるように1粒ずつシャーレに置床した。発芽試験は15℃もしくは20℃のインキュベータ内で10日間行い、発芽を調査した。成熟乾燥後の種子休眠性検定から、コムギ4A染色体に座乗するQTLが主働遺伝子Phs1として2.6cMの域内にマッピングされている(非特許文献13)。Phs1近傍のDNAマーカーがLeader型ホモに固定している63個体、春よ恋型に固定している65個体を選抜し、種子休眠性を評価した結果、Phs1がLeader型と春よ恋型で明瞭に種子休眠性(発芽率)の差異が認められた(図8)。
【0091】
(1)Phs1遺伝子のマッピングによる絞込み
(i)大規模連鎖解析
6倍体パンコムギ(Triticum aestivum L.)品種Chinese Springから作成したBACライブラリー(Allouis et al. (2003) Cereal Research Communications. 31:331-338)からAゲノム特異的プライマーを用いてクローンを選抜し、末端配列および内部配列を解読して新たなマーカーおよびBACスクリーニング用プライマーを作成した。単離された代表的なBACクロ−ンの位置関係を図9に示した。
【0092】
一方、Leader(強種子休眠性型)と春よ恋(弱種子休眠性型)間から作出したBC4F2集団(一部BC3F2個体含む)3,235個体を用いて、文献(非特許文献13)に記載の種子休眠性評価方法によりPhs1を0.19cMの域内に絞り込んだ(非特許文献14)。すなわち、マーカー間で組み換えを生じたF2個体からF3系統10〜20個体を展開し、春よ恋型およびLeader型に固定した個体を複数個体選抜した。選抜された個体の種子休眠性を評価し、分離が認められればF2のPhs1遺伝子型がヘテロ、春よ恋型(発芽率が高い)およびLeader型(発芽率が低い)に分離が認められなければ、F2のPhs1遺伝子型がそれぞれ、春よ恋ホモ、Leaderホモと判定しマッピングした。
【0093】
Phs1をカバーする連続的なBACコンティグの作成には至っていなかったが、CSBAC4からCSBAC6にわたる約180kbの領域がPhs1と共分離したことから、このBACクローンの塩基配列をショットガンシーケンス法により決定した。その結果、4種の遺伝子(No.1〜4)が想定された(図9)。遺伝子の推定にあたっては、RAP-DB(http://rapdb.dna.affrc.go.jp/)、NCBI(http://blast.ncbi.nlm.nih.gov/Blast.cgi)、GrainGenes (http://wheat.pw.usda.gov/GG2/index.shtml)のデータベースをサーベイした。
【0094】
(ii)オオムギ、イネとの比較ゲノム解析
イネのゲノム情報を通じて、オオムギゲノムとの比較解析を行った。オオムギSD2遺伝子近傍にマップされたマーカーとコムギPhs1近傍にマップされたマーカーのうち、イネの第3染色体に座乗するOs03g0860000(候補遺伝子No2に相当)、第6染色体に座乗するOs06g0473200(候補遺伝子No1に相当)が共通にマップされかつ同じ順序で並んでいた(図10)。このことから、コムギPhs1とオオムギSD2遺伝子は共通の遺伝子に制御されていることが強く示唆された。4種の候補遺伝子のうち、SD2遺伝子の有力な原因遺伝子と共通のMitogen-activated protein kinase kinase(MAPKK)遺伝子(候補遺伝子No1に相当)がPhs1の原因遺伝子と考えられ、コムギ「TaMEK-A1」と名づけた。なお、コムギ4ALは6倍体パンコムギへの進化の過程で逆位や5A染色体と7B染色体との部分的な相互転座が生じている(Devos et al. (1995) Theor Appl Genet 91:282-288)。Phs1近傍領域は5A末端領域が逆向きになっているため、オオムギ5HL染色体とはテロメア側とセントロメア側が逆になっている。
【0095】
(2)候補遺伝子TaMEK-A1の解析
(i)候補遺伝子TaMEK-A1の塩基配列決定
CSBAC4およびCSBAC6の内部塩基配列情報をもとにして、Leaderと春よ恋のゲノムDNAを鋳型にしたPCR産物の塩基配列を決定した。TaMEK-A1は、10のイントロンと11のエキソンからなる。ORF長は1572bpと推定され、推定翻訳産物は523残基からなるアミノ酸をコードする(図11、13)。
