
植物油脂の製造方法
【課題】植物体の光合成を継続させながら、種子以外の植物組織から、植物油脂を効率的に製造する方法の提供。
【解決手段】デンプンの生合成が抑制された変異植物の植物体を栽培し、種子以外の組織に蓄積した中性脂質を回収することを特徴とする植物油脂の製造方法。
【解決手段】デンプンの生合成が抑制された変異植物の植物体を栽培し、種子以外の組織に蓄積した中性脂質を回収することを特徴とする植物油脂の製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は植物油脂の製造方法に関する。
【背景技術】
【0002】
植物油脂は、マーガリン、ショートニング、ドレッシング、食用硬化油及び精製ラード等の食用、及び燃料用、潤滑油や界面活性剤原料などの工業用として世界で毎年1億トン以上が用いられており、世界的な人口増加などによりその需要は更に高まりつつある。一方、その生産量もパーム油及び大豆油を中心に、2001年から2008年までの間に約1.5倍に伸長してきているが、これらは例えばマレーシア及びインドネシアなどの主産地での植林量増加や品種改良などによるものや、特にヨーロッパにおけるバイオディーゼル用途の増加による補助金も含めた政策による影響も大きいと考えられる。
しかしながら、例えばパーム油やヤシ油は栽培可能地域が東南アジアなどの一部に限られていること、また生物多様性確保の問題から森林伐採による植林地の拡大化は望めず、単位植物体あたりの油脂生産量の増大などが望まれている。
【0003】
こうした背景から、植物の育種や栽培条件の改変による油脂生産量の増大を図る試みが成されている。古くから行われている交配によるもののほか、近年では遺伝子組換え技術を利用した育種例も報告されており、例えば、特許文献1及び非特許文献1には、脂肪酸鎖長伸長反応における伸長ユニットとなるマロニルCoA合成に関わるシロイヌナズナ由来のアセチルCoAカルボキシラーゼ(ACCase)遺伝子をナタネに導入することによりオレイン酸を中心に総脂肪酸量を向上させることが開示されている。また特許文献2及び非特許文献2には、トリアシルグリセロール(TAG)生合成の最終段階であるジアシルグリセロール(DAG)骨格へのアシル基転移反応を行うジアシルグリセロールアシルトランスフェラーゼ(DGAT)をシロイヌナズナで過剰発現させることで総脂質量を向上させることが報告されている。更に非特許文献3では特定の制御因子遺伝子を過剰発現させることによるTAGの高生産化、また特許文献3では、シロイヌナズナにおいて特定の色素合成に関与するカルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子、またはフラボン-3-ヒドラーゼ遺伝子のいずれかの機能を欠損した組換え植物体を用いるTAGの高生産化方法が開示されている。更に特許文献4には遺伝子組換え技術により脂肪酸分解系を欠損したシロイヌナズナを数日間、暗所処理することにより葉に油滴を蓄積させる方法が開示されている。
【0004】
また、非特許文献4では、シロイヌナズナの脂質輸送タンパクとして知られるTGD1の欠損株において葉へのTAG蓄積が報告されている。
しかし、これらの方法を用いた場合でも、暗所や乾燥などの条件が採用され、植物の光合成が阻害されること等により、植物油脂の高生産化効率は満足できるものではなかった。
【0005】
藻類における油脂の蓄積については、緑藻クラミドモナス(Chlamydomonas reinhardtii)のデンプン非蓄積株を窒素飢餓条件で培養することにより著量の油脂を蓄積すること(非特許文献5、非特許文献6)、デンプン合成系を欠損したADP-グルコースピロホスホリラーゼのChlamydomonas reinhardtii変異体は、弱光(100molm-2s-1)下、窒素飢餓の条件で油滴中にトリアシルグリセロールを蓄積し、脂質量は乾燥細胞重量の50%以上に達する(非特許文献7、非特許文献8)こと、が報告されている。
【0006】
一方、植物は、プラスチドにおいてデンプンを生合成する。デンプンは、多数のα−グルコースがα1−4、或いはα1−6グリコシド結合によって重合した高分子であり、1)グルコース−6−リン酸からグルコース−1−リン酸の生成工程、2)グルコース−1−リン酸からADPグルコースの生成工程、及び3)ADPグルコースのグルコース残基を、伸長中のアミロースやアミロペクチンの非還元末端のグルコース残基の4位の水酸基と脱水縮合して新たなα−1,4グルコシド結合を形成して取り込む工程、の3つのステップをキーとして合成されることが知られている。α-1,4グルコシド結合の繰り返しによって生じたグルコース残基の直鎖が、枝分かれ酵素によって一部切断され、その切断により生じた還元末端のグルコース残基の1位の水酸基と直鎖部分の中間のグルコース残基の6位の水酸基の間でα-1,6グルコシド結合が生じる。非特許文献9において、上記1)の工程に関与するホスホグルコムターゼ遺伝子(AtPGMp)を欠損したシロイヌナズナにて、その種子の脂質量が減少することが報告されている。
【0007】
しかしながら、植物体において、斯かるデンプンの生合成の欠損と組織への油脂蓄積量の増加との関係については、全く知られていない。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】米国特許第5925805号明細書
【特許文献2】国際公開第2000/036114号パンフレット
【特許文献3】特開2009-207421号公報
【特許文献4】国際公開第2008/068498号パンフレット
【非特許文献】
【0009】
【非特許文献1】Plant Physiology (1997) 11巻, 75頁
【非特許文献2】Plant Physiology (2001) 126巻, 861頁
【非特許文献3】Plant J. (2004) 40巻, 575頁
【非特許文献4】Plant Cell (2005) 17巻, 3094頁
【非特許文献5】19th International Symposium on Plant Lipids, Symposium Program(2010),40頁, P-20
【非特許文献6】Eukaryotic Cell (2009) 8巻, 1856頁
【非特許文献7】19th International Symposium on Plant Lipids, Symposium Program(2010),43頁, P-33
【非特許文献8】Metabolic Engineering (2010) 12巻, 387頁
【非特許文献9】Plant Physiology (2000) 122巻,1193頁
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明は、植物体の光合成を継続させながら、種子以外の植物組織から、植物油脂を効率的に製造する方法を提供することに関する。
【課題を解決するための手段】
【0011】
本発明者らは、デンプンの生合成が抑制された変異植物を用いることにより、光合成を継続させながら、中性脂質を含有する植物油脂を種子以外の植物組織に効率的に蓄積し得ることを見出した。
【0012】
すなわち、本発明は、以下の(1)〜(11)に係るものである。
(1)デンプンの生合成が抑制された変異植物の植物体を栽培し、種子以外の組織に蓄積した中性脂質を回収することを特徴とする植物油脂の製造方法。
(2)変異植物が、ホスホグルコムターゼ遺伝子、ADPグルコースピロホスホリラーゼ遺伝子及びデンプンシンターゼ遺伝子から選ばれる1種以上の遺伝子の機能を不活性化又は発現抑制したものである、上記(1)の植物油脂の製造方法。
(3)植物体を、リンが欠乏した状態で栽培する、上記(1)又は(2)の植物油脂の製造方法。
(4)リンが欠乏した状態での栽培が、組織が十分に生育した植物体を、リンが欠乏した媒体に移植するか或いは媒体をリンが欠乏した媒体に置換して栽培する、又は栽培過程の媒体において生じるリンの欠乏状態を維持しながら栽培するものである上記(3)の植物油脂の製造方法。
(5)種子以外の組織が、葉、茎、または根である、上記(1)〜(4)の植物油脂の製造方法。
