
植物細胞における高効率ペプチド生産
本発明は、組換えペプチドの生産のための改善された方法を提供する。特に、本発明は、植物細胞懸濁培養においてインビボで集合され得るウイルスカプシド融合タンパク質の形態の組換えペプチドの生産のための改善された方法を提供する。本発明はまた、非感染性ウイルスカプシド融合ペプチド生産を可能にするプラスミド、配列及び植物細胞を含む。

【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の相互参照)
本出願は、「植物細胞における高効率ペプチド生産」と題する、2004年2月27日出願の米国特許仮出願第60/548,744号の優先権を主張する。
【0002】
(政府権益の言明)
本出願は、国立衛生研究所、国立アレルギー及び感染症研究所(NIAID)、共同契約第1−U01−AI054641−01号との米国政府契約下にある。
【0003】
(技術分野)
本発明は、組換えペプチドの生産のための改善された方法を提供する。特に、本発明は、植物細胞懸濁培養においてインビボで集合され得るウイルスカプシド融合タンパク質の形態の組換えペプチドの生産のための改善された方法を提供する。本発明はまた、非感染性ウイルスカプシド融合ペプチド生産を可能にするプラスミド、配列及び植物細胞を含む。
【背景技術】
【0004】
細菌、酵母、植物、昆虫及び哺乳動物細胞発現系が、組換えペプチドを生産するために現在使用されており、成功の程度は様々である。異種ペプチドの生産のための発現系を創造する上での1つのゴールは、商業、治療及びワクチン適用において利用できる、広範囲で、柔軟性があり、効率的、経済的及び実用的なプラットフォームと方法を提供することである。例えばある種のペプチドの生産のためには、下流の再構築(reassembly)コストを取り除く又は軽減するために、インビボで効率良く且つ安価に大量の所望の最終産物を生産することができる発現系を得ることが理想である。
【0005】
現在、細菌は、豊富な量の組換えペプチドを生産するそれらの潜在能力の故に、組換えペプチドの生産のために最も広く使用されている発現系である。細菌は一般に、真核生物タンパク質をグリコシル化、アセチル化、アシル化、リン酸化又はγ−カルボキシル化せず、それ故、翻訳後修飾を必要とするある種の異種真核生物ペプチドをインビボで生産するそれらの能力には限界がある。細菌によって生産される真核生物ペプチドを修飾する付加的な工程は、所要時間を増大し、ペプチド生産の全体的収率を低下させると考えられ、細菌宿主発現系の利点の多くを色褪せさせる。それ故、細菌発現系の固有の限界を克服するために代替的な非細菌発現系が利用されてきた。
【0006】
異種ペプチドを生産する能力に関して分析された1つの特定宿主発現系は、全植物体及び植物細胞懸濁培養であった。植物の安全で安価な培養物は、ある種のペプチドのコスト効果的生産のための改善された代替的宿主を提供する。この代替物は、タンパク質が複雑で、グリコシル化を必要とし、ヒト又は動物感染性ウイルス及び細菌毒素不含である必要がある場合に特に魅力的である。
【0007】
全植物体は、比較的安価に膨大な量で成長するという利点を提供し、収集されたトランスジェニック又はウイルス感染植物の大量のバイオマスからの、所望組換えタンパク質の大きな収量の潜在的可能性を有する。その結果として、全植物体は、ヒト又は動物への投与を意図する生物薬剤タンパク質の商業生産のための発現系として開発されてきた。
【0008】
これらの系の固有の能力を利用するために、非細菌系における異種タンパク質の生産コストと収率を改善する試みに多大の時間と資源が費やされてきた。これらの領域の両方で進歩があったが、非細菌発現系における異種ペプチドの生産のためのさらなる方法とプラットフォームは有益であろう。
【0009】
ウイルス及びウイルス様粒子
宿主細胞発現系におけるペプチド生産を改善するための1つのアプローチは、対象組換えペプチドを生産するために感染性組換えウイルスの性質を利用することである。植物宿主系における感染性ウイルスの使用は特に周知である。例えばPorta & Lomonossoff, (2002) “Viruses as vectors for the expression of foreign sequences in plants, “Biotechnology and Genetic Enginering Reviews 19:245-291参照。
【0010】
最近の戦略は、ウイルス様粒子(VLP)構造における異種ペプチドの生産に集中している。一般に、カプシド形成ウイルスは、ウイルス核酸を含有するように集合されたタンパク質コート又は「カプシド」を含む。多くのウイルスは、個別に発現されたカプシドから「自己集合(self-assembly)」することができるカプシドを有し、カプシドは細胞内部で(「インビボ集合(in vivo assembly)」)及び単離と精製後に細胞の外側で(「インビトロ集合(in vitro assembly)」)発現される。理想的には、標的組換えペプチドを含むようにカプシドを改変し、組換えウイルスカプシド−ペプチド融合物を生成する。次に融合ペプチドを細胞において発現させ、理想的には、組換えウイルス又はウイルス様粒子を形成するようにインビボで構築させることができる。
【0011】
植物においてVLPに集合されたウイルスカプシド融合タンパク質による異種タンパク質の生産は、様々な程度の成功を収めている。例えば C Marusic et al., J Virol. 75 (18): 8434-39 (Sep 2001)(組換えウイルス粒子のインビボ形成を伴う、末端で抗原性6アミノ酸HIVペプチドに融合されたウイルスカプシドを発現する全ニコチアナ・ベンタミアナにおける感染性螺旋ジャガイモXウイルスの使用); FR Brennan et al., Vaccine 17 (15-16): 1846-57 (09 Apr 1999) (カウピー(ササゲ)全植物体(ビグナ・ユニキクラータ)における組換えウイルス粒子発現のインビボ形成を伴う、末端で抗原性黄色ブドウ球菌ペプチドに融合された感染性カウピーモザイクウイルス又は螺旋ジャガイモXウイルスの使用); C Porta et al.,Intervirology 39 (1-2): 79-84 (1996) (全植物体において抗原性HIV配列を含むキメラコートタンパク質を発現する感染性カウピーモザイクウイルス)参照。
【0012】
Lomonossoff & Johnsonへの米国特許第5,874,087号は、ウイルスカプシドが生物活性ペプチド、例えばホルモン、成長因子又は抗原性ペプチドを含むように工作されている、植物細胞又は全植物体における感染性植物ウイルスの生産を述べている。コモウイルス、トンブスウイルス、ソベモウイルス及びネポウイルス属から選択されるウイルスを、外来性ペプチドをコードする配列を含むように工作し、組換えウイルスを生産するために工作したウイルスのゲノム全体を発現させる。前記明細書は、改変ウイルスの増殖が発明の中心部分であることを強調する。
【0013】
チャップマン(Chapman)らへの米国特許第6,232,099号は、全植物体においてウイルスカプシドサブユニットに結合した異種タンパク質を生産するための感染性桿状ウイルスの使用を述べる。螺旋ウイルスとしても分類される桿状ウイルス、例えばジャガイモXウイルス(PVX)は、組換えウイルスカプシド−ペプチド融合物を創造するために対象組換えペプチドがウイルスのゲノムに挿入されている。次に、組換え感染性ウイルスを使用して全植物体の植物細胞に感染させ、ウイルスは植物細胞において活発に複製して、さらに他の細胞に感染し、最終的に宿主植物全体に感染する。最後に、組換えウイルスカプシド−ペプチド融合物を植物から精製する。
【0014】
チャップマン(Chapman)らはまた、限られた挿入物サイズが二十面体ウイルスによって許容されることを教示する。チャップマン(Chapman)らは、自らの主張を裏付けるのに、植物宿主細胞における発現のための二十面体ウイルス内の組換えペプチドの大きさを26アミノ酸の長さに限定する、国際公開公報第WO92/18618号を引用している。チャップマン(Chapman)らは、二十面体ウイルスのカプシドの内部挿入部位に存在するより大きなペプチドは、キメラウイルスの集合の失敗を導く、タンパク質の形状及び/又は他のカプシドと成功裏に相互作用するその能力の破壊をもたらし得ると理論付ける。
【0015】
カプロウスキー(Koprowski)らへの米国特許第6,042,832号は、抗原性ポリペプチドに融合した植物ウイルスカプシドタンパク質を含む融合カプシドタンパク質を述べる。生じる粒子は全植物体において生産される。
【0016】
カプシド融合タンパク質において異種タンパク質を生産するための感染性組換えウイルスの使用は、しかしながら、欠点がないわけではない。1つの特に厄介な面は、対象とする融合異種タンパク質を含まない基本的に野生型の復帰変異株、又は突然変異した望ましくない組換えカプシド融合タンパク質産物であるカプシドタンパク質を生じる、インビボで突然変異するこれらのウイルスの能力である。例えばPorta & Lomonossoff (1996) “Use of viral replicons for the expression of genes in plants, “ Molecular Biotechnology 5: 209- 221;Dolja et al. (1993) “Spontaneous mutagenesis of a plant potyvirus genome after insertion of a foreign gene,” J. Virol. 67 (10): 5968-5975; Dawson et al. (2001)”Assessment of recombinants that arise from the use of a TMV-based transient expression vector, “Virol. 284 (2): 182-189(接種した全植物体における異種挿入遺伝子の欠失を述べる)参照。全植物体におけるこれらのウイルスベクターの安定性の欠如は、全体的タンパク質産物の収率を潜在的に低下させ、カプシド−融合産物の不一致や不規則性を導き得る。そのような不規則性は、タンパク質産物の完全性がペプチドの特定所望物理化学的性質にとって必須である場合に特に厄介であり得る。
【発明の開示】
【発明が解決しようとする課題】
【0017】
植物における感染性組換えウイルスの固有の不安定性の結果として、商業的組換えタンパク質生産の分野において植物系型の利益を提供する効率的なペプチド生産系に対する需要がまだ存在する。
【0018】
加えて、組換えペプチドの生産のための全植物体の使用はまた、潜在的な問題を提示する。例えば長い発現時間、バッチごとの生成物収率の変動、封じ込めの問題、生産の初期段階に製造管理規則(Good Manufacturing Practice, GMP)を適用することの難しさ、農薬及び肥料による汚染の可能性、並びに害虫、病害及び微気候や土壌の相違による一定しない栽培条件は全て、組換えペプチドの生産のための潜在的に一貫しない宿主系を生じさせる。
【0019】
それ故、カプシド融合タンパク質を含むウイルス様粒子の生産のための安定で一貫した植物細胞発現系を提供することが本発明の1つの目的である。
【0020】
カプシド融合タンパク質を含むウイルス様粒子の生産のための安定な発現系における宿主細胞として使用のための植物細胞を提供することが本発明のもう1つの目的である。
【0021】
植物細胞懸濁培養を含む、植物細胞におけるカプシド融合タンパク質を含むウイルス様粒子の改善された生産のための方法を提供することが本発明のさらにもう1つの目的である。
【0022】
カプシド融合タンパク質を含むウイルス様粒子の生産のための植物細胞発現系において使用するための新規構築物及び核酸を提供することが本発明のさらにもう1つの目的である。
【課題を解決するための手段】
【0023】
本発明は、ウイルスカプシドと対象組換えペプチドを含む融合ペプチドをコードする非感染性プラスミドが宿主植物細胞のゲノムに安定に挿入され、懸濁植物細胞培養において発現される、組換えペプチドの生産のための方法を提供する。ウイルスカプシド−異種ペプチド融合産物は、インビボでウイルス様粒子として発現され得る。本発明は、感染性ウイルス因子の使用を必要としない;というよりもむしろ、カプシド−異種ペプチド融合産物をコードする非感染性核酸が植物細胞のゲノムに安定に挿入され、それを、発酵工程において培養して対象ペプチドを生産することができる。そのような工程は、異種ペプチドを含むカプシド融合タンパク質の発現のためにより変動が少なく、より安定な宿主系を生じさせる。
【0024】
全植物体において対象異種ペプチドを発現するために使用される、カプシド−異種ペプチド融合タンパク質を含む感染性ウイルスは、組換えカプシドタンパク質核酸における突然変異を生じさせる、全植物体での遺伝的不安定性を示し、発現される突然変異型カプシドは、ウイルス粒子へと集合することができないか又は突然変異した標的ペプチドを含むことが見出された。本発明は、増殖条件に対する厳密な制御、バッチごとの一貫性、高いレベルの封じ込め、及び製造管理規則(GMP)に従って組換えタンパク質を生産する能力という付加的な利益と共に、生じる異種ペプチドの高い安定性を提供する。
【0025】
カプシド−融合タンパク質産物は細胞内でウイルス様粒子を形成し得る。ウイルス様粒子は、回収されるペプチドの高純度を達成する改善された効率をもたらし得る。細胞において生産されるウイルス様粒子は、典型的には植物細胞に感染することができない。ウイルスカプシド配列は、栄養性及び非栄養性ウイルスに両方に由来することができ、栄養機能は発現宿主として使用される特定植物細胞によって決定される。1つの実施形態では、ウイルスカプシドタンパク質は、融合産物を発現するために使用される植物細胞に対して天然又は自然の栄養機能を示すウイルスに由来する。1つの実施形態では、ウイルスカプシドタンパク質は、融合産物を発現するために使用される植物細胞に対して天然又は自然の栄養機能を示さないウイルスに由来する。1つの実施形態では、細胞は、融合産物を生産するために使用される所望カプシドタンパク質配列以外の人工的に導入されたウイルスタンパク質を含まない。もう1つの実施形態では、細胞は、融合産物を生産するために使用される所望カプシドタンパク質配列以外の人工的に導入されたウイルスタンパク質又は核酸を含み、前記の付加的なウイルスタンパク質又は核酸は、核酸配列に感染性を付与しない。1つの実施形態では、ウイルスカプシドは、植物細胞発現宿主とは異なる科の生物に対して栄養機能を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、植物細胞発現宿主とは異なる属の生物に対して栄養機能を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、融合産物を発現するために使用される細胞とは異なる種の生物に対して親和性を有するウイルスに由来する。本発明の1つの実施形態では、カプシドは桿状植物ウイルスに由来する。特定実施形態では、カプシドは、タバコモザイクウイルス及びジャガイモXウイルス(PVX)から選択される群に由来する桿状ウイルスカプシドである。本発明の1つの実施形態では、カプシドタンパク質は二十面体ウイルスに由来する。特定実施形態では、カプシドは植物二十面体二十面体ウイルスに由来する。より特定の実施形態では、二十面体カプシドは、カウピーモザイクウイルス(ササゲモザイクウイルス)(Cowpea Mosaic Virus)、カウピークロロティックモトルウイルス(ササゲ退緑斑紋ウイルス)(Cowpea Clorotic Mottle Virus)及びアルファルファモザイクウイルスから選択される群に由来する。
【0026】
本発明はまた、ウイルスカプシドと組換えペプチドの融合タンパク質をコードする発現カセットを含む非感染性核酸構築物を含む植物細胞を提供する。ウイルスカプシドと組換えペプチドの融合ペプチドは、植物細胞において機能するプロモーター及びターミネーターに作動可能に連結されている。1つの詳細な実施形態では、核酸は植物細胞のゲノムに組み込まれ、この組込みは、カプシド融合タンパク質をコードする核酸の世代から世代への安定な遺伝と発現をもたらす。本発明の1つの詳細な実施形態では、カプシドタンパク質は二十面体ウイルスに由来する。1つの実施形態では、細胞はウイルス様粒子又は可溶性ケージ構造を生産する。1つの実施形態では、植物細胞は単子葉植物又は双子葉植物である。特定実施形態では、植物細胞はニコチアナ・タバカム(タバコ)である。代替的実施形態では、植物細胞はオリザ・サティバ(イネ科イネ)である。本発明の1つの実施形態では、ウイルスカプシドタンパク質に融合した組換えペプチドは、ヒト又は動物の治療のために有用な治療ペプチドである。1つの特定実施形態では、組換えペプチドは抗原性ペプチドである。特定実施形態では、抗原性ペプチドはグリコシル化抗原性ペプチドである。1つの実施形態では、抗原性ペプチドを含むカプシド−組換えペプチドウイルス様粒子を含有する植物細胞又は抽出物は、ワクチンとしてヒト又は動物に投与することができる。代替的実施形態では、抗原性ペプチドを含む精製カプシド−組換えペプチドウイルス様粒子は、ワクチンとしてヒト又は動物に投与することができる。1つの実施形態では、異種ペプチドは抗菌ペプチドである。もう1つの特定実施形態では、組換えペプチドは、遊離単量体形態であるとき植物宿主細胞に対して毒性であるペプチドである。1つの実施形態では、カプシドに融合した組換えペプチドは、少なくとも7、少なくとも8、少なくとも9、少なくとも10、少なくとも12、少なくとも15、少なくとも20、少なくとも25、少なくとも30、少なくとも35、少なくとも40、少なくとも45、少なくとも50、少なくとも55、少なくとも60、少なくとも65、少なくとも75、少なくとも85、少なくとも95、少なくとも99又は少なくとも100アミノ酸の長さである。
【0027】
本発明の1つの実施形態では、カプシドに融合した組換えペプチドは、所望標的ペプチドの少なくとも1個の単量体を含む。代替的実施形態では、組換えペプチドは、所望標的ペプチドの2個以上の単量体を含む。ある実施形態では、前記ペプチドは、コンカテマーペプチドとしてカプシドに作動可能に連結された少なくとも2個、少なくとも5個、少なくとも10個、少なくとも15個又は少なくとも20個の別個の単量体から成る。もう1つの実施形態では、コンカテマーペプチド内の個々の単量体は、切断可能なリンカー領域によって連結される。さらにもう1つの実施形態では、組換えペプチドをウイルスカプシドの少なくとも1つの表面ループに挿入する。1つの実施形態では、組換えペプチドを二十面体ウイルスカプシドの少なくとも1つの表面ループに挿入する。1つの実施形態では、少なくとも1個の単量体をウイルスカプシドタンパク質の2つ以上の表面ループに挿入する。さらにもう1つの実施形態では、組換えペプチドをウイルスカプシドの少なくとも1つの外側表面ループに挿入する。代替的実施形態では、組換えペプチドをウイルスカプシドの少なくとも1つの内側表面ループに挿入する。
【0028】
ウイルス様粒子の2つ以上のループを修飾することができる。1つの特定実施形態では、組換えペプチドをウイルス様粒子の少なくとも2つの表面ループ上で発現する。もう1つの実施形態では、少なくとも2個の異なるペプチドをウイルスカプシド、ケージ又はウイルス様粒子の少なくとも2つの表面ループに挿入する。もう1つの実施形態では、少なくとも3個の組換えペプチドをウイルス様粒子の少なくとも3つの表面ループに挿入する。表面ループ内の組換えペプチドは同じアミノ酸配列を有し得る。別の実施形態では、表面ループ内の組換えペプチドのアミノ酸配列は異なる。
【0029】
さらにもう1つの実施形態では、前記細胞は、野生型カプシド又は異なるカプシド−組換えペプチド融合ペプチドのいずれかをコードする少なくとも1個の付加的な核酸を含み、多数のカプシドは、インビボでキメラウイルス様粒子を生成するように集合され得る。
【0030】
本発明の1つの態様では、ウイルスカプシドと組換えペプチドの融合タンパク質を含む植物細胞が提供される。本発明の1つの詳細な実施形態では、植物細胞は単子葉植物である。代替的実施形態では、植物細胞は双子葉植物である。1つの実施形態では、カプシド−組換えペプチド融合タンパク質は、インビボでウイルス様粒子を形成するように集合する。
【0031】
本発明の1つの実施形態では、カプシドタンパク質−組換えペプチド融合ペプチドをコードする核酸構築物を含む植物細胞は、核形質転換によって得られる。1つの実施形態では、前記植物細胞は色素体(プラスチド(plastid))形質転換によって得られる。さらにもう1つの実施形態では、前記植物細胞は葉緑体形質転換によって得られる。
【0032】
本発明はさらに、ウイルスカプシドと組換えペプチドの融合タンパク質をコードする発現カセットを含む非感染性核酸構築物を提供する。本発明の1つの実施形態では、ウイルスカプシドと組換えペプチドの融合タンパク質をコードする発現カセットは、プロモーター及びターミネーターに作動可能に連結されている。本発明の1つの実施形態では、カプシドは植物ウイルスに由来する。本発明の1つの実施形態では、カプシドは二十面体植物ウイルスに由来する。特定実施形態では、カプシドは、カウピーモザイクウイルス、カウピークロロティックモトルウイルス及びアルファルファモザイクウイルスから選択される群に由来する二十面体ウイルスカプシドである。
【0033】
本発明の1つの実施形態では、組換えペプチドは、所望標的ペプチドの少なくとも1個の単量体を含む。代替的実施形態では、組換えペプチドは、所望標的ペプチドの2個以上の単量体を含む。さらにもう1つの実施形態では、組換えペプチドは、二十面体ウイルスカプシドの少なくとも1つの表面ループに挿入される。
【0034】
もう1つの実施形態では、前記核酸構築物は、植物細胞において機能する少なくとも1個のプロモーターを含む付加的な核酸配列を含む。もう1つの実施形態では、前記核酸構築物は、植物細胞において機能する少なくとも1個のプロモーター及び植物細胞において機能する少なくとも1個のターミネーターを含む付加的な核酸配列を含む。1つの実施形態では、プロモーター及びターミネーター配列に作動可能に連結された選択マーカーをコードする核酸配列が、前記核酸構築物に含まれる。代替的実施形態では、プロモーター及びターミネーター配列に作動可能に連結された選択マーカーは、別個の核酸構築物上で提供される。もう1つの実施形態では、前記核酸構築物は、ウイルスRNAの3’非翻訳領域(3’UTR)に由来する付加的な核酸配列を含む。さらにもう1つの実施形態では、核酸構築物は、ウイルスRNAに由来する少なくとも1つのカプシド形成シグナルを含む。さらにもう1つの実施形態では、非感染性核酸構築物は、ウイルス核酸に感染性を付与しない、ウイルスRNAに由来する付加的な核酸配列を含む。
【0035】
1つの態様では、本発明は、
a)植物細胞を提供すること;
b)少なくとも1個の異種ペプチドと少なくとも1個のウイルスカプシドタンパク質から成る融合ペプチドをコードする非感染性核酸を提供すること;
c)前記植物細胞を形質転換し、前記核酸がその後植物細胞のゲノムに組み込まれること;
d)前記植物細胞を発酵工程において懸濁植物細胞培養中で増殖させ、前記植物細胞において前記核酸を発現させること;
e)前記植物細胞における発現が、前記融合ペプチドのウイルス様粒子へのインビボ集合を提供すること;及び
d)前記ウイルス様粒子を単離すること
を含む、組換えペプチドを生産するための方法を提供する。
【0036】
1つの実施形態では、前記方法は、e)ウイルスカプシドタンパク質から組換えペプチドを分離するために融合産物を切断すること、をさらに含む。さらにもう1つの実施形態では、前記方法は、f)工程(e)の後に、組換えペプチドを単離すること、をさらに提供する。例えば図1参照。本発明の1つの実施形態では、植物細胞は、オリザ・サティバ及びニコチアナ・タバカムから成る群より選択される。
【0037】
組換えペプチドとウイルスカプシドタンパク質をコードする核酸は、プロモーター配列及びターミネーター配列に作動可能に連結される。
【0038】
1つの実施形態では、前記方法は、野生型カプシド又は異なるカプシド−組換えペプチド融合ペプチドをコードするもう1つ別の核酸を同時発現することを含み、前記カプシドは、インビボでキメラウイルス様粒子を生成するように集合される。
【0039】
本発明のもう1つの態様では、
a)植物細胞培地中で増殖することができる植物細胞;
b)少なくとも1個の組換えペプチドと少なくとも1個のウイルスカプシドを含む融合ペプチドをコードし、植物細胞においてゲノムに組み込まれて発現することができる、非感染性核酸;及び
c)増殖培地
を含む、組換えペプチドの生産のための発現系が提供される。
【0040】
融合ペプチドをコードする核酸は、プロモーター配列及びターミネーター配列に作動可能に連結され得る。発現されたとき、融合ペプチドは、細胞内でウイルス様粒子に集合することができる。1つの実施形態では、前記プロモーターは植物プロモーターである。
【発明を実施するための最良の形態】
【0041】
本発明は、ウイルスカプシドと対象組換えペプチドを含む融合ペプチドをコードする非感染性プラスミドが宿主植物細胞のゲノム内に安定に挿入され、懸濁植物細胞培養中で発現される、組換えペプチドの生産のための方法を提供する。ウイルスカプシド−異種ペプチド融合産物は、インビボでウイルス様粒子として発現され得る。本発明はさらに、前記方法における使用のための植物細胞及び核酸構築物を提供する。詳細には、本発明は、ウイルスカプシドと組換えペプチドの融合ペプチドをコードする発現カセットを含む核酸構築物と共に、懸濁植物細胞培養中で増殖することができる植物細胞を提供する。前記融合ペプチドは、プロモーター配列及び終結配列に作動可能に連結され得る。1つの実施形態では、融合ペプチドの植物細胞における発現は、ウイルス様粒子又は可溶性ケージ構造を生成する。本発明はまた、植物細胞のゲノムに組み込むことができ、1つの実施形態では、ヒト及び動物の治療のために有用な治療ペプチドであり得る、ウイルスカプシドと組換えペプチドの融合ペプチドをコードすることができる核酸構築物を提供する。
【0042】
本発明はまた、プロモーター及び終結配列に作動可能に連結された組換えペプチドとウイルスカプシドの融合ペプチドをコードする発現カセットを含む植物細胞のゲノムに組み込むことができる核酸;懸濁植物細胞培養中で前記植物細胞において核酸を発現させ、前記植物細胞における発現が前記融合ペプチドのウイルス様粒子へのインビボ集合を提供すること;及び前記ウイルス様粒子を単離すること、を提供することによって懸濁植物細胞培養において組換えペプチドを生産するための方法を提供する。
【0043】
ここで使用する「感染性」という用語は、ウイルス粒子がその核酸を宿主に転移する能力又は宿主へのウイルス核酸の導入を意味し、その場合ウイルス核酸が複製され、ウイルスタンパク質が翻訳されて、核酸を宿主にさらに転移することができる新しいウイルス粒子が集合される。ここで使用する「非感染性」という用語は、宿主への導入後にウイルス由来核酸が宿主において複製できないことを意味し、その場合ウイルスタンパク質が翻訳され、宿主においてウイルス感染過程を開始することができない新しいウイルス様粒子が構築される。ここで使用する「ペプチド」という用語は、特定分子量に限定されず、タンパク質又はポリペプチドも包含し得る。
(I.組換え植物細胞)
【0044】
本発明は、ゲノム組込みが可能であり、プロモーター配列及び終結配列に作動可能に連結されたウイルスカプシドと組換えペプチドの融合ペプチドをコードする非感染性核酸構築物を含む植物細胞を提供する。前記細胞は組換えペプチドを生産するための方法において使用することができる。
【0045】
植物細胞は、プロモーター及び終結配列に作動可能に連結されたウイルスカプシドと組換えペプチドの融合ペプチドをコードする発現カセットを含み、宿主細胞のゲノムに安定に組み込まれる核酸構築物で天然植物細胞を形質転換することによって誘導できる。安定な形質転換は、ゲノム内への異種DNAの組込みに依存する。異種DNAは植物の核又は色素体に組み込まれ得る。核酸構築物は、植物のゲノムにランダムに挿入され得るか又は相同的組換えを通してゲノムの特定領域を指向し得る。前記植物細胞は、核形質転換、色素体形質転換及び葉緑体形質転換を含む、当技術分野において公知の何らかの手段によって入手できる。例えば米国特許第6,218,145号;欧州特許第012345149号;国際公開公報第WO0121782号;米国特許第6,515,206号;国際公開公報第WO99/10513号;米国特許第5,693,507号;国際公開公報第WO02055651号;国際公開公報第WO0170939号;米国特許第6,472,586号;国際公開公報第WO02057466号;米国特許第5,057,422号;国際公開公報第WO0120974号; Staub, J. M. and Maliga, P, (1992) “Long regions of homologous DNA are incorporated into the tobacco plastid genome by transformation,”
Plant Cell 4: 39-45 参照。
【0046】
ウイルスカプシド
1つの実施形態では、本発明は、宿主ゲノムに安定に組み込まれた非感染性プラスミドからのウイルスカプシドに融合したペプチドの発現によってペプチドを生産するための方法における使用のための植物細胞を提供する。前記発現は、細胞内で少なくとも1個のウイルス様粒子(VLP)の形成を生じさせ得る。
【0047】
ウイルスは、螺旋対称又は二十面体対称を有するものに分類できる。一般に認識されているカプシド形態は、二十面体(正二十面体(icosahedral proper)、等軸晶系、準等軸晶系、及び双生又は「双晶(twinned)」を含む)、多面体(球状、卵形及びレモン形を含む)、杆状(棒状又は弾丸状、及び紡錘状又は葉巻形を含む)、及び螺旋状(桿状、円柱状及びひも状を含む)を含み、いずれも尾部を有していてもよく及び/又は表面突起、例えば棘又はこぶを含んでいてもよい。
【0048】
(形態)
本発明の1つの実施形態では、カプシドのアミノ酸配列は、何らかの形態を有すると分類されるウイルスのカプシドから選択される。1つの実施形態では、カプシドは桿状ウイルスに由来する。特定実施形態では、カプシドは桿状植物ウイルスに由来する。より特定の実施形態では、カプシドは、タバコモザイクウイルス(TMV)及びジャガイモXウイルス(PVX)から選択される群に由来する桿状植物ウイルスカプシドである。TMVは、桿状粒子(300nm)を生じさせる独自のコードタンパク質(17.5kDa)でカプシド形成された単一プラス−センスゲノムRNA(6.5kb)から成る。ジャガイモXウイルスは、515nmの明瞭なモード長と13nmの幅を有するひも状で被包されていない、通常は屈曲性のウイルスである(Brands, 1964)。カプシド構造は3.4nmのピッチで基本螺旋を形成する(Verma et al., 1968)。
【0049】
1つの実施形態では、カプシドは二十面体形態を有する。1つの実施形態では、カプシドアミノ酸配列は、正二十面体である実在のカプシドから選択される。もう1つの特定実施形態では、カプシドアミノ酸配列は、二十面体ウイルスのカプシドから選択される。1つの特定実施形態では、カプシドアミノ酸配列は、二十面体植物ウイルスのカプシドから選択される。しかし、もう1つの実施形態では、ウイルスカプシドは、植物に対して感染性でない二十面体ウイルスに由来する。例えば1つの実施形態では、ウイルスは、哺乳動物に対して感染性のウイルスである。
【0050】
一般に、二十面体ウイルスのウイルスカプシドは、二十面体(立方)対称に配列された多数のタンパク質サブユニットから成る。天然二十面体カプシドは、例えばカプシドの各々の三角形の面を形成し、3個のサブユニットで構築することができ、完全なカプシドを形成する60個のサブユニットを生じさせ得る。この小さなウイルス構造の典型は、例えばバクテリオファージφX174である。多くの二十面体ウイルスカプシドは60個以上のサブユニットを含む。二十面体ウイルスの多くのカプシドは、逆平行の8本鎖β−バレル型折りたたみモチーフを含む。このモチーフは、一方の側に4本のβ鎖(BIDGと称される)及び他方の側に4本のβ鎖(CHEFと称される)を備えるくさび形ブロックを有する。また2つの保存されたα−ヘリックス(A及びBと称される)も存在し、1つはβCとβDの間、他方はβEとβFの間にある。
【0051】
エンベロープを持つウイルスは、膜を通して粒子が突出(出芽)することにより、その全面的な破壊を伴わずに感染細胞を出て行くことができ、その間に粒子は、細胞膜に由来する脂質エンベロープ内に被包される(例えばAJ Cann (ed. ) (2001) Principles of Molecular Virology (Academic Press); A Granoff and RG Webster (eds.) (1999) Encyclopedia of Virology (Academic Press); DLD Caspar (1980) Biophys. J. 32: 103; DLD Caspar and A Klug (1962) Cold Spring Harbor Symp. Quant. Biol. 27: 1; J Grimes et al. (1988) Nature 395: 470; JE Johnson (1996) Proc. Nat'l Acad. Sci. USA 93: 27; and J Johnson and J Speir (1997) J. Mol. Biol. 269: 665参照)。
【0052】
(ウイルス)
ウイルス分類学は、以下の分類群のカプシド形成粒子実体を承認する:
・グループIウイルス、すなわちdsDNAウイルス;
・グループIIウイルス、すなわちssDNAウイルス;
・グループIIIウイルス、すなわちdsRNAウイルス;
・グループIVウイルス、すなわちDNA段階を持たないssRNA(+)鎖ウイルス;
・グループVウイルス、すなわちssRNA(−)鎖ウイルス;
・グループVIウイルス、すなわちssRNA逆転写ウイルスである、RNAレトロイドウイルス;
・グループVIIウイルス、すなわちdsDNA逆転写ウイルスである、DNAレトロイドウイルス;
・デルタウイルス;
・ウイロイド;及び
・サテライト核酸及びプリオンを除く、サテライトファージ及びサテライトウイルス。
【0053】
これらの分類群の成員は当業者に周知であり、H. V. Van Regenmortel et al. (eds.), Virus Taxonomy: Seventh Report of the International Committee on Taxonomy of Viruses (2000) (Academic Press/Elsevier, Burlington Mass., USA); レスター大学微生物・免疫学部のウイルス分類学ウエブページhttp://wwwmicro.msb.le.ac.uk/3035/ Virusgroups.html;及び米国保健社会福祉省の国立衛生研究所・国立医学図書館・国立バイオテクノロジー情報センター(NCBI)(Washington, D. C. , USA)の分類学ブラウザのオンライン「ウイルス」及び「ウイロイド」部門http://www.ncbi.nlm.nih.gov/Taxonomy/tax.html.において総説されている。
【0054】
カプシドのアミノ酸配列は、これらの分類群のいずれかの成員のカプシドから選択され得る。これらの分類群の成員のカプシドについてのアミノ酸配列は、例えば、NCBIによって提供されるPubMedオンライン検索機能の「ヌクレオチド」(Genbank)、「タンパク質」及び「構造」部門http://www.ncbi.nlm.nih.gov/entrez/ query.fcgi.を含むがこれらに限定されないソースから入手し得る。
【0055】
1つの実施形態では、カプシドアミノ酸配列は、以下の宿主の少なくとも1つに特異的な分類群成員から選択される:酵母を含む真菌、植物、藻類を含む原生生物、無脊椎動物、脊椎動物、及びヒト。1つの実施形態では、カプシドアミノ酸配列は以下の分類群のいずれかの成員から選択される:グループI、グループII、グループIII、グループIV、グループV、グループVII、ウイロイド、及びサテライトウイルス。1つの実施形態では、カプシドアミノ酸配列は、上述した6つの宿主型の少なくとも1つに特異的なこれら7つの分類群のいずれかの成員から選択される。より詳細な実施形態では、カプシドアミノ酸配列は、グループII、グループIII、グループIV、グループVII、及びサテライトウイルスのいずれかの成員から;又はグループII、グループIV、グループVII、及びサテライトウイルスのいずれかから選択される。もう1つの実施形態では、ウイルスカプシドはグループIV又はグループVIIから選択される。
【0056】
ウイルスカプシド配列は、当該細胞に対して親和性でないウイルスに由来し得る。1つの実施形態では、当該細胞は、所望二十面体カプシド以外の特定選択ウイルスからのウイルスタンパク質を含まない。1つの実施形態では、ウイルスカプシドは、当該細胞とは異なる科の生物に対して親和性を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、当該細胞とは異なる属の生物に対して親和性を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、当該細胞とは異なる種の生物に対して親和性を有するウイルスに由来する。
【0057】
詳細な実施形態では、ウイルスカプシドはグループIVのウイルスから選択される。
【0058】
1つの実施形態では、ウイルスカプシドは桿状植物ウイルスから選択される。より特定の実施形態では、ウイルスカプシドは、タバコモザイクウイルス及びジャガイモXウイルスから成る群より選択される。
【0059】
1つの実施形態では、ウイルスカプシドは二十面体ウイルスから選択される。二十面体ウイルスは、乳頭腫ウイルス科、トティウイルス科、ジシストロウイルス科、ヘパドナウイルス科、トガウイルス科、ポリオーマウイルス科、ノダウイルス科、テクティウイルス科、レヴィウイルス科、ミクロウイルス科、サイフォウイルス科、ノダウイルス科、ピコルナウイルス科、パルボウイルス科、カルシウイルス科、テトラウイルス科、及びサテライトウイルスのいずれかの成員から選択され得る。
【0060】
特定実施形態では、前記配列は、少なくとも1つの植物宿主に特異的な分類群のいずれかの成員から選択される。1つの実施形態では、二十面体植物ウイルス種は、ブンヤウイルス科、レオウイルス科、ラブドウイルス科、ルテオウイルス科、ナノウイルス科、パルティティウイルス科、セキウイルス科、ティモウイルス科、ウルミアウイルス属、タバコ壊死サテライトウイルス、カリモウイルス科、ジェミニウイルス科、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科分類群のいずれかであるか又はそのいずれかの成員である、植物感染性ウイルス種である。1つの実施形態では、二十面体植物ウイルス種は、ルテオウイルス科、ナノウイルス科、パルティティウイルス科、セキウイルス科、ティモウイルス科、ウルミアウイルス属、タバコ壊死サテライトウイルス、カリモウイルス科、ジェミニウイルス科、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科分類群のいずれかであるか又はそのいずれかの成員である、植物感染性ウイルス種である。詳細な実施形態では、二十面体植物ウイルス種は、カリモウイルス科、ジェミニウイルス科、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科のいずれかであるか又はそのいずれかの成員である、植物感染性ウイルス種である。より特定の実施形態では、二十面体植物ウイルス種は、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科のいずれかであるか又はそのいずれかの成員である、植物感染性ウイルス種である。より特定の実施形態では、二十面体植物ウイルス種は、コモウイルス科又はブロモウイルス科の成員である植物感染性ウイルス種である。特定実施形態では、ウイルスカプシドは、カウピーモザイクウイルス又はカウピークロロティックモトルウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、ブロモウイルス科分類群の種に由来する。詳細な実施形態では、カプシドは、イラルウイルス属又はアルファモウイルス属に由来する。より詳細な実施形態では、カプシドは、タバコ条斑病ウイルス又はアルファルファモザイクウイルス(AMV)(AMV1又はAMV2を含む)に由来する。
【0061】
(VLP)
