
植物ホルモン・オーキシンの生合成阻害剤及び該阻害剤を有効成分として含有する植物化学調節剤、除草剤並びにその使用方法
【課題】本願発明は、内在性のオーキシンの合成阻害剤を提供することを第1の課題とする。更に本願発明は、上記オーキシン生合成阻害剤を使用する植物成長抑制剤または除草剤を提供することを第2の課題とする。
【解決手段】本願発明は、植物体にオーキシン阻害剤候補化合物を添加し、オーキシン誘導性遺伝子の発現を妨げるオーキシン生合成阻害剤の候補をスクーニングする方法、並びに候補化合物を添加後に植物体を破砕して内生オーキシン量を測定することにより、植物体内でオーキシン合成を実際に阻害している化合物を選抜する方法、並びにあらかじめ調整したトリプトファンデアミナーゼとトリプトファンの存在下でインドールピルビン酸の生成を阻害する化合物を選抜する方法を提供する。より具体的には、本願発明は、AVG、AOA、AOIBA、L-AOPP又はその類縁体からなるオーキシン生合成阻害剤、及びこれら化合物を有効成分とする植物成長制御剤、除草剤を包含する。
【解決手段】本願発明は、植物体にオーキシン阻害剤候補化合物を添加し、オーキシン誘導性遺伝子の発現を妨げるオーキシン生合成阻害剤の候補をスクーニングする方法、並びに候補化合物を添加後に植物体を破砕して内生オーキシン量を測定することにより、植物体内でオーキシン合成を実際に阻害している化合物を選抜する方法、並びにあらかじめ調整したトリプトファンデアミナーゼとトリプトファンの存在下でインドールピルビン酸の生成を阻害する化合物を選抜する方法を提供する。より具体的には、本願発明は、AVG、AOA、AOIBA、L-AOPP又はその類縁体からなるオーキシン生合成阻害剤、及びこれら化合物を有効成分とする植物成長制御剤、除草剤を包含する。
【発明の詳細な説明】
【技術分野】
【0001】
本願発明は、オーキシン(インドール-3-酢酸等)の生合成阻害剤を有効成分として含有する植物化学調節剤または除草剤に関する。更に、本願発明は、該植物化学調節剤を用いて、様々な植物種において、様々な生育段階、器官において植物の生育を人為的に制御する方法に関する。
【背景技術】
【0002】
オーキシンは、植物ホルモンとして最初に認識された物質で、植物の示す屈曲性に関与するホルモンとして、同定された。その後、主要な天然オーキシンとしては、インドール−3-酢酸(IAA)であることが明らかにされ、IAA以外にも、インドール-3-酪酸、4−クロロインドール酢酸、及びフェニル酢酸などが知られている。また合成オーキシンとしては、2,4,ジクロロフェノキシ酢酸(2,4-D)、α-ナフタレン酢酸(NAA)、2,6-ジクロロ安息香酸などが知られている(非特許文献1)。
【0003】
天然オーキシンであるIAAは不安定であり、植物体内では分解経路も存在するため、農業用途には、合成オーキシンが用いられていることが多い。例えば、水田での双子葉植物除草剤としては、4-(4-chloro-o-tolyloxy)butyric acid (MCPB)や2,4-Dが用いられている。また2,3,5-トリヨード安息香酸(TIBA)は、側芽の発芽、成長を高めることができる。さらに、組織培養には、2,4−Dやナフタレン酢酸(NAA)が用いられる。また、発根には、NAAやインドール酪酸が用いられている。また、例えば、トマトでは、4−クロロフェノキシ酢酸は、未受精のトマトでも着果促進する効果がある(非特許文献1)。
【0004】
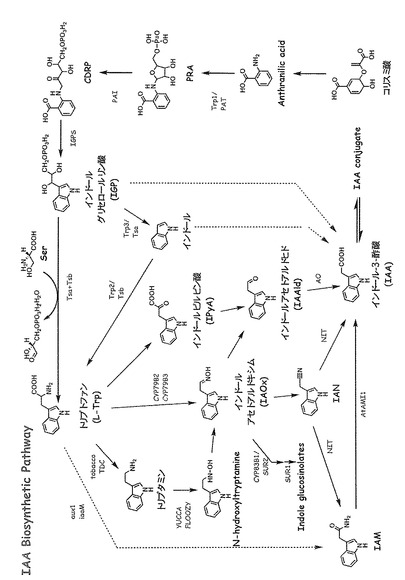
植物体内でオーキシンはコリスミ酸から合成され、以前は、L-トリプトファン(L-Trp)を経由して合成されると考えられていたが、現在では大きく分けて(1)L-Trpを経由する経路と、(2)L-Trpを経由しない経路の2つの経路が確認されている。L-Trpを経由する経路は、さらに4つ以上の枝に分岐しており、それぞれの経路は異なった酵素によって触媒されている(図1)。現在でも、どの経路が主要経路なのか、あるいはどのような役割分担があるのかは不明である。
【0005】
自身はほとんどオーキシン活性を示さないが、IAAなどのオーキシンと競合してオーキシンの結合部位を奪うことにより、オーキシン作用を拮抗的に阻害する抗オーキシンも合成されている。抗オーキシンとしては、2,4-ジクロロフェノキシイソ酪酸(PCIB)等が知られている。また、マレイン酸ヒドラジド、マレイン酸ヒドラジドコリンなども、オーキシンの作用を抑制し、細胞分裂を阻害すると共に、伸長を抑制することが知られている。タバコの芯止め後のえき芽発生抑制や、野菜の発芽抑制に用いられている(非特許文献2)。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】増田芳雄編著「植物ホルモン入門」オーム社(平成4年(1992年)7月30日発行)第9−17及び41−45頁
【非特許文献2】農薬ハンドブック1998年版編集委員会編「農薬ハンドブック1998年版」社団法人日本植物防除協会発行 第10版 平成10年(1998年)12月15日 第525-526頁
【発明の概要】
【発明が解決しようとする課題】
【0007】
これまで、抗オーキシンと呼ばれるオーキシンの信号を阻害するとされる物質(PCIBなど)や、オーキシン極性輸送阻害剤と呼ばれるTIBAなど、オーキシンの作用を制御する化合物としては知られていたが、内在性オーキシンの量を制御する(減少させる)物質は知られていなかった。また、モデル植物を中心に分子遺伝学が発達した今日に於いても、内生オーキシン量が減少する突然変異体は知られておらず、RNAi法やアンチセンス法など分子生物学的な手法を用いても内生オーキシンの量を抑制する技術は存在しなかった。本年4月になって、PLP酵素の変異体taa1/sav3/wei8が単離され、本遺伝子とそのファミリー遺伝子(TAR2)の2重変異体wei8 tar2変異体ではIAA内生量が半分以下に減少することが報告された(Cell 133: 177-191 (2008); Cell 133 164-176 (2008))。
【0008】
今日のオーキシン生合成経路に関する知見では、オーキシンは網目状の多様な経路を通して合成されており、そもそも一つの化合物(つまり一つの酵素反応の阻害)で、その生合成を強く阻害することは困難と考えられてきた。
【0009】
他方、植物の内在性のオーキシン合成量を調節できれば、更に効率的に植物成長を調節できる可能性がある。
【0010】
今日、国際市場で最も良く普及している除草剤グリホサートは、芳香族アミノ酸合成経路の阻害剤である(つまり、コリスミ酸の上流で代謝を阻害)。一方、本願発明で提供される化合物は、芳香族アミノ酸合成経路経路の下流末端に位置するオーキシンの生合成経路を阻害する。従って、既存の除草剤よりも制御する経路がより狭く、特異的であるため、本願発明によって提供される薬剤は、既存の除草剤よりも副作用が少なく、作用の選択性が高く、より効率的に(すなわち低濃度で)植物を除草する効果をもたらすことが期待できる。
【0011】
そこで、本願発明は、内在性のオーキシンの合成阻害剤を提供することを第1の課題とする。更に本願発明は、上記オーキシン生合成阻害剤を使用する植物成長制御剤または除草剤を提供することを第2の課題とする。
【課題を解決するための手段】
【0012】
本願発明者等は、その類まれなる洞察力により、オーキシン生合成阻害作用を奏する化合物を見出して、本願発明を完成させた。
【0013】
本願発明は、オーキシン誘導性遺伝子の発現を妨げるオーキシン生合成阻害剤候補をスクーニングする方法を提供する。これにはオーキシン応答性遺伝子に対するプローブを担持するマクロアレイを用いて、オーキシン誘導性遺伝子の発現を妨げる候補をスクーニングする方法を含む。さらに、本願発明は、実際に植物に前記オーキシン阻害剤候補を添加し、その後植物体を破砕して生体内のオーキシン量を測定することにより、オーキシン阻害剤の候補が植物体内でオーキシン合成を阻害しているオーキシン阻害剤候補をオーキシン阻害剤として選抜する、オーキシン生合成阻害剤のスクリーニング方法を提供する。さらに、オーキシン生合成経路のうち、L-TrpからIndole-3-pyrvic acid(IPyA)を合成する酵素抽出液に対して、前記オーキシン阻害剤候補とL-Trpを添加し、IPyAの生成を妨げる候補化合物をスクーニングする方法を含む。
【0014】
また、本願発明は、オーキシン生合成阻害剤を提供する。本願発明は、L-trpを経由してIAAを合成する経路で働く、ピリドキサルリン酸(PLP)要求性の酵素又はL-tryptophanを脱アミノ化してIndole-3-pyrvic acid(IPyA)に変換するトランスデアミナーゼを阻害することにより、オーキシン生合成を阻害する阻害剤を提供する。
【0015】
より具体的には、本願発明は、AVG(aminoethoxyvinylglycine、2-amino-4-(2-aminoethoxy)-3-butenoic acid(USP3869277))、AOA(aminooxyacetic acid)、2-aminooxy isobutyric acid hydlochloride (AOIBA)、L-aminooxyphenylpropionic acid (L-AOPP)を含有するオーキシン生合成阻害剤及びこれらを有効成分とする植物成長調節剤を包含する。
【0016】
本明細書は本願の優先権の基礎である日本国特許出願2007-152808号の明細書および/または図面に記載される内容を包含する。
【図面の簡単な説明】
【0017】
【図1】オーキシン合成経路を示す。オーキシンはコリスミ酸から合成され、L-Trpを経由する経路と、経由しない経路の2つの経路が確認されている。L-Trpを経由する経路は、4つ以上の枝に分岐しており、それぞれの経路は異なった酵素によって触媒される。それぞれの経路の中間体と生合成反応を触媒する酵素(または遺伝子)の名称(略号)を記す。
【図2】エチレン誘導性遺伝子(A)とオーキシン誘導性遺伝子(B)のAVG処理に対する挙動を示す。(A)エチレン応答性遺伝子の発現。(B)オーキシン応答性遺伝子の発現。
【図3】AVG処理後のシロイヌナズナ内生IAA量の変化を示す。
【図4】ACCとオーキシン生合成中間体を用いた、AVG処理からの遺伝子発現回復実験を示す。
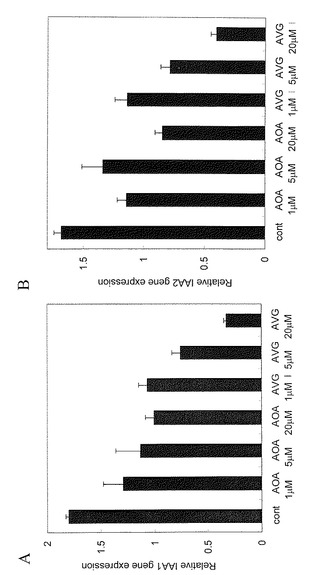
【図5】AVG, AOAがオーキシン応答性遺伝子の発現を抑制する。
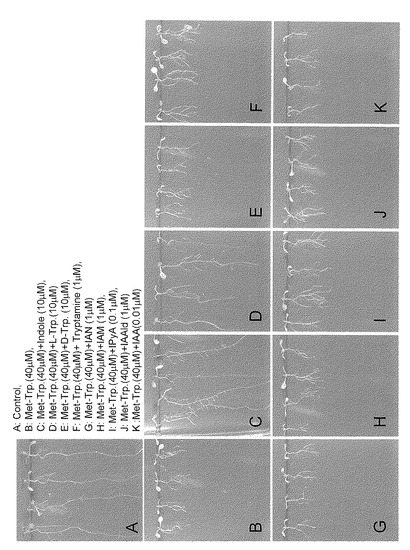
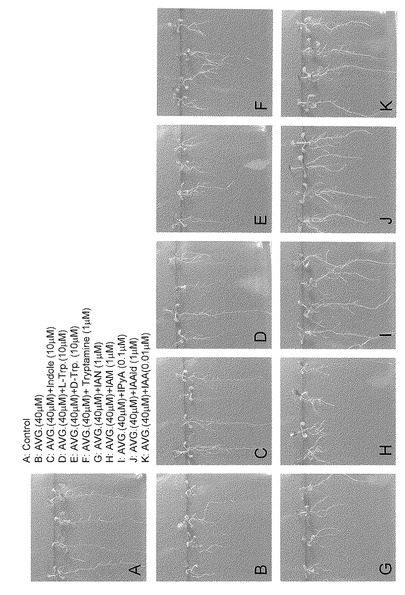
【図6】オーキシンとその生合成中間体を用いた、αメチルトリプトファン(Met-Trp)処理からの根の伸長回復実験を示す。
【図7】オーキシンと生合成中間体を用いた、AVG処理からの根の伸長回復実験を示す。
