
水産養殖用経口投与免疫賦活剤
【課題】 加水分解又は分解を受けることが無く、しかも、投与されるべき微生物により完全に吸収される、経口投与に適した新規な免疫賦活剤を提供する。
【解決手段】 本発明の水産養殖用の経口投与免疫製品は、マイクロカプセル化サイトカインと、サイトカインを保護するための腸溶性保護ポリマーからなる。サイトカインは魚、貝又は甲殻類のサイトカインであり、好ましくは、宿主微生物に過剰に発現する腫瘍壊死因子α(TNFα)のような組換サイトカインである。
【解決手段】 本発明の水産養殖用の経口投与免疫製品は、マイクロカプセル化サイトカインと、サイトカインを保護するための腸溶性保護ポリマーからなる。サイトカインは魚、貝又は甲殻類のサイトカインであり、好ましくは、宿主微生物に過剰に発現する腫瘍壊死因子α(TNFα)のような組換サイトカインである。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は水産養殖用の経口投与可能な免疫賦活剤に関する。
【背景技術】
【0002】
水産養殖は、採取漁業によりカバーできない水産食品の成長要求をカバーする唯一の手段であると思われる。世界の水産養殖生産量は過去50年間の間に、1950年代初頭の100万トン未満の生産量から2004年までの5940万トンまでに増大している。この生産量レベルはUSドルで703億ドルの価値に相当する。全世界の生産量の69.6%は中国により、21.9%は他のアジア及び太平洋諸国により、3.9%は西ヨーロッパ諸国により、0.4%は中央及び東ヨーロッパ諸国により生産された。環境に関連して、2004年度における海中養殖(海水水産養殖)による養殖生産量は3020万トンであり、これは養殖全体の50.9%に相当する。真水による水産養殖は2580万トン(43.4%)であり、残りは340万トン(5.7%)であり、これらは汽水環境における生産に由来する。ここに列挙したデータは全てFisheries Technical Paper 2006, State of World Aquaculture (FAO)に基づく。
【0003】
ヨーロッパにおける水産養殖の生産量は全世界の生産量の僅か3%である。しかし、アトランティックサーモン、トラウト(鱒)、シーバス(鱸)、ヨーロッパヘダイ(gilthead sesbream)、ターボット(鮃又は鰈などの扁平な魚、例えば、砂ヒラメ)、及びムラサキイガイなどのような幾つかの種の生産量では全世界でトップである。2002年度の生産量は130万トン超であり、32億8000万ユーロの価値があった。地中海諸国における最も重要な種はヨーロッパヘダイ、シーバス、ターボットである。2002年度におけるヨーロッパヘダイ及びシーバスの生産量は181000トンであり、主な生産国はギリシャ、トルコ、スペイン、イタリヤ、及びフランスである。2002年度におけるターボットの生産量は5320トンであり、その75%はスペインで生産された。これらのデータは全てFAOのデータに基づく。
【0004】
魚類の病気は水産養殖の発展の妨げとなり、世界中の多くの国々における経済発展に対する苛酷な打撃の原因となる。病気は多くの場合大量死を起こし、重大な生産量低下の原因となる。水産養殖の損失はまた、食品利用可能性の減退、収入喪失及び雇用喪失なども起こし、これら全てが付随的な社会的重大性を伴う。前記の種を襲う最も一般的な病気及びこれらの原因となる病原体(バクテリア又はウイルス)はビブリオ症(Photobacterium damselae subsp. damselae, Vibrio spp.),パスツレラ症(Photobacterium damselae subp. pasteurella),発光桿菌症(Photobacterium damselae subp. pasteurella.),滑走細菌症(Tenacibaculum maritimum),粘液桿菌症(Flexibacter maritimus),非定型エロモナス症(Aeromonas salmonicida),連鎖球菌感染症(Streptococcus parauberis),冬不全症候群(Pseudomonas anguilliseptica),ウイルス性神経壊死症(Nodavirus),リンホシスチス症(Iridoviridae),膨張腸症候群(ウイルス様粒子),伝染性膵臓壊死症(IPNV),伝染性造血器壊死症(IHNV)又はウイルス性出血性敗血症(VHS)などである。これらのデータは全て、Cultured Aquatic Species Information programme(FAO)に基づく。水産養殖場における種の養殖状態(すなわち、高密度の養殖魚類又は水産養殖環境等)は固体間で伝達される伝染性疾患病原体にとって好適である。更に、養魚場で養殖中の魚類が受けるストレスは病原体による感染を助長する免疫系の低下を起こすであろう。従って、多くの研究努力が魚類の病気を予防するためのワクチン又は免疫賦活剤などの製品の開発に捧げられてきた。また、病気の予防は環境影響も低減する。なぜなら、水産養殖における抗生剤の大量使用が避けられるからである。幾つかの製品が魚のワクチン接種又は免疫システムの賦活に使用されてきた。実際、ビブリオ症、発光桿菌症、非定型エロモナス症、滑走細菌症、冬不全症候群又は連鎖球菌感染症等のような通常の細菌性魚類疾患及び(Toranzoら,2005及びSommersetら,2005により報告されたIPNV又はIHNV等のような通常のウイルス性魚類疾患に使用可能なワクチン類が存在する。
【0005】
ワクチン接種には、浸漬、注入及び経口投与からなる3種類の一般的方法が存在する。ワクチン接種には腹腔内注射が最も効果的な方法である。なぜなら、腹腔内注射は投与量を自在にコントロールでき、しかも、優れた免疫応答をもたらすアジュバントの使用が可能だからである。しかし、腹腔内注射法には欠点がある。例えば、魚類の鎮静及び注射操作は魚類にストレスを与え、細心の注意を払いながらワクチンを投与しないと魚類を損傷してしまう。更に、小型魚類にはこの方法ではワクチン接種できない。浸漬方法は養殖場で頻繁に使用されている。なぜなら、簡単であり、しかも、迅速だからである。しかし、投与量の厳密なコントロールが出来ず、一貫性のある免疫系の抑制を有する魚類にはストレスが掛かり過ぎる。ワクチンの経口投与では魚の操作は不要であり、給餌を始めたばかりの幼魚を含めて全てのサイズの魚類のワクチン接種に適した方法である。幼魚にもワクチン接種できるこの方法は極めて重要である。なぜなら、これら幼魚は病原体に非常に感染し易いからである。経口投与法にも幾つかの欠点がある。例えば、投与量の厳密なコントロールができないこと、及び免疫化のために多量の抗原が必要なことなどである。経口投与法の最も重要な障害は、酸性環境又はプロテアーゼ活性により抗原が腸内でしばしば不活化し、その結果、腸による活性抗原吸収が抑制されることである。このため、効果的な経口投与を行うには抗原を保護することが必要である。すなわち、腸内における抗原分解を避けるために抗原をカプセル化することが必要である(Hartら,1988; Quentel及びVigneulle, 1997)。抗原を分解から保護するための別の方法(例えば、リポソーム内への閉じ込め又はアルギン酸塩マイクロカプセル化など)は幾つかの有望な結果を例証している(Ireら,2005; Mauriceら,2004)。
【0006】
魚類の病気に対する耐性を高めるために、水産養殖において免疫賦活剤が使用されている。化学特性及び作用モードにより、多数の様々な種類の免疫賦活剤が知られている。例えば、細菌(LPS)の構造要素による、細菌及び菌糸菌由来の様々なβ−1,3グルカン製品、酵母の細胞壁由来のβ−1,3/1,6グルカン、様々な生物源由来の複合炭水化物構造構造体(グリカン類)、ある種の動物の、又は魚類蛋白の酵素加水分解物により産生されたエキス(抽出物)内に存在するペプチド類、ヌクレオチド類、合成製品類(レバミソール)及びビタミンC(Raa,1996により研究された)などである。また、使用された幾つかの免疫賦活剤は特定の抗体反応を高める(Sakai,1999により研究された)。幾つかの状態は水産養殖における免疫賦活剤の使用に好適である。例えば、魚類の操作、水温変化、高濃度病原体曝露、ワクチンアジュバント及び全ての高ストレス状況などである。ヨーロッパヘダイ(gilthead sesbream)免疫系における免疫賦活剤の効果に関して多数の文献が発行されている。例えば、出芽酵母(Saccharomyces cerevisiae)に関するOrtunoらの2002年の論文及びRodriguezらの2003年の論文、キチン質に関するEstebanらの2002年の論文、Estebanらの2001年の論文及びCuestaらの2003年の論文、ビタミンC及びEに関するCuestaらの2002年の論文及びOrtunoらの2003年の論文及びレバミソール(levamisole)に関するMuleroらの1998年の論文及びCuestaらの2002年の論文などである。魚類に有効であることが知られている免疫賦活剤のうち、グルカン、キチン質及びレバミソールは食細胞活性を高め、一方、酵母グルカン及びビタミンCは補体活性を活性化する。免疫賦活剤としてのB-1,3/1,6グルカン投与に関する幾つかの研究は、オヒョウ(halibut)幼魚の発達段階の重大な局面でのオヒョウ幼魚の生存率を高めることに関して優れた結果を示すことがOttesn, Lunda及びEngstadらの1999年の論文に発表され、細菌感染症に対する耐性を高めることがRaaらの1990年の論文及びRobertsenらの1990年の論文に発表され、また、ワクチンの効果を高めることがRaaらの1990年の論文、Rorstad, Assjord及びRobertsenの1993年の論文に発表されている。
【0007】
サイトカイン(Cytokine)は細胞成長、細胞活性、炎症、免疫、組織修復、繊維増多及び形態形成などのような重量な生物学的プロセスの量を調節するので、“免疫調節剤”の一般的定義に属するタンパク質類である。サイトカインは幾つかの特性を共有するタンパク質類の異なるグループであり、先天的及び適応的免疫応答の両仲介エフェクター相の発生と作用にとって必須のものである。過去数年にわたり、非常に多数のサイトカインがクローンとして発生され、幾つかの魚属において配列決定された(例えば、Birdらの2002年の論文参照)。サイトカインはその機能により下記の4種類のグループに分類することができる。
(a)先天的免疫を仲介するサイトカイン類。このグループの魚類における最良の特性を有するサイトカインはタイプIインターフェロン(IFN)であり、これにはIFNαとIFNβ、インターロイキン1(IL-1)ファミリー(IL-1α、IL-1β、IL-1レセプター拮抗体及びIL-18を含む)及び腫瘍壊死因子(TNF)ファミリー(TNFα、TNFβ、Ltβ及びFas配位子を含む。)などが含まれる。これらのうち、TNFαが免疫系の調節機能を有する最も重要な一員である。
(b)造血を調整するサイトカイン、
(c)リンパ球を調節するサイトカイン及び
(d)非特定エフェクター細胞を制御するサイトカイン。
【0008】
これらのグループにおいて、最も優れた特性を有するサイトカインは、グループ(b)のケースではインターロイキン2(IL-2)であり、グループ(c)では形質転換成長因子ベータ(TGFβ)である。
【0009】
TNFαは急性炎症中にモノサイト/マクロファージにより産生される炎症性サイトカインであり、細胞内の異なった範囲内の前兆イベントに応答する義務がある。TNFαは、寄生感染症、細菌感染症及びウイルス感染症に対する抵抗性における重要なメディエータとしての役割を果たす(Czarnieckiの1993年の論文、Goldfeld及びTsaiの1996年の論文及びSteinshamnらの1996年の論文参照)。TNFαはその他の重要な治療作用も有する。例えば、耐腫瘍性(Vilcek及びLeeの1991年の論文参照)、睡眠調整(Kruegerらの1998年の論文参照)及び胚成長(Wride及びSandersの1995年の論文参照)。TNFαは細胞膜レセプター,TNFR-1又はTNFR-2にトリマーとして結合することによりこれらの多くの作用を発揮する(Dembicらの1990年の論文及びLoetscherらの1990年の論文参照)。これらの両レセプターは殆どの細胞内に非常に大量に存在する(Loetscherらの1991年の論文及びSchoenfeldらの1991年の論文参照)。
【0010】
TNFαmRNAは広範な種類の細胞内で転写されるものと思われる。従って、転写後レベルでは著しく抑制される(Hanらの1990年の論文参照)。TNFαは、膜結合形と可溶性形の2つの形で存在する。各形とも恐らく、明確な生理学的役割を有する(Wattsらの1990年の論文参照)。TNFαは26kDa膜結合前駆体として明示され、これはディスインテグリン及び金属蛋白分解酵素(metalloproteinase)(TACE:TNFα変換酵素)によりタンパク質切断され、17kDaC−末端活性形が生成される(Blackらの1997年の論文及びMossらの1997年の論文参照)。
【0011】