本発明のウイルスカプシドは、ここで述べる宿主細胞において非感染性である。1つの実施形態では、ウイルス科の発現の間又は発現後に宿主細胞においてウイルス様粒子(VLP)又はケージ構造が形成される。1つの実施形態では、VLP又はケージ構造はまた、対象とするペプチドも含み、特定実施形態では、対象ペプチドがVLPの表面に発現される。前記発現系は、典型的にはウイルスの感染性を可能にする付加的なウイルスタンパク質を含まない。典型的実施形態では、発現系は、宿主細胞、及び1又はそれ以上のウイルスカプシド及び作動可能に連結された対象ペプチドをコードするベクターを含む。前記ベクターは、典型的には付加的なウイルスタンパク質を含まない。本発明は、ウイルスカプシドがある種の宿主細胞においてより大きな度合で形成され、組換えペプチドのより効率的な回収を可能にするという発見に由来する。
【0062】
1つの実施形態では、VLP又はケージ構造は、3から約1,000カプシドを含む、カプシドの多量体集合物である。1つの実施形態では、VLP又はケージ構造は、少なくとも30、少なくとも50、少なくとも60、少なくとも90又は少なくとも120カプシドを含む。もう1つの実施形態では、各々のVLP又はケージ構造は、少なくとも150カプシド、少なくとも160、少なくとも170又は少なくとも180カプシドを含む。
【0063】
1つの実施形態では、VLPは二十面体構造として発現される。もう1つの実施形態では、VLPは、そのカプシド配列が由来する天然ウイルスと同じ形状で発現される。別の実施形態では、しかしながら、VLPは天然ウイルスと同じ形状を持たない。ある実施形態では、例えば、多数のカプシドで形成されるが天然型VLPを形成しない粒子中で構造が生成される。例えばわずか3個のウイルスカプシドのケージ構造が形成され得る。別の実施形態では、約6、9、12、15、18、21、24、27、30、33、36、39、42、45、48、51、54、57又は60カプシドのケージ構造が形成され得る。
【0064】
1つの実施形態では、カプシドの少なくとも1個は、少なくとも1個の対象ペプチドを含む。1つの実施形態では、ペプチドは、VLPの少なくとも1つの内部ループ内で又は少なくとも1つの外側表面ループにおいて発現される。
【0065】
ウイルスカプシドの2つ以上のループを修飾することができる。1つの特定実施形態では、組換えペプチドは、ウイルス様粒子の少なくとも2つの表面ループ上で発現される。もう1つの実施形態では、少なくとも2個の異なるペプチドをウイルスカプシド、ケージ又はウイルス様粒子の少なくとも2つの表面ループに挿入する。もう1つの実施形態では、少なくとも3個の組換えペプチドをウイルス様粒子の少なくとも3つの表面ループに挿入する。表面ループ内の組換えペプチドは同じアミノ酸配列を有し得る。別の実施形態では、表面ループ内の組換えペプチドのアミノ酸配列は異なる。
【0066】
ある実施形態では、VLPの集合を改善するために宿主細胞を修飾することができる。宿主細胞を、例えば、発現されたウイルスカプシドからのVLPの形成を促進するシャペロンタンパク質を含むように改変することができる。もう1つの実施形態では、宿主細胞を、VLPの調節された形成を促進するためにカプシドの発現をより効率的に調節するリプレッサータンパク質を含むように改変する。
【0067】
ウイルスカプシド又はタンパク質をコードする核酸配列はまた、VLPの形成を変化させるように付加的に修飾することができる(例えばBrumfield, et al. (2004) J. Gen. Virol. 85: 1049-1053参照)。例えばこれらの修飾は、集合されたタンパク質ケージ内の隣接サブユニットの内部、外部又はその間の界面を変化させるように設計される。これを実現するために、突然変異誘発プライマーを使用して、(i)N末端の塩基性残基(例えばK、R)を酸性グルタミン酸に置き換えることによってウイルス核酸結合領域の内表面電荷を変化させる(Douglas et al., 2002b);(ii)N末端から内部残基を欠失させる(CCMVにおいて、通常は残基4−37);(iii)11アミノ酸ペプチド細胞標的配列をコードするcDNA(Graf et al., 1987)を表面露出ループに挿入する;及び(iv)金属結合部位を変化させることによってウイルスサブユニットの間の相互作用を修飾する(CCMVにおいて、残基81/148突然変異体)ことができる。
【0068】
組換えペプチドの大きさ
1つの実施形態では、ウイルスカプシド配列に作動可能に連結されたペプチドは、少なくとも2個のアミノ酸を含む。もう1つの実施形態では、前記ペプチドは、少なくとも3、少なくとも4、少なくとも5又は少なくとも6アミノ酸の長さである。別の実施形態では、ペプチドは少なくとも7アミノ酸の長さである。ペプチドはまた、少なくとも8、少なくとも9、少なくとも10、少なくとも11、12、13、14、15、16、17、18、19、20、30、45、50、60、65、75、85、95、96、99又はそれ以上のアミノ酸の長さである。1つの実施形態では、コードされるペプチドは少なくとも25kDである。
【0069】
1つの実施形態では、ペプチドは、2から約300アミノ酸、又は約5から約250アミノ酸、又は約5から約200アミノ酸、又は約5から約150アミノ酸、又は約5から約100アミノ酸を含む。もう1つの実施形態では、ペプチドは、約10から約140アミノ酸、又は約10から約120アミノ酸、又は約10から約100アミノ酸を含む。
【0070】
1つの実施形態では、ウイルスカプシド配列に作動可能に連結されたペプチド又はタンパク質は、約500個のアミノ酸を含む。1つの実施形態では、ペプチドは500個未満のアミノ酸を含む。もう1つの実施形態では、ペプチドは、約300個までのアミノ酸、又は約250個まで、又は約200個まで、又は約180個まで、又は約160個まで、又は約150個まで、又は約140個まで、又は約120個まで、又は約110個まで、又は約100個まで、又は約90個まで、又は約80個まで、又は約70個まで、又は約60個まで、又は約50個まで、又は約40個まで、又は約30個までのアミノ酸を含む。
【0071】
1つの実施形態では、カプシドに融合した組換えペプチドは、少なくとも7、少なくとも8、少なくとも9、少なくとも10、少なくとも12、少なくとも15、少なくとも20、少なくとも25、少なくとも30、少なくとも35、少なくとも40、少なくとも45、少なくとも50、少なくとも55、少なくとも60、少なくとも65、少なくとも75、少なくとも85、少なくとも95、少なくとも99、又は少なくとも100アミノ酸である。
【0072】
本発明の1つの実施形態では、組換えペプチドは、所望標的ペプチドの少なくとも1個の単量体を含む。代替的実施形態では、組換えペプチドは、所望標的ペプチドの2個以上の単量体を含む。ある実施形態では、ペプチドは、コンカテマーペプチドとしてカプシドに作動可能に連結された、少なくとも2個、少なくとも5個、少なくとも10個、少なくとも15個又は少なくとも20個の別個の単量体から成る。もう1つの実施形態では、コンカテマーペプチド内の個々の単量体は、切断可能なリンカー領域によって連結される。さらにもう1つの実施形態では、組換えペプチドを、二十面体ウイルス様粒子の少なくとも1つの表面ループに挿入する。1つの実施形態では、少なくとも1個の単量体をウイルス様粒子の表面ループに挿入する。
【0073】
(分類)
ウイルスカプシドに融合する対象ペプチドは、ウイルスに由来しない、及び場合により、当該細胞と同じ種に由来しない、異種タンパク質であり得る。
【0074】
ウイルスカプシドに融合する対象ペプチドは、機能性ペプチド:構造ペプチド;抗原性ペプチド;毒性ペプチド、抗菌ペプチド、それらのフラグメント;それらの前駆体;前記のいずれかの組合せ;及び/又は前記のいずれかのコンカテマーであり得る。本発明の1つの実施形態では、組換えペプチドは、ワクチン戦略において使用される抗原性ペプチドを含む、ヒト及び動物の治療のために有用な治療ペプチドである。特定実施形態では、抗原性ペプチドは、インビボでグリコシル化される。
【0075】
機能性ペプチドは、例えば:生物活性ペプチド(すなわち生物学的実体、例えば生物、細胞、培養物、組織、器官又は細胞小器官、における又はその生物学的機能又は活性の開始、増強、延長、減衰、終結又は防止を及ぼす、惹起する又はさもなければ生じさせるペプチド);触媒ペプチド;ミクロ構造及びナノ構造活性ペプチド(すなわちその中で又はそれに関連して、作用、例えば運動、エネルギー変換、を実施する工作されたミクロ又はナノ構造の一部を形成するペプチド);及び刺激剤ペプチド(例えばペプチド着香料、着色料、着臭剤、フェロモン、誘引物質、抑制物質及び忌避物質)を含むが、これらに限定されない。
【0076】
生物活性ペプチドは、例えば:免疫活性ペプチド(例えば抗原性ペプチド、アレルゲンペプチド、ペプチド免疫調節剤(immunoregulator)、ペプチド免疫調節剤(immunomodulator):シグナル伝達ペプチド(signaling and signal transduction peptides)(例えばペプチドホルモン、サイトカイン及び神経伝達物質;受容体;アゴニスト及びアンタゴニストペプチド;ペプチドターゲティング及び分泌シグナルペプチド);及び生物阻害性ペプチド(例えば毒性、生物致死性又は生物抑制性ペプチド、例えばペプチド毒素及び抗菌ペプチド)を含むが、これらに限定されない。
【0077】
構造ペプチドは、例えば:ペプチドアプタマー、折りたたみペプチド(例えばもう1つ別の分子の物理的立体配座の形成又は維持を促進する又は誘導するペプチド);接着促進ペプチド(例えば接着ペプチド、細胞接着促進ペプチド);界面ペプチド(例えばペプチド界面活性剤及び乳化剤);ミクロ構造及びナノ構造構築ペプチド(すなわち工作されたミクロ又はナノ構造の一部を形成する構造ペプチド);及びプレ活性化ペプチド(例えばプレ、プロ及びプレ−プロタンパク質及びペプチドのリーダーペプチド;インテイン)を含むが、これらに限定されない。
【0078】
触媒ペプチドは、例えばアポB RNA編集シチジンデアミナーゼペプチド;グルタミニル−tRNAシンテターゼの触媒ペプチド:アスパラギン酸トランスカルバモイラーゼの触媒ペプチド;植物1型リボソーム不活性化ペプチド;ウイルス触媒ペプチド、例えば口蹄疫ウイルス[FMDV−2A]触媒ペプチド;マトリックスメタロプロテイナーゼペプチド;及び触媒メタロオリゴペプチドを含む。
【0079】
ペプチドはまた、ペプチドエピトープ、ハプテン又は関連ペプチド(例えば抗原性ウイルスペプチド;ウイルス関連ペプチド、例えばHIV関連ペプチド、肝炎関連ペプチド;抗体イディオタイプドメイン;細胞表面ペプチド;抗原性ヒト、動物、原生生物、植物、真菌、細菌及び/又は古細菌ペプチド;アレルゲンペプチド及びアレルゲン脱感作ペプチド)であり得る。
【0080】
ペプチドはまた、ペプチド免疫調節剤(peptide immunoregulators or immunomodulators)(例えばインターフェロン、インターロイキン、ペプチド免疫抑制剤及び免疫増強剤);抗体ペプチド(例えば一本鎖抗体;一本鎖抗体フラグメント及び構築物、例えば一本鎖Fv分子;抗体軽鎖分子、抗体重鎖分子、ドメイン欠失抗体軽鎖又は重鎖分子;一本鎖抗体ドメイン及び分子、例えばCH1、CH1−3、CH3、CH1−4、CH4、VHCH1、CL、CDR1又はFR1−CDR1−FR2ドメイン;パラトープペプチド;ミクロ抗体);別の結合ペプチド(例えばペプチドアプタマー、細胞内及び細胞表面受容体タンパク質、受容体フラグメント;抗腫瘍壊死因子ペプチド)であり得る。
【0081】
ペプチドはまた、酵素基質ペプチド又は酵素阻害剤ペプチド(例えばカスパーゼ基質及び阻害剤、プロテインキナーゼ基質及び阻害剤、蛍光共鳴エネルギー転移ペプチド酵素基質)であり得る。
【0082】
ペプチドはまた、細胞表面受容体ペプチドリガンド、アゴニスト及びアンタゴニスト(例えばセルレイン、ダイノルフィン、オレキシン、下垂体アデニル酸シクラーゼ活性化ペプチド、腫瘍壊死因子ペプチド;合成ペプチドリガンド、アゴニスト及びアンタゴニスト);ペプチドホルモン(例えば内分泌、パラ分泌及び自己分泌ホルモン、例えばアミリン、アンギオテンシン、ブラジキニン、カルシトニン、心臓興奮性神経ペプチド、カゾモルフィン、コレシストキニン、コルチコトロピン及びコルチコトロピン関連ペプチド、分化因子、エンドルフィン、エンドセリン、エンケファリン、エリスロポエチン、エキセンディン、卵胞刺激ホルモン、ガラニン、ガストリン、グルカゴン及びグルカゴン様ペプチド、ゴナドトロピン、成長ホルモン及び成長因子、インスリン、カリジン、キニン、レプチン、脂肪親和性ホルモン、黄体形成ホルモン、メラニン細胞刺激ホルモン、メラトニン、ナトリウム排泄増加性ペプチド、ニューロキニン、ニューロメジン、ノシセプチン、オステオカルシン、オキシトシン(すなわちオシトシン)、副甲状腺ホルモン、プレイオトロフィン、プロラクチン、レラキシン、セクレチン、セロトニン、睡眠誘発ペプチド、ソマトメジン、サイモポイエチン、甲状腺刺激ホルモン、チロトロピン、ウロテンシン、血管作用性小腸ペプチド、バソプレシン);ペプチドサイトカイン、ケモカイン、ウイロカイン(virokine)及びウイロセプター(viroceptor)ホルモン放出及び放出抑制ペプチド(例えばコルチコトロピン放出ホルモン、コルチスタチン、卵胞刺激ホルモン放出因子、胃抑制性ペプチド、ガストリン放出ペプチド、ゴナドトロピン放出ホルモン、成長ホルモン放出ホルモン、黄体形成ホルモン放出ホルモン、メラノトロピン放出ホルモン、メラノトロピン放出抑制因子;ノシスタチン、パンクリアスタチン、プロラクチン放出ペプチド、プロラクチン放出抑制因子;ソマトスタチン;甲状腺刺激ホルモン放出ホルモン);ペプチド神経伝達物質又はチャネルブロッカー(例えばボンベシン、神経ペプチドY、ニューロテンシン、サブスタンスP);ペプチド毒素、毒素前駆体ペプチド又は毒素ペプチド部分であり得る。ある実施形態では、ペプチド毒素はD−アミノ酸を含まない。毒素前駆体ペプチドは、D−アミノ酸を含まないもの及び/又は翻訳後修飾によって、例えばアミノ酸にD−立体配置を導入することができるD立体配置誘導剤(例えばペプチドイソメラーゼ又はエピメラーゼ又はラセマーゼ又はトランスアミノアーゼ)の作用によって、及び/又は環状ペプチド構造を形成することができる環化剤(例えばペプチドチオエステラーゼ又はペプチドリガーゼ、例えばトランススプライシングタンパク質又はインテイン)の作用によって、天然毒素構造に変換されていないものであり得る。
【0083】
毒素ペプチド部分は、ペプチド含有毒素の線状又は前環化オリゴ及びポリペプチド部分であり得る。ペプチド毒素の例は、例えばアガトキシン、アマトキシン、カリブドトキシン、クロロトキシン、コノトキシン、デンドロトキシン、インセクトトキシン、マルガトキシン、マスト細胞脱顆粒ペプチド、サポリン、サラフォトキシン;及び細菌体外毒素、例えば炭疽毒素、ボツリヌス毒素、ジフテリア毒素及び破傷風毒素を含む。
【0084】
ペプチドはまた、代謝及び消化関連ペプチド(例えばコレシストキニン−パンクレオザイミンペプチド、ペプチドyy、膵ペプチド、モチリン);細胞接着調節又は仲介ペプチド、細胞外マトリックスペプチド(例えば付着因子、セレクチン、ラミニン);神経保護又は髄鞘形成促進ペプチド;凝集阻害ペプチド(例えば細胞又は血小板凝集阻害剤ペプチド、アミロイド形成又は沈着阻害剤ペプチド);連結ペプチド(例えば心臓血管連結神経ペプチド、iga連結ペプチド);又はその他のペプチド(例えばアグーチ関連ペプチド、アミロイドペプチド、骨関連ペプチド、細胞透過性ペプチド、コナントキシン、コントリファン、コンチュラキン、ミエリン塩基性タンパク質等)であり得る。
【0085】
ペプチドはまた、インビボで翻訳後修飾され得る。特定実施形態では、ペプチドは、単独で又は組合せとして、グリコシル化、アセチル化、アシル化、リン酸化又はγ−カルボキシル化され得る。
【0086】
ある実施形態では、対象ペプチドは、選択されるウイルスカプシドに対して外因性である。ペプチドは、天然又は合成配列のいずれでもよい(及びそれらのコード配列は天然又は合成ヌクレオチド配列のいずれでもよい)。それ故、例えばアミノ酸の天然、修飾天然及び完全人工配列が包含される。これらのアミノ酸配列をコードする核酸分子の配列は、同様に、天然、修飾天然及び完全人工核酸配列であり得、例えばその核酸分子を得るために使用される(すなわち人為作用によって適用される)1又はそれ以上の合理的又はランダム突然変異及び/又は組換え及び/又は合成及び/又は選択工程の結果であり得る。
【0087】
コード配列は、使用可能な場合は、標的ペプチドについての天然コード配列であり得るが、より典型的には、例えば宿主の種のコドン使用頻度を反映する遺伝子を合成することによって、選択発現宿主細胞における使用のために選択された、改善された又は最適化されたコード配列である。本発明の1つの実施形態では、宿主の種はニコチアナ・タバカムであり、シグナル配列及びペプチド配列の両方を設計するときにはニコチアナ・タバカムのコドン使用頻度を考慮する。本発明の代替的実施形態では、宿主の種はオリザ・サティバであり、シグナル配列及びペプチド配列の両方を設計するときにはオリザ・サティバのコドン使用頻度を考慮する。
【0088】
(抗原性ペプチド(ペプチドエピトープ))
1つの実施形態では、抗原性ペプチドはウイルスカプシドによる発現を通して生産される。抗原性ペプチドは、感染因子、寄生生物、癌細胞及び他の病原因子を含む、ヒト又は動物病原因子の抗原性ペプチドであるものから選択することができる。そのような病原因子はまた、ビルレンス因子及び病原因子、例えばそれらの因子の体外毒素、内毒素等を含む。病原因子は何らかのレベルのビルレンスを示し得る、すなわちそれらは、例えば毒性、無毒性、偽毒性、半毒性等であり得る。1つの実施形態では、抗原性ペプチドは、病原因子からのエピトープアミノ酸配列を含む。1つの実施形態では、エピトープアミノ酸配列は、少なくとも1個のそのような因子の表面ペプチドの少なくとも一部であるものを含む。1つの実施形態では、カプシド−組換えペプチドウイルス様粒子は、ヒト又は動物適用におけるワクチンとして使用することができる。
【0089】
2個以上の抗原性ペプチドを選択してもよく、その場合、生じるウイルス様粒子は多数の異なる抗原性ペプチドを提示し得る。多抗原性ペプチド様式の特定実施形態では、様々な抗原性ペプチドは全て同じ病原因子からの複数のエピトープから選択される。多抗原性ペプチド様式の特定実施形態では、様々な抗原性ペプチドは全て、密接に関連する病原因子、例えば同じ種又は同じ属の異なる種の異なる株、亜種、次亜種、病原型、血清型又は遺伝子型(genovar)から選択される。
【0090】
1つの実施形態では、病原因子は以下の群の少なくとも1つに属する:病原性:バチルス属種、例えば炭疽菌;バルトネラ属種、例えばB.クインタナ;ブルセラ属種;ブルクホルデリア属種、例えばB.シュードマレイ(類鼻疽菌);カンピロバクター種;クロストリジウム属種、例えば破傷風菌、ボツリヌス菌;コクシエラ属種、例えばC.バーネティー;エドワードシエラ属種、例えばE.ターダ;エンテロバクター属種、例えばE.クロアカ;エンテロコッカス属種、例えばE.フェカリス、E.フェシウム;エシェリキア属種、例えば大腸菌;フランシセラ属種、例えば野兎病菌(F. tularensis);ヘモフィルス属種、例えばインフルエンザ菌;クレブシエラ属種、例えば肺炎杆菌;レジオネラ属種;リステリア属種、例えばリステリア菌(L. monocytogenes);髄膜炎菌及び淋菌、例えばナイセリア属種;モラクセラ属種;ミコバクテリウム属種、例えばらい菌、ヒト型結核菌;肺炎球菌、例えば肺炎双球菌;シュードモナス属種、例えば緑膿菌;リケッチア属種、例えば発疹チフスリケッチア、斑点熱リケッチア、発疹熱リケッチア;サルモネラ属菌、例えば腸チフス菌;ブドウ球菌属種、例えば黄色ブドウ球菌;A型連鎖球菌属及び溶血連鎖球菌属を含む、連鎖球菌属種、例えば肺炎連鎖球菌、化膿連鎖球菌;ストレプトミセス属種;赤痢菌属種;ビブリオ属種、例えばコレラ菌;及びエルジニア属種、例えばペスト菌、エルジニア小腸結腸炎菌(E.enterocolitica)を含むが、これらに限定されない細菌及びマイコプラスマ因子。病原性:アルテルナリア属種;アスペルギルス属種;ブラストミセス属種、例えばB.デルマチチジス;カンジダ属種、例えばカンジダアルビカンス;クラドスポリウム属種;コクシジオイデス属種、例えばC.イミチス;クリプトコッカス属種、例えばC.ネオフォルマンス;ヒストプラスマ属種、例えばH.カプスラーツム;及びスポロトリクス属種、例えばS.シェンキイ、を含むが、これらに限定されない真菌及び酵母因子。
【0091】
1つの実施形態では、病原因子は、病原性:アカントアメーバ属種、アメーバ属種、ネグレリア属種、エントアメーバ属種、例えば赤痢アメーバを含むアメーバ;クリプトスポリジウム属種、例えばC.パルブム;シクロスポラ属種;エンセファリトゾーン属種、例えばE.インテスティナリス;エンテロシトゾーン属種;ジアルジア属種、例えばランブル鞭毛虫;イソスポラ属種;ミクロスポリジウム属種;プラスモディウム属種、例えば熱帯熱マラリア原虫、四日熱マラリア原虫、卵形マラリア原虫、三日熱マラリア原虫;トキソプラスマ属種、例えばトキソプラスマ原虫;及びトリパノソーマ属種、例えばトリパノソーマブルセイ、を含むが、これらに限定されない原生生物因子由来である。
【0092】
1つの実施形態では、病原因子は、病原性:回虫属種、例えば回虫;ドラクンクルス属種、例えばメジナ虫;オンコセルカ属種、例えば回旋糸状虫;住血吸虫属種;旋毛虫属種、例えば旋毛虫;及び鞭虫属種、例えばヒト鞭虫、を含むが、これらに限定されない寄生生物因子(例えば蠕虫寄生生物)由来である。
【0093】
もう1つの実施形態では、病原因子は、病原性:アデノウイルス;アレナウイルス、例えばラッサ熱ウイルス;アストロウイルス;ブンヤウイルス、例えばハンタウイルス、リフトバレー熱ウイルス;コロナウイルス、デルタウイルス;サイトメガロウイルス、エプスタイン−バーウイルス、ヘルペスウイルス、水痘ウイルス;フィロウイルス、例えばエボラウイルス、マルブルクウイルス;フラビウイルス、例えばデング熱ウイルス、西ナイル熱ウイルス、黄熱病ウイルス;肝炎ウイルス;インフルエンザウイルス;レンチウイルス、T細胞リンパ球向性ウイルス、他の白血病ウイルス;ノーウォークウイルス;パピローマウイルス、他の腫瘍ウイルス;パラミクソウイルス、例えば麻疹ウイルス、流行性耳下腺炎ウイルス、パラインフルエンザウイルス、肺炎ウイルス、センダイウイルス;パルボウイルス;ピコルナウイルス、例えばカルジオウイルス、コクサッキーウイルス、エコーウイルス、ポリオウイルス、ライノウイルス、他のエンテロウイルス;ポックスウイルス、例えば痘瘡ウイルス、ワクシニアウイルス、パラポックスウイルス;レオウイルス、例えばコルティウイルス、オルビウイルス、ロタウイルス;ラブドウイルス、例えばリッサウイルス、水泡性口内炎ウイルス;及びトガウイルス、例えば風疹ウイルス、シンドビスウイルス、西部脳炎ウイルス、を含むが、これらに限定されないウイルス因子由来である。
【0094】
1つの実施形態では、抗原性ペプチドは、イヌパルボウイルスペプチド、炭疽菌防御抗原(PA)抗原性ペプチド及び東部ウマ脳炎ウイルス抗原性ペプチドから成る群より選択される。特定実施形態では、抗原性ペプチドは、PA1(配列番号1)、PA2(配列番号2)、PA3(配列番号3)又はPA4(配列番号4)から選択されるアミノ酸配列を有する炭疽菌由来ペプチドである。
【0095】
(宿主細胞毒性ペプチド)
もう1つの特定実施形態では、組換えペプチドは、遊離単量体形態であるとき宿主細胞に対して毒性であるペプチドである。ある実施形態では、ウイルスカプシドと共に発現される対象ペプチドは宿主細胞毒性ペプチドである。宿主細胞毒性ペプチドは、それが発現される宿主細胞に対して、又は宿主細胞がその成員である細胞培養中又は生物中の他の細胞に対して、又は宿主細胞を提供する生物又は種の細胞に対して、生物抑制性、生物致死性又は毒性である生物阻害性ペプチドを指示する。1つの実施形態では、宿主細胞毒性ペプチドは、それが発現される宿主細胞に対して生物抑制性、生物致死性又は毒性である生物阻害性ペプチドである。宿主細胞毒性ペプチドの一部の例は、ペプチド毒素、抗菌ペプチド及び他の抗生物質ペプチドを含むが、これらに限定されない。
【0096】
(抗菌ペプチド)
もう1つの特定実施形態では、組換えペプチドは、抗菌性ペプチドであるペプチドである。抗菌ペプチドは、例えば抗細菌ペプチド、例えばマガイニン、βデフェンシン、一部のαデフェンシン;カテリシジン;ヒスタチン;抗真菌ペプチド;抗原生動物ペプチド;合成AMP;ペプチド抗生物質又はその線状又は前環化オリゴ又はポリペプチド部分;他の抗生物質ペプチド(例えば駆虫性ペプチド、溶血ペプチド、殺腫瘍性ペプチド);及び抗ウイルスペプチド(例えば一部のαデフェンシン;殺ウイルス性ペプチド;ウイルス感染を阻害するペプチド)を含む。1つの特定実施形態では、抗菌ペプチドは、D2A21抗菌ペプチドから成る群より選択される。
【0097】
(VLPを発現するときに使用するための細胞)
本発明のウイルスカプシド又はウイルスカプシド融合ペプチドの発現のための宿主として使用する細胞(「宿主細胞」とも称する)は、ウイルスカプシドが細胞の複製又は感染を許容しないものである。1つの実施形態では、ウイルスカプシドは、宿主細胞が由来する細胞の特定種に感染しないウイルスに由来する。例えば、1つの実施形態では、ウイルスカプシドは二十面体植物ウイルスに由来し、ウイルスの天然栄養宿主ではない植物種の宿主細胞において発現される。もう1つの実施形態では、ウイルス種は哺乳動物に感染し、発現系は植物宿主細胞を含む。代替的実施形態では、ウイルスは、カプシド−組換えペプチド融合物を発現するための宿主細胞として使用される特定植物種の栄養ウイルスである。
【0098】
植物宿主細胞は、維管束植物門、ユーフィロフィタ(Euphyllophyta)、種子植物門又は被子植物門からの植物細胞であり得る。特定実施形態では、植物細胞は、例えば:ヤシ科、例えばココヤシ属、アラブヤシ属;ヤマオイモ科、例えばヤムイモ属;イネ科(Gramineae(Poaceae))、例えばカラスムギ属、オオムギ属、イネ属、キビ属、コムギ属、トウモロコシ属;及びバショウ科、例えばバナナ、の成員を含むが、これらに限定されない単子葉植物である。
【0099】
代替的実施形態では、植物細胞は、例えば:ヒユ科、例えばアマランサス属、ベタ属、アカザ属;セリ科(Umbelliferae)、例えばダウクス属、パスティナカ属;アブラナ科(Cruciferae)、例えばアラビドプシス属、アブラナ属;キク科、例えばヒマワリ属;ヒルガオ科、例えばイポメア属;ウリ科、例えばスイカ属、キュウリ属、カボチャ属;トウダイグサ科;マメ科(Fabaceae)、例えばダイズ属、ヒラマメ属、ウマゴヤシ属、インゲンマメ属、エンドウ属、シャジクソウ属、ソラマメ属、ササゲ属;アマ科、例えばアマ属;及びナス科、例えばトウガラシ属、トマト属、タバコ属、ナス属を含むが、これらに限定されない双子葉植物の成員である。
【0100】
特定実施形態では、宿主細胞は、アラディポシス・タリアナ(Aradiposis thaliana)、日本産イチイ(タクスス・クスビダータ)、ニチニチソウ(カサランツス・ロセウス)、ニコチアナ・タバカム、オリザ・サティバ、リコペルシカム・エスキュレンタム、及びダイズ(グリシン・マックス)から成る群より選択される。
【0101】
(II.核酸構築物)
本発明はさらに、宿主細胞のゲノムに安定に組み込まれることができ、カプシドと組換えペプチドの融合ペプチドをコードする非感染性核酸構築物を提供する。融合ペプチドはプロモーター配列と終結配列に作動可能に連結され得る。1つの実施形態では、a)組換えペプチドをコードする核酸配列、及びb)ウイルスカプシドをコードする核酸配列、を含む植物宿主細胞を形質転換するときに使用するための核酸構築物が提供され、a)の核酸とb)の核酸は融合タンパク質を形成するように作動可能に連結されており、前記核酸構築物は植物宿主細胞のゲノムに安定に挿入され、植物細胞において発現される。
【0102】
ある実施形態では、ベクターは、多数のカプシド又は多数の対象ペプチドについての配列を含み得る。1つの実施形態では、ベクターは、少なくとも2個の異なるカプシド−ペプチドコード配列を含み得る。1つの実施形態では、コード配列は同じプロモーターに連結される。ある実施形態では、コード配列は、内部リボソーム結合部位によって分離される。他の実施形態では、コード配列は、細胞内でのウイルス様粒子の形成を可能にするリンカー配列によって連結される。もう1つの実施形態では、コード配列は異なるプロモーターに連結される。これらのプロモーターは、構成的であるか、又は同じか又は異なる誘導条件によって駆動され得る。もう1つの実施形態では、ゲノム内に挿入されることができ、異なるカプシド−ペプチドの組合せをコードする多数のベクターが提供される。多数のベクターは、構成的であるか又は同じ誘導条件によって駆動されるか又は異なる誘導条件によって駆動されるプロモーターを含み得る。1つの実施形態では、ウイルスカプシドは二十面体ウイルスカプシドに由来する。
【0103】
対象ペプチドについてのコード配列は、あらかじめ定められた部位でウイルスカプシド又はカプシドのコード配列に挿入することができる。前記ペプチドはまた、あらかじめ定められていない部位に挿入し、細胞をVLPの生産に関してスクリーニングすることができる。1つの実施形態では、VLPの形成の間にループとして発現されるように、ペプチドをカプシドコード配列に挿入する。1つの実施形態では、1個のペプチドコード配列をベクター内に含めるが、また別の実施形態では、多数の配列を含める。多数の配列は、コンカテマーの形態、例えば切断可能なリンカー配列によって連結されたコンカテマーの形態であり得る。
【0104】
ペプチドは、カプシド内の2つ以上の挿入部位に挿入し得る。それ故、ペプチドは、カプシドの2個以上の表面ループモチーフに挿入し得る;ペプチドはまた、所与のループモチーフ内の多数の部位に挿入し得る。挿入物の個々の機能性及び/又は構造ペプチド、及び/又はペプチド挿入物全体は、切断部位、すなわちタンパク質を切断する又は加水分解する因子がカプシド構造又は集合物の残りの部分からペプチドを分離するように作用し得る部位によって分離され得る。
【0105】
ペプチドは、外側に面したループ及び/又は内側に面したループ、すなわちそれぞれカプシドの中心から遠ざかる方向又は中心に向かう方向に面したカプシドのループ内に挿入され得る。カプシドの表面ループ内のいかなるアミノ酸又はペプチド結合も、ペプチドのための挿入部位として役立ち得る。典型的には、挿入部位は、ループのほぼ中央、すなわち折りたたまれたカプシドペプチドの三次構造の中心からほぼ最も遠位の位置で選択される。ペプチドコード配列は、選択されたループのこのほぼ中央に対応するカプシドコード配列の位置に作動可能に挿入され得る。これは、ペプチド挿入部位から下流で合成されるカプシドのペプチド配列のその部分についての読み枠の維持を含む。
【0106】
もう1つの実施形態では、ペプチドをカプシドのアミノ末端に挿入することができる。ペプチドは、上述した切断可能リンカーを含む、1又はそれ以上のリンカー配列を通してカプシドに連結することができる。さらにもう1つの実施形態では、ペプチドをカプシドのカルボキシ末端に挿入することができる。ペプチドを、化学的又は酵素的加水分解によって切断され得る1又はそれ以上のリンカー配列を通してカルボキシ末端に連結することができる。1つの実施形態では、ペプチド配列は、アミノ末端とカルボキシ末端の両方で、又は1つの末端と少なくとも1つの内部位置で、例えばその三次元立体配座においてカプシドの表面に発現される位置で、連結される。
【0107】
1つの実施形態では、ペプチドは、カウピークロロティックモザイクウイルスからのカプシドに挿入することができる。1つの特定実施形態では、ペプチドは、CCMVコートタンパク質のアミノ酸129で挿入できる。1つの実施形態では、ペプチド配列は、CCMVコートタンパク質のアミノ酸63、102、114、129又は160で挿入することができる。もう1つの実施形態では、ペプチド配列は、CCMVコートタンパク質のアミノ酸60、61、62又は63で挿入することができる。さらにもう1つの実施形態では、ペプチドは、CCMVタンパク質のアミノ酸129及びアミノ酸60−63の両方で挿入できる。
【0108】
特定実施形態では、本発明は、a)抗原性ペプチドをコードする核酸及びb)ウイルスカプシドをコードする核酸を含み、a)の核酸とb)の核酸が、細胞において発現されたとき融合タンパク質を形成するように作動可能に連結されている、核酸構築物を提供する。核酸構築物を構築する上で有用な他のカプシド及び組換えペプチドは上記に開示されている。
【0109】
(プロモーター)
1つの実施形態では、核酸構築物は、カプシド−組換えペプチド融合ペプチドをコードする核酸配列に作動可能に結合されたプロモーター配列を含む。作動可能な結合又は連結は、転写及び何らかの翻訳調節エレメントが、宿主細胞の作用により、調節エレメントが対象配列の発現を指令することができるように前記配列に共有結合している立体配置を指す。代替的実施形態では、カプシド融合タンパク質をコードする核酸構築物は、遺伝子捕獲戦略を利用して宿主細胞天然プロモーターとインフレームで挿入することができる。
【0110】
発酵工程では、ひとたび発酵が始まれば、発現系の効率を最大化するために高レベルの生産を得ることが理想的である。プロモーターは転写を開始させ、一般に転写開始部位の1−100ヌクレオチド上流に位置する。理想的には、プロモーターは大量の組換えペプチド蓄積を可能にするのに十分なほど強力である。
【0111】
本発明に従って使用されるプロモーターは、構成的プロモーター又は調節を受けるプロモーターであり得る。当技術分野で使用される有用な調節を受けるプロモーターの一般的な例は、エタノール誘導性プロモーター;鉄欠乏誘導性プロモーター;創傷誘導性プロモーター;及びホルモン誘導性、例えばオーキシン誘導性プロモーターを含む。有用な構成的プロモーターの一般的な例は、ユビキチンプロモーター、アクチンプロモーター、チューブリンプロモーター及びキャッサバ葉脈モザイクウイルスプロモーターを含む。本発明において有用な他の種類のプロモーターは、組織特異的プロモーター、例えばトウモロコシスクロースシンテターゼ1(Yang el al., 1990)、トウモロコシアルコールデヒドロゲナーゼ1(Vogel et al., 1989)、トウモロコシ光捕集複合体 (Simpson, 1986)、トウモロコシ熱ショックタンパク質 (Odell et al., 1985)、エンドウマメ小サブユニットRuBPカルボキシラーゼ (Poulsen et al., 1986; Cashmore et al., 1983)、Tiプラスミドマンノピンシンターゼ (Langridge et al., 1989)、Tiプラスミドノパリンシンターゼ (Langridge et al., 1989)、ペチュニアカルコンイソメラーゼ (Van Tunen et al., 1988)、インゲンマメグリシンリッチタンパク質1(Keller et al., 1989)、CaMV 35s転写産物 (Odell et al., 1985) 及びジャガイモパタチン(Wenzler et al., 1989)を含む。好ましいプロモーターは、カリフラワーモザイクウイルスCaMV 35S)プロモーター、イネα−アミラーゼRAmy3Dプロモーター、及びS−E9小サブユニットRuBPカルボキシラーゼプロモーターである。加えて、環境、ホルモン、化学物質及び/又は発生シグナルに応答して調節される多様な植物遺伝子プロモーターが、植物細胞における作動可能に連結された遺伝子の発現のために使用でき、(1)熱(Callis et al., Plant Physiol. 88:965, 1988)、(2)光(例えばエンドウマメrbcS−3Aプロモーター、Kuhlemeier et al., Plant Cell 1:471, 1989;トウモロコシrbcSプロモーター、 Schaffher and Sheen, Plant Cell 3:997, 1991;又はクロロフィルa/b結合タンパク質プロモーター、Simpson et al., EMBO J. 4:2723, 1985)、(3)ホルモン、例えばアブシジン酸 (Marcotte et al., Plant Cell 1:969, 1989)、(4)創傷(例えばwunI、Siebertz et al., Plant Cell 1:961, 1989); 又は(5)化学物質、例えばジャスモン酸メチル、サリチル酸又は薬害軽減剤、によって調節されるプロモーターを含む。また、(6)器官特異的プロモーター(例えば Roshal et al., EMBO J. 6:1155, 1987; Schernthaner et al., EMBO J. 7:1249, 1988; Bustos et al., Plant Cell 1:839, 1989)を使用することも有益である。本発明において有用なさらなるプロモーターは、国際公開公報第WO97/48819号に述べられているもの;米国特許第 5,591,605号に述べられているファゼオリンプロモーター; 米国特許第5,641,876号に述べられているイネアクチンプロモーター;国際公開公報第WO98/56921号に述べられているper5プロモーター;国際公開公報第WO00/12681号に述べられているγ−ゼインプロモーター;及び米国特許第6,825,006号、同第6,660,911号に述べられている他のプロモーター、及びShahmuradov et al. (2003) “PlantProm: a database of plant promoter sequences,” Nucleic Acid Res. 31(1): 114-117の中で述べられており、http://mendel.cs.rhul.ac.uk/mendel.php?topic=plantpromにおいて入手可能な、植物プロモーター配列データベースPlantPromに述べられている植物プロモーターを含む。さらなるプロモーターはまた、カリフラワーモザイクウイルス(CaMV)35Sプロモーター又はその強化型、ハイブリッド(ocs)3masプロモーター、及びトウモロコシ又はA.タリアナからのユビキチンプロモーターを含み得る。
【0112】
選択植物宿主細胞にネイティブなプロモーターのヌクレオチド配列を有するプロモーターも、標的ペプチドをコードする導入遺伝子の発現を制御するために使用できる。2個以上のプロモーターが、同じか又は異なる配列の、もう1つ別のプロモーターに共有結合している縦列(タンデム)プロモーターも使用し得る。
【0113】
ウイルスに由来するヌクレオチド配列を有するプロモーターも、標的ペプチドをコードする導入遺伝子の発現を指令するために使用できる。
【0114】
調節を受けるプロモーターは、プロモーターがその一部である遺伝子の転写を制御するためにプロモーター調節タンパク質を利用することができる。調節されるプロモーターをここで使用する場合は、対応するプロモーター調節タンパク質も、本発明に従った発現系の一部である。多くの調節されるプロモーター/プロモーター調節タンパク質の対が当技術分野において公知である。
【0115】
(他のエレメント)
他の調節エレメントを発現構築物に含めることができる。そのようなエレメントは、例えば転写エンハンサー配列、翻訳エンハンサー配列、他のプロモーター、アクチベーター、翻訳開始及び終結シグナル、転写ターミネーター、シストロニック調節因子、ポリシストロニック調節因子、発現されるペプチドの特定、分離、精製又は単離を容易にするタグ配列、例えばヌクレオチド配列「タグ」及び「タグ」ペプチドコード配列、例えばHisタグ、Flagタグ、T7タグ、Sタグ、HSVタグ、Bタグ、Strepタグ、ポリアルギニン、ポリシステイン、ポリフェニルアラニン、ポリアスパラギン酸、(Ala−Trp−Trp−Pro)n、チオレドキシン、β−ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、シクロマルトデキストリングルコノトランスフェラーゼ、CTP:CMP−3−デオキシ−D−マンノ−オクツロソネートシチジルトランスフェラーゼ、trpE又はtrpLE、アビジン、ストレプトアビジン、T7遺伝子10、T4gp55、ブドウ球菌プロテインA、連鎖球菌プロテインG、GST、DHFR、CBP、MBP、ガラクトース結合ドメイン、カルモジュリン結合ドメイン、GFP、KSI、c−myc、ompT、ompA、pelB、NusA、ユビキチン及びヘモシリンA、を含むが、これらに限定されない。
【0116】
1つの実施形態では、核酸構築物は、対象組換えペプチドについてのコード配列に隣接する、又はウイルスカプシドについてのコード配列に連結された、タグ配列をさらに含む。1つの実施形態では、タグ配列はタンパク質の精製を可能にする。タグ配列は、アフィニティータグ、例えばヘキサヒスチジンアフィニティータグであり得る。もう1つの実施形態では、アフィニティータグは、グルタチオン−S−トランスフェラーゼ分子であり得る。タグはまた、蛍光分子、例えばYFP又はGFP、又はそのような蛍光タンパク質の類似体であり得る。タグはまた、抗体分子の一部、又は精製のために有用な公知の結合パートナーについての公知の抗原又はリガンドであり得る。
