
植物の開花時期を制御する遺伝子、タンパク質およびその利用
【課題】ナンヨウアブラギリに代表されるトウダイグサ目植物などの産業上、非常に有用であると考えられる植物の開花時期を制御する機能を有するポリペプチドを提供する。
【解決手段】特定のアミノ酸配列を有するポリペプチドをコードするポリヌクレオチドを用いる。つまり、(1)前記特定のアミノ酸配列で示されるポリペプチド、(2)前記アミノ配列で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド、(3)前記アミノ酸配列で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド。
【解決手段】特定のアミノ酸配列を有するポリペプチドをコードするポリヌクレオチドを用いる。つまり、(1)前記特定のアミノ酸配列で示されるポリペプチド、(2)前記アミノ配列で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド、(3)前記アミノ酸配列で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、植物、特にナンヨウアブラギリ等のトウダイグサ目植物の開花時期を制御する遺伝子、タンパク質およびその利用に関するものである。
【背景技術】
【0002】
近年、食物としてのみではなくバイオ燃料としても、各種植物が注目を浴びつつある。莫大な数の人類の生活を支えるには莫大な量のこれら植物を必要とするが、耕作面積には限りがあり、それ故に、限られた面積の耕作地において最大限の収穫量を実現するための研究がなされつつある。そして、近年、開花時期などを人為的に制御することによって植物のライフサイクルを短縮し、これによって、植物の収穫量を増加させようとする試みもなされている。
【0003】
通常、植物の開花時期は、品種特有の内的要因の他、日長時間、気温および栄養状態などの外的要因によっても決定される。それ故に、従来から、外的要因を調節することによって、植物の開花時期が調節されてきた。しかしながら、外的要因を調節するためには特別な施設(例えば、光照射装置および/または温度調節装置など)を利用する必要があり、これらの設備費が高いとともに、照明費および/または空調費などが高くなるという問題点を有している。
【0004】
そこで、近年、植物の開花時期の制御に関わる遺伝子を同定し(例えば、非特許文献1参照)、当該遺伝子を用いて植物の開花時期を制御しようとする試みがなされている。
【0005】
例えば、特許文献1では、日長の制御を直接受ける遺伝子よりも下流で制御される遺伝子が検索され、当該遺伝子を用いれば開花時期を早めることが可能となり、その結果、発芽から種子取得までの期間を大幅に短縮できることが記載されている。更に具体的には、特許文献1には、花成遺伝子(FT遺伝子:Flowering Locus T遺伝子)をシロイヌナズナ内で強制的に発現させることにより、開花が早くなることが記載されている。
【0006】
FT遺伝子は、現在までに、数種類の植物において同定されている。例えば、シロイヌナズナ、イネ、コムギ、ライグラス、アカザ、ミカン、カボチャ、リンゴ、トマト、ブドウおよびポプラにおいて、花成遺伝子が同定されている(例えば、非特許文献2参照)。
【0007】
FT遺伝子は、日長などに応答して維管束師部での発現が増大し、FT遺伝子産物であるFTタンパク質が茎頂に移動して、茎頂で発現しているFDと呼ばれるbZIP型転写因子と相互作用することにより開花時期を促進することが知られている(例えば、非特許文献3参照)。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特開2000−139250号公報(公開日:2000年5月23日)
【非特許文献】
【0009】
【非特許文献1】Kobayashi Y et al, 1999, Science 286, 1960-1962
【非特許文献2】Igasaki T et al., 2008, Plant Cell Physiol. 49(3), 291-300
【非特許文献3】Kobayashi Y and Weigel D、2007、Genes Dev 21、2371-2384
【発明の概要】
【発明が解決しようとする課題】
【0010】
ナンヨウアブラギリに代表されるトウダイグサ目植物などの産業上、非常に有用であると考えられる植物においては花成遺伝子が単離されておらず、当該花成遺伝子を用いてこれらの植物の開花時期を制御したという報告はない。
【0011】
本発明は、上記従来の問題点に鑑みなされたものであって、ナンヨウアブラギリに代表されるトウダイグサ目植物などの産業上、非常に有用であると考えられる植物の開花時期を制御することを目的とする。
【課題を解決するための手段】
【0012】
シロイヌナズナなどのモデル生物に関しては、研究者の数も多く、遺伝子配列などの多くの情報が流通している。しかしながら、ナンヨウアブラギリに関しては、研究者の数も少なく、流通している遺伝子配列などの情報量も限られている状況にある。しかも、シロイヌナズナとナンヨウアブラギリとでは種間の距離も大きく、遺伝子配列のホモロジーも低いと考えられた。そこで本発明者は、複数の縮重プライマーを作成してFT遺伝子の単離を試みた。
【0013】
当初、ナンヨウアブラギリのFT遺伝子が、どの生育段階で、どの部位で、どのような気象条件で、どれだけ発現しているのか不明であり、cDNAを鋳型としてナンヨウアブラギリのFT遺伝子をクローニングしようとしても成功しなかった。そこで、まず、ゲノムDNAを鋳型としてナンヨウアブラギリのFT遺伝子の部分断片を取得し、当該部分断片の情報に基づいて、ナンヨウアブラギリのFT遺伝子の発現解析を行った。その結果、ナンヨウアブラギリのFT遺伝子が高発現する葉位および気象条件を特定することができた。
【0014】
このような、多くの試行錯誤の末にナンヨウアブラギリからFT遺伝子を単離することに成功し、これによって本発明を完成させるに至った。
【0015】
つまり、本発明は、以下の〔1〕〜〔9〕の発明を包含する。
【0016】
〔1〕:下記(1)〜(3)の何れかのポリペプチドをコードすることを特徴とするポリヌクレオチド。つまり、
(1)配列番号14で示されるポリペプチド、
(2)配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド、
(3)配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド。
【0017】
〔2〕:下記(4)〜(10)の何れかであることを特徴とするポリヌクレオチド。つまり、
(4)配列番号11、12または13で示されるポリヌクレオチド、
(5)配列番号11、12または13で示されるポリヌクレオチドのアンチセンス配列をコードするポリヌクレオチド、
(6)配列番号17および配列番号18に示すプライマーセットによって増幅されるポリヌクレオチドを含むポリヌクレオチド、
(7)配列番号19および配列番号20に示すプライマーセットによって増幅されるポリヌクレオチドを含むポリヌクレオチド、
(8)上記(4)〜(7)の何れかに記載されるポリヌクレオチドにおいて、1つ以上30以下の塩基が置換、欠失、挿入および/または付加されたポリヌクレオチドからなり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド、
(9)上記(4)〜(7)の何れかに記載されるポリヌクレオチドと相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド、
(10)上記(4)〜(7)の何れかに記載されるポリヌクレオチドとの相同性が84%以上であり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド。
【0018】
〔3〕:バラ下綱植物由来であることを特徴とする〔1〕に記載のポリヌクレオチド。
【0019】
〔4〕:〔1〕〜〔3〕の何れかに記載のポリヌクレオチドを含有することを特徴とする組換えベクター。
【0020】
〔5〕:上記組換えベクターが、プラスミドベクター、ウイルスベクター、ファージベクターまたはコスミドベクターであることを特徴とする〔4〕に記載の組換えベクター。
【0021】
〔6〕:下記(11)〜(13)の何れかのポリペプチド。つまり、
(11)配列番号14で示されるポリペプチド、
(12)配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド、
(13)配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド。
【0022】
〔7〕:〔1〕〜〔3〕の何れかに記載のポリヌクレオチド、〔4〕若しくは〔5〕に記載の組換えベクター、または、〔6〕に記載のポリペプチドを植物に導入する工程を含むことを特徴とする植物の開花時期の制御方法。
【0023】
〔8〕:〔1〕〜〔3〕の何れかに記載のポリヌクレオチド、〔4〕若しくは〔5〕に記載の組換えベクター、または、〔6〕に記載のポリペプチドを植物に導入する工程を含むことを特徴とする形質転換植物の製造方法。
【0024】
〔9〕:〔1〕〜〔3〕の何れかに記載のポリヌクレオチド、〔4〕若しくは〔5〕に記載の組換えベクター、または、〔6〕に記載のポリペプチドが導入されていることを特徴とする形質転換植物。
【発明の効果】
【0025】
トウダイグサ目植物のナンヨウアブラギリ等の、産業上、有用であると考えられる植物の開花時期を制御(早期化または遅延化)することができるという効果を奏する。
【図面の簡単な説明】
【0026】
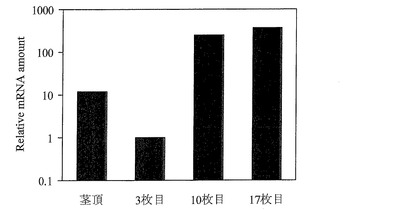
【図1】実施例において、JcFT遺伝子のmRNA量をリアルタイムPCRによって定量した結果を示すグラフである。
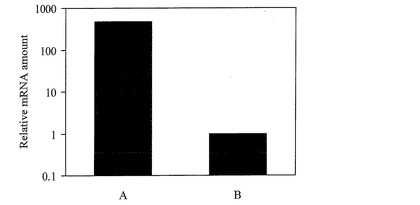
【図2】実施例において、JcFT遺伝子のmRNA量をリアルタイムPCRによって定量した結果を示すグラフである。
【図3】実施例において、pRI−35S−To71sGFP−CRが導入されている組換えシロイヌナズナの写真である。
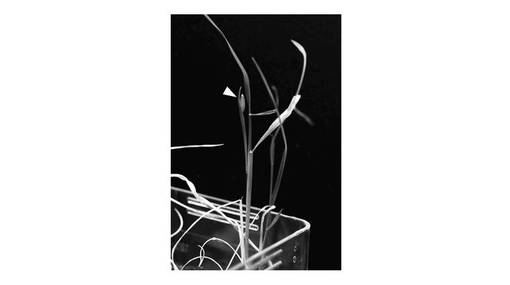
【図4】実施例において、pRH−2x35S−faiJcFT−CRが導入されている組換えイネの写真である(三角の矢印は花芽の位置を示す)。
【発明を実施するための形態】
【0027】
本発明の実施の形態について詳細に説明すれば以下のとおりであるが、本発明はこれに限定されるものではない。なお、本明細書中に記載された非特許文献および特許文献の全てが、本明細書中において参考として援用される。
【0028】
本明細書中で使用される場合、用語「ポリペプチド」は、「ペプチド」または「タンパク質」と交換可能に使用される。また、本明細書中で使用される場合、用語「ポリヌクレオチド」は、「遺伝子」、「核酸」または「核酸分子」と交換可能に使用され、ヌクレオチドの重合体が意図される。本明細書中で使用される場合、用語「塩基配列」は、「核酸配列」または「ヌクレオチド配列」と交換可能に使用され、デオキシリボヌクレオチド(A、G、CおよびTと省略される)の配列として示される。
【0029】
〔1.ポリヌクレオチド〕
本実施の形態のポリヌクレオチドは、下記(1)〜(3)の何れかのポリペプチドをコードするポリヌクレオチドであり得る。つまり、
(1)配列番号14で示されるポリペプチド、
(2)配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド、
(3)配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド。
【0030】
なお、配列番号14で示されるポリペプチドとは、発明者らによって同定された、ナンヨウアブラギリ(Jatropha curucas)由来のFT遺伝子(JcFT遺伝子)にコードされているタンパク質(JcFTタンパク質)である。
【0031】
また、本実施の形態のポリヌクレオチドは、下記(4)〜(10)の何れかであり得る。つまり、
(4)配列番号11、12または13で示されるポリヌクレオチド、
(5)配列番号11、12または13で示されるポリヌクレオチドのアンチセンス配列をコードするポリヌクレオチド、
(6)配列番号17および配列番号18に示すプライマーセットによって増幅されるポリヌクレオチドを含むポリヌクレオチド、
(7)配列番号19および配列番号20に示すプライマーセットによって増幅されるポリヌクレオチドを含むポリヌクレオチド、
(8)上記(4)〜(7)の何れかに記載されるポリヌクレオチドにおいて、1つ以上30以下の塩基が置換、欠失、挿入および/または付加されたポリヌクレオチドからなり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド、
(9)上記(4)〜(7)の何れかに記載されるポリヌクレオチドと相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド、
(10)上記(4)〜(7)の何れかに記載されるポリヌクレオチドとの相同性が84%以上であり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド。
【0032】
なお、上記(6)および(7)に記載のポリヌクレオチドは、各プライマーセットによって増幅されるポリヌクレオチドを含むものであるが、各プライマーセットによって増幅されるポリヌクレオチド以外の部分の具体的な塩基配列は、特に限定されない。勿論、上記(6)および(7)に記載のポリヌクレオチドは、各プライマーセットによって増幅されるポリヌクレオチド自体であってもよい。
【0033】
配列番号11で示されるポリヌクレオチドとは、JcFT遺伝子のゲノムDNAであり、配列番号12で示されるポリヌクレオチドとは、JcFT遺伝子の完全長cDNAであり、配列番号13で示されるポリヌクレオチドとは、JcFT遺伝子の翻訳領域配列である。
【0034】
また、配列番号17および配列番号18に示すプライマーセットとは、JcFT遺伝子の翻訳領域に相当する塩基配列の一部(配列番号13の塩基番号1〜124で示される塩基配列)を増幅することが可能なプライマーセットであり、配列番号19および配列番号20に示すプライマーセットとは、JcFT遺伝子の完全長cDNAの一部(配列番号12の塩基番号1〜816で示される塩基配列)を増幅することが可能なプライマーセットである。なお、これらのプライマーの具体的な配列を以下に記載する。
【0035】
JcFTL−2F :5'−ATGCCTAGGGATCAATTTAGAGACC−3'(配列番号17)
JcFTL−2RC:5'−AGCCATTGTTAACCTCTCTGTGATT−3'(配列番号18)
JcFTL−3F :5'−ACGCGGGGATGATAATACGAGTGTAGC−3'(配列番号19)
JcFTL−3RC:5'−AGAGATTAATATTCAGTAAATTTGATAGCATTTGTGATC−3'(配列番号20)。
【0036】
上記4つのプライマーの塩基配列は、他の植物のFT遺伝子とは配列の相同性が低い。従って、これらのプライマーを用いれば、ナンヨウアブラギリ由来のFT遺伝子と他の植物由来のFT遺伝子とを、より明確に判別することができる。勿論、これらのプライマーセットによって増幅されるポリヌクレオチド自体を用いて、ナンヨウアブラギリ由来のFT遺伝子と他の植物由来のFT遺伝子とを判別することも可能である。
【0037】
また、配列番号19および配列番号20に示すプライマーセットによって増幅されるポリヌクレオチドにコードされるポリペプチドは、それ自体、植物の開花時期を早める作用をも備えている。
【0038】
本実施の形態のポリヌクレオチドを用いれば、植物内のFT遺伝子の発現量・発現時期などを調節することが可能となり、その結果、植物の開花時期を制御することができる。一例としては、FT遺伝子のセンス鎖に相当する本実施形態のポリヌクレオチドを用いれば、植物内におけるFT遺伝子の発現量の上昇、および/または、発現時期の早期化を実現することが可能となり、これによって、植物の開花時期を早めることができる。また、FT遺伝子のアンチセンス鎖に相当する本実施形態のポリヌクレオチドを用いれば、植物内におけるFT遺伝子の発現量の低下、および/または、発現時期の遅延化を実現することが可能となり、これによって、植物の開花時期を遅くすることが可能となる。なお、後述する〔4.植物の開花時期の制御方法〕を一読すれば、植物の開花時期を制御するための更に詳細な方法を理解できるであろう。
