
樹状細胞調節分子
本発明は、哺乳動物樹状細胞の分化および成熟化を調節(好ましくは阻害)する樹状細胞調節分子を提供する。本発明はまた、樹状細胞調節分子およびそのホモログおよび活性フラグメントを含有する医薬組成物、それに対する抗体、およびそれらの分子を利用する治療法およびスクリーニング法を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は樹状細胞(DC)調節分子に関する。特に、本発明は哺乳動物DC(特にヒトDC)の分化および成熟化を調節(好ましくは阻害)する分子に関する。それらの分子は節足動物の唾液、より具体的にはマダニの唾液から単離できる。本発明はまた、それらの分子の治療への使用、特に、自己免疫疾患、アレルギーおよび他の過敏性反応、移植片拒絶反応および移植片対ホスト病、感染性疾患(例えばマダニによって伝染するもの)、癌(例えば血液系腫瘍)、そして急性および慢性炎症性疾患(例えば上記の疾患に伴う炎症)の治療へのそれらの分子の使用にも関する。
【背景技術】
【0002】
哺乳動物(特にヒト)の免疫系には、2つの系統、すなわち自然免疫系および適応免疫系がある。自然免疫系の細胞は包括的様式で感染性病原体を認識および応答する。自然免疫系は感染に対する重要な即時的バリアであるが、外来物質、例えば侵入病原体に対して長期間に及ぶ特異的な防御を与えるものではない。これに対して適応免疫系の細胞は特定の外来物質を認識し、ホストにおいてそれらの特定物質に対する免疫学的記憶を誘導する。
【0003】
DCは病原体による感染直後に自然免疫系の成分と相互作用し、また、哺乳動物の適応免疫応答の中心的役割を果たす。DCは前駆細胞から分化して未成熟DCとなる。未成熟DCは全身にわたって存在し、他の免疫系細胞もこの役割に関わるが、それらは、主にT細胞の活性化を惹起する能力によって適応免疫系の発動に関与する主要な細胞型である。
【0004】
未成熟DCは、外来物質(例えば病原体表面)上の特定の化学シグナルを認識するパターン認識受容体(PRR)(例えばToll様受容体(TRL))によって、常に周辺環境から感染性物質(例えばウィルス、微生物、および寄生生物)をサンプリングしている。病原体のような物質が外来であると同定されると、未成熟DCはその物質またはそのフラグメントを取り込み、タンパク質および脂質抗原を分解してペプチドおよびグリコペプチドまたは脂質フラグメントとし、DC表面に提示する。
【0005】
外来物質の認識および/または他の細胞環境内のシグナル(例えば炎症性サイトカイン)に応答して、未成熟DCは「成熟化」と総称されるいくつかの変化を受け、成熟DCへの発達を開始する。成熟DCは主要組織適合遺伝子複合体(MHC)およびMHC関連分子(例えばCD1)(それぞれ、外来物質由来のペプチドおよびグリコペプチド、および脂質と結合する)の発現を亢進的に調節し、それらをDC表面上に提示させる。これと同時に、DCは共刺激分子として知られる細胞表面受容体(CD80、CD86、およびCD40など。Tリンパ球活性化のための補助受容体として作用する)の発現を亢進的に調節する。更にDCは走化性シグナルを受け、リンパ系組織(例えばリンパ節および/または脾臓)への移動を開始する。リンパ系組織に到達すると、DCは外来物質由来のペプチドおよびグリコペプチドまたは脂質フラグメントを提示し、好適な共刺激シグナルを伝達することによって、Tリンパ球を活性化する。それらの活性化Tリンパ球は、適応免疫応答の伝達に関与している。外来物質は病原体、アレルゲン、または、自己免疫応答の場合は身体によって異物と誤認される自己抗原でありうる。
【0006】
抗原提示および共刺激によるT細胞活性化の惹起の役割に加え、成熟DCはT細胞調節、例えばヘルパーT細胞のTh1、Th2、Th17、または調節性(Treg)細胞への分極化(polarisation)、細胞傷害性T細胞の活性化、および(例えば皮膚または腸および他の粘膜部位への)T細胞ホーミングの調節にも関与する。
【0007】
適応免疫応答においてDCが中心的役割を果たしていることから、治療を目的とするDCの機能調節に関心が向けられており、動物実験ではDCの調節物質が自己免疫疾患および他の炎症性疾患の治療に有用であり得ることが示唆されている(Subkleweら Human Immunology, 2007, 68(3), 147-155)。また、DC調節物質は癌の治療に有用であり得ることも示唆されている。治療を目的としてDC調節物質として作用する更なる分子を同定することは明らかに有益である。
【0008】
特に、DC調節活性、特に阻害活性を有する化合物の同定、並びに、自己免疫疾患、他の炎症性疾患、および癌の治療におけるその使用法の開発がなお必要とされている。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】Subkleweら Human Immunology, 2007, 68(3), 147-155
【発明の概要】
【課題を解決するための手段】
【0010】
発明の概要
従って、本発明は単離されたDC調節分子を提供し、該分子は哺乳動物DCの分化および成熟化を調節(好ましくは阻害)する。
【0011】
ある態様では、単離されたDC調節分子はヒトDCの分化および成熟化を調節(好ましくは阻害)する。本発明のDC調節分子は節足動物、特に吸血性節足動物から単離することができる。単離されたDC調節分子はタンパク質であってもよい。
【0012】
吸血性節足動物はそのホスト(哺乳動物(例えばヒト)など)に付着し、長期間にわたって摂食する。それらがホストに運搬する成分(唾液中の成分など)はホストの免疫応答を惹起する可能性がある。それらの応答は節足動物に有害でありえ、従って節足動物はそれらを抑制する必要があり得る。免疫の惹起においてDCが中心的役割を果たしていることを考慮すると、節足動物にとって、その機能を阻害する物質を生成させることは有益であり得る。
【0013】
吸血性節足動物、特にマダニは、ホストに種々の抗炎症および免疫調節成分を接種することによって、ホストの免疫系を阻害しうる(Ribeiroら, Infectious Agents and Disease, 1992, 4(3), 143-152)。
【0014】
以下のようないくつかの免疫調節分子がマダニ唾液中で同定されている:マクロファージ遊走阻止因子(MIF)のホモログ(Jaworskiら, Insect Molecular Biology, 2001, 10(4), 323-331)、ヒト・マクロファージ、単球、および好中球によって分泌される白血球エラスターゼ阻害剤のホモログ(Leboulleら, The Journal of Biological Chemistry, 2002, 277(12), 10083-10089)、グリコシル化されたタンパク質p36(マイトジェンによって作動されるマウス脾臓細胞のインビトロ増殖を阻害すると考えられる)(Bergmanら, Journal of Parasitology, 2000, 86, 516-525)、B細胞阻害タンパク質(BIP)(Hannierら, Immunology, 2004, 113, 401-408)、およびB細胞抑制因子(BIF)(Yuら, Biochemical and Biophysical Research Communications, 2006, 343, 585 - 590)。しかしながら、これらの分子の多くは明確な細胞標的を有さず、これらの分子のうちで哺乳動物DC(特にヒトDC)の分化および成熟化の両方に阻害効果を与えることが確認されているものは存在しない。
【0015】
Salp15はマダニ唾液中に存在するタンパク質であり、未成熟ヒトDCに作用することが明らかになっている(Anguitaら, Immunity, 2002, 16, 849-859、およびHoviusら, Vector borne and Zoonotic diseases, 2007, 7(3), 296-302)。しかしながら、免疫調節刺激物質の存在下で未成熟ヒトDCをSalp15と共にインキュベートすることを伴うアッセイでは、Salp15が共刺激分子(例えばCD86)の亢進的調節を阻害しないことが明らかにされた。従って、Salp15はヒトDCの成熟化を阻害しない。
【0016】
プロスタグランジンE2(PGE2)はマダニ唾液中に存在する非タンパク質分子であり、未成熟マウスDCの活性を調節しうるが、これらのマウスDCの成熟化に対しては最小限の影響しか及ぼさない(Sa-Nunesら, The Journal of Immunology, 2007, 179, 1497-1505)。PGE2はヒトDCの成熟化を促進する能力を有するが、ヒトDCの分化および成熟化を阻害するように作用するという証拠はない。
【0017】
また、マダニ唾液および唾液腺抽出物(SGE)がマウスDCの分化および成熟化を調節する能力を有しうることも示唆されている(Cavassaniら, Immunology, 2005, 114, 235-245、およびSkallovaら, Journal of Immunology, 2008, 180, 6186-6192)。しかしながら、これらの活性に関与する分子は単離されておらず、これまでのところ、マダニ唾液がヒトDCの分化および成熟化の両方を阻害する能力を有するという証拠はない。
【0018】
驚くべきことに、出願人は哺乳動物DC、特にヒトDCの分化および成熟化の両方を調節(好ましくは阻害)する分子を単離した。
【0019】
「単離された」という用語は、分子がもはやその天然での環境中にないことを伝えることを意図する。この用語には、その天然での環境から除去された分子、およびそれらと同一であるが合成されたものである分子も含まれる。本発明の単離された分子は一般に実質的に純粋である。「実質的に純粋」とは、組成物が少なくとも約50%の目的分子を含有することを意味する。ある態様では、組成物は少なくとも約60%、少なくとも約70%、少なくとも約80%、少なくとも約90%、少なくとも約95%、少なくとも約99%、またはそれ以上の分子を含有することができる。換言すれば、組成物は約50%未満の他の分子を含有してもよい。他の態様では、組成物は約40%未満、約30%未満、約20%未満、約10%未満、約5%未満、約1%未満、またはそれ未満の他の分子を含有してもよい。
【図面の簡単な説明】
【0020】
【図1】図1は、ヒトDCによるCD86発現の亢進的調節に対する給餌および未給餌のオスおよびメスRhipicephalus appendiculatusマダニ由来SGEの影響を示す。F=メス、M=オス、0/3/6=給餌日数。
【図2】図2は、CD1a+細胞による成熟マーカーの発現に対する3日間給餌メスRhipicephalus appendiculatusマダニ由来SGEの影響を示す。黒色のヒストグラムはSGEおよびLPSで処理したCD1a+細胞による成熟マーカーの発現を示し、灰色のヒストグラムはLPS(SGE未含有)で処理したCD1a+細胞による成熟マーカーの発現を示す。
【図3】図3は、LPSで処理したCD1a+細胞による成熟マーカーの発現およびLPS処理を行っていないCD1a+細胞による成熟マーカーの発現に対するQカラム分離したSGEの影響を示す。QFT=pH7でQカラムを通過した物質。QFR=pH7でQカラムに保持された物質。
【図4】図4は、A)LPS+IFNγおよびB)ポリ(I:C)+TNFαの存在下でのCD86亢進的調節に対するSGEまたはQFTの影響を示す。
【図5】図5は、QFTのDC調節作用に対するプロテイナーゼK処理の影響を示す。
【図6】図6は、DC調節活性に関するサイズ分画の結果を示す。活性は、LPS存在下でのCD86亢進的調節の阻害として評価した。
【図7】図7は、サイズ排除クロマトグラフィーを使用して得た最も活性の高いフラグメントでのポリアクリルアミドゲル電気泳動の結果を示す。
【図8】図8は、HPLC精製の結果を示す:A)HPLCカラムからの溶出プロフィール。B)HPLCから溶出した画分の活性プロフィール。
【図9】図9は、エドマン分解によってシークエンシングしたN-末端16アミノ酸を示す。
【図10】図10は、ジャパニンDNAの3'領域をクローニングするためのPCRの結果を示す。
【図11】図11は、ジャパニンDNAの3'領域をクローニングするためのPCRの結果を示す。
【図12】図12は、ジャパニンの3'領域のコンセンサス配列を示す。
【図13a】図13aはジャパニンの5'領域増幅のためのプライマー設計を示す。
【図13b】図13bはジャパニンの5'領域のPCR増幅結果を示す。「PCR」のレーンは未処理のPCR産物である。「精製PCR」と表示した2つの500bpのレーンは、カラム精製後の容量が異なる。
【図14】図14は、ジャパニンの5' cDNA配列を示す。
【図15】図15は、完全長ジャパニンcDNA、およびそれにコードされる176アミノ酸タンパク質を示す。
【図16】図16は、完全長ジャパニン配列中に位置する、可能性のある開裂部位を示す。
【図17】図17は、ジャパニン発現ベクターを含有する細胞から得た昆虫細胞上清のDC調節活性を示す。
【図18】図18は、ジャパニン含有昆虫細胞から回収した上清の硫酸アンモニウム沈殿後の、種々の画分中の活性の存在を示す。
【図19】図19は、ジャパニンの存在を示す、銀染色したSDS-PAGEゲルを示す。
【図20】図20は、ジャパニンを単離するために使用するHisタグの存在を確認するウェスタンブロットを示す。
【図21】図21は、ジャパニン-TEV-his上清のDC調節効果を示す。
【図22】図22は、精製したHisタグ・ジャパニンのDC調節活性を示す。
【図23】図23は、DCによるCD1aおよびCD14の発現に対するジャパニンの影響を示す。
【図24】図24は、MLRアッセイにおける、ジャパニンによって誘導されるT細胞増殖の低下を示す。
【図25】図25は、ジャパニンおよびDermacentor andersoni(D. andersoni E1244)から同定されたジャパニン・ホモログとの2配列間アラインメントを示す。
【図26】図26は、ジャパニンおよびRhipicephalus microplus(R. microplus RM-CK185494)から同定されたジャパニン・ホモログ間の2配列間アラインメントを示す。
【図27】図27は、ジャパニンおよびAmblyomma americanum(A. americanum CX766068)から同定されたジャパニン・ホモログ間の2配列間アラインメントを示す。
【図28】図28は、ジャパニンおよびRhipicephalus appendiculatus(R. appendiculatus CD796501)から同定されたジャパニン・ホモログ間の2配列間アラインメントを示す。
【図29】図29は、ジャパニンおよびRhipicephalus microplus(R. microplus CV443471)から同定されたジャパニン・ホモログ間の2配列間アラインメントを示す。
【図30a】図30aは、GC-MS分析で得た質量分析データを示す。
【図30b】図30bは、GC-MS分析で得た質量分析データを示す。
【図30c】図30cは、GC-MS分析で得た質量分析データを示す。
【図31】図31は、3H-コレステロールのジャパニンへの結合を示す。
【図32a】図32aは、蛍光標識したジャパニンの5日目単球由来樹状細胞への結合のFACS分析を示す。
【図32b】図32bは、蛍光標識したジャパニンの単球への結合のFACS分析を示す。
【図32c】図32cは、蛍光標識したジャパニンの骨髄由来樹状細胞への結合のFACS分析を示す。
【図32d】図32dは、蛍光標識したジャパニンの1日目単球由来樹状細胞への結合のFACS分析を示す。
【図32e】図32eは、蛍光標識したジャパニンのマンナン存在下での単球由来樹状細胞への結合のFACS分析を示す。
【図32f】図32fは、蛍光標識したジャパニンのEDTA存在下での単球由来樹状細胞への結合のFACS分析を示す。
【図33】図33は、ジャパニンのN-グリコシル化を示す。
【図34a】図34aは、LPSの存在下での単球由来樹状細胞成熟化の阻害に対するジャパニンの影響を示す。
【図34b】図34bは、IFNγの存在下での単球由来樹状細胞成熟化の阻害に対するジャパニンの影響を示す。
【図34c】図34cは、TNFαの存在下での単球由来樹状細胞成熟化の阻害に対するジャパニンの影響を示す。
【図34d】図34dは、可溶性CD40Lの存在下での単球由来樹状細胞成熟化の阻害に対するジャパニンの影響を示す。
【図34e】図34eは、IFNαの存在下での単球由来樹状細胞成熟化の阻害に対するジャパニンの影響を示す。
【図34f】図34fは、CD097(TLR7/8リガンド)の存在下での単球由来樹状細胞成熟化の阻害に対するジャパニンの影響を示す。
【図35】図35は、TNFαの樹状細胞分泌に対するジャパニンの影響を示す。
【発明を実施するための形態】
【0021】
本発明の分子の活性
本発明の分子は、哺乳動物DCの分化および成熟化の両方を「調節」すなわち改変、または「阻害」すなわち低減する。ある態様では、それらはヒトDCであることができる。DCの分化および成熟化の調節または阻害を評価する好適なアッセイを以下に記載する。当業者に明らかなように、本明細書に記載する分化および成熟化の調節または阻害の評価のためのマーカーは単に例として提供するものであり、制限を意図するものではない。ある態様では、本発明の分子はDCの分化および成熟化の両方を、例えば以下に記載するアッセイによる測定で少なくとも20%低減させる。更なる態様では、DCの分化および成熟化の両方の阻害は30%、40%、50%、60%、70%、80%、90%、またはそれ以上であってもよい。
【0022】
「DCの分化」とは、細胞前駆体、例えば骨髄由来前駆細胞または血中単球の未成熟DCへの発達を意味する。DC分化の調節、例えば阻害は、当該分野で公知の標準的なアッセイを用いて表現型的に、最終的には機能的に評価できる。
【0023】
前駆体の未成熟DCへの表現型分化の阻害は、前駆細胞が未成熟DCに分化する際に発現が変更される細胞マーカーを用いて評価することができる。例えば、図23および付記する説明に記載するように、単球はCD14陽性およびCD1陰性であるDC前駆体である。未成熟DCはCD14陰性およびCD1陽性である。従って、未成熟DCへの単球の分化はCD14の減少およびCD1の増加によって検出することができる。本発明の分子による前駆細胞の未成熟DCへの分化の阻害は、CD14陽性およびCD1陰性である前駆細胞の持続的な存在によって検出してもよい。
【0024】
前駆細胞および発達したDCの機能的相違は、前駆細胞と発達したDCをそれらの活性に基づいて識別するアッセイを用いて評価できる。例えば、前駆細胞とは異なり、発達したDCは、特に後述のような刺激の後、インビトロアッセイにおいてT細胞増殖を惹起する能力を有する。一般的なT細胞増殖アッセイには同種異系混合リンパ球反応(MLR)および酸化的有糸分裂誘発がある。
【0025】
「DCの成熟化」とは、前駆細胞が分化して未成熟DCになった後に起こる過程を意味する。特にこの用語は、分化した未成熟DCが刺激物質に遭遇し、成熟DCに変換する際に起こる変化に関連する。刺激物質はPRR(例えばTLR)を介して感知される感染性物質(例えば病原体)の成分、サイトカイン受容体を通じて作用するある種のサイトカイン、および/または他の細胞型の特殊化された細胞表面分子(例えば活性化T細胞のCD154)であってもよい。未成熟DCの成熟化に伴う変化には、一般に、共刺激分子(例えばCD80およびCD86)発現の亢進的調節、およびDC表面上での病原体由来成分からの(一般的にはペプチド-MHCおよび脂質-CD1複合体としての)抗原提示がある。未成熟DCの成熟化は二次リンパ組織へのDCの遊走を伴う。
【0026】
本発明の分子は、未成熟DCの成熟化に伴うこれらの変化のいずれを調節または阻害するように作用することができる。従って、本発明の分子がDCの成熟化を阻害する能力は、CD86および/またはCD80および/またはMHC分子の発現を低下させる能力によって評価することができる。本発明の分子がDCの成熟化を阻害する能力は、TNFαの発現および/または分泌を低下させる能力によって評価してもよい。本発明の分子が未成熟DCの成熟化を阻害する能力は、必要により、ポリ(I:C)、LPS、またはIFNγ刺激後に評価してもよい。本発明の分子が未成熟DCの成熟化を阻害する能力は、必要により、CD40L、IFNα、またはTLR7もしくはTLR8リガンド(例えばCL097)刺激後に評価してもよい。未成熟DCの成熟化の阻害を評価するための実験法は実施例に記載する。未成熟DCの成熟化の阻害を評価するための更なる方法は当業者に公知である。
【0027】
上記のように、本発明の分子は前駆細胞の未成熟DCへの分化を調節(好ましくは阻害)し、その後の未成熟DCの成熟DCへの成熟化を調節(好ましくは阻害)するように作用する。このようなDCの分化および成熟化の両方の調節または阻害は、全体として免疫系に対して阻害的調節作用を有すると考えられるが、詳細については後述する。
【0028】
抗原提示細胞によって活性化されるまで、Tリンパ球は「ナイーブ」であると称される。各Tリンパ球は特定の抗原に特異的であり、この同族の抗原を提示している「特殊化された」抗原提示細胞(例えばDC)によってしか活性化されない。通常型の(conventional)Tリンパ球は、α鎖およびβ鎖を含むヘテロ二量体構造を持つT細胞受容体(TCR)を介して抗原-MHC複合体を認識する。しかしながら、共刺激の非存在下でのTCRによるシグナル伝達は、抗原無反応もしくはアネルギー状態、または活性化不全および細胞死を引き起こす。従って、成熟化過程で未成熟DCの表面上で亢進的に調節され、Tリンパ球表面上で受容体分子(例えばCD28(CD80およびCD86の場合)またはCD154(CD40の場合))によって認識される共刺激分子は、Tリンパ球の活性化に不可欠である。
【0029】
本発明のある観点では、上記の分子によって提供されるDCの分化および成熟化の阻害により、Tリンパ球活性化が低下する。「Tリンパ球活性化」とは、ヘルパーT細胞(Th1、Th2、Th17、またはTreg細胞など)の活性化、場合によっては、細胞傷害性T細胞の活性化(多くの場合、先行のヘルパーT細胞の活性化に依存する)を意味する。
【0030】
T細胞活性化の低下は当該分野で公知の方法によって評価することができる。制限ではなく一例として、T細胞活性化は以下によって評価することができる:サイトカイン分泌(例えばインターロイキン(IL)-2生成)もしくはDCによって惹起されるT細胞増殖(例えば同種異系MLRまたは酸化的有糸分裂誘発)のインビトロアッセイ、または抗原再刺激前および後にex vivoで単離されたT細胞と同様のアッセイを用いる正常またはトランスジェニック動物におけるモデル抗原(例えばオボアルブミン)に対するT細胞応答のin vivoアッセイ。
【0031】
ある態様では、本発明の分子は、DC分化および成熟化阻害分子の非存在下で、Tリンパ球活性化を標準のアッセイに比較して少なくとも約20%低下させる。更なる態様では、DC分化および成熟化の阻害によって、Tリンパ球活性化が少なくとも約30%、40%、50%、60%、70%、80%、90%、またはそれ以上低減される。
【0032】
本発明のある観点では、上記の分子によって提供されるDC分化および成熟化の調節または阻害により、Tリンパ球制御が調節される。特に、本発明の分子はTリンパ球のTh1、Th2、Th17、調節性T(Treg)、または濾胞性ヘルパーT(Thf)細胞への分極化を調節することができる。本発明の分子によるTリンパ球分極化の調節は、インビトロアッセイで、一般的に種々のタイプのCD4およびCD8 Tリンパ球に関係するTリンパ球誘導型サイトカインの測定によって評価することができる。これらには、Th1細胞ではIFN-γ;Th2細胞ではIL-4、IL-5、およびIL-13;Th17細胞ではIL-17;そしてTreg細胞ではIL-10およびTGFβがある。各タイプのCD4細胞は、それぞれT-bet、GATA-3、ROR-γ-t、およびFoxP3の発現の測定、またはThf細胞ではBcl6発現の測定によってアッセイすることもできる。あるいはまた、種々の細胞の表現型を、例えばケモカイン受容体およびそれらが発現する他の表現型マーカーの測定によって評価することもできる。
【0033】
Tリンパ球は哺乳動物の免疫応答の主な促進物質の一つである。従って、上記のような本発明の分子によるTリンパ球活性化の低下またはそれらTリンパ球の分極化の変更により、免疫応答の全体的な調節、特に免疫応答に関係するサイトカインのレベルの変化が起こる。例えば、本発明の分子は一般に免疫抑制効果を有しうる。
【0034】
当業者に明白なように、免疫応答の調節、例えば免疫抑制効果は、種々の方法のいずれかを用いて測定できる。全体的な免疫応答の低下または調節は、TLR刺激に応答して最も迅速に生成される炎症促進性サイトカイン、例えばインターロイキン-1および腫瘍壊死因子α(TNFα)、インターフェロンα、インターフェロンβ、または一般に感染直後に生成されるIL-6もしくはIL-12のようなサイトカインのレベル低下を観察することによって測定できる。本発明の分子は抗炎症性サイトカイン、例えばIL-10またはTGF-βのレベル増加も誘発しうる。
【0035】
ある態様では、本発明の分子は、DCの分化および成熟化を阻害する分子の非存在下での標準アッセイに比較して約20%まで、炎症促進性サイトカインのレベルを低下させるか、または抗炎症サイトカインのレベルを増加させる。更なる態様では、DC分化および成熟化の阻害により、少なくとも約30%、40%、50%、60%、70%、80%、90%、またはそれ以上、炎症促進性サイトカインのレベルが低下するか、または抗炎症サイトカインのレベルが増加する。
【0036】
免疫抑制効果は種々の他の方法、例えば炎症の局所的低下、または生成させた抗原特異的TおよびB細胞プールのサイズもしくは活性の低下によって評価してもよい。
【0037】
本発明の分子を単離しうる節足動物
本発明の分子は節足動物から単離することができる。「節足動物」は節足動物門に属する動物と定義され、それらには昆虫、甲殻類、およびクモ形類動物がある。節足動物は、分節した体とキチンを成分とする硬い外骨格を特徴とする。
【0038】
本発明のある観点では、本発明の分子は吸血性節足動物から単離することができる。「吸血性節足動物」という用語は、好適なホストから血液を摂取する全ての節足動物を含む。これには昆虫、マダニ(ticks)、シラミ、ノミ、およびダニ(mites)が含まれる。これらは一般に吸血性(blood feeding)節足動物として知られ、これらの2つの用語は本明細書において同義に使用される。
【0039】
本発明の更なる観点では、単離された吸血性節足動物はマダニであってもよい。「マダニ」という用語はマダニ上科(Ixodoidea)の小型クモ型類の通称であり、吸血性節足動物に包含される。マダニは外部寄生虫であり、哺乳動物、鳥類、および爬虫類の血液に依存して生息する。
【0040】
世界中に約900種のマダニが生息している。種々のマダニ種はその主な生息地および地理的分布によってキャラクタライズされる。ほとんどのマダニ種は種々のホスト種(ヒトを含む)に依存して生息できる。上記のように、節足動物、特にマダニは、ホストに種々の抗炎症および免疫調節成分を接種することによって、ホストの免疫系を阻害しうる。
【0041】
本発明の単離されたDC調節分子は任意の既知のマダニ種、例えばIxodinae、Bothriocrotoninae、Amblyomminae、Haemaphysalinae、Rhipicephalinae、Hyalomminae、Nuttalliellidae、Argasinae、Otobinae、Antricolinae、Nothoaspinae 、およびOrnithodorinae群に含まれる種、例えば以下のマダニ種のいずれかから単離することができる:Rhipicephalus appendiculatus、Rhipicephalus sanguineus、Rhipicephalus bursa、Amblyomma americanum、Amblyomma cajennense、Amblyommahebraeum、Amblyomma variegatum、Rhipicephalus(Boophilus)microplus、Rhipicephalus(Boophilus)annulatus、Rhipicephalus(Boophilus)decoloratus、Dermacentor reticulatus、Dermacentor andersoni、Dermacentor marginatus、Dermacentor variabilis、Haemaphysalis inermis、Haemaphysalis leachii、 Haemaphysalis punctata、Hyalomma anatolicum anatolicum、Hyalomma dromedarii、Hyalomma marginatum marginatum、Ixodes ricinus、Ixodes persulcatus、Ixodes scapularis、Ixodes hexagonus、Argas persicus、Argas reflexus、Ornithodoros erraticus、Ornithodoros moubata moubata、Ornithodoros moubata porcinus、およびOrnithodoros savignyi。
【0042】
本発明のタンパク質
本発明の分子はタンパク質であってもよい。実施例3から18に詳述するように、出願人はマダニ種Rhipicephalus appendiculatusから哺乳動物DCの分化および成熟化を阻害する節足動物タンパク質を同定および単離した。本明細書においてはこのタンパク質をジャパニン(Japanin)と称するが、そのアミノ酸配列を図15およびSEQ ID NO:2に示す。
【0043】
従って、本発明のある観点では、本発明の分子は以下を含むことができる:
i)SEQ ID NO:2のアミノ酸配列を含むタンパク質;
ii)i)のタンパク質のホモログ;
iii)上記i)のタンパク質もしくは上記ii)のホモログの活性フラグメント;または
iv)i)、ii)、もしくはiii)の機能的同等物。
【0044】
本発明の分子はSEQ ID NO:2のアミノ酸配列から成るタンパク質またはその活性フラグメントであってもよい。
【0045】
「機能的同等物」という用語は、本明細書では、上記のアッセイを用いて、SEQ ID NO:2のアミノ酸配列を含む完全長ジャパニン・タンパク質に匹敵する方法でDCの分化および成熟化を調節(好ましくは阻害)する能力を有する分子を表す。これには合成タンパク質、タンパク質の合成変異体、匹敵する活性を有する異なる配列のタンパク質分子、匹敵する活性を有する天然に存在する非タンパク質分子、および匹敵する活性を有する合成非タンパク質分子がある。
【0046】
特に、ジャパニン分子の三次構造または活性部位(単数または複数)を模倣するように設計した合成分子も機能的同等物であるとみなされる。ある態様では、機能的同等物は、上記のようにDCの分化および成熟化を調節(好ましくは阻害)する能力を有し、それによってTリンパ球活性化の低下またはTリンパ球分極化の調節が起こる。更なる態様では、機能的同等物は、上記のようにDCの分化および成熟化を調節(好ましくは阻害)する能力を有し、それによって免疫応答の抑制が起こる。
【0047】
グリコシル化
実施例26に示すように、ジャパニンは、おそらく2つの部位で、グリコシル化されると考えられる。アスパラギン結合型グリコシル化のためのコンセンサス配列の一部であるアスパラギン残基はジャパニン(SEQ ID NO:2)アミノ酸配列の59および155位に位置する。これらは、リーダーペプチド配列が無い成熟タンパク質の35および131位に相当する。
【0048】
従って、本発明のタンパク質は1つまたはそれ以上の位置でグリコシル化されていてもよい。ある態様では、タンパク質は1、2、3、またはそれ以上の位置でグリコシル化されていてもよい。本発明のタンパク質はN-グリコシル化されているが、タンパク質は更に1つまたはそれ以上の位置でO-グリコシル化されていてもよい。
【0049】
本発明のタンパク質は天然でグリコシル化されていてもよい。これは、タンパク質が天然物を単離したものである場合、またはタンパク質を組換え技術によって、タンパク質が天然で生成される生体のグリコシル化パターンと酷似したホスト細胞内で生成させる場合に特に当てはまる。あるいはまた、本発明のタンパク質を化学的にグリコシル化する必要もありうる。これは、タンパク質を化学的に合成する場合、またはタンパク質を組換え技術によって、天然とは異なるタンパク質グリコシル化パターンを有する生体で生成させる場合に特に当てはまる。また、更なるグリコシル化がなされてタンパク質の特性が改変または改善されてもよい。
【0050】
リポカインの構造および脂質結合性
実施例11に示すように、出願人はジャパニンがリポカリンであることを発見した。リポカリンは類似した折りたたみ構造を有するタンパク質のファミリーである。特徴的なリポカリンの折りたたみは8つのストランドから成る逆平行βバレルであり、内部にリガンド結合部位を形成する。リポカリンは一般に少なくとも2つ、多くの場合は4つのシステイン残基を間隔をあけて有する。システイン残基は内部にジスルフィド結合部位を形成し、これによってリポカリンの折りたたみが安定化されている可能性がある。本発明のある観点では、分子はリポカリンであってもよい。従って、本発明の範囲内に含まれるホモログ、フラグメント、および機能的同等物は、通常マダニ・リポカリン中に存在するCys/Try X Leu Trpモチーフを含有してもよい。
【0051】
リポカリンの構造が推測通りだとすると、ジャパニン・タンパク質は脂質または脂質様分子と結合(associated)しうる。「脂質」という用語は、有機溶媒に可溶であるが水に不溶である疎水性または両親媒性分子を含むことを意図する。これには脂質、油、トリアシルグリセロール、糖脂質、リン脂質およびステロイド、脂肪酸アシル、脂肪酸、グリセロ脂質、グリセロリン脂質、スフィンゴ脂質、サッカロ脂質(saccharolipid)、ポリケチド、ステロール脂質、およびプレノール脂質がある。「脂質様分子」という用語は、脂質と類似または同一の特性を有するいずれの分子も包含する。また「脂質様分子」という用語は非脂質分子と複合体を形成した脂質も包含する。これには糖脂質、リン脂質、ホスホ糖脂質、標識された脂質、およびアセチル化された脂質がある。
【0052】
in vivoにおいて、本発明のタンパク質は小胞体内での折りたたみ中もしくはその直後に、またはその位置から排出された後の段階で、脂質分子に結合する可能性がある。
【0053】
本発明の範囲内で、本発明のタンパク質はその生成の際に脂質と結合してもよい。例えば本発明のタンパク質が天然の供与源から単離されるか、または組換えによって生成される場合、タンパク質はいずれの介入も必要とせずに、自動的に脂質と結合してもよい。あるいはまた、脂質を本発明のタンパク質を含有する組成物に添加し、脂質およびタンパク質間で複合体を形成させてもよい。特にタンパク質を化学的に合成する場合、タンパク質を含有する組成物に脂質を外因的に添加し、複合体を形成させてもよい。上記のように、本発明のタンパク質は脂質と「結合」または「複合体を形成」してもよい。本明細書では、これらの用語は、本発明の脂質およびタンパク質間のあらゆる種類の接触に関連して同義に用いられる。特に、本発明のタンパク質および脂質間は相互作用してもよい。ある態様では、相互作用は単純に構造的なものであってもよい。すなわち、脂質がタンパク質内の結合ポケットに隙間無くはめ込まれていてもよい。別の態様では、脂質は任意の引力によってタンパク質と物理的に相互作用してもよい。それらの引力には静電相互作用、疎水性相互作用、親水性相互作用、ファンデルワールス力、水素結合、および共有結合性相互作用がある。脂質およびタンパク質間の相互作用は、構造的相互作用および引力が組み合わさったものであってもよい。
【0054】
ある態様では、本発明のタンパク質は脂質と結合してもよい。別の態様では、脂質はステロイドまたはステロール、例えばコレステロールであってもよい。別の態様では、脂質はコレステロールの代謝物、例えばビタミンD3であってもよい。実施例23に示すように、ジャパニンはコレステロールに結合することが明らかになっている。ある態様では、本発明のタンパク質はコレステロールの代謝物に結合してもよい。特に、本発明はジャパニンおよび脂質(例えばコレステロールまたはコレステロールの代謝物)を含有する、またはそれらから成る複合体を提供する。
【0055】
ジャパニン・タンパク質の機能的同等物、ホモログ、およびフラグメントは、脂質または他の疎水性分子(単数または複数)(例えば脂質様分子)と結合してもよい。特に、ジャパニン・タンパク質の機能的同等物、ホモログ、およびフラグメントはコレステロールまたはコレステロールの代謝物と結合してもよい。
【0056】
ジャパニンはコレステロールに結合することが明らかになっているので、出願人は、脂質を保持するがジャパニンの生体活性を有さないように遺伝子操作した分子は有用な特性を有しうると考える。従って、本発明は脂質に結合し、DC表面上の受容体を標的とするキャリア分子を含む。それらのキャリア分子は、それ自身は生体活性を有さない。ある態様では、ジャパニンをその生体活性を阻害するように遺伝子操作することによって、キャリア分子を生成してもよい。別の態様では、細胞受容体への結合によって、キャリア分子が保持する脂質に生体機能が施与されてもよい。従って「機能的同等物」という用語はキャリア分子を含む。
【0057】
受容体結合
実施例24に示すように、ジャパニンはC型レクチン細胞表面受容体に結合すると考えられる。これは、ジャパニンが、標的細胞の表面上にある受容体に結合し、阻害の原因となる内部細胞シグナル伝達経路を惹起させることによって、樹状細胞の分化および成熟化を調節(好ましくは阻害)するように機能することを示唆している。
【0058】
ある態様では、タンパク質は標的細胞(例えばDC)の外表面上の受容体に結合することができる。別の態様では、タンパク質は二価陽イオン依存性受容体、特にC型レクチン受容体に結合することができる。ある態様では、タンパク質は受容体に結合し、受容体の天然のリガンドを模倣してもよい。当業者に明白なように、タンパク質のいずれの部分が受容体に結合してもよい。特に、タンパク質がグリコシル化される、そして/または脂質分子に結合する場合、それは炭化水素部分または関連脂質であって、それが標的細胞上の受容体に結合してもよい。
【0059】
ある態様では、本発明のタンパク質および受容体を含有する、またはそれらから成る複合体は本発明に包含される。別の態様では、複合体は本発明のタンパク質、受容体(例えば二価陽イオン依存性受容体(C型レクチン受容体など))、および脂質(例えばコレステロールまたはコレステロールの代謝物)を含有するか、またはそれらから成ってもよい。
【0060】
ホモログおよびフラグメント
上記のように、本発明はアミノ酸配列SEQ ID NO:2で示すジャパニン・タンパク質のホモログおよび活性フラグメントを含む。本発明はこれらのホモログおよびフラグメントの機能的同等物も含む。
【0061】
「ホモログ」という用語は、SEQ ID NO:2に示すジャパニン配列のパラログおよびオルソログでDCの分化および成熟化を調節(好ましくは阻害)する能力を保有するものへの言及を含むことを意図する。ホモログは、上記のようにDCの分化および成熟化を調節(好ましくは阻害)し、それによってTリンパ球活性化の低下またはTリンパ球分極化の調節を起こす能力を有することができる。更なる態様では、ホモログは、上記のようにDCの分化および成熟化を阻害し、それによって免疫応答の阻害を起こす能力を有してもよい。別の態様では、ホモログは、脂質(例えばコレステロールまたはコレステロールの代謝物)に結合する能力および/または膜結合型受容体(例えば二価陽イオン依存性受容体(C型レクチン受容体など))に結合する能力を有してもよい。
【0062】
ホモログは以下のような、Rhipicephalus appendiculatus以外のマダニ種由来であってもよい:Rhipicephalus sanguineus、Rhipicephalus bursa、Amblyommaamericanum、Amblyomma cajennense、Amblyomma hebraeum、Ambylomma variegatum、Rhicephalus(Boophilus)microplus、Rhicephalus(Boophilus)annulatus、Rhicephalus(Boophilus)decoloratus、Dermacentor reticulatus、Dermacentor andersoni、Dermacentor marginatus、Dermacentor variabilis、Haemaphysalis inermis、Haemaphysalis leachii、 Haemaphysalis punctata、Hyalomma anatolicum anatolicum、Hyalomma dromedarii、Hyalomma marginatum marginatum、Ixodes ricinus、Ixodes persulcatus、Ixodes scapularis、Ixodes hexagonus、Argas persicus、Argas reflexus、Ornithodoros erraticus、Ornithodoros moubata moubata、Ornithodoros moubata porcinus、およびOrnithodoros savignyi。ホモログは蚊種(例えばCulex、Anopheles、およびAedes genera、特にCulex quinquefasciatus、Aedes aegypti、およびAnopheles gambiae);ノミ種(例えばCtenocephalides felis(ネコノミ));ウマバエ;スナバエ;ブユ;ツェツェバエ;シラミ;ダニ由来のものであってもよい。
【0063】
一般に、ホモログは任意の既知のマダニ種、例えばIxodinae、Bothriocrotoninae、Amblyomminae、Haemaphysalinae、Rhipicephalinae(Hyalomminaeなど)、Nuttalliellidae、Argasinae、Otobinae、Antricolinae、およびOrnithodorinaeの群に含まれるものに由来してもよい。
【0064】