【図8】In vitroでのL-トリプトファンからIPyAへの変換酵素(コムギ由来)に対するAVGの抑制作用。
【図9】シロイヌナズナ芽生の生育へのAVG、AOAの効果。
【図10】In vitro でのL-トリプトファンからIPyAへの変換酵素(シロイヌナズナ由来)に対するAVG, L-AOPPの効果。
【図11】In vitro でのL-トリプトファンからIPyAへの変換酵素(シロイヌナズナ由来)に対するAOA, AOIBAの効果。
【図12】L-AOPPとD-AOPPによるオーキシン応答性遺伝子の発現への影響。L、D-AOPP処理におけるIAA応答性遺伝子発現量
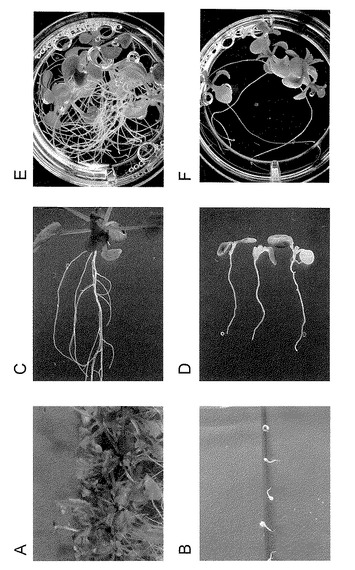
【図13】L-AOPPのシロイヌナズナ生育への影響を示す写真。
【図14】シロイヌナズナIAA内生量への影響。(A)阻害剤1h処理後のIAA内生量。(B)阻害剤1h処理後の1株あたりのIAA内生量。
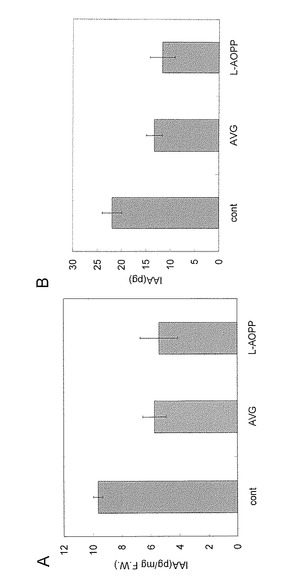
【図15】イネIAA内生量への影響。コシヒカリ阻害剤処理3h IAA内生量。
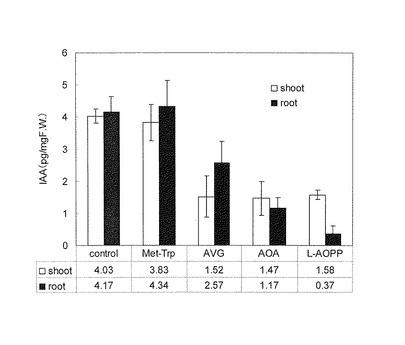
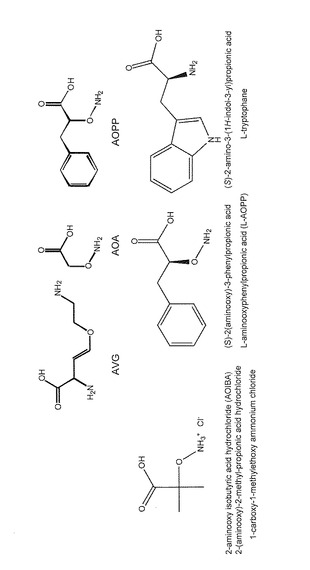
【図16】阻害剤とIAA前駆体の構造。
【発明を実施するための形態】
【0018】
1.はじめに
植物体内でのオーキシンの合成経路は複雑で、網目状であることが示されてきた (図1)。現在までのところ、どの経路が、内在性のオーキシンの主要な合成経路であるかは確認されていない。そのため、どの経路のどの酵素反応を阻害すれば、内在性オーキシン生合成を阻害し、それによって植物の成長を制御または抑制できるか明らかではない。
【0019】
2.遺伝子発現及びマイクロアレイを用いるオーキシン生合成阻害剤候補のスクリーニング方法
本願発明者等は、既に、オーキシンに応答して発現の影響を受ける遺伝子、つまりオーキシン誘導性遺伝子を多数分離している。
【0020】
本願発明者等は、このようなオーキシン誘導性遺伝子に対するプローブを担持するマクロアレイを用いて、オーキシン誘導性遺伝子の発現を妨げるオーキシン生合成阻害剤候補をスクーニングできることを見出した。このようなマイクロアレイとしては、以下の表1に記載の任意の遺伝子を適宜選択し、該遺伝子の塩基配列と同一の少なくとも20オリゴヌクレオチドをプローブとして、周知の任意のマイクロアレイ上に載せたものを使用できる。
【0021】
例えばシロイヌナズナにおいては、表1に掲げられる任意の遺伝子を、例えば、10種類以上選択し、各遺伝子に対するに対する20オリゴヌクレオチド以上からなるプローブを用いるマイクロアレイ及び当該マイクロアレイを用いるオーキシン生合成阻害剤候補のスクリーニング方法を包含する。ここで、マイクロアレイシステムとしては、たとえば、affymetrixのGeneChip systemを用いることができる。
【0022】
本マイクロアレイを用いて、例えば、オーキシン等対象化合物の存在下で発現が誘導される程度と、試験化合物の存在下で発現が抑制される程度を比較することで、オーキシン生合成阻害剤候補を選抜できる。具体的には、例えば、シロイヌナズナ(アラビドプシス)のようなモデル植物に、対象化合物としてオーキシンを与える場合の遺伝子発現のレベルを測定する。次にオーキシンを与えない場合での遺伝子発現の程度を測定して、これらを比較する。更に、該試験植物に試験化合物を与えた場合と与えない場合での遺伝子発現とを比較する。オーキシンの存在下で発現が増大する遺伝子群が、前記試験化合物の存在下で発現が減少する関係があるようであれば、当該試験化合物をオーキシン生合成阻害剤候補として選抜できる。
【0023】
一方、オーキシン応答性遺伝子のプロモーターを利用して人為的に作成したマーカー遺伝子(例えば、DR5-GUSやSAUR-GUSなど)を組み込んだ遺伝子組換え植物体に対して、候補化合物を処理し、マーカー遺伝子の発現レベルを測定することによっても候補化合物を選抜することができる。あるいは、野生型植物に候補化合物を処理し、処理後の植物からトータルRNAを抽出して、オーキシン応答性遺伝子の発現量を定法に従って測定することによって、オーキシン応答性遺伝子の発現を抑制する化合物の中から、候補を選抜することも可能である。
【0024】
【表1】

【0025】
本発明者等は、このようなマイクロアレイを用いたスクリーニング方法により、オーキシン生合成阻害剤候補として、AVGを同定した。
【0026】
3.IAA生合成や酵素活性を指標としたオーキシン生合成阻害剤のスクリーニング方法
植物体内でオーキシン合成を阻害する化合物は、実際に植物体に試験化合物又はオーキシン等の対象化合物を添加して、その後植物体を破砕して、質量分析装置等を用いて、生体内のオーキシン量を測定し、スクリーニングすることができる。
【0027】
また、上記2.で選ばれたオーキシン阻害剤候補が、植物体内でオーキシン合成を阻害している否かは、実際に植物体にオーキシン生合成阻害剤候補を添加して、その後植物体を破砕して、質量分析装置等を用いて、生体内のオーキシン量を測定し確認することもできる。この方法により、本願発明者らは、AVG及びその類縁体がオーキシン生合成阻害剤であることを確認した。
【0028】
さらに、オーキシン生合成経路の中間体をAVGと同時に植物に投与することで、当該化合物がAVGによる成育阻害を回復するか否かを検討した。これにより、本願発明者等は、AVGの成育阻害の作用点が、植物体内でのオーキシンの生合成経路上に存在することを確認した。さらに、植物より抽出した酵素活性をin vitroにおいて評価する系を確立した。この酵素活性の評価系を用いてAVGの作用点を調べ、オーキシン生合成を阻害するターゲットの1つは、ピリドキサルリン酸(PLP)要求性の酵素、又はL-tryptophanを脱アミノ化してIndole-3-pyrvic acid(IPyA)に変換するトランスデアミナーゼ、より好適なオーキシン生合成を阻害するターゲットとしては、L-tryptophanを脱アミノ化してIndole-3-pyrvic acid(IPyA)に変換するピリドキサルリン酸(PLP)要求性のトランスデアミナーゼであることを見出した。従って、これら酵素活性の阻害剤をin vitroでスクリーニングすることによっても、オーキシン合成阻害剤を探索できる。
【0029】
4.ピリドキサルリン酸(PLP)要求性の酵素の阻害剤又はトリプトファンアミノ基転移酵素の阻害剤を有効成分として含有するオーキシン生合成阻害剤
本願発明のオーキシン生合成阻害剤には、AVG及びその類縁体からなるピリドキサルリン酸要求性酵素の阻害剤を包含する。
【0030】
また、本願発明は、ピリドキサルリン酸(PLP)要求性の酵素(トランスデアミナーゼ、デカルボキシラーゼ、C-Sライエース、ラセマーゼ等を含む)の阻害剤を用いて、オーキシンの生合成を阻害することが可能であることを初めて示す。特に、PLP要求性酵素の阻害剤として知られる化合物の中から、前記3.の方法によってオーキシンの生合成を阻害する化合物を新たに選抜することが可能である。PLP要求性酵素の阻害剤としては、例えば、Annual Review of Biochemistry 73:383-415 (2004)に記載の化合物が知られている。本発明者等はこのようなスクリーニングを行うことにより、ACC合成酵素(PLP要求性酵素)の阻害剤として知られているAOA(aminooxyacetic acid)がオーキシン生合成を阻害できることを見いだした。よって、AOAの類縁体も本願発明のオーキシン生合成阻害剤に包含する。
【0031】
また、本願発明は、トランスデアミナーゼ(例えば、Trpアミノトランスフェラーゼ、フェニルアラニンアンモニアリアーゼ)の阻害剤を用いて、オーキシンの生合成を阻害することが可能であることを初めて示す。特に、トランスアミナーゼ阻害剤として知られる化合物の中から、前記3.の方法によってオーキシンの生合成を阻害する化合物を新たに選抜することが可能である。PLPを要求しないトランスアミラーゼとしては、例えばフェニルアラニンアンモニアリアーゼ(PAL)がある。PALの阻害剤としては、例えばPhytochemistry68:407-415 (2007)やPhytochemistry62:415-422 (2003)に記載の化合物、具体的には、1-amino-3-phenylpropylphosphonate、1-amino-2-(4-fluorophenyl)ethylphosphonic acid、(R)-1-amino-2-phenylwthylphosphonic acid、1-amino-2-phenylethyl-phosphonic acidが知られている。本発明者等はこのようなスクリーニングを行うことにより、PALの阻害剤として知られているL-AOPPがオーキシン生合成を阻害できることを見いだした。よって、L-AOPPの類縁体も本願発明のオーキシン生合成阻害剤に包含する。
【0032】
具体的には、下記(式I)又は(式II)から選ばれる化合物を有効成分として含有するオーキシン生合成阻害剤が挙げられる。
【0033】
【化1】
【0034】
但し、R1はカルボキシル基又はホスホン基を意味する。R2は水素原子もしくはメチル基を示す。R3はアミノ基、又はアミノオキシ基を示す。R4は、水素又はR5-CH2-であって、ここで、R5は、炭素原子が塩素又はメチル基で修飾されていてもよい、インドール環、ナフタレン環、ベンゼン環、ピリジン環、ピロール環を示す。
【0035】
【化2】
【0036】
R6は、アミノ基を有していても良い炭素数1−4のアルキル基を示す。
【0037】
更に具体的には、
【化3】
【0038】
但し、R1はカルボキシル基、又はホスホン基を意味する。R2は水素原子もしくはメチル基を示す。R3はアミノ基、又はアミノオキシ基(-O-NH2)を示す。
【0039】
不斉炭素の立体構造は、R1がカルボキシル基の場合にはS体がより好ましく、R1がホスホン基の場合にはR体が好ましい。
【0040】
R5は炭素原子が塩素又はメチル基で置換されていてもよい、インドール環、ナフタレン環、ベンゼン環、ピリジン環、ピロール環を示す。具体的には、R5としては、
【化4】
【化5】
が挙げられる。
【0041】
好適には、R1がカルボキシル基のものとしては、L-AOPPや
【化6】
が、R1がホスホン基のものとしては、1-amino-2-phenylethyl-phosphonic acidや
【化7】
がそれぞれ挙げられる。
【0042】
また、好適には、R3がアミノ基のものとしては、 1-amino-2-(1H-indol-3-yl)ethylphosphonic acidや1-amino-2-phenylethyl-phosphonic acid、R3がアミノオキシ基のものとしては、AOPPや2-(aminooxy)-3-(1H-indol-3-yl)propanoic acidが挙げられる。