【0096】
また、春よ恋の種子胚由来RNAからSuperScriptIII逆転写酵素(インビトロジェン)およびSMART RACE cDNA Amplification Kit(クローンテック)を用いて調製したcDNAから、TaMEK-A1に相当する完全長cDNAを得た。得られたcDNAを鋳型にして得られたTaMEK-A1遺伝子に相当するPCR産物の塩基配列を決定した。
【0097】
春よ恋およびLeaderの塩基配列の解読方法は以下の通りである。ゲノムDNAを鋳型にしてPCRを行い、目的領域に含まれる産物のみをクローニングして、シーケンスを決定した。クローニングにあたってはPCR産物をpGEM T-Easy Vector(プロメガ)にライゲーションし、大腸菌XL10-Goldに形質転換した(ストラタジーン)。コロニーPCRにより、組換え型クローンを選び、インサートDNAのシーケンスを決定した。また、染色体欠失系統を利用しても目的の産物を確認できなかった場合は、マッピング集団の両親のそれぞれについてPCR産物を得て、クローニングし、6〜8クローンを無作為に選んでシーケンスを決定した。Aゲノムの祖先種であるTriticum urartuのDNAを鋳型とするPCR産物も同時にシーケンスすることにより、Aゲノムシーケンスを同定した。得られたシーケンスのアライメントとグルーピングは、Sequencher ver. 4.1.2(Gene Code)を用いて行った。以下の(ii)、(iii)の実験も同様に行った。アミノ酸配列の解析は、BLAST-p, tblastn(http://blast.ncbi.nlm.nih.gov/Blast.cgi)を用いて行った。
【0098】
(ii)TAMEK-A1遺伝子の塩基配列と推定されるアミノ酸配列の品種間差
4A染色体のPhs1近傍にQTLが報告されている14品種・系統を用いて、TaMEK-A1およびNo2〜4遺伝子の想定されるエキソンおよび5′側の上流域について、春よ恋とLeader間に見出された塩基配列変異を調査した。その結果、TaMEK-A1のエキソン内に認められたSNPだけが唯一休眠性の品種間差とSNPの変異とがよく一致した(図12)。
【0099】
なお、弱種子休眠性品種のハルユタカ(日本)、Munstertaler(スイス)、Janz、Cascades、Cunningham、Cranbrook(以上オーストラリア)のTaMEK-A1のアミノ酸配列は、春よ恋(日本)と同一であった。また、強種子休眠性品種・系統のきたもえ、OS21-5(日本)、AC Domain(カナダ)、SW95-50213、AUS1408、AUS1490(以上オーストラリア)のTaMEK-A1のアミノ酸配列はLeader(カナダ)と同一であった。
【0100】
春よ恋およびLeaderのゲノムDNAを鋳型にして、5′上流域を含めると全長8,892bpのTaMEK-A1配列を決定した。2654番目の塩基に位置するエキソンに認められたSNPはTaMEK-A1遺伝子の推定アミノ酸配列内に予測されるセリン/スレオニン型タンパク質リン酸化酵素の活性部位に隣接するアミノ酸の置換を伴う変異であった。すなわちLeaderは塩基がCであるため、アミノ酸でアスパラギン(N)に翻訳されるのに対して春よ恋は塩基がAであるため、アミノ酸がリシン(K)に翻訳される(図13、図14)。タンパク質モチーフの予測はScanProsite(http://au.expasy.org/tools/scanprosite/)を利用した。
【0101】
(iii)TaMEK1のゲノム間および種間の比較
TaMEK-A1遺伝子のアミノ酸置換を伴う1塩基多型(SNP)だけが14品種の休眠性の差異とよく一致した。そこで、TaMEK1遺伝子のコムギ同祖遺伝子の単離を試みたところ、3種の異なる配列を同定した(TaMEK-B1、TAMEK-D1)。これらが同祖遺伝子の候補としてSNPのハプロタイプを判定した結果、B、Dゲノム(春よ恋のDゲノム遺伝子は未単離)のSNPはいずれもAゲノムのLeader(強種子休眠性型)と同じハプロタイプ(DNAでC、アミノ酸でアスパラギン)であった(図15)。また、Aゲノム祖先種T.urartu、Dゲノム祖先種のAe.squarrosaデータベースに登録されているホモログのハプロタイプはいずれもLeader型であった。従って、これまでのところLeader型のアリルが野生型で、Phs1の座乗するAゲノム遺伝子の春よ恋(弱種子休眠性型)アリルが変異型と考えられた。
