
植物体の子実を増大させる遺伝子、並びにその利用
【課題】本発明は、植物体の子実粒(種子、頴花又は果実を含む)ひいては粒長の伸長に関する新規な遺伝子の単離・同定、並びに該遺伝子を利用した植物体の子実を増大させる育種方法を提供することを目的とする。
【解決手段】連鎖解析により、植物体の子実(種子、頴花又は果実を含む)の伸長ひいては収量の増加に関するGL5遺伝子の単離・同定に成功した。また、該遺伝子を利用して単子葉植物又は双子葉植物の子実を増大させることができることが明らかとなった。
【解決手段】連鎖解析により、植物体の子実(種子、頴花又は果実を含む)の伸長ひいては収量の増加に関するGL5遺伝子の単離・同定に成功した。また、該遺伝子を利用して単子葉植物又は双子葉植物の子実を増大させることができることが明らかとなった。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、植物体の子実(種子、頴花又は果実を含む)を増大させる機能を有するイネ由来のタンパク質をコードするDNA、および該DNAを用いた子実収量(種子、頴花又は果実の収量のことをいう)の増加方法、子実収量の検出方法、子実収量が増加する植物体の育種方法に関する。
【背景技術】
【0002】
世界人口が爆発的に増え作物(例えば穀物)の生産増が求められている。世界の年間人口増加率1.4%に対し、穀物生産増加率は1.0%と人口増加率に比べ低く、世界人口が80億を突破する2025年には作物需要は50%上昇すると予想され、食糧不足も一層加速していると予想される。作物生産量を上昇させるためには、収量増加と関与する遺伝子の特定と特定した遺伝子を利用した効率的な穀物育種が必須である。
【0003】
イネにおいて子実の粒サイズはシンクのキャパシティーを示す値であり、収量に直接関わる。これまでに、イネにおいて種子(粒)の大きさに関わる遺伝子として、GW2(非特許文献1)及びGS3(非特許文献2)がポジショナルクローニング法により単離されている。しかし、これらの遺伝子は収量増に貢献しない。また、plastchron 1遺伝子を発現させ粒を拡大したトランスジェニック植物とその利用方法(特許文献1)、改変した3量体Gタンパク質αサブユニット遺伝子を利用し巨大粒を結実させる方法(特許文献2)、及びイネの粒長を制御するLk3遺伝子およびその利用(特許文献3)が特許出願されている。さらに、本願発明者らによって、粒重ひいては収量を増加させる遺伝子(TGW6)が単離されている(特許文献4)。
【0004】
しかしながら、イネの種子を増大させるには、現在までに単離された遺伝子だけでは十分ではなかった。また、種を超えて子実を増大させる方法は明らかではなかった。
なお、本出願の発明に関連する先行技術文献情報を以下に示す。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2005-204621
【特許文献2】特開2004-033199
【特許文献3】再表2006-112327
【特許文献4】特願2008-292285
【非特許文献】
【0006】
【非特許文献1】Song X.J. et al., (2007) Nature Genet. 39, 623-630
【非特許文献2】Fan C. et al., (2006) Theor. Appl. Genet. 112, 1164-1171
【非特許文献3】Shomura A et al. (2008) Nat. Genet. 40, 1023-1028.
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明は、このような状況に鑑みてなされたものであり、その目的は、植物の子実粒形(頴花・果実・種子を含む)ひいては粒長の伸長に関する新規な遺伝子の単離・同定、並びに該遺伝子を利用した植物の子実(頴花・果実・種子を含む)の粒サイズを増加させる育種方法を提供することにある。
【課題を解決するための手段】
【0008】
発明者らは、これまで植物体の作物収量増加のため、単子葉植物のモデルであるイネを用いて直接的に収量増加を担う遺伝子、すなわち粒長の伸長に関する遺伝子の探索を試みてきた。粒長は複数の遺伝子の相互作用による量的形質として支配されている。
【0009】
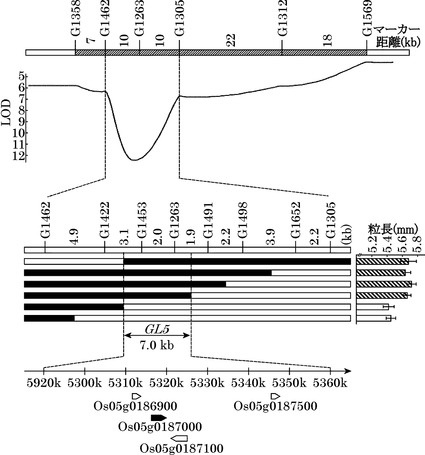
発明者らは、まず、コシヒカリと、コシヒカリを遺伝的背景にGL5領域をカサラス型染色体に置換した準同質遺伝子系統NIL(GL5)との戻し交雑後代を用いて、連鎖解析を行った。GL5遺伝子座近傍のSNPマーカーを用いて連鎖解析を行った結果、GL5遺伝子座をSNPマーカーG1422とSNPマーカーG1491との間、約7kbpの範囲に特定した(図1)。次に、イネアノテーションプロジェクトにより公開された遺伝子予測データベースにより、遺伝子の発現量・発現部位からGL5遺伝子を1つの遺伝子(Os05g0187000)に絞り込み、本遺伝子は20Sプロテアソームのβ2サブユニットをコードしていることが明らかとなった(図2)。
【0010】
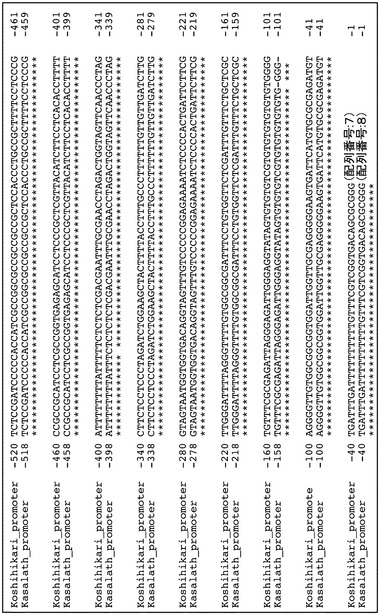
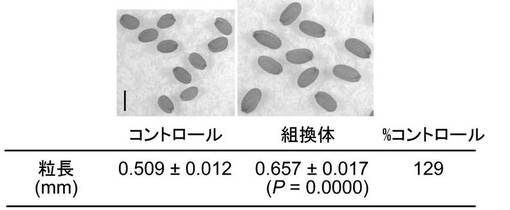
コシヒカリとカサラスのcDNA塩基配列解析の結果、GL5候補遺伝子(OsPBB1)のコーディング領域に2カ所のSNPを有していたが、これらはいずれもアミノ酸配列に変化を生じさせず、コシヒカリとカサラスのGL5候補遺伝子のアミノ酸配列は同一であることが明らかとなった(図3)。また、OsPBB1の推定プロモーター領域にもいくつかの変異が存在することが明らかとなった(図4)。
【0011】
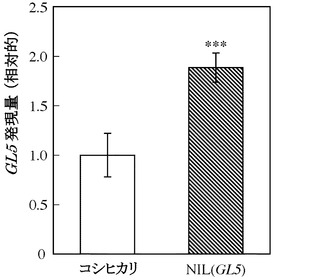
次に、OsPBB1遺伝子の発現量を比較したところ、コシヒカリに比べてNIL(GL5)は、発現量が約2.8倍高いことが明らかとなった(図5)。
【0012】
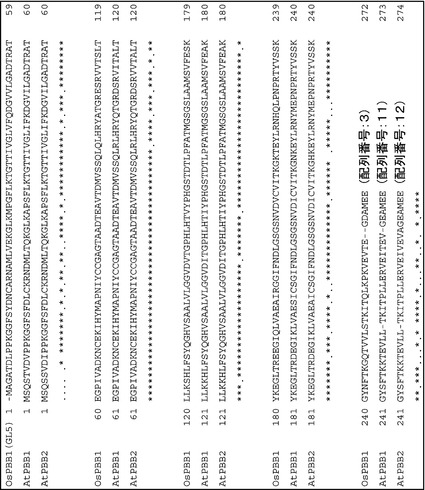
次に、コシヒカリと同等のOsPBB1発現量を持つ日本晴にOsPBB1を過剰発現させる形質転換体によりOsPBB1の機能証明を行ったところ、OsPBB1の過剰発現は玄米の長さを有意に増加させることが明らかとなった(図6)。
【0013】
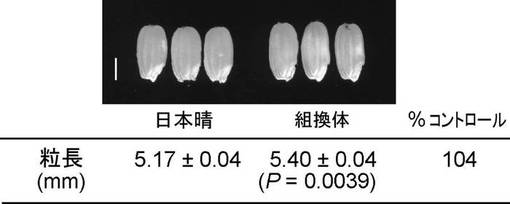
GL5遺伝子(OsPBB1)の高発現がイネの粒長を増大させることが判明したことから、さらに他の植物種における同遺伝子高発現の効果を調べた。その結果、OsPBB1の高発現は、シロイヌナズナにおいても種子サイズを増大させることが明らかとなった(図8)。これによりGL5は単子葉植物であるイネのみならず双子葉植物であるシロイヌナズナにおいても種子サイズを増大させる効果があり、幅広い植物種で同様の効果を有する事が示唆された。
【0014】
本発明におけるGL5遺伝子は粒長、粒長の伸長を介し収量特性を向上させる遺伝子として初めて特定されたものであり、GL5遺伝子を利用することで収量増、生産性の向上に利用できる。即ち、本発明者らは、植物体の粒形の増大に関与する新たな遺伝子を単離することに成功した。
【0015】
本発明は、より具体的には以下の〔1〕〜〔19〕を提供するものである。
〔1〕下記(a)から(d)のいずれかに記載のDNAを含む、植物体の子実(種子、頴花又は果実を含む)の大きさを増大させた植物体。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
〔2〕植物体が単子葉植物又は双子葉植物である、〔1〕に記載の植物体。
〔3〕下記(a)から(d)のいずれかに記載のDNAが発現するように導入されたベクター。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
〔4〕〔3〕 に記載のベクターが導入された宿主細胞。
〔5〕〔4〕に記載のベクターが導入された植物細胞。
〔6〕〔5〕に記載の植物細胞を含む形質転換植物体。
〔7〕〔6〕に記載の形質転換植物体の子孫またはクローンである、形質転換植物体。
〔8〕〔6〕又は〔7〕に記載の形質転換植物体の繁殖材料。
〔9〕下記(a)から(d)のいずれかに記載のDNAを植物体の細胞内で過剰発現させる工程を含む、植物体の子実(種子、頴花又は果実を含む)の大きさを増大させる方法。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
〔10〕植物体が単子葉植物又は双子葉植物である、〔9〕に記載の方法。
〔11〕被検植物体について、配列番号:1に記載のDNA領域に相当する部位、またはその周辺配列に存在する変異を含むDNAマーカーを検出することを特徴とする、被検植物体の子実(種子、頴花又は果実を含む)の収量を検出する方法。
〔12〕変異が一塩基多型である、〔11〕に記載の方法。
〔13〕以下の工程(a)及び(b)を含む、〔11〕に記載の方法。
(a)被検植物における配列番号:1に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域に相当する部位、またはその周辺配列に存在する多型部位の塩基種を決定する工程、及び
(b)(a)で決定された多型部位の塩基種において、配列番号:1またはその周辺配列と異なるアレルが検出された場合に、被検植物は子実(種子、頴花又は果実を含む)の収量が多いと判定する工程
〔14〕多型部位が、配列番号:1に記載の塩基配列における、32位、129位、161位、557位、839位、843位、976位又は1699位から選択される少なくとも一つの多型部位に相当する部位である、〔13〕に記載の方法。
〔15〕多型部位の塩基種の変異が、配列番号:1に記載の塩基配列における32位の塩基種のGからTへの変異、129位の塩基種のAからGへの変異、161位の塩基種のTからGへの変異、557位の塩基種のTからCへの変異、839位の塩基種のTの欠損、843位の塩基種のGの欠損、976位の塩基種のCからAへの変異、又は1699位の塩基種のCからTへの変異から選択される少なくとも一つの変異である場合には、子実(種子、頴花又は果実を含む)が増大すると判定する、〔14〕に記載の方法。
〔16〕配列番号:1に記載のDNAとストリンジェントな条件で特異的にハイブリダイズし、少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドを含む、被検植物の子実(種子、頴花又は果実を含む)の収量を検出するためのプライマー。
〔17〕〔14〕または〔15〕に記載の多型部位を含む領域を増幅するための、〔16〕に記載のプライマー。
〔18〕配列番号:1に記載のDNAとストリンジェントな条件で特異的にハイブリダイズし、少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドを含む、被検植物体の子実(種子、頴花又は果実を含む)の収量を検出するためのプローブ。
〔19〕〔14〕または〔15〕に記載の多型部位を含む領域に特異的にハイブリダイズする、〔18〕に記載のプローブ。
【発明の効果】
【0016】
連鎖解析により、植物の子実粒(種子、頴花又は果実を含む)の伸長ひいては収量の増加に関する遺伝子の単離・同定に成功した。また、該遺伝子を利用した植物の子実粒(種子、頴花又は果実を含む)の長さを増加させる育種手法も見出した。本発明は、植物の品種改良等の分野において有用である。
【図面の簡単な説明】
【0017】
【図1】粒長に関与するQTLの遺伝子特定結果を示す図である。
【図2】GL5に類似する遺伝子の系統樹を示す図である。
【図3】コシヒカリ、カサラスのGL5遺伝子のアミノ酸配列比較を示す図である。
【図4−1】コシヒカリ、カサラスのGL5遺伝子のプロモーター配列比較(前半部分)を示す図である。
【図4−2】コシヒカリ、カサラスのGL5遺伝子のプロモーター配列比較(後半部分)を示す図である。
【図5】コシヒカリとNIL(GL5)におけるGL5遺伝子発現量の比較を示すグラフである。
【図6】日本晴にGL5遺伝子を過剰発現させた形質転換体イネの玄米粒サイズを示す図である。
【図7】イネ、シロイヌナズナのGL5に類似する遺伝子のアミノ酸配列比較を示す図である。
【図8】シロイヌナズナにGL5遺伝子を過剰発現させた形質転換体シロイヌナズナの種子粒サイズを示す図である。
【発明を実施するための形態】
【0018】
本発明は、植物体の子実(種子、頴花又は果実を含む)の収量に関する遺伝子GL5を含む植物体、および該遺伝子を用いた子実収量(種子、頴花又は果実の収量のことをいう)の増加方法、子実収量の検出方法、子実収量が増加する植物体の育種方法を提供する。
【0019】
本発明において子実の収量の増加とは、具体的には種子、頴花又は果実を増大させることによってもたらされる。本発明において子実が増大するとは、子実の大きさが増加することをいい、該遺伝子の発現量が低い植物体の子実と比較して、それぞれ30%以上、好ましくは20%以上、より好ましくは15%、10%、5%以上のサイズの増加をいう。
【0020】
本発明者らにより作物の子実収量との関係が明らかにされた、イネカサラスのGL5遺伝子のゲノムDNAの塩基配列を配列番号:4に、cDNAの塩基配列を配列番号:5に、これら遺伝子がコードするタンパク質のアミノ酸配列を配列番号:6に示す。
【0021】
また、イネコシヒカリのGL5遺伝子のゲノムDNAの塩基配列を配列番号:1に、cDNAの塩基配列を配列番号:2に、これら遺伝子がコードするタンパク質のアミノ酸配列を配列番号:3に示す。
【0022】
本発明は、植物体の子実を増大させる機能を有するイネ由来のGL5遺伝子を含むベクターならびに形質転換植物細胞を提供する。
【0023】
本発明のGL5遺伝子としては、具体的には、植物体の子実を増大させる機能を有するイネ由来のタンパク質をコードする、下記(a)から(d)のいずれかに記載のDNAが含まれる。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
【0024】
本発明において、本発明の遺伝子が由来する植物体としては、特に制限はないが、例えばイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビ等の穀物、ダイズ、エンドウ、ピーナッツ等の豆類、カボチャ、トマト等のいわゆる実のなる野菜類、ゴマ、ナタネ、ヒマワリ等の油料作物、シロイヌナズナ等が挙げられる。本発明の遺伝子が由来する植物として、より好ましくは、イネを挙げることができる。
【0025】
本発明の「GL5遺伝子」は、「GL5タンパク質」をコードしうるものであれば、その形態に特に制限はなく、「GL5遺伝子」には、cDNAの他、ゲノムDNA、化学合成DNAなども含まれる。また、GL5遺伝子はGL5タンパク質をコードするものであれば、遺伝暗号の縮重に基づく任意の塩基配列を有するDNAが含まれる。
【0026】
本発明の遺伝子のコード領域は、例えば、配列番号:5に記載の塩基配列における217位〜1035位の領域を挙げることができる。
【0027】
ゲノムDNAおよびcDNAの調製は、当業者にとって常套手段を利用して行うことが可能である。ゲノムDNAは、例えば、植物からゲノムDNAを抽出し、ゲノミックライブラリー(ベクターとしては、プラスミド、ファージ、コスミド、BAC、PACなどが利用できる)を作成し、これを展開して、GL5遺伝子(例えば、配列番号:4または5のいずれかに記載のDNA)を基に調製したプローブを用いてコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより当該クローンを得、調製することが可能である。また、GL5遺伝子に特異的なプライマーを作成し、これを利用したPCRをおこなうことによって調製することも可能である。また、cDNAは、例えば、植物から抽出したmRNAを基にcDNAを合成し、これをλZAP等のベクターに挿入してcDNAライブラリーを作成し、これを展開して、上記と同様にコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより、また、PCRを行うことにより調製することが可能である。
【0028】
さらに、GL5遺伝子は広く穀物植物に存在すると考えられるため、GL5遺伝子には、種々の植物に存在する相同遺伝子も含まれる。ここで「相同遺伝子」とは、種々の植物において、イネにおける活性型GL5遺伝子産物と機能的に同等なタンパク質をコードする遺伝子を指す。このようなタンパク質には、例えば、活性型GL5タンパク質の変異体、アレル、バリアント、ホモログ、活性型GL5タンパク質の部分ペプチド、または、他のタンパク質との融合タンパク質などが挙げられるが、これらに限定されない。
【0029】
本発明における活性型GL5タンパク質の変異体としては、配列番号:6に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなる天然由来のタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質のN末側活性断片と機能的に同等なタンパク質を挙げることが出来る。また、配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズする天然由来のDNAよりコードされるタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質のN末側活性断片と機能的に同等なタンパク質も、活性型GL5タンパク質の変異体として挙げることができる。
