
植物用耐病性増強剤
【課題】ポリアミンをネクロトロフの性質を有する病原に対する抵抗性誘導剤として開発し、その剤の施用によって、植物がもっているポリアミンシグナル伝達経路、もしくはオーキシンシグナル伝達経路を活性化させ、ネクロトロフ病原が生成する植物毒素の働きを抑え、病原の侵入と病斑拡大を抑制する。
【解決手段】ポリアミン、リン酸塩、及びカルシウム塩のうち、少なくとも1つ以上の物質を用いて、植物を処理し、植物のポリアミンシグナル伝達経路及びオーキシンシグナル伝達経路のうち1つ以上の経路を活性化させることにより、植物の耐病性を増強する。
【解決手段】ポリアミン、リン酸塩、及びカルシウム塩のうち、少なくとも1つ以上の物質を用いて、植物を処理し、植物のポリアミンシグナル伝達経路及びオーキシンシグナル伝達経路のうち1つ以上の経路を活性化させることにより、植物の耐病性を増強する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、生体物質であるポリアミン、及びリン酸塩とカルシュウム塩で植物を処理し、植物のポリアミンシグナル伝達経路、もしくはオーキシンシグナル伝達経路を活性化し、植物が本来もっている耐病性を増強することに関する。
【背景技術】
【0002】
高等植物では、様々な病原の攻撃に対して、クチクラ層や細胞壁の強化、抗菌性物質の生産など、物理的手段から生化学的な手段に至る様々な抵抗性機構を発達させてきたが、植物の抵抗性応答の中でもっとも特徴的なのが過敏感反応(hypersensitive response : HR)である。HRは、感染した細胞の自発的な死(プログラム化された細胞死:PCD)を起こし病原の増殖を阻止するとともに、ファイトアレキシンと呼ばれる抗菌物質、病原特異的タンパク質(pathogenesis-related:PR)、及び植物のストレスホルモンであるサリチル酸(SA)、ジャスモン酸(JA)、エチレン(ET)などの生成蓄積を引き起こす。また、これらの生成蓄積によって、感染していない部位において次の感染を防ぐための全身獲得抵抗性(systemic acquired resistance : SAR)が誘導される。
【0003】
このようなHRの誘導は、多くの場合、宿主が抵抗性R遺伝子の産物を通じて病原特有の分子を認識することから開始され、植物病原の認識が、SA、JA、ETなどの植物ホルモン生合成の引き金を引く。これらホルモンシステムのバランス及び相互作用は、特定の病原と病害に対する抵抗性の発現に中枢的な役割をもっており、これらシグナル間の幅広いクロストークは、様々な病原のタイプに応じて宿主の適用すべき抵抗性機構を決定している。
【0004】
他方、ポリアミンは、第1級アミノ基を2つ以上有する脂肪族炭化水素の総称で、すべての生物に普遍的に存在する天然物質である。ジアミンのプトレシン、トリアミンのスペルミジン、テトラアミンのスペルミンが主要なものである。ポリアミンの基本的な機能として、核酸と融合してDNAの安定化とRNAの転写の効率化と制御、及びカチオン成分として細胞のイオンバランスの維持に関わっていることが知られている。また、細胞分裂やDNA複製のような細胞内過程、形態形成、さらには病害、乾燥、低・高温などの環境ストレスなどにも役割をもっていることを示唆するおびただしい研究報告がなされている。
【0005】
ポリアミンが、何故以上のような多岐にわたる複雑な生理活性を示すのかを考える際に、高度好熱性菌におけるポリアミンの働きをみていくことが極めて示唆的である。高度好熱性菌は、通常の生物には見られない特有のポリアミンが含まれていることが知られており、アミン数が多いポリアミンをより多くもった好熱性菌ほど高温に適応できることが明瞭に示されている。特に、テトラ、ペンタ、ヘキサアミンは核酸の立体構造の安定化を通じてタンパク質合成を含む高分子合成を高温でも可能なものにしていると考えられている(五十嵐一衛著「神秘の物質−ポリアミン」)。一方では、古細菌を含む好熱性菌や極限環境微生物の研究が進行し、生命の起源に近いとされている、つまりすべての生物の共通祖先に近い生物群は、好熱性を示すものが多く見られることが次第に明らかになっている。原始の単細胞生物は、過酷な環境に適応するためポリアミンのもつ化学的性質を、生命の誕生・進化のごく初期の段階で生命維持装置の1つとして取り込み、進化が進むにつれ、ポリアミンの機能も多面性が発揮されるようになってきたという考え方が成立する。
【0006】
ポリアミンの多面的な機能の1つとして、植物の病害抵抗性へのかかわりを挙げることができる。抵抗性N遺伝子をもったタバコ植物体にタバコモザイクウイルスを感染させると、ポリアミンは細胞間隙に蓄積した後、ポリアミン酸化酵素(PAO)で分解される過程で過酸化水素が生成され、病害抵抗性のひとつである過敏感細胞死が誘導されることが証明された(Hiroshi Yodaら2003)。また、HRには2つの主要な経路がある。1つは、特殊な宿主タンパク質による病原の認識を通じて、いわゆる遺伝子対遺伝子と称される病原と宿主の特異的な関係を有するhost HRであり、もう1つは、感染に対して通常の生化学的変化で抵抗性を誘導するnonhost HRの2つである。ポリアミンが双方のHRにかかわっているかどうかを検証するため、タバコ植物体に代表的なnonhost病原Pseudomonas cichoriiに感染させると、ポリアミン代謝にかかわっている酵素をコードしている6つの遺伝子の転写が同時的に誘導され、細胞間隙におけるポリアミンの蓄積、過酸化水素の生成、感染部位における過敏感細胞死が観察された。また、ウイルスで誘導される遺伝子サイレンシング方法によってPAOの活性を低下させたタバコ植物体は、病原細菌の生長を伴った過酸化水素生成の抑制と可視的な過敏感細胞死の消失をもたらした(Hiroshi Yodaら2009)。このことは、ポリアミンが植物防御に普遍的にかかわっていることを示唆している。
【0007】
ポリアミンが植物防御にもつ主要な役割は、以上に述べたように、細胞壁にあるPAOによる分解を通じて、シグナル分子として多数の遺伝子発現を制御する活性があることが証明されている過酸化水素(R.Desikanら2001)を生成させることに求められる。病原が侵入したときに、抵抗性R遺伝子産物を通じて病原の認知がなされ、その後の防御反応の誘導には、原形質膜状のNADPHオキシダーゼの働きによって生じる活性酸素種(ROS)が関与することが知られている。ROSには、スーパーオキサイドアニオン、一重項酸素、ヒドロキシルラジカル、さらにはスーパーオキサイドディスミューターゼの作用によって生じる過酸化水素などの多数の分子種が含まれる。このようなラジカル分子が生体膜近辺に急速に生成される現象は、オキシダティブバーストと呼ばれている。HR時におけるROSの生成は、NADPHオキシダーゼの働きによる急速でかつ一時的なものと、しばらく時間が経過して発生するがより持続的なものの2つの相があることが知られている。HRを誘導するためには、この2つの相のROS生成が必要とされることが近年明らかになっているが、前項の知見は、ポリアミンが後者のROS生成の普遍的な源泉であることを示唆している。
【0008】
ポリアミンの植物防御へのかかわりを明確にする上で、見落としてはならない報告がある。ポリアミン類の1つであるスペルミンは、タバコ植物においてサリチル酸とは独立して病原特異性タンパク質を誘導すること、さらにタバコモザイクウイルスに対する抵抗性を付与することが示され、病原体感染時のシグナル分子の1つとして考えられている。さらに、このスペルミンの作用に着目した研究では、病害抵抗性に関連しているMAPキナーゼであるサルチル酸誘導性タンパク質キナーゼ(SIPK)と傷害誘導性タンパク質キナーゼ(WIPK)を共に活性化させることが明らかになった。また、スペルミンによるMAPキナーゼの活性化はカルシュウム依存的であり、抗酸化剤の前処理によって強く抑制され、スペルミンシグナル伝達経路には活性酸素種とカルシュウムが関与していることが示唆されている(高橋ら2005−植物生理学会講演要旨集)。
【0009】
MAPキナーゼ(MAPK)は、タンパク質にリン酸基を付与するタンパク質キナーゼ(PK)の一種である。細胞は、その機能を維持するため、タンパク質のリン酸化と脱リン酸化を繰り返し行っている。このリン酸化及び脱リン酸化を通じて、酵素活性や他のタンパク質との相互作用が調節されており、MAPKは植物の病害を含めた環境ストレス応答反応に重要な酵素であり、とりわけSIPK及びWIPKは植物と病原の相互作用における防御反応の1つの収束的なポイントであることが認識されてきている。
【0010】
ところで、ジャガイモそうか病の病原であるStreptomyces scabieiは、バレイショの塊茎に侵入し重篤な病斑を形成して、商品価値を著しく低下させる病害であるが、ニンジン、ダイコン、サツマイモなどの根菜類にはほとんど発病させない。バレイショの塊茎肥大については、多くの研究者によって環境及び植物ホルモンの面から研究されている。実生バレイショにおける腋芽の伸長方向は、その着生部位によって異なり、一般的に上位節の腋芽は上方に、下位節の腋芽は下方に伸びてストロンとなり、地面に達した後、多くの場合、地中でイモを形成することが知られている。このような腋芽の伸長方向に植物ホルモンがどのようにかかわっているのかを、実生バレイショの切除処理と切除部に生長調節剤を塗布して調べると、腋芽の伸長方向は主として頂芽部及び若い葉でつくられる内生オーキシンによって制御されていることが示唆されている(池田ら1976−日本作物学会記事)。つまり、バレイショは、本来は腋芽になる幼芽組織を、オーキシンの制御でストロンと塊茎種子を形成する繁殖器官につくり変えていくという特殊なプロセスを進化させたといえる。S.scabieiのバレイショ塊茎への特異的な感染様式と種子塊茎をつくる特殊な器官であるストロンを形成する分子機構に、親和的な相互作用の要因があるという可能性が考えられ、オーキシンシグナル伝達経路との関連性があるのではないのかという疑問が生じる。
【0011】
事実、近年の英国での連続した研究で、オーキシンアナログのバレイショへの葉面散布はそうか病を有意的に減少させることが証明されている(McIntoshら1981〜1988)。さらに、オーストラリアのグループはこの英国での研究をもとに、2,4−Dのそうか病抑制活性を確認するとともに、塊茎の詳細な生理学的実験で、これまでそうか病菌感染に決定的と考えられている皮目数、皮目の外部容積、及び周皮の薄さと構造は、2,4−D処理による実質的な変化がなく、これらの構造変化を通じた病害抑制機構の可能性を否定し、オーキシン処理は、病原が産生するそうか病誘発に必須な植物毒素サクストミンAの毒性を抑制することに、病害抑制の機作を求めている(R.S. Teggら2008)。
【0012】
S.scabiei の植物毒素サクストミンAは、典型的な過敏感細胞死ではないが、アポトーシスの特徴である核DNAの断片化などを伴った過敏感細胞死に似た細胞死を誘導することが知られている。サクストミンAはセルロース生合成を阻害すると報告されているが、セルロース合成阻害剤イソキサベンもシロイヌナズナ懸濁培養細胞のプログラム化された細胞死(PCD)を誘導することが認められ、セルロース合成阻害など植物細胞壁の組成と構成における急速な変化を起こすことで、細胞のPCD を誘発すると示唆されている(Isabella Duvalら2005)。
【0013】
一方、無病育苗土を詰めた紙筒にマイクロチューバーあるいは無病種イモ切断塊茎を播種し、苗ステージごとにそうか病菌接種源6倍希釈の汚染土に裸移植し、その後の発病を調べると、塊茎ステージが進むに従って発病も減少し、塊茎肥大に伴ってそうか病に対する抵抗性が増加することが確認されているが(内藤ら1998)、S.scabieiの感染は、塊茎肥大のごく初期の段階から始まり、肥大の後期段階になると抵抗性が増強されることを意味している。前に記述した英国での研究報告では、オーキシンアナログの葉面散布は、生育の遅い段階よりも早い段階がより効果的であることが確認されており、塊茎肥大に伴って増強されるそうか病抵抗性にはオーキシンシグナル伝達経路がかかわっている可能性が大きい。
【0014】
植物病原は、これらの生活史からしばしばバイオトロフ、ネクロトロフ、及び生活史に応じて双方の性質を有するセミバイオトロフに分類される。バイオトロフとは生きた宿主の組織から栄養を摂るタイプ(死体栄養性)であり、一方、バイオトロフとは宿主の組織を殺しその残渣から栄養を摂るタイプ(生体栄養性)である。真性バイオトロフの典型であるErysiphe orontiiなどの糸状菌は、宿主細胞に細胞死を起こすことなく表皮細胞に感染し、吸器を発達させ胞子形成を十分に行うが、その一方、ネクロトロフと考えられているBotrytis cinereaなどの糸状菌は、宿主細胞の死を促進させそうな植物毒素を生産し、感染の非常に早い段階で宿主細胞を死に至らしめ、組織にダメージを拡大していくことが知られている。
【0015】
以上の知見を考え合わせると、ジャガイモそうか病の病原であるS.scabiei は真性ネクロトロフと分類でき、S.scabiei に対する抵抗性誘導にはオーキシンシグナル伝達経路が介在していることが示唆される。
【0016】
多くの植物病原は、植物との相互作用の中でオーキシンを生成させ、宿主のオーキシンシグナリングを強めることで病徴を進行させることが知られており、オーキシンが病原の攻撃に対する抵抗性に貢献することは、長い間、疑問視されてきた。
【0017】
