
植物細胞に分子性物質を移行させる方法
細胞壁を含む植物細胞に目的の分子を導入するための方法が提供される。植物を遺伝子改変するまたは他の形で改変するため、および細胞壁を含む植物細胞において病害を治療または予防するための方法が、提供される。
【発明の詳細な説明】
【技術分野】
【0001】
優先権主張
本出願は、METHODS FOR TRANSFERRING MOLECULAR SUBSTANCES INTO PLANT CELLSについての、2007年10月5日出願の米国仮特許出願第60/978,059号出願日の利益を主張する。
【背景技術】
【0002】
ナノ粒子は、細胞へのDNA送達における使用について開発されてきた独自の特性を有する。調査された全てのナノ粒子の中で金(Au)ナノ粒子は、それらの細胞毒性の低さおよび生物学的に重要な種々のリガンドでの機能化の容易さから送達のための優れた候補になるようである。Auナノ粒子の慣用の合成では、負に荷電した(例えばクエン酸コーティング)表面を生じる。塩基が部分的に非コイル状になるように十分に可動性である場合があるプラスミドDNAは金ナノ粒子(「GNP」)にさらされ得る。これらの部分的に非コイル状である条件下でDNA主鎖上の負電荷は、塩基と金ナノ粒子との間の誘引性ファンデルワールス力がプラスミドDNAの金ナノ粒子の表面への付着を生じるのに十分であるように十分に離れている場合がある。
【0003】
金属ナノ粒子に加えて、サイズ範囲3〜5nm内の半導体ナノ粒子(例えば量子ドット)(「QD」)も、細胞内に分子を送達するための担体として使用されている。DNAおよびタンパク質はQD表面に付着するリガンドに連結できる(Patolsky,F.ら、J.Am.Chem.Soc.125,13918(2003)を参照されたい)。カルボン酸またはアミンで被覆されたQDは、標準的な生体コンジュゲーションプロトコールを使用することによってチオール基、(例えばDubertret B.ら、Science 298,1759(2002)、Akerman,M.E.,W.C.W.Chan,P.Laakkonen,S.N.Bhatia,E.Ruoslahti,Proc.Natl.Acad.Sci.U.S.A.99,12617(2002)、Mitchell,G.P.,C.A.Mirkin,R.L.Letsinger,J.Am.Chem.Soc.121,8122(1999)を参照されたい)またはN−ヒドロキシサクシニミル(NHS)エステル基(例えばPinaud,F.,D.King,H.−P.Moore,S.Weiss,J.Am.Chem.Soc.126,6115(2004)、Bruchez,M..,M.Moronne,P.Gin,S.Weiss,A.P.Alivisatos,Science 281,2013(1998)を参照されたい)を含む分子と交差結合できる。代替法は、ストレプトアビジンで被覆されたQDとビオチン化タンパク質、オリゴまたは抗体のコンジュゲーションである(例えばDahan M.ら、Science 302,442(2003)、Pinaud,F.,D.King,H.−P.Moore,S.Weiss,J.Am.Chem.Soc.126,6115(2004)、Dahan M.ら、Science 302.442(2003)、Wu.X.Y.,ら、Nature Biotechnol.21,41(2003)、Jaiswal,J.K.,H.Mattoussi,J.M.Mauro,S.M.Simon,Nature Biotechnol.21,47(2003)およびMansson,A.ら、Biochem.Biophys.Res.Commun.314,529(2004)を参照されたい。
【0004】
ナノ粒子は、プラスミドDNAを種々の動物細胞に送達するために使用されている。DNAで被覆されたナノ粒子を細胞壁を有さない細胞と共にインキュベートすると、細胞がナノ粒子を取り込み、DNAにコードされた任意の遺伝子の発現を開始することが見出されている。通常は細胞壁を有する細胞へのナノ粒子の送達が望ましい場合は、植物のプロトプラストへの粒子の添加の前に細胞壁が取り除かれる(Torney,F.ら、Nature Nanotechnol.2,(2007)を参照されたい。植物細胞において、細胞壁は外来的に添加された分子の送達に対する障壁として存在する。遺伝子銃(微粒子銃)、微量注入法、電気穿孔法およびアグロバクテリウムなどの多数の侵襲的方法は、これらの壁を有する植物細胞への遺伝子および小分子の送達を達成するために使用されているが、タンパク質の送達は微量注入法によってだけ達成されている。植物細胞の細胞壁の存在下での小分子およびタンパク質の送達は、いまだ探究されておらず、in vitroおよびin vivo操作のための無傷の植物細胞/組織または器官で展開される実用化技術を開発するために有益である。
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明は、ナノ粒子を使用して、細胞壁を有する細胞に分子性物質を非侵襲的に送達する方法に関する。
【課題を解決するための手段】
【0006】
以下の実施形態は、典型的かつ例示的であるシステム、手段および方法と共に記載され、範囲を限定しない。
【0007】
本発明によれば、細胞壁を含む植物細胞に目的の分子を導入する方法であって、細胞壁を有する植物細胞とナノ粒子および目的の分子とを接触させて置くステップと、ナノ粒子および目的の分子の細胞への取り込みを可能にするステップとを含む方法が提供される。
【0008】
細胞壁を有する植物細胞に目的の分子を導入する方法であって、細胞壁を有する植物細胞とナノ粒子および病害を治療できる分子とを接触させて置くステップと、ナノ粒子および病害を治療できる分子の細胞への取り込みを可能にするステップとを含む方法がさらに提供される。
【0009】
上に記載の例示的態様および実施形態に加えて、さらなる態様および実施形態が以下の記載を考慮することにおいて明らかになるであろう。
【図面の簡単な説明】
【0010】
【図1】共焦点画像化システムを取り付けた微分干渉顕微鏡を使用して観察したBY2の単一細胞を示す写真である(パネルA)。色素体(アミロプラスト)を強調するためにI2KIで染色されたBY2バリアント由来の単一細胞の光学顕微鏡観察を示す図である(パネルB)。
【図2】図2パネルAは光学顕微鏡で観察した最少培地および5%二酸化炭素中で維持されたタバコ(NT1)の光独立栄養細胞を示す図であり、顕著な葉緑体が見られる。図2パネルB蛍光顕微鏡下で赤色の自己蛍光を発する活性葉緑体を有して観察される同様のNT1細胞を示す図である。
【図3】SAMSAフルオレセイン単独で、およびSAMSAフルオレセインで被覆されたGNPで処置したBY2懸濁凝集物を示す図である。図3パネルAは、SAMSAフルオレセイン単独で処置した細胞のDIC画像を示し、図3パネルBは、同じ細胞の蛍光画像を示す。図3パネルCはSAMSAフルオレセインで被覆されたGNPで処置した細胞のDIC画像を示し、図3パネルDは、SAMSAフルオレセインで被覆されたGNPで処置した細胞の蛍光画像を示す。核(Nu)および細胞壁(CW)の位置が示されている。
【図4】SAMSAフルオレセインで被覆されたGNPで処置した単一細胞を高拡大率で示す図である。パネルBは、核における多数のGNPの存在を示す。パネルAは、パネルBにおいて示されたのと同じ核の異なる焦点平面での明視野観察を示す。
【図5】SAMSAフルオレセインで被覆されたGNPで処置した光独立栄養細胞を示す図である。パネルAは、細胞壁の内側に並ぶ多量の葉緑体を有する3〜4個の細胞クラスター中のガラス質細胞を示す。パネルBは、葉緑体中のナノ粒子の蓄積を示す。パネルCおよびDは、蛍光顕微鏡を使用するより高い拡大率での単一の葉緑体を示す。ナノ粒子は、葉緑体の膜薄層形成において観察され、赤色の自己蛍光を発する葉緑素色素中に分散している。
【図6】ナノ粒子を含む細胞の反射顕微鏡および蛍光顕微鏡の画像である。図6のパネルAは、GNPが優先的に観察される反射画像を示す。パネルBは、赤色の自己蛍光を発する葉緑体の背景中で蛍光を発する粒子を示す。反射および蛍光のマージした画像をパネルCに示し、黄色蛍光を発する粒子は、葉緑体の境界内にある。
【図7】本発明の実施形態により考えられる1つの形質転換スキームを図示する。
【図8】処置の2時間後に反射顕微鏡を通して観察されるGFPの細胞内部移行を示す図である。パネルAおよびA1は、DIC顕微鏡下での未処置対照細胞(パネルA)およびDIC顕微鏡下で観察されるGFPテザーAu−NP処置細胞(パネルA1)を示す;パネルBおよびB1は、反射顕微鏡下での対照細胞(パネルB)および、反射顕微鏡下で観察されるGFPテザーAu−NP処置細胞(パネルB1)を示し、反射されたAu−NPにより粒子内部移行を示している;パネルCおよびC1は、対照細胞のDIC顕微鏡と反射顕微鏡とを重ね合わせた画像(パネルC)および、処置細胞のDIC顕微鏡と反射顕微鏡とを重ね合わせた画像(パネルC1)を示す;パネルDおよびD1は、背景に粒子が観察されない対照細胞の反射反転画像(パネルD)および、非常に明白な粒子の内部移行を示す処置細胞の反射反転画像(パネルD1)を示す。
【図9】単一細胞でのSAMSA染料で被覆されたGNPの内部移行を示す図である。パネルAは、蛍光を示す細胞壁および培地を有するが、染料の内部移行を示さない、蛍光染色した単一細胞を示し、パネルBは、DIC顕微鏡下での単一細胞を示し、パネルCは、粒子および1時間までの長時間UV光にさらしたプラズマライズ細胞と共にだけ内部移行したフルオレセインでの、サイトゾルおよび核へのナノ粒子(GNP 150nm)の内部移行を示す位相差画像を示す。
【図10】単一細胞を混合する前の、蛍光を発するGFP分子を有するAu−GFPコンジュゲートを示す図である。パネルA(FITC)、B(明視野)、C(反射)、D(パネルA+B+C):蛍光顕微鏡を通して観察されるGFP蛍光発光Au−GNP、インキュベーション2時間後だが細胞を混合する前。同様の蛍光発光粒子が核で反射を示す粒子について観察できた(図8を参照されたい)。
【図11】ナノ粒子(GNP90nm)が介在したBY2−E細胞系へのGFPの細胞内部移行を示す図である。パネルAは活性細胞質糸(cytoplasmic strand)を有する分裂対照細胞を示し(位相差画像)、パネルBは、FITCフィルターを通して検査した時の、パネルAと同じ細胞を示し、そこで、細胞質中の非緑色色素体由来および分裂核に関連するこれらの色素体由来の自己蛍光が周辺部を形成する;細胞質糸および細胞の周辺部に近い細胞質は自己蛍光を示さない;パネルCは、GNP(FITC)に付着していないGFPで処置した対照BY2細胞を示し、該細胞はGFP取り込みを示さず、GFPは細胞を取り囲んでいるが内部移行していない;パネルDは、FITCフィルターを通して観察される、パネルBでの対照と比較してGFPの内部移行を示す細胞質周辺部、細胞質糸および核を伴うGNP介在GFP内部移行を示す。
【図12】細胞のインキュベーションの2時間後にGNP介在YFP内部移行を示すBY2−E単一細胞株を示す図である。パネルA(FITC)、B(ローダミン)、C(DIC)、D(パネルA+B)、E(パネルA+B+C)、F(反射画像反転):蛍光顕微鏡を通して観察されたYFP内部移行。黄色矢印は、サイトゾルにYFPを有する生きた単一細胞での内部移行(核での拡散および濃縮)を示す。橙色矢印は、細胞内の収縮したプロトプラスト塊が細胞へのYFPの内部移行を示唆する強い蛍光を示すプラズマライズ細胞での内部移行を示す。この細胞は、近接して位置する生細胞と同じ共焦点面だが、他の生細胞の下に見出される。高レベルの粒子およびYFP蛍光が蓄積している細胞は、蛍光顕微鏡下での長時間の検査において細胞死を示す。
【図13】および
【図14】は、PATおよびYFP増幅遺伝子産物が増幅されたゲル画像を示す図である。
【図15】ペプチドのQDへの付着を確認するためにQD−ペプチドコンジュゲートについて実施された電気泳動を示す図である。
【図16】プラスミドpDAB3831を示す図である。
【発明を実施するための形態】
【0011】
以下の記載および表において、多数の用語が使用される。明細書および特許請求の範囲の明確かつ一貫した理解を提供するために、そのような用語に与えられる範囲を含めて以下の定義が提供される。
【0012】
戻し交雑 戻し交雑は、育種家がハイブリッド後代をもとの親(例えばF1ハイブリッドの親の遺伝型の内の1つを有する第1世代ハイブリッドF1)の内の1つと繰り返し交雑させる工程であり得る。
【0013】
胚 胚は、成熟種子中に含有される小植物体であり得る。
【0014】
ナノ粒子 通常100nm未満である、少なくとも1つのナノスケールの大きさを有する顕微鏡的粒子。本発明における使用に適するナノ粒子は、1nm〜0.4umの大きさを有し得る。量子ドットは、1nm〜10nmの、好ましくは2〜4nmのメジアン径を有し得る。ナノ粒子は、金ナノ粒子、金被覆ナノ粒子、多孔性ナノ粒子、メソ多孔性ナノ粒子、シリカナノ粒子、ポリマーナノ粒子、タングステンナノ粒子、ゼラチンナノ粒子、ナノシェル、ナノコア、ナノスフェア、ナノロッド、磁性ナノ粒子およびそれらの組合せから選択され得る。
【0015】
量子ドット 量子ドットは、3つ全ての空間方向における伝導帯電子、価電子帯正孔または励起子(伝導帯電子と価電子帯正孔との束縛された対)の動きを限定する半導体ナノ構造である。限定は、静電ポテンシャル(外部電極、不純物添加、歪み、不純物によって作り出される)、異なる半導体物質の間の境界面の存在(例えばコア−シェルナノ結晶系)、半導体表面の存在(例えば半導体ナノ結晶)またはこれらの組合せによる場合がある。量子ドットは、不連続の量子化されたエネルギースペクトルを有し得る。対応する波動関数は、量子ドットに空間的に局在するが、結晶格子の複数のピリオドに及ぶ。量子ドットは、少数の限定された数の(ほぼ1〜100程度)の伝導帯電子、価電子帯正孔または励起子(すなわち限定された数の素電荷)を含む。
【0016】
グリフォセート耐性 グリフォセート投与に対する耐性は、その投与量までのグリフォセートでの植物の生存能力(すなわち植物が枯らされない)を意味する。いくつかの場合において、耐性植物は一時的に黄変または他にいくつかのグリフォセート誘発性障害(例えば分けつ過剰および/または生育阻害)を呈するが、回復する。
【0017】
安定化 安定化は、同じ品種の自殖植物の一世代から次世代へ繁殖により伝えられる植物の特徴を意味する。
【0018】
取り込み 取り込みは、例えば金または量子ドットであるナノ粒子などの粒子の細胞壁または細胞膜を超える移動であって、粒子が取り込まれる細胞以外の何かによって粒子に与えられるモーメントの結果として単独に生じるのではない移動を意味する。粒子に与えられるモーメントの結果として単独で細胞壁または細胞膜を超える粒子の移動を生じる装置または方法の非限定的例は、微粒子銃、遺伝子銃、微量注入法および/またはインペールフェクション(impalefection)技術である。
【0019】
本発明の実施形態によれば、細胞壁を含む植物細胞に目的の分子を導入する方法であって、含有するナノ粒子および目的の分子を植物細胞と接触させて置くステップと、植物細胞壁を越えるナノ粒子の取り込みを可能にするステップとを含む方法が提供され得る。本発明の具体的な態様において、ナノ粒子は、任意のナノ粒子であってよく、目的の分子を可逆的または不可逆的に含有し、目的の分子で被覆され、または他の形で目的の分子と結合し、かつ/または目的の分子を保有し得る。特定の実施形態において、目的の分子は、細胞壁を有する植物細胞と接触する前にまたは細胞壁を有する植物細胞へのナノ粒子の導入と同時にナノ粒子に導入され得る。本発明の実施形態において使用され得るナノ粒子の例として、それだけに限られないが、金、量子ドット、金被覆ナノ粒子、多孔性ナノ粒子、メソ多孔性ナノ粒子、シリカナノ粒子、ポリマーナノ粒子、タングステンナノ粒子、ゼラチンナノ粒子、ナノシェル、ナノコア、ナノスフェア、ナノロッド、磁性ナノ粒子および/またはそれらの組合せが挙げられる。
【0020】
本発明の実施形態によれば、細胞壁を有する植物細胞は、無傷かつ完全な細胞壁を有する任意の植物細胞であってよい。細胞壁を有する細胞の例として、それだけに限られないが、藻類、タバコ、ニンジン、トウモロコシ、セイヨウアブラナ、ナタネ、ワタ、パーム、ピーナッツ、ダイズ、サトウキビ、オリザ種(Oryza sp)、シロイヌナズナ種(Arabidopsis sp)およびトウゴマ種(Ricinus sp)が、好ましくはタバコ、ニンジントウモロコシ、ワタ、セイヨウアブラナ、ダイズおよびサトウキビが、より好ましくはタバコおよびニンジンが挙げられる。本発明の実施形態は、任意の組織または、それだけに限られないが、胚、分裂組織細胞、カルス、花粉、葉、葯、根、根端、花、種子、さや、茎および組織培養物が挙げられるそれらが見出されるあらゆる箇所由来の細胞壁を有する細胞を含み得る。
【0021】
本発明の実施形態において目的の分子は、本発明により植物細胞に送達され得る任意の分子であり得る。目的の分子または目的の分子の構成要素は、それだけに限られないが、核酸、DNA、RNA、RNAi分子、遺伝子、プラスミド、コスミド、YAC、BAC、ポリペプチド、酵素、ホルモン、糖ペプチド、糖類、脂肪、シグナル伝達ペプチド、抗体、ビタミン、メッセンジャー、セカンドメッセンジャー、アミノ酸、cAMP、薬剤、除草剤、殺菌剤、抗生物質および/またはそれらの組合せを含み得る。
【0022】
本発明の実施形態は、病害の予防または治療のための方法を含む。非限定的例の実施形態は、殺菌剤、抗生物質および/または他の薬剤のそれを必要とする細胞への本発明の方法を使用する送達を含む。
【0023】
本発明の具体的な実施形態において、ナノ粒子の表面は、例えば標的化された取り込みを可能にするように、またはナノ粒子の表面への他の物質の可逆的もしくは不可逆的結合を可能にするように機能化され得る。非限定的な例として、ナノ粒子(例えば金ナノ粒子または量子ドット)の表面は、例えばさらに機能化または誘導体化され得るアルカンチオレートである自己組織化単分子層で機能化され得る。さらなる非限定的例においてナノ粒子の表面は、それ自体がさらに機能化または誘導体化され得るリンカーで誘導体化され得る。一実施形態においてナノ粒子は、PEG化され得る、他の実施形態においてナノ粒子は、1つまたは複数のコア(活性もしくは不活性)、ステリック被覆(活性もしくは不活性)、切断可能な結合、および/または標的化分子もしくはリガンドを含み得るまたはそれで多機能化され得る。
【0024】
本発明の態様において、ナノ粒子は、細胞の種々の部分に取り込まれ得る。ナノ粒子が取り込まれ得る位置の例として、それだけに限られないが、サイトゾル、核、液胞膜、色素体、エチオプラスト、有色体、白色体、エライオプラスト、プロテイノプラスト、アミロプラスト、葉緑体、および二重膜の内腔が挙げられる。本発明の他の実施形態において細胞壁を有する細胞へのナノ粒子取り込みは、シンプラスト的またはアポプラスト的経路を介して生じ得る。
【0025】
本発明の追加的実施形態は、植物細胞が本発明の方法を介して導入された1つまたは複数の核酸を有する、遺伝子改変された植物細胞およびそれらを生成する方法を含む。実施形態の一例において、目的の遺伝子および選択可能なマーカーを含むプラスミドは、本発明によるナノ粒子を介して細胞壁を有する植物細胞に導入され得る。さらなる実施形態において、目的の遺伝子および/または選択可能なマーカーを安定に組み込まれている安定形質転換体が、選択され得る。代替の実施形態においてこのとき目的の遺伝子を含む植物細胞は、目的の分子を含む他の細胞を生成するために増殖され得る。他の実施形態においてこのとき目的の分子を含む植物細胞は、目的の分子を含む植物体全体を再生するために使用できる再生可能な細胞であり得る。
【0026】
他の態様において、本発明は、組織培養における使用のための目的の分子を含む再生可能な植物細胞を作製する方法を提供する。組織培養は、好ましくは、再生可能な細胞と実質的に同じ遺伝子型を有する植物を再生可能な細胞として、再生することができる。そのような組織培養における再生可能な細胞は、胚、プロトプラスト、分裂組織細胞、カルス、花粉、葉、葯、根、根端、花、種子、さやまたは茎であり得る。さらに本発明の実施形態は、本発明の組織培養から再生された植物を提供する。
【0027】
代替として、本発明は、細胞壁を有する植物細胞に所望の形質を導入する方法であって、植物細胞に所望の形質を提供できるナノ粒子および目的の分子を細胞と接触させて置くステップと、細胞壁を越えるナノ粒子の取り込みを可能にするステップとを含む方法を提供する。所望の形質の例として、それだけに限られないが、雄性不稔、除草剤耐性、虫害耐性、ならびに細菌性病害、真菌性病害および/またはウイルス性病害への耐性から選択される形質が挙げられる。
【0028】
本発明のさらなる態様は、所望の形質または目的の分子を含む安定化した植物系の産生方法であって、所望の形質または目的の分子が植物細胞壁を越えるナノ粒子の取り込みによって最初に導入され得る方法を提供する。安定化した植物系の産生方法は、当業者に十分に既知であり、それだけに限られないが、自家受粉、戻し交雑、ハイブリッド産生、個体群への交雑などの技術を含み得る。細胞壁を越えるナノ粒子の取り込みによって植物細胞(またはその祖先)に最初に導入される所望の形質または目的の分子を含む全ての植物および植物細胞は、本発明の範囲内である。有利には、細胞壁を越えるナノ粒子の取り込みによって植物または細胞(もしくはその前身)に最初に導入される所望の形質または目的の分子を含む植物細胞は、優れた特徴を有する第一世代(F1)ハイブリッド細胞、種子および/または植物を産生するために他のさまざまな植物細胞との交雑に使用され得る。
【0029】
目的の分子が1つまたは複数の遺伝子を含む実施形態において、(1つまたは複数の)遺伝子は、優性または劣性アレルであり得る。例として(1つまたは複数の)遺伝子は、除草剤耐性、虫害耐性、細菌耐性への耐性、真菌耐性、ウイルス性病害耐性、雄の妊性、雄性不稔、増強された栄養価および産業利用などの形質を付与する。
【0030】
特定のタンパク質またはRNA産物(例えばRNAi)をコードする遺伝子の単離および特徴付けを可能にしている分子生物学的技術の到来により植物生物学の分野の科学者らは、特定のやり方で細胞の形質を変えるために外来遺伝子、または天然もしくは内在遺伝子(おそらく異なるプロモーターによって駆動される)の追加的もしくは改変バージョンを含有し、かつ発現するために細胞のゲノムを操作することに強い関心を発展させた。そのような外来の追加的および/または改変遺伝子は、本明細書において総称として「導入遺伝子」と称される。最近の15から20年間にわたってトランスジェニック細胞を産生するためのいくつかの方法が開発されており、特定の実施形態において、本発明は、細胞壁を越えるナノ粒子の取り込みを介して細胞壁を有する細胞に導入遺伝子を導入することにより形質転換型の細胞およびそれらを産生する方法に関する。本発明の実施形態において導入遺伝子は、発現ベクターに含有され得る。
【0031】
細胞形質転換は、特定の細胞で機能する発現ベクターの構築に関与し得る。そのようなベクターは、調節エレメント(例えばプロモーター)の制御下の、またはそれに機能的に結合した遺伝子を含むDNAを含み得る。発現ベクターは、1つまたは複数のそのように機能的に結合した遺伝子/調節エレメントの組合せを含有できる。(1つまたは複数の)ベクターは、プラスミドの形態であってよく、細胞壁を有する植物細胞の遺伝物質に(1つまたは複数の)導入遺伝子を組み込むための本明細書において記載の形質転換方法を使用して形質転換された細胞を提供するために単独でまたは他のプラスミドとの組合せで使用され得る。
【0032】
ナノ粒子を介する取り込みのための発現ベクター:マーカー遺伝子
