
毛成長休止期短縮剤
【課題】毛成長サイクルの休止期を規程する内在性因子を決定し、当該内在性因子の活性もしくは発現を阻害する物質のスクリーニング、毛成長休止期短縮剤又は抜け毛防止剤の提供。
【解決手段】毛包に内在するFGF18は毛の成長サイクルの休止期を規程し、脱落期を開始させ、さらにヘアサイクルドメインを規定する制御因子である。FGF18活性阻害物質又はFGF18遺伝子発現阻害物質をスクリーニングして、毛成長休止期短縮剤、抜け毛防止剤及び毛並み調整剤の候補とする方法。また、FGF18の部分ペプチド、抗FGF18抗体などのFGF18活性阻害物質及び/又は発現抑制物質を有効成分として含有する毛成長休止期短縮剤、抜け毛防止剤及び毛並み調整剤。
【解決手段】毛包に内在するFGF18は毛の成長サイクルの休止期を規程し、脱落期を開始させ、さらにヘアサイクルドメインを規定する制御因子である。FGF18活性阻害物質又はFGF18遺伝子発現阻害物質をスクリーニングして、毛成長休止期短縮剤、抜け毛防止剤及び毛並み調整剤の候補とする方法。また、FGF18の部分ペプチド、抗FGF18抗体などのFGF18活性阻害物質及び/又は発現抑制物質を有効成分として含有する毛成長休止期短縮剤、抜け毛防止剤及び毛並み調整剤。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、毛の成長サイクルにおける休止期を短縮すること及び脱落期の開始を遅らせることによる抜け毛の予防又は治療を目的とするものであり、毛髪が成長休止することを規定している物質の阻害剤及びそのスクリーニング方法に関する。
【背景技術】
【0002】
抜け毛、脱毛は遺伝的な要素が大きいといわれるものの、加齢に伴って加速されるため、男性のみならず女性においても悩む人は多い。脱毛症には加齢男性で多く発症する男性型脱毛症、女性で問題となる事の多いびまん性脱毛症、男女ともに頻度の高い円形脱毛症、さらに抗癌剤など細胞毒性のある薬剤投与によって引き起こされる脱毛症などがある。
これらの問題を解決すべく、従来から発毛効果、育毛効果のある物質として多数の物質が見出され提案されており、既に商品化されている物質も多いが、科学的な臨床実験で発毛効果が確認された育毛剤として「男性型脱毛症診療ガイドライン(2010版)」で推奨された薬剤は、血管拡張作用によって発毛を促すミノキシジル(商品名ロゲイン、リアップ)及び頭皮における男性ホルモン作用を抑制する内服薬(商品名プロペシア)のみである。
男性型脱毛症(AGA)の場合、その主要な原因の1つは、男性ホルモンであるテストステロンが5αリダクターゼの作用でジヒドロテストステロン(DHT)である。DHTが乳毛頭細胞上の男性ホルモン受容体(AR)に結合すると、それによって毛包の矮小化プロセスが始まると説明されているが、その分子機構の詳細は依然として不明である。一説によればDHTによる毛包の矮小化には、細胞増殖因子関連分子であるTGF-β1を介した細胞のアポトーシスが関与しているとされる。通常の毛髪成長サイクルによれば2〜6年は維持されるはずの毛包細胞がアポトーシスにより死滅してしまえば、毛髪は次々と休止期に移行することになる。したがって、テストステロンをDHTに変換する反応を抑制する内服薬の服用は、男性ホルモン過多の傾向にある男性の男性型脱毛症の予防には効果的であるとしても、DHTを抑制してしまうことの副作用を懸念する見方もあり、また男性ホルモン過多が原因となる典型的なAGA以外の脱毛症には効果がない。
ところで、上記のように説明されている男性型脱毛症以外の、びまん性脱毛症、男女ともに頻度の高い円形脱毛症、さらに抗癌剤などの薬剤投与によって引き起こされる脱毛症などにおいては、脱毛症の発症のメカニズムも多様であり、そこに関与する分子も、上述の男性型脱毛症におけるTGF-β1などの細胞増殖因子関連分子、円形脱毛症における免疫細胞関連サイトカインやケモカイン、毛包細胞自体を死滅させてしまう抗癌剤、などの他に、様々な分子があり、それら分子を産生・受容する多種の細胞が関与する。
したがって、根本的な脱毛防止のためには毛髪の成長サイクルにおける詳細な検討が欠かせない。
【0003】
毛の成長サイクル(図1)は、毛を作り出す器官である毛包自体の周期的な変化に規定されている。すなわち毛包は、成長期、退行期、休止期、という3つの相をこの順に繰り返す。このうち成長期には、毛包の先端が皮膚よりも深部の皮下脂肪層にまで到達すると共に、大きく太くなりつつ、長い毛軸を形成し、これを体外に長く伸ばす。これが外観的な毛の成長である。退行期は、毛包の構成細胞がアポトーシスを起こすことに伴い、毛包が退縮し、毛包の先端は皮下脂肪層よりも皮膚表面に近い真皮内部にまで移動する。次に来る休止期においては、毛包の構成細胞はほとんど増殖もアポトーシスも起こさず、小さい状態で静止すると考えられている。これら退行期、休止期において、外観的な毛の成長は止まるが、毛が抜けるわけではない。毛が抜けることは、脱毛期と称される独立の相で独立の制御により起こると考えられているが、脱毛期は、上記の三相と完全に無関係ではなく、休止期後期や次の成長期の途中で起こることが多いとされている。しかし病的な脱毛などでは、これよりも早く起こることもあり、その場合には、外観的にまったく毛の生えていない毛包が長期間存在することになる。この脱毛期を制御する分子メカニズムは、これまでほとんどわかっていない。これらの理由により、毛髪の成長周期を説明する場合に、成長期、退行期、休止期の三相は必ず説明の対象となるが、脱毛期は省略される場合も多い。動物の種によって毛の成長サイクルの各相の持続時間は異なり、同一種内でも、個体、性別、体の部位や毛の種類によっても異なるが、その成長のサイクルは、いずれも図1に示される各相を経由している。ヒトの頭髪の場合は個人差、年齢差はあるが、一般に、成長期は女性で4〜8年間、男性で3〜6年間、退行期は1〜2週間、休止期は3ヶ月間程度とされている。毛の研究に多用されるモデル動物であるマウスの場合、その背部皮膚に存在する毛包においては、成長期は18〜19日間、退行期は2日間、休止期は3〜5週間程度である。
一般的な脱毛症とは、外観的に毛が無い状態、または毛が非常に細く短い状態を意味するので、その原因を毛髪の成長サイクルから見た場合に、脱毛は毛の成長が不十分なこと、または、脱毛が正常よりも早期にまたは過剰に起こっていること、または、前の周期で形成された毛が脱落した後も次の毛の成長が始まらないこと、といえるので、その原因を、(1)成長期の短縮、(2)休止期の延長、(3)脱毛期の早期開始などにわけることができる。毛の成長サイクルにおけるそれぞれの期間を規定する因子として、従来成長期を維持する細胞増殖因子Wntファミリータンパク質(非特許文献1)、退行期を誘導する細胞増殖因子FGF5(非特許文献2)が知られていたが、休止期の維持や制御に係る因子や脱毛期の制御に関わる因子に関する詳細な研究はほとんどなされていなかった。
したがって、脱毛症患者それぞれの脱毛症の原因に個別に対応するためには、毛の成長サイクルの各相で作用する因子の働きを特定する必要がある。また、成長サイクル全体で見れば比較的長時間を要する休止期間を短縮することができれば、完全な抜け毛が起こる前に新しい毛髪が生えることになり、脱毛自体の防止が可能となり、毛髪全体の量も増えいわゆる薄毛予防ともなる。さらに、解明の進んでいない脱毛期を制御する因子が解明できれば、直接的に脱毛時期を遅らせることができることになり、脱毛症患者にとっての大きな福音となる。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】WO2008/102782
【特許文献2】WO2008/102783
【特許文献3】特開平4-224522号公報
【非特許文献】
【0005】
【非特許文献1】Shimizu,H, et al.,J.Invest.Dermatol.122,239-245(2004)
【非特許文献2】Hebert,J.M., et al.,Cell78,1017-1025(1994)
【非特許文献4】Kawano,M., et al.,J. Invest.Dermatol.124,877-885(2005)
【非特許文献5】Ohbayashi,N., et al.,Genes & Dev.16,870-879(2002)
【非特許文献6】Tarutani,M., et al.Proc Natl Acad Sci USA 94,7400-7405(1997)
【非特許文献7】Ornitz DM,Itoh N:Fibroblast growth factors.Genome Biol 2:REVIEWS3005,2001
【非特許文献8】du Cros DL:Fibroblast growth factor and epidermal growth factor in hair development.J Invest Dermatol 101:106S-113S.1993
【非特許文献9】du Cros DL, Isaacs K, Moore GP:Distribution of acidic and basic fibroblast growth factorsin ovine skin during follicle morphogenesis. J Cell Sci 105:667-674,1993
【非特許文献10】Hebert JM,R osenquist T,Gotz J,Martin GR:FGF5 as a regulator of the hair growth cycle:Evidence from targeted and spontaneous mutations.Cell 78:1017-1025,1994
【非特許文献11】Danilenko DM,Ring BD,Yanagihara D,Benson W,Wiemann B,Starnes CO,Pierce GF:Keratinocyte growth factor is an important endogenous mediator of hair follicle growth,development, and differentiation.American J Pathol 147:145-154,1995
【非特許文献12】Marchese C,Chedid M,Dirsch OR, et al:Modulation of keratinocyte growth factor and its receptor in reepithelializing human skin.J Exp Med 182:1369-1376,1995
【非特許文献13】Guo L,Degenstein L,Fuchs E:Keratinocyte growth factor is required for hair development but not for wound healing.Genes Dev 10:165-175,1996
【非特許文献14】Rosenquist TA,Martin GR:Fibroblast growth factor signalling in the hair growth cycle:Expression of the fibroblast growth factor receptor and ligand genes in the murine hair follicle.Developmental Dynamics 205:379-386,1996
【非特許文献15】Petho-Schramm A,Muller HJ,Paus R:FGF5 and the murine hair cycle.Arch Dermatol Res 288:264-266,1996
【非特許文献16】Mitsui S,Ohuchi A,Hotta M,Tsuboi R,Ogawa H:Genes for a range of growth factors and cyclin-dependent kinase inhibitors are expressed by isolated human hair follicles.Br J Dermatol 137:693-698,1997
【非特許文献17】Ortega S,Ittmann M,Tsang SH,Ehrlich M,Basilico C:Neuronal defects and delayed wound healing in mice lacking fibroblast growth factor-2.Proc Natl Acad Sci USA 95:5672-5677,1998
【非特許文献18】Suzuki S,Kato T,Takimoto H,et al:Localization of rat FGF-5 protein in skin macrophage-like cells and FGF-5S protein in hair follicle:Possible involvement of two Fgf-5 gene products in hair growth cycle regulation.J Invest Dermatol 111:963-972,1998
【非特許文献19】Suzuki S,Ota Y,Ozawa K,Imamura T:Dual-mode regulation of hair growth cycle by two Fgf-5 gene products.J Invest Dermatol 114:456-463,2000
【非特許文献20】Nakatake Y, Hoshikawa M, Asaki T, Kassai Y, Itoh N:Identification of a novel fibroblast growth factor,FGF-22,preferentially expressed in the inner root sheath of the hair follicle.Biochem Biophys Acta 1517:460-463,2001
【非特許文献21】Stenn KS,Paus R:Controls of hair follicle cycling.Physiol Rev 81:449-494,2001
【非特許文献22】Beyer TA, Werner S, Dickson C, Grose R:Fibroblast growth factor 22 and its potential role during skin development and repair.Exp Cell Res 287:228-236,2003
【非特許文献23】Kawano M, Suzuki S, Suzuki M, Oki J, Imamura T:Bulge- and basal layer-specific expression of fibroblast growth factor 13(FHF-2) in mouse skin.J Invest Dermatol 122:1084-1090,2004
【非特許文献24】Rendl, M., Polak, L. & Fuchs, E.BMP signaling in dermal papilla cells is required for their hair follicle-inductive properties.Genes & Dev.22,543-557(2008)
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、毛成長サイクルにおける休止期を長期間に制御する内在性因子を決定し、その作用機構を解明すると共に、脱毛期を制御する内在性因子を決定し、当該内在性因子の活性もしくは発現を阻害する物質をスクリーニングすることで、新規な毛成長休止期短縮剤、脱毛抑制剤を提供することを目的としている。
また、そのためのスクリーニング方法も本発明の他の目的である。
【課題を解決するための手段】
【0007】
マウス及びヒトの皮膚組織及び毛包細胞には、22種類の繊維芽細胞増殖因子(FGF)(非特許文献7)のうちで、FGF1、FGF2、FGF5、FGF7、FGF10、FGF13及びFGF22が発現しており、毛髪成長及び皮膚再生を制御していることが報告されている(非特許文献8〜23、特許文献3参照)。これらの文献内容から見て、各種FGFには皮膚細胞の増殖及び分化に重要な役割を担っていることが示唆されるが、毛包の成長を促進及び休止する作用、それに伴う毛髪成長期及び休止期制御作用及び脱毛促進作用に対して、FGF群の個々の因子がそれぞれどのように関与するのかといった知見は依然として多くは不明のままである。
上記の背景にあって発明者らは以前、FGF18が毛包を含む皮膚で高いレベルで発現することを見出し、毛成長周期の休止期で特に高い発現レベルであることを報告した(非特許文献4)。そして、FGF18をマウス休止期皮膚毛包において、抜毛により強制的に毛包の成長期を誘導したあとに、FGF18を背部皮下に持続的に存在させることで、毛の成長を著しく阻害することを見いだし、FGF18及びその活性化物質からなる毛髪成長阻害剤、及びFGF18阻害物質からなる発毛剤について特許出願している(特許文献1,2)。このように、毛包細胞に外部から導入したFGF18遺伝子産物の毛成長に及ぼす影響については解明されてきているが、毛包細胞に内在しているFGF18遺伝子が毛包細胞内でどのようなメカニズムで発現し、毛の成長周期にどのように係わっているのか、という遺伝子機能についてはよくわかっていなかった。
内在遺伝子の機能を直接的に解明する手法として遺伝子ノックアウト技術があるが、FGF18は、軟骨細胞や骨芽細胞の増殖分化や肺の形成など、個体形成や生命維持に不可欠な因子であるため、通常の全身ノックアウトを行うと、個体の機能的形成が正常に起こらず、胎生期または出生直後に死亡することが知られている(非特許文献5)。
そこで、本発明者らは、皮膚とその付属器官においては毛包で選択的に高いレベルで発現しているFGF18遺伝子を毛包特異的にノックアウトすることを想起し、毛包細胞がケラチン細胞の1種であることから、ケラチン5陽性細胞特異的な遺伝子ノックアウト法を、フォスファチジルイノシトールグリカンアンカー合成に関わるPig-a遺伝子に対して適用した技術(非特許文献6)を参考にし、ケラチン5陽性細胞特異的にFgf18遺伝子をノックアウトしたマウスを得た。ここで、ケラチン5は、ケラチン14と同様に、皮膚と毛包など皮膚付属器官の中では表皮、毛包、皮脂腺など多くの細胞で発現しているがFgf18を高いレベルで発現するのは、この中で毛包だけであるため、得られたコンディショナルノックアウトマウスは、実質的に「毛包特異的Fgf18遺伝子欠損マウス」と表現することができる。
毛包特異的FGF18欠損マウスは、健康に育ち、生殖能力も備えており、外観的に検知しうる障害を有さないが、驚くべきことに、毛の成長サイクルの進行の状態が、野生型と著しく異なることを確認した。すなわち、通常の野生型マウスでは3〜5週間以上も続く毛の成長サイクルの休止期が、毛包特異的FGF18欠損マウスでは約1週間で終了し、一周期に要する期間も約3週間にまで短縮されていた。また、毛包内のクラブヘアの脱落が防止され、脱落期の開始時期が遅くなった結果、毛包あたりの毛軸数が全体で2倍以上増加した。しかも、毛成長周期の進行が体表上の「毛周期ドメイン」構造の影響を受けにくく極めてスムーズに進行し、加齢が進むにつれ複数の縞模様をなすような毛成長フェーズの繰り返しが個体皮膚に現れる。このため、毛包特異的FGF18欠損マウスを用いれば、野生型のマウスにおいては同一個体皮膚表面で再現性良く行うことが極めて困難であった毛髪関連疾患や各種皮膚疾患の予防、治療物質あるいは発毛や脱毛を促進・抑制する物質の評価・スクリーニングを行うことができることを見出し、本出願と同日付で特許出願した。
【0008】
そしてこれらの結果から見て、毛包内の内在性FGF18には、毛の成長サイクルの休止期を延長させることで、毛の成長サイクル自体を延長させる作用と共に、毛包内のクラブヘアを脱落させるという脱毛期開始作用があるといえることから、FGF18こそが休止期延長制御因子であり、かつ脱毛促進因子であることが明らかとなった。
したがって、毛包内の内在性FGF18遺伝子の発現を抑える物質又はFGF18蛋白質の活性を抑える物質を毛包に投与することで、毛の成長サイクルの休止期を短縮して毛の成長サイクル全体の速度を速めることができ、また脱毛、抜け毛を防止することができる。つまり、毛包内の内在性FGF18遺伝子の発現を抑える物質又はFGF18蛋白質の活性を抑える物質を検索することで、毛の成長サイクルの休止期短縮剤であり、脱毛防止剤を提供できることを見出した。
以上の知見を得て、本発明を完成させた。
【0009】
すなわち、本発明は以下を包含する。
〔1〕 FGF18の活性阻害物質及び/又は発現抑制物質を有効成分として含有する毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔2〕 前記FGF18の活性阻害物質がFGF18の部分ペプチドであることを特徴とする前記〔1〕に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔3〕 前記FGF18の活性阻害物質が抗FGF18抗体であることを特徴とする前記〔1〕に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔4〕 前記FGF18の活性阻害物質がゴーヤー抽出物であることを特徴とする前記〔1〕に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔5〕 前記FGF18の活性阻害物質が、FGF18の活性を阻害する活性を有するFGF18の部分ペプチドをコードするcDNAを組み込んだ発現ベクターであることを特徴とする前記〔1〕に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔6〕 前記FGF18の発現抑制物質が、FGF18の発現を阻害する活性を有するsiRNAを組み込んだ発現ベクターであることを特徴とする前記〔1〕に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔7〕 他の毛髪成長促進剤、発毛促進剤、又は脱毛抑制剤を更に含むことを特徴とする前記〔1〕ないし〔6〕のいずれかに記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔8〕 前記〔1〕ないし〔4〕のいずれかに記載のFGF18の活性阻害物質をスクリーニングし、被検物質を毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤の候補とする方法であって、以下の(a)〜(c)の工程を含む方法;
(a)FGFR1c、FGFR2c、FGFR3c及びFGFR4から選ばれた少なくとも1つのFGF受容体遺伝子を用いて遺伝子操作により細胞表面に強制的に発現させ、当該細胞を培養する工程、
(b)(a)工程で得られた、細胞表面にFGF受容体を有する細胞系にFGF18とともに披検物質を接触させる工程、
(c)(b)工程において、FGF18の細胞増殖促進活性を阻害する活性を観察した披検物質を選択する工程
〔9〕 前記〔8〕に記載のFGF受容体がFGFR3cである前記〔8〕に記載のスクリーニング方法。
〔10〕 前記〔8〕に記載のFGF受容体がFGFR4である前記〔8〕に記載のスクリーニング方法。
〔11〕 前記〔8〕に記載のFGF受容体を細胞表面に強制的に発現させるための細胞がマウスIL-3依存性Ba/F3細胞株である前記〔8〕ないし〔10〕のいずれかに記載のスクリーニング方法。
〔12〕 前記〔1〕に記載のFGF18の発現抑制物質をスクリーニングし、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤の候補とするための方法であって、下記の(a)〜(d)の工程を含む方法;
(a)FGF18遺伝子が観察可能な程度発現する動物培養細胞又は実験動物を用意する工程、
(b)被検物質を、(a)の動物培養細胞に接触させるか又は実験動物に接触もしくは投与する工程、
(c)(b)工程後の動物培養細胞又は実験動物におけるFGF18遺伝子の発現をモニターする工程、
(d)FGF18遺伝子の発現を阻害する機能を有する被検物質を選択する工程。
〔13〕 前記(c)工程において、前記(b)工程後の実験動物又は動物培養細胞から、mRNAを抽出し、発現されたFGF18のmRNA量を解析することによりFGF18遺伝子の発現をモニターし、前記(d)工程において、披検物質を作用させない場合と比較して低いレベルのFGF18のmRNA量を観察した系を選択することにより、FGF18遺伝子の発現を阻害する機能を有する被検物質を選択することを特徴とする、前記〔12〕に記載の方法。
