
欠陥インフルエンザウイルス粒子
【課題】条件欠陥インフルエンザウイルス粒子を得るための方法によって得られたインフルエンザウイルス粒子の提供。
【解決手段】7個の異なるインフルエンザ核酸セグメントを有しそして酸性ポリメラーゼを本質的にコードする一つのインフルエンザ核酸セグメントを欠失する欠陥インフルエンザウイルス粒子、並びに機能性ポリメラーゼを産生できないが、しかしウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを妨げない該遺伝子構築物を含む発現プラスミドを用いて複数の細胞をトランスフェクションし、ヘルパーウイルスを使用しないことを特徴とするインフルエンザウイルス粒子の生産方法。
【解決手段】7個の異なるインフルエンザ核酸セグメントを有しそして酸性ポリメラーゼを本質的にコードする一つのインフルエンザ核酸セグメントを欠失する欠陥インフルエンザウイルス粒子、並びに機能性ポリメラーゼを産生できないが、しかしウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを妨げない該遺伝子構築物を含む発現プラスミドを用いて複数の細胞をトランスフェクションし、ヘルパーウイルスを使用しないことを特徴とするインフルエンザウイルス粒子の生産方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、インフルエンザウイルスおよび流感に対するワクチン接種の分野に関する。
【背景技術】
【0002】
インフルエンザウイルス(オルトミクソウイルス科)は、分節ゲノムを有するエンベロープ付マイナス鎖RNAウイルスである(非特許文献1)。それらは二つの属に分けられる。それらの核タンパク質およびマトリックスタンパク質の間の著しい抗原性差異に基づいて、一方はA型およびB型インフルエンザを含み、他はC型インフルエンザから成る。3種のウイルス型は、病原性およびゲノム構成でも異なる。A型は広範囲の温血動物に見られるが、一方BおよびC型は、主としてヒト病原体である。A型インフルエンザウイルスは、ビリオンの表面から突出する赤血球凝集素(HA)およびNA表面糖タンパク質の抗原性特性化によりさらに細分される。現在15のHAおよびNAサブタイプがある。A型インフルエンザウイルスは、鳥類、ブタ、ウマ、ヒト、およびその他の哺乳動物を含む広範囲の動物に感染する。水鳥は、すべての既知のA型インフルエンザサブタイプの天然の貯蔵庫となり、そして多分、ヒト汎流行インフルエンザ株の遺伝物質の起源であろう。
【0003】
関連するパラミクソウイルスとは異なり、インフルエンザウイルスは分節RNAゲノムを有する。A型およびB型インフルエンザウイルスは同様の構造を有し、一方C型はさらに分岐している。A型およびB型ウイルスは、それぞれ少なくとも1個のタンパク質をコードする8個の分離した遺伝子セグメント(分節)をそれぞれ含み、C型は7個の分離したセグメントを含み、A型およびB型のセグメント4および6を結合している。A型およびB型インフルエンザウイルスは、3種のタンパク質:HA、NAおよび基質2(M2)の突起により覆われている。C型インフルエンザウイルスは、ただ1種の表面糖タンパク質を有する。それぞれのインフルエンザRNAセグメントは、核タンパク質(NP)により包膜化されてリボヌクレオチド核タンパク質(RNP)複合体を形成する。3個のポリメラーゼタンパク質がRNP複合体の1個の末端と会合する。RNPは、一体部分としてマトリックスタンパク質(基質1)を有する膜により包囲されている。エンベロープのリン脂質部分は、細胞宿主膜から誘導される。ウイルス粒子中には非構造タンパク質2(NS2)も見いだされる。
【0004】
インフルエンザウイルスの命名法に関する世界保健機構(WHO)ガイドラインは下記である。最初にウイルスの型(A、B、またはC)、次いで宿主(ヒト以外の場合)、単離の場所、単離番号、および単離の年(斜線で分離)を指定する。A型インフルエンザに対しては、HAおよびNAサブタイプを括弧内に記載する。例えば、2000および2001のシーズンでの最近の三価ワクチン内中含まれる株は:A/パナマ/2007/99(H3N2)、A/ニュー・カレドニア/20/99(H1N1)、およびB/山梨/16/98である。1977年以後、2種のインフルエンザAサブタイプ:H1N1およびH3N2がヒト間で同時に流行している。
【0005】
インフルエンザウイルスのRNAポリメラーゼが複合体プルーフリーディング活性を持たないので、インフルエンザウイルスは複製の間に点変異を蓄積する。表面糖タンパク質の抗原性部分内のアミノ酸を変化させる変異は、すでに存在する免疫性を避けることをそれに可能とさせることにより、ウイルス株に選択的利益を与える。HA分子は、一定の宿主細胞上の受容体に結合することにより感染を開始する。HAタンパク質に対する抗体は、受容体結合を防止しそして同一株による再感染を防止するために非常に有効である。HAは、現在流行しているHA遺伝子の変異が抗体結合を破壊する抗原ドリフトによるか、またはウイルスが新しいサブタイプのHAを取得する抗原シフトのいずれかにより、以前
取得した免疫性を避けることができる。抗原ドリフト圧力は、HA分子間で非平等であり、正に選択された変化は、主としてHAタンパク質の球状頭部上に起きる。それらの変化も、NAよりもHA中でより広範囲に蓄積される。他のインフルエンザタンパク質内の変化はさらに遅く起きる。同様に、抗原性ドリフト圧力は、ヒト適合インフルエンザ株中で最大であり、ブタおよびウマ適合株中では中間であり、そして鳥類適合株中では最低である。
【0006】
インフルエンザウイルスは分節ゲノムを有するので、同じ宿主内での2種の異なる株の同時感染は、親の遺伝子セグメントの異なる組合せを含む新規の再構築されたインフルエンザ株の産生に導くことがある。15種のHAサブタイプが野生鳥類中に存在することが知られており、そしてヒトには先例がないHAの起源を提供する。抗原シフトによる新規サブタイプを有するインフルエンザ株のヒト流行の発生は、1957年および1968年の少なくとも二回のインフルエンザ汎流行の原因であり、1918年のインフルエンザ汎流行の原因である可能性が高い。汎流行インフルエンザウイルスの発生に関するすべての知識に沿って考えると、汎流行性株は現在流行しているものとは抗原的に異なるHAを有するに違いない;このHAは、ヒト内で60〜70年間は流行できなかったものである;そしてウイルスはヒトからヒトに伝播できなければならない。1957年および1968年の双方で、汎流行はHAのシフトからもたらされ、そして両方の場合に、汎流行株のHAは、鳥類株と密接に関連していた。汎流行のための絶対的な要求の一つは、HAが変化しなければならないことであるけれども、ウイルスの残部が変化可能または必須である範囲は知られていない。1957年および1968年の汎流行ウイルスのみが直接研究に利用でき、1918年汎流行インフルエンザウイルスは、分子考古学を用いて特性が研究されている。1957年には、3個の遺伝子が鳥類様遺伝子:HA、NA、およびポリメラーゼ複合体(PB1)のサブユニットにより置換された。1968年では、HAおよびPB1のみが置換された。
【0007】
インフルエンザ感染の特定診断は、ウイルス単離、赤血球凝集阻害(HI)試験、免疫アッセイによる抗原検出、血清学的試験、分泌物中のNA活性の証明、または分子に基づくアッセイにより行うことができる。試料は、痰、鼻咽頭スワブ、または緩衝塩溶液のうがいにより得られる鼻咽頭洗浄物として採取できる。インフルエンザ診断のための標準は、培養後の免疫学的特性化であった。血清学的分析は、それが急性および回復期血清の双方の採取を必要とするので、インフルエンザ感染に対しては、正確ではあるがしかし遡及的な方法となる。
【0008】
インフルエンザウイルスは、孵化鶏卵または多数の組織培養システム中で培養できる。トリプシンの添加(HAの切断活性化のため)は、Madin−Darbyイヌ腎(MDCK)細胞およびその他の系統中でインフルエンザウイルス繁殖を可能とする。ワクチン生産の最初の方法は、卵内のままでのインフルエンザウイルスの培養である。細胞系統の培養物は、ヒトインフルエンザウイルス(A型およびB型の同胞)の一次単離のために一般的に使用される。多数のヒトインフルエンザウイルスが孵化卵内の尿膜腔内で直接培養できる。一部のインフルエンザAおよびB型ウイルスは、羊膜腔での初期培養、引き続いて尿膜腔に順応させることを要する。培養物単離の後、大部分のインフルエンザ単離物は免疫アッセイまたは免疫蛍光法を用いて厳格に分離できる。インフルエンザウイルスのHA分子は、エントリーを増すためにウイルスの呼吸細胞の表面上のサリチル酸残基に結合する。
【0009】
インフルエンザ株は、イン・ビトロで赤血球を凝集させるインフルエンザウイルスの能力を利用して抗原的に特性化できる。抗HA抗体は、凝集を阻害できる。従って、赤血球凝集阻害(HI)アッセイはインフルエンザ株を特性化するために使用される標準的方法の一つである。HIアッセイは、試料株が最近のワクチン株に免疫学的に関連(すなわち
交差反応性)するかどうかを決定するために使用される。一般的にフェレット内で産生される類型決定血清(typing sera)を一連の二倍希釈ウエルに加え、そして実験者が赤血球の懸濁物に対する凝集物を観察してウエルを採点する。大部分の場合に、血清のパネルがワクチンおよび対照試料に対して試料血清を一致させるために使用され、そして該当するインフルエンザシーズンの間に、大多数の使用株がHIアッセイにより一致させることに成功する。
【0010】
WHOはガイドラインを提供し、そしてWHO協力センター(Collaborating Center)は個別のウイルス株の抗原的特性の同定に関するガイダンスを提供する。試料株は、免疫学的系統、例えばA/モスクワ/10/99(H3N2)、A/ニュー・カレドニア/20/99(H1N1)、およびB/北京/184/93様ウイルス、に従って分類される。HIアッセイで特性化できなかった試料株に対して、実験者は、それらをフェレットに接種して株特異性抗血清を生産させなければならない。新しい抗血清が準備されると、上記のようにしてHIアッセイを再度行う。新血清が交差反応性に有意のギャップ(通常試料とワクチン株との間の4倍の差異として定義される)を示す場合には、それを通常の実験室パネル内に組入れ、そして新規の流行株を探索するために使用される。このようにして、HIアッセイは、ワクチン株選択のためのインフルエンザウイルス監視の努力に非常に重要でありそして抗原ドリフトを評価するために最も一般的に使用される方法である。
【0011】
インフルエンザウイルスは、個別の遺伝子セグメントの配列比較により一般的に特性化でき、そしてこの場合にもWHOガイドラインおよびWHO協力センターがインフルエンザゲノムを含んでなるRNAセグメントの個別同一性の同定に関するガイダンスを提供している;核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、酸ポリメラーゼ(PA)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするA型およびB型インフルエンザウイルス核酸セグメント、および核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素−ノイラミニダーゼ様糖タンパク質(HN)、マトリックスタンパク質(M1およびM2)、および非構造タンパク質(NS1およびNS2)をコードするC型ウイルス核酸セグメント。抗原分析のため、核酸配列比較のため、およびワクチンウイルスの同定のための参照株の要求は、WHO Collaborating Centre for Reference and Research on Influenza,45 Poplar Road,Parkville,Victoria 3052,Australia(ファックス:+61 3 9389 1881,ウエブサイト:http://www.influenza centre,org);WHO Collaborating Centre for Reference and Research on Influenza,National Institute of
Infectious Diseases,Gakuen 4−7−1,Musashi−Murayama,Tokyo 208−0011,Japan(ファックス:+81
42 5610812または+81 42 5652498);WHO Collaborating Center for Surveillance,Epidemio
logy and Control of Influenza,Centers for Disease Control and Prevention,1600 Clifton Road,Mail Stop G16,Atlanta,GA 30333,United States of America(ファックス:+1 404 639 23 34);またはWHO Collaborating Centre for Reference and Research on Influenza, National Institute for Medical Research,The Ridgeway,Mill Hill,London NW7 1AA,En
gland(ファックス:+44 208 906 4477)。最新の疫学的情報は、WHOのウエブサイト:http://www.who.int/influenzeおよび地理学的情報システム、FluNet、http://www.who.int/flunetから入手できる。
【0012】
インフルエンザのインパクトおよびその予防の健康および経済的利益の意識は増大しており、そして過去10年間はワクチン接種の使用および利益が見られそして多数の抗インフルエンザ医薬がますます増えている。多数の国でより長い期待寿命の結果として、より多数の人々に合併症の危険があり、インフルエンザ流行の間の健康管理システムに対する負荷がさらに広く認識され、そしてさらに頻繁な国際旅行がウイルスの拡散の機会を与え、一方新製品の導入は疾患の予防および処置に対する選択肢を増やしている。約50の国々が、政府出資の国内インフルエンザ免疫プログラムを有しそしてワクチンはその他の多数の国々でも利用できる。ワクチンの使用のための特定の指針はさまざまであるが、しかし一般的に、高齢者および既存の慢性病的条件のために重篤な疾患の危険が上昇している6カ月以上の人々への毎年の接種を含む。一部の国では、医療リスクが高い人々へのインフルエンザの拡散を低減するためにワクチンが使用される。メンバーステイト(Member State)は、それらの全般的な公衆衛生優先事項の範囲内で、インフルエンザ予防活動の利益を考慮する必要がある。
【0013】
不活化ワクチンは、それらが全ウイルス粒子、部分的に破壊されたウイルス粒子(スプリットワクチン)または精製されたエンベロープ抗原(サブユニットワクチン)を含むかどうかに依存して、数種のタイプに分類される。いくらかのサブユニットワクチンは、アジュバントまたは送達システムと複合される。
【0014】
いくつかの国は一定の標的群に対して生弱毒インフルエンザワクチンを許可している。ロシア連邦では、1ワクチンの2種の異なる製剤が、健康な成人および小児に使用されており、別の生ワクチンが広範に試験されている。しかし、生弱毒ワクチンがさらに広範に入手可能となるまで、それはインフルエンザ予防のためにまだ一般的には推奨されない。
【0015】
2種の抗ウイルス剤がインフルエンザの予防および処置のために開発されている。M2阻害剤、アマンタジン(amantadine)およびリマンタジン(rimantadine)は、A型インフルエンザウイルスの処置に限定され、そして感染の予防に有効であると報告されている。両方の製品はいくらかの副作用を起こすが。著しい神経学的副作用はアマンタジンの方が大きい。ノイラミニダーゼ阻害剤、例えばザナミビル(zanamivir)およびオセルタミビル(oseltamivir)は、A型およびB型インフルエンザの処置のために多数の国で最近許可され、そして予防に有効であることが報告されている。双方の抗ウイルス剤を投与された患者内で、耐性変異体が検出されている。これは現在までは重要な公衆衛生問題として考慮されていないけれども、それらの薬剤がさらに大きいスケールで使用されると、状況は変化するであろう。
【0016】
WHOは、全世界的な国際監視プログラムを、82カ国に存在する110箇所の国内インフルエンザセンターおよび4箇所のインフルエンザ参照および研究のためのWHO協力センター(アトランタ(米国)、ロンドン(英国)、メルボルン(オーストラリア)および東京(日本)に位置する)の協力を得て維持している。それらのセンターは、流行ポテンシャルと共に発生する株の早期警告システムを提供する。インフルエンザワクチンが現在流行している株を含まない場合、インフルエンザワクチンの効力が低下するので、このシステムは重要である。WHOはワクチン組成の推奨を発行し、それは世界保健機構により出版されるWeekly Epidemiological Record(例えば非特許文献2参照)中に見ることができ、2月には北半球で使用されるワクチン、9月には南半球で使用されるワクチンに関して公開されている。インフルエンザは赤道付近の地域
では季節的パターンがあまりはっきりしないので、疫学的考慮が、それらの推奨事項(2月または9月のどちらか)が赤道付近の国々に使用されるワクチンに適合するかに影響する。
【0017】
協力センターは、各国センターから提出されたインフルエンザ単離物の抗原および遺伝子分析を行う。抗原変化の証拠が観察されると、変種の疫学的重要性を評価するために疫学的データを用いてこれを精査する。ワクチン接種の前後に採取したヒト血清のパネルを使用して、代表的な単離物を現在のワクチン株と比較して、現在のワクチンにそれらのウイルスに対する防御が期待できるかどうかを評価する。WHOの年間ワクチン推奨を公表に続いて、高度増殖株を開発しそしてワクチン生産のための種ウイルスの生成を支援するための参照ウイルスとして生産業者に提供する。インフルエンザワクチンの安全性および効力の試験は、ウイルス不活化、微生物の無菌性、ウイルスを破壊するために使用した化学薬品の測定および推奨する抗原濃度の確認を含む。ワクチンはWHOの要求に合致すべきことが推奨されるが、しかし、各国の規制機関はその国内で使用される特定のワクチンウイルスを承認するべきである。国の公共保健機関は、ワクチンの使用に関する推奨に責任を持つべきである。また、WHOはインフルエンザの予防に関する推奨を発行している(非特許文献3参照)。
【0018】
現在のワクチンがナイーブな個体を防御しないこと、すなわちそれまで流感感染に遭遇していない多数の個体が危険にさらされた場合に、インフルエンザの汎流行発生の場合に直ちに重要となるという事実はすでに証明されている。ウイルスは一般に細胞に入り宿主細胞表面受容体に付着し、そしてそれらのウイルス核酸を脱外被し、次いでウイルスゲノムの複製を行うことによりそれらはそのライフサイクルを開始する。ウイルスタンパク質および遺伝子の新しいコピーが合成された後、それらの成分は後代ビリオンに構築され、次いで細胞から出る。構築段階の間に、後代ウイルスは、細胞質内に存在するウイルスおよび細胞核酸の大きいプールからそのゲノム核酸を効率的に選択しなければならない。ビリオンへのウイルスゲノムのパッケージングは、典型的には、ウイルス核酸内のシス作用性配列のウイルス成分、いわゆる「パケージング・シグナル」による認識を含む。かかるシグナルの定義は、ウイルスライフサイクルを理解するために重要でありそして他者タンパク質の発現のためのウイルスベクターを構築するために使用できる情報を我々に提供する。実際、他者タンパク質の発現のための遺伝子送達ベクターのベヒクルとしてのレトロウイルスの利用は、それらのvRNAの後代ビリオン内へのパッケージングのプロセスの良く確立された知識に大部分が帰せられる。
【0019】
他のRNAウイルスのゲノムパッケージングシグナルは、良く理解されておらず、他者遺伝子の発現および送達のためのベクターとしてのそれらの使用に関する進歩を妨げている。例えばA型インフルエンザウイルスは、エンベロープされたマイナス鎖RNAウイルスであり、その分節化ゲノムは、核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、酸性ポリメラーゼ(PA)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)、および非構造タンパク質(NS1およびNS2)のコーディング容量を有する。
【0020】
このウイルスは、エンベロープ上に2種の膜貫通糖タンパク質、赤血球凝集素(HA)およびノイラミニダーゼ(NA)を有する。HAタンパク質は、宿主細胞表面上のシアル酸含有受容体に結合し、そして受容体媒介エンドサイトーシスの後にエンドソーム膜とのウイルスエンベロープの融合を媒介する。対照的に、NAタンパク質はシアリルオリゴサッカリドからシアル酸を除去し、従って新規に構築されたビリオンを細胞表面から離しそしてウイルス粒子の自己凝集を防止することにより、感染の後期に重要な役割を果たす。エンベロープ内で、8個の異なるウイルスRNA(vRNA)セグメントを含んでなるウイルスゲノムは、核タンパク質(NP)およびポリメラーゼタンパク質(PA、PB1、
およびPB2)と緊密に連結し、リボヌクレオタンパク質複合体を形成する。すべての8個(またはC型の場合には、すべての7種)の機能性遺伝子セグメントは、感染性ウイルスを産生するために必要である。ポリメラーゼ遺伝子内の種々の変種が記載されており(特許文献1、特許文献2、特許文献3、非特許文献4)それらはポリメラーゼ活性を変化させるかまたは別の様式でポリメラーゼを改変するがしかしかかる複製が不能な変異ポリメラーゼを含むウイルスを与えるウイルスRNAの合成におけるその機能は放さない。特許文献1中では、変異PA遺伝子を有する感染性ウイルスが産生され、それにより変異遺伝子を有する感染性ウイルスを産生および回収する選択システムの利益を示す。特許文献2中では、生弱毒ワクチンウイルス産生に関係する望ましい性質、例えば温度感受性またはその他の形の弱毒化を有するインフルエンザウイルスの産生および回収を可能とするポリメラーゼ遺伝子内の変異を有する感染性ウイルスを産生する方法が示されている。特許文献3中では、種々のポリメラーゼの特異性を改変しそして活性を低下させるであろうポリメラーゼ遺伝子セグメントが示唆されている。しかし、そのポリメラーゼ活性を全て喪失したウイルスを再構築することは別にして、それらはウイルス再構築には使用されなかった。さらに、上記の適用のいずれでも、複製するそれらの能力を失った欠陥粒子は産生されなかった。A型ウイルスが高い多重度の感染で継代されると、1個もしくはそれ以上の機能性遺伝子セグメントを欠失した欠陥ウイルス粒子が産生される。かかるウイルス粒子中では、1個もしくはそれ以上の機能性遺伝子が、インフルエンザウイルスポリメラーゼにより作成されたエラーにより、欠陥干渉性(DI)遺伝子セグメントと置換される。高い多重度の感染により、そして1個を越えるウイルス粒子による細胞の感染により、DI RNAを含むウイルスの欠陥は、欠損した機能性遺伝子の変化のないコピーを含むウイルスにより相補される。最近、酸性ポリメラーゼ遺伝子内の一定の変異が、欠陥遺伝子を有するウイルス粒子の生成の効率を上昇できることが示された(非特許文献4)。それらの実験における欠陥ウイルス粒子の生成および相補の双方が無作為的にかかる実験中で起きることが認識されたことは重要である。この無作為プロセスは、実際の適用におけるDI RNAおよび条件つき欠陥ウイルス粒子の使用を制限する。さらに、欠陥ウイルス粒子がそれらの公開された方法を用いて産生されると、所望の条件つき欠陥ウイルスの外に野生型複製適格ウイルスが産生される。かかる複製適格ウイルスは、完全に野生型(ヘルパーウイルス)であるかまたはヘルパーウイルスと欠陥ウイルスとの遺伝子混合からもたらされる再構築物のいずれかであろう。無作為的または特異的機構のいずれかを介するインフルエンザウイルスの遺伝子セグメントのパッケージングプロセスは、多年にわたって論議されている。双方の選択肢に数件の証拠が記載されている。無作為パッケージングの証拠は、凝集したウイルス粒子が非凝集ウイルス粒子よりも高い感染性を有しそして細胞培養物が低い感染モード(moi)で感染された場合には、一部の感染細胞は一つのセグメントの発現を欠失するというものであり、両方共に全てのインフルエンザウイルスゲノムは含まないビリオンがあることを示唆する。無作為パッケージングの別の証拠は、9個のセグメントを有するインフルエンザウイルスが実験的に産生されたことである。
【0021】
特異性パッケージングプロセスのための一つの論拠は、すべての遺伝子セグメントがウイルス原料内に等量で存在するけれども、それらは、生産細胞内では異なる量で存在するというものである。さらに、欠陥干渉性(DI)粒子が生成された場合に、DIvRNAは、それが誘導されたセグメントを置換する(欠陥干渉性粒子とは、遺伝子セグメントの一つが大きい内部欠失を有するウイルス粒子である。それらの粒子は、ウイルスが高いmoiで継代された場合に生じる)。最後に、ビリオン形成の効率は、遺伝子セグメントの数が増えると増加する。
【先行技術文献】
【特許文献】
【0022】
【特許文献1】国際特許出願公開(WO)第2004/094466号明細書
【特許文献2】国際特許出願公開(WO)第2003/091401号明細書
【特許文献3】米国特許(US)第5578473号明細書
【非特許文献】
【0023】