【0039】
本実施の形態のポリヌクレオチドは、DNAの形態(例えば、cDNAもしくはゲノムDNA)、またはRNA(例えば、mRNA)の形態であり得る。DNAまたはRNAは二本鎖であっても、一本鎖であってもよい。一本鎖DNAまたはRNAは、コード鎖(センス鎖)であっても、非コード鎖(アンチセンス鎖)であってもよい。
【0040】
また、本実施の形態のポリヌクレオチドは、化学的に合成されたものであってもよく、コードするタンパク質の発現が向上するように、コドンユーセージ(Codon usage)が改変されたものであってもよい。勿論、本実施の形態のポリヌクレオチドは、自然界から単離されたものであってもよい。
【0041】
自然界から単離されたポリヌクレオチドを本実施の形態のポリヌクレオチドとして用いる場合には、当該ポリヌクレオチドの由来は特に限定されないが、例えば、バラ下綱植物、トウダイグサ目植物、トウダイグサ科植物、または、ナンヨウアブラギリ植物に由来するポリヌクレオチドであることが好ましい。これらの中では、バラ下綱植物由来であることが好ましく、トウダイグサ目植物由来であることが更に好ましく、トウダイグサ科植物由来であることが更に好ましく、ナンヨウアブラギリ植物由来であることが最も好ましいといえる。
【0042】
本実施の形態のポリヌクレオチドは、配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなるポリペプチドをコードしたものであり得る。1または数個のアミノ酸が置換、欠失、挿入および/または付加される部位は、置換、欠失、挿入および/または付加後のポリペプチドが植物の開花時期を制御する機能を有していれば、該アミノ酸配列中のどの部位であってもよい。ここで「1または数個のアミノ酸残基」とは、具体的には10個以内の範囲のアミノ酸残基数であり、好ましくは6個以内の範囲のアミノ酸残基である。
【0043】
置換などの変異が生じる場合には、アミノ酸側鎖の性質が保存されている別のアミノ酸に置換されることが好ましい。例えばアミノ酸側鎖の性質としては、疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)、親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)、脂肪族側鎖などの特徴的な側鎖を有するアミノ酸(G、A、V、L、I、P)、水酸基含有側鎖を有するアミノ酸(S、T、Y)、硫黄原子含有側鎖を有するアミノ酸(C、M)、カルボン酸及びアミド含有側鎖を有するアミノ酸(D、N、E、Q)、塩基含有側鎖を有するアミノ酸(R、K、H)、芳香族含有側鎖などの特徴的な側鎖を有するアミノ酸(H、F、Y、W)を挙げることができる(括弧内はいずれもアミノ酸の一文字標記を表す)。アミノ酸側鎖の性質が同じアミノ酸間の置換であれば、ポリペプチドが備える生物学的活性を維持することができる。
【0044】
また、本実施の形態のポリヌクレオチドは、上記(4)〜(7)の何れかに記載されるポリヌクレオチドにおいて、1つ以上30以下の塩基が置換、欠失、挿入および/または付加されたポリヌクレオチドからなるポリヌクレオチドであり得る。塩基が置換、欠失、挿入および/または付加される部位は、置換、欠失、挿入および/または付加後のポリヌクレオチドが植物の開花時期を制御する機能を有していれば、該塩基配列中のどの部位であってもよい。
【0045】
以上のようなポリヌクレオチドは、特定のポリヌクレオチド内の塩基を置換、欠失、挿入および/または付加することによって得ることが可能である。ポリヌクレオチドの塩基を変換する具体的な方法としては、例えば市販のキット(Transformer Site−Directed Mutagenesis Kit;Clonetech製,QuickChange Site Directed Mutagenesis Kit;Stratagene製など)の使用、またはポリメラーゼ連鎖反応法(PCR)の利用が挙げられる。これらの方法は当業者に公知である。
【0046】
本実施の形態のポリヌクレオチドは、上記(4)〜(7)の何れかに記載されるポリヌクレオチドに対して相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズするものであり得る。
【0047】
ここで、「ストリンジェントな条件」とは、相同性が高い核酸同士、例えば完全にマッチしたハイブリッドの融解温度(Tm値)から15℃、好ましくは10℃低い温度までの範囲の温度でハイブリダイズする条件をいう。具体例としては、一般的なハイブリダイゼーション用緩衝液中で、68℃、20時間の条件でハイブリダイズする条件をいう。
【0048】
更に具体的には、「ストリンジェントな条件」とは、いわゆる塩基配列に特異的な2本鎖のポリヌクレオチドが形成され、非特異的な2本鎖のポリヌクレオチドが形成されない条件をいう。換言すれば、相同性が高い核酸同士が、例えば完全にマッチしたハイブリッドの融解温度(Tm値)から15℃、好ましくは10℃、更に好ましくは5℃低い温度までの範囲の温度でハイブリダイズする条件ともいえる。例えば、一般的なハイブリダイゼーション用緩衝液中で、68℃、20時間の条件でハイブリダイズする条件を挙げることができる。より具体的には、0.9M NaCl、0.09M sodium citrate、0.01M sodium phosphate pH6.8、1mM EDTA pH8.0、0.5% SDS、1×デンハルト溶液からなる緩衝液中で、60〜68℃、好ましくは65℃、さらに好ましくは68℃の条件下で16〜24時間ハイブリダイズさせた後、さらに0.3M NaCl、0.03M sodium citrate、1% SDSからなる緩衝液中で、60〜68℃、好ましくは65℃、さらに好ましくは68℃の条件下で15分間の洗浄を2回行う条件を挙げることができる。当業者であれば、Molecular Cloning(Sambrook, J. et al., Molecular Cloning :a Laboratory Manual 2nd ed., Cold Spring Harbor Laboratory Press, 10 Skyline Drive Plainview, NY (1989))等を参照することにより、こうした遺伝子を容易に取得することができる。
【0049】
本実施の形態のポリヌクレオチドは、配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチドをコードするポリヌクレオチドであり得る。なお、配列番号14で示されるポリペプチドとの相同性は、95%以上であることが更に好ましいといえる。また、本実施の形態のポリヌクレオチドは、上記(4)〜(7)の何れかに記載されるポリヌクレオチドとの相同性が84%以上であり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチドであり得る。なお、上記(4)〜(7)の何れかに記載されるポリヌクレオチドとの相同性は、90%以上であることが更に好ましいといえる。上記構成によれば各種植物の開花時期を好適に制御することができるが、その中でも、バラ下綱植物の開花時期を更に好適に、トウダイグサ目植物の開花時期を更に好適に、トウダイグサ科植物の開花時期を更に好適に、ナンヨウアブラギリ植物の開花時期を最も好適に制御することができる。
【0050】
本発明において「相同性」とは、2つの塩基配列または2つのアミノ酸配列間の相同性が意図される。相同性は、比較対象の配列の全領域にわたって最適な状態にアラインメントされた2つの配列を比較することにより決定される。ここで、比較対象の塩基配列またはアミノ酸配列を最適な状態にアラインメントするために、付加または欠失(例えば、ギャップ等)を許容してもよい。このような配列相同性は、例えば、FASTA(Pearson & Lipman、1988、PNAS、4、2444-2448)、BLAST(Altschul et al、1990、Journal of Molecular Biol.、215、403-410)、CLUSTAL W(Thompson et al、1994、Nucleic Acid Research、22、4673-4680)等のプログラムを用いて算出することができる。
【0051】
上記のプログラムは、例えば、DNA Data Bank of Japan(国立遺伝学研究所生命情報・DDBJ研究センター Center for Information Biology and DNA Data Bank of Japan; CIB/DDBJ)内で運営される国際DNAデータバンクのWEBページ(http://www.ddbj.nig.ac.jp/Welcome-e.html)等において、一般的に利用可能である。また、配列の相同性は市販の配列解析ソフトウェアを用いて求めることもできる。具体的には、配列の相同性は、DNASYS Pro ver.2.06(Hitachi Software Engineering)のSmith−Watermanプログラムにより相同性解析を行ってアラインメントすることにより算出することができる。
【0052】
本実施の形態のポリヌクレオチドは、後述する本発明のポリペプチドをコードするポリヌクレオチドのみからなるものであってもよいが、その他の塩基配列が付加されていてもよい。付加される塩基配列としては限定されないが、標識(例えば、ヒスチジンタグ、Mycタグ、およびFLAGタグなど)、本発明のポリペプチドと融合タンパク質を構成し得るポリペプチド(例えば、GSTおよびMBPなど)、プロモーター配列(例えば、植物由来プロモーター配列、酵母由来プロモーター配列、ファージ由来プロモーター配列、および大腸菌由来プロモーター配列など)、およびシグナル配列(例えば、小胞体移行シグナル配列、および分泌配列など)をコードする塩基配列などが挙げられる。これらの塩基配列が付加される部位は特に限定されるものではなく、例えば、ポリヌクレオチドの5’末端であっても、3’末端でもあってもよい。
【0053】
〔2.組換えベクター〕
本実施の形態の組換えベクターは、上述した本発明のポリヌクレオチドを含有するものである。
【0054】
本実施の形態の組み換えベクターを用いれば、植物内のFT遺伝子の発現量を調節することが可能となり、その結果、植物の開花時期を制御することができる。
【0055】
本実施の形態の組換えベクターとしては特に限定されないが、例えば、プラスミドベクター、ウイルスベクター、ファージベクターまたはコスミドベクターであることが好ましく、これらの中では、プラスミドベクターまたはウイルスベクターであることが更に好ましい。上記構成によれば、所望の宿主を形質転換することができる。
【0056】
本実施の形態の組換えベクターの更に具体的な構成としては特に限定されないが、発現制御配列を含むことが好ましい。なお、発現制御配列とは、所望の遺伝子の発現を制御するための塩基配列が意図される。当該発現制御配列としては特に限定されないが、例えば、プロモーター配列、エンハンサー配列、ターミネーター配列、5’非翻訳領域配列、3’非翻訳領域配列、開始コドン配列、イントロンのスプライシングシグナル配列、翻訳フレーム維持配列および停止コドン配列などを挙げることが可能である。そして、本実施の形態の組換えベクターは、これら具体的な発現制御配列を1つだけ含むことも可能であるし、複数種類を含むことも可能である。なお、複数種類の発現制御配列を含む場合には、その組み合わせは特に限定されない。
【0057】
上記「プロモーター配列」とは、遺伝子の転写を開始するために十分な最小の塩基配列が意図される。プロモーター配列としては、構成的プロモーター、組織特異的プロモーター、または、特定の刺激によって転写が誘導される誘導性プロモーターなどを用いることが可能であるが、これらに限定されない。組換えベクターの使用目的に応じて、適宜選択することが好ましい。
【0058】
上記構成的プロモーターとしては特に限定されないが、例えば、CaMV 35Sプロモーター(Benfey PN & Chua NH、1990、Science 250、959-966)、PG10−90(特開09-131187)、ユビキチンプロモーター(国際公開01/094394)、アクチンプロモーター(国際公開00/070067)等を用いることが好ましい。
【0059】
上記組織特異的プロモーターとしては特に限定されないが、例えば、ダイズ種子グリシニンプロモーター(特開06-189777)、プロラミンプロモーター(国際公開2004/056993)、インゲンマメ種子ファゼオリンプロモーター(国際公開91/013993)、ナタネ種子ナピンプロモーター(国際公開91/013972)、シロイヌナズナSultr2;2プロモーター(Takahashi H et al、2000、Plant J 23、171-82)、アグロバクテリウムrolCプロモーター(Almon E et al、1997、Physiol 115、1599-1607)等を用いることが好ましい。
【0060】
上記誘導性プロモーターとしては特に限定されないが、例えば、銅イオン誘導性プロモーター(国際公開08/111661)、ステロイドホルモン誘導性プロモーター(米国特許第6063985号)、エタノール誘導性システム(国際公開93/21334)、テトラサイクリン誘導性システム(例えば、Weinmann P et al、1994、Plant J. 5、559-569)、除草剤セーフナー誘導性プロモーター(Hershey et al、1991、Plant Mol. Biol.、17、679)、熱誘導性プロモーター(米国特許第5447858号)、低温誘導性プロモーター(米国特許第5847102号)、植物病原体の攻撃により誘導されるプロモーター(米国特許第5942662号)等を用いることが好ましい。
【0061】
上記「ターミネーター配列」とは、遺伝子の転写を終結させ、mRNAを安定化させるためのポリアデニン配列を付加するために十分な最小の配列が意図される。上記ターミネーター配列としては特に限定されないが、例えば、NOSターミネーター、CR16ターミネーター(特開2000-166577)、ダイズ種子グリシニンターミネーター(特開06-189777)等を用いることが好ましい。
【0062】
〔3.ポリペプチド〕
本実施の形態のポリペプチドは、下記(11)〜(13)の何れかのポリペプチドであり得る。つまり、
(11)配列番号14で示されるポリペプチド、
(12)配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド、
(13)配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド。
【0063】
なお、配列番号14で示されるポリペプチドとは、発明者らによって同定された、ナンヨウアブラギリ(Jatropha curucas)由来のFT遺伝子(JcFT遺伝子)にコードされているタンパク質(JcFTタンパク質)である。
【0064】
本実施の形態のポリペプチドを用いれば、植物内のFT遺伝子の発現量を調節することが可能となり、その結果、植物の開花時期を制御することができる。
【0065】
本実施の形態のポリペプチドは、配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチドであり得るが、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加される部位は、置換、欠失、挿入および/または付加後のタンパク質が植物の開花時期を制御する機能を有していれば、該アミノ酸配列中のどの部位であってもよい。ここで「1または数個のアミノ酸残基」とは、具体的には10個以内の範囲のアミノ酸残基数であり、好ましくは6個以内の範囲のアミノ酸残基である。
【0066】
置換などの変異が生じる場合には、アミノ酸側鎖の性質が保存されている別のアミノ酸に置換されることが好ましい。例えばアミノ酸側鎖の性質としては、疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)、親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)、脂肪族側鎖などの特徴的な側鎖を有する疎水性アミノ酸(G、A、V、L、I、P)、水酸基含有側鎖を有するアミノ酸(S、T、Y)、硫黄原子含有側鎖を有するアミノ酸(C、M)、カルボン酸及びアミド含有側鎖を有するアミノ酸(D、N、E、Q)、塩基含有側鎖を有するアミノ酸(R、K、H)、芳香族含有側鎖などの特徴的な側鎖を有するアミノ酸(H、F、Y、W)を挙げることができる(括弧内はいずれもアミノ酸の一文字標記を表す)。アミノ酸側鎖の性質が同じアミノ酸間の置換であれば、ポリペプチドが備える生物学的活性を維持することができる。
