
構成的植物プロモーター
【課題】植物、細胞、組織または器官を、一または複数の生物的および/または非生物的ストレスに晒したときに、特定のプロモーター活性が有意に低下しない植物、植物細胞、組織または器官の創製。
【解決手段】ゲノムに統合されたキメラ遺伝子を含む、トランスジェニック植物または植物細胞または植物組織または器官であって、前記キメラ遺伝子が、同種または異種の核酸配列に動作可能に連結された構成的プロモーターを含み、前記プロモーターが、特定核酸配列の群から選択されることを特徴とする、トランスジェニック植物または植物細胞または植物組織または器官。
【解決手段】ゲノムに統合されたキメラ遺伝子を含む、トランスジェニック植物または植物細胞または植物組織または器官であって、前記キメラ遺伝子が、同種または異種の核酸配列に動作可能に連結された構成的プロモーターを含み、前記プロモーターが、特定核酸配列の群から選択されることを特徴とする、トランスジェニック植物または植物細胞または植物組織または器官。
【発明の詳細な説明】
【発明の分野】
【0001】
本発明は、強力な構成的植物プロモーターおよびその使用の分野に関する。プロモーターは、植物または植物細胞において同種または異種のタンパク質を発現させるため、または活性な核酸分子、たとえばセンスおよび/またはアンチセンスRNAを発現させるために使用することができる。プロモーター活性を有する核酸配列、並びにキメラ遺伝子、ベクター、およびこれらを含む組換え(トランスジェニック)細胞および生物が提供される。また、プロモーターを含むトランスジェニック細胞または生物、とりわけ植物および植物細胞を作成する方法が提供される。更に、細胞質ゾルのシステインシンターゼタンパク質およびこれをコードする核酸配列が提供される。
【発明の背景】
【0002】
多くの植物プロモーターが、当該技術分野で公知であり、これらは、トランスジェニック植物または植物細胞においてタンパク質またはペプチドを発現させるため、または遺伝子または遺伝子ファミリーの発現を停止させる(silencing)ための有用なツールである。これら植物プロモーターには、構成的プロモーター、誘導性プロモーター、発生調節プロモーター、組織優先的または組織特異的プロモーターが含まれる。一般に使用される構成的プロモーターの例は、以下のとおりである:カリフラワーモザイクウイルス(CaMV)の単離体CM 1841 (Gardner et al., 1981, Nucleic Acids Research 9, 2871-2887)、CabbB-S (Franck et al., 1980, Cell 21, 285-294)、およびCabbB-JI (Hull and Howell, 1987, Virology 86, 482-493)の35Sプロモーターまたは増強(enhanced)35Sプロモーター(“35Sプロモーター”);Odell et al. (1985, Nature 313, 810-812) またはUS5164316に記載される35Sプロモーター、ユビキチンファミリー由来のプロモーター (たとえばトウモロコシのユビキチンプロモーター、Christensen et al., 1992, Plant Mol. Biol. 18, 675-689, EP 0 342 926、Cornejo et al. 1993, Plant Mol. Biol. 23, 567-581参照)、gos2プロモーター (de Pater et al., 1992 Plant J. 2, 837-844)、emuプロモーター (Last et al., 1990, Theor. Appl. Genet. 81,581-588)、アラビドプシス(Arabidopsis)のアクチンプロモーター、たとえばAn et al. (1996, Plant J. 10, 107-121)に記載されるプロモーター、イネのアクチンプロモーター、たとえばZhang et al.(1991, The Plant Cell 3, 1155-1165)に記載されるプロモーターおよびUS 5,641,876に記載されるプロモーターまたはWO 00/70067に記載されるようなイネのアクチン2プロモーター;キャッサバ葉脈モザイクウイルスのプロモーター (WO 97/48819, Verdaguer et al. 1998, Plant Mol. Biol. 37, 1055-1067)、サブタレニアン・クローバー萎縮ウイルス由来のpPLEXシリーズのプロモーター (WO 96/06932、とりわけS7プロモーター)、アルコールデヒドロゲナーゼプロモーター、たとえばpAdh1S (GenBankアクセッション番号X04049、X00581)、US 6 051 753およびEP 426 641に記載されるゴマノハグサモザイクウイルスプロモーター、ヒストン遺伝子プロモーター、たとえばアラビドプシス(Arabidopsis)由来のPh4a748プロモーター (Plant Mol. Biol. 8: 179-191)、CoYMV (ツユクサ黄色斑紋ウイルス)プロモーター (Medberry et al. 1992, The Plant Cell 4:185-192)など。
【0003】
しかし、公知の構成的プロモーターの共通の欠点は、これらが、器官または発生で調節される発現、またはストレス誘導性の発現変化など、しばしば活性の変動を示すことである。本発明者らは、トランスジェニック植物においてCaMV 35Sプロモーターの活性が、非生物的ストレス、とりわけトランスジェニック植物をスペインの農場で生育させたときに引き起こされる熱ストレスに感受性を示すことを見出した。したがって、一または複数の生物的および/または非生物的ストレスに感受性を示さない新規な構成的プロモーターを提供する必要がある。加えて、トランスジェニック植物の生成において植物由来のプロモーターを使用することが、調節の観点から望ましい。
【0004】
加えて、植物に感染可能なウイルスに由来するウイルス性プロモーターは、ウイルスの植物への感染が、トランスジェニックプロモーターのサイレンシングを引き起こし得るため、宿主植物種の形質転換にとってあまり好ましくない(Seemanpillai et al., 2003, Mol Plant Microbe Interact. 16(5): 429-38; Al-Kaff et al., 2000, Nat Biotechnol. 18: 995-9)。幾つかの同一プロモーターの使用は、サイレンシングという結果を招き得るため、遺伝子スタッキングアプローチのためには、異なる構成的プロモーターも必要である(Yang et al., 2005, Plant Mol Biol. 58: 351-66)。
【発明の概要】
【0005】
ゲノムに統合されたキメラ遺伝子を含む、トランスジェニック植物または植物細胞または植物組織または器官であって、前記キメラ遺伝子が、同種または異種の核酸配列に動作可能に連結された構成的プロモーターを含み、前記プロモーターが、
(a)配列番号1または配列番号2の核酸配列、または配列番号6〜9の何れか一つ、またはCBSに(Centraalbureau voor Schimmelcultures、Uppsalalaan 8, 3584 CT Utrecht, The Netherlandsに、2006年8月3日、それぞれアクセッション番号CBS 120175およびCBS 120176で)寄託されたベクターpKG8135またはpKG8137にクローニングされたプロモーター配列;
(b)配列番号1または配列番号2の機能的断片、または(a)に記載の配列の何れかの機能的断片;
(c)配列番号1または2と少なくとも70%の配列同一性を備えた核酸配列、または(a)に記載の配列の何れかと少なくとも70%の配列同一性を備えた核酸配列;
(d)(c)の核酸配列の機能的断片
の群から選択されることを特徴とする、トランスジェニック植物または植物細胞または植物組織または器官が提供される。
【0006】
また、植物細胞に導入された際にプロモーター活性を有する単離された核酸配列であって、
(a)配列番号1または配列番号2の核酸配列、または配列番号6〜9の何れか一つ、またはCBSに(Centraalbureau voor Schimmelcultures、Uppsalalaan 8, 3584 CT Utrecht, The Netherlandsに、2006年8月3日、それぞれアクセッション番号CBS 120175およびCBS 120176で)寄託されたベクターpKG8135またはpKG8137にクローニングされたプロモーター配列;
(b)配列番号1または配列番号2の断片、または(a)に記載の配列の何れかの断片;
(c)配列番号1または2と少なくとも70%の配列同一性を備えた核酸配列、または(a)に記載の配列の何れかと少なくとも70%の配列同一性を備えた核酸配列;
(d)(c)の核酸配列の断片
から選択される配列を含む核酸配列が提供される。
【0007】
また、上記配列を含む、ベクター、キメラ遺伝子および宿主細胞も、本発明の一つの態様である。
【0008】
別の態様において、トランスジェニック植物または植物細胞または植物組織または器官におけるセンスおよび/またはアンチセンス核酸配列の構成的発現のための、細胞質ゾルの植物システインシンターゼ遺伝子のプロモーターの使用が提供される。
【0009】
更に、トランスジェニック植物または植物細胞を作成するための方法であって、
(a)転写される核酸配列に動作可能に連結され、必要に応じて更に3’UTR核酸配列に連結された上述のプロモーターを含むキメラ遺伝子、またはかかるキメラ遺伝子を含むベクターを生成する工程;
(b)前記キメラ遺伝子またはベクターを用いて植物または植物細胞を形質転換する工程;および必要に応じて、
(c)トランスジェニック植物または植物細胞を再生させる工程
を含む方法が提供される。
【0010】
更に別の態様において、配列番号3または4のヌクレオチド配列を含む単離された核酸分子が提供される。
【一般的な定義】
【0011】
「核酸配列」(または核酸分子)の用語は、一本鎖または二本鎖の形態のDNAまたはRNA分子、とりわけ、本発明のプロモーター活性を有するDNAまたはタンパク質もしくはタンパク質断片をコードするDNAを指す。「単離された核酸配列」は、自然環境から単離され自然環境にもはや存在しない核酸配列、たとえば、細菌宿主細胞または植物の核もしくはプラスチドのゲノムに存在する当該核酸配列を指す。
【0012】
「タンパク質」または「ポリペプチド」の用語は、交換可能に使用され、具体的な作用の仕方、大きさ、3次元構造、由来に関係なく、アミノ酸の鎖からなる分子を指す。よって、タンパク質の「断片」または「部分」は、「タンパク質」と呼ばれてもよい。「単離されたタンパク質」は、自然環境にもはや存在しないタンパク質、たとえばインビトロ、または組換え細菌もしくは植物の宿主細胞に存在するタンパク質を指すために使用される。
【0013】
「遺伝子」の用語は、適切な転写調節領域(たとえばプロモーター)に動作可能に連結された、細胞でRNA分子(たとえばmRNA)に転写される領域(転写領域)を含むDNA配列を意味する。よって、遺伝子は、動作可能に連結された幾つかの配列、たとえば、プロモーター、たとえば翻訳開始に関与する配列などを含む5’非翻訳リーダー配列(これは、5’UTRとも呼ばれ、翻訳開始コドンの上流に存在する転写されるmRNA配列に相当する)、(タンパク質)コード領域(cDNAまたはゲノムDNA)、並びにたとえば転写終結部位およびポリアデニル化部位(たとえばAAUAAAまたはその変異体)などを含む3’非翻訳配列(これは3’非翻訳領域または3’UTRとも呼ばれる)を含み得る。
【0014】
「キメラ遺伝子」(または組換え遺伝子)は、ある生物種に自然の状態で通常見出されない任意の遺伝子、とりわけ自然の状態では互いに関連がない一または複数の部分の核酸配列が存在する遺伝子を指す。たとえば、プロモーターは、転写される領域の一部または全部と、または別の調節領域と、自然の状態で関連がない。「キメラ遺伝子」の用語は、プロモーターまたは転写調節配列が、一または複数のセンス配列(たとえばコード配列)、またはアンチセンス(センス鎖のリバース相補鎖)または逆方向反復配列(センスおよびアンチセンス、転写によりRNA転写産物は二本鎖RNAを形成する)に動作可能に連結された発現構造体を含むと理解される。
【0015】
「3’UTR」または「3’非翻訳配列」(しばしば3’非翻訳領域または3’端とも呼ばれる)は、たとえば転写終結部位および(全てではないがほとんどの真核生物のmRNAで)ポリアデニル化シグナル(たとえばAAUAAAまたはその変異体)などを含む、遺伝子のコード配列の下流に見出される核酸配列を指す。mRNA転写産物は、転写終結後、ポリアデニル化シグナルの下流で切断され、ポリ(A)テールが付加され、これは、mRNAの細胞質(ここで翻訳が起こる)への輸送に関与する。
【0016】
「遺伝子の発現」は、適切な調節領域、とりわけプロモーターに動作可能に連結されたDNA領域が、生物学的に活性なRNA、すなわち生物学的に活性なタンパク質またはペプチド(または活性なペプチドの断片)に翻訳され得るRNA、または(たとえば転写後の遺伝子サイレンシングまたはRNAiにおいて)それ自体で活性なRNAに転写されるプロセスを指す。ある態様において活性なタンパク質は、リプレッサードメインが存在するために優性のネガティブな機能を有するタンパク質を指す。コード配列は、好ましくは、センスの向きにあり、所望の生物学的に活性なタンパク質またはペプチド、または活性なペプチド断片をコードする。遺伝子サイレンシングのアプローチにおいて、DNA配列は、好ましくは、アンチセンスDNAまたは逆方向反復DNAの形態で存在し、ターゲット遺伝子の短い配列をアンチセンスの向き、またはセンスとアンチセンスの向きで含む。「異所性発現」は、その遺伝子が通常発現されない組織での発現を指す。
【0017】
「転写調節配列」は、転写調節配列に動作可能に連結された核酸配列の転写速度を調節することが可能な核酸配列と本明細書で規定される。よって、本明細書で規定される転写調節配列は、転写の開始に必要な配列エレメント(プロモーターエレメント)、転写を維持し調節するのに必要な配列エレメント、たとえばアテニュエーターまたはエンハンサー、サイレンサーなど、あらゆるものを含む。ほとんどの場合、コード配列の上流(5’)転写調節配列を指すが、コード配列の下流(3’)に見出される調節配列も、この規定に包含される。
【0018】
本明細書で使用される「プロモーター」の用語は、一または複数の遺伝子の転写をコントロールするために機能する核酸断片であって、遺伝子の転写開始部位の転写方向に対して上流(5’)に位置し(転写開始は、配列の+1位と呼ばれ、それに対して上流のヌクレオチドは、マイナスの数を用いて呼ばれ)、DNA依存性RNAポリメラーゼの結合部位、転写開始部位、および任意の他のDNAドメイン(cis作用性配列)(たとえば、転写因子結合部位、リプレッサーおよびアクチベータータンパク質結合部位、およびプロモーターに由来する転写量を調節するために直接的もしくは間接的に作用することが当業者に公知の任意の他のヌクレオチド配列を含むがこれらに限定されないもの)の存在により構造的に同定される核酸断片を指す。転写開始(+1)の上流の真核生物のcis作用性配列の例には、TATAボックス(一般に転写開始のおよそ−20から−30位)、CAATボックス(一般に転写開始に対しておよそ−75位)、5’エンハンサーまたはサイレンサーエレメントなどが含まれる。「構成的」プロモーターは、ほとんどの組織(または器官)で、ほとんどの生理的および発生的条件下において活性なプロモーターである。より好ましくは、構成的プロモーターは、全ての主要な器官、たとえば少なくとも葉、茎、根、種子、果実および花で、本質的に全ての生理的および発生的条件下において活性である。最も好ましくは、構成的プロモーターは、全ての器官で、ほとんどの(好ましくは全ての)生理的および発生的条件下において活性である。「誘導性」プロモーターは、生理的(たとえばある種の化合物の外からの適用により)または発生的に調節されるプロモーターである。「組織特異的」プロモーターは、特定のタイプの組織または細胞でのみ活性化である。したがって、プロモーター活性は、当該プロモーターが、プロモーターの下流(3’)に動作可能に連結された核酸配列を転写する環境を参照することにより説明することができる。したがって、「構成的活性を有するプロモーター」、または植物もしくは植物細胞で「構成的」なプロモーターは、植物またはほとんどの組織(または器官)の植物細胞で、ほとんどの生理的および発生的条件下において転写を行う核酸配列を指す。「一または複数の生物的および/または非生物的ストレスに対して感受性を示さない」プロモーター、またはその活性が「一または複数の生物的および/または非生物的ストレス条件に晒されたときに低下しない」プロモーターは、正常な生理的および発生的条件下においてプロモーター活性を有し、かつ生物的および/または非生物的ストレスが、当該プロモーターを含む生物(たとえば植物)または細胞または組織または器官に与えられたときに、その活性が定量的に低下しないか、または少なくとも有意に低下しない核酸配列を指す。
【0019】
「ストレス」は、植物または植物細胞に作用し、植物の収量ロスおよび/または品質ロスという結果を招き得るが、植物に致命的でない、物理学的、化学的または生物学的由来の条件または圧力を指す。
【0020】
「非ストレス条件」は、本明細書において、生理機能および発生が正常であるかまたは最適である条件を指す。
【0021】
「生物的ストレス」は、生物的(生きている)作用物質、たとえば真菌類、ウイルス、生物のようなマイコプラズマ、昆虫、細菌、線虫など(すなわち、とりわけ植物の害虫および病原体)により引き起こされるストレスを指す。
【0022】
「非生物的ストレス」は、非生物的(生きていない)作用物質、たとえば温度ストレス(低温/凍結、熱)、塩度(塩)、風、金属、日長(光周期)、水ストレス(たとえば過少もしくは過剰の水アベイラビリティ、すなわち干ばつ、脱水、水浸しなど)、傷害、照射などにより引き起こされるストレスを指す。
【0023】
本明細書で使用される「動作可能に連結された」の用語は、機能的な関係でのポリヌクレオチドエレメントの連結を指す。核酸は、別の核酸配列と機能的な関係に置かれたときに「動作可能に連結される」。たとえば、プロモーター、または転写調節配列は、コード配列の転写に影響を及ぼす場合、コード配列に動作可能に転結される。動作可能に連結されるということは、連結されるDNA配列が、典型的には隣接していることを意味し、二つのタンパク質コード領域をつなげる必要がある場合、「キメラタンパク質」を産生するようにリーディングフレームで隣接していることを意味する。「キメラタンパク質」または「ハイブリッドタンパク質」は、天然にはそういうものとして見出されないが、機能的タンパク質を形成するようにつながれ、つながれたドメインの機能性(たとえばDNA結合ドメインまたは優性ネガティブ機能につながる機能ドメインの抑制)を示す、種々のタンパク質「ドメイン」(またはモチーフ)から構成されるタンパク質である。キメラタンパク質は、天然に存在する二以上のタンパク質の融合タンパク質であってもよい。本明細書で使用される「ドメイン」の用語は、当該ドメインの機能的特性を少なくとも備えた新たなハイブリッドタンパク質を提供するための、別のタンパク質に移転可能な特定の構造または機能を備えたタンパク質の任意の部分またはドメインを意味する。
【0024】
「ターゲットペプチド」の用語は、細胞内オルガネラ、たとえばプラスチド、好ましくは葉緑体、ミトコンドリア、または細胞外空間(分泌シグナルペプチド)へタンパク質を向かわせるアミノ酸配列を指す。ターゲットペプチドをコードする核酸配列は、タンパク質のアミノ末端(N-末端)をコードする核酸配列に(フレームで)融合され得る。
【0025】
「核酸構築物」または「ベクター」は、本明細書において、組換えDNA技術の使用により得られ、外因性DNAを宿主細胞に運搬するために使用される、人工の核酸分子を意味すると理解される。ベクターの骨格は、たとえば、バイナリーまたはスーパーバイナリーベクター(たとえばUS 5,591,616、US2002138879およびWO 95/06722参照)、コインテグレート(co-integrate)ベクターまたはT-DNAベクターとすることができ、これらは、当該技術分野で公知であり、本明細書の他の箇所に記載されており、ベクターの骨格に、キメラ遺伝子が統合されるか、あるいは適切な転写調節配列/プロモーターが既に存在する場合には、所望の核酸配列のみ(たとえばコード配列、アンチセンスまたは逆方向反復配列)が、転写調節配列/プロモーターの下流に統合される。ベクターは、通常、分子クローニングでの使用を容易にするための更なる遺伝的エレメント、たとえば選択マーカー、マルチクローニングサイトなどを含む(下記参照)。
【0026】
「宿主細胞」または「組換え宿主細胞」または「形質転換細胞」は、所望のタンパク質をコードするキメラ遺伝子、またはターゲット遺伝子/遺伝子ファミリーのサイレンシングのためのアンチセンスRNAまたは逆方向反復RNA(またはヘアピンRNA)を転写により産生する核酸配列をとりわけ含む、少なくとも一の核酸分子を細胞に導入した結果生じる、新たな個々の細胞(生物)を指す用語である。宿主細胞は、好ましくは植物細胞であるが、細菌細胞、真菌細胞(酵母細胞を含む)などであってもよい。宿主細胞は、染色体外(エピソーム)複製分子として核酸構築物を含有してもよいし、あるいはより好ましくは、宿主細胞の核もしくはプラスチドのゲノムに統合されたキメラ遺伝子を含んでいてもよい。「選択マーカー」の用語は、当業者によく知られた用語であり、発現した際に、当該選択マーカーを含有する一または複数の細胞を選択するために使用することができる任意の遺伝的構成要素(entity)を記載するために本明細書で使用される。選択マーカー遺伝子産物は、たとえば、抗生物質に対する抵抗性、あるいはより好ましくは、除草剤に対する抵抗性または別の選択可能な特性、たとえば表現型の形質(たとえばの色素形成の変化)または栄養上の要件などを付与する。「レポーター」の用語は、可視的マーカー、たとえば緑色蛍光タンパク質(GFP)、eGFP、ルシフェラーゼ、GUSなどを指すために主に使用される。
【0027】
遺伝子またはタンパク質の「オルソログ(ortholog)」の用語は、本明細書において、当該遺伝子またはタンパク質と同じ機能を有するが、当該遺伝子を有する種が分岐した時点(すなわち、当該遺伝子が、種分化により共通の祖先から進化した時点)から配列が(通常)相違している、別の種に見出される相同な遺伝子またはタンパク質を指す。よって、ある植物種に由来する遺伝子のオロソログは、両方の配列の比較(たとえば配列全体または特定のドメインにわたるパーセンテージ配列同一性)および機能分析に基いて、他の植物種で同定され得る。
【0028】
「同種」および「異種」の用語は、核酸またはアミノ酸の配列と、その宿主細胞または生物、とりわけトランスジェニック生物との間の関係を指す。よって、同種の配列は、宿主種に天然に見出されるが(たとえば、トマト遺伝子で形質転換されたトマト植物)、異種の配列は、宿主細胞に天然に見出されない(たとえば、ポテト植物由来の配列で形質転換されたトマト植物)。「相同体(homolog)」または「相同な」の用語は、文脈に応じて、共通の祖先の配列に由来する子孫の配列を択一的に指し得る(たとえば、これらはオルソログであり得る)。
【0029】
「ストリンジェントなハイブリダイゼーション条件」は、所定のヌクレオチド配列と実質的に同一のヌクレオチド配列を同定するために使用することができる。ハイブリダイゼーション条件のストリンジェンシーは、配列に依存し、様々な環境で異なる。一般に、ストリンジェントな条件は、規定のイオン強度およびpHにおける特定配列の熱融点(Tm)より約5℃下げるように選択される。Tmは、ターゲット配列の50%が、完全一致のプローブとハイブリダイズする(規定のイオン強度およびpHの下での)温度である。典型的には、塩濃度がpH 7において約0.02モルであり、温度が少なくとも60℃であるストリンジェントな条件が選択される。塩濃度の低下および/または温度の増加は、ストリンジェンシーを増大させる。RNA−DNAハイブリダイゼーション(たとえば100 ntのプローブを用いたノーザンブロット)のためのストリンジェントな条件は、たとえば、63℃で20分間の0.2×SSC中での少なくとも1回の洗浄を含む条件、または同等の条件である。DNA−DNAハイブリダイゼーション(たとえば100 ntのプローブを用いたサザンブロット)のためのストリンジェントな条件は、たとえば、少なくとも50℃、通常約55℃の温度で20分間の0.2×SSC中での少なくとも1回の洗浄(通常2回)を含む条件、または同等の条件である。Sambrook et al. (1989)およびSambrook and Russell (2001)も参照されたい。
【0030】
「配列同一性」および「配列類似性」は、二つの配列の長さに依存して、グローバルまたはローカルアラインメントアルゴリズムを用いて、二つのペプチドまたは二つのヌクレオチドの配列を整列させること(alignment)により決定することができる。同様の長さの配列は、好ましくは、全長にわたって最適に配列を整列させるグローバルアラインメントアルゴリズム(たとえばNeedleman Wunsch)を用いて整列させるが、実質的に異なる長さの配列は、好ましくは、ローカルアラインメントアルゴリズム(たとえばSmith Waterman)を用いて整列させる。(デフォルトパラメーターを用いてたとえばプログラムGAPまたはBESTFITにより最適に整列させた際の)配列が、(以下で規定される)配列同一性の最小パーセンテージを少なくとも共有する場合、配列は、「実質的に同一である」または「本質的に同様である」ということができる。GAPは、Needleman and Wunschのグローバルアラインメントアルゴリズムを使用して、二つの配列を完全な長さ(全長)にわたって整列させ、一致の数を最大にし、ギャップの数を最小にする。二つの配列が同様の長さを有する場合、配列同一性を決定するためにグローバルアラインメントが適切に使用される。一般に、GAPデフォルトパラメーターが使用され、ギャップ生成ペナルティー=50(ヌクレオチド)/8(タンパク質)であり、ギャップ伸長ペナルティー=3(ヌクレオチド)/2(タンパク質)である。ヌクレオチドについて、使用されるデフォルトスコアリングマトリクスは、nwsgapdnaであり、タンパク質について、デフォルトスコアリングマトリクスは、Blosum62である(Henikoff & Henikoff, 1992, PNAS 89, 915-919)。配列アラインメントおよびパーセンテージ配列同一性のスコアは、コンピュータープログラム、たとえばGCG Wisconsin Package, Version 10.3(Accelrys Inc., 9685 Scranton Road, San Diego, CA 92121-3752 USAから入手可能)を用いて、あるいはオープンソースソフトウェア、たとえばEmbossWIN version 2.10.0に含まれるプログラム“needle”(グローバルNeedleman Wunschアルゴリズムを使用)もしくは“water”(ローカルSmith Watermanアルゴリズムを使用)を用いて、上記GAPと同じパラメーターを用いて、あるいはデフォルトセッティング(“needle”および“water”の何れについても、また、タンパク質およびDNAアラインメントの何れについても、デフォルトギャップオープニングペナルティーは10.0であり、デフォルトギャップ伸長ペナルティーは0.5であり; デフォルトスコアリングマトリクスは、タンパク質についてはBlossum62であり、DNAについてはDNAFullである)を用いて決定され得る。配列が、実質的に異なる全長を有している場合、たとえばSmith Watermanアルゴリズムを使用したローカルアラインメントが好ましい。あるいは、パーセンテージ類似性または同一性は、たとえばFASTA、BLASTなどのアルゴリズムを用いて公共データベースに対してサーチすることにより決定してもよい。
