
歯周病の予防に有用なポルフィロモナス・ジンジバリスポリペプチド
本発明は、歯周病の予防及び治療において用いられ得るヘム利用能により調節されると同定されるポルフィロモナス・ジンジバリスタンパク質を基礎にしたワクチン組成物及び方法に関する。特に2つの特定のインターナリン様ポルフィロモナス・ジンジバリスタンパク質、即ち宿主細胞によるポルフィロモナス・ジンジバリスのインターナリゼーションに関与するPG0350及びPG1374、細胞表面リポタンパク質であるとされる仮説的タンパク質PG1019、並びにアルキルヒドロペルオキシドレダクターゼタンパク質PG0618は、歯周病の予防及び治療のための有用な標的と同定された。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、歯周病の予防及び治療に有用なポルフィロモナス・ジンジバリス(P. gingivalis)タンパク質の組成物及び単離方法に関する。より詳細には、本発明は、歯周病の予防及び治療に用いられ得るヘム利用能により調節されることが同定されたポルフィロモナス・ジンジバリスタンパク質を基礎にしたワクチン組成物及び方法に関する。
【背景技術】
【0002】
歯周病は、歯を支持する歯茎及び骨に影響を及ぼす慢性細菌感染である。歯周病は、プラーク(歯の上に絶えず生じる粘着性バイオフィルム)中の細菌が歯茎を炎症させるようになると開始する。歯周病は、歯肉組織(歯茎);歯根膜(セメント質及び歯槽骨中に埋め込まれる結合組織);セメント質(歯根を覆っている石化結合組織);並びに歯槽骨(骨槽窩)に影響を及ぼし得る。疾患の進行によって、歯根膜の破壊、歯槽骨損失並びに結合組織性付着の根尖側移動が起こり得る。進行型歯周病は、細菌プラークを保有する歯周ポケットの形成、歯の進行性弛緩及び結果的損失を生じ得る。歯周病は、歯周炎に進み得る歯肉炎を包含する。慢性歯周炎は、歯肉下歯垢中の特定細菌に関連した歯の支持組織の炎症性疾患である。当該疾患は、約35%の有歯成人に影響を及ぼし、そして西欧諸国における歯牙損失の一主因である(1)。歯肉下歯垢の正常口腔内微生物相の一成員であるポルフィロモナス・ジンジバリスは、この疾患の進行における主要な日和見病原体の1つとして関連付けられてきた(2)。
【0003】
ポルフィロモナス・ジンジバリスは、エネルギー産生のためのアミノ酸の発酵に頼る黒色色素性、糖非分解性のグラム陰性嫌気性球桿菌である(3)。大半の細菌と同様に、ポルフィロモナス・ジンジバリスは、それが、配位第一鉄原子を有するプロトポルフィリンIX環(PPIX)から成る分子をヘムの形態で優先的に獲得する鉄に対する必須増殖要件を有する(4)。鉄供給源としてのヘムのこの利用は、ポルフィロモナス・ジンジバリスがde novoでPPIXを合成できないことを反映し得る(5)。ヘムはヘモグロビンから優先的に得られ、そしておそらくはTonB連結外膜受容体HmuRと協力して(8)、細胞表面Arg及びLys特異的プロテイナーゼ/アドヘシン複合体の活性により獲得される(4,6,7)。シデロフォアを用いて鉄を得る好気性細菌又は通性細菌と違って、ポルフィロモナス・ジンジバリスはシデロフォアを産生せず、そして通常はシデロフォア媒介性鉄獲得と関連する第二鉄レダクターゼ活性を欠く(9,10)。ポルフィロモナス・ジンジバリスは、酸化的攻撃から細胞を保護するのに役立つ固有のカタラーゼ活性を有するμ−オキソビス−ヘムの形態でヘムをその表面に貯蔵する(11)。口腔の微量栄養素制限環境内の多数且つ多様な細菌と競合可能であり得る(12)ポルフィロモナス・ジンジバリスに関しては、それはそれ自体を定着しなければならないだけでなく、多数の宿主防御を忌避するか又は克服しなければならない。
【0004】
歯周病の開始及び進行は、疾患の部位での出血と関連し、それにより高レベルのヘモグロビンを提供する。したがって、ポルフィロモナス・ジンジバリスが歯肉下歯垢中で定着し、増殖する、そして疾患を開始するメカニズムの理解を助けるために、貧(ヘム制限)微量栄養素条件から富(ヘム過剰)微量栄養素条件への移行中の相対的タンパク質存在量の変化を測定することは重要である。
【0005】
多数のタンパク質がヘム制限下での増殖と関連付けられてきたが(9,13)、しかしポルフィロモナス・ジンジバリスプロテオーム又はヘム制限中のプロテオームへの変化に関する広範な研究は報告されていない。
【0006】
歯周病の予防及び治療に有用である組成物を開発するに際しては、疾患過程の初期段階を妨害し、防止する作用物質を同定することが望ましい。
【0007】
ここで本発明者らは、歯周病の予防及び治療のための適切な標的として用いられ得るヘム利用能により調節される特定のポルフィロモナス・ジンジバリスタンパク質を同定するための方法を開発した。
【発明の概要】
【課題を解決するための手段】
【0008】
本発明者らは、ポルフィロモナス・ジンジバリスの代謝、病原性及び宿主細胞の侵襲に関与するヘム利用能により調節されるポルフィロモナス・ジンジバリスタンパク質の同定方法を首尾よく開発した。特に2つの特定のインターナリン様ポルフィロモナス・ジンジバリスタンパク質、即ち宿主細胞によるポルフィロモナス・ジンジバリスのインターナリゼーションに関与するPG0350及びPG1374、細胞表面リポタンパク質であるとされる仮説的タンパク質PG1019、並びにアルキルヒドロペルオキシドレダクターゼタンパク質PG0618は、歯周病の予防及び治療のための有用な標的と同定された。
【0009】
本発明の第一の態様は、単離抗原性ポルフィロモナス・ジンジバリスポリペプチドであって、以下の:
(i)配列番号1のアミノ酸配列を有するPG0350タンパク質;
(ii)配列番号2のアミノ酸配列を有するPG1374タンパク質;
(iii)配列番号3のアミノ酸配列を有するPG1019タンパク質;
(iv)配列番号4のアミノ酸配列を有するPG0618タンパク質;
(v)配列番号1若しくは配列番号2若しくは配列番号3若しくは配列番号4のアミノ酸配列と少なくとも85%同一であるアミノ酸配列;又は
(vi)配列番号1若しくは配列番号2若しくは配列番号3若しくは配列番号4の連続アミノ酸配列と同一の少なくとも10個のアミノ酸を含むアミノ酸配列
から成る群から選択されるポリペプチドである。
【0010】
本発明の第二の態様は、被験体においてポルフィロモナス・ジンジバリスに向けられる免疫応答を惹起するのに用いるためのワクチン組成物であって、有効量の本発明の第一の態様の少なくとも1つのポリペプチド、並びに薬学的に許容される担体を含む組成物である。
【0011】
本発明の第三の態様は、歯周病に関して被験体を予防するか又は治療する方法であって、本発明によるワクチン組成物を被験体に投与することを包含する方法である。
【0012】
本発明の第四の態様は、本発明の第一の態様のポリペプチドに対して惹起される抗体である。好ましくは抗体は、本発明のポリペプチドと特異的に結合する。
【0013】
本発明の第五の態様は、歯周病の予防又は治療に有用な組成物であって、本発明の第四の態様の抗体並びに薬学的に許容される担体を含む組成物である。
【0014】
本発明の第六の態様は、歯周病の進行に関与するポルフィロモナス・ジンジバリスポリペプチドの同定方法であって、以下の:
a)ヘム制限条件下で増殖されるポルフィロモナス・ジンジバリスにより産生されるポリペプチド又はそのペプチドの相対量を測定する工程;並びに
b)工程a)より高いヘム条件下で増殖されるポルフィロモナス・ジンジバリスにより産生されるポリペプチド又はそのペプチドの相対量を測定する工程
を包含する方法であり、工程b)と比較した場合の工程a)で検出されるポリペプチド又はそのペプチド断片の量の増大がポリペプチドが歯周病の進行に関与するということを示す方法である。
【0015】
本発明の第七の態様では、各鎖中に少なくとも19塩基対の二本鎖領域を含む干渉RNA分子であって、上記二本鎖領域の鎖のうちの1つが配列番号5又は配列番号6又は配列番号7又は配列番号8の領域と相補的である干渉RNA分子が提供される。
【0016】
本発明の第八の態様では、被験体における歯周病の治療のための薬剤の製造における本発明の第一の態様の少なくとも1つのポリペプチドの使用が提供される。
【図面の簡単な説明】
【0017】
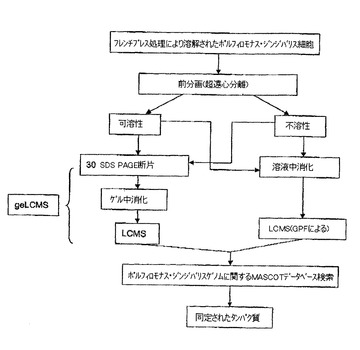
【図1】ヘム制限下で増殖されるポルフィロモナス・ジンジバリスからのタンパク質を同定するために用いられる組合せ戦略の概略図である。超遠心分離を用いて溶解細胞を可溶性及び不溶性画分に前分画し、その後、これら2つの画分を分析した。分離及び分析手順は、2つの主な方法、即ち気相分別によるLCMSとgeLCMSとから成る。
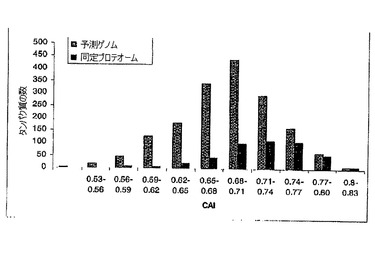
【図2】INCA(14)を用いて算定される同定ポルフィロモナス・ジンジバリスプロテオソーム及び予測ポルフィロモナス・ジンジバリスゲノムのコドン使用頻度指数(CAI)分布を示すグラフである。リボソームシンターゼ及びtRNAシンターゼは高度発現遺伝子として定義された。コドンが100個未満の遺伝子は除外し、全算定遺伝子を1685個とした。
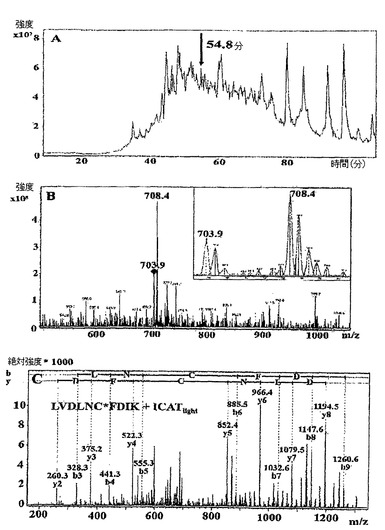
【図3】単一ペプチドの検出に基づいたPG0390の分析及び同定を示す図である。(A)総イオンクロマトグラム。(B)54.8分での質量スペクトル。差込図はm/z 703.9及び708.4での拡大ICATペプチドイオン対(比 1:2)(L/H)を示す。(C)前駆体m/z 703.9に関する生成物イオンスペクトル。このペプチドイオンは、配列LVDLNC*FDIK(MASCOTスコア=40;C*はICAT修飾システインを意味する)を有すると同定された。
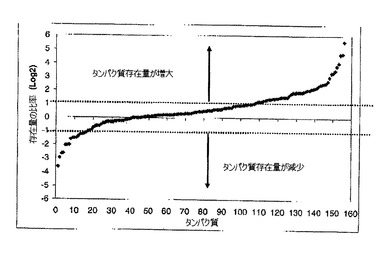
【図4】ヘム過剰を上回るヘム制限の比率を基礎にしたタンパク質存在量の分布を示す図である(Log2スケールでのすべての一単位は2倍変化を示す)。
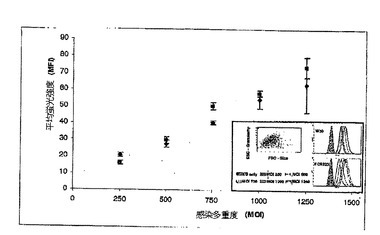
【図5】ポルフィロモナス・ジンジバリスW50(◆)及びECR312(■)によるKB細胞への結合を示す図である。2つの生物学的複製(n=6)を用いて、検定を実行した。差込図は、前方及び側方散乱特性を基礎にした生KB細胞のゲーティングを示し(左上)、5つのピークは、250〜1250のポルフィロモナス・ジンジバリス:KB細胞比でのKB細胞との結合ポルフィロモナス・ジンジバリスW50(右上)、並びにECR312(右下)のFITC蛍光を表わす。
【図6】PG0350タンパク質のアミノ酸配列(配列表中にも示されるように配列番号1と呼ばれる)を示す図である。
【図7】PG1374タンパク質のアミノ酸配列(配列表中にも示されるように配列番号2と呼ばれる)を示す図である。
【図8】PG1019タンパク質のアミノ酸配列(配列表中にも示されるように配列番号3と呼ばれる)を示す図である。
【図9】PG0618タンパク質のアミノ酸配列(配列表中にも示されるように配列番号4と呼ばれる)を示す図である。
【発明を実施するための形態】
【0018】
本発明は、歯周病の予防及び治療のための有用な標的としてのヘム利用能により調節されるポルフィロモナス・ジンジバリスタンパク質の同定を有利に提供する。好ましくは本発明は、歯周病の予防及び治療のための有用な標的としてのヘム制限により上方調節されるポルフィロモナス・ジンジバリスタンパク質の同定を提供する。
【0019】
特に本発明は、単離抗原性ポルフィロモナス・ジンジバリスポリペプチドであって、以下の:
(i)配列番号1のアミノ酸配列を有するPG0350タンパク質;
(ii)配列番号2のアミノ酸配列を有するPG1374タンパク質;
(iii)配列番号3のアミノ酸配列を有するPG1019タンパク質;
(iv)配列番号4のアミノ酸配列を有するPG0618タンパク質;
(v)配列番号1若しくは配列番号2若しくは配列番号3若しくは配列番号4のアミノ酸配列と少なくとも85%同一であるアミノ酸配列;又は
(vi)配列番号1若しくは配列番号2若しくは配列番号3若しくは配列番号4の連続アミノ酸配列と同一の少なくとも10個のアミノ酸を含むアミノ酸配列
から成る群から選択されるポリペプチドを提供する。
【0020】
好ましくは、単離抗原性ポリペプチドは、配列番号1又は配列番号2又は配列番号3又は配列番号4のアミノ酸配列と90%、95%、96%、97%、98%、99%又は100%同一である。
【0021】
好ましくは、単離抗原性ポリペプチドは、配列番号1又は配列番号2又は配列番号3又は配列番号4の連続アミノ酸配列と同一である少なくとも20個、30個、40個、50個、60個、70個、80個、90個又は100個のアミノ酸を含むアミノ酸配列を含む。
【0022】
好ましい一実施形態では、抗原性ポリペプチドは、PG1374又はPG0350又はPG1019又はPG0618タンパク質の親水性表面露出領域を構成するアミノ酸配列を含む。
【0023】
「ペプチド、タンパク質及びポリペプチド」という用語は、本明細書中では互換的に用いられる。本発明のポリペプチドは、組換えポリペプチド、例えば融合ポリペプチドを含み得る。融合ポリペプチドの製造方法は、当業者に周知である。
【0024】
当業者により十分に理解されるように、配列表中に示されたアミノ酸配列に改変がなされ得る。これらの改変は、アミノ酸残基の欠失、挿入又は置換であり得る。改変ポリペプチドは、天然(即ち、天然供給源から精製又は単離される)又は合成(例えばコード化DNA上での部位特異的突然変異誘発)であり得る。配列表に示された配列と少なくとも85%、好ましくは少なくとも90%、95%、96%、97%、98%又は99%の同一性を有するこのような改変ポリペプチドは本発明の範囲内である、と意図される。これらの改変ポリペプチドに対して産生される抗体はさらにまた、配列表に示された配列のうちの1つを有するポリペプチドと結合するであろう。
【0025】
保存的置換の概念は当業者により十分に理解されるが、一方、分かり易くするためには、保存的置換は以下に記述されるものである:
Gly、Ala、Val、Ile、Leu、Met;
Asp、Glu、Ser;
Asn、Gln;
Ser、Thr;
Lys、Arg、His;
Phe、Tyr、Trp、His;及び
Pro、Nα−アルキル(alkal)アミノ酸。
【0026】
本発明の実行は、別記しない限り、当業者に周知の化学、分子生物学、微生物学、組換えDNA及び免疫学の従来の技法を用いる。このような技法は、Perbal, A Practical Guide to Molecular Cloning, John Wiley and Sons (1984)(15)、Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbour Laboratory Press (1989)(16)、Brown (editor), Essential Molecular Biology: A Practical Approach, Volumes 1 and 2, IRL Press (1991)(17)、Glover & Hames (editors), DNA Cloning: A Practical Approach, Volumes 1-4, IRL Press (1995 and 1996)(18)及びAusubel et al., (Editors), Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience (1988、今日までのすべての最新情報を含む)(19)のような出典の文献全体に記載され、説明されている。これらのテキストの開示内容は、参照により本明細書中で援用される。
【0027】
「単離ポリペプチド」とは、本明細書中で用いる場合、それが天然に生じるか、或いはそれを用いてポリペプチド又はペプチドが合成的に合成され得る他のタンパク質、脂質及び核酸から分離されたポリペプチドを指す。好ましくはポリペプチドは、それを精製するために用いられる物質、例えば抗体又はゲルマトリックス、例えばポリアクリルアミドからも分離される。好ましくはポリペプチドは、精製調製物の乾燥重量の少なくとも10%、20%、50%、70%及び80%を構成する。好ましくは調製物は、タンパク質シーケンシングを可能にするのに十分な量のポリペプチドを含有する(即ち、少なくとも1mg、10mg又は100mg)。
【0028】
本明細書中に記載される単離ポリペプチドは、標準技法、例えばカラムクロマトグラフィー(タンパク質産物と相互作用する種々のマトリックス、例えばイオン交換マトリックス、疎水性マトリックス等を使用する)、アフィニティークロマトグラフィー(タンパク質又はタンパク質と結合するその他のリガンドに特異的な抗体を利用する)により精製され得る。
【0029】
「抗原性ポリペプチド」とは、本明細書中で用いる場合、検出可能な抗原−抗体複合体を形成するのに十分に高い親和性を有する特定の抗体と結合可能なポリペプチド、その類似体又は断片のような部分である。好ましくは抗原性ポリペプチドは、宿主動物中で体液性及び/又は細胞性の免疫応答を引き出すことが可能な免疫原性構成成分を含む。
【0030】
「連続アミノ酸配列」とは、本明細書中で用いる場合、アミノ酸の連続伸長を指す。
【0031】
2つのアミノ酸配列が指定パーセンテージ限度内に入るか否かを決定するに際して、配列の並列比較又は多重アラインメントを実行することが必要である、ということに当業者は気付くだろう。このような比較又はアラインメントでは、アラインメントを実施するために用いられるアルゴリズムによって、非同一残基の位置決めにおいて差異が生じる。本発明の状況では、2つ以上のアミノ酸配列間のパーセンテージ同一性又は類似性への言及は、当業者に既知の任意の標準アルゴリズムを用いて測定されるような上記の配列間のそれぞれ同一の及び類似の残基の数を指すと解釈される。例えばアミノ酸配列同一性又は類似性は、GAPプログラムを用いて算定され得るし、及び/又はComputer Genetics Group, Inc., University Research Park, Madison, Wisconsin, United States of America(20)のPILEUPプログラムを用いて整列され得る。GAPプログラムは、Needleman and Wunsch(21)のアルゴリズムを利用して、同一の/類似の残基の数を最大化し、そしてアラインメント中の配列ギャップの数及び長さを最小化する。代替的に又はさらに、2つより多くのアミノ酸配列が比較される場合、Clustal Wプログラム(22)が用いられる。
【0032】
本発明は、被験体においてポルフィロモナス・ジンジバリスに向けられる免疫応答を惹起するのに用いるためのワクチン組成物であって、有効量の本発明の第一の態様の少なくとも1つのポリペプチド並びに薬学的に許容される担体を含む組成物も提供する。
【0033】
本発明のワクチン組成物は、好ましくは、ポルフィロモナス・ジンジバリスに対する免疫を付与するために用いられ得る少なくとも1つの抗原を含む抗原性ポリペプチドを含む。本発明の方法により治療される被験体は、ヒト、ヒツジ、畜牛、ウマ、ウシ、ブタ、家禽、イヌ及びネコから成る群から選択され得るが、これらに限定されない。好ましくは、被験体はヒトである。ポルフィロモナス・ジンジバリスに対して向けられる免疫応答は、被験体の免疫系が特定の抗原性ポリペプチドに対する抗体を産生する場合、被験体において達成される。
【0034】
ワクチン組成物は、好ましくは、ポルフィロモナス・ジンジバリスに対する免疫を誘導し、それにより歯周病を予防するために被験体に投与される。「有効量」という用語は、本明細書中で用いる場合、ポルフィロモナス・ジンジバリスに対する免疫応答を引き出すのに十分な用量を意味する。これは、被験体並びにポルフィロモナス・ジンジバリス感染のレベルによって変わり、そして終局的には担当する科学者、医者又は獣医により決定される。
【0035】