幾つかの哺乳類のTNFα情報は完全に又は部分的に配列決定されている。このような哺乳類は例えば、ヒト(Pnnicaらの1984年の論文参照)、チンパンジー(Kutsuiらの2002年の論文参照)、マウス(Shiraiらの1988年の論文参照)、イヌ(Zuckerらの1994年の論文参照)又はネコ(McGrawらの1990年の論文参照)などである。また、幾つかの魚類のTNFα情報又はTNFα情報断片も配列決定されている。このような魚類はクロマダイ(black seabream)(Caiらの2003年の論文参照)、ニジマス(Laingらの2001年の論文参照)、コイ(Saeijらの2003年の論文参照)又はゼブラフィッシュ(Phelanらの2003年の論文参照)若しくはヨーロッパヘダイ(Garcia-Castilloらの2002年の論文参照)などである。
【0012】
ヨーロッパヘダイのTNFα情報は4個のエクソンと3個のイントロン及び1244pbの長さを有する。cDNAは1359pbからなり、これは、762pbのオープン読み枠(open reading frame;ORF)、142pbの5’非翻訳領域(UTR)及び455pbの3’UTRを含む。推定タンパク質は253個のアミノ酸を有し、概算分子量は28kDaであり、その他の魚種TNFαとの、特にレセプターとの相互作用に必要なC−末端エクストリームにおける高い配列相同性を与える。
【0013】
TNFαタンパク質は37及び54残基との間のトランスメンブラン領域と、TACEタンパク質切断に付随する保存配列Thr-Leuを包含する(Garcia-Castilloらの2002年の論文参照)。従って、253アミノ酸膜結合タンパク質(proTNFα)と167アミノ酸可溶性タンパク質(sbTNFα)とを区別することができる。
【0014】
発現研究は、肝臓、エラ、血液、頭腎、腹腔侵出物、脳及び脾臓などのような試験されたヨーロッパヘダイの全ての組織でTNFαの構成的発現を明らかにした。これは、この分子の主要源の一つのマクロファージである。
【0015】
特許文献1(米国特許第5871751号明細書)は、魚類の細菌性腎臓病菌(Renibacterium salmoninarum)による感染を受けやすい魚類の治療用ワクチン及び治療方法を開示している。このワクチンは無傷の細胞表面結合タンパク質p57を欠いた死滅微生物からなり、かつ、経口投与のための腸溶剤皮が施されている。腸溶剤皮は胃内における溶解又は分解を受け付けないが、腸内の高pH環境を通過したら溶解されるポリマー剤皮からなる。開示されたワクチンの好ましい実施態様は、球形糖微小球を用いて生成され、無傷の細胞表面結合タンパク質p57を欠いた死滅微生物からなる第1層で被覆し、次いで、魚類の胃内で溶解又は分解を受け付けない材料からなる腸溶剤皮の第2層で被覆されている。
【0016】
特許文献2(国際公開第WO03/020040A1号公報)は、抗原を含有する単細胞微生物をカプセル封入した多細胞生物からなるワクチンを開示している。このワクチンは予防接種すべき水生動物(例えば、魚類)に給餌するのに使用される。特許文献2によれば、単細胞微生物中に発現される抗原は2段階給餌により水生動物に送達される。すなわち、(抗原を含有する)単細胞微生物を多細胞生物に給餌する第1段階と、多細胞生物を水生動物に給餌する第2段階である。特許文献2によれば、単細胞微生物と多細胞生物の両方の同時存在が常に必要である。発現されるべき抗原は、誘発免疫反応が目標病原体に対するものであるか否かに依存する。特許文献2には、細菌性抗原として緑膿菌外毒素A(PE)、単細胞微生物として大腸菌(E. coli)、多細胞生物としてアルテミア・ノープリウス幼生(Artemia nauplii)が開示されている。特許文献2は経口投与ワクチンの製造を開示しているので、選択された抗原は、このワクチンが向けられなければならない目標病原体に厳密に依存する。
【0017】
特許文献3(スペイン特許公開第2189508A1号公報)は、ヨーロッパヘダイ(Sparus aurata L.)の1L-1β組換え体の製造方法及びワクチンアジュバント並びに市販魚類における免疫賦活剤としての使用法を開示している。特許文献3による1L-1β組換え体の製造はこれを大腸菌発現ベクターにクローニングすることにより行われる。このようにして得られた組換え体タンパク質は精製され、そして、魚類の大量養殖における免疫賦活剤及びワクチンアジュバントとして使用される。
【先行技術文献】
【特許文献】
【0018】
【特許文献1】米国特許第5871751号明細書
【特許文献2】国際公開第WO03/020040A1号公報
【特許文献3】スペイン特許公開第2189508A1号公報
【発明の概要】
【発明が解決しようとする課題】
【0019】
本発明の目的は、加水分解又は分解を受けることが無く、しかも、投与されるべき微生物により完全に吸収される、経口投与に適した免疫賦活剤を提供することである。
【0020】
本発明の別の目的は、目標対象物に直接投与することができる免疫賦活剤を提供することである。
【0021】
また、本発明の他の目的は、初期投与量に対して大量に投与される微生物により吸収され得る免疫賦活剤を提供することである。
【0022】
本発明の更に別の目的は、特定の病原体又は特定の病原体群に対して特別に送達されない免疫賦活剤を提供することである。
【0023】
本発明の更に他の目的は、経口投与可能な免疫賦活剤の製造方法及び水産養殖における免疫応答の活性化のための免疫賦活剤の使用法を提供することである。
【課題を解決するための手段】
【0024】
前記課題を解決するために、本発明は、少なくともマイクロカプセル化された組換魚類サイトカイン、特に腫瘍壊死因子α(TNFα)からなる経口投与可能な免疫賦活剤及びマイクロカプセル化された組換魚類サイトカインからなる経口投与可能な免疫賦活剤の製造方法及び水産養殖における免疫応答の活性化のためのその使用法を提供する。
【0025】
本発明の免疫賦活剤の特徴は、免疫賦活剤として作用する活性分子(すなわち、サイトカイン及び特定の腫瘍壊死因子α(TNFα))が、免疫応答の活性化が望まれる同じ魚種に属するサイトカインであるということである。この特徴は非常に重要である。なぜなら、当該魚種に既に属する特定の一組の免疫賦活剤を正に高める特定の魚種の免疫応答を活性化させることができるからである。この事実は次のことを意味する。すなわち、当該魚種に任意の方法で投与される外部因子、化合物類又は製品は存在せず、その結果、得られる結果が優れた利益を上げるばかりか、安全なアプローチ法としても優れている。前記のように、本発明によれば、免疫賦活剤として作用する活性分子は魚類自体から得られたサイトカイン、腫瘍壊死因子α(TNFα)である。従って、本発明の免疫賦活剤が投与されるとき、外来物質は魚類に導入されない。その結果、副作用の可能性は避けられる。
【0026】
本発明によれば、特定のサイトカインをコードする遺伝子は魚類の組織から単離され、適当な宿主微生物中でサイトカイン発現用の発現ベクターにクローンされる。宿主細胞は最適なサイトカイン発現及び高バイオマス産生に適した溶媒を用いてバイオリアクター中で培養される。本発明の好ましい実施態様によれば、宿主微生物は酵母(イースト)であり、好ましくはメタノール資化酵母(Pichia pastoris)及び出芽酵母(Saccharomyces cerevisiae)であり、最も好ましくは メタノール資化酵母である。宿主微生物としての酵母の選択は偶然ではない。なぜなら、酵母は真核微生物であり、細胞の外部及び内部の両方に特定のサイトカインを発現できるからである。これにより、発酵方法及び斯くして得られた生成物の精製に多くの効果をもたらす組換タンパク質の発現方法を決定することができる。次いで、組換タンパク質(単独で得られたもの又は宿主微生物の内部で得られたものの何れか)をマイクロカプセル化処理し、必要に応じて精製処理工程を経て、投与可能なマイクロカプセル化タンパク質が得られる。本明細書で使用する「マイクロカプセル化方法」という用語は、組換サイトカインに少なくとも部分的に被覆(この被覆は腸内分解からサイトカインを保護するのに適している)を施すのに好適な全ての方法を意味する。
【0027】
実際、免疫賦活剤を経口投与する場合、免疫賦活剤の腸内分解を避けるため、及び腸管により完全吸収を可能にするため、効率的な保護方法が必要である。マイクロカプセル化は、液体又は固体の極めて小さな液滴又は粒子を高分子材料で包囲又は被覆し、微粒子を得る技術的方法である。マイクロカプセル化技術は幾つかの生物学的及び化学的システムで汎用されており、莫大な量の製品を製造することができる。本発明によれば、所望の及び特定のサイトカイン(特に、TNFα)のマイクロカプセル化製剤を成功裡に得るために使用される主要技術は噴霧乾燥による微粉化である。噴霧乾燥による微粉化の技術では、活性分子は溶融又はポリマー溶液内に溶解又は懸濁されており、そして、乾燥粒子内に閉じ込められる。本発明によれば、腸内の産生pHから組換タンパク質の微粒子を保護するのに使用された最も有用なポリマーは例えば、酢酸フタル酸セルロース、フタル酸ヒドロキシプロピルセルロース、カルボキシメチルセルロース、メタクリル酸コポリマー(例えば、Eudragit L-30及びLollicoatなどのようなメタクリル酸コポリマーLS)などから選択される。その他のポリマー類、例えば、マルトデキストリン、キトサン、ゼラチン、スターチ又はアラビアゴムなどから選択されるポリマー類も使用できる。
【発明の効果】
【0028】
本発明では、微生物中で発現され、次いで、腸内の酸性環境及びプロテアーゼ活性からタンパク質を保護するためにポリマーと混合されるサイトカインの一例としてTNFαを使用する。組換タンパク質、すなわちTNFαは、宿主微生物が細胞外にその過剰発現を可能にするとき、そのまま使用することができる。効果的な別法として、宿主微生物がタンパク質を内部的に過剰発現する場合、同じ微生物をマイクロカプセル化し、そのまま使用することができるマイクロカプセル化微生物が得られると共に、前記のような全ての効果が発揮される。過剰発現したタンパク質を噴霧乾燥により乾燥し、魚類の健康に有効な量だけ魚類の餌に添加する。本発明による製品は、感染により魚類の疾患を予防し、何らかの高ストレス環境中の魚類を保護するため、及び、ワクチンのアジュバントとして、水産養殖市場で桁外れの重要性を有する。
【0029】
本発明によれば、免疫賦活剤が成魚の治療に差し向けられる場合、マイクロカプセル化処理により得られる利点により、免疫賦活剤は常にそのまま投与することができる。これに対して、免疫賦活剤が幼魚又は稚魚の治療に差し向けられる場合、例えば、アルテミア・ノープリウス幼生(Artemia nauplii)などのような多細胞生物に免疫賦活剤を給餌し、次いで、幼魚又は稚魚群により容易に摂食可能な、この多細胞生物を幼魚又は稚魚に給餌することができる。何れの場合も、マイクロカプセル化された免疫賦活剤は製品の主要特性及び活性を安全に維持することができ、水産養殖に多大な利益をもたらす。
【0030】
一例として、ヨーロッパヘダイの組換sbTNFαは精製され、そして、その生物活性は生体内及び試験管内で評価された。組換タンパク質はアフィニティークロマトグラフィー精製及び免疫ブロット法における抗ポリヒスチジンmAb検出用にN−末端エクストリーム(extreme)に6個のヒスチジン類を含有する。腹膜内注射は、(a)腹膜侵出及び頭腎(HK)白血球の呼吸性バースト(respiratory burst)の呼び水、(b)注射部位に対する食細胞顆粒白血球の急速な固体増加及び(c)HKにおける顆粒白血球形成の誘発を生じた。sbTNFαは試験管内でHK細胞の強力な分芽増殖を誘発することができた。
【0031】
本発明による別の効果は、TNFαサイトカインを発現する宿主微生物それ自体が免疫賦活剤物質の追加源として機能出来ることである。細菌の幾つかの構造的要素類、酵母の細胞壁からのグルカン類又は様々な生物源からの複合糖質構造(グリカン類)は実際、免疫賦活剤として使用できる。
【0032】
水産養殖において非常に重要性の高い幾つかの魚種のTNFαを用いて行われる下記の実施例により本発明を更に詳細に説明する。しかし、その他の魚種のTNFα及びその他のサイトカインも本発明を実施するのに使用できる。また、本発明は合成により抽出又は生成されたサイトカインにも適用できる。
【図面の簡単な説明】
【0033】
【図1】遺伝子構成に使用されるプライマー類を示す。クローニング用のEcoRi及びBamHi制限部位はイタリック体で示されている。
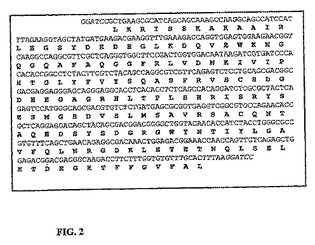
【図2】ヨーロッパヘダイのsbTNFαを含有するDNA断片及び演繹タンパク質配列を示す。
【図3】ヨーロッパヘダイのHis6-sbTNFαを含有するDNA断片及びメタノール資化酵母における発現用演繹タンパク質配列を示す。図中の下線部は6個のCAT/Cコドン類及びヒスチジン類を示す。メタノール資化酵母におけるタンパク質発現用コンセンサス配列は強調されている。EcoRi及びBamHi制限部位はイタリック体で示されている。
【図4】シーバスのHis6-sbTNFαを含有するDNA断片及びメタノール資化酵母における発現用演繹タンパク質配列を示す。図中の下線部は6個のCAT/Cコドン類及びヒスチジン類を示す。メタノール資化酵母におけるタンパク質発現用コンセンサス配列は強調されている。EcoRi及びBamHi制限部位はイタリック体で示されている。