【0067】
また、本実施の形態のポリペプチドは、配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチドであり得るが、より好ましくは、配列番号14で示されるポリペプチドとの相同性が95%以上である。
【0068】
本実施の形態のポリペプチドは、例えば、自然界から単離した当該ポリペプチドを産生する生物を用いて生産されてもよいし、遺伝子組み換え技術を用いて生産されてもよいし、またはアミノ酸合成機などを用いて化学合成されてもよい。
【0069】
遺伝子組み換え技術において、好適に用いられる各種組換えタンパク質発現系は、例えば、大腸菌発現系、酵母発現系、昆虫細胞発現系、哺乳類細胞発現系、および無細胞発現系を用いてもよく、これらに限定されない。
【0070】
本実施の形態のポリペプチドは、例えば、分子間架橋および/または分子内架橋(例えば、ジスルフィド結合など)が施されたもの、化学修飾(例えば、糖鎖、リン酸もしくはその他の官能基など)が施されたもの、標識(例えば、ヒスチジンタグなど)が付与されたもの、または本発明のポリペプチドと融合タンパク質を構成し得るポリペプチド(例えば、ストレプトアビジン、シトクロム、およびGFPなど)が連結されたものなどが含まれるが、特にこれらに限定されない。さらに、本実施の形態のポリペプチドには、植物の開花時期を制御する機能が実質的に維持される限り、数種のタンパク質の断片を組み合わせて構成されたキメラタンパク質も含まれる。
【0071】
〔4.植物の開花時期の制御方法〕
本実施の形態の植物の開花時期の制御方法は、本発明のポリヌクレオチド、本発明の組換えベクター、または、本発明のポリペプチドを植物に導入する工程を含む方法である。
【0072】
本発明のポリヌクレオチド、組換えベクターおよびポリペプチドに関しては既に説明したので、ここでは、その説明を省略する。
【0073】
上記植物(換言すれば、開花時期が制御される植物)としては特に限定されず、単子葉植物であっても双子葉植物であっても良い。具体的には、イネ科植物、バラ下綱植物、トウダイグサ目植物、トウダイグサ科植物、または、ナンヨウアブラギリ植物であることが好ましい。これらの中では、バラ下綱植物由来であることが好ましく、トウダイグサ目植物由来であることが更に好ましく、トウダイグサ科植物由来であることが更に好ましく、ナンヨウアブラギリ植物由来であることが最も好ましいといえる。
【0074】
また、上記植物としては、例えば、ダイズ、エンドウ、インゲン、アルファルファ、ミヤコグサ、クローバ、ピーナッツ、スイートピー、クルミ、チャ、ワタ、コショウ、キュウリ、スイカ、カボチャ、メロン、ダイコン、ナタネ、キャノーラ、テンサイ、レタス、キャベツ、ブロッコリー、カリフラワー、シロイヌナズナ、タバコ、ナス、ジャガイモ、サツマイモ、サトイモ、キクイモ、トマト、ホウレンソウ、アスパラガス、ニンジン、アマ、ゴマ、エンダイブ、キク、フウロウソウ、キンギョソウ、カーネーション、ナデシコ、ニチニチソウ、ブバルディア、カスミソウ、ガーベラ、トルコキキョウ、チューリップ、ストック、スターチス、シクラメン、ユキノシタ、ノースポール、スミレ、バラ、サクラ、リンゴ、ナシ、ブドウ、イチゴ、ウメ、アーモンド、ミカン、レモン、バナナ、マンゴー、パパイヤ、キウイ、コーヒー、ボケ、サツキ、ツツジ、ポインセチア、キャッサバ、アブラヤシ、ココヤシ、オリーブ、リンドウ、コスモス、アサガオ、ヒマワリ、イチョウ、スギ、ヒノキ、ポプラ、マツ、セコイア、オーク、ヤナギ、ユーカリ、ケナフ、スイレン、トチュウ、ブナ、ヒマ、タケ、サトウキビ、イネ、コムギ、オオムギ、ライムギ、エンバク、トウモロコシ、ソルガム、シバ、トールフェスキュー、スイッチグラス、ススキ、ネギ、タマネギ、ニンニク、ユリ、オニユリ、ラン、グラジオラス、またはパイナップルを挙げることも可能であるが、これらに限定されない。
【0075】
本発明のポリヌクレオチド、本発明の組換えベクター、または、本発明のポリペプチドを上記植物に導入する方法は特に限定されない。例えば、アグロバクテリウム法、パーティクルガン法、エレクトロポレーション法、リン酸カルシウム法、インジェクション法、またはウイルスベクター法などの公知方法を用いることが可能である。
【0076】
本実施の形態の植物の開花時期の制御方法によれば、植物の開花時期を早期化したり(換言すれば、開花の促進)、あるいは、植物の開花時期を遅延化する(換言すれば、開花の抑制)ことが可能となる。以下に、開花時期を早期化する場合の更に具体的な方法、および、開花時期を遅延化する場合の更に具体的な方法の一例に関して説明するが、本発明はこれらに限定されない。
【0077】
<4−1>開花時期の早期化
植物の開花時期の早期化は、例えば、以下の(A)〜(C)の方法によって実現することが可能である。
【0078】
(A)FT遺伝子が植物細胞中で発現可能となるように、任意の発現制御配列とFT遺伝子とを、任意のベクターに連結する。当該ベクターを植物細胞に導入して形質転換植物を得る。当該形質転換植物において開花が促進される。なお、FT遺伝子が植物細胞中で発現可能であれば、任意の発現制御配列や任意のベクターは必ずしも必要ではなく、形質転換植物を取得する必要もない。FT遺伝子が植物細胞中で発現可能となるよう、宿主植物に応じて,FT遺伝子を植物細胞へ導入すればよい。
【0079】
(B)植物のゲノムDNAに任意の塩基配列を組み込むことにより、植物に内在するFT遺伝子の発現を促進する。任意の塩基配列としては、好ましくは、任意の発現制御配列を挙げることができる。更に具体的には、任意の発現制御配列としては、プロモーター配列およびエンハンサー配列などを挙げることが可能である。当該発現制御配列が機能し得るように植物のゲノムDNA内に組み込まれ、植物に内在するFT遺伝子の発現を促進し、これによって植物の開花が促進される。
【0080】
(C)FT遺伝子がコードするタンパク質を、例えば、師管液中に注入し、植物細胞中に導入する。FT遺伝子がコードするタンパク質は、茎頂や腋芽に移動し、開花を促進する。FT遺伝子がコードするタンパク質は、組換え微生物を用いて生産してもよいし、野生型の植物あるいは形質転換植物から抽出してもよい。或は、FT遺伝子がコードするタンパク質が蓄積している植物(例えば、形質転換植物)を台木として接木してもよい。
【0081】
<4−2>開花時期の遅延化
植物の開花時期の遅延化は、例えば、以下の(D)〜(I)の方法によって実現することが可能である。
【0082】
(D)FT遺伝子のアンチセンス配列が植物細胞中で発現可能となるよう、任意の発現制御配列とFT遺伝子のアンチセンス配列とを、任意のベクターに連結する。当該ベクターを植物細胞に導入して形質転換植物を得る。当該形質転換植物において開花時期が遅延する。なお、FT遺伝子のアンチセンス配列が植物細胞中で発現可能であれば、任意の発現制御配列や任意のベクターは必ずしも必要はない。FT遺伝子のアンチセンス配列が植物細胞中で発現可能となるよう、宿主植物に応じて、FT遺伝子のアンチセンス配列を植物細胞へ導入すればよい。
【0083】
本明細書において「アンチセンス配列」とは、特定のmRNA分子の少なくとも一部に相補的なDNA分子またはRNA分子である。植物細胞中で、アンチセンス配列は、対応するmRNAにハイブリダイズして二本鎖分子を形成し、これによって、mRNAの翻訳を阻害する。
【0084】
(E)VIGS(Virus Induced Gene Silencing)を誘発するウイルスベクターにFT遺伝子の全長あるいはFT遺伝子の一部を組み込み、当該ベクターを植物細胞に導入することによって内在性のFT遺伝子の発現を抑制し、これによって、開花時期を遅延させる。
【0085】
(F)FT遺伝子の1アミノ酸または複数のアミノ酸を別のアミノ酸に置換することでFT遺伝子を花成抑制遺伝子に転換することが可能である(Hanzawa et al、2005、PNAS、102、7748-7753)。当該置換のように優性ネガティブ変異が導入された花成抑制遺伝子が植物細胞中で発現可能となるよう、任意の発現制御配列と花成抑制遺伝子とを、任意のベクターに連結する。当該ベクターを植物細胞に導入して形質転換植物を得る。当該形質転換植物では、開花時期が遅延する。なお、花成抑制遺伝子が植物細胞中で発現可能であれば、任意の発現制御配列や任意のベクターは、必ずしも必要ではない。花成抑制遺伝子が植物細胞中で発現可能となるよう、宿主植物に応じて、花成抑制遺伝子植物細胞へ導入すればよい。
【0086】
本明細書において「優性ネガティブ変異」とは、本来の機能を損なう形で変異が導入されており、かつ野生型のタンパク質の活性に対して優性的に働き、野生型の表現型を打ち消して負の表現型を示すことができる変異である。
【0087】
(G)植物のゲノムDNA中に任意の塩基配列を組み込むことにより、植物に内在するFT遺伝子の発現を抑制する。任意の塩基配列は、植物内在のFT遺伝子の発現を抑制し得るように、植物のゲノムDNA中に組み込まれる必要がある。具体的には、例えば、植物に内在するFT遺伝子内への任意の塩基配列の挿入、または、植物に内在するFT遺伝子の発現に関与する発現制御配列内への任意の塩基配列の挿入が挙げられるが、これらに限定されない。
【0088】
(H)リボザイムや三重鎖形成オリゴヌクレオチドを植物細胞中に導入する。リボザイムによってFT遺伝子のmRNAを切断したり、三重鎖形成オリゴヌクレオチドによってFT遺伝子の転写や翻訳を阻害したりすることによって、開花時期を遅延する。
【0089】
なお、本明細書において「リボザイム」とは、一本鎖RNAを特異的に切断する能力を有するRNA分子が意図される。リボザイムをコードする塩基配列を遺伝子工学的に改変することにより、RNA分子内の特定の塩基配列を認識して当該塩基配列を切断する分子を作製することができる(Ceeh et al、1988、J. Amer. Med. Assn.、260、3030)。
【0090】
また、本明細書において「三重鎖形成オリゴヌクレオチド」とは、二重らせんDNAの周りに巻き付いて三重らせんを形成し、遺伝子の転写を停止させる分子である。特定の塩基配列を認識するように、三重鎖形成オリゴヌクレオチドを設計することが可能である(Maher et al、1991、Antisense Res. and Dev.、1、227)。
【0091】
(I)優性ネガティブ変異が導入された花成抑制遺伝子がコードするタンパク質を、例えば、師管液中に注入し、植物細胞中に導入する。花成抑制遺伝子がコードするタンパク質は、茎頂や腋芽に移動し、開花時期を遅延する。花成抑制遺伝子がコードするタンパク質は、組換え微生物を用いて生産してもよいし、形質転換植物から抽出してもよい。或は花成抑制遺伝子がコードするタンパク質が蓄積している植物(例えば、形質転換植物)を台木として接木してもよい。
【0092】
上述した開花時期を早期化する方法と開花時期を遅延化する方法とは、各々単独で実施されることも可能であり、可能な範囲で自由に組み合わせられることも可能である。具体的には、例えば、植物に内在するFT遺伝子へ任意の塩基配列を挿入してFT遺伝子を機能的に欠損させた植物に、外来性のFT遺伝子(または、花成抑制遺伝子)が植物細胞中で発現可能となるよう、新たにFT遺伝子(または、花成抑制遺伝子)を導入してもよい。また、FT遺伝子(または、花成抑制遺伝子)をすでに導入した植物に、追加的にFT遺伝子(または、花成抑制遺伝子)を導入しても良い。
【0093】
また、具体的にナンヨウアブラギリの開花時期の制御には、各種誘導性プロモーター、具体的には例えば、銅イオン誘導性プロモーター(例えば、国際公開08/111661)に本発明のポリヌクレオチドを連結し、トウダイグサ目植物、具体的には例えば、ナンヨウアブラギリに導入し、当該形質転換植物に銅イオンを接触させることで開花時期を制御することが好ましい。上記構成によれば、例えば、JcFTタンパク質とのアミノ酸配列の相同性が90%未満のシロイヌナズナFT遺伝子由来のDNA断片を上記プロモーターに連結して導入した場合と比較して、開花時期の制御を効果的に行うことができるとともに、形態異常などの望ましくない表現型の出現を抑制することができる。
〔5.形質転換植物の製造方法〕
本実施の形態の形質転換植物の製造方法は、本発明のポリヌクレオチド、本発明の組換えベクター、または、本発明のポリペプチドを植物に導入する工程を含む方法である。つまり、本実施の形態の形質転換植物の製造方法は、既に説明した本発明の植物の開花時期の制御方法と実質的に同じ方法に基づいて実施することが可能である。
【0094】
本発明のポリヌクレオチド、組換えベクターおよびポリペプチドに関しては既に説明したので、ここでは、その説明を省略する。
【0095】
また、上記植物の具体的な構成、当該植物へ本発明のポリヌクレオチドなどを導入する具体的な構成については〔4.植物の開花時期の制御方法〕にて既に説明したので、ここではその説明を省略する。
【0096】
〔6.形質転換植物〕
本実施の形態の形質転換植物は、本発明のポリヌクレオチド、本発明の組換えベクター、または、本発明のポリペプチドが導入されているものである。
【0097】
本実施の形態の形質転換植物は、〔5.形質転換植物の製造方法〕にて説明した方法に基づいて製造することができる。
【0098】
本実施の形態の形質転換植物では、開花時期が制御されている。当該制御としては、開花時期の早期化、または開花時期の遅延化などを挙げることができるが、これらに限定されない。
【0099】
例えば、開花時期を早期化すれば、当該形質転換植物のライフサイクルを短くすることができる。つまり、短い時間で当該植物(例えば、種子、果実など)を収穫することが可能になる。その結果、例えば、品種改良を効率よく行うことが可能になる。
【0100】
逆に、開花時期を遅延化すれば、当該形質転換植物のライフサイクルを長くすることができる。つまり、時間をかけて十分に生育した当該植物(例えば、種子、果実など)を収穫することが可能になる。その結果、例えば、葉菜類や根菜類の商品価値を高めることが可能になる。
【0101】
本発明によれば、開花時期を早期化したり遅延化したりすることができるので、栽培環境、播種時期および栽培地などに幅をもたせることが可能となる。そして、その結果、生産性を向上させることが可能になる。
【実施例】
【0102】
以下、実施例に基づいて本発明を更に詳細に説明するが、本発明はこれら実施例によって限定されるものではない。
【0103】
〔実施例1.JcFT遺伝子断片の単離〕
DNeasy Plant mini kit(キアゲン)を用いて、ナンヨウアブラギリ(Jatropha curucas)の本葉からゲノムDNAを調製した。当該ゲノムDNAを鋳型として、4種類の縮重プライマー(FTD−1F、FTD−1RC、FTD−2F、FTD−2RC)を用いてPCR反応を行った。
【0104】
FTD−1F :5'−GACCCCTTYACAAGRTCYATYTCYCTGAGGGT−3' (配列番号1)
FTD−1RC:5'−CARAGTGTAGAAGGTCCKRAGRTC−3' (配列番号2)
FTD−2F :5'−CAAGAGATTGTGTGYTAYGARAGYCCAMGGCCAAC−3'(配列番号3)
FTD−2RC:5'−CGGAGCCRCYYTCCCTYTGRCA−3' (配列番号4)
混合塩基はIUBコードに従い、M(A or C)、R(A or G)、W(A or T/U)、S(C or G)、Y(C or T/U)、K(G or T/U)、V(A or C or G)、H(A or C or T/U)、D(A or G or T/U)、B(C or G or T/U)、N(A or C or G or T/U)で示した。
【0105】
PCR産物をアガロースゲルにて電気泳動した後、得られた増幅断片をアガロースゲルから切り出し、MagExtractor(東洋紡)を用いて上記増幅断片を精製した。精製された増幅断片を、ZeroBlunt TOPO PCR Cloning kit(インビトロジェン)を用いてpCR4Blunt−TOPO vectorにクローニングした。
【0106】
当該ベクターを用いて大腸菌DH5αを形質転換して、薬剤選抜されたコロニーをピックアップした。ピックアップされた大腸菌をLB培地(0.5%酵母エキス、1.0%バクトトリプトン、0.5%食塩)で培養し、QIAprep spin miniprep kit(キアゲン)を用いてプラスミドを調製した。
【0107】
M13 reverseプライマー、BigDye terminator v3.1(ABI)、3100 Genetic Analyzer(ABI)を用いて、上記プラスミド内に挿入された増幅断片の塩基配列を解析した。