本明細書に記載する単離されたジャパニン・タンパク質配列のホモログの同定法は、当業者に公知である。例えばホモログは、公的および私的な配列データベースのホモロジー検索によって同定することができる。公的なデータベースを使用してもよいが、特に公的データベースに掲載されていないデータが含まれる場合は、私的または市販のデータベースも同様に有用である。主要なデータベースは主要なヌクレオチドまたはアミノ酸配列データ預託のサイトであり、公的または市販のものがある。主な公的データベースの例として以下がある:GenBankデータベース(http://www.ncbi.nlm.nih.gov)、EMBLデータベース(http://www.ebi.ac.uk/)、DDBJデータベース(http://www.ddbj.nig.ac.jp/)、SWISS-PROTタンパク質データベース(http://expasy.hcuge.ch/)、PIR(http://pir.georgetown.edu/)、TrEMBL(http://www.ebi.ac. uk/)、TIGRデータベース(例えばhttp://www.tigr.org/tdb/index.html)、NRL-3Dデータベース(http://www.nbrfa.georgetown.edu、Protein Data Base(http://www.rcsb.org/pdb)、NRDBデータベース(ftp://ncbi.nlm.nih.gov/pub/nrdb/README)、OWLデータベース(http://www.biochem.ucl.ac.uk/bsm/dbbrowser/OWL/)、および二次的なデータベース、PROSITE(http://expasy.hcuge.ch/sprot/prosite.html)、PRINTS(http://iupab.leeds.ac.uk/bmb5dp/prints.html)、Profiles(http://ulrec3.unil.ch/software/PFSCAN_form.html)、Pfam (http://www.sanger.ac.uk/software/pfam)、Identify(http://dna.stanford. edu/identify/)、およびBlocks(http://www.blocks.fhcrc.org)データベース。市販のデータベースまたは私的データベースの例としてPathoGenome(Genome Therapeutics社)およびPathoSeq(Incyte Pharmaceuticals社)。
【0065】
一般的に、2つのポリペプチド間の同一性が(好ましくは特定の領域にわたって)30%より高ければ、それらは機能的同等物であることを示しており、従って2つのタンパク質はホモログであることを示していると見なされる。ある態様では、ホモログであるタンパク質はSEQ ID NO:2のジャパニン・タンパク質配列と60%を超える配列同一性を有する。他の態様では、ホモログは単離された節足動物タンパク質配列、SEQ ID NO:2と70%、80%、90%、95%、98%、または99%を超える同一性を有する。本明細書の記述では、同一性%の測定はNCBI(the National Center for Biotechnology Information; http://www.ncbi.nlm.nih.gov/) のデフォルト・パラメータ([Blosum 62 matrix; gap open penalty=11 and gap extension penalty=1])を用い、BLAST(バーション2.1.3)を使用して行う。
【0066】
ジャパニン・タンパク質配列(SEQ ID NO:2)のホモログには、野生型タンパク質配列が示すDC分化および成熟化の調節(好ましくは阻害)を保持している限り、野生型配列に比較してアミノ酸置換、挿入、または欠失を含む変異体も含まれる。変異体は、上記のようにDCの分化および成熟化を調節(好ましくは阻害)し、それによってTリンパ球活性化の低下またはTリンパ球分極化の調節を起こす能力を有することができる。更なる態様では、変異体は、上記のようにDCの分化および成熟化を調節(好ましくは阻害)し、それによって免疫応答を阻害する能力を有してもよい。
【0067】
従って変異体には、タンパク質の機能または活性に有害な影響を及ぼさない、保存的アミノ酸置換を含有するタンパク質も含まれる。この用語には天然の生物学的変異体(例えば、本発明の単離された節足動物タンパク質誘導の供与源である種内に含まれる対立遺伝子変異体または地理的変異体も包含されることが意図される。野生型タンパク質配列に比較してDCの分化および成熟化の調節または阻害活性が向上された変異体は、タンパク質配列中の特定残基の系統的または部位特異的変異によって設計してもよい。
【0068】
実施例20に記載するように、出願人はDermacentor andersoni中に含まれるジャパニン・ホモログを同定した。実施例21に記載するように、出願人は、Rhipicephalus(Boophilus)microplus(2つのホモログ)、Amblyomma americanum、およびRhipicephalus appendiculatusからそれぞれジャパニンのホモログを同定した。配列を図25-29に示すが、これはSEQ ID NO:4、6、8、10、および12に相当する。
【0069】
従って、本発明の更なる観点では、本発明の単離された分子は以下を含むことができる:
i)SEQ ID NO:4、6、8、10、もしくは12のいずれか1つのアミノ酸配列を含有するタンパク質
ii)i)のタンパク質のホモログ;
iii)上記i)のタンパク質もしくは上記ii)のホモログの活性フラグメント;または、
iv)i)、ii)、もしくはiii)の機能的同等物。
【0070】
理論に拘束されることは望まないが、これらの配列は完全長配列でなくてもよいと想定される。従って、本発明では、更なるアミノ酸がSEQ ID NO:4、6、8、10、または12のアミノ酸配列を含有する分子のN-末端および/またはC-末端に存在してもよい。
【0071】
本発明は単離されたジャパニン分子の「活性フラグメント」および単離されたジャパニン分子のホモログも提供する。完全長ジャパニン分子の哺乳動物DC分化および成熟を調節または阻害する能力を有する任意のフラグメントがこの定義に包含される。
【0072】
それらのフラグメントとしては、本明細書でSEQ ID NO:2、4、6、8、10、および12として定義される単離された節足動物タンパク質のフラグメントだけでなく、上記のように、このタンパク質のホモログのフラグメントも包含される。それらのホモログのフラグメントは一般にSEQ ID NO:2、4、6、8、10、および12の単離された節足動物タンパク質のフラグメントと60%を超える同一性を有するが、より好ましいホモログのフラグメントはSEQ ID NO:2、4、6、8、10、および12の単離された節足動物タンパク質のフラグメントとそれぞれ70%、80%、90%、95%、98%、または99%を超える同一性を示す。
【0073】
これらのSEQ ID NO:2、4、6、8、10、および12の単離された節足動物タンパク質の活性フラグメントおよびそのホモログのフラグメントはDCの分化および成熟化を調節(好ましくは阻害)する。ある態様では、SEQ ID NO:2、4、6、8、10、および12の単離された節足動物タンパク質のフラグメントおよびそのホモログのフラグメントは、上記のようにDCの分化および成熟化を調節(好ましくは阻害)し、それによってTリンパ球活性化の低下またはTリンパ球分極化の調節を起こす能力を有する。更なる態様では、SEQ ID NO:2、4、6、8、10、および12の単離された節足動物タンパク質のフラグメントおよびそのホモログのフラグメントは、上記のようにDCの分化および成熟化を調節(好ましくは阻害)し、それによって免疫応答を阻害する能力を有する。もちろん、DCの分化および成熟化を調節または阻害する活性が向上されたフラグメントを野生型配列の系統的変異または断片化によって合理的に設計し、その後公的な活性アッセイを行ってもよい。
【0074】
ある態様では、ホモログは脂質(例えばコレステロールまたはコレステロールの代謝物)との結合能、および/または膜受容体(例えば二価陽イオン依存性受容体(C型レクチン受容体など))との結合能を有してもよい。
【0075】
ある態様では、上記の単離された節足動物タンパク質のフラグメントは少なくとも約100アミノ酸長であってもよい。更なる態様では、上記の単離された節足動物タンパク質のフラグメントは少なくとも約90、少なくとも約80、少なくとも約70、少なくとも約60、少なくとも約50、少なくとも約40、少なくとも約30、少なくとも約20、少なくとも約10、または少なくとも約5アミノ酸長であってもよい。
【0076】
抗体
本発明はまた、本発明の分子、特に上記のジャパニン・タンパク質、そのホモログ、フラグメント、および機能的同等物に結合する抗体も提供する。抗体は分子の検出のための試薬として使用することができる。これは、下記のように、分子のDCの分化および成熟化を調節または阻害する活性を中和し、そのために治療の目的に有用である抗体であってもよい。本発明のこの観点には、上記のような本発明の範囲に包含される任意の機能的同等物、ホモログ、およびタンパク質フラグメントに結合する抗体も含まれる。
【0077】
本発明には、本発明のタンパク質の炭化水素部分に結合する抗体も含まれる。特に、本発明には天然でジャパニンに結合する炭化水素部分の1つまたはそれ以上に結合する抗体も含まれる。
【0078】
「アンチカリン」も本発明の範囲内に含まれる。これらは特定のタンパク質エピトープを認識および結合するようにリポカリンを遺伝子操作して作製した分子である。ある態様では、アンチカリンはペプチド、グリコペプチド、または糖脂質の形態を取ってもよい。本明細書では、アンチカリンは「抗体」という用語の範囲に包含される。アンチカリンは非免疫グロブリン誘導型分子ではあるが、抗体と同様の方法でタンパク質エピトープを認識する。
【0079】
ポリクローナル抗体が所望される場合、選択された哺乳動物、例えばマウス、ウサギ、ヤギ、またはウマを本発明の分子、例えばジャパニン・タンパク質、フラグメント、ホモログ、または機能的同等物で免疫化することができる。必要により、分子をキャリアタンパク質にコンジュゲートさせてもよい。一般的に使用されるキャリアにはウシ血清アルブミン、サイログロブリン、およびキーホールリンペットヘモシアニンがある。その後、カップリングした分子を用いて動物を免疫化する。免疫化動物の血清を回収し、既知の方法、例えば免疫親和性クロマトグラフィーで処理する。
【0080】
本発明の分子に対するモノクローナル抗体も当業者によって容易に生成される。ハイブリドーマ法を用いてモノクローナル抗体を生成する一般的な方法は公知である(例えばKohler, G.およびMilstein, C., Nature 256: 495-497 (1975);Kozborら, Immunology Today 4: 72 (1983);Coleら,Monoclonal Antibodies and Cancer Therapy,77-96, Alan R. Liss社(1985)参照)。
【0081】
本明細書で使用する「抗体」という用語は、抗体のフラグメント(例えばFab、F(ab')2、およびFvフラグメントでDC調節分子に特異的に結合するものを含む。更に「抗体」という用語は、本発明の分子、特にジャパニン・タンパク質、そのホモログおよびフラグメントに特異的なキメラおよびヒト化抗体分子を含む。キメラ抗体は、非ヒト可変領域をヒト定常領域に結合または融合させた抗体である(例えばLiuら, Proc. Natl. Acad. Sci. USA, 84, 3439 (1987)参照)。本明細書に記載する「ヒト化抗体」という用語は、非ヒト・ドナー抗体の重鎖および/または軽鎖の可変ドメイン中にあるCDRアミノ酸および選択された他のアミノ酸でヒト抗体中の同等のアミノ酸を置換した抗体分子をいう。従って、ヒト化抗体はヒト抗体と酷似しているが、ドナー抗体の結合能を有する(Jonesら, Nature, 321, 522 (1986);Verhoeyenら, Science, 239, 1534 (1988);Kabatら, J. Immunol., 147, 1709 (1991);Queenら, Proc. Natl Acad. Sci. USA, 86, 10029 (1989);Gormanら, Proc. Natl Acad. Sci. USA, 88, 34181 (1991);およびHodgsonら, Bio/Technology, 9, 421 (1991)参照)。
【0082】
ある態様では、抗体に(例えば検出を容易にするための)標識部分を結合させルのが望ましい。標識は酵素、放射能標識、ビオチンなどの化合物、または蛍光色素であってもよい。
【0083】
融合タンパク質
本発明は、1つまたはそれ以上のペプチド、ポリペプチド、または他の分子に遺伝子的に融合するか、または化学的に結合した本発明の分子、特にジャパニン・タンパク質、そのホモログ、フラグメント、または機能的同等物を含有する融合タンパク質を含む。更なるペプチドもしくはポリペプチドまたは分子の目的は、タンパク質の検出、発現、分離、もしくは精製を容易にすることであるか、またはタンパク質に所望の更なる特性を与えることであってもよい。有望な融合パートナーの例として、β-ガラクトシダーゼ、グルタチオン-S-トランスフェラーゼ、ルシフェラーゼ、ポリヒスチジン・タグ、T7ポリメラーゼ・フラグメント、および分泌シグナルペプチドがある。融合パートナーはin vivoにおける分子の寿命を延長するものであってもよい(例えばFcフラグメント)。融合タンパク質の例を実施例15-18に記載する。
【0084】
他の有望な融合パートナーには有望な生物医薬品、例えば特定の疾患を治療するための薬剤として使用するために開発されたタンパク質または他の分子がある。更なる有望な融合パートナーには、免疫系内で本発明の分子を細胞(例えばDC)にターゲッティングさせる抗原がある。例えば、融合パートナーは、in vivoにおいて分子に融合してこれをDCに運搬しうる自己もしくは外来抗原またはアレルゲンであってもよい。更なる融合パートナーは、DCの異なる細胞表面成分に結合してDCへの運搬を容易にする分子であってもよい。ある態様では、複数の融合パートナーが含まれてもよい。それらの抗原の例は以下に、より詳細に記載する。
【0085】
核酸
本発明は本発明の分子をコードする核酸配列を含有する核酸分子も含む。DNA分子、RNA分子、およびDNA-RNA分子混合物も「核酸分子」という用語に包含されることが意図される。更に、ゲノムDNA、cDNA分子、mRNA分子、そして改変された塩基を含有するRNAおよびDNA分子もこの定義に含まれる。当業者に明白なように、遺伝子コードの縮重により、単離されたタンパク質、タンパク質フラグメント、またはその機能的同等物の所定のタンパク質配列をコードする能力のある多くの異なる核酸配列が存在するが、それらは本発明の範囲内に含まれる。本発明には、融合タンパク質(例えば上記の融合タンパク質)をコードする核酸分子も含まれる。
【0086】
本発明のある観点では、本発明の分子をコードする核酸配列を含有する核酸分子はSEQ ID NO:1またはその縮重配列を含有するか、またはそれから成ってもよい。本発明の更なる観点では、核酸分子はSEQ ID NO:3、5、7、9、もしくは11、またはその縮重配列のいずれかを含有してもよい。縮重配列の一例はSEQ ID NO:32の核酸分子であり、これはSEQ ID NO:3の核酸分子と同じ、SEQ ID NO:4のDermacentor andersoniタンパク質をコードする。
【0087】
また本発明は、本発明の分子、特に上記のような単離された節足動物タンパク質、そのホモログ、フラグメント、または機能的同等物をコードする核酸配列を含有する核酸分子に高ストリンジェンシー・ハイブリダイゼーション条件下でハイブリダイズするアンチセンス核酸分子も提供する。高ストリンジェンシー・ハイブリダイゼーション条件として以下がある:50% ホルムアミド、5X SSC(150mM NaCl、15mM クエン酸三ナトリウム、)、50mM リン酸ナトリウム(pH7.6)、5x デンハルト溶液、10% 硫酸デキストラン、および20μg/ml 変性断片化サケ精子DNAを含有する溶液中、42℃で一晩インキュベートした後、フィルターを0.1X SSC中、約65℃で洗浄する。アンチセンス核酸分子にはアンチセンスDNAオリゴヌクレオチド、およびRNAオリゴヌクレオチド(siRNAなど)がある。
【0088】
また本発明には、単離された節足動物タンパク質、そのホモログ、フラグメント、もしくは機能的同等物をコードする核酸配列を含有する核酸配列、または高ストリンジェンシー・ハイブリダイゼーション条件下で該核酸分子にハイブリダイズするアンチセンス核酸分子を含有するベクターも含まれる。該ベクターにはクローニングおよび発現ベクターが含まれる。それらの発現ベクターには、本発明の核酸分子とインフレームで結合した好適な転写および翻訳制御配列、例えばエンハンサー要素、プロモーター/オペレーター領域、ターミネーション停止配列、mRNA安定化配列、開始もしくは停止コドン、またはリボソーム結合部位が組み込まれてもよい。これらの制御配列は例として提供するものであり、制限を意図するものではない。
【0089】
更に、組換えタンパク質の場合、特定のホストから分泌させるのが便宜であり得る。従って、それらのベクターの更なる成分として、分泌、シグナリング、およびプロセシング配列をコードする核酸配列が含まれてもよい。
【0090】
本発明にかかるベクターにはプラスミドおよびウィルス(バクテリオファージおよび真核ウィルス)、並びに他の直鎖または環状DNAキャリア、例えば転位因子または相同組換え技術を使用するものが包含される。特に好適なウィルスベクターにはバキュロウィルス、レトロウィルス、アデノウィルス、およびワクシニアウィルスに基づくベクターがある。
【0091】
また本発明には、本発明の分子、特にDCの分化および成熟化を調節(好ましくは阻害)する節足動物タンパク質、ホモログ、フラグメント、または機能的同等物をコードするベクター、核酸分子、またはアンチセンス核酸を含有するホスト細胞が包含される。本発明の範囲内で、任意の型のホスト細胞を利用してもよい。ある態様では、ホスト細胞は原核ホスト細胞であってもよい。この態様では、原核ホスト細胞はE. coliホスト細胞であってもよい。別の態様では、ホスト細胞は真核ホスト細胞であってもよい。この態様では、ホスト細胞は真核酵母細胞であってもよい。更なる態様では、ホスト細胞は哺乳動物ホスト細胞であってもよい。更に別の態様では、ホスト細胞は昆虫細胞であってもよく、この態様では、発現系はバキュロウィルス発現系であってもよい。
【0092】
種々の技術を用いて本発明のベクターまたは核酸をホスト細胞に導入することができる。好適な形質転換またはトランスフェクション法は文献に十分報告されている(Sambrookら, 1989;Ausubelら, 1991;Spector, GoldmanおよびLeinwald, 1998)。真核細胞では、発現系は系の必要性に従って、一過性(例えばエピソーム系)または恒久的(染色体への組込み)のいずれであってもよい。
【0093】
本発明の更なる態様では、本発明の分子、特にDCの分化および成熟化を調節(好ましくは阻害)する、単離された節足動物タンパク質、ホモログ、フラグメント、または機能的同等物の調製法を提供し、該方法は以下を含む:
i)本発明の分子、特に本発明にかかるDCの分化および成熟化を調節または阻害する節足動物タンパク質、ホモログ、フラグメント、または機能的同等物をコードする核酸配列を含有するベクターを含有するホスト細胞を、該タンパク質が発現される条件下で培養すること;および
ii)そのようにして生成された該タンパク質を回収すること。
【0094】
本発明のこの観点において、タンパク質発現に必要な条件はホスト細胞系、ベクター、およびその後のタンパク質回収法によって異なる。実施例に、本発明の単離されたタンパク質の生成および回収のための特定の方法を開示する。それらの条件の変更は当業者に明白である。
【0095】
医薬組成物
同定されているDCの分化および成熟化の調節(好ましくは阻害)における本発明の分子の活性から、本発明の分子、タンパク質、核酸、アンチセンス核酸、ベクター、ホスト細胞、および抗体を治療に使用することが意図される。
【0096】
本発明は単離されたDC調節分子、例えばDCの分化および成熟化を調節(好ましくは阻害)する節足動物タンパク質もしくはその機能的同等物、それら分子をコードする核酸、該核酸を含有するベクター、該ベクターを含有するホスト細胞、または該分子に結合する抗体、および医薬的に許容されるキャリアを含有する医薬組成物を提供する。
【0097】
本明細書で使用する「医薬的に許容されるキャリア」という用語には、賦形剤がそれ自体毒性効果を与えず、医薬組成物を投与する個体に有害な抗体産生を惹起しないことを条件に、遺伝子、ポリペプチド、抗体、リポソーム、多糖、ポリ乳酸、ポリグルコール酸、および不活性ウィルス粒子、または、任意の他の薬剤が包含される。医薬的に許容されるキャリアは、液体、例えば見ず、生理食塩水、グリセロール、エタノール、または補助物質、例えば湿潤剤、乳化剤、pH緩衝剤などを更に含有してもよい。賦形剤により医薬組成物を錠剤、ピル、糖衣錠、カプセル、液体、ジェル、シロップ、スラリー、懸濁液に製剤し、患者の摂取を容易にしてもよい。医薬的に許容されるキャリアについての総説はRemington's Pharmaceutical Sciences(Mack Pub.社, ニュージャージー州 1991)に記載されている。
【0098】
ある態様では、医薬組成物はタンパク質と相互作用する1つまたはそれ以上の脂質分子を含有することができる。ある特定の態様では、この脂質分子はコレステロールまたはコレステロールの代謝物、例えばビタミンD3であってもよい。別の態様では、医薬組成物はジャパニンおよび脂質(例えばコレステロールまたはコレステロールの代謝物)を含有する、またはそれらから成る複合体を含有してもよい。更なる態様では、医薬組成物はジャパニン、脂質(例えばコレステロールまたはコレステロールの代謝物)、および受容体(例えば二価陽イオン依存性受容体(C型レクチン受容体など))を含有する、またはそれらから成る複合体を含有してもよい。
【0099】
本発明のある態様では、医薬組成物は1つまたはそれ以上の更なる治療薬を含有することができる。本発明のこの観点には、当業者が本発明の分子と共投与するのに有益であると見なしうる、いずれの更なる治療薬も包含される。特に、該更なる治療薬は抗炎症剤、免疫調節剤、免疫抑制剤、サイトカイン、サイトカイン模倣タンパク質、またはサイトカイン結合タンパク質を含有してもよい。特定の態様では、1つまたはそれ以上の更なる治療薬は抗炎症剤を含んでもよい。
【0100】
治療法
本発明は治療に使用するための単離された分子、例えば哺乳動物DCの分化および成熟化を調節(好ましくは阻害)するタンパク質、それらタンパク質をコードする核酸、アンチセンス核酸、該核酸もしくはアンチセンス核酸を含有するベクター、該ベクターを含有するホスト細胞、該タンパク質もしくは分子に結合する抗体、または該分子、タンパク質、核酸、ベクター、ホスト細胞、もしくは抗体を含有する医薬組成物を提供する。
【0101】
本明細書で使用する「治療」という用語は、ヒトまたは動物患者の利益のための、本明細書に記載するタンパク質、分子、核酸、ベクター、ホスト細胞、抗体、または医薬組成物の使用を包含する。特にこの用語には治療処置、予防的処置、診断、およびワクチン接種も包含される。このリストは例証のためだけに提供するものであり、制限を意図するものではない。
【0102】
本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物は、いずれの動物の治療に使用してもよい。ある態様では、動物は哺乳動物であってもよい。更なる態様では、哺乳動物はウシ、ブタ、ヒツジ、ネコ、イヌ、またはウサギであってもよい。更なる態様では、哺乳動物はヒトであってもよい。
【0103】
別の態様では、哺乳動物DCの分化および成熟化を調節(好ましくは阻害)する単離された分子、例えば哺乳動物DCの分化および成熟化を調節(好ましくは阻害)する単離された節足動物タンパク質、それらタンパク質をコードする核酸、それらタンパク質をコードする核酸に結合するアンチセンス核酸、該核酸もしくはアンチセンス核酸を含有するベクター、該ベクターを含有するホスト細胞、該タンパク質もしくは分子に結合する抗体、または該タンパク質、分子、核酸、ベクター、ホスト細胞、もしくは抗体を含有する医薬組成物の、DC活性に関係する疾患の治療のための使用法を提供する。
【0104】
また、本発明はDCに関係する疾患に罹患した動物の治療法を提供し、該方法は該動物にDCの分化および成熟化を調節(好ましくは阻害)する分子、例えば哺乳動物DCの分化および成熟化を調節(好ましくは阻害)する単離された節足動物タンパク質、それらタンパク質をコードする核酸、それらタンパク質をコードする核酸に結合するアンチセンス核酸、該核酸もしくはアンチセンス核酸を含有するベクター、該ベクターを含有するホスト細胞、該タンパク質もしくは分子に結合する抗体、または該タンパク質、分子、核酸、ベクター、ホスト細胞、もしくは抗体を含有する組成物を投与することを含む。
【0105】
本発明の範囲内で、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物は、多くの投与方法のうちの任意の1つまたはそれ以上を用いて患者に投与することができる。それらの投与方法は当該分野で公知であり、非経口注入(例えば静脈内、皮下、腹腔内、筋肉内、または組織の間質空間への)または直腸内、経口、膣内、局所、経皮、皮内、クモ膜下腔内、経鼻、点眼、耳内、肺内、もしくは他の粘膜投与がある。ナノパッチを使用して本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物の経皮投与を行ってもよい。遺伝子銃を使用して本発明の核酸、ベクター、または医薬組成物を投与してもよい。正確な投与方法は治療すべき疾患または症状によって異なる。
【0106】
ある態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を用いて、自己免疫疾患、アレルギーもしくは他の過敏性疾患、移植片拒絶反応および移植片対ホスト病、および急性および慢性炎症性疾患の治療または予防を行うことができる。
【0107】
自己免疫疾患には、限定されるわけではないが以下がある:無酸症、自己免疫性慢性活動性肝炎、アジソン病、円形脱毛症、筋萎縮性側索硬化症(ALS、ルー・ゲーリック病)、強直性脊椎炎、抗GBM腎炎または抗TBM腎炎、抗リン脂質抗体症候群、再生不良性貧血、関節炎、喘息、アトピー性アレルギー、アトピー性皮膚炎、自己免疫性アジソン病、自己免疫性溶血性貧血、自己免疫性肝炎、自己免疫性内耳疾患(AIED)、自己免疫性リンパ球増殖性症候群(ALPS)、自己免疫性血小板減少性紫斑病(ATP)、Balo病、ベーチェット病、ベルジェ病(IgA腎症)、水疱性類天疱瘡、心筋症、セリアック病、セリアック・スプルー皮膚炎、慢性疲労性免疫不全症候群(CFIDS)、慢性疲労免疫機能障害症候群(CFIDS)、慢性炎症性脱髄性多発性神経障害、チャーグ・ストラウス症候群、瘢痕性類天疱瘡、コーガン症候群、寒冷凝集素症、大腸炎、頭部動脈炎、クレスト症候群、クローン病、クッシング病、デゴス病、皮膚炎、皮膚筋炎、若年性皮膚筋炎、デビック病、I型糖尿病、円板上ループス、ダウリング・デゴス病、ドレスラー症候群、好酸球性筋膜炎、後天性表皮水疱症、本態性混合型クリオグロブリン血症、エバンス症候群、線維筋痛、線維筋炎、線維化生肺胞炎、胃炎、巨細胞性動脈炎、糸球体腎炎、グッドパスチャー症候群、グレーブス病、ギラン・バレー症候群、橋本甲状腺炎、溶血性貧血、ヘノッホ・シェーンライン紫斑病、肝炎、ヒューズ(Hughes)症候群、特発性副腎萎縮、特発性肺線維症、特発性血小板減少性紫斑病(ITP)、IgA腎症、炎症性脱髄性多発性神経障害、インスリン依存性糖尿病(I型)、過敏性腸症候群、若年性関節炎、川崎病、扁平苔癬、ルー・ゲーリック病、ルポイド肝炎、ライム病、メニエル病、混合性結合組織病、多発性骨髄腫、多発性硬化症、重症筋無力症、筋炎、眼部瘢痕性類天疱瘡、骨粗鬆症、扁平部炎、尋常性天疱瘡、悪性貧血、結節性多発性動脈炎、多発性軟骨炎、多腺性症候群、多腺性自己免疫症候群、リウマチ性多発性筋痛(PMR)、多発性筋炎、および皮膚筋炎、原発性無ガンマグロブリン血症、原発性胆汁性肝硬変、原発性硬化性胆管炎、乾癬、レイノー現象、ライター症候群、リウマチ性関節炎、サルコイドーシス、強膜炎、強皮症、シェーグレン症候群、スティッキー・ブラッド(sticky blood)症候群、スティッフマン症候群、スティル病、シデナム舞踏病、全身性エリテマトーデス(SLE)、高安動脈炎、側頭動脈炎/巨細胞性動脈炎、潰瘍性大腸炎、ブドウ膜炎、脈管炎、白斑、ウェグナー肉芽腫症、およびウィルソン症候群。
【0108】
アレルギーまたは過敏性障害は、Gell-Coombs分類のI型、II型、III 型、またはIV型を含む任意の既知のアレルギーまたは過敏性障害、および、一般的ではないがV型過敏性障害であってもよい。それらの障害には、限定されるわけではないが以下がある:アトピー、喘息、胎児赤芽球症、グッドパスチャー症候群、自己免疫性溶血性貧血、血清病、アルツス反応、全身性エリテマトーデス、接触性皮膚炎、ツベルクリン皮膚試験、慢性移植片拒絶反応、グレーブス病、重症筋無力症、全身性アナフィラキシー、局所性アナフィラキシー、アレルギー性鼻炎、結膜炎、胃腸炎、湿疹、輸血反応、新生児溶血性疾患、リウマチ性関節炎、糸球体腎炎、接触性皮膚炎、アトピー性皮膚炎、結節性病変、薬剤性溶血性貧血、ループス腎炎、アスペルギルス症、多発性動脈炎、多発性筋炎、強皮症、過敏性肺炎、ウェグナー肉芽腫症、I型糖尿病、蕁麻疹/血管浮腫、または甲状腺の炎症。アレルギーまたは過敏性障害は感染性疾患に関係しうるが、それらには、限定されるわけではないが結核、ハンセン病、ブラストミセス症、ヒストプラズマ症、トキソプラズマ症、リーシュマニア症、または他の感染がある。治療しうるアレルギーには、限定されるわけではないが花粉(例えば樺の木、ブタクサ、ナタネ)、食物(例えばナッツ、卵、またはシーフード)、薬物(例えばペニシリンまたはサリチル酸塩)、昆虫(例えばハチもしくはスズメバチ毒またはチリダニ)、または獣毛、および合成品(例えばラテックス)に対するアレルギー反応がある。治療しうる他の炎症性疾患にはアテローム性動脈硬化または他の心血管疾患、アルツハイマー病、血管炎、筋炎、脳炎、再灌流障害、2型糖尿病、脂肪肝疾患、および創傷治癒(例えば炎症期、血管新生段階、線維増殖および上皮形成、そして組織修復期)がある。
【0109】
ある態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を用いて、治療または診断処置の過程で体液または組織が合成または非哺乳動物由来物質と接触することから生じる有害症状の治療または予防を行うことができる。それらの処置は一時的または恒久的であってもよく、それらには、限定されるわけではないが体外循環、例えば腎臓または肝臓透析、腹膜透析、心肺バイパスおよび血液濾過、血管、膀胱、くも膜下腔内、または他の管腔臓器のいずれかに挿入した留置カテーテル、プロテーゼ(人工関節、心臓弁、血管内ステント、CSFシャント、人工血管、および冠動脈血管形成カテーテルなど)の使用がある。
【0110】
本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物は免疫抑制効果を有すると考えられ、これは移植片拒絶反応の予防に有用であり得る。移植片は同じ個体間の同型移植片、同じ種の異なるメンバー間の同種移植片、または異なる種間の異種移植片であってもよい。本発明の分子は種々の移植片の拒絶反応を予防するのに有用であり得るが、それらには、限定されるわけではないが心臓、肺、心肺、腎臓、肝臓、膵臓、腸、手、角膜、皮膚移植(顔面再移植および顔面移植など)、ランゲルハンス島、骨髄移植、輸血、血管、心臓弁、骨、および皮膚がある。分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を用いて、骨髄移植後の移植片対ホスト病を予防してもよい。
【0111】
理論に拘束されることは望まないが、ジャパニン・タンパク質は癌に関与するシグナル伝達経路の阻害に関係しうると推測される。本発明の更なる態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を癌の治療に用いることができる。本発明は癌に罹患した動物の治療法を提供し、該方法は、該動物に上記のような本発明の分子、タンパク質、核酸、ベクター、抗体、または医薬組成物を投与することを含む。それらの治療法は、癌における免疫反応の再分極化または調節を伴ってもよい。
【0112】
特に、癌は血液系腫瘍、例えばリンパ腫もしくは白血病、または多発性骨髄腫であってもよい。本発明に従って治療できる白血病には急性リンパ芽球性白血病(ALL)、急性骨髄性白血病(AML)、慢性骨髄性白血病(CML)、慢性リンパ芽球性白血病(CLL)、およびヘアリー細胞白血病がある。本発明に従って治療できるリンパ腫にはホジキン病および非ホジキンリンパ腫がある。関係する障害を治療してもよく、それらには例えば骨髄異形成症候群(MDS)(最終的にALLになりうる)、骨髄増殖性疾患(例えば真性赤血球増加症、本態性血小板血症、または骨髄線維症)、および軽鎖疾患によるアミロイドがある。
【0113】
更なる態様では、癌は癌腫、肉腫、または芽細胞腫であってもよい。本発明はあらゆる臓器の癌(限定されるわけではないが乳房、肺、卵巣、膵臓、精巣、皮膚、結腸、脳、肝臓、または子宮頸部など)および黒色腫の治療を企図する。治療または予防しうる癌には組織球腫、特にイヌ皮膚組織球腫がある。
【0114】
本発明の更なる態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を感染性疾患の治療に用いることができる。特に、それらの治療は病原体、例えばウィルス、細菌、および原虫、例えばHIV、TB、およびマラリアの原因物質、並びに他の寄生生物による感染に対する免疫応答の再分極化または調節を伴ってもよい。
【0115】
吸血性節足動物、例えばマダニは、感染性疾患の原因物質(例えばマダニ媒介性脳炎ウィルス、クリミア・コンゴ出血熱ウィルス、ナイロビ・ヒツジ病ウィルス、ブルグドルフェリ(Borrelia burgdorferi。ライム病の原因物質)、およびタイレリア・バルバ(Theileria parva。東海岸熱の原因物質))の供与源である。ジャパニンはマダニ媒介性疾患の伝染を促進するように作用すると考えられる。従って、ジャパニン・タンパク質、そのホモログ、フラグメント、および機能的同等物は、動物に接種して免疫応答を惹起し、マダニ媒介性疾患の治療または予防を行うのに有用である。従って、本発明の更なる態様では、節足動物媒介性感染性疾患の伝染の予防法、または節足動物媒介性感染性疾患の治療法を提供し、該方法は本発明の分子(例えばタンパク質)、または本発明の核酸分子、ベクター、ホスト細胞、抗体、もしくは医薬組成物を動物に投与することを含む。本発明はまた、節足動物媒介性感染性疾患の伝染の予防、または節足動物媒介性感染性疾患の治療に使用するための、本発明の分子(例えばタンパク質)、または本発明の核酸分子、ベクター、ホスト細胞、抗体、もしくは医薬組成物も提供する。節足動物は吸血性節足動物であってもよい。節足動物媒介性疾患はライム病、マダニ媒介性脳炎、クリミア・コンゴ出血熱、ナイロビ・ヒツジ病、または東海岸熱であってもよい。
【0116】
ジャパニン・タンパク質、そのホモログ、フラグメント、および機能的同等物は吸血性節足動物そのもの、並びにそれらが運搬する疾患に対するワクチンとしても有用でありうる。従って本発明は更に、動物への吸血性節足動物(マダニであってもよい)に対するワクチン接種の方法を提供し、該方法は本発明の分子(例えばタンパク質)、または本発明の核酸分子、ベクター、ホスト細胞、抗体、もしくは医薬組成物を動物に投与することを含む。本発明はまた、動物への吸血性節足動物(マダニであってもよい)に対するワクチン接種に有用な本発明の分子(例えばタンパク質)、または本発明の核酸分子、ベクター、ホスト細胞、抗体、もしくは医薬組成物を提供する。
【0117】
上記のように、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を1つまたはそれ以上の更なる治療物質、例えば抗炎症剤、免疫調節剤、免疫抑制剤、サイトカイン、サイトカイン模倣タンパク質、サイトカイン結合タンパク質、または上記の障害のいずれかを治療するために開発された別の生物医薬品と共に投与するのが有益であり得る。
【0118】
本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、または抗体を、in vivoでそれらをDCにターゲッティングさせる抗原と共に投与して、所望しない免疫応答に関係するDCの分化および成熟化を調節または阻害することが有益であり得る。この態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を疾病関連要素と共に患者に投与し、本発明のタンパク質、分子、核酸、ベクター、ホスト細胞、抗体、または医薬組成物の好適なDCへのターゲッティングを促進することができる。本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、または抗体が既にDCへの特異的結合によってDCにターゲッティングされている態様では、疾病関連要素は不要である。
【0119】
「疾病関連要素」という用語は、患者における疾病と関係するいずれの成分も包含することを意図する。疾病には上記のように、自己免疫疾患、アレルギーおよび他の過敏性反応、移植片拒絶反応および移植片対ホスト病、感染性疾患(例えばマダニによって伝染するもの)、癌(例えば血液系腫瘍)、そして急性および慢性炎症性疾患がある。従って「疾病関連要素」には以下が包含される:i)感染性物質、例えばウィルス、微生物、寄生生物、および微生物毒に関係する成分;ii)アレルギーに関係する非自己分子であるアレルゲン;iii)アレルギー以外の過敏性反応に関係する非自己成分;iv)自己免疫疾患に関係する自己成分;v)同種の遺伝的に異なるメンバー由来(アロ抗原)または異なる種由来(異種抗原)の移植抗原;およびvi)腫瘍関連抗原および腫瘍特異的抗原。
【0120】
「疾病関連要素」という用語は、これらの疾病関連要素のフラグメントおよび誘導体も包含する。それらの誘導体には、無毒化された物質、合成ミモトープ、および抗原で、その構造中に置換、付加、または欠失を含有するもので、本発明のタンパク質、分子、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を好適なDCに指向させるように作用する能力を有するものが含まれる。
【0121】
疾病関連要素を有する動物を提供することにより、疾病に関係するDC分化および成熟化の調節または阻害の特異性が向上される。このように特定のDCを標的とすることは、全体的な免疫応答を阻害する必要を無くし、結果として副作用が低減しうるため、有益である。
【0122】
本発明のある観点では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を疾病関連要素と別々に投与してもよい。この観点では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物、および疾病関連要素を連続的に投与してもよい。別の態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を疾病関連要素の前に投与してもよい。
【0123】
更なる態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を疾病関連要素の後に投与してもよい。更なる態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物および疾病関連要素を同時に投与してもよい。本発明のこの観点では、本発明のタンパク質、分子、または抗体は疾病関連要素に結合して、例えば融合タンパク質の形態であってもよい。
【0124】
従って本発明は、本発明にかかるDC分化および成熟化を調節または阻害する分子および疾病関連要素を含有する融合タンパク質も提供する。それらの融合タンパク質をコードする核酸も提供する。
【0125】
本発明はまた、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物、および疾病関連要素、および医薬的に許容されるキャリアを含有する医薬組成物を提供する。
【0126】
上記の方法は、本発明の分子を動物に投与して、動物においてin vivoでDCの分化および成熟化を調節(好ましくは阻害)することを含む。分子は単独で投与してもよく、または分子をDCにターゲッティングさせる更なる物質(例えば疾病原因要素)と共に投与してもよい。
【0127】
上記の疾病を治療する別の方法は、本発明の分子を目的とする治療にex vivoで使用するものである。この方法は、本発明の分子をインビトロでDCに運搬してDCを調節し、調節されたDCを治療を必要とする動物に運搬することを伴う。
【0128】
更なる観点では、本発明はDCを調節する方法を提供し、該方法は上記のように、DCを本発明の分子、例えばジャパニン・タンパク質、そのホモログ、フラグメント、および機能的同等物と接触させることを含む。この方法によって生成される、調節されたDCも提供する。
【0129】
また本発明は、それを必要とする動物においてDCに関係する障害を治療または予防する方法も提供し、該方法はこの方法で生成される調節されたDCを動物に投与することを含む。
【0130】
DC分化を調節する分子は、上記のようにDC分化および成熟化を調節(好ましくは阻害)する任意の分子(例えばジャパニン・タンパク質、そのホモログ、フラグメント、および機能的同等物)であってもよい。これらの分子をコードする核酸分子を、核酸分子を含有するベクターとして使用してもよい。
【0131】
DCは治療を必要とする動物から直接単離してもよい。あるいはまた、DC前駆体(例えば骨髄前駆細胞の単球)を動物から単離し、これを使用してDCを生成させてもよい。本発明のこの観点では、DCまたはDC前駆体は、本発明の分子での処理後に調節されたDCを導入する動物にとって、自己移植であってもよく、または同種異系であってもよい。
【0132】
