
植物体における製薬に重要な蛋白質の一過性産生
本発明は、真核細胞システムにおけるバイオ製薬蛋白質及び他の有用な蛋白質を産生する迅速で、用途の広い方法に関する。本発明は、アグロバクテリウムにより作製した遺伝子を生長した植物宿主へ導入し、次いで所定の蛋白質を回収することによる、モノクローナル抗体及び他の製薬に重要な蛋白質の効果的且つ安価な一過性産生方法を特徴とするものである。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願)
本出願は、2002年12月23日に出願された米国仮出願第60/436,403号の優先権を主張するものであり、その全体が参照により本明細書に取り込まれる。
(技術分野)
【0002】
本発明は、植物における高レベル一過性蛋白質産生についての方法及びキットに関する。
【背景技術】
【0003】
製薬に重要な組換え蛋白質の発現は、通常、微生物又は哺乳類の宿主を用いて実施される。微生物のシステムはクローニング及び形質転換細胞を産生する速さの点でしばしば利点を提供する。一般に、異種の遺伝子産物の収量を高くすることができるが、蓄積する産物はしばしば生物学的な活性がなく、活性のある物質を得るにはコストのかかる困難なリフォールディングが要求される。真核生物に較べて、多くの翻訳後修飾も細菌では異なっており、ある種の蛋白質は原核生物システムでは適切に発現させることができない。
【0004】
製薬に重要な蛋白質の発現に異種システムを使用することの主な問題点の一つは、遺伝子のクローニングから、適切な動物モデルで使用するのに十分な量の機能的な蛋白質の産生までに必要とされる時間である。ほとんどのシステムで、この時間は数ヶ月から一年又はそれ以上になる。時間が消費されることに加えて、(例えば、COS又はCHO細胞の一過性のトランスフェクションによる、又は、昆虫細胞培養のバキュロウイルス感染による等の)細胞培養物における組換え蛋白質の一過性産生は、通常、特殊な装置を必要とし、産生に必要とされる細胞やウイルスを取扱うには特別な技術が要求される。加えて、より高価で特殊な装置と訓練が要求されるスケールアップの過程が導入されない限り、得られる蛋白質は通常非常に少量である。(ナノグラムからマイクログラム量)。
【0005】
1980年代後半から、作物に於ける外来又は異種蛋白質のコストパーフォーマンスのよい発現の例が多数開示されている。特に、モノクローナル抗体や他の製薬に重要な蛋白質の産生のための発現システムとして、近年、植物が浮上してきた。植物は、蛋白質のグリコシル化に主な相違があるが、動物細胞の経路に非常に良く似た蛋白質合成経路を有している(Fischer and Emans,Transgenic Res.9:279−299(2000);Cabanes−Macheteau,et al.,Glycobiology 9:365−72(1999))。さらに、所定の蛋白質のなかには植物体内に高レベルで蓄積されるものがあることが示されてきた。さらに、植物由来の抗体はハイブリドーマによって産生される抗体と機能的に同等である(Fischer and Emans,上記,(2000))。最終的には、植物由来の抗体や他の蛋白質は、ヒト又は動物の病原体、すなわち、共精製された血液感染性の病原体及び他のシステムで精製された組換え蛋白質に付随する腫瘍形成性の配列を含まない(Fischer and Emans,上記,(2000)。
【0006】
組換え蛋白質はトランスジェニック植物体内で安定的に合成された遺伝子から産生できる。異種遺伝子から蛋白質を産生するその他の方法は、一過性発現システムを使用することである。遺伝子クローニングのアプローチ、プラスミドの構築、プロモーター等を開発するために、これに適当なシステムがいくつか用いられてきた。特に、プロトプラストのエレクトロポレーションは、微粒子銃及びある程度のウイルスベクターと同様に、広く使用されてきた。
【0007】
微粒子銃は通常少数の細胞にのみに達するものであるが、転写が成就するにはDNAは細胞の核に到達しなければならない(Christou,Plant Mol.Biol.35:197−203(1996);Fischer and Emans,上記(2000))。浸潤によって送り込まれるアグロバクテリウム(agro−infiltration)の使用は、非常に多くの細胞に外来遺伝子を送り込むことができる。さらに、所定の遺伝子を収容しているT−DNAは、いくつかの細菌蛋白質の助けにより活発に核へと移入する(Kapilaら,Plant Sci.122:101−108(1997);Fischer and Emans,上記(2000))。微粒子銃及びagro−infiltrationともに処理後3−5日で異種蛋白質の発現が見られる一方、ウイルスによる送達は2から4週間かかる。微粒子銃の使用は一過性の発現にはあまり効率的ではないが、トランスジェニック穀物の再生には非常に重要である(Christou,上記(1996);Fischer and Emans,上記(2000))。
【0008】
修飾されたウイルスベクターでの感染は、殆どの植物細胞中にウイルスを組織的に広めることになる。導入された遺伝子は、ウイルスRNAレプリカーゼによって細胞質で転写され所定の蛋白質に翻訳される。標的の遺伝子は、ウイルスの複製中に高レベルに増加するので組換えウイルスベクターにより高いレベルで発現される(Porta and Lomonossoff,Mol.Biotechnol.5:209−21(1996))。しかしながら、通常このシステムは60−70Kdより小さい分子量の蛋白質に制限される。
【0009】
一過性発現のためのagro−infiltrationは多くの利点を有する。その方法は数日で所定の蛋白質を産生し、安定性と機能という特徴において満足できる蛋白質を多量に生ずる(Fischer and Emans,上記(2000))。安定な形質転換植物体を必要とすることなく、組換え蛋白質を数十ミリグラム産生するために、agro−infiltrationをスケールアップできることが提案されてきた。しかしながら、報告された(Fischer and Emans,上記(2000))発現レベルでは、より多量(例えば10−100mgs)の蛋白質が必要とされるモデル動物を使ったインビボでのより大規模な研究には実用的ではない。
【0010】
【非特許文献1】Artsaenkoら,Plant J.8:745−50(1995)
【非特許文献2】Banjokoら,Plant Physiol.107:1201−08(1995)
【非特許文献3】Baumら,Mol.Plant−Microbe Interact.5:382−87(1996)
【非特許文献4】Binetら,Plant Science 79:87−94(1991)
【非特許文献5】Borisjukら,Nature Biotechnology 17:466−69(1999)
【非特許文献6】Brownら,Nucleic Acid Res.17:8991(1989)
【非特許文献7】Cabanes−Macheteauら,Glycobiology 9:365−72(1999)
【非特許文献8】Christensenら,Plant Molec.Biol.12:619−632(1989)
【非特許文献9】Christou,P.Plant Mol Biol.35:197−203(1996)
【非特許文献10】Conrad&Fiedler,Plant Mol.Biol.38:101−09(1998)
【非特許文献11】Dennisら,Nucleic Acid Res.12:3983(1984)
【非特許文献12】DeWildeら,Plant Sci.114:231−41(1996)
【非特許文献13】Duringら,Plant Molecular Biology 15:287−93(1990)
【発明の開示】
【発明が解決しようとする課題】
【0011】
一過性発現のためのアグロバクテリウム浸潤の元来のシステムはKapilaによって記述され(Kapilaら,上記(1997))、植物体組織が有する病気に対する抵抗性にとって有用であると考えられていた蛋白質の機能性を、迅速に試験するために開発された。この適用においては、植物体組織全体をバイオアッセイに使用することができるので、蛋白質が精製され又は特徴づけされる必要はない。このシステムは後に製薬に重要な蛋白質の発現に使用された(Vaqueroら,Mol.Biotechnol.5:209−21(1996))。ヒト癌胚抗原に対するキメラ抗体及び同じ抗原に対する組換え一本鎖抗体が作製され、精製され、生化学的な解析がなされ、動物由来の蛋白質に類似するものであることが示された。しかしながら、このシステムからの産物は比較的少ないものであった。
【課題を解決するための手段】
【0012】
(発明の要約)
本発明は、植物栽培設備の必要のない、すでに生長した商業的に入手可能な植物体で蛋白質を産生するといった実質的に改良された工程を提供する。本発明の方法は、動物の試験、多様なアッセイによる解析、結晶構造の特徴づけ、蛋白質修飾のアッセイ等に適した規模で、最小限の時間と費用により、生物学的に機能的な蛋白質を非常に迅速に提供する。本発明の方法で、少なくとも約1mg、少なくとも約5mgまた少なくとも約10mgの蛋白質を1回で産生することができる。
【0013】
本発明の方法は、繰り返し試験するのに必要とされるミリグラム量を提供する規模まで容易かつ迅速に拡大することができ、製薬に用いる蛋白質を機能的に評価するのに使用することができる。
【0014】
本発明は、モノクローナル抗体及び他の製薬に重要な蛋白質の一過性発現のための方法とキットを提供する。この方法は、特に、ウイルスの制御配列を伴って又は伴わずに、プラスミドベクター上に所定の組み換え遺伝子を作製するAgrobacterium tumefaciensによって減圧浸潤されたレタスでの蛋白質産生に成功した。
【0015】
一態様では、本発明は、非常に迅速にkgあたり10−50mg/kgの蛋白質を産生するのに使用できる、拡大可能な細菌/植物ハイブリッド発現システムを提供する。一つの好ましい態様では、1又はそれ以上の異種遺伝子がアグロバクテリウムの無毒化された又は毒性の株を用いて植物体組織に送達される。好ましくは、植物体組織はすでに生長した植物(例えば、商店で入手できる植物等)から得られる。植物体組織の特に好ましい供給元はレタスである。本明細書に開示される方法は、細菌システムにおける迅速な遺伝子クローニングと操作の利点と、植物の栽培及びトランスジェニック植物材料の取り扱いの必要なしに、適当なターゲッティング、プロセッシング、修飾及び会合に必要な環境を提供する真核細胞環境で蛋白質が産生されることの利点を併せ持つ。
【0016】
一態様では、異種遺伝子又は所定の遺伝子を有する発現構成体を作製するアグロバクテリウムの細胞は、植物体組織の細胞及び/又は細胞外空間での一過性発現のために、1又は複数の異種遺伝子を植物体組織へ送達するのに使用される。一般的に、適当な発現構成体は以下を含む:少なくとも1つのT−DNA境界配列、発現制御配列(例えば、誘導可能な及び/又は組織特異的な、又は構成性であるかもしれないプロモーター)、及び発現制御配列に作動可能に結合された所定の遺伝子。一態様では、発現構成体は、発現構成体を含むベクターがアグロバクテリウム中で複製するのに適した1又はそれ以上の複製起点、少なくとも1つの複製起点を含むベクターの一部である。
【0017】
発現構成体を含むアグロバクテリウム細胞の培養物は、界面活性剤の存在下、植物体組織に浸潤される。好ましくは、浸潤は減圧状態で起こる。発現構成体が複数の植物細胞中で蛋白質を発現することができる適当な条件下で植物体組織をインキュベーションした後、蛋白質は細胞から分離される。本発明の方法は、ベクターを含むアグロバクテリウムと植物体組織を接触させ、植物体組織をベクターで浸潤し、約500μg−500mgの収量を得るために異種蛋白質を発現させるという、一連の操作を1回のみ必要とする。しかしながら、agro−infiltration及び精製の操作は手順をスケールアップするために追加で実施してもよい。本発明の方法の一回の操作で、より多くの植物体組織を使用するのが更に好ましい。
【0018】
使用されるアグロバクテリウムは、野生型(例えば、毒性の)又は無毒化されたものであってもよい。それぞれが異なる遺伝子を発現する多くのアグロバクテリウム株が、複数の個々の蛋白質又はヘテロ多量体蛋白質(例えば、抗体)を産生するために、又は、代謝経路、化学合成経路又は情報伝達経路等の経路を再生するために、使用され得る。あるいは又は付加的に、単一のアグロバクテリウム株が異なる異種遺伝子からなる複数の配列を含んでいるかもしれない。異なる異種遺伝子は、単一の核酸分子(例えば、単一のベクター)中に含まれているかもしれないし、異なるベクターで提供されるかもしれない。一態様では、少なくとも1つのアグロバクテリウム株はAgrobacterium tumefaciensを包含する。
【0019】
本発明は異種蛋白質を発現するためのハイスループットシステムを提供するもので、当当該システムは蛋白質の機能に関して所定の遺伝子及び/又は蛋白質の変異による効果を決定するのに使用することができる。一態様では、莫大な数の様々な異種遺伝子を評価するために、標準的な分子生物学的手法(例えば、ランダム突然変異誘発により又はコンビナトリアルクローニングにより等)を用いて、細菌中に複数の発現構成体が作製される。その細菌は、実質的に同一であり(例えば、約50%以上同一、好ましくは75%以上同一、より好ましくは約90%又は95%以上同一)、生物学的活性について迅速にスクリーニングされるために、本発明に従った一過性発現システムを用いて産生されるかもしれない蛋白質をコードする核酸を含んでいる。一態様では、複数の発現構成体は、約100以上、約500以上、約1×103以上、約1×104以上、約1×105以上、約1×106以上、約1×107以上、約1×108以上又は約1×109以上の様々なコード配列を包含する。
【0020】
複数の発現構成体は、異種ポリペプチドのコード配列にランダム又はややランダムな変異を含む配列のライブラリーを包含することができる。ライブラリーはE.coliに基づくライブラリー(すなわち、個々のライブラリー要素がE.coli中でクローン化され複製される)であっても良いし、アグロバクテリウムに基づくライブラリー(すなわち、個々のライブラリー要素がアグロバクテリウム中でクローン化され複製される)であっても良いし、それらの組み合わせ(すなわち、クローニングは最初E.coli中で実施され、ライブラリー要素がさらなる複製及び/又はクローニングのためにアグロバクテリウム細胞中に続いて導入されるかもしれない)であっても良い。ライブラリーの個々の要素は、例えば、大規模な産生及び/又はトランスジェニック植物の産生に適したポリペプチドを同定するために、植物体組織で一過性の発現をした後、ポリペプチドの機能について試験される。
【0021】
ある好ましい態様では、本発明の方法は、最適な活性及び/又は結合のために、複数のサブユニット蛋白質複合体の相互作用しているサブユニット要素(ISEs)の機能を最適化するために使用される。例えば、抗体の結合を最適化するために、抗体の可変領域の複数の変異体が細菌中に標準的な分子生物学的手法(例えば、ランダム突然変異誘発により又はコンビナトリアルクローニングにより等)を用いて作製される。好ましくは、約1×103以上、約1×104以上、約1×105以上、約1×106以上、約1×107以上、約1×108以上又は約1×109以上の様々な発現構成体が作製される。適当な可変領域の配列は抗体の軽鎖(LC)又は重鎖(HC)又は軽鎖及び重鎖の両者を包含する。これらの配列を含む変異体ポリペプチドは、選択された抗原に対する特異的な結合について評価される。変異体ポリペプチドは完全長の抗体又はそれらの抗原結合フラグメント(scFv、Fab’等)を含んでいても良い。異なる変異体は上述のアグロバクテリウムベクター中に容易にクローン化され、HC及びLCの全ての組合せが迅速に試験され得る。ある好ましい態様では、試験は並行して行われる。
【0022】
本発明の方法は、生長している植物体に於ける蛋白質の発現に最も適した発現ベクターを、あらかじめスクリーニングするために使用することができる。一態様では、本発明の方法は、最適な蛋白質発現を提供する、対照となる発現配列及び/又は翻訳配列の変異体を迅速にスクリーニングするのに使用される。あるいは又は付加的に、変異体配列は、増加した安定性又は他の望ましい製薬上の特性を有した蛋白質をコードする配列を同定するためにスクリーニングされる。
【0023】
本発明は、本発明の方法を実施するのに有用なキットも提供する。一態様では、本発明に従ったキットは、少なくともA.tumefaciens等のアグロバクテリウム種で発現するのに適したクローニング/発現ベクター、及び植物種から所望の異種蛋白質を浸潤、抽出及び/又は精製するための1又はそれ以上の構成要素を含む。別の態様では、キットはさらに1又はそれ以上の細菌種(例えば、E.coli及びA.tumefaciens等)を含む。さらなる態様では、キットは変異体異種配列をコードする核酸を含む複数の発現構成体を包含する。
【0024】
(発明の詳細な説明)
本発明は、商業的に入手可能な生長した植物体に於いて蛋白質の産生を可能にする方法を提供し、異種蛋白質において、バイオアッセイで利用するために必要な量を短期間で調達しなければならないという問題に新規な解決法を提供する。本発明の方法は、少なくとも約50μg−100mgの蛋白質を必要とするアッセイ、例えば、薬剤スクリーニング、蛋白質の特徴付け、結合アッセイ及び動物試験等のアッセイでの使用に対して、生物学的に活性な異種蛋白質を提供する。
【0025】
本発明の一態様では、本発明の方法は、異種蛋白質又は生物学的に活性な当該フラグメントをコードする発現構成体を植物体組織に導入すること、及び、植物体組織で当該蛋白質を一過性に発現させることを包含する。コード配列は、植物体組織の細胞及び/又は細胞外空間におけるコード配列の転写を制御することができる発現制御配列に作動可能に結合される。好ましくは、発現構成体は、大きな腫瘤誘導(“Ti”)プラスミドのT−DNA由来の少なくとも1つのT境界配列を包含する。また、好ましくは、発現構成体は、少なくともAgrobacterium tumefaciens等のアグロバクテリウム種の細胞で複製することができるベクター中に含まれる。一態様では、植物体組織はすでに生長した植物(例えば、商店で入手できる植物等)由来の葉組織を含む。好ましくは、植物は比較的大きな(例えば、少なくとも一方向が約3インチより大きな)葉からなり、例えば植物はレタス(Lactuca sativa)である。
【0026】
(定義)
以下の定義は、以下に記載される説明中で使用される特別な用語に対して与えられる。
明細書及び請求項で使用される時、単数形“a”“an”及び“the”は、文脈が別の方法で明らかに指示しない限りは、複数への言及も包含する。例えば、用語“a cell”は、複数の細胞の混合物も含む複数の細胞を包含する。用語“a protein”は複数の蛋白質を包含する。
【0027】
本明細書で使用される「植物細胞」は植物細胞、又は、分離された又は一部分離された細胞を包含する。「植物体組織」は分化した及び未分化の植物組織を包含し、根、シュート、葉、花粉及び種子を含むがそれらに限定されない。
【0028】
本明細書で使用される時、「植物材料」は、植物材料が処理されたことにより生じた派生物を包含し、食物生産物、食物原料、栄養補助食品、抽出物、濃縮物、丸剤、トローチ剤、かみ砕ける物、粉末、粉ミルク、シロップ、キャンディー、ウエハース、カプセル及び錠剤を含むがそれらに限定されない。
【0029】
本明細書で使用される時、「複数のサブユニット蛋白質」は複合体を形成するために互いに結合する1より多くの独立したポリペプチド又は蛋白質鎖を含む蛋白質であり、ここで少なくとも2つの独立したポリペプチドは異なる遺伝子によってコードされる。ある好ましい態様では、複数のサブユニット蛋白質は、少なくとも抗体の免疫学的に活性な部分を含み、よって特異的に抗原と結合できる。例えば、複数のサブユニット蛋白質は抗体分子の重鎖及び軽鎖又はその一部を包含することができる。複数の抗原結合部位が、多価抗体を作成するために、異なる構造遺伝子によってコードされ得る。
【0030】
製薬生産物の場合には、「実質的に純粋」なる用語は、一般的に、少なくとも純度90%、より好ましくは少なくとも純度95%及びさらにより好ましくは少なくとも純度98%の生産物をいう。
【0031】
「組織液(interstitial fluid)」は、細胞膜すなわち細胞表面の膜によって囲まれていない植物の全ての領域から得られる抽出液を意味する。この用語は、顕著な細胞溶解なしにこの処理によって細胞膜から放出されるかもしれない分子を含む、植物の細胞内に存在しない液体、物質、領域又は空間の全てを含むことになっている(ここで、細胞内は細胞質膜内に含まれる中身として定義される)。この用語の類義語は、「外質(exoplasm)」「apoplasm」「間質液」「細胞外液」及び「溢液」を含むがそれらに限定されない。
【0032】
「プロモーター」なる用語は、遺伝子の転写の開始を指示する、遺伝子の5’末端にある核酸配列をいう。一般的に、プロモーター配列は、それが作動可能に結合する遺伝子の発現を制御するのに必要であるが、常に十分なわけではない。プロモーター/異種遺伝子の組合せからなる構築物では、遺伝子はプロモーターに十分近接して、プロモーターに対して適した方向で配置されるので、遺伝子の発現はプロモーター配列によって調節される。プロモーターは、それが天然の位置で調節している遺伝子とそれ自身の間の距離に近似したある一定の距離を転写開始点からおいて、遺伝子の上流に優先的に配置される。公知のように、この距離におけるいくらかの多様性はプロモーター機能を損なわずに許容され得る。本明細書で使用される時、「作動可能に結合」なる用語は、コード領域の転写がプロモーターによって調節され制御されるように、当該プロモーターが当該コード領域に連結されることを意味する。コード領域に作動可能に結合しているプロモーターの意味は、当業界で周知である。
【0033】
本明細書で使用される時、「発現制御配列」はプロモーターを含み、1又はそれ以上のエンハンサー配列、転写終結配列、ポリA配列、3’又は5’非翻訳配列、イントロン配列、リボゾーム結合部位、及び、植物細胞で遺伝子の発現を安定化するか又は他の方法で制御する他の配列を含むかもしれないが、それらに限定されない。発現制御配列は内因性(すなわち、植物宿主で天然に見出される)又は外因性(植物宿主で天然に見出されない)であってもよい。外因性の発現配列は、選択された条件下にてそれらが植物細胞中で機能的である限り、植物の配列であってもなくてもよい。
【0034】
「異種遺伝子」又は「異種コード配列」は、形質転換又は処理された植物にとって外因性又はそこでは天然に見出されない遺伝子であり、「異種ポリペプチド」又はその生物学的に活性なフラグメントをコードする遺伝子である。異種遺伝子配列は、ウイルス、原核生物及び真核生物の配列を含む。原核生物のコード配列は、微生物の配列(例えば、ワクチンとして投与されるかもしれない抗原作製のため、ウイルスの配列もこの目的に使用されるかもしれない)を含むがそれに限定されない。真核生物のコード配列は、哺乳動物の配列を含むが、非哺乳動物、他の植物からの配列を含んでいてもよく、リーダー又は分泌シグナル配列、ターゲッティング配列等を含むがそれらに限定されない。ある好ましい態様では、異種遺伝子の核酸はヒトの蛋白質をコードする。「異種遺伝子」又は「異種コード配列」なる用語は、自然に発生した、突然変異させた、変異した、化学合成された、ゲノムの、cDNAの、又はこれらの配列のいかなる組合せをも包含するがそれらに限定されない。「遺伝子」への言及は、生物学的に活性な蛋白質をコードする完全長の遺伝子又はそのフラグメントを包含する。
【0035】
本明細書で使用される時、「蛋白質」なる用語は、一般的に、遺伝子によってコードされるアミノ酸配列全体、その処理された又は修飾された形又はその生物学的に活性なフラグメント(例えば、ポリペプチド又はペプチド)に言及するために使用される。
【0036】
「融合蛋白質」は、その配列の組合せは、単一の蛋白質として天然では発現されることのない、ポリペプチド中で結合した少なくとも2つの異なるアミノ酸配列を含む蛋白質である。
【0037】
本明細書で使用される時、「T DNA境界」は、A.tumefaciens又はA.rhizogenes株、又はそれらの修飾された又は突然変異された株等のアグロバクテリウム株が有する毒性遺伝子産物によって認識され得る約25ヌクレオチド長の配列を含むDNAフラグメントを意味し、それが連結しているDNA配列を真核生物細胞、好ましくは植物細胞へ移入させる能力のあるDNAフラグメントをいう。この定義は、野生型Tiプラスミド由来の全ての自然に発生したT−DNA境界及びその機能的ないかなる派生物をも含むがそれらに限定されず、化学的に合成されたT−DNA境界も包含する。一態様では、本発明に従った発現構成体のコード配列及び発現制御配列は2つのT−DNA境界の間に存在する。
【0038】
植物種