【0108】
その結果、FTD−1FおよびFTD−1RCを用いてPCR反応を行うことによって、JcFT遺伝子の第1エクソン内の塩基配列(144bp)の情報が得られ、FTD−2FおよびFTD−2RCを用いてPCR反応を行うことによって、JcFT遺伝子の第4エクソン内の塩基配列(205bp)の情報が得られた。
【0109】
得られた配列情報に基づいて2種類の特異的プライマー(JcFTL−1FおよびJcFTL−1RC)を設計した。
【0110】
JcFTL−1F :5'−TATAATCACAGAGAGGTTAACAATGGCTGTGAGCTCAAAC−3'(配列番号5)
JcFTL−1RC:5'−CTGACGCCACCCTGGTGGATACACGGTCTG−3'(配列番号6)
設計した2種類の特異的プライマー(JcFTL−1FおよびJcFTL−1RC)を用いて、ゲノムDNAを鋳型にしてPCR反応を行った。PCR産物を前述と同様にクローニングして、増幅断片の塩基配列を解析した。その結果、JcFT遺伝子の第1イントロン(149bp)、第2エクソン(62bp)、第2イントロン(2575bp)、第3エクソン(41bp)、第3イントロン(108bp)の塩基配列に関する情報が得られた。
【0111】
〔実施例2.ナンヨウアブラギリにおけるJcFT遺伝子の発現解析〕
培土に播種後2ヶ月〜4ヶ月のナンヨウアブラギリの茎頂および本葉から、RNeasy Plant mini kit(キアゲン)を用いて全RNAを調製した。
【0112】
ReverTra Ace qPCR RT Kit(TOYOBO)を用いて、調製した全RNAに基づいてcDNAを合成した。7500 Fast Real−time PCR装置(Applied Biosystems)を用いて、合成されたcDNAを鋳型として、SYBR green法に基づくリアルタイムPCRを行うことにより、mRNAの定量を行った。
【0113】
JcFT遺伝子のmRNAの定量には、2種の特異的プライマー(JcFT−1F、JcFT−1RC)を用いた。内部標準としては、ナンヨウアブラギリの5.8S rRNA(GenBank Accession Number AM774639)を用いた。5.8S rRNA遺伝子のmRNAの定量には、2種の特異的プライマー(Jc5.8−1F、Jc5.8−1RC)を用いた。
【0114】
JcFT−1F :5'−GACCCTAATCTCAGAGAATACTTGCA−3'(配列番号7)
JcFT−1RC :5'−CCAAAAGTTACCCCAGTAGTTGCT−3' (配列番号8)
Jc5.8−1F :5'−CTTGGTGTGAATTGCAGAATCC−3' (配列番号9)
Jc5.8−1RC:5'−GGCTTCGGGCGCAACT−3' (配列番号10)
花芽分化の起こりやすい気象条件A(明期平均温度32℃、明期平均湿度50%、暗期平均温度26℃、暗期平均湿度80%、日長12時間明期/12時間暗期)で栽培したナンヨウアブラギリ(反復4個体)の上位10枚目の本葉および17枚目の本葉におけるJcFT遺伝子の発現は、茎頂と比較して平均20倍程度、上位3枚目の本葉と比較して平均250倍程度上昇していた(図1参照)。
【0115】
また、花芽分化の起こりやすい気象条件A(明期平均温度32℃、明期平均湿度50%、暗期平均温度26℃、暗期平均湿度80%、日長12時間明期/12時間暗期)で栽培したナンヨウアブラギリ(反復6個体)の上位10枚目の本葉におけるJcFT遺伝子の発現は、花芽分化の起こりにくい気象条件B(明期平均温度40℃、明期平均湿度10%、暗期平均温度20℃、暗期平均湿度40%、日長12時間明期/12時間暗期)で栽培したナンヨウアブラギリ(反復6個体)の上位10枚目の本葉におけるJcFT遺伝子の発現と比較して、平均500倍程度上昇していた(図2参照)。これらの発現解析により、JcFT遺伝子が高発現する葉位および気象条件を特定した。
【0116】
なお、図1は、JcFT遺伝子のmRNA量をリアルタイムPCRにより定量した結果を示す図である。当該図面では、上位3枚目における反復4個体の平均値を1として、茎頂および上位10枚目、上位17枚目における反復4個体のそれぞれの平均値を相対値で示した。また、図2は、JcFT遺伝子のmRNA量をリアルタイムPCRにより定量した結果を示す図である。当該図面では、花芽分化の起こりにくい気象条件Bで栽培した反復6個体の平均値を1として、花芽分化の起こりやすい気象条件Aで栽培した反復6個体の平均値を相対値で示した。
【0117】
〔実施例3.JcFT遺伝子の単離〕
花芽分化の起こりやすい気象条件で栽培したナンヨウアブラギリ(Jatropha curucas)の上位10枚目の本葉から、RNeasy Plant mini kit(キアゲン)を用いて全RNAを調製した。
【0118】
SMART RACE cDNA Amplification kit(クロンテック)およびPrimeScript Reverse Transcriptase(タカラバイオ)を用いて、調製した全RNAに基づいて5'−RACEおよび3'−RACE用のFirst−Strand cDNAを合成した。
【0119】
1種の特異的プライマー(JcFTL−1RC)、およびSMART RACE cDNA Amplification kit(クロンテック)付属のプライマーミックス(UPM)を用いて、5'−RACE用に合成したcDNAを鋳型として5'−RACE反応を行った。また、1種の特異的プライマー(JcFTL−1F)、およびSMART RACE cDNA Amplification kit(クロンテック)付属のプライマーミックス(UPM)を用いて、3'−RACE用に合成したcDNAを鋳型として3'−RACE反応を行った。
【0120】
各RACE反応産物を、前述と同様にそれぞれクローニングし塩基配列を解析した。その結果、JcFT遺伝子の5'非翻訳領域(71bp)、第1エクソン(204bp)、第4エクソン(224bp)、3'非翻訳領域(246bp)の5'末端および3'末端の配列情報が得られた。
【0121】
このようにして、JcFT遺伝子全長に関する情報(例えば、ゲノムDNAの塩基配列(配列番号11参照)、完全長cDNAの塩基配列(配列番号12参照)、翻訳領域の塩基配列(配列番号13参照)、タンパク質のアミノ酸配列(配列番号14参照))を取得した。
【0122】
〔実施例4.JcFT遺伝子発現カセットの構築〕
p35S−ACE1/VP16AD−CR(WO2008111661A2参照)を制限酵素HindIIIおよびEcoRIにて処理し、当該プラスミドから転写因子遺伝子発現カセットを切り出した。制限酵素HindIIIおよびEcoRIにて処理したpRI909(タカラバイオ)に前述の転写因子遺伝子発現カセットをライゲーションし、pRI−35S−ACE1/VP16AD−CRを得た。
【0123】
pMRE4/35S(−46)−To71sGFP(WO2008111661A2参照)を制限酵素XbaIおよびSacIにて処理し、当該プラスミドからTo71sGFP遺伝子を切り出した。制限酵素XbaIおよびSacIにて処理したpRI−35S−ACE1/VP16AD−CRにTo71sGFP遺伝子をライゲーションし、pRI−35S−To71sGFP−CRを得た。
【0124】
また、ナンヨウアブラギリ(Jatropha curucas)の本葉から、RNeasy Plant mini kit(キアゲン)を用いて全RNAを調製した。ReverTra Ace qPCR RT kit(TOYOBO)を用いて、調製した全RNAに基づいてcDNAを合成した。合成したcDNAを鋳型として、2種類の特異的プライマー(BamJFT−1FおよびSpeJFT−1RC)を用いてPCR反応を行った。
【0125】
BamJFT−1F :5'−ATGGATCCAACAATGCCTAGGGATCAATTTAGAGACC−3'(配列番号15)
SpeJFT−1RC:5'−ATACTAGTTCACCGTCTCCGTCCTCCGGTG−3'(配列番号16)
PCR産物をBlunting Kination Ligation kit(タカラバイオ)を用いて平滑末端化およびリン酸化した後、当該PCR産物を制限酵素BamHIにて処理した。制限酵素SacIで処理した後、Blunting Kination Ligation kitを用いて平滑末端化およびリン酸化し、更に制限酵素BamHIで処理したpRI−35S−To71sGFP−CRに、前述のPCR産物をライゲーションし、pRI−35S−To71JcFT−CRを得た。
【0126】
〔実施例5.組換えシロイヌナズナにおけるJcFT遺伝子の機能評価〕
実施例4で作製されたpRI−35S−To71sGFP−CRおよびpRI−35S−To71JcFT−CRをアグロバクテリウム(Agrobacterium tumefaciens strain C58C1)に導入した。当該アグロバクテリウムを50mg/Lカナマイシン、100mg/Lアンピシリン、100mg/Lリファンピシンを含むLB寒天培地(0.5%酵母エキス、1.0%バクトトリプトン、0.5%食塩、1%寒天)で培養し、薬剤耐性コロニーを選抜することにより、組換えアグロバクテリウムを得た。
【0127】
得られた組換えアグロバクテリウムを、「モデル植物ラボマニュアル」(岩渕雅樹他編集、2000年、シュプリンガー・フェアラーク東京株式会社、ISBN 4-431-70881-2 C3045)に記載されている方法に準じて、シロイヌナズナ(Arabidopsis thaliana ecotype Columbia)に感染させることにより、シロイヌナズナへの遺伝子導入を行った。
【0128】
遺伝子導入されたシロイヌナズナから採取されたT1種子を、20mg/Lベンレート、200mg/Lクラフォラン、25mg/Lカナマイシンを含む改変MS寒天培地(MS無機塩類、B5ビタミン、1%ショ糖、0.8%寒天)に播種して生育させた後、カナマイシンに耐性を示す植物個体を選抜した。pRI−35S−To71JcFT−CRが導入されている組換えシロイヌナズナは、播種後30日目に寒天培地上で開花が認められた(図3参照)。
【0129】
選抜された植物個体を予め培土が入れられたポットに移植し、人工気象器内で生育させることにより、T2種子を得た。得られたT2種子を25mg/Lカナマイシンを含む改変MS寒天培地(MS無機塩類、B5ビタミン、2%ショ糖、0.8%寒天)に播種して生育させた後、χ2検定に基づいた5%の有意水準において、カナマイシンに耐性を示す植物個体が3:1の割合で出現するラインを選抜した。尚、植物個体を生育させるときの培養条件は、明期23時間、暗期1時間、23〜25℃とした。
【0130】
選抜されたラインについて、T2種子を、25mg/Lの濃度にてカナマイシンを含む改変MS寒天培地(MS無機塩類、B5ビタミン、2%ショ糖、0.8%寒天)に播種した。その結果、pRI−35S−To71JcFT−CRが導入されている組換えシロイヌナズナは、pRI−35S−To71sGFP−CRが導入されている組換えシロイヌナズナと比較して早期に開花が誘導された。
【0131】
播種後25日目に、開花時期の指標となるロゼット葉の数をカウントした。その結果、pRI−35S−To71JcFT−CRが導入されている組換えシロイヌナズナのロゼット葉は4.3±0.6枚(n=24)であるのに対して、pRI−35S−To71sGFP−CRが導入されている組換えシロイヌナズナのロゼット葉は5.5±0.7枚(n=39)であった。培養条件は、明期12時間、暗期12時間、23〜25℃とした。
【0132】
〔実施例6.組換えイネにおけるJcFT遺伝子の機能評価〕
pRI909(タカラバイオ)のカナマイシン耐性NPTII遺伝子をハイグロマイシン耐性APH4遺伝子に置換したpRH909を作製した。
【0133】
pRH909に、2x35Sプロモーター(Liu et al(2002)Plant J. 30、415-429)および翻訳エンハンサー配列fai(Mori et al(2006)Plant Biotech. 23、55-61)、JcFT遺伝子、CRターミネーターを挿入し、pRH−2x35S−faiJcFT−CRを得た。得られたpRH−2x35S−faiJcFT−CRをアグロバクテリウム(Agrobacterium tumefaciens strain LBA4404)に導入した。当該アグロバクテリウムを50mg/Lカナマイシン、100mg/Lストレプトマイシンを含むLB寒天培地(0.5%酵母エキス、1.0%バクトトリプトン、0.5%食塩、1%寒天)で培養し、薬剤耐性コロニーを選抜することにより、組換えアグロバクテリウムを得た。
【0134】
得られた組換えアグロバクテリウムを、Toki et al (2006)Plant J. 47、969-976に記載されている方法に準じて、イネ(Oryza sativa subsp. japonica cv. Nipponbare)に感染させることにより、イネへの遺伝子導入を行った。ハイグロマイシンに耐性を示す再生個体を取得した。pRH−2x35S−faiJcFT−CRが導入されている組換えイネは早期に花芽が形成された(図4参照)。尚、植物個体を生育させるときの培養条件は、明期23時間、暗期1時間、23〜25℃とした。
【0135】
なお本発明は、以上説示した各構成に限定されるものではなく、特許請求の範囲に示した範囲で種々の変更が可能であり、異なる実施形態や実施例にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態や実施例についても本発明の技術的範囲に含まれる。
【産業上の利用可能性】
【0136】
本発明は、開花時期が制御された植物を製造する目的に使用することができる。
【技術分野】
【0001】
本発明は、植物、特にナンヨウアブラギリ等のトウダイグサ目植物の開花時期を制御する遺伝子、タンパク質およびその利用に関するものである。
【背景技術】
【0002】
近年、食物としてのみではなくバイオ燃料としても、各種植物が注目を浴びつつある。莫大な数の人類の生活を支えるには莫大な量のこれら植物を必要とするが、耕作面積には限りがあり、それ故に、限られた面積の耕作地において最大限の収穫量を実現するための研究がなされつつある。そして、近年、開花時期などを人為的に制御することによって植物のライフサイクルを短縮し、これによって、植物の収穫量を増加させようとする試みもなされている。
【0003】
通常、植物の開花時期は、品種特有の内的要因の他、日長時間、気温および栄養状態などの外的要因によっても決定される。それ故に、従来から、外的要因を調節することによって、植物の開花時期が調節されてきた。しかしながら、外的要因を調節するためには特別な施設(例えば、光照射装置および/または温度調節装置など)を利用する必要があり、これらの設備費が高いとともに、照明費および/または空調費などが高くなるという問題点を有している。
【0004】
そこで、近年、植物の開花時期の制御に関わる遺伝子を同定し(例えば、非特許文献1参照)、当該遺伝子を用いて植物の開花時期を制御しようとする試みがなされている。
【0005】
例えば、特許文献1では、日長の制御を直接受ける遺伝子よりも下流で制御される遺伝子が検索され、当該遺伝子を用いれば開花時期を早めることが可能となり、その結果、発芽から種子取得までの期間を大幅に短縮できることが記載されている。更に具体的には、特許文献1には、花成遺伝子(FT遺伝子:Flowering Locus T遺伝子)をシロイヌナズナ内で強制的に発現させることにより、開花が早くなることが記載されている。
【0006】
FT遺伝子は、現在までに、数種類の植物において同定されている。例えば、シロイヌナズナ、イネ、コムギ、ライグラス、アカザ、ミカン、カボチャ、リンゴ、トマト、ブドウおよびポプラにおいて、花成遺伝子が同定されている(例えば、非特許文献2参照)。
【0007】
FT遺伝子は、日長などに応答して維管束師部での発現が増大し、FT遺伝子産物であるFTタンパク質が茎頂に移動して、茎頂で発現しているFDと呼ばれるbZIP型転写因子と相互作用することにより開花時期を促進することが知られている(例えば、非特許文献3参照)。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特開2000−139250号公報(公開日:2000年5月23日)
【非特許文献】
【0009】
【非特許文献1】Kobayashi Y et al, 1999, Science 286, 1960-1962
【非特許文献2】Igasaki T et al., 2008, Plant Cell Physiol. 49(3), 291-300
