
植物細胞においてDNA配列を発現させるための人工プロモーター
本発明は、組換えDNAのキメラ分子を含むことを特徴とする人工プロモーターに関し、そのプロモーターは任意のクラスの植物細胞にいったん導入された場合、その3’末端に融合した任意のDNA分子の高い発現レベルを促進する。本発明のプロモーター分子の基本的な遺伝子エレメントは、コンセンサスTATAボックスを有するプロモーター核、それに続くエキソン/イントロン/エキソン領域及び翻訳活性強化エレメントであり、それらのすべてが人工的に産生される。器官又は組織に特異的時間応答能を有した発現をもたらすために、転写発現調節エレメントをプロモーターの上流に挿入できる。設計された人工遺伝子エレメントを植物細胞における任意の活性プロモーター及び任意のDNA配列の間に、後者の転写/翻訳レベルを増加させるために機能的に挿入できる。

【発明の詳細な説明】
【技術分野】
【0001】
本発明はバイオテクノロジーに関し、さらに詳細には植物遺伝子工学に関する。詳細には、キメラDNA構築物を提供し、それらの構築物は双子葉及び単子葉植物の細胞においてそれらの構築物に融合した任意のヌクレオチド配列の高い転写/翻訳プロモーター活性を示し、さらに高発現レベルの目的の遺伝子及びDNA配列を有するトランスジェニック植物を得ることを可能にする。
【背景技術】
【0002】
植物遺伝子工学は、新しいバイオテクノロジー産物の基礎研究及び商業生産に非常に生産力の高いことが実証された技術である。(Hammond J.、Curr.Top.Microbiol.Immunol 1999、240:1〜19頁;Simoens C.及びVan Montagu M.、Reproduction Update 1995、1:523〜542頁)。
【0003】
分子生物学的手法により遺伝子操作された植物に導入された遺伝子又はDNA配列の強度及び時間的又は空間的特異性に関して十分な発現を保証するプロモーターシグナルの選択は、植物遺伝子工学が成功するために非常に重要である。このような理由から、必要な導入遺伝子のそれぞれの発現を確実にできるプロモーター及びシグナルの探索に多数の努力が過去20年間に捧げられてきた。このように、種々の起源(植物、ウイルス、アグロバクテリウム(Agrobacterium)のTi又はRi、或いはキメラ)のプロモーターが評価され、トランスジェニック植物の産生に採用されてきた。
【0004】
植物遺伝子操作にさらに広く使用された構成性プロモーターは、カリフラワーモザイクウイルス(CaMV)35S ARNプロモーター(Odell J.T;Nagy F;Chua N.H.、Nature 1985、313:810〜812頁)、A.ツメファシエンス(A.tumefaciens)Tiプラスミド由来ノパリンシンテターゼ遺伝子(nos)プロモーター(An G;Costa M.A;Mitra A;Ha S;Marton L.、Plant Phisiol.1986、88:547〜552頁)、イネアクチン−1遺伝子プロモーター(McElroy D;Zhang W;Cao J;Wu R.、Plant Cell 1990、2:163〜171頁)及びトウモロコシユビキチン−1遺伝子プロモーター(Christensen A.H;Sharrock R.A;Quail P.H.、Plant Mol.Biol.1992、18:675〜689頁)であった。しかし、どの植物クラスにおいても発現レベルが十分に高くないことが主な原因となって、これらの天然発現系にはそれぞれ限界がある。例えばプロモーターの発現が双子葉植物の細胞において低く、単子葉類ではほとんど検出できない一方で、最も広く使用されているプロモーターであるCaMV 35Sの発現は単子葉植物よりもタバコ細胞においてずっと強い(Topfer R;Maas C;Horicke−Grandpierre C;Schell J;Steinbiss H.H.、Methods Enzymol.1993、217:67〜78頁;Mitsuhara I;Ugaki M;Hirochika H;Ohshima M;Murakami T;Gotoh Yら、Plant Cell Physiol.1996、37:49〜59頁)。同様にイネアクチン−1及びトウモロコシユビキチン−1プロモーターは単子葉植物の細胞において下流の遺伝子の転写の促進に極めて効率的であるが、タバコ細胞におけるそのプロモーター活性は低い(Schledzewski K;Mendel R.R.、Transgenic Research 1994、3:249〜225頁)。
【0005】
トランスジェニック植物における異種タンパク質の発現レベルを増加させるために、天然プロモーターを転写又は翻訳エンハンサーと組み合わせた多様なキメラプロモーターが設計された。これらのエンハンサーエレメントの中で、タバコモザイクウイルス(TMV)のオメガ翻訳エンハンサー(Gallie D.R;Sleat D.E;Watts J.W;Turner P.C;Wilson T.M.A.、Nucleic Acids Res.1987、15:3257〜3273頁)、タバコエッチウイルス(TEV)の翻訳エンハンサー(Carrington J.C;Freed D.D.、J.Virol.1990、64:1590〜1597頁)、オクトピンシンターゼ由来プロモーター転写エンハンサー(Fromm H;Katagiri F;Chua N.H.、Plant Cell 1989、1:977〜984頁)、マンノピンシンターゼ由来(Comai L;Moran P;Maslyar D.、Plant Mol Biol.1990、15:373〜381頁)及びCaMV 35Sプロモーター由来(Kay R;Chan A;Daly M;McPherson J.、Science 1987、236:1299〜1302頁)、並びに天然エキソン及びイントロン、例えばトウモロコシアルコールデヒドロゲナーゼイントロン1(Callis J;Fromm M;Walbot V.、Genes Devel 1987、1:1183〜1200頁;Last D.I;Brettell R.I.S;Chamberlaine D.A;Chaudhury A.M;Larkin P.Jら、Theor Appl.Gen.1991、81:581〜588頁)、トウモロコシスクロースシンターゼ由来第一エキソン/イントロン(Maas C;Laufs J;Grant S;Korfhage C;Werr W.、Plant Mol.Biol.1991、16:199〜207頁)、イネアクチン−1遺伝子由来第一エキソン/イントロン(McElroy D;Blowers A.D;Jenes B;Wu R.、Mol.Gen.Genet.1991、231:150〜160頁)などを挙げることができる。それにより、2X35S、Mac、Emuなどのプロモーターが生じ(EP0459643、EP0651812)、それらは双子葉又は単子葉類の主に特定のクラスの植物細胞において強力である(Schledzewski K;Mendel R.R.、Transgenic Research 1994、3:249〜255頁)。
【0006】
双子葉及び単子葉植物両方の細胞において遺伝子を発現させるために採用できる強力なプロモーターの開発は、これまでも現在も多くの研究室に関係する問題である。それは、多様なクラスの植物を形質転換するための独特な遺伝子構築物を手に入れるという科学的挑戦やそれが意味する節約のためだけではなく、バイオテクノロジー産物の生産及び商品化をさらに容易にする特有の発現系を手に入れるためでもある。特許出願W09943838において請求された合成プロモーターは、5’末端を35Sプロモーター、トウモロコシユビキチン−1プロモーター及びオクトピンシンターゼプロモーターからの転写エンハンサー配列に融合した、高いGC含量(64%以上)を有するTATAボックスから転写開始部位までの配列を請求した。他方、双子葉及び単子葉植物における発現を探索するだけではなく、配列相同性に依存した遺伝子サイレンシングを避けるために(Park Y.D;Papp I;Moscone E;Iglesias V;Vaucheret H;Matzke A;Matzke M.A.、Plant J.1996、9:183〜194頁)、特許出願WO0058485は2つのウイルスゲノム、すなわちツユクサ黄色斑紋ウイルス(CoYMV)及びキャッサバ葉脈モザイクウイルス(CsVMV)からの配列並びに35Sプロモーターからのエンハンサー配列の組合せに由来する人工プロモーターを請求している。
【0007】
種々の遺伝子エレメントがヌクレオチド配列の転写又は翻訳を増強することを可能にするメカニズムはまだ明らかではない。例えば、多くのRNAウイルスからのリーダー配列が、5’末端に融合したキャップ(m7G(5’)ppp(5’)N)の存在と無関係に種々のメッセンジャーRNA(mRNA)の翻訳を増強できることが報告された(Sleat D.E;Wilson T.M.A.1992.「Plant virus genomes as sources of novel functions for genetic manipulations」、T.M.A.Wilson & J.W.Davies編、Genetic engineering with plant viruses、CRC Press,Inc.、55〜113頁;Gallie D.R;Sleat D.E;Watts J.W;Turner P.C;Wilson T.M.、Nucleic Acids Res.1987、15:8693〜8711頁)。しかし、これらすべてのウイルスリーダーのRNA二次構造が複雑でないことを除いて、翻訳エンハンサーの性質を表すために応答するヌクレオチド配列で別の共通エレメントは決定されていない。
【0008】
具体的には、TMVオメガ断片の翻訳増強は、繰り返し配列である少なくとも1つのコピーのオクタマーACATTTAC及び重大なモチーフと考えられる25塩基の(CAA)n領域の存在によることが報告された(2つのコピーの(CAA)n領域は高いエンハンサーレベルを付与するために十分である)(Gallie D.R;Walbot V.、Nucleic Acids Res.1992、20:4631〜4638)。しかし、ジャガイモXウイルス(PVX)リーダーからの富CA領域である28塩基は「それ自体では」翻訳エンハンサー活性を示さなかった(Pooggin M.M;Skryabin K.G.、Mol.Gen.Genet.1992、234:329〜331頁)が、PVXリーダーのCA領域に存在するCCACCペンタヌクレオチドは、18S rRNAの3’末端と対形成相互作用を有する可能性があることが報告されている(Tomashevskaya O.L;Solovyev A.G;Karpova O.V;Fedorkin O.N;Rodionova N.P;Morozov S.Y;Atabekov J.G.、J.Gen.Virol.1993、74:2717〜2724頁)。一部のウイルスリーダーは翻訳エンハンサー活性に関与する配列エレメントを有することが測定され、その配列エレメントには、ジャガイモウイルスS(PVS)様のカルラウイルスリーダーに保存されているCCTTTAGGTT配列(Turner R;Bate N;Tewell D;Foster G.D.、Arch.Virol.1994、134:321〜333頁)及びアルファルファモザイクウイルス(AlMV)のRNA3リーダーにおける27塩基繰り返し領域にみられ、最適な翻訳レベルに達するために2つを必要とするいわゆる内部制御領域2型(ICR2)モチーフ(GGTTCGANTCC)などがある(van der Vossen E.A.G;Neeleman L;Bol J.F.、Nucleic Acids Res.1993、21:1361〜1367頁)。
【0009】
TEVリーダーの場合、それぞれヌクレオチド28〜65及び66〜118のCIRE−1及びCIRE−2と称する2つの領域は、この148bpのウイルスリーダーの翻訳増強性を担っていることが明らかにされた(Niepel M;Gallie D.R.、J.Virol.1999、73:9080〜9088頁)。しかし、CIRE領域内部でこれらのウイルスリーダーのエンハンサー活性に重大とみなされる特異的エレメントは規定されていない。
【発明の開示】
【発明が解決しようとする課題】
【0010】
上に述べたように、天然起源のイントロン及びその隣接配列は、特にそのイントロンが遺伝子の5’末端近くである場合に、種々の遺伝子発現系を増強するためにも広く採用されてきた。しかし、イントロンの起源、エキソンフランキング領域及び細胞の種類のような要因に依存した遺伝子発現のイントロン介在性増強(IME)が報告されている。発現の強いIMEは主に単子葉植物の細胞で観察されたが、双子葉類ではIMEは通常2〜5倍を超えない。IMEの分子メカニズムは完全には開示されていない(Simpson G.G;Filipowicz W.、Plant Mol.Biol.1996、32:1〜41頁;Schuler M.A.1998.「Plant pre−MRNA splicing」、J.Bailey−Serres & D.R.Gallie編、A look beyond transcription:mechanisms determining MRNA stability and translation in plants、American Society of Plant Physiologists、1〜19頁;Lorkovic Z.J;Kirk D.A.W;Lambermon M.H.L;Filipowicz W.、Trends in Plant Science.2000、5:160〜167頁)。
【0011】
単子葉及び双子葉植物の細胞の間で観察された発現のIMEの変動は、異なるクラスの植物細胞における十分なプレmRNAプロセシングに要する要求性の公知の差が原因であり得る。実際に双子葉植物の細胞ではなく単子葉植物の細胞では、イントロン配列における富AUセグメントの存在はプロセシングに欠かせないわけではなく、単子葉植物の細胞は高GC含量(50%を超える)及び複雑な二次構造(ヘアピンループ)を有してイントロンをプロセシングでき、このことは双子葉植物の細胞が複雑な二次構造を有するイントロンをプロセシングできないことを示している(Goodall G.J;Filipowicz W.、The EMBO Journal 1991、10:2635〜2644頁;Lorkovic Z.J;Kirk D.A.W;Lambermon M.H.L;Filipowicz W.、Trends in Plant Science.2000、5:160〜167頁)。これらの理由はヌクレオチド配列の発現を人工的に増強するためにIMEを採用している現行の系がどうしてクラス特異的であるかを少なくとも部分的に説明している。
【課題を解決するための手段】
【0012】
本特許出願において提起された発現プロモーターの配列は、以下の特有の特徴を提供する。1)双子葉及び単子葉植物の細胞において活性であるので、普遍的に機能的であり、高発現レベルの目的遺伝子及び目的DNA配列を有する任意のクラスのトランスジェニック植物を得ることを可能にする、2)人工的に集合させた遺伝子エレメントの組合せに基づき、IMEによるだけでなく翻訳も促進してmRNAレベルを増加させる、3)本プロモーターが天然又はウイルス遺伝子と同一の配列を有する長鎖DNA断片を欠くことは、RNAが介在する相同遺伝子サイレンシングのリスク及びin plantaの相同組換えの結果としての新しいウイルス品種又はウイルス株が出現する可能性を最小限にする、4)TATAボックスから転写開始部位までの配列のGC含量は必ずしも高くなくともよい、5)我々のプロモーター配列の汎用性は5’末端に転写調節エレメントを挿入することを可能にし、このことは発現に時間、器官又は組織特異性を付与する、6)それを有する人工遺伝子エレメントを、植物細胞において活性な任意のプロモーター及び任意のDNA配列の間にも機能的に挿入でき、その転写/翻訳を増加できる。
【発明を実施するための最良の形態】
【0013】
任意のクラスの植物細胞においてmRNAの蓄積を大きく増強する活性を有するキメラエキソン/イントロン/エキソン領域の設計、及びそれの人工翻訳エンハンサーへの機能的組み込みは、本特許出願の2つの本質的要素を構成している。それは、これらのエレメントにより植物細胞に任意の目的DNA配列を効率的に発現することが可能になるからである。
【0014】
我々が領域、分子又はDNA配列が人工又はキメラであると称する場合、我々はそれがin vitroで設計及び合成されたため、その配列の小断片は天然起源を有し得るが、その配列は同一の一次DNA構造を有する遺伝子エレメントではまったくないことを言及していると明らかにしておくことは重要である。
【0015】
発現のIMEを促進できる、対応する隣接エキソン配列を有するイントロンを設計するために、我々はどの配列モチーフ及び遺伝子構成要素が報告された転写エンハンサー活性を有する植物イントロンに共通であるかを研究した。同時に、GC含量には依存せずに双子葉及び単子葉植物においてこのイントロンの十分で効率的なプロセシングを達成するという挑戦を決意しなければならなかった。
【0016】
転写エンハンサーとして広く使用されているプロモーターであるイネアクチン−1、トウモロコシユビキチン−1及びスクロースシンターゼ−1のイントロン配列を比較した場合、それらのすべてに共通の繰り返し配列モチーフを検出できる(図1)。これらのイントロンが付与する遺伝子発現のIMEにこれらのモチーフが果たす責務はまったく実証されないが、これらの領域、及び多様な植物プロモーターにおけるTATAボックスの5’領域においてCTCCモチーフ(又はその相同配列であるCTC、TCC及びTC)の保存レベルが高いことは、それがRNAポリメラーゼII活性を促進できる転写因子の結合に有利であり得るという可能性を示している。同時に、第一エキソン(翻訳されないmRNA非翻訳リーダーとして残る領域)及び報告された翻訳エンハンサー活性を有するウイルスリーダーにおける富C及びA配列の存在度並びに保存は、そのような配列が生じるMRNAの安定性及びそれが翻訳される能力を促進できることを示している。
【0017】
上に説明した理論のうちどれも完全な科学的実証をもたない。このように、特定の繰り返し配列モチーフを含む隣接するエキソン配列を有する人工イントロンの構築は、mRNAの高い転写レベル及び蓄積を促進する領域を生じることは明白ではないが、我々の研究の結果はこれを示していることを強調するのは評価される。
【0018】