【図5】ターボットのHis6-sbTNFαを含有するDNA断片及びメタノール資化酵母における発現用演繹タンパク質配列を示す。図中の下線部は6個のCAT/Cコドン類及びヒスチジン類を示す。メタノール資化酵母におけるタンパク質発現用コンセンサス配列は強調されている。EcoRi及びBamHi制限部位はイタリック体で示されている。
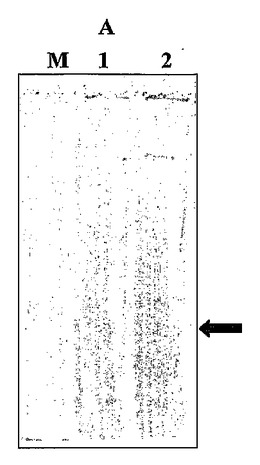
【図6】AはSDS-PAGE(ドデシル硫酸ナトリウム-ポリアクリルアミドゲル電気泳動)による泳動結果を示し、Bは抗ポリヒスチジンmAbによるウエスタン・ブロット法の分析結果を示す。図中の矢線は(1)大腸菌、(2)出芽酵母及び(3)メタノール資化酵母で発現されたヨーロッパヘダイ(Sparus aurata)のHis6-sbTNFαを示す。
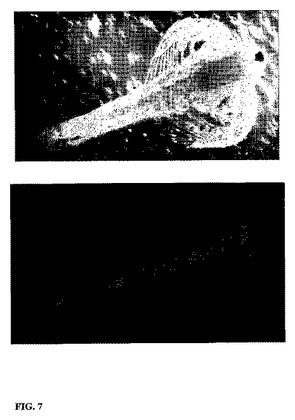
【図7】染色組換酵母を60分間給餌した後のアルテミア・ノープリウス幼生(Artemia nauplii)の顕微鏡観察図であり、Aはアルテミア・ノープリウス幼生を示し、BはDTAFを有する蛍光染色酵母による消化管内部を示す。
【図8】頭腎白血球により産生されたルミノール依存性化学発光による呼吸性バースト活性を示すグラフである。縦軸は、化学発光(相対光単位/分)
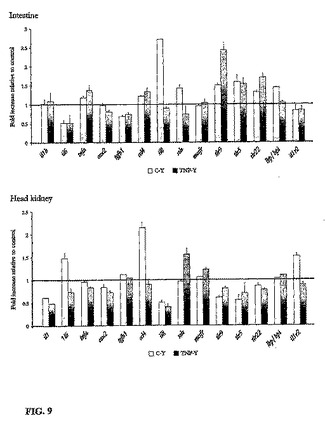
【図9】頭腎及び腸管における幾つかの炎症性遺伝子のmRNAレベルを示すグラフである。上のグラフは腸管に、下のグラフは頭腎に関する。縦軸はどちらも、対照に対する増大率である。
【発明を実施するための形態】
【0034】
以下、実施例により本発明を更に詳細に説明する。
【実施例1】
【0035】
sbTNFαをpET15bにクローニングする大腸菌内におけるヨーロッパヘダイTNFαの発現
FE4及びRE5(図1参照)によるsbTNFαの増幅のためのPCRにおけるテンプレート(鋳型)として、LPS刺激頭腎cDNAを使用した。両方のプライマーとも、プラスミドpET15b(Novagen社製)の同じ部位でPCR生成物を後クローニングするためのBamHI制限部位を有する。増幅はcDNAテンプレート、各dNTP50μM、プライマー0.2mM、MgCl2含有1X buffer PLUS及びEco Taq PULS DNAポリメラーゼ(Ecogen社製)1単位を含有するサンプル中で行った。サイクル反応はエッペンドルフ・マスターサイクラー・グラジエント内で、95℃・2分間を1サイクル、95℃・45秒間を25サイクル、60℃・45秒間を25サイクル及び72℃・30秒間を25サイクル続いて72℃・10分間を1サイクルの順で行った。PCR生成物をQIAquick・PCR精製キット(Qiagen社製)で精製し、T4DNAリガーゼ(New England Biolabs社製)1単位を使用し、インサート:プラスミド(3:1)の関係でプラスミドpGEM-T Easy(Promega社製)と室温で16時間かけてライゲーションさせた。ライゲーション混合物を形質転換大腸菌DH5αコンピテントセル(形質転換受容性細胞)に対して使用し、そして、アンピシリンとX-Gal(Sigma社製)を含有するLBプレート内に展延させた。このプレートを37℃でインキュベーションし、幾つかの生成白色コロニーを選び出し、QIAprep・Spin・Miniprepキット(Qiagen社製)によるプラスミド単離及びBamHIによる消化を通じてインサートの存在を試験した。インサート(BamHI端部を有するsbTNFα,図2参照)を含有する特定のプラスミドはpVP81と命名された。このプラスミド及び500ngのpET15bを、pVP81のsbTNFα遊離のため及びpET15b線状化のために10単位のBamHIで消化させた。インサート及び直線プラスミドの両方とも、QIAquickゲル抽出キット(Qiagen社製)によりアガロース低融点ゲル(Pronadisa社製)中で電気泳動することにより分離した後、精製し、そして、1単位のT4DNAリガーゼ(New England Biolabs社製)を使用し、室温で16時間かけてライゲーションさせた。ライゲーション混合物を形質転換大腸菌DH5αコンピテントセルに対して使用し、そして、アンピシリンを含有するLBプレート内に展延させた。このプレートを37℃でインキュベーションし、幾つかの生成コロニーのプラスミドを単離し、BamHI消化によるインサート遊離及びPvull消化によるインサート配向について試験した。選択されたプラスミドの配列をABI・Prism377遺伝子分析器(CIB,CSIC社製)で決定した。
【0036】
大腸菌の形質転換及び発現測定
大腸菌BL21(DE3)コンピテントセルをsbTNFα含有プラスミドpET15bで形質転換し、得られた混合物をアンピシリン含有LBプレート内に展延した。幾つかの生成コロニーをLB-アンピシリン媒体中で一晩培養した。新鮮なLB-アンピシリン中に希釈した後、培養物を37℃でOD600=0.8まで成長させ、1mMのイソプロピル-D-チオガラクトシド(IPTG; Applichem社製)を用いて0.25〜4時間、25℃又は37℃で誘導した。誘導培養物0.1mlを遠心分離(1400rpm)することにより全細胞抽出物中のタンパク質発現をチェックし、細胞ペレットを、SDS-PAGE及び抗ポリヒスチジンmAb(Sigma社製)を用いるウエスタン・ブロット法による分析のためにSDS-ローディングバッファー液中で煮沸することにより溶解させた。
【実施例2】
【0037】
出芽酵母におけるヨーロッパヘダイsbTNFαの発現
His6-sbTNFαのp424-GPDへのクローニング
613pbのHis6-sbTNFαを含む断片の増幅のためにTNF-ECOF及びTNF-ECORプライマー(図1参照)と共にPCRにおけるテンプレートとしてsbTNFα含有プラスミドpET15bを使用した。両方のプライマーとも、プラスミドp424-GPD(ATCC87357)の同じ部位内でPCR生成物をクローニングするためのEcoRI制限部位を含有する。増幅は、テンプレートを5ng、MgCl2を2.5mM、各dNTPを50μM、プライマーを0.2mM、1X・High・Speec添加剤、1X buffer PLUS及びEco・Taq・PLUS・DNAポリメラーゼを2単位含有するサンプル中で行った。サイクル反応はSmartサイクラーII(Cepheid社製)中で、95℃・5分間を1サイクル、95℃・30秒間を30サイクル、62℃・45秒間を30サイクル及び72℃・2分間を30サイクル、続いて72℃・10分間を1サイクルの順で行った。臭化エチジウムを0.5μg/ml含有する0.8%アガロースゲル(Pronadisa社製)中で電気泳動することによりPCR生成物を分離し、QIAquickゲル抽出キット(Qiagen社製)で精製した。精製断片及びp424-GPD250ngをEcoRI(Fermentas社製)10単位により37℃で消化させ、その後、T4DNAリガーゼ(Fermentas社製)5単位を用いて22℃でライゲーションさせた。ライゲーション反応生成物を大腸菌DH5αコンピテントセルに形質転換させ、アンピシリン100μg/mlを有するLBプレート内でプラスミド含有コロニーを選別した。2X・PCR・Master・Mix(Fermentas社製)及びPRO-GPD及びTER-GPDプライマー(図1参照)を用いるコロニーPCRにより組換体を識別した。得られたHis6-sbTNFα含有プラスミドp424-GPDをQIAprep・Spin・Miniprepキット(Qiagen社製)で精製し、EcoRIによる消化後のインサート遊離及び消化Smal及びNcolによるインサート配向について試験した。また、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学)によりプラスミドの配列を決定した。
【0038】
出芽酵母の形質転換及び発現測定
形質転換に使用された出芽細胞菌株はEGY48(Invitrogen社製)であった。形質転換用のEGY48を作製するために、YPD培地(2%ペプトン、1%酵母エキス、2%グルコース)中でOD600=0.5まで成長させた培養物100mlの細胞を2500rpmで4℃で5分間遠心分離した。滅菌水で洗浄後、細胞を1X・LiAc-TE(0.1M酢酸リチウム, 10mM Tris pH7.5,1mMEDTA pH8.0)に再懸濁させた。形質転換のために、濃度0.3μg/μlのDNA10μlを細胞50μl及びDNA担体50μgと混合した(サーモンのDNAを予め煮沸しておいてから使用した)。続いて、この混合物を1X・LiAc-TE-PEG(1X・LiAc-TEは前記のものと同一であり、PEGは40%)混合物に添加し、30℃で30分間インキュベーションし、その後、DMSOを10%まで添加し、42℃で10分間インキュベーションした。細胞をMMSS+SDC-Trp(グルコース2%、酵母窒素塩基0.17%、硫酸アンモニウム0.25%、1Mソルビトール及びSDC-Trpを含有する最少培地)プレートに展延し、コロニーが出現するまで30℃で数日間インキュベーションした。SDC(完全合成デキストロース, Synthetic Dextrose Complete)はアデニン、アルギニン、ヒスチジン、トリプトファン及びウラシルを20mg/l,ロイシン、リジン、チロシン及びイソロイシンを30mg/l,フェニルアラニンを50mg/l,スレオニンを200mg/l,バリンを150mg/lそれぞれ含有する。
【0039】
His6-sbTNFα発現を試験するために、幾つかのコロニーをYPD培地又はMM+SDC-Trp(グルコース2%、酵母窒素塩基0.17%、硫酸アンモニウム0.25%及びSDC-Trpを含有する最少培地)で数日間培養した。タンパク質発現をチェックするために、全細胞抽出物を、30秒間で10パルスの周期でBranson社製の超音波ホモジナイザーにより又は1分間で8パルスの周期でガラスビーズを用いるビードビーターにより粉砕した。粉砕処理完了後、細胞残留物を遠心分離により回収し、SDS-PAGE及び抗ポリヒスチジンmAb(Sigma社製)を用いるウエスタン・ブロット法による分析のためにSDS-ローディングバッファー液中で煮沸することにより上澄液のHis6-sbTNFαを測定した。
【実施例3】
【0040】
アルコール資化酵母におけるヨーロッパヘダイsbTNFαの発現
His6-sbTNFαのpPCICZAへのクローニング
618pbのHis6-sbTNFαを含む断片の増幅のためにTNF-ECOF及びTNF-ECORプライマー(図1参照)と共にPCRにおけるテンプレートとしてsbTNFα含有プラスミドpET15bを使用した。両方のプライマーとも、プラスミドpPCICZA(Invitrogen社製)の同じ部位内でPCR生成物をクローニングするためのEcoRI制限部位を含有する。増幅は、テンプレートを5ng、MgCl2を2.5mM、各dNTPを50μM、プライマーを0.2mM、1X・High・Speec添加剤、1Xbuffer PLUS及びEco・Taq・PLUS・DNAポリメラーゼを2単位含有するサンプル中で行った。サイクル反応はSmartサイクラーII(Cepheid社製)中で、95℃・5分間を1サイクル、95℃・30秒間を30サイクル、55℃・45秒間を30サイクル及び72℃・2分間を30サイクル、続いて72℃・10分間を1サイクルの順で行った。臭化エチジウムを0.5μg/ml含有する0.8%アガロースゲル(Pronadisa社製)中で電気泳動することによりPCR生成物を分離し、QIAquickゲル抽出キット(Qiagen社製)で精製した。精製断片及びpPICZA200ngをEcoRI(Fermentas社製)10単位と共に37℃で消化させ、その後、T4DNAリガーゼ(Fermentas社製)5単位を用いて22℃でライゲーションさせた。ライゲーション反応生成物を大腸菌TOP10F'コンピテントセルに形質転換させ、ゼオシン(Invitrogen社製)25μg/mlを有するLBローソルトプレート内でプラスミド含有コロニーを選別した。2X・PCR・Master・Mix(Fermentas社製)及びPIC5及PIC3プライマー(図1参照)を用いるコロニーPCRにより組換体を識別した。得られたHis6-sbTNFα含有プラスミドpPCICZAをQIAprep・Spin・Miniprepキット(Qiagen社製)で精製し、EcoRIによる消化後のインサート遊離及びXholによる消化を通じてインサート配向について試験した。また、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学)によりプラスミドの配列を決定した。組換プラスミドにおいて、sbTNFαのスターコドンATGは、酵母における外来遺伝子発現(Romanosらの1992年の論文参照)のために、酵母のコンセンサス配列AAAAATGTCT(図3参照)中に存在する。このコンセンサス配列はプライマーTNF-ECOPP中に含まれる。
【0041】
アルコール資化酵母のエレクトロポレーション及び発現測定