【非特許文献3】Kobayashi Y and Weigel D、2007、Genes Dev 21、2371-2384
【発明の概要】
【発明が解決しようとする課題】
【0010】
ナンヨウアブラギリに代表されるトウダイグサ目植物などの産業上、非常に有用であると考えられる植物においては花成遺伝子が単離されておらず、当該花成遺伝子を用いてこれらの植物の開花時期を制御したという報告はない。
【0011】
本発明は、上記従来の問題点に鑑みなされたものであって、ナンヨウアブラギリに代表されるトウダイグサ目植物などの産業上、非常に有用であると考えられる植物の開花時期を制御することを目的とする。
【課題を解決するための手段】
【0012】
シロイヌナズナなどのモデル生物に関しては、研究者の数も多く、遺伝子配列などの多くの情報が流通している。しかしながら、ナンヨウアブラギリに関しては、研究者の数も少なく、流通している遺伝子配列などの情報量も限られている状況にある。しかも、シロイヌナズナとナンヨウアブラギリとでは種間の距離も大きく、遺伝子配列のホモロジーも低いと考えられた。そこで本発明者は、複数の縮重プライマーを作成してFT遺伝子の単離を試みた。
【0013】
当初、ナンヨウアブラギリのFT遺伝子が、どの生育段階で、どの部位で、どのような気象条件で、どれだけ発現しているのか不明であり、cDNAを鋳型としてナンヨウアブラギリのFT遺伝子をクローニングしようとしても成功しなかった。そこで、まず、ゲノムDNAを鋳型としてナンヨウアブラギリのFT遺伝子の部分断片を取得し、当該部分断片の情報に基づいて、ナンヨウアブラギリのFT遺伝子の発現解析を行った。その結果、ナンヨウアブラギリのFT遺伝子が高発現する葉位および気象条件を特定することができた。
【0014】
このような、多くの試行錯誤の末にナンヨウアブラギリからFT遺伝子を単離することに成功し、これによって本発明を完成させるに至った。
【0015】
つまり、本発明は、以下の〔1〕〜〔9〕の発明を包含する。
【0016】
〔1〕:下記(1)〜(3)の何れかのポリペプチドをコードすることを特徴とするポリヌクレオチド。つまり、
(1)配列番号14で示されるポリペプチド、
(2)配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド、
(3)配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド。
【0017】
〔2〕:下記(4)〜(10)の何れかであることを特徴とするポリヌクレオチド。つまり、
(4)配列番号11、12または13で示されるポリヌクレオチド、
(5)配列番号11、12または13で示されるポリヌクレオチドのアンチセンス配列をコードするポリヌクレオチド、
(6)配列番号17および配列番号18に示すプライマーセットによって増幅されるポリヌクレオチドを含むポリヌクレオチド、
(7)配列番号19および配列番号20に示すプライマーセットによって増幅されるポリヌクレオチドを含むポリヌクレオチド、
(8)上記(4)〜(7)の何れかに記載されるポリヌクレオチドにおいて、1つ以上30以下の塩基が置換、欠失、挿入および/または付加されたポリヌクレオチドからなり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド、
(9)上記(4)〜(7)の何れかに記載されるポリヌクレオチドと相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド、
(10)上記(4)〜(7)の何れかに記載されるポリヌクレオチドとの相同性が84%以上であり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド。
【0018】
〔3〕:バラ下綱植物由来であることを特徴とする〔1〕に記載のポリヌクレオチド。
【0019】
〔4〕:〔1〕〜〔3〕の何れかに記載のポリヌクレオチドを含有することを特徴とする組換えベクター。
【0020】
〔5〕:上記組換えベクターが、プラスミドベクター、ウイルスベクター、ファージベクターまたはコスミドベクターであることを特徴とする〔4〕に記載の組換えベクター。
【0021】
〔6〕:下記(11)〜(13)の何れかのポリペプチド。つまり、
(11)配列番号14で示されるポリペプチド、
(12)配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド、
(13)配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド。
【0022】
〔7〕:〔1〕〜〔3〕の何れかに記載のポリヌクレオチド、〔4〕若しくは〔5〕に記載の組換えベクター、または、〔6〕に記載のポリペプチドを植物に導入する工程を含むことを特徴とする植物の開花時期の制御方法。
【0023】
〔8〕:〔1〕〜〔3〕の何れかに記載のポリヌクレオチド、〔4〕若しくは〔5〕に記載の組換えベクター、または、〔6〕に記載のポリペプチドを植物に導入する工程を含むことを特徴とする形質転換植物の製造方法。
【0024】
〔9〕:〔1〕〜〔3〕の何れかに記載のポリヌクレオチド、〔4〕若しくは〔5〕に記載の組換えベクター、または、〔6〕に記載のポリペプチドが導入されていることを特徴とする形質転換植物。
【発明の効果】
【0025】
トウダイグサ目植物のナンヨウアブラギリ等の、産業上、有用であると考えられる植物の開花時期を制御(早期化または遅延化)することができるという効果を奏する。
【図面の簡単な説明】
【0026】
【図1】実施例において、JcFT遺伝子のmRNA量をリアルタイムPCRによって定量した結果を示すグラフである。
【図2】実施例において、JcFT遺伝子のmRNA量をリアルタイムPCRによって定量した結果を示すグラフである。
【図3】実施例において、pRI−35S−To71sGFP−CRが導入されている組換えシロイヌナズナの写真である。
【図4】実施例において、pRH−2x35S−faiJcFT−CRが導入されている組換えイネの写真である(三角の矢印は花芽の位置を示す)。
【発明を実施するための形態】
【0027】
本発明の実施の形態について詳細に説明すれば以下のとおりであるが、本発明はこれに限定されるものではない。なお、本明細書中に記載された非特許文献および特許文献の全てが、本明細書中において参考として援用される。
【0028】
本明細書中で使用される場合、用語「ポリペプチド」は、「ペプチド」または「タンパク質」と交換可能に使用される。また、本明細書中で使用される場合、用語「ポリヌクレオチド」は、「遺伝子」、「核酸」または「核酸分子」と交換可能に使用され、ヌクレオチドの重合体が意図される。本明細書中で使用される場合、用語「塩基配列」は、「核酸配列」または「ヌクレオチド配列」と交換可能に使用され、デオキシリボヌクレオチド(A、G、CおよびTと省略される)の配列として示される。
【0029】
〔1.ポリヌクレオチド〕
本実施の形態のポリヌクレオチドは、下記(1)〜(3)の何れかのポリペプチドをコードするポリヌクレオチドであり得る。つまり、
(1)配列番号14で示されるポリペプチド、
(2)配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド、
(3)配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド。
【0030】
なお、配列番号14で示されるポリペプチドとは、発明者らによって同定された、ナンヨウアブラギリ(Jatropha curucas)由来のFT遺伝子(JcFT遺伝子)にコードされているタンパク質(JcFTタンパク質)である。
【0031】
また、本実施の形態のポリヌクレオチドは、下記(4)〜(10)の何れかであり得る。つまり、
(4)配列番号11、12または13で示されるポリヌクレオチド、
(5)配列番号11、12または13で示されるポリヌクレオチドのアンチセンス配列をコードするポリヌクレオチド、
(6)配列番号17および配列番号18に示すプライマーセットによって増幅されるポリヌクレオチドを含むポリヌクレオチド、
(7)配列番号19および配列番号20に示すプライマーセットによって増幅されるポリヌクレオチドを含むポリヌクレオチド、
(8)上記(4)〜(7)の何れかに記載されるポリヌクレオチドにおいて、1つ以上30以下の塩基が置換、欠失、挿入および/または付加されたポリヌクレオチドからなり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド、
(9)上記(4)〜(7)の何れかに記載されるポリヌクレオチドと相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド、
(10)上記(4)〜(7)の何れかに記載されるポリヌクレオチドとの相同性が84%以上であり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド。
【0032】
なお、上記(6)および(7)に記載のポリヌクレオチドは、各プライマーセットによって増幅されるポリヌクレオチドを含むものであるが、各プライマーセットによって増幅されるポリヌクレオチド以外の部分の具体的な塩基配列は、特に限定されない。勿論、上記(6)および(7)に記載のポリヌクレオチドは、各プライマーセットによって増幅されるポリヌクレオチド自体であってもよい。
【0033】
配列番号11で示されるポリヌクレオチドとは、JcFT遺伝子のゲノムDNAであり、配列番号12で示されるポリヌクレオチドとは、JcFT遺伝子の完全長cDNAであり、配列番号13で示されるポリヌクレオチドとは、JcFT遺伝子の翻訳領域配列である。
【0034】
また、配列番号17および配列番号18に示すプライマーセットとは、JcFT遺伝子の翻訳領域に相当する塩基配列の一部(配列番号13の塩基番号1〜124で示される塩基配列)を増幅することが可能なプライマーセットであり、配列番号19および配列番号20に示すプライマーセットとは、JcFT遺伝子の完全長cDNAの一部(配列番号12の塩基番号1〜816で示される塩基配列)を増幅することが可能なプライマーセットである。なお、これらのプライマーの具体的な配列を以下に記載する。
【0035】
JcFTL−2F :5'−ATGCCTAGGGATCAATTTAGAGACC−3'(配列番号17)
JcFTL−2RC:5'−AGCCATTGTTAACCTCTCTGTGATT−3'(配列番号18)
JcFTL−3F :5'−ACGCGGGGATGATAATACGAGTGTAGC−3'(配列番号19)
JcFTL−3RC:5'−AGAGATTAATATTCAGTAAATTTGATAGCATTTGTGATC−3'(配列番号20)。
【0036】
上記4つのプライマーの塩基配列は、他の植物のFT遺伝子とは配列の相同性が低い。従って、これらのプライマーを用いれば、ナンヨウアブラギリ由来のFT遺伝子と他の植物由来のFT遺伝子とを、より明確に判別することができる。勿論、これらのプライマーセットによって増幅されるポリヌクレオチド自体を用いて、ナンヨウアブラギリ由来のFT遺伝子と他の植物由来のFT遺伝子とを判別することも可能である。
【0037】
また、配列番号19および配列番号20に示すプライマーセットによって増幅されるポリヌクレオチドにコードされるポリペプチドは、それ自体、植物の開花時期を早める作用をも備えている。
【0038】
本実施の形態のポリヌクレオチドを用いれば、植物内のFT遺伝子の発現量・発現時期などを調節することが可能となり、その結果、植物の開花時期を制御することができる。一例としては、FT遺伝子のセンス鎖に相当する本実施形態のポリヌクレオチドを用いれば、植物内におけるFT遺伝子の発現量の上昇、および/または、発現時期の早期化を実現することが可能となり、これによって、植物の開花時期を早めることができる。また、FT遺伝子のアンチセンス鎖に相当する本実施形態のポリヌクレオチドを用いれば、植物内におけるFT遺伝子の発現量の低下、および/または、発現時期の遅延化を実現することが可能となり、これによって、植物の開花時期を遅くすることが可能となる。なお、後述する〔4.植物の開花時期の制御方法〕を一読すれば、植物の開花時期を制御するための更に詳細な方法を理解できるであろう。
【0039】
本実施の形態のポリヌクレオチドは、DNAの形態(例えば、cDNAもしくはゲノムDNA)、またはRNA(例えば、mRNA)の形態であり得る。DNAまたはRNAは二本鎖であっても、一本鎖であってもよい。一本鎖DNAまたはRNAは、コード鎖(センス鎖)であっても、非コード鎖(アンチセンス鎖)であってもよい。
【0040】
また、本実施の形態のポリヌクレオチドは、化学的に合成されたものであってもよく、コードするタンパク質の発現が向上するように、コドンユーセージ(Codon usage)が改変されたものであってもよい。勿論、本実施の形態のポリヌクレオチドは、自然界から単離されたものであってもよい。
【0041】
自然界から単離されたポリヌクレオチドを本実施の形態のポリヌクレオチドとして用いる場合には、当該ポリヌクレオチドの由来は特に限定されないが、例えば、バラ下綱植物、トウダイグサ目植物、トウダイグサ科植物、または、ナンヨウアブラギリ植物に由来するポリヌクレオチドであることが好ましい。これらの中では、バラ下綱植物由来であることが好ましく、トウダイグサ目植物由来であることが更に好ましく、トウダイグサ科植物由来であることが更に好ましく、ナンヨウアブラギリ植物由来であることが最も好ましいといえる。
【0042】
本実施の形態のポリヌクレオチドは、配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなるポリペプチドをコードしたものであり得る。1または数個のアミノ酸が置換、欠失、挿入および/または付加される部位は、置換、欠失、挿入および/または付加後のポリペプチドが植物の開花時期を制御する機能を有していれば、該アミノ酸配列中のどの部位であってもよい。ここで「1または数個のアミノ酸残基」とは、具体的には10個以内の範囲のアミノ酸残基数であり、好ましくは6個以内の範囲のアミノ酸残基である。
【0043】
置換などの変異が生じる場合には、アミノ酸側鎖の性質が保存されている別のアミノ酸に置換されることが好ましい。例えばアミノ酸側鎖の性質としては、疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)、親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)、脂肪族側鎖などの特徴的な側鎖を有するアミノ酸(G、A、V、L、I、P)、水酸基含有側鎖を有するアミノ酸(S、T、Y)、硫黄原子含有側鎖を有するアミノ酸(C、M)、カルボン酸及びアミド含有側鎖を有するアミノ酸(D、N、E、Q)、塩基含有側鎖を有するアミノ酸(R、K、H)、芳香族含有側鎖などの特徴的な側鎖を有するアミノ酸(H、F、Y、W)を挙げることができる(括弧内はいずれもアミノ酸の一文字標記を表す)。アミノ酸側鎖の性質が同じアミノ酸間の置換であれば、ポリペプチドが備える生物学的活性を維持することができる。
【0044】
また、本実施の形態のポリヌクレオチドは、上記(4)〜(7)の何れかに記載されるポリヌクレオチドにおいて、1つ以上30以下の塩基が置換、欠失、挿入および/または付加されたポリヌクレオチドからなるポリヌクレオチドであり得る。塩基が置換、欠失、挿入および/または付加される部位は、置換、欠失、挿入および/または付加後のポリヌクレオチドが植物の開花時期を制御する機能を有していれば、該塩基配列中のどの部位であってもよい。
【0045】
以上のようなポリヌクレオチドは、特定のポリヌクレオチド内の塩基を置換、欠失、挿入および/または付加することによって得ることが可能である。ポリヌクレオチドの塩基を変換する具体的な方法としては、例えば市販のキット(Transformer Site−Directed Mutagenesis Kit;Clonetech製,QuickChange Site Directed Mutagenesis Kit;Stratagene製など)の使用、またはポリメラーゼ連鎖反応法(PCR)の利用が挙げられる。これらの方法は当業者に公知である。
【0046】
本実施の形態のポリヌクレオチドは、上記(4)〜(7)の何れかに記載されるポリヌクレオチドに対して相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズするものであり得る。
【0047】