しかし、最近、ネクロトロフ糸状菌に対する抵抗性にオーキシンが関与しているという、間接的ではあるが確かな証拠が出されている。例えば、植物細菌性病原であるErwinia amylova由来のタンパク質エリシターである精製されたharpinを、抵抗性N遺伝子をもつタバコ葉にオーキシンアナログである2.4−Dと併せて浸潤させると、対照葉には細胞死が生じたこととは対照的に、オーキシンは完全に細胞死を抑制でき、また、harpinを浸潤させた6時間後に2.4−Dを施用すると、細胞死の進行を回復させることができた(Suresh Gopalan2007)。一方、シロイヌナズナのオーキシンシグナル伝達経路に変異性があり、オーキシン促進的なSCF(Skp-Cullin-F-box)ユビキチン化経路に欠陥をもつ、axr1、axr2及びaxr6は、ネクロトロフの糸状菌であるPlectosphaerella cucumerina及びBotrytis cinereaに対し感受性の増加を示していることが証明された。また、これら変異体は、通常SCF-ユビキチン/プロテアーゼ機構による除去のターゲットになっているオーキシン転写抑制物質AXR3の安定化が、P. cucumeriaの感染時に生じていることが示された。これらの結果から、オーキシンシグナル伝達経路はネクロトロフ糸状菌であるP. cucumeria及びB. cinereaに対しての抵抗性に重要であることが示唆されている(Francisco Llorenteら2008)。
【0018】
さらに、オーキシン受容体であるTIR1の3次元構造が解明され、TIR1のLRRs(ロイシン−リッチリピート)にあるオーキシン結合ポケットの構造を安定化する共因子として作用するイノシトールヘキサキスフォスフェート(フィチン酸:InsP6)分子が、TIR1と共役しオーキシンのシグナル転換を促進する可能性が指摘された(Xu Tanら2007)。他方では、低フィチン植物として開発された、イノシトール3フォスフェート合成(IPS:フィチン酸合成における最初のステップを触媒する)のアンチセンス遺伝子配列を構造的に発現するバレイショ組み換え植物体は、フィチン酸レベルを低下させ、非病原性ジャガイモYウイルス及び病原性タバコモザイクウイルス(TMV)に対する抵抗性が低下し、一方、シロイヌナズナにおいて、IPS遺伝子に変異性があるatips2はInsP6を枯渇しており、糸状菌病原であるB. cinerea、及び細菌病原であるPseudomonas病原性・非病原性のレースに対する感受性を高めた。また、変異体atpis2及びatpik1は、SAを蓄積させることができない植物体と同じように、感染に対して過敏な感受性を示したが、これら昂進された感受性はSAレベルの低下によるものではなかった。さらに、これもInsP6が枯渇している変異体atips1は、病原に対する抵抗性を低下させない。これら変異体に見られるInsP6合成の撹乱は、ウイルス、細菌、糸状菌の3つの幅広い病原の感染に対して感受性の昂進をもたらし、植物病原に対する防御を調節する新たなリンクがあることを想定しなければならない事象が明らかにされた(Alex M. Murphyら2008)。
【0019】
前項で示した数々の知見は、生長ホルモンであるオーキシンは、植物防御に関しネガティブな効果しかもたないとされてきた従来の考え方を覆すに十分である。とりわけ、InsP6分子は、様々な病原に対しての抵抗性誘導の重要な鍵を担っている証拠が示され、その作用モードは、オーキシンシグナル伝達経路を転換し、多様な病原に対して、込み入った防御反応の誘導を幅広く調節しているという考え方を導く。InsP6分子の作用モードが解明されていくと、植物防御反応におけるオーキシンシグナル伝達経路の役割が明らかになり、これまでの考え方に大きな変更を強いるような新たな知見が加えられる可能性がある。
【0020】
ポリアミンの多面的な機能をもっていることのもう1つの例として、マメ科植物と共生して窒素固定を行う根粒菌の根への侵入・着生をポリアミンが制御していることを示唆する報告(J.Terakadoら2006)を挙げることができる。ダイズ変異体En6500は根粒菌を根に過剰に着生させ、地下部との光合成産物の競合によるもと考えられるが、地上部の極端な矮性症状を示す。このEn6500の葉中のスペルミジン及びスペルミンの含有量はその親株であるエンレイのそれと比較するといつも低く、前駆体であるプトレシンからのスペルミジン及びスペルミンの合成が抑制されていた。En6500の葉にスペルミジン及びスペルミンを散布すると、根粒菌の着生制御機能をとりもどすとともに根の生長を抑制する事実が示されている。この報告に示された事実は、ポリアミンの直接的な効果ではなく、若枝におけるプトレシン、スペルミジン、スペルミンなどの構成もしくはその含量レベルの顕著な変化が葉中の生化学的な変化を生じさせ、その結果として、根における生化学的、植物生理的もしくは生態的な重要な変化を誘導する可能性を示唆したものである。
【0021】
さらに、根粒菌とマメ科植物の根への着生に関して、ポリアミンに関するものではないが、もう1つの興味深い知見がある。根粒菌を着生させないダイズ変異体T201の培地上に微量の2,4−Dを添加すると、通常のダイズ根粒と同じレベルの窒素固定活性を示す根粒が誘導されるという事実が示された(赤尾ら1995)。培地上に微量の2,4−Dを施用すると、根粒菌の根への侵入を容易にするという知見である。
【0022】
オーキシンは主根の伸長を抑制し、側根と不定根の形成を促進することは、古くから知られている。ダイズ変異体En6500は、主根の長さが短く側根数が異常に多いというオーキシン過剰の表現型を示す。おそらく、何らかの変異性によってポリアミン代謝に欠陥があり、外生ポリアミンに感受性を示す。一方、ダイズ変異体T201は、おそらくオーキシンの生成を含めたシグナル伝達経路もしくは輸送に異状があるオーキシン欠乏型の変異体で、外生的なオーキシンに感受性を示す。En6500の親株であるエンレイは、生育初期の葉中おいてはプトレシンの含量が高く、生殖生長に移行する段階になると、スペルミジン及びスペルミンの含量が高くなり、葉中のポリアミン組成の変動を通じて、根の生態的及び生理的な変化を起こし、おそらく根の伸長、特に側根の伸長が抑制され、根粒菌の侵入・着生を制御すると考えられる。
【0023】
Terakadoらが示唆した知見、及び赤尾らが示した知見、さらにはオーキシンの働きに関するこれまでの知見を総合的に考え合わせれば、ポリアミンがオーキシンの生成や輸送に影響を与えるという報告は見出せないものの、PAOで分解される過程で生成される過酸化水素によってオーキシンシグナル伝達経路が活性化される可能性がある。さらに、葉中のポリアミン組成の変動は、ダイズの根において、オーキシン受容のモードを変え、異なるオーキシ反応因子(ARF)が発現し、根における生化学的、植物生理的もしくは生態的な重要な変化を誘導するという仮説が成立する。
【0024】
前にも触れたが、バレイショはオーキシンによって制御されたストロン形成を行うが、この機構には、進化論的に考えると、側根もしくは不定根をつくりだす分子機構と同じものが用いられていることが類推できる。もし、この類推と前項の仮説が正しいとすれば、バレイショの葉にポリアミンを散布することによって、ダイズの根において根粒菌の着生を制御するように、ジャガイモそうか病菌の侵入を抑制できる可能性がある。
【0025】
最近の研究報告では、キュウリにおいて、主根を切断して得られた実生の外植片で過酸化水素が生成され、それが不定根の形成に先立ってオーキシンシグナルを仲介することが証明された(Shiweng Liら2007)。これは、シグナル分子として多様な遺伝子の発現を制御していることが証明されている過酸化水素が、内生オーキシンの生成を誘導することを示唆するもので、ポリアミンが、PAOよって分解される過程で生成される過酸化水素はオーキシン生成を促進するというわれわれの見解を支持している。
【0026】
以上、ポリアミン処理は、様々な文献に示された知見を総合的に検討し、植物が本来もっているオーキシンシグナル伝達経路を介した耐病性を増強できる可能性があることを示してきた。しかし、先端的な研究報告から得られた知見を単純に積み上げれば、ポリアミンの散布によってあるタイプの病原に対して実用性がある抵抗性を誘導できることが証明されるというものではない。その証拠に、ポリアミンの植物体への直接散布が、病害に対しての抑制効果を示した報告もなければ、ポリアミンがオーキシン活性を高めるという事実を示した報告も皆無である。
【0027】
ポリアミンはオーキシンシグナル伝達経路を活性化させオーキシンの働きを変えるという考え方に基づいた実験は、ジャガイモそうか病で例証するように、植物細胞にPCDを誘発するネクロトロフ病原などのあるタイプの病害に対して、植物に実用性がある耐病性を増強できることを示したデータをもたらした。
【先行技術文献】
【特許文献】
【0028】
【特許文献1】特許公開2004−242505 環境ストレス抵抗性を増強した植物の作出方法
【特許文献2】特許公開2007−000021 ポリアミン代謝制御による環境ストレス耐性を改良した植物、及びその作出方法
【特許文献3】特許公開2005−237387 種々の環境ストレス耐性を改良した植物、その作出方法並びにポリアミン代謝関連酵素遺伝子
【特許文献4】特許公開2004−242510 生産性を改良した植物、及び作出方法
【特許文献5】特許公開2004−180588 環境ストレス抵抗性を改良した植物及びその作出方法
【特許文献6】WO 02/023974 種々の環境ストレス耐性を改良した植物、その作出方法並びにポリアミン代謝関連酵素遺伝子
【非特許文献】
【0029】
【非特許文献1】五十嵐一衛著「神秘の物質−ポリアミン」共立出版(株) p.26〜27
【非特許文献2】Hiroshi Yoda et al., 「Induction of Hypersensitive Cell Death by Hydrogen Peroxide Produced through polyamine Degradation in Tobacco Plants.」Plant Physiology, August 2003 Vol.132, p. 1973〜1981
【非特許文献3】Hiroshi Yoda et al., 「Polyamine as a common source of hydrogen peroxide in host and nonhosthypersensitive response during pathogen infection.」Plant Mol Biol 2009 70:103〜112
【非特許文献4】Radhika Desikan et al., 「Regulation of the Arabidopsis Transcriptome by Oxidative Stress.」Plant Physiology, September 2001, Vol.127,p.159〜172
【非特許文献5】高橋ら「植物の抵抗性におけるスペルミンのシグナル伝達経路」、日本ポリアミン研究会−第19回研究発表会− 講演要旨集p.18
【非特許文献6】池田ら「ジャガイモの匐枝の生長に関する研究」日本作物学会記事 45(2) : 314-321 (1976)
【非特許文献7】McIntosh A. H. et al., 「Decreased Severity of Potato Commonc Scab after Foliar Sprays of 3,5-D Acid, a Possible Antipathogenic Agent.」Annals of Applied Biology 1981 Vol.99, p.275〜281
【非特許文献8】Robert S. Tegg et al., 「Auxin-Induced Resistance to Common Scab Disease of Potato Linked to Inhibition of Thaxtomin A Toxicity.」Plant Disease 2008 Vol.92 No.9, p.1321〜1328
【非特許文献9】Isabelle Duval et. al., 「Thaxtomin A induces programmed cell death in Arabidopsis thaliana suspension-cultured cells.」 Planta Vol.222, No.5 2005/11
【非特許文献10】内藤ら「紙筒移植栽培によるジャガイモそうか病回避の可能性」、日本植物病理学会報1998, 64(6), pp.580
【非特許文献11】Suresh Gopalan 「Reversal of an immunity associated plant cell death program by the growth regulators auxin.」Nature Precedings: Posted 18 Nov 2007
【非特許文献12】Francisco Llorenteet al., 「Repression of the AuxinResponse Pathway Increase Arabidopsis Susceptibility to NecrotrophicFungi.」Molecular Plant Vol. 1, No.3, p.496〜509, May 2008
【非特許文献13】Xu Tan et al., 「Mechanism of auxin perception by the TIR1 ubiquitinligarse」Nature 446, 640〜645, (5 April 2007)