【0030】
また、本発明における活性型GL5タンパク質の変異体としては、配列番号:6に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなる天然由来のタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質と機能的に同等なタンパク質を挙げることが出来る。また、配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズする天然由来のDNAよりコードされるタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質と機能的に同等なタンパク質も、活性型GL5タンパク質の変異体として挙げることができる。
【0031】
本発明において、変異するアミノ酸数は特に制限されないが、通常、30アミノ酸以内であり、好ましくは15アミノ酸以内であり、さらに好ましくは5アミノ酸以内(例えば、3アミノ酸以内)であると考えられる。変異するアミノ酸残基においては、アミノ酸側鎖の性質が保存されている別のアミノ酸に変異されることが望ましい(これは、保存的アミノ酸置換として知られている)。例えばアミノ酸側鎖の性質としては、疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)および親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)の二種類に大別することができる。また、その側鎖の構造に基づいて、脂肪族側鎖を有するアミノ酸(G、A、V、L、I、P)、水酸基含有側鎖を有するアミノ酸(S、T、Y)、硫黄原子含有側鎖を有するアミノ酸(C、M)、カルボン酸及びアミド含有側鎖を有するアミノ酸(D、N、E、Q)、塩基含有側鎖を有するアミノ酸(R、K、H)、芳香族含有側鎖を有するアミノ酸(H、F、Y、W)などのようにアミノ酸を分類することもできる。さらに、例えば、変異マトリクス(mutational matrix)によってアミノ酸を分類することも周知である(Taylor 1986, J, Theor. Biol. 119, 205-218; Sambrook, J. et al., Molecular Cloning 3rd ed. A7.6-A7.9, Cold Spring Harbor Lab. Press, 2001)。この分類を以下に要約すると、脂肪族アミノ酸(L、I、V)、芳香族アミノ酸(H、W、Y、F)、荷電アミノ酸(D、E、R、K、H)、正荷電アミノ酸(R、K、H)、負荷電アミノ酸(D、E)、疎水性アミノ酸(H、W、Y、F、M、L、I、V、C、A、G、T、K)、極性アミノ酸(T、S、N、D、E、Q、R、K、H、W、Y)、小型アミノ酸(P、V、C、A、G、T、S、N、D)、微小アミノ酸(A、G、S)および大型(非小型)アミノ酸(Q、E、R、K、H、W、Y、F、M、L、I)が挙げられる(括弧内はいずれもアミノ酸の一文字標記を表す)。
【0032】
あるアミノ酸配列に対する1又は複数個のアミノ酸残基の欠失、付加及び/又は他のアミノ酸による置換により修飾されたアミノ酸配列を有するポリペプチドがその生物学的活性を維持することはすでに知られている。さらに、標的アミノ酸残基は、共通した性質をできるだけ多く有するアミノ酸残基に変異させることがより好ましい。
【0033】
本発明において「機能的に同等」とは、対象となるタンパク質が、活性型GL5タンパク質と同等の生物学的機能や生化学的機能を有することを指す。本発明において、活性型GL5タンパク質の生物学的機能や生化学的機能としては、例えば穀物の種子を増大させる機能を挙げることができる。生物学的な性質には発現する部位の特異性や、発現量等も含まれる。
【0034】
相同遺伝子を単離するための当業者によく知られた方法としては、ハイブリダイゼーション技術(Southern, E. M., Journal of Molecular Biology, Vol. 98, 503, 1975)やポリメラーゼ連鎖反応(PCR)技術(Saiki, R. K., et al. Science, vol. 230, 1350-1354, 1985, Saiki, R. K. et al. Science, vol.239, 487-491,1988)が挙げられる。即ち、当業者にとっては、GL5遺伝子の塩基配列(例えば、配列番号:4又は5のいずれかに記載のDNA)もしくはその一部をプローブとして、またGL5遺伝子に特異的にハイブリダイズするオリゴヌクレオチドをプライマーとして、種々の植物からGL5遺伝子の相同遺伝子を単離することは通常行いうることである。
【0035】
このような相同遺伝子をコードするDNAを単離するためには、通常ストリンジェントな条件下でハイブリダイゼーション反応を行なう。ストリンジェントなハイブリダイゼーション条件は当業者であれば、適宜選択することができる。一例を示せば、25%ホルムアミド、より厳しい条件では50%ホルムアミド、4xSSC、50mM Hepes pH7.0、10×デンハルト溶液、20μg/ml変性サケ精子DNAを含むハイブリダイゼーション溶液中、42℃で一晩プレハイブリダイゼーションを行った後、標識したプローブを添加し、42℃で一晩保温することによりハイブリダイゼーションを行う。その後の洗浄における洗浄液および温度条件は、「1xSSC、0.1% SDS、37℃」程度で、より厳しい条件としては「0.5xSSC、0.1% SDS、42℃」程度で、さらに厳しい条件としては「0.2xSSC、0.1% SDS、65℃」程度で実施することができる。このようにハイブリダイゼーションの洗浄の条件が厳しくなるほどプローブ配列と高い相同性を有するDNAの単離を期待しうる。但し、上記SSC、SDSおよび温度の条件の組み合わせは例示であり、当業者であれば、ハイブリダイゼーションのストリンジェンシーを決定する上記若しくは他の要素(例えば、プローブ濃度、プローブの長さ、ハイブリダイゼーション反応時間など)を適宜組み合わせることにより、上記と同様のストリンジェンシーを実現することが可能である。
【0036】
単離されたDNAの相同性は、アミノ酸配列全体で、少なくとも50%以上、さらに好ましくは70%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。配列の相同性は、BLASTN(核酸レベル)やBLASTX(アミノ酸レベル)のプログラム(Altschul et al. J. Mol. Biol., 215: 403-410, 1990)を利用して決定することができる。該プログラムは、Karlin及びAltschulによるアルゴリズムBLAST (Proc. Natl. Acad. Sci. USA, 87:2264-2268, 1990, Proc. Natl. Acad. Sci. USA, 90: 5873-5877, 1993) に基づいている。BLASTNによって塩基配列を解析する場合には、パラメーターは例えばscore = 100、wordlength =12とする。また、BLASTXによってアミノ酸配列を解析する場合には、パラメーターは例えばscore = 50、wordlength = 3とする。また、Gapped BLASTプログラムを用いて、アミノ酸配列を解析する場合は、Altschulら(Nucleic Acids Res. 25: 3389-3402, 1997)に記載されているように行うことができる。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である。
【0037】
本発明のベクターとしては、形質転換体作製のために細胞内で本発明のDNAを発現させるベクター、例えば形質転換植物体作製のために植物細胞内で本発明のDNAを発現させるためのベクターが含まれる。植物細胞の形質転換に用いられるベクターとしては、該細胞内で挿入遺伝子を発現させることが可能なものであれば特に制限はない。例えばプラスミド、ファージ、またはコスミドなどを例示することができる。
【0038】
また上記「植物細胞」には、種々の形態の植物細胞、例えば、懸濁培養細胞、プロトプラスト、葉の切片、カルス等が含まれる。
【0039】
本発明のベクターは、本発明のDNAを恒常的または誘導的に発現させるためのプロモーターを含有してもよい。
【0040】
当業者においては、所望のDNAを有するベクターを、一般的な遺伝子工学技術によって、適宜、作製することが可能である。通常、市販の種々のベクターを利用することができる。
【0041】
本発明のベクターは、宿主細胞内において本発明のDNAを保持したり、発現させるためにも有用である。
【0042】
本発明におけるDNAは、通常、適当なベクターへ担持(挿入)され、宿主細胞へ導入される。即ち本発明は、本発明のDNAまたはベクターを保持する宿主細胞を提供する。該ベクターとしては、挿入したDNAを安定に保持するものであれば特に制限されず、例えば宿主に大腸菌を用いるのであれば、クローニング用ベクターとしてpBluescriptベクター(Stratagene社製)などが好ましいが、市販の種々のベクターを利用することができる。本発明のDNAを内在性遺伝子を有する細胞内に導入および発現させる目的としてベクターを用いる場合には、特に発現ベクターが有用である。発現ベクターとしては、試験管内、大腸菌内、培養細胞内、生物個体内でDNAを発現するベクターであれば特に制限されないが、例えば、試験管内発現であればpBESTベクター(プロメガ社製)、大腸菌であればpETベクター(Invitrogen社製)、培養細胞であればpME18S-FL3ベクター(GenBank Accession No. AB009864)、生物個体であればpME18Sベクター(Mol Cell Biol. 8:466-472(1988))、植物個体であればpBINPLUSベクター(van Engelen, F.A. et al., (1995). pBINPLUS: an improved plant transformation vector based on pBIN19. Transgenic Res. 4, 288-290.)などを例示することができる。ベクターへの本発明のDNAの挿入は、常法(Molecular Cloning, 5.61-5.63)により、例えば、制限酵素サイトを用いたリガーゼ反応により行うことができる。
【0043】
上記宿主細胞としては特に制限はなく、目的に応じて種々の宿主細胞が用いられる。本発明のDNAを発現させるための細胞としては、例えば、細菌細胞(例:ストレプトコッカス、スタフィロコッカス、大腸菌、ストレプトミセス、枯草菌)、昆虫細胞(例:ドロソフィラS2、スポドプテラSF9)、動物細胞(例:CHO、COS、HeLa、C127、3T3、BHK、HEK293、Bowes メラノーマ細胞)および植物細胞を例示することができる。
【0044】
また、生体内で本発明のDNAを発現させる方法としては、本発明のDNAを適当なベクターに組み込み、例えば、ポリエチレングリコール法、エレクトロポレーション法、アグロバクテリウム法、リポソーム法、カチオニックリポソーム法、リン酸カルシウム沈殿法、電気パルス穿孔法(エレクトロポーレーション)(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons.Section 9.1-9.9)、リポフェクション法(GIBCO-BRL社製)、マイクロインジェクション法、パーティクルガン法などの当業者に公知の方法により生体内に導入する方法などが挙げられる。
【0045】
植物体内への投与は、ex vivo法であっても、in vivo法であってもよい。
【0046】
また、植物体内へ本発明のDNAを導入する場合、DNAは、マイクロインジェクション法、エレクトロポレーション法、ポリエチレングリコール法等を用いて、植物細胞に直接導入することもできるが、植物への遺伝子導入用プラスミドに組込み、これをベクターとして、植物感染能のあるウイルスあるいは細菌を介して、間接的に植物細胞に導入することもできる。かかるウイルスとしては、例えば代表的なウイルスとして、カリフラワーモザイクウイルス、タバコモザイクウイルス、ジェミニウイルス等が挙げられ、細菌としては、アグロバクテリウム等が挙げられる。アグロバクテリウム法により、植物への遺伝子導入を行う場合には、市販のプラスミドを用いることができる。このようなベクターを用いて、植物体内へ本発明のDNAを導入する場合の方法としては、好ましくは、アグロバクテリウムを介して遺伝子を導入するリーフディスク法(Jorgensen, R.A. et al., (1996). Chalcone synthase cosuppression phenotypes in petunia flowers: comparison of sense vs. antisense constructs and single-copy vs. complex T-DNA sequences. Plant Mol. Biol. 31, 957-973.)が挙げられる。
【0047】
なおこれら上述の形質転換方法は、宿主となる植物などの種類(例えば単子葉植物又は双子葉植物)に応じて適宜選択することが好ましい。
【0048】
本発明において「植物」とは、特に制限されないが、例えばイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビ等の穀物、ダイズ、エンドウ、ピーナッツ等の豆類、カボチャ、トマト等のいわゆる実のなる野菜類、ゴマ、ナタネ、ヒマワリ等の油料作物、シロイヌナズナ等を挙げることができる。
【0049】
また、本発明は、本発明のDNAまたは本発明のベクターを保持する植物細胞を提供する。さらに本発明は、本発明の植物細胞を含む形質転換植物体を提供する。本発明のDNAまたは本発明のベクターが導入される細胞には、形質転換植物体作製のための植物細胞が含まれる。植物細胞としては特に制限はない。
【0050】
本発明の植物細胞には、培養細胞の他、植物体中の細胞も含まれる。また、プロトプラスト、苗条原基、多芽体、毛状根も含まれる。
【0051】
形質転換植物細胞からの植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。例えば、形質転換植物体を作出する手法については、ポリエチレングリコールによりプロトプラストへ遺伝子導入し植物体を再生させる方法、電気パルスによりプロトプラストへ遺伝子導入し植物体を再生させる方法、パーティクルガン法により細胞へ遺伝子を直接導入し、植物体を再生させる方法、およびアグロバクテリウムを介して遺伝子を導入し、植物体を再生させる方法などを挙げることができるが、特に制限されるものではない。いくつかの技術については既に確立し、本願発明の技術分野において広く用いられている。本発明においては、これらの方法を好適に用いることができる。
【0052】
本発明のDNAを含むベクターの導入により形質転換した植物細胞を効率的に選択するために、上記組み換えベクターは、適当な選抜マーカー遺伝子を含む、もしくは選抜マーカー遺伝子を含むプラスミドベクターと共に植物細胞へ導入することが好ましい。この目的に使用される選抜マーカー遺伝子は、例えば抗生物質カナマイシンまたはゲンタマイシンに耐性であるネオマイシンホスホトランスフェラーゼ遺伝子、ハイグロマイシンに耐性であるハイグロマイシンホスホトランスフェラーゼ遺伝子、および除草剤ホスフィノスリシンに耐性であるアセチルトランスフェラーゼ遺伝子等が挙げられる。
【0053】
組み換えベクターを導入した植物細胞は、導入された選抜マーカー遺伝子の種類に従って適当な選抜用薬剤を含む公知の選抜用培地に置床し培養する。これにより形質転換された植物培養細胞を得ることができる。
【0054】
形質転換された植物細胞は、再分化させることにより植物体を再生させることが可能である。再分化の方法は植物細胞の種類により異なるが、例えばイネであればFujimuraら(Plant Tissue Culture Lett. 2:74 (1995))の方法が挙げられ、トウモロコシであればShillitoら(Bio/Technology 7:581 (1989))の方法やGorden-Kammら(Plant Cell 2:603(1990))の方法が挙げられる。
【0055】
なお、このように再生され、かつ栽培した形質転換植物体中の導入された外来DNAの存在は、公知のPCR法やサザンハイブリダイゼーション法によって、または植物体中のDNAの塩基配列を解析することによって確認することができる。
【0056】
この場合、形質転換植物体からのDNAの抽出は、公知のJ.Sambrookらの方法(Molecular Cloning、第2版、Cold Spring Harbor Laboratory Press, 1989)に準じて実施することができる。
【0057】