【発明の効果】
【0010】
本発明において、毛包内の内在性FGF18遺伝子の発現を抑える物質又はFGF18蛋白質の活性を抑える物質を検索し、これらの物質を毛包に投与することで、毛の成長サイクルの休止期を短縮することができ、かつ毛の成長サイクル全体の速度を速めることができる。また、脱毛期を遅らせて、抜け毛自体を防止することができる。つまり、完全な抜け毛が起こる前に新しい毛髪が生えることになり、しかも脱毛自体の防止が可能となるため、毛髪全体の量が増えて、いわゆる薄毛予防ともなる。
【図面の簡単な説明】
【0011】
【図1】毛成長周期 図中、「anagen(成長期)」は毛包の成長と毛軸形成が進行する時期、「catagen(退行期)」は毛包が退縮する時期、「telogen(休止期)」は毛包の活動が休止する時期を表し、「anagen」が「propagating anagen」及び「autonomous anagen」に、「telogen」が「refractory telogen」及び「competent telogen」に分けられることを示す。なお、外観的に毛軸が脱落する「脱毛期(exogen)」は、退行期や休止期と直接連結するのではなく、その毛軸が完成した成長期の次の休止期又はその次の成長期の最中に、独立した相として起こるとされている。
【図2】FGF18遺伝子ノックアウト模式図 a:マウスFGF18遺伝子ゲノム第3エキソン付近の構造 図中、太い黒い線は第3エキソンを、線部分はイントロン構造を示す。EcoRIは、制限酵素EcoRIの位置を示す。 b:ターゲティングベクター構造ならびにゲノム相同組換えの概略図。 図中、「KIprobe」はサザンブロット法による組換え判定・遺伝子型判定などに用いるDNAプローブであり、その相同位置を太線で示す。ボックスで囲んだPGK-Neorは、neomycin耐性遺伝子のカセットを示し、黒三角はloxP配列を示し、灰色三角はFRT配列を示す。 c:bの配列からFlp recombinaseによりFRT配列で囲まれたneomycin耐性遺伝子のカセットが除去された様子を示す。 d:cの配列からCre recombinaseによりloxP配列で囲まれたFgf18遺伝子の第3エクソンが除去された様子を示す。 e:サザンブロット解析の結果の一例。hetero、homo、及び野生型の解析結果を示す。
【図3】毛包特異的FGF18欠損マウス aとbは、毛包特異的FGF18欠損マウス(K5Cretg;Fgf18flox/flox:ホモノックアウトマウス)において速やかに繰り返される毛成長周期(1) a:若齢における毛包特異的FGF18欠損マウス(ホモノックアウトマウス:下段)及び対照のヘテロノックアウトマウス(K5Cretg;Fgf18+/flox:上段)のそれぞれ代表的な一個体について、背部皮膚の毛成長周期の変遷を経時的に観察したもの。各写真とも、生えている毛をバリカンで短く刈ってから撮影しており、皮膚の色が毛成長周期状の相を反映している。 b:毛包特異的FGF18欠損マウス(ホモノックアウトマウス:左)及び対照の野生型マウス(C57BL/6:右)のそれぞれ代表的成体一個体について、それらの背部皮膚における発毛パターンの一例を示したものである。 cとdは、毛包特異的FGF18欠損マウス(K5Cretg;Fgf18flox/flox:ホモノックアウトマウス)において速やかに繰り返される毛成長周期(2) c:毛包特異的FGF18欠損マウス(ホモノックアウトマウス)の背部皮膚における毛成長周期の変化(Y軸)を、マウスの日齢(X軸)に対してプロットしたものである。 d:加齢の進んだ毛包特異的FGF18欠損マウス(ホモノックアウトマウス)の背部皮膚における毛成長周期の変遷を、概ね1週間毎に観察したものであり、各写真とも、観察日前の1週間の毛成長を反映している。
【図4】ヘアサイクルドメインの例示 図はマウスの背部皮膚に存在するヘアサイクルドメインを表す。マウス背部皮膚には、図中の縦横線で仕切られた区切りのようにヘアサイクルドメインが存在し、それぞれのドメイン毎に毛周期の制御が異なることが経験的に知られている。ただし、図中の縦横線の数や位置は例示であり、実際は様々なパターンがある。
【図5】毛包特異的FGF18欠損マウス aとbは、加齢の進んだ毛包特異的FGF18欠損マウス(ホモノックアウトマウス)におけるクラブヘアの不完全な脱落(1) a:左:加齢の進んだ毛包特異的FGF18欠損マウス(ホモノックアウトマウス)において、背部から指で抜去した毛の状態を示したものである。右:対照として左と同齢のヘテロノックアウトマウスから抜去したクラブヘアーの状態。 b:成長期毛包周囲における毛包の状態。なお、図中バーは200μmを表す。 cとdは、加齢の進んだ毛包特異的FGF18欠損マウスにおけるクラブヘアの不完全な脱落(2) c:休止期毛包周囲における毛包の状態。なお、図中バーは200μmを表す。 d:cの枠囲み部分の拡大図 eとfは、加齢の進んだ毛包特異的FGF18欠損マウスにおけるクラブヘアの不完全な脱落(3) e:成長期毛包を含む皮膚の切片の免疫染色写真。ケラチン15とPCNA抗原に対する二重染色である。PCNA陽性の成長期毛包のバルジ領域に寄り添うクラブヘアの毛包が観察される。 f:eの視野の一部のケラチン15のシグナルのみを表したものである。ケラチン15は皮膚幹細胞に特徴的な抗原であり、成長期毛包のバルジ領域とクラブヘアを包む袋状構造がいずれもケラチン15陽性で連続的な構造体を成していることが分かる。 gとhは、加齢の進んだ毛包特異的FGF18欠損マウスにおけるクラブヘアの不完全な脱落(4) g:加齢の進んだ毛包特異的FGF18欠損マウス(ホモノックアウトマウス)背部皮膚の成長期毛包周囲の輪切り切片の蛍光観察像。毛軸のタンパク質を染める蛍光色素チオフラビンTで染色してある。なお、図中バーは100μmを表す。 h:eと同じ試料の拡大観察像図。位相差観察像とマージしてある。なお、図中バーは100μmを表す。矢印はメラニンを含む成長期毛包に生える毛軸、それ以外はクラブヘアーを示す。 iとjは、加齢の進んだ毛包特異的FGF18欠損マウスにおけるクラブヘアの不完全な脱落(5) i:対照としてe,fと同齢のヘテロノックアウトマウス背部皮膚の成長期毛包周囲の輪切り切片の蛍光観察像。毛軸のタンパク質を染める蛍光色素チオフラビンTで染色してある。なお、図中バーは200μmを表す。 j:gと同じ試料の拡大観察像。位相差観察像とマージしてある。なお、図中バーは100μmを表す。矢印はメラニンを含む成長期毛包に生える毛軸、それ以外はクラブヘアーを示す。
【図6】FGF18がFGF受容体(FGFR)発現細胞の増殖を促進する効果が、FGF18部分ペプチドにより抑制されることを示す図である。FGFR3c発現細胞(R3c)、FGFR4発現細胞(R4)を用いた解析結果を示す。 A:試験物質を1μg/mlで加えた場合の結果 <対照> カラム1:FGF18以外の試料添加なし。 <R3c> カラム2:(d4)。カラム3:(d16)。カラム4:(d18)。カラム5:(d22)。カラム6:(d37)。カラム7:(d48)。カラム8:(d77)。カラム9:(d95)。 <R4> カラム10:(d22)。カラム11:(d37)。カラム12:(d48)。カラム13:(d67)。カラム14:(d77)。カラム15:(d95)。 B:試験物質を100ng/mlで加えた場合の結果 <対照> カラム1:FGF18以外の試料添加なし。 <R3c> カラム2:(d4)。カラム3:(d12)。カラム4:(d16)。カラム5:(d18)。カラム6:(d37)。カラム7:(d48)。カラム8:(d67)。カラム9:(d95)。 <R4> カラム10:(d37)。カラム11:(d67)。
【図7】ゴーヤー熱水抽出液のR4/BaF3 細胞増殖阻害作用:FGF18 存在下でFGFR4/BaF3 細胞をゴーヤー熱水抽出液とともに培養することにより、ゴーヤー熱水抽出液0.083%、0.83%及び8.3%の添加によるFGF18のFGFR4/BaF3細胞の増殖促進作用に対する阻害効果を示す。
【図8】ゴーヤー熱水抽出液のin vivo 解析:ゴーヤー熱水抽出液のエタノール溶液(49.5%エタノール、1%グリセロールを含有)を毛周期の休止期にあるC3H/He マウス背部皮膚に11日間(5日目、及び6日目を除く)に渡って毎日塗布し、第10日、14日、17日、21日、24日目のマウス背部の発毛状態を発毛スコアーで示す。
【発明を実施するための形態】
【0012】
以下、本発明を詳細に説明する。
1.毛包内の毛の成長サイクル
毛の成長サイクル(図1)は、毛を作り出す器官である毛包自体の周期的な変化に規定されている。すなわち毛包は、成長期(anagen)、退行期(catagen)、休止期(telogen)、という3つの相をこの順に繰り返す。成長期(anagen)は、さらに「propagating anagen」及び「autonomous anagen」に、休止期(telogen)は「refractory telogen」及び「competent telogen」に分けられるとされる。このうちの成長期には、毛包の先端が皮膚よりも深部の皮下脂肪層にまで到達すると共に、大きく太くなりつつ、長い毛軸を形成し、これを体外に長く伸ばす。これが外観的な毛の成長である。退行期は、毛包の構成細胞がアポトーシスを起こすことに伴い、毛包が退縮し、毛包の先端は皮下脂肪層よりも皮膚表面に近い真皮内部にまで移動する。次に来る休止期においては、毛包の構成細胞はほとんど増殖もアポトーシスも起こさず、小さい状態で静止すると考えられている。有色体毛を持つマウスでは、成長期の初期にはメラニン色素が合成され、皮膚が青くなっているのを視認することができる。したがって、皮膚の外側から青い色を見て毛包の成長周期の進行を評価することもできる。また、成長期に皮膚を切開して裏側から見ると、メラニン色素を多量に含んだ毛包が高い密度で並ぶこととなるため、皮膚の裏側が黒なっているのを視認することができる。これに対して休止期においては、皮膚の裏側は白いままであることを視認することができる。一般に、マウスの実験系では、成長期は脱毛後1〜19日目の期間であり、退行期は20〜21日目の期間である。また、脱毛後、21〜22日目に休止期に入り、休止期は3〜5週間以上続くことが知られている。本発明前に、休止期(telogen)のうち最初の短期間である「refractory telogen」の制御因子として、細胞増殖因子の一群であるBMP(bone morphogenetic protein)シグナルが関わっていることが報告された(非特許文献24)。しかし、休止期全体のうち大きな割合を占め、「refractory telogen」に続く時期である「competent telogen」の制御因子についてはまったく知られていなかった。本発明においてはじめて「competent telogen」の制御因子がFgf18因子であることを見出した。従って実質的に休止期を延長する制御因子がFgf18因子であることを、本発明においてはじめて見出した。なお、Fgf18因子が「refractory telogen」の制御因子としても機能を果たしている可能性は存在し、今後の検証が待たれる。
これら退行期、休止期において、外観的な毛の成長は止まるが、毛が抜けるわけではなく、毛が抜けることは、脱毛期(exogen)と称される独立の相で独立の制御により起こると考えられているが、この脱毛期を制御する分子メカニズムは、これまでほとんどわかっていなかった。本発明において、はじめてこの脱毛期開始を制御する因子の1つがFgf18であることを見出した。
【0013】
2.毛包特異的FGF18遺伝子ノックアウトマウスの製造について
本発明の「毛包特異的FGF18遺伝子ノックアウトマウス」は、以下のようにして作成される。
マウスFgf18遺伝子におけるエクソン3を、Cre-loxP法を用いてケラチン5陽性細胞特異的にノックアウトする。エクソン3を欠失させることで、Fgf18蛋白質の分泌シグナルの一部とその下流が発現されず、Fgf18蛋白質の生物活性が失われる。FGF18遺伝子をケラチン5陽性細胞特異的に欠損する方法は非特許文献6を参考に行った。
マウス由来Fgf18遺伝子のエクソン3を含むゲノム領域の塩基配列は、GenBankなど公的データベースから取得できる(Entrez Gene;Gene ID:14172,updated on 21-Jul-2010;Official Symbol:Fgf18 provided by MGI;Official Full Name:fibroblast growth factor 18 provided by MGI;Primary source:MGI:1277980)ので、その上流に、2つのFRT配列で挟まれたネオマイシン耐性遺伝子カセットを挿入し、このネオマイシンカセットとエクソン3が2つのloxP配列で挟まれたターゲティングベクターを作製した(図1)。このターゲティングベクターを用い、マウスES細胞に対してFgf18遺伝子エクソン3アリルをターゲティングした。この細胞をC57BL/6マウスの胚盤胞に注入したものをICRマウスの子宮内に移植して発生させ、得られたキメラの産子をC57BL/6マウスと交配して得られた産子の中から、導入したターゲット遺伝子を有する個体を選択した(F1)。F1の個体をFlpe トランスジェニックマウス個体(理研BRCから分譲:RIKEN RBRC01834;非特許文献7)と交配して、FRT配列で挟まれたネオマイシン耐性遺伝子カセットを欠失した個体を得た(F2)。次にF2マウスをK5Creトランスジェニックマウス(熊本大学CARDから分譲:CARD ID323;非特許文献6)と交配させてケラチン5陽性細胞特異的にFgf18遺伝子エクソン3を除去した。得られたK5Cretg; Fgf18+/floxマウス同士を交配しK5Cretg; Fgf18flox/floxマウスを得た。
ところでケラチン5は、ケラチン14と同様に、皮膚と毛包など皮膚付属器官の中では表皮、毛包、皮脂腺など多くの細胞で発現している。ところがFgf18を高いレベルで発現するのは、この中で毛包だけである。従って、上記のようにして得られたコンディショナルノックアウトマウスは、実質的に毛包特異的にFgf18遺伝子をノックアウトしたマウスと表現することができる。
【0014】
3.毛包特異的Fgf18遺伝子ノックアウトマウスの特性
(3−1)全体の特徴
FGF18遺伝子はマウスなど哺乳動物では骨、心筋、胎児肺、その他の組織に発現し、個体形成や生命維持に重要な働きをしていると考えられる。しかし、皮膚と毛包など皮膚付属器官の中でFgf18を高いレベルで発現するのは、毛包だけである。そして、ケラチン5遺伝子を高いレベルで発現するのは、皮膚の他では、骨、舌、気管支など、ごく一部の組織であるため、上記のようにして得られたコンディショナルノックアウトマウスにおいて、天然型のマウスに比べて実質的にFgf18遺伝子の生理的機能が失われていると言えるのは毛包のみである。
本発明で得られた毛包特異的Fgf18遺伝子ノックアウトマウスは、個体の健康状態はすこぶる良く、具体的に観察された特性は以下の通りである。
1)毛の成長周期のうち休止期が1/3〜1/5に短縮されたため、毛の成長周期全体の進行が早い。
2)背部皮膚での毛成長周期が帯状に整然と進行する。その際の進行速度が首に近い前方部が最も早く尾部後方部が最も遅いが、その進行速度の速まり方が帯状に整然と進行するため、剃毛を繰り返すと複数の横縞を形成する。その際の横縞数は加齢に比例する。
3)毛包細胞数は増加しないが、毛包からの脱毛速度が遅くなり、結果的に毛包内の毛軸の本数が多く、皮膚表面の全体の毛軸数は野生型の2倍以上となる。
【0015】
(3−2)毛成長周期の進行が早い特性
本発明の毛包特異的Fgf18遺伝子ノックアウトマウスは野生型に比べ、遙かに早い周期で毛成長が起きる。特に、毛成長周期のうち、休止期が短く(図3c)約1週間で終了し、次の成長期に入ることが示されている。これに対し、野生型マウスの例では、図3cの48日齢頃から始まる休止期が通常3〜5週間続くことが良く知られている。その結果として、一サイクルに要する期間も野生型マウスが6週間から8週間程度またはそれ以上であるのに対して、約3週間にまで短縮されていた。
すなわち、毛包内の内在性Fgf遺伝子の発現が阻害されたことで、毛成長サイクルの休止期が短縮し、同時に毛成長サイクル自体の速度が速まったことがわかる。
【0016】
(3−3)毛成長周期相の固有の表現型
従来からマウスの背部皮膚での毛成長周期が首に近い前方部が最も早く尾部後方部が最も遅い傾向にあることは知られていた。しかし、毛の成長周期の開始時期は背部皮膚の複数の位置で始まるため、野生型マウスの背部皮膚を剃毛すると、図2b右にあるように、背部皮膚全体的にランダムに発毛が始まる。このことの説明のために、マウスの背部皮膚において「ヘアサイクルドメイン」と呼ばれるブロック状に分かれて毛成長周期を決定する制御系の存在を想定することが一般的であった(図4)。マウスの背部皮膚には図4の縦横線で仕切られた区切りのようにヘアサイクルドメインが存在し、それぞれのドメイン毎に毛周期の制御が異なることが経験的に知られている。ただし、図4における縦横線の数や位置は例示であり、実際には様々なパターンがある。
一方、本発明の毛包特異的Fgf18遺伝子ノックアウトマウスは、このようなヘアサイクルドメインを示さず、ブロック状に分かれて毛成長周期を決定する制御系を決定づける因子が失われた可能性が大きく、毛成長周期が首に近い前方部が最も早く尾部後方部が最も遅いという本来の毛成長周期の進行速度の傾向がそのまま発現し、その結果、加齢が進むに従って各相の毛包がシマウマのストライプのように整然と配列した様相を示す。
すなわち、毛包内の内在性FGF遺伝子の発現が阻害されたことで、ヘアサイクルドメインのうちでも、特にマウスの体軸に対して直交する方向のドメイン間の境界線を失わせたことがわかる。
【0017】
(3−4)毛包内の毛軸の本数が多い特性
本発明の毛包特異的FGF18遺伝子ノックアウトマウスは、皮膚の単位面積あたりの毛包数は増加しないが、毛成長周期の進行が早まるにもかかわらず、脱毛期の開始時期が遅くなり、前周期の産物としての毛の脱落が起こりにくい。毛包内では、成長期の毛包に生えている成長期毛軸に寄り添い、脱落していないクラブヘアが多数存在している状態となる。すなわち、毛包内の内在性FGF遺伝子の発現が阻害されたことで、脱毛期の開始時期が遅くなり、クラブヘアの脱落が防止できたことがわかる。
【0018】
4.毛包細胞におけるFGF18遺伝子及び遺伝子産物であるFGF18蛋白の作用について
Fgf18遺伝子のノックアウトマウスの表現型を野生型のマウスと比較解析したことではじめて、毛包細胞内の内在性FGF18遺伝子、またはその発現産物であるFGF18の毛包における制御作用が解明できてきた。FGF18には具体的には以下の3つの作用があると推定される。
1)FGF18は毛包の休止期を維持する機能を有する。すなわち、毛包内の内在性FGF18は、毛の成長周期の休止期を延長させる制御因子であると考えられる。
2)FGF18はクラブヘアを皮膚から脱落させる機能を有する。すなわち、毛包内の内在性FGF18は、今まで未解明であった毛の脱落期を開始させる制御因子であると考えられる。
3)FGF18は動物の体軸(仮にX軸とする)に対して直交する方向(仮にY軸とする)を共有するヘアサイクルドメイン間の境界線を決定づける機能を有する。すなわち、皮膚表面のヘアサイクルドメインを決定する制御因子、すなわち毛周期の開始時期を制御する制御因子の1つであると考えられる。
【0019】
5.FGF18活性阻害物質
上記4.に示すように、毛包内で発現する内在性FGF18には1)〜3)の作用があることから、当該内在性FGF18の作用を阻害又は抑制する物質、すなわち「FGF18活性阻害物質」には以下の作用効果が期待できる。
1)毛髪の成長サイクルの休止期を短縮させると共にその結果毛髪の成長サイクル全体の速度を速める作用を持つと考えられるから、毛髪成長サイクルの休止期短縮剤又は毛髪成長サイクル促進剤としての作用効果を有する。
2)毛髪の脱落期を遅延させる作用があるといえるから、脱毛抑制剤、抜け毛防止剤又は抜け毛予防剤などとしての作用効果を有する。
3)毛の成長サイクルの開始時期について、体軸と直交する方向でのばらつきを抑え、体軸方向のみの整然とした成長サイクルでの発毛を促すため、毛並みをそろえる作用効果がある。
つまり、本発明は、毛髪成長サイクルの休止期短縮剤もしくは毛髪成長サイクル促進剤(あわせて「毛成長休止期短縮剤」という)、脱毛防止剤、抜け毛防止剤もしくは抜け毛予防剤(あわせて「抜け毛防止剤」という)、又は「毛並み調整剤」に係るものである。
本発明に係る「毛成長休止期短縮剤」、「抜け毛防止剤」又は「毛並み調整剤」の作用を有するFGF18活性阻害物質としては、例えばFGF18の部分断片であっても良い。
FGF18は、ヒトとマウスでは産生細胞の細胞質で207アミノ酸のポリペプチドとして合成され、それが細胞外に分泌される際にN末端のシグナルペプチドが切断され、181アミノ酸より成る分泌体として生理作用を発揮する。そして、7種類あるFGF受容体サブクラス(FGFR1c、FGFR1b、FGFR2c、FGFR2b、FGFR3c、FGFR3b、FGFR4)のうち少なくとも4つ、すなわちFGFR1c、FGFR2c、FGFR3c、FGFR4と反応する。
本発明において「FGF18」というとき、ヒト由来に限定されず、例えば、他の哺乳動物由来のFGF18を使用することができる。他の哺乳動物としては、マウス、ラット、ニワトリ、七面鳥、ウシ、ブタ、ヒツジ及びウサギ等を挙げることができるが、これらに限定されない。例えば、配列番号1に示すヒト由来FGF18の塩基配列に基づいて作製したプローブを定法に従って用いれば、ヒトを除く他の哺乳動物からFGF18をコードする遺伝子を単離することができる。
シグナル配列除去後のヒト由来FGF18の塩基配列及びアミノ酸配列をそれぞれ配列番号1及び2に示す。シグナル配列除去後のマウス由来FGF18の塩基配列及びアミノ酸配列をそれぞれ配列番号3及び14に示す。これら配列番号2及び14に示すアミノ酸配列を比較すると判るように、哺乳動物においてFGF18は非常に高い相同性を有しており、哺乳動物におけるFGF18の機能はほぼ同様であると理解することができる。
【0020】
ところで、一般にアンタゴニストとは、受容体には結合はするが、その受容体を刺激する生体物質(リガンド)とは異なり生体反応を起こさないかまたは比較的弱い生体反応しか起こさず、また本来結合すべき生体内物質と受容体の結合を阻害し、生体応答反応を引き起こさないかまたは減弱させる物質をいう。FGF18は、上述のように、7種類あるFGF受容体サブクラスのうちの少なくともFGFR1c、FGFR2c、FGFR3c、FGFR4と反応するので、本発明において「FGF18アンタゴニスト作用」というときは、FGFR1c、FGFR2c、FGFR3c、FGFR4のいずれかの受容体を発現している細胞に対して、FGF18による応答を引き起こさないかまたは減弱させる物質をいう。FGF18アンタゴニストがFGF18の受容体への結合を妨げることになってFGF18に起因する毛髪成長サイクルにおける休止期の延長作用、毛髪の脱落期開始作用などを阻害し、結果的に毛髪成長サイクルの促進効果、毛髪脱落防止効果を有することになる。
FGF18アンタゴニスト作用を有するFGF18部分ペプチドの場合としてヒトFGF18の場合を例にとれば、配列番号2に示すアミノ酸配列における32〜151番目の領域は、FGFファミリーで共通性の高いコア配列と呼ばれる領域であって、この領域だけで構築される3次元構造は受容体との基本的な結合能を有しているが完全な活性を持たないと考えられることから、当該領域に対応する領域のアミノ酸配列からなる部分ペプチドは、FGF18アンタゴニスト作用を有すると考えられる部分ペプチドである。当該領域に対応するコア配列を含むFGF18部分ペプチドであって、FGFR4などの受容体への結合能は十分有しており、かつFGF18応答を引き起こすほどに十分な長さを有していない場合は、FGF18アンタゴニスト作用を有する。