【0043】
また、具体的には、2-amino-4-methoxy-3-butenoic acid (AMB)やリゾビトキシン(2-amino-4-(2-amino-3-hydroxypropoxy)-3-butenoic acid)を包含する。特に、L-tryptophanの類縁体、2-amino-4-alkoxy-3-butenoic acidの構造を持つ化合物とそれらの塩は有効である。図17参照。
【0044】
5.植物成長制御剤
本願発明のオーキシン生合成阻害剤は、根の伸長の阻害を始め、芽生えの成長抑制などの植物成長抑制剤、植物成長制御剤、又は除草剤として使用することができる。例えば、オーキシン生合成阻害剤は、タルク、クレー、でんぷん、水等の各種担体と混合して、固体製剤としても、又は液体製剤としても使用することができる。液体製剤としては、適宜の担体を用いて、液剤、乳剤、マイクロエマルション、サスポエマルション、油剤、油性フロアブルなどの液体製剤化をすることができる。固体製剤としては、粉末剤、粒剤、顆粒剤、錠剤、水和剤、顆粒水和剤などとして用いることができる。更に、農薬等の製剤上使用される補助剤、例えば展着剤、乳化剤、着色剤等を必要に応じて添加することができる。
【実施例】
【0045】
[実施例1]
オーキシン誘導性遺伝子とエチレン誘導性遺伝子のAVG処理に対する挙動
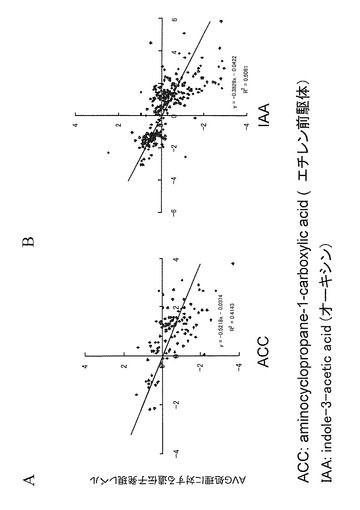
野生型シロイヌナズナ(コロンビア)の種子を4℃で48時間低温処理した後、1/2MS(Murashige&Skoog) 1%Sucroseを含む液体培地で7日間培養した。このシロイヌナズナの芽生えを10μMのAVG、10μMのエチレン前駆体ACC(アミノシクロプロパンカルボン酸)、又は1μMのIAAで3時間処理した。このシロイヌナズナの芽生えからトータルRNAを精製した後、Affymetrix社のプロトコールに従ってGeneChipを用いて遺伝子の発現レベルを測定した。結果を図2に示す。縦軸はAVG処理に対する遺伝子の発現応答を無処理のコントロールに対する比の対数(Signal Log Ratio)で示す。横軸はそれぞれACC処理(図2A)とIAA処理(図2B)に対する遺伝子発現の応答を無処理のコントロールに対するSignal Log Ratioで示す。AVGはエチレン生合成の阻害剤として知られており、ACC処理とAVG処理をエチレン誘導性遺伝子を対象としてプロットして(図2A)比較すると、既知のように負の相関が見られた。一方、IAA処理とAVG処理を、オーキシン誘導性遺伝子をプロットして(図2B)比較すると負の相関が見られた。この結果はAVGがこれまで考えられていた、エチレン抑制作用だけでなく、オーキシン抑制作用も持っていることを示唆している。なお、使用したAVGは、以下の実施例も含む全てS体である。
【0046】
[実施例2]
AVG処理後のシロイヌナズナ内生IAA量の変化
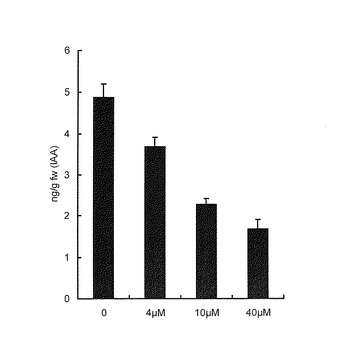
野生型シロイヌナズナ(コロンビア)の種子を4℃で48時間低温処理した後、1/2MS 1%Sucroseを含む液体培地で7日間培養した。このシロイヌナズナの芽生えにAVG4、10,又は40μMを24時間処理した。このシロイヌナズナの芽生えをメタノールで抽出後、内部標準物質として13C6-IAAを添加した。抽出液を濃縮後、N(CH3)2 HPLCカラムで精製し、IAA溶出画分を分取。IAA画分をトリメチルシリル化後、GC-MSで定量分析した。結果を図3に示す。AVG処理によりシロイヌナズナ内生IAA量は処理したAVGの濃度に依存して減少した。
【0047】
[実施例3]
ACCとオーキシン関連化合物を用いた、 AVG処理からの遺伝子発現回復実験
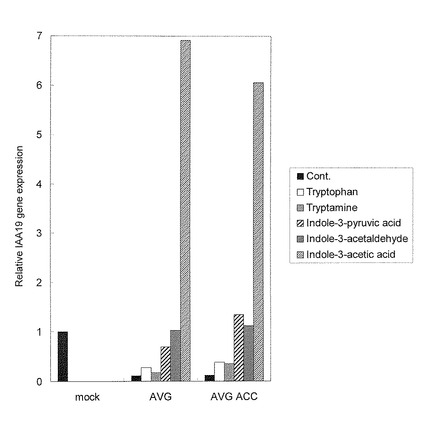
野生型シロイヌナズナ(Col.)の種子を4℃で48時間低温処理した後、 1/2MS 1%Sucroseを含む液体培地で7日間培養した。このシロイヌナズナの芽生えに40μM AVGと10μMのエチレン生合成前駆体ACC、IAA、IAA生合成前駆体IPyA(Indole-3-pyruvic acid)、又はIAAld(Indole-3-acetaldehyde)を24時間処理した。これらシロイヌナズナの芽生えからRNAを精製した後、Taqmanプローブを用いた定量PCR法(ABI PRISM7500)を用いてオーキシン作用の指標となる遺伝子の一つである、IAA19/ At3g15540遺伝子の発現レベルを測定した。プライマーはフォワード(GAGCATGGATGGTGTGCCTTAT)、リバースプライマー(TTCGCAGTTGTCACCATCTTTC)、TaqManプローブ(FAM-ATAAGCTCTTCGGTTTCCGTGGCATCG-TAMRA)を用い、測定は文献に記載の方法によって行った(PlantPhysiol131:287-297)。結果を図4に示す。
【0048】
縦軸は対照区を1とした遺伝子発現レベルの相対値を示す。 IAA19遺伝子の発現レベルはIPyAを経るオーキシン生合成の前駆体(IPyA, IAAld)で対照区レベルまで回復した。この傾向はACCをさらに加えても変化しなかった。この結果は、AVGはIAAの生合成を阻害することにより、オーキシン応答性遺伝子の発現を抑制することを示す。また、AVGによるオーキシン応答性遺伝子の発現抑制効果はエチレンの生合成阻害を介した間接作用ではないことを示す。
【0049】
[実施例4]
AVG、AOA処理後のオーキシン誘導生遺伝子の発現変化
野生型シロイヌナズナ(コロンビア)の種子を4℃で48時間低温処理した後、 1/2MS 1%Sucroseを含む液体培地で7日間培養した。このシロイヌナズナの芽生えに、AVGまたはAOAを1、5,又は20μMで24時間処理した。AOAはAVGと同様にピリドキサルリン酸依存性酵素の作用を阻害することで、エチレンの生合成を阻害することが知られている。処理後のシロイヌナズナの芽生えからRNAを精製した後、Taqman プローブを用いた定量PCR法を用いてオーキシン作用の指標となるIAA1、IAA2遺伝子の発現レベルを測定した(図5)。縦軸は対照区を1とした遺伝子発現レベルの相対値を示す。 IAA1/ At4g14560、IAA2/ At3g23030遺伝子共にAVG処理では濃度に依存してその発現量が減少した。同様にAOA処理によってもオーキシン誘導生遺伝子の発現は減少したが、その作用はAVGよりもやや弱かった。
【0050】
[実施例5]
オーキシンとその関連化合物を用いた、 Met-Trp処理又はAVG処理からの根の伸長回復実験
野生型シロイヌナズナ(Col.)の種子を4℃で48時間低温処理した後、1/2MS 1%Sucroseを含む寒天培地で2日間培養した。このシロイヌナズナの芽生をαメチルトリプトファン(Met-Trp)を含む1/2MS 1%Sucrose寒天培地に移植し垂直に6日間生育させた。Met-TrpはL-Trpの合成経路を上流で阻害するが、Met-Trpによる生育阻害をIAAとその生合成前駆体が回復できるかどうか、根の伸長回復を指標に観察した。結果を図6に示す。
【0051】
対照区(A)と比較してMet-Trp処理により根の伸長が著しく阻害された(B)。この根の伸長阻害作用はインドール(Indole)とトリプトファン(Trp)で回復するが(C、D)、他のオーキシン前駆物質では回復しなかった(E-K)。この結果はMet-Trpがトリプトファンやインドールの上流でこれらの生合成を阻害していることを示している。
【0052】
同様に、発芽2日後のシロイヌナズナの芽生えをAVGとオーキシンとその前駆物質を含む1/2MS 1%Sucrose寒天培地培地に垂直に並べ、6日後根の伸長回復を観察した。結果を図7に示す。対照区(A)と比較してAVG処理により根の伸長が著しく阻害された(B)。この根の伸長阻害作用はインドールとL-トリプトファン(C、D) 、IPyAを経るオーキシン合成経路の前駆体で回復するが(I-J)、IPyAを経由しない他の前駆体では回復しなかった(E-H)。またインドールやL-トリプトファンの回復と比較してインドールピルビン酸を経るオーキシン合成経路の化合物での回復は顕著であった。この結果はAVGがインドールからIAAが合成される途中のステップでオーキシン合成を阻害していることを示している。一方、インドールや、L-トリプトファンでも根の伸長が部分的に回復するのは、IAAの合成経路が複数有り、インドールやL-トリプトファンがAVGによって阻害されない経路を経由してIAAに変換されるからであると考えられる。
【0053】
[実施例6]
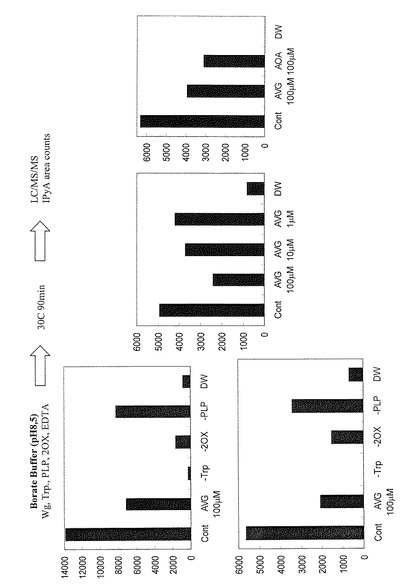
In vitroでのL-トリプトファンからIPyAの変換に対するAVG, AOAの抑制作用
コムギ胚芽抽質物(Wg)と5mM L-トリプトファン、0.2mMピリドキサルリン酸(PLP)、0.5mM 2オキソグルタル酸(20X)、及び4mM EDTAを、ホウ酸緩衝液中で30℃で90分間反応した。産物をMeOHを含むイミダゾールバッファー中でヒドロキシアミンによりオキシムに誘導体化する。溶液を塩酸酸性にした後、酢酸エチルで抽出、OASIS MCXカラムで精製し、生成したIPyAの量をLC/MS/MSを用いて測定した(LC: HP 1100 Series HPLC System (Hewlett Packard/Agilent Technologies), MS: QSTAR Pulsar (PE Sciex/Applied Biosystems))。結果を図8に示す。なお、図8中、−TrpはL-トリプトファンを添加しなかったことを、−2OXは2オキソグルタル酸(20X)を添加しなかったことを、更に―PLPはピリドキサルリン酸(PLP)を添加しなかったことを、それぞれ示す。L-トリプトファンから生じるIPyAの量は、PLP, 2OXの添加に依存して増加した。またその量はAVGまたはAOAを加えることにより減少した。このことはトリプトファンからインドールピルビン酸へのオーキシン生合成ステップがPLP酵素によって触媒されており、その活性をAVG、AOAが阻害していることを示している。
【0054】
[実施例7]
シロイヌナズナ芽生えの生育へのAVG、AOAの効果
野生型シロイヌナズナ(コロンビア)の種子を4℃で48時間処理した後、1/2MS1%SucroseにAVGを10μM (B)、2μM (D)含む寒天培地で培養した。結果を図9に示す。(A)(C)は対照区。21℃で培養し、14日目 (C)(D)と30日目(A)(B)に観察をおこなった。2μM AVG処理により発芽後の芽生えの育成が阻害された(D)。また、10μMでは、芽生えは発芽直後に白化して枯死した。根の伸長方向は、重力に従わず、方向が定まっていない。
【0055】