(対応するエキソンを有する)異なるイントロンの間のすでに言及した比較研究から、我々が遺伝子発現のIMEに関連性があると考えたモチーフに富むイネアクチン−1及びトウモロコシユビキチン−1のイントロン/エキソン配列断片を組み合わせた人工エキソン/イントロン/エキソン領域を設計することを決意した。この目標を達成するために、双子葉植物の細胞にも遺伝子発現の増加を起こすことができるためには、生じる人工イントロンがこれらのクラスの植物で効率的にプロセシングされなければならないことを考慮に入れなければならなかった。それにもかかわらず、最初の物質として我々が使用したイントロン配列は高いGC含量及び豊富なヘアピンループを有する複雑な二次構造を有し、そのAGアクセプターの3’スプライシング部位の配列は分岐点のコンセンサス配列とは幾分異なることから、これらのイントロンは双子葉植物の細胞でプロセシングが困難なおそれがあることを我々は発見した。
【0019】
我々が設計したエキソン/イントロン/エキソンの二次構造を簡単にしていかなるクラスの植物細胞においてもプロセシングできるようにするために、我々はその配列にいくつかの規則的な変化を作り、配列のプロセシングを活性化するUUUUUAU様配列を挿入することを決意した(Gniadkowski M;Hemmings−Mieszczak M;Klahre U;Liu H.X;Filipowicz W.、Nucleic Acids Res.1996、24:619〜627頁)。さらに、我々のキメラ配列を第二エキソンに融合させ、双子葉類(例えばタバコ)における効率的なプロセシングを利用してトウモロコシアクチン−1遺伝子の第二イントロン(IVS2)に挿入した(Goodall G.J;Filipowicz W.、The EMBO Journal 1991、10:2635〜2644頁)。それぞれの人工エキソン/イントロン/エキソン変異体の推定二次構造をPCFOLD4.0プログラムを用いたコンピュータ法で検討した(Zuker M.、Meth.Enzymology 1989、180:262〜288頁)。我々が創作した人工エキソン/イントロン/エキソン配列を本時点からARTと称する。
【0020】
すでに言及したように、本特許出願の第二の関連構成要素は、遺伝子発現レベルを増加させるためにキメラエキソン/イントロンの下流に融合させた人工翻訳エンハンサーである。
【0021】
いくつかのウイルスのリーダーで形成した配列及び二次構造の分析から人工翻訳エンハンサーを設計した。この分析から我々は翻訳エンハンサーには3つの本質的な構成要素があると結論する:1)複雑性の低い二次構造、2)富C及びA配列セグメント、3)コンセンサスHCAYYY配列(H=C又はU又はA;Y=C又はU、表1参照)と83%までの相同性を有し、露出し、繰り返しが頻繁で、かつ/又は低融解温度の尾部を有するヘアピン構造をとるモチーフ。
【0022】
表1.いくつかのRNAウイルスのリーダー断片において構造が保存された配列(H=C/U/A、Y=C/U)
【表1】
【0023】
上に指摘した前提から、我々はそれぞれヘアピンループ構造をとるHCAYYY様配列を45塩基の富C及びA配列に挿入した人工翻訳エンハンサーを設計した。この人工翻訳エンハンサーは、その調製に採用された理論的前提を提供したRNAウイルスのリーダーと55%を超えない相同性を有し、6個を超えるヌクレオチドと100%の相同性を有する配列セグメントはまったく存在しない。この理由から、我々の翻訳エンハンサーは以前に報告又は保護された(EP0270611)いかなる翻訳エンハンサーの誘導体でもなく、それらから直接誘導された配列も有さないことを我々は断言できる。
【0024】
操作及び目的遺伝子の融合をさらに容易にするために、我々の翻訳エンハンサーに制限部位を付加した。最終的に、我々が創作した人工エキソン/イントロンに新しい翻訳エンハンサーを融合する前に、キメラ遺伝子の発現を増強する能力がTMVオメガ断片と比べて同じであることを示し、そのキメラ遺伝子の機能性はin vivoであった。我々が創作した人工翻訳エンハンサーはEurekaと称する(図2)。
【0025】
本特許において請求されたプロモーター配列を構築するために、コンセンサスTATAボックスにより形成されるコアプロモーター(Joshi C.P.、Nucleic Acids Res.1987、15:6643〜6653頁)を最初に設計し、それをCaMV 35Sの(転写開始部位から)−24〜−4領域に融合させ、次に転写開始部位並びに富C及びA領域をもたらすアクチン−1の−5〜+27プロモーター領域に融合させた。転写開始部位からの+26〜+72であるトウモロコシユビキチン−1領域を下流に融合させ、富AC領域及び富TC領域をもたらし、第一人工エキソンを得、それをトウモロコシアクチン−1のIVS2イントロンの5’スプライス部位前の12塩基であるトウモロコシアクチン−1第二エキソン及びIVS2イントロン自体と連結した。RNAの成熟に影響し得る推定二次構造を避けるために、コンピュータ法の予測にしたがってこの結合の周辺に塩基の変化、付加又は欠失を作製した。我々が設計した人工イントロンは、IVS2イントロンの最初の54塩基が、転写開始部位からの塩基+89〜+126に対応するトウモロコシユビキチン−1の第一イントロンの5’領域からの37塩基に融合し、次にイネアクチン−1の第一イントロンからの375塩基(この転写開始部位から位置+103〜+477)が続き、トウモロコシユビキチン−1のイントロンの3’末端からの33塩基(転写開始部位から位置+1051〜1083)に融合し、アクチン−1のIVS2イントロンの後半(3’プロセシング開始部位から位置−52〜+5)並びに制限部位及び翻訳開始コンセンサス配列を含む29塩基のキメラ配列と連結したものから構成される(Lutcke H.A;Chow K.C;Mickel F.S;Moss K.A;Kern H.F;Scheele G.A.、The EMBO Journal.1987、6:43〜48頁)。我々が創作した人工エキソン/イントロン/エキソンであるARTの配列を図3に示す。構築された人工エキソン/イントロン/エキソンの効率的なプロセシングを、タバコ細胞及びイネ細胞の両方における一過性発現によりいったん試験してから、その3’末端に翻訳エンハンサーEurekaを融合した。図4に本発明のプロモーター配列物(PARTEプロモーター)の最終構造がみられる。
【0026】
我々が設計したエンハンサーエレメントであるARTは、通常使用されているイネアクチン−1遺伝子の第一エキソン/イントロン/エキソンよりも遺伝子発現エンハンサーとして高い効率を示したことを強調しなければならない。Eureka断片はARTの活性の追加のエンハンサーである。
【0027】
本研究において任意のクラスのトランスジェニック植物細胞において任意のDNA配列の発現を増強する、2つの非常に効率の高い人工遺伝子エレメントを初めて成し遂げることができ、このことは我々が基づいている理論的指針の妥当性を実証している。AT含量が52%を下回る人工プロモーターが双子葉植物の細胞において十分にプロセシングされ、発現の高いIMEを促進することも最初である。ARNウイルスのリーダーとの相同性が低い、完全に人工の、高効率翻訳エンハンサーの構築も新規である。
【0028】
種々の転写エンハンサー配列を本発明のプロモーター配列物の5’に融合した。このように、図5に示すようにイネアクチン−1領域の(転写開始部位から)−43〜−310がプロモーターPARTEの5’末端に融合され、プロモーター領域APARTEが形成し、これにas−1様転写エンハンサーエレメント(Benfey P.N;Chua N.H.、Science、1990、250:959〜966頁)及びトウモロコシユビキチン−1プロモーターの5’領域からの556塩基断片(転写開始部位から−299〜−855)も有する、U3ARTEプロモーターを最終的に得、その構造を図6に示す。
【0029】
記載された遺伝子エレメントから多くのプロモーター配列変異体を構築し(図7参照)、採用されたすべてのエンハンサー領域及びアクチベーター領域の遺伝子発現にわたり相乗効果をもたらすことから、それらのすべてにおいてin vivoアッセイによりその機能性が実証された。
【0030】
転写エンハンサーとして我々の構築物に採用されたas−1エレメント(図6参照)は、オクトピンシンターゼのパリンドロームエンハンサーと50%未満の相同性を有することから革新的な設計を有し(Ellis J.G;Llewellyn D.J;Walker J.C;Dennis E.S;Peacock W.J.、EMBO J.1987、6:3203〜3208頁;EP0278659)、Ellisら(Bouchez D;Tokuhisa J.G;Llewellyn D.J;Dennis E.S;Ellis J.G.、EMBO J.1989、8:4197〜4204頁;米国特許第5837849号)によりなされた研究において請求されたこの種の配列変異体のいずれとも同一でなく(同一性85%未満)、それにみられるTGACGモチーフは独特なフランク配列に関連して見出される。
【0031】
イネアクチン−1遺伝子はすでに記載され、種々の遺伝子の発現に使用されたが(McElroy D;Zhang W;Cao J;Wu R.、Plant Cell 1990、2:163〜171頁;W09109948)、我々の研究はその5’領域の転写エンハンサー活性及び異種発現エンハンサーとしてのその使用を明らかにしていることを強調しておくことが重要である。同様に、トウモロコシユビキチン−1の5’転写エンハンサー領域の使用が請求された(EP0342926)が、我々が採用した556塩基の断片はこのエンハンサーの機能性に必須であると規定された「熱ショック」エレメント(このプロモーター配列の位置−188〜−214の間にみられる)を含まないことから、この断片は独特であり我々が使用したユビキチン配列の転写エンハンサー活性は不要とならない。
【0032】
PARTEプロモーターはイネgluB−1遺伝子の転写開始部位から−31〜−245領域に対応する小さな214塩基断片にも融合され(Takaiwa F;Oono K;Kato A.、Plant Mol.Biol.1991、16:49〜58頁)、新しいプロモーター領域であるGARTE(図8)が形成した。一過性発現アッセイは、この新しい人工プロモーターGARTEが種子内乳に高効率でDNA配列を発現することを実証している。
【0033】
このように、これらのキメラ5’転写エンハンサーの機能的組合せが新規性を有し、植物が属するクラスとは無関係にその植物の細胞におけるDNA配列の高い発現レベルの達成を可能にして、遺伝子エレメントの産生に極めて有用であると結論する。
【0034】
種々のプロモーターからの他の5’調節因子領域を、高い発現レベルを達成するために、かつ/或いは発現に時間特異性、器官特異性又は組織特異性を付与するために本発明の物体に明らかにcys融合できる。
【0035】
遺伝子工学技術の経験者にとっては、本明細書に記載した事柄により、植物細胞においてART及びEurekaエンハンサーを任意の転写プロモーターエレメントと組み合わせて使用して、単子葉又は双子葉植物の細胞において任意のDNA配列の転写/翻訳を増強することが可能となる。同様に分子生物学技術の経験者は、我々の観察から得られたART及びEurekaの設計に我々を導いた理論的指針を採用し、植物細胞において新しいDNA発現エンハンサー配列を構築することができる。
【0036】
過去20年間における植物遺伝子工学の高まる開発を考慮すると、任意の遺伝子及び転写終結配列と連結した本発明のプロモーター物を植物細胞遺伝子形質転換ベクターに挿入でき、確立した効率的な方法の使用により目的遺伝子を発現できるトランスジェニック植物を得ることができることは明白である。
【0037】
本特許出願において、遺伝子形質転換ベクターはそれに予め挿入された任意のDNA断片を植物細胞に導入するための運搬媒体として役立つ(精製されたか、或いは細菌細胞又はウイルス内部に含まれる)DNA分子を称する。
【0038】
微生物の寄託
プラスミドpC−EURGUSint、pC−ARTEGUSint、pGARTEGUSint及びpC−U3ARTEGUSintを微生物保護のためのブダペスト条約に基づいてベルギー共同微生物プラスミド保存機関(the Belgian Coordinated Collection of Microorganism, Plasmid Collection)(BCCM/LMBP)、ゲント大学、「Fiers−Schell−Van Montagu」ビル、Technologiepark927、B−9052 Gent−Zwijnaarde、ベルギーに寄託した。プラスミドpC−EURGUSint、pC−ARTEGUSint、pGARTEGUSintの寄託番号はそれぞれLMBP4727、LMBP4725、LMBP4728、寄託日は2003年5月19日であり、pC−U3ARTEGUSintの寄託番号は4791で寄託日は2003年11月25日である。
【実施例】
【0039】
実施例1:植物細胞にDNA配列を発現させるための新しいキメラ系の構成エレメントの構築
正確なクローニングを容易にするために種々のII型制限エンドヌクレアーゼの制限部位に対する接着末端を有するようにすべての合成DNA断片を創作した。
【0040】
a)Eureka翻訳エンハンサーのクローニング
翻訳エンハンサーEUREKA(配列番号1)に対応する86塩基対(bp)のDNA断片を、合成DNA断片の設計に含めた制限酵素PstI及びSacIに対する接着末端を利用して予め両酵素で分解したベクターpBluescriptII SK(Stratagene、米国)にクローニングした。生じたプラスミドをpBS−Eurekaと名付けた。
【0041】
b)人工エキソン/イントロン/エキソン領域ARTの集合
人工エキソン/イントロン/エキソン領域であるARTを、設計したDNA断片のクローニング及び前後の集合により構築した。まず、コアプロモーター、第一エキソン及び人工イントロンの一部を含むP35AcUと名付けたDNA合成断片(配列番号2)を、EcoRI及びSpeI制限酵素で分解したpBluescriptII SKベクターにクローニングし、プラスミドpBS−AcUを得た。その後に、そのプラスミドをSpeI及びSacIで分解し、人工イントロンの一部をコードするDNA合成断片I−U/Ac(配列番号3)をそれに挿入した。このようにしてpBS−AcUAcプラスミドを得た。
【0042】
次に、人工イントロンの末端を保有するDNA合成断片I−Ac/U(配列番号4)を、制限酵素BamHI及びSacIで分解したpBS−AcUAcプラスミドに挿入し、プラスミドpBS−AcUAcUを産生させた。
【0043】
断片IniT(配列番号5)を、SpeI/SacIで分解したpBS−AcUAcUに挿入したときに、プラスミドpBS−ARTを一致させる人工エキソン/イントロン/エキソンであるARTが完成した(配列番号6)。pBluescriptIIベクターの制限部位EcoRI及びSacIの間の一次構造を配列番号7の配列に示す。
【0044】
c)PARTEプロモーターの構築
本発明のプロモーター配列物(PARTEプロモーター)を構築するために、コアプロモーター及び(3’領域を有さない)エキソン/イントロン/エキソン領域ARTを含むDNA断片を、XhoI/PstI分解によりpBS−ARTプラスミドから得、それを同酵素で分解したpBS−Eurekaプラスミドに挿入した。このように、我々はプラスミドpPARTE(図4、図7D)を実現し、そのEcoRI及びSacI部位の間の配列を配列番号9の配列に示す。
【0045】
実施例2:植物細胞におけるEureka及びARTエンハンサーエレメントの機能性の実証
a)タバコ細胞における翻訳エンハンサーEurekaの機能性
タバコ及びイネ細胞におけるEurekaのエンハンサー力を証明するために、一連の補助遺伝子構築物を作製した。
【0046】
SnaBI部位に挿入されたジャガイモST−LS1遺伝子のIV2イントロンを有するレポーター遺伝子uidA(GUSint)を、プラスミドpUC−GUSintのNcoI/SacI分解により得(図9)、プラスミドpBS−Eurekaの同部位にクローニングし、ベクターPbs−EURGUSintを生成させた。これをさらに制限酵素PstI及びSalIで分解し、S−1ヌクレアーゼで処理してプラスミドpBS−ΔEURGUSintを得た。それをXhoI/KpnIで分解してGUSint遺伝子に融合したEurekaエンハンサーを含むDNA断片を得て、それをpBPFΩ(オメガ)7ベクター(図10)に挿入した。このように、我々はpBPF−EURGUSintベクター(図7A)を得た。そのベクターはCaMV 35Sプロモーター(1.3kb版)、Eurekaエンハンサー及びアグロバクテリウムツメファシエンス(Agrobacterium tumefaciens)のnos遺伝子転写終結シグナル(tNOS)の制御下でGUSint遺伝子を発現する。
【0047】
構築物pBPF−EURGUSintの発現を評価するための対照として、SalI/クレノウ及びKpnI分解によりプラスミドpUC−GUSintから得られたGUSint遺伝子を、pBPFΩ(オメガ)7ベクターのSmaI及びKpnI部位の間にクローニングすることでプラスミドpBPFΩ(オメガ)−GUSintを構築した。このプラスミドは、Eurekaの代わりにGUSintを制御している翻訳エンハンサーΩ(オメガ)が存在することを除き、pBPF−EURGUSintに類似している。別の対照プラスミドを、XhoI−NcoI分解によりプラスミドpBPFΩ(オメガ)−GUSintのエンハンサーオメガを除き、クレノウにより処理し、プラスミドの自己連結を行うことにより構築し、pBPF−GUSintベクターを得た。
【0048】
プラスミドpBPFΩ(オメガ)−GUSint、pBPF−GUSint、及びpBPF−EURGUSintをHindIIIで分解して、植物においてGUSintを発現させるためのカセットを得た。これらをHindIIIで分解したバイナリーベクターpCAMBIA2300にクローニングし、それぞれバイナリーベクターpC−Ω(オメガ)7GUSint、pC−GUSint及びpC−EURGUSintを生成させた。
【0049】
得られたバイナリープラスミドをA.ツメファシエンスLBA4404株に導入し、我々は、Anら(An G.、Plant Physiol.1985、79:568〜570頁)が記載したプロトコルを幾分修正したものにしたがってNT1タバコ細胞において一過性発現実験によりエンハンサーEurekaの機能性のアッセイを始めた。タバコ細胞を、各バイナリーベクターを保有するアグロバクテリウムと4日間共培養した後で、細胞を回収し、Jefferson(Jefferson R.A.1988.「Plant reporter genes:the GUS gene fusion system」、J.K.Setlow編、Genetic Engineering、第10巻、Plenum Publishing Corporation、247〜263頁)が記載したように処理し、β−グルクロニダーゼ(GUS)活性を測定した。各実験を3回繰り返し、各回に各構築物につき5回の繰り返しを行った。結果を以下の表に示す。