実際に実験的に確かめた結果からは、図3に示されるように、配列番号14において、N末端から翻訳開始のために導入されたメチオニンを除く16個欠失した場合でもアンタゴニスト作用を及ぼし、好ましくは22個、より好ましくは77個、最も好ましくは95個のアミノ酸を欠失した部分ペプチドはさらに強いFGF18アンタゴニスト作用を有するということができる。すなわち、少なくともN末側のアミノ酸配列については、95個まで欠失しても受容体結合能を失わず、むしろアンタゴニスト作用が強まっているといえるから、N末側のアミノ酸配列は32番目から始まる上記コア配列の全てがFGF受容体の結合に必須の配列ではないことは明らかであり、N末側アミノ酸をさらに除去した場合であってもFGF18アンタゴニストであるといえる。また、このことからみて、C末側についても、コア配列のC末側に相当する、C末端から31番目までのアミノ酸を欠失した場合の部分ペプチドにFGF18アンタゴニスト作用があることは明らかである上に、さらにC末側アミノ酸配列を大きく欠失させた場合も、受容体結合能を完全には失わないことが十分期待できるから、C末端から31個以上の、例えば43個、57個、67個、82個、94個、108個、113個、125個などのアミノ酸を欠失した部分ペプチドも強いFGF18のアンタゴニスト作用のある部分ペプチドであると考えられる。そして、これはN末側とC末側を同時に除去しても当該アンタゴニスト作用が保たれることはもちろんであるから、少なくとも32〜151番目のアミノ酸配列を含有するペプチドのみならず、むしろ好ましくは77〜151、より好ましくは95〜151番目のアミノ酸配列を含むペプチドもFGF18アンタゴニスト作用を有するということができる。
【0021】
また、FGF18又はFGF18反応性受容体に結合する抗体、すなわち、抗FGF18抗体や抗FGFR3c抗体、抗FGFR4抗体も同様にFGF18とその受容体との結合を阻害するので、FGF18による毛包内の毛成長サイクルの休止期を長期に亘って保持する作用、毛髪の脱落の開始促進作用又は毛の成長サイクルスタート時期をヘアサイクルドメイン毎に規定する作用を阻害することになる。
そこで、本発明では、上記FGF18アンタゴニストも含めて、FGF18とその受容体との結合を阻害する物質など、FGF18の受容体を介しての作用を阻害する物質を「FGF18の活性阻害物質」という。
このような、FGF18活性阻害作用を有する物質は、FGF18とは無関係な物質群から選択された物質であっても良く、下記2.のスクリーニング方法を適用することにより、簡単にFGF18活性阻害作用を有する物質を取得することができる。本願の実施例においても、当該スクリーニング方法により、FGF18の活性阻害作用があるゴーヤー抽出物を選択し、当該ゴーヤー抽出物に、FGF18の毛包成長抑制作用を阻害し、かつ毛包成長作用があることを確認した。このように、本発明におけるFGF18の活性阻害物質は、FGF18の受容体との結合を阻害することでFGF18による毛包の毛成長サイクルの休止期を長期に亘って保持する作用と共に、毛髪の脱落の開始促進作用を阻害するので、毛成長休止期短縮作用、抜け毛防止作用があるが、短期間の観察だと毛髪の成長促進作用として観察される。
これらのFGF18の活性阻害物質が、ペプチド、タンパク質もしくは糖タンパク質の場合、これらペプチドなど自身を含む製剤として投与してもよいことはもちろんであるが、当該ペプチドなどをコードするDNAを組み込んだ発現ベクターの状態で投与することもできる。たとえば、FGF18部分ペプチドをコードするDNAを組み込んだ発現ベクターを有効成分とする製剤も、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤として用いることができる。
そして、これらのFGF18の活性阻害物質は、単独で、もしくは併用して、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤として用いることができる。
【0022】
6.FGF18活性阻害物質のスクリーニング方法
FGF18の活性阻害物質をスクリーニングし、被検物質を毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤の候補とする方法は、具体的には、以下の(a)〜(c)の工程を含むものである。
(a)FGFR1c、FGFR2c、FGFR3c及びFGFR4から選ばれた少なくとも1つのFGF受容体遺伝子を用いて遺伝子操作により細胞表面に強制的に発現させ、当該細胞を培養する工程、
(b)(a)工程で得られた、細胞表面にFGF受容体を有する細胞系にFGF18とともに披検物質を接触させる工程、
(c)(b)工程において、FGF18の細胞増殖促進活性を阻害する活性を観察した披検物質を選択する工程
ここで、FGF受容体遺伝子としては、FGF18が結合し、かつ細胞表面に受容体を有する細胞に対するFGF18の細胞増殖作用が確認されているFGFR1c、FGFR2c、FGFR3c及びFGFR4の受容体遺伝子の少なくとも1つを用いる。好ましくは、FGFR3cまたはFGFR4遺伝子を用いる。(b)工程で、被検物質を細胞に接触させる場合に、典型的には、細胞培養液に被検物質を直接投与するが、特に被検物質がタンパク質の場合などは、当該被検物質をコードする遺伝子をFGF受容体発現細胞中に導入することも可能である。
また、FGF受容体を細胞表面に強制的に発現させるための細胞は、培養可能な細胞であればどのような細胞でもよいが、好ましくはマウスIL-3依存性Ba/F3細胞株である。
スクリーニングに際しては、48時間以上、例えば72時間程度培養し、FGF18を単独で添加した場合に比べ、FGF受容体発現細胞の細胞増殖能が、5%以上、好ましくは10%以上低下していることを目安にスクリーニングすることが好ましい。
そして、FGF受容体遺伝子を導入していない親細胞は対照試験に用いることができ、披検物質を用いて(b)工程で添加したFGF18をIL-3に置き換えて同様の操作を行い、披検物質がこれら親細胞に対してはIL-3の細胞増殖促進活性を阻害しないことを確認する工程を設けることが好ましい。
【0023】
7.FGF18の発現抑制物質及びそのスクリーニング方法
FGF18は毛包内に存在する内在因子であるので、毛包細胞内でのFGF18遺伝子の転写及び翻訳を阻害する物質、すなわちFGF18遺伝子の発現を低下させる物質も毛包細胞中のFGF18濃度を低下させて、FGF18による毛包内の毛成長サイクルの休止期を長期に亘って保持する作用、毛髪の脱落の開始促進作用又は毛の成長サイクルスタート時期をヘアサイクルドメイン毎に規定する作用を阻害することができるから、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤毛髪成長促進作用及び発毛作用を有する。
このようなFGF18遺伝子の発現を阻害する物質は、FGF18遺伝子に対するsiRNAまたはその発現ベクターとして公知の方法で設計することができ、その活性は、動物培養細胞又は実験動物を用いてFGF18遺伝子の発現を阻害するか否かをモニターすることで確認することができる。
また別に、このようなFGF18遺伝子の発現を阻害する物質は、動物培養細胞又は実験動物を用いてFGF18遺伝子の発現を阻害するか否かをモニターすることでスクリーニングすることができる。
【0024】
具体的には、先ず、FGF18遺伝子を観察可能な程度に発現する動物培養細胞又は実験動物を用意し、被検物質を当該動物培養細胞もしくは実験動物に接触させるか又は実験動物に投与する。なお、実験動物とは、ヒトを除く、例えば、マウス、ラット、ニワトリ、七面鳥、ウシ、ブタ、ヒツジ及びウサギ等を意味する。被検物質としては、何ら限定されないが、例えば、植物抽出液、ペプチド、タンパク質、非ペプチド性化合物、低分子化合物、合成化合物、発酵生産物、細胞抽出液、動物組織抽出液等が挙げられる。これらの物質は新規な物質であってもよいし、公知の物質であってもよい。被検物質を細胞もしくは動物に接触させる場合に、典型的には、被検物質を細胞培養液に添加するか又は実験動物に投与するが、特に被検物質がタンパク質の場合などは、当該被検物質をコードする遺伝子をFGF受容体発現細胞中に導入することも可能である。
次に、当該動物培養細胞又は実験動物におけるFGF18遺伝子の発現をモニターする。動物培養細胞又は実験動物におけるFGF18遺伝子の発現は、例えば、FGF18抗体を用いたELISA等の常法を用いて解析するか、あるいは該細胞内又は実験動物内におけるFGF18遺伝子のmRNA量を定量的逆転写PCR法やノーザンブロット法等により解析するといった方法によりモニターすることができる。
これらいずれかの解析により、被検物質の非存在下で培養された動物培養細胞内におけるFGF18遺伝子の発現量と比べて、被検物質の存在下で培養された動物培養細胞内又は実験動物内におけるFGF18遺伝子の発現量が低下すれば、当該被検物質は毛髪成長促進或いは発毛といった機能を有する可能性があると判断できる。具体的には、被検物質を作用させない場合と比較して、mRNA量が0.8倍以下、好ましくは0.7倍以下、より好ましくは0.5倍以下に減少すれば、確実にFGF18の発現抑制物質であるといえる。培養角化細胞や培養真皮細胞、培養毛乳頭細胞でのFGF18のmRNA発現量は、培養条件や細胞の種類により様々であるので、上記の方法などにより個別に測定し、その数値が0.8倍以下に減少することを目安にスクリーニングすればよい。
このような工程を経てスクリーニングされたFGF18の発現抑制物質は、単独で、もしくは併用して、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤として用いることができる。
【0025】
8.毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤
上記「FGF18の活性阻害物質」及び/又は「FGF18の発現抑制物質」の項で説明した物質は、例えば、皮膚適用向けに適合化された溶液、クリーム、軟膏、ゲル、ローシヨン、シャンプー又はエアゾールといった形態で製剤化され、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤として提供される。なお、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤は、ヒトのみならず愛玩動物、毛を利用する家畜、実験動物などに適用した場合に、特に効果がある。
特に、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤は、局所投与用に適合化された薬理学上許容されるキャリアと共にFGF18活性阻害物質及び/又は発現抑制物質を含んだ医薬組成物の形で投与される。FGF18活性阻害物質及び/又は発現抑制物質を含有する毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤は、薬学上許容されるキャリア中に通常約0.01〜約100μg/日/cm2、好ましくは約0.1〜約10μg/日/cm2の活性化合物を含有している。言い換えると、FGF18の濃度は薬学上許容されるキャリア中で通常約0.01〜約100μg/日/cm2、好ましくは約0.1〜約10μg/日/cm2の活性化合物である。
さらにまた、局所投与用に適合化された薬理学上許容されるキャリアとしては、特に限定されないが、親水ワセリン又はポリエチレングリコール軟膏のような軟膏、キサンゴムのようなゴム等のペースト、アルコール、水性又は緩衝液のような溶液、水酸化アルミニウム又はアルギン酸ナトリウムゲルのようなゲル、ヒト又は動物アルブミンのようなアルブミン、ヒト又は動物コラーゲンのようなコラーゲン、アルキルセルロース、ヒドロキシアルキルセルロース及びアルキルヒドロキシアルキルセルロースのようなセルロース、メチルセルロース、ヒドロキシエチルセルロース、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース及びヒドロキシプロピルセルロース、プルロニックF−127で例示されるプルロニック(Pluronic(商標))ポリオ−ルのようなポリマ−;テトロニック1508のようなテトロニック(tetronic)、アルギン酸ナトリウムのようなアルギン酸塩を挙げることができる。
本発明に係るFGF18の活性阻害物質としては、FGF18部分ペプチドなどのFGF18の活性阻害タンパク質、ペプチドをコードするDNAを組み込んだ発現ベクターを用いることができるが、その場合は、通常の遺伝子治療の形態で提供することができる。発現ベクターとしては、動物細胞においてFGF18部分ペプチドなどを発現させるためのプロモーターなどの配列を備えるが、特に限定されるものではない。発現ベクターとしては、例えば、プラスミドベクター、ウイルスベクターなどが利用可能であるが、これらに限定されない。
【0026】
また、本発明に係る毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤は、FGF18発現抑制物質を有効成分として含有するものであってもよい。すなわち、本発明に係る毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤は、FGF18に対するsiRNA発現ベクターを用いた遺伝子治療として提供することができる。発現ベクターとしては、動物細胞においてsiRNAを発現するためのプロモーターなどの配列を備えるが、特に限定されるものではない。発現ベクターとしては、例えば、プラスミドベクター、ウイルスベクターなどが利用可能であるが、これらに限定されない。また、FGF18発現抑制物質は、FGF18に対するsiRNA発現ベクターに限定されない。
遺伝子治療用の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤を細胞内に導入する方法としては、ウイルスベクターを利用した遺伝子導入方法、非ウイルス性の遺伝子導入方法(日経サイエンス,1994年4月号,20-45頁、実験医学増刊,12(15)(1994)、実験医学別冊「遺伝子治療 の基礎技術」,羊土社(1996))のいずれの方法も適用することができる。
ウイルスベクターによる遺伝子導入方法としては、例えばレトロウイルス、アデノウイルス、アデノ関連ウイルス、ヘルペスウイルス、ワクシニアウイルス、ポックスウイルス、ポリオウイルス、シンビスウイルス等のDNAウイルス又はRNAウイルスに、TR4あるいは変異TR4をコードするDNAを組み込んで導入する方法が挙げられる。このうち、レトロウイルス、アデノウイルス、アデノ関連ウイルス、ワクシニアウイルスを用いた方法が、特に好ましい。非ウイルス性の遺伝子導入方法としては、発現プラスミドを直接筋肉内に投与する方法(DNAワクチン法)、リポソーム法、リポフェクション法、マイクロインジェクション法、リン酸カルシウム法、エレクトロポレーション法等が挙げられ、特にDNAワクチン法、リポソーム法が好ましい。
【0027】
9.インビトロ毛髪再生系への応用
FGF18の活性阻害物質及び/又は発現抑制物質を用いて、再生された皮膚組織における毛髪のインビトロ再生系を構築することができる。ここで、皮膚組織とは、分離した皮膚幹細胞を培養することによって得られる、皮膚の各種の細胞からなる組織を意味する。皮膚の各種の細胞とは、特に限定されないが、皮膚表皮の上皮細胞、皮膚上皮基底層の細胞、毛胞を構成する各種細胞、真皮の細胞及び脂肪細胞等を意味する。皮膚組織を再生する際に使用される細胞は、異種細胞、同種他家細胞及び同種自家細胞のいずれであっても良い。
先ず、皮膚幹細胞からの皮膚の各種細胞への分化を制御する方法としては、特に確立された技術がないため、本発明においては特に限定されない。例えば、自然分化(spontaneous differentiation)が起きた段階において異なる発現を示す増殖因子受容体を利用し、それぞれに対応するリガンド増殖因子を培地中に加えることにより、異なった分化方向を有する細胞を選択的に増幅することができる。異なった分化方向を有する細胞を選択的に増幅した後、皮膚組織を作製することができる。
人工皮膚組織の作製方法としては、特に定まった技術はないため、本発明においては限定されないが、例えば、上皮細胞だけを培養し層状に仕上げる方法、繊維芽細胞など真皮の構成細胞により真皮層を形成した後、上皮細胞を重層させて一体化する方法、その後上皮細胞の表面を空気に暴露することにより表皮化を促進させる方法、真皮層の変わりに、生物分解が可能な成分により形成された層状フィルムを用いる方法、など、様々な方法をとることができる。また、本発明は、ヒトから採取した皮膚組織から皮膚幹細胞を分離し、分離した皮膚幹細胞を用いて皮膚組織を作製するといった、いわゆる再生医療における皮膚組織の作製方法も適用される。このとき、新たに作製した皮膚組織は、採取したヒトと同一人に対して治療のために戻すことを前提としても良いし、採取したヒトと異なる他人に対して治療のために移植することを前提するものであってもよい。
このような皮膚組織の作製方法において、培地中にFGF18の活性阻害物質及び/又は発現抑制物質を適当な時期に添加することによって、再生した皮膚組織において毛包の成長サイクルの休止期を短縮し、その結果毛成長サイクル自体の速度を早くすることができ、当該皮膚組織における毛髪の抜け毛を防止することができる。さらに、皮膚組織の作製方法において、培地中にFGF18活性阻害物質及び/又は発現抑制物質を適当な時期に添加することによって、皮膚細胞の増殖を促進することができ、皮膚組織全体の体積の増大を図ることができる。
【0028】
10.ゴーヤー抽出物
本発明の前記9.のスクリーニング方法により得られたFGF18活性阻害物質はゴーヤー抽出物であり、実施例に示すようにその発毛促進効果も確認された。
ゴーヤー抽出物の原料となるゴーヤーは、ウリ科ツルレイシ属の植物で、学名はMomordica charantiaであり、ニガウリ、または、ツルレイシとも呼ばれている。
ゴーヤーからその抽出物を得るに当たっては、植物体の全部位が使用可能であり、各部位を単独に、或いは適宜混合して抽出原料に用いることができる。また、原料となる植物体の性状は、乾燥状態のもの、非乾燥状態のもの、いずれも用いることができる。
【0029】
ゴーヤーから、本発明に用いる抽出物を得るには、これらの原料を必要により細切ないし粉砕し、適当な溶媒を用いて、一般に用いられる抽出法で抽出すれば良い。抽出に用いられる溶媒は、特に限定されないが、例えば水、無水或いは含水有機溶媒を例示することができる。無水或いは含水有機溶媒としては、1価アルコール、多価アルコールまたはその誘導体、ケトン、エステル、エーテル、石油エーテル、脂肪族炭化水素、またはハロゲン化合物、芳香族炭化水素より選択された一種または二種以上を例示することができる。具体的な溶媒としては、水、メタノール、エタノール、ブタノール、アセトン、酢酸エチルエステルが例示され、これらは一種または二種以上を組み合わせて用いることができる。このうち、特に水、或いはメタノールやエタノール等の1価アルコールを用いることが好ましい。抽出に当たって、上記溶媒の使用量は特に限定されないが、抽出原料であるゴーヤーに対して、0.1-1000重量倍、好ましくは1-100重量倍、更に好ましくは2-50重量倍である。
【0030】
上記溶媒による抽出法は、常法に従って行なうことができる。例えば抽出温度については、室温程度の温度で抽出しても良く、また用いられる溶媒の沸点付近の温度で抽出しても良い。また、抽出操作についても、ゴーヤーの乾燥粉砕物もしくは粉砕物を室温下で1〜30日間浸漬することにより抽出する方法や、抽出溶媒の沸点程度の温度において還流抽出する方法等を用いることができる。
【0031】
下記の実施例において示されるように、本発明のゴーヤーの抽出物からなる物質は、FGFR4遺伝子を発現したBa/F3細胞に対して、FGF18による細胞増殖を阻害するFGF18阻害活性を有する。実施例では、FGFレセプターの1つであるFGFR4を遺伝子発現操作により細胞表面に強制的に発現させた細胞を用いたときに、FGF18の存在下で、FGF18による細胞増殖に対して、ゴーヤー抽出物の添加により濃度依存的に阻害することが示された。ゴーヤー抽出物のこの作用は、同時に行われたFGF18非存在下の細胞増殖に対しては増殖阻害作用を示さなかったことから、細胞障害作用ではなく、FGF18活性阻害作用であることが示された。
すなわち、本発明のゴーヤーの抽出物からなる物質は、FGF18による細胞増殖を阻害するFGF18活性阻害物質である。
【実施例】
【0032】
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
なお、遺伝子操作的手法として、特に断らない限りモレキュラー・クローニング((Molecular Cloning - A laboratory Manual、Sambrook
and Russell社刊)に記載された公知の方法を用いた。
【0033】
〔実施例1〕 Fgf18遺伝子毛包特異的欠損(ノックアウト)マウス作製
マウスFgf18遺伝子を含むBACクローンを129マウスBACクローンライブラリーから取得した。このクローンを用い、Fgf18遺伝子のエクソン3の上流にloxP-FRT[PGK-Neor]FRT配列を導入し、下流にloxP配列を導入した、エクソン3を欠損するためのターゲティングベクターを構築した(図2)。なお、当該ターゲティングベクター作製は、「マウスFgf18遺伝子のエクソン3欠失用のターゲティングベクター」として、フェニックスバイオ社に外注した。
129マウス由来のES細胞にこのベクターを導入し、細胞集団の中から、Fgf18遺伝子がこの配列によって組み換えられた(ターゲットされたと称する)細胞を選択し、クローン化した。そのうちの一つのES細胞株をトランスジェニックマウスの作出に用いた。C57BL/6マウス(濃褐色体毛、日本SLC社)の胚盤胞にこの細胞を注入し、さらにこの胚盤胞を、偽妊娠状態にした雌のICRマウス(白色体毛、日本SLC社)の子宮内に導入した。マウスが分娩した仔について、体毛の色からキメラマウスを区別した。これらの操作を63個の胚盤胞について行い、キメラマウス仔が16個体得られた。これらのマウスを飼育し、性成熟後、C57BL/6マウスと交配し、35匹の仔が得られた。これらのゲノムを分析したところ、15匹の個体がターゲットされたFgf18遺伝子を含んでいることが示された(これをF1とする)。
次に[PGK-Neor]カセットを除去するため、F1マウスをFlpe トランスジェニックマウスと交配し、仔マウスが67個体得られた。これらについてFgf18遺伝子アリルを分析したところ、12個体について、目的の配列(導入されたloxP-FRT[PGK-Neor]FRT-Exon3-loxPをloxP-FRT-Exon3-loxPとして)を有していることが確認された(F2)。
ここで用いたFlpe トランスジェニックマウスは、理研BRCから入手した(RIKEN RBRC01834; 非特許文献8)。
次に毛包特異的にFgf18遺伝子エクソン3を除去する目的で、F2マウスをさらにK5-Creトランスジェニックマウスと交配した。得られた仔マウスは4週齢に達した時点で離乳させ、皮膚を含む尾の一部を切除し、Fgf18遺伝子の遺伝子型とCreの発現を確認した。離乳した37匹の個体のうち、6匹の K5Cretg;Fgf18+/floxマウス(メス1匹、オス5匹)が得られた。K5Cretg;Fgf18+/floxマウス同士を交配することにより、29匹のマウスが得られた。これらの遺伝子型を解析したところ、一匹の個体はK5Cretg;Fgf18flox/flox(以降、これをFgf18ホモ欠失体と称する)であることが示された。同様にしてK5Cretg;Fgf18+/flox マウスコロニーを交配することで、Fgf18ホモ欠失体を多数得ることが出来た。
以下、このFgf18ホモ欠失体をFgf18遺伝子毛包特異的欠損動物と称する。
ここで用いたK5Creトランスジェニックマウスは、九州大学CARDから入手した (CARD ID323;非特許文献9)。
これらのマウス皮膚においてFgf18第3エキソンが欠落しているDNAが存在することは、サザンブロットで確認された。その例を示す。
なお、K5Cretg;Fgf18+/floxマウス同士の交配において生じる仔の遺伝型は、通常のメンデルの法則に従うものだった。
【0034】
〔実施例2〕 Fgf18遺伝子毛包特異的欠損(ノックアウト)マウスにおける休止期の短縮と速やかに繰り返される毛成長周期