本発明のワクチン組成物は、ヒト又は動物被験体に投与するのに適した適切な薬学的に許容される担体、例えば希釈剤及び/又はアジュバントを含む。ワクチンは、ポルフィロモナス・ジンジバリスに対する特定の免疫応答を惹起するために、好ましくは、鼻噴霧により、或いは注射により、経口的に送達するための適切なアジュバントを含む。本発明のワクチンは、本発明の抗原性ポリペプチドをコードする組換え核酸配列にも基づき得るが、この場合、当該核酸配列は、適切なベクター中に組入れられて、ベクターを含有する適切な形質転換宿主(例えば大腸菌、枯草菌、サッカロミセス・セレビシエ(saccharomyces cerevisiae)、COS細胞、CHO細胞及びHeLa細胞)中で発現される。ワクチンは、本明細書中に示されるような組換えDNA法を用いて製造され得るし、或いは本発明に記載されるアミノ酸配列から化学的に合成され得る。付加的には、本発明によれば、抗原性ポリペプチドを用いて、ポルフィロモナス・ジンジバリスにより引き起こされる歯周病及び感染に対する受動免疫化のために有用なポルフィロモナス・ジンジバリス抗血清を生成し得る。
【0036】
当業者に既知の種々のアジュバントは、ワクチン処方物と関連して一般的に用いられる。アジュバントは、免疫応答を調整することにより、そしてワクチン抗原が単独で投与された場合より少量のワクチン抗原又はより少ない用量を用いて、より永続的な且つより高レベルの免疫を獲得するに際して、助けとなる。アジュバントの例としては、不完全フロイントアジュバント(IFA)、アジュバント65(ピーナツ油、モノオレイン酸マンニド及びモノステアリン酸アルミニウムを含有)、オイルエマルジョン、Ribiアジュバント、プルロニック・ポリオール、ポリアミン、Avridine、Quil A、サポニン、MPL、QS−21及びミネラルゲル、例えばアルミニウム塩が挙げられる。その他の例としては、水中油型エマルジョン、例えばSAF−1、SAF−0、MF59、SeppicのISA720、並びにその他の粒状アジュバント、例えばISCOM及びISCOMマトリックスが挙げられる。アジュバントの他の例の広範なしかし包括的な一覧は、Cox and Coulter 1992(23)に列挙されている。アジュバントのほかに、ワクチンは、必要に応じて、従来の薬学的に許容される担体、賦形剤、充填剤、緩衝剤又は希釈剤を含み得る。アジュバントを含有する1回又は複数回用量のワクチンが、歯周病を防止するために予防的に、或いはすでに存在する歯周病を治療するために治療的に投与され得る。
【0037】
別の好ましいワクチン組成物において、調製物は、粘膜アジュバントと組合され、そして口又は鼻経路を介して投与される。粘膜アジュバントの例は、コレラ毒素及び易熱性大腸菌毒素、これらの毒素の非毒性Bサブユニット、低減毒性を示すこれらの毒素の遺伝子突然変異体である。抗原性ポリペプチドを経口的又は経鼻的に送達するために利用され得るその他の方法としては、消化管又は鼻腔からのマイクロスフェアの取り込みを手助けするための、そしてタンパク質の分解を防ぐための、マイクロカプセル封入による生分解性ポリマー(例えばアクリレート又はポリエステル)の粒子中へのポリペプチドの組入れが挙げられる。リポソーム、ISCOM、ヒドロゲルは、粘膜免疫系への抗原性ポリペプチドの送達のためのLTB、CTB又はレクチン(マンナン、キチン及びキトサン)のような標的分子の組入れによりさらに増強され得る他の可能な方法の例である。ワクチン並びに粘膜アジュバント又は送達系のほかに、ワクチンは、従来の薬学的に許容される担体、賦形剤、充填剤、コーティング、分散媒質、抗菌剤及び抗真菌剤、緩衝剤又は希釈剤を必要に応じて含み得る。
【0038】
この実施形態の別の方式は、ポルフィロモナス・ジンジバリスにより引き起こされる感染に対して防御するために用いられる生組換えウイルスワクチン、組換え細菌ワクチン、組換え弱毒化細菌ワクチン、又は不活性化組換えウイルスワクチンを提供する。ワクチンウイルスは、他の生物体由来のワクチン抗原を発現するよう工学処理される感染性ウイルスの当該技術分野における最も良く知られた例である。それ自体が疾患を引き起こさないよう弱毒化されるか又はそうでなければ処理される組換え生ワクシニアウイルスは、宿主を免疫化するために用いられる。宿主内の組換えウイルスのその後の複製は、抗原性ポリペプチドのようなワクチン抗原による免疫系の連続的刺激を提供し、それにより長期継続性免疫を提供する。
【0039】
他の生ワクチンベクターとしては、アデノウイルス、サイトメガロウイルス、好ましくはポックスウイルス、例えばワクシニア(24)、並びに弱毒化サルモネラ菌株(25〜28)が挙げられる。生ワクチンは、免疫系を連続的に刺激し、実質的に長期継続性の免疫を付与し得るため、特に有利である。免疫応答がその後のポルフィロモナス・ジンジバリス感染に対して防御的である場合、生ワクチンそれ自体はポルフィロモナス・ジンジバリスに対する防御ワクチンで用いられ得る。特に、生ワクチンは、口腔の共生棲息生物である細菌を基礎にし得る。この細菌は、組換え不活性化ポリペプチドを保有するベクターで形質転換されて、次に、口腔、特に口腔粘膜にコロニー形成するために用いられ得る。一旦口腔粘膜にコロニーが形成されると、組換えタンパク質の発現は、粘膜関連リンパ系組織を刺激して、中和抗体を産生する。例えば分子生物学技法を用いて、ポリペプチドをコードする遺伝子は、エピトープの発現を可能にするが、ワクシニアウイルスベクターの増殖又は複製に負の影響を及ぼさない部位でウイルスゲノムDNA中に挿入され得る。その結果生じる組換えウイルスは、ワクチン処方物中の免疫原として用いられ得る。同一方法を用いて、不活化組換えウイルスワクチン処方物を構築し得るが、但し、組換えウイルスは、例えば免疫原として用いる前に、そして発現免疫原の免疫原性に実質的に影響を及ぼすことなく、当該技術分野で既知の化学的手段により、不活性化される。
【0040】
能動免疫化に代わり得るものとして、免疫化は受動的、即ち本発明のポリペプチドに対する抗体を含有する精製免疫グロブリンの投与を包含する免疫化であり得る。
【0041】
本発明の方法及び組成物に用いられる抗原性ポリペプチドは、適切な賦形剤、例えば乳化剤、界面活性剤、安定剤、染料、浸透強化剤、酸化防止剤、水、塩溶液、アルコール、ポリエチレングリコール、ゼラチン、ラクトース、ステアリン酸マグネシウム及びケイ酸と組合せられ得る。抗原性ポリペプチドは、好ましくは滅菌水溶液として処方される。本発明のワクチン組成物は、歯周病のための現行治療を補足するために用いられ得る。
【0042】
本発明の第三の態様は、歯周病に関する被験体の予防又は治療方法であって、本発明によるワクチン組成物を被験体に投与することを包含する方法である。
【0043】
本発明の方法では、被験体は、歯周病のための予防的処置を含めて治療される。歯周病は、単純歯肉炎症から、歯を支持する柔組織及び骨に対する大きな損傷を生じる重篤な疾患まで多岐にわたる。歯周病は歯肉炎及び歯周炎を包含する。細菌、主にグラム陰性種、例えばポルフィロモナス・ジンジバリスは、「歯肉炎」と呼ばれる歯肉の炎症を引き起こす。歯肉炎では、歯肉は赤色になり、腫脹し、そして容易に出血し得る。歯肉炎が治療されない場合、それは「歯周炎」(「歯の周囲の炎症」を意味する)に進行し得る。歯周炎において、歯肉は歯から離れて、感染された「ポケット」を形成する。プラークが広がり、歯肉線より下に増殖すると、身体の免疫系は細菌と戦う。治療されない場合、歯を支持する骨、歯肉及び結合組織は破壊される。歯は、最終的には弛緩するようになり、除去されねばならない。
【0044】
本発明の第四の態様は、本発明の第一の態様のポリペプチドに対して産生される抗体である。好ましくは抗体は、本発明のポリペプチドに対して特異的に向けられる。
【0045】
本明細書中では、「抗体」という用語は最も広い意味で用いられ、そして詳細には、モノクローナル抗体、ポリクローナル抗体、多重特異性抗体(例えば二重特異性抗体)、キメラ抗体、ダイアボディ(diabodies)、トリアボディ(triabodies)及び抗体断片を包含する。本発明の抗体は、好ましくは、他のポリペプチドの抗原と交差反応することなく、本明細書中に上記されたような抗原性ポリペプチドと特異的に結合することができる。
【0046】
「〜と特異的に結合する」という用語は、本明細書中で用いる場合、例えばBIAcore(商標)表面プラズモン共鳴系及びBIAcore(商標)動的評価ソフトウェア(例えばバージョン2.1)を用いて表面プラズモン共鳴分析により測定した場合に1μM以下の解離定数(Kd)を有する抗体の免疫グロブリン可変領域による抗原の結合を指すよう意図される。特異的結合相互作用に関する親和性又は解離定数(Kd)は、好ましくは約500nM〜約50pM、さらに好ましくは約500nM以下、さらに好ましくは約300nM以下、そして好ましくは少なくとも約300nM〜約50pM、約200nM〜約50pM、さらに好ましくは少なくとも約100nM〜約50pM、約75nM〜約50pM、約10nM〜約50pMである。
【0047】
抗体の抗原結合機能は、全長抗体の断片により実行され得る、ということが示されている。抗体の結合断片の例としては、(i)Fab断片:VLドメイン、VHドメイン、CLドメイン及びCH1ドメインから成る一価断片;(ii)F(ab’)2断片:ヒンジ部でジスルフィド架橋により連結される2つのFab断片を含む二価断片;(iii)VHドメイン及びCIIIドメインから成るFd断片;(iv)抗体の単一アームのVLドメイン及びVHドメインから成るFv断片;(v)VHドメイン又はVLドメインから成るdAb断片;並びに(vi)単離相補性決定領域(CDR)が挙げられる。さらに、Fv断片の2つのドメインであるVL及びVHは、別個の遺伝子によりコードされるが、それらは、VL領域及びVH領域が対合して一価分子(1本鎖Fv(scFv)として既知である)を形成する単一タンパク質鎖としてそれらを作製させる合成リンカーにより、組換え法を用いて接合され得る。他の形態の1本鎖抗体、例えばダイアボディ又はトリアボディも包含される。ダイアボディは、VHドメイン及びVLドメインが単一ポリペプチド鎖上で、しかし同一鎖上の2つのドメイン間の対合を可能にするには短すぎるリンカーを用いて発現され、それにより当該ドメインを別の鎖の相補的ドメインと対合させて、2つの抗原結合部位を作製する二価二重特異性抗体である。
【0048】
当該技術分野で既知の種々の手法も、本発明の抗原性ポリペプチドと結合し得るモノクローナル抗体及びポリクローナル抗体、並びに種々の組換え体及び合成抗体の産生のために用いられ得る。さらに、宿主種によって免疫学的応答を増大するために用いられ得る種々のアジュバントは当業者に既知であり、例としては、フロイント(完全及び不完全)、ミネラルゲル、例えば水酸化アルミニウム、表面活性物質、例えばリゾレシチン、プルロニックポリオール、ポリアニオン、ペプチド、オイルエマルジョン、ジニトロフェノール、並びに潜在的に有用なヒトアジュバント、例えばカルメット・ゲラン桿菌(BCG)及びコリネバクテリウム・パルブム(Corynebacterium parvum)が挙げられるが、これらに限定されない。抗体及び抗体断片は、標準技法(例えば発酵器を用いて組織培養物又は無血清中で)により大量に産生され、そしてアフィニティーカラム、例えばプロテインA(例えばネズミMabに関して)、プロテインG(例えばラットMabに関して)又はMEP HYPERCEL(例えばIgM及びIgG Mabに関して)を用いて精製され得る。
【0049】
組換えヒト又はヒト化バージョンのモノクローナル抗体は、ヒト療法的用途のための好ましい一実施形態である。ヒト化抗体は、文献(29,30)中の手法に従って調製され得る。ヒト化モノクローナル抗体の産生のための近年記載された「遺伝子変換突然変異誘発」戦略も、ヒト化抗体の産生に用いられ得る(31)。代替的には、重鎖領域及び軽鎖領域の無作為組合せの組換え相ライブラリーを生成するための技法は、組換え抗体を調製するために用いられ得る(32)。
【0050】
本発明は、歯周病の予防又は治療に有用な組成物であって、本発明の第一の態様のポルフィロモナス・ジンジバリスポリペプチドのアンタゴニスト並びに薬学的に許容される担体を含み、当該アンタゴニストがポルフィロモナス・ジンジバリス感染を阻害する組成物も提供する。
【0051】
本明細書中で用いる場合、「アンタゴニスト」という用語は、対象のポリペプチドの生物学的活性を阻害する核酸、ペプチド、抗体、リガンド又はその他の化学的実体を指す。特定のタンパク質の適切なアンタゴニストを試験し、選択する技法は当業者には既知であり、このような技法は結合検定を包含し得る。PG0350、PG1374、PG1019及びPG0618の可能なアンタゴニストは、好ましくは、これらのタンパク質と宿主細胞又は他の基質との結合を阻害する抗体(モノクローナル又はポリクローナル)であり、或いはそれらは、これらのタンパク質の結合を妨げるタンパク質又はペプチドであり得る。
【0052】
本発明の抗体及びアンタゴニストは、多数の用途を有し、例えばそれらは、歯垢の制御並びに虫歯及び歯周病に関連する病原体の抑制のための口腔ケア製品(練り歯磨き及び洗口液)における抗菌防腐剤として用いられ得る。本発明の抗体及びアンタゴニストは、薬学的調製物(例えば、局所及び全身抗感染薬)にも用いられ得る。
【0053】
本発明の第六の態様では、歯周病の進行に関与するポルフィロモナス・ジンジバリスポリペプチドの同定方法であって、以下の:
a)ヘム制限条件下で増殖されるポルフィロモナス・ジンジバリスにより産生されるポリペプチド又はそのペプチドの相対量を測定する工程;並びに
b)工程a)より高いヘム条件下で増殖されるポルフィロモナス・ジンジバリスにより産生されるポリペプチド又はそのペプチドの相対量を測定する工程
を包含する方法であり、工程b)と比較した場合の工程a)で検出されるポリペプチド又はそのペプチド断片の量の増大がポリペプチドが歯周病の進行に関与するということを示す方法が提供される。
【0054】
ヘム制限条件下でポルフィロモナス・ジンジバリスを増殖させるためには、ヘミンの濃度は約0.1μg/ml〜約0.5μg/mlである、というのが好ましい。ヘム制限条件は、ポルフィロモナス・ジンジバリス細胞の細胞密度が本発明の方法の工程(b)の増殖条件下で観察されるものより有意に低い場合、達成される。工程(b)のより高いヘム条件は、好ましくは、5μg/mlを超えるヘミン濃度を用いて達成される。
【0055】
ヘム制限条件及び高ヘム条件におけるポリペプチドの相対量の比較は、好ましくは、鑑別プロテオミクス手法を用いることにより決定され得る。好ましくはポリペプチドの量は、当該技術分野で一般に用いられる定性的プロテオーム分析、例えば溶液中消化及びゲル中消化並びにLC−MS/MSの組合せ戦略、並びにMSと組合せた安定同位体標識戦略(ICAT)を用いた分析(これらに限定されない)により測定される。
【0056】
本発明の第六の態様で明示される方法に従って同定される単離抗原性ポルフィロモナス・ジンジバリスポリペプチドは、歯周病を治療し、予防するための標的として用いられ得る。特に、単離ポリペプチドは、ポルフィロモナス・ジンジバリス、例えばポルフィロモナス・ジンジバリス感染、例えば歯周病に対するワクチン組成物を開発するために用いられ得る。
【0057】
本発明は、本発明の第一の態様のポリペプチドをコードするmRNA分子に対して標的にされる干渉RNA分子も提供する。したがって本発明の第七の態様では、各鎖中に少なくとも19塩基対の二本鎖領域を含む干渉RNA分子であって、二本鎖領域の鎖のうちの1つが配列番号5又は配列番号6又は配列番号7又は配列番号8の領域と相補的である干渉RNA分子が提供される。
【0058】
いわゆるRNA干渉又はRNAiは周知であり、RNAiに関するさらなる情報は、Hannon (2002) Nature 418: 244-251及びMcManus & Sharp (2002) Nature Reviews: Genetics 3(10): 737-747(これらの記載内容は参照により本明細書中で援用される)に提供されている。
【0059】
本発明は、siRNA安定性を増強し、in vivoでのsiRNAの安定性を向上し、それらの使用を支持するsiRNAの化学的修飾(複数可)も意図する(例えばShen et al, (2006) Gene Therapy 13: 225-234参照)。これらの修飾は、センス鎖オリゴヌクレオチドの5’末端及び3’末端での逆方向脱塩基部分、並びにアンチセンス鎖の3’末端での最後の2つのヌクレオチド間の単一ホスホルチオエート結合を含み得る。
【0060】
干渉RNAの二本鎖領域は、二本鎖領域の各鎖中に少なくとも20塩基対、好ましくは少なくとも25塩基対、最も好ましくは少なくとも30塩基対を含む、というのが好ましい。本発明は、歯周病に関する被験体の治療方法であって、本発明の干渉RNA分子のうちの少なくとも1つを被験体に投与することを包含する方法も提供する。
【0061】
本発明の性質がより明白に理解され得るために、ここで、その好ましい形態を以下の実施例を参照しながら説明する。
【実施例】
【0062】
[実施例1]
1. 材料及び方法
1.1 細菌株及び化学物質
ポルフィロモナス・ジンジバリスW50(ATCC 53978)を、the Centre for Oral Health Science, The University of Melbourneの培養コレクションから得た。用いられる化学物質は超高純度であったが、但し、MS作業に関しては、LC MS等級試薬(Sigma, Reidel-de Haen)を用いた。
【0063】
1.2 ポルフィロモナス・ジンジバリスの増殖及び採取
400mLの作業容積を有するBioflo 110発酵器/バイオリアクター(New Brunswick Scientific)を用いて、ポルフィロモナス・ジンジバリスW50を連続培養で増殖させた。増殖培地は、5mg/mLのフィルター滅菌塩酸システイン、5.0μg/mLのヘミン(ヘム過剰)又は0.1μg/mLのヘミン(ヘム制限)を補足された37g/mLの脳心臓注入培地(Oxoid)であった。同一培地(ヘム過剰)中で増殖させたポルフィロモナス・ジンジバリスの24時間バッチ培養(100mL)を培養容器に接種することにより増殖を開始した。24時間のバッチ培養増殖後、培地貯蔵槽ポンプを作動させて、培地流を調整して、0.1 h−1(平均生成時間(MGT)6.9時間)の希釈率を生じた。容器の温度を、pH7.4±0.1で37℃に維持した。95%N2中の5%CO2で、培養を連続的にガス処理した。定常状態増殖中に細胞を採取し、5000gで30分間、洗浄緩衝液(50mMのトリス−HCl(pH8.0)、150mMのNaCl、5mMのMgCl2)で3回洗浄し、138MPaでフレンチプレス細胞破砕機(SLM, AMINCO)に3回通して破砕した。次に溶解細胞を2000gで30分間遠心分離して、非破壊細胞を除去し、その後、100000gで超遠心分離して、可溶性画分(上清)及び膜画分を生じた。全画分を氷上で実行した。
【0064】
1.3 非定量的プロテオーム分析のための試料の調製
2つの方法、即ち、トリプシンによる溶液中消化とその後の気相分別(GPF)によるLCMS、並びにゲル中消化とその後の組合せ戦略の一部としてのLCMS(geLC−MS)を用いて、非定量的プロテオーム分析を実行した(図1)。溶液中消化法に関しては、タンパク質を95℃で3分間煮沸し、TCA(16%)を用いて沈殿させて、可溶化緩衝液(8Mの尿素、50mMのトリス−HCl(pH8.3)、5mMのEDTA、0.05%SDS)中に再懸濁した。BCAタンパク質試薬(Pierce)を用いてタンパク質濃度を測定し、2μg/μLに調整した。1mMのDTTを用いて30分間、還元を実行し、10mMのヨードアセトアミドを用いて60分間、アルキル化した。溶液を水で希釈して最終濃度を1Mの尿素とした後、消化した。消化は、1:100(w/w)トリプシン対タンパク質の比率で、37℃で16時間、シーケンシング等級修飾トリプシン(Promega)を用いて実行した。蟻酸を添加して最終濃度を1%(v/v)とすることにより、消化を終結させた。次に、Sep−Pak C18カートリッジ(Waters)を用いてペプチドを脱塩し、真空遠心分離機(Thermosavant)を用いて乾燥し、そして0.1%TFA中5%アセトニトリル中に再懸濁した。2μgと等価のペプチドの量を、各LC−MS/MS分析のために注入した。
【0065】