【非特許文献1】Taubenberger and Layne,Molecular Diagnosis、第6巻、第4号、2001
【非特許文献2】第9号、2004、79、88ページまたはhttp://www.who.int/wer
【非特許文献3】WER第35号、2002、281−288ページ
【非特許文献4】Fodor et al.,J.Virol.77,5017−5020.2003
【発明の概要】
【0024】
欠陥インフルエンザウイルス粒子(例えばMena I.et al.,J.Virol.70:5016−24(1996);Neumann G.et al.,J.Virol.74:547−51(2000))は、ワクチン候補として有用であり、それはそれらがHAおよびNA以外のウイルスタンパク質に対する抗体を誘導するからであり、そして、それらが宿主細胞に入ることができる場合には、それらが、体液反応に加えてウイルスに対する細胞免疫反応を誘導できるからである(例えばヘルパーT細胞、細胞毒性T細胞)。これまで、欠陥インフルエンザウイルス粒子の生産は、トランスフェクションにより達成され(Mena I.et al.,J.Virol.70:5016−24(1996);Neumann G.et al.,J.Virol.74:547−51(2000))、かかる粒子を大量に生産する可能性を低下する。この方法とは別の方法では、条件つき欠陥であるウイルス粒子を生産し、それらの一定の生産システム内での複製を可能とするが、しかし正常の細胞または生産システム内では可能としないものであろう。この目的で、生産システムの細胞は、欠陥インフルエンザウイルス粒子のトランス−相補を可能とするインフルエンザウイルス遺伝子または遺伝子産物の1種もしくはそれ以上の生産を可能とするように変性される。本発明は、欠陥インフルエンザウイルス粒子の一定のトランス相補(transcomplementation)を最初に開示する。実験室内で、インフルエンザウイルス粒子のトランス相補は、欠陥干渉性インフルエンザウイルスが、欠陥干渉性遺伝子セグメントの野生型バージョンを有するウイルスにより同じ細胞内で相補される場合に観察される。トランス相補の「天然システム」は、一定の条件つき欠陥インフルエンザウイルス粒子を生産するために有用ではない。第一に、このシステムは、完全に感染性のウイルスの望ましくない産生をもたらすであろう少なくとも1つの(部分的)複製適格ウイルスによる一つの(部分的)欠陥ウイルスの相補を必要とする。第二に、欠陥干渉性粒子の産生は種々の遺伝子セグメントに無作為的に起きるので、一定の条件つき欠陥ウイルス粒子を産生することは不可能である。
【0025】
条件つき欠陥インフルエンザウイルス粒子は、理論的には、それらの全遺伝子セグメントまたは部分の欠失に基づくことができる。全遺伝子セグメントを欠失することにより一定の条件つき欠陥ウイルス粒子を産生(およびコードされた遺伝子産物をイントランス(in trans)で産生)する能力は、インフルエンザウイルスゲノムのパッケージングがすべての8セグメントの存在に依存するならば、限定されるであろうし、これは多く論議されている問題である(本明細書の別の箇所参照)。パッケージングプロセスがすべての8セグメントの存在を必要とする場合に、すべての遺伝子セグメントが全長として存在する必要であるかどうかは知られていず、それは条件つき欠陥ウイルス粒子の生産をさらに複雑にする。本発明はそれらの問題を解決する。
【0026】
本発明は、インフルエンザポリメラーゼをコードする核酸を内部欠失することにより誘導されて本明細書中で提供される内部欠失を有する遺伝子構築物、例えばpΔPB2、pΔPB1、pΔPAまたはpDIPAであって、ここで、該遺伝子構築物はウイルスRN
Aをコピーまたは合成できる機能性ポリメラーゼを産生できなくなるものを用い、そしてインフルエンザウイルスをコードする相補性インフルエンザウイルス核酸セグメント、例えばA/WSN/33をコードする7個の相補性構築物(HW181−188、Hoffmann et al.,2000)を用い、そしてかかる細胞内に該ポリメラーゼを発現できる発現プラスミド、例えば本明細書中に記載のHMG−PB2、HMG−PB1、HMG−PAを用いて、適切な第一細胞(単数または複数)、例えば293T細胞をトランスフェクションし、そして適当な時点、例えばトランスフェクションの後10〜50、好ましくは約20〜30時間以内に該第一細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取する第一工程、および細胞内に該ポリメラーゼを発現できる発現プラスミドを用いて適切な第二細胞(単数または複数)例えばMDCK細胞をトランスフェクションする第二工程、および該第一細胞から得た少なくとも1個のウイルス粒子を含んでなる上清を用いて該第二細胞(単数または複数)をトランスフェクションする第三工程、および適切な時点、例えばトランスフェクションの24〜96、好ましくは48〜72時間後に該第一細胞(単数または複数)の上清から少なくとも1個(内部欠失を有する遺伝子セグメントをパッケージングされたために産生されたウイルスがウイルスRNAのコピーまたは合成が可能な機能性ポリメラーゼを発現する遺伝子セグメントを欠失するウイルスが産生されるので条件つき欠陥である)のウイルス粒子を採取することを含んでなる第四工程を含んでなる条件つき欠陥インフルエンザウイルス粒子を得る方法を提供する。
【0027】
好ましくは、機能性タンパク質を産生不能ではあるが、しかしウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを阻止するほどは大きくない内部欠失である。好ましくは、それらの欠失は5’および3’非コード領域からそれぞれ数えられる。A型インフルエンザでは、かかる好ましい欠失は、例えばPAタンパク質ではヌクレオチド58と75の間であってそれらは包含されない位置にある5’ヌクレオチドから初まり、そしてヌクレオチド27と50の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一1タンパク質ではヌクレオチド43と75の間であってそれらは包含されない位置にある5’ヌクレオチドから初まり、そしてヌクレオチド24と50の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一2タンパク質ではヌクレオチド34と50の間であってそれらは包含されない位置にある5’ヌクレオチドから初まり、そしてヌクレオチド27と50の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる。さらに好ましくは、それらの欠失は、PAタンパク質ではヌクレオチド58と100の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と100の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一1タンパク質ではヌクレオチド43と100の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド24と100の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一2タンパク質ではヌクレオチド34と100の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と100の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる。もっとさらに好ましくは、それらの欠失は、PAタンパク質ではヌクレオチド58と150の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と150の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一1タンパク質ではヌクレオチド43と150の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド24と150の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一2タンパク質ではヌクレオチド34と150の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と150の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる。もっとさらに好ましくは、それらの欠失は、PAタンパク質ではヌクレオチド58と175の間であってそれらは包含されない位置にある5’ヌクレオ
チドから始まり、そしてヌクレオチド27と175の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一1タンパク質ではヌクレオチド43と175の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド24と175の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一2タンパク質ではヌクレオチド34と175の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と175の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる。最も好ましくは、それらの欠失は、PAタンパク質ではヌクレオチド58と207の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と194の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一1タンパク質ではヌクレオチド43と246の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド24と197の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一2タンパク質ではヌクレオチド34と234の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と209の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる。
【0028】
本明細書中で、相補性セグメントとは、例えばA型インフルエンザウイルスの8個の遺伝子セグメントの完全な組に導くセグメントとして定義される。従って、セグメント1が欠陥セグメントを産生するために以前に使用された場合には、相補性(非欠陥)セグメントは、セグメント2、3、4、5、6、7および8である。2が欠陥の場合には、相補セグメントはセグメント1、3、4、5、6、7および8である。以下同様。
【0029】
有利には、本発明はヘルパーウイルスが必要でなくまたは存在しない方法をもたらす。
【0030】
本発明は、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)および塩基性ポリメラーゼ2(PB2)の群から選択されたポリメラーゼをコードする機能性インフルエンザウイルス核酸セグメントを欠失する単離された条件つき欠陥インフルエンザウイルス粒子(本明細書中では条件つきの欠陥インフルエンザウイルス粒子とも呼ぶ)を提供し、かかる粒子はウイルスRNAをコピーまたは合成するポリメラーゼを生成する供給源として生成または役立つことができず、それにより機能ポリメラーゼを用いてトランス相補された細胞内での複製ウイルス粒子の生成を条件つきで可能とするだけである。さらに、本発明は、機能性インフルエンザウイルスポリメラーゼを用いるトランス相補により細胞を提供することを含んでなる条件つき欠陥インフルエンザウイルス粒子を得る方法を提供する。
【0031】
好ましい態様では、本発明による粒子は、粒子自体内では欠失している類似核酸セグメントを相補された細胞内で複製し、例えば、機能性インフルエンザウイルス核酸PAセグメントを欠失する粒子は少なくとも機能性インフルエンザウイルス核酸PAセグメントを備えた細胞内で複製し、機能性インフルエンザウイルス核酸PB1セグメントを欠失する粒子は少なくとも機能性インフルエンザウイルス核酸PB1セグメントを備えた細胞内で複製し、機能性インフルエンザウイルス核酸PB2セグメントを欠失する粒子は少なくとも機能性インフルエンザウイルス核酸PBセグメントを備えた細胞内でそれぞれ複製する。好ましい態様では、本発明は、ウイルス糖タンパク質をコードするインフルエンザウイルス核酸セグメントを有し、さらに好ましくは、核タンパク質(NP)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)、および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する本発明による粒子を提供する。一つの態様では、A型インフルエンザウイルスから誘導されたインフルエンザウイルス核酸セグメントを有する本発明による粒子が提供される。また、インフルエンザペプチドをコードしない核酸も備えている本発明に
よる粒子が提供される。また本発明は本発明による粒子を含んでなる単離された細胞を提供し、該細胞は野生型インフルエンザウイルスまたはヘルパーウイルスを含まないがしかし好ましくはインフルエンザウイルスポリメラーゼまたはそれをコードする遺伝子セグメントを備えるかまたは相補されている。好ましい態様では、かかる細胞はトランス相補された293TまたはMDCK細胞である。一つの態様では、本発明は機能性インフルエンザウイルス核酸PAセグメントを欠失した粒子を含んでなる単離された細胞を提供し、該細胞は野生型インフルエンザウイルスまたはヘルパーウイルスを含まないがしかし少なくとも機能性インフルエンザウイルス核酸PAセグメントまたは機能性PAを備えているかまたは相補されている。別の態様では、本発明は、機能性インフルエンザウイルス核酸PB1セグメントを欠失した粒子を含んでなる単離された細胞を提供し、該細胞は野生型インフルエンザウイルスまたはヘルパーウイルスを含まないがしかし少なくとも機能性インフルエンザウイルス核酸PB1セグメントまたは機能性PB1を備えている。さらに別の態様では、本発明は、機能性インフルエンザウイルス核酸PB2セグメントを欠失した粒子を含んでなる単離された細胞を提供し、該細胞は野生型インフルエンザウイルスまたはヘルパーウイルスを含まないがしかし好ましくは少なくとも機能性インフルエンザウイルス核酸PB2セグメントまたは機能性PB2を備えているかまたは相補されている。さらに、本発明は、本発明による粒子または本発明による細胞から誘導された細胞もしくは物質を含んでなる組成物、およびインフルエンザウイルスによる患者(subject)の感染に対する免疫学的防御を生成することを目的とする製薬学的組成物の製造のためのかかる組成物の使用を提供する。これによって、本発明は免疫学的防御を必要とする患者に本発明による組成物を提供することを含んでなる、インフルエンザウイルスの患者の感染に対する免疫学的防御を生成するための方法を提供する。また、本発明はインフルエンザペプチドをコードしない核酸の細胞への送達を目的とする組成物の生産のための本発明によるインフルエンザウイルス粒子の使用を提供する。また、本発明は、インフルエンザペプチドをコードしない核酸の患者の細胞への送達を目的とする製薬学的組成物の生産のための本発明による粒子の使用、および該細胞または患者へ本発明による粒子を提供することを含んでなる細胞または患者へのインフルエンザペプチドをコードしない核酸の送達のための方法を提供する。
【0032】
本発明は、天然のゲノムと比較すると一つの機能性インフルエンザウイルスセグメントを欠失する、すなわち:野生型またはヘルパーAもしくはB型ウイルスと比較して、7個(8個ではなくて)の異なる機能性インフルエンザウイルス核酸セグメントを有するか、または野生型またはヘルパーC型ウイルスと比較して、6個(7個ではなくて)の異なるインフルエンザウイルス核酸セグメントを有する条件つき欠陥インフルエンザウイルス粒子を提供する。本明細書中で「条件つき欠陥」の用語が使用される場合には、それはウイルスの遺伝子セグメントの一つがそれから発現されると非機能性タンパク質をもたらす大きい内部欠失を有するウイルス粒子を含み、それらに限定はされない。例えばA型インフルエンザウイルスのすべての8個の遺伝子セグメントおよびそれらによりコードされるすべてのタンパク質は、感染性ウイルスの生産に必要である。従って、欠陥遺伝子セグメントを含むウイルスはそれら自体が欠陥である:それは、すべてのウイルスタンパク質がビリオン内に存在するので細胞に感染できそして複製を一廻りできるが(このタンパク質は例えばウイルスが生産された場合に発現プラスミドにより産生される)しかしウイルスタンパク質の一つがウイルスにより産生できないので感染細胞内に感染性ウイルス粒子は産生されない。しかし、欠陥遺伝子セグメントにより通常発現されるタンパク質を発現する細胞が感染されると、すべてのウイルスタンパク質が存在するので欠陥ウイルスはそれらの細胞内で複製ができる。従って、それらのウイルスは、条件つき欠陥である:それらは、正しい状態の細胞が提供されない限り複製できない(この場合に、遺伝子セグメント内の欠失のためにウイルスによりコードされないウイルスタンパク質を細胞は発現する)。
【0033】
さらに、本発明はポリメラーゼをコードする機能性インフルエンザウイルス核酸セグメ
ントを欠失する条件つき欠陥インフルエンザウイルス粒子を提供する。本明細書中で、機能性インフルエンザウイルス核酸セグメントは、複製性ウイルスの生成を可能としそのために必要な機能性インフルエンザタンパク質をコードする核酸を含んでなる。例えば、A型インフルエンザウイルスは、8個の分節ゲノムを有するマイナス鎖RNAウイルスである。8個の遺伝子セグメントは11個のタンパク質をコードする;遺伝子セグメント1−8は、塩基性ポリメラーゼ2(PB2)、塩基性ポリメラーゼ1(PB1)およびPB1−ORF2(F2)、酸性ポリメラーゼ(PA)、赤血球凝集素(HA)、核タンパク質(NP)、ノイラミニダーゼ(NA)、マトリックスタンパク質1および2(M1、M2)および非構造タンパク質1および2(NS1、NS2)をそれぞれコードする。8個の遺伝子セグメントのコード領域は、ウイルスRNA合成に必要な非コード領域(NCR)に隣接している。ウイルスゲノムRNAの5’および3’末端のそれぞれ端部13および12ヌクレオチドは、すべてのA型インフルエンザウイルスセグメントのなかでも保存されそして部分的に相補的であり、ウイルスポリメラーゼ複合体により認識される二次構造を形成する。NCRは、8個の遺伝子セグメントの間で保存はされないがしかし異なるインフルエンザウイルス間では比較的保存される60個までの追加のヌクレオチドを含んでいるらしい。コード領域内のNCRおよび隣接配列は、効率的なウイルスゲノムパッケージングのために必要らしい。従って、機能性インフルエンザウイルス核酸セグメントは、複製可能なウイルスの生成を可能とする機能性インフルエンザタンパク質のためのコーディングポテンシャルを有する配列(セグメント当たりに1または2個のオープンリーディングフレーム)、mRNAの転写に必要なNCR、ウイルスRNA(vRNA)およびウイルスRNAに相補的なRNA(cRNA)およびNCR内に存在しそしてコード配列に隣接するパッケージングシグナルから成る。
【0034】
一つのインフルエンザウイルス核酸を欠失する該条件つき欠陥インフルエンザウイルス粒子が機能性ポリメラーゼを、それがPA、PB1、PB2であってもコードするセグメントを欠失することは好ましい。さらに、ワクチンの目的で、該粒子がウイルス糖タンパク質(単数または複数)をコードするインフルエンザウイルス核酸セグメント(単数または複数)を有することは好ましい。
【0035】
一つの態様では、本発明は7個の異なるA型インフルエンザ核酸セグメントを有するA型インフルエンザ粒子を提供する。本発明による欠陥インフルエンザウイルス粒子は、相補されていなくても、ただ一回であっても適切な宿主動物または細胞内で複製できる。適切に相補された細胞内では、本発明による粒子はさらに多くの回数複製できる。ワクチンおよび遺伝子送達の目的で、欠陥粒子は正常のトランス相補されていない細胞内では無限に複製はできず、そのため、宿主から宿主へのワクチンウイルスの拡散のリスクを低減しそして野生型ウイルスへの復帰の危険が低減されることは大きい利点である。
【0036】
7個の遺伝子セグメントのみを含みそして一回の複製のみを行うことができるか、または欠陥遺伝子セグメントがトランス相補された場合に複数回の複製ができる欠陥A型インフルエンザウイルスが逆遺伝子学を用いて生産できるのは最初のことである。一つの態様では、本発明は、本質的に酸性ポリメラーゼ(PA)をコードするインフルエンザ核酸セグメントを欠失する条件つき欠陥インフルエンザウイルス粒子を提供する。PAのトランス相補と同様に、他のインフルエンザウイルス遺伝子のトランス相補も考えられる。しかし、他のインフルエンザウイルスタンパク質の発現レベルと比較してPA発現レベルは重要性が低いので、PAは欠失されるポリメラーゼ群の好ましい遺伝子セグメントであり、PB2およびPB1欠失ウイルスも同様に生産でき、そしてNP欠失ウイルスはトランス相補できなかった。好ましい態様では、本発明は、7個の異なるA型インフルエンザ核酸セグメントを有しそして本質的に酸性ポリメラーゼをコードするA型インフルエンザ核酸セグメントを欠失する条件つき欠陥A型インフルエンザウイルス粒子を提供する。ワクチンの目的で、本発明による好ましい条件つき欠陥A型インフルエンザウイルス粒子は、赤
血球凝集素(HA)およびノイラミニダーゼ(NA)タンパク質を本質的にコードするA型インフルエンザウイルス核酸セグメントを有し、それらのタンパク質は防御性を与えるために免疫学的に最も関係が深い。ワクチン内に包含するために適切な遺伝子セグメントを選択するために、ワクチン使用のためにWHOにより推奨されたウイルスから遺伝子セグメントを選択するのが好ましい。勿論、HAおよびNAサブタイプは、ワクチン化しようとするインフルエンザ変種のHAおよびNAサブタイプに依存して変化できる。核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)、および非構造タンパク質(NS1およびNS2)を本質的にコードし、すなわちそれぞれの遺伝子セグメントから機能性タンパク質が発現されることを本明細書中で特定して指定するものを本質的にコードするA型インフルエンザウイルス核酸セグメントを有する本発明による条件つき欠陥インフルエンザウイルス粒子を生成することが最も好ましい。かかる粒子は、機能性PAまたはPAをコードする機能性遺伝子セグメントを備えた単離された細胞内に特定して提供される。別の態様では、核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)、および非構造タンパク質(NS1およびNS2)を本質的にコードし、すなわちそれぞれの遺伝子セグメントから機能性タンパク質が発現されることを本明細書中で特定して指定するものを本質的にコードするA型インフルエンザ核酸セグメントを有する本発明による条件つき欠陥インフルエンザウイルス粒子が提供される。かかる粒子は、機能性PB1またはPB1をコードする機能性遺伝子セグメントを備えた単離された細胞内に特定して提供される。別の態様では、核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)、および非構造タンパク質(NS1およびNS2)を本質的にコードし、すなわちそれぞれの遺伝子セグメントから機能性タンパク質が発現されることを本明細書中で特定して指定するものを本質的にコードするA型インフルエンザウイルス核酸セグメントを有する本発明による条件つき欠陥インフルエンザウイルス粒子が提供される。かかる粒子は、機能性PB2またはPB2をコードする機能性遺伝子セグメントを備えた単離された細胞内に特定して提供される。別の態様では、本発明は、インフルエンザペプチドをコードせず、例えば免疫反応を発現するために有用な他者タンパク質またはペプチドをコードする核酸を追加して備えるか、または細胞内の細胞もしくは病原体の機能に干渉できる核酸を備える本発明による粒子を提供する。
【0037】
さらに、本発明は、本発明によるインフルエンザウイルス粒子を含んでなる細胞を提供する。粒子が所要のポリメラーゼを本質的にコードする遺伝子セグメントを備えていなかった場合には、このようにして相補された細胞内の欠陥インフルエンザウイルス粒子の複数回の複製を可能とすることに適合する機能性のインフルエンザポリメラーゼウイルスを細胞が備えるように考慮することが有効である。
【0038】
また、本発明は、本発明による欠陥インフルエンザウイルス粒子または本発明による細胞もしくは細胞から誘導される物質を含んでなる組成物を提供し、かかる組成物は、例えば、インフルエンザウイルスによる患者の感染に対する免疫学的防御を生成することを目的とする製薬学的組成物の生産のために使用できる。また、本発明は、防御が必要な患者にかかる組成物を提供することを含んでなる、インフルエンザウイルスによる患者の感染に対する免疫学的防御を生成するための方法を提供する。ワクチンまたは免疫原的組成物としての本発明による粒子の使用の外に、かかる組成物は好ましくはワクチンとして、すなわちウイルス粒子、またはかかる粒子から誘導されたウイルスタンパク質(スプリットワクチン)を適当な製薬学的担体、例えば塩溶液またはアジュバント(例えばアルミニウム塩またはその他の一般的に使用される賦形剤(例えばhttp://www.cdc.gov/nip/publications/pink/Appendices/A/E