DCに関係する障害は上記の疾病のいずれであってもよく、それらには自己免疫疾患、アレルギーおよび他の過敏性反応、移植片拒絶反応および移植片対ホスト病、感染性疾患(例えばマダニによって伝染するもの)、癌(例えば血液系腫瘍)、そして急性および慢性炎症性疾患がある。この方法を使用して移植ドナーから調節されたDCを生成し、これを移植に先立って所定の移植レシピエントに投与し、移植片に対して不応答とし、それによって免疫抑制剤投与の必要を低減することができる。
【0133】
本発明のこの観点では、上記のようにDCを疾病関連要素と接触させ、DCの分化および成熟化を調節(好ましくは阻害)する分子を好適なDCにターゲッティングさせることができる。あるいはまた、調節されたDCを上記のような疾病関連要素と共に動物に投与してもよい。
【0134】
スクリーニング法
ジャパニンの同族受容体を同定することにより、受容体をスクリーニング法に使用し、可能性のあるジャパニンのアゴニストおよびアンタゴニストを同定し、治療または他の使用可能性を有する化合物を同定することが可能となる。
【0135】
可能性のあるアゴニストまたはアンタゴニスト化合物は、例えば細胞、無細胞標品、化学ライブラリー、または天然産物混合物から単離することができる。それらのスクリーニング法の好適な総説については、例えばColiganら, Current Protocols in Immunology 1(2):Chapter 5 (1991)を参照されたい。
【0136】
良好なアンタゴニストである可能性が最も高い化合物は、ジャパニンによって誘導される生物学的作用を誘導させずにジャパニンの同族受容体に結合し、ジャパニンの機能を競合的に阻害する分子である。上記のように、ジャパニンの同族受容体は二価陽イオン依存性受容体であるC型レクチン受容体であることが考えられ、これはジャパニンが結合すると細胞内シグナル伝達系を惹起し、それによって樹状細胞の分化および成熟化の阻害が起こる。可能性のあるアンタゴニストには、シグナル伝達経路を活性化することなく受容体に結合する、または負のシグナル伝達経路を活性化することによって受容体に結合する小型有機分子、ペプチド、ポリペプチド、および抗体がある。特に、好適かつ有望なアンタゴニストには炭化水素部分、および、受容体への結合親和性を保持したままその機能を低減するように操作した組換えジャパニン分子がある。更なる好適な化合物にはジャパニンに対する抗体、およびジャパニンに結合した炭化水素部分に対する抗体、および標的特異性を遺伝子操作できるアンチカリンがある。
【0137】
良好なアゴニストとして機能する可能性の最も高い化合物は、ジャパニンの同族受容体に結合し、ジャパニンと同じ細胞内シグナル伝達経路を惹起し、それによってジャパニンと同様の方法で樹状細胞の分化および成熟化を調節(好ましくは阻害)するように機能する化合物である。好適かつ有望なアゴニストの例には、受容体を活性化する能力および/または受容体に対する結合親和性が増加するように操作したジャパニン分子がある。
【0138】
それらのスクリーニング法に使用する同族受容体(二価陽イオン依存性受容体、例えばC型レクチン)は溶液中に含有されるか、固相支持体に固定されるか、細胞表面上に担持されるか、または細胞内に存在してもよい。一般に、それらのスクリーニング法は受容体を発現する好適な細胞または細胞膜を使用し、それらを試験化合物と接触させて結合または機能的反応の刺激もしくは阻害を観察することを伴う。その後、試験化合物と接触させた細胞の機能的反応を、試験化合物と接触させていないコントロール細胞と比較する。それらのアッセイにより、好適な検出系を用いて、試験化合物がジャパニンによる受容体活性化によって生成されるのと同様のシグナルを生成するか否かを評価してもよい。ジャパニンは細胞表面二価陽イオン依存性受容体(例えばC型レクチン受容体)に結合し、細胞内シグナル伝達経路を惹起することによって機能すると信じられるため、スクリーニング法が最も効果的に機能するのは、細胞表面上の受容体を使用し、化合物の受容体への結合によって細胞内シグナルの生成をモニタリングすることを伴う場合であると考えられる。
【0139】
ある態様では、ジャパニンのアゴニストまたはアンタゴニスト化合物の同定法は以下を含む:
(a)受容体への結合が可能な条件下で、その表面上に二価陽イオン依存性受容体(例えばC型レクチン受容体)を発現する細胞をスクリーニングを行う化合物と接触させること(受容体は化合物の結合に応答して検出可能なシグナルを生成する能力を有する);および
(b)化合物と受容体の相互作用から生成されるシグナルのレベルを測定することによって化合物が受容体に結合するか否か、また、受容体を活性化もしくは阻害するか否かを測定する。
【0140】
ある態様では、スクリーニングを行う化合物をリガンドの存在下で受容体と接触させることができる。それらのリガンドは脂質、例えばコレステロールまたはコレステロールの代謝物(例えばビタミンD3)であってもよい。
【0141】
別の態様では、ジャパニンのアンタゴニスト化合物の同定法は以下を含む:
(a)ジャパニンがその同族受容体に結合できる条件下で、その表面上に二価陽イオン依存性受容体(例えばC型レクチン受容体)を発現する細胞をジャパニンと接触させること(受容体はジャパニンの結合に応答して検出可能なシグナルを生成する能力を有する)
(b)ジャパニンと受容体の相互作用から生成されるシグナルのレベルを測定すること;
(c)スクリーニングを行う化合物を受容体への結合が可能な条件下で添加すること;および
(d)化合物と受容体の相互作用から生成されるシグナルのレベル変化を測定することによって、ジャパニン結合の際の化合物の影響を測定すること。
【0142】
ある態様では、上記のようなジャパニンのホモログまたは機能的同等物を、上記のスクリーニング法に用いることができる。
【0143】
更なる態様では、スクリーニングを行う化合物および/またはジャパニンをリガンドの存在下で受容体と接触させることができる。それらのリガンドは、脂質、例えばコレステロールまたはコレステロールの代謝物であってもよい。
【0144】
上記の条件は、培養液の存在、生理学的濃度のCa2+を含有する溶液の存在、および/またはpH 7-8であることを含んでもよい。
【0145】
上記の検出可能なシグナルには細胞内リン酸化、核局在化、遺伝子発現、および/またはサイトカイン放出がある。
【0146】
上記の方法のある態様では、単純な結合アッセイを使用し、C型レクチン受容体を保有する表面への試験化合物の接着を、試験化合物に直接もしくは間接的に結合する標識によって、または標識された競合物質との競合を伴うアッセイによって検出する。別の態様では、競合薬剤スクリーニング・アッセイを使用し、二価陽イオン依存性受容体(例えばC型レクチン受容体)との結合能を有する中和抗体を試験化合物と結合に関して特異的に競合させる。この方法では、抗体を使用して受容体と特異的な結合親和性を有する試験化合物の存在を検出することができる。
【0147】
当業者は、ジャパニンの同族受容体に作用する化合物を同定するためのアッセイを考案することができる。受容体との好適な結合親和性を有する化合物のハイスループット・スクリーニングを提供するのに使用してもよい技術については、国際特許出願WO84/03564を参照されたい。この方法では、多数の異なる小型試験化合物を固相支持体上に合成し、その後、受容体と反応させ、洗浄することができる。ポリペプチドを固定化する一つの方法は、非中和抗体の使用である。その後、当該分野で公知の方法によって、結合された受容体を検出してもよい。精製された受容体分子をプレート上に直接コーティングし、上記の薬剤スクリーニング法に用いてもよい。
【0148】
本発明は、上記の方法によって同定されたアゴニスト、アンタゴニスト、および他の化合物も包含する。これらのアゴニスト、アンタゴニスト、および他の化合物は、上記の医薬組成物の一部を成してもよく、上記の治療法に使用してもよい。
【0149】
ここで、本発明の種々の観点および態様について、実施例によってより詳細に記載する。認識されるように、本発明の範囲を逸脱することなく、詳細の改変を行ってもよい。
【実施例】
【0150】
試料と方法
マダニ
Rhipicephalus appendiculatusマダニはthe Centre of Ecology and Hydrology(オックスフォード)で飼育した。給餌は、ガーゼでカバーした保持チャンバー(retaining chambers)中で、それらを剪毛したモルモットの背上に配して行った。
【0151】
唾液腺抽出物の調製
給餌の1-6日後、モルモットからマダニを注意深く分離し、顕微鏡下でその唾液腺を切除した。唾液腺を冷リン酸緩衝液(PBS;Oxoid社)で軽く洗浄し、1.5mLの微小遠心管に移し、使用まで-70℃で保存した。Dounceホモジナイザーを用い、PBS中で唾液腺をホモジナイズして唾液腺抽出物(SGE)を調製した。ホモジネートを≧10000gで3分間遠心分離し、上清を回収して使用まで-20℃で保存した。
【0152】
細胞培養
細胞培養液および補足物質は、特に記載しない限りPAA社のものを使用した。それらにはウシ胎仔血清(FCS)があり、全試験を通してバッチ#A04304-0511を使用した。哺乳動物細胞の培養は37℃、5% CO2で行った。昆虫細胞の培養はSf900 II培養液(Invitrogen社)中、28℃で行った。共トランスフェクション培養を除いて、液体培養は三角フラスコ中、100-130rpmで軌道撹拌して行った。
【0153】
樹状細胞
ヒト樹状細胞(DC)を、健常成人ドナーから単離した末梢血単球から生成した。簡潔に記載すると、バフィーコート(National blood service、ブリストル)をCa2+/Mg2+未含有ハンクス緩衝液(HBSS)と1:2(v/v)で混合し、Lymphoprep(Axis Shield社)上に注意深く積層し、800gで30分間(22℃)遠心分離した。HBSS/バフィーコート混合液とLymphoprep間の接触面に生成された末梢血単核細胞(PBMC)層を注意深く回収し、HBSSで3回洗浄し、血小板を除去した。一時的接着または磁気ビーズによる陰性選択のいずれかによって、単球をPBMCから単離した。
【0154】
一時的接着による単球の単離では、PBMCペレットをRPMI-5(5% ヒトAB+血清(National blood service)、2mM L-グルタミン、100U/ml ペニシリン、および100μg/ml ストレプトマイシンを補足したRPMI 1640)に再懸濁した。PBMCを100mm細胞培養用ペトリ皿(BD Biosciences社)中に、1×107 細胞/ml、10ml/プレートで播種し、37℃、5% CO2で45分間インキュベートした。その後、プレートをHBSSで3回洗浄して接着していない細胞を除去し、10mlのRPMI-5をプレートに添加した。培養1日後、ヒトGM-CSF(John Radcliffe病院薬局)を1000U/mlの濃度で添加し、ヒトIL-4(Peprotech社)を20ng/mlの濃度で添加した。培養3日後および5日後、総容量の3分の1をフレッシュに調製したGM-CSF + IL-4を補足したRPMI-5で置換した。
【0155】
陰性選択(つまり非単球の除去)による単球の単離では、PBMCペレットをCa2+/Mg2+未含有HBSSに再懸濁した後、Dynal Monocyte Negative Isolationキット(Invitrogen社)を使用し、製造者の説明書に従って単球を単離した。精製した単球をDC-RPMI(RPMI 1640に10% FCS、2mM L-グルタミン、100U/ml ペニシリン、100μg/ml ストレプトマイシン、1000U/ml ヒトGM-CSF、および1000U/ml ヒトIL-4(どちらもGentaur社)に5x105/mlで再懸濁した。3日後、培養液の3分の1を除去して遠心分離し、ペレットを同量のフレッシュの培養液(3000U/mlの各サイトカインを含有)に再懸濁した後、再び培養液に添加した。5日後、HBSS/2% FCSで洗浄した後、等張食塩水に混合した5.5%ヘタスターチ(John Radcliffe病院薬局の「Voluven」)/4.8% DMSO(Sigma社、Hybrimaxグレード)/3.8% FCSで再懸濁し、その後、温度制御凍結装置(1度/分)に入れ、-80℃で細胞を凍結させた。
【0156】
DC調節活性のスクリーニング
上記のようにFCS含有培養液中で生成し、凍結したDCを用いて、DC調節活性の慣例的なスクリーニングを行った。DCを解凍した後、96ウェル平底Primariaプレート(BD Biosciences社)中、スクリーニングするサンプルを添加したDC-RPMI中で培養した。24時間培養後、ポリ(I:C)(25μg/ml)またはLPS(200ng/ml)を添加してDCを刺激したが、刺激剤の選択は各バッチのDCの異なる刺激剤に対する反応性によって決定した。更に18-24時間培養した後、CD1a+ 細胞によるCD86発現レベルをフローサイトメトリーで評価した。
【0157】
フローサイトメトリー
フローサイトメトリーはFACSortフローサイトメトリー(Becton Dickson社)を使用し、CellQuest Pro(Becton Dickinson社)で制御して行った。データ解析はCellQuest ProおよびFloJo(Treestar Software社)を用いて行った。
【0158】
PCR
PCRはDNAEngineサーモサイクラー(Biorad社)を用いて行った。プライマーはMWG Biosciences製(HPSFクォリティー)、dNTPはBioline製であった。他の試薬は本文に記載する通りである。
【0159】
DNAシークエンシング
DNAシークエンシングはGeneservice社(オックスフォード)で、BigDye v3.1化学を使用して実施した。
【0160】
E. coli形質転換
化学的コンピテントE. coliのヒートショック形質転換には、50μlのバクテリアを氷上で解凍し、氷上でDNAと共に5分間インキュベートし、42℃で40秒間熱処理した後、氷上に戻した。次いで、250μlのSOC培養液(室温)を添加し、細胞を37℃で、200rpmで軌道撹拌しながら45分から1時間30分インキュベートした後、50μlアリコートを100μg/mlアンピシリン含有LB寒天プレート上に播種した。
【0161】
実施例1 SGEによるLPS誘導型CD86亢進的調節の抑制
前記のように、一時的接着により単球を単離し、GM-CSF + IL-4と共に培養してDCを生成させた。5日間培養後、オスまたはメスR. appendiculatusマダニ由来のSGEを培養液に添加した。SGEは給餌していないマダニ、または3もしくは6日間給餌を行ったマダニ由来のいずれかであり、最終濃度50μg/mlのマダニ由来タンパク質となるように添加した。合計6日間の培養後、DCをLPS(50ng/ml)で処理し、7日間の培養後、抗CD1aおよび抗CD86で染色し、フローサイトメトリーで分析した。図1にCD86+細胞数を、CD1a+細胞%xこれらの細胞のCD86染色の幾何平均蛍光強度(geometric mean fluorescence intensity;GMFI)で示す。図1に示すように、3日間の給餌を行ったメスRhipicephalus appendiculatusマダニ由来SGEはヒトDCによる共刺激受容体CD86のLPS誘導型亢進的調節を抑制し、未給餌のメス、6日間給餌したメス、またはオス(給餌および未給餌)のマダニ由来のSGEでは抑制しないことが明らかになった。この実験を2回行い、同様の結果を得た。SGEを最終濃度がml当たり1つの唾液腺から誘導される物質となるように添加した場合にも、同様の結果が得られた。
【0162】
実施例2 SGEによるCD80およびMHCクラスIIのLPS誘導型亢進的調節の抑制
前記のように、一時的接着により単球を単離し、GM-CSF + IL-4と共に培養してDCを生成させた。5日間培養後、3日間給餌したメスR. appendiculatusマダニ由来のSGEをいくつかの培養液に添加し、最終濃度が培養液ml当たり、1つの唾液腺から誘導された物質となるようにした。合計6日間の培養後、DCをLPS(50ng/ml)で処理し、7日間の培養後、抗CD1a、抗CD80、抗CD86、および抗HLA-DRで染色し、フローサイトメトリーで分析した。図2に、SGEの存在下(黒色)および非存在下(灰色)におけるLPS処理したCD1a+細胞による成熟マーカーの発現を示す。図2に示すように、3日間給餌したメスRhipicephalus appendiculatus唾液腺抽出物はLPSに応答してCD86による亢進的調節を阻害するだけでなく、CD80およびMHCクラスIIの亢進的調節も阻害した。この実験を2回行った。
【0163】
実施例3 SGEの活性成分のQカラム分画
3日間給餌したメスR. appendiculatusマダニ由来の唾液腺112検体から得たSGEを50mM リン酸ナトリウム(pH7.0)で1:10に希釈した後、予め同じバッファーで平衡化しておいたHi-Trap Qセファロースイオン交換カラム(Amersham)に適用した。未結合の物質(Qカラム・フロースルー=QFT)を回収し、カラムを2カラム容量の50mM リン酸ナトリウム(pH7.0)で洗浄した後、50mM リン酸ナトリウム、1M NaCl(pH7.0)をカラムに適用して、結合した物質(Qカラム結合画分=QBF)を溶出した。Vivaspin 6 5kDa MWCO遠心濃縮器(Sartorius)(タンパク質の非特異的吸着を予防するためにγ-グロブリンで前処理したもの)を使用して、QFTおよびQFRを最終容量500μlまで濃縮した。
【0164】
上記のように、一時的接着によって単球を単離し、GM-CSF + IL-4と共に培養してDCを生成させた。5日間培養後、3日間給餌したメスR. appendiculatusマダニ由来のSGE、または上記で生成したQFTもしくはQBFを培養的に添加した。それぞれの最終濃度が培養液ml当たり、1つの唾液腺から誘導された物質となるようにした。合計6日間の培養後、DCをLPS(50ng/ml)で処理し、7日間の培養後、抗CD1aおよび抗CD86で染色し、フローサイトメトリーで分析した。図3にCD86+細胞数を、CD1a+細胞%xこれらの細胞のCD86染色の幾何学的平均蛍光強度(GMFI)で示す。エラーバーは2重複培養液間の範囲を表す。
【0165】
図4は、DC調節活性はLPS誘導型成熟化(おそらくTLR4を通じて作用)に限定されず、25μg/ml ポリ(I:C)(TLR3リガンド)または50ng/ml IFN-γでの処理にも応答してCD86亢進的調節を阻害した(図4a)。しかしながら、QFTは100ng/ml TNF-α誘導型CD86亢進的調節に対してほとんど、または全く影響を与えなかった(図4b)。この理由は現在のところ明らかではない。
【0166】
実施例4 QFTのDC調節活性に対するプロテイナーゼKの影響
上記のように調製した凍結された5日目DCを解凍し、最終濃度0.2唾液腺相当量/mlのQFTと共に培養した。この実験では、QFTの一部を後述のようにプロテイナーゼKで前処理した。一日後、DCをポリ(I:C)(25μg/ml)で処理し、更に1日培養した後、抗CD1aおよび抗CD86で染色し、フローサイトメトリーで分析した。
【0167】
プロテイナーゼK処理は、QFTを、50mM Na2HPO4(pH7.0)に混合した150μg/ml プロテイナーゼK(Sigma社)と共に50℃で2時間インキュベートして行った。その後、98℃で10分間加熱して、酵素を失活させた。プロテアーゼのみのコントロール[プロテイナーゼK+ポリ(I:C)]は、同じ反応をQFTを用いずに行って得られた産物を用いて行い、プロテアーゼ未含有コントロール[QFT+ポリ(I:C)]に用いたQFTも同様に処理したところ、DC調節活性の抑止は活性成分の熱変性によるものではないことが明らかになった。2つの更なるコントロールを実施し、加熱不活性化したプロテイナーゼKをQFT存在下または非存在下でインキュベートし([不活化プロテイナーゼK+ポリ(I:C)]および[QFT+不活化プロテイナーゼK+ポリ(I:C)])、プロテイナーゼKの(熱に不安定な)タンパク質分解能がDC調節活性の抑止に必要であることを確認した。そのため、プロテイナーゼKを98℃で10分間前処理し、その後、50℃でインキュベートした。図5に示すように、プロテイナーゼK処理によってQFTのDC調節能は抑止され、これがタンパク質性であることが確認された。結果は2重複ウェルの平均値±2 S.Eで示す。
【0168】
実施例5 QFTのサイズ分画
350検体の唾液腺を3日間給餌したメスR. appendiculatusマダニから切除し、前記のようにこれを用いてQFTを調製した。8つの独立した実験で、このQFTをSuperdex 75カラムでゲル濾過し、サイズ分画した。各実験で、約40画分をカラムから回収し、上記のように、解凍した5日目DCと共に培養してDC調節活性をスクリーニングした。24時間後に200ng/ml LPSを添加してDCを刺激し、更に24時間後、フローサイトメトリーでCD1a+細胞のCD86発現を測定した。
【0169】
8つのうち、1つの代表的な分画から得た結果を図6に示す。初期スクリーニングは1唾液腺相当量/mlで行い、活性があると推測される画分の2次スクリーニングは0.04唾液腺相当量/mlで行った。全ての分画で、画分12および13が最も高い活性を有すると同定された。
【0170】
各分画からの最も活性の高い画分をプールし、低塩バッファー(50mM HEPES、pH8.3)に対して透析した後、還元的条件下、4-12% Bis-Trisポリアクリルアミドゲル(Invitrogen社)で泳動した。比較のために、SGE、QFT、および2つのQカラム結合画分をプールした活性画分と並行して行った。結果を図7に示す。SGEおよびQFTはDC調節活性を有したが、Q結合画分は有さなかった。約20kDaのバンドがQFTおよびプール画分中に存在し、2つの不活性サンプル中には存在しなかったことから、このバンドが活性タンパク質であると考えられる。出願人はこのタンパク質を「ジャパニン(Japanin)」と命名した。
【0171】
QFTのゲル濾過クロマトグラフィーで得た最も活性の高い画分(上記参照)をプールし、低塩バッファーに対して透析した後、HPLC(C4カラム)を用いて分画した。アセトニトリルの増加グラジエント溶出によって23画分をカラムから回収し、上記のように解凍した5日目DCと共に培養してDC調節活性についてスクリーニングした。24時間後に25μg/ml LPSを添加してDCを刺激し、更に24時間後、フローサイトメトリーでCD1a+細胞のCD86発現を測定した。
【0172】
実施例6 HPLCによるDC調節活性の単離
各画分は1:200に希釈して評価し(画分の容量によって、2.6から7.9唾液腺相当量/mlの濃度となる)、インプットした物質(プールした活性ゲル濾過画分)およびQFTはコントロールとしてそれぞれ1.8および0.2唾液腺相当量/mlで含有させた。画分23は約100%アセトニトリルであるため、その作用はQFTの存在下および非存在下で評価し、HPLC溶媒による直接的DC細胞調節効果、あるいは逆に、溶媒がそれらの効果をマスキングする能力を除外した。図8にHPLCの結果を示す。図8aに溶出プロファイルを示し、図8bは各画分に関する活性(DCによるDC86の亢進的調節の阻害として測定)を示す。
【0173】
実施例7 エドマン分解
HPLC画分19が最も効果の高いDC調節活性を有することを確認した後、これを用いてエドマン分解シークエンシングを行い、16残基のN-末端配列を得た:(Thr)Pro Ser Met Pro Ala Ile Asn Thr Gln Thr Leu Tyr Leu Ala(Arg)(括弧内の残基の同定は暫定)。エドマン分解の読み取り結果を図9に示す。このN-末端配列を有するタンパク質を以下、「ジャパニン」と称する。
【0174】
実施例8 ジャパニンDNA 3'領域のPCR増幅
N-末端配列を用いてジャパニン DNAのPCR増幅のための外側順方向プライマーをポリ(dT)逆方向プライマーと組み合わせて設計した。配列「M P A I N T Q」に対する4つの変性プライマー・セットを使用した。それら4つは非常に類似しているが、縮重を低減するために個別に使用する。
【化1】
【0175】
PCRのテンプレートを得るために、1日間給餌したメスR. appendiculatus唾液腺からcDNAを生成した。Trizol試薬(Invitrogen社)を使用し、製造者の説明書に従って30検体の唾液腺からRNAを単離し、その後、1/3容量の8M 塩化リチウムを添加して水相から沈殿させた。冷75%エタノールで洗浄した後、RNAを5μlのRNase未含有水に再溶解した。
【0176】
ImPromII逆転写酵素(Promega社)を使用し、製造者の説明書に従って、40μl反応で逆転写を行った。反応液は4μgのRNAを含有し、2.5mMのMgCl2濃度、0.5mMの総dNTP濃度であった。逆転写を開始させるためにオリゴ(dT)15を使用し、0.1μg/mlを添加した。反応は42℃で1時間行い、その後70℃で15分間、加熱不活性化を行った。
【0177】
このcDNAを用いるPCRは、Taq DNAポリメラーゼ(New England Biosciences社)を用い、1X Thermopolバッファー(New England Biosciences社)(62.5Mの各dNTP、250nM変性プライマー、および3.25μM オリゴ(dT)20-V含有)中で行った。テンプレートcDNAは1:40に希釈して使用した。初期変性段階は94℃で1分間、次いで94℃で30秒間(変性)/45℃で30秒間(アニーリング)/72℃で60秒間(伸長)を5サイクル、その後、94℃/50℃で30秒間/72℃で60秒間を30サイクル、最後に72℃で更に5分間とした。陽性コントロールは既知のマダニ・タンパク質の順方向プライマーを使用し、同じ条件下で行った(RH1-PE)。生成物を1.8%アガロースゲルで泳動した。結果を図10に示す。
【0178】
順方向プライマー2および4で増幅した、より大きい生成物は約600bpであり、それらが約22kDaのタンパク質をコードしていることを示唆している(平均アミノ酸残基質量を110Daと仮定)。これはSDS-PAGEゲルで約20kDaのバンドが活性画分中で同定されたことと十分一致している。このcDNAをクローニングする前に、ネステッドPCRを実施してこれがN-末端配列と一致するか否かを確認した。
【0179】
配列「Asn Thr Gln Thr Leu Tyr Leu Ala」に対して内側プライマーを設計した。これはN-末端配列中にあるが、上記で使用したプライマーの結合位置より下流である:
【化2】
標準外コード「I」はニュートラル塩基としてのイノシンの導入を表す。
【0180】
順方向プライマー2および4にコードされるプライマーをオリゴ(dT)20-Vと共に1:20希釈で用いて増幅したDNAをテンプレートとした。反応条件は上記の通りであるが、アニーリング温度は35サイクル全てで50℃とし、72℃での伸長段階を60秒間から40秒間に短縮した。生成物を1.8%アガロースゲルで泳動した。
【0181】
これらのPCRにより、予想されたサイズ(すなわち約600bp)のバンドが生成され(図11)、N-末端タンパク質配列をコードするcDNAが特異的に増幅されることを強く示唆している。
【0182】
実施例9 ジャパニンcDNAの3'領域のコンセンサス配列の同定
ジャパニンの3'配列を得るために、外側順方向プライマー2および4を用いてDNAを増幅し、pCR2.1にクローニングし、シークエンシングを行った。
【0183】
上記のように、外側順方向プライマー2および4と、逆方向プライマーとしてオリゴ(dT)20-Vを併用し、40μl反応で行った。反応液をアガロースゲルで泳動し、約600bp DNAバンドを切り出し、QIAquickゲル抽出キット(Qiagen社)を使用し、製造者の説明書に従って精製した(30μlの溶出バッファーでカラムから溶出)。
【0184】
各精製産物6μlをpCR2.1にライゲートするため、10μl反応で50ngのpCR2.1-TA(Invitrogen社)およびT4 DNAリガーゼ(NEB Biosciences社)と共に14℃で一晩インキュベートした。ライゲーション混合液を用いてTOP10 E. coliコンピテントセルに形質転換し、挿入断片を含有するコロニーを、順方向プライマー2および4とオリゴ(dT)20-Vを併用するPCRスクリーニングで同定した。QIAprep Spinキット(Qiagen社)を使用し、製造者の説明書に従って、陽性コロニーの5ml培養液から挿入断片を含有するpCR2.1 DNAを単離した。
【0185】
4つの挿入断片含有プラスミドのシークエンシングによりジャパニンcDNAの3'領域のコンセンサス配列を構築することが可能となった。これを図12に示す。
【0186】
実施例10 ジャパニンの5'領域のシークエンシング
5' RACE(Rapid Amplification of cDNA Ends:cDNA末端迅速増幅)法を使用してジャパニンの5'領域を組み込んだ約500bpの生成物を増幅した。
【0187】
5' RACEは既知の3' cDNA配列(この場合、新たに得たジャパニン 3'配列)を使用して遺伝子特異的プライマーの設計の情報を得る。これらのプライマーを使用して遺伝子特異的逆転写を行い、ネステッドPCRを用いてcDNAの5'領域を増幅する。後者の場合、順方向プライマーは実験的に導入したオリゴヌクレオチドのキャップ領域に特異的であり、逆方向プライマーは遺伝子特異的逆方向プライマーである。使用される種々のジャパニン特異的プライマーの相対的な位置を図13aに示す。
【0188】
RNAqueous-4PCRキット(AmbionRNA社)を使用し、製造者の説明書に従って、2日間給餌したメスR. appendiculatusマダニ唾液腺からRNAを抽出した。カラム溶出物からRNAを沈殿させ、20μl溶出バッファーに再溶解した。
【0189】
GSP1Bプライマーを用いて遺伝子特異的逆転写を行った。ImProm II RT酵素(Promega社)を、1μgのRNAを含有する20μl反応で、製造者の説明書に従って使用した。この反応でMgCl2濃度は2.5mM、総dNTP濃度は0.5mM、プライマー濃度は125nMであった。RT反応を48℃で1時間行った後、70℃で15分間加熱不活性化を行った。その後、1μlのRNase混合液(Invitrogen社。5' RACE Systemキットより)を添加し、室温で30分間インキュベートしてRNAを除去した。
【0190】
SNAPカラム(Invitrogen社。5' RACE Systemキットより)を製造者の説明書に従って使用し、生成したcDNAから酵素、プライマー、およびヌクレオチドを除去した。50μLのヌクレアーゼ未含有水でcDNAを溶出した。
【0191】
15μlのcDNAへのテール付加を、25μl反応でTdT酵素およびdCTP(Invitrogen社。5’RACE Systemキットより)を用い、製造者の説明書に従って行った。
【0192】
ポリ(dC)テールを付加したcDNA 1μlを20μl反応でネステッドPCRのテンプレートとして使用した。増幅の第1ラウンドを、GSP2AプライマーをAAPプライマー(Invitrogen社。5’RACE Systemキットより)と併用して行い、生成物をゲル精製し、これをテンプレートとして使用し、GSP3プライマーとAUAPプライマー(Invitrogen社。5’RACE Systemキットより)を併用して増幅の第2ラウンドを行った。
【0193】
いずれのPCRもTaq DNAポリメラーゼ(New England Biosciences社)を使用し、2mM Mg2+を含有する1X Thermopolバッファー(New England Biosciences社)に62.5μMの各dNTP(Bioline社)および250μMの各プライマーを添加したものを用いて行った。GSP2A/AAP PCRは初期変性段階を94℃で1分間行った後、94℃で30秒間/66℃で30秒間/72℃で40秒間を35サイクル、そして最後に72℃で更に5分間とした。生成物20μlをアガロースゲルで泳動し、約650bpのバンドを切り出し、QIAquickゲル精製キット(Qiagen社)を使用して製造者の説明書に従って抽出した。精製した生成物を1:1000の希釈率で第2ラウンドの増幅のテンプレートとして使用した。
【0194】
GSP3/AUAP PCRは、アニーリング温度が66℃ではなく68℃とした以外は、同様である。シークエンシングのために十分なDNAを得るため、150μlのGSP3/AUAP PCRを行った。生成物75μlをアガロースゲルで泳動し、約500bpのバンドを切り出し、QIAquickゲル精製キット(Qiagen社)を使用して製造者の説明書に従って抽出した。30μlの溶出バッファーでDNAをカラムから溶出し、サンプルを未精製のPCR産物と並行してゲルで泳動して回収を確認し、濃度を算出し(約100ng/μl)、その後、残りのサンプルでシークエンシングを行った(図13b)。
【0195】
AUAPプライマーを用いてゲル精製したPCR産物のシークエンシングを行い、ジャパニン cDNAの5'配列を得た(図14)。
【化3】
【0196】
実施例11 完全長ジャパニンのクローニングのためのプライマー設計
5'および3' cDNA配列がクローニングされたことにより、ジャパニンcDNAの推定される完全長配列(176残基ペプチドをコード)を構築することが可能となった(図15)。ジャパニン配列中の残基配置および他の既知の分子との配列類似性は、ジャパニンがリポカリンであることを示唆している。
【0197】
神経ネットワーク分析は、最初の24アミノ酸が分泌のためのシグナル配列であることを示唆している。それらの後に続く配列はThr Pro Ser Met Pro Ala Ile Asn Thr Gln Thr Leu Tyr Leu Alaであり、DC調節HPLC画分から得たN-末端配列と一致する。これによって、正確なcDNAがシークエンシングされたことが確認された。
【0198】
ネステッドPCR法を用いて完全長cDNAのクローニングを行った。いずれのPCRラウンドも、ポリメラーゼによって生じる変異導入を最小限にするために、ハイフィデリティDNAポリメラーゼを使用した。推定される完全長cDNA配列に基づき、シグナル配列を含有するプライマーを設計し、第2ラウンドのプライマーには、サブクローニングを容易にするために、順方向および逆方向プライマーの5'末端にそれぞれBamHIおよびNotI制限部位を導入した。制限酵素は直鎖状核酸の最末端を切断することはできないことが報告されているので、制限部位の前に5個の更なるヌクレオチドを付加した。
【化4】
【0199】
いずれのPCRラウンドもPhusion HotStart DNAポリメラーゼ(New England Biosciences社)を使用し、50μMの各dNTP(Bioline社)および250nMの各プライマーを添加した1X HFバッファー(New England Biosciences社)中で行った。
【0200】
前記と同様、第1ラウンドのPCRは順方向プライマー5および逆方向プライマー3を使用し、1日給餌を行ったメスR. appendiculatusの唾液腺から生成されたcDNAをテンプレートとして行った。初期変性段階は98℃で30秒間、その後、98℃で10秒間/64.6℃で30秒間/72℃で30秒間を35サイクル、そして72℃で更に1分間とした。この反応生成物を1:100000の希釈率で第2ラウンドの増幅のテンプレートとして使用した。
【0201】
第2ラウンドのPCRは順方向プライマー6および逆方向プライマー4を使用して行った。初期変性段階は98℃で30秒間、その後、98℃で10秒間/41℃で30秒間/72℃で30秒間を2サイクル、98℃で10秒間/72℃で30秒間を20サイクル、そして72℃で更に5分間とした。
【0202】
実施例12 ジャパニンのクローニング
5' BamHI部位および3' NotI部位を付加した完全長ジャパニンをコードするDNAを上記のようにPCRで増幅し、BamHIおよびNotIで切断し、同様に処理したpBacPAK8ベクターにライゲートした。ライゲートしたDNAを用いてTOP10 E. coliに形質転換した後、個々のコロニーを増殖およびミニプレップし、それらのプラスミドDNAをシークエンシングした。
【0203】
完全長ジャパニン(5' BamHI部位および3' NotI部位を有する)をコードするDNAを増幅するための反応(50μl)を上記のように行った。生成物35μlをQIAquick PCR精製キット(Qiagen社)で処理してプライマーおよびヌクレオチドを除去し、バッファー強度およびその後の制限酵素バッファーpHに対する影響を低減するために水で0.33Xに希釈した溶出バッファー30μlでプラスミドを溶出した。
【0204】
増幅されたDNAをBamHIおよびNotIで消化した(BSAを添加した1X BamHIバッファー中、37℃で1時間インキュベーション)。その後、酵素を除去するため、QIAquick PCR精製キット(Qiagen社)によってDNAを精製し、30μlの溶出バッファーで溶出した。酵素およびバッファーは全てNew England Biosciences社製であった。
【0205】
pBacPAK8プラスミドを同様に消化し、その後、切断したマルチクローニング部位フラグメントが確実に除去されるよう、QIAquickゲル抽出キットを用いてゲルを精製した。
【0206】
10μlの反応液(1X T4 DNAリガーゼバッファー(New England Biosciences社)中に約60ngの切断したpBacPAK8、約5ngの切断したジャパニンPCR産物、および1μl T4 DNAリガーゼ(New England Biosciences社)を含有)中で、ジャパニンDNAをpBacPAK8にライゲートさせた。
【0207】
3μlのライゲーション反応液を用いて、50μlの化学的コンピテントセルTOP10 E. coliに形質転換した。アンピシリンを含有するLB寒天培地上で一晩培養した後、独立したコロニーを5mlのLB培地(+アンピシリン)に接種して一晩培養し、QIAprep Spinキット(Qiagen社)を使用し、製造者の説明書に従って、プラスミドDNAを単離した。
【0208】
Bac1およびBac2プライマーを用いてシークエンシングを行った。得られた配列から、推定される完全長cDNA配列が正確であることが確認され、組換えバキュロウィルス(pBacPAK8-ジャパニン)作製のための変異未含有クローンの選別が可能となった。
【0209】
実施例13 昆虫細胞培養液中でのジャパニンの発現
flashBac系を用いてジャパニン発現組換えバキュロウィルスを作製し、これを用いてSf9昆虫細胞をジャパニン移入ベクターおよび必須遺伝子欠損を有する変異ウィルスで共トランスフェクトした。強力なプロモーターの制御下で、ベクターおよびウィルス間の相同組換えにより必須遺伝子の機能が回復され、同時に、ジャパニン配列がウィルスに挿入された。これにより、生存可能ウィルスは全てジャパニンDNAを含有するということが確認される。増幅後、組換えウィルスを用いて新たなSf9細胞に感染させ、培養液上清を回収し、DC調節活性についてスクリーニングを行った。図17に示すように、上清がDC調節活性を有することが明らかとなり、これは機能性ジャパニンが生成され、培養液中に分泌されたことを示しているが、この活性をアンマスクするために、まず上清を100kDA MWCOフィルターに通過させてウィルス粒子を除去する必要があった。おそらくこれはウィルス粒子の高度な刺激作用がジャパニンの阻害作用を上回るか、または回避するためである。
【0210】
log期のSf9細胞を1.1×106細胞/ウェルの密度で6ウェルプレートに接着させた後、Cellfectinトランスフェクション試薬(Invitrogen社)を使用し、flashBacゴールド・バキュロウィルスDNA(Oxford Expression Technologies社)およびpBacPAK8-ジャパニンの混合物でトランスフェクションを行った。500ngのプラスミドDNAを、1mlのSf900 II培養液に混合した0.5μlのflashBacゴールドDNAおよび5μlのCellfectinと混合し、室温で25分間インキュベートして複合体を形成させた。培養液を接着Sf9細胞から除去し、DNA/Cellfectin複合体で置換し、細胞を一晩インキュベートした後、更に1mlのSf900 II培養液を添加し、更に4日間インキュベーションを継続した。この時点でウィルスを含有する上清を回収し、4℃の暗所に保存した。
【0211】
次いで、このようにして得た少量のウィルス・ストックをより多量のSf9細胞培養液に播種し、ウィルスを増幅した。0.5mlのウィルス含有上清をlog期にあるSf9細胞の撹拌培養液250ml(細胞密度:約1.5X106/ml)に添加した。その後培養液を5日間インキュベートし、上清を回収し、4℃の暗所に保存した。
【0212】
増幅したウィルス・ストックを用いてSf9細胞を多重感染させ、タンパク質を発現させた。25mlのウィルス・ストックをlog期のSf9細胞培養液250ml(8×105細胞/ml)に添加した。その後、培養液を3日間インキュベートし、遠心分離によって上清を回収した。
【0213】
ウィルス粒子を除去するために、上清サンプル5mlを100kDa MWCO Vivaspin 6遠心濃縮キット(Sartorius社)に通過させた。このサンプルを通常の方法でDC調節活性についてスクリーニングし、QFT(陽性コントロール)、未濾過のバキュロウィルス/ジャパニン上清、および無関係のタンパク質を発現するバキュロウィルス-Sf9細胞培養液上清についても並行して行った。結果は、明らかに上清中にDC調節活性が存在することを示したが、これは未濾過上清中のウィルス粒子の存在によってマスキングされ、おそらくこれは粒子そのものがDCに対して圧倒的な刺激を与えるためであると考えられる。この結果により、「ジャパニン」としてクローニングされたタンパク質が実際に予測されたDC調節特性を有し、Sf9細胞によって活性型として生成されることが確認された。
【0214】
実施例14 ジャパニンを含有する昆虫細胞の上清から単離されたタンパク質の沈殿
ポリエチレングリコール(PEG)または硫酸アンモニウムのいずれかを添加して、バキュロウィルス/ジャパニン上清からタンパク質を沈殿させた。硫酸アンモニウムで沈殿させたタンパク質を、Superdex 75カラムを用いるゲル濾過によって更に分画した。図18に示すように、DC調節活性は硫酸アンモニウム沈殿させたタンパク質中には保持されるが、PEG沈殿させたタンパク質中には保持されず、また、プールしたSuperdex画分22-38(タンパク質の大部分を含有)中にも存在することが明らかになった。その後の画分22-38のスクリーニングにより、プールした画分23+24が最も活性が高いことが明らかになった。