アルコール資化酵母のエレクトロポレーションの前に、後のアルコール資化酵母のゲノムDNAにおける組換及び組込みのために、His6-sbTNFα含有プラスミドpPICZAを37℃でSacl又はPmelによる消化を通じて線状化させた。線状プラスミドの全量5μgをMiniElute・Reaction・Cleanupキット(Qiagen社製)で精製し、滅菌水10μlで溶離した。エレクトロポレーションに使用したアルコール資化酵母菌株はX-33であった。エレクトロポレーション用のX-33を作製するために、OD600=1.5の培養物100mlの細胞を1500xgで4℃で5分間遠心分離した。滅菌水で2回洗浄後、細胞を1Mのソルビトール中に再懸濁させ、前記のように遠心分離し、そして最後に1Mのソルビトール100ml中に再懸濁させた。エレクトロポレーションを行うために、DNA10μlを細胞80μlと混合し、氷冷された0.2cmのエレクトロポレーション・キュベット(BioRad社製)に移送した。5分間インキュベーションした後、MicroPulserエレクトロポレーター(BioRad社製)により25μF、1.5kV及び400Ωの条件でパルスを印加した。その直後に、氷冷された1Mソルビトール溶液1mlをキュベットに添加し、得られた混合物を30℃で1時間インキュベーションした。細胞をゼオシン(Invitrogen社製)100μg/mlを有するYPDSプレート上に展延し、コロニーが出現するまで30℃で数日間インキュベーションした。PIC5及びPIC3プライマーと2X・PCRマスターミックス(Fermentas社製)を用いるPCR法によりインサートの存在に関して幾つかの形質転換体を分析し、His6-sbTNFα発現に関して測定した。特定のコロニーにおいて、ゲノムDNA精製キット(Fermentas社製)を用いてゲノムDNAを単離し、そして、His6-sbTNFα含有DNA断片をEcoTaq・PULS・DNAポリメラーゼ(Ecogen社製)で増幅し、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学)によりその配列を決定した。
【0042】
His6-sbTNFαを有する組換アルコール資化酵母菌株をグリセロール含有最少培地25ml中で30℃、250rpmで、培養物がOD600〜6に達するまで、メタノール導入前にバイオマスを発生させるために成長させた。3000xgで5分間遠心分離することにより細胞を回収し、発現を誘発させるために、OD600〜1に達するまで、ペレットを0.5%メタノール含有最少培地200ml中に再懸濁させた。前記に述べたように培養物をインキュベーションし、発現誘導を維持するために24時間毎に0.5%メタノールを添加した。幾つかの時点でサンプルを回収し、タンパク質発現を分析した。タンパク質発現をチェックするために、全細胞抽出物を30秒間で10パルスの周期でBranson社製の超音波ホモジナイザーにより又は1分間で8パルスの周期でガラスビーズを用いるビードビーターにより粉砕した。粉砕処理完了後、細胞残留物を遠心分離により回収し、SDS-PAGE及び抗ポリヒスチジンmAb(Sigma社製)を用いるウエスタン・ブロット法による分析のためにSDS-ローディングバッファー液中で煮沸することにより上澄液のHis6-sbTNFαを測定した。
【実施例4】
【0043】
大腸菌におけるシーバスsbTNFαの発現
sbTNFαのpET15bへのクローニング
FE4及びRE5(図1参照)によるsbTNFαの増幅のためのPCRにおけるテンプレート(鋳型)として、シーバスの肝臓から得られたcDNAを使用した。両方のプライマーとも、プラスミドpET15b(Novagen社製)の同じ部位でPCR生成物を後クローニングするためのBamHI制限部位を有する。増幅はcDNAテンプレート、各dNTP50μM、プライマー0.2mM、MgCl2含有1X buffer PLUS及びEco Taq PULS DNAポリメラーゼ(Ecogen社製)1単位を含有するサンプル中で行った。サイクル反応はSmartサイクラー(Cepheid社製)内で、95℃・5分間を1サイクル、95℃・45秒間を30サイクル、55℃・45秒間を30サイクル及び72℃・90秒間を30サイクル続いて72℃・10分間を1サイクルの順で行った。0.8%アガロースゲル(Pronadisa社製)中で電気泳動することにより分離した後、PCR生成物をQIAquick・PCR精製キット(Qiagen社製)で精製した。精製断片及びプラスミドpET15bをBamHIを10単位使用し、37℃で2時間かけて消化し、PCR精製キットによる前記PCR精製物をT4DNAリガーゼ(Fermentas社製)1単位を用いて22℃でライゲーションさせた。ライゲーション混合物を形質転換大腸菌DH5αコンピテントセル(形質転換受容性細胞)に対して使用し、そして、アンピシリン含有LBプレート内に展延させた。このプレートを37℃でインキュベーションし、2X・PCRマスターミックス(fermentas社製)及びプライマーT7F及びT7R(図1参照)を用いるコロニーPCR法により組換体を識別した。幾つかの生成コロニーのプラスミドをQIAprep・Spin・Miniprepキット(Qiagen社製)で単離し、BamHI消化によるインサート遊離及びPvull消化によるインサート配向について試験した。組換プラスミドの配列はABI・Prism3130遺伝子分析器(SACE,Murcia大学製)で決定した。
【0044】
大腸菌の形質転換及び発現測定
シーバスHis6-sbTNFαの大腸菌の形質転換及び発現測定は実施例1に述べた方法と同様な方法で実施した。
【実施例5】
【0045】
アルコール資化酵母におけるシーバスsbTNFαの発現
His6-sbTNFαのpPICZAへのクローニング
シーバスのHis6-sbTNFαのpPICZAへのクローニングは実施例3に述べた方法と同様な同様な方法で行った。シーバスのHis6-sbTNFαを含有する生成プラスミドpPICZAの配列をABI・Prism3130遺伝子分析器(SACE,Murcia大学製)で決定した。実施例3で述べたように、sbTNFαのスターコドンATGは、酵母における外来遺伝子発現(Romanosらの1992年の論文参照)のために酵母のコンセンサス配列AAAAATGTCT(図4参照)中に存在する。このコンセンサス配列はプライマーTNF-ECOPP中に含まれる。
【0046】
アルコール資化酵母のエレクトロポレーション及び発現測定
エレクトロポレーションは実施例3に述べた方法と同様な方法で行った。しかし、プラスミドの線状化はPmel(Fermentas社製)で行った。選択されたコロニーにおいて、ゲノムDNA精製キット(Fermentas社製)でDNAを単離し、His6-sbTNFα含有DNA断片をEcoTaq・PULS・DNAポリメラーゼ(Ecogen社製)で増幅し、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学製)によりその配列を決定した。
【0047】
発現測定も実施例3に述べた方法と同様な方法で行った。
【実施例6】
【0048】
大腸菌におけるターボットsbTNFαの発現
His6-sbTNFαのpET15bへのクローニング
プライマーTNFRO-BAMF及びTNFRO-BAMR(図1参照)を用いるsbTNFα増幅のためのPCR法におけるテンプレートとしてターボットの肝臓から得られたcDNAを使用した。両方のプライマーとも、プラスミドpET15b(Novagen社製)の同じ部位でPCR生成物を後クローニングするためのBamHI制限部位を有する。増幅はcDNAテンプレート、各dNTP50μM、プライマー0.2mM、MgCl2含有1X buffer PLUS及びEco Taq PULS DNAポリメラーゼ(Ecogen社製)1単位を含有するサンプル中で行った。サイクル反応はSmartサイクラー(Cepheid社製)内で、95℃・5分間を1サイクル、95℃・45秒間を30サイクル、55℃・45秒間を30サイクル及び72℃・90秒間を30サイクル続いて72℃・10分間を1サイクルの順で行った。0.8%アガロースゲル(Pronadisa社製)で電気泳動することにより分離した後、PCR生成物をQIAquick・PCR精製キット(Qiagen社製)で精製した。精製断片及びプラスミドpET15bをBamHI10単位により37℃で2時間かけて消化し、PCR精製キットによる前記PCR精製物をT4DNAリガーゼ(Fermentas社製)1単位を用いて22℃でライゲーションさせた。ライゲーション混合物を形質転換大腸菌DH5αコンピテントセル(形質転換受容性細胞)に対して使用し、そして、アンピシリン含有LBプレート内に展延させた。このプレートを37℃でインキュベーションし、2X・PCRマスターミックス(fermentas社製)を用いるコロニーPCR法により組換体を識別した。幾つかの生成コロニーのプラスミドを単離し、BamHI消化によるインサート遊離及びPvull消化によるインサート配向について試験した。選択されたプラスミドの配列をABI・Prism377遺伝子分析器(CIB,CSIC社製)で決定した。
【0049】
大腸菌の形質転換及び発現測定
大腸菌の形質転換及びターボットHis6-sbTNFαの発現測定は実施例1に述べた方法と同様な方法で実施した。
【実施例7】
【0050】
アルコール資化酵母におけるターボットsbTNFαの発現
His6-sbTNFαのpPICZAへのクローニング
ターボットのHis6-sbTNFαのpPICZAへのクローニングは実施例3に述べた方法と同様な同様な方法で行った。ターボットのHis6-sbTNFαを含有する生成プラスミドpPICZAの配列をABI・Prism3130遺伝子分析器(SACE,Murcia大学製)で決定した。実施例3で述べたように、sbTNFαのスターコドンATGは、酵母における外来遺伝子発現(Romanosらの1992年の論文参照)のために酵母のコンセンサス配列AAAAATGTCT(図5参照)中に存在する。このコンセンサス配列はプライマーTNF-ECOPP中に含まれる。
【0051】
アルコール資化酵母のエレクトロポレーション及び発現測定
エレクトロポレーションは実施例3に述べた方法と同様な方法で行った。しかし、プラスミドの線状化はPmel(Fermentas社製)で行った。選択されたコロニーにおいて、ゲノムDNA精製キット(Fermentas社製)でDNAを単離し、His6-sbTNFα含有DNA断片をEcoTaq・PULS・DNAポリメラーゼ(Ecogen社製)で増幅し、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学製)によりその配列を決定した。
【0052】
発現測定も実施例3に述べた方法と同様な方法で行った。
【実施例8】
【0053】
クロマトグラフ法によるsbTNFαの精製
大腸菌、出芽酵母又はアルコール資化酵母において発現されたHis6-sbTNFαは、AKTA・Explorer・FPLC(GE Healthcare社製)を用いてアフィニティクロマトグラフ法により精製した。精製はNi2+による固定金属イオンアフィニティクロマトグラフ法(ヒスチジン標識タンパク質の精製方法)によりHisTrap・FFカラム(GE Healthcare社製)を用いて行った。280nmにおける吸収をUNICORN5.10により測定した。容量1mlのカラムを、少なくとも5カラム容量のバインディングバッファー(20mM燐酸ナトリウム、0.5M塩化ナトリウム、5mMイミダゾール,pH7.4)を流速1ml/分で平衡化させた。次いで、成長培養物のサンプルを超音波又はビーズビーターで粉砕し、更に遠心分離してペレットを除去し、明澄な上澄液を得た。実施例2で述べたように、この直後に、この上澄液2mLをカラム内に装填した。吸収が安定したベースラインに達するまで(10〜15カラム容量)、カラムをバインディングバッファーで洗浄した。最後に、カラムに結合したタンパク質を溶出するために、溶出バッファー(20mM燐酸ナトリウム、0.5M塩化ナトリウム、5mMイミダゾール,pH7.4)の量を増大させる20〜25カラム容量の直線勾配を適用した。自動画分収集器を使用し、溶出液1mlの画分を収集した。His6-sbTNFαの存在について、SDS-PAGE及び抗ポリヒスチジンmAb(Sigma社製)を用いる免疫ブロット法によりタンパク質を含有する画分を試験した。選択された画分を合わせ、Bradfold(Sigma社製)によりHis6-sbTNFαの量を測定した。
【0054】
必要に応じて、トロンビン−アガロース(Recom-T,Sigma社製)を用いてHis6タグ(標識)を除去した。
【実施例9】
【0055】
SDS-PAGE及び免疫ブロット法によるsbTNFαの検出
12.5%アクリルアミド/ビサクリルアミドを用いるSDS-ポリアクリルミドゲル電気泳動法(SDS-PAGE)によりHis6-sbTNFαを有するサンプルを分析した。ゲルをcoomassie(登録商標)0.1%で30分間染色し、His6-sbTNFαに対応する21kDaのバンドを区別した。
【0056】