再生させた植物体中に存在する本発明のDNAよりなる外来遺伝子を、PCR法を用いて解析する場合には、上記のように再生植物体から抽出したDNAを鋳型として増幅反応を行う。また、本発明のDNA、あるいは本発明により改変されたDNAの塩基配列に従って適当に選択された塩基配列をもつ合成したオリゴヌクレオチドをプライマーとして用い、これらを混合させた反応液中において増幅反応を行うこともできる。増幅反応においては、DNAの変性、アニーリング、伸張反応を数十回繰り返すと、本発明のDNA配列を含むDNA断片の増幅生成物を得ることができる。増幅生成物を含む反応液を例えばアガロース電気泳動にかけると、増幅された各種のDNA断片が分画されて、そのDNA断片が本発明のDNAに対応することを確認することが可能である。
【0058】
一旦、ゲノム内に本発明のDNAが導入された形質転換植物体が得られれば、該植物体から有性生殖または無性生殖により子孫を得ることが可能である。また、該植物体やその子孫あるいはクローンから繁殖材料(例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラスト等)を得て、それらを基に該植物体を量産することも可能である。本発明には、本発明のDNAまたはベクターが導入された植物細胞、該細胞を含む植物体、該植物体の子孫およびクローン、並びに該植物体、その子孫、およびクローンの繁殖材料が含まれる。
【0059】
このようにして作出された植物体は通常の穀物に比べて子実収量が増加することが期待される。
【0060】
上述のように、本発明のDNAもしくはベクターを植物細胞へ導入し、該植物細胞から植物体を再生させる工程を含む、形質転換植物体の製造方法もまた本発明に含まれる。
【0061】
本発明の子実収量が増加した植物体もしくはその種子は、育種法によっても作出することが可能である。
【0062】
上記育種法としては、例えば、本発明のDNAを有する品種と交雑させることを特徴とする一般的な育種法(交雑育種法等)を挙げることができる。該方法によって、子実が増大した植物体もしくはその種子を作出することができる。
【0063】
育種法によって本発明の植物体もしくは種子を作製する際には、公知の種々の文献を参照して適宜実施することができる(細胞工学別冊・植物細胞工学シリーズ15「モデル植物の実験プロトコール」秀潤社、2001年、加藤鎌司著「2.1 イネ・コムギの交配」p.6-9; 高牟礼逸朗、佐野芳雄著「2. 交配法」p.46-48)。
【0064】
本発明の上記育種方法の好ましい態様としては、以下の(a)および(b)に記載の工程を含む方法である。
(a)子実が増大した植物と任意の機能を有する植物とが交配された品種を作製する工程、
(b)工程(a)で作製された植物の子実の収量を検出する工程
【0065】
本発明の植物体を育種する方法のより具体的な例としては、以下の(a)および(b)に記載の工程を含む方法が挙げられる。
(a)本発明のDNAを有する植物と交雑させる工程、
(b)前記DNAを有する植物改変体を選抜する工程
【0066】
本発明の上記植物体の作成方法を育種法によって実施する場合には、さらに具体的には、以下のような工程を含む方法を挙げることができる。
(a)植物Aと、本発明のDNAを有する他の植物Bを交雑させ、F1を作出する工程、
(b)前記F1と前記植物Aを交雑させる工程、
(c)前記DNAを有する植物を選抜する工程、
(d)工程(c)によって選抜された植物と、前記植物Aを交雑させる工程
【0067】
上記方法においては、本発明のDNAを有する植物Bと、子実収量を増加させたい植物もしくは子実を増大させたい植物(これら植物を「植物A」と記載する。)を交雑し、Bのもつ本発明のDNAが受け継がれ、かつ植物Aに近い個体を選抜し、これに植物Aによる交雑を重ねていく「戻し交雑」を行って、Bが有する本発明のDNAの形質を意図的に導入する。その際、一般的にゲノム育種に利用されるDNAマーカーを利用して本発明のDNAを有する植物を選抜することにより、上記「戻し交雑」による置換を効率的に行うことが可能である。その結果、育種期間の短縮に繋がり、また、余分なゲノム領域の混入を正確に除くことができる。通常、「戻し交雑」では、本発明のDNAと非常に強く連鎖する他のDNAに依存する形質がどうしても排除できないという現象が問題となることがあるが、本発明のDNAの近傍に存在するDNAマーカーを利用することにより、所望の植物の正確な選抜が可能となる。
【0068】
上記方法においては、必要に応じて、本発明のDNA以外のゲノム全域が目的の遺伝形質でホモ固定するまで、繰り返して行うことができる。即ち、本発明の好ましい態様のおいては、上記工程(d)によって交雑された個体について、一般的なDNAマーカーを利用して、本発明のDNAを有し、かつ、ゲノム構造が植物Aに近い植物個体を選抜することができる。さらに、この選抜された植物個体は、必要に応じて、「戻し交雑」(イネ品種Aと交雑)させることができる。
【0069】
特にDNAマーカーを利用したゲノム育種方法では、置換率の高い個体を選抜して次の交雑に進むことができるため、世代を進めるほどに選抜効率が良くなる。また、本方法では、少ない個体数を扱えば済むので、省スペースでの育種が可能になる。さらに、温室や人工気象室を利用して1年に複数回もの交雑が可能になる。
【0070】
上記工程(c)において、DNAマーカーを用いて選抜するとは、当該DNAマーカーを特徴付ける塩基配列(例えば、多型等)についての塩基種の情報を基に、選抜を行うことを言う。例えば、本発明のDNAの近傍に多型変異が存在する場合、当該多型変異と同一の多型変異を有する個体を選抜すること等を言う。
【0071】
本発明の育種方法は、好ましくは、DNAマーカーを利用した「ゲノム育種」方法である。該「ゲノム育種」は「マーカー育種」とも呼ばれる。
【0072】
本発明の育種方法において利用可能なDNAマーカーは、特に制限されず、一般的に知られている種々のDNAマーカーを好適に用いることができる。例えば、RFLP(制限酵素断片長多型)マーカー、SSR(単純反復配列)マーカー、SNP(一塩基多型)マーカー等を例示することができる。
【0073】
本発明者らは、カサラス由来のGL5遺伝子をもつイネが種子を増大させる機能を有することを見出した。従って、コシヒカリ型とカサラス型のGL5遺伝子を判別することで被検植物の種子の収量を検出することができる。言い換えれば、コシヒカリ型に対するカサラス型の変異部位をDNAマーカーとすることで、上記育種方法に使用することができる。
【0074】
上記の知見に基づき、本発明は、被検植物について、配列番号:1に記載のDNA領域(例えば配列番号:1に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域)に相当する部位、またはその周辺配列に存在する変異を含むDNAマーカーを検出することを特徴とする、被検植物の種子の収量を検出する方法を提供する。
【0075】
本発明において、配列番号:1に記載のDNA領域に「相当する部位」とは、種々の植物において、当該植物に存在する上記遺伝子の相同遺伝子における対応する部位をいう。
【0076】
本発明において「子実の収量を検出」とは、子実のサイズが増大するか否かを判定するための検査が含まれる。本発明の方法においては、上記配列番号:1に記載のDNA領域に相当する部位、またはその周辺配列において、配列番号:4と同型の変異が検出された場合には、子実のサイズは増大すると判定される。
【0077】
本発明の方法により、被検植物における変異を検出することで、実際に子実を観察しなくても、被検植物の子実のサイズが増大するか否かを判定することができる。
【0078】
本発明における「周辺配列」とは、通常、該遺伝子の近傍の染色体上の領域を指す。近傍とは、特に制限されるものではないが、通常、本発明の多型部位を含むDNA領域である。
【0079】
上記本発明の検出方法における「変異」の位置は、予め規定することは困難であるが、通常、上記遺伝子のORF中、あるいは上記遺伝子の発現を制御する領域(例えば、プロモーター領域、エンハンサー領域等)中に存在するが、これらに限定されるものではない。また、この「変異」とは、上記遺伝子の発現量を変化させる、mRNAの安定性等の性質を変化させる、あるいは上記遺伝子によってコードされるタンパク質の有する活性を変化させるような変異であることが多いが、特に制限されない。本発明の変異としては、例えば、塩基の付加、欠失、置換、挿入変異等を挙げることができる。
【0080】
本発明者らは、被検植物における、植物の子実収量を増加させる機能を有する植物由来のタンパク質をコードする配列番号:1のDNA領域において、植物の子実収量に対して有意に関連する多型変異を見出すことに成功した。
【0081】
本発明の子実収量を検出する方法における「多型部位」は、上記DNAのいずれかに記載の塩基配列もしくは該塩基配列の近傍DNA領域に存在する多型であれば、特に制限されない。具体的には、本発明の子実収量を検出する方法に利用可能な多型部位として、配列番号:1に記載の塩基における32位、129位、161位、557位、839位、843位、976位又は1699位から選択される少なくとも一つの多型部位に相当する部位を挙げることができる(なお、本明細書においては、これらの多型部位を単に『本発明の多型部位』と記載する場合がある)。
【0082】
上記表中に示した多型部位の塩基種は配列表に示した配列に対して相補鎖側にある塩基種を示している場合があるが、本明細書において記載された前後配列を用いれば異同を確認することは当業者にとって容易であり、検出を行うにあたってはプラス鎖とマイナス鎖のどちらを調べても必然的にもう一方の結果を決定することができる。
【0083】
本発明の好ましい態様においては、上述した多型部位における塩基種の変異が、配列番号:配列番号:1に記載の塩基配列における32位の塩基種のGからTへの変異、129位の塩基種のAからGへの変異、161位の塩基種のTからGへの変異、557位の塩基種のTからCへの変異、839位の塩基種のTの欠損、843位の塩基種のGの欠損、976位の塩基種のCからAへの変異、又は1699位の塩基種のCからTへの変異から選択される少なくとも一つの変異である場合に、子実が増大すると判定される。
【0084】
以上のように、本発明により、子実収量に関連する遺伝子上の領域が明らかになったことにより、当業者に過度の負担を強いることなく、子実収量の検出を行うことができる。
【0085】
本発明の多型部位における塩基種の決定は、当業者においては種々の方法によって行うことができる。一例を示せば、本発明の多型部位を含むDNAの塩基配列を直接決定することによって行うことができる。
【0086】
本発明の検出方法に供する「被検植物」としては、特に制限されないが、好ましくはイネ、さらに好ましくはイネのコシヒカリ品種、カサラス品種が挙げられる。
【0087】
本発明の検出方法に供する被検試料は、通常、予め被検植物から取得された生体試料であることが好ましい。生体試料としては、例えばDNA試料を挙げることができる。本発明におけるDNA試料は、例えば被検植物の組織または細胞等から抽出した染色体DNA、あるいはRNAを基に調製することができる。
【0088】
即ち本発明は、通常、被検者由来の生体試料(予め被検植物から取得された生体試料)を被検試料として検査に供する方法である。
【0089】
当業者においては、公知の技術を用いて、適宜、生体試料の調製を行うことができる。例えば、DNA試料は、本発明の多型部位を含むDNAにハイブリダイズするプライマーを用いて、染色体DNA、あるいはRNAを鋳型としたPCR等によって調製することができる。
【0090】
本方法においては、次いで、単離したDNAの塩基配列を決定する。単離したDNAの塩基配列の決定は、当業者においては、DNAシークエンサー等を用いて容易に実施することができる。
【0091】
予め塩基のバリエーションが明らかにされている多型部位について、その塩基種を決定するための様々な方法が公知である。本発明の塩基種の決定方法は、特に限定されない。例えば、PCR法を応用した解析方法として、TaqMan PCR法、AcycloPrime法、およびMALDI-TOF/MS法等が実用化されている。またPCRに依存しない塩基種の決定法としてInvader法やRCA法が知られている。更にDNAアレイを使って塩基種を決定することもできる。ここに述べた方法は、いずれも本発明における多型部位の塩基種の決定に応用できる。
【0092】
これらの方法はいずれも多量のサンプルを高速にジェノタイピングするために開発された方法である。MALDI-TOF/MSを除けば、通常、いずれの方法にも何らかの形で標識プローブなどを用意する必要がある。これに対して、標識プローブなどに頼らない塩基種決定法も古くから行われている。このような方法の一つとして、例えば、制限酵素断片長多型(Restriction Fragment Length Polymorphism/RFLP)を利用した方法やPCR-RFLP法等が挙げられる。
【0093】
RFLPは、制限酵素の認識部位の変異、あるいは制限酵素処理によって生じるDNA断片内における塩基の挿入または欠失が、制限酵素処理後に生じる断片の大きさの変化として検出できることを利用している。検出対象となる多型を含む塩基配列を認識する制限酵素が存在すれば、RFLPの原理によって多型部位の塩基を知ることができる。
【0094】
また、CAPS (Cleaved Amplifeid Polymorphic Sequence)マーカーあるいはプライマーに変異を導入し、制限酵素サイトを作り出すdCAPS (derived CAPS)マーカーを使用することもできる。dCAPSマーカーは、PCRのプライマーにテンプレートのDNAとミスマッチを起こして、PCR産物上に制限酵素サイトを作り出すという手法である(特開2003-259898)。
【0095】
標識プローブを必要としない方法として、DNAの二次構造の変化を指標として塩基の違いを検出する方法も公知である。PCR-SSCPでは、1本鎖DNAの二次構造がその塩基配列の相違を反映することを利用している(Cloning and polymerase chain reaction-single-strand conformation polymorphism analysis of anonymous Alu repeats on chromosome 11. Genomics. 1992 Jan 1; 12(1): 139-146.、Detection of p53 gene mutations in human brain tumors by single-strand conformation polymorphism analysis of polymerase chain reaction products. Oncogene. 1991 Aug 1; 6(8): 1313-1318.、Multiple fluorescence-based PCR-SSCP analysis with postlabeling.、PCR Methods Appl. 1995 Apr 1; 4(5): 275-282.)。PCR-SSCP法は、PCR産物を1本鎖DNAに解離させ、非変性ゲル上で分離する工程により実施される。ゲル上の移動度は、1本鎖DNAの二次構造によって変動するので、もしも多型部位における塩基の相違があれば、移動度の違いとして検出することができる。
【0096】
その他、標識プローブを必要としない方法として、例えば、変性剤濃度勾配ゲル(denaturant gradient gel electrophoresis:DGGE法)等を例示することができる。DGGE法は、変性剤の濃度勾配のあるポリアクリルアミドゲル中で、DNA断片の混合物を泳動し、それぞれの不安定性の違いによってDNA断片を分離する方法である。ミスマッチのある不安定なDNA断片が、ゲル中のある変性剤濃度の部分まで移動すると、ミスマッチ周辺のDNA配列はその不安定さのために、部分的に1本鎖へと解離する。部分的に解離したDNA断片の移動度は、非常に遅くなり、解離部分のない完全な二本鎖DNAの移動度と差がつくことから、両者を分離することができる。
【0097】
更にDNAアレイを使って塩基種を決定することもできる(細胞工学別冊「DNAマイクロアレイと最新PCR法」,秀潤社,2000.4/20発行,pp97-103「オリゴDNAチップによるSNPの解析」,梶江慎一)。DNAアレイは、同一平面上に配置した多数のプローブに対してサンプルDNA(あるいはRNA)をハイブリダイズさせ、当該平面をスキャンすることによって、各プローブに対するハイブリダイズが検出される。多くのプローブに対する反応を同時に観察することができることから、例えば、多数の多型部位について同時に解析するには、DNAアレイは有用である。
【0098】
上記の方法以外にも、特定部位の塩基を検出するために、アリル特異的オリゴヌクレオチド(Allele Specific Oligonucleotide/ASO)ハイブリダイゼーション法が利用できる。アリル特異的オリゴヌクレオチド(ASO)は、検出すべき多型部位が存在する領域にハイブリダイズする塩基配列で構成される。ASOを試料DNAにハイブリダイズさせるとき、多型によって多型部位にミスマッチが生じるとハイブリッド形成の効率が低下する。ミスマッチは、サザンブロット法や、特殊な蛍光試薬がハイブリッドのギャップにインターカレーションすることにより消光する性質を利用した方法等によって検出することができる。また、リボヌクレアーゼAミスマッチ切断法によって、ミスマッチを検出することもできる。