(6)植物体が被子植物である、上記(1)〜(5)の植物油脂の製造方法。
(7)植物体が裸子植物である、上記(1)〜(5)の植物油脂の製造方法。
(8)植物体が双子葉植物である、上記(1)〜(5)の植物油脂の製造方法。
(9)植物体がシロイヌナズナである、上記(1)〜(5)の植物油脂の製造方法。
(10)デンプンの生合成が抑制された変異植物の植物体を栽培することを特徴とする、種子以外の組織への中性脂質の蓄積方法。
(11)リンが欠乏した状態で栽培する、上記(10)の中性脂質の蓄積方法。
【発明の効果】
【0013】
本発明の植物油脂の製造方法によれば、植物の光合成能力を停止させることなく、かつ種子の成熟を待つことなく早期に中性脂質を種子以外の植物組織に蓄積し得ることから植物油脂を効率良く生産することができる。また更に、種子以外の植物組織に蓄積した脂質中に脂溶性成分を溶解させることにより、植物ステロール類やポリフェノール類等の機能性成分を効率良く生産すること等も期待される。
【図面の簡単な説明】
【0014】
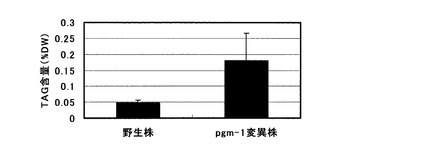
【図1】1mM可溶性リン酸を含む培地を用いた栽培におけるシロイヌナズナ地上部(葉・茎)へのTAG蓄積量を示すグラフ。
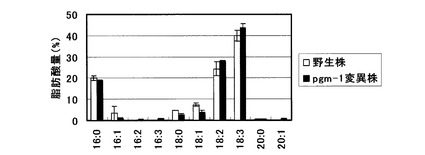
【図2】1mM可溶性リン酸を含む培地を用いた栽培におけるシロイヌナズナ地上部(葉・茎)の脂肪酸組成を示すグラフ。
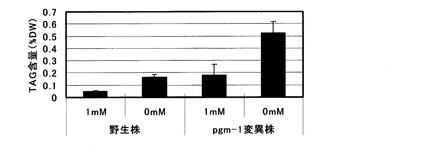
【図3】可溶性リン酸を含まない培地、及び1mM可溶性リン酸を含む培地を用いた栽培におけるシロイヌナズナ地上部(葉・茎)のTAG蓄積量を示すグラフ。
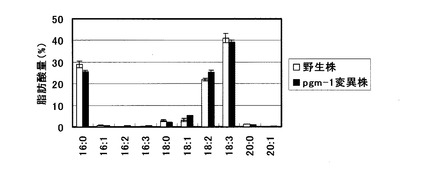
【図4】可溶性リン酸を含まない培地を用いた栽培におけるシロイヌナズナ地上部(葉・茎)の脂肪酸組成を示すグラフ。
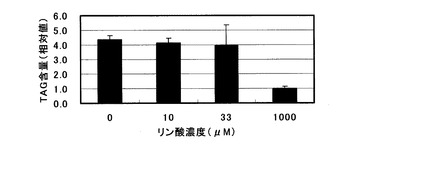
【図5】各種可溶性リン酸濃度の培地を用いた栽培におけるシロイヌナズナ地上部(葉・茎)へのTAG蓄積量を示すグラフ。
【発明を実施するための形態】
【0015】
本発明の植物油脂の製造方法は、デンプンの生合成が抑制された変異植物の植物体を栽培し、種子以外の組織に蓄積した中性脂質を回収するものである。
本発明において、用いられる植物としては、陸上植物であればその種類は特に限定されないが、好適には生育速度や得られる植物体量(バイオマス量)などから種子植物を用いることが効率的生産に有利である。種子植物のうち被子植物としては、例えばヤシ科やイネ科等の単子葉植物、あるいはマメ科、アブラナ科、キク科、トウダイグサ科、ゴマ科、モクセイ科、ミソハギ科、シソ科、セリ科、アカザ科、アオイ科等の双子葉植物、また裸子植物としては、例えばマツ科、イチョウ科等が挙げられる。
【0016】
より具体的な植物種の例としては、ヤシ科のココヤシ(Cocos nucifera)、パームヤシ(Elaeis guineensi, Elaeis oleifera)等、イネ科のイネ(Oryza sativa, Oryza glaberrima)、トウモロコシ(Zea mays)、ミスカンザス(Miscanthus giganteus)等、マメ科のダイズ(Glycine max)等、アブラナ科のナタネ(Brassica rapa, Brassica napus)、シロイヌナズナ(Arabidopsis thaliana)、ナガミノアマナズナ(Camelina sativa)、ハクサイ(Brassica rapa var. glabra)、キャベツ(Brassica oleracea var. capitata)、コマツナ(Brassica rapa var. peruviridis)、ミズナ(Brassica rapa var. nipposinica)、クレソン(Nasturtium officinale)等、キク科のヒマワリ(Helianthus annuus)、ベニバナ(Carthamus tinctorius)、レタス(Lactuca sativa)等、トウダイグサ科のヒマ(Ricinus communis)、ヤトロファ(Jatropha curcas)、ゴマ科のゴマ(Sesamum indicum)等、モクセイ科のオリーブ(Olea europea)等、ミソハギ科のクフェア(Cuphea hyssopifolia)等、シソ科のアオジソ(Perilla frutescens var. crispa)、アカジソ(Perilla frutescen var. crispa)、バジル(Ocimum basilicum L.)等、セリ科のミツバ(Cryptotaenia japonica)、コリアンダー(Coriandrum sativum L.)、パセリ(Petroselium crispum)等、アカザ科のホウレンソウ(Spinacia oleracea)等、ナス科のタバコ(Nicotiana tabacum)等が挙げられる。このうち、被子植物が好ましく、より好適には双子葉植物、更に好適にはアブラナ科植物が挙げられ、このうちシロイヌナズナがより好ましい。
【0017】
本発明の変異植物は、上記植物において、デンプンの生合成が抑制された変異体を意味する。具体的には、デンプンの生合成が抑制され、植物組織、例えば葉におけるデンプン蓄積が本来(野生型)よりも低下している植物、例えば、葉におけるデンプン蓄積量が、野生型の0〜50%程度、好ましくは0〜20%程度まで低下している植物が挙げられる。
デンプンの生合成を抑制する方法としては、例えば、デンプンの生合成のキーステップ、すなわちグルコース−6−リン酸からグルコース−1−リン酸の生成、グルコース−1−リン酸からADPグルコースの生成、及びADPグルコースからデンプンの生成に関与する酵素の構造遺伝子に対する、欠失或いは挿入変異による機能の不活性化の他、当該遺伝子の発現に関わる領域に対する、欠失或いは挿入変異による遺伝子発現の抑制(不活性化等)が挙げられる。
【0018】
デンプンの生合成に関わる酵素としては、グルコース−6−リン酸からグルコース−1−リン酸の生成に関与するホスホグルコムターゼ(Phosphoglucomutase(PGM))、グルコース−1−リン酸からADPグルコースを生成に関与するADPグルコースピロホスホリラーゼ(ADP-glucose pyrophosphorylase(AGPase)、及びADPグルコースからデンプンを合成するデンプンシンターゼ(Starch synthase(SS))が挙げられ、その構造遺伝子としては、PGM遺伝子、APL遺伝子、ADG遺伝子及びsoluble glycogen synthase-related遺伝子が挙げられる。
【0019】
本発明の変異植物において、機能の不活性化又は発現が制御される遺伝子としては、これらの何れでもよいが、具体的には、シロイヌナズナ由来のPGM遺伝子(AGIコード:AT5G51820(配列番号1))又は当該遺伝子の塩基配列に対して、80%以上、好ましくは88%以上、より好ましくは90%以上、さらに好ましくは98%の同一性を有する塩基配列からなり、且つホスホグルコムターゼ活性を有するタンパク質をコードする遺伝子、シロイヌナズナ由来のAPL1遺伝子(AGIコード:AT5G19220(配列番号2))又は当該遺伝子の塩基配列に対して、80%以上、好ましくは88%以上、より好ましくは90%以上、さらに好ましくは98%の同一性を有する塩基配列からなり、且つADPグルコースピロホスホリラーゼ活性を有するタンパク質をコードする遺伝子、シロイヌナズナ由来のADG1遺伝子(AGIコード:AT5G48300(配列番号3))又は当該遺伝子の塩基配列に対して、80%以上、好ましくは88%以上、より好ましくは90%以上、さらに好ましくは98%の同一性を有する塩基配列からなり、且つADPグルコースピロホスホリラーゼ活性を有するタンパク質をコードする遺伝子、シロイヌナズナ由来のsoluble glycogen synthase-related遺伝子(AGIコード:AT5G65685(配列番号4))又は当該遺伝子の塩基配列に対して、80%以上、好ましくは88%以上、より好ましくは90%以上、さらに好ましくは98%の同一性を有する塩基配列からなり、且つデンプンシンターゼ活性を有するタンパク質をコードする遺伝子が挙げられる。
