
植物への病原ウイルス感染を防除する組換えALSV
【課題】植物病原性ウイルスを対象として、それぞれに干渉効果を発揮する組換えウイルスを作製でき、また増殖の過程での強毒化の危険性のない、新しい植物病原ウイルス感染防除手段を提供する。
【解決手段】1種または2種以上の植物病原ウイルスのゲノムRNAまたはそのcDNAの一部断片を保有する組換えリンゴ小球型潜在ウイルス(ALSV)。および該組換えウイルスを幼苗に接種する病原ウイルス感染防除方法と、接種された抵抗性植物。
【解決手段】1種または2種以上の植物病原ウイルスのゲノムRNAまたはそのcDNAの一部断片を保有する組換えリンゴ小球型潜在ウイルス(ALSV)。および該組換えウイルスを幼苗に接種する病原ウイルス感染防除方法と、接種された抵抗性植物。
【発明の詳細な説明】
【技術分野】
【0001】
本願発明は、農作物等の植物への病原類する感染に対する防除剤の成分等として有用な組換えALSVに関するものである。
【背景技術】
【0002】
1.リンゴ小球形潜在ウイルス
リンゴ小球形潜在ウイルス(Apple latent spherical virus:ALSV)は2分節1本鎖RNAゲノム(RNA1、RNA2)と3種類の外被タンパク質(Vp25、Vp20、Vp24)からなる径25nmの球形ウイルスである(非特許文献1)。ALSVはリンゴから分離されたウイルスであるが、比較的広い宿主範囲を持つウイルスで、Chenopodium quinoa(キノア)には退緑症状を示すが、モデル植物のシロイヌナズナをはじめ、Nicotiana属植物(タバコ、ベンサミアナタバコ、オキシデンタリス、グルチノーザ)、トマト、ウリ科植物(キュウリ、メロン、ズッキーニ、ヘチマ)、マメ科植物(ダイズ、アズキ、エンドウ)、バラ科果樹植物(リンゴ、セイヨウナシ、ニホンナシ、モモ)など多くの植物種に無病徴感染する。本願発明者らは既に、ALSVの感染性cDNAクローンを構築し、RNA2の細胞間移行タンパク質(movement protein:MP)とVp25の間に外来遺伝子導入サイトを付加したALSVベクターによる草本植物やリンゴでの外来遺伝子の発現に成功している(例えば、非特許文献2)。また、GFP(green fluorescent protein)を発現するタバコにGFPを組み込んだALSVベクターを接種すると、タバコで発現しているGFPのサイレンシングが誘導され、接種葉ではGFP蛍光が円状に消失し、やがて上位葉全体でGFP蛍光が消失することが明らかにされた(例えば、非特許文献3)。さらに、植物の内在性遺伝子であるphytoene desaturase(PDS)遺伝子やsulfer(SU)遺伝子の一部を組み込んだALSV感染により、タバコ、シロイヌナズナ、ダイズ、キュウリなどにおいてウイルス誘導ジーンサイレンシング(virus induced gene silencing:VIGS)が誘導され、ALSVが感染植物において全身的なVIGSを誘導することが明らかにされた(例えば、非特許文献4)。
【0003】
2.RNAサイレンシング
RNAサイレンシングは1990年に植物で初めて報告された真核生物に普遍的に保存されている配列特異的な遺伝子不活化機構であり(非特許文献5)、主にdouble-stranded RNA(dsRNA)により誘導される。RNAウイルスが感染すると、RNA依存RNAポリメラーゼ(RdRp)の働きでdsRNAが合成され、これが2本鎖RNA分解酵素であるDicer(植物の場合Dicer-like:DCL)によって21から25 塩基(nucleotide:nt)のsmall interfering RNA(siRNA)に分解される(非特許文献6、7)。siRNAはRNA-induced silencing complex(RISC)に取り込まれ、siRNAと相補的な塩基配列のRNAはRISCにより分解される。また、siRNAは隣接する細胞へ移行し、RdRpの働きによってsiRNAと相補的なRNAがdsRNAに変換され、サイレンシングは全身的に拡大する。免疫機構を持たない植物において、RNAサイレンシングはウイルスに対する防御機構として重要な役割を持っている(非特許文献8)。1995年、植物内在性遺伝子を組み込んだTMV感染によりその遺伝子のRNAサイレンシングが誘導されることが報告され(非特許文献9)、その後VIGSは逆遺伝学的手法として広く利用されている。
【0004】
3.植物の病原ウイルスとその対策
近年、ズッキーニ黄斑モザイクウイルス(Zucchini yellow mosaic virus:ZYMV)の感染によるウリ科作物の被害が世界で問題となり、日本国内においてもキュウリにおける被害が深刻化している。ZYMVはPotyvirus属に属し、サイズが約9,600ntの1本鎖RNAゲノムを持つ、長さ約750nmのひも状ウイルスである。アブラムシによる非永続伝搬によって蔓延し、特にウリ科植物(キュウリ、カボチャ、メロン、スイカなど)において激しいモザイク、葉および果実の奇形、植物体の萎縮等の症状を引き起こすことが知られている(非特許文献10、11)。
【0005】
キュウリモザイク病はZYMVと、Potyvirus属のカボチャモザイクウイルス(Watermelon mosaic virus 2:WMV2)およびCucumovirus属のキュウリモザイクウイルス(Cucumber mosaic virus:CMV)の3種が病原ウイルスとされ、これらウイルスの単独および重複感染によって発生する(非特許文献11)。CMVはCucumovirus属のタイプ種で、3分節の1本鎖RNAゲノム(RNA1、RNA2、RNA3)およびサブゲノムRNA(RNA4)を持つ径29nmの球状ウイルスである。宿主や病原性の異なる系統が多数存在し、なかにはサテライトRNAを有するものも存在する。CMVはアブラムシによって非永続伝搬され、その宿主範囲は極めて広く1,000種以上の植物に感染する(非特許文献10)。
【0006】
ZYMVおよびCMVによる農作物の被害に対し、抵抗性品種の利用やアブラムシ防除に加えて、病原性の弱いZYMVやCMV(弱毒ZYMV、弱毒CMV)を用いた干渉効果を利用した防除が行われている(非特許文献12、13)。干渉効果は、あるウイルスに感染した植物はその後同種のウイルスに感染しない現象で、その発見は古く1929年にタバコモザイクウイルスで最初に報告された。干渉効果の機構については、ウイルスの外被タンパク質(coat protein:CP)を介する干渉効果とRNAを介する干渉効果の2つが主なものと考えられている(非特許文献14、15)。
【0007】
ZYMVやCMVに対する防除手段としての弱毒ウイルスに関する発明としては、特許文献1、2が知られており、特許文献3には、同一ウイルスグループ内の2種以上の弱毒ウイルスを混合した混合弱毒型ウイルスが開示されている。また、ウイルスのサテライトRNAを当該ウイルスに組込んで弱毒ウイルスとする技術として特許文献4、5が知られている。さらに市販品として、日本では、生物農薬として製剤化された弱毒ZYMVが販売され、キュウリで利用されている(非特許文献7、12)。また、トマトではサテライトRNAを有する弱毒CMVを感染させた苗が販売されている(非特許文献16)。
【0008】
ただし、病原ウイルスと異なるベクターウイルスに病原ウイルス遺伝子を組込んだ組換えウイルスを病原ウイルス感染の防除手段として用いる技術は全く知られていない。
【特許文献1】特開平5-68540号公報
【特許文献2】特開平10-203901号公報
【特許文献3】特開2000-264806号公報
【特許文献4】特開平5-3789号公報
【特許文献5】特開平11-279013号公報
【非特許文献1】Li,C., Yoshikawa,N., Takahashi,T., Ito,T., Yoshida,K. and Koganezawa,H. (2000). Nucleotide sequence and genome organization of apple latent spherical virus:a new virus classified into the family Comoviridae. J.Gen.Virol.81:541-547.
【非特許文献2】Li,C., Sasaki,N., Isogai,M. and Yoshikawa,N. (2004). Stable expression of foreign proteins in herbaceous and apple plants using Apple latent spherical virus RNA2 vectors. Arch. Virol. 149:1541-1558.
【非特許文献3】Yaegashi,H., Yamatsuta,T., Takahashi,T., Li,C., Isogai,M., Kobori,T., Ohki,S. and Yoshikawa,N. (2007). Characterization of virus-induced gene silencing in tobacco plants infected with apple latent spherical virus. Arch. Virol. 152:1839-1849.
【非特許文献4】Igarasi,A., Yamagata,K., Sugai,T., Takahashi,Y., Sugawara,E., Tamura,A., Yaegashi,H., Yamagishi,N., Takahashi,T., Isogai,M., Takahashi,H. and Yoshikawa,N. (2009). Apple latent spherical virus vectors for reliable and effective virus-induced gene silencing among abroad range of plants including tobacco, tomato, Arabidopsis thaliana, cucurbits, and legumes. Virology386:407-416.
【非特許文献6】Hamiltom,A., Voinnet,O., Chappell,L. and Baulcombe,D. (2002). Two classes of short interfering RNA in RNA silencing. EMBO. J.21:4671-4679.
【非特許文献7】Kosaka,Y. and Fukunishi,T. (1997). Multiple Inoculation with Three Attenuated Viruses for the Control of Cucumber Virus Disease. Plant Dis.81:733-738.
【非特許文献8】Voinnet O. (2001). RNA silencing as a plant immune system against viruses. Trends Genet. 17:449-459.
【非特許文献9】Kumagai,M.H., Donson,J., Della-Cioppa,G., Harvey,D., Hanley,K. and Grill,L.K. (1995). Cytoplasmic inhibition of carotenoid biosynthesis with virus-derived RNA. Proc. Natl. Acad. Sci. USA 92:1679-1683.
【非特許文献10】畑中正一編 (1997). ウイルス学. 朝倉書店. pp.427-434, 461-466.
【非特許文献11】岸國平編. (1988). 作物病害事典. 全国農村教育協会. pp.321-322
【非特許文献12】Kosaka,Y., Ryang,Bo-Song., Kobori,T., Shiomi,H., Yasuhara,H. and Kataoka,M. (2006). Effectiveness of an Attenuated Zucchini yellow mosaic virus Isolate for Cross-Protecting Cucumber. Plant Dis. 90:67-72.
【非特許文献13】Wang,Wei-Qin., Natsuaki,T. and Kosaka,Y. (2006). Comparison of the nucleotide and amino acid sequences of parental and attenuated isolates of Zucchini yellow mosaic virus. J. Gen. Plant Pathol. 72:52-56.
【非特許文献14】Beachy,R.N. (1999). Coat-protein-mediated resistance to tobacco mosaic virus:discovery mechanisms and exploitation. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 354:659-664.
【非特許文献15】Lin,S-S., Henriques,R., Wu,H-W., Niu,Q-W., Yen,S-D. and Chua,N-H. (2007). Strategies and mechanisms of plant virus resistance. Plant Biotechnol. Rep. 1:125-134.
【非特許文献16】津田和久, 小坂能尚, 小堀崇, 塩見寛, 六角啓一, 片岡光信. (2005). キュウリモザイクウイルス弱毒株CM95を接種したトマトの収量及びビタミンC含量に及ぼす肥培管理の影響. 日植病報 71:1-5.
【発明の概要】
【発明が解決しようとする課題】
【0009】
前記のとおり、植物の病原ウイルスの感染に対して、弱毒ウイルスの干渉効果を利用した防除が知られているが、弱毒ウイルスを用いた防除法は、弱毒ウイルスが利用できる一部のウイルスに限られており、弱毒ウイルスの自然界からの分離や変異誘導による作製が困難なウイルスには応用の難しい技術である。また、異なるウイルスに対して、弱毒ウイルスによる干渉効果は発揮されないため、各ウイルスごとに弱毒ウイルスが必要とされているが、弱毒ウイルスの作製には長い時間と多くの労力を必要とする。
【0010】
またサテライトRNAの挿入による弱毒ウイルスの場合には、親株ウイルスは病原性であり、植物体内での増殖の過程で強毒ウイルスに回帰する危険性は皆無ではない。
【0011】
本願発明は、以上のとおりの事情に鑑みてなされたものであり、理論的にはあらゆる病原性ウイルスを対象として、それぞれに干渉効果を発揮するウイルスを作製でき、また増殖の過程での強毒化の危険性のない新しい手段を提供することを課題としている。
【課題を解決するための手段】
【0012】
前記課題を解決するための発明として、1種または2種以上の植物病原ウイルスのゲノムRNAまたはそのcDNAの一部断片を保有する組換えリンゴ小球型潜在ウイルス(ALSV)を提供する。
【0013】
この組換えALSVにおいては、植物病原ウイルスがズッキーニ黄斑モザイクウイルス(ZYMV)、キュウリモザイクウイルス(CMV)、またはダイズモザイクウイルス(SMV)であることを好ましい態様としている。
【0014】
さらにこの出願は、前記の組換えALSVを植物の幼苗に感染させることを特徴とする植物病原ウイルスの防除方法と、前記の組換えALSVが幼苗期に接種された病原ウイルス抵抗性植物を提供する。
【発明の効果】
【0015】
本願発明では、植物病原ウイルスのゲノムRNAまたはそのcDNAの一部断片を組込んだ組換えALSVによって当該病原ウイルスの感染を防除する。対象となる病原ウイルスに制限はなく、あらゆる病原ウイルスの感染を防除することができる組換えALSVの作製が理論的には可能である。組換えALSVの作成は、当該技術分野の通常の知識を有するものであれば、本願明細書の記載に基づき容易に行なうことができ、従来の弱毒ウイルスに比して、極めて少ない時間と労力で作成することができる。
【0016】
また、ALSVは無病徴感染ウイルスであり、このALSVに挿入する病原ウイルスのゲノムRNAまたはそのcDNAの一部断片は、それ自体では病原性を持たないものであるため、組換えALSVが植物体内で毒性を獲得する危険性はなく、安全に使用することができる。
【図面の簡単な説明】
【0017】
【図1】ALSVの感染性cDNAクローンとALSVベクターの構成を示す。pEALSR1:ALSV RNA1の感染性cDNAクローン、pEALSR2L5R5:ALSV RNA2の感染性cDNAクローン、P35S:35S promoter、Tnos:nopaline synthase terminator、Q/G:タンパク質の切断部位、PRO-co:protease cofactor、HEL:NTP-binding helicase、C-PRO:cystein protease、POL:RNA polymerase、MP:movement protein、VP25, VP20, VP24:capsid protein。
【図2】実施例1においてZYMV遺伝子断片を導入したALSVベクターの構築工程を示す。
【図3】実施例1においてリーフディスクを採種した場所を示す。
【図4】実施例1の結果を示す。第1本葉二次接種区のキュウリ(品種:青大胡瓜)のZYMVによる病徴(21dpi)である。写真左上:(A)第5本葉、写真左下:(B)第6本葉、写真右:(C)植物全体。(A)(B)は左上から右下の順に、(C)は左から順に、それぞれ、1:非接種区、2:ALSVのみ区、3:ZYMVのみ区、4:wtALSV+ZYMV区、5:ALSV-Z:CP200+ZYMV区。
【図5】実施例1の結果を示す。ZYMVのみ区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図6】実施例1の結果を示す。wtALSV+ZYMV区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図7】実施例1の結果を示す。ALSV-Z:CP200+ZYMV区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図8】実施例1の結果である。第1本葉二次接種区のキュウリ(品種:青大胡瓜)におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図9】実施例1の結果である。第2本葉二次接種区のキュウリ(品種:青大胡瓜)におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図10】実施例1の結果である。第3本葉二次接種区のキュウリ(品種:青大胡瓜)におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図11】実施例1の結果である。異なるZYMV遺伝子断片を導入したALSVベクターを一次接種したキュウリ(品種:青大胡瓜)の各試験区における病徴(21dpi)である。写真左上:(A)第5本葉、写真左下:(B)第6本葉、写真右:(C)植物全体。(A)(B)は左上から右下の順に、(C)は左から順に、それぞれ、1:非接種区、2:ALSVのみ区、3:ZYMVのみ区、4:wtALSV+ZYMV区、5:ALSV-Z:P1+ZYMV区、6:ALSV-Z:P3+ZYMV区、7:ALSV-Z:CP200+ZYMV区。
【図12】実施例1の結果を示す。ALSV-Z:P3+ZYMV区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図13】実施例1の結果を示す。ALSV-Z:P1+ZYMV区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図14】実施例1の結果である。異なるZYMV遺伝子断片を導入したALSVベクターを一次接種したキュウリ(品種:青大胡瓜)の第1本葉二次接種区におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図15】実施例1の結果である。キュウリ(品種:鈴成四葉)の第1本葉二次接種区におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図16】実施例1の結果である。キュウリ(品種:つばさ)の第1本葉二次接種区におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図17】実施例1の結果である。長さの異なるZYMV遺伝子断片を導入したALSVベクターを一次接種したキュウリ(品種:青大胡瓜)の各試験区における病徴(21dpi)である。写真左上:(A)第5本葉、写真左下:(B)第6本葉、写真右:(C)植物全体。(A)(B)は左上から右下の順に、(C)は左から順に、それぞれ、1:非接種区、2:ALSVのみ区、3:ZYMVのみ区、4:wtALSV+ZYMV区、5:ALSV-Z:CP100+ZYMV区、6: ALSV-Z:CP200+ZYMV区。
【図18】実施例1の結果を示す。ALSV-Z:CP100+ZYMV区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図19】実施例1の結果である。長さの異なるZYMV遺伝子断片を導入したALSVベクターを一次接種したキュウリ(品種:青大胡瓜)の第1本葉二次接種区におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図20】実施例1の結果である。弱毒ZYMVおよび異なるZYMV遺伝子断片を導入した各種ALSVを一次接種したキュウリ(品種:青大胡瓜)の各試験区における病徴(21dpi)である。(A)第5本葉、(B)植物全体。(A)(B)は左から順に、それぞれ、1:非接種区、2:ALSVのみ区、3:ZYMVのみ区、4:wtALSV+ZYMV区、5:ALSV-Z:P1+ZYMV区、6:ALSV-Z:P3+ZYMV区、7:ALSV-Z:CP200+ZYMV区、8:ZYMV 2002+ZYMV区。
【図21】実施例1の結果を示す。ZYMV 2002 +ZYMV区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図22】実施例1の結果である。弱毒ZYMVおよび異なるZYMV遺伝子断片を導入した各種ALSVを一次接種したキュウリ(品種:青大胡瓜)の第1本葉二次接種区におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図23】実施例2におけるCMV-Y遺伝子のクローニングとCMV-Y遺伝子断片の構築工程。
【図24】実施例2におけるCMV-Y遺伝子断片を導入したALSVベクターの構築工程。
【図25】実施例2の結果である。各CMV-Y遺伝子断片導入ALSVベクターを一次接種したタバコにおけるCMV-Yの病徴(9dpi)。左上から右下の順に、非接種区、CMV-Yのみ区、ALSVのみ区、wtALSV+CMV-Y区、ALSV-C:1a+CMV-Y区、ALSV-C:2a+CMV-Y区、ALSV-C:CP+CMV-Y区。
【図26】実施例2の結果である。異なるCMV-Y遺伝子断片を導入した各種ALSVを一次接種したタバコのCMV-YおよびALSV蓄積量を直接ELISA法により解析した結果。
【図27】実施例2の結果である。各CMV-Y遺伝子断片導入ALSVベクターを一次接種したキュウリ(品種:青大胡瓜)におけるCMV-Yの病徴(18dpi)。(A)は植物全体、(B)は第6本葉、(A)は左から順に、(B)は左上から右下の順に、1:非接種区、2:ALSVのみ区、3:CMV-42CMのみ区、4:wtALSV+CMV-42CM区、5:ALSV-C:1a+CMV-42CM区、6:ALSV-C:2a+CMV-42CM区、7:ALSV-C:CP+CMV-42CM区。
【図28】実施例2の結果である。各CMV-Y遺伝子断片導入ALSVベクターを一次接種したキュウリ(品種:青大胡瓜)第1本葉二次接種区におけるCMV-42CMおよびALSV蓄積量を直接ELISA法により解析した結果。
【発明を実施するための形態】
【0018】
本願発明の組換えALSVは、植物病原ウイルスのゲノムRNAまたはそのcDNAの一部断片が挿入されていることを特徴とする。RNAサイレンシングの原理に従い、この組換えALSVが複製すると、挿入断片の配列を含むsiRNAが合成され、それに相補的な病原ウイルスゲノムの領域を破壊する。
【0019】
ALSVは、リンゴから単離したものを使用することもできるが、好ましくは、特許文献2に記載されているALSVのRNA1感染性cDNAクローン(pEALSR1)、RNA2感染性cDNAクローン(pEALSR2L5R5)などのALSVベクターを使用することができる(図1参照)。
【0020】
植物病原ウイルスとしては、実施例に示したキュウリモザイクウイルス(CMV)、ズッキーニ黄斑モザイクウイルス(ZYMV)、ダイズモザイクウイルス(SMV)の他、原則的にあらゆるウイルスを対象とすることができる。例えば、CMVやZYMVと同様にアブラムシ類により媒介されるカボチャモザイクウイルス(WMV-2)、ソラマメウイルトウイルス(BBWV)、カブモザイクウイルス(TuMV)など;アザミウマ類により媒介されるトマト黄化えそウイルス(TSWV)、インパチエンスネクロティックスポットウイルス(INSV)、アイリスイエロースポットウイルス(IYSV)など;コナジラミ類により媒介されるキュウリ黄化ウイルス(CuYV)など;土壌伝染するトマトモザイクウイルス(ToMV)などである。
【0021】
本願発明の組換えALSVは、これらの病原ウイルスの1種または2種以上(例えば3種)の各々のゲノムRNAまたはcDNAの一部断片を組合わせて(例えば、融合ポリヌクレオチドとして)ALSVベクターに挿入したものであってもよい。このような2種以上の病原ウイルスに対する組換えALSVは、いわゆる混合ワクチン様の機能を有し、同一植物個体への2種以上の病原ウイルスの感染を防御することができる。例えば、実施例に示したような、キュウリに対するズッキーニ黄斑モザイクウイルス(ZYMV)とキュウリモザイクウイルス(CMV)の感染防御である。
【0022】
ALSVへの挿入断片は、これらの病原ウイルスのゲノムRNA、もしくはそのcDNAから調製する。挿入断片は病原ウイルスゲノムの任意の領域から選択することができるが、好ましくは、病原ウイルスの生存、増殖、あるいはその病原性に関わる遺伝子領域等から選択することができる。例えば、実施例に示したように、ズッキーニ黄斑モザイクウイルス(ZYMV)の場合はP1、HC-Pro、P3、Cl、Nla、Nlb、CP(好ましくは、P3、Cl、Nla、Nlb、CP)、キュウリモザイクウイルス(CMV)の場合は1a、2a、CP(好ましくは2a)、ダイズモザイクウイルス(SMV)の場合はCPをそれぞれコードするゲノムRNAの一部領域またはこの領域に対応するcDNAである。
【0023】
挿入断片のサイズは、例えば、ALSVベクターに挿入した場合に、その植物個体への感染能、あるいは植物個体内での増殖能に影響を及ぼすことのないサイズの範囲内で任意に選択することができる。また、挿入断片のサイズは、挿入断片の配列を含むsiRNAが病原ウイルスゲノムを効果的に破壊し、かつその挿入断片が安定的に維持される範囲を選択する。すなわち、具体的には、挿入断片のサイズは例えば100〜300bp、好ましくは150〜200bp程度とすることができる。
【0024】
なお、この挿入断片がコードするペプチドが、ALSVペプチドとの融合ペプチドとして発現するように(すなわち、一部断片の挿入によって停止コドンが出現しないように)、フレームを一致させることが必要である。ALSVベクターを使用する場合、そのクローニング部位に一致した制限酵素で前記の一部断片を調製し、市販のライゲーションキット等を用いて一部断片をALSVベクターに挿入する。次いで、この組換えALSVベクターをキノアに感染させ、感染葉からウイルスを単離して組換えALSVを作成する。
【0025】
この組換えALSVは、そのまま植物に接種して病原ウイルスの感染防除に使用することができる。あるいは、安定剤等を含む緩衝液中で凍結乾燥させて感染防除剤として製品化することもできる(例えば、特許文献2など)。
【0026】
組換えALSV、または溶解した感染防除剤は、植物の幼苗の葉(例えば、キュウリの場合は子葉展開期の子葉、タバコの場合は5葉期の第3から第5本葉など)にカーボランダム法など公知の方法で接種することができる。
【0027】