ここで、「ストリンジェントな条件」とは、相同性が高い核酸同士、例えば完全にマッチしたハイブリッドの融解温度(Tm値)から15℃、好ましくは10℃低い温度までの範囲の温度でハイブリダイズする条件をいう。具体例としては、一般的なハイブリダイゼーション用緩衝液中で、68℃、20時間の条件でハイブリダイズする条件をいう。
【0048】
更に具体的には、「ストリンジェントな条件」とは、いわゆる塩基配列に特異的な2本鎖のポリヌクレオチドが形成され、非特異的な2本鎖のポリヌクレオチドが形成されない条件をいう。換言すれば、相同性が高い核酸同士が、例えば完全にマッチしたハイブリッドの融解温度(Tm値)から15℃、好ましくは10℃、更に好ましくは5℃低い温度までの範囲の温度でハイブリダイズする条件ともいえる。例えば、一般的なハイブリダイゼーション用緩衝液中で、68℃、20時間の条件でハイブリダイズする条件を挙げることができる。より具体的には、0.9M NaCl、0.09M sodium citrate、0.01M sodium phosphate pH6.8、1mM EDTA pH8.0、0.5% SDS、1×デンハルト溶液からなる緩衝液中で、60〜68℃、好ましくは65℃、さらに好ましくは68℃の条件下で16〜24時間ハイブリダイズさせた後、さらに0.3M NaCl、0.03M sodium citrate、1% SDSからなる緩衝液中で、60〜68℃、好ましくは65℃、さらに好ましくは68℃の条件下で15分間の洗浄を2回行う条件を挙げることができる。当業者であれば、Molecular Cloning(Sambrook, J. et al., Molecular Cloning :a Laboratory Manual 2nd ed., Cold Spring Harbor Laboratory Press, 10 Skyline Drive Plainview, NY (1989))等を参照することにより、こうした遺伝子を容易に取得することができる。
【0049】
本実施の形態のポリヌクレオチドは、配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチドをコードするポリヌクレオチドであり得る。なお、配列番号14で示されるポリペプチドとの相同性は、95%以上であることが更に好ましいといえる。また、本実施の形態のポリヌクレオチドは、上記(4)〜(7)の何れかに記載されるポリヌクレオチドとの相同性が84%以上であり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチドであり得る。なお、上記(4)〜(7)の何れかに記載されるポリヌクレオチドとの相同性は、90%以上であることが更に好ましいといえる。上記構成によれば各種植物の開花時期を好適に制御することができるが、その中でも、バラ下綱植物の開花時期を更に好適に、トウダイグサ目植物の開花時期を更に好適に、トウダイグサ科植物の開花時期を更に好適に、ナンヨウアブラギリ植物の開花時期を最も好適に制御することができる。
【0050】
本発明において「相同性」とは、2つの塩基配列または2つのアミノ酸配列間の相同性が意図される。相同性は、比較対象の配列の全領域にわたって最適な状態にアラインメントされた2つの配列を比較することにより決定される。ここで、比較対象の塩基配列またはアミノ酸配列を最適な状態にアラインメントするために、付加または欠失(例えば、ギャップ等)を許容してもよい。このような配列相同性は、例えば、FASTA(Pearson & Lipman、1988、PNAS、4、2444-2448)、BLAST(Altschul et al、1990、Journal of Molecular Biol.、215、403-410)、CLUSTAL W(Thompson et al、1994、Nucleic Acid Research、22、4673-4680)等のプログラムを用いて算出することができる。
【0051】
上記のプログラムは、例えば、DNA Data Bank of Japan(国立遺伝学研究所生命情報・DDBJ研究センター Center for Information Biology and DNA Data Bank of Japan; CIB/DDBJ)内で運営される国際DNAデータバンクのWEBページ(http://www.ddbj.nig.ac.jp/Welcome-e.html)等において、一般的に利用可能である。また、配列の相同性は市販の配列解析ソフトウェアを用いて求めることもできる。具体的には、配列の相同性は、DNASYS Pro ver.2.06(Hitachi Software Engineering)のSmith−Watermanプログラムにより相同性解析を行ってアラインメントすることにより算出することができる。
【0052】
本実施の形態のポリヌクレオチドは、後述する本発明のポリペプチドをコードするポリヌクレオチドのみからなるものであってもよいが、その他の塩基配列が付加されていてもよい。付加される塩基配列としては限定されないが、標識(例えば、ヒスチジンタグ、Mycタグ、およびFLAGタグなど)、本発明のポリペプチドと融合タンパク質を構成し得るポリペプチド(例えば、GSTおよびMBPなど)、プロモーター配列(例えば、植物由来プロモーター配列、酵母由来プロモーター配列、ファージ由来プロモーター配列、および大腸菌由来プロモーター配列など)、およびシグナル配列(例えば、小胞体移行シグナル配列、および分泌配列など)をコードする塩基配列などが挙げられる。これらの塩基配列が付加される部位は特に限定されるものではなく、例えば、ポリヌクレオチドの5’末端であっても、3’末端でもあってもよい。
【0053】
〔2.組換えベクター〕
本実施の形態の組換えベクターは、上述した本発明のポリヌクレオチドを含有するものである。
【0054】
本実施の形態の組み換えベクターを用いれば、植物内のFT遺伝子の発現量を調節することが可能となり、その結果、植物の開花時期を制御することができる。
【0055】
本実施の形態の組換えベクターとしては特に限定されないが、例えば、プラスミドベクター、ウイルスベクター、ファージベクターまたはコスミドベクターであることが好ましく、これらの中では、プラスミドベクターまたはウイルスベクターであることが更に好ましい。上記構成によれば、所望の宿主を形質転換することができる。
【0056】
本実施の形態の組換えベクターの更に具体的な構成としては特に限定されないが、発現制御配列を含むことが好ましい。なお、発現制御配列とは、所望の遺伝子の発現を制御するための塩基配列が意図される。当該発現制御配列としては特に限定されないが、例えば、プロモーター配列、エンハンサー配列、ターミネーター配列、5’非翻訳領域配列、3’非翻訳領域配列、開始コドン配列、イントロンのスプライシングシグナル配列、翻訳フレーム維持配列および停止コドン配列などを挙げることが可能である。そして、本実施の形態の組換えベクターは、これら具体的な発現制御配列を1つだけ含むことも可能であるし、複数種類を含むことも可能である。なお、複数種類の発現制御配列を含む場合には、その組み合わせは特に限定されない。
【0057】
上記「プロモーター配列」とは、遺伝子の転写を開始するために十分な最小の塩基配列が意図される。プロモーター配列としては、構成的プロモーター、組織特異的プロモーター、または、特定の刺激によって転写が誘導される誘導性プロモーターなどを用いることが可能であるが、これらに限定されない。組換えベクターの使用目的に応じて、適宜選択することが好ましい。
【0058】
上記構成的プロモーターとしては特に限定されないが、例えば、CaMV 35Sプロモーター(Benfey PN & Chua NH、1990、Science 250、959-966)、PG10−90(特開09-131187)、ユビキチンプロモーター(国際公開01/094394)、アクチンプロモーター(国際公開00/070067)等を用いることが好ましい。
【0059】
上記組織特異的プロモーターとしては特に限定されないが、例えば、ダイズ種子グリシニンプロモーター(特開06-189777)、プロラミンプロモーター(国際公開2004/056993)、インゲンマメ種子ファゼオリンプロモーター(国際公開91/013993)、ナタネ種子ナピンプロモーター(国際公開91/013972)、シロイヌナズナSultr2;2プロモーター(Takahashi H et al、2000、Plant J 23、171-82)、アグロバクテリウムrolCプロモーター(Almon E et al、1997、Physiol 115、1599-1607)等を用いることが好ましい。
【0060】
上記誘導性プロモーターとしては特に限定されないが、例えば、銅イオン誘導性プロモーター(国際公開08/111661)、ステロイドホルモン誘導性プロモーター(米国特許第6063985号)、エタノール誘導性システム(国際公開93/21334)、テトラサイクリン誘導性システム(例えば、Weinmann P et al、1994、Plant J. 5、559-569)、除草剤セーフナー誘導性プロモーター(Hershey et al、1991、Plant Mol. Biol.、17、679)、熱誘導性プロモーター(米国特許第5447858号)、低温誘導性プロモーター(米国特許第5847102号)、植物病原体の攻撃により誘導されるプロモーター(米国特許第5942662号)等を用いることが好ましい。
【0061】
上記「ターミネーター配列」とは、遺伝子の転写を終結させ、mRNAを安定化させるためのポリアデニン配列を付加するために十分な最小の配列が意図される。上記ターミネーター配列としては特に限定されないが、例えば、NOSターミネーター、CR16ターミネーター(特開2000-166577)、ダイズ種子グリシニンターミネーター(特開06-189777)等を用いることが好ましい。
【0062】
〔3.ポリペプチド〕
本実施の形態のポリペプチドは、下記(11)〜(13)の何れかのポリペプチドであり得る。つまり、
(11)配列番号14で示されるポリペプチド、
(12)配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド、
(13)配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド。
【0063】
なお、配列番号14で示されるポリペプチドとは、発明者らによって同定された、ナンヨウアブラギリ(Jatropha curucas)由来のFT遺伝子(JcFT遺伝子)にコードされているタンパク質(JcFTタンパク質)である。
【0064】
本実施の形態のポリペプチドを用いれば、植物内のFT遺伝子の発現量を調節することが可能となり、その結果、植物の開花時期を制御することができる。
【0065】
本実施の形態のポリペプチドは、配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチドであり得るが、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加される部位は、置換、欠失、挿入および/または付加後のタンパク質が植物の開花時期を制御する機能を有していれば、該アミノ酸配列中のどの部位であってもよい。ここで「1または数個のアミノ酸残基」とは、具体的には10個以内の範囲のアミノ酸残基数であり、好ましくは6個以内の範囲のアミノ酸残基である。
【0066】
置換などの変異が生じる場合には、アミノ酸側鎖の性質が保存されている別のアミノ酸に置換されることが好ましい。例えばアミノ酸側鎖の性質としては、疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)、親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)、脂肪族側鎖などの特徴的な側鎖を有する疎水性アミノ酸(G、A、V、L、I、P)、水酸基含有側鎖を有するアミノ酸(S、T、Y)、硫黄原子含有側鎖を有するアミノ酸(C、M)、カルボン酸及びアミド含有側鎖を有するアミノ酸(D、N、E、Q)、塩基含有側鎖を有するアミノ酸(R、K、H)、芳香族含有側鎖などの特徴的な側鎖を有するアミノ酸(H、F、Y、W)を挙げることができる(括弧内はいずれもアミノ酸の一文字標記を表す)。アミノ酸側鎖の性質が同じアミノ酸間の置換であれば、ポリペプチドが備える生物学的活性を維持することができる。
【0067】
また、本実施の形態のポリペプチドは、配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチドであり得るが、より好ましくは、配列番号14で示されるポリペプチドとの相同性が95%以上である。
【0068】
本実施の形態のポリペプチドは、例えば、自然界から単離した当該ポリペプチドを産生する生物を用いて生産されてもよいし、遺伝子組み換え技術を用いて生産されてもよいし、またはアミノ酸合成機などを用いて化学合成されてもよい。
【0069】
遺伝子組み換え技術において、好適に用いられる各種組換えタンパク質発現系は、例えば、大腸菌発現系、酵母発現系、昆虫細胞発現系、哺乳類細胞発現系、および無細胞発現系を用いてもよく、これらに限定されない。
【0070】
本実施の形態のポリペプチドは、例えば、分子間架橋および/または分子内架橋(例えば、ジスルフィド結合など)が施されたもの、化学修飾(例えば、糖鎖、リン酸もしくはその他の官能基など)が施されたもの、標識(例えば、ヒスチジンタグなど)が付与されたもの、または本発明のポリペプチドと融合タンパク質を構成し得るポリペプチド(例えば、ストレプトアビジン、シトクロム、およびGFPなど)が連結されたものなどが含まれるが、特にこれらに限定されない。さらに、本実施の形態のポリペプチドには、植物の開花時期を制御する機能が実質的に維持される限り、数種のタンパク質の断片を組み合わせて構成されたキメラタンパク質も含まれる。
【0071】
〔4.植物の開花時期の制御方法〕
本実施の形態の植物の開花時期の制御方法は、本発明のポリヌクレオチド、本発明の組換えベクター、または、本発明のポリペプチドを植物に導入する工程を含む方法である。
【0072】
本発明のポリヌクレオチド、組換えベクターおよびポリペプチドに関しては既に説明したので、ここでは、その説明を省略する。
【0073】
上記植物(換言すれば、開花時期が制御される植物)としては特に限定されず、単子葉植物であっても双子葉植物であっても良い。具体的には、イネ科植物、バラ下綱植物、トウダイグサ目植物、トウダイグサ科植物、または、ナンヨウアブラギリ植物であることが好ましい。これらの中では、バラ下綱植物由来であることが好ましく、トウダイグサ目植物由来であることが更に好ましく、トウダイグサ科植物由来であることが更に好ましく、ナンヨウアブラギリ植物由来であることが最も好ましいといえる。
【0074】
また、上記植物としては、例えば、ダイズ、エンドウ、インゲン、アルファルファ、ミヤコグサ、クローバ、ピーナッツ、スイートピー、クルミ、チャ、ワタ、コショウ、キュウリ、スイカ、カボチャ、メロン、ダイコン、ナタネ、キャノーラ、テンサイ、レタス、キャベツ、ブロッコリー、カリフラワー、シロイヌナズナ、タバコ、ナス、ジャガイモ、サツマイモ、サトイモ、キクイモ、トマト、ホウレンソウ、アスパラガス、ニンジン、アマ、ゴマ、エンダイブ、キク、フウロウソウ、キンギョソウ、カーネーション、ナデシコ、ニチニチソウ、ブバルディア、カスミソウ、ガーベラ、トルコキキョウ、チューリップ、ストック、スターチス、シクラメン、ユキノシタ、ノースポール、スミレ、バラ、サクラ、リンゴ、ナシ、ブドウ、イチゴ、ウメ、アーモンド、ミカン、レモン、バナナ、マンゴー、パパイヤ、キウイ、コーヒー、ボケ、サツキ、ツツジ、ポインセチア、キャッサバ、アブラヤシ、ココヤシ、オリーブ、リンドウ、コスモス、アサガオ、ヒマワリ、イチョウ、スギ、ヒノキ、ポプラ、マツ、セコイア、オーク、ヤナギ、ユーカリ、ケナフ、スイレン、トチュウ、ブナ、ヒマ、タケ、サトウキビ、イネ、コムギ、オオムギ、ライムギ、エンバク、トウモロコシ、ソルガム、シバ、トールフェスキュー、スイッチグラス、ススキ、ネギ、タマネギ、ニンニク、ユリ、オニユリ、ラン、グラジオラス、またはパイナップルを挙げることも可能であるが、これらに限定されない。
【0075】
本発明のポリヌクレオチド、本発明の組換えベクター、または、本発明のポリペプチドを上記植物に導入する方法は特に限定されない。例えば、アグロバクテリウム法、パーティクルガン法、エレクトロポレーション法、リン酸カルシウム法、インジェクション法、またはウイルスベクター法などの公知方法を用いることが可能である。