【0020】
尚、塩基配列の同一性は、例えば、リップマン−パーソン法(Lipman-Pearson法;Science,227,1435(1985))によって計算される。具体的には、遺伝情報処理ソフトウェアGenetyx−Win(Ver.5.1.1;ソフトウェア開発)のホモロジー解析(Search Homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
【0021】
当該酵素の構造遺伝子、或いは当該遺伝子発現に関わる領域への欠失変異を行う方法としては、メタンスルホン酸エチルやニトロソグアニジン等の突然変異誘起剤を用いる方法やγ線等を照射する方法が挙げられる。これらの欠失方法により生じたランダムな変異体の集団から、デンプン蓄積の抑制を指標として目的とする変異体を選抜すれば良い。
【0022】
また、当該酵素の構造遺伝子、或いは遺伝子発現に関わる領域への挿入変異を行う方法としては、アグロバクテリウム形質転換法によるTiプラスミド上のT-DNA領域の挿入や、トランスポゾンを用いた方法等が挙げられる。これらの挿入方法により生じたランダムな変異体の集団から、デンプン蓄積の抑制を指標として目的とする変異体を選抜すれば良い。
【0023】
デンプン蓄積の抑制の判定は、例えば、エタノールに浸漬する等の方法により脱色した植物の葉をヨウ素溶液に浸し、デンプンの蓄積を示す紫色を呈さないことを確認すれば良い。
その他、遺伝子発現を不活性化する方法として、対象とする遺伝子の配列情報を利用するアンチセンス法やRNA干渉法等が挙げられる。
【0024】
本発明において、栽培に用いられる「植物体」としては、根、茎、及び葉等の植物組織が生育した植物体を意味し、好適には、十分な植物体量(バイオマス量)を有する植物体が挙げられる。
【0025】
当該植物体の栽培は、土壌、水耕液(培養液)、或いは固体培地等を用いて行うことができる。また屋外での日照、及び屋内における人工照明等のいずれも利用可能であり、光量や照射時間も特に制限されないが、植物体固有の最適な温度、湿度、pH、光照射条件で栽培を行えばよい。
例えば、植物体としてシロイヌナズナを用いる場合、栽培は、18〜25℃、光強度30〜70μE/cm2、照射時間6〜24時間/日の範囲で行うことが好ましい。
尚、当該植物体を得るための、種子からの生育栽培も、植物固有の最適な温度、湿度、pH、光照射条件で栽培を行えばよい。シロイヌナズナを用いる場合、上記と同様の条件で栽培すればよい。
【0026】
本発明の植物体の栽培は、中性脂質の蓄積促進の点で、リンが欠乏した状態で栽培するのがより好ましい。
ここで、「リンが欠乏した状態で栽培する」とは、例えば、1)組織が十分に生育した植物体をリンが欠乏した媒体に移植して栽培すること、2)媒体をリンが欠乏した媒体に置換して当該植物体を栽培すること、3)栽培過程の媒体において生じるリンの欠乏状態を維持しながら栽培すること等が挙げられる。
【0027】
「リンが欠乏した状態」とは、媒体中にリンを含まないか又は媒体中のリン濃度が極めて低い状態を意味する。具体的には、中性脂質の蓄積促進の点から、媒体中のリン濃度がゼロに近いことが望ましいが、ゼロでなくても通常の栽培で用いるリン酸濃度の1/30未満、好適には1/100以下、より好適には1/300以下、更に好ましくは1/1000以下である。
例えば、後記表1に示した培地を用いて水耕栽培や寒天培地での栽培を行う場合、極めて低い濃度のリンを含む媒体のリン濃度は、33μM未満であり、好ましくは10μM以下、更に好適には3.3μM以下が望ましく、最適には1μM以下、或いは0.33μM以下とすることができる。
【0028】
リンが欠乏した状態での栽培は、通常の温度、湿度、pH、光照射条件で栽培を行えばよい。このリン欠乏条件における栽培期間は特に限定されないが、中性脂質蓄積の点から、好適には数日〜数週間、特には3日間〜3週間、最適には1〜2週間である。
例えば、植物体としてシロイヌナズナを用いる場合、18〜25℃、光強度30〜70μE/cm2、照射時間6〜24時間/日の範囲であれば良い。
【0029】
リンが欠乏した状態での栽培において、媒体の調製方法は特に限定されないが、一般にリン欠乏土壌と言われる土壌、例えば可溶性リン酸が100mg/100g以下、更には50mg/100g以下の土壌、或いはこの様な土壌にNK化成肥料等のリンを含まない肥料の施肥を行うか、或いはこの様な土壌にNK化成肥料等のリンを含まない肥料に少量のリン酸等のリン肥料を加えた肥料の施肥を行うことにより調製することができる。
また水耕液や固体培地を用いる場合は、リン以外の必要栄養素を適当に配合した培養液、或いは培地を用いることができる。リン以外の必要栄養素は特に限定されないが、硝酸カリウム、硝酸アンモニウム、硫酸アンモニウム、硝酸カルシウム、硝酸ナトリウム、塩化カリウム。塩化カルシウム、硫酸マグネシウム、硫酸ナトリウム、硫酸鉄(III)、塩化鉄(III)、硫酸鉄(III)、エチレンジアミン四酢酸二ナトリウム、エチレンジ
アミン四酢酸鉄ナトリウム、硫酸マンガン、硫酸亜鉛、ホウ酸、硫酸銅、モリブデン酸ナトリウム、三酸化モリブデン、ヨウ化カリウム、塩化コバルト、塩化アルミニウム、塩化ニッケル、ミオイノシトール、チアミン塩酸塩、ピリドキシン塩酸塩、ニコチン酸、葉酸、ビオチン、グリシン等を含むことができる。また固体培地のゲル化剤としては、特に限定されないが、寒天、ゼラチン、ゲルライト(和光純薬社製)等を用いることができる。
【0030】
「リンが欠乏した状態」での栽培の一態様としては、例えば、MS培地(Physiologia Plantarum (1962) 15巻, 473頁)で、組織が十分に生育した植物体を表1に示した組成をもつ培地に移すか又はMS培地を表1に示した培地に置換して、リンを含まないか又はリン濃度が極めて低い状態、例えば33μM未満、好ましくは10μM以下、更に好適には3.3μM以下、最適には1μM以下、或いは0.33μM以下で更に数日間〜数週間の栽培をすることが挙げられる。
【0031】
また別の一態様として、栽培過程において生じるリンの欠乏状態を維持しながら栽培することが挙げられ、この場合には、植物体を適当な期間栽培してリン濃度が極めて低くなってから、更に数日〜数週間の栽培を継続すればよい。この場合に用いる媒体中のリン濃度は、植物体が十分に生育した後に媒体中のリン濃度が上記に示した極めて低い濃度またはゼロとなる様に、初期濃度を調整することにより行えばよい。
【0032】
斯くして、植物体の根、葉、或いは茎等に蓄積した中性脂質を植物体から回収する方法は、特に限定されず、例えば、植物体の根、葉、或いは茎等を粉砕、圧搾すること、或いは適当な溶剤により抽出する方法等で行うことができる。より具体的には例えば、植物体の根、葉、或いは茎等を粉砕後、ノルマルヘキサンにより抽出する方法、或いはBligh and Dyer法(Can. J. Biochem. Physiol. (1959) 37巻, 911頁)などの方法を用いることで効率的に植物油脂を抽出することができ、植物油脂に含まれる形で中性脂質を回収することが可能である。斯かる植物油脂は、そのまま中性脂質として用いることもできるが、一般的な脱ガム、脱酸、脱色、脱臭等の精製の後に用いることもでき、その方法は特に限定されない。更に植物油脂から中性脂質を分離・回収することもでき、その方法についても特に限定されない。より具体的に例えば、薄層クロマトグラフィーによる分離およびシリカゲルプレートからの回収、高速液体クロマトグラフィーを用いた分離・回収などが挙げられる。
【0033】
斯くして得られる中性脂質はその大半がTAGであるが、少量のDAG等が含まれることもある。また中性脂質中の脂肪酸鎖長は植物体に固有な脂質生合成能力にほぼ依存することが多い。
【実施例】
【0034】
以下、実施例により本発明を詳細に説明する。なお、植物組織中の油脂成分の分析は、下記に記載の方法により行った。
(1)抽出及び前処理