以下、実施例を示して本願発明をさらに詳細かつ具体的に説明するが、本願発明は以下の例によって限定されるものではない。
【実施例1】
【0028】
ZYMVの遺伝子断片を導入したALSVベクターによる干渉効果
1:材料と方法
1-1:実験材料
1-1-1:供試植物
Chenopodium quinoa(以下キノア)をALSV増殖用植物として用いた。また干渉効果の検定にはCucumis sativus(以下キュウリ)のZYMV抵抗性を持たない3品種(青大胡瓜、鈴成四葉、つばさ)を供試した。
1-1-2.供試感染性クローン
供試感染性クローンは、非特許文献2等に記載されているALSVのRNA1感染性cDNAクローン(pEALSR1)とRNA2感染性cDNAクローン(pEALSR2L5R5)(図1)、および宇都宮大学農学部教授夏秋知英博士より分譲された、強毒株ZYMV isolate Z5-1(accession number:AB188115)の感染性cDNAクローン(p35S-Z5vector+GFP-FL)を供試した。また、(株)微生物化学研究所梁宝成博士より分譲された、弱毒株のZYMV isolate 2002(以下ZYMV 2002、accession number:AB188116)を供試した。
1-2:ZYMV遺伝子断片のALSVベクターへの導入
1-2-1:ZYMV遺伝子断片のクローニング
p35S-Z5vector+GFP-FLを鋳型とし、既知のZYMV isolate Z5-1のP1遺伝子、P3遺伝子およびCP遺伝子配列をもとに設計したプライマーを用いて、P1遺伝子(210 nt)の増幅にはZ:P1-Xho(+)とZ:P1-Bam(-)、P3遺伝子の増幅(189 nt)にはZ:P3-Xho(+)とZ:P3-Bam(-)、CP遺伝子の増幅(225 nt)にはZ:CP-Xho(+)とZ:CP-Bam(-)、CP遺伝子(117 nt)の増幅にはZ:CP-Xho(+)とZ:CP100-Bam(-)の組み合わせを用いた(表1:配列番号1−7)。
【0029】
【表1】

【0030】
2μlの鋳型DNA溶液(1ng/μl)を0.2ml容PCRチューブにとり、滅菌水を35μl、10×Ex TaqBuffer(TaKaRa)を5μl、2.5mM dNTP Mixture(TaKaRa)を5μl、10μMの各遺伝子断片増幅プライマーを各1μl、TaKaRa Ex TaqTM(TaKaRa)を1μl加えて混合し、TaKaRa PCR Thermal Cycler Dice VersionIII Model TP600(TaKaRa)を使用して94℃で5分間処理後、〔94℃、30秒→57℃、30秒→72℃、60秒〕という反応を30サイクル行い、その後72℃で5分間処理し、最後に4℃で5分間処理しPCRを終了した。得られたPCR産物は以下の方法で電気泳動を行った。Agarose S(ニッポンジーン)を0.15g、TAE[40mM Tris,40mM 酢酸,1mM EDTA(pH8.0)]を15ml、エチジウムブロマイドを0.6μl用いて作成した1%アガロースゲルをTAEで泳動槽内を満たしたMupid-2plus(ADVANCE)に設置し、2μlのPCR産物に1μlの10×Loading Buffer(TaKaRa)を混合し、アガロースゲルのウェルにアプライして電気泳動を行い、設計したサイズの断片であることを確認した。
【0031】
サイズの確認を行ったPCR産物はMonoFas DNA精製キットI(GL Sciences)を用いて以下のように精製した。1%アガロースを用いて泳動した20μl分のPCR産物のバンドをメスで切り取り、1.5ml容マイクロチューブに入れ、ゲルの重量を量った。ゲルの重量と等量のBuffer Aを加え60℃で10分間保温し、ゲルを完全に溶解した。コレクションチューブに取り付けたスピンカラムに溶解したゲルサンプル溶液を入れ、10,000rpm(4℃)で30秒遠心した。遠心後、スピンカラムに500μlのBuffer Bを入れ、10,000rpm(4℃)で30秒遠心した。遠心後、スピンカラムを1.5ml容マイクロチューブに付け替え、20μlのBuffer Cを入れ10,000rpm(4℃)で60秒遠心し、得られた溶液をZYMVの各遺伝子断片とした。
【0032】
各遺伝子断片はpGEM-T Easy Vector System(Promega)を用いて、TAクローニングを以下のように行った。1.5ml容マイクロチューブに4μlのP1遺伝子断片をとり、2×Rapid Ligation Bufferを5μl、pGEM-T Easy Vectorを1μl、T4 DNA Ligaseを1μl加えて混合後、室温で60分間静置しライゲーション反応を行った。ライゲーション反応を行ったサンプル溶液10μlを、あらかじめ氷上で解凍しておいた100μlの大腸菌DH5αのコンピテントセルに加え、穏やかに混合後氷上で20分間静置した。静置後、42℃のウォーターバスで45秒間ヒートショック処理を行い、ただちに氷上で2分間静置した。静置後、サンプルに1mlのSOC培地(pH7.5)[1Lあたりbacto tryptone 20g,bacto yeast extract 5g,NaCl 0.5g,1M MgCl2 10ml,1M MgSO410ml]を加え、37℃のシェイカーで60分間振盪培養した。培養後、0.15%の寒天を含むLB培地(pH7.5)[1Lあたりbacto tryptone 10g,bacto yeast extract 5g,NaCl 10g,アンピシリン(25mg/ml) 2ml]のプレートにサンプル50μlをスプレッドし、サンプルを塗り広げた。サンプルを塗り広げたプレートは37℃のインキュベーターに14~16時間静置した。
【0033】
プレートに形成されたコロニーをスモールスケールで培養し、煮沸法を用いた以下の方法でプラスミドの抽出を行った。オートクレーブ滅菌済みの爪楊枝を用いて、試験管内に分注した2mlのLB培地に植菌し、37℃に設定したシェイカーで8時間振盪培養した。培養液を1.5ml容マイクロチューブに移し、14,000rpm(室温)で1分間遠心した。遠心後、上清をアスピレーターで除去し、350μlのSTET[0.1M NaCl,10mM Tris-HCl(pH8.0),1mM EDTA(pH8.0),5% TritonX-100]を加えてボルテックスミキサーで沈殿を懸濁した。リゾチーム溶液[10mM Tris-HCl(pH8.0),10mg/mlリゾチーム]を25μl加え、3秒間ボルテックスミキサーで混合した。混合後、マイクロチューブを40秒間煮沸し、直ちに氷中に移して5分間冷却した。冷却後、14,000rpm(室温)で10分間遠心し、沈殿をオートクレーブ滅菌済みの爪楊枝で除去した。上清に40μlの3M酢酸ナトリウム(pH5.2)と420μlのイソプロピルアルコールを加えて混合し、-20℃の冷凍庫内に5分間静置した。静置後、14,000rpm(4℃)で遠心し、上清を除去した。マイクロチューブ内の沈殿に1mlの70%エタノールを加え、14,000rpm(4℃)で2分間遠心し、上清を除去後、減圧乾燥機でマイクロチューブ内の沈殿を2分間乾燥した。乾燥した沈殿を50μlのRNase溶液[10mM Tris-HCl(pH8.0),1mM EDTA(pH8.0),20μg/ml DNase free RNase ]懸濁した。懸濁後、37℃のインキュベーターで20分間静置し、プラスミド溶液とした。
【0034】
抽出したプラスミド溶液を2μl用いて、1%アガロースゲルで電気泳動を行い、インサートを保持していると考えられるプラスミドの選抜を行った。選抜したプラスミドはEcoRIを用いた制限酵素処理を以下の方法で行い、インサートの保持を確認した。マイクロチューブに2μlのプラスミドをとり、滅菌水を6.8μl、10×H Buffer(TaKaRa)を1μl、EcoRI(TaKaRa)を0.2μl加えて混合し、37℃のインキュベーターで3時間静置した。静置後、1%アガロースゲルで電気泳動を行い、インサートの保持を確認し、TAクローニングを終了した。
1-2-2:ZYMV遺伝子断片のシークエンス
TAクローニングによって得られたプラスミドは以下の方法で精製した。46μlのプラスミドに154μlのTE [10mM Tris-HCl(pH8.0),1mM EDTA(pH8.0)]、100μlのTE飽和フェノール、100μlのクロロホルムを加えボルテックスミキサーで5分間混合し、14,000rpm(4℃)で5分間遠心した。遠心後、200μlの水層を新しいマイクロチューブに移し、200μlのクロロホルムを加えボルテックスミキサーで混合し、14,000rpm(4℃)で5分間遠心した。150μlの水層を新しいマイクロチューブに移し、3M酢酸ナトリウム(pH5.2)を15μl、99%エタノールを375μl加えボルテックスミキサーで2分間混合し、-20℃の冷凍庫で20分間静置した。14,000rpm(4℃)で20分間遠心して上清を除去後、沈殿に1mlの70%エタノールを加え、14,000rpm(4℃)で5分間遠心した。遠心後、上清を除去し、減圧乾燥機で2分間乾燥後、50μlのTEに懸濁した。50μlのPEG溶液(13% polyethylene glycol 8000,1.6M NaCl)を加えて混合し、氷上に1時間静置した。静置後、14,000rpm(4℃)で20分間遠心し、沈殿に500μlの70%エタノールを加え、14,000rpm(4℃)で2分間遠心した。上清を除去し、沈殿を減圧乾燥機で2分間乾燥した。沈殿を20μlの滅菌水で懸濁し、NanoDrop ND-1000 Spectrophotometer(旭テクノグラス)を用いて吸光度(波長260nm)を測り、濃度を300ng/μlに調整し、精製プラスミドとした。
【0035】
精製プラスミドは以下の方法でシークエンス分析を行った。シークエンス分析に供試する泳動試料はBigDye Terminator v3.1 Cycle Sequencing Kit(Applied Biosystems)を用いて以下のように作成した。0.2ml容PCRチューブに1μlの精製プラスミドをとり、滅菌水を4μl、5×BigDye Sequencing Bufferを1μl、プライマーACUNI6745(+)を2μl、Ready reaction mixを2μl加えて混合し、TaKaRa PCR Thermal Cycler Dice VersionIII Model TP600(TaKaRa)を使用して94℃で1分間処理後、〔94℃、30秒→50℃、15秒→60℃、4分〕という反応を30サイクル行い、その後60℃で3分間処理し、最後に4℃で5分間処理し、PCRを終了した。PCR後、1.5ml容マイクロチューブに試料全量をとり、滅菌水を7.25μl、3M酢酸ナトリウムを1.5μl、99%エタノールを31.25μl加えてボルテックスミキサーで1分間混合し、アルミホイルで包んで遮光し、室温で10分間静置した。静置後、14,000rpm(4℃)で20分間遠心し、沈殿に1mlの70%エタノールを加え14,000rpm(4℃)で5分間遠心した。遠心後、減圧乾燥機で沈殿を乾燥し、20μlのHi-Di Formamidoに懸濁した。これを2分間煮沸し、ただちに氷上で5分間静置したものを泳動試料とした。
【0036】
試料の泳動はABI PRISM 3100-Avant Genetic Analyzerのマニュアルに従い行った。20μlの泳動試料をMicroAmp 96-well Reaction Plateのwellにのせ、プレートアセンブリを組み立て、オートサンプラーに設置した。キャピラリー長は80cm、ポリマーはPOP-4、泳動緩衝液は滅菌水で10倍に希釈した10×Genetic Analyzer Buffer with EDTAを用いた。次にプレートレコードを作成し、サンプル名、泳動条件および解析条件などを設定した。Dye SetはZ、Mobility FileはDT3100POP4{BDv3}v1.mob、BioLIMS Projectは3100_Project1、Run ModuleはLongseq80_POP4DefaultModule、Analysis ModuleはBC-3100APOP4_80cm_SeqOffFtOff.sazにそれぞれ設定し、泳動を行った。泳動終了後、ABI PRISM 3100-Avant Genetic Analyzerのマニュアルに従い、解析データを塩基配列に変換した。得られた塩基配列データは配列情報解析ソフトウェアのDNASIS Pro ver 2.09.000.001(HitachiSoft)を用いて解析を行い、作成したZYMV遺伝子断片が鋳型と同一であることを確認した。
1-2-3:ZYMV遺伝子断片のALSVベクターへの導入
配列の確認をした各ZYMV遺伝子断片はALSR2L5R5に導入するため、遺伝子断片の精製、BamHIとXhoIを用いた制限酵素処理を以下のように行った。
【0037】
各ZYMV遺伝子断片28μlにTEを72μl、TE飽和フェノールを50μl、クロロホルムを50μl加えボルテックスミキサーで3分間混合し、14,000rpm(4℃)で10分間遠心した。水層100μlを新しいマイクロチューブに移し、3M酢酸ナトリウム(pH5.2)を10μl、99%エタノールを250μl加えボルテックスミキサーで2分間混合し、-80℃の冷凍庫で15分間静置した。14,000rpm(4℃)で15分間遠心し、上清を除去し、沈殿に70%エタノールを1ml加え、14,000rpm(4℃)で5分間遠心した。上清を除去し、減圧乾燥機で沈殿を乾燥した。17μlの滅菌水に懸濁し、10×K Buffer(TaKaRa)を2μl、BamHI(TaKaRa)を0.5μl、XhoI(TaKaRa)を0.5μl加え穏やかに混合し、37℃のインキュベーターで12時間静置した。ALSR2L5R5も同様の方法で制限酵素処理を行った。
【0038】
制限酵素処理を行った各種ZYMV遺伝子断片およびALSR2L5R5はMonoFas DNA精製キットIを用いて前記1-2-1の方法で精製し、得られた溶液をそれぞれ精製Z:P1遺伝子、精製Z:P3遺伝子、精製Z:CP200遺伝子、精製Z:CP100遺伝子(ZYMVインサートDNA)および精製ALSR2L5R5(ベクターDNA)とした。
【0039】
各種ZYMVインサートDNAとベクターDNAにDNA Ligation Kit Ver.2.1(TaKaRa)を用いてライゲーション反応を以下の方法で行った。マイクロチューブに4μlの各種インサートDNAをとり、それぞれにベクターDNAを1μlとI液を5μlずつ加え、穏やかに混合し、16℃のウォーターバスで4時間ライゲーション反応を行った。各ライゲーション産物を用いて前記1-2-1と同様の方法で、大腸菌DH5αの形質転換とスモールスケール培養および煮沸法によるプラスミドの抽出を行った。抽出したプラスミドは1%アガロースで電気泳動し、インサートを保持していると考えられるものを選抜した。選抜したプラスミドはBamHIとXhoIを用いた制限酵素処理を以下の方法で行い、インサートの保持を確認した。マイクロチューブに2μlのプラスミドをとり、滅菌水を6.6μl、10×K Bufferを1μl、BamHIを0.2μl、XhoIを0.2μl加えて混合し、37℃のインキュベーターで3時間静置した。静置後、1%アガロースゲルで電気泳動を行い、各プラスミドのインサートの保持を確認した。
【0040】
インサートを保持したクローンはラージスケール培養を行い、QIAGEN Plasmid Midi Kit(QIAGEN)を用いて、遠心機にはRPR16ローター(HITACHI)を設置した高速冷却遠心機himac CR20G(HITACHI)を使用し、以下の方法で精製した。500ml容の坂口フラスコに100mlのLB培地を分注し、コロニーをオートクレーブ滅菌済みの爪楊枝で植菌し、37℃のシェイカーを用いて180rpmで14~16時間振盪培養した。培養液を50ml容の遠心管に移し、7,500rpm(4℃)で15分間遠心した。上清を除去し、残りの培養液を移して7,500rpm(4℃)で15分間遠心した。上清を除去し、沈殿を4mlのBuffer P1に懸濁した。懸濁液に4mlのBuffer P2を加えて転倒混和し室温で4分間静置した。直ちに4mlのBuffer P3を加えて転倒混和し、氷上で20分間静置した。13,000rpm(4℃)で30分間遠心し、上清を新しい50ml容の遠心管に移し、さらに13,000rpm(4℃)で15分間遠心した。4mlのBuffer QBTでカラムを平衡化したQIAGEN-tip 100に遠心上清を移し、自然落下でカラムにプラスミドDNAを吸着させた。QIAGEN-tip 100を10mlのBuffer QCで2回洗浄し、QIAGEN-tip 100を新しい50ml容スミロンチューブに設置し、5mlのBuffer QFでDNAを溶出した。溶出したDNA溶液にイソプロピルアルコールを3.5ml加えて混合し、直ちに15ml容のCOREXチューブに移し、11,000rpm(4℃)で30分間遠心した。上清を除去し、沈殿に2mlの70%エタノールを加え11,000rpm(4℃)で10分間遠心した。上清を除去し、減圧乾燥機で沈殿を乾燥した。沈殿を200μlのTEに懸濁し、NanoDrop ND-1000 Spectrophotometerを用いて吸光度(波長260nm)を測り、濃度を1μg/μlに調整した。各プラスミドはそれぞれALSVR2L5R5-Z:P1、ALSVR2L5R5-Z:P3、ALSVR2L5R5-Z:CP:200、ALSVR2L5R5-Z:CP:100とした(図2)。
1-2-4:ALSVベクターの接種とウイルス化
精製した各プラスミド(ALSVR2L5R5-Z:P1、ALSVR2L5R5-Z:P3、ALSVR2L5R5-Z:CP200、ALSVR2L5R5-Z:CP100)は、前記1-2-3の方法で精製したpEALSR1と等量ずつ混合し、DNA溶液とした。作製したDNA溶液を7葉期のキノアの第3から第6本葉に本葉1枚につき8μlずつ、カーボランダム法で接種した。接種後、退緑症状の現れた上葉をサンプリングし、2倍容のALSV磨砕緩衝液[0.1M Tris-HCl(pH7.8),0.1M NaCl,5mM MgCl2]で磨砕し、8葉期のキノアの第3から第8本葉にカーボランダム法で接種した。接種後、退緑症状の現れた上葉をサンプリングし、各ウイルスをそれぞれALSV-Z:P1、ALSV-Z:P3、ALSV-Z:CP200、ALSV-Z:CP100とし、以後の接種試験の接種源とした。また、ALSVR2L5R5も同様の方法でウイルス化し、得たウイルスをwtALSVとし、以後の接種試験の接種源とした。
1-3:キュウリへの接種法と一次接種および二次接種
1-3-1:キュウリへの一次接種
キュウリへの一次接種は各ALSV(ALSV-Z:P1、ALSV-Z:P3、ALSV-Z:CP200、ALSV-Z:CP100、wtALSV)およびZYMV 2002を用いて以下の方法で行った。各ALSV感染キノア葉を、それぞれ2倍容のALSV磨砕緩衝液で磨砕した。各ALSV磨砕液と、ZYMV 2002感染カボチャ葉を2倍容の0.1M リン酸緩衝液(pH7.0)で磨砕して調整したZYMV磨砕液を、子葉展開期のキュウリの子葉2枚にカーボランダム法で接種した。キュウリは25℃の長日条件で育成した
1-3-2:キュウリへの二次接種
一次接種したキュウリに、p35S-Z5vector+GFP-FL を以下の方法で金粒子にコーティングし、Helios Gene Gunシステム(BIO-RAD)を用いて二次接種した。1.5ml容マイクロチューブに径1.6μmの金粒子(BIO-RAD)を10mg量りとり、50μlの50%グリセロールを加えてボルテックスミキサーで混合した。超音波洗浄機で2分間ソニケーションを行い、ボルテックスミキサーで混合しながら25μlのp35S-Z5vector+GFP-FLを加え、次に150μlの1M CaCl2を加え、さらに150μlの0.05Mスペルミジンを加えて混合した。室温で15分間静置し、上清をピペットマンで除去した。沈殿を崩さないように1mlの99%エタノールを加えて洗浄し、ピペットマンで上清を除去した。同様の洗浄を3回行い、洗浄後、沈殿に500μlの99%エタノールを加え、指ではじいて懸濁し、金粒子の分散を確認した。懸濁した金粒子を全て15ml容チューブに移し、2.5mlの99%エタノールを加え混合した。調整した金粒子懸濁液はHelios Gene Gun簡易操作マニュアルに従い、以下の方法でゴールドコート用チューブにコーティングし、接種を行った。チュービングプレップステーションで0.35-0.40LPM(liters per minute)のN2ガスを用いてチューブ内を20分間乾燥したゴールドコート用チューブに、シリンジ付き懸濁液ローディング用チューブを取り付け、このシリンジを用いて金粒子懸濁液をゴールドコート用チューブに装填し、5分間静置した。静置後、シリンジ付き懸濁液ローディング用チューブを取り除き、エタノール除去用シリンジ付きチューブを取り付け、ゴールドコート用チューブ内のエタノールを全て除去した。除去後、直ちにゴールドコート用チューブを180°回転し、チュービングプレップステーションの回転スイッチを作動した。ゴールドコート用チューブ内に0.35-0.40LPMのN2ガスを通気しチューブ内を乾燥した。乾燥したゴールドコート用チューブをチュービングプレップステーションから取り出し、チュービングカッターを用いて切断し、サンプルカートリッジとした。サンプルカートリッジをHelios Gene Gunのカートリッジホルダーに取り付け、1葉期のキュウリの第1本葉に180PSI(pounds per square inch)で1 shot接種し、第1本葉二次接種区とした。上記の方法で2葉期の第2本葉のみに二次接種したものを第2本葉二次接種区、3葉期の第3本葉のみに二次接種したものを第3本葉二次接種区とした。二次接種後、キュウリは一次接種後と同様に25℃の長日条件で育成した。
【0041】
なお、二次接種の接種源であるp35S-Z5vector+GFP-FLは、使用する培地をLB培地から2×YT培地(pH7.5)[1Lあたりbacto tryptone 16g,bacto yeast extract 10g,NaCl 5g,アンピシリン(25mg/ml) 2ml]に変更したラージスケール培養で前記1-2-3と同様の方法で精製し、NanoDrop ND-1000 Spectrophotometerで濃度を測定後、400ng/μlに調整し、-80℃で保存しておいたものを使用した。
1-4:病徴観察とウイルス検定
1-4-1:病徴の観察
病徴の観察は二次接種後毎日行い、二次接種後21日目にデジタルカメラFinePix S1Pro(FUJIFILM)を用いて病徴の撮影を行った。
1-4-2:GFP蛍光の観察
GFP蛍光の観察は二次接種後7日目、14日目、21日目に蛍光顕微鏡システムVBシリーズ(KEYENCE)と顕微鏡デジタルカメラDP70(OLYMPUS)を用いて行い、同時に撮影した。
1-4-3:ウエスタンブロット法
一次接種後7日目にキュウリの子葉2枚を採取し、各一次接種ウイルスの感染を以下の方法で確認した。採取した子葉2枚の重量を量り、3倍容のALSV磨砕緩衝液とともに乳鉢と乳棒で磨砕し、100μlの磨砕液を1.5ml容マイクロチューブに移した。100μlの2×サンプルバッファー[0.1M Tris-HCl(pH8.0),4% SDS,30% スクロース,0.05% BPB,2% メルカプトエタノール]を加え、ボルテックスミキサーで1分間混合し、その後5分間煮沸し、直ちに氷上で5分間静置した。静置後、14,000rpm(4℃)で5分間遠心し、上清を泳動試料とした。泳動試料は10μl使用し、stacking gel(5%アクリルアミドゲル)とrunning gel(12.5%アクリルアミドゲル)および泳動バッファー[0.6%Tris,28%グリシン,0.1%SDS]を用いて泳動(160V,20mA,90分間,室温)を行った。泳動後、転写バッファー[0.1M Tris,0.192M グリシン,20%メタノール]を用いてセミドライエレクトロブロッティングを行い、PVDF膜(MILIPOLE)にタンパク質の転写を行った。転写後、PVDF膜をブロッキング液[0.02M Tris-HCl(pH7.5),0.5M NaCl,2% Tween-20,2%スキムミルク,2%PVP]に浸し、37℃のインキュベーターで60分間静置した。静置後、TTBS[0.02M Tris-HCl(pH7.5),0.5M NaCl,2% Tween-20]で25倍希釈したALSVおよびZYMVに対する吸収抗体を用いて、一次抗体処理を37℃のインキュベーター内で60分間行った。一次抗体処理後、PVDF膜をTTBSで満たしたタッパーに浸し、シーソーで5分間洗浄した。TTBSを新しいものに交換し、同様の方法でさらに2回洗浄を行った。洗浄したPVDF膜はTTBSで2000倍に希釈したアルカリフォスファターゼ標識坑ウサギ免疫グロブリンヤギ血清(Cell Signaling TECHNOLOGY)用いて、二次抗体処理を37℃のインキュベーター内で60分間行った。二次抗体処理後、PVDF膜をTTBSで3回洗浄し、発色基質液[0.2M Tris-HCl(pH8.2),3mg/ml Fast Red TR salt,1mg/ml Naphthol AS-MX phosphate]を用いて発色処理を行った。十分に発色した後、PVDF膜を蒸留水で洗浄し、反応を停止した。なお、吸収抗体はあらかじめ以下の方法で作成した。まず、5倍容のTTBSで磨砕した供試植物の磨砕液を3,000rpm(4℃)で5分間遠心し、上清を粗汁液とした。1.5ml容のマイクロチューブに10μlの一次抗体(各種ウイルス抗血清)をとり、粗汁液90μlを加えて混合し、37℃で60分間静置した。静置後、14,000rpm(4℃)で5分間遠心し、上清100μlを新しい1.5ml容のマイクロチューブに移した。上清に磨砕液を100μl加えて混合し、37℃で60分間静置した。静置後、14,000rpm(4℃)で5分間遠心し、上清200μlを新しい1.5ml容のマイクロチューブに移した。上清に磨砕液を200μl加えて混合し37℃で60分間静置した。14,000rpm(4℃)で5分間遠心し、上清400μlを新しい1.5ml容のマイクロチューブに移した。上清に400μlの100%グリセロールを加えボルテックスミキサーで十分混合し吸収抗体とした。吸収抗体は-20℃の冷凍庫で保存し適宜使用した。
1-4-4:ELISA法
ALSVとZYMVの蓄積量は直接ELISA法を用い、GFPの蓄積量は間接ELISA法を用いて以下の方法で定量した。
【0042】
検定試料は二次接種後21日目にコルクボーラーを用いて次のように採取した。第1本葉接種区では第3から第6本葉の各本葉を、第2本葉接種区では第4から第7本葉の各本葉を、第3本葉接種区では第5から第8本葉の各本葉をそれぞれ採取し、採取した本葉の基部、中心部、周辺部から太い葉脈を避けて直径1cmのリーフディスクを2枚ずつくりぬき、本葉1枚から計6枚のリーフディスクを採取した(図3)。採取した6枚のリーフディスクは1つのステンレスビーズSUB-50(TOMY)とともに2ml容凍結保存チューブ(アシスト)に入れ、液体窒素で凍結し、Micro Smash MS100-R(TOMY)を用いて2,500rpmで30秒間破砕した。破砕後、1.8mlのPBST[137mM NaCl,8.1mM Na2HPO4,2.68mM KCl,1.47mM KH2PO4,0.05% Tween20]を加え、Micro Smash MS100-Rを用いて2,500rpmで30秒間混合した。その後、14,000rpm(4℃)で5分間遠心し、得られた上清を検定試料とした。
【0043】
ALSVの定量は直接ELISA法を用いて以下の方法で行った。ELISAプレートのウェルにコーティングバッファー[0.05M炭酸緩衝液(pH9.6)]で1000倍に希釈したALSV抗体溶液を150μl入れ、プレートカバーを被せ37℃のインキュベーターで2時間静置した。溶液を捨て、PBSTでウェルを3回洗浄し、ブロッキング溶液[137mM NaCl,8.1mM Na2HPO4,2.68mM KCl,1.47mM KH2PO4,2%スキムミルク]をウェルに150μl入れ、プレートカバーを被せ37℃のインキュベーターで1時間静置した。溶液を捨て、PBSTでウェルを3回洗浄し、1検定試料当り2つのウェルに150μlずつ検定試料を入れ、プレートカバーを被せ4℃の冷蔵庫に一晩静置した。溶液を捨て、PBSTでウェルを10回洗浄し、PBSTで1000倍に希釈した酵素標識をしたALSV抗体溶液をウェルに150μl入れ、プレートカバーを被せ37℃のインキュベーターで3時間静置した。溶液を捨て、PBSTでウェルを10回洗浄し、基質液[10%ジエタノールアミン(pH9.8),0.67mg/ml p-ニトロフェニルリン酸]を150μl入れ、室温に静置して色を呈するまで反応させた。反応後、モデル550マイクロプレートリーダー(BIO-RAD)でウェルの吸光度を測定し、ALSVの蓄積量とした。