Fgf18遺伝子毛包特異的欠損マウスは、生存・成長し、生殖可能であり、健康状態も外観的に良好であった。その背部皮膚に生えている体毛を、短く刈ることにより、特徴的な毛成長サイクルの波を示すことが分かった。図3aに示すのは、Fgf18遺伝子毛包特異的欠損マウスの代表的な個体(ホモノックアウト体) [K5Cretg;Fgf18flox/flox]と、その対象としてFgf18遺伝子の1対2本のうち1本のみが破壊されている対照群の代表的な個体(ヘテロノックアウト体) [K5Cretg;Fgf18+/flox]を、32日齢以降、写真撮影したものである。尚、これらの写真撮影に先行して体毛をバリカンで刈ってあるので、体表の色調がわかるようになっている。32日齢では、背部皮膚はいずれのマウスも初めの生理的成長期にあり、黒い色を示している。37日齢になると、首に近い部分がピンクがかった色になり、休止期に入ったことを示す。40日齢では、背部皮膚全体が休止期である。ホモノックアウト体の場合、47日齢で、次の成長期に入ったことが、皮膚が青みがかった着色を示すことでわかる(図3a)。これに対して、対照のヘテロノックアウト体では、休止期がずっと長く続く(図3a)。
Fgf18遺伝子毛包特異的欠損マウス(68日齢)の背部皮膚における毛成長周期は、前位(首に近い位置)から後位(尾に近い位置)に向かってスムーズに進捗し。毛包の形態形成(図3cのMで示すサイクル)に加えて既に2回のサイクル(図3cの1,2)がまわり、3回目のサイクル(図3cの3)に入ったことが見て取れる(Fig. 3b 左)。 これに対して野生型のマウス(89日齢)では、次の成長期の開始の場所や時期は、個体によって多様であり、一個体の背部皮膚の中においてさえも、「毛成長ドメイン」によって異なる相を示す (Fig. 3b 右,及び図4)。従って、Fgf18遺伝子毛包特異的欠損マウスにおいては、マウスの齢に対する関数として、一般的な毛成長周期における各相の変遷をFig.2aと同様にして示すことが出来る(Fig. 3C)。
ここでは、マウスの首に近い背部皮膚(Fig.3bで点線囲みの部分)における毛成長周期の相を、皮膚の色及び前週の毛成長履歴によって判断し、単純化のために成長期または休止期として表している。その結果、Fgf18遺伝子毛包特異的欠損マウスにおいては、休止期はわずか1週間程度しか持続しないこと、各成長周期は休止期を含めた全体として約3週間であることがわかった。これに対して野生型マウスでは通常、休止期は3から5週間、あるいはより長い期間持続する。
図3dでは、加齢の進んだFgf18遺伝子毛包特異的欠損マウスの代表的な3個体を示す。これらのマウスは背部皮膚の毛を刈ってから1週間飼育し、その一週間に生えた毛を含めた姿を撮影している。特徴的な表現型として、このように加齢が進んだFgf18遺伝子毛包特異的欠損マウスにおいては、同じ成長周期相の毛包が、外観上ストライプを形成するように並んだ。さらに特徴として、ストライプの数は、マウスの日齢に依存して、加齢が進むほど増えていくことが分かった。加齢が進んだマウスにおいて、毛成長周期が一回回るために必要な期間は、前位においては約3週間であったが(Fig. 3d) 、後位においては、約4週間だった (Fig. 3d)。したがって、加齢が進んだFgf18遺伝子毛包特異的欠損マウスにおいて、この前位と後位の間の成長周期長の差が、ストライプ間の距離を縮めていることが理解できる。
【0035】
〔実施例3〕 Fgf18遺伝子毛包特異的欠損マウスにおける不完全な生理的脱毛
休止期の毛包に生えている毛は、それ以前の毛成長周期で完成された毛であり、クラブヘアー(club hair)と称する。一般的にマウスにおいては、休止期毛包に生えているクラブヘアーをグローブをはめた指先でつまみ、緩やかに引き抜くことが出来る。Fgf18遺伝子毛包特異的欠損マウス(ホモノックアウトマウス)においては、このようにして引き抜いた毛が、複数の毛の束になっていることを発見した(図5a、左)。
対象のヘテロノックアウトマウスにおいては、そのような束はまれであった(図5a、右)。引き抜いた毛の長さを、それぞれの群で200本以上測定すると、ホモノックアウトマウス(6.40+/-0.53mm)でもヘテロノックアウトマウス(6.38+/-0.65mm)でも、ほとんど同じであった。さらに毛の種類(guard,awl, and zigzag hairs)の分布にも差異は認められなかった。これらのことから、Fgf18遺伝子毛包特異的欠損マウスにおいて、毛成長周期の成長期は正常に進行していることが強く示唆された。
毛周期が成長期である特徴を示す皮膚の3.7mm2領域内の全毛包とそこに存在する毛軸数を、加齢の進んだFgf18遺伝子毛包特異的欠損マウス(ホモノックアウトマウス)及び対象のヘテロノックアウトマウスにおいて、その背部皮膚の毛包の数及びそこに存在する毛軸数を計測した。(表1)
【0036】
表1:毛包細胞が保持する毛軸数

この結果から見て、Fgf18遺伝子毛包特異的欠損マウスの毛包数は対照群と変わらないものの、それぞれの毛包内に保持される毛軸数の数が多いため、全体で毛軸数が2倍強となっていることが理解できる。
【0037】
さらに、加齢の進んだFgf18遺伝子毛包特異的欠損マウス(ホモノックアウトマウス)及び対象のヘテロノックアウトマウスにおいて、その背部皮膚の毛包の状態を拡大して観察した。すると、成長期の毛包に生えている成長期毛軸に寄り添って、脱落していないクラブヘアーが多数存在していることが明らかになった(図5c)。さらにフルフォーカスイメージングで皮膚を観察したところ、脱落していないクラブヘアーを包む袋様の構造は、成長中の毛軸のバルジ領域付近に一体化したように寄り添っていることが確認された。
また、加齢の進んだFgf18遺伝子毛包特異的欠損マウスにおいて、その背部皮膚の毛包を輪切りにした切片を作成し、チオフラビン T (黄色) 及び DAPI (赤色)で染色し、観察した。すると、ホモノックアウトマウスにおいては、それぞれの毛包にはメラニンに富む成長期毛軸と、多くは3〜6本の、メラニンのないクラブヘアーが共存していることが示された (図5f)。これに対して、対象のヘテロノックアウトマウスにおいては、成長期毛包に含まれるクラブヘアーの本数は多くは1〜2本であった。その分布は、表1に示されるとおりである。これらの観察により、Fgf18遺伝子毛包特異的欠損マウス(ホモノックアウトマウス)においては、クラブヘアーの正常な脱落(脱毛期)のためのメカニズムが不完全になっていることが示唆された。なお、そうではあっても、毛包あたりの残存毛軸数が、完了したと考えられる毛成長周期の数(10 to 12 cycles)よりも少ないことから、 不完全とはいってもある程度の毛軸の脱落は起きていると考えられた。以上より、FGF18活性を欠失させることにより、クラブヘアーの脱毛が阻害されることが初めて示された。
【0038】
〔実施例4〕 FGF18の部分ペプチドによるFGF18活性の阻害
FGF18の部分ペプチドとして、マウスFGF18のアミノ酸配列に対応する配列番号14のN末端から翻訳開始のために導入されたメチオニンを除く4番目まで〜95番目までのアミノ酸を除去した部分ポリペプチド(d4〜d95:メチオニンを除くN末端から除去したアミノ酸数で表す。)を作製した。
なお、それぞれの部分ポリペプチドのアミノ酸配列(塩基配列)は、以下の配列番号に対応する。
d4:配列番号15(配列番号4)
d12:配列番号16(配列番号5)
d16:配列番号17(配列番号6)
d18:配列番号18(配列番号7)
d22:配列番号19(配列番号8)
d37:配列番号20(配列番号9)
d48:配列番号21(配列番号10)
d67:配列番号22(配列番号11)
d77:配列番号23(配列番号12)
d95:配列番号24(配列番号13)
FGF18は4種類のFGFレセプターFGFR1c, FGFR2c, FGFR3c, FGFR4 を刺激するが、それらを刺激する強さは異なり、また、これらの受容体への刺激の総和としての結果として、毛成長サイクルの休止期を長期間保持し、脱毛の開始時期を早め、かつ毛成長サイクルの開始をドメイン毎に制御すると考えられる。
文献の教示にしたがって、マウスIL-3依存性proB細胞であるBa/F3細胞株(理研BRCより入手)に、遺伝子発現操作によりFGFレセプターであるFGFR1c, FGFR2c, FGFR3c, FGFR4のいずれかを導入して、FGFRが細胞表面に強制的に発現されている細胞を作成した(Ornitz,DM.,Xu,J.,Colvin,JS.,McEwen,DG.,MacArthur,CA.,Coulier,F.,Gao,G. and Goldfarb,M.,1996.Receptor specificity of the fibroblast growth factor family.J.Biol.Chem.271(25):15292-15297;Yoneda,A.,Asada,M.,Oda,Y.,Suzuki,M. and Imamura,T.,2000.Engineering of an FGF-proteoglycan fusion protein with heparin-independent,mitogenic activity.Nat.Biotec.18(6):641-644)。
【0039】
これらの細胞を用い、それぞれのレセプターを30 ng/mlのFGF18で刺激している際に、それぞれのFGF18の部分ペプチドを1μg/ml(図3A)または100ng/ml(図3B)の濃度で共存させ、FGF18で刺激された細胞のDNA合成(これを100% とする)を阻害するかどうかアンタゴニスト活性を調べた。すなわち、FGF18による細胞増殖刺激効果を抑制する活性を解析した。その結果を、図3 に示す。各カラムとテスト試料の対応は以下の通りである。
(図6A) カラム1:FGF18以外の試料添加なし(対照)。 カラム2:(d4)。カラム3 :(d16)。カラム4:(d18)。カラム5:(d22)。カラム6:(d37) 。カラム7:(d48) 。カラム8 :(d77)。カラム9:(d95) 。カラム10:(d22) 。カラム11:(d37)。カラム12:(d48)。カラム13:(d67)。カラム14:(d77)。カラム15:(d95)。
(図6B) カラム1:FGF18以外の試料添加なし(対照)。カラム2:(d4)。カラム3:(d12)。カラム4:(d16)。カラム5:(d18)。カラム6:(d37)。カラム7:(d48)。カラム8:(d67)。カラム9:(d95)。カラム10:(d37)。カラム11:(d67)。
図6Aから、披検物質を1μg/mlの濃度で試験すると、d16, d22, d37, d48, d77, d95などがFGFR3c/BaF3細胞上でFGF18の活性を強く阻害することが分かる。またd22, d48, d77, d95などがFGFR4/BaF3細胞上でFGF18の活性を強く阻害することが分かる。他の部分ペプチドも図に示すように様々な阻害効果を示す。
図6Bから、披検物質を100ng/mlの濃度で試験すると、d16, d95などがFGFR3c/BaF3細胞上でFGF18の活性を強く阻害することが分かる。
これらのFGF18の活性阻害物質となるFGF18の部分ペプチドを用いることで、毛成長制御におけるFGF18による毛成長サイクルの休止期制御作用、脱毛開始の制御作用を解除することができるから、毛成長休止期を短縮し、抜け毛を防止し、毛並みをそろえることができる。
また、同様に、マウスFGF18のアミノ酸配列に対応する配列番号14のC末端から25番目まで〜125番目までのアミノ酸を除去した部分ポリペプチド(dc25〜dc125:C末端から除去したアミノ酸数で表す。)を作製した。
なお、それぞれの部分ポリペプチドのアミノ酸配列(塩基配列)は、以下の配列番号に対応する。
dc25:配列番号34(配列番号25)
dc43:配列番号35(配列番号26)
dc57:配列番号36(配列番号27)
dc67:配列番号37(配列番号28)
dc82:配列番号38(配列番号29)
dc94:配列番号39(配列番号30)
dc108:配列番号40(配列番号31)
dc113:配列番号41(配列番号32)
dc125:配列番号42(配列番号33)
これらのFGF18部分ペプチドのうちdc25を除く部分ペプチドはN末端を除去した上記部分ペプチドと同様のFGF18の活性阻害作用を有するため、FGF18による毛成長サイクルの休止期制御作用、脱毛開始の制御作用を解除することができるから、毛成長休止期を短縮し、抜け毛を防止し、毛並みをそろえることができる。
【0040】
〔実施例5〕 FGFR4発現細胞を用いたFGF18活性阻害物質のスクリーニング
作製したFGFR4発現細胞(R4/Ba/F3細胞)を、FGF18存在下で、被験液として種々の植物抽出液と共に培養した。陽性対照としては市販のFGF18蛋白質を用いた。一定時間培養した後の細胞数を、Cell Counting Kit-8(株式会社同仁化学研究所製造、和光純薬工業株式会社販売)を用いて、WST-8ホルマザンの産生量に伴う450 nmの発色を測定して求めた。
その結果、ゴーヤーの抽出物が、FGF18活性阻害物質として作用し、FGF18によるFGFR4/BaF3細胞の増殖を阻害することを見いだした。
【0041】
〔実施例6〕 本発明のFGF18活性阻害物質の細胞増殖阻害活性
乾燥したゴーヤー1.5gに、蒸留水30mlを加え、15分間煮沸した。以上の様にして得られた抽出液をろ紙でろ過してろ液を取り、ゴーヤーの熱水抽出液を得た。乾燥したゴーヤー1.5gに、70%エタノール水溶液30mlを加え、室温で7日間浸漬抽出した。この様にして得られた抽出液をろ紙でろ過し、ゴーヤーの70%エタノール抽出液を得た。
【0042】
実施例5と同様にして、FGFR4/BaF3細胞を用いて本発明のFGF18活性阻害物質の活性測定を行った。具体的には、測定は次の様にして行った。96穴の細胞培養用プレートの各ウエルに、1ウエル当たり50μlの10% FBS、1% Antibiotic G-418 Sulfate(プロメガ株式会社、V7983)を含むRPMI1640培地を加えた。次いで、各試料を水で希釈して調製した種々の濃度の被験液 10μlを添加した後、10%FBS、1% Antibiotic G-418 Sulfateを含むRPMI1640培地中に5×104個のFGFR4/BaF3細胞が懸濁した細胞懸濁液50μlを添加し、軽く撹拌した。更に、heparin/10% FBS/1% Antibiotic G-418 Sulfate/RPMI1640培地、10μl(heparin終濃度5μg/ml)、及びFGF18(ぺプロテック株式会社、100-28)溶液 10μl(FGF18終濃度3ng/ml)を加え、5%CO2存在下、37℃の炭酸ガスインキュベーターで72時間培養した。FGFR4/BaF3細胞の増殖は、72時間培養後の各ウエルに、Cell Counting Kit-8(株式会社同仁化学研究所製造、和光純薬工業株式会社販売)/PBS溶液10μlを加えた後、更に3時間培養し、WST-8ホルマザンの産生量に伴う450nmの発色を測定して求めた。
【0043】
測定に際しては、陽性対照としては、FGF18(ぺプロテック株式会社、100-28)溶液 10μl(FGF18終濃度3ng/ml)を、また陰性対照としては、被験液作製時に使用した水またはエタノール溶液(エタノール溶液の終濃度は1%以下になるように調製)10μl を用いて測定した。細胞増殖阻害率(%)の算定は、以下の式(A)により算出した。
〔式(A)〕
細胞増殖阻害率(%)=100×{(FGF18及び水またはエタノール溶液添加時の吸光度)−(FGF18及び被験液添加時の吸光度)}/FGF18及び水またはエタノール溶液添加時の吸光度−水またはエタノール溶液添加時の吸光度)
【0044】
ゴーヤー熱水抽出液についての結果を図4に示す。FGF18の存在下で、終濃度8.3%のゴーヤー熱水抽出液を被験液とした場合、発色量が減少し、ゴーヤー熱水抽出液はFGF18によるFGFR4/BaF3細胞の増殖を阻害することが確認された。一方、FGF18無添加で被験液添加時の吸光度は、同様にFGF18無添加で水またはエタノール溶液添加時の吸光度に比べてほとんど減少することは無く、被験液による細胞障害作用はほとんど認められなかった。
【0045】
〔実施例7〕 本発明のFGF18活性阻害物質の in vivo 解析
FGF18活性阻害物質の in vivo での作用を見るために、毛周期の休止期にあるC3H/He マウスで試験した。
乾燥したゴーヤー1.1gに、蒸留水15mlを加え、20分間煮沸した。抽出液が室温に戻った後、ろ紙でろ過し、ろ液については同量のエタノールを添加することによって、ゴーヤー熱水抽出液の50%エタノール溶液を作製した。更に0.45μmのマイレクスHV滅菌フィルター(ミリポア社)を通過させ、その後、グリセロールを1%になるように添加し、被験液とした。この様にして得られたゴーヤー熱水抽出液(49.5%エタノール、1%グリセロールを含有)について、毛周期の休止期にあるC3H/He マウス背部皮膚に塗布した。7週齢のC3H/He 雄マウスに麻酔をし、背部毛をバリカンにより優しく剃毛した。剃毛後、被験液としてゴーヤー熱水抽出液(49.5%エタノール、1%グリセロールを含有)、又は水−エタノール−グリセロール溶液(49.5%水、49.5%エタノール、1%グリセロールを含有)200μlをそれぞれ1群5匹のマウス背部皮膚に塗布した(第0日)。同様にして、11日間(5日目、及び6日目を除く)に渡って毎日塗布し、第10日、14日、17日、21日、24日目のマウス背部の発毛状態を目視により観察し、発毛スコアーを得た。発毛スコアーは、1)色素沈着:1点、2)短かな毛並み:2点、3)通常の毛並み:3点とし、それぞれの発毛状態の面積が全剃毛面積に対して占める割合を%で示し、以下の式で求めた。本算出法によると、全剃毛面積が通常の毛並みに回復した場合、100点となる。
発毛スコアー=(色素沈着面積の割合(%)×1+短かな毛並み面積の割合(%)×2+通常毛並み面積の割合(%)×3)/3
第一回の塗布後、第10日、14日、17日、21日、24日目におけるマウス背部の発毛状態を観察した結果、図5に示す通り、ゴーヤー抽出液塗布群は、陰性対照群(水−エタノール−グリセロール溶液(49.5%水、49.5%エタノール、1%グリセロールを含有)塗布群)に比較して特に21日目以降に発毛スコアーが高く、発毛が有意に促進された。
以上の様に、FGF18活性阻害物質であるゴーヤー熱水抽出液にはマウス背部皮膚表面でのin vivo解析においてもFGF18本来の活性を阻害する作用があることが実証された。この実験では、24日目までのデータしか取得していないので、「発毛促進効果」として観察されるが、さらに観察を続ければ、当該効果は毛成長サイクルの休止期が短縮されたこと及び脱毛が開始しないことによる毛量全体の増量効果に基づくものであることが判明するといえる。
【0046】
〔実施例8〕 ゴーヤー熱水抽出物を用いたヘアシャンプー
本発明であるゴーヤー熱水抽出物を用いたヘアシャンプーの処方及び製造法を示す。
以下の処方、及び製法に従ってヘアシャンプーを製造した。
(処方)
成 分 重量%
1.実施例5で得られた希釈液 0.1
2.ラウリルエーテル硫酸ナトリウムエタノール 20
3.ラウリル硫酸ナトリウム 10
4.1,3ブチレングリコール 1
5.香料 適量
6.精製水 残量(全量で100とする)
(製法)
処方成分を80℃に加熱し、攪拌混合した後、攪拌冷却し本発明のヘアシャンプーを製造した。
【0047】
〔実施例9〕 ゴーヤー熱水抽出物を用いたヘアリキッド
本発明であるゴーヤー熱水抽出物を用いたヘアリキッドの処方及び製造法を示す。
以下の処方、及び製法に従ってヘアリキッドを製造した。
(処 方)
成 分 重量%
1.実施例5で得られた希釈液 0.1
2.エタノール 40
3.グリセリン 1
4.香料 適量
5.精製水 残量(全量で100とする)
(製法)
処方成分を加えて攪拌溶解した後、精製水を加えてヘアリキッドを製造した。
【0048】
〔実施例10〕
本発明であるゴーヤー熱水抽出物を用いたヘアクリームの処方及び製造法を示す。
以下の処方、及び製法に従ってヘアクリームを製造した。
(処方)
成 分 重量%
1.実施例5で得られた希釈液 0.1
2.流動パラフィン 40
3.ワセリン 1
4.セトステアリルアルコール 1
5.メチルポリシロキサン 1
6.パラオキシ安息香酸メチル 0.2
7.プロピレングリコール 5
8.香料 適量
9.精製水 残量(全量で100とする)
(製法)
処方成分を攪拌混合し、本発明のヘアクリームを製造した。
【産業上の利用可能性】
【0049】
本発明により、効果的な毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤が提供でき、これらを配合することにより、毛髪成長促進活性を有するシャンプー、ヘアリキッドなどの他、脱毛症治療用の医薬組成物などが提供でき、また非ヒト動物などで、毛の成長サイクルが早まった実験動物や毛並みのそろった愛玩動物が提供できる。
【技術分野】
【0001】
本発明は、毛の成長サイクルにおける休止期を短縮すること及び脱落期の開始を遅らせることによる抜け毛の予防又は治療を目的とするものであり、毛髪が成長休止することを規定している物質の阻害剤及びそのスクリーニング方法に関する。
【背景技術】
【0002】
抜け毛、脱毛は遺伝的な要素が大きいといわれるものの、加齢に伴って加速されるため、男性のみならず女性においても悩む人は多い。脱毛症には加齢男性で多く発症する男性型脱毛症、女性で問題となる事の多いびまん性脱毛症、男女ともに頻度の高い円形脱毛症、さらに抗癌剤など細胞毒性のある薬剤投与によって引き起こされる脱毛症などがある。
これらの問題を解決すべく、従来から発毛効果、育毛効果のある物質として多数の物質が見出され提案されており、既に商品化されている物質も多いが、科学的な臨床実験で発毛効果が確認された育毛剤として「男性型脱毛症診療ガイドライン(2010版)」で推奨された薬剤は、血管拡張作用によって発毛を促すミノキシジル(商品名ロゲイン、リアップ)及び頭皮における男性ホルモン作用を抑制する内服薬(商品名プロペシア)のみである。
男性型脱毛症(AGA)の場合、その主要な原因の1つは、男性ホルモンであるテストステロンが5αリダクターゼの作用でジヒドロテストステロン(DHT)である。DHTが乳毛頭細胞上の男性ホルモン受容体(AR)に結合すると、それによって毛包の矮小化プロセスが始まると説明されているが、その分子機構の詳細は依然として不明である。一説によればDHTによる毛包の矮小化には、細胞増殖因子関連分子であるTGF-β1を介した細胞のアポトーシスが関与しているとされる。通常の毛髪成長サイクルによれば2〜6年は維持されるはずの毛包細胞がアポトーシスにより死滅してしまえば、毛髪は次々と休止期に移行することになる。したがって、テストステロンをDHTに変換する反応を抑制する内服薬の服用は、男性ホルモン過多の傾向にある男性の男性型脱毛症の予防には効果的であるとしても、DHTを抑制してしまうことの副作用を懸念する見方もあり、また男性ホルモン過多が原因となる典型的なAGA以外の脱毛症には効果がない。
ところで、上記のように説明されている男性型脱毛症以外の、びまん性脱毛症、男女ともに頻度の高い円形脱毛症、さらに抗癌剤などの薬剤投与によって引き起こされる脱毛症などにおいては、脱毛症の発症のメカニズムも多様であり、そこに関与する分子も、上述の男性型脱毛症におけるTGF-β1などの細胞増殖因子関連分子、円形脱毛症における免疫細胞関連サイトカインやケモカイン、毛包細胞自体を死滅させてしまう抗癌剤、などの他に、様々な分子があり、それら分子を産生・受容する多種の細胞が関与する。
したがって、根本的な脱毛防止のためには毛髪の成長サイクルにおける詳細な検討が欠かせない。
【0003】
毛の成長サイクル(図1)は、毛を作り出す器官である毛包自体の周期的な変化に規定されている。すなわち毛包は、成長期、退行期、休止期、という3つの相をこの順に繰り返す。このうち成長期には、毛包の先端が皮膚よりも深部の皮下脂肪層にまで到達すると共に、大きく太くなりつつ、長い毛軸を形成し、これを体外に長く伸ばす。これが外観的な毛の成長である。退行期は、毛包の構成細胞がアポトーシスを起こすことに伴い、毛包が退縮し、毛包の先端は皮下脂肪層よりも皮膚表面に近い真皮内部にまで移動する。次に来る休止期においては、毛包の構成細胞はほとんど増殖もアポトーシスも起こさず、小さい状態で静止すると考えられている。これら退行期、休止期において、外観的な毛の成長は止まるが、毛が抜けるわけではない。毛が抜けることは、脱毛期と称される独立の相で独立の制御により起こると考えられているが、脱毛期は、上記の三相と完全に無関係ではなく、休止期後期や次の成長期の途中で起こることが多いとされている。しかし病的な脱毛などでは、これよりも早く起こることもあり、その場合には、外観的にまったく毛の生えていない毛包が長期間存在することになる。この脱毛期を制御する分子メカニズムは、これまでほとんどわかっていない。これらの理由により、毛髪の成長周期を説明する場合に、成長期、退行期、休止期の三相は必ず説明の対象となるが、脱毛期は省略される場合も多い。動物の種によって毛の成長サイクルの各相の持続時間は異なり、同一種内でも、個体、性別、体の部位や毛の種類によっても異なるが、その成長のサイクルは、いずれも図1に示される各相を経由している。ヒトの頭髪の場合は個人差、年齢差はあるが、一般に、成長期は女性で4〜8年間、男性で3〜6年間、退行期は1〜2週間、休止期は3ヶ月間程度とされている。毛の研究に多用されるモデル動物であるマウスの場合、その背部皮膚に存在する毛包においては、成長期は18〜19日間、退行期は2日間、休止期は3〜5週間程度である。
