
植物体の着色制御遺伝子、およびその利用
【課題】植物体の着色を制御する遺伝子の提供を課題とする。より詳しくは、植物体もしくはその種子の着色を制御する遺伝子、および、該遺伝子を発現し種子が着色した植物体(形質転換植物体)の製造方法、さらに、該遺伝子の発現を改変することにより、種子の色を制御する方法の提供を課題とする。
【解決手段】植物体の着色を制御する機能を有するRcおよびRd遺伝子を同定することに成功した。これらの遺伝子を植物体内において発現させることにより、例えば、白米を赤色米・褐色米等へ改変することができる。また、本発明の方法によって、タンニン類が蓄積された植物体もしくはその種子を作出することが可能である。タンニン類が蓄積されたコメは、機能性食品として有用である。さらに、これら遺伝子の発現を抑制させることにより、有色米を白米へ改変することも可能である。
【解決手段】植物体の着色を制御する機能を有するRcおよびRd遺伝子を同定することに成功した。これらの遺伝子を植物体内において発現させることにより、例えば、白米を赤色米・褐色米等へ改変することができる。また、本発明の方法によって、タンニン類が蓄積された植物体もしくはその種子を作出することが可能である。タンニン類が蓄積されたコメは、機能性食品として有用である。さらに、これら遺伝子の発現を抑制させることにより、有色米を白米へ改変することも可能である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、植物体もしくはその種子の着色を制御する遺伝子、および、該遺伝子を発現し着色した植物体(形質転換植物体)もしくはその種子の製造方法、該遺伝子(DNA配列)を利用して選抜育種される植物体及び種子、さらに、該遺伝子の発現を改変することにより、種子の色を制御する方法に関する。
【背景技術】
【0002】
赤色あるいは赤褐色を呈する赤米といわれるイネ種子には、カテキン、タンニンなどが蓄積することが知られているが、約50年以上前にその染色体上の遺伝子座が同定されているものの、未だに遺伝子レベルでの決定はなされていない。
なお、本出願の発明に関連する先行技術文献情報を以下に示す。
【先行技術文献】
【非特許文献】
【0003】
【非特許文献1】Kinoshita, T. & Takahashi, M. (1991) J. Fac. Agr. Hokkaido Univ. 65, 1-61.
【非特許文献2】Kinoshita, T. (1995) Rice Genetics Newsletter 12, 9-153.
【非特許文献3】Chen, M. & L. J. Bennetzen (1996) Plant Molecular Biology 32, 999-1001.
【非特許文献4】Nakai, K., Inagaki, Y., Nagata, H., Miyazaki, C., & Iida, S. (1998) Plant Biotechnology 15, 221-225.
【発明の概要】
【発明が解決しようとする課題】
【0004】
植物体の着色を制御する遺伝子の提供を課題とする。より詳しくは、植物体もしくはその種子の着色を制御する遺伝子、および、該遺伝子を発現し種子が着色した植物体(形質転換植物体)の製造方法、該遺伝子(DNA配列)を利用して選抜育種される植物体及び種子、さらに、該遺伝子の発現を改変することにより、植物体もしくはその種子の色を制御する方法の提供を課題とする。
【課題を解決するための手段】
【0005】
本発明者らは、上記の課題を解決するために、イネの赤米色素着色に関与するRcとRd遺伝子の同定を試みた。
【0006】
Rc遺伝子については、本発明者らによって同定されたいくつかの候補遺伝子を逆遺伝学的な手法で日本晴に導入し、表現型を白色から赤褐色に改変させる機能を有する遺伝子を原因遺伝子として特定した。その構造的な特徴からRc遺伝子はbHLHドメインを持つ転写因子であるMYC遺伝子であることが初めて明らかとなった。本発明者らによって同定された遺伝子の機能は、これまでのところ明らかになっておらず、MYC遺伝子が赤米関係の色素発現に関与していることが本発明者らによって初めて証明された。
【0007】
Rc遺伝子が存在すると考えられた染色体上の遺伝子座の領域は、非常に広範な領域(2.3Mbp)であり、その中から実際に植物体の着色を制御する原因遺伝子を同定することは非常に困難であった。即ち、上記領域の中において、アミノ酸が100個以上の長さのタンパク質をコードすると考えられる遺伝子の数は200以上にものぼり、その中から、着色を制御する原因遺伝子を同定することは極めて困難なことである。該原因遺伝子を同定することは、当業者であっても過度の困難性を伴うことであり、本発明者らによって原因遺伝子が実際に同定されたことは非常に大きな成果と言える。
【0008】
本発明者らによって同定された遺伝子のRc(MYC)遺伝子は、これまでのところ機能が全く知られておらず、本発明者らによって植物体の着色を制御する機能を有することが初めて見出された。
【0009】
またRd遺伝子は、本発明者らによる鋭意研究の結果、遺伝子DFRであることが分った。DFRの遺伝子構造については3種の多型が認められた。本発明者らはDFR抗体を作成し、タンパク質レベルでの発現を解析したところ、種子特異的なalternative translation initiationにより複数のペプチドとして翻訳されていることを初めて証明した。
【0010】
さらに本発明者らは、Rdが酵素タンパク質(DFR)をコードしており、その着色に関わる発現はRcである調節遺伝子MYCにより制御されていることを見出した。DFRタンパク質の翻訳は種子特異的なalternative translation initiationにより制御されている。その結果、全長のDFRを有するイネでは種子が赤色を呈し、DFRのORFが短い構造のイネにおいては種子が赤褐色を呈すること、さらにRcが機能しない場合は種子が白色を呈することが新たに見出された。
【0011】
より詳細な解析により、rd型においては、DFR遺伝子からalternative translation initiationにより下流から翻訳され、通常の46 kDaの翻訳産物ではなく、33 kDaおよび22 kDaの翻訳産物が生成されることが示された。さらに、alternative translation initiation機構によって翻訳される33 kDaおよび22 kDaの産物が、DFRの機能を相補し得ることを、本発明者らは初めて見出した。即ち、通常の全長DFRタンパク質に加え、より分子量の小さい翻訳産物についても、植物体を着色させる機能、もしくはタンニン類を蓄積させる機能を有することが判明した。
【0012】
上述の如く本発明者らは、植物体もしくはその種子の着色を制御する遺伝子を単離することに成功し本発明を完成させた。該遺伝子を利用することにより、植物体、特に種子の着色を制御することが可能である。
【0013】
本発明は、植物体もしくはその種子の着色を制御する遺伝子、および、該遺伝子を発現し種子が着色した植物体(形質転換植物体)の製造方法、該遺伝子(DNA配列)を利用して選抜育種される植物体及び種子、さらに、該遺伝子の発現を改変することにより、植物体、特に種子の色を制御する方法に関し、より具体的には、
〔1〕 植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能を有する植物由来のタンパク質をコードする、下記(a)〜(e)のいずれかに記載のDNA、
(a)配列番号:3または6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:1または4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:2または5に記載の塩基配列からなるDNA
(d)配列番号:3または6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:1または4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
〔2〕 植物が単子葉植物である、〔1〕に記載のDNA、
〔3〕 単子葉植物がイネである、〔2〕に記載のDNA、
〔4〕 〔1〕〜〔3〕のいずれかに記載のDNAによりコードされるタンパク質、
〔5〕 〔1〕〜〔3〕のいずれかに記載のDNAを含むベクター、
〔6〕 〔5〕に記載のベクターが導入された宿主細胞、
〔7〕 〔5〕に記載のベクターが導入された植物細胞、
〔8〕 〔7〕に記載の植物細胞を含む形質転換植物体、
〔9〕 〔8〕に記載の形質転換植物体の子孫またはクローンである、形質転換植物体、
〔10〕 〔8〕または〔9〕に記載の形質転換植物体の繁殖材料、
〔11〕 〔6〕に記載の宿主細胞を培養し、該細胞またはその培養上清から組換えタンパク質を回収する工程を含む、〔4〕に記載のタンパク質の製造方法、
〔12〕 〔4〕に記載のタンパク質に結合する抗体、
〔13〕 〔1〕〜〔3〕のいずれかに記載のDNAの塩基配列またはその相補配列に相補的な少なくとも15の連続する塩基を含むポリヌクレオチド、
〔14〕 下記(a)〜(c)のいずれかを有効成分として含有する、植物体着色化剤、
(a)〔1〕〜〔3〕のいずれかに記載のDNA
(b)〔4〕に記載のタンパク質
(c)〔5〕に記載のベクター
〔15〕 下記(a)〜(c)のいずれかを有効成分として含有する、タンニン類蓄積剤、
(a)〔1〕〜〔3〕のいずれかに記載のDNA
(b)〔4〕に記載のタンパク質
(c)〔5〕に記載のベクター
〔16〕 下記(a)〜(c)のいずれかを有効成分として含有する、植物体脱色化剤、
(a)〔1〕〜〔3〕のいずれかに記載のDNAの転写産物またはその一部に対するアンチセンス核酸
(b)〔1〕〜〔3〕のいずれかに記載のDNAの転写産物を特異的に開裂するリボザイム活性を有する核酸
(c)〔1〕〜〔3〕のいずれかに記載のDNAの発現をRNAi効果による阻害作用を有する核酸
〔17〕 〔1〕〜〔3〕のいずれかに記載のDNA、または、〔5〕に記載のベクターを植物細胞へ導入し、該植物細胞から植物体を再生させる工程を含む、形質転換植物体の製造方法、
〔18〕 〔1〕〜〔3〕のいずれかに記載のDNAを植物体の細胞内で発現させる工程を含む、植物体もしくはその種子を着色させる、または植物体もしくはその種子にタンニン類を蓄積させる方法、
〔19〕 〔1〕〜〔3〕のいずれかに記載のDNAを植物体の細胞内で発現させる工程を含む、着色した、もしくはタンニン類が蓄積した植物体もしくはその種子の製造方法、
〔20〕 〔1〕〜〔3〕のいずれかに記載のDNA、または〔5〕に記載のベクターを植物細胞へ導入する工程を含む、〔18〕または〔19〕に記載の方法、
〔21〕 下記工程(a)および(b)を含む、〔18〕または〔19〕に記載の方法、
(a)〔1〕〜〔3〕のいずれかに記載のDNAを有する植物と交雑させる工程を含む、
(b)前記DNAを有する植物改変体を選抜する工程
〔22〕 着色した種子、もしくはタンニン類が蓄積した種子を有する植物を育種する方法であって、下記工程(a)〜(d)を含む育種方法、
(a)植物Aと、〔1〕〜〔3〕のいずれかに記載のDNAを有する他の植物Bを交雑させ、F1を作出する工程
(b)前記F1と前記植物Aを交雑させる工程
(c)前記DNAを有する植物を選抜する工程
(d)工程(c)によって選抜された植物と、前記植物Aを交雑させる工程
〔23〕 前記工程(c)において、植物ゲノム中の〔1〕〜〔3〕のいずれかに記載のDNA配列、またはその周辺配列に存在するDNAマーカーを利用して選抜されることを特徴とする、〔22〕に記載の方法、
〔24〕 植物に内在する遺伝子であって、植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能を有するタンパク質をコードする遺伝子の発現を抑制する工程を含む、脱色された植物体もしくはその種子の製造方法、
〔25〕 植物がイネである、〔17〕〜〔24〕のいずれかに記載の方法、
〔26〕 〔18〕〜〔25〕のいずれかに記載の方法によって取得される植物体、またはその種子、
〔27〕 人為的に作製された植物体もしくはその種子であって、〔1〕〜〔3〕のいずれかに記載のDNAを有し着色されていることを特徴とする植物体もしくはその種子、
〔28〕 人為的に作製された植物体もしくはその種子であって、〔1〕〜〔3〕のいずれかに記載のDNAを有しタンニン類が蓄積していることを特徴とする植物体もしくはその種子、
〔29〕 植物に内在する遺伝子であって、植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能を有するタンパク質をコードする遺伝子の発現が人為的に抑制され、脱色されていることを特徴とする植物体もしくはその種子、
〔30〕 植物がイネである、〔27〕〜〔29〕のいずれかに記載の植物体もしくはその種子、を提供するものである。
【発明の効果】
【0014】
本発明の方法により、RcもしくはRd遺伝子を植物体内において発現させることにより、植物もしくはその種子(例えば、コメ等)を人為的に着色させる手法が確立された。一般種子への遺伝子組換え作物(Genetically Modified Organisms; GMO)混入などの社会的不安に答えるため、GMO米を人為的に着色させることで対応することができる。本発明の利点のひとつは、例えば、コメの果皮を着色させることによりGMOのモニタリングが可能になる。必要に応じ、精米により最終的に着色部分を除去することも可能である。
【0015】
また、本発明においてDNAマーカーを利用する育種法により、良食味米を数年で着色米に育種できる。ごく短期間に「コシヒカリカテキン」「カテキンヒトメボレ」「カテキンマンゲツモチ」のような良食味着色米あるいは機能性良食味米系統が育種することが可能である。食味については全く未開拓の着色米であるが、本発明の技術を用いることにより良食味着色米が開発できる。本発明の方法によって作出される植物は、抗酸化機能の高いカテキン、タンニンを含むことから、例えば、機能性米として付加価値を有するコメを効率的に作出することが可能である。
【図面の簡単な説明】
【0016】
【図1】DFRとタンニン合成・アントシアニン合成の関係を示す図である。酵素の略称は以下の通りである。F3H,flavanone 3-hydroxylase; DFR,dihydroflavonol-4-reductase; ANS,anthocyanin synthase; LAR,leucoananthocyanidin reductase; ANR,anthocyanidin reductase; BAN,BANYULS; GT,anthocyanidin glucosyl transferase
【図2】イネの玄米色に関わる遺伝子の位置を示す図である。水平な線は、マーカーの位置を表す。(A)Rc,Rdに関する初期の座乗位置の情報(Nagao, S. & Takahashi, M. (1963) Agr., Hokkaido Univ., Sapporo 53, 76-131.)。第一染色体上の遺伝マーカーとして、Aはanthocyanin activator, Pnはpurple nodeをそれぞれ示す。第7染色体上の遺伝マーカーとして、d6はlop-leaved dwarf, gはrecessive long empty glumesをそれぞれ示す。(B)イネゲノムプロジェクト終了後のRc,Rdの座乗位置の情報(IRGSP 2005 Nature)R1807,R1582,R2374,C808は(http://rgp.dna.affrc.go.jp/publicdata/geneticmap2000/)に記載されている分子マーカーである。(C)今回の研究によるRc,Rdの位置(本発明)。Rd座はDFR遺伝子をコードしており、染色体上での位置は第1染色体のAP004317の68929 bp−70543 bpである。Rc座は、MYC related proteinをコードしており、染色体上での位置は第7染色体のAP005098 150211 bp−156639 bpである。
【図3】RcRdの遺伝子型とその表現型を示す写真である。写真上部は玄米、下部は籾である。(A)RcRdの遺伝子型をもつカサラスの玄米色。(B)Rcrdの遺伝子型をもつtRc(NIL)の玄米色。(C)rcrdの遺伝子型を持つ日本晴の玄米色。rcRdの遺伝子型を持つ玄米色も白色になる。(Nagao, S. (1951) Advances in Genetics 4, 181-212.)
【図4】イネのDFR遺伝子の構造を示す図である。水平の黒い四角はDFRのエクソンを示す。黒い四角の間の水平の線は、それぞれ第1イントロン、第2イントロンをしめす。イネのDFR遺伝子のPCRには、OsDFRF4とOsDFRR4を用いた。イネDFRの第1エクソン、第1イントロン、第2エクソンの塩基配列の決定には、OsDFRR5を用いた。
【図5】イネDFRのmRNAの構造を示す図である。黒の線はORFを示す。イネDFRのRT-PCRには、OsDFRF8とOsDFRR5を用いた。
【図6】pGEX4T-3ベクターに連結したDFR(pGEX4T-3 DFR)を示す図である。pGEX4T-3をベースに構築した。AmpRはアンピシリン耐性遺伝子、Ptacはtacプロモーター、GSTはグルタチオン-S-トランスフェラーゼを示す。BamHI、EcoRIは、ベクターを作成する際に用いた制限酵素である。
【図7】35S::Murasaki DFRベクターの構造を示す図である。pCAMBIA1301(ベクター部分:11.8kb)をベースに構築した。LB,RBはレフトボーダー、ライトボーダーをそれぞれ示す。T35S,P35Sはカリフラワーモザイクウィルス由来の強発現用プロモーターである。MCSはマルチクローニングサイトを示す。NosTはノパリンシンターゼ由来のターミネーターを示す。HygR,KanRは、それぞれハイグロマイシン、カナマイシンの耐性遺伝子である。Murasakiine DFRは紫稲のDFRのゲノム配列(イントロン含む)である。
【図8】35S::Koshi DFRベクターの構造を示す図である。pCAMBIA1301(ベクター部分:11.8kb)をベースに構築した。LB,RBはレフトボーダー、ライトボーダーをそれぞれ示す。T35S,P35Sはカリフラワーモザイクウィルス由来の強発現用プロモーターである。MCSはマルチクローニングサイトを示す。NosTはノパリンシンターゼ由来のターミネーターを示す。HygR,KanRは、それぞれハイグロマイシン、カナマイシンの耐性遺伝子である。Koshihikari DFRはコシヒカリのDFRのゲノム配列(イントロン含む)である。
【図9】AsDFRベクターの構造を示す図である。pCAMBIA1301(ベクター部分:11.8kb)をベースに構築した。LB,RBはレフトボーダー、ライトボーダーをそれぞれ示す。T35S,P35Sはカリフラワーモザイクウィルス由来の強発現用プロモーターである。MCSはマルチクローニングサイトを示す。NosTはノパリンシンターゼ由来のターミネーターを示す。HygR,KanRは、それぞれハイグロマイシン、カナマイシンの耐性遺伝子である。Murasakiine DFRは紫稲のDFRのcDNA配列(イントロン含まない)である。Antisense法により、DFRの発現抑制をするベクターである。
【図10】pRc1-1301、pRc2-1301、pRc3-1301ベクターの構造を示す図である。pCAMBIA1301(ベクター部分:11.8kb)をベースに構築した。LB,RBはレフトボーダー、ライトボーダーをそれぞれ示す。T35S,P35Sはカリフラワーモザイクウィルス由来の強発現用プロモーターである。MCSはマルチクローニングサイトを示す。NosTはノパリンシンターゼ由来のターミネーターを示す。HygR,KanRは、それぞれハイグロマイシン、カナマイシンの耐性遺伝子である。Gus geneはGus遺伝子を示す。promoterはそれぞれRc1,Rc2,Rc3の由来のプロモーターを示す。TはそれぞれRc1,Rc2,Rc3の由来のターミネーターを示す。
【図11】DFRを遺伝子導入した個体の表現型を示す写真である。(A)35S::Murasaki DFRを遺伝子導入した個体の表現型。一番上は発芽後7日目の葉におけるDFR抗体を用いた免疫染色の写真を表す。二段目は出穂後7日目の穂を表す。三段目は完熟種子を表す。(B) As DFRを遺伝子導入した個体の表現型。一番上は発芽後7日目の葉におけるDFR抗体を用いた免疫染色の写真を表す。二段目は出穂後7日目の穂を表す。三段目は完熟種子を表す。(C) tRc (Rcrd)の表現型。二段目は出穂後7日目の穂を表す。三段目は完熟種子を表す。
【図12】DFR遺伝子の多型を示す図である。黒い四角はエクソンを示す。水平の線はイントロンを示す。白抜きの丸は翻訳開始コドンを示す。白抜きの四角は翻訳終止コドンを示す。
【図13】イネDFRのRT-PCRを示す写真である。DFRのPCRは35サイクル行った。アクチンのPCRは30サイクル行った。右側の数字は増幅断片の大きさである。
【図14】DFRの発現の比較を示す図及び写真である。(A)日本晴、カサラスのDFRの構造。水平な黒の四角はエクソンを表す。水平な黒の線はイントロンを表す。白抜きの丸はコザックコンセンサスを持つ開始コドンを表す(ANNATGN or GNNATGG)。上向きの三角はイントロンの位置を示す。白抜きの四角は翻訳終止コドンの位置を表す。(B)OsDFRの構造から予想されるORF(Alternative translation initiation)。水平な線は予想されるORFを示し、右の数値は予想されるタンパク質の分子量を表す。(C)、(D)は、登熟期の穂、発芽後7日目の葉におけるDFRの免疫染色を示す写真である。(C)は、GST-DFR(ポジティブコントロール); カサラス; 日本晴; コシヒカリから全タンパク質を抽出し、Anti-DFR抗体によって免疫染色した。右の数値はタンパク質の分子量を示す。(D)はGST-DFR(ポジティブコントロール); 35S::DFR遺伝子導入個体; カサラス; 日本晴; コシヒカリから全タンパク質を抽出し、Anti-DFR抗体によって免疫染色した。右の数値はタンパク質の分子量を示す。
【図15A−C】RiceGAASプログラムを用いたRcのアノテーションを示す図である。(A)Rcの7番染色体上での位置を示す。(B)Rcが座乗すると考えられるR1807とR1582の間に存在するMYC type protein,MYB related protein。白丸は分子マーカーを示す。矢印は、MYC type protein(predicted gene)、MYB related protein(predicted gene)をそれぞれ示す。(C)Rcの精密なマッピング。白丸は分子マーカーを示す。一番下の段は、Rice Genome research programの日本晴クローンを示す。矢印は、データベースサーチから予想されたRc遺伝子の位置を示している。黒色の四角は、Rcの相補実験に用いた領域である。
【図15D】Rc(本発明)とTT8(Nesi, N., Debeaujon, I., Jond, C., Pelletier, G., Caboche, M. & Lepiniec, L. (2001) Plant Cell 12, 1863-1878.)の予想アミノ酸配列のアラインメントを示す図である。同一のアミノ酸は黒の四角で示し、類似のアミノ酸は囲みで示す。水平な黒の線は、MYC type proteinに特有のbHLHドメインを示す。
【図16】Rcの相補実験結果を示す写真である。写真上部は玄米、下部は籾である。(A)RcRdの遺伝子型を持つカサラス。(B)Non-transformant。(C)rcrdの遺伝子型を持つ日本晴。(D)Rc-MYC-16 (rcrdの遺伝子型を持つ日本晴にRc-MYCを相補した植物のNO.16)。(E)Rc-MYC-23 (rcrdの遺伝子型を持つ日本晴にRc-MYCを相補した植物のNO.23)。(F)Rc-MYC-13(rcrdの遺伝子型を持つ日本晴にRc-MYCを相補した植物のNO.13)。(G)Rcrdの遺伝子型を持つtRc。(H)Rc-MYB1 (rcrdの遺伝子型を持つ日本晴にRc-MYB1を相補した植物)。(I)Rc-MYB2 (rcrdの遺伝子型を持つ日本晴にRc-MYB2を相補した植物)。
【図17】RcとRdによる玄米色の制御モデルを示す図である。
【図18】Rc-MYCの構造(カサラス)を示す図である。箱はエクソンを、水平の線はイントロンをそれぞれ示す。上向きの黒い三角は日本晴における欠失を、下向きの白い三角は日本晴における挿入変異をそれぞれ示す。
【図19】Rc-MYCの第7エクソンに存在する14 bpの欠失変異の多型解析を示す図である。用いたサンプルは、玄米色が赤いRcRdの遺伝子型をもつO. sativa からIndicaタイプのKasalath(カサラス)、Japonicaタイプのトボシ(TOBOSHI)、アカマイアサヒ(AKAMAIASAH)、アカマイ(AKAMAI)、宝満新田稲(HOUMAN)、ダネマ(DANEMA)O.rufipogonからは、DAMA/THAI/86/148 (O.R.D)とCOL/THAILAND/1990/MAFF/181 (O.R.C)を用いた。玄米色が赤褐色のRcrdの遺伝子型を持つものについては、NILであるtRcとCOL/KAGOSHIMA/1963 MAJIRI/1187 (col-kago)を用いた。玄米色が白いrcrdの遺伝子型を持つものについては、日本晴(nipponbare)、コシヒカリ(Koshi)、台中65号(T65)、ドウゴワセ(DOUGO)、ミトニシキ(MITONISHIKI)、フサクモチ(FUSAKUMOCHI)、シミズシラズ(SHIMIZU)を用いた。着色(赤及び赤褐)と白色の関係は欠失のある・なしと一致することから、当該遺伝子の着色への関与が図16の結果に加えて証明された。
【図20】Rc-MYCの系統解析を示す図である。bHLHドメインを持つ、アミノ酸配列をClastal Xを用いてアラインメントした。系統樹の作成、Boots strap valueはClastal X上のNJ法、最節約法を用いて計算した。それぞれIN1,Lc, R-S, B-Peruはトウモロコシ、Raはイネ、TT8はアラビドプシス、JAF13はペチュニア、DELILAは金魚草のアントシアニン合成系制御因子である。
【図21】本発明のRc(MYC)およびRd(DFR)遺伝子の近傍に存在し、本発明の育種方法に利用可能なDNAマーカーの具体例を示す図である。
【発明を実施するための形態】
【0017】
本発明者らによって、植物体を着色させる機能、または、植物体へタンニン類を蓄積させる機能を有する遺伝子が同定された。
【0018】
本発明の上記遺伝子の好ましい態様としては、イネのRc遺伝子、またはRd遺伝子が挙げられる。該Rc遺伝子によってコードされるタンパク質は、プロアントシアニジン合成系支配因子であり、転写制御因子としての機能を有する。該Rc遺伝子は、「MYC遺伝子」、「Rc-MYC遺伝子」、または「Myc related protein gene」等と表記される場合がある。
【0019】
一方、Rd遺伝子は、「DFR遺伝子」あるいは「Dihydroflavonol 4-reductase gene」と表記される場合がある。その発現は転写因子であるRc(MYC)によって、制御されていることが明らかになった。また、本発明者らによって、Rd(DFR)タンパク質の翻訳は、Alternative translation initiationによって制御されていることが見出された。全長DFRを有する植物(例えば、イネ等)では、種子が赤色を呈し、DFRのORFが短い構造の植物(例えば、イネ等)では種子が赤褐色を呈し、Rdが機能しない場合には種子が白色を呈する。
【0020】
後述の実施例において示すように、rd型のDFRは、Alternative translation initiationにより下流のATGから翻訳され、この翻訳産物はDFRの機能を相補する機能を有することが、本発明者らによって初めて見出された。従って、本発明においてRd遺伝子とは、全長のDFRタンパク質をコードするDNAに限定されず、DFRの機能を相補し得るDFRの下流のATGから翻訳される産物をコードするDNAが含まれる。
【0021】
例えば、本発明のRd(DFR)タンパク質には、図14に示される46 kDa(配列番号:6)、33 kDa(配列番号:7)、22 kDa(配列番号:8)に相当するタンパク質が挙げられる。該33 kDa、22 kDaに相当するタンパク質は、Rd(DFR)遺伝子からAlternative translation initiation機構によって翻訳されるタンパク質と考えられる。
【0022】
また、上記タンパク質以外であっても、DFRの機能を相補し得るようなDFRタンパク質変異体もしくはDFRタンパク質の部分断片ペプチドもまた、本発明のRd(DFR)タンパク質に含まれる。
【0023】
本発明のRcおよびRdタンパク質は、イネ由来のタンパク質であることが好ましいが、その由来する植物種は特に制限されず、イネ以外の植物におけるRcもしくはRdと同等なタンパク質(RcもしくはRdのホモログ・オルソログ等)も本発明における「Rcタンパク質」もしくは「Rdタンパク質」に含まれる。例えば、イネRc(MYC)もしくはRd(DFR)に相当するタンパク質を有する植物、または、イネRcもしくはRdタンパク質が実質的に機能し得る植物であれば、本発明を実施することが可能である。
【0024】
本発明において「植物」とは、特に制限されないが、好ましくは単子葉植物であり、より好ましくは、イネ科植物である。本発明における植物として具体的には、イネ、トウモロコシ、コムギ、オオムギ等を例示することができる。(これらのRcおよびRdタンパク質、および該タンパク質をコードする遺伝子を本明細書においてそれぞれ、「本発明のタンパク質」、「本発明の遺伝子」と記載する場合あり。)
【0025】
Rc(MYC)遺伝子のゲノムDNA配列を配列番号:1に、Rc(MYC)遺伝子のcDNA配列を配列番号:2に、該DNAによってコードされるタンパク質のアミノ酸配列を配列番号:3に記載する。また、Rd(DFR)遺伝子のゲノムDNA配列を配列番号:4に、Rd(DFR)遺伝子のcDNA配列を配列番号:5に、該DNAによってコードされるタンパク質のアミノ酸配列を配列番号:6に記載する。(イネRc(MYC)のGenBankのアクセッション番号はAB247503である。また、イネRd(DFR)のGenBankのアクセッション番号はAB003496である。)
【0026】
ただし、本発明におけるRcもしくはRd遺伝子は、必ずしも、配列表に具体的に記載された配列からなるDNAに限定されない。また、本発明におけるRcもしくはRdタンパク質は、必ずしも、配列表に具体的に記載されたアミノ酸配列からなるタンパク質に限定されない。
【0027】
上記以外のタンパク質であっても、例えば配列表に記載された配列と高い相同性(通常70%以上、好ましくは80%以上、より好ましくは90%以上、最も好ましくは95%以上)を有し、かつ、本発明のタンパク質が有する機能(例えば、種子を着色させる機能、または、種子へタンニン類を蓄積させる機能等)を持つタンパク質は、本発明のタンパク質に含まれる。
【0028】
上記タンパク質とは、例えば、配列番号:3または6〜8のいずれかに記載のアミノ酸配列において、1以上のアミノ酸が付加、欠失、置換、挿入されたアミノ酸配列からなるタンパク質であって、通常変化するアミノ酸数が30アミノ酸以内、好ましくは10アミノ酸以内、より好ましくは5アミノ酸以内、最も好ましくは3アミノ酸以内である。
【0029】
本発明における「Rc遺伝子」もしくは「Rd遺伝子」には、例えば、配列番号:2もしくは5に記載の塩基配列からなるDNAに対応する他の植物における内在性の遺伝子(イネのRcもしくはRd遺伝子のホモログ等)が含まれる。
【0030】
また、配列番号:1、2、4もしくは5に記載の塩基配列からなるDNAに対応する他の植物の内在性のDNAは、一般的に、配列番号:1、2、4もしくは5に記載のDNAと高い相同性を有する。高い相同性とは、50%以上、好ましくは70%以上、さらに好ましくは80%以上、より好ましくは90%以上(例えば、95%以上、さらには96%、97%、98%または99%以上)の相同性を意味する。この相同性は、mBLASTアルゴリズム(Altschul et al. (1990) Proc. Natl. Acad. Sci. USA 87: 2264-8; Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90: 5873-7)によって決定することができる。また、該DNAは、生体内から単離した場合、配列番号:1、2、4もしくは5に記載のDNAとストリンジェントな条件下でハイブリダイズすると考えられる。ここで「ストリンジェントな条件」としては、例えば「2×SSC、0.1%SDS、50℃」、「2×SSC、0.1%SDS、42℃」、「1×SSC、0.1%SDS、37℃」、よりストリンジェントな条件として「2×SSC、0.1%SDS、65℃」、「0.5×SSC、0.1%SDS、42℃」および「0.2×SSC、0.1%SDS、65℃」の条件を挙げることができる。当業者においては、他の植物における本発明の遺伝子に相当する内在性の遺伝子を、本発明の遺伝子の塩基配列を基に適宜取得することが可能である。なお、本明細書においては、イネ以外の植物におけるRcタンパク質(遺伝子)もしくはRdタンパク質(遺伝子)に相当するタンパク質(遺伝子)、または、これらのタンパク質と機能的に同等なタンパク質(遺伝子)を、単に「本発明のタンパク質(遺伝子)」と記載する場合がある。
【0031】
本発明のタンパク質は、天然のタンパク質のほか、遺伝子組み換え技術を利用した組換えタンパク質として調製することができる。天然のタンパク質は、例えば本発明のタンパク質が発現していると考えられる細胞(組織)の抽出液に対し、本発明のタンパク質に対する抗体を用いたアフィニティークロマトグラフィーを用いる方法により調製することが可能である。一方、組換えタンパク質は、本発明のタンパク質をコードするDNAで形質転換した細胞を培養することにより調製することが可能である。
【0032】
本発明の好ましい態様としては、植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能を有する植物由来のタンパク質をコードする、下記(a)〜(e)のいずれかに記載のDNAを提供する。
(a)配列番号:3または6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:1または4に記載の塩基配列のコード領域を含むDNA
(c)配列番号:2または5に記載の塩基配列からなるDNA