【0044】
ZYMVの定量は直接ELISA法を用いて以下の方法で行った。ELISAプレートのウェルにコーティングバッファーで500倍に希釈したZYMV抗体(日本植物防疫協会研究所)溶液を150μl入れ、プレートカバーを被せ37℃のインキュベーターで3時間静置した。溶液を捨て、PBSTでウェルを3回洗浄し、ブロッキング溶液をウェルに150μl入れ、プレートカバーを被せ37℃のインキュベーターで1時間静置した。溶液を捨て、PBSTでウェルを3回洗浄し、1検定試料当り2つのウェルに150μlずつ検定試料を入れ、プレートカバーを被せ4℃の冷蔵庫に一晩静置した。溶液を捨て、PBSTでウェルを10回洗浄し、PBSTで1000倍に希釈した酵素標識をしたZYMV抗体(日本植物防疫協会研究所)溶液をウェルに150μl入れ、プレートカバーを被せ37℃のインキュベーターで4時間静置した。溶液を捨て、PBSTでウェルを10回洗浄し、基質液[10%ジエタノールアミン(pH9.8),1mg/ml p-ニトロフェニルリン酸]を150μl入れ、室温に静置して色を呈するまで反応させた。反応後、モデル550マイクロプレートリーダーでウェルの吸光度を測定し、ZYMVの蓄積量とした。
【0045】
GFPの定量は以下の方法で行った。ELISAプレートのウェルに1検定試料当り2つのウェルに150μlずつ検定試料を入れ、プレートカバーを被せ37℃のインキュベーターに2時間静置した。溶液を捨て、PBSTでウェルを5回洗浄し、ブロッキング溶液をウェルに150μl入れ、プレートカバーを被せ37℃のインキュベーターで1時間静置した。溶液を捨て、PBSTでウェルを5回洗浄し、PBSTで1000倍に希釈したGFP抗体(Clontech)を150μlいれ、プレートカバーを被せ37℃の冷蔵庫に一晩静置した。溶液を捨て、PBSTでウェルを10回洗浄し、PBSTで2000倍に希釈したアルカリフォスファターゼ標識坑ウサギ免疫グロブリンヤギ血清をウェルに150μl入れ、プレートカバーを被せ37℃のインキュベーターで3時間静置した。溶液を捨て、PBSTでウェルを10回洗浄し、基質液[10%ジエタノールアミン(pH9.8),1mg/ml p-ニトロフェニルリン酸]を150μl入れ、室温に静置して色を呈するまで反応させた。反応後、モデル550マイクロプレートリーダーでウェルの吸光度を測定し、GFPの蓄積量とした。
1-4-5:RNA抽出法とRT-PCR法
二次接種後21日目にキュウリの最上位展開葉を採取し、-80℃の冷凍庫で凍結し検定試料とした。0.1gの検定試料を乳鉢と乳棒で磨砕し、1mlのTriPure Isolation Reagent(Roche)を加えさらに磨砕した。磨砕液1mlを1.5ml容マイクロチューブに移し、室温で5分間静置した。静置後、200μlのクロロホルムを加えボルテックスミキサーで20秒間混合し、室温で10分間静置した。静置後、14,000rpm(4℃)で15分間遠心し、500μlの水層を新しい1.5ml容マイクロチューブに移した。水飽和フェノールを250μl、クロロホルムを250μl加えボルテックスミキサーで5分間混合し、14,000rpm(4℃)で5分間遠心した。遠心後、500μlの水層を新しいマイクロチューブに移し、420μlのイソプロピルアルコールを加えボルテックスミキサーで混合し、室温で10分間静置した。静置後、14,000rpm(4℃)で10分間遠心し、上清を除去し70%エタノールを1.4ml加えボルテックスミキサーで5分間混合した。14,000rpm(4℃)で5分間遠心し、上清を除去後、沈殿を20μlの滅菌水に懸濁し、NanoDrop ND-1000 SpectrophotometerでRNA濃度を測定し、500ng/μlに調整しRNA溶液とした。
【0046】
逆転写反応は以下の方法で行った。0.2ml容PCRチューブにRNA溶液を2μl、滅菌水を6μl、5×RT Buffer(TOYOBO)を4μl、2.5mM dNTP mixture(TaKaRa)を8μl、10μM Oligo(dT)12プライマーを1μl、RNase Inhibitor(Wako)を0.5μl、ReverTra Ace(TOYOBO)を0.5μl 混合し、TaKaRa PCR Thermal Cycler Dice VersionIII Model TP600(TaKaRa)を使用して42℃で50分間処理し、その後95℃で5分間処理し、最後に4℃で5分間処理し、獲られた産物をRT産物とした。
【0047】
PCRにはALSV-RNA2ベクターの外来遺伝子導入サイトを含む配列を増幅できる10μM R2ALS 1363(+)[5’-GCGAGGCACTCCTTA-3’:配列番号8]と10μM R2ALS 1511(-)[5’-GCAAGGTGGTCGTGA-3’:配列番号9]をそれぞれプラス鎖プライマーおよびマイナス鎖プライマーとして用いた。1μlのRT産物を0.2ml容PCRチューブにとり、滅菌水を4.9μl、10×Ex Taq Buffer(TaKaRa)を1μl、2.5mM dNTP Mixture(TaKaRa)を1μl、10μM R2ALS 1363(+)と10μM R2ALS 1511(-)を各1μl、TaKaRa Ex TaqTM(TaKaRa)を0.1μl加えて混合し、TaKaRa PCR Thermal Cycler Dice VersionIII Model TP600(TaKaRa)を使用して94℃で5分間処理後、〔94℃、30秒→58℃、30秒→72℃、60秒〕という反応を30サイクル行い、その後72℃で5分間処理し、最後に4℃で5分間処理しPCRを終了した。得られたPCR産物2μlに1μlの10×Loading Buffer(TaKaRa)を混合し、1%アガロースゲルを用いた電気泳動を行い、各種ALSVのインサート配列の保持を確認した。
2:結果
2-1:ZYMV遺伝子を導入したALSVベクターのZYMVに対する干渉効果
ZYMV遺伝子断片を導入したALSVベクターのZYMVに対する干渉効果を調べるため、wtALSVおよびZYMVのCP遺伝子断片を導入したALSV(ALSV-Z:CP200)を一次接種したキュウリ(青大胡瓜)におけるZYMVの病徴、GFP蛍光、ウイルス量を調べた。
【0048】
p35S-Z5vector+GFP-FLのみを接種した試験区(ZYMVのみ区)では、接種後12日前後に第3本葉の基部周辺で退緑斑点や脈間退緑などの病徴が観察された。その後展開した第4本葉で葉のモザイク症状が見られ、第5本葉以上の葉では葉全面で激しいモザイク症状が観察された(図4A)。さらに、第6本葉以上の葉では激しいモザイク症状とともに葉の奇形や萎縮が観察され(図4B)、植物体全体も矮化を示した(図4C)。wtALSVを一次接種後にp35S-Z5vector+GFP-FLを二次接種した試験区(wtALSV+ZYMV区)では、第3から第5本葉での病徴の現れ方はZYMVのみ区とほとんど同じであったが、症状はZYMVのみ区に比べ軽く、植物体全体の矮化は観察されなかった(図4)。これに対して、ALSV-Z:CP200を一次接種後にp35S-Z5vector+GFP-FLを二次接種した試験区(ALSV-Z:CP200+ZYMV区)では、非接種区およびALSVのみを接種した試験区(ALSVのみ区)と同様に、全ての上葉において病徴が全く観察されなかった(図4)。
【0049】
蛍光顕微鏡でGFP蛍光を観察したところ、ZYMVのみ区では、第1本葉の接種部位周辺でスポット状の蛍光や葉脈上に蛍光が観察された。第2本葉では葉脈で観察された蛍光が葉の展開とともに拡大した。第3本葉から第5本葉では葉の全体が蛍光で埋め尽くされていた。第6本葉では葉全体で蛍光が観察されたが、葉の所々に蛍光のない領域が観察された(図5)。第7葉より上位の葉では蛍光の観察される領域と観察されない領域がモザイク状に混在していた。wtALSV+ZYMV区では各葉位でZYMVのみ区とほぼ同じ蛍光が観察された(図6)。これに対して、ALSV-Z:CP200+ZYMV区の第1葉では、ZYMVのみ区やwtALSV+ZYMV区と同様に、接種部位周辺でスポット状の蛍光や葉脈上に蛍光が観察されたが、第2本葉では葉脈で観察された蛍光の拡がる範囲がZYMV区やwtALSV+ZYMV区に比べ狭い領域に限定されていた。さらに、第3本葉では数箇所に蛍光が観察されるのみで、第4葉より上位の葉では蛍光が観察されなかった(図7)。
【0050】
続いて、ZYMVの定量を行った結果、第3本葉から第6本葉全てでwtALSV+ZYMV区ではZYMVのみ区と同程度のZYMVが蓄積していたのに対し、ALSV-Z:CP200+ZYMV区ではZYMVの蓄積は認められなかった(図8)。また、ALSVを定量したところ、wtALSV+ZYMV区ではALSVのみ区に比べ約2倍のELISA値を示し、ALSV-Z:CP200+ZYMV区ではALSVのみ区に比べやや減少していた(図8)。一次接種に用いたALSV-Z:CP200のインサートチェックを行ったところ、全てのキュウリ個体でALSVのインサートが保持されていることが確認された。
【0051】
第2本葉二次接種区の各試験区における病徴は第1本葉二次接種区のものとほぼ同じで、病徴の現れる葉位が1枚ずつ上位にずれていた。
【0052】
GFP蛍光の観察では、ALSV-Z:CP200+ZYMV区で接種葉の第2本葉のみで接種部位周辺でスポット状の蛍光や葉脈の一部に蛍光が観察され、第3本葉以降に展開した本葉では蛍光が観察されなかった。これに対し、ZYMVのみ区やwtALSV+ZYMV区では、第1本葉二次接種区の各試験区と同様に葉全体でGFP蛍光が観察された。
【0053】
直接ELISA法によりZYMVの蓄積を調べたところ、各試験区で第1本葉二次接種区とほとんど同じようにZYMVが蓄積していた(図9)。一次接種に用いたALSV-Z:CP200のインサートチェックでは、全てのキュウリ個体でALSVのインサートは保持されていた。
【0054】
第3本葉二次接種区の各試験区の病徴の様子やGFP蛍光の分布では、ALSV-Z:CP200+ZYMV区において接種葉を含む全ての本葉で蛍光が観察されなかったこと意外は、第1本葉二次接種区とほぼ同じであった。ただし、病徴やGFP蛍光は2枚上位にずれて観察された。
【0055】
直接ELISA法によるZYMVおよびALSVの定量では、ALSV-Z:CP200+ZYMV区で、ALSVのみ区より若干多くALSVが蓄積していることが明らかになり、それ以外の試験区では第1本葉二次接種区および第2本葉二次接種区とほぼ同じ結果が得られた(図10)。ALSV-Z:CP200のインサート保持も確認された。
【0056】
予備試験で行ったp35S-Z5vector+GFP-FL接種によるZYMVの病徴観察をキュウリ3品種(青大胡瓜、鈴成四葉、つばさ)で比較したところ、青大胡瓜が最も激しい症状を示し、鈴成四葉では軽度の症状が、つばさではほとんど病徴が現れなかった。よって、以後の全ての試験では青大胡瓜を用いることにした。
2-2:異なるZYMV遺伝子断片を導入したALSVベクターの干渉効果
ZYMV遺伝子の異なる領域を組み込んだALSVベクターによって誘導される干渉効果を比較するため、ALSV-Z:P1、ALSV-Z:P3およびALSV-Z:CP200をキュウリの子葉に一次接種し、第1本葉にp35S-Z5vector+GFP-FLを二次接種した。
【0057】
ALSV-Z:CP200+ZYMV区では前記2-1に示した通りである。ALSV-Z:P3を一次接種後p35S-Z5vector+GFP-FLを二次接種した試験区(ALSV-Z:P3+ZYMV区)では、ALSV-Z:CP200+ZYMV区と同様に病徴は観察されず(図11)、蛍光は第1本葉に接種部位周辺でスポット状の蛍光と葉脈上の蛍光が観察された。第2本葉では葉脈上に観察された蛍光が狭い範囲で拡大した。第3本葉および第4本葉では数箇所で蛍光が観察されるのみであり、第5本葉より上位の葉では蛍光が観察されなかった(図12)。一方、ALSV-Z:P1を一次接種後にp35S-Z5vector+GFP-FLを二次接種した試験区(ALSV-Z:P1+ZYMV区)では、第3本葉に葉の周辺部に弱い退緑症状が観察され、この症状はその後に展開した上葉全てで観察された。退緑症状は観察されたものの、ZYMVのみ区に観察された激しいモザイク症状や葉の矮化、植物体全体の矮化は観察されなかった(図11)。GFP蛍光について見ると、第1本葉煮接種部位周辺でスポット状の蛍光と葉脈上の蛍光が観察され、第2葉本では葉脈上の蛍光が狭い範囲で拡大した。また、第3本葉より上葉では、葉脈の蛍光や葉脈の周辺部でのスポット状の蛍光、葉の周辺部や先端部における蛍光が観察された(図13)。
【0058】
直接ELISA法によりZYMV量を測定したところ、ALSV-Z:CP200+ZYMV区とALSV-Z:P3+ZYMV区では第3本葉から第6本葉全てにおいて、ZYMVの蓄積は認められなかった。一方、ALSV-Z:P1+ZYMV区ではZYMVのみ区やwtALSV+ZYMV区の約半分程度蓄積していた(図14)。ALSVは、wtALSV+ZYMV区でALSVのみ区の約2倍蓄積していたが、ALSV-Z:P1+ZYMV区、ALSV-Z:P3+ZYMV区およびALSV-Z:CP200+ZYMV区では、ALSVのみ区よりもやや多い程度であった(図14)。一次接種に用いたZYMVの各遺伝子断片を導入したALSV(ALSV-Z:P1 、ALSV-Z:P3、ALSV-Z:CP200)のインサートチェックを行ったところ、全てのキュウリ個体でALSVのインサートが保持されていた。
【0059】
なお、ZYMVの他の遺伝子領域(HC-Pro、Cl、Nla、Nlb)の断片(180-201 nt)についても干渉効果を同様に試験した。その結果、Cl、NlaおよびNlbの各断片は、前記のCP200断片およびP3断片と同様の効果を示した。一方、HC-Pro断片の効果は前記P1断片の効果とほぼ同一であった。
2-3:異なるキュウリ品種における干渉効果
青大胡瓜とは異なる2つの品種(鈴成四葉、つばさ)においてALSV-Z:CP200がZYMVに対する干渉効果を誘導するか検討した。
【0060】
鈴成四葉では、ZYMVのみ区やwtALSV+ZYMV区で、退緑症状のみが現れ、激しいモザイクや葉の萎縮、植物体の矮化は観察されなかった。また、ALSV-Z:CP200+ZYMV区では、病徴が全く観察されなかった。つばさでは、全ての試験区でZYMVの症状は観察されなかった。
【0061】
鈴成四葉のZYMVのみ区やwtALSV+ZYMV区においては、第1本葉から第5本葉で青大胡瓜の第1本葉二次接種区場合と同様のGFP蛍光の分布が観察され。第6本葉では、基部において蛍光のない領域が一部観察されたが、それ以外では蛍光が観察された。第7葉以降に展開した葉ではスポット状の蛍光が葉の周辺部から中心部に多く観察され、基部では蛍光が観察されなかった。ALSV-Z:CP200+ZYMV区では、第1本葉の接種部位周辺部にスポット状の蛍光のみが僅かに観察され、第2本葉においても数個の蛍光スポットが観察されるのみであった。第3本葉より上位の葉では蛍光は観察されなかった。
【0062】
つばさのZYMVのみ区やwtALSV+ZYMV区では、第1本葉から第5本葉で青大胡瓜や鈴成四葉の場合と同様のGFP蛍光の分布が観察され。第6本葉では基部において蛍光のない領域が一部観察されたが、それ以外では蛍光が観察された。第7本葉より上位の葉ではスポット状の蛍光が葉の周辺部を縁取るように観察され、それ以外の部分では蛍光が観察されなかった。ALSV-Z:CP200+ZYMV区では第1本葉の接種部位周辺でスポット状の蛍光が僅かに観察されるのみで、第2本葉においても数個の蛍光スポットが観察されるのみだった。第3本葉より上葉では蛍光は観察されなかった。
【0063】
直接ELISA法でZYMVを定量したところ、鈴成四葉およびつばさのALSV-Z:CP200+ZYMV区では第3本葉から第6本葉でZYMVが蓄積しないことが明らかになった(図15、図16)。また、鈴成四葉およびつばさの両品種において、ALSVのみ区に比べwtALSV+ZYMV区ではALSVの蓄積量が増加し、ALSV-Z:CP200+ZYMV区では若干減少していた。(図15、図16)。ALSV-Z:CP200のインサートチェックを行ったところ、両品種のキュウリ個体でALSVのインサートは保持されていた。
2-4:異なる長さのZYMV遺伝子断片を導入したALSVベクターの干渉効果
ALSVベクターに導入したZYMV遺伝子断片の長さが干渉効果へ影響するかどうか調べるため、約100 ntと約200 ntのZYMV-CP遺伝子断片を挿入したALSV(ALSV-Z:CP100、ALSV-Z:CP200)の干渉効果を調べた。
【0064】
ALSV-Z:CP200+ZYMV区における病徴は前記2-1の第1本葉二次接種区と同様で、全ての上葉に病徴が観察されなかったのに対し、ALSV-Z:CP100を一次接種後p35S-Z5vector+GFP-FLを二次接種接種した試験区(ALSV-Z:CP100+ZYMV区)では、葉の一部に弱い退緑の症状が観察された(図17)。
【0065】
ALSV-Z:CP200+ZYMV区では、第1本葉の接種部位周辺でスポット状のGFP蛍光と葉脈上の蛍光が観察され、第2本葉で葉脈上の蛍光が狭い範囲で拡がった。第3本葉では数箇所で蛍光が観察されるのみで、第4本葉より上位の葉では蛍光が観察されず、前記2-1の第1本葉二次接種区と同様であった。これに対し、ALSV-Z:CP100+ZYMV区では、第1本葉の二次接種部位周辺でスポット状の蛍光と葉脈上の蛍光が観察された。第2本葉では、葉脈上に蛍光が観察され、葉の展開とともに蛍光も拡大した。第3本葉では、葉の全体が蛍光で埋め尽くされていることが観察された。第4本葉では、スポット状の蛍光が葉全体で観察され、特に葉の周辺部で多数観察された。第5本葉では、葉の周辺部の葉脈上に蛍光が観察された。第6本葉より上位の葉では、葉の先端部でのみ蛍光が観察された(図18)。
【0066】
直接ELISA法でZYMVを定量したところ、ALSV-Z:CP200+ZYMV区の第3本葉から第6本葉でZYMVが蓄積されないのに対し、ALSV-Z:CP100+ZYMV区では、ZYMVのみ区やwtALSV+ZYMY区に比べやや少ないがZYMVが蓄積していることが明らかになった(図19)。ALSVの定量をしたところ、ALSVのみ区に比べwtALSV+ZYMV区では蓄積量が増加し、ALSV-Z:CP200+ZYMV区では同程度蓄積していた。これに対して、ALSV-Z:CP100+ZYMV区では、ALSVのみ区と比較すると、第3本葉から第5本葉ではALSVの蓄積量が増加していたが、第6本葉では同程度であった(図19)。一次接種に用いた異なる長さのZYMVのCP遺伝子断片を導入したALSV(ALSV-Z:CP100 、ALSV-Z:CP200)のインサートチェックを行ったところ、全てのキュウリ個体でALSVのインサートが保持されていた。
【0067】
なお、同様にして、ZYMV-CP遺伝子の66 nt断片と33 nt断片をそれぞれ挿入したALSVの干渉効果を調べた。その結果、挿入遺伝子断片のサイズが小さくなるにつれて干渉効果が弱まることが確認された。
2-5:ZYMV遺伝子断片導入ALSVと弱毒系統ZYMVの干渉効果の比較
ウイルス製剤として市販されているZYMV 2002 による干渉効果と、ZYMV遺伝子断片を導入した3種のALSVによる干渉効果を比較するために、各試験区の病徴、GFP蛍光の分布およびウイルス蓄積を調べた。
【0068】
ZYMV 2002を一次接種後p35S-Z5vector+GFP-FLを二次接種した試験区(ZYMV 2002+ZYMV区)では接種葉および上葉において病徴は観察されなかった(図20)。
【0069】
また、ALSV-Z:P1+ZYMV区、ALSV-Z:P3+ZYMV区およびALSV-Z:CP200+ZYMV区では前記2-2のようにそれぞれ接種葉および上葉においてGFP蛍光が観察されたのに対し、ZYMV 2002+ZYMV区では接種葉および上葉においてGFP蛍光は観察されなかった(図21)。
【0070】
GFP抗体を用いた間接ELISA法で強毒ZYMVの蓄積を調べたところ、ZYMVのみ区とwtALSV+ZYMV区においてGFPが蓄積したのに対し、ALSV-Z:P3+ZYMV区、ALSV-Z:CP200+ZYMV区およびZYMV 2002+ZYMV区では、GFPが全く蓄積せず、ALSV-Z:P1+ZYMV区では僅かにGFPが蓄積した(図22)。一次接種に用いたZYMVの各遺伝子断片を導入したALSV(ALSV-Z:P1 、ALSV-Z:P3、ALSV-Z:CP200)のインサートチェックを行ったところ、全てのキュウリ個体でALSVのインサートが保持されていた。
3:考察
ZYMV遺伝子断片を導入したALSVをキュウリに接種すると、VIGSが誘導されZYMVに対する防御体制をとると予想された。先ず、ALSVベクター接種後の時間が干渉効果の誘導にどのように影響するか調べるため、ZYMVのCP遺伝子を導入したALSV(ALSV-Z:CP200)を接種後6日目、9日目および12日目に、それぞれ第1本葉、第2本葉および第3本葉に強毒ZYMV株の感染性クローンを接種し、干渉効果の誘導を調べた。その結果、3つの条件(第1本葉二次接種区、第2本葉二次接種区、第3本葉二次接種区)全てにおいて、上位葉でのZYMVの蓄積は認められなかった。また、GFP蛍光を観察したところ、第1本葉二次接種区では第1本葉から第3本葉でGFP蛍光が観察されたが、第2本葉二次接種区の接種葉でのみGFP蛍光が観察され、第3本葉二次接種区では接種葉においてもGFP蛍光が観察されなかった。以上よりALSV-Z:CP200接種後の日数が経過するほどZYMVの拡がりを抑えることが明らかになった。この結果は、ALSVベクターによるサイレンシングの誘導がALSVの存在する領域でのみ誘導され、常にALSV増殖後にVIGSが誘導されるという観察結果と合わせて考えると、第1本葉および第2本葉に二次接種した試験区では、まだALSVが分布していないため接種葉および上葉ではZYMVの増殖と移行が起こったものと考えられる。これに対し、第3本葉に二次接種した試験区では既にALSVが分布し、VIGSが誘導されていたため、ZYMVの増殖が抑えられGFP蛍光が観察されなかったと考えられた。第1本葉に接種した試験区では第3本葉のごく一部に蛍光が観察されたものの、直接ELISA法を用いたZYMV量の測定ではウイルスは検出されないレベルであり、また全ての試験区で病徴が観察されなかった。以上より、ZYMV遺伝子断片を導入したALSVが感染したキュウリにおいて、接種から6日以上経過すればZYMVに対する干渉効果が誘導され、ZYMVの増殖を抑制できることを示している。
【0071】
次に、ALSVベクターに導入するZYMV遺伝子領域の違いが干渉効果の誘導に影響を与えるか調べるため、3種のZYMV遺伝子導入ALSV(ALSV-Z:P1、ALSV-Z:P3、ALSV-Z:CP200)間における干渉効果を比較した。ALSV-Z:P1接種区では弱い病徴やGFP蛍光が観察されZYMVが蓄積したのに対し、ALSV-Z:P3およびALSV-Z:CP200接種区では、病徴は観察されず、また一部GFP蛍光が観察されたもののZYMVの蓄積は認められなかった。また、Cl、NlaおよびNlbの各断片も、前記のCP200断片およびP3断片と同様の効果を示した。
【0072】
キュウリ品種の違いが干渉効果に影響を与えるか調べるためALSV-Z:CP200を用いて比較試験を行ったところ、誘導されるワクチン効果に品種間差は認められなかった。よって、品種の違いは干渉効果に影響を与えないと考えられ、3品種以外の品種においてもALSVベクターによる干渉効果の誘導が可能であると考えられた。
【0073】
ALSVベクターに導入する遺伝子断片の長さが干渉効果の誘導に影響を与えるか調べるため、ALSV-Z:CP200 とALSV-Z:CP100による干渉効果を比較した。ALSV-Z:CP100を一次接種した試験区ではZYMVが蓄積され、軽微な病徴やGFP蛍光が観察されたことから、誘導される干渉効果がALSV-Z:CP200に比べ弱くなったと考えられた。この結果より、タバコにおいてPDS遺伝子のサイレンシングを誘導する際、異なるサイズのPDS遺伝子断片や同一のサイズで異なる部分のPDS遺伝子断片をALSVベクターに導入すると、サイレンシングの程度に違いが認められる報告(例えば、非特許文献4)と一致するものと考えられた。なお、挿入遺伝子のサイズをさらに小さくしたALSV-Z:CP66、ALSV-Z:CP33の試験でも、遺伝子サイズが小さくなるにつれて干渉効果が低減することが確認された。これらのことから、ALSVベクターを用いてあるウイルスに対して干渉効果を誘導する際、ALSVベクターに導入する遺伝子のサイズや部位によって、誘導される干渉効果が異なる可能性がある。そのため、複数のウイルス遺伝子断片を導入したALSVを作出し、その干渉効果について比較検討することが好ましい。
【0074】
市販されている弱毒ZYMVと、ZYMV遺伝子導入ALSV(ALSV-Z:P1、ALSV-Z:P3、ALSV-Z:CP200)の干渉効果の比較試験では、GFP蛍光の分布に差が認められた。すなわち、弱毒ZYMV接種区ではZYMVのGFP蛍光は全く観察されなかった。これは、弱毒ウイルスとALSVによる干渉効果の機構の差があるためと考えられる。つまり、ALSVベクターでは、導入遺伝子断片のサイレンシングによって干渉効果を誘導するRNA介在性機構であるため、ALSVが分布していない領域ではサイレンシングの誘導がなされていないため、強毒株のZYMVが増殖したと考えられる。これに対し、弱毒ZYMVでは、同種のウイルスによる干渉効果であるため、タンパク質介在性干渉効果が強毒ZYMVの増殖を抑え、蛍光が全く観察されなかったものと考えられた。さらに、ALSVベクターとZYMVでは感染葉でのウイルスの拡がり方が異なり、ALSVは葉の基部から先端部に向けてその範囲を拡げるように拡がるのに対し、ZYMVは葉脈全体に先ず拡がり、その後葉脈から染み出すように拡がるといった移行の違いも干渉効果に影響しているのかもしれない。ALSVを一次接種後にZYMVを二次接種した区では、ALSVの蓄積量が増加したのは、ZYMVとの重複感染によるsynergismが起こっていたものと考えられる(例えば、非特許文献7)。また、wtALSV接種区でも弱い干渉効果すなわち、植物体の矮化が認められなかったことは、このsynergismが影響しているのかもしれない。
【0075】
以上をまとめると、ALSVベクターにZYMVの遺伝子断片を組み込むことによってZYMVに対する干渉効果が誘導された。しかし、導入した遺伝子断片の種類、サイズ、位置によって干渉効果に強弱が見られ、実際にウイルスに対する干渉効果の誘導ベクターとして利用する場合には、ウイルス遺伝子断片導入ALSVを複数作製し、その効果を比較することが好ましい。
【実施例2】
【0076】
キュウリモザイクウイルス(CMV)の遺伝子断片を導入したALSVベクターによる干渉効果
1:材料と方法
1-1:実験材料
1-1-1:供試植物
キノア、Nicotiana tabacum cv Xanthi nc(タバコ)、とキュウリ(青大胡瓜)を供試した。
1-1-2:供試ウイルス
供試ウイルスは東北大学農学部准教授高橋英樹博士より分譲されたCucumber mosaic virus strain Y(以下CMV-Y)および、独立行政法人農業生物資源研究所より入手したCMV isolate 42CM(以下CMV-42CM、MAFF番号104087)である。
1-2:CMV遺伝子断片のALSVベクターへの導入
1-2-1:CMV遺伝子断片のクローニング
実施例1の1-4-5と同様の方法CMV-Y感染タバコ葉からでRNA抽出を行い、抽出したRNAを鋳型とし、既知のCMV-Y遺伝子配列をもとに設計したマイナス鎖のプライマーCMVY 1a(-)、CMVY 2a(-)、CMVY CP(-)を用いて(表2:配列番号10−15)、CMV-YのRNA1(accession number:D12537)、RNA2(accession number:D12538)、RNA3(accession number:D12499)に対する逆転写反応を以下の方法で行った。
【0077】
【表2】
【0078】
