
標的遺伝子の誘導性変異誘発
本発明は概して、B細胞の天然の変異誘発能力を利用し、多様化のプロセスを制御下に置くことによってこれらの能力を増強する、標的遺伝子の変異誘発に関する。本発明は、抗体遺伝子のような発現遺伝子に点変異および他のタイプの多様化を迅速かつ誘導的に発生させるための方法を提供する。この方法は、例えば、親和性または特異性が高い抗体を産生するB細胞クローンを同定するための選択と併用することができる。多様化プロセスは、変異誘発法間で調整、加速、停止、および切り替えなどをすることができる。本発明による多様化の調整は誘導性かつ可逆性である。本発明は、変異免疫グロブリンおよび他のポリペプチドのレパートリーを迅速かつ実行可能に開発する手段を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本出願は、その内容全体が参照により本出願に組み入れられる、2007年5月31日に出願された米国特許仮出願第60/932,672号の恩典を主張する。
【0002】
連邦政府による資金提供を受けた研究に関する記載
本発明は、国立衛生研究所(National Institutes of Health)によって授与されたR01 GM041712のもと、米国政府の支援を受けて行った。米国政府は、本発明において一定の権利を有する。
【0003】
発明の技術分野
本発明は概して、B細胞の天然の変異誘発能力を利用し、多様化のプロセスを制御下に置くことによってこれらの能力を増強する、標的遺伝子の変異誘発に関する。本発明は、抗体遺伝子のような発現遺伝子に点変異および他のタイプの多様化を迅速かつ誘導的に発生させるための方法を提供する。この方法は、例えば、親和性または特異性が高い抗体を産生するB細胞クローンを同定するための選択と併用することができる。多様化プロセスは、変異誘発法間で調整、加速、停止、および切り替えなどをすることができる。本発明による多様化の調整は誘導的かつ可逆的である。本発明は、変異免疫グロブリンおよび他のポリペプチドのレパートリーを迅速かつ実行可能に開発する手段を提供する。
【背景技術】
【0004】
発明の背景
抗体は、ヒトにおいて感染に対抗する重要な防御を提供する分子である。これらは、感染症から癌に至る様々な疾患の治療において治療物質として使用される。これらはまた、臨床的実験室および研究実験室で日々実施される多種多様な試験において診断試薬として使用される。
【0005】
抗体の特異性および親和性は、抗体をコードする遺伝子内の特定の領域を標的とする変異プロセスによってインビボで改変される。抗体は、免疫グロブリン(Ig)重鎖遺伝子および軽鎖遺伝子と呼ばれる2つの遺伝子にコードされる。Ig遺伝子にコードされるポリペプチドの重鎖および軽鎖は相互に作用して四量体分子を形成し、これは受容体として細胞表面で発現される。抗体分子は二価性である:一方のドメインは抗原を認識し、他方は身体からの抗原の除去を促進する。可変(V)領域と呼ばれる認識ドメインは、天然抗体中の重鎖ポリペプチドおよび軽鎖ポリペプチドが相互作用して作り出される。実際には、抗体によって配列は様々である。V領域の一次配列(ならびにその結果、三次元構造および抗原特異性)の多様性は、不可逆的な遺伝的変化を引き起こすことによってV領域配列を改変するプロセスの結果である。これらの変化は、B細胞発達の間にプログラムされており、また、B細胞を活性化する環境シグナルに応答して体内で誘導されることもある。いくつかの遺伝的メカニズムがこの多様性に寄与している。同じメカニズムの2つの副経路が、体細胞超変異および遺伝子変換と呼ばれる2つの異なる変異誘発結果をもたらす((Maizels, 2005)に総説がある)。体細胞超変異は、点変異を挿入する。体細胞超変異は、本質的に任意の変異が生じることを可能にするという利点を与え、したがって、変異したV領域を集めたものは、種々の起こり得る変異の本質的なサンプルとなる。遺伝子変換は、関連しているが同一ではない配列による「鋳型式の」変異を挿入する。遺伝子変換は、進化的選択によって最適化され得る、ゲノム中に既に保存されている情報に基づいてレパートリーを作り出すという利点を与える。
【0006】
改変された抗原受容体は選択の標的である。Igは細胞表面で発現されるため、生理学的環境または培養細胞内での所望の特異性または親和性を有するB細胞のクローン選択が可能である。細胞表面Igは、抗Igで染色した細胞をフローサイトメトリーに供することによって容易に検出することができる。特定の化合物へのIg分子の結合は、それらの化合物の蛍光性誘導体との相互作用として検出し、フローサイトメトリーによって解析することができ、また、フローサイトメトリーによる選別の際に、特定の化合物に結合するB細胞を回収することもできる。特定の化合物に結合するB細胞はまた、それらの化合物を担持する固体支持体上で選択することもできる。逆に、固体支持体に結合させることによって、望まれない結合特異性を有するB細胞を除去することも可能である。結合と遊離の反復サイクルにより、高親和性の結合を得るために富化することが可能になる。
【0007】
変異および遺伝子ターゲティングがDT40 B細胞株で起こる。DT40は、培養で容易に増殖するニワトリB細胞株である。DT40は、免疫グロブリン重鎖遺伝子および軽鎖遺伝子の構成的変異誘発を実施するために、多くの事例において文書に記録されている(Reynaud et al., 1987; ThompsonおよびNeiman, 1987; Reynaud et al., 1989)。変異誘発はV領域を標的としている。変異は、通常、「偽V」遺伝子と呼ばれ、各機能的V領域の上流に直線状アレイとして存在する、関連した非機能的V遺伝子セグメントをコピーすることによって鋳型式となる。鋳型式であることは、変異したV標的と偽V遺伝子の内の1つとが共有する配列の区域(tract)として明らかである。DT40の遺伝的改変は、鋳型式変異誘発から非鋳型式変異誘発への切り替えを引き起こし、ヒトB細胞においてV領域の配列を改変する体細胞超変異と本質的に同一の経路によって、体細胞超変異をもたらすことが示された(Sale et al., 2001)。また、DT40は非常に効率的な相同組換えまたは遺伝子ターゲティングを支援することも多くの事例において文書に記録されている(Buerstedde et al., 2002; Sale, 2004)。これにより、特定の遺伝子もしくはゲノム領域が改変もしくは除去されるか、または1つの遺伝的領域が別の領域で置換されている派生体を作製することが可能になる。
【0008】
標的化変異誘発のための現在利用可能なアプローチによってもたらされる制約および難題があるため、代替の方法および構築物を開発する必要が当技術分野において存在する。本発明はこの必要を満たし、他の関連した利点をさらに提供する。
【発明の概要】
【0009】
本発明は、標的配列を多様化するための材料および方法を提供することにより、これらおよび他の必要を満たす。本発明は、標的遺伝子の多様化を可逆的に誘導することを可能にするように改変したB細胞を提供する。この細胞は、関心対象の標的遺伝子に機能的に連結されたシス調節エレメントを含む。次いで、多様化を調整する因子を、シス調節エレメントに結合する係留因子(tethering factor)に融合させ、それによって、標的遺伝子の発現を制御する領域に多様化因子を係留することができる。B細胞は、ニワトリDT40 B細胞または他の脊椎動物B細胞でよく、いくつかの態様においては、ヒトB細胞、またはヒト化免疫グロブリン(Ig)遺伝子(ヒトIgHおよびIgLでニワトリIgHおよびIgLを置換している)を含むニワトリDT40 B細胞が好ましい。
【0010】
典型的には、標的遺伝子は、プロモーターおよびコード領域を含む。1つの態様において、標的遺伝子は、Ig遺伝子エンハンサーおよびコード領域を含む免疫グロブリン(Ig)遺伝子を含む。Ig遺伝子は、IgL遺伝子および/またはIgH遺伝子の全体または一部分でよい。コード領域は、Ig遺伝子に先天的なものでもよく、または異種遺伝子でもよい。いくつかの態様において、標的遺伝子は、多様化のための非Ig標的ドメイン、ならびに、膜貫通ドメインおよび細胞質尾部を含む、B細胞表面で遺伝子産物を提示することを可能にするドメインであるか、またはそれを含む。本発明のB細胞中の標的遺伝子のコード領域は、コード領域全体を含む必要はない。いくつかの態様において、ある特定の領域またはドメインが多様化のために標的とされ、コード領域は、関心対象の領域またはドメインを含む部分のみを任意でコードしてよい。
【0011】
シス調節エレメントは、標的遺伝子の多様化および/または発現を制御するランディングパッドを領域中に提供する。このランディングパッドは、DNAのこの領域に対して配列特異的な様式で係留因子が結合できる場所である。典型的な態様において、シス調節エレメントは、重合(polymerized)ラクトースオペレーター(LacO)である。1つの態様において、エレメントは、約80〜100個繰り返したLacOを含む。別の態様において、シス調節エレメントは、単独または多量体化したテトラサイクリンオペレーター(TetO)である。係留因子(配列特異的DNA結合タンパク質)がDNAに結合し、係留因子に融合された多様化因子をコード領域の十分近くに届けることができ、その結果、コード領域の多様化が可逆的に調節可能になる場所を提供するというランディングパッド機能をエレメントが果たす限り、様々な分子をシス調節エレメントとして使用することができる。
【0012】
係留因子は、配列特異的な様式でシス調節エレメントに結合する因子である。LacOがシス調節エレメントとして働く態様において、LacリプレッサーLacIは係留因子として働くことができ、シス調節エレメントLacOへの結合は、イソプロピル-β-D-チオ-ガラクトシド(IPTG)によって調節され得る。IPTGの不在下では、LacIはLacOに結合し、多様化は多様化因子の存在によって加速される(またはそうでなければ調節される)。IPTGは、多様化因子の活性の停止または低減が望ましい事象において添加され得る。TetOがシス調節エレメントとして働く態様において、TetRは適切な係留因子であり得、多様化因子の活性はテトラサイクリンまたはドキシサイクリンによって調節され得る。
【0013】
いくつかの態様において、多様化因子は、転写調節因子、ヘテロクロマチン関連タンパク質、ヒストンシャペロン、クロマチン再構築因子、核膜孔複合体の構成要素、遺伝子調節因子、またはそれらの組合せである。多様化因子として働くことができる他の分子には、DNA修復因子、DNA複製因子、リゾルベース、ヘリカーゼ、細胞周期調節因子、ユビキチン化因子、SUMO化因子、またはそれらの組合せが含まれるがこれらに限定されるわけではない。1つの態様において、転写調節因子は、VP16またはE47である。多様化因子として使用するための典型的なヘテロクロマチン関連タンパク質はHP1である。代表的なヒストンシャペロンはHIRAである。
【0014】
また、ポリペプチドをコードするポリヌクレオチド配列の多様化を通して、関心対象のポリペプチドの変異配列を有するポリペプチドのレパートリーを作製する方法も提供される。典型的には、この方法は、多様化因子の発現を可能にする条件で本発明のB細胞を培養する段階を含み、B細胞の標的遺伝子は関心対象のポリペプチドのコード領域を含み、それによって、コード領域の多様化が可能になる。この方法は、複数の変異ポリペプチドおよび所望のレパートリーが得られるまで、B細胞の増殖を可能にする条件下で培養物を維持する段階をさらに含んでよい。
【0015】
別の態様において、本発明は、最適化された関心対象のポリペプチドを産生するB細胞を作製する方法を提供する。この方法は、多様化因子の発現を可能にする条件で本発明のB細胞を培養する段階を含み、B細胞の標的遺伝子は関心対象のポリペプチドのコード領域を含み、B細胞は関心対象のポリペプチドをその表面で発現する。この方法は、関心対象のポリペプチドに特異的に結合するリガンドに結合する細胞を選択することにより、関心対象のポリペプチドをB細胞表面で発現する細胞を培養物から選択する段階をさらに含む。関心対象のポリペプチドに特異的に結合するリガンドに対して所望の親和性を有する細胞を選択するまで、これらの培養段階および選択段階を繰り返すことができる。関心対象のポリペプチドがIg、例えばIgL、IgH、または両方である態様において、リガンドは、抗原に相当する、組換えまたは他の手段によって作製されたポリペプチドでよい。例えば、磁気活性化細胞選択(MACS)による選択を容易にするために、リガンドを固体支持体に結合または連結することができる。別の例において、蛍光活性化細胞選別(FACS)を可能にするために、リガンドを蛍光タグに結合または連結することができる。
【0016】
本発明の方法は、培養物に調節分子を添加する段階をさらに含んでよく、調節分子は、シス調節エレメントへの係留因子の結合を調整し、それによって、コード領域の多様化を調整する。上記の例において、IPTG、テトラサイクリン、およびドキシサイクリンが、調節分子として働く。当業者は、多様化活性を調節するために個々の係留因子と共に使用できる他の調節分子を知っている。
【0017】
調節分子は、コード領域の多様化を様々な方法で調整することができる。例えば、いくつかの態様において、調節分子は、調整を行うために培養物に添加される。個々の調節分子がシス調節エレメントへの係留因子の結合を増加させる因子であるか、または減少させる因子であるかどうか、および結合の個々の変化が、多様化活性を増大させる効果を有するか、または低減させる効果を有するかどうかによって、調整の結果、多様化を低減もしくは停止するか、または多様化を促進もしくは加速することができる。他の態様において、調整は、調整の低減であれ停止であれ、促進であれ加速であれ、調節分子を培養物から除去または取り除くことによって行われる。同様に、多様化の調整は、B細胞に遺伝子を添加することもしくはB細胞から遺伝子を取り除くことによって、またはB細胞における遺伝子の発現を増大させることもしくは減少させることによって行うこともできる。当業者は、上記に説明した利用可能な変更(permutation)すべてを容易に理解することができ、これらはそれぞれ、B細胞における調節分子のレベルまたは存在を改変し、その結果として、シス調節エレメントへの多様化因子の係留を改変し、それによって、多様化活性を改変する効果を有する。
【0018】
また、本発明の方法を実施するために使用され得るキットも提供される。このキットは、本発明のB細胞ならびに対応する係留因子および多様化因子を発現する融合構築物を含む。例えば、B細胞は、プロモーターおよびコード領域を含む標的遺伝子に機能的に連結されたシス調節エレメントを含む。キットは、1種または複数種の融合構築物が中に保存された1つまたは複数の容器をさらに含む。各融合構築物は、B細胞で発現され得、かつ、多様化因子に融合された係留因子をコードするポリヌクレオチドを含み、この係留因子は、B細胞のシス調節エレメントに特異的に結合する。B細胞は、複数の融合構築物と共に使用するために複数のシス調節エレメントを含んでよい。
【図面の簡単な説明】
【0019】
【図1】抗体の体細胞超変異の概略図。上のパネル:Ig分子の相補性決定領域(CDR)が抗原に接触する。下のパネル:変異は、CDRをコードするゲノム領域に集中している。
【図2】DT40ポリLacO λ細胞におけるIg遺伝子多様化に対する、係留されたシス調節因子の効果を示す棒グラフ。IPTGの存在下および不在下でのクローンの多様化速度を比較した。
【図3】B細胞クローンの増殖中に抗体特異性が進化する方法を示す概略図。抗体は細胞表面で発現される。最初に多重特異性を示す集団(左上)を選択し、続いて、所望の特異性(細い二重円、中央)およびより高い親和性(太い円、右下)を富化する。
【図4】E2Aが、再配列されたIgλ座位と結合することの説明。図4A:ニワトリB細胞リンパ腫株DT40の再配列されたIgλ座位および再配列されていないIgλ座位の概略図。プロモーター(P)領域、リーダー(L)領域、可変(V)領域、連結(J)領域、および定常(Cλ)領域、J-Cイントロン中の推定のマトリックス結合領域(M)、3’-エンハンサー(E-3’)、ならびに遺伝子変換のための鋳型である、上流の非機能的偽可変領域の最も近位のもの(ψV1)および遠位のもの(ψV25)を示す。再配列された対立遺伝子および再配列されていない対立遺伝子は、矢印で示すプライマーを用いたPCRによって容易に区別することができる。図4B:対照の卵白アルブミン遺伝子アンプリコン(Ova)と比べた、DT40細胞の再配列されたλR座位および再配列されていないλU座位におけるE2A富化のChIP解析。富化倍率を下に示す。NTC、鋳型無しの対照。
【図5】E2Aは、シスで作用してIgλ遺伝子多様化を調節する。図5A:ポリLacOタグ付きの再配列されたIgλ座位の概略図。ポリLacOはψV17〜20の間に組み込まれる。他の概念は図4Aと同じである。図5B:DT40細胞およびDT40ポリLacO-λR細胞の細胞周期プロファイル。図5C:DT40細胞およびDT40ポリLacO-λR細胞によるsIgM喪失変異体の蓄積。6週間のクローン増殖後、各株に由来するサブクローン24個におけるsIgM喪失変異体の出現率をフローサイトメトリーによって定量した。図に示した出現率は、DT40に対して標準化した。sIgM喪失の平均出現率はそれぞれ0.8%および0.9%であった。図5D:DT40ポリLacO-λR GFP-LacI細胞およびDT40ポリLacO-λR E47-LacI細胞の細胞周期プロファイル。図5E:DT40ポリLacO-λR GFP-LacIトランスフェクタントおよびDT40ポリLacO-λR E47-LacIトランスフェクタントにおけるIgλ、AID、またはβ-アクチンmRNAの発現のRT-PCR解析。三角形は、cDNA鋳型の30倍、10倍、3倍、および1倍の相対濃度を示す。図5F:100μM IPTGの不在下または存在下で3週間培養した、独立したクローンDT40ポリLacO-λR GFP-LacIトランスフェクタント(n=13)およびDT40ポリLacO-λR E47-LacIトランスフェクタント(n=19)のsIgM平均喪失量。値は、IPTG無しで培養したDT40ポリLacO-λR GFP-LacI細胞に対して標準化した。
【図6】核の半径は細胞周期と相関している。図6A:G1期およびG2/M期の富化された細胞の代表的な画像。G1細胞(左)およびG2/M細胞(右)をHoechst 33342(10μM; Molecular Probes)で染色し、次いで、DNA含有量に基づいて選別した。スケールバー:10μm。図6B:DT40ポリLacO-λR細胞の代表的な細胞周期プロファイル。代表的なプロファイル内の水平な棒として、平均半径±標準偏差を示す。点線の縦線は、実験解析で使用したG1およびG2のカットオフ値を示す:G1、r<4μm;G2、r>5.2μm。
【図7】E2Aは、細胞周期のG1期にλRに局在化する。図7A:DT40ポリLacO-λR GFP-LacI細胞におけるλRおよびE2Aの共局在化の代表的な画像。DAPI染色によって決定される核周囲長の輪郭を白い点線で描いている。スケールバー:5μm。図7B:DT40ポリLacO-λR GFP-LacI細胞において細胞周期の各段階で起こるλR/E2A共局在化の割合。図7C:DT40ポリLacO-λR RFP-LacI細胞におけるλRおよび活性Pol II(P*-Pol II)の共局在化の代表的な画像。記号は図7Aと同じ。図7D:DT40ポリLacO-λR RFP-LacI細胞において細胞周期の各段階で起こるλR/P*-Pol II共局在化の割合。
【図8】Id1発現は、G1期のλR/E2A共局在化を阻害する。図8A:DT40ポリLacO-λR RFP-LacI細胞(左)およびDT40ポリLacO-λR RFP-LacI Id1細胞(右)におけるId1発現を分析するウェスタンブロット。左にマーカーポリペプチドのサイズ(kDa)を示す。図8B:DT40ポリLacO-λR RFP-LacI細胞およびDT40ポリLacO-λR RFP-LacI Id1細胞の細胞周期プロファイル。図8C:6週間培養した、独立したクローンDT40ポリLacO-λR RFP-LacI Id1トランスフェクタント(n=6)のsIgM平均喪失量。値は、DT40ポリLacO-λR RFP-LacI細胞に対して標準化した。図8D:Id1発現により、IgλまたはAIDの転写物レベルは変化しない。DT40ポリLacO-λR RFP-LacI細胞およびDT40ポリLacO-λR RFP-LacI Id1細胞中のIgλ、AID、または対照β-アクチンmRNAの発現のRT-PCR解析。図8E:DT40ポリLacO-λR RFP-LacI細胞およびId1を安定に発現する派生体(それぞれId1-および+)におけるλR/E2A共局在化の細胞周期依存性に対するId1発現の影響。細胞周期の各段階における共局在化の比率(%)を示す。図8F:DT40ポリLacO-λR RFP-LacI細胞およびId1を安定に発現する派生体におけるλR/P*-Pol II共局在化の共局在化細胞周期依存性に対するId1発現の影響。詳細は図8Eに同じ。
【図9】DT40 Igλ座位におけるクロマチン改変。図9A:再配列されたニワトリIgλ座位の概略図。25ψVλ領域および再配列されたVλR遺伝子(リーダー、L;可変領域、Vλ-Jλ;定常領域、Cλ領域)を示す。図9B:ヒストンH3およびH4のN末端アセチル化(AcH3およびAcH4)を分析した、代表的なクロマチン免疫沈降実験の概略。調べた部位は次のものであった:ψVλアレイの約1kb上流の領域(フランク)、ψVλ5とVλの間の領域、ψVλ1、ψVλ5、ψVλ13、ψVλ18、ψVλ24、ψVλ25、ならびに再配列されたVλR対立遺伝子および再配列されていないVλu対立遺伝子。詳細に関しては材料および方法を参照されたい。棒は標準偏差を示す。
【図10】DT40ポリLacO-λRにおけるψVλアレイへのGFP-LacIの可逆的係留。図10A:DT40の再配列されたニワトリIgλ座位の概略図。重合ラクトースオペレーター(ポリLacO)がψVλ17〜20の間に挿入されている。記号は図9と同じ。図10B:DT40 GFP-LacIトランスフェクタント、およびIPTGの不在下で培養するか(左)または100μM IPTGの存在下で一晩培養した(右)DT40ポリLacO-VλR GFP-LacIトランスフェクタントの蛍光画像。
【図11】HP1が係留されると、活性クロマチンに特徴的な改変が減少する。図11A:単一のDT40ポリLacO-VλR LacI-HP1トランスフェクタントの代表的な蛍光画像。抗LacI抗体で染色(左);DAPIで染色(中央);または合体した画像(右)。図11B:DT40ポリLacO-VλR GFP-LacIトランスフェクタントおよびDT40ポリLacO-VλR LacI-HP1トランスフェクタントのψVλ17Rにおけるアセチル化H3(AcH3)およびアセチル化H4(AcH4)の富化。ChIPに続いて、OvaプライマーおよびψVλ17Rプライマーを用いて二重PCRを実施した。富化は、全DNAインプット対照に対し、段階的に増量した鋳型DNAを4回別々に増幅した結果の±標準偏差として表す。図11C:DT40ポリLacO-VλR GFP-LacIトランスフェクタントおよびDT40ポリLacO-VλR LacI-HP1トランスフェクタントのψVλ17R(パネルBから)およびpol εプロモーターにおけるAcH3およびAcH4の富化を示すヒストグラム。棒は標準偏差を示す。
【図12】HP1の係留は、Igλ発現に影響を及ぼさない。図12A:DT40ポリLacO-VλR GFP-LacIトランスフェクタントおよびDT40ポリLacO-VλR LacI-HP1トランスフェクタントのVλRにおけるAcH3およびAcH4の相対的富化。富化の値は、対照のDT40ポリLacO-VλR GFP-LacIに対して標準化した。棒は、段階的に増量した鋳型DNAを4回別々に増幅した結果の標準偏差を示す。図12B:250μM IPTGの存在下および不在下で培養したDT40ポリLacO-λR GFP-LacIトランスフェクタント(GFP-LacI)およびDT40ポリLacO-λR LacI-HP1トランスフェクタント(LacI-HP1)における細胞表面IgM(sIgM)の相対強度。sIgMレベルは、マウス抗ニワトリIgM抗体による染色の強度を測定することによって定量し、DT40ポリLacO-λR GFP-LacIトランスフェクタントにおけるレベルに対して標準化した。詳細は図12Aに同じ。図12C:DT40ポリLacO-λR GFP-LacIトランスフェクタント(GFP-LacI)およびDT40ポリLacO-λR LacI-HP1トランスフェクタント(LacI-HP1)におけるVλR転写物の相対的レベル。転写物レベルはRT-PCRによって定量し、DT40ポリLacO-λR GFP-LacIトランスフェクタントにおけるレベルに対して標準化した。詳細は図12Aに同じ。
【図13】HP1が係留されると、ψVλアレイの全体に渡ってヒストンアセチル化が減少する。図13A:250μM IPTGの存在下および不在下で3日間培養したDT40ポリLacO-VλR LacI-HP1細胞株に由来するクロマチンにおけるヒストンH3のN末端アセチル化を分析した、クロマチン免疫沈降実験の概略。ChIP富化の値(実施例4を参照されたい)は、DT40ポリLacO-VλR GFP-LacI細胞に由来するクロマチンを並行して分析して得た値に対して標準化した。棒は、段階的に増量した鋳型DNAを4回別々に増幅した結果の標準偏差を示す。図13B:250μM IPTGの存在下および不在下で3日間培養したDT40ポリLacO-VλR LacI-HP1細胞株に由来するクロマチンにおけるヒストンH4のN末端アセチル化を分析した、クロマチン免疫沈降実験の概略。詳細は図13Aに同じ。
【図14】HP1の係留によって促進された、非鋳型式変異および鋳型式変異。図14A:独立したDT40ポリLacO-VλR GFP-LacI(n=27)およびDT40ポリLacO-VλR LacI-HP1(n=16)のパネルのsIgM喪失変動アッセイ法。図は、各融合構築物に関して少なくとも2個の独立したトランスフェクタントから得て組み合わせたデータを示す。多様化速度の中央値を下に示す。図14B:単一細胞PCRによって解析した、DT40ポリLacO-VλR GFP-LacIトランスフェクタント(n=78)およびDT40ポリLacO-VλR LacI-HP1トランスフェクタント(n=36)に由来する独特な変異を有するVλ領域の配列解析の概略。2つの独立したトランスフェクタントから配列を集めた。
【図15】変異したDT40ポリLacO-λR LacI-HP1クローンの配列アラインメント。多様化したDT40ポリLacO-λR LacI-HP1細胞に由来する14種の独特な変異Vλ領域の配列。青い四角は、遺伝子変換区域を囲み、赤色の円は点変異を示し、黒の点線の四角は非鋳型式挿入を示し、オレンジ色の三角は欠失を示す。DT40領域の親配列(SEQ ID NO: 31)を提示し、変異部位を示す。
【図16】Ig遺伝子変換は、ψVλアレイへのVP16の係留によって加速される。図16A:DT40の再配列されたニワトリIgλ座位の概略図。重合ラクトースオペレーター(ポリLacO)がψVλ17〜20の間に挿入されている。リーダー、L;再配列された可変領域、Vλ-Jλ;定常領域、Cλ。図16B:図に示すトランスフェクタントにおけるGFP-LacIまたはGFP-LacI-VP16の発現のウェスタンブロット解析。抗LacI抗体を用いてブロッティングし、対照のβ-アクチンと比べて発現を分析した。図16C:DT40ポリLacO-λ GFP-LacIトランスフェクタントおよびDT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントの細胞周期プロファイル。図16D:DT40ポリLacO-λ GFP-LacIトランスフェクタントおよびDT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントにおけるVλ転写物レベルのRT-PCRによる比較定量。DT40ポリLacO-λ GFP-LacIに対して標準化した。図16E:DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントの蛍光画像。矢印は、Igλに結合されたGFP-LacI-VP16を示す。核をDAPIで対比染色した(青色)。
【図17】ψVλアレイへのVP16の係留により、AcH3レベルおよびAcH4レベルは上昇し、Ig遺伝子変換は加速する。図17A:DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントのIgλにおけるアセチル化H3(AcH3)の富化。ChIPに続いて、OvaプライマーおよびψVλ17Rプライマーを用いて二重PCRを実施した。富化は、全DNAインプット対照に対し、段階的に増量した鋳型DNAを4回別々に増幅した結果の±標準偏差として表す。図17B:DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントのIgλにおけるアセチル化H4(AcH4)の富化。記号は図17Aと同じ。図17C:独立したDT40ポリLacO-λ GFP-VP16トランスフェクタントのパネルのsIgM喪失量中央値。DT40ポリLacO-Vλ GFP-LacIトランスフェクタントに対して標準化した。図は、各融合構築物に関して少なくとも2個の独立したトランスフェクタントから得て組み合わせたデータを示す。対照のDT40ポリLacO-Vλ GFP-LacIトランスフェクタントと比べた増加倍率を下に示す。
【図18】H3.3ヒストン変異体は、DT40 Igλ座位で豊富である。図18A:DT40の再配列されたニワトリIgλ座位の概略図。記号は図16Aと同じ。図18B:H3.3ヒストン変異体を分析する、代表的なクロマチン免疫沈降実験の概略。調べた部位は次のものであった:ψVλアレイの約1kb上流の領域(フランク)、ψVλ5とVλの間の領域、ψVλ1、ψVλ5、ψVλ13、ψVλ18、ψVλ24、ψVλ25、ならびに再配列されたVλR対立遺伝子。棒は標準偏差を示す。
【図19】Ig遺伝子変換は、ψVλアレイへのHIRAの係留によって加速される。図19A:図に示すトランスフェクタントにおけるGFP-LacIまたはHIRA-LacIの発現のウェスタンブロット解析。抗LacI抗体を用いてブロッティングし、対照のβ-アクチンと比べて発現を分析した。図19B:DT40ポリLacO-λ GFP-LacIトランスフェクタントおよびDT40ポリLacO-λ HIRA-LacIトランスフェクタントの細胞周期プロファイル。図19C:DT40ポリLacO-λ GFP-LacIトランスフェクタントおよびDT40ポリLacO-λ HIRA-LacIトランスフェクタントにおけるVλ転写物レベルのRT-PCRによる比較定量。DT40ポリLacO-λ GFP-LacIに対して標準化した。図19E:DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントの蛍光画像。矢印は、Igλに結合されたGFP-LacI-VP16を示す。核をDAPIで対比染色した(青色)。
【図20】DT40ポリLacO-λにおけるHIRA係留により、H3.3ヒストン変異体は富化され、遺伝子変換が加速する。図20A:H3.3-FLAGを発現するDT40ポリLacO-λ細胞およびDT40ポリLacO-λ HIRA-LacI細胞のψVλ17RにおけるH3.3(緑)およびpan H3(青)の富化。DT40ポリLacO-λ HIRA-LacIの2つの独立した株の結果を示す。DT40ポリLacO-λの富化は、値「1」に対して標準化する。棒は、段階的に増量した鋳型DNAを4回別々に増幅した結果の±標準偏差として示す。図20B:DT40ポリLacO-GFP-LacI細胞およびDT40ポリLacO-λ HIRA-LacI細胞の代表的な集団中のsIgM+細胞のフローサイトメトリー。図20C:独立したDT40ポリLacO-λ HIRA-LacIトランスフェクタントおよびDT40 HIRAトランスフェクタントのパネルのsIgM喪失量中央値。DT40ポリLacO-λ GFP-LacIトランスフェクタントに対して標準化した。図は、各融合構築物に関して少なくとも2個の独立したトランスフェクタントから得て組み合わせたデータを示す。対照のDT40ポリLacO-λ GFP-LacIトランスフェクタントと比べた増加倍率を下に示す。
【図21】係留されたHIRAおよびVP16の異なるクロマチン効果。図21A:小球菌ヌクレアーゼ(MNアーゼ(MNase))でクロマチンを消化した後の、DT40ポリLacO-λ GFP-LacIトランスフェクタント、DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタント、およびDT40ポリLacO-λ HIRA-LacIトランスフェクタントのポリLacO領域のサザンブロット解析。MNアーゼ無し、(-)。右の数字はヌクレオソーム多量体を示す。図20B:ヒストン沈着の増加が、クロマチンの「はしご状(laddering)」パターンの強調に関与し得る方法のモデル。矢印先端部は、存在し得るMNアーゼ切断部位を示す。黒い矢印先端部は、限定的な範囲の切断可能なDNAを示し、灰色の印先端部は、予測される約150bpの切断産物から逸脱する広い範囲の切断可能なDNAを示唆する。
【図22】単一のDT40ポリLacO-λ GFP-LacI-VP16細胞に由来する変異したV領域の配列。単一のsIgM-細胞からV領域を増幅し、次いで配列決定した。中に色を塗っていない青い四角は、長区域の遺伝子変換事象を囲み、青色に塗った四角は、短区域の遺伝子変換事象を囲み、赤い円は点変異を示し、黒い点線の四角は挿入を示し、ダイヤ型(carat)は欠失を示す。
【図23】単一のDT40ポリLacO-λ HIRA-LacI細胞に由来する変異したV領域の配列。記号は図22と同じ。
【発明を実施するための形態】
【0020】
発明の詳細な説明
本発明は、標的免疫グロブリン遺伝子の変異誘発が誘導可能であるB細胞株の開発に基づいている。免疫グロブリンは細胞表面で発現されるため、特定の特異性、高い親和性、および/または新しい活性を有する免疫グロブリン分子を発現するクローンの選択が可能である。これは、標的部位で免疫グロブリン遺伝子および同様に非免疫グロブリン遺伝子を用いて実施することができる。
【0021】
定義
本出願において使用される科学用語および技術用語はすべて、別段の指定が無い限り、当技術分野において一般に使用される意味を有する。本出願において使用される場合、以下の単語または語句は、指定の意味を有する。
【0022】
本明細書において使用される場合、「ポリペプチド」は、タンパク質、タンパク質断片、およびペプチドを含み、天然供給源から単離されるか、組換え技術によって作製されるか、または化学合成されるかを問わない。本発明のペプチドは、典型的には少なくとも約6個のアミノ酸を含む。
【0023】
本明細書において使用される場合、標的遺伝子の「多様化」とは、標的遺伝子の配列または構造の変化または変異を意味する。多様化は、点変異、鋳型式変異、DNA欠失、およびDNA挿入をもたらし得る、体細胞超変異、遺伝子変換、およびクラススイッチ組換えといった生物学的プロセスを含む。本発明の多様化因子は、これらの多様化方法のいずれかを誘導、強化、または調節することができる。
【0024】
「変異」とは、ポリヌクレオチド配列の改変であり、1つもしくは複数のヌクレオチド塩基の改変、もしくは配列への1つもしくは複数のヌクレオチドの挿入、もしくは配列からの1つもしくは複数のヌクレオチドの欠失のいずれか、またはこれらの組合せを特徴とする。
【0025】
本明細書において使用される場合、「シス調節エレメント」とは、遺伝子の発現または多様化を制御する領域中に位置するDNA配列である。係留因子は、DNAのこの領域に配列特異的な様式で結合する。代表的なシス調節エレメントにはLacOおよびTetOが含まれるが、それらに限定されるわけではない。
【0026】
本明細書において使用される場合、「係留因子」とは、配列特異的な様式でシス調節エレメントに結合する分子である。係留因子の1つの例は「リプレッサー」、すなわち、調節遺伝子によって合成され、オペレーター座位に結合して、そのオペロンの転写を妨害するタンパク質である。例示的な係留因子にはLacIおよびTetRが含まれるが、それらに限定されるわけではない。
【0027】
本明細書において使用される場合、「多様化因子」とは、多様化または超変異を加速させるか、または調節する分子を意味する。代表的な多様化因子には、最初は転写調節因子として同定されたいくつか(例えば、VP16もしくはE47)、ヘテロクロマチン関連タンパク質(例えば、HP1)、ヒストンシャペロン(HIRA)、クロマチン再構築因子、核膜孔複合体の構成要素(NUP153)、遺伝子調節因子、またはそれらの組合せが含まれる。多様化因子として働くことができる他の分子には、DNA修復因子、DNA複製因子、リゾルベース、ヘリカーゼ、細胞周期調節因子、ユビキチン化因子、SUMO化因子、またはそれらの組合せが含まれるがこれらに限定されるわけではない。
【0028】
本明細書において使用される場合、「プロモーター」とは、一般にコード領域の上流(5')側にあり、転写の開始およびレベルを少なくともある程度制御するDNA領域を意味する。本明細書における「プロモーター」への言及は、最も広い文脈で理解されるべきであり、TATAボックスプロモーターまたは非TATAボックスプロモーターを含む、古典的なゲノム遺伝子の転写調節配列、ならびに、発達的刺激および/もしくは環境刺激に応答して、または組織特異的もしくは細胞型特異的な様式で遺伝子発現を改変するその他の調節エレメント(すなわち、活性化配列、エンサンサー、およびサイレンサー)を含む。プロモーターは、発現を調節する相手である構造遺伝子の上流または5'側に通常は位置するが、必ずしもそうとは限らない。さらに、プロモーターを含む調節エレメントは、遺伝子の転写開始部位から2kb以内に通常位置するが、それらはより遠く(many kb)離れていてもよい。プロモーターは、細胞における発現をさらに促進するために、かつ/または機能的に結合されている構造遺伝子の発現の時期もしくは誘導能を改変するために、開始部位からさらに遠位に位置するその他の特異的調節エレメントを含んでよい。
【0029】
本明細書において使用される場合、「機能的に結合される」または「機能的に連結される」などは、機能的な関係にあるポリヌクレオチドエレメントの結合の仕方を意味する。核酸は、別の核酸配列との機能的関係に置かれている場合、「機能的に連結され」ている。例えば、プロモーターまたはエンハンサーは、コード配列の転写に影響を及ぼす場合、コード配列に機能的に連結されている。「機能的に連結される」とは、連結されている核酸配列が典型的には隣接しており、2つのタンパク質コード領域を結合する必要がある場合は、隣接し、かつリーディングフレーム中にあることを意味する。転写可能なポリヌクレオチドにプロモーターを「機能的に連結する」とは、転写可能なポリヌクレオチド(例えば、タンパク質をコードするポリヌクレオチドまたは他の転写物)をプロモーターの調節的制御下に置くことを意味し、プロモーターは、次いでそのポリヌクレオチドの転写および任意で翻訳を制御する。
【0030】
「核酸」または「ポリヌクレオチド」という用語は、単鎖または二本鎖いずれかの形態のデオキシリボヌクレオチドポリマーまたはリボヌクレオチドポリマーを意味し、別段の制限がない限り、天然に存在するヌクレオチドと同様の様式で核酸にハイブリダイズする、天然ヌクレオチドの公知の類似体も包含する。
【0031】
本明細書において使用される場合、「1つの(a)」または「1つの(an)」は、特に別段の指示がない限り、少なくとも1つを意味する。
【0032】
B細胞
本発明は、標的遺伝子の多様化の可逆的誘導を可能にするように改変されたB細胞を提供する。この細胞は、関心対象の標的遺伝子に機能的に連結されたシス調節エレメントを含む。次いで、多様化を調整する因子を、シス調節エレメントに結合する係留因子に融合させ、それによって、標的遺伝子の発現を制御する領域に多様化因子を係留することができる。B細胞は、ニワトリDT40 B細胞または他の脊椎動物B細胞でよく、いくつかの態様においては、ヒトB細胞またはヒト化免疫グロブリン(Ig)遺伝子(ヒトIgHおよびIgLでニワトリIgHおよびIgLを置換している)を含むニワトリDT40 B細胞が好ましい。
【0033】
B細胞は、天然の抗体産生者であるため、改善された抗体ならびに改善された非免疫グロブリンタンパク質およびポリペプチドの両方を産生させるための魅力的な細胞となっている。DT40 B細胞は、超変異および選択の反復サイクルによって特異的かつ高親和性の抗体を進化させるための有効な開始点である(Cumbers et al., 2002; Seo et al., 2005)。DT40細胞には、この目的のために試験された他の媒体(vehicle)よりも優れた利点がいくつかある。DT40は、培養状態でそのIg遺伝子を構成的に多様化し、発現された抗体分子のCDRに変異を導く。DT40はヒトB細胞株よりも速く増殖する(世代時間は24時間に対して10〜12時間)。細胞は、特殊な因子もフィーダー層も添加せずに、限界希釈によって容易にクローニングされるため、クローン集団を容易に単離することができる。DT40は効率的な相同遺伝子ターゲティングを実施し(Sale, 2004)、したがって、特定の座位を意のままに置換して、超変異を調節する因子を操作できるようにすることができる。
【0034】
本発明は、高親和性抗体を作製するための新規なプラットホームを提供する。1つの態様において、抗体を進化させるための媒体はB細胞株DT40であり、これは、抗体を天然に産生し、かつ、免疫グロブリン遺伝子超変異を加速させ誘導性にするために操作された。他のB細胞と同様に、DT40は細胞表面で抗体を発現するため、蛍光活性化細胞選別または磁気活性化細胞選別によって、高親和性および最適な特異性に関して簡便にクローン選択することが可能である。DT40細胞株において、超変異は、生理学的環境でのIg遺伝子超変異への、何百万年もの脊椎動物の進化を通して完成された同じ経路によって実施される。この高度に保存された経路は、抗原と接触する可変(V)領域のサブドメインである相補性決定領域(CDR)に優先的に(ただし排他的ではない)変異を導く(図1)。
【0035】