発現ベクターは、マーカーを含有する形質転換細胞がネガティブ選択(すなわち選択可能なマーカー遺伝子を含有しない細胞の増殖阻害)またはポジティブ選択(すなわち遺伝子マーカーによってコードされる産物についての選別)のいずれかによって回収されるようにする調節エレメント(例えばプロモーター)に機能的に結合した少なくとも1つの遺伝子マーカーを含むことができる。形質転換のための多数の選択可能なマーカー遺伝子が形質転換の技術分野において十分に既知であり、例えば、抗生物質もしくは除草剤であり得る選択的化学物質を代謝的に解毒する酵素をコードする遺伝子、または阻害剤に非感受性であり得る改変された標的をコードする遺伝子を含む。いくつかのポジティブ選択方法も当技術分野において既知である。
【0033】
植物の形質転換に適した慣用される1つの選択可能なマーカー遺伝子としてカナマイシンへの耐性を付与する、植物調節シグナルの制御下でのネオマイシンホスホトランスフェラーゼII(nptII)遺伝子が挙げられる。例えばFraleyら、Proc.Natl.Acad.Sci.U.S.A.,80:4803(1983)を参照されたい。他の慣用される選択可能なマーカー遺伝子は、抗生物質ハイグロマイシンへの耐性を付与するハイグロマイシンホスホトランスフェラーゼ遺伝子であり得る。例えばVanden Elzenら、Plant Mol.Biol.,5:299(1985)を参照されたい。
【0034】
抗生物質への耐性を付与する細菌由来の追加的な選択可能なマーカー遺伝子として、ゲンタマイシンアセチルトランスフェラーゼ、ストレプトマイシンホスホトランスフェラーゼ、アミノグリコシド−3’−アデニルトランスフェラーゼおよびブレオマイシン耐性決定因子が挙げられる。Hayfordら、Plant Physiol.86:1216(1988)、Jonesら、Mol.Gen.Genet.,210:86(1987)、Svabら、Plant Mol.Biol.14:197(1990)、Hilleら、Plant Mel.Biol.7:171(1986)を参照されたい。他の選択可能なマーカー遺伝子はグリフォセート、グルフォシネートまたはブロモキシニルなどの除草剤への耐性を付与する。Comaiら、Nature 317:741〜744(1985)、Gordon−Kammら、Plant Cell 2:603〜618(1990)およびStalkerら、Science 242:419〜423(1988)を参照されたい。
【0035】
植物の形質転換に適する他の選択可能なマーカー遺伝子は、細菌由来ではない。これらの遺伝子として、例えばマウスジヒドロ葉酸還元酵素、植物5−エノールピルビルシキミ酸−3−リン酸合成酵素および植物アセト乳酸合成酵素が挙げられる。Eichholtzら、Somatic Cell Mol.Genet.13:67(1987)、Shahら、Science 233:478(1986)、Charestら、Plant Cell Rep.8:643(1990)を参照されたい。
【0036】
植物の形質転換に適するマーカー遺伝子の他のクラスは、抗生物質などの毒性物質への耐性についての形質転換細胞の直接的な遺伝的選択よりも形質転換されたと仮定される植物細胞の選別を必要とする。これらの遺伝子は、特定の組織における遺伝子発現の空間分布を定量または可視化するために特に有用であり、遺伝子発現の調査のためにそれらが遺伝子または遺伝子調節配列に融合され得ることからしばしばレポーター遺伝子と称される。形質転換細胞を選別するために慣用される遺伝子として、β−グルクロニダーゼ(GUS)、β−ガラクトシダーゼ、ルシフェラーゼおよびクロラムフェニコールアセチルトランスフェラーゼが挙げられる。Jefferson,R.A.,Plant Mol.Biol.Rep.5:387(1987)、Teeriら、EMBO J.8:343(1989)、Konczら、Proc.Nalt.Acad.Sci U.S.A.84:131(1987)、DeBlockら、EMBO J.3:1681(1984)を参照されたい。
【0037】
近年、植物組織の破壊を必要とせずにGUS活性を可視化するためのin vivoでの方法が利用可能になっている。Molecular Probes publication 2908、Imagene Green.TM.、1〜4ページ(1993)およびNalewayら、J.Cell Biol.115:151a(1991)。しかしGUS活性を可視化するためのこれらのin vivo法は、低感度、蛍光の高いバックグラウンドおよび選択可能なマーカーとしのルシフェラーゼ遺伝子の使用に伴う制限から形質転換細胞を回収するために有用であるとは証明されていない。
【0038】
最近になり、蛍光タンパク質をコードする遺伝子(例えばGFP、EGFP、EBFP、ECFPおよびYFP)が、原核細胞および真核細胞における遺伝子発現のためのマーカーとして利用されている。Chalfieら、Science 263:802(1994)を参照されたい。蛍光タンパク質および蛍光タンパク質の変異体も選択可能なマーカーとして使用できる。
【0039】
ナノ粒子を介する取り込みのための発現ベクター:プロモーター
発現ベクターに含まれる遺伝子は、調節エレメント、例えばプロモーターを含むヌクレオチド配列によって駆動されねばならない。プロモーターのいくつかの種類は、単独またはプロモーターとの組合せで使用できる他の調節エレメントと同様に形質転換の技術分野においていまや十分に既知である。
【0040】
本明細書において使用する「プロモーター」は、転写の開始点より上流であってよく、RNAポリメラーゼおよび転写を開始する他のタンパク質の認識および結合に関与できるDNAの領域に関連する。「植物プロモーター」は、植物細胞において転写を開始できるプロモーターであり得る。発生調節下のプロモーターの例として、葉、根、種子、繊維、木部導管、仮導管または厚壁組織などの特定の組織において転写を優先的に開始するプロモーターが挙げられる。そのようなプロモーターは、「組織優先」プロモーターと称される。特定の組織においてだけ転写を開始するプロモーターは、「組織特異的」と称される。「細胞型」特異的プロモーターは、1つまたは複数の器官、例えば根または葉の維管束細胞での特定の細胞型において主に発現を駆動する。「誘導可能な」プロモーターは、環境制御下にある場合があるプロモーターであり得る。誘導可能なプロモーターによる転写に影響し得る環境条件の例として、嫌気性条件または光の存在が挙げられる。組織特異的、組織優先、細胞型特異的および誘導可能なプロモーターは、「非構成的」プロモーターのクラスを構成する。「構成的」プロモーターは、ほとんどの環境条件下で活性であってよいプロモーターであり得る。
【0041】
A.誘導可能なプロモーター
誘導可能なプロモーターは、細胞での発現のための遺伝子に機能的に結合され得る。場合により、誘導可能なプロモーターは、細胞での発現のための遺伝子に機能的に結合され得るシグナル配列をコードするヌクレオチド配列に機能的に結合され得る。誘導可能なプロモーターにより転写速度は、誘導剤に反応して増大する。
【0042】
誘導可能なプロモーターは、本発明において使用され得る。Wardら、Plant Mol.Biol.22:361〜366(1993)を参照されたい。例示的な誘導可能なプロモーターとして、それだけに限られないが、銅に反応するACEI系由来のもの(Mettら、PNAS 90:4567〜4571(1993))、ベンゼンスルフホンアミド除草剤緩和剤に反応するトウモロコシ由来のIn2遺伝子(Hersheyら、Mol.Gen Genetics 227:229〜237(1991)およびGatzら、Mol.Gen Genetics 243:32〜38(1994))、ならびにTn10由来のTet抑制因子(Gatzら、Mol.Gen Genetics 227:229〜237(1991))が挙げられる。特に有用な誘導可能なプロモーターは、植物が通常は反応しない誘導剤に反応するプロモーターであり得る。例示的な誘導可能なプロモーターは、ステロイドホルモン遺伝子由来の誘導可能なプロモーターであることができ、その転写活性はグルココルチコステロイドホルモンによって誘導され得る。Schenaら、Proc.Natl.Acad.Sci.U.S.A.88:0421(1991)。
【0043】
B.構成的プロモーター
構成的プロモーターは、細胞での発現のための遺伝子に機能的に結合され得る、または構成的プロモーターは細胞での発現のための遺伝子に機能的に結合され得るシグナル配列をコードするヌクレオチド配列に機能的に結合され得る。
【0044】
さまざまな構成的プロモーターが、本発明において利用され得る。例示的な構成的プロモーターとして、それだけに限られないが、CaMV由来の35Sプロモーターなどの植物ウイルス由来のプロモーター(Odellら、Nature 313:810〜812(1985))、イネアクチン遺伝子由来プロモーター(McElroyら、Plant Cell 2:163〜171(1990))、ユビキチン(Christensenら、Plant Mol.Biol.12:619〜632(1989)およびChristensenら、Plant Mol.Biol.18:675〜689(1992))、pEMU(Lastら、Theor.Appl.Genet.81:581〜588(1991))、MAS(Veltenら、EMBO J.3:2723〜2730(1984))、ならびにトウモロコシH3ヒストン(Lepetitら、Mol.Gen.Genetics 231:276〜285(1992)およびAtanassovaら、Plant Journal 2(3):291〜300(1992))が挙げられる。ALSプロモーター、Xbal/NcoI断片5’からセイヨウアブラナ(Brassica napus)ALS3構造遺伝子(または前記Xbal/NcoI断片と類似のヌクレオチド配列)は、特に有用な構成的プロモーターである。PCT出願WO96/30530を参照されたい。
【0045】
C.組織特異的または組織優先プロモーター
組織特異的プロモーターは、細胞での発現のための遺伝子に機能的に結合され得る。場合により組織特異的プロモーターは、細胞での発現のための遺伝子に機能的に結合され得るシグナル配列をコードするヌクレオチド配列に機能的に結合され得る。組織特異的プロモーターに機能的に結合した目的の遺伝子で形質転換した植物は、特定の組織において導入遺伝子のタンパク質産物を排他的または優先的に産生できる。
【0046】
任意の組織特異的または組織優先プロモーターは、本発明において利用され得る。例示的な組織特異的または組織優先プロモーターとして、それだけに限られないが、ファゼオリン遺伝子由来のものなどの根優先プロモーター(Muraiら、Science 23:476〜482(1983)およびSengupta−Gopalanら、Proc.Natl.Acd.Sci.U.S.A.82:3320〜3324(1985))、cabまたはrubisco由来のものなどの葉特異的および光誘導プロモーター(Simpsonら、EMBO J.4(11):2723〜2729(1985)およびTimkoら、Nature 318:579〜582(1985))、LAT52由来のものなどの葯特異的プロモーター(Twellら、Mol.Gen.Genetics 217:240〜245(1989))、Zm13由来のものなどの花粉特異的プロモーター(Guerreroら、Mol.Gen.Genetics 244:161〜168(1993))またはapg由来のものなどの小胞子優先プロモーター(Twellら、Sex.Plant Reprod.6:217〜224(1993))が挙げられる。
【0047】
導入遺伝子によって産生されるタンパク質の、葉緑体、空胞、ペルオキシソーム、グリオキシソーム、細胞壁もしくはミトコンドリアなどの細胞内区画への輸送またはアポプラストへの分泌は、目的のタンパク質をコードする遺伝子のシグナル配列から5’および/または3’領域をコードするヌクレオチド配列に機能的に結合する手段によって達成され得る。構造遺伝子の5’および/または3’末端での標的化配列は、タンパク質合成およびプロセシングの間に、コードされたタンパク質がどこで最終的に区画化され得るかを決定できる。代替として、そのような細胞内区画を標的化するタンパク質は、所望の細胞内区画に目的の分子で被覆されたナノ粒子を方向付けるために直接ナノ粒子に結合され得る。
【0048】
シグナル配列の存在は、細胞内小器官もしくは細胞内区画のいずれかにまたアポプラストへの分泌にポリペプチドを方向付ける。多数のシグナル配列が当技術分野において既知である。例えばBeckerら、Plant Mol.Biol.20:49(1992)、Close,P.S.,Master’s Thesis.Iowa State University(1993)、Knox,C.ら、「Structure and Organization of Two Divergent Alpha−Amylase Genes from Barley」、Plant Mol.Biol.9:3〜17(1987)、Lernerら、Plant Physiol.91:124〜129(1989)、Fontesら、Plant Cell 3:483〜496(1991)、Matsuokaら、Proc.Natl.Acad.Sci.88:834(1991)、Gouldら、J.Cell.Biol.108:1657(1989)、Creissenら、Plant J.2:129(1991)、Kalderonら、A short amino acid sequence able to specify nuclear location,Cell 39:499〜509(1984)、Steifelら、Expression of a maize cell wall hydroxyproline−rich glycoprotein gene in early leaf and root vascular differentiation,Plant Cell 2:785〜793(1990)を参照されたい。
【0049】
外来タンパク質遺伝子および農学的遺伝子
本発明によるトランスジェニック植物で外来タンパク質は、商業的な量で産生され得る。したがって、当技術分野において十分に理解されている形質転換された植物の選択および繁殖のための技術は、従来の手段で収穫される複数のトランスジェニック植物を産出し、それにより外来タンパク質は目的の組織から、またはバイオマス全体から抽出され得る。植物バイオマスからのタンパク質抽出は、例えばHeneyおよびOrr,Anal,Biochem.114:92〜6(1981)によって論じられた既知の方法によって達成され得る。
【0050】
本発明の態様において外来タンパク質の商業的産生のために提供されるトランスジェニック植物は、細胞または植物であり得る。他の態様において目的のバイオマスは種子であり得る。高いレベルの発現を示す比較的少数のトランスジェニック植物について遺伝子地図は、組み込まれたDNA分子の染色体上のおよその位置を同定する従来型のRFLP、PCRおよびSSR分析を主に介して作成され得る。この点に関する例示的な方法について、GlickおよびThompson,Methods in Plant Molecular Biology and Biotechnology CRC Press,Boca Raton 269:284(1993)を参照されたい。染色体上の位置に関する地図情報は、対象のトランスジェニック植物の所有権の保護において有用であり得る。認可されていない繁殖が行われ、他の生殖質との交雑物が作製され得る場合、組込み領域の地図は、疑われる植物についての同様の地図と、後者が対象植物と共通の起源を有するかどうかを決定するために比較され得る。地図の比較は、ハイブリダイゼーション、RFLP、PCR、SSRおよび配列決定を含み、その全ては従来型の技術である。
【0051】
同様に農業的な遺伝子は、形質転換された細胞またはそれらの後代において発現され得る。より具体的には植物は、農業的目的での種々の表現型を発現するために本発明の方法を介して遺伝的に操作され得る。この点において使用され得る例示的な遺伝子として、それだけに限られないが、以下に分類されるものが挙げられる。
【0052】
1.有害生物または病害への耐性を付与し、かつ以下をコードする遺伝子:
A)植物の病害耐性遺伝子。植物の防御は、植物における病害耐性遺伝子(R)の産物と病原体中の対応する非病原性(Avr)遺伝子の産物との間の特定の相互作用によってしばしば活性化される。植物種は、特定の病原体株に耐性である植物を操作するためにクローン化された耐性遺伝子で形質転換され得る。例えばJonesら、Science 266:789(1994)(クラドスポリウム・フルバム(Cladosporium fulvum)への耐性のためのトマトCf−9遺伝子のクローニング)、Martinら、Science 262:1432(1993)(シュードモナス・シリンゲ(Pseudomonas syringae)病原型への耐性のためのトマトPto遺伝子。トマトはプロテインキナーゼをコードする)、Mindrinosら、Cell 78:1089(1994)(シュードモナス・シリンゲへの耐性のためのアラビドプス(Arabidops)はRSP2遺伝子であり得る)を参照されたい。
【0053】
B)ダイズシストセンチュウ(soybean cyst nematode)などの有害生物への耐性を付与する遺伝子。例えば、PCT出願WO96/30517、PCT出願WO93/19181を参照されたい。
【0054】
C)バチルス・チューリンゲンシスタンパク質、その誘導体またはそれをモデル化した合成ポリペプチド。例えばBt δ−エンドトキシン遺伝子のクローニングおよびヌクレオチド配列を開示するGeiserら、Gene 48:109(1986)を参照されたい。さらにδ−エンドトキシン遺伝子をコードするDNA分子は、例えばATCC登録番号40098、67136、31995および31998でAmerican Type Culture Collection,Manassas,Vaから購入できる。
【0055】
D)レクチン。例えば、いくつかのクンシラン(Clivia miniata)マンノース結合レクチン遺伝子のヌクレオチド配列を開示しているVan Dammeら、Plant Molec.Biol.24:25(1994)による開示を参照されたい。
【0056】
E)アビジンなどのビタミン結合タンパク質。PCT出願US93/06487を参照されたい。出願は、害虫に対する殺幼虫剤(larvicide)としてのアビジンおよびアビジン相同体の使用を教示している。
【0057】
F)酵素阻害剤、例えばプロテアーゼもしくはプロテイナーゼ阻害剤またはアミラーゼ阻害剤。例えばAbeら、J.Biol.Chem.262:16793(1987)(イネシステインプロテイナーゼ阻害剤のヌクレオチド配列)、Huubら、Plant Molec.Biol.21:985(1993)(タバコプロテイナーゼ阻害剤IをコードするcDNAのヌクレオチド配列)、Sumitaniら、Biosci.Biotech.Biochem.57:1243(1993)(ストレプトマイセス・ニトロスポレウス(Streptomyces nitrosporeus)α−アミラーゼ阻害剤のヌクレオチド配列)および米国特許第5,494,813号(HepherおよびAtkinson、1996年2月27日発行)を参照されたい。
【0058】
G)エクジステロイドまたは幼若ホルモン、それらのバリアント、それらに基づく模倣物、またはそれらのアンタゴニストもしくはアゴニストなどの昆虫特異的ホルモンまたはフェロモン。例えばHammockら、Nature 344:458(1990)による、クローン化された幼若ホルモンエステラーゼ、幼若ホルモンの不活性化因子のバキュロウイルス発現の開示、を参照されたい。
【0059】
H)感染した有害生物の生理機能を発現によって破壊する昆虫特異的ペプチドまたは神経ペプチド。例えばRegan,J.Biol.Chem.269:9(1994)(発現クローニングは昆虫利尿ホルモン受容体をコードするDNAを産出する)およびPrattら、Biochem.Biophys.Res.Comm.163:1243(1989)(アロスタチン(allostatin)はディプロプテラ・パンタタ(Diploptera puntata)において同定され得る。)の開示を参照されたい。昆虫特異的な麻痺性神経毒をコードする遺伝子を開示しているTomalskiらの米国特許第5,266,317号も参照されたい。
【0060】
I)天然でヘビ、スズメバチまたは任意の他の生物によって産生される昆虫特異的毒液。例えばサソリ昆虫毒性ペプチドをコードする遺伝子の植物における異種性発現の開示についてPangら、Gene 116:165(1992)を参照されたい。
【0061】
J)モノテルペン、セスキテルペン、ステロイド、ヒドロキサム酸、フェニルプロパノイド誘導体または殺虫活性を有する他の非タンパク質性分子の高度集積に関連する酵素。
【0062】
K)生物学的に活性な分子、例えば天然または合成での、解糖系酵素、タンパク質分解酵素、脂肪分解酵素、ヌクレアーゼ、シクラーゼ、トランスアミナーゼ、エステラーゼ、ヒドロラーゼ、ホスファターゼ、キナーゼ、ホスホリラーゼ、ポリメラーゼ、エラスターゼ、キチナーゼおよびグルカナーゼ、の翻訳後修飾を含む修飾に関与する酵素。カルラーゼ(callase)遺伝子のヌクレオチド配列を開示しているScottらによるPCT出願WO93/02197を参照されたい。キチナーゼをコードする配列を含有するDNA分子は、例えばATCCから登録番号39637および67152で得ることができる。タバコイモムシキチナーゼをコードするcDNAのヌクレオチド配列を教示しているKramerら、Insect Biochem.Molec.Biol.23:691(1993)および、パセリubi4−2ポリユビキチン遺伝子のヌクレオチド配列を提供しているKawalleckら、Plant Molec.Biol.21:673(1993)も参照されたい。
【0063】
L)シグナル伝達を刺激する分子。例えばBotellaらPlant Molec.Biol.24:757(1994)によるリョクトウカルモジュリンcDNAクローンについてのヌクレオチド配列の開示、およびトウモロコシカルモジュリンcDNAクローンのヌクレオチド配列を提供しているGriessら、Plant Physiol.104:1467(1994)を参照されたい。
【0064】
M)疎水性モーメントペプチド。PCT出願WO95/16776(真菌性植物病原菌を抑制するタキプレシンのペプチド誘導体の開示)およびPCT出願WO95/18855(病害への耐性を付与する合成抗菌性ペプチドを教示)を参照されたい。
【0065】
N)膜透過酵素、チャネル形成剤またはチャネル遮断剤。例えば青枯病菌(Pseudomonas solanacearum)に耐性のトランスジェニックタバコ植物を提供するセクロピン−β溶解性ペプチド類似物の異種発現についてのJaynesら、Plant Sci 89:43(1993)の開示を参照されたい。
【0066】
O)ウイルス性侵襲性タンパク質またはそれら由来の複合毒素。例えば、形質転換された植物細胞におけるウイルス被覆タンパク質の蓄積は、被覆タンパク質遺伝子が由来し得るウイルスによって、および関連するウイルスよってもたらされるウイルス感染および/または病害の進行への耐性を与える。Beachyら、Ann.rev.Phytopathol.28:451(1990)を参照されたい。被覆タンパク質介在耐性は形質転換植物に、アルファルファモザイクウイルス、キュウリモザイクウイルス、タバコ条斑病ウイルス、ジャガイモウイルスX、ジャガイモウイルスY、タバコエッチウイルス、タバコ茎壊疽ウイルスおよびタバコモザイクウイルスに対して与えられている。同著者。
【0067】