0.2ml容PCRチューブにRNA溶液を2μl、滅菌水を6μl、5×RT Bufferを4μl、2.5mM dNTP mixture(TaKaRa)を8μl、10μM のマイナス鎖プライマーを1μl、RNase Inhibitorを0.5μl、ReverTra Aceを0.5μl 混合し、TaKaRa PCR Thermal Cycler Dice VersionIII Model TP600を使用して42℃で50分間処理し、その後95℃で5分間処理し、最後に4℃で5分間処理し、獲られた産物をRT産物とした。
【0079】
得られたRT産物を鋳型とし、1a遺伝子(750 nt)の増幅にはCMVY 1a(+)とCMVY 1a(-)、2a遺伝子(663 nt)の増幅にはCMVY 2a(+)とCMVY 2a(-)、CP遺伝子(558 nt)の増幅にはCMVY CP(+)とCMVY CP(-)の組み合わせ(第2表)のプライマーを用いたPCRによる各遺伝子断片の増幅とPCR産物のサイズ確認、PCR産物のMonoFas DNA精製キットIを用いた精製を、実施例1の1-2-1と同様の方法で行い、精製した遺伝子断片をそれぞれC:1a遺伝子断片、C:2a遺伝子断片、C:CP遺伝子断片とした。
【0080】
精製した3種の遺伝子断片を用いたpGEM-T Easy Vector SystemによるTAクローニングと、そのライゲーション産物を用いた大腸菌DH5αの形質転換、スモールスケール培養と煮沸法によるプラスミド抽出、抽出プラスミドの1%アガロースゲルを用いた電気泳動による選抜とEcoRIを用いた制限酵素処理によるインサート保持の確認は、実施例1の1-2-1と同様の方法で行った。
1-2-2:CMV遺伝子断片のシークエンス
TAクローニングによって作製したプラスミドは、実施例1の1-2-2と同様の方法でプラスミドの精製とシークエンス解析を行った。配列を確認したプラスミドをそれぞれpGEM-C:1a、pGEM-C:2a、pGEM-C:CPとした(図23)。
1-2-3: CMV遺伝子断片のALSVベクターへの導入
pGEM-C:1aを鋳型にCMV-Y 1a Xho(+)とCMV-Y 1a Bam(-)、pGEM-C:2aを鋳型にCMV-Y 2a Xho(+)とCMV-Y 2a Bam(-)、pGEM-C:CPを鋳型にCMV-Y CP Xho(+)とCMV-Y CP Bam(-)の組み合わせ(表3:配列番号16−21)で、実施例1の1-2-1と同様の方法でPCRを行った。PCR産物のBamHIとXhoIを用いた制限酵素処理、MonoFas DNA精製キットIを用いた精製はII-1-(2)-3)と同様の方法で行った。各種CMVインサートDNAとベクターDNAをDNA Ligation Kit Ver.2.1を用いて行い大腸菌DH5αを形質転換後、スモールスケール培養し、煮沸法によってプラスミドを抽出した。
【0081】
【表3】
【0082】
各CMVインサートDNAを保持するクローンは、それぞれ1μg/μlの濃度に調整し、得られたプラスミドをALSVR2L5R5-C:1a:、 ALSVR2L5R5-C:2a、ALSVR2L5R5-C:CPとした(図24)。
1-2-4:ALSVベクターの接種とウイルス化
精製した各プラスミド(ALSVR2L5R5-C:1a、ALSVR2L5R5-C:2a、ALSVR2L5R5-C:CP)は、実施例1の1-2-4と同様の方法でウイルス化を行い、各プラスミドの接種によって得たウイルスをそれぞれALSV-C:1a、ALSV-C:2a、ALSV-C:CPとし、以後の試験の接種源とした。
1-3:供試植物への接種法と一次接種および二次接種
1-3-1:一次接種
タバコへの一次接種は以下のとおり行った。wtALSV、ALSV-C:1a、ALSV-C:2a、ALSV-C:CPにそれぞれ感染したキノア葉を2倍容のALSV磨砕緩衝液で磨砕した。作製した各ALSV磨砕液を5葉期のタバコの第3から第5本葉の3枚に、カーボランダム法で接種した。タバコは25℃の長日条件で育成した。
【0083】
キュウリへの一次接種はII-(3)-1)と同様の方法で行い、25℃の長日条件で育成した。
1-3-2:二次接種
タバコでの二次接種にはCMV-Yを用いて以下の通り行った。CMV-Y感染タバコ葉を2倍容の0.1Mリン酸緩衝液(pH7.0)で磨砕し、第7から第9本葉の3枚にカーボランダム法で二次接種した。二次接種後、タバコは一次接種後と同様に25℃の長日条件で育成した。
【0084】
キュウリへの二次接種にはCMV-42CMを用いて以下の方法で接種した。CMV-42CM感染タバコ葉を2倍容の0.1Mリン酸緩衝液(pH7.0)で磨砕し、第1本葉1枚にカーボランダム法で二次接種した。二次接種後、キュウリは一次接種後と同様に25℃の長日条件で育成した。
【0085】
なお、二次接種に用いたCMV-YとCMV-42CMは、あらかじめ0.1Mリン酸緩衝液(pH7.0)を用いてタバコに接種し、病徴の現れたタバコの上葉を採取し、-80℃で凍結保存したものを使用した。
1-4:病徴観察とウイルス検定
1-4-1:病徴の観察
病徴の観察は二次接種後毎日行った。タバコは二次接種後9日目に、キュウリは二次接種後18日目にデジタルカメラFinePix S1Proを用いて病徴を撮影した。
1-4-2:ウエスタンブロット法
一次接種後7日目に、タバコでは第3本葉から第5本葉を採取し、キュウリでは子葉2枚を採取し、II-1-(4)-3)と同様の方法でウエスタンブロットを行い、一次接種したALSVの感染を確認した。
1-4-3:ELISA法
タバコにおけるCMV-YとALSVの蓄積量は以下のように検定試料を作製し、直接ELISA法で定量した。二次接種後9日目に第12本葉から第15本葉を採取し、乳鉢と乳棒を用いて20倍容のPBSTで磨砕し、磨砕液1mlを1.5ml容マイクロチューブに移した。14,000rpm(4℃)で5分間遠心し、得られた上清を検定試料とした。上記の方法で作製した検定試料を用いて、実施例1の1-4-4の直接ELISA法でウイルスの定量をした。
【0086】
キュウリにおけるCMV-42CMとALSVの蓄積量は以下のように検定試料を作製し、実施例1の1-4-4の直接ELISA法で定量した。二次接種後18日目に第3本葉から第6本葉のそれぞれからII-1-(4)-4)と同様にリーフディスクを6枚採取し、検定試料を作製した。
【0087】
直接ELISA法によるCMV-YおよびCMV-42CMの定量には、CMV抗体(日本植物防疫協会研究所)と酵素標識をしたCMV抗体(日本植物防疫協会研究所)を使用した。
1-4-4:RNA抽出法とRT-PCR法
タバコでは二次接種後9日目にタバコの最上位葉を採取し、キュウリでは二次接種後18日目にキュウリの最上位葉を採取し、それぞれ-80℃で凍結し、検定試料とした。RNA抽出と逆転写反応およびPCRは全て実施例1の1-4-5と同様の方法で行い、ALSVのインサート配列の保持を確認した。
2:結果
2-1:タバコにおけるCMV-Y遺伝子断片を導入したALSVベクターの干渉効果
CMV-Y遺伝子断片を導入したALSVベクターの接種によって、タバコでCMV-Yに対する干渉効果の誘導が可能かどうか調べるため、wtALSVおよびCMV-Yの各遺伝子断片を導入したALSV(ALSV-C:1a 、ALSV-C:2a、ALSV-C:CP)を一次接種したタバコにCMV-Yを二次接種し、CMV-Yに対する干渉効果の誘導を調べた。
【0088】
ALSVの一次接種を行わずCMV-Yのみを二次接種した試験区(CMV-Yのみ区)やwtALSVを一次接種後CMV-Yの二次接種を行った試験区(wtALSV+CMV-Y区)では、二次接種後約3日で全個体の接種葉で黄斑が観察され、約7日には上葉に激しいモザイク症状が観察された。これに対して、ALSV-C:2aを一次接種後にCMV-Yのを二次接種した試験区(ALSV-C:2a+CMV-Y区)では、全ての個体で病徴は観察されなかった。ALSV-C:1aを一次接種後にCMV-Yを二次接種した試験区(ALSV-C:1a+CMV-Y区)では、6個体中3個体の上葉に弱いモザイク症状が観察され、残りの3個体では病徴が観察されなかった。一方、ALSV-C:CPを一次接種後にCMV-Yを二次接種した試験区(ALSV-C:CP+CMV-Y区)では6個体全ての上葉に弱いモザイク症状が観察された(図25、表4)。
【0089】
【表4】
【0090】
CMV-Yの蓄積を調べたところ、ALSV-C:2a+CMV-Y区ではCMV-Yの蓄積は認められなかった。これに対して、ALSV-C:CP+CMV-Y区ではCMV-Yのみ区やwtALSV+CMV-Y区と同程度蓄積していた。ALSV-C:1a+CMV-Y区では、CMV-Yのみ区やwtALSV+CMV-Y区の3分の1程度蓄積していた(図26)。ALSVの定量ではALSV-C:1a+CMV-Y区とALSV-C:2a+CMV-Y区ではALSVのみ区と同程度蓄積し、wtALSV+CMV-Y区ではALSVのみ区に比べ蓄積量が若干増加していることが確認された。一方、ALSV-C:CP+CMV-Y区では、ALSVのみ区の半分から4分の1程度少なく蓄積していることが確認された(図26)。一次接種に用いたCMV-Yの各遺伝子断片を導入したALSV(ALSV-C:1a 、ALSV-C:2a、ALSV-C:CP)のインサートチェックを行ったところ、全てのキュウリ個体でALSVのインサートが保持されていた。
2-2:キュウリにおけるCMV-Y遺伝子断片を導入したALSVベクターの干渉効果
wtALSVおよびCMV-Yの各遺伝子断片を導入したALSV(ALSV-C:1a 、ALSV-C:2a、ALSV-C:CP)を一次接種したキュウリにおけるCMV-42CMに対する干渉効果を調べた。
【0091】
CMV-42CMのみ二次接種した試験区(CMV-42CMのみ区)やwtALSVを一次接種後にCMV-42CMを二次接種した試験区(wtALSV+CMV-42CM区)では、二次接種後約3日で全ての個体の第2本葉で退緑斑が観察され、その後展開した上葉で退緑斑が観察された。特に、第6本葉から第8本葉ではモザイク症状が観察された。これに対し、ALSV-C:2aを一次接種後にCMV-42CMを二次接種した試験区(ALSV-C:2a+CMV-42CM区)では、全個体で病徴が観察されなかった。ALSV-C:1aを一次接種後にCMV-42CMを二次接種した試験区(ALSV-C:2a+CMV-42CM区)では、6個体中2個体で接種葉に退緑斑が観察され、第2本葉から第4本葉で退緑斑観察された。第5本葉より上の葉では病徴が観察されなかった。また、残り4個体では病徴が観察されなかった。ALSV-C:CPを一次接種後にCMV-42CMを二次接種した試験区(ALSV-C:CP+CMV-42CM区)では6個体全てにCMV-42CMのみ区やwtALSV+CMV-42CM区と同様の病徴が観察された(図27)。
【0092】
CMV-42CMの定量の結果、ALSV-C:2a+CMV-42CM区ではCMV-42CMの蓄積は認められなかった。これに対し、ALSV-C:CP+CMV-42CM区では、CMV-42CMのみ区やwtALSV+CMV-42CM区と同程度CMV-42CMが蓄積し、ALSV-C:1a+CMV-42CM区では、約半分のZYMVが蓄積していた(図28)。また、ALSV-C:1a+CMV-42CM区とALSV-C:2a+CMV-42CM区では、ALSVのみ区と同程度のALSVが蓄積し、wtALSV+CMV-42CM区ではALSVのみに比べ蓄積量が増加していた。一方、ALSV-C:CP+CMV-42CM区では、ALSVのみ区より少なく蓄積していた(図28)。一次接種に用いたCMV-Yの各遺伝子断片を導入したALSV(ALSV-C:1a 、ALSV-C:2a、ALSV-C:CP)のインサートチェックの結果、全てのALSVがインサートを保持していた。
3:考察
タバコおよびキュウリの両植物のALSV-C:2a を接種した試験区では、CMV-YおよびCMV-42CMに対する干渉効果が誘導され、病徴やウイルスの蓄積は確認されなかった。これに対し、ALSV-C:1aおよびALSV-C:CPを接種した試験区では穏かな病徴が観察され、CMV-YおよびCMV-42CMの蓄積が確認された。この干渉効果の違いはALSVベクターに導入したそれぞれの遺伝子断片の違いであると考えられる。
【0093】
2a遺伝子断片を組み込んだALSV-C:2a ではCMV-YおよびCMV-42CMの病徴やウイルスの蓄積が全く確認されなかったことから、ALSVベクターによる干渉効果の誘導はタバコとキュウリの両植物においても可能であることが示され、ALSVベクターの宿主植物において干渉効果誘導ベクターとしての利用が期待される。
【0094】
ALSVに導入した各CMV-Y遺伝子断片領域とそれに対応するCMV-42CMの遺伝子領域の塩基配列の相同性を比較したところ、1a遺伝子では90.3%、2a遺伝子では98.1%、CP遺伝子では97.9%相同であった。
【0095】
本実施例2では、既知のCMV-Y遺伝子を基に構築した2a遺伝子断片を導入したALSVによって、2つのCMV(CMV-YとCMV-42CM)に対して干渉効果を誘導することができた。この結果から、塩基配列の相同性が保存されている遺伝子領域の断片を導入したALSVベクターを宿主に接種することで、異なる系統や分離株に対する干渉効果を誘導可能であることが示された。
【0096】
実施例1の結果と合わせて考察すると、ALSVベクターは異なる属のウイルスにおいて干渉効果の誘導に成功したことから、ALSVベクターに導入する遺伝子断片を変えることで、多数のウイルスに対する干渉効果を誘導する防除剤(植物ワクチン)の作出が短期間で可能であることが示された。
【実施例3】
【0097】
SMVの遺伝子断片を導入したALSVベクターによる干渉効果
SMVの外被タンパク質(CP)をコードするゲノム配列の一部(201 nt)を挿入したALSV(SMVCP-ALSV)を作出し、SMVに対する干渉効果を試験した。SMVCP-ALSVをダイズ品種(Jackおよび丹波黒)の子葉に1次接種後、展開した初生葉もしくは第一本葉にSMVをカーボランダム法により2次接種(チャレンジ接種)した。
【0098】
その結果、SMV単独接種区では上葉にモザイクと縮葉症状を示したのに対し、SMVCP-ALSVを1次接種した区では病徴はほとんど観察されなかった。また、SMV単独接種区と比較するとSMVの増殖量が著しく低下しており、SMVCP-ALSVによる干渉効果が生じていることが確認された。また、上葉におけるSMVの分布をティッシュブロット分析により解析したところ、SMV単独接種個体ではSMVが葉全面にモザイク状に分布しているのに対し、SMVCP-ALSVを1次接種した個体ではウイルスが葉脈とその近傍のみに局在していることが明らかとなった。
【実施例4】
【0099】
ZYMVとCMVの遺伝子断片を導入したALSVベクターによる干渉効果
実施例1と同様のZYMV遺伝子断片(150 nt)と実施例2のCMV遺伝子断片(150 nt)を融合した形で挿入したALSV(ZC-ALSV)を作出し、ZYMVとCMVに対する干渉効果を試験した。ZC-ALSVをキュウリ子葉に1次接種後、5〜6日後に第1本葉にZYMVとCMVをそれぞれ単独で、または混合し2次接種(チャレンジ接種)した。
【0100】
その結果、対象区では上葉にZYMVの奇形を伴う激しいモザイク(ZYMV2次接種区)や激しいモザイク(CMV2次接種区)、激しいモザイクと矮化症状(混合2次接種区)が現れたのに対して、ZC-ALSV1次接種区ではZYMV単独、CMV単独、ZYMVとCMV混合のいずれの2次接種区に対しても干渉効果を示し、明瞭な病徴は認められず、また各ウイルスの蓄積も抑制されていた。
【0101】
以上の結果から、2種類のウイルスを同時に抑制するALSV混合ワクチン株の作出が可能であることが確認された。
【技術分野】
【0001】
本願発明は、農作物等の植物への病原類する感染に対する防除剤の成分等として有用な組換えALSVに関するものである。
【背景技術】
【0002】
1.リンゴ小球形潜在ウイルス
リンゴ小球形潜在ウイルス(Apple latent spherical virus:ALSV)は2分節1本鎖RNAゲノム(RNA1、RNA2)と3種類の外被タンパク質(Vp25、Vp20、Vp24)からなる径25nmの球形ウイルスである(非特許文献1)。ALSVはリンゴから分離されたウイルスであるが、比較的広い宿主範囲を持つウイルスで、Chenopodium quinoa(キノア)には退緑症状を示すが、モデル植物のシロイヌナズナをはじめ、Nicotiana属植物(タバコ、ベンサミアナタバコ、オキシデンタリス、グルチノーザ)、トマト、ウリ科植物(キュウリ、メロン、ズッキーニ、ヘチマ)、マメ科植物(ダイズ、アズキ、エンドウ)、バラ科果樹植物(リンゴ、セイヨウナシ、ニホンナシ、モモ)など多くの植物種に無病徴感染する。本願発明者らは既に、ALSVの感染性cDNAクローンを構築し、RNA2の細胞間移行タンパク質(movement protein:MP)とVp25の間に外来遺伝子導入サイトを付加したALSVベクターによる草本植物やリンゴでの外来遺伝子の発現に成功している(例えば、非特許文献2)。また、GFP(green fluorescent protein)を発現するタバコにGFPを組み込んだALSVベクターを接種すると、タバコで発現しているGFPのサイレンシングが誘導され、接種葉ではGFP蛍光が円状に消失し、やがて上位葉全体でGFP蛍光が消失することが明らかにされた(例えば、非特許文献3)。さらに、植物の内在性遺伝子であるphytoene desaturase(PDS)遺伝子やsulfer(SU)遺伝子の一部を組み込んだALSV感染により、タバコ、シロイヌナズナ、ダイズ、キュウリなどにおいてウイルス誘導ジーンサイレンシング(virus induced gene silencing:VIGS)が誘導され、ALSVが感染植物において全身的なVIGSを誘導することが明らかにされた(例えば、非特許文献4)。
【0003】
2.RNAサイレンシング
RNAサイレンシングは1990年に植物で初めて報告された真核生物に普遍的に保存されている配列特異的な遺伝子不活化機構であり(非特許文献5)、主にdouble-stranded RNA(dsRNA)により誘導される。RNAウイルスが感染すると、RNA依存RNAポリメラーゼ(RdRp)の働きでdsRNAが合成され、これが2本鎖RNA分解酵素であるDicer(植物の場合Dicer-like:DCL)によって21から25 塩基(nucleotide:nt)のsmall interfering RNA(siRNA)に分解される(非特許文献6、7)。siRNAはRNA-induced silencing complex(RISC)に取り込まれ、siRNAと相補的な塩基配列のRNAはRISCにより分解される。また、siRNAは隣接する細胞へ移行し、RdRpの働きによってsiRNAと相補的なRNAがdsRNAに変換され、サイレンシングは全身的に拡大する。免疫機構を持たない植物において、RNAサイレンシングはウイルスに対する防御機構として重要な役割を持っている(非特許文献8)。1995年、植物内在性遺伝子を組み込んだTMV感染によりその遺伝子のRNAサイレンシングが誘導されることが報告され(非特許文献9)、その後VIGSは逆遺伝学的手法として広く利用されている。
【0004】
3.植物の病原ウイルスとその対策
近年、ズッキーニ黄斑モザイクウイルス(Zucchini yellow mosaic virus:ZYMV)の感染によるウリ科作物の被害が世界で問題となり、日本国内においてもキュウリにおける被害が深刻化している。ZYMVはPotyvirus属に属し、サイズが約9,600ntの1本鎖RNAゲノムを持つ、長さ約750nmのひも状ウイルスである。アブラムシによる非永続伝搬によって蔓延し、特にウリ科植物(キュウリ、カボチャ、メロン、スイカなど)において激しいモザイク、葉および果実の奇形、植物体の萎縮等の症状を引き起こすことが知られている(非特許文献10、11)。
【0005】
キュウリモザイク病はZYMVと、Potyvirus属のカボチャモザイクウイルス(Watermelon mosaic virus 2:WMV2)およびCucumovirus属のキュウリモザイクウイルス(Cucumber mosaic virus:CMV)の3種が病原ウイルスとされ、これらウイルスの単独および重複感染によって発生する(非特許文献11)。CMVはCucumovirus属のタイプ種で、3分節の1本鎖RNAゲノム(RNA1、RNA2、RNA3)およびサブゲノムRNA(RNA4)を持つ径29nmの球状ウイルスである。宿主や病原性の異なる系統が多数存在し、なかにはサテライトRNAを有するものも存在する。CMVはアブラムシによって非永続伝搬され、その宿主範囲は極めて広く1,000種以上の植物に感染する(非特許文献10)。
【0006】
ZYMVおよびCMVによる農作物の被害に対し、抵抗性品種の利用やアブラムシ防除に加えて、病原性の弱いZYMVやCMV(弱毒ZYMV、弱毒CMV)を用いた干渉効果を利用した防除が行われている(非特許文献12、13)。干渉効果は、あるウイルスに感染した植物はその後同種のウイルスに感染しない現象で、その発見は古く1929年にタバコモザイクウイルスで最初に報告された。干渉効果の機構については、ウイルスの外被タンパク質(coat protein:CP)を介する干渉効果とRNAを介する干渉効果の2つが主なものと考えられている(非特許文献14、15)。
【0007】
ZYMVやCMVに対する防除手段としての弱毒ウイルスに関する発明としては、特許文献1、2が知られており、特許文献3には、同一ウイルスグループ内の2種以上の弱毒ウイルスを混合した混合弱毒型ウイルスが開示されている。また、ウイルスのサテライトRNAを当該ウイルスに組込んで弱毒ウイルスとする技術として特許文献4、5が知られている。さらに市販品として、日本では、生物農薬として製剤化された弱毒ZYMVが販売され、キュウリで利用されている(非特許文献7、12)。また、トマトではサテライトRNAを有する弱毒CMVを感染させた苗が販売されている(非特許文献16)。
【0008】
ただし、病原ウイルスと異なるベクターウイルスに病原ウイルス遺伝子を組込んだ組換えウイルスを病原ウイルス感染の防除手段として用いる技術は全く知られていない。
【特許文献1】特開平5-68540号公報
【特許文献2】特開平10-203901号公報
【特許文献3】特開2000-264806号公報
【特許文献4】特開平5-3789号公報
【特許文献5】特開平11-279013号公報
【非特許文献1】Li,C., Yoshikawa,N., Takahashi,T., Ito,T., Yoshida,K. and Koganezawa,H. (2000). Nucleotide sequence and genome organization of apple latent spherical virus:a new virus classified into the family Comoviridae. J.Gen.Virol.81:541-547.
【非特許文献2】Li,C., Sasaki,N., Isogai,M. and Yoshikawa,N. (2004). Stable expression of foreign proteins in herbaceous and apple plants using Apple latent spherical virus RNA2 vectors. Arch. Virol. 149:1541-1558.
【非特許文献3】Yaegashi,H., Yamatsuta,T., Takahashi,T., Li,C., Isogai,M., Kobori,T., Ohki,S. and Yoshikawa,N. (2007). Characterization of virus-induced gene silencing in tobacco plants infected with apple latent spherical virus. Arch. Virol. 152:1839-1849.
【非特許文献4】Igarasi,A., Yamagata,K., Sugai,T., Takahashi,Y., Sugawara,E., Tamura,A., Yaegashi,H., Yamagishi,N., Takahashi,T., Isogai,M., Takahashi,H. and Yoshikawa,N. (2009). Apple latent spherical virus vectors for reliable and effective virus-induced gene silencing among abroad range of plants including tobacco, tomato, Arabidopsis thaliana, cucurbits, and legumes. Virology386:407-416.
【非特許文献6】Hamiltom,A., Voinnet,O., Chappell,L. and Baulcombe,D. (2002). Two classes of short interfering RNA in RNA silencing. EMBO. J.21:4671-4679.
【非特許文献7】Kosaka,Y. and Fukunishi,T. (1997). Multiple Inoculation with Three Attenuated Viruses for the Control of Cucumber Virus Disease. Plant Dis.81:733-738.
【非特許文献8】Voinnet O. (2001). RNA silencing as a plant immune system against viruses. Trends Genet. 17:449-459.
【非特許文献9】Kumagai,M.H., Donson,J., Della-Cioppa,G., Harvey,D., Hanley,K. and Grill,L.K. (1995). Cytoplasmic inhibition of carotenoid biosynthesis with virus-derived RNA. Proc. Natl. Acad. Sci. USA 92:1679-1683.
【非特許文献10】畑中正一編 (1997). ウイルス学. 朝倉書店. pp.427-434, 461-466.
【非特許文献11】岸國平編. (1988). 作物病害事典. 全国農村教育協会. pp.321-322
【非特許文献12】Kosaka,Y., Ryang,Bo-Song., Kobori,T., Shiomi,H., Yasuhara,H. and Kataoka,M. (2006). Effectiveness of an Attenuated Zucchini yellow mosaic virus Isolate for Cross-Protecting Cucumber. Plant Dis. 90:67-72.
【非特許文献13】Wang,Wei-Qin., Natsuaki,T. and Kosaka,Y. (2006). Comparison of the nucleotide and amino acid sequences of parental and attenuated isolates of Zucchini yellow mosaic virus. J. Gen. Plant Pathol. 72:52-56.
【非特許文献14】Beachy,R.N. (1999). Coat-protein-mediated resistance to tobacco mosaic virus:discovery mechanisms and exploitation. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 354:659-664.
【非特許文献15】Lin,S-S., Henriques,R., Wu,H-W., Niu,Q-W., Yen,S-D. and Chua,N-H. (2007). Strategies and mechanisms of plant virus resistance. Plant Biotechnol. Rep. 1:125-134.
【非特許文献16】津田和久, 小坂能尚, 小堀崇, 塩見寛, 六角啓一, 片岡光信. (2005). キュウリモザイクウイルス弱毒株CM95を接種したトマトの収量及びビタミンC含量に及ぼす肥培管理の影響. 日植病報 71:1-5.