geLC−MS法に関しては、25μgのタンパク質をプレキャストNovex 12%トリス−HClグリシンゲル(Invitrogen)上で分離し、クーマシー・ブリリアントブルーG−250(Sigma)で一晩染色した。ゲルを30の個々の断片に分けて、これを次に切り出して、約1mm3の立方体に切断した。50%エタノール及び25mMの重炭酸アンモニウム(ABC)緩衝液の溶液で3回、脱色を実行して、その後、100%エタノールで脱水した。25mMのABC中の10mMのDTTとともに、56℃で60分間、脱水ゲル立方体をインキュベートすることにより、還元を実行した。次に還元溶液を25mMのABC中55mMのヨードアセトアミドに取り替えて、45分間インキュベートした。ゲル立方体を50mMのABC中で2回洗浄し、100%エタノールで脱水した。30μLの修飾シーケンシング等級トリプシンを、25mMのABC緩衝液及び1mMのCaCl2中に5μg/mLの濃度で添加し、4℃で30分間インキュベートした。過剰量のトリプシン溶液を除去し、15μLの25mMのABC緩衝液を添加した。消化を37℃で一晩実行し、TFAを添加して最終濃度0.1%(v/v)とすることにより、終結させた。次に上清をエッペンドルフ管に移した。ゲル片に、0.1%TFA中の50%エタノールを50μL添加し、15分間超音波処理した。当該プロセスを反復し、1つのゲル断片由来の全上清をプールして、真空遠心機を用いて乾燥して約10μLとした。
【0066】
1.4 定量的ICAT分析のための試料の調製
タンパク質の標識化及び分離は、開裂性ICAT試薬(Applied Biosystems)を用いるgeLC−MS/MSアプローチ(33)を基礎にした。TCA(16%)を用いてタンパク質を先ず沈殿させて、6Mの尿素、5mMのEDTA、0.05%SDS及び50mMのトリス−HCl(pH8.3)を用いて可溶化した。BCAタンパク質試薬を用いてタンパク質濃度を測定し、1mg/mlに調整した。各増殖条件からのタンパク質100μgを、別々に、2μLの50mMのトリス(2−カルボキシ−エチル)ホスフィン塩酸塩を用いて、37℃で1時間、還元した。次にヘム制限増殖条件からの還元タンパク質をICATheavy試薬で、そしてヘム過剰増殖条件からのタンパク質をICATlight試薬でアルキル化した。次に2つの試料を合わせ、プレキャストNovex 10%NUPAGEゲル(Invitrogen)上でSDS−PAGEに付した。ゲルを、SimplyBlue(商標)SafeStain(Invitrogen)を用いて5分間染色し、その後、水で脱色した。次に、ゲルレーンを、ゲルの上部から染色前面まで、20の断片に切り出した。
【0067】
切断断片をさらにさいの目に切って1mm3の立方体として、一晩ゲル内消化し、上記の手順に従って2回抽出した。プールした上清を減圧下で乾燥して約50μLとし、その後、500μLのアフィニティーローディングバッファーと混合した後、メーカーの使用説明書(Applied Biosystems)に従ってアフィニティーカラム上に載せた。溶離ペプチドを乾燥し、ビオチンタグを、37℃で2時間、ニートTFAで切断し、その後、減圧下で乾燥した。乾燥試料を、0.1%TFA中の5%アセトニトリル 35μL中に懸濁した。
【0068】
1.5 液体クロマトグラフィー及び質量分析
UltiMate Nano LCシステム(LC Packings - Dionex)と接続されたEsquire HCTイオントラップ質量分光計(Bruker Daltonics)を用いて、MSを実行した。LC Packings逆相カラム(C18 PepMap 100、内径75μm×15cm、3μm、100Å)を用いて分離を達成し、以下のアセトニトリル勾配を用いて0.1%蟻酸で溶出した:0分〜5分(0%)、5分〜10分(0%〜10%)、10分〜100分(10%〜50%)、100分〜120分(50%〜80%)、120分〜130分(80%〜100%)。
【0069】
LC出力を、ナノスプレーイオン源と直接接続した。100ミリ秒の最大蓄積時間で、300〜1500のm/z範囲で、100000のイオン荷電制御下で、MS獲得を実施した。GPFを用いる場合、3つの付加的m/z範囲(300〜800、700〜1200及び1100〜1500)を用いて前駆体イオンに関して選択し、そして各m/z範囲を二重反復実験で実行して、同定されるペプチドの数を増大した。MS/MS獲得を100〜3000m/zの質量範囲全体で得て、初期完全プロテオーム分析に関して10個までの前駆体で、そして2分の能動除去時間を有する最強多重荷電イオンに関してはICAT分析のために3個の前駆体で実施した。
【0070】
1.6 非定量的プロテオーム分析のためのタンパク質同定
化合物検出閾値 10000並びにシグナル対ノイズ閾値 5で、Apexピークファインダーアルゴリズムを用いるDataAnalysis 3.2(Bruker Daltonics)を用いて、ピーク一覧を作成した。エクスポートされたデータに関して、+2及び+3という全体的負荷制限を設定した。ゲノム研究所(TIGR)ウェブサイト(www.tigr.org)から得られるポルフィロモナス・ジンジバリスデータベースに対して問い合わされるMS/MSデータに関するMASCOT検索エンジン(MASCOT2.1.02、Matrix Science)を用いて、タンパク質同定を達成した。以下の判定基準を用いて、整合ペプチドをさらに評価した:i)最大0.05のp値に対応する確率ベースのMowseスコアを有するペプチドは、当該スコアが−logX10(log(P))でありそしてPが観察された整合が無作為事象である確率である場合、ポジティブに同定されるとみなされた;ii)特定タンパク質の同定に1つのペプチドだけが用いられ、そしてMASCOTスコアが30より低い場合、スペクトルの手動的立証が実施された。
【0071】
1.7 ICATに関するタンパク質同定
特に単一ペプチドヒットを有するものに関するICAT標識化タンパク質の同定における信頼を増大するために、付加的フィルターを以下のように適用した:i)ICAT対の重ペプチド及び軽ペプチドは、それらの抽出イオンクロマトグラムから測定されるような溶離ピークと密接に示されていなければならない;ii)単一の独特のペプチドを有するタンパク質に関しては、このペプチドは(例えば、異なるSDS−PAGE画分で、又は軽ICAT型及び重ICAT型の両方で)1回より多く同定されていなければならない;或いはiii)単一ペプチドが(ii)の判定基準を満たさない場合、MASCOTスコアは25以上でなければならず、期待値≦0.01及びMS/MSスペクトルは、強いイオンが説明されている「b」又は「y」型イオンの連続シリーズを示していなければならない。
【0072】
1.8 擬陽性の概算
擬陽性割当のレベルを別々に概算するために、タンパク質数、アミノ酸のサイズ及び分布に関する標準データベースに対してサイズで当該データベースを同定するよう、各タンパク質に関するアミノ酸配列の順序を逆にすることにより、ポルフィロモナス・ジンジバリスの逆データベースを作製した(34)。したがって擬陽性率は、NR/NF(ここで、NR=逆データベースで同定されるペプチドの数(逆データベースに関する閾値を上回るペプチドのMASCOTスコア)、並びにNF=標準データベースで同定されるペプチドの数(標準データベースに関する閾値を上回るペプチドのMASCOTスコア))と概算された。包括的プロテオーム分析(NF=18375ペプチド)及び定量的ICAT分析(NF=530ペプチド)から、擬陽性を測定した。
【0073】
1.9 相対的存在量の定量
同位体的に重い13C対軽い12C ICAT標識化ペプチドの比率を、DataAnalysis(Bruker Daltonics)からのスクリプトを用いて測定し、単一MSスペクトルでのモノアイソトピックピーク強度の測定値(シグナル強度及びピーク面積)に基づいて、手動で立証した。定量のために用いられる親イオンの最小イオン計数は2000であったが、96%を超える重及び軽前駆体イオンは10000を超えた。貧解析スペクトルの場合、親イオンの再構成抽出イオンクロマトグラム(EIC)の面積から、当該比率を決定した。単一親タンパク質由来の多ペプチドに関して平均を算定し、外れ値を、Grubb検定を用いて除外した(α=0.05)。
【0074】
1.10 ゲノム分析
CELLO(http://cello.life.nctu.edu.tw)(35)を用いてポルフィロモナス・ジンジバリスタンパク質の、そしてTIGRから得られる配列に基づいたTMHMM2.0(www.cbs.dtu.dk/services/TMHMM-2.0)を用いて膜貫通へリックスの細胞内局在性を予測した。理論的プロテオームと比較した場合の同定されたタンパク質の相対的発現レベルを概算するために、高度発現遺伝子と定義されているリボソームタンパク質及びtRNAシンターゼを伴うプログラムINteractive Codon Analysis 1.12a(http://www.bioinfo-hr.org/inca)(14)を用いて、genebank(ftp://ftp.ncbi.nih.gov/genbank/genomes/Bacteria/Porphyromonas_gingivalis_W83/)からのポルフィロモナス・ジンジバリスのコーディング配列に基づきCAI値を算定した。Microbesonlineのウェブサイト(http://microbesonline.org)(36)から、オペロン予測を実行した。
【0075】
1.11 ECR312突然変異体の構築
ポルフィロモナス・ジンジバリスW50オープンリーディングフレームPG1374は、潜在的に、ポルフィロモナス・ジンジバリスW83ゲノムを基礎にして免疫反応性47KDa抗原(PG97)をコードする(www.tigr.org)。ポルフィロモナス・ジンジバリスPG1374突然変異体を構築するために、プライマーECR312ApaI−For(5’−AGAGGGCCCTAGCAATCATTGCATTGCT−3’)及びECR312AatII−Rev(5’−TGCGACGTCGTGTTACCAATAGAGGATT−3’)を用いたPCRにより、W50ゲノムDNAから、フランキングApaI及びAatII制限部位(下線付き)を有するPG1374遺伝子の672bp上流断片を生成した。この断片を、サブクローン化ermFカセット(37)を含有するpAL30、pGem(登録商標)T−easy(Promega)上のAatII及びBamHI部位中でクローン化して、pAL36を作製した。同様に、フランキングPstI及びNdeI制限部位を有するPG1374の下流の565bp断片を、ECR312PstI−For2(5’−TGACTGCAGGCTTTCGACCTTGGATCTT−3’)及びECR312NdeI−Rev2(5’−TCGCATATGAAGAAATAAGTGCCGTCGG−3’)プライマーを用いて増幅して、pAL36中のPstI及びNdeI制限部位中にクローン化した。その結果生じたプラスミド(PG1374オープンリーディングフレームの上流及び下流断片が側面に位置するermFカセットを有する)(pAL36.1と呼ばれる)をScaIで直線化し、以前記載されたように(38)ポルフィロモナス・ジンジバリスW50中で形質転換した。10μg/mLのエリスロマイシンを含有するHBAプレート上で、嫌気性条件下で37℃で7日間のインキュベーション後、形質転換細胞を選択した。PCR分析によりDNA組込みの確認を実施して、その結果生じた突然変異体をECR312と名付けた。
【0076】
1.12 抗生物質防御侵襲検定
W50及びECR312の侵襲効率を比較するために、24ウエル細胞培養プレート中で、以前記載されたように(39)抗生物質防御検定を実行した。要するに、一定数のポルフィロモナス・ジンジバリス細胞をKB単層に侵襲させた(各ウエル中に約105個の細胞)。ゲンタマイシン(300μg/mL)及びメトロニダゾール(200μg/mL)とともに1時間さらにインキュベートすることにより、非侵襲細胞又は接着細胞を殺した。次に侵襲細菌のコロニー形成単位を、ウマ血液寒天プレート上で数えた。
【0077】
1.13 細胞結合検定
以前記載されたように(40)、細胞結合検定を実行した。要するに、ポルフィロモナス・ジンジバリスを先ず、中対数期に増殖させて、約2.9×109細胞/mLの細胞密度とした。次に細胞を洗浄し、その後、500μgのフルオレセインイソチオシアネート(FITC)(Invitrogen)で標識し、500μLのDMSO中に再懸濁した後、振盪しながら37℃で45分間インキュベートした。インキュベーション後、細胞をさらに洗浄し、不完全アール最小必須培地(JRH Biosciences)中に再懸濁して、FACSCaliberフローサイトメーター(Becton Dickinson, San Jose, CA)を用いた細胞計数に基づいて、ポルフィロモナス・ジンジバリス細胞を調整した。FITCの緑色発光を、525nmフィルター(FL1)を用いて測定した。CellQuestソフトウェア(Becton Dickinson, San Jose, CA)を用いて、多重パラメータデータを分析した。全測定を二重反復実験で実行し、FITC蛍光の定量に関しては、平均蛍光強度(MFI)値を用いた。
【0078】
野生型ポルフィロモナス・ジンジバリス及びECR312の結合を、37℃で40分間、5%CO2大気でKB細胞上に200μLの細胞懸濁液を接種することにより、並行して実行した。インキュベーション後、KB細胞及び細菌を含有する上清を、1.5mL管に移した。次に、残りの結合細胞を、37℃で5分間、200μLのトリプシン−EDTA混合物(JRH Bioscience)を用いてウエルから剥がし、対応する収集上清とともにプールした。次に500μLの完全EMEMを添加してトリプシンを不活性化し、その後、3回洗浄して、最後に1mLのPBS中に懸濁した。以前記載されたようにフローサイトメーターで、結合細胞を計数した。
【0079】
2. 結果及び考察
2.1 連続培養中のポルフィロモナス・ジンジバリスの増殖
過剰ヘムを含有する富栄養培地中で連続培養で増殖した場合、ポルフィロモナス・ジンジバリスW50は、2.03±0.04mgの細胞乾燥重量/mLの接種後約48時間で定常状態細胞密度を達成した。増殖培地中のヘミンの濃度が5.0μg/mlから0.1μg/mlに低減された場合、0.99±0.20mgの細胞乾燥重量/mLという有意に低い定常状態細胞密度が達成されたが、これは、ヘム利用能が増殖を制限していた、ということを実証する。ヘムが培養物に添加し戻された場合、細胞密度に及ぼすヘム制限の作用は軽減された。
【0080】
ヘム制限中のポルフィロモナス・ジンジバリスの増殖は、6.9時間のポルフィロモナス・ジンジバリス平均世代時間を有するケモスタットを用いた従来の研究(41,42)と一致した。ポルフィロモナス・ジンジバリスがPPIXを合成できないため、この必須栄養素はヘモグロビン及び血漿タンパク質を含むその他のヘムのタンパク質分解により獲得されると考えられ(9)、したがってヘムにおける欠損は有意に低い細胞密度に反映された。
【0081】
2.2 ヘム制限下で増殖されるポルフィロモナス・ジンジバリスのプロテオーム分析
2つの異なるアプローチにより、ヘム制限下で増殖されるポルフィロモナス・ジンジバリスのプロテオームを広範に分析した。溶液中消化とその後の気相分別を伴うLCMSを用いて、344種のタンパク質を同定した。geLCMSアプローチでは、385種のタンパク質を同定したが、一方、247種のタンパク質は両方のアプローチにより見出された。組合せ戦略を用いて、ポルフィロモナス・ジンジバリス逆データベースに対する検索から算定される0.4%という概算擬陽性率を伴って、総計478種のタンパク質を同定した(表1参照)。個々のLCMS実行から(異なるm/z範囲又はSDS PAGE帯域から)の2以上の独特のペプチドにより、又は2以上の同一ペプチドにより、全タンパク質の77.0%を同定した。478種の同定タンパク質は、全ゲノム分析(43)により同定される1988個の割当タンパク質コード遺伝子のうちの約25%を表わす。総予測プロテオームの4分の1を同定したが、この数字は、いずれかの増殖条件中に発現される遺伝子の実数を考慮する場合、より高くなる。緑膿菌及び枯草菌では、単一増殖条件中に転写される総ORFのパーセンテージは、それぞれ60%及び40%であると概算された(44、45)。それらの質量分光計のデューティーサイクル限界を調べることにより、ORFを予測されたポルフィロモナス・ジンジバリスの約60%がそれらの増殖条件下で発現される、ということをZhangと共同研究者ら(46)は概算した。したがってこれらの数字に基づいて、本発明の増殖条件下で発現されたポルフィロモナス・ジンジバリスタンパク質の41%〜62%が同定されている。478種の同定タンパク質の機能的分類を、表1に示す。
【0082】
【表1】

【0083】
今日まで、多元的プロテオミクス手法を用いて全体的にポルフィロモナス・ジンジバリスタンパク質を同定するための試みは、1つだけ報告されている。1014ポルフィロモナス・ジンジバリスATCC 33277タンパク質がこの菌株がケラチノサイト増殖培地中で培養され、同一培地が分泌された上皮細胞構成成分に曝露された場合に同定されたZhangと共同研究者らの研究と比較して、本発明者らは、ヘム制限下での連続培養中に発現される478種のポルフィロモナス・ジンジバリスW50タンパク質を同定した。用いられた菌株、増殖条件、並びにMSデータの処理は2つの試験で異なったので、2つのデータセット間で直接比較を行なうことは困難である。それにもかかわらず、本発明の方法では、75のタンパク質を独自に同定した。
【0084】
細胞内タンパク質局在化を予測するためにCELLOを用いて、この試験で同定されたタンパク質のほとんどが、細胞質(予測された1350のタンパク質のうち347)から、その後、ペリプラズム空間(48/113)、外膜(47/154)、内膜(24/256)及び細胞外(12/35)由来であると予測された。予測通り、低パーセンテージの予測内膜タンパク質を同定した。内膜タンパク質の予測信頼度をさらに増大するために、膜貫通隠れマルコフモデル(TMHMM)を用いた。TMHMMアプローチを用いて、2を超える膜貫通ドメイン(TMD)を有すると予測された242のタンパク質のうちの20が同定され、そして10を超えるTMDを有すると予測された44のタンパク質のうちの5つが同定された。注目すべきは、10を超える膜貫通ドメインを有する12の膜タンパク質のすべてが溶液中消化法から同定された。
【0085】
2.3 同定タンパク質の有意性
多数の細菌中での高度発現遺伝子は、コドン使用に関する強い組成の偏りを有することが多い。コドン使用頻度指数(CAI)を用いて、そのコドン配列を基礎にした遺伝子の発現レベルを予測することができ、CAI値が高いということは発現が増大したことを示す(47)。100を超えるコドンを有するポルフィロモナス・ジンジバリスゲノム中の1685遺伝子のうち、ほぼ92%が0.62〜0.80のCAI値を有する(図2)。この範囲は真核生物と比較して狭いが(48)、しかしそれは、予測遺伝子の89%が0.35〜0.50のCAI値を有する好熱性サーモアナエロバクター・テングコンゲンシス(Thermoanaerobacter tengcongensis)と類似する。この試験における478の同定タンパク質をコードする遺伝子のCAI値は、理論的プロテオームと類似の分布を有するが、より高いタンパク質存在量の検出への偏りがあった。この偏りにも拘わらず、非常に低いCAIを有する遺伝子によりコードされる多数のタンパク質が同定された。この結果は、明らかに、細胞中のタンパク質存在量の大きなダイナミックレンジという問題を例示し、このことは、すべてのタンパク質を一度に検出することが現在のところ不可能であることを示す。
【0086】
同定タンパク質の最高パーセンテージを有するタンパク質の機能的クラスは、エネルギー代謝に関与するもの(表1)、典型的には発酵(95%、CAI 0.70〜0.80)、解糖(82%、CAI 0.71〜0.83)、並びにアミノ酸及びアミンの代謝(81%、CAI 0.71〜0.84)に関与するものである。これは、細菌細胞中で非常に豊富であることが示されているエネルギー代謝のような基礎代謝機能に関与する必須タンパク質として大いに期待された。これらのタンパク質のほぼ90%が細胞質中に存在すると予測され、このことがさらにまた膜タンパク質と比較して検出を容易にさせる、ということは最も重要である。最少を示したタンパク質の機能的クラスは、仮説的タンパク質の転位及び調節機能に関与するものである。
【0087】