一過性発現に対するアグロバクテリウム浸潤の初期の適用は、ポプラ及びPhaseolus(Kapila,et al.,1997)に基づいており、そらから後にたばこ(Vaquero,et al.,1999)へ応用された。ポプラ及びPhaseolusの両者は、即座に本発明で使用される植物体組織の適当な供給源となる。他の適当な植物は、レタス、アルファルファ、インゲンマメ、ホウレンソウ、タンポポ、アカチコリー、キバナスズシロ、エンダイブ、キクチシャ、チコリー、アーティチョーク、トウモロコシ、ポテト、イネ、ダイズ、Crucifera(例えば、Brassica、アラビドプシス(シロイヌナズナ))、ウキクサ、トウモロコシ、ポテト、イネ、ダイズ、ホウレンソウ、トマト及びタバコを含むがそれらに限定されない。Brassica科の例となる植物は、B.oleracea(例えば、キャベツ、コラード、カリフラワー、ブロッコリー、芽キャベツ、ケール、コールラビ)、B.campestris(例えば、ハクサイ、ターサイ、Chinese cabbage、celery cabbage、Siberian kale、カブ、カラシナ、ナタネ、スウェーデンカブ及びハツカダイコン)、Brassica juncea(例えば、Brown and Indian Mastard)、Brassica carinata(例えば、Abyssinian Mastard)、Brassica napus(スウェーデンカブ、Swede、Swede Turnip、Siberian kale、Hanover Salad、カノーラ)、Brassica nigra(例えば、Black Mastard)、Rorippa nasturtiumaquatkum(例えば、Water Cress)等を含むがそれらに限定されない。植物材料の特に好ましい供給源はレタスである。適したレタス植物は、Butterhead、Crisphead及びLeaf lettuce(例えば、Oak leaf、Salad Bowl、frilly Red Leaf及びcrinkly Green Leaf)を含むがそれらに限定されない。レタスのさらなる種類は当業界で公知であり、例えばhttp://www.thompson−morgan.com/seeds/us/list_lettuce_2.htmlに記載される。
【0039】
好ましくは、適する植物は一年中商業的に入手可能であり、発現制御配列に作動可能に結合したレポーター遺伝子(例えば、GUS等)の高レベルな一過性発現を維持することができる植物である。本明細書で使用される時、「高レベルな一過性発現」は、植物体組織量キログラム当たり、少なくとも約250μg、少なくとも約500μg、少なくとも約750μg、少なくとも約1mg、少なくとも約2mg、少なくとも約3mg、少なくとも約4mg、少なくとも約5mg、少なくとも約10mg、少なくとも約15mg、少なくとも約25mg、少なくとも約50mg、少なくとも約75mg、少なくとも約100mg、少なくとも約150mg、少なくとも約200mg又は少なくとも約500mgを発現する能力をいう。本明細書で使用される時、「一過性」は、適した植物体組織から蛋白質を分離することができる程十分長い期間をいう。好ましくは、蛋白質の発現は、植物体組織に発現構成体を導入後、少なくとも約1日、少なくとも約2日、少なくとも約3日、少なくとも約4日、少なくとも約5日以内に適当な高いレベルとなる。一態様では、植物体組織に発現構成体を導入後、適当な高いレベルが3−7日以内に、より好ましくは3−5日以内に得られる。
【0040】
適する植物体組織は、一般に、植物のいかなる部分でもあり得る。ある好ましい態様では、植物体組織は葉組織である。一態様では、植物体組織は、少なくとも一方向が少なくとも約3インチの葉からなる植物由来の葉組織である。しかしながら、葉の大きさは制限されず、一態様では本発明の方法はアラビドプシス(シロイヌナズナ)で蛋白質を一過性に発現させるために使用される。
【0041】
別の態様では、異種蛋白質を消化するプロテアーゼのレベルがほとんど又は全くない細胞、例えば、異種蛋白質を発現する核酸の導入から少なくとも蛋白質が植物体組織から分離される頃までの期間に、植物体に於いて発現される異種蛋白質の約5%より少なく、約1%より少なく、約0.1%より少ない量が消化されるような細胞からなる組織が選択される。プロテアーゼレベルは、異種蛋白質発現のウエスタンブロット解析を含む、当業界でごく普通の方法を用いてアッセイすることができる。
【0042】
本発明の特別な利点は、少なくとも約1日、少なくとも約2日、少なくとも約5日、少なくとも約1週間又は少なくとも約2週間、収穫され貯蔵された植物を含む、すでに生長した植物を植物体組織の供給源として使用できる点である。従って、植物体組織はいかなる一般的な食料品店でも得ることができる。
【0043】
レタスは、本発明に従った方法で使用される葉組織の特に適した供給源である。なぜなら、レタスは容易に入手でき、異種蛋白質の発現、安定性及び機能を高いレベルで提供するからである。さらに、レタス細胞は異種蛋白質を認識するプロテアーゼを量的にほとんど含んでいない。使用に適した様々なレタスの内、特に好ましいのは赤い葉の表現型を有するレタスである。レタスで観察される異種蛋白質の高レベルな発現に加えて、葉の多いレタスを使用することの利点は、特別に設計された装置の必要がなく葉を簡単に操作できる様式で植物が生長している点にある。
【0044】
トウモロコシは、安定なトランスジェニック植物から製薬に用いられる蛋白質を産生するために、最もよく使用される作物の一つである。トウモロコシでは、異種蛋白質は種子で産生され貯蔵される。しかしながら、トウモロコシは形質転換するのが難しく、一世代の時間が長く、室内で種子を産生するのが難しい。トウモロコシは、一過性発現システムから大いなる利益を得ることができる作物である。トウモロコシを形質転換するためにアグロバクテリウムを使用することは今や本当に一般的であるので、トウモロコシの種子由来の異種蛋白質の有用性/機能を迅速に評価し確認するために、トウモロコシ胚を処理することによって、本発明に従った一過性の発現システムを使用し、トウモロコシの種子で作られる蛋白質の産生量を高めることができる。
【0045】
発現カセット
本発明の好ましい実施形態では、野生型レタス、突然変異又は修飾された変異体レタス(例えば、トランスジェニックレタス等)が処理され、所望のDNA構築物から所定の遺伝子を発現する。このような構築物は、遺伝子の転写及び最終的な蛋白質の翻訳を容易にするために、プロモーター及び/又は他の調節要素(すなわち、発現制御配列)に作動可能に結合した、所望の蛋白質をコードする核酸配列を少なくとも含んでいる。
【0046】
一態様では、発現構成体は、5’から3’方向に作動可能に結合された以下のもの、プロモーター、遺伝子及びターミネーターを含むよう巧みに処理されている。別の態様では、遺伝子構築物は、1つの共通のプラスミド又は植物に同時トランスフェクションされるプラスミド上に作動可能に結合した複数のコード領域を含む(このような同時トランスフェクションされる構築物は、本明細書で使用される時には「遺伝子構築物」なる用語に一括して包含される)。同義遺伝子は分離されたシストロンとして又はポリシストロン性のユニットの部分としてコードされていてもよい。さらなる態様では、遺伝子構築物は1又はそれ以上のIRES要素を包含する。
【0047】
遺伝子構築物が選択可能なマーカーを含んでいる必要はなく、形態学的に正常で安定なトランスジェニック植物の作製で要求されるように、DNA構築物が「腫瘤誘導」遺伝子を欠いていることも必要とされない。
【0048】
蛋白質
本発明によって産生される蛋白質については予想される制限はないが、管理され、そして再生産が可能であるという条件下で特定の産物を産生する必要性があることから、特に適した蛋白質の範疇がある。特にこれには、医薬品の製造及び品質管理に関する基準や承認された方法がその生産過程で使用されなければならない製薬及び/診断のための全種類の蛋白質が含まれる。
【0049】
それらの産生物はヒトに直接、経口摂取、注射又は塗布(例えば、局所的な投与)するために使用されるので、蛋白質もまた、栄養補助食品及び薬用化粧品としての有用性があるので発現させてもよい。同様に規定された獣医向けの産物の産生に有用な蛋白質もまた発現させてもよい。
【0050】
産生されてもよい蛋白質の例は以下のものを含むがそれらに限定されない。成長因子(例えば、血小板由来増殖因子、インシュリン様増殖因子等)、受容体、リガンド、情報伝達分子、キナーゼ、酵素、ホルモン、腫瘍抑制遺伝子産物、血液凝固蛋白質、細胞周期蛋白質、テロメアーゼ、代謝蛋白質、神経蛋白質、心臓の蛋白質、特異的な疾患状態で定義される蛋白質、抗体、T細胞受容体(TCR)、主要組織適合性複合体(MHC)、抗原、病気に対する抵抗性を与える蛋白質、抗菌蛋白質、インターフェロン及びサイトカイン。
【0051】
一態様では、保護免疫反応を誘導する配列を含んでいる配列をコードする(例えば、ワクチン製剤に含まれるような)抗原である。このような適する抗原は、以下のものを含むがそれらに限定されない。微生物抗原(ウイルス抗原、細菌抗原、菌類の抗原、寄生虫抗原等)、多細胞生物(多細胞寄生虫等)由来の抗原、アレルゲン及びヒト又は動物の病理に関係のある抗原(例えば、癌、自己免疫疾患等)。ある好ましい態様では、ウイルス抗原は、以下のものを含むがそれらに限定されない。HIV抗原、天然痘に保護免疫反応を与える抗原(例えば、ワクシニアウイルス抗原)、炭疽菌抗原、狂犬病抗原等。ワクチン抗原は、例えば、1の発現構成体に、任意で1又はそれ以上のリンカー配列で分離された状態で、繰り返し、異なる又は同じ抗原性をコードする配列を含んでいる多価ペプチド又はポリペプチドとしてコードされ得る。
【0052】
植物は、また、化学合成又は工業的工程のための酵素経路を再現するために1又はそれ以上の遺伝子を発現するのに使用されるかもしれない。
【0053】
一態様では、所望の蛋白質をコードする核酸配列が選択される。そこでは、核酸配列は、レタス又は所望の蛋白質の大規模な生産に最終的に使用されるかもしれない植物によって好まれるコドン、ただし、そのコドンの選択が一過性システムで有用なレベル以下に発現量を減少させない場合、そのコドンを提供するように設計される。いくつかの植物におけるコドン使用の指標は入手可能であり、例えば、Wadaら,“Codon Usage Tabulated From The GenBank Genetic Sequence Data,”Nucleic Acids Reseach 19(Supp.):1981−1986(1991)に記載されている。
【0054】
以下にさらに記載するように、一態様では、本発明は複数の組換え蛋白質を発現するための方法を提供する。このような蛋白質は、独立した構築物の同時浸潤によって発現されるかもしれないし、以下にさらに記載するポリシストロン性の発現ユニットから発現されるかもしれない。このような蛋白質は、その天然の状態で、生物学的に活性になるために複数の構造遺伝子の協調した発現を必要とする蛋白質を含むことができる。一態様では、蛋白質は、活性を有するために複数のサブユニットの会合が必要とされる。別の態様では、蛋白質は成熟していない形で生産され、プロセッシング、例えば、活性化するために、1又はそれ以上のさらなる蛋白質によって、蛋白質分解での切断又は修飾(例えば、リン酸化、グリコシル化、リボシル化、アセチル化、ファルネシル化等)されることを必要とする。
【0055】
このような蛋白質の限定されない例としては、T細胞受容体、MHC分子、他の免疫グロブリンスーパーファミリーの蛋白質(フラグメント及び一本鎖変異体を含む)、核酸結合蛋白質(例えば、複製因子、翻訳因子等)、酵素、アブザイム、受容体(特に可溶性の受容体)、成長因子、細胞膜蛋白質、分化因子、ヘモグロビン様蛋白質、多重結合キナーゼ等のヘテロ二量体又はヘテロ多量体蛋白質を含む。
【0056】
本発明の好ましい態様では、発現カセットはヒト蛋白質(すなわち、ヒトで発現する蛋白質)をコードし、又は、他の状態で非ヒト蛋白質に含まれているヒトポリペプチド領域を含む蛋白質をコードする。
【0057】
ある特に好ましい態様では、発現カセットは、モノクローナル抗体の1又はそれ以上の遺伝子をコードする。このような遺伝子は、マウス、ヒト及び/又は他の動物から得ることができる。あるいは、例えば、抗体分子の構成要素である重鎖又は軽鎖をコードする遺伝子のキメラ又は修飾された形で、それらを合成することもできる。構築物中のコード領域の順番、例えば、重鎖それから軽鎖又は軽鎖それから重鎖は、重要ではない。重鎖又は軽鎖ポリペプチド(例えば、重鎖可変又は軽鎖可変ドメインポリペプチド等)をコードする遺伝子は、IgA、IgD、IgE、IgG又はIgMを産生する細胞から得ることができる。免疫グロブリン可変領域遺伝子をクローニングすることができるゲノムDNAのフラグメントを調製する方法は、当業界で周知である。例えば、Herrmannら,Methods in Enzymol.152:180−183(1987);Frischauf,Methods in Enzymol.152:183−190(1987);Frischauf,Methods in Enzymol.152:199−212(1987)参照。ある好ましい態様では、以下に記載するように、このような遺伝子はポリシストロン性ユニットの部分としてコードされる。
【0058】
調節要素
特定の構築物を作成するのに適した調節要素は、発現される組換え蛋白質の種類に基づいて選択されるだろう。一般に、浸潤される植物体組織において高いレベルで発現する能力が望まれる。
【0059】
植物のプロモーター
使用される遺伝子構築物は、生物学的に活性な蛋白質フラグメントをコードしている遺伝子全体又はその一部からなる、遺伝的な材料全てを含んでいるかもしれない。好ましくは、コード配列は発現制御配列に作動可能に結合している。発現制御配列はプロモーター、エンハンサー、IRES要素等の配列を含む。発現制御配列は、発現を誘導するための外部からの刺激として特定の栄養素又は薬剤の付加、温度変化等を要求することもできるし、植物体組織の浸潤及び/又は培養中に迅速に及び/又は自然発生的にコードされる蛋白質を発現するよう設計することもできる。
【0060】
このように、構成的又は調節されたプロモーターは所望の蛋白質をコードする遺伝子の発現を制御するかもしれない。調節されたプロモーターは、環境から情報を受け取るかもしれないし、化学的な誘導因子や抑制因子によって制御可能であるかもしれない。このような因子は天然のもの又は合成物由来であってもよく、プロモーターは天然のもの又は設計されたものであってもよい。プロモーターはキメラ、すなわち、2又はそれ以上の異なる天然又は合成されたプロモーター由来の配列要素を使ってできたものでもあり得る。
【0061】
好ましくは、構築物中で使用されるプロモーターは遺伝子の高い発現レベルを生じ、植物体組織重量(例えば、葉組織生物量)kg当たり少なくとも約250μg、少なくとも約500μg、少なくとも約750μg、少なくとも約1mg、少なくとも約2mg、少なくとも約3mg、少なくとも約4mg、少なくとも約5mg、少なくとも約10mg、少なくとも約15mg、少なくとも約25mg、少なくとも約50mg、少なくとも約75mg、少なくとも約100mg、少なくとも約150mg、少なくとも約200mg又は少なくとも約500mgの蛋白質の蓄積を可能にする。
【0062】
本発明では、アラビドプシス(シロイヌナズナ)アクチン2プロモーター、OCS3(MAS)プロモーター、CaMV 35Sプロモーター及びゴマノハグサモザイクウイルス34Sプロモーターが好まれる。しかしながら、他の構成的及び誘導可能なプロモーターを使用することもできる。例えば、ユビキチンプロモーターは植物体での使用のためにいくつかの種からクローニングされた(例えば、ヒマワリ(Binetら,Plant Science 79:87−94(1991))及びトウモロコシ(Christensenら,Plant Molec.Biol.12:619−632(1989))。さらに有用なプロモーターはトウモロコシ由来のU2及びU5 snRNAプロモーター(Brownら,Nucleic Acid Res.17:8991(1989))及びアルコールデヒドロゲナーゼ由来のプロモーター(Dennisら,Nucleic Acid Res.12:3983(1984))である。
【0063】
別の態様では、調節されたプロモーターは遺伝子に作動可能に結合される。調節されたプロモーターは、外部からの影響によって(例えば、化学薬品、光、温度等の外部因子の適用によって等)調節されるプロモーター、又は、植物における調節された発生上の変化等の内的なきっかけによって調節されるプロモーターを含むがそれらに限定されない。調節されたプロモーターは、収穫の時期又はその付近において特異的に所望の遺伝子の高レベルな発現を誘導するのに有用である。これは、所望の蛋白質が植物の成長を制限したり別の方法で抑制したり又は所望の蛋白質がいくつかの方法では不安定である場合に、特に有用である。発現構成体が一過性発現アッセイにおいてだけでなくトランスジェニック植物の生産においても使用されることが期待される場合には、このようなプロモーターが望まれるかもしれない。
【0064】
異なる植物体組織で移入された遺伝子の発現を制御する植物のプロモーターは当業者に公知である(Gasser&Fraley,Science 244:1293−99(1989))。カリフラワーモザイクウイルス35Sプロモーター(CaMV)及びCaMVプロモーターの増強された誘導体(Odellら,Nature,3(13):810(1985))、アクチンプロモーター(McElroyら,Plant Cell 2:163−71(1990))、AdhIプロモーター(Frommら,Bio/Technology 8:833−39(1990)、Kyozukaら,Mol.Gen.Genet.228:40−48(1991))、ユビキチンプロモーター、ゴマノハグサモザイクウイルスプロモーター、マンノピンシンターゼプロモーター、ノパリンシンターゼプロモーター、オクトピンシンターゼプロモーター及びそれらの誘導体が構成的プロモーターとみなされる。調節されたプロモーターは、光で誘導可能なもの(例えば、リブロースビスリン酸カルボキシラーゼ小サブユニットのプロモーター)、熱ショックプロモーター、硝酸塩及び他の化学的に誘導可能なプロモーター(例えば、米国特許第5,364,780号、第5,364,780号、第5,777,200号参照)として記述される。
【0065】
組織特異的なプロモーターは、植物の特定の部分で蛋白質を発現させる理由がある時に使用される。葉特異的なプロモーターは35Sエンハンサーが先行するC4PPDKプロモーター(Sheen,EMBO,12:3497−505(1993))又は葉での発現に特異的ないかなる他のプロモーターを包含していてもよい。
【0066】
一般に、植物において発現可能である、あらゆる遺伝的な構築物が本発明の方法で使用されるのに適している。特定のプロモーターが発現させる組換え蛋白質の種類を考慮して選択されるかもしれない。
【0067】
本発明にとって特に興味深いのは、OCSエンハンサー及び核となるMASプロモーターの複数ユニット由来の合成プロモーターの使用であり、ここでOCS及びMASの要素はアグロバクテリウム由来である(Gelvinら,米国特許第5,955,646号)。このプロモーターは、後に行われる浸潤及び植物ホルモン2,4−Dでの処理、このホルモンはより高い濃度で除草剤として使用されるが、に対して特に強いことが示されている。
【0068】
標的化配列
好ましい実施形態では、発現産物は、細胞膜、細胞外空間又は細胞小器官、例えばクロロプラスト等のプラスチドのような植物細胞の特異的な場所へと向けられる。好ましい実施形態では、発現産物は、細胞外空間へと向けられ、その結果、細胞内液の分離に基づく精製を可能にする。例えば、米国特許第6,096,546号、米国特許第6,284,875号及び国際公開第0,009,725号公報参照。
【0069】
蛋白質は、標的化配列の効力によって特異的な細胞内の一部又は細胞外の場所へと向けることができる。ある場合には、アミノ酸配列がポリペプチドのアミノ末端部分として合成され、移動又は局在化する過程の後又はその間に、プロテアーゼによって切断される。例えば、真核生物における蛋白質分泌経路のモデルは、リボソームのmRNAへの結合及び翻訳の開始に続いて、発生しようとしているポリペプチド鎖が出現する。それが分泌される運命にある蛋白質ならば、出現した蛋白質のアミノ末端は、シグナル認識粒子(SRP)、それはmRNA、リボソーム及びSRP複合体が小胞体(ER)に付着する間、翻訳の一時的な失速を引き起こすが、それによって認識される。付着した後、ポリペプチド鎖は今や翻訳されると同時にERルーメンへと移動するが、翻訳は再開する。
【0070】
液胞中に局在化するため又は分泌のために内膜システムへと標的化される蛋白質についてのシグナル配列は、植物及び動物で類似している。シグナルペプチドは、標準的な技術に従った本発明での使用(例えば、あらゆる他の遺伝子でフレームに合わせてクローニングするために適した末端に調製される)に適合しているかもしれない。
【0071】
一態様では、所望の蛋白質をコードする発現カセットは、所望の蛋白質をコードする配列にフレームを合わせて融合したシグナル配列を含む。ある好ましい態様では、シグナル配列は分泌経路に遺伝子の発現産物を向けることができる配列である。
【0072】
抗体は正常に分泌される蛋白質であるので、分泌過程は成熟した抗体分子の産生に重要な役割を演じている。植物ではこれを達成するために、天然の状態での哺乳動物のシグナルペプチドをコードする領域を有するか、植物の分泌シグナルペプチドが所定の遺伝子のシグナルペプチドとして置換され、所定の遺伝子に作動可能に結合した融合物として、遺伝子が合成又は他の方法で(例えば、クローニング)得られる。シグナルペプチドと蛋白質の融合は植物によってプロセッシングされるはずであり、結果として生じる蛋白質のアミノ末端はヒト宿主で生じるものと同一である。しかしながら、クロロプラストへの標的化も期待される。
【0073】
好ましい実施形態では、カルレティキュリン由来のシグナル配列(Borisjukら,Nature Biotechnology 17:466−69(1999))が使用される。より好ましい実施形態はトマト由来のズブチラーゼ(subtilase)配列を使用する(Janzikら,2000)。これらの植物シグナルペプチドは植物のapoplastic空間へと外来蛋白質を向かわせるのに有効であることが示されてきた(例えば、Janzikら,2000参照)。他の植物蛋白質シグナルペプチドはオオムギに対して記述されるシグナルペプチドも使用されるかもしれない(α−アミラーゼ、Duringら,Plant Molecular Biology 15:287−93(1990)、Schillbergら,Transgenic Research 8:255−63(1999))。
【0074】
蛋白質を植物の内膜システムへ標的化することは、その成熟形に到達するためにアミノ末端のプロセッシングを通常要求する蛋白質に対して、本発明の好ましい実施形態である。なぜなら、それは蛋白質アミノ末端の正確な成熟化を提供するからである。さらに、内膜システムの特異的な領域への局在化は、もし、所定の蛋白質がさらなる標的化情報を含むように作られているか作られるならば、達成することができる(例えば、Vossら,Mol.Breeding 1:39−50(1995)、Duringら,Plant Mol.Biol.15:281−93(1990)、Baumら,Mol.Plant−Microbe Interact.9:382−87(1996)、DeWildeら,Plant Sci.114:231−41(1996)、Maら,Immunology 24:131−38(1994)、Schoutenら,Plant Mol.Biol.30:781−93(1996)、Firekら,Plant Mol.Biol.23:861−70(1993)、Artsaenkoら,Plant J.8:745−50(1995)、Conrad&Fiedler,Plant Mol.Biol.38:101−09(1998)参照)。
【0075】
プラスチド(例えば、クロロプラスト及びミトコンドリア)等の細胞小器官への標的化は所望のアミノ末端の成熟を達成するために利点を有している。なぜなら、これらの局所いずれかへの標的化は、続いて切断が起こるアミノ末端シグナル配列によって指揮されるからである。好ましい実施形態では、シグナルペプチドは発現産物をプラスチド(例えば、クロロプラスト)又は他の細胞小器官へと向ける。ある例は、アルファルファのリブロースビスリン酸カルボキシラーゼ小サブユニットの通過ペプチドである(Khoudiら,Gene 197:343−5(1997))。ペルオキシソーム標的化配列は、植物のC−末端標的化トリペプチドSKL等の蛋白質をペルオキシソームへ標的化できる、N−末端、内部又はC−末端のいかなるペプチド配列にもあてはまる(Banjokoら,Plant Physiol.107:1201−08(1995))。
【0076】
さらに、又は特異的な細胞内局所に蛋白質を向かわせるためのものに代えて、一態様では、「エピトープタグ」及び/又は部位特異的な切断部位が融合蛋白質を作成するために付加される。このタグの有用性は、それらが簡便な精製メカニズムを提供できることにある。例えば、ストレプトアビジンに結合するためにビオチン由来の重要なアミノ酸配列を含む小さなペプチドが、所定の遺伝子の5’末端に付けられる。そして新たに合成された蛋白質は、基本的にビオチン:ストレプトアビジン結合に基づく多くの知られた方法によって捕捉することができる。「ビオチン様」ペプチドが蛋白質から除去することが望まれるならば、プロテアーゼ認識部位も含むことが可能である。プロテアーゼ認識部位は「エピトープタグ」配列の下流で、所望の蛋白質の成熟体をコードする配列の直前に挿入することができる。当業者は、エピトープタグ及びプロテアーゼ(Xa因子、タバコEtchウイルスプロテアーゼ、エンテロキナーゼ等)について多くの選択肢があり、好ましい部位やプロテアーゼの選択は問題となっている特異的な蛋白質のアミノ酸及びDNA配列に依存するかもしれないことを理解するであろう。
【0077】