【発明の概要】
【発明が解決しようとする課題】
【0009】
前記のとおり、植物の病原ウイルスの感染に対して、弱毒ウイルスの干渉効果を利用した防除が知られているが、弱毒ウイルスを用いた防除法は、弱毒ウイルスが利用できる一部のウイルスに限られており、弱毒ウイルスの自然界からの分離や変異誘導による作製が困難なウイルスには応用の難しい技術である。また、異なるウイルスに対して、弱毒ウイルスによる干渉効果は発揮されないため、各ウイルスごとに弱毒ウイルスが必要とされているが、弱毒ウイルスの作製には長い時間と多くの労力を必要とする。
【0010】
またサテライトRNAの挿入による弱毒ウイルスの場合には、親株ウイルスは病原性であり、植物体内での増殖の過程で強毒ウイルスに回帰する危険性は皆無ではない。
【0011】
本願発明は、以上のとおりの事情に鑑みてなされたものであり、理論的にはあらゆる病原性ウイルスを対象として、それぞれに干渉効果を発揮するウイルスを作製でき、また増殖の過程での強毒化の危険性のない新しい手段を提供することを課題としている。
【課題を解決するための手段】
【0012】
前記課題を解決するための発明として、1種または2種以上の植物病原ウイルスのゲノムRNAまたはそのcDNAの一部断片を保有する組換えリンゴ小球型潜在ウイルス(ALSV)を提供する。
【0013】
この組換えALSVにおいては、植物病原ウイルスがズッキーニ黄斑モザイクウイルス(ZYMV)、キュウリモザイクウイルス(CMV)、またはダイズモザイクウイルス(SMV)であることを好ましい態様としている。
【0014】
さらにこの出願は、前記の組換えALSVを植物の幼苗に感染させることを特徴とする植物病原ウイルスの防除方法と、前記の組換えALSVが幼苗期に接種された病原ウイルス抵抗性植物を提供する。
【発明の効果】
【0015】
本願発明では、植物病原ウイルスのゲノムRNAまたはそのcDNAの一部断片を組込んだ組換えALSVによって当該病原ウイルスの感染を防除する。対象となる病原ウイルスに制限はなく、あらゆる病原ウイルスの感染を防除することができる組換えALSVの作製が理論的には可能である。組換えALSVの作成は、当該技術分野の通常の知識を有するものであれば、本願明細書の記載に基づき容易に行なうことができ、従来の弱毒ウイルスに比して、極めて少ない時間と労力で作成することができる。
【0016】
また、ALSVは無病徴感染ウイルスであり、このALSVに挿入する病原ウイルスのゲノムRNAまたはそのcDNAの一部断片は、それ自体では病原性を持たないものであるため、組換えALSVが植物体内で毒性を獲得する危険性はなく、安全に使用することができる。
【図面の簡単な説明】
【0017】
【図1】ALSVの感染性cDNAクローンとALSVベクターの構成を示す。pEALSR1:ALSV RNA1の感染性cDNAクローン、pEALSR2L5R5:ALSV RNA2の感染性cDNAクローン、P35S:35S promoter、Tnos:nopaline synthase terminator、Q/G:タンパク質の切断部位、PRO-co:protease cofactor、HEL:NTP-binding helicase、C-PRO:cystein protease、POL:RNA polymerase、MP:movement protein、VP25, VP20, VP24:capsid protein。
【図2】実施例1においてZYMV遺伝子断片を導入したALSVベクターの構築工程を示す。
【図3】実施例1においてリーフディスクを採種した場所を示す。
【図4】実施例1の結果を示す。第1本葉二次接種区のキュウリ(品種:青大胡瓜)のZYMVによる病徴(21dpi)である。写真左上:(A)第5本葉、写真左下:(B)第6本葉、写真右:(C)植物全体。(A)(B)は左上から右下の順に、(C)は左から順に、それぞれ、1:非接種区、2:ALSVのみ区、3:ZYMVのみ区、4:wtALSV+ZYMV区、5:ALSV-Z:CP200+ZYMV区。
【図5】実施例1の結果を示す。ZYMVのみ区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図6】実施例1の結果を示す。wtALSV+ZYMV区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図7】実施例1の結果を示す。ALSV-Z:CP200+ZYMV区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図8】実施例1の結果である。第1本葉二次接種区のキュウリ(品種:青大胡瓜)におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図9】実施例1の結果である。第2本葉二次接種区のキュウリ(品種:青大胡瓜)におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図10】実施例1の結果である。第3本葉二次接種区のキュウリ(品種:青大胡瓜)におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図11】実施例1の結果である。異なるZYMV遺伝子断片を導入したALSVベクターを一次接種したキュウリ(品種:青大胡瓜)の各試験区における病徴(21dpi)である。写真左上:(A)第5本葉、写真左下:(B)第6本葉、写真右:(C)植物全体。(A)(B)は左上から右下の順に、(C)は左から順に、それぞれ、1:非接種区、2:ALSVのみ区、3:ZYMVのみ区、4:wtALSV+ZYMV区、5:ALSV-Z:P1+ZYMV区、6:ALSV-Z:P3+ZYMV区、7:ALSV-Z:CP200+ZYMV区。
【図12】実施例1の結果を示す。ALSV-Z:P3+ZYMV区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図13】実施例1の結果を示す。ALSV-Z:P1+ZYMV区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図14】実施例1の結果である。異なるZYMV遺伝子断片を導入したALSVベクターを一次接種したキュウリ(品種:青大胡瓜)の第1本葉二次接種区におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図15】実施例1の結果である。キュウリ(品種:鈴成四葉)の第1本葉二次接種区におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図16】実施例1の結果である。キュウリ(品種:つばさ)の第1本葉二次接種区におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図17】実施例1の結果である。長さの異なるZYMV遺伝子断片を導入したALSVベクターを一次接種したキュウリ(品種:青大胡瓜)の各試験区における病徴(21dpi)である。写真左上:(A)第5本葉、写真左下:(B)第6本葉、写真右:(C)植物全体。(A)(B)は左上から右下の順に、(C)は左から順に、それぞれ、1:非接種区、2:ALSVのみ区、3:ZYMVのみ区、4:wtALSV+ZYMV区、5:ALSV-Z:CP100+ZYMV区、6: ALSV-Z:CP200+ZYMV区。
【図18】実施例1の結果を示す。ALSV-Z:CP100+ZYMV区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図19】実施例1の結果である。長さの異なるZYMV遺伝子断片を導入したALSVベクターを一次接種したキュウリ(品種:青大胡瓜)の第1本葉二次接種区におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図20】実施例1の結果である。弱毒ZYMVおよび異なるZYMV遺伝子断片を導入した各種ALSVを一次接種したキュウリ(品種:青大胡瓜)の各試験区における病徴(21dpi)である。(A)第5本葉、(B)植物全体。(A)(B)は左から順に、それぞれ、1:非接種区、2:ALSVのみ区、3:ZYMVのみ区、4:wtALSV+ZYMV区、5:ALSV-Z:P1+ZYMV区、6:ALSV-Z:P3+ZYMV区、7:ALSV-Z:CP200+ZYMV区、8:ZYMV 2002+ZYMV区。
【図21】実施例1の結果を示す。ZYMV 2002 +ZYMV区の第1本葉から第6本葉のGFP蛍光の分布(14dpi)である。
【図22】実施例1の結果である。弱毒ZYMVおよび異なるZYMV遺伝子断片を導入した各種ALSVを一次接種したキュウリ(品種:青大胡瓜)の第1本葉二次接種区におけるZYMVおよびALSV蓄積量を直接ELISA法により解析した結果。
【図23】実施例2におけるCMV-Y遺伝子のクローニングとCMV-Y遺伝子断片の構築工程。
【図24】実施例2におけるCMV-Y遺伝子断片を導入したALSVベクターの構築工程。
【図25】実施例2の結果である。各CMV-Y遺伝子断片導入ALSVベクターを一次接種したタバコにおけるCMV-Yの病徴(9dpi)。左上から右下の順に、非接種区、CMV-Yのみ区、ALSVのみ区、wtALSV+CMV-Y区、ALSV-C:1a+CMV-Y区、ALSV-C:2a+CMV-Y区、ALSV-C:CP+CMV-Y区。
【図26】実施例2の結果である。異なるCMV-Y遺伝子断片を導入した各種ALSVを一次接種したタバコのCMV-YおよびALSV蓄積量を直接ELISA法により解析した結果。
【図27】実施例2の結果である。各CMV-Y遺伝子断片導入ALSVベクターを一次接種したキュウリ(品種:青大胡瓜)におけるCMV-Yの病徴(18dpi)。(A)は植物全体、(B)は第6本葉、(A)は左から順に、(B)は左上から右下の順に、1:非接種区、2:ALSVのみ区、3:CMV-42CMのみ区、4:wtALSV+CMV-42CM区、5:ALSV-C:1a+CMV-42CM区、6:ALSV-C:2a+CMV-42CM区、7:ALSV-C:CP+CMV-42CM区。
【図28】実施例2の結果である。各CMV-Y遺伝子断片導入ALSVベクターを一次接種したキュウリ(品種:青大胡瓜)第1本葉二次接種区におけるCMV-42CMおよびALSV蓄積量を直接ELISA法により解析した結果。
【発明を実施するための形態】
【0018】
本願発明の組換えALSVは、植物病原ウイルスのゲノムRNAまたはそのcDNAの一部断片が挿入されていることを特徴とする。RNAサイレンシングの原理に従い、この組換えALSVが複製すると、挿入断片の配列を含むsiRNAが合成され、それに相補的な病原ウイルスゲノムの領域を破壊する。
【0019】
ALSVは、リンゴから単離したものを使用することもできるが、好ましくは、特許文献2に記載されているALSVのRNA1感染性cDNAクローン(pEALSR1)、RNA2感染性cDNAクローン(pEALSR2L5R5)などのALSVベクターを使用することができる(図1参照)。
【0020】
植物病原ウイルスとしては、実施例に示したキュウリモザイクウイルス(CMV)、ズッキーニ黄斑モザイクウイルス(ZYMV)、ダイズモザイクウイルス(SMV)の他、原則的にあらゆるウイルスを対象とすることができる。例えば、CMVやZYMVと同様にアブラムシ類により媒介されるカボチャモザイクウイルス(WMV-2)、ソラマメウイルトウイルス(BBWV)、カブモザイクウイルス(TuMV)など;アザミウマ類により媒介されるトマト黄化えそウイルス(TSWV)、インパチエンスネクロティックスポットウイルス(INSV)、アイリスイエロースポットウイルス(IYSV)など;コナジラミ類により媒介されるキュウリ黄化ウイルス(CuYV)など;土壌伝染するトマトモザイクウイルス(ToMV)などである。
【0021】
本願発明の組換えALSVは、これらの病原ウイルスの1種または2種以上(例えば3種)の各々のゲノムRNAまたはcDNAの一部断片を組合わせて(例えば、融合ポリヌクレオチドとして)ALSVベクターに挿入したものであってもよい。このような2種以上の病原ウイルスに対する組換えALSVは、いわゆる混合ワクチン様の機能を有し、同一植物個体への2種以上の病原ウイルスの感染を防御することができる。例えば、実施例に示したような、キュウリに対するズッキーニ黄斑モザイクウイルス(ZYMV)とキュウリモザイクウイルス(CMV)の感染防御である。
【0022】
ALSVへの挿入断片は、これらの病原ウイルスのゲノムRNA、もしくはそのcDNAから調製する。挿入断片は病原ウイルスゲノムの任意の領域から選択することができるが、好ましくは、病原ウイルスの生存、増殖、あるいはその病原性に関わる遺伝子領域等から選択することができる。例えば、実施例に示したように、ズッキーニ黄斑モザイクウイルス(ZYMV)の場合はP1、HC-Pro、P3、Cl、Nla、Nlb、CP(好ましくは、P3、Cl、Nla、Nlb、CP)、キュウリモザイクウイルス(CMV)の場合は1a、2a、CP(好ましくは2a)、ダイズモザイクウイルス(SMV)の場合はCPをそれぞれコードするゲノムRNAの一部領域またはこの領域に対応するcDNAである。
【0023】
挿入断片のサイズは、例えば、ALSVベクターに挿入した場合に、その植物個体への感染能、あるいは植物個体内での増殖能に影響を及ぼすことのないサイズの範囲内で任意に選択することができる。また、挿入断片のサイズは、挿入断片の配列を含むsiRNAが病原ウイルスゲノムを効果的に破壊し、かつその挿入断片が安定的に維持される範囲を選択する。すなわち、具体的には、挿入断片のサイズは例えば100〜300bp、好ましくは150〜200bp程度とすることができる。
【0024】
なお、この挿入断片がコードするペプチドが、ALSVペプチドとの融合ペプチドとして発現するように(すなわち、一部断片の挿入によって停止コドンが出現しないように)、フレームを一致させることが必要である。ALSVベクターを使用する場合、そのクローニング部位に一致した制限酵素で前記の一部断片を調製し、市販のライゲーションキット等を用いて一部断片をALSVベクターに挿入する。次いで、この組換えALSVベクターをキノアに感染させ、感染葉からウイルスを単離して組換えALSVを作成する。
【0025】
この組換えALSVは、そのまま植物に接種して病原ウイルスの感染防除に使用することができる。あるいは、安定剤等を含む緩衝液中で凍結乾燥させて感染防除剤として製品化することもできる(例えば、特許文献2など)。
【0026】
組換えALSV、または溶解した感染防除剤は、植物の幼苗の葉(例えば、キュウリの場合は子葉展開期の子葉、タバコの場合は5葉期の第3から第5本葉など)にカーボランダム法など公知の方法で接種することができる。
【0027】
以下、実施例を示して本願発明をさらに詳細かつ具体的に説明するが、本願発明は以下の例によって限定されるものではない。
【実施例1】
【0028】
ZYMVの遺伝子断片を導入したALSVベクターによる干渉効果
1:材料と方法
1-1:実験材料
1-1-1:供試植物
Chenopodium quinoa(以下キノア)をALSV増殖用植物として用いた。また干渉効果の検定にはCucumis sativus(以下キュウリ)のZYMV抵抗性を持たない3品種(青大胡瓜、鈴成四葉、つばさ)を供試した。
1-1-2.供試感染性クローン
供試感染性クローンは、非特許文献2等に記載されているALSVのRNA1感染性cDNAクローン(pEALSR1)とRNA2感染性cDNAクローン(pEALSR2L5R5)(図1)、および宇都宮大学農学部教授夏秋知英博士より分譲された、強毒株ZYMV isolate Z5-1(accession number:AB188115)の感染性cDNAクローン(p35S-Z5vector+GFP-FL)を供試した。また、(株)微生物化学研究所梁宝成博士より分譲された、弱毒株のZYMV isolate 2002(以下ZYMV 2002、accession number:AB188116)を供試した。
1-2:ZYMV遺伝子断片のALSVベクターへの導入
1-2-1:ZYMV遺伝子断片のクローニング
p35S-Z5vector+GFP-FLを鋳型とし、既知のZYMV isolate Z5-1のP1遺伝子、P3遺伝子およびCP遺伝子配列をもとに設計したプライマーを用いて、P1遺伝子(210 nt)の増幅にはZ:P1-Xho(+)とZ:P1-Bam(-)、P3遺伝子の増幅(189 nt)にはZ:P3-Xho(+)とZ:P3-Bam(-)、CP遺伝子の増幅(225 nt)にはZ:CP-Xho(+)とZ:CP-Bam(-)、CP遺伝子(117 nt)の増幅にはZ:CP-Xho(+)とZ:CP100-Bam(-)の組み合わせを用いた(表1:配列番号1−7)。
【0029】
【表1】
【0030】
2μlの鋳型DNA溶液(1ng/μl)を0.2ml容PCRチューブにとり、滅菌水を35μl、10×Ex TaqBuffer(TaKaRa)を5μl、2.5mM dNTP Mixture(TaKaRa)を5μl、10μMの各遺伝子断片増幅プライマーを各1μl、TaKaRa Ex TaqTM(TaKaRa)を1μl加えて混合し、TaKaRa PCR Thermal Cycler Dice VersionIII Model TP600(TaKaRa)を使用して94℃で5分間処理後、〔94℃、30秒→57℃、30秒→72℃、60秒〕という反応を30サイクル行い、その後72℃で5分間処理し、最後に4℃で5分間処理しPCRを終了した。得られたPCR産物は以下の方法で電気泳動を行った。Agarose S(ニッポンジーン)を0.15g、TAE[40mM Tris,40mM 酢酸,1mM EDTA(pH8.0)]を15ml、エチジウムブロマイドを0.6μl用いて作成した1%アガロースゲルをTAEで泳動槽内を満たしたMupid-2plus(ADVANCE)に設置し、2μlのPCR産物に1μlの10×Loading Buffer(TaKaRa)を混合し、アガロースゲルのウェルにアプライして電気泳動を行い、設計したサイズの断片であることを確認した。
【0031】
サイズの確認を行ったPCR産物はMonoFas DNA精製キットI(GL Sciences)を用いて以下のように精製した。1%アガロースを用いて泳動した20μl分のPCR産物のバンドをメスで切り取り、1.5ml容マイクロチューブに入れ、ゲルの重量を量った。ゲルの重量と等量のBuffer Aを加え60℃で10分間保温し、ゲルを完全に溶解した。コレクションチューブに取り付けたスピンカラムに溶解したゲルサンプル溶液を入れ、10,000rpm(4℃)で30秒遠心した。遠心後、スピンカラムに500μlのBuffer Bを入れ、10,000rpm(4℃)で30秒遠心した。遠心後、スピンカラムを1.5ml容マイクロチューブに付け替え、20μlのBuffer Cを入れ10,000rpm(4℃)で60秒遠心し、得られた溶液をZYMVの各遺伝子断片とした。
【0032】
各遺伝子断片はpGEM-T Easy Vector System(Promega)を用いて、TAクローニングを以下のように行った。1.5ml容マイクロチューブに4μlのP1遺伝子断片をとり、2×Rapid Ligation Bufferを5μl、pGEM-T Easy Vectorを1μl、T4 DNA Ligaseを1μl加えて混合後、室温で60分間静置しライゲーション反応を行った。ライゲーション反応を行ったサンプル溶液10μlを、あらかじめ氷上で解凍しておいた100μlの大腸菌DH5αのコンピテントセルに加え、穏やかに混合後氷上で20分間静置した。静置後、42℃のウォーターバスで45秒間ヒートショック処理を行い、ただちに氷上で2分間静置した。静置後、サンプルに1mlのSOC培地(pH7.5)[1Lあたりbacto tryptone 20g,bacto yeast extract 5g,NaCl 0.5g,1M MgCl2 10ml,1M MgSO410ml]を加え、37℃のシェイカーで60分間振盪培養した。培養後、0.15%の寒天を含むLB培地(pH7.5)[1Lあたりbacto tryptone 10g,bacto yeast extract 5g,NaCl 10g,アンピシリン(25mg/ml) 2ml]のプレートにサンプル50μlをスプレッドし、サンプルを塗り広げた。サンプルを塗り広げたプレートは37℃のインキュベーターに14~16時間静置した。
【0033】
プレートに形成されたコロニーをスモールスケールで培養し、煮沸法を用いた以下の方法でプラスミドの抽出を行った。オートクレーブ滅菌済みの爪楊枝を用いて、試験管内に分注した2mlのLB培地に植菌し、37℃に設定したシェイカーで8時間振盪培養した。培養液を1.5ml容マイクロチューブに移し、14,000rpm(室温)で1分間遠心した。遠心後、上清をアスピレーターで除去し、350μlのSTET[0.1M NaCl,10mM Tris-HCl(pH8.0),1mM EDTA(pH8.0),5% TritonX-100]を加えてボルテックスミキサーで沈殿を懸濁した。リゾチーム溶液[10mM Tris-HCl(pH8.0),10mg/mlリゾチーム]を25μl加え、3秒間ボルテックスミキサーで混合した。混合後、マイクロチューブを40秒間煮沸し、直ちに氷中に移して5分間冷却した。冷却後、14,000rpm(室温)で10分間遠心し、沈殿をオートクレーブ滅菌済みの爪楊枝で除去した。上清に40μlの3M酢酸ナトリウム(pH5.2)と420μlのイソプロピルアルコールを加えて混合し、-20℃の冷凍庫内に5分間静置した。静置後、14,000rpm(4℃)で遠心し、上清を除去した。マイクロチューブ内の沈殿に1mlの70%エタノールを加え、14,000rpm(4℃)で2分間遠心し、上清を除去後、減圧乾燥機でマイクロチューブ内の沈殿を2分間乾燥した。乾燥した沈殿を50μlのRNase溶液[10mM Tris-HCl(pH8.0),1mM EDTA(pH8.0),20μg/ml DNase free RNase ]懸濁した。懸濁後、37℃のインキュベーターで20分間静置し、プラスミド溶液とした。
【0034】
抽出したプラスミド溶液を2μl用いて、1%アガロースゲルで電気泳動を行い、インサートを保持していると考えられるプラスミドの選抜を行った。選抜したプラスミドはEcoRIを用いた制限酵素処理を以下の方法で行い、インサートの保持を確認した。マイクロチューブに2μlのプラスミドをとり、滅菌水を6.8μl、10×H Buffer(TaKaRa)を1μl、EcoRI(TaKaRa)を0.2μl加えて混合し、37℃のインキュベーターで3時間静置した。静置後、1%アガロースゲルで電気泳動を行い、インサートの保持を確認し、TAクローニングを終了した。
1-2-2:ZYMV遺伝子断片のシークエンス