ポルフィロモナス・ジンジバリスの完全ゲノムはシーケンシングされているが、細胞機能に関する多数の重大な問題は依然として解決されていない。したがって翻訳遺伝子産物を同定するプロテオーム試験は、機能的ゲノムへの付加的洞察を提供するのに役立つ。例えばポルフィロモナス・ジンジバリスは糖非分解性であることが既知であるが(3)、しかしゲノムは、解糖経路の全酵素に関する推定ORFを含有する(43)。この経路が十分に利用されないことは、グルコキナーゼ(glucose kinase:グルコースキナーゼ)遺伝子が挿入エレメントにより妨害される原因となっている(43)。この知見と一致して、グルコキナーゼ、或いは別の解糖特異的酵素であるホスホフルクトキナーゼは同定されなかった。それに反して、糖新生に関与するすべての酵素が見出され、このことは、多糖生合成のようなプロセスに必要なグルコースがこの経路により得られ得る、ということを示唆した。
【0088】
2.4 ICAを用いて測定した場合のヘム制限に対するポルフィロモナス・ジンジバリスの応答
ヘム制限に対するポルフィロモナス・ジンジバリス応答の定量的ICAT分析を実行するために、溶液中ICAT法が強妨害三価イオン(strong interfering triply charged ions)の存在のため要求を満たさなかったので、geLCMSアプローチを選択した。これらの三価イオンの存在により、極少数のタンパク質同一性が生じた。したがって、SDS−PAGEによりICAT標識化可溶性及び不溶性タンパク質画分を別々に分離し、各ゲルレーンをゲル中トリプシン消化のために20断片に分けて、その後アフィニティー精製及びLCMS処理した。全体で142のタンパク質を同定した。逆データベースとの整合は得られなかったが、これは、低レベルの擬陽性同一性を示した。両画分中で検出されたタンパク質を考慮すると、多くて0.05のp値に対応する確率ベースのMowseスコアを有する2以上の独特のペプチドの存在に基づいて、53のタンパク質(34.0%)が同定され、2以上の異なる画分或いは両ICAT標識状態(それらのうちの58が25以上のMASCOTスコアを有する)から同定される1つの独特のペプチドの存在に基づいて60のタンパク質(38.5%)が同定され、そして25以上のMASCOTスコア、0.01以下の期待値、「b」又は「y」型イオンの連続シリーズを有する単一独自ペプチドに基づいて43のタンパク質(27.5%)が同定された(手動で妨害するのは強イオンのためである)。単一独自ペプチドの分析に基づいたタンパク質同一性の一例を、図3に示す。
【0089】
同定されたタンパク質のうち、103は可溶性画分中に見出され、53は不溶性画分中に見出され、そして14のタンパク質は両画分中に見出された。ヘム過剰からヘム制限への環境条件の変化に応答して、同定タンパク質のうちの70は存在量における少なくとも2倍の変化を示した(図4)。これらのうち、53のタンパク質の存在量は2倍より多く増大し、そして17のタンパク質の存在量はヘム制限中に2分の1未満に低減した。本発明のデータの再現性を査定するために、そして不溶性画分に関するタンパク質同一性を増大するために、ヘム制限及び過剰増殖中の異なる日に、ケモスタットから細胞を採取した。次に、異なる抽出方法とその後のICAT標識化を、変法プロトコル(示されていない)で反復した。得られたデータは、表2に示した選択タンパク質に関して類似の存在量比率を示すタンパク質に関して再現性があった(例えばPG0695 L/H=1.1、再標識化=1.3;PG0350 L/H=3.2、再標識化=2.6;PG0159 L/H=2.0、再標識化=2.2;PG0232 L/H=0.4、再標識化=0.4)。データをさらに検証するために、オペロンを形成する遺伝子によりコードされると予測される同定タンパク質の相対的存在量を比較した(50)。タンパク質の5つの群が、予測オペロンにより、又は特定の遺伝子座で分類される遺伝子によりコードされることが判明した(表2 影付)。各々の場合、コード化タンパク質の存在量は同様であると思われた。予測オペロンのうちの1つは、外膜タンパク質Omp40(PG0694)及びOmp41(PG0695)をコードし、その存在量は1.1の比率(ヘム制限/ヘム過剰、 L/E)で不変であった。これらのタンパク質は、グラム陰性細菌のOmpA様ポリンとの高い配列類似性を有し(51)、そして外膜とペプチドグリカン層との間の物理的連結を提供すると考えられる。これらの構造タンパク質は、環境ヘムレベルの変化を伴って存在量を変えるとは予期されない。残り4つの予測オペロンは、グルタメート又はアスパルテート異化作用と関連付けられることが判明した。
【0090】
【表2】
【0091】
2.5 宿主細胞侵襲関連タンパク質
宿主細胞の侵襲に関与すると思われる3つのタンパク質、インターナリン関連タンパク質(PG0350)、免疫反応性47kDaタンパク質(PG1374)及びエンドペプチダーゼPepO(PG0159)は、ヘム制限中の存在量がより高かった(表2)。抗生物質防御侵襲検定中、機能的PG1374を欠くポルフィロモナス・ジンジバリスは、野生型と比較して、上皮細胞中への約50%低い侵襲能力を有した(W50、32625±2582cfu/mL、ECR312、16250±1089cfu/mL;p<0.01、スチューデントT検定)。別個の結合検定において、野生型W50と比較した場合、PG1374突然変異体の付着において有意差は認められない(図5)。
【0092】
PG1374及びPG0350は、リステリア・モノサイトジェネス(L. monocytogenes)インターナリンタンパク質InlJと類似の富ロイシン反復ドメインを有するシステイン含有タンパク質の新規のクラスに属する(52)。リステリア・モノサイトジェネスにおいて、インターナリンファミリーの少なくとも15の成員が存在し、そしてすべてが、シグナルペプチド、N末端富ロイシン反復ドメインとその後の保存反復間領域から成る或る種の構造的特徴を共有することが判明している。これらのタンパク質の多くが、細胞侵襲過程に関与する(53)。多数のインターナリンがなぜ存在するかは実証されていないが、それらは、異なる細胞型への向性を付与するよう意図される(54)。
【0093】
最新の研究におけるヘム制限中のPG1374及びPG0350のより高い存在量(それぞれ6.5倍増及び3.2倍増)は、これら2つのタンパク質の発現が低ヘム増殖条件中に刺激されることを示唆する。配列情報及び予測構造から、インターナリンLRR残基の半分より多くが外側に面し、そして可変性であって、このことは、それらが異なるインターナリンクラスに特異的なタンパク質−タンパク質相互作用表面に関するものであることを示唆する(55)。PG1374及びPG0350はともに、シグナルペプチドを有し、そして約80残基の保存C末端ドメイン(CTD)は別として、有意の配列類似性を有しない34までの細胞表面に位置する外膜タンパク質の新規のクラスの一部である(56)。さらに、PG1374は、ヒト歯周炎患者からの血清で精査される場合、強免疫原性であって(57)、このことは、PG1374が細胞表面タンパク質相互作用に関与することをさらに示唆する。
【0094】
歯肉上皮細胞中へのポルフィロモナス・ジンジバリスのインターナリゼーションのプロセスは、フィムブリエ並びに種々の細胞表面プロテイナーゼにより媒介される付着及び侵襲の協調プロセスを包含すると考えられる(58〜60)。機能的推定インターナリン(PG0350)を欠くポルフィロモナス・ジンジバリス33277突然変異体は、野生型細菌と類似の侵襲特質を表示することが示されたが、バイオフィルム形成能力は低減された(46、61)。類似の侵襲は菌株33277による上皮細胞侵襲にも関与するフィムブリエの存在のためであると考えられたが、この突然変異体の類似の侵襲性は、ポルフィロモナス・ジンジバリスゲノムにおいてコードされた第二の推定インターナリンタンパク質(PG1374)の存在のためである、という可能性もあった。これら2つの推定インターナリンタンパク質のダブルノックアウトは、それらの考え得る協同的侵襲的役割を潜在的に解明する。
【0095】
ECR312による上皮細胞侵襲の50%低減並びに細胞結合において差異がないことは、PG1374が欠損したポルフィロモナス・ジンジバリスによる上皮細胞中への観察された侵襲低減がより少ない付着のためでなく、侵襲プロセスで実質的に欠損する、ということを明らかに実証する(図5)。細菌侵襲は、多数のタンパク質及び受容体が関与する高度複合プロセスであることが示されている(62)。ポルフィロモナス・ジンジバリス侵襲に関与する多数の因子の関連性は、フロリダ大学のLamontのグループにより実証されており、例としては、ハロ酸デハロゲナーゼ、エンドペプチダーゼ、陽イオン輸送ATPアーゼ、及びATP結合カセット輸送体が挙げられる(60、63)。したがって上皮細胞侵襲関連タンパク質としてのPG1374の発見は、細菌病原体によるこの複合宿主細胞侵襲プロセスに関与するタンパク質の一覧に加える。
【0096】
多数の細菌病原体に関しては、鉄利用能が病原性及び侵襲プロセスに影響を及ぼす、ということが十分に確立されているが、ポルフィロモナス・ジンジバリス侵襲遺伝子の発現に及ぼすヘムの影響についてはほとんど分かっていない。本発明者らは、μ−オキソビス・ヘム形態の鉄PPIXの保護層を欠くためであると思われる酸化ストレスからのポルフィロモナス・ジンジバリスの高死亡率のために、ヘム制限細胞に関する結合及び侵襲検定を実施できなかった(64)〜(66)。しかしながらこの試験において、推定侵襲関連タンパク質PcpO、PG0350及びPG1374のレベルはヘム制限下で増大し、そしてPG1374は細胞侵襲に関与する、ということを本発明者らは示した。
【0097】
上記のこれらの実験は、環境条件の変化に対するポルフィロモナス・ジンジバリスの応答の最初の定量的プロテオーム分析であり、完全プロテオーム分析と組合された安定同位体標識アプローチの有用性を実証する。ポルフィロモナス・ジンジバリスは、酸化ストレス応答、病原性及び宿主細胞の侵襲と関連付けられる多数のタンパク質の存在量により必須微量栄養素の制限に応答する。
【0098】
2.6 細胞表面に位置するタンパク質PG1019
ポルフィロモナス・ジンジバリス仮説的タンパク質PG1019は、細菌がヘム制限下で増殖された場合、25倍より豊富に存在することが観察された。バイオインフォマティック分析は、PG1019が、予測オペロン中の推定外膜受容体タンパク質(PG1020)をコードする遺伝子の直ぐ上流に位置する遺伝子によりコードされるリポタンパク質である、ということを示唆する。既知のポルフィロモナス・ジンジバリスTonB連結外膜受容体とのPG1020の多重アラインメント(示されていない)は、TonB連結受容体の特質の1つである推定TonBボックス(残基118〜126)(67)、並びにSimpson及び共同研究者ら(68)がTonBボックスIV領域と呼ぶ保存領域(残基236〜272)の存在を示す。TonB連結外膜受容体は、多数の鉄、鉄錯体及びその他の微量栄養素取込み系と関連付けられている。ヘム制限増殖条件下でのPG1019の高存在量は、このタンパク質が細胞中への鉄/鉄錯体の輸送に関与するTon−B連結系に対するアクセサリータンパク質であることと一致するが、これは未だ実証されていない。プロテオームデータのほかに、機能feoB1遺伝子を欠く突然変異体(ポルフィロモナス・ジンジバリスFB1)と比較されるポルフィロモナス・ジンジバリスW50のカスタムポルフィロモナス・ジンジバリスDNAマイクロアレイを用いるトランスクリプトーム(transcriptomic)分析を実施した。野生型W50及びFB1突然変異体をともに、ヘム過剰条件で連続培養で増殖させた。ポルフィロモナス・ジンジバリスFB1は、野生型W50の細胞鉄含量の約半分を有した(69)。したがって転写の増大を示す遺伝子は、細胞内又は環境鉄含量の低減に応答して上方調節されると思われる。PG1019及びPG1020はともに、野生型と比較して、FB1突然変異体において類似のレベルに有意に上方調節された。生物学的複製においてP<0.05の有意レベルでPG1019は2.46のLog2増大を示し、そしてPG1020は2.33のLog2増大を示し、これは、これらの遺伝子がオペロン中に存在する、というさらなる証拠である。さらに、ポルフィロモナス・ジンジバリスW50の別個のトランスクリプトームDNAマイクロアレイ分析は、ヘム過剰増殖中にPG1019遺伝子の発現がほとんど又は全く認められない、ということを示した。
【0099】
2.7 アルキルヒドロペルオキシドレダクターゼタンパク質AhpC(PG0618)
ポルフィロモナス・ジンジバリスにおけるペルオキシド耐性に重要な役割を果たすことが示されているペルオキシド除去酵素であるアルキルヒドロペルオキシドレダクターゼタンパク質AhpC(PG0618、表2)を用いて、ヘム過剰からヘム制限への移行中のポルフィロモナス・ジンジバリスのタンパク質存在量における最も実質的な変化を観察した(65)。ポルフィロモナス・ジンジバリスにおいて、細胞表面に酸素を有するμ−オキソビス・ヘム形態の鉄PPIXの層の形成は、その固有のカタラーゼ様活性のために酸化緩衝剤として作用すると考えられる(11)。この層は、鉄及びPPIXの細胞表面貯蔵にも役立ち得る(70)。ヘム制限中、鉄PPIXのこの供給源の枯渇は、酸化ストレスに対する感受性増大を生じることが示された(64)。したがってヘム制限中のアルキルヒドロペルオキシドレダクターゼの存在量の実質的増大は、μ−オキソビスヘム層の非存在/低減により引き起こされる酸化ストレス増大に応答し得た。OxyR(酸素感受性転写活性剤)も、嫌気性増殖中のアルキルヒドロペルオキシドの発現において関与するが(71)、この場合、ポルフィロモナス・ジンジバリスOxyR−突然変異体は、遺伝子発現の16分の1の低減を示す。さらに近年、Duran-Pinedo及び共同研究者らも、RprY応答レギュレーターによるaphC発現の正の調節を実証した。したがって存在量の実質的増大は、ヘム利用能がRprY及びOxyR制御遺伝子発現において関与し得る、ということを示唆する。興味深いことに、このタンパク質の非常に高いコドン使用頻度指数(CAI)値(0.838)は、このタンパク質が、このようなストレスに応答して迅速誘導のために細胞中で高度に発現され得る、ということを示唆する。
【0100】
本明細書全体を通して、「〜を含む」という単語、或いは「〜を含む(単数)(comprises)」又は「〜から成る(comprising)」という変形は、記述された素子、整数又は工程、或いは素子、整数又は工程の群を包含することを意味するが、任意の他の素子、整数又は工程、或いは素子、整数又は工程の群の排除を意味しないと理解される。
【0101】
本明細書中で言及された出版物はすべて、参照により本明細書中で援用される。本明細書中に包含されている文書、行為、材料、装置、論文等についての任意の考察は、単に、本発明に一情況を提供するためにある。これらの事柄のいずれか又はすべてが従来技術基礎の一部を構成する、或いは本出願の各特許請求の優先日前にどこかに存在したので、本発明に関連する分野における共通の一般知識であった、ということを認めるものと解釈されるべきでない。
【0102】
広範に記載したような本発明の精神又は範囲を逸脱しない限り、特定の実施形態に示したように、多数の変更及び/又は修正が本発明に対して成され得る、と当業者は理解する。したがって本発明の実施形態は、すべての点において例示であって、本発明を限定するものではない、と考えられるべきである。
引用文献
【0103】
【表3】
【技術分野】
【0001】
本発明は、歯周病の予防及び治療に有用なポルフィロモナス・ジンジバリス(P. gingivalis)タンパク質の組成物及び単離方法に関する。より詳細には、本発明は、歯周病の予防及び治療に用いられ得るヘム利用能により調節されることが同定されたポルフィロモナス・ジンジバリスタンパク質を基礎にしたワクチン組成物及び方法に関する。
【背景技術】
【0002】
歯周病は、歯を支持する歯茎及び骨に影響を及ぼす慢性細菌感染である。歯周病は、プラーク(歯の上に絶えず生じる粘着性バイオフィルム)中の細菌が歯茎を炎症させるようになると開始する。歯周病は、歯肉組織(歯茎);歯根膜(セメント質及び歯槽骨中に埋め込まれる結合組織);セメント質(歯根を覆っている石化結合組織);並びに歯槽骨(骨槽窩)に影響を及ぼし得る。疾患の進行によって、歯根膜の破壊、歯槽骨損失並びに結合組織性付着の根尖側移動が起こり得る。進行型歯周病は、細菌プラークを保有する歯周ポケットの形成、歯の進行性弛緩及び結果的損失を生じ得る。歯周病は、歯周炎に進み得る歯肉炎を包含する。慢性歯周炎は、歯肉下歯垢中の特定細菌に関連した歯の支持組織の炎症性疾患である。当該疾患は、約35%の有歯成人に影響を及ぼし、そして西欧諸国における歯牙損失の一主因である(1)。歯肉下歯垢の正常口腔内微生物相の一成員であるポルフィロモナス・ジンジバリスは、この疾患の進行における主要な日和見病原体の1つとして関連付けられてきた(2)。
【0003】
ポルフィロモナス・ジンジバリスは、エネルギー産生のためのアミノ酸の発酵に頼る黒色色素性、糖非分解性のグラム陰性嫌気性球桿菌である(3)。大半の細菌と同様に、ポルフィロモナス・ジンジバリスは、それが、配位第一鉄原子を有するプロトポルフィリンIX環(PPIX)から成る分子をヘムの形態で優先的に獲得する鉄に対する必須増殖要件を有する(4)。鉄供給源としてのヘムのこの利用は、ポルフィロモナス・ジンジバリスがde novoでPPIXを合成できないことを反映し得る(5)。ヘムはヘモグロビンから優先的に得られ、そしておそらくはTonB連結外膜受容体HmuRと協力して(8)、細胞表面Arg及びLys特異的プロテイナーゼ/アドヘシン複合体の活性により獲得される(4,6,7)。シデロフォアを用いて鉄を得る好気性細菌又は通性細菌と違って、ポルフィロモナス・ジンジバリスはシデロフォアを産生せず、そして通常はシデロフォア媒介性鉄獲得と関連する第二鉄レダクターゼ活性を欠く(9,10)。ポルフィロモナス・ジンジバリスは、酸化的攻撃から細胞を保護するのに役立つ固有のカタラーゼ活性を有するμ−オキソビス−ヘムの形態でヘムをその表面に貯蔵する(11)。口腔の微量栄養素制限環境内の多数且つ多様な細菌と競合可能であり得る(12)ポルフィロモナス・ジンジバリスに関しては、それはそれ自体を定着しなければならないだけでなく、多数の宿主防御を忌避するか又は克服しなければならない。
【0004】
歯周病の開始及び進行は、疾患の部位での出血と関連し、それにより高レベルのヘモグロビンを提供する。したがって、ポルフィロモナス・ジンジバリスが歯肉下歯垢中で定着し、増殖する、そして疾患を開始するメカニズムの理解を助けるために、貧(ヘム制限)微量栄養素条件から富(ヘム過剰)微量栄養素条件への移行中の相対的タンパク質存在量の変化を測定することは重要である。
【0005】
多数のタンパク質がヘム制限下での増殖と関連付けられてきたが(9,13)、しかしポルフィロモナス・ジンジバリスプロテオーム又はヘム制限中のプロテオームへの変化に関する広範な研究は報告されていない。
【0006】
歯周病の予防及び治療に有用である組成物を開発するに際しては、疾患過程の初期段階を妨害し、防止する作用物質を同定することが望ましい。
【0007】
ここで本発明者らは、歯周病の予防及び治療のための適切な標的として用いられ得るヘム利用能により調節される特定のポルフィロモナス・ジンジバリスタンパク質を同定するための方法を開発した。
【発明の概要】
【課題を解決するための手段】
【0008】
本発明者らは、ポルフィロモナス・ジンジバリスの代謝、病原性及び宿主細胞の侵襲に関与するヘム利用能により調節されるポルフィロモナス・ジンジバリスタンパク質の同定方法を首尾よく開発した。特に2つの特定のインターナリン様ポルフィロモナス・ジンジバリスタンパク質、即ち宿主細胞によるポルフィロモナス・ジンジバリスのインターナリゼーションに関与するPG0350及びPG1374、細胞表面リポタンパク質であるとされる仮説的タンパク質PG1019、並びにアルキルヒドロペルオキシドレダクターゼタンパク質PG0618は、歯周病の予防及び治療のための有用な標的と同定された。
【0009】
本発明の第一の態様は、単離抗原性ポルフィロモナス・ジンジバリスポリペプチドであって、以下の:
(i)配列番号1のアミノ酸配列を有するPG0350タンパク質;
(ii)配列番号2のアミノ酸配列を有するPG1374タンパク質;