【0099】
上記オリゴヌクレオチドのうち、配列番号:1に記載の塩基配列における、32位、129位、161位、557位、839位、843位、976位又は1699位から選択される少なくとも一つの多型部位に相当する部位のうち、いずれかの多型部位を含むDNAにハイブリダイズし、少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドは、子実収量を検出するための試薬(検査薬)として利用できる。これは遺伝子発現を指標とする検査、または遺伝子多型を指標とする検査に使用される。
【0100】
該オリゴヌクレオチドは、本発明の上記多型部位のいずれかの多型部位を含むDNAに特異的にハイブリダイズするものである。ここで「特異的にハイブリダイズする」とは、通常のハイブリダイゼーション条件下、好ましくはストリンジェントなハイブリダイゼーション条件下(例えば、サムブルックら,Molecular Cloning,Cold Spring Harbour Laboratory Press,New York,USA,第2版1989に記載の条件)において、他のタンパク質をコードするDNAとクロスハイブリダイゼーションを有意に生じないことを意味する。特異的なハイブリダイズが可能であれば、該オリゴヌクレオチドは、検出する遺伝子もしくは該遺伝子の近傍DNA領域における、上記植物の子実を増大させる機能を有する植物由来のタンパク質をコードするDNA塩基配列に対し、完全に相補的である必要はない。
【0101】
該オリゴヌクレオチドは、上記本発明の検査方法におけるプローブやプライマーとして用いることができる。該オリゴヌクレオチドをプライマーとして用いる場合、その長さは、通常15bp〜100bpであり、好ましくは17bp〜30bpである。プライマーは、本発明の上記配列番号:1に記載の塩基配列における、32位、129位、161位、557位、839位、843位、976位又は1699位から選択される少なくとも一つの多型部位に相当する部位のうち、いずれかの多型部位を含むDNAの少なくとも一部を増幅しうるものであれば、特に制限されない。
【0102】
本発明は、本発明の多型部位を含む領域を増幅するためのプライマー、および多型部位を含むDNA領域にハイブリダイズするプローブを提供する。
【0103】
本発明において、多型部位を含む領域を増幅するためのプライマーには、多型部位を含むDNAを鋳型として、多型部位に向かって相補鎖合成を開始することができるプライマーも含まれる。該プライマーは、多型部位を含むDNAにおける、多型部位の3'側に複製開始点を与えるためのプライマーと表現することもできる。プライマーがハイブリダイズする領域と多型部位との間隔は任意である。両者の間隔は、多型部位の塩基の解析手法に応じて、好適な塩基数を選択することができる。たとえば、DNAチップによる解析のためのプライマーであれば、多型部位を含む領域として、20〜500、通常50〜200塩基の長さの増幅産物が得られるようにプライマーをデザインすることができる。当業者においては、多型部位を含む周辺DNA領域についての塩基配列情報を基に、解析手法に応じたプライマーをデザインすることができる。本発明のプライマーを構成する塩基配列は、ゲノムの塩基配列に対して完全に相補的な塩基配列のみならず、適宜改変することができる。
【0104】
本発明のプライマーには、ゲノムの塩基配列に相補的な塩基配列に加え、任意の塩基配列を付加することができる。例えば、IIs型の制限酵素を利用した多型の解析方法のためのプライマーにおいては、IIs型制限酵素の認識配列を付加したプライマーが利用される。このような、塩基配列を修飾したプライマーは、本発明のプライマーに含まれる。更に、本発明のプライマーは、修飾することができる。例えば、蛍光物質や、ビオチンまたはジゴキシンのような結合親和性物質で標識したプライマーが各種のジェノタイピング方法において利用される。これらの修飾を有するプライマーも本発明に含まれる。
【0105】
本発明は、上記プライマーを有効成分として含有する、本発明の多型部位の検査薬またはキットも提供する。
【0106】
一方本発明において、多型部位を含む領域にハイブリダイズするプローブとは、多型部位を含む領域の塩基配列を有するポリヌクレオチドとハイブリダイズすることができるプローブを言う。より具体的には、プローブの塩基配列中に多型部位を含むプローブは本発明のプローブとして好ましい。あるいは、多型部位における塩基の解析方法によっては、プローブの末端が多型部位に隣接する塩基に対応するように、デザインされる場合もある。従って、プローブ自身の塩基配列には多型部位が含まれないが、多型部位に隣接する領域に相補的な塩基配列を含むプローブも、本発明における望ましいプローブとして示すことができる。
【0107】
言いかえれば、ゲノムDNA上の本発明の多型部位、または多型部位に隣接する部位にハイブリダイズすることができるプローブは、本発明のプローブとして好ましい。本発明のプローブには、プライマーと同様に、塩基配列の改変、塩基配列の付加、あるいは修飾が許される。例えば、Invader法に用いるプローブは、フラップを構成するゲノムとは無関係な塩基配列が付加される。このようなプローブも、多型部位を含む領域にハイブリダイズする限り、本発明のプローブに含まれる。本発明のプローブを構成する塩基配列は、ゲノムにおける本発明の多型部位の周辺DNA領域の塩基配列をもとに、解析方法に応じてデザインすることができる。
【0108】
本発明のプライマーまたはプローブは、それを構成する塩基配列をもとに、任意の方法によって合成することができる。本発明のプライマーまたはプローブの、ゲノムDNAに相補的な塩基配列の長さは、通常15〜100、一般に15〜50、通常15〜30である。与えられた塩基配列に基づいて、当該塩基配列を有するオリゴヌクレオチドを合成する手法は公知である。更に、オリゴヌクレオチドの合成において、蛍光色素やビオチンなどで修飾されたヌクレオチド誘導体を利用して、オリゴヌクレオチドに任意の修飾を導入することもできる。あるいは、合成されたオリゴヌクレオチドに、蛍光色素などを結合する方法も公知である。
【0109】
本発明のプローブの具体的な例としては、それぞれ配列番号:1に記載の塩基配列における32位の塩基種のGからTへの変異、129位の塩基種のAからGへの変異、161位の塩基種のTからGへの変異、557位の塩基種のTからCへの変異、839位の塩基種のTの欠損、843位の塩基種のGの欠損、976位の塩基種のCからAへの変異、又は1699位の塩基種のCからTへの変異から選択される少なくとも一つの変異いずれかの多型部位に相当する多型部位を含む領域にハイブリダイズするプローブであって、少なくとも15ヌクレオチドの鎖長を有するプローブが挙げられる。
【0110】
本発明はまた、本発明の子実収量を検出する方法に使用するための試薬(検査薬)を提供する。本発明の試薬は、前記本発明のプライマーおよび/またはプローブを含む。子実収量の検出においては上記、本発明の多型部位のいずれかに記載の多型部位を含むDNAを増幅するためのプライマーおよび/またはプローブを用いる。
【0111】
本発明の試薬には、塩基種の決定方法に応じて、各種の酵素、酵素基質、および緩衝液などを組み合わせることができる。酵素としては、DNAポリメラーゼ、DNAリガーゼ、あるいはIIs制限酵素などの、上記の塩基種決定方法として例示した各種の解析方法に必要な酵素を示すことができる。緩衝液は、これらの解析に用いる酵素の活性の維持に好適な緩衝液が、適宜選択される。更に、酵素基質としては、例えば、相補鎖合成用の基質等が用いられる。
【0112】
さらに、本発明における試薬の別の態様は、本発明の多型部位を含むDNAとハイブリダイズするヌクレオチドプローブが固定された固相からなる、子実収量を検出するための試薬である。
【0113】
これらは本発明の多型部位を指標とする検査に使用される。これらの調製方法に関しては、当業者に公知の方法で行なうことができる。
【0114】
なお、本明細書において引用された全ての先行技術文献は、参照として本明細書に組み入れられる。
【実施例】
【0115】
以下、本発明を実施例によりさらに具体的に説明するが本発明はこれら実施例に制限されるものではない。
【0116】
〔実施例1〕GL5遺伝子のポジショナルクローニング
連鎖解析によりGL5遺伝子の特定を行った。解析に用いた集団は、コシヒカリとコシヒカリを遺伝的背景にGL5領域をカサラス型染色体に置換した準同質遺伝子系統NIL(GL5)との戻し交雑後代を用いた。GL5領域がヘテロ型となった個体の自殖後代2,000個体から、GL5領域を挟み込む一塩基多型(SNP)マーカーG1358とSNPマーカーG1569を利用してGL5遺伝子座近傍で組換えが生じた個体を選抜した。GL5遺伝子座近傍のSNPマーカーを用いて連鎖解析を行った結果、GL5遺伝子座をSNPマーカーG1422とSNPマーカーG1491との間、約7kbpの範囲に特定した(図1)。
【0117】
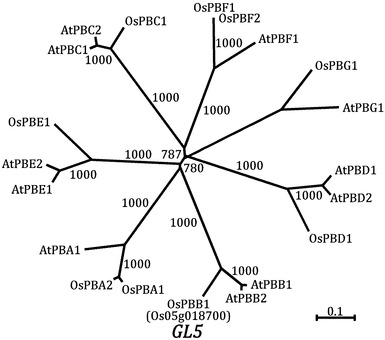
イネアノテーションプロジェクトにより公開された遺伝子予測データベース(http://rapdb.dna.affrc.go.jp/)により、SNPマーカーG1422とSNPマーカーG1491との間には、4つの遺伝子の存在が予測された。この4つの遺伝子の中から、遺伝子の発現量・発現部位からGL5遺伝子を1つの遺伝子(Os05g0187000)に絞り込んだ。この遺伝子は、20Sプロテアソームのβ2サブユニットをコードしていた。20Sプロテアソームのβ2サブユニットをコードする遺伝子は、イネには1つ(OsPBB1)しか存在しない(図2)。
【0118】
〔実施例2〕候補遺伝子の塩基配列と発現解析
コシヒカリとカサラスのcDNA塩基配列解析の結果、GL5候補遺伝子(OsPBB1)のコーディング領域に2カ所のSNP〔33位(配列番号:1の976位に相当)のCがA、267位(配列番号:1の1699位に相当)のCがT〕を有していたが、これらはいずれもアミノ酸配列に変化を生じさせず、コシヒカリとカサラスのGL5候補遺伝子のアミノ酸配列は同一であった(図3)。
【0119】
OsPBB1の推定プロモーター領域(1kbp)には、SNPが3ヵ所〔-912位(配列番号:1の32位に相当)のGがT、-815位(配列番号:1の129位に相当)のAがG、-783位(配列番号:1の161位に相当)のTがG〕、一塩基欠損挿入が2カ所〔-105位(配列番号:1の839位に相当)のTが欠損、-101位(配列番号:1の843位に相当)のGが欠損〕あった(図4-1及び4-2)。そこでOsPBB1遺伝子の発現量を解析した。コシヒカリおよびNIL(GL5)から全長10 cmの幼穂を採取し、酸性フェノール法で抽出した全RNAを鋳型とし、WAKO社のAMV逆転写酵素反応によりcDNAを得た。得られたcDNAを鋳型としてプライマー(5’-GGCAATTCGTGGTGGTATCT-3’、配列番号:9および5’-ACCTTCGGCTTCAGTTGTGT-3’、配列番号:10)を用いてリアルタイムPCR法によりOsPBB1遺伝子の発現量を比較した。その結果、コシヒカリに比べてNIL(GL5)は、発現量が約2.8倍高かった(図5)。
【0120】
〔実施例3〕GL5候補遺伝子(OsPBB1)の機能証明
OsPBB1の発現量がコシヒカリとNIL(GL5)の粒長を決定すると考えられた事から、コシヒカリと同等のOsPBB1発現量を持つ日本晴にOsPBB1を過剰発現させる形質転換体によりOsPBB1の機能証明を行った。
【0121】
OsPBB1の完全長cDNA(AK103126)をトウモロコシ由来ユビキチン1プロモーターとノパリン合成酵素ターミネーターの間に連結させ過剰発現させた形質転換体(BM349)をイネFOXハンティングシステム(Nakamura et al. (2007) Plant Mol. Biol. 65, 357-371)より選抜した。この形質転換体T2世代植物から収穫した充実した種子を25度で2週間乾燥後、覆っている内穎と外穎を取り除き、玄米を得た。この玄米の粒長は有意(P=0.0039)に増加しており、OsPBB1の過剰発現は玄米の長さを有意に増加させた(図6)。これにより、OsPBB1がGL5遺伝子であることが証明された。
【0122】
〔実施例4〕他の植物種におけるGL5の効果
GL5遺伝子(OsPBB1)の高発現がイネの粒長を増大させることが判明したことから、他の植物種における同遺伝子高発現の効果を調べた。シロイヌナズナ(Arabidopsis thaliana)は、OsPBB1のホモログ遺伝子としてAtPBB1(配列番号:11)およびAtPBB2(配列番号:12)の2つの遺伝子を持ち、OspBB1とのアミノ酸レベルでの相同性は両者とも82.7%と高い相同性を持つ(図2、図7)。そこでイネ由来OsPBB1の高発現がシロイヌナズナにおいても種子サイズを増加させると仮説し、実証を試みた。
【0123】
イネ由来OsPBB1の完全長cDNA(AK103126)の遺伝子コーディング領域をXbaIサイトを連結したプライマー(5’-CGCATCTAGACCGCGCGGGATGGCCGGAGC-3’、配列番号:13)とSacIサイトを連結したプライマー(5’-GTAAGAGCTCAAAATCACTCCTCCATTGCA-3’、配列番号:14)で増幅し、増幅産物をXbaIおよびSacIで処理してインサートを得た。このインサートをpSTARH301GバイナリーベクターのXbaI、SacIサイトにライゲーションして、OsPBB1の完全長cDNAをカリフラワーモザイクウイルス由来35Sプロモーターとノパリン合成酵素ターミネーターとの間に連結させたバイナリーベクター(pSTARH301G/GL5)を得た。pSTARH301G/GL5をエレクトロポレーション法によりアグロバクテリウム(Agrobacterium tumefaciens EHA101)へ導入し、さらにこのアグロバクテリウムを減圧浸潤法によりシロイヌナズナへ感染させ形質転換体T1種子を得た。この種子をハイグロマイシン50mg/lを含有するGM寒天培地で形質転換スクリーニングを行った。得られた形質転換体(T1世代)からT2種子を得て、再びハイグロマイシンを含む培地に播種し、3:1の分離比を示す系統を選択し、導入遺伝子が1遺伝子座に存在すると考えられる形質転換体を選抜した。この種子のサイズを測定したところ、OsPBB1のcDNAを含まないコントロールベクター系統に比べて種子の長径が有意に増加していた(図8)。OsPBB1の高発現は、シロイヌナズナにおいても種子サイズを増大させることが明らかとなった。これによりGL5は単子葉植物であるイネのみならず双子葉植物であるシロイヌナズナにおいても種子サイズを増大させる効果があり、幅広い植物種で同様の効果を有する事が示唆された。
【技術分野】
【0001】
本発明は、植物体の子実(種子、頴花又は果実を含む)を増大させる機能を有するイネ由来のタンパク質をコードするDNA、および該DNAを用いた子実収量(種子、頴花又は果実の収量のことをいう)の増加方法、子実収量の検出方法、子実収量が増加する植物体の育種方法に関する。
【背景技術】
【0002】
世界人口が爆発的に増え作物(例えば穀物)の生産増が求められている。世界の年間人口増加率1.4%に対し、穀物生産増加率は1.0%と人口増加率に比べ低く、世界人口が80億を突破する2025年には作物需要は50%上昇すると予想され、食糧不足も一層加速していると予想される。作物生産量を上昇させるためには、収量増加と関与する遺伝子の特定と特定した遺伝子を利用した効率的な穀物育種が必須である。
【0003】
イネにおいて子実の粒サイズはシンクのキャパシティーを示す値であり、収量に直接関わる。これまでに、イネにおいて種子(粒)の大きさに関わる遺伝子として、GW2(非特許文献1)及びGS3(非特許文献2)がポジショナルクローニング法により単離されている。しかし、これらの遺伝子は収量増に貢献しない。また、plastchron 1遺伝子を発現させ粒を拡大したトランスジェニック植物とその利用方法(特許文献1)、改変した3量体Gタンパク質αサブユニット遺伝子を利用し巨大粒を結実させる方法(特許文献2)、及びイネの粒長を制御するLk3遺伝子およびその利用(特許文献3)が特許出願されている。さらに、本願発明者らによって、粒重ひいては収量を増加させる遺伝子(TGW6)が単離されている(特許文献4)。
【0004】
しかしながら、イネの種子を増大させるには、現在までに単離された遺伝子だけでは十分ではなかった。また、種を超えて子実を増大させる方法は明らかではなかった。
なお、本出願の発明に関連する先行技術文献情報を以下に示す。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2005-204621
【特許文献2】特開2004-033199
【特許文献3】再表2006-112327
【特許文献4】特願2008-292285
【非特許文献】
【0006】
【非特許文献1】Song X.J. et al., (2007) Nature Genet. 39, 623-630
【非特許文献2】Fan C. et al., (2006) Theor. Appl. Genet. 112, 1164-1171
【非特許文献3】Shomura A et al. (2008) Nat. Genet. 40, 1023-1028.