【非特許文献14】Alex M. Murpry et al., 「A role for inositol hexakisphosphate in the maintenance of basal resistance to plant pathogens」The Plant Journal (2008) 56, 638〜652
【非特許文献15】Junko Terakado et al., 「Production Shoot-applied polyamine suppress nodule formation in soybean」Journal of Plant Physiology 163 (2005) 497〜505
【非特許文献16】赤尾ら「2,4-D誘導根粒の形成過程と根粒菌の感染様式」 日本作物学会記事 64巻(別1号) 1995
【非特許文献17】Shiweng Li et al., 「Hydorogen peroxide involvement in formation and development of adventitious roots in cucumber」Plant Growth Regulation, Vol.52, No.2, 2007/6
【発明の概要】
【発明が解決しようとする課題】
【0030】
本発明は、ポリアミンの外生的な処理は、植物のオーキシンシグナル伝達経路、もしくはそれとは独立したポリアミンシグナル伝達経路を活性化させ、ネクロトロフ病原であるジャガイモそうか病菌に対する抵抗性を誘導できるという事実に基づくものであり、宿主細胞にPCDもしくは壊死を引き起こすタイプの病原に対する植物の耐病性を増強できると考えられる。しかし、その適用範囲をジャガイモそうか病以外に広げて確定するには、今後の試験研究に待たねばならない。したがって、以下では、ジャガイモそうか病に限定し、どのような課題を解決できるか、また、どのような方法を用い、どのような効果があるのかについて例示する。
【0031】
ジャガイモそうか病は、バレイショを連作することでStreptomyces scabieiを中心とした病原性のある放線菌が土壌中での生息密度を高め、植物体が塊茎を形成するごく初期のステージから塊茎細胞に侵入し、塊茎肥大とともにかさぶた状の病斑を拡大させるバレイショの世界的な重要病害である。1891年にタクスターによって初めて報告された大変古い病害であり、その後世界各地で確認されている。わが国でも、北海道で1906年北海道農会報に記述されたのが最も古い記録のようであるが、主要なバレイショ生産地では代表的な土壌伝染性病害の1つとして、また、難防除病害として広く知られている。とりわけ、わが国において、西南暖地を代表するバレイショ産地では、長年にわたり年二作に及ぶ連作を続けた結果、そうか病はバレイショ連作障害の中心をなすものとして大きな問題となっている。
【0032】
そうか病に感染すると、ジャガイモの表面にかさぶた状の激しい病斑を形成し、食用、食品加工用としての商品価値が著しく低下する。そうか病による経済的な被害は、少なくとも、生産額の数パーセントに上ると考えられている。
【0033】
世界中のバレイショ生産地では、そうか病による経済的損失を最小化するため様々な対策が用いられているが、いかなる対策も経済性がある効果的で安定的な病害抑制に成り得ていない(Plant Disease/Vol.92 No.9 1321-1328)。クロルピクリンによる全面的な土壌消毒、S.scabieiの土壌中における活性を低下させるため、土壌を酸性側に維持する、また堆肥等の有機物を投入しないなどの土壌管理、あるいは病原菌の侵入する時期に潅水を行うなどの防除対策が行われているが、これらの対策は、環境への負荷、土壌生産性の低下、潅水時期の判断が難しいなどの理由から、安定的な防除対策とはいえない。また、拮抗微生物や抗菌物質を利用した生物的防除法があるが、効果に安定性がないため、生産現場において十分な普及をみていない。
【0034】
本発明は、病原とバレイショ植物体の相互作用に着目し、幅広い生理的活性をもっている生体物質のポリアミン、及びリン酸塩とカルシュウム塩で植物を処理することによって、オーキシンシグナル伝達経路、もしくはポリアミンシグナル伝達経路を活性化させ、植物がもともともっている抵抗性を増強することによって、病原菌のバレイショ塊茎細胞への侵入と病斑拡大を抑制し、防除困難と考えられてきたジャガイモそうか病の安定的な防除法を提供するものである。
【0035】
生産地においては、ジャガイモそうか病を抑制する土壌管理は、バレイショ作の低生産性を余儀なくされている。安定した防除法が確立されると、堆肥や石灰資材の投入による望ましい土壌管理が可能になり、とりわけバレイショ連作地帯では収量性と品質の双方とも大幅に改善され、土地生産性が飛躍的に向上する。
【課題を解決するための手段】
【0036】
スペルミジン、スペルミンなどのポリアミン、もしくはスペルミジン三塩酸塩、スペルミン四塩酸塩などのポリアミン化合物からなる物質群のうち、少なくともいずれか1つ以上の物質を水に溶かし、最大10μモル/ml以下の濃度で散布液を調製し、植え付けた種子が出芽して5日後を第1回の散布日にして、以降5〜10日間隔で、該調製液を、1植物体当たり2〜20ml、回数にして1〜4回程度の葉面散布を行う。
【0037】
ポリアミンを処理する前に、リン酸カルシュウムを水に溶かし、該水溶液を出芽して間もないバレイショの株元に1株当たりリン酸カルシュウムが最大3〜10グラムになるよう施用するか、リン酸カルシュウムの0.3パーセント程度の水溶液に種イモを2時間程度浸漬処理し、その後十分に風乾させた後に植え付けるなどの前処理を行うと、ポリアミンの作用性が増幅され、防除効果が高まる。
【0038】
そうか病菌は塊茎形成のごく初期(米粒以下の大きさ)から侵入するので、出芽後の散布適期を逃すと防除効果は極端に劣る。適期を逃さないことが本手段のもっとも大きなポイントである。
【発明の効果】
【0039】
本発明は、病原菌が病原性を発揮するために必須な植物毒素サクストミンAの毒性を抑制し、病原菌のバレイショ塊茎細胞への侵入と病斑拡大を抑制することを特徴とするまったく新しい手法による防除対策である。
【0040】
バレイショは、塊茎肥大のステージが進むにつれ病害に対する抵抗性を発現させることが知られているが、その抵抗性発現には、植物ホルモンである内生オーキシンがかかわっていることはほぼ確実である。外生的なポリアミン及びリン酸カルシュウムによる処理は、葉中のポリアミン組成を変化させるとともに、ポリアミンシグナル伝達経路を活性化させ、塊茎肥大の後期に発現する抵抗性を肥大の初期に前倒しにして誘導させる働きがあり、そうか病菌の塊茎細胞への侵入と病斑拡大を抑制する。その機構は、オーキシンは病原菌が生成する毒素サクストミンAの宿主細胞に細胞死を引き起こす働きを抑制する、あるいはそれとは独立したポリアミンシグナル伝達経路を通じたファイトアレキシンを生成させることなどに求められるだろうが、詳細な機構は不明であり、今後の究明を待たなければならない。
【0041】
いずれにしても、リン酸カルシュウムを併用したポリアミン処理は、実施例に示すように、そうか病の多発条件下において、そうか病の発生を70〜80パーセント程度に抑えることができ、すでに実用化技術として利用できるレベルまで達している。
【0042】
本発明は、これまでの防除対策とは異なり、病原菌の毒素サクストミンAの活性を阻害し、病原菌の侵入と病斑拡大を抑制するものであるから、普遍性のある効果が期待できる。
【0043】
本発明は、通常の人工的な合成物ではなくポリアミンという生体物質を用いること、またポリアミンが塊茎内に蓄積して直接的に毒素サクストミンAに作用するのではなく、オーキシンシグナル伝達経路、もしくはポリアミンシグナル伝達経路を活性化させ、サクストミンAの毒性を抑制し病原菌の侵入と病斑拡大を抑制すること、要するに生体機能に基づいた抵抗性を増強することを特徴とするものである。したがって、食の安全性についても何ら問題がない。また、オーキシンアナログである2,4−Dなどを用いると、収量の低下、奇形果の発生、塊茎での残留毒性などの問題が生じるが、ポリアミンを用いた場合、そのようなデメリットは生じない。
【図面の簡単な説明】
【0044】
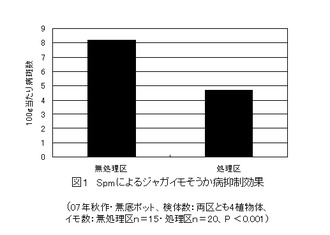
【図1】バレイショの植え付け25日後にスペルミン(Spm)を葉面に散布し、ジャガイモそうか病の抑制効果を示したものである。(実施事例1)
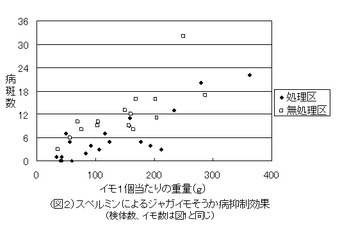
【図2】図1のもとになったデータをジャガイモ1個の重量と病斑数でプロットし、その分散状況を示した図である。(実施事例1)
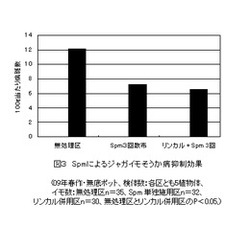
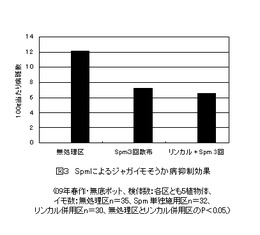
【図3】Spmを散布する前にリン酸カルシュウム(リンカル)を土壌に施用し、併用処理の効果を示した図である。(実施事例2)
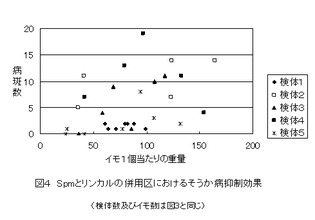
【図4】図1のもとになったデータのうち、Spmとリンカルの併用処理区だけのデータをとりだし、ジャガイモ1個の重量と病斑数でプロットし、分散状況を示した図である。(実施事例2)
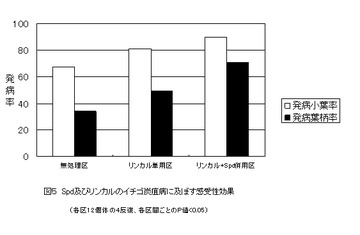
【図5】スペルミジン(Spd)とリンカルの併用処理が、イチゴ炭疽病の発病度合いにどのような影響を与えるかを示した図である。(実施事例3)
【発明を実施するための形態】
【0045】
本発明は、ポリアミンの生理活性機能を利用し、葉面散布もしくは土壌処理によって植物の生理を制御し、ジャガイモそうか病に代表されるネクロトロフ(死体栄養性)、もしくは植物細胞に壊死を起こす性質を有する病原に対する抵抗性を誘導し、これら病原が誘発する病害を抑制するものである。したがって、ポリアミンもしくはポリアミン塩酸塩などの化合物を、農薬取締法に基づいて殺菌剤もしくは植物生長調節剤として農薬登録を行い、農薬の製造・販売を経て、バレイショ生産者が農薬として散布して発明を実施することになる。
【実施例1】
【0046】
(図1及び図2の関連)
土壌中の微生物相は、温度、湿度、有機物の種類や量などの環境因子で大きく変り、通常の圃場でそうか病の多発条件をつくることは困難である。このため、圃場に直径30cm、深さ40cm塩化ビニール製の無底ポットを埋め込み、クロルピクリンで消毒した後、ポットに一定の病原汚染土を均質に混ぜること(接種)で、人工的に安定したそうか病の多発条件をつくり、そうか病の抵抗性品種検定等の効果判定が可能になった。
【0047】
この実験システムを用い、種イモを植え付け、1)植え付け25日後に、市販されているポリアミン化合物であるスペルミン4塩酸塩を用い、展着剤としてトリトン0.5mg/mlを加えて調整したスペルミン(Spm)2μM/ml水溶液を1植物体当たり20ml散布処理したSpm散布区、2)比較対照としての無処理区の2つの区を設定した。それぞれの区は5検体で反復なし。
【0048】
それぞれの区の検体ごとに収穫し、それぞれのイモごとに重量と病斑数をカウントし、処理区と無処理区のそれぞれの発病度を比較できるように、イモ1個ごとに100g当たりの病斑数に換算した。その1個ごとのデータを用いて、処理区と無処理のデータ分散状況を検定するとP値は0.001未満となり、完璧な有意差がとれた。図1は、処理区と無処理区の区ごとの平均値を示したもので、Spmによるジャガイモそうか病の抑制効果が確認できる。ただし、それぞれの区で出芽時期が遅れた1検体を、生育時期のずれによる降雨及び気温などの気象要因の違いの影響を受けている可能性があるので除外し、生育が揃っているそれぞれの区の4検体のデータを用いて、図を作成している。
【0049】
図2は、それぞれのイモの重量と病斑数でプロットし、処理区と無処理区の分散状況を示したものである。無処理区では、重量と病斑数に比較的強い相関が見られるが、その一方で、処理区ではN字状の連続性のある分散パターンを示し、重量200g前後の領域にSpmの強い作用領域が認められるとともに、200gより軽いイモの病斑数は、無処理区と比較して概して少ないことが認められる。また、イモの重量の違いはストロン形成の早晩によって異なるものと考えられ、重量を示したX軸は時間軸を表しているとも考えられ、無処理区のN字状のパターンは、Spmによる処理時期が多少遅かったことを意味している。
【0050】
以上の結果から、1)Spmによるそうか病の抑制効果は確認できるが、実用化のためには処理方法をさらに検討し、抑制率を高める必要があること、2)図2の分布パターンから判断し、処理時期を早める、処理効果には強いものがあるが効果が長続きしないので複数回の処理が必要とされる、ことなどが示唆される。
【実施例2】
【0051】
(図3〜4の関連)
Spmのシグナル伝達経路を増幅するにはMAPKを活性化させることが重要と考えられるので、Spmの処理前に土壌にリン酸カルシュウム水溶液を処理し、植物体中のリン酸とカルシュウムレベルを高めておくと、MAPKの活性化が期待できるのではないかと考えた。このため、1)出芽後5日、10日、20日の3回、実施例1と同じスぺルミン2μM/ml水溶液を1植物体当たり20ml散布処理したSpm単用区、2)Spmを処理する前に出芽後1〜2日後にリン酸カルシュウム(リンカル)3%水溶液を1ポット当たり100mlを土壌に施用したリンカル併用区、及び3)比較対照としての無処理区の3つの区を設けた。それぞれの区は5検体の反復なし。
【0052】