【0050】
表2.タバコ細胞における翻訳エンハンサーEurekaの機能性の実証
【表2】
【0051】
表2に示す結果にみられるように、Eurekaのエンハンサー活性はTMVオメガリーダー配列のエンハンサー活性と有意差をもたない。このことは完全に人工の効率的な翻訳エンハンサーを最初に実現したことを実証している。このことは、C及びAに富む配列の領域をモチーフHCAYYY(H=C/T/A、Y=C/T)と相同の配列と組み合わせることにより3’方向下流に融合したDNA配列の翻訳を増強させる重大な性質を有する遺伝子エレメントを構築することが可能であることを示し、我々の理論的指針も確認している。
【0052】
b)タバコ細胞における人工エキソン/イントロン/エキソンARTの機能性
タバコ細胞におけるARTエレメントの機能性を試験するために、プラスミドpUC−GUSintからまずGUSint断片を得、それをNcoI−SacIで分解し、同酵素で分解したpBS−ARTにクローニングしてプラスミドpBS−ARTGUSintを得た。次に、このプラスミドをSalI−BglIIで分解しS1ヌクレアーゼで処理してpBS−ΔARTGUSintを得、それをXhoI−KpnIで分解してARTGUSint断片を得、同酵素で分解したベクターpBPFΩ(オメガ)7−GUSintにクローニングし、プラスミドpBPFARTGUSint(図7B)を得た。ここで、GUSint発現はCaMV 35Sプロモーター(1.3kb版)、人工エキソン/イントロン/エキソン領域であるART、及びA.ツメファシエンスnos遺伝子転写終結シグナル(tNOS)の制御下にある。
【0053】
プラスミドpBS−ΔARTGUSintからXhoI/PstI分解によりエレメントARTを含むバンドを得、それを同酵素で分解したpBS−EURGUSintベクターに融合させプラスミドpBS−ARTEGUSintを得た。これからXhoI/KpnI分解によりGUSint遺伝子を遺伝子エレメントART及びEurekaのシグナル下に置くためのDNA断片を得、同様に分解したpBPFΩ(オメガ)7ベクターに導入しpBPFARTEGUSintプラスミドを産生させた(図7C)。
【0054】
プラスミドpBPFARTGUSint及びpBPFARTEGUSintをHindIIIで分解し、植物にGUSintを発現させるためのカセットを得た。これらをバイナリーベクターpCAMBIA2300にクローニングし、バイナリープラスミドpC−ARTGUSint及びpC−ARTEGUSintをそれぞれ生じさせた。
【0055】
得られたバイナリープラスミドをアグロバクテリウムツメファシエンスLBA4404株に導入した後に、我々は本実施例の(a)項に述べたようにNT1タバコ細胞を用いた一過性発現アッセイにおいて翻訳エンハンサーEurekaの機能性のアッセイを始めた。得られた結果を表3に示す。
【0056】
表3.タバコ細胞における遺伝子エレメントART及びEurekaの機能性の実証
【表3】
【0057】
タバコ細胞におけるARTの機能性評価の結果は、人工イントロンが双子葉植物の細胞において正確にプロセシングされ、発現エンハンサー活性を保持することを明らかにした。我々の実験結果は構築物pBPFARTEGUSintにおけるART及びEurekaエレメントの間の相互作用から得られた発現レベルに及ぼす正の相乗作用も示し、ここで公知の天然プロモーターCaMV 35Sの発現能が5倍に増加する。
【0058】
設計された人工遺伝子エレメント(ART及びEureka)を、植物細胞において活性な任意のプロモーター(この場合CaMV 35Sプロモーター)及び他の任意のDNA配列(我々の場合GUSint)の間に機能的に挿入でき、その転写/翻訳を増加できることも証明される。
【0059】
c)イネ細胞におけるエンハンサーエレメントEureka及びARTの機能性
単子葉植物の細胞における我々の人工エンハンサーの機能性を証明するために新しい構築物のセットを作製した。まず、5’末端を人工エキソン/イントロンARTに融合したuidA(gus)遺伝子を保有するプラスミドpBPFARTGUSintからのXhoI〜KpnI断片を、同制限酵素で分解したpBPFA19−リンカーベクター(図11)に挿入して、プラスミドpBPFA19ARTGUSintを形成させた。このプラスミドにおいて、pBPFA19−リンカーに存在するイネアクチン−1のエキソン/イントロン/エキソン領域は、人工エレメントARTに置き換わり、他の調節エレメントとして(4重のオクトピンシンターゼas−1様エンハンサーはCaMV 35Sプロモーター(400bp版)に融合している)キメラプロモーターA19及びtNOS転写終結シグナルを残す。
【0060】
同様に、プラスミドpBS−ARTEGUSintからのXhoI〜KpnIのバンドをpBPFA19−リンカーベクターにクローニングして構築物pBPFA19ARTEGUSintを得た。対照として使用した構築物pBPFA19GUSintを、プラスミドpUC−GUSintからGUSint断片をNcoI/SacIで分解したpBPFA19−リンカーベクターにクローニングすることにより作製した。使用した別の対照プラスミドはpBPFΩ(オメガ)−GUSintであった。
【0061】
イネ細胞においてART及びEurekaが遺伝子発現を増強する能力の定性評価を、インディカPerla品種のイネのカルスにおける一過性発現をアッセイすることにより実施した。次亜塩素酸ナトリウム及びアルコールで予め滅菌した成熟した種子からカルスを得、遮光してN6−2培地すなわちN6塩類及びビタミン類(Chu C.Cら、Scientia Sinic 1975、18:659頁)、0.1g/Lミオイノシトール、1g/Lカゼイン加水分解物、2mg/L 2,4D、30g/Lスクロース、3g/L Phytagel、pH5.7)中で21日間培養した。マイクロプロジェクタイルを用いたボンバードメント法により形質転換を実施した。ボンバードメントの前に、浸透圧前処理用に0.4Mマンニトールを補充したN6−2培地にカルスを継代した。出版されたプロトコルにしたがってボンバードメントには1μmの球状金粒子(BioRad)をマイクロプロジェクタイルとして使用した(Russell D.R.、Wallace K.M.、Bathe J.H.ら、Plant Cell Rep.1993、12:165〜169頁)。PDS−1000/Heシステム(BioRad)を採用して形質転換を実施した。ボンバードメントには30個のカルスを平板中央に置き、条件は圧力1100psi、距離6cm、平板1枚あたり1回発射した。ボンバードメント後にカルスを遮光下で同じ浸透圧培地中で16時間維持してから、マンニトールを有さないN6−2培地で2日間継代培養した。組織化学法によりX−Glucを用いてGUS活性を明らかにする(Jefferson R.A.1988.「Plant reporter genes:the GUS gene fusion system」、J.K.Setlow編、Genetic Engineering、第10巻、Plenum Publishing Corporation、247〜263頁)。立体顕微鏡で各カルスにおける青い点及び帯域を計数することにより評価を実施した(図12)。表4は繰り返しをそれぞれ3回行った4回の実験から得られた結果を示す。
【0062】
表4.イネ細胞におけるART及びEurekaの機能性の比較実証
【表4】
【0063】
見て取れるように、これらの実験結果は単子葉植物の細胞における遺伝子発現エンハンサーエレメントとしてのART及びEurekaの機能性も確認している。我々のアッセイにおいて人工エキソン/イントロン/エキソンART(pA19ARTGUSint)により発生するIME活性は、容認された遺伝子発現エンハンサー能を有する遺伝子エレメントであるイネアクチン−1遺伝子の第一エキソン/イントロン/エキソン(pBPFA19GUSint)で観察された活性よりも高かったことを強調しておくのは重要である。さらに、我々の実験において構築物pA19ARTGUSint及びpA19ARTEGUSintに対して得られた結果の間の有意差を評価することはできないが、pA19ARTEGUSintのボンバードメントを行ったカルスのX−Gluc組織化学染色後の青く着色した帯域の大きさ及び強度から(図12)、最終構築物中にEureka断片が存在すると発現が大きく増加することは図12において注目に値する。
【0064】
結論として、本実施例は任意の種類の植物細胞における遺伝子発現エンハンサーとしての人工遺伝子エレメントART及びEurekaの機能性を示している。そのうえ、これらのエンハンサーエレメントは効率が高く、それらが融合したプロモーターに依存せずに発現レベルを増加させることが実証された。最後に、ART及びEurekaは下流の遺伝子の発現をなお相乗的に増強させるために組み合わせることができることも実証された。
【0065】
ART及びEurekaを植物に活性な任意のプロモーター(例えばA19)及び任意のDNA配列(GUSint遺伝子)の間に機能的に挿入してその転写/翻訳を増加できることを新たに示す。
【0066】
実施例3:種々の5’転写エンハンサー領域を採用したPARTE発現系変異体
a)PARTEプロモーターへのイネアクチン−1の5’転写アクチベーター領域の付加
イネアクチン−1遺伝子からPARTEプロモーターに5’転写エンハンサー領域をcys−融合するために、pPARTEプラスミドを酵素EcoRI及びEcoRVで分解し、これらの酵素で連結する両極端を有する合成DNA断片En−Ac1(イネアクチン−1遺伝子の転写開始部位から−43〜−221、配列番号10)をそれに挿入した。得られたプラスミドpA1PARTEをEcoRV及びHindIIIで分解して、合成DNA断片En−Ac2(イネアクチン−1の転写開始部位から−226〜−310、配列番号11)を挿入し、アクチン−1遺伝子プロモーターの5’アクチベーター領域を完成させプラスミドpAPARTE(図5、図7E)を産生させた。このプラスミドの制限部位HindIII及びSacIの間のヌクレオチド配列を配列番号12に示す。
【0067】
b)APARTEプロモーターへのas−1様転写エンハンサー配列の付加
プラスミドpA1PARTEをNruI及びSalIで分解し、ASP(配列番号13)と称するDNA合成断片をそれに挿入し、構築物pASPΔA1PARTEを産生させる。ASPは上記の制限酵素に対する接着末端を保有しas−1−様転写エンハンサー配列のために成分化する。このプラスミドをSalIで分解し、S1ヌクレアーゼで処理してからPstIで分解し約900bpのDNA断片を得、それをその後NruI及びPstIで分解したベクターpA1PARTEにクローニングし、最終的にプラスミドpASPA1PARTEを得た。そのプラスミドはイネアクチン−1の5’転写活性化領域のNruI部位に挿入されたASPエンハンサーを有する。それをEcoRV−XhoIで分解することによって、第二のASPエンハンサーをプラスミドpASPA1PARTEに挿入し、ベクターp2A1PARTE(配列番号14、制限部位KpnI及びSacIの間のヌクレオチド配列を示す)を生成させたが、図7Fにその構造を示す。
【0068】
pA1PARTEについて上に解説したように、En−Ac2断片もpASPA1PARTEに挿入し構築物pASPAPARTEを得た。ここでそれをEcoRV−SalIで分解することにより第二ASPエンハンサーを挿入し、最終的にベクターp2APARTE(図7G)を得た。SalI及びSacI部位の間のヌクレオチド配列を配列番号15に示す。
【0069】
c)U3ARTEプロモーターの構築
まず、プライマーOli−U1(配列番号16)及びOli−U2(配列番号17)を用いたポリメラーゼ連鎖反応(PCR)によってトウモロコシubi−1遺伝子の転写開始部位から−299〜−680の領域に対応する約395bpのDNA断片を増幅させた。増幅した断片を制限酵素KpnI及びXhoI(両部位はプライマー内に含まれていた)で分解し、同様にプロセシングしたpBluescriptII SKベクターにクローニングし、構築物pBS−Ubi1を得た。その後、トウモロコシubi−1遺伝子の−680〜−855(配列番号18)のために成文化している(codify)合成DNA断片(En−U2)をNcoI/KpnIで分解したpBS−Ubi1ベクターにクローニングし、構築物pBS−Ubi2を生成させた。その構築物はトウモロコシユビキチン−1遺伝子5’アクチベーター領域(−299〜−855、配列番号19)を含む。
【0070】
トウモロコシユビキチン−1遺伝子からクローニングされた「熱ショック」ボックスを含まない5’転写アクチベーター領域をXhoI−KpnI分解によりプラスミドpBS−Ubi2から得、ベクターのSalI−KpnI分解によりプロモーター2APARTEの5’にcis−挿入し、いわゆる構築物pU3ARTEを得た(図6、図7H)。部位KpnI及びSacIの間のベクターpU3ARTEの配列を配列番号20に示す。
【0071】
d)GARTEプロモーターの構築
本発明物の適応性を実証するために、我々は器官、組織又は発生特異性を有するプロモーターからの5’調節領域を融合させることを決意し、これによりすでに言及した特徴を有する高い発現レベルが付与された。このように、グルテリンの器官特異的発現を制御するイネgluB−1遺伝子5’調節領域をPARTEプロモーターのcys及び5’に融合した。この目標を達成するために、gluB−1転写開始部位から−31〜−245領域(配列番号21)に対応する214塩基対の断片(GLU)及び都合のよいクローニング部位を合成し、EcoRI及びXhoIで分解したpPARTEプラスミドにそれを挿入して、pGARTEベクターを産生させ(図8)、XhoI及びSacI部位の間のその一次構造を配列番号22に示す。
【0072】
実施例4:タバコ及びイネ細胞におけるPARTE発現系の種々の変異体の機能性
単子葉及び双子葉植物の細胞において本発明の各プロモーター変異体物の発現レベルを評価するために、バイナリーベクターpC−ARTE−GUSintにおいてGUSintの発現を制御しているCaMV 35SプロモーターをSmaI−SpeI分解により欠失させ、その場所に構築物pAPARTE、p2A1PARTE、p2APARTE及びpU3ARTEからのKpnI/S1ヌクレアーゼ−SpeI断片を挿入し、新しいバイナリーベクターpC−APARTEGUSint、pC−2A1PARTEGUSint、pC−2APARTEGUSint及びpC−U3ARTEGUSintを産生させた。
【0073】
a)タバコでのアッセイ
バイナリーベクターpC−APARTEGUSint、pC−2A1PARTEGUSint、pC−2APARTEGUSint及びpC−U3ARTEGUSintをA.ツメファシエンス細胞に導入し、実施例2の(a)項に記載したようにNT1タバコ細胞における一過性発現アッセイ実験を実施した。使用した対照プラスミドpC−GUSintは、GUSの発現がCaMV 35Sプロモーター(1.3kb版)及びtNOSターミネーターに制御されている。実験を2回実施し(各処理について5回の繰り返し)、結果を以下の表に示す。
【0074】
表5.タバコ細胞におけるPARTE発現系の種々の変異体の機能性
【表5】
【0075】
我々の結果が本発明物における種々のプロモーター変異体の機能性の確証となり、PARTEに融合した5’転写調節領域に応じてその活性を調整することが可能であることも示していることは明らかである。双子葉植物の細胞において、我々の遺伝子構築物を用いて我々は天然プロモーターCaMV 35Sにより制御されている場合に達成されたものを上回る発現レベルに達したことを強調しなければならない。
【0076】
b)イネを用いたアッセイ
PARTEプロモーターの種々の変異体の活性の一過性評価を実施するために、実施例2(c)項に記載したようにバイナリーベクターpC−APARTEGUSint、pC−2A1PARTEGUSint、pC−2APARTEGUSint及びpC−U3ARTEGUSintを用いてイネカルスにボンバードメントを行った。対照プラスミドpActl−F(McElroy D;Zhang W;Cao J;Wu R.、Plant Cell 1990、2:163〜171頁)は、イネアクチン−1遺伝子プロモーター及びtNOSターミネーターの制御下でgus遺伝子を発現する。発現カセットをKpnI−XbaI分解によりこれらのプラスミドから抽出し、同制限酵素で分解したバイナリープラスミドpCAMBIA2300に挿入し、ベクターpC−Act1Fを産生させた。
【0077】
評価する各構築物について3回の繰り返しでボンバードメント実験を3回実施した。得られた結果を以下の表に示す。
【0078】
表6.イネ細胞におけるPARTEプロモーター発現系の種々の変異体の機能性
【表6】
【0079】
本表に示した結果は、単子葉植物の細胞におけるPARTEプロモーターの種々の変異体がイネアクチン−1遺伝子の天然プロモーターを上回る発現レベルに達することから、それらの変異体の機能性を証明している。したがって、本発明物は、その制御下でcysに配置されたDNA配列の高い発現レベルを達成するための効率の高い遺伝子ツールとしての有用性が確認される。
【0080】
c)イネ種子のアッセイ
イネ細胞内乳におけるGARTEプロモーターの組織特異性を評価するために、GUSint遺伝子に融合しnosターミネーター(tNOS)を有するEureka断片を含む、pBPFARTEGUSintベクターからの約2.5KbのSalI/クレノウ−PstI断片を、XbaI/クレノウ−PstIで分解したpGARTEベクターにクローニングした結果、構築物pGARTEGUSintが生じた。
【0081】
pGARTEGUSintにおけるGARTEプロモーターをXhoI−NcoI分解することにより、pGEM−T−GluB−1プラスミドのSalI−NcoI分解から得られた種子内乳特異的で高効率のイネグルテリンB−1プロモーターに置き換えた対照プラスミドも構築した。このように、我々はpGluGUSintを得た。
【0082】