一般的な脱毛症とは、外観的に毛が無い状態、または毛が非常に細く短い状態を意味するので、その原因を毛髪の成長サイクルから見た場合に、脱毛は毛の成長が不十分なこと、または、脱毛が正常よりも早期にまたは過剰に起こっていること、または、前の周期で形成された毛が脱落した後も次の毛の成長が始まらないこと、といえるので、その原因を、(1)成長期の短縮、(2)休止期の延長、(3)脱毛期の早期開始などにわけることができる。毛の成長サイクルにおけるそれぞれの期間を規定する因子として、従来成長期を維持する細胞増殖因子Wntファミリータンパク質(非特許文献1)、退行期を誘導する細胞増殖因子FGF5(非特許文献2)が知られていたが、休止期の維持や制御に係る因子や脱毛期の制御に関わる因子に関する詳細な研究はほとんどなされていなかった。
したがって、脱毛症患者それぞれの脱毛症の原因に個別に対応するためには、毛の成長サイクルの各相で作用する因子の働きを特定する必要がある。また、成長サイクル全体で見れば比較的長時間を要する休止期間を短縮することができれば、完全な抜け毛が起こる前に新しい毛髪が生えることになり、脱毛自体の防止が可能となり、毛髪全体の量も増えいわゆる薄毛予防ともなる。さらに、解明の進んでいない脱毛期を制御する因子が解明できれば、直接的に脱毛時期を遅らせることができることになり、脱毛症患者にとっての大きな福音となる。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】WO2008/102782
【特許文献2】WO2008/102783
【特許文献3】特開平4-224522号公報
【非特許文献】
【0005】
【非特許文献1】Shimizu,H, et al.,J.Invest.Dermatol.122,239-245(2004)
【非特許文献2】Hebert,J.M., et al.,Cell78,1017-1025(1994)
【非特許文献4】Kawano,M., et al.,J. Invest.Dermatol.124,877-885(2005)
【非特許文献5】Ohbayashi,N., et al.,Genes & Dev.16,870-879(2002)
【非特許文献6】Tarutani,M., et al.Proc Natl Acad Sci USA 94,7400-7405(1997)
【非特許文献7】Ornitz DM,Itoh N:Fibroblast growth factors.Genome Biol 2:REVIEWS3005,2001
【非特許文献8】du Cros DL:Fibroblast growth factor and epidermal growth factor in hair development.J Invest Dermatol 101:106S-113S.1993
【非特許文献9】du Cros DL, Isaacs K, Moore GP:Distribution of acidic and basic fibroblast growth factorsin ovine skin during follicle morphogenesis. J Cell Sci 105:667-674,1993
【非特許文献10】Hebert JM,R osenquist T,Gotz J,Martin GR:FGF5 as a regulator of the hair growth cycle:Evidence from targeted and spontaneous mutations.Cell 78:1017-1025,1994
【非特許文献11】Danilenko DM,Ring BD,Yanagihara D,Benson W,Wiemann B,Starnes CO,Pierce GF:Keratinocyte growth factor is an important endogenous mediator of hair follicle growth,development, and differentiation.American J Pathol 147:145-154,1995
【非特許文献12】Marchese C,Chedid M,Dirsch OR, et al:Modulation of keratinocyte growth factor and its receptor in reepithelializing human skin.J Exp Med 182:1369-1376,1995
【非特許文献13】Guo L,Degenstein L,Fuchs E:Keratinocyte growth factor is required for hair development but not for wound healing.Genes Dev 10:165-175,1996
【非特許文献14】Rosenquist TA,Martin GR:Fibroblast growth factor signalling in the hair growth cycle:Expression of the fibroblast growth factor receptor and ligand genes in the murine hair follicle.Developmental Dynamics 205:379-386,1996
【非特許文献15】Petho-Schramm A,Muller HJ,Paus R:FGF5 and the murine hair cycle.Arch Dermatol Res 288:264-266,1996
【非特許文献16】Mitsui S,Ohuchi A,Hotta M,Tsuboi R,Ogawa H:Genes for a range of growth factors and cyclin-dependent kinase inhibitors are expressed by isolated human hair follicles.Br J Dermatol 137:693-698,1997
【非特許文献17】Ortega S,Ittmann M,Tsang SH,Ehrlich M,Basilico C:Neuronal defects and delayed wound healing in mice lacking fibroblast growth factor-2.Proc Natl Acad Sci USA 95:5672-5677,1998
【非特許文献18】Suzuki S,Kato T,Takimoto H,et al:Localization of rat FGF-5 protein in skin macrophage-like cells and FGF-5S protein in hair follicle:Possible involvement of two Fgf-5 gene products in hair growth cycle regulation.J Invest Dermatol 111:963-972,1998
【非特許文献19】Suzuki S,Ota Y,Ozawa K,Imamura T:Dual-mode regulation of hair growth cycle by two Fgf-5 gene products.J Invest Dermatol 114:456-463,2000
【非特許文献20】Nakatake Y, Hoshikawa M, Asaki T, Kassai Y, Itoh N:Identification of a novel fibroblast growth factor,FGF-22,preferentially expressed in the inner root sheath of the hair follicle.Biochem Biophys Acta 1517:460-463,2001
【非特許文献21】Stenn KS,Paus R:Controls of hair follicle cycling.Physiol Rev 81:449-494,2001
【非特許文献22】Beyer TA, Werner S, Dickson C, Grose R:Fibroblast growth factor 22 and its potential role during skin development and repair.Exp Cell Res 287:228-236,2003
【非特許文献23】Kawano M, Suzuki S, Suzuki M, Oki J, Imamura T:Bulge- and basal layer-specific expression of fibroblast growth factor 13(FHF-2) in mouse skin.J Invest Dermatol 122:1084-1090,2004
【非特許文献24】Rendl, M., Polak, L. & Fuchs, E.BMP signaling in dermal papilla cells is required for their hair follicle-inductive properties.Genes & Dev.22,543-557(2008)
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、毛成長サイクルにおける休止期を長期間に制御する内在性因子を決定し、その作用機構を解明すると共に、脱毛期を制御する内在性因子を決定し、当該内在性因子の活性もしくは発現を阻害する物質をスクリーニングすることで、新規な毛成長休止期短縮剤、脱毛抑制剤を提供することを目的としている。
また、そのためのスクリーニング方法も本発明の他の目的である。
【課題を解決するための手段】
【0007】
マウス及びヒトの皮膚組織及び毛包細胞には、22種類の繊維芽細胞増殖因子(FGF)(非特許文献7)のうちで、FGF1、FGF2、FGF5、FGF7、FGF10、FGF13及びFGF22が発現しており、毛髪成長及び皮膚再生を制御していることが報告されている(非特許文献8〜23、特許文献3参照)。これらの文献内容から見て、各種FGFには皮膚細胞の増殖及び分化に重要な役割を担っていることが示唆されるが、毛包の成長を促進及び休止する作用、それに伴う毛髪成長期及び休止期制御作用及び脱毛促進作用に対して、FGF群の個々の因子がそれぞれどのように関与するのかといった知見は依然として多くは不明のままである。
上記の背景にあって発明者らは以前、FGF18が毛包を含む皮膚で高いレベルで発現することを見出し、毛成長周期の休止期で特に高い発現レベルであることを報告した(非特許文献4)。そして、FGF18をマウス休止期皮膚毛包において、抜毛により強制的に毛包の成長期を誘導したあとに、FGF18を背部皮下に持続的に存在させることで、毛の成長を著しく阻害することを見いだし、FGF18及びその活性化物質からなる毛髪成長阻害剤、及びFGF18阻害物質からなる発毛剤について特許出願している(特許文献1,2)。このように、毛包細胞に外部から導入したFGF18遺伝子産物の毛成長に及ぼす影響については解明されてきているが、毛包細胞に内在しているFGF18遺伝子が毛包細胞内でどのようなメカニズムで発現し、毛の成長周期にどのように係わっているのか、という遺伝子機能についてはよくわかっていなかった。
内在遺伝子の機能を直接的に解明する手法として遺伝子ノックアウト技術があるが、FGF18は、軟骨細胞や骨芽細胞の増殖分化や肺の形成など、個体形成や生命維持に不可欠な因子であるため、通常の全身ノックアウトを行うと、個体の機能的形成が正常に起こらず、胎生期または出生直後に死亡することが知られている(非特許文献5)。
そこで、本発明者らは、皮膚とその付属器官においては毛包で選択的に高いレベルで発現しているFGF18遺伝子を毛包特異的にノックアウトすることを想起し、毛包細胞がケラチン細胞の1種であることから、ケラチン5陽性細胞特異的な遺伝子ノックアウト法を、フォスファチジルイノシトールグリカンアンカー合成に関わるPig-a遺伝子に対して適用した技術(非特許文献6)を参考にし、ケラチン5陽性細胞特異的にFgf18遺伝子をノックアウトしたマウスを得た。ここで、ケラチン5は、ケラチン14と同様に、皮膚と毛包など皮膚付属器官の中では表皮、毛包、皮脂腺など多くの細胞で発現しているがFgf18を高いレベルで発現するのは、この中で毛包だけであるため、得られたコンディショナルノックアウトマウスは、実質的に「毛包特異的Fgf18遺伝子欠損マウス」と表現することができる。
毛包特異的FGF18欠損マウスは、健康に育ち、生殖能力も備えており、外観的に検知しうる障害を有さないが、驚くべきことに、毛の成長サイクルの進行の状態が、野生型と著しく異なることを確認した。すなわち、通常の野生型マウスでは3〜5週間以上も続く毛の成長サイクルの休止期が、毛包特異的FGF18欠損マウスでは約1週間で終了し、一周期に要する期間も約3週間にまで短縮されていた。また、毛包内のクラブヘアの脱落が防止され、脱落期の開始時期が遅くなった結果、毛包あたりの毛軸数が全体で2倍以上増加した。しかも、毛成長周期の進行が体表上の「毛周期ドメイン」構造の影響を受けにくく極めてスムーズに進行し、加齢が進むにつれ複数の縞模様をなすような毛成長フェーズの繰り返しが個体皮膚に現れる。このため、毛包特異的FGF18欠損マウスを用いれば、野生型のマウスにおいては同一個体皮膚表面で再現性良く行うことが極めて困難であった毛髪関連疾患や各種皮膚疾患の予防、治療物質あるいは発毛や脱毛を促進・抑制する物質の評価・スクリーニングを行うことができることを見出し、本出願と同日付で特許出願した。
【0008】
そしてこれらの結果から見て、毛包内の内在性FGF18には、毛の成長サイクルの休止期を延長させることで、毛の成長サイクル自体を延長させる作用と共に、毛包内のクラブヘアを脱落させるという脱毛期開始作用があるといえることから、FGF18こそが休止期延長制御因子であり、かつ脱毛促進因子であることが明らかとなった。
したがって、毛包内の内在性FGF18遺伝子の発現を抑える物質又はFGF18蛋白質の活性を抑える物質を毛包に投与することで、毛の成長サイクルの休止期を短縮して毛の成長サイクル全体の速度を速めることができ、また脱毛、抜け毛を防止することができる。つまり、毛包内の内在性FGF18遺伝子の発現を抑える物質又はFGF18蛋白質の活性を抑える物質を検索することで、毛の成長サイクルの休止期短縮剤であり、脱毛防止剤を提供できることを見出した。
以上の知見を得て、本発明を完成させた。
【0009】
すなわち、本発明は以下を包含する。
〔1〕 FGF18の活性阻害物質及び/又は発現抑制物質を有効成分として含有する毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔2〕 前記FGF18の活性阻害物質がFGF18の部分ペプチドであることを特徴とする前記〔1〕に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔3〕 前記FGF18の活性阻害物質が抗FGF18抗体であることを特徴とする前記〔1〕に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔4〕 前記FGF18の活性阻害物質がゴーヤー抽出物であることを特徴とする前記〔1〕に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔5〕 前記FGF18の活性阻害物質が、FGF18の活性を阻害する活性を有するFGF18の部分ペプチドをコードするcDNAを組み込んだ発現ベクターであることを特徴とする前記〔1〕に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔6〕 前記FGF18の発現抑制物質が、FGF18の発現を阻害する活性を有するsiRNAを組み込んだ発現ベクターであることを特徴とする前記〔1〕に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔7〕 他の毛髪成長促進剤、発毛促進剤、又は脱毛抑制剤を更に含むことを特徴とする前記〔1〕ないし〔6〕のいずれかに記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
〔8〕 前記〔1〕ないし〔4〕のいずれかに記載のFGF18の活性阻害物質をスクリーニングし、被検物質を毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤の候補とする方法であって、以下の(a)〜(c)の工程を含む方法;
(a)FGFR1c、FGFR2c、FGFR3c及びFGFR4から選ばれた少なくとも1つのFGF受容体遺伝子を用いて遺伝子操作により細胞表面に強制的に発現させ、当該細胞を培養する工程、
(b)(a)工程で得られた、細胞表面にFGF受容体を有する細胞系にFGF18とともに披検物質を接触させる工程、
(c)(b)工程において、FGF18の細胞増殖促進活性を阻害する活性を観察した披検物質を選択する工程
〔9〕 前記〔8〕に記載のFGF受容体がFGFR3cである前記〔8〕に記載のスクリーニング方法。
〔10〕 前記〔8〕に記載のFGF受容体がFGFR4である前記〔8〕に記載のスクリーニング方法。
〔11〕 前記〔8〕に記載のFGF受容体を細胞表面に強制的に発現させるための細胞がマウスIL-3依存性Ba/F3細胞株である前記〔8〕ないし〔10〕のいずれかに記載のスクリーニング方法。
〔12〕 前記〔1〕に記載のFGF18の発現抑制物質をスクリーニングし、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤の候補とするための方法であって、下記の(a)〜(d)の工程を含む方法;
(a)FGF18遺伝子が観察可能な程度発現する動物培養細胞又は実験動物を用意する工程、
(b)被検物質を、(a)の動物培養細胞に接触させるか又は実験動物に接触もしくは投与する工程、
(c)(b)工程後の動物培養細胞又は実験動物におけるFGF18遺伝子の発現をモニターする工程、
(d)FGF18遺伝子の発現を阻害する機能を有する被検物質を選択する工程。
〔13〕 前記(c)工程において、前記(b)工程後の実験動物又は動物培養細胞から、mRNAを抽出し、発現されたFGF18のmRNA量を解析することによりFGF18遺伝子の発現をモニターし、前記(d)工程において、披検物質を作用させない場合と比較して低いレベルのFGF18のmRNA量を観察した系を選択することにより、FGF18遺伝子の発現を阻害する機能を有する被検物質を選択することを特徴とする、前記〔12〕に記載の方法。
【発明の効果】
【0010】
本発明において、毛包内の内在性FGF18遺伝子の発現を抑える物質又はFGF18蛋白質の活性を抑える物質を検索し、これらの物質を毛包に投与することで、毛の成長サイクルの休止期を短縮することができ、かつ毛の成長サイクル全体の速度を速めることができる。また、脱毛期を遅らせて、抜け毛自体を防止することができる。つまり、完全な抜け毛が起こる前に新しい毛髪が生えることになり、しかも脱毛自体の防止が可能となるため、毛髪全体の量が増えて、いわゆる薄毛予防ともなる。
【図面の簡単な説明】
【0011】
【図1】毛成長周期 図中、「anagen(成長期)」は毛包の成長と毛軸形成が進行する時期、「catagen(退行期)」は毛包が退縮する時期、「telogen(休止期)」は毛包の活動が休止する時期を表し、「anagen」が「propagating anagen」及び「autonomous anagen」に、「telogen」が「refractory telogen」及び「competent telogen」に分けられることを示す。なお、外観的に毛軸が脱落する「脱毛期(exogen)」は、退行期や休止期と直接連結するのではなく、その毛軸が完成した成長期の次の休止期又はその次の成長期の最中に、独立した相として起こるとされている。
【図2】FGF18遺伝子ノックアウト模式図 a:マウスFGF18遺伝子ゲノム第3エキソン付近の構造 図中、太い黒い線は第3エキソンを、線部分はイントロン構造を示す。EcoRIは、制限酵素EcoRIの位置を示す。 b:ターゲティングベクター構造ならびにゲノム相同組換えの概略図。 図中、「KIprobe」はサザンブロット法による組換え判定・遺伝子型判定などに用いるDNAプローブであり、その相同位置を太線で示す。ボックスで囲んだPGK-Neorは、neomycin耐性遺伝子のカセットを示し、黒三角はloxP配列を示し、灰色三角はFRT配列を示す。 c:bの配列からFlp recombinaseによりFRT配列で囲まれたneomycin耐性遺伝子のカセットが除去された様子を示す。 d:cの配列からCre recombinaseによりloxP配列で囲まれたFgf18遺伝子の第3エクソンが除去された様子を示す。 e:サザンブロット解析の結果の一例。hetero、homo、及び野生型の解析結果を示す。
【図3】毛包特異的FGF18欠損マウス aとbは、毛包特異的FGF18欠損マウス(K5Cretg;Fgf18flox/flox:ホモノックアウトマウス)において速やかに繰り返される毛成長周期(1) a:若齢における毛包特異的FGF18欠損マウス(ホモノックアウトマウス:下段)及び対照のヘテロノックアウトマウス(K5Cretg;Fgf18+/flox:上段)のそれぞれ代表的な一個体について、背部皮膚の毛成長周期の変遷を経時的に観察したもの。各写真とも、生えている毛をバリカンで短く刈ってから撮影しており、皮膚の色が毛成長周期状の相を反映している。 b:毛包特異的FGF18欠損マウス(ホモノックアウトマウス:左)及び対照の野生型マウス(C57BL/6:右)のそれぞれ代表的成体一個体について、それらの背部皮膚における発毛パターンの一例を示したものである。 cとdは、毛包特異的FGF18欠損マウス(K5Cretg;Fgf18flox/flox:ホモノックアウトマウス)において速やかに繰り返される毛成長周期(2) c:毛包特異的FGF18欠損マウス(ホモノックアウトマウス)の背部皮膚における毛成長周期の変化(Y軸)を、マウスの日齢(X軸)に対してプロットしたものである。 d:加齢の進んだ毛包特異的FGF18欠損マウス(ホモノックアウトマウス)の背部皮膚における毛成長周期の変遷を、概ね1週間毎に観察したものであり、各写真とも、観察日前の1週間の毛成長を反映している。
【図4】ヘアサイクルドメインの例示 図はマウスの背部皮膚に存在するヘアサイクルドメインを表す。マウス背部皮膚には、図中の縦横線で仕切られた区切りのようにヘアサイクルドメインが存在し、それぞれのドメイン毎に毛周期の制御が異なることが経験的に知られている。ただし、図中の縦横線の数や位置は例示であり、実際は様々なパターンがある。
【図5】毛包特異的FGF18欠損マウス aとbは、加齢の進んだ毛包特異的FGF18欠損マウス(ホモノックアウトマウス)におけるクラブヘアの不完全な脱落(1) a:左:加齢の進んだ毛包特異的FGF18欠損マウス(ホモノックアウトマウス)において、背部から指で抜去した毛の状態を示したものである。右:対照として左と同齢のヘテロノックアウトマウスから抜去したクラブヘアーの状態。 b:成長期毛包周囲における毛包の状態。なお、図中バーは200μmを表す。 cとdは、加齢の進んだ毛包特異的FGF18欠損マウスにおけるクラブヘアの不完全な脱落(2) c:休止期毛包周囲における毛包の状態。なお、図中バーは200μmを表す。 d:cの枠囲み部分の拡大図 eとfは、加齢の進んだ毛包特異的FGF18欠損マウスにおけるクラブヘアの不完全な脱落(3) e:成長期毛包を含む皮膚の切片の免疫染色写真。ケラチン15とPCNA抗原に対する二重染色である。PCNA陽性の成長期毛包のバルジ領域に寄り添うクラブヘアの毛包が観察される。 f:eの視野の一部のケラチン15のシグナルのみを表したものである。ケラチン15は皮膚幹細胞に特徴的な抗原であり、成長期毛包のバルジ領域とクラブヘアを包む袋状構造がいずれもケラチン15陽性で連続的な構造体を成していることが分かる。 gとhは、加齢の進んだ毛包特異的FGF18欠損マウスにおけるクラブヘアの不完全な脱落(4) g:加齢の進んだ毛包特異的FGF18欠損マウス(ホモノックアウトマウス)背部皮膚の成長期毛包周囲の輪切り切片の蛍光観察像。毛軸のタンパク質を染める蛍光色素チオフラビンTで染色してある。なお、図中バーは100μmを表す。 h:eと同じ試料の拡大観察像図。位相差観察像とマージしてある。なお、図中バーは100μmを表す。矢印はメラニンを含む成長期毛包に生える毛軸、それ以外はクラブヘアーを示す。 iとjは、加齢の進んだ毛包特異的FGF18欠損マウスにおけるクラブヘアの不完全な脱落(5) i:対照としてe,fと同齢のヘテロノックアウトマウス背部皮膚の成長期毛包周囲の輪切り切片の蛍光観察像。毛軸のタンパク質を染める蛍光色素チオフラビンTで染色してある。なお、図中バーは200μmを表す。 j:gと同じ試料の拡大観察像。位相差観察像とマージしてある。なお、図中バーは100μmを表す。矢印はメラニンを含む成長期毛包に生える毛軸、それ以外はクラブヘアーを示す。