実施例1と同じように、検体ごとに収穫し、イモ1個ごとにそれぞれの重量の測定と病斑数をカウントした。図3は、それぞれの区の100g当たり病斑数の平均値を表したものである。Spmとリンカルの併用区と無処理区の間では、P値<0.05の有意差がとれ、リンカルはSpmのシグナル伝達経路を増幅することが確認できる。しかし、単用区及びリンカル併用区とも処理効果の個体差が大きいことが見出された。無処理区、Spm単用区、及びリンカル併用区における1検体における100g当たりの病斑数の最大値と最小値の幅は、それぞれ14.5〜9.6、14.4〜2.3、10.9〜2.0となっており、処理区おけるSpmの効果が顕著な検体とそうでない検体とに別れている。
【0053】
図4は、リンカル併用区におけるイモの重量と病斑数でプロットした分散パターンを示している。無処理区の100g当たり病斑数データ群とリンカル併用区のそれぞれの検体のデータ群間の分散状況を検定すると、P値は、検体4(0.98)>検体2(0.91)>検体3(0.22)>検体5(0.03)>検体1(0.02)となる。無処理区の分散パターンとほとんど変らない検体から有意的に病斑数が減少した検体の両極端に分かれている。本来ならば、検体1程度の病斑数の減少が期待できるにもかかわらず、何かの要因がそれを妨害したと考えなければならない。
【0054】
実施例2は2009年の春作に行ったが、それと同じ時期に、野外ではなく温室でも同じようなジャガイモそうか病抑制試験を行った。このときの気温推移は、処理時期の前後に、最高気温では8.6℃、平均気温では6.4℃と大幅な気温上昇があり、その時期の最高気温は通常であれば19℃程度であるが、そのときの最高気温は、バレイショの生育適温とされる22〜23℃を越え24.9℃を記録した。温室においては、重要な感染時期と考えられる塊茎形成と肥大の初期段階で、植物体を2週間程度35℃前後の高温に数時間継続的に遭遇させる結果になった。
【0055】
植物体がこのような高温ストレス条件下に置かれると、Spm効果が見出せないだけでなく、そうか病に対して本来発動されるべき抵抗性も消失し、通常は塊茎の表層だけにとどまる病斑部は塊茎内部に奥深く広がり、発病度合いを評価する通常の方法が適用できないほどの激しい病徴を示した。抵抗性R遺伝子の働きには温度感応性があり、28℃を超える高温では抵抗性が消失することが知られているが、それに似た現象が起きたことになる。
【0056】
キュウリを高温順化させるため38℃に遭遇させると、ポリアミン代謝に変化が起こり、葉にスペルミジン及びスペルミンが徐々に蓄積され、また、高温に遭遇させない非順化葉にスペルミジンまたはスペルミンを葉面散布すると、光合成器官の熱安定性が高まることが知られている(Tachibanaら2003)。ポリアミンは、プトレシンを前駆体として、順次、スペルミジン、スペルミンとアミン数が多いポリアミンが合成される。アミン数が多いポリアミンをより多くもった好熱性菌ほど高温に適応できることが知られているが、植物においても同様で、このような高温ストレス時には、ポリアミン酸化酵素(PAO)が活性をなくし、よりアミン数の多いポリアミンの蓄積が起きると考えられる。
【0057】
前にも触れたが、Hiroshi Yodaらは、PAOの活性を低下させたタバコ植物体は、病原菌の生長を伴った過酸化水素生成の抑制と可視的な過敏感細胞死の消失を観察したが、われわれも同様に、高温に遭遇したバレイショ植物体は、おそらくPAO活性を喪失し、スペルミジン及びスペルミンの分解による過酸化水素の発生が抑制された結果と考えられる病害抵抗性の消失を観察した。
【0058】
野外では温室ほどの高温ではないが、急激な気温上昇と生育適温を超える温度に遭遇することによって、植物のポリアミン代謝が変化し、PAO活性レベルが低下する個体が現れてくるとともに、微妙な温度域になるので、高温に過剰に反応せず活性レベルを低下させない個体もあり、そこにSpm効果に大きな振れが生じた原因があると考えられる。
【0059】
バレイショの種イモを植え付け、その後の降雨が少なく乾燥状態になると、早く出芽がするものと、まとまった雨を待って遅く出芽するものの両極端に分かれる現象が観察される。植物の環境ストレスに対する応答反応は、すべての個体が一律的に反応よりも、個体差をもたせた方が環境に柔軟に対応でき、種が生き残る機会を多くするだろうから、植物体の環境ストレス応答における個体差は、もともとプログラム化された応答システムとして埋め込まれているのかも知れない。
【0060】
Spmとリンカルの併用処理区における顕著な効果が見出せる検体の病徴と無処理区のそれとを比較すると、病斑数の減少だけでなく病斑のサイズが低下していることが確認でき、Spmの効果には、病原の侵入抑制と侵入を受けても病斑拡大を抑制する2つの効果がある。
【実施例3】
【0061】
(図5の関連)
スペルミジン(Spd)とリン酸カルシュウム(リンカル)の処理が、他の病害に対してどのような効果をもつかを調べるため、イチゴ炭疽病菌懸濁液をイチゴ苗に接種し、炭疽病の発病度合いをみてみた。ポリポット苗で育成したイチゴ苗に、1)リンカル3%水溶液を各ポットに50mlを施用したリンカル単用区、2)同じようにリンカル水溶液を処理した2日後にイチゴ苗に展着剤としてトリトンを加えたSpd水溶液2μモル/mlを20ml葉面散布したSpd併用区、3)比較対照としての無処理区の3区構成。Spd散布処理21日後に炭疽病菌の懸濁液(1×104個/mlの7.6ml/鉢で接種。病徴は複葉(1本の葉柄に3枚の小葉がついた葉)における発病小葉率及び発病葉柄率で評価。それぞれ12個体の4反復。
【0062】
図5は、この実施例の結果を示している。各区とも最終的な枯死率は変らなかったが、初期感染時において、小葉及び葉柄における病斑形成に有意差が生じた。無処理<リンカル単用区<リンカルとSpd 併用区の順に、小葉及び葉柄ともに発病率が有意的に高まった。
【0063】
イチゴ炭疽病の葉細胞への侵入と病斑形成は、旧葉よりも新葉で多く見られ、新葉の炭疽病に対する感受性は旧葉よりも高いことが知られている。つまり、オーキシン活性が高い若い組織ほど炭疽病に感受性を示すと考えてよい。多くの植物病原は、植物との相互作用の中でオーキシンを生成させ、宿主のオーキシンシグナリングを強めることで病徴を進行させることが知られているが、イチゴ炭疽病菌もそのようなタイプの1つであり、おそらくその抵抗性にはSAシグナル伝達経路がかかわっている。
【0064】
植物防御におけるオーキシンとSA との対立的関係は、近年、多数の研究報告の中で証明されている。例えば、オーキシンはSA シグナル伝達経路において中心的な役割をもっているPR-1遺伝子の十分な発現を阻害することが証明されている(Dong Wangら2007)。これまで得られている知見を総合して考えると、本実施例で得られたデータは、リンカルとSpdで処理したイチゴ植物体は、炭疽病に対する感受性が昂進されているので、リンカル及びSpd 処理は植物体のオーキシンシグナル伝達経路を活性化させるという証拠を提供している。このようなオーキシンとSA との対立的関係は、遺伝子の発現レベルをみるアプローチ法や変異体を用いた実験で証明されているが、実際のイチゴ苗を用いた防除試験において、そのことが裏付けられたことになる。
【0065】
この結果をもとに、Spdがジャガイモそうか病に対してどのような効果があるかを検証するための実験を行ったが、Spdの抑制効果はなく、むしろ病徴を助長する傾向が認められた。一方、Terakadoらは、Spdにはダイズ変異体En6500の根の伸長を抑制する効果があることを指摘しているが、バレイショを用いた本実施例でも、側根伸長の抑制によるものと考えられる生育遅延が認められた。背景技術の0023項で立てた仮説は正しいとしても、根において生じた生理的、生化学的、生態的な変化は、根粒菌の侵入は抑制できても、そうか病菌の塊茎細胞への侵入を抑制することができなかったことになる。しかし、Spdは、オーキシンの受容モードを変化させ、異なるオーキシン反応を誘導していることはほぼ間違いはない。
【0066】
そうか病菌の塊茎細胞への侵入と病斑拡大を抑制する効果をもつのは、Spmに限られ、SpmはSpdと比較してオーキシンの働きを変えていくより高次の機能があると考えざるを得ない。
【0067】
Spmによる植物防御効果には、1)オーキシンの受容モードをさらに変化させ、細胞壁の物理的性質を強化し病原の植物毒素が誘発するPCDを抑制する、2)ある種のMAPKはシロイヌナズナのファイトアレキシン(抗菌物質)であるカマレキシン生成を促進することが証明されており、4つのオーキシン(IAA)生合成経路の1つにおいて、IAAとカマレキシンは前駆体を共有しているので、Spmはある種のMAPKを活性化させIAA生成をカマレキシン生成に転換するなどの分子機構が想定されるが、これらの同定は、今後の基礎研究による解明を待たねばならない。
【0068】
前に触れた高橋らは、Spmはサリチル酸(SA)とは独立して病原特異的タンパク質(PR-1)を発現させ、タバコモザイクウィルスに対する抵抗性を付与できることを証明し、また、Hiroshi Yodaらは、host HR及びnonhost HRの双方の誘導に、ポリアミンがポリアミン酸化酵素(PAO)で分解される過程で生成される過酸化水素が必要であることを示し、ポリアミンが植物防御に普遍的にかかわっていることを示唆した。さらに、最近の報告では、オーキシンが植物防御ホルモンとしてネクロトロフ病原に対する抵抗性に貢献していることを示唆している。これらの知見を総合すると、ポリアミンがジャガイモそうか病を抑制できるという本事例のデータは、1つの例証に止まらず、少なくとも、ポリアミンがネクロトロフの性質をもつ、もしくは植物細胞に壊死を起こしきれる病原に対して植物の抵抗性を誘導できるという一般性を証明したものである。
【0069】
オーキシンは、生長ホルモンとして植物の生長と発達に重要な役割をもっており、一方、防御反応の活性化は、植物生長に有害な結果を及ぼすと考えられている。人間にとって有益な収量性や品質にかかわる形質を伸ばす方向で行われてきた人為的な育種選抜は、植物ホルモン、とりわけオーキシンの生長と発達促進的なシグナリングを増幅し、その反面、植物体の限られた資源が耐病性の構築に向けられなくなり、防御反応の低下をもたらす過程でもあったと考えられる。生長と発達に促進的な栽培種と耐病性を多くもっている野生種の表現型の比較は、そのことを物語っている。
【0070】
以上の実施例が示したデータ、及び背景技術に示した数々の知見を考え合わせると、耐病性を強化するキー的物質であるポリアミン、リン酸塩、カルシュウム塩を外生的に補給することで、あるタイプの病原に対する栽培種の耐病性を増強できるという考え方を導く。
【産業上の利用可能性】
【0071】
SA反応経路を通じてバイオトロフ(生体栄養性)病原に対する病害抵抗性を付与する薬剤はすでに開発され、その一部は、実用技術として古くから利用され、近年、適用範囲も広くなっている。しかし、ネクロトロフ(死体栄養性)の性質を有する病原に対する抵抗性を付与できる実用的な薬剤はまだ開発されていない。
【0072】
本発明で植物に実用的な抵抗性を付与できる可能性がある病害は、じゃがいもそうか病のようなネクロトロフの性質をもった病原で、宿主細胞にPCDを誘発できる、あるいは壊死を引き起こす毒素を生成する病原が引き起こす病害である。例えば、ジャガイモそうか病のような土壌伝染性病害は、クロルピクリン等による全面的な土壌消毒、あるいは輪作などの耕種的防除法しか通用しない難防除病害である。ジャガイモそうか病病原の宿主範囲は限られているが、それ以外の大半の病原は、宿主範囲が広い。連作によって土壌中の病原菌の密度を高め、顕著な連作障害を引き起こすことによって、世界各地で野菜などの様々な産地が衰退を余儀なくされた事例は枚挙に暇がない。
【0073】
本発明をもとに、このような病害に対する抵抗性を作物に付与できる技術開発ができれば、作物選択の自由度を高めるとともに、各地域の農業生産性の向上に大きく貢献する。
【0074】
農薬使用に伴う食品の安全性問題、あるいは、農薬が与える環境や生態系への負荷、さらには薬剤耐性や抵抗性を増強させた病害虫の多発などが、例えば、土壌消毒剤である臭化メチル使用禁止等々の事例があるように、生産現場での従来の農薬使用に様々な制約を与えるようになった。本発明をもとに新たな技術開発ができれば、様々な局面で農業生産の大きな進展をもたらすであろう。
【技術分野】
【0001】
本発明は、生体物質であるポリアミン、及びリン酸塩とカルシュウム塩で植物を処理し、植物のポリアミンシグナル伝達経路、もしくはオーキシンシグナル伝達経路を活性化し、植物が本来もっている耐病性を増強することに関する。
【背景技術】
【0002】
高等植物では、様々な病原の攻撃に対して、クチクラ層や細胞壁の強化、抗菌性物質の生産など、物理的手段から生化学的な手段に至る様々な抵抗性機構を発達させてきたが、植物の抵抗性応答の中でもっとも特徴的なのが過敏感反応(hypersensitive response : HR)である。HRは、感染した細胞の自発的な死(プログラム化された細胞死:PCD)を起こし病原の増殖を阻止するとともに、ファイトアレキシンと呼ばれる抗菌物質、病原特異的タンパク質(pathogenesis-related:PR)、及び植物のストレスホルモンであるサリチル酸(SA)、ジャスモン酸(JA)、エチレン(ET)などの生成蓄積を引き起こす。また、これらの生成蓄積によって、感染していない部位において次の感染を防ぐための全身獲得抵抗性(systemic acquired resistance : SAR)が誘導される。