(d)配列番号:3または6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(e)配列番号:1または4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
【0033】
また、上記DNAによってコードされるタンパク質もまた、本発明に含まれる。
本発明において、「植物の種子」とは、例えば、植物がイネの場合には、「コメ」、「玄米」等を指す。
【0034】
本発明において「着色」とは、通常、植物体もしくはその種子において、元の色を別の色へ変化させることを言う。一例を示せば、種子の全体あるいは一部を、赤色、赤褐色、褐色、または紫黒色等へ着色させることを指す。より具体的には、植物がイネである場合には、白色米(白米)を赤色米(赤米)、赤褐色米、褐色米、または紫黒色米等へ着色することを言う。(左記のように白米以外の有色のコメを、本発明において「有色米」と表記する場合がある。)
【0035】
また、本発明において「着色を制御する」とは、上述のように、植物体もしくはその種子の全体またはその一部の色を、他の色へ変化させることを言う。例えば、植物がイネの場合、上述のように、白米を、赤色米等の有色米へ変化させることを言うが、反対に、有色米を白色米へ変化させる場合も、本発明における「着色を制御する」ことに含まれる。
【0036】
本発明において着色が制御される植物体の部位は、特に制限されないが、例えば、種子、葉、節間、茎、えい(もみ)等を挙げることができる。
【0037】
本発明における植物がイネの場合には、例えば、イネの種子(コメ)の種皮および/または果皮が着色される。また、本発明における植物がコムギ、オオムギ、トウモロコシ等の場合には、種子の果皮部位が着色される。
【0038】
本発明のおける植物の着色部位は、必ずしも上述の部位に限定されるものではなく、植物の節間、種子の胚乳等を着色させることも可能である。
【0039】
また、「タンニン(tannin)」とは、一般的には、広く植物界に分布する多数のフェノール性ヒドロキシル基を持つ芳香族化合物の総称であり、「タンニン酸(tannic acid)」、あるいは「ガロタンニン酸(gallotannic acid)」とも呼ばれる。
【0040】
本発明における「タンニン類」には、例えば、「タンニン」、「カテキン(catechin)」、「アントシアニン(anthocyanin)」、「プロアントシアニジン(proanthocyanidin)」等が含まれる。上記カテキンは、「3,3',4',5,7-ペンタヒドロキシフラバン(3,3',4',5,7-pentahydroxyflavan)」とも呼ばれる。また、「カテキン」には、所謂「カテキン類」に属する化合物、例えば、エピカテキン等も含まれる。また、タンニン類が各種化合物と反応して生成される物質もまた、本発明における「タンニン類」に含まれる。上記「アントシアニン」は、紫色の色素であり、上記「プロアントシアニジン」は赤褐色もしくは赤色の色素であるものと考えられている。
【0041】
赤米品種である「紅衣」の玄米には、100g中に0.29gのタンニン、1.6mgのカテキン、0.09gのアントシアンが含まれている。従って、本発明の方法によって白米にタンニン類を蓄積させる場合、コメ中に蓄積されるタンニン類の量(含有量)としては、例えば、上記の量を例示することができる。
【0042】
本発明において「タンニン類を蓄積させる」とは、通常、植物体において元々タンニン類が存在しない部位にタンニン類を生成・蓄積させること、または、植物体においてタンニン類が存在する部位において、タンニン類の量(含有量)を増加させることを言う。
【0043】
本明細書におけるタンパク質とは、複数のアミノ酸からなる重合体を意味し、そのアミノ酸の長さは特に制限されない。従って、本発明のタンパク質には、所謂「ポリペプチド」、および「オリゴペプチド」も含まれる。本発明のタンパク質は、天然に存在する状態から修飾されていないもの、および修飾されているものの双方を含む。修飾としては、アセチル化、アシル化、ADP-リボシル化、アミド化、フラビンの共有結合、ヘム部分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトールの共有結合、架橋、環化、ジスルフィド結合の形成、脱メチル化、共有架橋の形成、シスチンの形成、ピログルタメートの形成、ホルミル化、γ-カルボキシル化、グリコシル化、GPIアンカー形成、ヒドロキシル化、ヨウ素化、メチル化、ミリストイル化、酸化、タンパク質分解処理、リン酸化、プレニル化、ラセミ化、セレノイル化、硫酸化、アルギニル化のようなタンパク質へのアミノ酸の転移RNA媒介付加、ユビキチン化等が含まれる。
【0044】
本発明のポリペプチドは、そのアミノ酸配列に従って、一般的な化学合成法により製造することが可能であり、該方法には、通常の液相法および固相法によるペプチド合成法が包含される。かかるペプチド合成法は、より詳しくはアミノ酸配列の情報に基づいて、各アミノ酸を1個ずつ逐次合成させて鎖を延長していくステップワイズエロンゲーション法と、アミノ酸数個からなるフラグメントを予め合成し、次いで各フラグメントをカップリング反応させるフラグメント・コンデンセーション法を包含し、本発明のタンパク質の合成は、いずれの方法を用いてもよい。
【0045】
このようなペプチド合成法にて用いられる縮合法も、各種方法に従って行うことができる。その具体例としては、例えばアジド法、混合酸無水物法、DCC法、活性エステル法、酸化還元法、DPPA(ジフェニルホスホリルアジド)法、ウッドワード法等を例示できる。
【0046】
これら各種方法に利用できる溶媒もまた、一般的に使用されるものを適宜利用することができる。その例としては、例えばジメチルホルムアミド(DMF)、ジメチルスルホキシド(DMSO)、ヘキサホスホロアミド、ジオキサン、テトラヒドロフラン(THF)、酢酸エチル等及びこれらの混合溶媒等を挙げることができる。なお、上記ペプチド合成反応に際して、反応に関与しないアミノ酸およびペプチドにおけるカルボキシル基は、一般にはエステル化により、例えばメチルエステル、エチルエステル、第三級ブチルエステル等の低級アルキルエステル、例えばベンジルエステル、P−メトキシベンジルエステル、P−ニトロベンジルエステルアラルキルエステル等として保護することができる。また、側鎖に官能基を有するアミノ酸、例えばTyrの水酸基は、アセチル基、ベンジル基、ベンジルオキシカルボニル基、第三級ブチル基等で保護されてもよいが、必ずしもかかる保護は必須ではない。また、例えば、Argのグアニジノ基は、ニトロ基、トシル基、2−メトキシベンゼンスルホニル基、メチシレン−2−スルホニル基、ベンジルオキシカルボニル基、イソボルニルオキシカルボニル基、アダマンチルオキシカルボニル基等の適当な保護基により保護することができる。
【0047】
上記のようにして得ることが可能な本発明のタンパク質は、通常の方法に従って、例えばイオン交換樹脂、分配クロマトグラフィー、ゲルクロマトグラフィー、アフィニティークロマトグラフィー、高速液体クロマトグラフィー(HPLC)、向流分配法等のペプチド化学の分野で汎用されている方法に従って、適宜、精製を行うことができる。
【0048】
本発明のタンパク質は、例えば、配列番号:3もしくは6〜8に記載のポリペプチド、または配列番号:2もしくは5に記載のDNA核酸分子を合成し、次いで適当な発現ベクターへ導入した後、宿主細胞内において発現させる遺伝子工学的手法によっても取得することができる。
【0049】
本発明のタンパク質には、例えば、Rc(MYC)もしくはRd(DFR)タンパク質と機能的に同等なタンパク質(ポリペプチド)が含まれる。ここで「機能的に同等」とは、対象となるタンパク質がRc(MYC)もしくはRd(DFR)タンパク質と同様の(同等の)生物学的あるいは生化学的機能(活性)を有することを指す。このような機能としては、例えば、植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能等を挙げることができる。
【0050】
あるDNAが植物体を着色させる機能、またはタンニン類を蓄積させる機能を有するタンパク質をコードするか否かは以下のようにして評価することができる。最も一般的な方法としては、該DNAが導入された植物を栽培し、植物体の色、または植物体中に含まれるタンニン類の量を評価する方法である。
【0051】
対象となるタンパク質(ポリペプチド)が、Rc(MYC)もしくはRd(DFR)タンパク質と同等の生物学的あるいは生化学的な機能(活性)を有しているか否かは、例えば、該タンパク質を発現させた植物もしくはその種子におけるタンニン類の量を測定することにより適宜評価することも可能である。
【0052】
あるポリペプチドと機能的に同等なポリペプチドを調製するための、当業者によく知られた方法としては、例えばポリペプチド中のアミノ酸配列に変異を導入する方法が挙げられる。具体的には当業者であれば部位特異的変異誘発法(Hashimoto-Gotoh, T. et al. (1995) Gene 152, 271-275、Zoller, MJ, and Smith, M.(1983) Methods Enzymol. 100, 468-500、Kramer, W. et al. (1984) Nucleic Acids Res. 12, 9441-9456、Kramer W, and Fritz HJ(1987) Methods. Enzymol. 154, 350-367、Kunkel,TA(1985) Proc Natl Acad Sci USA. 82, 488-492、Kunkel (1988) Methods Enzymol. 85, 2763-2766)などを用いて、配列番号:4に記載のアミノ酸配列に適宜変異を導入することにより、該ポリペプチドと機能的に同等なポリペプチドを調製することができる。また、ポリペプチド中のアミノ酸の変異は自然に生じることもある。このように、人工的か自然に生じたものかを問わず、本発明者らにより同定されたRcもしくはRdタンパク質(配列番号:3または6〜8)のアミノ酸配列において1もしくは複数のアミノ酸配列が変異したアミノ酸配列を有し、該ポリペプチドと機能的に同等なポリペプチドは、本発明のタンパク質(ポリペプチド)に含まれる。
【0053】
上記変異体における、変異するアミノ酸数は、本発明のタンパク質の有する機能が保持される限り制限はないが、通常15アミノ酸以内であり、好ましくは10アミノ酸以内であり、より好ましくは5アミノ酸以内であり、さらに好ましくは1〜4アミノ酸である。
【0054】
変異するアミノ酸残基としては、アミノ酸側鎖の性質が保存されている別のアミノ酸に変異されることが望ましい。例えばアミノ酸側鎖の性質としては、疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)、親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)、脂肪族側鎖を有するアミノ酸(G、A、V、L、I、P)、水酸基含有側鎖を有するアミノ酸(S、T、Y)、硫黄原子含有側鎖を有するアミノ酸(C、M)、カルボン酸及びアミド含有側鎖を有するアミノ酸(D、N、E、Q)、塩基含有側鎖を有するアミノ離(R、K、H)、芳香族含有側鎖を有するアミノ酸(H、F、Y、W)を挙げることができる(括弧内はいずれもアミノ酸の一文字表記を表す)。
【0055】
あるアミノ酸配列に対する1または複数個のアミノ酸残基の欠失、付加及び/又は他のアミノ酸による置換により修飾されたアミノ酸配列を有するポリペプチドがその生物学的機能(活性)を維持し得ることはすでに知られている(Mark, D. F. et al., Proc. Natl. Acad. Sci. USA (1984) 81, 5662-5666 、Zoller, M. J. & Smith, M. Nucleic Acids Research (1982) 10, 6487-6500 、Wang, A. et al., Science 224, 1431-1433 、Dalbadie-McFarland, G. et al., Proc. Natl. Acad. Sci. USA (1982) 79, 6409-6413)。
【0056】
具体的なアミノ酸配列(例えば、配列番号:3または6〜8)が開示された場合においては、当業者であれば、これらアミノ酸配列を基に、適宜アミノ酸が改変された配列からなるポリペプチドを作製し、当該ポリペプチドについて、上述の機能を有するか否かを評価し、本発明のタンパク質(ポリペプチド)を適宜選択することが可能である。
【0057】
本発明におけるRdタンパク質は、そのN末領域が欠損している場合であっても本発明のタンパク質の有する機能を相補し得ることが明らかとなった。従って、Rdタンパク質の改変体としては、例えば、配列番号:7もしくは8で示されるアミノ酸領域を含み、N末領域において変異もしくは欠失を有するRdタンパク質の改変体(変異体、部分ペプチド等)を例示することができる。
【0058】
本発明のタンパク質のアミノ酸配列に複数個のアミノ酸残基が付加されたタンパク質には、これらタンパク質を含む融合ポリペプチドが含まれる。融合ポリペプチドは、これらポリペプチドと他のペプチド又はポリペプチドとが融合したものである。融合ポリペプチドを作製する方法は、本発明のタンパク質(例えば、配列番号:3または6〜8)をコードするDNA(例えば、配列番号:2または5)と他のペプチド又はポリペプチドをコードするDNAをフレームが一致するように連結してこれを発現ベクターに導入し、宿主で発現させればよく、当業者に公知の手法を用いることができる。本発明のタンパク質との融合に付される他のペプチド又はポリペプチドは、特に制限されない。
【0059】
本発明のポリペプチドとの融合に付される他のペプチドとしては、例えば、GST(グルタチオン−S−トランスフェラーゼ)、イムノグロブリン定常領域、β−ガラクトシダーゼ、MBP(マルトース結合タンパク質)等が挙げられる。市販されているこれらペプチドまたはポリペプチドをコードするポリヌクレオチドを本発明のポリペプチドをコードするポリヌクレオチドと融合させ、これにより調製された融合ポリヌクレオチドを発現させることにより、融合ポリペプチドを調製することができる。
【0060】
またあるポリペプチドと機能的に同等なポリペプチドを調製する当業者によく知られた他の方法としては、ハイブリダイゼーション技術(Sambrook,J et al., Molecular Cloning 2nd ed., 9.47-9.58, Cold Spring Harbor Lab. press, 1989)を利用する方法が挙げられる。即ち、当業者であれば、本発明のタンパク質をコードするDNA(配列番号:2または5に記載の塩基配列)もしくはその一部をもとに、同種または異種植物由来のDNA試料から、これと相同性の高いDNAを単離して、該DNAから本発明のタンパク質と機能的に同等なポリペプチドを単離することも通常行いうることである。
【0061】
本発明には、本発明のタンパク質をコードするDNAとハイブリダイズするDNAによってコードされるタンパク質であって、本発明のタンパク質と機能的に同等なタンパク質が含まれる。このようなタンパク質としては、例えばイネあるいは他の植物のホモログ(例えば、トウモロコシ、コムギ、オオムギ等の植物に由来するタンパク質)が挙げられる。
【0062】
本発明のタンパク質と機能的に同等なタンパク質をコードするDNAを単離するためのハイブリダイゼーションの条件は、当業者であれば適宜選択することができる。ハイブリダイゼーションの条件としては、例えば、低ストリンジェントな条件が挙げられる。低ストリンジェントな条件とは、ハイブリダイゼーション後の洗浄において、例えば42℃、0.1×SSC、0.1%SDSの条件であり、好ましくは50℃、0.1×SSC、0.1%SDSの条件である。より好ましいハイブリダイゼーションの条件としては、高ストリンジェントな条件が挙げられる。高ストリンジェントな条件とは、例えば65℃、5×SSC及び0.1%SDSの条件である。これらの条件において、温度を上げる程に高い相同性を有するDNAが効率的に得られることが期待できる。但し、ハイブリダイゼーションのストリンジェンシーに影響する要素としては温度や塩濃度など複数の要素が考えられ、当業者であればこれら要素を適宜選択することで同様のストリンジェンシーを実現することが可能である。
【0063】
また、ハイブリダイゼーションにかえて、遺伝子増幅技術(PCR)(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley&Sons Section 6.1-6.4)を用いて、本発明のタンパク質をコードするDNA(例えば、配列番号:2または5)の一部を基にプライマーを設計し、本発明者らにより同定されたタンパク質をコードするDNAと相同性の高いDNA断片を単離し、該DNAを基に本発明者らにより同定されたタンパク質と機能的に同等なタンパク質を取得することも可能である。
【0064】
本発明のタンパク質は「成熟」ポリペプチドの形であっても、融合ポリペプチドのような、より大きいポリペプチドの一部であってもよい。本発明のタンパク質には、リーダー配列、プロ配列、多重ヒスチジン残基のような精製に役立つ配列、または組換え生産の際の安定性を確保する付加的配列などが含まれていてもよい。
【0065】
これらハイブリダイゼーション技術や遺伝子増幅技術により単離されるDNAによってコードされる、本発明のタンパク質と機能的に同等なタンパク質は、通常、本発明のタンパク質(例えば、配列番号:3または6〜8)とアミノ酸配列において高い相同性を有する。本発明のタンパク質には、RcもしくはRdタンパク質と機能的に同等であり、かつ該タンパク質のアミノ酸配列と高い相同性を有するタンパク質も含まれる。高い相同性とは、アミノ酸レベルにおいて、通常、少なくとも50%以上の同一性、好ましくは75%以上の同一性、さらに好ましくは85%以上の同一性、さらに好ましくは95%以上(例えば、96%以上、97%以上、98%以上、99%以上)の同一性を指す。タンパク質の相同性を決定するには、文献(Wilbur, W. J. and Lipman, D. J. Proc. Natl. Acad. Sci. USA (1983) 80, 726-730)に記載のアルゴリズムに従えばよい。
【0066】
アミノ酸配列の同一性は、例えば、Karlin and Altschul によるアルゴリズムBLAST (Proc. Natl. Acad. Sci. USA 87:2264-2268, 1990、Proc. Natl. Acad. Sci. USA 90:5873-5877, 1993)によって決定することができる。このアルゴリズムに基づいて、BLASTXと呼ばれるプログラムが開発されている(Altschul et al. J. Mol. Biol.215: 403-410, 1990)。BLASTXによってアミノ酸配列を解析する場合には、パラメーターは例えば、score = 50、wordlength = 3とする。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である(http://www.ncbi.nlm.nih.gov.)。
【0067】
本発明のタンパク質は、当業者に公知の方法により、組み換えポリペプチドとして、また天然のポリペプチドとして調製することが可能である。組み換えポリペプチドであれば、例えば、本発明のタンパク質をコードするDNA(例えば、配列番号:2または5に記載のDNA)を、適当な発現ベクターに組み込み、これを適当な宿主細胞に導入して得た形質転換体を回収し、抽出物を得た後、イオン交換、逆相、ゲル濾過などのクロマトグラフィー、あるいは本発明のタンパク質に対する抗体をカラムに固定したアフィニティークロマトグラフィーにかけることにより、または、さらにこれらのカラムを複数組み合わせることにより精製し、調製することが可能である。
【0068】
また、本発明のタンパク質をグルタチオンS-トランスフェラーゼタンパク質との融合ポリペプチドとして、あるいはヒスチジンを複数付加させた組み換えポリペプチドとして宿主細胞(例えば、植物細胞や微生物細胞等)内で発現させた場合には、発現させた組み換えポリペプチドはグルタチオンカラムあるいはニッケルカラムを用いて精製することができる。融合ポリペプチドの精製後、必要に応じて融合ポリペプチドのうち、目的のポリペプチド以外の領域を、トロンビンまたはファクターXaなどにより切断し、除去することも可能である。
【0069】
天然のタンパク質であれば、当業者に周知の方法、例えば本発明のタンパク質を発現している組織や細胞の抽出物に対し、本発明のタンパク質と親和性を有する抗体が結合したアフィニティーカラムを作用させて精製することにより単離することができる。抗体はポリクローナル抗体であっても、モノクローナル抗体であってもよい。
【0070】
また、本発明のタンパク質は例えば、本発明のタンパク質を認識する抗体の作製等に利用することが可能である。
【0071】
本発明のDNAは、本発明のタンパク質をコードし得るものであればいかなる形態でもよい。即ち、mRNAから合成されたcDNAであるか、ゲノムDNAであるか、化学合成DNAであるかなどを問わない。また、本発明のタンパク質をコードしうる限り、遺伝暗号の縮重に基づく任意の塩基配列を有するDNAが含まれる。
【0072】
本発明のDNAは、当業者に公知の方法により調製することができる。例えば、本発明のタンパク質を発現している細胞よりcDNAライブラリーを作製し、本発明のDNA(例えば、配列番号:2または5に記載の塩基配列)の一部をプローブとしてハイブリダイゼーションを行うことにより調製できる。cDNAライブラリーは、例えば、文献(Sambrook, J. et al., Molecular Cloning、Cold Spring Harbor Laboratory Press (1989))に記載の方法により調製してもよく、あるいは市販のDNAライブラリーを用いてもよい。また、本発明のタンパク質を発現している細胞よりRNAを調製し、逆転写酵素によりcDNAを合成後、本発明のDNA(例えば、配列番号:2または5に記載の塩基配列)に基づいてオリゴDNAを合成し、これをプライマーとして用いてPCR反応を行い、本発明のタンパク質をコードするcDNAを増幅させることにより調製することも可能である。
【0073】
また、得られたcDNAの塩基配列を決定することにより、それがコードする翻訳領域を決定でき、本発明のタンパク質のアミノ酸配列を得ることができる。また、得られたcDNAをプローブとしてゲノムDNAライブラリーをスクリーニングすることにより、ゲノムDNAを単離することができる。
【0074】
具体的には、次のようにすればよい。まず、本発明のタンパク質を発現する細胞、組織、器官からmRNAを単離する。mRNAの単離は、公知の方法、例えば、グアニジン超遠心法(Chirgwin, J. M. et al., Biochemistry (1979) 18, 5294-5299)、AGPC法(Chomczynski, P. and Sacchi, N., Anal. Biochem. (1987) 162, 156-159)等により全RNAを調製し、mRNA Purification Kit (Pharmacia) 等を使用して全RNAからmRNAを精製する。また、QuickPrep mRNA Purification Kit (Pharmacia) を用いることによりmRNAを直接調製することもできる。
【0075】
得られたmRNAから逆転写酵素を用いてcDNAを合成する。cDNAの合成は、AMV Reverse Transcriptase First-strand cDNA Synthesis Kit (生化学工業)等を用いて行うこともできる。また、本明細書に記載されたプライマー等を用いて、5'-Ampli FINDER RACE Kit (Clontech製)およびポリメラーゼ連鎖反応 (polymerase chain reaction ; PCR)を用いた5'-RACE法(Frohman, M. A. et al., Proc. Natl. Acad. Sci. U.S.A. (1988) 85, 8998-9002 ; Belyavsky, A. et al., Nucleic Acids Res. (1989) 17, 2919-2932) に従い、cDNAの合成および増幅を行うことができる。得られたPCR産物から目的とするDNA断片を調製し、ベクターDNAと連結する。さらに、これより組換えベクターを作製し、大腸菌等に導入してコロニーを選択して所望の組換えベクターを調製する。目的とするDNAの塩基配列は、公知の方法、例えば、ジデオキシヌクレオチドチェインターミネーション法により確認することができる。
【0076】
また、本発明のDNAの作成においては、発現に使用する宿主のコドン使用頻度を考慮し、より発現効率の高い塩基配列を設計することができる(Grantham, R. et al., Nucelic Acids Research (1981) 9, r43-74)。また、本発明のDNAは、市販のキットや公知の方法によって改変することができる。改変としては、例えば、制限酵素による消化、合成オリゴヌクレオチドや適当なDNAフラグメントの挿入、リンカーの付加、開始コドン(ATG)及び/又は終止コドン(TAA、TGA、又はTAG)の挿入等が挙げられる。
【0077】
また本発明は、本発明のDNAが挿入されたベクターを提供する。本発明のベクターとしては、組み換えタンパク質の生産に用いる上記ベクターの他、形質転換植物体作製のために植物細胞内で本発明のDNAを発現させるためのベクターも含まれる。このようなベクターとしては、好ましくは、植物細胞で転写可能なプロモーター配列と転写産物の安定化に必要なポリアデニレーション部位を含むターミネーター配列を含む。植物細胞の形質転換に用いられるベクターとしては、該細胞内で挿入遺伝子を発現させることが可能なものであれば特に制限はない。例えば、植物細胞内での恒常的な遺伝子発現を行うためのプロモーター(例えば、カリフラワーモザイクウイルスの35Sプロモーター)を有するベクターや外的な刺激により誘導的に活性化されるプロモーターを有するベクターを用いることも可能である。上記「植物細胞」には、種々の形態の植物細胞、例えば、懸濁培養細胞、プロトプラスト、葉の切片、カルス等が含まれる。
【0078】
本発明のベクターは、本発明のタンパク質を恒常的または誘導的に発現させるためのプロモーターを含有してもよい。恒常的に発現させるためのプロモーターとしては、例えば、カリフラワーモザイクウイルスの35Sプロモーター、イネのアクチンプロモーター、トウモロコシのユビキチンプロモーター等が挙げられる。
【0079】
また、誘導的に発現させるためのプロモーターとしては、例えば細菌・ウイルスの感染や侵入、低温、高温、乾燥、紫外線の照射、特定の化合物の散布などの外因によって発現することが知られているプロモーターなどが挙げられる。このようなプロモーターとしては、例えば、細菌・ウイルスの感染や侵入によって発現するイネキチナーゼ遺伝子のプロモーターやタバコのPRタンパク質遺伝子のプロモーター、低温によって誘導されるイネの「lip19」遺伝子のプロモーター、高温によって誘導されるイネの「hsp80」遺伝子と「hsp72」遺伝子のプロモーター、乾燥によって誘導されるシロイヌナズナの「rab16」遺伝子のプロモーター、紫外線の照射によって誘導されるパセリのカルコン合成酵素遺伝子のプロモーター、嫌気的条件で誘導されるトウモロコシのアルコールデヒドロゲナーゼ遺伝子のプロモーターなどが挙げられる。また、イネキチナーゼ遺伝子のプロモーターとタバコのPRタンパク質遺伝子のプロモーターはサリチル酸などの特定の化合物によって、「rab16」は植物ホルモンのアブシジン酸の散布によっても誘導される。
【0080】
当業者においては、所望のDNAを有するベクターを、一般的な遺伝子工学技術によって、適宜、作製することが可能である。通常、市販の種々のベクターを利用することができる。
【0081】
本発明のベクターは、宿主細胞内において本発明のDNAを保持したり、本発明のタンパク質を発現させるためにも有用である。
【0082】
本発明におけるDNAは、通常、適当なベクターへ担持(挿入)され、宿主細胞へ導入される。該ベクターとしては、挿入したDNAを安定に保持するものであれば特に制限されず、例えば宿主に大腸菌を用いるのであれば、クローニング用ベクターとしてpBluescriptベクター(Stratagene社製)などが好ましいが、市販の種々のベクターを利用することができる。本発明のタンパク質を生産する目的としてベクターを用いる場合には、特に発現ベクターが有用である。発現ベクターとしては、試験管内、大腸菌内、培養細胞内、植物個体内でポリペプチドを発現するベクターであれば特に制限されないが、例えば、試験管内発現であればpBESTベクター(プロメガ社製)、大腸菌であればpETベクター(Invitrogen社製)、培養細胞であればpME18S-FL3ベクター(GenBank Accession No. AB009864)、生物個体であればpME18Sベクター(Mol Cell Biol. 8:466-472(1988))などを例示することができる。ベクターへの本発明の核酸の挿入は、常法により、例えば、制限酵素サイトを用いたリガーゼ反応により行うことができる。
【0083】
上記宿主細胞としては特に制限はなく、目的に応じて種々の宿主細胞が用いられる。本発明のタンパク質を発現させるための細胞としては、例えば、細菌細胞(例:ストレプトコッカス、スタフィロコッカス、大腸菌、ストレプトミセス、枯草菌)、昆虫細胞(例:ドロソフィラS2、スポドプテラSF9)、動物細胞(例:CHO、COS、HeLa、C127、3T3、BHK、HEK293、Bowes メラノーマ細胞)および植物細胞を例示することができる。宿主細胞へのベクター導入は、例えば、リン酸カルシウム沈殿法、電気パルス穿孔法(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons.Section 9.1-9.9)、リポフェクション法(GIBCO-BRL社製)、マイクロインジェクション法などの公知の方法で行うことが可能である。
【0084】
宿主細胞において発現したタンパク質を小胞体の内腔に、細胞周辺腔に、または細胞外の環境に分泌させるために、適当な分泌シグナルを目的のタンパク質に組み込むことができる。これらのシグナルは目的のタンパク質に対して内因性であっても、異種シグナルであってもよい。
【0085】
上記製造方法におけるタンパク質の回収は、本発明のタンパク質が培地に分泌される場合は、培地を回収する。本発明のタンパク質が細胞内に産生される場合は、その細胞をまず溶解し、その後にタンパク質を回収する。
【0086】
組換え細胞培養物から本発明のタンパク質を回収し精製するには、硫酸アンモニウムまたはエタノール沈殿、酸抽出、アニオンまたはカチオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィーおよびレクチンクロマトグラフィーを含めた公知の方法を用いることができる。
【0087】
また、植物の生体内で本発明のDNAを発現させる方法としては、本発明のDNAを適当なベクターに組み込み、例えば、エレクトロポレーション法、アグロバクテリウム法、リポソーム法、カチオニックリポソーム法などにより生体内に導入する方法などが挙げられる。ベクターへの本発明のDNAの挿入などの一般的な遺伝子操作は、常法に従って行うことが可能である(Molecular Cloning, 5.61-5.63)。植物体内への投与は、ex vivo法であっても、in vivo法であってもよい。また、植物体内へ本発明のDNAを導入する方法としては、好ましくは、アグロバクテリウムを介して遺伝子を導入する方法が挙げられる。
【0088】
また、後述する手法で、本発明のDNAが導入された形質転換植物体を作成し、該植物体から本発明のタンパク質を調製することも可能である。
【0089】
また、上述のようにして取得される組換えタンパク質を用いれば、これに結合する抗体を調製することができる。例えば、ポリクローナル抗体は、精製した本発明のタンパク質もしくはその一部のペプチドをウサギなどの免疫動物に免疫し、一定期間の後に血液を採取し、血ぺいを除去することにより調製することが可能である。また、モノクローナル抗体は、上記タンパク質若しくはペプチドで免疫した動物の抗体産生細胞と骨腫瘍細胞とを融合させ、目的とする抗体を産生する単一クローンの細胞(ハイブリドーマ)を単離し、該細胞から抗体を得ることにより調製することができる。これにより得られた抗体は、本発明のタンパク質の精製や検出などに利用することが可能である。本発明には、本発明のタンパク質に結合する抗体が含まれる。これらの抗体を用いることにより、植物体における本発明のタンパク質の発現部位の判別、または、植物種が本発明のタンパク質を発現するか否かの判別を行うことが可能である。
【0090】
本発明のDNAを利用して、着色した、もしくはタンニン類が蓄積した形質転換植物体を作製する場合には、本発明のタンパク質をコードするDNAを適当なベクターに挿入して、これを植物細胞に導入し、これにより得られた形質転換植物細胞を再生させる。本発明者等により単離されたタンパク質は、植物体もしくはその種子を着色させる、またはタンニン類を蓄積させる機能を有するが、本発明のタンパク質を任意の植物種(イネ品種等)に導入し発現させることによりそれらの植物体もしくはその種子を着色させたり、植物体もしくはその種子へタンニン類を蓄積させたりすることが可能である。この形質転換に要する期間は、従来のような交雑による遺伝子移入に比較して極めて短期間であり、また、他の形質の変化を伴わない点で有利である。
【0091】
上述のように、本発明のDNAを植物体の細胞内で発現させる工程を含む、植物体もしくはその種子を着色させる、または植物体もしくはその種子にタンニン類を蓄積させる方法もまた本発明に含まれる。該方法によって、着色された、もしくはタンニン類が蓄積した植物体もしくはその種子が作出される。従って本発明は、本発明のDNAを植物体の細胞内で発現させる工程を含む、着色した、もしくは、タンニン類が蓄積した植物体もしくはその種子の製造方法を提供する。
【0092】
より具体的には、本発明の方法によって、例えば、白米を、赤米または赤褐色米等の有色米へ改変することが可能である。
【0093】
本発明の方法によって着色可能な白米品種は、特に制限されず、任意の白米を着色させることが可能である。具体的には、日本晴、コシヒカリ、台中65号、あきたこまち等の品種を例示することができるが、これらの品種に限定されない。
【0094】