(iii)配列番号3のアミノ酸配列を有するPG1019タンパク質;
(iv)配列番号4のアミノ酸配列を有するPG0618タンパク質;
(v)配列番号1若しくは配列番号2若しくは配列番号3若しくは配列番号4のアミノ酸配列と少なくとも85%同一であるアミノ酸配列;又は
(vi)配列番号1若しくは配列番号2若しくは配列番号3若しくは配列番号4の連続アミノ酸配列と同一の少なくとも10個のアミノ酸を含むアミノ酸配列
から成る群から選択されるポリペプチドである。
【0010】
本発明の第二の態様は、被験体においてポルフィロモナス・ジンジバリスに向けられる免疫応答を惹起するのに用いるためのワクチン組成物であって、有効量の本発明の第一の態様の少なくとも1つのポリペプチド、並びに薬学的に許容される担体を含む組成物である。
【0011】
本発明の第三の態様は、歯周病に関して被験体を予防するか又は治療する方法であって、本発明によるワクチン組成物を被験体に投与することを包含する方法である。
【0012】
本発明の第四の態様は、本発明の第一の態様のポリペプチドに対して惹起される抗体である。好ましくは抗体は、本発明のポリペプチドと特異的に結合する。
【0013】
本発明の第五の態様は、歯周病の予防又は治療に有用な組成物であって、本発明の第四の態様の抗体並びに薬学的に許容される担体を含む組成物である。
【0014】
本発明の第六の態様は、歯周病の進行に関与するポルフィロモナス・ジンジバリスポリペプチドの同定方法であって、以下の:
a)ヘム制限条件下で増殖されるポルフィロモナス・ジンジバリスにより産生されるポリペプチド又はそのペプチドの相対量を測定する工程;並びに
b)工程a)より高いヘム条件下で増殖されるポルフィロモナス・ジンジバリスにより産生されるポリペプチド又はそのペプチドの相対量を測定する工程
を包含する方法であり、工程b)と比較した場合の工程a)で検出されるポリペプチド又はそのペプチド断片の量の増大がポリペプチドが歯周病の進行に関与するということを示す方法である。
【0015】
本発明の第七の態様では、各鎖中に少なくとも19塩基対の二本鎖領域を含む干渉RNA分子であって、上記二本鎖領域の鎖のうちの1つが配列番号5又は配列番号6又は配列番号7又は配列番号8の領域と相補的である干渉RNA分子が提供される。
【0016】
本発明の第八の態様では、被験体における歯周病の治療のための薬剤の製造における本発明の第一の態様の少なくとも1つのポリペプチドの使用が提供される。
【図面の簡単な説明】
【0017】
【図1】ヘム制限下で増殖されるポルフィロモナス・ジンジバリスからのタンパク質を同定するために用いられる組合せ戦略の概略図である。超遠心分離を用いて溶解細胞を可溶性及び不溶性画分に前分画し、その後、これら2つの画分を分析した。分離及び分析手順は、2つの主な方法、即ち気相分別によるLCMSとgeLCMSとから成る。
【図2】INCA(14)を用いて算定される同定ポルフィロモナス・ジンジバリスプロテオソーム及び予測ポルフィロモナス・ジンジバリスゲノムのコドン使用頻度指数(CAI)分布を示すグラフである。リボソームシンターゼ及びtRNAシンターゼは高度発現遺伝子として定義された。コドンが100個未満の遺伝子は除外し、全算定遺伝子を1685個とした。
【図3】単一ペプチドの検出に基づいたPG0390の分析及び同定を示す図である。(A)総イオンクロマトグラム。(B)54.8分での質量スペクトル。差込図はm/z 703.9及び708.4での拡大ICATペプチドイオン対(比 1:2)(L/H)を示す。(C)前駆体m/z 703.9に関する生成物イオンスペクトル。このペプチドイオンは、配列LVDLNC*FDIK(MASCOTスコア=40;C*はICAT修飾システインを意味する)を有すると同定された。
【図4】ヘム過剰を上回るヘム制限の比率を基礎にしたタンパク質存在量の分布を示す図である(Log2スケールでのすべての一単位は2倍変化を示す)。
【図5】ポルフィロモナス・ジンジバリスW50(◆)及びECR312(■)によるKB細胞への結合を示す図である。2つの生物学的複製(n=6)を用いて、検定を実行した。差込図は、前方及び側方散乱特性を基礎にした生KB細胞のゲーティングを示し(左上)、5つのピークは、250〜1250のポルフィロモナス・ジンジバリス:KB細胞比でのKB細胞との結合ポルフィロモナス・ジンジバリスW50(右上)、並びにECR312(右下)のFITC蛍光を表わす。
【図6】PG0350タンパク質のアミノ酸配列(配列表中にも示されるように配列番号1と呼ばれる)を示す図である。
【図7】PG1374タンパク質のアミノ酸配列(配列表中にも示されるように配列番号2と呼ばれる)を示す図である。
【図8】PG1019タンパク質のアミノ酸配列(配列表中にも示されるように配列番号3と呼ばれる)を示す図である。
【図9】PG0618タンパク質のアミノ酸配列(配列表中にも示されるように配列番号4と呼ばれる)を示す図である。
【発明を実施するための形態】
【0018】
本発明は、歯周病の予防及び治療のための有用な標的としてのヘム利用能により調節されるポルフィロモナス・ジンジバリスタンパク質の同定を有利に提供する。好ましくは本発明は、歯周病の予防及び治療のための有用な標的としてのヘム制限により上方調節されるポルフィロモナス・ジンジバリスタンパク質の同定を提供する。
【0019】
特に本発明は、単離抗原性ポルフィロモナス・ジンジバリスポリペプチドであって、以下の:
(i)配列番号1のアミノ酸配列を有するPG0350タンパク質;
(ii)配列番号2のアミノ酸配列を有するPG1374タンパク質;
(iii)配列番号3のアミノ酸配列を有するPG1019タンパク質;
(iv)配列番号4のアミノ酸配列を有するPG0618タンパク質;
(v)配列番号1若しくは配列番号2若しくは配列番号3若しくは配列番号4のアミノ酸配列と少なくとも85%同一であるアミノ酸配列;又は
(vi)配列番号1若しくは配列番号2若しくは配列番号3若しくは配列番号4の連続アミノ酸配列と同一の少なくとも10個のアミノ酸を含むアミノ酸配列
から成る群から選択されるポリペプチドを提供する。
【0020】
好ましくは、単離抗原性ポリペプチドは、配列番号1又は配列番号2又は配列番号3又は配列番号4のアミノ酸配列と90%、95%、96%、97%、98%、99%又は100%同一である。
【0021】
好ましくは、単離抗原性ポリペプチドは、配列番号1又は配列番号2又は配列番号3又は配列番号4の連続アミノ酸配列と同一である少なくとも20個、30個、40個、50個、60個、70個、80個、90個又は100個のアミノ酸を含むアミノ酸配列を含む。
【0022】
好ましい一実施形態では、抗原性ポリペプチドは、PG1374又はPG0350又はPG1019又はPG0618タンパク質の親水性表面露出領域を構成するアミノ酸配列を含む。
【0023】
「ペプチド、タンパク質及びポリペプチド」という用語は、本明細書中では互換的に用いられる。本発明のポリペプチドは、組換えポリペプチド、例えば融合ポリペプチドを含み得る。融合ポリペプチドの製造方法は、当業者に周知である。
【0024】
当業者により十分に理解されるように、配列表中に示されたアミノ酸配列に改変がなされ得る。これらの改変は、アミノ酸残基の欠失、挿入又は置換であり得る。改変ポリペプチドは、天然(即ち、天然供給源から精製又は単離される)又は合成(例えばコード化DNA上での部位特異的突然変異誘発)であり得る。配列表に示された配列と少なくとも85%、好ましくは少なくとも90%、95%、96%、97%、98%又は99%の同一性を有するこのような改変ポリペプチドは本発明の範囲内である、と意図される。これらの改変ポリペプチドに対して産生される抗体はさらにまた、配列表に示された配列のうちの1つを有するポリペプチドと結合するであろう。
【0025】
保存的置換の概念は当業者により十分に理解されるが、一方、分かり易くするためには、保存的置換は以下に記述されるものである:
Gly、Ala、Val、Ile、Leu、Met;
Asp、Glu、Ser;
Asn、Gln;
Ser、Thr;
Lys、Arg、His;
Phe、Tyr、Trp、His;及び
Pro、Nα−アルキル(alkal)アミノ酸。
【0026】
本発明の実行は、別記しない限り、当業者に周知の化学、分子生物学、微生物学、組換えDNA及び免疫学の従来の技法を用いる。このような技法は、Perbal, A Practical Guide to Molecular Cloning, John Wiley and Sons (1984)(15)、Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbour Laboratory Press (1989)(16)、Brown (editor), Essential Molecular Biology: A Practical Approach, Volumes 1 and 2, IRL Press (1991)(17)、Glover & Hames (editors), DNA Cloning: A Practical Approach, Volumes 1-4, IRL Press (1995 and 1996)(18)及びAusubel et al., (Editors), Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience (1988、今日までのすべての最新情報を含む)(19)のような出典の文献全体に記載され、説明されている。これらのテキストの開示内容は、参照により本明細書中で援用される。
【0027】
「単離ポリペプチド」とは、本明細書中で用いる場合、それが天然に生じるか、或いはそれを用いてポリペプチド又はペプチドが合成的に合成され得る他のタンパク質、脂質及び核酸から分離されたポリペプチドを指す。好ましくはポリペプチドは、それを精製するために用いられる物質、例えば抗体又はゲルマトリックス、例えばポリアクリルアミドからも分離される。好ましくはポリペプチドは、精製調製物の乾燥重量の少なくとも10%、20%、50%、70%及び80%を構成する。好ましくは調製物は、タンパク質シーケンシングを可能にするのに十分な量のポリペプチドを含有する(即ち、少なくとも1mg、10mg又は100mg)。
【0028】
本明細書中に記載される単離ポリペプチドは、標準技法、例えばカラムクロマトグラフィー(タンパク質産物と相互作用する種々のマトリックス、例えばイオン交換マトリックス、疎水性マトリックス等を使用する)、アフィニティークロマトグラフィー(タンパク質又はタンパク質と結合するその他のリガンドに特異的な抗体を利用する)により精製され得る。
【0029】
「抗原性ポリペプチド」とは、本明細書中で用いる場合、検出可能な抗原−抗体複合体を形成するのに十分に高い親和性を有する特定の抗体と結合可能なポリペプチド、その類似体又は断片のような部分である。好ましくは抗原性ポリペプチドは、宿主動物中で体液性及び/又は細胞性の免疫応答を引き出すことが可能な免疫原性構成成分を含む。
【0030】
「連続アミノ酸配列」とは、本明細書中で用いる場合、アミノ酸の連続伸長を指す。
【0031】
2つのアミノ酸配列が指定パーセンテージ限度内に入るか否かを決定するに際して、配列の並列比較又は多重アラインメントを実行することが必要である、ということに当業者は気付くだろう。このような比較又はアラインメントでは、アラインメントを実施するために用いられるアルゴリズムによって、非同一残基の位置決めにおいて差異が生じる。本発明の状況では、2つ以上のアミノ酸配列間のパーセンテージ同一性又は類似性への言及は、当業者に既知の任意の標準アルゴリズムを用いて測定されるような上記の配列間のそれぞれ同一の及び類似の残基の数を指すと解釈される。例えばアミノ酸配列同一性又は類似性は、GAPプログラムを用いて算定され得るし、及び/又はComputer Genetics Group, Inc., University Research Park, Madison, Wisconsin, United States of America(20)のPILEUPプログラムを用いて整列され得る。GAPプログラムは、Needleman and Wunsch(21)のアルゴリズムを利用して、同一の/類似の残基の数を最大化し、そしてアラインメント中の配列ギャップの数及び長さを最小化する。代替的に又はさらに、2つより多くのアミノ酸配列が比較される場合、Clustal Wプログラム(22)が用いられる。
【0032】
本発明は、被験体においてポルフィロモナス・ジンジバリスに向けられる免疫応答を惹起するのに用いるためのワクチン組成物であって、有効量の本発明の第一の態様の少なくとも1つのポリペプチド並びに薬学的に許容される担体を含む組成物も提供する。
【0033】
本発明のワクチン組成物は、好ましくは、ポルフィロモナス・ジンジバリスに対する免疫を付与するために用いられ得る少なくとも1つの抗原を含む抗原性ポリペプチドを含む。本発明の方法により治療される被験体は、ヒト、ヒツジ、畜牛、ウマ、ウシ、ブタ、家禽、イヌ及びネコから成る群から選択され得るが、これらに限定されない。好ましくは、被験体はヒトである。ポルフィロモナス・ジンジバリスに対して向けられる免疫応答は、被験体の免疫系が特定の抗原性ポリペプチドに対する抗体を産生する場合、被験体において達成される。
【0034】
ワクチン組成物は、好ましくは、ポルフィロモナス・ジンジバリスに対する免疫を誘導し、それにより歯周病を予防するために被験体に投与される。「有効量」という用語は、本明細書中で用いる場合、ポルフィロモナス・ジンジバリスに対する免疫応答を引き出すのに十分な用量を意味する。これは、被験体並びにポルフィロモナス・ジンジバリス感染のレベルによって変わり、そして終局的には担当する科学者、医者又は獣医により決定される。
【0035】
本発明のワクチン組成物は、ヒト又は動物被験体に投与するのに適した適切な薬学的に許容される担体、例えば希釈剤及び/又はアジュバントを含む。ワクチンは、ポルフィロモナス・ジンジバリスに対する特定の免疫応答を惹起するために、好ましくは、鼻噴霧により、或いは注射により、経口的に送達するための適切なアジュバントを含む。本発明のワクチンは、本発明の抗原性ポリペプチドをコードする組換え核酸配列にも基づき得るが、この場合、当該核酸配列は、適切なベクター中に組入れられて、ベクターを含有する適切な形質転換宿主(例えば大腸菌、枯草菌、サッカロミセス・セレビシエ(saccharomyces cerevisiae)、COS細胞、CHO細胞及びHeLa細胞)中で発現される。ワクチンは、本明細書中に示されるような組換えDNA法を用いて製造され得るし、或いは本発明に記載されるアミノ酸配列から化学的に合成され得る。付加的には、本発明によれば、抗原性ポリペプチドを用いて、ポルフィロモナス・ジンジバリスにより引き起こされる歯周病及び感染に対する受動免疫化のために有用なポルフィロモナス・ジンジバリス抗血清を生成し得る。
【0036】
当業者に既知の種々のアジュバントは、ワクチン処方物と関連して一般的に用いられる。アジュバントは、免疫応答を調整することにより、そしてワクチン抗原が単独で投与された場合より少量のワクチン抗原又はより少ない用量を用いて、より永続的な且つより高レベルの免疫を獲得するに際して、助けとなる。アジュバントの例としては、不完全フロイントアジュバント(IFA)、アジュバント65(ピーナツ油、モノオレイン酸マンニド及びモノステアリン酸アルミニウムを含有)、オイルエマルジョン、Ribiアジュバント、プルロニック・ポリオール、ポリアミン、Avridine、Quil A、サポニン、MPL、QS−21及びミネラルゲル、例えばアルミニウム塩が挙げられる。その他の例としては、水中油型エマルジョン、例えばSAF−1、SAF−0、MF59、SeppicのISA720、並びにその他の粒状アジュバント、例えばISCOM及びISCOMマトリックスが挙げられる。アジュバントの他の例の広範なしかし包括的な一覧は、Cox and Coulter 1992(23)に列挙されている。アジュバントのほかに、ワクチンは、必要に応じて、従来の薬学的に許容される担体、賦形剤、充填剤、緩衝剤又は希釈剤を含み得る。アジュバントを含有する1回又は複数回用量のワクチンが、歯周病を防止するために予防的に、或いはすでに存在する歯周病を治療するために治療的に投与され得る。
【0037】
別の好ましいワクチン組成物において、調製物は、粘膜アジュバントと組合され、そして口又は鼻経路を介して投与される。粘膜アジュバントの例は、コレラ毒素及び易熱性大腸菌毒素、これらの毒素の非毒性Bサブユニット、低減毒性を示すこれらの毒素の遺伝子突然変異体である。抗原性ポリペプチドを経口的又は経鼻的に送達するために利用され得るその他の方法としては、消化管又は鼻腔からのマイクロスフェアの取り込みを手助けするための、そしてタンパク質の分解を防ぐための、マイクロカプセル封入による生分解性ポリマー(例えばアクリレート又はポリエステル)の粒子中へのポリペプチドの組入れが挙げられる。リポソーム、ISCOM、ヒドロゲルは、粘膜免疫系への抗原性ポリペプチドの送達のためのLTB、CTB又はレクチン(マンナン、キチン及びキトサン)のような標的分子の組入れによりさらに増強され得る他の可能な方法の例である。ワクチン並びに粘膜アジュバント又は送達系のほかに、ワクチンは、従来の薬学的に許容される担体、賦形剤、充填剤、コーティング、分散媒質、抗菌剤及び抗真菌剤、緩衝剤又は希釈剤を必要に応じて含み得る。
【0038】
この実施形態の別の方式は、ポルフィロモナス・ジンジバリスにより引き起こされる感染に対して防御するために用いられる生組換えウイルスワクチン、組換え細菌ワクチン、組換え弱毒化細菌ワクチン、又は不活性化組換えウイルスワクチンを提供する。ワクチンウイルスは、他の生物体由来のワクチン抗原を発現するよう工学処理される感染性ウイルスの当該技術分野における最も良く知られた例である。それ自体が疾患を引き起こさないよう弱毒化されるか又はそうでなければ処理される組換え生ワクシニアウイルスは、宿主を免疫化するために用いられる。宿主内の組換えウイルスのその後の複製は、抗原性ポリペプチドのようなワクチン抗原による免疫系の連続的刺激を提供し、それにより長期継続性免疫を提供する。
【0039】
他の生ワクチンベクターとしては、アデノウイルス、サイトメガロウイルス、好ましくはポックスウイルス、例えばワクシニア(24)、並びに弱毒化サルモネラ菌株(25〜28)が挙げられる。生ワクチンは、免疫系を連続的に刺激し、実質的に長期継続性の免疫を付与し得るため、特に有利である。免疫応答がその後のポルフィロモナス・ジンジバリス感染に対して防御的である場合、生ワクチンそれ自体はポルフィロモナス・ジンジバリスに対する防御ワクチンで用いられ得る。特に、生ワクチンは、口腔の共生棲息生物である細菌を基礎にし得る。この細菌は、組換え不活性化ポリペプチドを保有するベクターで形質転換されて、次に、口腔、特に口腔粘膜にコロニー形成するために用いられ得る。一旦口腔粘膜にコロニーが形成されると、組換えタンパク質の発現は、粘膜関連リンパ系組織を刺激して、中和抗体を産生する。例えば分子生物学技法を用いて、ポリペプチドをコードする遺伝子は、エピトープの発現を可能にするが、ワクシニアウイルスベクターの増殖又は複製に負の影響を及ぼさない部位でウイルスゲノムDNA中に挿入され得る。その結果生じる組換えウイルスは、ワクチン処方物中の免疫原として用いられ得る。同一方法を用いて、不活化組換えウイルスワクチン処方物を構築し得るが、但し、組換えウイルスは、例えば免疫原として用いる前に、そして発現免疫原の免疫原性に実質的に影響を及ぼすことなく、当該技術分野で既知の化学的手段により、不活性化される。
【0040】
能動免疫化に代わり得るものとして、免疫化は受動的、即ち本発明のポリペプチドに対する抗体を含有する精製免疫グロブリンの投与を包含する免疫化であり得る。
【0041】
本発明の方法及び組成物に用いられる抗原性ポリペプチドは、適切な賦形剤、例えば乳化剤、界面活性剤、安定剤、染料、浸透強化剤、酸化防止剤、水、塩溶液、アルコール、ポリエチレングリコール、ゼラチン、ラクトース、ステアリン酸マグネシウム及びケイ酸と組合せられ得る。抗原性ポリペプチドは、好ましくは滅菌水溶液として処方される。本発明のワクチン組成物は、歯周病のための現行治療を補足するために用いられ得る。
【0042】