野生型シロイヌナズナ(コロンビア)の種子を4℃で72時間処理した後、1/2MS 1%Sucroseに100μM AOAを含む液体培地(F)で21℃で6日間培養した。(E)は対照区。AOAの添加により、芽生えは地上部、地下部共に生育が抑えられたが、その作用はAVGよりも緩やかであった(図9)。
【0056】
[実施例8]
In vitro でのL-トリプトファンからIPyAの変換酵素(シロイヌナズナ由来)に対するAVG, L-AOPPの効果
シロイヌナズナの芽生えを1/2MS 1%sucroseの培地で3日間生育させた。この芽生えを-80℃で凍結して粉砕し、抽出バッファー(40mM HEPES-OKH (pH7.6), 10 mM potassimu chloride, 5 mM magnesium chloride, 2 mM calcium chloride, 4 mM dithiothreitol)でタンパク質を抽出した。これを20000g10分間遠心したのち、Sephadex G-25カラムを用いてゲル濾過を行い、高分子画分を粗酵素抽出液として得た。これを用いて、L-トリプトファンからインドールピルビン酸への酵素反応を行った。
【0057】
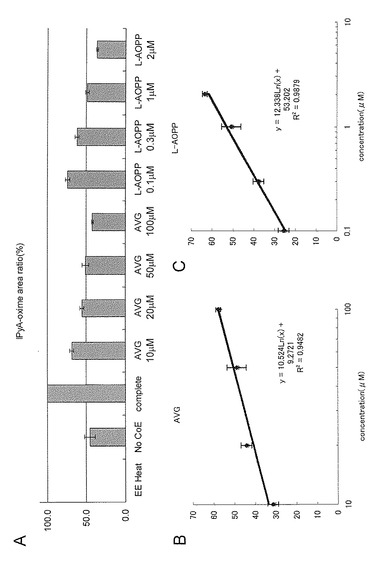
反応は、抽出タンパク質76μg/ml, 0.1M borate buffer(pH 8.5), 0.25 mM L-トリプトファン、0.5mM 2-oxoglutaric acid、0.05 mM PLPの存在下で、35℃1時間暗所でインキュベートした。 インドールピルビン酸の生成量は実施例6に従って定量した。結果を図10に示す。コントロールとして、シロイヌナズナの抽出酵素を95℃5分間処理したものを用いた(EE Heat)。また、反応からPLPと2オキソグルタル酸を抜いた際に出来るIPyAの量も示した(No CoE)。抽出液を熱処理すると活性は検出されず、PLPと2オキソグルタル酸を除いた際は活性が50%以下となることから、この反応は、PLPと2オキソグルタルに依存した酵素反応であることがわかる。
【0058】
次ぎに、阻害剤無し(complete)、AVG (10, 20, 50, 100 μM) 又はL-AOPP (0.1, 0.3, 1, 2 μM)を加えて反応を行った。
【0059】
縦軸はIPyAの生成量を相対値で示す。図10Aには3回繰り返しの平均値を示す。AVG, L-AOPPは共にシロイヌナズナのIPyA合成酵素の活性を阻害した。
この結果を基に、IC50を算出したところ、AVGは47.9 μM, L-AOPPは0.77 μMとなり、L-AOPPはAVGよりも2桁程度高活性であった(図10 B,C)。
【0060】
[実施例9]
In vitro でのL-トリプトファンからIPyAの変換酵素(シロイヌナズナ由来)に対するAOA, AOIBAの効果
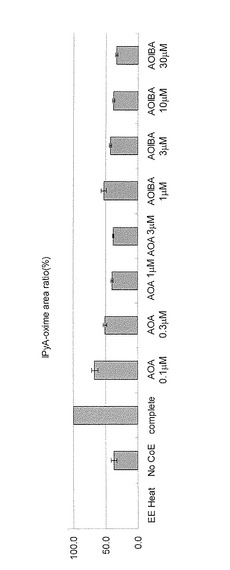
実施例8と同様にシロイヌナズナの芽生えから粗酵素抽出液を生成し、酵素反応を行った。反応条件は実施例8と同一とした。コントロールとして、シロイヌナズナの抽出酵素を95℃5分間処理したもの(EE Heat)と、反応からPLPと2オキソグルタル酸を除いた際のIPyAの生成量を示した(No CoE)。インドールピルビン酸は実施例6に従って定量した。結果を図11に示す。縦軸はインドールピルビン酸の生成量を相対値で示す。グラフは3回繰り返しの平均値を示す。
【0061】
次に、阻害剤無し(complete)とAOIBA (1, 3, 10, 30 μM) ,又はAOA (0.1, 0.3, 1, 3 μM)を加えて反応を行った。
【0062】
AOIBA, AOAは共にシロイヌナズナのIPyA合成酵素活性を阻害した。結果を基に、IC50を算出したところ、AOAは0.4 μM, AOIBAは1.26 μMとなり、いずれもAVGより高活性であった。
【0063】
[実施例10]
L-AOPPとD-AOPPによるオーキシン応答性遺伝子の発現への影響
AOPPがインドールピルビン酸合成酵素を阻害する際の立体構造の特異性を明らかにするために、L-AOPPとD-AOPPのオーキシン応答性遺伝子発現に対する影響を調べた。
シロイヌナズナを1/2MS、1%sucroseを含む液体培地で7日間生育させ、L-AOPP (1, 3, 10, 30 μM)とD-AOPP( 1, 3, 10, 30 μM)で3 h処理し、植物体をサンプリングして、RNAを精製した。実施例3の方法に従って、IAA19/ At3g15540およびIAA1/ At4g14560遺伝子の発現レベルを定量PCRを用いて測定した。結果を図12に示す。D-AOPPはIAA遺伝子の発現を3μMから抑制した。一方、D-AOPPはIAA1遺伝子の発現を30 μMで阻害したが、IAA19遺伝子の発現は阻害しなかった。このことから、S体はR体よりもIAA遺伝子発現の阻害により効果的であると言える。S体の立体構造はL-トリプトファンの立体構造と相同の関係があり、これら化合物がL-トリプトファンのアナログとして働いていること、つまり、オーキシン生合成阻害にはS体がより効果的であることが示された(実験は2回繰り返しの平均値である。)。
【0064】
[実施例11]
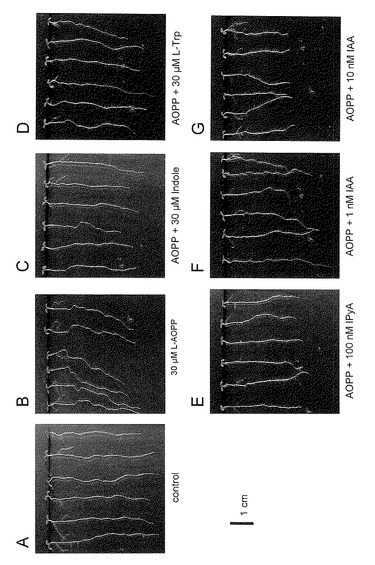
L-AOPPのシロイヌナズナ生育への影響
シロイヌナズナをL-AOPPを含む1%シュークロース1/2MS培地で生育させたが、生育阻害は見られなかった。これはL-AOPPがMS培地中で不安定であることが原因であった。そこで、MS培地成分を含まない糖寒天でL-AOPPのシロイヌナズナの生育への影響を調べた。シロイヌナズナ芽生えを4日間1%シュークロースを含む1/2MS培地で主根の長さが約1cmまで育てた。これらの幼植物体を薬剤を添加した1%シュークロースの寒天に移植し、4日間21℃で培養した。結果を図13及び図14に示す。
【0065】
30 μMのL-AOPPの存在下ではシロイヌナズナの根は左側に傾斜し、かつ波打って伸長した。これは根の重力屈性が正常に働いていないことを示しているとされている。これに対して、インドール、 Lトリプトファン, インドールピルビン酸を加えた場合には、根の重力屈性は回復した。また、1 nM IAAを添加した場合には重力屈性はほぼ回復したのに対し、10 nM IAAを添加した場合には、根の伸長に若干阻害が見られるが、根の重力屈性は完全に回復した。このことから、L-AOPPは重力屈性を制御するオーキシンの生合成を阻害し、この阻害はトリプトファンを経由するIAAの中間体とIAAで回復することが示された。なお、L-AOPPを50μM以上の濃度で処理すると主根の伸長が阻害され、この阻害はIAA、IPyAで回復した。
【0066】
[実施例12]
シロイヌナズナIAA内生量への影響
シロイヌナズナの芽生えを1/2MS寒天培地で6日間22℃連続光下で培養した。生育がそろっている芽生えを20株ずつ1/2MS液体培地(5ml)に移植し、24時間培養し、30 μMのAVGまたはL-AOPPで1時間処理した。内部標準としてd5-IAAを添加して、MeOHで抽出し、オアシスHLB, MCXカラムで精製し、LC-MS/MSでIAA量を定量した。LC-MS/MSによる定量分析は実施例6に記載の方法に従って行った。結果は図15に示し、3回の独立した実験結果の平均と標準誤差で、縦軸はIAA量を示す。左が生重量1g当たりのIAA量を示し、右が芽生え1本あたりのIAA量を示す。この結果、AVGとL-AOPPはどちらもIAAの内生量を大きく減少させた。
【0067】
[実施例13]
イネIAA内生量への影響
イネ(コシヒカリ)の種を滅菌し、25℃3日間吸水させて発芽させた。これを寒天上に播種し、28℃で2日間培養した。
【0068】
生育がそろった芽生えを10 mlの水に10本ずつ移して、30 μMの阻害剤Met-Trp, AVG, AOA, L-AOPPの存在下で28℃3時間振とう培養した。培養後、芽生えを地上部、根、種子に分け、地上部と地下部のそれぞれに100 pg/mgFW(Fresh Weight、他の図面中のFWも同じ。)となるようにd5-IAAを内部標準物質として加え、実施例12に従って、IAAの量をLC-MS/MSを用いて定量した。結果を図16に示す。グラフの縦軸は生重量1mgあたりのIAA量を示す。この実験を3回繰り返した平均値と標準誤差を示している。トリプトファンの生合成を阻害するMet-TrpはIAAの内生量に影響を与えなかった。一方、AVG, AOA, L-AOPPはIAAの内生量を減少させた。
【産業上の利用可能性】
【0069】
本願発明は、初めて、オーキシン生合成阻害剤及びそのスクリーニング方法を提供するというきわめて優れた効果を奏するものである。これにより従来不可能であった、植物体内でのオーキシン量を制御することが可能となり、今後まったく新たな植物成長調節剤、除草剤の開発が可能となる。
【0070】
本願発明は、植物成長調節剤、除草剤及びその製造分野で利用することができる。
本明細書で引用した全ての刊行物、特許および特許出願をそのまま参考として本明細書にとり入れるものとする。
【技術分野】
【0001】
本願発明は、オーキシン(インドール-3-酢酸等)の生合成阻害剤を有効成分として含有する植物化学調節剤または除草剤に関する。更に、本願発明は、該植物化学調節剤を用いて、様々な植物種において、様々な生育段階、器官において植物の生育を人為的に制御する方法に関する。
【背景技術】
【0002】
オーキシンは、植物ホルモンとして最初に認識された物質で、植物の示す屈曲性に関与するホルモンとして、同定された。その後、主要な天然オーキシンとしては、インドール−3-酢酸(IAA)であることが明らかにされ、IAA以外にも、インドール-3-酪酸、4−クロロインドール酢酸、及びフェニル酢酸などが知られている。また合成オーキシンとしては、2,4,ジクロロフェノキシ酢酸(2,4-D)、α-ナフタレン酢酸(NAA)、2,6-ジクロロ安息香酸などが知られている(非特許文献1)。
【0003】
天然オーキシンであるIAAは不安定であり、植物体内では分解経路も存在するため、農業用途には、合成オーキシンが用いられていることが多い。例えば、水田での双子葉植物除草剤としては、4-(4-chloro-o-tolyloxy)butyric acid (MCPB)や2,4-Dが用いられている。また2,3,5-トリヨード安息香酸(TIBA)は、側芽の発芽、成長を高めることができる。さらに、組織培養には、2,4−Dやナフタレン酢酸(NAA)が用いられる。また、発根には、NAAやインドール酪酸が用いられている。また、例えば、トマトでは、4−クロロフェノキシ酢酸は、未受精のトマトでも着果促進する効果がある(非特許文献1)。
【0004】