前記のようにSDS-PAGEの後に免疫ブロット法を実施することによりHis6-sbTNFαを検出するために、ゲルをTBS(20mM Tris-HCl,150mM NaCl,0.1% Tween20)中で100Vで1時間かけてニトロセルロース膜(Sigma社製)に移行させた。この膜を5%BSAを有するTTBS(10mM Tris-HCl,100mM NaCl,0.1% Tween20)と共にインキュベーションし、4℃でタンパク質をブロックした。膜をTTBSで洗浄後、これをTTBS+5%BSA+1:1000抗ポリヒスチジンmAb(Sigma社製)と共に2時間インキュベーションした。次いで、ECL(Amersham Biosciences社製)で染色する前に、この膜をTTBS及びTBSで再び洗浄した。His6-sbTNFαに対応する21kDaの単一のバンドが現れた(図6参照)。
【実施例10】
【0057】
発酵により酵母の培養
発酵による形質転換酵母の培養をバイオリアクターシステム(Applicon社製)で行った。
【0058】
一例として、アルコール資化酵母菌株をYPGly培地を有するフラスコ内で、培養物のOD600が30超に達するまで、培養した。次いで、成長培養物をMMG(グリセロール含有最少培地)含有バイオリアクターシステム内で接種した。培養条件は次の通りであった。温度30℃、pH5.00(NH3を添加することにより調節した)、撹拌速度500−1250rpm、酸素濃度30%、撹拌しながらのカスケード30%。これらのパラメータはBioXpert・V2により測定した。発酵は、培養物のOD600が50〜80に達するまで、バッチ法で行った。培養物のOD600が50〜80に達した直後に、発酵を半回分培養法に変更し、グリセロール含有飼料の給餌を10時間続けた(培養物のOD600は260〜280に達した)。最後に、メタノール含有飼料の給餌を40時間続けた。結局、培養物のOD600は380〜420に達した。
【0059】
組換TNF His6-sbTNFαの発現分析は免疫ブロット法(実施例9参照)により行った。
【実施例11】
【0060】
組換酵母のマイクロカプセル化
発酵が所望のバイオマスに達し、かつ、組換TNF・His6-sbTNFαの最適発現が得られた後、培養液を無菌的に採取し、20%マロデキストリン及び10%保護ポリマーKollicoat(登録商標)・MAE100P(BASF社製)を用いて製剤化した。
【0061】
次いで、製剤化された懸濁液を噴霧乾燥機(Buchi社製)から噴出させた。入口温度は120℃、出口温度は85℃に制御した。
【0062】
組換酵母の最終マイクロカプセルを分析し、細胞生存性の不存在及びTNF His6-sbTNFαの存在を免疫ブロット法(実施例10参照)により測定した。
【実施例12】
【0063】
マイクロカプセル化組換酵母によるアルテミア・ノープリウス幼生(Artemia nauplii)の富化
滅菌完全海水500mlが入ったフラスコ内でアルテミア・ノープリウス幼生を孵化させた。フラスコ内の海水は水槽用空気ポンプに接続された空気ホースにより曝気し、往復振盪水浴中で28℃に維持した。孵化してから24時間後に、ノープリウス幼生を富化システムに採取した。このシステムは、容量100mlの複数個のフラスコからなり、各フラスコには新鮮な海水が入れられており、各フラスコは全て同じ水浴中に配置され、そして、それぞれ別々に曝気されている。全ての実験について、海水0.5mlのサンプルを採取することにより、孵化率及びノープリウス幼生の個数を見積もった。
【0064】
マイクロカプセル化組換酵母による富化アルテミア・ノープリウス幼生の製造最適時間は、DTAFによる酵母の染色により評価した。次いで、アルテミア・ノープリウス幼生に給餌するために染色酵母を使用した。給餌後、0、30、45、60及び120分の各時点でアルテミア・ノープリウス幼生を殺した。富化アルテミア・ノープリウス幼生は蛍光鏡検法により観察した。
【0065】
結果を図7に示す。図示されているように、給餌開始後、初めのうちはマイクロカプセル化組換酵母の量の増加は口部領域で観察され、その後、腸管全体がマイクロカプセル化組換酵母で満たされ、個々の細胞は認められなかった。
【0066】
富化アルテミア・ノープリウス幼生の製造最適時間は60分であった。
【実施例13】
【0067】
アルコール資化酵母におけるヨーロッパヘダイsbTNFαの発現及び分泌
His6-sbTNFαのpPICZαAへのクローニング
615pbのHis6-sbTNFα含有断片の増幅のために、TNF-ECOF及びTNF-ECOR(図1参照)によるPCR法におけるテンプレートとしてsbTNFα含有プラスミドpET15bを使用した。両方のプライマーとも、プラスミドpPICZαAの同じ部位内でPCR生成物をクローニングするためのEcoRI制限部位を含有する。増幅は、テンプレートを5ng、MgCl2を2.5mM、各dNTPを50μM、プライマーを0.2mM、1X・High・Speec添加剤、1X buffer PLUS及びEco・Taq・PLUS・DNAポリメラーゼ(Ecogen社製)を2単位含有するサンプル中で行った。サイクル反応はSmartサイクラーII(Cepheid社製)中で、95℃・5分間を1サイクル、95℃・30秒間を30サイクル、55℃・45秒間を30サイクル及び72℃・2分間を30サイクル、続いて72℃・10分間を1サイクルの順で行った。臭化エチジウムを0.5μg/ml含有する0.8%アガロースゲル(Pronadisa社製)中で電気泳動することによりPCR生成物を分離し、QIAquickゲル抽出キット(Qiagen社製)で精製した。精製断片及びpPICZαA200ngをEcoRI(Fermentas社製)10単位により37℃で消化させ、その後、T4DNAリガーゼ(Fermentas社製)5単位を用いて22℃でライゲーションさせた。ライゲーション反応生成物を大腸菌TOP10F'コンピテントセルに形質転換させ、ゼオシン(Invitrogen社製)25μg/mlを有するLBプレート内でプラスミド含有コロニーを選別した。2X・PCR・Master・Mix(Fermentas社製)及びPICα及びPIC3プライマーを用いるコロニーPCRにより組換体を識別した。得られたHis6-sbTNFα含有プラスミドpPICZαAをQIAprep・Spin・Miniprepキット(Qiagen社製)で精製し、EcoRIによる消化後のインサート遊離及び消化Smal及びNcolによるインサート配向について試験した。また、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学)によりプラスミドの配列を決定した。
【0068】
アルコール資化酵母のエレクトロポレーション及び発現測定
アルコール資化酵母のエレクトロポレーションの前に、後のアルコール資化酵母のゲノムDNAにおける組換及び組込みのために、His6-sbTNFα含有プラスミドpPICZαAを37℃でSacl又はPmelによる消化を通じて線状化させた。線状プラスミドの全量5μgをMiniElute・Reaction・Cleanupキット(Qiagen社製)で精製し、滅菌水10μlで溶離した。エレクトロポレーションに使用したアルコール資化酵母菌株はX-33であった。エレクトロポレーション用のX-33を作製するために、OD600=1.5の培養物100mlの細胞を1500xgで4℃で5分間遠心分離した。滅菌水で2回洗浄後、細胞を1Mのソルビトール中に再懸濁させ、前記のように遠心分離し、そして最後に1Mのソルビトール100ml中に再懸濁させた。エレクトロポレーションを行うために、DNA10μlを細胞80μlと混合し、氷冷された0.2cmのエレクトロポレーション・キュベット(BioRad社製)に移送した。5分間インキュベーションした後、MicroPulserエレクトロポレーター(BioRad社製)により25μF、1.5kV及び400Ωの条件でパルスを印加した。その直後に、氷冷された1Mソルビトール溶液1mlをキュベットに添加し、得られた混合物を30℃で1時間インキュベーションした。細胞をゼオシン(Invitrogen社製)100μg/mlを有するYPDSプレート上に展延し、コロニーが出現するまで30℃で数日間インキュベーションした。PIC5及びPIC3プライマーと2X・PCRマスターミックス(Fermentas社製)を用いるPCR法によりインサートの存在に関して幾つかの形質転換体を分析し、His6-sbTNFα発現に関して測定した。特定のコロニーにおいて、ゲノムDNA精製キット(Fermentas社製)を用いてゲノムDNAを単離し、そして、His6-sbTNFα含有DNA断片をEcoTaq・PULS・DNAポリメラーゼ(Ecogen社製)で増幅し、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学)によりその配列を決定した。
【0069】
His6-sbTNFαを有する組換アルコール資化酵母菌株をグリセロール含有最少培地25ml中で30℃、250rpmで、培養物がOD600〜6に達するまで、メタノール導入前にバイオマスを発生させるために成長させた。3000xgで5分間遠心分離することにより細胞を回収し、発現を誘発させるために、OD600〜1に達するまで、ペレットを0.5%メタノール含有最少培地200ml中に再懸濁させた。前記に述べたように培養物をインキュベーションし、発現誘導を維持するために24時間毎に0.5%メタノールを添加した。幾つかの時点でサンプルを回収し、タンパク質発現を分析した。タンパク質発現をチェックするために、SDS-PAGE及び抗ポリヒスチジンmAb(Sigma社製)を用いるウエスタン・ブロット法による分析のためにSDS-ローディングバッファー液中で煮沸することにより上澄液のHis6-sbTNFαを測定した。
【実施例14】
【0070】
マイクロカプセル化組換酵母の効力
体重120gの健康なヨーロッパヘダイ(Sparus aurata L.)(硬骨魚類,スズキ目,タイ科)のサンプルを容量2m3の遊泳海水水槽内で飼育した。海水には20%水槽容量/時の流量で酸素6ppmを溶解させた。水槽は常温及び日長に維持され、市販のペレット飼料(スペインのブルゴスに所在するTrouvit社製)を1日に2回給餌した。試験群には、基礎飼料に対照酵母又は成熟(活性)sbTNFα過剰発現マイクロカプセル化酵母を0.1、1又は10%追加した餌を2日毎に給餌した。全てのサンプリングタイム(処置から2日、4日、6日及び10日後)時点で、標本魚の体重を測定し、頭腎及び腸管を採取し、光学顕微鏡で検査し、かつ、遺伝子発現を調査した。ここに述べた試験法は、実験動物の使用に関する欧州連合理事会(European Union Council)(86/609/EU)のガイドライン及びスペインのMurcia大学の生命倫理委員会のガイドラインに従っている。呼吸性バースト活性は、異なる刺激時間後の頭腎ロイコサイト懸濁液により生成されたルミノール依存性化学ルミネッセンス(Sepulcreらの2007年の論文参照)として測定した。全ての試験は5匹の魚と3重のサンプルを用いて行った。結果の有効性を確保するために、同じ試験を2回繰り返した。SPSS・13.0.1統計ソフトウエアパッケージを全ての統計分析に使用した。試験群間の差を決定するために、データは平方偏差(ANOVA)及びテューキー(Tukey)多重範囲検定により分析した。
【0071】
魚の食欲、成長速度、遊泳行動及び外的形態は処置により影響を受けなかった。また、試験中の魚の死亡は認められなかった。更に、10日間にわたってsbTNFα含有酵母を給餌された魚の腸管内には、食細胞(すなわち、好酸性顆粒又はマクロファージ)の浸潤ではない病理組織学的病変は認められなかった。酵母単独の投与は、頭腎ロイコサイトの呼吸性バーストとして測定した場合、魚の免疫状態に殆ど作用を及ぼさなかった。これと際立って対称的に、過剰発現sbTNFα含有マイクロカプセル化酵母を0.1及び1%含有する魚類飼料は、治療開始から4日後には、投与量に依存して増大する呼吸性バーストを示した(図8参照)。驚くべきことに、TLR9の例外を除いて、酵母の投与は頭腎及び腸管における幾つかの炎症性遺伝子のmRNAレベルに対して殆ど影響力を有していなかった。TLR9の発現は顕著に上方制御された(図9参照)。
【0072】
前記の結果は、10日間の免疫賦活剤の投与は魚類に何らの副作用も及ぼさないことを示す。呼吸性バーストは魚類の免疫状態の指標として広く使用されているので(Muleroらの1988年の論文、Garcia-Castilloらの2004年の論文、Chaves-Pozoらの2005年の論文及びSepulcreらの2007年の論文参照)、TNFα含有酵母を有する魚類飼料におけるこの免疫応答の活性化は、本発明の免疫賦活剤が非常に有効であり、その結果、非生物的ストレス(すなわち、魚類操作により惹起されるストレス)及び生物的ストレス(すなわち、病原体により惹起されるストレス)に対して魚類を保護することを示している。更に、本発明の免疫賦活剤の投与による腸管内のTLR9の発現の上方制御(すなわち、過剰発現)も、魚類の腸管がウイルス及び細菌感染に対して良好に保護されていることを示唆している。なぜなら、この受容体(レセプター)はこれら病原体の認識の主要成分だからである(Ishii及びAkiraの2006年の論文参照)。総括的に、前記の結果も全て、組換sbTNFαは、生来の免疫細胞の系統的活性化及びTLR9の発現量の増大により、経口ワクチンの優れたアジュバントであることを示している。
【0073】
以上の説明は、本発明の一実施例に関するもので、この技術分野の当業者であれば、本発明の種々の変形例を考え得るが、それらはいずれも本発明の技術的範囲に包含される。