【0215】
氷上で撹拌しながら、上清にPEGを最終濃度30%(w/v)まで徐々に添加した。同様に硫酸アンモニウムを70%(w/v)まで添加した。
【0216】
PEG沈殿したタンパク質を50mM HEPES(pH7.2)に再溶解し、Qカラムを用いて分画した。A280のリアルタイム測定により、画分4が分離したタンパク質ピークを含有することが示唆されたため、Superdex 75を用いるゲル濾過によってこの画分を更に分画した。このカラムから大部分のタンパク質が画分22および23に溶出された。
【0217】
硫酸アンモニウム沈殿したタンパク質を50mM HEPES(pH7.2)に再溶解し、5kDa MWCO Vivaspin 6遠心濃縮キット(Sartorius社)を使用してx15に濃縮した。Superdex 75カラムを用いるゲル濾過によって、濃縮されたタンパク質を分画した。A280のリアルタイム測定により、画分22-28が大部分のタンパク質を含有することが明らかになった。
【0218】
これらのサンプルについて、通常の方法でDC調節活性のスクリーニングを行ったところ(それぞれ1:100希釈)、活性は硫酸アンモニウム沈殿させたタンパク質中には保持されたが、PEG沈殿させたタンパク質中には検出されないことが明らかとなった。この事から予測されるように、選択されたPEG沈殿タンパク質のQカラム結合画分は活性を示さなかったが、このQカラム結合画分からのSuperdex画分22+23と共にインキュベートした後にはCD86発現のわずかな低下が観察された:この結果は明確なものではなく、更なる検討は行わなかった。反対に、硫酸アンモニウム沈殿したタンパク質は濃縮およびゲル濾過クロマトグラフィー後もその活性を保持し、その後のゲル濾過画分の比較から、画分23+24が最も高い活性を有することが明らかとなり、従って最も高濃度のジャパニンを含有することが想定される。
【0219】
実施例15 hisタグ・ジャパニンのクローニング
3段階ネステッドPCRを用い、pBacPAK8-ジャパニンをテンプレートとして、C末端に6残基のポリヒスチジン融合タグ(hisタグ)を付加したジャパニンを再クローニングした。ネステッドPCR産物を制限酵素で消化し、同様に制限酵素消化したpBacPAK8プラスミドへのライゲーションを行った。hisタグと天然のタンパク質配列の間に4つの更なる残基(グルタミン-グリシン-グリシン-セリン)が導入されるようにプライマーを設計した。これは、天然配列にhisタグが近接することによる立体障害を防止し、また、TEVプロテアーゼの共通開裂部位を導入する(これによって、タンパク質分解によるタグの除去が容易になる可能性がある)ことを目的とする。
【化5】
【0220】
第1段階のPCRは、50μMの各dNTP(Bioline社)、500nMの各プライマー、および10pg/μlのテンプレート・プラスミドを添加した1X HFバッファー(New England Biosciences社)中、Phusion DNAポリメラーゼ(New England Biosciences社)を用いて行った。初期変性段階は98℃で30秒間、その後、98℃で10秒間(変性)/70℃で30秒間(アニーリング)/72℃で15秒間(伸長)を15サイクル、そして最後に72℃で更に5分間とした。使用したプライマーは順方向プライマー7および逆方向プライマー5であった。このように行った20μlの反応液の産物をQIAquick PCR精製キットで精製し(30μl溶出バッファーで溶出)、プライマーおよびヌクレオチドを除去した。
【0221】
第2段階のPCRも同じ方法で行った。ただし、アニーリング温度を69℃とし、精製した第1段階のPCR産物を蒸留水で1:10に希釈してテンプレートとし、逆方向プライマーとして逆方向プライマー6を使用した。このように行った20μlの反応液の産物をQIAquick PCR精製キットで精製し(30μl溶出バッファーで溶出)、プライマーおよびヌクレオチドを除去した。
【0222】
第3段階のPCRは第2段階のPCRと同様に行った。ただし、精製した第2段階のPCR産物を蒸留水で1:10に希釈してテンプレートとし、逆方向プライマーとして逆方向プライマー7を使用した。このように行った50μlの反応液の産物をQIAquick PCR精製キットで精製し(30μlの0.5X溶出バッファーで溶出)、プライマーおよびヌクレオチドを除去した。反応から得たサンプルをアガロースゲルで泳動したところ、PCR産物の濃度は約20ng/μlと試算された。
【0223】
精製した第3段階のPCR産物をBamHIおよびNotIで消化した(BSAを添加した1X BamHIバッファーを含有する50μlの反応液中、37℃で20分間インキュベート)。バッファーおよび酵素はNew England Biosciences社のものを使用した。反応液の産物をQIAquick PCR精製キットで精製し(30μlの溶出バッファーで溶出)、酵素および切り出されたフラグメントを除去した。
【0224】
約1:1のモル比としてライゲーションを最適化するため、制限酵素消化産物約4.5ngを前記のBamHI/NotI消化およびCIP処理したpBacPAK8(50ng)とライゲートさせた。ライゲーションはT4 DNAリガーゼ(New England Biosciences社)を用い、T4 DNAリガーゼ・バッファー(New England Biosciences社)を含有する10μl反応液中、室温で15分間行った。ライゲーション混合液を用い、TOP10 E.coliコンピテントセルに形質転換した。分離したコロニーを5mlの液体LB培地に接種し、QIAprep Spinキット(Qiagen)を製造者の説明書に従って使用し、プラスミドDNAを単離した。Bac1およびBac2プライマーを用いてシークエンシングを行い、ジャパニン融合タンパク質をコードする挿入断片の存在を確認した。以下、このプラスミドをpBacPAK8-Jap-TEV-hisと称する。
【0225】
実施例16 hisタグ・ジャパニンの単離
pBacPAK8-Jap-TEV-hisプラスミドDNAを用い、flashBacゴールドシステムを使用して、前記の未標識組換えジャパニンの生成と同様に、組換えバキュロウィルスを調製した。次いで、これらの組換えバキュロウィルスを用いて、前記のようにSf9細胞の発現培養液250mlに感染させた。その後、水酸化アンモニウムを用いて培養液上清からタンパク質を沈殿させ、ポリヒスチジン・モチーフを結合させるIMACカラムを用いて精製した。SDS-PAGEゲルの銀染色により、組換え特異的タンパク質が約20kDaに存在することが明らかとなり(図19)、ウェスタン・ブロットにより、実際にこのタンパク質にhisタグが付加されていることが確認された(図20)。これらの結果は、hisタグ・ジャパニンがうまく発現されたことを示している。
【0226】
前記の未標識組換えジャパニンの生成と同様に、3日目のpBacPAK8-Jap-TEV-his発現培養液250mlの上清に硫酸アンモニウムを70%(w/v)まで添加し、タンパク質を沈殿させた。沈殿させたタンパク質を60mlの40mM Na2HPO4/300mM NaCl/10%グリセロール(pH8)(結合バッファー)に再溶解し、あらかじめ1mlのTalon樹脂(Clontech社)を充填しておいたカラムに負荷した。その後、カラムを20mlの結合バッファーで2回洗浄し、6mlの40mM Na2HPO4/100mM NaCl/300mMイミダゾール(pH8)(溶出バッファー)で溶出した。溶出されたタンパク質を5000MWCO Vivaspin 6(Sartorius)でX10に濃縮した。
【0227】
0.5μlのサンプルを4-12%ポリアクリルアミドBis-Trisゲル(Invitrogen社のNuPageプレキャスト・ゲル)で泳動した後、SilverXpressキット(Invitrogen社)を用い、製造者の説明書に従って銀染色を行った。2つの組換えタンパク質上清には約20kDaに2つの主要なバンドが明らかに存在したが、陰性コントロール精製物(野生型バキュロウィルスに感染させた細胞からの上清を用いて並行して行った)には存在しなかった。
【0228】
ウェスタン・ブロット分析のために、6.5μlのサンプルを4-12%ポリアクリルアミドBis-Trisゲル(Invitrogen社のNuPageプレキャスト・ゲル)で泳動した後、ウェット式ブロッティング(10%のメタノールを添加したNuPageトランスファー・バッファー中、30Vの定電圧を1時間適用)によって45μm孔のニトロセルロース膜(Biorad社)に転写した。転写完了後、膜を蒸留水で洗浄し、Trisバッファー/0.1% Tween 20(TBST)で5分間洗浄した後、ブロッキング試薬(#B6429、Sigma社)中、室温で1時間インキュベートした。ブロッキング後、膜をTBSTで軽く洗浄し、ブロッキング試薬で1:1000に希釈した抗ペンタ-his抗体(Qiagen社)と共に4℃で一晩インキュベートした。その後、TBSTで5分間の洗浄を5回行い、10%脱脂粉乳(Marvel社)/TBSで1:20000に希釈したロバ抗マウス-HRP(Jackson Immunoresearch社)中、室温で1時間インキュベートし、TBSTで5分間の洗浄を更に7回行った。最後に抗体を可視化するために、膜をECL基質(Amersham社)で製造者の説明書に従って処理し、これにX線フィルムを10秒間暴露させた。カラムから溶出された最初の4つの1ml画分中に約20kDaのバンドが明らかに存在し、予測されたサイズのhisタグ・タンパク質の存在が確認された。
【0229】
実施例17 ジャパニン-TEV-his上清由来サンプルのCD調節活性
前記のようにジャパニン-TEV-his上清由来サンプルについてDC調節活性のスクリーニングを、サンプルを1:100希釈して行った。タグを付加していない組換えジャパニン由来のSuperdex画分23+24を陽性コントロール、野生型バキュロウィルスに感染させたSf9細胞培養液由来のサンプルを陰性コントロールとして、DC調節活性についてのスクリーニングを行った。図21に示すように、ジャパニン-TEV-his上清から硫酸アンモニウムを用いて沈殿させたタンパク質に活性があり、活性組換え融合タンパク質が生成されたことを示している。活性はジャパニン-TEV-his上清由来の10X濃縮したTalonカラム結合タンパク質中にも存在し、活性タンパク質が実際にTalon樹脂に結合することが確認された。10X濃縮Talon溶出物がバルクの硫酸アンモニウムで沈殿させたタンパク質に比較して活性が低いことは必ずしも活性の低下を示しているものではなく、Talon溶出バッファーの刺激作用を反映している可能性がある。
【0230】
実施例18 hisタグ・ジャパニンの精製
Talonカラム溶出物をSuperdex75(ゲル濾過)カラムに通過させて、ポリヒスチジン・タグ・ジャパニンを更に精製した。銀染色したSDS-PAGEゲル上の約20kDaのバンドの存在によってジャパニンを含有する画分を同定し、プールした。プールした画分中のタンパク質濃度を、280nmの吸光度およびexpasy.orgのProtParamツールによって成熟ジャパニン配列から予測される吸光係数から算出した。
【0231】
精製したタンパク質のDC調節活性に関するアッセイを、前記のようにポリ(I:C)を24時間後に細胞に添加し、25ng/mlから1.6μg/mlまでの種々の濃度で行った。図22は、100ng/ml以上の濃度で最大活性に達することを示しており、これまでに行った1回の実験では、<25ng/mlで50%活性に達しうることが示唆された。
【0232】
実施例19 DC分化に対するジャパニンの影響
前記のように、GM-CSFおよびIL-4との培養によって、ヒト単球からDCを生成させた。いくつかの培養液には、200ng/mlの組換えジャパニンを更に添加した。3日目から6日目まで、日毎に培養液のCD14およびCD1a発現を分析した。
【0233】
分化に対するジャパニンの影響が、ジャパニンそのものに対する影響ではなく、組換えジャパニンの内毒素の混入によるものであるという反論もありうるため、ジャパニンの内毒素含有量の評価をLALアッセイを用いて行い、約0.540EU/μg(およそ、ジャパニン1μg当たり0.054ngのE. coli LPSに相当)であることが明らかとなった。
【0234】
図23に見られるように、200ng/mlのジャパニンによって分化培養物の生育は大きく改変され、約50%の単球がDCへの分化の痕跡であるCD1aの亢進的調節およびCD14の抑制的調節を行うことができなかった。9pg/mlまたは40ng/mlのいずれかのE. coli LPS(それぞれ、200ng/mlで使用した場合の組換えジャパニンの内毒素含量、およびその内毒素含量の>4000倍に相当)を添加したコントロールにより、いずれの濃度も分化に大きな影響を与えないことから、これはジャパニンの内毒素混入の副作用ではないということが明らかとなった。
【0235】
実施例20 T細胞増殖アッセイ-混合白血球反応(MLR)
混合白血球反応(MLR)を用いて、特異的抗原(この場合、同種異系MHC)を提示するmoDCに応答したT細胞増殖に対するジャパニンの影響を評価した。この系においては、ジャパニンは顕著なT細胞増殖阻害を示した。
【0236】
上記のように調製した、凍結された5日目の単球由来DCを解凍し、200ng/mlの組換えジャパニンの存在下または非存在下で更に2日間培養した。
【0237】
CD3 MACSマイクロビーズ(Miltenyi Biotech社)を製造者の説明書に従って用い、前記のように、Lymphoprepを用いて行うPBMCの初期分画によって同種異系T細胞をバフィーコートから単離した。
【0238】
1x105 T細胞/ウェルを、段階的に希釈した放射線照射樹状細胞と共に96ウェル丸底プレート(Corning社)に播種した。細胞培養液はRPMI 1640に10% FCS + 2mM L-グルタミン、100U/ml ペニシリン、および100μg/ml ストレプトマイシンを添加したもので、最終容量は200μl/ウェルとした。ジャパニンと共に2日間インキュベーションしておいたDCを含むウェルには、更に200ng/mlのジャパニンを追加した。DC未含有のT細胞を含むコントロールも含めた。
【0239】
MLR培養液を4日間インキュベートした後、0.5μCi/ウェルの3H-チミジンで瞬間標識した。次いで、これらを更に16-18時間インキュベートした後、自動細胞回収装置を用いてガラス繊維フィルター上に回収し、フィルターに結合した放射性DNAをシンチレーション・カウンターで定量した。
【0240】
この実験を3回行ったところ、いずれの実験でも、DCをジャパニンで前処理しMLR培養液中にもジャパニンを含有させるとT細胞の増殖が低減されることが明らかになった。結果の例を図24に示す。
【0241】
実施例21 ジャパニン・ホモログの検索(Dermacentor andersoni)
EMBL Expressed Sequence Tag(EST)データベースのBLAST検索で、Dermacentor andersoni由来のジャパニン・ホモログを同定した。このホモログは現在、D. andersoni E1244(EBI ID = EG363153)と称されている。SignalPを用いてDA-E1244のシグナルペプチド部分と考えられる部分(残基1-17)を同定し、成熟ジャパニン配列および予測される成熟DA-E1244配列を比較した。BLOSUM62マトリクスを用いて実施したEMBOSS2配列間アラインメントにより、2つのタンパク質間で30.5%の同一性および50.3%の類似性が確認された(図25)。データベース中に存在するヌクレオチド配列にはSEQ ID NO:32があり、推定上のコードされるタンパク質はSEQ ID NO:4である。
【0242】
このジャパニンとの高度なホモロジーは、DA-E1244がジャパニン様生体活性を有することを強く示唆しており、このため、組換えタンパク質を生成するための研究に着手している。
【0243】
このために、DA-E1244の完全長アミノ酸配列をコードするDNAを合成遺伝子として生成し(SEQ ID NO:3)、クローニング・ベクターに導入し、pCR4TOPO-DA-E1244_optとした(これはEntelechon社(レーゲンスブルク)による契約に従って行われた)。合成されたDNA配列はATGスタート・コドンで始まり、コザック・コンセンサス配列(真核細胞系における効率的な翻訳の必須条件である)に一致する上流配列は導入しなかった。これを改善するために、DA-1244を増幅し、産物の5'末端にBamHI認識部位(“GGATCC”)およびコザック対応(Kozak-compliant)配列(“TCCAAA”)を、3'末端にNotI認識部位(“GCGGCCGC”)を付加するようにプライマーを設計した。「余分の」塩基を5'および3'末端の両方に付加し、制限酵素部位が産物の末端に位置して制限酵素消化を阻害することがないようにした。
【化6】
【0244】
Phusion HotStart DNAポリメラーゼ(New England Biosciences社)を用い、50μMの各dNTP(Bioline社)および250μMの各プライマーを添加した1X HFバッファー(New England Biosciences社)中、20μl反応でPCRを行った。pCR4TOPO-DA-E1244_optプラスミドをテンプレートとして使用し、反応液中に1ng/μlの濃度で含有させた。初期変性段階は98℃で30秒間、次いで98℃で10秒間/69℃で30秒間/72℃で15秒間を15サイクル、最後に72℃で更に5分間とした。
【0245】
完了した反応液の5μlをアガロースゲルで泳動して期待されるサイズ(600bp)の産物の存在を確認し、残りはQIAquickカラム(Qiagen社)を製造者の説明書に従って用いて精製し、BamHIバッファー(NEB社)中、BamHIおよびNotI(共にNEB社)で消化し、再度QIAquickカラムで精製した。
【0246】
その後、切断および精製したDA-1244 PCR産物を、BamHI/NotI消化および仔ウシ腸ホスファターゼ処理を行ったpBacPAK8(バキュロウイルス・トランスファー・ベクター)にライゲートし、ライゲーション反応液を用いてTOP10 E.coliコンピテントセルに形質転換した。
【0247】
形質転換したE.coliの独立したコロニーを採取し、培養液中で増殖し、QIAprepミニプレップ・キットを用いてDNAを抽出した。シーケンシングを行い(Bac1およびBac2プライマーを使用)、変異が導入されることなくDA-E1244がクローニングされたことを確認した。
【0248】
好適なクローンを同定し(pBacPAK8-E1244)、組換えバキュロウィルスの作製に使用した。
【0249】
実施例22 更なるジャパニン・ホモログの検索
EMBL Expressed Sequence Tag(EST)データベースのBLAST検索で、ジャパニン・ホモログがRhipicephalus microplus(2つのホモログ)、Amblyomma americanum、およびRhipicephalusappendiculatusから同定された。現在これらのホモログはそれぞれ、R. microplusCK185494、A.americanum CX766068、R. appendiculatus CD796501、およびR. microplus CV436349と称されている。これらのタンパク質の配列アラインメントおよび同一性%の図を図26-29に示す。
【0250】
R. microplus CV436349は、これらの同定されたホモログのうち、唯一、シグナルペプチドを含有する。他の3つの同定されたホモログにシグナル配列が存在しないことに関しては、3つの説明が可能である:i)それらは分泌タンパク質ではない;ii)配列の一部が欠失している;またはiii)非標準的分泌が関与している。
【0251】
R.microplus CK185494およびA. americanum CX766068はいずれもジャパニンと実質的なホモロジーを有するリポカリンである。しかしながら、これらの配列はいずれもシグナル配列を有さず、上記のように、それらが分泌タンパク質ではないか、あるいは配列の一部が欠失していることを示唆している。
【0252】
これらのジャパニン・ホモログはジャパニンとの配列同一性がDermacentor andersoniから同定されたホモログより低いが、その機能はジャパニンに匹敵すると考えられる。
【0253】
実施例23 ジャパニン・リガンドとしてのコレステロールの同定
ジャパニンはリポカリンであると報告されており、これは脂質リガンドに結合しうる可能性を示唆している。OmCI(脂肪酸結合マダニ・リポカリン)の結晶構造に基づいて仮定されるジャパニンの構造モデルの構築により、ジャパニン中に疎水性のオープン結合ポケットが存在することが示唆され、更にこの見解を支持している(未掲載)。
【0254】
これを更に検討するため、実施例13に記載するように組換えジャパニンを昆虫細胞培養液中で生成し、前記のように金属アフィニティークロマトグラフィーおよびゲル濾過に連続的に施与して精製した。この精製組換えタンパク質400μgを用い、下記のようにBligh & Dyer法で脂質を抽出した後、ガスクロマトグラフィー質量分析(GC-MS)を行った。
【0255】
Bligh & Dyer脂質抽出法
3.75mlのクロロホルム:メタノール(1:2)を0.5mlのタンパク質サンプル(または0.5mlのバッファーコントロール)に添加した。この混合液を10-15分間撹拌し、更に1.25mlのクロロホルムを添加し、ボルテックスで1分間混合した。その後、1.25mlの超純粋を添加し、更に1分間ボルテックスを行った。得られたサンプルを遠心分離し、脂質を含有する下層を残し、上層を廃棄した。これを窒素雰囲気下で乾燥し、500μlのジクロロメタンに再懸濁した。
【0256】
ガスクロマトグラフィー/電子衝撃質量分析(GC/EI-MS)
組換えジャパニンまたはバッファー・ブランクから抽出したサンプル1μlを、キャピラリー・ガスクロマトグラフが組み込まれたPerkin Elmer Turbomass四重極質量分析計に注入した。以下の条件を使用した:
ガスクロマトグラフィー:カラム=DB-5。注入=オンカラム。注入温度=40℃。温度グラジエント=40℃、1分間。その後、8℃/分で325℃まで(10分間保持)。キャリアガス=ヘリウム。
質量分析:イオン化電圧=70eV。イオン化モード=スキャニング。MS分解能=ユニット。
【0257】
質量分析から得られたデータは、ジャパニン・サンプルで33.1分にピークを示し、これはバッファー・ブランクには存在しなかった(図30a)。このピークからの平均分布(図30b)をNISTライブラリーの分布と比較することにより、これがコレステロールであることが同定された。これを確認するために、コレステロール標準品を同じ条件下で処理したところ、33.1分にピークが得られ、平均分布は一致した(図30c)。
【0258】
実施例24 組換えジャパニンは遊離コレステロールに結合する
組換えオリゴヒスチジン・タグ・ジャパニンをNi-NTA磁気ビーズに固定するため、0.5μgのタンパク質を、ビーズを含有させた500μlのバッファーA(120mM NaCl、0.02% Tween、5%グリセロール、40mMリン酸二ナトリウム)中、室温で2時間インキュベートした。コントロールサンプルにはタンパク質を含有させなかった。タンパク質でコーティングされたビーズを500μlのバッファーAで3回洗浄した後、0.2μlの3H-コレステロールを含有するバッファーB(6M グアニジン、120mM NaCl、0.02% Tween、5%グリセロール、40mMリン酸二ナトリウム)50μlを添加した(変性バッファーは、起こりうる、放射能標識されたコレステロールを有するタンパク質に結合した細胞培養液由来のコールドリガンドの交換を促進するために使用した)。5分後、バッファーBを除去し、更に0.2μlの3H-コレステロールを含有させたバッファーAを500μl添加した。その後、室温で3時間インキュベートした。氷冷したバッファーAでビーズを洗浄し(500μlで1回、50μlで2回)、未結合のコレステロールを除去した。その後、これを0.5Mのイミダゾールを含有するバッファーA、100μlに再懸濁して、タンパク質をビーズから溶出させた。図31の洗浄1および2は50μlの洗浄であり、各サンプルの右側のバーはビーズ/タンパク質に結合した放射活性の量を示す。
【0259】
図31に示されるように、これらの結果は明らかに3H-コレステロールがジャパニンに結合することを示している。変性/リフォールディングがタンパク質の結合に必要であるか否かはまだ明らかではなく、結合の強度および特異性も確認する必要が残されている。
【0260】
実施例25 ジャパニンは樹状細胞上のC型レクチン細胞表面受容体に結合する
ジャパニンが樹状細胞成熟化を阻害する能力は、その樹状細胞表面への結合能を暗示するものである。これは、ジャパニンとの膜受容体特異的相互作用を介して起こる可能性が最も高いと考えられるが、ジャパニンが非特異的な(おそらく結合したコレステロールと血漿膜との相互作用を伴う)方法で細胞に結合および侵入し、その後、細胞内シグナル伝達経路に細胞型特異的に作用する作用機構も考えられる。
【0261】
ジャパニンが特異的に樹状細胞表面に結合するか否かを検討し、任意の相互作用の性質の検討を可能にするために、市販のキットを使用して、ジャパニンを蛍光色素Alexa 488で標識した。この蛍光標識したジャパニン、500ng/mlと共に細胞を4℃で30-60分間インキュベートした後、洗浄し、フローサイトメトリーでジャパニン結合を可視化した。
【0262】
ジャパニンが単球由来樹状細胞に特異的に結合することが図32aで明らかであり、ジャパニン-Alexa 488(32a-fの塗りつぶしのヒストグラム)は5日目の単球に由来する樹状細胞(前記のように作製)に結合するが、コントロール・リポカリンとして使用したムバチン(moubatin)-Alexa 488は結合しない(図32a-fの点線のヒストグラム)ことを示している。更に、ジャパニンは単球にも(図32b)、マウス骨髄由来樹状細胞にも(図32c)結合しない。
【0263】
先に証明されたジャパニンが単球の樹状細胞への分化を阻害する能力を考慮すると、ジャパニンが単球に結合しないことは予想外のことである。このことから、ジャパニンは、明らかに結合しない細胞型に対して、いかにして作用できるのかという疑問が浮上する。この問題に取り組むため、ジャパニン-Alexa 488(32a-fの塗りつぶしのヒストグラム)またはムバチン-Alexa 488(図32a-fの点線のヒストグラム)を1日目の単球から作製した樹状細胞と共にインキュベートした(図32d)。ジャパニンは、5日目単球由来樹状細胞より程度は低いが、1日目の単球由来樹状細胞に結合することが明らかになった。これは、ジャパニン結合受容体の亢進的調節が樹状細胞分化の非常に早い段階で開始され、そのためジャパニンはこの初期段階で細胞に作用してその分化を停止させるという可能性を示唆している。
【0264】
ジャパニン-樹状細胞相互作用の性質を検討するため、マンナンおよびEDTAの影響を試験した。1mg/mlのマンナンの存在により、単球由来樹状細胞へのジャパニン-Alexa 488の結合は大幅に低減されたが(図32e。塗りつぶしのヒストグラムはマンナン非存在下でのジャパニン-Alexa 488結合、白抜きのヒストグラムはマンナン存在下での結合、そして点線のヒストグラムはコントロール・タンパク質の結合を示す)、0.5mM EDTAの存在下ではこれが完全に阻害された(図32f。白抜きのヒストグラムがEDTA存在下での結合を示す以外は、図32dと同様)。考え合わせると、これらの知見はジャパニンが単球由来樹状細胞上のC型レクチン細胞表面受容体に結合することを強く示唆している。
【0265】
実施例26 ジャパニンはN-グリコシル化される
NetNGlyc 1.0(Center for Biological Sequence Analysis、デンマーク工科大学)の使用から、ジャパニンが1つの可能性の高い、そしてもう1つの可能性を有するN-グリコシル化部位を含有することが示唆される(図33a)。また、前記のように、ある程度のグリコシル化の存在が、ジャパニンとC型レクチン受容体との相互作用から示唆されている。
【0266】
N-グリコシル化の存在を確認するために、精製した組換えジャパニン(前記のように作製)をPNGase F(ほとんどの型のN-グリコシル化を阻害する酵素)で37℃、16時間処理した。次いでPNGase F処理をしたジャパニンを模擬処理および未処理のジャパニンと並行してSDS-PAGEゲルで泳動し、抗hisタグ・ウェスタンブロッティングで可視化した。図33bに示すように、PNGase F処理では、模擬処理および未処理のジャパニンにも見られる2つのバンドに加えて更なるより小さいバンドが存在した。これはより大きな2つのバンドの少なくとも一方、おそらくは両方がN-グリコシル化されたジャパニンであることを示している。
【0267】
実施例27 組換えジャパニンは多くの多様な刺激に反応して樹状細胞成熟化を阻害する
前記のように、組換えジャパニンはポリ(I:C)(TLR3刺激)に応答して単球由来樹状細胞の成熟化を阻害し、3日間給餌したメスR. appendiculatusマダニ由来のジャパニン含有Qカラム・フロースルーはLPS、TLR4刺激、およびIFNγ(γ-インターフェロン受容体を介して作用する)に応答して単球由来樹状細胞の成熟化を阻害するが、TNFR1を介して作用する可溶性TNFαには応答しない。これらの知見は、(前記と同様の方法に従って)これらの実験を反復して得られたが、これには精製組換えジャパニン(前記のように作製)を用い、LPS(図34a)、IFNγ(図34b)、TNFα(図34c)、可溶性CD40L(図34d)、IFNα(図34e)、またはCL097(TLR7/8リガンド)(図34f)で刺激を行った。TNFα以外のこれら全ての刺激で惹起される樹状細胞の成熟化はジャパニンで阻害される。TNFαで誘導される成熟化に対する有意な作用は観察されなかったが、最低限の阻害は起こりうる。これらの知見から、ジャパニンが、多くの異なる受容体および下流シグナル伝達系を介して作用する広範な刺激に応答して、樹状細胞の成熟化を阻害する能力を有することが確認される。
【0268】
実施例28 組換えジャパニンは刺激に応答して樹状細胞のTNFα分泌を阻害する
共刺激分子およびMHCクラスIIの亢進的調節に加えて、樹状細胞は炎症性刺激にも種々のサイトカインを生成することによって応答する。ジャパニンが樹状細胞成熟化を阻害する、または何らかの方法で変更する能力を有するか否かを評価するために、2つの刺激、LPSおよびIFNγを合わせたものに応答する前炎症性サイトカイン、TNFαの単球由来樹状細胞生成に対するジャパニンの影響を評価した。
【0269】
ヒト単球由来樹状細胞を前記のように作製した。5日目にそれらを回収し、前記のように、FCS、GCSF、およびIL4を含有するフレッシュの培養液に5x105細胞/mlの密度で再懸濁した。次いでそれらを組織培養用24ウェル・プレート中、精製組換えジャパニン(500ng/ml)の存在下または非存在下で培養し、24時間後、組換えヒトIFNγ(Peprotech社)の刺激カクテルおよび超高純度E.coli 011:B4 LPS(Alexis Biochemicals社)をいくつかのウェルに添加した(最終濃度:20ng/ml IFNγ+200ng/ml LPS)。更に48時間後、培養液上清を回収し、遠心分離して細胞およびデブリを除去した。その後、ELISAキット(Insight Biotechnology社)を製造者の説明書に従って使用し、TNFα濃度を測定した。
【0270】
ジャパニンは刺激カクテルに応答してTNFαの樹状細胞分泌を低減することが明らかになった(図35)。
【技術分野】
【0001】
本発明は樹状細胞(DC)調節分子に関する。特に、本発明は哺乳動物DC(特にヒトDC)の分化および成熟化を調節(好ましくは阻害)する分子に関する。それらの分子は節足動物の唾液、より具体的にはマダニの唾液から単離できる。本発明はまた、それらの分子の治療への使用、特に、自己免疫疾患、アレルギーおよび他の過敏性反応、移植片拒絶反応および移植片対ホスト病、感染性疾患(例えばマダニによって伝染するもの)、癌(例えば血液系腫瘍)、そして急性および慢性炎症性疾患(例えば上記の疾患に伴う炎症)の治療へのそれらの分子の使用にも関する。
【背景技術】
【0002】
哺乳動物(特にヒト)の免疫系には、2つの系統、すなわち自然免疫系および適応免疫系がある。自然免疫系の細胞は包括的様式で感染性病原体を認識および応答する。自然免疫系は感染に対する重要な即時的バリアであるが、外来物質、例えば侵入病原体に対して長期間に及ぶ特異的な防御を与えるものではない。これに対して適応免疫系の細胞は特定の外来物質を認識し、ホストにおいてそれらの特定物質に対する免疫学的記憶を誘導する。
【0003】
DCは病原体による感染直後に自然免疫系の成分と相互作用し、また、哺乳動物の適応免疫応答の中心的役割を果たす。DCは前駆細胞から分化して未成熟DCとなる。未成熟DCは全身にわたって存在し、他の免疫系細胞もこの役割に関わるが、それらは、主にT細胞の活性化を惹起する能力によって適応免疫系の発動に関与する主要な細胞型である。
【0004】
未成熟DCは、外来物質(例えば病原体表面)上の特定の化学シグナルを認識するパターン認識受容体(PRR)(例えばToll様受容体(TRL))によって、常に周辺環境から感染性物質(例えばウィルス、微生物、および寄生生物)をサンプリングしている。病原体のような物質が外来であると同定されると、未成熟DCはその物質またはそのフラグメントを取り込み、タンパク質および脂質抗原を分解してペプチドおよびグリコペプチドまたは脂質フラグメントとし、DC表面に提示する。
【0005】
外来物質の認識および/または他の細胞環境内のシグナル(例えば炎症性サイトカイン)に応答して、未成熟DCは「成熟化」と総称されるいくつかの変化を受け、成熟DCへの発達を開始する。成熟DCは主要組織適合遺伝子複合体(MHC)およびMHC関連分子(例えばCD1)(それぞれ、外来物質由来のペプチドおよびグリコペプチド、および脂質と結合する)の発現を亢進的に調節し、それらをDC表面上に提示させる。これと同時に、DCは共刺激分子として知られる細胞表面受容体(CD80、CD86、およびCD40など。Tリンパ球活性化のための補助受容体として作用する)の発現を亢進的に調節する。更にDCは走化性シグナルを受け、リンパ系組織(例えばリンパ節および/または脾臓)への移動を開始する。リンパ系組織に到達すると、DCは外来物質由来のペプチドおよびグリコペプチドまたは脂質フラグメントを提示し、好適な共刺激シグナルを伝達することによって、Tリンパ球を活性化する。それらの活性化Tリンパ球は、適応免疫応答の伝達に関与している。外来物質は病原体、アレルゲン、または、自己免疫応答の場合は身体によって異物と誤認される自己抗原でありうる。
【0006】
抗原提示および共刺激によるT細胞活性化の惹起の役割に加え、成熟DCはT細胞調節、例えばヘルパーT細胞のTh1、Th2、Th17、または調節性(Treg)細胞への分極化(polarisation)、細胞傷害性T細胞の活性化、および(例えば皮膚または腸および他の粘膜部位への)T細胞ホーミングの調節にも関与する。
【0007】
適応免疫応答においてDCが中心的役割を果たしていることから、治療を目的とするDCの機能調節に関心が向けられており、動物実験ではDCの調節物質が自己免疫疾患および他の炎症性疾患の治療に有用であり得ることが示唆されている(Subkleweら Human Immunology, 2007, 68(3), 147-155)。また、DC調節物質は癌の治療に有用であり得ることも示唆されている。治療を目的としてDC調節物質として作用する更なる分子を同定することは明らかに有益である。
【0008】
特に、DC調節活性、特に阻害活性を有する化合物の同定、並びに、自己免疫疾患、他の炎症性疾患、および癌の治療におけるその使用法の開発がなお必要とされている。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】Subkleweら Human Immunology, 2007, 68(3), 147-155
【発明の概要】
【課題を解決するための手段】
【0010】
発明の概要
従って、本発明は単離されたDC調節分子を提供し、該分子は哺乳動物DCの分化および成熟化を調節(好ましくは阻害)する。
【0011】
ある態様では、単離されたDC調節分子はヒトDCの分化および成熟化を調節(好ましくは阻害)する。本発明のDC調節分子は節足動物、特に吸血性節足動物から単離することができる。単離されたDC調節分子はタンパク質であってもよい。
【0012】
吸血性節足動物はそのホスト(哺乳動物(例えばヒト)など)に付着し、長期間にわたって摂食する。それらがホストに運搬する成分(唾液中の成分など)はホストの免疫応答を惹起する可能性がある。それらの応答は節足動物に有害でありえ、従って節足動物はそれらを抑制する必要があり得る。免疫の惹起においてDCが中心的役割を果たしていることを考慮すると、節足動物にとって、その機能を阻害する物質を生成させることは有益であり得る。
【0013】
吸血性節足動物、特にマダニは、ホストに種々の抗炎症および免疫調節成分を接種することによって、ホストの免疫系を阻害しうる(Ribeiroら, Infectious Agents and Disease, 1992, 4(3), 143-152)。
【0014】
以下のようないくつかの免疫調節分子がマダニ唾液中で同定されている:マクロファージ遊走阻止因子(MIF)のホモログ(Jaworskiら, Insect Molecular Biology, 2001, 10(4), 323-331)、ヒト・マクロファージ、単球、および好中球によって分泌される白血球エラスターゼ阻害剤のホモログ(Leboulleら, The Journal of Biological Chemistry, 2002, 277(12), 10083-10089)、グリコシル化されたタンパク質p36(マイトジェンによって作動されるマウス脾臓細胞のインビトロ増殖を阻害すると考えられる)(Bergmanら, Journal of Parasitology, 2000, 86, 516-525)、B細胞阻害タンパク質(BIP)(Hannierら, Immunology, 2004, 113, 401-408)、およびB細胞抑制因子(BIF)(Yuら, Biochemical and Biophysical Research Communications, 2006, 343, 585 - 590)。しかしながら、これらの分子の多くは明確な細胞標的を有さず、これらの分子のうちで哺乳動物DC(特にヒトDC)の分化および成熟化の両方に阻害効果を与えることが確認されているものは存在しない。
【0015】
Salp15はマダニ唾液中に存在するタンパク質であり、未成熟ヒトDCに作用することが明らかになっている(Anguitaら, Immunity, 2002, 16, 849-859、およびHoviusら, Vector borne and Zoonotic diseases, 2007, 7(3), 296-302)。しかしながら、免疫調節刺激物質の存在下で未成熟ヒトDCをSalp15と共にインキュベートすることを伴うアッセイでは、Salp15が共刺激分子(例えばCD86)の亢進的調節を阻害しないことが明らかにされた。従って、Salp15はヒトDCの成熟化を阻害しない。
【0016】
プロスタグランジンE2(PGE2)はマダニ唾液中に存在する非タンパク質分子であり、未成熟マウスDCの活性を調節しうるが、これらのマウスDCの成熟化に対しては最小限の影響しか及ぼさない(Sa-Nunesら, The Journal of Immunology, 2007, 179, 1497-1505)。PGE2はヒトDCの成熟化を促進する能力を有するが、ヒトDCの分化および成熟化を阻害するように作用するという証拠はない。
【0017】
また、マダニ唾液および唾液腺抽出物(SGE)がマウスDCの分化および成熟化を調節する能力を有しうることも示唆されている(Cavassaniら, Immunology, 2005, 114, 235-245、およびSkallovaら, Journal of Immunology, 2008, 180, 6186-6192)。しかしながら、これらの活性に関与する分子は単離されておらず、これまでのところ、マダニ唾液がヒトDCの分化および成熟化の両方を阻害する能力を有するという証拠はない。
【0018】
驚くべきことに、出願人は哺乳動物DC、特にヒトDCの分化および成熟化の両方を調節(好ましくは阻害)する分子を単離した。
【0019】
「単離された」という用語は、分子がもはやその天然での環境中にないことを伝えることを意図する。この用語には、その天然での環境から除去された分子、およびそれらと同一であるが合成されたものである分子も含まれる。本発明の単離された分子は一般に実質的に純粋である。「実質的に純粋」とは、組成物が少なくとも約50%の目的分子を含有することを意味する。ある態様では、組成物は少なくとも約60%、少なくとも約70%、少なくとも約80%、少なくとも約90%、少なくとも約95%、少なくとも約99%、またはそれ以上の分子を含有することができる。換言すれば、組成物は約50%未満の他の分子を含有してもよい。他の態様では、組成物は約40%未満、約30%未満、約20%未満、約10%未満、約5%未満、約1%未満、またはそれ未満の他の分子を含有してもよい。
【図面の簡単な説明】
【0020】
【図1】図1は、ヒトDCによるCD86発現の亢進的調節に対する給餌および未給餌のオスおよびメスRhipicephalus appendiculatusマダニ由来SGEの影響を示す。F=メス、M=オス、0/3/6=給餌日数。
【図2】図2は、CD1a+細胞による成熟マーカーの発現に対する3日間給餌メスRhipicephalus appendiculatusマダニ由来SGEの影響を示す。黒色のヒストグラムはSGEおよびLPSで処理したCD1a+細胞による成熟マーカーの発現を示し、灰色のヒストグラムはLPS(SGE未含有)で処理したCD1a+細胞による成熟マーカーの発現を示す。
【図3】図3は、LPSで処理したCD1a+細胞による成熟マーカーの発現およびLPS処理を行っていないCD1a+細胞による成熟マーカーの発現に対するQカラム分離したSGEの影響を示す。QFT=pH7でQカラムを通過した物質。QFR=pH7でQカラムに保持された物質。