【0102】
(iv)遺伝子発現
開花後40日目の種子胚からRNeasy Plant mini kit(キアゲン)を用いてRNAを抽出し、SuperScriptIII逆転写酵素(インビトロジェン)を用いてcDNAを合成した。cDNAを鋳型にしてTaMEK-A1遺伝子のゲノム特異的なプライマーを用いたRT-PCRにより遺伝子発現の解析を行った。その結果、春よ恋、LeaderともにTaMEK-A1遺伝子は種子胚で発現していることが明らかになった(図16)。
【0103】
(v)TaMEK-A1の判定方法
TaMEK-A1のエキソン内に見出されたSNPをもとにdCAPS(derived cleaved amplified polymorphic sequence)マーカーを作成した。ゲノムDNAに対してプライマー配列A(フォワードTAAAGCCAGCAAATTTACTGGTCGA/配列番号:9、リバースGCTCATATTTAAGGAATATCAAAAGAAC/配列番号:10)を用いたPCRを行い(アニーリング温度55℃)、制限酵素SalIで処理した場合に春よ恋型の塩基Cタイプのみが切断される。一方、プライマー配列B(フォワードTCTAAAGCCAGCAAATTTACTGGGTAA/配列番号:11、リバースGCTCATATTTAAGGAATATCAAAAGAAC/配列番号:12)を用いたPCR産物(アニーリング温度55℃)を制限酵素DraIで処理するとLeader型のDNAのみが切断される。制限酵素処理したPCR産物をアガロースゲルで検出することができる。この2種を組み合わせることにより、Phs1遺伝子のタイプを簡易に判定できる。
【0104】
PCRの条件は以下の通りである。ゲノムDNAはDneasy Plant mini kit(キアゲン)およびMagAttract96 DNA Plant Core kit(キアゲン)を用いてDNAを抽出した。次に鋳型DNA、2.5mM MgCl2、1.5mM dNTP、1.5μM プライマー 1unitのTaqポリメラーゼを混合して反応液とした。PCRは94℃3分後、「94℃40秒55℃40秒72℃1分」を30サイクル繰り返し、最後に72℃7分伸長反応を行った。PCR反応液を制限酵素で処理後、アガロースゲルで電気泳動しSYBR green I(Cambrex Bio Science)で染色した後、多型を検出した。
【産業上の利用可能性】
【0105】
上記した通り、本発明により、植物の種子休眠性の程度を支配する新規な遺伝子SD2/Phs1が同定され、同定されたSD2/Phs1遺伝子を利用して、植物の種子休眠性の程度の判定や種子休眠性が改変された植物の作出・育種が可能となった。本発明における種子休眠性の程度の判定は、それを支配する遺伝子を標的としており、しかも、育成早期の個体(例えば、種子)を用いて実施することが可能である。このため、本発明の判定方法を利用すれば、種子休眠性が改変された品種を特異的かつ効率的に育種することが可能である。
【0106】
こうして作出・育種された弱種子休眠性の植物は、休眠が深くて発芽が著しく遅延する場合や乾燥地域で降水量が少ないために発芽が遅延する場合に、十分かつ早期に発芽個体を確保できる点で有用である。また、強種子休眠性の植物は、降雨の多い地域で穂発芽による種子の品質劣化を防げる点で有用である。従って、本発明は、特に農作物の収穫量の向上及び品質劣化による経済的損失の軽減に大きく貢献することができる。
【配列表フリーテキスト】
【0107】
配列番号9〜12
<223> 人工的に合成されたプライマーの配列
【特許請求の範囲】
【請求項1】
植物の種子休眠性の形質を弱める活性を有するタンパク質をコードする、下記(a)〜(d)のいずれかに記載のDNA。
(a)配列番号:2または4に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:1または3に記載の塩基配列のコード領域を含むDNA
(c)配列番号:2または4に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号:1または3に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNA
【請求項2】
植物の種子休眠性の形質を強める活性を有するタンパク質をコードする、下記(a)〜(d)のいずれかに記載のDNA。
(a)配列番号:6または8に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:5または7に記載の塩基配列のコード領域を含むDNA
(c)配列番号:6または8に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号:5または7に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNA
【請求項3】
植物の種子休眠性の形質を強める活性を有する、下記(a)〜(c)のいずれかに記載のDNA。
(a)請求項1に記載のDNAの転写産物と相補的な二重鎖RNAをコードするDNA
(b)請求項1に記載のDNAの転写産物と相補的なアンチセンスRNAをコードするDNA
(c)請求項1に記載のDNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA
【請求項4】
請求項1〜3のいずれかに記載のDNAを含むベクター。
【請求項5】
請求項1〜3のいずれかに記載のDNAが導入された植物細胞。
【請求項6】
請求項5に記載の細胞を含む植物植物体。
【請求項7】
請求項6に記載の植物体の子孫またはクローンである、植物植物体。
【請求項8】
請求項6または7に記載の植物植物体の繁殖材料。
【請求項9】
植物に請求項1に記載のDNAを導入する工程を含む、種子休眠性の形質が弱められた植物の作出方法。
【請求項10】
植物における請求項1に記載のDNAの発現または機能を抑制することを特徴とする、種子休眠性の形質が強められた植物の作出方法。
【請求項11】
植物に請求項2または3に記載のDNAを導入する工程を含む、種子休眠性の形質が強められた植物の作出方法。
【請求項12】
請求項1に記載のDNA、または該DNAが挿入されたベクターを含む、植物の種子休眠性の形質を弱めるための薬剤。
【請求項13】
請求項2もしくは3に記載のDNA、または該DNAが挿入されたベクターを含む、植物の種子休眠性の形質を強めるための薬剤。
【請求項14】
植物における種子休眠性の程度を判定する方法であって、被検植物における請求項1もしくは2に記載のDNAまたはその発現制御領域の塩基配列を解析し、対照の塩基配列と比較することを特徴とする方法。
【請求項15】
弱い種子休眠性の形質の植物を育種する方法であって、
(a)弱い種子休眠性の形質の植物品種と任意の植物品種とを交配させる工程、
(b)工程(a)における交配により得られた個体における種子休眠性の程度を、請求項14に記載の方法により判定する工程、および
(c)弱い種子休眠性を有すると判定された品種を選抜する工程、を含む方法。
【請求項16】
強い種子休眠性の形質の植物を育種する方法であって、
(a)強い種子休眠性の形質の植物品種と任意の植物品種とを交配させる工程、
(b)工程(a)における交配により得られた個体における種子休眠性の程度を、請求項14に記載の方法により判定する工程、および
(c)強い種子休眠性を有すると判定された品種を選抜する工程、を含む方法。
【請求項1】
植物の種子休眠性の形質を弱める活性を有するタンパク質をコードする、下記(a)〜(d)のいずれかに記載のDNA。
(a)配列番号:2または4に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:1または3に記載の塩基配列のコード領域を含むDNA
(c)配列番号:2または4に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号:1または3に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNA
【請求項2】
植物の種子休眠性の形質を強める活性を有するタンパク質をコードする、下記(a)〜(d)のいずれかに記載のDNA。
(a)配列番号:6または8に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:5または7に記載の塩基配列のコード領域を含むDNA