xcipient.pdf.)と混合して処方される。本発明による条件つき欠陥インフルエンザウイルス粒子も、他者遺伝子送達のためおよび他者タンパク質の発現のための候補ベクターでもあり、それは機能性遺伝子が例えば5’および3’PA配列の間に挿入できるからである。本発明が、可能ならば他者もしくは宿主核酸セグメントまたはそれらのフラグメントを備えた、条件つき欠陥インフルエンザウイルス粒子を得るための方法を提供することを考慮して、適当な第一細胞(単数または複数)を、インフルエンザタンパク質をコードする核酸を内部欠失することにより誘導された1個もしくはそれ以上の遺伝子構築物であって、ここで該遺伝子構築物は機能性タンパク質を産生できす、そしてウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを妨げないものを用い、そしてインフルエンザウイルスをコードする相補性インフルエンザウイルス核酸セグメントを用い、そして該細胞内に該タンパク質を発現できる1個もしくはそれ以上の発現プラスミドを用いてトランスフェクションし、そしてトランスフェクションの後適当な時点で該第一細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取する第一工程、および細胞内に該タンパク質を発現できる1個もしくはそれ以上の発現プラスミドを用いて適切な第二細胞(単数または複数)をトランスフェクションする第二工程、および該第一細胞から得られた少なくとも1個のウイルス粒子を含んでなる上清を用いて該第二細胞(単数または複数)に感染させる第三工程、および感染の後の適当な時点で該第二細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取することを含んでなる第四工程を含んでなる。これと共に、本発明は、適切な細胞(単数または複数)を、インフルエンザポリメラーゼをコードする核酸を内部欠失することにより誘導された1個もしくはそれ以上の遺伝子構築物であって、ここで、該遺伝子構築物は機能性ポリメラーゼを産生できないが、しかしウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを妨げないものを用い、そしてインフルエンザウイルスをコードする相補性インフルエンザウイルス核酸セグメントを用い、そして該細胞内に該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いてトランスフェクションし、そしてトランスフェクションの後適当な時点で該細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取する工程を含んでなる条件付欠陥インフルエンザウイルス粒子を得るための方法を提供する。かかる条件付欠陥インフルエンザウイルス粒子を得るための方法は、細胞内にインフルエンザポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いて適切な細胞(単数または複数)をトランスフェクションする第一工程、および条件つき欠陥インフルエンザウイルス粒子を含んでなる上清を用いて該細胞(単数または複数)に感染させる第二工程、および感染の後の適当な時点で該細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取することを含んでなる第三工程を含んでなるか、または条件つき欠陥インフルエンザウイルス粒子を得る方法は、適当な第一細胞(単数または複数)を、インフルエンザポリメラーゼをコードする核酸を内部欠失することにより誘導された1個もしくはそれ以上の遺伝子構築物であって、ここで該遺伝子構築物は機能性ポリメラーゼを産生できないが、しかしウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを妨げないものを用い、そしてインフルエンザウイルスをコードする相補性インフルエンザウイルス核酸セグメントを用い、そして該細胞内に該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いてトランスフェクションし、そしてトランスフェクションの後適当な時点で該第一細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取する第一工程、および細胞内に該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いて適切な第二細胞(単数または複数)をトランスフェクションする第二工程、および該第一細胞から得られた少なくとも1個のウイルス粒子を含んでなる上清を用いて該第二細胞(単数または複数)に感染させる第三工程、および感染の後の適当な時点で該第二細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取することを含んでなる第四工程を含んでなる方法である。該方法において、該ポリメラーゼは、例えば酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)であることができる。好ましくは、本発明は、内部欠失が、PAタンパク質では非コード領域から数えてヌ
クレオチド58と207の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして非コード領域から数えてヌクレオチド27と194の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、あるいはPB1タンパク質では非コード領域から数えてヌクレオチド43と246の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして非コード領域から数えてヌクレオチド24と197の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、あるいはPB2タンパク質では非コード領域から数えてヌクレオチド34と234の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして非コード領域から数えてヌクレオチド27と209の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる、インフルエンザポリメラーゼをコードする核酸を内部欠失することから内部欠失がもたらされる方法を提供する。別の変法では、他者フラグメントがこの内部欠失内に挿入される。さらに、本発明は、条件つき欠陥インフルエンザウイルス粒子を含んでなる上清を用いて感染される細胞(単数または複数)が、非機能性ポリメラーゼ、例えば酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)および本明細書中に記載の方法により得られるインフルエンザ粒子をすでに発現している方法を提供する。例えば、遺伝子構築物および核酸セグメントを用いてトランスフェクションされた細胞(単数または複数)が非機能性ポリメラーゼをすでに発現することが本明細書中で提供される。具体的には、本発明は、セグメントが機能性インフルエンザポリメラーゼを産生できないようにするが、しかしウイルスの遺伝子セグメントのウイルス粒子内へのパッケージングは妨げないセグメント内の内部欠失を有する1個もしくはそれ以上の核酸セグメントを含んでなるインフルエンザウイルス粒子であって、ここで、ポリメラーゼが酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択されるものを提供する。内部欠失が:PAタンパク質では非コード領域から数えてヌクレオチド58と207の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして非コード領域から数えてヌクレオチド27と194の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB1タンパク質では非コード領域から数えてヌクレオチド43と246の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして非コード領域から数えてヌクレオチド24と197の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB2タンパク質では非コード領域から数えてヌクレオチド34と234の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして非コード領域から数えてヌクレオチド27と209の間であってそれらは包含されない位置にある3’ヌクレオチドで終わると好ましい。好ましい態様では、本発明はウイルス糖タンパク質をコードするインフルエンザウイルス核酸セグメントを有する本発明による粒子を提供する。本発明は、核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する本発明による粒子、または核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する粒子、または核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する粒子を提供する。具体的には、本発明は、A型インフルエンザウイルスから誘導されるインフルエンザウイルス核酸セグメントを有する本発明による粒子を提供する。本発明は、インフルエンザペプチドをコードしない核酸を備える本発明による粒子も提供する。さらに、本発明は、本発明による粒子を含んでなる細胞、具体的には、ポリメラーゼは酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(P
B2)から選択される1個もしくはそれ以上のインフルエンザウイルスポリメラーゼを備えた細胞を提供する。それに加えて、本発明は、本発明による粒子または本発明による細胞もしくは細胞から誘導された物質を含んでなる組成物、インフルエンザウイルスによる患者の感染に対する免疫学的防御を生成することを目的とする製薬学的組成物の生産のためのかかる組成物の使用、およびかかる組成物を防御を必要とする患者に提供することを含んでなるインフルエンザウイルスへの患者の感染に対する免疫学的防御を生成する方法を提供する。さらに、本発明は、インフルエンザペプチドをコードしない核酸を細胞に送達することを目的とする組成物の生産のための本発明による粒子の使用、およびインフルエンザペプチドをコードしない核酸を患者の細胞に送達することを目的とする製薬学的組成物の生産のための本発明による粒子の使用を提供する。かかる核酸(本明細書中では他者核酸と称する)は、適合する抗原性エピトープもしくはタンパク質をコードする他者遺伝子もしくは遺伝子フラグメントをコードしてもよく、または細胞内で核酸転写を干渉できるヌクレオチドの鎖をコードしてもよい。一つの態様では、本発明は、インフルエンザペプチドをコードしない核酸をある細胞または患者の細胞に送達することを目的とする組成物の生産のための本発明によるA型インフルエンザウイルス粒子の使用を提供する。さらに、本発明は、本発明による他者核酸を備えた欠陥インフルエンザウイルス粒子を細胞または患者が備えることを含んでなる、該細胞または該患者に、インフルエンザペプチドをコードしない核酸を送達する方法を提供する。
【図面の簡単な説明】
【0039】
【図1】条件つき欠陥A型インフルエンザウイルスの生産および増殖。最初に、293T細胞を、A/PR/8/34、pHMG−PAおよび適合する場合には、pΔPAもしくはpDIPAをコードする7個の二方向プラスミドを用いてトランスフェクションした。トランスフェクションの48時間後に、トランスフェクションされた細胞の上清を採取しそしてMDCK細胞および24時間前にHMG−PAを用いてトランスフェクションされたMDCK細胞を接種するために使用した。ウイルス複製にポジティブなMDCK−PA細胞の上清をMDCKおよびMDCK−PA細胞上で4回継代した。
【図2】条件つき欠陥ウイルス粒子の生成に使用した構築物。上図は、野生型PA遺伝子セグメントを示す。非コード領域(NCR)、および開始コドンが示されている。pΔPAは、A/WSN/33のPAを含む二方向プラスミドであるpHW183(9)のStulを用いる消化、次いで再結合して構築された。pDIPAは、A/PR/8/34のPA遺伝子セグメントの5’194および3’207ntをpSP72中にクローニングして構築された。次いで、挿入物は、二方向逆遺伝学ベクターに転移された。pΔPB1およびpΔPB2は、本文記載のようにして構築された。
【図3】上清rPR8−7、rPR8−ΔPAおよびrPR8−DIPA内のPA遺伝子セグメントの存在のRT−PCR分析。MDCK−PA継代4上清を22μmフィルターを通しそして遠心分離により濃縮した。続いて、RNAを単離しそしてPAセグメントの非コード領域を目標とするプライマーを用いてRT−PCRを行った。野生型A/PR/8/34から単離されたRNAを対照として使用した。レーン1:rPR8−7、レーン2:rPR8−ΔPA、レーン3:rPR8−DIPA、レーン4:野生型A/PR/8/34。マーカーサイズは左に記入されている。
【図4】欠失されたpΔPA構築物の追加の大きい部分、pΔPA−2、pΔPA−3、pΔPA−4、pΔPA−5をもたらす。
【0040】
詳細な説明
【実施例1】
【0041】
組換えDNAから欠陥A型インフルエンザウイルス粒子の生成
A型インフルエンザウイルスは、ネガティブ・センスの分節化ウイルスである。ゲノムは8個の遺伝子セグメントから成る。すべての8個の機能性遺伝子セグメントは、感染性ウイルス、すなわちインフルエンザウイルス複製に通常適切と考えられる細胞内で無限もしくは少なくとも数回の複製が可能な複製可能ウイルスを産生することを要求される。無作為的または特異的機構のいずれかを介するA型インフルエンザウイルスの遺伝子セグメントのパッケージングプロセスが長年の間論議されている。双方の意見の証拠が記載されている。無作為的パッケージングの証拠は、凝集したウイルス粒子が非凝集ウイルス粒子よりも高い感染性を有すること(6)および細胞培養物が低いmoiで感染された場合に、一部の感染細胞は一つのセグメントの発現を欠失すること(8)であり、双方共にすべてのインフルエンザウイルスゲノムは含まないビリオンがあることを示唆する。無作為的パッケージングのその他の証拠は、9個のセグメントを有するインフルエンザウイルスが実験的に産生されたことである(4)。BancroftおよびParslowは、ビリオン中のパッケージングのために同じ遺伝子セグメントに由来するvRNA間に競合がなかったことを見いだした(1)。
【0042】
特異的パッケージングプロセスのための一つの論議は、すべての遺伝子セグメントがウイルス原料中に等量で存在するけれども、それらは生産細胞中では異なる量で存在するというものである(10)。さらに、欠陥干渉性(DI)粒子が生成した場合に、DI vRNAはそれが誘導されたセグメントを置換する(3)(欠陥干渉性粒子は遺伝子セグメントの一つが大きい内部欠失を有するウイルス粒子である。それらの粒子は、ウイルスが高いmoiで継代された場合に起き、そしてポリメラーゼ酸性タンパク質のR638A変異によっても起きると考えられる〔Fodor et al;J.Virol.77:5017−5020,2003〕)。最後に、ビリオン形成の効率は、遺伝子セグメントの数が増えると増加する(5)。フジイら(Fujii et al.)は、ビリオン内のセグメントの効率的な組込みに必要なNAセグメントの領域も示し、その後同じグループはHAおよびNSの領域がウイルス粒子内へのパッケージングに重要であることを示した。〔Fujii,2005#256;Watanabe,2003#184〕。
【0043】
ここに、我々は特異性パッケージングの証拠を提出する。7個の機能性遺伝子セグメントのみを含むウイルス粒子を産生するために、我々はウイルス産生を止めることなくどの遺伝子セグメントを省くことができるかを決定する必要がある。ワクチンとしての複製欠失ウイルスの使用を考慮して、HAおよびNAは省かれず、そしてMAもNSも省かれないが、それはこの場合に2個の分離した発現プラスミドが必要なためである。我々はポリメラーゼ遺伝子が欠失したウイルスを生産した。我々は、欠失された遺伝子セグメントが発現プラスミドを用いてトランス相補されなかった場合にはウイルスを産生できなかった(表1、2および3、rPR8−7ntc)。7個の遺伝子セグメントおよび通常、非常に低い力価で欠失遺伝子セグメントにより発現されるタンパク質を発現するプラスミドのトランスフェクションによりウイルスが産生できた(表1、2および3、rPR8−7)。従って、インフルエンザウイルスA/WSN/33の遺伝子セグメント1、2および3の欠失変異体は、1032、528および1120ヌクレオチドの内部欠失をそれぞれ包含して産生された。それらの欠失変異体は、pΔPB2、pΔPB1およびpΔPAと命名された。図2参照。293T細胞は、以前の記載(de Wit,E.,M.I.Sproken,T.M.Bestebroer,G.F.Rimmelzwaan,A.D.Osterhaus,and R.A.Fouchier,2004,Efficient generation and growth of influenza virus A/PR/8/34 from eight cDNA fragments,Virus Res.103:155−61)のようにして、それらの欠失遺伝子セグメントそれぞれ1個およびA/PR/88.34をコードする7個の相補二方向構築物(de Wit et al,1994)および適切な発現プラスミドを用いてトランスフェクションされた。上清はトランスフェクションの48時間後に採取された。引き続いて、
MDCK細胞が、発現プラスミドHMG−PB2、HMG−PB1またはHMG−PAの一つを用いて以前の記載(2)のようにしてトランスフェクションされた。それらのトランスフェクションされた細胞は、トランスフェクションされた293T細胞の相当する上清を用いて接種された(実験手順の説明に関しては図1参照)。それらのMDCK細胞内のウイルス複製は、HAアッセイにより決定された。最初に、非トランスフェクションMDCK細胞内にはウイルス複製はなかった。ウイルス複製は相当する上清を接種されたHMG−PB2、HMG−PB1またはHMG−PAのいずれかを用いてトランスフェクションされたMDCK細胞内に示された。次いで、我々は、インフルエンザ配列データベース(www.flu.lanl.gov、アクセッション番号K00867)から得られたインフルエンザウイルスA/PR/8/34の欠陥干渉性PA vRNAの配列に基づいて欠陥PA遺伝子セグメントをクローンした。PAの5’207ntおよび3’194ntをPCR増幅しそして以前に記載したようにして(De Wit et al.,2004)変性したpHW2000(7)から誘導された二方向転写ベクター内にクローンした。得られたプラスミドはpDIPAと呼ばれた。図2)参照のこと。293T細胞をpDIPA,HMG−PAおよびインフルエンザウイルスA/PR/8/34の残りの遺伝子セグメントをコードする7個の二方向構築物を用いてトランスフェクションした(図2参照)。トランスフェクションの48時間後に上清を採取しそして引き続いて、24時間前にHMG−PAを用いてトランスフェクションされたMDCK細胞にこの上清を接種した。接種の72時間後にそれらのMDECK細胞の上清についてHAアッセイを行うと陽性であることがわかり、それはそれらの細胞内でのウイルス複製を示した。非トランスフェクションMDCK細胞の接種もHAアッセイにより測定するとウイルス産生をもたらさなかった。非トランスフェクションもしくはHMG−PAを用いてトランスフェクションしたMDCK細胞上でのPA欠損ウイルス粒子を含む上清の引き続く継代は、同様の結果に導いた(表1)。継代4まで、ウイルスはHMG−PAを用いてトランスフェクションされたMDCK細胞内で産生された。MDCKp4の上清は、ウイルス力価の指数を得るために血清で順列的に希釈され、それは約104TCID50/mlであることが示された。
【0044】
使用された方法の工程は以下である:293T細胞を構築物pΔPB2、pΔPB1、pΔPA、pΔNA、A/PR/8/34をコードする7個の相補構築物(De Wit
et al,2004)の一つおよびHMG−PB2、HMG−PB1、HMG−PA(発現プラスミドは、例えばPleschka,S.,R.Jaskunas,O.G.Engelhardt,T.Zurcher,P.Palese,and A.Garcia−Sastre,1996,A plasmid−based reverse genetics system for influenza A virus.J.Virol 70:4188−92;A.Garcia−Sastre and P.Paleseから入手)の一つを用いてトランスフェクションした(トランスフェクション・プロトコールはDe Wit et al,2004参照)。トランスフェクションの48時間後に、トランスフェクションされた293T細胞の上清を採取した。ウイルスが産生された場合には、それらは上清中に存在する。同時に、MDCK細胞を、発現プラスミドHMG−PB2、HMG−PB1、PMG−PAの一つ(使用した欠損変異体に応じて、すなわちpΔPB2を用いた場合には、MDCK細胞はここでHMG−PB2を用いてトランスフェクションされる)を用いてトランスフェクションした(トランスフェクションプロトコールはBasler et al.,2000参照)が、それはそれらが内部欠失を有する遺伝子セグメントをパッケージングしたので、産生されたウイルスがこのタンパク質を発現する遺伝子セグメントを欠失しているからである。トランスフェクションの24時間後に、トランスフェクションされたMDCK細胞を、トランスフェクションされた293T細胞から得た上清を用いて接種する。293T上清中にウイルスが存在する場合には、このウイルスはトランスフェクションMDCK細胞中で複製しそしてさらなるウイルスが産生される。この上清は、接種の72時間後に再び採取できる。
【0045】
機能性PA遺伝子セグメントをもたらす発生PAまたはDIPAの組換えが起きていないことを証明するために、MDCKp4の上清からRNAを単離した。最初に、上清を22μMフィルターを通しそして遠心分離により濃縮した。次いで、RNAを分離しそしてPAセグメントの非コード領域を目的とするプライマーを用いてRT−PCRを行った。PA vRNAに特異性のプライマーを用いて行ったRT−PCRは、ΔPAおよびDIPAが複数回の継代の間も安定のままであることを示した。DIPAウイルス粒子で感染されたMDCK細胞の上清中に、約400bpの明確なバンドが現れ、ΔPAを含むウイルスを用いて感染されたMDCK細胞の上清中には、1100bpのバンドが現れる。野生型A/PR/8/34を用いて感染されたMDCK細胞の上清中には、約2300ntのバンドが見られる(図3)。それらの結果は、ΔPAPR8遺伝子上清が安定にビリオン内にパッケージングされていることを示す。
【0046】
PB2を欠失する細胞を産生するために、インフルエンザウイルスA/PR/8/34の遺伝子セグメント2、3、4、5、6、7および8をコードする7個の二方向構築物(Hoffmann,E.G.,G.Neumann,Y.Kawaoka,G.Hobom,and R.G.Webster,2000,A DNA transfection system for generation of influenza A virus from eight plasmids.Proc.Natl.Acad
Sci U S A 97:6108−13)を用いて293T細胞をトランスフェクションすると(de Wit,E.,I.Sproken,T.M.Bestebroer,G.F.Rimmelzeaan,A.D.Osterhaus,and R.A.Fouchier,2004,Efficient Generation and growth of influenza virus A/PR/8/34 from eight cDNA fragments.Virus Res.103:155−61)、vRNAおよびmRNAの発現をもたらした。A/PR/8/34のPB2を発現するプラスミド、pHMG−PB2(Pleschka,S.,R.Jaskunas,O.G.Engelhardt,T.Zurcher,P.Palese,and A.Garcia−Sastre,1996,A plasmid−based reverse genetics system for influenza A virus,J.Virol.70:4188−92)を共トランスフェクションした。対照として、A/PR/8/34をコードする7個の二方向構築物のみをトランスフェクションし、pHMG−PB2を除外した。上清をトランスフェクションの48時間後に採取し、MDCK細胞または24時間前に100ml皿内でpHMG−PB2(MDCK−PB2)を用いてトランスフェクションされたMDCK細胞中に接種した。接種の3日後に、接種されたMDCK細胞の上清を、ウイルス産生に関する指示としてシチメンチョウ赤血球を用いて血球凝集活性を試験した。pHMG−PB2を除いて7個の遺伝子セグメントのみを用いてトランスフェクションされた293T細胞の上清を用いて接種された細胞中にウイルスは検出されなかった(rPR8−7ntc、表2)。7個の遺伝子セグメントおよびpHMG−PB2を用いてトランスフェクションされた293T細胞の上清を用いてトランスフェクションされたMDCK−PB2細胞の上清は陽性であった。引き続いて、rPR8−7上清をMDCKおよびMDCK−PB2細胞内で継代した。rPR8−7はMDCK−PB2細胞内で複製したが、しかしMDCK細胞内ではしなかった(表2)。我々は次いでインフルエンザウイルスA/WSN/33の遺伝子のセグメント1の1032nt欠失変異体を生成させると、344アミノ酸欠失をもたらした(pΔPB2、図2)。ΔPB2を含む組換えウイルス(rPR8−ΔPB2)を上記のようにして生産した(図1)。MDCK細胞中にはウイルスは検出できなかったが、rPR8−ΔPB2を用いて接種されたMDCK−PB2細胞中にはウイルスが検出された。rPR8−ΔPB2を継代した後、MDCK細胞内にウイルス産生の証拠はなく、MDCK−PB2細胞とは対照的であった(表2)。