これまで、抗体選択のためにDT40(および他の培養B細胞株)を使用することは、超変異の速度が非常に遅く、生理学的超変異の約0.1%〜1%の速度であるため、制限されていた。超変異を加速させるために、超変異の分子メカニズムについての本発明者らの現在の高度な理解を利用して、重要な調節部位および調節因子が操作された。
【0036】
ニワトリDT40 B細胞は多くの利点を与えるものの、いくつかの態様において、ヒトB細胞を使用することが望ましい場合がある。あるいは、ニワトリDT40 B細胞と共にヒト化Ig遺伝子を使用してもよい。DT40免疫グロブリン遺伝子をヒト化することによって、DT40プラットホームにおいて作製した抗体を治療に直接使用できるなど、治療学のためのこのプラットホームの有用性を広げることができる。
【0037】
ヒト化抗体遺伝子の有用性に関する十分な記録があり、最近再検討されたように、ヒト化するためのいくつかの有効なアプローチがある(WaldmannおよびMorris, 2006; AlmagroおよびFransson, 2008)。ヒト化は、ニワトリIg遺伝子をヒトIg遺伝子で置換することによって行われ、これは、高効率の相同遺伝子ターゲティングを利用することにより、DT40において容易に実施される。これらの置換は、下記の実施例で説明する、シス調節エレメントを挿入するための手順に類似しているが、重鎖座位および軽鎖座位の別個の部分を改変するように設計された異なるターゲティング構築物を用いる手順によって実施される。置換によって、V(D)J領域とC領域を交換することにより、完全にヒト化した抗体、またはキメラ抗体(C領域はヒト化されているがV領域はされていない)を産生するDT40派生体を作製することができる。これらの置換は、隣接したシス調節エレメントを改変することも、それらが超変異を加速させる能力に影響を及ぼすこともない。超変異を促進する保存されたメカニズムは、ヒト化配列のCDRに変異誘発を導く。したがって、ヒト化した株は、細胞培養においてヒトモノクローナルの発達を加速させるために使用することができ、治療目的または診断目的のいずれかに有用な抗体を迅速に作製するための二重プラットホームを提供する。
【0038】
さらに、C領域の置換によって抗体エフェクター機能を最適化することもできる。抗体に基づく免疫療法は、療法のための強力なアプローチであるが、このアプローチはこれまで、有用なエフェクター特性を有する特定の抗体の入手しやすさにある程度制限されていた(Hung et al., 2008; Liu et al., 2008)。抗体の定常(C)領域は、エフェクター機能を決定する。DT40媒体における相同遺伝子ターゲティングによって、天然または人工いずれかのヒトC領域を置換して、所望のエフェクター機能を有する抗体を作製することができる。
【0039】
標的遺伝子
典型的には、標的遺伝子は、プロモーターおよびコード領域を含む。本発明のB細胞中の標的遺伝子のコード領域は、関心対象の任意のタンパク質またはペプチドをコードするものでよく、完全なコード領域を含む必要はない。いくつかの態様において、特定の領域またはドメインが多様化のための標的とされ、コード領域は、関心対象の領域またはドメインを含む部分のみを任意でコードしてよい。
【0040】
1つの態様において、標的遺伝子は、Ig遺伝子のエンハンサーおよびコード領域を含む免疫グロブリン(Ig)遺伝子を含む。Ig遺伝子は、IgL遺伝子および/またはIgH遺伝子の全体または一部分でよい。コード領域は、Ig遺伝子に先天的なものでもよく、または異種遺伝子でもよい。いくつかの態様において、標的遺伝子は、多様化のための非Ig標的ドメイン、ならびに、膜貫通ドメインおよび細胞質尾部を含む、B細胞表面で遺伝子産物を提示することを可能にするドメインであるか、またはそれらを含む。
【0041】
シス調節エレメント
シス調節エレメントは、標的遺伝子の発現を制御するランディングパッドを領域中に提供する。このランディングパッドは、DNAのこの領域に対して配列特異的な様式で係留因子が結合できる場所を提供する。係留因子(配列特異的DNA結合タンパク質)がDNAに結合し、係留因子に融合された多様化因子をコード領域の十分近くに届けることができ、その結果、コード領域の多様化が可逆的に調節可能になる場所を提供するというランディングパッド機能をエレメントが果たす限り、様々な分子をシス調節エレメントとして使用することができる。典型的な態様において、シス調節エレメントは、重合ラクトースオペレーター(LacO)である。1つの態様において、エレメントは、約80〜100個繰り返したLacOを含む。別の態様において、シス調節エレメントは、テトラサイクリンオペレーター(TetO)である。
【0042】
係留因子および多様化因子
係留因子は、配列特異的な様式でシス調節エレメントに結合する因子である。いくつかの態様において、多様化の調節は、シス調節エレメントへの調節可能な結合を特徴とする係留因子を用いることによって実現される。LacOがシス調節エレメントとして働く態様において、LacリプレッサーLacIは、係留因子として働くことができ、シス調節エレメントLacOへの結合は、イソプロピル-β-D-チオ-ガラクトシド(IPTG)によって調節され得る。IPTGの不在下では、LacIはLacOに結合し、多様化は、多様化因子の存在によって加速される(またはそうでなければ調節される)。IPTGは、多様化因子の活性の停止または低減が望ましい事象において添加され得、それによって、変異誘発プロセスが可逆的プロセスになる。TetOがシス調節エレメントとして働く態様において、TetRは適切な係留因子であり得、多様化因子の活性はテトラサイクリンまたはドキシサイクリンによって調節され得る。
【0043】
多様化因子は、B細胞における変異誘発の調整を可能にする因子である。下記の実施例は、いくつかの調節因子を係留および遊離する効果を説明する。1つはヘテロクロマチンタンパク質HP1であり、超変異を5.6倍に加速した(実施例4)。もう1つはE47-LacI(E47は、E2A調節因子の1つのアイソフォームであり、B細胞の発達に不可欠である)であり、超変異を4.5倍に加速した(実施例3)。クロマチン構造体の他の2つの改変因子(modifier)は、超変異を8.4倍および11.0倍に加速した(図2)。さらに、係留部位は、発現されるIg重鎖座位を標的としていた。IgHおよびIgLにおける超変異の二重調節により、超変異を25倍から100倍に加速して、新しい抗体の迅速な作製を可能にすることができる。
【0044】
多様化している免疫グロブリン遺伝子は、細胞周期の過程で核内部を移動し、この移動は多様化と相関関係があり、多様化に必要である。(下記の実施例で説明する方法を用いて)DT40ポリLacO-λ細胞においてNP153-LacIを発現させることにより核膜孔に係留すると、多様化が5.7倍に加速する。この結果から、他の核膜孔タンパク質および核中で遺伝子の位置を決定または調節する他の因子を含む、遺伝子の位置を調節する他の因子もまた多様化を調節することが予測される。
【0045】
クロマチン構造も同様に多様化を調節する。実施例4のデータは、LacIに融合されたヘテロクロマチンタンパク質HP1の発現により多様化が加速すること、およびこの効果がIPTGによって可逆的であることを示す。この知見は、ヒストンシャペロンであるHIRAにも適用された。
【0046】
また、遺伝子の発現または機能の調節因子も、多様化因子として働く。下記の実施例3は、調節因子E2A(E47アイソフォームとして、LacI融合物として発現される)の発現により多様化が加速すること、およびこの効果がIPTGによって可逆的であることを示す。ヘルペスウイルス由来のVP16調節ドメインは多様化を加速させ、この効果はIPTGによって可逆的である。
【0047】
脱アミノもまた、変異誘発を加速させるが、これは誘導性ではない。AIDは、Ig遺伝子多様化を開始するB細胞特異的DNAデアミナーゼである。下記の実施例2は、DT40ポリLacO-λにAID (AID-LacI)を係留することにより変異誘発が促進されるが、IPTGはこの効果を抑えないことを示す。したがって、変異誘発の影響(affect)はゲノムワイドであるため、この因子は、さらに改変することなしには不適切であると思われる。
【0048】
いくつかの態様において、多様化因子は、転写調節因子、ヘテロクロマチン関連タンパク質、ヒストンシャペロン、クロマチン再構築因子、核膜孔複合体の構成要素、遺伝子調節因子、またはそれらの組合せである。多様化因子として働くことができる他の分子には、DNA修復因子、DNA複製因子、リゾルベース、ヘリカーゼ、細胞周期調節因子、ユビキチン化因子、SUMO化因子、またはそれらの組合せが含まれるがこれらに限定されるわけではない。1つの態様において、転写調節因子は、VP16またはE47である。多様化因子として使用するための典型的なヘテロクロマチン関連タンパク質はHP1である。代表的なヒストンシャペロンはHIRAである。
【0049】
(表1)代表的な多様化因子
【0050】
融合構築物
シス調節エレメントへの多様化因子の係留は、係留因子への多様化因子の融合を通して達成される。因子のこの組合せをコードする融合構築物は、本発明のB細胞中に存在し、本発明のB細胞によって発現されてよく、また、所望の時点および特定の目的のために望ましい因子を選択する際にB細胞に添加するために、別々に提供されてもよい。
【0051】
一般に、融合構築物は標準技術を用いて調製してよい。例えば、これらのペプチド構成要素をコードするDNA配列を別々に組み立て、適切な発現ベクターに連結してよい。連結されるDNA配列は、適切な転写調節エレメントまたは翻訳調節エレメントに機能的に連結される。一方のペプチド構成要素をコードするDNA配列の3'末端が、ペプチドリンカーを用いてまたは用いずに、第2のペプチド構成要素をコードするDNA配列の5'末端に連結され、その結果、これらの配列のリーディングフレームは協調する。これにより、両方の構成要素ペプチドの生物活性を保持している単一の融合タンパク質に翻訳されることが可能になる。
【0052】
ペプチドリンカー配列を使用して、第1のペプチド構成要素と第2のペプチド構成要素を、各ペプチドが折り畳まって二次構造および三次構造を形成するのを確実にするのに充分な距離だけ離れさせてもよい。このようなペプチドリンカー配列は、当技術分野において周知の標準技術を用いて融合タンパク質中に組み入れられる。適切なペプチドリンカー配列は、次の因子に基づいて選択することができる:(1)可撓性の拡張された立体構造をとれること;(2)第1のペプチドおよび第2のペプチド上の機能的領域と相互作用し得る二次構造をとれないこと;ならびに(3)ペプチドの機能的領域と反応する可能性がある疎水性残基または荷電残基が無いこと。好ましいプチドリンカー配列は、Gly残基、Asn残基、およびSer残基を含む。ThrおよびAlaなど他のほぼ中性のアミノ酸もまた、リンカー配列中で使用されてよい。
【0053】
本発明の方法および用途
本発明は、ポリペプチドをコードするポリヌクレオチド配列の多様化を通して、関心対象のポリペプチドの変異配列を有するポリペプチドのレパートリーを作製する方法を提供する。典型的には、この方法は、多様化因子の発現を可能にする条件で本発明のB細胞を培養する段階を含み、B細胞の標的遺伝子は関心対象のポリペプチドのコード領域を含み、それによって、コード領域の多様化が可能になる。この方法は、複数の変異ポリペプチドおよび所望のレパートリーが得られるまで、B細胞の増殖を可能にする条件下で培養物を維持する段階をさらに含んでよい。B細胞は外表面でポリペプチドを発現するため、レパートリーの性質および範囲を決定することができる。次いで、所望の特性を有するポリペプチドを選択するためにレパートリーを使用することができる。
【0054】
別の態様において、本発明は、最適化された関心対象のポリペプチドを産生するB細胞を作製する方法を提供する。この方法は、多様化因子の発現を可能にする条件で本発明のB細胞を培養する段階を含み、B細胞の標的遺伝子は関心対象のポリペプチドのコード領域を含み、B細胞は関心対象のポリペプチドをその表面で発現する。この方法は、関心対象のポリペプチドに特異的に結合するリガンドに結合する細胞を選択することにより、関心対象のポリペプチドをB細胞表面で発現する細胞を培養物から選択する段階をさらに含む。関心対象のポリペプチドに特異的に結合するリガンドに対して所望の親和性を有する細胞を選択するまで、これらの培養段階および選択段階を繰り返すことができる。B細胞クローンの増殖中の抗体特異性の進化を図3に例示する。最初に多重特異性を示す集団(左上)を選択し、続いて、所望の特異性(中央)およびより高い親和性(右下)を富化する。
【0055】
関心対象のポリペプチドがIg、例えばIgL、IgH、または両方である態様において、リガンドは、抗原に相当する、組換えまたは他の手段によって作製されたポリペプチドでよい。例えば、磁気活性化細胞選択(MACS)による選択を容易にするために、リガンドを固体支持体に結合または連結することができる。別の例において、蛍光活性化細胞選別(FACS)を可能にするために、リガンドを蛍光タグに結合または連結することができる。当業者は、細胞を標識し選択する他の方法が公知であり、本方法において使用できることを理解している。
【0056】
本発明の方法は、培養物に調節分子を添加する段階をさらに含んでよく、調節分子は、シス調節エレメントへの係留因子の結合を調整し、それによって、コード領域の多様化を調整する。上記の例において、IPTG、テトラサイクリン、およびドキシサイクリンが、調節分子として働く。当業者は、多様化活性を調節するために個々の係留因子と共に使用できる他の調節分子を知っている。
【0057】
調節分子は、コード領域の多様化を様々な方法で調整することができる。例えば、いくつかの態様において、調節分子は、調整を行うために培養物に添加される。個々の調節分子がシス調節エレメントへの係留因子の結合を増加させる因子であるか、または減少させる因子であるかどうか、および結合の個々の変化が、多様化活性を増大させる効果を有するか、または低減させる効果を有するかどうかによって、調整の結果、多様化を低減もしくは停止するか、または多様化を促進もしくは加速化することができる。他の態様において、調整は、調整の低減であれ停止であれ、促進であれ加速であれ、調節分子を培養物から除去または取り除くことによって行われる。同様に、多様化の調整は、B細胞に遺伝子を添加することもしくはB細胞から遺伝子を取り除くことによって、またはB細胞における遺伝子の発現を増大させることもしくは減少させることによって行うこともできる。当業者は、上記に説明した利用可能な変更すべてを容易に理解することができ、これらはそれぞれ、B細胞における調節分子のレベルまたは存在を改変し、その結果として、シス調節エレメントへの多様化因子の係留を改変し、それによって、多様化活性を改変する効果を有する。
【0058】
本発明の方法のその他の用途には、IgλにおけるIg遺伝子標的の誘導性超変異が含まれる。DT40ポリLacO-λ LacI-HP1細胞株(下記の実施例4を参照されたい)は、Vλ偽Vアレイの下流の任意のIg遺伝子配列を改変するために使用することができる。Ig座位のゲノム構造は、V領域の変異誘発を促進するが、Ig分子の定常領域の変異誘発は促進しないように進化した。Ig座位におけるIg遺伝子配列の改変は、これをうまく利用して、変異誘発による産生物が機能的な定常領域を確実に保持するようにする。DT40において遺伝子を置換することは容易であるため、その遺伝子に有用な変化を起こすことができる。例えば、ニワトリ遺伝子(可変領域および定常領域)をヒト抗体遺伝子(重鎖または軽鎖のいずれか)で置換して、治療的用途のある抗体を作製することができる。また、これを用いて、他の種のための治療的抗体を作製することもできる。これにより、ヘテロ二量体抗体の一方の鎖のみまたは単鎖抗体の単鎖のいずれかに変異誘発するための迅速な系が提供される。
【0059】
このアプローチの用途には、高親和性Igを産生するB細胞クローンを作製するための超変異、特異的エピトープの認識を保持しつつ、抗体の交差反応性を改変するための超変異、特定の化合物に対して高親和性を有するV領域配列を同定するための超変異が含まれる。これは、単一のIg座位のみ、または両方で実施することができる。
【0060】
また、誘導性超変異はIgHで実施することもできる。DT40のIgH座位は、Igλと同様に、効率的な変異誘発部位である。類似技術によって作製された派生細胞株では、IgHおよびIgλの両方に、または各対立遺伝子に別々に変異誘発することが可能である。DT40ポリLacO-λポリLacO-H LacI-HP1派生体を作り出すために、IgHにポリLacOを挿入すると、H鎖遺伝子およびL鎖遺伝子の両方における超変異が可能になる。培地にIPTGを添加すると、鋳型式事象を減少させ、非鋳型式超変異を増加させる阻害効果がいくらかもたらされる。したがって、IPTGの存在および不在を変更することによって、抗体の両方の鎖をコードする遺伝子の鋳型式変異誘発および非鋳型式変異誘発を行ったり来たり切り替えることができる。
【0061】
一方の対立遺伝子におけるLacリプレッサー(LacI)のIPTGによる調節と他方の対立遺伝子におけるTetリプレッサー(TetR)のテトラサイクリンによる調節とを組み合わせた派生体を作製することができる。このTetR派生体では、IgH対立遺伝子およびIgL対立遺伝子で、かつIgL座位およびIgH座位で多様化を調節するための異なるメカニズムを同時に用いて、多様化を独立に調節することが可能になる。
【0062】
本発明はまた、T細胞受容体を選択するための媒体も提供する。T細胞に基づいた免疫療法には大きな可能性がある(BlattmanおよびGreenberg, 2004)。T細胞受容体の特異性および親和性は、CDR接触によって決定されている(Chlewicki et al., 2005)。特異性または高親和性のT細胞受容体の加速された誘導性選択は、Ig座位をT細胞受容体(V領域または遺伝子全体)で置換することによって改変したDT40ポリLacO媒体において実施することができる。
【0063】
触媒的Igの作製は、本発明の別の局面である。標的Igは触媒のためにも使用され得るため、本発明のIgに関連した方法は、結合および認識のためのIg作製だけに限定されない。酵素反応の遷移状態を模倣する安定な分子を開発した後、DT40ポリLacO-λ LacI-HP1細胞を用いて、結合し実際の化学的遷移状態を安定化させる抗体を得ることができる。中間体に結合できるIgを産生するクローンを同定した後、この系を再び用いて、培養状態での実際の基質に対するIgの触媒活性を求めてスクリーニングすることができる。この系において何らかの活性が実証された後で、変異誘発を通してIg座位をさらに進化させることによって活性の最適化を進めることができる。したがって、本発明は、動物の免疫化(時間のかかる段階)、ハイブリドーマ技術による不死化、および触媒活性を示す抗体を得るためにハイブリドーマをスクリーニングしなければならない非効率な後段階(later)を要求しない。
【0064】
IgλまたはIgHにおける非Ig遺伝子標的の誘導性超変異は、本発明の別の局面である。Ig座位のゲノム構造は、プロモーターの1〜1.5kb下流の変異誘発を促進するように進化した。この系を利用して、遺伝子の短い領域を変異させることができる。表面のタンパク質発現に基づいたクローン選択は、表面発現を媒介するエレメントを発現する遺伝子の一部分に関心対象の領域を融合することによって組み入れることができる。表面発現のための例示的なエレメントには、B細胞表面で発現されるタンパク質に由来するシグナルペプチド、膜貫通ドメイン、および細胞質尾部が含まれる(Chou et al., 1999; Liao et al., 2001)。
【0065】
本発明はまた、認識アレイを作製するために使用することもできる。広範囲の抗原に対する親和性を有する受容体を内部に持つ細胞を進化させることができると、認識アレイを開発することが可能になる。この技術を、DT40における受容体刺激に由来する細胞内応答/シグナル伝達(エクオリンによるCa2+の測定値(Rider et al., 2003)またはレポーター遺伝子転写の使用など)と組み合わせると、有用なバイオセンサーが作り出される。多様化されたクローンをアレイまたは96ウェルプレートにスポットし、試料に曝露させる。各試料は、刺激の「指紋」を生じるはずである。これらのアレイにより、生物学的/医学的試料、環境的試料、および化学的試料の定性的比較が可能になると思われる。あらゆる形態の化合物が抗原特性を有し得るため、2Dゲルのような比較技術と同様に、解析はタンパク質解析に限定される必要はない。さらに、これらのアレイにより、その存在を前もって知らずとも、構成要素が同定される。
【0066】
キット
本明細書において説明する方法で使用する場合、キットもまた、本発明の範囲内である。このようなキットは、バイアルおよびチューブなど1つまたは複数の容器(各容器は、本発明において使用される個別の要素(例えば、細胞、構築物)の内の1つを含む)を入れるように仕切られたキャリア、パッケージ、または容器を含んでよい。
【0067】
典型的には、キットは、本発明のB細胞ならびに対応する係留因子および多様化因子を発現する融合構築物を含む。例えば、B細胞は、プロモーターおよびコード領域を含む標的遺伝子に機能的に連結されたシス調節エレメントを含む。キットは、1種または複数種の融合構築物が中に保存された1つまたは複数の容器をさらに含む。各融合構築物は、B細胞で発現され得、かつ、多様化因子に融合された係留因子をコードするポリヌクレオチドを含み、この係留因子は、B細胞のシス調節エレメントに特異的に結合する。B細胞は、複数の融合構築物と共に使用するために複数のシス調節エレメントを含んでよい。
【0068】
典型的には、本発明のキットは、前述の容器、ならびに緩衝剤、希釈剤、フィルター、針、シリンジ、および使用に関する指示が書かれた添付文書を含む、商業的観点かつ使用者の観点から望ましい材料を含む1つまたは他の容器を含む。さらに、組成物が特定の治療的用途または非治療的用途のために使用されることを示し、かつ、使用する際の指示も示してよいラベルを容器の表面に貼ってもよい。指示およびまたは他の情報は、キットと共に含まれる挿入物に含まれてもよい。
【0069】
実施例
以下の実施例は、本発明を例示し、かつ当業者がそれを作製および使用するのを補助するために提示される。これらの実施例は、本発明の範囲を別な具合に限定することを決して意図しない。
【0070】
実施例1:DT40 B細胞株における変異およびターゲティング
本実施例は、2段階で準備される本発明の態様を例示する。
(1)DT40ポリLacO-λ
本発明者らは、Igλ座位でのシスな調節を可能にする遺伝子エレメントを含む、ニワトリB細胞株DT40のこの派生体を最初に構築した。DT40ポリLacO-λでは、重合ラクトースオペレーター(LacO)が、Igλ軽鎖座位の偽Vλアレイのすぐ上流に挿入されている。これは、StraightおよびBelmontによって開発された系を拡張したものであり、その中のポリLacOは、20bpのLacO結合部位の約80個の繰り返しからなる(Straight et al., 1996; Belmont, 2001)。本発明者らは、これが鋳型式変異誘発の正常なプロセスに影響を及ぼさないことを示した。DT40ポリLacO-λの構築は、下記の実施例2で詳細に説明する。
【0071】
LacOには、ラクトースリプレッサー(LacI)が非常に高い親和性(<10-12M)で結合する。このメカニズムは、大腸菌(E. coli)においてラクトースを代謝するためにオペロンの発現を調節し、また、様々な細胞の状況においても使用され得る。本発明者らは、蛍光タンパク質(GFP、RFP、YFP、またはCFP)に融合されたLacIを発現する安定なトランスフェクタントを作製し、かつ、GFP-LacI、RFP-LacI、YFP-LacI、またはCFP-LacIを安定に発現する正常に増殖中のDT40ポリLacの核において単一のスポットとして画像化され得る結合されたタンパク質を免疫蛍光顕微鏡検査法によって示すことにより、LacIがDT40ポリLacO-λ中のポリLacOに結合することを示した。
【0072】
IPTGは、インビトロおよび培養細胞中の両方で、LacOからのLacIの遊離を引き起こす小分子である。DT40ポリLacO-λ GFP-LacIをわずか10μMのIPTGと共に培養すると、ポリLacOからのGFP-LacIの遊離が起こる。IPTGと共に培養しても、細胞増殖は影響を受けない。
【0073】
(2)DT40ポリLacO-λ LacI-HP1
本発明者らは、クロマチン構造の改変因子であるキイロショウジョウバエ(Drosophila melanogaster)HP1(L. Wallrath(University of Iowa)から入手)がポリLacOに係留されると体細胞超変異への切り替えが起こる、DT40ポリLacO-λのこの派生体を次に構築した。係留は、非常に十分に特徴付けられているシス調節因子およびトランス調節因子のペアである大腸菌LacO/LacIを用いて実現した。HP1は、ヘテロクロマチン遺伝子サイレンシングおよびヘテロクロマチンの伝播において機能する、十分に特徴付けられている非ヒストンヘテロクロマチンタンパク質であり、ショウジョウバエでは、係留されたLacI-HP1が、クロマチン構造の閉鎖およびlacOリピートに隣接したレポーター遺伝子の不活性化を促進することが示されている(Li et al., 2003; DanzerおよびWallrath, 2004)。本発明者らは、HP1の係留により、ChIPによって測定されるように、開放状態から非許容状態へとドナー配列が変化すること、および遺伝子変換ではなく体細胞超変異による変異誘発をVλが受けるようになることを実証した。これは、鋳型式変異誘発から体細胞超変異への切り替えを引き起こすことが示されている、偽Vλアレイの欠失と機能的に等価である(Arakawa et al., 2004)。DT40 ポリLacO-λLacI-HP1のさらなる特徴付けは、下記の実施例4において実証する。
【0074】
(3)DT40ポリLacO-λ VP16-LacI
次に、本発明者らは、ポリLacOにVP16-LacIを係留するために、発現プラスミドp3’ss-EGFP-VP16(A. Belmont(University of Illinois)から入手)を使用した。VP16は、強力な転写活性化因子であり、ヒストンアセチルトランスフェラーゼ複合体を動員することが実証されている(Tumbar et al., 1999)。本発明者らは、VP16を係留すると、ChIPによって測定されるように、許容性のより高いクロマチン構造が作り出されること、および野生型のレベルを上回る遺伝子変換の刺激による変異誘発をVλが受けるようになることを実証した。
【0075】
実施例2:遺伝子を核膜孔に係留することによる、変異誘発の加速
本発明者らは、多様化している免疫グロブリン遺伝子が、細胞周期の過程で核内部を移動すること、この移動は多様化経路の段階と相関関係があること、および多様化は核周辺で開始することを発見した。活性化誘導型デアミナーゼ(AID)は、多様化を開始する酵素であり、核外輸送シグナルを有し、核膜孔を介して核の外へと絶えず輸送されるため、その濃度は核周辺で高くなり得る。このことから、核膜孔に遺伝子を接近させることにより、多様化を加速させることが可能であり得ることが示唆された。本発明者らは、Lacリプレッサー(LacI)への核膜孔タンパク質Nup153の融合物を用いて、DT40ポリLacOのIgH座位を核膜孔に係留することにより、これが事実であることを示した。本発明者らは、これにより、クローンの多様化速度が5.7倍に加速されることを見出した。対照実験により、ポリLacOへの係留が変異誘発の加速にとって不可欠であることが示された。この方法は、標的の変異誘発を促進するために、AID核外移行シグナルおよび遺伝子局在化に依存する一般的な方法である。したがって、この方法を拡張して、非Ig遺伝子を孔に係留することにより、B細胞における非Ig遺伝子の変異誘発を促進することができる。さらに、非B細胞でAIDを発現させることにより、それらの細胞における遺伝子の変異誘発を促進するように拡張することもできる。
【0076】
実施例3:E2Aは、細胞周期のG1期にシスで作用してIg遺伝子多様化を促進する
本実施例は、核におけるIg遺伝子多様化の細胞周期に依存的な調節を説明する。再配列されたIg遺伝子は、DNAデアミナーゼAIDによって開始される配列および構造の多様化を経験する。多様化が起こるためにはIg遺伝子が転写されなければならないが、シス活性化のためにその他の必要条件があるかどうかは明らかにされていない。本実施例は、クロマチン免疫沈降法によって、調節因子E2Aが、構成的なIg遺伝子多様化を実行するニワトリDT40 B細胞株中の再配列されたIgλR遺伝子と結合することを示す。再配列されたλR遺伝子を重合ラクトースオペレーターによってタグ化したDT40派生体を直接画像化することにより、本発明者らは、λR/E2A共局在化が細胞周期のG1期に最も顕著であることを示す。本発明者らは、E2AアンタゴニストId1の発現により、G1期のλR/E2A共局在化が妨害され多様化が減退するが、λR(以下、IgλR遺伝子をIgλまたはλと呼ぶことがある)の転写は減退しないことをさらに示す。したがって、E2Aは、シスで作用してIg遺伝子多様化を促進し、G1期は、E2A作用のための決定的に重要な期間である。
【0077】
Ig座位で起こるゲノム配列および構造の調節された変化は、これらの遺伝子へのDNA損傷のターゲティングおよび正確な修復の回避の両方を反映している。体細胞超変異、クラススイッチ組換え、および遺伝子変はすべて、B細胞特異的酵素である活性化誘導型デアミナーゼ(AID)によって開始される(5〜8)。AIDはシトシンを脱アミノしてウラシルにし、単鎖DNAを好むことが明白である(9〜11)。転写は多様化するための必要条件であり、これは、AIDが単鎖基質を好むことを反映している可能性がある。DNA中のウラシルはよく起こる損傷であり、高度に保存された効率的な経路によって正確に修復することができる(12)。しかしながら、Ig座位は正確な修復を回避し、誤りがちな経路による修復を受けることができる(13)。
【0078】
E2Aは、bHLHタンパク質のEファミリーのメンバーであり、リンパ球発達の多くの局面の決定的に重要な調節因子である(14〜17)。Eタンパク質は二量体化してEボックスモチーフCANNTGに結合し、それらの機能は、Eタンパク質とヘテロ二量体化してDNA結合を妨害するIdタンパク質によって弱められる。E2Aは、活性化されたマウスB細胞において誘導され、クラススイッチ組換え(18)ならびにAIDをコードする遺伝子の発現(19)を調節する。ニワトリB細胞において、E2A遺伝子が不活性化されると、Igλ遺伝子の多様化が減退するが転写は減退しない(20、21)。一方、逆に、E47(E2Aにコードされる機能的に等価なアイソフォーム2つの内の1つ)が異所発現されると、Igλ遺伝子の多様化が促進されるが、Igλ転写レベルは影響を受けない(22)。
【0079】
E2Aがシスの部位に結合することによってIg遺伝子多様化を調節する可能性は、マウスにおいて、多量体化したEボックスは超変異を刺激するがIg導入遺伝子の転写は刺激しないという証拠に基づいて最初に示唆された(23)。この可能性は、ニワトリB細胞において、多量体化したEボックスはIg遺伝子多様化を促進できるが転写は促進できないことの証明によってさらに裏付けられた(24)。しかしながら、E2AがIg遺伝子に直接作用して多様化を促進するかという疑問を完全に解消することは、いくつかの理由によって困難であった。Eボックスは、特定の状況においてのみ、E2A依存的調節のための部位として機能し、したがって、Eボックスの存在は、ある部位でのE2A機能を保証するものではない。Eボックスモチーフのあいまいな一致(loose consensus)および頻繁な出現により、各個別部位の変異解析は妨げられる。いくつかの部位で、E2Aはタンパク質-DNA相互作用ではなくタンパク質-タンパク質相互作用によって動員され、したがって、Eボックスは、常にE2A依存的調節の必要条件であるとは限らない(25)。
【0080】
本発明者らは以下に、再配列されたIgλ対立遺伝子が、重合ラクトースオペレーターでタグ化された、構成的に多様化するニワトリB細胞株DT40の派生体(DT40ポリLacO-λR)を利用する実験において、E2AがIg遺伝子でシスに作用して多様化を促進することを証明した。クロマチン免疫沈降法(ChIP)により、本発明者らは、E2Aが、親株DT40において、再配列されたIgλ対立遺伝子とは結合するが再配列されていないIgλ対立遺伝子とは結合しないことを示す。本実施例では、DT40ポリLacO-λR細胞において、E47をλRに効果的に係留するE47-LacI融合タンパク質が発現されると多様化が加速すること、およびE47-LacI発現の刺激効果が、IPTGと共に培養された細胞では明らかではなく、したがって、シスに結合することが多様化を促進するのに必要であることを実証する。DT40ポリLacO-λR GFP-LacI細胞中の再配列されたλR遺伝子を直接画像化することにより、本発明者らは、λR/E2A共局在化がG1期に顕著であること、およびE2AアンタゴニストであるId1の発現により、多様化が減退し、具体的にはG1期のλR/E2A共局在化が減少するが、λ転写レベルも活性な転写場所へのλRの局在化も影響を受けないことを示す。これらの結果から、E2AがG1期にシスで作用してIg遺伝子多様化を促進することが示される。
【0081】
材料および方法
細胞培養、トランスフェクション、sIgM喪失アッセイ法、および細胞周期の解析
以前に説明されたようにして、ニワトリ嚢リンパ腫株DT40およびその派生体DT40ポリLacO-λRを維持し、トランスフェクトした(26、 27)。S003 E47プラスミド(28)(Cornelis Murre(University of California, San Diego, California)によって提供された)に由来するE47 cDNAを、p3’SS-GFP-LacIプラスミド(Andrew Belmont(University of Illinois, Urbana, Illinois)によって提供された)中にサブクローニングすることによって、E47-LacI発現構築物を作製した。Id1発現構築物(29)は、Barbara Christy (University of Texas, San Antonio, Texas)によって提供された。sIgM喪失アッセイ法を以前に説明されているようにして実施し(26、30)、Rソフトウェアパッケージ(http://www.r-project.org)を用い、マンホイットニーU検定によって結果を比較した。DNA含有量に基づいて細胞周期プロファイルを得るために、指数関数的に増殖中の細胞1×106個を0.1% Triton X-100中に懸濁し、200μg/mlのRNase Aおよび50μg/mlのヨウ化プロピジウムで処理し、以前に説明されたようにして解析した(26)。
【0082】
ChIP解析
以前に説明されているようにして、クロマチンを調製し、免疫沈降した(27、31、32)。その際、抗E2A抗体(ab11176; Abcam)または対照IgGを使用した。VλRおよびVλUに対する以前に説明されたプライマー(27)ならびに卵白アルブミン(Ova)に対するプライマー
を使用し、FastStart Taq DNAポリメラーゼ(Roche)を用いて半定量的PCRを実施した。ImageQuantソフトウェア(Amersham)を用いてPCR産物を定量した。対照IgGの場合の比率に対して標準化した、関心対象のアンプリコンとOvaアンプリコンの比率として富化を算出した。例えば、富化VλR=(抗E2A[VλR/Ova])/(IgG[VλR/Ova])。
【0083】
RT-PCRおよびウェスタンブロット法
RT-PCRアッセイ法のために、説明されているようにしてプライマーを用いてAIDおよびb-アクチンを増幅し(7)、プライマー
を用いてIgλ転写物を増幅した。ウェスタンブロット法のために、DT40ポリLacO-λR RFP-LacIおよびDT40ポリLacO-λR RFP-LacI Id1に由来する全細胞溶解物(50μg)を分離し、抗Id1抗体(JC-FL; Santa Cruz)を用い、FluorChem HD2 (Alpha Innotech)によってId1タンパク質を検出した。
【0084】
蛍光顕微鏡検査および画像解析
ポリLacOを画像化するために、GFP-LacI発現構築物p3’SS-GFP-LacI(Andrew Belmont(University of Illinois, Urbana, Illinois)から入手)(SV40核移行シグナルを含むように操作され、四量体形成に必要な配列を欠いているLacIをコードする)(33)、またはその派生体RFP-LacI(GFPがRFP(DsRed単量体;Clontech)で置き換えられている)をDT40ポリLacO-IR細胞にトランスフェクトした。免疫染色のために、Cytospin3(800rpm、4分;Shandon)を用いてスライドガラス上に細胞(約3×105個)を載せ、2%パラホルムアルデヒドで20分間固定し、以前に説明されたようにして染色した(26)。使用した一次抗体は、抗E2A(ab11176、1:200;Abcam)、Ser5でリン酸化された抗Pol II C末端ドメイン(ab5131、1:500;Abcam)であった。Alexa Fluor 488または594を結合させた抗IgG(Molecular Probes)を二次抗体として使用した。DeltaVision顕微鏡検査システム(Applied Precision)を用いて蛍光画像を獲得し、softWoRxソフトウェア(Applied Precision)およびImarisソフトウェア(Bitplane)を用いて処理および解析を行った。蛍光シグナルは、時折、完全にではなく部分的に重なることがあり、これは、ポリLacO-タグとVλ領域の間にかなりの距離(約17kb)があることを反映している可能性がある;どちらの配置も共局在化として記録した。共局在化の割合をピアソンのカイ2乗検定を用いて解析した。少なくとも2回の独立した直径測定値を平均し、2で割って、核半径を算出した。平均核半径の細胞周期依存性を各細胞株に関して独立に決定し、比較的不変であることが判明した。核半径と細胞周期の相関関係を示すために用いた標準値は、次のとおりであった:G1、r<4μm;G2、r≧5.2μm。
【0085】
結果
E2Aは、再配列されたIgλ遺伝子と結合するが、再配列されていないIgλ遺伝子とは結合しない
Ig遺伝子多様化におけるE2Aの重要性を示すかなりの証拠があるものの、この因子がIg遺伝子と直接結合することは示されていなかった。E2AとIgλの結合を試験するために、本発明者らは抗E2A抗体を用いて、ニワトリDT40 B細胞株からクロマチンを免疫沈降させた。この株は嚢リンパ腫に由来し、遺伝子変換によってIg重鎖遺伝子および軽鎖遺伝子の両方の構成的多様化を実施する。11kbのニワトリλ軽鎖座位の探索により、E2AのコンセンサスCANNTGに対する50個を超えるマッチ、VλとψVλ1(上流の偽遺伝子の内で最も近位のもの)の間の領域に17個、J-Cイントロンのマトリックス結合領域(MAR)に2個、および3’エンハンサーに6個が確認された。DT40 B細胞において、機能的対立遺伝子は、VセグメントとJセグメントを連結する1.8kb領域を欠失するVJ組換えをB細胞発達初期に経験するが、不活性なλ対立遺伝子は再配列されていないため(図4A)、2つの対立遺伝子をPCRによって容易に区別することができる。クロマチン免疫沈降(ChIP)後、再配列されたおよび再配列されていないλ対立遺伝子の回収物を、対照遺伝子アルブミンと比べて分析した。これにより、E2Aは、再配列されたVλR対立遺伝子において17.6倍に富化されているが、再配列されていないVλU対立遺伝子では富化されていないことが示された(図4B)。したがって、E2Aは、再配列されたVλR対立遺伝子と直接結合する。
【0086】
E2Aはシスに作用してIgλ多様化を調節する
E2Aがシスに結合して多様化を促進するに違いないかを問うために、本発明者らは、DT40の派生体であるDT40ポリLacO-λR(重合ラクトースオペレーターが相同遺伝子ターゲティングによってψVλアレイ中に挿入されており(図5A)、ラクトースリプレッサー(LacI)との融合物として発現される因子は、再配列されたλR対立遺伝子に係留され、IPTGとの培養によって遊離されることが可能である)を利用した(27)。細胞周期分布およびクローンのIg遺伝子変換速度は、DT40ポリLacO-λRおよび野生型DT40において同程度であった(図5B、5C)。LacIに融合されたE2AアイソフォームE47を発現するプラスミド(E47-LacI)またはLacIに融合された緑色蛍光タンパク質を発現する対照プラスミド(GFP-LacI)をDT40ポリLacO-λR細胞に安定にトランスフェクトした。細胞周期分布は、GFP-LacIトランスフェクタントおよびE47-LacIトランスフェクタントにおいて同程度であったが、後者の株の培養物は、いくつかのサブG1(アポトーシス性)細胞を含んだ(図5D)。E47-LacIトランスフェクタントにおいてIgλ転写物のレベルは変わっていなかったことから(図5E)、ニワトリB細胞においてE2AはIg遺伝子発現を調節しないことを示す、公開されている結果が確認された(21、22)。AID転写物のレベルは、E47-LacIトランスフェクタントの方が約3倍高かった(図5E)。E47の異所発現に応答したAID発現の同様の増大が、他の研究者によって観察されている(22)。
【0087】
E2Aが、Igλへの結合を介して多様化を直接的に調節するかを問うために、本発明者らは、IPTGの存在下および不在下で独立したE47-LacI(n=19)トランスフェクタントまたはGFP-LacIトランスフェクタント(n=13)を培養し、sIgM喪失変動アッセイ法を用いて、クローンの多様化速度を決定した(26、27、30)。このアッセイ法では、遺伝子変換、点変異、欠失、または挿入によってそれらが起こるかどうかに関わらず、不活性化変異を記録し、それによって、変異誘発の結果とは無関係な起因事象を定量する。この解析により、E47-LacIトランスフェクタントのクローン多様化速度が、GFP-LacI対照と比べて4.5倍速いことが示された(P=0.019、マンホイットニーU検定、図5F)。さらに、E47-LacIをポリLacOから遊離させるIPTGと共に培養すると、DT40ポリLacO-λR E47-LacI細胞の多様化速度はバックグラウンドレベルにほぼ戻ったが、GFP-LacI対照には影響を及ぼさなかった(図5F)。したがって、E47-LacIは、シスに作用することによって多様化を促進する。