【0117】
本発明は、カプシド−組換えペプチドコード配列に加えて、それに作動可能に連結された以下の調節エレメント:プロモーター、転写ターミネーター、翻訳開始及び終結シグナルを含み得る。
【0118】
翻訳及び転写エレメント、及び本発明において有用な他のエレメントのさらなる例は、JD Watson et al. (eds. ), Recombinant DNA, pp. 273-92 (1992) (Scientific American Books, W. H. Freeman and Co. , New York, NY, USA); and I Mitsuhara et al.,”Efficient promoter cassettes for enhanced expression of foreign genes in dicotyledonous and monocotyledonous plants,”Plant Cell Physiol. 37(1) : 49-59 (1996)に述べられている。
【0119】
1つの特定実施形態では、核酸構築物は、ウイルスカプシドタンパク質を導くために使用されるウイルスゲノム核酸配列の3’非翻訳領域(3’UTR)に由来する付加的な核酸配列を含む。1つの特定実施形態では、3’UTRは、野生型ウイルスにおける翻訳の調節又はカプシド形成において役割を果たす配列を含む。1つの実施形態では、3’UTR配列は上流シュードノットドメイン(UPD)である。例えばLeathers et al.(1993)”A phylogenetically conserved sequence within viral 3' untranslated RNA pseudolmots regulate stranslation,”Mol. Cell Biol. 13 (9): 5331-5347参照。特定実施形態では、3’UTRは、ウイルスゲノムのウイルスカプシドコード領域の3’UTRに由来する。より特定の実施形態では、3’UTRは、カウピークロロティックモザイクウイルスRNA3 3’非翻訳領域に由来する。1つの実施形態では、3’UTR配列はカプシド形成シグナルを含む。
【0120】
代替的実施形態では、核酸構築物は、ウイルス核酸配列に由来する少なくとも1個のカプシド形成シグナルを含む。カプシド形成シグナルは、ウイルスゲノム配列の5’UTR、3’UTR又は内部配列に由来し得る。1つの実施形態では、核酸構築物はVLPにおいてカプシド形成される。
【0121】
1つの実施形態では、非感染性核酸構築物はまた、ウイルスカプシドタンパク質配列以外のウイルス又は非ウイルス核酸配列に由来する他の配列も含む。
【0122】
(ベクター)
宿主細胞ゲノムに安定に組込み、カプシド−組換えペプチド融合ペプチドを発現するとき植物細胞において使用するための有用な発現ベクターは、機能的プロモーターを有する作動可能な読み枠内に、適切な翻訳開始及び終結シグナルと共にカプシドペプチドに融合した所望標的ペプチドをコードする構造DNA配列を挿入することによって構築される。ベクターはまた、宿主細胞の選択を可能にする1又はそれ以上の表現型選択マーカー又はスクリーニングマーカーを含み得る。あるいは、選択又はスクリーニングマーカーは別のプラスミド上で提供され得る。
【0123】
極めて多様なベクター及び/又は担体が、発現のために組換えカプシドをコードする核酸で標的植物宿主細胞を形質転換するために有用であることが当技術分野において公知であり、これらのいずれもが本発明に従った遺伝子を発現するために使用し得る。例えばIJ Goderis IJ et al.,”A set of modular plant transformation vectors allowing flexible insertion of up to six expression units,”Plant Mol. Biol. 50(1): 17-27 (2002); AH Christensen & PH Quail,”Ubiquitin promoter-based vectors for high-level expression of selectable and/or screenable marker genes in monocotyledonousplants,”Transgenic Res. 5 (3): 213-18 (1996); JD Jones et al., “Effective vectors for transformation, expression of heterologous genes, and assaying transposon excision in transgenic plants,”Transgenic Res. 1 (6): 285-97 (1992); AP Gleave AP,”A versatile binary vector system with a T-DNA organisational structure conducive to efficient integration of cloned DNA into the plant genome,”Plant Mol. Biol. 20 (6): 1203-07 (1992)参照。
【0124】
それ故、プラスミド、トランスポゾン、ゲノムDNA、ゲノムRNA、植物人工染色体及び他の核酸ベクターが使用し得る。使用し得る一部のベクターの例は、pRT101、pRT101−MCS、pRT104、pRT104.24LS、pRT104.N−myc、pRT104.C−myc、pRT104.N−3HA、pRT104−N3HAdAsp、pRT104.N−6HA、pRT104−N6HAdAsp、pRT104.C−3HA、pRT104−GST、pRT104 NES(A2)、pRT103−3HA、pRTd35S−Luci(−)、pRTd35S−Luci−NES、pRTd35S−GFP、pRTd35S−GFP.SacI、pRTdS GFP.BglII、pRTd35S−Ds−Red、pRTd35S−プロフィリン、pBIGal4DBD、pRT−3HAxHsfA2DBD、pRT−3HAxGal4DBD、pUC、pIL−TAB、pET、pME、pBBR及びpROKIIを含む。ベクターを細胞に導入するためにパーティクルガンを使用するときは、いかなるDNAプラスミド又はDNA分子も本発明において使用し得る。
【0125】
特定実施形態では、ベクターはpIL−Tabである。より特定の実施形態では、ベクターは、カウピークロロティックモザイクウイルス(CCMV)からのコートタンパク質をコードするpIL−Tabである。特定実施形態では、ベクターはpIL−Tabであり、異種ペプチドは、PA1、PA2、PA3又はPA4から成る群より選択される。さらなる実施形態では、ベクターは、pDOW2160(配列番号5)、pDOW2161(配列番号6)、pDOW2162(配列番号7)、pDOW2163(配列番号8)、pDOW2169(配列番号9)、pDOW2170(配列番号10)、pDOW2171(配列番号11)及びpDOW2172(配列番号12)から成る群より選択される。
【0126】
あるいは又はその上に、担体、例えばリポソーム、デンドリマー、カチオン性ポリマー及びカチオン性ポリマー−脂質複合体が使用でき、前記全てが細胞又はプロトプラストへの核酸の送達のために広く公表されている方法において使用されている。あるいは、裸のDNA又は裸のRNAも植物宿主に送達され得る。
【0127】
ベクター、担体又は裸の核酸は、そのために有効であることが当技術分野で公知の何らかの方法によって植物細胞又はプロトプラストに直接形質転換され得る。例えば、組換えカプシドをコードする核酸で植物標的宿主を形質転換するために以下の方法のいずれかを使用し得る:アグロバクテリウム属;V. Vasil et al., Bio/Technology 9, 743 (1991)によって述べられているようなマイクロパーティクルガン法;例えばMC Chupeau et al., Bio/Technology 7,503 (1989)によってレタスに関して述べられているようなプロトプラストの電気穿孔法;例えば炭化ケイ素ウィスカー、ガラス又はカーボランダムを使用する表面研磨法;例えばA. Deshayes et al., EMBO J. 4, p.2731-2737 (1985)によって述べられているようなプロトプラストとのリポソーム融合;細胞−細胞(プロトプラスト−プロトプラスト)融合;及び内因性植物ウイルス感染、すなわち選択植物標的宿主がその天然宿主であるタイプのウイルスであり、ウイルスが組換えカプシド−タンパク質をコードする核酸を含むように工作されている(組換え体が作られるカプシド−タンパク質−核酸が、植物標的宿主がその天然宿主ではないウイルスから選択される)、工作されたベクターによる植物標的宿主の形質転換;例えば I.Potrykus et al., Mol. Gen. Genetics, 197, 183-188 によって述べられているようなポリエチレングリコールを介した形質転換;及び例えばR Griesbach, Biotechnology 3, p. 348-350およびCK Shewmaker Mol. Gen. Genetics, 202 p. 179-185 (1986)によって述べられているような微量注入法。
【0128】
(III.植物細胞におけるカプシド融合産物の発現)
本発明はまた、組換えペプチドを生産するための方法を提供する。その方法は、
a)植物細胞を提供すること;
b)組換えペプチドと二十面体カプシドの融合ペプチドをコードする発現カセットを含む非感染性核酸を提供すること;
c)前記植物において前記核酸を発現し、前記細胞における発現が前記融合ペプチドのウイルス様粒子へのインビボ集合を提供すること;及び
d)前記ウイルス様粒子を単離すること
を含む。
【0129】
融合ペプチドは、プロモーター配列及びターミネーターに作動可能に連結される。1つの実施形態では、前記方法は、e)ウイルスカプシドタンパク質から組換えペプチドを切断すること、をさらに含む。さらにもう1つの実施形態では、前記方法は、f)工程eから組換えペプチドを単離すること、をさらに含む。例えば図1参照。
【0130】
ペプチドは、カプシドペプチド内で1コピーペプチド挿入物として発現され得る(すなわちペプチドに関してモノシストロニックである組換えカプシドペプチドコード配列から個別挿入物として発現される)か、又は2、3又は多コピーペプチド挿入物として発現され得る(すなわちペプチドに関してポリシストロニックである組換えカプシドペプチドコード配列からコンカテマー挿入物として発現される;コンカテマー挿入物は、同じ外来性対象ペプチドの多数のコピーを含み得るか又は異なる外来性対象ペプチドのコピーを含み得る)。コンカテマーは、ホモ又はヘテロコンカテマーであり得る。
【0131】
1つの実施形態では、単離されたウイルス様粒子をワクチン戦略においてヒト又は動物に投与することができる。
【0132】
もう1つの実施形態では、核酸構築物を、野生型カプシドをコードするもう1つ別の核酸と共発現することができる。特定実施形態では、共発現されたカプシド/カプシド−組換えペプチド融合粒子は、キメラウイルス様粒子を形成するようにインビボで集合する。キメラVLPは、少なくとも2個の異なる核酸構築物によってコードされるカプシド又はカプシド−ペプチド融合物を含むウイルス様粒子である。
【0133】
さらにもう1つの実施形態では、核酸構築物を、異なるカプシド−組換えペプチド融合粒子をコードするもう1つ別の核酸と共発現することができる。特定実施形態では、共発現されたカプシド融合粒子は、キメラウイルス様粒子を形成するようにインビボで集合する。
【0134】
さらにもう1つの実施形態では、異なるペプチド、例えばシャペロンタンパク質を発現するように設計された第二核酸を、融合ペプチドをコードする核酸と同時に発現することができる。
【0135】
本発明のために有用な植物細胞、カプシド及び組換えペプチドは上記に述べられている。
【0136】
1つの実施形態では、発現されるウイルスカプシド−異種ペプチド融合産物は、細胞において可溶性形態で発現される。
【0137】
1つの実施形態では、発現されるウイルスカプシド−異種ペプチド融合産物は、細胞内でVLPに構築される。
【0138】
別の実施形態では、発現される対象ウイルスカプシド−異種ペプチド融合産物の一部は、細胞において不溶性凝集物として形成される。1つの実施形態では、対象ペプチドを不溶性凝集物から復元することができる。
【0139】
(対象ペプチドの切断)
1つの実施形態では、前記方法は、e)カプシドから組換えペプチドを分離するために前記融合産物を切断すること、をさらに提供する。
【0140】
切断可能な連結配列は、ウイルスタンパク質と組換えペプチドの間に含まれ得る。そのような配列を切断することができる物質の例は、化学試薬、例えば酸(塩酸、ギ酸)、CNBr、ヒドロキシルアミン(アスパラギン−グリシンに関して)、2−ニトロ−5−チオシアノベンゾエート、O−ヨードソベンゾエート、及び酵素物質、例えばエンドペプチダーゼ、エンドプロテアーゼ、トリプシン、クロストリパイン及びブドウ球菌プロテアーゼを含むが、これらに限定されない。
【0141】
切断可能な連結配列は当技術分野において周知である。本発明では、Asp−Proなどのジペプチド切断配列を含む、切断物質によって認識されるいかなる切断可能連結配列も使用できる。
【0142】
(発現)
本発明の方法は、最適には、所望配列及び立体配座での、宿主植物細胞における所望カプシド−融合産物の生産上昇を導く。生産上昇は、選択的に、生産されるタンパク質のグラム当り又は宿主タンパク質のグラム当り又は生産される総組換えペプチドのパーセンテージとしての、所望ペプチドのレベル上昇であり得る。生産上昇はまた、組換え体のグラム当り又は宿主細胞タンパク質のグラム当りで生産される回収可能なペプチド、例えば可溶性タンパク質のレベル上昇であり得る。生産上昇はまた、タンパク質の総レベル上昇と活性又は可溶性レベル上昇の何らかの組合せであり得る。
【0143】
組換えタンパク質の改善された発現は、カプシド融合タンパク質としてのタンパク質の発現を通してであり得、その後VLPに挿入することができる。ある実施形態では、少なくとも60、少なくとも70、少なくとも80、少なくとも90、少なくとも100、少なくとも110、少なくとも120、少なくとも130、少なくとも140、少なくとも150、少なくとも160、少なくとも170、又は少なくとも180コピーの対象ペプチドが各々のVLPにおいて発現される。VLPを生産し、宿主細胞の細胞質、ペリプラズム又は細胞外液から回収することができる。
【0144】
もう1つの実施形態では、ペプチドは細胞において不溶性であり得る。ある実施形態では、可溶性又は不溶性ペプチドは、多数のカプシドから形成されるが天然型VLPを形成しない粒子において生産される。例えばわずか3個のウイルスカプシドのケージ構造が形成され得る。ある実施形態では、カプシド構造は2コピー以上の対象ペプチドを含み、ある実施形態では、少なくとも10、少なくとも20又は少なくとも30コピーを含む。ある実施形態では、ペプチドは植物細胞の液胞において形成される。
【0145】
ペプチド又はウイルスカプシド配列はまた、1又はそれ以上のターゲティング配列又は精製を助けるための配列を含み得る。これらはアフィニティータグであり得る。これらはまた、カプシドのVLPへの構築を指令するターゲティング配列であり得る。
【0146】
(細胞増殖)
ベクターによる植物宿主細胞の形質転換は、当技術分野で公知のいかなる形質転換法を用いて実施してもよく、植物宿主細胞を無傷細胞(intact cells)として又はプロトプラストとして形質転換し得る。例示的な形質転換法は上記に述べられている。
【0147】
ここで使用する、「発酵」という用語は、文字通りの発酵を用いる実施形態と、他の非発酵培養法を用いる実施形態の両方を包含する。発酵はいかなる規模で実施してもよい。当技術分野で一般的に知られるいかなる方法も発酵において使用し得る。例えばHellwig et al. (2004) “Plant cell cultures for the production of recombinant proteins, “Nature Biotech 22 (11): 1415-1422; Sajc et al. (2000) “Bioreactors for plant engineering : an outlook for further research, ”Biochem Engin. J. 4: 89-99参照。1つの実施形態では、発酵培地は、何らかの生育可能植物細胞培地から選択され得る。場合により、培地への添加物は、例えばPVP、BSA、NaCl、ブレフェルジンA、還元マンガン、プルロニック消泡剤、ポリエチレングリコール、ゼラチン、プロテアーゼ阻害剤、酸化還元補因子及びジメチルスルホキシドを含み得る。
【0148】
本発明に従った発現系は、いかなる発酵方式でも培養することができる。例えばバッチ、フェドバッチ、半連続、及び連続発酵方式がここで使用し得る。
【0149】
本発明に従った発現系は、いかなる規模(すなわち容量)の発酵においても導入遺伝子発現のために有用である。それ故、例えばマイクロリットル規模、センチリットル規模及びデシリットル規模の発酵容量を使用し得る;及び1リットル又はそれ以上の規模の発酵容量が使用できる。1つの実施形態では、発酵容量は1リットル又はそれ以上である。もう1つの実施形態では、発酵容量は、5リットル又はそれ以上、10リットル、15リットル、20リットル、25リットル、50リットル、75リットル、100リットル、200リットル、500リットル、1,000リットル、2,000リットル、5,000リットル、10,000リットル又は50,000リットルである。
【0150】
本発明では、形質転換宿主細胞の増殖、培養及び/又は発酵は、宿主細胞の生存を許容する温度範囲内、好ましくは約4℃から約55℃(両端を含む)の範囲内の温度で実施される。
【0151】
(細胞密度)(充填湿細胞(Packed wet cell))
本発明に従った植物細胞発現系は、約20g/L(2%充填細胞量)から550g/L(55%充填細胞量)より多くまでの細胞密度又は匹敵する充填湿細胞密度を提供することができる。1つの実施形態では、充填細胞量は2%−約60%である。もう1つの実施形態では、充填細胞量は、2%、5%、10%、15%、20%、25%、35%、40%、50%、55%、60%、70%又は85%である。
【0152】
(VLP又は対象ペプチドの単離)
ある実施形態では、本発明は、ウイルスカプシドとの連結及び共発現を通して発現の間ペプチドを保護することにより、対象ペプチドの回収率を改善するための方法を提供する。ある実施形態では、ウイルスカプシド融合は、細胞溶解産物から容易に分離することができるVLPを形成する。
【0153】
本発明のタンパク質は、PEG沈殿、硫酸アンモニウム又はエタノール沈殿、酸抽出、陰イオン又は陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ニッケルクロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィー、逆相クロマトグラフィー、レクチンクロマトグラフィー、分取電気泳動、界面活性剤可溶化、カラムクロマトグラフィーのような物質による選択的沈殿、免疫精製法等を含むが、これらに限定されない、当技術分野において周知の標準手法によって単離し、実質的純度に精製し得る。例えば確立された分子接着特性を有するタンパク質は、リガンドに可逆的に融合することができる。適切なリガンドを用いて、タンパク質を選択的に精製カラムに吸着させ、その後比較的純粋な形態でカラムから遊離することができる。次に、酵素又は他の作用によって融合タンパク質を取り出すことができる。加えて、免疫アフィニティーカラム又はNi−NTAカラムを用いてタンパク質を精製することができる。一般的な手法は、例えばR. Scopes, Protein Purification: Principles and Practice, Springer-Verlag: N. Y. (1982); Deutscher, Guide to Protein Purification, Academic Press (1990);米国特許第4,511,503号; S. Roe, Protein Purification Techniques: A Practical Approach (Practical Approach Series), Oxford Press (2001); D. Bollag, et al., Protein Methods, Wiley-Lisa, Inc. (1996); AK Patra et al., Protein Expr Purif, 18 (2): p/182-92 (2000);およびR. Mukhija, et al., Gene 165 (2): p. 303-6 (1995)において詳述されている。また、例えばAusubel, et al.(1987 and periodic supplements); Deutscher (1990) “Guide to Protein Purification, “Methods in Enzymology vol.182及びこのシリーズの他の巻; Coligan, et al. (1996 and periodic Supplements) Current Protocols in Protein Science Wiley/Greene, NY ;及びタンパク質精製産物の使用に関する製造者の文献、例えばPharmacia,Piscataway,N.J.又はBio−Rad,Richmond,Calif.も参照のこと。組換え手法の併用は、適切なセグメント、例えばFLAG配列又はプロテアーゼ−除去可能配列によって融合できる等価物、への融合を可能にする。また、例えばHochuli (1989) Chemische Industrie 12: 69-70; Hochuli (1990) “Purification of Recombinant Proteins with Metal Chelate Absorbent”in Setlow (ed.) Genetic Engineering, Principle and Methods 12: 87-98, Plenum Press, NY;およびCrowe, et al.(1992) QIAexpress : The High Level Expression & Protein Purification System QUIAGEN, Inc., Chatsworth, Calif.も参照のこと。
【0154】
同様に、ウイルス様粒子又はケージ様構造も、当技術分野で周知の標準手法によって単離し、実質的純度に精製することができる。VLPの単離のための手法は、上述したものに加えて、沈殿法、例えばポリエチレングリコール又は塩沈殿法を含む。分離手法は、陰イオン又は陽イオン交換クロマトグラフィー、サイズ排除クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ニッケルクロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィー、逆相クロマトグラフィー、レクチンクロマトグラフィー、分取電気泳動、免疫精製法、遠心分離、超遠心分離、密度勾配遠心分離(例えばスクロース又は塩化セシウム(CsCl)勾配で)、サイズ排除フィルターを通しての限外ろ過、及び当技術分野で公知の何らかの他のタンパク質単離法を含む。
【0155】
本発明はまた、活性組換えペプチドの回収率を改善することができる。活性タンパク質のレベルは、例えば特定されたペプチドと親ペプチド、ペプチド変異体、セグメント置換ペプチド及び/又は残基置換ペプチドの間の相互作用を従来の何らかのインビトロ又はインビボアッセイで測定することによって測定できる。そこで、インビトロアッセイは、特定されたタンパク質と対象ペプチドの間、例えば酵素と基質の間、ホルモンとホルモン受容体の間、抗体と抗原の間等の、何らかの検出可能な相互作用を測定するために使用できる。そのような検出は、比色変化、放射能の変化、溶解度の変化、ゲル電気泳動及び/又はゲル排除法によって測定したときの分子量の変化等の測定を含み得る。インビボアッセイは、生理的作用、例えば体重増加、電解質バランスの変化、血液凝固時間の変化、血餅溶解の変化及び抗原応答の誘導を検出するアッセイを含むが、これらに限定されない。一般に、特定されたペプチドと対象ペプチドの間の相互作用の変化を検出するための可変パラメータが存在する限り、いかなるインビボアッセイも使用することができる。例えば米国特許第5,834,250号参照。
【0156】
発現されたタンパク質の検出は、当技術分野で公知の方法によって達成され、例えば放射免疫測定法、ウエスタンブロット法又は免疫沈降法を含む。
【0157】
初期塩分画は、望ましくない宿主細胞タンパク質(又は細胞培養培地に由来するタンパク質)の多くを対象組換えタンパク質から分離することができる。1つのそのような例は硫酸アンモニウムであり得る。硫酸アンモニウムは、タンパク質混合物中の水分量を有効に減少させることによってタンパク質を沈殿させる。タンパク質はその後、それらの溶解度に基づいて沈殿する。タンパク質がより疎水性であるほど、より低い硫酸アンモニウム濃度で沈殿する可能性が高い。典型的なプロトコールは、生じる硫酸アンモニウム濃度が20−30%になるように飽和硫酸アンモニウムをタンパク質溶液に添加することを含む。この濃度は最も疎水性のタンパク質を沈殿させる。次に沈殿物を廃棄し(対象タンパク質が疎水性でない限り)、対象タンパク質を沈殿させることが既知である濃度まで硫酸アンモニウムを上清に添加する。次に沈殿物を緩衝液に溶解し、必要に応じて、透析又はダイアフィルトレーションのいずれかを通して過剰の塩を除去する。タンパク質の溶解度に基づく他の方法、例えば低温エタノール沈殿は当業者に周知であり、複雑なタンパク質混合物を分画するために使用できる。
【0158】
組換えタンパク質の分子量を利用して、異なる細孔径の膜(例えばアミコン又はミリポア膜)を通す限外ろ過を用いてより大きい及びより小さいサイズのタンパク質から単離することができる。第一段階として、タンパク質混合物を、対象タンパク質の分子量よりも低い分子量カットオフ値を有する細孔径の膜を通して限外ろ過することができる。限外ろ過の保持物質を、次に、対象タンパク質の分子量よりも大きい分子量カットオフ値を有する膜に対して限外ろ過することができる。組換えタンパク質は膜を通過してろ液中に入る。その後ろ液をクロマトグラフィーに供することができる。
【0159】
組換えタンパク質はまた、その大きさ、正味表面電荷、疎水性、及びリガンドに対する親和性に基づいて他のタンパク質から分離することができる。加えて、タンパク質に対して惹起される抗体をカラムマトリックスに結合(固定化)し、タンパク質を免疫精製することができる。これらの方法は全て当技術分野において周知である。クロマトグラフィー手法が、いかなる規模でも及び多くの異なる製造者(例えばPharmacia Biotech)からの装置を使用して実施できることは当業者には明白である。
【0160】
(復元及びリフォールディング)
二次及び三次タンパク質構造立体配座を生成するために不溶性タンパク質を復元又はリフォールディングすることができる。タンパク質リフォールディング工程は、必要に応じて、組換え産物の立体配置を完成させるときに使用できる。リフォールディング及び復元は、タンパク質の解離/会合を促進する当技術分野で公知の物質を用いて実施することができる。例えばタンパク質をジチオトレイトールと共にインキュベートし、次に酸化グルタチオンニナトリウム塩と共にインキュベートして、さらにその後リフォールディング剤、例えば尿素を含む緩衝液と共にインキュベートすることができる。
【0161】
組換えタンパク質はまた、例えばリン酸緩衝食塩水(PBS)又は50mM 酢酸ナトリウム、pH6緩衝液プラス200mM NaClに対して透析することによっても復元できる。あるいは、タンパク質を、カラム、例えばNi NTAカラムに固定化して、プロテアーゼ阻害因子を含有する、500mM NaCl、20%グリセロール及び20mM トリス/HCl pH7.4中の6M−1M 尿素直線勾配を使用することによって再生できる。復元は1.5時間又はそれ以上の期間で実施できる。復元後、250mM イミダゾールの添加によってタンパク質を溶出することができる。イミダゾールは、PBS又は50mM 酢酸ナトリウム、pH6緩衝液プラス200mM NaClに対する最終透析工程によって除去することができる。精製タンパク質は、室温、4℃で保存するか又は−20℃から−80℃で凍結して保存することができる。
【0162】
他の方法は、例えばMH Lee et al., Protein Expr. Purif., 25(1) : p. 166-73 (2002), W. K. Cho et al. , J. Biotechnology, 77 (2-3): p. 169-78 (2000), Ausubel, et al. (1987 and periodic supplements), Deutscher (1990) “Guide to Protein Purification, “Methods in Enzymology vol. 182, and other volumes in this series, Coligan, et al. (1996 and periodic Supplements) Current Protocols in Protein Science Wiley/Greene, NY, S. Roe, Protein Purification Techniques: A Practical Approach (Practical Approach Series), Oxford Press (2001); D. Bollag, et al. , Protein Methods, Wiley-Lisa, Inc. (1996)に述べられているものを含む。
【0163】
(活性ペプチド分析)
活性タンパク質は、その配列が由来する天然ペプチドの活性の少なくとも20%、30%又は40%、好ましくは少なくとも50%、60%又は70%、最も好ましくは少なくとも80%、90%又は95%の比活性を有し得る。さらに、基質特異性(Kcat/Km)は、場合により天然ペプチドと実質的に同様である。典型的には、Kcat/Kmは、天然ペプチドの少なくとも30%、40%又は50%、より好ましくは少なくとも60%、70%、80%又は90%である。タンパク質及びペプチドの活性及び基質特異性(Kcat/Km)を検定する方法及び定量手段は当業者に周知である。
【0164】
本発明に従って生産される組換えペプチドの活性は、当技術分野で公知の何らかのタンパク質特異的な慣例的又は標準的インビトロ又はインビボアッセイによって測定することができる。組換えタンパク質が、同じ又は類似の生理的条件下で天然ペプチドにおいて一般に認められる活性と実質的に類似又は同等の活性を示すかどうかを判定するために、植物細胞宿主が生産する組換えペプチドの活性を、対応する天然タンパク質の活性と比較することができる。
【0165】
組換えペプチドの活性を、これまでに確立された天然ペプチド標準活性と比較することができる。あるいは、組換えペプチドの活性を、天然ペプチドと同時又は実質的に同時の比較アッセイにおいて測定することができる。例えばインビトロアッセイを使用して、組換えペプチドと標的の間、例えば発現される酵素と基質の間、発現されるホルモンとホルモン受容体の間、発現される抗体と抗原の間等の、検出可能な相互作用を測定することができる。そのような検出は、比色変化、増殖変化、細胞死、細胞反発、放射能の変化、溶解度の変化、ゲル電気泳動及び/又はゲル排除法によって測定したときの分子量の変化、リン酸化能、抗体特異性アッセイ、例えばELISAアッセイ等の測定を含み得る。加えて、インビボアッセイは、天然ペプチドの生理的作用、例えば抗原応答の誘導と比較して、植物宿主細胞が生産するペプチドの生理的作用を検出するアッセイを含むが、これらに限定されない。一般に、いかなるインビトロ又はインビボアッセイも、そのような活性が検定可能である限り、天然ペプチドとの比較分析を可能にする組換えペプチドの活性を測定するために使用できる。あるいは、本発明において生産されるペプチドは、ペプチドと、通常そのペプチドと相互作用する分子、例えば基質又は天然タンパク質が通常相互作用するシグナル経路の成分、との間の相互作用を刺激する又は阻害する能力に関して検定することができる。そのようなアッセイは、典型的には、ペプチドが標的分子と相互作用することを許容する条件下でタンパク質と基質分子を組み合わせる工程を含み、タンパク質と標的分子の相互作用の生化学的結果を検出することができる。
【0166】
ペプチド活性を測定するために使用できるアッセイは、例えばRalph, P. J., et al. (1984) J. Immunol. 132:1858 or Saiki et al. (1981) J. Immunol. 127: 1044, Steward, W. E.II (1980) The Interferon Systems. Springer-Verlag, Vienna and New York, Broxmeyer, H. E., et al. (1982) Blood 60: 595, “Molecular Cloning: A Laboratory Manual”, 2d ed., Cold Spring Harbor Laboratory Press, Sambrook, J., E. F. Fritsch and T. Maniatis eds. , 1989, and “Methods in Enzymology: Guide to Molecular Cloning Techniques”, Academic Press, Berger, S. L. and A. R. Kimmel eds., 1987, AK Patra et al., Protein Expr Purif, 18 (2):p/182-92 (2000), Kodama et al., J. Biochem. 99: 1465-1472 (1986); Stewart et al., Proc.Nat'1 Acad. Sci. USA 90: 5209- 5213 (1993); (Lombillo et al., J. Cell Biol. 128: 107-115 (1995); (Vale et al., Cell 42: 39-50 (1985) に述べられている。
【実施例】
【0167】
コートタンパク質をコードする核酸のウイルスソース
以下の実施例では、カウピークロロティックモトルウイルス(CCMV)を所望組換えペプチドの発現のためのコートタンパク質のソースとして使用した。CCMVは、ブロモウイルス科ブロモウイルス群の成員である。ブロモウイルスは、4成分のプラスセンス一本鎖RNAゲノムを有する直径25−28nmの二十面体ウイルスである。RNA1及びRNA2はレプリカーゼ酵素をコードする。RNA3は、植物宿主内のウイルス運動に関与するタンパク質をコードする。RNA4(RNA3に由来するサブゲノムRNA)、すなわちsgRNA4は、20kDaのコートタンパク質(CP)(配列番号13)(表1)をコードする。各々のCCMV粒子は、180コピーのCCMV CPを含む。CCMV CPをコードする例示的DNA配列を配列番号14(表2)に示す。
【表1】
【表2】
【0168】
CCMVの結晶構造を解明した。この構造は、粒子の溶解度と動力学にとって決定的に重要であると思われるコートタンパク質相互作用のより明瞭な像を提供し、挿入部位の合理的設計を導くのに有用であった。これまでの試験は、CCMVコートタンパク質を、粒子を形成するそれらの能力を妨げることなく異種ペプチドを担持するように遺伝的に修飾できることを明らかにした。多くの適切な挿入部位が特定されている。合計約180コピーまでの異種ペプチド単位(個々のペプチド単位として又はコンカテマー単位内で)を、CCMV CP内の単一挿入部位でCCMV粒子に挿入することができる。現在までにCCMV CP内で特定された挿入部位は、様々な長さのペプチドを収容することができる。
【0169】
実験材料及び方法
異なる記載がない限り、分子生物学の分野において公知の標準手法、ベクター、制御配列エレメント及び他の発現系エレメントを、核酸操作、形質転換及び発現のために使用する。そのような標準手法、ベクター及びエレメントは、例えばAusubel et al. (eds.), Current Protocols in Molecular Biology (1995) (John Wiley & Sons); Sambrook, Fritsch, & Maniatis (eds.), Molecular Cloning (1989) (Cold Spring Harbor Laboratory Press, NY) ; Berger & Kimmel, Methods in Enzymology 152: Guide to Molecular Cloning Techniques(1987) (Academic Press); and Bukhari et al. (eds. ), DNA Insertion Elements, Plasmids and Episomes (1977) (Cold Spring Harbor Laboratory Press, NY)に見出すことができる。
【0170】
異なる記載がない限り、PCR反応は、以下のプロトコールに従ってPTC225サーモサイクラー(MJ Research,South San Francisco,CA,USA)を使用して実施した:
【表3】
【0171】
(実施例1)CCMV RNA1、RNA2及びキメラRNA3を接種したカウピー全植物体におけるCCMVウイルス粒子内での抗原性ペプチドの生産
カウピークロロティックモトルウイルス(CCMV)を、4個の異なる抗原性ペプチドの1個を含むように工作したカプシドタンパク質(CP)と共に使用して、炭疽菌抗原性ペプチドの発現を全植物体において実施した。
【0172】
CCMV RNA1のヌクレオチド配列を有するDNA及びCCMV RNA2のヌクレオチド配列を有するDNAを、T7プロモーターの下流でその制御下に、及びユニークXbaI部位の上流で、クローニングベクターpUC19に各々別々にサブクローニングした。これはプラスミドpDOW2122(CCMV RNA1)及びpDOW2123(CCMV RNA2)を生産した。
【0173】
5つのBamHI制限酵素切断部位を含むように工作した、CCMV RNA3のヌクレオチド配列を有するDNAを、組換えカプシドタンパク質をコードする核酸の生産のためにさらに工作した。各々が、4個の外来性ペプチド(炭疽菌防御抗原PAからの4個の異なる抗原性ペプチド)の異なる1個をコードする4個のDNA分子を、各々合成オリゴヌクレオチドのSOE(オーバーラップ伸長によるスプライシング)(splicing-by-overlap-extension)によって合成した。生じた核酸はBamHI認識部位末端を含んだ。各々のPA DNAフラグメントをBamHI制限酵素で制限し、CCMVコートタンパク質コード配列内の5つの異なる工作制限工切断部位の1つでコートタンパク質に独立して挿入した:コドン63のBamHI、コドン102のBamHI、コドン114のBamHI、コドン129のBamHI、コドン160のBamHI。
【0174】
4個の異なるペプチドは、PA1(配列番号15によってコードされる配列番号1)、PA2(配列番号16によってコードされる配列番号2)、PA3(配列番号17によってコードされる配列番号3)及びPA4(配列番号18によってコードされる配列番号4)であった。
【表4】
【0175】
これらの各々を、BamHI制限酵素で消化して、その後脱リン酸化しておいた5個のプラスミド:pDOW2125(pUC−CCMV−RNA3−CP63BamHI)、pDOW2126(pUC−CCMV−RNA3−102BamHI)、pDOW2127(pUC−CCMV−RNA3−CP114BamHI)、pDOW2128(pUC−CCMV−RNA3−CP129BamHI)及びpDOW2129(pUC−CCMV−RNA3−CP160BamHI)、の各々に挿入した。
【0176】
これは、「キメラRNA3」についての合計20の異なる型のDNAを生産した。プラスミドpDOW2135(pUC−CCMV−RNA3−CP63BamHI−PA1)、pDOW2139(pUC−CCMV−RNA3−102BamHI−PA1)、pDOW2143(pUC−CCMV−RNA3−CP114BamHI−PA1)、pDOW2147(pUC−CCMV−RNA3−CP129BamHI−PA1)及びpDOW2151(pUC−CCMV−RNA3−CP160BamHI−PA1)は、63、102、114、129及び160の位置にPA1挿入物を有するキメラRNA3の例である。野生型CCMVコートタンパク質をコードするRNA3及びBamHI制限部位を含むが挿入物を含まない工作したCCMVコートタンパク質をコードするRNAを対照として使用した。各々の修飾RNA3構築物を、T7プロモーターの下流でその制御下に、及びユニークXbaI部位の上流で、クローニングベクターpUC19に別々にサブクローニングした。