上記のように、プロモーター、エンハンサー、IRES要素及びシグナル配列等の調節要素の選択は、一般に、発現させる蛋白質の種類に依存するであろう。例えば、一態様では、IgGを作成する目的で好まれるいくつかの構築物は、5’OCS3MASプロモーター、(いずれかの生物の)ズブチラーゼ(subtilase)シグナルペプチド、IgG重鎖遺伝子の成熟部分に対するコード領域、翻訳終結シグナル、転写終結及びポリアデニル酸配列を有する構築物と共に、上記のものに類似の要素を含み、重鎖遺伝子が軽鎖遺伝子と置換された第2の構築物を含むだろう(すなわち、本明細書中で“binary”又は“dual”ベクターとして言及される2つのベクター)。あるいは、別の好ましい実施形態では、重鎖及び軽鎖遺伝子は同じDNA構築物上に存在する。さらに別の実施形態では、重鎖及び軽鎖遺伝子は、IRES要素によって分離されているが同じDNA構築物上の同じプロモーターから発現される。
【0078】
他の配列
発現構成体は発現ベクターの一部であってもよく、細菌の複製開始点(アグロバクテリウム及び/又はE.coliの複製開始点)、アグロバクテリウム及び/又は植物細胞等の細菌中で機能的であるレポーター遺伝子(例えば、GUS、GFP、EGFP、BFP、β−ガラクトシダーゼ及びそれらの修飾体)及び選択可能なマーカー遺伝子(例えば、抗生物質耐性遺伝子等)等のさらなる所望の配列を包含することができる。このために、本発明の方法で使用される外来DNAは、マーカー遺伝子を包含していてもよく、そのマーカー遺伝子の発現により、クローニングの開始段階で、非形質転換細胞から形質転換細胞の分離が可能となる。このようなマーカー遺伝子は、一般に、形質転換細胞を非形質転換細胞から表現型により区別することを可能とする蛋白質をコードしている。植物体に於いては、選択可能なマーカー遺伝子は、このように、ホスフィノトリシン等のグルタミンシンセターゼ阻害剤を含む除草剤等の除草剤に抵抗性を与える蛋白質をコードしていてもよい(例えば、欧州特許第0242236号、欧州特許第0242246号、De Blockら,1987,EMBO J.6:2513−2518)。しかしながら、マーカー遺伝子が、発現構成体/ベクターが導入された植物体組織から異種蛋白質を分離するのに必要とされないことは、本発明に従った一過性の蛋白質生産方法の利点である。
【0079】
異種蛋白質をコードする配列に融合できる、さらなる配列は以下のものを含むがそれらに限定されない。(例えば、Martinら,EMBO J.13(22):5303−5309(1994)に記載されるような)高次コイル配列、minibody配列、すなわち最低限の抗体相補性領域からなる配列(例えば、Bianchiら,J.Mol.Biol.236(2):649−59(1994)参照)、安定化配列、二量体化配列、リンカー配列、ミリスチル化配列(例えば、米国特許公開第2002/0146710号に記載される)、(例えば、免疫付着因子を産生するための)Fc領域等。
【0080】
発現構成体のライブラリー
一態様では、複数の発現構成体が、本発明に従った一過性の蛋白質発現システムにおいて変異体蛋白質配列の発現及び試験をするために、実質的に同一のコード配列(例えば、約50%以上同一、約75%以上同一、約90、95%又は99%以上同一)を含んで作成される。
【0081】
一態様では、構築物のコード領域は無作為化されている。構築物は、十分に無作為化されることもできるし、無作為化に偏りがあるようにすることもできる(すなわち、無作為か又は1又はそれ以上の選択された場所である)。一態様では、複数の発現構成体のうち1の発現構成体によってコードされる少なくとも1の蛋白質が所望の生物学的活性(例えば、抗原等の特定の結合パートナーに結合する能力等)を有するように、十分に多様な個体群を含む、発現構成体のライブラリーが作成される。別の態様では、発現構成体の数は、約100以上、約500以上、約1x103以上、約1x104以上、約1x105以上、約1x106以上、約1x107以上、約1x108以上又は約1x109以上の変異体コード配列を包含する。好ましくは、ライブラリーの多様性は、約1x107−1x109の異なる変異体コード配列を包含するような多様性である。蛋白質の1又はそれ以上のアミノ酸は一度に無作為化されてもよい。一態様では、約1、約2、約3、約4、約5又は約6、又はそれ以上のアミノ酸が一度に無作為化される。一態様では、“n”アミノ酸を含む蛋白質に対して、nアミノ酸のそれぞれ1つずつが独立して無作為化され、蛋白質は活性について試験される。他の領域が不変である一方、異種蛋白質の1又は複数の特定領域が多様化するかもしれないように(1の抗原結合部位又は特定の1アミノ酸等)、無作為化は偏りがあってもよい。
【0082】
蛋白質の方向付けられた進化のために核酸配列を突然変異させる方法は、例えば以下に記載されている。Leungら,Technique 1:11−15(1989)、Cadwell and Joyce,PCR Methods Appl.2:28−33(1992)、Shafikhaniら,Biotechniques 23:304−310(1997)、Wanら,Proc.Natl.Acad.Sci.U.S.A.95:12825−12831(1998)、You and Arnold,Protein Eng.9:77−83(1996)、Cherryら,Nat.Biotechnol.17:379−384(1999)、Tukeyら,J Immunol Methods 270(2):247−57(2002)、Choら,Mol.Biol.297(2):309−19(2000)。
【0083】
植物体組織サンプルは、以下に詳述するように、複数の発現構成体で浸潤することができ、所望の種類及び/又はレベルの生物学的活性を有する蛋白質を発現する組織/細胞を高い処理量で選択することができる。発現構成体は、所望の種類/レベルの活性を示すサンプル中の1又はそれ以上の細胞から回収することができ、活性の種類/レベルに関連する変異体配列を同定するために配列決定又は他の方法で特徴付けすることができる。構築物は、構築物の配列が決定される前又は後に、結果を確認するために、1又はそれ以上の植物体組織サンプルに再び導入することができる。偏りのある無作為化の2回目は、蛋白質の改変されなかった部位及び/又は増強された特性で蛋白質を同定するために(例えば、特定の結合パートナーに対する増強された結合親和性を有する蛋白質)選択された領域を変化させるのに使用されるかもしれない。
【0084】
一態様では、本発明の方法は、抗体の生成に関与している自然の選択過程をまねるために使用される。すなわち、特定の抗原に結合する発現構成体を同定し、構築物を突然変異させ、
特定の抗原に対する最も高い親和性を提供する構築物を同定するために、第2の選択工程を行う。
【0085】
アグロバクテリウムによる形質転換及び培養物の調製
アグロバクテリウムによる植物の形質転換及び安定なトランスジェニック植物の作成におけるその使用は、十分に証明されてきた。植物細胞とT−DNA境界配列を含むアグロバクテリウム細胞の相互作用の結果、植物核へ蛋白質と複合体を形成したアグロバクテリウム T−DNAの1本鎖コピーが移入する。安定な形質転換のために、T−DNAは核のDNAへ融合する。
【0086】
その過程は明らかに非常に効果的であるが、T−DNAの融合されなかったコピーは、一過性に転写されることができ、T−DNA遺伝子及び同時に移入された他のいかなる遺伝子も短期間は発現されることになる。一過性発現は、DNAの融合又は植物の再生に依存しないので、使用されるかもしれないが、無毒化したベクター(すなわち、もはや腫瘤を産生する遺伝子を含まないベクター)を使用する必要なしにより、毒性のあるアグロバクテリウム株を使用することが可能である。
【0087】
適した無毒化ベクターは、左境界及び左内側ホモロジー(LIH)配列は部分はそのままである一方、T−DNAの右境界は、サイトカイニン及びオーキシンをコードする植物ホルモン遺伝子と共に取り除かれ、細菌のカナマイシン耐性遺伝子によって置換されているSEVシリーズ、及び、植物ホルモン遺伝子が切除されpBR322ベクター配列の一部によって置換されており、左及び右境界配列は、Tiプラスミドのノパリンシンターゼ遺伝子と同様に、保存されているpGVシリーズを包含する。中間のベクターがヘルパー配列で結合されて使用されるかもしれない。いくつかの好ましい態様では、binaryベクターが当業界で公知であり、例えば、米国特許第4,940,838号、欧州特許公報第120516号、米国特許第5,464,763号に記載されている。
【0088】
適するアグロバクテリウム株は、野生型株(例えば、Agrobacterium tumefaciens)、又は、例えば、vir遺伝子の発現及び/又はその誘導が、突然変異又はキメラのvirA又はvirG遺伝子の存在によって改変されているアグロバクテリウム株等の1又はそれ以上の遺伝子が形質転換効率を高めるために突然変異されている株を包含する(例えば、Chen and Winans,1991,J.Bacteriol.173:1139−1144及びScheeren−Grootら,1994,J.Bacteriol.176:6418−6246)。別の実施形態では、例えば、米国特許第6,483,013号に記載されるような、pTiBo542由来のスーパーvirG遺伝子等の余分なvirG遺伝子コピーを含むアグロバクテリウム株が、好ましくは、複数コピーのプラスミドと結合された。
【0089】
他の適する株は、以下のものを含むがそれらに限定されない。A.tumefaciens C58C1(Van Larebekeら,Nature 252:169−170(1974))、A136(Watsonら,J.Bacteriol 123:255−264(1975))、LBA4011(Klapwijkら,J.Bacteriol 141:128−136(1980))、LBA4404(Hoekemaら,Nature 303:179−180(1983))、EHA101(Hoodら,J.Bac.168:1291−1301(1986))、EHA105(Hoodら,Trans Res.2:208−218(1993))、AGL1(Lazoら,Bio/Technology 9:963−967(1991))、A281(Hoodら,上記(1986))。
【0090】
一態様では、本発明は、アグロバクテリウムを処理する単純化された方法を提供する。好ましくは、アグロバクテリウム株は適する培養培地中で600nmでの光学密度(O.D.)2.5−3.5となるように培養される。細胞は一般にO.D.が2.5となるように希釈される。アグロバクテリウム細胞は、植物体組織の体積当たりおよそ1−3体積の培養培地中のアグロバクテリウム懸濁液を用いて、直接(すなわち、初期の濃縮工程又は遠心分離工程なしに)、植物体組織と接触される。ある実施形態では、植物材料1kg当たり約4Lより少ない量の細菌培養液が、少なくとも約250μg、少なくとも約500μg、少なくとも約750μg、少なくとも約1mg、少なくとも約2mg、少なくとも約3mg、少なくとも約4mg、少なくとも約5mg、少なくとも約10mg、少なくとも約15mg、少なくとも約25mg、少なくとも約50mg、少なくとも約75mg、少なくとも約100mg、少なくとも約150mg、少なくとも約200mg又は少なくとも約500mgの異種蛋白質を提供する。
【0091】
本発明の方法は、より少ない工程で実施することができるので、使用される他の方法と比較して、より少ない人的資源と15−20倍減少した生産コストを要する。
【0092】
減圧浸潤
ある特に好ましい実施形態では、界面活性剤が、植物体組織から異種蛋白質の収量を高めるために、アグロバクテリウム懸濁液に添加される。一態様では、アグロバクテリウム細胞又はその一部は、発現構成体/発現ベクターを発現するために、宿主の植物体組織に浸潤される。好ましくは、この工程は減圧状態で実施される。
【0093】
本明細書で使用される時、「界面活性剤」なる用語は、一般に疎水性部分及び親水性部分からなる表面活性化剤をいう(例えば、Bhairi,A Guide to the Properties and Uses of Detergents in Biological Systems,Calbiochem−Novabiochem Corp.1997参照)。界面活性剤は、それらが1又はそれ以上の荷電した官能基を含むか否かによって、陰イオン性、非イオン性、両性イオン又は陽イオン性と分類されるかもしれない。陰イオン性界面活性剤は、陰性に荷電した官能基を含み、正味の陰性電荷を有する。非イオン性界面活性剤は、荷電していない極性官能基を含み、電荷を有さない。これらの界面活性剤は、一般に、アルキレン酸化物と、アルキルフェノール、第1級又は第2級アルコールとの反応生成物、又は、アミン酸化物、ホスフィン酸化物又はジアルキルスルホキシドである。
【0094】
例となる非イオン性界面活性剤は以下のものを含むが、それらに限定されない。トクチルフェノキシポリエトキシエタノール(TritonX−100)、ポリオキシエチレンソルビタンモノラウリン酸(Tween 20)、ポリオキシエチレンソルビタンモノラウリン酸(Tween 21)、ポリオキシエチレンソルビタンモノパルミチン酸(Tween 40)、ポリオキシエチレンソルビタンモノステアリン酸(Tween 60)、ポリオキシエチレンソルビタンモノオレイン酸(Tween 80)、ポリオキシエチレンソルビタンモノトリオレイン酸(Tween 85)、オクチルフェノキシポリエトキシエタノール(IGEPAL CA−630/NP−40)、トリエチレングリコールモノラウリルエーテル(Brij 30)及びソルビタンモノラウリン酸(Span 20)。
【0095】
両性イオン界面活性剤は、陽性に荷電した官能基と陰性に荷電した官能基の両者を含み、正味の電荷は有さない。適する両性イオン界面活性剤は以下のものを含むがそれらに限定されない。カルボキシベタイン、スルホベタイン(スルタインとしても知られる)、アミドベタイン及びスルホアミドベタイン等のベタイン、これらはC8−C18、好ましくは、C10−C18のアルキルベタイン、スルホベタイン、アミドベタイン及びスルホアミドベタインを含んでいてもよく、例えば、ラウリルアミドプロピルベタイン(LAB)型ベタイン、n−アルキルジメチルアンモニオメタンカルボキシレート(DAMC)、n−アルキルジメチルアンモニオエタンカルボキシレート(DAEC)及びn−アルキルジメチルアンモニオプロパンカルボキシレート(DAPC)、n−アルキルスルタイン、n−アルキルジメチルアンモニオアルキルスルホン酸、N−アルキルジメチルアンモニオエタンスルホン酸(DAES)、n−アルキルジメチルアンモニオプロパンスルホン酸(DAPS)及びn−アルキルジメチルアンモニオブタンスルホン酸(DABS)、ヘキサデシルジメチルアンモニオプロパンスルホン酸、n−アルキルアミドメタンジメチルアンモニオメタンカルボン酸、n−アルキルアミドメタンジメチルアンモニオエタンカルボン酸、ラウリルアミドプロピルベタイン(LAB)、n−アルキルアミドメタンジメチルアンモニオメタンスルホン酸、n−アルキルアミドエタンジメチルアンモニオエタンスルホン酸及びn−アルキルアミドプロパンジメチルアンモニオプロパンスルホン酸、3−[(3−コールアミドプロピル)ジメチルアンモニオ]−1−プロパンスルホン酸(CHAPS)及び3−[(3−コールアミドプロピル)ジメチルアンモニオ]−2−ヒドロキシ−1−プロパンスルホン酸(CHAPSO)、リン脂質(例えば、ホスファチジルエタノールアミン、ホスファチジルグリセロール、ホスファチジルイノシトール、ジアシルホスファチジルコリン、ジ−O−アルキルホスファチジルコリン、リゾホスファチジルコリン、リゾホスファチジルエタノールアミン、リゾホスファチジルグリセロール、リゾホスファチジルイノシトール)、飽和及び不飽和脂肪酸誘導体(例えば、エチルエステル、プロピルエステル、コレステロールエステル、コエンザイムAエステル、ニトロフェニルエステル、ナフチルエステル、モノグリセリド、ジグリセリド及びトリグリセリド、脂肪族アルコール、脂肪族アルコール酢酸塩等)、リポ多糖、糖脂質、スフィンゴ脂質(例えば、セラミド、セレブロシド、ガラクトシルジグリセリド、ガングリオシド、ラクトセレブロシド、リゾスルファチド等)。
【0096】
「陽イオン性界面活性剤」は浸潤の条件下で陽性に荷電した官能基を有する。適する陽イオン性界面活性剤は、第4級又は第3級アミンを含むがそれらに限定されない。例となる第4級アミン界面活性剤は、以下のものを含むがそれらに限定されない。塩化セチルピリジニウム、臭化セチルトリメチルアンモニウム(CTAB;Calbiochem#B22633又はAldrich#85582−0)、塩化セチルトリメチルアンモニウム(CTACl;Aldrich#29273−7)、臭化ドデシルトリメチルアンモニウム(DTAB;Sigma#D−8638)、塩化ドデシルトリメチルアンモニウム(DTACl)、臭化オクチルトリメチルアンモニウム、臭化テトラデシルトリメチルアンモニウム(TTAB)、塩化テトラデシルトリメチルアンモニウム(TTACl)、臭化ドデシルエチルジメチルアンモニウム(DEDTAB)、臭化デシルトリメチルアンモニウム(DlOTAB)、臭化ドデシルトリフェニルホスホニウム(DTPB)、臭化オクタデシルイルトリメチルアンモニウム、塩化ステアロアルコニウム、塩化オレアルコニウム、塩化セトリモニウム、アルキルトリメチルアンモニウムメトサルフェート、パルミトアミドプロピルトリメチルクロリド、quaternium84(Mackernium NLE;Mcintyre Group,Ltd.)及び小麦脂質エポキシド(Mackernium WLE;Mcintyre Group,Ltd.)。例となる第3級アミン界面活性剤は、以下のものを含むがそれらに限定されない。オクチルジメチルアミン、デシルジメチルアミン、ドデシルジメチルアミン、テトラデシルジメチルアミン、ヘキサデシルジメチルアミン、オクチルデシルジメチルアミン、オクチルデシルメチルアミン、ジデシルメチルアミン、ドデシルメチルアミン、塩化トリアセチルアンモニウム、塩化セトリモニウム及び塩化アルキルジメチルベンジルアンモニウム。陽イオン界面活性剤のさらなる種類は、ホスホニウム、イミダゾリン及びエチル化アミン基を含むが、それらに限定されない。
【0097】
陰イオン性界面活性剤は、一般に、有機硫酸エステル及びスルホン酸エステルの水溶解アルカリ金属塩である。これらは、以下のものを含むがそれらに限定されない。ラウリン酸カリウム、硫酸ラウリルナトリウム、硫酸ドデシルナトリウム、硫酸アルキルポリオキシエチレン、アルギン酸ナトリウム、スルホコハク酸ジオクチルナトリウム、ホスファチジルコリン、ホスファチジルグリセロール、ホスファチジルイノシン、ホスファチジルセリン、ホスファチジン酸及びそれらの塩、グリセリルエステル、カルボキシルメチルセルロースナトリウム、コール酸及び他の胆汁酸(例えば、コール酸、デオキシコール酸、グリココール酸、タウロコール酸、グリコデオキシコール酸)及びそれらの塩(例えば、デオキシコール酸ナトリウム等)。
【0098】
エタノール、1−プロパノール及び1−ブタノール等の短鎖アルコールのような共力剤(co−surfactant)がさらに使用されてもよい。
上記界面活性剤のいずれかを組み合わせて使用してもよい。特に上記に列挙されていない界面活性剤も本発明の範囲に包含される。
【0099】
使用される界面活性剤の量は、界面活性剤及び処理される植物体組織(すなわち、葉の表面を覆っているワックスの厚さ等)の種類によって様々であろう。一般に、界面活性剤は、アグロバクテリウム懸濁液の体積の0.005%から約1%の範囲の濃度で使用される。好ましくは0.005%から約0.5%より好ましくは約0.005%から約0.05%の範囲の濃度である。一般に、イオン性界面活性剤は非イオン性界面活性剤より低い濃度で使用されるだろう。
ある好ましい態様では、Tween20等の非イオン性界面活性剤が使用される。
【0100】
界面活性剤を混合することに加えて、蛋白質発現を増加させるために、減圧衝撃を増大させるための浸透圧による衝撃工程を追加することができる。それゆえに、一態様では、ショ糖等の浸透圧衝撃剤が、蛋白質発現を増加させるために使用される。浸透圧衝撃剤の適する濃度は、20g/Lから100g/Lの範囲である。植物体組織がレタスである、一態様では、ショ糖が約60g/L使用される。
【0101】
例となる方法では、組み換えアグロバクテリウム培養物(すなわち、本発明に従った発現構成体を含んでいる)を、宿主及びベクター上に見出される耐性によって選択するために、適当な抗生物質を補充した修飾YEB培地(酵母抽出物6g/L、ペプトン5g/L、硫酸マグネシウム2mM及びショ糖5g/L)でおよそ2日間、生長させる。一過性発現に向けて、細胞を生長させるために、開始時のアグロバクテリウム培養物を、新たな修飾YEB培地で1:50に希釈する。抗生物質、50mMのリン酸カリウム緩衝液(pH5.8)及び20μMのアセトシリンゴンを添加する。28℃で18−24時間培養後、細胞は600nmでの吸光度(光学密度又はO.D.ともいわれる)2.5−3.5に達する。細胞を、好ましくは、もし必要であれば同じ培地を用いて、600nmでの吸光度2.5まで希釈する。
【0102】
細胞に、その後、ショ糖及びアセトシリンゴンを、最大で220μMのアセトシリンゴンと60g/Lのショ糖となるように補充する。懸濁液を、22℃で約1時間培養し、その後、浸潤に使用する。
【0103】
細胞を、再懸濁工程の必要がないように、いかなる遠心分離又は濃縮工程もなしに、直接、減圧条件下で浸潤する。この修正は、対数期にある培養物から細胞を遠心分離し、それらをムラシゲ・スクーグ培地(MS培地)に再懸濁する必要(Kapilaら,上記(1997))をなくし、新たに成長したアグロバクテリウム懸濁液で直接減圧浸潤することを可能にする。O.D.600nmが0.7−0.8(Kapilaら,1997)であるのに代えて、O.D.600nmが2.5−3.5までアグロバクテリウム細胞を成長させること、修飾YEB培地及びMESに代えてリン酸カリウム緩衝液を使用することは、発現レベル又は浸潤効率を変化させなかったが、処理の中でこの部分に要求されるコストと努力を顕著に減少させた。
【0104】
例えば、植物体組織がレタス由来であるような、一態様では、植物材料は、ビーカー中で、100μg/mlの2,4−D、0.005%のTween20と共に、あらかじめ培養されたアグロバクテリウム懸濁液に浸される。ビーカーは減圧乾燥器中に置かれ、減圧(29インチの水柱又は約7kPaに等しい)が20分間適用され、減圧の急速な解除による減圧衝撃が続く。レタス葉球全体の使用は、以前に記載されるような(Kapilaら.,上記(1997)、Vaqueroら,上記(1999))組織化されていない葉を集めたものと比べて、著しく簡便である。一般に、レタスの葉球全体は、葉の生物量に比較してより多くの発現レベルを与えると思われる。本発明の方法は、多数の葉からなるレタス葉球すなわちより多量の葉を一斉に処理することによって、容易に規模を大きくすることができる。同じアグロバクテリウム細胞懸濁液が、発現効率を顕著に減少させることなく、生産コストと労力をより減少させる減圧浸潤に、少なくとも2度使用することができる。
【0105】
一態様では、レタスの葉球全体、およそ300−400gが使用される。別の態様では、分離された葉が、界面活性剤及び/又は浸透圧剤の量及び組合せを最適化するために、予備的に使用される。
【0106】
好ましくは、減圧浸潤に続いてレタスの葉球が室温で約3−7日間光を受けて培養され、蛋白質抽出のためにホモジナイズされる。モノクローナル抗体及び他の製薬に重要な蛋白質が植物ホモジネートから適当な精製手順を用いて精製される。
【0107】
他の従来技術の方法と比べて、本過程は単純であり、より少ない工程からなり、異種蛋白質の発現が劇的に増加するようになる。
【0108】
蛋白質の分離
採収した後、蛋白質の分離が当業界で日常的に使用されている方法を用いて実施されるかもしれない。例えば、生物量の少なくとも一部はホモジナイズされ、組換え蛋白質が抽出され、さらに精製されるかもしれない。抽出は、適する溶媒にホモジネートを浸すか又は沈めることを包含するかもしれない。蛋白質は、例えば、米国特許第6,284,875号に記載されるような、減圧浸潤方法によって、植物の組織液からも分離されるかもしれない。
【0109】
精製方法は、免疫親和性による精製、蛋白質又は蛋白質複合体の特異的な大きさ、電気泳動移動度、生物学的な活性、及び/又は分離される異種蛋白質の正味の電荷に基づく又は蛋白質中のタグとなる分子の存在に基づく精製手順を含むがそれらに限定されない。
【0110】
分離された蛋白質の特徴は、免疫アッセイによって又は当業界で公知の他の方法によって導くことができる。例えば、蛋白質が実質的に分離されると、ウエスタンブロッティングによって又はクーマシーブルー染色によってSDS−PAGEゲル上で蛋白質が解析できる。分離された蛋白質は、生物学的な活性をアッセイするため、蛋白質の構造を(例えば、結晶化アッセイで)特徴付けるため、疾患の非動物ヒトモデルで有効性試験を行うため、最適な蛋白質活性及び/又は最適な製薬特性についてスクリーニングするため等に使用できる。
【0111】
本明細書で引用した全ての特許及び非特許文献は、本発明の属する分野における当業者の技術レベルを示すものである。全てのこれら刊行物及び特許出願は、個々の刊行物又は特許出願が、明確にそれぞれ、本明細書に参照として取り込まれるものとして示されているように、参照として本明細書に取り込まれる。
(実施例)
本発明は、実用的な実施例の方法によってしめされるが、これらの実施例は説明を目的としたものであり、本発明をなんら限定するものではない。
【実施例1】
【0112】
新規で迅速な蛋白質発現システムによるhOATの生産
本明細書に記載される方法は、上記のKapilaら、(1997)の教示による一過性発現システムを著しく改変するものであり、結果として劇的に改良された、低コストで、またより少ない工程で高品質な産物を発現させるものである。