総脂質の抽出は、Bligh and Dyer法(Can. J. Biochem. Physiol. (1959) 37巻, 911頁)に基づき行った。総脂質からTAGの分離は、薄層クロマトグラフィー(TLC Silica gel 60, 20x20 cm, メルク, 製品コード1.05721.0009、展開溶媒組成は、ヘキサン:ジエチルエーテル:酢酸=160:40:4(vol/vol))により行い、プレートからTAGのスポットをかきとって含量の測定を行った。
【0035】
(2)中性脂質量の測定
15:0脂肪酸を内部標準試料として、TAGをメタノリシス処理した。具体的には、ネジ栓付きガラス試験管内で、TAGを含むシリカゲル粉末に100 μlの1 mM 15:0ヘキサン溶液 (pentadecanoic acid, シグマ, P-6125)および350μlの5% 塩化水素メタノール溶液(和光, 089-03971)を添加して85℃で1時間処理した。メタノリシス処理後、ヘキサンで脂肪酸メチルエステルを回収し、窒素ガスで乾固後、60μlのヘキサンで回収し、そのうち3μlをガスクロマトグラフィーで解析した。ガスクロマトグラフィー(島津、GC-2014, カラムULBON HR-SS-10 (25 m, 0.25 mm ID)、カラム温度180℃、気化室および検出器250℃、入口圧(kPa) 68.2、カラム流量(ml/min) 0.53、スプリット比68.8、計測時間15分)を用いて分離・定量を行った。
【0036】
(3)植物組織乾燥重量の測定
凍結乾燥処理を20時間行った後、重量を測定し、乾燥重量とした。
【0037】
実施例1 リン酸含有培地における中性脂質の生産性
シロイヌナズナ(Arabidopsis thaliana)Col-0株(野生株)及びpgm-1変異株の種子35個体をMS寒天培地(Physiologia Plantarum (1962) 15巻, 473頁)に播種し、22℃、光強度40〜70μE/cm2、照射時間24時間/日の条件で10日間栽培した。尚、pgm-1遺伝子変異株はArabidopsis Biological Resource Center保有のストックCS210(参考文献:Plant Physiology (1985) 79巻, 11頁、Proceedings of the National Academy of Sciences of the United States of America (1989) 86巻, 5830頁,Plant Physiology (2000)122巻,1193頁)を購入し、葉におけるデンプン蓄積が抑制されていることを確認した個体から収穫した種子を使用した。本pgm-1変異株はAGIコードAT5G51820の遺伝子が変異により不活性化している。
【0038】
生育した植物体をMS寒天培地から注意深く引き抜き、可溶性リン酸(KH2PO4)を1mM含む表1の培地に35個体を移植した。これらを上記と同条件で更に10日間栽培した。栽培後の植物体を寒天培地から引き抜き、地上部(葉、茎)および根に切り分け、地上部の重量測定を行った後、液体窒素下すり鉢で粉砕し、Bligh and Dyer法で脂肪抽出を行った。この後、薄層クロマトグラフィーによりTAGを分離・精製後、塩酸メタノールでメタノリシス処理をしてガスクロマトグラフィーを用いて脂質分析を行った。また栽培した一部の個体は凍結乾燥し、乾燥重量を測定した。
【0039】
【表1】

【0040】
この結果、図1に示した様に、pgm-1変異株の地上部では、乾燥重量あたり野生株を有意に上回るTAG蓄積向上効果が認められた。また、得られた各株におけるTAGの脂肪酸組成分析を行ったところ、図2に示す様に脂肪酸組成の大きな変化は認められなかった。
【0041】
実施例2 リン酸非含有培地における中性脂質の生産性
シロイヌナズナ野生株及びpgm-1変異株の種子70個体をMS寒天培地(Physiologia Plantarum (1962) 15巻, 473頁)に播種し、22℃、光強度40〜70μE/cm2、照射時間24時間/日の条件で10日間栽培した。
生育した植物体をMS寒天培地から注意深く引き抜き、可溶性リン酸(KH2PO4)を含まない(0mM)、上記表1の培地に35個体を移植した。またそれぞれ35個体を1mM可溶性リン酸を含む表1の培地に移植し、これらを上記と同条件で更に10日間栽培した。
栽培後の植物体を寒天培地から引き抜き、地上部(葉、茎)および根に切り分け、地上部の重量測定を行った後、液体窒素下すり鉢で粉砕し、Bligh and Dyer法で脂肪抽出を行った。この後、薄層クロマトグラフィーによりTAGを分離・精製後、塩酸メタノールでメタノリシス処理をしてガスクロマトグラフィーを用いて脂質分析を行った。
【0042】
この結果、図3に示した様に、野生株及びpgm-1変異株共に、可溶性リン酸0mMの培地で栽培した地上部では1mM可溶性リン酸を含む培地で栽培した場合の約3倍のTAG蓄積向上効果が認められた。またpgm-1変異株はリン酸濃度に関わらず野生株の約3倍のTAG蓄積向上効果を示した。即ちこれらの結果より、pgm-1変異株の使用と可溶性リン酸0mMの培地での栽培を組み合わせることで、相乗的なTAG蓄積向上効果が得られることが明らかとなった。尚、得られた各株におけるTAGの脂肪酸組成分析を行ったところ、図4に示す様に脂肪酸組成の大きな変化は認められなかった。
【0043】
実施例3
シロイヌナズナ(Arabidopsis thaliana)野生株の種子96個体をMS寒天培地(Physiologia Plantarum (1962) 15巻, 473頁)に播種し、22℃、光強度40〜70μE/cm2、照射時間24時間/日の条件で10日間栽培した。
生育した植物体をMS寒天培地から注意深く引き抜き、可溶性リン酸(KH2PO4)の濃度が0mM、0.01mM、0.033mM、或いは1mMである上記表1の培地にそれぞれ24個体を移植し、これらを上記と同条件で更に10日間栽培した。
栽培後の植物体を寒天培地から引き抜き、地上部(葉、茎)および根に切り分け、地上部の重量測定を行った後、液体窒素下すり鉢で粉砕し、Bligh and Dyer法で脂肪抽出を行った。この後、薄層クロマトグラフィーによりTAGを分離・精製後、塩酸メタノールでメタノリシス処理をしてガスクロマトグラフィーを用いて脂質分析を行った。
【0044】
この結果、図5に示した様に、可溶性リン酸0mM、0.01mM(10μM)、及び0.033mM(33μM)の培地で栽培したシロイヌナズナ地上部では、1mM(1000μM))可溶性リン酸を含む培地で栽培した場合のそれぞれ約4倍のTAG蓄積向上効果が認められた。
【技術分野】
【0001】
本発明は植物油脂の製造方法に関する。
【背景技術】
【0002】
植物油脂は、マーガリン、ショートニング、ドレッシング、食用硬化油及び精製ラード等の食用、及び燃料用、潤滑油や界面活性剤原料などの工業用として世界で毎年1億トン以上が用いられており、世界的な人口増加などによりその需要は更に高まりつつある。一方、その生産量もパーム油及び大豆油を中心に、2001年から2008年までの間に約1.5倍に伸長してきているが、これらは例えばマレーシア及びインドネシアなどの主産地での植林量増加や品種改良などによるものや、特にヨーロッパにおけるバイオディーゼル用途の増加による補助金も含めた政策による影響も大きいと考えられる。
しかしながら、例えばパーム油やヤシ油は栽培可能地域が東南アジアなどの一部に限られていること、また生物多様性確保の問題から森林伐採による植林地の拡大化は望めず、単位植物体あたりの油脂生産量の増大などが望まれている。
【0003】