(c)配列番号:6または8に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号:5または7に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNA
【請求項3】
植物の種子休眠性の形質を強める活性を有する、下記(a)〜(c)のいずれかに記載のDNA。
(a)請求項1に記載のDNAの転写産物と相補的な二重鎖RNAをコードするDNA
(b)請求項1に記載のDNAの転写産物と相補的なアンチセンスRNAをコードするDNA
(c)請求項1に記載のDNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA
【請求項4】
請求項1〜3のいずれかに記載のDNAを含むベクター。
【請求項5】
請求項1〜3のいずれかに記載のDNAが導入された植物細胞。
【請求項6】
請求項5に記載の細胞を含む植物植物体。
【請求項7】
請求項6に記載の植物体の子孫またはクローンである、植物植物体。
【請求項8】
請求項6または7に記載の植物植物体の繁殖材料。
【請求項9】
植物に請求項1に記載のDNAを導入する工程を含む、種子休眠性の形質が弱められた植物の作出方法。
【請求項10】
植物における請求項1に記載のDNAの発現または機能を抑制することを特徴とする、種子休眠性の形質が強められた植物の作出方法。
【請求項11】
植物に請求項2または3に記載のDNAを導入する工程を含む、種子休眠性の形質が強められた植物の作出方法。
【請求項12】
請求項1に記載のDNA、または該DNAが挿入されたベクターを含む、植物の種子休眠性の形質を弱めるための薬剤。
【請求項13】
請求項2もしくは3に記載のDNA、または該DNAが挿入されたベクターを含む、植物の種子休眠性の形質を強めるための薬剤。
【請求項14】
植物における種子休眠性の程度を判定する方法であって、被検植物における請求項1もしくは2に記載のDNAまたはその発現制御領域の塩基配列を解析し、対照の塩基配列と比較することを特徴とする方法。
【請求項15】
弱い種子休眠性の形質の植物を育種する方法であって、
(a)弱い種子休眠性の形質の植物品種と任意の植物品種とを交配させる工程、
(b)工程(a)における交配により得られた個体における種子休眠性の程度を、請求項14に記載の方法により判定する工程、および
(c)弱い種子休眠性を有すると判定された品種を選抜する工程、を含む方法。
【請求項16】
強い種子休眠性の形質の植物を育種する方法であって、
(a)強い種子休眠性の形質の植物品種と任意の植物品種とを交配させる工程、
(b)工程(a)における交配により得られた個体における種子休眠性の程度を、請求項14に記載の方法により判定する工程、および
(c)強い種子休眠性を有すると判定された品種を選抜する工程、を含む方法。
【図1】


【図2】
【図3】
【図4】

【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【公開番号】特開2012−115178(P2012−115178A)
【公開日】平成24年6月21日(2012.6.21)
【国際特許分類】
【出願番号】特願2010−266305(P2010−266305)
【出願日】平成22年11月30日(2010.11.30)
【国等の委託研究の成果に係る記載事項】(出願人による申告)農林水産省、平成22年度、新農業展開ゲノムプロジェクト(麦類の遺伝子単離と機能解明)委託事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【出願人】(390022954)ホクレン農業協同組合連合会 (14)
【Fターム(参考)】
【公開日】平成24年6月21日(2012.6.21)
【国際特許分類】
【出願日】平成22年11月30日(2010.11.30)
【国等の委託研究の成果に係る記載事項】(出願人による申告)農林水産省、平成22年度、新農業展開ゲノムプロジェクト(麦類の遺伝子単離と機能解明)委託事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【出願人】(390022954)ホクレン農業協同組合連合会 (14)
【Fターム(参考)】
[ Back to top ]