TAクローニングによって得られたプラスミドは以下の方法で精製した。46μlのプラスミドに154μlのTE [10mM Tris-HCl(pH8.0),1mM EDTA(pH8.0)]、100μlのTE飽和フェノール、100μlのクロロホルムを加えボルテックスミキサーで5分間混合し、14,000rpm(4℃)で5分間遠心した。遠心後、200μlの水層を新しいマイクロチューブに移し、200μlのクロロホルムを加えボルテックスミキサーで混合し、14,000rpm(4℃)で5分間遠心した。150μlの水層を新しいマイクロチューブに移し、3M酢酸ナトリウム(pH5.2)を15μl、99%エタノールを375μl加えボルテックスミキサーで2分間混合し、-20℃の冷凍庫で20分間静置した。14,000rpm(4℃)で20分間遠心して上清を除去後、沈殿に1mlの70%エタノールを加え、14,000rpm(4℃)で5分間遠心した。遠心後、上清を除去し、減圧乾燥機で2分間乾燥後、50μlのTEに懸濁した。50μlのPEG溶液(13% polyethylene glycol 8000,1.6M NaCl)を加えて混合し、氷上に1時間静置した。静置後、14,000rpm(4℃)で20分間遠心し、沈殿に500μlの70%エタノールを加え、14,000rpm(4℃)で2分間遠心した。上清を除去し、沈殿を減圧乾燥機で2分間乾燥した。沈殿を20μlの滅菌水で懸濁し、NanoDrop ND-1000 Spectrophotometer(旭テクノグラス)を用いて吸光度(波長260nm)を測り、濃度を300ng/μlに調整し、精製プラスミドとした。
【0035】
精製プラスミドは以下の方法でシークエンス分析を行った。シークエンス分析に供試する泳動試料はBigDye Terminator v3.1 Cycle Sequencing Kit(Applied Biosystems)を用いて以下のように作成した。0.2ml容PCRチューブに1μlの精製プラスミドをとり、滅菌水を4μl、5×BigDye Sequencing Bufferを1μl、プライマーACUNI6745(+)を2μl、Ready reaction mixを2μl加えて混合し、TaKaRa PCR Thermal Cycler Dice VersionIII Model TP600(TaKaRa)を使用して94℃で1分間処理後、〔94℃、30秒→50℃、15秒→60℃、4分〕という反応を30サイクル行い、その後60℃で3分間処理し、最後に4℃で5分間処理し、PCRを終了した。PCR後、1.5ml容マイクロチューブに試料全量をとり、滅菌水を7.25μl、3M酢酸ナトリウムを1.5μl、99%エタノールを31.25μl加えてボルテックスミキサーで1分間混合し、アルミホイルで包んで遮光し、室温で10分間静置した。静置後、14,000rpm(4℃)で20分間遠心し、沈殿に1mlの70%エタノールを加え14,000rpm(4℃)で5分間遠心した。遠心後、減圧乾燥機で沈殿を乾燥し、20μlのHi-Di Formamidoに懸濁した。これを2分間煮沸し、ただちに氷上で5分間静置したものを泳動試料とした。
【0036】
試料の泳動はABI PRISM 3100-Avant Genetic Analyzerのマニュアルに従い行った。20μlの泳動試料をMicroAmp 96-well Reaction Plateのwellにのせ、プレートアセンブリを組み立て、オートサンプラーに設置した。キャピラリー長は80cm、ポリマーはPOP-4、泳動緩衝液は滅菌水で10倍に希釈した10×Genetic Analyzer Buffer with EDTAを用いた。次にプレートレコードを作成し、サンプル名、泳動条件および解析条件などを設定した。Dye SetはZ、Mobility FileはDT3100POP4{BDv3}v1.mob、BioLIMS Projectは3100_Project1、Run ModuleはLongseq80_POP4DefaultModule、Analysis ModuleはBC-3100APOP4_80cm_SeqOffFtOff.sazにそれぞれ設定し、泳動を行った。泳動終了後、ABI PRISM 3100-Avant Genetic Analyzerのマニュアルに従い、解析データを塩基配列に変換した。得られた塩基配列データは配列情報解析ソフトウェアのDNASIS Pro ver 2.09.000.001(HitachiSoft)を用いて解析を行い、作成したZYMV遺伝子断片が鋳型と同一であることを確認した。
1-2-3:ZYMV遺伝子断片のALSVベクターへの導入
配列の確認をした各ZYMV遺伝子断片はALSR2L5R5に導入するため、遺伝子断片の精製、BamHIとXhoIを用いた制限酵素処理を以下のように行った。
【0037】
各ZYMV遺伝子断片28μlにTEを72μl、TE飽和フェノールを50μl、クロロホルムを50μl加えボルテックスミキサーで3分間混合し、14,000rpm(4℃)で10分間遠心した。水層100μlを新しいマイクロチューブに移し、3M酢酸ナトリウム(pH5.2)を10μl、99%エタノールを250μl加えボルテックスミキサーで2分間混合し、-80℃の冷凍庫で15分間静置した。14,000rpm(4℃)で15分間遠心し、上清を除去し、沈殿に70%エタノールを1ml加え、14,000rpm(4℃)で5分間遠心した。上清を除去し、減圧乾燥機で沈殿を乾燥した。17μlの滅菌水に懸濁し、10×K Buffer(TaKaRa)を2μl、BamHI(TaKaRa)を0.5μl、XhoI(TaKaRa)を0.5μl加え穏やかに混合し、37℃のインキュベーターで12時間静置した。ALSR2L5R5も同様の方法で制限酵素処理を行った。
【0038】
制限酵素処理を行った各種ZYMV遺伝子断片およびALSR2L5R5はMonoFas DNA精製キットIを用いて前記1-2-1の方法で精製し、得られた溶液をそれぞれ精製Z:P1遺伝子、精製Z:P3遺伝子、精製Z:CP200遺伝子、精製Z:CP100遺伝子(ZYMVインサートDNA)および精製ALSR2L5R5(ベクターDNA)とした。
【0039】
各種ZYMVインサートDNAとベクターDNAにDNA Ligation Kit Ver.2.1(TaKaRa)を用いてライゲーション反応を以下の方法で行った。マイクロチューブに4μlの各種インサートDNAをとり、それぞれにベクターDNAを1μlとI液を5μlずつ加え、穏やかに混合し、16℃のウォーターバスで4時間ライゲーション反応を行った。各ライゲーション産物を用いて前記1-2-1と同様の方法で、大腸菌DH5αの形質転換とスモールスケール培養および煮沸法によるプラスミドの抽出を行った。抽出したプラスミドは1%アガロースで電気泳動し、インサートを保持していると考えられるものを選抜した。選抜したプラスミドはBamHIとXhoIを用いた制限酵素処理を以下の方法で行い、インサートの保持を確認した。マイクロチューブに2μlのプラスミドをとり、滅菌水を6.6μl、10×K Bufferを1μl、BamHIを0.2μl、XhoIを0.2μl加えて混合し、37℃のインキュベーターで3時間静置した。静置後、1%アガロースゲルで電気泳動を行い、各プラスミドのインサートの保持を確認した。
【0040】
インサートを保持したクローンはラージスケール培養を行い、QIAGEN Plasmid Midi Kit(QIAGEN)を用いて、遠心機にはRPR16ローター(HITACHI)を設置した高速冷却遠心機himac CR20G(HITACHI)を使用し、以下の方法で精製した。500ml容の坂口フラスコに100mlのLB培地を分注し、コロニーをオートクレーブ滅菌済みの爪楊枝で植菌し、37℃のシェイカーを用いて180rpmで14~16時間振盪培養した。培養液を50ml容の遠心管に移し、7,500rpm(4℃)で15分間遠心した。上清を除去し、残りの培養液を移して7,500rpm(4℃)で15分間遠心した。上清を除去し、沈殿を4mlのBuffer P1に懸濁した。懸濁液に4mlのBuffer P2を加えて転倒混和し室温で4分間静置した。直ちに4mlのBuffer P3を加えて転倒混和し、氷上で20分間静置した。13,000rpm(4℃)で30分間遠心し、上清を新しい50ml容の遠心管に移し、さらに13,000rpm(4℃)で15分間遠心した。4mlのBuffer QBTでカラムを平衡化したQIAGEN-tip 100に遠心上清を移し、自然落下でカラムにプラスミドDNAを吸着させた。QIAGEN-tip 100を10mlのBuffer QCで2回洗浄し、QIAGEN-tip 100を新しい50ml容スミロンチューブに設置し、5mlのBuffer QFでDNAを溶出した。溶出したDNA溶液にイソプロピルアルコールを3.5ml加えて混合し、直ちに15ml容のCOREXチューブに移し、11,000rpm(4℃)で30分間遠心した。上清を除去し、沈殿に2mlの70%エタノールを加え11,000rpm(4℃)で10分間遠心した。上清を除去し、減圧乾燥機で沈殿を乾燥した。沈殿を200μlのTEに懸濁し、NanoDrop ND-1000 Spectrophotometerを用いて吸光度(波長260nm)を測り、濃度を1μg/μlに調整した。各プラスミドはそれぞれALSVR2L5R5-Z:P1、ALSVR2L5R5-Z:P3、ALSVR2L5R5-Z:CP:200、ALSVR2L5R5-Z:CP:100とした(図2)。
1-2-4:ALSVベクターの接種とウイルス化
精製した各プラスミド(ALSVR2L5R5-Z:P1、ALSVR2L5R5-Z:P3、ALSVR2L5R5-Z:CP200、ALSVR2L5R5-Z:CP100)は、前記1-2-3の方法で精製したpEALSR1と等量ずつ混合し、DNA溶液とした。作製したDNA溶液を7葉期のキノアの第3から第6本葉に本葉1枚につき8μlずつ、カーボランダム法で接種した。接種後、退緑症状の現れた上葉をサンプリングし、2倍容のALSV磨砕緩衝液[0.1M Tris-HCl(pH7.8),0.1M NaCl,5mM MgCl2]で磨砕し、8葉期のキノアの第3から第8本葉にカーボランダム法で接種した。接種後、退緑症状の現れた上葉をサンプリングし、各ウイルスをそれぞれALSV-Z:P1、ALSV-Z:P3、ALSV-Z:CP200、ALSV-Z:CP100とし、以後の接種試験の接種源とした。また、ALSVR2L5R5も同様の方法でウイルス化し、得たウイルスをwtALSVとし、以後の接種試験の接種源とした。
1-3:キュウリへの接種法と一次接種および二次接種
1-3-1:キュウリへの一次接種
キュウリへの一次接種は各ALSV(ALSV-Z:P1、ALSV-Z:P3、ALSV-Z:CP200、ALSV-Z:CP100、wtALSV)およびZYMV 2002を用いて以下の方法で行った。各ALSV感染キノア葉を、それぞれ2倍容のALSV磨砕緩衝液で磨砕した。各ALSV磨砕液と、ZYMV 2002感染カボチャ葉を2倍容の0.1M リン酸緩衝液(pH7.0)で磨砕して調整したZYMV磨砕液を、子葉展開期のキュウリの子葉2枚にカーボランダム法で接種した。キュウリは25℃の長日条件で育成した
1-3-2:キュウリへの二次接種
一次接種したキュウリに、p35S-Z5vector+GFP-FL を以下の方法で金粒子にコーティングし、Helios Gene Gunシステム(BIO-RAD)を用いて二次接種した。1.5ml容マイクロチューブに径1.6μmの金粒子(BIO-RAD)を10mg量りとり、50μlの50%グリセロールを加えてボルテックスミキサーで混合した。超音波洗浄機で2分間ソニケーションを行い、ボルテックスミキサーで混合しながら25μlのp35S-Z5vector+GFP-FLを加え、次に150μlの1M CaCl2を加え、さらに150μlの0.05Mスペルミジンを加えて混合した。室温で15分間静置し、上清をピペットマンで除去した。沈殿を崩さないように1mlの99%エタノールを加えて洗浄し、ピペットマンで上清を除去した。同様の洗浄を3回行い、洗浄後、沈殿に500μlの99%エタノールを加え、指ではじいて懸濁し、金粒子の分散を確認した。懸濁した金粒子を全て15ml容チューブに移し、2.5mlの99%エタノールを加え混合した。調整した金粒子懸濁液はHelios Gene Gun簡易操作マニュアルに従い、以下の方法でゴールドコート用チューブにコーティングし、接種を行った。チュービングプレップステーションで0.35-0.40LPM(liters per minute)のN2ガスを用いてチューブ内を20分間乾燥したゴールドコート用チューブに、シリンジ付き懸濁液ローディング用チューブを取り付け、このシリンジを用いて金粒子懸濁液をゴールドコート用チューブに装填し、5分間静置した。静置後、シリンジ付き懸濁液ローディング用チューブを取り除き、エタノール除去用シリンジ付きチューブを取り付け、ゴールドコート用チューブ内のエタノールを全て除去した。除去後、直ちにゴールドコート用チューブを180°回転し、チュービングプレップステーションの回転スイッチを作動した。ゴールドコート用チューブ内に0.35-0.40LPMのN2ガスを通気しチューブ内を乾燥した。乾燥したゴールドコート用チューブをチュービングプレップステーションから取り出し、チュービングカッターを用いて切断し、サンプルカートリッジとした。サンプルカートリッジをHelios Gene Gunのカートリッジホルダーに取り付け、1葉期のキュウリの第1本葉に180PSI(pounds per square inch)で1 shot接種し、第1本葉二次接種区とした。上記の方法で2葉期の第2本葉のみに二次接種したものを第2本葉二次接種区、3葉期の第3本葉のみに二次接種したものを第3本葉二次接種区とした。二次接種後、キュウリは一次接種後と同様に25℃の長日条件で育成した。
【0041】
なお、二次接種の接種源であるp35S-Z5vector+GFP-FLは、使用する培地をLB培地から2×YT培地(pH7.5)[1Lあたりbacto tryptone 16g,bacto yeast extract 10g,NaCl 5g,アンピシリン(25mg/ml) 2ml]に変更したラージスケール培養で前記1-2-3と同様の方法で精製し、NanoDrop ND-1000 Spectrophotometerで濃度を測定後、400ng/μlに調整し、-80℃で保存しておいたものを使用した。
1-4:病徴観察とウイルス検定
1-4-1:病徴の観察
病徴の観察は二次接種後毎日行い、二次接種後21日目にデジタルカメラFinePix S1Pro(FUJIFILM)を用いて病徴の撮影を行った。
1-4-2:GFP蛍光の観察
GFP蛍光の観察は二次接種後7日目、14日目、21日目に蛍光顕微鏡システムVBシリーズ(KEYENCE)と顕微鏡デジタルカメラDP70(OLYMPUS)を用いて行い、同時に撮影した。
1-4-3:ウエスタンブロット法
一次接種後7日目にキュウリの子葉2枚を採取し、各一次接種ウイルスの感染を以下の方法で確認した。採取した子葉2枚の重量を量り、3倍容のALSV磨砕緩衝液とともに乳鉢と乳棒で磨砕し、100μlの磨砕液を1.5ml容マイクロチューブに移した。100μlの2×サンプルバッファー[0.1M Tris-HCl(pH8.0),4% SDS,30% スクロース,0.05% BPB,2% メルカプトエタノール]を加え、ボルテックスミキサーで1分間混合し、その後5分間煮沸し、直ちに氷上で5分間静置した。静置後、14,000rpm(4℃)で5分間遠心し、上清を泳動試料とした。泳動試料は10μl使用し、stacking gel(5%アクリルアミドゲル)とrunning gel(12.5%アクリルアミドゲル)および泳動バッファー[0.6%Tris,28%グリシン,0.1%SDS]を用いて泳動(160V,20mA,90分間,室温)を行った。泳動後、転写バッファー[0.1M Tris,0.192M グリシン,20%メタノール]を用いてセミドライエレクトロブロッティングを行い、PVDF膜(MILIPOLE)にタンパク質の転写を行った。転写後、PVDF膜をブロッキング液[0.02M Tris-HCl(pH7.5),0.5M NaCl,2% Tween-20,2%スキムミルク,2%PVP]に浸し、37℃のインキュベーターで60分間静置した。静置後、TTBS[0.02M Tris-HCl(pH7.5),0.5M NaCl,2% Tween-20]で25倍希釈したALSVおよびZYMVに対する吸収抗体を用いて、一次抗体処理を37℃のインキュベーター内で60分間行った。一次抗体処理後、PVDF膜をTTBSで満たしたタッパーに浸し、シーソーで5分間洗浄した。TTBSを新しいものに交換し、同様の方法でさらに2回洗浄を行った。洗浄したPVDF膜はTTBSで2000倍に希釈したアルカリフォスファターゼ標識坑ウサギ免疫グロブリンヤギ血清(Cell Signaling TECHNOLOGY)用いて、二次抗体処理を37℃のインキュベーター内で60分間行った。二次抗体処理後、PVDF膜をTTBSで3回洗浄し、発色基質液[0.2M Tris-HCl(pH8.2),3mg/ml Fast Red TR salt,1mg/ml Naphthol AS-MX phosphate]を用いて発色処理を行った。十分に発色した後、PVDF膜を蒸留水で洗浄し、反応を停止した。なお、吸収抗体はあらかじめ以下の方法で作成した。まず、5倍容のTTBSで磨砕した供試植物の磨砕液を3,000rpm(4℃)で5分間遠心し、上清を粗汁液とした。1.5ml容のマイクロチューブに10μlの一次抗体(各種ウイルス抗血清)をとり、粗汁液90μlを加えて混合し、37℃で60分間静置した。静置後、14,000rpm(4℃)で5分間遠心し、上清100μlを新しい1.5ml容のマイクロチューブに移した。上清に磨砕液を100μl加えて混合し、37℃で60分間静置した。静置後、14,000rpm(4℃)で5分間遠心し、上清200μlを新しい1.5ml容のマイクロチューブに移した。上清に磨砕液を200μl加えて混合し37℃で60分間静置した。14,000rpm(4℃)で5分間遠心し、上清400μlを新しい1.5ml容のマイクロチューブに移した。上清に400μlの100%グリセロールを加えボルテックスミキサーで十分混合し吸収抗体とした。吸収抗体は-20℃の冷凍庫で保存し適宜使用した。
1-4-4:ELISA法
ALSVとZYMVの蓄積量は直接ELISA法を用い、GFPの蓄積量は間接ELISA法を用いて以下の方法で定量した。
【0042】
検定試料は二次接種後21日目にコルクボーラーを用いて次のように採取した。第1本葉接種区では第3から第6本葉の各本葉を、第2本葉接種区では第4から第7本葉の各本葉を、第3本葉接種区では第5から第8本葉の各本葉をそれぞれ採取し、採取した本葉の基部、中心部、周辺部から太い葉脈を避けて直径1cmのリーフディスクを2枚ずつくりぬき、本葉1枚から計6枚のリーフディスクを採取した(図3)。採取した6枚のリーフディスクは1つのステンレスビーズSUB-50(TOMY)とともに2ml容凍結保存チューブ(アシスト)に入れ、液体窒素で凍結し、Micro Smash MS100-R(TOMY)を用いて2,500rpmで30秒間破砕した。破砕後、1.8mlのPBST[137mM NaCl,8.1mM Na2HPO4,2.68mM KCl,1.47mM KH2PO4,0.05% Tween20]を加え、Micro Smash MS100-Rを用いて2,500rpmで30秒間混合した。その後、14,000rpm(4℃)で5分間遠心し、得られた上清を検定試料とした。
【0043】
ALSVの定量は直接ELISA法を用いて以下の方法で行った。ELISAプレートのウェルにコーティングバッファー[0.05M炭酸緩衝液(pH9.6)]で1000倍に希釈したALSV抗体溶液を150μl入れ、プレートカバーを被せ37℃のインキュベーターで2時間静置した。溶液を捨て、PBSTでウェルを3回洗浄し、ブロッキング溶液[137mM NaCl,8.1mM Na2HPO4,2.68mM KCl,1.47mM KH2PO4,2%スキムミルク]をウェルに150μl入れ、プレートカバーを被せ37℃のインキュベーターで1時間静置した。溶液を捨て、PBSTでウェルを3回洗浄し、1検定試料当り2つのウェルに150μlずつ検定試料を入れ、プレートカバーを被せ4℃の冷蔵庫に一晩静置した。溶液を捨て、PBSTでウェルを10回洗浄し、PBSTで1000倍に希釈した酵素標識をしたALSV抗体溶液をウェルに150μl入れ、プレートカバーを被せ37℃のインキュベーターで3時間静置した。溶液を捨て、PBSTでウェルを10回洗浄し、基質液[10%ジエタノールアミン(pH9.8),0.67mg/ml p-ニトロフェニルリン酸]を150μl入れ、室温に静置して色を呈するまで反応させた。反応後、モデル550マイクロプレートリーダー(BIO-RAD)でウェルの吸光度を測定し、ALSVの蓄積量とした。
【0044】
ZYMVの定量は直接ELISA法を用いて以下の方法で行った。ELISAプレートのウェルにコーティングバッファーで500倍に希釈したZYMV抗体(日本植物防疫協会研究所)溶液を150μl入れ、プレートカバーを被せ37℃のインキュベーターで3時間静置した。溶液を捨て、PBSTでウェルを3回洗浄し、ブロッキング溶液をウェルに150μl入れ、プレートカバーを被せ37℃のインキュベーターで1時間静置した。溶液を捨て、PBSTでウェルを3回洗浄し、1検定試料当り2つのウェルに150μlずつ検定試料を入れ、プレートカバーを被せ4℃の冷蔵庫に一晩静置した。溶液を捨て、PBSTでウェルを10回洗浄し、PBSTで1000倍に希釈した酵素標識をしたZYMV抗体(日本植物防疫協会研究所)溶液をウェルに150μl入れ、プレートカバーを被せ37℃のインキュベーターで4時間静置した。溶液を捨て、PBSTでウェルを10回洗浄し、基質液[10%ジエタノールアミン(pH9.8),1mg/ml p-ニトロフェニルリン酸]を150μl入れ、室温に静置して色を呈するまで反応させた。反応後、モデル550マイクロプレートリーダーでウェルの吸光度を測定し、ZYMVの蓄積量とした。
【0045】
GFPの定量は以下の方法で行った。ELISAプレートのウェルに1検定試料当り2つのウェルに150μlずつ検定試料を入れ、プレートカバーを被せ37℃のインキュベーターに2時間静置した。溶液を捨て、PBSTでウェルを5回洗浄し、ブロッキング溶液をウェルに150μl入れ、プレートカバーを被せ37℃のインキュベーターで1時間静置した。溶液を捨て、PBSTでウェルを5回洗浄し、PBSTで1000倍に希釈したGFP抗体(Clontech)を150μlいれ、プレートカバーを被せ37℃の冷蔵庫に一晩静置した。溶液を捨て、PBSTでウェルを10回洗浄し、PBSTで2000倍に希釈したアルカリフォスファターゼ標識坑ウサギ免疫グロブリンヤギ血清をウェルに150μl入れ、プレートカバーを被せ37℃のインキュベーターで3時間静置した。溶液を捨て、PBSTでウェルを10回洗浄し、基質液[10%ジエタノールアミン(pH9.8),1mg/ml p-ニトロフェニルリン酸]を150μl入れ、室温に静置して色を呈するまで反応させた。反応後、モデル550マイクロプレートリーダーでウェルの吸光度を測定し、GFPの蓄積量とした。
1-4-5:RNA抽出法とRT-PCR法