GARTEプロモーターの評価及びGluB1のプロモーターとの比較を、Y−S.Hwang(Hwang Y−S;McCullar Cass;Huang N.、Plant Science.2001、161:1107〜1116頁)にしたがって、温室栽培されたイネ品種インディカPerlaの穂の穎果(ear cariopsis)から単離した未熟な内乳(受粉後8〜9日)にボンバードメントを行うことによって実施した。内乳におけるGUS活性を測定するための蛍光アッセイを、Jefferson(Jefferson R.A.1988.「Plant reporter genes:the GUS gene fusion system」、J.K.Setlow(編)、Genetic Engineering、第10巻、Plenum Publishing Corporation、247〜263頁)にしたがって実施し、プラスミドDNAで覆った金微粒子を用いたボンバードメントを行った24時間後に評価した。それぞれ5回の繰り返しを行った独立した2回の実験で得られたGUS活性の結果を表7に示す。
【0083】
表7.イネ種子内乳におけるGARTEプロモーターの機能性
【表7】
【0084】
これらの結果は、我々が設計した人工エレメントに基づくキメラプロモーターGARTEが種子内乳において遺伝子を発現する効率が高いことを確認している。GARTEプロモーターに挿入されたGLU配列は発現に特異性を付与できるが、それは「それ自体は」高レベルを保証せず、プロモーターを一致させる他のエレメントにも依存する(Takaiwa F;Yamanouchi U;Yoshihara T;Washida H;Tanabe F;Kato A;Yamada K.、Plant Mol Biol.1996、30:1207〜1221頁)。
【0085】
示された結果は、本発明のエレメント物上流に調節領域を挿入すると、発生、器官又は組織特異性を有する任意のDNA配列の発現を効率的に導くためにそのエレメント物を使用できることを再び支持している。分子生物学の経験者には、GARTEプロモーターに挿入されたGluB−1プロモーター調節配列を、生物によるストレス(例えば病原体の攻撃)、非生物因子(例えば創傷、極端な高温又は低温、塩分、乾燥、一部の化学物質の存在)、酸化ストレス、種々の器官及び組織発生段階などに応答する調節配列とうまく置き換えられることは明白である。
【0086】
本発明の調節領域物下でクローニングされたDNA配列を植物細胞に導入でき、公知の生物学的又は物理化学的形質転換法により安定挿入できること、並びにこれらの遺伝修飾された細胞から、DNA配列が融合したプロモーター変異体にしたがってその配列が都合よく発現する受精能のある植物を再生させることが可能であることも明らかである。このように、本発明は害虫、疾病、多様なストレスに対する抵抗レベルがさらに大きく、農業の収穫量がさらに大きいか、又は使用の中でとりわけ医学又は工業応用を有する化合物を高効率で産生するトランスジェニック植物の製造に貢献する潜在性を示している。
【図面の簡単な説明】
【0087】
【図1】転写開始部位からのイネアクチン−1(Act)、トウモロコシユビキチン−1(Ubi)及びトウモロコシスクロースシンターゼ(Shrun)遺伝子配列を示す図である。大文字は第一エキソンを、小文字は第一イントロンの5’領域を示し、繰り返し共通配列モチーフの部分に下線を付す。
【図2】Eureka人工翻訳エンハンサー配列を示す図であり、関連のあるエレメント及び制限エンドヌクレアーゼ認識部位を示す。
【図3】ARTエキソン/イントロン/エキソン人工配列を示す図であり、その構成要素の断片のそれぞれの起点を示す(小文字:人工イントロン、UUUUUAU様配列を作り出すために挿入した塩基に下線2本を付す、制限エンドヌクレアーゼのいくつかの関連性のある認識部位に下線1本で印を付ける)。
【図4】本発明物(PARTEプロモーター)の一次構造を示す図であり、ARTエキソン/イントロン/エキソン領域(イントロン塩基は小文字、エキソンは大文字)及び人工翻訳エンハンサーEUREKAに融合したコアプロモーター(斜体小文字)を示す。制限エンドヌクレアーゼのいくつかの関連性のある認識部位に下線を付し、TATAボックスに下線2本を付し、翻訳開始コドンを太字で示す。
【図5】APARTEプロモーターの一次構造を示す図であり、PARTEプロモーター(プロモーターは斜体小文字、イントロンは小文字、エキソンは大文字)に融合したイネアクチン−1の5’調節領域(斜体大文字で転写開始部位から−43〜−310領域)を示す。制限エンドヌクレアーゼのいくつかの関連性のある認識部位に下線1本で印を付す。TATAボックスに下線2本を付し、翻訳開始コドンを太字で示す。
【図6】U3ARTEプロモーターの一次構造を示す図であり、その構成エレメントであるトウモロコシubi−1遺伝子の転写開始部位から−299〜−855領域を大文字で、as−1様転写エンハンサーを太字大文字で、イネact−1遺伝子の転写開始部位から−43〜−310領域を斜体大文字で、PARTEプロモーターを小文字で示す(TATAボックスに下線2本を付し、ARTイントロンを斜体で示し、翻訳開始部位に下線1本を付す)。
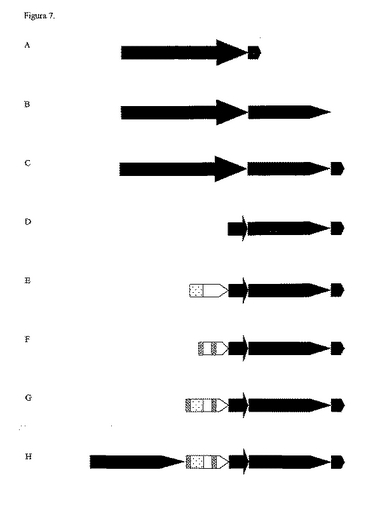
【図7】本発明のエンハンサーエレメント物を有するプロモーター変異体を示す図である。A:35SEureka; B:35SART; C:35SARTE; D:PARTE; E:APARTE; F:2A1PARTE; G:2APARTE; H:U3ARTE. れんが模様部分:35S プロモーター(1.3Kb); チェック柄部分:翻訳エンハンサー Eureka; 縦縞模様部分:ART エキソン/イントロン/エキソン; 格子柄部分:人工プロモーターコア; 白抜き部分:イネアクチン−1 遺伝子5’活性化領域(−43〜−221); 点(疎)模様部分:イネアクチン−1 遺伝子5’活性化領域(−226〜−310); 斜め縞模様部分:ASP(as−1−様エンハンサー); 点(密)模様部分:トウモロコシユビキチン−1 プロモーター5’活性化領域(−299〜−855)。
【図8】GARTEプロモーターの一次構造を示す図であり、その構成エレメントである、転写開始部位から−31〜−245のイネgluB−1遺伝子領域を斜体大文字で示す、PARTEプロモーター(プロモーターを斜体小文字で、イントロンを小文字で、エキソンを大文字で示し、いくつかの関連性のある制限部位に下線を付し、TATAボックスに下線2本を付し、翻訳開始コドンは太字)を示す。
【図9】pUC−GUSintの地図を示す図である。
【図10】pBPFΩ(オメガ)7の地図を示す図である。
【図11】pBPFA19−リンカーの地図を示す図である。
【図12】促進したマイクロプロジェクタイルを用いたボンバードメントにより誘導されたGUSint遺伝子を保有する種々の遺伝子構築物の一過性発現によりイネ細胞におけるART及びEUREKAエレメントの機能性のX−Gluc組織化学染色による比較実証を示す図である。
【技術分野】
【0001】
本発明はバイオテクノロジーに関し、さらに詳細には植物遺伝子工学に関する。詳細には、キメラDNA構築物を提供し、それらの構築物は双子葉及び単子葉植物の細胞においてそれらの構築物に融合した任意のヌクレオチド配列の高い転写/翻訳プロモーター活性を示し、さらに高発現レベルの目的の遺伝子及びDNA配列を有するトランスジェニック植物を得ることを可能にする。
【背景技術】
【0002】
植物遺伝子工学は、新しいバイオテクノロジー産物の基礎研究及び商業生産に非常に生産力の高いことが実証された技術である。(Hammond J.、Curr.Top.Microbiol.Immunol 1999、240:1〜19頁;Simoens C.及びVan Montagu M.、Reproduction Update 1995、1:523〜542頁)。
【0003】
分子生物学的手法により遺伝子操作された植物に導入された遺伝子又はDNA配列の強度及び時間的又は空間的特異性に関して十分な発現を保証するプロモーターシグナルの選択は、植物遺伝子工学が成功するために非常に重要である。このような理由から、必要な導入遺伝子のそれぞれの発現を確実にできるプロモーター及びシグナルの探索に多数の努力が過去20年間に捧げられてきた。このように、種々の起源(植物、ウイルス、アグロバクテリウム(Agrobacterium)のTi又はRi、或いはキメラ)のプロモーターが評価され、トランスジェニック植物の産生に採用されてきた。
【0004】
植物遺伝子操作にさらに広く使用された構成性プロモーターは、カリフラワーモザイクウイルス(CaMV)35S ARNプロモーター(Odell J.T;Nagy F;Chua N.H.、Nature 1985、313:810〜812頁)、A.ツメファシエンス(A.tumefaciens)Tiプラスミド由来ノパリンシンテターゼ遺伝子(nos)プロモーター(An G;Costa M.A;Mitra A;Ha S;Marton L.、Plant Phisiol.1986、88:547〜552頁)、イネアクチン−1遺伝子プロモーター(McElroy D;Zhang W;Cao J;Wu R.、Plant Cell 1990、2:163〜171頁)及びトウモロコシユビキチン−1遺伝子プロモーター(Christensen A.H;Sharrock R.A;Quail P.H.、Plant Mol.Biol.1992、18:675〜689頁)であった。しかし、どの植物クラスにおいても発現レベルが十分に高くないことが主な原因となって、これらの天然発現系にはそれぞれ限界がある。例えばプロモーターの発現が双子葉植物の細胞において低く、単子葉類ではほとんど検出できない一方で、最も広く使用されているプロモーターであるCaMV 35Sの発現は単子葉植物よりもタバコ細胞においてずっと強い(Topfer R;Maas C;Horicke−Grandpierre C;Schell J;Steinbiss H.H.、Methods Enzymol.1993、217:67〜78頁;Mitsuhara I;Ugaki M;Hirochika H;Ohshima M;Murakami T;Gotoh Yら、Plant Cell Physiol.1996、37:49〜59頁)。同様にイネアクチン−1及びトウモロコシユビキチン−1プロモーターは単子葉植物の細胞において下流の遺伝子の転写の促進に極めて効率的であるが、タバコ細胞におけるそのプロモーター活性は低い(Schledzewski K;Mendel R.R.、Transgenic Research 1994、3:249〜225頁)。
【0005】
トランスジェニック植物における異種タンパク質の発現レベルを増加させるために、天然プロモーターを転写又は翻訳エンハンサーと組み合わせた多様なキメラプロモーターが設計された。これらのエンハンサーエレメントの中で、タバコモザイクウイルス(TMV)のオメガ翻訳エンハンサー(Gallie D.R;Sleat D.E;Watts J.W;Turner P.C;Wilson T.M.A.、Nucleic Acids Res.1987、15:3257〜3273頁)、タバコエッチウイルス(TEV)の翻訳エンハンサー(Carrington J.C;Freed D.D.、J.Virol.1990、64:1590〜1597頁)、オクトピンシンターゼ由来プロモーター転写エンハンサー(Fromm H;Katagiri F;Chua N.H.、Plant Cell 1989、1:977〜984頁)、マンノピンシンターゼ由来(Comai L;Moran P;Maslyar D.、Plant Mol Biol.1990、15:373〜381頁)及びCaMV 35Sプロモーター由来(Kay R;Chan A;Daly M;McPherson J.、Science 1987、236:1299〜1302頁)、並びに天然エキソン及びイントロン、例えばトウモロコシアルコールデヒドロゲナーゼイントロン1(Callis J;Fromm M;Walbot V.、Genes Devel 1987、1:1183〜1200頁;Last D.I;Brettell R.I.S;Chamberlaine D.A;Chaudhury A.M;Larkin P.Jら、Theor Appl.Gen.1991、81:581〜588頁)、トウモロコシスクロースシンターゼ由来第一エキソン/イントロン(Maas C;Laufs J;Grant S;Korfhage C;Werr W.、Plant Mol.Biol.1991、16:199〜207頁)、イネアクチン−1遺伝子由来第一エキソン/イントロン(McElroy D;Blowers A.D;Jenes B;Wu R.、Mol.Gen.Genet.1991、231:150〜160頁)などを挙げることができる。それにより、2X35S、Mac、Emuなどのプロモーターが生じ(EP0459643、EP0651812)、それらは双子葉又は単子葉類の主に特定のクラスの植物細胞において強力である(Schledzewski K;Mendel R.R.、Transgenic Research 1994、3:249〜255頁)。
【0006】
双子葉及び単子葉植物両方の細胞において遺伝子を発現させるために採用できる強力なプロモーターの開発は、これまでも現在も多くの研究室に関係する問題である。それは、多様なクラスの植物を形質転換するための独特な遺伝子構築物を手に入れるという科学的挑戦やそれが意味する節約のためだけではなく、バイオテクノロジー産物の生産及び商品化をさらに容易にする特有の発現系を手に入れるためでもある。特許出願W09943838において請求された合成プロモーターは、5’末端を35Sプロモーター、トウモロコシユビキチン−1プロモーター及びオクトピンシンターゼプロモーターからの転写エンハンサー配列に融合した、高いGC含量(64%以上)を有するTATAボックスから転写開始部位までの配列を請求した。他方、双子葉及び単子葉植物における発現を探索するだけではなく、配列相同性に依存した遺伝子サイレンシングを避けるために(Park Y.D;Papp I;Moscone E;Iglesias V;Vaucheret H;Matzke A;Matzke M.A.、Plant J.1996、9:183〜194頁)、特許出願WO0058485は2つのウイルスゲノム、すなわちツユクサ黄色斑紋ウイルス(CoYMV)及びキャッサバ葉脈モザイクウイルス(CsVMV)からの配列並びに35Sプロモーターからのエンハンサー配列の組合せに由来する人工プロモーターを請求している。
【0007】
種々の遺伝子エレメントがヌクレオチド配列の転写又は翻訳を増強することを可能にするメカニズムはまだ明らかではない。例えば、多くのRNAウイルスからのリーダー配列が、5’末端に融合したキャップ(m7G(5’)ppp(5’)N)の存在と無関係に種々のメッセンジャーRNA(mRNA)の翻訳を増強できることが報告された(Sleat D.E;Wilson T.M.A.1992.「Plant virus genomes as sources of novel functions for genetic manipulations」、T.M.A.Wilson & J.W.Davies編、Genetic engineering with plant viruses、CRC Press,Inc.、55〜113頁;Gallie D.R;Sleat D.E;Watts J.W;Turner P.C;Wilson T.M.、Nucleic Acids Res.1987、15:8693〜8711頁)。しかし、これらすべてのウイルスリーダーのRNA二次構造が複雑でないことを除いて、翻訳エンハンサーの性質を表すために応答するヌクレオチド配列で別の共通エレメントは決定されていない。
【0008】
具体的には、TMVオメガ断片の翻訳増強は、繰り返し配列である少なくとも1つのコピーのオクタマーACATTTAC及び重大なモチーフと考えられる25塩基の(CAA)n領域の存在によることが報告された(2つのコピーの(CAA)n領域は高いエンハンサーレベルを付与するために十分である)(Gallie D.R;Walbot V.、Nucleic Acids Res.1992、20:4631〜4638)。しかし、ジャガイモXウイルス(PVX)リーダーからの富CA領域である28塩基は「それ自体では」翻訳エンハンサー活性を示さなかった(Pooggin M.M;Skryabin K.G.、Mol.Gen.Genet.1992、234:329〜331頁)が、PVXリーダーのCA領域に存在するCCACCペンタヌクレオチドは、18S rRNAの3’末端と対形成相互作用を有する可能性があることが報告されている(Tomashevskaya O.L;Solovyev A.G;Karpova O.V;Fedorkin O.N;Rodionova N.P;Morozov S.Y;Atabekov J.G.、J.Gen.Virol.1993、74:2717〜2724頁)。一部のウイルスリーダーは翻訳エンハンサー活性に関与する配列エレメントを有することが測定され、その配列エレメントには、ジャガイモウイルスS(PVS)様のカルラウイルスリーダーに保存されているCCTTTAGGTT配列(Turner R;Bate N;Tewell D;Foster G.D.、Arch.Virol.1994、134:321〜333頁)及びアルファルファモザイクウイルス(AlMV)のRNA3リーダーにおける27塩基繰り返し領域にみられ、最適な翻訳レベルに達するために2つを必要とするいわゆる内部制御領域2型(ICR2)モチーフ(GGTTCGANTCC)などがある(van der Vossen E.A.G;Neeleman L;Bol J.F.、Nucleic Acids Res.1993、21:1361〜1367頁)。
【0009】
TEVリーダーの場合、それぞれヌクレオチド28〜65及び66〜118のCIRE−1及びCIRE−2と称する2つの領域は、この148bpのウイルスリーダーの翻訳増強性を担っていることが明らかにされた(Niepel M;Gallie D.R.、J.Virol.1999、73:9080〜9088頁)。しかし、CIRE領域内部でこれらのウイルスリーダーのエンハンサー活性に重大とみなされる特異的エレメントは規定されていない。
【発明の開示】
【発明が解決しようとする課題】
【0010】