P)昆虫特異的抗体またはそれら由来の免疫毒素。このように昆虫の消化管において不可欠な代謝機能を標的とする抗体は、影響される酵素を不活性化し、昆虫を殺す。参照、Taylorら、Abstract#497,Seventh Int’l Symposium on Molecular Plant−Microbe Interactions(Edinburgh,Scotland)(1994)(1本鎖抗体断片の産生を介するトランスジェニックタバコにおける酵素の不活性化)。
【0068】
Q)ウイルス特異的抗体。例えば組換え抗体遺伝子を発現するトランスジェニック植物はウイルス攻撃から防御されることを示している、Tavladorakiら、Nature 366:469(1993)を参照されたい。
【0069】
R)病原体または寄生生物によって天然で産生される発生抑制タンパク質。例えば真菌エンドα−1,4−Dポリガラクツロナーゼは、植物細胞壁ホモ−α−1,4−D−ガラクツロナーゼを可溶化することによって真菌のコロニー形成および植物栄養素の放出を促進する。Lambら、Bio/Technology 10:1436(1992)を参照されたい。マメエンドポリガラクツロナーゼ阻害タンパク質をコードする遺伝子のクローニングおよび特徴付けはToubartら、Plant J.2:367(1992)によって記載され得る。
【0070】
S)植物によって天然で産生される発生抑制タンパク質。例えばLogemannら、Bio/Technology 10:305(1992)は、オオムギリボソーム不活性化遺伝子を発現するトランスジェニック植物は真菌性病害への耐性が増大していることを示している。
【0071】
2.除草剤への耐性を付与する遺伝子
A)成長点または分裂組織を阻害する、イミダゾリノンまたはスルホニル尿素などの除草剤。この分類の例示的な遺伝子は、例えばLeeら、EMBO J.7:1241(1988)およびMikiら、Theor.Appl.Genet.80:449(1990)によってそれぞれ記載のとおり変異体ALSおよびAHAS酵素をコードする。
【0072】
B)グリフォセート(例えば変異体5−エノールピルビルシキミ酸−3−リン酸合成酵素(EPSP)遺伝子によって(組換え核酸の導入および/または種々の形態の天然EPSP遺伝子のin vivo変異誘発を介して)、aroA遺伝子およびグリフォセートアセチルトランスフェラーゼ(GAT)遺伝子、それぞれによって耐性が付与される)、グルフォシネートなどの他のホスホノ化合物(ストレプトマイセス・ハイグロスコピカス(Streptomyces hygroscopicus)およびストレプトマイセス・ビリジクロモゲネス(Streptomyces viridichromogenes)を含むストレプトマイセス種由来のホスフィノトリシンアセチルトランスフェラーゼ(PAT)遺伝子)およびピリジノキシまたはフェノキシプロプリオン酸ならびにシクロヘキソン類(ACCase阻害剤をコードする遺伝子)、例えば植物にグリフォセート耐性を付与できるEPSPの形態のヌクレオチド配列を開示しているShanらの米国特許第4,940,835号およびBarryらの米国特許第6,248,876号を参照されたい。変異体aroA遺伝子をコードするDNA分子は、ATCC登録番号39256で得ることができ、変異体遺伝子のヌクレオチド配列はComaiの米国特許第4,769,061号において開示され得る。Kumadaらの欧州特許出願第0333033号およびGoodmanらの米国特許第4,975,374号は、L−ホスホリノトリシンなどの除草剤への耐性を付与するグルタミン合成酵素遺伝子のヌクレオチド配列を開示している。PAT遺伝子のヌクレオチド配列は、PAT活性をコードするキメラbar遺伝子を発現するトランスジェニック植物の産生を記載しているLeemansらの欧州特許出願第0242246号、DeGreefら、Bio/Technology 7:61(1989)によって提供され得る。フェノキシプロプリオン酸ならびに、セトキシジムおよびハロキシホップなどのシクロヘキソン類への耐性を付与する遺伝子の例として、Marshallら、Theor.Appl.Genet.83:435(1992)によって記載のAcc1−S1、Acc1−S2およびAcc1−S3遺伝子が挙げられる。グリフォセート耐性を付与できるGAT遺伝子は、CastleらのWO2005012515に記載されている。2,4−D、fopおよびピリジロキシオーキシン除草剤への耐性を付与する遺伝子は、Dow AgroSciences LLCに譲渡されたWO2005107437に記載されている。
【0073】
C)トリアジン(psbAおよびgs+遺伝子)またはベンゾニトリル(ニトリラーゼ遺伝子)などの光合成を阻害する除草剤。Przibilaら、Plant Cell 3:169(1991)は変異体psbA遺伝子をコードするプラスミドでのクラミドモナスの形質転換を記載している。ニトリラーゼ遺伝子のヌクレオチド配列は、Stalkerの米国特許第4,810,648号に記載されており、これらの遺伝子を含有するDNA分子はATCC登録番号53435、67441および67442で入手できる。グルタチオンS−トランスフェラーゼをコードするDNAのクローニングおよび発現は、Hayesら、Biochem.J.285:173(1992)によって記載されている。
【0074】
3.次のような付加価値のある特質を付与するまたはそれに寄与する遺伝子
A)例えば、植物のステアリン酸含有量を増加させるためにステアリルACP不飽和化酵素のアンチセンス遺伝子で植物を形質転換することによる、改変された脂肪酸代謝。Knultzonら、Proc.Natl.Acad.Sci.U.S.A.89:2624(1992)を参照されたい。
【0075】
B)フィチン酸含有量の減少−1)フィターゼをコードする遺伝子の導入は、フィチン酸の分解を増強し、形質転換植物の遊離リン酸を増やす。例えば、クロコウジカビ(Aspergillus niger)フィターゼ遺伝子のヌクレオチド配列の開示についてのVan Hartingsveldtら、Gene 127:87(1993)を参照されたい。2)フィターゼ含有量を減少させる遺伝子は導入され得る。例えばトウモロコシにおいて、これはクローニングおよびそれに次ぐ、フィチン酸の低いレベルによって特徴付けられるトウモロコシ変異体に関与し得る単一アレルに関連するDNAの再導入によって達成され得る。Raboyら、Maydica 35:383(1990)を参照されたい。
【0076】
C)例えば、デンプンの分枝様式を変化させる酵素をコードする遺伝子で植物を形質転換することによって生じた改変された炭水化物組成物。Shirozaら、J.Bacteol.170:810(1988)(連鎖球菌変異体フルクトース転移酵素遺伝子のヌクレオチド配列)、Steinmetzら、Mol.Gen.Genet.20:220(1985)(バチルス・サブティル(Bacillus subtil)はレバンスクラーゼ遺伝子のヌクレオチド配列であり得る)、Penら、Bio/Technology 10:292(1992)(バチルス・リケニフォン(Bacillus lichenifonn)を発現するトランスジェニック植物の産生物はα−アミラーゼであり得る)、Elliotら、Plant Molec.Biol.21:515(1993)(トマトインベルターゼ遺伝子のヌクレオチド配列)、Sogaardら、J.Biol.Chem.268:22480(1993)(オオムギα−アミラーゼ遺伝子の部位特異的ミュータジェンズ(mutagens)であり得る)およびFisherら、Plant.Physiol.102:1045(1993)(トウモロコシ胚乳デンプン分枝酵素II)を参照されたい。
【実施例】
【0077】
本発明は、例示の方法により提示され、いかなるやり方においても本発明を限定することを意図しない以下の実施例においてさらに記述される。
【実施例1】
【0078】
単一細胞植物材料の調製
BY2細胞およびNT1細胞の両方を使用した。BY2細胞は、非緑色、高増殖性タバコ細胞系である。NY1細胞は、タバコから単離された光独立栄養細胞である。形質転換の3から4日前に、1週間経過した懸濁培養物を、NT1またはBY2培養物の2mlを、250mLフラスコ中の50nM DAS−PMTI−1(微小管阻害剤)および0.5〜0.1%(v/v)DMSOを含有するNT1BまたはLSBY2培地40mlに移すことによって新鮮培地に継代した。単一細胞を微小管阻害剤処置の4日後または7日後のいずれかで回収した。使用したBY2単一細胞は、生細胞を計数するためにBeckmanフローサイトメーターにより処理した。平均径10.43umおよび容積593.8μm3であった1mlあたり658250個の生細胞があった。図1で観察されるとおり、全ての細胞は単一細胞であった(図1における対は端が重なっている)。細胞を共焦点画像化システムを取り付けた微分干渉(DIC)顕微鏡を使用して検査した(パネルA)。パネルBは、色素体(アミロプラスト)を強調するためにI2KIで染色したBY2細胞(EP12%培地で馴化および維持した培養物)由来の単一細胞の光学顕微鏡像を示す。ここで観察されるようにBY2細胞の単一細胞は、細胞の細胞質全体に分布した多数の色素体(アミロプラスト)を含む。
【0079】
図2、パネルAは、最少培地および5%二酸化炭素で維持した、顕著な葉緑体を有するタバコ(NT1)の光学顕微鏡での光独立栄養細胞を示す。これらの細胞を3.0OD600での懸濁物1mlを移すことによって14日ごとに1回継代した。図2、パネルBは蛍光顕微鏡下で観察した同様のNT1細胞を示し、赤色に自己蛍光を発している活性葉緑体が観察できる。
【0080】
上に記載する細胞型を形質転換のための標的細胞として使用した。緑色細胞(NT1細胞)は、クラスター化する細胞がほとんど無く、かつ透明であることから葉緑体中にナノ粒子を追跡するための最適な細胞型である。加えて細胞は、赤色の自己蛍光を発する非常に顕著な葉緑体を有する(図2、パネルBに観察されるとおり)。
【実施例2】
【0081】
ナノ粒子の調製および細胞の処置
培養物中で細胞が蛍光色素を取り込むかどうかを決定するために、BY2細胞の単一細胞のおよび多細胞性の標準的な凝集懸濁培養物を使用した。細胞懸濁培養物をMolecular ProbesからのSAMSAフルオレセイン(5−((2−(および−3)−S−(アセチルメルカプト)サクシノイル)アミノ)フルオレセイン)に、ナノ粒子の非存在下で20分間さらし、次いで蛍光顕微鏡下で観察する前に短時間洗浄した。
【0082】
金ナノ粒子(GNP)を製品技術指針(probes.invitrogen.com/media/pis/mp00685.pdfでワールドワイドウェブにおいて入手できる)によりSAMSAフルオレセインで被覆した。金−フルオレセインコンジュゲートを本明細書以下に記載の方法を使用して調製した。SAMSAフルオレセイン1mgを0.1M NaOH 100μlに溶解し、チオールを保護しているアセチル基を除去するために15分間ボルテックスした。この活性化SAMSAを次いで150nm金コロイド(1mlあたり粒子約109個)100μlと混合した。得られた溶液を次いで反応の確実な完了のために1時間インキュベートした。次いで1M HCl 50μLを溶液を中和するために加えた。3000RPMで30分間遠心分離し、上清を除去した。得られた黄色沈殿を0.1M PBS 200μLに再懸濁し、橙色溶液を得た。この精製ステップを、遊離SAMSAフルオレセインを確実に除去するために2回繰り返した。SAMSAの金への付着様式は、主にチオール結合を介する。顕著な静電反発力(SAMSAはpH>7で2価陰イオン性である)により、SAMSAは全体が金コロイド表面に垂直に位置している。粒子は、DICおよび多光子共焦点顕微鏡下で観察した場合にいかなるバックグラウンドも示さない明確な蛍光を示した。被覆した金ナノ粒子20および40μlをBY2/NT1タバコ懸濁液または光独立栄養タバコ細胞500μlに移し、暗所で20分間インキュベートした。
【0083】
インキュベーション後、細胞懸濁液50μl一定分量を顕微鏡の灌流スライドに載せ、粒子を追跡するために顕微鏡下で観察した。加えて、試料の一定分量を20分間のインキュベーションの2〜20時間後に顕微鏡観察のために調製した。
【実施例3】
【0084】
フルオレセインで被覆されたナノ粒子のBY2/NT1細胞凝集物ならびに単一の光独立栄養タバコ細胞の核および色素体における送達および蓄積
SAMSAフルオレセイン単独でおよびSAMSAフルオレセインで被覆されたGNPで処置したBY2/NT1懸濁凝集物を、DIC、明視野および蛍光顕微鏡を使用する低拡大率下および高拡大率下で検査した。図3、パネルAはSAMSAフルオレセイン単独で処置した細胞のDIC画像を示し、一方図3、パネルBは同じ細胞の蛍光画像を示す。図3、パネルCはSAMSAフルオレセインで被覆されたGNPで処置した細胞のDIC画像を示し、一方図3パネルDはSAMSAフルオレセインで被覆されたGNPで処置した細胞の蛍光画像を示す。図3、パネルBにおいて明確に観察されるとおり、SAMSAフルオレセイン単独で処置した細胞の細胞壁だけがフルオレセインで染色され、他のバッククラウンド蛍光はわずかしか観察されなかった。これは、細胞がナノ粒子の非存在下ではSAMSAフルオレセインを取り込まなかったことを示唆する。
【0085】
図3、パネルDで見られるとおり、対照的にSAMSAフルオレセインで被覆されたGNPは、細胞中および核(Nu)中へと追跡された。SAMSAフルオレセインで被覆されたGNPが空胞を除く細胞の全ての区画において見出されたことは、DIC観察から明らかであった。核区画に加えて空胞を裏打ちする細胞質糸もSAMSAフルオレセインで被覆されたGNPを有した。ナノ粒子は、細胞壁を越えるそれらの輸送を妨げられないようである。したがってSAMSAフルオレセインで被覆されたGNPの蓄積は、アポプラストの連続体ではなくシンプラストの連続体においてであると考えられる。SAMSAフルオレセインで被覆されたGNPで処置した単一細胞の高拡大率下でのさらなる検査は、核中の多数のGNPの存在を示し、GNPはこれらの小器官に優先的に蓄積されると考えられた(図4、パネルB)。図4、パネルAは、パネルBと同じ核の異なる焦点面での明視野画像を示す。
【0086】
図5は、SAMSAフルオレセインで被覆されたGNPで処置した光独立栄養細胞を示す。パネルAは、細胞壁の内側を満たしている大きな葉緑体を有する細胞3〜4個のクラスターでの非常に透明な細胞を示す。パネルBは、葉緑体中のナノ粒子の蓄積を示す。パネルCおよびDは、蛍光顕微鏡を使用する、より高拡大率での単一の葉緑体を示す。ナノ粒子は、葉緑体の膜薄層形成において観察され、赤色自己蛍光を発しているクロロフィル色素中に散在している。
【0087】
したがって、即時的に明視野および蛍光の顕微鏡で追跡した生存光独立栄養細胞は、ナノ粒子が膜および葉緑体マトリックスの両方に蓄積していることを示した。粒子は、葉緑体の二重膜の内腔でも追跡され得た。
【0088】
葉緑体中の粒子を追跡するために実施した実験は、粒子が色素体に蓄積するらしいことを明らかにしたが、不十分な解像度のため光学顕微鏡を使用することによって葉緑体包膜中の粒子の存在を可視的に同定することは困難であった。したがって反射顕微鏡および蛍光顕微鏡を使用して粒子を追加的に追跡し、粒子を明確に位置付けるために画像をマージした、図6示すとおり。図6、パネルAは、GNPが優先的に見られる反射画像を示す。この写真は葉緑体中のナノ粒子の存在を示すだけでなく、葉緑体中のナノ粒子の多量の蓄積も示し、活発な取り込みを示唆している。パネルBは、赤色自己蛍光を発する葉緑体のバックグラウンド中での蛍光発光粒子を示す。マージした反射および蛍光での画像をパネルCに示す、ここで黄色蛍光を発している粒子は葉緑体の境界内であり、色素体中の粒子の存在を確認している。
【実施例4】
【0089】
核形質転換のためのDNA付着GNP送達
DNAで被覆されたGNPを2経路、すなわち非特異的相互作用および特異的相互作用(PEGをプラットホームとして使用する)を介して合成し、BY2/NT1細胞とインキュベートした。非特異的相互作用のために、3%マンニトール 9mLを細胞懸濁物1mLに加え、次いで1000rpmで5分間遠心分離した。次いで上清をデカントし、細胞を3%マンニトール300μlに再懸濁した。直径150nmの金ナノ粒子(BBI Internationalから入手できる(EM.GC150))50μlおよびYFPをコードするプラスミドDNA(pDAB3831)(図16)(配列番号1および2)50μgを再懸濁した細胞に加え、混合物を20分間インキュベートした。インキュベーション後、3%マンニトール20mLを溶液に加え、得られた混合物を1000rpmで5分間遠心分離した。次いで上清をデカントし、細胞を増殖培地3mLに再懸濁した。次いで再懸濁した細胞を選択プレートに移す少なくとも48時間前にマイクロウェルに移した。特異的相互作用(PEG経路)のために、大過剰量のチオールリガンドを使用した:粒子1個あたり100単分子膜、1個のチオール分子によって占有される表面積がおよそ0.20nmであると仮定することにより推定した。この計算を使用して、SH−PEG(3)−OCH3 2mgをクエン酸GNP溶液に加えた。混合物を直ちに室温で20時間撹拌し、その間に溶液の色はわずかに濃くなった。次いでTHF 3容量を反応混合物に加え、得られた溶液を13K rpmで4℃、30分間遠心分離した。上清を除去し、沈殿を超純水(18MΩ.cm)10mLに再溶解し、THF 30mLを加え、同じ条件で2度目の遠心分離を実施した。次いで沈殿を超純水(18MΩ.cm)に再溶解し、室温に置いた。形質転換実験用に、プラスミドDNAをH3CO−PEG−SH−GNPに被覆するために、精製プラスミドDNA 1mgを金粒子10mgと水50ml中で2時間、23°でインキュベートした(Torney,F.ら、Nature Nanotechnol.2,(2007)を参照されたい)。
【0090】
考えられる1つの形質転換計画の図画表示を図7で概説する。形質転換用に、YFPレポーター遺伝子を含むプラスミドDNA、pDAB3831を使用した。BY2細胞を上に記載のとおり処置し、懸濁液をマイクロウェルプレートでゆっくり撹拌しながら48〜72時間インキュベートした。懸濁液の50μl一定分量を合計0.5〜1ml混合物から採取し、レポーター遺伝子の発現を観察するために蛍光顕微鏡下で検査した。YFPレポーター遺伝子を含有するプラスミドで形質転換したBY2細胞は、YFPの一過的発現を示した。
【実施例5】
【0091】
核形質転換のためのDNA付着PEG化量子ドット送達
細胞侵入評価調査のためのQDのPEG機能化。この手順は、Dubertret B.ら、Science 298,1759(2002)から採用した。TOPO(トリオクチルホスフィンオキシド)で被覆されたCdSe/ZnS QD(Ocean nanotechnology,Cat#QSO0630−0010)2mgをPEG−PE(1,2−ジアシル−sn−グリセロ−3−ホスホエタノールアミン−N−[メトキシ−ポリ(エチレングリコール)])(Avanti lipids,Cat#880160).015g(5.5μmol)とクロロホルム中に懸濁し、溶媒の蒸発および水での安定化が続いた。PEGコンジュゲート化を細胞毒性からの完全な防御を確実にするために実施した。
【0092】
プラスミドDNAとのQDコンジュゲーション。TOPO(トリオクチルホスフィンオキシド)で被覆されたQD(Ocean nanotechnology,Cat#QSO0630−0010)2mgをHS−PEG−OCH3(Prochimia,Cat#TH014−01)4mgと、一晩、約60〜70℃で懸濁した。真空オーブンで溶媒を除去した。次いで残渣を水(18M)1mLに懸濁した。最後のステップは赤色残渣の橙色、光学的に透明の、透き通った溶液への変化を伴う。形質転換実験用にプラスミドDNAをH3CO−PEG−SH−QDに被覆するために、精製プラスミドDNA(pDAB3831)0.02mgを得られたQDコンジュゲートと水2ml中で2時間、23°、暗所でインキュベートした(Torney,F.ら、Nature Nanotechnol.2,(2007))。
【0093】
タバコ無傷細胞とのQDのインキュベーション。細胞系での実験を、LSBY2培地中、25℃で維持したBright Yellow(BY2)タバコ単一細胞系を使用して実施した。これらの単一細胞系は、IDM#64901で概説されたのと同じ方法によって産生される。1〜3μL/mLの濃度を24ウェルマイクロタイタープレート中の細胞500μlに加え、細胞を分析する前に振盪機でおだやかに20分間回転させた。
【実施例6】
【0094】
無傷植物細胞への蛍光タンパク質のナノ粒子介在形質導入および細胞内部移行ならびに利用可能性。
植物細胞へのタンパク質のナノ粒子介在形質導入および細胞内部移行を検査するための材料は、サイズ直径150nmの金コロイド(BBI International,GC150)、5−((2−(および−3)−S(アセチルメルカプト)サクシノイル)アミノ)フルオレセイン(SAMSA fluorescein:Invitrogen,A−685)、サイズ80および90nmのカルボン酸被覆金コロイドのナノ粒子(TedPella,32019)、スルホ−NHS(N−ヒドロキシスルホサクシニミド)、EDC(1−エチル−3−[3−ジメチルアミノプロピル]カルボジイミド塩酸塩)、(Pierce Bitoechnology,24510,22980,)、MES(2−[N−モルホリノ]エタンスルホン酸)(Fisher Scientific,AC32776−1000)、リン酸緩衝食塩溶液緩衝剤包(Sigma,P5368−10PAK)、ヒストジンタグ付きGFP(Evrogen、励起最大−482nm、発光最大−502nm、FP611)、turbo YFP(Evrogen、励起最大−525nm、発光最大−538nm、FP611)およびヨウ化プロピジウム(Sigma−P4864)、フルオレセイン二酢酸(Sigma,F7378)を含む。
【0095】
細胞培養物(BY2−Eタバコ単一細胞):BY2細胞およびNT1細胞の両方を使用した。BY2/NT1細胞は、非緑色、高増殖性タバコ細胞系である。実験の3から4日前に、1週間経過した懸濁培養物を、NT1またはBY2培養物2mlを、250mLフラスコ中のDAS−医薬用MTI−1(PMTI−1)(微小管阻害剤)1μl、1〜3%グリセロールおよび0.05〜0.1%(v/v)DMSOを含有するNT1BまたはLSBY2培地40mlに移すことによって新鮮培地に継代した。単一細胞を微小管阻害剤処置の3.5日後または7日後のいずれかで回収した。使用したBY2単一細胞は、生細胞を計数するためにBeckmanフローサイトメーターにより処理した。平均径10.43um(容積593.8μm3)〜50.42um(容積67079.05μm3)で1mlあたり658250個の生細胞があった。これらの培養物中の生細胞は、PMTI−1処置後に全て単一細胞であった。細胞を共焦点蛍光画像化システムを取り付けた微分干渉(DIC)顕微鏡を使用して検査した。
【0096】
ナノ粒子コンジュゲート:金−フルオレセインコンジュゲート、金−ヒスチジンタグ付きGFPコンジュゲート、および金−YFPコンジュゲートを合成した。
【0097】