【0088】
E2Aは細胞周期のG1期にIgλRに局在化する
ヒトB細胞において、細胞周期のG1期の受容体架橋は、体細胞超変異を開始して、同定可能な変異を90分以内に生じさせることができる(34)。したがって、E2AがIgλで作用する細胞周期の段階を決定することが関心対象であった。再配列され、多様化しているλR遺伝子は、GFP-LacIを発現するDT40ポリLacO-λR細胞において明るい点として容易に画像化することができる(27)。しかしながら、細胞をHoechst 33342で染色し、DNA含有量に基づいて選別してG1段階、S段階、またはG2/Mの細胞を富化すると、タグ化された遺伝子に由来するはっきりした蛍光シグナルを提示している細胞の割合は、未選別の細胞で通常観察される90〜95%から、約45%に減少した。したがって、シグナルのこのような減少は結果を偏らせ得るため、本発明者らは、異なるアプローチによって細胞周期段階を決定した。Hoechst 33342で染色し選別した細胞の解析により、G1期の核のサイズはG2/M期の細胞よりもかなり小さいことが示された(例えば、図6A)。したがって、本発明者らは、Hoechst 33342で染色しDNA含有量に基づいて選別した、指数関数的に増殖中のDT40ポリLacO-λR集団に由来するG1細胞(n=55)およびG2細胞(n=55)の核半径を測定することにより、核半径(r)を用いて細胞周期の段階を明らかにできるかを問うた。G1細胞の平均核半径は3.8±0.3μmであり、G2細胞の平均核半径は5.4±0.5μmであった(図6B)。核半径によって決定したG1細胞:S細胞:G2細胞の比率(3:6:1)および染色によって決定した比率(2.7:5.5:1.7)を比較して、このアプローチをさらに確認した。したがって、G1細胞はr<4μm、G2細胞はr>5.2μmであることが実験で確認された。
【0089】
λR/E2Aの共局在化は、抗E2A抗体で染色したDT40ポリLacO-λ GFP-LacI細胞のデコンボリューション顕微鏡解析によって容易に同定された(例えば、図7A)。λR/E2A共局在化は、非同期細胞(n=227)の26%で明らかであった。共局在化の細胞周期分布の解析により、λR/E2A共局在化の45%がG1期の細胞で起こり、38%がS期の細胞で起こり、17%がG2期の細胞で起こることが示された(図7B)。したがって、G1期細胞の割合(25%)と比べて、G1期におけるλR/E2A共局在化(45%)は明らかに過剰であった(P<0.0001、カイ2乗検定)。
【0090】
本発明者らはまた、λR転写の細胞周期依存性も決定して、RNAポリメラーゼIIのC末端ドメインのリン酸化Ser5(P*-Pol II)(伸長しているPol II分子に特徴的な改変)に対する抗体で染色することによって、活性な転写場所を同定した(35)。DT40ポリLacO-λR RFP-LacI細胞の非同期細胞集団において、核の全体にわたって多数の活性な転写場所を同定することができ、λR/P*-Pol II共局在化が、19%の細胞で容易に観察された(n=392;例えば、図7C)。λR/P*-Pol II共局在化の細胞周期分布の解析により、共局在化の21%がG1期の細胞で起こり、58%がS期の細胞で起こり、21%がG2期の細胞で起こることが示された(図7D)。これは、細胞周期分布に類似している。したがって、λRは細胞周期の全体を通して転写されるが、λR/E2A共局在化はG1期で顕著である。
【0091】
Id1発現はIg遺伝子多様化およびG1期のλR/E2A共局在化を阻害する
G1期のλR/E2A共局在化が多様化にとって決定的に重要であるかを問うために、本発明者らは、これらの共局在化に対するId発現の影響を測定した。IdはE2Aに拮抗し、DT40 B細胞におけるId1またはId3の発現は、Ig遺伝子多様化を減弱させることが以前に示されている(22)。本発明者らは、安定なDT40ポリLacO-λR RFP-LacI Id1トランスフェクタントを作製し、ウェスタンブロット法によってId1発現を確認し(図8A)、Id1発現によって細胞周期プロファイルが変化しないことを示した(図8B)。本発明者らは、Id1発現により、クローンのIg遺伝子多様化速度が低下する(P<0.001、マンホイットニーU検定;図8C)が、Igλ転写物のレベルにもAID転写物のレベルにも影響を及ぼさないことを確認した(図8D)。次いで、本発明者らは、抗E2A抗体で染色することにより、Id1発現派生体および親株におけるλR/E2A共局在化を比較した。λR/E2A共局在化は、DT40ポリLacO-λR RFP-LacI細胞での26%に対し、非同期DT40ポリLacO-λR RFP-LacI Id1細胞(n=90)では13%で明らかであった(P=0.0030、カイ2乗検定)。
【0092】
Id1発現が細胞周期の特定の段階における共局在化に影響を及ぼすかどうかを判定するために、Id1トランスフェクタントにおけるλR/E2A共局在化を、核半径および細胞周期に関して定量した。これにより、DT40ポリLacO-λR RFP-LacI Id1細胞において、共局在化の10%がG1期の細胞で起こり、58%がS期の細胞で起こり、32%がG2期の細胞で起こることが示された(図8E)。したがって、Id1発現により、G1期のλR/E2A共局在化の割合が有意に減少し、親株での45%からId1トランスフェクタントでは10%となった(P<0.0001、カイ2乗検定;図8E)。Id1トランスフェクタントでのλR/E2A共局在化の90%は細胞周期のG1期より後に起こった。Id1トランスフェクタントで明らかな多様化の減弱と総合すると、これにより、G1期が、E2Aが多様化を促進する決定的に重要な期間であることが示される。
【0093】
λR/P*-Pol II共局在化は、活性な転写場所に対する抗体で染色したDT40ポリLacO-λR RFP-LacI Id1細胞(n =290)の18%で同定され、親株と類似していた(19%、P=0.80、カイ2乗検定;図7D)。また、λR/P*-Pol II共局在化の細胞周期プロファイルも、Id1トランスフェクタントと親株とで類似していた(図8F)。λR/P*-Pol II共局在化に対するId1発現の影響が無いことは、DT40ポリLacO-λR RFP-LacI Id1トランスフェクタントのIgλ転写物レベルが低下しないことと一致している(図8D)。したがって、Id1発現は、λRとE2Aの共局在化の細胞周期分布に影響を及ぼすが、P*-Pol IIとの共局在化の細胞周期分布には影響を及ぼさない。
【0094】
考察
これらの結果から、E2Aはシスに作用してIg遺伝子多様化を促進するに違いないこと、およびG1期が、このプロセスでE2Aが機能する決定的に重要な期間であることが示される。これらの実験では、ポリLacOでタグ化し、GFP-LacIまたはRFP-LacIに結合させることによって画像化したλ遺伝子を検査した。これは、遺伝子多様化を研究するための強力なアプローチを提供する。タグ化された座位は、固定した細胞の90%超で視認できるため、多様化に関与する因子との共局在化の解析が可能になる。潜在的な調節因子を係留し、IPTGとの培養によって遊離させることができるため、他の標的から独立してIgλ座位における因子の効果を研究することが可能になる。これは、大きくかつ複雑な調節階層の上位で機能するE2Aのような因子にとって特に有用である(36)。
【0095】
これらの結果から、Ig座位との物理的結合により、E2AがIg遺伝子多様化を直接的に調節することが確かめられる。ChIPにより、E2AがDT40 B細胞中の再配列されたλR対立遺伝子と結合することの明白な証拠が提供された。E2Aがシスに機能するに違いないということは、DT40ポリLacO-λR細胞においてE2A(E47-Lac融合物)をIgλ対立遺伝子に係留した結果起こる多様化の加速が、LacOからLacIを遊離させるIPTGと共に培養した細胞では明らかではないことを示すことによって確かめられた。
【0096】
E2Aは転写調節因子として最もよく知られているが、Igλ転写物のレベルはE47の異所発現によって変わらなかったため、多様化におけるE2Aの機能は、Igλでの転写活性化を反映したものではない。このことは、ニワトリおよびマウスB細胞において多様化に対するE2Aの影響を検査した他の研究者による結果を裏付ける(20、22、23)。さらに、G1期で最も顕著であるλRとE2Aの共局在化は、 細胞周期分布に類似している活性な転写場所へのλRの細胞周期分布とは違っていることから、λRが細胞周期の全体を通して転写されることが示唆される。多様化および転写におけるE2A機能の独立性は、再配列されたλRとE2Aの共局在化および活性な転写場所に対するId1発現の影響を対比することによってさらに裏付けられる。Id1発現により多様化は減退し、また、λRとE2Aの共局在化も減少した。一方、Id1発現は、活性な転写場所へのλRの局在化の数にも細胞周期分布にも影響を及ぼさなかった。λRを転写場所に動員するのにE2Aは必要とされないということは、Igλ転写に対するId1発現の影響が無いことと一致している。
【0097】
E2Aは、AID発現を調節し、これはIg遺伝子多様化を間接的に刺激する。AID遺伝子は、マウスB細胞においてE2Aによる転写調節の標的であることが示されていた(19)。本発明者らおよび他の研究者ら(22)は、E2Aの異所発現により、DT40ニワトリB細胞におけるAID転写物レベルが上昇することを示した。これは、IPTGと共に培養したDT40ポリLacO-λR E47-LacI細胞において明らかな中等度の多様化加速に寄与するか、または原因である可能性がある。E2Aの除去によってニワトリB細胞中のAID転写物レベルは低下しないことが報告されているものの(21)、E2Aと機能が重複した(redundant)因子が、E2Aの不在下で最低レベルのAID発現を保証している可能性がある。
【0098】
これらの結果から、G1期が、IgλでE2Aが機能する決定的に重要な期間であると特定される。本発明者らは、細胞周期の他の段階と比べてG1期のλR/E2A共局在化が過剰であることを観察し、Id1発現によってG1期の共局在化が特異的に減少し、多様化速度が低下することを示した。Idタンパク質はEタンパク質とヘテロ二量体化して、DNA結合を阻害する(14)。G1の間の共局在化がId1発現によって特異的に影響されることは、細胞周期の間のE2AとIgλの結合の様式が異なることを示している可能性がある。S期およびG2期では、E2Aは、DNAへの直接結合によってではなく、他のタンパク質との相互作用を介して動員され得る。
【0099】
その他の種類の証拠により、多様化がG1期に開始されるという考えが裏付けされる。ヒトBL2細胞株での体細胞超変異は、G1期のみに行われるインビトロ刺激によって誘導することができ、点変異は、刺激から90分以内、細胞がまだG1期にある間に最初に明らかになる(34)。クラススイッチ組換えのために活性化されたマウスB細胞において、スイッチ組換え経路の関与物であるNBS1またはγ-H2AXとのIgH共局在化は、G1期で顕著であり(37)、S領域でのDNA切断はG1期に検出され得る(38)。DNA切断はまた、超変異しているヒトB細胞株において細胞周期の後期にも確認されるが(39)、これらはAID依存性であることが判明した(40)。
【0100】
E2Aは、G1期に機能して、細胞周期の後期、またはさらに、後続の細胞周期の間に起こる事象のために座位を準備することができる。最近、E2AはヒストンH4アセチル化の維持に関係があるとされており(21)、娘細胞でのAID攻撃または効果的な多様化に都合の良い局所的なクロマチン環境を確立するようにE2Aが機能することが可能である。
【0101】
参考文献
【0102】
実施例4:クロマチン構造は遺伝子変換を調節する
本実施例は、ニワトリB細胞株DT40をモデルとして用いて、クロマチン構造が修復用のドナーとしての相同配列の使用に寄与する方法を示す。DT40において、免疫グロブリン遺伝子は、偽遺伝子ドナーを鋳型とする遺伝子変換による、調節された配列多様化を経験する。本発明者らは、Vλ偽遺伝子アレイが活性クロマチンに関連したヒストン改変を特徴とすることを見出した。本発明者らは、Lacリプレッサーとの融合物として発現されるヘテロクロマチンタンパク質HP1が、偽Vλドナーアレイ内部に組み込まれた重合ラクトースオペレーターに係留される調節可能な実験系を用いて、遺伝子変換のためにクロマチン構造が重要であることを直接的に実証した。HP1が係留されると、偽Vλドナーアレイ内部でのヒストンアセチル化が減少し、Vλ多様化の結果が変わり、その結果、鋳型式変異よりも非鋳型式変異が優勢になった。したがって、クロマチン構造は、相同性の指示に従った修復を調節する。これらの結果から、ヒストン改変が、反復配列間の組換えを妨害することにより、ゲノム安定性の維持に寄与し得ることが示唆される。
【0103】
本実施例では次の略語を使用する:AcH3、アセチル化ヒストンH3;AcH4、アセチル化ヒストンH4;AID、活性化誘導型デアミナーゼ;DSB、二本鎖切断;GFP、緑色蛍光タンパク質;Ig、免疫グロブリン;MRN、MRE11/RAD50/NBS1;NHEJ、非相同末端結合;V、可変性;UNG、ウラシルDNAグリコシラーゼ;AP、脱塩基;ポリLacO、重合ラクトースオペレーター;ChIP、クロマチン免疫沈降法;Ova、卵白アルブミン;LOH、ヘテロ接合性の喪失。
【0104】
材料および方法
クロマチン免疫沈降法(ChIP)
以前に説明されたようにしてChIPを実施した[48、89]。すべての実験において、安定にトランスフェクトされた少なくとも2つの独立な株に由来する少なくとも2つのクロマチン調製物を解析した。図面は1つの代表的な実験を示し、4回の別々の増幅の解析結果を用いて標準偏差を算出した。鋳型DNAの段階希釈物を4回別々に増幅して、測定される産生物強度が直線範囲内であることを確かめた。実験アンプリコンの富化を、二重PCRによって同じチューブ中で増幅した卵白アルブミン(Ova)遺伝子に由来する内部対照アンプリコンの富化に対して標準化し、特異的抗体を用いたChIPの際の富化は、対照の全インプットDNAを用いてChIPを実施した並行実験に対して標準化した。Ova内部対照アンプリコンを含めることによって、本発明者らは、IP効率、バックグラウンドから残った影響、およびゲル添加の差異に関して標準化することができた。富化=[(ψVλ/Ova)Ab]/[(ψVλ/Ova)インプット]。追加の対照として、対照の全インプット中の実験アンプリコンと対照アンプリコンの比率を、多特異性IgGを用いた対照ChIPと比較した。いずれの場合も、インプット対照およびIgG対照の富化は本質的に等しかった。代表的な実験のデータを提示する。標準偏差は、鋳型DNAの段階希釈物を4回別々に増幅した結果から算出した。
【0105】
使用した抗体は、Upstate (Lake Placid, NY)社製の抗AcH3(06-599)、抗AcH4(06-866)、およびジメチル化H3(K4)(07-030)であった。ChIP用のPCRプライマーは次のものであった:
【0106】
構築物、トランスフェクション、および細胞培養
pAIDPuro(H. Arakawa(Munich, Germany)から入手)中のAIDをL. Wallrath (University of Iowa, Iowa City)によって提供された構築物に由来するLacI-HP1aで置換して、ニワトリβ-アクチンプロモーターの下流にLacI-HP1を配置することによって、LacI-HP1発現プラスミドを作製した。GFP-LacI発現プラスミド(p3’ss-EGFP-LacI)は、A. Belmont(University of Illinois, Urbana)によって提供された。細胞培養およびトランスフェクションを、以前に説明されたようにして実施した[47]。転写されたVλRの17kb上流のψVλ17〜ψVλ20の間の領域を標的とするように設計されたアームが隣接する約3.8 kbの重合ラクトースオペレーター(ポリLacO)を有する構築物を用いて、相同遺伝子ターゲティングによってDT40ポリLacO-λRを作製した。手短に言えば、相同な組み込み体をPCRによって同定し、Cre発現によって選択マーカーを欠失させた。DT40嚢リンパ腫は、1つのIgλ対立遺伝子のみが再配列されており、2つの親染色体(chromome)が、ψVλ17付近の多型性に基づいて区別されるB細胞に由来する。これにより、本発明者らは、再配列された対立遺伝子または再配列されていない対立遺伝子が標的とされたかどうかをPCRによって判定することができた。対照実験を行って、DT40ポリLacO-λR細胞、DT40ポリLacO-λR GFP-LacI細胞、およびDT40ポリLacO-λR LacI-HP1細胞の細胞周期分布が類似していること、ならびに最高500μMのIPTGと共に7日間細胞を培養しても、DT40ポリLacO-λRGFP-LacI対照細胞の増殖速度にもψVλ17Rにおけるクロマチン改変にも影響は無いことを確かめた。Vλ配列を解析するためのオリゴヌクレオチドは以前に説明されている[47]。
【0107】
蛍光画像法
蛍光画像法のために、細胞(2×105個)をサイトスピンでスライドガラス上に載せ、2%パラホルムアルデヒドで20分間固定し、0.1%NP-40で15分間透過処理し、以前に説明されたようにして染色した[90]。一次染色には抗LacIモノクローナル抗体(1:500希釈;Upstate)を用いた。二次抗体は、ロバ抗マウスIgG Alexa Fluor 594 (1:2000; Molecular Probes, Eugene, OR)であった。核を可視化するために、細胞をDAPI(Sigma, Saint Louis, MO)で染色した。DeltaVision顕微鏡検査システム(Applied Precision)を用いて蛍光画像を獲得し、softWoRxソフトウェア(Applied Precision)を用いて処理した。
【0108】
RT-PCR
TRIzol試薬(Invitrogen)を用いてRNAを細胞から回収し、PreAnalytiXカラム(Qiagen)を用いて精製した。鋳型の希釈(1:1300)後にVλ転写物を増幅し、未希釈の試料からβ-アクチンを増幅した。Vλを増幅するためのプライマーは、
であった。β-アクチンを増幅するためのプライマーは、以前に説明されている[36]。
【0109】
sIgM喪失変異体の定量および配列解析
変異したV領域におけるフレームシフトまたはナンセンス変異の結果として生じ、蓄積したsIgM喪失変異体を測定する、sIgM喪失変異体のアッセイ法を用いて、IgV領域の多様化を定量した[47、50]。手短に言えば、フローサイトメトリーとそれに続く限界希釈クローニングによってsIgM+細胞を単離し、4週間増殖させた。sIgM-細胞の割合を定量するために、約1×106個の細胞を抗ニワトリIgM-RPE(Southern Biotechnology Associates, Birmingham, AL)で染色し、CellQuestソフトウェア(BD Biosciences)を用いたFACScanによって解析した。
【0110】
単一細胞PCRおよび配列解析を、以前に説明されたようにして実施した[47]。手短に言えば、sIgM-細胞を選別し、個々のウェルに等分し、Vλ領域を増幅および配列決定し、それらの配列をψVλドナーと比較して、変異が鋳型式であるか、または非鋳型式であるかを判定した。鋳型式変異の判定基準は、ドナーとレシピエントにおいて9個の連続した塩基が完全に一致しなければならないというものであった。配列は、独立にトランスフェクトされた2つの株に由来した。特有の配列のみを、変異の分類のために含めた。
【0111】
結果
VλおよびψVλドナー鋳型における許容性クロマチン構造
DT40 B細胞において、一方のIgλ対立遺伝子の機能的なVλ遺伝子は再配列され発現され、他方は再配列されず発現されない。本発明者らは、再配列された対立遺伝子(VλR)および再配列されていない対立遺伝子(Vλu)ならびにψVλアレイにおけるクロマチン構造をクロマチン免疫沈降法(ChIP)によって特徴付けた。ChIPは、ヒストンH3およびH4のN末端におけるリジンアセチル化に特異的な抗体を用いて実施した。回収されたDNAを二重PCR反応で増幅した。回収量は、B細胞では発現されない卵白アルブミン(Ova)遺伝子に由来するアンプリコンに対して標準化し、富化は、対照の全DNAインプットに対して標準化した(詳細については、材料および方法を参照されたい)。VλRおよびVλuはゲノム構造が異なるため、特異的プライマーを用いたPCRによってそれらを区別することができる。ChIPにより、再配列されたVλR遺伝子においてアセチル化ヒストンH3およびH4(AcH3およびAcH4)がかなり富化していることが実証された。典型的な実験において、AcH3のVλRにおける富化率は80倍を超え、AcH4の富化率は30倍を超えた(図9B)。一方、再配列されていないVλU対立遺伝子におけるAcH3およびAcH4のレベルは、VλRでのレベルよりはるかに低く(それぞれ、16分の1および7分の1)、インプットDNAと比べて数倍にしか富化しなかった。
【0112】
7箇所の部位(ψVλ1とVλ遺伝子の間の領域、ψVλ1、ψVλ5、ψVλ13、ψVλ18、ψVλ24、ψVλ25を含む)および上流のフランキング領域を調べるプライマーを用いた増幅によって、ψVλアレイ内のクロマチン構造を分析した。(多型性が足りないために、DT40中の2つのIgλ対立遺伝子におけるψVλアレイはPCRによって容易に区別することができない)。印象的なことには、本発明者らは、ψVλアレイの全体にわたってAcH3およびAcH4がかなり富化していることを観察した(図9B)。VλRから遠位の部位が、近位部位よりも低レベルの富化を一貫して示すということはなく、富化は、転写されたVλR遺伝子からの距離に比例していなかった(図9B)。したがって、ψVλアレイ内のアセチル化ヒストンの富化は、転写されたVλR遺伝子から上流部位へのクロマチン改変の段階的な伝播を単に示すわけではない。この座位のクロマチン構造が非均一であることから、ψVλアレイのクロマチン構造を調節するシスエレメントの存在が示唆される。
【0113】
DT40ポリLacO-λRにおけるψVλアレイへのLacI融合タンパク質の可逆的係留
クロマチン構造の局所的改変は、適切な融合タンパク質としてDNA結合部位に調節因子を係留することによって実現することができる。この戦略は、例えば、ショウジョウバエにおいて、ラクトースリプレッサーとの融合物(LacI-HP1)として発現されるヘテロクロマチンタンパク質HP1aが、クロマチン構造の閉鎖およびLacOリピートに隣接したレポーター遺伝子の不活性化を促進する[61、62]ことを示すため、ならびに脊椎動物のG9aヒストンメチルトランスフェラーゼがV(D)Jミニ遺伝子レポーター内のGAL4結合部位に係留されると、その構築物の非相同的媒介による組換えが減退する[63]ことを示すために使用された。重合ラクトースオペレーター(ポリLacO)を、転写されたVλRの17kb上流のψVλ17〜ψVλ20の間に相同遺伝子ターゲティングによって挿入したDT40派生体である細胞株DT40ポリLacO-λR(図10A)。ポリLacO挿入物の長さは3.8kbであり、20量体のオペレーターのコピー約100個からなる[64]。この細胞株を用いて、内因性座位内の生理学的プロセスにおける相同組換えに対する係留された調節因子の影響を分析し、導入遺伝子レポーターの必要性を回避することが可能である。対照実験により、ポリLacOタグが、細胞増殖にも細胞周期にもIg遺伝子多様化にも影響を及ぼさないことが示された。
【0114】
ラクトースリプレッサーに融合された高感度緑色蛍光タンパク質(GFP-LacI)を安定に発現するDT40ポリLacO-λR GFP-LacIにおいて、タグ化されたλR対立遺伝子は、蛍光顕微鏡によって直接画像化することができ、各細胞中に明瞭な斑点として現れる(図10B、左)。LacIがLacOに結合するのを妨げる100μM IPTGと共に一晩培養した後は明るい斑点がはっきりとしないように、係留は可逆的である(図10B、右)。
【0115】
HP1が係留されると、ψVλにおける活性クロマチンに特徴的な改変が減少する
ψVλアレイにおけるクロマチン構造を操作するために、本発明者らは、LacIに融合されたキイロショウジョウバエHP1タンパク質(LacI-HP1)を発現する、DT40ポリLacO-λRの安定なトランスフェクタントを作製した。HP1は、ヘテロクロマチンの遺伝子サイレンシング、ヘテロクロマチンの伝播、およびヒストン脱アセチル化において機能する非ヒストンヘテロクロマチンタンパク質である[58〜60]。係留されたHP1は、隣接した遺伝子でのクロマチン構造の閉鎖を促進することが示されている[61、62、66〜68]。DT40ポリLacO-λR LacI-HP1トランスフェクタントを抗LacI抗体で染色することにより、LacI-HP1が、挟動原体ヘテロクロマチンに対応するDAPI密集領域と共局在して(図11A)、ヘテロクロマチンの機能的マーカーとして挙動することが示された[65]。
【0116】
LacI-HP1が係留されるとクロマチン構造が変化するかを問うために、本発明者らは、ψVλ17におけるクロマチン改変を分析した。DT40ポリLacOを構築する間に配列多型性が生じるため、これは、再配列された対立遺伝子および再配列されていない対立遺伝子を、特異的PCRプライマーを用いて区別できるψVλアレイ中の唯一の部位である。ChIP後、標的とする再配列された対立遺伝子(ψVλ17R)に特異的なPCRプライマーを用いてDNAを増幅した。ψVλ17Rの富化を、内部対照としての発現されない卵白アルブミン遺伝子(Ova)と比較し、全インプットDNA中のψVλ17R:Ovaの富化率に対して標準化した(材料および方法を参照されたい)。対照のDT40ポリLacO-λR GFP-LacIのψVλ17RにおけるAcH3およびAcH4の富化率はそれぞれ2.2倍および5.9倍であった(図11B、C)。これらの富化レベルは、DT40において実証されたものと類似している(図9B)。(親DT40株の調査でのψVλにおける改変の解析は、両方の対立遺伝子を必ず含み、再配列された対立遺伝子における活性化改変を過小評価する場合があることに留意されたい。一方、ψVλ17Rにおける改変の解析では、活性な対立遺伝子のみを調べる)。AcH3およびAcH4は、DT40ポリLacO-λR LacI-HP1トランスフェクタントのψVλ17Rでは富化されておらず(それぞれ0.6倍および1.0倍;図11B、C)、HP1を介したサイレンシングと一致していた。HP1は、ヒストンH3のリジン9を改変するヒストンメチルトランスフェラーゼを動員することによってサイレンシングを行うことができる[66〜68]が、この改変とは無関係にサイレンシングを促進する場合もある[61]。ジおよびトリメチル化H3(K9)のいずれかに対する抗体を用いたChIPでは、H3 K9-Me改変の明らかな富化が示されなかった(データ不掲載)。ヒストンH3のリジン4のメチル化(ジMeK4)は転写と関連しており、一般に、アセチル化と時間的に重複した分布を示す[69、70]。ψVλ17RのジMeK4(H3)の分析により、この改変が、DT40ポリLacO-λR GFP-LacI細胞では18.9倍に富化されているが、DT40ポリLacO-λR LacI-HP1細胞ではバックグラウンドレベルであることが実証された(図11B、C)。
【0117】
HP1はヘテロクロマチンの維持および伝播を促進する[66]。係留されたHP1によって促進されるクロマチン構造の変化が染色体全体に伝播しないことを確認するために、本発明者らは、第15染色体上のλ軽鎖座位近くの別の部位、すなわちDNA polεの触媒サブユニットをコードする遺伝子を検査した。DNA polεは、遍在的に発現され、真核生物の染色体複製に不可欠であり[71]、Igλから約2.1Mbに位置する遺伝子にコードされている。本発明者らは、対照のDT40ポリLacO-λR GFP-LacIと比べて、DT40ポリLacO-λR LacI-HP1トランスフェクタントのpolεプロモーター領域のAcH3富化に差がないことを見出した(polε/Ova富化はそれぞれ8.5倍および8.4倍であった;図3C)。同様に、対照のDT40ポリLacO-λR GFP-LacIと比べて、DT40ポリLacO-λR LacI-HP1トランスフェクタントのpolεプロモーター領域のAcH4にも差がなかった(polε/Ova富化はそれぞれ1.9倍および1.7倍であった;図11C)。したがって、ψVλにLacI-HP1が係留されると、クロマチン構造が局所的に改変されて、ψVλ17Rにおける開放型クロマチンに特徴的なAcH3、AcH4、およびジMeK4(H3)改変が減少し、クロマチンが許容性の低い状態を取るようになった。
【0118】
HP1の係留は、Vλ遺伝子発現に影響を及ぼさない
本発明者らは、DT40ポリLacO-λR LacI-HP1細胞および対照のDT40ポリLacO-λR GFP-LacIトランスフェクタントにおけるこれらの改変を比較することによって、HP1の係留が、発現されたVλRにおけるAcH3レベルおよびAcH4レベルにどのように影響を及ぼすかを問うた(図12A)。HP1が係留されると、AcH3レベルおよびAcH4レベルはそれぞれ対照レベルの約40%および20%に低下した。これが遺伝子発現に影響を及ぼすかを問うために、本発明者らは、表面IgM(sIgM)発現およびVλ転写物レベルの両方を分析した。マウス抗ニワトリIgMで細胞を染色することにより、IPTGの存在下または不在下のいずれかで培養したDT40ポリLacO-λR GFP-LacI株およびDT40ポリLacO-λR LacI-HP1株のsIgM発現は類似していることが示された(図12B)。DT40ポリLacO-λR GFP-LacI細胞およびDT40ポリLacO-λR LacI-HP1細胞から回収したRNAの定量的RT-PCRによって、Vλ転写物レベルを分析し、対照としてのβ-アクチンに対して標準化した(図12C)。2種の細胞株のVλ転写物レベルに有意な差は観察されず、転写はψVλアレイ内でのHP1係留の影響を受けないことが実証された。したがって、LacI-HP1の係留により、下流のIg遺伝子の発現は影響を受けなかったが、VλRにおけるAcH3レベルおよびAcH4レベルは低下した。したがって、Vλに特徴的な非常に高いAcH3レベルおよびAcH4レベル(図9B、図12A)は、高レベルの遺伝子発現を維持するために不可欠ではない。
【0119】
HP1が係留されると、局所的なクロマチン構造が変化する
LacI-HP1のクロマチンへの影響がどれくらい大規模であるかを評価するために、本発明者らは、Igλ軽鎖座位の全体にわたって、図9で検査した同じアンプリコン(フランク中の1つ、ψVλアレイ中の6つを含む)、ならびに発現されたVλにおけるAcH3レベルおよびAcH4レベルを検査した。図11Bのような免疫沈降およびインプット条件のψVλ17R:Ova比率を比較することによって、改変のレベルを決定した。調査した部位におけるAcH3改変は、対照の同じ部位におけるレベルの24%〜63%の範囲であった(図13A、黒い棒)。これらの部位すべてに渡るH3アセチル化の平均レベルは、対照のポリLacO-λR GFP-LacIの38%であった。250μM IPTGと共に3日間、DT40ポリLacO-λR LacI-HP1トランスフェクタントを培養すると、調査した8箇所の部位すべてでH3アセチル化が増加した(図13A、黒い棒と白い棒を比較されたい)。IPTG培養の影響はいくらか変動したが、大半の部位で、IPTG培養により、対照のDT40ポリLacO-λR GFP-LacI細胞でのレベルの少なくとも45%までAcH3のレベルが回復し、平均は80%を上回った。したがって、DT40ポリLacO-λR LacI-HP1細胞のψVλ17Rにおけるクロマチン改変は、LacI-HP1の係留に直接起因し、大部分は可逆的であった。
【0120】
同じ8箇所の部位のH4アセチル化を調査した(図13B、黒い棒)。AcH4改変は対照のレベルの18%〜42%の範囲であることが判明した。平均レベルは、対照の細胞株の29%であった。IPTGと共に3日間培養すると、調査した8箇所の部位すべてでH4アセチル化が増加し(図13B、黒い棒と白い棒を比較されたい)、対照のDT40ポリLacO-λR GFP-LacI細胞でのレベルの少なくとも57%までH4アセチル化のレベルが回復し、平均は80%を上回った。さらに、IPTGは、LacI-HP1の影響を少なくともある程度取り消すことができる。
【0121】
これらの結果から、ψVλアレイにおいて観察されるクロマチン改変が、HP1の係留に起因することが示される。さらに、これらの改変が可逆的であるということから、活性なメカニズムが、ψVλでのクロマチン改変因子の係留によって課されるヒストン改変を元に戻すことが示される。
【0122】
HP1が係留されると、鋳型式変異誘発が減退する
LacI-HP1の係留によってψVλにおけるクロマチン構造を操作できるため(図11〜13)、本発明者らは、クロマチン構造がIg遺伝子変換に影響するかどうか、およびその方法を直接問うことができた。本発明者らは、sIgM喪失変異体アッセイ法を用いて、係留されたLacI-HP1が、再配列されたVλR遺伝子のクローン配列多様化速度に影響を及ぼすか判定した。この変動アッセイ法では、構造的に完全なsIgMをもはや発現しない変異体細胞の割合を測定し、したがって、遺伝子変換または点変異誘発のいずれかに起因する変異事象を記録する[47、50]。sIgM+細胞を限界希釈法でクローニングすることによってDT40ポリLacO-λR GFP-LacIおよびDT40ポリLacO-λR LacI-HP1の独立したクローン派生体を樹立し、4週間培養し、次いで抗IgM抗体で染色した細胞をフローサイトメトリーに供して、各集団中のsIgM-細胞の割合を決定した。sIgM喪失率の中央値は、DT40ポリLacO-λR GFP-LacI細胞の場合は0.5%、およびDT40ポリLacO-λR LacI-HP1細胞の場合は2.8%であった(図14A)。これは、対照のGFP-LacIと比べて、LacI-HP1トランスフェクタントのクローン多様化速度が5.6倍に加速したことに対応する。
【0123】
ニワトリB細胞におけるIg遺伝子多様化は、主に遺伝子変換(鋳型式変異)によって起こるが、例えば、不可欠な因子の欠如によって遺伝子変換が減退している場合には、修復により、非鋳型式変異がかなりの割合で生じ得る[50〜55]。遺伝子変換のためのψVλ鋳型は、再配列された遺伝子と約80%同一であり、遺伝子変換によって修復されるかなりの割合のDNA損傷は、配列の改変をまったく経験しないため、これは、典型的にはクローン多様化速度の上昇を伴う。一方、変異誘発性ポリメラーゼによる修復の方が、DNA配列を改変する可能性が高い。HP1の係留によってどのように多様化が加速するかを決定するために、本発明者らは、DT40ポリLacO-λR GFP-LacIトランスフェクタントおよびDT40ポリLacO-λR LacI-HP1トランスフェクタントからsIgM-細胞だけ(single)を選別し、発現されたVλ領域を単一細胞PCRによって増幅し、これらの領域を配列決定した。配列変化が、2つまたはそれ以上の変異を含む区域内にあり、その区域がドナーψVλ配列の少なくとも9bpに厳密に一致している場合には、その配列変化を「鋳型式」と分類し、配列変化が単一の塩基変化のみからなるが、ドナーψVλ配列の少なくとも9bpに一致した場合には、その配列変化を「あいまい」と分類した。点変異、欠失、および挿入からなる非鋳型式事象もまた、記録した。対照のDT40ポリLacO-λR GFP-LacIトランスフェクタントでは、70個の独特な変異のうち54個の鋳型式事象および2個のあいまい事象が記録された。したがって、大半の事象(77%)は鋳型式であり、低比率の事象(20%)が点変異であった(図14B、左;図15)。印象的なことには、DT40ポリLacO-λR LacI-HP1細胞では、点変異が優勢であり(59%)、欠失(8%)および挿入(14%)を伴った。一方、36個の独特な変異のうちで、ただ1個の明らかな鋳型式事象および6個のあいまいな事象が記録された(図14B、右;図15)。したがって、変異の3%しか、明らかに鋳型式ではなく、さらに、潜在的な鋳型式変異であるあいまいクラスを含んでも、鋳型式の割合は変異の19%以下しか占めることができなかった。統計学的比較により、対照のDT40ポリLacO-λR GFP-LacI細胞およびDT40ポリLacO-λR LacI-HP1トランスフェクタントにおける明らかな鋳型式変異の割合の差(77%対3%)は、有意度が高い(P=7.1×10-7;フィッシャーの正確確率検定)ことが示された。対照細胞(3%)およびHP1トランスフェクタント(17%)におけるあいまいな潜在的鋳型式変異の割合の差もまた、有意である(P=0.05;フィッシャーの正確確率検定)。このことから、この範疇のいくつかの変異は、非許容性のドナークロマチンによって課される遺伝子変換区域の長さに対する制約の結果として起こり得ることが示唆される。したがって、HP1が係留されると、鋳型式変異が減退することにより、クローン変異誘発速度が加速した。
【0124】
考察
ニワトリIg座位における遺伝子変換は、再配列され転写されたV遺伝子を標的とする損傷を相同性の指示に従って修復するための鋳型として上流のψVドナーのアレイを使用する。本実施例では、活性なIg遺伝子変換を実施するニワトリB細胞において、ドナーψVλアレイ内部のクロマチンが、開放型のクロマチン構造と相関がある改変であるアセチル化H3およびアセチル化H4の富化を特徴とすることを示す。本発明者らは、ψVλドナーアレイにヘテロクロマチンタンパク質HP1が係留されると、クロマチン構造が局所的に変化して、開放型クロマチンに特徴的なAcH3、AcH4、およびジMeK4(H3)改変が減少することを示すことによって、Ig遺伝子変換のために許容性クロマチン構造が重要であることを直接的に実証した。これらの変化は、閉鎖型クロマチンに特徴的なMe-K9(H3)改変を伴わなかったが、遺伝子変換に対する許容性が低い状態をその領域に取らせた。HP1の係留は、IgVλ変異範囲の大幅な変化を伴い、その結果、鋳型式変異は少数派となり、点変異が優勢になった。重要なことには、変異誘発に対するこの影響は、クロマチン構造の変化と相関関係があるが、座位の発現の変化とは相関関係がなかった。したがって、クロマチン構造は、内因的に発生したDNA損傷で遺伝子変換が起こるかどうかを指令することができる。
【0125】
複雑なクロマチンランドスケープ内部での遺伝子変換のメカニズム
Vλでの遺伝子変換は、ψVλ領域を鋳型として用いて、切断部(break)の3'末端で新しいDNA合成を開始した結果として生じる。遺伝子変換は、ドナーDNAとレシピエントDNAの対合、ならびに相同性の指示に従った修復を行う因子のドナーへの接近を必要とする。野生型DT40のψVλアレイに特徴的な高レベルのH3アセチル化およびH4アセチル化は、トランス作用性因子へのψVλ遺伝子の接近容易性を高め、また、配列対合に都合が良い三次元構造物を作り出すと思われる、弛緩したクロマチン構造の証拠である。
【0126】
ψVλドナーアレイ内部に係留されたHP1は、Vλ遺伝子発現に影響を及ぼすことなく、再配列されたVλRにおける遺伝子変換を減退させた。HP1の係留によって引き起こされるクロマチン変化により、修復因子の接近の妨害およびドナー鋳型に対する侵入鎖によって遺伝子変換が減退し得る。また、係留されたHP1は、ドナー配列およびレシピエント配列を並べるのに必要なループ形成のようなDNA修復経路のメカニズムに影響を及ぼす、より大きな染色体構築にも寄与し得る。LacI-HP1トランスフェクタントにおいて蓄積した点変異は、邪魔された組み換え修復に典型的であり、組換えに不可欠なトランス作用性因子[49〜55]またはψVドナーアレイの一部もしくは全部[56]のいずれかを欠いた細胞に特徴的である。HP1は、2つの方法で、すなわち、ヒストンメチルトランスフェラーゼと組むことによって[66]、およびヒストンデアセチラーゼを動員することによって[60]、クロマチン構造およびヘテロクロマチン遺伝子サイレンシングを調節する。HP1が係留されると、ψVλ内部の非許容性クロマチン構造に特徴的な改変が引き起こされた。
【0127】
体細胞超変異およびクラススイッチ組換えを経験する活性に転写された哺乳動物Ig遺伝子におけるヒストンアセチル化が実証されているが、ヒストンアセチル化が多様化のターゲティングに寄与するかどうかはまだこれから解決しなければならない[72〜76]。ヒストンアセチル化と遺伝子変換の関連は、ヒストンデアセチラーゼ阻害剤であるトリコスタチンAでDT40細胞を処理すると、VλRにおける遺伝子変換の増大を伴うゲノムワイドなヒストン脱アセチル化が促進されることを示す実験によって示唆された[77]。しかしながら、トリコスタチンAの効果はゲノムワイドであり、特異的ではないことから、これらの結果の解釈は複雑である。DT40ポリLacO-λR細胞株では、クロマチン構造を局所的に操作して、この複雑さを回避することができる。さらに、本発明者らは、LacI-HP1融合タンパク質を係留する効果の大部分が、IPTGと培養すると取り消され、したがって、活性なメカニズムがψVλにおけるクロマチン改変を決定するに違いないことを実証することができた。
【0128】
相同組換えを調査する場合、DT40ポリLacO-λR B細胞株は、Ig遺伝子変換が内因性座位内の生理学的プロセスであり、導入遺伝子レポーターの必要性を回避するというさらなる利点を有する。
【0129】
クロマチン構造、ゲノム安定性、老化、および遺伝子療法
相同組換えの結果に対するクロマチン構造の重要性は、ゲノム安定性を通常は維持するメカニズムを理解するのに関わりがある。脊椎動物のゲノムには全体に分布する膨大な数の反復エレメントがあり、これらのエレメント間の組換えは、ゲノムの不安定性につながり得る[78]。ヒトゲノムには、約100万個のAluエレメントがあり、Aluエレメント間の組換えは、腫瘍形成および遺伝病を招く重複の原因となり得る[79、80]。抑圧的な改変を有するヒストンは、反復エレメントで豊富である[81]。これらの改変は、転写抑制を維持することは疑いない。本発明者らの結果から、これらが組換えの抑圧にも寄与し得ることが示唆される。