【0031】
本明細書および特許請求の範囲において、動詞「含むこと(to comprise)」およびその語形変化は、非限定的な意味で使用され、そのワードの後につづく項目を含むが、具体的に挙げられていない項目を排除しないことを意味する。加えて、不定冠詞“a”または“an”によるエレメントの指示は、その文脈が、一つおよび唯一のエレメントが存在することを明らかに要求しない限り、二つ以上のエレメントが存在する可能性を排除しない。よって、不定冠詞“a”または“an”は、通常「少なくとも一つ」を意味する。更に、本明細書で「配列」を参照する場合、ある配列のサブユニットを備えた実際の物理的分子(たとえばアミノ酸)を指すと理解される。
【0032】
本発明に従って“植物(plant)”または“植物(plants)”(すなわち複数の植物)を参照する場合はいつも、別途示さない限り、植物の部分(細胞、組織または器官、種子、切断または収穫された部分、葉、芽生え、花、花粉、果実、茎、根、カルス、プロトプラストなど)、親の際立った特徴(たとえば導入遺伝子の存在)を保持する当該植物の子孫またはクローン繁殖体、たとえば自家受粉または交配により得られた種子、たとえば(二つの近交親系統の交配により得られた)ハイブリッド種子、これに由来するハイブリッド植物および植物部分も包含すると理解される。
【詳細な説明】
【0033】
スペインの農場試験でCaMV 35Sプロモーター(Franck et al., 1980, Cell 21, 285-294に記載されるシングル35Sプロモーター)を含むトランスジェニック植物を試験したところ、35Sプロモーターの活性は、高い夏の温度に影響を受けることが見出された。老齢の葉では、発現の全損失が起こった。したがって、植物の細胞および器官で構成的な活性を示し、一または複数の生物的および/または非生物的ストレスに本質的に感受性を示さない植物プロモーターは、トランスジェニック植物、とりわけ発生の間または成熟期に一または複数のストレス条件に晒され得る植物における核酸配列の制御発現にとって望ましいため、かかるプロモーターを単離するためのプログラムを本発明者らは開始した。cDNA-AFLPを用いて、強力で構成的なプロモーターの遺伝子をトマト植物から単離し、これは、ストレス条件、たとえば干ばつストレス、低温ストレス、熱ストレス、病原体ストレス(CMV感染)、日長変動、照射(たとえばUV誘発ストレス)、水ストレス(水撒き過多および水撒き過少)などの下において、強力で構成的な転写プロファイルを保持していた(実施例参照)。対応するプロモーターを単離することにより、強力で構成的な発現を植物に付与した(または付与することが可能な)プロモーターの部類が見出された。プロモーター強度は、CaMV 35Sプロモーターのプロモーター強度と少なくとも同等であり、幾つかの宿主種(トマト)ではそれより有意に高かった。更に、プロモーターは、これらを含む植物または植物部分が、種々の生物的および/または非生物的ストレス条件を受けたときに、活性の低下を示さない(すなわち、強力で構成的なままである)。これらプロモーターは、本明細書において「AA6プロモーター」と呼ばれる。
【0034】
核酸配列、キメラ遺伝子およびベクター
一つの態様において、植物細胞でプロモーター活性を有する単離された核酸配列(好ましくはゲノムまたは合成DNA配列)が提供され、これは、植物および植物の組織または器官において強力で構成的な転写活性を示す。プロモーターは、好ましくは植物の全ての器官で活性であるが、少なくとも主要な器官で活性であり、プロモーター強度は、35S(Franck et al., 同上に記載されるCaMV 35Sプロモーター)のプロモーター強度と少なくともほとんど同等である(またはそれより強力である)。好ましくは、この核酸配列のプロモーター活性は、トランスジェニック植物、または植物組織または器官が、一または複数の非生物的および/または生物的ストレスを受けたときに、低下しないか、または少なくとも有意に低下しない。この点において有意な低下は、非ストレス条件下での同一の組織または器官における活性と比較して、1%以上(たとえば2%、3%、5%、10%など、100%まで)のプロモーター活性の統計的に有意な(定量的)低下を指す。よって、好ましくは、プロモーターは、ストレス条件下において(好ましくは全ての器官、とりわけ少なくとも主要な植物器官で)強力で構成的なままである。
【0035】
一つの態様において、配列番号1(“3kb”プロモーター)または配列番号2(“5kb”プロモーター)、または配列番号6〜9の何れか一つ(配列番号6および8は“3kb”プロモーターであり、配列番号7および9は“5kb”プロモーターであり、配列番号1および2と比較して、幾つかの不明ヌクレオチドまたは僅かに異なる配列を有する)、またはpKG8135 (CBS120175)またはpKG8137 (CBS120176)にクローニングされたプロモーター配列、または少なくとも植物においてプロモーター活性を有するこれらの何れかの活性(機能的)断片、たとえば配列番号1または2、配列番号6〜9、またはpKG8135 (CBS120175)またはpKG8137 (CBS120176)内のプロモーターの少なくとも200、300、400、500、600、800、900、1000、1200、1500、2000、2500、2800、2900、3000、3500、4000、4500またはそれ以上の連続ヌクレオチドの断片を含む、構成的AA6プロモーターが提供される。「活性断片」または「機能的断片」、または「プロモーター活性を有する断片」は、植物細胞、器官および植物において、好ましくは、配列番号1および2、配列番号6〜9、またはpKG8135 (CBS120175)またはpKG8137 (CBS120176)内のプロモーターと少なくとも同じ組織および器官において構成的転写を与えることができる核酸断片を指す。これは、以下に記載するとおり、好ましくはレポーター遺伝子に動作可能に連結されたかかる断片で植物細胞を形質転換し、プロモーター活性を定性的(空間的−時間的転写)および/または定量的にアッセイすることにより試験することができる。
【0036】
本明細書において配列番号1を参照する場合、配列番号6または8の何れか、またはpKG8135 (CBS120175)に存在する配列も参照していること、並びにその文章は、これらプロモーター配列の何れかを参照するように読むことができることが理解される。同様に、本明細書において配列番号2を参照する場合、配列番号7または9の何れか、またはpKG8137 (CBS120175)に存在する配列も参照していること、並びにその文章は、これらプロモーター配列の何れかを指すように読むことができることが理解される。
【0037】
一つの態様において、プロモーター断片の強度は、定量的に、配列番号1および/または2の強度(したがって、35Sの強度)とほとんど同一であるか、またはそれよりも高い。配列番号1および2は、翻訳開始コドン(mRNA上のAUGまたはDNA上のATG)のすぐ上流の配列であるため、活性断片は、配列番号1および/または2の5’端での欠失により好ましくは作成される。したがって、断片は、好ましくは、配列番号1または2の3’領域の(上述のとおり)少なくとも200、300、400、500等のヌクレオチドを含む。明らかに、DNA断片は、幾つかの手法、たとえばデノボDNA合成、または制限酵素、または末端ヌクレアーゼなどを用いて作成され得る。
【0038】
しかし、ある種のシス作用性エレメント(たとえばエンハンサー配列)の除去は、低いプロモーター活性という結果を招き得る。したがって、様々な態様において、プロモーター断片の強度は、配列番号1および/または2の強度より(したがって、35Sの強度より)定量的に低い。幾つかのアプリケーションにとって、強度の低下したプロモーターが好ましい。当業者であれば、当該技術分野で公知の手法を用いて、全長プロモーターおよび任意のプロモーター断片の活性を容易に決定し、その強度および組織特異性を、35Sまたは本明細書で提供されるプロモーターのものと比較することができる。たとえば、Medberry et al. (Plant Cell, 1992, Vol. 4: 185-192)は、一過性発現アッセイを用いてプロモーター強度を比較する方法を記載する。
【0039】
配列番号1および2のプロモーター、およびその機能的断片は、好ましくは、プロモーターを含む植物または植物細胞、組織または器官が晒され得る少なくとも一つの(好ましくは複数の、最も好ましくはあらゆる)生物的および/または非生物的ストレスに感受性を示さない(下記参照)。よって、活性は、ストレス条件に晒されている間、構成的で強力なままである。
【0040】
また、上記AA6プロモーターの「変異体」およびかかる変異体の機能的断片も提供される。これら変異体には、配列番号1および/または2と本質的に類似している核酸配列(および上述のとおり、これら変異体配列の機能的断片)であって、構成的プロモーター活性を有している、すなわち、植物、植物細胞、組織または器官で、最も好ましくは配列番号1および2と少なくとも同じ組織および器官で構成的転写を提供することができる核酸配列が含まれる。配列番号1および/または2と「本質的に類似している」配列は、Needleman and Wunsch Pairwiseアラインメント (GCGのプログラム“GAP”またはEmbosswin, version 2.10.0の“needle”)を用いて、ギャップ生成ペナルティー = 5およびギャップ伸長ペナルティー = 3で、配列番号1および/または配列番号2と(全長にわたって)少なくとも約50、55、60、65、70、75、80、85、90、95、96、97、98、99%またはそれ以上の核酸配列同一性を備え、かつ植物または植物細胞においてプロモーター活性を有する核酸配列である。好ましい態様において、変異体(および機能的断片)の活性は、組織および器官の全てにおいて強力であり、すなわち、定量的に配列番号1または2と少なくとも同等に強力である(またはそれより強力である)。更なる態様において、これら変異体(および機能的断片)の活性は、一または複数の生物的および/または非生物的ストレスに感受性を示さず、すなわち強力で構成的なままである。
【0041】
本明細書で提供される核酸配列の変異体または機能的断片を同定、合成または単離するために多くの方法が使用可能であることは明らかであり、たとえば核酸ハイブリダイゼーション、PCR技術、インシリコ(in silico)分析、および核酸合成などが挙げられる。たとえば、核酸ハイブリダイゼーションは、ストリンジェントまたは適度にストリンジェントなハイブリダイゼーション条件下で、配列番号1または2、またはこれらの断片とハイブリダイズするDNA配列を他の植物種または品種で同定するために使用することができる。あるいは、配列データベースを、たとえばBLAST、FASTAなどの公知のアルゴリズムを用いて、変異体配列についてインシリコで(in silico)スクリーニングすることができる。このようにして、他の植物種またはトマトの他品種から、または他の生物全体から変異体配列を単離することは実現可能である。以下に記載されるとおり、トマトの他品種または他の植物種、とりわけナス属(Solanum)の種で見出される同じ遺伝子(細胞質ゾルのシステインシンターゼ遺伝子およびそのオルソログ)の他の対立遺伝子のプロモーターが、本明細書にとりわけ含まれる。たとえば、cDNAライブラリーは、一または複数の植物種、一または複数の品種、または一つの種または品種の異なる組織から構築され得る。cDNAライブラリーを、(たとえば、配列番号4(トマトシステインシンターゼcDNA)、またはその断片もしくは変異体に由来するプローブまたはプライマーを用いて)システインシンターゼcDNAについてスクリーニングしてもよい。同様に、実施例に記載されるとおり、ディファレンシャルディスプレイ法(たとえばcDNA-AFLP)を使用して、システインシンターゼ転写産物を同定してもよい。TAIL-PCR(Liu et al. 1995, Genomics 25(3):674-81; Liu et al. 2005, Methods Mol Biol. 286:341-8)、Linker-PCR、またはInverse PCR (IPCR)などの方法を使用して、遺伝子の上流転写調節領域を単離してもよい。
【0042】
核酸配列(または変異体の断片)が構成的プロモーター活性を有するかどうか、すなわち、全ての器官において構成的転写を与えることができるかどうか、その活性が「強力」であるかどうか、並びに、トランスジェニック細胞、組織、器官または生物(とりわけ植物または植物細胞)が晒され得る少なくとも一つの(好ましくは複数の、最も好ましくはあらゆる)生物的および/または非生物的ストレスに、その核酸配列の活性が感受性を示さないかどうかを、種々の方法を用いて決定することができる。一般に、定性的方法と定量的方法を区別することができる。定性的方法(たとえば組織学的GUS染色)は、プロモーターの空間的−時間的活性(プロモーターが、ある組織または器官で、あるいはある環境/発生条件の下で、活性であるか否か?)を決定するために使用され、定量的方法(たとえば蛍光GUSアッセイ)は、コントロールと比較した活性レベルを定量する。適切なコントロールは、たとえば、空のベクターで形質転換された植物(ネガティブコントロール)またはCaMV 35Sなどの他のプロモーターを含む構築物で形質転換された植物、または非トランスジェニック植物である。
【0043】
相対的または絶対的活性をテストし、必要に応じて定量するために、クローニングまたは合成された核酸分子、たとえば配列番号1、2、またはその変異体、またはこれらの何れかの断片を、公知の核酸配列(たとえば、gusAなどのレポーター遺伝子、または特定のタンパク質をコードする任意の遺伝子)に動作可能に連結し、これを使用して、公知の手法で植物細胞を形質転換してもよい。幾つかの態様において、細胞は、安定な様式で形質転換される必要はなく、すなわち、一過性発現アッセイを使用して(たとえばプロトプラストトランスフェクションまたはアグロインフィルトレーション)、プロモーターが細胞、組織または器官で活性であるかどうか、並びにどの程度までプロモーターが転写を駆動するかを決定してもよい。プロモーターの活性は、たとえば、下流核酸配列のRNA転写産物のレベルを検出することによりアッセイする(そして必要に応じて定量する)ことができる。これは、定量的方法、たとえば定量的RT-PCRまたは他のPCRベースの方法などを用いて行われてもよい。あるいは、レポータータンパク質またはレポータータンパク質の活性をアッセイし、定量してもよい。たとえば、レポーター遺伝子がgus遺伝子である場合、実施例に記載されるとおり、蛍光GUSアッセイを使用してもよい。このようにして、正常な生理的(非ストレス)条件下で維持された形質転換植物または植物細胞の定量的プロモーター活性レベルを、一または複数の生物的または非生物的ストレスに晒された植物または植物細胞のレベルと比較することができる。また、相対的または絶対的活性レベルを、構成的コントロールプロモーター、たとえば35Sプロモーター、ダブル35Sプロモーター、または配列番号1および/または2と比較することができる。好ましくは平均プロモーター活性レベルを決定し、統計的方法を用いて比較することが理解される。
【0044】
よって、本発明のプロモーターが、ある時間に、どの組織または器官において活性であるか(空間的−時間的活性)は、たとえば、植物または植物細胞をプロモーター−レポーター遺伝子構築物で形質転換し、種々の組織を種々の発生段階の間に、RNA転写産物またはレポータータンパク質(またはその活性)について分析することによりテストすることができる。単純なテストは、たとえば、組織学的GUS染色を採用し、これによる青色の視覚的評価は、種々の組織における種々の発生段階での活性を示す。
【0045】
既述のとおり、プロモーター活性は構成的であることが好ましく、好ましくは、植物および植物細胞、とりわけ配列が導入された宿主種または品種で強力である。構成的活性は、プロモーターに動作可能に連結された任意の核酸配列の転写産物が、好ましくは、ほとんどの組織(または器官)で、ほとんどの(正常な、非ストレス)生理的および発生条件下で産生されることを意味する。一つの態様において、本発明のプロモーターは、実施例で試験された組織または器官、たとえば葉(若い葉および老齢の葉)、根、花、種子、茎(主要な茎)、果実(たとえば未熟な果実および熟した果実)、発芽した種子などで少なくとも活性である。
【0046】
好ましくは、本発明のプロモーターは、すべての植物種、双子葉植物種および単子葉植物種の両方において、強力で構成的な活性を提供する。たとえば、配列番号1および配列番号2は、種々の植物種、たとえばトマト、タバコ、アブラナ属(Brassica)、メロンおよびレタスなどにおいて、強力で構成的な発現を提供することが見出された(実施例参照)。
【0047】
本発明のAA6プロモーター(断片または変異体を含む)の強度(定量的活性)は、下流(3’)に連結された核酸配列の発現を駆動する能力の観点から、種々の公知の方法を用いて定量的に決定することができる。たとえば、転写された転写産物(mRNA)の量は、定量的RT-PCRまたはノーザンブロッティングを用いて定量することができる。好ましくは、プロモーター強度は、正常な(非ストレス)条件下において、CaMV 35S(Franck et al., 同上)のプロモーター強度と少なくとも本質的に同等である。よって、「強力」とは、プロモーター強度が、正常な非ストレス条件下において、35Sのプロモーター強度と好ましくは少なくともほぼ同一であり、より好ましくはそれより強力であることを意味する。最も好ましくは、種々の組織および器官における平均定量的プロモーター活性は、CaMV 35Sプロモーターの活性と少なくとも同等であるか、あるいはCaMV 35Sプロモーターの平均活性より少なくとも5%、10%、20%、30%、35%、40%、45%、50%、55%、60%、70%、75%またはそれ以上高い。同じコピー数および接合状態レベルの形質転換体を比較すべきであることが理解され、たとえば導入遺伝子については半接合または同型接合である。好ましくは、シングルコピーの形質転換体を同定し、比較する。
【0048】
AA6プロモーターの発現は、ストレス条件下において、少なくとも本明細書の他の箇所に記載されるストレスから選択される一または複数のストレスの下で、構成的で強力なままである。
【0049】
よって、プロモーターを含む植物組織または器官または植物が、干ばつストレス、熱ストレス、水ストレス(過剰の水および過少の水の両方)、病原体ストレス(たとえばCMVなどのウイルス感染、真菌感染、細菌感染など)、害虫ストレス(たとえば昆虫の摂食)、傷害、塩ストレス、照射ストレスなどの一つまたは好ましくは幾つかから少なくとも選択されるストレス条件に晒されたときに、本発明のプロモーターの強度は、好ましくは、本質的に未変化のままであるか、あるいは少なくとも低下しない(または有意に低下しない)。再度、定量的テストを、これを決定するために使用することができる。たとえば、プロモーターを含む組換え植物を、正常な温度環境から温かい環境(たとえば約27℃から約50℃まで)に移し、種々の組織におけるプロモーター活性を、正常な温度条件と温かい温度条件の下での同じ組織の活性と比較してもよい。
【0050】
また、上記AA6プロモーターの変異体には、植物のシステインシンターゼ遺伝子、とりわけ細胞質ゾルのシステインシンターゼ遺伝子の任意の単離された核酸プロモーター(すなわち、システインシンターゼ酵素、とりわけ細胞質ゾルのアイソフォームをコードする植物遺伝子のプロモーター)であって、植物において強力で構成的な活性を示し、好ましくは、一または複数の生物的および/または非生物的ストレスに本質的に感受性を示さないものが含まれる。既述のとおり、他の植物のシステインシンターゼ遺伝子も、一または複数のストレス条件下で構成的発現を保持する強力で構成的なプロモーターを有していると考えられる。植物のシステインシンターゼ遺伝子は、O-アセチル-L-セリン[チオール]-リアーゼ(EC 4.2.99.8)とも称される酵素システインシンターゼをコードする遺伝子である。幾つかのシステインシンターゼ遺伝子がクローニングされているが(たとえば実施例で言及されるTC162833参照)、トランスジェニック植物における同型接合または異型接合配列の強力で構成的な発現を駆動するためのプロモーターの有用性については記載されていない。よって、かかる公知のシステインシンターゼ遺伝子または今のところ未知のシステインシンターゼ遺伝子のプロモーターを単離し、それらの活性についてスクリーニングすることができる。たとえば、cDNA-AFLP、他のPCRベースの方法またはノーザンハイブリダイゼーションを使用して、他のシステインシンターゼ遺伝子、とりわけ、構成的そして一または複数のストレス条件下においてもシステインシンターゼmRNAsを内因的に産生する遺伝子を単離または同定することができる。その後、所望の発現パターンを備えたシステインシンターゼ遺伝子を選択し、それらのプロモーターを公知の方法を用いてクローニングすることができる。好ましい態様において、プロモーターは、ナス科(Solanaceae)、たとえばナス属(Solanum)(再分類されたトマト属(Lycopersicon)の種を含む)、タバコ属(Nicotiana)、トウガラシ属(Capsicum)、ペチュニア属(Petunia)、コフィア属(Coffea)の種などに属する植物に由来するシステインシンターゼ遺伝子から得られる。
【0051】
本発明のプロモーターは、好ましくは、シロイヌナズナ(Arabidopsis thaliana)などの草植物のプロモーターではなく、好ましくは、Gutierrez-Alcala et al. (J. of Exp. Botany 56, p24872494) に記載されるシロイヌナズナのOASA1遺伝子のプロモーター、またはその断片ではない。
【0052】
(配列番号5のシステインシンターゼ酵素をコードする)配列番号4で提供されるシステインシンターゼ遺伝子の他に、他のシステインシンターゼ遺伝子を同定し、それらのプロモーターを単離することができる。上述のとおり種々の方法が使用され得る。
【0053】
よって、本発明の一つの態様において、植物および/または植物細胞で構成的活性を有し、かつ植物または植物細胞、組織または器官が、たとえば既述のストレスなどの一または複数の生物的および/または非生物的ストレスに晒されたときにその活性が低下しないか、または有意に低下しない、植物(細胞質ゾル)のシステインシンターゼ遺伝子のプロモーターが提供される。
【0054】
かかるプロモーターは、以下のものを含む:(a)植物から入手可能であり、植物のシステインシンターゼ遺伝子の発現を駆動する任意のプロモーター、とりわけ配列番号5のタンパク質をコードする任意の遺伝子のプロモーター;(b)配列番号5に対して(全長にわたって)少なくとも30、40、50、60、70、80、90、95、99%またはそれ以上のアミノ酸同一性を有するシステインシンターゼ酵素をコードする植物遺伝子のプロモーター;(c)システインシンターゼ酵素をコードし、その核酸配列が、配列番号4に対して(全長にわたって)少なくとも30、40、50、60、70、80、90、95、98、99%またはそれ以上のヌクレオチド配列同一性を含む、核酸配列の発現を駆動する植物プロモーター。
【0055】
別の態様において、組換え細胞または生物、とりわけ植物細胞または植物における同型接合または異型接合の核酸配列の構成的な過剰発現のための、植物のシステインシンターゼ遺伝子のプロモーターの使用が提供される。この使用は、既述のとおり、同型接合または異型接合の核酸配列にプロモーターを動作可能に連結し、植物または植物細胞を形質転換することを含む。
【0056】
上述の焦点は、植物および植物細胞における本発明のプロモーターの使用にあるが、他の細胞および生物、たとえば任意の原核または真核細胞または生物、たとえば細菌、真菌類(Pichia、Hansenulaなどの酵母を含む)、哺乳類、ヒト細胞または細胞株などにおいて同型接合または異型接合の核酸配列を発現させるためにプロモーターを使用することも本発明の態様である。
【0057】
本発明のキメラ遺伝子およびベクター
本発明の一つの態様において、プロモーター活性を有する上記核酸配列の何れかを使用して、キメラ遺伝子およびこれらを含むベクターが、キメラ遺伝子の宿主細胞への転移および動作可能に連結された異型接合または同型接合の核酸配列の宿主細胞(たとえば細胞、組織、器官または形質転換細胞に由来する生物全体)での発現のために作成される。
【0058】