金−フルオレセインコンジュゲートの合成:金−フルオレセインコンジュゲートは既に記載されている方法(Cannone,F.,G.Chiricoら、(2006)、Quenching and Blinking of Fluorescence of a Single Dye Molecule Bound to Gold Nanoparticles.J.Phys.Chem.B 110(33):16491〜16498)を使用することにより調製した。SAMSAフルオレセイン1mgを0.1M NaOH 100μlに溶解し、チオールを保護しているアセチル基を除去するために15分間ボルテックスした。この活性化SAMSAを150nm金コロイド(1mlあたり粒子約109個)100μlと混合した。この溶液を次いで反応を確実に完了させるために1時間インキュベートした。インキュベーション後、1M HCl 50μLを溶液を中和するために加えた。3000RPMで30分間遠心分離し、上清を除去した。得られた黄色沈殿を0.1M PBS 200μLに再懸濁し、橙色溶液を得た。この精製ステップを、遊離SAMSAフルオレセインの除去を確実にするために2回繰り返した。SAMSAの金への付着様式は、主にチオール結合を介する。顕著な静電反発力(SAMSAはpH>7で2価陰イオン性である)により、SAMSAは金コロイド表面に垂直に位置していると考えられている(Cannoneら、2006)。
【0098】
金−ヒスチジンタグ付きGFPおよび金−YFPコンジュゲートの合成:金−タンパク質コンジュゲートを、2種のタンパク質の連続的カップリングを例示したGrabarek(Grabarek,Z.およびJ.Gergely(1990)、Zero−length cross linking procedure with the use of active esters.Analytical Biochemistry 185(1):131〜135)によって記載された手順をわずかに変更して使用して合成した。90nmカルボン酸被覆金コロイド溶液(1mlあたり粒子約109個)0.25mlを3000RPMで10分間遠心分離した。上清を捨てた後、赤色沈殿を活性化緩衝液(0.1M MES、0.5M NaCl、pH6.0)1mlに懸濁した。EDC 0.4mgおよびスルホNHS 1.1mgをこの溶液に加え、室温で15分間ボルテックスした。次いでタンパク質(ヒスチジンタグ付きGFPまたはturbo YFP)9μlを加え、タンパク質と金とが完全に反応するように得られた溶液を2時間、暗所、室温でインキュベートした。この反応に使用した金コロイドとタンパク質との割合は、金コロイド上に存在するカルボン酸の数を得ることによって決定した。最初に金コロイド上に存在するカルボキシル基の数を、金粒子(球体と仮定)1個の表面積をカルボキシル基1個で占有される表面で割ることによって算出した(0.20nm2 Kimura,K.;Takashima,S.:Ohshima,H.Journal of Physical Chemistry B 2002,106,7260〜7266)。金コロイド溶液全体に存在するカルボキシル基の総数を得るために、この結果に存在する金コロイドの総数を掛けた。これを所与の量のタンパク質に存在するアミノ基の数と等しくした。これらの金コロイドは、金に存在するカルボン酸とタンパク質に存在するアミノ基との間のアミド結合の形成を介してタンパク質に付着する(Grabarek,Z.およびJ.Gergely(1990).Zero−length cross linking procedure with the use of active esters.Analytical Biochemistry 185(1):131〜135)。タンパク質分子およそ127285個が金ナノ粒子1個に繋ぎ止められている。
【0099】
細胞処置 3種の個々の試料を検査用に以下のとおり調製した:
金取り込みおよび細胞生存率の経時変化 以下の試料を24ウェル滅菌プレートに調製した。
i)単一BY2−E細胞500μl(対照)、
ii)単一BY2−E細胞500μl+GNP 20μl+フルオレセイン二酢酸(FDA)25μl+ヨウ化プロピジウム25μl;および
iii)他の処置は、細胞とのGNP 40、60、80μl、および上に記載の細胞生死判別染色を含む。処置した細胞(Ref)は、蛍光顕微鏡下で5、20、120分および最終的に18〜20時間後に検査した。
【0100】
金−SAMSAフルオレセイン処置 以下の試料を24ウェル滅菌プレートに調製した。
i)単一BY2−E細胞500μl(対照)、
ii)単一BY2−E細胞500μl+SAMSA−フルオレセイン20μl(対照);および
iii)単一BY2−E細胞500μl+Au−SAMSA−フルオレセイン20μl。
処置した細胞を、顕微鏡検査を実施する前に20分間、暗所、室温でインキュベートした。
【0101】
金−ヒスチジンタグ付きGFP処置 以下の試料を24ウェル滅菌プレートに調製した。
i)単一BY2−E細胞500μl(対照)、
ii)単一BY2−E細胞500μl+ヒスチジンタグ付きGFP 10μl(対照)、
iii)単一BY2−E細胞500μl+Au−ヒスチジンタグ付きGFP 20μl。
処置した細胞を、顕微鏡検査を実施する前に2時間、暗所、室温でインキュベートした。
【0102】
顕微鏡:Au−SAMSAフルオレセインおよびAu−ヒスチジンタグ付きGFPでの単一細胞実験の位相差顕微鏡法および蛍光顕微鏡法をLeica inverted fluorescence microscope(DAS)を使用して実施した。全ての実験は拡大率20×で実施した。FITC(フルオレセインイソチオシアネート)およびGFPフィルターをSAMSAフルオレセインおよびGFP単一細胞処置についてそれぞれ使用した。
【0103】
微分画像コントラスト(DIC)、共焦点および反射顕微鏡:これらの調査はUIUC(University of Illinois at Urbana Champaign)microscopy centerにおいてZeiss倒立顕微鏡で実施した。これら全ての方法について拡大率は63×に保った。共焦点用にFITC、GFPおよびYFPのフィルターをさまざまな細胞処置のために使用した。反射調査用には二色性鏡を透明なガラススライドに置き換え、発光フィルターを外した。
【0104】
画像取得:懸濁培養したタバコ細胞をX−Cite 120照明システムにつないだapotome光学切片システムを取り付けたZeiss Axiovert M 200顕微鏡(Carl Zeiss microimaging,Obercohen,Germany)を使用して画像化した。金粒子を635/20励起フィルターを通した水銀照明を使用して反射画像化設定下で画像化し、UVを遮断するためのGG420ガラス、KG5(1Rブロッカー)、50/50ビームスプリッターならびに励起および発光平行偏光板からなるIGS偏光フィルターセット(33001、Chroma Technology Corp.,Rockingham,VTから入手できる)を使用して画像化した。同時に、DIC/透過光画像を標準的DIC光学を使用して取得し、GFP−DNA被覆金粒子中のGFP(疑似カラー緑色)をバンドパスFITCフィルター(HQ480/40励起フィルター、Q505LP二色性鏡およびHQ535/50発光)で取得した。細胞を、高解像度画像化のために厚さ500ミクロンのチャンバーカバーガラスセット(Grace Bio−labs,Bend,OR)に分注した。大部分の画像は、細胞の大きさにより63×1.4 NA Planapochromat対物レンズまたは40×1.4でNA Planapochromat対物レンズで取得した(Carl Zeiss Microimaging,Obercohen,Germanyから入手できる)。露光時間は、各チャネル(すなわち、DIC、反射および/またはFITC)について設定し、ディメンジョン1388×1034ピクセルの高解像度Axiocam MRmモノクロカメラ(Carl Zeiss Obercohen,Germanyから入手できる)と共にAxiovision 4.6プログラムを使用して連続的に露光した。必要な場合には、解像度は1024×1024に設定し、およそ150〜250フレームを含む2〜5分間にわたる粒子の動態を解像するために可能な最高速度で経時的に連続な画像を得た。画像をAxiovision 4.6ギャラリーモジュールまたはAdobe Photoshop(Adobe Systems,San Jose,CA)のいずれかで作成した。
【0105】
経時変化およびGNP内部移行調査:粒子の取り込みおよびGNP濃度の細胞の正常性および生存率への影響を評価するために、経時的実験をクエン酸機能性付与GNP(直径90nm)とインキュベートしたBY2−E単一細胞系で実施した。種々の濃度のGNP(20、40、60、80μl)をこの実験に使用した。粒子は細胞との混合後5分間以内で内部移行され、粒子の蓄積は、細胞のサイトゾルおよび核におけるレベルの上昇を示すために2時間まで要した。検査した濃度中で高レベルの細胞生存率および細胞の活力は、FDAおよびPI(生/死細胞染色)手順で調査したとおり20μl処置で観察された。処置した全ての試料において細胞の平均生存率は98%に近いが、検査した最高濃度においてであり、FDAで染色された核は80μl処置では見られなかった。しかしこれらの染色されなかった核は、PIに反応せず、したがって細胞死ではないことを示唆していた。この結果は、最高濃度の粒子が、処置の20時間後に細胞が休眠する場合があるが、まだ生きている程度までの内部分布を生じ得ることを示唆する。
【0106】
【化1】

【0107】
反射顕微鏡追跡調査:金タンパク質(GFP/YFP)コンジュゲートで処置した単一BY2/NT1タバコ細胞での反射調査は、細胞内の金ナノ粒子の存在を示す。これを同様の条件下で暗くなって見えた未処置の対照単一BY2細胞と比較した、図8に示すとおり。明るい反射を発している単独の金ナノ粒子が観察された、図8に示すとおり。これは、これらの壁を有するBY2細胞による金ナノ粒子の取り込みの明らかな証拠である。
【0108】
金−SAMSAフルオレセイン実験:DASで実施した位相差実験は、対照単一細胞について観察されたシルバーコントラストと比較して、処置細胞についての細胞内腔および核の鮮黄色の染色を明らかにした。多数の細胞についてプラズマリシスの条件下でも細胞膜が細胞壁から離れ、図9に示すとおり細胞の部分的なまたは完全な原形質分離を示唆するそれらの間の空間が生じた。SAMSAフルオレセイン単独で処置した単一細胞の共焦点蛍光実験下で観察した場合そのような細胞は、細胞質および核に蛍光を示すが未処置の対照細胞においては暗く見えた。SAMSAフルオレセイン染色単独で処置した細胞もいくらかの壁での蛍光を示したが細胞内には示さなかった。これは、SAMSAフルオレセインがそれ自体は細胞内に内部移行されず、金ナノ粒子がその取り込みのための担体の様に作用することを意味する。
【0109】
金−ヒスチジンタグ付きGFP実験:GNPを介する未処置細胞へのタンパク質の送達を確立するために本発明者らは蛍光顕微鏡を使用してGNPへのGFPの付着を確認した、図10に示すとおり。ヒスチジンタグ付きGFPで処置したBY2細胞の蛍光画像は、中央部に蛍光を示さない暗い細胞での細胞外蛍光を示す。これは、ヒスチジンタグ付きGFPがAu粒子を伴わずに細胞に添加される対照処置は、粒子を内部移行させないことを示唆する。細胞内へのタンパク質の取り込みを支持する証拠は、i)処置細胞の内部での蛍光の蛍光強度の増大、ii)対照細胞における暗い糸と比較して処置細胞における蛍光発光している細胞質糸(図11を参照されたい)。同様の観察結果がYFPテザーGNPにおいても見られ、未処置細胞での植物細胞へのこれらの蛍光タンパク質の明確な内部移行を示唆していた(図12を参照されたい)。
【0110】
プラズマライジング/染色、またはプログラム細胞死(PCD)様の細胞学的特徴を示す細胞に由来する単一細胞でのある一定レベルのバックグラウンド反射および自己蛍光が存在する。そのようなバックグラウンドの問題からタンパク質が内部移行した細胞を描写し、かつタンパク質内部移行を直接的な証拠で明確に証明するために個々の粒子または粒子の凝集レベルを注目および追跡する詳細な反射顕微鏡調査を実施した。この調査の結果は、タンパク質を伴う粒子の細胞内および核内への内部移行を明確に示した。しかし蛍光タンパク質を伴う粒子の数が増加して蓄積した細胞は、顕微鏡下で観察した場合にプラズマライズする傾向を有した。GFPまたはYFPのいずれかに繋ぎ止められた多量のGNPの蓄積によるタンパク質濃度の上昇が毒性レベルに達するか、または顕微鏡下での長時間の観察がそのような細胞に結果的に有害効果を有するROSを誘導する可能性がある。
【実施例7】
【0111】
シロイヌナズナ(Arabidopsis thaliana)栽培種コロンビア(Columbia)のT1後代における導入遺伝子のゲノムへの組込みのための分子分析および立証
シロイヌナズナトランスジェニック植物由来のゲノムDNAを6週齢の葉全体の材料からDNeasy Plant Mini kitを製造者の説明書(Qiagen Inc)に従って使用して抽出した。以下のYFPおよびPAT PCRプライマーを、野外噴霧レベルの4〜5×の除草剤Finaleに耐性のT1実生由来の鋳型ゲノムDNAを使用するPCR反応において使用した。
YFP
順方向プライマー:5’―TGTTCCACGGCAAGATCCCCTACG―3’(配列番号3)
逆方向プライマー:5’―TATTCATCTGGGTGTGATCGGCCA―3’(配列番号4)
PAT
順方向プライマー:5’―GGAGAGGAGACCAGTTGAGATTAG―3’(配列番号5)
逆方向プライマー:5’―AGATCTGGGTAACTGGCCTAACTG―3’(配列番号6)
【0112】
PATおよびYFP(Yellow Florescent tag、Evrogen)遺伝子産物についてのPCRは、ゲノム鋳型DNA 100ng、1×ExTaq反応緩衝液(TaKaRa Bio)、0.2mM dNTP、各プライマー10pmolおよび0.025単位/μL ExTaqを含有する混合物の合計反応容量50μL中で増幅した。以下のPCR条件を使用した:96℃、5分間を1サイクルおよび以下のPCRプログラム:94℃で15秒間、65℃で30秒間、72℃で1分間を31サイクルおよび産物合成を完了するための72℃で7分間の最終伸長を実施した。ゲルの画像をBio Rad Gel imaging Systemを使用して得た(図13および14)。増幅した断片をゲル精製キット(Qiagen Inc)を製造者の説明書に従って使用してゲル精製した。
【0113】
PCR断片をPAT順方向プライマーおよびYFP順方向を使用して、advanced Sanger sequencing technology(MWG Biotechnologies,Inc)で使用して配列決定し、配列をSequencherソフトウェアを使用して分析した。
【0114】
結果は、PATおよびYFP配列はナノ粒子を通じて送達され、量子ドットはDNA送達を介在することを示し、したがってT1植物のゲノムDNAにおける導入遺伝子の安定なゲノムへの組込みの明確な証拠を提供している。
【実施例8】
【0115】
JTNT1タバコ単一細胞壁を越えるQDの送達の促進
細胞壁を越えるQDの非侵襲性送達を検査した実施例7において論じられた手順に基づいて、いくつかのペプチドを表面に機能化した。細胞透過性ペプチド(CPP)/タンパク質導入ユニット(PTD)付着決定を以下に記載のとおりゲル電気泳動を介して実施した。
【0116】
ゲル電気泳動をQD−ペプチドコンジュゲートについて、ペプチドのQDへの付着を確認するために実施した。使用した試料は、QD−アミン(対照)、QD−アミン−R9、QD−アミン−Zein、QD−アミン−Pep1およびQD−アミン−MPGであった。R9、Zein、Pep1およびMPGはペプチドである。2%(w/v)アガロースゲルでTBE(1×、pH8)緩衝液中、120V、1時間実施した。QD−アミン−ペプチドは、電極の陰性端に向かって移動し、QDに付着したペプチドの強く陽性の性質の付着を示したが、QD−アミンは静止したままであり、塩基性Phでのゲル緩衝液の中和効果によるアミン基の弱い陽電荷を示した、図15に示すとおり(レーン1:QD−アミン、2:QD アミン−R9、3:QD−アミンΥ−Zein、4:QD アミン−Pep1、5:QD−アミン−MPG)。
【0117】
ペプチドを細胞への内部移行について検査し、細胞および区画内の粒子の内部移行レベルを追跡するための方法として細胞内QDの発光を使用した。無傷の壁を有するJTNT1単一細胞を標的細胞としてこれらの実験において使用した。表1は、試料の処置を示す。細胞を顕微鏡下で追跡した。
【0118】
顕微鏡法は、試料の調製後1分間以内にSpinning Disk Confocal microscope(Andor Technology Revolution System)で行った。励起フィルターは488nmに設定し、発光フィルターは650nm超に設定した。
【0119】
表1に示すとおり、対照プロトプラストおよびJTNT1は、QDについて使用した発光波長で自己蛍光を示さなかった。顕著な細片(破損細胞の一部)がこれらの試料のそれぞれにおいて観察された。試料4、5、6および7は、単一細胞またはプロトプラスト内にQDの内部移行を示さなかった。試料8および9は、壁がある単一細胞およびプロトプラストの両方において核の周辺でのQDの明確な存在を示した。これは、核局在化シグナル(NLS)を有するQD上の細胞透過性ペプチドの存在によるものであった。しかしQDに繋ぎ止められたy−Zeinを有する試料番号6および7は、細胞内へのQD内部移行を示さなかった。これは細胞透過性ペプチド(CPP)またはタンパク質導入ユニット(PTD)によってQDが細胞に取り込まれたことを示唆し、アミンによってのみ機能化され、CPP/PTDでは機能化されなかったQDと同様にy−Zeinは内部移行されなかった。
【0120】
【表1】
【0121】
このデータは、無傷の機能性細胞の細胞壁を越えてQDを取り込む核局在化シグナル(NLS)を有するCPP/PTDに繋ぎ止められた量子ドットの細胞内部移行のSpinning Disk Confocal microscope(Andor Technology Revolution System)を介しての証拠を実証している。QDの核局在化は、酵素処理によって細胞壁を取り除かれたプロトプラストを基とする細胞の内部移行において観察されたとおり、試料9および細胞壁の非存在時に細胞壁を越えるQDの核局在化が可能である。したがって細胞壁の単なる存在は量子ドットの内部移行を妨げず、無傷の細胞壁を有する植物細胞における粒子の侵入がCPP/PTDで非侵襲的に実証されることを証明する。
【0122】
本発明は特定の実施形態で記載されているが、本発明は本開示の精神および範囲の中でさらに変更され得る。したがって本出願は、その一般的原理を使用する本発明の任意の変更、使用または適用に及ぶことを意図する。さらに本出願は、本発明に関連し、かつ添付の特許請求の範囲およびその等価物の限定内にある当技術分野での既知または慣用の実施内にある、本開示からのそのような逸脱を包含することを意図する。
【技術分野】
【0001】
優先権主張
本出願は、METHODS FOR TRANSFERRING MOLECULAR SUBSTANCES INTO PLANT CELLSについての、2007年10月5日出願の米国仮特許出願第60/978,059号出願日の利益を主張する。
【背景技術】
【0002】
ナノ粒子は、細胞へのDNA送達における使用について開発されてきた独自の特性を有する。調査された全てのナノ粒子の中で金(Au)ナノ粒子は、それらの細胞毒性の低さおよび生物学的に重要な種々のリガンドでの機能化の容易さから送達のための優れた候補になるようである。Auナノ粒子の慣用の合成では、負に荷電した(例えばクエン酸コーティング)表面を生じる。塩基が部分的に非コイル状になるように十分に可動性である場合があるプラスミドDNAは金ナノ粒子(「GNP」)にさらされ得る。これらの部分的に非コイル状である条件下でDNA主鎖上の負電荷は、塩基と金ナノ粒子との間の誘引性ファンデルワールス力がプラスミドDNAの金ナノ粒子の表面への付着を生じるのに十分であるように十分に離れている場合がある。
【0003】
金属ナノ粒子に加えて、サイズ範囲3〜5nm内の半導体ナノ粒子(例えば量子ドット)(「QD」)も、細胞内に分子を送達するための担体として使用されている。DNAおよびタンパク質はQD表面に付着するリガンドに連結できる(Patolsky,F.ら、J.Am.Chem.Soc.125,13918(2003)を参照されたい)。カルボン酸またはアミンで被覆されたQDは、標準的な生体コンジュゲーションプロトコールを使用することによってチオール基、(例えばDubertret B.ら、Science 298,1759(2002)、Akerman,M.E.,W.C.W.Chan,P.Laakkonen,S.N.Bhatia,E.Ruoslahti,Proc.Natl.Acad.Sci.U.S.A.99,12617(2002)、Mitchell,G.P.,C.A.Mirkin,R.L.Letsinger,J.Am.Chem.Soc.121,8122(1999)を参照されたい)またはN−ヒドロキシサクシニミル(NHS)エステル基(例えばPinaud,F.,D.King,H.−P.Moore,S.Weiss,J.Am.Chem.Soc.126,6115(2004)、Bruchez,M..,M.Moronne,P.Gin,S.Weiss,A.P.Alivisatos,Science 281,2013(1998)を参照されたい)を含む分子と交差結合できる。代替法は、ストレプトアビジンで被覆されたQDとビオチン化タンパク質、オリゴまたは抗体のコンジュゲーションである(例えばDahan M.ら、Science 302,442(2003)、Pinaud,F.,D.King,H.−P.Moore,S.Weiss,J.Am.Chem.Soc.126,6115(2004)、Dahan M.ら、Science 302.442(2003)、Wu.X.Y.,ら、Nature Biotechnol.21,41(2003)、Jaiswal,J.K.,H.Mattoussi,J.M.Mauro,S.M.Simon,Nature Biotechnol.21,47(2003)およびMansson,A.ら、Biochem.Biophys.Res.Commun.314,529(2004)を参照されたい。
【0004】
ナノ粒子は、プラスミドDNAを種々の動物細胞に送達するために使用されている。DNAで被覆されたナノ粒子を細胞壁を有さない細胞と共にインキュベートすると、細胞がナノ粒子を取り込み、DNAにコードされた任意の遺伝子の発現を開始することが見出されている。通常は細胞壁を有する細胞へのナノ粒子の送達が望ましい場合は、植物のプロトプラストへの粒子の添加の前に細胞壁が取り除かれる(Torney,F.ら、Nature Nanotechnol.2,(2007)を参照されたい。植物細胞において、細胞壁は外来的に添加された分子の送達に対する障壁として存在する。遺伝子銃(微粒子銃)、微量注入法、電気穿孔法およびアグロバクテリウムなどの多数の侵襲的方法は、これらの壁を有する植物細胞への遺伝子および小分子の送達を達成するために使用されているが、タンパク質の送達は微量注入法によってだけ達成されている。植物細胞の細胞壁の存在下での小分子およびタンパク質の送達は、いまだ探究されておらず、in vitroおよびin vivo操作のための無傷の植物細胞/組織または器官で展開される実用化技術を開発するために有益である。
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明は、ナノ粒子を使用して、細胞壁を有する細胞に分子性物質を非侵襲的に送達する方法に関する。
【課題を解決するための手段】
【0006】
以下の実施形態は、典型的かつ例示的であるシステム、手段および方法と共に記載され、範囲を限定しない。
【0007】
本発明によれば、細胞壁を含む植物細胞に目的の分子を導入する方法であって、細胞壁を有する植物細胞とナノ粒子および目的の分子とを接触させて置くステップと、ナノ粒子および目的の分子の細胞への取り込みを可能にするステップとを含む方法が提供される。
【0008】