こうした背景から、植物の育種や栽培条件の改変による油脂生産量の増大を図る試みが成されている。古くから行われている交配によるもののほか、近年では遺伝子組換え技術を利用した育種例も報告されており、例えば、特許文献1及び非特許文献1には、脂肪酸鎖長伸長反応における伸長ユニットとなるマロニルCoA合成に関わるシロイヌナズナ由来のアセチルCoAカルボキシラーゼ(ACCase)遺伝子をナタネに導入することによりオレイン酸を中心に総脂肪酸量を向上させることが開示されている。また特許文献2及び非特許文献2には、トリアシルグリセロール(TAG)生合成の最終段階であるジアシルグリセロール(DAG)骨格へのアシル基転移反応を行うジアシルグリセロールアシルトランスフェラーゼ(DGAT)をシロイヌナズナで過剰発現させることで総脂質量を向上させることが報告されている。更に非特許文献3では特定の制御因子遺伝子を過剰発現させることによるTAGの高生産化、また特許文献3では、シロイヌナズナにおいて特定の色素合成に関与するカルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子、またはフラボン-3-ヒドラーゼ遺伝子のいずれかの機能を欠損した組換え植物体を用いるTAGの高生産化方法が開示されている。更に特許文献4には遺伝子組換え技術により脂肪酸分解系を欠損したシロイヌナズナを数日間、暗所処理することにより葉に油滴を蓄積させる方法が開示されている。
【0004】
また、非特許文献4では、シロイヌナズナの脂質輸送タンパクとして知られるTGD1の欠損株において葉へのTAG蓄積が報告されている。
しかし、これらの方法を用いた場合でも、暗所や乾燥などの条件が採用され、植物の光合成が阻害されること等により、植物油脂の高生産化効率は満足できるものではなかった。
【0005】
藻類における油脂の蓄積については、緑藻クラミドモナス(Chlamydomonas reinhardtii)のデンプン非蓄積株を窒素飢餓条件で培養することにより著量の油脂を蓄積すること(非特許文献5、非特許文献6)、デンプン合成系を欠損したADP-グルコースピロホスホリラーゼのChlamydomonas reinhardtii変異体は、弱光(100molm-2s-1)下、窒素飢餓の条件で油滴中にトリアシルグリセロールを蓄積し、脂質量は乾燥細胞重量の50%以上に達する(非特許文献7、非特許文献8)こと、が報告されている。
【0006】
一方、植物は、プラスチドにおいてデンプンを生合成する。デンプンは、多数のα−グルコースがα1−4、或いはα1−6グリコシド結合によって重合した高分子であり、1)グルコース−6−リン酸からグルコース−1−リン酸の生成工程、2)グルコース−1−リン酸からADPグルコースの生成工程、及び3)ADPグルコースのグルコース残基を、伸長中のアミロースやアミロペクチンの非還元末端のグルコース残基の4位の水酸基と脱水縮合して新たなα−1,4グルコシド結合を形成して取り込む工程、の3つのステップをキーとして合成されることが知られている。α-1,4グルコシド結合の繰り返しによって生じたグルコース残基の直鎖が、枝分かれ酵素によって一部切断され、その切断により生じた還元末端のグルコース残基の1位の水酸基と直鎖部分の中間のグルコース残基の6位の水酸基の間でα-1,6グルコシド結合が生じる。非特許文献9において、上記1)の工程に関与するホスホグルコムターゼ遺伝子(AtPGMp)を欠損したシロイヌナズナにて、その種子の脂質量が減少することが報告されている。
【0007】
しかしながら、植物体において、斯かるデンプンの生合成の欠損と組織への油脂蓄積量の増加との関係については、全く知られていない。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】米国特許第5925805号明細書
【特許文献2】国際公開第2000/036114号パンフレット
【特許文献3】特開2009-207421号公報
【特許文献4】国際公開第2008/068498号パンフレット
【非特許文献】
【0009】
【非特許文献1】Plant Physiology (1997) 11巻, 75頁
【非特許文献2】Plant Physiology (2001) 126巻, 861頁
【非特許文献3】Plant J. (2004) 40巻, 575頁
【非特許文献4】Plant Cell (2005) 17巻, 3094頁
【非特許文献5】19th International Symposium on Plant Lipids, Symposium Program(2010),40頁, P-20
【非特許文献6】Eukaryotic Cell (2009) 8巻, 1856頁
【非特許文献7】19th International Symposium on Plant Lipids, Symposium Program(2010),43頁, P-33
【非特許文献8】Metabolic Engineering (2010) 12巻, 387頁
【非特許文献9】Plant Physiology (2000) 122巻,1193頁
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明は、植物体の光合成を継続させながら、種子以外の植物組織から、植物油脂を効率的に製造する方法を提供することに関する。
【課題を解決するための手段】
【0011】
本発明者らは、デンプンの生合成が抑制された変異植物を用いることにより、光合成を継続させながら、中性脂質を含有する植物油脂を種子以外の植物組織に効率的に蓄積し得ることを見出した。
【0012】
すなわち、本発明は、以下の(1)〜(11)に係るものである。
(1)デンプンの生合成が抑制された変異植物の植物体を栽培し、種子以外の組織に蓄積した中性脂質を回収することを特徴とする植物油脂の製造方法。
(2)変異植物が、ホスホグルコムターゼ遺伝子、ADPグルコースピロホスホリラーゼ遺伝子及びデンプンシンターゼ遺伝子から選ばれる1種以上の遺伝子の機能を不活性化又は発現抑制したものである、上記(1)の植物油脂の製造方法。
(3)植物体を、リンが欠乏した状態で栽培する、上記(1)又は(2)の植物油脂の製造方法。
(4)リンが欠乏した状態での栽培が、組織が十分に生育した植物体を、リンが欠乏した媒体に移植するか或いは媒体をリンが欠乏した媒体に置換して栽培する、又は栽培過程の媒体において生じるリンの欠乏状態を維持しながら栽培するものである上記(3)の植物油脂の製造方法。
(5)種子以外の組織が、葉、茎、または根である、上記(1)〜(4)の植物油脂の製造方法。
(6)植物体が被子植物である、上記(1)〜(5)の植物油脂の製造方法。
(7)植物体が裸子植物である、上記(1)〜(5)の植物油脂の製造方法。
(8)植物体が双子葉植物である、上記(1)〜(5)の植物油脂の製造方法。
(9)植物体がシロイヌナズナである、上記(1)〜(5)の植物油脂の製造方法。
(10)デンプンの生合成が抑制された変異植物の植物体を栽培することを特徴とする、種子以外の組織への中性脂質の蓄積方法。
(11)リンが欠乏した状態で栽培する、上記(10)の中性脂質の蓄積方法。
【発明の効果】
【0013】
本発明の植物油脂の製造方法によれば、植物の光合成能力を停止させることなく、かつ種子の成熟を待つことなく早期に中性脂質を種子以外の植物組織に蓄積し得ることから植物油脂を効率良く生産することができる。また更に、種子以外の植物組織に蓄積した脂質中に脂溶性成分を溶解させることにより、植物ステロール類やポリフェノール類等の機能性成分を効率良く生産すること等も期待される。
【図面の簡単な説明】
【0014】
【図1】1mM可溶性リン酸を含む培地を用いた栽培におけるシロイヌナズナ地上部(葉・茎)へのTAG蓄積量を示すグラフ。
【図2】1mM可溶性リン酸を含む培地を用いた栽培におけるシロイヌナズナ地上部(葉・茎)の脂肪酸組成を示すグラフ。
【図3】可溶性リン酸を含まない培地、及び1mM可溶性リン酸を含む培地を用いた栽培におけるシロイヌナズナ地上部(葉・茎)のTAG蓄積量を示すグラフ。
【図4】可溶性リン酸を含まない培地を用いた栽培におけるシロイヌナズナ地上部(葉・茎)の脂肪酸組成を示すグラフ。
【図5】各種可溶性リン酸濃度の培地を用いた栽培におけるシロイヌナズナ地上部(葉・茎)へのTAG蓄積量を示すグラフ。
【発明を実施するための形態】
【0015】
本発明の植物油脂の製造方法は、デンプンの生合成が抑制された変異植物の植物体を栽培し、種子以外の組織に蓄積した中性脂質を回収するものである。