用語「又は」に関して、例えば「A又はB」は、「Aのみ」、「Bのみ」ならず、「AとBの両方」を選択することも含む。特に記載のない限り、装置又は手段の数は、単数か複数かを問わない。
【技術分野】
【0001】
本発明は水産養殖用の経口投与可能な免疫賦活剤に関する。
【背景技術】
【0002】
水産養殖は、採取漁業によりカバーできない水産食品の成長要求をカバーする唯一の手段であると思われる。世界の水産養殖生産量は過去50年間の間に、1950年代初頭の100万トン未満の生産量から2004年までの5940万トンまでに増大している。この生産量レベルはUSドルで703億ドルの価値に相当する。全世界の生産量の69.6%は中国により、21.9%は他のアジア及び太平洋諸国により、3.9%は西ヨーロッパ諸国により、0.4%は中央及び東ヨーロッパ諸国により生産された。環境に関連して、2004年度における海中養殖(海水水産養殖)による養殖生産量は3020万トンであり、これは養殖全体の50.9%に相当する。真水による水産養殖は2580万トン(43.4%)であり、残りは340万トン(5.7%)であり、これらは汽水環境における生産に由来する。ここに列挙したデータは全てFisheries Technical Paper 2006, State of World Aquaculture (FAO)に基づく。
【0003】
ヨーロッパにおける水産養殖の生産量は全世界の生産量の僅か3%である。しかし、アトランティックサーモン、トラウト(鱒)、シーバス(鱸)、ヨーロッパヘダイ(gilthead sesbream)、ターボット(鮃又は鰈などの扁平な魚、例えば、砂ヒラメ)、及びムラサキイガイなどのような幾つかの種の生産量では全世界でトップである。2002年度の生産量は130万トン超であり、32億8000万ユーロの価値があった。地中海諸国における最も重要な種はヨーロッパヘダイ、シーバス、ターボットである。2002年度におけるヨーロッパヘダイ及びシーバスの生産量は181000トンであり、主な生産国はギリシャ、トルコ、スペイン、イタリヤ、及びフランスである。2002年度におけるターボットの生産量は5320トンであり、その75%はスペインで生産された。これらのデータは全てFAOのデータに基づく。
【0004】
魚類の病気は水産養殖の発展の妨げとなり、世界中の多くの国々における経済発展に対する苛酷な打撃の原因となる。病気は多くの場合大量死を起こし、重大な生産量低下の原因となる。水産養殖の損失はまた、食品利用可能性の減退、収入喪失及び雇用喪失なども起こし、これら全てが付随的な社会的重大性を伴う。前記の種を襲う最も一般的な病気及びこれらの原因となる病原体(バクテリア又はウイルス)はビブリオ症(Photobacterium damselae subsp. damselae, Vibrio spp.),パスツレラ症(Photobacterium damselae subp. pasteurella),発光桿菌症(Photobacterium damselae subp. pasteurella.),滑走細菌症(Tenacibaculum maritimum),粘液桿菌症(Flexibacter maritimus),非定型エロモナス症(Aeromonas salmonicida),連鎖球菌感染症(Streptococcus parauberis),冬不全症候群(Pseudomonas anguilliseptica),ウイルス性神経壊死症(Nodavirus),リンホシスチス症(Iridoviridae),膨張腸症候群(ウイルス様粒子),伝染性膵臓壊死症(IPNV),伝染性造血器壊死症(IHNV)又はウイルス性出血性敗血症(VHS)などである。これらのデータは全て、Cultured Aquatic Species Information programme(FAO)に基づく。水産養殖場における種の養殖状態(すなわち、高密度の養殖魚類又は水産養殖環境等)は固体間で伝達される伝染性疾患病原体にとって好適である。更に、養魚場で養殖中の魚類が受けるストレスは病原体による感染を助長する免疫系の低下を起こすであろう。従って、多くの研究努力が魚類の病気を予防するためのワクチン又は免疫賦活剤などの製品の開発に捧げられてきた。また、病気の予防は環境影響も低減する。なぜなら、水産養殖における抗生剤の大量使用が避けられるからである。幾つかの製品が魚のワクチン接種又は免疫システムの賦活に使用されてきた。実際、ビブリオ症、発光桿菌症、非定型エロモナス症、滑走細菌症、冬不全症候群又は連鎖球菌感染症等のような通常の細菌性魚類疾患及び(Toranzoら,2005及びSommersetら,2005により報告されたIPNV又はIHNV等のような通常のウイルス性魚類疾患に使用可能なワクチン類が存在する。
【0005】
ワクチン接種には、浸漬、注入及び経口投与からなる3種類の一般的方法が存在する。ワクチン接種には腹腔内注射が最も効果的な方法である。なぜなら、腹腔内注射は投与量を自在にコントロールでき、しかも、優れた免疫応答をもたらすアジュバントの使用が可能だからである。しかし、腹腔内注射法には欠点がある。例えば、魚類の鎮静及び注射操作は魚類にストレスを与え、細心の注意を払いながらワクチンを投与しないと魚類を損傷してしまう。更に、小型魚類にはこの方法ではワクチン接種できない。浸漬方法は養殖場で頻繁に使用されている。なぜなら、簡単であり、しかも、迅速だからである。しかし、投与量の厳密なコントロールが出来ず、一貫性のある免疫系の抑制を有する魚類にはストレスが掛かり過ぎる。ワクチンの経口投与では魚の操作は不要であり、給餌を始めたばかりの幼魚を含めて全てのサイズの魚類のワクチン接種に適した方法である。幼魚にもワクチン接種できるこの方法は極めて重要である。なぜなら、これら幼魚は病原体に非常に感染し易いからである。経口投与法にも幾つかの欠点がある。例えば、投与量の厳密なコントロールができないこと、及び免疫化のために多量の抗原が必要なことなどである。経口投与法の最も重要な障害は、酸性環境又はプロテアーゼ活性により抗原が腸内でしばしば不活化し、その結果、腸による活性抗原吸収が抑制されることである。このため、効果的な経口投与を行うには抗原を保護することが必要である。すなわち、腸内における抗原分解を避けるために抗原をカプセル化することが必要である(Hartら,1988; Quentel及びVigneulle, 1997)。抗原を分解から保護するための別の方法(例えば、リポソーム内への閉じ込め又はアルギン酸塩マイクロカプセル化など)は幾つかの有望な結果を例証している(Ireら,2005; Mauriceら,2004)。
【0006】
魚類の病気に対する耐性を高めるために、水産養殖において免疫賦活剤が使用されている。化学特性及び作用モードにより、多数の様々な種類の免疫賦活剤が知られている。例えば、細菌(LPS)の構造要素による、細菌及び菌糸菌由来の様々なβ−1,3グルカン製品、酵母の細胞壁由来のβ−1,3/1,6グルカン、様々な生物源由来の複合炭水化物構造構造体(グリカン類)、ある種の動物の、又は魚類蛋白の酵素加水分解物により産生されたエキス(抽出物)内に存在するペプチド類、ヌクレオチド類、合成製品類(レバミソール)及びビタミンC(Raa,1996により研究された)などである。また、使用された幾つかの免疫賦活剤は特定の抗体反応を高める(Sakai,1999により研究された)。幾つかの状態は水産養殖における免疫賦活剤の使用に好適である。例えば、魚類の操作、水温変化、高濃度病原体曝露、ワクチンアジュバント及び全ての高ストレス状況などである。ヨーロッパヘダイ(gilthead sesbream)免疫系における免疫賦活剤の効果に関して多数の文献が発行されている。例えば、出芽酵母(Saccharomyces cerevisiae)に関するOrtunoらの2002年の論文及びRodriguezらの2003年の論文、キチン質に関するEstebanらの2002年の論文、Estebanらの2001年の論文及びCuestaらの2003年の論文、ビタミンC及びEに関するCuestaらの2002年の論文及びOrtunoらの2003年の論文及びレバミソール(levamisole)に関するMuleroらの1998年の論文及びCuestaらの2002年の論文などである。魚類に有効であることが知られている免疫賦活剤のうち、グルカン、キチン質及びレバミソールは食細胞活性を高め、一方、酵母グルカン及びビタミンCは補体活性を活性化する。免疫賦活剤としてのB-1,3/1,6グルカン投与に関する幾つかの研究は、オヒョウ(halibut)幼魚の発達段階の重大な局面でのオヒョウ幼魚の生存率を高めることに関して優れた結果を示すことがOttesn, Lunda及びEngstadらの1999年の論文に発表され、細菌感染症に対する耐性を高めることがRaaらの1990年の論文及びRobertsenらの1990年の論文に発表され、また、ワクチンの効果を高めることがRaaらの1990年の論文、Rorstad, Assjord及びRobertsenの1993年の論文に発表されている。
【0007】
サイトカイン(Cytokine)は細胞成長、細胞活性、炎症、免疫、組織修復、繊維増多及び形態形成などのような重量な生物学的プロセスの量を調節するので、“免疫調節剤”の一般的定義に属するタンパク質類である。サイトカインは幾つかの特性を共有するタンパク質類の異なるグループであり、先天的及び適応的免疫応答の両仲介エフェクター相の発生と作用にとって必須のものである。過去数年にわたり、非常に多数のサイトカインがクローンとして発生され、幾つかの魚属において配列決定された(例えば、Birdらの2002年の論文参照)。サイトカインはその機能により下記の4種類のグループに分類することができる。
(a)先天的免疫を仲介するサイトカイン類。このグループの魚類における最良の特性を有するサイトカインはタイプIインターフェロン(IFN)であり、これにはIFNαとIFNβ、インターロイキン1(IL-1)ファミリー(IL-1α、IL-1β、IL-1レセプター拮抗体及びIL-18を含む)及び腫瘍壊死因子(TNF)ファミリー(TNFα、TNFβ、Ltβ及びFas配位子を含む。)などが含まれる。これらのうち、TNFαが免疫系の調節機能を有する最も重要な一員である。
(b)造血を調整するサイトカイン、
(c)リンパ球を調節するサイトカイン及び
(d)非特定エフェクター細胞を制御するサイトカイン。
【0008】
これらのグループにおいて、最も優れた特性を有するサイトカインは、グループ(b)のケースではインターロイキン2(IL-2)であり、グループ(c)では形質転換成長因子ベータ(TGFβ)である。
【0009】
TNFαは急性炎症中にモノサイト/マクロファージにより産生される炎症性サイトカインであり、細胞内の異なった範囲内の前兆イベントに応答する義務がある。TNFαは、寄生感染症、細菌感染症及びウイルス感染症に対する抵抗性における重要なメディエータとしての役割を果たす(Czarnieckiの1993年の論文、Goldfeld及びTsaiの1996年の論文及びSteinshamnらの1996年の論文参照)。TNFαはその他の重要な治療作用も有する。例えば、耐腫瘍性(Vilcek及びLeeの1991年の論文参照)、睡眠調整(Kruegerらの1998年の論文参照)及び胚成長(Wride及びSandersの1995年の論文参照)。TNFαは細胞膜レセプター,TNFR-1又はTNFR-2にトリマーとして結合することによりこれらの多くの作用を発揮する(Dembicらの1990年の論文及びLoetscherらの1990年の論文参照)。これらの両レセプターは殆どの細胞内に非常に大量に存在する(Loetscherらの1991年の論文及びSchoenfeldらの1991年の論文参照)。
【0010】
TNFαmRNAは広範な種類の細胞内で転写されるものと思われる。従って、転写後レベルでは著しく抑制される(Hanらの1990年の論文参照)。TNFαは、膜結合形と可溶性形の2つの形で存在する。各形とも恐らく、明確な生理学的役割を有する(Wattsらの1990年の論文参照)。TNFαは26kDa膜結合前駆体として明示され、これはディスインテグリン及び金属蛋白分解酵素(metalloproteinase)(TACE:TNFα変換酵素)によりタンパク質切断され、17kDaC−末端活性形が生成される(Blackらの1997年の論文及びMossらの1997年の論文参照)。
【0011】
幾つかの哺乳類のTNFα情報は完全に又は部分的に配列決定されている。このような哺乳類は例えば、ヒト(Pnnicaらの1984年の論文参照)、チンパンジー(Kutsuiらの2002年の論文参照)、マウス(Shiraiらの1988年の論文参照)、イヌ(Zuckerらの1994年の論文参照)又はネコ(McGrawらの1990年の論文参照)などである。また、幾つかの魚類のTNFα情報又はTNFα情報断片も配列決定されている。このような魚類はクロマダイ(black seabream)(Caiらの2003年の論文参照)、ニジマス(Laingらの2001年の論文参照)、コイ(Saeijらの2003年の論文参照)又はゼブラフィッシュ(Phelanらの2003年の論文参照)若しくはヨーロッパヘダイ(Garcia-Castilloらの2002年の論文参照)などである。
【0012】