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明は、このような状況に鑑みてなされたものであり、その目的は、植物の子実粒形(頴花・果実・種子を含む)ひいては粒長の伸長に関する新規な遺伝子の単離・同定、並びに該遺伝子を利用した植物の子実(頴花・果実・種子を含む)の粒サイズを増加させる育種方法を提供することにある。
【課題を解決するための手段】
【0008】
発明者らは、これまで植物体の作物収量増加のため、単子葉植物のモデルであるイネを用いて直接的に収量増加を担う遺伝子、すなわち粒長の伸長に関する遺伝子の探索を試みてきた。粒長は複数の遺伝子の相互作用による量的形質として支配されている。
【0009】
発明者らは、まず、コシヒカリと、コシヒカリを遺伝的背景にGL5領域をカサラス型染色体に置換した準同質遺伝子系統NIL(GL5)との戻し交雑後代を用いて、連鎖解析を行った。GL5遺伝子座近傍のSNPマーカーを用いて連鎖解析を行った結果、GL5遺伝子座をSNPマーカーG1422とSNPマーカーG1491との間、約7kbpの範囲に特定した(図1)。次に、イネアノテーションプロジェクトにより公開された遺伝子予測データベースにより、遺伝子の発現量・発現部位からGL5遺伝子を1つの遺伝子(Os05g0187000)に絞り込み、本遺伝子は20Sプロテアソームのβ2サブユニットをコードしていることが明らかとなった(図2)。
【0010】
コシヒカリとカサラスのcDNA塩基配列解析の結果、GL5候補遺伝子(OsPBB1)のコーディング領域に2カ所のSNPを有していたが、これらはいずれもアミノ酸配列に変化を生じさせず、コシヒカリとカサラスのGL5候補遺伝子のアミノ酸配列は同一であることが明らかとなった(図3)。また、OsPBB1の推定プロモーター領域にもいくつかの変異が存在することが明らかとなった(図4)。
【0011】
次に、OsPBB1遺伝子の発現量を比較したところ、コシヒカリに比べてNIL(GL5)は、発現量が約2.8倍高いことが明らかとなった(図5)。
【0012】
次に、コシヒカリと同等のOsPBB1発現量を持つ日本晴にOsPBB1を過剰発現させる形質転換体によりOsPBB1の機能証明を行ったところ、OsPBB1の過剰発現は玄米の長さを有意に増加させることが明らかとなった(図6)。
【0013】
GL5遺伝子(OsPBB1)の高発現がイネの粒長を増大させることが判明したことから、さらに他の植物種における同遺伝子高発現の効果を調べた。その結果、OsPBB1の高発現は、シロイヌナズナにおいても種子サイズを増大させることが明らかとなった(図8)。これによりGL5は単子葉植物であるイネのみならず双子葉植物であるシロイヌナズナにおいても種子サイズを増大させる効果があり、幅広い植物種で同様の効果を有する事が示唆された。
【0014】
本発明におけるGL5遺伝子は粒長、粒長の伸長を介し収量特性を向上させる遺伝子として初めて特定されたものであり、GL5遺伝子を利用することで収量増、生産性の向上に利用できる。即ち、本発明者らは、植物体の粒形の増大に関与する新たな遺伝子を単離することに成功した。
【0015】
本発明は、より具体的には以下の〔1〕〜〔19〕を提供するものである。
〔1〕下記(a)から(d)のいずれかに記載のDNAを含む、植物体の子実(種子、頴花又は果実を含む)の大きさを増大させた植物体。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
〔2〕植物体が単子葉植物又は双子葉植物である、〔1〕に記載の植物体。
〔3〕下記(a)から(d)のいずれかに記載のDNAが発現するように導入されたベクター。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
〔4〕〔3〕 に記載のベクターが導入された宿主細胞。
〔5〕〔4〕に記載のベクターが導入された植物細胞。
〔6〕〔5〕に記載の植物細胞を含む形質転換植物体。
〔7〕〔6〕に記載の形質転換植物体の子孫またはクローンである、形質転換植物体。
〔8〕〔6〕又は〔7〕に記載の形質転換植物体の繁殖材料。
〔9〕下記(a)から(d)のいずれかに記載のDNAを植物体の細胞内で過剰発現させる工程を含む、植物体の子実(種子、頴花又は果実を含む)の大きさを増大させる方法。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
〔10〕植物体が単子葉植物又は双子葉植物である、〔9〕に記載の方法。
〔11〕被検植物体について、配列番号:1に記載のDNA領域に相当する部位、またはその周辺配列に存在する変異を含むDNAマーカーを検出することを特徴とする、被検植物体の子実(種子、頴花又は果実を含む)の収量を検出する方法。
〔12〕変異が一塩基多型である、〔11〕に記載の方法。
〔13〕以下の工程(a)及び(b)を含む、〔11〕に記載の方法。
(a)被検植物における配列番号:1に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域に相当する部位、またはその周辺配列に存在する多型部位の塩基種を決定する工程、及び
(b)(a)で決定された多型部位の塩基種において、配列番号:1またはその周辺配列と異なるアレルが検出された場合に、被検植物は子実(種子、頴花又は果実を含む)の収量が多いと判定する工程
〔14〕多型部位が、配列番号:1に記載の塩基配列における、32位、129位、161位、557位、839位、843位、976位又は1699位から選択される少なくとも一つの多型部位に相当する部位である、〔13〕に記載の方法。
〔15〕多型部位の塩基種の変異が、配列番号:1に記載の塩基配列における32位の塩基種のGからTへの変異、129位の塩基種のAからGへの変異、161位の塩基種のTからGへの変異、557位の塩基種のTからCへの変異、839位の塩基種のTの欠損、843位の塩基種のGの欠損、976位の塩基種のCからAへの変異、又は1699位の塩基種のCからTへの変異から選択される少なくとも一つの変異である場合には、子実(種子、頴花又は果実を含む)が増大すると判定する、〔14〕に記載の方法。
〔16〕配列番号:1に記載のDNAとストリンジェントな条件で特異的にハイブリダイズし、少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドを含む、被検植物の子実(種子、頴花又は果実を含む)の収量を検出するためのプライマー。
〔17〕〔14〕または〔15〕に記載の多型部位を含む領域を増幅するための、〔16〕に記載のプライマー。
〔18〕配列番号:1に記載のDNAとストリンジェントな条件で特異的にハイブリダイズし、少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドを含む、被検植物体の子実(種子、頴花又は果実を含む)の収量を検出するためのプローブ。
〔19〕〔14〕または〔15〕に記載の多型部位を含む領域に特異的にハイブリダイズする、〔18〕に記載のプローブ。
【発明の効果】
【0016】
連鎖解析により、植物の子実粒(種子、頴花又は果実を含む)の伸長ひいては収量の増加に関する遺伝子の単離・同定に成功した。また、該遺伝子を利用した植物の子実粒(種子、頴花又は果実を含む)の長さを増加させる育種手法も見出した。本発明は、植物の品種改良等の分野において有用である。
【図面の簡単な説明】
【0017】
【図1】粒長に関与するQTLの遺伝子特定結果を示す図である。
【図2】GL5に類似する遺伝子の系統樹を示す図である。
【図3】コシヒカリ、カサラスのGL5遺伝子のアミノ酸配列比較を示す図である。
【図4−1】コシヒカリ、カサラスのGL5遺伝子のプロモーター配列比較(前半部分)を示す図である。
【図4−2】コシヒカリ、カサラスのGL5遺伝子のプロモーター配列比較(後半部分)を示す図である。
【図5】コシヒカリとNIL(GL5)におけるGL5遺伝子発現量の比較を示すグラフである。
【図6】日本晴にGL5遺伝子を過剰発現させた形質転換体イネの玄米粒サイズを示す図である。
【図7】イネ、シロイヌナズナのGL5に類似する遺伝子のアミノ酸配列比較を示す図である。
【図8】シロイヌナズナにGL5遺伝子を過剰発現させた形質転換体シロイヌナズナの種子粒サイズを示す図である。
【発明を実施するための形態】
【0018】
本発明は、植物体の子実(種子、頴花又は果実を含む)の収量に関する遺伝子GL5を含む植物体、および該遺伝子を用いた子実収量(種子、頴花又は果実の収量のことをいう)の増加方法、子実収量の検出方法、子実収量が増加する植物体の育種方法を提供する。
【0019】
本発明において子実の収量の増加とは、具体的には種子、頴花又は果実を増大させることによってもたらされる。本発明において子実が増大するとは、子実の大きさが増加することをいい、該遺伝子の発現量が低い植物体の子実と比較して、それぞれ30%以上、好ましくは20%以上、より好ましくは15%、10%、5%以上のサイズの増加をいう。
【0020】
本発明者らにより作物の子実収量との関係が明らかにされた、イネカサラスのGL5遺伝子のゲノムDNAの塩基配列を配列番号:4に、cDNAの塩基配列を配列番号:5に、これら遺伝子がコードするタンパク質のアミノ酸配列を配列番号:6に示す。
【0021】
また、イネコシヒカリのGL5遺伝子のゲノムDNAの塩基配列を配列番号:1に、cDNAの塩基配列を配列番号:2に、これら遺伝子がコードするタンパク質のアミノ酸配列を配列番号:3に示す。
【0022】
本発明は、植物体の子実を増大させる機能を有するイネ由来のGL5遺伝子を含むベクターならびに形質転換植物細胞を提供する。
【0023】
本発明のGL5遺伝子としては、具体的には、植物体の子実を増大させる機能を有するイネ由来のタンパク質をコードする、下記(a)から(d)のいずれかに記載のDNAが含まれる。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
【0024】
本発明において、本発明の遺伝子が由来する植物体としては、特に制限はないが、例えばイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビ等の穀物、ダイズ、エンドウ、ピーナッツ等の豆類、カボチャ、トマト等のいわゆる実のなる野菜類、ゴマ、ナタネ、ヒマワリ等の油料作物、シロイヌナズナ等が挙げられる。本発明の遺伝子が由来する植物として、より好ましくは、イネを挙げることができる。
【0025】
本発明の「GL5遺伝子」は、「GL5タンパク質」をコードしうるものであれば、その形態に特に制限はなく、「GL5遺伝子」には、cDNAの他、ゲノムDNA、化学合成DNAなども含まれる。また、GL5遺伝子はGL5タンパク質をコードするものであれば、遺伝暗号の縮重に基づく任意の塩基配列を有するDNAが含まれる。
【0026】
本発明の遺伝子のコード領域は、例えば、配列番号:5に記載の塩基配列における217位〜1035位の領域を挙げることができる。
【0027】
ゲノムDNAおよびcDNAの調製は、当業者にとって常套手段を利用して行うことが可能である。ゲノムDNAは、例えば、植物からゲノムDNAを抽出し、ゲノミックライブラリー(ベクターとしては、プラスミド、ファージ、コスミド、BAC、PACなどが利用できる)を作成し、これを展開して、GL5遺伝子(例えば、配列番号:4または5のいずれかに記載のDNA)を基に調製したプローブを用いてコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより当該クローンを得、調製することが可能である。また、GL5遺伝子に特異的なプライマーを作成し、これを利用したPCRをおこなうことによって調製することも可能である。また、cDNAは、例えば、植物から抽出したmRNAを基にcDNAを合成し、これをλZAP等のベクターに挿入してcDNAライブラリーを作成し、これを展開して、上記と同様にコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより、また、PCRを行うことにより調製することが可能である。
【0028】
さらに、GL5遺伝子は広く穀物植物に存在すると考えられるため、GL5遺伝子には、種々の植物に存在する相同遺伝子も含まれる。ここで「相同遺伝子」とは、種々の植物において、イネにおける活性型GL5遺伝子産物と機能的に同等なタンパク質をコードする遺伝子を指す。このようなタンパク質には、例えば、活性型GL5タンパク質の変異体、アレル、バリアント、ホモログ、活性型GL5タンパク質の部分ペプチド、または、他のタンパク質との融合タンパク質などが挙げられるが、これらに限定されない。
【0029】
本発明における活性型GL5タンパク質の変異体としては、配列番号:6に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなる天然由来のタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質のN末側活性断片と機能的に同等なタンパク質を挙げることが出来る。また、配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズする天然由来のDNAよりコードされるタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質のN末側活性断片と機能的に同等なタンパク質も、活性型GL5タンパク質の変異体として挙げることができる。
【0030】
また、本発明における活性型GL5タンパク質の変異体としては、配列番号:6に記載のアミノ酸配列において1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなる天然由来のタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質と機能的に同等なタンパク質を挙げることが出来る。また、配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズする天然由来のDNAよりコードされるタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質と機能的に同等なタンパク質も、活性型GL5タンパク質の変異体として挙げることができる。
【0031】
本発明において、変異するアミノ酸数は特に制限されないが、通常、30アミノ酸以内であり、好ましくは15アミノ酸以内であり、さらに好ましくは5アミノ酸以内(例えば、3アミノ酸以内)であると考えられる。変異するアミノ酸残基においては、アミノ酸側鎖の性質が保存されている別のアミノ酸に変異されることが望ましい(これは、保存的アミノ酸置換として知られている)。例えばアミノ酸側鎖の性質としては、疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)および親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)の二種類に大別することができる。また、その側鎖の構造に基づいて、脂肪族側鎖を有するアミノ酸(G、A、V、L、I、P)、水酸基含有側鎖を有するアミノ酸(S、T、Y)、硫黄原子含有側鎖を有するアミノ酸(C、M)、カルボン酸及びアミド含有側鎖を有するアミノ酸(D、N、E、Q)、塩基含有側鎖を有するアミノ酸(R、K、H)、芳香族含有側鎖を有するアミノ酸(H、F、Y、W)などのようにアミノ酸を分類することもできる。さらに、例えば、変異マトリクス(mutational matrix)によってアミノ酸を分類することも周知である(Taylor 1986, J, Theor. Biol. 119, 205-218; Sambrook, J. et al., Molecular Cloning 3rd ed. A7.6-A7.9, Cold Spring Harbor Lab. Press, 2001)。この分類を以下に要約すると、脂肪族アミノ酸(L、I、V)、芳香族アミノ酸(H、W、Y、F)、荷電アミノ酸(D、E、R、K、H)、正荷電アミノ酸(R、K、H)、負荷電アミノ酸(D、E)、疎水性アミノ酸(H、W、Y、F、M、L、I、V、C、A、G、T、K)、極性アミノ酸(T、S、N、D、E、Q、R、K、H、W、Y)、小型アミノ酸(P、V、C、A、G、T、S、N、D)、微小アミノ酸(A、G、S)および大型(非小型)アミノ酸(Q、E、R、K、H、W、Y、F、M、L、I)が挙げられる(括弧内はいずれもアミノ酸の一文字標記を表す)。
【0032】
あるアミノ酸配列に対する1又は複数個のアミノ酸残基の欠失、付加及び/又は他のアミノ酸による置換により修飾されたアミノ酸配列を有するポリペプチドがその生物学的活性を維持することはすでに知られている。さらに、標的アミノ酸残基は、共通した性質をできるだけ多く有するアミノ酸残基に変異させることがより好ましい。
【0033】
本発明において「機能的に同等」とは、対象となるタンパク質が、活性型GL5タンパク質と同等の生物学的機能や生化学的機能を有することを指す。本発明において、活性型GL5タンパク質の生物学的機能や生化学的機能としては、例えば穀物の種子を増大させる機能を挙げることができる。生物学的な性質には発現する部位の特異性や、発現量等も含まれる。
【0034】
相同遺伝子を単離するための当業者によく知られた方法としては、ハイブリダイゼーション技術(Southern, E. M., Journal of Molecular Biology, Vol. 98, 503, 1975)やポリメラーゼ連鎖反応(PCR)技術(Saiki, R. K., et al. Science, vol. 230, 1350-1354, 1985, Saiki, R. K. et al. Science, vol.239, 487-491,1988)が挙げられる。即ち、当業者にとっては、GL5遺伝子の塩基配列(例えば、配列番号:4又は5のいずれかに記載のDNA)もしくはその一部をプローブとして、またGL5遺伝子に特異的にハイブリダイズするオリゴヌクレオチドをプライマーとして、種々の植物からGL5遺伝子の相同遺伝子を単離することは通常行いうることである。