【0047】
PB1を欠失するウイルスも生産した。293T細胞を、インフルエンザウイルスA/PR/8/34の遺伝子セグメント1、3、4、5、6、7および8をコードする7個の二方向構築物を用いてトランスフェクションすると、vRNAおよびmRNAの発現をもたらした。A/PR/8/34のPB1を発現するプラスミド、pHMG−PB1を共トランスフェクションした。対照として、pHMG−PB1を除いてA/PR/8/34をコードする7個の二方向構築物のみをトランスフェクションした。トランスフェクションの48時間後に上清を採取しそしてMDCK細胞または24時間前に100mm皿内でpHMG−PB1を用いてトランスフェクションされたMDCK細胞(MDCK−PB1)内に接種した(2)(図1)。接種の3日後に、接種されたMDCK細胞の上清を、ウイルス産生の指示としてシチメンチョウ赤血球を用いて血球凝集活性を試験した。pHMG−PB1を除いて7個の遺伝子セグメントのみを用いてトランスフェクションされた293T細胞の上清を用いて接種された細胞中にウイルスは検出されなかった(rPR8−7ntc、表3)。7個の遺伝子セグメントおよびpHMG−PB1を用いてトランスフェクションされた293T細胞の上清を用いて接種されたMDCK−PB1細胞の上清は陽性であった。引き続いて、rPR8−7上清をMDCKおよびMDCK−PB1細胞内で継代した。rPR8−7はMDCK−PB1細胞内で複製したが、しかしMDCK細胞内ではしなかった(表3)。我々は次いでインフルエンザウイルスA/WSN/33の遺伝子セグメント2の528nt欠失変異体を生成させると、178アミノ酸欠失をもたらした(pΔPB1、図2)。ΔPB1を含む組換えウイルス(rPR8−ΔPB1)を上記のようにして生産した(図1)。MDCK細胞中にはウイルスは検出できなかったが、rPR8−ΔPB1を用いて接種されたMDCK−PB1細胞中にはウイルスが検出された。rPR8−ΔPB1を継代した後、MDCK細胞内にウイルス産生の証拠はなく、MDCK−PB1細胞とは対照的であった(表3)。
【0048】
このように、我々は、上記のようにしてpΔPB2、pΔPB1またはpΔPA/pDIPA構築物をもたらしそしてRNAポリメラーゼII駆動PB2、PB1またはPA発現プラスミドを用いてトランス相補して、セグメント1、2または3を欠失するウイルスを産生できる。ここに記載する条件つき欠陥ウイルスは、トランス相補されていない細胞中ではただ一周の複製が可能であるが、しかしそれはトランス相補された細胞中では繁殖できる。これは、7個の機能性遺伝子セグメントのみを含み複製一回のみが可能、または欠失遺伝子セグメントがトランス相補された場合には複数回の複製が可能な逆遺伝学を用いて欠陥ウイルスが生産された最初である。
【0049】
この方法で生産された欠陥ウイルス粒子は、それらが感染性ウイルスを生産することなく一回の複製を行えるので、ワクチンの候補である。この一回複製の結果は、ワクチンが体液的および細胞的免疫反応の双方を誘導することである。それらの欠陥粒子は通常の細胞内では複製しないけれども、生産目的では欠陥タンパク質を発現する細胞系統内で大量のウイルスが増殖可能である。我々が示したように、複数回の複製はウイルスの遺伝子型に影響しない。ワクチンとしての欠陥ウイルス粒子の使用の外に、それらは遺伝子送達および他者タンパク質の発現のための候補ベクターでもあり、それというのも機能性遺伝子は5’および3’PA、PB2またはPB1配列の間に挿入できるからである。これはWatanabeら(11)によっても示されており、彼らはHAおよびNAを他者遺伝子で置換しそしてそれでもウイルス生産が可能なことも示した。
【0050】
pΔPAのさらなる末端短縮
さらに、pΔPA構築物のより大きい部分を欠失させると、pΔPA−2、pΔPA−3、pΔPA−4、pΔPA−5がもたらされる(図4)。293T細胞は、以前の記載(de Wit et al.,2004)のようにそれらの欠失された遺伝子セグメントのそれぞれ一つおよびA/PR/8/34をコードする7個の相補二方向性構築物(D
e Wit et al.,2004)およびPAを発現する発現プラスミドを用いてトランスフェクションされた。トランスフェクションの48時間後に上清を採取した。引き続いて、MDCK細胞を発現プラスミドHMG−PAを用いて以前の記載(Basler,C.F.,et al.,2000,Proc Natl Acad Sci U S
A 97:12289−94)のようにしてトランスフェクションした。それらのトランスフェクションされた細胞および非トランスフェクション細胞を、トランスフェクションされた293T細胞の上清を用いて接種した。それらのMDCKおよびMDCK−PA細胞内のウイルス複製は、HAアッセイにより決定した。非トランスフェクションMDCK細胞内では複製はなかった。ウイルス複製は、上清のいずれか一つを用いて接種された、HMG−PAを用いてトランスフェクションされたMDCK細胞内に見られた。それらの構築物からもたらされたすべてのvRNAをビリオン内にパッケージングした(表4)。
【0051】
【表1】

【0052】
【表2】
【0053】
【表3】
【0054】
【表4】
【実施例2】
【0055】
欠陥組換えウイルスを用いるワクチン接種
機能性PA、PB1もしくはPB2遺伝子を欠失する条件つき欠陥組換えウイルスを、高処理量ウイルスバックボーン(例えばワクチン株A/PR/8/34から誘導)に基づいて関連する流行性ウイルス(例えばA/モスクワ/10/99)のHAおよびNA遺伝子を用いて、本明細書中に記載のようにして生産する。条件つき欠陥ウイルスはトランスフェクションにより生産され、その際、ポリメラーゼタンパク質発現はトランス相補を介して達成される。次いで関連ポリメラーゼを安定して発現する適切な細胞基質、例えばMDCK細胞またはVero細胞内でウイルスを増幅する。ウイルス上清は、10分間、1000xgで遠心分離して清澄化する。ウイルスを20〜60%のスクロース勾配中で遠心分離して濃縮および精製し、ペレット化し、そしてリン酸緩衝食塩水(PBS)中に再懸濁する。ウイルス調製物の純度および品質は、クーマシー・ブリリアント・ブルーを用いて染色した12.5%SDSポリアクリルアミド・ゲルを用いて確認されそして条件つき欠陥ウイルスのウイルス力価は、MDCK細胞および関連するポリメラーゼを発現するMDCK細胞の感染および抗核タンパク質モノクローナル抗体を用いる染色により決定される。1x10E5 50%組織培養感染量(TCID−50)を用いて気管内または鼻腔内にネブライザーを用いてマウスに接種する。ワクチン接種の前後に採取した血清試料中のインフルエンザウイルスのHA、NAおよび内部タンパク質に対する抗体力価は、赤血球凝集阻害アッセイ、ノイラニミダーゼ阻害アッセイ、ELISA、またはウイルス中和アッセイを用いて決定される。ワクチン接種された動物内の抗原特異性細胞免疫反応は、フローサイトメトリー、CD4およびCD8陽性細胞のテトラマー染色、細胞溶解活性、T細胞増殖などにより細胞内サイトカイン発現を測定して定量化される。ワクチン接種
および対照動物を、ワクチン接種の6週間後にインフルエンザウイルスA/モスクワ/10/99または異種ウイルス単離物の1x10E6 TCID−50を用いて負荷する。負荷の後、鼻または咽頭スワブ試料を10日間、毎日動物から採取し、そして感染した動物により分泌されたウイルスの量が定量PCR分析またはウイルス滴定により決定される。得られたワクチン誘導体液免疫は抗体力価の上昇を定量することにより、得られたワクチン誘導細胞免疫はヘルパーおよび細胞毒性T細胞反応の上昇を定量することにより、そして免疫性の全体的レベルは負荷ウイルスを用いる感染に対する防御を確認することにより検出される。
【0056】
【表5】
【0057】
<本発明の特徴または態様>
限定されるものではないが、本発明の主たる特徴または態様を以下に記載する。
【0058】
態様1:条件欠陥インフルエンザウイルス粒子を得るための方法であって、インフルエンザポリメラーゼをコードする核酸を内部欠失することにより誘導された1個もしくはそれ以上の遺伝子構築物であって、機能性ポリメラーゼを産生できないが、しかしウイルス粒
子内へのウイルスの遺伝子セグメントのパッケージングを妨げない該遺伝子構築物を用い、かつ、インフルエンザウイルスをコードするが該インフルエンザポリメラーゼをコードしない相補性インフルエンザウイルス核酸セグメントを用い、かつ、該細胞内で該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いて、適当な単一もしくは複数の細胞をトランスフェクションする工程、およびトランスフェクションの後適当な時点で該単一もしくは複数の細胞の上清から少なくとも1個のウイルス粒子を採取する工程を含むが、ヘルパーウイルスを使用しない、ことを特徴とする方法。
【0059】
態様2:条件欠陥インフルエンザウイルス粒子を得るための方法であって、適切な単一もしくは複数の細胞を該細胞内でインフルエンザポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いてトランスフェクションする第一工程、および機能性インフルエンザポリメラーゼを産生できない請求項1に記載の条件欠陥インフルエンザウイルス粒子を含む上清を用いて該単一もしくは複数の細胞に感染させる第二工程、および感染の後の適当な時点で該単一もしくは複数の細胞の上清から少なくとも1個のウイルス粒子を採取することを含む第三工程を含むが、ヘルパーウイルスを使用しない、ことを特徴とする方法。
【0060】
態様3:条件欠陥インフルエンザウイルス粒子を得るための方法であって、
インフルエンザポリメラーゼをコードする核酸を内部欠失することにより誘導された1個もしくはそれ以上の遺伝子構築物であって、機能性ポリメラーゼを産生できないが、しかしウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを妨げない該遺伝子構築物を用い、かつ、インフルエンザウイルスをコードするが、該インフルエンザポリメラーゼをコードしない相補性インフルエンザウイルス核酸セグメントを用い、かつ、該細胞内で該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いて適当な第一の単一もしくは複数の細胞をトランスフェクションし、かつ、トランスフェクションの後適当な時点で該第一の単一もしくは複数の細胞の上清から少なくとも1個のウイルス粒子を採取する第一工程、および適切な第二の単一もしくは複数の細胞を該細胞内で該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いてトランスフェクションする第二工程、および該第一細胞から得られた少なくとも1個のウイルス粒子を含む上清を用いて該第二の単一もしくは複数の細胞に感染させる第三工程、および感染の後の適当な時点で該第二の単一もしくは複数の細胞の上清から少なくとも1個のウイルス粒子を採取することを含んでなる第四工程を含んでなるが、ヘルパーウイルスを使用しない、ことを特徴とする方法。
【0061】
態様4:インフルエンザポリメラーゼをコードする核酸の内部欠失することからもたらされる内部欠失が、PAタンパク質では5’非コード領域から数えてヌクレオチド58と207の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域のから数えてヌクレオチド27と194の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB1タンパク質では5’非コード領域から数えてヌクレオチド43と246の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド24と197の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB2タンパク質では5’非コード領域から数えてヌクレオチド34と234の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド27と209の間であってそれらは包含されない位置にある3’ヌクレオチドで終わることを特徴とする態様1および3のいずれかに記載の方法。
【0062】
態様5:条件欠陥インフルエンザウイルス粒子を含んでなる上清を用いて感染される単一もしくは複数の細胞が条件欠陥インフルエンザウイルス粒子内で非機能性であるポリメラ
ーゼに相当するインフルエンザポリメラーゼをすでに発現している、態様2〜4のいずれか1項に記載の方法。
【0063】
態様6:ポリメラーゼが酸性ポリメラーゼ(PA)である、態様1〜5のいずれか1項に記載の方法。
【0064】
態様7:ポリメラーゼが塩基性ポリメラーゼ1(PB1)である、態様1〜5のいずれか1項に記載の方法。
【0065】
態様8:ポリメラーゼが塩基性ポリメラーゼ2(PB2)である、態様1〜5のいずれか1項に記載の方法。
【0066】
態様9:態様1〜8のいずれかに記載の方法により得られるインフルエンザウイルス粒子。
【0067】
態様10:機能性インフルエンザポリメラーゼを産生できないようにし、かつ、複製について条件欠陥性のウイルス粒子をもたらすが、しかしインフルエンザウイルスの遺伝子セグメントのウイルス粒子内へのパッケージングは妨げないセグメント内の内部欠失を有する1個もしくはそれ以上の核酸セグメントを含んでなるインフルエンザウイルス粒子であって、ここで、該ポリメラーゼが酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択されることを特徴とするインフルエンザウイルス粒子。
【0068】
態様11:機能性インフルエンザポリメラーゼを産生できないようにし、かつ、複製について条件欠陥性のウイルス粒子をもたらすが、しかしインフルエンザウイルスの遺伝子セグメントのウイルス粒子内へのパッケージングは妨げないセグメント内の内部欠失を有する1個もしくはそれ以上の核酸セグメントを含んでなるインフルエンザウイルス粒子であって、ここで、ポリメラーゼが酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択され、かつ、ここで内部欠失が、PAタンパク質では5’非コード領域から数えてヌクレオチド58と207の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド27と194の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB1タンパク質では5’非コード領域から数えてヌクレオチド43と246の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド24と197の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB2タンパク質では5’非コード領域から数えてヌクレオチド34と234の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド27と209の間であってそれらは包含されない位置にある3’ヌクレオチドで終わることを特徴とするインフルエンザウイルス粒子。
【0069】
態様12:ウイルス糖タンパク質をコードするインフルエンザウイルス核酸セグメントを有する、態様9〜11のいずれか1項に記載の粒子。
【0070】
態様13:核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する、態様9〜12のいずれか1項に記載の粒子。
【0071】
態様14:核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する、態様9〜12のいずれか1項に記載の粒子。
【0072】
態様15:核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する、態様9〜12のいずれか1項に記載の粒子。
【0073】
態様16:A型インフルエンザウイルスから誘導されたインフルエンザウイルス核酸セグメントを有する、態様9〜15のいずれか1項に記載の粒子。
【0074】
態様17:インフルエンザペプチドをコードしない核酸を備えている、態様9〜16のいずれか1項に記載の粒子。
【0075】
態様18:態様9〜17のいずれか1項に記載の粒子を含むが、ヘルパーウイルスを含まないことを特徴とする細胞。
【0076】
態様19:ポリメラーゼが、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択される、1個もしくはそれ以上のインフルエンザウイルスポリメラーゼを発現する態様18記載の細胞。
【0077】
態様20:態様9〜17のいずれか1項に記載の粒子または態様18もしくは19記載の細胞もしくは細胞から誘導される物質を含んでなる組成物。
【0078】
態様21:インフルエンザウイルスへの患者の感染に対する免疫学的防御を生ぜしめる目的とする製薬学的組成物の生産のための、態様20記載の組成物の使用。
【0079】
態様22:有効成分として、態様9〜17のいずれか1項に記載の粒子または態様18もしくは19記載の細胞もしくは細胞から誘導される物質を含んでなるインフルエンザウイルスへの患者の感染に対する免疫学的防御を生ぜしめるための製薬学的製剤。
【0080】
態様23:細胞に対してインフルエンザペプチドをコードしない核酸の送達を目的とする組成物の生産のための、態様17記載の粒子の使用。
【0081】
態様24:対照者の細胞へのインフルエンザペプチドをコードしない核酸の送達を目的とする製薬学的組成物の生産のための、態様17記載の粒子の使用。
【0082】
態様25:態様17記載の欠陥インフルエンザウイルス粒子を細胞に提供することを含んでなる、該細胞へのインフルエンザペプチドをコードしない核酸のインビトロでの送達の方法。
【0083】
態様26:有効成分として、態様17記載の欠陥インフルエンザウイルス粒子を含んでなる、該患者へインフルエンザペプチドをコードしない核酸を送達するための製薬学的製剤。
【産業上の利用可能性】
【0084】
本発明により提供される条件欠陥インフルエンザウイルス粒子は、通常の条件下では、
生体内で複製、増殖されないので、インフルエンザの予防または治療に効果的に使用できる。したがって、例えば、医薬製造業で利用できる。
【技術分野】
【0001】
本発明は、インフルエンザウイルスおよび流感に対するワクチン接種の分野に関する。
【背景技術】
【0002】
インフルエンザウイルス(オルトミクソウイルス科)は、分節ゲノムを有するエンベロープ付マイナス鎖RNAウイルスである(非特許文献1)。それらは二つの属に分けられる。それらの核タンパク質およびマトリックスタンパク質の間の著しい抗原性差異に基づいて、一方はA型およびB型インフルエンザを含み、他はC型インフルエンザから成る。3種のウイルス型は、病原性およびゲノム構成でも異なる。A型は広範囲の温血動物に見られるが、一方BおよびC型は、主としてヒト病原体である。A型インフルエンザウイルスは、ビリオンの表面から突出する赤血球凝集素(HA)およびNA表面糖タンパク質の抗原性特性化によりさらに細分される。現在15のHAおよびNAサブタイプがある。A型インフルエンザウイルスは、鳥類、ブタ、ウマ、ヒト、およびその他の哺乳動物を含む広範囲の動物に感染する。水鳥は、すべての既知のA型インフルエンザサブタイプの天然の貯蔵庫となり、そして多分、ヒト汎流行インフルエンザ株の遺伝物質の起源であろう。
【0003】
関連するパラミクソウイルスとは異なり、インフルエンザウイルスは分節RNAゲノムを有する。A型およびB型インフルエンザウイルスは同様の構造を有し、一方C型はさらに分岐している。A型およびB型ウイルスは、それぞれ少なくとも1個のタンパク質をコードする8個の分離した遺伝子セグメント(分節)をそれぞれ含み、C型は7個の分離したセグメントを含み、A型およびB型のセグメント4および6を結合している。A型およびB型インフルエンザウイルスは、3種のタンパク質:HA、NAおよび基質2(M2)の突起により覆われている。C型インフルエンザウイルスは、ただ1種の表面糖タンパク質を有する。それぞれのインフルエンザRNAセグメントは、核タンパク質(NP)により包膜化されてリボヌクレオチド核タンパク質(RNP)複合体を形成する。3個のポリメラーゼタンパク質がRNP複合体の1個の末端と会合する。RNPは、一体部分としてマトリックスタンパク質(基質1)を有する膜により包囲されている。エンベロープのリン脂質部分は、細胞宿主膜から誘導される。ウイルス粒子中には非構造タンパク質2(NS2)も見いだされる。
【0004】
インフルエンザウイルスの命名法に関する世界保健機構(WHO)ガイドラインは下記である。最初にウイルスの型(A、B、またはC)、次いで宿主(ヒト以外の場合)、単離の場所、単離番号、および単離の年(斜線で分離)を指定する。A型インフルエンザに対しては、HAおよびNAサブタイプを括弧内に記載する。例えば、2000および2001のシーズンでの最近の三価ワクチン内中含まれる株は:A/パナマ/2007/99(H3N2)、A/ニュー・カレドニア/20/99(H1N1)、およびB/山梨/16/98である。1977年以後、2種のインフルエンザAサブタイプ:H1N1およびH3N2がヒト間で同時に流行している。
【0005】
インフルエンザウイルスのRNAポリメラーゼが複合体プルーフリーディング活性を持たないので、インフルエンザウイルスは複製の間に点変異を蓄積する。表面糖タンパク質の抗原性部分内のアミノ酸を変化させる変異は、すでに存在する免疫性を避けることをそれに可能とさせることにより、ウイルス株に選択的利益を与える。HA分子は、一定の宿主細胞上の受容体に結合することにより感染を開始する。HAタンパク質に対する抗体は、受容体結合を防止しそして同一株による再感染を防止するために非常に有効である。HAは、現在流行しているHA遺伝子の変異が抗体結合を破壊する抗原ドリフトによるか、またはウイルスが新しいサブタイプのHAを取得する抗原シフトのいずれかにより、以前
取得した免疫性を避けることができる。抗原ドリフト圧力は、HA分子間で非平等であり、正に選択された変化は、主としてHAタンパク質の球状頭部上に起きる。それらの変化も、NAよりもHA中でより広範囲に蓄積される。他のインフルエンザタンパク質内の変化はさらに遅く起きる。同様に、抗原性ドリフト圧力は、ヒト適合インフルエンザ株中で最大であり、ブタおよびウマ適合株中では中間であり、そして鳥類適合株中では最低である。
【0006】
インフルエンザウイルスは分節ゲノムを有するので、同じ宿主内での2種の異なる株の同時感染は、親の遺伝子セグメントの異なる組合せを含む新規の再構築されたインフルエンザ株の産生に導くことがある。15種のHAサブタイプが野生鳥類中に存在することが知られており、そしてヒトには先例がないHAの起源を提供する。抗原シフトによる新規サブタイプを有するインフルエンザ株のヒト流行の発生は、1957年および1968年の少なくとも二回のインフルエンザ汎流行の原因であり、1918年のインフルエンザ汎流行の原因である可能性が高い。汎流行インフルエンザウイルスの発生に関するすべての知識に沿って考えると、汎流行性株は現在流行しているものとは抗原的に異なるHAを有するに違いない;このHAは、ヒト内で60〜70年間は流行できなかったものである;そしてウイルスはヒトからヒトに伝播できなければならない。1957年および1968年の双方で、汎流行はHAのシフトからもたらされ、そして両方の場合に、汎流行株のHAは、鳥類株と密接に関連していた。汎流行のための絶対的な要求の一つは、HAが変化しなければならないことであるけれども、ウイルスの残部が変化可能または必須である範囲は知られていない。1957年および1968年の汎流行ウイルスのみが直接研究に利用でき、1918年汎流行インフルエンザウイルスは、分子考古学を用いて特性が研究されている。1957年には、3個の遺伝子が鳥類様遺伝子:HA、NA、およびポリメラーゼ複合体(PB1)のサブユニットにより置換された。1968年では、HAおよびPB1のみが置換された。
【0007】
インフルエンザ感染の特定診断は、ウイルス単離、赤血球凝集阻害(HI)試験、免疫アッセイによる抗原検出、血清学的試験、分泌物中のNA活性の証明、または分子に基づくアッセイにより行うことができる。試料は、痰、鼻咽頭スワブ、または緩衝塩溶液のうがいにより得られる鼻咽頭洗浄物として採取できる。インフルエンザ診断のための標準は、培養後の免疫学的特性化であった。血清学的分析は、それが急性および回復期血清の双方の採取を必要とするので、インフルエンザ感染に対しては、正確ではあるがしかし遡及的な方法となる。