【0130】
ヘテロ接合性消失(LOH)は、対立遺伝子部位における相同体間の不均等な有糸分裂組換の結果として生じる。LOHのメカニズムは、腫瘍形成を招く腫瘍抑制遺伝子機能の消失をもたらすため、特に関心対象である[82]。最近の実験で、S.セレビシエ(cerevisiae)におけるLOH[83]およびショウジョウバエ生殖細胞におけるレポーター遺伝子[84]の年齢依存的増加、ならびにマウス膵臓細胞における相同組換えの増加[85]が実証された。年齢に関連したLOHを説明するために提唱されたメカニズムには、老化に伴う高比率のDNA損傷、細胞周期分布の変化、および相同性依存的修復経路の不活性化が含まれる。これらの結果から、クロマチン構造の弛緩が老化に付随して起こり、老化細胞における相同組換えのゲノムワイドな増加を促進し得るという別の可能性が示唆される。この可能性は、ショウジョウバエに関する最近の解析[86]、ならびにハッチンソン・ギルフォード早老症候群に関与するラミンAの変異が、H3メチル化のゲノムワイドな減少を招くという最近の証拠[87]によって裏付けられている。
【0131】
クロマチン構造が相同組換えを調節するという知見には、実用的な派生問題もある。不完全な標的遺伝子を置換するために完全なドナー遺伝子を提供することにより、相同性依存的修復に対する細胞の能力を利用して遺伝子療法を促進する戦略を開発することに現在かなりの労力が向けられている[88]。これらの結果から、ドナーの許容的構造が、治療的用途のためのドナー遺伝子を開発する際の重要な設計パラメーターになるであろうことが示唆される。
【0132】
参考文献
【0133】
実施例5:クロマチンの異なる改変によって加速する遺伝子変換
本実施例は、修復の効率がドナーのクロマチン構造によって決定されることを実証する。この解析は、ニワトリDT40 B細胞株の転写され、多様化しているVλ遺伝子のすぐ上流に挿入されたシス調節部位(重合ラクトースオペレーター、ポリLacO)を利用する。データから、活性化因子VP16またはヒストンシャペロンHIRAのいずれかを係留すると、局所的なクロマチン構造が変化し、遺伝子変換が約10倍に加速することが示される。これら2種の因子の機能上の結果は類似しているが、クロマチン構造に対するそれらの影響は異なる。VP16の係留により、アセチル化ヒストンH3およびH4の局所レベルが上昇するのに対し、HIRAの係留により、ヌクレオソームの密度が上昇する。したがって、異なるクロマチン改変によって、類似した機能上の結果が実現され得る。
【0134】
材料および方法
クロマチン免疫沈降法(ChIP)
以前に説明されたようにしてChIPを実施した(Cummings et al., 2007)。すべての実験において、安定にトランスフェクトされた少なくとも2つの独立な株に由来する少なくとも2つのクロマチン調製物を解析した。図面は1つの代表的な実験を示し、4回の別々の増幅の解析結果を用いて標準偏差を算出した。鋳型DNAの段階希釈物を4回別々に増幅して、測定される産生物強度が直線範囲内であることを確かめた。実験アンプリコンの富化を、二重PCRによって同じチューブ中で増幅した卵白アルブミン(Ova)遺伝子に由来する内部対照アンプリコンの富化に対して標準化し、特異的抗体を用いたChIPの際の富化は、対照の全インプットDNAを用いてChIPを実施した並行実験に対して標準化した。Ova内部対照アンプリコンを含めることによって、本発明者らは、IP効率、バックグラウンドから残った影響、およびゲル添加の差異に関して標準化することができた。富化=[(ψVλ/Ova)Ab]/[(ψVλ/Ova)インプット]。追加の対照として、対照の全インプット中の実験アンプリコンと対照アンプリコンの比率を、多特異性IgGを用いた対照ChIPと比較した。いずれの場合も、インプット対照およびIgG対照における富化は本質的に等しかった。代表的な実験のデータを提示する。標準偏差は、鋳型DNAの段階希釈物を4回別々に増幅した結果から算出した。
【0135】
使用した抗体は、Upstate (Lake Placid, NY)社製の抗H3 CT-pan、抗AcH3(06-599)、抗AcH4(06-866)、およびジメチル化H3(K4)(07-030)であった。
【0136】
ChIP用のPCRプライマーは次のものであった:
【0137】
MNアーゼ消化およびサザンブロット法
核を調製し、以前に説明されているようにして、0、3、7、15、30、および60ユニットのMNアーゼを用いて消化を行った(Prioleau et al., 1999)。MNアーゼ消化後、フェノール:クロロホルム:イソアミルアルコールでDNAを3回抽出し、エタノールを用いて沈殿させた。20マイクログラムのDNAをゲルに添加し、電気泳動によって分離させ、サザンハイブリダイゼーション用に移した。ポリLacOリピートを含む長さ約500bpの断片を鋳型として用いて、ランダムプライミングによってlacOプローブを標識した。
【0138】
蛍光画像法
蛍光画像法のために、細胞(2×105個)をサイトスピンでスライドガラス上に載せ、2%パラホルムアルデヒドで15分間固定した。核を可視化するために、細胞をDAPI(Sigma, Saint Louis, MO)で染色した。DeltaVision顕微鏡検査システム(Applied Precision)を用いて蛍光画像を獲得し、softWoRxソフトウェア(Applied Precision)を用いて処理した。
【0139】
RT-PCR
TRIzol試薬(Invitrogen)を用いてRNAを細胞から回収した。鋳型の希釈後にVλ転写物およびβ-アクチン転写物を増幅した。Vλを増幅するためのプライマーは、
であった。β-アクチンを増幅するためのプライマーは、以前に説明されている(Arakawa et al., 2002)。
【0140】
sIgM喪失変異体の定量および配列解析
変異したV領域におけるフレームシフトまたはナンセンス変異の結果として生じ、蓄積したsIgM喪失変異体を測定する、sIgM喪失変異体のアッセイ法を用いて、IgV領域の多様化を定量した(Sale et al., 2001; Yabuki et al., 2005; Cummings et al., 2007)。手短に言えば、フローサイトメトリーとそれに続く限界希釈クローニングによってsIgM+細胞を単離し、4週間増殖させた。sIgM-細胞の割合を定量するために、約1×106個の細胞を抗ニワトリIgM-RPE(Southern Biotechnology Associates, Birmingham, AL)で染色し、CellQuestソフトウェア(BD Biosciences)を用いたFACScanによって解析した。
【0141】
単一細胞PCRおよび配列解析を、以前に説明されたようにして実施した(Yabuki et al., 2005; Cummings et al., 2007)。手短に言えば、sIgM-細胞を選別し、個々のウェルに等分し、Vλ領域を増幅および配列決定し、それらの配列をψVλドナーと比較して、変異が鋳型式であるか、または非鋳型式であるかを判定した。鋳型式変異の判定基準は、ドナーとレシピエントにおいて9個の連続した塩基が完全に一致しなければならないというものであった。配列は、独立にトランスフェクトされた2つの株に由来した。特有の配列のみを、変異の分類のために含めた。
【0142】
結果
VP16が係留されると遺伝子変換が加速する
抑圧的なドナークロマチン構造により遺伝子変換が減退するならば、活性化改変によって遺伝子変換が促進される可能性がある。この可能性を検証するために、本発明者らは、本発明者らの実験室で構築したDT40ポリLacO-λ細胞株を利用した。この細胞株では、重合ラクトースオペレーター(ポリLacO)が、ψVλアレイのψVλ17〜ψVλ20の間(発現されたVλ遺伝子の17kb上流)に挿入されている(図16A;実施例4)。これにより、本発明者らは、ラクトースリプレッサーとの融合物として発現される、係留された調節因子の効果を分析することができる。対照実験により、ポリLacOタグが、細胞増殖にも細胞周期プロファイルにもIg遺伝子多様化にも影響を及ぼさないことが示された。
【0143】
ψVλドナー内部のヒストンアセチル化の喪失は、Vλにおける遺伝子変換の減少と相関している(実施例4)。したがって、本発明者らは、クロマチンの弛緩およびヒストンアセチルトランスフェラーゼの動員と関連付けられている、ヘルペスウイルス由来の強力なトランス活性化因子であるVP16の影響を試験した(Tumbar et al., 1999; Carpenter et al., 2005)。本発明者らは、GFP-LacI-VP16を安定に発現するDT40ポリLacO-λRトランスフェクタントを作製した。ウェスタンブロット法によってタンパク質発現を確認した(図16B)。細胞周期プロファイル(図16C)およびλ転写レベル(図16D)は、対照DT40ポリLacO-λ GFP-LacIトランスフェクタントと比べて変わっていなかった。DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントの蛍光画像法では、GFP-LacI-VP16発現およびポリLacOへの結合の証拠である単一の緑色の斑点が核内部で示された(図16E)。
【0144】
本発明者らは、ポリLacO-λ GFP-LacI-VP16細胞および対照DT40ポリLacO-λ GFP-LacI細胞におけるヒストンアセチル化を分析した。ψVλ1とVλ遺伝子の間のプロモーター領域(Vλpro)、ψVλ1、ψVλ5、ψVλ13、ψVλ24、ψVλ25を含む7箇所の部位に特異的なプライマーを用いた増幅によって、ψVλアレイを分析した。(多型性を欠くために、DT40中の2つのIgλ対立遺伝子におけるψVλアレイはPCRによって容易に区別することができない)。さらに、本発明者らは、(1)DT40ポリLacO-λの構築中に生じる配列多型性の寄与により、再配列された対立遺伝子および再配列されていない対立遺伝子を特異的PCRプライマーを用いて区別できる、ψVλアレイ中の唯一の部位であるψVλ17、(2)Vλの再配列された対立遺伝子(VλR)、(3)Vλの再配列されていない対立遺伝子(VλU)、ならびに(4)染色体内対照であるPolε遺伝子、におけるクロマチン改変も分析した。ψVλ部位の増幅は、発現されない卵白アルブミン遺伝子(Ova)と同時に(duplex)実施した。したがって、Ovaは内部対照として機能し、免疫沈降法によって得たψVλ:Ova富化率を、全インプットDNAから得たψVλ:Ovaの比率に対して標準化した(材料および方法を参照されたい)。VP16が係留されると、対照DT40ポリLacO-λ GFP-LacI細胞と比べて、DT40ポリLacO-λGFP-LacI-VP16トランスフェクタントのAcH3レベル(図17A)およびAcH4レベル(図17B)の両方が上昇した。
【0145】
本発明者らは、sIgM喪失変異体アッセイ法を用いて、VP16の係留によって、クローンVλ配列多様化速度が変化するかを問うた。この変動アッセイ法では、構造的に完全なsIgMをもはや発現しない変異体細胞の割合を測定し、したがって、遺伝子変換、点変異、挿入、または欠失に起因する変異事象を記録する(Sale et al., 2001; Yabuki et al., 2005)。遺伝子変換は、ニワトリB細胞におけるIg遺伝子多様化の主な経路であるが、(例えば、不可欠な組換え因子の欠如によって)遺伝子変換が減退している場合には(Sale et al., 2001; Niedzwiedz et al., 2004; Hatanaka et al., 2005; Kawamoto et al., 2005; McIlwraith et al., 2005; Yamamoto et al., 2005)、他の修復結果、特に点変異が主になると考えられる。したがって、多様化は、sIgM-からsIgM+への機能獲得アッセイ法よりはむしろ、遺伝子変換のみを採点するこの機能喪失アッセイ法によって最もうまくモニターされる(Saribasak et al., 2006)。
【0146】
sIgM+細胞を限界希釈法でクローニングすることによってDT40ポリLacO-λR GFP-LacIトランスフェクタントおよびDT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントの独立したクローン派生体を樹立し、4週間培養した後、抗IgM抗体で染色した細胞をフローサイトメトリーに供して、各集団中のsIgM-細胞の割合を決定した。sIgM-細胞比率(%)の中央値の比較により、DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントでは、DT40ポリLacO-λ GFP-LacI対照細胞と比べて、多様化が8.4倍に増大していることが示された(図17B)。単一細胞PCRによって増幅したVλ領域の配列解析により、対照細胞およびVP16がIgλに係留された細胞の両方において、大半の多様化は遺伝子変換によることが示された(図22)。
【0147】
ヒストンH3.3は、ψVλドナー遺伝子において豊富である
ヒストン改変だけでなく、ヌクレオソームのヒストン組成の局所的な変化もまた、クロマチン構造の調節に寄与し得る。クロマチン活性化に関連し、複製とは無関係に沈着する1つのヒストン変異体はH3.3である。ヒストンH3.3は、活性化に関連した翻訳後修飾を含む主要なH3様変異体であり(McKittrick et al., 2004; Henikoff, 2008)、したがって、H3.3がψVλアレイで豊富であるかを問うことは関心対象であった。ChIPによってH3をH3.3と区別するために、本発明者らは、FLAGタグ付きH3.3(H3.3-FLAG)を安定に発現するDT40派生体を作製した。
【0148】
抗FLAG抗体を用いたChIPによるH3.3沈着の特徴付けにより、発現されたVλR遺伝子におけるH3.3変異体の富化が実証された。ψVλアレイ内のクロマチン構造を、再配列され発現されたVλ遺伝子(ψVλ1、ψVλ5、ψVλ13、ψVλ24、ψVλ25、ψVλ17R、VλRを含む)の上流の部位(図18A)、および再配列された対立遺伝子VλU;ならびにIgλと同じ染色体上の別の遺伝子Polεの増幅によって分析した。典型的な実験では、H3.3はVλRにおいてインプットDNAと比べて4倍超に富化されていたが、印象的なことには、本発明者らは、ψVλアレイの全体に渡ってH3.3がかなり富化されており、ψVλ5およびVλproにおいて最も富化されていることを観察した(図18B)。近位部位が遠位部位より高レベルの富化を一貫して示すことはなかったため、ψVλアレイ内でのH3.3の富化は、転写されたVλR遺伝子から上流部位への変異体の段階的な伝播を単に示すわけではない。この座位のクロマチン構造が非均一であることから、座位の発現および/または多様化を調節し得るシスエレメントの存在が、特にψVλ5領域において示唆される。
【0149】
HIRAが係留されると、ヒストン沈着が局所的に増加する
HIRAは、ヒストンシャペロンであり、その機能の内の1つは、ヒストン変異体H3.3を含むヌクレオソームを組み立てることである(Ray-Gallet et al., 2002; Tagami et al., 2004)。したがって、ψVλアレイにおけるH3.3の富化から、VP16の係留による効果と同様に、HIRAの係留によってIg遺伝子多様化が加速し得ることが示唆される。これを検証するために、本発明者らは、安定なDT40ポリLacO-λ HIRA-LacIトランスフェクタントを作製し、ウェスタンブロット法によって、タンパク質発現が同程度のレベルであることを確認し(図19A)、かつ、係留が細胞周期プロファイル(図19B)にもλ転写物レベル(図19C)にも影響を及ぼさないことを示した。ChIPアッセイ法によるAcH3レベルおよびAcH4レベルの分析では、HIRAまたはGFPがIgλに係留された細胞において差は示されなかった。
【0150】
HIRAの係留によりH3.3レベルが上昇するかを問うために、本発明者らは、DT40ポリLacO-λ FLAG-H3.3 HIRA-LacIトランスフェクタントを作製し、次いで、多型性があるため、係留されたIgλのみを解析することが可能であるψVλ17RにおけるFLAG-H3.3の富化を測定した。これらの実験では、2つの独立したHIRA-LacIトランスフェクタントのそれぞれで、対照株と比べて中程度(1.4倍)のFLAG-H3.3富化が示された(図20A)。さらに、汎的な(pan-)H3抗体(H3、およびH3.3を含む他の変異体を検出する)がChIPのために使用される場合、同程度の富化が明らかであった。したがって、HIRAは、H3.3を含むヒストンの正味の富化を引き起こすと思われ、したがって、ψVλでのヌクレオソーム密度に影響を及ぼした。
【0151】
HIRAの係留はIg遺伝子変換を促進する
HIRAの係留がクローンのIg遺伝子多様化速度に影響を及ぼすかを問うために、本発明者らは、sIgM+細胞を限界希釈法でクローニングすることによってDT40ポリLacO-λ HIRA-LacIトランスフェクタンの独立したクローン派生体を樹立し、4週間培養した後、各集団中のsIgM-細胞の割合を決定した。sIgM-細胞の割合は、対照細胞よりもHIRA-LacIトランスフェクタントの方が明らかに大きかった(例えば、図20B)。sIgM-細胞比率(%)の中央値の比較により、DT40ポリLacO-λ HIRA-LacI細胞では、DT40ポリLacO-λ GFP-LacI対照細胞と比べて、多様化が11倍に増大していることが示された(図20C)。HIRAを安定に発現するDT40のトランスフェクタントでは、多様化の中程度の加速(1.7倍)しか明らかではなかったため、この結果は単にHIRAの過剰発現を反映したものではなかった(図20C)。単一細胞PCRによって増幅したVλ領域の配列解析により、対照細胞およびHIRAがIgλに係留された細胞において、大半の多様化は遺伝子変換によることが示された(図23)。
【0152】
短区域の鋳型式変異の割合は、VP16またはHIRAの係留によって増加する
VP16およびHIRAが係留されると、クローンの遺伝子多様化速度は同じ程度加速し(それぞれ8.4倍および11倍)、どちらの場合も遺伝子変換が顕著であった。わずかな差によって変異範囲を区別し得るかどうかを確かめるため、DT40ポリLacO-λ HIRA-LacIトランスフェクタントおよびDT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントにおける独立した変異事象を、対照のDT40ポリLacO-λ GFP-LacIトランスフェクタントと比較した。配列変化がドナーψVλ配列の少なくとも9bpに厳密に一致している区域内に存在し、かつ、生殖系列配列と比べて1つ、2つ、またはそれ以上の一塩基変化を含む場合、その配列変化を遺伝子変換として分類した。一塩基の違いおよび2つまたはそれ以上の違いを含む事象は、「短」区域遺伝子変換および「長」区域遺伝子変換と別々に名付けた。これは、前者のクラスの一部の変異が、ψVλドナー配列と偶然に一致した点変異に原理的には起因し得るためである。点変異、欠失、および挿入からなる非鋳型式事象もまた、記録した。
【0153】
DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントにおいて、74%の事象は鋳型式であったが、かなりの比率の鋳型式事象(変異事象全体の39%または29%)は、長区域遺伝子変換ではなく短区域遺伝子変換であった(表1、図22)。また、かなりの割合で挿入もあった(0%に対して13%)。DT40ポリLacO-λR HIRA-LacI細胞において解析した事象でも、遺伝子変換事象が同様に顕著であった(87%)(表1、図23)。大半の遺伝子変換事象は長区域であったが、少数派であるが有意な事象(変異事象全体の24%または21%)は短区域遺伝子変換であった。残りの事象は、点変異(11%)および挿入(2%)であった。本発明者らは以前に、対照のDT40ポリLacO-λ GFP-LacIトランスフェクタントでの変異誘発事象を特徴付けし、大半の変異が長区域遺伝子変換事象に起因し(77%)、一方、ごく一部(3%)が、短区域変換事象またはドナー配列に偶然に一致した点変異のいずれかであることを発見した(実施例4:表1に要約した)。残りの変異(20%)は、偽遺伝子ドナーのいずれにも対応しなかったため、明らかに点変異であった。
【0154】
(表1)遺伝子変換に対するHP1およびVP16の係留の影響
DT40ポリLacO-λ GFP-LacIトランスフェクタント(n=71;(実施例4)、DT40ポリLacO-λ GFP-LacI-VP16(n=31)トランスフェクタント、およびDT40ポリLacO-λ HIRA-LacIトランスフェクタント(n=137)に由来する、特有の変異を有するVλ領域の配列の要約。データは、少なくとも2回の独立したトランスフェクションから導き出した。
【0155】
したがって、VP16およびHIRAの係留による多様化の加速は、鋳型式変異のレベル上昇に起因する。どちらの場合も、大半の鋳型区域は、2つまたはそれ以上の変異を含んだ。しかし、約3分の1の区域は、単一の変異のみを含んだ。一方、GFP-LacI対照細胞における本質的にすべての遺伝子変換事象は、少なくとも2つの変異を含んだ。このことから、ドナークロマチン構造の変化は、遺伝子変換を加速させるだけでなく、遺伝子変換区域の広さを制限する可能性があること、または、加速した遺伝子変換が、変換区域の長さの伸長に寄与する因子の量を調整する(titrate)可能性があることが示唆される。
【0156】
HIRAが係留されるとヌクレオソーム密度が増加する
上記の結果から、係留されたVP16およびHIRAがIg遺伝子多様化の結果に与える影響は類似していたが、それぞれの局所的影響は異なっていたことが示される:VP16はAcH3およびAcH4を増加させたのに対し、HIRAはH3.3および全体的なヒストン搭載(loading)を増加させた。これらの異なる因子の添加がどのようにクロマチンに影響を及ぼし得るかを確かめるために、本発明者らは、小球菌ヌクレアーゼ(MNアーゼ)を用いてクロマチン構造を探索した。核を採取し、様々な量のMNアーゼで処理し、ゲノムDNAを精製、分離、およびブロットし、次いで、LacOリピートに相当な標識DNAを用いて探索した。DT40ポリLacO-λ GFP-LacI細胞およびDT40ポリLacO-λ GFP-LacI-VP16細胞のMNアーゼ消化パターンは、このアッセイ法では同様と思われた(図21)。高レベルのMNアーゼで消化すると、明らかな制限生成物(clear limit product)が得られた。これは、ヌクレオソームに接触するDNA領域(約150bp)であり、したがって、MNアーゼから保護されている。個々のヌクレオソーム間の間隔は変動し得るため、部分的消化の際に産生されるヌクレオソーム多量体のサイズにはわずかな差が生じ、かつ、ゲル中でよりゆっくりと移動する特徴的にぼやけたバンドが生じる。
【0157】
一方、DT40ポリLacO HIRA-LacI細胞に由来するクロマチンのMNアーゼ消化では、異なるパターンが明らかになった:対照では認められない示差的なはしご状が観察された(図21、右)。対照およびVP16発現細胞では、4量体よりも大きいサイズではしごがぼやけたのに対し、輪郭のはっきりした一連の消化生成物が、制限消化の際に生じる1量体から、最高で8量体およびそれ以上に及んだ。このことから、ヒストンシャペロンHIRAの係留により、リンカーサイズの均一性が高まったことが示唆される。これは、局所的ヌクレオソーム密度が増加した場合に起こると思われ、ヒストンシャペロンとしてのHIRAの公知の役割、ならびに、HIRAの係留によってH3.3およびH3の局所的富化が増大したという証拠と一致する(図18C)。
【0158】
考察
本実施例では、遺伝子変換がクロマチン構造の変化によって加速し得ること、およびこのために少なくとも2つの異なる手段があることを示した。ニワトリDT40 B細胞株のψVλアレイに活性化因子VP16またはヒストンシャペロンHIRAのいずれかを係留すると、遺伝子変換の速さが1桁速くなった。VP16を係留すると、AcH3およびAcH4のレベルが上昇したのに対し、HIRAを係留すると、それらのクロマチンマークは影響を受けなかったが、一つにはH3.3の搭載によってヌクレオソーム密度が増加した。クロマチン状態の異なる変化が、相同組換えを同様に可能にするということから、組換えの際のDNA使用を調節するために利用可能な重複した(redundant)経路が細胞にはあることが示される。
【0159】
VP16およびHIRAの係留がIg遺伝子多様化に与える影響は、ヘテロクロマチンタンパク質HP1の係留による影響とは異なっている。HP1を係留すると、点変異がほぼ3倍に増加し、鋳型式変異の減少を伴った(実施例4)。これらの差異から、ドナークロマチン構造は、相同性の指示に従ったDNA修復を活性化または抑制される場合があり、修復結果に影響を与えることが示される。
【0160】
現在、相同性の指示に従った対立遺伝子修復は、S期に顕著であり、鋳型用の姉妹染色分体に依存すると考えられている。クロマチン構造の局所的改変が、相同性の指示に従った修復のための領域を活性化するという可能性から、調節は、もっと複雑であり得、相同なドナーの存在だけでなく、修復の助けとなるクロマチン構造も必要とし得ることが示唆される。
【0161】
HIRAまたはVP16が係留されると、鋳型式変異を促進することにより、クローンの多様化速度が加速した。どちらの場合も、単一の鋳型塩基の変化のみが変異区域中で明らかである鋳型式変異の割合の増加が明らかであった。これらの「短区域」変異は、VP16トランスフェクタントおよびHIR1トランスフェクタントにおける全変異のそれぞれ29%および21%しか含まなかったが、この範疇の変異は、GFP-LacIトランスフェクタントでは本質的に無かった(3%)。
【0162】
長区域の遺伝子変換を促進するのに必要な因子が、VP16またはHIRAのいずれかがポリLacOに係留された細胞中の遺伝子の増加を支援するのに充分なほど豊富ではない場合、短い遺伝子変換区域が生じ得る。あるいは、この範疇の変異の増加は、ヌクレオソームの位置およびリンカーの長さの両方に影響を及ぼし得るこれらの因子を係留することによって引き起こされたクロマチン構造の変化を反映している可能性もある。
【0163】
HIRAを係留すると、ヌクレオソーム密度が増加する。この密度増加は、短くなったリンカー長を反映している可能性が高い。リンカー領域が遺伝子変換のために好ましい鋳型である場合、HIRAトランスフェクタント中のこれらの領域の短縮は、遺伝子変換区域の長さの短縮の一因となり得る。あるいは、ヌクレオソーム長の短縮により、クロマチンは組換えに対して活性であるがψV間の非相同領域は接近しにくくなるように、ヌクレオソームを再調整(re-phase)することができる。
【0164】
偽遺伝子におけるHIRAの役割は、ヒストン搭載がDNA修復に影響を及ぼす方法に関する新規な例である。多くの研究により、デノボのH3.3沈着に必要とされる転写が発見されているため(AhmadおよびHenikoff, 2002a; AhmadおよびHenikoff, 2002b; Janicki et al., 2004; Wirbelauer et al., 2005)、転写されていない偽遺伝子におけるHIRAの役割は、特に興味深い。これらの結果から、現在、転写はH3.3搭載の必要条件ではないことが示唆される。
【0165】
これらの結果により、クロマチンの制約が組換えに対して及ぼす制約に関する本発明者らの理解は深まる。さらに、これらは、遺伝子ターゲティングおよび/または遺伝子療法のために使用される分子に必要なクロマチン化への洞察も与える。
【0166】
参考文献
【0167】
実施例6:メソテリンおよびHE4に対する診断用抗体の作製
本実施例では、本発明を使用して、十分に有効性を確認された卵巣癌バイオマーカーであるメソテリンおよびHE4に対する抗体をどのように開発できるかを例示する。同定された抗体は、新しい貴重な診断試薬であり、有用な冗長性を臨床試験に追加すると考えられる。本実施例では、他のバイオマーカーを認識する診断用抗体を選択するためのプラットホームとしてのDT40ポリLacOの有用性もさらに確認した。
【0168】
抗体開発のための媒体としてのDT40ポリLacO:2種の卵巣癌バイオマーカー、すなわちメソテリン(Scholler et al., 1999; Frierson et al., 2003)およびHE4(Hellstrom et al., 2003)に対する新しい抗体の同定。
【0169】
メソテリンは、卵巣癌を含む多くの起源の癌細胞において高発現される上皮分化マーカーである(Robinson et al., 2003)。HE4(別名WFDC2)は、乳清酸性タンパク質(WAP)ファミリーのメンバーであり、このファミリーのメンバーは高レベルで分泌され、癌に関連しているが、機能は理解されていない(Bouchard et al., 2006)。本発明者らは、組換えメソテリンおよびHE4を得た。これらのタンパク質は、選択またはELISAアッセイ法用のストレプトアビジン結合ビーズまたはプレートへの結合を促進するビオチン融合物として酵母において発現される(Scholler et al., 2006; Bergan et al., 2007)。本発明者らはまた、新しく同定された抗体の親和性を最初に比較するための標準を与える、メソテリンおよびHE4(Scholler et al., 2008)に対する単鎖抗体(scFv)も得た。
【0170】
メソテリンおよびHE4に対する高親和性抗体を同定するために、超変異および各組換え抗原の認識に関するクローン選択を反復的に実施する。超変異を加速させる条件下でクローンを増殖させ、メソテリンまたはHE4に高親和性で結合するクローンを磁気活性化細胞選別によって選択し、上清の抗体親和性をELISAアッセイ法によって分析する。高親和性抗体を産生する集団をさらに選択して、親和性および特異性を増大させる(図3)。
【0171】
選択は、それぞれ細胞108個(培養物100mlに相当する)からなる3つの集団において開始することができる。各集団は、限界希釈によって単離した単細胞に由来してよく、次いで、超変異を加速させる条件下で培養してよい。ビーズに単独で結合する「粘着性」細胞の富化を最小限にするために、所望の特異性を富化する前に、ビオチン結合ビーズで各集団を最初に清掃する。所望の特異性の富化は、磁気活性化細胞選別(MACS)によって便宜的に実行する。MACSを用いることにより、非常に大きな試料サイズ(細胞108個)から極めて迅速に富化することが可能であり、開始試料の<0.1%を含む集団を100倍に富化することを常に実現する(Volna et al., 2007)。選択された集団を増殖させ、同じプロトコールを用いて再選択する。
【0172】
他の研究者(例えば、Cumbers et al., 2002)の結果に基づき、本発明者らは、各「抗原」に特異的な抗体を産生する細胞が、反復的な超変異および選択の2〜3回目(2〜3週目)またはその前、およびおそらくはそれ以前に回収されると予想する。ELISAアッセイ法によって細胞プールの抗体親和性を試験することができ、親和性が1〜10nMの範囲である場合、限界希釈クローニングを実施し、高親和性抗体を産生するクローンを同定する。限界希釈クローニングは、オペレーターからリプレッサーを遊離させて超変異を減速させるIPTGを含む培地中で実施する。これらのクローン集団から産生された抗体を、後述するようにしてさらに特徴付ける。
【0173】
以下の3つの基準を用いて、このアプローチが成功したかを確かめることができる。
1.親和性および特異性
選択過程で抗体親和性が増大することを実証することによって、抗体開発の有効性を確認する。それ以上の超変異を最小限にするためのIPTGの存在下、ELISAのバックグラウンドを最小限にするためにIgMを枯渇させた培地中で48時間培養した細胞の上清希釈物をELISAアッセイ法に供することにより、抗体親和性を最初に測定する。抗体が抗原を認識し、それが提示されている組換え骨格を認識しないよう徹底するために、ビオチンおよびストレプトアビジンを用いて反応性を試験する。両方の組換え抗原に対する抗メソテリン抗体および抗HE4抗体を並行して比較することが、交差反応性の試験として役立つ。親和性を、メソテリンまたはHE4を対象とする対照scFv抗体と比較する。高親和性のクローン集団の樹立後、Biacore社の表面プラズモン共鳴によってKdを定量的に測定する。
【0174】
2.変異誘発のターゲティング
クローン選択の過程で抗体遺伝子を超変異させる細胞メカニズムにより、抗原に接触するV領域のサブドメインであるCDR中に変異が集中する。本発明者らは、加速した超変異および選択によって単離した高親和性クローンは、CDRの変異を同様に集中させると予想する。これを試験するには、単一細胞PCRを用いて、親和性選択後に個々のB細胞のVH領域およびVL領域を増幅し、VH領域およびVL領域を配列決定し、これらの配列を選択されていない集団の配列と比較することができる。
【0175】
3. IPTGと共に培養した際の特異性および親和性の維持
抗体作製用のこのプラットホームの力は、高親和性クローンが作製された際に超変異を制限する能力にある程度依存する。これを試験するには、IPTGの存在下および不在下でクローン集団を培養し、蛍光タグ付きのメソテリンまたはHE4に結合された細胞をフローサイトメトリーに供することにより、増殖した培養物の平均親和性を比較することができる。親和性は、IPTGを含む培養物では不変のままであると予想されるが、IPTGを欠く培養物では増大または減少すると予想される。したがって、蛍光シグナルの分布は、IPTGを含む培養物の方が広いと考えられる。
【0176】
参考文献
【0177】
本出願の全体を通して、様々な刊行物が参照される。これらの刊行物全体の開示は、本発明が関連する現況技術をより完全に説明するために、参照により本出願の本明細書に組み入れられる。
【0178】
前述の内容から、本発明の特定の態様が、例証のために本明細書において説明されるが、本発明の精神および範囲から逸脱することなく様々な修正を実施し得ることが、理解されると考えられる。したがって、本発明は、添付の特許請求の範囲による場合を除いて、限定されない。
【技術分野】
【0001】
本出願は、その内容全体が参照により本出願に組み入れられる、2007年5月31日に出願された米国特許仮出願第60/932,672号の恩典を主張する。
【0002】
連邦政府による資金提供を受けた研究に関する記載
本発明は、国立衛生研究所(National Institutes of Health)によって授与されたR01 GM041712のもと、米国政府の支援を受けて行った。米国政府は、本発明において一定の権利を有する。
【0003】
発明の技術分野
本発明は概して、B細胞の天然の変異誘発能力を利用し、多様化のプロセスを制御下に置くことによってこれらの能力を増強する、標的遺伝子の変異誘発に関する。本発明は、抗体遺伝子のような発現遺伝子に点変異および他のタイプの多様化を迅速かつ誘導的に発生させるための方法を提供する。この方法は、例えば、親和性または特異性が高い抗体を産生するB細胞クローンを同定するための選択と併用することができる。多様化プロセスは、変異誘発法間で調整、加速、停止、および切り替えなどをすることができる。本発明による多様化の調整は誘導的かつ可逆的である。本発明は、変異免疫グロブリンおよび他のポリペプチドのレパートリーを迅速かつ実行可能に開発する手段を提供する。
【背景技術】
【0004】
発明の背景
抗体は、ヒトにおいて感染に対抗する重要な防御を提供する分子である。これらは、感染症から癌に至る様々な疾患の治療において治療物質として使用される。これらはまた、臨床的実験室および研究実験室で日々実施される多種多様な試験において診断試薬として使用される。
【0005】
抗体の特異性および親和性は、抗体をコードする遺伝子内の特定の領域を標的とする変異プロセスによってインビボで改変される。抗体は、免疫グロブリン(Ig)重鎖遺伝子および軽鎖遺伝子と呼ばれる2つの遺伝子にコードされる。Ig遺伝子にコードされるポリペプチドの重鎖および軽鎖は相互に作用して四量体分子を形成し、これは受容体として細胞表面で発現される。抗体分子は二価性である:一方のドメインは抗原を認識し、他方は身体からの抗原の除去を促進する。可変(V)領域と呼ばれる認識ドメインは、天然抗体中の重鎖ポリペプチドおよび軽鎖ポリペプチドが相互作用して作り出される。実際には、抗体によって配列は様々である。V領域の一次配列(ならびにその結果、三次元構造および抗原特異性)の多様性は、不可逆的な遺伝的変化を引き起こすことによってV領域配列を改変するプロセスの結果である。これらの変化は、B細胞発達の間にプログラムされており、また、B細胞を活性化する環境シグナルに応答して体内で誘導されることもある。いくつかの遺伝的メカニズムがこの多様性に寄与している。同じメカニズムの2つの副経路が、体細胞超変異および遺伝子変換と呼ばれる2つの異なる変異誘発結果をもたらす((Maizels, 2005)に総説がある)。体細胞超変異は、点変異を挿入する。体細胞超変異は、本質的に任意の変異が生じることを可能にするという利点を与え、したがって、変異したV領域を集めたものは、種々の起こり得る変異の本質的なサンプルとなる。遺伝子変換は、関連しているが同一ではない配列による「鋳型式の」変異を挿入する。遺伝子変換は、進化的選択によって最適化され得る、ゲノム中に既に保存されている情報に基づいてレパートリーを作り出すという利点を与える。
【0006】
改変された抗原受容体は選択の標的である。Igは細胞表面で発現されるため、生理学的環境または培養細胞内での所望の特異性または親和性を有するB細胞のクローン選択が可能である。細胞表面Igは、抗Igで染色した細胞をフローサイトメトリーに供することによって容易に検出することができる。特定の化合物へのIg分子の結合は、それらの化合物の蛍光性誘導体との相互作用として検出し、フローサイトメトリーによって解析することができ、また、フローサイトメトリーによる選別の際に、特定の化合物に結合するB細胞を回収することもできる。特定の化合物に結合するB細胞はまた、それらの化合物を担持する固体支持体上で選択することもできる。逆に、固体支持体に結合させることによって、望まれない結合特異性を有するB細胞を除去することも可能である。結合と遊離の反復サイクルにより、高親和性の結合を得るために富化することが可能になる。
【0007】
変異および遺伝子ターゲティングがDT40 B細胞株で起こる。DT40は、培養で容易に増殖するニワトリB細胞株である。DT40は、免疫グロブリン重鎖遺伝子および軽鎖遺伝子の構成的変異誘発を実施するために、多くの事例において文書に記録されている(Reynaud et al., 1987; ThompsonおよびNeiman, 1987; Reynaud et al., 1989)。変異誘発はV領域を標的としている。変異は、通常、「偽V」遺伝子と呼ばれ、各機能的V領域の上流に直線状アレイとして存在する、関連した非機能的V遺伝子セグメントをコピーすることによって鋳型式となる。鋳型式であることは、変異したV標的と偽V遺伝子の内の1つとが共有する配列の区域(tract)として明らかである。DT40の遺伝的改変は、鋳型式変異誘発から非鋳型式変異誘発への切り替えを引き起こし、ヒトB細胞においてV領域の配列を改変する体細胞超変異と本質的に同一の経路によって、体細胞超変異をもたらすことが示された(Sale et al., 2001)。また、DT40は非常に効率的な相同組換えまたは遺伝子ターゲティングを支援することも多くの事例において文書に記録されている(Buerstedde et al., 2002; Sale, 2004)。これにより、特定の遺伝子もしくはゲノム領域が改変もしくは除去されるか、または1つの遺伝的領域が別の領域で置換されている派生体を作製することが可能になる。
【0008】
標的化変異誘発のための現在利用可能なアプローチによってもたらされる制約および難題があるため、代替の方法および構築物を開発する必要が当技術分野において存在する。本発明はこの必要を満たし、他の関連した利点をさらに提供する。
【発明の概要】
【0009】
本発明は、標的配列を多様化するための材料および方法を提供することにより、これらおよび他の必要を満たす。本発明は、標的遺伝子の多様化を可逆的に誘導することを可能にするように改変したB細胞を提供する。この細胞は、関心対象の標的遺伝子に機能的に連結されたシス調節エレメントを含む。次いで、多様化を調整する因子を、シス調節エレメントに結合する係留因子(tethering factor)に融合させ、それによって、標的遺伝子の発現を制御する領域に多様化因子を係留することができる。B細胞は、ニワトリDT40 B細胞または他の脊椎動物B細胞でよく、いくつかの態様においては、ヒトB細胞、またはヒト化免疫グロブリン(Ig)遺伝子(ヒトIgHおよびIgLでニワトリIgHおよびIgLを置換している)を含むニワトリDT40 B細胞が好ましい。
【0010】
典型的には、標的遺伝子は、プロモーターおよびコード領域を含む。1つの態様において、標的遺伝子は、Ig遺伝子エンハンサーおよびコード領域を含む免疫グロブリン(Ig)遺伝子を含む。Ig遺伝子は、IgL遺伝子および/またはIgH遺伝子の全体または一部分でよい。コード領域は、Ig遺伝子に先天的なものでもよく、または異種遺伝子でもよい。いくつかの態様において、標的遺伝子は、多様化のための非Ig標的ドメイン、ならびに、膜貫通ドメインおよび細胞質尾部を含む、B細胞表面で遺伝子産物を提示することを可能にするドメインであるか、またはそれを含む。本発明のB細胞中の標的遺伝子のコード領域は、コード領域全体を含む必要はない。いくつかの態様において、ある特定の領域またはドメインが多様化のために標的とされ、コード領域は、関心対象の領域またはドメインを含む部分のみを任意でコードしてよい。
【0011】
シス調節エレメントは、標的遺伝子の多様化および/または発現を制御するランディングパッドを領域中に提供する。このランディングパッドは、DNAのこの領域に対して配列特異的な様式で係留因子が結合できる場所である。典型的な態様において、シス調節エレメントは、重合(polymerized)ラクトースオペレーター(LacO)である。1つの態様において、エレメントは、約80〜100個繰り返したLacOを含む。別の態様において、シス調節エレメントは、単独または多量体化したテトラサイクリンオペレーター(TetO)である。係留因子(配列特異的DNA結合タンパク質)がDNAに結合し、係留因子に融合された多様化因子をコード領域の十分近くに届けることができ、その結果、コード領域の多様化が可逆的に調節可能になる場所を提供するというランディングパッド機能をエレメントが果たす限り、様々な分子をシス調節エレメントとして使用することができる。