また、本発明は、本発明のベクターが導入された形質転換細胞を提供する。本発明のベクターが導入される細胞には、組み換えタンパク質の生産に用いる上記した細胞の他に、形質転換植物体作製のための植物細胞が含まれる。植物細胞としては特に制限はなく、例えば、イネ、トウモロコシ、コムギ、オオムギなどの細胞が挙げられる。本発明の植物細胞には、培養細胞の他、植物体中の細胞も含まれる。また、プロトプラスト、苗条原基、多芽体、毛状根も含まれる。植物細胞へのベクターの導入は、ポリエチレングリコール法、電気穿孔法(エレクトロポーレーション)、アグロバクテリウムを介する方法、パーティクルガン法など当業者に公知の種々の方法を用いることができる。形質転換植物細胞からの植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。例えば、イネにおいては、形質転換植物体を作出する手法については、ポリエチレングリコールによりプロトプラストへ遺伝子導入し植物体を再生させる方法、電気パルスによりプロトプラストへ遺伝子導入し植物体を再生させる方法、パーティクルガン法により細胞へ遺伝子を直接導入し、植物体を再生させる方法、およびアグロバクテリウムを介して遺伝子を導入し、植物体を再生させる方法など、いくつかの技術が既に確立し、本願発明の技術分野において広く用いられている。本発明においては、これらの方法を好適に用いることができる。
【0095】
本発明のDNAを含むベクターの導入により形質転換した植物細胞を効率的に選択するために、上記組み換えベクターは、適当な選抜マーカー遺伝子を含む、もしくは選抜マーカー遺伝子を含むプラスミドベクターと共に植物細胞へ導入することが好ましい。この目的に使用される選抜マーカー遺伝子は、例えば抗生物質ハイグロマイシンに耐性であるハイグロマイシンホスホトランスフェラーゼ遺伝子、カナマイシンまたはゲンタマイシンに耐性であるネオマイシンホスホトランスフェラーゼ遺伝子、および除草剤ホスフィノスリシンに耐性であるアセチルトランスフェラーゼ遺伝子等が挙げられる。
【0096】
組み換えベクターを導入した植物細胞は、導入された選抜マーカー遺伝子の種類に従って適当な選抜用薬剤を含む公知の選抜用培地に置床し培養する。これにより形質転換された植物培養細胞を得ることができる。
【0097】
形質転換された植物細胞は、再分化させることにより植物体を再生させることが可能である。再分化の方法は植物細胞の種類により異なるが、例えば、イネであればFujimuraら(Plant Tissue Culture Lett. 2:74 (1995))の方法が挙げられ、トウモロコシであればShillitoら(Bio/Technology 7:581 (1989))の方法やGorden-Kammら(Plant Cell 2:603(1990))が挙げられる。
【0098】
一旦、ゲノム内に本発明のDNAが導入された形質転換植物体が得られれば、該植物体から有性生殖または無性生殖により子孫を得ることが可能である。また、該植物体やその子孫あるいはクローンから繁殖材料(例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラスト等)を得て、それらを基に該植物体を量産することも可能である。本発明には、本発明のDNAが導入された植物細胞、該細胞を含む植物体、該植物体の子孫およびクローン、並びに該植物体、その子孫、およびクローンの繁殖材料が含まれる。
【0099】
このようにして作出された植物体もしくはその種子の色は着色されることが期待される。
上述のように、本発明のDNAもしくはベクターを植物細胞へ導入し、該植物細胞から植物体を再生させる工程を含む、形質転換植物体の製造方法もまた本発明に含まれる。
【0100】
本発明の着色した植物体もしくはその種子、またはタンニン類が蓄積した植物体もしくはその種子は、育種法によっても作出することが可能である。
【0101】
上記育種法としては、例えば、本発明のDNAを有する品種と交雑させることを特徴とする一般的な育種法(交雑育種法等)を挙げることができる。該方法によって、着色した植物体もしくはその種子、またはタンニン類が蓄積した植物体もしくはその種子を作出することができる。
【0102】
育種法によって本発明の植物体もしくは種子を作製する際には、公知の種々の文献を参照して適宜実施することができる。(細胞工学別冊・植物細胞工学シリーズ15「モデル植物の実験プロトコール」秀潤社、2001年、加藤鎌司著「2.1 イネ・コムギの交配」p.6-9; 高牟礼逸朗、佐野芳雄著「2. 交配法」p.46-48)
【0103】
本発明の上記育種方法の好ましい態様としては、以下の工程(a)および(b)を含む方法である。
(a)本発明のDNAを有する植物と交雑させる工程を含む、
(b)前記DNAを有する植物改変体を選抜する工程
【0104】
本発明の上記方法を育種法によって実施する場合には、より具体的には、以下のような工程を含む方法を挙げることができる。
(a)植物Aと、本発明のDNAを有する他の植物Bを交雑させ、F1を作出する工程
(b)前記F1と前記植物Aを交雑させる工程
(c)前記DNAを有する植物を選抜する工程
(d)工程(c)によって選抜された植物と、前記植物Aを交雑させる工程
【0105】
上記方法においては、本発明のDNAを有する植物Bと、着色させたい植物もしくはタンニン類を蓄積させたい植物(これら植物を「植物A」と記載する。)を交雑し、Bのもつ本発明のDNAが受け継がれ、かつ植物Aに近い個体を選抜し、これに植物Aによる交雑を重ねていく「戻し交雑」を行って、Bが有する本発明のDNAの形質を意図的に導入する。その際、一般的にゲノム育種に利用されるDNAマーカーを利用して本発明のDNAを有する植物を選抜することにより、上記「戻し交雑」による置換を効率的に行うことが可能である。その結果、育種期間の短縮に繋がり、また、余分なゲノム領域の混入を正確に除くことができる。通常、「戻し交雑」では、本発明のDNAと非常に強く連鎖する他のDNAに依存する形質がどうしても排除できないという現象が問題となることがあるが、本発明のDNAの近傍に存在するDNAマーカーを利用することにより、所望の植物の正確な選抜が可能となる。
【0106】
上記方法においては、必要に応じて、本発明のDNA以外のゲノム全域が目的の遺伝形質でホモ固定するまで、繰り返して行うことができる。即ち、本発明の好ましい態様のおいては、上記工程(d)によって交雑された個体について、一般的なDNAマーカーを利用して、本発明のDNAを有し、かつ、ゲノム構造が植物Aに近い植物個体を選抜することができる。さらに、この選抜された植物個体は、必要に応じて、「戻し交雑」(イネ品種Aと交雑)させることができる。
【0107】
特にDNAマーカーを利用したゲノム育種方法では、置換率の高い個体を選抜して次の交雑に進むことができるため、世代を進めるほどに選抜効率が良くなる。また、本方法では、少ない個体数を扱えば済むので、省スペースでの育種が可能になる。さらに、温室や人工気象室を利用して1年に複数回もの交雑が可能になる。
【0108】
上記工程(c)において、DNAマーカーを用いて選抜するとは、当該DNAマーカーを特徴付ける塩基配列(例えば、多型等)についての塩基種の情報を基に、選抜を行うことを言う。例えば、本発明のDNAの近傍に多型変異が存在する場合、当該多型変異と同一の多型変異を有する個体を選抜すること等を言う。
【0109】
本発明の育種方法は、好ましくは、DNAマーカーを利用した「ゲノム育種」方法である。該「ゲノム育種」は「マーカー育種」とも呼ばれる。
【0110】
「ゲノム育種」とは、ゲノム全域に高密度で存在するDNAマーカーを利用し、交雑のたびに後代個体のゲノム全域についての染色体置換状況を図示したものを描写し、最も目的に適った染色体置換を有する個体を幼苗期に選抜して次の交雑に用いることを連続して行う育種方法である。従来の育種法では、次の交雑に用いる個体を選抜するために形質評価を用いていたため、多数の個体収穫期まで成長させる必要があり、1年に1回、あるいは2年に1回の交雑しかできず、また多数の個体を均一に栽培し正しく形質評価するには多大な労力と経験を要した。また、育種過程の最後に、染色体全域をホモ接合体とするために、複数回の自殖が必要であり、結果的に品種の完成までに10年以上を要することが多かった。また、この手法は長年の経験と勘に依存するものである。これに対して、ゲノム育種の戦略を用いれば、交雑して得られた多数の種子を発芽させ、幼苗期に選抜して少数の個体を育てて交雑することを繰り返し、形質評価も最終段階まで不要のため、温室や人工気象室を利用して1年に複数回もの交雑が可能である。さらに、染色体全域がホモ固定しているかどうかは選抜の過程で確認できるため、通常、交雑を終了した時点で品種が完成する。このため、開始から短期間で目的の植物を作出することが可能である。加えて、用いるDNAマーカーの密度を十分に高めることにより、形質転換に匹敵する精度で、目的の遺伝子領域のみを導入することも可能である。形質転換を用いないため、遺伝子組換え作物が受け入れられない国や地域でも問題なく栽培できる植物体が作製できる。
【0111】
上述のように、本発明の育種方法の好ましい態様としては、本発明のDNA配列、またはその周辺配列に存在するDNAマーカーを利用して選抜することを特徴とする方法である。
【0112】
本発明の育種方法において利用可能なDNAマーカーは、特に制限されず、一般的に知られている種々のDNAマーカーを好適に用いることができる。例えば、RFLP(制限酵素断片長多型)マーカー、SSR(単純反復配列)マーカー、SNP(一塩基多型)マーカー等を例示することができる。
【0113】
本発明の植物がイネの場合には、例えば、図21で示される各種マーカーを利用することが可能である。本発明のRcはR1807-R1582の2つのマーカーの間に存在し、E12196Sのマーカーの近傍に存在する。また、本発明のRd(DFR)はR2374とC808の間に存在する。従って、本発明の育種方法において利用可能なマーカーの好ましい具体例としては、これら4種のマーカーを挙げることができる。今回同定したDNA配列中にマーカーを作り出すことももちろん可能である。
【0114】
また、本発明者のRcもしくはRd遺伝子を植物内において発現させることによって、植物体もしくはその種子を着色可能であり、さらに、有色の植物体に内在するこれら遺伝子の発現を抑制することにより、脱色(白色化)された植物体もしくはその種子を作出することも可能である。
【0115】
即ち本発明は、植物に内在する遺伝子であって、植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能を有するタンパク質をコードする遺伝子(本発明のDNAからなる遺伝子)の発現を抑制する工程を含む、脱色された植物体もしくはその種子の製造方法を提供する。
【0116】
該植物体は、本発明のDNAからなる遺伝子の発現が人為的に抑制されていることを特徴とする、遺伝子改変植物である。該遺伝子改変植物は、例えば、「ノックアウト植物」、「ノックダウン植物」あるいは「トランスジェニック植物」と呼ばれる場合もある。
【0117】
本発明において「遺伝子の発現が人為的に抑制されている」には、例えば、(1)本発明のDNAからなる遺伝子における遺伝子対の一方または双方に、ヌクレオチドの挿入、欠失、置換等の遺伝子変異を有することによって該遺伝子の発現が抑制されている状態、(2)本発明の遺伝子の発現を抑制する機能を有する核酸(例えば、アンチセンスRNAまたはsiRNA等)の作用により遺伝子の発現が抑制されている状態、等を挙げることができる。
【0118】
本発明における「抑制」には、本発明の遺伝子の発現が完全に抑制されている場合、および、植物における本発明の遺伝子の発現量が他の植物における遺伝子の発現量と比較して有意に低下している場合等が含まれる。
【0119】
上記(1)には、本発明の遺伝子の遺伝子対の一方の遺伝子の発現のみが抑制されている場合も含まれる。本発明における遺伝子変異の存在する部位は、該遺伝子の発現が抑制されるような部位であれば特に制限されず、例えばエクソン部位、プロモーター部位等を挙げることができる。
【0120】
本発明の遺伝子ノックアウト植物は、当業者においては一般的に公知の遺伝子工学技術により作製することができる。例えば、以下のようにして遺伝子ノックアウト植物を作製することができる。まず、植物から本発明の遺伝子(植物がイネの場合には、RcもしくはRd遺伝子)のエクソン部分を含むDNAを単離し、このDNA断片に適当なマーカー遺伝子を挿入し、ターゲッティングベクターを構築する。このターゲッティングベクターをエレクトロポレーション法、アグロバクテリウム法などにより植物細胞に導入し、相同組み換えを生じた細胞株を選抜する。挿入するマーカー遺伝子としては、一般的な薬剤選択用遺伝子などが好ましい。薬剤選択用遺伝子を挿入した場合には、薬剤を含む培地で培養するだけで相同組み換えを生じた細胞株を選抜することができる。また、PCRおよびサザンブロットにより相同組み換え体の検定を行い、本発明の遺伝子が不活性化された細胞株を効率よく得ることもできる。
【0121】
本発明の上記ノックアウト植物(ノックダウン植物)は、例えば、本発明の遺伝子の発現を抑制する機能を有する核酸を植物へ導入することによって、作出することも可能である。
【0122】
本発明の遺伝子の発現を抑制する機能を有する核酸としては、以下の(a)〜(c)に記載の核酸を例示することができる。
【0123】
(a)本発明のDNAからなる遺伝子の転写産物またはその一部に対するアンチセンス核酸
(b)本発明のDNAからなる遺伝子の転写産物を特異的に開裂するリボザイム活性を有する核酸
(c)本発明のDNAからなる遺伝子の発現をRNAi効果による阻害作用を有する核酸
【0124】
本発明における「核酸」とはRNAまたはDNAを意味する。また、所謂PNA (peptide nucleic acid)等の化学合成核酸アナログも、本発明の核酸に含まれる。PNAは、核酸の基本骨格構造である五単糖・リン酸骨格を、グリシンを単位とするポリアミド骨格に置換したもので、核酸によく似た3次元構造を有する。
【0125】
特定の内在性遺伝子の発現を阻害する方法としては、アンチセンス技術を利用する方法が当業者によく知られている。アンチセンス核酸が標的遺伝子の発現を阻害する作用としては、以下のような複数の要因が存在する。即ち、三重鎖形成による転写開始阻害、RNAポリメラーゼによって局部的に開状ループ構造が作られた部位とのハイブリッド形成による転写阻害、合成の進みつつあるRNAとのハイブリッド形成による転写阻害、イントロンとエクソンとの接合点におけるハイブリッド形成によるスプライシング阻害、スプライソソーム形成部位とのハイブリッド形成によるスプライシング阻害、mRNAとのハイブリッド形成による核から細胞質への移行阻害、キャッピング部位やポリ(A)付加部位とのハイブリッド形成によるスプライシング阻害、翻訳開始因子結合部位とのハイブリッド形成による翻訳開始阻害、開始コドン近傍のリボソーム結合部位とのハイブリッド形成による翻訳阻害、mRNAの翻訳領域やポリソーム結合部位とのハイブリッド形成によるペプチド鎖の伸長阻害、および核酸とタンパク質との相互作用部位とのハイブリッド形成による遺伝子発現阻害などである。このようにアンチセンス核酸は、転写、スプライシングまたは翻訳など様々な過程を阻害することで、標的遺伝子の発現を阻害する(平島および井上, 新生化学実験講座2 核酸IV遺伝子の複製と発現, 日本生化学会編, 東京化学同人, 1993, 319-347.)。
【0126】
本発明で用いられるアンチセンス核酸は、上記のいずれの作用により本発明の遺伝子の発現を阻害してもよい。一つの態様としては、本発明の遺伝子のmRNAの5'端近傍の非翻訳領域に相補的なアンチセンス配列を設計すれば、遺伝子の翻訳阻害に効果的と考えられる。また、コード領域もしくは3'側の非翻訳領域に相補的な配列も使用することができる。このように、本発明の遺伝子の翻訳領域だけでなく非翻訳領域の配列のアンチセンス配列を含む核酸も、本発明で利用されるアンチセンス核酸に含まれる。使用されるアンチセンス核酸は、適当なプロモーターの下流に連結され、好ましくは3'側に転写終結シグナルを含む配列が連結される。このようにして調製された核酸は、公知の方法を用いることで、所望の植物へ形質転換できる。アンチセンス核酸の配列は、形質転換される植物が持つ内在性の本発明の遺伝子またはその一部と相補的な配列であることが好ましいが、遺伝子の発現を有効に抑制できる限りにおいて、完全に相補的でなくてもよい。転写されたRNAは、標的遺伝子の転写産物に対して好ましくは90%以上、最も好ましくは95%以上の相補性を有する。アンチセンス核酸を用いて標的遺伝子の発現を効果的に抑制するには、アンチセンス核酸の長さは少なくとも15塩基以上25塩基未満であることが好ましいが、本発明のアンチセンス核酸は、必ずしもこの長さに限定されない。
【0127】
また、本発明の遺伝子の発現の阻害は、リボザイム、またはリボザイムをコードするDNAを利用して行うことも可能である。リボザイムとは触媒活性を有するRNA分子を指す。リボザイムには種々の活性を有するものが存在するが、中でもRNAを切断する酵素としてのリボザイムに焦点を当てた研究により、RNAを部位特異的に切断するリボザイムの設計が可能となった。
【0128】
また、ヘアピン型リボザイムも本発明の目的に有用である。ヘアピン型リボザイムからも、標的特異的なRNA切断リボザイムを作出できることが示されている(Kikuchi, Y. & Sasaki, N., Nucl Acids Res, 1991, 19, 6751.、菊池洋, 化学と生物, 1992, 30, 112.)。このように、リボザイムを用いて本発明における遺伝子の転写産物を特異的に切断することで、該遺伝子の発現を阻害することができる。
【0129】
内在性遺伝子の発現の阻害は、さらに、標的遺伝子配列と同一もしくは類似した配列を有する二本鎖RNAを用いたRNA干渉(RNA interferance;RNAi)によっても行うことができる。本発明のRNAi効果による阻害作用を有する核酸は、一般的にsiRNAとも言われる。RNAiは、標的遺伝子のmRNAと相同な配列からなるセンスRNAとこれと相補的な配列からなるアンチセンスRNAとからなる二本鎖RNAを細胞等に導入することにより、標的遺伝子mRNAの破壊を誘導し、標的遺伝子の発現を抑制し得る現象である。このようにRNAiは、標的遺伝子の発現を抑制し得ることから、従来の煩雑で効率の低い相同組換えによる遺伝子破壊方法に代わる簡易な遺伝子ノックアウト方法として、または、遺伝子治療への応用可能な方法として注目を集めている。RNAiに用いるRNAは、本発明の遺伝子もしくは該遺伝子の部分領域と必ずしも完全に同一である必要はないが、完全な相同性を有することが好ましい。
【0130】
本発明の上記(c)の核酸の好ましい態様として、本発明の遺伝子に対してRNAi(RNA interference;RNA干渉)効果を有する二本鎖RNA(siRNA)を挙げることができる。より具体的には、配列番号:2または5に記載の塩基配列の部分配列に対するセンスRNAおよびアンチセンスRNAからなる二本鎖RNA(siRNA)を挙げることができる。
【0131】
なお、上記RNA分子において一方の端が閉じた構造の分子、例えば、ヘアピン構造を有するsiRNA(shRNA)も本発明に含まれる。即ち、分子内において二本鎖RNA構造を形成し得る一本鎖RNA分子もまた本発明に含まれる。
【0132】
本発明の上記「RNAi効果により抑制し得る二本鎖RNA」は、当業者においては、該二本鎖RNAの標的となる本発明の遺伝子の塩基配列を基に、適宜作製することができる。一例を示せば、配列番号:2または5に記載の塩基配列をもとに、本発明の二本鎖RNAを作製することができる。即ち、配列番号:2または5に記載の塩基配列をもとに、該配列の転写産物であるmRNAの任意の連続するRNA領域を選択し、この領域に対応する二本鎖RNAを作製することは、当業者においては、通常の試行の範囲内において適宜行い得ることである。また、該配列の転写産物であるmRNA配列から、より強いRNAi効果を有するsiRNA配列を選択することも、当業者においては、公知の方法によって適宜実施することが可能である。また、一方の鎖(例えば、配列番号:2または5に記載の塩基配列)が判明していれば、当業者においては容易に他方の鎖(相補鎖)の塩基配列を知ることができる。siRNAは、当業者においては市販の核酸合成機を用いて適宜作製することが可能である。また、所望のRNAの合成については、一般の合成受託サービスを利用することができる。
【0133】
さらに、本発明の上記RNAを発現し得るDNA(ベクター)もまた、本発明の遺伝子の発現を抑制し得る化合物の好ましい態様に含まれる。例えば、本発明の上記二本鎖RNAを発現し得るDNA(ベクター)は、該二本鎖RNAの一方の鎖をコードするDNA、および該二本鎖RNAの他方の鎖をコードするDNAが、それぞれ発現し得るようにプロモーターと連結した構造を有するDNAである。本発明の上記DNAは、当業者においては、一般的な遺伝子工学技術により、適宜作製することができる。より具体的には、本発明のRNAをコードするDNAを公知の種々の発現ベクターへ適宜挿入することによって、本発明の発現ベクターを作製することが可能である。
【0134】
本発明の上記ノックダウン植物は、上記核酸(アンチセンスRNAまたはsiRNA等)、あるいは該核酸を発現し得る構造のベクターを、所望の植物へ導入することによって作製することができる。
【0135】
また、本発明は、配列番号:1、2、4もしくは5に記載の塩基配列、またはその相補配列に相補的な少なくとも15の連続する塩基を含むポリヌクレオチドを提供する。ここで「相補配列」とは、A:T、G:Cの塩基対からなる2本鎖DNAの一方の鎖の配列に対する他方の鎖の配列を指す。また、「相補的」とは、少なくとも15個の連続したヌクレオチド領域で完全に相補配列である場合に限られず、少なくとも70% 、好ましくは少なくとも80% 、より好ましくは90% 、さらに好ましくは95% 以上の塩基配列の同一性を有すればよい。このようなDNAは、本発明のDNAの検出や、本発明のDNAを有する植物体(植物細胞)の選抜を行なうためのプローブとして、また、本発明のDNAの増幅を行うためのプライマーとして有用である。
【0136】
また本発明は、本発明の植物体もしくはその種子の製造方法、またはこれらの育種方法に用いるためのオリゴヌクレオチドを含む、試薬(例えば、植物体育種用試薬等)を提供する。
【0137】
本発明の試薬は、より具体的には、以下の(a)および(b)のオリゴヌクレオチドを含有する試薬である。
(a)本発明のDNAの全配列もしくはその一部の配列を増幅するためのオリゴヌクレオチドプライマー
(b)本発明のDNA領域とストリンジェントな条件でハイブリダイズし、少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドプローブ
【0138】
本発明のプライマー、またはプローブは、それを構成する塩基配列をもとに、任意の方法によって合成することができる。本発明のプライマーまたはプローブの、ゲノムDNAに相補的な塩基配列の長さは、通常15〜100、一般に15〜50、好ましくは15〜30である。与えられた塩基配列に基づいて、当該塩基配列を有するオリゴヌクレオチドを合成する手法は公知である。更に、オリゴヌクレオチドの合成において、蛍光色素やビオチンなどで修飾されたヌクレオチド誘導体を利用して、オリゴヌクレオチドに任意の修飾を導入することもできる。あるいは、合成されたオリゴヌクレオチドに、蛍光色素などを結合する方法も公知である。
【0139】
また本発明は、本発明の各種方法に使用するためのキット、例えば、本発明の植物育種方法に使用するための育種用キットを提供する。
【0140】
本発明のキットの好ましい態様においては、上記(a)または(b)の少なくとも1種のオリゴヌクレオチドを含む。本発明のキットには、適宜、陽性や陰性の標準試料、使用方法を記載した指示書等をパッケージしておくこともできる。
【0141】
本発明のDNAもしくはベクターは、例えば、着色された植物体、もしくは着色された種子の作出等に利用することが可能である。また、本発明のDNAもしくはベクターは、タンニン類が蓄積した植物体、もしくはタンニン類が蓄積した種子の作出に利用することも可能である。
【0142】
本発明のDNAもしくはベクターを、所望の植物体もしくはその種子において発現させることにより、着色された、あるいはタンニン類が蓄積した植物体もしくはその種子を作製することが可能である。
【0143】
従って本発明は、本発明のDNAもしくはベクターを有効成分とする、植物体着色剤を提供する。
【0144】
本発明における「植物体着色剤」とは、植物体もしくはその種子の全部またはその一部を着色させる作用を有する薬剤を言い、通常、植物体もしくはその種子を着色させることを用途とする、本発明のDNAもしくはベクターを有効成分とする物質、または組成物(混合物)を指す。
【0145】
また本発明は、本発明のDNAもしくはベクターを有効成分とする、タンニン類蓄積剤を提供する。
【0146】
本発明における「タンニン類蓄積剤」とは、植物体もしくはその種子の全部またはその一部へタンニン類を蓄積させる作用を有する薬剤を言い、通常、植物体もしくはその種子へタンニン類を蓄積させることを用途とする、本発明のDNAもしくはベクターを有効成分とする物質、または組成物(混合物)を指す。
【0147】
さらに本発明は、上記(a)〜(c)に記載の核酸を有効成分とする、植物体脱色化剤を提供する。
【0148】
本発明における「植物体脱色化剤」とは、植物体もしくはその種子の全部またはその一部を脱色させる作用を有する薬剤を言い、通常、植物体もしくはその種子を脱色させることを用途とする、本発明の上記核酸を有効成分とする物質、または組成物(混合物)を指す。
【0149】
上記「脱色」とは、例えば、赤色、赤褐色、褐色等の色から白色へ変化させること等を言う。植物がイネの場合には、その種子であるコメの色が、例えば、赤米から白米へ変化させること等を指す。
【0150】
本発明の薬剤においては、有効成分であるDNAまたはベクター以外に、例えば、滅菌水、生理食塩水、植物油、界面活性剤、脂質、溶解補助剤、緩衝剤、保存剤等が必要に応じて混合されていてもよい。
【0151】
また本発明は、本発明の上記製造方法、または育種方法によって作出される植物体もしくはその種子を提供する。
例えば、(1)人為的に作製された植物体もしくはその種子であって、本発明のDNAを有し着色されていることを特徴とする植物体もしくはその種子、(2)人為的に作製された植物体もしくはその種子であって、本発明のDNAを有しタンニン類が蓄積していることを特徴とする植物体もしくはその種子、(3)植物に内在する遺伝子であって、植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能を有するタンパク質をコードする遺伝子の発現が人為的に抑制され、脱色されていることを特徴とする植物体もしくはその種子、は本発明に含まれる。
【実施例】
【0152】
以下、本発明を実施例によりさらに具体的に説明するが本発明はこれら実施例に制限されるものではない。なお、遺伝子型の記号、学術用語はKinoshita and Takahashiの文献(Kinoshita, T. & Takahashi, M. (1991) J. Fac. Agr. Hokkaido Univ. 65, 1-61.)とKinoshitaの文献(Kinoshita, T. (1995) Rice Genetics Newsletter 12, 9-153.)を参考にした。
【0153】
〔実施例1〕 実験材料と方法
(a)資料植物及び核酸の単離
イネ日本晴品種、カサラス、コシヒカリ、tRc(NIL)、ムラサキイネ、及び形質転換イネは28℃の温室で栽培した。tRcはCI-69(赤米)にT-65を交雑させることによって作成した準同質遺伝系統(第7染色体のT-65(遺伝子型rcrd)が、第7染色体のCI-69(遺伝子型RcRd)に置換された準同質遺伝系統)である。ゲノムDNAは、発芽後10日目の若い葉から抽出した。DNA抽出には、DNeasy plant mini kit(Qiagen)を用い、キットに記載されているプロトコルに従った。葉のRNAは、発芽後10日目の若い葉から抽出した。穂のRNAは、開花後7日目の穂から抽出した。RNA抽出は、RNeasy plant mini kit(Qiagen)を用い、キットに記載されている方法に従った。
【0154】
(b)PCR,DNA 配列
OsDFRのゲノムDNAは、1UのLA Taq DNA polymerase(TaKaRa)によって増幅された。PCRに用いたプライマーは、OsDFR遺伝子の翻訳開始コドン1.7 kb上流のOsDFRF4(5'-CATGTGTGAGCATCTCTAAGTGACG-3'/配列番号:9)とDFR遺伝子の翻訳終止コドン0.5 kb 下流のOsDFRR4(5'-CTCTCTAGGAGCACGTGTAAAGG-3'/配列番号:10)を用いた(図4)。DFRのPCR産物のシーケンスは、CEQ Dye Terminator Cycle Sequencing Start KitおよびCEQ 2000XL DNA Analysis System(Beqman Coulter)を用い、これに記載されている方法に従った。シーケンスに用いたプライマーは、OsDFR遺伝子の第2エクソンに相当するOsDFRR5(5'-AGAAGTCGATGTCGCTCCAG-3'/配列番号:11)を用いた(図4)。
【0155】
(c)RT-PCR
PCRの基質に用いたcDNAの合成はSuperScriptTM 逆転写酵素とRandom Primers (Invitrogen)を用い、これに記載されている方法に従った。PCRはAmpli taq(Applied Biosystems)を用いて、94℃ 1分静置してのち、94℃15秒、60℃30秒、72℃30秒のサイクルを40回繰り返した。使用したプライマーは、OsDFR遺伝子の第1エクソンに対応するOsDFRF8(5'-GATTCGCGTGCGAATCCAAC-3'/配列番号:12)、及びOsDFR遺伝子の第2エクソンに対応するOsDFRR5(5'-AGAAGTCGATGTCGCTCCAG-3'/配列番号:13)を用いた(図5)。イネアクチン遺伝子(X15865)の転写産物の増幅には、イネアクチン遺伝子の第2エクソンに対応するRAc1cDNA-F プライマー(5'-TGTCATGGTCGGAATGGG-3'/配列番号:14)、及びイネアクチン遺伝子の第3エクソンに対応するRAc1cDNA-R プライマー(5'-TGCCAGGGAACATAGTGG -3'/配列番号:15)を用いた。PCR産物は1.8 % (w/v)のアガロースゲル電気泳動により分離・同定した。
【0156】
(d)タンパク質ブロット解析
穂の全タンパク質は、抽出Buffer(200 mM Tris-HCL.pH 6.8; 8 %(w/v) SDS; 40 % Glycerol; 5 mM 2-mercaptoethanol)で抽出した。抽出したサンプルは、4,000×gで5分間、4℃で遠心し、上静をSDS-PSGE running Bufferに溶かした。タンパクは12 %のポリアクリルアミドゲルで分離し、Immobillon PVDF membraneに転写した。
【0157】
DFRに対する抗血清は、ムラサキイネのOsDFRのORF全長に相当するcDNAをPCRにより増幅し、pGEX4T-3 vector(Amersham Pharmacia Biotech)に連結し、E.coli JM109株に遺伝子導入した(図6)。GST/DFRの融合タンパク質は、1 mMのIPTGを加えることによって誘導し、(Asano, T., Kusano, H., Okuda, T., Kubo, N., Shimada, H. & Kadowaki, K. (2001) Plant Cell Physial 43, 668-674.)に記載されている方法に従って精製した。精製された融合タンパク質は、2週間おきに6回ウサギに注射し、その血清を解析に用いた。
【0158】
(e)Rcのゲノムアノテーション
Rcのゲノムアノテーションによる候補の探索は、Rice GAAS(Rice Genome Automated Annotation System){ http://ricegaas.dna.affrc.go.jp/}を用いて行った。
【0159】
(f)形質転換植物の産出
ムラサキイネ型DFR、コシヒカリ型DFRの過剰発現体の作成についてはムラサキイネ、コシヒカリのOsDFRのORF全長に相当するゲノム領域を、インフレームでpCAMBIA1301 plasmidの35S promoterの下流にセンス方向に連結した(図7、図8)。またDFRのアンチセンス個体の作成についてはOsDFR cDNAのORFに相当する領域を、pCAMBIA1301 plasmidの35S promoterの下流にアンチセンス方向に連結した(図9)。ゲノムアノテーションにより得られたRcの3つの候補(Rc-MYB1、Rc-MYB2、Rc-MYC)は、予想されるプロモーター領域1.5 kbから予想されるポリA付加シグナルまでの約12 kbをpCAMBIA1301 plasmidのマルチクローニングサイトに連結した(図10)。Rc-MYB1、Rc-MYB2、Rc-MYCの遺伝子構築に使用したのは、それぞれカサラス由来の105659−107804 (AP005197)、 118440−126019 (AP003754)、 150211−156639 (AP005779) である。作成したプラスミドは、Agrobacterium tumefaciens strain EHA105に遺伝子導入した。アグロバクテリウム法によるイネの形質転換(DFRの形質転換は、tRc(NIL)に対して行い、Rcの形質転換は日本晴に対して行った)は、Patent No. WO01/06844に記載されている方法に従った。得られた形質転換植物は、温室で育てた。
【0160】
〔実施例2〕 Rd遺伝子の同定
(1)RcRd、Rcrd、rcRd及びrcrd遺伝子型の表現型
イネにおいて、タンニン類(CTs)による種皮の着色には、RcとRdの2つの支配因子が存在する(Nagao, S. (1951) Advances in Genetics 4, 181-212.)。Rcは、種皮の赤色及び赤褐色の着色に必須な因子である。Rdは、Rcが存在するときにのみRcと協調的に機能し、種皮の赤色の着色に必須な因子である。交雑実験の結果、RcRdの遺伝子型の玄米色は赤、Rcrdの玄米色は赤褐色(くぼみがあり、そこに濃い着色が見られる)、rcRd またrcrdの玄米色は白色になることが知られている(図3)。
【0161】
(2)DFRを伴うRd遺伝子座の分離