宿主細胞は、好ましくは植物細胞である。任意の植物が、適切な宿主であり、たとえば、単子葉植物または双子葉植物、たとえば、トウモロコシ(maize/corn)(Zeaの種、たとえばZ. mays、Z. diploperennis (chapule)、Zea luxurians (グアテマラのテオシンテ)、Zea mays subsp. huehuetenangensis (サンアントニオヒュースタのテオシンテ)、Z. mays subsp. mexicana (メキシコのテオシンテ)、Z. mays subsp. parviglumis (バルサスのテオシンテ)、Z. perennis (多年生テオシンテ) およびZ. ramosa、コムギ (Triticumの種)、オオムギ (たとえばHordeum vulgare)、カラスムギ (たとえばAvena sativa)、モロコシ (Sorghum bicolor)、ライムギ (Secale cereale)、ダイズ (Glycine spp、たとえばG. max)、ワタ (Gossypiumの種、たとえばG. hirsutum、G. barbadense)、Brassica spp. (たとえばB. napus、B. juncea、B. oleracea、B. rapaなど)、ヒマワリ (Helianthus annus)、タバコ (Nicotianaの種)、アルファルファ (Medicago sativa)、イネ (Oryzaの種、たとえばO. sativaインディカ栽培品種-グループまたはジャポニカ栽培品種-グループ)、まぐさ(forage grasses)、パールミレット (Pennisetum属の種、たとえばP. glaucum)、樹木の種、野菜の種、たとえばLycopersicon ssp (近年ナス属(Solanum)に属すると再分類された)、たとえばトマト (L. esculentum, syn. Solanum lycopersicum) たとえばチェリートマト、var. cerasiformeまたはカレントトマト、var. pimpinellifolium) またはツリートマト (S. betaceum, syn. Cyphomandra betaceae)、ジャガイモ (Solanum tuberosum) およびその他のナス属(Solanum)の種、たとえばナス (Solanum melongena)、ペピーノ (S. muricatum)、ココナ (S. sessiliflorum) およびナランヒージャ (S. quitoense);コショウ (Capsicum annuum、Capsicum frutescens)、エンドウ (たとえばPisum sativum)、マメ (たとえばPhaseolusの種)、ニンジン (Daucus carota)、Lactuca属の種 (たとえばLactuca sativa、Lactuca indica、Lactuca perennis)、キュウリ (Cucumis sativus)、メロン (Cucumis melo)、ズッキーニ (Cucurbita pepo)、スカッシュ (Cucurbita maxima、Cucurbita pepo、Cucurbita mixta)、パンプキン (Cucurbita pepo)、スイカ (Citrullus lanatus syn. Citrullus vulgaris)、多肉果の種 (グレープ、ピーチ、プラム、ストロベリー、マンゴー、メロン)、観賞植物種 (たとえばバラ、ペチュニア、キク、ユリ、チューリップ、ガーベラの種)、木本 (たとえばPopulus、Salix、Quercus、Eucalyptusの種)、繊維の種、たとえばアマ (Linum usitatissimum) およびアサ (Cannabis sativa)である。一つの態様において、野菜の種、とりわけナス属(Solanum)の種(トマト属(Lycopersicon)の種を含む)が好ましい。
【0059】
よって、たとえば以下の属の種が形質転換され得る:Cucurbita、Rosa、Vitis、Juglans、Fragaria、Lotus、Medicago、Onobrychis、Trifolium、Trigonella、Vigna、Citrus、Linum、Geranium、Manihot、Daucus、Arabidopsis、Brassica、Raphanus、Sinapis、Atropa、Capsicum、Datura、Cucumis、Hyoscyamus、Lycopersicon、Solanum、Nicotiana、Malus、Petunia、Digitalis、Majorana、Ciahorium、Helianthus、Lactuca、Bromus、Citrullus、Asparagus、Antirrhinum、Heterocallis、Nemesis、Pelargonium、Panieum、Pennisetum、Ranunculus、Senecio、Salpiglossis、Browaalia、Glycine、Pisum、Phaseolus、Gossypium、Glycine、Lolium、Festuca、Agrostis。更に好ましいのは、Cucurbita、Brassica、Lycopersicon、Solanum、OryzaおよびZeaの各々である。好ましいのは、Avena、Medicago、Capsicum、Nicotiana、Lactuca、Pisum、Cucumis、Cucurbita、Brassica、Solanum (Lycopersiconを含む)、OryzaおよびZeaの各々である。
【0060】
キメラ遺伝子、および宿主細胞のゲノムへのキメラ遺伝子の導入のためのベクターの構築は、一般に当該技術分野で公知である。キメラ遺伝子を作成するために、AA6プロモーター配列を、標準的な分子生物学技術を用いて、宿主細胞で転写される別の核酸配列に動作可能に連結する。プロモーター配列は、転写される核酸配列が、プロモーター配列の下流でベクターに簡単に挿入されるように、ベクターに既に存在していてもよい。その後、ベクターは、宿主細胞を形質転換するために使用され、キメラ遺伝子は、好ましくは、核ゲノム、またはプラスチド、ミトコンドリアまたは葉緑体のゲノムに挿入され、その結果、下流の核酸配列が、プロモーターの活性により発現される(たとえば、Mc Bride et al., 1995 Bio/Technology 13, 362; US 5,693,507)。
【0061】
したがって、キメラ遺伝子は、好ましくは上述のAA6プロモーターを含み、これは、同型接合または異型接合の核酸配列に動作可能に連結され、必要に応じてその後に3’非翻訳核酸配列(3’UTR)が続く。同型接合または異型接合の核酸配列は、タンパク質またはペプチドをコードする配列であってもよいし、活性なRNA分子、たとえば宿主細胞または生物における遺伝子または遺伝子ファミリーのサイレンシングに適したセンスおよび/またはアンチセンスRNA(dsRNA)に転写される配列であってもよい。
【0062】
AA6プロモーターを含むキメラ遺伝子は、単一の植物細胞の核ゲノムに慣用的なやり方で安定に挿入することができ、このように形質転換された植物細胞は、キメラ遺伝子の構成的発現により表現型の変化した形質転換植物を産生するために、慣用的なやり方で使用することができる。
【0063】
これに関して、アグロバクテリウムツメファシエンス(Agrobacterium tumefaciens)に含まれる、更なる核酸配列に動作可能に連結されたAA6プロモーター(または上述の変異体または断片)を含むT-DNAベクターは、植物細胞を形質転換するために使用することができ、その後、たとえばEP 0 116 718、EP 0 270 822、PCT公報 WO 84/02913および公開された欧州特許出願EP 0 242 246、およびGould et al. (1991, Plant Physiol. 95,426-434) に記載される手法を用いて、形質転換された植物細胞から形質転換植物を再生することができる。アグロバクテリウム媒介性の植物の形質転換は、当該技術分野で周知である。T-DNAベクターは、EP 0 120 561およびEP 0 120 515に記載されるバイナリーベクター、または相同組換えによりアグロバクテリウムTi-プラスミドに統合することができるEP 0 116 718に記載されるコインテグレート(co-integrate)ベクターの何れであってもよい。
【0064】
好ましいT-DNAベクターはそれぞれ、T-DNAのボーダー配列の間、または少なくとも右ボーダー配列の左側の位置に、転写される核酸配列に動作可能に連結されたAA6プロモーターを含有する。ボーダー配列は、Gielen et al. (1984, EMBO J 3,835-845) に記載される。当然、他のタイプのベクターを使用して、植物細胞を形質転換することができ、直接的な遺伝子導入(たとえばEP 0 223 247に記載される、またはUS 2005/055740およびWO 2004/092345に記載されるパーティクルまたはミクロプロジェクタイル(microprojectile)ボンバードメント)、花粉媒介性の形質転換(たとえばEP 0 270 356およびWO 85/01856に記載される)、プロトプラストの形質転換(たとえばUS 4,684,611に記載される)、植物ウイルス媒介性の形質転換、リポソーム媒介性の形質転換(たとえばUS 4,536,475に記載される)などの手法、並びにその他の方法、たとえばトウモロコシ (たとえば、US 6,140,553; Fromm et al., 1990, Bio/Technology 8, 833-839; Gordon-Kamm et al., 1990, The Plant Cell 2, 603-618) およびイネ (Shimamoto et al., 1989, Nature 338, 274-276; Datta et al. 1990, Bio/Technology 8, 736-740) のある系統を形質転換するために記載された方法、および一般に単子葉植物を形質転換するための方法 (WO 92/09696) が使用される。ワタの形質転換のためにはWO 00/71733も参照され、イネの形質転換のためにはWO 92/09696、WO 94/00977およびWO 95/06722に記載の方法も参照される。モロコシの形質転換のためには、たとえばJeoung JM et al. 2002, Hereditas 137: 20-8またはZhao ZY et al. 2000, Plant Mol Biol. 44: 789-98が参照される。トマトまたはタバコの形質転換のためには、An G. et al., 1986, Plant Physiol. 81: 301-305; Horsch R.B. et al., 1988, In: Plant Molecular Biology Manual A5, Dordrecht, Netherlands, Kluwer Academic Publishers. pp 1-9; Koornneef M. et al., 1986, In: Nevins D.J. and R.A. Jones, eds. Tomato Biotechnology, New York, NY, USA, Alan R. Liss, Inc. pp 169-178も参照される。同様に、形質転換された細胞からの形質転換植物の選択および再生は、当該技術分野で周知である。明らかに、様々な種に対して、更に単一種の様々な品種または栽培種に対しても、高い頻度で形質転換体を再生させるためにプロトコールを具体的に適合させる。
【0065】
核ゲノムの形質転換に加えて、プラスチドのゲノム、好ましくは葉緑体のゲノムの形質転換も本発明に含められる。プラスチドのゲノムの形質転換は、導入遺伝子の蔓延のリスクを低減できるという利点がある。プラスチドのゲノムの形質転換は、当該技術分野で公知のとおり行うことができ、たとえばSidorov VA et al. 1999, Plant J.19: 209-216またはLutz KA et al. 2004, Plant J. 37(6): 906-13が参照される。
【0066】
得られた形質転換植物は、慣用的な植物育種スキームで使用して、導入遺伝子を含有する多くの形質転換植物を産生することができる。シングルコピーの形質転換体は、たとえばサザンブロット分析またはPCRベースの方法またはInvader(登録商標)テクノロジーアッセイ(Third Wave Technologies, Inc.)を用いて選択することができる。形質転換された細胞および植物は、キメラ遺伝子の存在により非形質転換体と容易に区別することができる。導入遺伝子の挿入部位に隣接する植物DNAの配列も配列決定することができ、これにより、ルーチン使用のために「イベント特異的な」検出方法を開発することができる。たとえば、WO 01/41558が参照され、これは、たとえば統合された配列および隣接する(ゲノム)配列に基くエリートなイベント検出キット(たとえばPCR検出キット)を記載する。
【0067】
一つの態様において、転写され、(コード配列である場合)必要に応じて翻訳される核酸配列は、転写される配列が、適切な3’端の転写調節シグナル(“3’端”)(すなわち転写産物形成およびポリアデニル化シグナル)の上流(すなわち5’)に位置するように植物ゲノムに挿入される。ポリアデニル化および転写産物形成シグナルには、ノパリンシンターゼ遺伝子のもの(“3’nos”) (Depicker et al., 1982 J. Molec. Appl. Genetics 1, 561-573.)、オクトピンシンターゼ遺伝子のもの(“3’ocs”) (Gielen et al., 1984, EMBO J 3, 835-845)、およびT-DNA遺伝子7のもの (“3’遺伝子7”) (Velten and Schell, 1985, Nucleic Acids Research 13, 6981-6998) などが含まれ、これらは、形質転換された植物細胞において3’-非翻訳DNA配列として機能する。
【0068】
好ましい態様において、使用される3’端配列は、植物のシステインシンターゼ遺伝子のものであり、好ましくは、AA6プロモーターが入手可能であるシステインシンターゼ遺伝子に由来する。本明細書で提供される適切な3’端配列は、配列番号3、またはその断片もしくは変異体である。配列番号3の変異体には、配列番号3に対して少なくとも50、60、65、70、75、80、85、90、95、98、99%またはそれ以上の核酸配列同一性を含む核酸配列が含まれる。配列番号3の断片または配列番号3の変異体の断片には、配列番号3または配列番号3の変異体の少なくとも50、100、150、200、250、300またはそれ以上の連続ヌクレオチドを含む核酸配列が含まれる。これらの3’端配列は、それ自体で一つの態様でもあり、任意のキメラ遺伝子およびベクターを構築するために、すなわち様々なプロモーターと組合せて使用してもよい。
【0069】
発現される核酸配列は、一つの態様において、ハイブリッドタンパク質またはペプチドまたは融合タンパク質を含むタンパク質またはペプチドをコードする配列である。コード配列は、任意の由来、すなわち植物、真菌類(酵母を含む)、動物、細菌、合成、ウイルスなどであり得る。ターゲッティングペプチド、たとえば分泌シグナルペプチドまたはプラスチドターゲッティングシグナルをコードする配列を含んでいてもよい。また、コード配列を、選択またはスコア(scorable)マーカーをコードする遺伝子、たとえばカナマイシン抵抗性を付与するneo(またはnptII)遺伝子(EP 0 242 236)に読み取り枠を合わせて(in-frame)連結させて、細胞が、容易に検出可能な融合タンパク質を発現するようにしてもよい。任意の遺伝子のコード領域(cDNAまたはゲノムDNA)が使用され得るが、以下の遺伝子のコード領域の例が、好ましくは、本発明のAA6プロモーターに動作可能に連結される:1.ウイルスの核酸配列の逆方向反復配列 (たとえば、ウイルスのコートタンパク質遺伝子のセンスおよびアンチセンス配列;下記も参照);2.疾病シグナル伝達経路遺伝子または疾病抵抗性遺伝子;3.非生物的ストレス応答関連遺伝子 (たとえば、SHINE転写因子、またはCBF/DREB遺伝子);4.二次代謝産物生合成遺伝子、たとえば治療的および/または薬理的に重要な産物または産業上価値のある化合物の生産のための遺伝子。
【0070】
明らかに、他の遺伝子、たとえば、作物学的形質に影響を及ぼす遺伝子、たとえば除草剤抵抗性の遺伝子(たとえばEPSPS遺伝子、barまたはPAT遺伝子)、収量または品質の形質(たとえばタンパク質組成)に影響を及ぼす遺伝子なども、本発明のAA6プロモーターに動作可能に連結され得る。
【0071】
本発明のキメラ遺伝子またはベクターは、微生物、たとえば細菌(たとえば大腸菌(Escherichia coli)、シュードモナス属(Pseudomonas)、アグロバクテリウム属(Agrobacterium)、バチルス属(Bacillus)など)または真菌類または藻類または昆虫を形質転換するために使用することもできるし、あるいはこの遺伝子またはベクターを、ウイルスを操作するために使用してもよい。適切なクローニング媒体に組み込まれた本発明の核酸配列による細菌の形質転換は、慣用的なやり方で、好ましくはMaillon et al. (1989, FEMS Microbiol. Letters 60, 205-210.) およびWO 90/06999に記載される慣用的なエレクトロポレーション技術を用いて行うことができる。原核宿主細胞でコード配列を発現させるために、核酸配列のコドン使用頻度をそれに応じて最適化してもよい(同様に、植物細胞でコード配列を発現させるために、核酸配列のコドン使用頻度を公知のとおり最適化してもよい)。イントロン配列は除去すべきであり、最適な発現のための他の適応を公知のとおり行ってもよい。
【0072】
イネ科植物種などの単子葉植物、たとえばトウモロコシまたはイネにおいて核酸配列の高い発現を実現するために、イントロン、好ましくは単子葉植物のイントロンをキメラ遺伝子に付加することができる。たとえば、5’調節領域へのトウモロコシAdh1遺伝子のイントロンの挿入は、トウモロコシで発現を高めることが示されている(Callis et. al., 1987, Genes Develop. 1: 1183-1200)。同様に、US 5,859,347に記載されるHSP70のイントロンは、発現を高めるために使用され得る。このように、一または複数のイントロンは、本発明のプロモーター配列の何れかに、または5’UTRまたはコード配列に必要に応じて挿入され得る。
【0073】
別の態様において、AA6プロモーターは、遺伝子サイレンシングのためのキメラ遺伝子およびベクターを作成するために使用され、ここでAA6プロモーターは、ターゲット遺伝子(サイレンシングされる内在性遺伝子または遺伝子ファミリー)のセンスおよび/またはアンチセンス核酸配列に動作可能に連結される。更に別の態様において、ターゲット遺伝子は、侵入する植物病原体の遺伝子または遺伝子ファミリーであってもよい。たとえば、ウイルスのコートタンパク質遺伝子の逆方向反復配列は、ウイルス抵抗性植物を作成するために使用され得る。ウイルスのコートタンパク質遺伝子は、たとえばWO 96/21031に記載される。「遺伝子サイレンシング」は、一または複数のターゲット遺伝子の遺伝子発現のダウンレギュレーションまたは完全な阻害を指す。遺伝子発現を低下または廃止させるための抑制性RNAの使用は、当該技術分野で十分に確立されており、幾つかのレビューの主題である(たとえばBaulcombe, 1996, Plant Cell 8: 1833-1844; Stam et al., 1997, Plant Journal 12: 63-82; Depicker and Van Montagu, 1997, Curr. Opinion Cell Biol. 9: 373-382)。植物で遺伝子サイレンシングを達成するために利用可能な技術は幾つか存在し、たとえば、ターゲット遺伝子の全部または一部のアンチセンスRNAを産生するキメラ遺伝子(たとえばEP 0 140 308 B1、EP 0 240 208 B1およびEP 0 223 399 B1参照)、または(共抑制(co-suppression)とも称される)センスRNAを産生するキメラ遺伝子(EP 0 465 572 B1参照)がある。
【0074】
しかし、これまでに最も成功したアプローチは、ターゲット遺伝子のセンスRNAおよびアンチセンスRNAの両方(「逆方向反復配列」)の産生であり、これは、細胞内に二本鎖RNA(dsRNA)を形成し、ターゲット遺伝子をサイレンシングさせる。dsRNAの産生および遺伝子サイレンシングのための方法およびベクターは、EP 1 068 311、EP 983 370 A1、EP 1 042 462 A1、EP 1 071 762 A1およびEP 1 080 208 A1に記載される。
【0075】
したがって、本発明のベクターは、ターゲット遺伝子のセンスおよび/またはアンチセンスDNA断片に動作可能に連結されたAA6プロモーターを含んでいてもよい。ターゲット遺伝子配列の短い(センスおよびアンチセンス)区切り(stretch)、たとえば少なくとも約17、18、19、20、21、22、23、24または25ヌクレオチドのコードまたは非コード配列で十分であり得る。より長い配列、たとえば少なくとも約100、200、250、300、400、500、1000、1500ヌクレオチドまたはそれ以上の配列も頻繁に使用される。好ましくは、センスおよびアンチセンス断片は、スペーサー配列、たとえばイントロンにより分離されており、これにより、dsRNAの形成でループ(またはヘアピン)が形成される。ターゲット遺伝子の任意の区切りは、遺伝子サイレンシングベクター、およびターゲット遺伝子または遺伝子ファミリーがサイレンシングされたトランスジェニック植物を作成するために使用され得る。ヘアピン構築物を生成する簡便な方法は、ジェネリックベクター、たとえばpHANNIBALおよびpHELLSGATE、Gateway(登録商標)技術に基くベクターを使用することである (Wesley et al. 2004, Methods Mol Biol. 265: 117-30; Wesley et al. 2003, Methods Mol Biol. 236: 273-86およびHelliwell & Waterhouse 2003, Methods 30(4): 289-95参照、これら全ては参照により本明細書に組み込まれる)。
【0076】
ターゲット遺伝子の保存された核酸配列を選択することにより、宿主植物内のファミリーメンバーをサイレンシングすることができる。ターゲット遺伝子が、侵入する病原体の遺伝子または遺伝子ファミリーである場合、病原体のターゲット遺伝子がサイレンシングされ、植物は、病原体に抵抗性をもつようになる。ターゲット遺伝子核酸配列のセンスおよび/またはアンチセンスDNA断片に動作可能に連結されたAA6プロモーターを含み、ターゲット遺伝子サイレンシングの表現型を示すトランスジェニック植物も、本明細書に包含される。表現型は、遺伝子の機能に依存し、化学的変化であっても分子的変化であってもよく、肉眼で見えても見えなくてもよい。したがって、かかるキメラ遺伝子およびベクターは、遺伝子の機能を決定または検証するために使用することもできる。
【0077】
本発明のキメラ遺伝子は、宿主ゲノムに安定に導入してもよいし、エピソームユニットとして存在してもよい。
【0078】
本発明のトランスジェニック細胞および生物
上述の方法により入手可能なトランスジェニック細胞および生物、とりわけ植物、植物細胞、組織または器官が提供される。これらの細胞および生物は、本発明のAA6プロモーターの存在による、細胞またはゲノム内のキメラ遺伝子の存在により特徴づけられる。加えて、mRNA転写産物または翻訳されたタンパク質は、細胞または生物、たとえば植物細胞または植物の表現型を変化させ得る。
【0079】
AA6プロモーターは構成的であるが、ゲノム内の位置は、プロモーターの活性およびキメラ遺伝子の発現レベルに影響を及ぼすことができる。したがって、高い構成的なレベルのタンパク質、または(サイレンシング構築物を使用する場合)高い構成的なレベルのセンスおよび/またはアンチセンス転写産物を発現する形質転換体(“イベント”または“形質転換イベント”)は、たとえば、コピー数の分析(サザンブロット分析)、mRNA転写産物レベルの分析(たとえばノーザンブロット分析またはRT-PCR)により、あるいは核酸配列によりコードされるタンパク質の存在およびレベルの分析(たとえばSDS-PAGEとその後のウェスタンブロット分析;ELISA分析、免疫細胞学的アッセイなど)により選択することができる。また、形質転換体は、一または複数の生物的および/または非生物的ストレス条件の下での発現の安定性についてテストすることができ、一または複数の所望の条件の下で高い構成的発現を保持するイベントを同定し、更なる使用のために選択することができる。
【0080】
トランスジェニック植物は、伝統的な育種方法、たとえば交配、自家受粉、戻し交配などで使用することができる。形質転換体を自家受粉させることにより、導入遺伝子について同型接合である植物を生成することができる。育種の手順は、当該技術分野で公知であり、植物育種の標準テキストブック、たとえば、Allard, R.W., Principles of Plant Breeding (1960) New York, NY, Wiley, pp 485; Simmonds, N.W., Principles of Crop Improvement (1979), London, UK, Longman, pp 408; Sneep, J. et al., (1979) Tomato Breeding (p. 135-171) in: Breeding of Vegetable Crops, Mark J. Basset, (1986, editor), The Tomato crop: a scientific basis for improvement, by Atherton, J.G. & J. Rudich (editors), Plant Breeding Perspectives (1986); Fehr, Principles of Cultivar Development-Theory and Technique (1987) New York, NY, MacMillanに記載される。
【0081】
トランスジェニック細胞または生物は、たとえば組換えタンパク質の大規模生産のために、細胞培養(植物細胞培養、細菌または真菌細胞培養、たとえば酵母培養、ヒトまたは哺乳類細胞培養、昆虫細胞培養)において使用することもできる。一つの態様において、本発明のAA6プロモーターを含む細胞を含む細胞培養が提供される。
【0082】
本発明の方法および使用
また、トランスジェニック植物または植物細胞を作成するための方法であって、
(a)発現される核酸配列に動作可能に連結された本発明のAA6プロモーターおよび/またはAA6 3’UTRを含むキメラ遺伝子またはベクターを生成する工程;
(b)前記キメラ遺伝子またはベクターを用いて植物または植物細胞を形質転換する工程;および必要に応じて、