二次接種後21日目にキュウリの最上位展開葉を採取し、-80℃の冷凍庫で凍結し検定試料とした。0.1gの検定試料を乳鉢と乳棒で磨砕し、1mlのTriPure Isolation Reagent(Roche)を加えさらに磨砕した。磨砕液1mlを1.5ml容マイクロチューブに移し、室温で5分間静置した。静置後、200μlのクロロホルムを加えボルテックスミキサーで20秒間混合し、室温で10分間静置した。静置後、14,000rpm(4℃)で15分間遠心し、500μlの水層を新しい1.5ml容マイクロチューブに移した。水飽和フェノールを250μl、クロロホルムを250μl加えボルテックスミキサーで5分間混合し、14,000rpm(4℃)で5分間遠心した。遠心後、500μlの水層を新しいマイクロチューブに移し、420μlのイソプロピルアルコールを加えボルテックスミキサーで混合し、室温で10分間静置した。静置後、14,000rpm(4℃)で10分間遠心し、上清を除去し70%エタノールを1.4ml加えボルテックスミキサーで5分間混合した。14,000rpm(4℃)で5分間遠心し、上清を除去後、沈殿を20μlの滅菌水に懸濁し、NanoDrop ND-1000 SpectrophotometerでRNA濃度を測定し、500ng/μlに調整しRNA溶液とした。
【0046】
逆転写反応は以下の方法で行った。0.2ml容PCRチューブにRNA溶液を2μl、滅菌水を6μl、5×RT Buffer(TOYOBO)を4μl、2.5mM dNTP mixture(TaKaRa)を8μl、10μM Oligo(dT)12プライマーを1μl、RNase Inhibitor(Wako)を0.5μl、ReverTra Ace(TOYOBO)を0.5μl 混合し、TaKaRa PCR Thermal Cycler Dice VersionIII Model TP600(TaKaRa)を使用して42℃で50分間処理し、その後95℃で5分間処理し、最後に4℃で5分間処理し、獲られた産物をRT産物とした。
【0047】
PCRにはALSV-RNA2ベクターの外来遺伝子導入サイトを含む配列を増幅できる10μM R2ALS 1363(+)[5’-GCGAGGCACTCCTTA-3’:配列番号8]と10μM R2ALS 1511(-)[5’-GCAAGGTGGTCGTGA-3’:配列番号9]をそれぞれプラス鎖プライマーおよびマイナス鎖プライマーとして用いた。1μlのRT産物を0.2ml容PCRチューブにとり、滅菌水を4.9μl、10×Ex Taq Buffer(TaKaRa)を1μl、2.5mM dNTP Mixture(TaKaRa)を1μl、10μM R2ALS 1363(+)と10μM R2ALS 1511(-)を各1μl、TaKaRa Ex TaqTM(TaKaRa)を0.1μl加えて混合し、TaKaRa PCR Thermal Cycler Dice VersionIII Model TP600(TaKaRa)を使用して94℃で5分間処理後、〔94℃、30秒→58℃、30秒→72℃、60秒〕という反応を30サイクル行い、その後72℃で5分間処理し、最後に4℃で5分間処理しPCRを終了した。得られたPCR産物2μlに1μlの10×Loading Buffer(TaKaRa)を混合し、1%アガロースゲルを用いた電気泳動を行い、各種ALSVのインサート配列の保持を確認した。
2:結果
2-1:ZYMV遺伝子を導入したALSVベクターのZYMVに対する干渉効果
ZYMV遺伝子断片を導入したALSVベクターのZYMVに対する干渉効果を調べるため、wtALSVおよびZYMVのCP遺伝子断片を導入したALSV(ALSV-Z:CP200)を一次接種したキュウリ(青大胡瓜)におけるZYMVの病徴、GFP蛍光、ウイルス量を調べた。
【0048】
p35S-Z5vector+GFP-FLのみを接種した試験区(ZYMVのみ区)では、接種後12日前後に第3本葉の基部周辺で退緑斑点や脈間退緑などの病徴が観察された。その後展開した第4本葉で葉のモザイク症状が見られ、第5本葉以上の葉では葉全面で激しいモザイク症状が観察された(図4A)。さらに、第6本葉以上の葉では激しいモザイク症状とともに葉の奇形や萎縮が観察され(図4B)、植物体全体も矮化を示した(図4C)。wtALSVを一次接種後にp35S-Z5vector+GFP-FLを二次接種した試験区(wtALSV+ZYMV区)では、第3から第5本葉での病徴の現れ方はZYMVのみ区とほとんど同じであったが、症状はZYMVのみ区に比べ軽く、植物体全体の矮化は観察されなかった(図4)。これに対して、ALSV-Z:CP200を一次接種後にp35S-Z5vector+GFP-FLを二次接種した試験区(ALSV-Z:CP200+ZYMV区)では、非接種区およびALSVのみを接種した試験区(ALSVのみ区)と同様に、全ての上葉において病徴が全く観察されなかった(図4)。
【0049】
蛍光顕微鏡でGFP蛍光を観察したところ、ZYMVのみ区では、第1本葉の接種部位周辺でスポット状の蛍光や葉脈上に蛍光が観察された。第2本葉では葉脈で観察された蛍光が葉の展開とともに拡大した。第3本葉から第5本葉では葉の全体が蛍光で埋め尽くされていた。第6本葉では葉全体で蛍光が観察されたが、葉の所々に蛍光のない領域が観察された(図5)。第7葉より上位の葉では蛍光の観察される領域と観察されない領域がモザイク状に混在していた。wtALSV+ZYMV区では各葉位でZYMVのみ区とほぼ同じ蛍光が観察された(図6)。これに対して、ALSV-Z:CP200+ZYMV区の第1葉では、ZYMVのみ区やwtALSV+ZYMV区と同様に、接種部位周辺でスポット状の蛍光や葉脈上に蛍光が観察されたが、第2本葉では葉脈で観察された蛍光の拡がる範囲がZYMV区やwtALSV+ZYMV区に比べ狭い領域に限定されていた。さらに、第3本葉では数箇所に蛍光が観察されるのみで、第4葉より上位の葉では蛍光が観察されなかった(図7)。
【0050】
続いて、ZYMVの定量を行った結果、第3本葉から第6本葉全てでwtALSV+ZYMV区ではZYMVのみ区と同程度のZYMVが蓄積していたのに対し、ALSV-Z:CP200+ZYMV区ではZYMVの蓄積は認められなかった(図8)。また、ALSVを定量したところ、wtALSV+ZYMV区ではALSVのみ区に比べ約2倍のELISA値を示し、ALSV-Z:CP200+ZYMV区ではALSVのみ区に比べやや減少していた(図8)。一次接種に用いたALSV-Z:CP200のインサートチェックを行ったところ、全てのキュウリ個体でALSVのインサートが保持されていることが確認された。
【0051】
第2本葉二次接種区の各試験区における病徴は第1本葉二次接種区のものとほぼ同じで、病徴の現れる葉位が1枚ずつ上位にずれていた。
【0052】
GFP蛍光の観察では、ALSV-Z:CP200+ZYMV区で接種葉の第2本葉のみで接種部位周辺でスポット状の蛍光や葉脈の一部に蛍光が観察され、第3本葉以降に展開した本葉では蛍光が観察されなかった。これに対し、ZYMVのみ区やwtALSV+ZYMV区では、第1本葉二次接種区の各試験区と同様に葉全体でGFP蛍光が観察された。
【0053】
直接ELISA法によりZYMVの蓄積を調べたところ、各試験区で第1本葉二次接種区とほとんど同じようにZYMVが蓄積していた(図9)。一次接種に用いたALSV-Z:CP200のインサートチェックでは、全てのキュウリ個体でALSVのインサートは保持されていた。
【0054】
第3本葉二次接種区の各試験区の病徴の様子やGFP蛍光の分布では、ALSV-Z:CP200+ZYMV区において接種葉を含む全ての本葉で蛍光が観察されなかったこと意外は、第1本葉二次接種区とほぼ同じであった。ただし、病徴やGFP蛍光は2枚上位にずれて観察された。
【0055】
直接ELISA法によるZYMVおよびALSVの定量では、ALSV-Z:CP200+ZYMV区で、ALSVのみ区より若干多くALSVが蓄積していることが明らかになり、それ以外の試験区では第1本葉二次接種区および第2本葉二次接種区とほぼ同じ結果が得られた(図10)。ALSV-Z:CP200のインサート保持も確認された。
【0056】
予備試験で行ったp35S-Z5vector+GFP-FL接種によるZYMVの病徴観察をキュウリ3品種(青大胡瓜、鈴成四葉、つばさ)で比較したところ、青大胡瓜が最も激しい症状を示し、鈴成四葉では軽度の症状が、つばさではほとんど病徴が現れなかった。よって、以後の全ての試験では青大胡瓜を用いることにした。
2-2:異なるZYMV遺伝子断片を導入したALSVベクターの干渉効果
ZYMV遺伝子の異なる領域を組み込んだALSVベクターによって誘導される干渉効果を比較するため、ALSV-Z:P1、ALSV-Z:P3およびALSV-Z:CP200をキュウリの子葉に一次接種し、第1本葉にp35S-Z5vector+GFP-FLを二次接種した。
【0057】
ALSV-Z:CP200+ZYMV区では前記2-1に示した通りである。ALSV-Z:P3を一次接種後p35S-Z5vector+GFP-FLを二次接種した試験区(ALSV-Z:P3+ZYMV区)では、ALSV-Z:CP200+ZYMV区と同様に病徴は観察されず(図11)、蛍光は第1本葉に接種部位周辺でスポット状の蛍光と葉脈上の蛍光が観察された。第2本葉では葉脈上に観察された蛍光が狭い範囲で拡大した。第3本葉および第4本葉では数箇所で蛍光が観察されるのみであり、第5本葉より上位の葉では蛍光が観察されなかった(図12)。一方、ALSV-Z:P1を一次接種後にp35S-Z5vector+GFP-FLを二次接種した試験区(ALSV-Z:P1+ZYMV区)では、第3本葉に葉の周辺部に弱い退緑症状が観察され、この症状はその後に展開した上葉全てで観察された。退緑症状は観察されたものの、ZYMVのみ区に観察された激しいモザイク症状や葉の矮化、植物体全体の矮化は観察されなかった(図11)。GFP蛍光について見ると、第1本葉煮接種部位周辺でスポット状の蛍光と葉脈上の蛍光が観察され、第2葉本では葉脈上の蛍光が狭い範囲で拡大した。また、第3本葉より上葉では、葉脈の蛍光や葉脈の周辺部でのスポット状の蛍光、葉の周辺部や先端部における蛍光が観察された(図13)。
【0058】
直接ELISA法によりZYMV量を測定したところ、ALSV-Z:CP200+ZYMV区とALSV-Z:P3+ZYMV区では第3本葉から第6本葉全てにおいて、ZYMVの蓄積は認められなかった。一方、ALSV-Z:P1+ZYMV区ではZYMVのみ区やwtALSV+ZYMV区の約半分程度蓄積していた(図14)。ALSVは、wtALSV+ZYMV区でALSVのみ区の約2倍蓄積していたが、ALSV-Z:P1+ZYMV区、ALSV-Z:P3+ZYMV区およびALSV-Z:CP200+ZYMV区では、ALSVのみ区よりもやや多い程度であった(図14)。一次接種に用いたZYMVの各遺伝子断片を導入したALSV(ALSV-Z:P1 、ALSV-Z:P3、ALSV-Z:CP200)のインサートチェックを行ったところ、全てのキュウリ個体でALSVのインサートが保持されていた。
【0059】
なお、ZYMVの他の遺伝子領域(HC-Pro、Cl、Nla、Nlb)の断片(180-201 nt)についても干渉効果を同様に試験した。その結果、Cl、NlaおよびNlbの各断片は、前記のCP200断片およびP3断片と同様の効果を示した。一方、HC-Pro断片の効果は前記P1断片の効果とほぼ同一であった。
2-3:異なるキュウリ品種における干渉効果
青大胡瓜とは異なる2つの品種(鈴成四葉、つばさ)においてALSV-Z:CP200がZYMVに対する干渉効果を誘導するか検討した。
【0060】
鈴成四葉では、ZYMVのみ区やwtALSV+ZYMV区で、退緑症状のみが現れ、激しいモザイクや葉の萎縮、植物体の矮化は観察されなかった。また、ALSV-Z:CP200+ZYMV区では、病徴が全く観察されなかった。つばさでは、全ての試験区でZYMVの症状は観察されなかった。
【0061】
鈴成四葉のZYMVのみ区やwtALSV+ZYMV区においては、第1本葉から第5本葉で青大胡瓜の第1本葉二次接種区場合と同様のGFP蛍光の分布が観察され。第6本葉では、基部において蛍光のない領域が一部観察されたが、それ以外では蛍光が観察された。第7葉以降に展開した葉ではスポット状の蛍光が葉の周辺部から中心部に多く観察され、基部では蛍光が観察されなかった。ALSV-Z:CP200+ZYMV区では、第1本葉の接種部位周辺部にスポット状の蛍光のみが僅かに観察され、第2本葉においても数個の蛍光スポットが観察されるのみであった。第3本葉より上位の葉では蛍光は観察されなかった。
【0062】
つばさのZYMVのみ区やwtALSV+ZYMV区では、第1本葉から第5本葉で青大胡瓜や鈴成四葉の場合と同様のGFP蛍光の分布が観察され。第6本葉では基部において蛍光のない領域が一部観察されたが、それ以外では蛍光が観察された。第7本葉より上位の葉ではスポット状の蛍光が葉の周辺部を縁取るように観察され、それ以外の部分では蛍光が観察されなかった。ALSV-Z:CP200+ZYMV区では第1本葉の接種部位周辺でスポット状の蛍光が僅かに観察されるのみで、第2本葉においても数個の蛍光スポットが観察されるのみだった。第3本葉より上葉では蛍光は観察されなかった。
【0063】
直接ELISA法でZYMVを定量したところ、鈴成四葉およびつばさのALSV-Z:CP200+ZYMV区では第3本葉から第6本葉でZYMVが蓄積しないことが明らかになった(図15、図16)。また、鈴成四葉およびつばさの両品種において、ALSVのみ区に比べwtALSV+ZYMV区ではALSVの蓄積量が増加し、ALSV-Z:CP200+ZYMV区では若干減少していた。(図15、図16)。ALSV-Z:CP200のインサートチェックを行ったところ、両品種のキュウリ個体でALSVのインサートは保持されていた。
2-4:異なる長さのZYMV遺伝子断片を導入したALSVベクターの干渉効果
ALSVベクターに導入したZYMV遺伝子断片の長さが干渉効果へ影響するかどうか調べるため、約100 ntと約200 ntのZYMV-CP遺伝子断片を挿入したALSV(ALSV-Z:CP100、ALSV-Z:CP200)の干渉効果を調べた。
【0064】
ALSV-Z:CP200+ZYMV区における病徴は前記2-1の第1本葉二次接種区と同様で、全ての上葉に病徴が観察されなかったのに対し、ALSV-Z:CP100を一次接種後p35S-Z5vector+GFP-FLを二次接種接種した試験区(ALSV-Z:CP100+ZYMV区)では、葉の一部に弱い退緑の症状が観察された(図17)。
【0065】
ALSV-Z:CP200+ZYMV区では、第1本葉の接種部位周辺でスポット状のGFP蛍光と葉脈上の蛍光が観察され、第2本葉で葉脈上の蛍光が狭い範囲で拡がった。第3本葉では数箇所で蛍光が観察されるのみで、第4本葉より上位の葉では蛍光が観察されず、前記2-1の第1本葉二次接種区と同様であった。これに対し、ALSV-Z:CP100+ZYMV区では、第1本葉の二次接種部位周辺でスポット状の蛍光と葉脈上の蛍光が観察された。第2本葉では、葉脈上に蛍光が観察され、葉の展開とともに蛍光も拡大した。第3本葉では、葉の全体が蛍光で埋め尽くされていることが観察された。第4本葉では、スポット状の蛍光が葉全体で観察され、特に葉の周辺部で多数観察された。第5本葉では、葉の周辺部の葉脈上に蛍光が観察された。第6本葉より上位の葉では、葉の先端部でのみ蛍光が観察された(図18)。
【0066】
直接ELISA法でZYMVを定量したところ、ALSV-Z:CP200+ZYMV区の第3本葉から第6本葉でZYMVが蓄積されないのに対し、ALSV-Z:CP100+ZYMV区では、ZYMVのみ区やwtALSV+ZYMY区に比べやや少ないがZYMVが蓄積していることが明らかになった(図19)。ALSVの定量をしたところ、ALSVのみ区に比べwtALSV+ZYMV区では蓄積量が増加し、ALSV-Z:CP200+ZYMV区では同程度蓄積していた。これに対して、ALSV-Z:CP100+ZYMV区では、ALSVのみ区と比較すると、第3本葉から第5本葉ではALSVの蓄積量が増加していたが、第6本葉では同程度であった(図19)。一次接種に用いた異なる長さのZYMVのCP遺伝子断片を導入したALSV(ALSV-Z:CP100 、ALSV-Z:CP200)のインサートチェックを行ったところ、全てのキュウリ個体でALSVのインサートが保持されていた。
【0067】
なお、同様にして、ZYMV-CP遺伝子の66 nt断片と33 nt断片をそれぞれ挿入したALSVの干渉効果を調べた。その結果、挿入遺伝子断片のサイズが小さくなるにつれて干渉効果が弱まることが確認された。
2-5:ZYMV遺伝子断片導入ALSVと弱毒系統ZYMVの干渉効果の比較
ウイルス製剤として市販されているZYMV 2002 による干渉効果と、ZYMV遺伝子断片を導入した3種のALSVによる干渉効果を比較するために、各試験区の病徴、GFP蛍光の分布およびウイルス蓄積を調べた。
【0068】
ZYMV 2002を一次接種後p35S-Z5vector+GFP-FLを二次接種した試験区(ZYMV 2002+ZYMV区)では接種葉および上葉において病徴は観察されなかった(図20)。
【0069】
また、ALSV-Z:P1+ZYMV区、ALSV-Z:P3+ZYMV区およびALSV-Z:CP200+ZYMV区では前記2-2のようにそれぞれ接種葉および上葉においてGFP蛍光が観察されたのに対し、ZYMV 2002+ZYMV区では接種葉および上葉においてGFP蛍光は観察されなかった(図21)。
【0070】
GFP抗体を用いた間接ELISA法で強毒ZYMVの蓄積を調べたところ、ZYMVのみ区とwtALSV+ZYMV区においてGFPが蓄積したのに対し、ALSV-Z:P3+ZYMV区、ALSV-Z:CP200+ZYMV区およびZYMV 2002+ZYMV区では、GFPが全く蓄積せず、ALSV-Z:P1+ZYMV区では僅かにGFPが蓄積した(図22)。一次接種に用いたZYMVの各遺伝子断片を導入したALSV(ALSV-Z:P1 、ALSV-Z:P3、ALSV-Z:CP200)のインサートチェックを行ったところ、全てのキュウリ個体でALSVのインサートが保持されていた。
3:考察
ZYMV遺伝子断片を導入したALSVをキュウリに接種すると、VIGSが誘導されZYMVに対する防御体制をとると予想された。先ず、ALSVベクター接種後の時間が干渉効果の誘導にどのように影響するか調べるため、ZYMVのCP遺伝子を導入したALSV(ALSV-Z:CP200)を接種後6日目、9日目および12日目に、それぞれ第1本葉、第2本葉および第3本葉に強毒ZYMV株の感染性クローンを接種し、干渉効果の誘導を調べた。その結果、3つの条件(第1本葉二次接種区、第2本葉二次接種区、第3本葉二次接種区)全てにおいて、上位葉でのZYMVの蓄積は認められなかった。また、GFP蛍光を観察したところ、第1本葉二次接種区では第1本葉から第3本葉でGFP蛍光が観察されたが、第2本葉二次接種区の接種葉でのみGFP蛍光が観察され、第3本葉二次接種区では接種葉においてもGFP蛍光が観察されなかった。以上よりALSV-Z:CP200接種後の日数が経過するほどZYMVの拡がりを抑えることが明らかになった。この結果は、ALSVベクターによるサイレンシングの誘導がALSVの存在する領域でのみ誘導され、常にALSV増殖後にVIGSが誘導されるという観察結果と合わせて考えると、第1本葉および第2本葉に二次接種した試験区では、まだALSVが分布していないため接種葉および上葉ではZYMVの増殖と移行が起こったものと考えられる。これに対し、第3本葉に二次接種した試験区では既にALSVが分布し、VIGSが誘導されていたため、ZYMVの増殖が抑えられGFP蛍光が観察されなかったと考えられた。第1本葉に接種した試験区では第3本葉のごく一部に蛍光が観察されたものの、直接ELISA法を用いたZYMV量の測定ではウイルスは検出されないレベルであり、また全ての試験区で病徴が観察されなかった。以上より、ZYMV遺伝子断片を導入したALSVが感染したキュウリにおいて、接種から6日以上経過すればZYMVに対する干渉効果が誘導され、ZYMVの増殖を抑制できることを示している。
【0071】
次に、ALSVベクターに導入するZYMV遺伝子領域の違いが干渉効果の誘導に影響を与えるか調べるため、3種のZYMV遺伝子導入ALSV(ALSV-Z:P1、ALSV-Z:P3、ALSV-Z:CP200)間における干渉効果を比較した。ALSV-Z:P1接種区では弱い病徴やGFP蛍光が観察されZYMVが蓄積したのに対し、ALSV-Z:P3およびALSV-Z:CP200接種区では、病徴は観察されず、また一部GFP蛍光が観察されたもののZYMVの蓄積は認められなかった。また、Cl、NlaおよびNlbの各断片も、前記のCP200断片およびP3断片と同様の効果を示した。
【0072】
キュウリ品種の違いが干渉効果に影響を与えるか調べるためALSV-Z:CP200を用いて比較試験を行ったところ、誘導されるワクチン効果に品種間差は認められなかった。よって、品種の違いは干渉効果に影響を与えないと考えられ、3品種以外の品種においてもALSVベクターによる干渉効果の誘導が可能であると考えられた。
【0073】
ALSVベクターに導入する遺伝子断片の長さが干渉効果の誘導に影響を与えるか調べるため、ALSV-Z:CP200 とALSV-Z:CP100による干渉効果を比較した。ALSV-Z:CP100を一次接種した試験区ではZYMVが蓄積され、軽微な病徴やGFP蛍光が観察されたことから、誘導される干渉効果がALSV-Z:CP200に比べ弱くなったと考えられた。この結果より、タバコにおいてPDS遺伝子のサイレンシングを誘導する際、異なるサイズのPDS遺伝子断片や同一のサイズで異なる部分のPDS遺伝子断片をALSVベクターに導入すると、サイレンシングの程度に違いが認められる報告(例えば、非特許文献4)と一致するものと考えられた。なお、挿入遺伝子のサイズをさらに小さくしたALSV-Z:CP66、ALSV-Z:CP33の試験でも、遺伝子サイズが小さくなるにつれて干渉効果が低減することが確認された。これらのことから、ALSVベクターを用いてあるウイルスに対して干渉効果を誘導する際、ALSVベクターに導入する遺伝子のサイズや部位によって、誘導される干渉効果が異なる可能性がある。そのため、複数のウイルス遺伝子断片を導入したALSVを作出し、その干渉効果について比較検討することが好ましい。
【0074】