【図6】FGF18がFGF受容体(FGFR)発現細胞の増殖を促進する効果が、FGF18部分ペプチドにより抑制されることを示す図である。FGFR3c発現細胞(R3c)、FGFR4発現細胞(R4)を用いた解析結果を示す。 A:試験物質を1μg/mlで加えた場合の結果 <対照> カラム1:FGF18以外の試料添加なし。 <R3c> カラム2:(d4)。カラム3:(d16)。カラム4:(d18)。カラム5:(d22)。カラム6:(d37)。カラム7:(d48)。カラム8:(d77)。カラム9:(d95)。 <R4> カラム10:(d22)。カラム11:(d37)。カラム12:(d48)。カラム13:(d67)。カラム14:(d77)。カラム15:(d95)。 B:試験物質を100ng/mlで加えた場合の結果 <対照> カラム1:FGF18以外の試料添加なし。 <R3c> カラム2:(d4)。カラム3:(d12)。カラム4:(d16)。カラム5:(d18)。カラム6:(d37)。カラム7:(d48)。カラム8:(d67)。カラム9:(d95)。 <R4> カラム10:(d37)。カラム11:(d67)。
【図7】ゴーヤー熱水抽出液のR4/BaF3 細胞増殖阻害作用:FGF18 存在下でFGFR4/BaF3 細胞をゴーヤー熱水抽出液とともに培養することにより、ゴーヤー熱水抽出液0.083%、0.83%及び8.3%の添加によるFGF18のFGFR4/BaF3細胞の増殖促進作用に対する阻害効果を示す。
【図8】ゴーヤー熱水抽出液のin vivo 解析:ゴーヤー熱水抽出液のエタノール溶液(49.5%エタノール、1%グリセロールを含有)を毛周期の休止期にあるC3H/He マウス背部皮膚に11日間(5日目、及び6日目を除く)に渡って毎日塗布し、第10日、14日、17日、21日、24日目のマウス背部の発毛状態を発毛スコアーで示す。
【発明を実施するための形態】
【0012】
以下、本発明を詳細に説明する。
1.毛包内の毛の成長サイクル
毛の成長サイクル(図1)は、毛を作り出す器官である毛包自体の周期的な変化に規定されている。すなわち毛包は、成長期(anagen)、退行期(catagen)、休止期(telogen)、という3つの相をこの順に繰り返す。成長期(anagen)は、さらに「propagating anagen」及び「autonomous anagen」に、休止期(telogen)は「refractory telogen」及び「competent telogen」に分けられるとされる。このうちの成長期には、毛包の先端が皮膚よりも深部の皮下脂肪層にまで到達すると共に、大きく太くなりつつ、長い毛軸を形成し、これを体外に長く伸ばす。これが外観的な毛の成長である。退行期は、毛包の構成細胞がアポトーシスを起こすことに伴い、毛包が退縮し、毛包の先端は皮下脂肪層よりも皮膚表面に近い真皮内部にまで移動する。次に来る休止期においては、毛包の構成細胞はほとんど増殖もアポトーシスも起こさず、小さい状態で静止すると考えられている。有色体毛を持つマウスでは、成長期の初期にはメラニン色素が合成され、皮膚が青くなっているのを視認することができる。したがって、皮膚の外側から青い色を見て毛包の成長周期の進行を評価することもできる。また、成長期に皮膚を切開して裏側から見ると、メラニン色素を多量に含んだ毛包が高い密度で並ぶこととなるため、皮膚の裏側が黒なっているのを視認することができる。これに対して休止期においては、皮膚の裏側は白いままであることを視認することができる。一般に、マウスの実験系では、成長期は脱毛後1〜19日目の期間であり、退行期は20〜21日目の期間である。また、脱毛後、21〜22日目に休止期に入り、休止期は3〜5週間以上続くことが知られている。本発明前に、休止期(telogen)のうち最初の短期間である「refractory telogen」の制御因子として、細胞増殖因子の一群であるBMP(bone morphogenetic protein)シグナルが関わっていることが報告された(非特許文献24)。しかし、休止期全体のうち大きな割合を占め、「refractory telogen」に続く時期である「competent telogen」の制御因子についてはまったく知られていなかった。本発明においてはじめて「competent telogen」の制御因子がFgf18因子であることを見出した。従って実質的に休止期を延長する制御因子がFgf18因子であることを、本発明においてはじめて見出した。なお、Fgf18因子が「refractory telogen」の制御因子としても機能を果たしている可能性は存在し、今後の検証が待たれる。
これら退行期、休止期において、外観的な毛の成長は止まるが、毛が抜けるわけではなく、毛が抜けることは、脱毛期(exogen)と称される独立の相で独立の制御により起こると考えられているが、この脱毛期を制御する分子メカニズムは、これまでほとんどわかっていなかった。本発明において、はじめてこの脱毛期開始を制御する因子の1つがFgf18であることを見出した。
【0013】
2.毛包特異的FGF18遺伝子ノックアウトマウスの製造について
本発明の「毛包特異的FGF18遺伝子ノックアウトマウス」は、以下のようにして作成される。
マウスFgf18遺伝子におけるエクソン3を、Cre-loxP法を用いてケラチン5陽性細胞特異的にノックアウトする。エクソン3を欠失させることで、Fgf18蛋白質の分泌シグナルの一部とその下流が発現されず、Fgf18蛋白質の生物活性が失われる。FGF18遺伝子をケラチン5陽性細胞特異的に欠損する方法は非特許文献6を参考に行った。
マウス由来Fgf18遺伝子のエクソン3を含むゲノム領域の塩基配列は、GenBankなど公的データベースから取得できる(Entrez Gene;Gene ID:14172,updated on 21-Jul-2010;Official Symbol:Fgf18 provided by MGI;Official Full Name:fibroblast growth factor 18 provided by MGI;Primary source:MGI:1277980)ので、その上流に、2つのFRT配列で挟まれたネオマイシン耐性遺伝子カセットを挿入し、このネオマイシンカセットとエクソン3が2つのloxP配列で挟まれたターゲティングベクターを作製した(図1)。このターゲティングベクターを用い、マウスES細胞に対してFgf18遺伝子エクソン3アリルをターゲティングした。この細胞をC57BL/6マウスの胚盤胞に注入したものをICRマウスの子宮内に移植して発生させ、得られたキメラの産子をC57BL/6マウスと交配して得られた産子の中から、導入したターゲット遺伝子を有する個体を選択した(F1)。F1の個体をFlpe トランスジェニックマウス個体(理研BRCから分譲:RIKEN RBRC01834;非特許文献7)と交配して、FRT配列で挟まれたネオマイシン耐性遺伝子カセットを欠失した個体を得た(F2)。次にF2マウスをK5Creトランスジェニックマウス(熊本大学CARDから分譲:CARD ID323;非特許文献6)と交配させてケラチン5陽性細胞特異的にFgf18遺伝子エクソン3を除去した。得られたK5Cretg; Fgf18+/floxマウス同士を交配しK5Cretg; Fgf18flox/floxマウスを得た。
ところでケラチン5は、ケラチン14と同様に、皮膚と毛包など皮膚付属器官の中では表皮、毛包、皮脂腺など多くの細胞で発現している。ところがFgf18を高いレベルで発現するのは、この中で毛包だけである。従って、上記のようにして得られたコンディショナルノックアウトマウスは、実質的に毛包特異的にFgf18遺伝子をノックアウトしたマウスと表現することができる。
【0014】
3.毛包特異的Fgf18遺伝子ノックアウトマウスの特性
(3−1)全体の特徴
FGF18遺伝子はマウスなど哺乳動物では骨、心筋、胎児肺、その他の組織に発現し、個体形成や生命維持に重要な働きをしていると考えられる。しかし、皮膚と毛包など皮膚付属器官の中でFgf18を高いレベルで発現するのは、毛包だけである。そして、ケラチン5遺伝子を高いレベルで発現するのは、皮膚の他では、骨、舌、気管支など、ごく一部の組織であるため、上記のようにして得られたコンディショナルノックアウトマウスにおいて、天然型のマウスに比べて実質的にFgf18遺伝子の生理的機能が失われていると言えるのは毛包のみである。
本発明で得られた毛包特異的Fgf18遺伝子ノックアウトマウスは、個体の健康状態はすこぶる良く、具体的に観察された特性は以下の通りである。
1)毛の成長周期のうち休止期が1/3〜1/5に短縮されたため、毛の成長周期全体の進行が早い。
2)背部皮膚での毛成長周期が帯状に整然と進行する。その際の進行速度が首に近い前方部が最も早く尾部後方部が最も遅いが、その進行速度の速まり方が帯状に整然と進行するため、剃毛を繰り返すと複数の横縞を形成する。その際の横縞数は加齢に比例する。
3)毛包細胞数は増加しないが、毛包からの脱毛速度が遅くなり、結果的に毛包内の毛軸の本数が多く、皮膚表面の全体の毛軸数は野生型の2倍以上となる。
【0015】
(3−2)毛成長周期の進行が早い特性
本発明の毛包特異的Fgf18遺伝子ノックアウトマウスは野生型に比べ、遙かに早い周期で毛成長が起きる。特に、毛成長周期のうち、休止期が短く(図3c)約1週間で終了し、次の成長期に入ることが示されている。これに対し、野生型マウスの例では、図3cの48日齢頃から始まる休止期が通常3〜5週間続くことが良く知られている。その結果として、一サイクルに要する期間も野生型マウスが6週間から8週間程度またはそれ以上であるのに対して、約3週間にまで短縮されていた。
すなわち、毛包内の内在性Fgf遺伝子の発現が阻害されたことで、毛成長サイクルの休止期が短縮し、同時に毛成長サイクル自体の速度が速まったことがわかる。
【0016】
(3−3)毛成長周期相の固有の表現型
従来からマウスの背部皮膚での毛成長周期が首に近い前方部が最も早く尾部後方部が最も遅い傾向にあることは知られていた。しかし、毛の成長周期の開始時期は背部皮膚の複数の位置で始まるため、野生型マウスの背部皮膚を剃毛すると、図2b右にあるように、背部皮膚全体的にランダムに発毛が始まる。このことの説明のために、マウスの背部皮膚において「ヘアサイクルドメイン」と呼ばれるブロック状に分かれて毛成長周期を決定する制御系の存在を想定することが一般的であった(図4)。マウスの背部皮膚には図4の縦横線で仕切られた区切りのようにヘアサイクルドメインが存在し、それぞれのドメイン毎に毛周期の制御が異なることが経験的に知られている。ただし、図4における縦横線の数や位置は例示であり、実際には様々なパターンがある。
一方、本発明の毛包特異的Fgf18遺伝子ノックアウトマウスは、このようなヘアサイクルドメインを示さず、ブロック状に分かれて毛成長周期を決定する制御系を決定づける因子が失われた可能性が大きく、毛成長周期が首に近い前方部が最も早く尾部後方部が最も遅いという本来の毛成長周期の進行速度の傾向がそのまま発現し、その結果、加齢が進むに従って各相の毛包がシマウマのストライプのように整然と配列した様相を示す。
すなわち、毛包内の内在性FGF遺伝子の発現が阻害されたことで、ヘアサイクルドメインのうちでも、特にマウスの体軸に対して直交する方向のドメイン間の境界線を失わせたことがわかる。
【0017】
(3−4)毛包内の毛軸の本数が多い特性
本発明の毛包特異的FGF18遺伝子ノックアウトマウスは、皮膚の単位面積あたりの毛包数は増加しないが、毛成長周期の進行が早まるにもかかわらず、脱毛期の開始時期が遅くなり、前周期の産物としての毛の脱落が起こりにくい。毛包内では、成長期の毛包に生えている成長期毛軸に寄り添い、脱落していないクラブヘアが多数存在している状態となる。すなわち、毛包内の内在性FGF遺伝子の発現が阻害されたことで、脱毛期の開始時期が遅くなり、クラブヘアの脱落が防止できたことがわかる。
【0018】
4.毛包細胞におけるFGF18遺伝子及び遺伝子産物であるFGF18蛋白の作用について
Fgf18遺伝子のノックアウトマウスの表現型を野生型のマウスと比較解析したことではじめて、毛包細胞内の内在性FGF18遺伝子、またはその発現産物であるFGF18の毛包における制御作用が解明できてきた。FGF18には具体的には以下の3つの作用があると推定される。
1)FGF18は毛包の休止期を維持する機能を有する。すなわち、毛包内の内在性FGF18は、毛の成長周期の休止期を延長させる制御因子であると考えられる。
2)FGF18はクラブヘアを皮膚から脱落させる機能を有する。すなわち、毛包内の内在性FGF18は、今まで未解明であった毛の脱落期を開始させる制御因子であると考えられる。
3)FGF18は動物の体軸(仮にX軸とする)に対して直交する方向(仮にY軸とする)を共有するヘアサイクルドメイン間の境界線を決定づける機能を有する。すなわち、皮膚表面のヘアサイクルドメインを決定する制御因子、すなわち毛周期の開始時期を制御する制御因子の1つであると考えられる。
【0019】
5.FGF18活性阻害物質
上記4.に示すように、毛包内で発現する内在性FGF18には1)〜3)の作用があることから、当該内在性FGF18の作用を阻害又は抑制する物質、すなわち「FGF18活性阻害物質」には以下の作用効果が期待できる。
1)毛髪の成長サイクルの休止期を短縮させると共にその結果毛髪の成長サイクル全体の速度を速める作用を持つと考えられるから、毛髪成長サイクルの休止期短縮剤又は毛髪成長サイクル促進剤としての作用効果を有する。
2)毛髪の脱落期を遅延させる作用があるといえるから、脱毛抑制剤、抜け毛防止剤又は抜け毛予防剤などとしての作用効果を有する。
3)毛の成長サイクルの開始時期について、体軸と直交する方向でのばらつきを抑え、体軸方向のみの整然とした成長サイクルでの発毛を促すため、毛並みをそろえる作用効果がある。
つまり、本発明は、毛髪成長サイクルの休止期短縮剤もしくは毛髪成長サイクル促進剤(あわせて「毛成長休止期短縮剤」という)、脱毛防止剤、抜け毛防止剤もしくは抜け毛予防剤(あわせて「抜け毛防止剤」という)、又は「毛並み調整剤」に係るものである。
本発明に係る「毛成長休止期短縮剤」、「抜け毛防止剤」又は「毛並み調整剤」の作用を有するFGF18活性阻害物質としては、例えばFGF18の部分断片であっても良い。
FGF18は、ヒトとマウスでは産生細胞の細胞質で207アミノ酸のポリペプチドとして合成され、それが細胞外に分泌される際にN末端のシグナルペプチドが切断され、181アミノ酸より成る分泌体として生理作用を発揮する。そして、7種類あるFGF受容体サブクラス(FGFR1c、FGFR1b、FGFR2c、FGFR2b、FGFR3c、FGFR3b、FGFR4)のうち少なくとも4つ、すなわちFGFR1c、FGFR2c、FGFR3c、FGFR4と反応する。
本発明において「FGF18」というとき、ヒト由来に限定されず、例えば、他の哺乳動物由来のFGF18を使用することができる。他の哺乳動物としては、マウス、ラット、ニワトリ、七面鳥、ウシ、ブタ、ヒツジ及びウサギ等を挙げることができるが、これらに限定されない。例えば、配列番号1に示すヒト由来FGF18の塩基配列に基づいて作製したプローブを定法に従って用いれば、ヒトを除く他の哺乳動物からFGF18をコードする遺伝子を単離することができる。
シグナル配列除去後のヒト由来FGF18の塩基配列及びアミノ酸配列をそれぞれ配列番号1及び2に示す。シグナル配列除去後のマウス由来FGF18の塩基配列及びアミノ酸配列をそれぞれ配列番号3及び14に示す。これら配列番号2及び14に示すアミノ酸配列を比較すると判るように、哺乳動物においてFGF18は非常に高い相同性を有しており、哺乳動物におけるFGF18の機能はほぼ同様であると理解することができる。
【0020】
ところで、一般にアンタゴニストとは、受容体には結合はするが、その受容体を刺激する生体物質(リガンド)とは異なり生体反応を起こさないかまたは比較的弱い生体反応しか起こさず、また本来結合すべき生体内物質と受容体の結合を阻害し、生体応答反応を引き起こさないかまたは減弱させる物質をいう。FGF18は、上述のように、7種類あるFGF受容体サブクラスのうちの少なくともFGFR1c、FGFR2c、FGFR3c、FGFR4と反応するので、本発明において「FGF18アンタゴニスト作用」というときは、FGFR1c、FGFR2c、FGFR3c、FGFR4のいずれかの受容体を発現している細胞に対して、FGF18による応答を引き起こさないかまたは減弱させる物質をいう。FGF18アンタゴニストがFGF18の受容体への結合を妨げることになってFGF18に起因する毛髪成長サイクルにおける休止期の延長作用、毛髪の脱落期開始作用などを阻害し、結果的に毛髪成長サイクルの促進効果、毛髪脱落防止効果を有することになる。
FGF18アンタゴニスト作用を有するFGF18部分ペプチドの場合としてヒトFGF18の場合を例にとれば、配列番号2に示すアミノ酸配列における32〜151番目の領域は、FGFファミリーで共通性の高いコア配列と呼ばれる領域であって、この領域だけで構築される3次元構造は受容体との基本的な結合能を有しているが完全な活性を持たないと考えられることから、当該領域に対応する領域のアミノ酸配列からなる部分ペプチドは、FGF18アンタゴニスト作用を有すると考えられる部分ペプチドである。当該領域に対応するコア配列を含むFGF18部分ペプチドであって、FGFR4などの受容体への結合能は十分有しており、かつFGF18応答を引き起こすほどに十分な長さを有していない場合は、FGF18アンタゴニスト作用を有する。
実際に実験的に確かめた結果からは、図3に示されるように、配列番号14において、N末端から翻訳開始のために導入されたメチオニンを除く16個欠失した場合でもアンタゴニスト作用を及ぼし、好ましくは22個、より好ましくは77個、最も好ましくは95個のアミノ酸を欠失した部分ペプチドはさらに強いFGF18アンタゴニスト作用を有するということができる。すなわち、少なくともN末側のアミノ酸配列については、95個まで欠失しても受容体結合能を失わず、むしろアンタゴニスト作用が強まっているといえるから、N末側のアミノ酸配列は32番目から始まる上記コア配列の全てがFGF受容体の結合に必須の配列ではないことは明らかであり、N末側アミノ酸をさらに除去した場合であってもFGF18アンタゴニストであるといえる。また、このことからみて、C末側についても、コア配列のC末側に相当する、C末端から31番目までのアミノ酸を欠失した場合の部分ペプチドにFGF18アンタゴニスト作用があることは明らかである上に、さらにC末側アミノ酸配列を大きく欠失させた場合も、受容体結合能を完全には失わないことが十分期待できるから、C末端から31個以上の、例えば43個、57個、67個、82個、94個、108個、113個、125個などのアミノ酸を欠失した部分ペプチドも強いFGF18のアンタゴニスト作用のある部分ペプチドであると考えられる。そして、これはN末側とC末側を同時に除去しても当該アンタゴニスト作用が保たれることはもちろんであるから、少なくとも32〜151番目のアミノ酸配列を含有するペプチドのみならず、むしろ好ましくは77〜151、より好ましくは95〜151番目のアミノ酸配列を含むペプチドもFGF18アンタゴニスト作用を有するということができる。
【0021】
また、FGF18又はFGF18反応性受容体に結合する抗体、すなわち、抗FGF18抗体や抗FGFR3c抗体、抗FGFR4抗体も同様にFGF18とその受容体との結合を阻害するので、FGF18による毛包内の毛成長サイクルの休止期を長期に亘って保持する作用、毛髪の脱落の開始促進作用又は毛の成長サイクルスタート時期をヘアサイクルドメイン毎に規定する作用を阻害することになる。
そこで、本発明では、上記FGF18アンタゴニストも含めて、FGF18とその受容体との結合を阻害する物質など、FGF18の受容体を介しての作用を阻害する物質を「FGF18の活性阻害物質」という。
このような、FGF18活性阻害作用を有する物質は、FGF18とは無関係な物質群から選択された物質であっても良く、下記2.のスクリーニング方法を適用することにより、簡単にFGF18活性阻害作用を有する物質を取得することができる。本願の実施例においても、当該スクリーニング方法により、FGF18の活性阻害作用があるゴーヤー抽出物を選択し、当該ゴーヤー抽出物に、FGF18の毛包成長抑制作用を阻害し、かつ毛包成長作用があることを確認した。このように、本発明におけるFGF18の活性阻害物質は、FGF18の受容体との結合を阻害することでFGF18による毛包の毛成長サイクルの休止期を長期に亘って保持する作用と共に、毛髪の脱落の開始促進作用を阻害するので、毛成長休止期短縮作用、抜け毛防止作用があるが、短期間の観察だと毛髪の成長促進作用として観察される。
これらのFGF18の活性阻害物質が、ペプチド、タンパク質もしくは糖タンパク質の場合、これらペプチドなど自身を含む製剤として投与してもよいことはもちろんであるが、当該ペプチドなどをコードするDNAを組み込んだ発現ベクターの状態で投与することもできる。たとえば、FGF18部分ペプチドをコードするDNAを組み込んだ発現ベクターを有効成分とする製剤も、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤として用いることができる。
そして、これらのFGF18の活性阻害物質は、単独で、もしくは併用して、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤として用いることができる。
【0022】
6.FGF18活性阻害物質のスクリーニング方法
FGF18の活性阻害物質をスクリーニングし、被検物質を毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤の候補とする方法は、具体的には、以下の(a)〜(c)の工程を含むものである。
(a)FGFR1c、FGFR2c、FGFR3c及びFGFR4から選ばれた少なくとも1つのFGF受容体遺伝子を用いて遺伝子操作により細胞表面に強制的に発現させ、当該細胞を培養する工程、
(b)(a)工程で得られた、細胞表面にFGF受容体を有する細胞系にFGF18とともに披検物質を接触させる工程、
(c)(b)工程において、FGF18の細胞増殖促進活性を阻害する活性を観察した披検物質を選択する工程
ここで、FGF受容体遺伝子としては、FGF18が結合し、かつ細胞表面に受容体を有する細胞に対するFGF18の細胞増殖作用が確認されているFGFR1c、FGFR2c、FGFR3c及びFGFR4の受容体遺伝子の少なくとも1つを用いる。好ましくは、FGFR3cまたはFGFR4遺伝子を用いる。(b)工程で、被検物質を細胞に接触させる場合に、典型的には、細胞培養液に被検物質を直接投与するが、特に被検物質がタンパク質の場合などは、当該被検物質をコードする遺伝子をFGF受容体発現細胞中に導入することも可能である。
また、FGF受容体を細胞表面に強制的に発現させるための細胞は、培養可能な細胞であればどのような細胞でもよいが、好ましくはマウスIL-3依存性Ba/F3細胞株である。
スクリーニングに際しては、48時間以上、例えば72時間程度培養し、FGF18を単独で添加した場合に比べ、FGF受容体発現細胞の細胞増殖能が、5%以上、好ましくは10%以上低下していることを目安にスクリーニングすることが好ましい。
そして、FGF受容体遺伝子を導入していない親細胞は対照試験に用いることができ、披検物質を用いて(b)工程で添加したFGF18をIL-3に置き換えて同様の操作を行い、披検物質がこれら親細胞に対してはIL-3の細胞増殖促進活性を阻害しないことを確認する工程を設けることが好ましい。