植物体内でオーキシンはコリスミ酸から合成され、以前は、L-トリプトファン(L-Trp)を経由して合成されると考えられていたが、現在では大きく分けて(1)L-Trpを経由する経路と、(2)L-Trpを経由しない経路の2つの経路が確認されている。L-Trpを経由する経路は、さらに4つ以上の枝に分岐しており、それぞれの経路は異なった酵素によって触媒されている(図1)。現在でも、どの経路が主要経路なのか、あるいはどのような役割分担があるのかは不明である。
【0005】
自身はほとんどオーキシン活性を示さないが、IAAなどのオーキシンと競合してオーキシンの結合部位を奪うことにより、オーキシン作用を拮抗的に阻害する抗オーキシンも合成されている。抗オーキシンとしては、2,4-ジクロロフェノキシイソ酪酸(PCIB)等が知られている。また、マレイン酸ヒドラジド、マレイン酸ヒドラジドコリンなども、オーキシンの作用を抑制し、細胞分裂を阻害すると共に、伸長を抑制することが知られている。タバコの芯止め後のえき芽発生抑制や、野菜の発芽抑制に用いられている(非特許文献2)。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】増田芳雄編著「植物ホルモン入門」オーム社(平成4年(1992年)7月30日発行)第9−17及び41−45頁
【非特許文献2】農薬ハンドブック1998年版編集委員会編「農薬ハンドブック1998年版」社団法人日本植物防除協会発行 第10版 平成10年(1998年)12月15日 第525-526頁
【発明の概要】
【発明が解決しようとする課題】
【0007】
これまで、抗オーキシンと呼ばれるオーキシンの信号を阻害するとされる物質(PCIBなど)や、オーキシン極性輸送阻害剤と呼ばれるTIBAなど、オーキシンの作用を制御する化合物としては知られていたが、内在性オーキシンの量を制御する(減少させる)物質は知られていなかった。また、モデル植物を中心に分子遺伝学が発達した今日に於いても、内生オーキシン量が減少する突然変異体は知られておらず、RNAi法やアンチセンス法など分子生物学的な手法を用いても内生オーキシンの量を抑制する技術は存在しなかった。本年4月になって、PLP酵素の変異体taa1/sav3/wei8が単離され、本遺伝子とそのファミリー遺伝子(TAR2)の2重変異体wei8 tar2変異体ではIAA内生量が半分以下に減少することが報告された(Cell 133: 177-191 (2008); Cell 133 164-176 (2008))。
【0008】
今日のオーキシン生合成経路に関する知見では、オーキシンは網目状の多様な経路を通して合成されており、そもそも一つの化合物(つまり一つの酵素反応の阻害)で、その生合成を強く阻害することは困難と考えられてきた。
【0009】
他方、植物の内在性のオーキシン合成量を調節できれば、更に効率的に植物成長を調節できる可能性がある。
【0010】
今日、国際市場で最も良く普及している除草剤グリホサートは、芳香族アミノ酸合成経路の阻害剤である(つまり、コリスミ酸の上流で代謝を阻害)。一方、本願発明で提供される化合物は、芳香族アミノ酸合成経路経路の下流末端に位置するオーキシンの生合成経路を阻害する。従って、既存の除草剤よりも制御する経路がより狭く、特異的であるため、本願発明によって提供される薬剤は、既存の除草剤よりも副作用が少なく、作用の選択性が高く、より効率的に(すなわち低濃度で)植物を除草する効果をもたらすことが期待できる。
【0011】
そこで、本願発明は、内在性のオーキシンの合成阻害剤を提供することを第1の課題とする。更に本願発明は、上記オーキシン生合成阻害剤を使用する植物成長制御剤または除草剤を提供することを第2の課題とする。
【課題を解決するための手段】
【0012】
本願発明者等は、その類まれなる洞察力により、オーキシン生合成阻害作用を奏する化合物を見出して、本願発明を完成させた。
【0013】
本願発明は、オーキシン誘導性遺伝子の発現を妨げるオーキシン生合成阻害剤候補をスクーニングする方法を提供する。これにはオーキシン応答性遺伝子に対するプローブを担持するマクロアレイを用いて、オーキシン誘導性遺伝子の発現を妨げる候補をスクーニングする方法を含む。さらに、本願発明は、実際に植物に前記オーキシン阻害剤候補を添加し、その後植物体を破砕して生体内のオーキシン量を測定することにより、オーキシン阻害剤の候補が植物体内でオーキシン合成を阻害しているオーキシン阻害剤候補をオーキシン阻害剤として選抜する、オーキシン生合成阻害剤のスクリーニング方法を提供する。さらに、オーキシン生合成経路のうち、L-TrpからIndole-3-pyrvic acid(IPyA)を合成する酵素抽出液に対して、前記オーキシン阻害剤候補とL-Trpを添加し、IPyAの生成を妨げる候補化合物をスクーニングする方法を含む。
【0014】
また、本願発明は、オーキシン生合成阻害剤を提供する。本願発明は、L-trpを経由してIAAを合成する経路で働く、ピリドキサルリン酸(PLP)要求性の酵素又はL-tryptophanを脱アミノ化してIndole-3-pyrvic acid(IPyA)に変換するトランスデアミナーゼを阻害することにより、オーキシン生合成を阻害する阻害剤を提供する。
【0015】
より具体的には、本願発明は、AVG(aminoethoxyvinylglycine、2-amino-4-(2-aminoethoxy)-3-butenoic acid(USP3869277))、AOA(aminooxyacetic acid)、2-aminooxy isobutyric acid hydlochloride (AOIBA)、L-aminooxyphenylpropionic acid (L-AOPP)を含有するオーキシン生合成阻害剤及びこれらを有効成分とする植物成長調節剤を包含する。
【0016】
本明細書は本願の優先権の基礎である日本国特許出願2007-152808号の明細書および/または図面に記載される内容を包含する。
【図面の簡単な説明】
【0017】
【図1】オーキシン合成経路を示す。オーキシンはコリスミ酸から合成され、L-Trpを経由する経路と、経由しない経路の2つの経路が確認されている。L-Trpを経由する経路は、4つ以上の枝に分岐しており、それぞれの経路は異なった酵素によって触媒される。それぞれの経路の中間体と生合成反応を触媒する酵素(または遺伝子)の名称(略号)を記す。
【図2】エチレン誘導性遺伝子(A)とオーキシン誘導性遺伝子(B)のAVG処理に対する挙動を示す。(A)エチレン応答性遺伝子の発現。(B)オーキシン応答性遺伝子の発現。
【図3】AVG処理後のシロイヌナズナ内生IAA量の変化を示す。
【図4】ACCとオーキシン生合成中間体を用いた、AVG処理からの遺伝子発現回復実験を示す。
【図5】AVG, AOAがオーキシン応答性遺伝子の発現を抑制する。
【図6】オーキシンとその生合成中間体を用いた、αメチルトリプトファン(Met-Trp)処理からの根の伸長回復実験を示す。
【図7】オーキシンと生合成中間体を用いた、AVG処理からの根の伸長回復実験を示す。
【図8】In vitroでのL-トリプトファンからIPyAへの変換酵素(コムギ由来)に対するAVGの抑制作用。
【図9】シロイヌナズナ芽生の生育へのAVG、AOAの効果。
【図10】In vitro でのL-トリプトファンからIPyAへの変換酵素(シロイヌナズナ由来)に対するAVG, L-AOPPの効果。
【図11】In vitro でのL-トリプトファンからIPyAへの変換酵素(シロイヌナズナ由来)に対するAOA, AOIBAの効果。
【図12】L-AOPPとD-AOPPによるオーキシン応答性遺伝子の発現への影響。L、D-AOPP処理におけるIAA応答性遺伝子発現量
【図13】L-AOPPのシロイヌナズナ生育への影響を示す写真。
【図14】シロイヌナズナIAA内生量への影響。(A)阻害剤1h処理後のIAA内生量。(B)阻害剤1h処理後の1株あたりのIAA内生量。
【図15】イネIAA内生量への影響。コシヒカリ阻害剤処理3h IAA内生量。
【図16】阻害剤とIAA前駆体の構造。
【発明を実施するための形態】
【0018】
1.はじめに
植物体内でのオーキシンの合成経路は複雑で、網目状であることが示されてきた (図1)。現在までのところ、どの経路が、内在性のオーキシンの主要な合成経路であるかは確認されていない。そのため、どの経路のどの酵素反応を阻害すれば、内在性オーキシン生合成を阻害し、それによって植物の成長を制御または抑制できるか明らかではない。
【0019】
2.遺伝子発現及びマイクロアレイを用いるオーキシン生合成阻害剤候補のスクリーニング方法
本願発明者等は、既に、オーキシンに応答して発現の影響を受ける遺伝子、つまりオーキシン誘導性遺伝子を多数分離している。
【0020】
本願発明者等は、このようなオーキシン誘導性遺伝子に対するプローブを担持するマクロアレイを用いて、オーキシン誘導性遺伝子の発現を妨げるオーキシン生合成阻害剤候補をスクーニングできることを見出した。このようなマイクロアレイとしては、以下の表1に記載の任意の遺伝子を適宜選択し、該遺伝子の塩基配列と同一の少なくとも20オリゴヌクレオチドをプローブとして、周知の任意のマイクロアレイ上に載せたものを使用できる。
【0021】
例えばシロイヌナズナにおいては、表1に掲げられる任意の遺伝子を、例えば、10種類以上選択し、各遺伝子に対するに対する20オリゴヌクレオチド以上からなるプローブを用いるマイクロアレイ及び当該マイクロアレイを用いるオーキシン生合成阻害剤候補のスクリーニング方法を包含する。ここで、マイクロアレイシステムとしては、たとえば、affymetrixのGeneChip systemを用いることができる。
【0022】
本マイクロアレイを用いて、例えば、オーキシン等対象化合物の存在下で発現が誘導される程度と、試験化合物の存在下で発現が抑制される程度を比較することで、オーキシン生合成阻害剤候補を選抜できる。具体的には、例えば、シロイヌナズナ(アラビドプシス)のようなモデル植物に、対象化合物としてオーキシンを与える場合の遺伝子発現のレベルを測定する。次にオーキシンを与えない場合での遺伝子発現の程度を測定して、これらを比較する。更に、該試験植物に試験化合物を与えた場合と与えない場合での遺伝子発現とを比較する。オーキシンの存在下で発現が増大する遺伝子群が、前記試験化合物の存在下で発現が減少する関係があるようであれば、当該試験化合物をオーキシン生合成阻害剤候補として選抜できる。
【0023】
一方、オーキシン応答性遺伝子のプロモーターを利用して人為的に作成したマーカー遺伝子(例えば、DR5-GUSやSAUR-GUSなど)を組み込んだ遺伝子組換え植物体に対して、候補化合物を処理し、マーカー遺伝子の発現レベルを測定することによっても候補化合物を選抜することができる。あるいは、野生型植物に候補化合物を処理し、処理後の植物からトータルRNAを抽出して、オーキシン応答性遺伝子の発現量を定法に従って測定することによって、オーキシン応答性遺伝子の発現を抑制する化合物の中から、候補を選抜することも可能である。