(c)トランスジェニック植物(a transgenic plant or plants)を再生させる工程
を含む方法が提供される。
【0083】
更に、非ストレス条件下において強力で構成的なプロモーター活性を提供し、かつ一または複数の生物的および/または非生物的ストレスに晒されたときにプロモーター活性が本質的に変化しない(少なくとも低下しないか、または有意に低下しない)トランスジェニック植物が同定され得る。
【0084】
この植物は、慣用的な農業および育種の方法で使用され得る。とりわけ、植物は、一または複数の生物的および/または非生物的ストレス条件に植物を晒す環境において、プロモーターの活性を低下させることなく、生育させることができる。よって、たとえば、高温または低温、強風、高い塩、高い土壌汚染物質、高いまたは低い水レベル、干ばつ期(drought spells)、高い病気または害虫プレッシャーの領域、高い照射の領域などにおいて、プロモーター活性を低下させることなく、あるいは少なくとも有意に低下させることなく、トランスジェニック植物を生育させるための方法が提供される。
【0085】
配列
配列番号1:“3kb”(2986 bp) AA6プロモーター
配列番号2:“5kb”(5000 bp) AA6プロモーター
配列番号3:トマトのシステインシンターゼ遺伝子(AA6遺伝子)の3’UTR
配列番号4:トマトのシステインシンターゼcDNA (AA6 cDNA)
配列番号5:配列番号4によりコードされるタンパク質 (トマトの細胞質ゾルのシステインシンターゼ)
配列番号6:幾つかの不明ヌクレオチドを有する“3kb”AA6プロモーター
配列番号7:幾つかの不明ヌクレオチドを有する“5kb”AA6プロモーター
配列番号8:再度配列決定され、E.coliの寄託物CBS120175内にある、pKG8135の“3kb”AA6プロモーター
配列番号9:再度配列決定され、E.coliの寄託物CBS120176内にある、pKG8137の“5kb”AA6プロモーター。
【0086】
図面
図1:pKG1562のベクターマップ
図2:pKG1700のベクターマップ
図3:pKG8135、pKG8136およびpKG8137のベクターマップ。
【0087】
以下の非限定的な実施例は、本発明のAA6プロモーターの使用を記載する。実施例で別途記載しない限り、全ての組換えDNA技術は、Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press、およびSambrook and Russell (2001) Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Laboratory Press, NY; およびVolumes 1 and 2 of Ausubel et al. (1994) Current Protocols in Molecular Biology, Current Protocols, USAに記載されるとおり標準プロトコールに従って行われる。植物の分子レベル研究のための標準的な材料と方法は、Plant Molecular Biology Labfax (1993) by R.D.D. Croy, jointly published by BIOS Scientific Publications Ltd (UK) and Blackwell Scientific Publications, UKに記載される。
【実施例】
【0088】
例1−材料と方法
cDNAを用いたディファレンシャルディスプレイ分析は、Lycopersicon esculentum var. Moneybergについて実施した。
【0089】
ストレス条件下での発現テストは、L.esculentum var. Moneybergについて実施した。
【0090】
ベクターpKG8136、pKG8137およびpKG1700を用いた形質転換は、Nicotiana tabacum var. SR1およびL. esculentum var. RZ 52201で実施した。植物の形質転換は、以下の参考文献に記載されるとおり行った:
トマトおよびタバコの形質転換について:
An G., B.D. Watson and C.C. Chiang. 1986. Transformation of tobacco, tomato, potato, and Arabidopsis thaliana using a binary Ti vector system. Plant Physiol. 81: 301-305。
【0091】
タバコの形質転換について:Horsch R.B., J. Fry, N. Hoffman, J. Neidermeyer, S.G. Rogers and R.T. Fraley. 1988. Leaf disc transformation. In: Plant Molecular Biology Manual A5. Dordrecht, Netherlands, Kluwer Academic Publishers. pp 1-9。
【0092】
トマトの形質転換について:Koornneef M., M. Jongsma, R. Weide, P. Zabel and J. Hille. 1986. Transformation of tomato. In: Nevins D.J. and R.A. Jones, eds. Tomato Biotechnology. New York, NY, USA, Alan R. Liss, Inc. pp 169-178。
【0093】
5’および3’端RACEは、CLONTECH Laboratories IncからのSMARTTM RACE cDNA Amplification Kitを用いて実施した。
【0094】
PCR産物のクローニングは、Invitrogen BVからのOriginal TA Cloning(登録商標)Kitを用いて、プラスミドpCR(登録商標)2.1を使用して実施した。
【0095】
DNA配列決定は、BaseClear (P.O. Box 1336, 2302 BH, Leiden, The Netherlands) により実施した。
【0096】
形質転換ベクターは、アグロバクテリウムツメファシエンス(Agrobacterium tumefaciens)var. GV2260に組み込んだ。
【0097】
プロモーター活性の定性的分析は、組織学的GUSアッセイを用いて行った:
種々の植物の部分を、37℃で一晩、振盪しながら、大気中の酸素の存在下で、Xgluc基質とともに、リン酸緩衝液 (5-ブロモ-4-クロロ-インドリルグルクロニド、1 mg/ml、K2HPO4、40 mM、KH2PO4、10 mM、pH 7.4) 中でインキュベートした。サンプルは、エタノールで繰返し洗浄することにより脱色した。非トランスジェニック植物を、ネガティブコントロールとして使用した。
【0098】
プロモーター活性の定量的分析は、蛍光GUSアッセイを用いて行った:
トータルタンパク質サンプルは、若葉の材料から調製し;サンプルは、テストされる各植物の様々な部分からプールしたおよそ同じサイズおよび発生段階の葉片から調製した。新鮮な葉の材料を、リン酸緩衝液 (Na2HPO4、77.4 mM、NaH2PO4、22.6 mM) 中で金属ビーズを用いてすりつぶし、その後、遠心分離と上清の回収を行った。
【0099】
蛍光GUSアッセイ:
各上清の一部におけるタンパク質濃度を、Molecular Probes, IncからのNano Orange Kitを用いて測定した。タンパク質サンプルは、トータルタンパク質濃度を標準化するために希釈した。タンパク質サンプルの一部を、37℃で一晩、基質4-メチルウンベリフェリルβ-d-グルクロニド (MUG) (最終濃度 1 mg/ml) とともにインキュベートした。時間0と一晩インキュベーション後の蛍光測定を、反応混合物の一部を取り出し、Na2CO3溶液(最終濃度 1.1 M) を添加して反応を停止させ、その後、355nmでの励起により引き起こされる460nmでの発光を測定することにより行った。
【0100】
例2−プロモーター単離用遺伝子の選択
ディファレンシャル分析は、cDNA-AFLP(登録商標)(Volkmuth W., et al., 2003, Genome-Wide cDNA-AFLP(登録商標)Analysis of the Arabidopsis Transcriptome, OMICS, 7, 2; Vos, P. and Stanssens P., 2002, AFLP-based transcript profiling, Current Protocols in Molecular Biology, unit 25B.5., Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G. Smith, J.A., Struhl, K., John Wiley and Sons, New York; and Vos et al., 1995, AFLP: a new technique for DNA fingerprinting, Nucleic Acids Research 23: 4407-4414) を用いて、種々の発生段階におけるトマト植物材料(若葉、老葉、茎、根、インビトロの葉、インビトロの茎、インビトロの根、カルス、未熟および成熟した緑色の実、ブレーカー(breaker)段階の実、赤色の実、花、芽生え全体)から単離したcDNAについて実施した。発現プロファイルは、Taq1プライマーについては2つの選択ヌクレオチドを備え、Mse1プライマーについては3つの選択ヌクレオチドを備えた、全ての可能なTaq1/Mse1プライマーの組み合わせを用いて作成した。その後の遺伝子断片発現プロファイルは、Keygene N.V. 所有ソフトウェア (ImproveTM) を用いて作成し、データベースに保存した。全てのサンプル組織において強力で構成的な発現を示した、12のTaq1/Mse1プライマーの組み合わせに対応する13の候補転写産物を選択した。
【0101】
その後、候補遺伝子の発現を、種々の組織において、生物的および非生物的ストレスの下で、cDNA-AFLP(登録商標)により測定した。3週齢のトマト植物 (Moneyberg) を、以下の別個のストレス条件を用いて14日間生育させた:
キュウリモザイクウイルス(CMV)による感染、高温(昼37℃および夜25℃)、低温(昼10℃および夜5℃)、干ばつストレス(植物の死を回避する最小限の水、昼37℃および夜25℃)、並びに収穫2日前の傷害(ピンセットで葉を押しつぶす)。
【0102】
以下の材料を収穫した:若葉、老葉、茎、根および花芽。これらの材料からcDNAを作成し、ディファレンシャル分析を、cDNA-AFLP(登録商標)を使用して、選択された13の候補転写産物を増幅するのに必要な同一プライマーの組み合わせ(および少数の選択ヌクレオチドを備えたプライマーの組み合わせも使用)を用いて実施した(プライマー配列については下記参照)。上記ストレス条件の全ての下において強力で構成的な発現を提供し続けた7つの転写産物を選択した。
【0103】
システインシンターゼ遺伝子(AA6)断片を増幅する(+2/+2)AFLPプライマーのプライマー配列は、以下のとおりである:
【表A】

【0104】
候補cDNA AFLP断片を、ゲルから切り出し、精製し、クローニングし、配列決定を行った。その配列情報を、その後、RACE PCRのプライマーを設計するために使用した。
【0105】
5’および3’両端のSMART-RACE-PCR (Clontech) を実施し、7つの候補遺伝子の伸長遺伝子断片を作成し、これらをpCR2.1 (Invitrogen) にクローニングし、その後、配列決定を行った(BaseClear, P.O. Box 1336, 2302 BH, Leiden, The Netherlands)。伸長遺伝子断片は、公共のデータベースに由来する相同ESTsと共に組み立てを行った。これにより、5つの候補遺伝子について全長cDNA配列を得た。
【0106】
5つの遺伝子についてのコピー数の評価は、10種類の制限酵素で消化したトマトのゲノムDNAを用いたサザンブロッティングを使用し、プローブとしてcDNAの5’端断片を使用して実施した。同じプローブを、既存のMoneyberg BACライブラリー (Keygene N.V.)にもハイブリダイズさせた。
【0107】
全長cDNAの一つ (IS158-53と表示される; 配列番号4) は、トマトの細胞質ゾルのシステインシンターゼ遺伝子と非常に高い相同性を示した(UP|Q9FS27 (Q9FS27)に対するtomato|TC162833相同物)。TC162833および配列番号4は何れも、同一のアミノ酸配列(配列番号5)をコードするが、二つのヌクレオチドにおいて核酸レベルで異なる。配列番号4の207位のヌクレオチド‘C’は、TC162833で‘T’であり、配列番号4の372位のヌクレオチド‘A’は、TC162833で‘G’である。
【0108】
配列番号4は、生物的および非生物的ストレス条件下においても構成的に発現し、トマトのゲノムにおいて少ないコピー数(約2コピー)で存在し、Keygene N.V.トマトBACライブラリーのBAC 9と十分にハイブリダイズしたため、そのプロモーターの単離に着手した。
【0109】
全長cDNAがトマトのシステインシンターゼ遺伝子(配列番号4)と相同性を有する遺伝子のプロモーターを、以下に記載するとおり選択した。
【0110】
例3−AA6プロモーター配列の単離
システインシンターゼと指定されたトマト遺伝子の導出配列上に設計されたプライマー (プライマー配列については下記参照) を使用して、BAC 9由来のゲノムDNAを鋳型として用いたリンカーPCRを実施した。5’および3’配列は何れも、システインシンターゼのUTRについて得た。3’UTR配列 (配列番号3) は、MoneybergトマトのゲノムDNAを鋳型として用いた長距離PCRにより得た。
【0111】
システインシンターゼ配列上に設計されたプライマー:
プロモーター用 5’-GTTCGATGAGGACACTCTCGC-3
およびネスティッドプライマー 5’-CAATTAAAGTTGCTAAGCGTCCTGA-3’
ターミネーター用 5’-TCAGTTACATCCTTGGCAATTCC-3’
およびネスティッドプライマー 5’-CAGAGAACATGACTGTGGAGCC-3’。
【0112】
例4−形質転換ベクターの構築
システインシンターゼ5’上流領域の二つのAA6プロモーターDNA断片、5000bpのもの (配列番号2) と2986bpのもの (配列番号1) を、gusAコード領域と一緒に、内在性システインシンターゼターミネーター (配列番号3) またはnos ターミネーターの何れかを備えたプラスミドpKG1562 (ノパリンシンターゼプロモーターにより駆動されるnptIIを含有するベクター骨格) に連結した。
【0113】
構築物を指定した:
以下のものを含有するpKG8135 (図3参照):
2986 bpのシステインシンターゼ5’上流領域 (配列番号1): gusA : 3’nos
以下のものを含有するpKG8136 (図3参照):
5000 bpのシステインシンターゼ5’上流領域 (配列番号2): gusA : 3’nos
以下のものを含有するpKG8137 (図3参照):
5000 bpのシステインシンターゼ5’上流領域 (配列番号2): gusA : 3’システインシンターゼ (配列番号3)。
【0114】
コントロール構築物は、以下のとおり使用した(図2参照):
以下のものを含有するpKG1700: CaMV35Sプロモーター (Franck et al., 同上): gusA : 3’nos。
【0115】
使用した形質転換ベクターのベクターマップの詳細については、図1〜3が参照される。
【0116】
プラスミドpKG8135、pKG8136、pKG8137およびpKG1700を、アグロバクテリウムツメファシエンスに組み込んだ。タバコ (var. SR1) およびトマト (var. RZ 52201) を形質転換し、カナマイシン選択の下で再生させ、第一次再生体 (T0) を生育して結実させた。
【0117】
例5−形質転換体の発現分析
gusAの発現は、これらベクター内のAA6プロモーターまたはCaMV35Sプロモーターにより駆動されるため、植物で産生されるβ-D-グルクロニダーゼ (GUS) のレベルは、プロモーターの有効性を示す。GUSの存在についての組織学的染色アッセイは、基質として5-ブロモ-4-クロロ-インドリルグルクロニド (XGluc) を用いて実施した。
【0118】
染色実験は、(第一次形質転換体の自家受粉後に得られた) タバコおよびトマトの両方のT1植物の葉組織について、芽生えから開花植物までの幾つかの発生段階で実施した。加えて、GUSについての組織学的染色実験は、成熟したタバコおよびトマト系統に由来する様々な材料、たとえば花芽、開花している花、花茎 (タバコ)、花柄、頂端分裂組織の芽、葉、葉柄、主要な茎、根、種子および果実 (トマト)、並びに発芽している種子および芽生えについて実施した。
【0119】
AA6プロモーターの長い断片 (配列番号2) もしくは短い断片 (配列番号1) またはCaMV35SプロモーターによりgusAが駆動された場合、GUSの発現を、タバコおよびトマトの両方において、試験された全ての組織で検出した。
【0120】
視覚的 (染色) 比較により、タバコおよびトマトの両方において、AA6プロモーター断片並びにCaMV35Sプロモーターが機能していることが示された。
【0121】
GUS発現の定量的分析は、基質として4-メチルウンベリフェリルβ-d-グルクロニド (MUG) を用いた蛍光アッセイにより実施した。5000bp断片のAA6プロモーター (pKG8137) により駆動されるGUSの発現を、タバコおよびトマトの両方の若葉において、CaMV35Sプロモーター (pKG1700) により駆動されるGUSの発現と比較した。
【0122】
シングルコピーの植物を、サザンブロッティングを用いて選択した;HindIII制限酵素で消化したタバコおよびトマト植物由来のゲノムDNAを使用し、nptII断片をプローブとして使用し、そのプロセスを、制限酵素としてXbaIを用いて繰り返した。各T-DNA挿入物は、nptIIおよびgusAをそれぞれ一コピー含有するため、nptIIコピー数は、gusAコピー数を反映する。また、コピー数は、Invader(登録商標)アッセイ (Third Wave Technologies, Inc.)を用いて確認し、公知のトマトまたはタバコの内部コントロールと比較してnptII遺伝子の存在および相対量を検出した;このアッセイをT1世代に適用すると、半接合個体と同型接合個体との識別が可能である。
【0123】
トータルタンパク質抽出物を、タバコおよびトマト植物のpKG8137およびpKG1700半接合および同型接合シングルコピー形質転換体およびゼロコピーコントロールから作成した。タンパク質含量は、NanoOrangeTM Kit (Molecular Probes) を用いて測定し、その後、サンプル中のタンパク質濃度を、抽出バッファーでの希釈により標準化した。
【0124】
MUG基質の添加後、初期蛍光を測定した (励起フィルター、355nmおよび発光フィルター、460nm)。その後、サンプルをインキュベートし、切断されたメチルウンベリフェロン (MU) による蛍光を測定した。インキュベーション1時間あたりの蛍光の増加を計算した。(テストサンプルのトータルタンパク質含量に等しい) 非トランスジェニックタバコまたはトマトのトータルタンパク質抽出物のバックグラウンドにおいてGUS酵素 (Sigma-Aldrich Chemie B.V.) 活性を示すキャリブレーションカーブを作成した。トランスジェニックテストサンプルに存在するGUS酵素の濃度 (mU/トータル植物タンパク質抽出物mg) を、サンプルおよびキャリブレーションカーブからの蛍光データを用いて計算した。
【0125】
2回の実験の統計解析により、AA6プロモーター (pKG8137) およびCaMV35Sプロモーターにより駆動されるGUS発現のレベルは、タバコにおいて有意な差異はないが、トマトでは、同型接合シングルコピー植物においてAA6プロモーターによるGUSの発現は、CaMV35SプロモーターによるGUSの発現より79%高いことが示された(以下の表で示されるデータセット)。
【0126】
タバコでのGUSの発現
表1:半接合タバコ植物の系統ごとの平均GUS活性 (mU/トータル植物タンパク質抽出物mgで表示)、並びに標準偏差および平均の標準誤差 (SEM)
【表1】
【0127】
表2:同型接合タバコ植物の系統ごとの平均GUS活性 (mU/トータル植物タンパク質抽出物mgで表示)、並びに標準偏差および平均の標準誤差 (SEM)
【表2】
【0128】
以上示すとおり、AA6プロモーター (5kbプロモーター) の平均プロモーター活性は、(導入遺伝子について半接合および同型接合の植物の両方において)トランスジェニックタバコの葉において35Sプロモーターのものと本質的に同等である。
【0129】
トマトでのGUSの発現
表3:半接合トマト植物の系統ごとの平均GUS活性 (mU/トータル植物タンパク質抽出物mgで表示)、並びに標準偏差および平均の標準誤差 (SEM)
【表3】
【0130】
表4:同型接合トマト植物の系統ごとの平均GUS活性 (mU/トータル植物タンパク質抽出物mgで表示)、並びに標準偏差および平均の標準誤差 (SEM)
【表4】
【0131】
表4に示すとおり、AA6プロモーター (5kbプロモーター) の平均プロモーター活性は、(導入遺伝子について半接合および同型接合の植物の両方において)トランスジェニックトマトの葉における35Sプロモーターのものと比較して、トマトにおいて有意に高い(35S活性をスタンダードとした場合、データセット1では35Sより約52%高く、データセット2では35Sより約79%高い)。
【0132】
例6−他の植物種でのプロモーター活性
他の植物種においてAA6およびCaMV35Sプロモーターにより駆動されるGUS発現の比較をテストし、確認した。現在のところ、導入遺伝子のAA6プロモーター駆動発現についてテストを行った種のリストには、(上述のタバコおよびトマトに加えて)レタス、メロンおよびアブラナ属(Brassica)が含まれ、これらの各々において構成的発現が示されている。
【0133】
例7−“3kb”および“5kb”AA6プロモーターの再配列決定
ベクターpKG8135およびpKG8137 (図3) に存在する核酸配列を、再配列決定し、配列をチェックし、不明なヌクレオチドを解明した。
【0134】
不明な位置は、配列番号6 (“3kb”プロモーター) および配列番号7 (“5kb”プロモーター)の配列表に示し、ここでは、不明な位置において最も可能性のあるヌクレオチドも同定される (以下の表も参照)。
【0135】
不明なヌクレオチドのために、ごく僅かな配列の違いが存在する:配列番号1は、(‘ニードル’、ギャップオープニング = 10.0、ギャップ伸長 = 0.5; DNAFullマトリクスを用いて) 配列番号8および6とそれぞれ99.6%および99.7%の配列同一性を有する。配列番号2は、(‘ニードル’、ギャップオープニング = 10.0、ギャップ伸長 = 0.5; DNAFullマトリクスを用いて) 配列番号9および7とそれぞれ99.6%および99.8%の配列同一性を有する。
【表B】
【表C】
【図面の簡単な説明】
【0136】
【図1】pKG1562のベクターマップ。
【図2】pKG1700のベクターマップ。
【図3】pKG8135、pKG8136およびpKG8137のベクターマップ。
【発明の分野】
【0001】
本発明は、強力な構成的植物プロモーターおよびその使用の分野に関する。プロモーターは、植物または植物細胞において同種または異種のタンパク質を発現させるため、または活性な核酸分子、たとえばセンスおよび/またはアンチセンスRNAを発現させるために使用することができる。プロモーター活性を有する核酸配列、並びにキメラ遺伝子、ベクター、およびこれらを含む組換え(トランスジェニック)細胞および生物が提供される。また、プロモーターを含むトランスジェニック細胞または生物、とりわけ植物および植物細胞を作成する方法が提供される。更に、細胞質ゾルのシステインシンターゼタンパク質およびこれをコードする核酸配列が提供される。
【発明の背景】
【0002】
多くの植物プロモーターが、当該技術分野で公知であり、これらは、トランスジェニック植物または植物細胞においてタンパク質またはペプチドを発現させるため、または遺伝子または遺伝子ファミリーの発現を停止させる(silencing)ための有用なツールである。これら植物プロモーターには、構成的プロモーター、誘導性プロモーター、発生調節プロモーター、組織優先的または組織特異的プロモーターが含まれる。一般に使用される構成的プロモーターの例は、以下のとおりである:カリフラワーモザイクウイルス(CaMV)の単離体CM 1841 (Gardner et al., 1981, Nucleic Acids Research 9, 2871-2887)、CabbB-S (Franck et al., 1980, Cell 21, 285-294)、およびCabbB-JI (Hull and Howell, 1987, Virology 86, 482-493)の35Sプロモーターまたは増強(enhanced)35Sプロモーター(“35Sプロモーター”);Odell et al. (1985, Nature 313, 810-812) またはUS5164316に記載される35Sプロモーター、ユビキチンファミリー由来のプロモーター (たとえばトウモロコシのユビキチンプロモーター、Christensen et al., 1992, Plant Mol. Biol. 18, 675-689, EP 0 342 926、Cornejo et al. 1993, Plant Mol. Biol. 23, 567-581参照)、gos2プロモーター (de Pater et al., 1992 Plant J. 2, 837-844)、emuプロモーター (Last et al., 1990, Theor. Appl. Genet. 81,581-588)、アラビドプシス(Arabidopsis)のアクチンプロモーター、たとえばAn et al. (1996, Plant J. 10, 107-121)に記載されるプロモーター、イネのアクチンプロモーター、たとえばZhang et al.(1991, The Plant Cell 3, 1155-1165)に記載されるプロモーターおよびUS 5,641,876に記載されるプロモーターまたはWO 00/70067に記載されるようなイネのアクチン2プロモーター;キャッサバ葉脈モザイクウイルスのプロモーター (WO 97/48819, Verdaguer et al. 1998, Plant Mol. Biol. 37, 1055-1067)、サブタレニアン・クローバー萎縮ウイルス由来のpPLEXシリーズのプロモーター (WO 96/06932、とりわけS7プロモーター)、アルコールデヒドロゲナーゼプロモーター、たとえばpAdh1S (GenBankアクセッション番号X04049、X00581)、US 6 051 753およびEP 426 641に記載されるゴマノハグサモザイクウイルスプロモーター、ヒストン遺伝子プロモーター、たとえばアラビドプシス(Arabidopsis)由来のPh4a748プロモーター (Plant Mol. Biol. 8: 179-191)、CoYMV (ツユクサ黄色斑紋ウイルス)プロモーター (Medberry et al. 1992, The Plant Cell 4:185-192)など。