【0003】
このようなHRの誘導は、多くの場合、宿主が抵抗性R遺伝子の産物を通じて病原特有の分子を認識することから開始され、植物病原の認識が、SA、JA、ETなどの植物ホルモン生合成の引き金を引く。これらホルモンシステムのバランス及び相互作用は、特定の病原と病害に対する抵抗性の発現に中枢的な役割をもっており、これらシグナル間の幅広いクロストークは、様々な病原のタイプに応じて宿主の適用すべき抵抗性機構を決定している。
【0004】
他方、ポリアミンは、第1級アミノ基を2つ以上有する脂肪族炭化水素の総称で、すべての生物に普遍的に存在する天然物質である。ジアミンのプトレシン、トリアミンのスペルミジン、テトラアミンのスペルミンが主要なものである。ポリアミンの基本的な機能として、核酸と融合してDNAの安定化とRNAの転写の効率化と制御、及びカチオン成分として細胞のイオンバランスの維持に関わっていることが知られている。また、細胞分裂やDNA複製のような細胞内過程、形態形成、さらには病害、乾燥、低・高温などの環境ストレスなどにも役割をもっていることを示唆するおびただしい研究報告がなされている。
【0005】
ポリアミンが、何故以上のような多岐にわたる複雑な生理活性を示すのかを考える際に、高度好熱性菌におけるポリアミンの働きをみていくことが極めて示唆的である。高度好熱性菌は、通常の生物には見られない特有のポリアミンが含まれていることが知られており、アミン数が多いポリアミンをより多くもった好熱性菌ほど高温に適応できることが明瞭に示されている。特に、テトラ、ペンタ、ヘキサアミンは核酸の立体構造の安定化を通じてタンパク質合成を含む高分子合成を高温でも可能なものにしていると考えられている(五十嵐一衛著「神秘の物質−ポリアミン」)。一方では、古細菌を含む好熱性菌や極限環境微生物の研究が進行し、生命の起源に近いとされている、つまりすべての生物の共通祖先に近い生物群は、好熱性を示すものが多く見られることが次第に明らかになっている。原始の単細胞生物は、過酷な環境に適応するためポリアミンのもつ化学的性質を、生命の誕生・進化のごく初期の段階で生命維持装置の1つとして取り込み、進化が進むにつれ、ポリアミンの機能も多面性が発揮されるようになってきたという考え方が成立する。
【0006】
ポリアミンの多面的な機能の1つとして、植物の病害抵抗性へのかかわりを挙げることができる。抵抗性N遺伝子をもったタバコ植物体にタバコモザイクウイルスを感染させると、ポリアミンは細胞間隙に蓄積した後、ポリアミン酸化酵素(PAO)で分解される過程で過酸化水素が生成され、病害抵抗性のひとつである過敏感細胞死が誘導されることが証明された(Hiroshi Yodaら2003)。また、HRには2つの主要な経路がある。1つは、特殊な宿主タンパク質による病原の認識を通じて、いわゆる遺伝子対遺伝子と称される病原と宿主の特異的な関係を有するhost HRであり、もう1つは、感染に対して通常の生化学的変化で抵抗性を誘導するnonhost HRの2つである。ポリアミンが双方のHRにかかわっているかどうかを検証するため、タバコ植物体に代表的なnonhost病原Pseudomonas cichoriiに感染させると、ポリアミン代謝にかかわっている酵素をコードしている6つの遺伝子の転写が同時的に誘導され、細胞間隙におけるポリアミンの蓄積、過酸化水素の生成、感染部位における過敏感細胞死が観察された。また、ウイルスで誘導される遺伝子サイレンシング方法によってPAOの活性を低下させたタバコ植物体は、病原細菌の生長を伴った過酸化水素生成の抑制と可視的な過敏感細胞死の消失をもたらした(Hiroshi Yodaら2009)。このことは、ポリアミンが植物防御に普遍的にかかわっていることを示唆している。
【0007】
ポリアミンが植物防御にもつ主要な役割は、以上に述べたように、細胞壁にあるPAOによる分解を通じて、シグナル分子として多数の遺伝子発現を制御する活性があることが証明されている過酸化水素(R.Desikanら2001)を生成させることに求められる。病原が侵入したときに、抵抗性R遺伝子産物を通じて病原の認知がなされ、その後の防御反応の誘導には、原形質膜状のNADPHオキシダーゼの働きによって生じる活性酸素種(ROS)が関与することが知られている。ROSには、スーパーオキサイドアニオン、一重項酸素、ヒドロキシルラジカル、さらにはスーパーオキサイドディスミューターゼの作用によって生じる過酸化水素などの多数の分子種が含まれる。このようなラジカル分子が生体膜近辺に急速に生成される現象は、オキシダティブバーストと呼ばれている。HR時におけるROSの生成は、NADPHオキシダーゼの働きによる急速でかつ一時的なものと、しばらく時間が経過して発生するがより持続的なものの2つの相があることが知られている。HRを誘導するためには、この2つの相のROS生成が必要とされることが近年明らかになっているが、前項の知見は、ポリアミンが後者のROS生成の普遍的な源泉であることを示唆している。
【0008】
ポリアミンの植物防御へのかかわりを明確にする上で、見落としてはならない報告がある。ポリアミン類の1つであるスペルミンは、タバコ植物においてサリチル酸とは独立して病原特異性タンパク質を誘導すること、さらにタバコモザイクウイルスに対する抵抗性を付与することが示され、病原体感染時のシグナル分子の1つとして考えられている。さらに、このスペルミンの作用に着目した研究では、病害抵抗性に関連しているMAPキナーゼであるサルチル酸誘導性タンパク質キナーゼ(SIPK)と傷害誘導性タンパク質キナーゼ(WIPK)を共に活性化させることが明らかになった。また、スペルミンによるMAPキナーゼの活性化はカルシュウム依存的であり、抗酸化剤の前処理によって強く抑制され、スペルミンシグナル伝達経路には活性酸素種とカルシュウムが関与していることが示唆されている(高橋ら2005−植物生理学会講演要旨集)。
【0009】
MAPキナーゼ(MAPK)は、タンパク質にリン酸基を付与するタンパク質キナーゼ(PK)の一種である。細胞は、その機能を維持するため、タンパク質のリン酸化と脱リン酸化を繰り返し行っている。このリン酸化及び脱リン酸化を通じて、酵素活性や他のタンパク質との相互作用が調節されており、MAPKは植物の病害を含めた環境ストレス応答反応に重要な酵素であり、とりわけSIPK及びWIPKは植物と病原の相互作用における防御反応の1つの収束的なポイントであることが認識されてきている。
【0010】
ところで、ジャガイモそうか病の病原であるStreptomyces scabieiは、バレイショの塊茎に侵入し重篤な病斑を形成して、商品価値を著しく低下させる病害であるが、ニンジン、ダイコン、サツマイモなどの根菜類にはほとんど発病させない。バレイショの塊茎肥大については、多くの研究者によって環境及び植物ホルモンの面から研究されている。実生バレイショにおける腋芽の伸長方向は、その着生部位によって異なり、一般的に上位節の腋芽は上方に、下位節の腋芽は下方に伸びてストロンとなり、地面に達した後、多くの場合、地中でイモを形成することが知られている。このような腋芽の伸長方向に植物ホルモンがどのようにかかわっているのかを、実生バレイショの切除処理と切除部に生長調節剤を塗布して調べると、腋芽の伸長方向は主として頂芽部及び若い葉でつくられる内生オーキシンによって制御されていることが示唆されている(池田ら1976−日本作物学会記事)。つまり、バレイショは、本来は腋芽になる幼芽組織を、オーキシンの制御でストロンと塊茎種子を形成する繁殖器官につくり変えていくという特殊なプロセスを進化させたといえる。S.scabieiのバレイショ塊茎への特異的な感染様式と種子塊茎をつくる特殊な器官であるストロンを形成する分子機構に、親和的な相互作用の要因があるという可能性が考えられ、オーキシンシグナル伝達経路との関連性があるのではないのかという疑問が生じる。
【0011】
事実、近年の英国での連続した研究で、オーキシンアナログのバレイショへの葉面散布はそうか病を有意的に減少させることが証明されている(McIntoshら1981〜1988)。さらに、オーストラリアのグループはこの英国での研究をもとに、2,4−Dのそうか病抑制活性を確認するとともに、塊茎の詳細な生理学的実験で、これまでそうか病菌感染に決定的と考えられている皮目数、皮目の外部容積、及び周皮の薄さと構造は、2,4−D処理による実質的な変化がなく、これらの構造変化を通じた病害抑制機構の可能性を否定し、オーキシン処理は、病原が産生するそうか病誘発に必須な植物毒素サクストミンAの毒性を抑制することに、病害抑制の機作を求めている(R.S. Teggら2008)。
【0012】
S.scabiei の植物毒素サクストミンAは、典型的な過敏感細胞死ではないが、アポトーシスの特徴である核DNAの断片化などを伴った過敏感細胞死に似た細胞死を誘導することが知られている。サクストミンAはセルロース生合成を阻害すると報告されているが、セルロース合成阻害剤イソキサベンもシロイヌナズナ懸濁培養細胞のプログラム化された細胞死(PCD)を誘導することが認められ、セルロース合成阻害など植物細胞壁の組成と構成における急速な変化を起こすことで、細胞のPCD を誘発すると示唆されている(Isabella Duvalら2005)。
【0013】
一方、無病育苗土を詰めた紙筒にマイクロチューバーあるいは無病種イモ切断塊茎を播種し、苗ステージごとにそうか病菌接種源6倍希釈の汚染土に裸移植し、その後の発病を調べると、塊茎ステージが進むに従って発病も減少し、塊茎肥大に伴ってそうか病に対する抵抗性が増加することが確認されているが(内藤ら1998)、S.scabieiの感染は、塊茎肥大のごく初期の段階から始まり、肥大の後期段階になると抵抗性が増強されることを意味している。前に記述した英国での研究報告では、オーキシンアナログの葉面散布は、生育の遅い段階よりも早い段階がより効果的であることが確認されており、塊茎肥大に伴って増強されるそうか病抵抗性にはオーキシンシグナル伝達経路がかかわっている可能性が大きい。
【0014】
植物病原は、これらの生活史からしばしばバイオトロフ、ネクロトロフ、及び生活史に応じて双方の性質を有するセミバイオトロフに分類される。バイオトロフとは生きた宿主の組織から栄養を摂るタイプ(死体栄養性)であり、一方、バイオトロフとは宿主の組織を殺しその残渣から栄養を摂るタイプ(生体栄養性)である。真性バイオトロフの典型であるErysiphe orontiiなどの糸状菌は、宿主細胞に細胞死を起こすことなく表皮細胞に感染し、吸器を発達させ胞子形成を十分に行うが、その一方、ネクロトロフと考えられているBotrytis cinereaなどの糸状菌は、宿主細胞の死を促進させそうな植物毒素を生産し、感染の非常に早い段階で宿主細胞を死に至らしめ、組織にダメージを拡大していくことが知られている。
【0015】
以上の知見を考え合わせると、ジャガイモそうか病の病原であるS.scabiei は真性ネクロトロフと分類でき、S.scabiei に対する抵抗性誘導にはオーキシンシグナル伝達経路が介在していることが示唆される。
【0016】
多くの植物病原は、植物との相互作用の中でオーキシンを生成させ、宿主のオーキシンシグナリングを強めることで病徴を進行させることが知られており、オーキシンが病原の攻撃に対する抵抗性に貢献することは、長い間、疑問視されてきた。
【0017】
しかし、最近、ネクロトロフ糸状菌に対する抵抗性にオーキシンが関与しているという、間接的ではあるが確かな証拠が出されている。例えば、植物細菌性病原であるErwinia amylova由来のタンパク質エリシターである精製されたharpinを、抵抗性N遺伝子をもつタバコ葉にオーキシンアナログである2.4−Dと併せて浸潤させると、対照葉には細胞死が生じたこととは対照的に、オーキシンは完全に細胞死を抑制でき、また、harpinを浸潤させた6時間後に2.4−Dを施用すると、細胞死の進行を回復させることができた(Suresh Gopalan2007)。一方、シロイヌナズナのオーキシンシグナル伝達経路に変異性があり、オーキシン促進的なSCF(Skp-Cullin-F-box)ユビキチン化経路に欠陥をもつ、axr1、axr2及びaxr6は、ネクロトロフの糸状菌であるPlectosphaerella cucumerina及びBotrytis cinereaに対し感受性の増加を示していることが証明された。また、これら変異体は、通常SCF-ユビキチン/プロテアーゼ機構による除去のターゲットになっているオーキシン転写抑制物質AXR3の安定化が、P. cucumeriaの感染時に生じていることが示された。これらの結果から、オーキシンシグナル伝達経路はネクロトロフ糸状菌であるP. cucumeria及びB. cinereaに対しての抵抗性に重要であることが示唆されている(Francisco Llorenteら2008)。
【0018】