【0008】
インフルエンザウイルスは、孵化鶏卵または多数の組織培養システム中で培養できる。トリプシンの添加(HAの切断活性化のため)は、Madin−Darbyイヌ腎(MDCK)細胞およびその他の系統中でインフルエンザウイルス繁殖を可能とする。ワクチン生産の最初の方法は、卵内のままでのインフルエンザウイルスの培養である。細胞系統の培養物は、ヒトインフルエンザウイルス(A型およびB型の同胞)の一次単離のために一般的に使用される。多数のヒトインフルエンザウイルスが孵化卵内の尿膜腔内で直接培養できる。一部のインフルエンザAおよびB型ウイルスは、羊膜腔での初期培養、引き続いて尿膜腔に順応させることを要する。培養物単離の後、大部分のインフルエンザ単離物は免疫アッセイまたは免疫蛍光法を用いて厳格に分離できる。インフルエンザウイルスのHA分子は、エントリーを増すためにウイルスの呼吸細胞の表面上のサリチル酸残基に結合する。
【0009】
インフルエンザ株は、イン・ビトロで赤血球を凝集させるインフルエンザウイルスの能力を利用して抗原的に特性化できる。抗HA抗体は、凝集を阻害できる。従って、赤血球凝集阻害(HI)アッセイはインフルエンザ株を特性化するために使用される標準的方法の一つである。HIアッセイは、試料株が最近のワクチン株に免疫学的に関連(すなわち
交差反応性)するかどうかを決定するために使用される。一般的にフェレット内で産生される類型決定血清(typing sera)を一連の二倍希釈ウエルに加え、そして実験者が赤血球の懸濁物に対する凝集物を観察してウエルを採点する。大部分の場合に、血清のパネルがワクチンおよび対照試料に対して試料血清を一致させるために使用され、そして該当するインフルエンザシーズンの間に、大多数の使用株がHIアッセイにより一致させることに成功する。
【0010】
WHOはガイドラインを提供し、そしてWHO協力センター(Collaborating Center)は個別のウイルス株の抗原的特性の同定に関するガイダンスを提供する。試料株は、免疫学的系統、例えばA/モスクワ/10/99(H3N2)、A/ニュー・カレドニア/20/99(H1N1)、およびB/北京/184/93様ウイルス、に従って分類される。HIアッセイで特性化できなかった試料株に対して、実験者は、それらをフェレットに接種して株特異性抗血清を生産させなければならない。新しい抗血清が準備されると、上記のようにしてHIアッセイを再度行う。新血清が交差反応性に有意のギャップ(通常試料とワクチン株との間の4倍の差異として定義される)を示す場合には、それを通常の実験室パネル内に組入れ、そして新規の流行株を探索するために使用される。このようにして、HIアッセイは、ワクチン株選択のためのインフルエンザウイルス監視の努力に非常に重要でありそして抗原ドリフトを評価するために最も一般的に使用される方法である。
【0011】
インフルエンザウイルスは、個別の遺伝子セグメントの配列比較により一般的に特性化でき、そしてこの場合にもWHOガイドラインおよびWHO協力センターがインフルエンザゲノムを含んでなるRNAセグメントの個別同一性の同定に関するガイダンスを提供している;核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、酸ポリメラーゼ(PA)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするA型およびB型インフルエンザウイルス核酸セグメント、および核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素−ノイラミニダーゼ様糖タンパク質(HN)、マトリックスタンパク質(M1およびM2)、および非構造タンパク質(NS1およびNS2)をコードするC型ウイルス核酸セグメント。抗原分析のため、核酸配列比較のため、およびワクチンウイルスの同定のための参照株の要求は、WHO Collaborating Centre for Reference and Research on Influenza,45 Poplar Road,Parkville,Victoria 3052,Australia(ファックス:+61 3 9389 1881,ウエブサイト:http://www.influenza centre,org);WHO Collaborating Centre for Reference and Research on Influenza,National Institute of
Infectious Diseases,Gakuen 4−7−1,Musashi−Murayama,Tokyo 208−0011,Japan(ファックス:+81
42 5610812または+81 42 5652498);WHO Collaborating Center for Surveillance,Epidemio
logy and Control of Influenza,Centers for Disease Control and Prevention,1600 Clifton Road,Mail Stop G16,Atlanta,GA 30333,United States of America(ファックス:+1 404 639 23 34);またはWHO Collaborating Centre for Reference and Research on Influenza, National Institute for Medical Research,The Ridgeway,Mill Hill,London NW7 1AA,En
gland(ファックス:+44 208 906 4477)。最新の疫学的情報は、WHOのウエブサイト:http://www.who.int/influenzeおよび地理学的情報システム、FluNet、http://www.who.int/flunetから入手できる。
【0012】
インフルエンザのインパクトおよびその予防の健康および経済的利益の意識は増大しており、そして過去10年間はワクチン接種の使用および利益が見られそして多数の抗インフルエンザ医薬がますます増えている。多数の国でより長い期待寿命の結果として、より多数の人々に合併症の危険があり、インフルエンザ流行の間の健康管理システムに対する負荷がさらに広く認識され、そしてさらに頻繁な国際旅行がウイルスの拡散の機会を与え、一方新製品の導入は疾患の予防および処置に対する選択肢を増やしている。約50の国々が、政府出資の国内インフルエンザ免疫プログラムを有しそしてワクチンはその他の多数の国々でも利用できる。ワクチンの使用のための特定の指針はさまざまであるが、しかし一般的に、高齢者および既存の慢性病的条件のために重篤な疾患の危険が上昇している6カ月以上の人々への毎年の接種を含む。一部の国では、医療リスクが高い人々へのインフルエンザの拡散を低減するためにワクチンが使用される。メンバーステイト(Member State)は、それらの全般的な公衆衛生優先事項の範囲内で、インフルエンザ予防活動の利益を考慮する必要がある。
【0013】
不活化ワクチンは、それらが全ウイルス粒子、部分的に破壊されたウイルス粒子(スプリットワクチン)または精製されたエンベロープ抗原(サブユニットワクチン)を含むかどうかに依存して、数種のタイプに分類される。いくらかのサブユニットワクチンは、アジュバントまたは送達システムと複合される。
【0014】
いくつかの国は一定の標的群に対して生弱毒インフルエンザワクチンを許可している。ロシア連邦では、1ワクチンの2種の異なる製剤が、健康な成人および小児に使用されており、別の生ワクチンが広範に試験されている。しかし、生弱毒ワクチンがさらに広範に入手可能となるまで、それはインフルエンザ予防のためにまだ一般的には推奨されない。
【0015】
2種の抗ウイルス剤がインフルエンザの予防および処置のために開発されている。M2阻害剤、アマンタジン(amantadine)およびリマンタジン(rimantadine)は、A型インフルエンザウイルスの処置に限定され、そして感染の予防に有効であると報告されている。両方の製品はいくらかの副作用を起こすが。著しい神経学的副作用はアマンタジンの方が大きい。ノイラミニダーゼ阻害剤、例えばザナミビル(zanamivir)およびオセルタミビル(oseltamivir)は、A型およびB型インフルエンザの処置のために多数の国で最近許可され、そして予防に有効であることが報告されている。双方の抗ウイルス剤を投与された患者内で、耐性変異体が検出されている。これは現在までは重要な公衆衛生問題として考慮されていないけれども、それらの薬剤がさらに大きいスケールで使用されると、状況は変化するであろう。
【0016】
WHOは、全世界的な国際監視プログラムを、82カ国に存在する110箇所の国内インフルエンザセンターおよび4箇所のインフルエンザ参照および研究のためのWHO協力センター(アトランタ(米国)、ロンドン(英国)、メルボルン(オーストラリア)および東京(日本)に位置する)の協力を得て維持している。それらのセンターは、流行ポテンシャルと共に発生する株の早期警告システムを提供する。インフルエンザワクチンが現在流行している株を含まない場合、インフルエンザワクチンの効力が低下するので、このシステムは重要である。WHOはワクチン組成の推奨を発行し、それは世界保健機構により出版されるWeekly Epidemiological Record(例えば非特許文献2参照)中に見ることができ、2月には北半球で使用されるワクチン、9月には南半球で使用されるワクチンに関して公開されている。インフルエンザは赤道付近の地域
では季節的パターンがあまりはっきりしないので、疫学的考慮が、それらの推奨事項(2月または9月のどちらか)が赤道付近の国々に使用されるワクチンに適合するかに影響する。
【0017】
協力センターは、各国センターから提出されたインフルエンザ単離物の抗原および遺伝子分析を行う。抗原変化の証拠が観察されると、変種の疫学的重要性を評価するために疫学的データを用いてこれを精査する。ワクチン接種の前後に採取したヒト血清のパネルを使用して、代表的な単離物を現在のワクチン株と比較して、現在のワクチンにそれらのウイルスに対する防御が期待できるかどうかを評価する。WHOの年間ワクチン推奨を公表に続いて、高度増殖株を開発しそしてワクチン生産のための種ウイルスの生成を支援するための参照ウイルスとして生産業者に提供する。インフルエンザワクチンの安全性および効力の試験は、ウイルス不活化、微生物の無菌性、ウイルスを破壊するために使用した化学薬品の測定および推奨する抗原濃度の確認を含む。ワクチンはWHOの要求に合致すべきことが推奨されるが、しかし、各国の規制機関はその国内で使用される特定のワクチンウイルスを承認するべきである。国の公共保健機関は、ワクチンの使用に関する推奨に責任を持つべきである。また、WHOはインフルエンザの予防に関する推奨を発行している(非特許文献3参照)。
【0018】
現在のワクチンがナイーブな個体を防御しないこと、すなわちそれまで流感感染に遭遇していない多数の個体が危険にさらされた場合に、インフルエンザの汎流行発生の場合に直ちに重要となるという事実はすでに証明されている。ウイルスは一般に細胞に入り宿主細胞表面受容体に付着し、そしてそれらのウイルス核酸を脱外被し、次いでウイルスゲノムの複製を行うことによりそれらはそのライフサイクルを開始する。ウイルスタンパク質および遺伝子の新しいコピーが合成された後、それらの成分は後代ビリオンに構築され、次いで細胞から出る。構築段階の間に、後代ウイルスは、細胞質内に存在するウイルスおよび細胞核酸の大きいプールからそのゲノム核酸を効率的に選択しなければならない。ビリオンへのウイルスゲノムのパッケージングは、典型的には、ウイルス核酸内のシス作用性配列のウイルス成分、いわゆる「パケージング・シグナル」による認識を含む。かかるシグナルの定義は、ウイルスライフサイクルを理解するために重要でありそして他者タンパク質の発現のためのウイルスベクターを構築するために使用できる情報を我々に提供する。実際、他者タンパク質の発現のための遺伝子送達ベクターのベヒクルとしてのレトロウイルスの利用は、それらのvRNAの後代ビリオン内へのパッケージングのプロセスの良く確立された知識に大部分が帰せられる。
【0019】
他のRNAウイルスのゲノムパッケージングシグナルは、良く理解されておらず、他者遺伝子の発現および送達のためのベクターとしてのそれらの使用に関する進歩を妨げている。例えばA型インフルエンザウイルスは、エンベロープされたマイナス鎖RNAウイルスであり、その分節化ゲノムは、核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、酸性ポリメラーゼ(PA)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)、および非構造タンパク質(NS1およびNS2)のコーディング容量を有する。
【0020】
このウイルスは、エンベロープ上に2種の膜貫通糖タンパク質、赤血球凝集素(HA)およびノイラミニダーゼ(NA)を有する。HAタンパク質は、宿主細胞表面上のシアル酸含有受容体に結合し、そして受容体媒介エンドサイトーシスの後にエンドソーム膜とのウイルスエンベロープの融合を媒介する。対照的に、NAタンパク質はシアリルオリゴサッカリドからシアル酸を除去し、従って新規に構築されたビリオンを細胞表面から離しそしてウイルス粒子の自己凝集を防止することにより、感染の後期に重要な役割を果たす。エンベロープ内で、8個の異なるウイルスRNA(vRNA)セグメントを含んでなるウイルスゲノムは、核タンパク質(NP)およびポリメラーゼタンパク質(PA、PB1、
およびPB2)と緊密に連結し、リボヌクレオタンパク質複合体を形成する。すべての8個(またはC型の場合には、すべての7種)の機能性遺伝子セグメントは、感染性ウイルスを産生するために必要である。ポリメラーゼ遺伝子内の種々の変種が記載されており(特許文献1、特許文献2、特許文献3、非特許文献4)それらはポリメラーゼ活性を変化させるかまたは別の様式でポリメラーゼを改変するがしかしかかる複製が不能な変異ポリメラーゼを含むウイルスを与えるウイルスRNAの合成におけるその機能は放さない。特許文献1中では、変異PA遺伝子を有する感染性ウイルスが産生され、それにより変異遺伝子を有する感染性ウイルスを産生および回収する選択システムの利益を示す。特許文献2中では、生弱毒ワクチンウイルス産生に関係する望ましい性質、例えば温度感受性またはその他の形の弱毒化を有するインフルエンザウイルスの産生および回収を可能とするポリメラーゼ遺伝子内の変異を有する感染性ウイルスを産生する方法が示されている。特許文献3中では、種々のポリメラーゼの特異性を改変しそして活性を低下させるであろうポリメラーゼ遺伝子セグメントが示唆されている。しかし、そのポリメラーゼ活性を全て喪失したウイルスを再構築することは別にして、それらはウイルス再構築には使用されなかった。さらに、上記の適用のいずれでも、複製するそれらの能力を失った欠陥粒子は産生されなかった。A型ウイルスが高い多重度の感染で継代されると、1個もしくはそれ以上の機能性遺伝子セグメントを欠失した欠陥ウイルス粒子が産生される。かかるウイルス粒子中では、1個もしくはそれ以上の機能性遺伝子が、インフルエンザウイルスポリメラーゼにより作成されたエラーにより、欠陥干渉性(DI)遺伝子セグメントと置換される。高い多重度の感染により、そして1個を越えるウイルス粒子による細胞の感染により、DI RNAを含むウイルスの欠陥は、欠損した機能性遺伝子の変化のないコピーを含むウイルスにより相補される。最近、酸性ポリメラーゼ遺伝子内の一定の変異が、欠陥遺伝子を有するウイルス粒子の生成の効率を上昇できることが示された(非特許文献4)。それらの実験における欠陥ウイルス粒子の生成および相補の双方が無作為的にかかる実験中で起きることが認識されたことは重要である。この無作為プロセスは、実際の適用におけるDI RNAおよび条件つき欠陥ウイルス粒子の使用を制限する。さらに、欠陥ウイルス粒子がそれらの公開された方法を用いて産生されると、所望の条件つき欠陥ウイルスの外に野生型複製適格ウイルスが産生される。かかる複製適格ウイルスは、完全に野生型(ヘルパーウイルス)であるかまたはヘルパーウイルスと欠陥ウイルスとの遺伝子混合からもたらされる再構築物のいずれかであろう。無作為的または特異的機構のいずれかを介するインフルエンザウイルスの遺伝子セグメントのパッケージングプロセスは、多年にわたって論議されている。双方の選択肢に数件の証拠が記載されている。無作為パッケージングの証拠は、凝集したウイルス粒子が非凝集ウイルス粒子よりも高い感染性を有しそして細胞培養物が低い感染モード(moi)で感染された場合には、一部の感染細胞は一つのセグメントの発現を欠失するというものであり、両方共に全てのインフルエンザウイルスゲノムは含まないビリオンがあることを示唆する。無作為パッケージングの別の証拠は、9個のセグメントを有するインフルエンザウイルスが実験的に産生されたことである。
【0021】
特異性パッケージングプロセスのための一つの論拠は、すべての遺伝子セグメントがウイルス原料内に等量で存在するけれども、それらは、生産細胞内では異なる量で存在するというものである。さらに、欠陥干渉性(DI)粒子が生成された場合に、DIvRNAは、それが誘導されたセグメントを置換する(欠陥干渉性粒子とは、遺伝子セグメントの一つが大きい内部欠失を有するウイルス粒子である。それらの粒子は、ウイルスが高いmoiで継代された場合に生じる)。最後に、ビリオン形成の効率は、遺伝子セグメントの数が増えると増加する。
【先行技術文献】
【特許文献】
【0022】
【特許文献1】国際特許出願公開(WO)第2004/094466号明細書
【特許文献2】国際特許出願公開(WO)第2003/091401号明細書
【特許文献3】米国特許(US)第5578473号明細書
【非特許文献】
【0023】
【非特許文献1】Taubenberger and Layne,Molecular Diagnosis、第6巻、第4号、2001
【非特許文献2】第9号、2004、79、88ページまたはhttp://www.who.int/wer
【非特許文献3】WER第35号、2002、281−288ページ
【非特許文献4】Fodor et al.,J.Virol.77,5017−5020.2003
【発明の概要】
【0024】
欠陥インフルエンザウイルス粒子(例えばMena I.et al.,J.Virol.70:5016−24(1996);Neumann G.et al.,J.Virol.74:547−51(2000))は、ワクチン候補として有用であり、それはそれらがHAおよびNA以外のウイルスタンパク質に対する抗体を誘導するからであり、そして、それらが宿主細胞に入ることができる場合には、それらが、体液反応に加えてウイルスに対する細胞免疫反応を誘導できるからである(例えばヘルパーT細胞、細胞毒性T細胞)。これまで、欠陥インフルエンザウイルス粒子の生産は、トランスフェクションにより達成され(Mena I.et al.,J.Virol.70:5016−24(1996);Neumann G.et al.,J.Virol.74:547−51(2000))、かかる粒子を大量に生産する可能性を低下する。この方法とは別の方法では、条件つき欠陥であるウイルス粒子を生産し、それらの一定の生産システム内での複製を可能とするが、しかし正常の細胞または生産システム内では可能としないものであろう。この目的で、生産システムの細胞は、欠陥インフルエンザウイルス粒子のトランス−相補を可能とするインフルエンザウイルス遺伝子または遺伝子産物の1種もしくはそれ以上の生産を可能とするように変性される。本発明は、欠陥インフルエンザウイルス粒子の一定のトランス相補(transcomplementation)を最初に開示する。実験室内で、インフルエンザウイルス粒子のトランス相補は、欠陥干渉性インフルエンザウイルスが、欠陥干渉性遺伝子セグメントの野生型バージョンを有するウイルスにより同じ細胞内で相補される場合に観察される。トランス相補の「天然システム」は、一定の条件つき欠陥インフルエンザウイルス粒子を生産するために有用ではない。第一に、このシステムは、完全に感染性のウイルスの望ましくない産生をもたらすであろう少なくとも1つの(部分的)複製適格ウイルスによる一つの(部分的)欠陥ウイルスの相補を必要とする。第二に、欠陥干渉性粒子の産生は種々の遺伝子セグメントに無作為的に起きるので、一定の条件つき欠陥ウイルス粒子を産生することは不可能である。
【0025】
条件つき欠陥インフルエンザウイルス粒子は、理論的には、それらの全遺伝子セグメントまたは部分の欠失に基づくことができる。全遺伝子セグメントを欠失することにより一定の条件つき欠陥ウイルス粒子を産生(およびコードされた遺伝子産物をイントランス(in trans)で産生)する能力は、インフルエンザウイルスゲノムのパッケージングがすべての8セグメントの存在に依存するならば、限定されるであろうし、これは多く論議されている問題である(本明細書の別の箇所参照)。パッケージングプロセスがすべての8セグメントの存在を必要とする場合に、すべての遺伝子セグメントが全長として存在する必要であるかどうかは知られていず、それは条件つき欠陥ウイルス粒子の生産をさらに複雑にする。本発明はそれらの問題を解決する。
【0026】
本発明は、インフルエンザポリメラーゼをコードする核酸を内部欠失することにより誘導されて本明細書中で提供される内部欠失を有する遺伝子構築物、例えばpΔPB2、pΔPB1、pΔPAまたはpDIPAであって、ここで、該遺伝子構築物はウイルスRN
Aをコピーまたは合成できる機能性ポリメラーゼを産生できなくなるものを用い、そしてインフルエンザウイルスをコードする相補性インフルエンザウイルス核酸セグメント、例えばA/WSN/33をコードする7個の相補性構築物(HW181−188、Hoffmann et al.,2000)を用い、そしてかかる細胞内に該ポリメラーゼを発現できる発現プラスミド、例えば本明細書中に記載のHMG−PB2、HMG−PB1、HMG−PAを用いて、適切な第一細胞(単数または複数)、例えば293T細胞をトランスフェクションし、そして適当な時点、例えばトランスフェクションの後10〜50、好ましくは約20〜30時間以内に該第一細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取する第一工程、および細胞内に該ポリメラーゼを発現できる発現プラスミドを用いて適切な第二細胞(単数または複数)例えばMDCK細胞をトランスフェクションする第二工程、および該第一細胞から得た少なくとも1個のウイルス粒子を含んでなる上清を用いて該第二細胞(単数または複数)をトランスフェクションする第三工程、および適切な時点、例えばトランスフェクションの24〜96、好ましくは48〜72時間後に該第一細胞(単数または複数)の上清から少なくとも1個(内部欠失を有する遺伝子セグメントをパッケージングされたために産生されたウイルスがウイルスRNAのコピーまたは合成が可能な機能性ポリメラーゼを発現する遺伝子セグメントを欠失するウイルスが産生されるので条件つき欠陥である)のウイルス粒子を採取することを含んでなる第四工程を含んでなる条件つき欠陥インフルエンザウイルス粒子を得る方法を提供する。
【0027】
好ましくは、機能性タンパク質を産生不能ではあるが、しかしウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを阻止するほどは大きくない内部欠失である。好ましくは、それらの欠失は5’および3’非コード領域からそれぞれ数えられる。A型インフルエンザでは、かかる好ましい欠失は、例えばPAタンパク質ではヌクレオチド58と75の間であってそれらは包含されない位置にある5’ヌクレオチドから初まり、そしてヌクレオチド27と50の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一1タンパク質ではヌクレオチド43と75の間であってそれらは包含されない位置にある5’ヌクレオチドから初まり、そしてヌクレオチド24と50の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一2タンパク質ではヌクレオチド34と50の間であってそれらは包含されない位置にある5’ヌクレオチドから初まり、そしてヌクレオチド27と50の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる。さらに好ましくは、それらの欠失は、PAタンパク質ではヌクレオチド58と100の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と100の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一1タンパク質ではヌクレオチド43と100の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド24と100の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一2タンパク質ではヌクレオチド34と100の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と100の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる。もっとさらに好ましくは、それらの欠失は、PAタンパク質ではヌクレオチド58と150の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と150の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一1タンパク質ではヌクレオチド43と150の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド24と150の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一2タンパク質ではヌクレオチド34と150の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と150の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる。もっとさらに好ましくは、それらの欠失は、PAタンパク質ではヌクレオチド58と175の間であってそれらは包含されない位置にある5’ヌクレオ