本発明の第三の態様は、歯周病に関する被験体の予防又は治療方法であって、本発明によるワクチン組成物を被験体に投与することを包含する方法である。
【0043】
本発明の方法では、被験体は、歯周病のための予防的処置を含めて治療される。歯周病は、単純歯肉炎症から、歯を支持する柔組織及び骨に対する大きな損傷を生じる重篤な疾患まで多岐にわたる。歯周病は歯肉炎及び歯周炎を包含する。細菌、主にグラム陰性種、例えばポルフィロモナス・ジンジバリスは、「歯肉炎」と呼ばれる歯肉の炎症を引き起こす。歯肉炎では、歯肉は赤色になり、腫脹し、そして容易に出血し得る。歯肉炎が治療されない場合、それは「歯周炎」(「歯の周囲の炎症」を意味する)に進行し得る。歯周炎において、歯肉は歯から離れて、感染された「ポケット」を形成する。プラークが広がり、歯肉線より下に増殖すると、身体の免疫系は細菌と戦う。治療されない場合、歯を支持する骨、歯肉及び結合組織は破壊される。歯は、最終的には弛緩するようになり、除去されねばならない。
【0044】
本発明の第四の態様は、本発明の第一の態様のポリペプチドに対して産生される抗体である。好ましくは抗体は、本発明のポリペプチドに対して特異的に向けられる。
【0045】
本明細書中では、「抗体」という用語は最も広い意味で用いられ、そして詳細には、モノクローナル抗体、ポリクローナル抗体、多重特異性抗体(例えば二重特異性抗体)、キメラ抗体、ダイアボディ(diabodies)、トリアボディ(triabodies)及び抗体断片を包含する。本発明の抗体は、好ましくは、他のポリペプチドの抗原と交差反応することなく、本明細書中に上記されたような抗原性ポリペプチドと特異的に結合することができる。
【0046】
「〜と特異的に結合する」という用語は、本明細書中で用いる場合、例えばBIAcore(商標)表面プラズモン共鳴系及びBIAcore(商標)動的評価ソフトウェア(例えばバージョン2.1)を用いて表面プラズモン共鳴分析により測定した場合に1μM以下の解離定数(Kd)を有する抗体の免疫グロブリン可変領域による抗原の結合を指すよう意図される。特異的結合相互作用に関する親和性又は解離定数(Kd)は、好ましくは約500nM〜約50pM、さらに好ましくは約500nM以下、さらに好ましくは約300nM以下、そして好ましくは少なくとも約300nM〜約50pM、約200nM〜約50pM、さらに好ましくは少なくとも約100nM〜約50pM、約75nM〜約50pM、約10nM〜約50pMである。
【0047】
抗体の抗原結合機能は、全長抗体の断片により実行され得る、ということが示されている。抗体の結合断片の例としては、(i)Fab断片:VLドメイン、VHドメイン、CLドメイン及びCH1ドメインから成る一価断片;(ii)F(ab’)2断片:ヒンジ部でジスルフィド架橋により連結される2つのFab断片を含む二価断片;(iii)VHドメイン及びCIIIドメインから成るFd断片;(iv)抗体の単一アームのVLドメイン及びVHドメインから成るFv断片;(v)VHドメイン又はVLドメインから成るdAb断片;並びに(vi)単離相補性決定領域(CDR)が挙げられる。さらに、Fv断片の2つのドメインであるVL及びVHは、別個の遺伝子によりコードされるが、それらは、VL領域及びVH領域が対合して一価分子(1本鎖Fv(scFv)として既知である)を形成する単一タンパク質鎖としてそれらを作製させる合成リンカーにより、組換え法を用いて接合され得る。他の形態の1本鎖抗体、例えばダイアボディ又はトリアボディも包含される。ダイアボディは、VHドメイン及びVLドメインが単一ポリペプチド鎖上で、しかし同一鎖上の2つのドメイン間の対合を可能にするには短すぎるリンカーを用いて発現され、それにより当該ドメインを別の鎖の相補的ドメインと対合させて、2つの抗原結合部位を作製する二価二重特異性抗体である。
【0048】
当該技術分野で既知の種々の手法も、本発明の抗原性ポリペプチドと結合し得るモノクローナル抗体及びポリクローナル抗体、並びに種々の組換え体及び合成抗体の産生のために用いられ得る。さらに、宿主種によって免疫学的応答を増大するために用いられ得る種々のアジュバントは当業者に既知であり、例としては、フロイント(完全及び不完全)、ミネラルゲル、例えば水酸化アルミニウム、表面活性物質、例えばリゾレシチン、プルロニックポリオール、ポリアニオン、ペプチド、オイルエマルジョン、ジニトロフェノール、並びに潜在的に有用なヒトアジュバント、例えばカルメット・ゲラン桿菌(BCG)及びコリネバクテリウム・パルブム(Corynebacterium parvum)が挙げられるが、これらに限定されない。抗体及び抗体断片は、標準技法(例えば発酵器を用いて組織培養物又は無血清中で)により大量に産生され、そしてアフィニティーカラム、例えばプロテインA(例えばネズミMabに関して)、プロテインG(例えばラットMabに関して)又はMEP HYPERCEL(例えばIgM及びIgG Mabに関して)を用いて精製され得る。
【0049】
組換えヒト又はヒト化バージョンのモノクローナル抗体は、ヒト療法的用途のための好ましい一実施形態である。ヒト化抗体は、文献(29,30)中の手法に従って調製され得る。ヒト化モノクローナル抗体の産生のための近年記載された「遺伝子変換突然変異誘発」戦略も、ヒト化抗体の産生に用いられ得る(31)。代替的には、重鎖領域及び軽鎖領域の無作為組合せの組換え相ライブラリーを生成するための技法は、組換え抗体を調製するために用いられ得る(32)。
【0050】
本発明は、歯周病の予防又は治療に有用な組成物であって、本発明の第一の態様のポルフィロモナス・ジンジバリスポリペプチドのアンタゴニスト並びに薬学的に許容される担体を含み、当該アンタゴニストがポルフィロモナス・ジンジバリス感染を阻害する組成物も提供する。
【0051】
本明細書中で用いる場合、「アンタゴニスト」という用語は、対象のポリペプチドの生物学的活性を阻害する核酸、ペプチド、抗体、リガンド又はその他の化学的実体を指す。特定のタンパク質の適切なアンタゴニストを試験し、選択する技法は当業者には既知であり、このような技法は結合検定を包含し得る。PG0350、PG1374、PG1019及びPG0618の可能なアンタゴニストは、好ましくは、これらのタンパク質と宿主細胞又は他の基質との結合を阻害する抗体(モノクローナル又はポリクローナル)であり、或いはそれらは、これらのタンパク質の結合を妨げるタンパク質又はペプチドであり得る。
【0052】
本発明の抗体及びアンタゴニストは、多数の用途を有し、例えばそれらは、歯垢の制御並びに虫歯及び歯周病に関連する病原体の抑制のための口腔ケア製品(練り歯磨き及び洗口液)における抗菌防腐剤として用いられ得る。本発明の抗体及びアンタゴニストは、薬学的調製物(例えば、局所及び全身抗感染薬)にも用いられ得る。
【0053】
本発明の第六の態様では、歯周病の進行に関与するポルフィロモナス・ジンジバリスポリペプチドの同定方法であって、以下の:
a)ヘム制限条件下で増殖されるポルフィロモナス・ジンジバリスにより産生されるポリペプチド又はそのペプチドの相対量を測定する工程;並びに
b)工程a)より高いヘム条件下で増殖されるポルフィロモナス・ジンジバリスにより産生されるポリペプチド又はそのペプチドの相対量を測定する工程
を包含する方法であり、工程b)と比較した場合の工程a)で検出されるポリペプチド又はそのペプチド断片の量の増大がポリペプチドが歯周病の進行に関与するということを示す方法が提供される。
【0054】
ヘム制限条件下でポルフィロモナス・ジンジバリスを増殖させるためには、ヘミンの濃度は約0.1μg/ml〜約0.5μg/mlである、というのが好ましい。ヘム制限条件は、ポルフィロモナス・ジンジバリス細胞の細胞密度が本発明の方法の工程(b)の増殖条件下で観察されるものより有意に低い場合、達成される。工程(b)のより高いヘム条件は、好ましくは、5μg/mlを超えるヘミン濃度を用いて達成される。
【0055】
ヘム制限条件及び高ヘム条件におけるポリペプチドの相対量の比較は、好ましくは、鑑別プロテオミクス手法を用いることにより決定され得る。好ましくはポリペプチドの量は、当該技術分野で一般に用いられる定性的プロテオーム分析、例えば溶液中消化及びゲル中消化並びにLC−MS/MSの組合せ戦略、並びにMSと組合せた安定同位体標識戦略(ICAT)を用いた分析(これらに限定されない)により測定される。
【0056】
本発明の第六の態様で明示される方法に従って同定される単離抗原性ポルフィロモナス・ジンジバリスポリペプチドは、歯周病を治療し、予防するための標的として用いられ得る。特に、単離ポリペプチドは、ポルフィロモナス・ジンジバリス、例えばポルフィロモナス・ジンジバリス感染、例えば歯周病に対するワクチン組成物を開発するために用いられ得る。
【0057】
本発明は、本発明の第一の態様のポリペプチドをコードするmRNA分子に対して標的にされる干渉RNA分子も提供する。したがって本発明の第七の態様では、各鎖中に少なくとも19塩基対の二本鎖領域を含む干渉RNA分子であって、二本鎖領域の鎖のうちの1つが配列番号5又は配列番号6又は配列番号7又は配列番号8の領域と相補的である干渉RNA分子が提供される。
【0058】
いわゆるRNA干渉又はRNAiは周知であり、RNAiに関するさらなる情報は、Hannon (2002) Nature 418: 244-251及びMcManus & Sharp (2002) Nature Reviews: Genetics 3(10): 737-747(これらの記載内容は参照により本明細書中で援用される)に提供されている。
【0059】
本発明は、siRNA安定性を増強し、in vivoでのsiRNAの安定性を向上し、それらの使用を支持するsiRNAの化学的修飾(複数可)も意図する(例えばShen et al, (2006) Gene Therapy 13: 225-234参照)。これらの修飾は、センス鎖オリゴヌクレオチドの5’末端及び3’末端での逆方向脱塩基部分、並びにアンチセンス鎖の3’末端での最後の2つのヌクレオチド間の単一ホスホルチオエート結合を含み得る。
【0060】
干渉RNAの二本鎖領域は、二本鎖領域の各鎖中に少なくとも20塩基対、好ましくは少なくとも25塩基対、最も好ましくは少なくとも30塩基対を含む、というのが好ましい。本発明は、歯周病に関する被験体の治療方法であって、本発明の干渉RNA分子のうちの少なくとも1つを被験体に投与することを包含する方法も提供する。
【0061】
本発明の性質がより明白に理解され得るために、ここで、その好ましい形態を以下の実施例を参照しながら説明する。
【実施例】
【0062】
[実施例1]
1. 材料及び方法
1.1 細菌株及び化学物質
ポルフィロモナス・ジンジバリスW50(ATCC 53978)を、the Centre for Oral Health Science, The University of Melbourneの培養コレクションから得た。用いられる化学物質は超高純度であったが、但し、MS作業に関しては、LC MS等級試薬(Sigma, Reidel-de Haen)を用いた。
【0063】
1.2 ポルフィロモナス・ジンジバリスの増殖及び採取
400mLの作業容積を有するBioflo 110発酵器/バイオリアクター(New Brunswick Scientific)を用いて、ポルフィロモナス・ジンジバリスW50を連続培養で増殖させた。増殖培地は、5mg/mLのフィルター滅菌塩酸システイン、5.0μg/mLのヘミン(ヘム過剰)又は0.1μg/mLのヘミン(ヘム制限)を補足された37g/mLの脳心臓注入培地(Oxoid)であった。同一培地(ヘム過剰)中で増殖させたポルフィロモナス・ジンジバリスの24時間バッチ培養(100mL)を培養容器に接種することにより増殖を開始した。24時間のバッチ培養増殖後、培地貯蔵槽ポンプを作動させて、培地流を調整して、0.1 h−1(平均生成時間(MGT)6.9時間)の希釈率を生じた。容器の温度を、pH7.4±0.1で37℃に維持した。95%N2中の5%CO2で、培養を連続的にガス処理した。定常状態増殖中に細胞を採取し、5000gで30分間、洗浄緩衝液(50mMのトリス−HCl(pH8.0)、150mMのNaCl、5mMのMgCl2)で3回洗浄し、138MPaでフレンチプレス細胞破砕機(SLM, AMINCO)に3回通して破砕した。次に溶解細胞を2000gで30分間遠心分離して、非破壊細胞を除去し、その後、100000gで超遠心分離して、可溶性画分(上清)及び膜画分を生じた。全画分を氷上で実行した。
【0064】
1.3 非定量的プロテオーム分析のための試料の調製
2つの方法、即ち、トリプシンによる溶液中消化とその後の気相分別(GPF)によるLCMS、並びにゲル中消化とその後の組合せ戦略の一部としてのLCMS(geLC−MS)を用いて、非定量的プロテオーム分析を実行した(図1)。溶液中消化法に関しては、タンパク質を95℃で3分間煮沸し、TCA(16%)を用いて沈殿させて、可溶化緩衝液(8Mの尿素、50mMのトリス−HCl(pH8.3)、5mMのEDTA、0.05%SDS)中に再懸濁した。BCAタンパク質試薬(Pierce)を用いてタンパク質濃度を測定し、2μg/μLに調整した。1mMのDTTを用いて30分間、還元を実行し、10mMのヨードアセトアミドを用いて60分間、アルキル化した。溶液を水で希釈して最終濃度を1Mの尿素とした後、消化した。消化は、1:100(w/w)トリプシン対タンパク質の比率で、37℃で16時間、シーケンシング等級修飾トリプシン(Promega)を用いて実行した。蟻酸を添加して最終濃度を1%(v/v)とすることにより、消化を終結させた。次に、Sep−Pak C18カートリッジ(Waters)を用いてペプチドを脱塩し、真空遠心分離機(Thermosavant)を用いて乾燥し、そして0.1%TFA中5%アセトニトリル中に再懸濁した。2μgと等価のペプチドの量を、各LC−MS/MS分析のために注入した。
【0065】
geLC−MS法に関しては、25μgのタンパク質をプレキャストNovex 12%トリス−HClグリシンゲル(Invitrogen)上で分離し、クーマシー・ブリリアントブルーG−250(Sigma)で一晩染色した。ゲルを30の個々の断片に分けて、これを次に切り出して、約1mm3の立方体に切断した。50%エタノール及び25mMの重炭酸アンモニウム(ABC)緩衝液の溶液で3回、脱色を実行して、その後、100%エタノールで脱水した。25mMのABC中の10mMのDTTとともに、56℃で60分間、脱水ゲル立方体をインキュベートすることにより、還元を実行した。次に還元溶液を25mMのABC中55mMのヨードアセトアミドに取り替えて、45分間インキュベートした。ゲル立方体を50mMのABC中で2回洗浄し、100%エタノールで脱水した。30μLの修飾シーケンシング等級トリプシンを、25mMのABC緩衝液及び1mMのCaCl2中に5μg/mLの濃度で添加し、4℃で30分間インキュベートした。過剰量のトリプシン溶液を除去し、15μLの25mMのABC緩衝液を添加した。消化を37℃で一晩実行し、TFAを添加して最終濃度0.1%(v/v)とすることにより、終結させた。次に上清をエッペンドルフ管に移した。ゲル片に、0.1%TFA中の50%エタノールを50μL添加し、15分間超音波処理した。当該プロセスを反復し、1つのゲル断片由来の全上清をプールして、真空遠心機を用いて乾燥して約10μLとした。
【0066】
1.4 定量的ICAT分析のための試料の調製
タンパク質の標識化及び分離は、開裂性ICAT試薬(Applied Biosystems)を用いるgeLC−MS/MSアプローチ(33)を基礎にした。TCA(16%)を用いてタンパク質を先ず沈殿させて、6Mの尿素、5mMのEDTA、0.05%SDS及び50mMのトリス−HCl(pH8.3)を用いて可溶化した。BCAタンパク質試薬を用いてタンパク質濃度を測定し、1mg/mlに調整した。各増殖条件からのタンパク質100μgを、別々に、2μLの50mMのトリス(2−カルボキシ−エチル)ホスフィン塩酸塩を用いて、37℃で1時間、還元した。次にヘム制限増殖条件からの還元タンパク質をICATheavy試薬で、そしてヘム過剰増殖条件からのタンパク質をICATlight試薬でアルキル化した。次に2つの試料を合わせ、プレキャストNovex 10%NUPAGEゲル(Invitrogen)上でSDS−PAGEに付した。ゲルを、SimplyBlue(商標)SafeStain(Invitrogen)を用いて5分間染色し、その後、水で脱色した。次に、ゲルレーンを、ゲルの上部から染色前面まで、20の断片に切り出した。
【0067】
切断断片をさらにさいの目に切って1mm3の立方体として、一晩ゲル内消化し、上記の手順に従って2回抽出した。プールした上清を減圧下で乾燥して約50μLとし、その後、500μLのアフィニティーローディングバッファーと混合した後、メーカーの使用説明書(Applied Biosystems)に従ってアフィニティーカラム上に載せた。溶離ペプチドを乾燥し、ビオチンタグを、37℃で2時間、ニートTFAで切断し、その後、減圧下で乾燥した。乾燥試料を、0.1%TFA中の5%アセトニトリル 35μL中に懸濁した。
【0068】
1.5 液体クロマトグラフィー及び質量分析
UltiMate Nano LCシステム(LC Packings - Dionex)と接続されたEsquire HCTイオントラップ質量分光計(Bruker Daltonics)を用いて、MSを実行した。LC Packings逆相カラム(C18 PepMap 100、内径75μm×15cm、3μm、100Å)を用いて分離を達成し、以下のアセトニトリル勾配を用いて0.1%蟻酸で溶出した:0分〜5分(0%)、5分〜10分(0%〜10%)、10分〜100分(10%〜50%)、100分〜120分(50%〜80%)、120分〜130分(80%〜100%)。
【0069】
LC出力を、ナノスプレーイオン源と直接接続した。100ミリ秒の最大蓄積時間で、300〜1500のm/z範囲で、100000のイオン荷電制御下で、MS獲得を実施した。GPFを用いる場合、3つの付加的m/z範囲(300〜800、700〜1200及び1100〜1500)を用いて前駆体イオンに関して選択し、そして各m/z範囲を二重反復実験で実行して、同定されるペプチドの数を増大した。MS/MS獲得を100〜3000m/zの質量範囲全体で得て、初期完全プロテオーム分析に関して10個までの前駆体で、そして2分の能動除去時間を有する最強多重荷電イオンに関してはICAT分析のために3個の前駆体で実施した。
【0070】
1.6 非定量的プロテオーム分析のためのタンパク質同定
化合物検出閾値 10000並びにシグナル対ノイズ閾値 5で、Apexピークファインダーアルゴリズムを用いるDataAnalysis 3.2(Bruker Daltonics)を用いて、ピーク一覧を作成した。エクスポートされたデータに関して、+2及び+3という全体的負荷制限を設定した。ゲノム研究所(TIGR)ウェブサイト(www.tigr.org)から得られるポルフィロモナス・ジンジバリスデータベースに対して問い合わされるMS/MSデータに関するMASCOT検索エンジン(MASCOT2.1.02、Matrix Science)を用いて、タンパク質同定を達成した。以下の判定基準を用いて、整合ペプチドをさらに評価した:i)最大0.05のp値に対応する確率ベースのMowseスコアを有するペプチドは、当該スコアが−logX10(log(P))でありそしてPが観察された整合が無作為事象である確率である場合、ポジティブに同定されるとみなされた;ii)特定タンパク質の同定に1つのペプチドだけが用いられ、そしてMASCOTスコアが30より低い場合、スペクトルの手動的立証が実施された。
【0071】
1.7 ICATに関するタンパク質同定
特に単一ペプチドヒットを有するものに関するICAT標識化タンパク質の同定における信頼を増大するために、付加的フィルターを以下のように適用した:i)ICAT対の重ペプチド及び軽ペプチドは、それらの抽出イオンクロマトグラムから測定されるような溶離ピークと密接に示されていなければならない;ii)単一の独特のペプチドを有するタンパク質に関しては、このペプチドは(例えば、異なるSDS−PAGE画分で、又は軽ICAT型及び重ICAT型の両方で)1回より多く同定されていなければならない;或いはiii)単一ペプチドが(ii)の判定基準を満たさない場合、MASCOTスコアは25以上でなければならず、期待値≦0.01及びMS/MSスペクトルは、強いイオンが説明されている「b」又は「y」型イオンの連続シリーズを示していなければならない。