【0113】
【表1】

【0114】
高ショ糖存在下での前処理
植物発現ベクターの構築
所定の遺伝子を含むプラスミドベクターを標準的な分子生物学的手法を用いて構築した。基礎となる要素は、E.coli及びアグロバクテリウムの両者において複製を可能とする開始プラスミド、プロモーターによって制御される所定の遺伝子の側面に位置する右及び左T−DNA境界及び標的化配列を含む。図1A及び1Bに示されるプラスミドを作製にあたって、必要な要素を集めて組みあわせる。
【0115】
図1Aは、2,4−Dにて誘導可能なプロモーター(OCS)3Masプロモーター(Gelvinら,米国特許第5,955,646号)を含み、抗組織因子抗体(IgG1)の重鎖に翻訳上融合されたズブチリシン(subtilisin)分泌配列(Janzikら,Biol.Chem.275:5193−5199(2000))を制御する、プラスミドpSUNP1を示す。さらに、プラスミドはカナマイシン耐性に対する選択可能なマーカー及び本一過性発現システムには有用性のないbialphos耐性に対するBAR遺伝子を含む。
【0116】
図1Bに示される、プラスミドpSUNP2は、抗組織因子抗体の重鎖が同じ抗体の軽鎖(カッパー)によって置換された点を除いて、pSUNP1に類似している。
【0117】
形質転換及びアグロバクテリウムの調製
所望のバイナリーベクター作製用のAgrobacterium tumefaciens C58/C1培養物を、プラスミド及び適当な細菌を選択するために、適当な抗生物質(100μg/mLのカナマイシン、15μg/mLのリファンピシン、25μg/mLのゲンタマイシン)を含む、修飾YEB培地(6g/Lの酵母抽出物、5g/Lのペプトン、5g/Lのショ糖、2mMの硫酸マグネシウム)で28℃で2日間成長させた。この培養物を、抗生物質、50mMのリン酸カリウム緩衝液pH5.6、20μMのアセトシリンゴンを補充した修飾YEB培地で1:50に希釈し、およそ18−24時間、O.D.600nmがおよそ2.5−3.5になるまで培養した。必要に応じて、(220μMのアセトシリンゴン及び60g/Lのショ糖という最大量を与えるために)細菌細胞をO.D.600nmが2.4になるよう希釈し、55g/Lのショ糖及び200μMのアセトシリンゴンを補充した。懸濁液を室温(約22℃)にて1時間インキュベートし、100μgの2,4−D(2,4−ジクロロフェノキシ酢酸、Sigma)及び0.005%Tween20を使用前に添加した。
【0118】
比較のためのKapilaら,上記(1997)によって記述される方法では、YEB培地は、プラスミドを選択するための適当な抗生物質(100μg/mLのカナマイシン、15μg/mLのリファンピシン、25μg/mLのゲンタマイシン)と共に、5g/Lの牛肉抽出物、1g/Lの酵母抽出物、5g/Lのペプトン、5g/Lのショ糖、2mMの硫酸マグネシウムから成る。開始時の培養物を、抗生物質、10mMのMESpH5.6、20μMのアセトシリンゴンを補充したYEB培地で1:500に希釈し、およそ18−24時間、O.D.600nmがおよそ0.7−1になるまで培養した。培養物を細胞を回収するために遠心分離し、10mMのMESpH5.6、20g/Lのショ糖及び200μMのアセトシリンゴンを含むムラシゲ・スクーグ培地(MS培地)(Kapilaら,上記(1997))にO.D.600nmが2.4になるよう再懸濁した。懸濁液を室温(約22℃)にて1時間インキュベートし、100μgの2,4−Dを添加した。
【0119】
減圧浸潤及び処理されたレタスのインキュベーション
アグロバクテリウム懸濁液を減圧浸潤に直接使用した。軽鎖が1つのプラスミド上にあり重鎖が2つめのプラスミド上にある場合(デュアルベクターシステム(dual vector system))、2つのアグロバクテリウム培養物を調製し、浸潤前に等量ずつ混合しなければならない。この例では、カッパー軽鎖をコードする軽鎖ベクターはhOATと呼ばれる抗組織因子抗体に由来し、重鎖ベクターはhOATのIgG1重鎖をコードした。レタスの葉球全体を2Lビーカー中の1.2Lの懸濁液に浸し、減圧乾燥器中に置いた。減圧(7kPaに等しい)を20分間適用した後、減圧の急激な解除による減圧衝撃を続けた。レタスの葉を水で洗浄し、閉じた透明な箱内の湿ったペーパータオル上で室温にて16時間日光をあてながら3−4日間インキュベートした。3−4日間後、葉を基部から切り、蛋白質抽出のためにホモジナイズした。
【0120】
蛋白質の抽出及び精製
蛋白質を、葉の重量と等しい体積の緩衝液を用いて、緩衝液(100mMのTris−HCl、5mMのEDTA、pH8.0、使用前に1.5%不溶性PVPを添加)中で抽出した。葉をワーリングブレンダー中で高速で1分間ホモジナイズし、得られたホモジネートを10,000xgで15分間遠心分離した。沈降しなかった細胞破片を有する上清を12層の薄地の綿布で濾過し、20,000xgで15分間遠心分離した。濾液をrProteinA sepharose fast flow affinity column(5mL、樹脂はAmersham Pharmacia Biotech AB、Swedenより)に流速2mL/minでかけた。使用した洗浄緩衝液、溶出緩衝液はそれぞれ0.1Mの酢酸ナトリウム及び0.1Mの酢酸であった。蛋白質を、段階的な勾配pH法、すなわち2カラム容積に対して20%の0.1M酢酸、4カラム容積に対して40%、60%及び100%の0.1M酢酸を続けることによって、溶出した(図3)。ピーク画分を集め、1MのTris−HCl、pH8.0を用いてpHを8.0に調整した。精製した抗体を280nmのO.D.を用いて定量した。さらなる精製のために、Q Sepharose fast flow column(5mL、樹脂はAmersham Pharmacia Biotech AB、Swedenより)を20mMTris−HClpH8.5で平衡化し、同じ緩衝液でプロテインA精製した抗体の緩衝液を交換し、カラムにかけた。抗体を、塩段階的溶出法、すなわち2カラム容積に対して20mMのTris−HCl、pH8.0、1MのNaClの10%緩衝液、4カラム容積に対して50%緩衝液、2カラム容積に対して100%緩衝液を続けることによって、溶出した。ピーク画分を集め、280nmのO.D.を用いて定量した(図4)。緩衝液交換のために、Millipore Ultrafree Centrifugal filter device 15mL(Millipore Corporation、Bedford、MA)を少なくとも1時間0.1MのNaOHに浸した。Qセファロースカラムで精製した抗体の緩衝液の交換はPBSで行い、1000倍希釈以上を得た。緩衝液を交換した抗体をMillex−GV 0.22μm filter unit(Millipore Corporation、Bedford、MA)で濾過し、280nmのO.D.を用いて定量した。
【0121】
hOAT抗体をELISAアッセイを用いて定量した。1枚のプレートを調製するために、コーティング緩衝液(35mMのNaHCO3、15mMの炭酸水素ナトリウム、50mMのNaCl、pH9.0)11mL中に5.5μgの組換え組織因子(rTF)を溶解したコーティング溶液を調製する。100μlのコーティング溶液をそれぞれのウェルに移し、覆った状態で4℃で2週間まで貯蔵する。プレートを洗浄緩衝液(Kirkegaard&Perry)400μLで3回洗浄し、100μLのサンプルをそれぞれのウェルに移す。プレートを振動しながら室温にて1時間インキュベートし、その後、洗浄緩衝液で6回洗浄する。結合した抗体を、ペルオキシダーゼ結合ロバ抗ヒトIgG(H+L)(Jackson ImmunoResearch)を用いて室温にて10分間プレートをインキュベートし、洗浄緩衝液で6回洗浄することにより検出する。ABTS基質(BioFX)を(100μL)添加し、10分後反応を1%のSDS100μL添加することにより停止する。405nmにおける吸光度を測定する。本発明の実施例1の実施形態及びKapilaの方法、それぞれからの発現レベルの比較を表2に示す。
【0122】
【表2】
【0123】
SDS−PAGEを12%のトリスグリシンミニゲルでLaemmli(1970)によって記述されるように実施した。サンプルを、蛋白質抽出物をローディング緩衝液と混合し(4:1、v/v)、続けて70℃で10分間加熱することで調製した。蛋白質のバンドをクーマシーブルー(図5A)染色により検出し、又は電気泳動的にPVDF膜に転写した。膜を10%のスキムミルクを含む2XPBSの1:10希釈液で室温にて1時間ブロッキングした。洗浄後、膜を西洋ワサビペルオキシダーゼ結合抗ヒトIgG抗体(Binding site)と室温にて1時間インキュベートした。図5Bは、製造者の指示(Pierce)に従って強化化学発光ウエスタンブロット検出試薬で膜をインキュベートすることにより検出されるIgGの重鎖及び軽鎖を示している。
【実施例2】
【0124】
ショ糖濃度の最適化
ショ糖の濃度とショ糖による浸透圧衝撃の効果は、発現を増加させるための重要なパラメーターであろうと認識された。蛋白質の生産レベルにおけるショ糖濃度の効果を試験するため、ショ糖の濃度範囲を抗組織因子抗体の発現におけるその効果について試験した点を除いて、実施例1の手順に従った。図6の結果に基づき、ショ糖の最終濃度60g/Lを標準的な方法として選択した。
【実施例3】
【0125】
界面活性剤の最適化
界面活性剤の効果を特徴付けるため、使用する界面活性剤の種類と濃度の両者を多様化している点を除いて、実施例1に記載される手順に従った。
【0126】
【表3】
【0127】
表3は、異なる非イオン性及びイオン性界面活性剤の調査結果を示す。表は網羅的になってはいないが、このパラメーターの多様性から顕著な効果を示している。0.005%でTween−20は異種蛋白質の特に高い発現レベルを提供する。
【実施例4】
【0128】
一過性発現システムでの(OCS)3MASプロモーターによる発現における2,4−Dの影響
多くの異なるプロモーター(5’転写調節領域)が入手可能であり、植物における異種蛋白質の発現に広く用いられている。文献の調査及び限られた試験を、本発明の方法に適したプロモーターを決定するために行った。表4は、化学的な誘導物質2,4−Dを用いた合成プロモーターOCS3MASからのhOATの発現を示す。(OCS)3MASプロモーターは様々な因子、とりわけ創傷によって誘導されることが知られていたが、この研究はアグロバクテリウムを用いた遺伝子の一過性の送達も結果的に発現を誘導するものである。この研究は、実施例1で示されるように細菌の調製及び浸潤に関しては元来のKapilaの方法(MS方法)を用いて行われた、よって、他の実施例で見られるより発現レベルは低いものであった。この研究に基づき、最終濃度100μg/mLの2,4−Dが、……
【0129】
【表4】
【0130】
この限られた調査は単なる例を意味しており、Adh遺伝子又はeIF4等の「ハウスキーピング」遺伝子の報告されている強力なプロモーター又はRUBISCOの小サブユニットからの発現と比べて、非常に高い発現が嫌気的に誘導可能なプロモーターから得られると言えるかもしれない。
【実施例5】
【0131】
hOATの一過性発現におけるデュアルベクター(Dual vector)対2シストロン性ベクター(Bicistronic vector)の比較
この実施例では、(実施例1に記載されるような)デュアルベクターシステムの使用を単一のベクターからの重鎖及び軽鎖の発現(2シストロン性システム)と比較した。プラスミドpSUNP4(図2)は同じプラスミドのT−DNA領域に重鎖及び軽鎖を有するプラスミドベクターを表す。(OCS)3MASプロモーターが、ズブチリシン(subtilisin)をコードするシグナル配列と共に抗組織因子遺伝子の重鎖及び軽鎖の両者の発現を制御するために使用される。
【0132】
レタスを、デュアルベクター作製用の2つのアグロバクテリウム培養物又は2シストロン性ベクターpSUNP4作製用の単一のアグロバクテリウム培養物のいずれかで浸潤し、hOATの発現レベルを実施例1のように測定した。これら2つのベクターシステムを用いたhOATの発現レベルを表す結果は、表5に示される。
【0133】
【表5】
【実施例6】
【0134】
志賀毒素2に対する組換え抗体の一過性発現
この実施例では、一過性発現が他の抗体を生産するのに使用できることを示した。cαStx2は、腸管出血性大腸菌によって産生される志賀毒素2に結合し中和するキメラ抗体である。cαStx2重鎖及び軽鎖の遺伝子を、pSUNP1及びpSUNP2に類似したcαStx2ベクターを作成するために、デュアルベクターに導入した。これらの構築物を、実施例1に記載の方法を用いて、アグロバクテリウムに移入し、レタスをagro−infiltrationするために使用した。一過性発現後、プロテインA精製により得た抗体と共に、植物細胞からの粗抽出物をELISAにより解析した。
【0135】
ELISAを、Stx2抗原でmaxisorp96ウェルプレートをコートすることで実施したが、コートしたプレートはプラスチックフィルムで覆い、使用まで4℃で貯蔵した。抗体生産を測定するために、ウェルをコート溶液を取り除くため使用前に緩衝液で3回洗浄した。植物細胞抽出液又は精製された蛋白質溶液をコートしたウェルに添加した。室温で1時間後、ウェルを緩衝液で3回洗浄し、抗ヒトカッパ鎖−HRP抗体の希釈液を添加した。その後、プレートを室温で1時間インキュベートし、緩衝液で3回洗浄した。プローブ抗体の存在を検出するために、ABTS基質剤を添加し、室温で数分間インキュベートし、続けてABTS停止緩衝液を加えた。405nmの吸光度を自動プレートリーダーで測定した。
【0136】
ELISA解析によれば、一過性に発現された蛋白質は機能的に活性なcαStx2抗体を含むことが示された。
【0137】
当業者は通常の実験法のみで、本明細書中に記載されている特定の物質及び手順に対して多くの均等物を認識し、又は確かめることができるものである。このような均等物は本発明の範囲とみなされ、上述の特許請求の範囲により保護されるものである。
【0138】
本明細書の発明は、特定の実施態様を参照として記載されているが、これらの実施態様は単に本発明の原理の説明及び応用に過ぎないと理解されるべきである。それゆえに、多くの改変が説明のための実施態様に対してなされていてもよく、添付の請求項によって定義される、本発明の精神及び範囲から逸脱することなくその他の改良が工夫されてもよいものと理解されるべきである。
【0139】
特許、特許出願、国際出願及び本明細書中及び上記の参考文献は、それらの全体が本明細書中に取り込まれる。
【図面の簡単な説明】
【0140】
本発明の目的と特徴は、前述の詳細な説明と添付の図面を参照することにより、より良く理解することができるものである。
【図1A】図1Aは、本発明の一態様に従って植物体組織の同時浸潤に使用されたpSUNP1プラスミドの図である。プラスミドpSUNP1はOCS3MASプロモーターからhOATの重鎖を発現する。TRとTLの間に存在する全ての遺伝子を植物体の核へと導くT−DNAの境界が示される。これらのプラスミドのいずれかを有するアグロバクテリウムはレタス等の植物を同時浸潤するのに使用され、hOATの一過性発現を示した。
【図1B】図1Bは、本発明の一態様に従って植物組織の同時浸潤に使用されたpSUNP2プラスミドの図である。プラスミドpSUNP1はOCS3MASプロモーターからhOATの重鎖を発現する。TRとTLの間に存在する全ての遺伝子を植物核へと導くT−DNAの境界が示される。これらのプラスミドのいずれかを有するアグロバクテリウムはレタス等の植物を同時浸潤するのに使用され、hOATの一過性発現を示した。
【図2】図2は、植物体組織での一過性発現によって単一プラスミドからhOATの重鎖及び軽鎖の両者を発現するために使用可能な、2シストロン性の発現プラスミドpSUNP4の図である。
【図3】図3は、hOAT重鎖及び軽鎖の遺伝子を含むアグロバクテリウムで浸潤されたレタスから抽出された蛋白質の溶出プロフィールを示す。蛋白質はプロテインAカラムにかけられ溶出された。
【図4】図4は、本発明の一態様に従って分離され、Qセファロースカラムにかけられた蛋白質の溶出プロフィールを示す。かけられた蛋白質は、図3で使用されたプロテインAカラムからの溶出によって回収されたものであった。
【図5A】図5Aは、還元条件下で泳動されクーマシーブルー染色されたSDS−PAGEゲル及び本発明の一態様に従って得られたhOAT蛋白質の画分を示す。レーン1、分子量(Mr)標準;レーン2、精製されたCHO生産hOAT;レーン3、レタスで発現されプロテインAカラムから溶出されたhOAT。
【図5B】図5Bは、本発明の一態様に従って同定されSDS−PAGEで分離された蛋白質のウエスタンブロットを示す。プローブは抗H+L抗体。レーンA、市販のIgG;レーンB、プロテインAカラムから溶出され精製されたhOAT;レーンC、陰性コントロール−agro−infiltrationなしのレタスからの抽出物。
【図6】図6は、レタスでのhOAT発現レベルにおける、浸透圧衝撃に関して、ショ糖の様々な濃度効果を示す。hOATの発現は、抽出に用いられたレタス材料のキログラム当たり生産される(ELISAに基づく)抗体mgとして表される。
【技術分野】
【0001】
(関連出願)
本出願は、2002年12月23日に出願された米国仮出願第60/436,403号の優先権を主張するものであり、その全体が参照により本明細書に取り込まれる。
(技術分野)
【0002】
本発明は、植物における高レベル一過性蛋白質産生についての方法及びキットに関する。
【背景技術】
【0003】
製薬に重要な組換え蛋白質の発現は、通常、微生物又は哺乳類の宿主を用いて実施される。微生物のシステムはクローニング及び形質転換細胞を産生する速さの点でしばしば利点を提供する。一般に、異種の遺伝子産物の収量を高くすることができるが、蓄積する産物はしばしば生物学的な活性がなく、活性のある物質を得るにはコストのかかる困難なリフォールディングが要求される。真核生物に較べて、多くの翻訳後修飾も細菌では異なっており、ある種の蛋白質は原核生物システムでは適切に発現させることができない。
【0004】
製薬に重要な蛋白質の発現に異種システムを使用することの主な問題点の一つは、遺伝子のクローニングから、適切な動物モデルで使用するのに十分な量の機能的な蛋白質の産生までに必要とされる時間である。ほとんどのシステムで、この時間は数ヶ月から一年又はそれ以上になる。時間が消費されることに加えて、(例えば、COS又はCHO細胞の一過性のトランスフェクションによる、又は、昆虫細胞培養のバキュロウイルス感染による等の)細胞培養物における組換え蛋白質の一過性産生は、通常、特殊な装置を必要とし、産生に必要とされる細胞やウイルスを取扱うには特別な技術が要求される。加えて、より高価で特殊な装置と訓練が要求されるスケールアップの過程が導入されない限り、得られる蛋白質は通常非常に少量である。(ナノグラムからマイクログラム量)。
【0005】
1980年代後半から、作物に於ける外来又は異種蛋白質のコストパーフォーマンスのよい発現の例が多数開示されている。特に、モノクローナル抗体や他の製薬に重要な蛋白質の産生のための発現システムとして、近年、植物が浮上してきた。植物は、蛋白質のグリコシル化に主な相違があるが、動物細胞の経路に非常に良く似た蛋白質合成経路を有している(Fischer and Emans,Transgenic Res.9:279−299(2000);Cabanes−Macheteau,et al.,Glycobiology 9:365−72(1999))。さらに、所定の蛋白質のなかには植物体内に高レベルで蓄積されるものがあることが示されてきた。さらに、植物由来の抗体はハイブリドーマによって産生される抗体と機能的に同等である(Fischer and Emans,上記,(2000))。最終的には、植物由来の抗体や他の蛋白質は、ヒト又は動物の病原体、すなわち、共精製された血液感染性の病原体及び他のシステムで精製された組換え蛋白質に付随する腫瘍形成性の配列を含まない(Fischer and Emans,上記,(2000)。
【0006】
組換え蛋白質はトランスジェニック植物体内で安定的に合成された遺伝子から産生できる。異種遺伝子から蛋白質を産生するその他の方法は、一過性発現システムを使用することである。遺伝子クローニングのアプローチ、プラスミドの構築、プロモーター等を開発するために、これに適当なシステムがいくつか用いられてきた。特に、プロトプラストのエレクトロポレーションは、微粒子銃及びある程度のウイルスベクターと同様に、広く使用されてきた。
【0007】
微粒子銃は通常少数の細胞にのみに達するものであるが、転写が成就するにはDNAは細胞の核に到達しなければならない(Christou,Plant Mol.Biol.35:197−203(1996);Fischer and Emans,上記(2000))。浸潤によって送り込まれるアグロバクテリウム(agro−infiltration)の使用は、非常に多くの細胞に外来遺伝子を送り込むことができる。さらに、所定の遺伝子を収容しているT−DNAは、いくつかの細菌蛋白質の助けにより活発に核へと移入する(Kapilaら,Plant Sci.122:101−108(1997);Fischer and Emans,上記(2000))。微粒子銃及びagro−infiltrationともに処理後3−5日で異種蛋白質の発現が見られる一方、ウイルスによる送達は2から4週間かかる。微粒子銃の使用は一過性の発現にはあまり効率的ではないが、トランスジェニック穀物の再生には非常に重要である(Christou,上記(1996);Fischer and Emans,上記(2000))。
【0008】
修飾されたウイルスベクターでの感染は、殆どの植物細胞中にウイルスを組織的に広めることになる。導入された遺伝子は、ウイルスRNAレプリカーゼによって細胞質で転写され所定の蛋白質に翻訳される。標的の遺伝子は、ウイルスの複製中に高レベルに増加するので組換えウイルスベクターにより高いレベルで発現される(Porta and Lomonossoff,Mol.Biotechnol.5:209−21(1996))。しかしながら、通常このシステムは60−70Kdより小さい分子量の蛋白質に制限される。
【0009】
一過性発現のためのagro−infiltrationは多くの利点を有する。その方法は数日で所定の蛋白質を産生し、安定性と機能という特徴において満足できる蛋白質を多量に生ずる(Fischer and Emans,上記(2000))。安定な形質転換植物体を必要とすることなく、組換え蛋白質を数十ミリグラム産生するために、agro−infiltrationをスケールアップできることが提案されてきた。しかしながら、報告された(Fischer and Emans,上記(2000))発現レベルでは、より多量(例えば10−100mgs)の蛋白質が必要とされるモデル動物を使ったインビボでのより大規模な研究には実用的ではない。
【0010】
【非特許文献1】Artsaenkoら,Plant J.8:745−50(1995)
【非特許文献2】Banjokoら,Plant Physiol.107:1201−08(1995)
【非特許文献3】Baumら,Mol.Plant−Microbe Interact.5:382−87(1996)
【非特許文献4】Binetら,Plant Science 79:87−94(1991)
【非特許文献5】Borisjukら,Nature Biotechnology 17:466−69(1999)
【非特許文献6】Brownら,Nucleic Acid Res.17:8991(1989)
【非特許文献7】Cabanes−Macheteauら,Glycobiology 9:365−72(1999)
【非特許文献8】Christensenら,Plant Molec.Biol.12:619−632(1989)
【非特許文献9】Christou,P.Plant Mol Biol.35:197−203(1996)
【非特許文献10】Conrad&Fiedler,Plant Mol.Biol.38:101−09(1998)
【非特許文献11】Dennisら,Nucleic Acid Res.12:3983(1984)
【非特許文献12】DeWildeら,Plant Sci.114:231−41(1996)
【非特許文献13】Duringら,Plant Molecular Biology 15:287−93(1990)
【発明の開示】
【発明が解決しようとする課題】
【0011】
一過性発現のためのアグロバクテリウム浸潤の元来のシステムはKapilaによって記述され(Kapilaら,上記(1997))、植物体組織が有する病気に対する抵抗性にとって有用であると考えられていた蛋白質の機能性を、迅速に試験するために開発された。この適用においては、植物体組織全体をバイオアッセイに使用することができるので、蛋白質が精製され又は特徴づけされる必要はない。このシステムは後に製薬に重要な蛋白質の発現に使用された(Vaqueroら,Mol.Biotechnol.5:209−21(1996))。ヒト癌胚抗原に対するキメラ抗体及び同じ抗原に対する組換え一本鎖抗体が作製され、精製され、生化学的な解析がなされ、動物由来の蛋白質に類似するものであることが示された。しかしながら、このシステムからの産物は比較的少ないものであった。
【課題を解決するための手段】
【0012】
(発明の要約)
本発明は、植物栽培設備の必要のない、すでに生長した商業的に入手可能な植物体で蛋白質を産生するといった実質的に改良された工程を提供する。本発明の方法は、動物の試験、多様なアッセイによる解析、結晶構造の特徴づけ、蛋白質修飾のアッセイ等に適した規模で、最小限の時間と費用により、生物学的に機能的な蛋白質を非常に迅速に提供する。本発明の方法で、少なくとも約1mg、少なくとも約5mgまた少なくとも約10mgの蛋白質を1回で産生することができる。