【0024】
【表1】
【0025】
本発明者等は、このようなマイクロアレイを用いたスクリーニング方法により、オーキシン生合成阻害剤候補として、AVGを同定した。
【0026】
3.IAA生合成や酵素活性を指標としたオーキシン生合成阻害剤のスクリーニング方法
植物体内でオーキシン合成を阻害する化合物は、実際に植物体に試験化合物又はオーキシン等の対象化合物を添加して、その後植物体を破砕して、質量分析装置等を用いて、生体内のオーキシン量を測定し、スクリーニングすることができる。
【0027】
また、上記2.で選ばれたオーキシン阻害剤候補が、植物体内でオーキシン合成を阻害している否かは、実際に植物体にオーキシン生合成阻害剤候補を添加して、その後植物体を破砕して、質量分析装置等を用いて、生体内のオーキシン量を測定し確認することもできる。この方法により、本願発明者らは、AVG及びその類縁体がオーキシン生合成阻害剤であることを確認した。
【0028】
さらに、オーキシン生合成経路の中間体をAVGと同時に植物に投与することで、当該化合物がAVGによる成育阻害を回復するか否かを検討した。これにより、本願発明者等は、AVGの成育阻害の作用点が、植物体内でのオーキシンの生合成経路上に存在することを確認した。さらに、植物より抽出した酵素活性をin vitroにおいて評価する系を確立した。この酵素活性の評価系を用いてAVGの作用点を調べ、オーキシン生合成を阻害するターゲットの1つは、ピリドキサルリン酸(PLP)要求性の酵素、又はL-tryptophanを脱アミノ化してIndole-3-pyrvic acid(IPyA)に変換するトランスデアミナーゼ、より好適なオーキシン生合成を阻害するターゲットとしては、L-tryptophanを脱アミノ化してIndole-3-pyrvic acid(IPyA)に変換するピリドキサルリン酸(PLP)要求性のトランスデアミナーゼであることを見出した。従って、これら酵素活性の阻害剤をin vitroでスクリーニングすることによっても、オーキシン合成阻害剤を探索できる。
【0029】
4.ピリドキサルリン酸(PLP)要求性の酵素の阻害剤又はトリプトファンアミノ基転移酵素の阻害剤を有効成分として含有するオーキシン生合成阻害剤
本願発明のオーキシン生合成阻害剤には、AVG及びその類縁体からなるピリドキサルリン酸要求性酵素の阻害剤を包含する。
【0030】
また、本願発明は、ピリドキサルリン酸(PLP)要求性の酵素(トランスデアミナーゼ、デカルボキシラーゼ、C-Sライエース、ラセマーゼ等を含む)の阻害剤を用いて、オーキシンの生合成を阻害することが可能であることを初めて示す。特に、PLP要求性酵素の阻害剤として知られる化合物の中から、前記3.の方法によってオーキシンの生合成を阻害する化合物を新たに選抜することが可能である。PLP要求性酵素の阻害剤としては、例えば、Annual Review of Biochemistry 73:383-415 (2004)に記載の化合物が知られている。本発明者等はこのようなスクリーニングを行うことにより、ACC合成酵素(PLP要求性酵素)の阻害剤として知られているAOA(aminooxyacetic acid)がオーキシン生合成を阻害できることを見いだした。よって、AOAの類縁体も本願発明のオーキシン生合成阻害剤に包含する。
【0031】
また、本願発明は、トランスデアミナーゼ(例えば、Trpアミノトランスフェラーゼ、フェニルアラニンアンモニアリアーゼ)の阻害剤を用いて、オーキシンの生合成を阻害することが可能であることを初めて示す。特に、トランスアミナーゼ阻害剤として知られる化合物の中から、前記3.の方法によってオーキシンの生合成を阻害する化合物を新たに選抜することが可能である。PLPを要求しないトランスアミラーゼとしては、例えばフェニルアラニンアンモニアリアーゼ(PAL)がある。PALの阻害剤としては、例えばPhytochemistry68:407-415 (2007)やPhytochemistry62:415-422 (2003)に記載の化合物、具体的には、1-amino-3-phenylpropylphosphonate、1-amino-2-(4-fluorophenyl)ethylphosphonic acid、(R)-1-amino-2-phenylwthylphosphonic acid、1-amino-2-phenylethyl-phosphonic acidが知られている。本発明者等はこのようなスクリーニングを行うことにより、PALの阻害剤として知られているL-AOPPがオーキシン生合成を阻害できることを見いだした。よって、L-AOPPの類縁体も本願発明のオーキシン生合成阻害剤に包含する。
【0032】
具体的には、下記(式I)又は(式II)から選ばれる化合物を有効成分として含有するオーキシン生合成阻害剤が挙げられる。
【0033】
【化1】
【0034】
但し、R1はカルボキシル基又はホスホン基を意味する。R2は水素原子もしくはメチル基を示す。R3はアミノ基、又はアミノオキシ基を示す。R4は、水素又はR5-CH2-であって、ここで、R5は、炭素原子が塩素又はメチル基で修飾されていてもよい、インドール環、ナフタレン環、ベンゼン環、ピリジン環、ピロール環を示す。
【0035】
【化2】
【0036】
R6は、アミノ基を有していても良い炭素数1−4のアルキル基を示す。
【0037】
更に具体的には、
【化3】
【0038】
但し、R1はカルボキシル基、又はホスホン基を意味する。R2は水素原子もしくはメチル基を示す。R3はアミノ基、又はアミノオキシ基(-O-NH2)を示す。
【0039】
不斉炭素の立体構造は、R1がカルボキシル基の場合にはS体がより好ましく、R1がホスホン基の場合にはR体が好ましい。
【0040】
R5は炭素原子が塩素又はメチル基で置換されていてもよい、インドール環、ナフタレン環、ベンゼン環、ピリジン環、ピロール環を示す。具体的には、R5としては、
【化4】
【化5】
が挙げられる。
【0041】
好適には、R1がカルボキシル基のものとしては、L-AOPPや
【化6】
が、R1がホスホン基のものとしては、1-amino-2-phenylethyl-phosphonic acidや
【化7】
がそれぞれ挙げられる。
【0042】
また、好適には、R3がアミノ基のものとしては、 1-amino-2-(1H-indol-3-yl)ethylphosphonic acidや1-amino-2-phenylethyl-phosphonic acid、R3がアミノオキシ基のものとしては、AOPPや2-(aminooxy)-3-(1H-indol-3-yl)propanoic acidが挙げられる。
【0043】
また、具体的には、2-amino-4-methoxy-3-butenoic acid (AMB)やリゾビトキシン(2-amino-4-(2-amino-3-hydroxypropoxy)-3-butenoic acid)を包含する。特に、L-tryptophanの類縁体、2-amino-4-alkoxy-3-butenoic acidの構造を持つ化合物とそれらの塩は有効である。図17参照。
【0044】
5.植物成長制御剤
本願発明のオーキシン生合成阻害剤は、根の伸長の阻害を始め、芽生えの成長抑制などの植物成長抑制剤、植物成長制御剤、又は除草剤として使用することができる。例えば、オーキシン生合成阻害剤は、タルク、クレー、でんぷん、水等の各種担体と混合して、固体製剤としても、又は液体製剤としても使用することができる。液体製剤としては、適宜の担体を用いて、液剤、乳剤、マイクロエマルション、サスポエマルション、油剤、油性フロアブルなどの液体製剤化をすることができる。固体製剤としては、粉末剤、粒剤、顆粒剤、錠剤、水和剤、顆粒水和剤などとして用いることができる。更に、農薬等の製剤上使用される補助剤、例えば展着剤、乳化剤、着色剤等を必要に応じて添加することができる。
【実施例】
【0045】
[実施例1]
オーキシン誘導性遺伝子とエチレン誘導性遺伝子のAVG処理に対する挙動
野生型シロイヌナズナ(コロンビア)の種子を4℃で48時間低温処理した後、1/2MS(Murashige&Skoog) 1%Sucroseを含む液体培地で7日間培養した。このシロイヌナズナの芽生えを10μMのAVG、10μMのエチレン前駆体ACC(アミノシクロプロパンカルボン酸)、又は1μMのIAAで3時間処理した。このシロイヌナズナの芽生えからトータルRNAを精製した後、Affymetrix社のプロトコールに従ってGeneChipを用いて遺伝子の発現レベルを測定した。結果を図2に示す。縦軸はAVG処理に対する遺伝子の発現応答を無処理のコントロールに対する比の対数(Signal Log Ratio)で示す。横軸はそれぞれACC処理(図2A)とIAA処理(図2B)に対する遺伝子発現の応答を無処理のコントロールに対するSignal Log Ratioで示す。AVGはエチレン生合成の阻害剤として知られており、ACC処理とAVG処理をエチレン誘導性遺伝子を対象としてプロットして(図2A)比較すると、既知のように負の相関が見られた。一方、IAA処理とAVG処理を、オーキシン誘導性遺伝子をプロットして(図2B)比較すると負の相関が見られた。この結果はAVGがこれまで考えられていた、エチレン抑制作用だけでなく、オーキシン抑制作用も持っていることを示唆している。なお、使用したAVGは、以下の実施例も含む全てS体である。
【0046】
[実施例2]
AVG処理後のシロイヌナズナ内生IAA量の変化
野生型シロイヌナズナ(コロンビア)の種子を4℃で48時間低温処理した後、1/2MS 1%Sucroseを含む液体培地で7日間培養した。このシロイヌナズナの芽生えにAVG4、10,又は40μMを24時間処理した。このシロイヌナズナの芽生えをメタノールで抽出後、内部標準物質として13C6-IAAを添加した。抽出液を濃縮後、N(CH3)2 HPLCカラムで精製し、IAA溶出画分を分取。IAA画分をトリメチルシリル化後、GC-MSで定量分析した。結果を図3に示す。AVG処理によりシロイヌナズナ内生IAA量は処理したAVGの濃度に依存して減少した。
【0047】
[実施例3]
ACCとオーキシン関連化合物を用いた、 AVG処理からの遺伝子発現回復実験
野生型シロイヌナズナ(Col.)の種子を4℃で48時間低温処理した後、 1/2MS 1%Sucroseを含む液体培地で7日間培養した。このシロイヌナズナの芽生えに40μM AVGと10μMのエチレン生合成前駆体ACC、IAA、IAA生合成前駆体IPyA(Indole-3-pyruvic acid)、又はIAAld(Indole-3-acetaldehyde)を24時間処理した。これらシロイヌナズナの芽生えからRNAを精製した後、Taqmanプローブを用いた定量PCR法(ABI PRISM7500)を用いてオーキシン作用の指標となる遺伝子の一つである、IAA19/ At3g15540遺伝子の発現レベルを測定した。プライマーはフォワード(GAGCATGGATGGTGTGCCTTAT)、リバースプライマー(TTCGCAGTTGTCACCATCTTTC)、TaqManプローブ(FAM-ATAAGCTCTTCGGTTTCCGTGGCATCG-TAMRA)を用い、測定は文献に記載の方法によって行った(PlantPhysiol131:287-297)。結果を図4に示す。