上に述べたように、天然起源のイントロン及びその隣接配列は、特にそのイントロンが遺伝子の5’末端近くである場合に、種々の遺伝子発現系を増強するためにも広く採用されてきた。しかし、イントロンの起源、エキソンフランキング領域及び細胞の種類のような要因に依存した遺伝子発現のイントロン介在性増強(IME)が報告されている。発現の強いIMEは主に単子葉植物の細胞で観察されたが、双子葉類ではIMEは通常2〜5倍を超えない。IMEの分子メカニズムは完全には開示されていない(Simpson G.G;Filipowicz W.、Plant Mol.Biol.1996、32:1〜41頁;Schuler M.A.1998.「Plant pre−MRNA splicing」、J.Bailey−Serres & D.R.Gallie編、A look beyond transcription:mechanisms determining MRNA stability and translation in plants、American Society of Plant Physiologists、1〜19頁;Lorkovic Z.J;Kirk D.A.W;Lambermon M.H.L;Filipowicz W.、Trends in Plant Science.2000、5:160〜167頁)。
【0011】
単子葉及び双子葉植物の細胞の間で観察された発現のIMEの変動は、異なるクラスの植物細胞における十分なプレmRNAプロセシングに要する要求性の公知の差が原因であり得る。実際に双子葉植物の細胞ではなく単子葉植物の細胞では、イントロン配列における富AUセグメントの存在はプロセシングに欠かせないわけではなく、単子葉植物の細胞は高GC含量(50%を超える)及び複雑な二次構造(ヘアピンループ)を有してイントロンをプロセシングでき、このことは双子葉植物の細胞が複雑な二次構造を有するイントロンをプロセシングできないことを示している(Goodall G.J;Filipowicz W.、The EMBO Journal 1991、10:2635〜2644頁;Lorkovic Z.J;Kirk D.A.W;Lambermon M.H.L;Filipowicz W.、Trends in Plant Science.2000、5:160〜167頁)。これらの理由はヌクレオチド配列の発現を人工的に増強するためにIMEを採用している現行の系がどうしてクラス特異的であるかを少なくとも部分的に説明している。
【課題を解決するための手段】
【0012】
本特許出願において提起された発現プロモーターの配列は、以下の特有の特徴を提供する。1)双子葉及び単子葉植物の細胞において活性であるので、普遍的に機能的であり、高発現レベルの目的遺伝子及び目的DNA配列を有する任意のクラスのトランスジェニック植物を得ることを可能にする、2)人工的に集合させた遺伝子エレメントの組合せに基づき、IMEによるだけでなく翻訳も促進してmRNAレベルを増加させる、3)本プロモーターが天然又はウイルス遺伝子と同一の配列を有する長鎖DNA断片を欠くことは、RNAが介在する相同遺伝子サイレンシングのリスク及びin plantaの相同組換えの結果としての新しいウイルス品種又はウイルス株が出現する可能性を最小限にする、4)TATAボックスから転写開始部位までの配列のGC含量は必ずしも高くなくともよい、5)我々のプロモーター配列の汎用性は5’末端に転写調節エレメントを挿入することを可能にし、このことは発現に時間、器官又は組織特異性を付与する、6)それを有する人工遺伝子エレメントを、植物細胞において活性な任意のプロモーター及び任意のDNA配列の間にも機能的に挿入でき、その転写/翻訳を増加できる。
【発明を実施するための最良の形態】
【0013】
任意のクラスの植物細胞においてmRNAの蓄積を大きく増強する活性を有するキメラエキソン/イントロン/エキソン領域の設計、及びそれの人工翻訳エンハンサーへの機能的組み込みは、本特許出願の2つの本質的要素を構成している。それは、これらのエレメントにより植物細胞に任意の目的DNA配列を効率的に発現することが可能になるからである。
【0014】
我々が領域、分子又はDNA配列が人工又はキメラであると称する場合、我々はそれがin vitroで設計及び合成されたため、その配列の小断片は天然起源を有し得るが、その配列は同一の一次DNA構造を有する遺伝子エレメントではまったくないことを言及していると明らかにしておくことは重要である。
【0015】
発現のIMEを促進できる、対応する隣接エキソン配列を有するイントロンを設計するために、我々はどの配列モチーフ及び遺伝子構成要素が報告された転写エンハンサー活性を有する植物イントロンに共通であるかを研究した。同時に、GC含量には依存せずに双子葉及び単子葉植物においてこのイントロンの十分で効率的なプロセシングを達成するという挑戦を決意しなければならなかった。
【0016】
転写エンハンサーとして広く使用されているプロモーターであるイネアクチン−1、トウモロコシユビキチン−1及びスクロースシンターゼ−1のイントロン配列を比較した場合、それらのすべてに共通の繰り返し配列モチーフを検出できる(図1)。これらのイントロンが付与する遺伝子発現のIMEにこれらのモチーフが果たす責務はまったく実証されないが、これらの領域、及び多様な植物プロモーターにおけるTATAボックスの5’領域においてCTCCモチーフ(又はその相同配列であるCTC、TCC及びTC)の保存レベルが高いことは、それがRNAポリメラーゼII活性を促進できる転写因子の結合に有利であり得るという可能性を示している。同時に、第一エキソン(翻訳されないmRNA非翻訳リーダーとして残る領域)及び報告された翻訳エンハンサー活性を有するウイルスリーダーにおける富C及びA配列の存在度並びに保存は、そのような配列が生じるMRNAの安定性及びそれが翻訳される能力を促進できることを示している。
【0017】
上に説明した理論のうちどれも完全な科学的実証をもたない。このように、特定の繰り返し配列モチーフを含む隣接するエキソン配列を有する人工イントロンの構築は、mRNAの高い転写レベル及び蓄積を促進する領域を生じることは明白ではないが、我々の研究の結果はこれを示していることを強調するのは評価される。
【0018】
(対応するエキソンを有する)異なるイントロンの間のすでに言及した比較研究から、我々が遺伝子発現のIMEに関連性があると考えたモチーフに富むイネアクチン−1及びトウモロコシユビキチン−1のイントロン/エキソン配列断片を組み合わせた人工エキソン/イントロン/エキソン領域を設計することを決意した。この目標を達成するために、双子葉植物の細胞にも遺伝子発現の増加を起こすことができるためには、生じる人工イントロンがこれらのクラスの植物で効率的にプロセシングされなければならないことを考慮に入れなければならなかった。それにもかかわらず、最初の物質として我々が使用したイントロン配列は高いGC含量及び豊富なヘアピンループを有する複雑な二次構造を有し、そのAGアクセプターの3’スプライシング部位の配列は分岐点のコンセンサス配列とは幾分異なることから、これらのイントロンは双子葉植物の細胞でプロセシングが困難なおそれがあることを我々は発見した。
【0019】
我々が設計したエキソン/イントロン/エキソンの二次構造を簡単にしていかなるクラスの植物細胞においてもプロセシングできるようにするために、我々はその配列にいくつかの規則的な変化を作り、配列のプロセシングを活性化するUUUUUAU様配列を挿入することを決意した(Gniadkowski M;Hemmings−Mieszczak M;Klahre U;Liu H.X;Filipowicz W.、Nucleic Acids Res.1996、24:619〜627頁)。さらに、我々のキメラ配列を第二エキソンに融合させ、双子葉類(例えばタバコ)における効率的なプロセシングを利用してトウモロコシアクチン−1遺伝子の第二イントロン(IVS2)に挿入した(Goodall G.J;Filipowicz W.、The EMBO Journal 1991、10:2635〜2644頁)。それぞれの人工エキソン/イントロン/エキソン変異体の推定二次構造をPCFOLD4.0プログラムを用いたコンピュータ法で検討した(Zuker M.、Meth.Enzymology 1989、180:262〜288頁)。我々が創作した人工エキソン/イントロン/エキソン配列を本時点からARTと称する。
【0020】
すでに言及したように、本特許出願の第二の関連構成要素は、遺伝子発現レベルを増加させるためにキメラエキソン/イントロンの下流に融合させた人工翻訳エンハンサーである。
【0021】
いくつかのウイルスのリーダーで形成した配列及び二次構造の分析から人工翻訳エンハンサーを設計した。この分析から我々は翻訳エンハンサーには3つの本質的な構成要素があると結論する:1)複雑性の低い二次構造、2)富C及びA配列セグメント、3)コンセンサスHCAYYY配列(H=C又はU又はA;Y=C又はU、表1参照)と83%までの相同性を有し、露出し、繰り返しが頻繁で、かつ/又は低融解温度の尾部を有するヘアピン構造をとるモチーフ。
【0022】
表1.いくつかのRNAウイルスのリーダー断片において構造が保存された配列(H=C/U/A、Y=C/U)
【表1】
【0023】
上に指摘した前提から、我々はそれぞれヘアピンループ構造をとるHCAYYY様配列を45塩基の富C及びA配列に挿入した人工翻訳エンハンサーを設計した。この人工翻訳エンハンサーは、その調製に採用された理論的前提を提供したRNAウイルスのリーダーと55%を超えない相同性を有し、6個を超えるヌクレオチドと100%の相同性を有する配列セグメントはまったく存在しない。この理由から、我々の翻訳エンハンサーは以前に報告又は保護された(EP0270611)いかなる翻訳エンハンサーの誘導体でもなく、それらから直接誘導された配列も有さないことを我々は断言できる。
【0024】
操作及び目的遺伝子の融合をさらに容易にするために、我々の翻訳エンハンサーに制限部位を付加した。最終的に、我々が創作した人工エキソン/イントロンに新しい翻訳エンハンサーを融合する前に、キメラ遺伝子の発現を増強する能力がTMVオメガ断片と比べて同じであることを示し、そのキメラ遺伝子の機能性はin vivoであった。我々が創作した人工翻訳エンハンサーはEurekaと称する(図2)。
【0025】
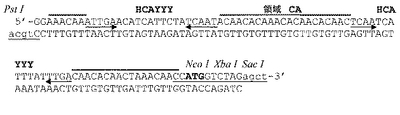
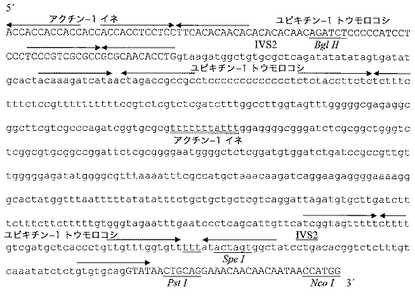
本特許において請求されたプロモーター配列を構築するために、コンセンサスTATAボックスにより形成されるコアプロモーター(Joshi C.P.、Nucleic Acids Res.1987、15:6643〜6653頁)を最初に設計し、それをCaMV 35Sの(転写開始部位から)−24〜−4領域に融合させ、次に転写開始部位並びに富C及びA領域をもたらすアクチン−1の−5〜+27プロモーター領域に融合させた。転写開始部位からの+26〜+72であるトウモロコシユビキチン−1領域を下流に融合させ、富AC領域及び富TC領域をもたらし、第一人工エキソンを得、それをトウモロコシアクチン−1のIVS2イントロンの5’スプライス部位前の12塩基であるトウモロコシアクチン−1第二エキソン及びIVS2イントロン自体と連結した。RNAの成熟に影響し得る推定二次構造を避けるために、コンピュータ法の予測にしたがってこの結合の周辺に塩基の変化、付加又は欠失を作製した。我々が設計した人工イントロンは、IVS2イントロンの最初の54塩基が、転写開始部位からの塩基+89〜+126に対応するトウモロコシユビキチン−1の第一イントロンの5’領域からの37塩基に融合し、次にイネアクチン−1の第一イントロンからの375塩基(この転写開始部位から位置+103〜+477)が続き、トウモロコシユビキチン−1のイントロンの3’末端からの33塩基(転写開始部位から位置+1051〜1083)に融合し、アクチン−1のIVS2イントロンの後半(3’プロセシング開始部位から位置−52〜+5)並びに制限部位及び翻訳開始コンセンサス配列を含む29塩基のキメラ配列と連結したものから構成される(Lutcke H.A;Chow K.C;Mickel F.S;Moss K.A;Kern H.F;Scheele G.A.、The EMBO Journal.1987、6:43〜48頁)。我々が創作した人工エキソン/イントロン/エキソンであるARTの配列を図3に示す。構築された人工エキソン/イントロン/エキソンの効率的なプロセシングを、タバコ細胞及びイネ細胞の両方における一過性発現によりいったん試験してから、その3’末端に翻訳エンハンサーEurekaを融合した。図4に本発明のプロモーター配列物(PARTEプロモーター)の最終構造がみられる。
【0026】
我々が設計したエンハンサーエレメントであるARTは、通常使用されているイネアクチン−1遺伝子の第一エキソン/イントロン/エキソンよりも遺伝子発現エンハンサーとして高い効率を示したことを強調しなければならない。Eureka断片はARTの活性の追加のエンハンサーである。
【0027】
本研究において任意のクラスのトランスジェニック植物細胞において任意のDNA配列の発現を増強する、2つの非常に効率の高い人工遺伝子エレメントを初めて成し遂げることができ、このことは我々が基づいている理論的指針の妥当性を実証している。AT含量が52%を下回る人工プロモーターが双子葉植物の細胞において十分にプロセシングされ、発現の高いIMEを促進することも最初である。ARNウイルスのリーダーとの相同性が低い、完全に人工の、高効率翻訳エンハンサーの構築も新規である。
【0028】
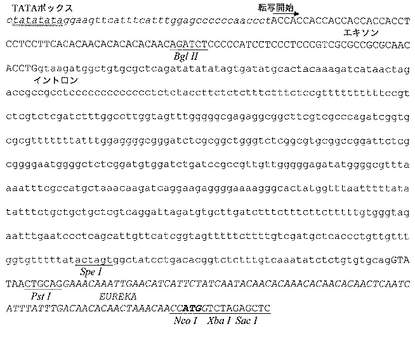
種々の転写エンハンサー配列を本発明のプロモーター配列物の5’に融合した。このように、図5に示すようにイネアクチン−1領域の(転写開始部位から)−43〜−310がプロモーターPARTEの5’末端に融合され、プロモーター領域APARTEが形成し、これにas−1様転写エンハンサーエレメント(Benfey P.N;Chua N.H.、Science、1990、250:959〜966頁)及びトウモロコシユビキチン−1プロモーターの5’領域からの556塩基断片(転写開始部位から−299〜−855)も有する、U3ARTEプロモーターを最終的に得、その構造を図6に示す。
【0029】
記載された遺伝子エレメントから多くのプロモーター配列変異体を構築し(図7参照)、採用されたすべてのエンハンサー領域及びアクチベーター領域の遺伝子発現にわたり相乗効果をもたらすことから、それらのすべてにおいてin vivoアッセイによりその機能性が実証された。
【0030】
転写エンハンサーとして我々の構築物に採用されたas−1エレメント(図6参照)は、オクトピンシンターゼのパリンドロームエンハンサーと50%未満の相同性を有することから革新的な設計を有し(Ellis J.G;Llewellyn D.J;Walker J.C;Dennis E.S;Peacock W.J.、EMBO J.1987、6:3203〜3208頁;EP0278659)、Ellisら(Bouchez D;Tokuhisa J.G;Llewellyn D.J;Dennis E.S;Ellis J.G.、EMBO J.1989、8:4197〜4204頁;米国特許第5837849号)によりなされた研究において請求されたこの種の配列変異体のいずれとも同一でなく(同一性85%未満)、それにみられるTGACGモチーフは独特なフランク配列に関連して見出される。
【0031】
イネアクチン−1遺伝子はすでに記載され、種々の遺伝子の発現に使用されたが(McElroy D;Zhang W;Cao J;Wu R.、Plant Cell 1990、2:163〜171頁;W09109948)、我々の研究はその5’領域の転写エンハンサー活性及び異種発現エンハンサーとしてのその使用を明らかにしていることを強調しておくことが重要である。同様に、トウモロコシユビキチン−1の5’転写エンハンサー領域の使用が請求された(EP0342926)が、我々が採用した556塩基の断片はこのエンハンサーの機能性に必須であると規定された「熱ショック」エレメント(このプロモーター配列の位置−188〜−214の間にみられる)を含まないことから、この断片は独特であり我々が使用したユビキチン配列の転写エンハンサー活性は不要とならない。
【0032】
PARTEプロモーターはイネgluB−1遺伝子の転写開始部位から−31〜−245領域に対応する小さな214塩基断片にも融合され(Takaiwa F;Oono K;Kato A.、Plant Mol.Biol.1991、16:49〜58頁)、新しいプロモーター領域であるGARTE(図8)が形成した。一過性発現アッセイは、この新しい人工プロモーターGARTEが種子内乳に高効率でDNA配列を発現することを実証している。
【0033】
このように、これらのキメラ5’転写エンハンサーの機能的組合せが新規性を有し、植物が属するクラスとは無関係にその植物の細胞におけるDNA配列の高い発現レベルの達成を可能にして、遺伝子エレメントの産生に極めて有用であると結論する。
【0034】
種々のプロモーターからの他の5’調節因子領域を、高い発現レベルを達成するために、かつ/或いは発現に時間特異性、器官特異性又は組織特異性を付与するために本発明の物体に明らかにcys融合できる。
【0035】
遺伝子工学技術の経験者にとっては、本明細書に記載した事柄により、植物細胞においてART及びEurekaエンハンサーを任意の転写プロモーターエレメントと組み合わせて使用して、単子葉又は双子葉植物の細胞において任意のDNA配列の転写/翻訳を増強することが可能となる。同様に分子生物学技術の経験者は、我々の観察から得られたART及びEurekaの設計に我々を導いた理論的指針を採用し、植物細胞において新しいDNA発現エンハンサー配列を構築することができる。
【0036】
過去20年間における植物遺伝子工学の高まる開発を考慮すると、任意の遺伝子及び転写終結配列と連結した本発明のプロモーター物を植物細胞遺伝子形質転換ベクターに挿入でき、確立した効率的な方法の使用により目的遺伝子を発現できるトランスジェニック植物を得ることができることは明白である。
【0037】
本特許出願において、遺伝子形質転換ベクターはそれに予め挿入された任意のDNA断片を植物細胞に導入するための運搬媒体として役立つ(精製されたか、或いは細菌細胞又はウイルス内部に含まれる)DNA分子を称する。
【0038】
微生物の寄託
プラスミドpC−EURGUSint、pC−ARTEGUSint、pGARTEGUSint及びpC−U3ARTEGUSintを微生物保護のためのブダペスト条約に基づいてベルギー共同微生物プラスミド保存機関(the Belgian Coordinated Collection of Microorganism, Plasmid Collection)(BCCM/LMBP)、ゲント大学、「Fiers−Schell−Van Montagu」ビル、Technologiepark927、B−9052 Gent−Zwijnaarde、ベルギーに寄託した。プラスミドpC−EURGUSint、pC−ARTEGUSint、pGARTEGUSintの寄託番号はそれぞれLMBP4727、LMBP4725、LMBP4728、寄託日は2003年5月19日であり、pC−U3ARTEGUSintの寄託番号は4791で寄託日は2003年11月25日である。