【0013】
本発明の方法は、繰り返し試験するのに必要とされるミリグラム量を提供する規模まで容易かつ迅速に拡大することができ、製薬に用いる蛋白質を機能的に評価するのに使用することができる。
【0014】
本発明は、モノクローナル抗体及び他の製薬に重要な蛋白質の一過性発現のための方法とキットを提供する。この方法は、特に、ウイルスの制御配列を伴って又は伴わずに、プラスミドベクター上に所定の組み換え遺伝子を作製するAgrobacterium tumefaciensによって減圧浸潤されたレタスでの蛋白質産生に成功した。
【0015】
一態様では、本発明は、非常に迅速にkgあたり10−50mg/kgの蛋白質を産生するのに使用できる、拡大可能な細菌/植物ハイブリッド発現システムを提供する。一つの好ましい態様では、1又はそれ以上の異種遺伝子がアグロバクテリウムの無毒化された又は毒性の株を用いて植物体組織に送達される。好ましくは、植物体組織はすでに生長した植物(例えば、商店で入手できる植物等)から得られる。植物体組織の特に好ましい供給元はレタスである。本明細書に開示される方法は、細菌システムにおける迅速な遺伝子クローニングと操作の利点と、植物の栽培及びトランスジェニック植物材料の取り扱いの必要なしに、適当なターゲッティング、プロセッシング、修飾及び会合に必要な環境を提供する真核細胞環境で蛋白質が産生されることの利点を併せ持つ。
【0016】
一態様では、異種遺伝子又は所定の遺伝子を有する発現構成体を作製するアグロバクテリウムの細胞は、植物体組織の細胞及び/又は細胞外空間での一過性発現のために、1又は複数の異種遺伝子を植物体組織へ送達するのに使用される。一般的に、適当な発現構成体は以下を含む:少なくとも1つのT−DNA境界配列、発現制御配列(例えば、誘導可能な及び/又は組織特異的な、又は構成性であるかもしれないプロモーター)、及び発現制御配列に作動可能に結合された所定の遺伝子。一態様では、発現構成体は、発現構成体を含むベクターがアグロバクテリウム中で複製するのに適した1又はそれ以上の複製起点、少なくとも1つの複製起点を含むベクターの一部である。
【0017】
発現構成体を含むアグロバクテリウム細胞の培養物は、界面活性剤の存在下、植物体組織に浸潤される。好ましくは、浸潤は減圧状態で起こる。発現構成体が複数の植物細胞中で蛋白質を発現することができる適当な条件下で植物体組織をインキュベーションした後、蛋白質は細胞から分離される。本発明の方法は、ベクターを含むアグロバクテリウムと植物体組織を接触させ、植物体組織をベクターで浸潤し、約500μg−500mgの収量を得るために異種蛋白質を発現させるという、一連の操作を1回のみ必要とする。しかしながら、agro−infiltration及び精製の操作は手順をスケールアップするために追加で実施してもよい。本発明の方法の一回の操作で、より多くの植物体組織を使用するのが更に好ましい。
【0018】
使用されるアグロバクテリウムは、野生型(例えば、毒性の)又は無毒化されたものであってもよい。それぞれが異なる遺伝子を発現する多くのアグロバクテリウム株が、複数の個々の蛋白質又はヘテロ多量体蛋白質(例えば、抗体)を産生するために、又は、代謝経路、化学合成経路又は情報伝達経路等の経路を再生するために、使用され得る。あるいは又は付加的に、単一のアグロバクテリウム株が異なる異種遺伝子からなる複数の配列を含んでいるかもしれない。異なる異種遺伝子は、単一の核酸分子(例えば、単一のベクター)中に含まれているかもしれないし、異なるベクターで提供されるかもしれない。一態様では、少なくとも1つのアグロバクテリウム株はAgrobacterium tumefaciensを包含する。
【0019】
本発明は異種蛋白質を発現するためのハイスループットシステムを提供するもので、当当該システムは蛋白質の機能に関して所定の遺伝子及び/又は蛋白質の変異による効果を決定するのに使用することができる。一態様では、莫大な数の様々な異種遺伝子を評価するために、標準的な分子生物学的手法(例えば、ランダム突然変異誘発により又はコンビナトリアルクローニングにより等)を用いて、細菌中に複数の発現構成体が作製される。その細菌は、実質的に同一であり(例えば、約50%以上同一、好ましくは75%以上同一、より好ましくは約90%又は95%以上同一)、生物学的活性について迅速にスクリーニングされるために、本発明に従った一過性発現システムを用いて産生されるかもしれない蛋白質をコードする核酸を含んでいる。一態様では、複数の発現構成体は、約100以上、約500以上、約1×103以上、約1×104以上、約1×105以上、約1×106以上、約1×107以上、約1×108以上又は約1×109以上の様々なコード配列を包含する。
【0020】
複数の発現構成体は、異種ポリペプチドのコード配列にランダム又はややランダムな変異を含む配列のライブラリーを包含することができる。ライブラリーはE.coliに基づくライブラリー(すなわち、個々のライブラリー要素がE.coli中でクローン化され複製される)であっても良いし、アグロバクテリウムに基づくライブラリー(すなわち、個々のライブラリー要素がアグロバクテリウム中でクローン化され複製される)であっても良いし、それらの組み合わせ(すなわち、クローニングは最初E.coli中で実施され、ライブラリー要素がさらなる複製及び/又はクローニングのためにアグロバクテリウム細胞中に続いて導入されるかもしれない)であっても良い。ライブラリーの個々の要素は、例えば、大規模な産生及び/又はトランスジェニック植物の産生に適したポリペプチドを同定するために、植物体組織で一過性の発現をした後、ポリペプチドの機能について試験される。
【0021】
ある好ましい態様では、本発明の方法は、最適な活性及び/又は結合のために、複数のサブユニット蛋白質複合体の相互作用しているサブユニット要素(ISEs)の機能を最適化するために使用される。例えば、抗体の結合を最適化するために、抗体の可変領域の複数の変異体が細菌中に標準的な分子生物学的手法(例えば、ランダム突然変異誘発により又はコンビナトリアルクローニングにより等)を用いて作製される。好ましくは、約1×103以上、約1×104以上、約1×105以上、約1×106以上、約1×107以上、約1×108以上又は約1×109以上の様々な発現構成体が作製される。適当な可変領域の配列は抗体の軽鎖(LC)又は重鎖(HC)又は軽鎖及び重鎖の両者を包含する。これらの配列を含む変異体ポリペプチドは、選択された抗原に対する特異的な結合について評価される。変異体ポリペプチドは完全長の抗体又はそれらの抗原結合フラグメント(scFv、Fab’等)を含んでいても良い。異なる変異体は上述のアグロバクテリウムベクター中に容易にクローン化され、HC及びLCの全ての組合せが迅速に試験され得る。ある好ましい態様では、試験は並行して行われる。
【0022】
本発明の方法は、生長している植物体に於ける蛋白質の発現に最も適した発現ベクターを、あらかじめスクリーニングするために使用することができる。一態様では、本発明の方法は、最適な蛋白質発現を提供する、対照となる発現配列及び/又は翻訳配列の変異体を迅速にスクリーニングするのに使用される。あるいは又は付加的に、変異体配列は、増加した安定性又は他の望ましい製薬上の特性を有した蛋白質をコードする配列を同定するためにスクリーニングされる。
【0023】
本発明は、本発明の方法を実施するのに有用なキットも提供する。一態様では、本発明に従ったキットは、少なくともA.tumefaciens等のアグロバクテリウム種で発現するのに適したクローニング/発現ベクター、及び植物種から所望の異種蛋白質を浸潤、抽出及び/又は精製するための1又はそれ以上の構成要素を含む。別の態様では、キットはさらに1又はそれ以上の細菌種(例えば、E.coli及びA.tumefaciens等)を含む。さらなる態様では、キットは変異体異種配列をコードする核酸を含む複数の発現構成体を包含する。
【0024】
(発明の詳細な説明)
本発明は、商業的に入手可能な生長した植物体に於いて蛋白質の産生を可能にする方法を提供し、異種蛋白質において、バイオアッセイで利用するために必要な量を短期間で調達しなければならないという問題に新規な解決法を提供する。本発明の方法は、少なくとも約50μg−100mgの蛋白質を必要とするアッセイ、例えば、薬剤スクリーニング、蛋白質の特徴付け、結合アッセイ及び動物試験等のアッセイでの使用に対して、生物学的に活性な異種蛋白質を提供する。
【0025】
本発明の一態様では、本発明の方法は、異種蛋白質又は生物学的に活性な当該フラグメントをコードする発現構成体を植物体組織に導入すること、及び、植物体組織で当該蛋白質を一過性に発現させることを包含する。コード配列は、植物体組織の細胞及び/又は細胞外空間におけるコード配列の転写を制御することができる発現制御配列に作動可能に結合される。好ましくは、発現構成体は、大きな腫瘤誘導(“Ti”)プラスミドのT−DNA由来の少なくとも1つのT境界配列を包含する。また、好ましくは、発現構成体は、少なくともAgrobacterium tumefaciens等のアグロバクテリウム種の細胞で複製することができるベクター中に含まれる。一態様では、植物体組織はすでに生長した植物(例えば、商店で入手できる植物等)由来の葉組織を含む。好ましくは、植物は比較的大きな(例えば、少なくとも一方向が約3インチより大きな)葉からなり、例えば植物はレタス(Lactuca sativa)である。
【0026】
(定義)
以下の定義は、以下に記載される説明中で使用される特別な用語に対して与えられる。
明細書及び請求項で使用される時、単数形“a”“an”及び“the”は、文脈が別の方法で明らかに指示しない限りは、複数への言及も包含する。例えば、用語“a cell”は、複数の細胞の混合物も含む複数の細胞を包含する。用語“a protein”は複数の蛋白質を包含する。
【0027】
本明細書で使用される「植物細胞」は植物細胞、又は、分離された又は一部分離された細胞を包含する。「植物体組織」は分化した及び未分化の植物組織を包含し、根、シュート、葉、花粉及び種子を含むがそれらに限定されない。
【0028】
本明細書で使用される時、「植物材料」は、植物材料が処理されたことにより生じた派生物を包含し、食物生産物、食物原料、栄養補助食品、抽出物、濃縮物、丸剤、トローチ剤、かみ砕ける物、粉末、粉ミルク、シロップ、キャンディー、ウエハース、カプセル及び錠剤を含むがそれらに限定されない。
【0029】
本明細書で使用される時、「複数のサブユニット蛋白質」は複合体を形成するために互いに結合する1より多くの独立したポリペプチド又は蛋白質鎖を含む蛋白質であり、ここで少なくとも2つの独立したポリペプチドは異なる遺伝子によってコードされる。ある好ましい態様では、複数のサブユニット蛋白質は、少なくとも抗体の免疫学的に活性な部分を含み、よって特異的に抗原と結合できる。例えば、複数のサブユニット蛋白質は抗体分子の重鎖及び軽鎖又はその一部を包含することができる。複数の抗原結合部位が、多価抗体を作成するために、異なる構造遺伝子によってコードされ得る。
【0030】
製薬生産物の場合には、「実質的に純粋」なる用語は、一般的に、少なくとも純度90%、より好ましくは少なくとも純度95%及びさらにより好ましくは少なくとも純度98%の生産物をいう。
【0031】
「組織液(interstitial fluid)」は、細胞膜すなわち細胞表面の膜によって囲まれていない植物の全ての領域から得られる抽出液を意味する。この用語は、顕著な細胞溶解なしにこの処理によって細胞膜から放出されるかもしれない分子を含む、植物の細胞内に存在しない液体、物質、領域又は空間の全てを含むことになっている(ここで、細胞内は細胞質膜内に含まれる中身として定義される)。この用語の類義語は、「外質(exoplasm)」「apoplasm」「間質液」「細胞外液」及び「溢液」を含むがそれらに限定されない。
【0032】
「プロモーター」なる用語は、遺伝子の転写の開始を指示する、遺伝子の5’末端にある核酸配列をいう。一般的に、プロモーター配列は、それが作動可能に結合する遺伝子の発現を制御するのに必要であるが、常に十分なわけではない。プロモーター/異種遺伝子の組合せからなる構築物では、遺伝子はプロモーターに十分近接して、プロモーターに対して適した方向で配置されるので、遺伝子の発現はプロモーター配列によって調節される。プロモーターは、それが天然の位置で調節している遺伝子とそれ自身の間の距離に近似したある一定の距離を転写開始点からおいて、遺伝子の上流に優先的に配置される。公知のように、この距離におけるいくらかの多様性はプロモーター機能を損なわずに許容され得る。本明細書で使用される時、「作動可能に結合」なる用語は、コード領域の転写がプロモーターによって調節され制御されるように、当該プロモーターが当該コード領域に連結されることを意味する。コード領域に作動可能に結合しているプロモーターの意味は、当業界で周知である。
【0033】
本明細書で使用される時、「発現制御配列」はプロモーターを含み、1又はそれ以上のエンハンサー配列、転写終結配列、ポリA配列、3’又は5’非翻訳配列、イントロン配列、リボゾーム結合部位、及び、植物細胞で遺伝子の発現を安定化するか又は他の方法で制御する他の配列を含むかもしれないが、それらに限定されない。発現制御配列は内因性(すなわち、植物宿主で天然に見出される)又は外因性(植物宿主で天然に見出されない)であってもよい。外因性の発現配列は、選択された条件下にてそれらが植物細胞中で機能的である限り、植物の配列であってもなくてもよい。
【0034】
「異種遺伝子」又は「異種コード配列」は、形質転換又は処理された植物にとって外因性又はそこでは天然に見出されない遺伝子であり、「異種ポリペプチド」又はその生物学的に活性なフラグメントをコードする遺伝子である。異種遺伝子配列は、ウイルス、原核生物及び真核生物の配列を含む。原核生物のコード配列は、微生物の配列(例えば、ワクチンとして投与されるかもしれない抗原作製のため、ウイルスの配列もこの目的に使用されるかもしれない)を含むがそれに限定されない。真核生物のコード配列は、哺乳動物の配列を含むが、非哺乳動物、他の植物からの配列を含んでいてもよく、リーダー又は分泌シグナル配列、ターゲッティング配列等を含むがそれらに限定されない。ある好ましい態様では、異種遺伝子の核酸はヒトの蛋白質をコードする。「異種遺伝子」又は「異種コード配列」なる用語は、自然に発生した、突然変異させた、変異した、化学合成された、ゲノムの、cDNAの、又はこれらの配列のいかなる組合せをも包含するがそれらに限定されない。「遺伝子」への言及は、生物学的に活性な蛋白質をコードする完全長の遺伝子又はそのフラグメントを包含する。
【0035】
本明細書で使用される時、「蛋白質」なる用語は、一般的に、遺伝子によってコードされるアミノ酸配列全体、その処理された又は修飾された形又はその生物学的に活性なフラグメント(例えば、ポリペプチド又はペプチド)に言及するために使用される。
【0036】
「融合蛋白質」は、その配列の組合せは、単一の蛋白質として天然では発現されることのない、ポリペプチド中で結合した少なくとも2つの異なるアミノ酸配列を含む蛋白質である。
【0037】
本明細書で使用される時、「T DNA境界」は、A.tumefaciens又はA.rhizogenes株、又はそれらの修飾された又は突然変異された株等のアグロバクテリウム株が有する毒性遺伝子産物によって認識され得る約25ヌクレオチド長の配列を含むDNAフラグメントを意味し、それが連結しているDNA配列を真核生物細胞、好ましくは植物細胞へ移入させる能力のあるDNAフラグメントをいう。この定義は、野生型Tiプラスミド由来の全ての自然に発生したT−DNA境界及びその機能的ないかなる派生物をも含むがそれらに限定されず、化学的に合成されたT−DNA境界も包含する。一態様では、本発明に従った発現構成体のコード配列及び発現制御配列は2つのT−DNA境界の間に存在する。
【0038】
植物種
一過性発現に対するアグロバクテリウム浸潤の初期の適用は、ポプラ及びPhaseolus(Kapila,et al.,1997)に基づいており、そらから後にたばこ(Vaquero,et al.,1999)へ応用された。ポプラ及びPhaseolusの両者は、即座に本発明で使用される植物体組織の適当な供給源となる。他の適当な植物は、レタス、アルファルファ、インゲンマメ、ホウレンソウ、タンポポ、アカチコリー、キバナスズシロ、エンダイブ、キクチシャ、チコリー、アーティチョーク、トウモロコシ、ポテト、イネ、ダイズ、Crucifera(例えば、Brassica、アラビドプシス(シロイヌナズナ))、ウキクサ、トウモロコシ、ポテト、イネ、ダイズ、ホウレンソウ、トマト及びタバコを含むがそれらに限定されない。Brassica科の例となる植物は、B.oleracea(例えば、キャベツ、コラード、カリフラワー、ブロッコリー、芽キャベツ、ケール、コールラビ)、B.campestris(例えば、ハクサイ、ターサイ、Chinese cabbage、celery cabbage、Siberian kale、カブ、カラシナ、ナタネ、スウェーデンカブ及びハツカダイコン)、Brassica juncea(例えば、Brown and Indian Mastard)、Brassica carinata(例えば、Abyssinian Mastard)、Brassica napus(スウェーデンカブ、Swede、Swede Turnip、Siberian kale、Hanover Salad、カノーラ)、Brassica nigra(例えば、Black Mastard)、Rorippa nasturtiumaquatkum(例えば、Water Cress)等を含むがそれらに限定されない。植物材料の特に好ましい供給源はレタスである。適したレタス植物は、Butterhead、Crisphead及びLeaf lettuce(例えば、Oak leaf、Salad Bowl、frilly Red Leaf及びcrinkly Green Leaf)を含むがそれらに限定されない。レタスのさらなる種類は当業界で公知であり、例えばhttp://www.thompson−morgan.com/seeds/us/list_lettuce_2.htmlに記載される。
【0039】
好ましくは、適する植物は一年中商業的に入手可能であり、発現制御配列に作動可能に結合したレポーター遺伝子(例えば、GUS等)の高レベルな一過性発現を維持することができる植物である。本明細書で使用される時、「高レベルな一過性発現」は、植物体組織量キログラム当たり、少なくとも約250μg、少なくとも約500μg、少なくとも約750μg、少なくとも約1mg、少なくとも約2mg、少なくとも約3mg、少なくとも約4mg、少なくとも約5mg、少なくとも約10mg、少なくとも約15mg、少なくとも約25mg、少なくとも約50mg、少なくとも約75mg、少なくとも約100mg、少なくとも約150mg、少なくとも約200mg又は少なくとも約500mgを発現する能力をいう。本明細書で使用される時、「一過性」は、適した植物体組織から蛋白質を分離することができる程十分長い期間をいう。好ましくは、蛋白質の発現は、植物体組織に発現構成体を導入後、少なくとも約1日、少なくとも約2日、少なくとも約3日、少なくとも約4日、少なくとも約5日以内に適当な高いレベルとなる。一態様では、植物体組織に発現構成体を導入後、適当な高いレベルが3−7日以内に、より好ましくは3−5日以内に得られる。
【0040】
適する植物体組織は、一般に、植物のいかなる部分でもあり得る。ある好ましい態様では、植物体組織は葉組織である。一態様では、植物体組織は、少なくとも一方向が少なくとも約3インチの葉からなる植物由来の葉組織である。しかしながら、葉の大きさは制限されず、一態様では本発明の方法はアラビドプシス(シロイヌナズナ)で蛋白質を一過性に発現させるために使用される。
【0041】
別の態様では、異種蛋白質を消化するプロテアーゼのレベルがほとんど又は全くない細胞、例えば、異種蛋白質を発現する核酸の導入から少なくとも蛋白質が植物体組織から分離される頃までの期間に、植物体に於いて発現される異種蛋白質の約5%より少なく、約1%より少なく、約0.1%より少ない量が消化されるような細胞からなる組織が選択される。プロテアーゼレベルは、異種蛋白質発現のウエスタンブロット解析を含む、当業界でごく普通の方法を用いてアッセイすることができる。
【0042】
本発明の特別な利点は、少なくとも約1日、少なくとも約2日、少なくとも約5日、少なくとも約1週間又は少なくとも約2週間、収穫され貯蔵された植物を含む、すでに生長した植物を植物体組織の供給源として使用できる点である。従って、植物体組織はいかなる一般的な食料品店でも得ることができる。
【0043】
レタスは、本発明に従った方法で使用される葉組織の特に適した供給源である。なぜなら、レタスは容易に入手でき、異種蛋白質の発現、安定性及び機能を高いレベルで提供するからである。さらに、レタス細胞は異種蛋白質を認識するプロテアーゼを量的にほとんど含んでいない。使用に適した様々なレタスの内、特に好ましいのは赤い葉の表現型を有するレタスである。レタスで観察される異種蛋白質の高レベルな発現に加えて、葉の多いレタスを使用することの利点は、特別に設計された装置の必要がなく葉を簡単に操作できる様式で植物が生長している点にある。
【0044】
トウモロコシは、安定なトランスジェニック植物から製薬に用いられる蛋白質を産生するために、最もよく使用される作物の一つである。トウモロコシでは、異種蛋白質は種子で産生され貯蔵される。しかしながら、トウモロコシは形質転換するのが難しく、一世代の時間が長く、室内で種子を産生するのが難しい。トウモロコシは、一過性発現システムから大いなる利益を得ることができる作物である。トウモロコシを形質転換するためにアグロバクテリウムを使用することは今や本当に一般的であるので、トウモロコシの種子由来の異種蛋白質の有用性/機能を迅速に評価し確認するために、トウモロコシ胚を処理することによって、本発明に従った一過性の発現システムを使用し、トウモロコシの種子で作られる蛋白質の産生量を高めることができる。
【0045】
発現カセット
本発明の好ましい実施形態では、野生型レタス、突然変異又は修飾された変異体レタス(例えば、トランスジェニックレタス等)が処理され、所望のDNA構築物から所定の遺伝子を発現する。このような構築物は、遺伝子の転写及び最終的な蛋白質の翻訳を容易にするために、プロモーター及び/又は他の調節要素(すなわち、発現制御配列)に作動可能に結合した、所望の蛋白質をコードする核酸配列を少なくとも含んでいる。
【0046】
一態様では、発現構成体は、5’から3’方向に作動可能に結合された以下のもの、プロモーター、遺伝子及びターミネーターを含むよう巧みに処理されている。別の態様では、遺伝子構築物は、1つの共通のプラスミド又は植物に同時トランスフェクションされるプラスミド上に作動可能に結合した複数のコード領域を含む(このような同時トランスフェクションされる構築物は、本明細書で使用される時には「遺伝子構築物」なる用語に一括して包含される)。同義遺伝子は分離されたシストロンとして又はポリシストロン性のユニットの部分としてコードされていてもよい。さらなる態様では、遺伝子構築物は1又はそれ以上のIRES要素を包含する。
【0047】
遺伝子構築物が選択可能なマーカーを含んでいる必要はなく、形態学的に正常で安定なトランスジェニック植物の作製で要求されるように、DNA構築物が「腫瘤誘導」遺伝子を欠いていることも必要とされない。
【0048】
蛋白質
本発明によって産生される蛋白質については予想される制限はないが、管理され、そして再生産が可能であるという条件下で特定の産物を産生する必要性があることから、特に適した蛋白質の範疇がある。特にこれには、医薬品の製造及び品質管理に関する基準や承認された方法がその生産過程で使用されなければならない製薬及び/診断のための全種類の蛋白質が含まれる。
【0049】
それらの産生物はヒトに直接、経口摂取、注射又は塗布(例えば、局所的な投与)するために使用されるので、蛋白質もまた、栄養補助食品及び薬用化粧品としての有用性があるので発現させてもよい。同様に規定された獣医向けの産物の産生に有用な蛋白質もまた発現させてもよい。
【0050】
産生されてもよい蛋白質の例は以下のものを含むがそれらに限定されない。成長因子(例えば、血小板由来増殖因子、インシュリン様増殖因子等)、受容体、リガンド、情報伝達分子、キナーゼ、酵素、ホルモン、腫瘍抑制遺伝子産物、血液凝固蛋白質、細胞周期蛋白質、テロメアーゼ、代謝蛋白質、神経蛋白質、心臓の蛋白質、特異的な疾患状態で定義される蛋白質、抗体、T細胞受容体(TCR)、主要組織適合性複合体(MHC)、抗原、病気に対する抵抗性を与える蛋白質、抗菌蛋白質、インターフェロン及びサイトカイン。
【0051】
一態様では、保護免疫反応を誘導する配列を含んでいる配列をコードする(例えば、ワクチン製剤に含まれるような)抗原である。このような適する抗原は、以下のものを含むがそれらに限定されない。微生物抗原(ウイルス抗原、細菌抗原、菌類の抗原、寄生虫抗原等)、多細胞生物(多細胞寄生虫等)由来の抗原、アレルゲン及びヒト又は動物の病理に関係のある抗原(例えば、癌、自己免疫疾患等)。ある好ましい態様では、ウイルス抗原は、以下のものを含むがそれらに限定されない。HIV抗原、天然痘に保護免疫反応を与える抗原(例えば、ワクシニアウイルス抗原)、炭疽菌抗原、狂犬病抗原等。ワクチン抗原は、例えば、1の発現構成体に、任意で1又はそれ以上のリンカー配列で分離された状態で、繰り返し、異なる又は同じ抗原性をコードする配列を含んでいる多価ペプチド又はポリペプチドとしてコードされ得る。
【0052】