チドから始まり、そしてヌクレオチド27と175の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一1タンパク質ではヌクレオチド43と175の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド24と175の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一2タンパク質ではヌクレオチド34と175の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と175の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる。最も好ましくは、それらの欠失は、PAタンパク質ではヌクレオチド58と207の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と194の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一1タンパク質ではヌクレオチド43と246の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド24と197の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB一2タンパク質ではヌクレオチド34と234の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そしてヌクレオチド27と209の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる。
【0028】
本明細書中で、相補性セグメントとは、例えばA型インフルエンザウイルスの8個の遺伝子セグメントの完全な組に導くセグメントとして定義される。従って、セグメント1が欠陥セグメントを産生するために以前に使用された場合には、相補性(非欠陥)セグメントは、セグメント2、3、4、5、6、7および8である。2が欠陥の場合には、相補セグメントはセグメント1、3、4、5、6、7および8である。以下同様。
【0029】
有利には、本発明はヘルパーウイルスが必要でなくまたは存在しない方法をもたらす。
【0030】
本発明は、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)および塩基性ポリメラーゼ2(PB2)の群から選択されたポリメラーゼをコードする機能性インフルエンザウイルス核酸セグメントを欠失する単離された条件つき欠陥インフルエンザウイルス粒子(本明細書中では条件つきの欠陥インフルエンザウイルス粒子とも呼ぶ)を提供し、かかる粒子はウイルスRNAをコピーまたは合成するポリメラーゼを生成する供給源として生成または役立つことができず、それにより機能ポリメラーゼを用いてトランス相補された細胞内での複製ウイルス粒子の生成を条件つきで可能とするだけである。さらに、本発明は、機能性インフルエンザウイルスポリメラーゼを用いるトランス相補により細胞を提供することを含んでなる条件つき欠陥インフルエンザウイルス粒子を得る方法を提供する。
【0031】
好ましい態様では、本発明による粒子は、粒子自体内では欠失している類似核酸セグメントを相補された細胞内で複製し、例えば、機能性インフルエンザウイルス核酸PAセグメントを欠失する粒子は少なくとも機能性インフルエンザウイルス核酸PAセグメントを備えた細胞内で複製し、機能性インフルエンザウイルス核酸PB1セグメントを欠失する粒子は少なくとも機能性インフルエンザウイルス核酸PB1セグメントを備えた細胞内で複製し、機能性インフルエンザウイルス核酸PB2セグメントを欠失する粒子は少なくとも機能性インフルエンザウイルス核酸PBセグメントを備えた細胞内でそれぞれ複製する。好ましい態様では、本発明は、ウイルス糖タンパク質をコードするインフルエンザウイルス核酸セグメントを有し、さらに好ましくは、核タンパク質(NP)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)、および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する本発明による粒子を提供する。一つの態様では、A型インフルエンザウイルスから誘導されたインフルエンザウイルス核酸セグメントを有する本発明による粒子が提供される。また、インフルエンザペプチドをコードしない核酸も備えている本発明に
よる粒子が提供される。また本発明は本発明による粒子を含んでなる単離された細胞を提供し、該細胞は野生型インフルエンザウイルスまたはヘルパーウイルスを含まないがしかし好ましくはインフルエンザウイルスポリメラーゼまたはそれをコードする遺伝子セグメントを備えるかまたは相補されている。好ましい態様では、かかる細胞はトランス相補された293TまたはMDCK細胞である。一つの態様では、本発明は機能性インフルエンザウイルス核酸PAセグメントを欠失した粒子を含んでなる単離された細胞を提供し、該細胞は野生型インフルエンザウイルスまたはヘルパーウイルスを含まないがしかし少なくとも機能性インフルエンザウイルス核酸PAセグメントまたは機能性PAを備えているかまたは相補されている。別の態様では、本発明は、機能性インフルエンザウイルス核酸PB1セグメントを欠失した粒子を含んでなる単離された細胞を提供し、該細胞は野生型インフルエンザウイルスまたはヘルパーウイルスを含まないがしかし少なくとも機能性インフルエンザウイルス核酸PB1セグメントまたは機能性PB1を備えている。さらに別の態様では、本発明は、機能性インフルエンザウイルス核酸PB2セグメントを欠失した粒子を含んでなる単離された細胞を提供し、該細胞は野生型インフルエンザウイルスまたはヘルパーウイルスを含まないがしかし好ましくは少なくとも機能性インフルエンザウイルス核酸PB2セグメントまたは機能性PB2を備えているかまたは相補されている。さらに、本発明は、本発明による粒子または本発明による細胞から誘導された細胞もしくは物質を含んでなる組成物、およびインフルエンザウイルスによる患者(subject)の感染に対する免疫学的防御を生成することを目的とする製薬学的組成物の製造のためのかかる組成物の使用を提供する。これによって、本発明は免疫学的防御を必要とする患者に本発明による組成物を提供することを含んでなる、インフルエンザウイルスの患者の感染に対する免疫学的防御を生成するための方法を提供する。また、本発明はインフルエンザペプチドをコードしない核酸の細胞への送達を目的とする組成物の生産のための本発明によるインフルエンザウイルス粒子の使用を提供する。また、本発明は、インフルエンザペプチドをコードしない核酸の患者の細胞への送達を目的とする製薬学的組成物の生産のための本発明による粒子の使用、および該細胞または患者へ本発明による粒子を提供することを含んでなる細胞または患者へのインフルエンザペプチドをコードしない核酸の送達のための方法を提供する。
【0032】
本発明は、天然のゲノムと比較すると一つの機能性インフルエンザウイルスセグメントを欠失する、すなわち:野生型またはヘルパーAもしくはB型ウイルスと比較して、7個(8個ではなくて)の異なる機能性インフルエンザウイルス核酸セグメントを有するか、または野生型またはヘルパーC型ウイルスと比較して、6個(7個ではなくて)の異なるインフルエンザウイルス核酸セグメントを有する条件つき欠陥インフルエンザウイルス粒子を提供する。本明細書中で「条件つき欠陥」の用語が使用される場合には、それはウイルスの遺伝子セグメントの一つがそれから発現されると非機能性タンパク質をもたらす大きい内部欠失を有するウイルス粒子を含み、それらに限定はされない。例えばA型インフルエンザウイルスのすべての8個の遺伝子セグメントおよびそれらによりコードされるすべてのタンパク質は、感染性ウイルスの生産に必要である。従って、欠陥遺伝子セグメントを含むウイルスはそれら自体が欠陥である:それは、すべてのウイルスタンパク質がビリオン内に存在するので細胞に感染できそして複製を一廻りできるが(このタンパク質は例えばウイルスが生産された場合に発現プラスミドにより産生される)しかしウイルスタンパク質の一つがウイルスにより産生できないので感染細胞内に感染性ウイルス粒子は産生されない。しかし、欠陥遺伝子セグメントにより通常発現されるタンパク質を発現する細胞が感染されると、すべてのウイルスタンパク質が存在するので欠陥ウイルスはそれらの細胞内で複製ができる。従って、それらのウイルスは、条件つき欠陥である:それらは、正しい状態の細胞が提供されない限り複製できない(この場合に、遺伝子セグメント内の欠失のためにウイルスによりコードされないウイルスタンパク質を細胞は発現する)。
【0033】
さらに、本発明はポリメラーゼをコードする機能性インフルエンザウイルス核酸セグメ
ントを欠失する条件つき欠陥インフルエンザウイルス粒子を提供する。本明細書中で、機能性インフルエンザウイルス核酸セグメントは、複製性ウイルスの生成を可能としそのために必要な機能性インフルエンザタンパク質をコードする核酸を含んでなる。例えば、A型インフルエンザウイルスは、8個の分節ゲノムを有するマイナス鎖RNAウイルスである。8個の遺伝子セグメントは11個のタンパク質をコードする;遺伝子セグメント1−8は、塩基性ポリメラーゼ2(PB2)、塩基性ポリメラーゼ1(PB1)およびPB1−ORF2(F2)、酸性ポリメラーゼ(PA)、赤血球凝集素(HA)、核タンパク質(NP)、ノイラミニダーゼ(NA)、マトリックスタンパク質1および2(M1、M2)および非構造タンパク質1および2(NS1、NS2)をそれぞれコードする。8個の遺伝子セグメントのコード領域は、ウイルスRNA合成に必要な非コード領域(NCR)に隣接している。ウイルスゲノムRNAの5’および3’末端のそれぞれ端部13および12ヌクレオチドは、すべてのA型インフルエンザウイルスセグメントのなかでも保存されそして部分的に相補的であり、ウイルスポリメラーゼ複合体により認識される二次構造を形成する。NCRは、8個の遺伝子セグメントの間で保存はされないがしかし異なるインフルエンザウイルス間では比較的保存される60個までの追加のヌクレオチドを含んでいるらしい。コード領域内のNCRおよび隣接配列は、効率的なウイルスゲノムパッケージングのために必要らしい。従って、機能性インフルエンザウイルス核酸セグメントは、複製可能なウイルスの生成を可能とする機能性インフルエンザタンパク質のためのコーディングポテンシャルを有する配列(セグメント当たりに1または2個のオープンリーディングフレーム)、mRNAの転写に必要なNCR、ウイルスRNA(vRNA)およびウイルスRNAに相補的なRNA(cRNA)およびNCR内に存在しそしてコード配列に隣接するパッケージングシグナルから成る。
【0034】
一つのインフルエンザウイルス核酸を欠失する該条件つき欠陥インフルエンザウイルス粒子が機能性ポリメラーゼを、それがPA、PB1、PB2であってもコードするセグメントを欠失することは好ましい。さらに、ワクチンの目的で、該粒子がウイルス糖タンパク質(単数または複数)をコードするインフルエンザウイルス核酸セグメント(単数または複数)を有することは好ましい。
【0035】
一つの態様では、本発明は7個の異なるA型インフルエンザ核酸セグメントを有するA型インフルエンザ粒子を提供する。本発明による欠陥インフルエンザウイルス粒子は、相補されていなくても、ただ一回であっても適切な宿主動物または細胞内で複製できる。適切に相補された細胞内では、本発明による粒子はさらに多くの回数複製できる。ワクチンおよび遺伝子送達の目的で、欠陥粒子は正常のトランス相補されていない細胞内では無限に複製はできず、そのため、宿主から宿主へのワクチンウイルスの拡散のリスクを低減しそして野生型ウイルスへの復帰の危険が低減されることは大きい利点である。
【0036】
7個の遺伝子セグメントのみを含みそして一回の複製のみを行うことができるか、または欠陥遺伝子セグメントがトランス相補された場合に複数回の複製ができる欠陥A型インフルエンザウイルスが逆遺伝子学を用いて生産できるのは最初のことである。一つの態様では、本発明は、本質的に酸性ポリメラーゼ(PA)をコードするインフルエンザ核酸セグメントを欠失する条件つき欠陥インフルエンザウイルス粒子を提供する。PAのトランス相補と同様に、他のインフルエンザウイルス遺伝子のトランス相補も考えられる。しかし、他のインフルエンザウイルスタンパク質の発現レベルと比較してPA発現レベルは重要性が低いので、PAは欠失されるポリメラーゼ群の好ましい遺伝子セグメントであり、PB2およびPB1欠失ウイルスも同様に生産でき、そしてNP欠失ウイルスはトランス相補できなかった。好ましい態様では、本発明は、7個の異なるA型インフルエンザ核酸セグメントを有しそして本質的に酸性ポリメラーゼをコードするA型インフルエンザ核酸セグメントを欠失する条件つき欠陥A型インフルエンザウイルス粒子を提供する。ワクチンの目的で、本発明による好ましい条件つき欠陥A型インフルエンザウイルス粒子は、赤
血球凝集素(HA)およびノイラミニダーゼ(NA)タンパク質を本質的にコードするA型インフルエンザウイルス核酸セグメントを有し、それらのタンパク質は防御性を与えるために免疫学的に最も関係が深い。ワクチン内に包含するために適切な遺伝子セグメントを選択するために、ワクチン使用のためにWHOにより推奨されたウイルスから遺伝子セグメントを選択するのが好ましい。勿論、HAおよびNAサブタイプは、ワクチン化しようとするインフルエンザ変種のHAおよびNAサブタイプに依存して変化できる。核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)、および非構造タンパク質(NS1およびNS2)を本質的にコードし、すなわちそれぞれの遺伝子セグメントから機能性タンパク質が発現されることを本明細書中で特定して指定するものを本質的にコードするA型インフルエンザウイルス核酸セグメントを有する本発明による条件つき欠陥インフルエンザウイルス粒子を生成することが最も好ましい。かかる粒子は、機能性PAまたはPAをコードする機能性遺伝子セグメントを備えた単離された細胞内に特定して提供される。別の態様では、核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)、および非構造タンパク質(NS1およびNS2)を本質的にコードし、すなわちそれぞれの遺伝子セグメントから機能性タンパク質が発現されることを本明細書中で特定して指定するものを本質的にコードするA型インフルエンザ核酸セグメントを有する本発明による条件つき欠陥インフルエンザウイルス粒子が提供される。かかる粒子は、機能性PB1またはPB1をコードする機能性遺伝子セグメントを備えた単離された細胞内に特定して提供される。別の態様では、核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)、および非構造タンパク質(NS1およびNS2)を本質的にコードし、すなわちそれぞれの遺伝子セグメントから機能性タンパク質が発現されることを本明細書中で特定して指定するものを本質的にコードするA型インフルエンザウイルス核酸セグメントを有する本発明による条件つき欠陥インフルエンザウイルス粒子が提供される。かかる粒子は、機能性PB2またはPB2をコードする機能性遺伝子セグメントを備えた単離された細胞内に特定して提供される。別の態様では、本発明は、インフルエンザペプチドをコードせず、例えば免疫反応を発現するために有用な他者タンパク質またはペプチドをコードする核酸を追加して備えるか、または細胞内の細胞もしくは病原体の機能に干渉できる核酸を備える本発明による粒子を提供する。
【0037】
さらに、本発明は、本発明によるインフルエンザウイルス粒子を含んでなる細胞を提供する。粒子が所要のポリメラーゼを本質的にコードする遺伝子セグメントを備えていなかった場合には、このようにして相補された細胞内の欠陥インフルエンザウイルス粒子の複数回の複製を可能とすることに適合する機能性のインフルエンザポリメラーゼウイルスを細胞が備えるように考慮することが有効である。
【0038】
また、本発明は、本発明による欠陥インフルエンザウイルス粒子または本発明による細胞もしくは細胞から誘導される物質を含んでなる組成物を提供し、かかる組成物は、例えば、インフルエンザウイルスによる患者の感染に対する免疫学的防御を生成することを目的とする製薬学的組成物の生産のために使用できる。また、本発明は、防御が必要な患者にかかる組成物を提供することを含んでなる、インフルエンザウイルスによる患者の感染に対する免疫学的防御を生成するための方法を提供する。ワクチンまたは免疫原的組成物としての本発明による粒子の使用の外に、かかる組成物は好ましくはワクチンとして、すなわちウイルス粒子、またはかかる粒子から誘導されたウイルスタンパク質(スプリットワクチン)を適当な製薬学的担体、例えば塩溶液またはアジュバント(例えばアルミニウム塩またはその他の一般的に使用される賦形剤(例えばhttp://www.cdc.gov/nip/publications/pink/Appendices/A/E
xcipient.pdf.)と混合して処方される。本発明による条件つき欠陥インフルエンザウイルス粒子も、他者遺伝子送達のためおよび他者タンパク質の発現のための候補ベクターでもあり、それは機能性遺伝子が例えば5’および3’PA配列の間に挿入できるからである。本発明が、可能ならば他者もしくは宿主核酸セグメントまたはそれらのフラグメントを備えた、条件つき欠陥インフルエンザウイルス粒子を得るための方法を提供することを考慮して、適当な第一細胞(単数または複数)を、インフルエンザタンパク質をコードする核酸を内部欠失することにより誘導された1個もしくはそれ以上の遺伝子構築物であって、ここで該遺伝子構築物は機能性タンパク質を産生できす、そしてウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを妨げないものを用い、そしてインフルエンザウイルスをコードする相補性インフルエンザウイルス核酸セグメントを用い、そして該細胞内に該タンパク質を発現できる1個もしくはそれ以上の発現プラスミドを用いてトランスフェクションし、そしてトランスフェクションの後適当な時点で該第一細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取する第一工程、および細胞内に該タンパク質を発現できる1個もしくはそれ以上の発現プラスミドを用いて適切な第二細胞(単数または複数)をトランスフェクションする第二工程、および該第一細胞から得られた少なくとも1個のウイルス粒子を含んでなる上清を用いて該第二細胞(単数または複数)に感染させる第三工程、および感染の後の適当な時点で該第二細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取することを含んでなる第四工程を含んでなる。これと共に、本発明は、適切な細胞(単数または複数)を、インフルエンザポリメラーゼをコードする核酸を内部欠失することにより誘導された1個もしくはそれ以上の遺伝子構築物であって、ここで、該遺伝子構築物は機能性ポリメラーゼを産生できないが、しかしウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを妨げないものを用い、そしてインフルエンザウイルスをコードする相補性インフルエンザウイルス核酸セグメントを用い、そして該細胞内に該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いてトランスフェクションし、そしてトランスフェクションの後適当な時点で該細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取する工程を含んでなる条件付欠陥インフルエンザウイルス粒子を得るための方法を提供する。かかる条件付欠陥インフルエンザウイルス粒子を得るための方法は、細胞内にインフルエンザポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いて適切な細胞(単数または複数)をトランスフェクションする第一工程、および条件つき欠陥インフルエンザウイルス粒子を含んでなる上清を用いて該細胞(単数または複数)に感染させる第二工程、および感染の後の適当な時点で該細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取することを含んでなる第三工程を含んでなるか、または条件つき欠陥インフルエンザウイルス粒子を得る方法は、適当な第一細胞(単数または複数)を、インフルエンザポリメラーゼをコードする核酸を内部欠失することにより誘導された1個もしくはそれ以上の遺伝子構築物であって、ここで該遺伝子構築物は機能性ポリメラーゼを産生できないが、しかしウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを妨げないものを用い、そしてインフルエンザウイルスをコードする相補性インフルエンザウイルス核酸セグメントを用い、そして該細胞内に該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いてトランスフェクションし、そしてトランスフェクションの後適当な時点で該第一細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取する第一工程、および細胞内に該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いて適切な第二細胞(単数または複数)をトランスフェクションする第二工程、および該第一細胞から得られた少なくとも1個のウイルス粒子を含んでなる上清を用いて該第二細胞(単数または複数)に感染させる第三工程、および感染の後の適当な時点で該第二細胞(単数または複数)の上清から少なくとも1個のウイルス粒子を採取することを含んでなる第四工程を含んでなる方法である。該方法において、該ポリメラーゼは、例えば酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)であることができる。好ましくは、本発明は、内部欠失が、PAタンパク質では非コード領域から数えてヌ
クレオチド58と207の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして非コード領域から数えてヌクレオチド27と194の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、あるいはPB1タンパク質では非コード領域から数えてヌクレオチド43と246の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして非コード領域から数えてヌクレオチド24と197の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、あるいはPB2タンパク質では非コード領域から数えてヌクレオチド34と234の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして非コード領域から数えてヌクレオチド27と209の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる、インフルエンザポリメラーゼをコードする核酸を内部欠失することから内部欠失がもたらされる方法を提供する。別の変法では、他者フラグメントがこの内部欠失内に挿入される。さらに、本発明は、条件つき欠陥インフルエンザウイルス粒子を含んでなる上清を用いて感染される細胞(単数または複数)が、非機能性ポリメラーゼ、例えば酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)および本明細書中に記載の方法により得られるインフルエンザ粒子をすでに発現している方法を提供する。例えば、遺伝子構築物および核酸セグメントを用いてトランスフェクションされた細胞(単数または複数)が非機能性ポリメラーゼをすでに発現することが本明細書中で提供される。具体的には、本発明は、セグメントが機能性インフルエンザポリメラーゼを産生できないようにするが、しかしウイルスの遺伝子セグメントのウイルス粒子内へのパッケージングは妨げないセグメント内の内部欠失を有する1個もしくはそれ以上の核酸セグメントを含んでなるインフルエンザウイルス粒子であって、ここで、ポリメラーゼが酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択されるものを提供する。内部欠失が:PAタンパク質では非コード領域から数えてヌクレオチド58と207の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして非コード領域から数えてヌクレオチド27と194の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB1タンパク質では非コード領域から数えてヌクレオチド43と246の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして非コード領域から数えてヌクレオチド24と197の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB2タンパク質では非コード領域から数えてヌクレオチド34と234の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして非コード領域から数えてヌクレオチド27と209の間であってそれらは包含されない位置にある3’ヌクレオチドで終わると好ましい。好ましい態様では、本発明はウイルス糖タンパク質をコードするインフルエンザウイルス核酸セグメントを有する本発明による粒子を提供する。本発明は、核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する本発明による粒子、または核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する粒子、または核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する粒子を提供する。具体的には、本発明は、A型インフルエンザウイルスから誘導されるインフルエンザウイルス核酸セグメントを有する本発明による粒子を提供する。本発明は、インフルエンザペプチドをコードしない核酸を備える本発明による粒子も提供する。さらに、本発明は、本発明による粒子を含んでなる細胞、具体的には、ポリメラーゼは酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(P