【0177】
3つのクラスのプラスミドの各々を大腸菌にクローニングした。プラスミドを単離し、XBaI制限酵素消化によって線状化して、次にMESSAGE mMACHINE T7 Kit(RNA転写キット、Ambion,Inc.,Austin,TX,USAより)を用いて各々1μgをインビトロでRNAに転写した。これは24個の異なるRNA種:RNA1について1個、RNA2について1個、キメラRNA3について20個、及びRNA3対照について2個、を生産した。
【0178】
カウピー植物に感染させるためにRNA1、RNA2、RNA3又はPA挿入物を含むキメラRNA3のカクテル混合物を使用した。カウピー植物を、カウピーカリフォルニアブラックアイNo.5種子(Ferry−Morse Seed Co.KY)から発芽させた。新芽を、Miracle−Gro鉢植え混合物(Miracle−Gro Lawn Products OH)と共に1つずつ6インチの鉢に移植した。2葉期(発芽後約7日目)にカウピー植物を感染させた。各々の植物の1枚の葉にカーボランダム(Carborundum)粉末400粗粒( Fisher Scientificカタログ番号409−21−2)の散粉を実施した。カーボランダム層にRNAカクテル混合物を適用した。手袋をはめた指で静かにこすって葉を削り取った。接種後7−14日で感染が確認された。
【0179】
意外にも、この全植物体系では、天然宿主植物を使用したにもかかわらず、キメラコートタンパク質は野生型に復帰するか、又は一部の場合は、所望外来性抗原ペプチドが部分的に欠失した。これはウイルス子孫の配列決定によって確認された。RNAを抽出し、ThermoScript RT−PCRシステム(Invitrogenカタログ番号11146−024)及びCCMVRNA4.R遺伝子特異的プライマー(配列番号19) (5’−CTC GAG CTA ATA CAC CGG AGT GAA AG−3’)を用いてDNAに逆転写した。cDNAを、プライマーCCMVRNA4.F(配列番号20)(5’−CTG CAG ATG TCT ACA GTC GGA ACA GG−3’)及びCCMVRNA4.Rを使用するPCRによってさらに増幅した。反応物を精製し、CCMV−CP−F1プライマー(配列番号21)(5’−AAC CCA TCG CTT CAG GCC AA−3’)で配列決定した。配列をシーケンサーソフトウエアバージョン4.0.5(Gene Codes Corporation)で分析した。例えばPA1挿入物を含むpDOW2128に由来するRNA3子孫を配列決定すると、 コートタンパク質は、PA1挿入物アミノ酸配列の代わりにRISDPLとして翻訳された、小さな認識されないヌクレオチド挿入物(5’−CGTATTTCTGATCCTCTC−3’)を含んだ。挿入したヌクレオチド配列はコートタンパク質の残りの部分をインフレームで保持した。工作CCMV RNA3は、全植物における発現時に不安定であり、容易に組換えを受け、全植物体系はキメラCCMVウイルス粒子上のエピトープの生産のために有用ではないと判定された。
【0180】
(実施例2)CCMV RNA1、RNA2及びキメラRNA3を接種したタバコ懸濁培養中のCCMVウイルス粒子における抗原性ペプチドの生産
同じCCMV−ペプチドをコードする構築物の発現を植物細胞懸濁培養において実施した。ニコチアナ・タバカムNT1細胞を、キメラCCMVコートタンパク質をコードするCCMV RNA3及びレプリカーゼ遺伝子をコードするCCMV RNA1及び2のRNA転写産物での電気穿孔によってトランスフェクトした。野生型CCMVコートタンパク質をコードするRNA3及び適切なBamHI制限部位を含むが挿入物を含まない工作したCCMVコートタンパク質をコードするRNA3を対照として使用した。
【0181】
24個の異なるRNA種:RNA1について1個、RNA2について1個、キメラRNA3について20個、及びRNA3対照について2個を、実施例1で述べたようなインビトロRNA転写によって得た。22個の異なる群において、3個の生じたRNA(RNA1、RNA2、及びRNA3の1個)の各々2μgを電気穿孔法によってタバコ細胞に形質転換した。
【0182】
以下のプロトコールを植物細胞トランスフェクションのために使用した:
1)培地の調製:
NTI培地(1リットル):
ムラシゲ&スクーグ基礎塩4.33g(Phyto Technology Laboratories KSカタログ番号M524)ミオ−イノシトール100mg(Sigmaカタログ番号I−3011)、塩酸チアミンの1mg/ml溶液1ml(Sigmaカタログ番号T−3902)、リン酸二水素カリウム(KH2PO4)180mg(Sigmaカタログ番号P−8416)、スクロース30g(Sigmaカタログ番号S−5390)、及び10mg/mlの2,4−D溶液200μl(Sigmaカタログ番号D−7299)を少量の水中で混合した。精製水を前記溶液に添加して容量を1リットルにした。pHを5.8に調整し、溶液を加圧滅菌した。
【0183】
マンニトール洗浄液0.4M:
マンニトール36.43g(Sigmaカタログ番号M−1902)を少量の精製水に添加した。精製水を添加して容量を500mlにし、pHを5.5に調整した。滅菌するために溶液をオートクレーブに入れた。
【0184】
酵素溶液:
0.4M マンニトール及び0.02M MESを少量の水中で混合した。精製水を添加して容量を500mlにし、pHを5.5に調整した。滅菌するために溶液をオートクレーブに入れ、溶液を4℃で保存した。使用前に、1%セルライシン(Carbiochemカタログ番号219466)及び0.3%マセラーゼペクチナーゼ(Carbiochemカタログ番号441201)を溶液に添加し、セルライシンとマセラーゼペクチナーゼが溶解するまで溶液を振とうした。
【0185】
電気穿孔緩衝液:
0.8%NaCl、0.02%KC1、0.02%KH2PO4及び0.11%Na2HPO4を少量の精製水に添加した。精製水を添加して溶液を100mlにし、pHを6.5に調整した。緩衝液を加圧滅菌し、4℃で保存した。
【0186】
2)タバコ細胞部分消化プロトコール:
タバコ細胞系NT1を電気穿孔の前に以下のように部分消化した。週に1回継代培養することによって強固な細胞系を維持し、細胞を、静かに振とうしながらNT1培地において24℃又は28℃で増殖させ、消化の3日前に、細胞懸濁液5mlをNT1培地50mlに継代培養して28℃でインキュベートした。細胞を50mlチューブにおいて800rpmで5分間遠心沈殿させた。酵素溶液を調製し、細胞をマンニトール洗浄液(約40ml)で洗った。細胞を800rpmで5分間遠心し、3容の酵素溶液を細胞に添加した。倒置して細胞を再懸濁し、10cmペトリ皿に注ぎ入れて移し、皿をアルミニウム箔に包んで60−120分間、室温で非常に緩やかに振とうした。次に細胞を50mlプラスチックチューブに戻し、800rpmで5分間遠心した。細胞をマンニトール洗浄液40mlで洗い、再び遠心して、電気穿孔緩衝液で洗浄し、遠心した。3容の電気穿孔緩衝液(総容量は通常20mlである)を細胞に添加した;細胞をアルミニウム箔に包んで4℃で保存した。
【0187】
部分消化した細胞の電気穿孔プロトコール:
消化した細胞1mlをアリコートに分けて電気穿孔キュベットに入れた(4mm間隔)。各々のRNA転写産物2μg−CCMV RNA1、RNA2及びキメラRNA3−をキュベットに添加し、5分間氷上に置いた。NT1平板培養培地10ml(NT1+0.4M マンニトール)を各々のペトリ皿に添加した。細胞を500μF、250Vで電気穿孔し、キュベットを氷上に戻した。次に細胞をペトリ皿に移し、暗所で振とうせずに室温でインキュベートした。トランスフェクション後48時間目に分析のために細胞を収集した。
【0188】
結果
キメラコートタンパク質の発現を、抗CCMVコートタンパク質ポリクローナル抗体を用いたウエスタンブロット法によって分析した。また、各々の培養物からRNAを抽出し、キメラCCMV RNA4をcDNAに逆転写して、それをPCRによって増幅した。PCR産物を実施例1で述べたように配列決定した。RNA1及び2だけでトランスフェクトした陰性対照を除いて、全ての試料がウエスタンブロット法で陽性であった。
【0189】
対照CCMVカプシドタンパク質と比較して、キメラコートタンパク質はより大きなサイズとゲル上でのより緩慢な運動性を有しており、キメラコートタンパク質がPA挿入物を含むことを示した。これは、ウイルスRNA子孫の配列決定によって確認された。結果は、20個のキメラ構築物のうち19個が、所望抗原ペプチド内に突然変異を含むことなく正しくキメラCCMVコートタンパク質を発現したことを明らかにした。この結果は、懸濁細胞において発現されたキメラCCMV RNA3が安定であることを示す。
【0190】
図2は、CCMV RNA1、RNA2、及びpDOW2125(CCMV63BamHI)、pDOW2126(CCMV102BamHI)、pDOW2127(CCMV−114BamHI)、pDOW2128(CCMV129BamHI)及びpDOW2129(CCMV160BamHI)から転写されたRNA3でトランスフェクトした細胞におけるCCMV CPの発現を示す。結果は、CP内の異なる位置に工作BamHI部位を有する全てのRNA3からCCMV CPが発現されることを明らかにする。
【0191】
63、102、114、129及び160BamHI部位でCCMVコートタンパク質に融合した全てのPAペプチド(PA1、PA2、PA3及びPA4)が、タバコ細胞において成功裏に生産された。図3及び図4は、CCMV RNA1、RNA2、及び63、102、114、129及び160位に4個のPAペプチド挿入物を有するCPを含むキメラRNA3でトランスフェクトした細胞におけるキメラCCMV CPの発現を示す。図3は、あらかじめ63、102又は114位に挿入されたPA1、PA2、PA3又はPA4ペプチドを有するCCMV CPの融合タンパク質が、63位でCCMV CPに挿入されたPA2ペプチドを除き、タバコ植物細胞において発現されることを明らかにする。図4は、129又は160位に挿入されたPA1、PA2、PA3又はPA4ペプチドを有するCCMV CPの融合タンパク質がタバコ植物細胞において発現されることを示す。
【0192】
(実施例3)キメラCCMV CPをコードする植物発現プラスミドでトランスフェクトしたタバコ懸濁培養中のCCMVウイルス様粒子における4つの抗原性ペプチドの生産
【0193】
1)ベクターの構築:
プラスミドpIL−Tab358を植物発現ベクターとして使用した。CCMV CP挿入のために選択した制限部位はXbaIとEcoRIであった。このプラスミドは、XbaI部位の上流にキャッサバ葉脈モザイクウイルスプロモーター及びEcoRI部位の下流にNosターミネーターを含む。前記ベクターは、XbaI及びEcoRIで消化し、脱リン酸化した後、挿入物と連結することによって作製した。
【0194】
2)挿入物の構築:
CCMV CP−PA融合物を、プライマーCCMV−CP−XbaI(配列番号22)及びCCMV−CP−EcoRI(配列番号23)を使用してpDOW2147(pUC−CCMV−RNA3−CP129BamHI−PA1)、pDOW2148(pUC−CCMV−RNA3−CP129BamHI−PA2)、pDOW2149(pUC−CCMV−RNA3−CP129BamHI−PA3)、pDOW2150(pUC−CCMV−RNA3−CP129BamHI−PA4)からPCRによって増幅し、pDOW2160(pIL−Tab−CCMV129BamHI−PA1)(配列番号5)、pDOW2161(pIL−Tab−CCMV129BamHI−PA2)(配列番号6)、pDOW2162(pIL−Tab−CCMV129BamHI−PA3)(配列番号7)及びpDOW2163(pIL−Tab−CCMV129BamHI−PA4)(配列番号8)を作製した。pDOW2160、pDOW2161、pDOW2162及びpDOW2163についてのプラスミド地図を図5、6、7及び8に示す。
【0195】
3)植物細胞トランスフェクション:
植物細胞をプラスミドpDOW2160、pDOW2161、pDOW2162及びpDOW2163 10μgでトランスフェクトした。植物細胞トランスフェクションは実施例2におけるように実施した。
【0196】
結果
129BamHI部位の4つのPAペプチド−CCMV CP融合物全てがタバコ細胞において成功裏に発現された。図9は、pDOW2160、pDOW2161、pDOW2162及びpDOW2163でトランスフェクトした細胞におけるキメラCCMV CPの発現を示す。対照(挿入物を含まないCCMVコートタンパク質)と比較して、4つのキメラコートタンパク質全てが、PA挿入物を有することを表すより緩慢な運動性を示した。
【0197】
(実施例4)キメラCCMV CP及び3’UTR(CCMV RNA3 3’非翻訳領域)をコードする植物発現プラスミドでトランスフェクトしたタバコ懸濁培養中のウイルス様粒子におけるCCMV CP融合物としての4つの抗原性ペプチドの生産
1)ベクターの構築:
プラスミドpIL−Tab358を植物発現ベクターとして使用した。CCMV CP挿入のために選択した制限部位はXbaIとEcoRIであった。このプラスミドは、XbaI部位の上流にキャッサバ葉脈モザイクウイルスプロモーター及びEcoRI部位の下流にNosターミネーターを含む。前記ベクターは、XbaI及びEcoRIで消化し、脱リン酸化した後、挿入物と連結することによって作製した。
【0198】
2)挿入物の構築:
RNA3の3’UTRを含むCCMV CP−PA融合物を、プライマーCCMV−CP−XbaI及びCCMV−CP−EcoRI−3’UTR(配列番号24)を使用してpDOW2147、pDOW2148、pDOW2149及びpDOW2150からPCRによって増幅し、pDOW2169(pIL−Tab−CP129BamHI−PA1−3’UTR)(配列番号9)、pDOW2170(pIL−Tab−CP129BamHI−PA2−3’UTR)(配列番号10)pDOW2171(pIL−Tab−CP129BamHI−PA3−3’UTR)(配列番号11)及びpDOW2172(pIL−Tab−CP129BamHI−PA4−3’UTR)(配列番号12)を作製した。
【0199】
3)植物細胞トランスフェクション:
植物細胞をプラスミドpDOW2169、pDOW2170、pDOW2171及びpDOW2172 10μgでトランスフェクトした。植物細胞トランスフェクションは実施例2におけるように実施した。
【0200】
結果
3’UTRを含む129BamHI部位の4つのPAペプチド−CCMV CP融合物全てがタバコ細胞において成功裏に発現された。図10は、pDOW2169、pDOW2170、pDOW2171及びpDOW2172でトランスフェクトした細胞におけるキメラCCMV CPの発現を示す。対照(挿入物を含まないCCMVコートタンパク質)と比較して、4つのキメラコートタンパク質全てが、PA挿入物を有することを表すより緩慢な運動性を示した。
【0201】
(実施例5)キメラCCMV CPをコードする植物発現プラスミドでトランスフェクトしたトランスジェニックタバコ懸濁培養中のウイルス様粒子におけるCCMV CP融合物としての4つの抗原性ペプチドの生産
タバコ細胞系NT1を、週に1回継代培養することによって維持した。細胞を、静かに振とうしながらNT1培地において24℃又は28℃で増殖させた。トランスフェクションの3日前に、細胞懸濁液5mlをNT1培地50mlに継代培養して28℃でインキュベートした。
【0202】
1)パーティクルガンによる植物細胞形質転換:
プラスミドpDOW2160、pDOW2161、pDOW2162、pDOW2163及びキャッサバ葉脈モザイクウイルスプロモーターによって駆動される植物選択マーカーPatを含む植物発現プラスミドpBBV(pBBV)を、マイクロパーティクルガン法によるNT1タバコ細胞形質転換のために使用した。使用したプラスミド比は、1:6 pBBV:pDOW2160、pBBV:pDOW2161、pBBV:pDOW2162又はpBBV:pDOW2163であった。合計5μgのプラスミドDNAは、6回のパーティクルガン法のために十分であった。Chen, L et al. Plant Cell Reports (1998) 18: 25-31 に述べられているBioradパーティクルガンプロトコールを形質転換のために使用した。
【0203】
2)プレーティング
パーティクルガンを打ち込んだ細胞を、非選択的NT1培地寒天プレートに4時間移した後、25μg/mlのグルフォシネート−アンモニウム(Sigmaカタログ番号45520)を含むNT1培地寒天プレートに移した。
【0204】
3)トランスジェニックカルスの選択
21日後、白色綿毛状細胞増殖を有していたカルスを、CP融合物の発現を調べるためのウエスタンブロット法及びプロモーター−CP融合物遺伝子−ターミネーターカセットの植物ゲノムへの組込みを調べるためのPCRによる分析のために選択した。キメラCCMV CPを発現するトランスジェニックカルスをNT1液体培地に移した。細胞を、静かに振とうしながらNT1培地において24℃又は28℃で増殖させ、週に1回継代培養した。
【0205】
結果
図11は、pDOW2160、pDOW2161、pDOW2162及びpDOW2163で安定に形質転換された細胞におけるキメラCCMV CPの発現を示す。CCMVに対するポリクローナル抗体によってCP融合物を検出した。4つのキメラコートタンパク質導入遺伝子全ての発現が検出された。
【0206】
(実施例6)キメラCCMV CPをコードする植物発現プラスミドでトランスフェクトしたトランスジェニックイネ懸濁培養中のCCMVウイルス様粒子におけるCCMV CP融合物としての4つの抗原性ペプチドの生産
イネ細胞系を、週に1回継代培養することによって維持した。細胞を、静かに振とうしながらNB培地(Li, L et al. Plant Cell Reports. (1993) 12: 250-255)において28℃で増殖させた。トランスフェクションの3日前に、細胞懸濁液5mlをNB培地50mlに継代培養して28℃でインキュベートした。
【0207】
1)パーティクルガンによる植物細胞形質転換:
プラスミドpDOW2160、pDOW2161、pDOW2162及びpDOW2163、及びキャッサバ葉脈モザイクウイルスプロモーターによって駆動される植物選択マーカーPatを含む植物発現プラスミドpBBVを、マイクロパーティクルガン法によるイネ細胞形質転換のために使用した。使用したプラスミド比は、1:6 pBBV:pDOW2160、pBBV:pDOW2161、pBBV:pDOW2162又はpBBV:pDOW2163であった。合計5μgのプラスミドDNAは、6回のパーティクルガン法のために十分であった。Chen, L et al. Plant Cell Reports (1998) 18: 25-31 に述べられているBioradパーティクルガンプロトコールを形質転換のために使用した。
【0208】
2)プレーティング
パーティクルガンを打ち込んだ細胞を、選択のために25μg/mlのグルフォシネート−アンモニウム(Sigmaカタログ番号45520)を含むNB培地寒天プレートに移した。
【0209】
3)トランスジェニックカルスの選択
21日後、白色綿毛状細胞増殖を有していたカルスを、CP融合物の発現を調べるためのウエスタンブロット法及びプロモーター−CP融合物遺伝子−ターミネーターカセットの植物ゲノムへの組込みを調べるためのPCRによる分析のために選択した。
【0210】
結果
図12は、CCMVに対するポリクローナル抗体によって検出したときの、pDOW2160、pDOW2161、pDOW2162及びpDOW2163で安定に形質転換された細胞におけるキメラCCMV CPの発現を示す。4つのキメラコートタンパク質導入遺伝子全ての発現が検出された。
【0211】
ゲノムDNAを抽出し、プロモーター−CP融合物遺伝子−ターミネーターカセットの植物ゲノムへの組込みをPCRによって調べるために以下のプロトコールを使用した。ゲノムDNAを抽出するためにQiagen DNeasy Plant Miniを製造者の指示に従って使用した。
【0212】
新鮮イネカルス約50−100mgを1.5mlチューブに入れた。即時DNA抽出のために組織を液体窒素中で凍結するか又は保存のために−80℃の冷凍庫に入れた。DNA抽出の前に、組織をミクロ乳鉢(micropestle)で摩砕することによって手動で破壊した。試料を氷上に置き、Qiagen DNeasy Plant Mini Kitに述べられているようにゲノムDNAを抽出した。Access Quick Master Mix 2x(Promegaカタログ番号A1720)を使用してPCR反応を以下のように設定した:
Master mix 25μl
CCMV−F(10mM)プライマー、配列番号25 1μl
Nos−term−R(10mM)プライマー、配列番号26 1μl
ゲノムDNA 1μl
水 22μl
合計 55μl
【0213】
ゲノムDNAからプロモーター−CP融合物遺伝子−ターミネーターカセットを増幅するために以下のPCRサイクルを使用した:
1サイクル 95℃ 2分間
35サイクル 95℃ 30秒間
55℃ 30秒間
70℃ 2分間
1サイクル 70℃ 5分間
4℃ 維持
【0214】
PCR反応物10μlを、EtBrで染色した1.2%アガロースゲル上で泳動させた。図13は、選択した個別イネカルスから増幅したPCR産物を示す。予測された大きさのPCR産物を含む試料を、安定形質転換及びキメラCP導入遺伝子の植物ゲノムへの組込みに関して陽性と評価した。
【0215】
キメラCCMV CPを発現するトランスジェニックカルスを、スケールアップ発酵に適した細胞懸濁培養を作製するために液体NB培地に移した。細胞を、静かに振とうしながらNB培地において28℃で増殖させ、週に1回継代培養した。
【0216】
VLP抽出:
以下のプロトコールに従って、振とうフラスコ培養試料を溶解し、生じた細胞溶解産物をPEG(ポリエチレングリコール)処理して、限外ろ過することにより、キメラVLPを沈殿させた:
【0217】
各々の振とうフラスコ培養物の50mLアリコートを遠心分離して細胞をペレット化した。ペレット化した細胞をウイルス緩衝液(0.2M 酢酸ナトリウム、pH5.2;10mM EDTA.0)中に緩衝液2容:ペレット1容の比率で再懸濁した。次に60秒間ずつ何度も混合し、混合と混合の間に2分間氷上で休ませることによって細胞を破壊した。生じたホモジネートを3層のチーズクロス(cheese cloth)を通してしぼり、その後4℃、15,000×Gで15分間遠心分離した。生じた上清を採取し、それらの容量を測定した。各々の上清に、10%の最終濃度までPEG8000を添加し、その溶液を氷上で1時間又は4℃で一晩インキュベートした。次に、溶液を4℃、15,000×Gで10分間遠心分離した。沈殿したペレットをウイルス緩衝液の1/10初期上清容量に再懸濁し、4℃で保存して、ポリクローナル抗CCMV抗体でのウエスタンブロット法によって分析した。
【0218】
図14は、キメラCCMV VLPが、pDOW2160で安定に形質転換した細胞から回収されたことを示す。キメラCP−PA1融合タンパク質は、再懸濁したPECペレット中で検出されたが、上清では検出されず、キメラCPがVLPに集合されたことを指示した。
【0219】
あるいは、再懸濁した試料を4℃、15,000×Gで10分間遠心分離し、上清を回収して、2回目のPEC沈殿に供した。PEG8000を15%の最終濃度まで添加し、4℃で2時間攪拌した。その溶液を15,000×Gで10分間遠心分離し、ペレットを少量のウイルス緩衝液に再懸濁した。再懸濁したVLP溶液を、カットオフ値が300K分子量のCentricon Plus−20に負荷し、4,000×Gで5分間遠心した。濃縮したVLP試料をポリクローナル抗CCMV抗体でのウエスタンブロット法によって分析した(図14参照)。キメラCP−PA1融合タンパク質は、再懸濁してサイズろ過した2回目のPECペレット中で検出されたが、上清では検出されず、キメラCPがVLPに構築されたことを示した。
【図面の簡単な説明】
【0220】
【図1】図1は、植物細胞内のウイルス様粒子(VLP)におけるペプチド単量体の生産についての概要図を示す。所望標的ペプチド(「I」)をコードする配列を、ウイルスコートタンパク質(「CP」)をコードする配列に挿入して、組換えウイルスコートタンパク質(「rCP」)をコードする遺伝子を構築し、それを、ベクターの一部として、植物細胞に形質転換し、組換えコートタンパク質(「rCP」)を形成するように発現させる。任意の切断部位(「*」)も示す。植物細胞を、rCP遺伝子についての植物発現プラスミドでトランスフェクト又は形質転換する。rCPは植物細胞においてVLPに集合することができる。ペプチド挿入物はVLP表面に提示される。キメラVLPを植物細胞から精製することができ、場合により、所望標的ペプチドを精製VLPから切断分離して、精製することができる。
【図2】図2は、感染性CCMV RNA1、RNA2、及びpDOW2124(CCMV−RNA3−CP)、pDOW2125(CCMV−RNA3−CP63BamHI)、pDOW2126(CCMV−RNA3−CP102BamHI)、pDOW2127(CCMV−RNA3−CP114BamHI)、pDOW2128(CCMV−RNA3−CP129BamHI)及びpDOW2129(CCMV−RNA3−CP160BamHI)から転写されたRNA3の1個でトランスフェクトしたタバコNT1細胞におけるCCMV CPの発現を示す。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1は緩衝液のみを示し、レーン2はRNA1及びRNA2のみを示し、レーン3は、RNA1及びRNA2+pDOW2124から転写される野生型RNA3を示し、レーン4は、RNA1及びRNA2+pDOW2125から転写されるRNA3を示し、レーン5は、RNA1及びRNA2+pDOW2126から転写されるRNA3を示し、レーン6は、RNA1及びRNA2+pDOW2127から転写されるRNA3を示し、レーン7は、RNA1及びRNA2+pDOW2128から転写されるRNA3を示し、レーン8は、RNA1及びRNA2+pDOW2129から転写されるRNA3を示す。
【図3】図3は、感染性CCMV RNA1、RNA2、及び63、102又は114位に抗原性ペプチドPA1、PA2、PA3又はPA4とのCCMV CP融合物を含む工作されたRNA3の1個でトランスフェクトしたタバコNT1細胞におけるCCMV CPの発現を示す。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1はサイズマーカーを示し、レーン2は緩衝液のみを示し、レーン3は、RNA1+RNA2+CP内の63位にPA1を有するキメラRNA3を示し、レーン4は、RNA1+RNA2+CP内の63位にPA2を有するキメラRNA3を示し、レーン5は、RNA1+RNA2+CP内の63位にPA3を有するキメラRNA3を示し、レーン6は、RNA1+RNA2+CP内の63位にPA4を有するキメラRNA3を示し、レーン7は、RNA1+RNA2+CP内の102位にPA1を有するキメラRNA3を示し、レーン8は、RNA1+RNA2+CP内の102位にPA2を有するキメラRNA3を示し、レーン9は、RNA1+RNA2+CP内の102位にPA3を有するキメラRNA3を示し、レーン10は、RNA1+RNA2+CP内の102位にPA4を有するキメラRNA3を示し、レーン11は、RNA1+RNA2+CP内の114位にPA1を有するキメラRNA3を示し、レーン12は、RNA1+RNA2+CP内の114位にPA2を有するキメラRNA3を示し、レーン13は、RNA1+RNA2+CP内の114位にPA3を有するキメラRNA3を示し、及びレーン14は、RNA1+RNA2+CP内の114位にPA4を有するキメラRNA3を示す。
【図4】図4は、感染性CCMV RNA1、RNA2、及び129又は160位に抗原性ペプチドPA1、PA2、PA3又はPA4とのCCMV CP融合物を含む工作されたRNA3の1個でトランスフェクトしたタバコNT1細胞におけるCCMV CPの発現を示す。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1はサイズマーカーを示し、レーン2は、RNA1+RNA2+CP内の129位にPA1を有するキメラRNA3を示し、レーン3は、RNA1+RNA2+CP内の129位にPA2を有するキメラRNA3を示し、レーン4は、RNA1+RNA2+CP内の129位にPA3を有するキメラRNA3を示し、レーン5は、RNA1+RNA2+CP内の129位にPA4を有するキメラRNA3を示し、レーン6は、RNA1+RNA2+CP内の160位にPA1を有するキメラRNA3を示し、レーン7は、RNA1+RNA2+CP内の160位にPA2を有するキメラRNA3を示し、レーン8は、RNA1+RNA2+CP内の160位にPA3を有するキメラRNA3を示し、レーン9は、RNA1+RNA2+CP内の160位にPA4を有するキメラRNA3を示し、レーン10は、RNA1+RNA2+野生型RNA3を示し、及びレーン11は緩衝液のみを示す。
【図5】図5は、pDOW2160のプラスミド地図を示す。
【図6】図6は、pDOW2161のプラスミド地図を示す。
【図7】図7は、pDOW2162のプラスミド地図を示す。
【図8】図8は、pDOW2163のプラスミド地図を示す。
【図9】図9は、非感染性プラスミドpDOW2160、pDOW2161、pDOW2162及びpDOW2163でトランスフェクトしたタバコNT1細胞におけるキメラCCMV CPの発現を示す。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1は緩衝液のみを示し、レーン2−6は、RNA1+RNA2+野生型RNA3(対照)を示し、レーン7はpDOW2160を示し、レーン8はpDOW2161を示し、レーン9はpDOW2162を示し、及びレーン10はpDOW2163を示す。
【図10】図10は、非感染性プラスミドpDOW2169、pDOW2170、pDOW2171及びpDOW2172でトランスフェクトしたタバコNT1細胞におけるキメラCCMV CPの発現を示す。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1はRNA1+RNA2+RNA3(対照)を示し、レーン2は緩衝液を示し、レーン3はpDOW2169を示し、レーン4はpDOW2170を示し、レーン5はpDOW2171を示し、及びレーン6はpDOW2172を示す。
【図11】図11は、非感染性プラスミドpDOW2160、pDOW2161、pDOW2162及びpDOW2163で安定に形質転換したタバコNT1細胞におけるキメラCCMV CPの発現を示す。21日間の選択後、白色綿毛状細胞増殖を有していたカルスを、CP発現を調べるためのウエスタンブロット法による分析のために選択した。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1は、非形質転換タバコ細胞(陰性対照)を示し、レーン2は、pDOW2160で形質転換したトランスジェニックタバコ細胞を示し、レーン3は、pDOW2161で形質転換したトランスジェニックタバコ細胞を示し、レーン4は、pDOW2162で形質転換したトランスジェニックタバコ細胞を示し、及びレーン5は、pDOW2163で形質転換したトランスジェニックタバコ細胞を示す。
【図12】図12は、非感染性プラスミドpDOW2160、pDOW2161、pDOW2162及びpDOW2163で安定に形質転換したイネ細胞におけるキメラCCMV CPの発現を示す。21日間の選択後、白色綿毛状細胞増殖を有していたカルスを、CP発現を調べるためのウエスタンブロット法による分析のために選択した。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1は野生型CCMV CP(陽性対照)を示し、レーン2はサイズラダーを示し、レーン3は非トランスジェニックイネ細胞(陰性対照)を示し、レーン4は、pDOW2160で形質転換したトランスジェニックイネ細胞を示し、レーン5は野生型CCMV CP(陽性対照)を示し、レーン6はサイズラダーを示し、レーン7は非トランスジェニックイネ細胞を示し、レーン8は、pDOW2161で形質転換したトランスジェニックイネ細胞を示し、レーン9は野生型CCMV CP(陽性対照)を示し、レーン10はサイズラダーを示し、レーン11は非トランスジェニックイネ細胞(陰性対照)を示し、レーン12は、pDOW2162で形質転換したトランスジェニックイネ細胞を示し、レーン13は野生型CCMV CP(陽性対照)を示し、レーン14はサイズラダーを示し、レーン15は非トランスジェニックイネ細胞(陰性対照)を示し、及びレーン16は、pDOW2163で形質転換したトランスジェニックイネ細胞を示す。
【図13】図13は、非感染性プラスミドpDOW2160、pDOW2161、pDOW2162及びpDOW2163で形質転換した選択個別イネカルスから増幅したPCR産物の検出を示す。選択個別イネカルスから増幅されたPCR産物を示す、EtBrで染色した1.2%アガロースゲル。21日間の選択後、白色綿毛状細胞増殖を有していたカルスを、植物ゲノムへのプロモーター−CP融合物遺伝子−ターミネーターカセットの組込みを調べるためのPCRによる分析のために選択した。レーン1はイネカルス46−5を示し、レーン2はイネカルス46−11を示し、レーン3はイネカルス46−17を示し、レーン4はサイズラダーを示し、レーン5はイネカルス47−6を示し、レーン6はイネカルス48−12を示し、レーン7はイネカルス48−18を示し、レーン8はイネカルス48−20を示し、レーン9はイネカルス49−11を示し、レーン10はイネカルス49−18を示し、及びレーン11は陰性対照(非トランスジェニックイネ細胞)を示す。
【図14】図14は、非感染性プラスミドpDOW2160で安定に形質転換したイネ細胞からPEG沈殿及び限外ろ過によって精製したキメラCCMV VLPのウエスタンブロット分析及び精製VLP試料におけるキメラCP−PA1融合タンパク質の検出を示す。試料をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1はサイズラダーを示し、レーン2は、CCMV−CP1融合に関してトランスジェニックのイネ懸濁細胞からの全細胞溶解産物を示し、レーン3は第一PEG上清を示し、レーン4は第一PEGペレットを示し、レーン5は第二PEGペレットを示し、レーン6は第二PEG上清を示す。
【図15】図15は、発酵による植物懸濁細胞でのキメラVLPの生産工程の一覧図を示す。
【技術分野】
【0001】
(関連出願の相互参照)
本出願は、「植物細胞における高効率ペプチド生産」と題する、2004年2月27日出願の米国特許仮出願第60/548,744号の優先権を主張する。
【0002】
(政府権益の言明)
本出願は、国立衛生研究所、国立アレルギー及び感染症研究所(NIAID)、共同契約第1−U01−AI054641−01号との米国政府契約下にある。
【0003】
(技術分野)
本発明は、組換えペプチドの生産のための改善された方法を提供する。特に、本発明は、植物細胞懸濁培養においてインビボで集合され得るウイルスカプシド融合タンパク質の形態の組換えペプチドの生産のための改善された方法を提供する。本発明はまた、非感染性ウイルスカプシド融合ペプチド生産を可能にするプラスミド、配列及び植物細胞を含む。
【背景技術】
【0004】
細菌、酵母、植物、昆虫及び哺乳動物細胞発現系が、組換えペプチドを生産するために現在使用されており、成功の程度は様々である。異種ペプチドの生産のための発現系を創造する上での1つのゴールは、商業、治療及びワクチン適用において利用できる、広範囲で、柔軟性があり、効率的、経済的及び実用的なプラットフォームと方法を提供することである。例えばある種のペプチドの生産のためには、下流の再構築(reassembly)コストを取り除く又は軽減するために、インビボで効率良く且つ安価に大量の所望の最終産物を生産することができる発現系を得ることが理想である。
【0005】
現在、細菌は、豊富な量の組換えペプチドを生産するそれらの潜在能力の故に、組換えペプチドの生産のために最も広く使用されている発現系である。細菌は一般に、真核生物タンパク質をグリコシル化、アセチル化、アシル化、リン酸化又はγ−カルボキシル化せず、それ故、翻訳後修飾を必要とするある種の異種真核生物ペプチドをインビボで生産するそれらの能力には限界がある。細菌によって生産される真核生物ペプチドを修飾する付加的な工程は、所要時間を増大し、ペプチド生産の全体的収率を低下させると考えられ、細菌宿主発現系の利点の多くを色褪せさせる。それ故、細菌発現系の固有の限界を克服するために代替的な非細菌発現系が利用されてきた。
【0006】
異種ペプチドを生産する能力に関して分析された1つの特定宿主発現系は、全植物体及び植物細胞懸濁培養であった。植物の安全で安価な培養物は、ある種のペプチドのコスト効果的生産のための改善された代替的宿主を提供する。この代替物は、タンパク質が複雑で、グリコシル化を必要とし、ヒト又は動物感染性ウイルス及び細菌毒素不含である必要がある場合に特に魅力的である。
【0007】
全植物体は、比較的安価に膨大な量で成長するという利点を提供し、収集されたトランスジェニック又はウイルス感染植物の大量のバイオマスからの、所望組換えタンパク質の大きな収量の潜在的可能性を有する。その結果として、全植物体は、ヒト又は動物への投与を意図する生物薬剤タンパク質の商業生産のための発現系として開発されてきた。
【0008】
これらの系の固有の能力を利用するために、非細菌系における異種タンパク質の生産コストと収率を改善する試みに多大の時間と資源が費やされてきた。これらの領域の両方で進歩があったが、非細菌発現系における異種ペプチドの生産のためのさらなる方法とプラットフォームは有益であろう。
【0009】
ウイルス及びウイルス様粒子
宿主細胞発現系におけるペプチド生産を改善するための1つのアプローチは、対象組換えペプチドを生産するために感染性組換えウイルスの性質を利用することである。植物宿主系における感染性ウイルスの使用は特に周知である。例えばPorta & Lomonossoff, (2002) “Viruses as vectors for the expression of foreign sequences in plants, “Biotechnology and Genetic Enginering Reviews 19:245-291参照。
【0010】
最近の戦略は、ウイルス様粒子(VLP)構造における異種ペプチドの生産に集中している。一般に、カプシド形成ウイルスは、ウイルス核酸を含有するように集合されたタンパク質コート又は「カプシド」を含む。多くのウイルスは、個別に発現されたカプシドから「自己集合(self-assembly)」することができるカプシドを有し、カプシドは細胞内部で(「インビボ集合(in vivo assembly)」)及び単離と精製後に細胞の外側で(「インビトロ集合(in vitro assembly)」)発現される。理想的には、標的組換えペプチドを含むようにカプシドを改変し、組換えウイルスカプシド−ペプチド融合物を生成する。次に融合ペプチドを細胞において発現させ、理想的には、組換えウイルス又はウイルス様粒子を形成するようにインビボで構築させることができる。
【0011】
植物においてVLPに集合されたウイルスカプシド融合タンパク質による異種タンパク質の生産は、様々な程度の成功を収めている。例えば C Marusic et al., J Virol. 75 (18): 8434-39 (Sep 2001)(組換えウイルス粒子のインビボ形成を伴う、末端で抗原性6アミノ酸HIVペプチドに融合されたウイルスカプシドを発現する全ニコチアナ・ベンタミアナにおける感染性螺旋ジャガイモXウイルスの使用); FR Brennan et al., Vaccine 17 (15-16): 1846-57 (09 Apr 1999) (カウピー(ササゲ)全植物体(ビグナ・ユニキクラータ)における組換えウイルス粒子発現のインビボ形成を伴う、末端で抗原性黄色ブドウ球菌ペプチドに融合された感染性カウピーモザイクウイルス又は螺旋ジャガイモXウイルスの使用); C Porta et al.,Intervirology 39 (1-2): 79-84 (1996) (全植物体において抗原性HIV配列を含むキメラコートタンパク質を発現する感染性カウピーモザイクウイルス)参照。
【0012】