植物は、また、化学合成又は工業的工程のための酵素経路を再現するために1又はそれ以上の遺伝子を発現するのに使用されるかもしれない。
【0053】
一態様では、所望の蛋白質をコードする核酸配列が選択される。そこでは、核酸配列は、レタス又は所望の蛋白質の大規模な生産に最終的に使用されるかもしれない植物によって好まれるコドン、ただし、そのコドンの選択が一過性システムで有用なレベル以下に発現量を減少させない場合、そのコドンを提供するように設計される。いくつかの植物におけるコドン使用の指標は入手可能であり、例えば、Wadaら,“Codon Usage Tabulated From The GenBank Genetic Sequence Data,”Nucleic Acids Reseach 19(Supp.):1981−1986(1991)に記載されている。
【0054】
以下にさらに記載するように、一態様では、本発明は複数の組換え蛋白質を発現するための方法を提供する。このような蛋白質は、独立した構築物の同時浸潤によって発現されるかもしれないし、以下にさらに記載するポリシストロン性の発現ユニットから発現されるかもしれない。このような蛋白質は、その天然の状態で、生物学的に活性になるために複数の構造遺伝子の協調した発現を必要とする蛋白質を含むことができる。一態様では、蛋白質は、活性を有するために複数のサブユニットの会合が必要とされる。別の態様では、蛋白質は成熟していない形で生産され、プロセッシング、例えば、活性化するために、1又はそれ以上のさらなる蛋白質によって、蛋白質分解での切断又は修飾(例えば、リン酸化、グリコシル化、リボシル化、アセチル化、ファルネシル化等)されることを必要とする。
【0055】
このような蛋白質の限定されない例としては、T細胞受容体、MHC分子、他の免疫グロブリンスーパーファミリーの蛋白質(フラグメント及び一本鎖変異体を含む)、核酸結合蛋白質(例えば、複製因子、翻訳因子等)、酵素、アブザイム、受容体(特に可溶性の受容体)、成長因子、細胞膜蛋白質、分化因子、ヘモグロビン様蛋白質、多重結合キナーゼ等のヘテロ二量体又はヘテロ多量体蛋白質を含む。
【0056】
本発明の好ましい態様では、発現カセットはヒト蛋白質(すなわち、ヒトで発現する蛋白質)をコードし、又は、他の状態で非ヒト蛋白質に含まれているヒトポリペプチド領域を含む蛋白質をコードする。
【0057】
ある特に好ましい態様では、発現カセットは、モノクローナル抗体の1又はそれ以上の遺伝子をコードする。このような遺伝子は、マウス、ヒト及び/又は他の動物から得ることができる。あるいは、例えば、抗体分子の構成要素である重鎖又は軽鎖をコードする遺伝子のキメラ又は修飾された形で、それらを合成することもできる。構築物中のコード領域の順番、例えば、重鎖それから軽鎖又は軽鎖それから重鎖は、重要ではない。重鎖又は軽鎖ポリペプチド(例えば、重鎖可変又は軽鎖可変ドメインポリペプチド等)をコードする遺伝子は、IgA、IgD、IgE、IgG又はIgMを産生する細胞から得ることができる。免疫グロブリン可変領域遺伝子をクローニングすることができるゲノムDNAのフラグメントを調製する方法は、当業界で周知である。例えば、Herrmannら,Methods in Enzymol.152:180−183(1987);Frischauf,Methods in Enzymol.152:183−190(1987);Frischauf,Methods in Enzymol.152:199−212(1987)参照。ある好ましい態様では、以下に記載するように、このような遺伝子はポリシストロン性ユニットの部分としてコードされる。
【0058】
調節要素
特定の構築物を作成するのに適した調節要素は、発現される組換え蛋白質の種類に基づいて選択されるだろう。一般に、浸潤される植物体組織において高いレベルで発現する能力が望まれる。
【0059】
植物のプロモーター
使用される遺伝子構築物は、生物学的に活性な蛋白質フラグメントをコードしている遺伝子全体又はその一部からなる、遺伝的な材料全てを含んでいるかもしれない。好ましくは、コード配列は発現制御配列に作動可能に結合している。発現制御配列はプロモーター、エンハンサー、IRES要素等の配列を含む。発現制御配列は、発現を誘導するための外部からの刺激として特定の栄養素又は薬剤の付加、温度変化等を要求することもできるし、植物体組織の浸潤及び/又は培養中に迅速に及び/又は自然発生的にコードされる蛋白質を発現するよう設計することもできる。
【0060】
このように、構成的又は調節されたプロモーターは所望の蛋白質をコードする遺伝子の発現を制御するかもしれない。調節されたプロモーターは、環境から情報を受け取るかもしれないし、化学的な誘導因子や抑制因子によって制御可能であるかもしれない。このような因子は天然のもの又は合成物由来であってもよく、プロモーターは天然のもの又は設計されたものであってもよい。プロモーターはキメラ、すなわち、2又はそれ以上の異なる天然又は合成されたプロモーター由来の配列要素を使ってできたものでもあり得る。
【0061】
好ましくは、構築物中で使用されるプロモーターは遺伝子の高い発現レベルを生じ、植物体組織重量(例えば、葉組織生物量)kg当たり少なくとも約250μg、少なくとも約500μg、少なくとも約750μg、少なくとも約1mg、少なくとも約2mg、少なくとも約3mg、少なくとも約4mg、少なくとも約5mg、少なくとも約10mg、少なくとも約15mg、少なくとも約25mg、少なくとも約50mg、少なくとも約75mg、少なくとも約100mg、少なくとも約150mg、少なくとも約200mg又は少なくとも約500mgの蛋白質の蓄積を可能にする。
【0062】
本発明では、アラビドプシス(シロイヌナズナ)アクチン2プロモーター、OCS3(MAS)プロモーター、CaMV 35Sプロモーター及びゴマノハグサモザイクウイルス34Sプロモーターが好まれる。しかしながら、他の構成的及び誘導可能なプロモーターを使用することもできる。例えば、ユビキチンプロモーターは植物体での使用のためにいくつかの種からクローニングされた(例えば、ヒマワリ(Binetら,Plant Science 79:87−94(1991))及びトウモロコシ(Christensenら,Plant Molec.Biol.12:619−632(1989))。さらに有用なプロモーターはトウモロコシ由来のU2及びU5 snRNAプロモーター(Brownら,Nucleic Acid Res.17:8991(1989))及びアルコールデヒドロゲナーゼ由来のプロモーター(Dennisら,Nucleic Acid Res.12:3983(1984))である。
【0063】
別の態様では、調節されたプロモーターは遺伝子に作動可能に結合される。調節されたプロモーターは、外部からの影響によって(例えば、化学薬品、光、温度等の外部因子の適用によって等)調節されるプロモーター、又は、植物における調節された発生上の変化等の内的なきっかけによって調節されるプロモーターを含むがそれらに限定されない。調節されたプロモーターは、収穫の時期又はその付近において特異的に所望の遺伝子の高レベルな発現を誘導するのに有用である。これは、所望の蛋白質が植物の成長を制限したり別の方法で抑制したり又は所望の蛋白質がいくつかの方法では不安定である場合に、特に有用である。発現構成体が一過性発現アッセイにおいてだけでなくトランスジェニック植物の生産においても使用されることが期待される場合には、このようなプロモーターが望まれるかもしれない。
【0064】
異なる植物体組織で移入された遺伝子の発現を制御する植物のプロモーターは当業者に公知である(Gasser&Fraley,Science 244:1293−99(1989))。カリフラワーモザイクウイルス35Sプロモーター(CaMV)及びCaMVプロモーターの増強された誘導体(Odellら,Nature,3(13):810(1985))、アクチンプロモーター(McElroyら,Plant Cell 2:163−71(1990))、AdhIプロモーター(Frommら,Bio/Technology 8:833−39(1990)、Kyozukaら,Mol.Gen.Genet.228:40−48(1991))、ユビキチンプロモーター、ゴマノハグサモザイクウイルスプロモーター、マンノピンシンターゼプロモーター、ノパリンシンターゼプロモーター、オクトピンシンターゼプロモーター及びそれらの誘導体が構成的プロモーターとみなされる。調節されたプロモーターは、光で誘導可能なもの(例えば、リブロースビスリン酸カルボキシラーゼ小サブユニットのプロモーター)、熱ショックプロモーター、硝酸塩及び他の化学的に誘導可能なプロモーター(例えば、米国特許第5,364,780号、第5,364,780号、第5,777,200号参照)として記述される。
【0065】
組織特異的なプロモーターは、植物の特定の部分で蛋白質を発現させる理由がある時に使用される。葉特異的なプロモーターは35Sエンハンサーが先行するC4PPDKプロモーター(Sheen,EMBO,12:3497−505(1993))又は葉での発現に特異的ないかなる他のプロモーターを包含していてもよい。
【0066】
一般に、植物において発現可能である、あらゆる遺伝的な構築物が本発明の方法で使用されるのに適している。特定のプロモーターが発現させる組換え蛋白質の種類を考慮して選択されるかもしれない。
【0067】
本発明にとって特に興味深いのは、OCSエンハンサー及び核となるMASプロモーターの複数ユニット由来の合成プロモーターの使用であり、ここでOCS及びMASの要素はアグロバクテリウム由来である(Gelvinら,米国特許第5,955,646号)。このプロモーターは、後に行われる浸潤及び植物ホルモン2,4−Dでの処理、このホルモンはより高い濃度で除草剤として使用されるが、に対して特に強いことが示されている。
【0068】
標的化配列
好ましい実施形態では、発現産物は、細胞膜、細胞外空間又は細胞小器官、例えばクロロプラスト等のプラスチドのような植物細胞の特異的な場所へと向けられる。好ましい実施形態では、発現産物は、細胞外空間へと向けられ、その結果、細胞内液の分離に基づく精製を可能にする。例えば、米国特許第6,096,546号、米国特許第6,284,875号及び国際公開第0,009,725号公報参照。
【0069】
蛋白質は、標的化配列の効力によって特異的な細胞内の一部又は細胞外の場所へと向けることができる。ある場合には、アミノ酸配列がポリペプチドのアミノ末端部分として合成され、移動又は局在化する過程の後又はその間に、プロテアーゼによって切断される。例えば、真核生物における蛋白質分泌経路のモデルは、リボソームのmRNAへの結合及び翻訳の開始に続いて、発生しようとしているポリペプチド鎖が出現する。それが分泌される運命にある蛋白質ならば、出現した蛋白質のアミノ末端は、シグナル認識粒子(SRP)、それはmRNA、リボソーム及びSRP複合体が小胞体(ER)に付着する間、翻訳の一時的な失速を引き起こすが、それによって認識される。付着した後、ポリペプチド鎖は今や翻訳されると同時にERルーメンへと移動するが、翻訳は再開する。
【0070】
液胞中に局在化するため又は分泌のために内膜システムへと標的化される蛋白質についてのシグナル配列は、植物及び動物で類似している。シグナルペプチドは、標準的な技術に従った本発明での使用(例えば、あらゆる他の遺伝子でフレームに合わせてクローニングするために適した末端に調製される)に適合しているかもしれない。
【0071】
一態様では、所望の蛋白質をコードする発現カセットは、所望の蛋白質をコードする配列にフレームを合わせて融合したシグナル配列を含む。ある好ましい態様では、シグナル配列は分泌経路に遺伝子の発現産物を向けることができる配列である。
【0072】
抗体は正常に分泌される蛋白質であるので、分泌過程は成熟した抗体分子の産生に重要な役割を演じている。植物ではこれを達成するために、天然の状態での哺乳動物のシグナルペプチドをコードする領域を有するか、植物の分泌シグナルペプチドが所定の遺伝子のシグナルペプチドとして置換され、所定の遺伝子に作動可能に結合した融合物として、遺伝子が合成又は他の方法で(例えば、クローニング)得られる。シグナルペプチドと蛋白質の融合は植物によってプロセッシングされるはずであり、結果として生じる蛋白質のアミノ末端はヒト宿主で生じるものと同一である。しかしながら、クロロプラストへの標的化も期待される。
【0073】
好ましい実施形態では、カルレティキュリン由来のシグナル配列(Borisjukら,Nature Biotechnology 17:466−69(1999))が使用される。より好ましい実施形態はトマト由来のズブチラーゼ(subtilase)配列を使用する(Janzikら,2000)。これらの植物シグナルペプチドは植物のapoplastic空間へと外来蛋白質を向かわせるのに有効であることが示されてきた(例えば、Janzikら,2000参照)。他の植物蛋白質シグナルペプチドはオオムギに対して記述されるシグナルペプチドも使用されるかもしれない(α−アミラーゼ、Duringら,Plant Molecular Biology 15:287−93(1990)、Schillbergら,Transgenic Research 8:255−63(1999))。
【0074】
蛋白質を植物の内膜システムへ標的化することは、その成熟形に到達するためにアミノ末端のプロセッシングを通常要求する蛋白質に対して、本発明の好ましい実施形態である。なぜなら、それは蛋白質アミノ末端の正確な成熟化を提供するからである。さらに、内膜システムの特異的な領域への局在化は、もし、所定の蛋白質がさらなる標的化情報を含むように作られているか作られるならば、達成することができる(例えば、Vossら,Mol.Breeding 1:39−50(1995)、Duringら,Plant Mol.Biol.15:281−93(1990)、Baumら,Mol.Plant−Microbe Interact.9:382−87(1996)、DeWildeら,Plant Sci.114:231−41(1996)、Maら,Immunology 24:131−38(1994)、Schoutenら,Plant Mol.Biol.30:781−93(1996)、Firekら,Plant Mol.Biol.23:861−70(1993)、Artsaenkoら,Plant J.8:745−50(1995)、Conrad&Fiedler,Plant Mol.Biol.38:101−09(1998)参照)。
【0075】
プラスチド(例えば、クロロプラスト及びミトコンドリア)等の細胞小器官への標的化は所望のアミノ末端の成熟を達成するために利点を有している。なぜなら、これらの局所いずれかへの標的化は、続いて切断が起こるアミノ末端シグナル配列によって指揮されるからである。好ましい実施形態では、シグナルペプチドは発現産物をプラスチド(例えば、クロロプラスト)又は他の細胞小器官へと向ける。ある例は、アルファルファのリブロースビスリン酸カルボキシラーゼ小サブユニットの通過ペプチドである(Khoudiら,Gene 197:343−5(1997))。ペルオキシソーム標的化配列は、植物のC−末端標的化トリペプチドSKL等の蛋白質をペルオキシソームへ標的化できる、N−末端、内部又はC−末端のいかなるペプチド配列にもあてはまる(Banjokoら,Plant Physiol.107:1201−08(1995))。
【0076】
さらに、又は特異的な細胞内局所に蛋白質を向かわせるためのものに代えて、一態様では、「エピトープタグ」及び/又は部位特異的な切断部位が融合蛋白質を作成するために付加される。このタグの有用性は、それらが簡便な精製メカニズムを提供できることにある。例えば、ストレプトアビジンに結合するためにビオチン由来の重要なアミノ酸配列を含む小さなペプチドが、所定の遺伝子の5’末端に付けられる。そして新たに合成された蛋白質は、基本的にビオチン:ストレプトアビジン結合に基づく多くの知られた方法によって捕捉することができる。「ビオチン様」ペプチドが蛋白質から除去することが望まれるならば、プロテアーゼ認識部位も含むことが可能である。プロテアーゼ認識部位は「エピトープタグ」配列の下流で、所望の蛋白質の成熟体をコードする配列の直前に挿入することができる。当業者は、エピトープタグ及びプロテアーゼ(Xa因子、タバコEtchウイルスプロテアーゼ、エンテロキナーゼ等)について多くの選択肢があり、好ましい部位やプロテアーゼの選択は問題となっている特異的な蛋白質のアミノ酸及びDNA配列に依存するかもしれないことを理解するであろう。
【0077】
上記のように、プロモーター、エンハンサー、IRES要素及びシグナル配列等の調節要素の選択は、一般に、発現させる蛋白質の種類に依存するであろう。例えば、一態様では、IgGを作成する目的で好まれるいくつかの構築物は、5’OCS3MASプロモーター、(いずれかの生物の)ズブチラーゼ(subtilase)シグナルペプチド、IgG重鎖遺伝子の成熟部分に対するコード領域、翻訳終結シグナル、転写終結及びポリアデニル酸配列を有する構築物と共に、上記のものに類似の要素を含み、重鎖遺伝子が軽鎖遺伝子と置換された第2の構築物を含むだろう(すなわち、本明細書中で“binary”又は“dual”ベクターとして言及される2つのベクター)。あるいは、別の好ましい実施形態では、重鎖及び軽鎖遺伝子は同じDNA構築物上に存在する。さらに別の実施形態では、重鎖及び軽鎖遺伝子は、IRES要素によって分離されているが同じDNA構築物上の同じプロモーターから発現される。
【0078】
他の配列
発現構成体は発現ベクターの一部であってもよく、細菌の複製開始点(アグロバクテリウム及び/又はE.coliの複製開始点)、アグロバクテリウム及び/又は植物細胞等の細菌中で機能的であるレポーター遺伝子(例えば、GUS、GFP、EGFP、BFP、β−ガラクトシダーゼ及びそれらの修飾体)及び選択可能なマーカー遺伝子(例えば、抗生物質耐性遺伝子等)等のさらなる所望の配列を包含することができる。このために、本発明の方法で使用される外来DNAは、マーカー遺伝子を包含していてもよく、そのマーカー遺伝子の発現により、クローニングの開始段階で、非形質転換細胞から形質転換細胞の分離が可能となる。このようなマーカー遺伝子は、一般に、形質転換細胞を非形質転換細胞から表現型により区別することを可能とする蛋白質をコードしている。植物体に於いては、選択可能なマーカー遺伝子は、このように、ホスフィノトリシン等のグルタミンシンセターゼ阻害剤を含む除草剤等の除草剤に抵抗性を与える蛋白質をコードしていてもよい(例えば、欧州特許第0242236号、欧州特許第0242246号、De Blockら,1987,EMBO J.6:2513−2518)。しかしながら、マーカー遺伝子が、発現構成体/ベクターが導入された植物体組織から異種蛋白質を分離するのに必要とされないことは、本発明に従った一過性の蛋白質生産方法の利点である。
【0079】
異種蛋白質をコードする配列に融合できる、さらなる配列は以下のものを含むがそれらに限定されない。(例えば、Martinら,EMBO J.13(22):5303−5309(1994)に記載されるような)高次コイル配列、minibody配列、すなわち最低限の抗体相補性領域からなる配列(例えば、Bianchiら,J.Mol.Biol.236(2):649−59(1994)参照)、安定化配列、二量体化配列、リンカー配列、ミリスチル化配列(例えば、米国特許公開第2002/0146710号に記載される)、(例えば、免疫付着因子を産生するための)Fc領域等。
【0080】
発現構成体のライブラリー
一態様では、複数の発現構成体が、本発明に従った一過性の蛋白質発現システムにおいて変異体蛋白質配列の発現及び試験をするために、実質的に同一のコード配列(例えば、約50%以上同一、約75%以上同一、約90、95%又は99%以上同一)を含んで作成される。
【0081】
一態様では、構築物のコード領域は無作為化されている。構築物は、十分に無作為化されることもできるし、無作為化に偏りがあるようにすることもできる(すなわち、無作為か又は1又はそれ以上の選択された場所である)。一態様では、複数の発現構成体のうち1の発現構成体によってコードされる少なくとも1の蛋白質が所望の生物学的活性(例えば、抗原等の特定の結合パートナーに結合する能力等)を有するように、十分に多様な個体群を含む、発現構成体のライブラリーが作成される。別の態様では、発現構成体の数は、約100以上、約500以上、約1x103以上、約1x104以上、約1x105以上、約1x106以上、約1x107以上、約1x108以上又は約1x109以上の変異体コード配列を包含する。好ましくは、ライブラリーの多様性は、約1x107−1x109の異なる変異体コード配列を包含するような多様性である。蛋白質の1又はそれ以上のアミノ酸は一度に無作為化されてもよい。一態様では、約1、約2、約3、約4、約5又は約6、又はそれ以上のアミノ酸が一度に無作為化される。一態様では、“n”アミノ酸を含む蛋白質に対して、nアミノ酸のそれぞれ1つずつが独立して無作為化され、蛋白質は活性について試験される。他の領域が不変である一方、異種蛋白質の1又は複数の特定領域が多様化するかもしれないように(1の抗原結合部位又は特定の1アミノ酸等)、無作為化は偏りがあってもよい。
【0082】
蛋白質の方向付けられた進化のために核酸配列を突然変異させる方法は、例えば以下に記載されている。Leungら,Technique 1:11−15(1989)、Cadwell and Joyce,PCR Methods Appl.2:28−33(1992)、Shafikhaniら,Biotechniques 23:304−310(1997)、Wanら,Proc.Natl.Acad.Sci.U.S.A.95:12825−12831(1998)、You and Arnold,Protein Eng.9:77−83(1996)、Cherryら,Nat.Biotechnol.17:379−384(1999)、Tukeyら,J Immunol Methods 270(2):247−57(2002)、Choら,Mol.Biol.297(2):309−19(2000)。
【0083】
植物体組織サンプルは、以下に詳述するように、複数の発現構成体で浸潤することができ、所望の種類及び/又はレベルの生物学的活性を有する蛋白質を発現する組織/細胞を高い処理量で選択することができる。発現構成体は、所望の種類/レベルの活性を示すサンプル中の1又はそれ以上の細胞から回収することができ、活性の種類/レベルに関連する変異体配列を同定するために配列決定又は他の方法で特徴付けすることができる。構築物は、構築物の配列が決定される前又は後に、結果を確認するために、1又はそれ以上の植物体組織サンプルに再び導入することができる。偏りのある無作為化の2回目は、蛋白質の改変されなかった部位及び/又は増強された特性で蛋白質を同定するために(例えば、特定の結合パートナーに対する増強された結合親和性を有する蛋白質)選択された領域を変化させるのに使用されるかもしれない。
【0084】
一態様では、本発明の方法は、抗体の生成に関与している自然の選択過程をまねるために使用される。すなわち、特定の抗原に結合する発現構成体を同定し、構築物を突然変異させ、
特定の抗原に対する最も高い親和性を提供する構築物を同定するために、第2の選択工程を行う。
【0085】
アグロバクテリウムによる形質転換及び培養物の調製
アグロバクテリウムによる植物の形質転換及び安定なトランスジェニック植物の作成におけるその使用は、十分に証明されてきた。植物細胞とT−DNA境界配列を含むアグロバクテリウム細胞の相互作用の結果、植物核へ蛋白質と複合体を形成したアグロバクテリウム T−DNAの1本鎖コピーが移入する。安定な形質転換のために、T−DNAは核のDNAへ融合する。
【0086】
その過程は明らかに非常に効果的であるが、T−DNAの融合されなかったコピーは、一過性に転写されることができ、T−DNA遺伝子及び同時に移入された他のいかなる遺伝子も短期間は発現されることになる。一過性発現は、DNAの融合又は植物の再生に依存しないので、使用されるかもしれないが、無毒化したベクター(すなわち、もはや腫瘤を産生する遺伝子を含まないベクター)を使用する必要なしにより、毒性のあるアグロバクテリウム株を使用することが可能である。
【0087】
適した無毒化ベクターは、左境界及び左内側ホモロジー(LIH)配列は部分はそのままである一方、T−DNAの右境界は、サイトカイニン及びオーキシンをコードする植物ホルモン遺伝子と共に取り除かれ、細菌のカナマイシン耐性遺伝子によって置換されているSEVシリーズ、及び、植物ホルモン遺伝子が切除されpBR322ベクター配列の一部によって置換されており、左及び右境界配列は、Tiプラスミドのノパリンシンターゼ遺伝子と同様に、保存されているpGVシリーズを包含する。中間のベクターがヘルパー配列で結合されて使用されるかもしれない。いくつかの好ましい態様では、binaryベクターが当業界で公知であり、例えば、米国特許第4,940,838号、欧州特許公報第120516号、米国特許第5,464,763号に記載されている。
【0088】
適するアグロバクテリウム株は、野生型株(例えば、Agrobacterium tumefaciens)、又は、例えば、vir遺伝子の発現及び/又はその誘導が、突然変異又はキメラのvirA又はvirG遺伝子の存在によって改変されているアグロバクテリウム株等の1又はそれ以上の遺伝子が形質転換効率を高めるために突然変異されている株を包含する(例えば、Chen and Winans,1991,J.Bacteriol.173:1139−1144及びScheeren−Grootら,1994,J.Bacteriol.176:6418−6246)。別の実施形態では、例えば、米国特許第6,483,013号に記載されるような、pTiBo542由来のスーパーvirG遺伝子等の余分なvirG遺伝子コピーを含むアグロバクテリウム株が、好ましくは、複数コピーのプラスミドと結合された。