B2)から選択される1個もしくはそれ以上のインフルエンザウイルスポリメラーゼを備えた細胞を提供する。それに加えて、本発明は、本発明による粒子または本発明による細胞もしくは細胞から誘導された物質を含んでなる組成物、インフルエンザウイルスによる患者の感染に対する免疫学的防御を生成することを目的とする製薬学的組成物の生産のためのかかる組成物の使用、およびかかる組成物を防御を必要とする患者に提供することを含んでなるインフルエンザウイルスへの患者の感染に対する免疫学的防御を生成する方法を提供する。さらに、本発明は、インフルエンザペプチドをコードしない核酸を細胞に送達することを目的とする組成物の生産のための本発明による粒子の使用、およびインフルエンザペプチドをコードしない核酸を患者の細胞に送達することを目的とする製薬学的組成物の生産のための本発明による粒子の使用を提供する。かかる核酸(本明細書中では他者核酸と称する)は、適合する抗原性エピトープもしくはタンパク質をコードする他者遺伝子もしくは遺伝子フラグメントをコードしてもよく、または細胞内で核酸転写を干渉できるヌクレオチドの鎖をコードしてもよい。一つの態様では、本発明は、インフルエンザペプチドをコードしない核酸をある細胞または患者の細胞に送達することを目的とする組成物の生産のための本発明によるA型インフルエンザウイルス粒子の使用を提供する。さらに、本発明は、本発明による他者核酸を備えた欠陥インフルエンザウイルス粒子を細胞または患者が備えることを含んでなる、該細胞または該患者に、インフルエンザペプチドをコードしない核酸を送達する方法を提供する。
【図面の簡単な説明】
【0039】
【図1】条件つき欠陥A型インフルエンザウイルスの生産および増殖。最初に、293T細胞を、A/PR/8/34、pHMG−PAおよび適合する場合には、pΔPAもしくはpDIPAをコードする7個の二方向プラスミドを用いてトランスフェクションした。トランスフェクションの48時間後に、トランスフェクションされた細胞の上清を採取しそしてMDCK細胞および24時間前にHMG−PAを用いてトランスフェクションされたMDCK細胞を接種するために使用した。ウイルス複製にポジティブなMDCK−PA細胞の上清をMDCKおよびMDCK−PA細胞上で4回継代した。
【図2】条件つき欠陥ウイルス粒子の生成に使用した構築物。上図は、野生型PA遺伝子セグメントを示す。非コード領域(NCR)、および開始コドンが示されている。pΔPAは、A/WSN/33のPAを含む二方向プラスミドであるpHW183(9)のStulを用いる消化、次いで再結合して構築された。pDIPAは、A/PR/8/34のPA遺伝子セグメントの5’194および3’207ntをpSP72中にクローニングして構築された。次いで、挿入物は、二方向逆遺伝学ベクターに転移された。pΔPB1およびpΔPB2は、本文記載のようにして構築された。
【図3】上清rPR8−7、rPR8−ΔPAおよびrPR8−DIPA内のPA遺伝子セグメントの存在のRT−PCR分析。MDCK−PA継代4上清を22μmフィルターを通しそして遠心分離により濃縮した。続いて、RNAを単離しそしてPAセグメントの非コード領域を目標とするプライマーを用いてRT−PCRを行った。野生型A/PR/8/34から単離されたRNAを対照として使用した。レーン1:rPR8−7、レーン2:rPR8−ΔPA、レーン3:rPR8−DIPA、レーン4:野生型A/PR/8/34。マーカーサイズは左に記入されている。
【図4】欠失されたpΔPA構築物の追加の大きい部分、pΔPA−2、pΔPA−3、pΔPA−4、pΔPA−5をもたらす。
【0040】
詳細な説明
【実施例1】
【0041】
組換えDNAから欠陥A型インフルエンザウイルス粒子の生成
A型インフルエンザウイルスは、ネガティブ・センスの分節化ウイルスである。ゲノムは8個の遺伝子セグメントから成る。すべての8個の機能性遺伝子セグメントは、感染性ウイルス、すなわちインフルエンザウイルス複製に通常適切と考えられる細胞内で無限もしくは少なくとも数回の複製が可能な複製可能ウイルスを産生することを要求される。無作為的または特異的機構のいずれかを介するA型インフルエンザウイルスの遺伝子セグメントのパッケージングプロセスが長年の間論議されている。双方の意見の証拠が記載されている。無作為的パッケージングの証拠は、凝集したウイルス粒子が非凝集ウイルス粒子よりも高い感染性を有すること(6)および細胞培養物が低いmoiで感染された場合に、一部の感染細胞は一つのセグメントの発現を欠失すること(8)であり、双方共にすべてのインフルエンザウイルスゲノムは含まないビリオンがあることを示唆する。無作為的パッケージングのその他の証拠は、9個のセグメントを有するインフルエンザウイルスが実験的に産生されたことである(4)。BancroftおよびParslowは、ビリオン中のパッケージングのために同じ遺伝子セグメントに由来するvRNA間に競合がなかったことを見いだした(1)。
【0042】
特異的パッケージングプロセスのための一つの論議は、すべての遺伝子セグメントがウイルス原料中に等量で存在するけれども、それらは生産細胞中では異なる量で存在するというものである(10)。さらに、欠陥干渉性(DI)粒子が生成した場合に、DI vRNAはそれが誘導されたセグメントを置換する(3)(欠陥干渉性粒子は遺伝子セグメントの一つが大きい内部欠失を有するウイルス粒子である。それらの粒子は、ウイルスが高いmoiで継代された場合に起き、そしてポリメラーゼ酸性タンパク質のR638A変異によっても起きると考えられる〔Fodor et al;J.Virol.77:5017−5020,2003〕)。最後に、ビリオン形成の効率は、遺伝子セグメントの数が増えると増加する(5)。フジイら(Fujii et al.)は、ビリオン内のセグメントの効率的な組込みに必要なNAセグメントの領域も示し、その後同じグループはHAおよびNSの領域がウイルス粒子内へのパッケージングに重要であることを示した。〔Fujii,2005#256;Watanabe,2003#184〕。
【0043】
ここに、我々は特異性パッケージングの証拠を提出する。7個の機能性遺伝子セグメントのみを含むウイルス粒子を産生するために、我々はウイルス産生を止めることなくどの遺伝子セグメントを省くことができるかを決定する必要がある。ワクチンとしての複製欠失ウイルスの使用を考慮して、HAおよびNAは省かれず、そしてMAもNSも省かれないが、それはこの場合に2個の分離した発現プラスミドが必要なためである。我々はポリメラーゼ遺伝子が欠失したウイルスを生産した。我々は、欠失された遺伝子セグメントが発現プラスミドを用いてトランス相補されなかった場合にはウイルスを産生できなかった(表1、2および3、rPR8−7ntc)。7個の遺伝子セグメントおよび通常、非常に低い力価で欠失遺伝子セグメントにより発現されるタンパク質を発現するプラスミドのトランスフェクションによりウイルスが産生できた(表1、2および3、rPR8−7)。従って、インフルエンザウイルスA/WSN/33の遺伝子セグメント1、2および3の欠失変異体は、1032、528および1120ヌクレオチドの内部欠失をそれぞれ包含して産生された。それらの欠失変異体は、pΔPB2、pΔPB1およびpΔPAと命名された。図2参照。293T細胞は、以前の記載(de Wit,E.,M.I.Sproken,T.M.Bestebroer,G.F.Rimmelzwaan,A.D.Osterhaus,and R.A.Fouchier,2004,Efficient generation and growth of influenza virus A/PR/8/34 from eight cDNA fragments,Virus Res.103:155−61)のようにして、それらの欠失遺伝子セグメントそれぞれ1個およびA/PR/88.34をコードする7個の相補二方向構築物(de Wit et al,1994)および適切な発現プラスミドを用いてトランスフェクションされた。上清はトランスフェクションの48時間後に採取された。引き続いて、
MDCK細胞が、発現プラスミドHMG−PB2、HMG−PB1またはHMG−PAの一つを用いて以前の記載(2)のようにしてトランスフェクションされた。それらのトランスフェクションされた細胞は、トランスフェクションされた293T細胞の相当する上清を用いて接種された(実験手順の説明に関しては図1参照)。それらのMDCK細胞内のウイルス複製は、HAアッセイにより決定された。最初に、非トランスフェクションMDCK細胞内にはウイルス複製はなかった。ウイルス複製は相当する上清を接種されたHMG−PB2、HMG−PB1またはHMG−PAのいずれかを用いてトランスフェクションされたMDCK細胞内に示された。次いで、我々は、インフルエンザ配列データベース(www.flu.lanl.gov、アクセッション番号K00867)から得られたインフルエンザウイルスA/PR/8/34の欠陥干渉性PA vRNAの配列に基づいて欠陥PA遺伝子セグメントをクローンした。PAの5’207ntおよび3’194ntをPCR増幅しそして以前に記載したようにして(De Wit et al.,2004)変性したpHW2000(7)から誘導された二方向転写ベクター内にクローンした。得られたプラスミドはpDIPAと呼ばれた。図2)参照のこと。293T細胞をpDIPA,HMG−PAおよびインフルエンザウイルスA/PR/8/34の残りの遺伝子セグメントをコードする7個の二方向構築物を用いてトランスフェクションした(図2参照)。トランスフェクションの48時間後に上清を採取しそして引き続いて、24時間前にHMG−PAを用いてトランスフェクションされたMDCK細胞にこの上清を接種した。接種の72時間後にそれらのMDECK細胞の上清についてHAアッセイを行うと陽性であることがわかり、それはそれらの細胞内でのウイルス複製を示した。非トランスフェクションMDCK細胞の接種もHAアッセイにより測定するとウイルス産生をもたらさなかった。非トランスフェクションもしくはHMG−PAを用いてトランスフェクションしたMDCK細胞上でのPA欠損ウイルス粒子を含む上清の引き続く継代は、同様の結果に導いた(表1)。継代4まで、ウイルスはHMG−PAを用いてトランスフェクションされたMDCK細胞内で産生された。MDCKp4の上清は、ウイルス力価の指数を得るために血清で順列的に希釈され、それは約104TCID50/mlであることが示された。
【0044】
使用された方法の工程は以下である:293T細胞を構築物pΔPB2、pΔPB1、pΔPA、pΔNA、A/PR/8/34をコードする7個の相補構築物(De Wit
et al,2004)の一つおよびHMG−PB2、HMG−PB1、HMG−PA(発現プラスミドは、例えばPleschka,S.,R.Jaskunas,O.G.Engelhardt,T.Zurcher,P.Palese,and A.Garcia−Sastre,1996,A plasmid−based reverse genetics system for influenza A virus.J.Virol 70:4188−92;A.Garcia−Sastre and P.Paleseから入手)の一つを用いてトランスフェクションした(トランスフェクション・プロトコールはDe Wit et al,2004参照)。トランスフェクションの48時間後に、トランスフェクションされた293T細胞の上清を採取した。ウイルスが産生された場合には、それらは上清中に存在する。同時に、MDCK細胞を、発現プラスミドHMG−PB2、HMG−PB1、PMG−PAの一つ(使用した欠損変異体に応じて、すなわちpΔPB2を用いた場合には、MDCK細胞はここでHMG−PB2を用いてトランスフェクションされる)を用いてトランスフェクションした(トランスフェクションプロトコールはBasler et al.,2000参照)が、それはそれらが内部欠失を有する遺伝子セグメントをパッケージングしたので、産生されたウイルスがこのタンパク質を発現する遺伝子セグメントを欠失しているからである。トランスフェクションの24時間後に、トランスフェクションされたMDCK細胞を、トランスフェクションされた293T細胞から得た上清を用いて接種する。293T上清中にウイルスが存在する場合には、このウイルスはトランスフェクションMDCK細胞中で複製しそしてさらなるウイルスが産生される。この上清は、接種の72時間後に再び採取できる。
【0045】
機能性PA遺伝子セグメントをもたらす発生PAまたはDIPAの組換えが起きていないことを証明するために、MDCKp4の上清からRNAを単離した。最初に、上清を22μMフィルターを通しそして遠心分離により濃縮した。次いで、RNAを分離しそしてPAセグメントの非コード領域を目的とするプライマーを用いてRT−PCRを行った。PA vRNAに特異性のプライマーを用いて行ったRT−PCRは、ΔPAおよびDIPAが複数回の継代の間も安定のままであることを示した。DIPAウイルス粒子で感染されたMDCK細胞の上清中に、約400bpの明確なバンドが現れ、ΔPAを含むウイルスを用いて感染されたMDCK細胞の上清中には、1100bpのバンドが現れる。野生型A/PR/8/34を用いて感染されたMDCK細胞の上清中には、約2300ntのバンドが見られる(図3)。それらの結果は、ΔPAPR8遺伝子上清が安定にビリオン内にパッケージングされていることを示す。
【0046】
PB2を欠失する細胞を産生するために、インフルエンザウイルスA/PR/8/34の遺伝子セグメント2、3、4、5、6、7および8をコードする7個の二方向構築物(Hoffmann,E.G.,G.Neumann,Y.Kawaoka,G.Hobom,and R.G.Webster,2000,A DNA transfection system for generation of influenza A virus from eight plasmids.Proc.Natl.Acad
Sci U S A 97:6108−13)を用いて293T細胞をトランスフェクションすると(de Wit,E.,I.Sproken,T.M.Bestebroer,G.F.Rimmelzeaan,A.D.Osterhaus,and R.A.Fouchier,2004,Efficient Generation and growth of influenza virus A/PR/8/34 from eight cDNA fragments.Virus Res.103:155−61)、vRNAおよびmRNAの発現をもたらした。A/PR/8/34のPB2を発現するプラスミド、pHMG−PB2(Pleschka,S.,R.Jaskunas,O.G.Engelhardt,T.Zurcher,P.Palese,and A.Garcia−Sastre,1996,A plasmid−based reverse genetics system for influenza A virus,J.Virol.70:4188−92)を共トランスフェクションした。対照として、A/PR/8/34をコードする7個の二方向構築物のみをトランスフェクションし、pHMG−PB2を除外した。上清をトランスフェクションの48時間後に採取し、MDCK細胞または24時間前に100ml皿内でpHMG−PB2(MDCK−PB2)を用いてトランスフェクションされたMDCK細胞中に接種した。接種の3日後に、接種されたMDCK細胞の上清を、ウイルス産生に関する指示としてシチメンチョウ赤血球を用いて血球凝集活性を試験した。pHMG−PB2を除いて7個の遺伝子セグメントのみを用いてトランスフェクションされた293T細胞の上清を用いて接種された細胞中にウイルスは検出されなかった(rPR8−7ntc、表2)。7個の遺伝子セグメントおよびpHMG−PB2を用いてトランスフェクションされた293T細胞の上清を用いてトランスフェクションされたMDCK−PB2細胞の上清は陽性であった。引き続いて、rPR8−7上清をMDCKおよびMDCK−PB2細胞内で継代した。rPR8−7はMDCK−PB2細胞内で複製したが、しかしMDCK細胞内ではしなかった(表2)。我々は次いでインフルエンザウイルスA/WSN/33の遺伝子のセグメント1の1032nt欠失変異体を生成させると、344アミノ酸欠失をもたらした(pΔPB2、図2)。ΔPB2を含む組換えウイルス(rPR8−ΔPB2)を上記のようにして生産した(図1)。MDCK細胞中にはウイルスは検出できなかったが、rPR8−ΔPB2を用いて接種されたMDCK−PB2細胞中にはウイルスが検出された。rPR8−ΔPB2を継代した後、MDCK細胞内にウイルス産生の証拠はなく、MDCK−PB2細胞とは対照的であった(表2)。
【0047】
PB1を欠失するウイルスも生産した。293T細胞を、インフルエンザウイルスA/PR/8/34の遺伝子セグメント1、3、4、5、6、7および8をコードする7個の二方向構築物を用いてトランスフェクションすると、vRNAおよびmRNAの発現をもたらした。A/PR/8/34のPB1を発現するプラスミド、pHMG−PB1を共トランスフェクションした。対照として、pHMG−PB1を除いてA/PR/8/34をコードする7個の二方向構築物のみをトランスフェクションした。トランスフェクションの48時間後に上清を採取しそしてMDCK細胞または24時間前に100mm皿内でpHMG−PB1を用いてトランスフェクションされたMDCK細胞(MDCK−PB1)内に接種した(2)(図1)。接種の3日後に、接種されたMDCK細胞の上清を、ウイルス産生の指示としてシチメンチョウ赤血球を用いて血球凝集活性を試験した。pHMG−PB1を除いて7個の遺伝子セグメントのみを用いてトランスフェクションされた293T細胞の上清を用いて接種された細胞中にウイルスは検出されなかった(rPR8−7ntc、表3)。7個の遺伝子セグメントおよびpHMG−PB1を用いてトランスフェクションされた293T細胞の上清を用いて接種されたMDCK−PB1細胞の上清は陽性であった。引き続いて、rPR8−7上清をMDCKおよびMDCK−PB1細胞内で継代した。rPR8−7はMDCK−PB1細胞内で複製したが、しかしMDCK細胞内ではしなかった(表3)。我々は次いでインフルエンザウイルスA/WSN/33の遺伝子セグメント2の528nt欠失変異体を生成させると、178アミノ酸欠失をもたらした(pΔPB1、図2)。ΔPB1を含む組換えウイルス(rPR8−ΔPB1)を上記のようにして生産した(図1)。MDCK細胞中にはウイルスは検出できなかったが、rPR8−ΔPB1を用いて接種されたMDCK−PB1細胞中にはウイルスが検出された。rPR8−ΔPB1を継代した後、MDCK細胞内にウイルス産生の証拠はなく、MDCK−PB1細胞とは対照的であった(表3)。
【0048】
このように、我々は、上記のようにしてpΔPB2、pΔPB1またはpΔPA/pDIPA構築物をもたらしそしてRNAポリメラーゼII駆動PB2、PB1またはPA発現プラスミドを用いてトランス相補して、セグメント1、2または3を欠失するウイルスを産生できる。ここに記載する条件つき欠陥ウイルスは、トランス相補されていない細胞中ではただ一周の複製が可能であるが、しかしそれはトランス相補された細胞中では繁殖できる。これは、7個の機能性遺伝子セグメントのみを含み複製一回のみが可能、または欠失遺伝子セグメントがトランス相補された場合には複数回の複製が可能な逆遺伝学を用いて欠陥ウイルスが生産された最初である。
【0049】
この方法で生産された欠陥ウイルス粒子は、それらが感染性ウイルスを生産することなく一回の複製を行えるので、ワクチンの候補である。この一回複製の結果は、ワクチンが体液的および細胞的免疫反応の双方を誘導することである。それらの欠陥粒子は通常の細胞内では複製しないけれども、生産目的では欠陥タンパク質を発現する細胞系統内で大量のウイルスが増殖可能である。我々が示したように、複数回の複製はウイルスの遺伝子型に影響しない。ワクチンとしての欠陥ウイルス粒子の使用の外に、それらは遺伝子送達および他者タンパク質の発現のための候補ベクターでもあり、それというのも機能性遺伝子は5’および3’PA、PB2またはPB1配列の間に挿入できるからである。これはWatanabeら(11)によっても示されており、彼らはHAおよびNAを他者遺伝子で置換しそしてそれでもウイルス生産が可能なことも示した。
【0050】
pΔPAのさらなる末端短縮
さらに、pΔPA構築物のより大きい部分を欠失させると、pΔPA−2、pΔPA−3、pΔPA−4、pΔPA−5がもたらされる(図4)。293T細胞は、以前の記載(de Wit et al.,2004)のようにそれらの欠失された遺伝子セグメントのそれぞれ一つおよびA/PR/8/34をコードする7個の相補二方向性構築物(D
e Wit et al.,2004)およびPAを発現する発現プラスミドを用いてトランスフェクションされた。トランスフェクションの48時間後に上清を採取した。引き続いて、MDCK細胞を発現プラスミドHMG−PAを用いて以前の記載(Basler,C.F.,et al.,2000,Proc Natl Acad Sci U S
A 97:12289−94)のようにしてトランスフェクションした。それらのトランスフェクションされた細胞および非トランスフェクション細胞を、トランスフェクションされた293T細胞の上清を用いて接種した。それらのMDCKおよびMDCK−PA細胞内のウイルス複製は、HAアッセイにより決定した。非トランスフェクションMDCK細胞内では複製はなかった。ウイルス複製は、上清のいずれか一つを用いて接種された、HMG−PAを用いてトランスフェクションされたMDCK細胞内に見られた。それらの構築物からもたらされたすべてのvRNAをビリオン内にパッケージングした(表4)。
【0051】
【表1】
【0052】
【表2】
【0053】
【表3】
【0054】
【表4】
【実施例2】
【0055】
欠陥組換えウイルスを用いるワクチン接種
機能性PA、PB1もしくはPB2遺伝子を欠失する条件つき欠陥組換えウイルスを、高処理量ウイルスバックボーン(例えばワクチン株A/PR/8/34から誘導)に基づいて関連する流行性ウイルス(例えばA/モスクワ/10/99)のHAおよびNA遺伝子を用いて、本明細書中に記載のようにして生産する。条件つき欠陥ウイルスはトランスフェクションにより生産され、その際、ポリメラーゼタンパク質発現はトランス相補を介して達成される。次いで関連ポリメラーゼを安定して発現する適切な細胞基質、例えばMDCK細胞またはVero細胞内でウイルスを増幅する。ウイルス上清は、10分間、1000xgで遠心分離して清澄化する。ウイルスを20〜60%のスクロース勾配中で遠心分離して濃縮および精製し、ペレット化し、そしてリン酸緩衝食塩水(PBS)中に再懸濁する。ウイルス調製物の純度および品質は、クーマシー・ブリリアント・ブルーを用いて染色した12.5%SDSポリアクリルアミド・ゲルを用いて確認されそして条件つき欠陥ウイルスのウイルス力価は、MDCK細胞および関連するポリメラーゼを発現するMDCK細胞の感染および抗核タンパク質モノクローナル抗体を用いる染色により決定される。1x10E5 50%組織培養感染量(TCID−50)を用いて気管内または鼻腔内にネブライザーを用いてマウスに接種する。ワクチン接種の前後に採取した血清試料中のインフルエンザウイルスのHA、NAおよび内部タンパク質に対する抗体力価は、赤血球凝集阻害アッセイ、ノイラニミダーゼ阻害アッセイ、ELISA、またはウイルス中和アッセイを用いて決定される。ワクチン接種された動物内の抗原特異性細胞免疫反応は、フローサイトメトリー、CD4およびCD8陽性細胞のテトラマー染色、細胞溶解活性、T細胞増殖などにより細胞内サイトカイン発現を測定して定量化される。ワクチン接種