さらに、オーキシン受容体であるTIR1の3次元構造が解明され、TIR1のLRRs(ロイシン−リッチリピート)にあるオーキシン結合ポケットの構造を安定化する共因子として作用するイノシトールヘキサキスフォスフェート(フィチン酸:InsP6)分子が、TIR1と共役しオーキシンのシグナル転換を促進する可能性が指摘された(Xu Tanら2007)。他方では、低フィチン植物として開発された、イノシトール3フォスフェート合成(IPS:フィチン酸合成における最初のステップを触媒する)のアンチセンス遺伝子配列を構造的に発現するバレイショ組み換え植物体は、フィチン酸レベルを低下させ、非病原性ジャガイモYウイルス及び病原性タバコモザイクウイルス(TMV)に対する抵抗性が低下し、一方、シロイヌナズナにおいて、IPS遺伝子に変異性があるatips2はInsP6を枯渇しており、糸状菌病原であるB. cinerea、及び細菌病原であるPseudomonas病原性・非病原性のレースに対する感受性を高めた。また、変異体atpis2及びatpik1は、SAを蓄積させることができない植物体と同じように、感染に対して過敏な感受性を示したが、これら昂進された感受性はSAレベルの低下によるものではなかった。さらに、これもInsP6が枯渇している変異体atips1は、病原に対する抵抗性を低下させない。これら変異体に見られるInsP6合成の撹乱は、ウイルス、細菌、糸状菌の3つの幅広い病原の感染に対して感受性の昂進をもたらし、植物病原に対する防御を調節する新たなリンクがあることを想定しなければならない事象が明らかにされた(Alex M. Murphyら2008)。
【0019】
前項で示した数々の知見は、生長ホルモンであるオーキシンは、植物防御に関しネガティブな効果しかもたないとされてきた従来の考え方を覆すに十分である。とりわけ、InsP6分子は、様々な病原に対しての抵抗性誘導の重要な鍵を担っている証拠が示され、その作用モードは、オーキシンシグナル伝達経路を転換し、多様な病原に対して、込み入った防御反応の誘導を幅広く調節しているという考え方を導く。InsP6分子の作用モードが解明されていくと、植物防御反応におけるオーキシンシグナル伝達経路の役割が明らかになり、これまでの考え方に大きな変更を強いるような新たな知見が加えられる可能性がある。
【0020】
ポリアミンの多面的な機能をもっていることのもう1つの例として、マメ科植物と共生して窒素固定を行う根粒菌の根への侵入・着生をポリアミンが制御していることを示唆する報告(J.Terakadoら2006)を挙げることができる。ダイズ変異体En6500は根粒菌を根に過剰に着生させ、地下部との光合成産物の競合によるもと考えられるが、地上部の極端な矮性症状を示す。このEn6500の葉中のスペルミジン及びスペルミンの含有量はその親株であるエンレイのそれと比較するといつも低く、前駆体であるプトレシンからのスペルミジン及びスペルミンの合成が抑制されていた。En6500の葉にスペルミジン及びスペルミンを散布すると、根粒菌の着生制御機能をとりもどすとともに根の生長を抑制する事実が示されている。この報告に示された事実は、ポリアミンの直接的な効果ではなく、若枝におけるプトレシン、スペルミジン、スペルミンなどの構成もしくはその含量レベルの顕著な変化が葉中の生化学的な変化を生じさせ、その結果として、根における生化学的、植物生理的もしくは生態的な重要な変化を誘導する可能性を示唆したものである。
【0021】
さらに、根粒菌とマメ科植物の根への着生に関して、ポリアミンに関するものではないが、もう1つの興味深い知見がある。根粒菌を着生させないダイズ変異体T201の培地上に微量の2,4−Dを添加すると、通常のダイズ根粒と同じレベルの窒素固定活性を示す根粒が誘導されるという事実が示された(赤尾ら1995)。培地上に微量の2,4−Dを施用すると、根粒菌の根への侵入を容易にするという知見である。
【0022】
オーキシンは主根の伸長を抑制し、側根と不定根の形成を促進することは、古くから知られている。ダイズ変異体En6500は、主根の長さが短く側根数が異常に多いというオーキシン過剰の表現型を示す。おそらく、何らかの変異性によってポリアミン代謝に欠陥があり、外生ポリアミンに感受性を示す。一方、ダイズ変異体T201は、おそらくオーキシンの生成を含めたシグナル伝達経路もしくは輸送に異状があるオーキシン欠乏型の変異体で、外生的なオーキシンに感受性を示す。En6500の親株であるエンレイは、生育初期の葉中おいてはプトレシンの含量が高く、生殖生長に移行する段階になると、スペルミジン及びスペルミンの含量が高くなり、葉中のポリアミン組成の変動を通じて、根の生態的及び生理的な変化を起こし、おそらく根の伸長、特に側根の伸長が抑制され、根粒菌の侵入・着生を制御すると考えられる。
【0023】
Terakadoらが示唆した知見、及び赤尾らが示した知見、さらにはオーキシンの働きに関するこれまでの知見を総合的に考え合わせれば、ポリアミンがオーキシンの生成や輸送に影響を与えるという報告は見出せないものの、PAOで分解される過程で生成される過酸化水素によってオーキシンシグナル伝達経路が活性化される可能性がある。さらに、葉中のポリアミン組成の変動は、ダイズの根において、オーキシン受容のモードを変え、異なるオーキシ反応因子(ARF)が発現し、根における生化学的、植物生理的もしくは生態的な重要な変化を誘導するという仮説が成立する。
【0024】
前にも触れたが、バレイショはオーキシンによって制御されたストロン形成を行うが、この機構には、進化論的に考えると、側根もしくは不定根をつくりだす分子機構と同じものが用いられていることが類推できる。もし、この類推と前項の仮説が正しいとすれば、バレイショの葉にポリアミンを散布することによって、ダイズの根において根粒菌の着生を制御するように、ジャガイモそうか病菌の侵入を抑制できる可能性がある。
【0025】
最近の研究報告では、キュウリにおいて、主根を切断して得られた実生の外植片で過酸化水素が生成され、それが不定根の形成に先立ってオーキシンシグナルを仲介することが証明された(Shiweng Liら2007)。これは、シグナル分子として多様な遺伝子の発現を制御していることが証明されている過酸化水素が、内生オーキシンの生成を誘導することを示唆するもので、ポリアミンが、PAOよって分解される過程で生成される過酸化水素はオーキシン生成を促進するというわれわれの見解を支持している。
【0026】
以上、ポリアミン処理は、様々な文献に示された知見を総合的に検討し、植物が本来もっているオーキシンシグナル伝達経路を介した耐病性を増強できる可能性があることを示してきた。しかし、先端的な研究報告から得られた知見を単純に積み上げれば、ポリアミンの散布によってあるタイプの病原に対して実用性がある抵抗性を誘導できることが証明されるというものではない。その証拠に、ポリアミンの植物体への直接散布が、病害に対しての抑制効果を示した報告もなければ、ポリアミンがオーキシン活性を高めるという事実を示した報告も皆無である。
【0027】
ポリアミンはオーキシンシグナル伝達経路を活性化させオーキシンの働きを変えるという考え方に基づいた実験は、ジャガイモそうか病で例証するように、植物細胞にPCDを誘発するネクロトロフ病原などのあるタイプの病害に対して、植物に実用性がある耐病性を増強できることを示したデータをもたらした。
【先行技術文献】
【特許文献】
【0028】
【特許文献1】特許公開2004−242505 環境ストレス抵抗性を増強した植物の作出方法
【特許文献2】特許公開2007−000021 ポリアミン代謝制御による環境ストレス耐性を改良した植物、及びその作出方法
【特許文献3】特許公開2005−237387 種々の環境ストレス耐性を改良した植物、その作出方法並びにポリアミン代謝関連酵素遺伝子
【特許文献4】特許公開2004−242510 生産性を改良した植物、及び作出方法
【特許文献5】特許公開2004−180588 環境ストレス抵抗性を改良した植物及びその作出方法
【特許文献6】WO 02/023974 種々の環境ストレス耐性を改良した植物、その作出方法並びにポリアミン代謝関連酵素遺伝子
【非特許文献】
【0029】
【非特許文献1】五十嵐一衛著「神秘の物質−ポリアミン」共立出版(株) p.26〜27
【非特許文献2】Hiroshi Yoda et al., 「Induction of Hypersensitive Cell Death by Hydrogen Peroxide Produced through polyamine Degradation in Tobacco Plants.」Plant Physiology, August 2003 Vol.132, p. 1973〜1981
【非特許文献3】Hiroshi Yoda et al., 「Polyamine as a common source of hydrogen peroxide in host and nonhosthypersensitive response during pathogen infection.」Plant Mol Biol 2009 70:103〜112
【非特許文献4】Radhika Desikan et al., 「Regulation of the Arabidopsis Transcriptome by Oxidative Stress.」Plant Physiology, September 2001, Vol.127,p.159〜172
【非特許文献5】高橋ら「植物の抵抗性におけるスペルミンのシグナル伝達経路」、日本ポリアミン研究会−第19回研究発表会− 講演要旨集p.18
【非特許文献6】池田ら「ジャガイモの匐枝の生長に関する研究」日本作物学会記事 45(2) : 314-321 (1976)
【非特許文献7】McIntosh A. H. et al., 「Decreased Severity of Potato Commonc Scab after Foliar Sprays of 3,5-D Acid, a Possible Antipathogenic Agent.」Annals of Applied Biology 1981 Vol.99, p.275〜281
【非特許文献8】Robert S. Tegg et al., 「Auxin-Induced Resistance to Common Scab Disease of Potato Linked to Inhibition of Thaxtomin A Toxicity.」Plant Disease 2008 Vol.92 No.9, p.1321〜1328
【非特許文献9】Isabelle Duval et. al., 「Thaxtomin A induces programmed cell death in Arabidopsis thaliana suspension-cultured cells.」 Planta Vol.222, No.5 2005/11
【非特許文献10】内藤ら「紙筒移植栽培によるジャガイモそうか病回避の可能性」、日本植物病理学会報1998, 64(6), pp.580
【非特許文献11】Suresh Gopalan 「Reversal of an immunity associated plant cell death program by the growth regulators auxin.」Nature Precedings: Posted 18 Nov 2007
【非特許文献12】Francisco Llorenteet al., 「Repression of the AuxinResponse Pathway Increase Arabidopsis Susceptibility to NecrotrophicFungi.」Molecular Plant Vol. 1, No.3, p.496〜509, May 2008
【非特許文献13】Xu Tan et al., 「Mechanism of auxin perception by the TIR1 ubiquitinligarse」Nature 446, 640〜645, (5 April 2007)
【非特許文献14】Alex M. Murpry et al., 「A role for inositol hexakisphosphate in the maintenance of basal resistance to plant pathogens」The Plant Journal (2008) 56, 638〜652
【非特許文献15】Junko Terakado et al., 「Production Shoot-applied polyamine suppress nodule formation in soybean」Journal of Plant Physiology 163 (2005) 497〜505
【非特許文献16】赤尾ら「2,4-D誘導根粒の形成過程と根粒菌の感染様式」 日本作物学会記事 64巻(別1号) 1995
【非特許文献17】Shiweng Li et al., 「Hydorogen peroxide involvement in formation and development of adventitious roots in cucumber」Plant Growth Regulation, Vol.52, No.2, 2007/6
【発明の概要】
【発明が解決しようとする課題】
【0030】
本発明は、ポリアミンの外生的な処理は、植物のオーキシンシグナル伝達経路、もしくはそれとは独立したポリアミンシグナル伝達経路を活性化させ、ネクロトロフ病原であるジャガイモそうか病菌に対する抵抗性を誘導できるという事実に基づくものであり、宿主細胞にPCDもしくは壊死を引き起こすタイプの病原に対する植物の耐病性を増強できると考えられる。しかし、その適用範囲をジャガイモそうか病以外に広げて確定するには、今後の試験研究に待たねばならない。したがって、以下では、ジャガイモそうか病に限定し、どのような課題を解決できるか、また、どのような方法を用い、どのような効果があるのかについて例示する。
【0031】