【0089】
他の適する株は、以下のものを含むがそれらに限定されない。A.tumefaciens C58C1(Van Larebekeら,Nature 252:169−170(1974))、A136(Watsonら,J.Bacteriol 123:255−264(1975))、LBA4011(Klapwijkら,J.Bacteriol 141:128−136(1980))、LBA4404(Hoekemaら,Nature 303:179−180(1983))、EHA101(Hoodら,J.Bac.168:1291−1301(1986))、EHA105(Hoodら,Trans Res.2:208−218(1993))、AGL1(Lazoら,Bio/Technology 9:963−967(1991))、A281(Hoodら,上記(1986))。
【0090】
一態様では、本発明は、アグロバクテリウムを処理する単純化された方法を提供する。好ましくは、アグロバクテリウム株は適する培養培地中で600nmでの光学密度(O.D.)2.5−3.5となるように培養される。細胞は一般にO.D.が2.5となるように希釈される。アグロバクテリウム細胞は、植物体組織の体積当たりおよそ1−3体積の培養培地中のアグロバクテリウム懸濁液を用いて、直接(すなわち、初期の濃縮工程又は遠心分離工程なしに)、植物体組織と接触される。ある実施形態では、植物材料1kg当たり約4Lより少ない量の細菌培養液が、少なくとも約250μg、少なくとも約500μg、少なくとも約750μg、少なくとも約1mg、少なくとも約2mg、少なくとも約3mg、少なくとも約4mg、少なくとも約5mg、少なくとも約10mg、少なくとも約15mg、少なくとも約25mg、少なくとも約50mg、少なくとも約75mg、少なくとも約100mg、少なくとも約150mg、少なくとも約200mg又は少なくとも約500mgの異種蛋白質を提供する。
【0091】
本発明の方法は、より少ない工程で実施することができるので、使用される他の方法と比較して、より少ない人的資源と15−20倍減少した生産コストを要する。
【0092】
減圧浸潤
ある特に好ましい実施形態では、界面活性剤が、植物体組織から異種蛋白質の収量を高めるために、アグロバクテリウム懸濁液に添加される。一態様では、アグロバクテリウム細胞又はその一部は、発現構成体/発現ベクターを発現するために、宿主の植物体組織に浸潤される。好ましくは、この工程は減圧状態で実施される。
【0093】
本明細書で使用される時、「界面活性剤」なる用語は、一般に疎水性部分及び親水性部分からなる表面活性化剤をいう(例えば、Bhairi,A Guide to the Properties and Uses of Detergents in Biological Systems,Calbiochem−Novabiochem Corp.1997参照)。界面活性剤は、それらが1又はそれ以上の荷電した官能基を含むか否かによって、陰イオン性、非イオン性、両性イオン又は陽イオン性と分類されるかもしれない。陰イオン性界面活性剤は、陰性に荷電した官能基を含み、正味の陰性電荷を有する。非イオン性界面活性剤は、荷電していない極性官能基を含み、電荷を有さない。これらの界面活性剤は、一般に、アルキレン酸化物と、アルキルフェノール、第1級又は第2級アルコールとの反応生成物、又は、アミン酸化物、ホスフィン酸化物又はジアルキルスルホキシドである。
【0094】
例となる非イオン性界面活性剤は以下のものを含むが、それらに限定されない。トクチルフェノキシポリエトキシエタノール(TritonX−100)、ポリオキシエチレンソルビタンモノラウリン酸(Tween 20)、ポリオキシエチレンソルビタンモノラウリン酸(Tween 21)、ポリオキシエチレンソルビタンモノパルミチン酸(Tween 40)、ポリオキシエチレンソルビタンモノステアリン酸(Tween 60)、ポリオキシエチレンソルビタンモノオレイン酸(Tween 80)、ポリオキシエチレンソルビタンモノトリオレイン酸(Tween 85)、オクチルフェノキシポリエトキシエタノール(IGEPAL CA−630/NP−40)、トリエチレングリコールモノラウリルエーテル(Brij 30)及びソルビタンモノラウリン酸(Span 20)。
【0095】
両性イオン界面活性剤は、陽性に荷電した官能基と陰性に荷電した官能基の両者を含み、正味の電荷は有さない。適する両性イオン界面活性剤は以下のものを含むがそれらに限定されない。カルボキシベタイン、スルホベタイン(スルタインとしても知られる)、アミドベタイン及びスルホアミドベタイン等のベタイン、これらはC8−C18、好ましくは、C10−C18のアルキルベタイン、スルホベタイン、アミドベタイン及びスルホアミドベタインを含んでいてもよく、例えば、ラウリルアミドプロピルベタイン(LAB)型ベタイン、n−アルキルジメチルアンモニオメタンカルボキシレート(DAMC)、n−アルキルジメチルアンモニオエタンカルボキシレート(DAEC)及びn−アルキルジメチルアンモニオプロパンカルボキシレート(DAPC)、n−アルキルスルタイン、n−アルキルジメチルアンモニオアルキルスルホン酸、N−アルキルジメチルアンモニオエタンスルホン酸(DAES)、n−アルキルジメチルアンモニオプロパンスルホン酸(DAPS)及びn−アルキルジメチルアンモニオブタンスルホン酸(DABS)、ヘキサデシルジメチルアンモニオプロパンスルホン酸、n−アルキルアミドメタンジメチルアンモニオメタンカルボン酸、n−アルキルアミドメタンジメチルアンモニオエタンカルボン酸、ラウリルアミドプロピルベタイン(LAB)、n−アルキルアミドメタンジメチルアンモニオメタンスルホン酸、n−アルキルアミドエタンジメチルアンモニオエタンスルホン酸及びn−アルキルアミドプロパンジメチルアンモニオプロパンスルホン酸、3−[(3−コールアミドプロピル)ジメチルアンモニオ]−1−プロパンスルホン酸(CHAPS)及び3−[(3−コールアミドプロピル)ジメチルアンモニオ]−2−ヒドロキシ−1−プロパンスルホン酸(CHAPSO)、リン脂質(例えば、ホスファチジルエタノールアミン、ホスファチジルグリセロール、ホスファチジルイノシトール、ジアシルホスファチジルコリン、ジ−O−アルキルホスファチジルコリン、リゾホスファチジルコリン、リゾホスファチジルエタノールアミン、リゾホスファチジルグリセロール、リゾホスファチジルイノシトール)、飽和及び不飽和脂肪酸誘導体(例えば、エチルエステル、プロピルエステル、コレステロールエステル、コエンザイムAエステル、ニトロフェニルエステル、ナフチルエステル、モノグリセリド、ジグリセリド及びトリグリセリド、脂肪族アルコール、脂肪族アルコール酢酸塩等)、リポ多糖、糖脂質、スフィンゴ脂質(例えば、セラミド、セレブロシド、ガラクトシルジグリセリド、ガングリオシド、ラクトセレブロシド、リゾスルファチド等)。
【0096】
「陽イオン性界面活性剤」は浸潤の条件下で陽性に荷電した官能基を有する。適する陽イオン性界面活性剤は、第4級又は第3級アミンを含むがそれらに限定されない。例となる第4級アミン界面活性剤は、以下のものを含むがそれらに限定されない。塩化セチルピリジニウム、臭化セチルトリメチルアンモニウム(CTAB;Calbiochem#B22633又はAldrich#85582−0)、塩化セチルトリメチルアンモニウム(CTACl;Aldrich#29273−7)、臭化ドデシルトリメチルアンモニウム(DTAB;Sigma#D−8638)、塩化ドデシルトリメチルアンモニウム(DTACl)、臭化オクチルトリメチルアンモニウム、臭化テトラデシルトリメチルアンモニウム(TTAB)、塩化テトラデシルトリメチルアンモニウム(TTACl)、臭化ドデシルエチルジメチルアンモニウム(DEDTAB)、臭化デシルトリメチルアンモニウム(DlOTAB)、臭化ドデシルトリフェニルホスホニウム(DTPB)、臭化オクタデシルイルトリメチルアンモニウム、塩化ステアロアルコニウム、塩化オレアルコニウム、塩化セトリモニウム、アルキルトリメチルアンモニウムメトサルフェート、パルミトアミドプロピルトリメチルクロリド、quaternium84(Mackernium NLE;Mcintyre Group,Ltd.)及び小麦脂質エポキシド(Mackernium WLE;Mcintyre Group,Ltd.)。例となる第3級アミン界面活性剤は、以下のものを含むがそれらに限定されない。オクチルジメチルアミン、デシルジメチルアミン、ドデシルジメチルアミン、テトラデシルジメチルアミン、ヘキサデシルジメチルアミン、オクチルデシルジメチルアミン、オクチルデシルメチルアミン、ジデシルメチルアミン、ドデシルメチルアミン、塩化トリアセチルアンモニウム、塩化セトリモニウム及び塩化アルキルジメチルベンジルアンモニウム。陽イオン界面活性剤のさらなる種類は、ホスホニウム、イミダゾリン及びエチル化アミン基を含むが、それらに限定されない。
【0097】
陰イオン性界面活性剤は、一般に、有機硫酸エステル及びスルホン酸エステルの水溶解アルカリ金属塩である。これらは、以下のものを含むがそれらに限定されない。ラウリン酸カリウム、硫酸ラウリルナトリウム、硫酸ドデシルナトリウム、硫酸アルキルポリオキシエチレン、アルギン酸ナトリウム、スルホコハク酸ジオクチルナトリウム、ホスファチジルコリン、ホスファチジルグリセロール、ホスファチジルイノシン、ホスファチジルセリン、ホスファチジン酸及びそれらの塩、グリセリルエステル、カルボキシルメチルセルロースナトリウム、コール酸及び他の胆汁酸(例えば、コール酸、デオキシコール酸、グリココール酸、タウロコール酸、グリコデオキシコール酸)及びそれらの塩(例えば、デオキシコール酸ナトリウム等)。
【0098】
エタノール、1−プロパノール及び1−ブタノール等の短鎖アルコールのような共力剤(co−surfactant)がさらに使用されてもよい。
上記界面活性剤のいずれかを組み合わせて使用してもよい。特に上記に列挙されていない界面活性剤も本発明の範囲に包含される。
【0099】
使用される界面活性剤の量は、界面活性剤及び処理される植物体組織(すなわち、葉の表面を覆っているワックスの厚さ等)の種類によって様々であろう。一般に、界面活性剤は、アグロバクテリウム懸濁液の体積の0.005%から約1%の範囲の濃度で使用される。好ましくは0.005%から約0.5%より好ましくは約0.005%から約0.05%の範囲の濃度である。一般に、イオン性界面活性剤は非イオン性界面活性剤より低い濃度で使用されるだろう。
ある好ましい態様では、Tween20等の非イオン性界面活性剤が使用される。
【0100】
界面活性剤を混合することに加えて、蛋白質発現を増加させるために、減圧衝撃を増大させるための浸透圧による衝撃工程を追加することができる。それゆえに、一態様では、ショ糖等の浸透圧衝撃剤が、蛋白質発現を増加させるために使用される。浸透圧衝撃剤の適する濃度は、20g/Lから100g/Lの範囲である。植物体組織がレタスである、一態様では、ショ糖が約60g/L使用される。
【0101】
例となる方法では、組み換えアグロバクテリウム培養物(すなわち、本発明に従った発現構成体を含んでいる)を、宿主及びベクター上に見出される耐性によって選択するために、適当な抗生物質を補充した修飾YEB培地(酵母抽出物6g/L、ペプトン5g/L、硫酸マグネシウム2mM及びショ糖5g/L)でおよそ2日間、生長させる。一過性発現に向けて、細胞を生長させるために、開始時のアグロバクテリウム培養物を、新たな修飾YEB培地で1:50に希釈する。抗生物質、50mMのリン酸カリウム緩衝液(pH5.8)及び20μMのアセトシリンゴンを添加する。28℃で18−24時間培養後、細胞は600nmでの吸光度(光学密度又はO.D.ともいわれる)2.5−3.5に達する。細胞を、好ましくは、もし必要であれば同じ培地を用いて、600nmでの吸光度2.5まで希釈する。
【0102】
細胞に、その後、ショ糖及びアセトシリンゴンを、最大で220μMのアセトシリンゴンと60g/Lのショ糖となるように補充する。懸濁液を、22℃で約1時間培養し、その後、浸潤に使用する。
【0103】
細胞を、再懸濁工程の必要がないように、いかなる遠心分離又は濃縮工程もなしに、直接、減圧条件下で浸潤する。この修正は、対数期にある培養物から細胞を遠心分離し、それらをムラシゲ・スクーグ培地(MS培地)に再懸濁する必要(Kapilaら,上記(1997))をなくし、新たに成長したアグロバクテリウム懸濁液で直接減圧浸潤することを可能にする。O.D.600nmが0.7−0.8(Kapilaら,1997)であるのに代えて、O.D.600nmが2.5−3.5までアグロバクテリウム細胞を成長させること、修飾YEB培地及びMESに代えてリン酸カリウム緩衝液を使用することは、発現レベル又は浸潤効率を変化させなかったが、処理の中でこの部分に要求されるコストと努力を顕著に減少させた。
【0104】
例えば、植物体組織がレタス由来であるような、一態様では、植物材料は、ビーカー中で、100μg/mlの2,4−D、0.005%のTween20と共に、あらかじめ培養されたアグロバクテリウム懸濁液に浸される。ビーカーは減圧乾燥器中に置かれ、減圧(29インチの水柱又は約7kPaに等しい)が20分間適用され、減圧の急速な解除による減圧衝撃が続く。レタス葉球全体の使用は、以前に記載されるような(Kapilaら.,上記(1997)、Vaqueroら,上記(1999))組織化されていない葉を集めたものと比べて、著しく簡便である。一般に、レタスの葉球全体は、葉の生物量に比較してより多くの発現レベルを与えると思われる。本発明の方法は、多数の葉からなるレタス葉球すなわちより多量の葉を一斉に処理することによって、容易に規模を大きくすることができる。同じアグロバクテリウム細胞懸濁液が、発現効率を顕著に減少させることなく、生産コストと労力をより減少させる減圧浸潤に、少なくとも2度使用することができる。
【0105】
一態様では、レタスの葉球全体、およそ300−400gが使用される。別の態様では、分離された葉が、界面活性剤及び/又は浸透圧剤の量及び組合せを最適化するために、予備的に使用される。
【0106】
好ましくは、減圧浸潤に続いてレタスの葉球が室温で約3−7日間光を受けて培養され、蛋白質抽出のためにホモジナイズされる。モノクローナル抗体及び他の製薬に重要な蛋白質が植物ホモジネートから適当な精製手順を用いて精製される。
【0107】
他の従来技術の方法と比べて、本過程は単純であり、より少ない工程からなり、異種蛋白質の発現が劇的に増加するようになる。
【0108】
蛋白質の分離
採収した後、蛋白質の分離が当業界で日常的に使用されている方法を用いて実施されるかもしれない。例えば、生物量の少なくとも一部はホモジナイズされ、組換え蛋白質が抽出され、さらに精製されるかもしれない。抽出は、適する溶媒にホモジネートを浸すか又は沈めることを包含するかもしれない。蛋白質は、例えば、米国特許第6,284,875号に記載されるような、減圧浸潤方法によって、植物の組織液からも分離されるかもしれない。
【0109】
精製方法は、免疫親和性による精製、蛋白質又は蛋白質複合体の特異的な大きさ、電気泳動移動度、生物学的な活性、及び/又は分離される異種蛋白質の正味の電荷に基づく又は蛋白質中のタグとなる分子の存在に基づく精製手順を含むがそれらに限定されない。
【0110】
分離された蛋白質の特徴は、免疫アッセイによって又は当業界で公知の他の方法によって導くことができる。例えば、蛋白質が実質的に分離されると、ウエスタンブロッティングによって又はクーマシーブルー染色によってSDS−PAGEゲル上で蛋白質が解析できる。分離された蛋白質は、生物学的な活性をアッセイするため、蛋白質の構造を(例えば、結晶化アッセイで)特徴付けるため、疾患の非動物ヒトモデルで有効性試験を行うため、最適な蛋白質活性及び/又は最適な製薬特性についてスクリーニングするため等に使用できる。
【0111】
本明細書で引用した全ての特許及び非特許文献は、本発明の属する分野における当業者の技術レベルを示すものである。全てのこれら刊行物及び特許出願は、個々の刊行物又は特許出願が、明確にそれぞれ、本明細書に参照として取り込まれるものとして示されているように、参照として本明細書に取り込まれる。
(実施例)
本発明は、実用的な実施例の方法によってしめされるが、これらの実施例は説明を目的としたものであり、本発明をなんら限定するものではない。
【実施例1】
【0112】
新規で迅速な蛋白質発現システムによるhOATの生産
本明細書に記載される方法は、上記のKapilaら、(1997)の教示による一過性発現システムを著しく改変するものであり、結果として劇的に改良された、低コストで、またより少ない工程で高品質な産物を発現させるものである。
【0113】
【表1】
【0114】
高ショ糖存在下での前処理
植物発現ベクターの構築
所定の遺伝子を含むプラスミドベクターを標準的な分子生物学的手法を用いて構築した。基礎となる要素は、E.coli及びアグロバクテリウムの両者において複製を可能とする開始プラスミド、プロモーターによって制御される所定の遺伝子の側面に位置する右及び左T−DNA境界及び標的化配列を含む。図1A及び1Bに示されるプラスミドを作製にあたって、必要な要素を集めて組みあわせる。
【0115】
図1Aは、2,4−Dにて誘導可能なプロモーター(OCS)3Masプロモーター(Gelvinら,米国特許第5,955,646号)を含み、抗組織因子抗体(IgG1)の重鎖に翻訳上融合されたズブチリシン(subtilisin)分泌配列(Janzikら,Biol.Chem.275:5193−5199(2000))を制御する、プラスミドpSUNP1を示す。さらに、プラスミドはカナマイシン耐性に対する選択可能なマーカー及び本一過性発現システムには有用性のないbialphos耐性に対するBAR遺伝子を含む。
【0116】
図1Bに示される、プラスミドpSUNP2は、抗組織因子抗体の重鎖が同じ抗体の軽鎖(カッパー)によって置換された点を除いて、pSUNP1に類似している。
【0117】
形質転換及びアグロバクテリウムの調製
所望のバイナリーベクター作製用のAgrobacterium tumefaciens C58/C1培養物を、プラスミド及び適当な細菌を選択するために、適当な抗生物質(100μg/mLのカナマイシン、15μg/mLのリファンピシン、25μg/mLのゲンタマイシン)を含む、修飾YEB培地(6g/Lの酵母抽出物、5g/Lのペプトン、5g/Lのショ糖、2mMの硫酸マグネシウム)で28℃で2日間成長させた。この培養物を、抗生物質、50mMのリン酸カリウム緩衝液pH5.6、20μMのアセトシリンゴンを補充した修飾YEB培地で1:50に希釈し、およそ18−24時間、O.D.600nmがおよそ2.5−3.5になるまで培養した。必要に応じて、(220μMのアセトシリンゴン及び60g/Lのショ糖という最大量を与えるために)細菌細胞をO.D.600nmが2.4になるよう希釈し、55g/Lのショ糖及び200μMのアセトシリンゴンを補充した。懸濁液を室温(約22℃)にて1時間インキュベートし、100μgの2,4−D(2,4−ジクロロフェノキシ酢酸、Sigma)及び0.005%Tween20を使用前に添加した。
【0118】
比較のためのKapilaら,上記(1997)によって記述される方法では、YEB培地は、プラスミドを選択するための適当な抗生物質(100μg/mLのカナマイシン、15μg/mLのリファンピシン、25μg/mLのゲンタマイシン)と共に、5g/Lの牛肉抽出物、1g/Lの酵母抽出物、5g/Lのペプトン、5g/Lのショ糖、2mMの硫酸マグネシウムから成る。開始時の培養物を、抗生物質、10mMのMESpH5.6、20μMのアセトシリンゴンを補充したYEB培地で1:500に希釈し、およそ18−24時間、O.D.600nmがおよそ0.7−1になるまで培養した。培養物を細胞を回収するために遠心分離し、10mMのMESpH5.6、20g/Lのショ糖及び200μMのアセトシリンゴンを含むムラシゲ・スクーグ培地(MS培地)(Kapilaら,上記(1997))にO.D.600nmが2.4になるよう再懸濁した。懸濁液を室温(約22℃)にて1時間インキュベートし、100μgの2,4−Dを添加した。
【0119】
減圧浸潤及び処理されたレタスのインキュベーション
アグロバクテリウム懸濁液を減圧浸潤に直接使用した。軽鎖が1つのプラスミド上にあり重鎖が2つめのプラスミド上にある場合(デュアルベクターシステム(dual vector system))、2つのアグロバクテリウム培養物を調製し、浸潤前に等量ずつ混合しなければならない。この例では、カッパー軽鎖をコードする軽鎖ベクターはhOATと呼ばれる抗組織因子抗体に由来し、重鎖ベクターはhOATのIgG1重鎖をコードした。レタスの葉球全体を2Lビーカー中の1.2Lの懸濁液に浸し、減圧乾燥器中に置いた。減圧(7kPaに等しい)を20分間適用した後、減圧の急激な解除による減圧衝撃を続けた。レタスの葉を水で洗浄し、閉じた透明な箱内の湿ったペーパータオル上で室温にて16時間日光をあてながら3−4日間インキュベートした。3−4日間後、葉を基部から切り、蛋白質抽出のためにホモジナイズした。
【0120】
蛋白質の抽出及び精製
蛋白質を、葉の重量と等しい体積の緩衝液を用いて、緩衝液(100mMのTris−HCl、5mMのEDTA、pH8.0、使用前に1.5%不溶性PVPを添加)中で抽出した。葉をワーリングブレンダー中で高速で1分間ホモジナイズし、得られたホモジネートを10,000xgで15分間遠心分離した。沈降しなかった細胞破片を有する上清を12層の薄地の綿布で濾過し、20,000xgで15分間遠心分離した。濾液をrProteinA sepharose fast flow affinity column(5mL、樹脂はAmersham Pharmacia Biotech AB、Swedenより)に流速2mL/minでかけた。使用した洗浄緩衝液、溶出緩衝液はそれぞれ0.1Mの酢酸ナトリウム及び0.1Mの酢酸であった。蛋白質を、段階的な勾配pH法、すなわち2カラム容積に対して20%の0.1M酢酸、4カラム容積に対して40%、60%及び100%の0.1M酢酸を続けることによって、溶出した(図3)。ピーク画分を集め、1MのTris−HCl、pH8.0を用いてpHを8.0に調整した。精製した抗体を280nmのO.D.を用いて定量した。さらなる精製のために、Q Sepharose fast flow column(5mL、樹脂はAmersham Pharmacia Biotech AB、Swedenより)を20mMTris−HClpH8.5で平衡化し、同じ緩衝液でプロテインA精製した抗体の緩衝液を交換し、カラムにかけた。抗体を、塩段階的溶出法、すなわち2カラム容積に対して20mMのTris−HCl、pH8.0、1MのNaClの10%緩衝液、4カラム容積に対して50%緩衝液、2カラム容積に対して100%緩衝液を続けることによって、溶出した。ピーク画分を集め、280nmのO.D.を用いて定量した(図4)。緩衝液交換のために、Millipore Ultrafree Centrifugal filter device 15mL(Millipore Corporation、Bedford、MA)を少なくとも1時間0.1MのNaOHに浸した。Qセファロースカラムで精製した抗体の緩衝液の交換はPBSで行い、1000倍希釈以上を得た。緩衝液を交換した抗体をMillex−GV 0.22μm filter unit(Millipore Corporation、Bedford、MA)で濾過し、280nmのO.D.を用いて定量した。
【0121】
hOAT抗体をELISAアッセイを用いて定量した。1枚のプレートを調製するために、コーティング緩衝液(35mMのNaHCO3、15mMの炭酸水素ナトリウム、50mMのNaCl、pH9.0)11mL中に5.5μgの組換え組織因子(rTF)を溶解したコーティング溶液を調製する。100μlのコーティング溶液をそれぞれのウェルに移し、覆った状態で4℃で2週間まで貯蔵する。プレートを洗浄緩衝液(Kirkegaard&Perry)400μLで3回洗浄し、100μLのサンプルをそれぞれのウェルに移す。プレートを振動しながら室温にて1時間インキュベートし、その後、洗浄緩衝液で6回洗浄する。結合した抗体を、ペルオキシダーゼ結合ロバ抗ヒトIgG(H+L)(Jackson ImmunoResearch)を用いて室温にて10分間プレートをインキュベートし、洗浄緩衝液で6回洗浄することにより検出する。ABTS基質(BioFX)を(100μL)添加し、10分後反応を1%のSDS100μL添加することにより停止する。405nmにおける吸光度を測定する。本発明の実施例1の実施形態及びKapilaの方法、それぞれからの発現レベルの比較を表2に示す。
【0122】
【表2】
【0123】
SDS−PAGEを12%のトリスグリシンミニゲルでLaemmli(1970)によって記述されるように実施した。サンプルを、蛋白質抽出物をローディング緩衝液と混合し(4:1、v/v)、続けて70℃で10分間加熱することで調製した。蛋白質のバンドをクーマシーブルー(図5A)染色により検出し、又は電気泳動的にPVDF膜に転写した。膜を10%のスキムミルクを含む2XPBSの1:10希釈液で室温にて1時間ブロッキングした。洗浄後、膜を西洋ワサビペルオキシダーゼ結合抗ヒトIgG抗体(Binding site)と室温にて1時間インキュベートした。図5Bは、製造者の指示(Pierce)に従って強化化学発光ウエスタンブロット検出試薬で膜をインキュベートすることにより検出されるIgGの重鎖及び軽鎖を示している。
【実施例2】
【0124】
ショ糖濃度の最適化