【0048】
縦軸は対照区を1とした遺伝子発現レベルの相対値を示す。 IAA19遺伝子の発現レベルはIPyAを経るオーキシン生合成の前駆体(IPyA, IAAld)で対照区レベルまで回復した。この傾向はACCをさらに加えても変化しなかった。この結果は、AVGはIAAの生合成を阻害することにより、オーキシン応答性遺伝子の発現を抑制することを示す。また、AVGによるオーキシン応答性遺伝子の発現抑制効果はエチレンの生合成阻害を介した間接作用ではないことを示す。
【0049】
[実施例4]
AVG、AOA処理後のオーキシン誘導生遺伝子の発現変化
野生型シロイヌナズナ(コロンビア)の種子を4℃で48時間低温処理した後、 1/2MS 1%Sucroseを含む液体培地で7日間培養した。このシロイヌナズナの芽生えに、AVGまたはAOAを1、5,又は20μMで24時間処理した。AOAはAVGと同様にピリドキサルリン酸依存性酵素の作用を阻害することで、エチレンの生合成を阻害することが知られている。処理後のシロイヌナズナの芽生えからRNAを精製した後、Taqman プローブを用いた定量PCR法を用いてオーキシン作用の指標となるIAA1、IAA2遺伝子の発現レベルを測定した(図5)。縦軸は対照区を1とした遺伝子発現レベルの相対値を示す。 IAA1/ At4g14560、IAA2/ At3g23030遺伝子共にAVG処理では濃度に依存してその発現量が減少した。同様にAOA処理によってもオーキシン誘導生遺伝子の発現は減少したが、その作用はAVGよりもやや弱かった。
【0050】
[実施例5]
オーキシンとその関連化合物を用いた、 Met-Trp処理又はAVG処理からの根の伸長回復実験
野生型シロイヌナズナ(Col.)の種子を4℃で48時間低温処理した後、1/2MS 1%Sucroseを含む寒天培地で2日間培養した。このシロイヌナズナの芽生をαメチルトリプトファン(Met-Trp)を含む1/2MS 1%Sucrose寒天培地に移植し垂直に6日間生育させた。Met-TrpはL-Trpの合成経路を上流で阻害するが、Met-Trpによる生育阻害をIAAとその生合成前駆体が回復できるかどうか、根の伸長回復を指標に観察した。結果を図6に示す。
【0051】
対照区(A)と比較してMet-Trp処理により根の伸長が著しく阻害された(B)。この根の伸長阻害作用はインドール(Indole)とトリプトファン(Trp)で回復するが(C、D)、他のオーキシン前駆物質では回復しなかった(E-K)。この結果はMet-Trpがトリプトファンやインドールの上流でこれらの生合成を阻害していることを示している。
【0052】
同様に、発芽2日後のシロイヌナズナの芽生えをAVGとオーキシンとその前駆物質を含む1/2MS 1%Sucrose寒天培地培地に垂直に並べ、6日後根の伸長回復を観察した。結果を図7に示す。対照区(A)と比較してAVG処理により根の伸長が著しく阻害された(B)。この根の伸長阻害作用はインドールとL-トリプトファン(C、D) 、IPyAを経るオーキシン合成経路の前駆体で回復するが(I-J)、IPyAを経由しない他の前駆体では回復しなかった(E-H)。またインドールやL-トリプトファンの回復と比較してインドールピルビン酸を経るオーキシン合成経路の化合物での回復は顕著であった。この結果はAVGがインドールからIAAが合成される途中のステップでオーキシン合成を阻害していることを示している。一方、インドールや、L-トリプトファンでも根の伸長が部分的に回復するのは、IAAの合成経路が複数有り、インドールやL-トリプトファンがAVGによって阻害されない経路を経由してIAAに変換されるからであると考えられる。
【0053】
[実施例6]
In vitroでのL-トリプトファンからIPyAの変換に対するAVG, AOAの抑制作用
コムギ胚芽抽質物(Wg)と5mM L-トリプトファン、0.2mMピリドキサルリン酸(PLP)、0.5mM 2オキソグルタル酸(20X)、及び4mM EDTAを、ホウ酸緩衝液中で30℃で90分間反応した。産物をMeOHを含むイミダゾールバッファー中でヒドロキシアミンによりオキシムに誘導体化する。溶液を塩酸酸性にした後、酢酸エチルで抽出、OASIS MCXカラムで精製し、生成したIPyAの量をLC/MS/MSを用いて測定した(LC: HP 1100 Series HPLC System (Hewlett Packard/Agilent Technologies), MS: QSTAR Pulsar (PE Sciex/Applied Biosystems))。結果を図8に示す。なお、図8中、−TrpはL-トリプトファンを添加しなかったことを、−2OXは2オキソグルタル酸(20X)を添加しなかったことを、更に―PLPはピリドキサルリン酸(PLP)を添加しなかったことを、それぞれ示す。L-トリプトファンから生じるIPyAの量は、PLP, 2OXの添加に依存して増加した。またその量はAVGまたはAOAを加えることにより減少した。このことはトリプトファンからインドールピルビン酸へのオーキシン生合成ステップがPLP酵素によって触媒されており、その活性をAVG、AOAが阻害していることを示している。
【0054】
[実施例7]
シロイヌナズナ芽生えの生育へのAVG、AOAの効果
野生型シロイヌナズナ(コロンビア)の種子を4℃で48時間処理した後、1/2MS1%SucroseにAVGを10μM (B)、2μM (D)含む寒天培地で培養した。結果を図9に示す。(A)(C)は対照区。21℃で培養し、14日目 (C)(D)と30日目(A)(B)に観察をおこなった。2μM AVG処理により発芽後の芽生えの育成が阻害された(D)。また、10μMでは、芽生えは発芽直後に白化して枯死した。根の伸長方向は、重力に従わず、方向が定まっていない。
【0055】
野生型シロイヌナズナ(コロンビア)の種子を4℃で72時間処理した後、1/2MS 1%Sucroseに100μM AOAを含む液体培地(F)で21℃で6日間培養した。(E)は対照区。AOAの添加により、芽生えは地上部、地下部共に生育が抑えられたが、その作用はAVGよりも緩やかであった(図9)。
【0056】
[実施例8]
In vitro でのL-トリプトファンからIPyAの変換酵素(シロイヌナズナ由来)に対するAVG, L-AOPPの効果
シロイヌナズナの芽生えを1/2MS 1%sucroseの培地で3日間生育させた。この芽生えを-80℃で凍結して粉砕し、抽出バッファー(40mM HEPES-OKH (pH7.6), 10 mM potassimu chloride, 5 mM magnesium chloride, 2 mM calcium chloride, 4 mM dithiothreitol)でタンパク質を抽出した。これを20000g10分間遠心したのち、Sephadex G-25カラムを用いてゲル濾過を行い、高分子画分を粗酵素抽出液として得た。これを用いて、L-トリプトファンからインドールピルビン酸への酵素反応を行った。
【0057】
反応は、抽出タンパク質76μg/ml, 0.1M borate buffer(pH 8.5), 0.25 mM L-トリプトファン、0.5mM 2-oxoglutaric acid、0.05 mM PLPの存在下で、35℃1時間暗所でインキュベートした。 インドールピルビン酸の生成量は実施例6に従って定量した。結果を図10に示す。コントロールとして、シロイヌナズナの抽出酵素を95℃5分間処理したものを用いた(EE Heat)。また、反応からPLPと2オキソグルタル酸を抜いた際に出来るIPyAの量も示した(No CoE)。抽出液を熱処理すると活性は検出されず、PLPと2オキソグルタル酸を除いた際は活性が50%以下となることから、この反応は、PLPと2オキソグルタルに依存した酵素反応であることがわかる。
【0058】
次ぎに、阻害剤無し(complete)、AVG (10, 20, 50, 100 μM) 又はL-AOPP (0.1, 0.3, 1, 2 μM)を加えて反応を行った。
【0059】
縦軸はIPyAの生成量を相対値で示す。図10Aには3回繰り返しの平均値を示す。AVG, L-AOPPは共にシロイヌナズナのIPyA合成酵素の活性を阻害した。
この結果を基に、IC50を算出したところ、AVGは47.9 μM, L-AOPPは0.77 μMとなり、L-AOPPはAVGよりも2桁程度高活性であった(図10 B,C)。
【0060】
[実施例9]
In vitro でのL-トリプトファンからIPyAの変換酵素(シロイヌナズナ由来)に対するAOA, AOIBAの効果
実施例8と同様にシロイヌナズナの芽生えから粗酵素抽出液を生成し、酵素反応を行った。反応条件は実施例8と同一とした。コントロールとして、シロイヌナズナの抽出酵素を95℃5分間処理したもの(EE Heat)と、反応からPLPと2オキソグルタル酸を除いた際のIPyAの生成量を示した(No CoE)。インドールピルビン酸は実施例6に従って定量した。結果を図11に示す。縦軸はインドールピルビン酸の生成量を相対値で示す。グラフは3回繰り返しの平均値を示す。
【0061】
次に、阻害剤無し(complete)とAOIBA (1, 3, 10, 30 μM) ,又はAOA (0.1, 0.3, 1, 3 μM)を加えて反応を行った。
【0062】
AOIBA, AOAは共にシロイヌナズナのIPyA合成酵素活性を阻害した。結果を基に、IC50を算出したところ、AOAは0.4 μM, AOIBAは1.26 μMとなり、いずれもAVGより高活性であった。
【0063】
[実施例10]
L-AOPPとD-AOPPによるオーキシン応答性遺伝子の発現への影響
AOPPがインドールピルビン酸合成酵素を阻害する際の立体構造の特異性を明らかにするために、L-AOPPとD-AOPPのオーキシン応答性遺伝子発現に対する影響を調べた。
シロイヌナズナを1/2MS、1%sucroseを含む液体培地で7日間生育させ、L-AOPP (1, 3, 10, 30 μM)とD-AOPP( 1, 3, 10, 30 μM)で3 h処理し、植物体をサンプリングして、RNAを精製した。実施例3の方法に従って、IAA19/ At3g15540およびIAA1/ At4g14560遺伝子の発現レベルを定量PCRを用いて測定した。結果を図12に示す。D-AOPPはIAA遺伝子の発現を3μMから抑制した。一方、D-AOPPはIAA1遺伝子の発現を30 μMで阻害したが、IAA19遺伝子の発現は阻害しなかった。このことから、S体はR体よりもIAA遺伝子発現の阻害により効果的であると言える。S体の立体構造はL-トリプトファンの立体構造と相同の関係があり、これら化合物がL-トリプトファンのアナログとして働いていること、つまり、オーキシン生合成阻害にはS体がより効果的であることが示された(実験は2回繰り返しの平均値である。)。