本発明において、用いられる植物としては、陸上植物であればその種類は特に限定されないが、好適には生育速度や得られる植物体量(バイオマス量)などから種子植物を用いることが効率的生産に有利である。種子植物のうち被子植物としては、例えばヤシ科やイネ科等の単子葉植物、あるいはマメ科、アブラナ科、キク科、トウダイグサ科、ゴマ科、モクセイ科、ミソハギ科、シソ科、セリ科、アカザ科、アオイ科等の双子葉植物、また裸子植物としては、例えばマツ科、イチョウ科等が挙げられる。
【0016】
より具体的な植物種の例としては、ヤシ科のココヤシ(Cocos nucifera)、パームヤシ(Elaeis guineensi, Elaeis oleifera)等、イネ科のイネ(Oryza sativa, Oryza glaberrima)、トウモロコシ(Zea mays)、ミスカンザス(Miscanthus giganteus)等、マメ科のダイズ(Glycine max)等、アブラナ科のナタネ(Brassica rapa, Brassica napus)、シロイヌナズナ(Arabidopsis thaliana)、ナガミノアマナズナ(Camelina sativa)、ハクサイ(Brassica rapa var. glabra)、キャベツ(Brassica oleracea var. capitata)、コマツナ(Brassica rapa var. peruviridis)、ミズナ(Brassica rapa var. nipposinica)、クレソン(Nasturtium officinale)等、キク科のヒマワリ(Helianthus annuus)、ベニバナ(Carthamus tinctorius)、レタス(Lactuca sativa)等、トウダイグサ科のヒマ(Ricinus communis)、ヤトロファ(Jatropha curcas)、ゴマ科のゴマ(Sesamum indicum)等、モクセイ科のオリーブ(Olea europea)等、ミソハギ科のクフェア(Cuphea hyssopifolia)等、シソ科のアオジソ(Perilla frutescens var. crispa)、アカジソ(Perilla frutescen var. crispa)、バジル(Ocimum basilicum L.)等、セリ科のミツバ(Cryptotaenia japonica)、コリアンダー(Coriandrum sativum L.)、パセリ(Petroselium crispum)等、アカザ科のホウレンソウ(Spinacia oleracea)等、ナス科のタバコ(Nicotiana tabacum)等が挙げられる。このうち、被子植物が好ましく、より好適には双子葉植物、更に好適にはアブラナ科植物が挙げられ、このうちシロイヌナズナがより好ましい。
【0017】
本発明の変異植物は、上記植物において、デンプンの生合成が抑制された変異体を意味する。具体的には、デンプンの生合成が抑制され、植物組織、例えば葉におけるデンプン蓄積が本来(野生型)よりも低下している植物、例えば、葉におけるデンプン蓄積量が、野生型の0〜50%程度、好ましくは0〜20%程度まで低下している植物が挙げられる。
デンプンの生合成を抑制する方法としては、例えば、デンプンの生合成のキーステップ、すなわちグルコース−6−リン酸からグルコース−1−リン酸の生成、グルコース−1−リン酸からADPグルコースの生成、及びADPグルコースからデンプンの生成に関与する酵素の構造遺伝子に対する、欠失或いは挿入変異による機能の不活性化の他、当該遺伝子の発現に関わる領域に対する、欠失或いは挿入変異による遺伝子発現の抑制(不活性化等)が挙げられる。
【0018】
デンプンの生合成に関わる酵素としては、グルコース−6−リン酸からグルコース−1−リン酸の生成に関与するホスホグルコムターゼ(Phosphoglucomutase(PGM))、グルコース−1−リン酸からADPグルコースを生成に関与するADPグルコースピロホスホリラーゼ(ADP-glucose pyrophosphorylase(AGPase)、及びADPグルコースからデンプンを合成するデンプンシンターゼ(Starch synthase(SS))が挙げられ、その構造遺伝子としては、PGM遺伝子、APL遺伝子、ADG遺伝子及びsoluble glycogen synthase-related遺伝子が挙げられる。
【0019】
本発明の変異植物において、機能の不活性化又は発現が制御される遺伝子としては、これらの何れでもよいが、具体的には、シロイヌナズナ由来のPGM遺伝子(AGIコード:AT5G51820(配列番号1))又は当該遺伝子の塩基配列に対して、80%以上、好ましくは88%以上、より好ましくは90%以上、さらに好ましくは98%の同一性を有する塩基配列からなり、且つホスホグルコムターゼ活性を有するタンパク質をコードする遺伝子、シロイヌナズナ由来のAPL1遺伝子(AGIコード:AT5G19220(配列番号2))又は当該遺伝子の塩基配列に対して、80%以上、好ましくは88%以上、より好ましくは90%以上、さらに好ましくは98%の同一性を有する塩基配列からなり、且つADPグルコースピロホスホリラーゼ活性を有するタンパク質をコードする遺伝子、シロイヌナズナ由来のADG1遺伝子(AGIコード:AT5G48300(配列番号3))又は当該遺伝子の塩基配列に対して、80%以上、好ましくは88%以上、より好ましくは90%以上、さらに好ましくは98%の同一性を有する塩基配列からなり、且つADPグルコースピロホスホリラーゼ活性を有するタンパク質をコードする遺伝子、シロイヌナズナ由来のsoluble glycogen synthase-related遺伝子(AGIコード:AT5G65685(配列番号4))又は当該遺伝子の塩基配列に対して、80%以上、好ましくは88%以上、より好ましくは90%以上、さらに好ましくは98%の同一性を有する塩基配列からなり、且つデンプンシンターゼ活性を有するタンパク質をコードする遺伝子が挙げられる。
【0020】
尚、塩基配列の同一性は、例えば、リップマン−パーソン法(Lipman-Pearson法;Science,227,1435(1985))によって計算される。具体的には、遺伝情報処理ソフトウェアGenetyx−Win(Ver.5.1.1;ソフトウェア開発)のホモロジー解析(Search Homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
【0021】
当該酵素の構造遺伝子、或いは当該遺伝子発現に関わる領域への欠失変異を行う方法としては、メタンスルホン酸エチルやニトロソグアニジン等の突然変異誘起剤を用いる方法やγ線等を照射する方法が挙げられる。これらの欠失方法により生じたランダムな変異体の集団から、デンプン蓄積の抑制を指標として目的とする変異体を選抜すれば良い。
【0022】
また、当該酵素の構造遺伝子、或いは遺伝子発現に関わる領域への挿入変異を行う方法としては、アグロバクテリウム形質転換法によるTiプラスミド上のT-DNA領域の挿入や、トランスポゾンを用いた方法等が挙げられる。これらの挿入方法により生じたランダムな変異体の集団から、デンプン蓄積の抑制を指標として目的とする変異体を選抜すれば良い。
【0023】
デンプン蓄積の抑制の判定は、例えば、エタノールに浸漬する等の方法により脱色した植物の葉をヨウ素溶液に浸し、デンプンの蓄積を示す紫色を呈さないことを確認すれば良い。
その他、遺伝子発現を不活性化する方法として、対象とする遺伝子の配列情報を利用するアンチセンス法やRNA干渉法等が挙げられる。
【0024】
本発明において、栽培に用いられる「植物体」としては、根、茎、及び葉等の植物組織が生育した植物体を意味し、好適には、十分な植物体量(バイオマス量)を有する植物体が挙げられる。
【0025】
当該植物体の栽培は、土壌、水耕液(培養液)、或いは固体培地等を用いて行うことができる。また屋外での日照、及び屋内における人工照明等のいずれも利用可能であり、光量や照射時間も特に制限されないが、植物体固有の最適な温度、湿度、pH、光照射条件で栽培を行えばよい。
例えば、植物体としてシロイヌナズナを用いる場合、栽培は、18〜25℃、光強度30〜70μE/cm2、照射時間6〜24時間/日の範囲で行うことが好ましい。
尚、当該植物体を得るための、種子からの生育栽培も、植物固有の最適な温度、湿度、pH、光照射条件で栽培を行えばよい。シロイヌナズナを用いる場合、上記と同様の条件で栽培すればよい。