【0012】
係留因子は、配列特異的な様式でシス調節エレメントに結合する因子である。LacOがシス調節エレメントとして働く態様において、LacリプレッサーLacIは係留因子として働くことができ、シス調節エレメントLacOへの結合は、イソプロピル-β-D-チオ-ガラクトシド(IPTG)によって調節され得る。IPTGの不在下では、LacIはLacOに結合し、多様化は多様化因子の存在によって加速される(またはそうでなければ調節される)。IPTGは、多様化因子の活性の停止または低減が望ましい事象において添加され得る。TetOがシス調節エレメントとして働く態様において、TetRは適切な係留因子であり得、多様化因子の活性はテトラサイクリンまたはドキシサイクリンによって調節され得る。
【0013】
いくつかの態様において、多様化因子は、転写調節因子、ヘテロクロマチン関連タンパク質、ヒストンシャペロン、クロマチン再構築因子、核膜孔複合体の構成要素、遺伝子調節因子、またはそれらの組合せである。多様化因子として働くことができる他の分子には、DNA修復因子、DNA複製因子、リゾルベース、ヘリカーゼ、細胞周期調節因子、ユビキチン化因子、SUMO化因子、またはそれらの組合せが含まれるがこれらに限定されるわけではない。1つの態様において、転写調節因子は、VP16またはE47である。多様化因子として使用するための典型的なヘテロクロマチン関連タンパク質はHP1である。代表的なヒストンシャペロンはHIRAである。
【0014】
また、ポリペプチドをコードするポリヌクレオチド配列の多様化を通して、関心対象のポリペプチドの変異配列を有するポリペプチドのレパートリーを作製する方法も提供される。典型的には、この方法は、多様化因子の発現を可能にする条件で本発明のB細胞を培養する段階を含み、B細胞の標的遺伝子は関心対象のポリペプチドのコード領域を含み、それによって、コード領域の多様化が可能になる。この方法は、複数の変異ポリペプチドおよび所望のレパートリーが得られるまで、B細胞の増殖を可能にする条件下で培養物を維持する段階をさらに含んでよい。
【0015】
別の態様において、本発明は、最適化された関心対象のポリペプチドを産生するB細胞を作製する方法を提供する。この方法は、多様化因子の発現を可能にする条件で本発明のB細胞を培養する段階を含み、B細胞の標的遺伝子は関心対象のポリペプチドのコード領域を含み、B細胞は関心対象のポリペプチドをその表面で発現する。この方法は、関心対象のポリペプチドに特異的に結合するリガンドに結合する細胞を選択することにより、関心対象のポリペプチドをB細胞表面で発現する細胞を培養物から選択する段階をさらに含む。関心対象のポリペプチドに特異的に結合するリガンドに対して所望の親和性を有する細胞を選択するまで、これらの培養段階および選択段階を繰り返すことができる。関心対象のポリペプチドがIg、例えばIgL、IgH、または両方である態様において、リガンドは、抗原に相当する、組換えまたは他の手段によって作製されたポリペプチドでよい。例えば、磁気活性化細胞選択(MACS)による選択を容易にするために、リガンドを固体支持体に結合または連結することができる。別の例において、蛍光活性化細胞選別(FACS)を可能にするために、リガンドを蛍光タグに結合または連結することができる。
【0016】
本発明の方法は、培養物に調節分子を添加する段階をさらに含んでよく、調節分子は、シス調節エレメントへの係留因子の結合を調整し、それによって、コード領域の多様化を調整する。上記の例において、IPTG、テトラサイクリン、およびドキシサイクリンが、調節分子として働く。当業者は、多様化活性を調節するために個々の係留因子と共に使用できる他の調節分子を知っている。
【0017】
調節分子は、コード領域の多様化を様々な方法で調整することができる。例えば、いくつかの態様において、調節分子は、調整を行うために培養物に添加される。個々の調節分子がシス調節エレメントへの係留因子の結合を増加させる因子であるか、または減少させる因子であるかどうか、および結合の個々の変化が、多様化活性を増大させる効果を有するか、または低減させる効果を有するかどうかによって、調整の結果、多様化を低減もしくは停止するか、または多様化を促進もしくは加速することができる。他の態様において、調整は、調整の低減であれ停止であれ、促進であれ加速であれ、調節分子を培養物から除去または取り除くことによって行われる。同様に、多様化の調整は、B細胞に遺伝子を添加することもしくはB細胞から遺伝子を取り除くことによって、またはB細胞における遺伝子の発現を増大させることもしくは減少させることによって行うこともできる。当業者は、上記に説明した利用可能な変更(permutation)すべてを容易に理解することができ、これらはそれぞれ、B細胞における調節分子のレベルまたは存在を改変し、その結果として、シス調節エレメントへの多様化因子の係留を改変し、それによって、多様化活性を改変する効果を有する。
【0018】
また、本発明の方法を実施するために使用され得るキットも提供される。このキットは、本発明のB細胞ならびに対応する係留因子および多様化因子を発現する融合構築物を含む。例えば、B細胞は、プロモーターおよびコード領域を含む標的遺伝子に機能的に連結されたシス調節エレメントを含む。キットは、1種または複数種の融合構築物が中に保存された1つまたは複数の容器をさらに含む。各融合構築物は、B細胞で発現され得、かつ、多様化因子に融合された係留因子をコードするポリヌクレオチドを含み、この係留因子は、B細胞のシス調節エレメントに特異的に結合する。B細胞は、複数の融合構築物と共に使用するために複数のシス調節エレメントを含んでよい。
【図面の簡単な説明】
【0019】
【図1】抗体の体細胞超変異の概略図。上のパネル:Ig分子の相補性決定領域(CDR)が抗原に接触する。下のパネル:変異は、CDRをコードするゲノム領域に集中している。
【図2】DT40ポリLacO λ細胞におけるIg遺伝子多様化に対する、係留されたシス調節因子の効果を示す棒グラフ。IPTGの存在下および不在下でのクローンの多様化速度を比較した。
【図3】B細胞クローンの増殖中に抗体特異性が進化する方法を示す概略図。抗体は細胞表面で発現される。最初に多重特異性を示す集団(左上)を選択し、続いて、所望の特異性(細い二重円、中央)およびより高い親和性(太い円、右下)を富化する。
【図4】E2Aが、再配列されたIgλ座位と結合することの説明。図4A:ニワトリB細胞リンパ腫株DT40の再配列されたIgλ座位および再配列されていないIgλ座位の概略図。プロモーター(P)領域、リーダー(L)領域、可変(V)領域、連結(J)領域、および定常(Cλ)領域、J-Cイントロン中の推定のマトリックス結合領域(M)、3’-エンハンサー(E-3’)、ならびに遺伝子変換のための鋳型である、上流の非機能的偽可変領域の最も近位のもの(ψV1)および遠位のもの(ψV25)を示す。再配列された対立遺伝子および再配列されていない対立遺伝子は、矢印で示すプライマーを用いたPCRによって容易に区別することができる。図4B:対照の卵白アルブミン遺伝子アンプリコン(Ova)と比べた、DT40細胞の再配列されたλR座位および再配列されていないλU座位におけるE2A富化のChIP解析。富化倍率を下に示す。NTC、鋳型無しの対照。
【図5】E2Aは、シスで作用してIgλ遺伝子多様化を調節する。図5A:ポリLacOタグ付きの再配列されたIgλ座位の概略図。ポリLacOはψV17〜20の間に組み込まれる。他の概念は図4Aと同じである。図5B:DT40細胞およびDT40ポリLacO-λR細胞の細胞周期プロファイル。図5C:DT40細胞およびDT40ポリLacO-λR細胞によるsIgM喪失変異体の蓄積。6週間のクローン増殖後、各株に由来するサブクローン24個におけるsIgM喪失変異体の出現率をフローサイトメトリーによって定量した。図に示した出現率は、DT40に対して標準化した。sIgM喪失の平均出現率はそれぞれ0.8%および0.9%であった。図5D:DT40ポリLacO-λR GFP-LacI細胞およびDT40ポリLacO-λR E47-LacI細胞の細胞周期プロファイル。図5E:DT40ポリLacO-λR GFP-LacIトランスフェクタントおよびDT40ポリLacO-λR E47-LacIトランスフェクタントにおけるIgλ、AID、またはβ-アクチンmRNAの発現のRT-PCR解析。三角形は、cDNA鋳型の30倍、10倍、3倍、および1倍の相対濃度を示す。図5F:100μM IPTGの不在下または存在下で3週間培養した、独立したクローンDT40ポリLacO-λR GFP-LacIトランスフェクタント(n=13)およびDT40ポリLacO-λR E47-LacIトランスフェクタント(n=19)のsIgM平均喪失量。値は、IPTG無しで培養したDT40ポリLacO-λR GFP-LacI細胞に対して標準化した。
【図6】核の半径は細胞周期と相関している。図6A:G1期およびG2/M期の富化された細胞の代表的な画像。G1細胞(左)およびG2/M細胞(右)をHoechst 33342(10μM; Molecular Probes)で染色し、次いで、DNA含有量に基づいて選別した。スケールバー:10μm。図6B:DT40ポリLacO-λR細胞の代表的な細胞周期プロファイル。代表的なプロファイル内の水平な棒として、平均半径±標準偏差を示す。点線の縦線は、実験解析で使用したG1およびG2のカットオフ値を示す:G1、r<4μm;G2、r>5.2μm。
【図7】E2Aは、細胞周期のG1期にλRに局在化する。図7A:DT40ポリLacO-λR GFP-LacI細胞におけるλRおよびE2Aの共局在化の代表的な画像。DAPI染色によって決定される核周囲長の輪郭を白い点線で描いている。スケールバー:5μm。図7B:DT40ポリLacO-λR GFP-LacI細胞において細胞周期の各段階で起こるλR/E2A共局在化の割合。図7C:DT40ポリLacO-λR RFP-LacI細胞におけるλRおよび活性Pol II(P*-Pol II)の共局在化の代表的な画像。記号は図7Aと同じ。図7D:DT40ポリLacO-λR RFP-LacI細胞において細胞周期の各段階で起こるλR/P*-Pol II共局在化の割合。
【図8】Id1発現は、G1期のλR/E2A共局在化を阻害する。図8A:DT40ポリLacO-λR RFP-LacI細胞(左)およびDT40ポリLacO-λR RFP-LacI Id1細胞(右)におけるId1発現を分析するウェスタンブロット。左にマーカーポリペプチドのサイズ(kDa)を示す。図8B:DT40ポリLacO-λR RFP-LacI細胞およびDT40ポリLacO-λR RFP-LacI Id1細胞の細胞周期プロファイル。図8C:6週間培養した、独立したクローンDT40ポリLacO-λR RFP-LacI Id1トランスフェクタント(n=6)のsIgM平均喪失量。値は、DT40ポリLacO-λR RFP-LacI細胞に対して標準化した。図8D:Id1発現により、IgλまたはAIDの転写物レベルは変化しない。DT40ポリLacO-λR RFP-LacI細胞およびDT40ポリLacO-λR RFP-LacI Id1細胞中のIgλ、AID、または対照β-アクチンmRNAの発現のRT-PCR解析。図8E:DT40ポリLacO-λR RFP-LacI細胞およびId1を安定に発現する派生体(それぞれId1-および+)におけるλR/E2A共局在化の細胞周期依存性に対するId1発現の影響。細胞周期の各段階における共局在化の比率(%)を示す。図8F:DT40ポリLacO-λR RFP-LacI細胞およびId1を安定に発現する派生体におけるλR/P*-Pol II共局在化の共局在化細胞周期依存性に対するId1発現の影響。詳細は図8Eに同じ。
【図9】DT40 Igλ座位におけるクロマチン改変。図9A:再配列されたニワトリIgλ座位の概略図。25ψVλ領域および再配列されたVλR遺伝子(リーダー、L;可変領域、Vλ-Jλ;定常領域、Cλ領域)を示す。図9B:ヒストンH3およびH4のN末端アセチル化(AcH3およびAcH4)を分析した、代表的なクロマチン免疫沈降実験の概略。調べた部位は次のものであった:ψVλアレイの約1kb上流の領域(フランク)、ψVλ5とVλの間の領域、ψVλ1、ψVλ5、ψVλ13、ψVλ18、ψVλ24、ψVλ25、ならびに再配列されたVλR対立遺伝子および再配列されていないVλu対立遺伝子。詳細に関しては材料および方法を参照されたい。棒は標準偏差を示す。
【図10】DT40ポリLacO-λRにおけるψVλアレイへのGFP-LacIの可逆的係留。図10A:DT40の再配列されたニワトリIgλ座位の概略図。重合ラクトースオペレーター(ポリLacO)がψVλ17〜20の間に挿入されている。記号は図9と同じ。図10B:DT40 GFP-LacIトランスフェクタント、およびIPTGの不在下で培養するか(左)または100μM IPTGの存在下で一晩培養した(右)DT40ポリLacO-VλR GFP-LacIトランスフェクタントの蛍光画像。
【図11】HP1が係留されると、活性クロマチンに特徴的な改変が減少する。図11A:単一のDT40ポリLacO-VλR LacI-HP1トランスフェクタントの代表的な蛍光画像。抗LacI抗体で染色(左);DAPIで染色(中央);または合体した画像(右)。図11B:DT40ポリLacO-VλR GFP-LacIトランスフェクタントおよびDT40ポリLacO-VλR LacI-HP1トランスフェクタントのψVλ17Rにおけるアセチル化H3(AcH3)およびアセチル化H4(AcH4)の富化。ChIPに続いて、OvaプライマーおよびψVλ17Rプライマーを用いて二重PCRを実施した。富化は、全DNAインプット対照に対し、段階的に増量した鋳型DNAを4回別々に増幅した結果の±標準偏差として表す。図11C:DT40ポリLacO-VλR GFP-LacIトランスフェクタントおよびDT40ポリLacO-VλR LacI-HP1トランスフェクタントのψVλ17R(パネルBから)およびpol εプロモーターにおけるAcH3およびAcH4の富化を示すヒストグラム。棒は標準偏差を示す。
【図12】HP1の係留は、Igλ発現に影響を及ぼさない。図12A:DT40ポリLacO-VλR GFP-LacIトランスフェクタントおよびDT40ポリLacO-VλR LacI-HP1トランスフェクタントのVλRにおけるAcH3およびAcH4の相対的富化。富化の値は、対照のDT40ポリLacO-VλR GFP-LacIに対して標準化した。棒は、段階的に増量した鋳型DNAを4回別々に増幅した結果の標準偏差を示す。図12B:250μM IPTGの存在下および不在下で培養したDT40ポリLacO-λR GFP-LacIトランスフェクタント(GFP-LacI)およびDT40ポリLacO-λR LacI-HP1トランスフェクタント(LacI-HP1)における細胞表面IgM(sIgM)の相対強度。sIgMレベルは、マウス抗ニワトリIgM抗体による染色の強度を測定することによって定量し、DT40ポリLacO-λR GFP-LacIトランスフェクタントにおけるレベルに対して標準化した。詳細は図12Aに同じ。図12C:DT40ポリLacO-λR GFP-LacIトランスフェクタント(GFP-LacI)およびDT40ポリLacO-λR LacI-HP1トランスフェクタント(LacI-HP1)におけるVλR転写物の相対的レベル。転写物レベルはRT-PCRによって定量し、DT40ポリLacO-λR GFP-LacIトランスフェクタントにおけるレベルに対して標準化した。詳細は図12Aに同じ。
【図13】HP1が係留されると、ψVλアレイの全体に渡ってヒストンアセチル化が減少する。図13A:250μM IPTGの存在下および不在下で3日間培養したDT40ポリLacO-VλR LacI-HP1細胞株に由来するクロマチンにおけるヒストンH3のN末端アセチル化を分析した、クロマチン免疫沈降実験の概略。ChIP富化の値(実施例4を参照されたい)は、DT40ポリLacO-VλR GFP-LacI細胞に由来するクロマチンを並行して分析して得た値に対して標準化した。棒は、段階的に増量した鋳型DNAを4回別々に増幅した結果の標準偏差を示す。図13B:250μM IPTGの存在下および不在下で3日間培養したDT40ポリLacO-VλR LacI-HP1細胞株に由来するクロマチンにおけるヒストンH4のN末端アセチル化を分析した、クロマチン免疫沈降実験の概略。詳細は図13Aに同じ。
【図14】HP1の係留によって促進された、非鋳型式変異および鋳型式変異。図14A:独立したDT40ポリLacO-VλR GFP-LacI(n=27)およびDT40ポリLacO-VλR LacI-HP1(n=16)のパネルのsIgM喪失変動アッセイ法。図は、各融合構築物に関して少なくとも2個の独立したトランスフェクタントから得て組み合わせたデータを示す。多様化速度の中央値を下に示す。図14B:単一細胞PCRによって解析した、DT40ポリLacO-VλR GFP-LacIトランスフェクタント(n=78)およびDT40ポリLacO-VλR LacI-HP1トランスフェクタント(n=36)に由来する独特な変異を有するVλ領域の配列解析の概略。2つの独立したトランスフェクタントから配列を集めた。
【図15】変異したDT40ポリLacO-λR LacI-HP1クローンの配列アラインメント。多様化したDT40ポリLacO-λR LacI-HP1細胞に由来する14種の独特な変異Vλ領域の配列。青い四角は、遺伝子変換区域を囲み、赤色の円は点変異を示し、黒の点線の四角は非鋳型式挿入を示し、オレンジ色の三角は欠失を示す。DT40領域の親配列(SEQ ID NO: 31)を提示し、変異部位を示す。
【図16】Ig遺伝子変換は、ψVλアレイへのVP16の係留によって加速される。図16A:DT40の再配列されたニワトリIgλ座位の概略図。重合ラクトースオペレーター(ポリLacO)がψVλ17〜20の間に挿入されている。リーダー、L;再配列された可変領域、Vλ-Jλ;定常領域、Cλ。図16B:図に示すトランスフェクタントにおけるGFP-LacIまたはGFP-LacI-VP16の発現のウェスタンブロット解析。抗LacI抗体を用いてブロッティングし、対照のβ-アクチンと比べて発現を分析した。図16C:DT40ポリLacO-λ GFP-LacIトランスフェクタントおよびDT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントの細胞周期プロファイル。図16D:DT40ポリLacO-λ GFP-LacIトランスフェクタントおよびDT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントにおけるVλ転写物レベルのRT-PCRによる比較定量。DT40ポリLacO-λ GFP-LacIに対して標準化した。図16E:DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントの蛍光画像。矢印は、Igλに結合されたGFP-LacI-VP16を示す。核をDAPIで対比染色した(青色)。
【図17】ψVλアレイへのVP16の係留により、AcH3レベルおよびAcH4レベルは上昇し、Ig遺伝子変換は加速する。図17A:DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントのIgλにおけるアセチル化H3(AcH3)の富化。ChIPに続いて、OvaプライマーおよびψVλ17Rプライマーを用いて二重PCRを実施した。富化は、全DNAインプット対照に対し、段階的に増量した鋳型DNAを4回別々に増幅した結果の±標準偏差として表す。図17B:DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントのIgλにおけるアセチル化H4(AcH4)の富化。記号は図17Aと同じ。図17C:独立したDT40ポリLacO-λ GFP-VP16トランスフェクタントのパネルのsIgM喪失量中央値。DT40ポリLacO-Vλ GFP-LacIトランスフェクタントに対して標準化した。図は、各融合構築物に関して少なくとも2個の独立したトランスフェクタントから得て組み合わせたデータを示す。対照のDT40ポリLacO-Vλ GFP-LacIトランスフェクタントと比べた増加倍率を下に示す。
【図18】H3.3ヒストン変異体は、DT40 Igλ座位で豊富である。図18A:DT40の再配列されたニワトリIgλ座位の概略図。記号は図16Aと同じ。図18B:H3.3ヒストン変異体を分析する、代表的なクロマチン免疫沈降実験の概略。調べた部位は次のものであった:ψVλアレイの約1kb上流の領域(フランク)、ψVλ5とVλの間の領域、ψVλ1、ψVλ5、ψVλ13、ψVλ18、ψVλ24、ψVλ25、ならびに再配列されたVλR対立遺伝子。棒は標準偏差を示す。
【図19】Ig遺伝子変換は、ψVλアレイへのHIRAの係留によって加速される。図19A:図に示すトランスフェクタントにおけるGFP-LacIまたはHIRA-LacIの発現のウェスタンブロット解析。抗LacI抗体を用いてブロッティングし、対照のβ-アクチンと比べて発現を分析した。図19B:DT40ポリLacO-λ GFP-LacIトランスフェクタントおよびDT40ポリLacO-λ HIRA-LacIトランスフェクタントの細胞周期プロファイル。図19C:DT40ポリLacO-λ GFP-LacIトランスフェクタントおよびDT40ポリLacO-λ HIRA-LacIトランスフェクタントにおけるVλ転写物レベルのRT-PCRによる比較定量。DT40ポリLacO-λ GFP-LacIに対して標準化した。図19E:DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントの蛍光画像。矢印は、Igλに結合されたGFP-LacI-VP16を示す。核をDAPIで対比染色した(青色)。
【図20】DT40ポリLacO-λにおけるHIRA係留により、H3.3ヒストン変異体は富化され、遺伝子変換が加速する。図20A:H3.3-FLAGを発現するDT40ポリLacO-λ細胞およびDT40ポリLacO-λ HIRA-LacI細胞のψVλ17RにおけるH3.3(緑)およびpan H3(青)の富化。DT40ポリLacO-λ HIRA-LacIの2つの独立した株の結果を示す。DT40ポリLacO-λの富化は、値「1」に対して標準化する。棒は、段階的に増量した鋳型DNAを4回別々に増幅した結果の±標準偏差として示す。図20B:DT40ポリLacO-GFP-LacI細胞およびDT40ポリLacO-λ HIRA-LacI細胞の代表的な集団中のsIgM+細胞のフローサイトメトリー。図20C:独立したDT40ポリLacO-λ HIRA-LacIトランスフェクタントおよびDT40 HIRAトランスフェクタントのパネルのsIgM喪失量中央値。DT40ポリLacO-λ GFP-LacIトランスフェクタントに対して標準化した。図は、各融合構築物に関して少なくとも2個の独立したトランスフェクタントから得て組み合わせたデータを示す。対照のDT40ポリLacO-λ GFP-LacIトランスフェクタントと比べた増加倍率を下に示す。
【図21】係留されたHIRAおよびVP16の異なるクロマチン効果。図21A:小球菌ヌクレアーゼ(MNアーゼ(MNase))でクロマチンを消化した後の、DT40ポリLacO-λ GFP-LacIトランスフェクタント、DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタント、およびDT40ポリLacO-λ HIRA-LacIトランスフェクタントのポリLacO領域のサザンブロット解析。MNアーゼ無し、(-)。右の数字はヌクレオソーム多量体を示す。図20B:ヒストン沈着の増加が、クロマチンの「はしご状(laddering)」パターンの強調に関与し得る方法のモデル。矢印先端部は、存在し得るMNアーゼ切断部位を示す。黒い矢印先端部は、限定的な範囲の切断可能なDNAを示し、灰色の印先端部は、予測される約150bpの切断産物から逸脱する広い範囲の切断可能なDNAを示唆する。
【図22】単一のDT40ポリLacO-λ GFP-LacI-VP16細胞に由来する変異したV領域の配列。単一のsIgM-細胞からV領域を増幅し、次いで配列決定した。中に色を塗っていない青い四角は、長区域の遺伝子変換事象を囲み、青色に塗った四角は、短区域の遺伝子変換事象を囲み、赤い円は点変異を示し、黒い点線の四角は挿入を示し、ダイヤ型(carat)は欠失を示す。
【図23】単一のDT40ポリLacO-λ HIRA-LacI細胞に由来する変異したV領域の配列。記号は図22と同じ。
【発明を実施するための形態】
【0020】
発明の詳細な説明
本発明は、標的免疫グロブリン遺伝子の変異誘発が誘導可能であるB細胞株の開発に基づいている。免疫グロブリンは細胞表面で発現されるため、特定の特異性、高い親和性、および/または新しい活性を有する免疫グロブリン分子を発現するクローンの選択が可能である。これは、標的部位で免疫グロブリン遺伝子および同様に非免疫グロブリン遺伝子を用いて実施することができる。
【0021】
定義
本出願において使用される科学用語および技術用語はすべて、別段の指定が無い限り、当技術分野において一般に使用される意味を有する。本出願において使用される場合、以下の単語または語句は、指定の意味を有する。
【0022】
本明細書において使用される場合、「ポリペプチド」は、タンパク質、タンパク質断片、およびペプチドを含み、天然供給源から単離されるか、組換え技術によって作製されるか、または化学合成されるかを問わない。本発明のペプチドは、典型的には少なくとも約6個のアミノ酸を含む。
【0023】
本明細書において使用される場合、標的遺伝子の「多様化」とは、標的遺伝子の配列または構造の変化または変異を意味する。多様化は、点変異、鋳型式変異、DNA欠失、およびDNA挿入をもたらし得る、体細胞超変異、遺伝子変換、およびクラススイッチ組換えといった生物学的プロセスを含む。本発明の多様化因子は、これらの多様化方法のいずれかを誘導、強化、または調節することができる。
【0024】
「変異」とは、ポリヌクレオチド配列の改変であり、1つもしくは複数のヌクレオチド塩基の改変、もしくは配列への1つもしくは複数のヌクレオチドの挿入、もしくは配列からの1つもしくは複数のヌクレオチドの欠失のいずれか、またはこれらの組合せを特徴とする。
【0025】
本明細書において使用される場合、「シス調節エレメント」とは、遺伝子の発現または多様化を制御する領域中に位置するDNA配列である。係留因子は、DNAのこの領域に配列特異的な様式で結合する。代表的なシス調節エレメントにはLacOおよびTetOが含まれるが、それらに限定されるわけではない。
【0026】
本明細書において使用される場合、「係留因子」とは、配列特異的な様式でシス調節エレメントに結合する分子である。係留因子の1つの例は「リプレッサー」、すなわち、調節遺伝子によって合成され、オペレーター座位に結合して、そのオペロンの転写を妨害するタンパク質である。例示的な係留因子にはLacIおよびTetRが含まれるが、それらに限定されるわけではない。
【0027】
本明細書において使用される場合、「多様化因子」とは、多様化または超変異を加速させるか、または調節する分子を意味する。代表的な多様化因子には、最初は転写調節因子として同定されたいくつか(例えば、VP16もしくはE47)、ヘテロクロマチン関連タンパク質(例えば、HP1)、ヒストンシャペロン(HIRA)、クロマチン再構築因子、核膜孔複合体の構成要素(NUP153)、遺伝子調節因子、またはそれらの組合せが含まれる。多様化因子として働くことができる他の分子には、DNA修復因子、DNA複製因子、リゾルベース、ヘリカーゼ、細胞周期調節因子、ユビキチン化因子、SUMO化因子、またはそれらの組合せが含まれるがこれらに限定されるわけではない。
【0028】
本明細書において使用される場合、「プロモーター」とは、一般にコード領域の上流(5')側にあり、転写の開始およびレベルを少なくともある程度制御するDNA領域を意味する。本明細書における「プロモーター」への言及は、最も広い文脈で理解されるべきであり、TATAボックスプロモーターまたは非TATAボックスプロモーターを含む、古典的なゲノム遺伝子の転写調節配列、ならびに、発達的刺激および/もしくは環境刺激に応答して、または組織特異的もしくは細胞型特異的な様式で遺伝子発現を改変するその他の調節エレメント(すなわち、活性化配列、エンサンサー、およびサイレンサー)を含む。プロモーターは、発現を調節する相手である構造遺伝子の上流または5'側に通常は位置するが、必ずしもそうとは限らない。さらに、プロモーターを含む調節エレメントは、遺伝子の転写開始部位から2kb以内に通常位置するが、それらはより遠く(many kb)離れていてもよい。プロモーターは、細胞における発現をさらに促進するために、かつ/または機能的に結合されている構造遺伝子の発現の時期もしくは誘導能を改変するために、開始部位からさらに遠位に位置するその他の特異的調節エレメントを含んでよい。
【0029】
本明細書において使用される場合、「機能的に結合される」または「機能的に連結される」などは、機能的な関係にあるポリヌクレオチドエレメントの結合の仕方を意味する。核酸は、別の核酸配列との機能的関係に置かれている場合、「機能的に連結され」ている。例えば、プロモーターまたはエンハンサーは、コード配列の転写に影響を及ぼす場合、コード配列に機能的に連結されている。「機能的に連結される」とは、連結されている核酸配列が典型的には隣接しており、2つのタンパク質コード領域を結合する必要がある場合は、隣接し、かつリーディングフレーム中にあることを意味する。転写可能なポリヌクレオチドにプロモーターを「機能的に連結する」とは、転写可能なポリヌクレオチド(例えば、タンパク質をコードするポリヌクレオチドまたは他の転写物)をプロモーターの調節的制御下に置くことを意味し、プロモーターは、次いでそのポリヌクレオチドの転写および任意で翻訳を制御する。
【0030】
「核酸」または「ポリヌクレオチド」という用語は、単鎖または二本鎖いずれかの形態のデオキシリボヌクレオチドポリマーまたはリボヌクレオチドポリマーを意味し、別段の制限がない限り、天然に存在するヌクレオチドと同様の様式で核酸にハイブリダイズする、天然ヌクレオチドの公知の類似体も包含する。
【0031】
本明細書において使用される場合、「1つの(a)」または「1つの(an)」は、特に別段の指示がない限り、少なくとも1つを意味する。
【0032】
B細胞
本発明は、標的遺伝子の多様化の可逆的誘導を可能にするように改変されたB細胞を提供する。この細胞は、関心対象の標的遺伝子に機能的に連結されたシス調節エレメントを含む。次いで、多様化を調整する因子を、シス調節エレメントに結合する係留因子に融合させ、それによって、標的遺伝子の発現を制御する領域に多様化因子を係留することができる。B細胞は、ニワトリDT40 B細胞または他の脊椎動物B細胞でよく、いくつかの態様においては、ヒトB細胞またはヒト化免疫グロブリン(Ig)遺伝子(ヒトIgHおよびIgLでニワトリIgHおよびIgLを置換している)を含むニワトリDT40 B細胞が好ましい。
【0033】
B細胞は、天然の抗体産生者であるため、改善された抗体ならびに改善された非免疫グロブリンタンパク質およびポリペプチドの両方を産生させるための魅力的な細胞となっている。DT40 B細胞は、超変異および選択の反復サイクルによって特異的かつ高親和性の抗体を進化させるための有効な開始点である(Cumbers et al., 2002; Seo et al., 2005)。DT40細胞には、この目的のために試験された他の媒体(vehicle)よりも優れた利点がいくつかある。DT40は、培養状態でそのIg遺伝子を構成的に多様化し、発現された抗体分子のCDRに変異を導く。DT40はヒトB細胞株よりも速く増殖する(世代時間は24時間に対して10〜12時間)。細胞は、特殊な因子もフィーダー層も添加せずに、限界希釈によって容易にクローニングされるため、クローン集団を容易に単離することができる。DT40は効率的な相同遺伝子ターゲティングを実施し(Sale, 2004)、したがって、特定の座位を意のままに置換して、超変異を調節する因子を操作できるようにすることができる。
【0034】
本発明は、高親和性抗体を作製するための新規なプラットホームを提供する。1つの態様において、抗体を進化させるための媒体はB細胞株DT40であり、これは、抗体を天然に産生し、かつ、免疫グロブリン遺伝子超変異を加速させ誘導性にするために操作された。他のB細胞と同様に、DT40は細胞表面で抗体を発現するため、蛍光活性化細胞選別または磁気活性化細胞選別によって、高親和性および最適な特異性に関して簡便にクローン選択することが可能である。DT40細胞株において、超変異は、生理学的環境でのIg遺伝子超変異への、何百万年もの脊椎動物の進化を通して完成された同じ経路によって実施される。この高度に保存された経路は、抗原と接触する可変(V)領域のサブドメインである相補性決定領域(CDR)に優先的に(ただし排他的ではない)変異を導く(図1)。
【0035】
これまで、抗体選択のためにDT40(および他の培養B細胞株)を使用することは、超変異の速度が非常に遅く、生理学的超変異の約0.1%〜1%の速度であるため、制限されていた。超変異を加速させるために、超変異の分子メカニズムについての本発明者らの現在の高度な理解を利用して、重要な調節部位および調節因子が操作された。
【0036】
ニワトリDT40 B細胞は多くの利点を与えるものの、いくつかの態様において、ヒトB細胞を使用することが望ましい場合がある。あるいは、ニワトリDT40 B細胞と共にヒト化Ig遺伝子を使用してもよい。DT40免疫グロブリン遺伝子をヒト化することによって、DT40プラットホームにおいて作製した抗体を治療に直接使用できるなど、治療学のためのこのプラットホームの有用性を広げることができる。
【0037】
ヒト化抗体遺伝子の有用性に関する十分な記録があり、最近再検討されたように、ヒト化するためのいくつかの有効なアプローチがある(WaldmannおよびMorris, 2006; AlmagroおよびFransson, 2008)。ヒト化は、ニワトリIg遺伝子をヒトIg遺伝子で置換することによって行われ、これは、高効率の相同遺伝子ターゲティングを利用することにより、DT40において容易に実施される。これらの置換は、下記の実施例で説明する、シス調節エレメントを挿入するための手順に類似しているが、重鎖座位および軽鎖座位の別個の部分を改変するように設計された異なるターゲティング構築物を用いる手順によって実施される。置換によって、V(D)J領域とC領域を交換することにより、完全にヒト化した抗体、またはキメラ抗体(C領域はヒト化されているがV領域はされていない)を産生するDT40派生体を作製することができる。これらの置換は、隣接したシス調節エレメントを改変することも、それらが超変異を加速させる能力に影響を及ぼすこともない。超変異を促進する保存されたメカニズムは、ヒト化配列のCDRに変異誘発を導く。したがって、ヒト化した株は、細胞培養においてヒトモノクローナルの発達を加速させるために使用することができ、治療目的または診断目的のいずれかに有用な抗体を迅速に作製するための二重プラットホームを提供する。
【0038】
さらに、C領域の置換によって抗体エフェクター機能を最適化することもできる。抗体に基づく免疫療法は、療法のための強力なアプローチであるが、このアプローチはこれまで、有用なエフェクター特性を有する特定の抗体の入手しやすさにある程度制限されていた(Hung et al., 2008; Liu et al., 2008)。抗体の定常(C)領域は、エフェクター機能を決定する。DT40媒体における相同遺伝子ターゲティングによって、天然または人工いずれかのヒトC領域を置換して、所望のエフェクター機能を有する抗体を作製することができる。
【0039】
標的遺伝子
典型的には、標的遺伝子は、プロモーターおよびコード領域を含む。本発明のB細胞中の標的遺伝子のコード領域は、関心対象の任意のタンパク質またはペプチドをコードするものでよく、完全なコード領域を含む必要はない。いくつかの態様において、特定の領域またはドメインが多様化のための標的とされ、コード領域は、関心対象の領域またはドメインを含む部分のみを任意でコードしてよい。
【0040】
1つの態様において、標的遺伝子は、Ig遺伝子のエンハンサーおよびコード領域を含む免疫グロブリン(Ig)遺伝子を含む。Ig遺伝子は、IgL遺伝子および/またはIgH遺伝子の全体または一部分でよい。コード領域は、Ig遺伝子に先天的なものでもよく、または異種遺伝子でもよい。いくつかの態様において、標的遺伝子は、多様化のための非Ig標的ドメイン、ならびに、膜貫通ドメインおよび細胞質尾部を含む、B細胞表面で遺伝子産物を提示することを可能にするドメインであるか、またはそれらを含む。
【0041】
シス調節エレメント
シス調節エレメントは、標的遺伝子の発現を制御するランディングパッドを領域中に提供する。このランディングパッドは、DNAのこの領域に対して配列特異的な様式で係留因子が結合できる場所を提供する。係留因子(配列特異的DNA結合タンパク質)がDNAに結合し、係留因子に融合された多様化因子をコード領域の十分近くに届けることができ、その結果、コード領域の多様化が可逆的に調節可能になる場所を提供するというランディングパッド機能をエレメントが果たす限り、様々な分子をシス調節エレメントとして使用することができる。典型的な態様において、シス調節エレメントは、重合ラクトースオペレーター(LacO)である。1つの態様において、エレメントは、約80〜100個繰り返したLacOを含む。別の態様において、シス調節エレメントは、テトラサイクリンオペレーター(TetO)である。
【0042】
係留因子および多様化因子
係留因子は、配列特異的な様式でシス調節エレメントに結合する因子である。いくつかの態様において、多様化の調節は、シス調節エレメントへの調節可能な結合を特徴とする係留因子を用いることによって実現される。LacOがシス調節エレメントとして働く態様において、LacリプレッサーLacIは、係留因子として働くことができ、シス調節エレメントLacOへの結合は、イソプロピル-β-D-チオ-ガラクトシド(IPTG)によって調節され得る。IPTGの不在下では、LacIはLacOに結合し、多様化は、多様化因子の存在によって加速される(またはそうでなければ調節される)。IPTGは、多様化因子の活性の停止または低減が望ましい事象において添加され得、それによって、変異誘発プロセスが可逆的プロセスになる。TetOがシス調節エレメントとして働く態様において、TetRは適切な係留因子であり得、多様化因子の活性はテトラサイクリンまたはドキシサイクリンによって調節され得る。
【0043】
多様化因子は、B細胞における変異誘発の調整を可能にする因子である。下記の実施例は、いくつかの調節因子を係留および遊離する効果を説明する。1つはヘテロクロマチンタンパク質HP1であり、超変異を5.6倍に加速した(実施例4)。もう1つはE47-LacI(E47は、E2A調節因子の1つのアイソフォームであり、B細胞の発達に不可欠である)であり、超変異を4.5倍に加速した(実施例3)。クロマチン構造体の他の2つの改変因子(modifier)は、超変異を8.4倍および11.0倍に加速した(図2)。さらに、係留部位は、発現されるIg重鎖座位を標的としていた。IgHおよびIgLにおける超変異の二重調節により、超変異を25倍から100倍に加速して、新しい抗体の迅速な作製を可能にすることができる。
【0044】
多様化している免疫グロブリン遺伝子は、細胞周期の過程で核内部を移動し、この移動は多様化と相関関係があり、多様化に必要である。(下記の実施例で説明する方法を用いて)DT40ポリLacO-λ細胞においてNP153-LacIを発現させることにより核膜孔に係留すると、多様化が5.7倍に加速する。この結果から、他の核膜孔タンパク質および核中で遺伝子の位置を決定または調節する他の因子を含む、遺伝子の位置を調節する他の因子もまた多様化を調節することが予測される。
【0045】
クロマチン構造も同様に多様化を調節する。実施例4のデータは、LacIに融合されたヘテロクロマチンタンパク質HP1の発現により多様化が加速すること、およびこの効果がIPTGによって可逆的であることを示す。この知見は、ヒストンシャペロンであるHIRAにも適用された。
【0046】
また、遺伝子の発現または機能の調節因子も、多様化因子として働く。下記の実施例3は、調節因子E2A(E47アイソフォームとして、LacI融合物として発現される)の発現により多様化が加速すること、およびこの効果がIPTGによって可逆的であることを示す。ヘルペスウイルス由来のVP16調節ドメインは多様化を加速させ、この効果はIPTGによって可逆的である。
【0047】
脱アミノもまた、変異誘発を加速させるが、これは誘導性ではない。AIDは、Ig遺伝子多様化を開始するB細胞特異的DNAデアミナーゼである。下記の実施例2は、DT40ポリLacO-λにAID (AID-LacI)を係留することにより変異誘発が促進されるが、IPTGはこの効果を抑えないことを示す。したがって、変異誘発の影響(affect)はゲノムワイドであるため、この因子は、さらに改変することなしには不適切であると思われる。
【0048】
いくつかの態様において、多様化因子は、転写調節因子、ヘテロクロマチン関連タンパク質、ヒストンシャペロン、クロマチン再構築因子、核膜孔複合体の構成要素、遺伝子調節因子、またはそれらの組合せである。多様化因子として働くことができる他の分子には、DNA修復因子、DNA複製因子、リゾルベース、ヘリカーゼ、細胞周期調節因子、ユビキチン化因子、SUMO化因子、またはそれらの組合せが含まれるがこれらに限定されるわけではない。1つの態様において、転写調節因子は、VP16またはE47である。多様化因子として使用するための典型的なヘテロクロマチン関連タンパク質はHP1である。代表的なヒストンシャペロンはHIRAである。
【0049】
(表1)代表的な多様化因子
【0050】
融合構築物