ジャガイモそうか病は、バレイショを連作することでStreptomyces scabieiを中心とした病原性のある放線菌が土壌中での生息密度を高め、植物体が塊茎を形成するごく初期のステージから塊茎細胞に侵入し、塊茎肥大とともにかさぶた状の病斑を拡大させるバレイショの世界的な重要病害である。1891年にタクスターによって初めて報告された大変古い病害であり、その後世界各地で確認されている。わが国でも、北海道で1906年北海道農会報に記述されたのが最も古い記録のようであるが、主要なバレイショ生産地では代表的な土壌伝染性病害の1つとして、また、難防除病害として広く知られている。とりわけ、わが国において、西南暖地を代表するバレイショ産地では、長年にわたり年二作に及ぶ連作を続けた結果、そうか病はバレイショ連作障害の中心をなすものとして大きな問題となっている。
【0032】
そうか病に感染すると、ジャガイモの表面にかさぶた状の激しい病斑を形成し、食用、食品加工用としての商品価値が著しく低下する。そうか病による経済的な被害は、少なくとも、生産額の数パーセントに上ると考えられている。
【0033】
世界中のバレイショ生産地では、そうか病による経済的損失を最小化するため様々な対策が用いられているが、いかなる対策も経済性がある効果的で安定的な病害抑制に成り得ていない(Plant Disease/Vol.92 No.9 1321-1328)。クロルピクリンによる全面的な土壌消毒、S.scabieiの土壌中における活性を低下させるため、土壌を酸性側に維持する、また堆肥等の有機物を投入しないなどの土壌管理、あるいは病原菌の侵入する時期に潅水を行うなどの防除対策が行われているが、これらの対策は、環境への負荷、土壌生産性の低下、潅水時期の判断が難しいなどの理由から、安定的な防除対策とはいえない。また、拮抗微生物や抗菌物質を利用した生物的防除法があるが、効果に安定性がないため、生産現場において十分な普及をみていない。
【0034】
本発明は、病原とバレイショ植物体の相互作用に着目し、幅広い生理的活性をもっている生体物質のポリアミン、及びリン酸塩とカルシュウム塩で植物を処理することによって、オーキシンシグナル伝達経路、もしくはポリアミンシグナル伝達経路を活性化させ、植物がもともともっている抵抗性を増強することによって、病原菌のバレイショ塊茎細胞への侵入と病斑拡大を抑制し、防除困難と考えられてきたジャガイモそうか病の安定的な防除法を提供するものである。
【0035】
生産地においては、ジャガイモそうか病を抑制する土壌管理は、バレイショ作の低生産性を余儀なくされている。安定した防除法が確立されると、堆肥や石灰資材の投入による望ましい土壌管理が可能になり、とりわけバレイショ連作地帯では収量性と品質の双方とも大幅に改善され、土地生産性が飛躍的に向上する。
【課題を解決するための手段】
【0036】
スペルミジン、スペルミンなどのポリアミン、もしくはスペルミジン三塩酸塩、スペルミン四塩酸塩などのポリアミン化合物からなる物質群のうち、少なくともいずれか1つ以上の物質を水に溶かし、最大10μモル/ml以下の濃度で散布液を調製し、植え付けた種子が出芽して5日後を第1回の散布日にして、以降5〜10日間隔で、該調製液を、1植物体当たり2〜20ml、回数にして1〜4回程度の葉面散布を行う。
【0037】
ポリアミンを処理する前に、リン酸カルシュウムを水に溶かし、該水溶液を出芽して間もないバレイショの株元に1株当たりリン酸カルシュウムが最大3〜10グラムになるよう施用するか、リン酸カルシュウムの0.3パーセント程度の水溶液に種イモを2時間程度浸漬処理し、その後十分に風乾させた後に植え付けるなどの前処理を行うと、ポリアミンの作用性が増幅され、防除効果が高まる。
【0038】
そうか病菌は塊茎形成のごく初期(米粒以下の大きさ)から侵入するので、出芽後の散布適期を逃すと防除効果は極端に劣る。適期を逃さないことが本手段のもっとも大きなポイントである。
【発明の効果】
【0039】
本発明は、病原菌が病原性を発揮するために必須な植物毒素サクストミンAの毒性を抑制し、病原菌のバレイショ塊茎細胞への侵入と病斑拡大を抑制することを特徴とするまったく新しい手法による防除対策である。
【0040】
バレイショは、塊茎肥大のステージが進むにつれ病害に対する抵抗性を発現させることが知られているが、その抵抗性発現には、植物ホルモンである内生オーキシンがかかわっていることはほぼ確実である。外生的なポリアミン及びリン酸カルシュウムによる処理は、葉中のポリアミン組成を変化させるとともに、ポリアミンシグナル伝達経路を活性化させ、塊茎肥大の後期に発現する抵抗性を肥大の初期に前倒しにして誘導させる働きがあり、そうか病菌の塊茎細胞への侵入と病斑拡大を抑制する。その機構は、オーキシンは病原菌が生成する毒素サクストミンAの宿主細胞に細胞死を引き起こす働きを抑制する、あるいはそれとは独立したポリアミンシグナル伝達経路を通じたファイトアレキシンを生成させることなどに求められるだろうが、詳細な機構は不明であり、今後の究明を待たなければならない。
【0041】
いずれにしても、リン酸カルシュウムを併用したポリアミン処理は、実施例に示すように、そうか病の多発条件下において、そうか病の発生を70〜80パーセント程度に抑えることができ、すでに実用化技術として利用できるレベルまで達している。
【0042】
本発明は、これまでの防除対策とは異なり、病原菌の毒素サクストミンAの活性を阻害し、病原菌の侵入と病斑拡大を抑制するものであるから、普遍性のある効果が期待できる。
【0043】
本発明は、通常の人工的な合成物ではなくポリアミンという生体物質を用いること、またポリアミンが塊茎内に蓄積して直接的に毒素サクストミンAに作用するのではなく、オーキシンシグナル伝達経路、もしくはポリアミンシグナル伝達経路を活性化させ、サクストミンAの毒性を抑制し病原菌の侵入と病斑拡大を抑制すること、要するに生体機能に基づいた抵抗性を増強することを特徴とするものである。したがって、食の安全性についても何ら問題がない。また、オーキシンアナログである2,4−Dなどを用いると、収量の低下、奇形果の発生、塊茎での残留毒性などの問題が生じるが、ポリアミンを用いた場合、そのようなデメリットは生じない。
【図面の簡単な説明】
【0044】
【図1】バレイショの植え付け25日後にスペルミン(Spm)を葉面に散布し、ジャガイモそうか病の抑制効果を示したものである。(実施事例1)
【図2】図1のもとになったデータをジャガイモ1個の重量と病斑数でプロットし、その分散状況を示した図である。(実施事例1)
【図3】Spmを散布する前にリン酸カルシュウム(リンカル)を土壌に施用し、併用処理の効果を示した図である。(実施事例2)
【図4】図1のもとになったデータのうち、Spmとリンカルの併用処理区だけのデータをとりだし、ジャガイモ1個の重量と病斑数でプロットし、分散状況を示した図である。(実施事例2)
【図5】スペルミジン(Spd)とリンカルの併用処理が、イチゴ炭疽病の発病度合いにどのような影響を与えるかを示した図である。(実施事例3)
【発明を実施するための形態】
【0045】
本発明は、ポリアミンの生理活性機能を利用し、葉面散布もしくは土壌処理によって植物の生理を制御し、ジャガイモそうか病に代表されるネクロトロフ(死体栄養性)、もしくは植物細胞に壊死を起こす性質を有する病原に対する抵抗性を誘導し、これら病原が誘発する病害を抑制するものである。したがって、ポリアミンもしくはポリアミン塩酸塩などの化合物を、農薬取締法に基づいて殺菌剤もしくは植物生長調節剤として農薬登録を行い、農薬の製造・販売を経て、バレイショ生産者が農薬として散布して発明を実施することになる。
【実施例1】
【0046】
(図1及び図2の関連)
土壌中の微生物相は、温度、湿度、有機物の種類や量などの環境因子で大きく変り、通常の圃場でそうか病の多発条件をつくることは困難である。このため、圃場に直径30cm、深さ40cm塩化ビニール製の無底ポットを埋め込み、クロルピクリンで消毒した後、ポットに一定の病原汚染土を均質に混ぜること(接種)で、人工的に安定したそうか病の多発条件をつくり、そうか病の抵抗性品種検定等の効果判定が可能になった。
【0047】
この実験システムを用い、種イモを植え付け、1)植え付け25日後に、市販されているポリアミン化合物であるスペルミン4塩酸塩を用い、展着剤としてトリトン0.5mg/mlを加えて調整したスペルミン(Spm)2μM/ml水溶液を1植物体当たり20ml散布処理したSpm散布区、2)比較対照としての無処理区の2つの区を設定した。それぞれの区は5検体で反復なし。
【0048】
それぞれの区の検体ごとに収穫し、それぞれのイモごとに重量と病斑数をカウントし、処理区と無処理区のそれぞれの発病度を比較できるように、イモ1個ごとに100g当たりの病斑数に換算した。その1個ごとのデータを用いて、処理区と無処理のデータ分散状況を検定するとP値は0.001未満となり、完璧な有意差がとれた。図1は、処理区と無処理区の区ごとの平均値を示したもので、Spmによるジャガイモそうか病の抑制効果が確認できる。ただし、それぞれの区で出芽時期が遅れた1検体を、生育時期のずれによる降雨及び気温などの気象要因の違いの影響を受けている可能性があるので除外し、生育が揃っているそれぞれの区の4検体のデータを用いて、図を作成している。
【0049】
図2は、それぞれのイモの重量と病斑数でプロットし、処理区と無処理区の分散状況を示したものである。無処理区では、重量と病斑数に比較的強い相関が見られるが、その一方で、処理区ではN字状の連続性のある分散パターンを示し、重量200g前後の領域にSpmの強い作用領域が認められるとともに、200gより軽いイモの病斑数は、無処理区と比較して概して少ないことが認められる。また、イモの重量の違いはストロン形成の早晩によって異なるものと考えられ、重量を示したX軸は時間軸を表しているとも考えられ、無処理区のN字状のパターンは、Spmによる処理時期が多少遅かったことを意味している。
【0050】
以上の結果から、1)Spmによるそうか病の抑制効果は確認できるが、実用化のためには処理方法をさらに検討し、抑制率を高める必要があること、2)図2の分布パターンから判断し、処理時期を早める、処理効果には強いものがあるが効果が長続きしないので複数回の処理が必要とされる、ことなどが示唆される。
【実施例2】
【0051】
(図3〜4の関連)
Spmのシグナル伝達経路を増幅するにはMAPKを活性化させることが重要と考えられるので、Spmの処理前に土壌にリン酸カルシュウム水溶液を処理し、植物体中のリン酸とカルシュウムレベルを高めておくと、MAPKの活性化が期待できるのではないかと考えた。このため、1)出芽後5日、10日、20日の3回、実施例1と同じスぺルミン2μM/ml水溶液を1植物体当たり20ml散布処理したSpm単用区、2)Spmを処理する前に出芽後1〜2日後にリン酸カルシュウム(リンカル)3%水溶液を1ポット当たり100mlを土壌に施用したリンカル併用区、及び3)比較対照としての無処理区の3つの区を設けた。それぞれの区は5検体の反復なし。
【0052】
実施例1と同じように、検体ごとに収穫し、イモ1個ごとにそれぞれの重量の測定と病斑数をカウントした。図3は、それぞれの区の100g当たり病斑数の平均値を表したものである。Spmとリンカルの併用区と無処理区の間では、P値<0.05の有意差がとれ、リンカルはSpmのシグナル伝達経路を増幅することが確認できる。しかし、単用区及びリンカル併用区とも処理効果の個体差が大きいことが見出された。無処理区、Spm単用区、及びリンカル併用区における1検体における100g当たりの病斑数の最大値と最小値の幅は、それぞれ14.5〜9.6、14.4〜2.3、10.9〜2.0となっており、処理区おけるSpmの効果が顕著な検体とそうでない検体とに別れている。
【0053】
図4は、リンカル併用区におけるイモの重量と病斑数でプロットした分散パターンを示している。無処理区の100g当たり病斑数データ群とリンカル併用区のそれぞれの検体のデータ群間の分散状況を検定すると、P値は、検体4(0.98)>検体2(0.91)>検体3(0.22)>検体5(0.03)>検体1(0.02)となる。無処理区の分散パターンとほとんど変らない検体から有意的に病斑数が減少した検体の両極端に分かれている。本来ならば、検体1程度の病斑数の減少が期待できるにもかかわらず、何かの要因がそれを妨害したと考えなければならない。
【0054】
実施例2は2009年の春作に行ったが、それと同じ時期に、野外ではなく温室でも同じようなジャガイモそうか病抑制試験を行った。このときの気温推移は、処理時期の前後に、最高気温では8.6℃、平均気温では6.4℃と大幅な気温上昇があり、その時期の最高気温は通常であれば19℃程度であるが、そのときの最高気温は、バレイショの生育適温とされる22〜23℃を越え24.9℃を記録した。温室においては、重要な感染時期と考えられる塊茎形成と肥大の初期段階で、植物体を2週間程度35℃前後の高温に数時間継続的に遭遇させる結果になった。
【0055】
植物体がこのような高温ストレス条件下に置かれると、Spm効果が見出せないだけでなく、そうか病に対して本来発動されるべき抵抗性も消失し、通常は塊茎の表層だけにとどまる病斑部は塊茎内部に奥深く広がり、発病度合いを評価する通常の方法が適用できないほどの激しい病徴を示した。抵抗性R遺伝子の働きには温度感応性があり、28℃を超える高温では抵抗性が消失することが知られているが、それに似た現象が起きたことになる。
【0056】