【図4】図4は、A)LPS+IFNγおよびB)ポリ(I:C)+TNFαの存在下でのCD86亢進的調節に対するSGEまたはQFTの影響を示す。
【図5】図5は、QFTのDC調節作用に対するプロテイナーゼK処理の影響を示す。
【図6】図6は、DC調節活性に関するサイズ分画の結果を示す。活性は、LPS存在下でのCD86亢進的調節の阻害として評価した。
【図7】図7は、サイズ排除クロマトグラフィーを使用して得た最も活性の高いフラグメントでのポリアクリルアミドゲル電気泳動の結果を示す。
【図8】図8は、HPLC精製の結果を示す:A)HPLCカラムからの溶出プロフィール。B)HPLCから溶出した画分の活性プロフィール。
【図9】図9は、エドマン分解によってシークエンシングしたN-末端16アミノ酸を示す。
【図10】図10は、ジャパニンDNAの3'領域をクローニングするためのPCRの結果を示す。
【図11】図11は、ジャパニンDNAの3'領域をクローニングするためのPCRの結果を示す。
【図12】図12は、ジャパニンの3'領域のコンセンサス配列を示す。
【図13a】図13aはジャパニンの5'領域増幅のためのプライマー設計を示す。
【図13b】図13bはジャパニンの5'領域のPCR増幅結果を示す。「PCR」のレーンは未処理のPCR産物である。「精製PCR」と表示した2つの500bpのレーンは、カラム精製後の容量が異なる。
【図14】図14は、ジャパニンの5' cDNA配列を示す。
【図15】図15は、完全長ジャパニンcDNA、およびそれにコードされる176アミノ酸タンパク質を示す。
【図16】図16は、完全長ジャパニン配列中に位置する、可能性のある開裂部位を示す。
【図17】図17は、ジャパニン発現ベクターを含有する細胞から得た昆虫細胞上清のDC調節活性を示す。
【図18】図18は、ジャパニン含有昆虫細胞から回収した上清の硫酸アンモニウム沈殿後の、種々の画分中の活性の存在を示す。
【図19】図19は、ジャパニンの存在を示す、銀染色したSDS-PAGEゲルを示す。
【図20】図20は、ジャパニンを単離するために使用するHisタグの存在を確認するウェスタンブロットを示す。
【図21】図21は、ジャパニン-TEV-his上清のDC調節効果を示す。
【図22】図22は、精製したHisタグ・ジャパニンのDC調節活性を示す。
【図23】図23は、DCによるCD1aおよびCD14の発現に対するジャパニンの影響を示す。
【図24】図24は、MLRアッセイにおける、ジャパニンによって誘導されるT細胞増殖の低下を示す。
【図25】図25は、ジャパニンおよびDermacentor andersoni(D. andersoni E1244)から同定されたジャパニン・ホモログとの2配列間アラインメントを示す。
【図26】図26は、ジャパニンおよびRhipicephalus microplus(R. microplus RM-CK185494)から同定されたジャパニン・ホモログ間の2配列間アラインメントを示す。
【図27】図27は、ジャパニンおよびAmblyomma americanum(A. americanum CX766068)から同定されたジャパニン・ホモログ間の2配列間アラインメントを示す。
【図28】図28は、ジャパニンおよびRhipicephalus appendiculatus(R. appendiculatus CD796501)から同定されたジャパニン・ホモログ間の2配列間アラインメントを示す。
【図29】図29は、ジャパニンおよびRhipicephalus microplus(R. microplus CV443471)から同定されたジャパニン・ホモログ間の2配列間アラインメントを示す。
【図30a】図30aは、GC-MS分析で得た質量分析データを示す。
【図30b】図30bは、GC-MS分析で得た質量分析データを示す。
【図30c】図30cは、GC-MS分析で得た質量分析データを示す。
【図31】図31は、3H-コレステロールのジャパニンへの結合を示す。
【図32a】図32aは、蛍光標識したジャパニンの5日目単球由来樹状細胞への結合のFACS分析を示す。
【図32b】図32bは、蛍光標識したジャパニンの単球への結合のFACS分析を示す。
【図32c】図32cは、蛍光標識したジャパニンの骨髄由来樹状細胞への結合のFACS分析を示す。
【図32d】図32dは、蛍光標識したジャパニンの1日目単球由来樹状細胞への結合のFACS分析を示す。
【図32e】図32eは、蛍光標識したジャパニンのマンナン存在下での単球由来樹状細胞への結合のFACS分析を示す。
【図32f】図32fは、蛍光標識したジャパニンのEDTA存在下での単球由来樹状細胞への結合のFACS分析を示す。
【図33】図33は、ジャパニンのN-グリコシル化を示す。
【図34a】図34aは、LPSの存在下での単球由来樹状細胞成熟化の阻害に対するジャパニンの影響を示す。
【図34b】図34bは、IFNγの存在下での単球由来樹状細胞成熟化の阻害に対するジャパニンの影響を示す。
【図34c】図34cは、TNFαの存在下での単球由来樹状細胞成熟化の阻害に対するジャパニンの影響を示す。
【図34d】図34dは、可溶性CD40Lの存在下での単球由来樹状細胞成熟化の阻害に対するジャパニンの影響を示す。
【図34e】図34eは、IFNαの存在下での単球由来樹状細胞成熟化の阻害に対するジャパニンの影響を示す。
【図34f】図34fは、CD097(TLR7/8リガンド)の存在下での単球由来樹状細胞成熟化の阻害に対するジャパニンの影響を示す。
【図35】図35は、TNFαの樹状細胞分泌に対するジャパニンの影響を示す。
【発明を実施するための形態】
【0021】
本発明の分子の活性
本発明の分子は、哺乳動物DCの分化および成熟化の両方を「調節」すなわち改変、または「阻害」すなわち低減する。ある態様では、それらはヒトDCであることができる。DCの分化および成熟化の調節または阻害を評価する好適なアッセイを以下に記載する。当業者に明らかなように、本明細書に記載する分化および成熟化の調節または阻害の評価のためのマーカーは単に例として提供するものであり、制限を意図するものではない。ある態様では、本発明の分子はDCの分化および成熟化の両方を、例えば以下に記載するアッセイによる測定で少なくとも20%低減させる。更なる態様では、DCの分化および成熟化の両方の阻害は30%、40%、50%、60%、70%、80%、90%、またはそれ以上であってもよい。
【0022】
「DCの分化」とは、細胞前駆体、例えば骨髄由来前駆細胞または血中単球の未成熟DCへの発達を意味する。DC分化の調節、例えば阻害は、当該分野で公知の標準的なアッセイを用いて表現型的に、最終的には機能的に評価できる。
【0023】
前駆体の未成熟DCへの表現型分化の阻害は、前駆細胞が未成熟DCに分化する際に発現が変更される細胞マーカーを用いて評価することができる。例えば、図23および付記する説明に記載するように、単球はCD14陽性およびCD1陰性であるDC前駆体である。未成熟DCはCD14陰性およびCD1陽性である。従って、未成熟DCへの単球の分化はCD14の減少およびCD1の増加によって検出することができる。本発明の分子による前駆細胞の未成熟DCへの分化の阻害は、CD14陽性およびCD1陰性である前駆細胞の持続的な存在によって検出してもよい。
【0024】
前駆細胞および発達したDCの機能的相違は、前駆細胞と発達したDCをそれらの活性に基づいて識別するアッセイを用いて評価できる。例えば、前駆細胞とは異なり、発達したDCは、特に後述のような刺激の後、インビトロアッセイにおいてT細胞増殖を惹起する能力を有する。一般的なT細胞増殖アッセイには同種異系混合リンパ球反応(MLR)および酸化的有糸分裂誘発がある。
【0025】
「DCの成熟化」とは、前駆細胞が分化して未成熟DCになった後に起こる過程を意味する。特にこの用語は、分化した未成熟DCが刺激物質に遭遇し、成熟DCに変換する際に起こる変化に関連する。刺激物質はPRR(例えばTLR)を介して感知される感染性物質(例えば病原体)の成分、サイトカイン受容体を通じて作用するある種のサイトカイン、および/または他の細胞型の特殊化された細胞表面分子(例えば活性化T細胞のCD154)であってもよい。未成熟DCの成熟化に伴う変化には、一般に、共刺激分子(例えばCD80およびCD86)発現の亢進的調節、およびDC表面上での病原体由来成分からの(一般的にはペプチド-MHCおよび脂質-CD1複合体としての)抗原提示がある。未成熟DCの成熟化は二次リンパ組織へのDCの遊走を伴う。
【0026】
本発明の分子は、未成熟DCの成熟化に伴うこれらの変化のいずれを調節または阻害するように作用することができる。従って、本発明の分子がDCの成熟化を阻害する能力は、CD86および/またはCD80および/またはMHC分子の発現を低下させる能力によって評価することができる。本発明の分子がDCの成熟化を阻害する能力は、TNFαの発現および/または分泌を低下させる能力によって評価してもよい。本発明の分子が未成熟DCの成熟化を阻害する能力は、必要により、ポリ(I:C)、LPS、またはIFNγ刺激後に評価してもよい。本発明の分子が未成熟DCの成熟化を阻害する能力は、必要により、CD40L、IFNα、またはTLR7もしくはTLR8リガンド(例えばCL097)刺激後に評価してもよい。未成熟DCの成熟化の阻害を評価するための実験法は実施例に記載する。未成熟DCの成熟化の阻害を評価するための更なる方法は当業者に公知である。
【0027】
上記のように、本発明の分子は前駆細胞の未成熟DCへの分化を調節(好ましくは阻害)し、その後の未成熟DCの成熟DCへの成熟化を調節(好ましくは阻害)するように作用する。このようなDCの分化および成熟化の両方の調節または阻害は、全体として免疫系に対して阻害的調節作用を有すると考えられるが、詳細については後述する。
【0028】
抗原提示細胞によって活性化されるまで、Tリンパ球は「ナイーブ」であると称される。各Tリンパ球は特定の抗原に特異的であり、この同族の抗原を提示している「特殊化された」抗原提示細胞(例えばDC)によってしか活性化されない。通常型の(conventional)Tリンパ球は、α鎖およびβ鎖を含むヘテロ二量体構造を持つT細胞受容体(TCR)を介して抗原-MHC複合体を認識する。しかしながら、共刺激の非存在下でのTCRによるシグナル伝達は、抗原無反応もしくはアネルギー状態、または活性化不全および細胞死を引き起こす。従って、成熟化過程で未成熟DCの表面上で亢進的に調節され、Tリンパ球表面上で受容体分子(例えばCD28(CD80およびCD86の場合)またはCD154(CD40の場合))によって認識される共刺激分子は、Tリンパ球の活性化に不可欠である。
【0029】
本発明のある観点では、上記の分子によって提供されるDCの分化および成熟化の阻害により、Tリンパ球活性化が低下する。「Tリンパ球活性化」とは、ヘルパーT細胞(Th1、Th2、Th17、またはTreg細胞など)の活性化、場合によっては、細胞傷害性T細胞の活性化(多くの場合、先行のヘルパーT細胞の活性化に依存する)を意味する。
【0030】
T細胞活性化の低下は当該分野で公知の方法によって評価することができる。制限ではなく一例として、T細胞活性化は以下によって評価することができる:サイトカイン分泌(例えばインターロイキン(IL)-2生成)もしくはDCによって惹起されるT細胞増殖(例えば同種異系MLRまたは酸化的有糸分裂誘発)のインビトロアッセイ、または抗原再刺激前および後にex vivoで単離されたT細胞と同様のアッセイを用いる正常またはトランスジェニック動物におけるモデル抗原(例えばオボアルブミン)に対するT細胞応答のin vivoアッセイ。
【0031】
ある態様では、本発明の分子は、DC分化および成熟化阻害分子の非存在下で、Tリンパ球活性化を標準のアッセイに比較して少なくとも約20%低下させる。更なる態様では、DC分化および成熟化の阻害によって、Tリンパ球活性化が少なくとも約30%、40%、50%、60%、70%、80%、90%、またはそれ以上低減される。
【0032】
本発明のある観点では、上記の分子によって提供されるDC分化および成熟化の調節または阻害により、Tリンパ球制御が調節される。特に、本発明の分子はTリンパ球のTh1、Th2、Th17、調節性T(Treg)、または濾胞性ヘルパーT(Thf)細胞への分極化を調節することができる。本発明の分子によるTリンパ球分極化の調節は、インビトロアッセイで、一般的に種々のタイプのCD4およびCD8 Tリンパ球に関係するTリンパ球誘導型サイトカインの測定によって評価することができる。これらには、Th1細胞ではIFN-γ;Th2細胞ではIL-4、IL-5、およびIL-13;Th17細胞ではIL-17;そしてTreg細胞ではIL-10およびTGFβがある。各タイプのCD4細胞は、それぞれT-bet、GATA-3、ROR-γ-t、およびFoxP3の発現の測定、またはThf細胞ではBcl6発現の測定によってアッセイすることもできる。あるいはまた、種々の細胞の表現型を、例えばケモカイン受容体およびそれらが発現する他の表現型マーカーの測定によって評価することもできる。
【0033】
Tリンパ球は哺乳動物の免疫応答の主な促進物質の一つである。従って、上記のような本発明の分子によるTリンパ球活性化の低下またはそれらTリンパ球の分極化の変更により、免疫応答の全体的な調節、特に免疫応答に関係するサイトカインのレベルの変化が起こる。例えば、本発明の分子は一般に免疫抑制効果を有しうる。
【0034】
当業者に明白なように、免疫応答の調節、例えば免疫抑制効果は、種々の方法のいずれかを用いて測定できる。全体的な免疫応答の低下または調節は、TLR刺激に応答して最も迅速に生成される炎症促進性サイトカイン、例えばインターロイキン-1および腫瘍壊死因子α(TNFα)、インターフェロンα、インターフェロンβ、または一般に感染直後に生成されるIL-6もしくはIL-12のようなサイトカインのレベル低下を観察することによって測定できる。本発明の分子は抗炎症性サイトカイン、例えばIL-10またはTGF-βのレベル増加も誘発しうる。
【0035】
ある態様では、本発明の分子は、DCの分化および成熟化を阻害する分子の非存在下での標準アッセイに比較して約20%まで、炎症促進性サイトカインのレベルを低下させるか、または抗炎症サイトカインのレベルを増加させる。更なる態様では、DC分化および成熟化の阻害により、少なくとも約30%、40%、50%、60%、70%、80%、90%、またはそれ以上、炎症促進性サイトカインのレベルが低下するか、または抗炎症サイトカインのレベルが増加する。
【0036】
免疫抑制効果は種々の他の方法、例えば炎症の局所的低下、または生成させた抗原特異的TおよびB細胞プールのサイズもしくは活性の低下によって評価してもよい。
【0037】
本発明の分子を単離しうる節足動物
本発明の分子は節足動物から単離することができる。「節足動物」は節足動物門に属する動物と定義され、それらには昆虫、甲殻類、およびクモ形類動物がある。節足動物は、分節した体とキチンを成分とする硬い外骨格を特徴とする。
【0038】
本発明のある観点では、本発明の分子は吸血性節足動物から単離することができる。「吸血性節足動物」という用語は、好適なホストから血液を摂取する全ての節足動物を含む。これには昆虫、マダニ(ticks)、シラミ、ノミ、およびダニ(mites)が含まれる。これらは一般に吸血性(blood feeding)節足動物として知られ、これらの2つの用語は本明細書において同義に使用される。
【0039】
本発明の更なる観点では、単離された吸血性節足動物はマダニであってもよい。「マダニ」という用語はマダニ上科(Ixodoidea)の小型クモ型類の通称であり、吸血性節足動物に包含される。マダニは外部寄生虫であり、哺乳動物、鳥類、および爬虫類の血液に依存して生息する。
【0040】
世界中に約900種のマダニが生息している。種々のマダニ種はその主な生息地および地理的分布によってキャラクタライズされる。ほとんどのマダニ種は種々のホスト種(ヒトを含む)に依存して生息できる。上記のように、節足動物、特にマダニは、ホストに種々の抗炎症および免疫調節成分を接種することによって、ホストの免疫系を阻害しうる。
【0041】
本発明の単離されたDC調節分子は任意の既知のマダニ種、例えばIxodinae、Bothriocrotoninae、Amblyomminae、Haemaphysalinae、Rhipicephalinae、Hyalomminae、Nuttalliellidae、Argasinae、Otobinae、Antricolinae、Nothoaspinae 、およびOrnithodorinae群に含まれる種、例えば以下のマダニ種のいずれかから単離することができる:Rhipicephalus appendiculatus、Rhipicephalus sanguineus、Rhipicephalus bursa、Amblyomma americanum、Amblyomma cajennense、Amblyommahebraeum、Amblyomma variegatum、Rhipicephalus(Boophilus)microplus、Rhipicephalus(Boophilus)annulatus、Rhipicephalus(Boophilus)decoloratus、Dermacentor reticulatus、Dermacentor andersoni、Dermacentor marginatus、Dermacentor variabilis、Haemaphysalis inermis、Haemaphysalis leachii、 Haemaphysalis punctata、Hyalomma anatolicum anatolicum、Hyalomma dromedarii、Hyalomma marginatum marginatum、Ixodes ricinus、Ixodes persulcatus、Ixodes scapularis、Ixodes hexagonus、Argas persicus、Argas reflexus、Ornithodoros erraticus、Ornithodoros moubata moubata、Ornithodoros moubata porcinus、およびOrnithodoros savignyi。
【0042】
本発明のタンパク質
本発明の分子はタンパク質であってもよい。実施例3から18に詳述するように、出願人はマダニ種Rhipicephalus appendiculatusから哺乳動物DCの分化および成熟化を阻害する節足動物タンパク質を同定および単離した。本明細書においてはこのタンパク質をジャパニン(Japanin)と称するが、そのアミノ酸配列を図15およびSEQ ID NO:2に示す。
【0043】
従って、本発明のある観点では、本発明の分子は以下を含むことができる:
i)SEQ ID NO:2のアミノ酸配列を含むタンパク質;
ii)i)のタンパク質のホモログ;
iii)上記i)のタンパク質もしくは上記ii)のホモログの活性フラグメント;または
iv)i)、ii)、もしくはiii)の機能的同等物。
【0044】
本発明の分子はSEQ ID NO:2のアミノ酸配列から成るタンパク質またはその活性フラグメントであってもよい。
【0045】
「機能的同等物」という用語は、本明細書では、上記のアッセイを用いて、SEQ ID NO:2のアミノ酸配列を含む完全長ジャパニン・タンパク質に匹敵する方法でDCの分化および成熟化を調節(好ましくは阻害)する能力を有する分子を表す。これには合成タンパク質、タンパク質の合成変異体、匹敵する活性を有する異なる配列のタンパク質分子、匹敵する活性を有する天然に存在する非タンパク質分子、および匹敵する活性を有する合成非タンパク質分子がある。
【0046】
特に、ジャパニン分子の三次構造または活性部位(単数または複数)を模倣するように設計した合成分子も機能的同等物であるとみなされる。ある態様では、機能的同等物は、上記のようにDCの分化および成熟化を調節(好ましくは阻害)する能力を有し、それによってTリンパ球活性化の低下またはTリンパ球分極化の調節が起こる。更なる態様では、機能的同等物は、上記のようにDCの分化および成熟化を調節(好ましくは阻害)する能力を有し、それによって免疫応答の抑制が起こる。
【0047】
グリコシル化
実施例26に示すように、ジャパニンは、おそらく2つの部位で、グリコシル化されると考えられる。アスパラギン結合型グリコシル化のためのコンセンサス配列の一部であるアスパラギン残基はジャパニン(SEQ ID NO:2)アミノ酸配列の59および155位に位置する。これらは、リーダーペプチド配列が無い成熟タンパク質の35および131位に相当する。
【0048】
従って、本発明のタンパク質は1つまたはそれ以上の位置でグリコシル化されていてもよい。ある態様では、タンパク質は1、2、3、またはそれ以上の位置でグリコシル化されていてもよい。本発明のタンパク質はN-グリコシル化されているが、タンパク質は更に1つまたはそれ以上の位置でO-グリコシル化されていてもよい。
【0049】
本発明のタンパク質は天然でグリコシル化されていてもよい。これは、タンパク質が天然物を単離したものである場合、またはタンパク質を組換え技術によって、タンパク質が天然で生成される生体のグリコシル化パターンと酷似したホスト細胞内で生成させる場合に特に当てはまる。あるいはまた、本発明のタンパク質を化学的にグリコシル化する必要もありうる。これは、タンパク質を化学的に合成する場合、またはタンパク質を組換え技術によって、天然とは異なるタンパク質グリコシル化パターンを有する生体で生成させる場合に特に当てはまる。また、更なるグリコシル化がなされてタンパク質の特性が改変または改善されてもよい。
【0050】
リポカインの構造および脂質結合性
実施例11に示すように、出願人はジャパニンがリポカリンであることを発見した。リポカリンは類似した折りたたみ構造を有するタンパク質のファミリーである。特徴的なリポカリンの折りたたみは8つのストランドから成る逆平行βバレルであり、内部にリガンド結合部位を形成する。リポカリンは一般に少なくとも2つ、多くの場合は4つのシステイン残基を間隔をあけて有する。システイン残基は内部にジスルフィド結合部位を形成し、これによってリポカリンの折りたたみが安定化されている可能性がある。本発明のある観点では、分子はリポカリンであってもよい。従って、本発明の範囲内に含まれるホモログ、フラグメント、および機能的同等物は、通常マダニ・リポカリン中に存在するCys/Try X Leu Trpモチーフを含有してもよい。
【0051】
リポカリンの構造が推測通りだとすると、ジャパニン・タンパク質は脂質または脂質様分子と結合(associated)しうる。「脂質」という用語は、有機溶媒に可溶であるが水に不溶である疎水性または両親媒性分子を含むことを意図する。これには脂質、油、トリアシルグリセロール、糖脂質、リン脂質およびステロイド、脂肪酸アシル、脂肪酸、グリセロ脂質、グリセロリン脂質、スフィンゴ脂質、サッカロ脂質(saccharolipid)、ポリケチド、ステロール脂質、およびプレノール脂質がある。「脂質様分子」という用語は、脂質と類似または同一の特性を有するいずれの分子も包含する。また「脂質様分子」という用語は非脂質分子と複合体を形成した脂質も包含する。これには糖脂質、リン脂質、ホスホ糖脂質、標識された脂質、およびアセチル化された脂質がある。
【0052】
in vivoにおいて、本発明のタンパク質は小胞体内での折りたたみ中もしくはその直後に、またはその位置から排出された後の段階で、脂質分子に結合する可能性がある。
【0053】
本発明の範囲内で、本発明のタンパク質はその生成の際に脂質と結合してもよい。例えば本発明のタンパク質が天然の供与源から単離されるか、または組換えによって生成される場合、タンパク質はいずれの介入も必要とせずに、自動的に脂質と結合してもよい。あるいはまた、脂質を本発明のタンパク質を含有する組成物に添加し、脂質およびタンパク質間で複合体を形成させてもよい。特にタンパク質を化学的に合成する場合、タンパク質を含有する組成物に脂質を外因的に添加し、複合体を形成させてもよい。上記のように、本発明のタンパク質は脂質と「結合」または「複合体を形成」してもよい。本明細書では、これらの用語は、本発明の脂質およびタンパク質間のあらゆる種類の接触に関連して同義に用いられる。特に、本発明のタンパク質および脂質間は相互作用してもよい。ある態様では、相互作用は単純に構造的なものであってもよい。すなわち、脂質がタンパク質内の結合ポケットに隙間無くはめ込まれていてもよい。別の態様では、脂質は任意の引力によってタンパク質と物理的に相互作用してもよい。それらの引力には静電相互作用、疎水性相互作用、親水性相互作用、ファンデルワールス力、水素結合、および共有結合性相互作用がある。脂質およびタンパク質間の相互作用は、構造的相互作用および引力が組み合わさったものであってもよい。
【0054】
ある態様では、本発明のタンパク質は脂質と結合してもよい。別の態様では、脂質はステロイドまたはステロール、例えばコレステロールであってもよい。別の態様では、脂質はコレステロールの代謝物、例えばビタミンD3であってもよい。実施例23に示すように、ジャパニンはコレステロールに結合することが明らかになっている。ある態様では、本発明のタンパク質はコレステロールの代謝物に結合してもよい。特に、本発明はジャパニンおよび脂質(例えばコレステロールまたはコレステロールの代謝物)を含有する、またはそれらから成る複合体を提供する。
【0055】
ジャパニン・タンパク質の機能的同等物、ホモログ、およびフラグメントは、脂質または他の疎水性分子(単数または複数)(例えば脂質様分子)と結合してもよい。特に、ジャパニン・タンパク質の機能的同等物、ホモログ、およびフラグメントはコレステロールまたはコレステロールの代謝物と結合してもよい。
【0056】
ジャパニンはコレステロールに結合することが明らかになっているので、出願人は、脂質を保持するがジャパニンの生体活性を有さないように遺伝子操作した分子は有用な特性を有しうると考える。従って、本発明は脂質に結合し、DC表面上の受容体を標的とするキャリア分子を含む。それらのキャリア分子は、それ自身は生体活性を有さない。ある態様では、ジャパニンをその生体活性を阻害するように遺伝子操作することによって、キャリア分子を生成してもよい。別の態様では、細胞受容体への結合によって、キャリア分子が保持する脂質に生体機能が施与されてもよい。従って「機能的同等物」という用語はキャリア分子を含む。
【0057】
受容体結合
実施例24に示すように、ジャパニンはC型レクチン細胞表面受容体に結合すると考えられる。これは、ジャパニンが、標的細胞の表面上にある受容体に結合し、阻害の原因となる内部細胞シグナル伝達経路を惹起させることによって、樹状細胞の分化および成熟化を調節(好ましくは阻害)するように機能することを示唆している。
【0058】
ある態様では、タンパク質は標的細胞(例えばDC)の外表面上の受容体に結合することができる。別の態様では、タンパク質は二価陽イオン依存性受容体、特にC型レクチン受容体に結合することができる。ある態様では、タンパク質は受容体に結合し、受容体の天然のリガンドを模倣してもよい。当業者に明白なように、タンパク質のいずれの部分が受容体に結合してもよい。特に、タンパク質がグリコシル化される、そして/または脂質分子に結合する場合、それは炭化水素部分または関連脂質であって、それが標的細胞上の受容体に結合してもよい。
【0059】
ある態様では、本発明のタンパク質および受容体を含有する、またはそれらから成る複合体は本発明に包含される。別の態様では、複合体は本発明のタンパク質、受容体(例えば二価陽イオン依存性受容体(C型レクチン受容体など))、および脂質(例えばコレステロールまたはコレステロールの代謝物)を含有するか、またはそれらから成ってもよい。
【0060】
ホモログおよびフラグメント
上記のように、本発明はアミノ酸配列SEQ ID NO:2で示すジャパニン・タンパク質のホモログおよび活性フラグメントを含む。本発明はこれらのホモログおよびフラグメントの機能的同等物も含む。
【0061】
「ホモログ」という用語は、SEQ ID NO:2に示すジャパニン配列のパラログおよびオルソログでDCの分化および成熟化を調節(好ましくは阻害)する能力を保有するものへの言及を含むことを意図する。ホモログは、上記のようにDCの分化および成熟化を調節(好ましくは阻害)し、それによってTリンパ球活性化の低下またはTリンパ球分極化の調節を起こす能力を有することができる。更なる態様では、ホモログは、上記のようにDCの分化および成熟化を阻害し、それによって免疫応答の阻害を起こす能力を有してもよい。別の態様では、ホモログは、脂質(例えばコレステロールまたはコレステロールの代謝物)に結合する能力および/または膜結合型受容体(例えば二価陽イオン依存性受容体(C型レクチン受容体など))に結合する能力を有してもよい。
【0062】
ホモログは以下のような、Rhipicephalus appendiculatus以外のマダニ種由来であってもよい:Rhipicephalus sanguineus、Rhipicephalus bursa、Amblyommaamericanum、Amblyomma cajennense、Amblyomma hebraeum、Ambylomma variegatum、Rhicephalus(Boophilus)microplus、Rhicephalus(Boophilus)annulatus、Rhicephalus(Boophilus)decoloratus、Dermacentor reticulatus、Dermacentor andersoni、Dermacentor marginatus、Dermacentor variabilis、Haemaphysalis inermis、Haemaphysalis leachii、 Haemaphysalis punctata、Hyalomma anatolicum anatolicum、Hyalomma dromedarii、Hyalomma marginatum marginatum、Ixodes ricinus、Ixodes persulcatus、Ixodes scapularis、Ixodes hexagonus、Argas persicus、Argas reflexus、Ornithodoros erraticus、Ornithodoros moubata moubata、Ornithodoros moubata porcinus、およびOrnithodoros savignyi。ホモログは蚊種(例えばCulex、Anopheles、およびAedes genera、特にCulex quinquefasciatus、Aedes aegypti、およびAnopheles gambiae);ノミ種(例えばCtenocephalides felis(ネコノミ));ウマバエ;スナバエ;ブユ;ツェツェバエ;シラミ;ダニ由来のものであってもよい。
【0063】
一般に、ホモログは任意の既知のマダニ種、例えばIxodinae、Bothriocrotoninae、Amblyomminae、Haemaphysalinae、Rhipicephalinae(Hyalomminaeなど)、Nuttalliellidae、Argasinae、Otobinae、Antricolinae、およびOrnithodorinaeの群に含まれるものに由来してもよい。
【0064】
本明細書に記載する単離されたジャパニン・タンパク質配列のホモログの同定法は、当業者に公知である。例えばホモログは、公的および私的な配列データベースのホモロジー検索によって同定することができる。公的なデータベースを使用してもよいが、特に公的データベースに掲載されていないデータが含まれる場合は、私的または市販のデータベースも同様に有用である。主要なデータベースは主要なヌクレオチドまたはアミノ酸配列データ預託のサイトであり、公的または市販のものがある。主な公的データベースの例として以下がある:GenBankデータベース(http://www.ncbi.nlm.nih.gov)、EMBLデータベース(http://www.ebi.ac.uk/)、DDBJデータベース(http://www.ddbj.nig.ac.jp/)、SWISS-PROTタンパク質データベース(http://expasy.hcuge.ch/)、PIR(http://pir.georgetown.edu/)、TrEMBL(http://www.ebi.ac. uk/)、TIGRデータベース(例えばhttp://www.tigr.org/tdb/index.html)、NRL-3Dデータベース(http://www.nbrfa.georgetown.edu、Protein Data Base(http://www.rcsb.org/pdb)、NRDBデータベース(ftp://ncbi.nlm.nih.gov/pub/nrdb/README)、OWLデータベース(http://www.biochem.ucl.ac.uk/bsm/dbbrowser/OWL/)、および二次的なデータベース、PROSITE(http://expasy.hcuge.ch/sprot/prosite.html)、PRINTS(http://iupab.leeds.ac.uk/bmb5dp/prints.html)、Profiles(http://ulrec3.unil.ch/software/PFSCAN_form.html)、Pfam (http://www.sanger.ac.uk/software/pfam)、Identify(http://dna.stanford. edu/identify/)、およびBlocks(http://www.blocks.fhcrc.org)データベース。市販のデータベースまたは私的データベースの例としてPathoGenome(Genome Therapeutics社)およびPathoSeq(Incyte Pharmaceuticals社)。
【0065】
一般的に、2つのポリペプチド間の同一性が(好ましくは特定の領域にわたって)30%より高ければ、それらは機能的同等物であることを示しており、従って2つのタンパク質はホモログであることを示していると見なされる。ある態様では、ホモログであるタンパク質はSEQ ID NO:2のジャパニン・タンパク質配列と60%を超える配列同一性を有する。他の態様では、ホモログは単離された節足動物タンパク質配列、SEQ ID NO:2と70%、80%、90%、95%、98%、または99%を超える同一性を有する。本明細書の記述では、同一性%の測定はNCBI(the National Center for Biotechnology Information; http://www.ncbi.nlm.nih.gov/) のデフォルト・パラメータ([Blosum 62 matrix; gap open penalty=11 and gap extension penalty=1])を用い、BLAST(バーション2.1.3)を使用して行う。
【0066】
ジャパニン・タンパク質配列(SEQ ID NO:2)のホモログには、野生型タンパク質配列が示すDC分化および成熟化の調節(好ましくは阻害)を保持している限り、野生型配列に比較してアミノ酸置換、挿入、または欠失を含む変異体も含まれる。変異体は、上記のようにDCの分化および成熟化を調節(好ましくは阻害)し、それによってTリンパ球活性化の低下またはTリンパ球分極化の調節を起こす能力を有することができる。更なる態様では、変異体は、上記のようにDCの分化および成熟化を調節(好ましくは阻害)し、それによって免疫応答を阻害する能力を有してもよい。
【0067】
従って変異体には、タンパク質の機能または活性に有害な影響を及ぼさない、保存的アミノ酸置換を含有するタンパク質も含まれる。この用語には天然の生物学的変異体(例えば、本発明の単離された節足動物タンパク質誘導の供与源である種内に含まれる対立遺伝子変異体または地理的変異体も包含されることが意図される。野生型タンパク質配列に比較してDCの分化および成熟化の調節または阻害活性が向上された変異体は、タンパク質配列中の特定残基の系統的または部位特異的変異によって設計してもよい。
【0068】
実施例20に記載するように、出願人はDermacentor andersoni中に含まれるジャパニン・ホモログを同定した。実施例21に記載するように、出願人は、Rhipicephalus(Boophilus)microplus(2つのホモログ)、Amblyomma americanum、およびRhipicephalus appendiculatusからそれぞれジャパニンのホモログを同定した。配列を図25-29に示すが、これはSEQ ID NO:4、6、8、10、および12に相当する。
【0069】
従って、本発明の更なる観点では、本発明の単離された分子は以下を含むことができる:
i)SEQ ID NO:4、6、8、10、もしくは12のいずれか1つのアミノ酸配列を含有するタンパク質
ii)i)のタンパク質のホモログ;
iii)上記i)のタンパク質もしくは上記ii)のホモログの活性フラグメント;または、
iv)i)、ii)、もしくはiii)の機能的同等物。
【0070】
理論に拘束されることは望まないが、これらの配列は完全長配列でなくてもよいと想定される。従って、本発明では、更なるアミノ酸がSEQ ID NO:4、6、8、10、または12のアミノ酸配列を含有する分子のN-末端および/またはC-末端に存在してもよい。
【0071】
本発明は単離されたジャパニン分子の「活性フラグメント」および単離されたジャパニン分子のホモログも提供する。完全長ジャパニン分子の哺乳動物DC分化および成熟を調節または阻害する能力を有する任意のフラグメントがこの定義に包含される。
【0072】
それらのフラグメントとしては、本明細書でSEQ ID NO:2、4、6、8、10、および12として定義される単離された節足動物タンパク質のフラグメントだけでなく、上記のように、このタンパク質のホモログのフラグメントも包含される。それらのホモログのフラグメントは一般にSEQ ID NO:2、4、6、8、10、および12の単離された節足動物タンパク質のフラグメントと60%を超える同一性を有するが、より好ましいホモログのフラグメントはSEQ ID NO:2、4、6、8、10、および12の単離された節足動物タンパク質のフラグメントとそれぞれ70%、80%、90%、95%、98%、または99%を超える同一性を示す。
【0073】
これらのSEQ ID NO:2、4、6、8、10、および12の単離された節足動物タンパク質の活性フラグメントおよびそのホモログのフラグメントはDCの分化および成熟化を調節(好ましくは阻害)する。ある態様では、SEQ ID NO:2、4、6、8、10、および12の単離された節足動物タンパク質のフラグメントおよびそのホモログのフラグメントは、上記のようにDCの分化および成熟化を調節(好ましくは阻害)し、それによってTリンパ球活性化の低下またはTリンパ球分極化の調節を起こす能力を有する。更なる態様では、SEQ ID NO:2、4、6、8、10、および12の単離された節足動物タンパク質のフラグメントおよびそのホモログのフラグメントは、上記のようにDCの分化および成熟化を調節(好ましくは阻害)し、それによって免疫応答を阻害する能力を有する。もちろん、DCの分化および成熟化を調節または阻害する活性が向上されたフラグメントを野生型配列の系統的変異または断片化によって合理的に設計し、その後公的な活性アッセイを行ってもよい。
【0074】
ある態様では、ホモログは脂質(例えばコレステロールまたはコレステロールの代謝物)との結合能、および/または膜受容体(例えば二価陽イオン依存性受容体(C型レクチン受容体など))との結合能を有してもよい。
【0075】
ある態様では、上記の単離された節足動物タンパク質のフラグメントは少なくとも約100アミノ酸長であってもよい。更なる態様では、上記の単離された節足動物タンパク質のフラグメントは少なくとも約90、少なくとも約80、少なくとも約70、少なくとも約60、少なくとも約50、少なくとも約40、少なくとも約30、少なくとも約20、少なくとも約10、または少なくとも約5アミノ酸長であってもよい。
【0076】
抗体
本発明はまた、本発明の分子、特に上記のジャパニン・タンパク質、そのホモログ、フラグメント、および機能的同等物に結合する抗体も提供する。抗体は分子の検出のための試薬として使用することができる。これは、下記のように、分子のDCの分化および成熟化を調節または阻害する活性を中和し、そのために治療の目的に有用である抗体であってもよい。本発明のこの観点には、上記のような本発明の範囲に包含される任意の機能的同等物、ホモログ、およびタンパク質フラグメントに結合する抗体も含まれる。
【0077】
本発明には、本発明のタンパク質の炭化水素部分に結合する抗体も含まれる。特に、本発明には天然でジャパニンに結合する炭化水素部分の1つまたはそれ以上に結合する抗体も含まれる。
【0078】
「アンチカリン」も本発明の範囲内に含まれる。これらは特定のタンパク質エピトープを認識および結合するようにリポカリンを遺伝子操作して作製した分子である。ある態様では、アンチカリンはペプチド、グリコペプチド、または糖脂質の形態を取ってもよい。本明細書では、アンチカリンは「抗体」という用語の範囲に包含される。アンチカリンは非免疫グロブリン誘導型分子ではあるが、抗体と同様の方法でタンパク質エピトープを認識する。
【0079】
ポリクローナル抗体が所望される場合、選択された哺乳動物、例えばマウス、ウサギ、ヤギ、またはウマを本発明の分子、例えばジャパニン・タンパク質、フラグメント、ホモログ、または機能的同等物で免疫化することができる。必要により、分子をキャリアタンパク質にコンジュゲートさせてもよい。一般的に使用されるキャリアにはウシ血清アルブミン、サイログロブリン、およびキーホールリンペットヘモシアニンがある。その後、カップリングした分子を用いて動物を免疫化する。免疫化動物の血清を回収し、既知の方法、例えば免疫親和性クロマトグラフィーで処理する。
【0080】
本発明の分子に対するモノクローナル抗体も当業者によって容易に生成される。ハイブリドーマ法を用いてモノクローナル抗体を生成する一般的な方法は公知である(例えばKohler, G.およびMilstein, C., Nature 256: 495-497 (1975);Kozborら, Immunology Today 4: 72 (1983);Coleら,Monoclonal Antibodies and Cancer Therapy,77-96, Alan R. Liss社(1985)参照)。
【0081】