細胞壁を有する植物細胞に目的の分子を導入する方法であって、細胞壁を有する植物細胞とナノ粒子および病害を治療できる分子とを接触させて置くステップと、ナノ粒子および病害を治療できる分子の細胞への取り込みを可能にするステップとを含む方法がさらに提供される。
【0009】
上に記載の例示的態様および実施形態に加えて、さらなる態様および実施形態が以下の記載を考慮することにおいて明らかになるであろう。
【図面の簡単な説明】
【0010】
【図1】共焦点画像化システムを取り付けた微分干渉顕微鏡を使用して観察したBY2の単一細胞を示す写真である(パネルA)。色素体(アミロプラスト)を強調するためにI2KIで染色されたBY2バリアント由来の単一細胞の光学顕微鏡観察を示す図である(パネルB)。
【図2】図2パネルAは光学顕微鏡で観察した最少培地および5%二酸化炭素中で維持されたタバコ(NT1)の光独立栄養細胞を示す図であり、顕著な葉緑体が見られる。図2パネルB蛍光顕微鏡下で赤色の自己蛍光を発する活性葉緑体を有して観察される同様のNT1細胞を示す図である。
【図3】SAMSAフルオレセイン単独で、およびSAMSAフルオレセインで被覆されたGNPで処置したBY2懸濁凝集物を示す図である。図3パネルAは、SAMSAフルオレセイン単独で処置した細胞のDIC画像を示し、図3パネルBは、同じ細胞の蛍光画像を示す。図3パネルCはSAMSAフルオレセインで被覆されたGNPで処置した細胞のDIC画像を示し、図3パネルDは、SAMSAフルオレセインで被覆されたGNPで処置した細胞の蛍光画像を示す。核(Nu)および細胞壁(CW)の位置が示されている。
【図4】SAMSAフルオレセインで被覆されたGNPで処置した単一細胞を高拡大率で示す図である。パネルBは、核における多数のGNPの存在を示す。パネルAは、パネルBにおいて示されたのと同じ核の異なる焦点平面での明視野観察を示す。
【図5】SAMSAフルオレセインで被覆されたGNPで処置した光独立栄養細胞を示す図である。パネルAは、細胞壁の内側に並ぶ多量の葉緑体を有する3〜4個の細胞クラスター中のガラス質細胞を示す。パネルBは、葉緑体中のナノ粒子の蓄積を示す。パネルCおよびDは、蛍光顕微鏡を使用するより高い拡大率での単一の葉緑体を示す。ナノ粒子は、葉緑体の膜薄層形成において観察され、赤色の自己蛍光を発する葉緑素色素中に分散している。
【図6】ナノ粒子を含む細胞の反射顕微鏡および蛍光顕微鏡の画像である。図6のパネルAは、GNPが優先的に観察される反射画像を示す。パネルBは、赤色の自己蛍光を発する葉緑体の背景中で蛍光を発する粒子を示す。反射および蛍光のマージした画像をパネルCに示し、黄色蛍光を発する粒子は、葉緑体の境界内にある。
【図7】本発明の実施形態により考えられる1つの形質転換スキームを図示する。
【図8】処置の2時間後に反射顕微鏡を通して観察されるGFPの細胞内部移行を示す図である。パネルAおよびA1は、DIC顕微鏡下での未処置対照細胞(パネルA)およびDIC顕微鏡下で観察されるGFPテザーAu−NP処置細胞(パネルA1)を示す;パネルBおよびB1は、反射顕微鏡下での対照細胞(パネルB)および、反射顕微鏡下で観察されるGFPテザーAu−NP処置細胞(パネルB1)を示し、反射されたAu−NPにより粒子内部移行を示している;パネルCおよびC1は、対照細胞のDIC顕微鏡と反射顕微鏡とを重ね合わせた画像(パネルC)および、処置細胞のDIC顕微鏡と反射顕微鏡とを重ね合わせた画像(パネルC1)を示す;パネルDおよびD1は、背景に粒子が観察されない対照細胞の反射反転画像(パネルD)および、非常に明白な粒子の内部移行を示す処置細胞の反射反転画像(パネルD1)を示す。
【図9】単一細胞でのSAMSA染料で被覆されたGNPの内部移行を示す図である。パネルAは、蛍光を示す細胞壁および培地を有するが、染料の内部移行を示さない、蛍光染色した単一細胞を示し、パネルBは、DIC顕微鏡下での単一細胞を示し、パネルCは、粒子および1時間までの長時間UV光にさらしたプラズマライズ細胞と共にだけ内部移行したフルオレセインでの、サイトゾルおよび核へのナノ粒子(GNP 150nm)の内部移行を示す位相差画像を示す。
【図10】単一細胞を混合する前の、蛍光を発するGFP分子を有するAu−GFPコンジュゲートを示す図である。パネルA(FITC)、B(明視野)、C(反射)、D(パネルA+B+C):蛍光顕微鏡を通して観察されるGFP蛍光発光Au−GNP、インキュベーション2時間後だが細胞を混合する前。同様の蛍光発光粒子が核で反射を示す粒子について観察できた(図8を参照されたい)。
【図11】ナノ粒子(GNP90nm)が介在したBY2−E細胞系へのGFPの細胞内部移行を示す図である。パネルAは活性細胞質糸(cytoplasmic strand)を有する分裂対照細胞を示し(位相差画像)、パネルBは、FITCフィルターを通して検査した時の、パネルAと同じ細胞を示し、そこで、細胞質中の非緑色色素体由来および分裂核に関連するこれらの色素体由来の自己蛍光が周辺部を形成する;細胞質糸および細胞の周辺部に近い細胞質は自己蛍光を示さない;パネルCは、GNP(FITC)に付着していないGFPで処置した対照BY2細胞を示し、該細胞はGFP取り込みを示さず、GFPは細胞を取り囲んでいるが内部移行していない;パネルDは、FITCフィルターを通して観察される、パネルBでの対照と比較してGFPの内部移行を示す細胞質周辺部、細胞質糸および核を伴うGNP介在GFP内部移行を示す。
【図12】細胞のインキュベーションの2時間後にGNP介在YFP内部移行を示すBY2−E単一細胞株を示す図である。パネルA(FITC)、B(ローダミン)、C(DIC)、D(パネルA+B)、E(パネルA+B+C)、F(反射画像反転):蛍光顕微鏡を通して観察されたYFP内部移行。黄色矢印は、サイトゾルにYFPを有する生きた単一細胞での内部移行(核での拡散および濃縮)を示す。橙色矢印は、細胞内の収縮したプロトプラスト塊が細胞へのYFPの内部移行を示唆する強い蛍光を示すプラズマライズ細胞での内部移行を示す。この細胞は、近接して位置する生細胞と同じ共焦点面だが、他の生細胞の下に見出される。高レベルの粒子およびYFP蛍光が蓄積している細胞は、蛍光顕微鏡下での長時間の検査において細胞死を示す。
【図13】および
【図14】は、PATおよびYFP増幅遺伝子産物が増幅されたゲル画像を示す図である。
【図15】ペプチドのQDへの付着を確認するためにQD−ペプチドコンジュゲートについて実施された電気泳動を示す図である。
【図16】プラスミドpDAB3831を示す図である。
【発明を実施するための形態】
【0011】
以下の記載および表において、多数の用語が使用される。明細書および特許請求の範囲の明確かつ一貫した理解を提供するために、そのような用語に与えられる範囲を含めて以下の定義が提供される。
【0012】
戻し交雑 戻し交雑は、育種家がハイブリッド後代をもとの親(例えばF1ハイブリッドの親の遺伝型の内の1つを有する第1世代ハイブリッドF1)の内の1つと繰り返し交雑させる工程であり得る。
【0013】
胚 胚は、成熟種子中に含有される小植物体であり得る。
【0014】
ナノ粒子 通常100nm未満である、少なくとも1つのナノスケールの大きさを有する顕微鏡的粒子。本発明における使用に適するナノ粒子は、1nm〜0.4umの大きさを有し得る。量子ドットは、1nm〜10nmの、好ましくは2〜4nmのメジアン径を有し得る。ナノ粒子は、金ナノ粒子、金被覆ナノ粒子、多孔性ナノ粒子、メソ多孔性ナノ粒子、シリカナノ粒子、ポリマーナノ粒子、タングステンナノ粒子、ゼラチンナノ粒子、ナノシェル、ナノコア、ナノスフェア、ナノロッド、磁性ナノ粒子およびそれらの組合せから選択され得る。
【0015】
量子ドット 量子ドットは、3つ全ての空間方向における伝導帯電子、価電子帯正孔または励起子(伝導帯電子と価電子帯正孔との束縛された対)の動きを限定する半導体ナノ構造である。限定は、静電ポテンシャル(外部電極、不純物添加、歪み、不純物によって作り出される)、異なる半導体物質の間の境界面の存在(例えばコア−シェルナノ結晶系)、半導体表面の存在(例えば半導体ナノ結晶)またはこれらの組合せによる場合がある。量子ドットは、不連続の量子化されたエネルギースペクトルを有し得る。対応する波動関数は、量子ドットに空間的に局在するが、結晶格子の複数のピリオドに及ぶ。量子ドットは、少数の限定された数の(ほぼ1〜100程度)の伝導帯電子、価電子帯正孔または励起子(すなわち限定された数の素電荷)を含む。
【0016】
グリフォセート耐性 グリフォセート投与に対する耐性は、その投与量までのグリフォセートでの植物の生存能力(すなわち植物が枯らされない)を意味する。いくつかの場合において、耐性植物は一時的に黄変または他にいくつかのグリフォセート誘発性障害(例えば分けつ過剰および/または生育阻害)を呈するが、回復する。
【0017】
安定化 安定化は、同じ品種の自殖植物の一世代から次世代へ繁殖により伝えられる植物の特徴を意味する。
【0018】
取り込み 取り込みは、例えば金または量子ドットであるナノ粒子などの粒子の細胞壁または細胞膜を超える移動であって、粒子が取り込まれる細胞以外の何かによって粒子に与えられるモーメントの結果として単独に生じるのではない移動を意味する。粒子に与えられるモーメントの結果として単独で細胞壁または細胞膜を超える粒子の移動を生じる装置または方法の非限定的例は、微粒子銃、遺伝子銃、微量注入法および/またはインペールフェクション(impalefection)技術である。
【0019】
本発明の実施形態によれば、細胞壁を含む植物細胞に目的の分子を導入する方法であって、含有するナノ粒子および目的の分子を植物細胞と接触させて置くステップと、植物細胞壁を越えるナノ粒子の取り込みを可能にするステップとを含む方法が提供され得る。本発明の具体的な態様において、ナノ粒子は、任意のナノ粒子であってよく、目的の分子を可逆的または不可逆的に含有し、目的の分子で被覆され、または他の形で目的の分子と結合し、かつ/または目的の分子を保有し得る。特定の実施形態において、目的の分子は、細胞壁を有する植物細胞と接触する前にまたは細胞壁を有する植物細胞へのナノ粒子の導入と同時にナノ粒子に導入され得る。本発明の実施形態において使用され得るナノ粒子の例として、それだけに限られないが、金、量子ドット、金被覆ナノ粒子、多孔性ナノ粒子、メソ多孔性ナノ粒子、シリカナノ粒子、ポリマーナノ粒子、タングステンナノ粒子、ゼラチンナノ粒子、ナノシェル、ナノコア、ナノスフェア、ナノロッド、磁性ナノ粒子および/またはそれらの組合せが挙げられる。
【0020】
本発明の実施形態によれば、細胞壁を有する植物細胞は、無傷かつ完全な細胞壁を有する任意の植物細胞であってよい。細胞壁を有する細胞の例として、それだけに限られないが、藻類、タバコ、ニンジン、トウモロコシ、セイヨウアブラナ、ナタネ、ワタ、パーム、ピーナッツ、ダイズ、サトウキビ、オリザ種(Oryza sp)、シロイヌナズナ種(Arabidopsis sp)およびトウゴマ種(Ricinus sp)が、好ましくはタバコ、ニンジントウモロコシ、ワタ、セイヨウアブラナ、ダイズおよびサトウキビが、より好ましくはタバコおよびニンジンが挙げられる。本発明の実施形態は、任意の組織または、それだけに限られないが、胚、分裂組織細胞、カルス、花粉、葉、葯、根、根端、花、種子、さや、茎および組織培養物が挙げられるそれらが見出されるあらゆる箇所由来の細胞壁を有する細胞を含み得る。
【0021】
本発明の実施形態において目的の分子は、本発明により植物細胞に送達され得る任意の分子であり得る。目的の分子または目的の分子の構成要素は、それだけに限られないが、核酸、DNA、RNA、RNAi分子、遺伝子、プラスミド、コスミド、YAC、BAC、ポリペプチド、酵素、ホルモン、糖ペプチド、糖類、脂肪、シグナル伝達ペプチド、抗体、ビタミン、メッセンジャー、セカンドメッセンジャー、アミノ酸、cAMP、薬剤、除草剤、殺菌剤、抗生物質および/またはそれらの組合せを含み得る。
【0022】
本発明の実施形態は、病害の予防または治療のための方法を含む。非限定的例の実施形態は、殺菌剤、抗生物質および/または他の薬剤のそれを必要とする細胞への本発明の方法を使用する送達を含む。
【0023】
本発明の具体的な実施形態において、ナノ粒子の表面は、例えば標的化された取り込みを可能にするように、またはナノ粒子の表面への他の物質の可逆的もしくは不可逆的結合を可能にするように機能化され得る。非限定的な例として、ナノ粒子(例えば金ナノ粒子または量子ドット)の表面は、例えばさらに機能化または誘導体化され得るアルカンチオレートである自己組織化単分子層で機能化され得る。さらなる非限定的例においてナノ粒子の表面は、それ自体がさらに機能化または誘導体化され得るリンカーで誘導体化され得る。一実施形態においてナノ粒子は、PEG化され得る、他の実施形態においてナノ粒子は、1つまたは複数のコア(活性もしくは不活性)、ステリック被覆(活性もしくは不活性)、切断可能な結合、および/または標的化分子もしくはリガンドを含み得るまたはそれで多機能化され得る。
【0024】
本発明の態様において、ナノ粒子は、細胞の種々の部分に取り込まれ得る。ナノ粒子が取り込まれ得る位置の例として、それだけに限られないが、サイトゾル、核、液胞膜、色素体、エチオプラスト、有色体、白色体、エライオプラスト、プロテイノプラスト、アミロプラスト、葉緑体、および二重膜の内腔が挙げられる。本発明の他の実施形態において細胞壁を有する細胞へのナノ粒子取り込みは、シンプラスト的またはアポプラスト的経路を介して生じ得る。
【0025】
本発明の追加的実施形態は、植物細胞が本発明の方法を介して導入された1つまたは複数の核酸を有する、遺伝子改変された植物細胞およびそれらを生成する方法を含む。実施形態の一例において、目的の遺伝子および選択可能なマーカーを含むプラスミドは、本発明によるナノ粒子を介して細胞壁を有する植物細胞に導入され得る。さらなる実施形態において、目的の遺伝子および/または選択可能なマーカーを安定に組み込まれている安定形質転換体が、選択され得る。代替の実施形態においてこのとき目的の遺伝子を含む植物細胞は、目的の分子を含む他の細胞を生成するために増殖され得る。他の実施形態においてこのとき目的の分子を含む植物細胞は、目的の分子を含む植物体全体を再生するために使用できる再生可能な細胞であり得る。
【0026】
他の態様において、本発明は、組織培養における使用のための目的の分子を含む再生可能な植物細胞を作製する方法を提供する。組織培養は、好ましくは、再生可能な細胞と実質的に同じ遺伝子型を有する植物を再生可能な細胞として、再生することができる。そのような組織培養における再生可能な細胞は、胚、プロトプラスト、分裂組織細胞、カルス、花粉、葉、葯、根、根端、花、種子、さやまたは茎であり得る。さらに本発明の実施形態は、本発明の組織培養から再生された植物を提供する。
【0027】
代替として、本発明は、細胞壁を有する植物細胞に所望の形質を導入する方法であって、植物細胞に所望の形質を提供できるナノ粒子および目的の分子を細胞と接触させて置くステップと、細胞壁を越えるナノ粒子の取り込みを可能にするステップとを含む方法を提供する。所望の形質の例として、それだけに限られないが、雄性不稔、除草剤耐性、虫害耐性、ならびに細菌性病害、真菌性病害および/またはウイルス性病害への耐性から選択される形質が挙げられる。
【0028】
本発明のさらなる態様は、所望の形質または目的の分子を含む安定化した植物系の産生方法であって、所望の形質または目的の分子が植物細胞壁を越えるナノ粒子の取り込みによって最初に導入され得る方法を提供する。安定化した植物系の産生方法は、当業者に十分に既知であり、それだけに限られないが、自家受粉、戻し交雑、ハイブリッド産生、個体群への交雑などの技術を含み得る。細胞壁を越えるナノ粒子の取り込みによって植物細胞(またはその祖先)に最初に導入される所望の形質または目的の分子を含む全ての植物および植物細胞は、本発明の範囲内である。有利には、細胞壁を越えるナノ粒子の取り込みによって植物または細胞(もしくはその前身)に最初に導入される所望の形質または目的の分子を含む植物細胞は、優れた特徴を有する第一世代(F1)ハイブリッド細胞、種子および/または植物を産生するために他のさまざまな植物細胞との交雑に使用され得る。
【0029】
目的の分子が1つまたは複数の遺伝子を含む実施形態において、(1つまたは複数の)遺伝子は、優性または劣性アレルであり得る。例として(1つまたは複数の)遺伝子は、除草剤耐性、虫害耐性、細菌耐性への耐性、真菌耐性、ウイルス性病害耐性、雄の妊性、雄性不稔、増強された栄養価および産業利用などの形質を付与する。
【0030】
特定のタンパク質またはRNA産物(例えばRNAi)をコードする遺伝子の単離および特徴付けを可能にしている分子生物学的技術の到来により植物生物学の分野の科学者らは、特定のやり方で細胞の形質を変えるために外来遺伝子、または天然もしくは内在遺伝子(おそらく異なるプロモーターによって駆動される)の追加的もしくは改変バージョンを含有し、かつ発現するために細胞のゲノムを操作することに強い関心を発展させた。そのような外来の追加的および/または改変遺伝子は、本明細書において総称として「導入遺伝子」と称される。最近の15から20年間にわたってトランスジェニック細胞を産生するためのいくつかの方法が開発されており、特定の実施形態において、本発明は、細胞壁を越えるナノ粒子の取り込みを介して細胞壁を有する細胞に導入遺伝子を導入することにより形質転換型の細胞およびそれらを産生する方法に関する。本発明の実施形態において導入遺伝子は、発現ベクターに含有され得る。
【0031】
細胞形質転換は、特定の細胞で機能する発現ベクターの構築に関与し得る。そのようなベクターは、調節エレメント(例えばプロモーター)の制御下の、またはそれに機能的に結合した遺伝子を含むDNAを含み得る。発現ベクターは、1つまたは複数のそのように機能的に結合した遺伝子/調節エレメントの組合せを含有できる。(1つまたは複数の)ベクターは、プラスミドの形態であってよく、細胞壁を有する植物細胞の遺伝物質に(1つまたは複数の)導入遺伝子を組み込むための本明細書において記載の形質転換方法を使用して形質転換された細胞を提供するために単独でまたは他のプラスミドとの組合せで使用され得る。
【0032】
ナノ粒子を介する取り込みのための発現ベクター:マーカー遺伝子
発現ベクターは、マーカーを含有する形質転換細胞がネガティブ選択(すなわち選択可能なマーカー遺伝子を含有しない細胞の増殖阻害)またはポジティブ選択(すなわち遺伝子マーカーによってコードされる産物についての選別)のいずれかによって回収されるようにする調節エレメント(例えばプロモーター)に機能的に結合した少なくとも1つの遺伝子マーカーを含むことができる。形質転換のための多数の選択可能なマーカー遺伝子が形質転換の技術分野において十分に既知であり、例えば、抗生物質もしくは除草剤であり得る選択的化学物質を代謝的に解毒する酵素をコードする遺伝子、または阻害剤に非感受性であり得る改変された標的をコードする遺伝子を含む。いくつかのポジティブ選択方法も当技術分野において既知である。
【0033】
植物の形質転換に適した慣用される1つの選択可能なマーカー遺伝子としてカナマイシンへの耐性を付与する、植物調節シグナルの制御下でのネオマイシンホスホトランスフェラーゼII(nptII)遺伝子が挙げられる。例えばFraleyら、Proc.Natl.Acad.Sci.U.S.A.,80:4803(1983)を参照されたい。他の慣用される選択可能なマーカー遺伝子は、抗生物質ハイグロマイシンへの耐性を付与するハイグロマイシンホスホトランスフェラーゼ遺伝子であり得る。例えばVanden Elzenら、Plant Mol.Biol.,5:299(1985)を参照されたい。
【0034】
抗生物質への耐性を付与する細菌由来の追加的な選択可能なマーカー遺伝子として、ゲンタマイシンアセチルトランスフェラーゼ、ストレプトマイシンホスホトランスフェラーゼ、アミノグリコシド−3’−アデニルトランスフェラーゼおよびブレオマイシン耐性決定因子が挙げられる。Hayfordら、Plant Physiol.86:1216(1988)、Jonesら、Mol.Gen.Genet.,210:86(1987)、Svabら、Plant Mol.Biol.14:197(1990)、Hilleら、Plant Mel.Biol.7:171(1986)を参照されたい。他の選択可能なマーカー遺伝子はグリフォセート、グルフォシネートまたはブロモキシニルなどの除草剤への耐性を付与する。Comaiら、Nature 317:741〜744(1985)、Gordon−Kammら、Plant Cell 2:603〜618(1990)およびStalkerら、Science 242:419〜423(1988)を参照されたい。
【0035】
植物の形質転換に適する他の選択可能なマーカー遺伝子は、細菌由来ではない。これらの遺伝子として、例えばマウスジヒドロ葉酸還元酵素、植物5−エノールピルビルシキミ酸−3−リン酸合成酵素および植物アセト乳酸合成酵素が挙げられる。Eichholtzら、Somatic Cell Mol.Genet.13:67(1987)、Shahら、Science 233:478(1986)、Charestら、Plant Cell Rep.8:643(1990)を参照されたい。
【0036】
植物の形質転換に適するマーカー遺伝子の他のクラスは、抗生物質などの毒性物質への耐性についての形質転換細胞の直接的な遺伝的選択よりも形質転換されたと仮定される植物細胞の選別を必要とする。これらの遺伝子は、特定の組織における遺伝子発現の空間分布を定量または可視化するために特に有用であり、遺伝子発現の調査のためにそれらが遺伝子または遺伝子調節配列に融合され得ることからしばしばレポーター遺伝子と称される。形質転換細胞を選別するために慣用される遺伝子として、β−グルクロニダーゼ(GUS)、β−ガラクトシダーゼ、ルシフェラーゼおよびクロラムフェニコールアセチルトランスフェラーゼが挙げられる。Jefferson,R.A.,Plant Mol.Biol.Rep.5:387(1987)、Teeriら、EMBO J.8:343(1989)、Konczら、Proc.Nalt.Acad.Sci U.S.A.84:131(1987)、DeBlockら、EMBO J.3:1681(1984)を参照されたい。
【0037】
近年、植物組織の破壊を必要とせずにGUS活性を可視化するためのin vivoでの方法が利用可能になっている。Molecular Probes publication 2908、Imagene Green.TM.、1〜4ページ(1993)およびNalewayら、J.Cell Biol.115:151a(1991)。しかしGUS活性を可視化するためのこれらのin vivo法は、低感度、蛍光の高いバックグラウンドおよび選択可能なマーカーとしのルシフェラーゼ遺伝子の使用に伴う制限から形質転換細胞を回収するために有用であるとは証明されていない。
【0038】
最近になり、蛍光タンパク質をコードする遺伝子(例えばGFP、EGFP、EBFP、ECFPおよびYFP)が、原核細胞および真核細胞における遺伝子発現のためのマーカーとして利用されている。Chalfieら、Science 263:802(1994)を参照されたい。蛍光タンパク質および蛍光タンパク質の変異体も選択可能なマーカーとして使用できる。
【0039】
ナノ粒子を介する取り込みのための発現ベクター:プロモーター
発現ベクターに含まれる遺伝子は、調節エレメント、例えばプロモーターを含むヌクレオチド配列によって駆動されねばならない。プロモーターのいくつかの種類は、単独またはプロモーターとの組合せで使用できる他の調節エレメントと同様に形質転換の技術分野においていまや十分に既知である。
【0040】