【0035】
このような相同遺伝子をコードするDNAを単離するためには、通常ストリンジェントな条件下でハイブリダイゼーション反応を行なう。ストリンジェントなハイブリダイゼーション条件は当業者であれば、適宜選択することができる。一例を示せば、25%ホルムアミド、より厳しい条件では50%ホルムアミド、4xSSC、50mM Hepes pH7.0、10×デンハルト溶液、20μg/ml変性サケ精子DNAを含むハイブリダイゼーション溶液中、42℃で一晩プレハイブリダイゼーションを行った後、標識したプローブを添加し、42℃で一晩保温することによりハイブリダイゼーションを行う。その後の洗浄における洗浄液および温度条件は、「1xSSC、0.1% SDS、37℃」程度で、より厳しい条件としては「0.5xSSC、0.1% SDS、42℃」程度で、さらに厳しい条件としては「0.2xSSC、0.1% SDS、65℃」程度で実施することができる。このようにハイブリダイゼーションの洗浄の条件が厳しくなるほどプローブ配列と高い相同性を有するDNAの単離を期待しうる。但し、上記SSC、SDSおよび温度の条件の組み合わせは例示であり、当業者であれば、ハイブリダイゼーションのストリンジェンシーを決定する上記若しくは他の要素(例えば、プローブ濃度、プローブの長さ、ハイブリダイゼーション反応時間など)を適宜組み合わせることにより、上記と同様のストリンジェンシーを実現することが可能である。
【0036】
単離されたDNAの相同性は、アミノ酸配列全体で、少なくとも50%以上、さらに好ましくは70%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。配列の相同性は、BLASTN(核酸レベル)やBLASTX(アミノ酸レベル)のプログラム(Altschul et al. J. Mol. Biol., 215: 403-410, 1990)を利用して決定することができる。該プログラムは、Karlin及びAltschulによるアルゴリズムBLAST (Proc. Natl. Acad. Sci. USA, 87:2264-2268, 1990, Proc. Natl. Acad. Sci. USA, 90: 5873-5877, 1993) に基づいている。BLASTNによって塩基配列を解析する場合には、パラメーターは例えばscore = 100、wordlength =12とする。また、BLASTXによってアミノ酸配列を解析する場合には、パラメーターは例えばscore = 50、wordlength = 3とする。また、Gapped BLASTプログラムを用いて、アミノ酸配列を解析する場合は、Altschulら(Nucleic Acids Res. 25: 3389-3402, 1997)に記載されているように行うことができる。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である。
【0037】
本発明のベクターとしては、形質転換体作製のために細胞内で本発明のDNAを発現させるベクター、例えば形質転換植物体作製のために植物細胞内で本発明のDNAを発現させるためのベクターが含まれる。植物細胞の形質転換に用いられるベクターとしては、該細胞内で挿入遺伝子を発現させることが可能なものであれば特に制限はない。例えばプラスミド、ファージ、またはコスミドなどを例示することができる。
【0038】
また上記「植物細胞」には、種々の形態の植物細胞、例えば、懸濁培養細胞、プロトプラスト、葉の切片、カルス等が含まれる。
【0039】
本発明のベクターは、本発明のDNAを恒常的または誘導的に発現させるためのプロモーターを含有してもよい。
【0040】
当業者においては、所望のDNAを有するベクターを、一般的な遺伝子工学技術によって、適宜、作製することが可能である。通常、市販の種々のベクターを利用することができる。
【0041】
本発明のベクターは、宿主細胞内において本発明のDNAを保持したり、発現させるためにも有用である。
【0042】
本発明におけるDNAは、通常、適当なベクターへ担持(挿入)され、宿主細胞へ導入される。即ち本発明は、本発明のDNAまたはベクターを保持する宿主細胞を提供する。該ベクターとしては、挿入したDNAを安定に保持するものであれば特に制限されず、例えば宿主に大腸菌を用いるのであれば、クローニング用ベクターとしてpBluescriptベクター(Stratagene社製)などが好ましいが、市販の種々のベクターを利用することができる。本発明のDNAを内在性遺伝子を有する細胞内に導入および発現させる目的としてベクターを用いる場合には、特に発現ベクターが有用である。発現ベクターとしては、試験管内、大腸菌内、培養細胞内、生物個体内でDNAを発現するベクターであれば特に制限されないが、例えば、試験管内発現であればpBESTベクター(プロメガ社製)、大腸菌であればpETベクター(Invitrogen社製)、培養細胞であればpME18S-FL3ベクター(GenBank Accession No. AB009864)、生物個体であればpME18Sベクター(Mol Cell Biol. 8:466-472(1988))、植物個体であればpBINPLUSベクター(van Engelen, F.A. et al., (1995). pBINPLUS: an improved plant transformation vector based on pBIN19. Transgenic Res. 4, 288-290.)などを例示することができる。ベクターへの本発明のDNAの挿入は、常法(Molecular Cloning, 5.61-5.63)により、例えば、制限酵素サイトを用いたリガーゼ反応により行うことができる。
【0043】
上記宿主細胞としては特に制限はなく、目的に応じて種々の宿主細胞が用いられる。本発明のDNAを発現させるための細胞としては、例えば、細菌細胞(例:ストレプトコッカス、スタフィロコッカス、大腸菌、ストレプトミセス、枯草菌)、昆虫細胞(例:ドロソフィラS2、スポドプテラSF9)、動物細胞(例:CHO、COS、HeLa、C127、3T3、BHK、HEK293、Bowes メラノーマ細胞)および植物細胞を例示することができる。
【0044】
また、生体内で本発明のDNAを発現させる方法としては、本発明のDNAを適当なベクターに組み込み、例えば、ポリエチレングリコール法、エレクトロポレーション法、アグロバクテリウム法、リポソーム法、カチオニックリポソーム法、リン酸カルシウム沈殿法、電気パルス穿孔法(エレクトロポーレーション)(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons.Section 9.1-9.9)、リポフェクション法(GIBCO-BRL社製)、マイクロインジェクション法、パーティクルガン法などの当業者に公知の方法により生体内に導入する方法などが挙げられる。
【0045】
植物体内への投与は、ex vivo法であっても、in vivo法であってもよい。
【0046】
また、植物体内へ本発明のDNAを導入する場合、DNAは、マイクロインジェクション法、エレクトロポレーション法、ポリエチレングリコール法等を用いて、植物細胞に直接導入することもできるが、植物への遺伝子導入用プラスミドに組込み、これをベクターとして、植物感染能のあるウイルスあるいは細菌を介して、間接的に植物細胞に導入することもできる。かかるウイルスとしては、例えば代表的なウイルスとして、カリフラワーモザイクウイルス、タバコモザイクウイルス、ジェミニウイルス等が挙げられ、細菌としては、アグロバクテリウム等が挙げられる。アグロバクテリウム法により、植物への遺伝子導入を行う場合には、市販のプラスミドを用いることができる。このようなベクターを用いて、植物体内へ本発明のDNAを導入する場合の方法としては、好ましくは、アグロバクテリウムを介して遺伝子を導入するリーフディスク法(Jorgensen, R.A. et al., (1996). Chalcone synthase cosuppression phenotypes in petunia flowers: comparison of sense vs. antisense constructs and single-copy vs. complex T-DNA sequences. Plant Mol. Biol. 31, 957-973.)が挙げられる。
【0047】
なおこれら上述の形質転換方法は、宿主となる植物などの種類(例えば単子葉植物又は双子葉植物)に応じて適宜選択することが好ましい。
【0048】
本発明において「植物」とは、特に制限されないが、例えばイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビ等の穀物、ダイズ、エンドウ、ピーナッツ等の豆類、カボチャ、トマト等のいわゆる実のなる野菜類、ゴマ、ナタネ、ヒマワリ等の油料作物、シロイヌナズナ等を挙げることができる。
【0049】
また、本発明は、本発明のDNAまたは本発明のベクターを保持する植物細胞を提供する。さらに本発明は、本発明の植物細胞を含む形質転換植物体を提供する。本発明のDNAまたは本発明のベクターが導入される細胞には、形質転換植物体作製のための植物細胞が含まれる。植物細胞としては特に制限はない。
【0050】
本発明の植物細胞には、培養細胞の他、植物体中の細胞も含まれる。また、プロトプラスト、苗条原基、多芽体、毛状根も含まれる。
【0051】
形質転換植物細胞からの植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。例えば、形質転換植物体を作出する手法については、ポリエチレングリコールによりプロトプラストへ遺伝子導入し植物体を再生させる方法、電気パルスによりプロトプラストへ遺伝子導入し植物体を再生させる方法、パーティクルガン法により細胞へ遺伝子を直接導入し、植物体を再生させる方法、およびアグロバクテリウムを介して遺伝子を導入し、植物体を再生させる方法などを挙げることができるが、特に制限されるものではない。いくつかの技術については既に確立し、本願発明の技術分野において広く用いられている。本発明においては、これらの方法を好適に用いることができる。
【0052】
本発明のDNAを含むベクターの導入により形質転換した植物細胞を効率的に選択するために、上記組み換えベクターは、適当な選抜マーカー遺伝子を含む、もしくは選抜マーカー遺伝子を含むプラスミドベクターと共に植物細胞へ導入することが好ましい。この目的に使用される選抜マーカー遺伝子は、例えば抗生物質カナマイシンまたはゲンタマイシンに耐性であるネオマイシンホスホトランスフェラーゼ遺伝子、ハイグロマイシンに耐性であるハイグロマイシンホスホトランスフェラーゼ遺伝子、および除草剤ホスフィノスリシンに耐性であるアセチルトランスフェラーゼ遺伝子等が挙げられる。
【0053】
組み換えベクターを導入した植物細胞は、導入された選抜マーカー遺伝子の種類に従って適当な選抜用薬剤を含む公知の選抜用培地に置床し培養する。これにより形質転換された植物培養細胞を得ることができる。
【0054】
形質転換された植物細胞は、再分化させることにより植物体を再生させることが可能である。再分化の方法は植物細胞の種類により異なるが、例えばイネであればFujimuraら(Plant Tissue Culture Lett. 2:74 (1995))の方法が挙げられ、トウモロコシであればShillitoら(Bio/Technology 7:581 (1989))の方法やGorden-Kammら(Plant Cell 2:603(1990))の方法が挙げられる。
【0055】
なお、このように再生され、かつ栽培した形質転換植物体中の導入された外来DNAの存在は、公知のPCR法やサザンハイブリダイゼーション法によって、または植物体中のDNAの塩基配列を解析することによって確認することができる。
【0056】
この場合、形質転換植物体からのDNAの抽出は、公知のJ.Sambrookらの方法(Molecular Cloning、第2版、Cold Spring Harbor Laboratory Press, 1989)に準じて実施することができる。
【0057】
再生させた植物体中に存在する本発明のDNAよりなる外来遺伝子を、PCR法を用いて解析する場合には、上記のように再生植物体から抽出したDNAを鋳型として増幅反応を行う。また、本発明のDNA、あるいは本発明により改変されたDNAの塩基配列に従って適当に選択された塩基配列をもつ合成したオリゴヌクレオチドをプライマーとして用い、これらを混合させた反応液中において増幅反応を行うこともできる。増幅反応においては、DNAの変性、アニーリング、伸張反応を数十回繰り返すと、本発明のDNA配列を含むDNA断片の増幅生成物を得ることができる。増幅生成物を含む反応液を例えばアガロース電気泳動にかけると、増幅された各種のDNA断片が分画されて、そのDNA断片が本発明のDNAに対応することを確認することが可能である。
【0058】
一旦、ゲノム内に本発明のDNAが導入された形質転換植物体が得られれば、該植物体から有性生殖または無性生殖により子孫を得ることが可能である。また、該植物体やその子孫あるいはクローンから繁殖材料(例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラスト等)を得て、それらを基に該植物体を量産することも可能である。本発明には、本発明のDNAまたはベクターが導入された植物細胞、該細胞を含む植物体、該植物体の子孫およびクローン、並びに該植物体、その子孫、およびクローンの繁殖材料が含まれる。
【0059】
このようにして作出された植物体は通常の穀物に比べて子実収量が増加することが期待される。
【0060】
上述のように、本発明のDNAもしくはベクターを植物細胞へ導入し、該植物細胞から植物体を再生させる工程を含む、形質転換植物体の製造方法もまた本発明に含まれる。
【0061】
本発明の子実収量が増加した植物体もしくはその種子は、育種法によっても作出することが可能である。
【0062】
上記育種法としては、例えば、本発明のDNAを有する品種と交雑させることを特徴とする一般的な育種法(交雑育種法等)を挙げることができる。該方法によって、子実が増大した植物体もしくはその種子を作出することができる。
【0063】
育種法によって本発明の植物体もしくは種子を作製する際には、公知の種々の文献を参照して適宜実施することができる(細胞工学別冊・植物細胞工学シリーズ15「モデル植物の実験プロトコール」秀潤社、2001年、加藤鎌司著「2.1 イネ・コムギの交配」p.6-9; 高牟礼逸朗、佐野芳雄著「2. 交配法」p.46-48)。
【0064】
本発明の上記育種方法の好ましい態様としては、以下の(a)および(b)に記載の工程を含む方法である。
(a)子実が増大した植物と任意の機能を有する植物とが交配された品種を作製する工程、
(b)工程(a)で作製された植物の子実の収量を検出する工程
【0065】
本発明の植物体を育種する方法のより具体的な例としては、以下の(a)および(b)に記載の工程を含む方法が挙げられる。
(a)本発明のDNAを有する植物と交雑させる工程、
(b)前記DNAを有する植物改変体を選抜する工程
【0066】
本発明の上記植物体の作成方法を育種法によって実施する場合には、さらに具体的には、以下のような工程を含む方法を挙げることができる。
(a)植物Aと、本発明のDNAを有する他の植物Bを交雑させ、F1を作出する工程、
(b)前記F1と前記植物Aを交雑させる工程、
(c)前記DNAを有する植物を選抜する工程、
(d)工程(c)によって選抜された植物と、前記植物Aを交雑させる工程
【0067】
上記方法においては、本発明のDNAを有する植物Bと、子実収量を増加させたい植物もしくは子実を増大させたい植物(これら植物を「植物A」と記載する。)を交雑し、Bのもつ本発明のDNAが受け継がれ、かつ植物Aに近い個体を選抜し、これに植物Aによる交雑を重ねていく「戻し交雑」を行って、Bが有する本発明のDNAの形質を意図的に導入する。その際、一般的にゲノム育種に利用されるDNAマーカーを利用して本発明のDNAを有する植物を選抜することにより、上記「戻し交雑」による置換を効率的に行うことが可能である。その結果、育種期間の短縮に繋がり、また、余分なゲノム領域の混入を正確に除くことができる。通常、「戻し交雑」では、本発明のDNAと非常に強く連鎖する他のDNAに依存する形質がどうしても排除できないという現象が問題となることがあるが、本発明のDNAの近傍に存在するDNAマーカーを利用することにより、所望の植物の正確な選抜が可能となる。
【0068】
上記方法においては、必要に応じて、本発明のDNA以外のゲノム全域が目的の遺伝形質でホモ固定するまで、繰り返して行うことができる。即ち、本発明の好ましい態様のおいては、上記工程(d)によって交雑された個体について、一般的なDNAマーカーを利用して、本発明のDNAを有し、かつ、ゲノム構造が植物Aに近い植物個体を選抜することができる。さらに、この選抜された植物個体は、必要に応じて、「戻し交雑」(イネ品種Aと交雑)させることができる。
【0069】
特にDNAマーカーを利用したゲノム育種方法では、置換率の高い個体を選抜して次の交雑に進むことができるため、世代を進めるほどに選抜効率が良くなる。また、本方法では、少ない個体数を扱えば済むので、省スペースでの育種が可能になる。さらに、温室や人工気象室を利用して1年に複数回もの交雑が可能になる。
【0070】
上記工程(c)において、DNAマーカーを用いて選抜するとは、当該DNAマーカーを特徴付ける塩基配列(例えば、多型等)についての塩基種の情報を基に、選抜を行うことを言う。例えば、本発明のDNAの近傍に多型変異が存在する場合、当該多型変異と同一の多型変異を有する個体を選抜すること等を言う。
【0071】
本発明の育種方法は、好ましくは、DNAマーカーを利用した「ゲノム育種」方法である。該「ゲノム育種」は「マーカー育種」とも呼ばれる。