Lomonossoff & Johnsonへの米国特許第5,874,087号は、ウイルスカプシドが生物活性ペプチド、例えばホルモン、成長因子又は抗原性ペプチドを含むように工作されている、植物細胞又は全植物体における感染性植物ウイルスの生産を述べている。コモウイルス、トンブスウイルス、ソベモウイルス及びネポウイルス属から選択されるウイルスを、外来性ペプチドをコードする配列を含むように工作し、組換えウイルスを生産するために工作したウイルスのゲノム全体を発現させる。前記明細書は、改変ウイルスの増殖が発明の中心部分であることを強調する。
【0013】
チャップマン(Chapman)らへの米国特許第6,232,099号は、全植物体においてウイルスカプシドサブユニットに結合した異種タンパク質を生産するための感染性桿状ウイルスの使用を述べる。螺旋ウイルスとしても分類される桿状ウイルス、例えばジャガイモXウイルス(PVX)は、組換えウイルスカプシド−ペプチド融合物を創造するために対象組換えペプチドがウイルスのゲノムに挿入されている。次に、組換え感染性ウイルスを使用して全植物体の植物細胞に感染させ、ウイルスは植物細胞において活発に複製して、さらに他の細胞に感染し、最終的に宿主植物全体に感染する。最後に、組換えウイルスカプシド−ペプチド融合物を植物から精製する。
【0014】
チャップマン(Chapman)らはまた、限られた挿入物サイズが二十面体ウイルスによって許容されることを教示する。チャップマン(Chapman)らは、自らの主張を裏付けるのに、植物宿主細胞における発現のための二十面体ウイルス内の組換えペプチドの大きさを26アミノ酸の長さに限定する、国際公開公報第WO92/18618号を引用している。チャップマン(Chapman)らは、二十面体ウイルスのカプシドの内部挿入部位に存在するより大きなペプチドは、キメラウイルスの集合の失敗を導く、タンパク質の形状及び/又は他のカプシドと成功裏に相互作用するその能力の破壊をもたらし得ると理論付ける。
【0015】
カプロウスキー(Koprowski)らへの米国特許第6,042,832号は、抗原性ポリペプチドに融合した植物ウイルスカプシドタンパク質を含む融合カプシドタンパク質を述べる。生じる粒子は全植物体において生産される。
【0016】
カプシド融合タンパク質において異種タンパク質を生産するための感染性組換えウイルスの使用は、しかしながら、欠点がないわけではない。1つの特に厄介な面は、対象とする融合異種タンパク質を含まない基本的に野生型の復帰変異株、又は突然変異した望ましくない組換えカプシド融合タンパク質産物であるカプシドタンパク質を生じる、インビボで突然変異するこれらのウイルスの能力である。例えばPorta & Lomonossoff (1996) “Use of viral replicons for the expression of genes in plants, “ Molecular Biotechnology 5: 209- 221;Dolja et al. (1993) “Spontaneous mutagenesis of a plant potyvirus genome after insertion of a foreign gene,” J. Virol. 67 (10): 5968-5975; Dawson et al. (2001)”Assessment of recombinants that arise from the use of a TMV-based transient expression vector, “Virol. 284 (2): 182-189(接種した全植物体における異種挿入遺伝子の欠失を述べる)参照。全植物体におけるこれらのウイルスベクターの安定性の欠如は、全体的タンパク質産物の収率を潜在的に低下させ、カプシド−融合産物の不一致や不規則性を導き得る。そのような不規則性は、タンパク質産物の完全性がペプチドの特定所望物理化学的性質にとって必須である場合に特に厄介であり得る。
【発明の開示】
【発明が解決しようとする課題】
【0017】
植物における感染性組換えウイルスの固有の不安定性の結果として、商業的組換えタンパク質生産の分野において植物系型の利益を提供する効率的なペプチド生産系に対する需要がまだ存在する。
【0018】
加えて、組換えペプチドの生産のための全植物体の使用はまた、潜在的な問題を提示する。例えば長い発現時間、バッチごとの生成物収率の変動、封じ込めの問題、生産の初期段階に製造管理規則(Good Manufacturing Practice, GMP)を適用することの難しさ、農薬及び肥料による汚染の可能性、並びに害虫、病害及び微気候や土壌の相違による一定しない栽培条件は全て、組換えペプチドの生産のための潜在的に一貫しない宿主系を生じさせる。
【0019】
それ故、カプシド融合タンパク質を含むウイルス様粒子の生産のための安定で一貫した植物細胞発現系を提供することが本発明の1つの目的である。
【0020】
カプシド融合タンパク質を含むウイルス様粒子の生産のための安定な発現系における宿主細胞として使用のための植物細胞を提供することが本発明のもう1つの目的である。
【0021】
植物細胞懸濁培養を含む、植物細胞におけるカプシド融合タンパク質を含むウイルス様粒子の改善された生産のための方法を提供することが本発明のさらにもう1つの目的である。
【0022】
カプシド融合タンパク質を含むウイルス様粒子の生産のための植物細胞発現系において使用するための新規構築物及び核酸を提供することが本発明のさらにもう1つの目的である。
【課題を解決するための手段】
【0023】
本発明は、ウイルスカプシドと対象組換えペプチドを含む融合ペプチドをコードする非感染性プラスミドが宿主植物細胞のゲノムに安定に挿入され、懸濁植物細胞培養において発現される、組換えペプチドの生産のための方法を提供する。ウイルスカプシド−異種ペプチド融合産物は、インビボでウイルス様粒子として発現され得る。本発明は、感染性ウイルス因子の使用を必要としない;というよりもむしろ、カプシド−異種ペプチド融合産物をコードする非感染性核酸が植物細胞のゲノムに安定に挿入され、それを、発酵工程において培養して対象ペプチドを生産することができる。そのような工程は、異種ペプチドを含むカプシド融合タンパク質の発現のためにより変動が少なく、より安定な宿主系を生じさせる。
【0024】
全植物体において対象異種ペプチドを発現するために使用される、カプシド−異種ペプチド融合タンパク質を含む感染性ウイルスは、組換えカプシドタンパク質核酸における突然変異を生じさせる、全植物体での遺伝的不安定性を示し、発現される突然変異型カプシドは、ウイルス粒子へと集合することができないか又は突然変異した標的ペプチドを含むことが見出された。本発明は、増殖条件に対する厳密な制御、バッチごとの一貫性、高いレベルの封じ込め、及び製造管理規則(GMP)に従って組換えタンパク質を生産する能力という付加的な利益と共に、生じる異種ペプチドの高い安定性を提供する。
【0025】
カプシド−融合タンパク質産物は細胞内でウイルス様粒子を形成し得る。ウイルス様粒子は、回収されるペプチドの高純度を達成する改善された効率をもたらし得る。細胞において生産されるウイルス様粒子は、典型的には植物細胞に感染することができない。ウイルスカプシド配列は、栄養性及び非栄養性ウイルスに両方に由来することができ、栄養機能は発現宿主として使用される特定植物細胞によって決定される。1つの実施形態では、ウイルスカプシドタンパク質は、融合産物を発現するために使用される植物細胞に対して天然又は自然の栄養機能を示すウイルスに由来する。1つの実施形態では、ウイルスカプシドタンパク質は、融合産物を発現するために使用される植物細胞に対して天然又は自然の栄養機能を示さないウイルスに由来する。1つの実施形態では、細胞は、融合産物を生産するために使用される所望カプシドタンパク質配列以外の人工的に導入されたウイルスタンパク質を含まない。もう1つの実施形態では、細胞は、融合産物を生産するために使用される所望カプシドタンパク質配列以外の人工的に導入されたウイルスタンパク質又は核酸を含み、前記の付加的なウイルスタンパク質又は核酸は、核酸配列に感染性を付与しない。1つの実施形態では、ウイルスカプシドは、植物細胞発現宿主とは異なる科の生物に対して栄養機能を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、植物細胞発現宿主とは異なる属の生物に対して栄養機能を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、融合産物を発現するために使用される細胞とは異なる種の生物に対して親和性を有するウイルスに由来する。本発明の1つの実施形態では、カプシドは桿状植物ウイルスに由来する。特定実施形態では、カプシドは、タバコモザイクウイルス及びジャガイモXウイルス(PVX)から選択される群に由来する桿状ウイルスカプシドである。本発明の1つの実施形態では、カプシドタンパク質は二十面体ウイルスに由来する。特定実施形態では、カプシドは植物二十面体二十面体ウイルスに由来する。より特定の実施形態では、二十面体カプシドは、カウピーモザイクウイルス(ササゲモザイクウイルス)(Cowpea Mosaic Virus)、カウピークロロティックモトルウイルス(ササゲ退緑斑紋ウイルス)(Cowpea Clorotic Mottle Virus)及びアルファルファモザイクウイルスから選択される群に由来する。
【0026】
本発明はまた、ウイルスカプシドと組換えペプチドの融合タンパク質をコードする発現カセットを含む非感染性核酸構築物を含む植物細胞を提供する。ウイルスカプシドと組換えペプチドの融合ペプチドは、植物細胞において機能するプロモーター及びターミネーターに作動可能に連結されている。1つの詳細な実施形態では、核酸は植物細胞のゲノムに組み込まれ、この組込みは、カプシド融合タンパク質をコードする核酸の世代から世代への安定な遺伝と発現をもたらす。本発明の1つの詳細な実施形態では、カプシドタンパク質は二十面体ウイルスに由来する。1つの実施形態では、細胞はウイルス様粒子又は可溶性ケージ構造を生産する。1つの実施形態では、植物細胞は単子葉植物又は双子葉植物である。特定実施形態では、植物細胞はニコチアナ・タバカム(タバコ)である。代替的実施形態では、植物細胞はオリザ・サティバ(イネ科イネ)である。本発明の1つの実施形態では、ウイルスカプシドタンパク質に融合した組換えペプチドは、ヒト又は動物の治療のために有用な治療ペプチドである。1つの特定実施形態では、組換えペプチドは抗原性ペプチドである。特定実施形態では、抗原性ペプチドはグリコシル化抗原性ペプチドである。1つの実施形態では、抗原性ペプチドを含むカプシド−組換えペプチドウイルス様粒子を含有する植物細胞又は抽出物は、ワクチンとしてヒト又は動物に投与することができる。代替的実施形態では、抗原性ペプチドを含む精製カプシド−組換えペプチドウイルス様粒子は、ワクチンとしてヒト又は動物に投与することができる。1つの実施形態では、異種ペプチドは抗菌ペプチドである。もう1つの特定実施形態では、組換えペプチドは、遊離単量体形態であるとき植物宿主細胞に対して毒性であるペプチドである。1つの実施形態では、カプシドに融合した組換えペプチドは、少なくとも7、少なくとも8、少なくとも9、少なくとも10、少なくとも12、少なくとも15、少なくとも20、少なくとも25、少なくとも30、少なくとも35、少なくとも40、少なくとも45、少なくとも50、少なくとも55、少なくとも60、少なくとも65、少なくとも75、少なくとも85、少なくとも95、少なくとも99又は少なくとも100アミノ酸の長さである。
【0027】
本発明の1つの実施形態では、カプシドに融合した組換えペプチドは、所望標的ペプチドの少なくとも1個の単量体を含む。代替的実施形態では、組換えペプチドは、所望標的ペプチドの2個以上の単量体を含む。ある実施形態では、前記ペプチドは、コンカテマーペプチドとしてカプシドに作動可能に連結された少なくとも2個、少なくとも5個、少なくとも10個、少なくとも15個又は少なくとも20個の別個の単量体から成る。もう1つの実施形態では、コンカテマーペプチド内の個々の単量体は、切断可能なリンカー領域によって連結される。さらにもう1つの実施形態では、組換えペプチドをウイルスカプシドの少なくとも1つの表面ループに挿入する。1つの実施形態では、組換えペプチドを二十面体ウイルスカプシドの少なくとも1つの表面ループに挿入する。1つの実施形態では、少なくとも1個の単量体をウイルスカプシドタンパク質の2つ以上の表面ループに挿入する。さらにもう1つの実施形態では、組換えペプチドをウイルスカプシドの少なくとも1つの外側表面ループに挿入する。代替的実施形態では、組換えペプチドをウイルスカプシドの少なくとも1つの内側表面ループに挿入する。
【0028】
ウイルス様粒子の2つ以上のループを修飾することができる。1つの特定実施形態では、組換えペプチドをウイルス様粒子の少なくとも2つの表面ループ上で発現する。もう1つの実施形態では、少なくとも2個の異なるペプチドをウイルスカプシド、ケージ又はウイルス様粒子の少なくとも2つの表面ループに挿入する。もう1つの実施形態では、少なくとも3個の組換えペプチドをウイルス様粒子の少なくとも3つの表面ループに挿入する。表面ループ内の組換えペプチドは同じアミノ酸配列を有し得る。別の実施形態では、表面ループ内の組換えペプチドのアミノ酸配列は異なる。
【0029】
さらにもう1つの実施形態では、前記細胞は、野生型カプシド又は異なるカプシド−組換えペプチド融合ペプチドのいずれかをコードする少なくとも1個の付加的な核酸を含み、多数のカプシドは、インビボでキメラウイルス様粒子を生成するように集合され得る。
【0030】
本発明の1つの態様では、ウイルスカプシドと組換えペプチドの融合タンパク質を含む植物細胞が提供される。本発明の1つの詳細な実施形態では、植物細胞は単子葉植物である。代替的実施形態では、植物細胞は双子葉植物である。1つの実施形態では、カプシド−組換えペプチド融合タンパク質は、インビボでウイルス様粒子を形成するように集合する。
【0031】
本発明の1つの実施形態では、カプシドタンパク質−組換えペプチド融合ペプチドをコードする核酸構築物を含む植物細胞は、核形質転換によって得られる。1つの実施形態では、前記植物細胞は色素体(プラスチド(plastid))形質転換によって得られる。さらにもう1つの実施形態では、前記植物細胞は葉緑体形質転換によって得られる。
【0032】
本発明はさらに、ウイルスカプシドと組換えペプチドの融合タンパク質をコードする発現カセットを含む非感染性核酸構築物を提供する。本発明の1つの実施形態では、ウイルスカプシドと組換えペプチドの融合タンパク質をコードする発現カセットは、プロモーター及びターミネーターに作動可能に連結されている。本発明の1つの実施形態では、カプシドは植物ウイルスに由来する。本発明の1つの実施形態では、カプシドは二十面体植物ウイルスに由来する。特定実施形態では、カプシドは、カウピーモザイクウイルス、カウピークロロティックモトルウイルス及びアルファルファモザイクウイルスから選択される群に由来する二十面体ウイルスカプシドである。
【0033】
本発明の1つの実施形態では、組換えペプチドは、所望標的ペプチドの少なくとも1個の単量体を含む。代替的実施形態では、組換えペプチドは、所望標的ペプチドの2個以上の単量体を含む。さらにもう1つの実施形態では、組換えペプチドは、二十面体ウイルスカプシドの少なくとも1つの表面ループに挿入される。
【0034】
もう1つの実施形態では、前記核酸構築物は、植物細胞において機能する少なくとも1個のプロモーターを含む付加的な核酸配列を含む。もう1つの実施形態では、前記核酸構築物は、植物細胞において機能する少なくとも1個のプロモーター及び植物細胞において機能する少なくとも1個のターミネーターを含む付加的な核酸配列を含む。1つの実施形態では、プロモーター及びターミネーター配列に作動可能に連結された選択マーカーをコードする核酸配列が、前記核酸構築物に含まれる。代替的実施形態では、プロモーター及びターミネーター配列に作動可能に連結された選択マーカーは、別個の核酸構築物上で提供される。もう1つの実施形態では、前記核酸構築物は、ウイルスRNAの3’非翻訳領域(3’UTR)に由来する付加的な核酸配列を含む。さらにもう1つの実施形態では、核酸構築物は、ウイルスRNAに由来する少なくとも1つのカプシド形成シグナルを含む。さらにもう1つの実施形態では、非感染性核酸構築物は、ウイルス核酸に感染性を付与しない、ウイルスRNAに由来する付加的な核酸配列を含む。
【0035】
1つの態様では、本発明は、
a)植物細胞を提供すること;
b)少なくとも1個の異種ペプチドと少なくとも1個のウイルスカプシドタンパク質から成る融合ペプチドをコードする非感染性核酸を提供すること;
c)前記植物細胞を形質転換し、前記核酸がその後植物細胞のゲノムに組み込まれること;
d)前記植物細胞を発酵工程において懸濁植物細胞培養中で増殖させ、前記植物細胞において前記核酸を発現させること;
e)前記植物細胞における発現が、前記融合ペプチドのウイルス様粒子へのインビボ集合を提供すること;及び
d)前記ウイルス様粒子を単離すること
を含む、組換えペプチドを生産するための方法を提供する。
【0036】
1つの実施形態では、前記方法は、e)ウイルスカプシドタンパク質から組換えペプチドを分離するために融合産物を切断すること、をさらに含む。さらにもう1つの実施形態では、前記方法は、f)工程(e)の後に、組換えペプチドを単離すること、をさらに提供する。例えば図1参照。本発明の1つの実施形態では、植物細胞は、オリザ・サティバ及びニコチアナ・タバカムから成る群より選択される。
【0037】
組換えペプチドとウイルスカプシドタンパク質をコードする核酸は、プロモーター配列及びターミネーター配列に作動可能に連結される。
【0038】
1つの実施形態では、前記方法は、野生型カプシド又は異なるカプシド−組換えペプチド融合ペプチドをコードするもう1つ別の核酸を同時発現することを含み、前記カプシドは、インビボでキメラウイルス様粒子を生成するように集合される。
【0039】
本発明のもう1つの態様では、
a)植物細胞培地中で増殖することができる植物細胞;
b)少なくとも1個の組換えペプチドと少なくとも1個のウイルスカプシドを含む融合ペプチドをコードし、植物細胞においてゲノムに組み込まれて発現することができる、非感染性核酸;及び
c)増殖培地
を含む、組換えペプチドの生産のための発現系が提供される。
【0040】
融合ペプチドをコードする核酸は、プロモーター配列及びターミネーター配列に作動可能に連結され得る。発現されたとき、融合ペプチドは、細胞内でウイルス様粒子に集合することができる。1つの実施形態では、前記プロモーターは植物プロモーターである。
【発明を実施するための最良の形態】
【0041】
本発明は、ウイルスカプシドと対象組換えペプチドを含む融合ペプチドをコードする非感染性プラスミドが宿主植物細胞のゲノム内に安定に挿入され、懸濁植物細胞培養中で発現される、組換えペプチドの生産のための方法を提供する。ウイルスカプシド−異種ペプチド融合産物は、インビボでウイルス様粒子として発現され得る。本発明はさらに、前記方法における使用のための植物細胞及び核酸構築物を提供する。詳細には、本発明は、ウイルスカプシドと組換えペプチドの融合ペプチドをコードする発現カセットを含む核酸構築物と共に、懸濁植物細胞培養中で増殖することができる植物細胞を提供する。前記融合ペプチドは、プロモーター配列及び終結配列に作動可能に連結され得る。1つの実施形態では、融合ペプチドの植物細胞における発現は、ウイルス様粒子又は可溶性ケージ構造を生成する。本発明はまた、植物細胞のゲノムに組み込むことができ、1つの実施形態では、ヒト及び動物の治療のために有用な治療ペプチドであり得る、ウイルスカプシドと組換えペプチドの融合ペプチドをコードすることができる核酸構築物を提供する。
【0042】
本発明はまた、プロモーター及び終結配列に作動可能に連結された組換えペプチドとウイルスカプシドの融合ペプチドをコードする発現カセットを含む植物細胞のゲノムに組み込むことができる核酸;懸濁植物細胞培養中で前記植物細胞において核酸を発現させ、前記植物細胞における発現が前記融合ペプチドのウイルス様粒子へのインビボ集合を提供すること;及び前記ウイルス様粒子を単離すること、を提供することによって懸濁植物細胞培養において組換えペプチドを生産するための方法を提供する。
【0043】
ここで使用する「感染性」という用語は、ウイルス粒子がその核酸を宿主に転移する能力又は宿主へのウイルス核酸の導入を意味し、その場合ウイルス核酸が複製され、ウイルスタンパク質が翻訳されて、核酸を宿主にさらに転移することができる新しいウイルス粒子が集合される。ここで使用する「非感染性」という用語は、宿主への導入後にウイルス由来核酸が宿主において複製できないことを意味し、その場合ウイルスタンパク質が翻訳され、宿主においてウイルス感染過程を開始することができない新しいウイルス様粒子が構築される。ここで使用する「ペプチド」という用語は、特定分子量に限定されず、タンパク質又はポリペプチドも包含し得る。
(I.組換え植物細胞)
【0044】
本発明は、ゲノム組込みが可能であり、プロモーター配列及び終結配列に作動可能に連結されたウイルスカプシドと組換えペプチドの融合ペプチドをコードする非感染性核酸構築物を含む植物細胞を提供する。前記細胞は組換えペプチドを生産するための方法において使用することができる。
【0045】
植物細胞は、プロモーター及び終結配列に作動可能に連結されたウイルスカプシドと組換えペプチドの融合ペプチドをコードする発現カセットを含み、宿主細胞のゲノムに安定に組み込まれる核酸構築物で天然植物細胞を形質転換することによって誘導できる。安定な形質転換は、ゲノム内への異種DNAの組込みに依存する。異種DNAは植物の核又は色素体に組み込まれ得る。核酸構築物は、植物のゲノムにランダムに挿入され得るか又は相同的組換えを通してゲノムの特定領域を指向し得る。前記植物細胞は、核形質転換、色素体形質転換及び葉緑体形質転換を含む、当技術分野において公知の何らかの手段によって入手できる。例えば米国特許第6,218,145号;欧州特許第012345149号;国際公開公報第WO0121782号;米国特許第6,515,206号;国際公開公報第WO99/10513号;米国特許第5,693,507号;国際公開公報第WO02055651号;国際公開公報第WO0170939号;米国特許第6,472,586号;国際公開公報第WO02057466号;米国特許第5,057,422号;国際公開公報第WO0120974号; Staub, J. M. and Maliga, P, (1992) “Long regions of homologous DNA are incorporated into the tobacco plastid genome by transformation,”
Plant Cell 4: 39-45 参照。
【0046】
ウイルスカプシド
1つの実施形態では、本発明は、宿主ゲノムに安定に組み込まれた非感染性プラスミドからのウイルスカプシドに融合したペプチドの発現によってペプチドを生産するための方法における使用のための植物細胞を提供する。前記発現は、細胞内で少なくとも1個のウイルス様粒子(VLP)の形成を生じさせ得る。
【0047】
ウイルスは、螺旋対称又は二十面体対称を有するものに分類できる。一般に認識されているカプシド形態は、二十面体(正二十面体(icosahedral proper)、等軸晶系、準等軸晶系、及び双生又は「双晶(twinned)」を含む)、多面体(球状、卵形及びレモン形を含む)、杆状(棒状又は弾丸状、及び紡錘状又は葉巻形を含む)、及び螺旋状(桿状、円柱状及びひも状を含む)を含み、いずれも尾部を有していてもよく及び/又は表面突起、例えば棘又はこぶを含んでいてもよい。
【0048】
(形態)
本発明の1つの実施形態では、カプシドのアミノ酸配列は、何らかの形態を有すると分類されるウイルスのカプシドから選択される。1つの実施形態では、カプシドは桿状ウイルスに由来する。特定実施形態では、カプシドは桿状植物ウイルスに由来する。より特定の実施形態では、カプシドは、タバコモザイクウイルス(TMV)及びジャガイモXウイルス(PVX)から選択される群に由来する桿状植物ウイルスカプシドである。TMVは、桿状粒子(300nm)を生じさせる独自のコードタンパク質(17.5kDa)でカプシド形成された単一プラス−センスゲノムRNA(6.5kb)から成る。ジャガイモXウイルスは、515nmの明瞭なモード長と13nmの幅を有するひも状で被包されていない、通常は屈曲性のウイルスである(Brands, 1964)。カプシド構造は3.4nmのピッチで基本螺旋を形成する(Verma et al., 1968)。
【0049】
1つの実施形態では、カプシドは二十面体形態を有する。1つの実施形態では、カプシドアミノ酸配列は、正二十面体である実在のカプシドから選択される。もう1つの特定実施形態では、カプシドアミノ酸配列は、二十面体ウイルスのカプシドから選択される。1つの特定実施形態では、カプシドアミノ酸配列は、二十面体植物ウイルスのカプシドから選択される。しかし、もう1つの実施形態では、ウイルスカプシドは、植物に対して感染性でない二十面体ウイルスに由来する。例えば1つの実施形態では、ウイルスは、哺乳動物に対して感染性のウイルスである。
【0050】
一般に、二十面体ウイルスのウイルスカプシドは、二十面体(立方)対称に配列された多数のタンパク質サブユニットから成る。天然二十面体カプシドは、例えばカプシドの各々の三角形の面を形成し、3個のサブユニットで構築することができ、完全なカプシドを形成する60個のサブユニットを生じさせ得る。この小さなウイルス構造の典型は、例えばバクテリオファージφX174である。多くの二十面体ウイルスカプシドは60個以上のサブユニットを含む。二十面体ウイルスの多くのカプシドは、逆平行の8本鎖β−バレル型折りたたみモチーフを含む。このモチーフは、一方の側に4本のβ鎖(BIDGと称される)及び他方の側に4本のβ鎖(CHEFと称される)を備えるくさび形ブロックを有する。また2つの保存されたα−ヘリックス(A及びBと称される)も存在し、1つはβCとβDの間、他方はβEとβFの間にある。
【0051】
エンベロープを持つウイルスは、膜を通して粒子が突出(出芽)することにより、その全面的な破壊を伴わずに感染細胞を出て行くことができ、その間に粒子は、細胞膜に由来する脂質エンベロープ内に被包される(例えばAJ Cann (ed. ) (2001) Principles of Molecular Virology (Academic Press); A Granoff and RG Webster (eds.) (1999) Encyclopedia of Virology (Academic Press); DLD Caspar (1980) Biophys. J. 32: 103; DLD Caspar and A Klug (1962) Cold Spring Harbor Symp. Quant. Biol. 27: 1; J Grimes et al. (1988) Nature 395: 470; JE Johnson (1996) Proc. Nat'l Acad. Sci. USA 93: 27; and J Johnson and J Speir (1997) J. Mol. Biol. 269: 665参照)。
【0052】
(ウイルス)
ウイルス分類学は、以下の分類群のカプシド形成粒子実体を承認する:
・グループIウイルス、すなわちdsDNAウイルス;
・グループIIウイルス、すなわちssDNAウイルス;
・グループIIIウイルス、すなわちdsRNAウイルス;
・グループIVウイルス、すなわちDNA段階を持たないssRNA(+)鎖ウイルス;
・グループVウイルス、すなわちssRNA(−)鎖ウイルス;
・グループVIウイルス、すなわちssRNA逆転写ウイルスである、RNAレトロイドウイルス;
・グループVIIウイルス、すなわちdsDNA逆転写ウイルスである、DNAレトロイドウイルス;
・デルタウイルス;
・ウイロイド;及び
・サテライト核酸及びプリオンを除く、サテライトファージ及びサテライトウイルス。
【0053】
これらの分類群の成員は当業者に周知であり、H. V. Van Regenmortel et al. (eds.), Virus Taxonomy: Seventh Report of the International Committee on Taxonomy of Viruses (2000) (Academic Press/Elsevier, Burlington Mass., USA); レスター大学微生物・免疫学部のウイルス分類学ウエブページhttp://wwwmicro.msb.le.ac.uk/3035/ Virusgroups.html;及び米国保健社会福祉省の国立衛生研究所・国立医学図書館・国立バイオテクノロジー情報センター(NCBI)(Washington, D. C. , USA)の分類学ブラウザのオンライン「ウイルス」及び「ウイロイド」部門http://www.ncbi.nlm.nih.gov/Taxonomy/tax.html.において総説されている。
【0054】
カプシドのアミノ酸配列は、これらの分類群のいずれかの成員のカプシドから選択され得る。これらの分類群の成員のカプシドについてのアミノ酸配列は、例えば、NCBIによって提供されるPubMedオンライン検索機能の「ヌクレオチド」(Genbank)、「タンパク質」及び「構造」部門http://www.ncbi.nlm.nih.gov/entrez/ query.fcgi.を含むがこれらに限定されないソースから入手し得る。
【0055】
1つの実施形態では、カプシドアミノ酸配列は、以下の宿主の少なくとも1つに特異的な分類群成員から選択される:酵母を含む真菌、植物、藻類を含む原生生物、無脊椎動物、脊椎動物、及びヒト。1つの実施形態では、カプシドアミノ酸配列は以下の分類群のいずれかの成員から選択される:グループI、グループII、グループIII、グループIV、グループV、グループVII、ウイロイド、及びサテライトウイルス。1つの実施形態では、カプシドアミノ酸配列は、上述した6つの宿主型の少なくとも1つに特異的なこれら7つの分類群のいずれかの成員から選択される。より詳細な実施形態では、カプシドアミノ酸配列は、グループII、グループIII、グループIV、グループVII、及びサテライトウイルスのいずれかの成員から;又はグループII、グループIV、グループVII、及びサテライトウイルスのいずれかから選択される。もう1つの実施形態では、ウイルスカプシドはグループIV又はグループVIIから選択される。
【0056】
ウイルスカプシド配列は、当該細胞に対して親和性でないウイルスに由来し得る。1つの実施形態では、当該細胞は、所望二十面体カプシド以外の特定選択ウイルスからのウイルスタンパク質を含まない。1つの実施形態では、ウイルスカプシドは、当該細胞とは異なる科の生物に対して親和性を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、当該細胞とは異なる属の生物に対して親和性を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、当該細胞とは異なる種の生物に対して親和性を有するウイルスに由来する。
【0057】
詳細な実施形態では、ウイルスカプシドはグループIVのウイルスから選択される。
【0058】
1つの実施形態では、ウイルスカプシドは桿状植物ウイルスから選択される。より特定の実施形態では、ウイルスカプシドは、タバコモザイクウイルス及びジャガイモXウイルスから成る群より選択される。
【0059】
1つの実施形態では、ウイルスカプシドは二十面体ウイルスから選択される。二十面体ウイルスは、乳頭腫ウイルス科、トティウイルス科、ジシストロウイルス科、ヘパドナウイルス科、トガウイルス科、ポリオーマウイルス科、ノダウイルス科、テクティウイルス科、レヴィウイルス科、ミクロウイルス科、サイフォウイルス科、ノダウイルス科、ピコルナウイルス科、パルボウイルス科、カルシウイルス科、テトラウイルス科、及びサテライトウイルスのいずれかの成員から選択され得る。
【0060】
特定実施形態では、前記配列は、少なくとも1つの植物宿主に特異的な分類群のいずれかの成員から選択される。1つの実施形態では、二十面体植物ウイルス種は、ブンヤウイルス科、レオウイルス科、ラブドウイルス科、ルテオウイルス科、ナノウイルス科、パルティティウイルス科、セキウイルス科、ティモウイルス科、ウルミアウイルス属、タバコ壊死サテライトウイルス、カリモウイルス科、ジェミニウイルス科、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科分類群のいずれかであるか又はそのいずれかの成員である、植物感染性ウイルス種である。1つの実施形態では、二十面体植物ウイルス種は、ルテオウイルス科、ナノウイルス科、パルティティウイルス科、セキウイルス科、ティモウイルス科、ウルミアウイルス属、タバコ壊死サテライトウイルス、カリモウイルス科、ジェミニウイルス科、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科分類群のいずれかであるか又はそのいずれかの成員である、植物感染性ウイルス種である。詳細な実施形態では、二十面体植物ウイルス種は、カリモウイルス科、ジェミニウイルス科、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科のいずれかであるか又はそのいずれかの成員である、植物感染性ウイルス種である。より特定の実施形態では、二十面体植物ウイルス種は、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科のいずれかであるか又はそのいずれかの成員である、植物感染性ウイルス種である。より特定の実施形態では、二十面体植物ウイルス種は、コモウイルス科又はブロモウイルス科の成員である植物感染性ウイルス種である。特定実施形態では、ウイルスカプシドは、カウピーモザイクウイルス又はカウピークロロティックモトルウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、ブロモウイルス科分類群の種に由来する。詳細な実施形態では、カプシドは、イラルウイルス属又はアルファモウイルス属に由来する。より詳細な実施形態では、カプシドは、タバコ条斑病ウイルス又はアルファルファモザイクウイルス(AMV)(AMV1又はAMV2を含む)に由来する。
【0061】
(VLP)
本発明のウイルスカプシドは、ここで述べる宿主細胞において非感染性である。1つの実施形態では、ウイルス科の発現の間又は発現後に宿主細胞においてウイルス様粒子(VLP)又はケージ構造が形成される。1つの実施形態では、VLP又はケージ構造はまた、対象とするペプチドも含み、特定実施形態では、対象ペプチドがVLPの表面に発現される。前記発現系は、典型的にはウイルスの感染性を可能にする付加的なウイルスタンパク質を含まない。典型的実施形態では、発現系は、宿主細胞、及び1又はそれ以上のウイルスカプシド及び作動可能に連結された対象ペプチドをコードするベクターを含む。前記ベクターは、典型的には付加的なウイルスタンパク質を含まない。本発明は、ウイルスカプシドがある種の宿主細胞においてより大きな度合で形成され、組換えペプチドのより効率的な回収を可能にするという発見に由来する。
【0062】
1つの実施形態では、VLP又はケージ構造は、3から約1,000カプシドを含む、カプシドの多量体集合物である。1つの実施形態では、VLP又はケージ構造は、少なくとも30、少なくとも50、少なくとも60、少なくとも90又は少なくとも120カプシドを含む。もう1つの実施形態では、各々のVLP又はケージ構造は、少なくとも150カプシド、少なくとも160、少なくとも170又は少なくとも180カプシドを含む。
【0063】
1つの実施形態では、VLPは二十面体構造として発現される。もう1つの実施形態では、VLPは、そのカプシド配列が由来する天然ウイルスと同じ形状で発現される。別の実施形態では、しかしながら、VLPは天然ウイルスと同じ形状を持たない。ある実施形態では、例えば、多数のカプシドで形成されるが天然型VLPを形成しない粒子中で構造が生成される。例えばわずか3個のウイルスカプシドのケージ構造が形成され得る。別の実施形態では、約6、9、12、15、18、21、24、27、30、33、36、39、42、45、48、51、54、57又は60カプシドのケージ構造が形成され得る。
【0064】
1つの実施形態では、カプシドの少なくとも1個は、少なくとも1個の対象ペプチドを含む。1つの実施形態では、ペプチドは、VLPの少なくとも1つの内部ループ内で又は少なくとも1つの外側表面ループにおいて発現される。
【0065】
ウイルスカプシドの2つ以上のループを修飾することができる。1つの特定実施形態では、組換えペプチドは、ウイルス様粒子の少なくとも2つの表面ループ上で発現される。もう1つの実施形態では、少なくとも2個の異なるペプチドをウイルスカプシド、ケージ又はウイルス様粒子の少なくとも2つの表面ループに挿入する。もう1つの実施形態では、少なくとも3個の組換えペプチドをウイルス様粒子の少なくとも3つの表面ループに挿入する。表面ループ内の組換えペプチドは同じアミノ酸配列を有し得る。別の実施形態では、表面ループ内の組換えペプチドのアミノ酸配列は異なる。
【0066】
ある実施形態では、VLPの集合を改善するために宿主細胞を修飾することができる。宿主細胞を、例えば、発現されたウイルスカプシドからのVLPの形成を促進するシャペロンタンパク質を含むように改変することができる。もう1つの実施形態では、宿主細胞を、VLPの調節された形成を促進するためにカプシドの発現をより効率的に調節するリプレッサータンパク質を含むように改変する。
【0067】
ウイルスカプシド又はタンパク質をコードする核酸配列はまた、VLPの形成を変化させるように付加的に修飾することができる(例えばBrumfield, et al. (2004) J. Gen. Virol. 85: 1049-1053参照)。例えばこれらの修飾は、集合されたタンパク質ケージ内の隣接サブユニットの内部、外部又はその間の界面を変化させるように設計される。これを実現するために、突然変異誘発プライマーを使用して、(i)N末端の塩基性残基(例えばK、R)を酸性グルタミン酸に置き換えることによってウイルス核酸結合領域の内表面電荷を変化させる(Douglas et al., 2002b);(ii)N末端から内部残基を欠失させる(CCMVにおいて、通常は残基4−37);(iii)11アミノ酸ペプチド細胞標的配列をコードするcDNA(Graf et al., 1987)を表面露出ループに挿入する;及び(iv)金属結合部位を変化させることによってウイルスサブユニットの間の相互作用を修飾する(CCMVにおいて、残基81/148突然変異体)ことができる。
【0068】
組換えペプチドの大きさ
1つの実施形態では、ウイルスカプシド配列に作動可能に連結されたペプチドは、少なくとも2個のアミノ酸を含む。もう1つの実施形態では、前記ペプチドは、少なくとも3、少なくとも4、少なくとも5又は少なくとも6アミノ酸の長さである。別の実施形態では、ペプチドは少なくとも7アミノ酸の長さである。ペプチドはまた、少なくとも8、少なくとも9、少なくとも10、少なくとも11、12、13、14、15、16、17、18、19、20、30、45、50、60、65、75、85、95、96、99又はそれ以上のアミノ酸の長さである。1つの実施形態では、コードされるペプチドは少なくとも25kDである。
【0069】
1つの実施形態では、ペプチドは、2から約300アミノ酸、又は約5から約250アミノ酸、又は約5から約200アミノ酸、又は約5から約150アミノ酸、又は約5から約100アミノ酸を含む。もう1つの実施形態では、ペプチドは、約10から約140アミノ酸、又は約10から約120アミノ酸、又は約10から約100アミノ酸を含む。
【0070】
1つの実施形態では、ウイルスカプシド配列に作動可能に連結されたペプチド又はタンパク質は、約500個のアミノ酸を含む。1つの実施形態では、ペプチドは500個未満のアミノ酸を含む。もう1つの実施形態では、ペプチドは、約300個までのアミノ酸、又は約250個まで、又は約200個まで、又は約180個まで、又は約160個まで、又は約150個まで、又は約140個まで、又は約120個まで、又は約110個まで、又は約100個まで、又は約90個まで、又は約80個まで、又は約70個まで、又は約60個まで、又は約50個まで、又は約40個まで、又は約30個までのアミノ酸を含む。
【0071】
1つの実施形態では、カプシドに融合した組換えペプチドは、少なくとも7、少なくとも8、少なくとも9、少なくとも10、少なくとも12、少なくとも15、少なくとも20、少なくとも25、少なくとも30、少なくとも35、少なくとも40、少なくとも45、少なくとも50、少なくとも55、少なくとも60、少なくとも65、少なくとも75、少なくとも85、少なくとも95、少なくとも99、又は少なくとも100アミノ酸である。
【0072】