シス調節エレメントへの多様化因子の係留は、係留因子への多様化因子の融合を通して達成される。因子のこの組合せをコードする融合構築物は、本発明のB細胞中に存在し、本発明のB細胞によって発現されてよく、また、所望の時点および特定の目的のために望ましい因子を選択する際にB細胞に添加するために、別々に提供されてもよい。
【0051】
一般に、融合構築物は標準技術を用いて調製してよい。例えば、これらのペプチド構成要素をコードするDNA配列を別々に組み立て、適切な発現ベクターに連結してよい。連結されるDNA配列は、適切な転写調節エレメントまたは翻訳調節エレメントに機能的に連結される。一方のペプチド構成要素をコードするDNA配列の3'末端が、ペプチドリンカーを用いてまたは用いずに、第2のペプチド構成要素をコードするDNA配列の5'末端に連結され、その結果、これらの配列のリーディングフレームは協調する。これにより、両方の構成要素ペプチドの生物活性を保持している単一の融合タンパク質に翻訳されることが可能になる。
【0052】
ペプチドリンカー配列を使用して、第1のペプチド構成要素と第2のペプチド構成要素を、各ペプチドが折り畳まって二次構造および三次構造を形成するのを確実にするのに充分な距離だけ離れさせてもよい。このようなペプチドリンカー配列は、当技術分野において周知の標準技術を用いて融合タンパク質中に組み入れられる。適切なペプチドリンカー配列は、次の因子に基づいて選択することができる:(1)可撓性の拡張された立体構造をとれること;(2)第1のペプチドおよび第2のペプチド上の機能的領域と相互作用し得る二次構造をとれないこと;ならびに(3)ペプチドの機能的領域と反応する可能性がある疎水性残基または荷電残基が無いこと。好ましいプチドリンカー配列は、Gly残基、Asn残基、およびSer残基を含む。ThrおよびAlaなど他のほぼ中性のアミノ酸もまた、リンカー配列中で使用されてよい。
【0053】
本発明の方法および用途
本発明は、ポリペプチドをコードするポリヌクレオチド配列の多様化を通して、関心対象のポリペプチドの変異配列を有するポリペプチドのレパートリーを作製する方法を提供する。典型的には、この方法は、多様化因子の発現を可能にする条件で本発明のB細胞を培養する段階を含み、B細胞の標的遺伝子は関心対象のポリペプチドのコード領域を含み、それによって、コード領域の多様化が可能になる。この方法は、複数の変異ポリペプチドおよび所望のレパートリーが得られるまで、B細胞の増殖を可能にする条件下で培養物を維持する段階をさらに含んでよい。B細胞は外表面でポリペプチドを発現するため、レパートリーの性質および範囲を決定することができる。次いで、所望の特性を有するポリペプチドを選択するためにレパートリーを使用することができる。
【0054】
別の態様において、本発明は、最適化された関心対象のポリペプチドを産生するB細胞を作製する方法を提供する。この方法は、多様化因子の発現を可能にする条件で本発明のB細胞を培養する段階を含み、B細胞の標的遺伝子は関心対象のポリペプチドのコード領域を含み、B細胞は関心対象のポリペプチドをその表面で発現する。この方法は、関心対象のポリペプチドに特異的に結合するリガンドに結合する細胞を選択することにより、関心対象のポリペプチドをB細胞表面で発現する細胞を培養物から選択する段階をさらに含む。関心対象のポリペプチドに特異的に結合するリガンドに対して所望の親和性を有する細胞を選択するまで、これらの培養段階および選択段階を繰り返すことができる。B細胞クローンの増殖中の抗体特異性の進化を図3に例示する。最初に多重特異性を示す集団(左上)を選択し、続いて、所望の特異性(中央)およびより高い親和性(右下)を富化する。
【0055】
関心対象のポリペプチドがIg、例えばIgL、IgH、または両方である態様において、リガンドは、抗原に相当する、組換えまたは他の手段によって作製されたポリペプチドでよい。例えば、磁気活性化細胞選択(MACS)による選択を容易にするために、リガンドを固体支持体に結合または連結することができる。別の例において、蛍光活性化細胞選別(FACS)を可能にするために、リガンドを蛍光タグに結合または連結することができる。当業者は、細胞を標識し選択する他の方法が公知であり、本方法において使用できることを理解している。
【0056】
本発明の方法は、培養物に調節分子を添加する段階をさらに含んでよく、調節分子は、シス調節エレメントへの係留因子の結合を調整し、それによって、コード領域の多様化を調整する。上記の例において、IPTG、テトラサイクリン、およびドキシサイクリンが、調節分子として働く。当業者は、多様化活性を調節するために個々の係留因子と共に使用できる他の調節分子を知っている。
【0057】
調節分子は、コード領域の多様化を様々な方法で調整することができる。例えば、いくつかの態様において、調節分子は、調整を行うために培養物に添加される。個々の調節分子がシス調節エレメントへの係留因子の結合を増加させる因子であるか、または減少させる因子であるかどうか、および結合の個々の変化が、多様化活性を増大させる効果を有するか、または低減させる効果を有するかどうかによって、調整の結果、多様化を低減もしくは停止するか、または多様化を促進もしくは加速化することができる。他の態様において、調整は、調整の低減であれ停止であれ、促進であれ加速であれ、調節分子を培養物から除去または取り除くことによって行われる。同様に、多様化の調整は、B細胞に遺伝子を添加することもしくはB細胞から遺伝子を取り除くことによって、またはB細胞における遺伝子の発現を増大させることもしくは減少させることによって行うこともできる。当業者は、上記に説明した利用可能な変更すべてを容易に理解することができ、これらはそれぞれ、B細胞における調節分子のレベルまたは存在を改変し、その結果として、シス調節エレメントへの多様化因子の係留を改変し、それによって、多様化活性を改変する効果を有する。
【0058】
本発明の方法のその他の用途には、IgλにおけるIg遺伝子標的の誘導性超変異が含まれる。DT40ポリLacO-λ LacI-HP1細胞株(下記の実施例4を参照されたい)は、Vλ偽Vアレイの下流の任意のIg遺伝子配列を改変するために使用することができる。Ig座位のゲノム構造は、V領域の変異誘発を促進するが、Ig分子の定常領域の変異誘発は促進しないように進化した。Ig座位におけるIg遺伝子配列の改変は、これをうまく利用して、変異誘発による産生物が機能的な定常領域を確実に保持するようにする。DT40において遺伝子を置換することは容易であるため、その遺伝子に有用な変化を起こすことができる。例えば、ニワトリ遺伝子(可変領域および定常領域)をヒト抗体遺伝子(重鎖または軽鎖のいずれか)で置換して、治療的用途のある抗体を作製することができる。また、これを用いて、他の種のための治療的抗体を作製することもできる。これにより、ヘテロ二量体抗体の一方の鎖のみまたは単鎖抗体の単鎖のいずれかに変異誘発するための迅速な系が提供される。
【0059】
このアプローチの用途には、高親和性Igを産生するB細胞クローンを作製するための超変異、特異的エピトープの認識を保持しつつ、抗体の交差反応性を改変するための超変異、特定の化合物に対して高親和性を有するV領域配列を同定するための超変異が含まれる。これは、単一のIg座位のみ、または両方で実施することができる。
【0060】
また、誘導性超変異はIgHで実施することもできる。DT40のIgH座位は、Igλと同様に、効率的な変異誘発部位である。類似技術によって作製された派生細胞株では、IgHおよびIgλの両方に、または各対立遺伝子に別々に変異誘発することが可能である。DT40ポリLacO-λポリLacO-H LacI-HP1派生体を作り出すために、IgHにポリLacOを挿入すると、H鎖遺伝子およびL鎖遺伝子の両方における超変異が可能になる。培地にIPTGを添加すると、鋳型式事象を減少させ、非鋳型式超変異を増加させる阻害効果がいくらかもたらされる。したがって、IPTGの存在および不在を変更することによって、抗体の両方の鎖をコードする遺伝子の鋳型式変異誘発および非鋳型式変異誘発を行ったり来たり切り替えることができる。
【0061】
一方の対立遺伝子におけるLacリプレッサー(LacI)のIPTGによる調節と他方の対立遺伝子におけるTetリプレッサー(TetR)のテトラサイクリンによる調節とを組み合わせた派生体を作製することができる。このTetR派生体では、IgH対立遺伝子およびIgL対立遺伝子で、かつIgL座位およびIgH座位で多様化を調節するための異なるメカニズムを同時に用いて、多様化を独立に調節することが可能になる。
【0062】
本発明はまた、T細胞受容体を選択するための媒体も提供する。T細胞に基づいた免疫療法には大きな可能性がある(BlattmanおよびGreenberg, 2004)。T細胞受容体の特異性および親和性は、CDR接触によって決定されている(Chlewicki et al., 2005)。特異性または高親和性のT細胞受容体の加速された誘導性選択は、Ig座位をT細胞受容体(V領域または遺伝子全体)で置換することによって改変したDT40ポリLacO媒体において実施することができる。
【0063】
触媒的Igの作製は、本発明の別の局面である。標的Igは触媒のためにも使用され得るため、本発明のIgに関連した方法は、結合および認識のためのIg作製だけに限定されない。酵素反応の遷移状態を模倣する安定な分子を開発した後、DT40ポリLacO-λ LacI-HP1細胞を用いて、結合し実際の化学的遷移状態を安定化させる抗体を得ることができる。中間体に結合できるIgを産生するクローンを同定した後、この系を再び用いて、培養状態での実際の基質に対するIgの触媒活性を求めてスクリーニングすることができる。この系において何らかの活性が実証された後で、変異誘発を通してIg座位をさらに進化させることによって活性の最適化を進めることができる。したがって、本発明は、動物の免疫化(時間のかかる段階)、ハイブリドーマ技術による不死化、および触媒活性を示す抗体を得るためにハイブリドーマをスクリーニングしなければならない非効率な後段階(later)を要求しない。
【0064】
IgλまたはIgHにおける非Ig遺伝子標的の誘導性超変異は、本発明の別の局面である。Ig座位のゲノム構造は、プロモーターの1〜1.5kb下流の変異誘発を促進するように進化した。この系を利用して、遺伝子の短い領域を変異させることができる。表面のタンパク質発現に基づいたクローン選択は、表面発現を媒介するエレメントを発現する遺伝子の一部分に関心対象の領域を融合することによって組み入れることができる。表面発現のための例示的なエレメントには、B細胞表面で発現されるタンパク質に由来するシグナルペプチド、膜貫通ドメイン、および細胞質尾部が含まれる(Chou et al., 1999; Liao et al., 2001)。
【0065】
本発明はまた、認識アレイを作製するために使用することもできる。広範囲の抗原に対する親和性を有する受容体を内部に持つ細胞を進化させることができると、認識アレイを開発することが可能になる。この技術を、DT40における受容体刺激に由来する細胞内応答/シグナル伝達(エクオリンによるCa2+の測定値(Rider et al., 2003)またはレポーター遺伝子転写の使用など)と組み合わせると、有用なバイオセンサーが作り出される。多様化されたクローンをアレイまたは96ウェルプレートにスポットし、試料に曝露させる。各試料は、刺激の「指紋」を生じるはずである。これらのアレイにより、生物学的/医学的試料、環境的試料、および化学的試料の定性的比較が可能になると思われる。あらゆる形態の化合物が抗原特性を有し得るため、2Dゲルのような比較技術と同様に、解析はタンパク質解析に限定される必要はない。さらに、これらのアレイにより、その存在を前もって知らずとも、構成要素が同定される。
【0066】
キット
本明細書において説明する方法で使用する場合、キットもまた、本発明の範囲内である。このようなキットは、バイアルおよびチューブなど1つまたは複数の容器(各容器は、本発明において使用される個別の要素(例えば、細胞、構築物)の内の1つを含む)を入れるように仕切られたキャリア、パッケージ、または容器を含んでよい。
【0067】
典型的には、キットは、本発明のB細胞ならびに対応する係留因子および多様化因子を発現する融合構築物を含む。例えば、B細胞は、プロモーターおよびコード領域を含む標的遺伝子に機能的に連結されたシス調節エレメントを含む。キットは、1種または複数種の融合構築物が中に保存された1つまたは複数の容器をさらに含む。各融合構築物は、B細胞で発現され得、かつ、多様化因子に融合された係留因子をコードするポリヌクレオチドを含み、この係留因子は、B細胞のシス調節エレメントに特異的に結合する。B細胞は、複数の融合構築物と共に使用するために複数のシス調節エレメントを含んでよい。
【0068】
典型的には、本発明のキットは、前述の容器、ならびに緩衝剤、希釈剤、フィルター、針、シリンジ、および使用に関する指示が書かれた添付文書を含む、商業的観点かつ使用者の観点から望ましい材料を含む1つまたは他の容器を含む。さらに、組成物が特定の治療的用途または非治療的用途のために使用されることを示し、かつ、使用する際の指示も示してよいラベルを容器の表面に貼ってもよい。指示およびまたは他の情報は、キットと共に含まれる挿入物に含まれてもよい。
【0069】
実施例
以下の実施例は、本発明を例示し、かつ当業者がそれを作製および使用するのを補助するために提示される。これらの実施例は、本発明の範囲を別な具合に限定することを決して意図しない。
【0070】
実施例1:DT40 B細胞株における変異およびターゲティング
本実施例は、2段階で準備される本発明の態様を例示する。
(1)DT40ポリLacO-λ
本発明者らは、Igλ座位でのシスな調節を可能にする遺伝子エレメントを含む、ニワトリB細胞株DT40のこの派生体を最初に構築した。DT40ポリLacO-λでは、重合ラクトースオペレーター(LacO)が、Igλ軽鎖座位の偽Vλアレイのすぐ上流に挿入されている。これは、StraightおよびBelmontによって開発された系を拡張したものであり、その中のポリLacOは、20bpのLacO結合部位の約80個の繰り返しからなる(Straight et al., 1996; Belmont, 2001)。本発明者らは、これが鋳型式変異誘発の正常なプロセスに影響を及ぼさないことを示した。DT40ポリLacO-λの構築は、下記の実施例2で詳細に説明する。
【0071】
LacOには、ラクトースリプレッサー(LacI)が非常に高い親和性(<10-12M)で結合する。このメカニズムは、大腸菌(E. coli)においてラクトースを代謝するためにオペロンの発現を調節し、また、様々な細胞の状況においても使用され得る。本発明者らは、蛍光タンパク質(GFP、RFP、YFP、またはCFP)に融合されたLacIを発現する安定なトランスフェクタントを作製し、かつ、GFP-LacI、RFP-LacI、YFP-LacI、またはCFP-LacIを安定に発現する正常に増殖中のDT40ポリLacの核において単一のスポットとして画像化され得る結合されたタンパク質を免疫蛍光顕微鏡検査法によって示すことにより、LacIがDT40ポリLacO-λ中のポリLacOに結合することを示した。
【0072】
IPTGは、インビトロおよび培養細胞中の両方で、LacOからのLacIの遊離を引き起こす小分子である。DT40ポリLacO-λ GFP-LacIをわずか10μMのIPTGと共に培養すると、ポリLacOからのGFP-LacIの遊離が起こる。IPTGと共に培養しても、細胞増殖は影響を受けない。
【0073】
(2)DT40ポリLacO-λ LacI-HP1
本発明者らは、クロマチン構造の改変因子であるキイロショウジョウバエ(Drosophila melanogaster)HP1(L. Wallrath(University of Iowa)から入手)がポリLacOに係留されると体細胞超変異への切り替えが起こる、DT40ポリLacO-λのこの派生体を次に構築した。係留は、非常に十分に特徴付けられているシス調節因子およびトランス調節因子のペアである大腸菌LacO/LacIを用いて実現した。HP1は、ヘテロクロマチン遺伝子サイレンシングおよびヘテロクロマチンの伝播において機能する、十分に特徴付けられている非ヒストンヘテロクロマチンタンパク質であり、ショウジョウバエでは、係留されたLacI-HP1が、クロマチン構造の閉鎖およびlacOリピートに隣接したレポーター遺伝子の不活性化を促進することが示されている(Li et al., 2003; DanzerおよびWallrath, 2004)。本発明者らは、HP1の係留により、ChIPによって測定されるように、開放状態から非許容状態へとドナー配列が変化すること、および遺伝子変換ではなく体細胞超変異による変異誘発をVλが受けるようになることを実証した。これは、鋳型式変異誘発から体細胞超変異への切り替えを引き起こすことが示されている、偽Vλアレイの欠失と機能的に等価である(Arakawa et al., 2004)。DT40 ポリLacO-λLacI-HP1のさらなる特徴付けは、下記の実施例4において実証する。
【0074】
(3)DT40ポリLacO-λ VP16-LacI
次に、本発明者らは、ポリLacOにVP16-LacIを係留するために、発現プラスミドp3’ss-EGFP-VP16(A. Belmont(University of Illinois)から入手)を使用した。VP16は、強力な転写活性化因子であり、ヒストンアセチルトランスフェラーゼ複合体を動員することが実証されている(Tumbar et al., 1999)。本発明者らは、VP16を係留すると、ChIPによって測定されるように、許容性のより高いクロマチン構造が作り出されること、および野生型のレベルを上回る遺伝子変換の刺激による変異誘発をVλが受けるようになることを実証した。
【0075】
実施例2:遺伝子を核膜孔に係留することによる、変異誘発の加速
本発明者らは、多様化している免疫グロブリン遺伝子が、細胞周期の過程で核内部を移動すること、この移動は多様化経路の段階と相関関係があること、および多様化は核周辺で開始することを発見した。活性化誘導型デアミナーゼ(AID)は、多様化を開始する酵素であり、核外輸送シグナルを有し、核膜孔を介して核の外へと絶えず輸送されるため、その濃度は核周辺で高くなり得る。このことから、核膜孔に遺伝子を接近させることにより、多様化を加速させることが可能であり得ることが示唆された。本発明者らは、Lacリプレッサー(LacI)への核膜孔タンパク質Nup153の融合物を用いて、DT40ポリLacOのIgH座位を核膜孔に係留することにより、これが事実であることを示した。本発明者らは、これにより、クローンの多様化速度が5.7倍に加速されることを見出した。対照実験により、ポリLacOへの係留が変異誘発の加速にとって不可欠であることが示された。この方法は、標的の変異誘発を促進するために、AID核外移行シグナルおよび遺伝子局在化に依存する一般的な方法である。したがって、この方法を拡張して、非Ig遺伝子を孔に係留することにより、B細胞における非Ig遺伝子の変異誘発を促進することができる。さらに、非B細胞でAIDを発現させることにより、それらの細胞における遺伝子の変異誘発を促進するように拡張することもできる。
【0076】
実施例3:E2Aは、細胞周期のG1期にシスで作用してIg遺伝子多様化を促進する
本実施例は、核におけるIg遺伝子多様化の細胞周期に依存的な調節を説明する。再配列されたIg遺伝子は、DNAデアミナーゼAIDによって開始される配列および構造の多様化を経験する。多様化が起こるためにはIg遺伝子が転写されなければならないが、シス活性化のためにその他の必要条件があるかどうかは明らかにされていない。本実施例は、クロマチン免疫沈降法によって、調節因子E2Aが、構成的なIg遺伝子多様化を実行するニワトリDT40 B細胞株中の再配列されたIgλR遺伝子と結合することを示す。再配列されたλR遺伝子を重合ラクトースオペレーターによってタグ化したDT40派生体を直接画像化することにより、本発明者らは、λR/E2A共局在化が細胞周期のG1期に最も顕著であることを示す。本発明者らは、E2AアンタゴニストId1の発現により、G1期のλR/E2A共局在化が妨害され多様化が減退するが、λR(以下、IgλR遺伝子をIgλまたはλと呼ぶことがある)の転写は減退しないことをさらに示す。したがって、E2Aは、シスで作用してIg遺伝子多様化を促進し、G1期は、E2A作用のための決定的に重要な期間である。
【0077】
Ig座位で起こるゲノム配列および構造の調節された変化は、これらの遺伝子へのDNA損傷のターゲティングおよび正確な修復の回避の両方を反映している。体細胞超変異、クラススイッチ組換え、および遺伝子変はすべて、B細胞特異的酵素である活性化誘導型デアミナーゼ(AID)によって開始される(5〜8)。AIDはシトシンを脱アミノしてウラシルにし、単鎖DNAを好むことが明白である(9〜11)。転写は多様化するための必要条件であり、これは、AIDが単鎖基質を好むことを反映している可能性がある。DNA中のウラシルはよく起こる損傷であり、高度に保存された効率的な経路によって正確に修復することができる(12)。しかしながら、Ig座位は正確な修復を回避し、誤りがちな経路による修復を受けることができる(13)。
【0078】
E2Aは、bHLHタンパク質のEファミリーのメンバーであり、リンパ球発達の多くの局面の決定的に重要な調節因子である(14〜17)。Eタンパク質は二量体化してEボックスモチーフCANNTGに結合し、それらの機能は、Eタンパク質とヘテロ二量体化してDNA結合を妨害するIdタンパク質によって弱められる。E2Aは、活性化されたマウスB細胞において誘導され、クラススイッチ組換え(18)ならびにAIDをコードする遺伝子の発現(19)を調節する。ニワトリB細胞において、E2A遺伝子が不活性化されると、Igλ遺伝子の多様化が減退するが転写は減退しない(20、21)。一方、逆に、E47(E2Aにコードされる機能的に等価なアイソフォーム2つの内の1つ)が異所発現されると、Igλ遺伝子の多様化が促進されるが、Igλ転写レベルは影響を受けない(22)。
【0079】
E2Aがシスの部位に結合することによってIg遺伝子多様化を調節する可能性は、マウスにおいて、多量体化したEボックスは超変異を刺激するがIg導入遺伝子の転写は刺激しないという証拠に基づいて最初に示唆された(23)。この可能性は、ニワトリB細胞において、多量体化したEボックスはIg遺伝子多様化を促進できるが転写は促進できないことの証明によってさらに裏付けられた(24)。しかしながら、E2AがIg遺伝子に直接作用して多様化を促進するかという疑問を完全に解消することは、いくつかの理由によって困難であった。Eボックスは、特定の状況においてのみ、E2A依存的調節のための部位として機能し、したがって、Eボックスの存在は、ある部位でのE2A機能を保証するものではない。Eボックスモチーフのあいまいな一致(loose consensus)および頻繁な出現により、各個別部位の変異解析は妨げられる。いくつかの部位で、E2Aはタンパク質-DNA相互作用ではなくタンパク質-タンパク質相互作用によって動員され、したがって、Eボックスは、常にE2A依存的調節の必要条件であるとは限らない(25)。
【0080】
本発明者らは以下に、再配列されたIgλ対立遺伝子が、重合ラクトースオペレーターでタグ化された、構成的に多様化するニワトリB細胞株DT40の派生体(DT40ポリLacO-λR)を利用する実験において、E2AがIg遺伝子でシスに作用して多様化を促進することを証明した。クロマチン免疫沈降法(ChIP)により、本発明者らは、E2Aが、親株DT40において、再配列されたIgλ対立遺伝子とは結合するが再配列されていないIgλ対立遺伝子とは結合しないことを示す。本実施例では、DT40ポリLacO-λR細胞において、E47をλRに効果的に係留するE47-LacI融合タンパク質が発現されると多様化が加速すること、およびE47-LacI発現の刺激効果が、IPTGと共に培養された細胞では明らかではなく、したがって、シスに結合することが多様化を促進するのに必要であることを実証する。DT40ポリLacO-λR GFP-LacI細胞中の再配列されたλR遺伝子を直接画像化することにより、本発明者らは、λR/E2A共局在化がG1期に顕著であること、およびE2AアンタゴニストであるId1の発現により、多様化が減退し、具体的にはG1期のλR/E2A共局在化が減少するが、λ転写レベルも活性な転写場所へのλRの局在化も影響を受けないことを示す。これらの結果から、E2AがG1期にシスで作用してIg遺伝子多様化を促進することが示される。
【0081】
材料および方法
細胞培養、トランスフェクション、sIgM喪失アッセイ法、および細胞周期の解析
以前に説明されたようにして、ニワトリ嚢リンパ腫株DT40およびその派生体DT40ポリLacO-λRを維持し、トランスフェクトした(26、 27)。S003 E47プラスミド(28)(Cornelis Murre(University of California, San Diego, California)によって提供された)に由来するE47 cDNAを、p3’SS-GFP-LacIプラスミド(Andrew Belmont(University of Illinois, Urbana, Illinois)によって提供された)中にサブクローニングすることによって、E47-LacI発現構築物を作製した。Id1発現構築物(29)は、Barbara Christy (University of Texas, San Antonio, Texas)によって提供された。sIgM喪失アッセイ法を以前に説明されているようにして実施し(26、30)、Rソフトウェアパッケージ(http://www.r-project.org)を用い、マンホイットニーU検定によって結果を比較した。DNA含有量に基づいて細胞周期プロファイルを得るために、指数関数的に増殖中の細胞1×106個を0.1% Triton X-100中に懸濁し、200μg/mlのRNase Aおよび50μg/mlのヨウ化プロピジウムで処理し、以前に説明されたようにして解析した(26)。
【0082】
ChIP解析
以前に説明されているようにして、クロマチンを調製し、免疫沈降した(27、31、32)。その際、抗E2A抗体(ab11176; Abcam)または対照IgGを使用した。VλRおよびVλUに対する以前に説明されたプライマー(27)ならびに卵白アルブミン(Ova)に対するプライマー
を使用し、FastStart Taq DNAポリメラーゼ(Roche)を用いて半定量的PCRを実施した。ImageQuantソフトウェア(Amersham)を用いてPCR産物を定量した。対照IgGの場合の比率に対して標準化した、関心対象のアンプリコンとOvaアンプリコンの比率として富化を算出した。例えば、富化VλR=(抗E2A[VλR/Ova])/(IgG[VλR/Ova])。
【0083】
RT-PCRおよびウェスタンブロット法
RT-PCRアッセイ法のために、説明されているようにしてプライマーを用いてAIDおよびb-アクチンを増幅し(7)、プライマー
を用いてIgλ転写物を増幅した。ウェスタンブロット法のために、DT40ポリLacO-λR RFP-LacIおよびDT40ポリLacO-λR RFP-LacI Id1に由来する全細胞溶解物(50μg)を分離し、抗Id1抗体(JC-FL; Santa Cruz)を用い、FluorChem HD2 (Alpha Innotech)によってId1タンパク質を検出した。
【0084】
蛍光顕微鏡検査および画像解析
ポリLacOを画像化するために、GFP-LacI発現構築物p3’SS-GFP-LacI(Andrew Belmont(University of Illinois, Urbana, Illinois)から入手)(SV40核移行シグナルを含むように操作され、四量体形成に必要な配列を欠いているLacIをコードする)(33)、またはその派生体RFP-LacI(GFPがRFP(DsRed単量体;Clontech)で置き換えられている)をDT40ポリLacO-IR細胞にトランスフェクトした。免疫染色のために、Cytospin3(800rpm、4分;Shandon)を用いてスライドガラス上に細胞(約3×105個)を載せ、2%パラホルムアルデヒドで20分間固定し、以前に説明されたようにして染色した(26)。使用した一次抗体は、抗E2A(ab11176、1:200;Abcam)、Ser5でリン酸化された抗Pol II C末端ドメイン(ab5131、1:500;Abcam)であった。Alexa Fluor 488または594を結合させた抗IgG(Molecular Probes)を二次抗体として使用した。DeltaVision顕微鏡検査システム(Applied Precision)を用いて蛍光画像を獲得し、softWoRxソフトウェア(Applied Precision)およびImarisソフトウェア(Bitplane)を用いて処理および解析を行った。蛍光シグナルは、時折、完全にではなく部分的に重なることがあり、これは、ポリLacO-タグとVλ領域の間にかなりの距離(約17kb)があることを反映している可能性がある;どちらの配置も共局在化として記録した。共局在化の割合をピアソンのカイ2乗検定を用いて解析した。少なくとも2回の独立した直径測定値を平均し、2で割って、核半径を算出した。平均核半径の細胞周期依存性を各細胞株に関して独立に決定し、比較的不変であることが判明した。核半径と細胞周期の相関関係を示すために用いた標準値は、次のとおりであった:G1、r<4μm;G2、r≧5.2μm。
【0085】
結果
E2Aは、再配列されたIgλ遺伝子と結合するが、再配列されていないIgλ遺伝子とは結合しない
Ig遺伝子多様化におけるE2Aの重要性を示すかなりの証拠があるものの、この因子がIg遺伝子と直接結合することは示されていなかった。E2AとIgλの結合を試験するために、本発明者らは抗E2A抗体を用いて、ニワトリDT40 B細胞株からクロマチンを免疫沈降させた。この株は嚢リンパ腫に由来し、遺伝子変換によってIg重鎖遺伝子および軽鎖遺伝子の両方の構成的多様化を実施する。11kbのニワトリλ軽鎖座位の探索により、E2AのコンセンサスCANNTGに対する50個を超えるマッチ、VλとψVλ1(上流の偽遺伝子の内で最も近位のもの)の間の領域に17個、J-Cイントロンのマトリックス結合領域(MAR)に2個、および3’エンハンサーに6個が確認された。DT40 B細胞において、機能的対立遺伝子は、VセグメントとJセグメントを連結する1.8kb領域を欠失するVJ組換えをB細胞発達初期に経験するが、不活性なλ対立遺伝子は再配列されていないため(図4A)、2つの対立遺伝子をPCRによって容易に区別することができる。クロマチン免疫沈降(ChIP)後、再配列されたおよび再配列されていないλ対立遺伝子の回収物を、対照遺伝子アルブミンと比べて分析した。これにより、E2Aは、再配列されたVλR対立遺伝子において17.6倍に富化されているが、再配列されていないVλU対立遺伝子では富化されていないことが示された(図4B)。したがって、E2Aは、再配列されたVλR対立遺伝子と直接結合する。
【0086】
E2Aはシスに作用してIgλ多様化を調節する
E2Aがシスに結合して多様化を促進するに違いないかを問うために、本発明者らは、DT40の派生体であるDT40ポリLacO-λR(重合ラクトースオペレーターが相同遺伝子ターゲティングによってψVλアレイ中に挿入されており(図5A)、ラクトースリプレッサー(LacI)との融合物として発現される因子は、再配列されたλR対立遺伝子に係留され、IPTGとの培養によって遊離されることが可能である)を利用した(27)。細胞周期分布およびクローンのIg遺伝子変換速度は、DT40ポリLacO-λRおよび野生型DT40において同程度であった(図5B、5C)。LacIに融合されたE2AアイソフォームE47を発現するプラスミド(E47-LacI)またはLacIに融合された緑色蛍光タンパク質を発現する対照プラスミド(GFP-LacI)をDT40ポリLacO-λR細胞に安定にトランスフェクトした。細胞周期分布は、GFP-LacIトランスフェクタントおよびE47-LacIトランスフェクタントにおいて同程度であったが、後者の株の培養物は、いくつかのサブG1(アポトーシス性)細胞を含んだ(図5D)。E47-LacIトランスフェクタントにおいてIgλ転写物のレベルは変わっていなかったことから(図5E)、ニワトリB細胞においてE2AはIg遺伝子発現を調節しないことを示す、公開されている結果が確認された(21、22)。AID転写物のレベルは、E47-LacIトランスフェクタントの方が約3倍高かった(図5E)。E47の異所発現に応答したAID発現の同様の増大が、他の研究者によって観察されている(22)。
【0087】
E2Aが、Igλへの結合を介して多様化を直接的に調節するかを問うために、本発明者らは、IPTGの存在下および不在下で独立したE47-LacI(n=19)トランスフェクタントまたはGFP-LacIトランスフェクタント(n=13)を培養し、sIgM喪失変動アッセイ法を用いて、クローンの多様化速度を決定した(26、27、30)。このアッセイ法では、遺伝子変換、点変異、欠失、または挿入によってそれらが起こるかどうかに関わらず、不活性化変異を記録し、それによって、変異誘発の結果とは無関係な起因事象を定量する。この解析により、E47-LacIトランスフェクタントのクローン多様化速度が、GFP-LacI対照と比べて4.5倍速いことが示された(P=0.019、マンホイットニーU検定、図5F)。さらに、E47-LacIをポリLacOから遊離させるIPTGと共に培養すると、DT40ポリLacO-λR E47-LacI細胞の多様化速度はバックグラウンドレベルにほぼ戻ったが、GFP-LacI対照には影響を及ぼさなかった(図5F)。したがって、E47-LacIは、シスに作用することによって多様化を促進する。
【0088】
E2Aは細胞周期のG1期にIgλRに局在化する
ヒトB細胞において、細胞周期のG1期の受容体架橋は、体細胞超変異を開始して、同定可能な変異を90分以内に生じさせることができる(34)。したがって、E2AがIgλで作用する細胞周期の段階を決定することが関心対象であった。再配列され、多様化しているλR遺伝子は、GFP-LacIを発現するDT40ポリLacO-λR細胞において明るい点として容易に画像化することができる(27)。しかしながら、細胞をHoechst 33342で染色し、DNA含有量に基づいて選別してG1段階、S段階、またはG2/Mの細胞を富化すると、タグ化された遺伝子に由来するはっきりした蛍光シグナルを提示している細胞の割合は、未選別の細胞で通常観察される90〜95%から、約45%に減少した。したがって、シグナルのこのような減少は結果を偏らせ得るため、本発明者らは、異なるアプローチによって細胞周期段階を決定した。Hoechst 33342で染色し選別した細胞の解析により、G1期の核のサイズはG2/M期の細胞よりもかなり小さいことが示された(例えば、図6A)。したがって、本発明者らは、Hoechst 33342で染色しDNA含有量に基づいて選別した、指数関数的に増殖中のDT40ポリLacO-λR集団に由来するG1細胞(n=55)およびG2細胞(n=55)の核半径を測定することにより、核半径(r)を用いて細胞周期の段階を明らかにできるかを問うた。G1細胞の平均核半径は3.8±0.3μmであり、G2細胞の平均核半径は5.4±0.5μmであった(図6B)。核半径によって決定したG1細胞:S細胞:G2細胞の比率(3:6:1)および染色によって決定した比率(2.7:5.5:1.7)を比較して、このアプローチをさらに確認した。したがって、G1細胞はr<4μm、G2細胞はr>5.2μmであることが実験で確認された。
【0089】
λR/E2Aの共局在化は、抗E2A抗体で染色したDT40ポリLacO-λ GFP-LacI細胞のデコンボリューション顕微鏡解析によって容易に同定された(例えば、図7A)。λR/E2A共局在化は、非同期細胞(n=227)の26%で明らかであった。共局在化の細胞周期分布の解析により、λR/E2A共局在化の45%がG1期の細胞で起こり、38%がS期の細胞で起こり、17%がG2期の細胞で起こることが示された(図7B)。したがって、G1期細胞の割合(25%)と比べて、G1期におけるλR/E2A共局在化(45%)は明らかに過剰であった(P<0.0001、カイ2乗検定)。
【0090】
本発明者らはまた、λR転写の細胞周期依存性も決定して、RNAポリメラーゼIIのC末端ドメインのリン酸化Ser5(P*-Pol II)(伸長しているPol II分子に特徴的な改変)に対する抗体で染色することによって、活性な転写場所を同定した(35)。DT40ポリLacO-λR RFP-LacI細胞の非同期細胞集団において、核の全体にわたって多数の活性な転写場所を同定することができ、λR/P*-Pol II共局在化が、19%の細胞で容易に観察された(n=392;例えば、図7C)。λR/P*-Pol II共局在化の細胞周期分布の解析により、共局在化の21%がG1期の細胞で起こり、58%がS期の細胞で起こり、21%がG2期の細胞で起こることが示された(図7D)。これは、細胞周期分布に類似している。したがって、λRは細胞周期の全体を通して転写されるが、λR/E2A共局在化はG1期で顕著である。
【0091】
Id1発現はIg遺伝子多様化およびG1期のλR/E2A共局在化を阻害する
G1期のλR/E2A共局在化が多様化にとって決定的に重要であるかを問うために、本発明者らは、これらの共局在化に対するId発現の影響を測定した。IdはE2Aに拮抗し、DT40 B細胞におけるId1またはId3の発現は、Ig遺伝子多様化を減弱させることが以前に示されている(22)。本発明者らは、安定なDT40ポリLacO-λR RFP-LacI Id1トランスフェクタントを作製し、ウェスタンブロット法によってId1発現を確認し(図8A)、Id1発現によって細胞周期プロファイルが変化しないことを示した(図8B)。本発明者らは、Id1発現により、クローンのIg遺伝子多様化速度が低下する(P<0.001、マンホイットニーU検定;図8C)が、Igλ転写物のレベルにもAID転写物のレベルにも影響を及ぼさないことを確認した(図8D)。次いで、本発明者らは、抗E2A抗体で染色することにより、Id1発現派生体および親株におけるλR/E2A共局在化を比較した。λR/E2A共局在化は、DT40ポリLacO-λR RFP-LacI細胞での26%に対し、非同期DT40ポリLacO-λR RFP-LacI Id1細胞(n=90)では13%で明らかであった(P=0.0030、カイ2乗検定)。
【0092】
Id1発現が細胞周期の特定の段階における共局在化に影響を及ぼすかどうかを判定するために、Id1トランスフェクタントにおけるλR/E2A共局在化を、核半径および細胞周期に関して定量した。これにより、DT40ポリLacO-λR RFP-LacI Id1細胞において、共局在化の10%がG1期の細胞で起こり、58%がS期の細胞で起こり、32%がG2期の細胞で起こることが示された(図8E)。したがって、Id1発現により、G1期のλR/E2A共局在化の割合が有意に減少し、親株での45%からId1トランスフェクタントでは10%となった(P<0.0001、カイ2乗検定;図8E)。Id1トランスフェクタントでのλR/E2A共局在化の90%は細胞周期のG1期より後に起こった。Id1トランスフェクタントで明らかな多様化の減弱と総合すると、これにより、G1期が、E2Aが多様化を促進する決定的に重要な期間であることが示される。
【0093】
λR/P*-Pol II共局在化は、活性な転写場所に対する抗体で染色したDT40ポリLacO-λR RFP-LacI Id1細胞(n =290)の18%で同定され、親株と類似していた(19%、P=0.80、カイ2乗検定;図7D)。また、λR/P*-Pol II共局在化の細胞周期プロファイルも、Id1トランスフェクタントと親株とで類似していた(図8F)。λR/P*-Pol II共局在化に対するId1発現の影響が無いことは、DT40ポリLacO-λR RFP-LacI Id1トランスフェクタントのIgλ転写物レベルが低下しないことと一致している(図8D)。したがって、Id1発現は、λRとE2Aの共局在化の細胞周期分布に影響を及ぼすが、P*-Pol IIとの共局在化の細胞周期分布には影響を及ぼさない。
【0094】
考察
これらの結果から、E2Aはシスに作用してIg遺伝子多様化を促進するに違いないこと、およびG1期が、このプロセスでE2Aが機能する決定的に重要な期間であることが示される。これらの実験では、ポリLacOでタグ化し、GFP-LacIまたはRFP-LacIに結合させることによって画像化したλ遺伝子を検査した。これは、遺伝子多様化を研究するための強力なアプローチを提供する。タグ化された座位は、固定した細胞の90%超で視認できるため、多様化に関与する因子との共局在化の解析が可能になる。潜在的な調節因子を係留し、IPTGとの培養によって遊離させることができるため、他の標的から独立してIgλ座位における因子の効果を研究することが可能になる。これは、大きくかつ複雑な調節階層の上位で機能するE2Aのような因子にとって特に有用である(36)。
【0095】
これらの結果から、Ig座位との物理的結合により、E2AがIg遺伝子多様化を直接的に調節することが確かめられる。ChIPにより、E2AがDT40 B細胞中の再配列されたλR対立遺伝子と結合することの明白な証拠が提供された。E2Aがシスに機能するに違いないということは、DT40ポリLacO-λR細胞においてE2A(E47-Lac融合物)をIgλ対立遺伝子に係留した結果起こる多様化の加速が、LacOからLacIを遊離させるIPTGと共に培養した細胞では明らかではないことを示すことによって確かめられた。
【0096】
E2Aは転写調節因子として最もよく知られているが、Igλ転写物のレベルはE47の異所発現によって変わらなかったため、多様化におけるE2Aの機能は、Igλでの転写活性化を反映したものではない。このことは、ニワトリおよびマウスB細胞において多様化に対するE2Aの影響を検査した他の研究者による結果を裏付ける(20、22、23)。さらに、G1期で最も顕著であるλRとE2Aの共局在化は、 細胞周期分布に類似している活性な転写場所へのλRの細胞周期分布とは違っていることから、λRが細胞周期の全体を通して転写されることが示唆される。多様化および転写におけるE2A機能の独立性は、再配列されたλRとE2Aの共局在化および活性な転写場所に対するId1発現の影響を対比することによってさらに裏付けられる。Id1発現により多様化は減退し、また、λRとE2Aの共局在化も減少した。一方、Id1発現は、活性な転写場所へのλRの局在化の数にも細胞周期分布にも影響を及ぼさなかった。λRを転写場所に動員するのにE2Aは必要とされないということは、Igλ転写に対するId1発現の影響が無いことと一致している。