本明細書で使用する「抗体」という用語は、抗体のフラグメント(例えばFab、F(ab')2、およびFvフラグメントでDC調節分子に特異的に結合するものを含む。更に「抗体」という用語は、本発明の分子、特にジャパニン・タンパク質、そのホモログおよびフラグメントに特異的なキメラおよびヒト化抗体分子を含む。キメラ抗体は、非ヒト可変領域をヒト定常領域に結合または融合させた抗体である(例えばLiuら, Proc. Natl. Acad. Sci. USA, 84, 3439 (1987)参照)。本明細書に記載する「ヒト化抗体」という用語は、非ヒト・ドナー抗体の重鎖および/または軽鎖の可変ドメイン中にあるCDRアミノ酸および選択された他のアミノ酸でヒト抗体中の同等のアミノ酸を置換した抗体分子をいう。従って、ヒト化抗体はヒト抗体と酷似しているが、ドナー抗体の結合能を有する(Jonesら, Nature, 321, 522 (1986);Verhoeyenら, Science, 239, 1534 (1988);Kabatら, J. Immunol., 147, 1709 (1991);Queenら, Proc. Natl Acad. Sci. USA, 86, 10029 (1989);Gormanら, Proc. Natl Acad. Sci. USA, 88, 34181 (1991);およびHodgsonら, Bio/Technology, 9, 421 (1991)参照)。
【0082】
ある態様では、抗体に(例えば検出を容易にするための)標識部分を結合させルのが望ましい。標識は酵素、放射能標識、ビオチンなどの化合物、または蛍光色素であってもよい。
【0083】
融合タンパク質
本発明は、1つまたはそれ以上のペプチド、ポリペプチド、または他の分子に遺伝子的に融合するか、または化学的に結合した本発明の分子、特にジャパニン・タンパク質、そのホモログ、フラグメント、または機能的同等物を含有する融合タンパク質を含む。更なるペプチドもしくはポリペプチドまたは分子の目的は、タンパク質の検出、発現、分離、もしくは精製を容易にすることであるか、またはタンパク質に所望の更なる特性を与えることであってもよい。有望な融合パートナーの例として、β-ガラクトシダーゼ、グルタチオン-S-トランスフェラーゼ、ルシフェラーゼ、ポリヒスチジン・タグ、T7ポリメラーゼ・フラグメント、および分泌シグナルペプチドがある。融合パートナーはin vivoにおける分子の寿命を延長するものであってもよい(例えばFcフラグメント)。融合タンパク質の例を実施例15-18に記載する。
【0084】
他の有望な融合パートナーには有望な生物医薬品、例えば特定の疾患を治療するための薬剤として使用するために開発されたタンパク質または他の分子がある。更なる有望な融合パートナーには、免疫系内で本発明の分子を細胞(例えばDC)にターゲッティングさせる抗原がある。例えば、融合パートナーは、in vivoにおいて分子に融合してこれをDCに運搬しうる自己もしくは外来抗原またはアレルゲンであってもよい。更なる融合パートナーは、DCの異なる細胞表面成分に結合してDCへの運搬を容易にする分子であってもよい。ある態様では、複数の融合パートナーが含まれてもよい。それらの抗原の例は以下に、より詳細に記載する。
【0085】
核酸
本発明は本発明の分子をコードする核酸配列を含有する核酸分子も含む。DNA分子、RNA分子、およびDNA-RNA分子混合物も「核酸分子」という用語に包含されることが意図される。更に、ゲノムDNA、cDNA分子、mRNA分子、そして改変された塩基を含有するRNAおよびDNA分子もこの定義に含まれる。当業者に明白なように、遺伝子コードの縮重により、単離されたタンパク質、タンパク質フラグメント、またはその機能的同等物の所定のタンパク質配列をコードする能力のある多くの異なる核酸配列が存在するが、それらは本発明の範囲内に含まれる。本発明には、融合タンパク質(例えば上記の融合タンパク質)をコードする核酸分子も含まれる。
【0086】
本発明のある観点では、本発明の分子をコードする核酸配列を含有する核酸分子はSEQ ID NO:1またはその縮重配列を含有するか、またはそれから成ってもよい。本発明の更なる観点では、核酸分子はSEQ ID NO:3、5、7、9、もしくは11、またはその縮重配列のいずれかを含有してもよい。縮重配列の一例はSEQ ID NO:32の核酸分子であり、これはSEQ ID NO:3の核酸分子と同じ、SEQ ID NO:4のDermacentor andersoniタンパク質をコードする。
【0087】
また本発明は、本発明の分子、特に上記のような単離された節足動物タンパク質、そのホモログ、フラグメント、または機能的同等物をコードする核酸配列を含有する核酸分子に高ストリンジェンシー・ハイブリダイゼーション条件下でハイブリダイズするアンチセンス核酸分子も提供する。高ストリンジェンシー・ハイブリダイゼーション条件として以下がある:50% ホルムアミド、5X SSC(150mM NaCl、15mM クエン酸三ナトリウム、)、50mM リン酸ナトリウム(pH7.6)、5x デンハルト溶液、10% 硫酸デキストラン、および20μg/ml 変性断片化サケ精子DNAを含有する溶液中、42℃で一晩インキュベートした後、フィルターを0.1X SSC中、約65℃で洗浄する。アンチセンス核酸分子にはアンチセンスDNAオリゴヌクレオチド、およびRNAオリゴヌクレオチド(siRNAなど)がある。
【0088】
また本発明には、単離された節足動物タンパク質、そのホモログ、フラグメント、もしくは機能的同等物をコードする核酸配列を含有する核酸配列、または高ストリンジェンシー・ハイブリダイゼーション条件下で該核酸分子にハイブリダイズするアンチセンス核酸分子を含有するベクターも含まれる。該ベクターにはクローニングおよび発現ベクターが含まれる。それらの発現ベクターには、本発明の核酸分子とインフレームで結合した好適な転写および翻訳制御配列、例えばエンハンサー要素、プロモーター/オペレーター領域、ターミネーション停止配列、mRNA安定化配列、開始もしくは停止コドン、またはリボソーム結合部位が組み込まれてもよい。これらの制御配列は例として提供するものであり、制限を意図するものではない。
【0089】
更に、組換えタンパク質の場合、特定のホストから分泌させるのが便宜であり得る。従って、それらのベクターの更なる成分として、分泌、シグナリング、およびプロセシング配列をコードする核酸配列が含まれてもよい。
【0090】
本発明にかかるベクターにはプラスミドおよびウィルス(バクテリオファージおよび真核ウィルス)、並びに他の直鎖または環状DNAキャリア、例えば転位因子または相同組換え技術を使用するものが包含される。特に好適なウィルスベクターにはバキュロウィルス、レトロウィルス、アデノウィルス、およびワクシニアウィルスに基づくベクターがある。
【0091】
また本発明には、本発明の分子、特にDCの分化および成熟化を調節(好ましくは阻害)する節足動物タンパク質、ホモログ、フラグメント、または機能的同等物をコードするベクター、核酸分子、またはアンチセンス核酸を含有するホスト細胞が包含される。本発明の範囲内で、任意の型のホスト細胞を利用してもよい。ある態様では、ホスト細胞は原核ホスト細胞であってもよい。この態様では、原核ホスト細胞はE. coliホスト細胞であってもよい。別の態様では、ホスト細胞は真核ホスト細胞であってもよい。この態様では、ホスト細胞は真核酵母細胞であってもよい。更なる態様では、ホスト細胞は哺乳動物ホスト細胞であってもよい。更に別の態様では、ホスト細胞は昆虫細胞であってもよく、この態様では、発現系はバキュロウィルス発現系であってもよい。
【0092】
種々の技術を用いて本発明のベクターまたは核酸をホスト細胞に導入することができる。好適な形質転換またはトランスフェクション法は文献に十分報告されている(Sambrookら, 1989;Ausubelら, 1991;Spector, GoldmanおよびLeinwald, 1998)。真核細胞では、発現系は系の必要性に従って、一過性(例えばエピソーム系)または恒久的(染色体への組込み)のいずれであってもよい。
【0093】
本発明の更なる態様では、本発明の分子、特にDCの分化および成熟化を調節(好ましくは阻害)する、単離された節足動物タンパク質、ホモログ、フラグメント、または機能的同等物の調製法を提供し、該方法は以下を含む:
i)本発明の分子、特に本発明にかかるDCの分化および成熟化を調節または阻害する節足動物タンパク質、ホモログ、フラグメント、または機能的同等物をコードする核酸配列を含有するベクターを含有するホスト細胞を、該タンパク質が発現される条件下で培養すること;および
ii)そのようにして生成された該タンパク質を回収すること。
【0094】
本発明のこの観点において、タンパク質発現に必要な条件はホスト細胞系、ベクター、およびその後のタンパク質回収法によって異なる。実施例に、本発明の単離されたタンパク質の生成および回収のための特定の方法を開示する。それらの条件の変更は当業者に明白である。
【0095】
医薬組成物
同定されているDCの分化および成熟化の調節(好ましくは阻害)における本発明の分子の活性から、本発明の分子、タンパク質、核酸、アンチセンス核酸、ベクター、ホスト細胞、および抗体を治療に使用することが意図される。
【0096】
本発明は単離されたDC調節分子、例えばDCの分化および成熟化を調節(好ましくは阻害)する節足動物タンパク質もしくはその機能的同等物、それら分子をコードする核酸、該核酸を含有するベクター、該ベクターを含有するホスト細胞、または該分子に結合する抗体、および医薬的に許容されるキャリアを含有する医薬組成物を提供する。
【0097】
本明細書で使用する「医薬的に許容されるキャリア」という用語には、賦形剤がそれ自体毒性効果を与えず、医薬組成物を投与する個体に有害な抗体産生を惹起しないことを条件に、遺伝子、ポリペプチド、抗体、リポソーム、多糖、ポリ乳酸、ポリグルコール酸、および不活性ウィルス粒子、または、任意の他の薬剤が包含される。医薬的に許容されるキャリアは、液体、例えば見ず、生理食塩水、グリセロール、エタノール、または補助物質、例えば湿潤剤、乳化剤、pH緩衝剤などを更に含有してもよい。賦形剤により医薬組成物を錠剤、ピル、糖衣錠、カプセル、液体、ジェル、シロップ、スラリー、懸濁液に製剤し、患者の摂取を容易にしてもよい。医薬的に許容されるキャリアについての総説はRemington's Pharmaceutical Sciences(Mack Pub.社, ニュージャージー州 1991)に記載されている。
【0098】
ある態様では、医薬組成物はタンパク質と相互作用する1つまたはそれ以上の脂質分子を含有することができる。ある特定の態様では、この脂質分子はコレステロールまたはコレステロールの代謝物、例えばビタミンD3であってもよい。別の態様では、医薬組成物はジャパニンおよび脂質(例えばコレステロールまたはコレステロールの代謝物)を含有する、またはそれらから成る複合体を含有してもよい。更なる態様では、医薬組成物はジャパニン、脂質(例えばコレステロールまたはコレステロールの代謝物)、および受容体(例えば二価陽イオン依存性受容体(C型レクチン受容体など))を含有する、またはそれらから成る複合体を含有してもよい。
【0099】
本発明のある態様では、医薬組成物は1つまたはそれ以上の更なる治療薬を含有することができる。本発明のこの観点には、当業者が本発明の分子と共投与するのに有益であると見なしうる、いずれの更なる治療薬も包含される。特に、該更なる治療薬は抗炎症剤、免疫調節剤、免疫抑制剤、サイトカイン、サイトカイン模倣タンパク質、またはサイトカイン結合タンパク質を含有してもよい。特定の態様では、1つまたはそれ以上の更なる治療薬は抗炎症剤を含んでもよい。
【0100】
治療法
本発明は治療に使用するための単離された分子、例えば哺乳動物DCの分化および成熟化を調節(好ましくは阻害)するタンパク質、それらタンパク質をコードする核酸、アンチセンス核酸、該核酸もしくはアンチセンス核酸を含有するベクター、該ベクターを含有するホスト細胞、該タンパク質もしくは分子に結合する抗体、または該分子、タンパク質、核酸、ベクター、ホスト細胞、もしくは抗体を含有する医薬組成物を提供する。
【0101】
本明細書で使用する「治療」という用語は、ヒトまたは動物患者の利益のための、本明細書に記載するタンパク質、分子、核酸、ベクター、ホスト細胞、抗体、または医薬組成物の使用を包含する。特にこの用語には治療処置、予防的処置、診断、およびワクチン接種も包含される。このリストは例証のためだけに提供するものであり、制限を意図するものではない。
【0102】
本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物は、いずれの動物の治療に使用してもよい。ある態様では、動物は哺乳動物であってもよい。更なる態様では、哺乳動物はウシ、ブタ、ヒツジ、ネコ、イヌ、またはウサギであってもよい。更なる態様では、哺乳動物はヒトであってもよい。
【0103】
別の態様では、哺乳動物DCの分化および成熟化を調節(好ましくは阻害)する単離された分子、例えば哺乳動物DCの分化および成熟化を調節(好ましくは阻害)する単離された節足動物タンパク質、それらタンパク質をコードする核酸、それらタンパク質をコードする核酸に結合するアンチセンス核酸、該核酸もしくはアンチセンス核酸を含有するベクター、該ベクターを含有するホスト細胞、該タンパク質もしくは分子に結合する抗体、または該タンパク質、分子、核酸、ベクター、ホスト細胞、もしくは抗体を含有する医薬組成物の、DC活性に関係する疾患の治療のための使用法を提供する。
【0104】
また、本発明はDCに関係する疾患に罹患した動物の治療法を提供し、該方法は該動物にDCの分化および成熟化を調節(好ましくは阻害)する分子、例えば哺乳動物DCの分化および成熟化を調節(好ましくは阻害)する単離された節足動物タンパク質、それらタンパク質をコードする核酸、それらタンパク質をコードする核酸に結合するアンチセンス核酸、該核酸もしくはアンチセンス核酸を含有するベクター、該ベクターを含有するホスト細胞、該タンパク質もしくは分子に結合する抗体、または該タンパク質、分子、核酸、ベクター、ホスト細胞、もしくは抗体を含有する組成物を投与することを含む。
【0105】
本発明の範囲内で、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物は、多くの投与方法のうちの任意の1つまたはそれ以上を用いて患者に投与することができる。それらの投与方法は当該分野で公知であり、非経口注入(例えば静脈内、皮下、腹腔内、筋肉内、または組織の間質空間への)または直腸内、経口、膣内、局所、経皮、皮内、クモ膜下腔内、経鼻、点眼、耳内、肺内、もしくは他の粘膜投与がある。ナノパッチを使用して本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物の経皮投与を行ってもよい。遺伝子銃を使用して本発明の核酸、ベクター、または医薬組成物を投与してもよい。正確な投与方法は治療すべき疾患または症状によって異なる。
【0106】
ある態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を用いて、自己免疫疾患、アレルギーもしくは他の過敏性疾患、移植片拒絶反応および移植片対ホスト病、および急性および慢性炎症性疾患の治療または予防を行うことができる。
【0107】
自己免疫疾患には、限定されるわけではないが以下がある:無酸症、自己免疫性慢性活動性肝炎、アジソン病、円形脱毛症、筋萎縮性側索硬化症(ALS、ルー・ゲーリック病)、強直性脊椎炎、抗GBM腎炎または抗TBM腎炎、抗リン脂質抗体症候群、再生不良性貧血、関節炎、喘息、アトピー性アレルギー、アトピー性皮膚炎、自己免疫性アジソン病、自己免疫性溶血性貧血、自己免疫性肝炎、自己免疫性内耳疾患(AIED)、自己免疫性リンパ球増殖性症候群(ALPS)、自己免疫性血小板減少性紫斑病(ATP)、Balo病、ベーチェット病、ベルジェ病(IgA腎症)、水疱性類天疱瘡、心筋症、セリアック病、セリアック・スプルー皮膚炎、慢性疲労性免疫不全症候群(CFIDS)、慢性疲労免疫機能障害症候群(CFIDS)、慢性炎症性脱髄性多発性神経障害、チャーグ・ストラウス症候群、瘢痕性類天疱瘡、コーガン症候群、寒冷凝集素症、大腸炎、頭部動脈炎、クレスト症候群、クローン病、クッシング病、デゴス病、皮膚炎、皮膚筋炎、若年性皮膚筋炎、デビック病、I型糖尿病、円板上ループス、ダウリング・デゴス病、ドレスラー症候群、好酸球性筋膜炎、後天性表皮水疱症、本態性混合型クリオグロブリン血症、エバンス症候群、線維筋痛、線維筋炎、線維化生肺胞炎、胃炎、巨細胞性動脈炎、糸球体腎炎、グッドパスチャー症候群、グレーブス病、ギラン・バレー症候群、橋本甲状腺炎、溶血性貧血、ヘノッホ・シェーンライン紫斑病、肝炎、ヒューズ(Hughes)症候群、特発性副腎萎縮、特発性肺線維症、特発性血小板減少性紫斑病(ITP)、IgA腎症、炎症性脱髄性多発性神経障害、インスリン依存性糖尿病(I型)、過敏性腸症候群、若年性関節炎、川崎病、扁平苔癬、ルー・ゲーリック病、ルポイド肝炎、ライム病、メニエル病、混合性結合組織病、多発性骨髄腫、多発性硬化症、重症筋無力症、筋炎、眼部瘢痕性類天疱瘡、骨粗鬆症、扁平部炎、尋常性天疱瘡、悪性貧血、結節性多発性動脈炎、多発性軟骨炎、多腺性症候群、多腺性自己免疫症候群、リウマチ性多発性筋痛(PMR)、多発性筋炎、および皮膚筋炎、原発性無ガンマグロブリン血症、原発性胆汁性肝硬変、原発性硬化性胆管炎、乾癬、レイノー現象、ライター症候群、リウマチ性関節炎、サルコイドーシス、強膜炎、強皮症、シェーグレン症候群、スティッキー・ブラッド(sticky blood)症候群、スティッフマン症候群、スティル病、シデナム舞踏病、全身性エリテマトーデス(SLE)、高安動脈炎、側頭動脈炎/巨細胞性動脈炎、潰瘍性大腸炎、ブドウ膜炎、脈管炎、白斑、ウェグナー肉芽腫症、およびウィルソン症候群。
【0108】
アレルギーまたは過敏性障害は、Gell-Coombs分類のI型、II型、III 型、またはIV型を含む任意の既知のアレルギーまたは過敏性障害、および、一般的ではないがV型過敏性障害であってもよい。それらの障害には、限定されるわけではないが以下がある:アトピー、喘息、胎児赤芽球症、グッドパスチャー症候群、自己免疫性溶血性貧血、血清病、アルツス反応、全身性エリテマトーデス、接触性皮膚炎、ツベルクリン皮膚試験、慢性移植片拒絶反応、グレーブス病、重症筋無力症、全身性アナフィラキシー、局所性アナフィラキシー、アレルギー性鼻炎、結膜炎、胃腸炎、湿疹、輸血反応、新生児溶血性疾患、リウマチ性関節炎、糸球体腎炎、接触性皮膚炎、アトピー性皮膚炎、結節性病変、薬剤性溶血性貧血、ループス腎炎、アスペルギルス症、多発性動脈炎、多発性筋炎、強皮症、過敏性肺炎、ウェグナー肉芽腫症、I型糖尿病、蕁麻疹/血管浮腫、または甲状腺の炎症。アレルギーまたは過敏性障害は感染性疾患に関係しうるが、それらには、限定されるわけではないが結核、ハンセン病、ブラストミセス症、ヒストプラズマ症、トキソプラズマ症、リーシュマニア症、または他の感染がある。治療しうるアレルギーには、限定されるわけではないが花粉(例えば樺の木、ブタクサ、ナタネ)、食物(例えばナッツ、卵、またはシーフード)、薬物(例えばペニシリンまたはサリチル酸塩)、昆虫(例えばハチもしくはスズメバチ毒またはチリダニ)、または獣毛、および合成品(例えばラテックス)に対するアレルギー反応がある。治療しうる他の炎症性疾患にはアテローム性動脈硬化または他の心血管疾患、アルツハイマー病、血管炎、筋炎、脳炎、再灌流障害、2型糖尿病、脂肪肝疾患、および創傷治癒(例えば炎症期、血管新生段階、線維増殖および上皮形成、そして組織修復期)がある。
【0109】
ある態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を用いて、治療または診断処置の過程で体液または組織が合成または非哺乳動物由来物質と接触することから生じる有害症状の治療または予防を行うことができる。それらの処置は一時的または恒久的であってもよく、それらには、限定されるわけではないが体外循環、例えば腎臓または肝臓透析、腹膜透析、心肺バイパスおよび血液濾過、血管、膀胱、くも膜下腔内、または他の管腔臓器のいずれかに挿入した留置カテーテル、プロテーゼ(人工関節、心臓弁、血管内ステント、CSFシャント、人工血管、および冠動脈血管形成カテーテルなど)の使用がある。
【0110】
本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物は免疫抑制効果を有すると考えられ、これは移植片拒絶反応の予防に有用であり得る。移植片は同じ個体間の同型移植片、同じ種の異なるメンバー間の同種移植片、または異なる種間の異種移植片であってもよい。本発明の分子は種々の移植片の拒絶反応を予防するのに有用であり得るが、それらには、限定されるわけではないが心臓、肺、心肺、腎臓、肝臓、膵臓、腸、手、角膜、皮膚移植(顔面再移植および顔面移植など)、ランゲルハンス島、骨髄移植、輸血、血管、心臓弁、骨、および皮膚がある。分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を用いて、骨髄移植後の移植片対ホスト病を予防してもよい。
【0111】
理論に拘束されることは望まないが、ジャパニン・タンパク質は癌に関与するシグナル伝達経路の阻害に関係しうると推測される。本発明の更なる態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を癌の治療に用いることができる。本発明は癌に罹患した動物の治療法を提供し、該方法は、該動物に上記のような本発明の分子、タンパク質、核酸、ベクター、抗体、または医薬組成物を投与することを含む。それらの治療法は、癌における免疫反応の再分極化または調節を伴ってもよい。
【0112】
特に、癌は血液系腫瘍、例えばリンパ腫もしくは白血病、または多発性骨髄腫であってもよい。本発明に従って治療できる白血病には急性リンパ芽球性白血病(ALL)、急性骨髄性白血病(AML)、慢性骨髄性白血病(CML)、慢性リンパ芽球性白血病(CLL)、およびヘアリー細胞白血病がある。本発明に従って治療できるリンパ腫にはホジキン病および非ホジキンリンパ腫がある。関係する障害を治療してもよく、それらには例えば骨髄異形成症候群(MDS)(最終的にALLになりうる)、骨髄増殖性疾患(例えば真性赤血球増加症、本態性血小板血症、または骨髄線維症)、および軽鎖疾患によるアミロイドがある。
【0113】
更なる態様では、癌は癌腫、肉腫、または芽細胞腫であってもよい。本発明はあらゆる臓器の癌(限定されるわけではないが乳房、肺、卵巣、膵臓、精巣、皮膚、結腸、脳、肝臓、または子宮頸部など)および黒色腫の治療を企図する。治療または予防しうる癌には組織球腫、特にイヌ皮膚組織球腫がある。
【0114】
本発明の更なる態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を感染性疾患の治療に用いることができる。特に、それらの治療は病原体、例えばウィルス、細菌、および原虫、例えばHIV、TB、およびマラリアの原因物質、並びに他の寄生生物による感染に対する免疫応答の再分極化または調節を伴ってもよい。
【0115】
吸血性節足動物、例えばマダニは、感染性疾患の原因物質(例えばマダニ媒介性脳炎ウィルス、クリミア・コンゴ出血熱ウィルス、ナイロビ・ヒツジ病ウィルス、ブルグドルフェリ(Borrelia burgdorferi。ライム病の原因物質)、およびタイレリア・バルバ(Theileria parva。東海岸熱の原因物質))の供与源である。ジャパニンはマダニ媒介性疾患の伝染を促進するように作用すると考えられる。従って、ジャパニン・タンパク質、そのホモログ、フラグメント、および機能的同等物は、動物に接種して免疫応答を惹起し、マダニ媒介性疾患の治療または予防を行うのに有用である。従って、本発明の更なる態様では、節足動物媒介性感染性疾患の伝染の予防法、または節足動物媒介性感染性疾患の治療法を提供し、該方法は本発明の分子(例えばタンパク質)、または本発明の核酸分子、ベクター、ホスト細胞、抗体、もしくは医薬組成物を動物に投与することを含む。本発明はまた、節足動物媒介性感染性疾患の伝染の予防、または節足動物媒介性感染性疾患の治療に使用するための、本発明の分子(例えばタンパク質)、または本発明の核酸分子、ベクター、ホスト細胞、抗体、もしくは医薬組成物も提供する。節足動物は吸血性節足動物であってもよい。節足動物媒介性疾患はライム病、マダニ媒介性脳炎、クリミア・コンゴ出血熱、ナイロビ・ヒツジ病、または東海岸熱であってもよい。
【0116】
ジャパニン・タンパク質、そのホモログ、フラグメント、および機能的同等物は吸血性節足動物そのもの、並びにそれらが運搬する疾患に対するワクチンとしても有用でありうる。従って本発明は更に、動物への吸血性節足動物(マダニであってもよい)に対するワクチン接種の方法を提供し、該方法は本発明の分子(例えばタンパク質)、または本発明の核酸分子、ベクター、ホスト細胞、抗体、もしくは医薬組成物を動物に投与することを含む。本発明はまた、動物への吸血性節足動物(マダニであってもよい)に対するワクチン接種に有用な本発明の分子(例えばタンパク質)、または本発明の核酸分子、ベクター、ホスト細胞、抗体、もしくは医薬組成物を提供する。
【0117】
上記のように、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を1つまたはそれ以上の更なる治療物質、例えば抗炎症剤、免疫調節剤、免疫抑制剤、サイトカイン、サイトカイン模倣タンパク質、サイトカイン結合タンパク質、または上記の障害のいずれかを治療するために開発された別の生物医薬品と共に投与するのが有益であり得る。
【0118】
本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、または抗体を、in vivoでそれらをDCにターゲッティングさせる抗原と共に投与して、所望しない免疫応答に関係するDCの分化および成熟化を調節または阻害することが有益であり得る。この態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を疾病関連要素と共に患者に投与し、本発明のタンパク質、分子、核酸、ベクター、ホスト細胞、抗体、または医薬組成物の好適なDCへのターゲッティングを促進することができる。本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、または抗体が既にDCへの特異的結合によってDCにターゲッティングされている態様では、疾病関連要素は不要である。
【0119】
「疾病関連要素」という用語は、患者における疾病と関係するいずれの成分も包含することを意図する。疾病には上記のように、自己免疫疾患、アレルギーおよび他の過敏性反応、移植片拒絶反応および移植片対ホスト病、感染性疾患(例えばマダニによって伝染するもの)、癌(例えば血液系腫瘍)、そして急性および慢性炎症性疾患がある。従って「疾病関連要素」には以下が包含される:i)感染性物質、例えばウィルス、微生物、寄生生物、および微生物毒に関係する成分;ii)アレルギーに関係する非自己分子であるアレルゲン;iii)アレルギー以外の過敏性反応に関係する非自己成分;iv)自己免疫疾患に関係する自己成分;v)同種の遺伝的に異なるメンバー由来(アロ抗原)または異なる種由来(異種抗原)の移植抗原;およびvi)腫瘍関連抗原および腫瘍特異的抗原。
【0120】
「疾病関連要素」という用語は、これらの疾病関連要素のフラグメントおよび誘導体も包含する。それらの誘導体には、無毒化された物質、合成ミモトープ、および抗原で、その構造中に置換、付加、または欠失を含有するもので、本発明のタンパク質、分子、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を好適なDCに指向させるように作用する能力を有するものが含まれる。
【0121】
疾病関連要素を有する動物を提供することにより、疾病に関係するDC分化および成熟化の調節または阻害の特異性が向上される。このように特定のDCを標的とすることは、全体的な免疫応答を阻害する必要を無くし、結果として副作用が低減しうるため、有益である。
【0122】
本発明のある観点では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を疾病関連要素と別々に投与してもよい。この観点では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物、および疾病関連要素を連続的に投与してもよい。別の態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を疾病関連要素の前に投与してもよい。
【0123】
更なる態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物を疾病関連要素の後に投与してもよい。更なる態様では、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物および疾病関連要素を同時に投与してもよい。本発明のこの観点では、本発明のタンパク質、分子、または抗体は疾病関連要素に結合して、例えば融合タンパク質の形態であってもよい。
【0124】
従って本発明は、本発明にかかるDC分化および成熟化を調節または阻害する分子および疾病関連要素を含有する融合タンパク質も提供する。それらの融合タンパク質をコードする核酸も提供する。
【0125】
本発明はまた、本発明の分子、タンパク質、核酸、ベクター、ホスト細胞、抗体、または医薬組成物、および疾病関連要素、および医薬的に許容されるキャリアを含有する医薬組成物を提供する。
【0126】
上記の方法は、本発明の分子を動物に投与して、動物においてin vivoでDCの分化および成熟化を調節(好ましくは阻害)することを含む。分子は単独で投与してもよく、または分子をDCにターゲッティングさせる更なる物質(例えば疾病原因要素)と共に投与してもよい。
【0127】
上記の疾病を治療する別の方法は、本発明の分子を目的とする治療にex vivoで使用するものである。この方法は、本発明の分子をインビトロでDCに運搬してDCを調節し、調節されたDCを治療を必要とする動物に運搬することを伴う。
【0128】
更なる観点では、本発明はDCを調節する方法を提供し、該方法は上記のように、DCを本発明の分子、例えばジャパニン・タンパク質、そのホモログ、フラグメント、および機能的同等物と接触させることを含む。この方法によって生成される、調節されたDCも提供する。
【0129】
また本発明は、それを必要とする動物においてDCに関係する障害を治療または予防する方法も提供し、該方法はこの方法で生成される調節されたDCを動物に投与することを含む。
【0130】
DC分化を調節する分子は、上記のようにDC分化および成熟化を調節(好ましくは阻害)する任意の分子(例えばジャパニン・タンパク質、そのホモログ、フラグメント、および機能的同等物)であってもよい。これらの分子をコードする核酸分子を、核酸分子を含有するベクターとして使用してもよい。
【0131】
DCは治療を必要とする動物から直接単離してもよい。あるいはまた、DC前駆体(例えば骨髄前駆細胞の単球)を動物から単離し、これを使用してDCを生成させてもよい。本発明のこの観点では、DCまたはDC前駆体は、本発明の分子での処理後に調節されたDCを導入する動物にとって、自己移植であってもよく、または同種異系であってもよい。
【0132】
DCに関係する障害は上記の疾病のいずれであってもよく、それらには自己免疫疾患、アレルギーおよび他の過敏性反応、移植片拒絶反応および移植片対ホスト病、感染性疾患(例えばマダニによって伝染するもの)、癌(例えば血液系腫瘍)、そして急性および慢性炎症性疾患がある。この方法を使用して移植ドナーから調節されたDCを生成し、これを移植に先立って所定の移植レシピエントに投与し、移植片に対して不応答とし、それによって免疫抑制剤投与の必要を低減することができる。
【0133】
本発明のこの観点では、上記のようにDCを疾病関連要素と接触させ、DCの分化および成熟化を調節(好ましくは阻害)する分子を好適なDCにターゲッティングさせることができる。あるいはまた、調節されたDCを上記のような疾病関連要素と共に動物に投与してもよい。
【0134】
スクリーニング法
ジャパニンの同族受容体を同定することにより、受容体をスクリーニング法に使用し、可能性のあるジャパニンのアゴニストおよびアンタゴニストを同定し、治療または他の使用可能性を有する化合物を同定することが可能となる。
【0135】
可能性のあるアゴニストまたはアンタゴニスト化合物は、例えば細胞、無細胞標品、化学ライブラリー、または天然産物混合物から単離することができる。それらのスクリーニング法の好適な総説については、例えばColiganら, Current Protocols in Immunology 1(2):Chapter 5 (1991)を参照されたい。
【0136】
良好なアンタゴニストである可能性が最も高い化合物は、ジャパニンによって誘導される生物学的作用を誘導させずにジャパニンの同族受容体に結合し、ジャパニンの機能を競合的に阻害する分子である。上記のように、ジャパニンの同族受容体は二価陽イオン依存性受容体であるC型レクチン受容体であることが考えられ、これはジャパニンが結合すると細胞内シグナル伝達系を惹起し、それによって樹状細胞の分化および成熟化の阻害が起こる。可能性のあるアンタゴニストには、シグナル伝達経路を活性化することなく受容体に結合する、または負のシグナル伝達経路を活性化することによって受容体に結合する小型有機分子、ペプチド、ポリペプチド、および抗体がある。特に、好適かつ有望なアンタゴニストには炭化水素部分、および、受容体への結合親和性を保持したままその機能を低減するように操作した組換えジャパニン分子がある。更なる好適な化合物にはジャパニンに対する抗体、およびジャパニンに結合した炭化水素部分に対する抗体、および標的特異性を遺伝子操作できるアンチカリンがある。
【0137】
良好なアゴニストとして機能する可能性の最も高い化合物は、ジャパニンの同族受容体に結合し、ジャパニンと同じ細胞内シグナル伝達経路を惹起し、それによってジャパニンと同様の方法で樹状細胞の分化および成熟化を調節(好ましくは阻害)するように機能する化合物である。好適かつ有望なアゴニストの例には、受容体を活性化する能力および/または受容体に対する結合親和性が増加するように操作したジャパニン分子がある。
【0138】
それらのスクリーニング法に使用する同族受容体(二価陽イオン依存性受容体、例えばC型レクチン)は溶液中に含有されるか、固相支持体に固定されるか、細胞表面上に担持されるか、または細胞内に存在してもよい。一般に、それらのスクリーニング法は受容体を発現する好適な細胞または細胞膜を使用し、それらを試験化合物と接触させて結合または機能的反応の刺激もしくは阻害を観察することを伴う。その後、試験化合物と接触させた細胞の機能的反応を、試験化合物と接触させていないコントロール細胞と比較する。それらのアッセイにより、好適な検出系を用いて、試験化合物がジャパニンによる受容体活性化によって生成されるのと同様のシグナルを生成するか否かを評価してもよい。ジャパニンは細胞表面二価陽イオン依存性受容体(例えばC型レクチン受容体)に結合し、細胞内シグナル伝達経路を惹起することによって機能すると信じられるため、スクリーニング法が最も効果的に機能するのは、細胞表面上の受容体を使用し、化合物の受容体への結合によって細胞内シグナルの生成をモニタリングすることを伴う場合であると考えられる。
【0139】
ある態様では、ジャパニンのアゴニストまたはアンタゴニスト化合物の同定法は以下を含む:
(a)受容体への結合が可能な条件下で、その表面上に二価陽イオン依存性受容体(例えばC型レクチン受容体)を発現する細胞をスクリーニングを行う化合物と接触させること(受容体は化合物の結合に応答して検出可能なシグナルを生成する能力を有する);および
(b)化合物と受容体の相互作用から生成されるシグナルのレベルを測定することによって化合物が受容体に結合するか否か、また、受容体を活性化もしくは阻害するか否かを測定する。
【0140】
ある態様では、スクリーニングを行う化合物をリガンドの存在下で受容体と接触させることができる。それらのリガンドは脂質、例えばコレステロールまたはコレステロールの代謝物(例えばビタミンD3)であってもよい。
【0141】
別の態様では、ジャパニンのアンタゴニスト化合物の同定法は以下を含む:
(a)ジャパニンがその同族受容体に結合できる条件下で、その表面上に二価陽イオン依存性受容体(例えばC型レクチン受容体)を発現する細胞をジャパニンと接触させること(受容体はジャパニンの結合に応答して検出可能なシグナルを生成する能力を有する)
(b)ジャパニンと受容体の相互作用から生成されるシグナルのレベルを測定すること;
(c)スクリーニングを行う化合物を受容体への結合が可能な条件下で添加すること;および
(d)化合物と受容体の相互作用から生成されるシグナルのレベル変化を測定することによって、ジャパニン結合の際の化合物の影響を測定すること。
【0142】
ある態様では、上記のようなジャパニンのホモログまたは機能的同等物を、上記のスクリーニング法に用いることができる。
【0143】
更なる態様では、スクリーニングを行う化合物および/またはジャパニンをリガンドの存在下で受容体と接触させることができる。それらのリガンドは、脂質、例えばコレステロールまたはコレステロールの代謝物であってもよい。
【0144】
上記の条件は、培養液の存在、生理学的濃度のCa2+を含有する溶液の存在、および/またはpH 7-8であることを含んでもよい。
【0145】
上記の検出可能なシグナルには細胞内リン酸化、核局在化、遺伝子発現、および/またはサイトカイン放出がある。
【0146】
上記の方法のある態様では、単純な結合アッセイを使用し、C型レクチン受容体を保有する表面への試験化合物の接着を、試験化合物に直接もしくは間接的に結合する標識によって、または標識された競合物質との競合を伴うアッセイによって検出する。別の態様では、競合薬剤スクリーニング・アッセイを使用し、二価陽イオン依存性受容体(例えばC型レクチン受容体)との結合能を有する中和抗体を試験化合物と結合に関して特異的に競合させる。この方法では、抗体を使用して受容体と特異的な結合親和性を有する試験化合物の存在を検出することができる。
【0147】
当業者は、ジャパニンの同族受容体に作用する化合物を同定するためのアッセイを考案することができる。受容体との好適な結合親和性を有する化合物のハイスループット・スクリーニングを提供するのに使用してもよい技術については、国際特許出願WO84/03564を参照されたい。この方法では、多数の異なる小型試験化合物を固相支持体上に合成し、その後、受容体と反応させ、洗浄することができる。ポリペプチドを固定化する一つの方法は、非中和抗体の使用である。その後、当該分野で公知の方法によって、結合された受容体を検出してもよい。精製された受容体分子をプレート上に直接コーティングし、上記の薬剤スクリーニング法に用いてもよい。
【0148】
本発明は、上記の方法によって同定されたアゴニスト、アンタゴニスト、および他の化合物も包含する。これらのアゴニスト、アンタゴニスト、および他の化合物は、上記の医薬組成物の一部を成してもよく、上記の治療法に使用してもよい。
【0149】
ここで、本発明の種々の観点および態様について、実施例によってより詳細に記載する。認識されるように、本発明の範囲を逸脱することなく、詳細の改変を行ってもよい。
【実施例】
【0150】
試料と方法
マダニ
Rhipicephalus appendiculatusマダニはthe Centre of Ecology and Hydrology(オックスフォード)で飼育した。給餌は、ガーゼでカバーした保持チャンバー(retaining chambers)中で、それらを剪毛したモルモットの背上に配して行った。
【0151】
唾液腺抽出物の調製
給餌の1-6日後、モルモットからマダニを注意深く分離し、顕微鏡下でその唾液腺を切除した。唾液腺を冷リン酸緩衝液(PBS;Oxoid社)で軽く洗浄し、1.5mLの微小遠心管に移し、使用まで-70℃で保存した。Dounceホモジナイザーを用い、PBS中で唾液腺をホモジナイズして唾液腺抽出物(SGE)を調製した。ホモジネートを≧10000gで3分間遠心分離し、上清を回収して使用まで-20℃で保存した。
【0152】
細胞培養
細胞培養液および補足物質は、特に記載しない限りPAA社のものを使用した。それらにはウシ胎仔血清(FCS)があり、全試験を通してバッチ#A04304-0511を使用した。哺乳動物細胞の培養は37℃、5% CO2で行った。昆虫細胞の培養はSf900 II培養液(Invitrogen社)中、28℃で行った。共トランスフェクション培養を除いて、液体培養は三角フラスコ中、100-130rpmで軌道撹拌して行った。
【0153】
樹状細胞
ヒト樹状細胞(DC)を、健常成人ドナーから単離した末梢血単球から生成した。簡潔に記載すると、バフィーコート(National blood service、ブリストル)をCa2+/Mg2+未含有ハンクス緩衝液(HBSS)と1:2(v/v)で混合し、Lymphoprep(Axis Shield社)上に注意深く積層し、800gで30分間(22℃)遠心分離した。HBSS/バフィーコート混合液とLymphoprep間の接触面に生成された末梢血単核細胞(PBMC)層を注意深く回収し、HBSSで3回洗浄し、血小板を除去した。一時的接着または磁気ビーズによる陰性選択のいずれかによって、単球をPBMCから単離した。
【0154】
一時的接着による単球の単離では、PBMCペレットをRPMI-5(5% ヒトAB+血清(National blood service)、2mM L-グルタミン、100U/ml ペニシリン、および100μg/ml ストレプトマイシンを補足したRPMI 1640)に再懸濁した。PBMCを100mm細胞培養用ペトリ皿(BD Biosciences社)中に、1×107 細胞/ml、10ml/プレートで播種し、37℃、5% CO2で45分間インキュベートした。その後、プレートをHBSSで3回洗浄して接着していない細胞を除去し、10mlのRPMI-5をプレートに添加した。培養1日後、ヒトGM-CSF(John Radcliffe病院薬局)を1000U/mlの濃度で添加し、ヒトIL-4(Peprotech社)を20ng/mlの濃度で添加した。培養3日後および5日後、総容量の3分の1をフレッシュに調製したGM-CSF + IL-4を補足したRPMI-5で置換した。
【0155】