【実施例】
【0039】
実施例1:植物細胞にDNA配列を発現させるための新しいキメラ系の構成エレメントの構築
正確なクローニングを容易にするために種々のII型制限エンドヌクレアーゼの制限部位に対する接着末端を有するようにすべての合成DNA断片を創作した。
【0040】
a)Eureka翻訳エンハンサーのクローニング
翻訳エンハンサーEUREKA(配列番号1)に対応する86塩基対(bp)のDNA断片を、合成DNA断片の設計に含めた制限酵素PstI及びSacIに対する接着末端を利用して予め両酵素で分解したベクターpBluescriptII SK(Stratagene、米国)にクローニングした。生じたプラスミドをpBS−Eurekaと名付けた。
【0041】
b)人工エキソン/イントロン/エキソン領域ARTの集合
人工エキソン/イントロン/エキソン領域であるARTを、設計したDNA断片のクローニング及び前後の集合により構築した。まず、コアプロモーター、第一エキソン及び人工イントロンの一部を含むP35AcUと名付けたDNA合成断片(配列番号2)を、EcoRI及びSpeI制限酵素で分解したpBluescriptII SKベクターにクローニングし、プラスミドpBS−AcUを得た。その後に、そのプラスミドをSpeI及びSacIで分解し、人工イントロンの一部をコードするDNA合成断片I−U/Ac(配列番号3)をそれに挿入した。このようにしてpBS−AcUAcプラスミドを得た。
【0042】
次に、人工イントロンの末端を保有するDNA合成断片I−Ac/U(配列番号4)を、制限酵素BamHI及びSacIで分解したpBS−AcUAcプラスミドに挿入し、プラスミドpBS−AcUAcUを産生させた。
【0043】
断片IniT(配列番号5)を、SpeI/SacIで分解したpBS−AcUAcUに挿入したときに、プラスミドpBS−ARTを一致させる人工エキソン/イントロン/エキソンであるARTが完成した(配列番号6)。pBluescriptIIベクターの制限部位EcoRI及びSacIの間の一次構造を配列番号7の配列に示す。
【0044】
c)PARTEプロモーターの構築
本発明のプロモーター配列物(PARTEプロモーター)を構築するために、コアプロモーター及び(3’領域を有さない)エキソン/イントロン/エキソン領域ARTを含むDNA断片を、XhoI/PstI分解によりpBS−ARTプラスミドから得、それを同酵素で分解したpBS−Eurekaプラスミドに挿入した。このように、我々はプラスミドpPARTE(図4、図7D)を実現し、そのEcoRI及びSacI部位の間の配列を配列番号9の配列に示す。
【0045】
実施例2:植物細胞におけるEureka及びARTエンハンサーエレメントの機能性の実証
a)タバコ細胞における翻訳エンハンサーEurekaの機能性
タバコ及びイネ細胞におけるEurekaのエンハンサー力を証明するために、一連の補助遺伝子構築物を作製した。
【0046】
SnaBI部位に挿入されたジャガイモST−LS1遺伝子のIV2イントロンを有するレポーター遺伝子uidA(GUSint)を、プラスミドpUC−GUSintのNcoI/SacI分解により得(図9)、プラスミドpBS−Eurekaの同部位にクローニングし、ベクターPbs−EURGUSintを生成させた。これをさらに制限酵素PstI及びSalIで分解し、S−1ヌクレアーゼで処理してプラスミドpBS−ΔEURGUSintを得た。それをXhoI/KpnIで分解してGUSint遺伝子に融合したEurekaエンハンサーを含むDNA断片を得て、それをpBPFΩ(オメガ)7ベクター(図10)に挿入した。このように、我々はpBPF−EURGUSintベクター(図7A)を得た。そのベクターはCaMV 35Sプロモーター(1.3kb版)、Eurekaエンハンサー及びアグロバクテリウムツメファシエンス(Agrobacterium tumefaciens)のnos遺伝子転写終結シグナル(tNOS)の制御下でGUSint遺伝子を発現する。
【0047】
構築物pBPF−EURGUSintの発現を評価するための対照として、SalI/クレノウ及びKpnI分解によりプラスミドpUC−GUSintから得られたGUSint遺伝子を、pBPFΩ(オメガ)7ベクターのSmaI及びKpnI部位の間にクローニングすることでプラスミドpBPFΩ(オメガ)−GUSintを構築した。このプラスミドは、Eurekaの代わりにGUSintを制御している翻訳エンハンサーΩ(オメガ)が存在することを除き、pBPF−EURGUSintに類似している。別の対照プラスミドを、XhoI−NcoI分解によりプラスミドpBPFΩ(オメガ)−GUSintのエンハンサーオメガを除き、クレノウにより処理し、プラスミドの自己連結を行うことにより構築し、pBPF−GUSintベクターを得た。
【0048】
プラスミドpBPFΩ(オメガ)−GUSint、pBPF−GUSint、及びpBPF−EURGUSintをHindIIIで分解して、植物においてGUSintを発現させるためのカセットを得た。これらをHindIIIで分解したバイナリーベクターpCAMBIA2300にクローニングし、それぞれバイナリーベクターpC−Ω(オメガ)7GUSint、pC−GUSint及びpC−EURGUSintを生成させた。
【0049】
得られたバイナリープラスミドをA.ツメファシエンスLBA4404株に導入し、我々は、Anら(An G.、Plant Physiol.1985、79:568〜570頁)が記載したプロトコルを幾分修正したものにしたがってNT1タバコ細胞において一過性発現実験によりエンハンサーEurekaの機能性のアッセイを始めた。タバコ細胞を、各バイナリーベクターを保有するアグロバクテリウムと4日間共培養した後で、細胞を回収し、Jefferson(Jefferson R.A.1988.「Plant reporter genes:the GUS gene fusion system」、J.K.Setlow編、Genetic Engineering、第10巻、Plenum Publishing Corporation、247〜263頁)が記載したように処理し、β−グルクロニダーゼ(GUS)活性を測定した。各実験を3回繰り返し、各回に各構築物につき5回の繰り返しを行った。結果を以下の表に示す。
【0050】
表2.タバコ細胞における翻訳エンハンサーEurekaの機能性の実証
【表2】
【0051】
表2に示す結果にみられるように、Eurekaのエンハンサー活性はTMVオメガリーダー配列のエンハンサー活性と有意差をもたない。このことは完全に人工の効率的な翻訳エンハンサーを最初に実現したことを実証している。このことは、C及びAに富む配列の領域をモチーフHCAYYY(H=C/T/A、Y=C/T)と相同の配列と組み合わせることにより3’方向下流に融合したDNA配列の翻訳を増強させる重大な性質を有する遺伝子エレメントを構築することが可能であることを示し、我々の理論的指針も確認している。
【0052】
b)タバコ細胞における人工エキソン/イントロン/エキソンARTの機能性
タバコ細胞におけるARTエレメントの機能性を試験するために、プラスミドpUC−GUSintからまずGUSint断片を得、それをNcoI−SacIで分解し、同酵素で分解したpBS−ARTにクローニングしてプラスミドpBS−ARTGUSintを得た。次に、このプラスミドをSalI−BglIIで分解しS1ヌクレアーゼで処理してpBS−ΔARTGUSintを得、それをXhoI−KpnIで分解してARTGUSint断片を得、同酵素で分解したベクターpBPFΩ(オメガ)7−GUSintにクローニングし、プラスミドpBPFARTGUSint(図7B)を得た。ここで、GUSint発現はCaMV 35Sプロモーター(1.3kb版)、人工エキソン/イントロン/エキソン領域であるART、及びA.ツメファシエンスnos遺伝子転写終結シグナル(tNOS)の制御下にある。
【0053】
プラスミドpBS−ΔARTGUSintからXhoI/PstI分解によりエレメントARTを含むバンドを得、それを同酵素で分解したpBS−EURGUSintベクターに融合させプラスミドpBS−ARTEGUSintを得た。これからXhoI/KpnI分解によりGUSint遺伝子を遺伝子エレメントART及びEurekaのシグナル下に置くためのDNA断片を得、同様に分解したpBPFΩ(オメガ)7ベクターに導入しpBPFARTEGUSintプラスミドを産生させた(図7C)。
【0054】
プラスミドpBPFARTGUSint及びpBPFARTEGUSintをHindIIIで分解し、植物にGUSintを発現させるためのカセットを得た。これらをバイナリーベクターpCAMBIA2300にクローニングし、バイナリープラスミドpC−ARTGUSint及びpC−ARTEGUSintをそれぞれ生じさせた。
【0055】
得られたバイナリープラスミドをアグロバクテリウムツメファシエンスLBA4404株に導入した後に、我々は本実施例の(a)項に述べたようにNT1タバコ細胞を用いた一過性発現アッセイにおいて翻訳エンハンサーEurekaの機能性のアッセイを始めた。得られた結果を表3に示す。
【0056】
表3.タバコ細胞における遺伝子エレメントART及びEurekaの機能性の実証
【表3】
【0057】
タバコ細胞におけるARTの機能性評価の結果は、人工イントロンが双子葉植物の細胞において正確にプロセシングされ、発現エンハンサー活性を保持することを明らかにした。我々の実験結果は構築物pBPFARTEGUSintにおけるART及びEurekaエレメントの間の相互作用から得られた発現レベルに及ぼす正の相乗作用も示し、ここで公知の天然プロモーターCaMV 35Sの発現能が5倍に増加する。
【0058】
設計された人工遺伝子エレメント(ART及びEureka)を、植物細胞において活性な任意のプロモーター(この場合CaMV 35Sプロモーター)及び他の任意のDNA配列(我々の場合GUSint)の間に機能的に挿入でき、その転写/翻訳を増加できることも証明される。
【0059】
c)イネ細胞におけるエンハンサーエレメントEureka及びARTの機能性
単子葉植物の細胞における我々の人工エンハンサーの機能性を証明するために新しい構築物のセットを作製した。まず、5’末端を人工エキソン/イントロンARTに融合したuidA(gus)遺伝子を保有するプラスミドpBPFARTGUSintからのXhoI〜KpnI断片を、同制限酵素で分解したpBPFA19−リンカーベクター(図11)に挿入して、プラスミドpBPFA19ARTGUSintを形成させた。このプラスミドにおいて、pBPFA19−リンカーに存在するイネアクチン−1のエキソン/イントロン/エキソン領域は、人工エレメントARTに置き換わり、他の調節エレメントとして(4重のオクトピンシンターゼas−1様エンハンサーはCaMV 35Sプロモーター(400bp版)に融合している)キメラプロモーターA19及びtNOS転写終結シグナルを残す。
【0060】
同様に、プラスミドpBS−ARTEGUSintからのXhoI〜KpnIのバンドをpBPFA19−リンカーベクターにクローニングして構築物pBPFA19ARTEGUSintを得た。対照として使用した構築物pBPFA19GUSintを、プラスミドpUC−GUSintからGUSint断片をNcoI/SacIで分解したpBPFA19−リンカーベクターにクローニングすることにより作製した。使用した別の対照プラスミドはpBPFΩ(オメガ)−GUSintであった。
【0061】
イネ細胞においてART及びEurekaが遺伝子発現を増強する能力の定性評価を、インディカPerla品種のイネのカルスにおける一過性発現をアッセイすることにより実施した。次亜塩素酸ナトリウム及びアルコールで予め滅菌した成熟した種子からカルスを得、遮光してN6−2培地すなわちN6塩類及びビタミン類(Chu C.Cら、Scientia Sinic 1975、18:659頁)、0.1g/Lミオイノシトール、1g/Lカゼイン加水分解物、2mg/L 2,4D、30g/Lスクロース、3g/L Phytagel、pH5.7)中で21日間培養した。マイクロプロジェクタイルを用いたボンバードメント法により形質転換を実施した。ボンバードメントの前に、浸透圧前処理用に0.4Mマンニトールを補充したN6−2培地にカルスを継代した。出版されたプロトコルにしたがってボンバードメントには1μmの球状金粒子(BioRad)をマイクロプロジェクタイルとして使用した(Russell D.R.、Wallace K.M.、Bathe J.H.ら、Plant Cell Rep.1993、12:165〜169頁)。PDS−1000/Heシステム(BioRad)を採用して形質転換を実施した。ボンバードメントには30個のカルスを平板中央に置き、条件は圧力1100psi、距離6cm、平板1枚あたり1回発射した。ボンバードメント後にカルスを遮光下で同じ浸透圧培地中で16時間維持してから、マンニトールを有さないN6−2培地で2日間継代培養した。組織化学法によりX−Glucを用いてGUS活性を明らかにする(Jefferson R.A.1988.「Plant reporter genes:the GUS gene fusion system」、J.K.Setlow編、Genetic Engineering、第10巻、Plenum Publishing Corporation、247〜263頁)。立体顕微鏡で各カルスにおける青い点及び帯域を計数することにより評価を実施した(図12)。表4は繰り返しをそれぞれ3回行った4回の実験から得られた結果を示す。
【0062】
表4.イネ細胞におけるART及びEurekaの機能性の比較実証
【表4】
【0063】
見て取れるように、これらの実験結果は単子葉植物の細胞における遺伝子発現エンハンサーエレメントとしてのART及びEurekaの機能性も確認している。我々のアッセイにおいて人工エキソン/イントロン/エキソンART(pA19ARTGUSint)により発生するIME活性は、容認された遺伝子発現エンハンサー能を有する遺伝子エレメントであるイネアクチン−1遺伝子の第一エキソン/イントロン/エキソン(pBPFA19GUSint)で観察された活性よりも高かったことを強調しておくのは重要である。さらに、我々の実験において構築物pA19ARTGUSint及びpA19ARTEGUSintに対して得られた結果の間の有意差を評価することはできないが、pA19ARTEGUSintのボンバードメントを行ったカルスのX−Gluc組織化学染色後の青く着色した帯域の大きさ及び強度から(図12)、最終構築物中にEureka断片が存在すると発現が大きく増加することは図12において注目に値する。
【0064】
結論として、本実施例は任意の種類の植物細胞における遺伝子発現エンハンサーとしての人工遺伝子エレメントART及びEurekaの機能性を示している。そのうえ、これらのエンハンサーエレメントは効率が高く、それらが融合したプロモーターに依存せずに発現レベルを増加させることが実証された。最後に、ART及びEurekaは下流の遺伝子の発現をなお相乗的に増強させるために組み合わせることができることも実証された。
【0065】
ART及びEurekaを植物に活性な任意のプロモーター(例えばA19)及び任意のDNA配列(GUSint遺伝子)の間に機能的に挿入してその転写/翻訳を増加できることを新たに示す。
【0066】
実施例3:種々の5’転写エンハンサー領域を採用したPARTE発現系変異体
a)PARTEプロモーターへのイネアクチン−1の5’転写アクチベーター領域の付加
イネアクチン−1遺伝子からPARTEプロモーターに5’転写エンハンサー領域をcys−融合するために、pPARTEプラスミドを酵素EcoRI及びEcoRVで分解し、これらの酵素で連結する両極端を有する合成DNA断片En−Ac1(イネアクチン−1遺伝子の転写開始部位から−43〜−221、配列番号10)をそれに挿入した。得られたプラスミドpA1PARTEをEcoRV及びHindIIIで分解して、合成DNA断片En−Ac2(イネアクチン−1の転写開始部位から−226〜−310、配列番号11)を挿入し、アクチン−1遺伝子プロモーターの5’アクチベーター領域を完成させプラスミドpAPARTE(図5、図7E)を産生させた。このプラスミドの制限部位HindIII及びSacIの間のヌクレオチド配列を配列番号12に示す。
【0067】
b)APARTEプロモーターへのas−1様転写エンハンサー配列の付加
プラスミドpA1PARTEをNruI及びSalIで分解し、ASP(配列番号13)と称するDNA合成断片をそれに挿入し、構築物pASPΔA1PARTEを産生させる。ASPは上記の制限酵素に対する接着末端を保有しas−1−様転写エンハンサー配列のために成分化する。このプラスミドをSalIで分解し、S1ヌクレアーゼで処理してからPstIで分解し約900bpのDNA断片を得、それをその後NruI及びPstIで分解したベクターpA1PARTEにクローニングし、最終的にプラスミドpASPA1PARTEを得た。そのプラスミドはイネアクチン−1の5’転写活性化領域のNruI部位に挿入されたASPエンハンサーを有する。それをEcoRV−XhoIで分解することによって、第二のASPエンハンサーをプラスミドpASPA1PARTEに挿入し、ベクターp2A1PARTE(配列番号14、制限部位KpnI及びSacIの間のヌクレオチド配列を示す)を生成させたが、図7Fにその構造を示す。
【0068】
pA1PARTEについて上に解説したように、En−Ac2断片もpASPA1PARTEに挿入し構築物pASPAPARTEを得た。ここでそれをEcoRV−SalIで分解することにより第二ASPエンハンサーを挿入し、最終的にベクターp2APARTE(図7G)を得た。SalI及びSacI部位の間のヌクレオチド配列を配列番号15に示す。
【0069】
c)U3ARTEプロモーターの構築