RcRdの玄米色は赤、Rcrdの玄米色は赤褐色、rcRd,rcrdの玄米色は白となることが遺伝学的解析により知られている(Nagao, S. & Takahashi, M. (1947) Jap. J. Genet. Supp. 1, 1-27.)。またRcRd、Rcrdの玄米には主要な色素であるプロアントシアニジンが存在することがNagaoら(Nagao, S., Takahashi, M. & Miyamoto, T. (1956) The Japanese Jourant of Genetics 32, 124-128.)によって確認された。イネの玄米の色調に影響を及ぼすことから、Rdはプロアントシアニジン合成に関与する遺伝子座であり、対立遺伝子であるrdは機能を欠失しているわけではなく、Rdに比べてプロアントシアニジン合成系における機能が弱いことが考えられる。ここでKinoshitaら(Kinoshita, T. & Takahashi, M. (1991) J. Fac. Agr. Hokkaido Univ. 65, 1-61.)の報告から、Rd座はA座の近傍に存在し、それは0.3 cM以内である(図2)。
【0162】
A座はイネのアントシアニン合成系のActivatorとして定義された遺伝子であって、現在までにA座がコードする遺伝子については明らかになっていない。イネのアントシアニンによる着色には色原素遺伝子であるC(Chromogen)、アントシアニン活性化因子であるA(Activator)、そして着色部位を規定するP(Purple)の3つの遺伝子が存在する。これら3つの因子が存在するときにイネの組織(玄米、葉、ラミナジョイント、茎、etc)にアントシアニンによる着色が起こる(表1)。
【0163】
【表1】

(*Nagaoら(Nagao, S. & Takahashi, M. (1963) Agr., Hokkaido Univ., Sapporo 53, 76-131.)およびKatoら(Kato, S., & Ishikawa. J. (1923) Jap. J. Genet. 1, 1-7.)によって決定された遺伝子座は、独立した研究であり、比較できない。)
【0164】
ここでCはトウモロコシのC1(MYB)のホモログであることが知られており(Reddy, V. S., Scheffler, B. E., Wienand, U., Wessler, S. R. & Reddy, A. R. (1998) Plant Molecular Biology 36, 497-498.; Saitoh, K., Onishi, K., Mikami, I., Thidar, K. & Sano, Y. (2004) Genetics 168, 997-1007.)、PはトウモロコシのR/B(bHLH)のホモログであることが近年明らかになった(Hu, J., Anderson, B. & Wessler, R. S. (1996) Genetics 142, 1021-1031.; Hu, J., Reddy, S. V. & Wessler, R. S. (2000) Plant Molecular Biology 42, 667-678.; Sakamoto, W., Ohmori., T., Kageyama, K., Miyazaki, C., Saito, A., Murata, M., Noda, K. & Maekawa, M. (2001) Plant Cell Physiol. 42, 982-991.)(表2)。
【0165】
【表2】
【0166】
このことはイネのA遺伝子がDFRをコードすることを示している。
【0167】
RdはA(DFR)の近傍に存在する(Nagao, S. & Takahashi, M. (1947) Jap. J. Genet. Supp. 1, 1-27.)。A(DFR)が存在するR2374(103.7 cM) −C808(106.2 cM)の間に存在する遺伝子をRice GAAS programを用いて解析した結果、この領域にDFR以外にタンニン合成に関与する遺伝子は存在しなかった。よってRdもAと同様にDFRをコードすることが示唆された。
【0168】
上述の結果から、Rd遺伝子とA遺伝子は同一であり、これらの遺伝子は同一のDFRタンパク質をコードしているという仮説が考えられた。この仮説を検証するために、準同質遺伝系統T-65(RcRd, A)とtRc系統(Rcrd, a)との間で、交配実験を行った。もし、A遺伝子とRd遺伝子が同一でないならば、AとRdとの間に起こる組換え体がF2において観察される。該組換え体は(Rcrd, A)、または(RcRd, A)の遺伝子型を有する。
【0169】
A遺伝子は、ふ先(apiculus)におけるアントシアニン色素の原因遺伝子であることから、(Rcrd, A)を有する植物は、種子の果皮が赤褐色となりアントシアニンによって着色されたふ先を有する。一方、(RcRd, a)を有する植物は、種子の果皮が赤色となり非着色のふ先を有する。
【0170】
実験の結果、F2において、672個体が種子の果皮が赤色となり、(RcRd, A)によるアントシアニンによって着色されたふ先が観察された。また232個体は種子の果皮が赤褐色となり、(Rcrd, a)による非着色のふ先が観察された。RdとAとの間の組換え体は観察されなかった。このことから、RdとAは遺伝学的に強く連鎖し、同一の遺伝子であることが示唆された。
【0171】
(3)Rcrd とゲノムOsDFRの相補性
tRc(Rcrd)にムラサキイネ由来のDFRを相補した結果、玄米色が赤褐から赤になった。RcRdの玄米色が赤であることを考えると、RdはDFRをコードすることが明らかになった(図11)。
【0172】
(4)DFRアレルの解析
Rd、rdの遺伝子型による種皮色の違いとDFR遺伝子の構造の関係を探るためにDNAシーケンスを行った。解析に用いたものはRcRdの遺伝子型を持つカサラス、Rcrd の遺伝子型を持つtRc、rcrdの遺伝子型を持つ日本晴とコシヒカリである。DFRの第1エクソンから第2エクソンまでの領域をgenomic DNAをテンプレートとしてPCRで増幅し、ダイレクトシーケンスを行った。その結果Rdを持つカサラスは、以前に単離されているイネのDFR遺伝子と同様な構造をとっていた(Chen, M. & L. J. Bennetzen (1996) Plant Molecular Biology 32, 999-1001.; Nakai, K., Inagaki, Y., Nagata, H., Miyazaki, C., & Iida, S. (1998) Plant Biotechnology 15, 221-225.)。一方、rdを持つtRcや日本晴のDFR遺伝子の構造には、第2エクソンに突然変異によって出来たと思われる翻訳終止コドン(TAG)が存在した。さらにrdを持つと思われていたコシヒカリのDFRの構造は、カサラスとも日本晴れとも異なり第1エクソンの開始コドンのすぐ下流に突然変異によってできたと思われる翻訳終止コドン(TAG)が存在した(図12)。以上のことから、RdはカサラスタイプのDFRをコードし、rdは第1および第2エクソンに翻訳終止コドンの存在するコシヒカリ及び日本晴タイプのDFRをコードすることが明らかになった。
【0173】
rdのDFR遺伝子の構造は、第2エクソンに終止コドンが存在しており、開始コドンから突然変異によって出来た翻訳終止コドンまでは、OsDFRが42 kDaをコードするのに対し4.5 kDaであり、機能を持っているとは考えにくい。またDFRはイネに1コピーしか存在しない(Nakai, K., Inagaki, Y., Nagata, H., Miyazaki, C., & Iida, S. (1998) Plant Biotechnology 15, 221-225.; International Rice Genome Sequence Project. (2005) Nature 436, 793-800.)。しかしRcrd のDFR遺伝子は日本晴タイプであり、ORFのN末側が欠損しているが、玄米色は赤褐色で、プロアントシアニジンによる着色であることが知られている(Nagao, S., Takahashi, M. & Miyamoto, T. (1956) The Japanese Jourant of Genetics 32, 124-128.)。従って、第2エクソンに終止コドンを持つ日本晴タイプのDFRに機能があることが考えられた。そこで、Rdの対立遺伝子であるDFRが転写しているか調べるために、RcRdの遺伝子型を持つカサラス、rcrdの遺伝子型を持つ日本晴、rcrd’の遺伝子型を持つコシヒカリに対して、RT-PCRを行った。その結果、カサラス、tRc、日本晴、コシヒカリとも開花後7日目の穂で特異的に発現しており、その発現レベルの強さは、カサラス>日本晴>コシヒカリ であった(図13)。
【0174】
次に本発明者らは、Rdの対立遺伝子であるDFRの翻訳の機構を調べるために、RcRdの遺伝子型を持つカサラス、rcrdの遺伝子型を持つ日本晴、rcrdの遺伝子型を持つコシヒカリに対して、開花後7日目の穂において免疫染色を行った。その結果驚くことに、カサラスは、Alternative translation initiation(Bab, I., Smith, E., Gavish, H., Malka, Attar-Namdar., Chorev, M., Yu-Chen, Chen., Muhlrad, A., Birnbaum, J. M., Stein, G. & Frenkel, B. (1999) The Journal OF BIOLOGICAL CHEMISTRY 274, 14474-14481.; Chabregas, M. S., Luche, D. D., Marie-Anne, Van, Sluys., Carlos, F. M. Menck., Marcio, C. Silva-Filho. (2002) Journal of Cell Science 116, 285-291.; Smith, E., Meyerrose, E. T., Kohler, T., Malka, Namdar-Attar., Bab, N., Lahat, O., Noh, T., Li, J., Karaman, W. M., Hacia, G. J., Chen, T. T., Nolta, A. J., Muller, R., Bab, I. & Frenkel, B. (2005) Nucleic Acids Research 33, 1298-1308.; Hanfrey, C., Elliott, A. K., Franceschetti, M., Mayer, J. M., Illingworth, C. & Michael, J. A. (2005) The Journal OF BIOLOGICAL CHEMISTRY, in press.; Kabeya, Y. & Sato, N. (2005) Plant Physiology 138, 369-382.)により46 kDa、33 kDa、22 kDaの3つのORFが存在した。また日本晴、コシヒカリにおいても33 kDa、22 kDaの2つのORFが存在した。この46 kDaのバンドは、予想されたタンパク質の大きさよりも大きいが、DFRのN末端側に塩基性のアミノ酸が多いため、バンドがシフトしていると考えられた(図4)。
【0175】
ここでOsDFRのORFには、最初の開始コドンを含め5個のKozak コンセンサス配列が存在する(Kozak, M. (2000) Genomics 70, 396-406.; Kozak, M. (2002) Gene 299, 1-34.)。46 kDa、22 kDaのバンドは比較的強く検出されているが、これは最も強いKozak コンセンサス配列を持つ GnnATGG (nはA,T,G,Cいずれかの塩基)を持つ、一番目と4番目のKozak コンセンサスだと考えられた。33 kDaのバンドの由来については、分子量の大きさから考えて、第2エクソンのKozak配列以外のATGから翻訳が開始されていると推察された(図4)。
【0176】
以上の結果より、rd型のDFRはAlternative translation initiation(Kozak, M. (2000) Genomics 70, 396-406.; Kozak, M. (2002) Gene 299, 1-34.)により下流のATGから翻訳され、DFRの機能を相補していることが示された。以上の結果よりrcrdの遺伝子型を持つ日本晴にRcを相補すれば、タンニン類による着色が起こることが示唆された。
【0177】
〔実施例3〕 Rc遺伝子の同定
(1)Rcのゲノムアノテーション
現在までに遺伝学的解析により、Rcは第7染色体に存在することが知られている(Nagao, S. & Takahashi, M. (1963) Agr., Hokkaido Univ., Sapporo 53, 76-131.)(図15A)。またイネゲノムプロジェクトの終了によりRcは、第7染色体上のR1807(42.6 cM)とR1582(47.7 cM)の二つの分子マーカーの間に存在することが分かった(IRGSP, 2005)(図5B)。この領域に存在するプロアントシアニジン合成系(図1)の酵素をRiceGAAS(http://ricegaas.dna.affrc.go.jp/)を用いて検索した。しかしこの領域にプロアントシアニジン合成系(図1)の酵素に対して、ホモロジーが高いものは存在しなかった。
【0178】
そこで次に、プロアントシアニジン合成系の転写制御因子に着目した。プロアントシアニジン合成系の転写制御因子としては、MYC type proteinをコードしているアラビドプシスのTT8(Transparent Testa 8)(Nesi, N., Debeaujon, I., Jond, C., Pelletier, G., Caboche, M. & Lepiniec, L. (2001) Plant Cell 12, 1863-1878.)、MYB related proteinをコードするアラビドプシスのTT2(Nesi, N., Jond, C., Debeaujon, I., Caboche, M. & Lepiniec, L. (2001) Plant Cell 13, 2099-2114.)などが知られている。そこでRcが存在する第7染色体上のR1807(42.6 cM)とR1582(47.7 cM)に存在するMYC type protein、MYB related proteinをコードする遺伝子についてRiceGAASを用いて検索した。
【0179】
その結果、MYB related proteinが2つ、MYC type proteinが1つ見つかった。MYB related proteinをコードする遺伝子の一つはRc-MYB1と名付け、IRGSPのAP005197の105659 bp - 107804 bpにORFが存在した。もう一つのMYB related proteinはRc-MYB2と名付け、IRGSPのAP003754の 118440 bp - 126019 bpにORFが存在した。またMYC type proteinはRc-MYCと名付け、そのORFはIRGSPのAP005779の150211 bp - 156639 bpに存在した(図15A,B,C)。Rc-MYB1とRc-MYB2については、アラビドプシスのプロアントシアニジン合成系の転写制御因子であるTT2(MYB related protein)との相同性はともに24 %(Identities)であった。一方Rc-MYCについては、アラビドプシスのプロアントシアニジン合成系の転写因子であるTT8(MYC type protein)との相同性が38 %(Identities)、その中の一部分のbHLHドメインでは相同性は高く73 %(Identities)であった(図15D)(ClustalW ver.1.82)。
【0180】
(2)rcrdとゲノム断片の相補性
RcRdの遺伝子型を持つカサラス(RcRd)のRc-MYB1、Rc-MYB2、Rc-MYC(図15)についてプロモーター領域である翻訳開始コドン上流1.5 kbからポリA付加シグナルまでの約12 kbをrcrdの遺伝子型を持つ日本晴(rcrd)に遺伝子導入した。それぞれ16個体の形質転換体が得られた。Rc-MYB1、Rc-MYB2を遺伝子導入した個体については葉、もみ、玄米ともプロアントシアニジンによる着色は見られなかった。一方、Rc-MYCを遺伝子導入した個体は、玄米にプロアントシアニジンによると思われる着色が見られた(図16)。着色が見られた形質転換体は10個体得られ、その表現型はRcrdの遺伝子型を持つtRcと非常に類似していた。
【0181】
以上の結果より、Rc-MYCがプロアントシアニジン合成系支配因子であるRcであることが示された。玄米以外の組織では着色は見られなかった。以上のことから、Rcは種子特異的に働く転写因子であることが示された。
【0182】
(3)Rc領域の解析
次に本発明者らはカサラス(RcRd)のRc-MYCの完全長cDNA配列を決定した。使用したRNAは開花後7日目の穂から抽出した。完全長のカサラスのRc-MYCの配列を決定した結果、カサラスのRc-MYCは673 a.a.のタンパク質をコードする8個のエクソンからなる遺伝子であることが分かった。
【0183】
次に本発明者らは、カサラス(RcRd)のRc-MYCのDNA配列と日本晴(rcrd)の遺伝子配列を比較してみることにした。その結果、rcを持つ日本晴には第7エクソンに14 bpの欠失変異、さらに第8エクソンに6 bpの挿入変異と9 bpの欠失変異が確認できた。日本晴の第7エクソンの14 bpの欠失変異は、フレームシフトを起こし、第7エクソンの欠失場所の9 bp下流に終止コドンを生じさせていた。その結果、日本晴のRc-MYCは478 a.a. のタンパク質をコードしており、カサラスのRc-MYCに比べて215 a.a. だけ短かった。DNAの結合に必要な bHLHドメインは第8エクソンにコードされている。よって日本晴のRc-MYCは第8エクソンまで欠失変異のため翻訳されないので、bHLHドメインを持たないことになる。そのために日本晴Rc-MYCは転写制御因子として機能できないと考えられた(図18)。
【0184】
Rc-MYCの第7エクソンの14 bp 欠失している領域について、シーケンスを用いた多型解析をRcRd(玄米色は赤)、Rcrd(玄米色は赤褐)、rcrd(玄米色は白)について行った。図19は、玄米色が赤色のRcRd、玄米色が赤褐色のRcrdには第7エクソンに欠失がなく、玄米色が白色のrcrdには第7エクソンに欠失があることを示している。図19の結果はプロアントシアニジンによる玄米の着色はRc-MYCによることを示唆している。
【0185】
遺伝学的解析、逆遺伝学的解析は日本晴の玄米色がRc-MYCの機能が欠失したために、白色になったことを明確に示した。
【0186】
(4)MYC型タンパク質の相同性解析
次にRc-MYCとそのホモログの遺伝距離を調べることにした。アミノ酸配列のアラインメントはClastalX ver1.83を用い、系統樹はclastalX上のNJ法を使用した。またBoots strap valueもClastalX上の機能を使用した。系統解析に使用したのは、トウモロコシのIN1(U57899), Lc(M26227), B-Peru(X57276), R-S(X15806), ペチュニアのJAF13(AF020545), キンギョソウのDELILA(M84913), アラビドプシスのTT8(AJ277509), イネのRa(U39860), Rc-MYC(this study)を用いた。系統解析の結果、イネのRc-MYCは、トウモロコシのアントシアニン合成系抑制因子として単離されているIntensifier(IN1)と最も相同性が高かった。しかしながら、Rc-MYCはタンニン類蓄積促進因子であるが、IN1はその遺伝子機能が失われるとタンニン類が蓄積するため、機能は正反対である。一方、アントシアニン合成系活性化因子として単離されたキンギョソウのDELILAやペチュニアのJAF13とは最も相同性が低かった(図20)。
【技術分野】
【0001】
本発明は、植物体もしくはその種子の着色を制御する遺伝子、および、該遺伝子を発現し着色した植物体(形質転換植物体)もしくはその種子の製造方法、該遺伝子(DNA配列)を利用して選抜育種される植物体及び種子、さらに、該遺伝子の発現を改変することにより、種子の色を制御する方法に関する。
【背景技術】
【0002】
赤色あるいは赤褐色を呈する赤米といわれるイネ種子には、カテキン、タンニンなどが蓄積することが知られているが、約50年以上前にその染色体上の遺伝子座が同定されているものの、未だに遺伝子レベルでの決定はなされていない。
なお、本出願の発明に関連する先行技術文献情報を以下に示す。
【先行技術文献】
【非特許文献】
【0003】
【非特許文献1】Kinoshita, T. & Takahashi, M. (1991) J. Fac. Agr. Hokkaido Univ. 65, 1-61.
【非特許文献2】Kinoshita, T. (1995) Rice Genetics Newsletter 12, 9-153.
【非特許文献3】Chen, M. & L. J. Bennetzen (1996) Plant Molecular Biology 32, 999-1001.
【非特許文献4】Nakai, K., Inagaki, Y., Nagata, H., Miyazaki, C., & Iida, S. (1998) Plant Biotechnology 15, 221-225.
【発明の概要】
【発明が解決しようとする課題】
【0004】
植物体の着色を制御する遺伝子の提供を課題とする。より詳しくは、植物体もしくはその種子の着色を制御する遺伝子、および、該遺伝子を発現し種子が着色した植物体(形質転換植物体)の製造方法、該遺伝子(DNA配列)を利用して選抜育種される植物体及び種子、さらに、該遺伝子の発現を改変することにより、植物体もしくはその種子の色を制御する方法の提供を課題とする。
【課題を解決するための手段】
【0005】
本発明者らは、上記の課題を解決するために、イネの赤米色素着色に関与するRcとRd遺伝子の同定を試みた。
【0006】
Rc遺伝子については、本発明者らによって同定されたいくつかの候補遺伝子を逆遺伝学的な手法で日本晴に導入し、表現型を白色から赤褐色に改変させる機能を有する遺伝子を原因遺伝子として特定した。その構造的な特徴からRc遺伝子はbHLHドメインを持つ転写因子であるMYC遺伝子であることが初めて明らかとなった。本発明者らによって同定された遺伝子の機能は、これまでのところ明らかになっておらず、MYC遺伝子が赤米関係の色素発現に関与していることが本発明者らによって初めて証明された。
【0007】
Rc遺伝子が存在すると考えられた染色体上の遺伝子座の領域は、非常に広範な領域(2.3Mbp)であり、その中から実際に植物体の着色を制御する原因遺伝子を同定することは非常に困難であった。即ち、上記領域の中において、アミノ酸が100個以上の長さのタンパク質をコードすると考えられる遺伝子の数は200以上にものぼり、その中から、着色を制御する原因遺伝子を同定することは極めて困難なことである。該原因遺伝子を同定することは、当業者であっても過度の困難性を伴うことであり、本発明者らによって原因遺伝子が実際に同定されたことは非常に大きな成果と言える。
【0008】
本発明者らによって同定された遺伝子のRc(MYC)遺伝子は、これまでのところ機能が全く知られておらず、本発明者らによって植物体の着色を制御する機能を有することが初めて見出された。
【0009】
またRd遺伝子は、本発明者らによる鋭意研究の結果、遺伝子DFRであることが分った。DFRの遺伝子構造については3種の多型が認められた。本発明者らはDFR抗体を作成し、タンパク質レベルでの発現を解析したところ、種子特異的なalternative translation initiationにより複数のペプチドとして翻訳されていることを初めて証明した。
【0010】
さらに本発明者らは、Rdが酵素タンパク質(DFR)をコードしており、その着色に関わる発現はRcである調節遺伝子MYCにより制御されていることを見出した。DFRタンパク質の翻訳は種子特異的なalternative translation initiationにより制御されている。その結果、全長のDFRを有するイネでは種子が赤色を呈し、DFRのORFが短い構造のイネにおいては種子が赤褐色を呈すること、さらにRcが機能しない場合は種子が白色を呈することが新たに見出された。
【0011】
より詳細な解析により、rd型においては、DFR遺伝子からalternative translation initiationにより下流から翻訳され、通常の46 kDaの翻訳産物ではなく、33 kDaおよび22 kDaの翻訳産物が生成されることが示された。さらに、alternative translation initiation機構によって翻訳される33 kDaおよび22 kDaの産物が、DFRの機能を相補し得ることを、本発明者らは初めて見出した。即ち、通常の全長DFRタンパク質に加え、より分子量の小さい翻訳産物についても、植物体を着色させる機能、もしくはタンニン類を蓄積させる機能を有することが判明した。
【0012】
上述の如く本発明者らは、植物体もしくはその種子の着色を制御する遺伝子を単離することに成功し本発明を完成させた。該遺伝子を利用することにより、植物体、特に種子の着色を制御することが可能である。
【0013】
本発明は、植物体もしくはその種子の着色を制御する遺伝子、および、該遺伝子を発現し種子が着色した植物体(形質転換植物体)の製造方法、該遺伝子(DNA配列)を利用して選抜育種される植物体及び種子、さらに、該遺伝子の発現を改変することにより、植物体、特に種子の色を制御する方法に関し、より具体的には、
〔1〕 植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能を有する植物由来のタンパク質をコードする、下記(a)〜(e)のいずれかに記載のDNA、
(a)配列番号:3または6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:1または4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:2または5に記載の塩基配列からなるDNA
(d)配列番号:3または6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:1または4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
〔2〕 植物が単子葉植物である、〔1〕に記載のDNA、
〔3〕 単子葉植物がイネである、〔2〕に記載のDNA、
〔4〕 〔1〕〜〔3〕のいずれかに記載のDNAによりコードされるタンパク質、
〔5〕 〔1〕〜〔3〕のいずれかに記載のDNAを含むベクター、
〔6〕 〔5〕に記載のベクターが導入された宿主細胞、
〔7〕 〔5〕に記載のベクターが導入された植物細胞、
〔8〕 〔7〕に記載の植物細胞を含む形質転換植物体、
〔9〕 〔8〕に記載の形質転換植物体の子孫またはクローンである、形質転換植物体、
〔10〕 〔8〕または〔9〕に記載の形質転換植物体の繁殖材料、
〔11〕 〔6〕に記載の宿主細胞を培養し、該細胞またはその培養上清から組換えタンパク質を回収する工程を含む、〔4〕に記載のタンパク質の製造方法、
〔12〕 〔4〕に記載のタンパク質に結合する抗体、
〔13〕 〔1〕〜〔3〕のいずれかに記載のDNAの塩基配列またはその相補配列に相補的な少なくとも15の連続する塩基を含むポリヌクレオチド、
〔14〕 下記(a)〜(c)のいずれかを有効成分として含有する、植物体着色化剤、
(a)〔1〕〜〔3〕のいずれかに記載のDNA
(b)〔4〕に記載のタンパク質
(c)〔5〕に記載のベクター
〔15〕 下記(a)〜(c)のいずれかを有効成分として含有する、タンニン類蓄積剤、
(a)〔1〕〜〔3〕のいずれかに記載のDNA
(b)〔4〕に記載のタンパク質
(c)〔5〕に記載のベクター
〔16〕 下記(a)〜(c)のいずれかを有効成分として含有する、植物体脱色化剤、
(a)〔1〕〜〔3〕のいずれかに記載のDNAの転写産物またはその一部に対するアンチセンス核酸
(b)〔1〕〜〔3〕のいずれかに記載のDNAの転写産物を特異的に開裂するリボザイム活性を有する核酸
(c)〔1〕〜〔3〕のいずれかに記載のDNAの発現をRNAi効果による阻害作用を有する核酸
〔17〕 〔1〕〜〔3〕のいずれかに記載のDNA、または、〔5〕に記載のベクターを植物細胞へ導入し、該植物細胞から植物体を再生させる工程を含む、形質転換植物体の製造方法、
〔18〕 〔1〕〜〔3〕のいずれかに記載のDNAを植物体の細胞内で発現させる工程を含む、植物体もしくはその種子を着色させる、または植物体もしくはその種子にタンニン類を蓄積させる方法、
〔19〕 〔1〕〜〔3〕のいずれかに記載のDNAを植物体の細胞内で発現させる工程を含む、着色した、もしくはタンニン類が蓄積した植物体もしくはその種子の製造方法、
〔20〕 〔1〕〜〔3〕のいずれかに記載のDNA、または〔5〕に記載のベクターを植物細胞へ導入する工程を含む、〔18〕または〔19〕に記載の方法、
〔21〕 下記工程(a)および(b)を含む、〔18〕または〔19〕に記載の方法、
(a)〔1〕〜〔3〕のいずれかに記載のDNAを有する植物と交雑させる工程を含む、
(b)前記DNAを有する植物改変体を選抜する工程
〔22〕 着色した種子、もしくはタンニン類が蓄積した種子を有する植物を育種する方法であって、下記工程(a)〜(d)を含む育種方法、
(a)植物Aと、〔1〕〜〔3〕のいずれかに記載のDNAを有する他の植物Bを交雑させ、F1を作出する工程
(b)前記F1と前記植物Aを交雑させる工程
(c)前記DNAを有する植物を選抜する工程
(d)工程(c)によって選抜された植物と、前記植物Aを交雑させる工程
〔23〕 前記工程(c)において、植物ゲノム中の〔1〕〜〔3〕のいずれかに記載のDNA配列、またはその周辺配列に存在するDNAマーカーを利用して選抜されることを特徴とする、〔22〕に記載の方法、
〔24〕 植物に内在する遺伝子であって、植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能を有するタンパク質をコードする遺伝子の発現を抑制する工程を含む、脱色された植物体もしくはその種子の製造方法、
〔25〕 植物がイネである、〔17〕〜〔24〕のいずれかに記載の方法、
〔26〕 〔18〕〜〔25〕のいずれかに記載の方法によって取得される植物体、またはその種子、
〔27〕 人為的に作製された植物体もしくはその種子であって、〔1〕〜〔3〕のいずれかに記載のDNAを有し着色されていることを特徴とする植物体もしくはその種子、
〔28〕 人為的に作製された植物体もしくはその種子であって、〔1〕〜〔3〕のいずれかに記載のDNAを有しタンニン類が蓄積していることを特徴とする植物体もしくはその種子、
〔29〕 植物に内在する遺伝子であって、植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能を有するタンパク質をコードする遺伝子の発現が人為的に抑制され、脱色されていることを特徴とする植物体もしくはその種子、
〔30〕 植物がイネである、〔27〕〜〔29〕のいずれかに記載の植物体もしくはその種子、を提供するものである。
【発明の効果】
【0014】
本発明の方法により、RcもしくはRd遺伝子を植物体内において発現させることにより、植物もしくはその種子(例えば、コメ等)を人為的に着色させる手法が確立された。一般種子への遺伝子組換え作物(Genetically Modified Organisms; GMO)混入などの社会的不安に答えるため、GMO米を人為的に着色させることで対応することができる。本発明の利点のひとつは、例えば、コメの果皮を着色させることによりGMOのモニタリングが可能になる。必要に応じ、精米により最終的に着色部分を除去することも可能である。
【0015】
また、本発明においてDNAマーカーを利用する育種法により、良食味米を数年で着色米に育種できる。ごく短期間に「コシヒカリカテキン」「カテキンヒトメボレ」「カテキンマンゲツモチ」のような良食味着色米あるいは機能性良食味米系統が育種することが可能である。食味については全く未開拓の着色米であるが、本発明の技術を用いることにより良食味着色米が開発できる。本発明の方法によって作出される植物は、抗酸化機能の高いカテキン、タンニンを含むことから、例えば、機能性米として付加価値を有するコメを効率的に作出することが可能である。
【図面の簡単な説明】
【0016】
【図1】DFRとタンニン合成・アントシアニン合成の関係を示す図である。酵素の略称は以下の通りである。F3H,flavanone 3-hydroxylase; DFR,dihydroflavonol-4-reductase; ANS,anthocyanin synthase; LAR,leucoananthocyanidin reductase; ANR,anthocyanidin reductase; BAN,BANYULS; GT,anthocyanidin glucosyl transferase
【図2】イネの玄米色に関わる遺伝子の位置を示す図である。水平な線は、マーカーの位置を表す。(A)Rc,Rdに関する初期の座乗位置の情報(Nagao, S. & Takahashi, M. (1963) Agr., Hokkaido Univ., Sapporo 53, 76-131.)。第一染色体上の遺伝マーカーとして、Aはanthocyanin activator, Pnはpurple nodeをそれぞれ示す。第7染色体上の遺伝マーカーとして、d6はlop-leaved dwarf, gはrecessive long empty glumesをそれぞれ示す。(B)イネゲノムプロジェクト終了後のRc,Rdの座乗位置の情報(IRGSP 2005 Nature)R1807,R1582,R2374,C808は(http://rgp.dna.affrc.go.jp/publicdata/geneticmap2000/)に記載されている分子マーカーである。(C)今回の研究によるRc,Rdの位置(本発明)。Rd座はDFR遺伝子をコードしており、染色体上での位置は第1染色体のAP004317の68929 bp−70543 bpである。Rc座は、MYC related proteinをコードしており、染色体上での位置は第7染色体のAP005098 150211 bp−156639 bpである。
【図3】RcRdの遺伝子型とその表現型を示す写真である。写真上部は玄米、下部は籾である。(A)RcRdの遺伝子型をもつカサラスの玄米色。(B)Rcrdの遺伝子型をもつtRc(NIL)の玄米色。(C)rcrdの遺伝子型を持つ日本晴の玄米色。rcRdの遺伝子型を持つ玄米色も白色になる。(Nagao, S. (1951) Advances in Genetics 4, 181-212.)