陰性選択(つまり非単球の除去)による単球の単離では、PBMCペレットをCa2+/Mg2+未含有HBSSに再懸濁した後、Dynal Monocyte Negative Isolationキット(Invitrogen社)を使用し、製造者の説明書に従って単球を単離した。精製した単球をDC-RPMI(RPMI 1640に10% FCS、2mM L-グルタミン、100U/ml ペニシリン、100μg/ml ストレプトマイシン、1000U/ml ヒトGM-CSF、および1000U/ml ヒトIL-4(どちらもGentaur社)に5x105/mlで再懸濁した。3日後、培養液の3分の1を除去して遠心分離し、ペレットを同量のフレッシュの培養液(3000U/mlの各サイトカインを含有)に再懸濁した後、再び培養液に添加した。5日後、HBSS/2% FCSで洗浄した後、等張食塩水に混合した5.5%ヘタスターチ(John Radcliffe病院薬局の「Voluven」)/4.8% DMSO(Sigma社、Hybrimaxグレード)/3.8% FCSで再懸濁し、その後、温度制御凍結装置(1度/分)に入れ、-80℃で細胞を凍結させた。
【0156】
DC調節活性のスクリーニング
上記のようにFCS含有培養液中で生成し、凍結したDCを用いて、DC調節活性の慣例的なスクリーニングを行った。DCを解凍した後、96ウェル平底Primariaプレート(BD Biosciences社)中、スクリーニングするサンプルを添加したDC-RPMI中で培養した。24時間培養後、ポリ(I:C)(25μg/ml)またはLPS(200ng/ml)を添加してDCを刺激したが、刺激剤の選択は各バッチのDCの異なる刺激剤に対する反応性によって決定した。更に18-24時間培養した後、CD1a+ 細胞によるCD86発現レベルをフローサイトメトリーで評価した。
【0157】
フローサイトメトリー
フローサイトメトリーはFACSortフローサイトメトリー(Becton Dickson社)を使用し、CellQuest Pro(Becton Dickinson社)で制御して行った。データ解析はCellQuest ProおよびFloJo(Treestar Software社)を用いて行った。
【0158】
PCR
PCRはDNAEngineサーモサイクラー(Biorad社)を用いて行った。プライマーはMWG Biosciences製(HPSFクォリティー)、dNTPはBioline製であった。他の試薬は本文に記載する通りである。
【0159】
DNAシークエンシング
DNAシークエンシングはGeneservice社(オックスフォード)で、BigDye v3.1化学を使用して実施した。
【0160】
E. coli形質転換
化学的コンピテントE. coliのヒートショック形質転換には、50μlのバクテリアを氷上で解凍し、氷上でDNAと共に5分間インキュベートし、42℃で40秒間熱処理した後、氷上に戻した。次いで、250μlのSOC培養液(室温)を添加し、細胞を37℃で、200rpmで軌道撹拌しながら45分から1時間30分インキュベートした後、50μlアリコートを100μg/mlアンピシリン含有LB寒天プレート上に播種した。
【0161】
実施例1 SGEによるLPS誘導型CD86亢進的調節の抑制
前記のように、一時的接着により単球を単離し、GM-CSF + IL-4と共に培養してDCを生成させた。5日間培養後、オスまたはメスR. appendiculatusマダニ由来のSGEを培養液に添加した。SGEは給餌していないマダニ、または3もしくは6日間給餌を行ったマダニ由来のいずれかであり、最終濃度50μg/mlのマダニ由来タンパク質となるように添加した。合計6日間の培養後、DCをLPS(50ng/ml)で処理し、7日間の培養後、抗CD1aおよび抗CD86で染色し、フローサイトメトリーで分析した。図1にCD86+細胞数を、CD1a+細胞%xこれらの細胞のCD86染色の幾何平均蛍光強度(geometric mean fluorescence intensity;GMFI)で示す。図1に示すように、3日間の給餌を行ったメスRhipicephalus appendiculatusマダニ由来SGEはヒトDCによる共刺激受容体CD86のLPS誘導型亢進的調節を抑制し、未給餌のメス、6日間給餌したメス、またはオス(給餌および未給餌)のマダニ由来のSGEでは抑制しないことが明らかになった。この実験を2回行い、同様の結果を得た。SGEを最終濃度がml当たり1つの唾液腺から誘導される物質となるように添加した場合にも、同様の結果が得られた。
【0162】
実施例2 SGEによるCD80およびMHCクラスIIのLPS誘導型亢進的調節の抑制
前記のように、一時的接着により単球を単離し、GM-CSF + IL-4と共に培養してDCを生成させた。5日間培養後、3日間給餌したメスR. appendiculatusマダニ由来のSGEをいくつかの培養液に添加し、最終濃度が培養液ml当たり、1つの唾液腺から誘導された物質となるようにした。合計6日間の培養後、DCをLPS(50ng/ml)で処理し、7日間の培養後、抗CD1a、抗CD80、抗CD86、および抗HLA-DRで染色し、フローサイトメトリーで分析した。図2に、SGEの存在下(黒色)および非存在下(灰色)におけるLPS処理したCD1a+細胞による成熟マーカーの発現を示す。図2に示すように、3日間給餌したメスRhipicephalus appendiculatus唾液腺抽出物はLPSに応答してCD86による亢進的調節を阻害するだけでなく、CD80およびMHCクラスIIの亢進的調節も阻害した。この実験を2回行った。
【0163】
実施例3 SGEの活性成分のQカラム分画
3日間給餌したメスR. appendiculatusマダニ由来の唾液腺112検体から得たSGEを50mM リン酸ナトリウム(pH7.0)で1:10に希釈した後、予め同じバッファーで平衡化しておいたHi-Trap Qセファロースイオン交換カラム(Amersham)に適用した。未結合の物質(Qカラム・フロースルー=QFT)を回収し、カラムを2カラム容量の50mM リン酸ナトリウム(pH7.0)で洗浄した後、50mM リン酸ナトリウム、1M NaCl(pH7.0)をカラムに適用して、結合した物質(Qカラム結合画分=QBF)を溶出した。Vivaspin 6 5kDa MWCO遠心濃縮器(Sartorius)(タンパク質の非特異的吸着を予防するためにγ-グロブリンで前処理したもの)を使用して、QFTおよびQFRを最終容量500μlまで濃縮した。
【0164】
上記のように、一時的接着によって単球を単離し、GM-CSF + IL-4と共に培養してDCを生成させた。5日間培養後、3日間給餌したメスR. appendiculatusマダニ由来のSGE、または上記で生成したQFTもしくはQBFを培養的に添加した。それぞれの最終濃度が培養液ml当たり、1つの唾液腺から誘導された物質となるようにした。合計6日間の培養後、DCをLPS(50ng/ml)で処理し、7日間の培養後、抗CD1aおよび抗CD86で染色し、フローサイトメトリーで分析した。図3にCD86+細胞数を、CD1a+細胞%xこれらの細胞のCD86染色の幾何学的平均蛍光強度(GMFI)で示す。エラーバーは2重複培養液間の範囲を表す。
【0165】
図4は、DC調節活性はLPS誘導型成熟化(おそらくTLR4を通じて作用)に限定されず、25μg/ml ポリ(I:C)(TLR3リガンド)または50ng/ml IFN-γでの処理にも応答してCD86亢進的調節を阻害した(図4a)。しかしながら、QFTは100ng/ml TNF-α誘導型CD86亢進的調節に対してほとんど、または全く影響を与えなかった(図4b)。この理由は現在のところ明らかではない。
【0166】
実施例4 QFTのDC調節活性に対するプロテイナーゼKの影響
上記のように調製した凍結された5日目DCを解凍し、最終濃度0.2唾液腺相当量/mlのQFTと共に培養した。この実験では、QFTの一部を後述のようにプロテイナーゼKで前処理した。一日後、DCをポリ(I:C)(25μg/ml)で処理し、更に1日培養した後、抗CD1aおよび抗CD86で染色し、フローサイトメトリーで分析した。
【0167】
プロテイナーゼK処理は、QFTを、50mM Na2HPO4(pH7.0)に混合した150μg/ml プロテイナーゼK(Sigma社)と共に50℃で2時間インキュベートして行った。その後、98℃で10分間加熱して、酵素を失活させた。プロテアーゼのみのコントロール[プロテイナーゼK+ポリ(I:C)]は、同じ反応をQFTを用いずに行って得られた産物を用いて行い、プロテアーゼ未含有コントロール[QFT+ポリ(I:C)]に用いたQFTも同様に処理したところ、DC調節活性の抑止は活性成分の熱変性によるものではないことが明らかになった。2つの更なるコントロールを実施し、加熱不活性化したプロテイナーゼKをQFT存在下または非存在下でインキュベートし([不活化プロテイナーゼK+ポリ(I:C)]および[QFT+不活化プロテイナーゼK+ポリ(I:C)])、プロテイナーゼKの(熱に不安定な)タンパク質分解能がDC調節活性の抑止に必要であることを確認した。そのため、プロテイナーゼKを98℃で10分間前処理し、その後、50℃でインキュベートした。図5に示すように、プロテイナーゼK処理によってQFTのDC調節能は抑止され、これがタンパク質性であることが確認された。結果は2重複ウェルの平均値±2 S.Eで示す。
【0168】
実施例5 QFTのサイズ分画
350検体の唾液腺を3日間給餌したメスR. appendiculatusマダニから切除し、前記のようにこれを用いてQFTを調製した。8つの独立した実験で、このQFTをSuperdex 75カラムでゲル濾過し、サイズ分画した。各実験で、約40画分をカラムから回収し、上記のように、解凍した5日目DCと共に培養してDC調節活性をスクリーニングした。24時間後に200ng/ml LPSを添加してDCを刺激し、更に24時間後、フローサイトメトリーでCD1a+細胞のCD86発現を測定した。
【0169】
8つのうち、1つの代表的な分画から得た結果を図6に示す。初期スクリーニングは1唾液腺相当量/mlで行い、活性があると推測される画分の2次スクリーニングは0.04唾液腺相当量/mlで行った。全ての分画で、画分12および13が最も高い活性を有すると同定された。
【0170】
各分画からの最も活性の高い画分をプールし、低塩バッファー(50mM HEPES、pH8.3)に対して透析した後、還元的条件下、4-12% Bis-Trisポリアクリルアミドゲル(Invitrogen社)で泳動した。比較のために、SGE、QFT、および2つのQカラム結合画分をプールした活性画分と並行して行った。結果を図7に示す。SGEおよびQFTはDC調節活性を有したが、Q結合画分は有さなかった。約20kDaのバンドがQFTおよびプール画分中に存在し、2つの不活性サンプル中には存在しなかったことから、このバンドが活性タンパク質であると考えられる。出願人はこのタンパク質を「ジャパニン(Japanin)」と命名した。
【0171】
QFTのゲル濾過クロマトグラフィーで得た最も活性の高い画分(上記参照)をプールし、低塩バッファーに対して透析した後、HPLC(C4カラム)を用いて分画した。アセトニトリルの増加グラジエント溶出によって23画分をカラムから回収し、上記のように解凍した5日目DCと共に培養してDC調節活性についてスクリーニングした。24時間後に25μg/ml LPSを添加してDCを刺激し、更に24時間後、フローサイトメトリーでCD1a+細胞のCD86発現を測定した。
【0172】
実施例6 HPLCによるDC調節活性の単離
各画分は1:200に希釈して評価し(画分の容量によって、2.6から7.9唾液腺相当量/mlの濃度となる)、インプットした物質(プールした活性ゲル濾過画分)およびQFTはコントロールとしてそれぞれ1.8および0.2唾液腺相当量/mlで含有させた。画分23は約100%アセトニトリルであるため、その作用はQFTの存在下および非存在下で評価し、HPLC溶媒による直接的DC細胞調節効果、あるいは逆に、溶媒がそれらの効果をマスキングする能力を除外した。図8にHPLCの結果を示す。図8aに溶出プロファイルを示し、図8bは各画分に関する活性(DCによるDC86の亢進的調節の阻害として測定)を示す。
【0173】
実施例7 エドマン分解
HPLC画分19が最も効果の高いDC調節活性を有することを確認した後、これを用いてエドマン分解シークエンシングを行い、16残基のN-末端配列を得た:(Thr)Pro Ser Met Pro Ala Ile Asn Thr Gln Thr Leu Tyr Leu Ala(Arg)(括弧内の残基の同定は暫定)。エドマン分解の読み取り結果を図9に示す。このN-末端配列を有するタンパク質を以下、「ジャパニン」と称する。
【0174】
実施例8 ジャパニンDNA 3'領域のPCR増幅
N-末端配列を用いてジャパニン DNAのPCR増幅のための外側順方向プライマーをポリ(dT)逆方向プライマーと組み合わせて設計した。配列「M P A I N T Q」に対する4つの変性プライマー・セットを使用した。それら4つは非常に類似しているが、縮重を低減するために個別に使用する。
【化1】
【0175】
PCRのテンプレートを得るために、1日間給餌したメスR. appendiculatus唾液腺からcDNAを生成した。Trizol試薬(Invitrogen社)を使用し、製造者の説明書に従って30検体の唾液腺からRNAを単離し、その後、1/3容量の8M 塩化リチウムを添加して水相から沈殿させた。冷75%エタノールで洗浄した後、RNAを5μlのRNase未含有水に再溶解した。
【0176】
ImPromII逆転写酵素(Promega社)を使用し、製造者の説明書に従って、40μl反応で逆転写を行った。反応液は4μgのRNAを含有し、2.5mMのMgCl2濃度、0.5mMの総dNTP濃度であった。逆転写を開始させるためにオリゴ(dT)15を使用し、0.1μg/mlを添加した。反応は42℃で1時間行い、その後70℃で15分間、加熱不活性化を行った。
【0177】
このcDNAを用いるPCRは、Taq DNAポリメラーゼ(New England Biosciences社)を用い、1X Thermopolバッファー(New England Biosciences社)(62.5Mの各dNTP、250nM変性プライマー、および3.25μM オリゴ(dT)20-V含有)中で行った。テンプレートcDNAは1:40に希釈して使用した。初期変性段階は94℃で1分間、次いで94℃で30秒間(変性)/45℃で30秒間(アニーリング)/72℃で60秒間(伸長)を5サイクル、その後、94℃/50℃で30秒間/72℃で60秒間を30サイクル、最後に72℃で更に5分間とした。陽性コントロールは既知のマダニ・タンパク質の順方向プライマーを使用し、同じ条件下で行った(RH1-PE)。生成物を1.8%アガロースゲルで泳動した。結果を図10に示す。
【0178】
順方向プライマー2および4で増幅した、より大きい生成物は約600bpであり、それらが約22kDaのタンパク質をコードしていることを示唆している(平均アミノ酸残基質量を110Daと仮定)。これはSDS-PAGEゲルで約20kDaのバンドが活性画分中で同定されたことと十分一致している。このcDNAをクローニングする前に、ネステッドPCRを実施してこれがN-末端配列と一致するか否かを確認した。
【0179】
配列「Asn Thr Gln Thr Leu Tyr Leu Ala」に対して内側プライマーを設計した。これはN-末端配列中にあるが、上記で使用したプライマーの結合位置より下流である:
【化2】
標準外コード「I」はニュートラル塩基としてのイノシンの導入を表す。
【0180】
順方向プライマー2および4にコードされるプライマーをオリゴ(dT)20-Vと共に1:20希釈で用いて増幅したDNAをテンプレートとした。反応条件は上記の通りであるが、アニーリング温度は35サイクル全てで50℃とし、72℃での伸長段階を60秒間から40秒間に短縮した。生成物を1.8%アガロースゲルで泳動した。
【0181】
これらのPCRにより、予想されたサイズ(すなわち約600bp)のバンドが生成され(図11)、N-末端タンパク質配列をコードするcDNAが特異的に増幅されることを強く示唆している。
【0182】
実施例9 ジャパニンcDNAの3'領域のコンセンサス配列の同定
ジャパニンの3'配列を得るために、外側順方向プライマー2および4を用いてDNAを増幅し、pCR2.1にクローニングし、シークエンシングを行った。
【0183】
上記のように、外側順方向プライマー2および4と、逆方向プライマーとしてオリゴ(dT)20-Vを併用し、40μl反応で行った。反応液をアガロースゲルで泳動し、約600bp DNAバンドを切り出し、QIAquickゲル抽出キット(Qiagen社)を使用し、製造者の説明書に従って精製した(30μlの溶出バッファーでカラムから溶出)。
【0184】
各精製産物6μlをpCR2.1にライゲートするため、10μl反応で50ngのpCR2.1-TA(Invitrogen社)およびT4 DNAリガーゼ(NEB Biosciences社)と共に14℃で一晩インキュベートした。ライゲーション混合液を用いてTOP10 E. coliコンピテントセルに形質転換し、挿入断片を含有するコロニーを、順方向プライマー2および4とオリゴ(dT)20-Vを併用するPCRスクリーニングで同定した。QIAprep Spinキット(Qiagen社)を使用し、製造者の説明書に従って、陽性コロニーの5ml培養液から挿入断片を含有するpCR2.1 DNAを単離した。
【0185】
4つの挿入断片含有プラスミドのシークエンシングによりジャパニンcDNAの3'領域のコンセンサス配列を構築することが可能となった。これを図12に示す。
【0186】
実施例10 ジャパニンの5'領域のシークエンシング
5' RACE(Rapid Amplification of cDNA Ends:cDNA末端迅速増幅)法を使用してジャパニンの5'領域を組み込んだ約500bpの生成物を増幅した。
【0187】
5' RACEは既知の3' cDNA配列(この場合、新たに得たジャパニン 3'配列)を使用して遺伝子特異的プライマーの設計の情報を得る。これらのプライマーを使用して遺伝子特異的逆転写を行い、ネステッドPCRを用いてcDNAの5'領域を増幅する。後者の場合、順方向プライマーは実験的に導入したオリゴヌクレオチドのキャップ領域に特異的であり、逆方向プライマーは遺伝子特異的逆方向プライマーである。使用される種々のジャパニン特異的プライマーの相対的な位置を図13aに示す。
【0188】
RNAqueous-4PCRキット(AmbionRNA社)を使用し、製造者の説明書に従って、2日間給餌したメスR. appendiculatusマダニ唾液腺からRNAを抽出した。カラム溶出物からRNAを沈殿させ、20μl溶出バッファーに再溶解した。
【0189】
GSP1Bプライマーを用いて遺伝子特異的逆転写を行った。ImProm II RT酵素(Promega社)を、1μgのRNAを含有する20μl反応で、製造者の説明書に従って使用した。この反応でMgCl2濃度は2.5mM、総dNTP濃度は0.5mM、プライマー濃度は125nMであった。RT反応を48℃で1時間行った後、70℃で15分間加熱不活性化を行った。その後、1μlのRNase混合液(Invitrogen社。5' RACE Systemキットより)を添加し、室温で30分間インキュベートしてRNAを除去した。
【0190】
SNAPカラム(Invitrogen社。5' RACE Systemキットより)を製造者の説明書に従って使用し、生成したcDNAから酵素、プライマー、およびヌクレオチドを除去した。50μLのヌクレアーゼ未含有水でcDNAを溶出した。
【0191】
15μlのcDNAへのテール付加を、25μl反応でTdT酵素およびdCTP(Invitrogen社。5’RACE Systemキットより)を用い、製造者の説明書に従って行った。
【0192】
ポリ(dC)テールを付加したcDNA 1μlを20μl反応でネステッドPCRのテンプレートとして使用した。増幅の第1ラウンドを、GSP2AプライマーをAAPプライマー(Invitrogen社。5’RACE Systemキットより)と併用して行い、生成物をゲル精製し、これをテンプレートとして使用し、GSP3プライマーとAUAPプライマー(Invitrogen社。5’RACE Systemキットより)を併用して増幅の第2ラウンドを行った。
【0193】
いずれのPCRもTaq DNAポリメラーゼ(New England Biosciences社)を使用し、2mM Mg2+を含有する1X Thermopolバッファー(New England Biosciences社)に62.5μMの各dNTP(Bioline社)および250μMの各プライマーを添加したものを用いて行った。GSP2A/AAP PCRは初期変性段階を94℃で1分間行った後、94℃で30秒間/66℃で30秒間/72℃で40秒間を35サイクル、そして最後に72℃で更に5分間とした。生成物20μlをアガロースゲルで泳動し、約650bpのバンドを切り出し、QIAquickゲル精製キット(Qiagen社)を使用して製造者の説明書に従って抽出した。精製した生成物を1:1000の希釈率で第2ラウンドの増幅のテンプレートとして使用した。
【0194】
GSP3/AUAP PCRは、アニーリング温度が66℃ではなく68℃とした以外は、同様である。シークエンシングのために十分なDNAを得るため、150μlのGSP3/AUAP PCRを行った。生成物75μlをアガロースゲルで泳動し、約500bpのバンドを切り出し、QIAquickゲル精製キット(Qiagen社)を使用して製造者の説明書に従って抽出した。30μlの溶出バッファーでDNAをカラムから溶出し、サンプルを未精製のPCR産物と並行してゲルで泳動して回収を確認し、濃度を算出し(約100ng/μl)、その後、残りのサンプルでシークエンシングを行った(図13b)。
【0195】
AUAPプライマーを用いてゲル精製したPCR産物のシークエンシングを行い、ジャパニン cDNAの5'配列を得た(図14)。
【化3】
【0196】
実施例11 完全長ジャパニンのクローニングのためのプライマー設計
5'および3' cDNA配列がクローニングされたことにより、ジャパニンcDNAの推定される完全長配列(176残基ペプチドをコード)を構築することが可能となった(図15)。ジャパニン配列中の残基配置および他の既知の分子との配列類似性は、ジャパニンがリポカリンであることを示唆している。
【0197】
神経ネットワーク分析は、最初の24アミノ酸が分泌のためのシグナル配列であることを示唆している。それらの後に続く配列はThr Pro Ser Met Pro Ala Ile Asn Thr Gln Thr Leu Tyr Leu Alaであり、DC調節HPLC画分から得たN-末端配列と一致する。これによって、正確なcDNAがシークエンシングされたことが確認された。
【0198】
ネステッドPCR法を用いて完全長cDNAのクローニングを行った。いずれのPCRラウンドも、ポリメラーゼによって生じる変異導入を最小限にするために、ハイフィデリティDNAポリメラーゼを使用した。推定される完全長cDNA配列に基づき、シグナル配列を含有するプライマーを設計し、第2ラウンドのプライマーには、サブクローニングを容易にするために、順方向および逆方向プライマーの5'末端にそれぞれBamHIおよびNotI制限部位を導入した。制限酵素は直鎖状核酸の最末端を切断することはできないことが報告されているので、制限部位の前に5個の更なるヌクレオチドを付加した。
【化4】
【0199】
いずれのPCRラウンドもPhusion HotStart DNAポリメラーゼ(New England Biosciences社)を使用し、50μMの各dNTP(Bioline社)および250nMの各プライマーを添加した1X HFバッファー(New England Biosciences社)中で行った。
【0200】
前記と同様、第1ラウンドのPCRは順方向プライマー5および逆方向プライマー3を使用し、1日給餌を行ったメスR. appendiculatusの唾液腺から生成されたcDNAをテンプレートとして行った。初期変性段階は98℃で30秒間、その後、98℃で10秒間/64.6℃で30秒間/72℃で30秒間を35サイクル、そして72℃で更に1分間とした。この反応生成物を1:100000の希釈率で第2ラウンドの増幅のテンプレートとして使用した。
【0201】
第2ラウンドのPCRは順方向プライマー6および逆方向プライマー4を使用して行った。初期変性段階は98℃で30秒間、その後、98℃で10秒間/41℃で30秒間/72℃で30秒間を2サイクル、98℃で10秒間/72℃で30秒間を20サイクル、そして72℃で更に5分間とした。
【0202】
実施例12 ジャパニンのクローニング
5' BamHI部位および3' NotI部位を付加した完全長ジャパニンをコードするDNAを上記のようにPCRで増幅し、BamHIおよびNotIで切断し、同様に処理したpBacPAK8ベクターにライゲートした。ライゲートしたDNAを用いてTOP10 E. coliに形質転換した後、個々のコロニーを増殖およびミニプレップし、それらのプラスミドDNAをシークエンシングした。
【0203】
完全長ジャパニン(5' BamHI部位および3' NotI部位を有する)をコードするDNAを増幅するための反応(50μl)を上記のように行った。生成物35μlをQIAquick PCR精製キット(Qiagen社)で処理してプライマーおよびヌクレオチドを除去し、バッファー強度およびその後の制限酵素バッファーpHに対する影響を低減するために水で0.33Xに希釈した溶出バッファー30μlでプラスミドを溶出した。
【0204】
増幅されたDNAをBamHIおよびNotIで消化した(BSAを添加した1X BamHIバッファー中、37℃で1時間インキュベーション)。その後、酵素を除去するため、QIAquick PCR精製キット(Qiagen社)によってDNAを精製し、30μlの溶出バッファーで溶出した。酵素およびバッファーは全てNew England Biosciences社製であった。
【0205】
pBacPAK8プラスミドを同様に消化し、その後、切断したマルチクローニング部位フラグメントが確実に除去されるよう、QIAquickゲル抽出キットを用いてゲルを精製した。
【0206】
10μlの反応液(1X T4 DNAリガーゼバッファー(New England Biosciences社)中に約60ngの切断したpBacPAK8、約5ngの切断したジャパニンPCR産物、および1μl T4 DNAリガーゼ(New England Biosciences社)を含有)中で、ジャパニンDNAをpBacPAK8にライゲートさせた。
【0207】
3μlのライゲーション反応液を用いて、50μlの化学的コンピテントセルTOP10 E. coliに形質転換した。アンピシリンを含有するLB寒天培地上で一晩培養した後、独立したコロニーを5mlのLB培地(+アンピシリン)に接種して一晩培養し、QIAprep Spinキット(Qiagen社)を使用し、製造者の説明書に従って、プラスミドDNAを単離した。
【0208】
Bac1およびBac2プライマーを用いてシークエンシングを行った。得られた配列から、推定される完全長cDNA配列が正確であることが確認され、組換えバキュロウィルス(pBacPAK8-ジャパニン)作製のための変異未含有クローンの選別が可能となった。
【0209】
実施例13 昆虫細胞培養液中でのジャパニンの発現
flashBac系を用いてジャパニン発現組換えバキュロウィルスを作製し、これを用いてSf9昆虫細胞をジャパニン移入ベクターおよび必須遺伝子欠損を有する変異ウィルスで共トランスフェクトした。強力なプロモーターの制御下で、ベクターおよびウィルス間の相同組換えにより必須遺伝子の機能が回復され、同時に、ジャパニン配列がウィルスに挿入された。これにより、生存可能ウィルスは全てジャパニンDNAを含有するということが確認される。増幅後、組換えウィルスを用いて新たなSf9細胞に感染させ、培養液上清を回収し、DC調節活性についてスクリーニングを行った。図17に示すように、上清がDC調節活性を有することが明らかとなり、これは機能性ジャパニンが生成され、培養液中に分泌されたことを示しているが、この活性をアンマスクするために、まず上清を100kDA MWCOフィルターに通過させてウィルス粒子を除去する必要があった。おそらくこれはウィルス粒子の高度な刺激作用がジャパニンの阻害作用を上回るか、または回避するためである。
【0210】
log期のSf9細胞を1.1×106細胞/ウェルの密度で6ウェルプレートに接着させた後、Cellfectinトランスフェクション試薬(Invitrogen社)を使用し、flashBacゴールド・バキュロウィルスDNA(Oxford Expression Technologies社)およびpBacPAK8-ジャパニンの混合物でトランスフェクションを行った。500ngのプラスミドDNAを、1mlのSf900 II培養液に混合した0.5μlのflashBacゴールドDNAおよび5μlのCellfectinと混合し、室温で25分間インキュベートして複合体を形成させた。培養液を接着Sf9細胞から除去し、DNA/Cellfectin複合体で置換し、細胞を一晩インキュベートした後、更に1mlのSf900 II培養液を添加し、更に4日間インキュベーションを継続した。この時点でウィルスを含有する上清を回収し、4℃の暗所に保存した。
【0211】
次いで、このようにして得た少量のウィルス・ストックをより多量のSf9細胞培養液に播種し、ウィルスを増幅した。0.5mlのウィルス含有上清をlog期にあるSf9細胞の撹拌培養液250ml(細胞密度:約1.5X106/ml)に添加した。その後培養液を5日間インキュベートし、上清を回収し、4℃の暗所に保存した。
【0212】
増幅したウィルス・ストックを用いてSf9細胞を多重感染させ、タンパク質を発現させた。25mlのウィルス・ストックをlog期のSf9細胞培養液250ml(8×105細胞/ml)に添加した。その後、培養液を3日間インキュベートし、遠心分離によって上清を回収した。
【0213】
ウィルス粒子を除去するために、上清サンプル5mlを100kDa MWCO Vivaspin 6遠心濃縮キット(Sartorius社)に通過させた。このサンプルを通常の方法でDC調節活性についてスクリーニングし、QFT(陽性コントロール)、未濾過のバキュロウィルス/ジャパニン上清、および無関係のタンパク質を発現するバキュロウィルス-Sf9細胞培養液上清についても並行して行った。結果は、明らかに上清中にDC調節活性が存在することを示したが、これは未濾過上清中のウィルス粒子の存在によってマスキングされ、おそらくこれは粒子そのものがDCに対して圧倒的な刺激を与えるためであると考えられる。この結果により、「ジャパニン」としてクローニングされたタンパク質が実際に予測されたDC調節特性を有し、Sf9細胞によって活性型として生成されることが確認された。
【0214】
実施例14 ジャパニンを含有する昆虫細胞の上清から単離されたタンパク質の沈殿
ポリエチレングリコール(PEG)または硫酸アンモニウムのいずれかを添加して、バキュロウィルス/ジャパニン上清からタンパク質を沈殿させた。硫酸アンモニウムで沈殿させたタンパク質を、Superdex 75カラムを用いるゲル濾過によって更に分画した。図18に示すように、DC調節活性は硫酸アンモニウム沈殿させたタンパク質中には保持されるが、PEG沈殿させたタンパク質中には保持されず、また、プールしたSuperdex画分22-38(タンパク質の大部分を含有)中にも存在することが明らかになった。その後の画分22-38のスクリーニングにより、プールした画分23+24が最も活性が高いことが明らかになった。
【0215】
氷上で撹拌しながら、上清にPEGを最終濃度30%(w/v)まで徐々に添加した。同様に硫酸アンモニウムを70%(w/v)まで添加した。
【0216】
PEG沈殿したタンパク質を50mM HEPES(pH7.2)に再溶解し、Qカラムを用いて分画した。A280のリアルタイム測定により、画分4が分離したタンパク質ピークを含有することが示唆されたため、Superdex 75を用いるゲル濾過によってこの画分を更に分画した。このカラムから大部分のタンパク質が画分22および23に溶出された。
【0217】
硫酸アンモニウム沈殿したタンパク質を50mM HEPES(pH7.2)に再溶解し、5kDa MWCO Vivaspin 6遠心濃縮キット(Sartorius社)を使用してx15に濃縮した。Superdex 75カラムを用いるゲル濾過によって、濃縮されたタンパク質を分画した。A280のリアルタイム測定により、画分22-28が大部分のタンパク質を含有することが明らかになった。
【0218】
これらのサンプルについて、通常の方法でDC調節活性のスクリーニングを行ったところ(それぞれ1:100希釈)、活性は硫酸アンモニウム沈殿させたタンパク質中には保持されたが、PEG沈殿させたタンパク質中には検出されないことが明らかとなった。この事から予測されるように、選択されたPEG沈殿タンパク質のQカラム結合画分は活性を示さなかったが、このQカラム結合画分からのSuperdex画分22+23と共にインキュベートした後にはCD86発現のわずかな低下が観察された:この結果は明確なものではなく、更なる検討は行わなかった。反対に、硫酸アンモニウム沈殿したタンパク質は濃縮およびゲル濾過クロマトグラフィー後もその活性を保持し、その後のゲル濾過画分の比較から、画分23+24が最も高い活性を有することが明らかとなり、従って最も高濃度のジャパニンを含有することが想定される。
【0219】
実施例15 hisタグ・ジャパニンのクローニング
3段階ネステッドPCRを用い、pBacPAK8-ジャパニンをテンプレートとして、C末端に6残基のポリヒスチジン融合タグ(hisタグ)を付加したジャパニンを再クローニングした。ネステッドPCR産物を制限酵素で消化し、同様に制限酵素消化したpBacPAK8プラスミドへのライゲーションを行った。hisタグと天然のタンパク質配列の間に4つの更なる残基(グルタミン-グリシン-グリシン-セリン)が導入されるようにプライマーを設計した。これは、天然配列にhisタグが近接することによる立体障害を防止し、また、TEVプロテアーゼの共通開裂部位を導入する(これによって、タンパク質分解によるタグの除去が容易になる可能性がある)ことを目的とする。
【化5】
【0220】
第1段階のPCRは、50μMの各dNTP(Bioline社)、500nMの各プライマー、および10pg/μlのテンプレート・プラスミドを添加した1X HFバッファー(New England Biosciences社)中、Phusion DNAポリメラーゼ(New England Biosciences社)を用いて行った。初期変性段階は98℃で30秒間、その後、98℃で10秒間(変性)/70℃で30秒間(アニーリング)/72℃で15秒間(伸長)を15サイクル、そして最後に72℃で更に5分間とした。使用したプライマーは順方向プライマー7および逆方向プライマー5であった。このように行った20μlの反応液の産物をQIAquick PCR精製キットで精製し(30μl溶出バッファーで溶出)、プライマーおよびヌクレオチドを除去した。
【0221】
第2段階のPCRも同じ方法で行った。ただし、アニーリング温度を69℃とし、精製した第1段階のPCR産物を蒸留水で1:10に希釈してテンプレートとし、逆方向プライマーとして逆方向プライマー6を使用した。このように行った20μlの反応液の産物をQIAquick PCR精製キットで精製し(30μl溶出バッファーで溶出)、プライマーおよびヌクレオチドを除去した。
【0222】
第3段階のPCRは第2段階のPCRと同様に行った。ただし、精製した第2段階のPCR産物を蒸留水で1:10に希釈してテンプレートとし、逆方向プライマーとして逆方向プライマー7を使用した。このように行った50μlの反応液の産物をQIAquick PCR精製キットで精製し(30μlの0.5X溶出バッファーで溶出)、プライマーおよびヌクレオチドを除去した。反応から得たサンプルをアガロースゲルで泳動したところ、PCR産物の濃度は約20ng/μlと試算された。
【0223】
精製した第3段階のPCR産物をBamHIおよびNotIで消化した(BSAを添加した1X BamHIバッファーを含有する50μlの反応液中、37℃で20分間インキュベート)。バッファーおよび酵素はNew England Biosciences社のものを使用した。反応液の産物をQIAquick PCR精製キットで精製し(30μlの溶出バッファーで溶出)、酵素および切り出されたフラグメントを除去した。
【0224】
約1:1のモル比としてライゲーションを最適化するため、制限酵素消化産物約4.5ngを前記のBamHI/NotI消化およびCIP処理したpBacPAK8(50ng)とライゲートさせた。ライゲーションはT4 DNAリガーゼ(New England Biosciences社)を用い、T4 DNAリガーゼ・バッファー(New England Biosciences社)を含有する10μl反応液中、室温で15分間行った。ライゲーション混合液を用い、TOP10 E.coliコンピテントセルに形質転換した。分離したコロニーを5mlの液体LB培地に接種し、QIAprep Spinキット(Qiagen)を製造者の説明書に従って使用し、プラスミドDNAを単離した。Bac1およびBac2プライマーを用いてシークエンシングを行い、ジャパニン融合タンパク質をコードする挿入断片の存在を確認した。以下、このプラスミドをpBacPAK8-Jap-TEV-hisと称する。
【0225】
実施例16 hisタグ・ジャパニンの単離
pBacPAK8-Jap-TEV-hisプラスミドDNAを用い、flashBacゴールドシステムを使用して、前記の未標識組換えジャパニンの生成と同様に、組換えバキュロウィルスを調製した。次いで、これらの組換えバキュロウィルスを用いて、前記のようにSf9細胞の発現培養液250mlに感染させた。その後、水酸化アンモニウムを用いて培養液上清からタンパク質を沈殿させ、ポリヒスチジン・モチーフを結合させるIMACカラムを用いて精製した。SDS-PAGEゲルの銀染色により、組換え特異的タンパク質が約20kDaに存在することが明らかとなり(図19)、ウェスタン・ブロットにより、実際にこのタンパク質にhisタグが付加されていることが確認された(図20)。これらの結果は、hisタグ・ジャパニンがうまく発現されたことを示している。
【0226】
前記の未標識組換えジャパニンの生成と同様に、3日目のpBacPAK8-Jap-TEV-his発現培養液250mlの上清に硫酸アンモニウムを70%(w/v)まで添加し、タンパク質を沈殿させた。沈殿させたタンパク質を60mlの40mM Na2HPO4/300mM NaCl/10%グリセロール(pH8)(結合バッファー)に再溶解し、あらかじめ1mlのTalon樹脂(Clontech社)を充填しておいたカラムに負荷した。その後、カラムを20mlの結合バッファーで2回洗浄し、6mlの40mM Na2HPO4/100mM NaCl/300mMイミダゾール(pH8)(溶出バッファー)で溶出した。溶出されたタンパク質を5000MWCO Vivaspin 6(Sartorius)でX10に濃縮した。
【0227】
0.5μlのサンプルを4-12%ポリアクリルアミドBis-Trisゲル(Invitrogen社のNuPageプレキャスト・ゲル)で泳動した後、SilverXpressキット(Invitrogen社)を用い、製造者の説明書に従って銀染色を行った。2つの組換えタンパク質上清には約20kDaに2つの主要なバンドが明らかに存在したが、陰性コントロール精製物(野生型バキュロウィルスに感染させた細胞からの上清を用いて並行して行った)には存在しなかった。
【0228】
ウェスタン・ブロット分析のために、6.5μlのサンプルを4-12%ポリアクリルアミドBis-Trisゲル(Invitrogen社のNuPageプレキャスト・ゲル)で泳動した後、ウェット式ブロッティング(10%のメタノールを添加したNuPageトランスファー・バッファー中、30Vの定電圧を1時間適用)によって45μm孔のニトロセルロース膜(Biorad社)に転写した。転写完了後、膜を蒸留水で洗浄し、Trisバッファー/0.1% Tween 20(TBST)で5分間洗浄した後、ブロッキング試薬(#B6429、Sigma社)中、室温で1時間インキュベートした。ブロッキング後、膜をTBSTで軽く洗浄し、ブロッキング試薬で1:1000に希釈した抗ペンタ-his抗体(Qiagen社)と共に4℃で一晩インキュベートした。その後、TBSTで5分間の洗浄を5回行い、10%脱脂粉乳(Marvel社)/TBSで1:20000に希釈したロバ抗マウス-HRP(Jackson Immunoresearch社)中、室温で1時間インキュベートし、TBSTで5分間の洗浄を更に7回行った。最後に抗体を可視化するために、膜をECL基質(Amersham社)で製造者の説明書に従って処理し、これにX線フィルムを10秒間暴露させた。カラムから溶出された最初の4つの1ml画分中に約20kDaのバンドが明らかに存在し、予測されたサイズのhisタグ・タンパク質の存在が確認された。
【0229】
実施例17 ジャパニン-TEV-his上清由来サンプルのCD調節活性
前記のようにジャパニン-TEV-his上清由来サンプルについてDC調節活性のスクリーニングを、サンプルを1:100希釈して行った。タグを付加していない組換えジャパニン由来のSuperdex画分23+24を陽性コントロール、野生型バキュロウィルスに感染させたSf9細胞培養液由来のサンプルを陰性コントロールとして、DC調節活性についてのスクリーニングを行った。図21に示すように、ジャパニン-TEV-his上清から硫酸アンモニウムを用いて沈殿させたタンパク質に活性があり、活性組換え融合タンパク質が生成されたことを示している。活性はジャパニン-TEV-his上清由来の10X濃縮したTalonカラム結合タンパク質中にも存在し、活性タンパク質が実際にTalon樹脂に結合することが確認された。10X濃縮Talon溶出物がバルクの硫酸アンモニウムで沈殿させたタンパク質に比較して活性が低いことは必ずしも活性の低下を示しているものではなく、Talon溶出バッファーの刺激作用を反映している可能性がある。
【0230】
実施例18 hisタグ・ジャパニンの精製
Talonカラム溶出物をSuperdex75(ゲル濾過)カラムに通過させて、ポリヒスチジン・タグ・ジャパニンを更に精製した。銀染色したSDS-PAGEゲル上の約20kDaのバンドの存在によってジャパニンを含有する画分を同定し、プールした。プールした画分中のタンパク質濃度を、280nmの吸光度およびexpasy.orgのProtParamツールによって成熟ジャパニン配列から予測される吸光係数から算出した。