本明細書において使用する「プロモーター」は、転写の開始点より上流であってよく、RNAポリメラーゼおよび転写を開始する他のタンパク質の認識および結合に関与できるDNAの領域に関連する。「植物プロモーター」は、植物細胞において転写を開始できるプロモーターであり得る。発生調節下のプロモーターの例として、葉、根、種子、繊維、木部導管、仮導管または厚壁組織などの特定の組織において転写を優先的に開始するプロモーターが挙げられる。そのようなプロモーターは、「組織優先」プロモーターと称される。特定の組織においてだけ転写を開始するプロモーターは、「組織特異的」と称される。「細胞型」特異的プロモーターは、1つまたは複数の器官、例えば根または葉の維管束細胞での特定の細胞型において主に発現を駆動する。「誘導可能な」プロモーターは、環境制御下にある場合があるプロモーターであり得る。誘導可能なプロモーターによる転写に影響し得る環境条件の例として、嫌気性条件または光の存在が挙げられる。組織特異的、組織優先、細胞型特異的および誘導可能なプロモーターは、「非構成的」プロモーターのクラスを構成する。「構成的」プロモーターは、ほとんどの環境条件下で活性であってよいプロモーターであり得る。
【0041】
A.誘導可能なプロモーター
誘導可能なプロモーターは、細胞での発現のための遺伝子に機能的に結合され得る。場合により、誘導可能なプロモーターは、細胞での発現のための遺伝子に機能的に結合され得るシグナル配列をコードするヌクレオチド配列に機能的に結合され得る。誘導可能なプロモーターにより転写速度は、誘導剤に反応して増大する。
【0042】
誘導可能なプロモーターは、本発明において使用され得る。Wardら、Plant Mol.Biol.22:361〜366(1993)を参照されたい。例示的な誘導可能なプロモーターとして、それだけに限られないが、銅に反応するACEI系由来のもの(Mettら、PNAS 90:4567〜4571(1993))、ベンゼンスルフホンアミド除草剤緩和剤に反応するトウモロコシ由来のIn2遺伝子(Hersheyら、Mol.Gen Genetics 227:229〜237(1991)およびGatzら、Mol.Gen Genetics 243:32〜38(1994))、ならびにTn10由来のTet抑制因子(Gatzら、Mol.Gen Genetics 227:229〜237(1991))が挙げられる。特に有用な誘導可能なプロモーターは、植物が通常は反応しない誘導剤に反応するプロモーターであり得る。例示的な誘導可能なプロモーターは、ステロイドホルモン遺伝子由来の誘導可能なプロモーターであることができ、その転写活性はグルココルチコステロイドホルモンによって誘導され得る。Schenaら、Proc.Natl.Acad.Sci.U.S.A.88:0421(1991)。
【0043】
B.構成的プロモーター
構成的プロモーターは、細胞での発現のための遺伝子に機能的に結合され得る、または構成的プロモーターは細胞での発現のための遺伝子に機能的に結合され得るシグナル配列をコードするヌクレオチド配列に機能的に結合され得る。
【0044】
さまざまな構成的プロモーターが、本発明において利用され得る。例示的な構成的プロモーターとして、それだけに限られないが、CaMV由来の35Sプロモーターなどの植物ウイルス由来のプロモーター(Odellら、Nature 313:810〜812(1985))、イネアクチン遺伝子由来プロモーター(McElroyら、Plant Cell 2:163〜171(1990))、ユビキチン(Christensenら、Plant Mol.Biol.12:619〜632(1989)およびChristensenら、Plant Mol.Biol.18:675〜689(1992))、pEMU(Lastら、Theor.Appl.Genet.81:581〜588(1991))、MAS(Veltenら、EMBO J.3:2723〜2730(1984))、ならびにトウモロコシH3ヒストン(Lepetitら、Mol.Gen.Genetics 231:276〜285(1992)およびAtanassovaら、Plant Journal 2(3):291〜300(1992))が挙げられる。ALSプロモーター、Xbal/NcoI断片5’からセイヨウアブラナ(Brassica napus)ALS3構造遺伝子(または前記Xbal/NcoI断片と類似のヌクレオチド配列)は、特に有用な構成的プロモーターである。PCT出願WO96/30530を参照されたい。
【0045】
C.組織特異的または組織優先プロモーター
組織特異的プロモーターは、細胞での発現のための遺伝子に機能的に結合され得る。場合により組織特異的プロモーターは、細胞での発現のための遺伝子に機能的に結合され得るシグナル配列をコードするヌクレオチド配列に機能的に結合され得る。組織特異的プロモーターに機能的に結合した目的の遺伝子で形質転換した植物は、特定の組織において導入遺伝子のタンパク質産物を排他的または優先的に産生できる。
【0046】
任意の組織特異的または組織優先プロモーターは、本発明において利用され得る。例示的な組織特異的または組織優先プロモーターとして、それだけに限られないが、ファゼオリン遺伝子由来のものなどの根優先プロモーター(Muraiら、Science 23:476〜482(1983)およびSengupta−Gopalanら、Proc.Natl.Acd.Sci.U.S.A.82:3320〜3324(1985))、cabまたはrubisco由来のものなどの葉特異的および光誘導プロモーター(Simpsonら、EMBO J.4(11):2723〜2729(1985)およびTimkoら、Nature 318:579〜582(1985))、LAT52由来のものなどの葯特異的プロモーター(Twellら、Mol.Gen.Genetics 217:240〜245(1989))、Zm13由来のものなどの花粉特異的プロモーター(Guerreroら、Mol.Gen.Genetics 244:161〜168(1993))またはapg由来のものなどの小胞子優先プロモーター(Twellら、Sex.Plant Reprod.6:217〜224(1993))が挙げられる。
【0047】
導入遺伝子によって産生されるタンパク質の、葉緑体、空胞、ペルオキシソーム、グリオキシソーム、細胞壁もしくはミトコンドリアなどの細胞内区画への輸送またはアポプラストへの分泌は、目的のタンパク質をコードする遺伝子のシグナル配列から5’および/または3’領域をコードするヌクレオチド配列に機能的に結合する手段によって達成され得る。構造遺伝子の5’および/または3’末端での標的化配列は、タンパク質合成およびプロセシングの間に、コードされたタンパク質がどこで最終的に区画化され得るかを決定できる。代替として、そのような細胞内区画を標的化するタンパク質は、所望の細胞内区画に目的の分子で被覆されたナノ粒子を方向付けるために直接ナノ粒子に結合され得る。
【0048】
シグナル配列の存在は、細胞内小器官もしくは細胞内区画のいずれかにまたアポプラストへの分泌にポリペプチドを方向付ける。多数のシグナル配列が当技術分野において既知である。例えばBeckerら、Plant Mol.Biol.20:49(1992)、Close,P.S.,Master’s Thesis.Iowa State University(1993)、Knox,C.ら、「Structure and Organization of Two Divergent Alpha−Amylase Genes from Barley」、Plant Mol.Biol.9:3〜17(1987)、Lernerら、Plant Physiol.91:124〜129(1989)、Fontesら、Plant Cell 3:483〜496(1991)、Matsuokaら、Proc.Natl.Acad.Sci.88:834(1991)、Gouldら、J.Cell.Biol.108:1657(1989)、Creissenら、Plant J.2:129(1991)、Kalderonら、A short amino acid sequence able to specify nuclear location,Cell 39:499〜509(1984)、Steifelら、Expression of a maize cell wall hydroxyproline−rich glycoprotein gene in early leaf and root vascular differentiation,Plant Cell 2:785〜793(1990)を参照されたい。
【0049】
外来タンパク質遺伝子および農学的遺伝子
本発明によるトランスジェニック植物で外来タンパク質は、商業的な量で産生され得る。したがって、当技術分野において十分に理解されている形質転換された植物の選択および繁殖のための技術は、従来の手段で収穫される複数のトランスジェニック植物を産出し、それにより外来タンパク質は目的の組織から、またはバイオマス全体から抽出され得る。植物バイオマスからのタンパク質抽出は、例えばHeneyおよびOrr,Anal,Biochem.114:92〜6(1981)によって論じられた既知の方法によって達成され得る。
【0050】
本発明の態様において外来タンパク質の商業的産生のために提供されるトランスジェニック植物は、細胞または植物であり得る。他の態様において目的のバイオマスは種子であり得る。高いレベルの発現を示す比較的少数のトランスジェニック植物について遺伝子地図は、組み込まれたDNA分子の染色体上のおよその位置を同定する従来型のRFLP、PCRおよびSSR分析を主に介して作成され得る。この点に関する例示的な方法について、GlickおよびThompson,Methods in Plant Molecular Biology and Biotechnology CRC Press,Boca Raton 269:284(1993)を参照されたい。染色体上の位置に関する地図情報は、対象のトランスジェニック植物の所有権の保護において有用であり得る。認可されていない繁殖が行われ、他の生殖質との交雑物が作製され得る場合、組込み領域の地図は、疑われる植物についての同様の地図と、後者が対象植物と共通の起源を有するかどうかを決定するために比較され得る。地図の比較は、ハイブリダイゼーション、RFLP、PCR、SSRおよび配列決定を含み、その全ては従来型の技術である。
【0051】
同様に農業的な遺伝子は、形質転換された細胞またはそれらの後代において発現され得る。より具体的には植物は、農業的目的での種々の表現型を発現するために本発明の方法を介して遺伝的に操作され得る。この点において使用され得る例示的な遺伝子として、それだけに限られないが、以下に分類されるものが挙げられる。
【0052】
1.有害生物または病害への耐性を付与し、かつ以下をコードする遺伝子:
A)植物の病害耐性遺伝子。植物の防御は、植物における病害耐性遺伝子(R)の産物と病原体中の対応する非病原性(Avr)遺伝子の産物との間の特定の相互作用によってしばしば活性化される。植物種は、特定の病原体株に耐性である植物を操作するためにクローン化された耐性遺伝子で形質転換され得る。例えばJonesら、Science 266:789(1994)(クラドスポリウム・フルバム(Cladosporium fulvum)への耐性のためのトマトCf−9遺伝子のクローニング)、Martinら、Science 262:1432(1993)(シュードモナス・シリンゲ(Pseudomonas syringae)病原型への耐性のためのトマトPto遺伝子。トマトはプロテインキナーゼをコードする)、Mindrinosら、Cell 78:1089(1994)(シュードモナス・シリンゲへの耐性のためのアラビドプス(Arabidops)はRSP2遺伝子であり得る)を参照されたい。
【0053】
B)ダイズシストセンチュウ(soybean cyst nematode)などの有害生物への耐性を付与する遺伝子。例えば、PCT出願WO96/30517、PCT出願WO93/19181を参照されたい。
【0054】
C)バチルス・チューリンゲンシスタンパク質、その誘導体またはそれをモデル化した合成ポリペプチド。例えばBt δ−エンドトキシン遺伝子のクローニングおよびヌクレオチド配列を開示するGeiserら、Gene 48:109(1986)を参照されたい。さらにδ−エンドトキシン遺伝子をコードするDNA分子は、例えばATCC登録番号40098、67136、31995および31998でAmerican Type Culture Collection,Manassas,Vaから購入できる。
【0055】
D)レクチン。例えば、いくつかのクンシラン(Clivia miniata)マンノース結合レクチン遺伝子のヌクレオチド配列を開示しているVan Dammeら、Plant Molec.Biol.24:25(1994)による開示を参照されたい。
【0056】
E)アビジンなどのビタミン結合タンパク質。PCT出願US93/06487を参照されたい。出願は、害虫に対する殺幼虫剤(larvicide)としてのアビジンおよびアビジン相同体の使用を教示している。
【0057】
F)酵素阻害剤、例えばプロテアーゼもしくはプロテイナーゼ阻害剤またはアミラーゼ阻害剤。例えばAbeら、J.Biol.Chem.262:16793(1987)(イネシステインプロテイナーゼ阻害剤のヌクレオチド配列)、Huubら、Plant Molec.Biol.21:985(1993)(タバコプロテイナーゼ阻害剤IをコードするcDNAのヌクレオチド配列)、Sumitaniら、Biosci.Biotech.Biochem.57:1243(1993)(ストレプトマイセス・ニトロスポレウス(Streptomyces nitrosporeus)α−アミラーゼ阻害剤のヌクレオチド配列)および米国特許第5,494,813号(HepherおよびAtkinson、1996年2月27日発行)を参照されたい。
【0058】
G)エクジステロイドまたは幼若ホルモン、それらのバリアント、それらに基づく模倣物、またはそれらのアンタゴニストもしくはアゴニストなどの昆虫特異的ホルモンまたはフェロモン。例えばHammockら、Nature 344:458(1990)による、クローン化された幼若ホルモンエステラーゼ、幼若ホルモンの不活性化因子のバキュロウイルス発現の開示、を参照されたい。
【0059】
H)感染した有害生物の生理機能を発現によって破壊する昆虫特異的ペプチドまたは神経ペプチド。例えばRegan,J.Biol.Chem.269:9(1994)(発現クローニングは昆虫利尿ホルモン受容体をコードするDNAを産出する)およびPrattら、Biochem.Biophys.Res.Comm.163:1243(1989)(アロスタチン(allostatin)はディプロプテラ・パンタタ(Diploptera puntata)において同定され得る。)の開示を参照されたい。昆虫特異的な麻痺性神経毒をコードする遺伝子を開示しているTomalskiらの米国特許第5,266,317号も参照されたい。
【0060】
I)天然でヘビ、スズメバチまたは任意の他の生物によって産生される昆虫特異的毒液。例えばサソリ昆虫毒性ペプチドをコードする遺伝子の植物における異種性発現の開示についてPangら、Gene 116:165(1992)を参照されたい。
【0061】
J)モノテルペン、セスキテルペン、ステロイド、ヒドロキサム酸、フェニルプロパノイド誘導体または殺虫活性を有する他の非タンパク質性分子の高度集積に関連する酵素。
【0062】
K)生物学的に活性な分子、例えば天然または合成での、解糖系酵素、タンパク質分解酵素、脂肪分解酵素、ヌクレアーゼ、シクラーゼ、トランスアミナーゼ、エステラーゼ、ヒドロラーゼ、ホスファターゼ、キナーゼ、ホスホリラーゼ、ポリメラーゼ、エラスターゼ、キチナーゼおよびグルカナーゼ、の翻訳後修飾を含む修飾に関与する酵素。カルラーゼ(callase)遺伝子のヌクレオチド配列を開示しているScottらによるPCT出願WO93/02197を参照されたい。キチナーゼをコードする配列を含有するDNA分子は、例えばATCCから登録番号39637および67152で得ることができる。タバコイモムシキチナーゼをコードするcDNAのヌクレオチド配列を教示しているKramerら、Insect Biochem.Molec.Biol.23:691(1993)および、パセリubi4−2ポリユビキチン遺伝子のヌクレオチド配列を提供しているKawalleckら、Plant Molec.Biol.21:673(1993)も参照されたい。
【0063】
L)シグナル伝達を刺激する分子。例えばBotellaらPlant Molec.Biol.24:757(1994)によるリョクトウカルモジュリンcDNAクローンについてのヌクレオチド配列の開示、およびトウモロコシカルモジュリンcDNAクローンのヌクレオチド配列を提供しているGriessら、Plant Physiol.104:1467(1994)を参照されたい。
【0064】
M)疎水性モーメントペプチド。PCT出願WO95/16776(真菌性植物病原菌を抑制するタキプレシンのペプチド誘導体の開示)およびPCT出願WO95/18855(病害への耐性を付与する合成抗菌性ペプチドを教示)を参照されたい。
【0065】
N)膜透過酵素、チャネル形成剤またはチャネル遮断剤。例えば青枯病菌(Pseudomonas solanacearum)に耐性のトランスジェニックタバコ植物を提供するセクロピン−β溶解性ペプチド類似物の異種発現についてのJaynesら、Plant Sci 89:43(1993)の開示を参照されたい。
【0066】
O)ウイルス性侵襲性タンパク質またはそれら由来の複合毒素。例えば、形質転換された植物細胞におけるウイルス被覆タンパク質の蓄積は、被覆タンパク質遺伝子が由来し得るウイルスによって、および関連するウイルスよってもたらされるウイルス感染および/または病害の進行への耐性を与える。Beachyら、Ann.rev.Phytopathol.28:451(1990)を参照されたい。被覆タンパク質介在耐性は形質転換植物に、アルファルファモザイクウイルス、キュウリモザイクウイルス、タバコ条斑病ウイルス、ジャガイモウイルスX、ジャガイモウイルスY、タバコエッチウイルス、タバコ茎壊疽ウイルスおよびタバコモザイクウイルスに対して与えられている。同著者。
【0067】
P)昆虫特異的抗体またはそれら由来の免疫毒素。このように昆虫の消化管において不可欠な代謝機能を標的とする抗体は、影響される酵素を不活性化し、昆虫を殺す。参照、Taylorら、Abstract#497,Seventh Int’l Symposium on Molecular Plant−Microbe Interactions(Edinburgh,Scotland)(1994)(1本鎖抗体断片の産生を介するトランスジェニックタバコにおける酵素の不活性化)。
【0068】
Q)ウイルス特異的抗体。例えば組換え抗体遺伝子を発現するトランスジェニック植物はウイルス攻撃から防御されることを示している、Tavladorakiら、Nature 366:469(1993)を参照されたい。
【0069】
R)病原体または寄生生物によって天然で産生される発生抑制タンパク質。例えば真菌エンドα−1,4−Dポリガラクツロナーゼは、植物細胞壁ホモ−α−1,4−D−ガラクツロナーゼを可溶化することによって真菌のコロニー形成および植物栄養素の放出を促進する。Lambら、Bio/Technology 10:1436(1992)を参照されたい。マメエンドポリガラクツロナーゼ阻害タンパク質をコードする遺伝子のクローニングおよび特徴付けはToubartら、Plant J.2:367(1992)によって記載され得る。
【0070】
S)植物によって天然で産生される発生抑制タンパク質。例えばLogemannら、Bio/Technology 10:305(1992)は、オオムギリボソーム不活性化遺伝子を発現するトランスジェニック植物は真菌性病害への耐性が増大していることを示している。
【0071】
2.除草剤への耐性を付与する遺伝子
A)成長点または分裂組織を阻害する、イミダゾリノンまたはスルホニル尿素などの除草剤。この分類の例示的な遺伝子は、例えばLeeら、EMBO J.7:1241(1988)およびMikiら、Theor.Appl.Genet.80:449(1990)によってそれぞれ記載のとおり変異体ALSおよびAHAS酵素をコードする。
【0072】
B)グリフォセート(例えば変異体5−エノールピルビルシキミ酸−3−リン酸合成酵素(EPSP)遺伝子によって(組換え核酸の導入および/または種々の形態の天然EPSP遺伝子のin vivo変異誘発を介して)、aroA遺伝子およびグリフォセートアセチルトランスフェラーゼ(GAT)遺伝子、それぞれによって耐性が付与される)、グルフォシネートなどの他のホスホノ化合物(ストレプトマイセス・ハイグロスコピカス(Streptomyces hygroscopicus)およびストレプトマイセス・ビリジクロモゲネス(Streptomyces viridichromogenes)を含むストレプトマイセス種由来のホスフィノトリシンアセチルトランスフェラーゼ(PAT)遺伝子)およびピリジノキシまたはフェノキシプロプリオン酸ならびにシクロヘキソン類(ACCase阻害剤をコードする遺伝子)、例えば植物にグリフォセート耐性を付与できるEPSPの形態のヌクレオチド配列を開示しているShanらの米国特許第4,940,835号およびBarryらの米国特許第6,248,876号を参照されたい。変異体aroA遺伝子をコードするDNA分子は、ATCC登録番号39256で得ることができ、変異体遺伝子のヌクレオチド配列はComaiの米国特許第4,769,061号において開示され得る。Kumadaらの欧州特許出願第0333033号およびGoodmanらの米国特許第4,975,374号は、L−ホスホリノトリシンなどの除草剤への耐性を付与するグルタミン合成酵素遺伝子のヌクレオチド配列を開示している。PAT遺伝子のヌクレオチド配列は、PAT活性をコードするキメラbar遺伝子を発現するトランスジェニック植物の産生を記載しているLeemansらの欧州特許出願第0242246号、DeGreefら、Bio/Technology 7:61(1989)によって提供され得る。フェノキシプロプリオン酸ならびに、セトキシジムおよびハロキシホップなどのシクロヘキソン類への耐性を付与する遺伝子の例として、Marshallら、Theor.Appl.Genet.83:435(1992)によって記載のAcc1−S1、Acc1−S2およびAcc1−S3遺伝子が挙げられる。グリフォセート耐性を付与できるGAT遺伝子は、CastleらのWO2005012515に記載されている。2,4−D、fopおよびピリジロキシオーキシン除草剤への耐性を付与する遺伝子は、Dow AgroSciences LLCに譲渡されたWO2005107437に記載されている。
【0073】
C)トリアジン(psbAおよびgs+遺伝子)またはベンゾニトリル(ニトリラーゼ遺伝子)などの光合成を阻害する除草剤。Przibilaら、Plant Cell 3:169(1991)は変異体psbA遺伝子をコードするプラスミドでのクラミドモナスの形質転換を記載している。ニトリラーゼ遺伝子のヌクレオチド配列は、Stalkerの米国特許第4,810,648号に記載されており、これらの遺伝子を含有するDNA分子はATCC登録番号53435、67441および67442で入手できる。グルタチオンS−トランスフェラーゼをコードするDNAのクローニングおよび発現は、Hayesら、Biochem.J.285:173(1992)によって記載されている。
【0074】
3.次のような付加価値のある特質を付与するまたはそれに寄与する遺伝子
A)例えば、植物のステアリン酸含有量を増加させるためにステアリルACP不飽和化酵素のアンチセンス遺伝子で植物を形質転換することによる、改変された脂肪酸代謝。Knultzonら、Proc.Natl.Acad.Sci.U.S.A.89:2624(1992)を参照されたい。