【0072】
本発明の育種方法において利用可能なDNAマーカーは、特に制限されず、一般的に知られている種々のDNAマーカーを好適に用いることができる。例えば、RFLP(制限酵素断片長多型)マーカー、SSR(単純反復配列)マーカー、SNP(一塩基多型)マーカー等を例示することができる。
【0073】
本発明者らは、カサラス由来のGL5遺伝子をもつイネが種子を増大させる機能を有することを見出した。従って、コシヒカリ型とカサラス型のGL5遺伝子を判別することで被検植物の種子の収量を検出することができる。言い換えれば、コシヒカリ型に対するカサラス型の変異部位をDNAマーカーとすることで、上記育種方法に使用することができる。
【0074】
上記の知見に基づき、本発明は、被検植物について、配列番号:1に記載のDNA領域(例えば配列番号:1に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域)に相当する部位、またはその周辺配列に存在する変異を含むDNAマーカーを検出することを特徴とする、被検植物の種子の収量を検出する方法を提供する。
【0075】
本発明において、配列番号:1に記載のDNA領域に「相当する部位」とは、種々の植物において、当該植物に存在する上記遺伝子の相同遺伝子における対応する部位をいう。
【0076】
本発明において「子実の収量を検出」とは、子実のサイズが増大するか否かを判定するための検査が含まれる。本発明の方法においては、上記配列番号:1に記載のDNA領域に相当する部位、またはその周辺配列において、配列番号:4と同型の変異が検出された場合には、子実のサイズは増大すると判定される。
【0077】
本発明の方法により、被検植物における変異を検出することで、実際に子実を観察しなくても、被検植物の子実のサイズが増大するか否かを判定することができる。
【0078】
本発明における「周辺配列」とは、通常、該遺伝子の近傍の染色体上の領域を指す。近傍とは、特に制限されるものではないが、通常、本発明の多型部位を含むDNA領域である。
【0079】
上記本発明の検出方法における「変異」の位置は、予め規定することは困難であるが、通常、上記遺伝子のORF中、あるいは上記遺伝子の発現を制御する領域(例えば、プロモーター領域、エンハンサー領域等)中に存在するが、これらに限定されるものではない。また、この「変異」とは、上記遺伝子の発現量を変化させる、mRNAの安定性等の性質を変化させる、あるいは上記遺伝子によってコードされるタンパク質の有する活性を変化させるような変異であることが多いが、特に制限されない。本発明の変異としては、例えば、塩基の付加、欠失、置換、挿入変異等を挙げることができる。
【0080】
本発明者らは、被検植物における、植物の子実収量を増加させる機能を有する植物由来のタンパク質をコードする配列番号:1のDNA領域において、植物の子実収量に対して有意に関連する多型変異を見出すことに成功した。
【0081】
本発明の子実収量を検出する方法における「多型部位」は、上記DNAのいずれかに記載の塩基配列もしくは該塩基配列の近傍DNA領域に存在する多型であれば、特に制限されない。具体的には、本発明の子実収量を検出する方法に利用可能な多型部位として、配列番号:1に記載の塩基における32位、129位、161位、557位、839位、843位、976位又は1699位から選択される少なくとも一つの多型部位に相当する部位を挙げることができる(なお、本明細書においては、これらの多型部位を単に『本発明の多型部位』と記載する場合がある)。
【0082】
上記表中に示した多型部位の塩基種は配列表に示した配列に対して相補鎖側にある塩基種を示している場合があるが、本明細書において記載された前後配列を用いれば異同を確認することは当業者にとって容易であり、検出を行うにあたってはプラス鎖とマイナス鎖のどちらを調べても必然的にもう一方の結果を決定することができる。
【0083】
本発明の好ましい態様においては、上述した多型部位における塩基種の変異が、配列番号:配列番号:1に記載の塩基配列における32位の塩基種のGからTへの変異、129位の塩基種のAからGへの変異、161位の塩基種のTからGへの変異、557位の塩基種のTからCへの変異、839位の塩基種のTの欠損、843位の塩基種のGの欠損、976位の塩基種のCからAへの変異、又は1699位の塩基種のCからTへの変異から選択される少なくとも一つの変異である場合に、子実が増大すると判定される。
【0084】
以上のように、本発明により、子実収量に関連する遺伝子上の領域が明らかになったことにより、当業者に過度の負担を強いることなく、子実収量の検出を行うことができる。
【0085】
本発明の多型部位における塩基種の決定は、当業者においては種々の方法によって行うことができる。一例を示せば、本発明の多型部位を含むDNAの塩基配列を直接決定することによって行うことができる。
【0086】
本発明の検出方法に供する「被検植物」としては、特に制限されないが、好ましくはイネ、さらに好ましくはイネのコシヒカリ品種、カサラス品種が挙げられる。
【0087】
本発明の検出方法に供する被検試料は、通常、予め被検植物から取得された生体試料であることが好ましい。生体試料としては、例えばDNA試料を挙げることができる。本発明におけるDNA試料は、例えば被検植物の組織または細胞等から抽出した染色体DNA、あるいはRNAを基に調製することができる。
【0088】
即ち本発明は、通常、被検者由来の生体試料(予め被検植物から取得された生体試料)を被検試料として検査に供する方法である。
【0089】
当業者においては、公知の技術を用いて、適宜、生体試料の調製を行うことができる。例えば、DNA試料は、本発明の多型部位を含むDNAにハイブリダイズするプライマーを用いて、染色体DNA、あるいはRNAを鋳型としたPCR等によって調製することができる。
【0090】
本方法においては、次いで、単離したDNAの塩基配列を決定する。単離したDNAの塩基配列の決定は、当業者においては、DNAシークエンサー等を用いて容易に実施することができる。
【0091】
予め塩基のバリエーションが明らかにされている多型部位について、その塩基種を決定するための様々な方法が公知である。本発明の塩基種の決定方法は、特に限定されない。例えば、PCR法を応用した解析方法として、TaqMan PCR法、AcycloPrime法、およびMALDI-TOF/MS法等が実用化されている。またPCRに依存しない塩基種の決定法としてInvader法やRCA法が知られている。更にDNAアレイを使って塩基種を決定することもできる。ここに述べた方法は、いずれも本発明における多型部位の塩基種の決定に応用できる。
【0092】
これらの方法はいずれも多量のサンプルを高速にジェノタイピングするために開発された方法である。MALDI-TOF/MSを除けば、通常、いずれの方法にも何らかの形で標識プローブなどを用意する必要がある。これに対して、標識プローブなどに頼らない塩基種決定法も古くから行われている。このような方法の一つとして、例えば、制限酵素断片長多型(Restriction Fragment Length Polymorphism/RFLP)を利用した方法やPCR-RFLP法等が挙げられる。
【0093】
RFLPは、制限酵素の認識部位の変異、あるいは制限酵素処理によって生じるDNA断片内における塩基の挿入または欠失が、制限酵素処理後に生じる断片の大きさの変化として検出できることを利用している。検出対象となる多型を含む塩基配列を認識する制限酵素が存在すれば、RFLPの原理によって多型部位の塩基を知ることができる。
【0094】
また、CAPS (Cleaved Amplifeid Polymorphic Sequence)マーカーあるいはプライマーに変異を導入し、制限酵素サイトを作り出すdCAPS (derived CAPS)マーカーを使用することもできる。dCAPSマーカーは、PCRのプライマーにテンプレートのDNAとミスマッチを起こして、PCR産物上に制限酵素サイトを作り出すという手法である(特開2003-259898)。
【0095】
標識プローブを必要としない方法として、DNAの二次構造の変化を指標として塩基の違いを検出する方法も公知である。PCR-SSCPでは、1本鎖DNAの二次構造がその塩基配列の相違を反映することを利用している(Cloning and polymerase chain reaction-single-strand conformation polymorphism analysis of anonymous Alu repeats on chromosome 11. Genomics. 1992 Jan 1; 12(1): 139-146.、Detection of p53 gene mutations in human brain tumors by single-strand conformation polymorphism analysis of polymerase chain reaction products. Oncogene. 1991 Aug 1; 6(8): 1313-1318.、Multiple fluorescence-based PCR-SSCP analysis with postlabeling.、PCR Methods Appl. 1995 Apr 1; 4(5): 275-282.)。PCR-SSCP法は、PCR産物を1本鎖DNAに解離させ、非変性ゲル上で分離する工程により実施される。ゲル上の移動度は、1本鎖DNAの二次構造によって変動するので、もしも多型部位における塩基の相違があれば、移動度の違いとして検出することができる。
【0096】
その他、標識プローブを必要としない方法として、例えば、変性剤濃度勾配ゲル(denaturant gradient gel electrophoresis:DGGE法)等を例示することができる。DGGE法は、変性剤の濃度勾配のあるポリアクリルアミドゲル中で、DNA断片の混合物を泳動し、それぞれの不安定性の違いによってDNA断片を分離する方法である。ミスマッチのある不安定なDNA断片が、ゲル中のある変性剤濃度の部分まで移動すると、ミスマッチ周辺のDNA配列はその不安定さのために、部分的に1本鎖へと解離する。部分的に解離したDNA断片の移動度は、非常に遅くなり、解離部分のない完全な二本鎖DNAの移動度と差がつくことから、両者を分離することができる。
【0097】
更にDNAアレイを使って塩基種を決定することもできる(細胞工学別冊「DNAマイクロアレイと最新PCR法」,秀潤社,2000.4/20発行,pp97-103「オリゴDNAチップによるSNPの解析」,梶江慎一)。DNAアレイは、同一平面上に配置した多数のプローブに対してサンプルDNA(あるいはRNA)をハイブリダイズさせ、当該平面をスキャンすることによって、各プローブに対するハイブリダイズが検出される。多くのプローブに対する反応を同時に観察することができることから、例えば、多数の多型部位について同時に解析するには、DNAアレイは有用である。
【0098】
上記の方法以外にも、特定部位の塩基を検出するために、アリル特異的オリゴヌクレオチド(Allele Specific Oligonucleotide/ASO)ハイブリダイゼーション法が利用できる。アリル特異的オリゴヌクレオチド(ASO)は、検出すべき多型部位が存在する領域にハイブリダイズする塩基配列で構成される。ASOを試料DNAにハイブリダイズさせるとき、多型によって多型部位にミスマッチが生じるとハイブリッド形成の効率が低下する。ミスマッチは、サザンブロット法や、特殊な蛍光試薬がハイブリッドのギャップにインターカレーションすることにより消光する性質を利用した方法等によって検出することができる。また、リボヌクレアーゼAミスマッチ切断法によって、ミスマッチを検出することもできる。
【0099】
上記オリゴヌクレオチドのうち、配列番号:1に記載の塩基配列における、32位、129位、161位、557位、839位、843位、976位又は1699位から選択される少なくとも一つの多型部位に相当する部位のうち、いずれかの多型部位を含むDNAにハイブリダイズし、少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドは、子実収量を検出するための試薬(検査薬)として利用できる。これは遺伝子発現を指標とする検査、または遺伝子多型を指標とする検査に使用される。
【0100】
該オリゴヌクレオチドは、本発明の上記多型部位のいずれかの多型部位を含むDNAに特異的にハイブリダイズするものである。ここで「特異的にハイブリダイズする」とは、通常のハイブリダイゼーション条件下、好ましくはストリンジェントなハイブリダイゼーション条件下(例えば、サムブルックら,Molecular Cloning,Cold Spring Harbour Laboratory Press,New York,USA,第2版1989に記載の条件)において、他のタンパク質をコードするDNAとクロスハイブリダイゼーションを有意に生じないことを意味する。特異的なハイブリダイズが可能であれば、該オリゴヌクレオチドは、検出する遺伝子もしくは該遺伝子の近傍DNA領域における、上記植物の子実を増大させる機能を有する植物由来のタンパク質をコードするDNA塩基配列に対し、完全に相補的である必要はない。
【0101】
該オリゴヌクレオチドは、上記本発明の検査方法におけるプローブやプライマーとして用いることができる。該オリゴヌクレオチドをプライマーとして用いる場合、その長さは、通常15bp〜100bpであり、好ましくは17bp〜30bpである。プライマーは、本発明の上記配列番号:1に記載の塩基配列における、32位、129位、161位、557位、839位、843位、976位又は1699位から選択される少なくとも一つの多型部位に相当する部位のうち、いずれかの多型部位を含むDNAの少なくとも一部を増幅しうるものであれば、特に制限されない。
【0102】
本発明は、本発明の多型部位を含む領域を増幅するためのプライマー、および多型部位を含むDNA領域にハイブリダイズするプローブを提供する。
【0103】
本発明において、多型部位を含む領域を増幅するためのプライマーには、多型部位を含むDNAを鋳型として、多型部位に向かって相補鎖合成を開始することができるプライマーも含まれる。該プライマーは、多型部位を含むDNAにおける、多型部位の3'側に複製開始点を与えるためのプライマーと表現することもできる。プライマーがハイブリダイズする領域と多型部位との間隔は任意である。両者の間隔は、多型部位の塩基の解析手法に応じて、好適な塩基数を選択することができる。たとえば、DNAチップによる解析のためのプライマーであれば、多型部位を含む領域として、20〜500、通常50〜200塩基の長さの増幅産物が得られるようにプライマーをデザインすることができる。当業者においては、多型部位を含む周辺DNA領域についての塩基配列情報を基に、解析手法に応じたプライマーをデザインすることができる。本発明のプライマーを構成する塩基配列は、ゲノムの塩基配列に対して完全に相補的な塩基配列のみならず、適宜改変することができる。
【0104】
本発明のプライマーには、ゲノムの塩基配列に相補的な塩基配列に加え、任意の塩基配列を付加することができる。例えば、IIs型の制限酵素を利用した多型の解析方法のためのプライマーにおいては、IIs型制限酵素の認識配列を付加したプライマーが利用される。このような、塩基配列を修飾したプライマーは、本発明のプライマーに含まれる。更に、本発明のプライマーは、修飾することができる。例えば、蛍光物質や、ビオチンまたはジゴキシンのような結合親和性物質で標識したプライマーが各種のジェノタイピング方法において利用される。これらの修飾を有するプライマーも本発明に含まれる。
【0105】
本発明は、上記プライマーを有効成分として含有する、本発明の多型部位の検査薬またはキットも提供する。
【0106】
一方本発明において、多型部位を含む領域にハイブリダイズするプローブとは、多型部位を含む領域の塩基配列を有するポリヌクレオチドとハイブリダイズすることができるプローブを言う。より具体的には、プローブの塩基配列中に多型部位を含むプローブは本発明のプローブとして好ましい。あるいは、多型部位における塩基の解析方法によっては、プローブの末端が多型部位に隣接する塩基に対応するように、デザインされる場合もある。従って、プローブ自身の塩基配列には多型部位が含まれないが、多型部位に隣接する領域に相補的な塩基配列を含むプローブも、本発明における望ましいプローブとして示すことができる。
【0107】
言いかえれば、ゲノムDNA上の本発明の多型部位、または多型部位に隣接する部位にハイブリダイズすることができるプローブは、本発明のプローブとして好ましい。本発明のプローブには、プライマーと同様に、塩基配列の改変、塩基配列の付加、あるいは修飾が許される。例えば、Invader法に用いるプローブは、フラップを構成するゲノムとは無関係な塩基配列が付加される。このようなプローブも、多型部位を含む領域にハイブリダイズする限り、本発明のプローブに含まれる。本発明のプローブを構成する塩基配列は、ゲノムにおける本発明の多型部位の周辺DNA領域の塩基配列をもとに、解析方法に応じてデザインすることができる。
【0108】
本発明のプライマーまたはプローブは、それを構成する塩基配列をもとに、任意の方法によって合成することができる。本発明のプライマーまたはプローブの、ゲノムDNAに相補的な塩基配列の長さは、通常15〜100、一般に15〜50、通常15〜30である。与えられた塩基配列に基づいて、当該塩基配列を有するオリゴヌクレオチドを合成する手法は公知である。更に、オリゴヌクレオチドの合成において、蛍光色素やビオチンなどで修飾されたヌクレオチド誘導体を利用して、オリゴヌクレオチドに任意の修飾を導入することもできる。あるいは、合成されたオリゴヌクレオチドに、蛍光色素などを結合する方法も公知である。
【0109】