【0231】
精製したタンパク質のDC調節活性に関するアッセイを、前記のようにポリ(I:C)を24時間後に細胞に添加し、25ng/mlから1.6μg/mlまでの種々の濃度で行った。図22は、100ng/ml以上の濃度で最大活性に達することを示しており、これまでに行った1回の実験では、<25ng/mlで50%活性に達しうることが示唆された。
【0232】
実施例19 DC分化に対するジャパニンの影響
前記のように、GM-CSFおよびIL-4との培養によって、ヒト単球からDCを生成させた。いくつかの培養液には、200ng/mlの組換えジャパニンを更に添加した。3日目から6日目まで、日毎に培養液のCD14およびCD1a発現を分析した。
【0233】
分化に対するジャパニンの影響が、ジャパニンそのものに対する影響ではなく、組換えジャパニンの内毒素の混入によるものであるという反論もありうるため、ジャパニンの内毒素含有量の評価をLALアッセイを用いて行い、約0.540EU/μg(およそ、ジャパニン1μg当たり0.054ngのE. coli LPSに相当)であることが明らかとなった。
【0234】
図23に見られるように、200ng/mlのジャパニンによって分化培養物の生育は大きく改変され、約50%の単球がDCへの分化の痕跡であるCD1aの亢進的調節およびCD14の抑制的調節を行うことができなかった。9pg/mlまたは40ng/mlのいずれかのE. coli LPS(それぞれ、200ng/mlで使用した場合の組換えジャパニンの内毒素含量、およびその内毒素含量の>4000倍に相当)を添加したコントロールにより、いずれの濃度も分化に大きな影響を与えないことから、これはジャパニンの内毒素混入の副作用ではないということが明らかとなった。
【0235】
実施例20 T細胞増殖アッセイ-混合白血球反応(MLR)
混合白血球反応(MLR)を用いて、特異的抗原(この場合、同種異系MHC)を提示するmoDCに応答したT細胞増殖に対するジャパニンの影響を評価した。この系においては、ジャパニンは顕著なT細胞増殖阻害を示した。
【0236】
上記のように調製した、凍結された5日目の単球由来DCを解凍し、200ng/mlの組換えジャパニンの存在下または非存在下で更に2日間培養した。
【0237】
CD3 MACSマイクロビーズ(Miltenyi Biotech社)を製造者の説明書に従って用い、前記のように、Lymphoprepを用いて行うPBMCの初期分画によって同種異系T細胞をバフィーコートから単離した。
【0238】
1x105 T細胞/ウェルを、段階的に希釈した放射線照射樹状細胞と共に96ウェル丸底プレート(Corning社)に播種した。細胞培養液はRPMI 1640に10% FCS + 2mM L-グルタミン、100U/ml ペニシリン、および100μg/ml ストレプトマイシンを添加したもので、最終容量は200μl/ウェルとした。ジャパニンと共に2日間インキュベーションしておいたDCを含むウェルには、更に200ng/mlのジャパニンを追加した。DC未含有のT細胞を含むコントロールも含めた。
【0239】
MLR培養液を4日間インキュベートした後、0.5μCi/ウェルの3H-チミジンで瞬間標識した。次いで、これらを更に16-18時間インキュベートした後、自動細胞回収装置を用いてガラス繊維フィルター上に回収し、フィルターに結合した放射性DNAをシンチレーション・カウンターで定量した。
【0240】
この実験を3回行ったところ、いずれの実験でも、DCをジャパニンで前処理しMLR培養液中にもジャパニンを含有させるとT細胞の増殖が低減されることが明らかになった。結果の例を図24に示す。
【0241】
実施例21 ジャパニン・ホモログの検索(Dermacentor andersoni)
EMBL Expressed Sequence Tag(EST)データベースのBLAST検索で、Dermacentor andersoni由来のジャパニン・ホモログを同定した。このホモログは現在、D. andersoni E1244(EBI ID = EG363153)と称されている。SignalPを用いてDA-E1244のシグナルペプチド部分と考えられる部分(残基1-17)を同定し、成熟ジャパニン配列および予測される成熟DA-E1244配列を比較した。BLOSUM62マトリクスを用いて実施したEMBOSS2配列間アラインメントにより、2つのタンパク質間で30.5%の同一性および50.3%の類似性が確認された(図25)。データベース中に存在するヌクレオチド配列にはSEQ ID NO:32があり、推定上のコードされるタンパク質はSEQ ID NO:4である。
【0242】
このジャパニンとの高度なホモロジーは、DA-E1244がジャパニン様生体活性を有することを強く示唆しており、このため、組換えタンパク質を生成するための研究に着手している。
【0243】
このために、DA-E1244の完全長アミノ酸配列をコードするDNAを合成遺伝子として生成し(SEQ ID NO:3)、クローニング・ベクターに導入し、pCR4TOPO-DA-E1244_optとした(これはEntelechon社(レーゲンスブルク)による契約に従って行われた)。合成されたDNA配列はATGスタート・コドンで始まり、コザック・コンセンサス配列(真核細胞系における効率的な翻訳の必須条件である)に一致する上流配列は導入しなかった。これを改善するために、DA-1244を増幅し、産物の5'末端にBamHI認識部位(“GGATCC”)およびコザック対応(Kozak-compliant)配列(“TCCAAA”)を、3'末端にNotI認識部位(“GCGGCCGC”)を付加するようにプライマーを設計した。「余分の」塩基を5'および3'末端の両方に付加し、制限酵素部位が産物の末端に位置して制限酵素消化を阻害することがないようにした。
【化6】
【0244】
Phusion HotStart DNAポリメラーゼ(New England Biosciences社)を用い、50μMの各dNTP(Bioline社)および250μMの各プライマーを添加した1X HFバッファー(New England Biosciences社)中、20μl反応でPCRを行った。pCR4TOPO-DA-E1244_optプラスミドをテンプレートとして使用し、反応液中に1ng/μlの濃度で含有させた。初期変性段階は98℃で30秒間、次いで98℃で10秒間/69℃で30秒間/72℃で15秒間を15サイクル、最後に72℃で更に5分間とした。
【0245】
完了した反応液の5μlをアガロースゲルで泳動して期待されるサイズ(600bp)の産物の存在を確認し、残りはQIAquickカラム(Qiagen社)を製造者の説明書に従って用いて精製し、BamHIバッファー(NEB社)中、BamHIおよびNotI(共にNEB社)で消化し、再度QIAquickカラムで精製した。
【0246】
その後、切断および精製したDA-1244 PCR産物を、BamHI/NotI消化および仔ウシ腸ホスファターゼ処理を行ったpBacPAK8(バキュロウイルス・トランスファー・ベクター)にライゲートし、ライゲーション反応液を用いてTOP10 E.coliコンピテントセルに形質転換した。
【0247】
形質転換したE.coliの独立したコロニーを採取し、培養液中で増殖し、QIAprepミニプレップ・キットを用いてDNAを抽出した。シーケンシングを行い(Bac1およびBac2プライマーを使用)、変異が導入されることなくDA-E1244がクローニングされたことを確認した。
【0248】
好適なクローンを同定し(pBacPAK8-E1244)、組換えバキュロウィルスの作製に使用した。
【0249】
実施例22 更なるジャパニン・ホモログの検索
EMBL Expressed Sequence Tag(EST)データベースのBLAST検索で、ジャパニン・ホモログがRhipicephalus microplus(2つのホモログ)、Amblyomma americanum、およびRhipicephalusappendiculatusから同定された。現在これらのホモログはそれぞれ、R. microplusCK185494、A.americanum CX766068、R. appendiculatus CD796501、およびR. microplus CV436349と称されている。これらのタンパク質の配列アラインメントおよび同一性%の図を図26-29に示す。
【0250】
R. microplus CV436349は、これらの同定されたホモログのうち、唯一、シグナルペプチドを含有する。他の3つの同定されたホモログにシグナル配列が存在しないことに関しては、3つの説明が可能である:i)それらは分泌タンパク質ではない;ii)配列の一部が欠失している;またはiii)非標準的分泌が関与している。
【0251】
R.microplus CK185494およびA. americanum CX766068はいずれもジャパニンと実質的なホモロジーを有するリポカリンである。しかしながら、これらの配列はいずれもシグナル配列を有さず、上記のように、それらが分泌タンパク質ではないか、あるいは配列の一部が欠失していることを示唆している。
【0252】
これらのジャパニン・ホモログはジャパニンとの配列同一性がDermacentor andersoniから同定されたホモログより低いが、その機能はジャパニンに匹敵すると考えられる。
【0253】
実施例23 ジャパニン・リガンドとしてのコレステロールの同定
ジャパニンはリポカリンであると報告されており、これは脂質リガンドに結合しうる可能性を示唆している。OmCI(脂肪酸結合マダニ・リポカリン)の結晶構造に基づいて仮定されるジャパニンの構造モデルの構築により、ジャパニン中に疎水性のオープン結合ポケットが存在することが示唆され、更にこの見解を支持している(未掲載)。
【0254】
これを更に検討するため、実施例13に記載するように組換えジャパニンを昆虫細胞培養液中で生成し、前記のように金属アフィニティークロマトグラフィーおよびゲル濾過に連続的に施与して精製した。この精製組換えタンパク質400μgを用い、下記のようにBligh & Dyer法で脂質を抽出した後、ガスクロマトグラフィー質量分析(GC-MS)を行った。
【0255】
Bligh & Dyer脂質抽出法
3.75mlのクロロホルム:メタノール(1:2)を0.5mlのタンパク質サンプル(または0.5mlのバッファーコントロール)に添加した。この混合液を10-15分間撹拌し、更に1.25mlのクロロホルムを添加し、ボルテックスで1分間混合した。その後、1.25mlの超純粋を添加し、更に1分間ボルテックスを行った。得られたサンプルを遠心分離し、脂質を含有する下層を残し、上層を廃棄した。これを窒素雰囲気下で乾燥し、500μlのジクロロメタンに再懸濁した。
【0256】
ガスクロマトグラフィー/電子衝撃質量分析(GC/EI-MS)
組換えジャパニンまたはバッファー・ブランクから抽出したサンプル1μlを、キャピラリー・ガスクロマトグラフが組み込まれたPerkin Elmer Turbomass四重極質量分析計に注入した。以下の条件を使用した:
ガスクロマトグラフィー:カラム=DB-5。注入=オンカラム。注入温度=40℃。温度グラジエント=40℃、1分間。その後、8℃/分で325℃まで(10分間保持)。キャリアガス=ヘリウム。
質量分析:イオン化電圧=70eV。イオン化モード=スキャニング。MS分解能=ユニット。
【0257】
質量分析から得られたデータは、ジャパニン・サンプルで33.1分にピークを示し、これはバッファー・ブランクには存在しなかった(図30a)。このピークからの平均分布(図30b)をNISTライブラリーの分布と比較することにより、これがコレステロールであることが同定された。これを確認するために、コレステロール標準品を同じ条件下で処理したところ、33.1分にピークが得られ、平均分布は一致した(図30c)。
【0258】
実施例24 組換えジャパニンは遊離コレステロールに結合する
組換えオリゴヒスチジン・タグ・ジャパニンをNi-NTA磁気ビーズに固定するため、0.5μgのタンパク質を、ビーズを含有させた500μlのバッファーA(120mM NaCl、0.02% Tween、5%グリセロール、40mMリン酸二ナトリウム)中、室温で2時間インキュベートした。コントロールサンプルにはタンパク質を含有させなかった。タンパク質でコーティングされたビーズを500μlのバッファーAで3回洗浄した後、0.2μlの3H-コレステロールを含有するバッファーB(6M グアニジン、120mM NaCl、0.02% Tween、5%グリセロール、40mMリン酸二ナトリウム)50μlを添加した(変性バッファーは、起こりうる、放射能標識されたコレステロールを有するタンパク質に結合した細胞培養液由来のコールドリガンドの交換を促進するために使用した)。5分後、バッファーBを除去し、更に0.2μlの3H-コレステロールを含有させたバッファーAを500μl添加した。その後、室温で3時間インキュベートした。氷冷したバッファーAでビーズを洗浄し(500μlで1回、50μlで2回)、未結合のコレステロールを除去した。その後、これを0.5Mのイミダゾールを含有するバッファーA、100μlに再懸濁して、タンパク質をビーズから溶出させた。図31の洗浄1および2は50μlの洗浄であり、各サンプルの右側のバーはビーズ/タンパク質に結合した放射活性の量を示す。
【0259】
図31に示されるように、これらの結果は明らかに3H-コレステロールがジャパニンに結合することを示している。変性/リフォールディングがタンパク質の結合に必要であるか否かはまだ明らかではなく、結合の強度および特異性も確認する必要が残されている。
【0260】
実施例25 ジャパニンは樹状細胞上のC型レクチン細胞表面受容体に結合する
ジャパニンが樹状細胞成熟化を阻害する能力は、その樹状細胞表面への結合能を暗示するものである。これは、ジャパニンとの膜受容体特異的相互作用を介して起こる可能性が最も高いと考えられるが、ジャパニンが非特異的な(おそらく結合したコレステロールと血漿膜との相互作用を伴う)方法で細胞に結合および侵入し、その後、細胞内シグナル伝達経路に細胞型特異的に作用する作用機構も考えられる。
【0261】
ジャパニンが特異的に樹状細胞表面に結合するか否かを検討し、任意の相互作用の性質の検討を可能にするために、市販のキットを使用して、ジャパニンを蛍光色素Alexa 488で標識した。この蛍光標識したジャパニン、500ng/mlと共に細胞を4℃で30-60分間インキュベートした後、洗浄し、フローサイトメトリーでジャパニン結合を可視化した。
【0262】
ジャパニンが単球由来樹状細胞に特異的に結合することが図32aで明らかであり、ジャパニン-Alexa 488(32a-fの塗りつぶしのヒストグラム)は5日目の単球に由来する樹状細胞(前記のように作製)に結合するが、コントロール・リポカリンとして使用したムバチン(moubatin)-Alexa 488は結合しない(図32a-fの点線のヒストグラム)ことを示している。更に、ジャパニンは単球にも(図32b)、マウス骨髄由来樹状細胞にも(図32c)結合しない。
【0263】
先に証明されたジャパニンが単球の樹状細胞への分化を阻害する能力を考慮すると、ジャパニンが単球に結合しないことは予想外のことである。このことから、ジャパニンは、明らかに結合しない細胞型に対して、いかにして作用できるのかという疑問が浮上する。この問題に取り組むため、ジャパニン-Alexa 488(32a-fの塗りつぶしのヒストグラム)またはムバチン-Alexa 488(図32a-fの点線のヒストグラム)を1日目の単球から作製した樹状細胞と共にインキュベートした(図32d)。ジャパニンは、5日目単球由来樹状細胞より程度は低いが、1日目の単球由来樹状細胞に結合することが明らかになった。これは、ジャパニン結合受容体の亢進的調節が樹状細胞分化の非常に早い段階で開始され、そのためジャパニンはこの初期段階で細胞に作用してその分化を停止させるという可能性を示唆している。
【0264】
ジャパニン-樹状細胞相互作用の性質を検討するため、マンナンおよびEDTAの影響を試験した。1mg/mlのマンナンの存在により、単球由来樹状細胞へのジャパニン-Alexa 488の結合は大幅に低減されたが(図32e。塗りつぶしのヒストグラムはマンナン非存在下でのジャパニン-Alexa 488結合、白抜きのヒストグラムはマンナン存在下での結合、そして点線のヒストグラムはコントロール・タンパク質の結合を示す)、0.5mM EDTAの存在下ではこれが完全に阻害された(図32f。白抜きのヒストグラムがEDTA存在下での結合を示す以外は、図32dと同様)。考え合わせると、これらの知見はジャパニンが単球由来樹状細胞上のC型レクチン細胞表面受容体に結合することを強く示唆している。
【0265】
実施例26 ジャパニンはN-グリコシル化される
NetNGlyc 1.0(Center for Biological Sequence Analysis、デンマーク工科大学)の使用から、ジャパニンが1つの可能性の高い、そしてもう1つの可能性を有するN-グリコシル化部位を含有することが示唆される(図33a)。また、前記のように、ある程度のグリコシル化の存在が、ジャパニンとC型レクチン受容体との相互作用から示唆されている。
【0266】
N-グリコシル化の存在を確認するために、精製した組換えジャパニン(前記のように作製)をPNGase F(ほとんどの型のN-グリコシル化を阻害する酵素)で37℃、16時間処理した。次いでPNGase F処理をしたジャパニンを模擬処理および未処理のジャパニンと並行してSDS-PAGEゲルで泳動し、抗hisタグ・ウェスタンブロッティングで可視化した。図33bに示すように、PNGase F処理では、模擬処理および未処理のジャパニンにも見られる2つのバンドに加えて更なるより小さいバンドが存在した。これはより大きな2つのバンドの少なくとも一方、おそらくは両方がN-グリコシル化されたジャパニンであることを示している。
【0267】
実施例27 組換えジャパニンは多くの多様な刺激に反応して樹状細胞成熟化を阻害する
前記のように、組換えジャパニンはポリ(I:C)(TLR3刺激)に応答して単球由来樹状細胞の成熟化を阻害し、3日間給餌したメスR. appendiculatusマダニ由来のジャパニン含有Qカラム・フロースルーはLPS、TLR4刺激、およびIFNγ(γ-インターフェロン受容体を介して作用する)に応答して単球由来樹状細胞の成熟化を阻害するが、TNFR1を介して作用する可溶性TNFαには応答しない。これらの知見は、(前記と同様の方法に従って)これらの実験を反復して得られたが、これには精製組換えジャパニン(前記のように作製)を用い、LPS(図34a)、IFNγ(図34b)、TNFα(図34c)、可溶性CD40L(図34d)、IFNα(図34e)、またはCL097(TLR7/8リガンド)(図34f)で刺激を行った。TNFα以外のこれら全ての刺激で惹起される樹状細胞の成熟化はジャパニンで阻害される。TNFαで誘導される成熟化に対する有意な作用は観察されなかったが、最低限の阻害は起こりうる。これらの知見から、ジャパニンが、多くの異なる受容体および下流シグナル伝達系を介して作用する広範な刺激に応答して、樹状細胞の成熟化を阻害する能力を有することが確認される。
【0268】
実施例28 組換えジャパニンは刺激に応答して樹状細胞のTNFα分泌を阻害する
共刺激分子およびMHCクラスIIの亢進的調節に加えて、樹状細胞は炎症性刺激にも種々のサイトカインを生成することによって応答する。ジャパニンが樹状細胞成熟化を阻害する、または何らかの方法で変更する能力を有するか否かを評価するために、2つの刺激、LPSおよびIFNγを合わせたものに応答する前炎症性サイトカイン、TNFαの単球由来樹状細胞生成に対するジャパニンの影響を評価した。
【0269】
ヒト単球由来樹状細胞を前記のように作製した。5日目にそれらを回収し、前記のように、FCS、GCSF、およびIL4を含有するフレッシュの培養液に5x105細胞/mlの密度で再懸濁した。次いでそれらを組織培養用24ウェル・プレート中、精製組換えジャパニン(500ng/ml)の存在下または非存在下で培養し、24時間後、組換えヒトIFNγ(Peprotech社)の刺激カクテルおよび超高純度E.coli 011:B4 LPS(Alexis Biochemicals社)をいくつかのウェルに添加した(最終濃度:20ng/ml IFNγ+200ng/ml LPS)。更に48時間後、培養液上清を回収し、遠心分離して細胞およびデブリを除去した。その後、ELISAキット(Insight Biotechnology社)を製造者の説明書に従って使用し、TNFα濃度を測定した。
【0270】
ジャパニンは刺激カクテルに応答してTNFαの樹状細胞分泌を低減することが明らかになった(図35)。
【特許請求の範囲】
【請求項1】
樹状細胞(DC)調節分子であって、該分子が哺乳動物樹状細胞の分化および成熟化を調節する、上記分子。
【請求項2】
該分子が哺乳動物DCの分化および成熟化を阻害する、請求項1記載の樹状細胞(DC)調節分子。
【請求項3】
該分子がヒトDCの分化および成熟化を調節または阻害する、請求項1または2記載のDC調節分子。
【請求項4】
吸血性節足動物から単離されたものである、請求項1から3のいずれかに記載のDC調節分子。
【請求項5】
該分子がタンパク質である、請求項1から4のいずれかに記載のDC調節分子。
【請求項6】
該分子がTリンパ球の活性化を阻害する、請求項1から5のいずれかに記載のDC調節分子。
【請求項7】
該分子がTリンパ球の分極化を調節する、請求項1から6のいずれかに記載のDC調節分子。
【請求項8】
該分子が免疫抑制剤である、請求項1から7のいずれかに記載のDC調節分子。
【請求項9】
該分子がグリコシル化されている、請求項5から8のいずれかに記載のDC調節分子。
【請求項10】
分子がマダニから単離されたものである、請求項4から9のいずれかに記載のDC調節分子。
【請求項11】
マダニがIxodes、Bothriocrotoninae、Amblyomminae、Haemaphysalinae、Rhipicephalinae(Hyalomminaeを含む)、Nuttalliellidae、Argasinae、Otobinae、Antricolinae、Nothhoaspinae、およびOrnithodorinaeの群から選択される、請求項10記載のDC調節分子。
【請求項12】
リポカリンである、請求項1−11のいずれかに記載のDC調節分子。
【請求項13】
分子が脂質と複合体を形成している、請求項12記載のDC調節分子。
【請求項14】
脂質がステロイドまたはステロールである、請求項13記載のDC調節分子。
【請求項15】
脂質がコレステロールまたはコレステロールの代謝物、例えばビタミンD3である、請求項14記載のDC調節分子。
【請求項16】
DCの外膜上に存在する受容体に結合する、請求項1−15のいずれかに記載のDC調節分子。
【請求項17】
C型レクチン受容体に結合する、請求項16記載のDC調節分子。
【請求項18】
該分子が、
i)SEQ ID NO:2のアミノ酸配列を含有するタンパク質;
ii)i)のタンパク質と少なくとも60%の同一性を有するタンパク質のホモログ;
iii)上記i)のタンパク質もしくは上記ii)のホモログの活性フラグメント;または、
iv)i)、ii)、もしくはiii)の機能的同等物
を含有する、請求項1−17のいずれかに記載のDC調節分子。
【請求項19】
該分子が、
i)SEQ ID NO:2、4、6、8、10、もしくは12のいずれかのアミノ酸配列を含有するタンパク質;
ii)i)のタンパク質と少なくとも60%の同一性を有するタンパク質ホモログ;
iii)上記i)のタンパク質もしくは上記ii)のホモログの活性フラグメント;または、
iv)i)、ii)、もしくはiii)の機能的同等物
を含有する、請求項1−18のいずれかに記載のDC調節分子。
【請求項20】
請求項1−19のいずれかに記載のDC調節分子をコードする核酸配列を含有する核酸分子。
【請求項21】
SEQ ID NO:1、3、5、7、9、または11のいずれかを含有する、請求項20記載の核酸分子。
【請求項22】
高ストリンジェンシー・ハイブリダイゼーション条件下で請求項20または21記載の核酸分子とハイブリダイズする核酸分子。
【請求項23】
請求項20から22のいずれかの核酸配列を含有するベクター。
【請求項24】
請求項23記載のベクターまたは請求項20から22のいずれかに記載の核酸分子を含有するホスト細胞。
【請求項25】
請求項1から19のいずれかに記載のDC調節分子の調製法であって、該タンパク質が発現される条件下で請求項24記載のホスト細胞を培養すること、およびそれによって生成される該タンパク質を回収することを含む上記方法。
【請求項26】
請求項1から19のいずれかに記載のDC調節分子に結合する抗体。
【請求項27】
該DCを請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、または請求項26記載の抗体と接触させることを含む、DCの調節法。
【請求項28】
請求項27に記載される方法によって生成される調節されたDC。
【請求項29】
請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、または請求項28記載のDC、および医薬的に許容されるキャリアを含有する医薬組成物。
【請求項30】
1つまたはそれ以上の更なる治療物質を更に含有する、請求項29記載の医薬組成物。
【請求項31】
1つまたはそれ以上の更なる治療物質が抗炎症剤、免疫調節剤、免疫抑制剤、サイトカイン、サイトカイン模倣タンパク質、またはサントカイン結合タンパク質を含有する、請求項30記載の医薬組成物。
【請求項32】
1つまたはそれ以上の治療物質が疾病関連要素を含有する、請求項29から30のいずれかに記載の医薬組成物。
【請求項33】
治療に使用するための、請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から32のいずれかに記載の医薬組成物。
【請求項34】
自己免疫疾患、移植片拒絶反応、急性および慢性炎症性疾患、アレルギー、または過敏性反応の治療または予防に使用するための、請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から32のいずれかに記載の医薬組成物。
【請求項35】
節足動物媒介性疾患を含む感染性疾患の治療または予防に使用するための、請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から32のいずれかに記載の医薬組成物。
【請求項36】
癌の治療または予防に使用するための、請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から30のいずれかに記載の医薬組成物。
【請求項37】
自己免疫疾患、移植片拒絶反応、急性および慢性炎症性疾患、アレルギー、または過敏症に罹患した動物の治療法であって、該動物に請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から32のいずれかに記載の医薬組成物を投与することを含む上記方法。
【請求項38】
節足動物媒介性疾患を含む感染性疾患に罹患した動物の治療法であって、該動物に請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から32のいずれかに記載の医薬組成物を投与することを含む上記方法。
【請求項39】
癌に罹患した動物の治療法であって、該動物に請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から30のいずれかに記載の医薬組成物を投与することを含む上記方法。
【請求項40】
DC調節分子、核酸、ベクター、ホスト細胞、抗体、DC、または医薬組成物を疾病関連要素と併せて投与する、請求項37から39のいずれかに記載の方法、または請求項31から34のいずれかに記載のDC調節分子、核酸、ベクター、ホスト細胞、抗体、DC、もしくは医薬組成物。
【請求項41】
疾病関連要素が、感染性物質に関係する成分;アレルゲン;アレルギー以外の過敏症に関係する非自己成分;自己免疫疾患に関係する自己成分;移植抗原;および腫瘍抗原から選択される、請求項38記載の方法または請求項38記載のDC調節分子、核酸、ベクター、ホスト細胞、抗体、DC、もしくは医薬組成物。
【請求項42】
請求項1から19のいずれかに記載の調節分子のアゴニストまたはアンタゴニストを同定するための方法であって、
(a)その表面上に受容体を発現する細胞を、スクリーニングすべき化合物と、受容体に結合できる条件下で接触させ(ここで、受容体は化合物の結合に応答して検出可能なシグナルを提供する能力を有する);および
(b)化合物と受容体の相互作用から生成されるシグナルのレベルを測定することによって、化合物が受容体に結合するか否か、および受容体を活性化または阻害するか否かを確認する、
ことを含む上記方法。
【請求項1】
樹状細胞(DC)調節分子であって、該分子が哺乳動物樹状細胞の分化および成熟化を調節する、上記分子。
【請求項2】
該分子が哺乳動物DCの分化および成熟化を阻害する、請求項1記載の樹状細胞(DC)調節分子。
【請求項3】
該分子がヒトDCの分化および成熟化を調節または阻害する、請求項1または2記載のDC調節分子。
【請求項4】
吸血性節足動物から単離されたものである、請求項1から3のいずれかに記載のDC調節分子。
【請求項5】
該分子がタンパク質である、請求項1から4のいずれかに記載のDC調節分子。
【請求項6】
該分子がTリンパ球の活性化を阻害する、請求項1から5のいずれかに記載のDC調節分子。
【請求項7】
該分子がTリンパ球の分極化を調節する、請求項1から6のいずれかに記載のDC調節分子。
【請求項8】
該分子が免疫抑制剤である、請求項1から7のいずれかに記載のDC調節分子。
【請求項9】
該分子がグリコシル化されている、請求項5から8のいずれかに記載のDC調節分子。
【請求項10】
分子がマダニから単離されたものである、請求項4から9のいずれかに記載のDC調節分子。
【請求項11】
マダニがIxodes、Bothriocrotoninae、Amblyomminae、Haemaphysalinae、Rhipicephalinae(Hyalomminaeを含む)、Nuttalliellidae、Argasinae、Otobinae、Antricolinae、Nothhoaspinae、およびOrnithodorinaeの群から選択される、請求項10記載のDC調節分子。
【請求項12】
リポカリンである、請求項1−11のいずれかに記載のDC調節分子。
【請求項13】
分子が脂質と複合体を形成している、請求項12記載のDC調節分子。
【請求項14】
脂質がステロイドまたはステロールである、請求項13記載のDC調節分子。
【請求項15】
脂質がコレステロールまたはコレステロールの代謝物、例えばビタミンD3である、請求項14記載のDC調節分子。
【請求項16】
DCの外膜上に存在する受容体に結合する、請求項1−15のいずれかに記載のDC調節分子。
【請求項17】
C型レクチン受容体に結合する、請求項16記載のDC調節分子。
【請求項18】
該分子が、
i)SEQ ID NO:2のアミノ酸配列を含有するタンパク質;
ii)i)のタンパク質と少なくとも60%の同一性を有するタンパク質のホモログ;
iii)上記i)のタンパク質もしくは上記ii)のホモログの活性フラグメント;または、
iv)i)、ii)、もしくはiii)の機能的同等物
を含有する、請求項1−17のいずれかに記載のDC調節分子。
【請求項19】
該分子が、
i)SEQ ID NO:2、4、6、8、10、もしくは12のいずれかのアミノ酸配列を含有するタンパク質;
ii)i)のタンパク質と少なくとも60%の同一性を有するタンパク質ホモログ;
iii)上記i)のタンパク質もしくは上記ii)のホモログの活性フラグメント;または、
iv)i)、ii)、もしくはiii)の機能的同等物
を含有する、請求項1−18のいずれかに記載のDC調節分子。
【請求項20】
請求項1−19のいずれかに記載のDC調節分子をコードする核酸配列を含有する核酸分子。
【請求項21】
SEQ ID NO:1、3、5、7、9、または11のいずれかを含有する、請求項20記載の核酸分子。
【請求項22】
高ストリンジェンシー・ハイブリダイゼーション条件下で請求項20または21記載の核酸分子とハイブリダイズする核酸分子。
【請求項23】
請求項20から22のいずれかの核酸配列を含有するベクター。
【請求項24】
請求項23記載のベクターまたは請求項20から22のいずれかに記載の核酸分子を含有するホスト細胞。
【請求項25】
請求項1から19のいずれかに記載のDC調節分子の調製法であって、該タンパク質が発現される条件下で請求項24記載のホスト細胞を培養すること、およびそれによって生成される該タンパク質を回収することを含む上記方法。
【請求項26】
請求項1から19のいずれかに記載のDC調節分子に結合する抗体。
【請求項27】
該DCを請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、または請求項26記載の抗体と接触させることを含む、DCの調節法。
【請求項28】
請求項27に記載される方法によって生成される調節されたDC。
【請求項29】
請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、または請求項28記載のDC、および医薬的に許容されるキャリアを含有する医薬組成物。
【請求項30】
1つまたはそれ以上の更なる治療物質を更に含有する、請求項29記載の医薬組成物。
【請求項31】
1つまたはそれ以上の更なる治療物質が抗炎症剤、免疫調節剤、免疫抑制剤、サイトカイン、サイトカイン模倣タンパク質、またはサントカイン結合タンパク質を含有する、請求項30記載の医薬組成物。
【請求項32】
1つまたはそれ以上の治療物質が疾病関連要素を含有する、請求項29から30のいずれかに記載の医薬組成物。
【請求項33】
治療に使用するための、請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から32のいずれかに記載の医薬組成物。
【請求項34】
自己免疫疾患、移植片拒絶反応、急性および慢性炎症性疾患、アレルギー、または過敏性反応の治療または予防に使用するための、請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から32のいずれかに記載の医薬組成物。
【請求項35】
節足動物媒介性疾患を含む感染性疾患の治療または予防に使用するための、請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から32のいずれかに記載の医薬組成物。
【請求項36】
癌の治療または予防に使用するための、請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から30のいずれかに記載の医薬組成物。
【請求項37】
自己免疫疾患、移植片拒絶反応、急性および慢性炎症性疾患、アレルギー、または過敏症に罹患した動物の治療法であって、該動物に請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から32のいずれかに記載の医薬組成物を投与することを含む上記方法。
【請求項38】
節足動物媒介性疾患を含む感染性疾患に罹患した動物の治療法であって、該動物に請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から32のいずれかに記載の医薬組成物を投与することを含む上記方法。
【請求項39】
癌に罹患した動物の治療法であって、該動物に請求項1から19のいずれかに記載のDC調節分子、請求項20から22のいずれかに記載の核酸、請求項23記載のベクター、請求項24記載のホスト細胞、請求項26記載の抗体、請求項28記載のDC、または請求項29から30のいずれかに記載の医薬組成物を投与することを含む上記方法。
【請求項40】
DC調節分子、核酸、ベクター、ホスト細胞、抗体、DC、または医薬組成物を疾病関連要素と併せて投与する、請求項37から39のいずれかに記載の方法、または請求項31から34のいずれかに記載のDC調節分子、核酸、ベクター、ホスト細胞、抗体、DC、もしくは医薬組成物。
【請求項41】
疾病関連要素が、感染性物質に関係する成分;アレルゲン;アレルギー以外の過敏症に関係する非自己成分;自己免疫疾患に関係する自己成分;移植抗原;および腫瘍抗原から選択される、請求項38記載の方法または請求項38記載のDC調節分子、核酸、ベクター、ホスト細胞、抗体、DC、もしくは医薬組成物。
【請求項42】
請求項1から19のいずれかに記載の調節分子のアゴニストまたはアンタゴニストを同定するための方法であって、
(a)その表面上に受容体を発現する細胞を、スクリーニングすべき化合物と、受容体に結合できる条件下で接触させ(ここで、受容体は化合物の結合に応答して検出可能なシグナルを提供する能力を有する);および
(b)化合物と受容体の相互作用から生成されるシグナルのレベルを測定することによって、化合物が受容体に結合するか否か、および受容体を活性化または阻害するか否かを確認する、
ことを含む上記方法。
【図1】

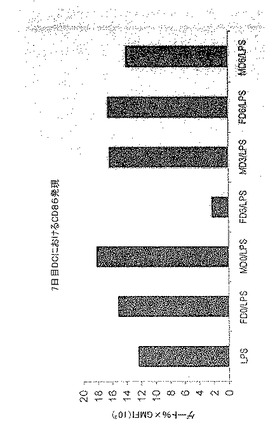
【図2】
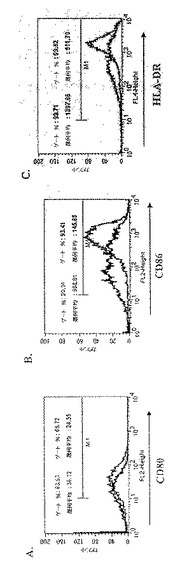
【図3】
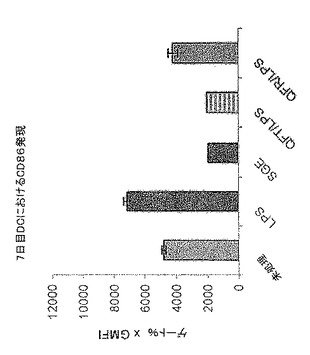
【図4】
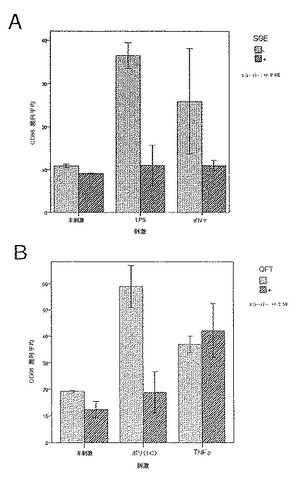
【図5】
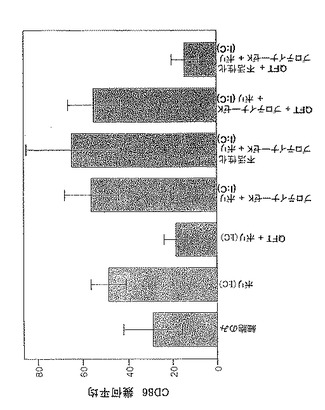
【図6】
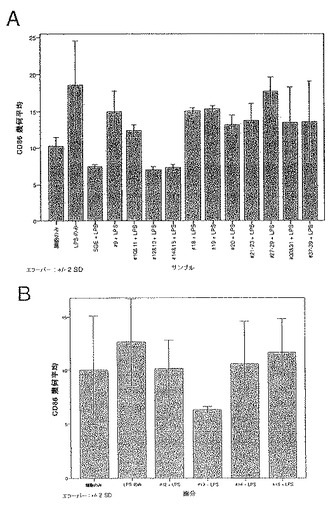
【図7】
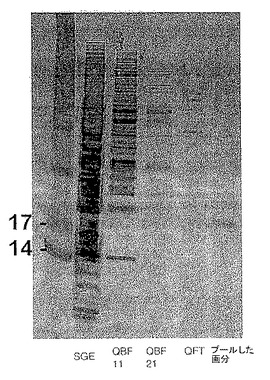
【図8】
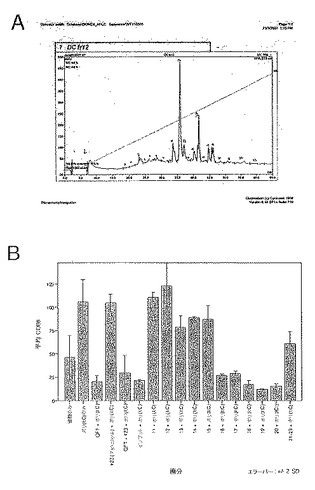
【図9】

【図10】
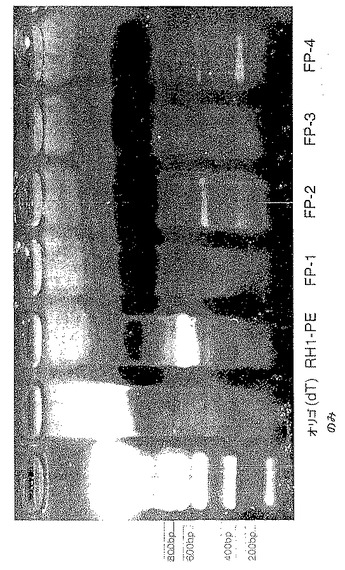
【図11】
【図12】

【図13a】
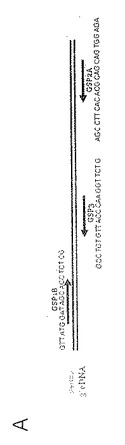
【図13b】

【図14】

【図15】

【図16】

【図17】
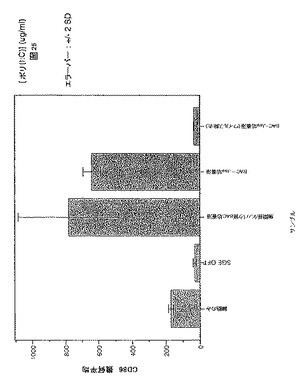
【図18】
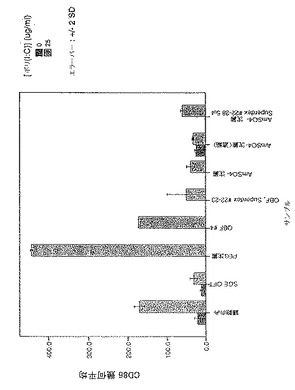
【図19】

【図20】
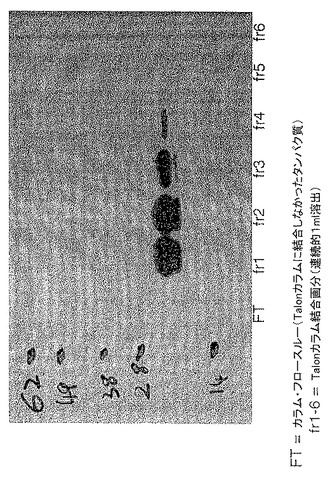
【図21】
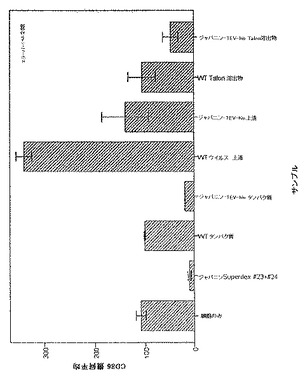
【図22】
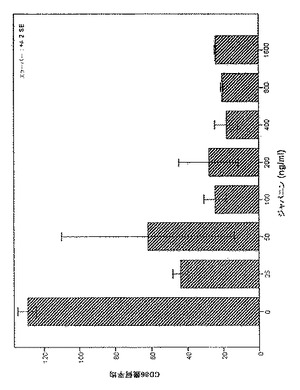
【図23】
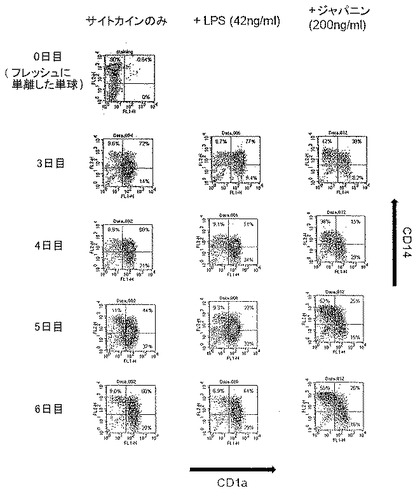
【図24】
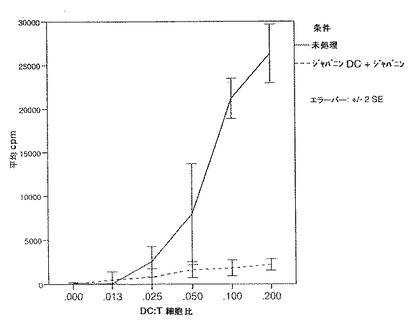
【図25】

【図26】

【図27】

【図28】
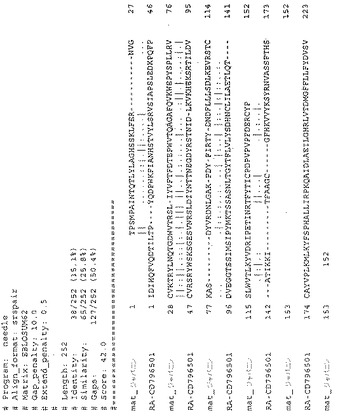
【図29】

【図30a】
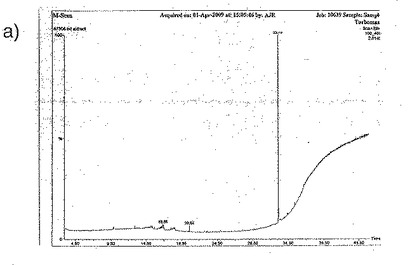
【図30b】
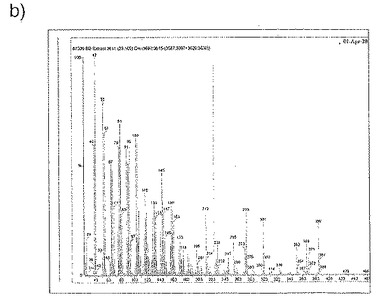
【図30c】
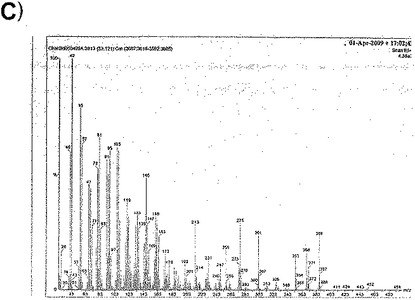
【図31】
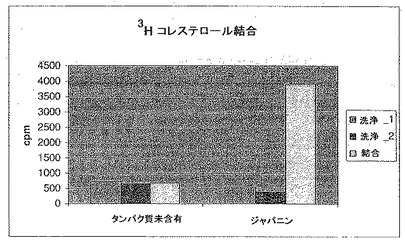
【図32a】
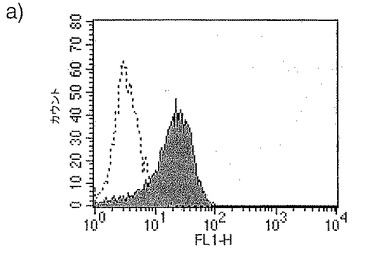
【図32b】
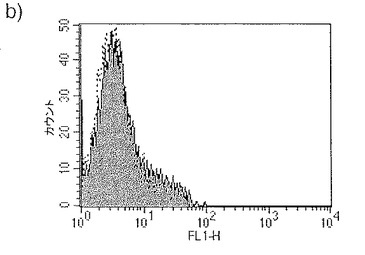
【図32c】
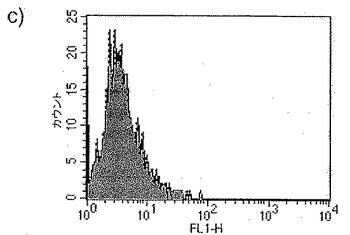
【図32d】
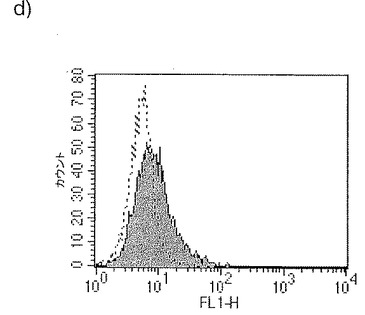
【図32e】
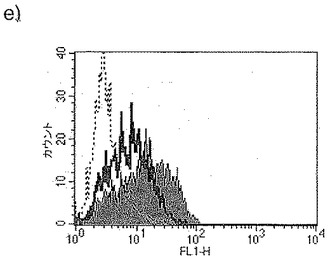
【図32f】
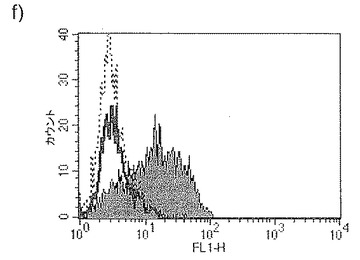
【図33】
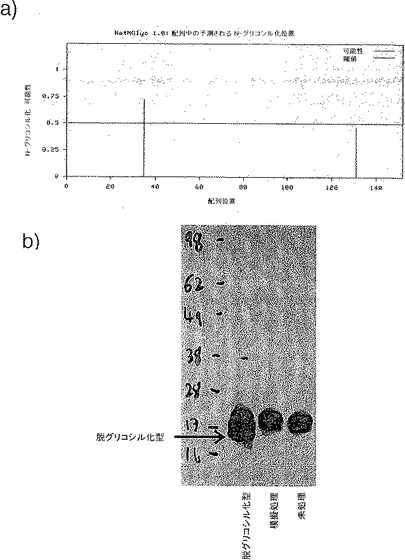
【図34a】
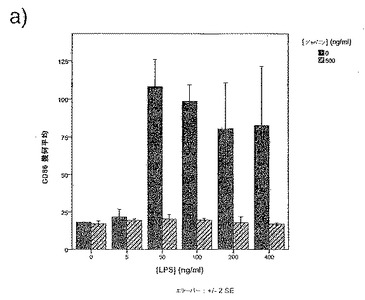
【図34b】
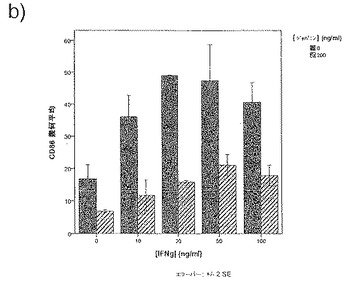
【図34c】
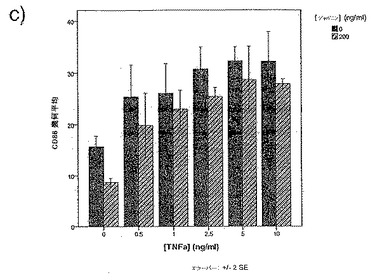
【図34d】
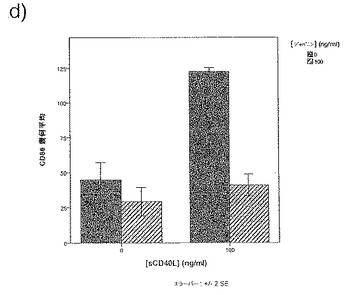
【図34e】
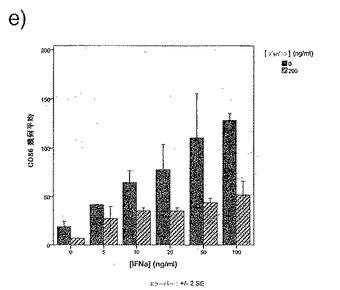
【図34f】

【図35】
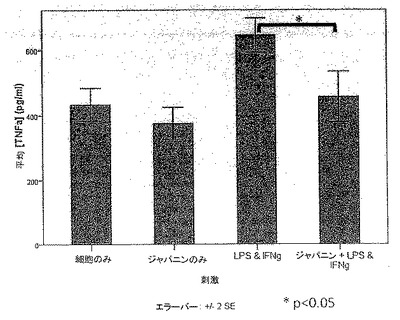
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13a】
【図13b】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30a】
【図30b】
【図30c】
【図31】
【図32a】
【図32b】
【図32c】
【図32d】
【図32e】
【図32f】
【図33】
【図34a】
【図34b】
【図34c】
【図34d】
【図34e】
【図34f】
【図35】
【公表番号】特表2012−502625(P2012−502625A)
【公表日】平成24年2月2日(2012.2.2)
【国際特許分類】
【出願番号】特願2011−526558(P2011−526558)
【出願日】平成21年9月16日(2009.9.16)
【国際出願番号】PCT/GB2009/002219
【国際公開番号】WO2010/032008
【国際公開日】平成22年3月25日(2010.3.25)
【出願人】(511065196)
【Fターム(参考)】
【公表日】平成24年2月2日(2012.2.2)
【国際特許分類】
【出願日】平成21年9月16日(2009.9.16)
【国際出願番号】PCT/GB2009/002219
【国際公開番号】WO2010/032008
【国際公開日】平成22年3月25日(2010.3.25)
【出願人】(511065196)
【Fターム(参考)】
[ Back to top ]