【0075】
B)フィチン酸含有量の減少−1)フィターゼをコードする遺伝子の導入は、フィチン酸の分解を増強し、形質転換植物の遊離リン酸を増やす。例えば、クロコウジカビ(Aspergillus niger)フィターゼ遺伝子のヌクレオチド配列の開示についてのVan Hartingsveldtら、Gene 127:87(1993)を参照されたい。2)フィターゼ含有量を減少させる遺伝子は導入され得る。例えばトウモロコシにおいて、これはクローニングおよびそれに次ぐ、フィチン酸の低いレベルによって特徴付けられるトウモロコシ変異体に関与し得る単一アレルに関連するDNAの再導入によって達成され得る。Raboyら、Maydica 35:383(1990)を参照されたい。
【0076】
C)例えば、デンプンの分枝様式を変化させる酵素をコードする遺伝子で植物を形質転換することによって生じた改変された炭水化物組成物。Shirozaら、J.Bacteol.170:810(1988)(連鎖球菌変異体フルクトース転移酵素遺伝子のヌクレオチド配列)、Steinmetzら、Mol.Gen.Genet.20:220(1985)(バチルス・サブティル(Bacillus subtil)はレバンスクラーゼ遺伝子のヌクレオチド配列であり得る)、Penら、Bio/Technology 10:292(1992)(バチルス・リケニフォン(Bacillus lichenifonn)を発現するトランスジェニック植物の産生物はα−アミラーゼであり得る)、Elliotら、Plant Molec.Biol.21:515(1993)(トマトインベルターゼ遺伝子のヌクレオチド配列)、Sogaardら、J.Biol.Chem.268:22480(1993)(オオムギα−アミラーゼ遺伝子の部位特異的ミュータジェンズ(mutagens)であり得る)およびFisherら、Plant.Physiol.102:1045(1993)(トウモロコシ胚乳デンプン分枝酵素II)を参照されたい。
【実施例】
【0077】
本発明は、例示の方法により提示され、いかなるやり方においても本発明を限定することを意図しない以下の実施例においてさらに記述される。
【実施例1】
【0078】
単一細胞植物材料の調製
BY2細胞およびNT1細胞の両方を使用した。BY2細胞は、非緑色、高増殖性タバコ細胞系である。NY1細胞は、タバコから単離された光独立栄養細胞である。形質転換の3から4日前に、1週間経過した懸濁培養物を、NT1またはBY2培養物の2mlを、250mLフラスコ中の50nM DAS−PMTI−1(微小管阻害剤)および0.5〜0.1%(v/v)DMSOを含有するNT1BまたはLSBY2培地40mlに移すことによって新鮮培地に継代した。単一細胞を微小管阻害剤処置の4日後または7日後のいずれかで回収した。使用したBY2単一細胞は、生細胞を計数するためにBeckmanフローサイトメーターにより処理した。平均径10.43umおよび容積593.8μm3であった1mlあたり658250個の生細胞があった。図1で観察されるとおり、全ての細胞は単一細胞であった(図1における対は端が重なっている)。細胞を共焦点画像化システムを取り付けた微分干渉(DIC)顕微鏡を使用して検査した(パネルA)。パネルBは、色素体(アミロプラスト)を強調するためにI2KIで染色したBY2細胞(EP12%培地で馴化および維持した培養物)由来の単一細胞の光学顕微鏡像を示す。ここで観察されるようにBY2細胞の単一細胞は、細胞の細胞質全体に分布した多数の色素体(アミロプラスト)を含む。
【0079】
図2、パネルAは、最少培地および5%二酸化炭素で維持した、顕著な葉緑体を有するタバコ(NT1)の光学顕微鏡での光独立栄養細胞を示す。これらの細胞を3.0OD600での懸濁物1mlを移すことによって14日ごとに1回継代した。図2、パネルBは蛍光顕微鏡下で観察した同様のNT1細胞を示し、赤色に自己蛍光を発している活性葉緑体が観察できる。
【0080】
上に記載する細胞型を形質転換のための標的細胞として使用した。緑色細胞(NT1細胞)は、クラスター化する細胞がほとんど無く、かつ透明であることから葉緑体中にナノ粒子を追跡するための最適な細胞型である。加えて細胞は、赤色の自己蛍光を発する非常に顕著な葉緑体を有する(図2、パネルBに観察されるとおり)。
【実施例2】
【0081】
ナノ粒子の調製および細胞の処置
培養物中で細胞が蛍光色素を取り込むかどうかを決定するために、BY2細胞の単一細胞のおよび多細胞性の標準的な凝集懸濁培養物を使用した。細胞懸濁培養物をMolecular ProbesからのSAMSAフルオレセイン(5−((2−(および−3)−S−(アセチルメルカプト)サクシノイル)アミノ)フルオレセイン)に、ナノ粒子の非存在下で20分間さらし、次いで蛍光顕微鏡下で観察する前に短時間洗浄した。
【0082】
金ナノ粒子(GNP)を製品技術指針(probes.invitrogen.com/media/pis/mp00685.pdfでワールドワイドウェブにおいて入手できる)によりSAMSAフルオレセインで被覆した。金−フルオレセインコンジュゲートを本明細書以下に記載の方法を使用して調製した。SAMSAフルオレセイン1mgを0.1M NaOH 100μlに溶解し、チオールを保護しているアセチル基を除去するために15分間ボルテックスした。この活性化SAMSAを次いで150nm金コロイド(1mlあたり粒子約109個)100μlと混合した。得られた溶液を次いで反応の確実な完了のために1時間インキュベートした。次いで1M HCl 50μLを溶液を中和するために加えた。3000RPMで30分間遠心分離し、上清を除去した。得られた黄色沈殿を0.1M PBS 200μLに再懸濁し、橙色溶液を得た。この精製ステップを、遊離SAMSAフルオレセインを確実に除去するために2回繰り返した。SAMSAの金への付着様式は、主にチオール結合を介する。顕著な静電反発力(SAMSAはpH>7で2価陰イオン性である)により、SAMSAは全体が金コロイド表面に垂直に位置している。粒子は、DICおよび多光子共焦点顕微鏡下で観察した場合にいかなるバックグラウンドも示さない明確な蛍光を示した。被覆した金ナノ粒子20および40μlをBY2/NT1タバコ懸濁液または光独立栄養タバコ細胞500μlに移し、暗所で20分間インキュベートした。
【0083】
インキュベーション後、細胞懸濁液50μl一定分量を顕微鏡の灌流スライドに載せ、粒子を追跡するために顕微鏡下で観察した。加えて、試料の一定分量を20分間のインキュベーションの2〜20時間後に顕微鏡観察のために調製した。
【実施例3】
【0084】
フルオレセインで被覆されたナノ粒子のBY2/NT1細胞凝集物ならびに単一の光独立栄養タバコ細胞の核および色素体における送達および蓄積
SAMSAフルオレセイン単独でおよびSAMSAフルオレセインで被覆されたGNPで処置したBY2/NT1懸濁凝集物を、DIC、明視野および蛍光顕微鏡を使用する低拡大率下および高拡大率下で検査した。図3、パネルAはSAMSAフルオレセイン単独で処置した細胞のDIC画像を示し、一方図3、パネルBは同じ細胞の蛍光画像を示す。図3、パネルCはSAMSAフルオレセインで被覆されたGNPで処置した細胞のDIC画像を示し、一方図3パネルDはSAMSAフルオレセインで被覆されたGNPで処置した細胞の蛍光画像を示す。図3、パネルBにおいて明確に観察されるとおり、SAMSAフルオレセイン単独で処置した細胞の細胞壁だけがフルオレセインで染色され、他のバッククラウンド蛍光はわずかしか観察されなかった。これは、細胞がナノ粒子の非存在下ではSAMSAフルオレセインを取り込まなかったことを示唆する。
【0085】
図3、パネルDで見られるとおり、対照的にSAMSAフルオレセインで被覆されたGNPは、細胞中および核(Nu)中へと追跡された。SAMSAフルオレセインで被覆されたGNPが空胞を除く細胞の全ての区画において見出されたことは、DIC観察から明らかであった。核区画に加えて空胞を裏打ちする細胞質糸もSAMSAフルオレセインで被覆されたGNPを有した。ナノ粒子は、細胞壁を越えるそれらの輸送を妨げられないようである。したがってSAMSAフルオレセインで被覆されたGNPの蓄積は、アポプラストの連続体ではなくシンプラストの連続体においてであると考えられる。SAMSAフルオレセインで被覆されたGNPで処置した単一細胞の高拡大率下でのさらなる検査は、核中の多数のGNPの存在を示し、GNPはこれらの小器官に優先的に蓄積されると考えられた(図4、パネルB)。図4、パネルAは、パネルBと同じ核の異なる焦点面での明視野画像を示す。
【0086】
図5は、SAMSAフルオレセインで被覆されたGNPで処置した光独立栄養細胞を示す。パネルAは、細胞壁の内側を満たしている大きな葉緑体を有する細胞3〜4個のクラスターでの非常に透明な細胞を示す。パネルBは、葉緑体中のナノ粒子の蓄積を示す。パネルCおよびDは、蛍光顕微鏡を使用する、より高拡大率での単一の葉緑体を示す。ナノ粒子は、葉緑体の膜薄層形成において観察され、赤色自己蛍光を発しているクロロフィル色素中に散在している。
【0087】
したがって、即時的に明視野および蛍光の顕微鏡で追跡した生存光独立栄養細胞は、ナノ粒子が膜および葉緑体マトリックスの両方に蓄積していることを示した。粒子は、葉緑体の二重膜の内腔でも追跡され得た。
【0088】
葉緑体中の粒子を追跡するために実施した実験は、粒子が色素体に蓄積するらしいことを明らかにしたが、不十分な解像度のため光学顕微鏡を使用することによって葉緑体包膜中の粒子の存在を可視的に同定することは困難であった。したがって反射顕微鏡および蛍光顕微鏡を使用して粒子を追加的に追跡し、粒子を明確に位置付けるために画像をマージした、図6示すとおり。図6、パネルAは、GNPが優先的に見られる反射画像を示す。この写真は葉緑体中のナノ粒子の存在を示すだけでなく、葉緑体中のナノ粒子の多量の蓄積も示し、活発な取り込みを示唆している。パネルBは、赤色自己蛍光を発する葉緑体のバックグラウンド中での蛍光発光粒子を示す。マージした反射および蛍光での画像をパネルCに示す、ここで黄色蛍光を発している粒子は葉緑体の境界内であり、色素体中の粒子の存在を確認している。
【実施例4】
【0089】
核形質転換のためのDNA付着GNP送達
DNAで被覆されたGNPを2経路、すなわち非特異的相互作用および特異的相互作用(PEGをプラットホームとして使用する)を介して合成し、BY2/NT1細胞とインキュベートした。非特異的相互作用のために、3%マンニトール 9mLを細胞懸濁物1mLに加え、次いで1000rpmで5分間遠心分離した。次いで上清をデカントし、細胞を3%マンニトール300μlに再懸濁した。直径150nmの金ナノ粒子(BBI Internationalから入手できる(EM.GC150))50μlおよびYFPをコードするプラスミドDNA(pDAB3831)(図16)(配列番号1および2)50μgを再懸濁した細胞に加え、混合物を20分間インキュベートした。インキュベーション後、3%マンニトール20mLを溶液に加え、得られた混合物を1000rpmで5分間遠心分離した。次いで上清をデカントし、細胞を増殖培地3mLに再懸濁した。次いで再懸濁した細胞を選択プレートに移す少なくとも48時間前にマイクロウェルに移した。特異的相互作用(PEG経路)のために、大過剰量のチオールリガンドを使用した:粒子1個あたり100単分子膜、1個のチオール分子によって占有される表面積がおよそ0.20nmであると仮定することにより推定した。この計算を使用して、SH−PEG(3)−OCH3 2mgをクエン酸GNP溶液に加えた。混合物を直ちに室温で20時間撹拌し、その間に溶液の色はわずかに濃くなった。次いでTHF 3容量を反応混合物に加え、得られた溶液を13K rpmで4℃、30分間遠心分離した。上清を除去し、沈殿を超純水(18MΩ.cm)10mLに再溶解し、THF 30mLを加え、同じ条件で2度目の遠心分離を実施した。次いで沈殿を超純水(18MΩ.cm)に再溶解し、室温に置いた。形質転換実験用に、プラスミドDNAをH3CO−PEG−SH−GNPに被覆するために、精製プラスミドDNA 1mgを金粒子10mgと水50ml中で2時間、23°でインキュベートした(Torney,F.ら、Nature Nanotechnol.2,(2007)を参照されたい)。
【0090】
考えられる1つの形質転換計画の図画表示を図7で概説する。形質転換用に、YFPレポーター遺伝子を含むプラスミドDNA、pDAB3831を使用した。BY2細胞を上に記載のとおり処置し、懸濁液をマイクロウェルプレートでゆっくり撹拌しながら48〜72時間インキュベートした。懸濁液の50μl一定分量を合計0.5〜1ml混合物から採取し、レポーター遺伝子の発現を観察するために蛍光顕微鏡下で検査した。YFPレポーター遺伝子を含有するプラスミドで形質転換したBY2細胞は、YFPの一過的発現を示した。
【実施例5】
【0091】
核形質転換のためのDNA付着PEG化量子ドット送達
細胞侵入評価調査のためのQDのPEG機能化。この手順は、Dubertret B.ら、Science 298,1759(2002)から採用した。TOPO(トリオクチルホスフィンオキシド)で被覆されたCdSe/ZnS QD(Ocean nanotechnology,Cat#QSO0630−0010)2mgをPEG−PE(1,2−ジアシル−sn−グリセロ−3−ホスホエタノールアミン−N−[メトキシ−ポリ(エチレングリコール)])(Avanti lipids,Cat#880160).015g(5.5μmol)とクロロホルム中に懸濁し、溶媒の蒸発および水での安定化が続いた。PEGコンジュゲート化を細胞毒性からの完全な防御を確実にするために実施した。
【0092】
プラスミドDNAとのQDコンジュゲーション。TOPO(トリオクチルホスフィンオキシド)で被覆されたQD(Ocean nanotechnology,Cat#QSO0630−0010)2mgをHS−PEG−OCH3(Prochimia,Cat#TH014−01)4mgと、一晩、約60〜70℃で懸濁した。真空オーブンで溶媒を除去した。次いで残渣を水(18M)1mLに懸濁した。最後のステップは赤色残渣の橙色、光学的に透明の、透き通った溶液への変化を伴う。形質転換実験用にプラスミドDNAをH3CO−PEG−SH−QDに被覆するために、精製プラスミドDNA(pDAB3831)0.02mgを得られたQDコンジュゲートと水2ml中で2時間、23°、暗所でインキュベートした(Torney,F.ら、Nature Nanotechnol.2,(2007))。
【0093】
タバコ無傷細胞とのQDのインキュベーション。細胞系での実験を、LSBY2培地中、25℃で維持したBright Yellow(BY2)タバコ単一細胞系を使用して実施した。これらの単一細胞系は、IDM#64901で概説されたのと同じ方法によって産生される。1〜3μL/mLの濃度を24ウェルマイクロタイタープレート中の細胞500μlに加え、細胞を分析する前に振盪機でおだやかに20分間回転させた。
【実施例6】
【0094】
無傷植物細胞への蛍光タンパク質のナノ粒子介在形質導入および細胞内部移行ならびに利用可能性。
植物細胞へのタンパク質のナノ粒子介在形質導入および細胞内部移行を検査するための材料は、サイズ直径150nmの金コロイド(BBI International,GC150)、5−((2−(および−3)−S(アセチルメルカプト)サクシノイル)アミノ)フルオレセイン(SAMSA fluorescein:Invitrogen,A−685)、サイズ80および90nmのカルボン酸被覆金コロイドのナノ粒子(TedPella,32019)、スルホ−NHS(N−ヒドロキシスルホサクシニミド)、EDC(1−エチル−3−[3−ジメチルアミノプロピル]カルボジイミド塩酸塩)、(Pierce Bitoechnology,24510,22980,)、MES(2−[N−モルホリノ]エタンスルホン酸)(Fisher Scientific,AC32776−1000)、リン酸緩衝食塩溶液緩衝剤包(Sigma,P5368−10PAK)、ヒストジンタグ付きGFP(Evrogen、励起最大−482nm、発光最大−502nm、FP611)、turbo YFP(Evrogen、励起最大−525nm、発光最大−538nm、FP611)およびヨウ化プロピジウム(Sigma−P4864)、フルオレセイン二酢酸(Sigma,F7378)を含む。
【0095】
細胞培養物(BY2−Eタバコ単一細胞):BY2細胞およびNT1細胞の両方を使用した。BY2/NT1細胞は、非緑色、高増殖性タバコ細胞系である。実験の3から4日前に、1週間経過した懸濁培養物を、NT1またはBY2培養物2mlを、250mLフラスコ中のDAS−医薬用MTI−1(PMTI−1)(微小管阻害剤)1μl、1〜3%グリセロールおよび0.05〜0.1%(v/v)DMSOを含有するNT1BまたはLSBY2培地40mlに移すことによって新鮮培地に継代した。単一細胞を微小管阻害剤処置の3.5日後または7日後のいずれかで回収した。使用したBY2単一細胞は、生細胞を計数するためにBeckmanフローサイトメーターにより処理した。平均径10.43um(容積593.8μm3)〜50.42um(容積67079.05μm3)で1mlあたり658250個の生細胞があった。これらの培養物中の生細胞は、PMTI−1処置後に全て単一細胞であった。細胞を共焦点蛍光画像化システムを取り付けた微分干渉(DIC)顕微鏡を使用して検査した。
【0096】
ナノ粒子コンジュゲート:金−フルオレセインコンジュゲート、金−ヒスチジンタグ付きGFPコンジュゲート、および金−YFPコンジュゲートを合成した。
【0097】
金−フルオレセインコンジュゲートの合成:金−フルオレセインコンジュゲートは既に記載されている方法(Cannone,F.,G.Chiricoら、(2006)、Quenching and Blinking of Fluorescence of a Single Dye Molecule Bound to Gold Nanoparticles.J.Phys.Chem.B 110(33):16491〜16498)を使用することにより調製した。SAMSAフルオレセイン1mgを0.1M NaOH 100μlに溶解し、チオールを保護しているアセチル基を除去するために15分間ボルテックスした。この活性化SAMSAを150nm金コロイド(1mlあたり粒子約109個)100μlと混合した。この溶液を次いで反応を確実に完了させるために1時間インキュベートした。インキュベーション後、1M HCl 50μLを溶液を中和するために加えた。3000RPMで30分間遠心分離し、上清を除去した。得られた黄色沈殿を0.1M PBS 200μLに再懸濁し、橙色溶液を得た。この精製ステップを、遊離SAMSAフルオレセインの除去を確実にするために2回繰り返した。SAMSAの金への付着様式は、主にチオール結合を介する。顕著な静電反発力(SAMSAはpH>7で2価陰イオン性である)により、SAMSAは金コロイド表面に垂直に位置していると考えられている(Cannoneら、2006)。
【0098】
金−ヒスチジンタグ付きGFPおよび金−YFPコンジュゲートの合成:金−タンパク質コンジュゲートを、2種のタンパク質の連続的カップリングを例示したGrabarek(Grabarek,Z.およびJ.Gergely(1990)、Zero−length cross linking procedure with the use of active esters.Analytical Biochemistry 185(1):131〜135)によって記載された手順をわずかに変更して使用して合成した。90nmカルボン酸被覆金コロイド溶液(1mlあたり粒子約109個)0.25mlを3000RPMで10分間遠心分離した。上清を捨てた後、赤色沈殿を活性化緩衝液(0.1M MES、0.5M NaCl、pH6.0)1mlに懸濁した。EDC 0.4mgおよびスルホNHS 1.1mgをこの溶液に加え、室温で15分間ボルテックスした。次いでタンパク質(ヒスチジンタグ付きGFPまたはturbo YFP)9μlを加え、タンパク質と金とが完全に反応するように得られた溶液を2時間、暗所、室温でインキュベートした。この反応に使用した金コロイドとタンパク質との割合は、金コロイド上に存在するカルボン酸の数を得ることによって決定した。最初に金コロイド上に存在するカルボキシル基の数を、金粒子(球体と仮定)1個の表面積をカルボキシル基1個で占有される表面で割ることによって算出した(0.20nm2 Kimura,K.;Takashima,S.:Ohshima,H.Journal of Physical Chemistry B 2002,106,7260〜7266)。金コロイド溶液全体に存在するカルボキシル基の総数を得るために、この結果に存在する金コロイドの総数を掛けた。これを所与の量のタンパク質に存在するアミノ基の数と等しくした。これらの金コロイドは、金に存在するカルボン酸とタンパク質に存在するアミノ基との間のアミド結合の形成を介してタンパク質に付着する(Grabarek,Z.およびJ.Gergely(1990).Zero−length cross linking procedure with the use of active esters.Analytical Biochemistry 185(1):131〜135)。タンパク質分子およそ127285個が金ナノ粒子1個に繋ぎ止められている。
【0099】
細胞処置 3種の個々の試料を検査用に以下のとおり調製した:
金取り込みおよび細胞生存率の経時変化 以下の試料を24ウェル滅菌プレートに調製した。
i)単一BY2−E細胞500μl(対照)、
ii)単一BY2−E細胞500μl+GNP 20μl+フルオレセイン二酢酸(FDA)25μl+ヨウ化プロピジウム25μl;および
iii)他の処置は、細胞とのGNP 40、60、80μl、および上に記載の細胞生死判別染色を含む。処置した細胞(Ref)は、蛍光顕微鏡下で5、20、120分および最終的に18〜20時間後に検査した。
【0100】
金−SAMSAフルオレセイン処置 以下の試料を24ウェル滅菌プレートに調製した。
i)単一BY2−E細胞500μl(対照)、
ii)単一BY2−E細胞500μl+SAMSA−フルオレセイン20μl(対照);および
iii)単一BY2−E細胞500μl+Au−SAMSA−フルオレセイン20μl。
処置した細胞を、顕微鏡検査を実施する前に20分間、暗所、室温でインキュベートした。
【0101】
金−ヒスチジンタグ付きGFP処置 以下の試料を24ウェル滅菌プレートに調製した。
i)単一BY2−E細胞500μl(対照)、
ii)単一BY2−E細胞500μl+ヒスチジンタグ付きGFP 10μl(対照)、
iii)単一BY2−E細胞500μl+Au−ヒスチジンタグ付きGFP 20μl。
処置した細胞を、顕微鏡検査を実施する前に2時間、暗所、室温でインキュベートした。
【0102】
顕微鏡:Au−SAMSAフルオレセインおよびAu−ヒスチジンタグ付きGFPでの単一細胞実験の位相差顕微鏡法および蛍光顕微鏡法をLeica inverted fluorescence microscope(DAS)を使用して実施した。全ての実験は拡大率20×で実施した。FITC(フルオレセインイソチオシアネート)およびGFPフィルターをSAMSAフルオレセインおよびGFP単一細胞処置についてそれぞれ使用した。
【0103】
微分画像コントラスト(DIC)、共焦点および反射顕微鏡:これらの調査はUIUC(University of Illinois at Urbana Champaign)microscopy centerにおいてZeiss倒立顕微鏡で実施した。これら全ての方法について拡大率は63×に保った。共焦点用にFITC、GFPおよびYFPのフィルターをさまざまな細胞処置のために使用した。反射調査用には二色性鏡を透明なガラススライドに置き換え、発光フィルターを外した。