【図4】イネのDFR遺伝子の構造を示す図である。水平の黒い四角はDFRのエクソンを示す。黒い四角の間の水平の線は、それぞれ第1イントロン、第2イントロンをしめす。イネのDFR遺伝子のPCRには、OsDFRF4とOsDFRR4を用いた。イネDFRの第1エクソン、第1イントロン、第2エクソンの塩基配列の決定には、OsDFRR5を用いた。
【図5】イネDFRのmRNAの構造を示す図である。黒の線はORFを示す。イネDFRのRT-PCRには、OsDFRF8とOsDFRR5を用いた。
【図6】pGEX4T-3ベクターに連結したDFR(pGEX4T-3 DFR)を示す図である。pGEX4T-3をベースに構築した。AmpRはアンピシリン耐性遺伝子、Ptacはtacプロモーター、GSTはグルタチオン-S-トランスフェラーゼを示す。BamHI、EcoRIは、ベクターを作成する際に用いた制限酵素である。
【図7】35S::Murasaki DFRベクターの構造を示す図である。pCAMBIA1301(ベクター部分:11.8kb)をベースに構築した。LB,RBはレフトボーダー、ライトボーダーをそれぞれ示す。T35S,P35Sはカリフラワーモザイクウィルス由来の強発現用プロモーターである。MCSはマルチクローニングサイトを示す。NosTはノパリンシンターゼ由来のターミネーターを示す。HygR,KanRは、それぞれハイグロマイシン、カナマイシンの耐性遺伝子である。Murasakiine DFRは紫稲のDFRのゲノム配列(イントロン含む)である。
【図8】35S::Koshi DFRベクターの構造を示す図である。pCAMBIA1301(ベクター部分:11.8kb)をベースに構築した。LB,RBはレフトボーダー、ライトボーダーをそれぞれ示す。T35S,P35Sはカリフラワーモザイクウィルス由来の強発現用プロモーターである。MCSはマルチクローニングサイトを示す。NosTはノパリンシンターゼ由来のターミネーターを示す。HygR,KanRは、それぞれハイグロマイシン、カナマイシンの耐性遺伝子である。Koshihikari DFRはコシヒカリのDFRのゲノム配列(イントロン含む)である。
【図9】AsDFRベクターの構造を示す図である。pCAMBIA1301(ベクター部分:11.8kb)をベースに構築した。LB,RBはレフトボーダー、ライトボーダーをそれぞれ示す。T35S,P35Sはカリフラワーモザイクウィルス由来の強発現用プロモーターである。MCSはマルチクローニングサイトを示す。NosTはノパリンシンターゼ由来のターミネーターを示す。HygR,KanRは、それぞれハイグロマイシン、カナマイシンの耐性遺伝子である。Murasakiine DFRは紫稲のDFRのcDNA配列(イントロン含まない)である。Antisense法により、DFRの発現抑制をするベクターである。
【図10】pRc1-1301、pRc2-1301、pRc3-1301ベクターの構造を示す図である。pCAMBIA1301(ベクター部分:11.8kb)をベースに構築した。LB,RBはレフトボーダー、ライトボーダーをそれぞれ示す。T35S,P35Sはカリフラワーモザイクウィルス由来の強発現用プロモーターである。MCSはマルチクローニングサイトを示す。NosTはノパリンシンターゼ由来のターミネーターを示す。HygR,KanRは、それぞれハイグロマイシン、カナマイシンの耐性遺伝子である。Gus geneはGus遺伝子を示す。promoterはそれぞれRc1,Rc2,Rc3の由来のプロモーターを示す。TはそれぞれRc1,Rc2,Rc3の由来のターミネーターを示す。
【図11】DFRを遺伝子導入した個体の表現型を示す写真である。(A)35S::Murasaki DFRを遺伝子導入した個体の表現型。一番上は発芽後7日目の葉におけるDFR抗体を用いた免疫染色の写真を表す。二段目は出穂後7日目の穂を表す。三段目は完熟種子を表す。(B) As DFRを遺伝子導入した個体の表現型。一番上は発芽後7日目の葉におけるDFR抗体を用いた免疫染色の写真を表す。二段目は出穂後7日目の穂を表す。三段目は完熟種子を表す。(C) tRc (Rcrd)の表現型。二段目は出穂後7日目の穂を表す。三段目は完熟種子を表す。
【図12】DFR遺伝子の多型を示す図である。黒い四角はエクソンを示す。水平の線はイントロンを示す。白抜きの丸は翻訳開始コドンを示す。白抜きの四角は翻訳終止コドンを示す。
【図13】イネDFRのRT-PCRを示す写真である。DFRのPCRは35サイクル行った。アクチンのPCRは30サイクル行った。右側の数字は増幅断片の大きさである。
【図14】DFRの発現の比較を示す図及び写真である。(A)日本晴、カサラスのDFRの構造。水平な黒の四角はエクソンを表す。水平な黒の線はイントロンを表す。白抜きの丸はコザックコンセンサスを持つ開始コドンを表す(ANNATGN or GNNATGG)。上向きの三角はイントロンの位置を示す。白抜きの四角は翻訳終止コドンの位置を表す。(B)OsDFRの構造から予想されるORF(Alternative translation initiation)。水平な線は予想されるORFを示し、右の数値は予想されるタンパク質の分子量を表す。(C)、(D)は、登熟期の穂、発芽後7日目の葉におけるDFRの免疫染色を示す写真である。(C)は、GST-DFR(ポジティブコントロール); カサラス; 日本晴; コシヒカリから全タンパク質を抽出し、Anti-DFR抗体によって免疫染色した。右の数値はタンパク質の分子量を示す。(D)はGST-DFR(ポジティブコントロール); 35S::DFR遺伝子導入個体; カサラス; 日本晴; コシヒカリから全タンパク質を抽出し、Anti-DFR抗体によって免疫染色した。右の数値はタンパク質の分子量を示す。
【図15A−C】RiceGAASプログラムを用いたRcのアノテーションを示す図である。(A)Rcの7番染色体上での位置を示す。(B)Rcが座乗すると考えられるR1807とR1582の間に存在するMYC type protein,MYB related protein。白丸は分子マーカーを示す。矢印は、MYC type protein(predicted gene)、MYB related protein(predicted gene)をそれぞれ示す。(C)Rcの精密なマッピング。白丸は分子マーカーを示す。一番下の段は、Rice Genome research programの日本晴クローンを示す。矢印は、データベースサーチから予想されたRc遺伝子の位置を示している。黒色の四角は、Rcの相補実験に用いた領域である。
【図15D】Rc(本発明)とTT8(Nesi, N., Debeaujon, I., Jond, C., Pelletier, G., Caboche, M. & Lepiniec, L. (2001) Plant Cell 12, 1863-1878.)の予想アミノ酸配列のアラインメントを示す図である。同一のアミノ酸は黒の四角で示し、類似のアミノ酸は囲みで示す。水平な黒の線は、MYC type proteinに特有のbHLHドメインを示す。
【図16】Rcの相補実験結果を示す写真である。写真上部は玄米、下部は籾である。(A)RcRdの遺伝子型を持つカサラス。(B)Non-transformant。(C)rcrdの遺伝子型を持つ日本晴。(D)Rc-MYC-16 (rcrdの遺伝子型を持つ日本晴にRc-MYCを相補した植物のNO.16)。(E)Rc-MYC-23 (rcrdの遺伝子型を持つ日本晴にRc-MYCを相補した植物のNO.23)。(F)Rc-MYC-13(rcrdの遺伝子型を持つ日本晴にRc-MYCを相補した植物のNO.13)。(G)Rcrdの遺伝子型を持つtRc。(H)Rc-MYB1 (rcrdの遺伝子型を持つ日本晴にRc-MYB1を相補した植物)。(I)Rc-MYB2 (rcrdの遺伝子型を持つ日本晴にRc-MYB2を相補した植物)。
【図17】RcとRdによる玄米色の制御モデルを示す図である。
【図18】Rc-MYCの構造(カサラス)を示す図である。箱はエクソンを、水平の線はイントロンをそれぞれ示す。上向きの黒い三角は日本晴における欠失を、下向きの白い三角は日本晴における挿入変異をそれぞれ示す。
【図19】Rc-MYCの第7エクソンに存在する14 bpの欠失変異の多型解析を示す図である。用いたサンプルは、玄米色が赤いRcRdの遺伝子型をもつO. sativa からIndicaタイプのKasalath(カサラス)、Japonicaタイプのトボシ(TOBOSHI)、アカマイアサヒ(AKAMAIASAH)、アカマイ(AKAMAI)、宝満新田稲(HOUMAN)、ダネマ(DANEMA)O.rufipogonからは、DAMA/THAI/86/148 (O.R.D)とCOL/THAILAND/1990/MAFF/181 (O.R.C)を用いた。玄米色が赤褐色のRcrdの遺伝子型を持つものについては、NILであるtRcとCOL/KAGOSHIMA/1963 MAJIRI/1187 (col-kago)を用いた。玄米色が白いrcrdの遺伝子型を持つものについては、日本晴(nipponbare)、コシヒカリ(Koshi)、台中65号(T65)、ドウゴワセ(DOUGO)、ミトニシキ(MITONISHIKI)、フサクモチ(FUSAKUMOCHI)、シミズシラズ(SHIMIZU)を用いた。着色(赤及び赤褐)と白色の関係は欠失のある・なしと一致することから、当該遺伝子の着色への関与が図16の結果に加えて証明された。
【図20】Rc-MYCの系統解析を示す図である。bHLHドメインを持つ、アミノ酸配列をClastal Xを用いてアラインメントした。系統樹の作成、Boots strap valueはClastal X上のNJ法、最節約法を用いて計算した。それぞれIN1,Lc, R-S, B-Peruはトウモロコシ、Raはイネ、TT8はアラビドプシス、JAF13はペチュニア、DELILAは金魚草のアントシアニン合成系制御因子である。
【図21】本発明のRc(MYC)およびRd(DFR)遺伝子の近傍に存在し、本発明の育種方法に利用可能なDNAマーカーの具体例を示す図である。
【発明を実施するための形態】
【0017】
本発明者らによって、植物体を着色させる機能、または、植物体へタンニン類を蓄積させる機能を有する遺伝子が同定された。
【0018】
本発明の上記遺伝子の好ましい態様としては、イネのRc遺伝子、またはRd遺伝子が挙げられる。該Rc遺伝子によってコードされるタンパク質は、プロアントシアニジン合成系支配因子であり、転写制御因子としての機能を有する。該Rc遺伝子は、「MYC遺伝子」、「Rc-MYC遺伝子」、または「Myc related protein gene」等と表記される場合がある。
【0019】
一方、Rd遺伝子は、「DFR遺伝子」あるいは「Dihydroflavonol 4-reductase gene」と表記される場合がある。その発現は転写因子であるRc(MYC)によって、制御されていることが明らかになった。また、本発明者らによって、Rd(DFR)タンパク質の翻訳は、Alternative translation initiationによって制御されていることが見出された。全長DFRを有する植物(例えば、イネ等)では、種子が赤色を呈し、DFRのORFが短い構造の植物(例えば、イネ等)では種子が赤褐色を呈し、Rdが機能しない場合には種子が白色を呈する。
【0020】
後述の実施例において示すように、rd型のDFRは、Alternative translation initiationにより下流のATGから翻訳され、この翻訳産物はDFRの機能を相補する機能を有することが、本発明者らによって初めて見出された。従って、本発明においてRd遺伝子とは、全長のDFRタンパク質をコードするDNAに限定されず、DFRの機能を相補し得るDFRの下流のATGから翻訳される産物をコードするDNAが含まれる。
【0021】
例えば、本発明のRd(DFR)タンパク質には、図14に示される46 kDa(配列番号:6)、33 kDa(配列番号:7)、22 kDa(配列番号:8)に相当するタンパク質が挙げられる。該33 kDa、22 kDaに相当するタンパク質は、Rd(DFR)遺伝子からAlternative translation initiation機構によって翻訳されるタンパク質と考えられる。
【0022】
また、上記タンパク質以外であっても、DFRの機能を相補し得るようなDFRタンパク質変異体もしくはDFRタンパク質の部分断片ペプチドもまた、本発明のRd(DFR)タンパク質に含まれる。
【0023】
本発明のRcおよびRdタンパク質は、イネ由来のタンパク質であることが好ましいが、その由来する植物種は特に制限されず、イネ以外の植物におけるRcもしくはRdと同等なタンパク質(RcもしくはRdのホモログ・オルソログ等)も本発明における「Rcタンパク質」もしくは「Rdタンパク質」に含まれる。例えば、イネRc(MYC)もしくはRd(DFR)に相当するタンパク質を有する植物、または、イネRcもしくはRdタンパク質が実質的に機能し得る植物であれば、本発明を実施することが可能である。
【0024】
本発明において「植物」とは、特に制限されないが、好ましくは単子葉植物であり、より好ましくは、イネ科植物である。本発明における植物として具体的には、イネ、トウモロコシ、コムギ、オオムギ等を例示することができる。(これらのRcおよびRdタンパク質、および該タンパク質をコードする遺伝子を本明細書においてそれぞれ、「本発明のタンパク質」、「本発明の遺伝子」と記載する場合あり。)
【0025】
Rc(MYC)遺伝子のゲノムDNA配列を配列番号:1に、Rc(MYC)遺伝子のcDNA配列を配列番号:2に、該DNAによってコードされるタンパク質のアミノ酸配列を配列番号:3に記載する。また、Rd(DFR)遺伝子のゲノムDNA配列を配列番号:4に、Rd(DFR)遺伝子のcDNA配列を配列番号:5に、該DNAによってコードされるタンパク質のアミノ酸配列を配列番号:6に記載する。(イネRc(MYC)のGenBankのアクセッション番号はAB247503である。また、イネRd(DFR)のGenBankのアクセッション番号はAB003496である。)
【0026】
ただし、本発明におけるRcもしくはRd遺伝子は、必ずしも、配列表に具体的に記載された配列からなるDNAに限定されない。また、本発明におけるRcもしくはRdタンパク質は、必ずしも、配列表に具体的に記載されたアミノ酸配列からなるタンパク質に限定されない。
【0027】
上記以外のタンパク質であっても、例えば配列表に記載された配列と高い相同性(通常70%以上、好ましくは80%以上、より好ましくは90%以上、最も好ましくは95%以上)を有し、かつ、本発明のタンパク質が有する機能(例えば、種子を着色させる機能、または、種子へタンニン類を蓄積させる機能等)を持つタンパク質は、本発明のタンパク質に含まれる。
【0028】
上記タンパク質とは、例えば、配列番号:3または6〜8のいずれかに記載のアミノ酸配列において、1以上のアミノ酸が付加、欠失、置換、挿入されたアミノ酸配列からなるタンパク質であって、通常変化するアミノ酸数が30アミノ酸以内、好ましくは10アミノ酸以内、より好ましくは5アミノ酸以内、最も好ましくは3アミノ酸以内である。
【0029】
本発明における「Rc遺伝子」もしくは「Rd遺伝子」には、例えば、配列番号:2もしくは5に記載の塩基配列からなるDNAに対応する他の植物における内在性の遺伝子(イネのRcもしくはRd遺伝子のホモログ等)が含まれる。
【0030】
また、配列番号:1、2、4もしくは5に記載の塩基配列からなるDNAに対応する他の植物の内在性のDNAは、一般的に、配列番号:1、2、4もしくは5に記載のDNAと高い相同性を有する。高い相同性とは、50%以上、好ましくは70%以上、さらに好ましくは80%以上、より好ましくは90%以上(例えば、95%以上、さらには96%、97%、98%または99%以上)の相同性を意味する。この相同性は、mBLASTアルゴリズム(Altschul et al. (1990) Proc. Natl. Acad. Sci. USA 87: 2264-8; Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90: 5873-7)によって決定することができる。また、該DNAは、生体内から単離した場合、配列番号:1、2、4もしくは5に記載のDNAとストリンジェントな条件下でハイブリダイズすると考えられる。ここで「ストリンジェントな条件」としては、例えば「2×SSC、0.1%SDS、50℃」、「2×SSC、0.1%SDS、42℃」、「1×SSC、0.1%SDS、37℃」、よりストリンジェントな条件として「2×SSC、0.1%SDS、65℃」、「0.5×SSC、0.1%SDS、42℃」および「0.2×SSC、0.1%SDS、65℃」の条件を挙げることができる。当業者においては、他の植物における本発明の遺伝子に相当する内在性の遺伝子を、本発明の遺伝子の塩基配列を基に適宜取得することが可能である。なお、本明細書においては、イネ以外の植物におけるRcタンパク質(遺伝子)もしくはRdタンパク質(遺伝子)に相当するタンパク質(遺伝子)、または、これらのタンパク質と機能的に同等なタンパク質(遺伝子)を、単に「本発明のタンパク質(遺伝子)」と記載する場合がある。
【0031】
本発明のタンパク質は、天然のタンパク質のほか、遺伝子組み換え技術を利用した組換えタンパク質として調製することができる。天然のタンパク質は、例えば本発明のタンパク質が発現していると考えられる細胞(組織)の抽出液に対し、本発明のタンパク質に対する抗体を用いたアフィニティークロマトグラフィーを用いる方法により調製することが可能である。一方、組換えタンパク質は、本発明のタンパク質をコードするDNAで形質転換した細胞を培養することにより調製することが可能である。
【0032】
本発明の好ましい態様としては、植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能を有する植物由来のタンパク質をコードする、下記(a)〜(e)のいずれかに記載のDNAを提供する。
(a)配列番号:3または6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:1または4に記載の塩基配列のコード領域を含むDNA
(c)配列番号:2または5に記載の塩基配列からなるDNA
(d)配列番号:3または6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(e)配列番号:1または4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA
【0033】
また、上記DNAによってコードされるタンパク質もまた、本発明に含まれる。
本発明において、「植物の種子」とは、例えば、植物がイネの場合には、「コメ」、「玄米」等を指す。
【0034】
本発明において「着色」とは、通常、植物体もしくはその種子において、元の色を別の色へ変化させることを言う。一例を示せば、種子の全体あるいは一部を、赤色、赤褐色、褐色、または紫黒色等へ着色させることを指す。より具体的には、植物がイネである場合には、白色米(白米)を赤色米(赤米)、赤褐色米、褐色米、または紫黒色米等へ着色することを言う。(左記のように白米以外の有色のコメを、本発明において「有色米」と表記する場合がある。)
【0035】
また、本発明において「着色を制御する」とは、上述のように、植物体もしくはその種子の全体またはその一部の色を、他の色へ変化させることを言う。例えば、植物がイネの場合、上述のように、白米を、赤色米等の有色米へ変化させることを言うが、反対に、有色米を白色米へ変化させる場合も、本発明における「着色を制御する」ことに含まれる。
【0036】
本発明において着色が制御される植物体の部位は、特に制限されないが、例えば、種子、葉、節間、茎、えい(もみ)等を挙げることができる。
【0037】
本発明における植物がイネの場合には、例えば、イネの種子(コメ)の種皮および/または果皮が着色される。また、本発明における植物がコムギ、オオムギ、トウモロコシ等の場合には、種子の果皮部位が着色される。
【0038】
本発明のおける植物の着色部位は、必ずしも上述の部位に限定されるものではなく、植物の節間、種子の胚乳等を着色させることも可能である。
【0039】
また、「タンニン(tannin)」とは、一般的には、広く植物界に分布する多数のフェノール性ヒドロキシル基を持つ芳香族化合物の総称であり、「タンニン酸(tannic acid)」、あるいは「ガロタンニン酸(gallotannic acid)」とも呼ばれる。
【0040】
本発明における「タンニン類」には、例えば、「タンニン」、「カテキン(catechin)」、「アントシアニン(anthocyanin)」、「プロアントシアニジン(proanthocyanidin)」等が含まれる。上記カテキンは、「3,3',4',5,7-ペンタヒドロキシフラバン(3,3',4',5,7-pentahydroxyflavan)」とも呼ばれる。また、「カテキン」には、所謂「カテキン類」に属する化合物、例えば、エピカテキン等も含まれる。また、タンニン類が各種化合物と反応して生成される物質もまた、本発明における「タンニン類」に含まれる。上記「アントシアニン」は、紫色の色素であり、上記「プロアントシアニジン」は赤褐色もしくは赤色の色素であるものと考えられている。
【0041】
赤米品種である「紅衣」の玄米には、100g中に0.29gのタンニン、1.6mgのカテキン、0.09gのアントシアンが含まれている。従って、本発明の方法によって白米にタンニン類を蓄積させる場合、コメ中に蓄積されるタンニン類の量(含有量)としては、例えば、上記の量を例示することができる。
【0042】
本発明において「タンニン類を蓄積させる」とは、通常、植物体において元々タンニン類が存在しない部位にタンニン類を生成・蓄積させること、または、植物体においてタンニン類が存在する部位において、タンニン類の量(含有量)を増加させることを言う。
【0043】
本明細書におけるタンパク質とは、複数のアミノ酸からなる重合体を意味し、そのアミノ酸の長さは特に制限されない。従って、本発明のタンパク質には、所謂「ポリペプチド」、および「オリゴペプチド」も含まれる。本発明のタンパク質は、天然に存在する状態から修飾されていないもの、および修飾されているものの双方を含む。修飾としては、アセチル化、アシル化、ADP-リボシル化、アミド化、フラビンの共有結合、ヘム部分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトールの共有結合、架橋、環化、ジスルフィド結合の形成、脱メチル化、共有架橋の形成、シスチンの形成、ピログルタメートの形成、ホルミル化、γ-カルボキシル化、グリコシル化、GPIアンカー形成、ヒドロキシル化、ヨウ素化、メチル化、ミリストイル化、酸化、タンパク質分解処理、リン酸化、プレニル化、ラセミ化、セレノイル化、硫酸化、アルギニル化のようなタンパク質へのアミノ酸の転移RNA媒介付加、ユビキチン化等が含まれる。
【0044】
本発明のポリペプチドは、そのアミノ酸配列に従って、一般的な化学合成法により製造することが可能であり、該方法には、通常の液相法および固相法によるペプチド合成法が包含される。かかるペプチド合成法は、より詳しくはアミノ酸配列の情報に基づいて、各アミノ酸を1個ずつ逐次合成させて鎖を延長していくステップワイズエロンゲーション法と、アミノ酸数個からなるフラグメントを予め合成し、次いで各フラグメントをカップリング反応させるフラグメント・コンデンセーション法を包含し、本発明のタンパク質の合成は、いずれの方法を用いてもよい。
【0045】
このようなペプチド合成法にて用いられる縮合法も、各種方法に従って行うことができる。その具体例としては、例えばアジド法、混合酸無水物法、DCC法、活性エステル法、酸化還元法、DPPA(ジフェニルホスホリルアジド)法、ウッドワード法等を例示できる。
【0046】
これら各種方法に利用できる溶媒もまた、一般的に使用されるものを適宜利用することができる。その例としては、例えばジメチルホルムアミド(DMF)、ジメチルスルホキシド(DMSO)、ヘキサホスホロアミド、ジオキサン、テトラヒドロフラン(THF)、酢酸エチル等及びこれらの混合溶媒等を挙げることができる。なお、上記ペプチド合成反応に際して、反応に関与しないアミノ酸およびペプチドにおけるカルボキシル基は、一般にはエステル化により、例えばメチルエステル、エチルエステル、第三級ブチルエステル等の低級アルキルエステル、例えばベンジルエステル、P−メトキシベンジルエステル、P−ニトロベンジルエステルアラルキルエステル等として保護することができる。また、側鎖に官能基を有するアミノ酸、例えばTyrの水酸基は、アセチル基、ベンジル基、ベンジルオキシカルボニル基、第三級ブチル基等で保護されてもよいが、必ずしもかかる保護は必須ではない。また、例えば、Argのグアニジノ基は、ニトロ基、トシル基、2−メトキシベンゼンスルホニル基、メチシレン−2−スルホニル基、ベンジルオキシカルボニル基、イソボルニルオキシカルボニル基、アダマンチルオキシカルボニル基等の適当な保護基により保護することができる。
【0047】
上記のようにして得ることが可能な本発明のタンパク質は、通常の方法に従って、例えばイオン交換樹脂、分配クロマトグラフィー、ゲルクロマトグラフィー、アフィニティークロマトグラフィー、高速液体クロマトグラフィー(HPLC)、向流分配法等のペプチド化学の分野で汎用されている方法に従って、適宜、精製を行うことができる。
【0048】
本発明のタンパク質は、例えば、配列番号:3もしくは6〜8に記載のポリペプチド、または配列番号:2もしくは5に記載のDNA核酸分子を合成し、次いで適当な発現ベクターへ導入した後、宿主細胞内において発現させる遺伝子工学的手法によっても取得することができる。
【0049】
本発明のタンパク質には、例えば、Rc(MYC)もしくはRd(DFR)タンパク質と機能的に同等なタンパク質(ポリペプチド)が含まれる。ここで「機能的に同等」とは、対象となるタンパク質がRc(MYC)もしくはRd(DFR)タンパク質と同様の(同等の)生物学的あるいは生化学的機能(活性)を有することを指す。このような機能としては、例えば、植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能等を挙げることができる。
【0050】
あるDNAが植物体を着色させる機能、またはタンニン類を蓄積させる機能を有するタンパク質をコードするか否かは以下のようにして評価することができる。最も一般的な方法としては、該DNAが導入された植物を栽培し、植物体の色、または植物体中に含まれるタンニン類の量を評価する方法である。
【0051】
対象となるタンパク質(ポリペプチド)が、Rc(MYC)もしくはRd(DFR)タンパク質と同等の生物学的あるいは生化学的な機能(活性)を有しているか否かは、例えば、該タンパク質を発現させた植物もしくはその種子におけるタンニン類の量を測定することにより適宜評価することも可能である。
【0052】
あるポリペプチドと機能的に同等なポリペプチドを調製するための、当業者によく知られた方法としては、例えばポリペプチド中のアミノ酸配列に変異を導入する方法が挙げられる。具体的には当業者であれば部位特異的変異誘発法(Hashimoto-Gotoh, T. et al. (1995) Gene 152, 271-275、Zoller, MJ, and Smith, M.(1983) Methods Enzymol. 100, 468-500、Kramer, W. et al. (1984) Nucleic Acids Res. 12, 9441-9456、Kramer W, and Fritz HJ(1987) Methods. Enzymol. 154, 350-367、Kunkel,TA(1985) Proc Natl Acad Sci USA. 82, 488-492、Kunkel (1988) Methods Enzymol. 85, 2763-2766)などを用いて、配列番号:4に記載のアミノ酸配列に適宜変異を導入することにより、該ポリペプチドと機能的に同等なポリペプチドを調製することができる。また、ポリペプチド中のアミノ酸の変異は自然に生じることもある。このように、人工的か自然に生じたものかを問わず、本発明者らにより同定されたRcもしくはRdタンパク質(配列番号:3または6〜8)のアミノ酸配列において1もしくは複数のアミノ酸配列が変異したアミノ酸配列を有し、該ポリペプチドと機能的に同等なポリペプチドは、本発明のタンパク質(ポリペプチド)に含まれる。
【0053】
上記変異体における、変異するアミノ酸数は、本発明のタンパク質の有する機能が保持される限り制限はないが、通常15アミノ酸以内であり、好ましくは10アミノ酸以内であり、より好ましくは5アミノ酸以内であり、さらに好ましくは1〜4アミノ酸である。
【0054】
変異するアミノ酸残基としては、アミノ酸側鎖の性質が保存されている別のアミノ酸に変異されることが望ましい。例えばアミノ酸側鎖の性質としては、疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)、親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)、脂肪族側鎖を有するアミノ酸(G、A、V、L、I、P)、水酸基含有側鎖を有するアミノ酸(S、T、Y)、硫黄原子含有側鎖を有するアミノ酸(C、M)、カルボン酸及びアミド含有側鎖を有するアミノ酸(D、N、E、Q)、塩基含有側鎖を有するアミノ離(R、K、H)、芳香族含有側鎖を有するアミノ酸(H、F、Y、W)を挙げることができる(括弧内はいずれもアミノ酸の一文字表記を表す)。
【0055】
あるアミノ酸配列に対する1または複数個のアミノ酸残基の欠失、付加及び/又は他のアミノ酸による置換により修飾されたアミノ酸配列を有するポリペプチドがその生物学的機能(活性)を維持し得ることはすでに知られている(Mark, D. F. et al., Proc. Natl. Acad. Sci. USA (1984) 81, 5662-5666 、Zoller, M. J. & Smith, M. Nucleic Acids Research (1982) 10, 6487-6500 、Wang, A. et al., Science 224, 1431-1433 、Dalbadie-McFarland, G. et al., Proc. Natl. Acad. Sci. USA (1982) 79, 6409-6413)。
【0056】
具体的なアミノ酸配列(例えば、配列番号:3または6〜8)が開示された場合においては、当業者であれば、これらアミノ酸配列を基に、適宜アミノ酸が改変された配列からなるポリペプチドを作製し、当該ポリペプチドについて、上述の機能を有するか否かを評価し、本発明のタンパク質(ポリペプチド)を適宜選択することが可能である。
【0057】