【0003】
しかし、公知の構成的プロモーターの共通の欠点は、これらが、器官または発生で調節される発現、またはストレス誘導性の発現変化など、しばしば活性の変動を示すことである。本発明者らは、トランスジェニック植物においてCaMV 35Sプロモーターの活性が、非生物的ストレス、とりわけトランスジェニック植物をスペインの農場で生育させたときに引き起こされる熱ストレスに感受性を示すことを見出した。したがって、一または複数の生物的および/または非生物的ストレスに感受性を示さない新規な構成的プロモーターを提供する必要がある。加えて、トランスジェニック植物の生成において植物由来のプロモーターを使用することが、調節の観点から望ましい。
【0004】
加えて、植物に感染可能なウイルスに由来するウイルス性プロモーターは、ウイルスの植物への感染が、トランスジェニックプロモーターのサイレンシングを引き起こし得るため、宿主植物種の形質転換にとってあまり好ましくない(Seemanpillai et al., 2003, Mol Plant Microbe Interact. 16(5): 429-38; Al-Kaff et al., 2000, Nat Biotechnol. 18: 995-9)。幾つかの同一プロモーターの使用は、サイレンシングという結果を招き得るため、遺伝子スタッキングアプローチのためには、異なる構成的プロモーターも必要である(Yang et al., 2005, Plant Mol Biol. 58: 351-66)。
【発明の概要】
【0005】
ゲノムに統合されたキメラ遺伝子を含む、トランスジェニック植物または植物細胞または植物組織または器官であって、前記キメラ遺伝子が、同種または異種の核酸配列に動作可能に連結された構成的プロモーターを含み、前記プロモーターが、
(a)配列番号1または配列番号2の核酸配列、または配列番号6〜9の何れか一つ、またはCBSに(Centraalbureau voor Schimmelcultures、Uppsalalaan 8, 3584 CT Utrecht, The Netherlandsに、2006年8月3日、それぞれアクセッション番号CBS 120175およびCBS 120176で)寄託されたベクターpKG8135またはpKG8137にクローニングされたプロモーター配列;
(b)配列番号1または配列番号2の機能的断片、または(a)に記載の配列の何れかの機能的断片;
(c)配列番号1または2と少なくとも70%の配列同一性を備えた核酸配列、または(a)に記載の配列の何れかと少なくとも70%の配列同一性を備えた核酸配列;
(d)(c)の核酸配列の機能的断片
の群から選択されることを特徴とする、トランスジェニック植物または植物細胞または植物組織または器官が提供される。
【0006】
また、植物細胞に導入された際にプロモーター活性を有する単離された核酸配列であって、
(a)配列番号1または配列番号2の核酸配列、または配列番号6〜9の何れか一つ、またはCBSに(Centraalbureau voor Schimmelcultures、Uppsalalaan 8, 3584 CT Utrecht, The Netherlandsに、2006年8月3日、それぞれアクセッション番号CBS 120175およびCBS 120176で)寄託されたベクターpKG8135またはpKG8137にクローニングされたプロモーター配列;
(b)配列番号1または配列番号2の断片、または(a)に記載の配列の何れかの断片;
(c)配列番号1または2と少なくとも70%の配列同一性を備えた核酸配列、または(a)に記載の配列の何れかと少なくとも70%の配列同一性を備えた核酸配列;
(d)(c)の核酸配列の断片
から選択される配列を含む核酸配列が提供される。
【0007】
また、上記配列を含む、ベクター、キメラ遺伝子および宿主細胞も、本発明の一つの態様である。
【0008】
別の態様において、トランスジェニック植物または植物細胞または植物組織または器官におけるセンスおよび/またはアンチセンス核酸配列の構成的発現のための、細胞質ゾルの植物システインシンターゼ遺伝子のプロモーターの使用が提供される。
【0009】
更に、トランスジェニック植物または植物細胞を作成するための方法であって、
(a)転写される核酸配列に動作可能に連結され、必要に応じて更に3’UTR核酸配列に連結された上述のプロモーターを含むキメラ遺伝子、またはかかるキメラ遺伝子を含むベクターを生成する工程;
(b)前記キメラ遺伝子またはベクターを用いて植物または植物細胞を形質転換する工程;および必要に応じて、
(c)トランスジェニック植物または植物細胞を再生させる工程
を含む方法が提供される。
【0010】
更に別の態様において、配列番号3または4のヌクレオチド配列を含む単離された核酸分子が提供される。
【一般的な定義】
【0011】
「核酸配列」(または核酸分子)の用語は、一本鎖または二本鎖の形態のDNAまたはRNA分子、とりわけ、本発明のプロモーター活性を有するDNAまたはタンパク質もしくはタンパク質断片をコードするDNAを指す。「単離された核酸配列」は、自然環境から単離され自然環境にもはや存在しない核酸配列、たとえば、細菌宿主細胞または植物の核もしくはプラスチドのゲノムに存在する当該核酸配列を指す。
【0012】
「タンパク質」または「ポリペプチド」の用語は、交換可能に使用され、具体的な作用の仕方、大きさ、3次元構造、由来に関係なく、アミノ酸の鎖からなる分子を指す。よって、タンパク質の「断片」または「部分」は、「タンパク質」と呼ばれてもよい。「単離されたタンパク質」は、自然環境にもはや存在しないタンパク質、たとえばインビトロ、または組換え細菌もしくは植物の宿主細胞に存在するタンパク質を指すために使用される。
【0013】
「遺伝子」の用語は、適切な転写調節領域(たとえばプロモーター)に動作可能に連結された、細胞でRNA分子(たとえばmRNA)に転写される領域(転写領域)を含むDNA配列を意味する。よって、遺伝子は、動作可能に連結された幾つかの配列、たとえば、プロモーター、たとえば翻訳開始に関与する配列などを含む5’非翻訳リーダー配列(これは、5’UTRとも呼ばれ、翻訳開始コドンの上流に存在する転写されるmRNA配列に相当する)、(タンパク質)コード領域(cDNAまたはゲノムDNA)、並びにたとえば転写終結部位およびポリアデニル化部位(たとえばAAUAAAまたはその変異体)などを含む3’非翻訳配列(これは3’非翻訳領域または3’UTRとも呼ばれる)を含み得る。
【0014】
「キメラ遺伝子」(または組換え遺伝子)は、ある生物種に自然の状態で通常見出されない任意の遺伝子、とりわけ自然の状態では互いに関連がない一または複数の部分の核酸配列が存在する遺伝子を指す。たとえば、プロモーターは、転写される領域の一部または全部と、または別の調節領域と、自然の状態で関連がない。「キメラ遺伝子」の用語は、プロモーターまたは転写調節配列が、一または複数のセンス配列(たとえばコード配列)、またはアンチセンス(センス鎖のリバース相補鎖)または逆方向反復配列(センスおよびアンチセンス、転写によりRNA転写産物は二本鎖RNAを形成する)に動作可能に連結された発現構造体を含むと理解される。
【0015】
「3’UTR」または「3’非翻訳配列」(しばしば3’非翻訳領域または3’端とも呼ばれる)は、たとえば転写終結部位および(全てではないがほとんどの真核生物のmRNAで)ポリアデニル化シグナル(たとえばAAUAAAまたはその変異体)などを含む、遺伝子のコード配列の下流に見出される核酸配列を指す。mRNA転写産物は、転写終結後、ポリアデニル化シグナルの下流で切断され、ポリ(A)テールが付加され、これは、mRNAの細胞質(ここで翻訳が起こる)への輸送に関与する。
【0016】
「遺伝子の発現」は、適切な調節領域、とりわけプロモーターに動作可能に連結されたDNA領域が、生物学的に活性なRNA、すなわち生物学的に活性なタンパク質またはペプチド(または活性なペプチドの断片)に翻訳され得るRNA、または(たとえば転写後の遺伝子サイレンシングまたはRNAiにおいて)それ自体で活性なRNAに転写されるプロセスを指す。ある態様において活性なタンパク質は、リプレッサードメインが存在するために優性のネガティブな機能を有するタンパク質を指す。コード配列は、好ましくは、センスの向きにあり、所望の生物学的に活性なタンパク質またはペプチド、または活性なペプチド断片をコードする。遺伝子サイレンシングのアプローチにおいて、DNA配列は、好ましくは、アンチセンスDNAまたは逆方向反復DNAの形態で存在し、ターゲット遺伝子の短い配列をアンチセンスの向き、またはセンスとアンチセンスの向きで含む。「異所性発現」は、その遺伝子が通常発現されない組織での発現を指す。
【0017】
「転写調節配列」は、転写調節配列に動作可能に連結された核酸配列の転写速度を調節することが可能な核酸配列と本明細書で規定される。よって、本明細書で規定される転写調節配列は、転写の開始に必要な配列エレメント(プロモーターエレメント)、転写を維持し調節するのに必要な配列エレメント、たとえばアテニュエーターまたはエンハンサー、サイレンサーなど、あらゆるものを含む。ほとんどの場合、コード配列の上流(5’)転写調節配列を指すが、コード配列の下流(3’)に見出される調節配列も、この規定に包含される。
【0018】
本明細書で使用される「プロモーター」の用語は、一または複数の遺伝子の転写をコントロールするために機能する核酸断片であって、遺伝子の転写開始部位の転写方向に対して上流(5’)に位置し(転写開始は、配列の+1位と呼ばれ、それに対して上流のヌクレオチドは、マイナスの数を用いて呼ばれ)、DNA依存性RNAポリメラーゼの結合部位、転写開始部位、および任意の他のDNAドメイン(cis作用性配列)(たとえば、転写因子結合部位、リプレッサーおよびアクチベータータンパク質結合部位、およびプロモーターに由来する転写量を調節するために直接的もしくは間接的に作用することが当業者に公知の任意の他のヌクレオチド配列を含むがこれらに限定されないもの)の存在により構造的に同定される核酸断片を指す。転写開始(+1)の上流の真核生物のcis作用性配列の例には、TATAボックス(一般に転写開始のおよそ−20から−30位)、CAATボックス(一般に転写開始に対しておよそ−75位)、5’エンハンサーまたはサイレンサーエレメントなどが含まれる。「構成的」プロモーターは、ほとんどの組織(または器官)で、ほとんどの生理的および発生的条件下において活性なプロモーターである。より好ましくは、構成的プロモーターは、全ての主要な器官、たとえば少なくとも葉、茎、根、種子、果実および花で、本質的に全ての生理的および発生的条件下において活性である。最も好ましくは、構成的プロモーターは、全ての器官で、ほとんどの(好ましくは全ての)生理的および発生的条件下において活性である。「誘導性」プロモーターは、生理的(たとえばある種の化合物の外からの適用により)または発生的に調節されるプロモーターである。「組織特異的」プロモーターは、特定のタイプの組織または細胞でのみ活性化である。したがって、プロモーター活性は、当該プロモーターが、プロモーターの下流(3’)に動作可能に連結された核酸配列を転写する環境を参照することにより説明することができる。したがって、「構成的活性を有するプロモーター」、または植物もしくは植物細胞で「構成的」なプロモーターは、植物またはほとんどの組織(または器官)の植物細胞で、ほとんどの生理的および発生的条件下において転写を行う核酸配列を指す。「一または複数の生物的および/または非生物的ストレスに対して感受性を示さない」プロモーター、またはその活性が「一または複数の生物的および/または非生物的ストレス条件に晒されたときに低下しない」プロモーターは、正常な生理的および発生的条件下においてプロモーター活性を有し、かつ生物的および/または非生物的ストレスが、当該プロモーターを含む生物(たとえば植物)または細胞または組織または器官に与えられたときに、その活性が定量的に低下しないか、または少なくとも有意に低下しない核酸配列を指す。
【0019】
「ストレス」は、植物または植物細胞に作用し、植物の収量ロスおよび/または品質ロスという結果を招き得るが、植物に致命的でない、物理学的、化学的または生物学的由来の条件または圧力を指す。
【0020】
「非ストレス条件」は、本明細書において、生理機能および発生が正常であるかまたは最適である条件を指す。
【0021】
「生物的ストレス」は、生物的(生きている)作用物質、たとえば真菌類、ウイルス、生物のようなマイコプラズマ、昆虫、細菌、線虫など(すなわち、とりわけ植物の害虫および病原体)により引き起こされるストレスを指す。
【0022】
「非生物的ストレス」は、非生物的(生きていない)作用物質、たとえば温度ストレス(低温/凍結、熱)、塩度(塩)、風、金属、日長(光周期)、水ストレス(たとえば過少もしくは過剰の水アベイラビリティ、すなわち干ばつ、脱水、水浸しなど)、傷害、照射などにより引き起こされるストレスを指す。
【0023】
本明細書で使用される「動作可能に連結された」の用語は、機能的な関係でのポリヌクレオチドエレメントの連結を指す。核酸は、別の核酸配列と機能的な関係に置かれたときに「動作可能に連結される」。たとえば、プロモーター、または転写調節配列は、コード配列の転写に影響を及ぼす場合、コード配列に動作可能に転結される。動作可能に連結されるということは、連結されるDNA配列が、典型的には隣接していることを意味し、二つのタンパク質コード領域をつなげる必要がある場合、「キメラタンパク質」を産生するようにリーディングフレームで隣接していることを意味する。「キメラタンパク質」または「ハイブリッドタンパク質」は、天然にはそういうものとして見出されないが、機能的タンパク質を形成するようにつながれ、つながれたドメインの機能性(たとえばDNA結合ドメインまたは優性ネガティブ機能につながる機能ドメインの抑制)を示す、種々のタンパク質「ドメイン」(またはモチーフ)から構成されるタンパク質である。キメラタンパク質は、天然に存在する二以上のタンパク質の融合タンパク質であってもよい。本明細書で使用される「ドメイン」の用語は、当該ドメインの機能的特性を少なくとも備えた新たなハイブリッドタンパク質を提供するための、別のタンパク質に移転可能な特定の構造または機能を備えたタンパク質の任意の部分またはドメインを意味する。
【0024】
「ターゲットペプチド」の用語は、細胞内オルガネラ、たとえばプラスチド、好ましくは葉緑体、ミトコンドリア、または細胞外空間(分泌シグナルペプチド)へタンパク質を向かわせるアミノ酸配列を指す。ターゲットペプチドをコードする核酸配列は、タンパク質のアミノ末端(N-末端)をコードする核酸配列に(フレームで)融合され得る。
【0025】
「核酸構築物」または「ベクター」は、本明細書において、組換えDNA技術の使用により得られ、外因性DNAを宿主細胞に運搬するために使用される、人工の核酸分子を意味すると理解される。ベクターの骨格は、たとえば、バイナリーまたはスーパーバイナリーベクター(たとえばUS 5,591,616、US2002138879およびWO 95/06722参照)、コインテグレート(co-integrate)ベクターまたはT-DNAベクターとすることができ、これらは、当該技術分野で公知であり、本明細書の他の箇所に記載されており、ベクターの骨格に、キメラ遺伝子が統合されるか、あるいは適切な転写調節配列/プロモーターが既に存在する場合には、所望の核酸配列のみ(たとえばコード配列、アンチセンスまたは逆方向反復配列)が、転写調節配列/プロモーターの下流に統合される。ベクターは、通常、分子クローニングでの使用を容易にするための更なる遺伝的エレメント、たとえば選択マーカー、マルチクローニングサイトなどを含む(下記参照)。
【0026】
「宿主細胞」または「組換え宿主細胞」または「形質転換細胞」は、所望のタンパク質をコードするキメラ遺伝子、またはターゲット遺伝子/遺伝子ファミリーのサイレンシングのためのアンチセンスRNAまたは逆方向反復RNA(またはヘアピンRNA)を転写により産生する核酸配列をとりわけ含む、少なくとも一の核酸分子を細胞に導入した結果生じる、新たな個々の細胞(生物)を指す用語である。宿主細胞は、好ましくは植物細胞であるが、細菌細胞、真菌細胞(酵母細胞を含む)などであってもよい。宿主細胞は、染色体外(エピソーム)複製分子として核酸構築物を含有してもよいし、あるいはより好ましくは、宿主細胞の核もしくはプラスチドのゲノムに統合されたキメラ遺伝子を含んでいてもよい。「選択マーカー」の用語は、当業者によく知られた用語であり、発現した際に、当該選択マーカーを含有する一または複数の細胞を選択するために使用することができる任意の遺伝的構成要素(entity)を記載するために本明細書で使用される。選択マーカー遺伝子産物は、たとえば、抗生物質に対する抵抗性、あるいはより好ましくは、除草剤に対する抵抗性または別の選択可能な特性、たとえば表現型の形質(たとえばの色素形成の変化)または栄養上の要件などを付与する。「レポーター」の用語は、可視的マーカー、たとえば緑色蛍光タンパク質(GFP)、eGFP、ルシフェラーゼ、GUSなどを指すために主に使用される。
【0027】
遺伝子またはタンパク質の「オルソログ(ortholog)」の用語は、本明細書において、当該遺伝子またはタンパク質と同じ機能を有するが、当該遺伝子を有する種が分岐した時点(すなわち、当該遺伝子が、種分化により共通の祖先から進化した時点)から配列が(通常)相違している、別の種に見出される相同な遺伝子またはタンパク質を指す。よって、ある植物種に由来する遺伝子のオロソログは、両方の配列の比較(たとえば配列全体または特定のドメインにわたるパーセンテージ配列同一性)および機能分析に基いて、他の植物種で同定され得る。
【0028】
「同種」および「異種」の用語は、核酸またはアミノ酸の配列と、その宿主細胞または生物、とりわけトランスジェニック生物との間の関係を指す。よって、同種の配列は、宿主種に天然に見出されるが(たとえば、トマト遺伝子で形質転換されたトマト植物)、異種の配列は、宿主細胞に天然に見出されない(たとえば、ポテト植物由来の配列で形質転換されたトマト植物)。「相同体(homolog)」または「相同な」の用語は、文脈に応じて、共通の祖先の配列に由来する子孫の配列を択一的に指し得る(たとえば、これらはオルソログであり得る)。
【0029】
「ストリンジェントなハイブリダイゼーション条件」は、所定のヌクレオチド配列と実質的に同一のヌクレオチド配列を同定するために使用することができる。ハイブリダイゼーション条件のストリンジェンシーは、配列に依存し、様々な環境で異なる。一般に、ストリンジェントな条件は、規定のイオン強度およびpHにおける特定配列の熱融点(Tm)より約5℃下げるように選択される。Tmは、ターゲット配列の50%が、完全一致のプローブとハイブリダイズする(規定のイオン強度およびpHの下での)温度である。典型的には、塩濃度がpH 7において約0.02モルであり、温度が少なくとも60℃であるストリンジェントな条件が選択される。塩濃度の低下および/または温度の増加は、ストリンジェンシーを増大させる。RNA−DNAハイブリダイゼーション(たとえば100 ntのプローブを用いたノーザンブロット)のためのストリンジェントな条件は、たとえば、63℃で20分間の0.2×SSC中での少なくとも1回の洗浄を含む条件、または同等の条件である。DNA−DNAハイブリダイゼーション(たとえば100 ntのプローブを用いたサザンブロット)のためのストリンジェントな条件は、たとえば、少なくとも50℃、通常約55℃の温度で20分間の0.2×SSC中での少なくとも1回の洗浄(通常2回)を含む条件、または同等の条件である。Sambrook et al. (1989)およびSambrook and Russell (2001)も参照されたい。
【0030】
「配列同一性」および「配列類似性」は、二つの配列の長さに依存して、グローバルまたはローカルアラインメントアルゴリズムを用いて、二つのペプチドまたは二つのヌクレオチドの配列を整列させること(alignment)により決定することができる。同様の長さの配列は、好ましくは、全長にわたって最適に配列を整列させるグローバルアラインメントアルゴリズム(たとえばNeedleman Wunsch)を用いて整列させるが、実質的に異なる長さの配列は、好ましくは、ローカルアラインメントアルゴリズム(たとえばSmith Waterman)を用いて整列させる。(デフォルトパラメーターを用いてたとえばプログラムGAPまたはBESTFITにより最適に整列させた際の)配列が、(以下で規定される)配列同一性の最小パーセンテージを少なくとも共有する場合、配列は、「実質的に同一である」または「本質的に同様である」ということができる。GAPは、Needleman and Wunschのグローバルアラインメントアルゴリズムを使用して、二つの配列を完全な長さ(全長)にわたって整列させ、一致の数を最大にし、ギャップの数を最小にする。二つの配列が同様の長さを有する場合、配列同一性を決定するためにグローバルアラインメントが適切に使用される。一般に、GAPデフォルトパラメーターが使用され、ギャップ生成ペナルティー=50(ヌクレオチド)/8(タンパク質)であり、ギャップ伸長ペナルティー=3(ヌクレオチド)/2(タンパク質)である。ヌクレオチドについて、使用されるデフォルトスコアリングマトリクスは、nwsgapdnaであり、タンパク質について、デフォルトスコアリングマトリクスは、Blosum62である(Henikoff & Henikoff, 1992, PNAS 89, 915-919)。配列アラインメントおよびパーセンテージ配列同一性のスコアは、コンピュータープログラム、たとえばGCG Wisconsin Package, Version 10.3(Accelrys Inc., 9685 Scranton Road, San Diego, CA 92121-3752 USAから入手可能)を用いて、あるいはオープンソースソフトウェア、たとえばEmbossWIN version 2.10.0に含まれるプログラム“needle”(グローバルNeedleman Wunschアルゴリズムを使用)もしくは“water”(ローカルSmith Watermanアルゴリズムを使用)を用いて、上記GAPと同じパラメーターを用いて、あるいはデフォルトセッティング(“needle”および“water”の何れについても、また、タンパク質およびDNAアラインメントの何れについても、デフォルトギャップオープニングペナルティーは10.0であり、デフォルトギャップ伸長ペナルティーは0.5であり; デフォルトスコアリングマトリクスは、タンパク質についてはBlossum62であり、DNAについてはDNAFullである)を用いて決定され得る。配列が、実質的に異なる全長を有している場合、たとえばSmith Watermanアルゴリズムを使用したローカルアラインメントが好ましい。あるいは、パーセンテージ類似性または同一性は、たとえばFASTA、BLASTなどのアルゴリズムを用いて公共データベースに対してサーチすることにより決定してもよい。
【0031】
本明細書および特許請求の範囲において、動詞「含むこと(to comprise)」およびその語形変化は、非限定的な意味で使用され、そのワードの後につづく項目を含むが、具体的に挙げられていない項目を排除しないことを意味する。加えて、不定冠詞“a”または“an”によるエレメントの指示は、その文脈が、一つおよび唯一のエレメントが存在することを明らかに要求しない限り、二つ以上のエレメントが存在する可能性を排除しない。よって、不定冠詞“a”または“an”は、通常「少なくとも一つ」を意味する。更に、本明細書で「配列」を参照する場合、ある配列のサブユニットを備えた実際の物理的分子(たとえばアミノ酸)を指すと理解される。
【0032】
本発明に従って“植物(plant)”または“植物(plants)”(すなわち複数の植物)を参照する場合はいつも、別途示さない限り、植物の部分(細胞、組織または器官、種子、切断または収穫された部分、葉、芽生え、花、花粉、果実、茎、根、カルス、プロトプラストなど)、親の際立った特徴(たとえば導入遺伝子の存在)を保持する当該植物の子孫またはクローン繁殖体、たとえば自家受粉または交配により得られた種子、たとえば(二つの近交親系統の交配により得られた)ハイブリッド種子、これに由来するハイブリッド植物および植物部分も包含すると理解される。
【詳細な説明】
【0033】
スペインの農場試験でCaMV 35Sプロモーター(Franck et al., 1980, Cell 21, 285-294に記載されるシングル35Sプロモーター)を含むトランスジェニック植物を試験したところ、35Sプロモーターの活性は、高い夏の温度に影響を受けることが見出された。老齢の葉では、発現の全損失が起こった。したがって、植物の細胞および器官で構成的な活性を示し、一または複数の生物的および/または非生物的ストレスに本質的に感受性を示さない植物プロモーターは、トランスジェニック植物、とりわけ発生の間または成熟期に一または複数のストレス条件に晒され得る植物における核酸配列の制御発現にとって望ましいため、かかるプロモーターを単離するためのプログラムを本発明者らは開始した。cDNA-AFLPを用いて、強力で構成的なプロモーターの遺伝子をトマト植物から単離し、これは、ストレス条件、たとえば干ばつストレス、低温ストレス、熱ストレス、病原体ストレス(CMV感染)、日長変動、照射(たとえばUV誘発ストレス)、水ストレス(水撒き過多および水撒き過少)などの下において、強力で構成的な転写プロファイルを保持していた(実施例参照)。対応するプロモーターを単離することにより、強力で構成的な発現を植物に付与した(または付与することが可能な)プロモーターの部類が見出された。プロモーター強度は、CaMV 35Sプロモーターのプロモーター強度と少なくとも同等であり、幾つかの宿主種(トマト)ではそれより有意に高かった。更に、プロモーターは、これらを含む植物または植物部分が、種々の生物的および/または非生物的ストレス条件を受けたときに、活性の低下を示さない(すなわち、強力で構成的なままである)。これらプロモーターは、本明細書において「AA6プロモーター」と呼ばれる。