【0104】
画像取得:懸濁培養したタバコ細胞をX−Cite 120照明システムにつないだapotome光学切片システムを取り付けたZeiss Axiovert M 200顕微鏡(Carl Zeiss microimaging,Obercohen,Germany)を使用して画像化した。金粒子を635/20励起フィルターを通した水銀照明を使用して反射画像化設定下で画像化し、UVを遮断するためのGG420ガラス、KG5(1Rブロッカー)、50/50ビームスプリッターならびに励起および発光平行偏光板からなるIGS偏光フィルターセット(33001、Chroma Technology Corp.,Rockingham,VTから入手できる)を使用して画像化した。同時に、DIC/透過光画像を標準的DIC光学を使用して取得し、GFP−DNA被覆金粒子中のGFP(疑似カラー緑色)をバンドパスFITCフィルター(HQ480/40励起フィルター、Q505LP二色性鏡およびHQ535/50発光)で取得した。細胞を、高解像度画像化のために厚さ500ミクロンのチャンバーカバーガラスセット(Grace Bio−labs,Bend,OR)に分注した。大部分の画像は、細胞の大きさにより63×1.4 NA Planapochromat対物レンズまたは40×1.4でNA Planapochromat対物レンズで取得した(Carl Zeiss Microimaging,Obercohen,Germanyから入手できる)。露光時間は、各チャネル(すなわち、DIC、反射および/またはFITC)について設定し、ディメンジョン1388×1034ピクセルの高解像度Axiocam MRmモノクロカメラ(Carl Zeiss Obercohen,Germanyから入手できる)と共にAxiovision 4.6プログラムを使用して連続的に露光した。必要な場合には、解像度は1024×1024に設定し、およそ150〜250フレームを含む2〜5分間にわたる粒子の動態を解像するために可能な最高速度で経時的に連続な画像を得た。画像をAxiovision 4.6ギャラリーモジュールまたはAdobe Photoshop(Adobe Systems,San Jose,CA)のいずれかで作成した。
【0105】
経時変化およびGNP内部移行調査:粒子の取り込みおよびGNP濃度の細胞の正常性および生存率への影響を評価するために、経時的実験をクエン酸機能性付与GNP(直径90nm)とインキュベートしたBY2−E単一細胞系で実施した。種々の濃度のGNP(20、40、60、80μl)をこの実験に使用した。粒子は細胞との混合後5分間以内で内部移行され、粒子の蓄積は、細胞のサイトゾルおよび核におけるレベルの上昇を示すために2時間まで要した。検査した濃度中で高レベルの細胞生存率および細胞の活力は、FDAおよびPI(生/死細胞染色)手順で調査したとおり20μl処置で観察された。処置した全ての試料において細胞の平均生存率は98%に近いが、検査した最高濃度においてであり、FDAで染色された核は80μl処置では見られなかった。しかしこれらの染色されなかった核は、PIに反応せず、したがって細胞死ではないことを示唆していた。この結果は、最高濃度の粒子が、処置の20時間後に細胞が休眠する場合があるが、まだ生きている程度までの内部分布を生じ得ることを示唆する。
【0106】
【化1】
【0107】
反射顕微鏡追跡調査:金タンパク質(GFP/YFP)コンジュゲートで処置した単一BY2/NT1タバコ細胞での反射調査は、細胞内の金ナノ粒子の存在を示す。これを同様の条件下で暗くなって見えた未処置の対照単一BY2細胞と比較した、図8に示すとおり。明るい反射を発している単独の金ナノ粒子が観察された、図8に示すとおり。これは、これらの壁を有するBY2細胞による金ナノ粒子の取り込みの明らかな証拠である。
【0108】
金−SAMSAフルオレセイン実験:DASで実施した位相差実験は、対照単一細胞について観察されたシルバーコントラストと比較して、処置細胞についての細胞内腔および核の鮮黄色の染色を明らかにした。多数の細胞についてプラズマリシスの条件下でも細胞膜が細胞壁から離れ、図9に示すとおり細胞の部分的なまたは完全な原形質分離を示唆するそれらの間の空間が生じた。SAMSAフルオレセイン単独で処置した単一細胞の共焦点蛍光実験下で観察した場合そのような細胞は、細胞質および核に蛍光を示すが未処置の対照細胞においては暗く見えた。SAMSAフルオレセイン染色単独で処置した細胞もいくらかの壁での蛍光を示したが細胞内には示さなかった。これは、SAMSAフルオレセインがそれ自体は細胞内に内部移行されず、金ナノ粒子がその取り込みのための担体の様に作用することを意味する。
【0109】
金−ヒスチジンタグ付きGFP実験:GNPを介する未処置細胞へのタンパク質の送達を確立するために本発明者らは蛍光顕微鏡を使用してGNPへのGFPの付着を確認した、図10に示すとおり。ヒスチジンタグ付きGFPで処置したBY2細胞の蛍光画像は、中央部に蛍光を示さない暗い細胞での細胞外蛍光を示す。これは、ヒスチジンタグ付きGFPがAu粒子を伴わずに細胞に添加される対照処置は、粒子を内部移行させないことを示唆する。細胞内へのタンパク質の取り込みを支持する証拠は、i)処置細胞の内部での蛍光の蛍光強度の増大、ii)対照細胞における暗い糸と比較して処置細胞における蛍光発光している細胞質糸(図11を参照されたい)。同様の観察結果がYFPテザーGNPにおいても見られ、未処置細胞での植物細胞へのこれらの蛍光タンパク質の明確な内部移行を示唆していた(図12を参照されたい)。
【0110】
プラズマライジング/染色、またはプログラム細胞死(PCD)様の細胞学的特徴を示す細胞に由来する単一細胞でのある一定レベルのバックグラウンド反射および自己蛍光が存在する。そのようなバックグラウンドの問題からタンパク質が内部移行した細胞を描写し、かつタンパク質内部移行を直接的な証拠で明確に証明するために個々の粒子または粒子の凝集レベルを注目および追跡する詳細な反射顕微鏡調査を実施した。この調査の結果は、タンパク質を伴う粒子の細胞内および核内への内部移行を明確に示した。しかし蛍光タンパク質を伴う粒子の数が増加して蓄積した細胞は、顕微鏡下で観察した場合にプラズマライズする傾向を有した。GFPまたはYFPのいずれかに繋ぎ止められた多量のGNPの蓄積によるタンパク質濃度の上昇が毒性レベルに達するか、または顕微鏡下での長時間の観察がそのような細胞に結果的に有害効果を有するROSを誘導する可能性がある。
【実施例7】
【0111】
シロイヌナズナ(Arabidopsis thaliana)栽培種コロンビア(Columbia)のT1後代における導入遺伝子のゲノムへの組込みのための分子分析および立証
シロイヌナズナトランスジェニック植物由来のゲノムDNAを6週齢の葉全体の材料からDNeasy Plant Mini kitを製造者の説明書(Qiagen Inc)に従って使用して抽出した。以下のYFPおよびPAT PCRプライマーを、野外噴霧レベルの4〜5×の除草剤Finaleに耐性のT1実生由来の鋳型ゲノムDNAを使用するPCR反応において使用した。
YFP
順方向プライマー:5’―TGTTCCACGGCAAGATCCCCTACG―3’(配列番号3)
逆方向プライマー:5’―TATTCATCTGGGTGTGATCGGCCA―3’(配列番号4)
PAT
順方向プライマー:5’―GGAGAGGAGACCAGTTGAGATTAG―3’(配列番号5)
逆方向プライマー:5’―AGATCTGGGTAACTGGCCTAACTG―3’(配列番号6)
【0112】
PATおよびYFP(Yellow Florescent tag、Evrogen)遺伝子産物についてのPCRは、ゲノム鋳型DNA 100ng、1×ExTaq反応緩衝液(TaKaRa Bio)、0.2mM dNTP、各プライマー10pmolおよび0.025単位/μL ExTaqを含有する混合物の合計反応容量50μL中で増幅した。以下のPCR条件を使用した:96℃、5分間を1サイクルおよび以下のPCRプログラム:94℃で15秒間、65℃で30秒間、72℃で1分間を31サイクルおよび産物合成を完了するための72℃で7分間の最終伸長を実施した。ゲルの画像をBio Rad Gel imaging Systemを使用して得た(図13および14)。増幅した断片をゲル精製キット(Qiagen Inc)を製造者の説明書に従って使用してゲル精製した。
【0113】
PCR断片をPAT順方向プライマーおよびYFP順方向を使用して、advanced Sanger sequencing technology(MWG Biotechnologies,Inc)で使用して配列決定し、配列をSequencherソフトウェアを使用して分析した。
【0114】
結果は、PATおよびYFP配列はナノ粒子を通じて送達され、量子ドットはDNA送達を介在することを示し、したがってT1植物のゲノムDNAにおける導入遺伝子の安定なゲノムへの組込みの明確な証拠を提供している。
【実施例8】
【0115】
JTNT1タバコ単一細胞壁を越えるQDの送達の促進
細胞壁を越えるQDの非侵襲性送達を検査した実施例7において論じられた手順に基づいて、いくつかのペプチドを表面に機能化した。細胞透過性ペプチド(CPP)/タンパク質導入ユニット(PTD)付着決定を以下に記載のとおりゲル電気泳動を介して実施した。
【0116】
ゲル電気泳動をQD−ペプチドコンジュゲートについて、ペプチドのQDへの付着を確認するために実施した。使用した試料は、QD−アミン(対照)、QD−アミン−R9、QD−アミン−Zein、QD−アミン−Pep1およびQD−アミン−MPGであった。R9、Zein、Pep1およびMPGはペプチドである。2%(w/v)アガロースゲルでTBE(1×、pH8)緩衝液中、120V、1時間実施した。QD−アミン−ペプチドは、電極の陰性端に向かって移動し、QDに付着したペプチドの強く陽性の性質の付着を示したが、QD−アミンは静止したままであり、塩基性Phでのゲル緩衝液の中和効果によるアミン基の弱い陽電荷を示した、図15に示すとおり(レーン1:QD−アミン、2:QD アミン−R9、3:QD−アミンΥ−Zein、4:QD アミン−Pep1、5:QD−アミン−MPG)。
【0117】
ペプチドを細胞への内部移行について検査し、細胞および区画内の粒子の内部移行レベルを追跡するための方法として細胞内QDの発光を使用した。無傷の壁を有するJTNT1単一細胞を標的細胞としてこれらの実験において使用した。表1は、試料の処置を示す。細胞を顕微鏡下で追跡した。
【0118】
顕微鏡法は、試料の調製後1分間以内にSpinning Disk Confocal microscope(Andor Technology Revolution System)で行った。励起フィルターは488nmに設定し、発光フィルターは650nm超に設定した。
【0119】
表1に示すとおり、対照プロトプラストおよびJTNT1は、QDについて使用した発光波長で自己蛍光を示さなかった。顕著な細片(破損細胞の一部)がこれらの試料のそれぞれにおいて観察された。試料4、5、6および7は、単一細胞またはプロトプラスト内にQDの内部移行を示さなかった。試料8および9は、壁がある単一細胞およびプロトプラストの両方において核の周辺でのQDの明確な存在を示した。これは、核局在化シグナル(NLS)を有するQD上の細胞透過性ペプチドの存在によるものであった。しかしQDに繋ぎ止められたy−Zeinを有する試料番号6および7は、細胞内へのQD内部移行を示さなかった。これは細胞透過性ペプチド(CPP)またはタンパク質導入ユニット(PTD)によってQDが細胞に取り込まれたことを示唆し、アミンによってのみ機能化され、CPP/PTDでは機能化されなかったQDと同様にy−Zeinは内部移行されなかった。
【0120】
【表1】
【0121】
このデータは、無傷の機能性細胞の細胞壁を越えてQDを取り込む核局在化シグナル(NLS)を有するCPP/PTDに繋ぎ止められた量子ドットの細胞内部移行のSpinning Disk Confocal microscope(Andor Technology Revolution System)を介しての証拠を実証している。QDの核局在化は、酵素処理によって細胞壁を取り除かれたプロトプラストを基とする細胞の内部移行において観察されたとおり、試料9および細胞壁の非存在時に細胞壁を越えるQDの核局在化が可能である。したがって細胞壁の単なる存在は量子ドットの内部移行を妨げず、無傷の細胞壁を有する植物細胞における粒子の侵入がCPP/PTDで非侵襲的に実証されることを証明する。
【0122】
本発明は特定の実施形態で記載されているが、本発明は本開示の精神および範囲の中でさらに変更され得る。したがって本出願は、その一般的原理を使用する本発明の任意の変更、使用または適用に及ぶことを意図する。さらに本出願は、本発明に関連し、かつ添付の特許請求の範囲およびその等価物の限定内にある当技術分野での既知または慣用の実施内にある、本開示からのそのような逸脱を包含することを意図する。
【特許請求の範囲】
【請求項1】
細胞壁を有する植物細胞に目的の分子を導入する方法であって、
細胞壁を有する植物細胞を提供するステップと、
ナノ粒子を目的の分子で被覆するステップと、
細胞壁を有する細胞と被覆されたナノ粒子とを互いに接触させて置くステップと、
細胞壁を含む細胞へのナノ粒子および目的の分子の取り込みを可能にするステップと
を含む方法。
【請求項2】
目的の分子でナノ粒子を被覆するステップが、ナノ粒子の表面での非共有結合的吸着により目的の分子を固定化するステップを含む、請求項1に記載の方法。
【請求項3】
ナノ粒子に目的の分子を吸着させるステップをさらに含む、請求項1に記載の方法。
【請求項4】
細胞壁を含む植物細胞の区画へのナノ粒子の取り込みを可能にするステップをさらに含む、請求項1に記載の方法。
【請求項5】
細胞内区画を標的化するタンパク質でナノ粒子を被覆するステップをさらに含む、請求項4に記載の方法。
【請求項6】
区画が、サイトゾル、核、液胞膜、色素体、エチオプラスト、有色体、白色体、エライオプラスト、プロテイノプラスト、アミロプラスト、葉緑体、および二重膜の内腔からなる群から選択される、請求項4に記載の方法。
【請求項7】
細胞壁を含む植物細胞がタバコ、ニンジン、トウモロコシ、セイヨウアブラナ、ナタネ、ワタ、パーム、ピーナッツ、ダイズ、オリザ種(Oryza sp)、シロイヌナズナ種(Arabidopsis sp)、トウゴマ種(Ricinus sp)およびサトウキビの細胞からなる群から選択される、請求項1に記載の方法。
【請求項8】
植物細胞が、胚、分裂組織、カルス、花粉、葉、葯、根、根端、花、種子、さやおよび茎からなる群から選択される組織由来である、請求項1に記載の方法。
【請求項9】
ナノ粒子が、金ナノ粒子、金被覆ナノ粒子、多孔性ナノ粒子、メソ多孔性ナノ粒子、シリカナノ粒子、ポリマーナノ粒子、タングステンナノ粒子、ゼラチンナノ粒子、ナノシェル、ナノコア、ナノスフェア、ナノロッド、磁性ナノ粒子およびそれらの組合せからなる群から選択される、請求項1に記載の方法。
【請求項10】
ナノ粒子の表面を誘導体化するステップをさらに含む、請求項1に記載の方法。
【請求項11】
目的の分子が、核酸、DNA、RNA、RNAi分子、遺伝子、プラスミド、コスミド、YAC、BAC、ポリペプチド、酵素、ホルモン、糖ペプチド、糖類、脂肪、シグナル伝達ペプチド、抗体、ビタミン、メッセンジャー、セカンドメッセンジャー、アミノ酸、cAMP、薬剤、除草剤、殺菌剤、抗生物質およびそれらの組合せからなる群から選択される構成要素を含む、請求項1に記載の方法。
【請求項12】
目的の分子が遺伝子を含む、請求項10に記載の方法。
【請求項13】
遺伝子が外来タンパク質遺伝子、農学的遺伝子、またはマーカー遺伝子である、請求項11に記載の方法。
【請求項14】
目的の分子が安定に組み込まれている細胞を選択するステップをさらに含む、請求項11に記載の方法。
【請求項15】
選択された細胞が再生可能な細胞である、請求項13に記載の方法。
【請求項16】
再生可能な細胞から植物を再生するステップをさらに含む、請求項14に記載の方法。
【請求項17】
遺伝子を発現させる方法であって、
細胞壁を有する植物細胞を提供するステップと、
ナノ粒子を遺伝子で被覆するステップと、
細胞壁を有する植物細胞と被覆されたナノ粒子とを互いに接触させて置くステップと、
細胞壁を含む植物細胞へのナノ粒子および目的の分子の取り込みを可能にするステップと、
遺伝子を発現させるステップと
を含む方法。
【請求項18】
遺伝子が葉緑体で発現される、請求項16に記載の方法。
【請求項19】
遺伝子を発現する細胞を選択するステップをさらに含む、請求項16に記載の方法。
【請求項20】
植物細胞に分子性物質を移行させるための方法であって、
ナノ粒子をプラスミドDNAで被覆するステップと、
被覆されたナノ粒子と細胞壁を有する無傷の植物細胞とを、植物細胞への被覆されたナノ粒子の取り込みを可能にする条件下で接触させるステップと
を含む方法。
【請求項21】
ナノ粒子が金を含む、請求項19に記載の方法。
【請求項1】
細胞壁を有する植物細胞に目的の分子を導入する方法であって、
細胞壁を有する植物細胞を提供するステップと、
ナノ粒子を目的の分子で被覆するステップと、
細胞壁を有する細胞と被覆されたナノ粒子とを互いに接触させて置くステップと、
細胞壁を含む細胞へのナノ粒子および目的の分子の取り込みを可能にするステップと
を含む方法。
【請求項2】
目的の分子でナノ粒子を被覆するステップが、ナノ粒子の表面での非共有結合的吸着により目的の分子を固定化するステップを含む、請求項1に記載の方法。
【請求項3】
ナノ粒子に目的の分子を吸着させるステップをさらに含む、請求項1に記載の方法。
【請求項4】
細胞壁を含む植物細胞の区画へのナノ粒子の取り込みを可能にするステップをさらに含む、請求項1に記載の方法。
【請求項5】
細胞内区画を標的化するタンパク質でナノ粒子を被覆するステップをさらに含む、請求項4に記載の方法。
【請求項6】
区画が、サイトゾル、核、液胞膜、色素体、エチオプラスト、有色体、白色体、エライオプラスト、プロテイノプラスト、アミロプラスト、葉緑体、および二重膜の内腔からなる群から選択される、請求項4に記載の方法。
【請求項7】
細胞壁を含む植物細胞がタバコ、ニンジン、トウモロコシ、セイヨウアブラナ、ナタネ、ワタ、パーム、ピーナッツ、ダイズ、オリザ種(Oryza sp)、シロイヌナズナ種(Arabidopsis sp)、トウゴマ種(Ricinus sp)およびサトウキビの細胞からなる群から選択される、請求項1に記載の方法。
【請求項8】
植物細胞が、胚、分裂組織、カルス、花粉、葉、葯、根、根端、花、種子、さやおよび茎からなる群から選択される組織由来である、請求項1に記載の方法。
【請求項9】
ナノ粒子が、金ナノ粒子、金被覆ナノ粒子、多孔性ナノ粒子、メソ多孔性ナノ粒子、シリカナノ粒子、ポリマーナノ粒子、タングステンナノ粒子、ゼラチンナノ粒子、ナノシェル、ナノコア、ナノスフェア、ナノロッド、磁性ナノ粒子およびそれらの組合せからなる群から選択される、請求項1に記載の方法。
【請求項10】
ナノ粒子の表面を誘導体化するステップをさらに含む、請求項1に記載の方法。
【請求項11】
目的の分子が、核酸、DNA、RNA、RNAi分子、遺伝子、プラスミド、コスミド、YAC、BAC、ポリペプチド、酵素、ホルモン、糖ペプチド、糖類、脂肪、シグナル伝達ペプチド、抗体、ビタミン、メッセンジャー、セカンドメッセンジャー、アミノ酸、cAMP、薬剤、除草剤、殺菌剤、抗生物質およびそれらの組合せからなる群から選択される構成要素を含む、請求項1に記載の方法。
【請求項12】
目的の分子が遺伝子を含む、請求項10に記載の方法。
【請求項13】
遺伝子が外来タンパク質遺伝子、農学的遺伝子、またはマーカー遺伝子である、請求項11に記載の方法。
【請求項14】
目的の分子が安定に組み込まれている細胞を選択するステップをさらに含む、請求項11に記載の方法。
【請求項15】
選択された細胞が再生可能な細胞である、請求項13に記載の方法。
【請求項16】
再生可能な細胞から植物を再生するステップをさらに含む、請求項14に記載の方法。
【請求項17】
遺伝子を発現させる方法であって、
細胞壁を有する植物細胞を提供するステップと、
ナノ粒子を遺伝子で被覆するステップと、
細胞壁を有する植物細胞と被覆されたナノ粒子とを互いに接触させて置くステップと、
細胞壁を含む植物細胞へのナノ粒子および目的の分子の取り込みを可能にするステップと、
遺伝子を発現させるステップと
を含む方法。
【請求項18】
遺伝子が葉緑体で発現される、請求項16に記載の方法。
【請求項19】
遺伝子を発現する細胞を選択するステップをさらに含む、請求項16に記載の方法。
【請求項20】
植物細胞に分子性物質を移行させるための方法であって、
ナノ粒子をプラスミドDNAで被覆するステップと、
被覆されたナノ粒子と細胞壁を有する無傷の植物細胞とを、植物細胞への被覆されたナノ粒子の取り込みを可能にする条件下で接触させるステップと
を含む方法。
【請求項21】
ナノ粒子が金を含む、請求項19に記載の方法。
【図1】

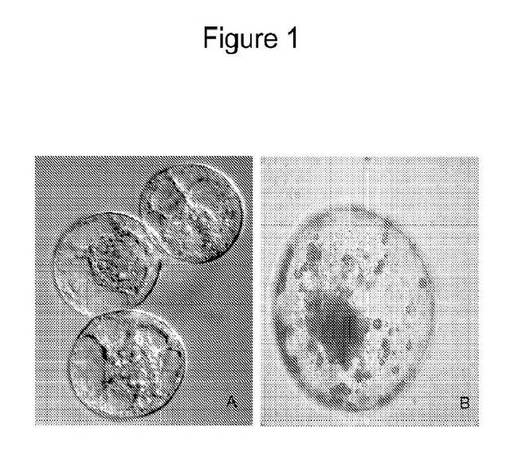
【図2】

【図3】
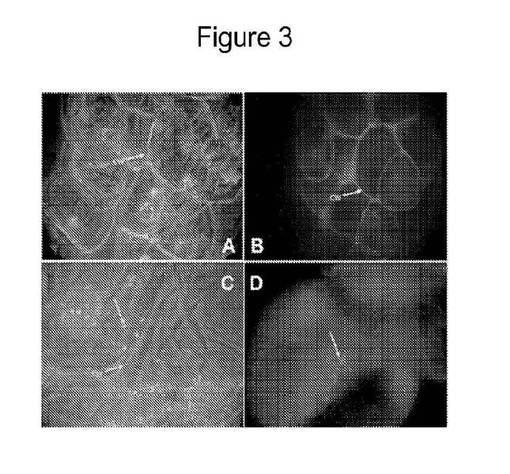
【図4】
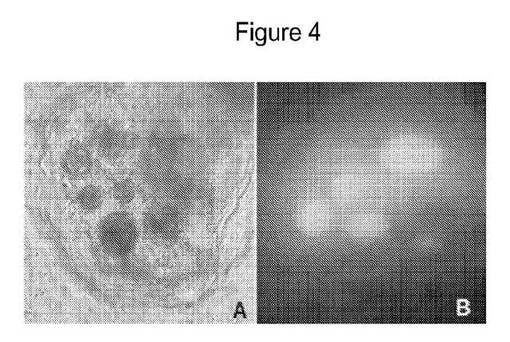
【図5】
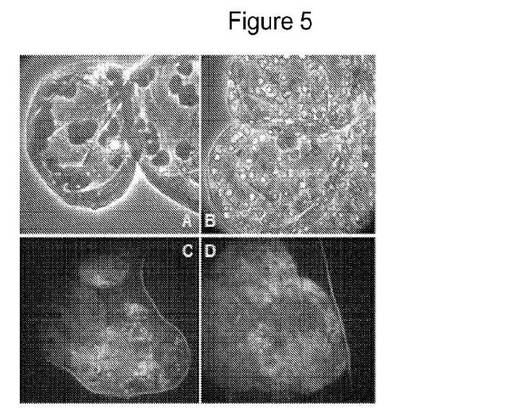
【図6】
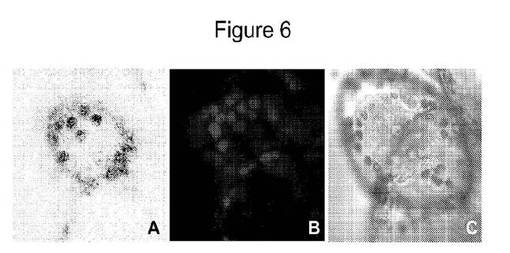
【図7】
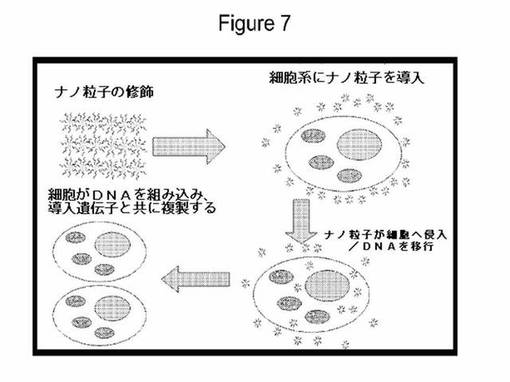
【図8】
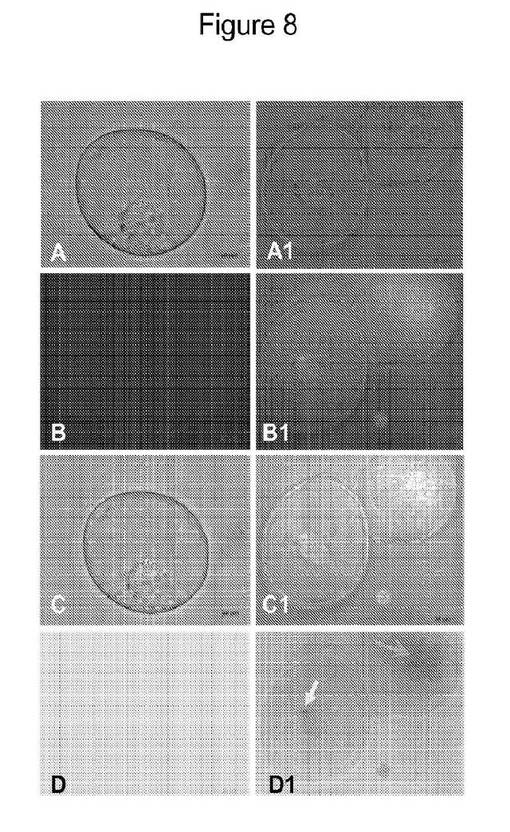
【図9】
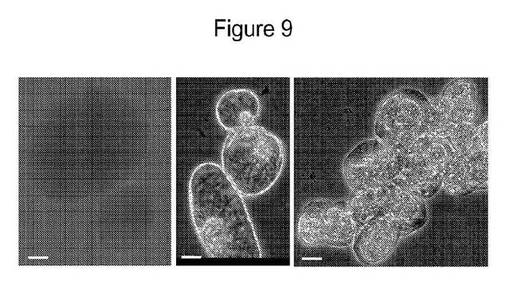
【図10】

【図11】
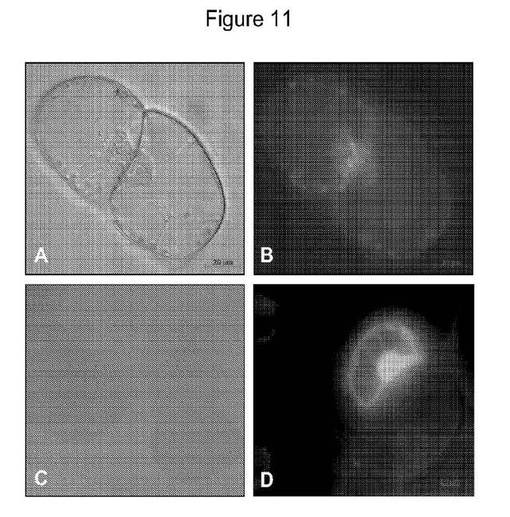
【図12】
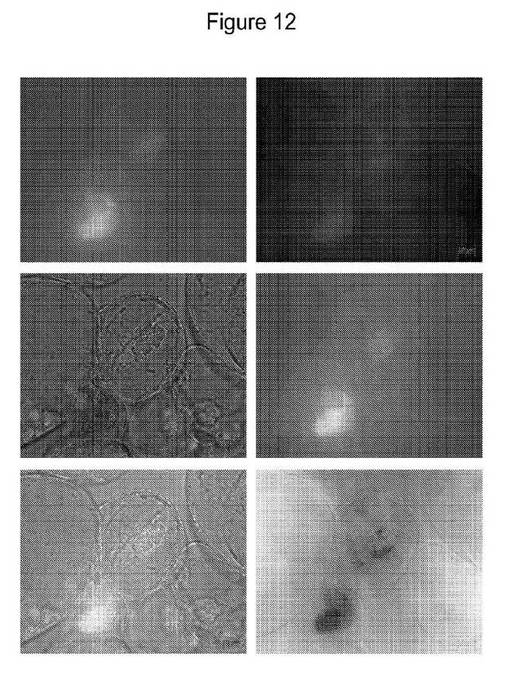
【図13】
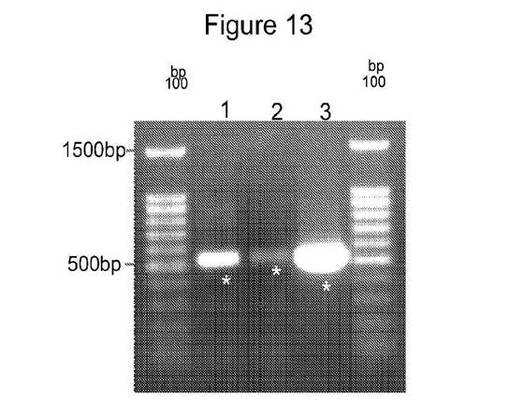
【図14】
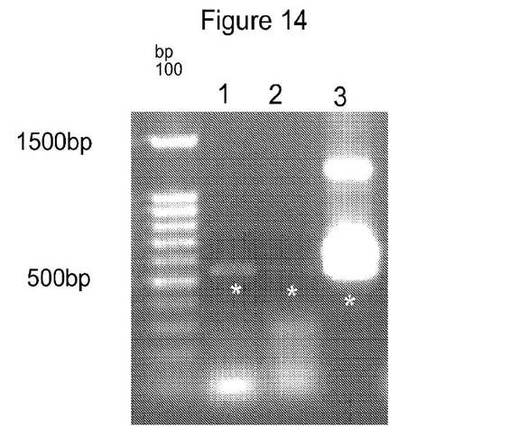
【図15】

【図16】
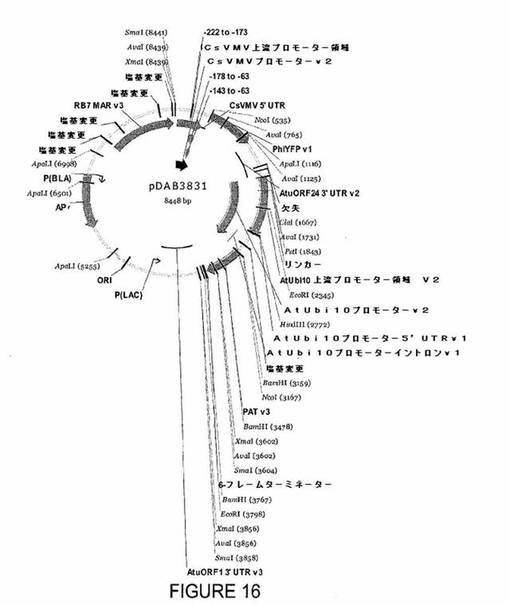
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【公表番号】特表2010−539989(P2010−539989A)
【公表日】平成22年12月24日(2010.12.24)
【国際特許分類】
【出願番号】特願2010−528195(P2010−528195)
【出願日】平成20年10月3日(2008.10.3)
【国際出願番号】PCT/US2008/078860
【国際公開番号】WO2009/046384
【国際公開日】平成21年4月9日(2009.4.9)
【出願人】(501035309)ダウ アグロサイエンシィズ エルエルシー (197)
【Fターム(参考)】
【公表日】平成22年12月24日(2010.12.24)
【国際特許分類】
【出願日】平成20年10月3日(2008.10.3)
【国際出願番号】PCT/US2008/078860
【国際公開番号】WO2009/046384
【国際公開日】平成21年4月9日(2009.4.9)
【出願人】(501035309)ダウ アグロサイエンシィズ エルエルシー (197)
【Fターム(参考)】
[ Back to top ]