本発明の1つの実施形態では、組換えペプチドは、所望標的ペプチドの少なくとも1個の単量体を含む。代替的実施形態では、組換えペプチドは、所望標的ペプチドの2個以上の単量体を含む。ある実施形態では、ペプチドは、コンカテマーペプチドとしてカプシドに作動可能に連結された、少なくとも2個、少なくとも5個、少なくとも10個、少なくとも15個又は少なくとも20個の別個の単量体から成る。もう1つの実施形態では、コンカテマーペプチド内の個々の単量体は、切断可能なリンカー領域によって連結される。さらにもう1つの実施形態では、組換えペプチドを、二十面体ウイルス様粒子の少なくとも1つの表面ループに挿入する。1つの実施形態では、少なくとも1個の単量体をウイルス様粒子の表面ループに挿入する。
【0073】
(分類)
ウイルスカプシドに融合する対象ペプチドは、ウイルスに由来しない、及び場合により、当該細胞と同じ種に由来しない、異種タンパク質であり得る。
【0074】
ウイルスカプシドに融合する対象ペプチドは、機能性ペプチド:構造ペプチド;抗原性ペプチド;毒性ペプチド、抗菌ペプチド、それらのフラグメント;それらの前駆体;前記のいずれかの組合せ;及び/又は前記のいずれかのコンカテマーであり得る。本発明の1つの実施形態では、組換えペプチドは、ワクチン戦略において使用される抗原性ペプチドを含む、ヒト及び動物の治療のために有用な治療ペプチドである。特定実施形態では、抗原性ペプチドは、インビボでグリコシル化される。
【0075】
機能性ペプチドは、例えば:生物活性ペプチド(すなわち生物学的実体、例えば生物、細胞、培養物、組織、器官又は細胞小器官、における又はその生物学的機能又は活性の開始、増強、延長、減衰、終結又は防止を及ぼす、惹起する又はさもなければ生じさせるペプチド);触媒ペプチド;ミクロ構造及びナノ構造活性ペプチド(すなわちその中で又はそれに関連して、作用、例えば運動、エネルギー変換、を実施する工作されたミクロ又はナノ構造の一部を形成するペプチド);及び刺激剤ペプチド(例えばペプチド着香料、着色料、着臭剤、フェロモン、誘引物質、抑制物質及び忌避物質)を含むが、これらに限定されない。
【0076】
生物活性ペプチドは、例えば:免疫活性ペプチド(例えば抗原性ペプチド、アレルゲンペプチド、ペプチド免疫調節剤(immunoregulator)、ペプチド免疫調節剤(immunomodulator):シグナル伝達ペプチド(signaling and signal transduction peptides)(例えばペプチドホルモン、サイトカイン及び神経伝達物質;受容体;アゴニスト及びアンタゴニストペプチド;ペプチドターゲティング及び分泌シグナルペプチド);及び生物阻害性ペプチド(例えば毒性、生物致死性又は生物抑制性ペプチド、例えばペプチド毒素及び抗菌ペプチド)を含むが、これらに限定されない。
【0077】
構造ペプチドは、例えば:ペプチドアプタマー、折りたたみペプチド(例えばもう1つ別の分子の物理的立体配座の形成又は維持を促進する又は誘導するペプチド);接着促進ペプチド(例えば接着ペプチド、細胞接着促進ペプチド);界面ペプチド(例えばペプチド界面活性剤及び乳化剤);ミクロ構造及びナノ構造構築ペプチド(すなわち工作されたミクロ又はナノ構造の一部を形成する構造ペプチド);及びプレ活性化ペプチド(例えばプレ、プロ及びプレ−プロタンパク質及びペプチドのリーダーペプチド;インテイン)を含むが、これらに限定されない。
【0078】
触媒ペプチドは、例えばアポB RNA編集シチジンデアミナーゼペプチド;グルタミニル−tRNAシンテターゼの触媒ペプチド:アスパラギン酸トランスカルバモイラーゼの触媒ペプチド;植物1型リボソーム不活性化ペプチド;ウイルス触媒ペプチド、例えば口蹄疫ウイルス[FMDV−2A]触媒ペプチド;マトリックスメタロプロテイナーゼペプチド;及び触媒メタロオリゴペプチドを含む。
【0079】
ペプチドはまた、ペプチドエピトープ、ハプテン又は関連ペプチド(例えば抗原性ウイルスペプチド;ウイルス関連ペプチド、例えばHIV関連ペプチド、肝炎関連ペプチド;抗体イディオタイプドメイン;細胞表面ペプチド;抗原性ヒト、動物、原生生物、植物、真菌、細菌及び/又は古細菌ペプチド;アレルゲンペプチド及びアレルゲン脱感作ペプチド)であり得る。
【0080】
ペプチドはまた、ペプチド免疫調節剤(peptide immunoregulators or immunomodulators)(例えばインターフェロン、インターロイキン、ペプチド免疫抑制剤及び免疫増強剤);抗体ペプチド(例えば一本鎖抗体;一本鎖抗体フラグメント及び構築物、例えば一本鎖Fv分子;抗体軽鎖分子、抗体重鎖分子、ドメイン欠失抗体軽鎖又は重鎖分子;一本鎖抗体ドメイン及び分子、例えばCH1、CH1−3、CH3、CH1−4、CH4、VHCH1、CL、CDR1又はFR1−CDR1−FR2ドメイン;パラトープペプチド;ミクロ抗体);別の結合ペプチド(例えばペプチドアプタマー、細胞内及び細胞表面受容体タンパク質、受容体フラグメント;抗腫瘍壊死因子ペプチド)であり得る。
【0081】
ペプチドはまた、酵素基質ペプチド又は酵素阻害剤ペプチド(例えばカスパーゼ基質及び阻害剤、プロテインキナーゼ基質及び阻害剤、蛍光共鳴エネルギー転移ペプチド酵素基質)であり得る。
【0082】
ペプチドはまた、細胞表面受容体ペプチドリガンド、アゴニスト及びアンタゴニスト(例えばセルレイン、ダイノルフィン、オレキシン、下垂体アデニル酸シクラーゼ活性化ペプチド、腫瘍壊死因子ペプチド;合成ペプチドリガンド、アゴニスト及びアンタゴニスト);ペプチドホルモン(例えば内分泌、パラ分泌及び自己分泌ホルモン、例えばアミリン、アンギオテンシン、ブラジキニン、カルシトニン、心臓興奮性神経ペプチド、カゾモルフィン、コレシストキニン、コルチコトロピン及びコルチコトロピン関連ペプチド、分化因子、エンドルフィン、エンドセリン、エンケファリン、エリスロポエチン、エキセンディン、卵胞刺激ホルモン、ガラニン、ガストリン、グルカゴン及びグルカゴン様ペプチド、ゴナドトロピン、成長ホルモン及び成長因子、インスリン、カリジン、キニン、レプチン、脂肪親和性ホルモン、黄体形成ホルモン、メラニン細胞刺激ホルモン、メラトニン、ナトリウム排泄増加性ペプチド、ニューロキニン、ニューロメジン、ノシセプチン、オステオカルシン、オキシトシン(すなわちオシトシン)、副甲状腺ホルモン、プレイオトロフィン、プロラクチン、レラキシン、セクレチン、セロトニン、睡眠誘発ペプチド、ソマトメジン、サイモポイエチン、甲状腺刺激ホルモン、チロトロピン、ウロテンシン、血管作用性小腸ペプチド、バソプレシン);ペプチドサイトカイン、ケモカイン、ウイロカイン(virokine)及びウイロセプター(viroceptor)ホルモン放出及び放出抑制ペプチド(例えばコルチコトロピン放出ホルモン、コルチスタチン、卵胞刺激ホルモン放出因子、胃抑制性ペプチド、ガストリン放出ペプチド、ゴナドトロピン放出ホルモン、成長ホルモン放出ホルモン、黄体形成ホルモン放出ホルモン、メラノトロピン放出ホルモン、メラノトロピン放出抑制因子;ノシスタチン、パンクリアスタチン、プロラクチン放出ペプチド、プロラクチン放出抑制因子;ソマトスタチン;甲状腺刺激ホルモン放出ホルモン);ペプチド神経伝達物質又はチャネルブロッカー(例えばボンベシン、神経ペプチドY、ニューロテンシン、サブスタンスP);ペプチド毒素、毒素前駆体ペプチド又は毒素ペプチド部分であり得る。ある実施形態では、ペプチド毒素はD−アミノ酸を含まない。毒素前駆体ペプチドは、D−アミノ酸を含まないもの及び/又は翻訳後修飾によって、例えばアミノ酸にD−立体配置を導入することができるD立体配置誘導剤(例えばペプチドイソメラーゼ又はエピメラーゼ又はラセマーゼ又はトランスアミノアーゼ)の作用によって、及び/又は環状ペプチド構造を形成することができる環化剤(例えばペプチドチオエステラーゼ又はペプチドリガーゼ、例えばトランススプライシングタンパク質又はインテイン)の作用によって、天然毒素構造に変換されていないものであり得る。
【0083】
毒素ペプチド部分は、ペプチド含有毒素の線状又は前環化オリゴ及びポリペプチド部分であり得る。ペプチド毒素の例は、例えばアガトキシン、アマトキシン、カリブドトキシン、クロロトキシン、コノトキシン、デンドロトキシン、インセクトトキシン、マルガトキシン、マスト細胞脱顆粒ペプチド、サポリン、サラフォトキシン;及び細菌体外毒素、例えば炭疽毒素、ボツリヌス毒素、ジフテリア毒素及び破傷風毒素を含む。
【0084】
ペプチドはまた、代謝及び消化関連ペプチド(例えばコレシストキニン−パンクレオザイミンペプチド、ペプチドyy、膵ペプチド、モチリン);細胞接着調節又は仲介ペプチド、細胞外マトリックスペプチド(例えば付着因子、セレクチン、ラミニン);神経保護又は髄鞘形成促進ペプチド;凝集阻害ペプチド(例えば細胞又は血小板凝集阻害剤ペプチド、アミロイド形成又は沈着阻害剤ペプチド);連結ペプチド(例えば心臓血管連結神経ペプチド、iga連結ペプチド);又はその他のペプチド(例えばアグーチ関連ペプチド、アミロイドペプチド、骨関連ペプチド、細胞透過性ペプチド、コナントキシン、コントリファン、コンチュラキン、ミエリン塩基性タンパク質等)であり得る。
【0085】
ペプチドはまた、インビボで翻訳後修飾され得る。特定実施形態では、ペプチドは、単独で又は組合せとして、グリコシル化、アセチル化、アシル化、リン酸化又はγ−カルボキシル化され得る。
【0086】
ある実施形態では、対象ペプチドは、選択されるウイルスカプシドに対して外因性である。ペプチドは、天然又は合成配列のいずれでもよい(及びそれらのコード配列は天然又は合成ヌクレオチド配列のいずれでもよい)。それ故、例えばアミノ酸の天然、修飾天然及び完全人工配列が包含される。これらのアミノ酸配列をコードする核酸分子の配列は、同様に、天然、修飾天然及び完全人工核酸配列であり得、例えばその核酸分子を得るために使用される(すなわち人為作用によって適用される)1又はそれ以上の合理的又はランダム突然変異及び/又は組換え及び/又は合成及び/又は選択工程の結果であり得る。
【0087】
コード配列は、使用可能な場合は、標的ペプチドについての天然コード配列であり得るが、より典型的には、例えば宿主の種のコドン使用頻度を反映する遺伝子を合成することによって、選択発現宿主細胞における使用のために選択された、改善された又は最適化されたコード配列である。本発明の1つの実施形態では、宿主の種はニコチアナ・タバカムであり、シグナル配列及びペプチド配列の両方を設計するときにはニコチアナ・タバカムのコドン使用頻度を考慮する。本発明の代替的実施形態では、宿主の種はオリザ・サティバであり、シグナル配列及びペプチド配列の両方を設計するときにはオリザ・サティバのコドン使用頻度を考慮する。
【0088】
(抗原性ペプチド(ペプチドエピトープ))
1つの実施形態では、抗原性ペプチドはウイルスカプシドによる発現を通して生産される。抗原性ペプチドは、感染因子、寄生生物、癌細胞及び他の病原因子を含む、ヒト又は動物病原因子の抗原性ペプチドであるものから選択することができる。そのような病原因子はまた、ビルレンス因子及び病原因子、例えばそれらの因子の体外毒素、内毒素等を含む。病原因子は何らかのレベルのビルレンスを示し得る、すなわちそれらは、例えば毒性、無毒性、偽毒性、半毒性等であり得る。1つの実施形態では、抗原性ペプチドは、病原因子からのエピトープアミノ酸配列を含む。1つの実施形態では、エピトープアミノ酸配列は、少なくとも1個のそのような因子の表面ペプチドの少なくとも一部であるものを含む。1つの実施形態では、カプシド−組換えペプチドウイルス様粒子は、ヒト又は動物適用におけるワクチンとして使用することができる。
【0089】
2個以上の抗原性ペプチドを選択してもよく、その場合、生じるウイルス様粒子は多数の異なる抗原性ペプチドを提示し得る。多抗原性ペプチド様式の特定実施形態では、様々な抗原性ペプチドは全て同じ病原因子からの複数のエピトープから選択される。多抗原性ペプチド様式の特定実施形態では、様々な抗原性ペプチドは全て、密接に関連する病原因子、例えば同じ種又は同じ属の異なる種の異なる株、亜種、次亜種、病原型、血清型又は遺伝子型(genovar)から選択される。
【0090】
1つの実施形態では、病原因子は以下の群の少なくとも1つに属する:病原性:バチルス属種、例えば炭疽菌;バルトネラ属種、例えばB.クインタナ;ブルセラ属種;ブルクホルデリア属種、例えばB.シュードマレイ(類鼻疽菌);カンピロバクター種;クロストリジウム属種、例えば破傷風菌、ボツリヌス菌;コクシエラ属種、例えばC.バーネティー;エドワードシエラ属種、例えばE.ターダ;エンテロバクター属種、例えばE.クロアカ;エンテロコッカス属種、例えばE.フェカリス、E.フェシウム;エシェリキア属種、例えば大腸菌;フランシセラ属種、例えば野兎病菌(F. tularensis);ヘモフィルス属種、例えばインフルエンザ菌;クレブシエラ属種、例えば肺炎杆菌;レジオネラ属種;リステリア属種、例えばリステリア菌(L. monocytogenes);髄膜炎菌及び淋菌、例えばナイセリア属種;モラクセラ属種;ミコバクテリウム属種、例えばらい菌、ヒト型結核菌;肺炎球菌、例えば肺炎双球菌;シュードモナス属種、例えば緑膿菌;リケッチア属種、例えば発疹チフスリケッチア、斑点熱リケッチア、発疹熱リケッチア;サルモネラ属菌、例えば腸チフス菌;ブドウ球菌属種、例えば黄色ブドウ球菌;A型連鎖球菌属及び溶血連鎖球菌属を含む、連鎖球菌属種、例えば肺炎連鎖球菌、化膿連鎖球菌;ストレプトミセス属種;赤痢菌属種;ビブリオ属種、例えばコレラ菌;及びエルジニア属種、例えばペスト菌、エルジニア小腸結腸炎菌(E.enterocolitica)を含むが、これらに限定されない細菌及びマイコプラスマ因子。病原性:アルテルナリア属種;アスペルギルス属種;ブラストミセス属種、例えばB.デルマチチジス;カンジダ属種、例えばカンジダアルビカンス;クラドスポリウム属種;コクシジオイデス属種、例えばC.イミチス;クリプトコッカス属種、例えばC.ネオフォルマンス;ヒストプラスマ属種、例えばH.カプスラーツム;及びスポロトリクス属種、例えばS.シェンキイ、を含むが、これらに限定されない真菌及び酵母因子。
【0091】
1つの実施形態では、病原因子は、病原性:アカントアメーバ属種、アメーバ属種、ネグレリア属種、エントアメーバ属種、例えば赤痢アメーバを含むアメーバ;クリプトスポリジウム属種、例えばC.パルブム;シクロスポラ属種;エンセファリトゾーン属種、例えばE.インテスティナリス;エンテロシトゾーン属種;ジアルジア属種、例えばランブル鞭毛虫;イソスポラ属種;ミクロスポリジウム属種;プラスモディウム属種、例えば熱帯熱マラリア原虫、四日熱マラリア原虫、卵形マラリア原虫、三日熱マラリア原虫;トキソプラスマ属種、例えばトキソプラスマ原虫;及びトリパノソーマ属種、例えばトリパノソーマブルセイ、を含むが、これらに限定されない原生生物因子由来である。
【0092】
1つの実施形態では、病原因子は、病原性:回虫属種、例えば回虫;ドラクンクルス属種、例えばメジナ虫;オンコセルカ属種、例えば回旋糸状虫;住血吸虫属種;旋毛虫属種、例えば旋毛虫;及び鞭虫属種、例えばヒト鞭虫、を含むが、これらに限定されない寄生生物因子(例えば蠕虫寄生生物)由来である。
【0093】
もう1つの実施形態では、病原因子は、病原性:アデノウイルス;アレナウイルス、例えばラッサ熱ウイルス;アストロウイルス;ブンヤウイルス、例えばハンタウイルス、リフトバレー熱ウイルス;コロナウイルス、デルタウイルス;サイトメガロウイルス、エプスタイン−バーウイルス、ヘルペスウイルス、水痘ウイルス;フィロウイルス、例えばエボラウイルス、マルブルクウイルス;フラビウイルス、例えばデング熱ウイルス、西ナイル熱ウイルス、黄熱病ウイルス;肝炎ウイルス;インフルエンザウイルス;レンチウイルス、T細胞リンパ球向性ウイルス、他の白血病ウイルス;ノーウォークウイルス;パピローマウイルス、他の腫瘍ウイルス;パラミクソウイルス、例えば麻疹ウイルス、流行性耳下腺炎ウイルス、パラインフルエンザウイルス、肺炎ウイルス、センダイウイルス;パルボウイルス;ピコルナウイルス、例えばカルジオウイルス、コクサッキーウイルス、エコーウイルス、ポリオウイルス、ライノウイルス、他のエンテロウイルス;ポックスウイルス、例えば痘瘡ウイルス、ワクシニアウイルス、パラポックスウイルス;レオウイルス、例えばコルティウイルス、オルビウイルス、ロタウイルス;ラブドウイルス、例えばリッサウイルス、水泡性口内炎ウイルス;及びトガウイルス、例えば風疹ウイルス、シンドビスウイルス、西部脳炎ウイルス、を含むが、これらに限定されないウイルス因子由来である。
【0094】
1つの実施形態では、抗原性ペプチドは、イヌパルボウイルスペプチド、炭疽菌防御抗原(PA)抗原性ペプチド及び東部ウマ脳炎ウイルス抗原性ペプチドから成る群より選択される。特定実施形態では、抗原性ペプチドは、PA1(配列番号1)、PA2(配列番号2)、PA3(配列番号3)又はPA4(配列番号4)から選択されるアミノ酸配列を有する炭疽菌由来ペプチドである。
【0095】
(宿主細胞毒性ペプチド)
もう1つの特定実施形態では、組換えペプチドは、遊離単量体形態であるとき宿主細胞に対して毒性であるペプチドである。ある実施形態では、ウイルスカプシドと共に発現される対象ペプチドは宿主細胞毒性ペプチドである。宿主細胞毒性ペプチドは、それが発現される宿主細胞に対して、又は宿主細胞がその成員である細胞培養中又は生物中の他の細胞に対して、又は宿主細胞を提供する生物又は種の細胞に対して、生物抑制性、生物致死性又は毒性である生物阻害性ペプチドを指示する。1つの実施形態では、宿主細胞毒性ペプチドは、それが発現される宿主細胞に対して生物抑制性、生物致死性又は毒性である生物阻害性ペプチドである。宿主細胞毒性ペプチドの一部の例は、ペプチド毒素、抗菌ペプチド及び他の抗生物質ペプチドを含むが、これらに限定されない。
【0096】
(抗菌ペプチド)
もう1つの特定実施形態では、組換えペプチドは、抗菌性ペプチドであるペプチドである。抗菌ペプチドは、例えば抗細菌ペプチド、例えばマガイニン、βデフェンシン、一部のαデフェンシン;カテリシジン;ヒスタチン;抗真菌ペプチド;抗原生動物ペプチド;合成AMP;ペプチド抗生物質又はその線状又は前環化オリゴ又はポリペプチド部分;他の抗生物質ペプチド(例えば駆虫性ペプチド、溶血ペプチド、殺腫瘍性ペプチド);及び抗ウイルスペプチド(例えば一部のαデフェンシン;殺ウイルス性ペプチド;ウイルス感染を阻害するペプチド)を含む。1つの特定実施形態では、抗菌ペプチドは、D2A21抗菌ペプチドから成る群より選択される。
【0097】
(VLPを発現するときに使用するための細胞)
本発明のウイルスカプシド又はウイルスカプシド融合ペプチドの発現のための宿主として使用する細胞(「宿主細胞」とも称する)は、ウイルスカプシドが細胞の複製又は感染を許容しないものである。1つの実施形態では、ウイルスカプシドは、宿主細胞が由来する細胞の特定種に感染しないウイルスに由来する。例えば、1つの実施形態では、ウイルスカプシドは二十面体植物ウイルスに由来し、ウイルスの天然栄養宿主ではない植物種の宿主細胞において発現される。もう1つの実施形態では、ウイルス種は哺乳動物に感染し、発現系は植物宿主細胞を含む。代替的実施形態では、ウイルスは、カプシド−組換えペプチド融合物を発現するための宿主細胞として使用される特定植物種の栄養ウイルスである。
【0098】
植物宿主細胞は、維管束植物門、ユーフィロフィタ(Euphyllophyta)、種子植物門又は被子植物門からの植物細胞であり得る。特定実施形態では、植物細胞は、例えば:ヤシ科、例えばココヤシ属、アラブヤシ属;ヤマオイモ科、例えばヤムイモ属;イネ科(Gramineae(Poaceae))、例えばカラスムギ属、オオムギ属、イネ属、キビ属、コムギ属、トウモロコシ属;及びバショウ科、例えばバナナ、の成員を含むが、これらに限定されない単子葉植物である。
【0099】
代替的実施形態では、植物細胞は、例えば:ヒユ科、例えばアマランサス属、ベタ属、アカザ属;セリ科(Umbelliferae)、例えばダウクス属、パスティナカ属;アブラナ科(Cruciferae)、例えばアラビドプシス属、アブラナ属;キク科、例えばヒマワリ属;ヒルガオ科、例えばイポメア属;ウリ科、例えばスイカ属、キュウリ属、カボチャ属;トウダイグサ科;マメ科(Fabaceae)、例えばダイズ属、ヒラマメ属、ウマゴヤシ属、インゲンマメ属、エンドウ属、シャジクソウ属、ソラマメ属、ササゲ属;アマ科、例えばアマ属;及びナス科、例えばトウガラシ属、トマト属、タバコ属、ナス属を含むが、これらに限定されない双子葉植物の成員である。
【0100】
特定実施形態では、宿主細胞は、アラディポシス・タリアナ(Aradiposis thaliana)、日本産イチイ(タクスス・クスビダータ)、ニチニチソウ(カサランツス・ロセウス)、ニコチアナ・タバカム、オリザ・サティバ、リコペルシカム・エスキュレンタム、及びダイズ(グリシン・マックス)から成る群より選択される。
【0101】
(II.核酸構築物)
本発明はさらに、宿主細胞のゲノムに安定に組み込まれることができ、カプシドと組換えペプチドの融合ペプチドをコードする非感染性核酸構築物を提供する。融合ペプチドはプロモーター配列と終結配列に作動可能に連結され得る。1つの実施形態では、a)組換えペプチドをコードする核酸配列、及びb)ウイルスカプシドをコードする核酸配列、を含む植物宿主細胞を形質転換するときに使用するための核酸構築物が提供され、a)の核酸とb)の核酸は融合タンパク質を形成するように作動可能に連結されており、前記核酸構築物は植物宿主細胞のゲノムに安定に挿入され、植物細胞において発現される。
【0102】
ある実施形態では、ベクターは、多数のカプシド又は多数の対象ペプチドについての配列を含み得る。1つの実施形態では、ベクターは、少なくとも2個の異なるカプシド−ペプチドコード配列を含み得る。1つの実施形態では、コード配列は同じプロモーターに連結される。ある実施形態では、コード配列は、内部リボソーム結合部位によって分離される。他の実施形態では、コード配列は、細胞内でのウイルス様粒子の形成を可能にするリンカー配列によって連結される。もう1つの実施形態では、コード配列は異なるプロモーターに連結される。これらのプロモーターは、構成的であるか、又は同じか又は異なる誘導条件によって駆動され得る。もう1つの実施形態では、ゲノム内に挿入されることができ、異なるカプシド−ペプチドの組合せをコードする多数のベクターが提供される。多数のベクターは、構成的であるか又は同じ誘導条件によって駆動されるか又は異なる誘導条件によって駆動されるプロモーターを含み得る。1つの実施形態では、ウイルスカプシドは二十面体ウイルスカプシドに由来する。
【0103】
対象ペプチドについてのコード配列は、あらかじめ定められた部位でウイルスカプシド又はカプシドのコード配列に挿入することができる。前記ペプチドはまた、あらかじめ定められていない部位に挿入し、細胞をVLPの生産に関してスクリーニングすることができる。1つの実施形態では、VLPの形成の間にループとして発現されるように、ペプチドをカプシドコード配列に挿入する。1つの実施形態では、1個のペプチドコード配列をベクター内に含めるが、また別の実施形態では、多数の配列を含める。多数の配列は、コンカテマーの形態、例えば切断可能なリンカー配列によって連結されたコンカテマーの形態であり得る。
【0104】
ペプチドは、カプシド内の2つ以上の挿入部位に挿入し得る。それ故、ペプチドは、カプシドの2個以上の表面ループモチーフに挿入し得る;ペプチドはまた、所与のループモチーフ内の多数の部位に挿入し得る。挿入物の個々の機能性及び/又は構造ペプチド、及び/又はペプチド挿入物全体は、切断部位、すなわちタンパク質を切断する又は加水分解する因子がカプシド構造又は集合物の残りの部分からペプチドを分離するように作用し得る部位によって分離され得る。
【0105】
ペプチドは、外側に面したループ及び/又は内側に面したループ、すなわちそれぞれカプシドの中心から遠ざかる方向又は中心に向かう方向に面したカプシドのループ内に挿入され得る。カプシドの表面ループ内のいかなるアミノ酸又はペプチド結合も、ペプチドのための挿入部位として役立ち得る。典型的には、挿入部位は、ループのほぼ中央、すなわち折りたたまれたカプシドペプチドの三次構造の中心からほぼ最も遠位の位置で選択される。ペプチドコード配列は、選択されたループのこのほぼ中央に対応するカプシドコード配列の位置に作動可能に挿入され得る。これは、ペプチド挿入部位から下流で合成されるカプシドのペプチド配列のその部分についての読み枠の維持を含む。
【0106】
もう1つの実施形態では、ペプチドをカプシドのアミノ末端に挿入することができる。ペプチドは、上述した切断可能リンカーを含む、1又はそれ以上のリンカー配列を通してカプシドに連結することができる。さらにもう1つの実施形態では、ペプチドをカプシドのカルボキシ末端に挿入することができる。ペプチドを、化学的又は酵素的加水分解によって切断され得る1又はそれ以上のリンカー配列を通してカルボキシ末端に連結することができる。1つの実施形態では、ペプチド配列は、アミノ末端とカルボキシ末端の両方で、又は1つの末端と少なくとも1つの内部位置で、例えばその三次元立体配座においてカプシドの表面に発現される位置で、連結される。
【0107】
1つの実施形態では、ペプチドは、カウピークロロティックモザイクウイルスからのカプシドに挿入することができる。1つの特定実施形態では、ペプチドは、CCMVコートタンパク質のアミノ酸129で挿入できる。1つの実施形態では、ペプチド配列は、CCMVコートタンパク質のアミノ酸63、102、114、129又は160で挿入することができる。もう1つの実施形態では、ペプチド配列は、CCMVコートタンパク質のアミノ酸60、61、62又は63で挿入することができる。さらにもう1つの実施形態では、ペプチドは、CCMVタンパク質のアミノ酸129及びアミノ酸60−63の両方で挿入できる。
【0108】
特定実施形態では、本発明は、a)抗原性ペプチドをコードする核酸及びb)ウイルスカプシドをコードする核酸を含み、a)の核酸とb)の核酸が、細胞において発現されたとき融合タンパク質を形成するように作動可能に連結されている、核酸構築物を提供する。核酸構築物を構築する上で有用な他のカプシド及び組換えペプチドは上記に開示されている。
【0109】
(プロモーター)
1つの実施形態では、核酸構築物は、カプシド−組換えペプチド融合ペプチドをコードする核酸配列に作動可能に結合されたプロモーター配列を含む。作動可能な結合又は連結は、転写及び何らかの翻訳調節エレメントが、宿主細胞の作用により、調節エレメントが対象配列の発現を指令することができるように前記配列に共有結合している立体配置を指す。代替的実施形態では、カプシド融合タンパク質をコードする核酸構築物は、遺伝子捕獲戦略を利用して宿主細胞天然プロモーターとインフレームで挿入することができる。
【0110】
発酵工程では、ひとたび発酵が始まれば、発現系の効率を最大化するために高レベルの生産を得ることが理想的である。プロモーターは転写を開始させ、一般に転写開始部位の1−100ヌクレオチド上流に位置する。理想的には、プロモーターは大量の組換えペプチド蓄積を可能にするのに十分なほど強力である。
【0111】
本発明に従って使用されるプロモーターは、構成的プロモーター又は調節を受けるプロモーターであり得る。当技術分野で使用される有用な調節を受けるプロモーターの一般的な例は、エタノール誘導性プロモーター;鉄欠乏誘導性プロモーター;創傷誘導性プロモーター;及びホルモン誘導性、例えばオーキシン誘導性プロモーターを含む。有用な構成的プロモーターの一般的な例は、ユビキチンプロモーター、アクチンプロモーター、チューブリンプロモーター及びキャッサバ葉脈モザイクウイルスプロモーターを含む。本発明において有用な他の種類のプロモーターは、組織特異的プロモーター、例えばトウモロコシスクロースシンテターゼ1(Yang el al., 1990)、トウモロコシアルコールデヒドロゲナーゼ1(Vogel et al., 1989)、トウモロコシ光捕集複合体 (Simpson, 1986)、トウモロコシ熱ショックタンパク質 (Odell et al., 1985)、エンドウマメ小サブユニットRuBPカルボキシラーゼ (Poulsen et al., 1986; Cashmore et al., 1983)、Tiプラスミドマンノピンシンターゼ (Langridge et al., 1989)、Tiプラスミドノパリンシンターゼ (Langridge et al., 1989)、ペチュニアカルコンイソメラーゼ (Van Tunen et al., 1988)、インゲンマメグリシンリッチタンパク質1(Keller et al., 1989)、CaMV 35s転写産物 (Odell et al., 1985) 及びジャガイモパタチン(Wenzler et al., 1989)を含む。好ましいプロモーターは、カリフラワーモザイクウイルスCaMV 35S)プロモーター、イネα−アミラーゼRAmy3Dプロモーター、及びS−E9小サブユニットRuBPカルボキシラーゼプロモーターである。加えて、環境、ホルモン、化学物質及び/又は発生シグナルに応答して調節される多様な植物遺伝子プロモーターが、植物細胞における作動可能に連結された遺伝子の発現のために使用でき、(1)熱(Callis et al., Plant Physiol. 88:965, 1988)、(2)光(例えばエンドウマメrbcS−3Aプロモーター、Kuhlemeier et al., Plant Cell 1:471, 1989;トウモロコシrbcSプロモーター、 Schaffher and Sheen, Plant Cell 3:997, 1991;又はクロロフィルa/b結合タンパク質プロモーター、Simpson et al., EMBO J. 4:2723, 1985)、(3)ホルモン、例えばアブシジン酸 (Marcotte et al., Plant Cell 1:969, 1989)、(4)創傷(例えばwunI、Siebertz et al., Plant Cell 1:961, 1989); 又は(5)化学物質、例えばジャスモン酸メチル、サリチル酸又は薬害軽減剤、によって調節されるプロモーターを含む。また、(6)器官特異的プロモーター(例えば Roshal et al., EMBO J. 6:1155, 1987; Schernthaner et al., EMBO J. 7:1249, 1988; Bustos et al., Plant Cell 1:839, 1989)を使用することも有益である。本発明において有用なさらなるプロモーターは、国際公開公報第WO97/48819号に述べられているもの;米国特許第 5,591,605号に述べられているファゼオリンプロモーター; 米国特許第5,641,876号に述べられているイネアクチンプロモーター;国際公開公報第WO98/56921号に述べられているper5プロモーター;国際公開公報第WO00/12681号に述べられているγ−ゼインプロモーター;及び米国特許第6,825,006号、同第6,660,911号に述べられている他のプロモーター、及びShahmuradov et al. (2003) “PlantProm: a database of plant promoter sequences,” Nucleic Acid Res. 31(1): 114-117の中で述べられており、http://mendel.cs.rhul.ac.uk/mendel.php?topic=plantpromにおいて入手可能な、植物プロモーター配列データベースPlantPromに述べられている植物プロモーターを含む。さらなるプロモーターはまた、カリフラワーモザイクウイルス(CaMV)35Sプロモーター又はその強化型、ハイブリッド(ocs)3masプロモーター、及びトウモロコシ又はA.タリアナからのユビキチンプロモーターを含み得る。
【0112】
選択植物宿主細胞にネイティブなプロモーターのヌクレオチド配列を有するプロモーターも、標的ペプチドをコードする導入遺伝子の発現を制御するために使用できる。2個以上のプロモーターが、同じか又は異なる配列の、もう1つ別のプロモーターに共有結合している縦列(タンデム)プロモーターも使用し得る。
【0113】
ウイルスに由来するヌクレオチド配列を有するプロモーターも、標的ペプチドをコードする導入遺伝子の発現を指令するために使用できる。
【0114】
調節を受けるプロモーターは、プロモーターがその一部である遺伝子の転写を制御するためにプロモーター調節タンパク質を利用することができる。調節されるプロモーターをここで使用する場合は、対応するプロモーター調節タンパク質も、本発明に従った発現系の一部である。多くの調節されるプロモーター/プロモーター調節タンパク質の対が当技術分野において公知である。
【0115】
(他のエレメント)
他の調節エレメントを発現構築物に含めることができる。そのようなエレメントは、例えば転写エンハンサー配列、翻訳エンハンサー配列、他のプロモーター、アクチベーター、翻訳開始及び終結シグナル、転写ターミネーター、シストロニック調節因子、ポリシストロニック調節因子、発現されるペプチドの特定、分離、精製又は単離を容易にするタグ配列、例えばヌクレオチド配列「タグ」及び「タグ」ペプチドコード配列、例えばHisタグ、Flagタグ、T7タグ、Sタグ、HSVタグ、Bタグ、Strepタグ、ポリアルギニン、ポリシステイン、ポリフェニルアラニン、ポリアスパラギン酸、(Ala−Trp−Trp−Pro)n、チオレドキシン、β−ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、シクロマルトデキストリングルコノトランスフェラーゼ、CTP:CMP−3−デオキシ−D−マンノ−オクツロソネートシチジルトランスフェラーゼ、trpE又はtrpLE、アビジン、ストレプトアビジン、T7遺伝子10、T4gp55、ブドウ球菌プロテインA、連鎖球菌プロテインG、GST、DHFR、CBP、MBP、ガラクトース結合ドメイン、カルモジュリン結合ドメイン、GFP、KSI、c−myc、ompT、ompA、pelB、NusA、ユビキチン及びヘモシリンA、を含むが、これらに限定されない。
【0116】
1つの実施形態では、核酸構築物は、対象組換えペプチドについてのコード配列に隣接する、又はウイルスカプシドについてのコード配列に連結された、タグ配列をさらに含む。1つの実施形態では、タグ配列はタンパク質の精製を可能にする。タグ配列は、アフィニティータグ、例えばヘキサヒスチジンアフィニティータグであり得る。もう1つの実施形態では、アフィニティータグは、グルタチオン−S−トランスフェラーゼ分子であり得る。タグはまた、蛍光分子、例えばYFP又はGFP、又はそのような蛍光タンパク質の類似体であり得る。タグはまた、抗体分子の一部、又は精製のために有用な公知の結合パートナーについての公知の抗原又はリガンドであり得る。
【0117】
本発明は、カプシド−組換えペプチドコード配列に加えて、それに作動可能に連結された以下の調節エレメント:プロモーター、転写ターミネーター、翻訳開始及び終結シグナルを含み得る。
【0118】
翻訳及び転写エレメント、及び本発明において有用な他のエレメントのさらなる例は、JD Watson et al. (eds. ), Recombinant DNA, pp. 273-92 (1992) (Scientific American Books, W. H. Freeman and Co. , New York, NY, USA); and I Mitsuhara et al.,”Efficient promoter cassettes for enhanced expression of foreign genes in dicotyledonous and monocotyledonous plants,”Plant Cell Physiol. 37(1) : 49-59 (1996)に述べられている。
【0119】
1つの特定実施形態では、核酸構築物は、ウイルスカプシドタンパク質を導くために使用されるウイルスゲノム核酸配列の3’非翻訳領域(3’UTR)に由来する付加的な核酸配列を含む。1つの特定実施形態では、3’UTRは、野生型ウイルスにおける翻訳の調節又はカプシド形成において役割を果たす配列を含む。1つの実施形態では、3’UTR配列は上流シュードノットドメイン(UPD)である。例えばLeathers et al.(1993)”A phylogenetically conserved sequence within viral 3' untranslated RNA pseudolmots regulate stranslation,”Mol. Cell Biol. 13 (9): 5331-5347参照。特定実施形態では、3’UTRは、ウイルスゲノムのウイルスカプシドコード領域の3’UTRに由来する。より特定の実施形態では、3’UTRは、カウピークロロティックモザイクウイルスRNA3 3’非翻訳領域に由来する。1つの実施形態では、3’UTR配列はカプシド形成シグナルを含む。
【0120】
代替的実施形態では、核酸構築物は、ウイルス核酸配列に由来する少なくとも1個のカプシド形成シグナルを含む。カプシド形成シグナルは、ウイルスゲノム配列の5’UTR、3’UTR又は内部配列に由来し得る。1つの実施形態では、核酸構築物はVLPにおいてカプシド形成される。
【0121】
1つの実施形態では、非感染性核酸構築物はまた、ウイルスカプシドタンパク質配列以外のウイルス又は非ウイルス核酸配列に由来する他の配列も含む。
【0122】
(ベクター)
宿主細胞ゲノムに安定に組込み、カプシド−組換えペプチド融合ペプチドを発現するとき植物細胞において使用するための有用な発現ベクターは、機能的プロモーターを有する作動可能な読み枠内に、適切な翻訳開始及び終結シグナルと共にカプシドペプチドに融合した所望標的ペプチドをコードする構造DNA配列を挿入することによって構築される。ベクターはまた、宿主細胞の選択を可能にする1又はそれ以上の表現型選択マーカー又はスクリーニングマーカーを含み得る。あるいは、選択又はスクリーニングマーカーは別のプラスミド上で提供され得る。
【0123】
極めて多様なベクター及び/又は担体が、発現のために組換えカプシドをコードする核酸で標的植物宿主細胞を形質転換するために有用であることが当技術分野において公知であり、これらのいずれもが本発明に従った遺伝子を発現するために使用し得る。例えばIJ Goderis IJ et al.,”A set of modular plant transformation vectors allowing flexible insertion of up to six expression units,”Plant Mol. Biol. 50(1): 17-27 (2002); AH Christensen & PH Quail,”Ubiquitin promoter-based vectors for high-level expression of selectable and/or screenable marker genes in monocotyledonousplants,”Transgenic Res. 5 (3): 213-18 (1996); JD Jones et al., “Effective vectors for transformation, expression of heterologous genes, and assaying transposon excision in transgenic plants,”Transgenic Res. 1 (6): 285-97 (1992); AP Gleave AP,”A versatile binary vector system with a T-DNA organisational structure conducive to efficient integration of cloned DNA into the plant genome,”Plant Mol. Biol. 20 (6): 1203-07 (1992)参照。
【0124】
それ故、プラスミド、トランスポゾン、ゲノムDNA、ゲノムRNA、植物人工染色体及び他の核酸ベクターが使用し得る。使用し得る一部のベクターの例は、pRT101、pRT101−MCS、pRT104、pRT104.24LS、pRT104.N−myc、pRT104.C−myc、pRT104.N−3HA、pRT104−N3HAdAsp、pRT104.N−6HA、pRT104−N6HAdAsp、pRT104.C−3HA、pRT104−GST、pRT104 NES(A2)、pRT103−3HA、pRTd35S−Luci(−)、pRTd35S−Luci−NES、pRTd35S−GFP、pRTd35S−GFP.SacI、pRTdS GFP.BglII、pRTd35S−Ds−Red、pRTd35S−プロフィリン、pBIGal4DBD、pRT−3HAxHsfA2DBD、pRT−3HAxGal4DBD、pUC、pIL−TAB、pET、pME、pBBR及びpROKIIを含む。ベクターを細胞に導入するためにパーティクルガンを使用するときは、いかなるDNAプラスミド又はDNA分子も本発明において使用し得る。
【0125】