【0076】
本実施の形態の植物の開花時期の制御方法によれば、植物の開花時期を早期化したり(換言すれば、開花の促進)、あるいは、植物の開花時期を遅延化する(換言すれば、開花の抑制)ことが可能となる。以下に、開花時期を早期化する場合の更に具体的な方法、および、開花時期を遅延化する場合の更に具体的な方法の一例に関して説明するが、本発明はこれらに限定されない。
【0077】
<4−1>開花時期の早期化
植物の開花時期の早期化は、例えば、以下の(A)〜(C)の方法によって実現することが可能である。
【0078】
(A)FT遺伝子が植物細胞中で発現可能となるように、任意の発現制御配列とFT遺伝子とを、任意のベクターに連結する。当該ベクターを植物細胞に導入して形質転換植物を得る。当該形質転換植物において開花が促進される。なお、FT遺伝子が植物細胞中で発現可能であれば、任意の発現制御配列や任意のベクターは必ずしも必要ではなく、形質転換植物を取得する必要もない。FT遺伝子が植物細胞中で発現可能となるよう、宿主植物に応じて,FT遺伝子を植物細胞へ導入すればよい。
【0079】
(B)植物のゲノムDNAに任意の塩基配列を組み込むことにより、植物に内在するFT遺伝子の発現を促進する。任意の塩基配列としては、好ましくは、任意の発現制御配列を挙げることができる。更に具体的には、任意の発現制御配列としては、プロモーター配列およびエンハンサー配列などを挙げることが可能である。当該発現制御配列が機能し得るように植物のゲノムDNA内に組み込まれ、植物に内在するFT遺伝子の発現を促進し、これによって植物の開花が促進される。
【0080】
(C)FT遺伝子がコードするタンパク質を、例えば、師管液中に注入し、植物細胞中に導入する。FT遺伝子がコードするタンパク質は、茎頂や腋芽に移動し、開花を促進する。FT遺伝子がコードするタンパク質は、組換え微生物を用いて生産してもよいし、野生型の植物あるいは形質転換植物から抽出してもよい。或は、FT遺伝子がコードするタンパク質が蓄積している植物(例えば、形質転換植物)を台木として接木してもよい。
【0081】
<4−2>開花時期の遅延化
植物の開花時期の遅延化は、例えば、以下の(D)〜(I)の方法によって実現することが可能である。
【0082】
(D)FT遺伝子のアンチセンス配列が植物細胞中で発現可能となるよう、任意の発現制御配列とFT遺伝子のアンチセンス配列とを、任意のベクターに連結する。当該ベクターを植物細胞に導入して形質転換植物を得る。当該形質転換植物において開花時期が遅延する。なお、FT遺伝子のアンチセンス配列が植物細胞中で発現可能であれば、任意の発現制御配列や任意のベクターは必ずしも必要はない。FT遺伝子のアンチセンス配列が植物細胞中で発現可能となるよう、宿主植物に応じて、FT遺伝子のアンチセンス配列を植物細胞へ導入すればよい。
【0083】
本明細書において「アンチセンス配列」とは、特定のmRNA分子の少なくとも一部に相補的なDNA分子またはRNA分子である。植物細胞中で、アンチセンス配列は、対応するmRNAにハイブリダイズして二本鎖分子を形成し、これによって、mRNAの翻訳を阻害する。
【0084】
(E)VIGS(Virus Induced Gene Silencing)を誘発するウイルスベクターにFT遺伝子の全長あるいはFT遺伝子の一部を組み込み、当該ベクターを植物細胞に導入することによって内在性のFT遺伝子の発現を抑制し、これによって、開花時期を遅延させる。
【0085】
(F)FT遺伝子の1アミノ酸または複数のアミノ酸を別のアミノ酸に置換することでFT遺伝子を花成抑制遺伝子に転換することが可能である(Hanzawa et al、2005、PNAS、102、7748-7753)。当該置換のように優性ネガティブ変異が導入された花成抑制遺伝子が植物細胞中で発現可能となるよう、任意の発現制御配列と花成抑制遺伝子とを、任意のベクターに連結する。当該ベクターを植物細胞に導入して形質転換植物を得る。当該形質転換植物では、開花時期が遅延する。なお、花成抑制遺伝子が植物細胞中で発現可能であれば、任意の発現制御配列や任意のベクターは、必ずしも必要ではない。花成抑制遺伝子が植物細胞中で発現可能となるよう、宿主植物に応じて、花成抑制遺伝子植物細胞へ導入すればよい。
【0086】
本明細書において「優性ネガティブ変異」とは、本来の機能を損なう形で変異が導入されており、かつ野生型のタンパク質の活性に対して優性的に働き、野生型の表現型を打ち消して負の表現型を示すことができる変異である。
【0087】
(G)植物のゲノムDNA中に任意の塩基配列を組み込むことにより、植物に内在するFT遺伝子の発現を抑制する。任意の塩基配列は、植物内在のFT遺伝子の発現を抑制し得るように、植物のゲノムDNA中に組み込まれる必要がある。具体的には、例えば、植物に内在するFT遺伝子内への任意の塩基配列の挿入、または、植物に内在するFT遺伝子の発現に関与する発現制御配列内への任意の塩基配列の挿入が挙げられるが、これらに限定されない。
【0088】
(H)リボザイムや三重鎖形成オリゴヌクレオチドを植物細胞中に導入する。リボザイムによってFT遺伝子のmRNAを切断したり、三重鎖形成オリゴヌクレオチドによってFT遺伝子の転写や翻訳を阻害したりすることによって、開花時期を遅延する。
【0089】
なお、本明細書において「リボザイム」とは、一本鎖RNAを特異的に切断する能力を有するRNA分子が意図される。リボザイムをコードする塩基配列を遺伝子工学的に改変することにより、RNA分子内の特定の塩基配列を認識して当該塩基配列を切断する分子を作製することができる(Ceeh et al、1988、J. Amer. Med. Assn.、260、3030)。
【0090】
また、本明細書において「三重鎖形成オリゴヌクレオチド」とは、二重らせんDNAの周りに巻き付いて三重らせんを形成し、遺伝子の転写を停止させる分子である。特定の塩基配列を認識するように、三重鎖形成オリゴヌクレオチドを設計することが可能である(Maher et al、1991、Antisense Res. and Dev.、1、227)。
【0091】
(I)優性ネガティブ変異が導入された花成抑制遺伝子がコードするタンパク質を、例えば、師管液中に注入し、植物細胞中に導入する。花成抑制遺伝子がコードするタンパク質は、茎頂や腋芽に移動し、開花時期を遅延する。花成抑制遺伝子がコードするタンパク質は、組換え微生物を用いて生産してもよいし、形質転換植物から抽出してもよい。或は花成抑制遺伝子がコードするタンパク質が蓄積している植物(例えば、形質転換植物)を台木として接木してもよい。
【0092】
上述した開花時期を早期化する方法と開花時期を遅延化する方法とは、各々単独で実施されることも可能であり、可能な範囲で自由に組み合わせられることも可能である。具体的には、例えば、植物に内在するFT遺伝子へ任意の塩基配列を挿入してFT遺伝子を機能的に欠損させた植物に、外来性のFT遺伝子(または、花成抑制遺伝子)が植物細胞中で発現可能となるよう、新たにFT遺伝子(または、花成抑制遺伝子)を導入してもよい。また、FT遺伝子(または、花成抑制遺伝子)をすでに導入した植物に、追加的にFT遺伝子(または、花成抑制遺伝子)を導入しても良い。
【0093】
また、具体的にナンヨウアブラギリの開花時期の制御には、各種誘導性プロモーター、具体的には例えば、銅イオン誘導性プロモーター(例えば、国際公開08/111661)に本発明のポリヌクレオチドを連結し、トウダイグサ目植物、具体的には例えば、ナンヨウアブラギリに導入し、当該形質転換植物に銅イオンを接触させることで開花時期を制御することが好ましい。上記構成によれば、例えば、JcFTタンパク質とのアミノ酸配列の相同性が90%未満のシロイヌナズナFT遺伝子由来のDNA断片を上記プロモーターに連結して導入した場合と比較して、開花時期の制御を効果的に行うことができるとともに、形態異常などの望ましくない表現型の出現を抑制することができる。
〔5.形質転換植物の製造方法〕
本実施の形態の形質転換植物の製造方法は、本発明のポリヌクレオチド、本発明の組換えベクター、または、本発明のポリペプチドを植物に導入する工程を含む方法である。つまり、本実施の形態の形質転換植物の製造方法は、既に説明した本発明の植物の開花時期の制御方法と実質的に同じ方法に基づいて実施することが可能である。
【0094】
本発明のポリヌクレオチド、組換えベクターおよびポリペプチドに関しては既に説明したので、ここでは、その説明を省略する。
【0095】
また、上記植物の具体的な構成、当該植物へ本発明のポリヌクレオチドなどを導入する具体的な構成については〔4.植物の開花時期の制御方法〕にて既に説明したので、ここではその説明を省略する。
【0096】
〔6.形質転換植物〕
本実施の形態の形質転換植物は、本発明のポリヌクレオチド、本発明の組換えベクター、または、本発明のポリペプチドが導入されているものである。
【0097】
本実施の形態の形質転換植物は、〔5.形質転換植物の製造方法〕にて説明した方法に基づいて製造することができる。
【0098】
本実施の形態の形質転換植物では、開花時期が制御されている。当該制御としては、開花時期の早期化、または開花時期の遅延化などを挙げることができるが、これらに限定されない。
【0099】
例えば、開花時期を早期化すれば、当該形質転換植物のライフサイクルを短くすることができる。つまり、短い時間で当該植物(例えば、種子、果実など)を収穫することが可能になる。その結果、例えば、品種改良を効率よく行うことが可能になる。
【0100】
逆に、開花時期を遅延化すれば、当該形質転換植物のライフサイクルを長くすることができる。つまり、時間をかけて十分に生育した当該植物(例えば、種子、果実など)を収穫することが可能になる。その結果、例えば、葉菜類や根菜類の商品価値を高めることが可能になる。
【0101】
本発明によれば、開花時期を早期化したり遅延化したりすることができるので、栽培環境、播種時期および栽培地などに幅をもたせることが可能となる。そして、その結果、生産性を向上させることが可能になる。
【実施例】
【0102】
以下、実施例に基づいて本発明を更に詳細に説明するが、本発明はこれら実施例によって限定されるものではない。
【0103】
〔実施例1.JcFT遺伝子断片の単離〕
DNeasy Plant mini kit(キアゲン)を用いて、ナンヨウアブラギリ(Jatropha curucas)の本葉からゲノムDNAを調製した。当該ゲノムDNAを鋳型として、4種類の縮重プライマー(FTD−1F、FTD−1RC、FTD−2F、FTD−2RC)を用いてPCR反応を行った。
【0104】
FTD−1F :5'−GACCCCTTYACAAGRTCYATYTCYCTGAGGGT−3' (配列番号1)
FTD−1RC:5'−CARAGTGTAGAAGGTCCKRAGRTC−3' (配列番号2)
FTD−2F :5'−CAAGAGATTGTGTGYTAYGARAGYCCAMGGCCAAC−3'(配列番号3)
FTD−2RC:5'−CGGAGCCRCYYTCCCTYTGRCA−3' (配列番号4)
混合塩基はIUBコードに従い、M(A or C)、R(A or G)、W(A or T/U)、S(C or G)、Y(C or T/U)、K(G or T/U)、V(A or C or G)、H(A or C or T/U)、D(A or G or T/U)、B(C or G or T/U)、N(A or C or G or T/U)で示した。
【0105】
PCR産物をアガロースゲルにて電気泳動した後、得られた増幅断片をアガロースゲルから切り出し、MagExtractor(東洋紡)を用いて上記増幅断片を精製した。精製された増幅断片を、ZeroBlunt TOPO PCR Cloning kit(インビトロジェン)を用いてpCR4Blunt−TOPO vectorにクローニングした。
【0106】
当該ベクターを用いて大腸菌DH5αを形質転換して、薬剤選抜されたコロニーをピックアップした。ピックアップされた大腸菌をLB培地(0.5%酵母エキス、1.0%バクトトリプトン、0.5%食塩)で培養し、QIAprep spin miniprep kit(キアゲン)を用いてプラスミドを調製した。
【0107】
M13 reverseプライマー、BigDye terminator v3.1(ABI)、3100 Genetic Analyzer(ABI)を用いて、上記プラスミド内に挿入された増幅断片の塩基配列を解析した。
【0108】
その結果、FTD−1FおよびFTD−1RCを用いてPCR反応を行うことによって、JcFT遺伝子の第1エクソン内の塩基配列(144bp)の情報が得られ、FTD−2FおよびFTD−2RCを用いてPCR反応を行うことによって、JcFT遺伝子の第4エクソン内の塩基配列(205bp)の情報が得られた。
【0109】
得られた配列情報に基づいて2種類の特異的プライマー(JcFTL−1FおよびJcFTL−1RC)を設計した。
【0110】
JcFTL−1F :5'−TATAATCACAGAGAGGTTAACAATGGCTGTGAGCTCAAAC−3'(配列番号5)
JcFTL−1RC:5'−CTGACGCCACCCTGGTGGATACACGGTCTG−3'(配列番号6)
設計した2種類の特異的プライマー(JcFTL−1FおよびJcFTL−1RC)を用いて、ゲノムDNAを鋳型にしてPCR反応を行った。PCR産物を前述と同様にクローニングして、増幅断片の塩基配列を解析した。その結果、JcFT遺伝子の第1イントロン(149bp)、第2エクソン(62bp)、第2イントロン(2575bp)、第3エクソン(41bp)、第3イントロン(108bp)の塩基配列に関する情報が得られた。
【0111】
〔実施例2.ナンヨウアブラギリにおけるJcFT遺伝子の発現解析〕
培土に播種後2ヶ月〜4ヶ月のナンヨウアブラギリの茎頂および本葉から、RNeasy Plant mini kit(キアゲン)を用いて全RNAを調製した。
【0112】
ReverTra Ace qPCR RT Kit(TOYOBO)を用いて、調製した全RNAに基づいてcDNAを合成した。7500 Fast Real−time PCR装置(Applied Biosystems)を用いて、合成されたcDNAを鋳型として、SYBR green法に基づくリアルタイムPCRを行うことにより、mRNAの定量を行った。
【0113】
JcFT遺伝子のmRNAの定量には、2種の特異的プライマー(JcFT−1F、JcFT−1RC)を用いた。内部標準としては、ナンヨウアブラギリの5.8S rRNA(GenBank Accession Number AM774639)を用いた。5.8S rRNA遺伝子のmRNAの定量には、2種の特異的プライマー(Jc5.8−1F、Jc5.8−1RC)を用いた。
【0114】
JcFT−1F :5'−GACCCTAATCTCAGAGAATACTTGCA−3'(配列番号7)
JcFT−1RC :5'−CCAAAAGTTACCCCAGTAGTTGCT−3' (配列番号8)
Jc5.8−1F :5'−CTTGGTGTGAATTGCAGAATCC−3' (配列番号9)
Jc5.8−1RC:5'−GGCTTCGGGCGCAACT−3' (配列番号10)
花芽分化の起こりやすい気象条件A(明期平均温度32℃、明期平均湿度50%、暗期平均温度26℃、暗期平均湿度80%、日長12時間明期/12時間暗期)で栽培したナンヨウアブラギリ(反復4個体)の上位10枚目の本葉および17枚目の本葉におけるJcFT遺伝子の発現は、茎頂と比較して平均20倍程度、上位3枚目の本葉と比較して平均250倍程度上昇していた(図1参照)。
【0115】
また、花芽分化の起こりやすい気象条件A(明期平均温度32℃、明期平均湿度50%、暗期平均温度26℃、暗期平均湿度80%、日長12時間明期/12時間暗期)で栽培したナンヨウアブラギリ(反復6個体)の上位10枚目の本葉におけるJcFT遺伝子の発現は、花芽分化の起こりにくい気象条件B(明期平均温度40℃、明期平均湿度10%、暗期平均温度20℃、暗期平均湿度40%、日長12時間明期/12時間暗期)で栽培したナンヨウアブラギリ(反復6個体)の上位10枚目の本葉におけるJcFT遺伝子の発現と比較して、平均500倍程度上昇していた(図2参照)。これらの発現解析により、JcFT遺伝子が高発現する葉位および気象条件を特定した。
【0116】
なお、図1は、JcFT遺伝子のmRNA量をリアルタイムPCRにより定量した結果を示す図である。当該図面では、上位3枚目における反復4個体の平均値を1として、茎頂および上位10枚目、上位17枚目における反復4個体のそれぞれの平均値を相対値で示した。また、図2は、JcFT遺伝子のmRNA量をリアルタイムPCRにより定量した結果を示す図である。当該図面では、花芽分化の起こりにくい気象条件Bで栽培した反復6個体の平均値を1として、花芽分化の起こりやすい気象条件Aで栽培した反復6個体の平均値を相対値で示した。