【0072】
1.8 擬陽性の概算
擬陽性割当のレベルを別々に概算するために、タンパク質数、アミノ酸のサイズ及び分布に関する標準データベースに対してサイズで当該データベースを同定するよう、各タンパク質に関するアミノ酸配列の順序を逆にすることにより、ポルフィロモナス・ジンジバリスの逆データベースを作製した(34)。したがって擬陽性率は、NR/NF(ここで、NR=逆データベースで同定されるペプチドの数(逆データベースに関する閾値を上回るペプチドのMASCOTスコア)、並びにNF=標準データベースで同定されるペプチドの数(標準データベースに関する閾値を上回るペプチドのMASCOTスコア))と概算された。包括的プロテオーム分析(NF=18375ペプチド)及び定量的ICAT分析(NF=530ペプチド)から、擬陽性を測定した。
【0073】
1.9 相対的存在量の定量
同位体的に重い13C対軽い12C ICAT標識化ペプチドの比率を、DataAnalysis(Bruker Daltonics)からのスクリプトを用いて測定し、単一MSスペクトルでのモノアイソトピックピーク強度の測定値(シグナル強度及びピーク面積)に基づいて、手動で立証した。定量のために用いられる親イオンの最小イオン計数は2000であったが、96%を超える重及び軽前駆体イオンは10000を超えた。貧解析スペクトルの場合、親イオンの再構成抽出イオンクロマトグラム(EIC)の面積から、当該比率を決定した。単一親タンパク質由来の多ペプチドに関して平均を算定し、外れ値を、Grubb検定を用いて除外した(α=0.05)。
【0074】
1.10 ゲノム分析
CELLO(http://cello.life.nctu.edu.tw)(35)を用いてポルフィロモナス・ジンジバリスタンパク質の、そしてTIGRから得られる配列に基づいたTMHMM2.0(www.cbs.dtu.dk/services/TMHMM-2.0)を用いて膜貫通へリックスの細胞内局在性を予測した。理論的プロテオームと比較した場合の同定されたタンパク質の相対的発現レベルを概算するために、高度発現遺伝子と定義されているリボソームタンパク質及びtRNAシンターゼを伴うプログラムINteractive Codon Analysis 1.12a(http://www.bioinfo-hr.org/inca)(14)を用いて、genebank(ftp://ftp.ncbi.nih.gov/genbank/genomes/Bacteria/Porphyromonas_gingivalis_W83/)からのポルフィロモナス・ジンジバリスのコーディング配列に基づきCAI値を算定した。Microbesonlineのウェブサイト(http://microbesonline.org)(36)から、オペロン予測を実行した。
【0075】
1.11 ECR312突然変異体の構築
ポルフィロモナス・ジンジバリスW50オープンリーディングフレームPG1374は、潜在的に、ポルフィロモナス・ジンジバリスW83ゲノムを基礎にして免疫反応性47KDa抗原(PG97)をコードする(www.tigr.org)。ポルフィロモナス・ジンジバリスPG1374突然変異体を構築するために、プライマーECR312ApaI−For(5’−AGAGGGCCCTAGCAATCATTGCATTGCT−3’)及びECR312AatII−Rev(5’−TGCGACGTCGTGTTACCAATAGAGGATT−3’)を用いたPCRにより、W50ゲノムDNAから、フランキングApaI及びAatII制限部位(下線付き)を有するPG1374遺伝子の672bp上流断片を生成した。この断片を、サブクローン化ermFカセット(37)を含有するpAL30、pGem(登録商標)T−easy(Promega)上のAatII及びBamHI部位中でクローン化して、pAL36を作製した。同様に、フランキングPstI及びNdeI制限部位を有するPG1374の下流の565bp断片を、ECR312PstI−For2(5’−TGACTGCAGGCTTTCGACCTTGGATCTT−3’)及びECR312NdeI−Rev2(5’−TCGCATATGAAGAAATAAGTGCCGTCGG−3’)プライマーを用いて増幅して、pAL36中のPstI及びNdeI制限部位中にクローン化した。その結果生じたプラスミド(PG1374オープンリーディングフレームの上流及び下流断片が側面に位置するermFカセットを有する)(pAL36.1と呼ばれる)をScaIで直線化し、以前記載されたように(38)ポルフィロモナス・ジンジバリスW50中で形質転換した。10μg/mLのエリスロマイシンを含有するHBAプレート上で、嫌気性条件下で37℃で7日間のインキュベーション後、形質転換細胞を選択した。PCR分析によりDNA組込みの確認を実施して、その結果生じた突然変異体をECR312と名付けた。
【0076】
1.12 抗生物質防御侵襲検定
W50及びECR312の侵襲効率を比較するために、24ウエル細胞培養プレート中で、以前記載されたように(39)抗生物質防御検定を実行した。要するに、一定数のポルフィロモナス・ジンジバリス細胞をKB単層に侵襲させた(各ウエル中に約105個の細胞)。ゲンタマイシン(300μg/mL)及びメトロニダゾール(200μg/mL)とともに1時間さらにインキュベートすることにより、非侵襲細胞又は接着細胞を殺した。次に侵襲細菌のコロニー形成単位を、ウマ血液寒天プレート上で数えた。
【0077】
1.13 細胞結合検定
以前記載されたように(40)、細胞結合検定を実行した。要するに、ポルフィロモナス・ジンジバリスを先ず、中対数期に増殖させて、約2.9×109細胞/mLの細胞密度とした。次に細胞を洗浄し、その後、500μgのフルオレセインイソチオシアネート(FITC)(Invitrogen)で標識し、500μLのDMSO中に再懸濁した後、振盪しながら37℃で45分間インキュベートした。インキュベーション後、細胞をさらに洗浄し、不完全アール最小必須培地(JRH Biosciences)中に再懸濁して、FACSCaliberフローサイトメーター(Becton Dickinson, San Jose, CA)を用いた細胞計数に基づいて、ポルフィロモナス・ジンジバリス細胞を調整した。FITCの緑色発光を、525nmフィルター(FL1)を用いて測定した。CellQuestソフトウェア(Becton Dickinson, San Jose, CA)を用いて、多重パラメータデータを分析した。全測定を二重反復実験で実行し、FITC蛍光の定量に関しては、平均蛍光強度(MFI)値を用いた。
【0078】
野生型ポルフィロモナス・ジンジバリス及びECR312の結合を、37℃で40分間、5%CO2大気でKB細胞上に200μLの細胞懸濁液を接種することにより、並行して実行した。インキュベーション後、KB細胞及び細菌を含有する上清を、1.5mL管に移した。次に、残りの結合細胞を、37℃で5分間、200μLのトリプシン−EDTA混合物(JRH Bioscience)を用いてウエルから剥がし、対応する収集上清とともにプールした。次に500μLの完全EMEMを添加してトリプシンを不活性化し、その後、3回洗浄して、最後に1mLのPBS中に懸濁した。以前記載されたようにフローサイトメーターで、結合細胞を計数した。
【0079】
2. 結果及び考察
2.1 連続培養中のポルフィロモナス・ジンジバリスの増殖
過剰ヘムを含有する富栄養培地中で連続培養で増殖した場合、ポルフィロモナス・ジンジバリスW50は、2.03±0.04mgの細胞乾燥重量/mLの接種後約48時間で定常状態細胞密度を達成した。増殖培地中のヘミンの濃度が5.0μg/mlから0.1μg/mlに低減された場合、0.99±0.20mgの細胞乾燥重量/mLという有意に低い定常状態細胞密度が達成されたが、これは、ヘム利用能が増殖を制限していた、ということを実証する。ヘムが培養物に添加し戻された場合、細胞密度に及ぼすヘム制限の作用は軽減された。
【0080】
ヘム制限中のポルフィロモナス・ジンジバリスの増殖は、6.9時間のポルフィロモナス・ジンジバリス平均世代時間を有するケモスタットを用いた従来の研究(41,42)と一致した。ポルフィロモナス・ジンジバリスがPPIXを合成できないため、この必須栄養素はヘモグロビン及び血漿タンパク質を含むその他のヘムのタンパク質分解により獲得されると考えられ(9)、したがってヘムにおける欠損は有意に低い細胞密度に反映された。
【0081】
2.2 ヘム制限下で増殖されるポルフィロモナス・ジンジバリスのプロテオーム分析
2つの異なるアプローチにより、ヘム制限下で増殖されるポルフィロモナス・ジンジバリスのプロテオームを広範に分析した。溶液中消化とその後の気相分別を伴うLCMSを用いて、344種のタンパク質を同定した。geLCMSアプローチでは、385種のタンパク質を同定したが、一方、247種のタンパク質は両方のアプローチにより見出された。組合せ戦略を用いて、ポルフィロモナス・ジンジバリス逆データベースに対する検索から算定される0.4%という概算擬陽性率を伴って、総計478種のタンパク質を同定した(表1参照)。個々のLCMS実行から(異なるm/z範囲又はSDS PAGE帯域から)の2以上の独特のペプチドにより、又は2以上の同一ペプチドにより、全タンパク質の77.0%を同定した。478種の同定タンパク質は、全ゲノム分析(43)により同定される1988個の割当タンパク質コード遺伝子のうちの約25%を表わす。総予測プロテオームの4分の1を同定したが、この数字は、いずれかの増殖条件中に発現される遺伝子の実数を考慮する場合、より高くなる。緑膿菌及び枯草菌では、単一増殖条件中に転写される総ORFのパーセンテージは、それぞれ60%及び40%であると概算された(44、45)。それらの質量分光計のデューティーサイクル限界を調べることにより、ORFを予測されたポルフィロモナス・ジンジバリスの約60%がそれらの増殖条件下で発現される、ということをZhangと共同研究者ら(46)は概算した。したがってこれらの数字に基づいて、本発明の増殖条件下で発現されたポルフィロモナス・ジンジバリスタンパク質の41%〜62%が同定されている。478種の同定タンパク質の機能的分類を、表1に示す。
【0082】
【表1】
【0083】
今日まで、多元的プロテオミクス手法を用いて全体的にポルフィロモナス・ジンジバリスタンパク質を同定するための試みは、1つだけ報告されている。1014ポルフィロモナス・ジンジバリスATCC 33277タンパク質がこの菌株がケラチノサイト増殖培地中で培養され、同一培地が分泌された上皮細胞構成成分に曝露された場合に同定されたZhangと共同研究者らの研究と比較して、本発明者らは、ヘム制限下での連続培養中に発現される478種のポルフィロモナス・ジンジバリスW50タンパク質を同定した。用いられた菌株、増殖条件、並びにMSデータの処理は2つの試験で異なったので、2つのデータセット間で直接比較を行なうことは困難である。それにもかかわらず、本発明の方法では、75のタンパク質を独自に同定した。
【0084】
細胞内タンパク質局在化を予測するためにCELLOを用いて、この試験で同定されたタンパク質のほとんどが、細胞質(予測された1350のタンパク質のうち347)から、その後、ペリプラズム空間(48/113)、外膜(47/154)、内膜(24/256)及び細胞外(12/35)由来であると予測された。予測通り、低パーセンテージの予測内膜タンパク質を同定した。内膜タンパク質の予測信頼度をさらに増大するために、膜貫通隠れマルコフモデル(TMHMM)を用いた。TMHMMアプローチを用いて、2を超える膜貫通ドメイン(TMD)を有すると予測された242のタンパク質のうちの20が同定され、そして10を超えるTMDを有すると予測された44のタンパク質のうちの5つが同定された。注目すべきは、10を超える膜貫通ドメインを有する12の膜タンパク質のすべてが溶液中消化法から同定された。
【0085】
2.3 同定タンパク質の有意性
多数の細菌中での高度発現遺伝子は、コドン使用に関する強い組成の偏りを有することが多い。コドン使用頻度指数(CAI)を用いて、そのコドン配列を基礎にした遺伝子の発現レベルを予測することができ、CAI値が高いということは発現が増大したことを示す(47)。100を超えるコドンを有するポルフィロモナス・ジンジバリスゲノム中の1685遺伝子のうち、ほぼ92%が0.62〜0.80のCAI値を有する(図2)。この範囲は真核生物と比較して狭いが(48)、しかしそれは、予測遺伝子の89%が0.35〜0.50のCAI値を有する好熱性サーモアナエロバクター・テングコンゲンシス(Thermoanaerobacter tengcongensis)と類似する。この試験における478の同定タンパク質をコードする遺伝子のCAI値は、理論的プロテオームと類似の分布を有するが、より高いタンパク質存在量の検出への偏りがあった。この偏りにも拘わらず、非常に低いCAIを有する遺伝子によりコードされる多数のタンパク質が同定された。この結果は、明らかに、細胞中のタンパク質存在量の大きなダイナミックレンジという問題を例示し、このことは、すべてのタンパク質を一度に検出することが現在のところ不可能であることを示す。
【0086】
同定タンパク質の最高パーセンテージを有するタンパク質の機能的クラスは、エネルギー代謝に関与するもの(表1)、典型的には発酵(95%、CAI 0.70〜0.80)、解糖(82%、CAI 0.71〜0.83)、並びにアミノ酸及びアミンの代謝(81%、CAI 0.71〜0.84)に関与するものである。これは、細菌細胞中で非常に豊富であることが示されているエネルギー代謝のような基礎代謝機能に関与する必須タンパク質として大いに期待された。これらのタンパク質のほぼ90%が細胞質中に存在すると予測され、このことがさらにまた膜タンパク質と比較して検出を容易にさせる、ということは最も重要である。最少を示したタンパク質の機能的クラスは、仮説的タンパク質の転位及び調節機能に関与するものである。
【0087】
ポルフィロモナス・ジンジバリスの完全ゲノムはシーケンシングされているが、細胞機能に関する多数の重大な問題は依然として解決されていない。したがって翻訳遺伝子産物を同定するプロテオーム試験は、機能的ゲノムへの付加的洞察を提供するのに役立つ。例えばポルフィロモナス・ジンジバリスは糖非分解性であることが既知であるが(3)、しかしゲノムは、解糖経路の全酵素に関する推定ORFを含有する(43)。この経路が十分に利用されないことは、グルコキナーゼ(glucose kinase:グルコースキナーゼ)遺伝子が挿入エレメントにより妨害される原因となっている(43)。この知見と一致して、グルコキナーゼ、或いは別の解糖特異的酵素であるホスホフルクトキナーゼは同定されなかった。それに反して、糖新生に関与するすべての酵素が見出され、このことは、多糖生合成のようなプロセスに必要なグルコースがこの経路により得られ得る、ということを示唆した。
【0088】
2.4 ICAを用いて測定した場合のヘム制限に対するポルフィロモナス・ジンジバリスの応答
ヘム制限に対するポルフィロモナス・ジンジバリス応答の定量的ICAT分析を実行するために、溶液中ICAT法が強妨害三価イオン(strong interfering triply charged ions)の存在のため要求を満たさなかったので、geLCMSアプローチを選択した。これらの三価イオンの存在により、極少数のタンパク質同一性が生じた。したがって、SDS−PAGEによりICAT標識化可溶性及び不溶性タンパク質画分を別々に分離し、各ゲルレーンをゲル中トリプシン消化のために20断片に分けて、その後アフィニティー精製及びLCMS処理した。全体で142のタンパク質を同定した。逆データベースとの整合は得られなかったが、これは、低レベルの擬陽性同一性を示した。両画分中で検出されたタンパク質を考慮すると、多くて0.05のp値に対応する確率ベースのMowseスコアを有する2以上の独特のペプチドの存在に基づいて、53のタンパク質(34.0%)が同定され、2以上の異なる画分或いは両ICAT標識状態(それらのうちの58が25以上のMASCOTスコアを有する)から同定される1つの独特のペプチドの存在に基づいて60のタンパク質(38.5%)が同定され、そして25以上のMASCOTスコア、0.01以下の期待値、「b」又は「y」型イオンの連続シリーズを有する単一独自ペプチドに基づいて43のタンパク質(27.5%)が同定された(手動で妨害するのは強イオンのためである)。単一独自ペプチドの分析に基づいたタンパク質同一性の一例を、図3に示す。
【0089】
同定されたタンパク質のうち、103は可溶性画分中に見出され、53は不溶性画分中に見出され、そして14のタンパク質は両画分中に見出された。ヘム過剰からヘム制限への環境条件の変化に応答して、同定タンパク質のうちの70は存在量における少なくとも2倍の変化を示した(図4)。これらのうち、53のタンパク質の存在量は2倍より多く増大し、そして17のタンパク質の存在量はヘム制限中に2分の1未満に低減した。本発明のデータの再現性を査定するために、そして不溶性画分に関するタンパク質同一性を増大するために、ヘム制限及び過剰増殖中の異なる日に、ケモスタットから細胞を採取した。次に、異なる抽出方法とその後のICAT標識化を、変法プロトコル(示されていない)で反復した。得られたデータは、表2に示した選択タンパク質に関して類似の存在量比率を示すタンパク質に関して再現性があった(例えばPG0695 L/H=1.1、再標識化=1.3;PG0350 L/H=3.2、再標識化=2.6;PG0159 L/H=2.0、再標識化=2.2;PG0232 L/H=0.4、再標識化=0.4)。データをさらに検証するために、オペロンを形成する遺伝子によりコードされると予測される同定タンパク質の相対的存在量を比較した(50)。タンパク質の5つの群が、予測オペロンにより、又は特定の遺伝子座で分類される遺伝子によりコードされることが判明した(表2 影付)。各々の場合、コード化タンパク質の存在量は同様であると思われた。予測オペロンのうちの1つは、外膜タンパク質Omp40(PG0694)及びOmp41(PG0695)をコードし、その存在量は1.1の比率(ヘム制限/ヘム過剰、 L/E)で不変であった。これらのタンパク質は、グラム陰性細菌のOmpA様ポリンとの高い配列類似性を有し(51)、そして外膜とペプチドグリカン層との間の物理的連結を提供すると考えられる。これらの構造タンパク質は、環境ヘムレベルの変化を伴って存在量を変えるとは予期されない。残り4つの予測オペロンは、グルタメート又はアスパルテート異化作用と関連付けられることが判明した。
【0090】
【表2】
【0091】
2.5 宿主細胞侵襲関連タンパク質
宿主細胞の侵襲に関与すると思われる3つのタンパク質、インターナリン関連タンパク質(PG0350)、免疫反応性47kDaタンパク質(PG1374)及びエンドペプチダーゼPepO(PG0159)は、ヘム制限中の存在量がより高かった(表2)。抗生物質防御侵襲検定中、機能的PG1374を欠くポルフィロモナス・ジンジバリスは、野生型と比較して、上皮細胞中への約50%低い侵襲能力を有した(W50、32625±2582cfu/mL、ECR312、16250±1089cfu/mL;p<0.01、スチューデントT検定)。別個の結合検定において、野生型W50と比較した場合、PG1374突然変異体の付着において有意差は認められない(図5)。
【0092】
PG1374及びPG0350は、リステリア・モノサイトジェネス(L. monocytogenes)インターナリンタンパク質InlJと類似の富ロイシン反復ドメインを有するシステイン含有タンパク質の新規のクラスに属する(52)。リステリア・モノサイトジェネスにおいて、インターナリンファミリーの少なくとも15の成員が存在し、そしてすべてが、シグナルペプチド、N末端富ロイシン反復ドメインとその後の保存反復間領域から成る或る種の構造的特徴を共有することが判明している。これらのタンパク質の多くが、細胞侵襲過程に関与する(53)。多数のインターナリンがなぜ存在するかは実証されていないが、それらは、異なる細胞型への向性を付与するよう意図される(54)。
【0093】