まず、プライマーOli−U1(配列番号16)及びOli−U2(配列番号17)を用いたポリメラーゼ連鎖反応(PCR)によってトウモロコシubi−1遺伝子の転写開始部位から−299〜−680の領域に対応する約395bpのDNA断片を増幅させた。増幅した断片を制限酵素KpnI及びXhoI(両部位はプライマー内に含まれていた)で分解し、同様にプロセシングしたpBluescriptII SKベクターにクローニングし、構築物pBS−Ubi1を得た。その後、トウモロコシubi−1遺伝子の−680〜−855(配列番号18)のために成文化している(codify)合成DNA断片(En−U2)をNcoI/KpnIで分解したpBS−Ubi1ベクターにクローニングし、構築物pBS−Ubi2を生成させた。その構築物はトウモロコシユビキチン−1遺伝子5’アクチベーター領域(−299〜−855、配列番号19)を含む。
【0070】
トウモロコシユビキチン−1遺伝子からクローニングされた「熱ショック」ボックスを含まない5’転写アクチベーター領域をXhoI−KpnI分解によりプラスミドpBS−Ubi2から得、ベクターのSalI−KpnI分解によりプロモーター2APARTEの5’にcis−挿入し、いわゆる構築物pU3ARTEを得た(図6、図7H)。部位KpnI及びSacIの間のベクターpU3ARTEの配列を配列番号20に示す。
【0071】
d)GARTEプロモーターの構築
本発明物の適応性を実証するために、我々は器官、組織又は発生特異性を有するプロモーターからの5’調節領域を融合させることを決意し、これによりすでに言及した特徴を有する高い発現レベルが付与された。このように、グルテリンの器官特異的発現を制御するイネgluB−1遺伝子5’調節領域をPARTEプロモーターのcys及び5’に融合した。この目標を達成するために、gluB−1転写開始部位から−31〜−245領域(配列番号21)に対応する214塩基対の断片(GLU)及び都合のよいクローニング部位を合成し、EcoRI及びXhoIで分解したpPARTEプラスミドにそれを挿入して、pGARTEベクターを産生させ(図8)、XhoI及びSacI部位の間のその一次構造を配列番号22に示す。
【0072】
実施例4:タバコ及びイネ細胞におけるPARTE発現系の種々の変異体の機能性
単子葉及び双子葉植物の細胞において本発明の各プロモーター変異体物の発現レベルを評価するために、バイナリーベクターpC−ARTE−GUSintにおいてGUSintの発現を制御しているCaMV 35SプロモーターをSmaI−SpeI分解により欠失させ、その場所に構築物pAPARTE、p2A1PARTE、p2APARTE及びpU3ARTEからのKpnI/S1ヌクレアーゼ−SpeI断片を挿入し、新しいバイナリーベクターpC−APARTEGUSint、pC−2A1PARTEGUSint、pC−2APARTEGUSint及びpC−U3ARTEGUSintを産生させた。
【0073】
a)タバコでのアッセイ
バイナリーベクターpC−APARTEGUSint、pC−2A1PARTEGUSint、pC−2APARTEGUSint及びpC−U3ARTEGUSintをA.ツメファシエンス細胞に導入し、実施例2の(a)項に記載したようにNT1タバコ細胞における一過性発現アッセイ実験を実施した。使用した対照プラスミドpC−GUSintは、GUSの発現がCaMV 35Sプロモーター(1.3kb版)及びtNOSターミネーターに制御されている。実験を2回実施し(各処理について5回の繰り返し)、結果を以下の表に示す。
【0074】
表5.タバコ細胞におけるPARTE発現系の種々の変異体の機能性
【表5】
【0075】
我々の結果が本発明物における種々のプロモーター変異体の機能性の確証となり、PARTEに融合した5’転写調節領域に応じてその活性を調整することが可能であることも示していることは明らかである。双子葉植物の細胞において、我々の遺伝子構築物を用いて我々は天然プロモーターCaMV 35Sにより制御されている場合に達成されたものを上回る発現レベルに達したことを強調しなければならない。
【0076】
b)イネを用いたアッセイ
PARTEプロモーターの種々の変異体の活性の一過性評価を実施するために、実施例2(c)項に記載したようにバイナリーベクターpC−APARTEGUSint、pC−2A1PARTEGUSint、pC−2APARTEGUSint及びpC−U3ARTEGUSintを用いてイネカルスにボンバードメントを行った。対照プラスミドpActl−F(McElroy D;Zhang W;Cao J;Wu R.、Plant Cell 1990、2:163〜171頁)は、イネアクチン−1遺伝子プロモーター及びtNOSターミネーターの制御下でgus遺伝子を発現する。発現カセットをKpnI−XbaI分解によりこれらのプラスミドから抽出し、同制限酵素で分解したバイナリープラスミドpCAMBIA2300に挿入し、ベクターpC−Act1Fを産生させた。
【0077】
評価する各構築物について3回の繰り返しでボンバードメント実験を3回実施した。得られた結果を以下の表に示す。
【0078】
表6.イネ細胞におけるPARTEプロモーター発現系の種々の変異体の機能性
【表6】
【0079】
本表に示した結果は、単子葉植物の細胞におけるPARTEプロモーターの種々の変異体がイネアクチン−1遺伝子の天然プロモーターを上回る発現レベルに達することから、それらの変異体の機能性を証明している。したがって、本発明物は、その制御下でcysに配置されたDNA配列の高い発現レベルを達成するための効率の高い遺伝子ツールとしての有用性が確認される。
【0080】
c)イネ種子のアッセイ
イネ細胞内乳におけるGARTEプロモーターの組織特異性を評価するために、GUSint遺伝子に融合しnosターミネーター(tNOS)を有するEureka断片を含む、pBPFARTEGUSintベクターからの約2.5KbのSalI/クレノウ−PstI断片を、XbaI/クレノウ−PstIで分解したpGARTEベクターにクローニングした結果、構築物pGARTEGUSintが生じた。
【0081】
pGARTEGUSintにおけるGARTEプロモーターをXhoI−NcoI分解することにより、pGEM−T−GluB−1プラスミドのSalI−NcoI分解から得られた種子内乳特異的で高効率のイネグルテリンB−1プロモーターに置き換えた対照プラスミドも構築した。このように、我々はpGluGUSintを得た。
【0082】
GARTEプロモーターの評価及びGluB1のプロモーターとの比較を、Y−S.Hwang(Hwang Y−S;McCullar Cass;Huang N.、Plant Science.2001、161:1107〜1116頁)にしたがって、温室栽培されたイネ品種インディカPerlaの穂の穎果(ear cariopsis)から単離した未熟な内乳(受粉後8〜9日)にボンバードメントを行うことによって実施した。内乳におけるGUS活性を測定するための蛍光アッセイを、Jefferson(Jefferson R.A.1988.「Plant reporter genes:the GUS gene fusion system」、J.K.Setlow(編)、Genetic Engineering、第10巻、Plenum Publishing Corporation、247〜263頁)にしたがって実施し、プラスミドDNAで覆った金微粒子を用いたボンバードメントを行った24時間後に評価した。それぞれ5回の繰り返しを行った独立した2回の実験で得られたGUS活性の結果を表7に示す。
【0083】
表7.イネ種子内乳におけるGARTEプロモーターの機能性
【表7】
【0084】
これらの結果は、我々が設計した人工エレメントに基づくキメラプロモーターGARTEが種子内乳において遺伝子を発現する効率が高いことを確認している。GARTEプロモーターに挿入されたGLU配列は発現に特異性を付与できるが、それは「それ自体は」高レベルを保証せず、プロモーターを一致させる他のエレメントにも依存する(Takaiwa F;Yamanouchi U;Yoshihara T;Washida H;Tanabe F;Kato A;Yamada K.、Plant Mol Biol.1996、30:1207〜1221頁)。
【0085】
示された結果は、本発明のエレメント物上流に調節領域を挿入すると、発生、器官又は組織特異性を有する任意のDNA配列の発現を効率的に導くためにそのエレメント物を使用できることを再び支持している。分子生物学の経験者には、GARTEプロモーターに挿入されたGluB−1プロモーター調節配列を、生物によるストレス(例えば病原体の攻撃)、非生物因子(例えば創傷、極端な高温又は低温、塩分、乾燥、一部の化学物質の存在)、酸化ストレス、種々の器官及び組織発生段階などに応答する調節配列とうまく置き換えられることは明白である。
【0086】
本発明の調節領域物下でクローニングされたDNA配列を植物細胞に導入でき、公知の生物学的又は物理化学的形質転換法により安定挿入できること、並びにこれらの遺伝修飾された細胞から、DNA配列が融合したプロモーター変異体にしたがってその配列が都合よく発現する受精能のある植物を再生させることが可能であることも明らかである。このように、本発明は害虫、疾病、多様なストレスに対する抵抗レベルがさらに大きく、農業の収穫量がさらに大きいか、又は使用の中でとりわけ医学又は工業応用を有する化合物を高効率で産生するトランスジェニック植物の製造に貢献する潜在性を示している。
【図面の簡単な説明】
【0087】
【図1】転写開始部位からのイネアクチン−1(Act)、トウモロコシユビキチン−1(Ubi)及びトウモロコシスクロースシンターゼ(Shrun)遺伝子配列を示す図である。大文字は第一エキソンを、小文字は第一イントロンの5’領域を示し、繰り返し共通配列モチーフの部分に下線を付す。
【図2】Eureka人工翻訳エンハンサー配列を示す図であり、関連のあるエレメント及び制限エンドヌクレアーゼ認識部位を示す。
【図3】ARTエキソン/イントロン/エキソン人工配列を示す図であり、その構成要素の断片のそれぞれの起点を示す(小文字:人工イントロン、UUUUUAU様配列を作り出すために挿入した塩基に下線2本を付す、制限エンドヌクレアーゼのいくつかの関連性のある認識部位に下線1本で印を付ける)。
【図4】本発明物(PARTEプロモーター)の一次構造を示す図であり、ARTエキソン/イントロン/エキソン領域(イントロン塩基は小文字、エキソンは大文字)及び人工翻訳エンハンサーEUREKAに融合したコアプロモーター(斜体小文字)を示す。制限エンドヌクレアーゼのいくつかの関連性のある認識部位に下線を付し、TATAボックスに下線2本を付し、翻訳開始コドンを太字で示す。
【図5】APARTEプロモーターの一次構造を示す図であり、PARTEプロモーター(プロモーターは斜体小文字、イントロンは小文字、エキソンは大文字)に融合したイネアクチン−1の5’調節領域(斜体大文字で転写開始部位から−43〜−310領域)を示す。制限エンドヌクレアーゼのいくつかの関連性のある認識部位に下線1本で印を付す。TATAボックスに下線2本を付し、翻訳開始コドンを太字で示す。
【図6】U3ARTEプロモーターの一次構造を示す図であり、その構成エレメントであるトウモロコシubi−1遺伝子の転写開始部位から−299〜−855領域を大文字で、as−1様転写エンハンサーを太字大文字で、イネact−1遺伝子の転写開始部位から−43〜−310領域を斜体大文字で、PARTEプロモーターを小文字で示す(TATAボックスに下線2本を付し、ARTイントロンを斜体で示し、翻訳開始部位に下線1本を付す)。
【図7】本発明のエンハンサーエレメント物を有するプロモーター変異体を示す図である。A:35SEureka; B:35SART; C:35SARTE; D:PARTE; E:APARTE; F:2A1PARTE; G:2APARTE; H:U3ARTE. れんが模様部分:35S プロモーター(1.3Kb); チェック柄部分:翻訳エンハンサー Eureka; 縦縞模様部分:ART エキソン/イントロン/エキソン; 格子柄部分:人工プロモーターコア; 白抜き部分:イネアクチン−1 遺伝子5’活性化領域(−43〜−221); 点(疎)模様部分:イネアクチン−1 遺伝子5’活性化領域(−226〜−310); 斜め縞模様部分:ASP(as−1−様エンハンサー); 点(密)模様部分:トウモロコシユビキチン−1 プロモーター5’活性化領域(−299〜−855)。
【図8】GARTEプロモーターの一次構造を示す図であり、その構成エレメントである、転写開始部位から−31〜−245のイネgluB−1遺伝子領域を斜体大文字で示す、PARTEプロモーター(プロモーターを斜体小文字で、イントロンを小文字で、エキソンを大文字で示し、いくつかの関連性のある制限部位に下線を付し、TATAボックスに下線2本を付し、翻訳開始コドンは太字)を示す。
【図9】pUC−GUSintの地図を示す図である。
【図10】pBPFΩ(オメガ)7の地図を示す図である。
【図11】pBPFA19−リンカーの地図を示す図である。
【図12】促進したマイクロプロジェクタイルを用いたボンバードメントにより誘導されたGUSint遺伝子を保有する種々の遺伝子構築物の一過性発現によりイネ細胞におけるART及びEUREKAエレメントの機能性のX−Gluc組織化学染色による比較実証を示す図である。
【特許請求の範囲】
【請求項1】
組換えDNA分子であることを特徴とする人工プロモーターであって、前記プロモーターが植物細胞においてその3’末端に融合した任意のDNA配列の発現を促進し、
a)5’転写調節エレメント、
b)それに続くTATAボックス、GC含量64%未満を有するヌクレオチド配列、及び転写開始部位を含む人工コアプロモーター、並びに
c)第一キメラエキソン、人工イントロン、及び第二キメラエキソンにより一致される、下流に挿入された任意の遺伝子の翻訳増強性を有する転写可能であるが翻訳可能ではない合成ヌクレオチド配列、
を含み、前記転写開始部位の3’末端が前記合成ヌクレオチド配列に融合し、前記合成イントロンが植物細胞においてそれに融合した遺伝子の発現を増強できるプロモーター。
【請求項2】
前記5’転写調節エレメントが人工である、組換えDNA分子であることを特徴とする請求項1記載の人工プロモーター。
【請求項3】
前記5’転写調節エレメントが植物細胞において遺伝子発現を自然に増強及び/又は調節するDNA配列と相同である、組換えDNA分子であることを特徴とする請求項1記載の人工プロモーター。
【請求項4】
5’転写調節エレメントがイネアクチン−1遺伝子に由来する請求項3記載の人工プロモーター。
【請求項5】
前記5’転写調節エレメントがイネアクチン−1遺伝子の転写開始部位から−43から−310の領域を含む請求項4記載の人工プロモーター。
【請求項6】
前記5’転写調節エレメントのヌクレオチド配列が配列番号10における配列又はその断片に対応する請求項5記載の人工プロモーター。
【請求項7】
前記5’転写調節エレメントのヌクレオチド配列が配列番号11における配列又はその断片に対応する請求項5記載の人工プロモーター。
【請求項8】
前記5’転写調節エレメントがトウモロコシユビキチン−1遺伝子に由来する請求項3記載の人工プロモーター。
【請求項9】
ヌクレオチド配列がトウモロコシユビキチン−1遺伝子の転写開始部位から−299から−855の領域を含む請求項8記載の人工プロモーター。
【請求項10】
前記5’転写調節エレメントが配列番号19における配列又はその断片に対応する請求項9記載の人工プロモーター。
【請求項11】
前記5’転写調節エレメントがas−1様転写エンハンサーである請求項2又は3に記載の人工プロモーター。
【請求項12】
as−1様転写エンハンサーのヌクレオチド配列が配列番号13におけるヌクレオチド7から26、又はその相補的配列に対応する配列断片と本質的に同一である、請求項11記載の人工プロモーター
【請求項13】
前記5’転写調節エレメントがウイルスプロモーターに由来する請求項3記載の人工プロモーター。
【請求項14】
前記5’転写調節エレメントがCaMV 35Sプロモーターに由来する請求項13記載の人工プロモーター。
【請求項15】
前記5’転写調節エレメントが発生特異性、器官特異性又は組織特異性を有して植物細胞における遺伝子発現を制御する請求項2又は3に記載の人工プロモーター。
【請求項16】
前記5’転写調節エレメントが種子における発現を制御する請求項15記載の人工プロモーター。
【請求項17】
前記5’転写調節エレメントがイネグルテリンB−1遺伝子に由来する請求項16記載の人工プロモーター。
【請求項18】
前記5’転写調節エレメントがイネグルテリンB−1遺伝子の転写開始部位から−31から−245の領域の断片を含む請求項17記載の人工プロモーター。
【請求項19】
前記5’転写調節エレメントが配列番号21における配列又はその断片に対応する請求項18記載の人工プロモーター。
【請求項20】
前記5’転写調節エレメントが生物又は非生物ストレス下にある植物細胞における遺伝子発現を制御する請求項15記載の人工プロモーター。
【請求項21】
前記5’転写調節エレメントが傷を有する植物組織における遺伝子発現を制御する請求項15記載の人工プロモーター。
【請求項22】
5’転写調節領域が異なる起源からの機能できる形で融合した2つ以上の調節エレメントを含み、前記調節エレメントが請求項2から21までに記載の性質のいずれかに対して個別に応答する請求項1記載の人工プロモーター。
【請求項23】
前記人工エキソン/イントロン/エキソン領域からの第一エキソンがC及びAリッチな(rich)配列モチーフを含む請求項1記載の人工プロモーター。
【請求項24】
前記人工エキソン/イントロン/エキソン領域からの第一エキソンが、モチーフCTCC並びに/又はその相同配列CTC、TCC及びTCの頻繁に繰り返す配列を含む請求項1記載の人工プロモーター。
【請求項25】
前記人工エキソン/イントロン/エキソン領域からのイントロンが、CTCCモチーフ並びに/又はその相同配列CTC、TCC及びTCの頻繁に繰り返す配列を含む請求項1記載の人工プロモーター。
【請求項26】
前記人工エキソン/イントロン/エキソン領域からのヌクレオチド配列が、配列番号6又はその断片に対応する請求項23〜25のいずれか一項に記載の人工プロモーター。
【請求項27】
前記人工エキソン/イントロン/エキソン領域からの第二エキソンが高いC及びA含量を有する配列モチーフを含む請求項1記載の人工プロモーター。
【請求項28】
前記人工エキソン/イントロン/エキソン領域からの第二エキソンがモチーフHCAYYY(H=C又はT又はA、Y=C又はT)と少なくとも83%相同の配列を含む請求項1記載の人工プロモーター。
【請求項29】
前記人工エキソン/イントロン/エキソン領域からの第二エキソンのヌクレオチド配列が配列番号1の配列に対応する請求項27又は28に記載の人工プロモーター。
【請求項30】
前記人工エキソン/イントロン/エキソン領域のヌクレオチド配列が配列番号8又はその断片に対応する請求項23〜29のいずれか一項に記載の人工プロモーター。