【0026】
本発明の植物体の栽培は、中性脂質の蓄積促進の点で、リンが欠乏した状態で栽培するのがより好ましい。
ここで、「リンが欠乏した状態で栽培する」とは、例えば、1)組織が十分に生育した植物体をリンが欠乏した媒体に移植して栽培すること、2)媒体をリンが欠乏した媒体に置換して当該植物体を栽培すること、3)栽培過程の媒体において生じるリンの欠乏状態を維持しながら栽培すること等が挙げられる。
【0027】
「リンが欠乏した状態」とは、媒体中にリンを含まないか又は媒体中のリン濃度が極めて低い状態を意味する。具体的には、中性脂質の蓄積促進の点から、媒体中のリン濃度がゼロに近いことが望ましいが、ゼロでなくても通常の栽培で用いるリン酸濃度の1/30未満、好適には1/100以下、より好適には1/300以下、更に好ましくは1/1000以下である。
例えば、後記表1に示した培地を用いて水耕栽培や寒天培地での栽培を行う場合、極めて低い濃度のリンを含む媒体のリン濃度は、33μM未満であり、好ましくは10μM以下、更に好適には3.3μM以下が望ましく、最適には1μM以下、或いは0.33μM以下とすることができる。
【0028】
リンが欠乏した状態での栽培は、通常の温度、湿度、pH、光照射条件で栽培を行えばよい。このリン欠乏条件における栽培期間は特に限定されないが、中性脂質蓄積の点から、好適には数日〜数週間、特には3日間〜3週間、最適には1〜2週間である。
例えば、植物体としてシロイヌナズナを用いる場合、18〜25℃、光強度30〜70μE/cm2、照射時間6〜24時間/日の範囲であれば良い。
【0029】
リンが欠乏した状態での栽培において、媒体の調製方法は特に限定されないが、一般にリン欠乏土壌と言われる土壌、例えば可溶性リン酸が100mg/100g以下、更には50mg/100g以下の土壌、或いはこの様な土壌にNK化成肥料等のリンを含まない肥料の施肥を行うか、或いはこの様な土壌にNK化成肥料等のリンを含まない肥料に少量のリン酸等のリン肥料を加えた肥料の施肥を行うことにより調製することができる。
また水耕液や固体培地を用いる場合は、リン以外の必要栄養素を適当に配合した培養液、或いは培地を用いることができる。リン以外の必要栄養素は特に限定されないが、硝酸カリウム、硝酸アンモニウム、硫酸アンモニウム、硝酸カルシウム、硝酸ナトリウム、塩化カリウム。塩化カルシウム、硫酸マグネシウム、硫酸ナトリウム、硫酸鉄(III)、塩化鉄(III)、硫酸鉄(III)、エチレンジアミン四酢酸二ナトリウム、エチレンジ
アミン四酢酸鉄ナトリウム、硫酸マンガン、硫酸亜鉛、ホウ酸、硫酸銅、モリブデン酸ナトリウム、三酸化モリブデン、ヨウ化カリウム、塩化コバルト、塩化アルミニウム、塩化ニッケル、ミオイノシトール、チアミン塩酸塩、ピリドキシン塩酸塩、ニコチン酸、葉酸、ビオチン、グリシン等を含むことができる。また固体培地のゲル化剤としては、特に限定されないが、寒天、ゼラチン、ゲルライト(和光純薬社製)等を用いることができる。
【0030】
「リンが欠乏した状態」での栽培の一態様としては、例えば、MS培地(Physiologia Plantarum (1962) 15巻, 473頁)で、組織が十分に生育した植物体を表1に示した組成をもつ培地に移すか又はMS培地を表1に示した培地に置換して、リンを含まないか又はリン濃度が極めて低い状態、例えば33μM未満、好ましくは10μM以下、更に好適には3.3μM以下、最適には1μM以下、或いは0.33μM以下で更に数日間〜数週間の栽培をすることが挙げられる。
【0031】
また別の一態様として、栽培過程において生じるリンの欠乏状態を維持しながら栽培することが挙げられ、この場合には、植物体を適当な期間栽培してリン濃度が極めて低くなってから、更に数日〜数週間の栽培を継続すればよい。この場合に用いる媒体中のリン濃度は、植物体が十分に生育した後に媒体中のリン濃度が上記に示した極めて低い濃度またはゼロとなる様に、初期濃度を調整することにより行えばよい。
【0032】
斯くして、植物体の根、葉、或いは茎等に蓄積した中性脂質を植物体から回収する方法は、特に限定されず、例えば、植物体の根、葉、或いは茎等を粉砕、圧搾すること、或いは適当な溶剤により抽出する方法等で行うことができる。より具体的には例えば、植物体の根、葉、或いは茎等を粉砕後、ノルマルヘキサンにより抽出する方法、或いはBligh and Dyer法(Can. J. Biochem. Physiol. (1959) 37巻, 911頁)などの方法を用いることで効率的に植物油脂を抽出することができ、植物油脂に含まれる形で中性脂質を回収することが可能である。斯かる植物油脂は、そのまま中性脂質として用いることもできるが、一般的な脱ガム、脱酸、脱色、脱臭等の精製の後に用いることもでき、その方法は特に限定されない。更に植物油脂から中性脂質を分離・回収することもでき、その方法についても特に限定されない。より具体的に例えば、薄層クロマトグラフィーによる分離およびシリカゲルプレートからの回収、高速液体クロマトグラフィーを用いた分離・回収などが挙げられる。
【0033】
斯くして得られる中性脂質はその大半がTAGであるが、少量のDAG等が含まれることもある。また中性脂質中の脂肪酸鎖長は植物体に固有な脂質生合成能力にほぼ依存することが多い。
【実施例】
【0034】
以下、実施例により本発明を詳細に説明する。なお、植物組織中の油脂成分の分析は、下記に記載の方法により行った。
(1)抽出及び前処理
総脂質の抽出は、Bligh and Dyer法(Can. J. Biochem. Physiol. (1959) 37巻, 911頁)に基づき行った。総脂質からTAGの分離は、薄層クロマトグラフィー(TLC Silica gel 60, 20x20 cm, メルク, 製品コード1.05721.0009、展開溶媒組成は、ヘキサン:ジエチルエーテル:酢酸=160:40:4(vol/vol))により行い、プレートからTAGのスポットをかきとって含量の測定を行った。
【0035】
(2)中性脂質量の測定
15:0脂肪酸を内部標準試料として、TAGをメタノリシス処理した。具体的には、ネジ栓付きガラス試験管内で、TAGを含むシリカゲル粉末に100 μlの1 mM 15:0ヘキサン溶液 (pentadecanoic acid, シグマ, P-6125)および350μlの5% 塩化水素メタノール溶液(和光, 089-03971)を添加して85℃で1時間処理した。メタノリシス処理後、ヘキサンで脂肪酸メチルエステルを回収し、窒素ガスで乾固後、60μlのヘキサンで回収し、そのうち3μlをガスクロマトグラフィーで解析した。ガスクロマトグラフィー(島津、GC-2014, カラムULBON HR-SS-10 (25 m, 0.25 mm ID)、カラム温度180℃、気化室および検出器250℃、入口圧(kPa) 68.2、カラム流量(ml/min) 0.53、スプリット比68.8、計測時間15分)を用いて分離・定量を行った。
【0036】
(3)植物組織乾燥重量の測定
凍結乾燥処理を20時間行った後、重量を測定し、乾燥重量とした。
【0037】
実施例1 リン酸含有培地における中性脂質の生産性
シロイヌナズナ(Arabidopsis thaliana)Col-0株(野生株)及びpgm-1変異株の種子35個体をMS寒天培地(Physiologia Plantarum (1962) 15巻, 473頁)に播種し、22℃、光強度40〜70μE/cm2、照射時間24時間/日の条件で10日間栽培した。尚、pgm-1遺伝子変異株はArabidopsis Biological Resource Center保有のストックCS210(参考文献:Plant Physiology (1985) 79巻, 11頁、Proceedings of the National Academy of Sciences of the United States of America (1989) 86巻, 5830頁,Plant Physiology (2000)122巻,1193頁)を購入し、葉におけるデンプン蓄積が抑制されていることを確認した個体から収穫した種子を使用した。本pgm-1変異株はAGIコードAT5G51820の遺伝子が変異により不活性化している。