【0097】
E2Aは、AID発現を調節し、これはIg遺伝子多様化を間接的に刺激する。AID遺伝子は、マウスB細胞においてE2Aによる転写調節の標的であることが示されていた(19)。本発明者らおよび他の研究者ら(22)は、E2Aの異所発現により、DT40ニワトリB細胞におけるAID転写物レベルが上昇することを示した。これは、IPTGと共に培養したDT40ポリLacO-λR E47-LacI細胞において明らかな中等度の多様化加速に寄与するか、または原因である可能性がある。E2Aの除去によってニワトリB細胞中のAID転写物レベルは低下しないことが報告されているものの(21)、E2Aと機能が重複した(redundant)因子が、E2Aの不在下で最低レベルのAID発現を保証している可能性がある。
【0098】
これらの結果から、G1期が、IgλでE2Aが機能する決定的に重要な期間であると特定される。本発明者らは、細胞周期の他の段階と比べてG1期のλR/E2A共局在化が過剰であることを観察し、Id1発現によってG1期の共局在化が特異的に減少し、多様化速度が低下することを示した。Idタンパク質はEタンパク質とヘテロ二量体化して、DNA結合を阻害する(14)。G1の間の共局在化がId1発現によって特異的に影響されることは、細胞周期の間のE2AとIgλの結合の様式が異なることを示している可能性がある。S期およびG2期では、E2Aは、DNAへの直接結合によってではなく、他のタンパク質との相互作用を介して動員され得る。
【0099】
その他の種類の証拠により、多様化がG1期に開始されるという考えが裏付けされる。ヒトBL2細胞株での体細胞超変異は、G1期のみに行われるインビトロ刺激によって誘導することができ、点変異は、刺激から90分以内、細胞がまだG1期にある間に最初に明らかになる(34)。クラススイッチ組換えのために活性化されたマウスB細胞において、スイッチ組換え経路の関与物であるNBS1またはγ-H2AXとのIgH共局在化は、G1期で顕著であり(37)、S領域でのDNA切断はG1期に検出され得る(38)。DNA切断はまた、超変異しているヒトB細胞株において細胞周期の後期にも確認されるが(39)、これらはAID依存性であることが判明した(40)。
【0100】
E2Aは、G1期に機能して、細胞周期の後期、またはさらに、後続の細胞周期の間に起こる事象のために座位を準備することができる。最近、E2AはヒストンH4アセチル化の維持に関係があるとされており(21)、娘細胞でのAID攻撃または効果的な多様化に都合の良い局所的なクロマチン環境を確立するようにE2Aが機能することが可能である。
【0101】
参考文献
【0102】
実施例4:クロマチン構造は遺伝子変換を調節する
本実施例は、ニワトリB細胞株DT40をモデルとして用いて、クロマチン構造が修復用のドナーとしての相同配列の使用に寄与する方法を示す。DT40において、免疫グロブリン遺伝子は、偽遺伝子ドナーを鋳型とする遺伝子変換による、調節された配列多様化を経験する。本発明者らは、Vλ偽遺伝子アレイが活性クロマチンに関連したヒストン改変を特徴とすることを見出した。本発明者らは、Lacリプレッサーとの融合物として発現されるヘテロクロマチンタンパク質HP1が、偽Vλドナーアレイ内部に組み込まれた重合ラクトースオペレーターに係留される調節可能な実験系を用いて、遺伝子変換のためにクロマチン構造が重要であることを直接的に実証した。HP1が係留されると、偽Vλドナーアレイ内部でのヒストンアセチル化が減少し、Vλ多様化の結果が変わり、その結果、鋳型式変異よりも非鋳型式変異が優勢になった。したがって、クロマチン構造は、相同性の指示に従った修復を調節する。これらの結果から、ヒストン改変が、反復配列間の組換えを妨害することにより、ゲノム安定性の維持に寄与し得ることが示唆される。
【0103】
本実施例では次の略語を使用する:AcH3、アセチル化ヒストンH3;AcH4、アセチル化ヒストンH4;AID、活性化誘導型デアミナーゼ;DSB、二本鎖切断;GFP、緑色蛍光タンパク質;Ig、免疫グロブリン;MRN、MRE11/RAD50/NBS1;NHEJ、非相同末端結合;V、可変性;UNG、ウラシルDNAグリコシラーゼ;AP、脱塩基;ポリLacO、重合ラクトースオペレーター;ChIP、クロマチン免疫沈降法;Ova、卵白アルブミン;LOH、ヘテロ接合性の喪失。
【0104】
材料および方法
クロマチン免疫沈降法(ChIP)
以前に説明されたようにしてChIPを実施した[48、89]。すべての実験において、安定にトランスフェクトされた少なくとも2つの独立な株に由来する少なくとも2つのクロマチン調製物を解析した。図面は1つの代表的な実験を示し、4回の別々の増幅の解析結果を用いて標準偏差を算出した。鋳型DNAの段階希釈物を4回別々に増幅して、測定される産生物強度が直線範囲内であることを確かめた。実験アンプリコンの富化を、二重PCRによって同じチューブ中で増幅した卵白アルブミン(Ova)遺伝子に由来する内部対照アンプリコンの富化に対して標準化し、特異的抗体を用いたChIPの際の富化は、対照の全インプットDNAを用いてChIPを実施した並行実験に対して標準化した。Ova内部対照アンプリコンを含めることによって、本発明者らは、IP効率、バックグラウンドから残った影響、およびゲル添加の差異に関して標準化することができた。富化=[(ψVλ/Ova)Ab]/[(ψVλ/Ova)インプット]。追加の対照として、対照の全インプット中の実験アンプリコンと対照アンプリコンの比率を、多特異性IgGを用いた対照ChIPと比較した。いずれの場合も、インプット対照およびIgG対照の富化は本質的に等しかった。代表的な実験のデータを提示する。標準偏差は、鋳型DNAの段階希釈物を4回別々に増幅した結果から算出した。
【0105】
使用した抗体は、Upstate (Lake Placid, NY)社製の抗AcH3(06-599)、抗AcH4(06-866)、およびジメチル化H3(K4)(07-030)であった。ChIP用のPCRプライマーは次のものであった:
【0106】
構築物、トランスフェクション、および細胞培養
pAIDPuro(H. Arakawa(Munich, Germany)から入手)中のAIDをL. Wallrath (University of Iowa, Iowa City)によって提供された構築物に由来するLacI-HP1aで置換して、ニワトリβ-アクチンプロモーターの下流にLacI-HP1を配置することによって、LacI-HP1発現プラスミドを作製した。GFP-LacI発現プラスミド(p3’ss-EGFP-LacI)は、A. Belmont(University of Illinois, Urbana)によって提供された。細胞培養およびトランスフェクションを、以前に説明されたようにして実施した[47]。転写されたVλRの17kb上流のψVλ17〜ψVλ20の間の領域を標的とするように設計されたアームが隣接する約3.8 kbの重合ラクトースオペレーター(ポリLacO)を有する構築物を用いて、相同遺伝子ターゲティングによってDT40ポリLacO-λRを作製した。手短に言えば、相同な組み込み体をPCRによって同定し、Cre発現によって選択マーカーを欠失させた。DT40嚢リンパ腫は、1つのIgλ対立遺伝子のみが再配列されており、2つの親染色体(chromome)が、ψVλ17付近の多型性に基づいて区別されるB細胞に由来する。これにより、本発明者らは、再配列された対立遺伝子または再配列されていない対立遺伝子が標的とされたかどうかをPCRによって判定することができた。対照実験を行って、DT40ポリLacO-λR細胞、DT40ポリLacO-λR GFP-LacI細胞、およびDT40ポリLacO-λR LacI-HP1細胞の細胞周期分布が類似していること、ならびに最高500μMのIPTGと共に7日間細胞を培養しても、DT40ポリLacO-λRGFP-LacI対照細胞の増殖速度にもψVλ17Rにおけるクロマチン改変にも影響は無いことを確かめた。Vλ配列を解析するためのオリゴヌクレオチドは以前に説明されている[47]。
【0107】
蛍光画像法
蛍光画像法のために、細胞(2×105個)をサイトスピンでスライドガラス上に載せ、2%パラホルムアルデヒドで20分間固定し、0.1%NP-40で15分間透過処理し、以前に説明されたようにして染色した[90]。一次染色には抗LacIモノクローナル抗体(1:500希釈;Upstate)を用いた。二次抗体は、ロバ抗マウスIgG Alexa Fluor 594 (1:2000; Molecular Probes, Eugene, OR)であった。核を可視化するために、細胞をDAPI(Sigma, Saint Louis, MO)で染色した。DeltaVision顕微鏡検査システム(Applied Precision)を用いて蛍光画像を獲得し、softWoRxソフトウェア(Applied Precision)を用いて処理した。
【0108】
RT-PCR
TRIzol試薬(Invitrogen)を用いてRNAを細胞から回収し、PreAnalytiXカラム(Qiagen)を用いて精製した。鋳型の希釈(1:1300)後にVλ転写物を増幅し、未希釈の試料からβ-アクチンを増幅した。Vλを増幅するためのプライマーは、
であった。β-アクチンを増幅するためのプライマーは、以前に説明されている[36]。
【0109】
sIgM喪失変異体の定量および配列解析
変異したV領域におけるフレームシフトまたはナンセンス変異の結果として生じ、蓄積したsIgM喪失変異体を測定する、sIgM喪失変異体のアッセイ法を用いて、IgV領域の多様化を定量した[47、50]。手短に言えば、フローサイトメトリーとそれに続く限界希釈クローニングによってsIgM+細胞を単離し、4週間増殖させた。sIgM-細胞の割合を定量するために、約1×106個の細胞を抗ニワトリIgM-RPE(Southern Biotechnology Associates, Birmingham, AL)で染色し、CellQuestソフトウェア(BD Biosciences)を用いたFACScanによって解析した。
【0110】
単一細胞PCRおよび配列解析を、以前に説明されたようにして実施した[47]。手短に言えば、sIgM-細胞を選別し、個々のウェルに等分し、Vλ領域を増幅および配列決定し、それらの配列をψVλドナーと比較して、変異が鋳型式であるか、または非鋳型式であるかを判定した。鋳型式変異の判定基準は、ドナーとレシピエントにおいて9個の連続した塩基が完全に一致しなければならないというものであった。配列は、独立にトランスフェクトされた2つの株に由来した。特有の配列のみを、変異の分類のために含めた。
【0111】
結果
VλおよびψVλドナー鋳型における許容性クロマチン構造
DT40 B細胞において、一方のIgλ対立遺伝子の機能的なVλ遺伝子は再配列され発現され、他方は再配列されず発現されない。本発明者らは、再配列された対立遺伝子(VλR)および再配列されていない対立遺伝子(Vλu)ならびにψVλアレイにおけるクロマチン構造をクロマチン免疫沈降法(ChIP)によって特徴付けた。ChIPは、ヒストンH3およびH4のN末端におけるリジンアセチル化に特異的な抗体を用いて実施した。回収されたDNAを二重PCR反応で増幅した。回収量は、B細胞では発現されない卵白アルブミン(Ova)遺伝子に由来するアンプリコンに対して標準化し、富化は、対照の全DNAインプットに対して標準化した(詳細については、材料および方法を参照されたい)。VλRおよびVλuはゲノム構造が異なるため、特異的プライマーを用いたPCRによってそれらを区別することができる。ChIPにより、再配列されたVλR遺伝子においてアセチル化ヒストンH3およびH4(AcH3およびAcH4)がかなり富化していることが実証された。典型的な実験において、AcH3のVλRにおける富化率は80倍を超え、AcH4の富化率は30倍を超えた(図9B)。一方、再配列されていないVλU対立遺伝子におけるAcH3およびAcH4のレベルは、VλRでのレベルよりはるかに低く(それぞれ、16分の1および7分の1)、インプットDNAと比べて数倍にしか富化しなかった。
【0112】
7箇所の部位(ψVλ1とVλ遺伝子の間の領域、ψVλ1、ψVλ5、ψVλ13、ψVλ18、ψVλ24、ψVλ25を含む)および上流のフランキング領域を調べるプライマーを用いた増幅によって、ψVλアレイ内のクロマチン構造を分析した。(多型性が足りないために、DT40中の2つのIgλ対立遺伝子におけるψVλアレイはPCRによって容易に区別することができない)。印象的なことには、本発明者らは、ψVλアレイの全体にわたってAcH3およびAcH4がかなり富化していることを観察した(図9B)。VλRから遠位の部位が、近位部位よりも低レベルの富化を一貫して示すということはなく、富化は、転写されたVλR遺伝子からの距離に比例していなかった(図9B)。したがって、ψVλアレイ内のアセチル化ヒストンの富化は、転写されたVλR遺伝子から上流部位へのクロマチン改変の段階的な伝播を単に示すわけではない。この座位のクロマチン構造が非均一であることから、ψVλアレイのクロマチン構造を調節するシスエレメントの存在が示唆される。
【0113】
DT40ポリLacO-λRにおけるψVλアレイへのLacI融合タンパク質の可逆的係留
クロマチン構造の局所的改変は、適切な融合タンパク質としてDNA結合部位に調節因子を係留することによって実現することができる。この戦略は、例えば、ショウジョウバエにおいて、ラクトースリプレッサーとの融合物(LacI-HP1)として発現されるヘテロクロマチンタンパク質HP1aが、クロマチン構造の閉鎖およびLacOリピートに隣接したレポーター遺伝子の不活性化を促進する[61、62]ことを示すため、ならびに脊椎動物のG9aヒストンメチルトランスフェラーゼがV(D)Jミニ遺伝子レポーター内のGAL4結合部位に係留されると、その構築物の非相同的媒介による組換えが減退する[63]ことを示すために使用された。重合ラクトースオペレーター(ポリLacO)を、転写されたVλRの17kb上流のψVλ17〜ψVλ20の間に相同遺伝子ターゲティングによって挿入したDT40派生体である細胞株DT40ポリLacO-λR(図10A)。ポリLacO挿入物の長さは3.8kbであり、20量体のオペレーターのコピー約100個からなる[64]。この細胞株を用いて、内因性座位内の生理学的プロセスにおける相同組換えに対する係留された調節因子の影響を分析し、導入遺伝子レポーターの必要性を回避することが可能である。対照実験により、ポリLacOタグが、細胞増殖にも細胞周期にもIg遺伝子多様化にも影響を及ぼさないことが示された。
【0114】
ラクトースリプレッサーに融合された高感度緑色蛍光タンパク質(GFP-LacI)を安定に発現するDT40ポリLacO-λR GFP-LacIにおいて、タグ化されたλR対立遺伝子は、蛍光顕微鏡によって直接画像化することができ、各細胞中に明瞭な斑点として現れる(図10B、左)。LacIがLacOに結合するのを妨げる100μM IPTGと共に一晩培養した後は明るい斑点がはっきりとしないように、係留は可逆的である(図10B、右)。
【0115】
HP1が係留されると、ψVλにおける活性クロマチンに特徴的な改変が減少する
ψVλアレイにおけるクロマチン構造を操作するために、本発明者らは、LacIに融合されたキイロショウジョウバエHP1タンパク質(LacI-HP1)を発現する、DT40ポリLacO-λRの安定なトランスフェクタントを作製した。HP1は、ヘテロクロマチンの遺伝子サイレンシング、ヘテロクロマチンの伝播、およびヒストン脱アセチル化において機能する非ヒストンヘテロクロマチンタンパク質である[58〜60]。係留されたHP1は、隣接した遺伝子でのクロマチン構造の閉鎖を促進することが示されている[61、62、66〜68]。DT40ポリLacO-λR LacI-HP1トランスフェクタントを抗LacI抗体で染色することにより、LacI-HP1が、挟動原体ヘテロクロマチンに対応するDAPI密集領域と共局在して(図11A)、ヘテロクロマチンの機能的マーカーとして挙動することが示された[65]。
【0116】
LacI-HP1が係留されるとクロマチン構造が変化するかを問うために、本発明者らは、ψVλ17におけるクロマチン改変を分析した。DT40ポリLacOを構築する間に配列多型性が生じるため、これは、再配列された対立遺伝子および再配列されていない対立遺伝子を、特異的PCRプライマーを用いて区別できるψVλアレイ中の唯一の部位である。ChIP後、標的とする再配列された対立遺伝子(ψVλ17R)に特異的なPCRプライマーを用いてDNAを増幅した。ψVλ17Rの富化を、内部対照としての発現されない卵白アルブミン遺伝子(Ova)と比較し、全インプットDNA中のψVλ17R:Ovaの富化率に対して標準化した(材料および方法を参照されたい)。対照のDT40ポリLacO-λR GFP-LacIのψVλ17RにおけるAcH3およびAcH4の富化率はそれぞれ2.2倍および5.9倍であった(図11B、C)。これらの富化レベルは、DT40において実証されたものと類似している(図9B)。(親DT40株の調査でのψVλにおける改変の解析は、両方の対立遺伝子を必ず含み、再配列された対立遺伝子における活性化改変を過小評価する場合があることに留意されたい。一方、ψVλ17Rにおける改変の解析では、活性な対立遺伝子のみを調べる)。AcH3およびAcH4は、DT40ポリLacO-λR LacI-HP1トランスフェクタントのψVλ17Rでは富化されておらず(それぞれ0.6倍および1.0倍;図11B、C)、HP1を介したサイレンシングと一致していた。HP1は、ヒストンH3のリジン9を改変するヒストンメチルトランスフェラーゼを動員することによってサイレンシングを行うことができる[66〜68]が、この改変とは無関係にサイレンシングを促進する場合もある[61]。ジおよびトリメチル化H3(K9)のいずれかに対する抗体を用いたChIPでは、H3 K9-Me改変の明らかな富化が示されなかった(データ不掲載)。ヒストンH3のリジン4のメチル化(ジMeK4)は転写と関連しており、一般に、アセチル化と時間的に重複した分布を示す[69、70]。ψVλ17RのジMeK4(H3)の分析により、この改変が、DT40ポリLacO-λR GFP-LacI細胞では18.9倍に富化されているが、DT40ポリLacO-λR LacI-HP1細胞ではバックグラウンドレベルであることが実証された(図11B、C)。
【0117】
HP1はヘテロクロマチンの維持および伝播を促進する[66]。係留されたHP1によって促進されるクロマチン構造の変化が染色体全体に伝播しないことを確認するために、本発明者らは、第15染色体上のλ軽鎖座位近くの別の部位、すなわちDNA polεの触媒サブユニットをコードする遺伝子を検査した。DNA polεは、遍在的に発現され、真核生物の染色体複製に不可欠であり[71]、Igλから約2.1Mbに位置する遺伝子にコードされている。本発明者らは、対照のDT40ポリLacO-λR GFP-LacIと比べて、DT40ポリLacO-λR LacI-HP1トランスフェクタントのpolεプロモーター領域のAcH3富化に差がないことを見出した(polε/Ova富化はそれぞれ8.5倍および8.4倍であった;図3C)。同様に、対照のDT40ポリLacO-λR GFP-LacIと比べて、DT40ポリLacO-λR LacI-HP1トランスフェクタントのpolεプロモーター領域のAcH4にも差がなかった(polε/Ova富化はそれぞれ1.9倍および1.7倍であった;図11C)。したがって、ψVλにLacI-HP1が係留されると、クロマチン構造が局所的に改変されて、ψVλ17Rにおける開放型クロマチンに特徴的なAcH3、AcH4、およびジMeK4(H3)改変が減少し、クロマチンが許容性の低い状態を取るようになった。
【0118】
HP1の係留は、Vλ遺伝子発現に影響を及ぼさない
本発明者らは、DT40ポリLacO-λR LacI-HP1細胞および対照のDT40ポリLacO-λR GFP-LacIトランスフェクタントにおけるこれらの改変を比較することによって、HP1の係留が、発現されたVλRにおけるAcH3レベルおよびAcH4レベルにどのように影響を及ぼすかを問うた(図12A)。HP1が係留されると、AcH3レベルおよびAcH4レベルはそれぞれ対照レベルの約40%および20%に低下した。これが遺伝子発現に影響を及ぼすかを問うために、本発明者らは、表面IgM(sIgM)発現およびVλ転写物レベルの両方を分析した。マウス抗ニワトリIgMで細胞を染色することにより、IPTGの存在下または不在下のいずれかで培養したDT40ポリLacO-λR GFP-LacI株およびDT40ポリLacO-λR LacI-HP1株のsIgM発現は類似していることが示された(図12B)。DT40ポリLacO-λR GFP-LacI細胞およびDT40ポリLacO-λR LacI-HP1細胞から回収したRNAの定量的RT-PCRによって、Vλ転写物レベルを分析し、対照としてのβ-アクチンに対して標準化した(図12C)。2種の細胞株のVλ転写物レベルに有意な差は観察されず、転写はψVλアレイ内でのHP1係留の影響を受けないことが実証された。したがって、LacI-HP1の係留により、下流のIg遺伝子の発現は影響を受けなかったが、VλRにおけるAcH3レベルおよびAcH4レベルは低下した。したがって、Vλに特徴的な非常に高いAcH3レベルおよびAcH4レベル(図9B、図12A)は、高レベルの遺伝子発現を維持するために不可欠ではない。
【0119】
HP1が係留されると、局所的なクロマチン構造が変化する
LacI-HP1のクロマチンへの影響がどれくらい大規模であるかを評価するために、本発明者らは、Igλ軽鎖座位の全体にわたって、図9で検査した同じアンプリコン(フランク中の1つ、ψVλアレイ中の6つを含む)、ならびに発現されたVλにおけるAcH3レベルおよびAcH4レベルを検査した。図11Bのような免疫沈降およびインプット条件のψVλ17R:Ova比率を比較することによって、改変のレベルを決定した。調査した部位におけるAcH3改変は、対照の同じ部位におけるレベルの24%〜63%の範囲であった(図13A、黒い棒)。これらの部位すべてに渡るH3アセチル化の平均レベルは、対照のポリLacO-λR GFP-LacIの38%であった。250μM IPTGと共に3日間、DT40ポリLacO-λR LacI-HP1トランスフェクタントを培養すると、調査した8箇所の部位すべてでH3アセチル化が増加した(図13A、黒い棒と白い棒を比較されたい)。IPTG培養の影響はいくらか変動したが、大半の部位で、IPTG培養により、対照のDT40ポリLacO-λR GFP-LacI細胞でのレベルの少なくとも45%までAcH3のレベルが回復し、平均は80%を上回った。したがって、DT40ポリLacO-λR LacI-HP1細胞のψVλ17Rにおけるクロマチン改変は、LacI-HP1の係留に直接起因し、大部分は可逆的であった。
【0120】
同じ8箇所の部位のH4アセチル化を調査した(図13B、黒い棒)。AcH4改変は対照のレベルの18%〜42%の範囲であることが判明した。平均レベルは、対照の細胞株の29%であった。IPTGと共に3日間培養すると、調査した8箇所の部位すべてでH4アセチル化が増加し(図13B、黒い棒と白い棒を比較されたい)、対照のDT40ポリLacO-λR GFP-LacI細胞でのレベルの少なくとも57%までH4アセチル化のレベルが回復し、平均は80%を上回った。さらに、IPTGは、LacI-HP1の影響を少なくともある程度取り消すことができる。
【0121】
これらの結果から、ψVλアレイにおいて観察されるクロマチン改変が、HP1の係留に起因することが示される。さらに、これらの改変が可逆的であるということから、活性なメカニズムが、ψVλでのクロマチン改変因子の係留によって課されるヒストン改変を元に戻すことが示される。
【0122】
HP1が係留されると、鋳型式変異誘発が減退する
LacI-HP1の係留によってψVλにおけるクロマチン構造を操作できるため(図11〜13)、本発明者らは、クロマチン構造がIg遺伝子変換に影響するかどうか、およびその方法を直接問うことができた。本発明者らは、sIgM喪失変異体アッセイ法を用いて、係留されたLacI-HP1が、再配列されたVλR遺伝子のクローン配列多様化速度に影響を及ぼすか判定した。この変動アッセイ法では、構造的に完全なsIgMをもはや発現しない変異体細胞の割合を測定し、したがって、遺伝子変換または点変異誘発のいずれかに起因する変異事象を記録する[47、50]。sIgM+細胞を限界希釈法でクローニングすることによってDT40ポリLacO-λR GFP-LacIおよびDT40ポリLacO-λR LacI-HP1の独立したクローン派生体を樹立し、4週間培養し、次いで抗IgM抗体で染色した細胞をフローサイトメトリーに供して、各集団中のsIgM-細胞の割合を決定した。sIgM喪失率の中央値は、DT40ポリLacO-λR GFP-LacI細胞の場合は0.5%、およびDT40ポリLacO-λR LacI-HP1細胞の場合は2.8%であった(図14A)。これは、対照のGFP-LacIと比べて、LacI-HP1トランスフェクタントのクローン多様化速度が5.6倍に加速したことに対応する。
【0123】
ニワトリB細胞におけるIg遺伝子多様化は、主に遺伝子変換(鋳型式変異)によって起こるが、例えば、不可欠な因子の欠如によって遺伝子変換が減退している場合には、修復により、非鋳型式変異がかなりの割合で生じ得る[50〜55]。遺伝子変換のためのψVλ鋳型は、再配列された遺伝子と約80%同一であり、遺伝子変換によって修復されるかなりの割合のDNA損傷は、配列の改変をまったく経験しないため、これは、典型的にはクローン多様化速度の上昇を伴う。一方、変異誘発性ポリメラーゼによる修復の方が、DNA配列を改変する可能性が高い。HP1の係留によってどのように多様化が加速するかを決定するために、本発明者らは、DT40ポリLacO-λR GFP-LacIトランスフェクタントおよびDT40ポリLacO-λR LacI-HP1トランスフェクタントからsIgM-細胞だけ(single)を選別し、発現されたVλ領域を単一細胞PCRによって増幅し、これらの領域を配列決定した。配列変化が、2つまたはそれ以上の変異を含む区域内にあり、その区域がドナーψVλ配列の少なくとも9bpに厳密に一致している場合には、その配列変化を「鋳型式」と分類し、配列変化が単一の塩基変化のみからなるが、ドナーψVλ配列の少なくとも9bpに一致した場合には、その配列変化を「あいまい」と分類した。点変異、欠失、および挿入からなる非鋳型式事象もまた、記録した。対照のDT40ポリLacO-λR GFP-LacIトランスフェクタントでは、70個の独特な変異のうち54個の鋳型式事象および2個のあいまい事象が記録された。したがって、大半の事象(77%)は鋳型式であり、低比率の事象(20%)が点変異であった(図14B、左;図15)。印象的なことには、DT40ポリLacO-λR LacI-HP1細胞では、点変異が優勢であり(59%)、欠失(8%)および挿入(14%)を伴った。一方、36個の独特な変異のうちで、ただ1個の明らかな鋳型式事象および6個のあいまいな事象が記録された(図14B、右;図15)。したがって、変異の3%しか、明らかに鋳型式ではなく、さらに、潜在的な鋳型式変異であるあいまいクラスを含んでも、鋳型式の割合は変異の19%以下しか占めることができなかった。統計学的比較により、対照のDT40ポリLacO-λR GFP-LacI細胞およびDT40ポリLacO-λR LacI-HP1トランスフェクタントにおける明らかな鋳型式変異の割合の差(77%対3%)は、有意度が高い(P=7.1×10-7;フィッシャーの正確確率検定)ことが示された。対照細胞(3%)およびHP1トランスフェクタント(17%)におけるあいまいな潜在的鋳型式変異の割合の差もまた、有意である(P=0.05;フィッシャーの正確確率検定)。このことから、この範疇のいくつかの変異は、非許容性のドナークロマチンによって課される遺伝子変換区域の長さに対する制約の結果として起こり得ることが示唆される。したがって、HP1が係留されると、鋳型式変異が減退することにより、クローン変異誘発速度が加速した。
【0124】
考察
ニワトリIg座位における遺伝子変換は、再配列され転写されたV遺伝子を標的とする損傷を相同性の指示に従って修復するための鋳型として上流のψVドナーのアレイを使用する。本実施例では、活性なIg遺伝子変換を実施するニワトリB細胞において、ドナーψVλアレイ内部のクロマチンが、開放型のクロマチン構造と相関がある改変であるアセチル化H3およびアセチル化H4の富化を特徴とすることを示す。本発明者らは、ψVλドナーアレイにヘテロクロマチンタンパク質HP1が係留されると、クロマチン構造が局所的に変化して、開放型クロマチンに特徴的なAcH3、AcH4、およびジMeK4(H3)改変が減少することを示すことによって、Ig遺伝子変換のために許容性クロマチン構造が重要であることを直接的に実証した。これらの変化は、閉鎖型クロマチンに特徴的なMe-K9(H3)改変を伴わなかったが、遺伝子変換に対する許容性が低い状態をその領域に取らせた。HP1の係留は、IgVλ変異範囲の大幅な変化を伴い、その結果、鋳型式変異は少数派となり、点変異が優勢になった。重要なことには、変異誘発に対するこの影響は、クロマチン構造の変化と相関関係があるが、座位の発現の変化とは相関関係がなかった。したがって、クロマチン構造は、内因的に発生したDNA損傷で遺伝子変換が起こるかどうかを指令することができる。
【0125】
複雑なクロマチンランドスケープ内部での遺伝子変換のメカニズム
Vλでの遺伝子変換は、ψVλ領域を鋳型として用いて、切断部(break)の3'末端で新しいDNA合成を開始した結果として生じる。遺伝子変換は、ドナーDNAとレシピエントDNAの対合、ならびに相同性の指示に従った修復を行う因子のドナーへの接近を必要とする。野生型DT40のψVλアレイに特徴的な高レベルのH3アセチル化およびH4アセチル化は、トランス作用性因子へのψVλ遺伝子の接近容易性を高め、また、配列対合に都合が良い三次元構造物を作り出すと思われる、弛緩したクロマチン構造の証拠である。
【0126】
ψVλドナーアレイ内部に係留されたHP1は、Vλ遺伝子発現に影響を及ぼすことなく、再配列されたVλRにおける遺伝子変換を減退させた。HP1の係留によって引き起こされるクロマチン変化により、修復因子の接近の妨害およびドナー鋳型に対する侵入鎖によって遺伝子変換が減退し得る。また、係留されたHP1は、ドナー配列およびレシピエント配列を並べるのに必要なループ形成のようなDNA修復経路のメカニズムに影響を及ぼす、より大きな染色体構築にも寄与し得る。LacI-HP1トランスフェクタントにおいて蓄積した点変異は、邪魔された組み換え修復に典型的であり、組換えに不可欠なトランス作用性因子[49〜55]またはψVドナーアレイの一部もしくは全部[56]のいずれかを欠いた細胞に特徴的である。HP1は、2つの方法で、すなわち、ヒストンメチルトランスフェラーゼと組むことによって[66]、およびヒストンデアセチラーゼを動員することによって[60]、クロマチン構造およびヘテロクロマチン遺伝子サイレンシングを調節する。HP1が係留されると、ψVλ内部の非許容性クロマチン構造に特徴的な改変が引き起こされた。
【0127】
体細胞超変異およびクラススイッチ組換えを経験する活性に転写された哺乳動物Ig遺伝子におけるヒストンアセチル化が実証されているが、ヒストンアセチル化が多様化のターゲティングに寄与するかどうかはまだこれから解決しなければならない[72〜76]。ヒストンアセチル化と遺伝子変換の関連は、ヒストンデアセチラーゼ阻害剤であるトリコスタチンAでDT40細胞を処理すると、VλRにおける遺伝子変換の増大を伴うゲノムワイドなヒストン脱アセチル化が促進されることを示す実験によって示唆された[77]。しかしながら、トリコスタチンAの効果はゲノムワイドであり、特異的ではないことから、これらの結果の解釈は複雑である。DT40ポリLacO-λR細胞株では、クロマチン構造を局所的に操作して、この複雑さを回避することができる。さらに、本発明者らは、LacI-HP1融合タンパク質を係留する効果の大部分が、IPTGと培養すると取り消され、したがって、活性なメカニズムがψVλにおけるクロマチン改変を決定するに違いないことを実証することができた。
【0128】
相同組換えを調査する場合、DT40ポリLacO-λR B細胞株は、Ig遺伝子変換が内因性座位内の生理学的プロセスであり、導入遺伝子レポーターの必要性を回避するというさらなる利点を有する。
【0129】
クロマチン構造、ゲノム安定性、老化、および遺伝子療法
相同組換えの結果に対するクロマチン構造の重要性は、ゲノム安定性を通常は維持するメカニズムを理解するのに関わりがある。脊椎動物のゲノムには全体に分布する膨大な数の反復エレメントがあり、これらのエレメント間の組換えは、ゲノムの不安定性につながり得る[78]。ヒトゲノムには、約100万個のAluエレメントがあり、Aluエレメント間の組換えは、腫瘍形成および遺伝病を招く重複の原因となり得る[79、80]。抑圧的な改変を有するヒストンは、反復エレメントで豊富である[81]。これらの改変は、転写抑制を維持することは疑いない。本発明者らの結果から、これらが組換えの抑圧にも寄与し得ることが示唆される。
【0130】
ヘテロ接合性消失(LOH)は、対立遺伝子部位における相同体間の不均等な有糸分裂組換の結果として生じる。LOHのメカニズムは、腫瘍形成を招く腫瘍抑制遺伝子機能の消失をもたらすため、特に関心対象である[82]。最近の実験で、S.セレビシエ(cerevisiae)におけるLOH[83]およびショウジョウバエ生殖細胞におけるレポーター遺伝子[84]の年齢依存的増加、ならびにマウス膵臓細胞における相同組換えの増加[85]が実証された。年齢に関連したLOHを説明するために提唱されたメカニズムには、老化に伴う高比率のDNA損傷、細胞周期分布の変化、および相同性依存的修復経路の不活性化が含まれる。これらの結果から、クロマチン構造の弛緩が老化に付随して起こり、老化細胞における相同組換えのゲノムワイドな増加を促進し得るという別の可能性が示唆される。この可能性は、ショウジョウバエに関する最近の解析[86]、ならびにハッチンソン・ギルフォード早老症候群に関与するラミンAの変異が、H3メチル化のゲノムワイドな減少を招くという最近の証拠[87]によって裏付けられている。
【0131】
クロマチン構造が相同組換えを調節するという知見には、実用的な派生問題もある。不完全な標的遺伝子を置換するために完全なドナー遺伝子を提供することにより、相同性依存的修復に対する細胞の能力を利用して遺伝子療法を促進する戦略を開発することに現在かなりの労力が向けられている[88]。これらの結果から、ドナーの許容的構造が、治療的用途のためのドナー遺伝子を開発する際の重要な設計パラメーターになるであろうことが示唆される。
【0132】
参考文献
【0133】
実施例5:クロマチンの異なる改変によって加速する遺伝子変換
本実施例は、修復の効率がドナーのクロマチン構造によって決定されることを実証する。この解析は、ニワトリDT40 B細胞株の転写され、多様化しているVλ遺伝子のすぐ上流に挿入されたシス調節部位(重合ラクトースオペレーター、ポリLacO)を利用する。データから、活性化因子VP16またはヒストンシャペロンHIRAのいずれかを係留すると、局所的なクロマチン構造が変化し、遺伝子変換が約10倍に加速することが示される。これら2種の因子の機能上の結果は類似しているが、クロマチン構造に対するそれらの影響は異なる。VP16の係留により、アセチル化ヒストンH3およびH4の局所レベルが上昇するのに対し、HIRAの係留により、ヌクレオソームの密度が上昇する。したがって、異なるクロマチン改変によって、類似した機能上の結果が実現され得る。
【0134】
材料および方法
クロマチン免疫沈降法(ChIP)
以前に説明されたようにしてChIPを実施した(Cummings et al., 2007)。すべての実験において、安定にトランスフェクトされた少なくとも2つの独立な株に由来する少なくとも2つのクロマチン調製物を解析した。図面は1つの代表的な実験を示し、4回の別々の増幅の解析結果を用いて標準偏差を算出した。鋳型DNAの段階希釈物を4回別々に増幅して、測定される産生物強度が直線範囲内であることを確かめた。実験アンプリコンの富化を、二重PCRによって同じチューブ中で増幅した卵白アルブミン(Ova)遺伝子に由来する内部対照アンプリコンの富化に対して標準化し、特異的抗体を用いたChIPの際の富化は、対照の全インプットDNAを用いてChIPを実施した並行実験に対して標準化した。Ova内部対照アンプリコンを含めることによって、本発明者らは、IP効率、バックグラウンドから残った影響、およびゲル添加の差異に関して標準化することができた。富化=[(ψVλ/Ova)Ab]/[(ψVλ/Ova)インプット]。追加の対照として、対照の全インプット中の実験アンプリコンと対照アンプリコンの比率を、多特異性IgGを用いた対照ChIPと比較した。いずれの場合も、インプット対照およびIgG対照における富化は本質的に等しかった。代表的な実験のデータを提示する。標準偏差は、鋳型DNAの段階希釈物を4回別々に増幅した結果から算出した。
【0135】
使用した抗体は、Upstate (Lake Placid, NY)社製の抗H3 CT-pan、抗AcH3(06-599)、抗AcH4(06-866)、およびジメチル化H3(K4)(07-030)であった。
【0136】
ChIP用のPCRプライマーは次のものであった:
【0137】
MNアーゼ消化およびサザンブロット法
核を調製し、以前に説明されているようにして、0、3、7、15、30、および60ユニットのMNアーゼを用いて消化を行った(Prioleau et al., 1999)。MNアーゼ消化後、フェノール:クロロホルム:イソアミルアルコールでDNAを3回抽出し、エタノールを用いて沈殿させた。20マイクログラムのDNAをゲルに添加し、電気泳動によって分離させ、サザンハイブリダイゼーション用に移した。ポリLacOリピートを含む長さ約500bpの断片を鋳型として用いて、ランダムプライミングによってlacOプローブを標識した。
【0138】
蛍光画像法
蛍光画像法のために、細胞(2×105個)をサイトスピンでスライドガラス上に載せ、2%パラホルムアルデヒドで15分間固定した。核を可視化するために、細胞をDAPI(Sigma, Saint Louis, MO)で染色した。DeltaVision顕微鏡検査システム(Applied Precision)を用いて蛍光画像を獲得し、softWoRxソフトウェア(Applied Precision)を用いて処理した。
【0139】
RT-PCR
TRIzol試薬(Invitrogen)を用いてRNAを細胞から回収した。鋳型の希釈後にVλ転写物およびβ-アクチン転写物を増幅した。Vλを増幅するためのプライマーは、
であった。β-アクチンを増幅するためのプライマーは、以前に説明されている(Arakawa et al., 2002)。
【0140】
sIgM喪失変異体の定量および配列解析
変異したV領域におけるフレームシフトまたはナンセンス変異の結果として生じ、蓄積したsIgM喪失変異体を測定する、sIgM喪失変異体のアッセイ法を用いて、IgV領域の多様化を定量した(Sale et al., 2001; Yabuki et al., 2005; Cummings et al., 2007)。手短に言えば、フローサイトメトリーとそれに続く限界希釈クローニングによってsIgM+細胞を単離し、4週間増殖させた。sIgM-細胞の割合を定量するために、約1×106個の細胞を抗ニワトリIgM-RPE(Southern Biotechnology Associates, Birmingham, AL)で染色し、CellQuestソフトウェア(BD Biosciences)を用いたFACScanによって解析した。
【0141】
単一細胞PCRおよび配列解析を、以前に説明されたようにして実施した(Yabuki et al., 2005; Cummings et al., 2007)。手短に言えば、sIgM-細胞を選別し、個々のウェルに等分し、Vλ領域を増幅および配列決定し、それらの配列をψVλドナーと比較して、変異が鋳型式であるか、または非鋳型式であるかを判定した。鋳型式変異の判定基準は、ドナーとレシピエントにおいて9個の連続した塩基が完全に一致しなければならないというものであった。配列は、独立にトランスフェクトされた2つの株に由来した。特有の配列のみを、変異の分類のために含めた。
【0142】
結果
VP16が係留されると遺伝子変換が加速する
抑圧的なドナークロマチン構造により遺伝子変換が減退するならば、活性化改変によって遺伝子変換が促進される可能性がある。この可能性を検証するために、本発明者らは、本発明者らの実験室で構築したDT40ポリLacO-λ細胞株を利用した。この細胞株では、重合ラクトースオペレーター(ポリLacO)が、ψVλアレイのψVλ17〜ψVλ20の間(発現されたVλ遺伝子の17kb上流)に挿入されている(図16A;実施例4)。これにより、本発明者らは、ラクトースリプレッサーとの融合物として発現される、係留された調節因子の効果を分析することができる。対照実験により、ポリLacOタグが、細胞増殖にも細胞周期プロファイルにもIg遺伝子多様化にも影響を及ぼさないことが示された。
【0143】
ψVλドナー内部のヒストンアセチル化の喪失は、Vλにおける遺伝子変換の減少と相関している(実施例4)。したがって、本発明者らは、クロマチンの弛緩およびヒストンアセチルトランスフェラーゼの動員と関連付けられている、ヘルペスウイルス由来の強力なトランス活性化因子であるVP16の影響を試験した(Tumbar et al., 1999; Carpenter et al., 2005)。本発明者らは、GFP-LacI-VP16を安定に発現するDT40ポリLacO-λRトランスフェクタントを作製した。ウェスタンブロット法によってタンパク質発現を確認した(図16B)。細胞周期プロファイル(図16C)およびλ転写レベル(図16D)は、対照DT40ポリLacO-λ GFP-LacIトランスフェクタントと比べて変わっていなかった。DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントの蛍光画像法では、GFP-LacI-VP16発現およびポリLacOへの結合の証拠である単一の緑色の斑点が核内部で示された(図16E)。