ショ糖の濃度とショ糖による浸透圧衝撃の効果は、発現を増加させるための重要なパラメーターであろうと認識された。蛋白質の生産レベルにおけるショ糖濃度の効果を試験するため、ショ糖の濃度範囲を抗組織因子抗体の発現におけるその効果について試験した点を除いて、実施例1の手順に従った。図6の結果に基づき、ショ糖の最終濃度60g/Lを標準的な方法として選択した。
【実施例3】
【0125】
界面活性剤の最適化
界面活性剤の効果を特徴付けるため、使用する界面活性剤の種類と濃度の両者を多様化している点を除いて、実施例1に記載される手順に従った。
【0126】
【表3】
【0127】
表3は、異なる非イオン性及びイオン性界面活性剤の調査結果を示す。表は網羅的になってはいないが、このパラメーターの多様性から顕著な効果を示している。0.005%でTween−20は異種蛋白質の特に高い発現レベルを提供する。
【実施例4】
【0128】
一過性発現システムでの(OCS)3MASプロモーターによる発現における2,4−Dの影響
多くの異なるプロモーター(5’転写調節領域)が入手可能であり、植物における異種蛋白質の発現に広く用いられている。文献の調査及び限られた試験を、本発明の方法に適したプロモーターを決定するために行った。表4は、化学的な誘導物質2,4−Dを用いた合成プロモーターOCS3MASからのhOATの発現を示す。(OCS)3MASプロモーターは様々な因子、とりわけ創傷によって誘導されることが知られていたが、この研究はアグロバクテリウムを用いた遺伝子の一過性の送達も結果的に発現を誘導するものである。この研究は、実施例1で示されるように細菌の調製及び浸潤に関しては元来のKapilaの方法(MS方法)を用いて行われた、よって、他の実施例で見られるより発現レベルは低いものであった。この研究に基づき、最終濃度100μg/mLの2,4−Dが、……
【0129】
【表4】
【0130】
この限られた調査は単なる例を意味しており、Adh遺伝子又はeIF4等の「ハウスキーピング」遺伝子の報告されている強力なプロモーター又はRUBISCOの小サブユニットからの発現と比べて、非常に高い発現が嫌気的に誘導可能なプロモーターから得られると言えるかもしれない。
【実施例5】
【0131】
hOATの一過性発現におけるデュアルベクター(Dual vector)対2シストロン性ベクター(Bicistronic vector)の比較
この実施例では、(実施例1に記載されるような)デュアルベクターシステムの使用を単一のベクターからの重鎖及び軽鎖の発現(2シストロン性システム)と比較した。プラスミドpSUNP4(図2)は同じプラスミドのT−DNA領域に重鎖及び軽鎖を有するプラスミドベクターを表す。(OCS)3MASプロモーターが、ズブチリシン(subtilisin)をコードするシグナル配列と共に抗組織因子遺伝子の重鎖及び軽鎖の両者の発現を制御するために使用される。
【0132】
レタスを、デュアルベクター作製用の2つのアグロバクテリウム培養物又は2シストロン性ベクターpSUNP4作製用の単一のアグロバクテリウム培養物のいずれかで浸潤し、hOATの発現レベルを実施例1のように測定した。これら2つのベクターシステムを用いたhOATの発現レベルを表す結果は、表5に示される。
【0133】
【表5】
【実施例6】
【0134】
志賀毒素2に対する組換え抗体の一過性発現
この実施例では、一過性発現が他の抗体を生産するのに使用できることを示した。cαStx2は、腸管出血性大腸菌によって産生される志賀毒素2に結合し中和するキメラ抗体である。cαStx2重鎖及び軽鎖の遺伝子を、pSUNP1及びpSUNP2に類似したcαStx2ベクターを作成するために、デュアルベクターに導入した。これらの構築物を、実施例1に記載の方法を用いて、アグロバクテリウムに移入し、レタスをagro−infiltrationするために使用した。一過性発現後、プロテインA精製により得た抗体と共に、植物細胞からの粗抽出物をELISAにより解析した。
【0135】
ELISAを、Stx2抗原でmaxisorp96ウェルプレートをコートすることで実施したが、コートしたプレートはプラスチックフィルムで覆い、使用まで4℃で貯蔵した。抗体生産を測定するために、ウェルをコート溶液を取り除くため使用前に緩衝液で3回洗浄した。植物細胞抽出液又は精製された蛋白質溶液をコートしたウェルに添加した。室温で1時間後、ウェルを緩衝液で3回洗浄し、抗ヒトカッパ鎖−HRP抗体の希釈液を添加した。その後、プレートを室温で1時間インキュベートし、緩衝液で3回洗浄した。プローブ抗体の存在を検出するために、ABTS基質剤を添加し、室温で数分間インキュベートし、続けてABTS停止緩衝液を加えた。405nmの吸光度を自動プレートリーダーで測定した。
【0136】
ELISA解析によれば、一過性に発現された蛋白質は機能的に活性なcαStx2抗体を含むことが示された。
【0137】
当業者は通常の実験法のみで、本明細書中に記載されている特定の物質及び手順に対して多くの均等物を認識し、又は確かめることができるものである。このような均等物は本発明の範囲とみなされ、上述の特許請求の範囲により保護されるものである。
【0138】
本明細書の発明は、特定の実施態様を参照として記載されているが、これらの実施態様は単に本発明の原理の説明及び応用に過ぎないと理解されるべきである。それゆえに、多くの改変が説明のための実施態様に対してなされていてもよく、添付の請求項によって定義される、本発明の精神及び範囲から逸脱することなくその他の改良が工夫されてもよいものと理解されるべきである。
【0139】
特許、特許出願、国際出願及び本明細書中及び上記の参考文献は、それらの全体が本明細書中に取り込まれる。
【図面の簡単な説明】
【0140】
本発明の目的と特徴は、前述の詳細な説明と添付の図面を参照することにより、より良く理解することができるものである。
【図1A】図1Aは、本発明の一態様に従って植物体組織の同時浸潤に使用されたpSUNP1プラスミドの図である。プラスミドpSUNP1はOCS3MASプロモーターからhOATの重鎖を発現する。TRとTLの間に存在する全ての遺伝子を植物体の核へと導くT−DNAの境界が示される。これらのプラスミドのいずれかを有するアグロバクテリウムはレタス等の植物を同時浸潤するのに使用され、hOATの一過性発現を示した。
【図1B】図1Bは、本発明の一態様に従って植物組織の同時浸潤に使用されたpSUNP2プラスミドの図である。プラスミドpSUNP1はOCS3MASプロモーターからhOATの重鎖を発現する。TRとTLの間に存在する全ての遺伝子を植物核へと導くT−DNAの境界が示される。これらのプラスミドのいずれかを有するアグロバクテリウムはレタス等の植物を同時浸潤するのに使用され、hOATの一過性発現を示した。
【図2】図2は、植物体組織での一過性発現によって単一プラスミドからhOATの重鎖及び軽鎖の両者を発現するために使用可能な、2シストロン性の発現プラスミドpSUNP4の図である。
【図3】図3は、hOAT重鎖及び軽鎖の遺伝子を含むアグロバクテリウムで浸潤されたレタスから抽出された蛋白質の溶出プロフィールを示す。蛋白質はプロテインAカラムにかけられ溶出された。
【図4】図4は、本発明の一態様に従って分離され、Qセファロースカラムにかけられた蛋白質の溶出プロフィールを示す。かけられた蛋白質は、図3で使用されたプロテインAカラムからの溶出によって回収されたものであった。
【図5A】図5Aは、還元条件下で泳動されクーマシーブルー染色されたSDS−PAGEゲル及び本発明の一態様に従って得られたhOAT蛋白質の画分を示す。レーン1、分子量(Mr)標準;レーン2、精製されたCHO生産hOAT;レーン3、レタスで発現されプロテインAカラムから溶出されたhOAT。
【図5B】図5Bは、本発明の一態様に従って同定されSDS−PAGEで分離された蛋白質のウエスタンブロットを示す。プローブは抗H+L抗体。レーンA、市販のIgG;レーンB、プロテインAカラムから溶出され精製されたhOAT;レーンC、陰性コントロール−agro−infiltrationなしのレタスからの抽出物。
【図6】図6は、レタスでのhOAT発現レベルにおける、浸透圧衝撃に関して、ショ糖の様々な濃度効果を示す。hOATの発現は、抽出に用いられたレタス材料のキログラム当たり生産される(ELISAに基づく)抗体mgとして表される。
【特許請求の範囲】
【請求項1】
界面活性剤の存在下アグロバクテリウム細胞で植物体組織を浸潤すること、そこで当該アグロバクテリウム細胞は当該植物体組織の細胞及び/又は細胞間に異種ポリペプチドを発現できる発現構成体を含み、及び
当該植物体組織を当該ポリペプチドを発現するのに適した条件下でインキュベートすること、
を包含する、植物体に於いて異種蛋白質を発現する方法。
【請求項2】
アグロバクテリウム細胞で植物体組織を浸潤すること、そこで当該アグロバクテリウム細胞は当該植物体組織の植物細胞及び/又は細胞間に異種ポリペプチドを発現できる発現構成体を含み、
当該植物体組織を当該ポリペプチドを発現するのに適した条件下でインキュベートすること、及び、
処理された1キログラムの植物体組織から約1mg以上の当該ポリペプチドを分離すること、
を包含する、植物体に於いて異種蛋白質を発現する方法。
【請求項3】
アグロバクテリウム細胞で植物体組織を浸潤すること、そこで当該アグロバクテリウム細胞は当該植物体組織の植物細胞及び/又は細胞間に異種ポリペプチドを発現できる発現構成体を含み、及び
当該植物体組織を当該ポリペプチドを発現するのに適した条件下でインキュベートすること、を包含し、
そこで、当該植物体組織は収穫後少なくとも約1週間である植物から得られる、植物体に於いて異種蛋白質を発現する方法。
【請求項4】
植物体組織の植物細胞及び/又は細胞間に異種ポリペプチドを発現できる発現構成体を含むアグロバクテリウム細胞を培養培地中で培養すること、
当該アグロバクテリウム細胞を含む培養培地を希釈工程あり又はなしに当該植物体組織と直接接触させること、
当該アグロバクテリウム細胞を含む当該培養培地で当該植物細胞を浸潤すること、及び、
当該植物体組織を当該ポリペプチドを発現するのに適した条件下でインキュベートすること、
を包含する、植物体に於いて異種蛋白質を発現する方法。
【請求項5】
前記異種ポリペプチドが前記植物体組織から分離される、請求項1から3及び5の何れか一項に記載の方法。
【請求項6】
前記ベクターが少なくとも1つのT−DNA境界を含む、請求項1から5の何れか一項に記載の方法。
【請求項7】
さらに、前記ベクターを少なくとも1つのアグロバクテリウム細胞に導入する、及び、当該細胞を前記植物体組織を浸潤するために十分な量の細胞が得られるまで培養する、工程を包含する、請求項1から5の何れか一項に記載の方法。
【請求項8】
前記植物体が、レタス、アルファルファ、インゲンマメ、ホウレンソウ、タンポポ、アカチコリー、キバナスズシロ、エンダイブ、キクチシャ、チコリー、アーティチョーク、トウモロコシ、ポテト、イネ、ダイズ、Crucifera、ウキクサ、トウモロコシ、ポテト、イネ、ダイズ、ホウレンソウ、トマト及びタバコからなる群より選択される、請求項1から5の何れか一項に記載の方法。
【請求項9】
前記アブラナ科(Crucifera)の植物が、ブラッシカ(アブラナ属)又はアラビドプシス(シロイヌナズナ)を包含する、請求項8に記載の方法。
【請求項10】
前記アグロバクテリウムが一晩培養され、使用前に600nmでの吸光度が約2.5まで希釈される、請求項1から4の何れか一項に記載の方法。
【請求項11】
前記アグロバクテリウムが減圧下に浸潤される、請求項1から4の何れか一項に記載の方法。
【請求項12】
前記浸潤が界面活性剤の存在下で行われる、請求項2から4の何れか一項に記載の方法。
【請求項13】
前記界面活性剤が、Tween−20、Tween−80、Triton X−100、NP−40、Silwet L−77からなる群より選択される、請求項1に記載の方法。
【請求項14】
前記界面活性剤が、Tween−20、Tween−80、Triton X−100、NP−40、Silwet L−77からなる群より選択される、請求項12に記載の方法。
【請求項15】
前記界面活性剤が、約0.005%のTween−20である、請求項1に記載の方法。
【請求項16】
前記界面活性剤が、約0.005%のTween−20である、請求項12に記載の方法。
【請求項17】
前記浸潤が、浸透圧衝撃を誘導するための薬剤の存在下で実施される、請求項1から5の何れか一項に記載の方法。
【請求項18】
前記浸透圧衝撃を誘導するための薬剤がショ糖である、請求項17に記載の方法。
【請求項19】
前記ショ糖が濃度約60g/Lである、請求項18に記載の方法。
【請求項20】
前記蛋白質が植物体の全部又は葉をすりつぶすことによって回収される、請求項1から4の何れか一項に記載の方法。
【請求項21】
前記蛋白質が前記植物体の細胞間液から回収される、請求項1から4の何れか一項に記載の方法。
【請求項22】
前記蛋白質が前記植物細胞の小胞体へ標的化されている、請求項1から4の何れか一項に記載の方法。
【請求項23】
同義遺伝子(multiple genes)がアグロバクテリウムによって前記植物に送達される、請求項1から4の何れか一項に記載の方法。
【請求項24】
複数のアグロバクテリウム株が組み合わされ、所望の植物に同時浸潤される、請求項1から4の何れか一項に記載の方法。
【請求項25】
同義遺伝子が前記アグロバクテリウム株によって送達される、請求項24に記載の方法。
【請求項26】
前記同義遺伝子が、会合してマルチサブユニット蛋白質を形成する蛋白質をコードする、請求項23に記載の方法。
【請求項27】
発現した前記蛋白質が免疫グロブリンスーパーファミリーの蛋白質を包含する、請求項1から4の何れか一項に記載の方法。
【請求項28】
前記蛋白質が、抗体、T細胞受容体及び主要組織適合性複合体、又はそれらの生物学的に機能的なフラグメント又は一本鎖誘導体である、請求項27に記載の方法。
【請求項29】
前記同義遺伝子が経路の一員を包含する蛋白質をコードする、請求項25に記載の方法。
【請求項30】
前記経路が化学合成経路又は情報伝達経路である、請求項29に記載の方法。
【請求項31】
前記蛋白質がグリコシル化されている、請求項1から4の何れか一項に記載の方法。
【請求項32】
すでに発現している前記発現構成体を得る、及び、それを植物体に安定に導入する、工程を更に包含する、請求項1から4の何れか一項に記載の方法。
【請求項33】
前記植物体組織がトランスジェニック植物由来である、請求項1から4の何れか一項に記載の方法。
【請求項34】
複数の発現構成体が、少なくとも約50%の相同性を有し、及び、1又はそれ以上の突然変異の存在によって互いに異なる、及び/又は、1又はそれ以上の変異体アミノ酸をコードする、コード配列を含み、当該複数の発現構成体を用いて複数の植物体組織サンプルにおいて請求項1から4の何れか一項に記載の方法を実施することを包含する、突然変異又は変異体の配列についてスクリーニングする方法。
【請求項35】
前記1又はそれ以上の突然変異が、欠失、挿入、置換又は転位を包含する、請求項34に記載の方法。
【請求項36】
1又はそれ以上の突然変異を含まず及び変異体アミノ酸配列をコードしない異種ポリペプチドと比較して、改良された特性を有する異種ポリペプチドをコードする発現構成体が同定されるものである、請求項34に記載の方法。
【請求項37】
前記改良された特性が増加した活性及び/又は安定性を包含する、請求項36に記載の方法。
【請求項38】
前記増加した活性が、異種蛋白質に特異的に結合する結合パートナーに対する異種蛋白質の増加した結合親和性を包含する、請求項37に記載の方法。
【請求項39】
前記蛋白質が免疫グロブリンスーパーファミリーの一員を包含する、請求項38に記載の方法。
【請求項40】
前記蛋白質が、抗体、T細胞受容体及び主要組織適合性複合体、又はそれらの生物学的に機能的なフラグメント又は一本鎖誘導体からなる群より選択される、請求項39に記載の方法。
【請求項41】
前記1又はそれ以上の突然変異が、ヒトではない動物由来の異種コード配列にヒト配列を挿入することを包含する、請求項33に記載の方法。
【請求項1】
界面活性剤の存在下アグロバクテリウム細胞で植物体組織を浸潤すること、そこで当該アグロバクテリウム細胞は当該植物体組織の細胞及び/又は細胞間に異種ポリペプチドを発現できる発現構成体を含み、及び
当該植物体組織を当該ポリペプチドを発現するのに適した条件下でインキュベートすること、
を包含する、植物体に於いて異種蛋白質を発現する方法。
【請求項2】
アグロバクテリウム細胞で植物体組織を浸潤すること、そこで当該アグロバクテリウム細胞は当該植物体組織の植物細胞及び/又は細胞間に異種ポリペプチドを発現できる発現構成体を含み、
当該植物体組織を当該ポリペプチドを発現するのに適した条件下でインキュベートすること、及び、
処理された1キログラムの植物体組織から約1mg以上の当該ポリペプチドを分離すること、
を包含する、植物体に於いて異種蛋白質を発現する方法。
【請求項3】
アグロバクテリウム細胞で植物体組織を浸潤すること、そこで当該アグロバクテリウム細胞は当該植物体組織の植物細胞及び/又は細胞間に異種ポリペプチドを発現できる発現構成体を含み、及び
当該植物体組織を当該ポリペプチドを発現するのに適した条件下でインキュベートすること、を包含し、
そこで、当該植物体組織は収穫後少なくとも約1週間である植物から得られる、植物体に於いて異種蛋白質を発現する方法。
【請求項4】
植物体組織の植物細胞及び/又は細胞間に異種ポリペプチドを発現できる発現構成体を含むアグロバクテリウム細胞を培養培地中で培養すること、
当該アグロバクテリウム細胞を含む培養培地を希釈工程あり又はなしに当該植物体組織と直接接触させること、
当該アグロバクテリウム細胞を含む当該培養培地で当該植物細胞を浸潤すること、及び、
当該植物体組織を当該ポリペプチドを発現するのに適した条件下でインキュベートすること、
を包含する、植物体に於いて異種蛋白質を発現する方法。
【請求項5】
前記異種ポリペプチドが前記植物体組織から分離される、請求項1から3及び5の何れか一項に記載の方法。
【請求項6】
前記ベクターが少なくとも1つのT−DNA境界を含む、請求項1から5の何れか一項に記載の方法。
【請求項7】
さらに、前記ベクターを少なくとも1つのアグロバクテリウム細胞に導入する、及び、当該細胞を前記植物体組織を浸潤するために十分な量の細胞が得られるまで培養する、工程を包含する、請求項1から5の何れか一項に記載の方法。
【請求項8】
前記植物体が、レタス、アルファルファ、インゲンマメ、ホウレンソウ、タンポポ、アカチコリー、キバナスズシロ、エンダイブ、キクチシャ、チコリー、アーティチョーク、トウモロコシ、ポテト、イネ、ダイズ、Crucifera、ウキクサ、トウモロコシ、ポテト、イネ、ダイズ、ホウレンソウ、トマト及びタバコからなる群より選択される、請求項1から5の何れか一項に記載の方法。
【請求項9】
前記アブラナ科(Crucifera)の植物が、ブラッシカ(アブラナ属)又はアラビドプシス(シロイヌナズナ)を包含する、請求項8に記載の方法。
【請求項10】
前記アグロバクテリウムが一晩培養され、使用前に600nmでの吸光度が約2.5まで希釈される、請求項1から4の何れか一項に記載の方法。
【請求項11】
前記アグロバクテリウムが減圧下に浸潤される、請求項1から4の何れか一項に記載の方法。
【請求項12】
前記浸潤が界面活性剤の存在下で行われる、請求項2から4の何れか一項に記載の方法。
【請求項13】
前記界面活性剤が、Tween−20、Tween−80、Triton X−100、NP−40、Silwet L−77からなる群より選択される、請求項1に記載の方法。
【請求項14】
前記界面活性剤が、Tween−20、Tween−80、Triton X−100、NP−40、Silwet L−77からなる群より選択される、請求項12に記載の方法。
【請求項15】
前記界面活性剤が、約0.005%のTween−20である、請求項1に記載の方法。
【請求項16】
前記界面活性剤が、約0.005%のTween−20である、請求項12に記載の方法。
【請求項17】
前記浸潤が、浸透圧衝撃を誘導するための薬剤の存在下で実施される、請求項1から5の何れか一項に記載の方法。
【請求項18】
前記浸透圧衝撃を誘導するための薬剤がショ糖である、請求項17に記載の方法。
【請求項19】
前記ショ糖が濃度約60g/Lである、請求項18に記載の方法。
【請求項20】
前記蛋白質が植物体の全部又は葉をすりつぶすことによって回収される、請求項1から4の何れか一項に記載の方法。
【請求項21】
前記蛋白質が前記植物体の細胞間液から回収される、請求項1から4の何れか一項に記載の方法。
【請求項22】
前記蛋白質が前記植物細胞の小胞体へ標的化されている、請求項1から4の何れか一項に記載の方法。
【請求項23】
同義遺伝子(multiple genes)がアグロバクテリウムによって前記植物に送達される、請求項1から4の何れか一項に記載の方法。
【請求項24】
複数のアグロバクテリウム株が組み合わされ、所望の植物に同時浸潤される、請求項1から4の何れか一項に記載の方法。
【請求項25】
同義遺伝子が前記アグロバクテリウム株によって送達される、請求項24に記載の方法。
【請求項26】
前記同義遺伝子が、会合してマルチサブユニット蛋白質を形成する蛋白質をコードする、請求項23に記載の方法。
【請求項27】
発現した前記蛋白質が免疫グロブリンスーパーファミリーの蛋白質を包含する、請求項1から4の何れか一項に記載の方法。
【請求項28】
前記蛋白質が、抗体、T細胞受容体及び主要組織適合性複合体、又はそれらの生物学的に機能的なフラグメント又は一本鎖誘導体である、請求項27に記載の方法。
【請求項29】
前記同義遺伝子が経路の一員を包含する蛋白質をコードする、請求項25に記載の方法。
【請求項30】
前記経路が化学合成経路又は情報伝達経路である、請求項29に記載の方法。
【請求項31】
前記蛋白質がグリコシル化されている、請求項1から4の何れか一項に記載の方法。
【請求項32】
すでに発現している前記発現構成体を得る、及び、それを植物体に安定に導入する、工程を更に包含する、請求項1から4の何れか一項に記載の方法。
【請求項33】
前記植物体組織がトランスジェニック植物由来である、請求項1から4の何れか一項に記載の方法。
【請求項34】
複数の発現構成体が、少なくとも約50%の相同性を有し、及び、1又はそれ以上の突然変異の存在によって互いに異なる、及び/又は、1又はそれ以上の変異体アミノ酸をコードする、コード配列を含み、当該複数の発現構成体を用いて複数の植物体組織サンプルにおいて請求項1から4の何れか一項に記載の方法を実施することを包含する、突然変異又は変異体の配列についてスクリーニングする方法。
【請求項35】
前記1又はそれ以上の突然変異が、欠失、挿入、置換又は転位を包含する、請求項34に記載の方法。
【請求項36】
1又はそれ以上の突然変異を含まず及び変異体アミノ酸配列をコードしない異種ポリペプチドと比較して、改良された特性を有する異種ポリペプチドをコードする発現構成体が同定されるものである、請求項34に記載の方法。
【請求項37】
前記改良された特性が増加した活性及び/又は安定性を包含する、請求項36に記載の方法。
【請求項38】
前記増加した活性が、異種蛋白質に特異的に結合する結合パートナーに対する異種蛋白質の増加した結合親和性を包含する、請求項37に記載の方法。
【請求項39】
前記蛋白質が免疫グロブリンスーパーファミリーの一員を包含する、請求項38に記載の方法。
【請求項40】
前記蛋白質が、抗体、T細胞受容体及び主要組織適合性複合体、又はそれらの生物学的に機能的なフラグメント又は一本鎖誘導体からなる群より選択される、請求項39に記載の方法。
【請求項41】
前記1又はそれ以上の突然変異が、ヒトではない動物由来の異種コード配列にヒト配列を挿入することを包含する、請求項33に記載の方法。
【図1A】

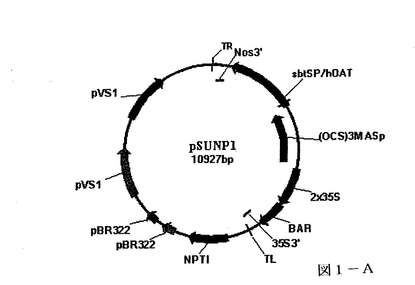
【図1B】
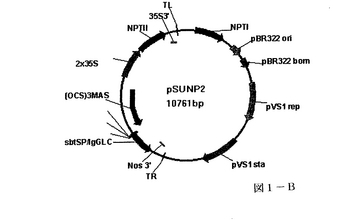
【図2】
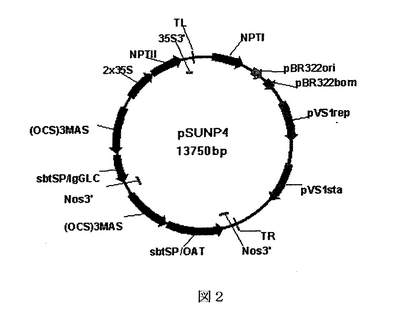
【図3】
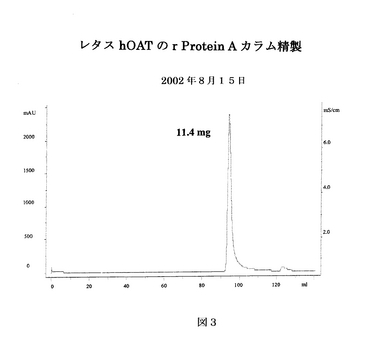
【図4】
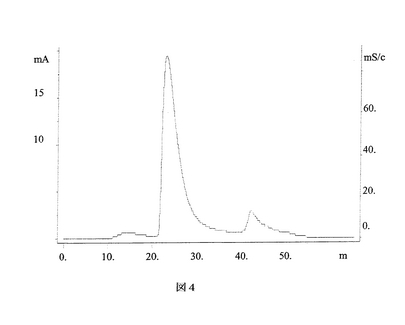
【図5A】
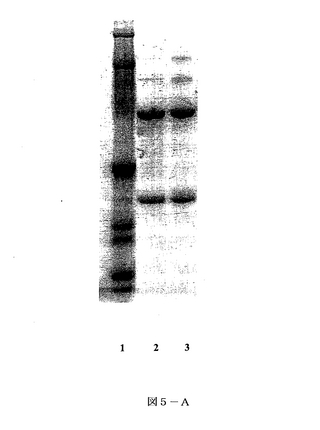
【図5B】

【図6】
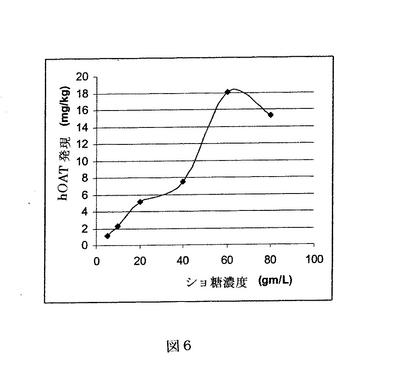
【図1B】
【図2】
【図3】
【図4】
【図5A】
【図5B】
【図6】
【公表番号】特表2006−518994(P2006−518994A)
【公表日】平成18年8月24日(2006.8.24)
【国際特許分類】
【出願番号】特願2005−516755(P2005−516755)
【出願日】平成15年12月17日(2003.12.17)
【国際出願番号】PCT/US2003/040451
【国際公開番号】WO2005/076766
【国際公開日】平成17年8月25日(2005.8.25)
【出願人】(504090190)アルター・バイオサイエンス・コーポレーション (8)
【Fターム(参考)】
【公表日】平成18年8月24日(2006.8.24)
【国際特許分類】
【出願日】平成15年12月17日(2003.12.17)
【国際出願番号】PCT/US2003/040451
【国際公開番号】WO2005/076766
【国際公開日】平成17年8月25日(2005.8.25)
【出願人】(504090190)アルター・バイオサイエンス・コーポレーション (8)
【Fターム(参考)】
[ Back to top ]