ヨーロッパヘダイのTNFα情報は4個のエクソンと3個のイントロン及び1244pbの長さを有する。cDNAは1359pbからなり、これは、762pbのオープン読み枠(open reading frame;ORF)、142pbの5’非翻訳領域(UTR)及び455pbの3’UTRを含む。推定タンパク質は253個のアミノ酸を有し、概算分子量は28kDaであり、その他の魚種TNFαとの、特にレセプターとの相互作用に必要なC−末端エクストリームにおける高い配列相同性を与える。
【0013】
TNFαタンパク質は37及び54残基との間のトランスメンブラン領域と、TACEタンパク質切断に付随する保存配列Thr-Leuを包含する(Garcia-Castilloらの2002年の論文参照)。従って、253アミノ酸膜結合タンパク質(proTNFα)と167アミノ酸可溶性タンパク質(sbTNFα)とを区別することができる。
【0014】
発現研究は、肝臓、エラ、血液、頭腎、腹腔侵出物、脳及び脾臓などのような試験されたヨーロッパヘダイの全ての組織でTNFαの構成的発現を明らかにした。これは、この分子の主要源の一つのマクロファージである。
【0015】
特許文献1(米国特許第5871751号明細書)は、魚類の細菌性腎臓病菌(Renibacterium salmoninarum)による感染を受けやすい魚類の治療用ワクチン及び治療方法を開示している。このワクチンは無傷の細胞表面結合タンパク質p57を欠いた死滅微生物からなり、かつ、経口投与のための腸溶剤皮が施されている。腸溶剤皮は胃内における溶解又は分解を受け付けないが、腸内の高pH環境を通過したら溶解されるポリマー剤皮からなる。開示されたワクチンの好ましい実施態様は、球形糖微小球を用いて生成され、無傷の細胞表面結合タンパク質p57を欠いた死滅微生物からなる第1層で被覆し、次いで、魚類の胃内で溶解又は分解を受け付けない材料からなる腸溶剤皮の第2層で被覆されている。
【0016】
特許文献2(国際公開第WO03/020040A1号公報)は、抗原を含有する単細胞微生物をカプセル封入した多細胞生物からなるワクチンを開示している。このワクチンは予防接種すべき水生動物(例えば、魚類)に給餌するのに使用される。特許文献2によれば、単細胞微生物中に発現される抗原は2段階給餌により水生動物に送達される。すなわち、(抗原を含有する)単細胞微生物を多細胞生物に給餌する第1段階と、多細胞生物を水生動物に給餌する第2段階である。特許文献2によれば、単細胞微生物と多細胞生物の両方の同時存在が常に必要である。発現されるべき抗原は、誘発免疫反応が目標病原体に対するものであるか否かに依存する。特許文献2には、細菌性抗原として緑膿菌外毒素A(PE)、単細胞微生物として大腸菌(E. coli)、多細胞生物としてアルテミア・ノープリウス幼生(Artemia nauplii)が開示されている。特許文献2は経口投与ワクチンの製造を開示しているので、選択された抗原は、このワクチンが向けられなければならない目標病原体に厳密に依存する。
【0017】
特許文献3(スペイン特許公開第2189508A1号公報)は、ヨーロッパヘダイ(Sparus aurata L.)の1L-1β組換え体の製造方法及びワクチンアジュバント並びに市販魚類における免疫賦活剤としての使用法を開示している。特許文献3による1L-1β組換え体の製造はこれを大腸菌発現ベクターにクローニングすることにより行われる。このようにして得られた組換え体タンパク質は精製され、そして、魚類の大量養殖における免疫賦活剤及びワクチンアジュバントとして使用される。
【先行技術文献】
【特許文献】
【0018】
【特許文献1】米国特許第5871751号明細書
【特許文献2】国際公開第WO03/020040A1号公報
【特許文献3】スペイン特許公開第2189508A1号公報
【発明の概要】
【発明が解決しようとする課題】
【0019】
本発明の目的は、加水分解又は分解を受けることが無く、しかも、投与されるべき微生物により完全に吸収される、経口投与に適した免疫賦活剤を提供することである。
【0020】
本発明の別の目的は、目標対象物に直接投与することができる免疫賦活剤を提供することである。
【0021】
また、本発明の他の目的は、初期投与量に対して大量に投与される微生物により吸収され得る免疫賦活剤を提供することである。
【0022】
本発明の更に別の目的は、特定の病原体又は特定の病原体群に対して特別に送達されない免疫賦活剤を提供することである。
【0023】
本発明の更に他の目的は、経口投与可能な免疫賦活剤の製造方法及び水産養殖における免疫応答の活性化のための免疫賦活剤の使用法を提供することである。
【課題を解決するための手段】
【0024】
前記課題を解決するために、本発明は、少なくともマイクロカプセル化された組換魚類サイトカイン、特に腫瘍壊死因子α(TNFα)からなる経口投与可能な免疫賦活剤及びマイクロカプセル化された組換魚類サイトカインからなる経口投与可能な免疫賦活剤の製造方法及び水産養殖における免疫応答の活性化のためのその使用法を提供する。
【0025】
本発明の免疫賦活剤の特徴は、免疫賦活剤として作用する活性分子(すなわち、サイトカイン及び特定の腫瘍壊死因子α(TNFα))が、免疫応答の活性化が望まれる同じ魚種に属するサイトカインであるということである。この特徴は非常に重要である。なぜなら、当該魚種に既に属する特定の一組の免疫賦活剤を正に高める特定の魚種の免疫応答を活性化させることができるからである。この事実は次のことを意味する。すなわち、当該魚種に任意の方法で投与される外部因子、化合物類又は製品は存在せず、その結果、得られる結果が優れた利益を上げるばかりか、安全なアプローチ法としても優れている。前記のように、本発明によれば、免疫賦活剤として作用する活性分子は魚類自体から得られたサイトカイン、腫瘍壊死因子α(TNFα)である。従って、本発明の免疫賦活剤が投与されるとき、外来物質は魚類に導入されない。その結果、副作用の可能性は避けられる。
【0026】
本発明によれば、特定のサイトカインをコードする遺伝子は魚類の組織から単離され、適当な宿主微生物中でサイトカイン発現用の発現ベクターにクローンされる。宿主細胞は最適なサイトカイン発現及び高バイオマス産生に適した溶媒を用いてバイオリアクター中で培養される。本発明の好ましい実施態様によれば、宿主微生物は酵母(イースト)であり、好ましくはメタノール資化酵母(Pichia pastoris)及び出芽酵母(Saccharomyces cerevisiae)であり、最も好ましくは メタノール資化酵母である。宿主微生物としての酵母の選択は偶然ではない。なぜなら、酵母は真核微生物であり、細胞の外部及び内部の両方に特定のサイトカインを発現できるからである。これにより、発酵方法及び斯くして得られた生成物の精製に多くの効果をもたらす組換タンパク質の発現方法を決定することができる。次いで、組換タンパク質(単独で得られたもの又は宿主微生物の内部で得られたものの何れか)をマイクロカプセル化処理し、必要に応じて精製処理工程を経て、投与可能なマイクロカプセル化タンパク質が得られる。本明細書で使用する「マイクロカプセル化方法」という用語は、組換サイトカインに少なくとも部分的に被覆(この被覆は腸内分解からサイトカインを保護するのに適している)を施すのに好適な全ての方法を意味する。
【0027】
実際、免疫賦活剤を経口投与する場合、免疫賦活剤の腸内分解を避けるため、及び腸管により完全吸収を可能にするため、効率的な保護方法が必要である。マイクロカプセル化は、液体又は固体の極めて小さな液滴又は粒子を高分子材料で包囲又は被覆し、微粒子を得る技術的方法である。マイクロカプセル化技術は幾つかの生物学的及び化学的システムで汎用されており、莫大な量の製品を製造することができる。本発明によれば、所望の及び特定のサイトカイン(特に、TNFα)のマイクロカプセル化製剤を成功裡に得るために使用される主要技術は噴霧乾燥による微粉化である。噴霧乾燥による微粉化の技術では、活性分子は溶融又はポリマー溶液内に溶解又は懸濁されており、そして、乾燥粒子内に閉じ込められる。本発明によれば、腸内の産生pHから組換タンパク質の微粒子を保護するのに使用された最も有用なポリマーは例えば、酢酸フタル酸セルロース、フタル酸ヒドロキシプロピルセルロース、カルボキシメチルセルロース、メタクリル酸コポリマー(例えば、Eudragit L-30及びLollicoatなどのようなメタクリル酸コポリマーLS)などから選択される。その他のポリマー類、例えば、マルトデキストリン、キトサン、ゼラチン、スターチ又はアラビアゴムなどから選択されるポリマー類も使用できる。
【発明の効果】
【0028】
本発明では、微生物中で発現され、次いで、腸内の酸性環境及びプロテアーゼ活性からタンパク質を保護するためにポリマーと混合されるサイトカインの一例としてTNFαを使用する。組換タンパク質、すなわちTNFαは、宿主微生物が細胞外にその過剰発現を可能にするとき、そのまま使用することができる。効果的な別法として、宿主微生物がタンパク質を内部的に過剰発現する場合、同じ微生物をマイクロカプセル化し、そのまま使用することができるマイクロカプセル化微生物が得られると共に、前記のような全ての効果が発揮される。過剰発現したタンパク質を噴霧乾燥により乾燥し、魚類の健康に有効な量だけ魚類の餌に添加する。本発明による製品は、感染により魚類の疾患を予防し、何らかの高ストレス環境中の魚類を保護するため、及び、ワクチンのアジュバントとして、水産養殖市場で桁外れの重要性を有する。
【0029】
本発明によれば、免疫賦活剤が成魚の治療に差し向けられる場合、マイクロカプセル化処理により得られる利点により、免疫賦活剤は常にそのまま投与することができる。これに対して、免疫賦活剤が幼魚又は稚魚の治療に差し向けられる場合、例えば、アルテミア・ノープリウス幼生(Artemia nauplii)などのような多細胞生物に免疫賦活剤を給餌し、次いで、幼魚又は稚魚群により容易に摂食可能な、この多細胞生物を幼魚又は稚魚に給餌することができる。何れの場合も、マイクロカプセル化された免疫賦活剤は製品の主要特性及び活性を安全に維持することができ、水産養殖に多大な利益をもたらす。
【0030】
一例として、ヨーロッパヘダイの組換sbTNFαは精製され、そして、その生物活性は生体内及び試験管内で評価された。組換タンパク質はアフィニティークロマトグラフィー精製及び免疫ブロット法における抗ポリヒスチジンmAb検出用にN−末端エクストリーム(extreme)に6個のヒスチジン類を含有する。腹膜内注射は、(a)腹膜侵出及び頭腎(HK)白血球の呼吸性バースト(respiratory burst)の呼び水、(b)注射部位に対する食細胞顆粒白血球の急速な固体増加及び(c)HKにおける顆粒白血球形成の誘発を生じた。sbTNFαは試験管内でHK細胞の強力な分芽増殖を誘発することができた。
【0031】
本発明による別の効果は、TNFαサイトカインを発現する宿主微生物それ自体が免疫賦活剤物質の追加源として機能出来ることである。細菌の幾つかの構造的要素類、酵母の細胞壁からのグルカン類又は様々な生物源からの複合糖質構造(グリカン類)は実際、免疫賦活剤として使用できる。
【0032】
水産養殖において非常に重要性の高い幾つかの魚種のTNFαを用いて行われる下記の実施例により本発明を更に詳細に説明する。しかし、その他の魚種のTNFα及びその他のサイトカインも本発明を実施するのに使用できる。また、本発明は合成により抽出又は生成されたサイトカインにも適用できる。
【図面の簡単な説明】
【0033】
【図1】遺伝子構成に使用されるプライマー類を示す。クローニング用のEcoRi及びBamHi制限部位はイタリック体で示されている。
【図2】ヨーロッパヘダイのsbTNFαを含有するDNA断片及び演繹タンパク質配列を示す。
【図3】ヨーロッパヘダイのHis6-sbTNFαを含有するDNA断片及びメタノール資化酵母における発現用演繹タンパク質配列を示す。図中の下線部は6個のCAT/Cコドン類及びヒスチジン類を示す。メタノール資化酵母におけるタンパク質発現用コンセンサス配列は強調されている。EcoRi及びBamHi制限部位はイタリック体で示されている。
【図4】シーバスのHis6-sbTNFαを含有するDNA断片及びメタノール資化酵母における発現用演繹タンパク質配列を示す。図中の下線部は6個のCAT/Cコドン類及びヒスチジン類を示す。メタノール資化酵母におけるタンパク質発現用コンセンサス配列は強調されている。EcoRi及びBamHi制限部位はイタリック体で示されている。
【図5】ターボットのHis6-sbTNFαを含有するDNA断片及びメタノール資化酵母における発現用演繹タンパク質配列を示す。図中の下線部は6個のCAT/Cコドン類及びヒスチジン類を示す。メタノール資化酵母におけるタンパク質発現用コンセンサス配列は強調されている。EcoRi及びBamHi制限部位はイタリック体で示されている。
【図6】AはSDS-PAGE(ドデシル硫酸ナトリウム-ポリアクリルアミドゲル電気泳動)による泳動結果を示し、Bは抗ポリヒスチジンmAbによるウエスタン・ブロット法の分析結果を示す。図中の矢線は(1)大腸菌、(2)出芽酵母及び(3)メタノール資化酵母で発現されたヨーロッパヘダイ(Sparus aurata)のHis6-sbTNFαを示す。