最新の研究におけるヘム制限中のPG1374及びPG0350のより高い存在量(それぞれ6.5倍増及び3.2倍増)は、これら2つのタンパク質の発現が低ヘム増殖条件中に刺激されることを示唆する。配列情報及び予測構造から、インターナリンLRR残基の半分より多くが外側に面し、そして可変性であって、このことは、それらが異なるインターナリンクラスに特異的なタンパク質−タンパク質相互作用表面に関するものであることを示唆する(55)。PG1374及びPG0350はともに、シグナルペプチドを有し、そして約80残基の保存C末端ドメイン(CTD)は別として、有意の配列類似性を有しない34までの細胞表面に位置する外膜タンパク質の新規のクラスの一部である(56)。さらに、PG1374は、ヒト歯周炎患者からの血清で精査される場合、強免疫原性であって(57)、このことは、PG1374が細胞表面タンパク質相互作用に関与することをさらに示唆する。
【0094】
歯肉上皮細胞中へのポルフィロモナス・ジンジバリスのインターナリゼーションのプロセスは、フィムブリエ並びに種々の細胞表面プロテイナーゼにより媒介される付着及び侵襲の協調プロセスを包含すると考えられる(58〜60)。機能的推定インターナリン(PG0350)を欠くポルフィロモナス・ジンジバリス33277突然変異体は、野生型細菌と類似の侵襲特質を表示することが示されたが、バイオフィルム形成能力は低減された(46、61)。類似の侵襲は菌株33277による上皮細胞侵襲にも関与するフィムブリエの存在のためであると考えられたが、この突然変異体の類似の侵襲性は、ポルフィロモナス・ジンジバリスゲノムにおいてコードされた第二の推定インターナリンタンパク質(PG1374)の存在のためである、という可能性もあった。これら2つの推定インターナリンタンパク質のダブルノックアウトは、それらの考え得る協同的侵襲的役割を潜在的に解明する。
【0095】
ECR312による上皮細胞侵襲の50%低減並びに細胞結合において差異がないことは、PG1374が欠損したポルフィロモナス・ジンジバリスによる上皮細胞中への観察された侵襲低減がより少ない付着のためでなく、侵襲プロセスで実質的に欠損する、ということを明らかに実証する(図5)。細菌侵襲は、多数のタンパク質及び受容体が関与する高度複合プロセスであることが示されている(62)。ポルフィロモナス・ジンジバリス侵襲に関与する多数の因子の関連性は、フロリダ大学のLamontのグループにより実証されており、例としては、ハロ酸デハロゲナーゼ、エンドペプチダーゼ、陽イオン輸送ATPアーゼ、及びATP結合カセット輸送体が挙げられる(60、63)。したがって上皮細胞侵襲関連タンパク質としてのPG1374の発見は、細菌病原体によるこの複合宿主細胞侵襲プロセスに関与するタンパク質の一覧に加える。
【0096】
多数の細菌病原体に関しては、鉄利用能が病原性及び侵襲プロセスに影響を及ぼす、ということが十分に確立されているが、ポルフィロモナス・ジンジバリス侵襲遺伝子の発現に及ぼすヘムの影響についてはほとんど分かっていない。本発明者らは、μ−オキソビス・ヘム形態の鉄PPIXの保護層を欠くためであると思われる酸化ストレスからのポルフィロモナス・ジンジバリスの高死亡率のために、ヘム制限細胞に関する結合及び侵襲検定を実施できなかった(64)〜(66)。しかしながらこの試験において、推定侵襲関連タンパク質PcpO、PG0350及びPG1374のレベルはヘム制限下で増大し、そしてPG1374は細胞侵襲に関与する、ということを本発明者らは示した。
【0097】
上記のこれらの実験は、環境条件の変化に対するポルフィロモナス・ジンジバリスの応答の最初の定量的プロテオーム分析であり、完全プロテオーム分析と組合された安定同位体標識アプローチの有用性を実証する。ポルフィロモナス・ジンジバリスは、酸化ストレス応答、病原性及び宿主細胞の侵襲と関連付けられる多数のタンパク質の存在量により必須微量栄養素の制限に応答する。
【0098】
2.6 細胞表面に位置するタンパク質PG1019
ポルフィロモナス・ジンジバリス仮説的タンパク質PG1019は、細菌がヘム制限下で増殖された場合、25倍より豊富に存在することが観察された。バイオインフォマティック分析は、PG1019が、予測オペロン中の推定外膜受容体タンパク質(PG1020)をコードする遺伝子の直ぐ上流に位置する遺伝子によりコードされるリポタンパク質である、ということを示唆する。既知のポルフィロモナス・ジンジバリスTonB連結外膜受容体とのPG1020の多重アラインメント(示されていない)は、TonB連結受容体の特質の1つである推定TonBボックス(残基118〜126)(67)、並びにSimpson及び共同研究者ら(68)がTonBボックスIV領域と呼ぶ保存領域(残基236〜272)の存在を示す。TonB連結外膜受容体は、多数の鉄、鉄錯体及びその他の微量栄養素取込み系と関連付けられている。ヘム制限増殖条件下でのPG1019の高存在量は、このタンパク質が細胞中への鉄/鉄錯体の輸送に関与するTon−B連結系に対するアクセサリータンパク質であることと一致するが、これは未だ実証されていない。プロテオームデータのほかに、機能feoB1遺伝子を欠く突然変異体(ポルフィロモナス・ジンジバリスFB1)と比較されるポルフィロモナス・ジンジバリスW50のカスタムポルフィロモナス・ジンジバリスDNAマイクロアレイを用いるトランスクリプトーム(transcriptomic)分析を実施した。野生型W50及びFB1突然変異体をともに、ヘム過剰条件で連続培養で増殖させた。ポルフィロモナス・ジンジバリスFB1は、野生型W50の細胞鉄含量の約半分を有した(69)。したがって転写の増大を示す遺伝子は、細胞内又は環境鉄含量の低減に応答して上方調節されると思われる。PG1019及びPG1020はともに、野生型と比較して、FB1突然変異体において類似のレベルに有意に上方調節された。生物学的複製においてP<0.05の有意レベルでPG1019は2.46のLog2増大を示し、そしてPG1020は2.33のLog2増大を示し、これは、これらの遺伝子がオペロン中に存在する、というさらなる証拠である。さらに、ポルフィロモナス・ジンジバリスW50の別個のトランスクリプトームDNAマイクロアレイ分析は、ヘム過剰増殖中にPG1019遺伝子の発現がほとんど又は全く認められない、ということを示した。
【0099】
2.7 アルキルヒドロペルオキシドレダクターゼタンパク質AhpC(PG0618)
ポルフィロモナス・ジンジバリスにおけるペルオキシド耐性に重要な役割を果たすことが示されているペルオキシド除去酵素であるアルキルヒドロペルオキシドレダクターゼタンパク質AhpC(PG0618、表2)を用いて、ヘム過剰からヘム制限への移行中のポルフィロモナス・ジンジバリスのタンパク質存在量における最も実質的な変化を観察した(65)。ポルフィロモナス・ジンジバリスにおいて、細胞表面に酸素を有するμ−オキソビス・ヘム形態の鉄PPIXの層の形成は、その固有のカタラーゼ様活性のために酸化緩衝剤として作用すると考えられる(11)。この層は、鉄及びPPIXの細胞表面貯蔵にも役立ち得る(70)。ヘム制限中、鉄PPIXのこの供給源の枯渇は、酸化ストレスに対する感受性増大を生じることが示された(64)。したがってヘム制限中のアルキルヒドロペルオキシドレダクターゼの存在量の実質的増大は、μ−オキソビスヘム層の非存在/低減により引き起こされる酸化ストレス増大に応答し得た。OxyR(酸素感受性転写活性剤)も、嫌気性増殖中のアルキルヒドロペルオキシドの発現において関与するが(71)、この場合、ポルフィロモナス・ジンジバリスOxyR−突然変異体は、遺伝子発現の16分の1の低減を示す。さらに近年、Duran-Pinedo及び共同研究者らも、RprY応答レギュレーターによるaphC発現の正の調節を実証した。したがって存在量の実質的増大は、ヘム利用能がRprY及びOxyR制御遺伝子発現において関与し得る、ということを示唆する。興味深いことに、このタンパク質の非常に高いコドン使用頻度指数(CAI)値(0.838)は、このタンパク質が、このようなストレスに応答して迅速誘導のために細胞中で高度に発現され得る、ということを示唆する。
【0100】
本明細書全体を通して、「〜を含む」という単語、或いは「〜を含む(単数)(comprises)」又は「〜から成る(comprising)」という変形は、記述された素子、整数又は工程、或いは素子、整数又は工程の群を包含することを意味するが、任意の他の素子、整数又は工程、或いは素子、整数又は工程の群の排除を意味しないと理解される。
【0101】
本明細書中で言及された出版物はすべて、参照により本明細書中で援用される。本明細書中に包含されている文書、行為、材料、装置、論文等についての任意の考察は、単に、本発明に一情況を提供するためにある。これらの事柄のいずれか又はすべてが従来技術基礎の一部を構成する、或いは本出願の各特許請求の優先日前にどこかに存在したので、本発明に関連する分野における共通の一般知識であった、ということを認めるものと解釈されるべきでない。
【0102】
広範に記載したような本発明の精神又は範囲を逸脱しない限り、特定の実施形態に示したように、多数の変更及び/又は修正が本発明に対して成され得る、と当業者は理解する。したがって本発明の実施形態は、すべての点において例示であって、本発明を限定するものではない、と考えられるべきである。
引用文献
【0103】
【表3】
【特許請求の範囲】
【請求項1】
単離抗原性ポルフィロモナス・ジンジバリスポリペプチドであって、以下の:
(i)配列番号1のアミノ酸配列を有するPG0350タンパク質;
(ii)配列番号2のアミノ酸配列を有するPG1374タンパク質;
(iii)配列番号3のアミノ酸配列を有するPG1019タンパク質;
(iv)配列番号4のアミノ酸配列を有するPG0618タンパク質;
(v)配列番号1若しくは配列番号2若しくは配列番号3若しくは配列番号4のアミノ酸配列と少なくとも85%同一であるアミノ酸配列;又は
(vi)配列番号1若しくは配列番号2若しくは配列番号3若しくは配列番号4の連続アミノ酸配列と同一の少なくとも10個のアミノ酸を含むアミノ酸配列
から成る群から選択される、単離抗原性ポルフィロモナス・ジンジバリスポリペプチド。
【請求項2】
配列番号1又は配列番号2又は配列番号3又は配列番号4の前記アミノ酸配列と90%、95%、96%、97%、98%、99%又は100%同一である、請求項1に記載のポリペプチド。
【請求項3】
配列番号1又は配列番号2又は配列番号3又は配列番号4の連続アミノ酸配列と同一である少なくとも20個、30個、40個、50個、60個、70個、80個、90個又は100個のアミノ酸の連続配列を含む、請求項1に記載のポリペプチド。
【請求項4】
被験体においてポルフィロモナス・ジンジバリスに向けられる免疫応答を惹起するのに用いるためのワクチン組成物であって、有効量の請求項1〜3のいずれか一項に記載の少なくとも1つのポリペプチド、並びに薬学的に許容される担体を含む、ワクチン組成物。
【請求項5】
歯周病に関して被験体を予防するか又は治療する方法であって、請求項4に記載のワクチン組成物を前記被験体に投与することを包含する、歯周病に関して被験体を予防するか又は治療する方法。
【請求項6】
被験体における歯周病の治療のための薬剤の製造における請求項1〜3のいずれか一項に記載の少なくとも1つのポリペプチドの使用。
【請求項7】
前記被験体がヒト、ヒツジ、畜牛、ウマ、ウシ、ブタ、家禽、イヌ及びネコから成る群から選択される、請求項9に記載の方法。
【請求項8】
前記被験体がヒトである、請求項10に記載の方法。
【請求項9】
請求項1〜3のいずれか一項に記載のポリペプチドと特異的に結合する抗体。
【請求項10】
ポリクローナル抗体である、請求項9に記載の抗体。
【請求項11】
モノクローナル抗体である、請求項9に記載の抗体。
【請求項12】
歯周病の予防又は治療に有用な組成物であって、請求項9〜11のいずれか一項に記載の抗体並びに薬学的に許容される担体を含む、歯周病の予防又は治療に有用な組成物。
【請求項13】
歯周病の進行に関与するポルフィロモナス・ジンジバリスポリペプチドの同定方法であって、以下の:
a)ヘム制限条件下で増殖されるポルフィロモナス・ジンジバリスにより産生されるポリペプチド又はそのペプチドの相対量を測定する工程;並びに
b)工程a)より高いヘム条件下で増殖されるポルフィロモナス・ジンジバリスにより産生されるポリペプチド又はそのペプチドの相対量を測定する工程
を包含する方法であり、工程b)と比較した場合の工程a)で検出される前記ポリペプチド又はそのペプチド断片の量の増大が前記ポリペプチドが歯周病の進行に関与するということを示す、歯周病の進行に関与するポルフィロモナス・ジンジバリスポリペプチドの同定方法。
【請求項14】
前記工程(a)のヘム制限条件でのヘミン濃度が約0.1μg/mL〜約0.5μg/mLである、請求項13に記載の方法。
【請求項15】
前記工程(b)のより高いヘム条件でのヘミン濃度が5μg/mLを超える、請求項13又は14に記載の方法。
【請求項16】
前記工程(a)のヘム制限条件における並びに前記工程(b)のより高いヘム条件でのポリペプチドの相対量が定性プロテオミクス手法を用いて測定される、請求項13〜15のいずれか一項に記載の方法。
【請求項17】
ポリペプチドの前記量が溶液中及びゲル中消化並びにLC−MS/MSの組合せ戦略を用いて、或いはMSと組合せた安定同位体標識戦略(ICAT)を用いた分析により測定される、請求項16に記載の方法。
【請求項18】
各鎖中に少なくとも19塩基対の二本鎖領域を含む干渉RNA分子であって、前記二本鎖領域の鎖のうちの1つが配列番号5又は配列番号6又は配列番号7又は配列番号8の領域と相補的である、各鎖中に少なくとも19塩基対の二本鎖領域を含む干渉RNA分子。
【請求項19】
前記干渉RNAの前記二本鎖領域が少なくとも20塩基対、好ましくは少なくとも25塩基対、最も好ましくは少なくとも30塩基対を前記二本鎖領域の各鎖中に含む、請求項18に記載の干渉RNA分子。
【請求項20】
歯周病に関する被験体の治療方法であって、請求項18又は請求項19に記載の少なくとも1つの干渉RNA分子を前記被験体に投与することを包含する、歯周病に関する被験体の治療方法。
【請求項1】
単離抗原性ポルフィロモナス・ジンジバリスポリペプチドであって、以下の:
(i)配列番号1のアミノ酸配列を有するPG0350タンパク質;
(ii)配列番号2のアミノ酸配列を有するPG1374タンパク質;
(iii)配列番号3のアミノ酸配列を有するPG1019タンパク質;
(iv)配列番号4のアミノ酸配列を有するPG0618タンパク質;
(v)配列番号1若しくは配列番号2若しくは配列番号3若しくは配列番号4のアミノ酸配列と少なくとも85%同一であるアミノ酸配列;又は
(vi)配列番号1若しくは配列番号2若しくは配列番号3若しくは配列番号4の連続アミノ酸配列と同一の少なくとも10個のアミノ酸を含むアミノ酸配列
から成る群から選択される、単離抗原性ポルフィロモナス・ジンジバリスポリペプチド。
【請求項2】
配列番号1又は配列番号2又は配列番号3又は配列番号4の前記アミノ酸配列と90%、95%、96%、97%、98%、99%又は100%同一である、請求項1に記載のポリペプチド。
【請求項3】
配列番号1又は配列番号2又は配列番号3又は配列番号4の連続アミノ酸配列と同一である少なくとも20個、30個、40個、50個、60個、70個、80個、90個又は100個のアミノ酸の連続配列を含む、請求項1に記載のポリペプチド。
【請求項4】
被験体においてポルフィロモナス・ジンジバリスに向けられる免疫応答を惹起するのに用いるためのワクチン組成物であって、有効量の請求項1〜3のいずれか一項に記載の少なくとも1つのポリペプチド、並びに薬学的に許容される担体を含む、ワクチン組成物。
【請求項5】
歯周病に関して被験体を予防するか又は治療する方法であって、請求項4に記載のワクチン組成物を前記被験体に投与することを包含する、歯周病に関して被験体を予防するか又は治療する方法。
【請求項6】
被験体における歯周病の治療のための薬剤の製造における請求項1〜3のいずれか一項に記載の少なくとも1つのポリペプチドの使用。
【請求項7】
前記被験体がヒト、ヒツジ、畜牛、ウマ、ウシ、ブタ、家禽、イヌ及びネコから成る群から選択される、請求項9に記載の方法。
【請求項8】
前記被験体がヒトである、請求項10に記載の方法。
【請求項9】
請求項1〜3のいずれか一項に記載のポリペプチドと特異的に結合する抗体。
【請求項10】
ポリクローナル抗体である、請求項9に記載の抗体。
【請求項11】
モノクローナル抗体である、請求項9に記載の抗体。
【請求項12】
歯周病の予防又は治療に有用な組成物であって、請求項9〜11のいずれか一項に記載の抗体並びに薬学的に許容される担体を含む、歯周病の予防又は治療に有用な組成物。
【請求項13】
歯周病の進行に関与するポルフィロモナス・ジンジバリスポリペプチドの同定方法であって、以下の:
a)ヘム制限条件下で増殖されるポルフィロモナス・ジンジバリスにより産生されるポリペプチド又はそのペプチドの相対量を測定する工程;並びに
b)工程a)より高いヘム条件下で増殖されるポルフィロモナス・ジンジバリスにより産生されるポリペプチド又はそのペプチドの相対量を測定する工程
を包含する方法であり、工程b)と比較した場合の工程a)で検出される前記ポリペプチド又はそのペプチド断片の量の増大が前記ポリペプチドが歯周病の進行に関与するということを示す、歯周病の進行に関与するポルフィロモナス・ジンジバリスポリペプチドの同定方法。
【請求項14】
前記工程(a)のヘム制限条件でのヘミン濃度が約0.1μg/mL〜約0.5μg/mLである、請求項13に記載の方法。
【請求項15】
前記工程(b)のより高いヘム条件でのヘミン濃度が5μg/mLを超える、請求項13又は14に記載の方法。
【請求項16】
前記工程(a)のヘム制限条件における並びに前記工程(b)のより高いヘム条件でのポリペプチドの相対量が定性プロテオミクス手法を用いて測定される、請求項13〜15のいずれか一項に記載の方法。
【請求項17】
ポリペプチドの前記量が溶液中及びゲル中消化並びにLC−MS/MSの組合せ戦略を用いて、或いはMSと組合せた安定同位体標識戦略(ICAT)を用いた分析により測定される、請求項16に記載の方法。
【請求項18】
各鎖中に少なくとも19塩基対の二本鎖領域を含む干渉RNA分子であって、前記二本鎖領域の鎖のうちの1つが配列番号5又は配列番号6又は配列番号7又は配列番号8の領域と相補的である、各鎖中に少なくとも19塩基対の二本鎖領域を含む干渉RNA分子。
【請求項19】
前記干渉RNAの前記二本鎖領域が少なくとも20塩基対、好ましくは少なくとも25塩基対、最も好ましくは少なくとも30塩基対を前記二本鎖領域の各鎖中に含む、請求項18に記載の干渉RNA分子。
【請求項20】
歯周病に関する被験体の治療方法であって、請求項18又は請求項19に記載の少なくとも1つの干渉RNA分子を前記被験体に投与することを包含する、歯周病に関する被験体の治療方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】

【図7】

【図8】

【図9】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2009−544279(P2009−544279A)
【公表日】平成21年12月17日(2009.12.17)
【国際特許分類】
【出願番号】特願2009−516822(P2009−516822)
【出願日】平成19年6月27日(2007.6.27)
【国際出願番号】PCT/AU2007/000890
【国際公開番号】WO2008/000028
【国際公開日】平成20年1月3日(2008.1.3)
【出願人】(508377141)オーラル ヘルス オーストラリア ピーティーワイ リミテッド (1)
【Fターム(参考)】
【公表日】平成21年12月17日(2009.12.17)
【国際特許分類】
【出願日】平成19年6月27日(2007.6.27)
【国際出願番号】PCT/AU2007/000890
【国際公開番号】WO2008/000028
【国際公開日】平成20年1月3日(2008.1.3)
【出願人】(508377141)オーラル ヘルス オーストラリア ピーティーワイ リミテッド (1)
【Fターム(参考)】
[ Back to top ]