【0023】
7.FGF18の発現抑制物質及びそのスクリーニング方法
FGF18は毛包内に存在する内在因子であるので、毛包細胞内でのFGF18遺伝子の転写及び翻訳を阻害する物質、すなわちFGF18遺伝子の発現を低下させる物質も毛包細胞中のFGF18濃度を低下させて、FGF18による毛包内の毛成長サイクルの休止期を長期に亘って保持する作用、毛髪の脱落の開始促進作用又は毛の成長サイクルスタート時期をヘアサイクルドメイン毎に規定する作用を阻害することができるから、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤毛髪成長促進作用及び発毛作用を有する。
このようなFGF18遺伝子の発現を阻害する物質は、FGF18遺伝子に対するsiRNAまたはその発現ベクターとして公知の方法で設計することができ、その活性は、動物培養細胞又は実験動物を用いてFGF18遺伝子の発現を阻害するか否かをモニターすることで確認することができる。
また別に、このようなFGF18遺伝子の発現を阻害する物質は、動物培養細胞又は実験動物を用いてFGF18遺伝子の発現を阻害するか否かをモニターすることでスクリーニングすることができる。
【0024】
具体的には、先ず、FGF18遺伝子を観察可能な程度に発現する動物培養細胞又は実験動物を用意し、被検物質を当該動物培養細胞もしくは実験動物に接触させるか又は実験動物に投与する。なお、実験動物とは、ヒトを除く、例えば、マウス、ラット、ニワトリ、七面鳥、ウシ、ブタ、ヒツジ及びウサギ等を意味する。被検物質としては、何ら限定されないが、例えば、植物抽出液、ペプチド、タンパク質、非ペプチド性化合物、低分子化合物、合成化合物、発酵生産物、細胞抽出液、動物組織抽出液等が挙げられる。これらの物質は新規な物質であってもよいし、公知の物質であってもよい。被検物質を細胞もしくは動物に接触させる場合に、典型的には、被検物質を細胞培養液に添加するか又は実験動物に投与するが、特に被検物質がタンパク質の場合などは、当該被検物質をコードする遺伝子をFGF受容体発現細胞中に導入することも可能である。
次に、当該動物培養細胞又は実験動物におけるFGF18遺伝子の発現をモニターする。動物培養細胞又は実験動物におけるFGF18遺伝子の発現は、例えば、FGF18抗体を用いたELISA等の常法を用いて解析するか、あるいは該細胞内又は実験動物内におけるFGF18遺伝子のmRNA量を定量的逆転写PCR法やノーザンブロット法等により解析するといった方法によりモニターすることができる。
これらいずれかの解析により、被検物質の非存在下で培養された動物培養細胞内におけるFGF18遺伝子の発現量と比べて、被検物質の存在下で培養された動物培養細胞内又は実験動物内におけるFGF18遺伝子の発現量が低下すれば、当該被検物質は毛髪成長促進或いは発毛といった機能を有する可能性があると判断できる。具体的には、被検物質を作用させない場合と比較して、mRNA量が0.8倍以下、好ましくは0.7倍以下、より好ましくは0.5倍以下に減少すれば、確実にFGF18の発現抑制物質であるといえる。培養角化細胞や培養真皮細胞、培養毛乳頭細胞でのFGF18のmRNA発現量は、培養条件や細胞の種類により様々であるので、上記の方法などにより個別に測定し、その数値が0.8倍以下に減少することを目安にスクリーニングすればよい。
このような工程を経てスクリーニングされたFGF18の発現抑制物質は、単独で、もしくは併用して、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤として用いることができる。
【0025】
8.毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤
上記「FGF18の活性阻害物質」及び/又は「FGF18の発現抑制物質」の項で説明した物質は、例えば、皮膚適用向けに適合化された溶液、クリーム、軟膏、ゲル、ローシヨン、シャンプー又はエアゾールといった形態で製剤化され、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤として提供される。なお、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤は、ヒトのみならず愛玩動物、毛を利用する家畜、実験動物などに適用した場合に、特に効果がある。
特に、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤は、局所投与用に適合化された薬理学上許容されるキャリアと共にFGF18活性阻害物質及び/又は発現抑制物質を含んだ医薬組成物の形で投与される。FGF18活性阻害物質及び/又は発現抑制物質を含有する毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤は、薬学上許容されるキャリア中に通常約0.01〜約100μg/日/cm2、好ましくは約0.1〜約10μg/日/cm2の活性化合物を含有している。言い換えると、FGF18の濃度は薬学上許容されるキャリア中で通常約0.01〜約100μg/日/cm2、好ましくは約0.1〜約10μg/日/cm2の活性化合物である。
さらにまた、局所投与用に適合化された薬理学上許容されるキャリアとしては、特に限定されないが、親水ワセリン又はポリエチレングリコール軟膏のような軟膏、キサンゴムのようなゴム等のペースト、アルコール、水性又は緩衝液のような溶液、水酸化アルミニウム又はアルギン酸ナトリウムゲルのようなゲル、ヒト又は動物アルブミンのようなアルブミン、ヒト又は動物コラーゲンのようなコラーゲン、アルキルセルロース、ヒドロキシアルキルセルロース及びアルキルヒドロキシアルキルセルロースのようなセルロース、メチルセルロース、ヒドロキシエチルセルロース、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース及びヒドロキシプロピルセルロース、プルロニックF−127で例示されるプルロニック(Pluronic(商標))ポリオ−ルのようなポリマ−;テトロニック1508のようなテトロニック(tetronic)、アルギン酸ナトリウムのようなアルギン酸塩を挙げることができる。
本発明に係るFGF18の活性阻害物質としては、FGF18部分ペプチドなどのFGF18の活性阻害タンパク質、ペプチドをコードするDNAを組み込んだ発現ベクターを用いることができるが、その場合は、通常の遺伝子治療の形態で提供することができる。発現ベクターとしては、動物細胞においてFGF18部分ペプチドなどを発現させるためのプロモーターなどの配列を備えるが、特に限定されるものではない。発現ベクターとしては、例えば、プラスミドベクター、ウイルスベクターなどが利用可能であるが、これらに限定されない。
【0026】
また、本発明に係る毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤は、FGF18発現抑制物質を有効成分として含有するものであってもよい。すなわち、本発明に係る毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤は、FGF18に対するsiRNA発現ベクターを用いた遺伝子治療として提供することができる。発現ベクターとしては、動物細胞においてsiRNAを発現するためのプロモーターなどの配列を備えるが、特に限定されるものではない。発現ベクターとしては、例えば、プラスミドベクター、ウイルスベクターなどが利用可能であるが、これらに限定されない。また、FGF18発現抑制物質は、FGF18に対するsiRNA発現ベクターに限定されない。
遺伝子治療用の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤を細胞内に導入する方法としては、ウイルスベクターを利用した遺伝子導入方法、非ウイルス性の遺伝子導入方法(日経サイエンス,1994年4月号,20-45頁、実験医学増刊,12(15)(1994)、実験医学別冊「遺伝子治療 の基礎技術」,羊土社(1996))のいずれの方法も適用することができる。
ウイルスベクターによる遺伝子導入方法としては、例えばレトロウイルス、アデノウイルス、アデノ関連ウイルス、ヘルペスウイルス、ワクシニアウイルス、ポックスウイルス、ポリオウイルス、シンビスウイルス等のDNAウイルス又はRNAウイルスに、TR4あるいは変異TR4をコードするDNAを組み込んで導入する方法が挙げられる。このうち、レトロウイルス、アデノウイルス、アデノ関連ウイルス、ワクシニアウイルスを用いた方法が、特に好ましい。非ウイルス性の遺伝子導入方法としては、発現プラスミドを直接筋肉内に投与する方法(DNAワクチン法)、リポソーム法、リポフェクション法、マイクロインジェクション法、リン酸カルシウム法、エレクトロポレーション法等が挙げられ、特にDNAワクチン法、リポソーム法が好ましい。
【0027】
9.インビトロ毛髪再生系への応用
FGF18の活性阻害物質及び/又は発現抑制物質を用いて、再生された皮膚組織における毛髪のインビトロ再生系を構築することができる。ここで、皮膚組織とは、分離した皮膚幹細胞を培養することによって得られる、皮膚の各種の細胞からなる組織を意味する。皮膚の各種の細胞とは、特に限定されないが、皮膚表皮の上皮細胞、皮膚上皮基底層の細胞、毛胞を構成する各種細胞、真皮の細胞及び脂肪細胞等を意味する。皮膚組織を再生する際に使用される細胞は、異種細胞、同種他家細胞及び同種自家細胞のいずれであっても良い。
先ず、皮膚幹細胞からの皮膚の各種細胞への分化を制御する方法としては、特に確立された技術がないため、本発明においては特に限定されない。例えば、自然分化(spontaneous differentiation)が起きた段階において異なる発現を示す増殖因子受容体を利用し、それぞれに対応するリガンド増殖因子を培地中に加えることにより、異なった分化方向を有する細胞を選択的に増幅することができる。異なった分化方向を有する細胞を選択的に増幅した後、皮膚組織を作製することができる。
人工皮膚組織の作製方法としては、特に定まった技術はないため、本発明においては限定されないが、例えば、上皮細胞だけを培養し層状に仕上げる方法、繊維芽細胞など真皮の構成細胞により真皮層を形成した後、上皮細胞を重層させて一体化する方法、その後上皮細胞の表面を空気に暴露することにより表皮化を促進させる方法、真皮層の変わりに、生物分解が可能な成分により形成された層状フィルムを用いる方法、など、様々な方法をとることができる。また、本発明は、ヒトから採取した皮膚組織から皮膚幹細胞を分離し、分離した皮膚幹細胞を用いて皮膚組織を作製するといった、いわゆる再生医療における皮膚組織の作製方法も適用される。このとき、新たに作製した皮膚組織は、採取したヒトと同一人に対して治療のために戻すことを前提としても良いし、採取したヒトと異なる他人に対して治療のために移植することを前提するものであってもよい。
このような皮膚組織の作製方法において、培地中にFGF18の活性阻害物質及び/又は発現抑制物質を適当な時期に添加することによって、再生した皮膚組織において毛包の成長サイクルの休止期を短縮し、その結果毛成長サイクル自体の速度を早くすることができ、当該皮膚組織における毛髪の抜け毛を防止することができる。さらに、皮膚組織の作製方法において、培地中にFGF18活性阻害物質及び/又は発現抑制物質を適当な時期に添加することによって、皮膚細胞の増殖を促進することができ、皮膚組織全体の体積の増大を図ることができる。
【0028】
10.ゴーヤー抽出物
本発明の前記9.のスクリーニング方法により得られたFGF18活性阻害物質はゴーヤー抽出物であり、実施例に示すようにその発毛促進効果も確認された。
ゴーヤー抽出物の原料となるゴーヤーは、ウリ科ツルレイシ属の植物で、学名はMomordica charantiaであり、ニガウリ、または、ツルレイシとも呼ばれている。
ゴーヤーからその抽出物を得るに当たっては、植物体の全部位が使用可能であり、各部位を単独に、或いは適宜混合して抽出原料に用いることができる。また、原料となる植物体の性状は、乾燥状態のもの、非乾燥状態のもの、いずれも用いることができる。
【0029】
ゴーヤーから、本発明に用いる抽出物を得るには、これらの原料を必要により細切ないし粉砕し、適当な溶媒を用いて、一般に用いられる抽出法で抽出すれば良い。抽出に用いられる溶媒は、特に限定されないが、例えば水、無水或いは含水有機溶媒を例示することができる。無水或いは含水有機溶媒としては、1価アルコール、多価アルコールまたはその誘導体、ケトン、エステル、エーテル、石油エーテル、脂肪族炭化水素、またはハロゲン化合物、芳香族炭化水素より選択された一種または二種以上を例示することができる。具体的な溶媒としては、水、メタノール、エタノール、ブタノール、アセトン、酢酸エチルエステルが例示され、これらは一種または二種以上を組み合わせて用いることができる。このうち、特に水、或いはメタノールやエタノール等の1価アルコールを用いることが好ましい。抽出に当たって、上記溶媒の使用量は特に限定されないが、抽出原料であるゴーヤーに対して、0.1-1000重量倍、好ましくは1-100重量倍、更に好ましくは2-50重量倍である。
【0030】
上記溶媒による抽出法は、常法に従って行なうことができる。例えば抽出温度については、室温程度の温度で抽出しても良く、また用いられる溶媒の沸点付近の温度で抽出しても良い。また、抽出操作についても、ゴーヤーの乾燥粉砕物もしくは粉砕物を室温下で1〜30日間浸漬することにより抽出する方法や、抽出溶媒の沸点程度の温度において還流抽出する方法等を用いることができる。
【0031】
下記の実施例において示されるように、本発明のゴーヤーの抽出物からなる物質は、FGFR4遺伝子を発現したBa/F3細胞に対して、FGF18による細胞増殖を阻害するFGF18阻害活性を有する。実施例では、FGFレセプターの1つであるFGFR4を遺伝子発現操作により細胞表面に強制的に発現させた細胞を用いたときに、FGF18の存在下で、FGF18による細胞増殖に対して、ゴーヤー抽出物の添加により濃度依存的に阻害することが示された。ゴーヤー抽出物のこの作用は、同時に行われたFGF18非存在下の細胞増殖に対しては増殖阻害作用を示さなかったことから、細胞障害作用ではなく、FGF18活性阻害作用であることが示された。
すなわち、本発明のゴーヤーの抽出物からなる物質は、FGF18による細胞増殖を阻害するFGF18活性阻害物質である。
【実施例】
【0032】
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
なお、遺伝子操作的手法として、特に断らない限りモレキュラー・クローニング((Molecular Cloning - A laboratory Manual、Sambrook
and Russell社刊)に記載された公知の方法を用いた。
【0033】
〔実施例1〕 Fgf18遺伝子毛包特異的欠損(ノックアウト)マウス作製
マウスFgf18遺伝子を含むBACクローンを129マウスBACクローンライブラリーから取得した。このクローンを用い、Fgf18遺伝子のエクソン3の上流にloxP-FRT[PGK-Neor]FRT配列を導入し、下流にloxP配列を導入した、エクソン3を欠損するためのターゲティングベクターを構築した(図2)。なお、当該ターゲティングベクター作製は、「マウスFgf18遺伝子のエクソン3欠失用のターゲティングベクター」として、フェニックスバイオ社に外注した。
129マウス由来のES細胞にこのベクターを導入し、細胞集団の中から、Fgf18遺伝子がこの配列によって組み換えられた(ターゲットされたと称する)細胞を選択し、クローン化した。そのうちの一つのES細胞株をトランスジェニックマウスの作出に用いた。C57BL/6マウス(濃褐色体毛、日本SLC社)の胚盤胞にこの細胞を注入し、さらにこの胚盤胞を、偽妊娠状態にした雌のICRマウス(白色体毛、日本SLC社)の子宮内に導入した。マウスが分娩した仔について、体毛の色からキメラマウスを区別した。これらの操作を63個の胚盤胞について行い、キメラマウス仔が16個体得られた。これらのマウスを飼育し、性成熟後、C57BL/6マウスと交配し、35匹の仔が得られた。これらのゲノムを分析したところ、15匹の個体がターゲットされたFgf18遺伝子を含んでいることが示された(これをF1とする)。
次に[PGK-Neor]カセットを除去するため、F1マウスをFlpe トランスジェニックマウスと交配し、仔マウスが67個体得られた。これらについてFgf18遺伝子アリルを分析したところ、12個体について、目的の配列(導入されたloxP-FRT[PGK-Neor]FRT-Exon3-loxPをloxP-FRT-Exon3-loxPとして)を有していることが確認された(F2)。
ここで用いたFlpe トランスジェニックマウスは、理研BRCから入手した(RIKEN RBRC01834; 非特許文献8)。
次に毛包特異的にFgf18遺伝子エクソン3を除去する目的で、F2マウスをさらにK5-Creトランスジェニックマウスと交配した。得られた仔マウスは4週齢に達した時点で離乳させ、皮膚を含む尾の一部を切除し、Fgf18遺伝子の遺伝子型とCreの発現を確認した。離乳した37匹の個体のうち、6匹の K5Cretg;Fgf18+/floxマウス(メス1匹、オス5匹)が得られた。K5Cretg;Fgf18+/floxマウス同士を交配することにより、29匹のマウスが得られた。これらの遺伝子型を解析したところ、一匹の個体はK5Cretg;Fgf18flox/flox(以降、これをFgf18ホモ欠失体と称する)であることが示された。同様にしてK5Cretg;Fgf18+/flox マウスコロニーを交配することで、Fgf18ホモ欠失体を多数得ることが出来た。
以下、このFgf18ホモ欠失体をFgf18遺伝子毛包特異的欠損動物と称する。
ここで用いたK5Creトランスジェニックマウスは、九州大学CARDから入手した (CARD ID323;非特許文献9)。
これらのマウス皮膚においてFgf18第3エキソンが欠落しているDNAが存在することは、サザンブロットで確認された。その例を示す。
なお、K5Cretg;Fgf18+/floxマウス同士の交配において生じる仔の遺伝型は、通常のメンデルの法則に従うものだった。
【0034】
〔実施例2〕 Fgf18遺伝子毛包特異的欠損(ノックアウト)マウスにおける休止期の短縮と速やかに繰り返される毛成長周期
Fgf18遺伝子毛包特異的欠損マウスは、生存・成長し、生殖可能であり、健康状態も外観的に良好であった。その背部皮膚に生えている体毛を、短く刈ることにより、特徴的な毛成長サイクルの波を示すことが分かった。図3aに示すのは、Fgf18遺伝子毛包特異的欠損マウスの代表的な個体(ホモノックアウト体) [K5Cretg;Fgf18flox/flox]と、その対象としてFgf18遺伝子の1対2本のうち1本のみが破壊されている対照群の代表的な個体(ヘテロノックアウト体) [K5Cretg;Fgf18+/flox]を、32日齢以降、写真撮影したものである。尚、これらの写真撮影に先行して体毛をバリカンで刈ってあるので、体表の色調がわかるようになっている。32日齢では、背部皮膚はいずれのマウスも初めの生理的成長期にあり、黒い色を示している。37日齢になると、首に近い部分がピンクがかった色になり、休止期に入ったことを示す。40日齢では、背部皮膚全体が休止期である。ホモノックアウト体の場合、47日齢で、次の成長期に入ったことが、皮膚が青みがかった着色を示すことでわかる(図3a)。これに対して、対照のヘテロノックアウト体では、休止期がずっと長く続く(図3a)。
Fgf18遺伝子毛包特異的欠損マウス(68日齢)の背部皮膚における毛成長周期は、前位(首に近い位置)から後位(尾に近い位置)に向かってスムーズに進捗し。毛包の形態形成(図3cのMで示すサイクル)に加えて既に2回のサイクル(図3cの1,2)がまわり、3回目のサイクル(図3cの3)に入ったことが見て取れる(Fig. 3b 左)。 これに対して野生型のマウス(89日齢)では、次の成長期の開始の場所や時期は、個体によって多様であり、一個体の背部皮膚の中においてさえも、「毛成長ドメイン」によって異なる相を示す (Fig. 3b 右,及び図4)。従って、Fgf18遺伝子毛包特異的欠損マウスにおいては、マウスの齢に対する関数として、一般的な毛成長周期における各相の変遷をFig.2aと同様にして示すことが出来る(Fig. 3C)。
ここでは、マウスの首に近い背部皮膚(Fig.3bで点線囲みの部分)における毛成長周期の相を、皮膚の色及び前週の毛成長履歴によって判断し、単純化のために成長期または休止期として表している。その結果、Fgf18遺伝子毛包特異的欠損マウスにおいては、休止期はわずか1週間程度しか持続しないこと、各成長周期は休止期を含めた全体として約3週間であることがわかった。これに対して野生型マウスでは通常、休止期は3から5週間、あるいはより長い期間持続する。
図3dでは、加齢の進んだFgf18遺伝子毛包特異的欠損マウスの代表的な3個体を示す。これらのマウスは背部皮膚の毛を刈ってから1週間飼育し、その一週間に生えた毛を含めた姿を撮影している。特徴的な表現型として、このように加齢が進んだFgf18遺伝子毛包特異的欠損マウスにおいては、同じ成長周期相の毛包が、外観上ストライプを形成するように並んだ。さらに特徴として、ストライプの数は、マウスの日齢に依存して、加齢が進むほど増えていくことが分かった。加齢が進んだマウスにおいて、毛成長周期が一回回るために必要な期間は、前位においては約3週間であったが(Fig. 3d) 、後位においては、約4週間だった (Fig. 3d)。したがって、加齢が進んだFgf18遺伝子毛包特異的欠損マウスにおいて、この前位と後位の間の成長周期長の差が、ストライプ間の距離を縮めていることが理解できる。
【0035】
〔実施例3〕 Fgf18遺伝子毛包特異的欠損マウスにおける不完全な生理的脱毛
休止期の毛包に生えている毛は、それ以前の毛成長周期で完成された毛であり、クラブヘアー(club hair)と称する。一般的にマウスにおいては、休止期毛包に生えているクラブヘアーをグローブをはめた指先でつまみ、緩やかに引き抜くことが出来る。Fgf18遺伝子毛包特異的欠損マウス(ホモノックアウトマウス)においては、このようにして引き抜いた毛が、複数の毛の束になっていることを発見した(図5a、左)。
対象のヘテロノックアウトマウスにおいては、そのような束はまれであった(図5a、右)。引き抜いた毛の長さを、それぞれの群で200本以上測定すると、ホモノックアウトマウス(6.40+/-0.53mm)でもヘテロノックアウトマウス(6.38+/-0.65mm)でも、ほとんど同じであった。さらに毛の種類(guard,awl, and zigzag hairs)の分布にも差異は認められなかった。これらのことから、Fgf18遺伝子毛包特異的欠損マウスにおいて、毛成長周期の成長期は正常に進行していることが強く示唆された。
毛周期が成長期である特徴を示す皮膚の3.7mm2領域内の全毛包とそこに存在する毛軸数を、加齢の進んだFgf18遺伝子毛包特異的欠損マウス(ホモノックアウトマウス)及び対象のヘテロノックアウトマウスにおいて、その背部皮膚の毛包の数及びそこに存在する毛軸数を計測した。(表1)
【0036】
表1:毛包細胞が保持する毛軸数
この結果から見て、Fgf18遺伝子毛包特異的欠損マウスの毛包数は対照群と変わらないものの、それぞれの毛包内に保持される毛軸数の数が多いため、全体で毛軸数が2倍強となっていることが理解できる。
【0037】
さらに、加齢の進んだFgf18遺伝子毛包特異的欠損マウス(ホモノックアウトマウス)及び対象のヘテロノックアウトマウスにおいて、その背部皮膚の毛包の状態を拡大して観察した。すると、成長期の毛包に生えている成長期毛軸に寄り添って、脱落していないクラブヘアーが多数存在していることが明らかになった(図5c)。さらにフルフォーカスイメージングで皮膚を観察したところ、脱落していないクラブヘアーを包む袋様の構造は、成長中の毛軸のバルジ領域付近に一体化したように寄り添っていることが確認された。