本発明のプローブの具体的な例としては、それぞれ配列番号:1に記載の塩基配列における32位の塩基種のGからTへの変異、129位の塩基種のAからGへの変異、161位の塩基種のTからGへの変異、557位の塩基種のTからCへの変異、839位の塩基種のTの欠損、843位の塩基種のGの欠損、976位の塩基種のCからAへの変異、又は1699位の塩基種のCからTへの変異から選択される少なくとも一つの変異いずれかの多型部位に相当する多型部位を含む領域にハイブリダイズするプローブであって、少なくとも15ヌクレオチドの鎖長を有するプローブが挙げられる。
【0110】
本発明はまた、本発明の子実収量を検出する方法に使用するための試薬(検査薬)を提供する。本発明の試薬は、前記本発明のプライマーおよび/またはプローブを含む。子実収量の検出においては上記、本発明の多型部位のいずれかに記載の多型部位を含むDNAを増幅するためのプライマーおよび/またはプローブを用いる。
【0111】
本発明の試薬には、塩基種の決定方法に応じて、各種の酵素、酵素基質、および緩衝液などを組み合わせることができる。酵素としては、DNAポリメラーゼ、DNAリガーゼ、あるいはIIs制限酵素などの、上記の塩基種決定方法として例示した各種の解析方法に必要な酵素を示すことができる。緩衝液は、これらの解析に用いる酵素の活性の維持に好適な緩衝液が、適宜選択される。更に、酵素基質としては、例えば、相補鎖合成用の基質等が用いられる。
【0112】
さらに、本発明における試薬の別の態様は、本発明の多型部位を含むDNAとハイブリダイズするヌクレオチドプローブが固定された固相からなる、子実収量を検出するための試薬である。
【0113】
これらは本発明の多型部位を指標とする検査に使用される。これらの調製方法に関しては、当業者に公知の方法で行なうことができる。
【0114】
なお、本明細書において引用された全ての先行技術文献は、参照として本明細書に組み入れられる。
【実施例】
【0115】
以下、本発明を実施例によりさらに具体的に説明するが本発明はこれら実施例に制限されるものではない。
【0116】
〔実施例1〕GL5遺伝子のポジショナルクローニング
連鎖解析によりGL5遺伝子の特定を行った。解析に用いた集団は、コシヒカリとコシヒカリを遺伝的背景にGL5領域をカサラス型染色体に置換した準同質遺伝子系統NIL(GL5)との戻し交雑後代を用いた。GL5領域がヘテロ型となった個体の自殖後代2,000個体から、GL5領域を挟み込む一塩基多型(SNP)マーカーG1358とSNPマーカーG1569を利用してGL5遺伝子座近傍で組換えが生じた個体を選抜した。GL5遺伝子座近傍のSNPマーカーを用いて連鎖解析を行った結果、GL5遺伝子座をSNPマーカーG1422とSNPマーカーG1491との間、約7kbpの範囲に特定した(図1)。
【0117】
イネアノテーションプロジェクトにより公開された遺伝子予測データベース(http://rapdb.dna.affrc.go.jp/)により、SNPマーカーG1422とSNPマーカーG1491との間には、4つの遺伝子の存在が予測された。この4つの遺伝子の中から、遺伝子の発現量・発現部位からGL5遺伝子を1つの遺伝子(Os05g0187000)に絞り込んだ。この遺伝子は、20Sプロテアソームのβ2サブユニットをコードしていた。20Sプロテアソームのβ2サブユニットをコードする遺伝子は、イネには1つ(OsPBB1)しか存在しない(図2)。
【0118】
〔実施例2〕候補遺伝子の塩基配列と発現解析
コシヒカリとカサラスのcDNA塩基配列解析の結果、GL5候補遺伝子(OsPBB1)のコーディング領域に2カ所のSNP〔33位(配列番号:1の976位に相当)のCがA、267位(配列番号:1の1699位に相当)のCがT〕を有していたが、これらはいずれもアミノ酸配列に変化を生じさせず、コシヒカリとカサラスのGL5候補遺伝子のアミノ酸配列は同一であった(図3)。
【0119】
OsPBB1の推定プロモーター領域(1kbp)には、SNPが3ヵ所〔-912位(配列番号:1の32位に相当)のGがT、-815位(配列番号:1の129位に相当)のAがG、-783位(配列番号:1の161位に相当)のTがG〕、一塩基欠損挿入が2カ所〔-105位(配列番号:1の839位に相当)のTが欠損、-101位(配列番号:1の843位に相当)のGが欠損〕あった(図4-1及び4-2)。そこでOsPBB1遺伝子の発現量を解析した。コシヒカリおよびNIL(GL5)から全長10 cmの幼穂を採取し、酸性フェノール法で抽出した全RNAを鋳型とし、WAKO社のAMV逆転写酵素反応によりcDNAを得た。得られたcDNAを鋳型としてプライマー(5’-GGCAATTCGTGGTGGTATCT-3’、配列番号:9および5’-ACCTTCGGCTTCAGTTGTGT-3’、配列番号:10)を用いてリアルタイムPCR法によりOsPBB1遺伝子の発現量を比較した。その結果、コシヒカリに比べてNIL(GL5)は、発現量が約2.8倍高かった(図5)。
【0120】
〔実施例3〕GL5候補遺伝子(OsPBB1)の機能証明
OsPBB1の発現量がコシヒカリとNIL(GL5)の粒長を決定すると考えられた事から、コシヒカリと同等のOsPBB1発現量を持つ日本晴にOsPBB1を過剰発現させる形質転換体によりOsPBB1の機能証明を行った。
【0121】
OsPBB1の完全長cDNA(AK103126)をトウモロコシ由来ユビキチン1プロモーターとノパリン合成酵素ターミネーターの間に連結させ過剰発現させた形質転換体(BM349)をイネFOXハンティングシステム(Nakamura et al. (2007) Plant Mol. Biol. 65, 357-371)より選抜した。この形質転換体T2世代植物から収穫した充実した種子を25度で2週間乾燥後、覆っている内穎と外穎を取り除き、玄米を得た。この玄米の粒長は有意(P=0.0039)に増加しており、OsPBB1の過剰発現は玄米の長さを有意に増加させた(図6)。これにより、OsPBB1がGL5遺伝子であることが証明された。
【0122】
〔実施例4〕他の植物種におけるGL5の効果
GL5遺伝子(OsPBB1)の高発現がイネの粒長を増大させることが判明したことから、他の植物種における同遺伝子高発現の効果を調べた。シロイヌナズナ(Arabidopsis thaliana)は、OsPBB1のホモログ遺伝子としてAtPBB1(配列番号:11)およびAtPBB2(配列番号:12)の2つの遺伝子を持ち、OspBB1とのアミノ酸レベルでの相同性は両者とも82.7%と高い相同性を持つ(図2、図7)。そこでイネ由来OsPBB1の高発現がシロイヌナズナにおいても種子サイズを増加させると仮説し、実証を試みた。
【0123】
イネ由来OsPBB1の完全長cDNA(AK103126)の遺伝子コーディング領域をXbaIサイトを連結したプライマー(5’-CGCATCTAGACCGCGCGGGATGGCCGGAGC-3’、配列番号:13)とSacIサイトを連結したプライマー(5’-GTAAGAGCTCAAAATCACTCCTCCATTGCA-3’、配列番号:14)で増幅し、増幅産物をXbaIおよびSacIで処理してインサートを得た。このインサートをpSTARH301GバイナリーベクターのXbaI、SacIサイトにライゲーションして、OsPBB1の完全長cDNAをカリフラワーモザイクウイルス由来35Sプロモーターとノパリン合成酵素ターミネーターとの間に連結させたバイナリーベクター(pSTARH301G/GL5)を得た。pSTARH301G/GL5をエレクトロポレーション法によりアグロバクテリウム(Agrobacterium tumefaciens EHA101)へ導入し、さらにこのアグロバクテリウムを減圧浸潤法によりシロイヌナズナへ感染させ形質転換体T1種子を得た。この種子をハイグロマイシン50mg/lを含有するGM寒天培地で形質転換スクリーニングを行った。得られた形質転換体(T1世代)からT2種子を得て、再びハイグロマイシンを含む培地に播種し、3:1の分離比を示す系統を選択し、導入遺伝子が1遺伝子座に存在すると考えられる形質転換体を選抜した。この種子のサイズを測定したところ、OsPBB1のcDNAを含まないコントロールベクター系統に比べて種子の長径が有意に増加していた(図8)。OsPBB1の高発現は、シロイヌナズナにおいても種子サイズを増大させることが明らかとなった。これによりGL5は単子葉植物であるイネのみならず双子葉植物であるシロイヌナズナにおいても種子サイズを増大させる効果があり、幅広い植物種で同様の効果を有する事が示唆された。
【特許請求の範囲】
【請求項1】
下記(a)から(d)のいずれかに記載のDNAを含む、植物体の子実(種子、頴花又は果実を含む)の大きさを増大させた植物体。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
【請求項2】
植物体が単子葉植物又は双子葉植物である、請求項1に記載の植物体。
【請求項3】
下記(a)から(d)のいずれかに記載のDNAが発現するように導入されたベクター。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
【請求項4】
請求項3に記載のベクターが導入された宿主細胞。
【請求項5】
請求項4に記載のベクターが導入された植物細胞。
【請求項6】
請求項5に記載の植物細胞を含む形質転換植物体。
【請求項7】
請求項6に記載の形質転換植物体の子孫またはクローンである、形質転換植物体。
【請求項8】
請求項6又は7に記載の形質転換植物体の繁殖材料。
【請求項9】
下記(a)から(d)のいずれかに記載のDNAを植物体の細胞内で過剰発現させる工程を含む、植物体の種子、頴花又は果実の大きさを増大させる方法。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
【請求項10】
植物体が単子葉植物又は双子葉植物である、請求項9に記載の方法。
【請求項11】
被検植物体について、配列番号:1に記載のDNA領域に相当する部位、またはその周辺配列に存在する変異を含むDNAマーカーを検出することを特徴とする、被検植物体の種子、頴花又は果実の収量を検出する方法。
【請求項12】
変異が一塩基多型である、請求項11に記載の方法。
【請求項13】
以下の工程(a)及び(b)を含む、請求項11に記載の方法。
(a)被検植物における配列番号:1に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域に相当する部位、またはその周辺配列に存在する多型部位の塩基種を決定する工程、及び
(b)(a)で決定された多型部位の塩基種において、配列番号:1またはその周辺配列と異なるアレルが検出された場合に、被検植物は子実(種子、頴花又は果実を含む)の収量が多いと判定する工程
【請求項14】
多型部位が、配列番号:1に記載の塩基配列における、32位、129位、161位、557位、839位、843位、976位又は1699位から選択される少なくとも一つの多型部位に相当する部位である、請求項13に記載の方法。
【請求項15】
多型部位の塩基種の変異が、配列番号:1に記載の塩基配列における32位の塩基種のGからTへの変異、129位の塩基種のAからGへの変異、161位の塩基種のTからGへの変異、557位の塩基種のTからCへの変異、839位の塩基種のTの欠損、843位の塩基種のGの欠損、976位の塩基種のCからAへの変異、又は1699位の塩基種のCからTへの変異から選択される少なくとも一つの変異である場合には、子実(種子、頴花又は果実を含む)が増大すると判定する、請求項14に記載の方法。
【請求項16】
配列番号:1に記載のDNAとストリンジェントな条件で特異的にハイブリダイズし、少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドを含む、被検植物の子実(種子、頴花又は果実を含む)の収量を検出するためのプライマー。
【請求項17】
請求項14または15に記載の多型部位を含む領域を増幅するための、請求項16に記載のプライマー。
【請求項18】
配列番号:1に記載のDNAとストリンジェントな条件で特異的にハイブリダイズし、少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドを含む、被検植物体の子実(種子、頴花又は果実を含む)の収量を検出するためのプローブ。
【請求項19】
請求項14または15に記載の多型部位を含む領域に特異的にハイブリダイズする、請求項18に記載のプローブ。
【請求項1】
下記(a)から(d)のいずれかに記載のDNAを含む、植物体の子実(種子、頴花又は果実を含む)の大きさを増大させた植物体。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
【請求項2】
植物体が単子葉植物又は双子葉植物である、請求項1に記載の植物体。
【請求項3】
下記(a)から(d)のいずれかに記載のDNAが発現するように導入されたベクター。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
【請求項4】
請求項3に記載のベクターが導入された宿主細胞。
【請求項5】
請求項4に記載のベクターが導入された植物細胞。
【請求項6】
請求項5に記載の植物細胞を含む形質転換植物体。
【請求項7】
請求項6に記載の形質転換植物体の子孫またはクローンである、形質転換植物体。
【請求項8】
請求項6又は7に記載の形質転換植物体の繁殖材料。
【請求項9】
下記(a)から(d)のいずれかに記載のDNAを植物体の細胞内で過剰発現させる工程を含む、植物体の種子、頴花又は果実の大きさを増大させる方法。
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4又は5に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA、又は
(d)配列番号:4又は5に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
【請求項10】
植物体が単子葉植物又は双子葉植物である、請求項9に記載の方法。
【請求項11】
被検植物体について、配列番号:1に記載のDNA領域に相当する部位、またはその周辺配列に存在する変異を含むDNAマーカーを検出することを特徴とする、被検植物体の種子、頴花又は果実の収量を検出する方法。
【請求項12】
変異が一塩基多型である、請求項11に記載の方法。
【請求項13】
以下の工程(a)及び(b)を含む、請求項11に記載の方法。
(a)被検植物における配列番号:1に記載の塩基配列のコード領域及び配列番号:1に記載の塩基配列のプロモーター領域に相当する部位、またはその周辺配列に存在する多型部位の塩基種を決定する工程、及び
(b)(a)で決定された多型部位の塩基種において、配列番号:1またはその周辺配列と異なるアレルが検出された場合に、被検植物は子実(種子、頴花又は果実を含む)の収量が多いと判定する工程
【請求項14】
多型部位が、配列番号:1に記載の塩基配列における、32位、129位、161位、557位、839位、843位、976位又は1699位から選択される少なくとも一つの多型部位に相当する部位である、請求項13に記載の方法。
【請求項15】
多型部位の塩基種の変異が、配列番号:1に記載の塩基配列における32位の塩基種のGからTへの変異、129位の塩基種のAからGへの変異、161位の塩基種のTからGへの変異、557位の塩基種のTからCへの変異、839位の塩基種のTの欠損、843位の塩基種のGの欠損、976位の塩基種のCからAへの変異、又は1699位の塩基種のCからTへの変異から選択される少なくとも一つの変異である場合には、子実(種子、頴花又は果実を含む)が増大すると判定する、請求項14に記載の方法。
【請求項16】
配列番号:1に記載のDNAとストリンジェントな条件で特異的にハイブリダイズし、少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドを含む、被検植物の子実(種子、頴花又は果実を含む)の収量を検出するためのプライマー。
【請求項17】
請求項14または15に記載の多型部位を含む領域を増幅するための、請求項16に記載のプライマー。
【請求項18】
配列番号:1に記載のDNAとストリンジェントな条件で特異的にハイブリダイズし、少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドを含む、被検植物体の子実(種子、頴花又は果実を含む)の収量を検出するためのプローブ。
【請求項19】
請求項14または15に記載の多型部位を含む領域に特異的にハイブリダイズする、請求項18に記載のプローブ。
【図1】


【図2】
【図3】

【図4−1】

【図4−2】
【図5】
【図7】
【図6】
【図8】
【図2】
【図3】
【図4−1】
【図4−2】
【図5】
【図7】
【図6】
【図8】
【公開番号】特開2011−101622(P2011−101622A)
【公開日】平成23年5月26日(2011.5.26)
【国際特許分類】
【出願番号】特願2009−258075(P2009−258075)
【出願日】平成21年11月11日(2009.11.11)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成20年度農林水産省プロジェクト「新農業展開ゲノムプロジェクト」(イネの量的形質遺伝子の同定)に関する委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【Fターム(参考)】
【公開日】平成23年5月26日(2011.5.26)
【国際特許分類】
【出願日】平成21年11月11日(2009.11.11)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成20年度農林水産省プロジェクト「新農業展開ゲノムプロジェクト」(イネの量的形質遺伝子の同定)に関する委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【Fターム(参考)】
[ Back to top ]