本発明におけるRdタンパク質は、そのN末領域が欠損している場合であっても本発明のタンパク質の有する機能を相補し得ることが明らかとなった。従って、Rdタンパク質の改変体としては、例えば、配列番号:7もしくは8で示されるアミノ酸領域を含み、N末領域において変異もしくは欠失を有するRdタンパク質の改変体(変異体、部分ペプチド等)を例示することができる。
【0058】
本発明のタンパク質のアミノ酸配列に複数個のアミノ酸残基が付加されたタンパク質には、これらタンパク質を含む融合ポリペプチドが含まれる。融合ポリペプチドは、これらポリペプチドと他のペプチド又はポリペプチドとが融合したものである。融合ポリペプチドを作製する方法は、本発明のタンパク質(例えば、配列番号:3または6〜8)をコードするDNA(例えば、配列番号:2または5)と他のペプチド又はポリペプチドをコードするDNAをフレームが一致するように連結してこれを発現ベクターに導入し、宿主で発現させればよく、当業者に公知の手法を用いることができる。本発明のタンパク質との融合に付される他のペプチド又はポリペプチドは、特に制限されない。
【0059】
本発明のポリペプチドとの融合に付される他のペプチドとしては、例えば、GST(グルタチオン−S−トランスフェラーゼ)、イムノグロブリン定常領域、β−ガラクトシダーゼ、MBP(マルトース結合タンパク質)等が挙げられる。市販されているこれらペプチドまたはポリペプチドをコードするポリヌクレオチドを本発明のポリペプチドをコードするポリヌクレオチドと融合させ、これにより調製された融合ポリヌクレオチドを発現させることにより、融合ポリペプチドを調製することができる。
【0060】
またあるポリペプチドと機能的に同等なポリペプチドを調製する当業者によく知られた他の方法としては、ハイブリダイゼーション技術(Sambrook,J et al., Molecular Cloning 2nd ed., 9.47-9.58, Cold Spring Harbor Lab. press, 1989)を利用する方法が挙げられる。即ち、当業者であれば、本発明のタンパク質をコードするDNA(配列番号:2または5に記載の塩基配列)もしくはその一部をもとに、同種または異種植物由来のDNA試料から、これと相同性の高いDNAを単離して、該DNAから本発明のタンパク質と機能的に同等なポリペプチドを単離することも通常行いうることである。
【0061】
本発明には、本発明のタンパク質をコードするDNAとハイブリダイズするDNAによってコードされるタンパク質であって、本発明のタンパク質と機能的に同等なタンパク質が含まれる。このようなタンパク質としては、例えばイネあるいは他の植物のホモログ(例えば、トウモロコシ、コムギ、オオムギ等の植物に由来するタンパク質)が挙げられる。
【0062】
本発明のタンパク質と機能的に同等なタンパク質をコードするDNAを単離するためのハイブリダイゼーションの条件は、当業者であれば適宜選択することができる。ハイブリダイゼーションの条件としては、例えば、低ストリンジェントな条件が挙げられる。低ストリンジェントな条件とは、ハイブリダイゼーション後の洗浄において、例えば42℃、0.1×SSC、0.1%SDSの条件であり、好ましくは50℃、0.1×SSC、0.1%SDSの条件である。より好ましいハイブリダイゼーションの条件としては、高ストリンジェントな条件が挙げられる。高ストリンジェントな条件とは、例えば65℃、5×SSC及び0.1%SDSの条件である。これらの条件において、温度を上げる程に高い相同性を有するDNAが効率的に得られることが期待できる。但し、ハイブリダイゼーションのストリンジェンシーに影響する要素としては温度や塩濃度など複数の要素が考えられ、当業者であればこれら要素を適宜選択することで同様のストリンジェンシーを実現することが可能である。
【0063】
また、ハイブリダイゼーションにかえて、遺伝子増幅技術(PCR)(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley&Sons Section 6.1-6.4)を用いて、本発明のタンパク質をコードするDNA(例えば、配列番号:2または5)の一部を基にプライマーを設計し、本発明者らにより同定されたタンパク質をコードするDNAと相同性の高いDNA断片を単離し、該DNAを基に本発明者らにより同定されたタンパク質と機能的に同等なタンパク質を取得することも可能である。
【0064】
本発明のタンパク質は「成熟」ポリペプチドの形であっても、融合ポリペプチドのような、より大きいポリペプチドの一部であってもよい。本発明のタンパク質には、リーダー配列、プロ配列、多重ヒスチジン残基のような精製に役立つ配列、または組換え生産の際の安定性を確保する付加的配列などが含まれていてもよい。
【0065】
これらハイブリダイゼーション技術や遺伝子増幅技術により単離されるDNAによってコードされる、本発明のタンパク質と機能的に同等なタンパク質は、通常、本発明のタンパク質(例えば、配列番号:3または6〜8)とアミノ酸配列において高い相同性を有する。本発明のタンパク質には、RcもしくはRdタンパク質と機能的に同等であり、かつ該タンパク質のアミノ酸配列と高い相同性を有するタンパク質も含まれる。高い相同性とは、アミノ酸レベルにおいて、通常、少なくとも50%以上の同一性、好ましくは75%以上の同一性、さらに好ましくは85%以上の同一性、さらに好ましくは95%以上(例えば、96%以上、97%以上、98%以上、99%以上)の同一性を指す。タンパク質の相同性を決定するには、文献(Wilbur, W. J. and Lipman, D. J. Proc. Natl. Acad. Sci. USA (1983) 80, 726-730)に記載のアルゴリズムに従えばよい。
【0066】
アミノ酸配列の同一性は、例えば、Karlin and Altschul によるアルゴリズムBLAST (Proc. Natl. Acad. Sci. USA 87:2264-2268, 1990、Proc. Natl. Acad. Sci. USA 90:5873-5877, 1993)によって決定することができる。このアルゴリズムに基づいて、BLASTXと呼ばれるプログラムが開発されている(Altschul et al. J. Mol. Biol.215: 403-410, 1990)。BLASTXによってアミノ酸配列を解析する場合には、パラメーターは例えば、score = 50、wordlength = 3とする。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である(http://www.ncbi.nlm.nih.gov.)。
【0067】
本発明のタンパク質は、当業者に公知の方法により、組み換えポリペプチドとして、また天然のポリペプチドとして調製することが可能である。組み換えポリペプチドであれば、例えば、本発明のタンパク質をコードするDNA(例えば、配列番号:2または5に記載のDNA)を、適当な発現ベクターに組み込み、これを適当な宿主細胞に導入して得た形質転換体を回収し、抽出物を得た後、イオン交換、逆相、ゲル濾過などのクロマトグラフィー、あるいは本発明のタンパク質に対する抗体をカラムに固定したアフィニティークロマトグラフィーにかけることにより、または、さらにこれらのカラムを複数組み合わせることにより精製し、調製することが可能である。
【0068】
また、本発明のタンパク質をグルタチオンS-トランスフェラーゼタンパク質との融合ポリペプチドとして、あるいはヒスチジンを複数付加させた組み換えポリペプチドとして宿主細胞(例えば、植物細胞や微生物細胞等)内で発現させた場合には、発現させた組み換えポリペプチドはグルタチオンカラムあるいはニッケルカラムを用いて精製することができる。融合ポリペプチドの精製後、必要に応じて融合ポリペプチドのうち、目的のポリペプチド以外の領域を、トロンビンまたはファクターXaなどにより切断し、除去することも可能である。
【0069】
天然のタンパク質であれば、当業者に周知の方法、例えば本発明のタンパク質を発現している組織や細胞の抽出物に対し、本発明のタンパク質と親和性を有する抗体が結合したアフィニティーカラムを作用させて精製することにより単離することができる。抗体はポリクローナル抗体であっても、モノクローナル抗体であってもよい。
【0070】
また、本発明のタンパク質は例えば、本発明のタンパク質を認識する抗体の作製等に利用することが可能である。
【0071】
本発明のDNAは、本発明のタンパク質をコードし得るものであればいかなる形態でもよい。即ち、mRNAから合成されたcDNAであるか、ゲノムDNAであるか、化学合成DNAであるかなどを問わない。また、本発明のタンパク質をコードしうる限り、遺伝暗号の縮重に基づく任意の塩基配列を有するDNAが含まれる。
【0072】
本発明のDNAは、当業者に公知の方法により調製することができる。例えば、本発明のタンパク質を発現している細胞よりcDNAライブラリーを作製し、本発明のDNA(例えば、配列番号:2または5に記載の塩基配列)の一部をプローブとしてハイブリダイゼーションを行うことにより調製できる。cDNAライブラリーは、例えば、文献(Sambrook, J. et al., Molecular Cloning、Cold Spring Harbor Laboratory Press (1989))に記載の方法により調製してもよく、あるいは市販のDNAライブラリーを用いてもよい。また、本発明のタンパク質を発現している細胞よりRNAを調製し、逆転写酵素によりcDNAを合成後、本発明のDNA(例えば、配列番号:2または5に記載の塩基配列)に基づいてオリゴDNAを合成し、これをプライマーとして用いてPCR反応を行い、本発明のタンパク質をコードするcDNAを増幅させることにより調製することも可能である。
【0073】
また、得られたcDNAの塩基配列を決定することにより、それがコードする翻訳領域を決定でき、本発明のタンパク質のアミノ酸配列を得ることができる。また、得られたcDNAをプローブとしてゲノムDNAライブラリーをスクリーニングすることにより、ゲノムDNAを単離することができる。
【0074】
具体的には、次のようにすればよい。まず、本発明のタンパク質を発現する細胞、組織、器官からmRNAを単離する。mRNAの単離は、公知の方法、例えば、グアニジン超遠心法(Chirgwin, J. M. et al., Biochemistry (1979) 18, 5294-5299)、AGPC法(Chomczynski, P. and Sacchi, N., Anal. Biochem. (1987) 162, 156-159)等により全RNAを調製し、mRNA Purification Kit (Pharmacia) 等を使用して全RNAからmRNAを精製する。また、QuickPrep mRNA Purification Kit (Pharmacia) を用いることによりmRNAを直接調製することもできる。
【0075】
得られたmRNAから逆転写酵素を用いてcDNAを合成する。cDNAの合成は、AMV Reverse Transcriptase First-strand cDNA Synthesis Kit (生化学工業)等を用いて行うこともできる。また、本明細書に記載されたプライマー等を用いて、5'-Ampli FINDER RACE Kit (Clontech製)およびポリメラーゼ連鎖反応 (polymerase chain reaction ; PCR)を用いた5'-RACE法(Frohman, M. A. et al., Proc. Natl. Acad. Sci. U.S.A. (1988) 85, 8998-9002 ; Belyavsky, A. et al., Nucleic Acids Res. (1989) 17, 2919-2932) に従い、cDNAの合成および増幅を行うことができる。得られたPCR産物から目的とするDNA断片を調製し、ベクターDNAと連結する。さらに、これより組換えベクターを作製し、大腸菌等に導入してコロニーを選択して所望の組換えベクターを調製する。目的とするDNAの塩基配列は、公知の方法、例えば、ジデオキシヌクレオチドチェインターミネーション法により確認することができる。
【0076】
また、本発明のDNAの作成においては、発現に使用する宿主のコドン使用頻度を考慮し、より発現効率の高い塩基配列を設計することができる(Grantham, R. et al., Nucelic Acids Research (1981) 9, r43-74)。また、本発明のDNAは、市販のキットや公知の方法によって改変することができる。改変としては、例えば、制限酵素による消化、合成オリゴヌクレオチドや適当なDNAフラグメントの挿入、リンカーの付加、開始コドン(ATG)及び/又は終止コドン(TAA、TGA、又はTAG)の挿入等が挙げられる。
【0077】
また本発明は、本発明のDNAが挿入されたベクターを提供する。本発明のベクターとしては、組み換えタンパク質の生産に用いる上記ベクターの他、形質転換植物体作製のために植物細胞内で本発明のDNAを発現させるためのベクターも含まれる。このようなベクターとしては、好ましくは、植物細胞で転写可能なプロモーター配列と転写産物の安定化に必要なポリアデニレーション部位を含むターミネーター配列を含む。植物細胞の形質転換に用いられるベクターとしては、該細胞内で挿入遺伝子を発現させることが可能なものであれば特に制限はない。例えば、植物細胞内での恒常的な遺伝子発現を行うためのプロモーター(例えば、カリフラワーモザイクウイルスの35Sプロモーター)を有するベクターや外的な刺激により誘導的に活性化されるプロモーターを有するベクターを用いることも可能である。上記「植物細胞」には、種々の形態の植物細胞、例えば、懸濁培養細胞、プロトプラスト、葉の切片、カルス等が含まれる。
【0078】
本発明のベクターは、本発明のタンパク質を恒常的または誘導的に発現させるためのプロモーターを含有してもよい。恒常的に発現させるためのプロモーターとしては、例えば、カリフラワーモザイクウイルスの35Sプロモーター、イネのアクチンプロモーター、トウモロコシのユビキチンプロモーター等が挙げられる。
【0079】
また、誘導的に発現させるためのプロモーターとしては、例えば細菌・ウイルスの感染や侵入、低温、高温、乾燥、紫外線の照射、特定の化合物の散布などの外因によって発現することが知られているプロモーターなどが挙げられる。このようなプロモーターとしては、例えば、細菌・ウイルスの感染や侵入によって発現するイネキチナーゼ遺伝子のプロモーターやタバコのPRタンパク質遺伝子のプロモーター、低温によって誘導されるイネの「lip19」遺伝子のプロモーター、高温によって誘導されるイネの「hsp80」遺伝子と「hsp72」遺伝子のプロモーター、乾燥によって誘導されるシロイヌナズナの「rab16」遺伝子のプロモーター、紫外線の照射によって誘導されるパセリのカルコン合成酵素遺伝子のプロモーター、嫌気的条件で誘導されるトウモロコシのアルコールデヒドロゲナーゼ遺伝子のプロモーターなどが挙げられる。また、イネキチナーゼ遺伝子のプロモーターとタバコのPRタンパク質遺伝子のプロモーターはサリチル酸などの特定の化合物によって、「rab16」は植物ホルモンのアブシジン酸の散布によっても誘導される。
【0080】
当業者においては、所望のDNAを有するベクターを、一般的な遺伝子工学技術によって、適宜、作製することが可能である。通常、市販の種々のベクターを利用することができる。
【0081】
本発明のベクターは、宿主細胞内において本発明のDNAを保持したり、本発明のタンパク質を発現させるためにも有用である。
【0082】
本発明におけるDNAは、通常、適当なベクターへ担持(挿入)され、宿主細胞へ導入される。該ベクターとしては、挿入したDNAを安定に保持するものであれば特に制限されず、例えば宿主に大腸菌を用いるのであれば、クローニング用ベクターとしてpBluescriptベクター(Stratagene社製)などが好ましいが、市販の種々のベクターを利用することができる。本発明のタンパク質を生産する目的としてベクターを用いる場合には、特に発現ベクターが有用である。発現ベクターとしては、試験管内、大腸菌内、培養細胞内、植物個体内でポリペプチドを発現するベクターであれば特に制限されないが、例えば、試験管内発現であればpBESTベクター(プロメガ社製)、大腸菌であればpETベクター(Invitrogen社製)、培養細胞であればpME18S-FL3ベクター(GenBank Accession No. AB009864)、生物個体であればpME18Sベクター(Mol Cell Biol. 8:466-472(1988))などを例示することができる。ベクターへの本発明の核酸の挿入は、常法により、例えば、制限酵素サイトを用いたリガーゼ反応により行うことができる。
【0083】
上記宿主細胞としては特に制限はなく、目的に応じて種々の宿主細胞が用いられる。本発明のタンパク質を発現させるための細胞としては、例えば、細菌細胞(例:ストレプトコッカス、スタフィロコッカス、大腸菌、ストレプトミセス、枯草菌)、昆虫細胞(例:ドロソフィラS2、スポドプテラSF9)、動物細胞(例:CHO、COS、HeLa、C127、3T3、BHK、HEK293、Bowes メラノーマ細胞)および植物細胞を例示することができる。宿主細胞へのベクター導入は、例えば、リン酸カルシウム沈殿法、電気パルス穿孔法(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons.Section 9.1-9.9)、リポフェクション法(GIBCO-BRL社製)、マイクロインジェクション法などの公知の方法で行うことが可能である。
【0084】
宿主細胞において発現したタンパク質を小胞体の内腔に、細胞周辺腔に、または細胞外の環境に分泌させるために、適当な分泌シグナルを目的のタンパク質に組み込むことができる。これらのシグナルは目的のタンパク質に対して内因性であっても、異種シグナルであってもよい。
【0085】
上記製造方法におけるタンパク質の回収は、本発明のタンパク質が培地に分泌される場合は、培地を回収する。本発明のタンパク質が細胞内に産生される場合は、その細胞をまず溶解し、その後にタンパク質を回収する。
【0086】
組換え細胞培養物から本発明のタンパク質を回収し精製するには、硫酸アンモニウムまたはエタノール沈殿、酸抽出、アニオンまたはカチオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィーおよびレクチンクロマトグラフィーを含めた公知の方法を用いることができる。
【0087】
また、植物の生体内で本発明のDNAを発現させる方法としては、本発明のDNAを適当なベクターに組み込み、例えば、エレクトロポレーション法、アグロバクテリウム法、リポソーム法、カチオニックリポソーム法などにより生体内に導入する方法などが挙げられる。ベクターへの本発明のDNAの挿入などの一般的な遺伝子操作は、常法に従って行うことが可能である(Molecular Cloning, 5.61-5.63)。植物体内への投与は、ex vivo法であっても、in vivo法であってもよい。また、植物体内へ本発明のDNAを導入する方法としては、好ましくは、アグロバクテリウムを介して遺伝子を導入する方法が挙げられる。
【0088】
また、後述する手法で、本発明のDNAが導入された形質転換植物体を作成し、該植物体から本発明のタンパク質を調製することも可能である。
【0089】
また、上述のようにして取得される組換えタンパク質を用いれば、これに結合する抗体を調製することができる。例えば、ポリクローナル抗体は、精製した本発明のタンパク質もしくはその一部のペプチドをウサギなどの免疫動物に免疫し、一定期間の後に血液を採取し、血ぺいを除去することにより調製することが可能である。また、モノクローナル抗体は、上記タンパク質若しくはペプチドで免疫した動物の抗体産生細胞と骨腫瘍細胞とを融合させ、目的とする抗体を産生する単一クローンの細胞(ハイブリドーマ)を単離し、該細胞から抗体を得ることにより調製することができる。これにより得られた抗体は、本発明のタンパク質の精製や検出などに利用することが可能である。本発明には、本発明のタンパク質に結合する抗体が含まれる。これらの抗体を用いることにより、植物体における本発明のタンパク質の発現部位の判別、または、植物種が本発明のタンパク質を発現するか否かの判別を行うことが可能である。
【0090】
本発明のDNAを利用して、着色した、もしくはタンニン類が蓄積した形質転換植物体を作製する場合には、本発明のタンパク質をコードするDNAを適当なベクターに挿入して、これを植物細胞に導入し、これにより得られた形質転換植物細胞を再生させる。本発明者等により単離されたタンパク質は、植物体もしくはその種子を着色させる、またはタンニン類を蓄積させる機能を有するが、本発明のタンパク質を任意の植物種(イネ品種等)に導入し発現させることによりそれらの植物体もしくはその種子を着色させたり、植物体もしくはその種子へタンニン類を蓄積させたりすることが可能である。この形質転換に要する期間は、従来のような交雑による遺伝子移入に比較して極めて短期間であり、また、他の形質の変化を伴わない点で有利である。
【0091】
上述のように、本発明のDNAを植物体の細胞内で発現させる工程を含む、植物体もしくはその種子を着色させる、または植物体もしくはその種子にタンニン類を蓄積させる方法もまた本発明に含まれる。該方法によって、着色された、もしくはタンニン類が蓄積した植物体もしくはその種子が作出される。従って本発明は、本発明のDNAを植物体の細胞内で発現させる工程を含む、着色した、もしくは、タンニン類が蓄積した植物体もしくはその種子の製造方法を提供する。
【0092】
より具体的には、本発明の方法によって、例えば、白米を、赤米または赤褐色米等の有色米へ改変することが可能である。
【0093】
本発明の方法によって着色可能な白米品種は、特に制限されず、任意の白米を着色させることが可能である。具体的には、日本晴、コシヒカリ、台中65号、あきたこまち等の品種を例示することができるが、これらの品種に限定されない。
【0094】
また、本発明は、本発明のベクターが導入された形質転換細胞を提供する。本発明のベクターが導入される細胞には、組み換えタンパク質の生産に用いる上記した細胞の他に、形質転換植物体作製のための植物細胞が含まれる。植物細胞としては特に制限はなく、例えば、イネ、トウモロコシ、コムギ、オオムギなどの細胞が挙げられる。本発明の植物細胞には、培養細胞の他、植物体中の細胞も含まれる。また、プロトプラスト、苗条原基、多芽体、毛状根も含まれる。植物細胞へのベクターの導入は、ポリエチレングリコール法、電気穿孔法(エレクトロポーレーション)、アグロバクテリウムを介する方法、パーティクルガン法など当業者に公知の種々の方法を用いることができる。形質転換植物細胞からの植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。例えば、イネにおいては、形質転換植物体を作出する手法については、ポリエチレングリコールによりプロトプラストへ遺伝子導入し植物体を再生させる方法、電気パルスによりプロトプラストへ遺伝子導入し植物体を再生させる方法、パーティクルガン法により細胞へ遺伝子を直接導入し、植物体を再生させる方法、およびアグロバクテリウムを介して遺伝子を導入し、植物体を再生させる方法など、いくつかの技術が既に確立し、本願発明の技術分野において広く用いられている。本発明においては、これらの方法を好適に用いることができる。
【0095】
本発明のDNAを含むベクターの導入により形質転換した植物細胞を効率的に選択するために、上記組み換えベクターは、適当な選抜マーカー遺伝子を含む、もしくは選抜マーカー遺伝子を含むプラスミドベクターと共に植物細胞へ導入することが好ましい。この目的に使用される選抜マーカー遺伝子は、例えば抗生物質ハイグロマイシンに耐性であるハイグロマイシンホスホトランスフェラーゼ遺伝子、カナマイシンまたはゲンタマイシンに耐性であるネオマイシンホスホトランスフェラーゼ遺伝子、および除草剤ホスフィノスリシンに耐性であるアセチルトランスフェラーゼ遺伝子等が挙げられる。
【0096】
組み換えベクターを導入した植物細胞は、導入された選抜マーカー遺伝子の種類に従って適当な選抜用薬剤を含む公知の選抜用培地に置床し培養する。これにより形質転換された植物培養細胞を得ることができる。
【0097】
形質転換された植物細胞は、再分化させることにより植物体を再生させることが可能である。再分化の方法は植物細胞の種類により異なるが、例えば、イネであればFujimuraら(Plant Tissue Culture Lett. 2:74 (1995))の方法が挙げられ、トウモロコシであればShillitoら(Bio/Technology 7:581 (1989))の方法やGorden-Kammら(Plant Cell 2:603(1990))が挙げられる。
【0098】
一旦、ゲノム内に本発明のDNAが導入された形質転換植物体が得られれば、該植物体から有性生殖または無性生殖により子孫を得ることが可能である。また、該植物体やその子孫あるいはクローンから繁殖材料(例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラスト等)を得て、それらを基に該植物体を量産することも可能である。本発明には、本発明のDNAが導入された植物細胞、該細胞を含む植物体、該植物体の子孫およびクローン、並びに該植物体、その子孫、およびクローンの繁殖材料が含まれる。
【0099】
このようにして作出された植物体もしくはその種子の色は着色されることが期待される。
上述のように、本発明のDNAもしくはベクターを植物細胞へ導入し、該植物細胞から植物体を再生させる工程を含む、形質転換植物体の製造方法もまた本発明に含まれる。
【0100】
本発明の着色した植物体もしくはその種子、またはタンニン類が蓄積した植物体もしくはその種子は、育種法によっても作出することが可能である。
【0101】
上記育種法としては、例えば、本発明のDNAを有する品種と交雑させることを特徴とする一般的な育種法(交雑育種法等)を挙げることができる。該方法によって、着色した植物体もしくはその種子、またはタンニン類が蓄積した植物体もしくはその種子を作出することができる。
【0102】
育種法によって本発明の植物体もしくは種子を作製する際には、公知の種々の文献を参照して適宜実施することができる。(細胞工学別冊・植物細胞工学シリーズ15「モデル植物の実験プロトコール」秀潤社、2001年、加藤鎌司著「2.1 イネ・コムギの交配」p.6-9; 高牟礼逸朗、佐野芳雄著「2. 交配法」p.46-48)
【0103】
本発明の上記育種方法の好ましい態様としては、以下の工程(a)および(b)を含む方法である。
(a)本発明のDNAを有する植物と交雑させる工程を含む、
(b)前記DNAを有する植物改変体を選抜する工程
【0104】
本発明の上記方法を育種法によって実施する場合には、より具体的には、以下のような工程を含む方法を挙げることができる。
(a)植物Aと、本発明のDNAを有する他の植物Bを交雑させ、F1を作出する工程
(b)前記F1と前記植物Aを交雑させる工程
(c)前記DNAを有する植物を選抜する工程
(d)工程(c)によって選抜された植物と、前記植物Aを交雑させる工程
【0105】
上記方法においては、本発明のDNAを有する植物Bと、着色させたい植物もしくはタンニン類を蓄積させたい植物(これら植物を「植物A」と記載する。)を交雑し、Bのもつ本発明のDNAが受け継がれ、かつ植物Aに近い個体を選抜し、これに植物Aによる交雑を重ねていく「戻し交雑」を行って、Bが有する本発明のDNAの形質を意図的に導入する。その際、一般的にゲノム育種に利用されるDNAマーカーを利用して本発明のDNAを有する植物を選抜することにより、上記「戻し交雑」による置換を効率的に行うことが可能である。その結果、育種期間の短縮に繋がり、また、余分なゲノム領域の混入を正確に除くことができる。通常、「戻し交雑」では、本発明のDNAと非常に強く連鎖する他のDNAに依存する形質がどうしても排除できないという現象が問題となることがあるが、本発明のDNAの近傍に存在するDNAマーカーを利用することにより、所望の植物の正確な選抜が可能となる。
【0106】
上記方法においては、必要に応じて、本発明のDNA以外のゲノム全域が目的の遺伝形質でホモ固定するまで、繰り返して行うことができる。即ち、本発明の好ましい態様のおいては、上記工程(d)によって交雑された個体について、一般的なDNAマーカーを利用して、本発明のDNAを有し、かつ、ゲノム構造が植物Aに近い植物個体を選抜することができる。さらに、この選抜された植物個体は、必要に応じて、「戻し交雑」(イネ品種Aと交雑)させることができる。
【0107】
特にDNAマーカーを利用したゲノム育種方法では、置換率の高い個体を選抜して次の交雑に進むことができるため、世代を進めるほどに選抜効率が良くなる。また、本方法では、少ない個体数を扱えば済むので、省スペースでの育種が可能になる。さらに、温室や人工気象室を利用して1年に複数回もの交雑が可能になる。
【0108】
上記工程(c)において、DNAマーカーを用いて選抜するとは、当該DNAマーカーを特徴付ける塩基配列(例えば、多型等)についての塩基種の情報を基に、選抜を行うことを言う。例えば、本発明のDNAの近傍に多型変異が存在する場合、当該多型変異と同一の多型変異を有する個体を選抜すること等を言う。
【0109】
本発明の育種方法は、好ましくは、DNAマーカーを利用した「ゲノム育種」方法である。該「ゲノム育種」は「マーカー育種」とも呼ばれる。
【0110】
「ゲノム育種」とは、ゲノム全域に高密度で存在するDNAマーカーを利用し、交雑のたびに後代個体のゲノム全域についての染色体置換状況を図示したものを描写し、最も目的に適った染色体置換を有する個体を幼苗期に選抜して次の交雑に用いることを連続して行う育種方法である。従来の育種法では、次の交雑に用いる個体を選抜するために形質評価を用いていたため、多数の個体収穫期まで成長させる必要があり、1年に1回、あるいは2年に1回の交雑しかできず、また多数の個体を均一に栽培し正しく形質評価するには多大な労力と経験を要した。また、育種過程の最後に、染色体全域をホモ接合体とするために、複数回の自殖が必要であり、結果的に品種の完成までに10年以上を要することが多かった。また、この手法は長年の経験と勘に依存するものである。これに対して、ゲノム育種の戦略を用いれば、交雑して得られた多数の種子を発芽させ、幼苗期に選抜して少数の個体を育てて交雑することを繰り返し、形質評価も最終段階まで不要のため、温室や人工気象室を利用して1年に複数回もの交雑が可能である。さらに、染色体全域がホモ固定しているかどうかは選抜の過程で確認できるため、通常、交雑を終了した時点で品種が完成する。このため、開始から短期間で目的の植物を作出することが可能である。加えて、用いるDNAマーカーの密度を十分に高めることにより、形質転換に匹敵する精度で、目的の遺伝子領域のみを導入することも可能である。形質転換を用いないため、遺伝子組換え作物が受け入れられない国や地域でも問題なく栽培できる植物体が作製できる。