また、加齢の進んだFgf18遺伝子毛包特異的欠損マウスにおいて、その背部皮膚の毛包を輪切りにした切片を作成し、チオフラビン T (黄色) 及び DAPI (赤色)で染色し、観察した。すると、ホモノックアウトマウスにおいては、それぞれの毛包にはメラニンに富む成長期毛軸と、多くは3〜6本の、メラニンのないクラブヘアーが共存していることが示された (図5f)。これに対して、対象のヘテロノックアウトマウスにおいては、成長期毛包に含まれるクラブヘアーの本数は多くは1〜2本であった。その分布は、表1に示されるとおりである。これらの観察により、Fgf18遺伝子毛包特異的欠損マウス(ホモノックアウトマウス)においては、クラブヘアーの正常な脱落(脱毛期)のためのメカニズムが不完全になっていることが示唆された。なお、そうではあっても、毛包あたりの残存毛軸数が、完了したと考えられる毛成長周期の数(10 to 12 cycles)よりも少ないことから、 不完全とはいってもある程度の毛軸の脱落は起きていると考えられた。以上より、FGF18活性を欠失させることにより、クラブヘアーの脱毛が阻害されることが初めて示された。
【0038】
〔実施例4〕 FGF18の部分ペプチドによるFGF18活性の阻害
FGF18の部分ペプチドとして、マウスFGF18のアミノ酸配列に対応する配列番号14のN末端から翻訳開始のために導入されたメチオニンを除く4番目まで〜95番目までのアミノ酸を除去した部分ポリペプチド(d4〜d95:メチオニンを除くN末端から除去したアミノ酸数で表す。)を作製した。
なお、それぞれの部分ポリペプチドのアミノ酸配列(塩基配列)は、以下の配列番号に対応する。
d4:配列番号15(配列番号4)
d12:配列番号16(配列番号5)
d16:配列番号17(配列番号6)
d18:配列番号18(配列番号7)
d22:配列番号19(配列番号8)
d37:配列番号20(配列番号9)
d48:配列番号21(配列番号10)
d67:配列番号22(配列番号11)
d77:配列番号23(配列番号12)
d95:配列番号24(配列番号13)
FGF18は4種類のFGFレセプターFGFR1c, FGFR2c, FGFR3c, FGFR4 を刺激するが、それらを刺激する強さは異なり、また、これらの受容体への刺激の総和としての結果として、毛成長サイクルの休止期を長期間保持し、脱毛の開始時期を早め、かつ毛成長サイクルの開始をドメイン毎に制御すると考えられる。
文献の教示にしたがって、マウスIL-3依存性proB細胞であるBa/F3細胞株(理研BRCより入手)に、遺伝子発現操作によりFGFレセプターであるFGFR1c, FGFR2c, FGFR3c, FGFR4のいずれかを導入して、FGFRが細胞表面に強制的に発現されている細胞を作成した(Ornitz,DM.,Xu,J.,Colvin,JS.,McEwen,DG.,MacArthur,CA.,Coulier,F.,Gao,G. and Goldfarb,M.,1996.Receptor specificity of the fibroblast growth factor family.J.Biol.Chem.271(25):15292-15297;Yoneda,A.,Asada,M.,Oda,Y.,Suzuki,M. and Imamura,T.,2000.Engineering of an FGF-proteoglycan fusion protein with heparin-independent,mitogenic activity.Nat.Biotec.18(6):641-644)。
【0039】
これらの細胞を用い、それぞれのレセプターを30 ng/mlのFGF18で刺激している際に、それぞれのFGF18の部分ペプチドを1μg/ml(図3A)または100ng/ml(図3B)の濃度で共存させ、FGF18で刺激された細胞のDNA合成(これを100% とする)を阻害するかどうかアンタゴニスト活性を調べた。すなわち、FGF18による細胞増殖刺激効果を抑制する活性を解析した。その結果を、図3 に示す。各カラムとテスト試料の対応は以下の通りである。
(図6A) カラム1:FGF18以外の試料添加なし(対照)。 カラム2:(d4)。カラム3 :(d16)。カラム4:(d18)。カラム5:(d22)。カラム6:(d37) 。カラム7:(d48) 。カラム8 :(d77)。カラム9:(d95) 。カラム10:(d22) 。カラム11:(d37)。カラム12:(d48)。カラム13:(d67)。カラム14:(d77)。カラム15:(d95)。
(図6B) カラム1:FGF18以外の試料添加なし(対照)。カラム2:(d4)。カラム3:(d12)。カラム4:(d16)。カラム5:(d18)。カラム6:(d37)。カラム7:(d48)。カラム8:(d67)。カラム9:(d95)。カラム10:(d37)。カラム11:(d67)。
図6Aから、披検物質を1μg/mlの濃度で試験すると、d16, d22, d37, d48, d77, d95などがFGFR3c/BaF3細胞上でFGF18の活性を強く阻害することが分かる。またd22, d48, d77, d95などがFGFR4/BaF3細胞上でFGF18の活性を強く阻害することが分かる。他の部分ペプチドも図に示すように様々な阻害効果を示す。
図6Bから、披検物質を100ng/mlの濃度で試験すると、d16, d95などがFGFR3c/BaF3細胞上でFGF18の活性を強く阻害することが分かる。
これらのFGF18の活性阻害物質となるFGF18の部分ペプチドを用いることで、毛成長制御におけるFGF18による毛成長サイクルの休止期制御作用、脱毛開始の制御作用を解除することができるから、毛成長休止期を短縮し、抜け毛を防止し、毛並みをそろえることができる。
また、同様に、マウスFGF18のアミノ酸配列に対応する配列番号14のC末端から25番目まで〜125番目までのアミノ酸を除去した部分ポリペプチド(dc25〜dc125:C末端から除去したアミノ酸数で表す。)を作製した。
なお、それぞれの部分ポリペプチドのアミノ酸配列(塩基配列)は、以下の配列番号に対応する。
dc25:配列番号34(配列番号25)
dc43:配列番号35(配列番号26)
dc57:配列番号36(配列番号27)
dc67:配列番号37(配列番号28)
dc82:配列番号38(配列番号29)
dc94:配列番号39(配列番号30)
dc108:配列番号40(配列番号31)
dc113:配列番号41(配列番号32)
dc125:配列番号42(配列番号33)
これらのFGF18部分ペプチドのうちdc25を除く部分ペプチドはN末端を除去した上記部分ペプチドと同様のFGF18の活性阻害作用を有するため、FGF18による毛成長サイクルの休止期制御作用、脱毛開始の制御作用を解除することができるから、毛成長休止期を短縮し、抜け毛を防止し、毛並みをそろえることができる。
【0040】
〔実施例5〕 FGFR4発現細胞を用いたFGF18活性阻害物質のスクリーニング
作製したFGFR4発現細胞(R4/Ba/F3細胞)を、FGF18存在下で、被験液として種々の植物抽出液と共に培養した。陽性対照としては市販のFGF18蛋白質を用いた。一定時間培養した後の細胞数を、Cell Counting Kit-8(株式会社同仁化学研究所製造、和光純薬工業株式会社販売)を用いて、WST-8ホルマザンの産生量に伴う450 nmの発色を測定して求めた。
その結果、ゴーヤーの抽出物が、FGF18活性阻害物質として作用し、FGF18によるFGFR4/BaF3細胞の増殖を阻害することを見いだした。
【0041】
〔実施例6〕 本発明のFGF18活性阻害物質の細胞増殖阻害活性
乾燥したゴーヤー1.5gに、蒸留水30mlを加え、15分間煮沸した。以上の様にして得られた抽出液をろ紙でろ過してろ液を取り、ゴーヤーの熱水抽出液を得た。乾燥したゴーヤー1.5gに、70%エタノール水溶液30mlを加え、室温で7日間浸漬抽出した。この様にして得られた抽出液をろ紙でろ過し、ゴーヤーの70%エタノール抽出液を得た。
【0042】
実施例5と同様にして、FGFR4/BaF3細胞を用いて本発明のFGF18活性阻害物質の活性測定を行った。具体的には、測定は次の様にして行った。96穴の細胞培養用プレートの各ウエルに、1ウエル当たり50μlの10% FBS、1% Antibiotic G-418 Sulfate(プロメガ株式会社、V7983)を含むRPMI1640培地を加えた。次いで、各試料を水で希釈して調製した種々の濃度の被験液 10μlを添加した後、10%FBS、1% Antibiotic G-418 Sulfateを含むRPMI1640培地中に5×104個のFGFR4/BaF3細胞が懸濁した細胞懸濁液50μlを添加し、軽く撹拌した。更に、heparin/10% FBS/1% Antibiotic G-418 Sulfate/RPMI1640培地、10μl(heparin終濃度5μg/ml)、及びFGF18(ぺプロテック株式会社、100-28)溶液 10μl(FGF18終濃度3ng/ml)を加え、5%CO2存在下、37℃の炭酸ガスインキュベーターで72時間培養した。FGFR4/BaF3細胞の増殖は、72時間培養後の各ウエルに、Cell Counting Kit-8(株式会社同仁化学研究所製造、和光純薬工業株式会社販売)/PBS溶液10μlを加えた後、更に3時間培養し、WST-8ホルマザンの産生量に伴う450nmの発色を測定して求めた。
【0043】
測定に際しては、陽性対照としては、FGF18(ぺプロテック株式会社、100-28)溶液 10μl(FGF18終濃度3ng/ml)を、また陰性対照としては、被験液作製時に使用した水またはエタノール溶液(エタノール溶液の終濃度は1%以下になるように調製)10μl を用いて測定した。細胞増殖阻害率(%)の算定は、以下の式(A)により算出した。
〔式(A)〕
細胞増殖阻害率(%)=100×{(FGF18及び水またはエタノール溶液添加時の吸光度)−(FGF18及び被験液添加時の吸光度)}/FGF18及び水またはエタノール溶液添加時の吸光度−水またはエタノール溶液添加時の吸光度)
【0044】
ゴーヤー熱水抽出液についての結果を図4に示す。FGF18の存在下で、終濃度8.3%のゴーヤー熱水抽出液を被験液とした場合、発色量が減少し、ゴーヤー熱水抽出液はFGF18によるFGFR4/BaF3細胞の増殖を阻害することが確認された。一方、FGF18無添加で被験液添加時の吸光度は、同様にFGF18無添加で水またはエタノール溶液添加時の吸光度に比べてほとんど減少することは無く、被験液による細胞障害作用はほとんど認められなかった。
【0045】
〔実施例7〕 本発明のFGF18活性阻害物質の in vivo 解析
FGF18活性阻害物質の in vivo での作用を見るために、毛周期の休止期にあるC3H/He マウスで試験した。
乾燥したゴーヤー1.1gに、蒸留水15mlを加え、20分間煮沸した。抽出液が室温に戻った後、ろ紙でろ過し、ろ液については同量のエタノールを添加することによって、ゴーヤー熱水抽出液の50%エタノール溶液を作製した。更に0.45μmのマイレクスHV滅菌フィルター(ミリポア社)を通過させ、その後、グリセロールを1%になるように添加し、被験液とした。この様にして得られたゴーヤー熱水抽出液(49.5%エタノール、1%グリセロールを含有)について、毛周期の休止期にあるC3H/He マウス背部皮膚に塗布した。7週齢のC3H/He 雄マウスに麻酔をし、背部毛をバリカンにより優しく剃毛した。剃毛後、被験液としてゴーヤー熱水抽出液(49.5%エタノール、1%グリセロールを含有)、又は水−エタノール−グリセロール溶液(49.5%水、49.5%エタノール、1%グリセロールを含有)200μlをそれぞれ1群5匹のマウス背部皮膚に塗布した(第0日)。同様にして、11日間(5日目、及び6日目を除く)に渡って毎日塗布し、第10日、14日、17日、21日、24日目のマウス背部の発毛状態を目視により観察し、発毛スコアーを得た。発毛スコアーは、1)色素沈着:1点、2)短かな毛並み:2点、3)通常の毛並み:3点とし、それぞれの発毛状態の面積が全剃毛面積に対して占める割合を%で示し、以下の式で求めた。本算出法によると、全剃毛面積が通常の毛並みに回復した場合、100点となる。
発毛スコアー=(色素沈着面積の割合(%)×1+短かな毛並み面積の割合(%)×2+通常毛並み面積の割合(%)×3)/3
第一回の塗布後、第10日、14日、17日、21日、24日目におけるマウス背部の発毛状態を観察した結果、図5に示す通り、ゴーヤー抽出液塗布群は、陰性対照群(水−エタノール−グリセロール溶液(49.5%水、49.5%エタノール、1%グリセロールを含有)塗布群)に比較して特に21日目以降に発毛スコアーが高く、発毛が有意に促進された。
以上の様に、FGF18活性阻害物質であるゴーヤー熱水抽出液にはマウス背部皮膚表面でのin vivo解析においてもFGF18本来の活性を阻害する作用があることが実証された。この実験では、24日目までのデータしか取得していないので、「発毛促進効果」として観察されるが、さらに観察を続ければ、当該効果は毛成長サイクルの休止期が短縮されたこと及び脱毛が開始しないことによる毛量全体の増量効果に基づくものであることが判明するといえる。
【0046】
〔実施例8〕 ゴーヤー熱水抽出物を用いたヘアシャンプー
本発明であるゴーヤー熱水抽出物を用いたヘアシャンプーの処方及び製造法を示す。
以下の処方、及び製法に従ってヘアシャンプーを製造した。
(処方)
成 分 重量%
1.実施例5で得られた希釈液 0.1
2.ラウリルエーテル硫酸ナトリウムエタノール 20
3.ラウリル硫酸ナトリウム 10
4.1,3ブチレングリコール 1
5.香料 適量
6.精製水 残量(全量で100とする)
(製法)
処方成分を80℃に加熱し、攪拌混合した後、攪拌冷却し本発明のヘアシャンプーを製造した。
【0047】
〔実施例9〕 ゴーヤー熱水抽出物を用いたヘアリキッド
本発明であるゴーヤー熱水抽出物を用いたヘアリキッドの処方及び製造法を示す。
以下の処方、及び製法に従ってヘアリキッドを製造した。
(処 方)
成 分 重量%
1.実施例5で得られた希釈液 0.1
2.エタノール 40
3.グリセリン 1
4.香料 適量
5.精製水 残量(全量で100とする)
(製法)
処方成分を加えて攪拌溶解した後、精製水を加えてヘアリキッドを製造した。
【0048】
〔実施例10〕
本発明であるゴーヤー熱水抽出物を用いたヘアクリームの処方及び製造法を示す。
以下の処方、及び製法に従ってヘアクリームを製造した。
(処方)
成 分 重量%
1.実施例5で得られた希釈液 0.1
2.流動パラフィン 40
3.ワセリン 1
4.セトステアリルアルコール 1
5.メチルポリシロキサン 1
6.パラオキシ安息香酸メチル 0.2
7.プロピレングリコール 5
8.香料 適量
9.精製水 残量(全量で100とする)
(製法)
処方成分を攪拌混合し、本発明のヘアクリームを製造した。
【産業上の利用可能性】
【0049】
本発明により、効果的な毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤が提供でき、これらを配合することにより、毛髪成長促進活性を有するシャンプー、ヘアリキッドなどの他、脱毛症治療用の医薬組成物などが提供でき、また非ヒト動物などで、毛の成長サイクルが早まった実験動物や毛並みのそろった愛玩動物が提供できる。
【特許請求の範囲】
【請求項1】
FGF18の活性阻害物質及び/又は発現抑制物質を有効成分として含有する毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項2】
前記FGF18の活性阻害物質がFGF18の部分ペプチドであることを特徴とする請求項1に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項3】
前記FGF18の活性阻害物質が抗FGF18抗体であることを特徴とする請求項1に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項4】
前記FGF18の活性阻害物質がゴーヤー抽出物であることを特徴とする請求項1に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項5】
前記FGF18の活性阻害物質が、FGF18の活性を阻害する活性を有するFGF18の部分ペプチドをコードするcDNAを組み込んだ発現ベクターであることを特徴とする請求項1に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項6】
前記FGF18の発現抑制物質が、FGF18の発現を阻害する活性を有するsiRNAを組み込んだ発現ベクターであることを特徴とする請求項1に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項7】
他の毛髪成長促進剤、発毛促進剤、又は脱毛抑制剤を更に含むことを特徴とする請求項1ないし6のいずれかに記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項8】
請求項1ないし4のいずれかに記載のFGF18の活性阻害物質をスクリーニングし、被検物質を毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤の候補とする方法であって、以下の(a)〜(c)の工程を含む方法。
(a)FGFR1c、FGFR2c、FGFR3c及びFGFR4から選ばれた少なくとも1つのFGF受容体遺伝子を用いて遺伝子操作により細胞表面に強制的に発現させ、当該細胞を培養する工程、
(b)(a)工程で得られた、細胞表面にFGF受容体を有する細胞系にFGF18とともに披検物質を接触させる工程、
(c)(b)工程において、FGF18の細胞増殖促進活性を阻害する活性を観察した披検物質を選択する工程
【請求項9】
請求項8に記載のFGF受容体がFGFR3cである請求項8に記載のスクリーニング方法。
【請求項10】
請求項8に記載のFGF受容体がFGFR4である請求項8に記載のスクリーニング方法。
【請求項11】
請求項8に記載のFGF受容体を細胞表面に強制的に発現させるための細胞がマウスIL-3依存性Ba/F3細胞株である請求項8ないし10のいずれかに記載のスクリーニング方法。
【請求項12】
請求項1に記載のFGF18の発現抑制物質をスクリーニングし、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤の候補とするための方法であって、下記の(a)〜(d)の工程を含む方法。
(a)FGF18遺伝子が観察可能な程度発現する動物培養細胞又は実験動物を用意する工程、
(b)被検物質を、(a)の動物培養細胞に接触させるか又は実験動物に接触もしくは投与する工程、
(c)(b)工程後の動物培養細胞又は実験動物におけるFGF18遺伝子の発現をモニターする工程、
(d)FGF18遺伝子の発現を阻害する機能を有する被検物質を選択する工程。
【請求項13】
前記(c)工程において、前記(b)工程後の実験動物又は動物培養細胞から、mRNAを抽出し、発現されたFGF18のmRNA量を解析することによりFGF18遺伝子の発現をモニターし、前記(d)工程において、披検物質を作用させない場合と比較して低いレベルのFGF18のmRNA量を観察した系を選択することにより、FGF18遺伝子の発現を阻害する機能を有する被検物質を選択することを特徴とする、請求項12に記載の方法。
【請求項1】
FGF18の活性阻害物質及び/又は発現抑制物質を有効成分として含有する毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項2】
前記FGF18の活性阻害物質がFGF18の部分ペプチドであることを特徴とする請求項1に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項3】
前記FGF18の活性阻害物質が抗FGF18抗体であることを特徴とする請求項1に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項4】
前記FGF18の活性阻害物質がゴーヤー抽出物であることを特徴とする請求項1に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項5】
前記FGF18の活性阻害物質が、FGF18の活性を阻害する活性を有するFGF18の部分ペプチドをコードするcDNAを組み込んだ発現ベクターであることを特徴とする請求項1に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項6】
前記FGF18の発現抑制物質が、FGF18の発現を阻害する活性を有するsiRNAを組み込んだ発現ベクターであることを特徴とする請求項1に記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項7】
他の毛髪成長促進剤、発毛促進剤、又は脱毛抑制剤を更に含むことを特徴とする請求項1ないし6のいずれかに記載の毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤。
【請求項8】
請求項1ないし4のいずれかに記載のFGF18の活性阻害物質をスクリーニングし、被検物質を毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤の候補とする方法であって、以下の(a)〜(c)の工程を含む方法。
(a)FGFR1c、FGFR2c、FGFR3c及びFGFR4から選ばれた少なくとも1つのFGF受容体遺伝子を用いて遺伝子操作により細胞表面に強制的に発現させ、当該細胞を培養する工程、
(b)(a)工程で得られた、細胞表面にFGF受容体を有する細胞系にFGF18とともに披検物質を接触させる工程、
(c)(b)工程において、FGF18の細胞増殖促進活性を阻害する活性を観察した披検物質を選択する工程
【請求項9】
請求項8に記載のFGF受容体がFGFR3cである請求項8に記載のスクリーニング方法。
【請求項10】
請求項8に記載のFGF受容体がFGFR4である請求項8に記載のスクリーニング方法。
【請求項11】
請求項8に記載のFGF受容体を細胞表面に強制的に発現させるための細胞がマウスIL-3依存性Ba/F3細胞株である請求項8ないし10のいずれかに記載のスクリーニング方法。
【請求項12】
請求項1に記載のFGF18の発現抑制物質をスクリーニングし、毛成長休止期短縮剤、抜け毛防止剤又は毛並み調整剤の候補とするための方法であって、下記の(a)〜(d)の工程を含む方法。
(a)FGF18遺伝子が観察可能な程度発現する動物培養細胞又は実験動物を用意する工程、
(b)被検物質を、(a)の動物培養細胞に接触させるか又は実験動物に接触もしくは投与する工程、
(c)(b)工程後の動物培養細胞又は実験動物におけるFGF18遺伝子の発現をモニターする工程、
(d)FGF18遺伝子の発現を阻害する機能を有する被検物質を選択する工程。
【請求項13】
前記(c)工程において、前記(b)工程後の実験動物又は動物培養細胞から、mRNAを抽出し、発現されたFGF18のmRNA量を解析することによりFGF18遺伝子の発現をモニターし、前記(d)工程において、披検物質を作用させない場合と比較して低いレベルのFGF18のmRNA量を観察した系を選択することにより、FGF18遺伝子の発現を阻害する機能を有する被検物質を選択することを特徴とする、請求項12に記載の方法。
【図6】

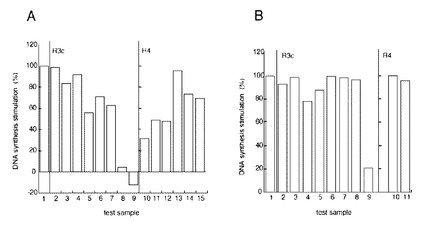
【図7】
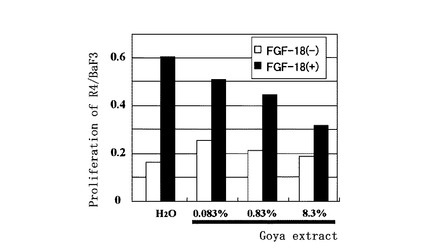
【図8】
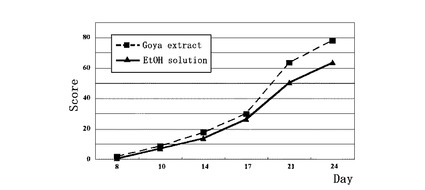
【図1】
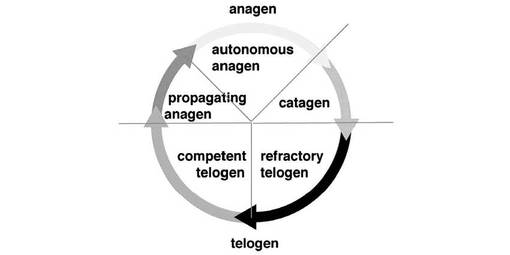
【図2】
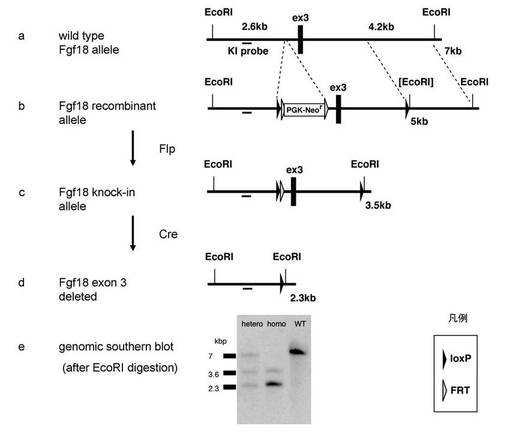
【図3】
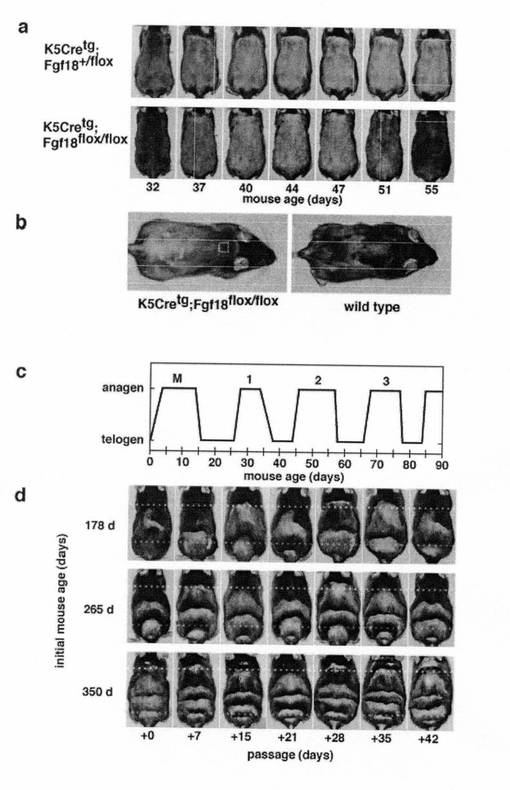
【図4】
【図5】
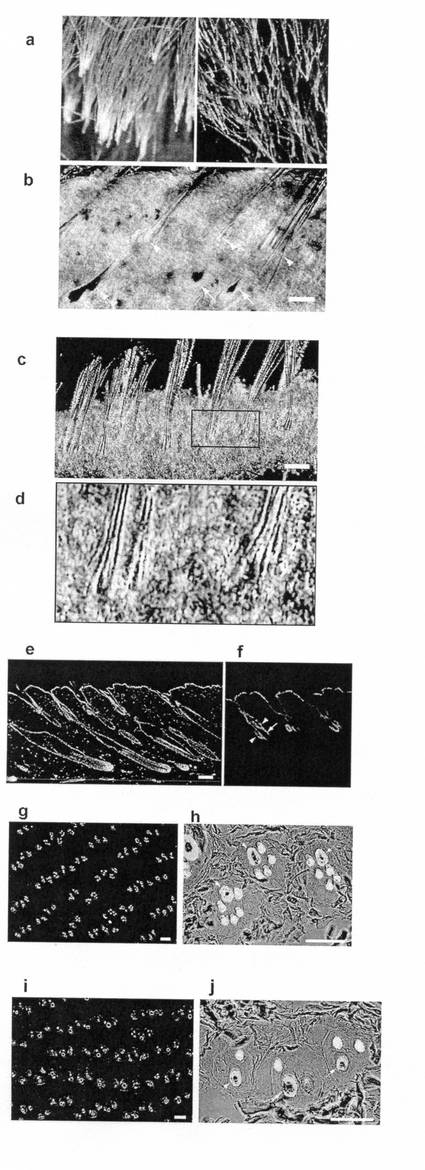
【図7】
【図8】
【図1】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−62294(P2012−62294A)
【公開日】平成24年3月29日(2012.3.29)
【国際特許分類】
【出願番号】特願2010−209749(P2010−209749)
【出願日】平成22年9月17日(2010.9.17)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成24年3月29日(2012.3.29)
【国際特許分類】
【出願日】平成22年9月17日(2010.9.17)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]