【0117】
〔実施例3.JcFT遺伝子の単離〕
花芽分化の起こりやすい気象条件で栽培したナンヨウアブラギリ(Jatropha curucas)の上位10枚目の本葉から、RNeasy Plant mini kit(キアゲン)を用いて全RNAを調製した。
【0118】
SMART RACE cDNA Amplification kit(クロンテック)およびPrimeScript Reverse Transcriptase(タカラバイオ)を用いて、調製した全RNAに基づいて5'−RACEおよび3'−RACE用のFirst−Strand cDNAを合成した。
【0119】
1種の特異的プライマー(JcFTL−1RC)、およびSMART RACE cDNA Amplification kit(クロンテック)付属のプライマーミックス(UPM)を用いて、5'−RACE用に合成したcDNAを鋳型として5'−RACE反応を行った。また、1種の特異的プライマー(JcFTL−1F)、およびSMART RACE cDNA Amplification kit(クロンテック)付属のプライマーミックス(UPM)を用いて、3'−RACE用に合成したcDNAを鋳型として3'−RACE反応を行った。
【0120】
各RACE反応産物を、前述と同様にそれぞれクローニングし塩基配列を解析した。その結果、JcFT遺伝子の5'非翻訳領域(71bp)、第1エクソン(204bp)、第4エクソン(224bp)、3'非翻訳領域(246bp)の5'末端および3'末端の配列情報が得られた。
【0121】
このようにして、JcFT遺伝子全長に関する情報(例えば、ゲノムDNAの塩基配列(配列番号11参照)、完全長cDNAの塩基配列(配列番号12参照)、翻訳領域の塩基配列(配列番号13参照)、タンパク質のアミノ酸配列(配列番号14参照))を取得した。
【0122】
〔実施例4.JcFT遺伝子発現カセットの構築〕
p35S−ACE1/VP16AD−CR(WO2008111661A2参照)を制限酵素HindIIIおよびEcoRIにて処理し、当該プラスミドから転写因子遺伝子発現カセットを切り出した。制限酵素HindIIIおよびEcoRIにて処理したpRI909(タカラバイオ)に前述の転写因子遺伝子発現カセットをライゲーションし、pRI−35S−ACE1/VP16AD−CRを得た。
【0123】
pMRE4/35S(−46)−To71sGFP(WO2008111661A2参照)を制限酵素XbaIおよびSacIにて処理し、当該プラスミドからTo71sGFP遺伝子を切り出した。制限酵素XbaIおよびSacIにて処理したpRI−35S−ACE1/VP16AD−CRにTo71sGFP遺伝子をライゲーションし、pRI−35S−To71sGFP−CRを得た。
【0124】
また、ナンヨウアブラギリ(Jatropha curucas)の本葉から、RNeasy Plant mini kit(キアゲン)を用いて全RNAを調製した。ReverTra Ace qPCR RT kit(TOYOBO)を用いて、調製した全RNAに基づいてcDNAを合成した。合成したcDNAを鋳型として、2種類の特異的プライマー(BamJFT−1FおよびSpeJFT−1RC)を用いてPCR反応を行った。
【0125】
BamJFT−1F :5'−ATGGATCCAACAATGCCTAGGGATCAATTTAGAGACC−3'(配列番号15)
SpeJFT−1RC:5'−ATACTAGTTCACCGTCTCCGTCCTCCGGTG−3'(配列番号16)
PCR産物をBlunting Kination Ligation kit(タカラバイオ)を用いて平滑末端化およびリン酸化した後、当該PCR産物を制限酵素BamHIにて処理した。制限酵素SacIで処理した後、Blunting Kination Ligation kitを用いて平滑末端化およびリン酸化し、更に制限酵素BamHIで処理したpRI−35S−To71sGFP−CRに、前述のPCR産物をライゲーションし、pRI−35S−To71JcFT−CRを得た。
【0126】
〔実施例5.組換えシロイヌナズナにおけるJcFT遺伝子の機能評価〕
実施例4で作製されたpRI−35S−To71sGFP−CRおよびpRI−35S−To71JcFT−CRをアグロバクテリウム(Agrobacterium tumefaciens strain C58C1)に導入した。当該アグロバクテリウムを50mg/Lカナマイシン、100mg/Lアンピシリン、100mg/Lリファンピシンを含むLB寒天培地(0.5%酵母エキス、1.0%バクトトリプトン、0.5%食塩、1%寒天)で培養し、薬剤耐性コロニーを選抜することにより、組換えアグロバクテリウムを得た。
【0127】
得られた組換えアグロバクテリウムを、「モデル植物ラボマニュアル」(岩渕雅樹他編集、2000年、シュプリンガー・フェアラーク東京株式会社、ISBN 4-431-70881-2 C3045)に記載されている方法に準じて、シロイヌナズナ(Arabidopsis thaliana ecotype Columbia)に感染させることにより、シロイヌナズナへの遺伝子導入を行った。
【0128】
遺伝子導入されたシロイヌナズナから採取されたT1種子を、20mg/Lベンレート、200mg/Lクラフォラン、25mg/Lカナマイシンを含む改変MS寒天培地(MS無機塩類、B5ビタミン、1%ショ糖、0.8%寒天)に播種して生育させた後、カナマイシンに耐性を示す植物個体を選抜した。pRI−35S−To71JcFT−CRが導入されている組換えシロイヌナズナは、播種後30日目に寒天培地上で開花が認められた(図3参照)。
【0129】
選抜された植物個体を予め培土が入れられたポットに移植し、人工気象器内で生育させることにより、T2種子を得た。得られたT2種子を25mg/Lカナマイシンを含む改変MS寒天培地(MS無機塩類、B5ビタミン、2%ショ糖、0.8%寒天)に播種して生育させた後、χ2検定に基づいた5%の有意水準において、カナマイシンに耐性を示す植物個体が3:1の割合で出現するラインを選抜した。尚、植物個体を生育させるときの培養条件は、明期23時間、暗期1時間、23〜25℃とした。
【0130】
選抜されたラインについて、T2種子を、25mg/Lの濃度にてカナマイシンを含む改変MS寒天培地(MS無機塩類、B5ビタミン、2%ショ糖、0.8%寒天)に播種した。その結果、pRI−35S−To71JcFT−CRが導入されている組換えシロイヌナズナは、pRI−35S−To71sGFP−CRが導入されている組換えシロイヌナズナと比較して早期に開花が誘導された。
【0131】
播種後25日目に、開花時期の指標となるロゼット葉の数をカウントした。その結果、pRI−35S−To71JcFT−CRが導入されている組換えシロイヌナズナのロゼット葉は4.3±0.6枚(n=24)であるのに対して、pRI−35S−To71sGFP−CRが導入されている組換えシロイヌナズナのロゼット葉は5.5±0.7枚(n=39)であった。培養条件は、明期12時間、暗期12時間、23〜25℃とした。
【0132】
〔実施例6.組換えイネにおけるJcFT遺伝子の機能評価〕
pRI909(タカラバイオ)のカナマイシン耐性NPTII遺伝子をハイグロマイシン耐性APH4遺伝子に置換したpRH909を作製した。
【0133】
pRH909に、2x35Sプロモーター(Liu et al(2002)Plant J. 30、415-429)および翻訳エンハンサー配列fai(Mori et al(2006)Plant Biotech. 23、55-61)、JcFT遺伝子、CRターミネーターを挿入し、pRH−2x35S−faiJcFT−CRを得た。得られたpRH−2x35S−faiJcFT−CRをアグロバクテリウム(Agrobacterium tumefaciens strain LBA4404)に導入した。当該アグロバクテリウムを50mg/Lカナマイシン、100mg/Lストレプトマイシンを含むLB寒天培地(0.5%酵母エキス、1.0%バクトトリプトン、0.5%食塩、1%寒天)で培養し、薬剤耐性コロニーを選抜することにより、組換えアグロバクテリウムを得た。
【0134】
得られた組換えアグロバクテリウムを、Toki et al (2006)Plant J. 47、969-976に記載されている方法に準じて、イネ(Oryza sativa subsp. japonica cv. Nipponbare)に感染させることにより、イネへの遺伝子導入を行った。ハイグロマイシンに耐性を示す再生個体を取得した。pRH−2x35S−faiJcFT−CRが導入されている組換えイネは早期に花芽が形成された(図4参照)。尚、植物個体を生育させるときの培養条件は、明期23時間、暗期1時間、23〜25℃とした。
【0135】
なお本発明は、以上説示した各構成に限定されるものではなく、特許請求の範囲に示した範囲で種々の変更が可能であり、異なる実施形態や実施例にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態や実施例についても本発明の技術的範囲に含まれる。
【産業上の利用可能性】
【0136】
本発明は、開花時期が制御された植物を製造する目的に使用することができる。
【特許請求の範囲】
【請求項1】
下記(1)〜(3)の何れかのポリペプチドをコードすることを特徴とするポリヌクレオチド。
(1)配列番号14で示されるポリペプチド
(2)配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド
(3)配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド
【請求項2】
下記(4)〜(10)の何れかであることを特徴とするポリヌクレオチド。
(4)配列番号11、12または13で示されるポリヌクレオチド
(5)配列番号11、12または13で示されるポリヌクレオチドのアンチセンス配列をコードするポリヌクレオチド
(6)配列番号17および配列番号18に示すプライマーセットによって増幅されるポリヌクレオチドを含むポリヌクレオチド
(7)配列番号19および配列番号20に示すプライマーセットによって増幅されるポリヌクレオチドを含むポリヌクレオチド
(8)上記(4)〜(7)の何れかに記載されるポリヌクレオチドにおいて、1つ以上30以下の塩基が置換、欠失、挿入および/または付加されたポリヌクレオチドからなり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド
(9)上記(4)〜(7)の何れかに記載されるポリヌクレオチドと相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド
(10)上記(4)〜(7)の何れかに記載されるポリヌクレオチドとの相同性が84%以上であり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド
【請求項3】
バラ下綱植物由来であることを特徴とする請求項1に記載のポリヌクレオチド。
【請求項4】
請求項1〜3の何れか1項に記載のポリヌクレオチドを含有することを特徴とする組換えベクター。
【請求項5】
上記組換えベクターが、プラスミドベクター、ウイルスベクター、ファージベクターまたはコスミドベクターであることを特徴とする請求項4に記載の組換えベクター。
【請求項6】
下記(11)〜(13)の何れかのポリペプチド。
(11)配列番号14で示されるポリペプチド
(12)配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド
(13)配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド
【請求項7】
請求項1〜3の何れか1項に記載のポリヌクレオチド、請求項4若しくは5に記載の組換えベクター、または、請求項6に記載のポリペプチドを植物に導入する工程を含むことを特徴とする植物の開花時期の制御方法。
【請求項8】
請求項1〜3の何れか1項に記載のポリヌクレオチド、請求項4若しくは5に記載の組換えベクター、または、請求項6に記載のポリペプチドを植物に導入する工程を含むことを特徴とする形質転換植物の製造方法。
【請求項9】
請求項1〜3の何れか1項に記載のポリヌクレオチド、請求項4若しくは5に記載の組換えベクター、または、請求項6に記載のポリペプチドが導入されていることを特徴とする形質転換植物。
【請求項1】
下記(1)〜(3)の何れかのポリペプチドをコードすることを特徴とするポリヌクレオチド。
(1)配列番号14で示されるポリペプチド
(2)配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド
(3)配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド
【請求項2】
下記(4)〜(10)の何れかであることを特徴とするポリヌクレオチド。
(4)配列番号11、12または13で示されるポリヌクレオチド
(5)配列番号11、12または13で示されるポリヌクレオチドのアンチセンス配列をコードするポリヌクレオチド
(6)配列番号17および配列番号18に示すプライマーセットによって増幅されるポリヌクレオチドを含むポリヌクレオチド
(7)配列番号19および配列番号20に示すプライマーセットによって増幅されるポリヌクレオチドを含むポリヌクレオチド
(8)上記(4)〜(7)の何れかに記載されるポリヌクレオチドにおいて、1つ以上30以下の塩基が置換、欠失、挿入および/または付加されたポリヌクレオチドからなり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド
(9)上記(4)〜(7)の何れかに記載されるポリヌクレオチドと相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド
(10)上記(4)〜(7)の何れかに記載されるポリヌクレオチドとの相同性が84%以上であり、且つ、植物の開花時期を制御する機能を有するポリヌクレオチド
【請求項3】
バラ下綱植物由来であることを特徴とする請求項1に記載のポリヌクレオチド。
【請求項4】
請求項1〜3の何れか1項に記載のポリヌクレオチドを含有することを特徴とする組換えベクター。
【請求項5】
上記組換えベクターが、プラスミドベクター、ウイルスベクター、ファージベクターまたはコスミドベクターであることを特徴とする請求項4に記載の組換えベクター。
【請求項6】
下記(11)〜(13)の何れかのポリペプチド。
(11)配列番号14で示されるポリペプチド
(12)配列番号14で示されるポリペプチドにおいて、1または数個のアミノ酸残基が置換、欠失、挿入および/または付加されたポリペプチドからなり、且つ、植物の開花時期を制御する機能を有するポリペプチド
(13)配列番号14で示されるポリペプチドとの相同性が90%以上であって、且つ、植物の開花時期を制御する機能を有するポリペプチド
【請求項7】
請求項1〜3の何れか1項に記載のポリヌクレオチド、請求項4若しくは5に記載の組換えベクター、または、請求項6に記載のポリペプチドを植物に導入する工程を含むことを特徴とする植物の開花時期の制御方法。
【請求項8】
請求項1〜3の何れか1項に記載のポリヌクレオチド、請求項4若しくは5に記載の組換えベクター、または、請求項6に記載のポリペプチドを植物に導入する工程を含むことを特徴とする形質転換植物の製造方法。
【請求項9】
請求項1〜3の何れか1項に記載のポリヌクレオチド、請求項4若しくは5に記載の組換えベクター、または、請求項6に記載のポリペプチドが導入されていることを特徴とする形質転換植物。
【図1】


【図2】
【図3】

【図4】
【図2】
【図3】
【図4】
【公開番号】特開2011−67192(P2011−67192A)
【公開日】平成23年4月7日(2011.4.7)
【国際特許分類】
【出願番号】特願2010−54992(P2010−54992)
【出願日】平成22年3月11日(2010.3.11)
【出願人】(000002093)住友化学株式会社 (8,981)
【Fターム(参考)】
【公開日】平成23年4月7日(2011.4.7)
【国際特許分類】
【出願日】平成22年3月11日(2010.3.11)
【出願人】(000002093)住友化学株式会社 (8,981)
【Fターム(参考)】
[ Back to top ]