【0111】
上述のように、本発明の育種方法の好ましい態様としては、本発明のDNA配列、またはその周辺配列に存在するDNAマーカーを利用して選抜することを特徴とする方法である。
【0112】
本発明の育種方法において利用可能なDNAマーカーは、特に制限されず、一般的に知られている種々のDNAマーカーを好適に用いることができる。例えば、RFLP(制限酵素断片長多型)マーカー、SSR(単純反復配列)マーカー、SNP(一塩基多型)マーカー等を例示することができる。
【0113】
本発明の植物がイネの場合には、例えば、図21で示される各種マーカーを利用することが可能である。本発明のRcはR1807-R1582の2つのマーカーの間に存在し、E12196Sのマーカーの近傍に存在する。また、本発明のRd(DFR)はR2374とC808の間に存在する。従って、本発明の育種方法において利用可能なマーカーの好ましい具体例としては、これら4種のマーカーを挙げることができる。今回同定したDNA配列中にマーカーを作り出すことももちろん可能である。
【0114】
また、本発明者のRcもしくはRd遺伝子を植物内において発現させることによって、植物体もしくはその種子を着色可能であり、さらに、有色の植物体に内在するこれら遺伝子の発現を抑制することにより、脱色(白色化)された植物体もしくはその種子を作出することも可能である。
【0115】
即ち本発明は、植物に内在する遺伝子であって、植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能を有するタンパク質をコードする遺伝子(本発明のDNAからなる遺伝子)の発現を抑制する工程を含む、脱色された植物体もしくはその種子の製造方法を提供する。
【0116】
該植物体は、本発明のDNAからなる遺伝子の発現が人為的に抑制されていることを特徴とする、遺伝子改変植物である。該遺伝子改変植物は、例えば、「ノックアウト植物」、「ノックダウン植物」あるいは「トランスジェニック植物」と呼ばれる場合もある。
【0117】
本発明において「遺伝子の発現が人為的に抑制されている」には、例えば、(1)本発明のDNAからなる遺伝子における遺伝子対の一方または双方に、ヌクレオチドの挿入、欠失、置換等の遺伝子変異を有することによって該遺伝子の発現が抑制されている状態、(2)本発明の遺伝子の発現を抑制する機能を有する核酸(例えば、アンチセンスRNAまたはsiRNA等)の作用により遺伝子の発現が抑制されている状態、等を挙げることができる。
【0118】
本発明における「抑制」には、本発明の遺伝子の発現が完全に抑制されている場合、および、植物における本発明の遺伝子の発現量が他の植物における遺伝子の発現量と比較して有意に低下している場合等が含まれる。
【0119】
上記(1)には、本発明の遺伝子の遺伝子対の一方の遺伝子の発現のみが抑制されている場合も含まれる。本発明における遺伝子変異の存在する部位は、該遺伝子の発現が抑制されるような部位であれば特に制限されず、例えばエクソン部位、プロモーター部位等を挙げることができる。
【0120】
本発明の遺伝子ノックアウト植物は、当業者においては一般的に公知の遺伝子工学技術により作製することができる。例えば、以下のようにして遺伝子ノックアウト植物を作製することができる。まず、植物から本発明の遺伝子(植物がイネの場合には、RcもしくはRd遺伝子)のエクソン部分を含むDNAを単離し、このDNA断片に適当なマーカー遺伝子を挿入し、ターゲッティングベクターを構築する。このターゲッティングベクターをエレクトロポレーション法、アグロバクテリウム法などにより植物細胞に導入し、相同組み換えを生じた細胞株を選抜する。挿入するマーカー遺伝子としては、一般的な薬剤選択用遺伝子などが好ましい。薬剤選択用遺伝子を挿入した場合には、薬剤を含む培地で培養するだけで相同組み換えを生じた細胞株を選抜することができる。また、PCRおよびサザンブロットにより相同組み換え体の検定を行い、本発明の遺伝子が不活性化された細胞株を効率よく得ることもできる。
【0121】
本発明の上記ノックアウト植物(ノックダウン植物)は、例えば、本発明の遺伝子の発現を抑制する機能を有する核酸を植物へ導入することによって、作出することも可能である。
【0122】
本発明の遺伝子の発現を抑制する機能を有する核酸としては、以下の(a)〜(c)に記載の核酸を例示することができる。
【0123】
(a)本発明のDNAからなる遺伝子の転写産物またはその一部に対するアンチセンス核酸
(b)本発明のDNAからなる遺伝子の転写産物を特異的に開裂するリボザイム活性を有する核酸
(c)本発明のDNAからなる遺伝子の発現をRNAi効果による阻害作用を有する核酸
【0124】
本発明における「核酸」とはRNAまたはDNAを意味する。また、所謂PNA (peptide nucleic acid)等の化学合成核酸アナログも、本発明の核酸に含まれる。PNAは、核酸の基本骨格構造である五単糖・リン酸骨格を、グリシンを単位とするポリアミド骨格に置換したもので、核酸によく似た3次元構造を有する。
【0125】
特定の内在性遺伝子の発現を阻害する方法としては、アンチセンス技術を利用する方法が当業者によく知られている。アンチセンス核酸が標的遺伝子の発現を阻害する作用としては、以下のような複数の要因が存在する。即ち、三重鎖形成による転写開始阻害、RNAポリメラーゼによって局部的に開状ループ構造が作られた部位とのハイブリッド形成による転写阻害、合成の進みつつあるRNAとのハイブリッド形成による転写阻害、イントロンとエクソンとの接合点におけるハイブリッド形成によるスプライシング阻害、スプライソソーム形成部位とのハイブリッド形成によるスプライシング阻害、mRNAとのハイブリッド形成による核から細胞質への移行阻害、キャッピング部位やポリ(A)付加部位とのハイブリッド形成によるスプライシング阻害、翻訳開始因子結合部位とのハイブリッド形成による翻訳開始阻害、開始コドン近傍のリボソーム結合部位とのハイブリッド形成による翻訳阻害、mRNAの翻訳領域やポリソーム結合部位とのハイブリッド形成によるペプチド鎖の伸長阻害、および核酸とタンパク質との相互作用部位とのハイブリッド形成による遺伝子発現阻害などである。このようにアンチセンス核酸は、転写、スプライシングまたは翻訳など様々な過程を阻害することで、標的遺伝子の発現を阻害する(平島および井上, 新生化学実験講座2 核酸IV遺伝子の複製と発現, 日本生化学会編, 東京化学同人, 1993, 319-347.)。
【0126】
本発明で用いられるアンチセンス核酸は、上記のいずれの作用により本発明の遺伝子の発現を阻害してもよい。一つの態様としては、本発明の遺伝子のmRNAの5'端近傍の非翻訳領域に相補的なアンチセンス配列を設計すれば、遺伝子の翻訳阻害に効果的と考えられる。また、コード領域もしくは3'側の非翻訳領域に相補的な配列も使用することができる。このように、本発明の遺伝子の翻訳領域だけでなく非翻訳領域の配列のアンチセンス配列を含む核酸も、本発明で利用されるアンチセンス核酸に含まれる。使用されるアンチセンス核酸は、適当なプロモーターの下流に連結され、好ましくは3'側に転写終結シグナルを含む配列が連結される。このようにして調製された核酸は、公知の方法を用いることで、所望の植物へ形質転換できる。アンチセンス核酸の配列は、形質転換される植物が持つ内在性の本発明の遺伝子またはその一部と相補的な配列であることが好ましいが、遺伝子の発現を有効に抑制できる限りにおいて、完全に相補的でなくてもよい。転写されたRNAは、標的遺伝子の転写産物に対して好ましくは90%以上、最も好ましくは95%以上の相補性を有する。アンチセンス核酸を用いて標的遺伝子の発現を効果的に抑制するには、アンチセンス核酸の長さは少なくとも15塩基以上25塩基未満であることが好ましいが、本発明のアンチセンス核酸は、必ずしもこの長さに限定されない。
【0127】
また、本発明の遺伝子の発現の阻害は、リボザイム、またはリボザイムをコードするDNAを利用して行うことも可能である。リボザイムとは触媒活性を有するRNA分子を指す。リボザイムには種々の活性を有するものが存在するが、中でもRNAを切断する酵素としてのリボザイムに焦点を当てた研究により、RNAを部位特異的に切断するリボザイムの設計が可能となった。
【0128】
また、ヘアピン型リボザイムも本発明の目的に有用である。ヘアピン型リボザイムからも、標的特異的なRNA切断リボザイムを作出できることが示されている(Kikuchi, Y. & Sasaki, N., Nucl Acids Res, 1991, 19, 6751.、菊池洋, 化学と生物, 1992, 30, 112.)。このように、リボザイムを用いて本発明における遺伝子の転写産物を特異的に切断することで、該遺伝子の発現を阻害することができる。
【0129】
内在性遺伝子の発現の阻害は、さらに、標的遺伝子配列と同一もしくは類似した配列を有する二本鎖RNAを用いたRNA干渉(RNA interferance;RNAi)によっても行うことができる。本発明のRNAi効果による阻害作用を有する核酸は、一般的にsiRNAとも言われる。RNAiは、標的遺伝子のmRNAと相同な配列からなるセンスRNAとこれと相補的な配列からなるアンチセンスRNAとからなる二本鎖RNAを細胞等に導入することにより、標的遺伝子mRNAの破壊を誘導し、標的遺伝子の発現を抑制し得る現象である。このようにRNAiは、標的遺伝子の発現を抑制し得ることから、従来の煩雑で効率の低い相同組換えによる遺伝子破壊方法に代わる簡易な遺伝子ノックアウト方法として、または、遺伝子治療への応用可能な方法として注目を集めている。RNAiに用いるRNAは、本発明の遺伝子もしくは該遺伝子の部分領域と必ずしも完全に同一である必要はないが、完全な相同性を有することが好ましい。
【0130】
本発明の上記(c)の核酸の好ましい態様として、本発明の遺伝子に対してRNAi(RNA interference;RNA干渉)効果を有する二本鎖RNA(siRNA)を挙げることができる。より具体的には、配列番号:2または5に記載の塩基配列の部分配列に対するセンスRNAおよびアンチセンスRNAからなる二本鎖RNA(siRNA)を挙げることができる。
【0131】
なお、上記RNA分子において一方の端が閉じた構造の分子、例えば、ヘアピン構造を有するsiRNA(shRNA)も本発明に含まれる。即ち、分子内において二本鎖RNA構造を形成し得る一本鎖RNA分子もまた本発明に含まれる。
【0132】
本発明の上記「RNAi効果により抑制し得る二本鎖RNA」は、当業者においては、該二本鎖RNAの標的となる本発明の遺伝子の塩基配列を基に、適宜作製することができる。一例を示せば、配列番号:2または5に記載の塩基配列をもとに、本発明の二本鎖RNAを作製することができる。即ち、配列番号:2または5に記載の塩基配列をもとに、該配列の転写産物であるmRNAの任意の連続するRNA領域を選択し、この領域に対応する二本鎖RNAを作製することは、当業者においては、通常の試行の範囲内において適宜行い得ることである。また、該配列の転写産物であるmRNA配列から、より強いRNAi効果を有するsiRNA配列を選択することも、当業者においては、公知の方法によって適宜実施することが可能である。また、一方の鎖(例えば、配列番号:2または5に記載の塩基配列)が判明していれば、当業者においては容易に他方の鎖(相補鎖)の塩基配列を知ることができる。siRNAは、当業者においては市販の核酸合成機を用いて適宜作製することが可能である。また、所望のRNAの合成については、一般の合成受託サービスを利用することができる。
【0133】
さらに、本発明の上記RNAを発現し得るDNA(ベクター)もまた、本発明の遺伝子の発現を抑制し得る化合物の好ましい態様に含まれる。例えば、本発明の上記二本鎖RNAを発現し得るDNA(ベクター)は、該二本鎖RNAの一方の鎖をコードするDNA、および該二本鎖RNAの他方の鎖をコードするDNAが、それぞれ発現し得るようにプロモーターと連結した構造を有するDNAである。本発明の上記DNAは、当業者においては、一般的な遺伝子工学技術により、適宜作製することができる。より具体的には、本発明のRNAをコードするDNAを公知の種々の発現ベクターへ適宜挿入することによって、本発明の発現ベクターを作製することが可能である。
【0134】
本発明の上記ノックダウン植物は、上記核酸(アンチセンスRNAまたはsiRNA等)、あるいは該核酸を発現し得る構造のベクターを、所望の植物へ導入することによって作製することができる。
【0135】
また、本発明は、配列番号:1、2、4もしくは5に記載の塩基配列、またはその相補配列に相補的な少なくとも15の連続する塩基を含むポリヌクレオチドを提供する。ここで「相補配列」とは、A:T、G:Cの塩基対からなる2本鎖DNAの一方の鎖の配列に対する他方の鎖の配列を指す。また、「相補的」とは、少なくとも15個の連続したヌクレオチド領域で完全に相補配列である場合に限られず、少なくとも70% 、好ましくは少なくとも80% 、より好ましくは90% 、さらに好ましくは95% 以上の塩基配列の同一性を有すればよい。このようなDNAは、本発明のDNAの検出や、本発明のDNAを有する植物体(植物細胞)の選抜を行なうためのプローブとして、また、本発明のDNAの増幅を行うためのプライマーとして有用である。
【0136】
また本発明は、本発明の植物体もしくはその種子の製造方法、またはこれらの育種方法に用いるためのオリゴヌクレオチドを含む、試薬(例えば、植物体育種用試薬等)を提供する。
【0137】
本発明の試薬は、より具体的には、以下の(a)および(b)のオリゴヌクレオチドを含有する試薬である。
(a)本発明のDNAの全配列もしくはその一部の配列を増幅するためのオリゴヌクレオチドプライマー
(b)本発明のDNA領域とストリンジェントな条件でハイブリダイズし、少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドプローブ
【0138】
本発明のプライマー、またはプローブは、それを構成する塩基配列をもとに、任意の方法によって合成することができる。本発明のプライマーまたはプローブの、ゲノムDNAに相補的な塩基配列の長さは、通常15〜100、一般に15〜50、好ましくは15〜30である。与えられた塩基配列に基づいて、当該塩基配列を有するオリゴヌクレオチドを合成する手法は公知である。更に、オリゴヌクレオチドの合成において、蛍光色素やビオチンなどで修飾されたヌクレオチド誘導体を利用して、オリゴヌクレオチドに任意の修飾を導入することもできる。あるいは、合成されたオリゴヌクレオチドに、蛍光色素などを結合する方法も公知である。
【0139】
また本発明は、本発明の各種方法に使用するためのキット、例えば、本発明の植物育種方法に使用するための育種用キットを提供する。
【0140】
本発明のキットの好ましい態様においては、上記(a)または(b)の少なくとも1種のオリゴヌクレオチドを含む。本発明のキットには、適宜、陽性や陰性の標準試料、使用方法を記載した指示書等をパッケージしておくこともできる。
【0141】
本発明のDNAもしくはベクターは、例えば、着色された植物体、もしくは着色された種子の作出等に利用することが可能である。また、本発明のDNAもしくはベクターは、タンニン類が蓄積した植物体、もしくはタンニン類が蓄積した種子の作出に利用することも可能である。
【0142】
本発明のDNAもしくはベクターを、所望の植物体もしくはその種子において発現させることにより、着色された、あるいはタンニン類が蓄積した植物体もしくはその種子を作製することが可能である。
【0143】
従って本発明は、本発明のDNAもしくはベクターを有効成分とする、植物体着色剤を提供する。
【0144】
本発明における「植物体着色剤」とは、植物体もしくはその種子の全部またはその一部を着色させる作用を有する薬剤を言い、通常、植物体もしくはその種子を着色させることを用途とする、本発明のDNAもしくはベクターを有効成分とする物質、または組成物(混合物)を指す。
【0145】
また本発明は、本発明のDNAもしくはベクターを有効成分とする、タンニン類蓄積剤を提供する。
【0146】
本発明における「タンニン類蓄積剤」とは、植物体もしくはその種子の全部またはその一部へタンニン類を蓄積させる作用を有する薬剤を言い、通常、植物体もしくはその種子へタンニン類を蓄積させることを用途とする、本発明のDNAもしくはベクターを有効成分とする物質、または組成物(混合物)を指す。
【0147】
さらに本発明は、上記(a)〜(c)に記載の核酸を有効成分とする、植物体脱色化剤を提供する。
【0148】
本発明における「植物体脱色化剤」とは、植物体もしくはその種子の全部またはその一部を脱色させる作用を有する薬剤を言い、通常、植物体もしくはその種子を脱色させることを用途とする、本発明の上記核酸を有効成分とする物質、または組成物(混合物)を指す。
【0149】
上記「脱色」とは、例えば、赤色、赤褐色、褐色等の色から白色へ変化させること等を言う。植物がイネの場合には、その種子であるコメの色が、例えば、赤米から白米へ変化させること等を指す。
【0150】
本発明の薬剤においては、有効成分であるDNAまたはベクター以外に、例えば、滅菌水、生理食塩水、植物油、界面活性剤、脂質、溶解補助剤、緩衝剤、保存剤等が必要に応じて混合されていてもよい。
【0151】
また本発明は、本発明の上記製造方法、または育種方法によって作出される植物体もしくはその種子を提供する。
例えば、(1)人為的に作製された植物体もしくはその種子であって、本発明のDNAを有し着色されていることを特徴とする植物体もしくはその種子、(2)人為的に作製された植物体もしくはその種子であって、本発明のDNAを有しタンニン類が蓄積していることを特徴とする植物体もしくはその種子、(3)植物に内在する遺伝子であって、植物体もしくはその種子を着色させる機能、または、植物体もしくはその種子へタンニン類を蓄積させる機能を有するタンパク質をコードする遺伝子の発現が人為的に抑制され、脱色されていることを特徴とする植物体もしくはその種子、は本発明に含まれる。
【実施例】
【0152】
以下、本発明を実施例によりさらに具体的に説明するが本発明はこれら実施例に制限されるものではない。なお、遺伝子型の記号、学術用語はKinoshita and Takahashiの文献(Kinoshita, T. & Takahashi, M. (1991) J. Fac. Agr. Hokkaido Univ. 65, 1-61.)とKinoshitaの文献(Kinoshita, T. (1995) Rice Genetics Newsletter 12, 9-153.)を参考にした。
【0153】
〔実施例1〕 実験材料と方法
(a)資料植物及び核酸の単離
イネ日本晴品種、カサラス、コシヒカリ、tRc(NIL)、ムラサキイネ、及び形質転換イネは28℃の温室で栽培した。tRcはCI-69(赤米)にT-65を交雑させることによって作成した準同質遺伝系統(第7染色体のT-65(遺伝子型rcrd)が、第7染色体のCI-69(遺伝子型RcRd)に置換された準同質遺伝系統)である。ゲノムDNAは、発芽後10日目の若い葉から抽出した。DNA抽出には、DNeasy plant mini kit(Qiagen)を用い、キットに記載されているプロトコルに従った。葉のRNAは、発芽後10日目の若い葉から抽出した。穂のRNAは、開花後7日目の穂から抽出した。RNA抽出は、RNeasy plant mini kit(Qiagen)を用い、キットに記載されている方法に従った。
【0154】
(b)PCR,DNA 配列
OsDFRのゲノムDNAは、1UのLA Taq DNA polymerase(TaKaRa)によって増幅された。PCRに用いたプライマーは、OsDFR遺伝子の翻訳開始コドン1.7 kb上流のOsDFRF4(5'-CATGTGTGAGCATCTCTAAGTGACG-3'/配列番号:9)とDFR遺伝子の翻訳終止コドン0.5 kb 下流のOsDFRR4(5'-CTCTCTAGGAGCACGTGTAAAGG-3'/配列番号:10)を用いた(図4)。DFRのPCR産物のシーケンスは、CEQ Dye Terminator Cycle Sequencing Start KitおよびCEQ 2000XL DNA Analysis System(Beqman Coulter)を用い、これに記載されている方法に従った。シーケンスに用いたプライマーは、OsDFR遺伝子の第2エクソンに相当するOsDFRR5(5'-AGAAGTCGATGTCGCTCCAG-3'/配列番号:11)を用いた(図4)。
【0155】
(c)RT-PCR
PCRの基質に用いたcDNAの合成はSuperScriptTM 逆転写酵素とRandom Primers (Invitrogen)を用い、これに記載されている方法に従った。PCRはAmpli taq(Applied Biosystems)を用いて、94℃ 1分静置してのち、94℃15秒、60℃30秒、72℃30秒のサイクルを40回繰り返した。使用したプライマーは、OsDFR遺伝子の第1エクソンに対応するOsDFRF8(5'-GATTCGCGTGCGAATCCAAC-3'/配列番号:12)、及びOsDFR遺伝子の第2エクソンに対応するOsDFRR5(5'-AGAAGTCGATGTCGCTCCAG-3'/配列番号:13)を用いた(図5)。イネアクチン遺伝子(X15865)の転写産物の増幅には、イネアクチン遺伝子の第2エクソンに対応するRAc1cDNA-F プライマー(5'-TGTCATGGTCGGAATGGG-3'/配列番号:14)、及びイネアクチン遺伝子の第3エクソンに対応するRAc1cDNA-R プライマー(5'-TGCCAGGGAACATAGTGG -3'/配列番号:15)を用いた。PCR産物は1.8 % (w/v)のアガロースゲル電気泳動により分離・同定した。
【0156】
(d)タンパク質ブロット解析
穂の全タンパク質は、抽出Buffer(200 mM Tris-HCL.pH 6.8; 8 %(w/v) SDS; 40 % Glycerol; 5 mM 2-mercaptoethanol)で抽出した。抽出したサンプルは、4,000×gで5分間、4℃で遠心し、上静をSDS-PSGE running Bufferに溶かした。タンパクは12 %のポリアクリルアミドゲルで分離し、Immobillon PVDF membraneに転写した。
【0157】
DFRに対する抗血清は、ムラサキイネのOsDFRのORF全長に相当するcDNAをPCRにより増幅し、pGEX4T-3 vector(Amersham Pharmacia Biotech)に連結し、E.coli JM109株に遺伝子導入した(図6)。GST/DFRの融合タンパク質は、1 mMのIPTGを加えることによって誘導し、(Asano, T., Kusano, H., Okuda, T., Kubo, N., Shimada, H. & Kadowaki, K. (2001) Plant Cell Physial 43, 668-674.)に記載されている方法に従って精製した。精製された融合タンパク質は、2週間おきに6回ウサギに注射し、その血清を解析に用いた。
【0158】
(e)Rcのゲノムアノテーション
Rcのゲノムアノテーションによる候補の探索は、Rice GAAS(Rice Genome Automated Annotation System){ http://ricegaas.dna.affrc.go.jp/}を用いて行った。
【0159】
(f)形質転換植物の産出
ムラサキイネ型DFR、コシヒカリ型DFRの過剰発現体の作成についてはムラサキイネ、コシヒカリのOsDFRのORF全長に相当するゲノム領域を、インフレームでpCAMBIA1301 plasmidの35S promoterの下流にセンス方向に連結した(図7、図8)。またDFRのアンチセンス個体の作成についてはOsDFR cDNAのORFに相当する領域を、pCAMBIA1301 plasmidの35S promoterの下流にアンチセンス方向に連結した(図9)。ゲノムアノテーションにより得られたRcの3つの候補(Rc-MYB1、Rc-MYB2、Rc-MYC)は、予想されるプロモーター領域1.5 kbから予想されるポリA付加シグナルまでの約12 kbをpCAMBIA1301 plasmidのマルチクローニングサイトに連結した(図10)。Rc-MYB1、Rc-MYB2、Rc-MYCの遺伝子構築に使用したのは、それぞれカサラス由来の105659−107804 (AP005197)、 118440−126019 (AP003754)、 150211−156639 (AP005779) である。作成したプラスミドは、Agrobacterium tumefaciens strain EHA105に遺伝子導入した。アグロバクテリウム法によるイネの形質転換(DFRの形質転換は、tRc(NIL)に対して行い、Rcの形質転換は日本晴に対して行った)は、Patent No. WO01/06844に記載されている方法に従った。得られた形質転換植物は、温室で育てた。
【0160】
〔実施例2〕 Rd遺伝子の同定
(1)RcRd、Rcrd、rcRd及びrcrd遺伝子型の表現型
イネにおいて、タンニン類(CTs)による種皮の着色には、RcとRdの2つの支配因子が存在する(Nagao, S. (1951) Advances in Genetics 4, 181-212.)。Rcは、種皮の赤色及び赤褐色の着色に必須な因子である。Rdは、Rcが存在するときにのみRcと協調的に機能し、種皮の赤色の着色に必須な因子である。交雑実験の結果、RcRdの遺伝子型の玄米色は赤、Rcrdの玄米色は赤褐色(くぼみがあり、そこに濃い着色が見られる)、rcRd またrcrdの玄米色は白色になることが知られている(図3)。
【0161】
(2)DFRを伴うRd遺伝子座の分離
RcRdの玄米色は赤、Rcrdの玄米色は赤褐色、rcRd,rcrdの玄米色は白となることが遺伝学的解析により知られている(Nagao, S. & Takahashi, M. (1947) Jap. J. Genet. Supp. 1, 1-27.)。またRcRd、Rcrdの玄米には主要な色素であるプロアントシアニジンが存在することがNagaoら(Nagao, S., Takahashi, M. & Miyamoto, T. (1956) The Japanese Jourant of Genetics 32, 124-128.)によって確認された。イネの玄米の色調に影響を及ぼすことから、Rdはプロアントシアニジン合成に関与する遺伝子座であり、対立遺伝子であるrdは機能を欠失しているわけではなく、Rdに比べてプロアントシアニジン合成系における機能が弱いことが考えられる。ここでKinoshitaら(Kinoshita, T. & Takahashi, M. (1991) J. Fac. Agr. Hokkaido Univ. 65, 1-61.)の報告から、Rd座はA座の近傍に存在し、それは0.3 cM以内である(図2)。
【0162】
A座はイネのアントシアニン合成系のActivatorとして定義された遺伝子であって、現在までにA座がコードする遺伝子については明らかになっていない。イネのアントシアニンによる着色には色原素遺伝子であるC(Chromogen)、アントシアニン活性化因子であるA(Activator)、そして着色部位を規定するP(Purple)の3つの遺伝子が存在する。これら3つの因子が存在するときにイネの組織(玄米、葉、ラミナジョイント、茎、etc)にアントシアニンによる着色が起こる(表1)。
【0163】
【表1】
(*Nagaoら(Nagao, S. & Takahashi, M. (1963) Agr., Hokkaido Univ., Sapporo 53, 76-131.)およびKatoら(Kato, S., & Ishikawa. J. (1923) Jap. J. Genet. 1, 1-7.)によって決定された遺伝子座は、独立した研究であり、比較できない。)
【0164】
ここでCはトウモロコシのC1(MYB)のホモログであることが知られており(Reddy, V. S., Scheffler, B. E., Wienand, U., Wessler, S. R. & Reddy, A. R. (1998) Plant Molecular Biology 36, 497-498.; Saitoh, K., Onishi, K., Mikami, I., Thidar, K. & Sano, Y. (2004) Genetics 168, 997-1007.)、PはトウモロコシのR/B(bHLH)のホモログであることが近年明らかになった(Hu, J., Anderson, B. & Wessler, R. S. (1996) Genetics 142, 1021-1031.; Hu, J., Reddy, S. V. & Wessler, R. S. (2000) Plant Molecular Biology 42, 667-678.; Sakamoto, W., Ohmori., T., Kageyama, K., Miyazaki, C., Saito, A., Murata, M., Noda, K. & Maekawa, M. (2001) Plant Cell Physiol. 42, 982-991.)(表2)。
【0165】
【表2】
【0166】
このことはイネのA遺伝子がDFRをコードすることを示している。
【0167】
RdはA(DFR)の近傍に存在する(Nagao, S. & Takahashi, M. (1947) Jap. J. Genet. Supp. 1, 1-27.)。A(DFR)が存在するR2374(103.7 cM) −C808(106.2 cM)の間に存在する遺伝子をRice GAAS programを用いて解析した結果、この領域にDFR以外にタンニン合成に関与する遺伝子は存在しなかった。よってRdもAと同様にDFRをコードすることが示唆された。
【0168】
上述の結果から、Rd遺伝子とA遺伝子は同一であり、これらの遺伝子は同一のDFRタンパク質をコードしているという仮説が考えられた。この仮説を検証するために、準同質遺伝系統T-65(RcRd, A)とtRc系統(Rcrd, a)との間で、交配実験を行った。もし、A遺伝子とRd遺伝子が同一でないならば、AとRdとの間に起こる組換え体がF2において観察される。該組換え体は(Rcrd, A)、または(RcRd, A)の遺伝子型を有する。
【0169】
A遺伝子は、ふ先(apiculus)におけるアントシアニン色素の原因遺伝子であることから、(Rcrd, A)を有する植物は、種子の果皮が赤褐色となりアントシアニンによって着色されたふ先を有する。一方、(RcRd, a)を有する植物は、種子の果皮が赤色となり非着色のふ先を有する。
【0170】