【0144】
本発明者らは、ポリLacO-λ GFP-LacI-VP16細胞および対照DT40ポリLacO-λ GFP-LacI細胞におけるヒストンアセチル化を分析した。ψVλ1とVλ遺伝子の間のプロモーター領域(Vλpro)、ψVλ1、ψVλ5、ψVλ13、ψVλ24、ψVλ25を含む7箇所の部位に特異的なプライマーを用いた増幅によって、ψVλアレイを分析した。(多型性を欠くために、DT40中の2つのIgλ対立遺伝子におけるψVλアレイはPCRによって容易に区別することができない)。さらに、本発明者らは、(1)DT40ポリLacO-λの構築中に生じる配列多型性の寄与により、再配列された対立遺伝子および再配列されていない対立遺伝子を特異的PCRプライマーを用いて区別できる、ψVλアレイ中の唯一の部位であるψVλ17、(2)Vλの再配列された対立遺伝子(VλR)、(3)Vλの再配列されていない対立遺伝子(VλU)、ならびに(4)染色体内対照であるPolε遺伝子、におけるクロマチン改変も分析した。ψVλ部位の増幅は、発現されない卵白アルブミン遺伝子(Ova)と同時に(duplex)実施した。したがって、Ovaは内部対照として機能し、免疫沈降法によって得たψVλ:Ova富化率を、全インプットDNAから得たψVλ:Ovaの比率に対して標準化した(材料および方法を参照されたい)。VP16が係留されると、対照DT40ポリLacO-λ GFP-LacI細胞と比べて、DT40ポリLacO-λGFP-LacI-VP16トランスフェクタントのAcH3レベル(図17A)およびAcH4レベル(図17B)の両方が上昇した。
【0145】
本発明者らは、sIgM喪失変異体アッセイ法を用いて、VP16の係留によって、クローンVλ配列多様化速度が変化するかを問うた。この変動アッセイ法では、構造的に完全なsIgMをもはや発現しない変異体細胞の割合を測定し、したがって、遺伝子変換、点変異、挿入、または欠失に起因する変異事象を記録する(Sale et al., 2001; Yabuki et al., 2005)。遺伝子変換は、ニワトリB細胞におけるIg遺伝子多様化の主な経路であるが、(例えば、不可欠な組換え因子の欠如によって)遺伝子変換が減退している場合には(Sale et al., 2001; Niedzwiedz et al., 2004; Hatanaka et al., 2005; Kawamoto et al., 2005; McIlwraith et al., 2005; Yamamoto et al., 2005)、他の修復結果、特に点変異が主になると考えられる。したがって、多様化は、sIgM-からsIgM+への機能獲得アッセイ法よりはむしろ、遺伝子変換のみを採点するこの機能喪失アッセイ法によって最もうまくモニターされる(Saribasak et al., 2006)。
【0146】
sIgM+細胞を限界希釈法でクローニングすることによってDT40ポリLacO-λR GFP-LacIトランスフェクタントおよびDT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントの独立したクローン派生体を樹立し、4週間培養した後、抗IgM抗体で染色した細胞をフローサイトメトリーに供して、各集団中のsIgM-細胞の割合を決定した。sIgM-細胞比率(%)の中央値の比較により、DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントでは、DT40ポリLacO-λ GFP-LacI対照細胞と比べて、多様化が8.4倍に増大していることが示された(図17B)。単一細胞PCRによって増幅したVλ領域の配列解析により、対照細胞およびVP16がIgλに係留された細胞の両方において、大半の多様化は遺伝子変換によることが示された(図22)。
【0147】
ヒストンH3.3は、ψVλドナー遺伝子において豊富である
ヒストン改変だけでなく、ヌクレオソームのヒストン組成の局所的な変化もまた、クロマチン構造の調節に寄与し得る。クロマチン活性化に関連し、複製とは無関係に沈着する1つのヒストン変異体はH3.3である。ヒストンH3.3は、活性化に関連した翻訳後修飾を含む主要なH3様変異体であり(McKittrick et al., 2004; Henikoff, 2008)、したがって、H3.3がψVλアレイで豊富であるかを問うことは関心対象であった。ChIPによってH3をH3.3と区別するために、本発明者らは、FLAGタグ付きH3.3(H3.3-FLAG)を安定に発現するDT40派生体を作製した。
【0148】
抗FLAG抗体を用いたChIPによるH3.3沈着の特徴付けにより、発現されたVλR遺伝子におけるH3.3変異体の富化が実証された。ψVλアレイ内のクロマチン構造を、再配列され発現されたVλ遺伝子(ψVλ1、ψVλ5、ψVλ13、ψVλ24、ψVλ25、ψVλ17R、VλRを含む)の上流の部位(図18A)、および再配列された対立遺伝子VλU;ならびにIgλと同じ染色体上の別の遺伝子Polεの増幅によって分析した。典型的な実験では、H3.3はVλRにおいてインプットDNAと比べて4倍超に富化されていたが、印象的なことには、本発明者らは、ψVλアレイの全体に渡ってH3.3がかなり富化されており、ψVλ5およびVλproにおいて最も富化されていることを観察した(図18B)。近位部位が遠位部位より高レベルの富化を一貫して示すことはなかったため、ψVλアレイ内でのH3.3の富化は、転写されたVλR遺伝子から上流部位への変異体の段階的な伝播を単に示すわけではない。この座位のクロマチン構造が非均一であることから、座位の発現および/または多様化を調節し得るシスエレメントの存在が、特にψVλ5領域において示唆される。
【0149】
HIRAが係留されると、ヒストン沈着が局所的に増加する
HIRAは、ヒストンシャペロンであり、その機能の内の1つは、ヒストン変異体H3.3を含むヌクレオソームを組み立てることである(Ray-Gallet et al., 2002; Tagami et al., 2004)。したがって、ψVλアレイにおけるH3.3の富化から、VP16の係留による効果と同様に、HIRAの係留によってIg遺伝子多様化が加速し得ることが示唆される。これを検証するために、本発明者らは、安定なDT40ポリLacO-λ HIRA-LacIトランスフェクタントを作製し、ウェスタンブロット法によって、タンパク質発現が同程度のレベルであることを確認し(図19A)、かつ、係留が細胞周期プロファイル(図19B)にもλ転写物レベル(図19C)にも影響を及ぼさないことを示した。ChIPアッセイ法によるAcH3レベルおよびAcH4レベルの分析では、HIRAまたはGFPがIgλに係留された細胞において差は示されなかった。
【0150】
HIRAの係留によりH3.3レベルが上昇するかを問うために、本発明者らは、DT40ポリLacO-λ FLAG-H3.3 HIRA-LacIトランスフェクタントを作製し、次いで、多型性があるため、係留されたIgλのみを解析することが可能であるψVλ17RにおけるFLAG-H3.3の富化を測定した。これらの実験では、2つの独立したHIRA-LacIトランスフェクタントのそれぞれで、対照株と比べて中程度(1.4倍)のFLAG-H3.3富化が示された(図20A)。さらに、汎的な(pan-)H3抗体(H3、およびH3.3を含む他の変異体を検出する)がChIPのために使用される場合、同程度の富化が明らかであった。したがって、HIRAは、H3.3を含むヒストンの正味の富化を引き起こすと思われ、したがって、ψVλでのヌクレオソーム密度に影響を及ぼした。
【0151】
HIRAの係留はIg遺伝子変換を促進する
HIRAの係留がクローンのIg遺伝子多様化速度に影響を及ぼすかを問うために、本発明者らは、sIgM+細胞を限界希釈法でクローニングすることによってDT40ポリLacO-λ HIRA-LacIトランスフェクタンの独立したクローン派生体を樹立し、4週間培養した後、各集団中のsIgM-細胞の割合を決定した。sIgM-細胞の割合は、対照細胞よりもHIRA-LacIトランスフェクタントの方が明らかに大きかった(例えば、図20B)。sIgM-細胞比率(%)の中央値の比較により、DT40ポリLacO-λ HIRA-LacI細胞では、DT40ポリLacO-λ GFP-LacI対照細胞と比べて、多様化が11倍に増大していることが示された(図20C)。HIRAを安定に発現するDT40のトランスフェクタントでは、多様化の中程度の加速(1.7倍)しか明らかではなかったため、この結果は単にHIRAの過剰発現を反映したものではなかった(図20C)。単一細胞PCRによって増幅したVλ領域の配列解析により、対照細胞およびHIRAがIgλに係留された細胞において、大半の多様化は遺伝子変換によることが示された(図23)。
【0152】
短区域の鋳型式変異の割合は、VP16またはHIRAの係留によって増加する
VP16およびHIRAが係留されると、クローンの遺伝子多様化速度は同じ程度加速し(それぞれ8.4倍および11倍)、どちらの場合も遺伝子変換が顕著であった。わずかな差によって変異範囲を区別し得るかどうかを確かめるため、DT40ポリLacO-λ HIRA-LacIトランスフェクタントおよびDT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントにおける独立した変異事象を、対照のDT40ポリLacO-λ GFP-LacIトランスフェクタントと比較した。配列変化がドナーψVλ配列の少なくとも9bpに厳密に一致している区域内に存在し、かつ、生殖系列配列と比べて1つ、2つ、またはそれ以上の一塩基変化を含む場合、その配列変化を遺伝子変換として分類した。一塩基の違いおよび2つまたはそれ以上の違いを含む事象は、「短」区域遺伝子変換および「長」区域遺伝子変換と別々に名付けた。これは、前者のクラスの一部の変異が、ψVλドナー配列と偶然に一致した点変異に原理的には起因し得るためである。点変異、欠失、および挿入からなる非鋳型式事象もまた、記録した。
【0153】
DT40ポリLacO-λ GFP-LacI-VP16トランスフェクタントにおいて、74%の事象は鋳型式であったが、かなりの比率の鋳型式事象(変異事象全体の39%または29%)は、長区域遺伝子変換ではなく短区域遺伝子変換であった(表1、図22)。また、かなりの割合で挿入もあった(0%に対して13%)。DT40ポリLacO-λR HIRA-LacI細胞において解析した事象でも、遺伝子変換事象が同様に顕著であった(87%)(表1、図23)。大半の遺伝子変換事象は長区域であったが、少数派であるが有意な事象(変異事象全体の24%または21%)は短区域遺伝子変換であった。残りの事象は、点変異(11%)および挿入(2%)であった。本発明者らは以前に、対照のDT40ポリLacO-λ GFP-LacIトランスフェクタントでの変異誘発事象を特徴付けし、大半の変異が長区域遺伝子変換事象に起因し(77%)、一方、ごく一部(3%)が、短区域変換事象またはドナー配列に偶然に一致した点変異のいずれかであることを発見した(実施例4:表1に要約した)。残りの変異(20%)は、偽遺伝子ドナーのいずれにも対応しなかったため、明らかに点変異であった。
【0154】
(表1)遺伝子変換に対するHP1およびVP16の係留の影響
DT40ポリLacO-λ GFP-LacIトランスフェクタント(n=71;(実施例4)、DT40ポリLacO-λ GFP-LacI-VP16(n=31)トランスフェクタント、およびDT40ポリLacO-λ HIRA-LacIトランスフェクタント(n=137)に由来する、特有の変異を有するVλ領域の配列の要約。データは、少なくとも2回の独立したトランスフェクションから導き出した。
【0155】
したがって、VP16およびHIRAの係留による多様化の加速は、鋳型式変異のレベル上昇に起因する。どちらの場合も、大半の鋳型区域は、2つまたはそれ以上の変異を含んだ。しかし、約3分の1の区域は、単一の変異のみを含んだ。一方、GFP-LacI対照細胞における本質的にすべての遺伝子変換事象は、少なくとも2つの変異を含んだ。このことから、ドナークロマチン構造の変化は、遺伝子変換を加速させるだけでなく、遺伝子変換区域の広さを制限する可能性があること、または、加速した遺伝子変換が、変換区域の長さの伸長に寄与する因子の量を調整する(titrate)可能性があることが示唆される。
【0156】
HIRAが係留されるとヌクレオソーム密度が増加する
上記の結果から、係留されたVP16およびHIRAがIg遺伝子多様化の結果に与える影響は類似していたが、それぞれの局所的影響は異なっていたことが示される:VP16はAcH3およびAcH4を増加させたのに対し、HIRAはH3.3および全体的なヒストン搭載(loading)を増加させた。これらの異なる因子の添加がどのようにクロマチンに影響を及ぼし得るかを確かめるために、本発明者らは、小球菌ヌクレアーゼ(MNアーゼ)を用いてクロマチン構造を探索した。核を採取し、様々な量のMNアーゼで処理し、ゲノムDNAを精製、分離、およびブロットし、次いで、LacOリピートに相当な標識DNAを用いて探索した。DT40ポリLacO-λ GFP-LacI細胞およびDT40ポリLacO-λ GFP-LacI-VP16細胞のMNアーゼ消化パターンは、このアッセイ法では同様と思われた(図21)。高レベルのMNアーゼで消化すると、明らかな制限生成物(clear limit product)が得られた。これは、ヌクレオソームに接触するDNA領域(約150bp)であり、したがって、MNアーゼから保護されている。個々のヌクレオソーム間の間隔は変動し得るため、部分的消化の際に産生されるヌクレオソーム多量体のサイズにはわずかな差が生じ、かつ、ゲル中でよりゆっくりと移動する特徴的にぼやけたバンドが生じる。
【0157】
一方、DT40ポリLacO HIRA-LacI細胞に由来するクロマチンのMNアーゼ消化では、異なるパターンが明らかになった:対照では認められない示差的なはしご状が観察された(図21、右)。対照およびVP16発現細胞では、4量体よりも大きいサイズではしごがぼやけたのに対し、輪郭のはっきりした一連の消化生成物が、制限消化の際に生じる1量体から、最高で8量体およびそれ以上に及んだ。このことから、ヒストンシャペロンHIRAの係留により、リンカーサイズの均一性が高まったことが示唆される。これは、局所的ヌクレオソーム密度が増加した場合に起こると思われ、ヒストンシャペロンとしてのHIRAの公知の役割、ならびに、HIRAの係留によってH3.3およびH3の局所的富化が増大したという証拠と一致する(図18C)。
【0158】
考察
本実施例では、遺伝子変換がクロマチン構造の変化によって加速し得ること、およびこのために少なくとも2つの異なる手段があることを示した。ニワトリDT40 B細胞株のψVλアレイに活性化因子VP16またはヒストンシャペロンHIRAのいずれかを係留すると、遺伝子変換の速さが1桁速くなった。VP16を係留すると、AcH3およびAcH4のレベルが上昇したのに対し、HIRAを係留すると、それらのクロマチンマークは影響を受けなかったが、一つにはH3.3の搭載によってヌクレオソーム密度が増加した。クロマチン状態の異なる変化が、相同組換えを同様に可能にするということから、組換えの際のDNA使用を調節するために利用可能な重複した(redundant)経路が細胞にはあることが示される。
【0159】
VP16およびHIRAの係留がIg遺伝子多様化に与える影響は、ヘテロクロマチンタンパク質HP1の係留による影響とは異なっている。HP1を係留すると、点変異がほぼ3倍に増加し、鋳型式変異の減少を伴った(実施例4)。これらの差異から、ドナークロマチン構造は、相同性の指示に従ったDNA修復を活性化または抑制される場合があり、修復結果に影響を与えることが示される。
【0160】
現在、相同性の指示に従った対立遺伝子修復は、S期に顕著であり、鋳型用の姉妹染色分体に依存すると考えられている。クロマチン構造の局所的改変が、相同性の指示に従った修復のための領域を活性化するという可能性から、調節は、もっと複雑であり得、相同なドナーの存在だけでなく、修復の助けとなるクロマチン構造も必要とし得ることが示唆される。
【0161】
HIRAまたはVP16が係留されると、鋳型式変異を促進することにより、クローンの多様化速度が加速した。どちらの場合も、単一の鋳型塩基の変化のみが変異区域中で明らかである鋳型式変異の割合の増加が明らかであった。これらの「短区域」変異は、VP16トランスフェクタントおよびHIR1トランスフェクタントにおける全変異のそれぞれ29%および21%しか含まなかったが、この範疇の変異は、GFP-LacIトランスフェクタントでは本質的に無かった(3%)。
【0162】
長区域の遺伝子変換を促進するのに必要な因子が、VP16またはHIRAのいずれかがポリLacOに係留された細胞中の遺伝子の増加を支援するのに充分なほど豊富ではない場合、短い遺伝子変換区域が生じ得る。あるいは、この範疇の変異の増加は、ヌクレオソームの位置およびリンカーの長さの両方に影響を及ぼし得るこれらの因子を係留することによって引き起こされたクロマチン構造の変化を反映している可能性もある。
【0163】
HIRAを係留すると、ヌクレオソーム密度が増加する。この密度増加は、短くなったリンカー長を反映している可能性が高い。リンカー領域が遺伝子変換のために好ましい鋳型である場合、HIRAトランスフェクタント中のこれらの領域の短縮は、遺伝子変換区域の長さの短縮の一因となり得る。あるいは、ヌクレオソーム長の短縮により、クロマチンは組換えに対して活性であるがψV間の非相同領域は接近しにくくなるように、ヌクレオソームを再調整(re-phase)することができる。
【0164】
偽遺伝子におけるHIRAの役割は、ヒストン搭載がDNA修復に影響を及ぼす方法に関する新規な例である。多くの研究により、デノボのH3.3沈着に必要とされる転写が発見されているため(AhmadおよびHenikoff, 2002a; AhmadおよびHenikoff, 2002b; Janicki et al., 2004; Wirbelauer et al., 2005)、転写されていない偽遺伝子におけるHIRAの役割は、特に興味深い。これらの結果から、現在、転写はH3.3搭載の必要条件ではないことが示唆される。
【0165】
これらの結果により、クロマチンの制約が組換えに対して及ぼす制約に関する本発明者らの理解は深まる。さらに、これらは、遺伝子ターゲティングおよび/または遺伝子療法のために使用される分子に必要なクロマチン化への洞察も与える。
【0166】
参考文献
【0167】
実施例6:メソテリンおよびHE4に対する診断用抗体の作製
本実施例では、本発明を使用して、十分に有効性を確認された卵巣癌バイオマーカーであるメソテリンおよびHE4に対する抗体をどのように開発できるかを例示する。同定された抗体は、新しい貴重な診断試薬であり、有用な冗長性を臨床試験に追加すると考えられる。本実施例では、他のバイオマーカーを認識する診断用抗体を選択するためのプラットホームとしてのDT40ポリLacOの有用性もさらに確認した。
【0168】
抗体開発のための媒体としてのDT40ポリLacO:2種の卵巣癌バイオマーカー、すなわちメソテリン(Scholler et al., 1999; Frierson et al., 2003)およびHE4(Hellstrom et al., 2003)に対する新しい抗体の同定。
【0169】
メソテリンは、卵巣癌を含む多くの起源の癌細胞において高発現される上皮分化マーカーである(Robinson et al., 2003)。HE4(別名WFDC2)は、乳清酸性タンパク質(WAP)ファミリーのメンバーであり、このファミリーのメンバーは高レベルで分泌され、癌に関連しているが、機能は理解されていない(Bouchard et al., 2006)。本発明者らは、組換えメソテリンおよびHE4を得た。これらのタンパク質は、選択またはELISAアッセイ法用のストレプトアビジン結合ビーズまたはプレートへの結合を促進するビオチン融合物として酵母において発現される(Scholler et al., 2006; Bergan et al., 2007)。本発明者らはまた、新しく同定された抗体の親和性を最初に比較するための標準を与える、メソテリンおよびHE4(Scholler et al., 2008)に対する単鎖抗体(scFv)も得た。
【0170】
メソテリンおよびHE4に対する高親和性抗体を同定するために、超変異および各組換え抗原の認識に関するクローン選択を反復的に実施する。超変異を加速させる条件下でクローンを増殖させ、メソテリンまたはHE4に高親和性で結合するクローンを磁気活性化細胞選別によって選択し、上清の抗体親和性をELISAアッセイ法によって分析する。高親和性抗体を産生する集団をさらに選択して、親和性および特異性を増大させる(図3)。
【0171】
選択は、それぞれ細胞108個(培養物100mlに相当する)からなる3つの集団において開始することができる。各集団は、限界希釈によって単離した単細胞に由来してよく、次いで、超変異を加速させる条件下で培養してよい。ビーズに単独で結合する「粘着性」細胞の富化を最小限にするために、所望の特異性を富化する前に、ビオチン結合ビーズで各集団を最初に清掃する。所望の特異性の富化は、磁気活性化細胞選別(MACS)によって便宜的に実行する。MACSを用いることにより、非常に大きな試料サイズ(細胞108個)から極めて迅速に富化することが可能であり、開始試料の<0.1%を含む集団を100倍に富化することを常に実現する(Volna et al., 2007)。選択された集団を増殖させ、同じプロトコールを用いて再選択する。
【0172】
他の研究者(例えば、Cumbers et al., 2002)の結果に基づき、本発明者らは、各「抗原」に特異的な抗体を産生する細胞が、反復的な超変異および選択の2〜3回目(2〜3週目)またはその前、およびおそらくはそれ以前に回収されると予想する。ELISAアッセイ法によって細胞プールの抗体親和性を試験することができ、親和性が1〜10nMの範囲である場合、限界希釈クローニングを実施し、高親和性抗体を産生するクローンを同定する。限界希釈クローニングは、オペレーターからリプレッサーを遊離させて超変異を減速させるIPTGを含む培地中で実施する。これらのクローン集団から産生された抗体を、後述するようにしてさらに特徴付ける。
【0173】
以下の3つの基準を用いて、このアプローチが成功したかを確かめることができる。
1.親和性および特異性
選択過程で抗体親和性が増大することを実証することによって、抗体開発の有効性を確認する。それ以上の超変異を最小限にするためのIPTGの存在下、ELISAのバックグラウンドを最小限にするためにIgMを枯渇させた培地中で48時間培養した細胞の上清希釈物をELISAアッセイ法に供することにより、抗体親和性を最初に測定する。抗体が抗原を認識し、それが提示されている組換え骨格を認識しないよう徹底するために、ビオチンおよびストレプトアビジンを用いて反応性を試験する。両方の組換え抗原に対する抗メソテリン抗体および抗HE4抗体を並行して比較することが、交差反応性の試験として役立つ。親和性を、メソテリンまたはHE4を対象とする対照scFv抗体と比較する。高親和性のクローン集団の樹立後、Biacore社の表面プラズモン共鳴によってKdを定量的に測定する。
【0174】
2.変異誘発のターゲティング
クローン選択の過程で抗体遺伝子を超変異させる細胞メカニズムにより、抗原に接触するV領域のサブドメインであるCDR中に変異が集中する。本発明者らは、加速した超変異および選択によって単離した高親和性クローンは、CDRの変異を同様に集中させると予想する。これを試験するには、単一細胞PCRを用いて、親和性選択後に個々のB細胞のVH領域およびVL領域を増幅し、VH領域およびVL領域を配列決定し、これらの配列を選択されていない集団の配列と比較することができる。
【0175】
3. IPTGと共に培養した際の特異性および親和性の維持
抗体作製用のこのプラットホームの力は、高親和性クローンが作製された際に超変異を制限する能力にある程度依存する。これを試験するには、IPTGの存在下および不在下でクローン集団を培養し、蛍光タグ付きのメソテリンまたはHE4に結合された細胞をフローサイトメトリーに供することにより、増殖した培養物の平均親和性を比較することができる。親和性は、IPTGを含む培養物では不変のままであると予想されるが、IPTGを欠く培養物では増大または減少すると予想される。したがって、蛍光シグナルの分布は、IPTGを含む培養物の方が広いと考えられる。
【0176】
参考文献
【0177】
本出願の全体を通して、様々な刊行物が参照される。これらの刊行物全体の開示は、本発明が関連する現況技術をより完全に説明するために、参照により本出願の本明細書に組み入れられる。
【0178】
前述の内容から、本発明の特定の態様が、例証のために本明細書において説明されるが、本発明の精神および範囲から逸脱することなく様々な修正を実施し得ることが、理解されると考えられる。したがって、本発明は、添付の特許請求の範囲による場合を除いて、限定されない。
【特許請求の範囲】
【請求項1】
以下を含む、標的遺伝子の多様化を可逆的に誘導するように改変されたB細胞:
(a)プロモーターおよびコード領域を含む標的遺伝子に機能的に連結されたシス調節エレメント;
(b)多様化因子に融合された係留因子をコードする融合構築物であって、該係留因子が前記シス調節エレメントに特異的に結合する、融合構築物。
【請求項2】
DT40ニワトリB細胞である、請求項1記載のB細胞。
【請求項3】
ヒトB細胞である、請求項1記載のB細胞。
【請求項4】
シス調節エレメントが重合(polymerized)ラクトースオペレーター(LacO)であり、係留因子がラクトースリプレッサー(LacI)である、前記請求項のいずれか一項記載のB細胞。
【請求項5】
シス調節エレメントがテトラサイクリンオペレーター(TetO)であり、係留因子がテトラサイクリンリプレッサー(TetR)である、請求項4記載のB細胞。
【請求項6】
多様化因子が、転写調節因子、ヘテロクロマチン関連タンパク質、ヒストンシャペロン、クロマチン再構築因子、核膜孔複合体の構成要素、遺伝子調節因子、またはそれらの組合せである、請求項5記載のB細胞。
【請求項7】
多様化因子が、DNA修復因子、DNA複製因子、リゾルベース、ヘリカーゼ、細胞周期調節因子、ユビキチン化因子、SUMO化因子、またはそれらの組合せである、請求項5または6記載のB細胞。
【請求項8】
転写調節因子がVP16またはE47である、請求項6記載のB細胞。
【請求項9】
ヘテロクロマチン関連タンパク質がHP1である、請求項6記載のB細胞。
【請求項10】
ヒストンシャペロンがHIRAである、請求項6記載のB細胞。
【請求項11】
標的遺伝子がヒトIg遺伝子である、前記請求項のいずれか一項記載のB細胞。
【請求項12】
標的遺伝子がIgL遺伝子である、前記請求項のいずれか一項記載のB細胞。
【請求項13】
標的遺伝子がIgH遺伝子である、前記請求項のいずれか一項記載のB細胞。
【請求項14】
標的遺伝子が、異種(heterologous)コード領域と、B細胞表面で標的遺伝子産物の提示を行うのに充分な膜貫通ドメインおよび細胞質尾部をコードする領域とを含む、前記請求項のいずれか一項記載のB細胞。
【請求項15】
以下の段階を含む、関心対象のポリペプチドの変異配列を有するポリペプチドのレパートリーを作製する方法:
(a)多様化因子の発現を可能にする条件下で請求項5〜14のいずれか一項記載のB細胞を培養する段階であって、該B細胞の標的遺伝子が、関心対象のポリペプチドのコード領域を含み、それによって、該コード領域の多様化が可能になる、段階;ならびに
(b)複数のB細胞および所望のレパートリーが得られるまで、前記B細胞の増殖を可能にする条件下で培養物を維持する段階。
【請求項16】
以下の段階を含む、最適化された関心対象のポリペプチドを産生するB細胞を作製する方法:
(a)多様化因子の発現を可能にする条件下で請求項5〜14のいずれか一項記載のB細胞を培養する段階であって、該B細胞の標的遺伝子が関心対象のポリペプチドのコード領域を含み、該B細胞が該関心対象のポリペプチドを該B細胞の表面で発現する、段階;
(b)前記B細胞表面で発現される前記関心対象のポリペプチドに特異的に結合するリガンドに結合する細胞を培養物から選択する段階;ならびに
(c)前記関心対象のポリペプチドに特異的に結合する前記リガンドに対して所望の親和性を有する細胞を選択するまで、段階(a)および段階(b)を繰り返す段階。
【請求項17】
関心対象のポリペプチドがIgである、請求項15または16記載の方法。
【請求項18】
Igが、IgL、IgH、または両方である、請求項17記載の方法。
【請求項19】
培養物に調節分子を添加する段階をさらに含み、該調節分子が、シス調節エレメントへの係留因子の結合を調整し、それによって、コード領域の多様化を調整する、請求項15または16記載の方法。
【請求項20】
培養物から調節分子を除去する段階をさらに含み、該調節分子が、シス調節エレメントへの係留因子の結合を調整し、それによって、コード領域の多様化を調整する、請求項15または16記載の方法。
【請求項21】
B細胞において調節遺伝子の発現を調整する段階をさらに含み、該調節遺伝子が多様化を調節し、それによって、関心対象のポリペプチドをコードする領域の多様化を調整する、請求項15または16記載の方法。
【請求項22】
以下を含むキット:
(a)プロモーターおよびコード領域を含む標的遺伝子に機能的に連結されたシス調節エレメントを含むB細胞;ならびに
(b)1つまたは複数の容器;ならびに
(c) (b)の容器中に保存された1種または複数種の融合構築物であって、各融合構築物が、前記B細胞で発現可能であり、かつ、多様化因子に融合された係留因子をコードし、該係留因子が、前記B細胞の前記シス調節エレメントに特異的に結合する、融合構築物。
【請求項23】
B細胞がDT40ニワトリB細胞である、請求項22記載のキット。
【請求項24】
B細胞がヒトB細胞である、請求項22記載のキット。
【請求項25】
シス調節エレメントが重合ラクトースオペレーター(LacO)であり、係留因子がラクトースリプレッサー(LacI)である、請求項22〜24のいずれか一項記載のキット。
【請求項26】
シス調節エレメントがテトラサイクリンオペレーター(TetO)であり、係留因子がテトラサイクリンリプレッサー(TetR)である、請求項22〜25のいずれか一項記載のキット。
【請求項27】
多様化因子が、転写調節因子、ヘテロクロマチン関連タンパク質、ヒストンシャペロン、クロマチン再構築因子、核膜孔複合体の構成要素、遺伝子調節因子、またはそれらの組合せである、請求項22〜26のいずれか一項記載のキット。
【請求項28】
多様化因子が、DNA修復因子、DNA複製因子、リゾルベース、ヘリカーゼ、細胞周期調節因子、ユビキチン化因子、SUMO化因子、またはそれらの組合せである、請求項22〜27のいずれか一項記載のキット。
【請求項29】
転写調節因子がVP16またはE47である、請求項27記載のキット。
【請求項30】
ヘテロクロマチン関連タンパク質がHP1である、請求項27記載のキット。
【請求項31】
ヒストンシャペロンがHIRAである、請求項27記載のキット。
【請求項32】
標的遺伝子がヒトIg遺伝子である、請求項22〜31のいずれか一項記載のキット。
【請求項33】
標的遺伝子がIgL遺伝子である、請求項22〜32のいずれか一項記載のキット。
【請求項34】
標的遺伝子がIgH遺伝子である、請求項22〜33のいずれか一項記載のキット。
【請求項35】
標的遺伝子が、異種コード領域と、B細胞表面で標的遺伝子産物の提示を行うのに充分な膜貫通ドメインおよび細胞質尾部をコードする領域とを含む、請求項22〜34のいずれか一項記載のキット。
【請求項1】
以下を含む、標的遺伝子の多様化を可逆的に誘導するように改変されたB細胞:
(a)プロモーターおよびコード領域を含む標的遺伝子に機能的に連結されたシス調節エレメント;
(b)多様化因子に融合された係留因子をコードする融合構築物であって、該係留因子が前記シス調節エレメントに特異的に結合する、融合構築物。
【請求項2】
DT40ニワトリB細胞である、請求項1記載のB細胞。
【請求項3】
ヒトB細胞である、請求項1記載のB細胞。
【請求項4】
シス調節エレメントが重合(polymerized)ラクトースオペレーター(LacO)であり、係留因子がラクトースリプレッサー(LacI)である、前記請求項のいずれか一項記載のB細胞。
【請求項5】
シス調節エレメントがテトラサイクリンオペレーター(TetO)であり、係留因子がテトラサイクリンリプレッサー(TetR)である、請求項4記載のB細胞。
【請求項6】
多様化因子が、転写調節因子、ヘテロクロマチン関連タンパク質、ヒストンシャペロン、クロマチン再構築因子、核膜孔複合体の構成要素、遺伝子調節因子、またはそれらの組合せである、請求項5記載のB細胞。
【請求項7】
多様化因子が、DNA修復因子、DNA複製因子、リゾルベース、ヘリカーゼ、細胞周期調節因子、ユビキチン化因子、SUMO化因子、またはそれらの組合せである、請求項5または6記載のB細胞。
【請求項8】
転写調節因子がVP16またはE47である、請求項6記載のB細胞。
【請求項9】
ヘテロクロマチン関連タンパク質がHP1である、請求項6記載のB細胞。
【請求項10】
ヒストンシャペロンがHIRAである、請求項6記載のB細胞。
【請求項11】
標的遺伝子がヒトIg遺伝子である、前記請求項のいずれか一項記載のB細胞。
【請求項12】
標的遺伝子がIgL遺伝子である、前記請求項のいずれか一項記載のB細胞。
【請求項13】
標的遺伝子がIgH遺伝子である、前記請求項のいずれか一項記載のB細胞。
【請求項14】
標的遺伝子が、異種(heterologous)コード領域と、B細胞表面で標的遺伝子産物の提示を行うのに充分な膜貫通ドメインおよび細胞質尾部をコードする領域とを含む、前記請求項のいずれか一項記載のB細胞。
【請求項15】
以下の段階を含む、関心対象のポリペプチドの変異配列を有するポリペプチドのレパートリーを作製する方法:
(a)多様化因子の発現を可能にする条件下で請求項5〜14のいずれか一項記載のB細胞を培養する段階であって、該B細胞の標的遺伝子が、関心対象のポリペプチドのコード領域を含み、それによって、該コード領域の多様化が可能になる、段階;ならびに
(b)複数のB細胞および所望のレパートリーが得られるまで、前記B細胞の増殖を可能にする条件下で培養物を維持する段階。
【請求項16】
以下の段階を含む、最適化された関心対象のポリペプチドを産生するB細胞を作製する方法:
(a)多様化因子の発現を可能にする条件下で請求項5〜14のいずれか一項記載のB細胞を培養する段階であって、該B細胞の標的遺伝子が関心対象のポリペプチドのコード領域を含み、該B細胞が該関心対象のポリペプチドを該B細胞の表面で発現する、段階;
(b)前記B細胞表面で発現される前記関心対象のポリペプチドに特異的に結合するリガンドに結合する細胞を培養物から選択する段階;ならびに
(c)前記関心対象のポリペプチドに特異的に結合する前記リガンドに対して所望の親和性を有する細胞を選択するまで、段階(a)および段階(b)を繰り返す段階。
【請求項17】
関心対象のポリペプチドがIgである、請求項15または16記載の方法。
【請求項18】
Igが、IgL、IgH、または両方である、請求項17記載の方法。
【請求項19】
培養物に調節分子を添加する段階をさらに含み、該調節分子が、シス調節エレメントへの係留因子の結合を調整し、それによって、コード領域の多様化を調整する、請求項15または16記載の方法。
【請求項20】
培養物から調節分子を除去する段階をさらに含み、該調節分子が、シス調節エレメントへの係留因子の結合を調整し、それによって、コード領域の多様化を調整する、請求項15または16記載の方法。
【請求項21】
B細胞において調節遺伝子の発現を調整する段階をさらに含み、該調節遺伝子が多様化を調節し、それによって、関心対象のポリペプチドをコードする領域の多様化を調整する、請求項15または16記載の方法。
【請求項22】
以下を含むキット:
(a)プロモーターおよびコード領域を含む標的遺伝子に機能的に連結されたシス調節エレメントを含むB細胞;ならびに
(b)1つまたは複数の容器;ならびに
(c) (b)の容器中に保存された1種または複数種の融合構築物であって、各融合構築物が、前記B細胞で発現可能であり、かつ、多様化因子に融合された係留因子をコードし、該係留因子が、前記B細胞の前記シス調節エレメントに特異的に結合する、融合構築物。
【請求項23】
B細胞がDT40ニワトリB細胞である、請求項22記載のキット。
【請求項24】
B細胞がヒトB細胞である、請求項22記載のキット。
【請求項25】
シス調節エレメントが重合ラクトースオペレーター(LacO)であり、係留因子がラクトースリプレッサー(LacI)である、請求項22〜24のいずれか一項記載のキット。
【請求項26】
シス調節エレメントがテトラサイクリンオペレーター(TetO)であり、係留因子がテトラサイクリンリプレッサー(TetR)である、請求項22〜25のいずれか一項記載のキット。
【請求項27】
多様化因子が、転写調節因子、ヘテロクロマチン関連タンパク質、ヒストンシャペロン、クロマチン再構築因子、核膜孔複合体の構成要素、遺伝子調節因子、またはそれらの組合せである、請求項22〜26のいずれか一項記載のキット。
【請求項28】
多様化因子が、DNA修復因子、DNA複製因子、リゾルベース、ヘリカーゼ、細胞周期調節因子、ユビキチン化因子、SUMO化因子、またはそれらの組合せである、請求項22〜27のいずれか一項記載のキット。
【請求項29】
転写調節因子がVP16またはE47である、請求項27記載のキット。
【請求項30】
ヘテロクロマチン関連タンパク質がHP1である、請求項27記載のキット。
【請求項31】
ヒストンシャペロンがHIRAである、請求項27記載のキット。
【請求項32】
標的遺伝子がヒトIg遺伝子である、請求項22〜31のいずれか一項記載のキット。
【請求項33】
標的遺伝子がIgL遺伝子である、請求項22〜32のいずれか一項記載のキット。
【請求項34】
標的遺伝子がIgH遺伝子である、請求項22〜33のいずれか一項記載のキット。
【請求項35】
標的遺伝子が、異種コード領域と、B細胞表面で標的遺伝子産物の提示を行うのに充分な膜貫通ドメインおよび細胞質尾部をコードする領域とを含む、請求項22〜34のいずれか一項記載のキット。
【図1】

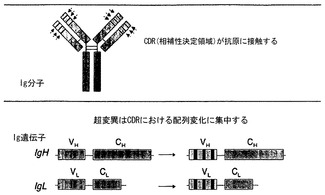
【図2】
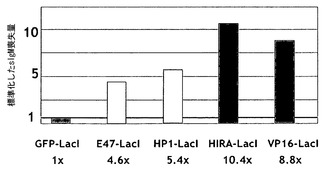
【図3】
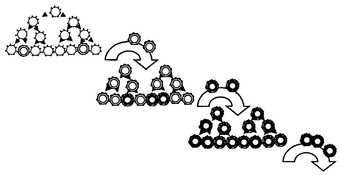
【図4】
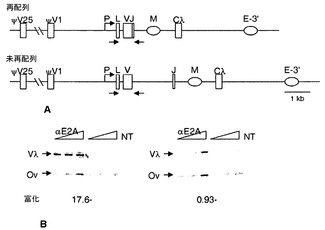
【図5】
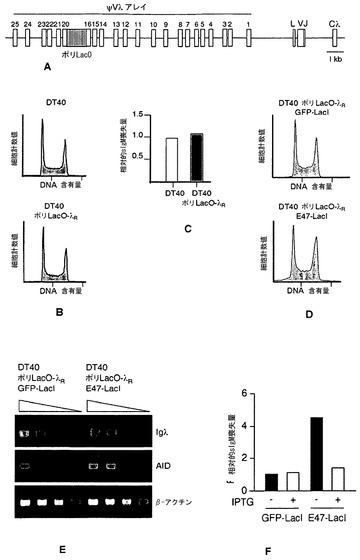
【図6】
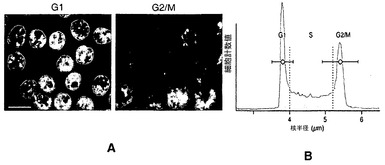
【図7】
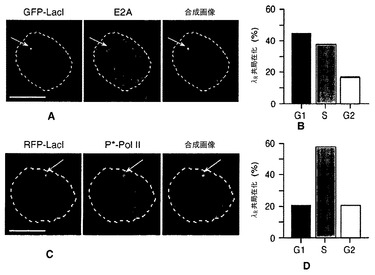
【図8】
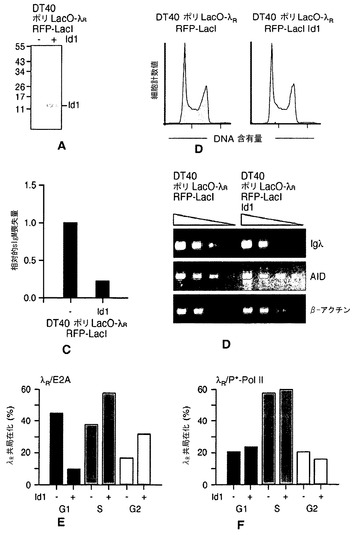
【図9】
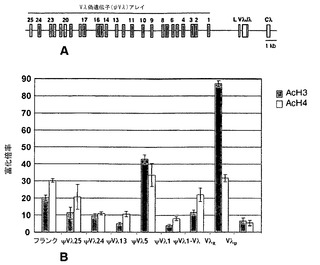
【図10】
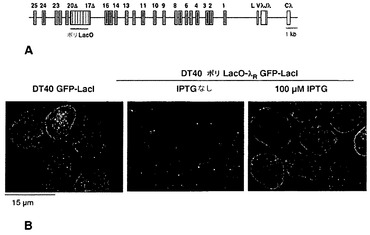
【図11】
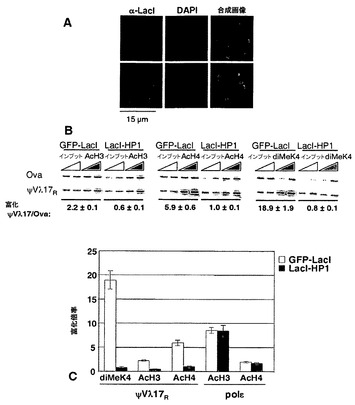
【図12】
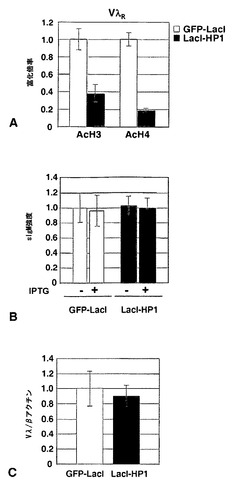
【図13】
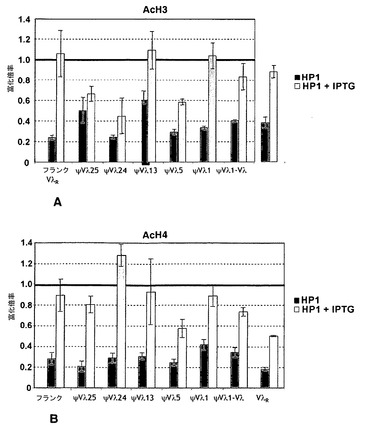
【図14】
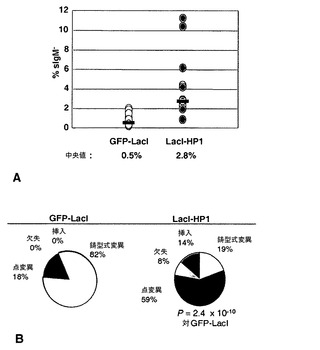
【図15−1】
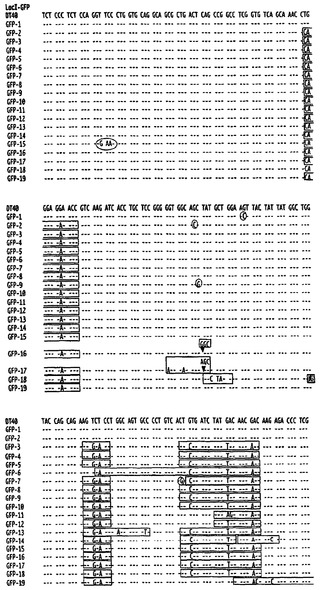
【図15−2】

【図15−3】

【図15−4】

【図16】
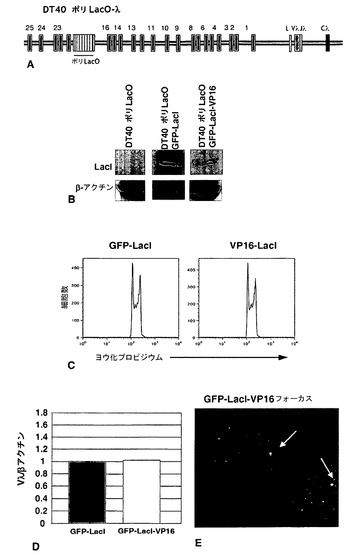
【図17】
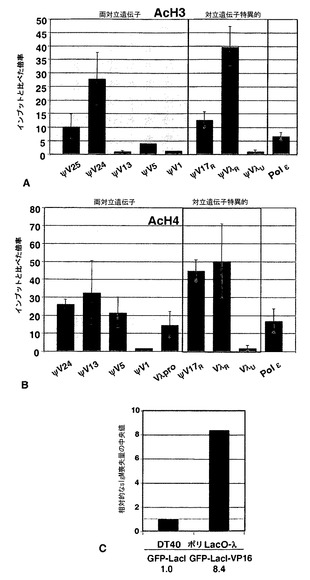
【図18】
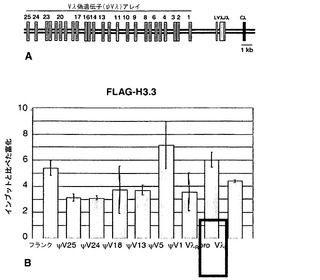
【図19】
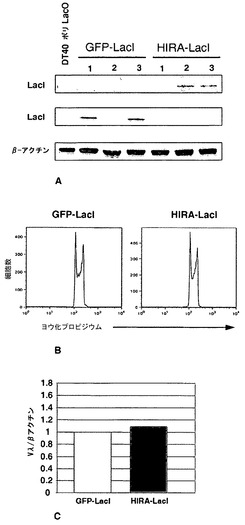
【図20】
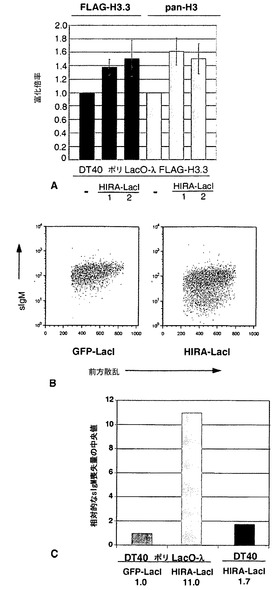
【図21】
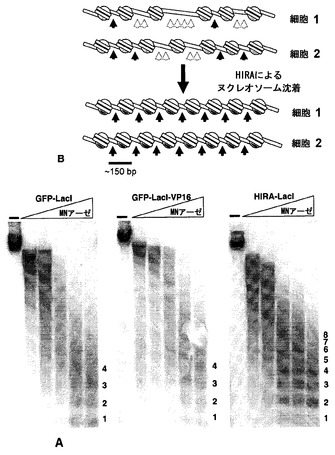
【図22−1】

【図22−2】
【図23−1】
【図23−2】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15−1】
【図15−2】
【図15−3】
【図15−4】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22−1】
【図22−2】
【図23−1】
【図23−2】
【公表番号】特表2010−528626(P2010−528626A)
【公表日】平成22年8月26日(2010.8.26)
【国際特許分類】
【出願番号】特願2010−510553(P2010−510553)
【出願日】平成20年6月2日(2008.6.2)
【国際出願番号】PCT/US2008/065528
【国際公開番号】WO2009/029315
【国際公開日】平成21年3月5日(2009.3.5)
【出願人】(507127439)ユニヴァーシティ オブ ワシントン (2)
【Fターム(参考)】
【公表日】平成22年8月26日(2010.8.26)
【国際特許分類】
【出願日】平成20年6月2日(2008.6.2)
【国際出願番号】PCT/US2008/065528
【国際公開番号】WO2009/029315
【国際公開日】平成21年3月5日(2009.3.5)
【出願人】(507127439)ユニヴァーシティ オブ ワシントン (2)
【Fターム(参考)】
[ Back to top ]