キュウリを高温順化させるため38℃に遭遇させると、ポリアミン代謝に変化が起こり、葉にスペルミジン及びスペルミンが徐々に蓄積され、また、高温に遭遇させない非順化葉にスペルミジンまたはスペルミンを葉面散布すると、光合成器官の熱安定性が高まることが知られている(Tachibanaら2003)。ポリアミンは、プトレシンを前駆体として、順次、スペルミジン、スペルミンとアミン数が多いポリアミンが合成される。アミン数が多いポリアミンをより多くもった好熱性菌ほど高温に適応できることが知られているが、植物においても同様で、このような高温ストレス時には、ポリアミン酸化酵素(PAO)が活性をなくし、よりアミン数の多いポリアミンの蓄積が起きると考えられる。
【0057】
前にも触れたが、Hiroshi Yodaらは、PAOの活性を低下させたタバコ植物体は、病原菌の生長を伴った過酸化水素生成の抑制と可視的な過敏感細胞死の消失を観察したが、われわれも同様に、高温に遭遇したバレイショ植物体は、おそらくPAO活性を喪失し、スペルミジン及びスペルミンの分解による過酸化水素の発生が抑制された結果と考えられる病害抵抗性の消失を観察した。
【0058】
野外では温室ほどの高温ではないが、急激な気温上昇と生育適温を超える温度に遭遇することによって、植物のポリアミン代謝が変化し、PAO活性レベルが低下する個体が現れてくるとともに、微妙な温度域になるので、高温に過剰に反応せず活性レベルを低下させない個体もあり、そこにSpm効果に大きな振れが生じた原因があると考えられる。
【0059】
バレイショの種イモを植え付け、その後の降雨が少なく乾燥状態になると、早く出芽がするものと、まとまった雨を待って遅く出芽するものの両極端に分かれる現象が観察される。植物の環境ストレスに対する応答反応は、すべての個体が一律的に反応よりも、個体差をもたせた方が環境に柔軟に対応でき、種が生き残る機会を多くするだろうから、植物体の環境ストレス応答における個体差は、もともとプログラム化された応答システムとして埋め込まれているのかも知れない。
【0060】
Spmとリンカルの併用処理区における顕著な効果が見出せる検体の病徴と無処理区のそれとを比較すると、病斑数の減少だけでなく病斑のサイズが低下していることが確認でき、Spmの効果には、病原の侵入抑制と侵入を受けても病斑拡大を抑制する2つの効果がある。
【実施例3】
【0061】
(図5の関連)
スペルミジン(Spd)とリン酸カルシュウム(リンカル)の処理が、他の病害に対してどのような効果をもつかを調べるため、イチゴ炭疽病菌懸濁液をイチゴ苗に接種し、炭疽病の発病度合いをみてみた。ポリポット苗で育成したイチゴ苗に、1)リンカル3%水溶液を各ポットに50mlを施用したリンカル単用区、2)同じようにリンカル水溶液を処理した2日後にイチゴ苗に展着剤としてトリトンを加えたSpd水溶液2μモル/mlを20ml葉面散布したSpd併用区、3)比較対照としての無処理区の3区構成。Spd散布処理21日後に炭疽病菌の懸濁液(1×104個/mlの7.6ml/鉢で接種。病徴は複葉(1本の葉柄に3枚の小葉がついた葉)における発病小葉率及び発病葉柄率で評価。それぞれ12個体の4反復。
【0062】
図5は、この実施例の結果を示している。各区とも最終的な枯死率は変らなかったが、初期感染時において、小葉及び葉柄における病斑形成に有意差が生じた。無処理<リンカル単用区<リンカルとSpd 併用区の順に、小葉及び葉柄ともに発病率が有意的に高まった。
【0063】
イチゴ炭疽病の葉細胞への侵入と病斑形成は、旧葉よりも新葉で多く見られ、新葉の炭疽病に対する感受性は旧葉よりも高いことが知られている。つまり、オーキシン活性が高い若い組織ほど炭疽病に感受性を示すと考えてよい。多くの植物病原は、植物との相互作用の中でオーキシンを生成させ、宿主のオーキシンシグナリングを強めることで病徴を進行させることが知られているが、イチゴ炭疽病菌もそのようなタイプの1つであり、おそらくその抵抗性にはSAシグナル伝達経路がかかわっている。
【0064】
植物防御におけるオーキシンとSA との対立的関係は、近年、多数の研究報告の中で証明されている。例えば、オーキシンはSA シグナル伝達経路において中心的な役割をもっているPR-1遺伝子の十分な発現を阻害することが証明されている(Dong Wangら2007)。これまで得られている知見を総合して考えると、本実施例で得られたデータは、リンカルとSpdで処理したイチゴ植物体は、炭疽病に対する感受性が昂進されているので、リンカル及びSpd 処理は植物体のオーキシンシグナル伝達経路を活性化させるという証拠を提供している。このようなオーキシンとSA との対立的関係は、遺伝子の発現レベルをみるアプローチ法や変異体を用いた実験で証明されているが、実際のイチゴ苗を用いた防除試験において、そのことが裏付けられたことになる。
【0065】
この結果をもとに、Spdがジャガイモそうか病に対してどのような効果があるかを検証するための実験を行ったが、Spdの抑制効果はなく、むしろ病徴を助長する傾向が認められた。一方、Terakadoらは、Spdにはダイズ変異体En6500の根の伸長を抑制する効果があることを指摘しているが、バレイショを用いた本実施例でも、側根伸長の抑制によるものと考えられる生育遅延が認められた。背景技術の0023項で立てた仮説は正しいとしても、根において生じた生理的、生化学的、生態的な変化は、根粒菌の侵入は抑制できても、そうか病菌の塊茎細胞への侵入を抑制することができなかったことになる。しかし、Spdは、オーキシンの受容モードを変化させ、異なるオーキシン反応を誘導していることはほぼ間違いはない。
【0066】
そうか病菌の塊茎細胞への侵入と病斑拡大を抑制する効果をもつのは、Spmに限られ、SpmはSpdと比較してオーキシンの働きを変えていくより高次の機能があると考えざるを得ない。
【0067】
Spmによる植物防御効果には、1)オーキシンの受容モードをさらに変化させ、細胞壁の物理的性質を強化し病原の植物毒素が誘発するPCDを抑制する、2)ある種のMAPKはシロイヌナズナのファイトアレキシン(抗菌物質)であるカマレキシン生成を促進することが証明されており、4つのオーキシン(IAA)生合成経路の1つにおいて、IAAとカマレキシンは前駆体を共有しているので、Spmはある種のMAPKを活性化させIAA生成をカマレキシン生成に転換するなどの分子機構が想定されるが、これらの同定は、今後の基礎研究による解明を待たねばならない。
【0068】
前に触れた高橋らは、Spmはサリチル酸(SA)とは独立して病原特異的タンパク質(PR-1)を発現させ、タバコモザイクウィルスに対する抵抗性を付与できることを証明し、また、Hiroshi Yodaらは、host HR及びnonhost HRの双方の誘導に、ポリアミンがポリアミン酸化酵素(PAO)で分解される過程で生成される過酸化水素が必要であることを示し、ポリアミンが植物防御に普遍的にかかわっていることを示唆した。さらに、最近の報告では、オーキシンが植物防御ホルモンとしてネクロトロフ病原に対する抵抗性に貢献していることを示唆している。これらの知見を総合すると、ポリアミンがジャガイモそうか病を抑制できるという本事例のデータは、1つの例証に止まらず、少なくとも、ポリアミンがネクロトロフの性質をもつ、もしくは植物細胞に壊死を起こしきれる病原に対して植物の抵抗性を誘導できるという一般性を証明したものである。
【0069】
オーキシンは、生長ホルモンとして植物の生長と発達に重要な役割をもっており、一方、防御反応の活性化は、植物生長に有害な結果を及ぼすと考えられている。人間にとって有益な収量性や品質にかかわる形質を伸ばす方向で行われてきた人為的な育種選抜は、植物ホルモン、とりわけオーキシンの生長と発達促進的なシグナリングを増幅し、その反面、植物体の限られた資源が耐病性の構築に向けられなくなり、防御反応の低下をもたらす過程でもあったと考えられる。生長と発達に促進的な栽培種と耐病性を多くもっている野生種の表現型の比較は、そのことを物語っている。
【0070】
以上の実施例が示したデータ、及び背景技術に示した数々の知見を考え合わせると、耐病性を強化するキー的物質であるポリアミン、リン酸塩、カルシュウム塩を外生的に補給することで、あるタイプの病原に対する栽培種の耐病性を増強できるという考え方を導く。
【産業上の利用可能性】
【0071】
SA反応経路を通じてバイオトロフ(生体栄養性)病原に対する病害抵抗性を付与する薬剤はすでに開発され、その一部は、実用技術として古くから利用され、近年、適用範囲も広くなっている。しかし、ネクロトロフ(死体栄養性)の性質を有する病原に対する抵抗性を付与できる実用的な薬剤はまだ開発されていない。
【0072】
本発明で植物に実用的な抵抗性を付与できる可能性がある病害は、じゃがいもそうか病のようなネクロトロフの性質をもった病原で、宿主細胞にPCDを誘発できる、あるいは壊死を引き起こす毒素を生成する病原が引き起こす病害である。例えば、ジャガイモそうか病のような土壌伝染性病害は、クロルピクリン等による全面的な土壌消毒、あるいは輪作などの耕種的防除法しか通用しない難防除病害である。ジャガイモそうか病病原の宿主範囲は限られているが、それ以外の大半の病原は、宿主範囲が広い。連作によって土壌中の病原菌の密度を高め、顕著な連作障害を引き起こすことによって、世界各地で野菜などの様々な産地が衰退を余儀なくされた事例は枚挙に暇がない。
【0073】
本発明をもとに、このような病害に対する抵抗性を作物に付与できる技術開発ができれば、作物選択の自由度を高めるとともに、各地域の農業生産性の向上に大きく貢献する。
【0074】
農薬使用に伴う食品の安全性問題、あるいは、農薬が与える環境や生態系への負荷、さらには薬剤耐性や抵抗性を増強させた病害虫の多発などが、例えば、土壌消毒剤である臭化メチル使用禁止等々の事例があるように、生産現場での従来の農薬使用に様々な制約を与えるようになった。本発明をもとに新たな技術開発ができれば、様々な局面で農業生産の大きな進展をもたらすであろう。
【特許請求の範囲】
【請求項1】
植物の生育過程、もしくは種子形態において、ポリアミン、リン酸塩、及びカルシュウム塩のうち、少なくとも1つ以上の物質を用いて、散布、土壌施用、及び種子浸漬のうち少なくとも1つ以上の方法で処理し、植物のポリアミンシグナル伝達経路及びオーキシンシグナル伝達経路のうち1つ以上の経路を活性化させることを特徴とする植物の耐病性を増強する方法。
【請求項2】
生育過程にある上記植物を部分的にポリアミン、リン酸塩、及びカルシュウム塩で処理する請求項1の植物の耐病性を増強する方法。
【請求項3】
処理される植物の部分が蕾、花、子房、果実、葉、茎、根のうち少なくとも1つ以上から選択される請求項2に記載された植物の耐病性を増強する方法。
【請求項4】
ポリアミンが、第1級アミノ基を2つ以上有する脂肪族炭化水素からなる群から少なくとも1つ以上選ばれたポリアミン、及び該ポリアミンの化合物である請求項1〜3のいずれかに記載された植物の耐病性を増強する方法。
【請求項5】
ポリアミンが、1,3-ジアミノプロパン、プトレシン、カダベリン、カルジン、スペルミジン、ホモスペルミジン、アミノプロピルカダベリン、テルミン、スペルミン、テルモスペルミン、カナバルミン、アミノペンチルノルスペルミジン、N,N-ビス(アミノプロピル)カダベリン、ホモスペルミン、カルドペンタミン、ホモカルドペンタミン、カルドヘキサミン、ホモカルドヘキサミンよりなる群から選ばれた少なくとも1種以上のポリアミン、及び該ポリアミンの化合物である請求項4に記載された植物の耐病性を増強する方法。
【請求項1】
植物の生育過程、もしくは種子形態において、ポリアミン、リン酸塩、及びカルシュウム塩のうち、少なくとも1つ以上の物質を用いて、散布、土壌施用、及び種子浸漬のうち少なくとも1つ以上の方法で処理し、植物のポリアミンシグナル伝達経路及びオーキシンシグナル伝達経路のうち1つ以上の経路を活性化させることを特徴とする植物の耐病性を増強する方法。
【請求項2】
生育過程にある上記植物を部分的にポリアミン、リン酸塩、及びカルシュウム塩で処理する請求項1の植物の耐病性を増強する方法。
【請求項3】
処理される植物の部分が蕾、花、子房、果実、葉、茎、根のうち少なくとも1つ以上から選択される請求項2に記載された植物の耐病性を増強する方法。
【請求項4】
ポリアミンが、第1級アミノ基を2つ以上有する脂肪族炭化水素からなる群から少なくとも1つ以上選ばれたポリアミン、及び該ポリアミンの化合物である請求項1〜3のいずれかに記載された植物の耐病性を増強する方法。
【請求項5】
ポリアミンが、1,3-ジアミノプロパン、プトレシン、カダベリン、カルジン、スペルミジン、ホモスペルミジン、アミノプロピルカダベリン、テルミン、スペルミン、テルモスペルミン、カナバルミン、アミノペンチルノルスペルミジン、N,N-ビス(アミノプロピル)カダベリン、ホモスペルミン、カルドペンタミン、ホモカルドペンタミン、カルドヘキサミン、ホモカルドヘキサミンよりなる群から選ばれた少なくとも1種以上のポリアミン、及び該ポリアミンの化合物である請求項4に記載された植物の耐病性を増強する方法。
【図1】


【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2011−132158(P2011−132158A)
【公開日】平成23年7月7日(2011.7.7)
【国際特許分類】
【出願番号】特願2009−292157(P2009−292157)
【出願日】平成21年12月24日(2009.12.24)
【出願人】(000214191)長崎県 (106)
【Fターム(参考)】
【公開日】平成23年7月7日(2011.7.7)
【国際特許分類】
【出願日】平成21年12月24日(2009.12.24)
【出願人】(000214191)長崎県 (106)
【Fターム(参考)】
[ Back to top ]