【0034】
核酸配列、キメラ遺伝子およびベクター
一つの態様において、植物細胞でプロモーター活性を有する単離された核酸配列(好ましくはゲノムまたは合成DNA配列)が提供され、これは、植物および植物の組織または器官において強力で構成的な転写活性を示す。プロモーターは、好ましくは植物の全ての器官で活性であるが、少なくとも主要な器官で活性であり、プロモーター強度は、35S(Franck et al., 同上に記載されるCaMV 35Sプロモーター)のプロモーター強度と少なくともほとんど同等である(またはそれより強力である)。好ましくは、この核酸配列のプロモーター活性は、トランスジェニック植物、または植物組織または器官が、一または複数の非生物的および/または生物的ストレスを受けたときに、低下しないか、または少なくとも有意に低下しない。この点において有意な低下は、非ストレス条件下での同一の組織または器官における活性と比較して、1%以上(たとえば2%、3%、5%、10%など、100%まで)のプロモーター活性の統計的に有意な(定量的)低下を指す。よって、好ましくは、プロモーターは、ストレス条件下において(好ましくは全ての器官、とりわけ少なくとも主要な植物器官で)強力で構成的なままである。
【0035】
一つの態様において、配列番号1(“3kb”プロモーター)または配列番号2(“5kb”プロモーター)、または配列番号6〜9の何れか一つ(配列番号6および8は“3kb”プロモーターであり、配列番号7および9は“5kb”プロモーターであり、配列番号1および2と比較して、幾つかの不明ヌクレオチドまたは僅かに異なる配列を有する)、またはpKG8135 (CBS120175)またはpKG8137 (CBS120176)にクローニングされたプロモーター配列、または少なくとも植物においてプロモーター活性を有するこれらの何れかの活性(機能的)断片、たとえば配列番号1または2、配列番号6〜9、またはpKG8135 (CBS120175)またはpKG8137 (CBS120176)内のプロモーターの少なくとも200、300、400、500、600、800、900、1000、1200、1500、2000、2500、2800、2900、3000、3500、4000、4500またはそれ以上の連続ヌクレオチドの断片を含む、構成的AA6プロモーターが提供される。「活性断片」または「機能的断片」、または「プロモーター活性を有する断片」は、植物細胞、器官および植物において、好ましくは、配列番号1および2、配列番号6〜9、またはpKG8135 (CBS120175)またはpKG8137 (CBS120176)内のプロモーターと少なくとも同じ組織および器官において構成的転写を与えることができる核酸断片を指す。これは、以下に記載するとおり、好ましくはレポーター遺伝子に動作可能に連結されたかかる断片で植物細胞を形質転換し、プロモーター活性を定性的(空間的−時間的転写)および/または定量的にアッセイすることにより試験することができる。
【0036】
本明細書において配列番号1を参照する場合、配列番号6または8の何れか、またはpKG8135 (CBS120175)に存在する配列も参照していること、並びにその文章は、これらプロモーター配列の何れかを参照するように読むことができることが理解される。同様に、本明細書において配列番号2を参照する場合、配列番号7または9の何れか、またはpKG8137 (CBS120175)に存在する配列も参照していること、並びにその文章は、これらプロモーター配列の何れかを指すように読むことができることが理解される。
【0037】
一つの態様において、プロモーター断片の強度は、定量的に、配列番号1および/または2の強度(したがって、35Sの強度)とほとんど同一であるか、またはそれよりも高い。配列番号1および2は、翻訳開始コドン(mRNA上のAUGまたはDNA上のATG)のすぐ上流の配列であるため、活性断片は、配列番号1および/または2の5’端での欠失により好ましくは作成される。したがって、断片は、好ましくは、配列番号1または2の3’領域の(上述のとおり)少なくとも200、300、400、500等のヌクレオチドを含む。明らかに、DNA断片は、幾つかの手法、たとえばデノボDNA合成、または制限酵素、または末端ヌクレアーゼなどを用いて作成され得る。
【0038】
しかし、ある種のシス作用性エレメント(たとえばエンハンサー配列)の除去は、低いプロモーター活性という結果を招き得る。したがって、様々な態様において、プロモーター断片の強度は、配列番号1および/または2の強度より(したがって、35Sの強度より)定量的に低い。幾つかのアプリケーションにとって、強度の低下したプロモーターが好ましい。当業者であれば、当該技術分野で公知の手法を用いて、全長プロモーターおよび任意のプロモーター断片の活性を容易に決定し、その強度および組織特異性を、35Sまたは本明細書で提供されるプロモーターのものと比較することができる。たとえば、Medberry et al. (Plant Cell, 1992, Vol. 4: 185-192)は、一過性発現アッセイを用いてプロモーター強度を比較する方法を記載する。
【0039】
配列番号1および2のプロモーター、およびその機能的断片は、好ましくは、プロモーターを含む植物または植物細胞、組織または器官が晒され得る少なくとも一つの(好ましくは複数の、最も好ましくはあらゆる)生物的および/または非生物的ストレスに感受性を示さない(下記参照)。よって、活性は、ストレス条件に晒されている間、構成的で強力なままである。
【0040】
また、上記AA6プロモーターの「変異体」およびかかる変異体の機能的断片も提供される。これら変異体には、配列番号1および/または2と本質的に類似している核酸配列(および上述のとおり、これら変異体配列の機能的断片)であって、構成的プロモーター活性を有している、すなわち、植物、植物細胞、組織または器官で、最も好ましくは配列番号1および2と少なくとも同じ組織および器官で構成的転写を提供することができる核酸配列が含まれる。配列番号1および/または2と「本質的に類似している」配列は、Needleman and Wunsch Pairwiseアラインメント (GCGのプログラム“GAP”またはEmbosswin, version 2.10.0の“needle”)を用いて、ギャップ生成ペナルティー = 5およびギャップ伸長ペナルティー = 3で、配列番号1および/または配列番号2と(全長にわたって)少なくとも約50、55、60、65、70、75、80、85、90、95、96、97、98、99%またはそれ以上の核酸配列同一性を備え、かつ植物または植物細胞においてプロモーター活性を有する核酸配列である。好ましい態様において、変異体(および機能的断片)の活性は、組織および器官の全てにおいて強力であり、すなわち、定量的に配列番号1または2と少なくとも同等に強力である(またはそれより強力である)。更なる態様において、これら変異体(および機能的断片)の活性は、一または複数の生物的および/または非生物的ストレスに感受性を示さず、すなわち強力で構成的なままである。
【0041】
本明細書で提供される核酸配列の変異体または機能的断片を同定、合成または単離するために多くの方法が使用可能であることは明らかであり、たとえば核酸ハイブリダイゼーション、PCR技術、インシリコ(in silico)分析、および核酸合成などが挙げられる。たとえば、核酸ハイブリダイゼーションは、ストリンジェントまたは適度にストリンジェントなハイブリダイゼーション条件下で、配列番号1または2、またはこれらの断片とハイブリダイズするDNA配列を他の植物種または品種で同定するために使用することができる。あるいは、配列データベースを、たとえばBLAST、FASTAなどの公知のアルゴリズムを用いて、変異体配列についてインシリコで(in silico)スクリーニングすることができる。このようにして、他の植物種またはトマトの他品種から、または他の生物全体から変異体配列を単離することは実現可能である。以下に記載されるとおり、トマトの他品種または他の植物種、とりわけナス属(Solanum)の種で見出される同じ遺伝子(細胞質ゾルのシステインシンターゼ遺伝子およびそのオルソログ)の他の対立遺伝子のプロモーターが、本明細書にとりわけ含まれる。たとえば、cDNAライブラリーは、一または複数の植物種、一または複数の品種、または一つの種または品種の異なる組織から構築され得る。cDNAライブラリーを、(たとえば、配列番号4(トマトシステインシンターゼcDNA)、またはその断片もしくは変異体に由来するプローブまたはプライマーを用いて)システインシンターゼcDNAについてスクリーニングしてもよい。同様に、実施例に記載されるとおり、ディファレンシャルディスプレイ法(たとえばcDNA-AFLP)を使用して、システインシンターゼ転写産物を同定してもよい。TAIL-PCR(Liu et al. 1995, Genomics 25(3):674-81; Liu et al. 2005, Methods Mol Biol. 286:341-8)、Linker-PCR、またはInverse PCR (IPCR)などの方法を使用して、遺伝子の上流転写調節領域を単離してもよい。
【0042】
核酸配列(または変異体の断片)が構成的プロモーター活性を有するかどうか、すなわち、全ての器官において構成的転写を与えることができるかどうか、その活性が「強力」であるかどうか、並びに、トランスジェニック細胞、組織、器官または生物(とりわけ植物または植物細胞)が晒され得る少なくとも一つの(好ましくは複数の、最も好ましくはあらゆる)生物的および/または非生物的ストレスに、その核酸配列の活性が感受性を示さないかどうかを、種々の方法を用いて決定することができる。一般に、定性的方法と定量的方法を区別することができる。定性的方法(たとえば組織学的GUS染色)は、プロモーターの空間的−時間的活性(プロモーターが、ある組織または器官で、あるいはある環境/発生条件の下で、活性であるか否か?)を決定するために使用され、定量的方法(たとえば蛍光GUSアッセイ)は、コントロールと比較した活性レベルを定量する。適切なコントロールは、たとえば、空のベクターで形質転換された植物(ネガティブコントロール)またはCaMV 35Sなどの他のプロモーターを含む構築物で形質転換された植物、または非トランスジェニック植物である。
【0043】
相対的または絶対的活性をテストし、必要に応じて定量するために、クローニングまたは合成された核酸分子、たとえば配列番号1、2、またはその変異体、またはこれらの何れかの断片を、公知の核酸配列(たとえば、gusAなどのレポーター遺伝子、または特定のタンパク質をコードする任意の遺伝子)に動作可能に連結し、これを使用して、公知の手法で植物細胞を形質転換してもよい。幾つかの態様において、細胞は、安定な様式で形質転換される必要はなく、すなわち、一過性発現アッセイを使用して(たとえばプロトプラストトランスフェクションまたはアグロインフィルトレーション)、プロモーターが細胞、組織または器官で活性であるかどうか、並びにどの程度までプロモーターが転写を駆動するかを決定してもよい。プロモーターの活性は、たとえば、下流核酸配列のRNA転写産物のレベルを検出することによりアッセイする(そして必要に応じて定量する)ことができる。これは、定量的方法、たとえば定量的RT-PCRまたは他のPCRベースの方法などを用いて行われてもよい。あるいは、レポータータンパク質またはレポータータンパク質の活性をアッセイし、定量してもよい。たとえば、レポーター遺伝子がgus遺伝子である場合、実施例に記載されるとおり、蛍光GUSアッセイを使用してもよい。このようにして、正常な生理的(非ストレス)条件下で維持された形質転換植物または植物細胞の定量的プロモーター活性レベルを、一または複数の生物的または非生物的ストレスに晒された植物または植物細胞のレベルと比較することができる。また、相対的または絶対的活性レベルを、構成的コントロールプロモーター、たとえば35Sプロモーター、ダブル35Sプロモーター、または配列番号1および/または2と比較することができる。好ましくは平均プロモーター活性レベルを決定し、統計的方法を用いて比較することが理解される。
【0044】
よって、本発明のプロモーターが、ある時間に、どの組織または器官において活性であるか(空間的−時間的活性)は、たとえば、植物または植物細胞をプロモーター−レポーター遺伝子構築物で形質転換し、種々の組織を種々の発生段階の間に、RNA転写産物またはレポータータンパク質(またはその活性)について分析することによりテストすることができる。単純なテストは、たとえば、組織学的GUS染色を採用し、これによる青色の視覚的評価は、種々の組織における種々の発生段階での活性を示す。
【0045】
既述のとおり、プロモーター活性は構成的であることが好ましく、好ましくは、植物および植物細胞、とりわけ配列が導入された宿主種または品種で強力である。構成的活性は、プロモーターに動作可能に連結された任意の核酸配列の転写産物が、好ましくは、ほとんどの組織(または器官)で、ほとんどの(正常な、非ストレス)生理的および発生条件下で産生されることを意味する。一つの態様において、本発明のプロモーターは、実施例で試験された組織または器官、たとえば葉(若い葉および老齢の葉)、根、花、種子、茎(主要な茎)、果実(たとえば未熟な果実および熟した果実)、発芽した種子などで少なくとも活性である。
【0046】
好ましくは、本発明のプロモーターは、すべての植物種、双子葉植物種および単子葉植物種の両方において、強力で構成的な活性を提供する。たとえば、配列番号1および配列番号2は、種々の植物種、たとえばトマト、タバコ、アブラナ属(Brassica)、メロンおよびレタスなどにおいて、強力で構成的な発現を提供することが見出された(実施例参照)。
【0047】
本発明のAA6プロモーター(断片または変異体を含む)の強度(定量的活性)は、下流(3’)に連結された核酸配列の発現を駆動する能力の観点から、種々の公知の方法を用いて定量的に決定することができる。たとえば、転写された転写産物(mRNA)の量は、定量的RT-PCRまたはノーザンブロッティングを用いて定量することができる。好ましくは、プロモーター強度は、正常な(非ストレス)条件下において、CaMV 35S(Franck et al., 同上)のプロモーター強度と少なくとも本質的に同等である。よって、「強力」とは、プロモーター強度が、正常な非ストレス条件下において、35Sのプロモーター強度と好ましくは少なくともほぼ同一であり、より好ましくはそれより強力であることを意味する。最も好ましくは、種々の組織および器官における平均定量的プロモーター活性は、CaMV 35Sプロモーターの活性と少なくとも同等であるか、あるいはCaMV 35Sプロモーターの平均活性より少なくとも5%、10%、20%、30%、35%、40%、45%、50%、55%、60%、70%、75%またはそれ以上高い。同じコピー数および接合状態レベルの形質転換体を比較すべきであることが理解され、たとえば導入遺伝子については半接合または同型接合である。好ましくは、シングルコピーの形質転換体を同定し、比較する。
【0048】
AA6プロモーターの発現は、ストレス条件下において、少なくとも本明細書の他の箇所に記載されるストレスから選択される一または複数のストレスの下で、構成的で強力なままである。
【0049】
よって、プロモーターを含む植物組織または器官または植物が、干ばつストレス、熱ストレス、水ストレス(過剰の水および過少の水の両方)、病原体ストレス(たとえばCMVなどのウイルス感染、真菌感染、細菌感染など)、害虫ストレス(たとえば昆虫の摂食)、傷害、塩ストレス、照射ストレスなどの一つまたは好ましくは幾つかから少なくとも選択されるストレス条件に晒されたときに、本発明のプロモーターの強度は、好ましくは、本質的に未変化のままであるか、あるいは少なくとも低下しない(または有意に低下しない)。再度、定量的テストを、これを決定するために使用することができる。たとえば、プロモーターを含む組換え植物を、正常な温度環境から温かい環境(たとえば約27℃から約50℃まで)に移し、種々の組織におけるプロモーター活性を、正常な温度条件と温かい温度条件の下での同じ組織の活性と比較してもよい。
【0050】
また、上記AA6プロモーターの変異体には、植物のシステインシンターゼ遺伝子、とりわけ細胞質ゾルのシステインシンターゼ遺伝子の任意の単離された核酸プロモーター(すなわち、システインシンターゼ酵素、とりわけ細胞質ゾルのアイソフォームをコードする植物遺伝子のプロモーター)であって、植物において強力で構成的な活性を示し、好ましくは、一または複数の生物的および/または非生物的ストレスに本質的に感受性を示さないものが含まれる。既述のとおり、他の植物のシステインシンターゼ遺伝子も、一または複数のストレス条件下で構成的発現を保持する強力で構成的なプロモーターを有していると考えられる。植物のシステインシンターゼ遺伝子は、O-アセチル-L-セリン[チオール]-リアーゼ(EC 4.2.99.8)とも称される酵素システインシンターゼをコードする遺伝子である。幾つかのシステインシンターゼ遺伝子がクローニングされているが(たとえば実施例で言及されるTC162833参照)、トランスジェニック植物における同型接合または異型接合配列の強力で構成的な発現を駆動するためのプロモーターの有用性については記載されていない。よって、かかる公知のシステインシンターゼ遺伝子または今のところ未知のシステインシンターゼ遺伝子のプロモーターを単離し、それらの活性についてスクリーニングすることができる。たとえば、cDNA-AFLP、他のPCRベースの方法またはノーザンハイブリダイゼーションを使用して、他のシステインシンターゼ遺伝子、とりわけ、構成的そして一または複数のストレス条件下においてもシステインシンターゼmRNAsを内因的に産生する遺伝子を単離または同定することができる。その後、所望の発現パターンを備えたシステインシンターゼ遺伝子を選択し、それらのプロモーターを公知の方法を用いてクローニングすることができる。好ましい態様において、プロモーターは、ナス科(Solanaceae)、たとえばナス属(Solanum)(再分類されたトマト属(Lycopersicon)の種を含む)、タバコ属(Nicotiana)、トウガラシ属(Capsicum)、ペチュニア属(Petunia)、コフィア属(Coffea)の種などに属する植物に由来するシステインシンターゼ遺伝子から得られる。
【0051】
本発明のプロモーターは、好ましくは、シロイヌナズナ(Arabidopsis thaliana)などの草植物のプロモーターではなく、好ましくは、Gutierrez-Alcala et al. (J. of Exp. Botany 56, p24872494) に記載されるシロイヌナズナのOASA1遺伝子のプロモーター、またはその断片ではない。
【0052】
(配列番号5のシステインシンターゼ酵素をコードする)配列番号4で提供されるシステインシンターゼ遺伝子の他に、他のシステインシンターゼ遺伝子を同定し、それらのプロモーターを単離することができる。上述のとおり種々の方法が使用され得る。
【0053】
よって、本発明の一つの態様において、植物および/または植物細胞で構成的活性を有し、かつ植物または植物細胞、組織または器官が、たとえば既述のストレスなどの一または複数の生物的および/または非生物的ストレスに晒されたときにその活性が低下しないか、または有意に低下しない、植物(細胞質ゾル)のシステインシンターゼ遺伝子のプロモーターが提供される。
【0054】
かかるプロモーターは、以下のものを含む:(a)植物から入手可能であり、植物のシステインシンターゼ遺伝子の発現を駆動する任意のプロモーター、とりわけ配列番号5のタンパク質をコードする任意の遺伝子のプロモーター;(b)配列番号5に対して(全長にわたって)少なくとも30、40、50、60、70、80、90、95、99%またはそれ以上のアミノ酸同一性を有するシステインシンターゼ酵素をコードする植物遺伝子のプロモーター;(c)システインシンターゼ酵素をコードし、その核酸配列が、配列番号4に対して(全長にわたって)少なくとも30、40、50、60、70、80、90、95、98、99%またはそれ以上のヌクレオチド配列同一性を含む、核酸配列の発現を駆動する植物プロモーター。
【0055】
別の態様において、組換え細胞または生物、とりわけ植物細胞または植物における同型接合または異型接合の核酸配列の構成的な過剰発現のための、植物のシステインシンターゼ遺伝子のプロモーターの使用が提供される。この使用は、既述のとおり、同型接合または異型接合の核酸配列にプロモーターを動作可能に連結し、植物または植物細胞を形質転換することを含む。
【0056】
上述の焦点は、植物および植物細胞における本発明のプロモーターの使用にあるが、他の細胞および生物、たとえば任意の原核または真核細胞または生物、たとえば細菌、真菌類(Pichia、Hansenulaなどの酵母を含む)、哺乳類、ヒト細胞または細胞株などにおいて同型接合または異型接合の核酸配列を発現させるためにプロモーターを使用することも本発明の態様である。
【0057】
本発明のキメラ遺伝子およびベクター
本発明の一つの態様において、プロモーター活性を有する上記核酸配列の何れかを使用して、キメラ遺伝子およびこれらを含むベクターが、キメラ遺伝子の宿主細胞への転移および動作可能に連結された異型接合または同型接合の核酸配列の宿主細胞(たとえば細胞、組織、器官または形質転換細胞に由来する生物全体)での発現のために作成される。
【0058】
宿主細胞は、好ましくは植物細胞である。任意の植物が、適切な宿主であり、たとえば、単子葉植物または双子葉植物、たとえば、トウモロコシ(maize/corn)(Zeaの種、たとえばZ. mays、Z. diploperennis (chapule)、Zea luxurians (グアテマラのテオシンテ)、Zea mays subsp. huehuetenangensis (サンアントニオヒュースタのテオシンテ)、Z. mays subsp. mexicana (メキシコのテオシンテ)、Z. mays subsp. parviglumis (バルサスのテオシンテ)、Z. perennis (多年生テオシンテ) およびZ. ramosa、コムギ (Triticumの種)、オオムギ (たとえばHordeum vulgare)、カラスムギ (たとえばAvena sativa)、モロコシ (Sorghum bicolor)、ライムギ (Secale cereale)、ダイズ (Glycine spp、たとえばG. max)、ワタ (Gossypiumの種、たとえばG. hirsutum、G. barbadense)、Brassica spp. (たとえばB. napus、B. juncea、B. oleracea、B. rapaなど)、ヒマワリ (Helianthus annus)、タバコ (Nicotianaの種)、アルファルファ (Medicago sativa)、イネ (Oryzaの種、たとえばO. sativaインディカ栽培品種-グループまたはジャポニカ栽培品種-グループ)、まぐさ(forage grasses)、パールミレット (Pennisetum属の種、たとえばP. glaucum)、樹木の種、野菜の種、たとえばLycopersicon ssp (近年ナス属(Solanum)に属すると再分類された)、たとえばトマト (L. esculentum, syn. Solanum lycopersicum) たとえばチェリートマト、var. cerasiformeまたはカレントトマト、var. pimpinellifolium) またはツリートマト (S. betaceum, syn. Cyphomandra betaceae)、ジャガイモ (Solanum tuberosum) およびその他のナス属(Solanum)の種、たとえばナス (Solanum melongena)、ペピーノ (S. muricatum)、ココナ (S. sessiliflorum) およびナランヒージャ (S. quitoense);コショウ (Capsicum annuum、Capsicum frutescens)、エンドウ (たとえばPisum sativum)、マメ (たとえばPhaseolusの種)、ニンジン (Daucus carota)、Lactuca属の種 (たとえばLactuca sativa、Lactuca indica、Lactuca perennis)、キュウリ (Cucumis sativus)、メロン (Cucumis melo)、ズッキーニ (Cucurbita pepo)、スカッシュ (Cucurbita maxima、Cucurbita pepo、Cucurbita mixta)、パンプキン (Cucurbita pepo)、スイカ (Citrullus lanatus syn. Citrullus vulgaris)、多肉果の種 (グレープ、ピーチ、プラム、ストロベリー、マンゴー、メロン)、観賞植物種 (たとえばバラ、ペチュニア、キク、ユリ、チューリップ、ガーベラの種)、木本 (たとえばPopulus、Salix、Quercus、Eucalyptusの種)、繊維の種、たとえばアマ (Linum usitatissimum) およびアサ (Cannabis sativa)である。一つの態様において、野菜の種、とりわけナス属(Solanum)の種(トマト属(Lycopersicon)の種を含む)が好ましい。
【0059】
よって、たとえば以下の属の種が形質転換され得る:Cucurbita、Rosa、Vitis、Juglans、Fragaria、Lotus、Medicago、Onobrychis、Trifolium、Trigonella、Vigna、Citrus、Linum、Geranium、Manihot、Daucus、Arabidopsis、Brassica、Raphanus、Sinapis、Atropa、Capsicum、Datura、Cucumis、Hyoscyamus、Lycopersicon、Solanum、Nicotiana、Malus、Petunia、Digitalis、Majorana、Ciahorium、Helianthus、Lactuca、Bromus、Citrullus、Asparagus、Antirrhinum、Heterocallis、Nemesis、Pelargonium、Panieum、Pennisetum、Ranunculus、Senecio、Salpiglossis、Browaalia、Glycine、Pisum、Phaseolus、Gossypium、Glycine、Lolium、Festuca、Agrostis。更に好ましいのは、Cucurbita、Brassica、Lycopersicon、Solanum、OryzaおよびZeaの各々である。好ましいのは、Avena、Medicago、Capsicum、Nicotiana、Lactuca、Pisum、Cucumis、Cucurbita、Brassica、Solanum (Lycopersiconを含む)、OryzaおよびZeaの各々である。