市販されている弱毒ZYMVと、ZYMV遺伝子導入ALSV(ALSV-Z:P1、ALSV-Z:P3、ALSV-Z:CP200)の干渉効果の比較試験では、GFP蛍光の分布に差が認められた。すなわち、弱毒ZYMV接種区ではZYMVのGFP蛍光は全く観察されなかった。これは、弱毒ウイルスとALSVによる干渉効果の機構の差があるためと考えられる。つまり、ALSVベクターでは、導入遺伝子断片のサイレンシングによって干渉効果を誘導するRNA介在性機構であるため、ALSVが分布していない領域ではサイレンシングの誘導がなされていないため、強毒株のZYMVが増殖したと考えられる。これに対し、弱毒ZYMVでは、同種のウイルスによる干渉効果であるため、タンパク質介在性干渉効果が強毒ZYMVの増殖を抑え、蛍光が全く観察されなかったものと考えられた。さらに、ALSVベクターとZYMVでは感染葉でのウイルスの拡がり方が異なり、ALSVは葉の基部から先端部に向けてその範囲を拡げるように拡がるのに対し、ZYMVは葉脈全体に先ず拡がり、その後葉脈から染み出すように拡がるといった移行の違いも干渉効果に影響しているのかもしれない。ALSVを一次接種後にZYMVを二次接種した区では、ALSVの蓄積量が増加したのは、ZYMVとの重複感染によるsynergismが起こっていたものと考えられる(例えば、非特許文献7)。また、wtALSV接種区でも弱い干渉効果すなわち、植物体の矮化が認められなかったことは、このsynergismが影響しているのかもしれない。
【0075】
以上をまとめると、ALSVベクターにZYMVの遺伝子断片を組み込むことによってZYMVに対する干渉効果が誘導された。しかし、導入した遺伝子断片の種類、サイズ、位置によって干渉効果に強弱が見られ、実際にウイルスに対する干渉効果の誘導ベクターとして利用する場合には、ウイルス遺伝子断片導入ALSVを複数作製し、その効果を比較することが好ましい。
【実施例2】
【0076】
キュウリモザイクウイルス(CMV)の遺伝子断片を導入したALSVベクターによる干渉効果
1:材料と方法
1-1:実験材料
1-1-1:供試植物
キノア、Nicotiana tabacum cv Xanthi nc(タバコ)、とキュウリ(青大胡瓜)を供試した。
1-1-2:供試ウイルス
供試ウイルスは東北大学農学部准教授高橋英樹博士より分譲されたCucumber mosaic virus strain Y(以下CMV-Y)および、独立行政法人農業生物資源研究所より入手したCMV isolate 42CM(以下CMV-42CM、MAFF番号104087)である。
1-2:CMV遺伝子断片のALSVベクターへの導入
1-2-1:CMV遺伝子断片のクローニング
実施例1の1-4-5と同様の方法CMV-Y感染タバコ葉からでRNA抽出を行い、抽出したRNAを鋳型とし、既知のCMV-Y遺伝子配列をもとに設計したマイナス鎖のプライマーCMVY 1a(-)、CMVY 2a(-)、CMVY CP(-)を用いて(表2:配列番号10−15)、CMV-YのRNA1(accession number:D12537)、RNA2(accession number:D12538)、RNA3(accession number:D12499)に対する逆転写反応を以下の方法で行った。
【0077】
【表2】
【0078】
0.2ml容PCRチューブにRNA溶液を2μl、滅菌水を6μl、5×RT Bufferを4μl、2.5mM dNTP mixture(TaKaRa)を8μl、10μM のマイナス鎖プライマーを1μl、RNase Inhibitorを0.5μl、ReverTra Aceを0.5μl 混合し、TaKaRa PCR Thermal Cycler Dice VersionIII Model TP600を使用して42℃で50分間処理し、その後95℃で5分間処理し、最後に4℃で5分間処理し、獲られた産物をRT産物とした。
【0079】
得られたRT産物を鋳型とし、1a遺伝子(750 nt)の増幅にはCMVY 1a(+)とCMVY 1a(-)、2a遺伝子(663 nt)の増幅にはCMVY 2a(+)とCMVY 2a(-)、CP遺伝子(558 nt)の増幅にはCMVY CP(+)とCMVY CP(-)の組み合わせ(第2表)のプライマーを用いたPCRによる各遺伝子断片の増幅とPCR産物のサイズ確認、PCR産物のMonoFas DNA精製キットIを用いた精製を、実施例1の1-2-1と同様の方法で行い、精製した遺伝子断片をそれぞれC:1a遺伝子断片、C:2a遺伝子断片、C:CP遺伝子断片とした。
【0080】
精製した3種の遺伝子断片を用いたpGEM-T Easy Vector SystemによるTAクローニングと、そのライゲーション産物を用いた大腸菌DH5αの形質転換、スモールスケール培養と煮沸法によるプラスミド抽出、抽出プラスミドの1%アガロースゲルを用いた電気泳動による選抜とEcoRIを用いた制限酵素処理によるインサート保持の確認は、実施例1の1-2-1と同様の方法で行った。
1-2-2:CMV遺伝子断片のシークエンス
TAクローニングによって作製したプラスミドは、実施例1の1-2-2と同様の方法でプラスミドの精製とシークエンス解析を行った。配列を確認したプラスミドをそれぞれpGEM-C:1a、pGEM-C:2a、pGEM-C:CPとした(図23)。
1-2-3: CMV遺伝子断片のALSVベクターへの導入
pGEM-C:1aを鋳型にCMV-Y 1a Xho(+)とCMV-Y 1a Bam(-)、pGEM-C:2aを鋳型にCMV-Y 2a Xho(+)とCMV-Y 2a Bam(-)、pGEM-C:CPを鋳型にCMV-Y CP Xho(+)とCMV-Y CP Bam(-)の組み合わせ(表3:配列番号16−21)で、実施例1の1-2-1と同様の方法でPCRを行った。PCR産物のBamHIとXhoIを用いた制限酵素処理、MonoFas DNA精製キットIを用いた精製はII-1-(2)-3)と同様の方法で行った。各種CMVインサートDNAとベクターDNAをDNA Ligation Kit Ver.2.1を用いて行い大腸菌DH5αを形質転換後、スモールスケール培養し、煮沸法によってプラスミドを抽出した。
【0081】
【表3】
【0082】
各CMVインサートDNAを保持するクローンは、それぞれ1μg/μlの濃度に調整し、得られたプラスミドをALSVR2L5R5-C:1a:、 ALSVR2L5R5-C:2a、ALSVR2L5R5-C:CPとした(図24)。
1-2-4:ALSVベクターの接種とウイルス化
精製した各プラスミド(ALSVR2L5R5-C:1a、ALSVR2L5R5-C:2a、ALSVR2L5R5-C:CP)は、実施例1の1-2-4と同様の方法でウイルス化を行い、各プラスミドの接種によって得たウイルスをそれぞれALSV-C:1a、ALSV-C:2a、ALSV-C:CPとし、以後の試験の接種源とした。
1-3:供試植物への接種法と一次接種および二次接種
1-3-1:一次接種
タバコへの一次接種は以下のとおり行った。wtALSV、ALSV-C:1a、ALSV-C:2a、ALSV-C:CPにそれぞれ感染したキノア葉を2倍容のALSV磨砕緩衝液で磨砕した。作製した各ALSV磨砕液を5葉期のタバコの第3から第5本葉の3枚に、カーボランダム法で接種した。タバコは25℃の長日条件で育成した。
【0083】
キュウリへの一次接種はII-(3)-1)と同様の方法で行い、25℃の長日条件で育成した。
1-3-2:二次接種
タバコでの二次接種にはCMV-Yを用いて以下の通り行った。CMV-Y感染タバコ葉を2倍容の0.1Mリン酸緩衝液(pH7.0)で磨砕し、第7から第9本葉の3枚にカーボランダム法で二次接種した。二次接種後、タバコは一次接種後と同様に25℃の長日条件で育成した。
【0084】
キュウリへの二次接種にはCMV-42CMを用いて以下の方法で接種した。CMV-42CM感染タバコ葉を2倍容の0.1Mリン酸緩衝液(pH7.0)で磨砕し、第1本葉1枚にカーボランダム法で二次接種した。二次接種後、キュウリは一次接種後と同様に25℃の長日条件で育成した。
【0085】
なお、二次接種に用いたCMV-YとCMV-42CMは、あらかじめ0.1Mリン酸緩衝液(pH7.0)を用いてタバコに接種し、病徴の現れたタバコの上葉を採取し、-80℃で凍結保存したものを使用した。
1-4:病徴観察とウイルス検定
1-4-1:病徴の観察
病徴の観察は二次接種後毎日行った。タバコは二次接種後9日目に、キュウリは二次接種後18日目にデジタルカメラFinePix S1Proを用いて病徴を撮影した。
1-4-2:ウエスタンブロット法
一次接種後7日目に、タバコでは第3本葉から第5本葉を採取し、キュウリでは子葉2枚を採取し、II-1-(4)-3)と同様の方法でウエスタンブロットを行い、一次接種したALSVの感染を確認した。
1-4-3:ELISA法
タバコにおけるCMV-YとALSVの蓄積量は以下のように検定試料を作製し、直接ELISA法で定量した。二次接種後9日目に第12本葉から第15本葉を採取し、乳鉢と乳棒を用いて20倍容のPBSTで磨砕し、磨砕液1mlを1.5ml容マイクロチューブに移した。14,000rpm(4℃)で5分間遠心し、得られた上清を検定試料とした。上記の方法で作製した検定試料を用いて、実施例1の1-4-4の直接ELISA法でウイルスの定量をした。
【0086】
キュウリにおけるCMV-42CMとALSVの蓄積量は以下のように検定試料を作製し、実施例1の1-4-4の直接ELISA法で定量した。二次接種後18日目に第3本葉から第6本葉のそれぞれからII-1-(4)-4)と同様にリーフディスクを6枚採取し、検定試料を作製した。
【0087】
直接ELISA法によるCMV-YおよびCMV-42CMの定量には、CMV抗体(日本植物防疫協会研究所)と酵素標識をしたCMV抗体(日本植物防疫協会研究所)を使用した。
1-4-4:RNA抽出法とRT-PCR法
タバコでは二次接種後9日目にタバコの最上位葉を採取し、キュウリでは二次接種後18日目にキュウリの最上位葉を採取し、それぞれ-80℃で凍結し、検定試料とした。RNA抽出と逆転写反応およびPCRは全て実施例1の1-4-5と同様の方法で行い、ALSVのインサート配列の保持を確認した。
2:結果
2-1:タバコにおけるCMV-Y遺伝子断片を導入したALSVベクターの干渉効果
CMV-Y遺伝子断片を導入したALSVベクターの接種によって、タバコでCMV-Yに対する干渉効果の誘導が可能かどうか調べるため、wtALSVおよびCMV-Yの各遺伝子断片を導入したALSV(ALSV-C:1a 、ALSV-C:2a、ALSV-C:CP)を一次接種したタバコにCMV-Yを二次接種し、CMV-Yに対する干渉効果の誘導を調べた。
【0088】
ALSVの一次接種を行わずCMV-Yのみを二次接種した試験区(CMV-Yのみ区)やwtALSVを一次接種後CMV-Yの二次接種を行った試験区(wtALSV+CMV-Y区)では、二次接種後約3日で全個体の接種葉で黄斑が観察され、約7日には上葉に激しいモザイク症状が観察された。これに対して、ALSV-C:2aを一次接種後にCMV-Yのを二次接種した試験区(ALSV-C:2a+CMV-Y区)では、全ての個体で病徴は観察されなかった。ALSV-C:1aを一次接種後にCMV-Yを二次接種した試験区(ALSV-C:1a+CMV-Y区)では、6個体中3個体の上葉に弱いモザイク症状が観察され、残りの3個体では病徴が観察されなかった。一方、ALSV-C:CPを一次接種後にCMV-Yを二次接種した試験区(ALSV-C:CP+CMV-Y区)では6個体全ての上葉に弱いモザイク症状が観察された(図25、表4)。
【0089】
【表4】
【0090】
CMV-Yの蓄積を調べたところ、ALSV-C:2a+CMV-Y区ではCMV-Yの蓄積は認められなかった。これに対して、ALSV-C:CP+CMV-Y区ではCMV-Yのみ区やwtALSV+CMV-Y区と同程度蓄積していた。ALSV-C:1a+CMV-Y区では、CMV-Yのみ区やwtALSV+CMV-Y区の3分の1程度蓄積していた(図26)。ALSVの定量ではALSV-C:1a+CMV-Y区とALSV-C:2a+CMV-Y区ではALSVのみ区と同程度蓄積し、wtALSV+CMV-Y区ではALSVのみ区に比べ蓄積量が若干増加していることが確認された。一方、ALSV-C:CP+CMV-Y区では、ALSVのみ区の半分から4分の1程度少なく蓄積していることが確認された(図26)。一次接種に用いたCMV-Yの各遺伝子断片を導入したALSV(ALSV-C:1a 、ALSV-C:2a、ALSV-C:CP)のインサートチェックを行ったところ、全てのキュウリ個体でALSVのインサートが保持されていた。
2-2:キュウリにおけるCMV-Y遺伝子断片を導入したALSVベクターの干渉効果
wtALSVおよびCMV-Yの各遺伝子断片を導入したALSV(ALSV-C:1a 、ALSV-C:2a、ALSV-C:CP)を一次接種したキュウリにおけるCMV-42CMに対する干渉効果を調べた。
【0091】
CMV-42CMのみ二次接種した試験区(CMV-42CMのみ区)やwtALSVを一次接種後にCMV-42CMを二次接種した試験区(wtALSV+CMV-42CM区)では、二次接種後約3日で全ての個体の第2本葉で退緑斑が観察され、その後展開した上葉で退緑斑が観察された。特に、第6本葉から第8本葉ではモザイク症状が観察された。これに対し、ALSV-C:2aを一次接種後にCMV-42CMを二次接種した試験区(ALSV-C:2a+CMV-42CM区)では、全個体で病徴が観察されなかった。ALSV-C:1aを一次接種後にCMV-42CMを二次接種した試験区(ALSV-C:2a+CMV-42CM区)では、6個体中2個体で接種葉に退緑斑が観察され、第2本葉から第4本葉で退緑斑観察された。第5本葉より上の葉では病徴が観察されなかった。また、残り4個体では病徴が観察されなかった。ALSV-C:CPを一次接種後にCMV-42CMを二次接種した試験区(ALSV-C:CP+CMV-42CM区)では6個体全てにCMV-42CMのみ区やwtALSV+CMV-42CM区と同様の病徴が観察された(図27)。
【0092】
CMV-42CMの定量の結果、ALSV-C:2a+CMV-42CM区ではCMV-42CMの蓄積は認められなかった。これに対し、ALSV-C:CP+CMV-42CM区では、CMV-42CMのみ区やwtALSV+CMV-42CM区と同程度CMV-42CMが蓄積し、ALSV-C:1a+CMV-42CM区では、約半分のZYMVが蓄積していた(図28)。また、ALSV-C:1a+CMV-42CM区とALSV-C:2a+CMV-42CM区では、ALSVのみ区と同程度のALSVが蓄積し、wtALSV+CMV-42CM区ではALSVのみに比べ蓄積量が増加していた。一方、ALSV-C:CP+CMV-42CM区では、ALSVのみ区より少なく蓄積していた(図28)。一次接種に用いたCMV-Yの各遺伝子断片を導入したALSV(ALSV-C:1a 、ALSV-C:2a、ALSV-C:CP)のインサートチェックの結果、全てのALSVがインサートを保持していた。
3:考察
タバコおよびキュウリの両植物のALSV-C:2a を接種した試験区では、CMV-YおよびCMV-42CMに対する干渉効果が誘導され、病徴やウイルスの蓄積は確認されなかった。これに対し、ALSV-C:1aおよびALSV-C:CPを接種した試験区では穏かな病徴が観察され、CMV-YおよびCMV-42CMの蓄積が確認された。この干渉効果の違いはALSVベクターに導入したそれぞれの遺伝子断片の違いであると考えられる。
【0093】
2a遺伝子断片を組み込んだALSV-C:2a ではCMV-YおよびCMV-42CMの病徴やウイルスの蓄積が全く確認されなかったことから、ALSVベクターによる干渉効果の誘導はタバコとキュウリの両植物においても可能であることが示され、ALSVベクターの宿主植物において干渉効果誘導ベクターとしての利用が期待される。
【0094】
ALSVに導入した各CMV-Y遺伝子断片領域とそれに対応するCMV-42CMの遺伝子領域の塩基配列の相同性を比較したところ、1a遺伝子では90.3%、2a遺伝子では98.1%、CP遺伝子では97.9%相同であった。
【0095】
本実施例2では、既知のCMV-Y遺伝子を基に構築した2a遺伝子断片を導入したALSVによって、2つのCMV(CMV-YとCMV-42CM)に対して干渉効果を誘導することができた。この結果から、塩基配列の相同性が保存されている遺伝子領域の断片を導入したALSVベクターを宿主に接種することで、異なる系統や分離株に対する干渉効果を誘導可能であることが示された。
【0096】
実施例1の結果と合わせて考察すると、ALSVベクターは異なる属のウイルスにおいて干渉効果の誘導に成功したことから、ALSVベクターに導入する遺伝子断片を変えることで、多数のウイルスに対する干渉効果を誘導する防除剤(植物ワクチン)の作出が短期間で可能であることが示された。
【実施例3】
【0097】
SMVの遺伝子断片を導入したALSVベクターによる干渉効果
SMVの外被タンパク質(CP)をコードするゲノム配列の一部(201 nt)を挿入したALSV(SMVCP-ALSV)を作出し、SMVに対する干渉効果を試験した。SMVCP-ALSVをダイズ品種(Jackおよび丹波黒)の子葉に1次接種後、展開した初生葉もしくは第一本葉にSMVをカーボランダム法により2次接種(チャレンジ接種)した。
【0098】
その結果、SMV単独接種区では上葉にモザイクと縮葉症状を示したのに対し、SMVCP-ALSVを1次接種した区では病徴はほとんど観察されなかった。また、SMV単独接種区と比較するとSMVの増殖量が著しく低下しており、SMVCP-ALSVによる干渉効果が生じていることが確認された。また、上葉におけるSMVの分布をティッシュブロット分析により解析したところ、SMV単独接種個体ではSMVが葉全面にモザイク状に分布しているのに対し、SMVCP-ALSVを1次接種した個体ではウイルスが葉脈とその近傍のみに局在していることが明らかとなった。
【実施例4】
【0099】
ZYMVとCMVの遺伝子断片を導入したALSVベクターによる干渉効果
実施例1と同様のZYMV遺伝子断片(150 nt)と実施例2のCMV遺伝子断片(150 nt)を融合した形で挿入したALSV(ZC-ALSV)を作出し、ZYMVとCMVに対する干渉効果を試験した。ZC-ALSVをキュウリ子葉に1次接種後、5〜6日後に第1本葉にZYMVとCMVをそれぞれ単独で、または混合し2次接種(チャレンジ接種)した。
【0100】
その結果、対象区では上葉にZYMVの奇形を伴う激しいモザイク(ZYMV2次接種区)や激しいモザイク(CMV2次接種区)、激しいモザイクと矮化症状(混合2次接種区)が現れたのに対して、ZC-ALSV1次接種区ではZYMV単独、CMV単独、ZYMVとCMV混合のいずれの2次接種区に対しても干渉効果を示し、明瞭な病徴は認められず、また各ウイルスの蓄積も抑制されていた。
【0101】
以上の結果から、2種類のウイルスを同時に抑制するALSV混合ワクチン株の作出が可能であることが確認された。
【特許請求の範囲】
【請求項1】
1種または2種以上の植物病原ウイルスのゲノムRNAまたはそのcDNAの一部断片を保有する組換えリンゴ小球型潜在ウイルス(ALSV)。
【請求項2】
植物病原ウイルスがズッキーニ黄斑モザイクウイルス(ZYMV)である請求項1の組換えALSV。
【請求項3】
植物病原ウイルスがキュウリモザイクウイルス(CMV)である請求項1の組換えALSV。
【請求項4】
植物病原ウイルスがダイズモザイクウイルス(SMV)である請求項1の組換えALSV。
【請求項5】
請求項1の組換えALSVを植物の幼苗に接種することを特徴とする植物病原ウイルスの感染防除方法。
【請求項6】
請求項1の組換えALSVが幼苗期に接種された病原ウイルス抵抗性植物。
【請求項1】
1種または2種以上の植物病原ウイルスのゲノムRNAまたはそのcDNAの一部断片を保有する組換えリンゴ小球型潜在ウイルス(ALSV)。
【請求項2】
植物病原ウイルスがズッキーニ黄斑モザイクウイルス(ZYMV)である請求項1の組換えALSV。
【請求項3】
植物病原ウイルスがキュウリモザイクウイルス(CMV)である請求項1の組換えALSV。
【請求項4】
植物病原ウイルスがダイズモザイクウイルス(SMV)である請求項1の組換えALSV。
【請求項5】
請求項1の組換えALSVを植物の幼苗に接種することを特徴とする植物病原ウイルスの感染防除方法。
【請求項6】
請求項1の組換えALSVが幼苗期に接種された病原ウイルス抵抗性植物。
【図23】

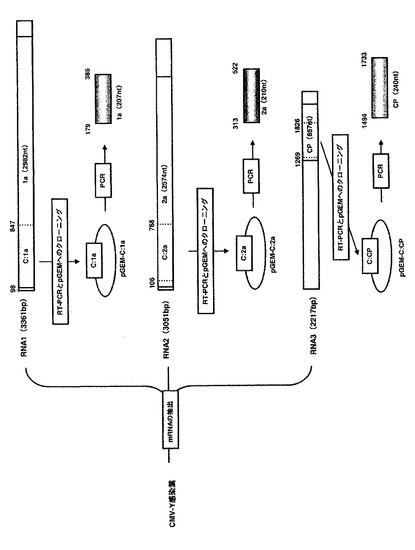
【図24】
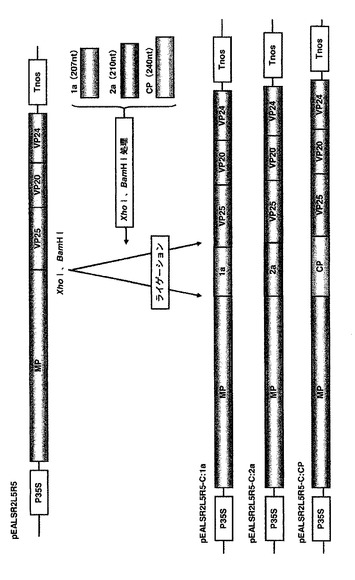
【図1】
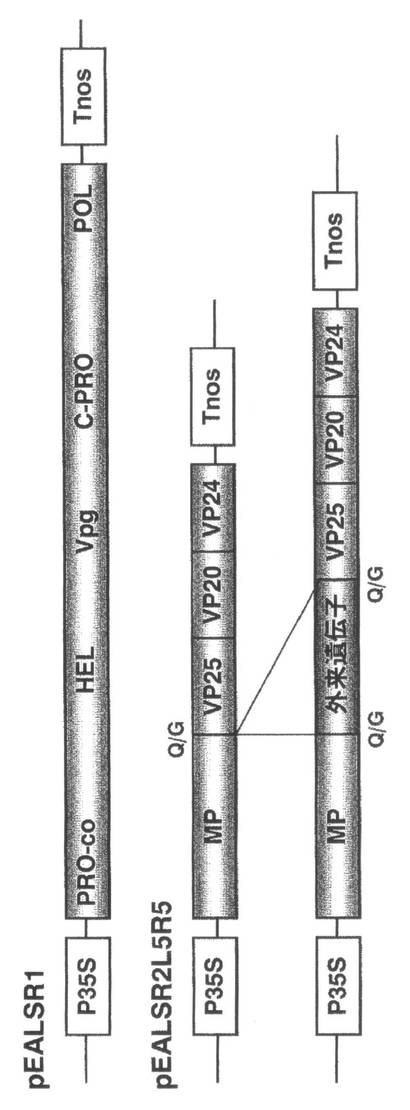
【図2】
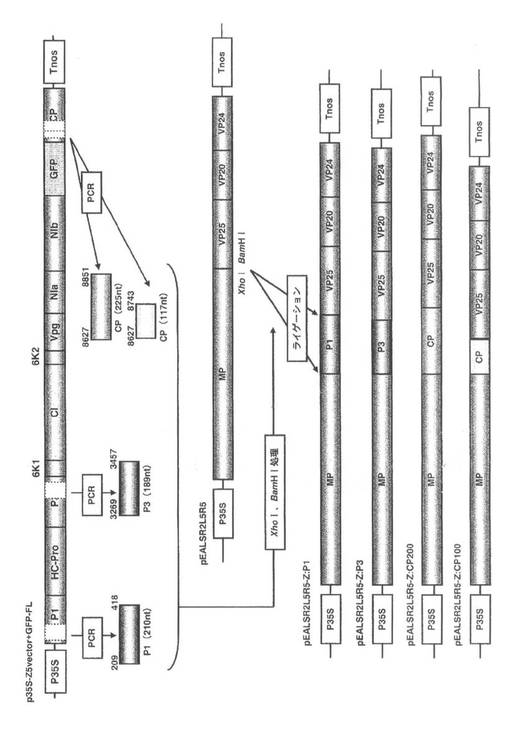
【図3】
【図4】
【図5】
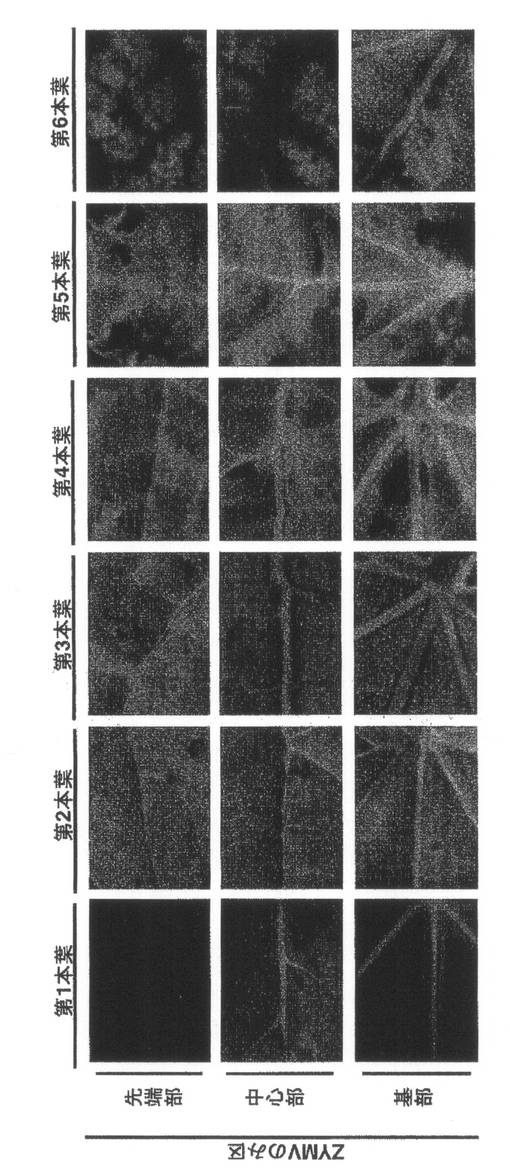
【図6】
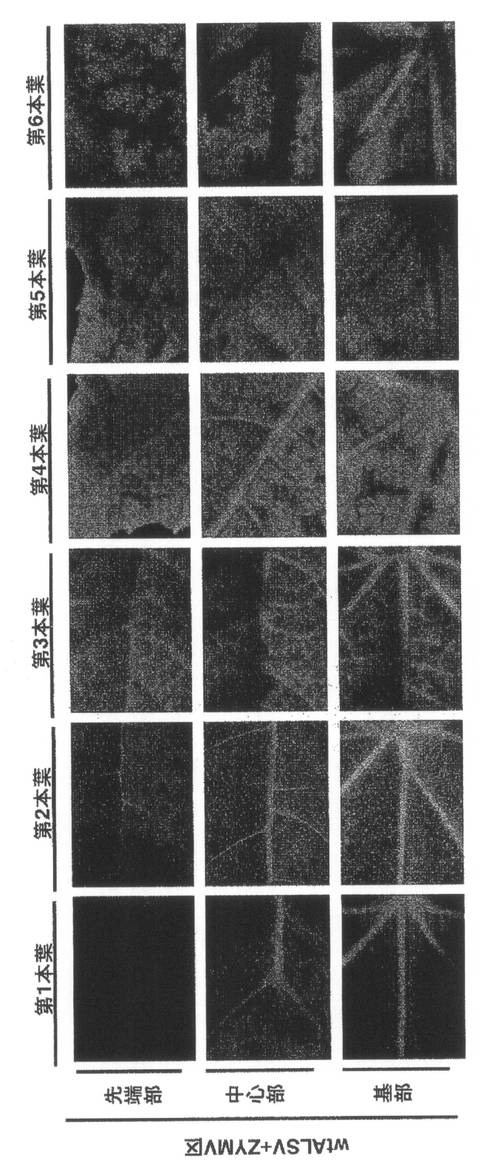
【図7】
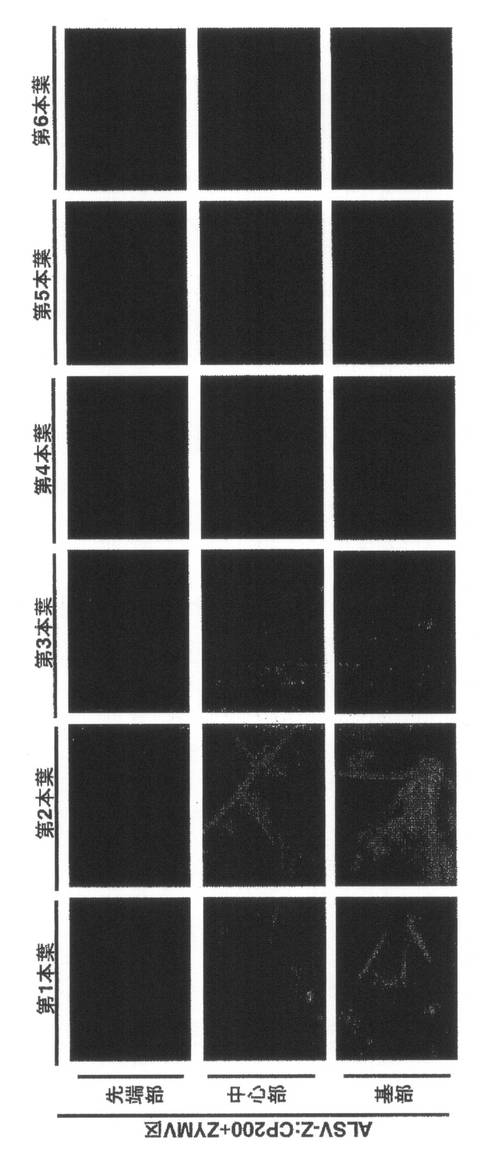
【図8】
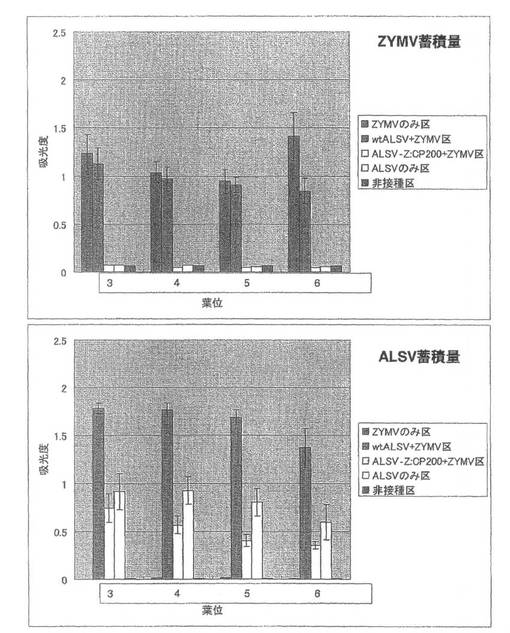
【図9】
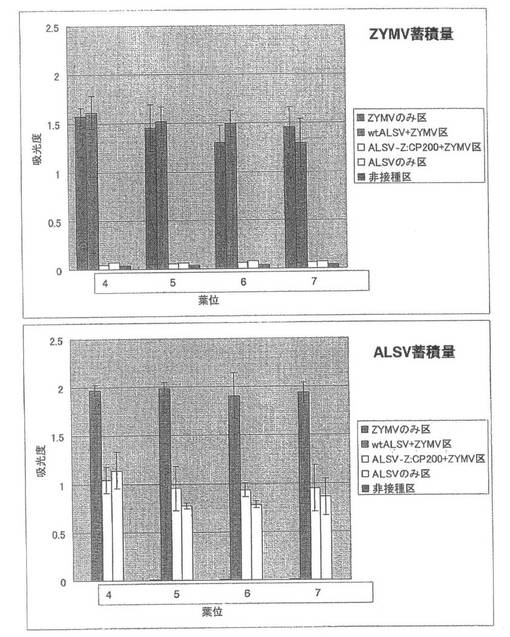
【図10】
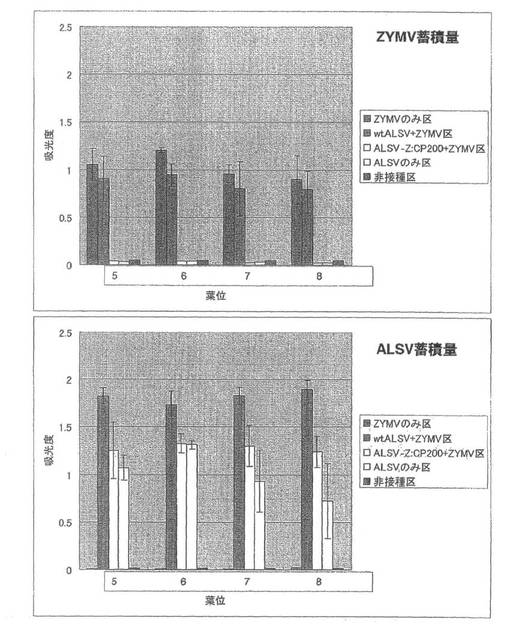
【図11】
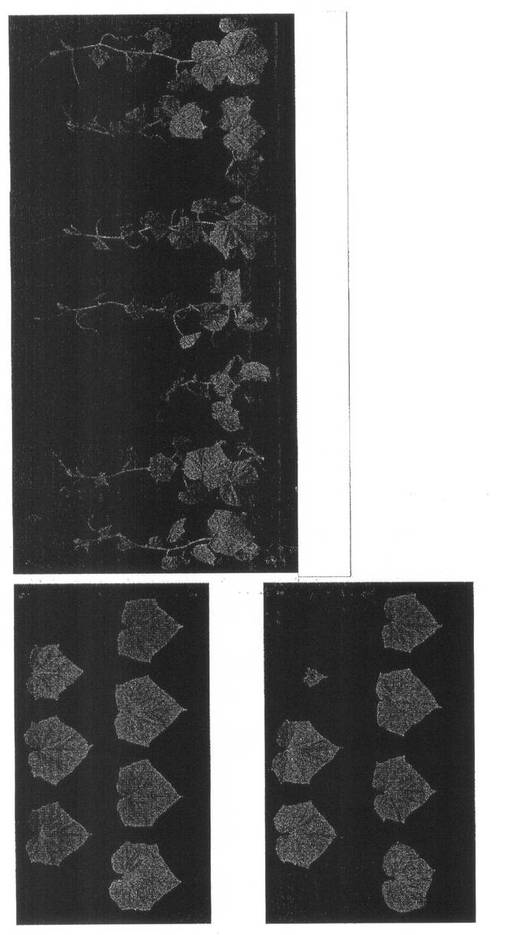
【図12】

【図13】

【図14】
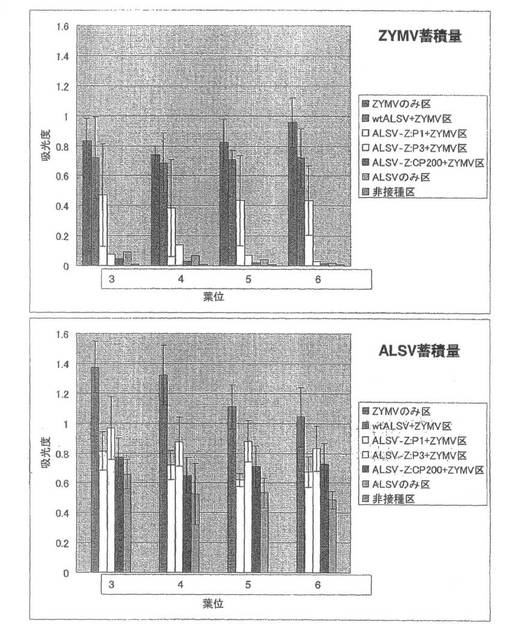
【図15】
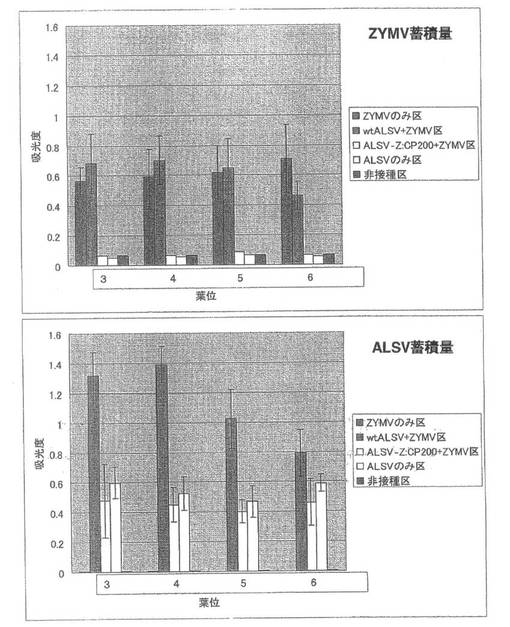
【図16】
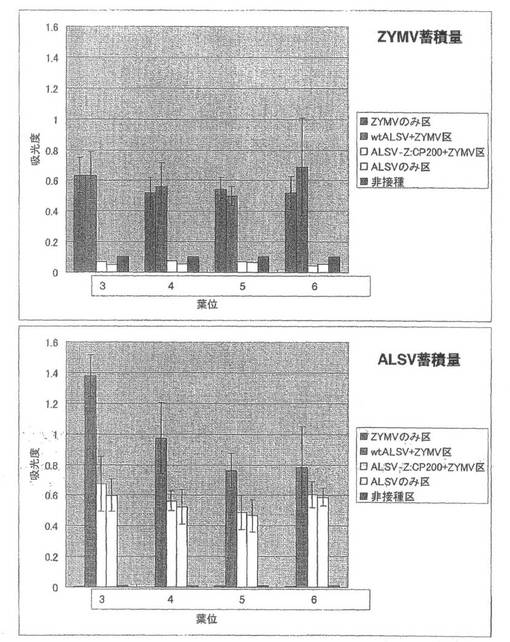
【図17】
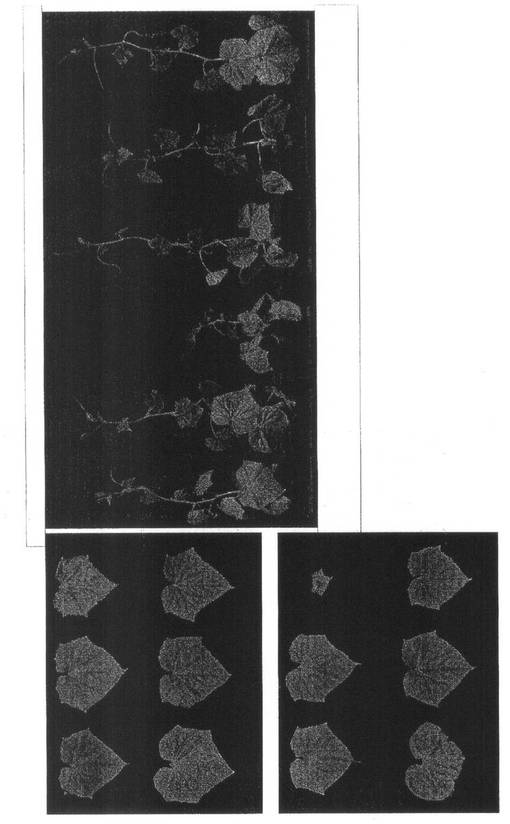
【図18】

【図19】
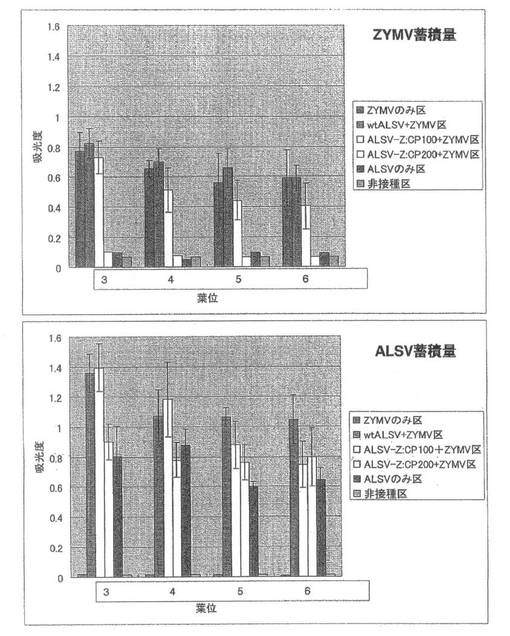
【図20】
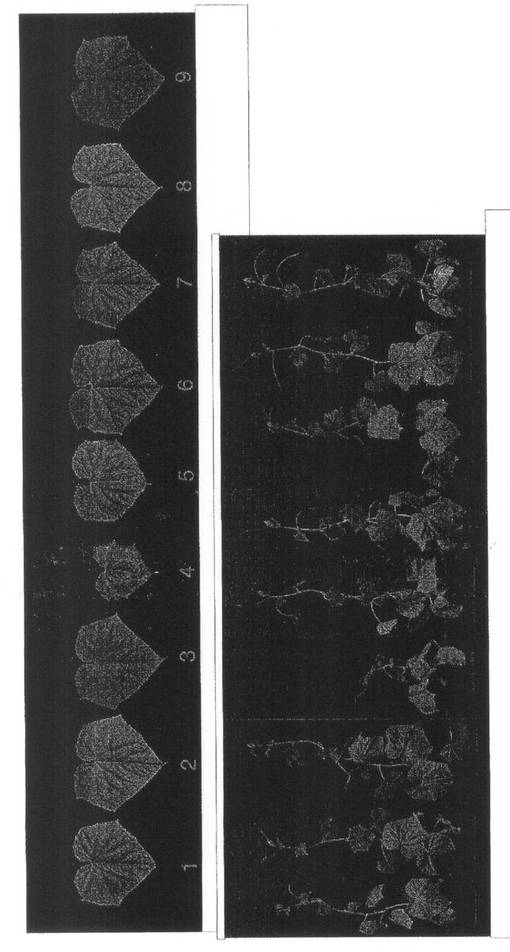
【図21】

【図22】
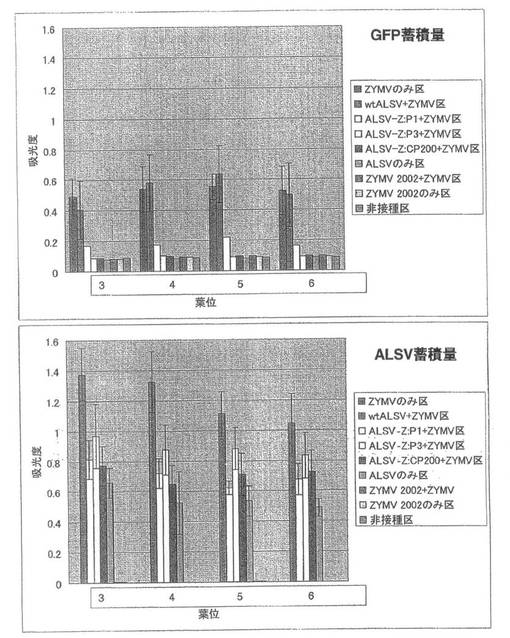
【図25】
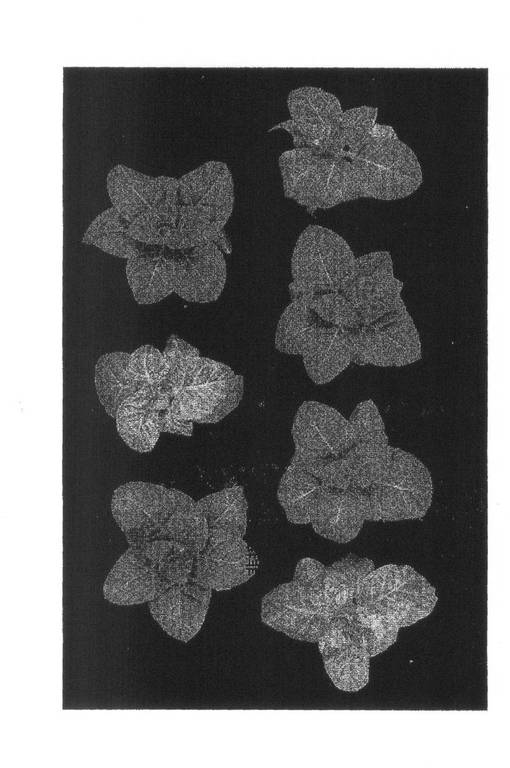
【図26】
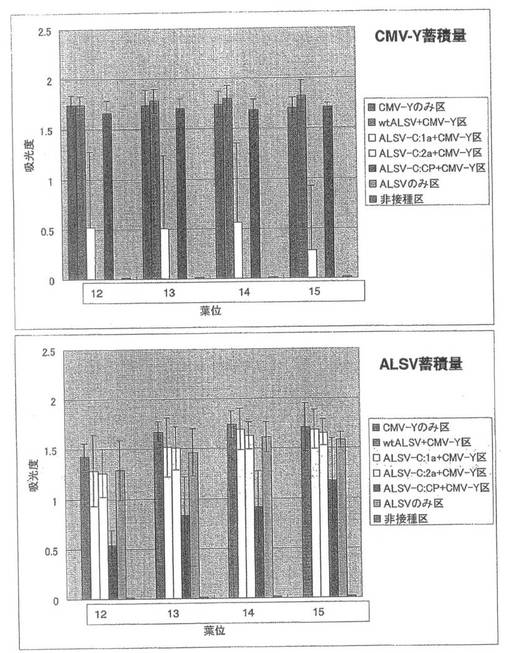
【図27】

【図28】
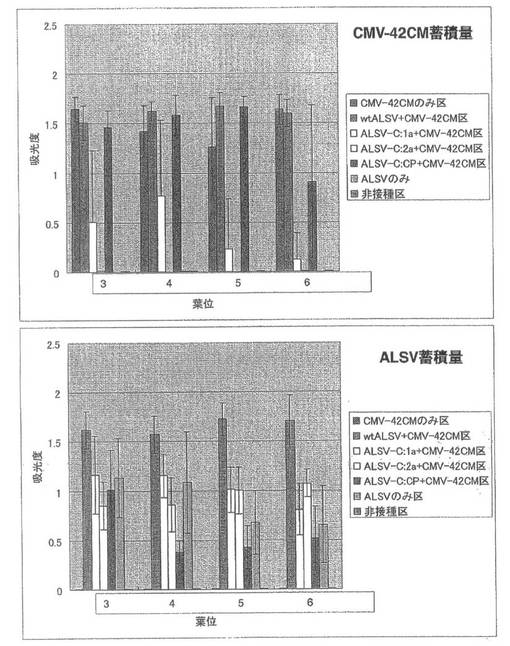
【図24】
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図25】
【図26】
【図27】
【図28】
【公開番号】特開2011−217744(P2011−217744A)
【公開日】平成23年11月4日(2011.11.4)
【国際特許分類】
【出願番号】特願2011−66704(P2011−66704)
【出願日】平成23年3月24日(2011.3.24)
【出願人】(504165591)国立大学法人岩手大学 (222)
【Fターム(参考)】
【公開日】平成23年11月4日(2011.11.4)
【国際特許分類】
【出願日】平成23年3月24日(2011.3.24)
【出願人】(504165591)国立大学法人岩手大学 (222)
【Fターム(参考)】
[ Back to top ]