【0038】
生育した植物体をMS寒天培地から注意深く引き抜き、可溶性リン酸(KH2PO4)を1mM含む表1の培地に35個体を移植した。これらを上記と同条件で更に10日間栽培した。栽培後の植物体を寒天培地から引き抜き、地上部(葉、茎)および根に切り分け、地上部の重量測定を行った後、液体窒素下すり鉢で粉砕し、Bligh and Dyer法で脂肪抽出を行った。この後、薄層クロマトグラフィーによりTAGを分離・精製後、塩酸メタノールでメタノリシス処理をしてガスクロマトグラフィーを用いて脂質分析を行った。また栽培した一部の個体は凍結乾燥し、乾燥重量を測定した。
【0039】
【表1】
【0040】
この結果、図1に示した様に、pgm-1変異株の地上部では、乾燥重量あたり野生株を有意に上回るTAG蓄積向上効果が認められた。また、得られた各株におけるTAGの脂肪酸組成分析を行ったところ、図2に示す様に脂肪酸組成の大きな変化は認められなかった。
【0041】
実施例2 リン酸非含有培地における中性脂質の生産性
シロイヌナズナ野生株及びpgm-1変異株の種子70個体をMS寒天培地(Physiologia Plantarum (1962) 15巻, 473頁)に播種し、22℃、光強度40〜70μE/cm2、照射時間24時間/日の条件で10日間栽培した。
生育した植物体をMS寒天培地から注意深く引き抜き、可溶性リン酸(KH2PO4)を含まない(0mM)、上記表1の培地に35個体を移植した。またそれぞれ35個体を1mM可溶性リン酸を含む表1の培地に移植し、これらを上記と同条件で更に10日間栽培した。
栽培後の植物体を寒天培地から引き抜き、地上部(葉、茎)および根に切り分け、地上部の重量測定を行った後、液体窒素下すり鉢で粉砕し、Bligh and Dyer法で脂肪抽出を行った。この後、薄層クロマトグラフィーによりTAGを分離・精製後、塩酸メタノールでメタノリシス処理をしてガスクロマトグラフィーを用いて脂質分析を行った。
【0042】
この結果、図3に示した様に、野生株及びpgm-1変異株共に、可溶性リン酸0mMの培地で栽培した地上部では1mM可溶性リン酸を含む培地で栽培した場合の約3倍のTAG蓄積向上効果が認められた。またpgm-1変異株はリン酸濃度に関わらず野生株の約3倍のTAG蓄積向上効果を示した。即ちこれらの結果より、pgm-1変異株の使用と可溶性リン酸0mMの培地での栽培を組み合わせることで、相乗的なTAG蓄積向上効果が得られることが明らかとなった。尚、得られた各株におけるTAGの脂肪酸組成分析を行ったところ、図4に示す様に脂肪酸組成の大きな変化は認められなかった。
【0043】
実施例3
シロイヌナズナ(Arabidopsis thaliana)野生株の種子96個体をMS寒天培地(Physiologia Plantarum (1962) 15巻, 473頁)に播種し、22℃、光強度40〜70μE/cm2、照射時間24時間/日の条件で10日間栽培した。
生育した植物体をMS寒天培地から注意深く引き抜き、可溶性リン酸(KH2PO4)の濃度が0mM、0.01mM、0.033mM、或いは1mMである上記表1の培地にそれぞれ24個体を移植し、これらを上記と同条件で更に10日間栽培した。
栽培後の植物体を寒天培地から引き抜き、地上部(葉、茎)および根に切り分け、地上部の重量測定を行った後、液体窒素下すり鉢で粉砕し、Bligh and Dyer法で脂肪抽出を行った。この後、薄層クロマトグラフィーによりTAGを分離・精製後、塩酸メタノールでメタノリシス処理をしてガスクロマトグラフィーを用いて脂質分析を行った。
【0044】
この結果、図5に示した様に、可溶性リン酸0mM、0.01mM(10μM)、及び0.033mM(33μM)の培地で栽培したシロイヌナズナ地上部では、1mM(1000μM))可溶性リン酸を含む培地で栽培した場合のそれぞれ約4倍のTAG蓄積向上効果が認められた。
【特許請求の範囲】
【請求項1】
デンプンの生合成が抑制された変異植物の植物体を栽培し、種子以外の組織に蓄積した中性脂質を回収することを特徴とする植物油脂の製造方法。
【請求項2】
変異植物が、ホスホグルコムターゼ遺伝子、ADPグルコースピロホスホリラーゼ遺伝子及びデンプンシンターゼ遺伝子から選ばれる1種以上の遺伝子の機能を不活性化又は発現抑制したものである、請求項1記載の植物油脂の製造方法。
【請求項3】
植物体を、リンが欠乏した状態で栽培する、請求項1又は2記載の植物油脂の製造方法。
【請求項4】
リンが欠乏した状態での栽培が、組織が十分に生育した植物体を、リンが欠乏した媒体に移植するか或いは媒体をリンが欠乏した媒体に置換して栽培する、又は栽培過程の媒体において生じるリンの欠乏状態を維持しながら栽培するものである請求項3記載の植物油脂の製造方法。
【請求項5】
種子以外の組織が、葉、茎、または根である、請求項1〜4のいずれか1項記載の植物油脂の製造方法。
【請求項6】
植物体が被子植物である、請求項1〜5のいずれか1項記載の植物油脂の製造方法。
【請求項7】
植物体が裸子植物である、請求項1〜5のいずれか1項記載の植物油脂の製造方法。
【請求項8】
植物体が双子葉植物である、請求項1〜5のいずれか1項記載の植物油脂の製造方法。
【請求項9】
植物体がシロイヌナズナである、請求項1〜5のいずれか1項記載の植物油脂の製造方法。
【請求項10】
デンプンの生合成が抑制された変異植物の植物体を栽培することを特徴とする、種子以外の組織への中性脂質の蓄積方法。
【請求項11】
リンが欠乏した状態で栽培する、請求項10記載の中性脂質の蓄積方法。
【請求項1】
デンプンの生合成が抑制された変異植物の植物体を栽培し、種子以外の組織に蓄積した中性脂質を回収することを特徴とする植物油脂の製造方法。
【請求項2】
変異植物が、ホスホグルコムターゼ遺伝子、ADPグルコースピロホスホリラーゼ遺伝子及びデンプンシンターゼ遺伝子から選ばれる1種以上の遺伝子の機能を不活性化又は発現抑制したものである、請求項1記載の植物油脂の製造方法。
【請求項3】
植物体を、リンが欠乏した状態で栽培する、請求項1又は2記載の植物油脂の製造方法。
【請求項4】
リンが欠乏した状態での栽培が、組織が十分に生育した植物体を、リンが欠乏した媒体に移植するか或いは媒体をリンが欠乏した媒体に置換して栽培する、又は栽培過程の媒体において生じるリンの欠乏状態を維持しながら栽培するものである請求項3記載の植物油脂の製造方法。
【請求項5】
種子以外の組織が、葉、茎、または根である、請求項1〜4のいずれか1項記載の植物油脂の製造方法。
【請求項6】
植物体が被子植物である、請求項1〜5のいずれか1項記載の植物油脂の製造方法。
【請求項7】
植物体が裸子植物である、請求項1〜5のいずれか1項記載の植物油脂の製造方法。
【請求項8】
植物体が双子葉植物である、請求項1〜5のいずれか1項記載の植物油脂の製造方法。
【請求項9】
植物体がシロイヌナズナである、請求項1〜5のいずれか1項記載の植物油脂の製造方法。
【請求項10】
デンプンの生合成が抑制された変異植物の植物体を栽培することを特徴とする、種子以外の組織への中性脂質の蓄積方法。
【請求項11】
リンが欠乏した状態で栽培する、請求項10記載の中性脂質の蓄積方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−153833(P2012−153833A)
【公開日】平成24年8月16日(2012.8.16)
【国際特許分類】
【出願番号】特願2011−15640(P2011−15640)
【出願日】平成23年1月27日(2011.1.27)
【出願人】(304021417)国立大学法人東京工業大学 (1,821)
【出願人】(000000918)花王株式会社 (8,290)
【Fターム(参考)】
【公開日】平成24年8月16日(2012.8.16)
【国際特許分類】
【出願日】平成23年1月27日(2011.1.27)
【出願人】(304021417)国立大学法人東京工業大学 (1,821)
【出願人】(000000918)花王株式会社 (8,290)
【Fターム(参考)】
[ Back to top ]