および対照動物を、ワクチン接種の6週間後にインフルエンザウイルスA/モスクワ/10/99または異種ウイルス単離物の1x10E6 TCID−50を用いて負荷する。負荷の後、鼻または咽頭スワブ試料を10日間、毎日動物から採取し、そして感染した動物により分泌されたウイルスの量が定量PCR分析またはウイルス滴定により決定される。得られたワクチン誘導体液免疫は抗体力価の上昇を定量することにより、得られたワクチン誘導細胞免疫はヘルパーおよび細胞毒性T細胞反応の上昇を定量することにより、そして免疫性の全体的レベルは負荷ウイルスを用いる感染に対する防御を確認することにより検出される。
【0056】
【表5】
【0057】
<本発明の特徴または態様>
限定されるものではないが、本発明の主たる特徴または態様を以下に記載する。
【0058】
態様1:条件欠陥インフルエンザウイルス粒子を得るための方法であって、インフルエンザポリメラーゼをコードする核酸を内部欠失することにより誘導された1個もしくはそれ以上の遺伝子構築物であって、機能性ポリメラーゼを産生できないが、しかしウイルス粒
子内へのウイルスの遺伝子セグメントのパッケージングを妨げない該遺伝子構築物を用い、かつ、インフルエンザウイルスをコードするが該インフルエンザポリメラーゼをコードしない相補性インフルエンザウイルス核酸セグメントを用い、かつ、該細胞内で該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いて、適当な単一もしくは複数の細胞をトランスフェクションする工程、およびトランスフェクションの後適当な時点で該単一もしくは複数の細胞の上清から少なくとも1個のウイルス粒子を採取する工程を含むが、ヘルパーウイルスを使用しない、ことを特徴とする方法。
【0059】
態様2:条件欠陥インフルエンザウイルス粒子を得るための方法であって、適切な単一もしくは複数の細胞を該細胞内でインフルエンザポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いてトランスフェクションする第一工程、および機能性インフルエンザポリメラーゼを産生できない請求項1に記載の条件欠陥インフルエンザウイルス粒子を含む上清を用いて該単一もしくは複数の細胞に感染させる第二工程、および感染の後の適当な時点で該単一もしくは複数の細胞の上清から少なくとも1個のウイルス粒子を採取することを含む第三工程を含むが、ヘルパーウイルスを使用しない、ことを特徴とする方法。
【0060】
態様3:条件欠陥インフルエンザウイルス粒子を得るための方法であって、
インフルエンザポリメラーゼをコードする核酸を内部欠失することにより誘導された1個もしくはそれ以上の遺伝子構築物であって、機能性ポリメラーゼを産生できないが、しかしウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを妨げない該遺伝子構築物を用い、かつ、インフルエンザウイルスをコードするが、該インフルエンザポリメラーゼをコードしない相補性インフルエンザウイルス核酸セグメントを用い、かつ、該細胞内で該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いて適当な第一の単一もしくは複数の細胞をトランスフェクションし、かつ、トランスフェクションの後適当な時点で該第一の単一もしくは複数の細胞の上清から少なくとも1個のウイルス粒子を採取する第一工程、および適切な第二の単一もしくは複数の細胞を該細胞内で該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いてトランスフェクションする第二工程、および該第一細胞から得られた少なくとも1個のウイルス粒子を含む上清を用いて該第二の単一もしくは複数の細胞に感染させる第三工程、および感染の後の適当な時点で該第二の単一もしくは複数の細胞の上清から少なくとも1個のウイルス粒子を採取することを含んでなる第四工程を含んでなるが、ヘルパーウイルスを使用しない、ことを特徴とする方法。
【0061】
態様4:インフルエンザポリメラーゼをコードする核酸の内部欠失することからもたらされる内部欠失が、PAタンパク質では5’非コード領域から数えてヌクレオチド58と207の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域のから数えてヌクレオチド27と194の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB1タンパク質では5’非コード領域から数えてヌクレオチド43と246の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド24と197の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB2タンパク質では5’非コード領域から数えてヌクレオチド34と234の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド27と209の間であってそれらは包含されない位置にある3’ヌクレオチドで終わることを特徴とする態様1および3のいずれかに記載の方法。
【0062】
態様5:条件欠陥インフルエンザウイルス粒子を含んでなる上清を用いて感染される単一もしくは複数の細胞が条件欠陥インフルエンザウイルス粒子内で非機能性であるポリメラ
ーゼに相当するインフルエンザポリメラーゼをすでに発現している、態様2〜4のいずれか1項に記載の方法。
【0063】
態様6:ポリメラーゼが酸性ポリメラーゼ(PA)である、態様1〜5のいずれか1項に記載の方法。
【0064】
態様7:ポリメラーゼが塩基性ポリメラーゼ1(PB1)である、態様1〜5のいずれか1項に記載の方法。
【0065】
態様8:ポリメラーゼが塩基性ポリメラーゼ2(PB2)である、態様1〜5のいずれか1項に記載の方法。
【0066】
態様9:態様1〜8のいずれかに記載の方法により得られるインフルエンザウイルス粒子。
【0067】
態様10:機能性インフルエンザポリメラーゼを産生できないようにし、かつ、複製について条件欠陥性のウイルス粒子をもたらすが、しかしインフルエンザウイルスの遺伝子セグメントのウイルス粒子内へのパッケージングは妨げないセグメント内の内部欠失を有する1個もしくはそれ以上の核酸セグメントを含んでなるインフルエンザウイルス粒子であって、ここで、該ポリメラーゼが酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択されることを特徴とするインフルエンザウイルス粒子。
【0068】
態様11:機能性インフルエンザポリメラーゼを産生できないようにし、かつ、複製について条件欠陥性のウイルス粒子をもたらすが、しかしインフルエンザウイルスの遺伝子セグメントのウイルス粒子内へのパッケージングは妨げないセグメント内の内部欠失を有する1個もしくはそれ以上の核酸セグメントを含んでなるインフルエンザウイルス粒子であって、ここで、ポリメラーゼが酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択され、かつ、ここで内部欠失が、PAタンパク質では5’非コード領域から数えてヌクレオチド58と207の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド27と194の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB1タンパク質では5’非コード領域から数えてヌクレオチド43と246の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド24と197の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、PB2タンパク質では5’非コード領域から数えてヌクレオチド34と234の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド27と209の間であってそれらは包含されない位置にある3’ヌクレオチドで終わることを特徴とするインフルエンザウイルス粒子。
【0069】
態様12:ウイルス糖タンパク質をコードするインフルエンザウイルス核酸セグメントを有する、態様9〜11のいずれか1項に記載の粒子。
【0070】
態様13:核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する、態様9〜12のいずれか1項に記載の粒子。
【0071】
態様14:核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する、態様9〜12のいずれか1項に記載の粒子。
【0072】
態様15:核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する、態様9〜12のいずれか1項に記載の粒子。
【0073】
態様16:A型インフルエンザウイルスから誘導されたインフルエンザウイルス核酸セグメントを有する、態様9〜15のいずれか1項に記載の粒子。
【0074】
態様17:インフルエンザペプチドをコードしない核酸を備えている、態様9〜16のいずれか1項に記載の粒子。
【0075】
態様18:態様9〜17のいずれか1項に記載の粒子を含むが、ヘルパーウイルスを含まないことを特徴とする細胞。
【0076】
態様19:ポリメラーゼが、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択される、1個もしくはそれ以上のインフルエンザウイルスポリメラーゼを発現する態様18記載の細胞。
【0077】
態様20:態様9〜17のいずれか1項に記載の粒子または態様18もしくは19記載の細胞もしくは細胞から誘導される物質を含んでなる組成物。
【0078】
態様21:インフルエンザウイルスへの患者の感染に対する免疫学的防御を生ぜしめる目的とする製薬学的組成物の生産のための、態様20記載の組成物の使用。
【0079】
態様22:有効成分として、態様9〜17のいずれか1項に記載の粒子または態様18もしくは19記載の細胞もしくは細胞から誘導される物質を含んでなるインフルエンザウイルスへの患者の感染に対する免疫学的防御を生ぜしめるための製薬学的製剤。
【0080】
態様23:細胞に対してインフルエンザペプチドをコードしない核酸の送達を目的とする組成物の生産のための、態様17記載の粒子の使用。
【0081】
態様24:対照者の細胞へのインフルエンザペプチドをコードしない核酸の送達を目的とする製薬学的組成物の生産のための、態様17記載の粒子の使用。
【0082】
態様25:態様17記載の欠陥インフルエンザウイルス粒子を細胞に提供することを含んでなる、該細胞へのインフルエンザペプチドをコードしない核酸のインビトロでの送達の方法。
【0083】
態様26:有効成分として、態様17記載の欠陥インフルエンザウイルス粒子を含んでなる、該患者へインフルエンザペプチドをコードしない核酸を送達するための製薬学的製剤。
【産業上の利用可能性】
【0084】
本発明により提供される条件欠陥インフルエンザウイルス粒子は、通常の条件下では、
生体内で複製、増殖されないので、インフルエンザの予防または治療に効果的に使用できる。したがって、例えば、医薬製造業で利用できる。
【特許請求の範囲】
【請求項1】
条件欠陥インフルエンザウイルス粒子を得るための方法によって得られたインフルエンザウイルス粒子であって、当該方法が、
インフルエンザポリメラーゼをコードする核酸を内部欠失することにより誘導された1個もしくはそれ以上の遺伝子構築物であって、機能性ポリメラーゼを産生できないが、しかしウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを妨げない該遺伝子構築物を用い、かつ、インフルエンザウイルスをコードするが該インフルエンザポリメラーゼをコードしない相補性インフルエンザウイルス核酸セグメントを用い、かつ、該細胞内で該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いて、適当な単一もしくは複数の細胞をトランスフェクションする工程、および
トランスフェクションの後適当な時点で該単一もしくは複数の細胞の上清から少なくとも1個のウイルス粒子を採取する工程を含むが、ヘルパーウイルスを使用しない、ことを特徴とする上記インフルエンザウイルス粒子。
【請求項2】
機能性インフルエンザポリメラーゼを産生できないようにし、かつ、複製について条件欠陥性のウイルス粒子をもたらすが、しかしインフルエンザウイルスの遺伝子セグメントのウイルス粒子内へのパッケージングは妨げないセグメント内の内部欠失を有する1個もしくはそれ以上の核酸セグメントを含んでなるインフルエンザウイルス粒子であって、ここで、該ポリメラーゼが酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択されることを特徴とするインフルエンザウイルス粒子。
【請求項3】
機能性インフルエンザポリメラーゼを産生できないようにし、かつ、複製について条件欠陥性のウイルス粒子をもたらすが、しかしインフルエンザウイルスの遺伝子セグメントのウイルス粒子内へのパッケージングは妨げないセグメント内の内部欠失を有する1個もしくはそれ以上の核酸セグメントを含んでなるインフルエンザウイルス粒子であって、ここで、ポリメラーゼが酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択され、かつ、ここで内部欠失が、
PAタンパク質では5’非コード領域から数えてヌクレオチド58と207の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド27と194の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、
PB1タンパク質では5’非コード領域から数えてヌクレオチド43と246の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド24と197の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、
PB2タンパク質では5’非コード領域から数えてヌクレオチド34と234の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド27と209の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる
ことを特徴とするインフルエンザウイルス粒子。
【請求項4】
ウイルス糖タンパク質をコードするインフルエンザウイルス核酸セグメントを有する、請求項1〜3のいずれか1項に記載の粒子。
【請求項5】
核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する、請求項1〜4のいずれか1項に記載の粒子。
【請求項6】
核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する、請求項1〜4のいずれか1項に記載の粒子。
【請求項7】
核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する、請求項1〜4のいずれか1項に記載の粒子。
【請求項8】
A型インフルエンザウイルスから誘導されたインフルエンザウイルス核酸セグメントを有する、請求項1〜7のいずれか1項に記載の粒子。
【請求項9】
インフルエンザペプチドをコードしない核酸を備えている、請求項1〜8のいずれか1項に記載の粒子。
【請求項10】
請求項1〜9のいずれか1項に記載の粒子を含むが、ヘルパーウイルスを含まないことを特徴とする細胞。
【請求項11】
ポリメラーゼが、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択される、1個もしくはそれ以上のインフルエンザウイルスポリメラーゼを発現する請求項10記載の細胞。
【請求項12】
請求項1〜9のいずれか1項に記載の粒子または請求項10もしくは11記載の細胞もしくは細胞から誘導される物質を含んでなる組成物。
【請求項13】
インフルエンザウイルスへの患者の感染に対する免疫学的防御を生ぜしめる目的とする製薬学的組成物の生産のための、請求項12記載の組成物の使用。
【請求項14】
有効成分として、請求項1〜9のいずれか1項に記載の粒子または請求項10もしくは11記載の細胞もしくは細胞から誘導される物質を含んでなるインフルエンザウイルスへの患者の感染に対する免疫学的防御を生ぜしめるための製薬学的製剤。
【請求項15】
細胞に対してインフルエンザペプチドをコードしない核酸の送達を目的とする組成物の生産のための、請求項9記載の粒子の使用。
【請求項16】
対照者の細胞へのインフルエンザペプチドをコードしない核酸の送達を目的とする製薬学的組成物の生産のための、請求項9記載の粒子の使用。
【請求項17】
請求項9記載の欠陥インフルエンザウイルス粒子を細胞に提供することを含んでなる、該細胞へのインフルエンザペプチドをコードしない核酸のインビトロでの送達の方法。
【請求項18】
有効成分として、請求項9記載の欠陥インフルエンザウイルス粒子を含んでなる、該患者へインフルエンザペプチドをコードしない核酸を送達するための製薬学的製剤。
【請求項1】
条件欠陥インフルエンザウイルス粒子を得るための方法によって得られたインフルエンザウイルス粒子であって、当該方法が、
インフルエンザポリメラーゼをコードする核酸を内部欠失することにより誘導された1個もしくはそれ以上の遺伝子構築物であって、機能性ポリメラーゼを産生できないが、しかしウイルス粒子内へのウイルスの遺伝子セグメントのパッケージングを妨げない該遺伝子構築物を用い、かつ、インフルエンザウイルスをコードするが該インフルエンザポリメラーゼをコードしない相補性インフルエンザウイルス核酸セグメントを用い、かつ、該細胞内で該ポリメラーゼを発現できる1個もしくはそれ以上の発現プラスミドを用いて、適当な単一もしくは複数の細胞をトランスフェクションする工程、および
トランスフェクションの後適当な時点で該単一もしくは複数の細胞の上清から少なくとも1個のウイルス粒子を採取する工程を含むが、ヘルパーウイルスを使用しない、ことを特徴とする上記インフルエンザウイルス粒子。
【請求項2】
機能性インフルエンザポリメラーゼを産生できないようにし、かつ、複製について条件欠陥性のウイルス粒子をもたらすが、しかしインフルエンザウイルスの遺伝子セグメントのウイルス粒子内へのパッケージングは妨げないセグメント内の内部欠失を有する1個もしくはそれ以上の核酸セグメントを含んでなるインフルエンザウイルス粒子であって、ここで、該ポリメラーゼが酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択されることを特徴とするインフルエンザウイルス粒子。
【請求項3】
機能性インフルエンザポリメラーゼを産生できないようにし、かつ、複製について条件欠陥性のウイルス粒子をもたらすが、しかしインフルエンザウイルスの遺伝子セグメントのウイルス粒子内へのパッケージングは妨げないセグメント内の内部欠失を有する1個もしくはそれ以上の核酸セグメントを含んでなるインフルエンザウイルス粒子であって、ここで、ポリメラーゼが酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択され、かつ、ここで内部欠失が、
PAタンパク質では5’非コード領域から数えてヌクレオチド58と207の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド27と194の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、
PB1タンパク質では5’非コード領域から数えてヌクレオチド43と246の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド24と197の間であってそれらは包含されない位置にある3’ヌクレオチドで終わり、
PB2タンパク質では5’非コード領域から数えてヌクレオチド34と234の間であってそれらは包含されない位置にある5’ヌクレオチドから始まり、そして3’非コード領域から数えてヌクレオチド27と209の間であってそれらは包含されない位置にある3’ヌクレオチドで終わる
ことを特徴とするインフルエンザウイルス粒子。
【請求項4】
ウイルス糖タンパク質をコードするインフルエンザウイルス核酸セグメントを有する、請求項1〜3のいずれか1項に記載の粒子。
【請求項5】
核タンパク質(NP)、塩基性ポリメラーゼ1(PB1)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する、請求項1〜4のいずれか1項に記載の粒子。
【請求項6】
核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ2(PB2)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する、請求項1〜4のいずれか1項に記載の粒子。
【請求項7】
核タンパク質(NP)、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)、赤血球凝集素(HA)、ノイラミニダーゼ(NA)、マトリックスタンパク質(M1およびM2)および非構造タンパク質(NS1およびNS2)をコードするインフルエンザウイルス核酸セグメントを有する、請求項1〜4のいずれか1項に記載の粒子。
【請求項8】
A型インフルエンザウイルスから誘導されたインフルエンザウイルス核酸セグメントを有する、請求項1〜7のいずれか1項に記載の粒子。
【請求項9】
インフルエンザペプチドをコードしない核酸を備えている、請求項1〜8のいずれか1項に記載の粒子。
【請求項10】
請求項1〜9のいずれか1項に記載の粒子を含むが、ヘルパーウイルスを含まないことを特徴とする細胞。
【請求項11】
ポリメラーゼが、酸性ポリメラーゼ(PA)、塩基性ポリメラーゼ1(PB1)または塩基性ポリメラーゼ2(PB2)の群から選択される、1個もしくはそれ以上のインフルエンザウイルスポリメラーゼを発現する請求項10記載の細胞。
【請求項12】
請求項1〜9のいずれか1項に記載の粒子または請求項10もしくは11記載の細胞もしくは細胞から誘導される物質を含んでなる組成物。
【請求項13】
インフルエンザウイルスへの患者の感染に対する免疫学的防御を生ぜしめる目的とする製薬学的組成物の生産のための、請求項12記載の組成物の使用。
【請求項14】
有効成分として、請求項1〜9のいずれか1項に記載の粒子または請求項10もしくは11記載の細胞もしくは細胞から誘導される物質を含んでなるインフルエンザウイルスへの患者の感染に対する免疫学的防御を生ぜしめるための製薬学的製剤。
【請求項15】
細胞に対してインフルエンザペプチドをコードしない核酸の送達を目的とする組成物の生産のための、請求項9記載の粒子の使用。
【請求項16】
対照者の細胞へのインフルエンザペプチドをコードしない核酸の送達を目的とする製薬学的組成物の生産のための、請求項9記載の粒子の使用。
【請求項17】
請求項9記載の欠陥インフルエンザウイルス粒子を細胞に提供することを含んでなる、該細胞へのインフルエンザペプチドをコードしない核酸のインビトロでの送達の方法。
【請求項18】
有効成分として、請求項9記載の欠陥インフルエンザウイルス粒子を含んでなる、該患者へインフルエンザペプチドをコードしない核酸を送達するための製薬学的製剤。
【図2】

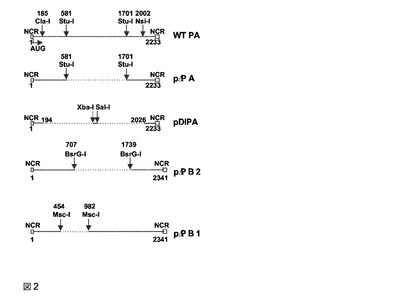
【図4】
【図1】
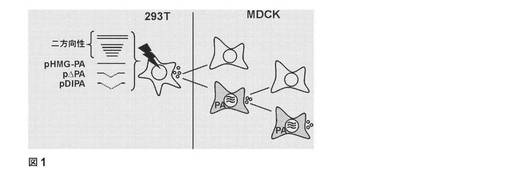
【図3】
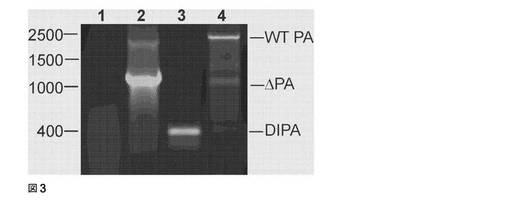
【図4】
【図1】
【図3】
【公開番号】特開2012−110339(P2012−110339A)
【公開日】平成24年6月14日(2012.6.14)
【国際特許分類】
【出願番号】特願2012−15022(P2012−15022)
【出願日】平成24年1月27日(2012.1.27)
【分割の表示】特願2007−540638(P2007−540638)の分割
【原出願日】平成17年11月8日(2005.11.8)
【出願人】(508282960)
【氏名又は名称原語表記】Abbott Biologicals B.V.
【出願人】(507154468)エラスムス・ユニバーシテイ・メデイカル・センター・ロツテルダム (4)
【Fターム(参考)】
【公開日】平成24年6月14日(2012.6.14)
【国際特許分類】
【出願日】平成24年1月27日(2012.1.27)
【分割の表示】特願2007−540638(P2007−540638)の分割
【原出願日】平成17年11月8日(2005.11.8)
【出願人】(508282960)
【氏名又は名称原語表記】Abbott Biologicals B.V.
【出願人】(507154468)エラスムス・ユニバーシテイ・メデイカル・センター・ロツテルダム (4)
【Fターム(参考)】
[ Back to top ]