実験の結果、F2において、672個体が種子の果皮が赤色となり、(RcRd, A)によるアントシアニンによって着色されたふ先が観察された。また232個体は種子の果皮が赤褐色となり、(Rcrd, a)による非着色のふ先が観察された。RdとAとの間の組換え体は観察されなかった。このことから、RdとAは遺伝学的に強く連鎖し、同一の遺伝子であることが示唆された。
【0171】
(3)Rcrd とゲノムOsDFRの相補性
tRc(Rcrd)にムラサキイネ由来のDFRを相補した結果、玄米色が赤褐から赤になった。RcRdの玄米色が赤であることを考えると、RdはDFRをコードすることが明らかになった(図11)。
【0172】
(4)DFRアレルの解析
Rd、rdの遺伝子型による種皮色の違いとDFR遺伝子の構造の関係を探るためにDNAシーケンスを行った。解析に用いたものはRcRdの遺伝子型を持つカサラス、Rcrd の遺伝子型を持つtRc、rcrdの遺伝子型を持つ日本晴とコシヒカリである。DFRの第1エクソンから第2エクソンまでの領域をgenomic DNAをテンプレートとしてPCRで増幅し、ダイレクトシーケンスを行った。その結果Rdを持つカサラスは、以前に単離されているイネのDFR遺伝子と同様な構造をとっていた(Chen, M. & L. J. Bennetzen (1996) Plant Molecular Biology 32, 999-1001.; Nakai, K., Inagaki, Y., Nagata, H., Miyazaki, C., & Iida, S. (1998) Plant Biotechnology 15, 221-225.)。一方、rdを持つtRcや日本晴のDFR遺伝子の構造には、第2エクソンに突然変異によって出来たと思われる翻訳終止コドン(TAG)が存在した。さらにrdを持つと思われていたコシヒカリのDFRの構造は、カサラスとも日本晴れとも異なり第1エクソンの開始コドンのすぐ下流に突然変異によってできたと思われる翻訳終止コドン(TAG)が存在した(図12)。以上のことから、RdはカサラスタイプのDFRをコードし、rdは第1および第2エクソンに翻訳終止コドンの存在するコシヒカリ及び日本晴タイプのDFRをコードすることが明らかになった。
【0173】
rdのDFR遺伝子の構造は、第2エクソンに終止コドンが存在しており、開始コドンから突然変異によって出来た翻訳終止コドンまでは、OsDFRが42 kDaをコードするのに対し4.5 kDaであり、機能を持っているとは考えにくい。またDFRはイネに1コピーしか存在しない(Nakai, K., Inagaki, Y., Nagata, H., Miyazaki, C., & Iida, S. (1998) Plant Biotechnology 15, 221-225.; International Rice Genome Sequence Project. (2005) Nature 436, 793-800.)。しかしRcrd のDFR遺伝子は日本晴タイプであり、ORFのN末側が欠損しているが、玄米色は赤褐色で、プロアントシアニジンによる着色であることが知られている(Nagao, S., Takahashi, M. & Miyamoto, T. (1956) The Japanese Jourant of Genetics 32, 124-128.)。従って、第2エクソンに終止コドンを持つ日本晴タイプのDFRに機能があることが考えられた。そこで、Rdの対立遺伝子であるDFRが転写しているか調べるために、RcRdの遺伝子型を持つカサラス、rcrdの遺伝子型を持つ日本晴、rcrd’の遺伝子型を持つコシヒカリに対して、RT-PCRを行った。その結果、カサラス、tRc、日本晴、コシヒカリとも開花後7日目の穂で特異的に発現しており、その発現レベルの強さは、カサラス>日本晴>コシヒカリ であった(図13)。
【0174】
次に本発明者らは、Rdの対立遺伝子であるDFRの翻訳の機構を調べるために、RcRdの遺伝子型を持つカサラス、rcrdの遺伝子型を持つ日本晴、rcrdの遺伝子型を持つコシヒカリに対して、開花後7日目の穂において免疫染色を行った。その結果驚くことに、カサラスは、Alternative translation initiation(Bab, I., Smith, E., Gavish, H., Malka, Attar-Namdar., Chorev, M., Yu-Chen, Chen., Muhlrad, A., Birnbaum, J. M., Stein, G. & Frenkel, B. (1999) The Journal OF BIOLOGICAL CHEMISTRY 274, 14474-14481.; Chabregas, M. S., Luche, D. D., Marie-Anne, Van, Sluys., Carlos, F. M. Menck., Marcio, C. Silva-Filho. (2002) Journal of Cell Science 116, 285-291.; Smith, E., Meyerrose, E. T., Kohler, T., Malka, Namdar-Attar., Bab, N., Lahat, O., Noh, T., Li, J., Karaman, W. M., Hacia, G. J., Chen, T. T., Nolta, A. J., Muller, R., Bab, I. & Frenkel, B. (2005) Nucleic Acids Research 33, 1298-1308.; Hanfrey, C., Elliott, A. K., Franceschetti, M., Mayer, J. M., Illingworth, C. & Michael, J. A. (2005) The Journal OF BIOLOGICAL CHEMISTRY, in press.; Kabeya, Y. & Sato, N. (2005) Plant Physiology 138, 369-382.)により46 kDa、33 kDa、22 kDaの3つのORFが存在した。また日本晴、コシヒカリにおいても33 kDa、22 kDaの2つのORFが存在した。この46 kDaのバンドは、予想されたタンパク質の大きさよりも大きいが、DFRのN末端側に塩基性のアミノ酸が多いため、バンドがシフトしていると考えられた(図4)。
【0175】
ここでOsDFRのORFには、最初の開始コドンを含め5個のKozak コンセンサス配列が存在する(Kozak, M. (2000) Genomics 70, 396-406.; Kozak, M. (2002) Gene 299, 1-34.)。46 kDa、22 kDaのバンドは比較的強く検出されているが、これは最も強いKozak コンセンサス配列を持つ GnnATGG (nはA,T,G,Cいずれかの塩基)を持つ、一番目と4番目のKozak コンセンサスだと考えられた。33 kDaのバンドの由来については、分子量の大きさから考えて、第2エクソンのKozak配列以外のATGから翻訳が開始されていると推察された(図4)。
【0176】
以上の結果より、rd型のDFRはAlternative translation initiation(Kozak, M. (2000) Genomics 70, 396-406.; Kozak, M. (2002) Gene 299, 1-34.)により下流のATGから翻訳され、DFRの機能を相補していることが示された。以上の結果よりrcrdの遺伝子型を持つ日本晴にRcを相補すれば、タンニン類による着色が起こることが示唆された。
【0177】
〔実施例3〕 Rc遺伝子の同定
(1)Rcのゲノムアノテーション
現在までに遺伝学的解析により、Rcは第7染色体に存在することが知られている(Nagao, S. & Takahashi, M. (1963) Agr., Hokkaido Univ., Sapporo 53, 76-131.)(図15A)。またイネゲノムプロジェクトの終了によりRcは、第7染色体上のR1807(42.6 cM)とR1582(47.7 cM)の二つの分子マーカーの間に存在することが分かった(IRGSP, 2005)(図5B)。この領域に存在するプロアントシアニジン合成系(図1)の酵素をRiceGAAS(http://ricegaas.dna.affrc.go.jp/)を用いて検索した。しかしこの領域にプロアントシアニジン合成系(図1)の酵素に対して、ホモロジーが高いものは存在しなかった。
【0178】
そこで次に、プロアントシアニジン合成系の転写制御因子に着目した。プロアントシアニジン合成系の転写制御因子としては、MYC type proteinをコードしているアラビドプシスのTT8(Transparent Testa 8)(Nesi, N., Debeaujon, I., Jond, C., Pelletier, G., Caboche, M. & Lepiniec, L. (2001) Plant Cell 12, 1863-1878.)、MYB related proteinをコードするアラビドプシスのTT2(Nesi, N., Jond, C., Debeaujon, I., Caboche, M. & Lepiniec, L. (2001) Plant Cell 13, 2099-2114.)などが知られている。そこでRcが存在する第7染色体上のR1807(42.6 cM)とR1582(47.7 cM)に存在するMYC type protein、MYB related proteinをコードする遺伝子についてRiceGAASを用いて検索した。
【0179】
その結果、MYB related proteinが2つ、MYC type proteinが1つ見つかった。MYB related proteinをコードする遺伝子の一つはRc-MYB1と名付け、IRGSPのAP005197の105659 bp - 107804 bpにORFが存在した。もう一つのMYB related proteinはRc-MYB2と名付け、IRGSPのAP003754の 118440 bp - 126019 bpにORFが存在した。またMYC type proteinはRc-MYCと名付け、そのORFはIRGSPのAP005779の150211 bp - 156639 bpに存在した(図15A,B,C)。Rc-MYB1とRc-MYB2については、アラビドプシスのプロアントシアニジン合成系の転写制御因子であるTT2(MYB related protein)との相同性はともに24 %(Identities)であった。一方Rc-MYCについては、アラビドプシスのプロアントシアニジン合成系の転写因子であるTT8(MYC type protein)との相同性が38 %(Identities)、その中の一部分のbHLHドメインでは相同性は高く73 %(Identities)であった(図15D)(ClustalW ver.1.82)。
【0180】
(2)rcrdとゲノム断片の相補性
RcRdの遺伝子型を持つカサラス(RcRd)のRc-MYB1、Rc-MYB2、Rc-MYC(図15)についてプロモーター領域である翻訳開始コドン上流1.5 kbからポリA付加シグナルまでの約12 kbをrcrdの遺伝子型を持つ日本晴(rcrd)に遺伝子導入した。それぞれ16個体の形質転換体が得られた。Rc-MYB1、Rc-MYB2を遺伝子導入した個体については葉、もみ、玄米ともプロアントシアニジンによる着色は見られなかった。一方、Rc-MYCを遺伝子導入した個体は、玄米にプロアントシアニジンによると思われる着色が見られた(図16)。着色が見られた形質転換体は10個体得られ、その表現型はRcrdの遺伝子型を持つtRcと非常に類似していた。
【0181】
以上の結果より、Rc-MYCがプロアントシアニジン合成系支配因子であるRcであることが示された。玄米以外の組織では着色は見られなかった。以上のことから、Rcは種子特異的に働く転写因子であることが示された。
【0182】
(3)Rc領域の解析
次に本発明者らはカサラス(RcRd)のRc-MYCの完全長cDNA配列を決定した。使用したRNAは開花後7日目の穂から抽出した。完全長のカサラスのRc-MYCの配列を決定した結果、カサラスのRc-MYCは673 a.a.のタンパク質をコードする8個のエクソンからなる遺伝子であることが分かった。
【0183】
次に本発明者らは、カサラス(RcRd)のRc-MYCのDNA配列と日本晴(rcrd)の遺伝子配列を比較してみることにした。その結果、rcを持つ日本晴には第7エクソンに14 bpの欠失変異、さらに第8エクソンに6 bpの挿入変異と9 bpの欠失変異が確認できた。日本晴の第7エクソンの14 bpの欠失変異は、フレームシフトを起こし、第7エクソンの欠失場所の9 bp下流に終止コドンを生じさせていた。その結果、日本晴のRc-MYCは478 a.a. のタンパク質をコードしており、カサラスのRc-MYCに比べて215 a.a. だけ短かった。DNAの結合に必要な bHLHドメインは第8エクソンにコードされている。よって日本晴のRc-MYCは第8エクソンまで欠失変異のため翻訳されないので、bHLHドメインを持たないことになる。そのために日本晴Rc-MYCは転写制御因子として機能できないと考えられた(図18)。
【0184】
Rc-MYCの第7エクソンの14 bp 欠失している領域について、シーケンスを用いた多型解析をRcRd(玄米色は赤)、Rcrd(玄米色は赤褐)、rcrd(玄米色は白)について行った。図19は、玄米色が赤色のRcRd、玄米色が赤褐色のRcrdには第7エクソンに欠失がなく、玄米色が白色のrcrdには第7エクソンに欠失があることを示している。図19の結果はプロアントシアニジンによる玄米の着色はRc-MYCによることを示唆している。
【0185】
遺伝学的解析、逆遺伝学的解析は日本晴の玄米色がRc-MYCの機能が欠失したために、白色になったことを明確に示した。
【0186】
(4)MYC型タンパク質の相同性解析
次にRc-MYCとそのホモログの遺伝距離を調べることにした。アミノ酸配列のアラインメントはClastalX ver1.83を用い、系統樹はclastalX上のNJ法を使用した。またBoots strap valueもClastalX上の機能を使用した。系統解析に使用したのは、トウモロコシのIN1(U57899), Lc(M26227), B-Peru(X57276), R-S(X15806), ペチュニアのJAF13(AF020545), キンギョソウのDELILA(M84913), アラビドプシスのTT8(AJ277509), イネのRa(U39860), Rc-MYC(this study)を用いた。系統解析の結果、イネのRc-MYCは、トウモロコシのアントシアニン合成系抑制因子として単離されているIntensifier(IN1)と最も相同性が高かった。しかしながら、Rc-MYCはタンニン類蓄積促進因子であるが、IN1はその遺伝子機能が失われるとタンニン類が蓄積するため、機能は正反対である。一方、アントシアニン合成系活性化因子として単離されたキンギョソウのDELILAやペチュニアのJAF13とは最も相同性が低かった(図20)。
【特許請求の範囲】
【請求項1】
下記(a)〜(f)のいずれかに記載のDNA、該DNAによりコードされるタンパク質、または、該DNAを含む発現ベクターを有効成分として含有する、イネ着色化剤。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項2】
下記(a)〜(f)のいずれかに記載のDNA、該DNAによりコードされるタンパク質、または、該DNAを含む発現ベクターを有効成分として含有する、イネへのタンニン類蓄積剤。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項3】
下記(a)〜(c)のいずれかを有効成分として含有する、イネ脱色化剤。
(a)下記(1)〜(6)のいずれかに記載のDNAの転写産物またはその一部に対するアンチセンス核酸
(b)下記(1)〜(6)のいずれかに記載のDNAの転写産物を特異的に開裂するリボザイム活性を有する核酸
(c)下記(1)〜(6)のいずれかに記載のDNAの発現をRNAi効果による阻害作用を有する核酸
(1)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(2)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(3)配列番号:5に記載の塩基配列からなるDNA。
(4)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(5)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(6)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項4】
下記(a)〜(f)のいずれかに記載のDNA、または、該DNAを含む発現ベクターをイネ細胞へ導入し、該イネ細胞からイネを再生させる工程を含む、着色した、もしくはタンニン類が蓄積した形質転換イネの製造方法。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項5】
下記(a)〜(f)のいずれかに記載のDNA、該DNAによりコードされるタンパク質、または、該DNAを含む発現ベクターをイネの細胞内で発現させる工程を含む、イネもしくはその種子を着色させる、またはイネもしくはその種子にタンニン類を蓄積させる方法。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項6】
下記(a)〜(f)のいずれかに記載のDNA、該DNAによりコードされるタンパク質、または、該DNAを含む発現ベクターを植物体の細胞内で発現させる工程を含む、着色した、もしくはタンニン類が蓄積したイネもしくはその種子の製造方法。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項7】
前記DNA、または前記ベクターをイネ細胞へ導入する工程を含む、請求項5または6に記載の方法。
【請求項8】
下記工程(a)および(b)を含む、請求項5または6に記載の方法。
(a)前記DNAを有するイネと交雑させる工程、
(b)前記DNAを有するイネ改変体を選抜する工程
【請求項9】
着色した種子、もしくはタンニン類が蓄積した種子を有するイネを育種する方法であって、下記工程(a)〜(d)を含む育種方法。
(a)イネAと、下記(1)〜(6)のいずれかに記載のDNAを有する他のイネBを交雑させ、F1を作出する工程
(1)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(2)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(3)配列番号:5に記載の塩基配列からなるDNA。
(4)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(5)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(6)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
(b)前記F1と前記イネAを交雑させる工程
(c)前記DNAを有するイネを選抜する工程
(d)工程(c)によって選抜されたイネと、前記イネAを交雑させる工程
【請求項10】
前記工程(c)において、植物ゲノム中の前記DNA配列、またはその周辺配列に存在するDNAマーカーを利用して選抜されることを特徴とする、請求項9に記載の方法。
【請求項11】
イネに内在する遺伝子であって、イネもしくはその種子を着色させる機能、または、イネもしくはその種子へタンニン類を蓄積させる機能を有するタンパク質をコードするDFR遺伝子の発現を抑制する工程を含む、脱色された植物体もしくはその種子の製造方法。
【請求項12】
請求項5〜11のいずれかに記載の方法によって取得されるイネ、またはその種子。
【請求項13】
人為的に作製されたイネもしくはその種子であって、下記(a)〜(f)のいずれかに記載のDNAを有し着色されていることを特徴とするイネもしくはその種子。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項14】
人為的に作製されたイネもしくはその種子であって、下記(a)〜(f)のいずれかに記載のDNAを有しタンニン類が蓄積していることを特徴とするイネもしくはその種子。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項15】
イネに内在する遺伝子であって、イネもしくはその種子を着色させる機能、または、イネもしくはその種子へタンニン類を蓄積させる機能を有するタンパク質をコードするDFR遺伝子の発現が人為的に抑制され、脱色されていることを特徴とするイネもしくはその種子。
【請求項1】
下記(a)〜(f)のいずれかに記載のDNA、該DNAによりコードされるタンパク質、または、該DNAを含む発現ベクターを有効成分として含有する、イネ着色化剤。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項2】
下記(a)〜(f)のいずれかに記載のDNA、該DNAによりコードされるタンパク質、または、該DNAを含む発現ベクターを有効成分として含有する、イネへのタンニン類蓄積剤。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項3】
下記(a)〜(c)のいずれかを有効成分として含有する、イネ脱色化剤。
(a)下記(1)〜(6)のいずれかに記載のDNAの転写産物またはその一部に対するアンチセンス核酸
(b)下記(1)〜(6)のいずれかに記載のDNAの転写産物を特異的に開裂するリボザイム活性を有する核酸
(c)下記(1)〜(6)のいずれかに記載のDNAの発現をRNAi効果による阻害作用を有する核酸
(1)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(2)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(3)配列番号:5に記載の塩基配列からなるDNA。
(4)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(5)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(6)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項4】
下記(a)〜(f)のいずれかに記載のDNA、または、該DNAを含む発現ベクターをイネ細胞へ導入し、該イネ細胞からイネを再生させる工程を含む、着色した、もしくはタンニン類が蓄積した形質転換イネの製造方法。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項5】
下記(a)〜(f)のいずれかに記載のDNA、該DNAによりコードされるタンパク質、または、該DNAを含む発現ベクターをイネの細胞内で発現させる工程を含む、イネもしくはその種子を着色させる、またはイネもしくはその種子にタンニン類を蓄積させる方法。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項6】
下記(a)〜(f)のいずれかに記載のDNA、該DNAによりコードされるタンパク質、または、該DNAを含む発現ベクターを植物体の細胞内で発現させる工程を含む、着色した、もしくはタンニン類が蓄積したイネもしくはその種子の製造方法。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項7】
前記DNA、または前記ベクターをイネ細胞へ導入する工程を含む、請求項5または6に記載の方法。
【請求項8】
下記工程(a)および(b)を含む、請求項5または6に記載の方法。
(a)前記DNAを有するイネと交雑させる工程、
(b)前記DNAを有するイネ改変体を選抜する工程
【請求項9】
着色した種子、もしくはタンニン類が蓄積した種子を有するイネを育種する方法であって、下記工程(a)〜(d)を含む育種方法。
(a)イネAと、下記(1)〜(6)のいずれかに記載のDNAを有する他のイネBを交雑させ、F1を作出する工程
(1)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(2)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(3)配列番号:5に記載の塩基配列からなるDNA。
(4)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(5)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(6)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
(b)前記F1と前記イネAを交雑させる工程
(c)前記DNAを有するイネを選抜する工程
(d)工程(c)によって選抜されたイネと、前記イネAを交雑させる工程
【請求項10】
前記工程(c)において、植物ゲノム中の前記DNA配列、またはその周辺配列に存在するDNAマーカーを利用して選抜されることを特徴とする、請求項9に記載の方法。
【請求項11】
イネに内在する遺伝子であって、イネもしくはその種子を着色させる機能、または、イネもしくはその種子へタンニン類を蓄積させる機能を有するタンパク質をコードするDFR遺伝子の発現を抑制する工程を含む、脱色された植物体もしくはその種子の製造方法。
【請求項12】
請求項5〜11のいずれかに記載の方法によって取得されるイネ、またはその種子。
【請求項13】
人為的に作製されたイネもしくはその種子であって、下記(a)〜(f)のいずれかに記載のDNAを有し着色されていることを特徴とするイネもしくはその種子。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項14】
人為的に作製されたイネもしくはその種子であって、下記(a)〜(f)のいずれかに記載のDNAを有しタンニン類が蓄積していることを特徴とするイネもしくはその種子。
(a)配列番号:6〜8のいずれかに記載のアミノ酸配列からなるタンパク質をコードするDNA。
(b)配列番号:4に記載の塩基配列のコード領域を含むDNA。
(c)配列番号:5に記載の塩基配列からなるDNA。
(d)配列番号:6〜8のいずれかに記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA。
(e)配列番号:4に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA。
(f)配列番号:5に記載の塩基配列からなるDNAと95%以上の同一性を示すDNA。
【請求項15】
イネに内在する遺伝子であって、イネもしくはその種子を着色させる機能、または、イネもしくはその種子へタンニン類を蓄積させる機能を有するタンパク質をコードするDFR遺伝子の発現が人為的に抑制され、脱色されていることを特徴とするイネもしくはその種子。
【図1】

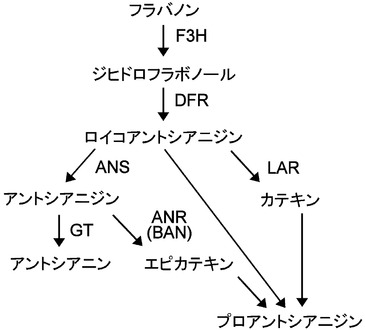
【図2】
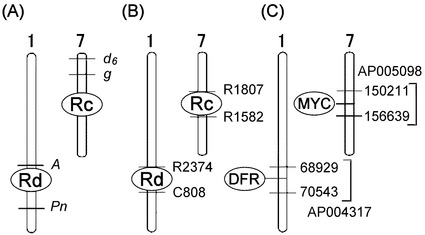
【図4】
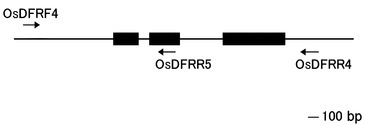
【図5】
【図6】
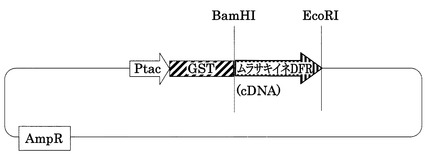
【図7】
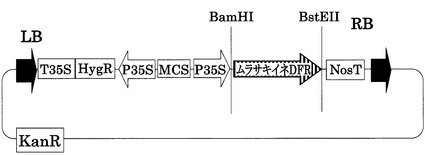
【図8】
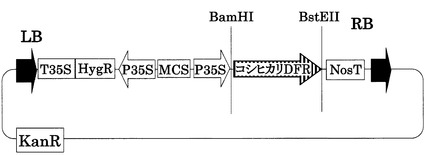
【図9】
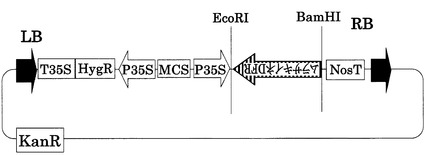
【図10】
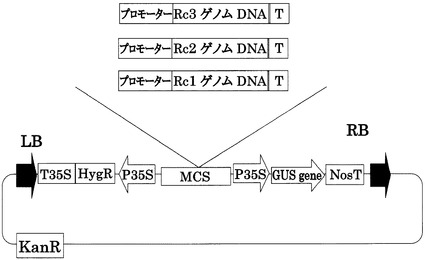
【図12】
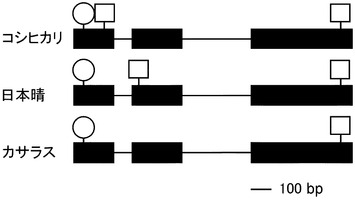
【図15A−C】
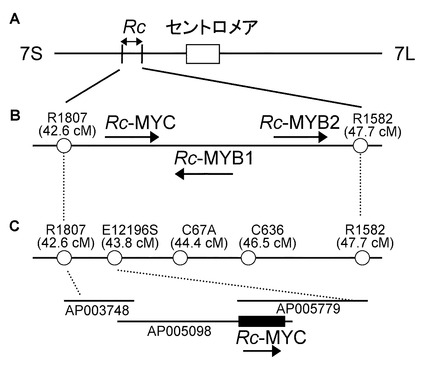
【図15D】
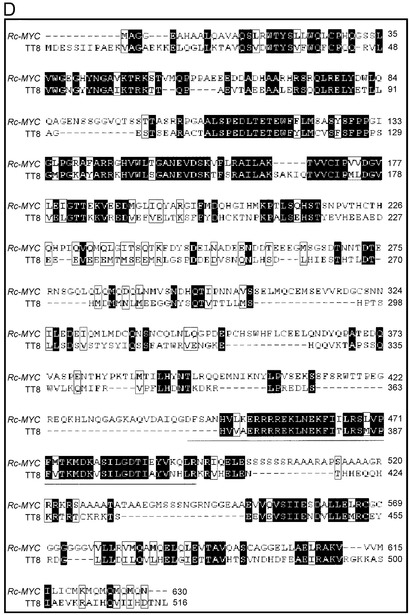
【図17】
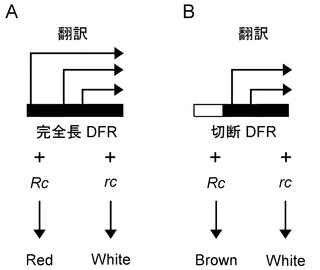
【図20】
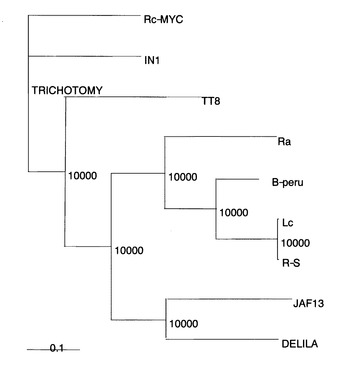
【図21】
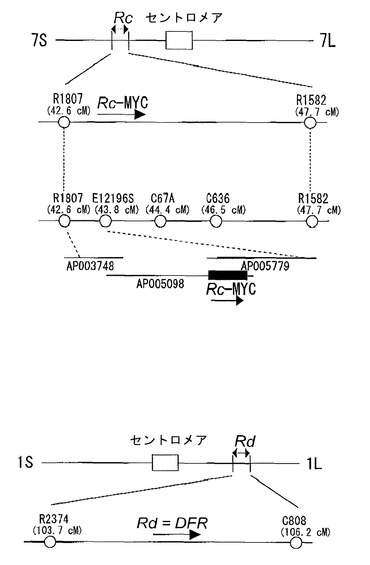
【図3】
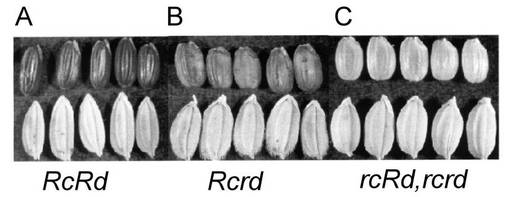
【図11】
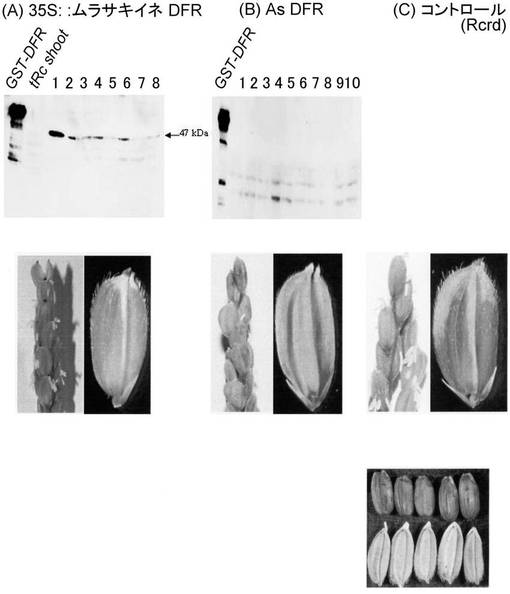
【図13】
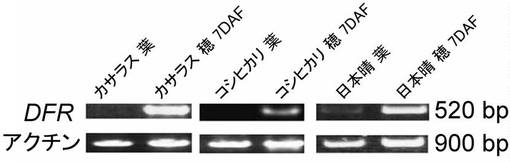
【図14】
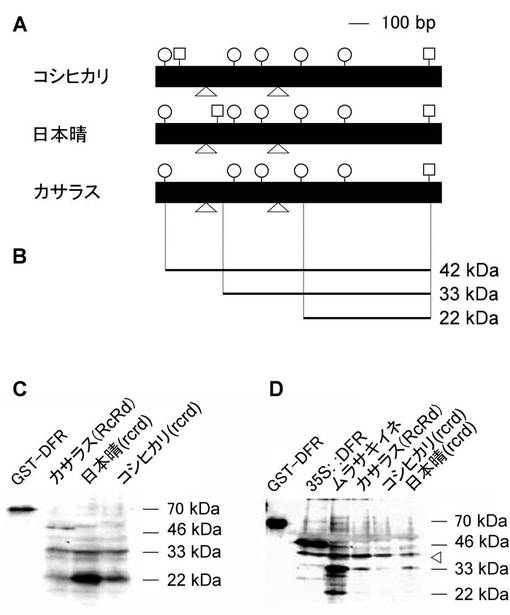
【図16】
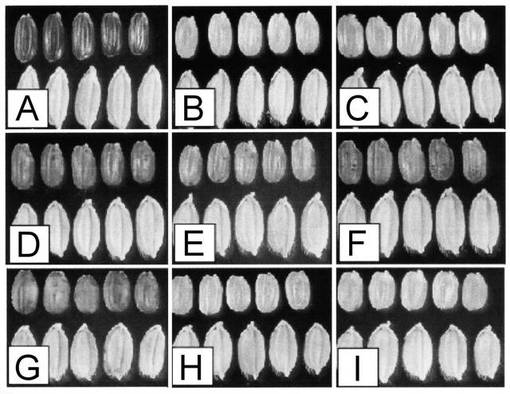
【図18】
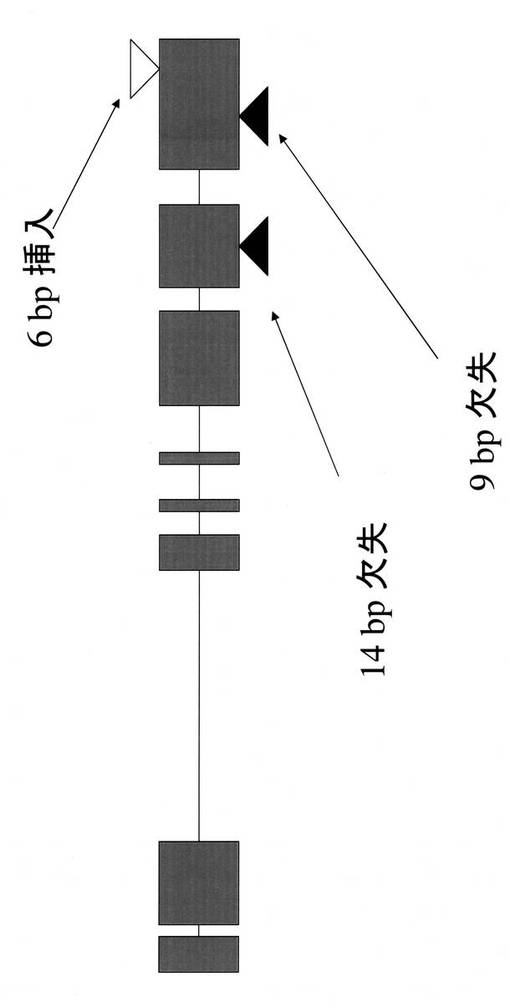
【図19】

【図2】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図12】
【図15A−C】
【図15D】
【図17】
【図20】
【図21】
【図3】
【図11】
【図13】
【図14】
【図16】
【図18】
【図19】
【公開番号】特開2012−24103(P2012−24103A)
【公開日】平成24年2月9日(2012.2.9)
【国際特許分類】
【出願番号】特願2011−234737(P2011−234737)
【出願日】平成23年10月26日(2011.10.26)
【分割の表示】特願2006−22132(P2006−22132)の分割
【原出願日】平成18年1月31日(2006.1.31)
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【出願人】(504147243)国立大学法人 岡山大学 (444)
【Fターム(参考)】
【公開日】平成24年2月9日(2012.2.9)
【国際特許分類】
【出願日】平成23年10月26日(2011.10.26)
【分割の表示】特願2006−22132(P2006−22132)の分割
【原出願日】平成18年1月31日(2006.1.31)
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【出願人】(504147243)国立大学法人 岡山大学 (444)
【Fターム(参考)】
[ Back to top ]