【請求項31】
前記人工エキソン/イントロン/エキソン領域のヌクレオチド配列が少なくとも部分的に配列番号20の配列に対応する請求項1〜30のいずれか一項に記載の人工プロモーター。
【請求項32】
植物において機能的な任意のプロモーターに融合した場合に前記プロモーターにより制御されるDNA配列の発現を増強するように貢献する請求項1〜31のいずれか一項に記載の人工プロモーターからのDNA断片。
【請求項33】
人工プロモーターの断片であって、前記断片に融合した遺伝子の翻訳を増強できる、請求項32記載の断片。
【請求項34】
モチーフHCAYYY(H=C又はT又はA、Y=C又はT)と少なくとも83%相同の配列を含む請求項33記載の人工プロモーターの断片。
【請求項35】
C及びAリッチな(rich)配列モチーフを有する請求項33記載の人工プロモーターの断片。
【請求項36】
ヌクレオチド配列が配列番号1における配列に対応する請求項33記載の人工プロモーターの断片。
【請求項37】
植物細胞においてCaMV 35Sプロモーターから産生されるmRNAの翻訳の増強に貢献する請求項33〜36のいずれか一項に記載の人工プロモーターの断片。
【請求項38】
エキソン/イントロン/エキソン領域に対応する請求項32記載の人工プロモーターの断片。
【請求項39】
第一エキソンがC及びAリッチな(rich)配列モチーフを含む請求項38記載の人工プロモーターの断片。
【請求項40】
前記第一エキソンがモチーフCTCC並びに/又はその相同配列CTC、TCC及びTCの頻繁に繰り返す配列を含む請求項38記載の人工プロモーターの断片。
【請求項41】
前記イントロンがモチーフCTCC並びに/又はその相同配列CTC、TCC及びTCの頻繁に繰り返す配列を含む請求項38記載の人工プロモーターの断片。
【請求項42】
ヌクレオチド配列が配列番号6における配列に対応する請求項38記載の人工プロモーターの断片。
【請求項43】
ヌクレオチド配列が配列番号8における配列に対応する請求項38記載の人工プロモーターの断片。
【請求項44】
植物細胞においてCaMV 35Sプロモーターの制御下で任意の遺伝子の発現の増強に貢献する請求項38〜43のいずれか一項に記載の人工プロモーターの断片。
【請求項45】
as−1様転写エンハンサーに対応する請求項32記載の人工プロモーターの断片。
【請求項46】
ヌクレオチド配列が配列番号13におけるヌクレオチド7から26又はその相補的配列に対応する断片の配列と本質的に同一である人工プロモーターの断片。
【請求項47】
5’転写調節エレメントに対応する請求項32記載の人工プロモーターの断片。
【請求項48】
前記5’転写調節エレメントがイネアクチン−1遺伝子に由来する請求項47記載の人工プロモーターの断片。
【請求項49】
ヌクレオチド配列がイネアクチン−1遺伝子の転写開始部位から−43から−310の断片を含む請求項48記載の人工プロモーターの断片。
【請求項50】
ヌクレオチド配列が配列番号10における配列又はその断片に対応する請求項49記載の人工プロモーターの断片。
【請求項51】
ヌクレオチド配列が配列番号11における配列又はその断片に対応する請求項49記載の人工プロモーターの断片。
【請求項52】
前記5’転写調節エレメントがトウモロコシユビキチン−1遺伝子に由来する請求項47記載の人工プロモーターの断片。
【請求項53】
ヌクレオチド配列がトウモロコシユビキチン−1遺伝子の転写開始部位から−299から−855の領域を含む請求項52記載の人工プロモーターの断片。
【請求項54】
前記5’転写調節エレメントが配列番号19における配列又はその断片に対応する請求項53記載の人工プロモーターの断片。
【請求項55】
請求項1から31までのいずれかに対応する人工プロモーターを含む、植物細胞におけるDNA配列の発現のためのカセット。
【請求項56】
請求項32から54までのいずれかに対応するDNA断片に機能的に融合した転写エンハンサーエレメントを含む、植物細胞におけるDNA配列の発現のためのカセット。
【請求項57】
請求項55又は56に記載の発現カセットの1つを含む、植物細胞の形質転換のためのDNAベクター。
【請求項58】
請求項57記載のベクターを保有する細菌細胞及びその子孫。
【請求項59】
請求項57記載のベクターで形質転換された植物細胞及びその子孫。
【請求項60】
請求項57記載のベクターにより導入された前記発現カセットにおいて人工プロモーターの制御下でDNA断片を発現する請求項59記載の植物細胞。
【請求項61】
請求項55又は56に記載の発現カセットがゲノムに完全に組み込まれた請求項59記載の植物細胞。
【請求項62】
請求項61記載の植物細胞から再生したトランスジェニック植物。
【請求項63】
請求項57に記載したベクターにより導入された発現カセットに含まれる人工プロモーターの制御下でDNA断片を発現する請求項62記載のトランスジェニック植物。
【請求項64】
請求項63記載のトランスジェニック植物の子孫。
【請求項65】
双子葉類である請求項64記載の植物。
【請求項66】
ナス科(Solanaceae)である請求項65記載の植物。
【請求項67】
タバコ、トマト又はジャガイモのうち1つの種に属する請求項66記載の植物。
【請求項68】
単子葉類である請求項64記載の植物。
【請求項69】
イネ科(Gramineae)である請求項68記載の植物。
【請求項70】
イネ、サトウキビ、トウモロコシ、コムギ又はオオムギのうちの1つの種に属する請求項69記載の植物。
【請求項71】
請求項57記載のベクターにより導入された発現カセットに含まれる人工プロモーターの制御下に位置するDNA断片の発現の結果としての、請求項60又は63に記載の細胞又は植物により産生される組換えタンパク質の精製又は使用。
【請求項1】
組換えDNA分子であることを特徴とする人工プロモーターであって、前記プロモーターが植物細胞においてその3’末端に融合した任意のDNA配列の発現を促進し、
a)5’転写調節エレメント、
b)それに続くTATAボックス、GC含量64%未満を有するヌクレオチド配列、及び転写開始部位を含む人工コアプロモーター、並びに
c)第一キメラエキソン、人工イントロン、及び第二キメラエキソンにより一致される、下流に挿入された任意の遺伝子の翻訳増強性を有する転写可能であるが翻訳可能ではない合成ヌクレオチド配列、
を含み、前記転写開始部位の3’末端が前記合成ヌクレオチド配列に融合し、前記合成イントロンが植物細胞においてそれに融合した遺伝子の発現を増強できるプロモーター。
【請求項2】
前記5’転写調節エレメントが人工である、組換えDNA分子であることを特徴とする請求項1記載の人工プロモーター。
【請求項3】
前記5’転写調節エレメントが植物細胞において遺伝子発現を自然に増強及び/又は調節するDNA配列と相同である、組換えDNA分子であることを特徴とする請求項1記載の人工プロモーター。
【請求項4】
5’転写調節エレメントがイネアクチン−1遺伝子に由来する請求項3記載の人工プロモーター。
【請求項5】
前記5’転写調節エレメントがイネアクチン−1遺伝子の転写開始部位から−43から−310の領域を含む請求項4記載の人工プロモーター。
【請求項6】
前記5’転写調節エレメントのヌクレオチド配列が配列番号10における配列又はその断片に対応する請求項5記載の人工プロモーター。
【請求項7】
前記5’転写調節エレメントのヌクレオチド配列が配列番号11における配列又はその断片に対応する請求項5記載の人工プロモーター。
【請求項8】
前記5’転写調節エレメントがトウモロコシユビキチン−1遺伝子に由来する請求項3記載の人工プロモーター。
【請求項9】
ヌクレオチド配列がトウモロコシユビキチン−1遺伝子の転写開始部位から−299から−855の領域を含む請求項8記載の人工プロモーター。
【請求項10】
前記5’転写調節エレメントが配列番号19における配列又はその断片に対応する請求項9記載の人工プロモーター。
【請求項11】
前記5’転写調節エレメントがas−1様転写エンハンサーである請求項2又は3に記載の人工プロモーター。
【請求項12】
as−1様転写エンハンサーのヌクレオチド配列が配列番号13におけるヌクレオチド7から26、又はその相補的配列に対応する配列断片と本質的に同一である、請求項11記載の人工プロモーター
【請求項13】
前記5’転写調節エレメントがウイルスプロモーターに由来する請求項3記載の人工プロモーター。
【請求項14】
前記5’転写調節エレメントがCaMV 35Sプロモーターに由来する請求項13記載の人工プロモーター。
【請求項15】
前記5’転写調節エレメントが発生特異性、器官特異性又は組織特異性を有して植物細胞における遺伝子発現を制御する請求項2又は3に記載の人工プロモーター。
【請求項16】
前記5’転写調節エレメントが種子における発現を制御する請求項15記載の人工プロモーター。
【請求項17】
前記5’転写調節エレメントがイネグルテリンB−1遺伝子に由来する請求項16記載の人工プロモーター。
【請求項18】
前記5’転写調節エレメントがイネグルテリンB−1遺伝子の転写開始部位から−31から−245の領域の断片を含む請求項17記載の人工プロモーター。
【請求項19】
前記5’転写調節エレメントが配列番号21における配列又はその断片に対応する請求項18記載の人工プロモーター。
【請求項20】
前記5’転写調節エレメントが生物又は非生物ストレス下にある植物細胞における遺伝子発現を制御する請求項15記載の人工プロモーター。
【請求項21】
前記5’転写調節エレメントが傷を有する植物組織における遺伝子発現を制御する請求項15記載の人工プロモーター。
【請求項22】
5’転写調節領域が異なる起源からの機能できる形で融合した2つ以上の調節エレメントを含み、前記調節エレメントが請求項2から21までに記載の性質のいずれかに対して個別に応答する請求項1記載の人工プロモーター。
【請求項23】
前記人工エキソン/イントロン/エキソン領域からの第一エキソンがC及びAリッチな(rich)配列モチーフを含む請求項1記載の人工プロモーター。
【請求項24】
前記人工エキソン/イントロン/エキソン領域からの第一エキソンが、モチーフCTCC並びに/又はその相同配列CTC、TCC及びTCの頻繁に繰り返す配列を含む請求項1記載の人工プロモーター。
【請求項25】
前記人工エキソン/イントロン/エキソン領域からのイントロンが、CTCCモチーフ並びに/又はその相同配列CTC、TCC及びTCの頻繁に繰り返す配列を含む請求項1記載の人工プロモーター。
【請求項26】
前記人工エキソン/イントロン/エキソン領域からのヌクレオチド配列が、配列番号6又はその断片に対応する請求項23〜25のいずれか一項に記載の人工プロモーター。
【請求項27】
前記人工エキソン/イントロン/エキソン領域からの第二エキソンが高いC及びA含量を有する配列モチーフを含む請求項1記載の人工プロモーター。
【請求項28】
前記人工エキソン/イントロン/エキソン領域からの第二エキソンがモチーフHCAYYY(H=C又はT又はA、Y=C又はT)と少なくとも83%相同の配列を含む請求項1記載の人工プロモーター。
【請求項29】
前記人工エキソン/イントロン/エキソン領域からの第二エキソンのヌクレオチド配列が配列番号1の配列に対応する請求項27又は28に記載の人工プロモーター。
【請求項30】
前記人工エキソン/イントロン/エキソン領域のヌクレオチド配列が配列番号8又はその断片に対応する請求項23〜29のいずれか一項に記載の人工プロモーター。
【請求項31】
前記人工エキソン/イントロン/エキソン領域のヌクレオチド配列が少なくとも部分的に配列番号20の配列に対応する請求項1〜30のいずれか一項に記載の人工プロモーター。
【請求項32】
植物において機能的な任意のプロモーターに融合した場合に前記プロモーターにより制御されるDNA配列の発現を増強するように貢献する請求項1〜31のいずれか一項に記載の人工プロモーターからのDNA断片。
【請求項33】
人工プロモーターの断片であって、前記断片に融合した遺伝子の翻訳を増強できる、請求項32記載の断片。
【請求項34】
モチーフHCAYYY(H=C又はT又はA、Y=C又はT)と少なくとも83%相同の配列を含む請求項33記載の人工プロモーターの断片。
【請求項35】
C及びAリッチな(rich)配列モチーフを有する請求項33記載の人工プロモーターの断片。
【請求項36】
ヌクレオチド配列が配列番号1における配列に対応する請求項33記載の人工プロモーターの断片。
【請求項37】
植物細胞においてCaMV 35Sプロモーターから産生されるmRNAの翻訳の増強に貢献する請求項33〜36のいずれか一項に記載の人工プロモーターの断片。
【請求項38】
エキソン/イントロン/エキソン領域に対応する請求項32記載の人工プロモーターの断片。
【請求項39】
第一エキソンがC及びAリッチな(rich)配列モチーフを含む請求項38記載の人工プロモーターの断片。
【請求項40】
前記第一エキソンがモチーフCTCC並びに/又はその相同配列CTC、TCC及びTCの頻繁に繰り返す配列を含む請求項38記載の人工プロモーターの断片。
【請求項41】
前記イントロンがモチーフCTCC並びに/又はその相同配列CTC、TCC及びTCの頻繁に繰り返す配列を含む請求項38記載の人工プロモーターの断片。
【請求項42】
ヌクレオチド配列が配列番号6における配列に対応する請求項38記載の人工プロモーターの断片。
【請求項43】
ヌクレオチド配列が配列番号8における配列に対応する請求項38記載の人工プロモーターの断片。
【請求項44】
植物細胞においてCaMV 35Sプロモーターの制御下で任意の遺伝子の発現の増強に貢献する請求項38〜43のいずれか一項に記載の人工プロモーターの断片。
【請求項45】
as−1様転写エンハンサーに対応する請求項32記載の人工プロモーターの断片。
【請求項46】
ヌクレオチド配列が配列番号13におけるヌクレオチド7から26又はその相補的配列に対応する断片の配列と本質的に同一である人工プロモーターの断片。
【請求項47】
5’転写調節エレメントに対応する請求項32記載の人工プロモーターの断片。
【請求項48】
前記5’転写調節エレメントがイネアクチン−1遺伝子に由来する請求項47記載の人工プロモーターの断片。
【請求項49】
ヌクレオチド配列がイネアクチン−1遺伝子の転写開始部位から−43から−310の断片を含む請求項48記載の人工プロモーターの断片。
【請求項50】
ヌクレオチド配列が配列番号10における配列又はその断片に対応する請求項49記載の人工プロモーターの断片。
【請求項51】
ヌクレオチド配列が配列番号11における配列又はその断片に対応する請求項49記載の人工プロモーターの断片。
【請求項52】
前記5’転写調節エレメントがトウモロコシユビキチン−1遺伝子に由来する請求項47記載の人工プロモーターの断片。
【請求項53】
ヌクレオチド配列がトウモロコシユビキチン−1遺伝子の転写開始部位から−299から−855の領域を含む請求項52記載の人工プロモーターの断片。
【請求項54】
前記5’転写調節エレメントが配列番号19における配列又はその断片に対応する請求項53記載の人工プロモーターの断片。
【請求項55】
請求項1から31までのいずれかに対応する人工プロモーターを含む、植物細胞におけるDNA配列の発現のためのカセット。
【請求項56】
請求項32から54までのいずれかに対応するDNA断片に機能的に融合した転写エンハンサーエレメントを含む、植物細胞におけるDNA配列の発現のためのカセット。
【請求項57】
請求項55又は56に記載の発現カセットの1つを含む、植物細胞の形質転換のためのDNAベクター。
【請求項58】
請求項57記載のベクターを保有する細菌細胞及びその子孫。
【請求項59】
請求項57記載のベクターで形質転換された植物細胞及びその子孫。
【請求項60】
請求項57記載のベクターにより導入された前記発現カセットにおいて人工プロモーターの制御下でDNA断片を発現する請求項59記載の植物細胞。
【請求項61】
請求項55又は56に記載の発現カセットがゲノムに完全に組み込まれた請求項59記載の植物細胞。
【請求項62】
請求項61記載の植物細胞から再生したトランスジェニック植物。
【請求項63】
請求項57に記載したベクターにより導入された発現カセットに含まれる人工プロモーターの制御下でDNA断片を発現する請求項62記載のトランスジェニック植物。
【請求項64】
請求項63記載のトランスジェニック植物の子孫。
【請求項65】
双子葉類である請求項64記載の植物。
【請求項66】
ナス科(Solanaceae)である請求項65記載の植物。
【請求項67】
タバコ、トマト又はジャガイモのうち1つの種に属する請求項66記載の植物。
【請求項68】
単子葉類である請求項64記載の植物。
【請求項69】
イネ科(Gramineae)である請求項68記載の植物。
【請求項70】
イネ、サトウキビ、トウモロコシ、コムギ又はオオムギのうちの1つの種に属する請求項69記載の植物。
【請求項71】
請求項57記載のベクターにより導入された発現カセットに含まれる人工プロモーターの制御下に位置するDNA断片の発現の結果としての、請求項60又は63に記載の細胞又は植物により産生される組換えタンパク質の精製又は使用。
【図1】


【図2】
【図3】
【図4】
【図5】

【図6】

【図7】
【図8】

【図9】
【図10】
【図11】
【図12】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2006−512067(P2006−512067A)
【公表日】平成18年4月13日(2006.4.13)
【国際特許分類】
【出願番号】特願2004−562477(P2004−562477)
【出願日】平成15年12月19日(2003.12.19)
【国際出願番号】PCT/CU2003/000018
【国際公開番号】WO2004/058979
【国際公開日】平成16年7月15日(2004.7.15)
【出願人】(304012895)セントロ デ インジエニエリア ジエネテイカ イ バイオテクノロジア (46)
【Fターム(参考)】
【公表日】平成18年4月13日(2006.4.13)
【国際特許分類】
【出願日】平成15年12月19日(2003.12.19)
【国際出願番号】PCT/CU2003/000018
【国際公開番号】WO2004/058979
【国際公開日】平成16年7月15日(2004.7.15)
【出願人】(304012895)セントロ デ インジエニエリア ジエネテイカ イ バイオテクノロジア (46)
【Fターム(参考)】
[ Back to top ]