【0060】
キメラ遺伝子、および宿主細胞のゲノムへのキメラ遺伝子の導入のためのベクターの構築は、一般に当該技術分野で公知である。キメラ遺伝子を作成するために、AA6プロモーター配列を、標準的な分子生物学技術を用いて、宿主細胞で転写される別の核酸配列に動作可能に連結する。プロモーター配列は、転写される核酸配列が、プロモーター配列の下流でベクターに簡単に挿入されるように、ベクターに既に存在していてもよい。その後、ベクターは、宿主細胞を形質転換するために使用され、キメラ遺伝子は、好ましくは、核ゲノム、またはプラスチド、ミトコンドリアまたは葉緑体のゲノムに挿入され、その結果、下流の核酸配列が、プロモーターの活性により発現される(たとえば、Mc Bride et al., 1995 Bio/Technology 13, 362; US 5,693,507)。
【0061】
したがって、キメラ遺伝子は、好ましくは上述のAA6プロモーターを含み、これは、同型接合または異型接合の核酸配列に動作可能に連結され、必要に応じてその後に3’非翻訳核酸配列(3’UTR)が続く。同型接合または異型接合の核酸配列は、タンパク質またはペプチドをコードする配列であってもよいし、活性なRNA分子、たとえば宿主細胞または生物における遺伝子または遺伝子ファミリーのサイレンシングに適したセンスおよび/またはアンチセンスRNA(dsRNA)に転写される配列であってもよい。
【0062】
AA6プロモーターを含むキメラ遺伝子は、単一の植物細胞の核ゲノムに慣用的なやり方で安定に挿入することができ、このように形質転換された植物細胞は、キメラ遺伝子の構成的発現により表現型の変化した形質転換植物を産生するために、慣用的なやり方で使用することができる。
【0063】
これに関して、アグロバクテリウムツメファシエンス(Agrobacterium tumefaciens)に含まれる、更なる核酸配列に動作可能に連結されたAA6プロモーター(または上述の変異体または断片)を含むT-DNAベクターは、植物細胞を形質転換するために使用することができ、その後、たとえばEP 0 116 718、EP 0 270 822、PCT公報 WO 84/02913および公開された欧州特許出願EP 0 242 246、およびGould et al. (1991, Plant Physiol. 95,426-434) に記載される手法を用いて、形質転換された植物細胞から形質転換植物を再生することができる。アグロバクテリウム媒介性の植物の形質転換は、当該技術分野で周知である。T-DNAベクターは、EP 0 120 561およびEP 0 120 515に記載されるバイナリーベクター、または相同組換えによりアグロバクテリウムTi-プラスミドに統合することができるEP 0 116 718に記載されるコインテグレート(co-integrate)ベクターの何れであってもよい。
【0064】
好ましいT-DNAベクターはそれぞれ、T-DNAのボーダー配列の間、または少なくとも右ボーダー配列の左側の位置に、転写される核酸配列に動作可能に連結されたAA6プロモーターを含有する。ボーダー配列は、Gielen et al. (1984, EMBO J 3,835-845) に記載される。当然、他のタイプのベクターを使用して、植物細胞を形質転換することができ、直接的な遺伝子導入(たとえばEP 0 223 247に記載される、またはUS 2005/055740およびWO 2004/092345に記載されるパーティクルまたはミクロプロジェクタイル(microprojectile)ボンバードメント)、花粉媒介性の形質転換(たとえばEP 0 270 356およびWO 85/01856に記載される)、プロトプラストの形質転換(たとえばUS 4,684,611に記載される)、植物ウイルス媒介性の形質転換、リポソーム媒介性の形質転換(たとえばUS 4,536,475に記載される)などの手法、並びにその他の方法、たとえばトウモロコシ (たとえば、US 6,140,553; Fromm et al., 1990, Bio/Technology 8, 833-839; Gordon-Kamm et al., 1990, The Plant Cell 2, 603-618) およびイネ (Shimamoto et al., 1989, Nature 338, 274-276; Datta et al. 1990, Bio/Technology 8, 736-740) のある系統を形質転換するために記載された方法、および一般に単子葉植物を形質転換するための方法 (WO 92/09696) が使用される。ワタの形質転換のためにはWO 00/71733も参照され、イネの形質転換のためにはWO 92/09696、WO 94/00977およびWO 95/06722に記載の方法も参照される。モロコシの形質転換のためには、たとえばJeoung JM et al. 2002, Hereditas 137: 20-8またはZhao ZY et al. 2000, Plant Mol Biol. 44: 789-98が参照される。トマトまたはタバコの形質転換のためには、An G. et al., 1986, Plant Physiol. 81: 301-305; Horsch R.B. et al., 1988, In: Plant Molecular Biology Manual A5, Dordrecht, Netherlands, Kluwer Academic Publishers. pp 1-9; Koornneef M. et al., 1986, In: Nevins D.J. and R.A. Jones, eds. Tomato Biotechnology, New York, NY, USA, Alan R. Liss, Inc. pp 169-178も参照される。同様に、形質転換された細胞からの形質転換植物の選択および再生は、当該技術分野で周知である。明らかに、様々な種に対して、更に単一種の様々な品種または栽培種に対しても、高い頻度で形質転換体を再生させるためにプロトコールを具体的に適合させる。
【0065】
核ゲノムの形質転換に加えて、プラスチドのゲノム、好ましくは葉緑体のゲノムの形質転換も本発明に含められる。プラスチドのゲノムの形質転換は、導入遺伝子の蔓延のリスクを低減できるという利点がある。プラスチドのゲノムの形質転換は、当該技術分野で公知のとおり行うことができ、たとえばSidorov VA et al. 1999, Plant J.19: 209-216またはLutz KA et al. 2004, Plant J. 37(6): 906-13が参照される。
【0066】
得られた形質転換植物は、慣用的な植物育種スキームで使用して、導入遺伝子を含有する多くの形質転換植物を産生することができる。シングルコピーの形質転換体は、たとえばサザンブロット分析またはPCRベースの方法またはInvader(登録商標)テクノロジーアッセイ(Third Wave Technologies, Inc.)を用いて選択することができる。形質転換された細胞および植物は、キメラ遺伝子の存在により非形質転換体と容易に区別することができる。導入遺伝子の挿入部位に隣接する植物DNAの配列も配列決定することができ、これにより、ルーチン使用のために「イベント特異的な」検出方法を開発することができる。たとえば、WO 01/41558が参照され、これは、たとえば統合された配列および隣接する(ゲノム)配列に基くエリートなイベント検出キット(たとえばPCR検出キット)を記載する。
【0067】
一つの態様において、転写され、(コード配列である場合)必要に応じて翻訳される核酸配列は、転写される配列が、適切な3’端の転写調節シグナル(“3’端”)(すなわち転写産物形成およびポリアデニル化シグナル)の上流(すなわち5’)に位置するように植物ゲノムに挿入される。ポリアデニル化および転写産物形成シグナルには、ノパリンシンターゼ遺伝子のもの(“3’nos”) (Depicker et al., 1982 J. Molec. Appl. Genetics 1, 561-573.)、オクトピンシンターゼ遺伝子のもの(“3’ocs”) (Gielen et al., 1984, EMBO J 3, 835-845)、およびT-DNA遺伝子7のもの (“3’遺伝子7”) (Velten and Schell, 1985, Nucleic Acids Research 13, 6981-6998) などが含まれ、これらは、形質転換された植物細胞において3’-非翻訳DNA配列として機能する。
【0068】
好ましい態様において、使用される3’端配列は、植物のシステインシンターゼ遺伝子のものであり、好ましくは、AA6プロモーターが入手可能であるシステインシンターゼ遺伝子に由来する。本明細書で提供される適切な3’端配列は、配列番号3、またはその断片もしくは変異体である。配列番号3の変異体には、配列番号3に対して少なくとも50、60、65、70、75、80、85、90、95、98、99%またはそれ以上の核酸配列同一性を含む核酸配列が含まれる。配列番号3の断片または配列番号3の変異体の断片には、配列番号3または配列番号3の変異体の少なくとも50、100、150、200、250、300またはそれ以上の連続ヌクレオチドを含む核酸配列が含まれる。これらの3’端配列は、それ自体で一つの態様でもあり、任意のキメラ遺伝子およびベクターを構築するために、すなわち様々なプロモーターと組合せて使用してもよい。
【0069】
発現される核酸配列は、一つの態様において、ハイブリッドタンパク質またはペプチドまたは融合タンパク質を含むタンパク質またはペプチドをコードする配列である。コード配列は、任意の由来、すなわち植物、真菌類(酵母を含む)、動物、細菌、合成、ウイルスなどであり得る。ターゲッティングペプチド、たとえば分泌シグナルペプチドまたはプラスチドターゲッティングシグナルをコードする配列を含んでいてもよい。また、コード配列を、選択またはスコア(scorable)マーカーをコードする遺伝子、たとえばカナマイシン抵抗性を付与するneo(またはnptII)遺伝子(EP 0 242 236)に読み取り枠を合わせて(in-frame)連結させて、細胞が、容易に検出可能な融合タンパク質を発現するようにしてもよい。任意の遺伝子のコード領域(cDNAまたはゲノムDNA)が使用され得るが、以下の遺伝子のコード領域の例が、好ましくは、本発明のAA6プロモーターに動作可能に連結される:1.ウイルスの核酸配列の逆方向反復配列 (たとえば、ウイルスのコートタンパク質遺伝子のセンスおよびアンチセンス配列;下記も参照);2.疾病シグナル伝達経路遺伝子または疾病抵抗性遺伝子;3.非生物的ストレス応答関連遺伝子 (たとえば、SHINE転写因子、またはCBF/DREB遺伝子);4.二次代謝産物生合成遺伝子、たとえば治療的および/または薬理的に重要な産物または産業上価値のある化合物の生産のための遺伝子。
【0070】
明らかに、他の遺伝子、たとえば、作物学的形質に影響を及ぼす遺伝子、たとえば除草剤抵抗性の遺伝子(たとえばEPSPS遺伝子、barまたはPAT遺伝子)、収量または品質の形質(たとえばタンパク質組成)に影響を及ぼす遺伝子なども、本発明のAA6プロモーターに動作可能に連結され得る。
【0071】
本発明のキメラ遺伝子またはベクターは、微生物、たとえば細菌(たとえば大腸菌(Escherichia coli)、シュードモナス属(Pseudomonas)、アグロバクテリウム属(Agrobacterium)、バチルス属(Bacillus)など)または真菌類または藻類または昆虫を形質転換するために使用することもできるし、あるいはこの遺伝子またはベクターを、ウイルスを操作するために使用してもよい。適切なクローニング媒体に組み込まれた本発明の核酸配列による細菌の形質転換は、慣用的なやり方で、好ましくはMaillon et al. (1989, FEMS Microbiol. Letters 60, 205-210.) およびWO 90/06999に記載される慣用的なエレクトロポレーション技術を用いて行うことができる。原核宿主細胞でコード配列を発現させるために、核酸配列のコドン使用頻度をそれに応じて最適化してもよい(同様に、植物細胞でコード配列を発現させるために、核酸配列のコドン使用頻度を公知のとおり最適化してもよい)。イントロン配列は除去すべきであり、最適な発現のための他の適応を公知のとおり行ってもよい。
【0072】
イネ科植物種などの単子葉植物、たとえばトウモロコシまたはイネにおいて核酸配列の高い発現を実現するために、イントロン、好ましくは単子葉植物のイントロンをキメラ遺伝子に付加することができる。たとえば、5’調節領域へのトウモロコシAdh1遺伝子のイントロンの挿入は、トウモロコシで発現を高めることが示されている(Callis et. al., 1987, Genes Develop. 1: 1183-1200)。同様に、US 5,859,347に記載されるHSP70のイントロンは、発現を高めるために使用され得る。このように、一または複数のイントロンは、本発明のプロモーター配列の何れかに、または5’UTRまたはコード配列に必要に応じて挿入され得る。
【0073】
別の態様において、AA6プロモーターは、遺伝子サイレンシングのためのキメラ遺伝子およびベクターを作成するために使用され、ここでAA6プロモーターは、ターゲット遺伝子(サイレンシングされる内在性遺伝子または遺伝子ファミリー)のセンスおよび/またはアンチセンス核酸配列に動作可能に連結される。更に別の態様において、ターゲット遺伝子は、侵入する植物病原体の遺伝子または遺伝子ファミリーであってもよい。たとえば、ウイルスのコートタンパク質遺伝子の逆方向反復配列は、ウイルス抵抗性植物を作成するために使用され得る。ウイルスのコートタンパク質遺伝子は、たとえばWO 96/21031に記載される。「遺伝子サイレンシング」は、一または複数のターゲット遺伝子の遺伝子発現のダウンレギュレーションまたは完全な阻害を指す。遺伝子発現を低下または廃止させるための抑制性RNAの使用は、当該技術分野で十分に確立されており、幾つかのレビューの主題である(たとえばBaulcombe, 1996, Plant Cell 8: 1833-1844; Stam et al., 1997, Plant Journal 12: 63-82; Depicker and Van Montagu, 1997, Curr. Opinion Cell Biol. 9: 373-382)。植物で遺伝子サイレンシングを達成するために利用可能な技術は幾つか存在し、たとえば、ターゲット遺伝子の全部または一部のアンチセンスRNAを産生するキメラ遺伝子(たとえばEP 0 140 308 B1、EP 0 240 208 B1およびEP 0 223 399 B1参照)、または(共抑制(co-suppression)とも称される)センスRNAを産生するキメラ遺伝子(EP 0 465 572 B1参照)がある。
【0074】
しかし、これまでに最も成功したアプローチは、ターゲット遺伝子のセンスRNAおよびアンチセンスRNAの両方(「逆方向反復配列」)の産生であり、これは、細胞内に二本鎖RNA(dsRNA)を形成し、ターゲット遺伝子をサイレンシングさせる。dsRNAの産生および遺伝子サイレンシングのための方法およびベクターは、EP 1 068 311、EP 983 370 A1、EP 1 042 462 A1、EP 1 071 762 A1およびEP 1 080 208 A1に記載される。
【0075】
したがって、本発明のベクターは、ターゲット遺伝子のセンスおよび/またはアンチセンスDNA断片に動作可能に連結されたAA6プロモーターを含んでいてもよい。ターゲット遺伝子配列の短い(センスおよびアンチセンス)区切り(stretch)、たとえば少なくとも約17、18、19、20、21、22、23、24または25ヌクレオチドのコードまたは非コード配列で十分であり得る。より長い配列、たとえば少なくとも約100、200、250、300、400、500、1000、1500ヌクレオチドまたはそれ以上の配列も頻繁に使用される。好ましくは、センスおよびアンチセンス断片は、スペーサー配列、たとえばイントロンにより分離されており、これにより、dsRNAの形成でループ(またはヘアピン)が形成される。ターゲット遺伝子の任意の区切りは、遺伝子サイレンシングベクター、およびターゲット遺伝子または遺伝子ファミリーがサイレンシングされたトランスジェニック植物を作成するために使用され得る。ヘアピン構築物を生成する簡便な方法は、ジェネリックベクター、たとえばpHANNIBALおよびpHELLSGATE、Gateway(登録商標)技術に基くベクターを使用することである (Wesley et al. 2004, Methods Mol Biol. 265: 117-30; Wesley et al. 2003, Methods Mol Biol. 236: 273-86およびHelliwell & Waterhouse 2003, Methods 30(4): 289-95参照、これら全ては参照により本明細書に組み込まれる)。
【0076】
ターゲット遺伝子の保存された核酸配列を選択することにより、宿主植物内のファミリーメンバーをサイレンシングすることができる。ターゲット遺伝子が、侵入する病原体の遺伝子または遺伝子ファミリーである場合、病原体のターゲット遺伝子がサイレンシングされ、植物は、病原体に抵抗性をもつようになる。ターゲット遺伝子核酸配列のセンスおよび/またはアンチセンスDNA断片に動作可能に連結されたAA6プロモーターを含み、ターゲット遺伝子サイレンシングの表現型を示すトランスジェニック植物も、本明細書に包含される。表現型は、遺伝子の機能に依存し、化学的変化であっても分子的変化であってもよく、肉眼で見えても見えなくてもよい。したがって、かかるキメラ遺伝子およびベクターは、遺伝子の機能を決定または検証するために使用することもできる。
【0077】
本発明のキメラ遺伝子は、宿主ゲノムに安定に導入してもよいし、エピソームユニットとして存在してもよい。
【0078】
本発明のトランスジェニック細胞および生物
上述の方法により入手可能なトランスジェニック細胞および生物、とりわけ植物、植物細胞、組織または器官が提供される。これらの細胞および生物は、本発明のAA6プロモーターの存在による、細胞またはゲノム内のキメラ遺伝子の存在により特徴づけられる。加えて、mRNA転写産物または翻訳されたタンパク質は、細胞または生物、たとえば植物細胞または植物の表現型を変化させ得る。
【0079】
AA6プロモーターは構成的であるが、ゲノム内の位置は、プロモーターの活性およびキメラ遺伝子の発現レベルに影響を及ぼすことができる。したがって、高い構成的なレベルのタンパク質、または(サイレンシング構築物を使用する場合)高い構成的なレベルのセンスおよび/またはアンチセンス転写産物を発現する形質転換体(“イベント”または“形質転換イベント”)は、たとえば、コピー数の分析(サザンブロット分析)、mRNA転写産物レベルの分析(たとえばノーザンブロット分析またはRT-PCR)により、あるいは核酸配列によりコードされるタンパク質の存在およびレベルの分析(たとえばSDS-PAGEとその後のウェスタンブロット分析;ELISA分析、免疫細胞学的アッセイなど)により選択することができる。また、形質転換体は、一または複数の生物的および/または非生物的ストレス条件の下での発現の安定性についてテストすることができ、一または複数の所望の条件の下で高い構成的発現を保持するイベントを同定し、更なる使用のために選択することができる。
【0080】
トランスジェニック植物は、伝統的な育種方法、たとえば交配、自家受粉、戻し交配などで使用することができる。形質転換体を自家受粉させることにより、導入遺伝子について同型接合である植物を生成することができる。育種の手順は、当該技術分野で公知であり、植物育種の標準テキストブック、たとえば、Allard, R.W., Principles of Plant Breeding (1960) New York, NY, Wiley, pp 485; Simmonds, N.W., Principles of Crop Improvement (1979), London, UK, Longman, pp 408; Sneep, J. et al., (1979) Tomato Breeding (p. 135-171) in: Breeding of Vegetable Crops, Mark J. Basset, (1986, editor), The Tomato crop: a scientific basis for improvement, by Atherton, J.G. & J. Rudich (editors), Plant Breeding Perspectives (1986); Fehr, Principles of Cultivar Development-Theory and Technique (1987) New York, NY, MacMillanに記載される。
【0081】
トランスジェニック細胞または生物は、たとえば組換えタンパク質の大規模生産のために、細胞培養(植物細胞培養、細菌または真菌細胞培養、たとえば酵母培養、ヒトまたは哺乳類細胞培養、昆虫細胞培養)において使用することもできる。一つの態様において、本発明のAA6プロモーターを含む細胞を含む細胞培養が提供される。
【0082】
本発明の方法および使用
また、トランスジェニック植物または植物細胞を作成するための方法であって、
(a)発現される核酸配列に動作可能に連結された本発明のAA6プロモーターおよび/またはAA6 3’UTRを含むキメラ遺伝子またはベクターを生成する工程;
(b)前記キメラ遺伝子またはベクターを用いて植物または植物細胞を形質転換する工程;および必要に応じて、
(c)トランスジェニック植物(a transgenic plant or plants)を再生させる工程
を含む方法が提供される。
【0083】
更に、非ストレス条件下において強力で構成的なプロモーター活性を提供し、かつ一または複数の生物的および/または非生物的ストレスに晒されたときにプロモーター活性が本質的に変化しない(少なくとも低下しないか、または有意に低下しない)トランスジェニック植物が同定され得る。
【0084】
この植物は、慣用的な農業および育種の方法で使用され得る。とりわけ、植物は、一または複数の生物的および/または非生物的ストレス条件に植物を晒す環境において、プロモーターの活性を低下させることなく、生育させることができる。よって、たとえば、高温または低温、強風、高い塩、高い土壌汚染物質、高いまたは低い水レベル、干ばつ期(drought spells)、高い病気または害虫プレッシャーの領域、高い照射の領域などにおいて、プロモーター活性を低下させることなく、あるいは少なくとも有意に低下させることなく、トランスジェニック植物を生育させるための方法が提供される。
【0085】
配列
配列番号1:“3kb”(2986 bp) AA6プロモーター
配列番号2:“5kb”(5000 bp) AA6プロモーター
配列番号3:トマトのシステインシンターゼ遺伝子(AA6遺伝子)の3’UTR
配列番号4:トマトのシステインシンターゼcDNA (AA6 cDNA)
配列番号5:配列番号4によりコードされるタンパク質 (トマトの細胞質ゾルのシステインシンターゼ)
配列番号6:幾つかの不明ヌクレオチドを有する“3kb”AA6プロモーター
配列番号7:幾つかの不明ヌクレオチドを有する“5kb”AA6プロモーター
配列番号8:再度配列決定され、E.coliの寄託物CBS120175内にある、pKG8135の“3kb”AA6プロモーター
配列番号9:再度配列決定され、E.coliの寄託物CBS120176内にある、pKG8137の“5kb”AA6プロモーター。
【0086】
図面
図1:pKG1562のベクターマップ
図2:pKG1700のベクターマップ
図3:pKG8135、pKG8136およびpKG8137のベクターマップ。
【0087】
以下の非限定的な実施例は、本発明のAA6プロモーターの使用を記載する。実施例で別途記載しない限り、全ての組換えDNA技術は、Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press、およびSambrook and Russell (2001) Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Laboratory Press, NY; およびVolumes 1 and 2 of Ausubel et al. (1994) Current Protocols in Molecular Biology, Current Protocols, USAに記載されるとおり標準プロトコールに従って行われる。植物の分子レベル研究のための標準的な材料と方法は、Plant Molecular Biology Labfax (1993) by R.D.D. Croy, jointly published by BIOS Scientific Publications Ltd (UK) and Blackwell Scientific Publications, UKに記載される。
【実施例】
【0088】
例1−材料と方法
cDNAを用いたディファレンシャルディスプレイ分析は、Lycopersicon esculentum var. Moneybergについて実施した。
【0089】
ストレス条件下での発現テストは、L.esculentum var. Moneybergについて実施した。
【0090】
ベクターpKG8136、pKG8137およびpKG1700を用いた形質転換は、Nicotiana tabacum var. SR1およびL. esculentum var. RZ 52201で実施した。植物の形質転換は、以下の参考文献に記載されるとおり行った:
トマトおよびタバコの形質転換について:
An G., B.D. Watson and C.C. Chiang. 1986. Transformation of tobacco, tomato, potato, and Arabidopsis thaliana using a binary Ti vector system. Plant Physiol. 81: 301-305。
【0091】
タバコの形質転換について:Horsch R.B., J. Fry, N. Hoffman, J. Neidermeyer, S.G. Rogers and R.T. Fraley. 1988. Leaf disc transformation. In: Plant Molecular Biology Manual A5. Dordrecht, Netherlands, Kluwer Academic Publishers. pp 1-9。
【0092】
トマトの形質転換について:Koornneef M., M. Jongsma, R. Weide, P. Zabel and J. Hille. 1986. Transformation of tomato. In: Nevins D.J. and R.A. Jones, eds. Tomato Biotechnology. New York, NY, USA, Alan R. Liss, Inc. pp 169-178。
【0093】
5’および3’端RACEは、CLONTECH Laboratories IncからのSMARTTM RACE cDNA Amplification Kitを用いて実施した。
【0094】
PCR産物のクローニングは、Invitrogen BVからのOriginal TA Cloning(登録商標)Kitを用いて、プラスミドpCR(登録商標)2.1を使用して実施した。
【0095】
DNA配列決定は、BaseClear (P.O. Box 1336, 2302 BH, Leiden, The Netherlands) により実施した。
【0096】
形質転換ベクターは、アグロバクテリウムツメファシエンス(Agrobacterium tumefaciens)var. GV2260に組み込んだ。
【0097】
プロモーター活性の定性的分析は、組織学的GUSアッセイを用いて行った:
種々の植物の部分を、37℃で一晩、振盪しながら、大気中の酸素の存在下で、Xgluc基質とともに、リン酸緩衝液 (5-ブロモ-4-クロロ-インドリルグルクロニド、1 mg/ml、K2HPO4、40 mM、KH2PO4、10 mM、pH 7.4) 中でインキュベートした。サンプルは、エタノールで繰返し洗浄することにより脱色した。非トランスジェニック植物を、ネガティブコントロールとして使用した。
【0098】
プロモーター活性の定量的分析は、蛍光GUSアッセイを用いて行った:
トータルタンパク質サンプルは、若葉の材料から調製し;サンプルは、テストされる各植物の様々な部分からプールしたおよそ同じサイズおよび発生段階の葉片から調製した。新鮮な葉の材料を、リン酸緩衝液 (Na2HPO4、77.4 mM、NaH2PO4、22.6 mM) 中で金属ビーズを用いてすりつぶし、その後、遠心分離と上清の回収を行った。
【0099】
蛍光GUSアッセイ:
各上清の一部におけるタンパク質濃度を、Molecular Probes, IncからのNano Orange Kitを用いて測定した。タンパク質サンプルは、トータルタンパク質濃度を標準化するために希釈した。タンパク質サンプルの一部を、37℃で一晩、基質4-メチルウンベリフェリルβ-d-グルクロニド (MUG) (最終濃度 1 mg/ml) とともにインキュベートした。時間0と一晩インキュベーション後の蛍光測定を、反応混合物の一部を取り出し、Na2CO3溶液(最終濃度 1.1 M) を添加して反応を停止させ、その後、355nmでの励起により引き起こされる460nmでの発光を測定することにより行った。
【0100】
例2−プロモーター単離用遺伝子の選択
ディファレンシャル分析は、cDNA-AFLP(登録商標)(Volkmuth W., et al., 2003, Genome-Wide cDNA-AFLP(登録商標)Analysis of the Arabidopsis Transcriptome, OMICS, 7, 2; Vos, P. and Stanssens P., 2002, AFLP-based transcript profiling, Current Protocols in Molecular Biology, unit 25B.5., Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G. Smith, J.A., Struhl, K., John Wiley and Sons, New York; and Vos et al., 1995, AFLP: a new technique for DNA fingerprinting, Nucleic Acids Research 23: 4407-4414) を用いて、種々の発生段階におけるトマト植物材料(若葉、老葉、茎、根、インビトロの葉、インビトロの茎、インビトロの根、カルス、未熟および成熟した緑色の実、ブレーカー(breaker)段階の実、赤色の実、花、芽生え全体)から単離したcDNAについて実施した。発現プロファイルは、Taq1プライマーについては2つの選択ヌクレオチドを備え、Mse1プライマーについては3つの選択ヌクレオチドを備えた、全ての可能なTaq1/Mse1プライマーの組み合わせを用いて作成した。その後の遺伝子断片発現プロファイルは、Keygene N.V. 所有ソフトウェア (ImproveTM) を用いて作成し、データベースに保存した。全てのサンプル組織において強力で構成的な発現を示した、12のTaq1/Mse1プライマーの組み合わせに対応する13の候補転写産物を選択した。
【0101】
その後、候補遺伝子の発現を、種々の組織において、生物的および非生物的ストレスの下で、cDNA-AFLP(登録商標)により測定した。3週齢のトマト植物 (Moneyberg) を、以下の別個のストレス条件を用いて14日間生育させた:
キュウリモザイクウイルス(CMV)による感染、高温(昼37℃および夜25℃)、低温(昼10℃および夜5℃)、干ばつストレス(植物の死を回避する最小限の水、昼37℃および夜25℃)、並びに収穫2日前の傷害(ピンセットで葉を押しつぶす)。
【0102】
以下の材料を収穫した:若葉、老葉、茎、根および花芽。これらの材料からcDNAを作成し、ディファレンシャル分析を、cDNA-AFLP(登録商標)を使用して、選択された13の候補転写産物を増幅するのに必要な同一プライマーの組み合わせ(および少数の選択ヌクレオチドを備えたプライマーの組み合わせも使用)を用いて実施した(プライマー配列については下記参照)。上記ストレス条件の全ての下において強力で構成的な発現を提供し続けた7つの転写産物を選択した。
【0103】
システインシンターゼ遺伝子(AA6)断片を増幅する(+2/+2)AFLPプライマーのプライマー配列は、以下のとおりである:
【表A】
【0104】
候補cDNA AFLP断片を、ゲルから切り出し、精製し、クローニングし、配列決定を行った。その配列情報を、その後、RACE PCRのプライマーを設計するために使用した。
【0105】
5’および3’両端のSMART-RACE-PCR (Clontech) を実施し、7つの候補遺伝子の伸長遺伝子断片を作成し、これらをpCR2.1 (Invitrogen) にクローニングし、その後、配列決定を行った(BaseClear, P.O. Box 1336, 2302 BH, Leiden, The Netherlands)。伸長遺伝子断片は、公共のデータベースに由来する相同ESTsと共に組み立てを行った。これにより、5つの候補遺伝子について全長cDNA配列を得た。
【0106】
5つの遺伝子についてのコピー数の評価は、10種類の制限酵素で消化したトマトのゲノムDNAを用いたサザンブロッティングを使用し、プローブとしてcDNAの5’端断片を使用して実施した。同じプローブを、既存のMoneyberg BACライブラリー (Keygene N.V.)にもハイブリダイズさせた。
【0107】
全長cDNAの一つ (IS158-53と表示される; 配列番号4) は、トマトの細胞質ゾルのシステインシンターゼ遺伝子と非常に高い相同性を示した(UP|Q9FS27 (Q9FS27)に対するtomato|TC162833相同物)。TC162833および配列番号4は何れも、同一のアミノ酸配列(配列番号5)をコードするが、二つのヌクレオチドにおいて核酸レベルで異なる。配列番号4の207位のヌクレオチド‘C’は、TC162833で‘T’であり、配列番号4の372位のヌクレオチド‘A’は、TC162833で‘G’である。
【0108】
配列番号4は、生物的および非生物的ストレス条件下においても構成的に発現し、トマトのゲノムにおいて少ないコピー数(約2コピー)で存在し、Keygene N.V.トマトBACライブラリーのBAC 9と十分にハイブリダイズしたため、そのプロモーターの単離に着手した。
【0109】
全長cDNAがトマトのシステインシンターゼ遺伝子(配列番号4)と相同性を有する遺伝子のプロモーターを、以下に記載するとおり選択した。
【0110】
例3−AA6プロモーター配列の単離
システインシンターゼと指定されたトマト遺伝子の導出配列上に設計されたプライマー (プライマー配列については下記参照) を使用して、BAC 9由来のゲノムDNAを鋳型として用いたリンカーPCRを実施した。5’および3’配列は何れも、システインシンターゼのUTRについて得た。3’UTR配列 (配列番号3) は、MoneybergトマトのゲノムDNAを鋳型として用いた長距離PCRにより得た。
【0111】
システインシンターゼ配列上に設計されたプライマー:
プロモーター用 5’-GTTCGATGAGGACACTCTCGC-3
およびネスティッドプライマー 5’-CAATTAAAGTTGCTAAGCGTCCTGA-3’
ターミネーター用 5’-TCAGTTACATCCTTGGCAATTCC-3’
およびネスティッドプライマー 5’-CAGAGAACATGACTGTGGAGCC-3’。
【0112】
例4−形質転換ベクターの構築
システインシンターゼ5’上流領域の二つのAA6プロモーターDNA断片、5000bpのもの (配列番号2) と2986bpのもの (配列番号1) を、gusAコード領域と一緒に、内在性システインシンターゼターミネーター (配列番号3) またはnos ターミネーターの何れかを備えたプラスミドpKG1562 (ノパリンシンターゼプロモーターにより駆動されるnptIIを含有するベクター骨格) に連結した。
【0113】
構築物を指定した:
以下のものを含有するpKG8135 (図3参照):
2986 bpのシステインシンターゼ5’上流領域 (配列番号1): gusA : 3’nos
以下のものを含有するpKG8136 (図3参照):
5000 bpのシステインシンターゼ5’上流領域 (配列番号2): gusA : 3’nos
以下のものを含有するpKG8137 (図3参照):
5000 bpのシステインシンターゼ5’上流領域 (配列番号2): gusA : 3’システインシンターゼ (配列番号3)。
【0114】
コントロール構築物は、以下のとおり使用した(図2参照):
以下のものを含有するpKG1700: CaMV35Sプロモーター (Franck et al., 同上): gusA : 3’nos。
【0115】
使用した形質転換ベクターのベクターマップの詳細については、図1〜3が参照される。
【0116】
プラスミドpKG8135、pKG8136、pKG8137およびpKG1700を、アグロバクテリウムツメファシエンスに組み込んだ。タバコ (var. SR1) およびトマト (var. RZ 52201) を形質転換し、カナマイシン選択の下で再生させ、第一次再生体 (T0) を生育して結実させた。
【0117】
例5−形質転換体の発現分析
gusAの発現は、これらベクター内のAA6プロモーターまたはCaMV35Sプロモーターにより駆動されるため、植物で産生されるβ-D-グルクロニダーゼ (GUS) のレベルは、プロモーターの有効性を示す。GUSの存在についての組織学的染色アッセイは、基質として5-ブロモ-4-クロロ-インドリルグルクロニド (XGluc) を用いて実施した。
【0118】
染色実験は、(第一次形質転換体の自家受粉後に得られた) タバコおよびトマトの両方のT1植物の葉組織について、芽生えから開花植物までの幾つかの発生段階で実施した。加えて、GUSについての組織学的染色実験は、成熟したタバコおよびトマト系統に由来する様々な材料、たとえば花芽、開花している花、花茎 (タバコ)、花柄、頂端分裂組織の芽、葉、葉柄、主要な茎、根、種子および果実 (トマト)、並びに発芽している種子および芽生えについて実施した。
【0119】
AA6プロモーターの長い断片 (配列番号2) もしくは短い断片 (配列番号1) またはCaMV35SプロモーターによりgusAが駆動された場合、GUSの発現を、タバコおよびトマトの両方において、試験された全ての組織で検出した。
【0120】
視覚的 (染色) 比較により、タバコおよびトマトの両方において、AA6プロモーター断片並びにCaMV35Sプロモーターが機能していることが示された。
【0121】
GUS発現の定量的分析は、基質として4-メチルウンベリフェリルβ-d-グルクロニド (MUG) を用いた蛍光アッセイにより実施した。5000bp断片のAA6プロモーター (pKG8137) により駆動されるGUSの発現を、タバコおよびトマトの両方の若葉において、CaMV35Sプロモーター (pKG1700) により駆動されるGUSの発現と比較した。
【0122】
シングルコピーの植物を、サザンブロッティングを用いて選択した;HindIII制限酵素で消化したタバコおよびトマト植物由来のゲノムDNAを使用し、nptII断片をプローブとして使用し、そのプロセスを、制限酵素としてXbaIを用いて繰り返した。各T-DNA挿入物は、nptIIおよびgusAをそれぞれ一コピー含有するため、nptIIコピー数は、gusAコピー数を反映する。また、コピー数は、Invader(登録商標)アッセイ (Third Wave Technologies, Inc.)を用いて確認し、公知のトマトまたはタバコの内部コントロールと比較してnptII遺伝子の存在および相対量を検出した;このアッセイをT1世代に適用すると、半接合個体と同型接合個体との識別が可能である。
【0123】
トータルタンパク質抽出物を、タバコおよびトマト植物のpKG8137およびpKG1700半接合および同型接合シングルコピー形質転換体およびゼロコピーコントロールから作成した。タンパク質含量は、NanoOrangeTM Kit (Molecular Probes) を用いて測定し、その後、サンプル中のタンパク質濃度を、抽出バッファーでの希釈により標準化した。
【0124】
MUG基質の添加後、初期蛍光を測定した (励起フィルター、355nmおよび発光フィルター、460nm)。その後、サンプルをインキュベートし、切断されたメチルウンベリフェロン (MU) による蛍光を測定した。インキュベーション1時間あたりの蛍光の増加を計算した。(テストサンプルのトータルタンパク質含量に等しい) 非トランスジェニックタバコまたはトマトのトータルタンパク質抽出物のバックグラウンドにおいてGUS酵素 (Sigma-Aldrich Chemie B.V.) 活性を示すキャリブレーションカーブを作成した。トランスジェニックテストサンプルに存在するGUS酵素の濃度 (mU/トータル植物タンパク質抽出物mg) を、サンプルおよびキャリブレーションカーブからの蛍光データを用いて計算した。
【0125】
2回の実験の統計解析により、AA6プロモーター (pKG8137) およびCaMV35Sプロモーターにより駆動されるGUS発現のレベルは、タバコにおいて有意な差異はないが、トマトでは、同型接合シングルコピー植物においてAA6プロモーターによるGUSの発現は、CaMV35SプロモーターによるGUSの発現より79%高いことが示された(以下の表で示されるデータセット)。
【0126】
タバコでのGUSの発現
表1:半接合タバコ植物の系統ごとの平均GUS活性 (mU/トータル植物タンパク質抽出物mgで表示)、並びに標準偏差および平均の標準誤差 (SEM)
【表1】
【0127】
表2:同型接合タバコ植物の系統ごとの平均GUS活性 (mU/トータル植物タンパク質抽出物mgで表示)、並びに標準偏差および平均の標準誤差 (SEM)
【表2】
【0128】
以上示すとおり、AA6プロモーター (5kbプロモーター) の平均プロモーター活性は、(導入遺伝子について半接合および同型接合の植物の両方において)トランスジェニックタバコの葉において35Sプロモーターのものと本質的に同等である。
【0129】
トマトでのGUSの発現
表3:半接合トマト植物の系統ごとの平均GUS活性 (mU/トータル植物タンパク質抽出物mgで表示)、並びに標準偏差および平均の標準誤差 (SEM)
【表3】
【0130】
表4:同型接合トマト植物の系統ごとの平均GUS活性 (mU/トータル植物タンパク質抽出物mgで表示)、並びに標準偏差および平均の標準誤差 (SEM)
【表4】
【0131】
表4に示すとおり、AA6プロモーター (5kbプロモーター) の平均プロモーター活性は、(導入遺伝子について半接合および同型接合の植物の両方において)トランスジェニックトマトの葉における35Sプロモーターのものと比較して、トマトにおいて有意に高い(35S活性をスタンダードとした場合、データセット1では35Sより約52%高く、データセット2では35Sより約79%高い)。
【0132】
例6−他の植物種でのプロモーター活性
他の植物種においてAA6およびCaMV35Sプロモーターにより駆動されるGUS発現の比較をテストし、確認した。現在のところ、導入遺伝子のAA6プロモーター駆動発現についてテストを行った種のリストには、(上述のタバコおよびトマトに加えて)レタス、メロンおよびアブラナ属(Brassica)が含まれ、これらの各々において構成的発現が示されている。
【0133】
例7−“3kb”および“5kb”AA6プロモーターの再配列決定
ベクターpKG8135およびpKG8137 (図3) に存在する核酸配列を、再配列決定し、配列をチェックし、不明なヌクレオチドを解明した。
【0134】
不明な位置は、配列番号6 (“3kb”プロモーター) および配列番号7 (“5kb”プロモーター)の配列表に示し、ここでは、不明な位置において最も可能性のあるヌクレオチドも同定される (以下の表も参照)。
【0135】
不明なヌクレオチドのために、ごく僅かな配列の違いが存在する:配列番号1は、(‘ニードル’、ギャップオープニング = 10.0、ギャップ伸長 = 0.5; DNAFullマトリクスを用いて) 配列番号8および6とそれぞれ99.6%および99.7%の配列同一性を有する。配列番号2は、(‘ニードル’、ギャップオープニング = 10.0、ギャップ伸長 = 0.5; DNAFullマトリクスを用いて) 配列番号9および7とそれぞれ99.6%および99.8%の配列同一性を有する。
【表B】
【表C】
【図面の簡単な説明】
【0136】
【図1】pKG1562のベクターマップ。
【図2】pKG1700のベクターマップ。
【図3】pKG8135、pKG8136およびpKG8137のベクターマップ。
【特許請求の範囲】
【請求項1】
ゲノムに統合されたキメラ遺伝子を含む、トランスジェニック植物または植物細胞または植物組織または器官であって、前記キメラ遺伝子が、同種または異種の核酸配列に動作可能に連結された構成的プロモーターを含み、前記プロモーターが、
(a)配列番号1または配列番号2の核酸配列;
(b)配列番号1または配列番号2の機能的断片;
(c)配列番号1または2と少なくとも70%の配列同一性を備えた核酸配列;
(d)(c)の核酸配列の機能的断片
の群から選択されることを特徴とする、トランスジェニック植物または植物細胞または植物組織または器官。
【請求項2】
前記植物、細胞、組織または器官を、一または複数の生物的および/または非生物的ストレスに晒したときに、前記プロモーターの活性が有意に低下しない、請求項1に記載の植物、植物細胞、組織または器官。
【請求項3】
植物細胞に導入された際にプロモーター活性を有する単離された核酸配列であって、
(a)配列番号1または配列番号2の核酸配列;
(b)配列番号1または配列番号2の断片;
(c)配列番号1または2と少なくとも70%の配列同一性を備えた核酸配列;
(d)(c)の核酸配列の断片
から選択される配列を含む核酸配列。
【請求項4】
請求項1の(b)または請求項3の(b)に記載の断片であって、配列番号1または配列番号2の少なくとも200の連続ヌクレオチドを含む断片。
【請求項5】
同種または異種の核酸配列に動作可能に連結され、必要に応じて3’UTR配列に連結された請求項3または4に記載の核酸配列を含むキメラ遺伝子。
【請求項6】
前記3’UTR配列が、配列番号3、または配列番号3の断片、または配列番号3と少なくとも70%の核酸同一性を含む核酸配列である、請求項5に記載のキメラ遺伝子。
【請求項7】
請求項3または4に記載の核酸配列、または請求項5または6に記載のキメラ遺伝子を含むベクター。
【請求項8】
請求項5または6に記載のキメラ遺伝子を含むトランスジェニック植物または植物細胞。
【請求項9】
トランスジェニック植物または植物細胞または植物組織または器官におけるセンスおよび/またはアンチセンス核酸配列の構成的発現のための、細胞質ゾルの植物システインシンターゼ遺伝子のプロモーターの使用。
【請求項10】
細胞質ゾルの植物システインシンターゼ遺伝子が配列番号5のタンパク質をコードする、請求項9に記載の使用。
【請求項11】
トランスジェニック植物または植物細胞を作成するための方法であって、
(a)請求項5または6に記載のキメラ遺伝子または請求項7に記載のベクターを生成する工程;
(b)前記キメラ遺伝子またはベクターを用いて植物または植物細胞を形質転換する工程;および必要に応じて、
(c)トランスジェニック植物を再生させる工程
を含む方法。
【請求項12】
配列番号3または4のヌクレオチド配列を含む単離された核酸分子。
【請求項1】
ゲノムに統合されたキメラ遺伝子を含む、トランスジェニック植物または植物細胞または植物組織または器官であって、前記キメラ遺伝子が、同種または異種の核酸配列に動作可能に連結された構成的プロモーターを含み、前記プロモーターが、
(a)配列番号1または配列番号2の核酸配列;
(b)配列番号1または配列番号2の機能的断片;
(c)配列番号1または2と少なくとも70%の配列同一性を備えた核酸配列;
(d)(c)の核酸配列の機能的断片
の群から選択されることを特徴とする、トランスジェニック植物または植物細胞または植物組織または器官。
【請求項2】
前記植物、細胞、組織または器官を、一または複数の生物的および/または非生物的ストレスに晒したときに、前記プロモーターの活性が有意に低下しない、請求項1に記載の植物、植物細胞、組織または器官。
【請求項3】
植物細胞に導入された際にプロモーター活性を有する単離された核酸配列であって、
(a)配列番号1または配列番号2の核酸配列;
(b)配列番号1または配列番号2の断片;
(c)配列番号1または2と少なくとも70%の配列同一性を備えた核酸配列;
(d)(c)の核酸配列の断片
から選択される配列を含む核酸配列。
【請求項4】
請求項1の(b)または請求項3の(b)に記載の断片であって、配列番号1または配列番号2の少なくとも200の連続ヌクレオチドを含む断片。
【請求項5】
同種または異種の核酸配列に動作可能に連結され、必要に応じて3’UTR配列に連結された請求項3または4に記載の核酸配列を含むキメラ遺伝子。
【請求項6】
前記3’UTR配列が、配列番号3、または配列番号3の断片、または配列番号3と少なくとも70%の核酸同一性を含む核酸配列である、請求項5に記載のキメラ遺伝子。
【請求項7】
請求項3または4に記載の核酸配列、または請求項5または6に記載のキメラ遺伝子を含むベクター。
【請求項8】
請求項5または6に記載のキメラ遺伝子を含むトランスジェニック植物または植物細胞。
【請求項9】
トランスジェニック植物または植物細胞または植物組織または器官におけるセンスおよび/またはアンチセンス核酸配列の構成的発現のための、細胞質ゾルの植物システインシンターゼ遺伝子のプロモーターの使用。
【請求項10】
細胞質ゾルの植物システインシンターゼ遺伝子が配列番号5のタンパク質をコードする、請求項9に記載の使用。
【請求項11】
トランスジェニック植物または植物細胞を作成するための方法であって、
(a)請求項5または6に記載のキメラ遺伝子または請求項7に記載のベクターを生成する工程;
(b)前記キメラ遺伝子またはベクターを用いて植物または植物細胞を形質転換する工程;および必要に応じて、
(c)トランスジェニック植物を再生させる工程
を含む方法。
【請求項12】
配列番号3または4のヌクレオチド配列を含む単離された核酸分子。
【図1】

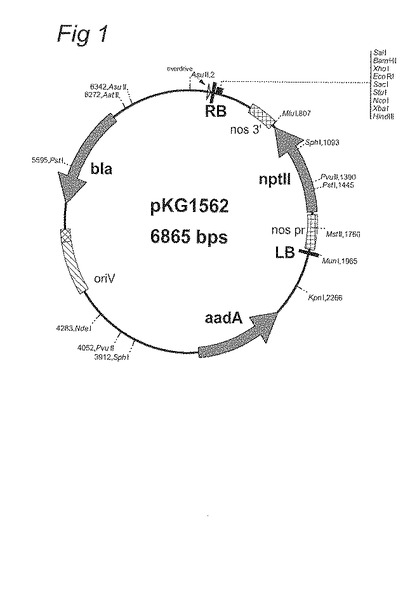
【図2】
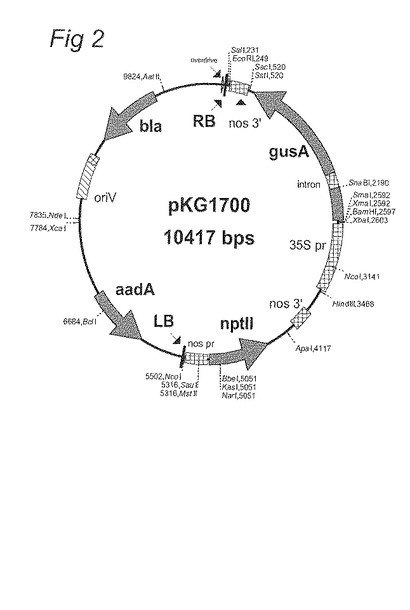
【図3】
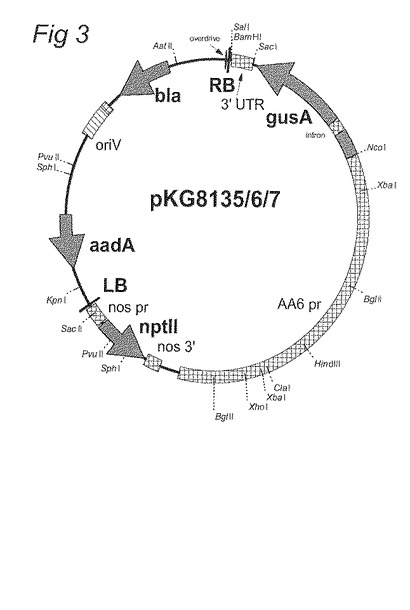
【図2】
【図3】
【公開番号】特開2013−55946(P2013−55946A)
【公開日】平成25年3月28日(2013.3.28)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−236995(P2012−236995)
【出願日】平成24年10月26日(2012.10.26)
【分割の表示】特願2008−545520(P2008−545520)の分割
【原出願日】平成18年12月12日(2006.12.12)
【出願人】(505477187)ケイヘーネ・エヌ・ブイ (10)
【Fターム(参考)】
【公開日】平成25年3月28日(2013.3.28)
【国際特許分類】
【出願番号】特願2012−236995(P2012−236995)
【出願日】平成24年10月26日(2012.10.26)
【分割の表示】特願2008−545520(P2008−545520)の分割
【原出願日】平成18年12月12日(2006.12.12)
【出願人】(505477187)ケイヘーネ・エヌ・ブイ (10)
【Fターム(参考)】
[ Back to top ]