特定実施形態では、ベクターはpIL−Tabである。より特定の実施形態では、ベクターは、カウピークロロティックモザイクウイルス(CCMV)からのコートタンパク質をコードするpIL−Tabである。特定実施形態では、ベクターはpIL−Tabであり、異種ペプチドは、PA1、PA2、PA3又はPA4から成る群より選択される。さらなる実施形態では、ベクターは、pDOW2160(配列番号5)、pDOW2161(配列番号6)、pDOW2162(配列番号7)、pDOW2163(配列番号8)、pDOW2169(配列番号9)、pDOW2170(配列番号10)、pDOW2171(配列番号11)及びpDOW2172(配列番号12)から成る群より選択される。
【0126】
あるいは又はその上に、担体、例えばリポソーム、デンドリマー、カチオン性ポリマー及びカチオン性ポリマー−脂質複合体が使用でき、前記全てが細胞又はプロトプラストへの核酸の送達のために広く公表されている方法において使用されている。あるいは、裸のDNA又は裸のRNAも植物宿主に送達され得る。
【0127】
ベクター、担体又は裸の核酸は、そのために有効であることが当技術分野で公知の何らかの方法によって植物細胞又はプロトプラストに直接形質転換され得る。例えば、組換えカプシドをコードする核酸で植物標的宿主を形質転換するために以下の方法のいずれかを使用し得る:アグロバクテリウム属;V. Vasil et al., Bio/Technology 9, 743 (1991)によって述べられているようなマイクロパーティクルガン法;例えばMC Chupeau et al., Bio/Technology 7,503 (1989)によってレタスに関して述べられているようなプロトプラストの電気穿孔法;例えば炭化ケイ素ウィスカー、ガラス又はカーボランダムを使用する表面研磨法;例えばA. Deshayes et al., EMBO J. 4, p.2731-2737 (1985)によって述べられているようなプロトプラストとのリポソーム融合;細胞−細胞(プロトプラスト−プロトプラスト)融合;及び内因性植物ウイルス感染、すなわち選択植物標的宿主がその天然宿主であるタイプのウイルスであり、ウイルスが組換えカプシド−タンパク質をコードする核酸を含むように工作されている(組換え体が作られるカプシド−タンパク質−核酸が、植物標的宿主がその天然宿主ではないウイルスから選択される)、工作されたベクターによる植物標的宿主の形質転換;例えば I.Potrykus et al., Mol. Gen. Genetics, 197, 183-188 によって述べられているようなポリエチレングリコールを介した形質転換;及び例えばR Griesbach, Biotechnology 3, p. 348-350およびCK Shewmaker Mol. Gen. Genetics, 202 p. 179-185 (1986)によって述べられているような微量注入法。
【0128】
(III.植物細胞におけるカプシド融合産物の発現)
本発明はまた、組換えペプチドを生産するための方法を提供する。その方法は、
a)植物細胞を提供すること;
b)組換えペプチドと二十面体カプシドの融合ペプチドをコードする発現カセットを含む非感染性核酸を提供すること;
c)前記植物において前記核酸を発現し、前記細胞における発現が前記融合ペプチドのウイルス様粒子へのインビボ集合を提供すること;及び
d)前記ウイルス様粒子を単離すること
を含む。
【0129】
融合ペプチドは、プロモーター配列及びターミネーターに作動可能に連結される。1つの実施形態では、前記方法は、e)ウイルスカプシドタンパク質から組換えペプチドを切断すること、をさらに含む。さらにもう1つの実施形態では、前記方法は、f)工程eから組換えペプチドを単離すること、をさらに含む。例えば図1参照。
【0130】
ペプチドは、カプシドペプチド内で1コピーペプチド挿入物として発現され得る(すなわちペプチドに関してモノシストロニックである組換えカプシドペプチドコード配列から個別挿入物として発現される)か、又は2、3又は多コピーペプチド挿入物として発現され得る(すなわちペプチドに関してポリシストロニックである組換えカプシドペプチドコード配列からコンカテマー挿入物として発現される;コンカテマー挿入物は、同じ外来性対象ペプチドの多数のコピーを含み得るか又は異なる外来性対象ペプチドのコピーを含み得る)。コンカテマーは、ホモ又はヘテロコンカテマーであり得る。
【0131】
1つの実施形態では、単離されたウイルス様粒子をワクチン戦略においてヒト又は動物に投与することができる。
【0132】
もう1つの実施形態では、核酸構築物を、野生型カプシドをコードするもう1つ別の核酸と共発現することができる。特定実施形態では、共発現されたカプシド/カプシド−組換えペプチド融合粒子は、キメラウイルス様粒子を形成するようにインビボで集合する。キメラVLPは、少なくとも2個の異なる核酸構築物によってコードされるカプシド又はカプシド−ペプチド融合物を含むウイルス様粒子である。
【0133】
さらにもう1つの実施形態では、核酸構築物を、異なるカプシド−組換えペプチド融合粒子をコードするもう1つ別の核酸と共発現することができる。特定実施形態では、共発現されたカプシド融合粒子は、キメラウイルス様粒子を形成するようにインビボで集合する。
【0134】
さらにもう1つの実施形態では、異なるペプチド、例えばシャペロンタンパク質を発現するように設計された第二核酸を、融合ペプチドをコードする核酸と同時に発現することができる。
【0135】
本発明のために有用な植物細胞、カプシド及び組換えペプチドは上記に述べられている。
【0136】
1つの実施形態では、発現されるウイルスカプシド−異種ペプチド融合産物は、細胞において可溶性形態で発現される。
【0137】
1つの実施形態では、発現されるウイルスカプシド−異種ペプチド融合産物は、細胞内でVLPに構築される。
【0138】
別の実施形態では、発現される対象ウイルスカプシド−異種ペプチド融合産物の一部は、細胞において不溶性凝集物として形成される。1つの実施形態では、対象ペプチドを不溶性凝集物から復元することができる。
【0139】
(対象ペプチドの切断)
1つの実施形態では、前記方法は、e)カプシドから組換えペプチドを分離するために前記融合産物を切断すること、をさらに提供する。
【0140】
切断可能な連結配列は、ウイルスタンパク質と組換えペプチドの間に含まれ得る。そのような配列を切断することができる物質の例は、化学試薬、例えば酸(塩酸、ギ酸)、CNBr、ヒドロキシルアミン(アスパラギン−グリシンに関して)、2−ニトロ−5−チオシアノベンゾエート、O−ヨードソベンゾエート、及び酵素物質、例えばエンドペプチダーゼ、エンドプロテアーゼ、トリプシン、クロストリパイン及びブドウ球菌プロテアーゼを含むが、これらに限定されない。
【0141】
切断可能な連結配列は当技術分野において周知である。本発明では、Asp−Proなどのジペプチド切断配列を含む、切断物質によって認識されるいかなる切断可能連結配列も使用できる。
【0142】
(発現)
本発明の方法は、最適には、所望配列及び立体配座での、宿主植物細胞における所望カプシド−融合産物の生産上昇を導く。生産上昇は、選択的に、生産されるタンパク質のグラム当り又は宿主タンパク質のグラム当り又は生産される総組換えペプチドのパーセンテージとしての、所望ペプチドのレベル上昇であり得る。生産上昇はまた、組換え体のグラム当り又は宿主細胞タンパク質のグラム当りで生産される回収可能なペプチド、例えば可溶性タンパク質のレベル上昇であり得る。生産上昇はまた、タンパク質の総レベル上昇と活性又は可溶性レベル上昇の何らかの組合せであり得る。
【0143】
組換えタンパク質の改善された発現は、カプシド融合タンパク質としてのタンパク質の発現を通してであり得、その後VLPに挿入することができる。ある実施形態では、少なくとも60、少なくとも70、少なくとも80、少なくとも90、少なくとも100、少なくとも110、少なくとも120、少なくとも130、少なくとも140、少なくとも150、少なくとも160、少なくとも170、又は少なくとも180コピーの対象ペプチドが各々のVLPにおいて発現される。VLPを生産し、宿主細胞の細胞質、ペリプラズム又は細胞外液から回収することができる。
【0144】
もう1つの実施形態では、ペプチドは細胞において不溶性であり得る。ある実施形態では、可溶性又は不溶性ペプチドは、多数のカプシドから形成されるが天然型VLPを形成しない粒子において生産される。例えばわずか3個のウイルスカプシドのケージ構造が形成され得る。ある実施形態では、カプシド構造は2コピー以上の対象ペプチドを含み、ある実施形態では、少なくとも10、少なくとも20又は少なくとも30コピーを含む。ある実施形態では、ペプチドは植物細胞の液胞において形成される。
【0145】
ペプチド又はウイルスカプシド配列はまた、1又はそれ以上のターゲティング配列又は精製を助けるための配列を含み得る。これらはアフィニティータグであり得る。これらはまた、カプシドのVLPへの構築を指令するターゲティング配列であり得る。
【0146】
(細胞増殖)
ベクターによる植物宿主細胞の形質転換は、当技術分野で公知のいかなる形質転換法を用いて実施してもよく、植物宿主細胞を無傷細胞(intact cells)として又はプロトプラストとして形質転換し得る。例示的な形質転換法は上記に述べられている。
【0147】
ここで使用する、「発酵」という用語は、文字通りの発酵を用いる実施形態と、他の非発酵培養法を用いる実施形態の両方を包含する。発酵はいかなる規模で実施してもよい。当技術分野で一般的に知られるいかなる方法も発酵において使用し得る。例えばHellwig et al. (2004) “Plant cell cultures for the production of recombinant proteins, “Nature Biotech 22 (11): 1415-1422; Sajc et al. (2000) “Bioreactors for plant engineering : an outlook for further research, ”Biochem Engin. J. 4: 89-99参照。1つの実施形態では、発酵培地は、何らかの生育可能植物細胞培地から選択され得る。場合により、培地への添加物は、例えばPVP、BSA、NaCl、ブレフェルジンA、還元マンガン、プルロニック消泡剤、ポリエチレングリコール、ゼラチン、プロテアーゼ阻害剤、酸化還元補因子及びジメチルスルホキシドを含み得る。
【0148】
本発明に従った発現系は、いかなる発酵方式でも培養することができる。例えばバッチ、フェドバッチ、半連続、及び連続発酵方式がここで使用し得る。
【0149】
本発明に従った発現系は、いかなる規模(すなわち容量)の発酵においても導入遺伝子発現のために有用である。それ故、例えばマイクロリットル規模、センチリットル規模及びデシリットル規模の発酵容量を使用し得る;及び1リットル又はそれ以上の規模の発酵容量が使用できる。1つの実施形態では、発酵容量は1リットル又はそれ以上である。もう1つの実施形態では、発酵容量は、5リットル又はそれ以上、10リットル、15リットル、20リットル、25リットル、50リットル、75リットル、100リットル、200リットル、500リットル、1,000リットル、2,000リットル、5,000リットル、10,000リットル又は50,000リットルである。
【0150】
本発明では、形質転換宿主細胞の増殖、培養及び/又は発酵は、宿主細胞の生存を許容する温度範囲内、好ましくは約4℃から約55℃(両端を含む)の範囲内の温度で実施される。
【0151】
(細胞密度)(充填湿細胞(Packed wet cell))
本発明に従った植物細胞発現系は、約20g/L(2%充填細胞量)から550g/L(55%充填細胞量)より多くまでの細胞密度又は匹敵する充填湿細胞密度を提供することができる。1つの実施形態では、充填細胞量は2%−約60%である。もう1つの実施形態では、充填細胞量は、2%、5%、10%、15%、20%、25%、35%、40%、50%、55%、60%、70%又は85%である。
【0152】
(VLP又は対象ペプチドの単離)
ある実施形態では、本発明は、ウイルスカプシドとの連結及び共発現を通して発現の間ペプチドを保護することにより、対象ペプチドの回収率を改善するための方法を提供する。ある実施形態では、ウイルスカプシド融合は、細胞溶解産物から容易に分離することができるVLPを形成する。
【0153】
本発明のタンパク質は、PEG沈殿、硫酸アンモニウム又はエタノール沈殿、酸抽出、陰イオン又は陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ニッケルクロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィー、逆相クロマトグラフィー、レクチンクロマトグラフィー、分取電気泳動、界面活性剤可溶化、カラムクロマトグラフィーのような物質による選択的沈殿、免疫精製法等を含むが、これらに限定されない、当技術分野において周知の標準手法によって単離し、実質的純度に精製し得る。例えば確立された分子接着特性を有するタンパク質は、リガンドに可逆的に融合することができる。適切なリガンドを用いて、タンパク質を選択的に精製カラムに吸着させ、その後比較的純粋な形態でカラムから遊離することができる。次に、酵素又は他の作用によって融合タンパク質を取り出すことができる。加えて、免疫アフィニティーカラム又はNi−NTAカラムを用いてタンパク質を精製することができる。一般的な手法は、例えばR. Scopes, Protein Purification: Principles and Practice, Springer-Verlag: N. Y. (1982); Deutscher, Guide to Protein Purification, Academic Press (1990);米国特許第4,511,503号; S. Roe, Protein Purification Techniques: A Practical Approach (Practical Approach Series), Oxford Press (2001); D. Bollag, et al., Protein Methods, Wiley-Lisa, Inc. (1996); AK Patra et al., Protein Expr Purif, 18 (2): p/182-92 (2000);およびR. Mukhija, et al., Gene 165 (2): p. 303-6 (1995)において詳述されている。また、例えばAusubel, et al.(1987 and periodic supplements); Deutscher (1990) “Guide to Protein Purification, “Methods in Enzymology vol.182及びこのシリーズの他の巻; Coligan, et al. (1996 and periodic Supplements) Current Protocols in Protein Science Wiley/Greene, NY ;及びタンパク質精製産物の使用に関する製造者の文献、例えばPharmacia,Piscataway,N.J.又はBio−Rad,Richmond,Calif.も参照のこと。組換え手法の併用は、適切なセグメント、例えばFLAG配列又はプロテアーゼ−除去可能配列によって融合できる等価物、への融合を可能にする。また、例えばHochuli (1989) Chemische Industrie 12: 69-70; Hochuli (1990) “Purification of Recombinant Proteins with Metal Chelate Absorbent”in Setlow (ed.) Genetic Engineering, Principle and Methods 12: 87-98, Plenum Press, NY;およびCrowe, et al.(1992) QIAexpress : The High Level Expression & Protein Purification System QUIAGEN, Inc., Chatsworth, Calif.も参照のこと。
【0154】
同様に、ウイルス様粒子又はケージ様構造も、当技術分野で周知の標準手法によって単離し、実質的純度に精製することができる。VLPの単離のための手法は、上述したものに加えて、沈殿法、例えばポリエチレングリコール又は塩沈殿法を含む。分離手法は、陰イオン又は陽イオン交換クロマトグラフィー、サイズ排除クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ニッケルクロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィー、逆相クロマトグラフィー、レクチンクロマトグラフィー、分取電気泳動、免疫精製法、遠心分離、超遠心分離、密度勾配遠心分離(例えばスクロース又は塩化セシウム(CsCl)勾配で)、サイズ排除フィルターを通しての限外ろ過、及び当技術分野で公知の何らかの他のタンパク質単離法を含む。
【0155】
本発明はまた、活性組換えペプチドの回収率を改善することができる。活性タンパク質のレベルは、例えば特定されたペプチドと親ペプチド、ペプチド変異体、セグメント置換ペプチド及び/又は残基置換ペプチドの間の相互作用を従来の何らかのインビトロ又はインビボアッセイで測定することによって測定できる。そこで、インビトロアッセイは、特定されたタンパク質と対象ペプチドの間、例えば酵素と基質の間、ホルモンとホルモン受容体の間、抗体と抗原の間等の、何らかの検出可能な相互作用を測定するために使用できる。そのような検出は、比色変化、放射能の変化、溶解度の変化、ゲル電気泳動及び/又はゲル排除法によって測定したときの分子量の変化等の測定を含み得る。インビボアッセイは、生理的作用、例えば体重増加、電解質バランスの変化、血液凝固時間の変化、血餅溶解の変化及び抗原応答の誘導を検出するアッセイを含むが、これらに限定されない。一般に、特定されたペプチドと対象ペプチドの間の相互作用の変化を検出するための可変パラメータが存在する限り、いかなるインビボアッセイも使用することができる。例えば米国特許第5,834,250号参照。
【0156】
発現されたタンパク質の検出は、当技術分野で公知の方法によって達成され、例えば放射免疫測定法、ウエスタンブロット法又は免疫沈降法を含む。
【0157】
初期塩分画は、望ましくない宿主細胞タンパク質(又は細胞培養培地に由来するタンパク質)の多くを対象組換えタンパク質から分離することができる。1つのそのような例は硫酸アンモニウムであり得る。硫酸アンモニウムは、タンパク質混合物中の水分量を有効に減少させることによってタンパク質を沈殿させる。タンパク質はその後、それらの溶解度に基づいて沈殿する。タンパク質がより疎水性であるほど、より低い硫酸アンモニウム濃度で沈殿する可能性が高い。典型的なプロトコールは、生じる硫酸アンモニウム濃度が20−30%になるように飽和硫酸アンモニウムをタンパク質溶液に添加することを含む。この濃度は最も疎水性のタンパク質を沈殿させる。次に沈殿物を廃棄し(対象タンパク質が疎水性でない限り)、対象タンパク質を沈殿させることが既知である濃度まで硫酸アンモニウムを上清に添加する。次に沈殿物を緩衝液に溶解し、必要に応じて、透析又はダイアフィルトレーションのいずれかを通して過剰の塩を除去する。タンパク質の溶解度に基づく他の方法、例えば低温エタノール沈殿は当業者に周知であり、複雑なタンパク質混合物を分画するために使用できる。
【0158】
組換えタンパク質の分子量を利用して、異なる細孔径の膜(例えばアミコン又はミリポア膜)を通す限外ろ過を用いてより大きい及びより小さいサイズのタンパク質から単離することができる。第一段階として、タンパク質混合物を、対象タンパク質の分子量よりも低い分子量カットオフ値を有する細孔径の膜を通して限外ろ過することができる。限外ろ過の保持物質を、次に、対象タンパク質の分子量よりも大きい分子量カットオフ値を有する膜に対して限外ろ過することができる。組換えタンパク質は膜を通過してろ液中に入る。その後ろ液をクロマトグラフィーに供することができる。
【0159】
組換えタンパク質はまた、その大きさ、正味表面電荷、疎水性、及びリガンドに対する親和性に基づいて他のタンパク質から分離することができる。加えて、タンパク質に対して惹起される抗体をカラムマトリックスに結合(固定化)し、タンパク質を免疫精製することができる。これらの方法は全て当技術分野において周知である。クロマトグラフィー手法が、いかなる規模でも及び多くの異なる製造者(例えばPharmacia Biotech)からの装置を使用して実施できることは当業者には明白である。
【0160】
(復元及びリフォールディング)
二次及び三次タンパク質構造立体配座を生成するために不溶性タンパク質を復元又はリフォールディングすることができる。タンパク質リフォールディング工程は、必要に応じて、組換え産物の立体配置を完成させるときに使用できる。リフォールディング及び復元は、タンパク質の解離/会合を促進する当技術分野で公知の物質を用いて実施することができる。例えばタンパク質をジチオトレイトールと共にインキュベートし、次に酸化グルタチオンニナトリウム塩と共にインキュベートして、さらにその後リフォールディング剤、例えば尿素を含む緩衝液と共にインキュベートすることができる。
【0161】
組換えタンパク質はまた、例えばリン酸緩衝食塩水(PBS)又は50mM 酢酸ナトリウム、pH6緩衝液プラス200mM NaClに対して透析することによっても復元できる。あるいは、タンパク質を、カラム、例えばNi NTAカラムに固定化して、プロテアーゼ阻害因子を含有する、500mM NaCl、20%グリセロール及び20mM トリス/HCl pH7.4中の6M−1M 尿素直線勾配を使用することによって再生できる。復元は1.5時間又はそれ以上の期間で実施できる。復元後、250mM イミダゾールの添加によってタンパク質を溶出することができる。イミダゾールは、PBS又は50mM 酢酸ナトリウム、pH6緩衝液プラス200mM NaClに対する最終透析工程によって除去することができる。精製タンパク質は、室温、4℃で保存するか又は−20℃から−80℃で凍結して保存することができる。
【0162】
他の方法は、例えばMH Lee et al., Protein Expr. Purif., 25(1) : p. 166-73 (2002), W. K. Cho et al. , J. Biotechnology, 77 (2-3): p. 169-78 (2000), Ausubel, et al. (1987 and periodic supplements), Deutscher (1990) “Guide to Protein Purification, “Methods in Enzymology vol. 182, and other volumes in this series, Coligan, et al. (1996 and periodic Supplements) Current Protocols in Protein Science Wiley/Greene, NY, S. Roe, Protein Purification Techniques: A Practical Approach (Practical Approach Series), Oxford Press (2001); D. Bollag, et al. , Protein Methods, Wiley-Lisa, Inc. (1996)に述べられているものを含む。
【0163】
(活性ペプチド分析)
活性タンパク質は、その配列が由来する天然ペプチドの活性の少なくとも20%、30%又は40%、好ましくは少なくとも50%、60%又は70%、最も好ましくは少なくとも80%、90%又は95%の比活性を有し得る。さらに、基質特異性(Kcat/Km)は、場合により天然ペプチドと実質的に同様である。典型的には、Kcat/Kmは、天然ペプチドの少なくとも30%、40%又は50%、より好ましくは少なくとも60%、70%、80%又は90%である。タンパク質及びペプチドの活性及び基質特異性(Kcat/Km)を検定する方法及び定量手段は当業者に周知である。
【0164】
本発明に従って生産される組換えペプチドの活性は、当技術分野で公知の何らかのタンパク質特異的な慣例的又は標準的インビトロ又はインビボアッセイによって測定することができる。組換えタンパク質が、同じ又は類似の生理的条件下で天然ペプチドにおいて一般に認められる活性と実質的に類似又は同等の活性を示すかどうかを判定するために、植物細胞宿主が生産する組換えペプチドの活性を、対応する天然タンパク質の活性と比較することができる。
【0165】
組換えペプチドの活性を、これまでに確立された天然ペプチド標準活性と比較することができる。あるいは、組換えペプチドの活性を、天然ペプチドと同時又は実質的に同時の比較アッセイにおいて測定することができる。例えばインビトロアッセイを使用して、組換えペプチドと標的の間、例えば発現される酵素と基質の間、発現されるホルモンとホルモン受容体の間、発現される抗体と抗原の間等の、検出可能な相互作用を測定することができる。そのような検出は、比色変化、増殖変化、細胞死、細胞反発、放射能の変化、溶解度の変化、ゲル電気泳動及び/又はゲル排除法によって測定したときの分子量の変化、リン酸化能、抗体特異性アッセイ、例えばELISAアッセイ等の測定を含み得る。加えて、インビボアッセイは、天然ペプチドの生理的作用、例えば抗原応答の誘導と比較して、植物宿主細胞が生産するペプチドの生理的作用を検出するアッセイを含むが、これらに限定されない。一般に、いかなるインビトロ又はインビボアッセイも、そのような活性が検定可能である限り、天然ペプチドとの比較分析を可能にする組換えペプチドの活性を測定するために使用できる。あるいは、本発明において生産されるペプチドは、ペプチドと、通常そのペプチドと相互作用する分子、例えば基質又は天然タンパク質が通常相互作用するシグナル経路の成分、との間の相互作用を刺激する又は阻害する能力に関して検定することができる。そのようなアッセイは、典型的には、ペプチドが標的分子と相互作用することを許容する条件下でタンパク質と基質分子を組み合わせる工程を含み、タンパク質と標的分子の相互作用の生化学的結果を検出することができる。
【0166】
ペプチド活性を測定するために使用できるアッセイは、例えばRalph, P. J., et al. (1984) J. Immunol. 132:1858 or Saiki et al. (1981) J. Immunol. 127: 1044, Steward, W. E.II (1980) The Interferon Systems. Springer-Verlag, Vienna and New York, Broxmeyer, H. E., et al. (1982) Blood 60: 595, “Molecular Cloning: A Laboratory Manual”, 2d ed., Cold Spring Harbor Laboratory Press, Sambrook, J., E. F. Fritsch and T. Maniatis eds. , 1989, and “Methods in Enzymology: Guide to Molecular Cloning Techniques”, Academic Press, Berger, S. L. and A. R. Kimmel eds., 1987, AK Patra et al., Protein Expr Purif, 18 (2):p/182-92 (2000), Kodama et al., J. Biochem. 99: 1465-1472 (1986); Stewart et al., Proc.Nat'1 Acad. Sci. USA 90: 5209- 5213 (1993); (Lombillo et al., J. Cell Biol. 128: 107-115 (1995); (Vale et al., Cell 42: 39-50 (1985) に述べられている。
【実施例】
【0167】
コートタンパク質をコードする核酸のウイルスソース
以下の実施例では、カウピークロロティックモトルウイルス(CCMV)を所望組換えペプチドの発現のためのコートタンパク質のソースとして使用した。CCMVは、ブロモウイルス科ブロモウイルス群の成員である。ブロモウイルスは、4成分のプラスセンス一本鎖RNAゲノムを有する直径25−28nmの二十面体ウイルスである。RNA1及びRNA2はレプリカーゼ酵素をコードする。RNA3は、植物宿主内のウイルス運動に関与するタンパク質をコードする。RNA4(RNA3に由来するサブゲノムRNA)、すなわちsgRNA4は、20kDaのコートタンパク質(CP)(配列番号13)(表1)をコードする。各々のCCMV粒子は、180コピーのCCMV CPを含む。CCMV CPをコードする例示的DNA配列を配列番号14(表2)に示す。
【表1】
【表2】
【0168】
CCMVの結晶構造を解明した。この構造は、粒子の溶解度と動力学にとって決定的に重要であると思われるコートタンパク質相互作用のより明瞭な像を提供し、挿入部位の合理的設計を導くのに有用であった。これまでの試験は、CCMVコートタンパク質を、粒子を形成するそれらの能力を妨げることなく異種ペプチドを担持するように遺伝的に修飾できることを明らかにした。多くの適切な挿入部位が特定されている。合計約180コピーまでの異種ペプチド単位(個々のペプチド単位として又はコンカテマー単位内で)を、CCMV CP内の単一挿入部位でCCMV粒子に挿入することができる。現在までにCCMV CP内で特定された挿入部位は、様々な長さのペプチドを収容することができる。
【0169】
実験材料及び方法
異なる記載がない限り、分子生物学の分野において公知の標準手法、ベクター、制御配列エレメント及び他の発現系エレメントを、核酸操作、形質転換及び発現のために使用する。そのような標準手法、ベクター及びエレメントは、例えばAusubel et al. (eds.), Current Protocols in Molecular Biology (1995) (John Wiley & Sons); Sambrook, Fritsch, & Maniatis (eds.), Molecular Cloning (1989) (Cold Spring Harbor Laboratory Press, NY) ; Berger & Kimmel, Methods in Enzymology 152: Guide to Molecular Cloning Techniques(1987) (Academic Press); and Bukhari et al. (eds. ), DNA Insertion Elements, Plasmids and Episomes (1977) (Cold Spring Harbor Laboratory Press, NY)に見出すことができる。
【0170】
異なる記載がない限り、PCR反応は、以下のプロトコールに従ってPTC225サーモサイクラー(MJ Research,South San Francisco,CA,USA)を使用して実施した:
【表3】
【0171】
(実施例1)CCMV RNA1、RNA2及びキメラRNA3を接種したカウピー全植物体におけるCCMVウイルス粒子内での抗原性ペプチドの生産
カウピークロロティックモトルウイルス(CCMV)を、4個の異なる抗原性ペプチドの1個を含むように工作したカプシドタンパク質(CP)と共に使用して、炭疽菌抗原性ペプチドの発現を全植物体において実施した。
【0172】
CCMV RNA1のヌクレオチド配列を有するDNA及びCCMV RNA2のヌクレオチド配列を有するDNAを、T7プロモーターの下流でその制御下に、及びユニークXbaI部位の上流で、クローニングベクターpUC19に各々別々にサブクローニングした。これはプラスミドpDOW2122(CCMV RNA1)及びpDOW2123(CCMV RNA2)を生産した。
【0173】
5つのBamHI制限酵素切断部位を含むように工作した、CCMV RNA3のヌクレオチド配列を有するDNAを、組換えカプシドタンパク質をコードする核酸の生産のためにさらに工作した。各々が、4個の外来性ペプチド(炭疽菌防御抗原PAからの4個の異なる抗原性ペプチド)の異なる1個をコードする4個のDNA分子を、各々合成オリゴヌクレオチドのSOE(オーバーラップ伸長によるスプライシング)(splicing-by-overlap-extension)によって合成した。生じた核酸はBamHI認識部位末端を含んだ。各々のPA DNAフラグメントをBamHI制限酵素で制限し、CCMVコートタンパク質コード配列内の5つの異なる工作制限工切断部位の1つでコートタンパク質に独立して挿入した:コドン63のBamHI、コドン102のBamHI、コドン114のBamHI、コドン129のBamHI、コドン160のBamHI。
【0174】
4個の異なるペプチドは、PA1(配列番号15によってコードされる配列番号1)、PA2(配列番号16によってコードされる配列番号2)、PA3(配列番号17によってコードされる配列番号3)及びPA4(配列番号18によってコードされる配列番号4)であった。
【表4】
【0175】
これらの各々を、BamHI制限酵素で消化して、その後脱リン酸化しておいた5個のプラスミド:pDOW2125(pUC−CCMV−RNA3−CP63BamHI)、pDOW2126(pUC−CCMV−RNA3−102BamHI)、pDOW2127(pUC−CCMV−RNA3−CP114BamHI)、pDOW2128(pUC−CCMV−RNA3−CP129BamHI)及びpDOW2129(pUC−CCMV−RNA3−CP160BamHI)、の各々に挿入した。
【0176】
これは、「キメラRNA3」についての合計20の異なる型のDNAを生産した。プラスミドpDOW2135(pUC−CCMV−RNA3−CP63BamHI−PA1)、pDOW2139(pUC−CCMV−RNA3−102BamHI−PA1)、pDOW2143(pUC−CCMV−RNA3−CP114BamHI−PA1)、pDOW2147(pUC−CCMV−RNA3−CP129BamHI−PA1)及びpDOW2151(pUC−CCMV−RNA3−CP160BamHI−PA1)は、63、102、114、129及び160の位置にPA1挿入物を有するキメラRNA3の例である。野生型CCMVコートタンパク質をコードするRNA3及びBamHI制限部位を含むが挿入物を含まない工作したCCMVコートタンパク質をコードするRNAを対照として使用した。各々の修飾RNA3構築物を、T7プロモーターの下流でその制御下に、及びユニークXbaI部位の上流で、クローニングベクターpUC19に別々にサブクローニングした。
【0177】
3つのクラスのプラスミドの各々を大腸菌にクローニングした。プラスミドを単離し、XBaI制限酵素消化によって線状化して、次にMESSAGE mMACHINE T7 Kit(RNA転写キット、Ambion,Inc.,Austin,TX,USAより)を用いて各々1μgをインビトロでRNAに転写した。これは24個の異なるRNA種:RNA1について1個、RNA2について1個、キメラRNA3について20個、及びRNA3対照について2個、を生産した。
【0178】
カウピー植物に感染させるためにRNA1、RNA2、RNA3又はPA挿入物を含むキメラRNA3のカクテル混合物を使用した。カウピー植物を、カウピーカリフォルニアブラックアイNo.5種子(Ferry−Morse Seed Co.KY)から発芽させた。新芽を、Miracle−Gro鉢植え混合物(Miracle−Gro Lawn Products OH)と共に1つずつ6インチの鉢に移植した。2葉期(発芽後約7日目)にカウピー植物を感染させた。各々の植物の1枚の葉にカーボランダム(Carborundum)粉末400粗粒( Fisher Scientificカタログ番号409−21−2)の散粉を実施した。カーボランダム層にRNAカクテル混合物を適用した。手袋をはめた指で静かにこすって葉を削り取った。接種後7−14日で感染が確認された。
【0179】
意外にも、この全植物体系では、天然宿主植物を使用したにもかかわらず、キメラコートタンパク質は野生型に復帰するか、又は一部の場合は、所望外来性抗原ペプチドが部分的に欠失した。これはウイルス子孫の配列決定によって確認された。RNAを抽出し、ThermoScript RT−PCRシステム(Invitrogenカタログ番号11146−024)及びCCMVRNA4.R遺伝子特異的プライマー(配列番号19) (5’−CTC GAG CTA ATA CAC CGG AGT GAA AG−3’)を用いてDNAに逆転写した。cDNAを、プライマーCCMVRNA4.F(配列番号20)(5’−CTG CAG ATG TCT ACA GTC GGA ACA GG−3’)及びCCMVRNA4.Rを使用するPCRによってさらに増幅した。反応物を精製し、CCMV−CP−F1プライマー(配列番号21)(5’−AAC CCA TCG CTT CAG GCC AA−3’)で配列決定した。配列をシーケンサーソフトウエアバージョン4.0.5(Gene Codes Corporation)で分析した。例えばPA1挿入物を含むpDOW2128に由来するRNA3子孫を配列決定すると、 コートタンパク質は、PA1挿入物アミノ酸配列の代わりにRISDPLとして翻訳された、小さな認識されないヌクレオチド挿入物(5’−CGTATTTCTGATCCTCTC−3’)を含んだ。挿入したヌクレオチド配列はコートタンパク質の残りの部分をインフレームで保持した。工作CCMV RNA3は、全植物における発現時に不安定であり、容易に組換えを受け、全植物体系はキメラCCMVウイルス粒子上のエピトープの生産のために有用ではないと判定された。
【0180】
(実施例2)CCMV RNA1、RNA2及びキメラRNA3を接種したタバコ懸濁培養中のCCMVウイルス粒子における抗原性ペプチドの生産
同じCCMV−ペプチドをコードする構築物の発現を植物細胞懸濁培養において実施した。ニコチアナ・タバカムNT1細胞を、キメラCCMVコートタンパク質をコードするCCMV RNA3及びレプリカーゼ遺伝子をコードするCCMV RNA1及び2のRNA転写産物での電気穿孔によってトランスフェクトした。野生型CCMVコートタンパク質をコードするRNA3及び適切なBamHI制限部位を含むが挿入物を含まない工作したCCMVコートタンパク質をコードするRNA3を対照として使用した。
【0181】
24個の異なるRNA種:RNA1について1個、RNA2について1個、キメラRNA3について20個、及びRNA3対照について2個を、実施例1で述べたようなインビトロRNA転写によって得た。22個の異なる群において、3個の生じたRNA(RNA1、RNA2、及びRNA3の1個)の各々2μgを電気穿孔法によってタバコ細胞に形質転換した。
【0182】
以下のプロトコールを植物細胞トランスフェクションのために使用した:
1)培地の調製:
NTI培地(1リットル):
ムラシゲ&スクーグ基礎塩4.33g(Phyto Technology Laboratories KSカタログ番号M524)ミオ−イノシトール100mg(Sigmaカタログ番号I−3011)、塩酸チアミンの1mg/ml溶液1ml(Sigmaカタログ番号T−3902)、リン酸二水素カリウム(KH2PO4)180mg(Sigmaカタログ番号P−8416)、スクロース30g(Sigmaカタログ番号S−5390)、及び10mg/mlの2,4−D溶液200μl(Sigmaカタログ番号D−7299)を少量の水中で混合した。精製水を前記溶液に添加して容量を1リットルにした。pHを5.8に調整し、溶液を加圧滅菌した。
【0183】
マンニトール洗浄液0.4M:
マンニトール36.43g(Sigmaカタログ番号M−1902)を少量の精製水に添加した。精製水を添加して容量を500mlにし、pHを5.5に調整した。滅菌するために溶液をオートクレーブに入れた。
【0184】
酵素溶液:
0.4M マンニトール及び0.02M MESを少量の水中で混合した。精製水を添加して容量を500mlにし、pHを5.5に調整した。滅菌するために溶液をオートクレーブに入れ、溶液を4℃で保存した。使用前に、1%セルライシン(Carbiochemカタログ番号219466)及び0.3%マセラーゼペクチナーゼ(Carbiochemカタログ番号441201)を溶液に添加し、セルライシンとマセラーゼペクチナーゼが溶解するまで溶液を振とうした。
【0185】
電気穿孔緩衝液:
0.8%NaCl、0.02%KC1、0.02%KH2PO4及び0.11%Na2HPO4を少量の精製水に添加した。精製水を添加して溶液を100mlにし、pHを6.5に調整した。緩衝液を加圧滅菌し、4℃で保存した。
【0186】
2)タバコ細胞部分消化プロトコール:
タバコ細胞系NT1を電気穿孔の前に以下のように部分消化した。週に1回継代培養することによって強固な細胞系を維持し、細胞を、静かに振とうしながらNT1培地において24℃又は28℃で増殖させ、消化の3日前に、細胞懸濁液5mlをNT1培地50mlに継代培養して28℃でインキュベートした。細胞を50mlチューブにおいて800rpmで5分間遠心沈殿させた。酵素溶液を調製し、細胞をマンニトール洗浄液(約40ml)で洗った。細胞を800rpmで5分間遠心し、3容の酵素溶液を細胞に添加した。倒置して細胞を再懸濁し、10cmペトリ皿に注ぎ入れて移し、皿をアルミニウム箔に包んで60−120分間、室温で非常に緩やかに振とうした。次に細胞を50mlプラスチックチューブに戻し、800rpmで5分間遠心した。細胞をマンニトール洗浄液40mlで洗い、再び遠心して、電気穿孔緩衝液で洗浄し、遠心した。3容の電気穿孔緩衝液(総容量は通常20mlである)を細胞に添加した;細胞をアルミニウム箔に包んで4℃で保存した。
【0187】
部分消化した細胞の電気穿孔プロトコール:
消化した細胞1mlをアリコートに分けて電気穿孔キュベットに入れた(4mm間隔)。各々のRNA転写産物2μg−CCMV RNA1、RNA2及びキメラRNA3−をキュベットに添加し、5分間氷上に置いた。NT1平板培養培地10ml(NT1+0.4M マンニトール)を各々のペトリ皿に添加した。細胞を500μF、250Vで電気穿孔し、キュベットを氷上に戻した。次に細胞をペトリ皿に移し、暗所で振とうせずに室温でインキュベートした。トランスフェクション後48時間目に分析のために細胞を収集した。
【0188】
結果
キメラコートタンパク質の発現を、抗CCMVコートタンパク質ポリクローナル抗体を用いたウエスタンブロット法によって分析した。また、各々の培養物からRNAを抽出し、キメラCCMV RNA4をcDNAに逆転写して、それをPCRによって増幅した。PCR産物を実施例1で述べたように配列決定した。RNA1及び2だけでトランスフェクトした陰性対照を除いて、全ての試料がウエスタンブロット法で陽性であった。
【0189】
対照CCMVカプシドタンパク質と比較して、キメラコートタンパク質はより大きなサイズとゲル上でのより緩慢な運動性を有しており、キメラコートタンパク質がPA挿入物を含むことを示した。これは、ウイルスRNA子孫の配列決定によって確認された。結果は、20個のキメラ構築物のうち19個が、所望抗原ペプチド内に突然変異を含むことなく正しくキメラCCMVコートタンパク質を発現したことを明らかにした。この結果は、懸濁細胞において発現されたキメラCCMV RNA3が安定であることを示す。
【0190】
図2は、CCMV RNA1、RNA2、及びpDOW2125(CCMV63BamHI)、pDOW2126(CCMV102BamHI)、pDOW2127(CCMV−114BamHI)、pDOW2128(CCMV129BamHI)及びpDOW2129(CCMV160BamHI)から転写されたRNA3でトランスフェクトした細胞におけるCCMV CPの発現を示す。結果は、CP内の異なる位置に工作BamHI部位を有する全てのRNA3からCCMV CPが発現されることを明らかにする。
【0191】
63、102、114、129及び160BamHI部位でCCMVコートタンパク質に融合した全てのPAペプチド(PA1、PA2、PA3及びPA4)が、タバコ細胞において成功裏に生産された。図3及び図4は、CCMV RNA1、RNA2、及び63、102、114、129及び160位に4個のPAペプチド挿入物を有するCPを含むキメラRNA3でトランスフェクトした細胞におけるキメラCCMV CPの発現を示す。図3は、あらかじめ63、102又は114位に挿入されたPA1、PA2、PA3又はPA4ペプチドを有するCCMV CPの融合タンパク質が、63位でCCMV CPに挿入されたPA2ペプチドを除き、タバコ植物細胞において発現されることを明らかにする。図4は、129又は160位に挿入されたPA1、PA2、PA3又はPA4ペプチドを有するCCMV CPの融合タンパク質がタバコ植物細胞において発現されることを示す。
【0192】
(実施例3)キメラCCMV CPをコードする植物発現プラスミドでトランスフェクトしたタバコ懸濁培養中のCCMVウイルス様粒子における4つの抗原性ペプチドの生産
【0193】
1)ベクターの構築:
プラスミドpIL−Tab358を植物発現ベクターとして使用した。CCMV CP挿入のために選択した制限部位はXbaIとEcoRIであった。このプラスミドは、XbaI部位の上流にキャッサバ葉脈モザイクウイルスプロモーター及びEcoRI部位の下流にNosターミネーターを含む。前記ベクターは、XbaI及びEcoRIで消化し、脱リン酸化した後、挿入物と連結することによって作製した。
【0194】
2)挿入物の構築:
CCMV CP−PA融合物を、プライマーCCMV−CP−XbaI(配列番号22)及びCCMV−CP−EcoRI(配列番号23)を使用してpDOW2147(pUC−CCMV−RNA3−CP129BamHI−PA1)、pDOW2148(pUC−CCMV−RNA3−CP129BamHI−PA2)、pDOW2149(pUC−CCMV−RNA3−CP129BamHI−PA3)、pDOW2150(pUC−CCMV−RNA3−CP129BamHI−PA4)からPCRによって増幅し、pDOW2160(pIL−Tab−CCMV129BamHI−PA1)(配列番号5)、pDOW2161(pIL−Tab−CCMV129BamHI−PA2)(配列番号6)、pDOW2162(pIL−Tab−CCMV129BamHI−PA3)(配列番号7)及びpDOW2163(pIL−Tab−CCMV129BamHI−PA4)(配列番号8)を作製した。pDOW2160、pDOW2161、pDOW2162及びpDOW2163についてのプラスミド地図を図5、6、7及び8に示す。
【0195】
3)植物細胞トランスフェクション:
植物細胞をプラスミドpDOW2160、pDOW2161、pDOW2162及びpDOW2163 10μgでトランスフェクトした。植物細胞トランスフェクションは実施例2におけるように実施した。
【0196】
結果
129BamHI部位の4つのPAペプチド−CCMV CP融合物全てがタバコ細胞において成功裏に発現された。図9は、pDOW2160、pDOW2161、pDOW2162及びpDOW2163でトランスフェクトした細胞におけるキメラCCMV CPの発現を示す。対照(挿入物を含まないCCMVコートタンパク質)と比較して、4つのキメラコートタンパク質全てが、PA挿入物を有することを表すより緩慢な運動性を示した。
【0197】
(実施例4)キメラCCMV CP及び3’UTR(CCMV RNA3 3’非翻訳領域)をコードする植物発現プラスミドでトランスフェクトしたタバコ懸濁培養中のウイルス様粒子におけるCCMV CP融合物としての4つの抗原性ペプチドの生産
1)ベクターの構築:
プラスミドpIL−Tab358を植物発現ベクターとして使用した。CCMV CP挿入のために選択した制限部位はXbaIとEcoRIであった。このプラスミドは、XbaI部位の上流にキャッサバ葉脈モザイクウイルスプロモーター及びEcoRI部位の下流にNosターミネーターを含む。前記ベクターは、XbaI及びEcoRIで消化し、脱リン酸化した後、挿入物と連結することによって作製した。
【0198】
2)挿入物の構築:
RNA3の3’UTRを含むCCMV CP−PA融合物を、プライマーCCMV−CP−XbaI及びCCMV−CP−EcoRI−3’UTR(配列番号24)を使用してpDOW2147、pDOW2148、pDOW2149及びpDOW2150からPCRによって増幅し、pDOW2169(pIL−Tab−CP129BamHI−PA1−3’UTR)(配列番号9)、pDOW2170(pIL−Tab−CP129BamHI−PA2−3’UTR)(配列番号10)pDOW2171(pIL−Tab−CP129BamHI−PA3−3’UTR)(配列番号11)及びpDOW2172(pIL−Tab−CP129BamHI−PA4−3’UTR)(配列番号12)を作製した。
【0199】
3)植物細胞トランスフェクション:
植物細胞をプラスミドpDOW2169、pDOW2170、pDOW2171及びpDOW2172 10μgでトランスフェクトした。植物細胞トランスフェクションは実施例2におけるように実施した。
【0200】
結果
3’UTRを含む129BamHI部位の4つのPAペプチド−CCMV CP融合物全てがタバコ細胞において成功裏に発現された。図10は、pDOW2169、pDOW2170、pDOW2171及びpDOW2172でトランスフェクトした細胞におけるキメラCCMV CPの発現を示す。対照(挿入物を含まないCCMVコートタンパク質)と比較して、4つのキメラコートタンパク質全てが、PA挿入物を有することを表すより緩慢な運動性を示した。
【0201】
(実施例5)キメラCCMV CPをコードする植物発現プラスミドでトランスフェクトしたトランスジェニックタバコ懸濁培養中のウイルス様粒子におけるCCMV CP融合物としての4つの抗原性ペプチドの生産
タバコ細胞系NT1を、週に1回継代培養することによって維持した。細胞を、静かに振とうしながらNT1培地において24℃又は28℃で増殖させた。トランスフェクションの3日前に、細胞懸濁液5mlをNT1培地50mlに継代培養して28℃でインキュベートした。
【0202】
1)パーティクルガンによる植物細胞形質転換:
プラスミドpDOW2160、pDOW2161、pDOW2162、pDOW2163及びキャッサバ葉脈モザイクウイルスプロモーターによって駆動される植物選択マーカーPatを含む植物発現プラスミドpBBV(pBBV)を、マイクロパーティクルガン法によるNT1タバコ細胞形質転換のために使用した。使用したプラスミド比は、1:6 pBBV:pDOW2160、pBBV:pDOW2161、pBBV:pDOW2162又はpBBV:pDOW2163であった。合計5μgのプラスミドDNAは、6回のパーティクルガン法のために十分であった。Chen, L et al. Plant Cell Reports (1998) 18: 25-31 に述べられているBioradパーティクルガンプロトコールを形質転換のために使用した。
【0203】
2)プレーティング
パーティクルガンを打ち込んだ細胞を、非選択的NT1培地寒天プレートに4時間移した後、25μg/mlのグルフォシネート−アンモニウム(Sigmaカタログ番号45520)を含むNT1培地寒天プレートに移した。
【0204】
3)トランスジェニックカルスの選択
21日後、白色綿毛状細胞増殖を有していたカルスを、CP融合物の発現を調べるためのウエスタンブロット法及びプロモーター−CP融合物遺伝子−ターミネーターカセットの植物ゲノムへの組込みを調べるためのPCRによる分析のために選択した。キメラCCMV CPを発現するトランスジェニックカルスをNT1液体培地に移した。細胞を、静かに振とうしながらNT1培地において24℃又は28℃で増殖させ、週に1回継代培養した。
【0205】
結果
図11は、pDOW2160、pDOW2161、pDOW2162及びpDOW2163で安定に形質転換された細胞におけるキメラCCMV CPの発現を示す。CCMVに対するポリクローナル抗体によってCP融合物を検出した。4つのキメラコートタンパク質導入遺伝子全ての発現が検出された。
【0206】
(実施例6)キメラCCMV CPをコードする植物発現プラスミドでトランスフェクトしたトランスジェニックイネ懸濁培養中のCCMVウイルス様粒子におけるCCMV CP融合物としての4つの抗原性ペプチドの生産
イネ細胞系を、週に1回継代培養することによって維持した。細胞を、静かに振とうしながらNB培地(Li, L et al. Plant Cell Reports. (1993) 12: 250-255)において28℃で増殖させた。トランスフェクションの3日前に、細胞懸濁液5mlをNB培地50mlに継代培養して28℃でインキュベートした。
【0207】
1)パーティクルガンによる植物細胞形質転換:
プラスミドpDOW2160、pDOW2161、pDOW2162及びpDOW2163、及びキャッサバ葉脈モザイクウイルスプロモーターによって駆動される植物選択マーカーPatを含む植物発現プラスミドpBBVを、マイクロパーティクルガン法によるイネ細胞形質転換のために使用した。使用したプラスミド比は、1:6 pBBV:pDOW2160、pBBV:pDOW2161、pBBV:pDOW2162又はpBBV:pDOW2163であった。合計5μgのプラスミドDNAは、6回のパーティクルガン法のために十分であった。Chen, L et al. Plant Cell Reports (1998) 18: 25-31 に述べられているBioradパーティクルガンプロトコールを形質転換のために使用した。
【0208】
2)プレーティング
パーティクルガンを打ち込んだ細胞を、選択のために25μg/mlのグルフォシネート−アンモニウム(Sigmaカタログ番号45520)を含むNB培地寒天プレートに移した。
【0209】
3)トランスジェニックカルスの選択
21日後、白色綿毛状細胞増殖を有していたカルスを、CP融合物の発現を調べるためのウエスタンブロット法及びプロモーター−CP融合物遺伝子−ターミネーターカセットの植物ゲノムへの組込みを調べるためのPCRによる分析のために選択した。
【0210】
結果
図12は、CCMVに対するポリクローナル抗体によって検出したときの、pDOW2160、pDOW2161、pDOW2162及びpDOW2163で安定に形質転換された細胞におけるキメラCCMV CPの発現を示す。4つのキメラコートタンパク質導入遺伝子全ての発現が検出された。
【0211】
ゲノムDNAを抽出し、プロモーター−CP融合物遺伝子−ターミネーターカセットの植物ゲノムへの組込みをPCRによって調べるために以下のプロトコールを使用した。ゲノムDNAを抽出するためにQiagen DNeasy Plant Miniを製造者の指示に従って使用した。
【0212】
新鮮イネカルス約50−100mgを1.5mlチューブに入れた。即時DNA抽出のために組織を液体窒素中で凍結するか又は保存のために−80℃の冷凍庫に入れた。DNA抽出の前に、組織をミクロ乳鉢(micropestle)で摩砕することによって手動で破壊した。試料を氷上に置き、Qiagen DNeasy Plant Mini Kitに述べられているようにゲノムDNAを抽出した。Access Quick Master Mix 2x(Promegaカタログ番号A1720)を使用してPCR反応を以下のように設定した:
Master mix 25μl
CCMV−F(10mM)プライマー、配列番号25 1μl
Nos−term−R(10mM)プライマー、配列番号26 1μl
ゲノムDNA 1μl
水 22μl
合計 55μl
【0213】
ゲノムDNAからプロモーター−CP融合物遺伝子−ターミネーターカセットを増幅するために以下のPCRサイクルを使用した:
1サイクル 95℃ 2分間
35サイクル 95℃ 30秒間
55℃ 30秒間
70℃ 2分間
1サイクル 70℃ 5分間
4℃ 維持
【0214】
PCR反応物10μlを、EtBrで染色した1.2%アガロースゲル上で泳動させた。図13は、選択した個別イネカルスから増幅したPCR産物を示す。予測された大きさのPCR産物を含む試料を、安定形質転換及びキメラCP導入遺伝子の植物ゲノムへの組込みに関して陽性と評価した。
【0215】
キメラCCMV CPを発現するトランスジェニックカルスを、スケールアップ発酵に適した細胞懸濁培養を作製するために液体NB培地に移した。細胞を、静かに振とうしながらNB培地において28℃で増殖させ、週に1回継代培養した。
【0216】
VLP抽出:
以下のプロトコールに従って、振とうフラスコ培養試料を溶解し、生じた細胞溶解産物をPEG(ポリエチレングリコール)処理して、限外ろ過することにより、キメラVLPを沈殿させた:
【0217】
各々の振とうフラスコ培養物の50mLアリコートを遠心分離して細胞をペレット化した。ペレット化した細胞をウイルス緩衝液(0.2M 酢酸ナトリウム、pH5.2;10mM EDTA.0)中に緩衝液2容:ペレット1容の比率で再懸濁した。次に60秒間ずつ何度も混合し、混合と混合の間に2分間氷上で休ませることによって細胞を破壊した。生じたホモジネートを3層のチーズクロス(cheese cloth)を通してしぼり、その後4℃、15,000×Gで15分間遠心分離した。生じた上清を採取し、それらの容量を測定した。各々の上清に、10%の最終濃度までPEG8000を添加し、その溶液を氷上で1時間又は4℃で一晩インキュベートした。次に、溶液を4℃、15,000×Gで10分間遠心分離した。沈殿したペレットをウイルス緩衝液の1/10初期上清容量に再懸濁し、4℃で保存して、ポリクローナル抗CCMV抗体でのウエスタンブロット法によって分析した。
【0218】
図14は、キメラCCMV VLPが、pDOW2160で安定に形質転換した細胞から回収されたことを示す。キメラCP−PA1融合タンパク質は、再懸濁したPECペレット中で検出されたが、上清では検出されず、キメラCPがVLPに集合されたことを指示した。
【0219】
あるいは、再懸濁した試料を4℃、15,000×Gで10分間遠心分離し、上清を回収して、2回目のPEC沈殿に供した。PEG8000を15%の最終濃度まで添加し、4℃で2時間攪拌した。その溶液を15,000×Gで10分間遠心分離し、ペレットを少量のウイルス緩衝液に再懸濁した。再懸濁したVLP溶液を、カットオフ値が300K分子量のCentricon Plus−20に負荷し、4,000×Gで5分間遠心した。濃縮したVLP試料をポリクローナル抗CCMV抗体でのウエスタンブロット法によって分析した(図14参照)。キメラCP−PA1融合タンパク質は、再懸濁してサイズろ過した2回目のPECペレット中で検出されたが、上清では検出されず、キメラCPがVLPに構築されたことを示した。
【図面の簡単な説明】
【0220】
【図1】図1は、植物細胞内のウイルス様粒子(VLP)におけるペプチド単量体の生産についての概要図を示す。所望標的ペプチド(「I」)をコードする配列を、ウイルスコートタンパク質(「CP」)をコードする配列に挿入して、組換えウイルスコートタンパク質(「rCP」)をコードする遺伝子を構築し、それを、ベクターの一部として、植物細胞に形質転換し、組換えコートタンパク質(「rCP」)を形成するように発現させる。任意の切断部位(「*」)も示す。植物細胞を、rCP遺伝子についての植物発現プラスミドでトランスフェクト又は形質転換する。rCPは植物細胞においてVLPに集合することができる。ペプチド挿入物はVLP表面に提示される。キメラVLPを植物細胞から精製することができ、場合により、所望標的ペプチドを精製VLPから切断分離して、精製することができる。
【図2】図2は、感染性CCMV RNA1、RNA2、及びpDOW2124(CCMV−RNA3−CP)、pDOW2125(CCMV−RNA3−CP63BamHI)、pDOW2126(CCMV−RNA3−CP102BamHI)、pDOW2127(CCMV−RNA3−CP114BamHI)、pDOW2128(CCMV−RNA3−CP129BamHI)及びpDOW2129(CCMV−RNA3−CP160BamHI)から転写されたRNA3の1個でトランスフェクトしたタバコNT1細胞におけるCCMV CPの発現を示す。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1は緩衝液のみを示し、レーン2はRNA1及びRNA2のみを示し、レーン3は、RNA1及びRNA2+pDOW2124から転写される野生型RNA3を示し、レーン4は、RNA1及びRNA2+pDOW2125から転写されるRNA3を示し、レーン5は、RNA1及びRNA2+pDOW2126から転写されるRNA3を示し、レーン6は、RNA1及びRNA2+pDOW2127から転写されるRNA3を示し、レーン7は、RNA1及びRNA2+pDOW2128から転写されるRNA3を示し、レーン8は、RNA1及びRNA2+pDOW2129から転写されるRNA3を示す。
【図3】図3は、感染性CCMV RNA1、RNA2、及び63、102又は114位に抗原性ペプチドPA1、PA2、PA3又はPA4とのCCMV CP融合物を含む工作されたRNA3の1個でトランスフェクトしたタバコNT1細胞におけるCCMV CPの発現を示す。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1はサイズマーカーを示し、レーン2は緩衝液のみを示し、レーン3は、RNA1+RNA2+CP内の63位にPA1を有するキメラRNA3を示し、レーン4は、RNA1+RNA2+CP内の63位にPA2を有するキメラRNA3を示し、レーン5は、RNA1+RNA2+CP内の63位にPA3を有するキメラRNA3を示し、レーン6は、RNA1+RNA2+CP内の63位にPA4を有するキメラRNA3を示し、レーン7は、RNA1+RNA2+CP内の102位にPA1を有するキメラRNA3を示し、レーン8は、RNA1+RNA2+CP内の102位にPA2を有するキメラRNA3を示し、レーン9は、RNA1+RNA2+CP内の102位にPA3を有するキメラRNA3を示し、レーン10は、RNA1+RNA2+CP内の102位にPA4を有するキメラRNA3を示し、レーン11は、RNA1+RNA2+CP内の114位にPA1を有するキメラRNA3を示し、レーン12は、RNA1+RNA2+CP内の114位にPA2を有するキメラRNA3を示し、レーン13は、RNA1+RNA2+CP内の114位にPA3を有するキメラRNA3を示し、及びレーン14は、RNA1+RNA2+CP内の114位にPA4を有するキメラRNA3を示す。
【図4】図4は、感染性CCMV RNA1、RNA2、及び129又は160位に抗原性ペプチドPA1、PA2、PA3又はPA4とのCCMV CP融合物を含む工作されたRNA3の1個でトランスフェクトしたタバコNT1細胞におけるCCMV CPの発現を示す。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1はサイズマーカーを示し、レーン2は、RNA1+RNA2+CP内の129位にPA1を有するキメラRNA3を示し、レーン3は、RNA1+RNA2+CP内の129位にPA2を有するキメラRNA3を示し、レーン4は、RNA1+RNA2+CP内の129位にPA3を有するキメラRNA3を示し、レーン5は、RNA1+RNA2+CP内の129位にPA4を有するキメラRNA3を示し、レーン6は、RNA1+RNA2+CP内の160位にPA1を有するキメラRNA3を示し、レーン7は、RNA1+RNA2+CP内の160位にPA2を有するキメラRNA3を示し、レーン8は、RNA1+RNA2+CP内の160位にPA3を有するキメラRNA3を示し、レーン9は、RNA1+RNA2+CP内の160位にPA4を有するキメラRNA3を示し、レーン10は、RNA1+RNA2+野生型RNA3を示し、及びレーン11は緩衝液のみを示す。
【図5】図5は、pDOW2160のプラスミド地図を示す。
【図6】図6は、pDOW2161のプラスミド地図を示す。
【図7】図7は、pDOW2162のプラスミド地図を示す。
【図8】図8は、pDOW2163のプラスミド地図を示す。
【図9】図9は、非感染性プラスミドpDOW2160、pDOW2161、pDOW2162及びpDOW2163でトランスフェクトしたタバコNT1細胞におけるキメラCCMV CPの発現を示す。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1は緩衝液のみを示し、レーン2−6は、RNA1+RNA2+野生型RNA3(対照)を示し、レーン7はpDOW2160を示し、レーン8はpDOW2161を示し、レーン9はpDOW2162を示し、及びレーン10はpDOW2163を示す。
【図10】図10は、非感染性プラスミドpDOW2169、pDOW2170、pDOW2171及びpDOW2172でトランスフェクトしたタバコNT1細胞におけるキメラCCMV CPの発現を示す。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1はRNA1+RNA2+RNA3(対照)を示し、レーン2は緩衝液を示し、レーン3はpDOW2169を示し、レーン4はpDOW2170を示し、レーン5はpDOW2171を示し、及びレーン6はpDOW2172を示す。
【図11】図11は、非感染性プラスミドpDOW2160、pDOW2161、pDOW2162及びpDOW2163で安定に形質転換したタバコNT1細胞におけるキメラCCMV CPの発現を示す。21日間の選択後、白色綿毛状細胞増殖を有していたカルスを、CP発現を調べるためのウエスタンブロット法による分析のために選択した。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1は、非形質転換タバコ細胞(陰性対照)を示し、レーン2は、pDOW2160で形質転換したトランスジェニックタバコ細胞を示し、レーン3は、pDOW2161で形質転換したトランスジェニックタバコ細胞を示し、レーン4は、pDOW2162で形質転換したトランスジェニックタバコ細胞を示し、及びレーン5は、pDOW2163で形質転換したトランスジェニックタバコ細胞を示す。
【図12】図12は、非感染性プラスミドpDOW2160、pDOW2161、pDOW2162及びpDOW2163で安定に形質転換したイネ細胞におけるキメラCCMV CPの発現を示す。21日間の選択後、白色綿毛状細胞増殖を有していたカルスを、CP発現を調べるためのウエスタンブロット法による分析のために選択した。植物細胞抽出物をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1は野生型CCMV CP(陽性対照)を示し、レーン2はサイズラダーを示し、レーン3は非トランスジェニックイネ細胞(陰性対照)を示し、レーン4は、pDOW2160で形質転換したトランスジェニックイネ細胞を示し、レーン5は野生型CCMV CP(陽性対照)を示し、レーン6はサイズラダーを示し、レーン7は非トランスジェニックイネ細胞を示し、レーン8は、pDOW2161で形質転換したトランスジェニックイネ細胞を示し、レーン9は野生型CCMV CP(陽性対照)を示し、レーン10はサイズラダーを示し、レーン11は非トランスジェニックイネ細胞(陰性対照)を示し、レーン12は、pDOW2162で形質転換したトランスジェニックイネ細胞を示し、レーン13は野生型CCMV CP(陽性対照)を示し、レーン14はサイズラダーを示し、レーン15は非トランスジェニックイネ細胞(陰性対照)を示し、及びレーン16は、pDOW2163で形質転換したトランスジェニックイネ細胞を示す。
【図13】図13は、非感染性プラスミドpDOW2160、pDOW2161、pDOW2162及びpDOW2163で形質転換した選択個別イネカルスから増幅したPCR産物の検出を示す。選択個別イネカルスから増幅されたPCR産物を示す、EtBrで染色した1.2%アガロースゲル。21日間の選択後、白色綿毛状細胞増殖を有していたカルスを、植物ゲノムへのプロモーター−CP融合物遺伝子−ターミネーターカセットの組込みを調べるためのPCRによる分析のために選択した。レーン1はイネカルス46−5を示し、レーン2はイネカルス46−11を示し、レーン3はイネカルス46−17を示し、レーン4はサイズラダーを示し、レーン5はイネカルス47−6を示し、レーン6はイネカルス48−12を示し、レーン7はイネカルス48−18を示し、レーン8はイネカルス48−20を示し、レーン9はイネカルス49−11を示し、レーン10はイネカルス49−18を示し、及びレーン11は陰性対照(非トランスジェニックイネ細胞)を示す。
【図14】図14は、非感染性プラスミドpDOW2160で安定に形質転換したイネ細胞からPEG沈殿及び限外ろ過によって精製したキメラCCMV VLPのウエスタンブロット分析及び精製VLP試料におけるキメラCP−PA1融合タンパク質の検出を示す。試料をSDS−PAGEゲルで泳動し、抗CCMVポリクローナル抗体でプローブした。レーン1はサイズラダーを示し、レーン2は、CCMV−CP1融合に関してトランスジェニックのイネ懸濁細胞からの全細胞溶解産物を示し、レーン3は第一PEG上清を示し、レーン4は第一PEGペレットを示し、レーン5は第二PEGペレットを示し、レーン6は第二PEG上清を示す。
【図15】図15は、発酵による植物懸濁細胞でのキメラVLPの生産工程の一覧図を示す。
【特許請求の範囲】
【請求項1】
a)融合ペプチドをコードする核酸配列を含む非感染性核酸で植物細胞を形質転換すること、但し前記融合ペプチドは少なくとも1個の対象異種ペプチドと少なくとも1個のウイルスカプシドタンパク質から成り、前記核酸は前記植物細胞ゲノムに組み込まれている;
b)前記植物細胞を発酵工程において懸濁植物細胞培養中で増殖させ、前記植物細胞において前記核酸を発現させること;
c)前記植物細胞における発現が、前記融合ペプチドのウイルス様粒子へのインビボ集合を提供すること;及び
d)前記ウイルス様粒子を単離すること
を含む、異種ペプチドを生産するための方法。
【請求項2】
前記融合ペプチドをコードする前記核酸が、プロモーター配列及びターミネーター配列に作動可能に連結されている、請求項1に記載の方法。
【請求項3】
前記核酸がウイルス3’UTRをさらに含む、請求項2に記載の方法。
【請求項4】
前記3’UTRがカプシド形成シグナルを含む、請求項3に記載の方法。
【請求項5】
前記核酸がカプシド形成シグナルをさらに含む、請求項2に記載の方法。
【請求項6】
前記ウイルスカプシドタンパク質が、植物細胞宿主に対して天然栄養機能を示さないウイルスに由来する、請求項1に記載の方法。
【請求項7】
前記ウイルスカプシドタンパク質が、植物細胞宿主に対して天然栄養機能を示すウイルスに由来する、請求項1に記載の方法。
【請求項8】
前記ウイルスカプシドタンパク質が二十面体植物ウイルスに由来する、請求項1に記載の方法。
【請求項9】
前記二十面体ウイルスが、カウピーモザイクウイルス(Cowpea Mosaic Virus)、カウピークロロティックモトルウイルス(Cowpea Chlorotic Mottle Virus)及びアルファルファモザイクウイルス(Alfalfa Mosaic Virus)から成るウイルスの群より選択される、請求項8に記載の方法。
【請求項10】
前記異種ペプチドが抗原性ペプチドである、請求項1に記載の方法。
【請求項11】
前記抗原性ペプチドが炭疽菌(Bacillus anthracis)ペプチドである、請求項10に記載の方法。
【請求項12】
前記抗原性ペプチドが、配列番号1−4から成る群より選択される、請求項11に記載の方法。
【請求項13】
前記核酸が、配列番号5−12から成る群より選択される、請求項1に記載の方法。
【請求項14】
前記ウイルス様粒子をワクチンとして動物に投与する、請求項1に記載の方法。
【請求項15】
前記植物細胞が双子葉植物である、請求項1に記載の方法。
【請求項16】
前記植物細胞がニコチアナ・タバカム(タバコ)(Nicotiana tabacum)である、請求項15に記載の方法。
【請求項17】
前記植物細胞が単子葉植物である、請求項1に記載の方法。
【請求項18】
前記植物細胞がオリザ・サティバ(イネ科イネ)(Oryza sativa)である、請求項17に記載の方法。
【請求項19】
前記核酸が、前記カプシドタンパク質以外のウイルスタンパク質をコードするウイルス配列を含まない、請求項1に記載の方法。
【請求項20】
融合ペプチドをコードする核酸配列を含む非感染性核酸を含む植物細胞であって、前記融合ペプチドは少なくとも1個の対象異種ペプチドと少なくとも1個のウイルスカプシドタンパク質から成り、前記核酸は前記植物細胞ゲノムに組み込まれている、前記植物細胞。
【請求項21】
前記植物細胞が前記融合ペプチドを発現する、請求項20に記載の植物細胞。
【請求項22】
前記融合ペプチドがインビボでウイルス様粒子に集合される、請求項21に記載の植物細胞。
【請求項23】
前記抗原性ペプチドが、配列番号1−4から成る群より選択される、請求項20に記載の植物細胞。
【請求項24】
前記核酸が、配列番号5−12から成る群より選択される、請求項20に記載の植物細胞。
【請求項25】
前記植物細胞がニコチアナ・タバカムである、請求項20に記載の植物細胞。
【請求項26】
前記植物細胞がオリザ・サティバである、請求項20に記載の植物細胞。
【請求項27】
前記核酸が3’UTRをさらに含む、請求項20に記載の植物細胞。
【請求項28】
前記核酸がウイルス3’UTRをさらに含む、請求項20に記載の植物細胞。
【請求項29】
前記3’UTRがカプシド形成シグナルを含む、請求項28に記載の植物細胞。
【請求項30】
前記核酸がカプシド形成シグナルをさらに含む、請求項20に記載の植物細胞。
【請求項31】
前記ウイルスカプシドタンパク質が、植物細胞宿主に対して天然栄養機能を示さないウイルスに由来する、請求項20に記載の植物細胞。
【請求項32】
前記ウイルスカプシドタンパク質が、植物細胞宿主に対して天然栄養機能を示すウイルスに由来する、請求項20に記載の植物細胞。
【請求項33】
前記ウイルスカプシドタンパク質が二十面体植物ウイルスに由来する、請求項20に記載の植物細胞。
【請求項34】
前記二十面体ウイルスが、カウピーモザイクウイルス、カウピークロロティックモトルウイルス及びアルファルファモザイクウイルスから成るウイルスの群より選択される、請求項33に記載の植物細胞。
【請求項35】
配列番号5−12から成る群より選択される配列を含む核酸構築物。
【請求項1】
a)融合ペプチドをコードする核酸配列を含む非感染性核酸で植物細胞を形質転換すること、但し前記融合ペプチドは少なくとも1個の対象異種ペプチドと少なくとも1個のウイルスカプシドタンパク質から成り、前記核酸は前記植物細胞ゲノムに組み込まれている;
b)前記植物細胞を発酵工程において懸濁植物細胞培養中で増殖させ、前記植物細胞において前記核酸を発現させること;
c)前記植物細胞における発現が、前記融合ペプチドのウイルス様粒子へのインビボ集合を提供すること;及び
d)前記ウイルス様粒子を単離すること
を含む、異種ペプチドを生産するための方法。
【請求項2】
前記融合ペプチドをコードする前記核酸が、プロモーター配列及びターミネーター配列に作動可能に連結されている、請求項1に記載の方法。
【請求項3】
前記核酸がウイルス3’UTRをさらに含む、請求項2に記載の方法。
【請求項4】
前記3’UTRがカプシド形成シグナルを含む、請求項3に記載の方法。
【請求項5】
前記核酸がカプシド形成シグナルをさらに含む、請求項2に記載の方法。
【請求項6】
前記ウイルスカプシドタンパク質が、植物細胞宿主に対して天然栄養機能を示さないウイルスに由来する、請求項1に記載の方法。
【請求項7】
前記ウイルスカプシドタンパク質が、植物細胞宿主に対して天然栄養機能を示すウイルスに由来する、請求項1に記載の方法。
【請求項8】
前記ウイルスカプシドタンパク質が二十面体植物ウイルスに由来する、請求項1に記載の方法。
【請求項9】
前記二十面体ウイルスが、カウピーモザイクウイルス(Cowpea Mosaic Virus)、カウピークロロティックモトルウイルス(Cowpea Chlorotic Mottle Virus)及びアルファルファモザイクウイルス(Alfalfa Mosaic Virus)から成るウイルスの群より選択される、請求項8に記載の方法。
【請求項10】
前記異種ペプチドが抗原性ペプチドである、請求項1に記載の方法。
【請求項11】
前記抗原性ペプチドが炭疽菌(Bacillus anthracis)ペプチドである、請求項10に記載の方法。
【請求項12】
前記抗原性ペプチドが、配列番号1−4から成る群より選択される、請求項11に記載の方法。
【請求項13】
前記核酸が、配列番号5−12から成る群より選択される、請求項1に記載の方法。
【請求項14】
前記ウイルス様粒子をワクチンとして動物に投与する、請求項1に記載の方法。
【請求項15】
前記植物細胞が双子葉植物である、請求項1に記載の方法。
【請求項16】
前記植物細胞がニコチアナ・タバカム(タバコ)(Nicotiana tabacum)である、請求項15に記載の方法。
【請求項17】
前記植物細胞が単子葉植物である、請求項1に記載の方法。
【請求項18】
前記植物細胞がオリザ・サティバ(イネ科イネ)(Oryza sativa)である、請求項17に記載の方法。
【請求項19】
前記核酸が、前記カプシドタンパク質以外のウイルスタンパク質をコードするウイルス配列を含まない、請求項1に記載の方法。
【請求項20】
融合ペプチドをコードする核酸配列を含む非感染性核酸を含む植物細胞であって、前記融合ペプチドは少なくとも1個の対象異種ペプチドと少なくとも1個のウイルスカプシドタンパク質から成り、前記核酸は前記植物細胞ゲノムに組み込まれている、前記植物細胞。
【請求項21】
前記植物細胞が前記融合ペプチドを発現する、請求項20に記載の植物細胞。
【請求項22】
前記融合ペプチドがインビボでウイルス様粒子に集合される、請求項21に記載の植物細胞。
【請求項23】
前記抗原性ペプチドが、配列番号1−4から成る群より選択される、請求項20に記載の植物細胞。
【請求項24】
前記核酸が、配列番号5−12から成る群より選択される、請求項20に記載の植物細胞。
【請求項25】
前記植物細胞がニコチアナ・タバカムである、請求項20に記載の植物細胞。
【請求項26】
前記植物細胞がオリザ・サティバである、請求項20に記載の植物細胞。
【請求項27】
前記核酸が3’UTRをさらに含む、請求項20に記載の植物細胞。
【請求項28】
前記核酸がウイルス3’UTRをさらに含む、請求項20に記載の植物細胞。
【請求項29】
前記3’UTRがカプシド形成シグナルを含む、請求項28に記載の植物細胞。
【請求項30】
前記核酸がカプシド形成シグナルをさらに含む、請求項20に記載の植物細胞。
【請求項31】
前記ウイルスカプシドタンパク質が、植物細胞宿主に対して天然栄養機能を示さないウイルスに由来する、請求項20に記載の植物細胞。
【請求項32】
前記ウイルスカプシドタンパク質が、植物細胞宿主に対して天然栄養機能を示すウイルスに由来する、請求項20に記載の植物細胞。
【請求項33】
前記ウイルスカプシドタンパク質が二十面体植物ウイルスに由来する、請求項20に記載の植物細胞。
【請求項34】
前記二十面体ウイルスが、カウピーモザイクウイルス、カウピークロロティックモトルウイルス及びアルファルファモザイクウイルスから成るウイルスの群より選択される、請求項33に記載の植物細胞。
【請求項35】
配列番号5−12から成る群より選択される配列を含む核酸構築物。
【図1】

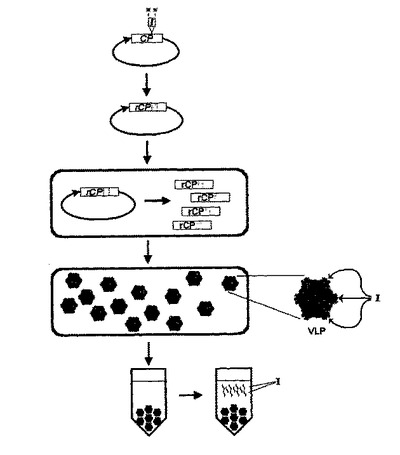
【図2】
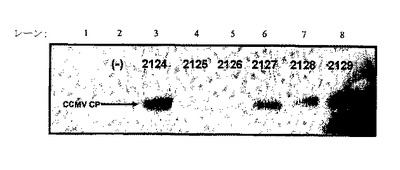
【図3】
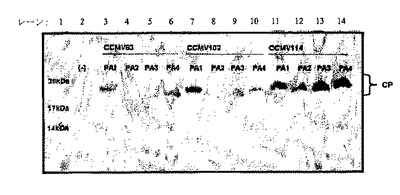
【図4】

【図5】
【図6】
【図7】
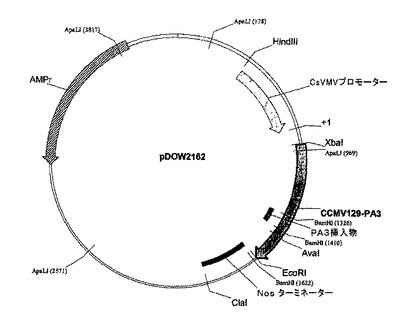
【図8】
【図9】
【図10】
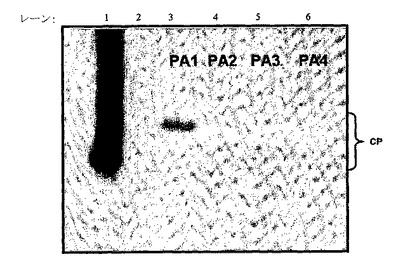
【図11】

【図12】
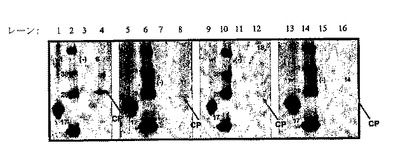
【図13】
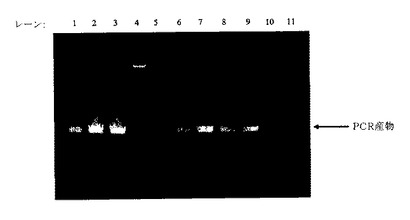
【図14】
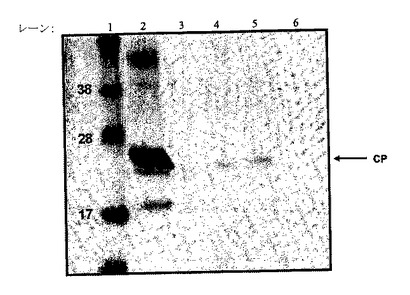
【図15】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公表番号】特表2007−528727(P2007−528727A)
【公表日】平成19年10月18日(2007.10.18)
【国際特許分類】
【出願番号】特願2007−500784(P2007−500784)
【出願日】平成17年2月28日(2005.2.28)
【国際出願番号】PCT/US2005/006342
【国際公開番号】WO2005/086667
【国際公開日】平成17年9月22日(2005.9.22)
【出願人】(590000020)ザ ダウ ケミカル カンパニー (24)
【氏名又は名称原語表記】THE DOW CHEMICAL COMPANY
【Fターム(参考)】
【公表日】平成19年10月18日(2007.10.18)
【国際特許分類】
【出願日】平成17年2月28日(2005.2.28)
【国際出願番号】PCT/US2005/006342
【国際公開番号】WO2005/086667
【国際公開日】平成17年9月22日(2005.9.22)
【出願人】(590000020)ザ ダウ ケミカル カンパニー (24)
【氏名又は名称原語表記】THE DOW CHEMICAL COMPANY
【Fターム(参考)】
[ Back to top ]