【0064】
[実施例11]
L-AOPPのシロイヌナズナ生育への影響
シロイヌナズナをL-AOPPを含む1%シュークロース1/2MS培地で生育させたが、生育阻害は見られなかった。これはL-AOPPがMS培地中で不安定であることが原因であった。そこで、MS培地成分を含まない糖寒天でL-AOPPのシロイヌナズナの生育への影響を調べた。シロイヌナズナ芽生えを4日間1%シュークロースを含む1/2MS培地で主根の長さが約1cmまで育てた。これらの幼植物体を薬剤を添加した1%シュークロースの寒天に移植し、4日間21℃で培養した。結果を図13及び図14に示す。
【0065】
30 μMのL-AOPPの存在下ではシロイヌナズナの根は左側に傾斜し、かつ波打って伸長した。これは根の重力屈性が正常に働いていないことを示しているとされている。これに対して、インドール、 Lトリプトファン, インドールピルビン酸を加えた場合には、根の重力屈性は回復した。また、1 nM IAAを添加した場合には重力屈性はほぼ回復したのに対し、10 nM IAAを添加した場合には、根の伸長に若干阻害が見られるが、根の重力屈性は完全に回復した。このことから、L-AOPPは重力屈性を制御するオーキシンの生合成を阻害し、この阻害はトリプトファンを経由するIAAの中間体とIAAで回復することが示された。なお、L-AOPPを50μM以上の濃度で処理すると主根の伸長が阻害され、この阻害はIAA、IPyAで回復した。
【0066】
[実施例12]
シロイヌナズナIAA内生量への影響
シロイヌナズナの芽生えを1/2MS寒天培地で6日間22℃連続光下で培養した。生育がそろっている芽生えを20株ずつ1/2MS液体培地(5ml)に移植し、24時間培養し、30 μMのAVGまたはL-AOPPで1時間処理した。内部標準としてd5-IAAを添加して、MeOHで抽出し、オアシスHLB, MCXカラムで精製し、LC-MS/MSでIAA量を定量した。LC-MS/MSによる定量分析は実施例6に記載の方法に従って行った。結果は図15に示し、3回の独立した実験結果の平均と標準誤差で、縦軸はIAA量を示す。左が生重量1g当たりのIAA量を示し、右が芽生え1本あたりのIAA量を示す。この結果、AVGとL-AOPPはどちらもIAAの内生量を大きく減少させた。
【0067】
[実施例13]
イネIAA内生量への影響
イネ(コシヒカリ)の種を滅菌し、25℃3日間吸水させて発芽させた。これを寒天上に播種し、28℃で2日間培養した。
【0068】
生育がそろった芽生えを10 mlの水に10本ずつ移して、30 μMの阻害剤Met-Trp, AVG, AOA, L-AOPPの存在下で28℃3時間振とう培養した。培養後、芽生えを地上部、根、種子に分け、地上部と地下部のそれぞれに100 pg/mgFW(Fresh Weight、他の図面中のFWも同じ。)となるようにd5-IAAを内部標準物質として加え、実施例12に従って、IAAの量をLC-MS/MSを用いて定量した。結果を図16に示す。グラフの縦軸は生重量1mgあたりのIAA量を示す。この実験を3回繰り返した平均値と標準誤差を示している。トリプトファンの生合成を阻害するMet-TrpはIAAの内生量に影響を与えなかった。一方、AVG, AOA, L-AOPPはIAAの内生量を減少させた。
【産業上の利用可能性】
【0069】
本願発明は、初めて、オーキシン生合成阻害剤及びそのスクリーニング方法を提供するというきわめて優れた効果を奏するものである。これにより従来不可能であった、植物体内でのオーキシン量を制御することが可能となり、今後まったく新たな植物成長調節剤、除草剤の開発が可能となる。
【0070】
本願発明は、植物成長調節剤、除草剤及びその製造分野で利用することができる。
本明細書で引用した全ての刊行物、特許および特許出願をそのまま参考として本明細書にとり入れるものとする。
【特許請求の範囲】
【請求項1】
試験化合物及び対照化合物で植物を処理し、オーキシン応答性遺伝子の発現レベルを測定し、オーキシン応答性遺伝子の発現を抑制する化合物の中からオーキシン生合成阻害剤候補をスクリーニングする方法において、オーキシン応答性遺伝子に対するプローブを担持したマイクロアレイを用い、試験化合物又は対照化合物で植物を処理し遺伝子発現を測定し、オーキシン応答性遺伝子群の発現を抑制する化合物をオーキシン生合成阻害剤候補として選択する、オーキシン生合成阻害剤候補をスクリーニングする方法。
【請求項2】
試験化合物として、ピリドキサルリン酸要求性酵素又はアミノ基転移酵素の阻害剤を用いる、請求項1のオーキシン生合成阻害剤候補をスクリーニングする方法。
【請求項3】
試験化合物及び対照化合物で植物を処理し、処理した植物から内生オーキシンを抽出し、その量を測定し、オーキシン合成を阻害するものをオーキシン生合成阻害剤として選択する、オーキシン生合成阻害剤をスクリーニングする方法。
【請求項4】
試験化合物として、ピリドキサルリン酸要求性酵素又はアミノ基転移酵素の阻害剤を用いる、請求項3記載のオーキシン生合成阻害剤をスクリーニングする方法。
【請求項5】
試験化合物及び対照化合物でトリプトファンを含有するトリプトファンデアミナーゼ抽出物を処理し、処理した抽出物中のインドールピルビン酸量を測定し、インドールピルビン酸の生成を阻害するものをオーキシン生合成阻害剤として選択する、オーキシン生合成阻害剤をスクリーニングする方法。
【請求項6】
試験化合物として、ピリドキサルリン酸要求性酵素又はアミノ基転移酵素の阻害剤を用いる、請求項5記載のオーキシン生合成阻害剤をスクリーニングする方法。
【請求項7】
下記式(I)又は式(II)から選ばれる化合物を有効成分として含有するオーキシン生合成阻害剤。
【化1】
但し、R1はカルボキシル基、又はホスホン基を意味する。R2は水素原子もしくはメチル基を示す。R3はアミノ基又はアミノオキシ基(−O-NH2)を示す。R4は、水素又はR5-CH2-であって、ここで、R5は炭素原子が塩素又はメチル基で修飾されていてもよい、インドール環、ナフタレン環、ベンゼン環、ピリジン環、ピロール環を示す。
【化2】
R6は、アミノ基を有していても良い炭素数1−4のアルキルを示す。
【請求項8】
AVG、AOA、AOIBA、AOPP又はそれらの類縁体を有効成分として含有するオーキシン生合成阻害剤。
【請求項9】
ピリドキサルリン酸要求性酵素の阻害剤又はトリプトファンアミノ基転移酵素阻害剤からなるオーキシン生合成阻害剤。
【請求項10】
オーキシン生合成阻害剤を有効成分として含有する植物成長制御剤または除草剤。
【請求項1】
試験化合物及び対照化合物で植物を処理し、オーキシン応答性遺伝子の発現レベルを測定し、オーキシン応答性遺伝子の発現を抑制する化合物の中からオーキシン生合成阻害剤候補をスクリーニングする方法において、オーキシン応答性遺伝子に対するプローブを担持したマイクロアレイを用い、試験化合物又は対照化合物で植物を処理し遺伝子発現を測定し、オーキシン応答性遺伝子群の発現を抑制する化合物をオーキシン生合成阻害剤候補として選択する、オーキシン生合成阻害剤候補をスクリーニングする方法。
【請求項2】
試験化合物として、ピリドキサルリン酸要求性酵素又はアミノ基転移酵素の阻害剤を用いる、請求項1のオーキシン生合成阻害剤候補をスクリーニングする方法。
【請求項3】
試験化合物及び対照化合物で植物を処理し、処理した植物から内生オーキシンを抽出し、その量を測定し、オーキシン合成を阻害するものをオーキシン生合成阻害剤として選択する、オーキシン生合成阻害剤をスクリーニングする方法。
【請求項4】
試験化合物として、ピリドキサルリン酸要求性酵素又はアミノ基転移酵素の阻害剤を用いる、請求項3記載のオーキシン生合成阻害剤をスクリーニングする方法。
【請求項5】
試験化合物及び対照化合物でトリプトファンを含有するトリプトファンデアミナーゼ抽出物を処理し、処理した抽出物中のインドールピルビン酸量を測定し、インドールピルビン酸の生成を阻害するものをオーキシン生合成阻害剤として選択する、オーキシン生合成阻害剤をスクリーニングする方法。
【請求項6】
試験化合物として、ピリドキサルリン酸要求性酵素又はアミノ基転移酵素の阻害剤を用いる、請求項5記載のオーキシン生合成阻害剤をスクリーニングする方法。
【請求項7】
下記式(I)又は式(II)から選ばれる化合物を有効成分として含有するオーキシン生合成阻害剤。
【化1】
但し、R1はカルボキシル基、又はホスホン基を意味する。R2は水素原子もしくはメチル基を示す。R3はアミノ基又はアミノオキシ基(−O-NH2)を示す。R4は、水素又はR5-CH2-であって、ここで、R5は炭素原子が塩素又はメチル基で修飾されていてもよい、インドール環、ナフタレン環、ベンゼン環、ピリジン環、ピロール環を示す。
【化2】
R6は、アミノ基を有していても良い炭素数1−4のアルキルを示す。
【請求項8】
AVG、AOA、AOIBA、AOPP又はそれらの類縁体を有効成分として含有するオーキシン生合成阻害剤。
【請求項9】
ピリドキサルリン酸要求性酵素の阻害剤又はトリプトファンアミノ基転移酵素阻害剤からなるオーキシン生合成阻害剤。
【請求項10】
オーキシン生合成阻害剤を有効成分として含有する植物成長制御剤または除草剤。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】

【図13】
【図14】
【図15】
【図16】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【公開番号】特開2013−67656(P2013−67656A)
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願番号】特願2013−207(P2013−207)
【出願日】平成25年1月4日(2013.1.4)
【分割の表示】特願2009−517932(P2009−517932)の分割
【原出願日】平成20年6月6日(2008.6.6)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【Fターム(参考)】
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願日】平成25年1月4日(2013.1.4)
【分割の表示】特願2009−517932(P2009−517932)の分割
【原出願日】平成20年6月6日(2008.6.6)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【Fターム(参考)】
[ Back to top ]