【図7】染色組換酵母を60分間給餌した後のアルテミア・ノープリウス幼生(Artemia nauplii)の顕微鏡観察図であり、Aはアルテミア・ノープリウス幼生を示し、BはDTAFを有する蛍光染色酵母による消化管内部を示す。
【図8】頭腎白血球により産生されたルミノール依存性化学発光による呼吸性バースト活性を示すグラフである。縦軸は、化学発光(相対光単位/分)
【図9】頭腎及び腸管における幾つかの炎症性遺伝子のmRNAレベルを示すグラフである。上のグラフは腸管に、下のグラフは頭腎に関する。縦軸はどちらも、対照に対する増大率である。
【発明を実施するための形態】
【0034】
以下、実施例により本発明を更に詳細に説明する。
【実施例1】
【0035】
sbTNFαをpET15bにクローニングする大腸菌内におけるヨーロッパヘダイTNFαの発現
FE4及びRE5(図1参照)によるsbTNFαの増幅のためのPCRにおけるテンプレート(鋳型)として、LPS刺激頭腎cDNAを使用した。両方のプライマーとも、プラスミドpET15b(Novagen社製)の同じ部位でPCR生成物を後クローニングするためのBamHI制限部位を有する。増幅はcDNAテンプレート、各dNTP50μM、プライマー0.2mM、MgCl2含有1X buffer PLUS及びEco Taq PULS DNAポリメラーゼ(Ecogen社製)1単位を含有するサンプル中で行った。サイクル反応はエッペンドルフ・マスターサイクラー・グラジエント内で、95℃・2分間を1サイクル、95℃・45秒間を25サイクル、60℃・45秒間を25サイクル及び72℃・30秒間を25サイクル続いて72℃・10分間を1サイクルの順で行った。PCR生成物をQIAquick・PCR精製キット(Qiagen社製)で精製し、T4DNAリガーゼ(New England Biolabs社製)1単位を使用し、インサート:プラスミド(3:1)の関係でプラスミドpGEM-T Easy(Promega社製)と室温で16時間かけてライゲーションさせた。ライゲーション混合物を形質転換大腸菌DH5αコンピテントセル(形質転換受容性細胞)に対して使用し、そして、アンピシリンとX-Gal(Sigma社製)を含有するLBプレート内に展延させた。このプレートを37℃でインキュベーションし、幾つかの生成白色コロニーを選び出し、QIAprep・Spin・Miniprepキット(Qiagen社製)によるプラスミド単離及びBamHIによる消化を通じてインサートの存在を試験した。インサート(BamHI端部を有するsbTNFα,図2参照)を含有する特定のプラスミドはpVP81と命名された。このプラスミド及び500ngのpET15bを、pVP81のsbTNFα遊離のため及びpET15b線状化のために10単位のBamHIで消化させた。インサート及び直線プラスミドの両方とも、QIAquickゲル抽出キット(Qiagen社製)によりアガロース低融点ゲル(Pronadisa社製)中で電気泳動することにより分離した後、精製し、そして、1単位のT4DNAリガーゼ(New England Biolabs社製)を使用し、室温で16時間かけてライゲーションさせた。ライゲーション混合物を形質転換大腸菌DH5αコンピテントセルに対して使用し、そして、アンピシリンを含有するLBプレート内に展延させた。このプレートを37℃でインキュベーションし、幾つかの生成コロニーのプラスミドを単離し、BamHI消化によるインサート遊離及びPvull消化によるインサート配向について試験した。選択されたプラスミドの配列をABI・Prism377遺伝子分析器(CIB,CSIC社製)で決定した。
【0036】
大腸菌の形質転換及び発現測定
大腸菌BL21(DE3)コンピテントセルをsbTNFα含有プラスミドpET15bで形質転換し、得られた混合物をアンピシリン含有LBプレート内に展延した。幾つかの生成コロニーをLB-アンピシリン媒体中で一晩培養した。新鮮なLB-アンピシリン中に希釈した後、培養物を37℃でOD600=0.8まで成長させ、1mMのイソプロピル-D-チオガラクトシド(IPTG; Applichem社製)を用いて0.25〜4時間、25℃又は37℃で誘導した。誘導培養物0.1mlを遠心分離(1400rpm)することにより全細胞抽出物中のタンパク質発現をチェックし、細胞ペレットを、SDS-PAGE及び抗ポリヒスチジンmAb(Sigma社製)を用いるウエスタン・ブロット法による分析のためにSDS-ローディングバッファー液中で煮沸することにより溶解させた。
【実施例2】
【0037】
出芽酵母におけるヨーロッパヘダイsbTNFαの発現
His6-sbTNFαのp424-GPDへのクローニング
613pbのHis6-sbTNFαを含む断片の増幅のためにTNF-ECOF及びTNF-ECORプライマー(図1参照)と共にPCRにおけるテンプレートとしてsbTNFα含有プラスミドpET15bを使用した。両方のプライマーとも、プラスミドp424-GPD(ATCC87357)の同じ部位内でPCR生成物をクローニングするためのEcoRI制限部位を含有する。増幅は、テンプレートを5ng、MgCl2を2.5mM、各dNTPを50μM、プライマーを0.2mM、1X・High・Speec添加剤、1X buffer PLUS及びEco・Taq・PLUS・DNAポリメラーゼを2単位含有するサンプル中で行った。サイクル反応はSmartサイクラーII(Cepheid社製)中で、95℃・5分間を1サイクル、95℃・30秒間を30サイクル、62℃・45秒間を30サイクル及び72℃・2分間を30サイクル、続いて72℃・10分間を1サイクルの順で行った。臭化エチジウムを0.5μg/ml含有する0.8%アガロースゲル(Pronadisa社製)中で電気泳動することによりPCR生成物を分離し、QIAquickゲル抽出キット(Qiagen社製)で精製した。精製断片及びp424-GPD250ngをEcoRI(Fermentas社製)10単位により37℃で消化させ、その後、T4DNAリガーゼ(Fermentas社製)5単位を用いて22℃でライゲーションさせた。ライゲーション反応生成物を大腸菌DH5αコンピテントセルに形質転換させ、アンピシリン100μg/mlを有するLBプレート内でプラスミド含有コロニーを選別した。2X・PCR・Master・Mix(Fermentas社製)及びPRO-GPD及びTER-GPDプライマー(図1参照)を用いるコロニーPCRにより組換体を識別した。得られたHis6-sbTNFα含有プラスミドp424-GPDをQIAprep・Spin・Miniprepキット(Qiagen社製)で精製し、EcoRIによる消化後のインサート遊離及び消化Smal及びNcolによるインサート配向について試験した。また、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学)によりプラスミドの配列を決定した。
【0038】
出芽酵母の形質転換及び発現測定
形質転換に使用された出芽細胞菌株はEGY48(Invitrogen社製)であった。形質転換用のEGY48を作製するために、YPD培地(2%ペプトン、1%酵母エキス、2%グルコース)中でOD600=0.5まで成長させた培養物100mlの細胞を2500rpmで4℃で5分間遠心分離した。滅菌水で洗浄後、細胞を1X・LiAc-TE(0.1M酢酸リチウム, 10mM Tris pH7.5,1mMEDTA pH8.0)に再懸濁させた。形質転換のために、濃度0.3μg/μlのDNA10μlを細胞50μl及びDNA担体50μgと混合した(サーモンのDNAを予め煮沸しておいてから使用した)。続いて、この混合物を1X・LiAc-TE-PEG(1X・LiAc-TEは前記のものと同一であり、PEGは40%)混合物に添加し、30℃で30分間インキュベーションし、その後、DMSOを10%まで添加し、42℃で10分間インキュベーションした。細胞をMMSS+SDC-Trp(グルコース2%、酵母窒素塩基0.17%、硫酸アンモニウム0.25%、1Mソルビトール及びSDC-Trpを含有する最少培地)プレートに展延し、コロニーが出現するまで30℃で数日間インキュベーションした。SDC(完全合成デキストロース, Synthetic Dextrose Complete)はアデニン、アルギニン、ヒスチジン、トリプトファン及びウラシルを20mg/l,ロイシン、リジン、チロシン及びイソロイシンを30mg/l,フェニルアラニンを50mg/l,スレオニンを200mg/l,バリンを150mg/lそれぞれ含有する。
【0039】
His6-sbTNFα発現を試験するために、幾つかのコロニーをYPD培地又はMM+SDC-Trp(グルコース2%、酵母窒素塩基0.17%、硫酸アンモニウム0.25%及びSDC-Trpを含有する最少培地)で数日間培養した。タンパク質発現をチェックするために、全細胞抽出物を、30秒間で10パルスの周期でBranson社製の超音波ホモジナイザーにより又は1分間で8パルスの周期でガラスビーズを用いるビードビーターにより粉砕した。粉砕処理完了後、細胞残留物を遠心分離により回収し、SDS-PAGE及び抗ポリヒスチジンmAb(Sigma社製)を用いるウエスタン・ブロット法による分析のためにSDS-ローディングバッファー液中で煮沸することにより上澄液のHis6-sbTNFαを測定した。
【実施例3】
【0040】
アルコール資化酵母におけるヨーロッパヘダイsbTNFαの発現
His6-sbTNFαのpPCICZAへのクローニング
618pbのHis6-sbTNFαを含む断片の増幅のためにTNF-ECOF及びTNF-ECORプライマー(図1参照)と共にPCRにおけるテンプレートとしてsbTNFα含有プラスミドpET15bを使用した。両方のプライマーとも、プラスミドpPCICZA(Invitrogen社製)の同じ部位内でPCR生成物をクローニングするためのEcoRI制限部位を含有する。増幅は、テンプレートを5ng、MgCl2を2.5mM、各dNTPを50μM、プライマーを0.2mM、1X・High・Speec添加剤、1Xbuffer PLUS及びEco・Taq・PLUS・DNAポリメラーゼを2単位含有するサンプル中で行った。サイクル反応はSmartサイクラーII(Cepheid社製)中で、95℃・5分間を1サイクル、95℃・30秒間を30サイクル、55℃・45秒間を30サイクル及び72℃・2分間を30サイクル、続いて72℃・10分間を1サイクルの順で行った。臭化エチジウムを0.5μg/ml含有する0.8%アガロースゲル(Pronadisa社製)中で電気泳動することによりPCR生成物を分離し、QIAquickゲル抽出キット(Qiagen社製)で精製した。精製断片及びpPICZA200ngをEcoRI(Fermentas社製)10単位と共に37℃で消化させ、その後、T4DNAリガーゼ(Fermentas社製)5単位を用いて22℃でライゲーションさせた。ライゲーション反応生成物を大腸菌TOP10F'コンピテントセルに形質転換させ、ゼオシン(Invitrogen社製)25μg/mlを有するLBローソルトプレート内でプラスミド含有コロニーを選別した。2X・PCR・Master・Mix(Fermentas社製)及びPIC5及PIC3プライマー(図1参照)を用いるコロニーPCRにより組換体を識別した。得られたHis6-sbTNFα含有プラスミドpPCICZAをQIAprep・Spin・Miniprepキット(Qiagen社製)で精製し、EcoRIによる消化後のインサート遊離及びXholによる消化を通じてインサート配向について試験した。また、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学)によりプラスミドの配列を決定した。組換プラスミドにおいて、sbTNFαのスターコドンATGは、酵母における外来遺伝子発現(Romanosらの1992年の論文参照)のために、酵母のコンセンサス配列AAAAATGTCT(図3参照)中に存在する。このコンセンサス配列はプライマーTNF-ECOPP中に含まれる。
【0041】
アルコール資化酵母のエレクトロポレーション及び発現測定
アルコール資化酵母のエレクトロポレーションの前に、後のアルコール資化酵母のゲノムDNAにおける組換及び組込みのために、His6-sbTNFα含有プラスミドpPICZAを37℃でSacl又はPmelによる消化を通じて線状化させた。線状プラスミドの全量5μgをMiniElute・Reaction・Cleanupキット(Qiagen社製)で精製し、滅菌水10μlで溶離した。エレクトロポレーションに使用したアルコール資化酵母菌株はX-33であった。エレクトロポレーション用のX-33を作製するために、OD600=1.5の培養物100mlの細胞を1500xgで4℃で5分間遠心分離した。滅菌水で2回洗浄後、細胞を1Mのソルビトール中に再懸濁させ、前記のように遠心分離し、そして最後に1Mのソルビトール100ml中に再懸濁させた。エレクトロポレーションを行うために、DNA10μlを細胞80μlと混合し、氷冷された0.2cmのエレクトロポレーション・キュベット(BioRad社製)に移送した。5分間インキュベーションした後、MicroPulserエレクトロポレーター(BioRad社製)により25μF、1.5kV及び400Ωの条件でパルスを印加した。その直後に、氷冷された1Mソルビトール溶液1mlをキュベットに添加し、得られた混合物を30℃で1時間インキュベーションした。細胞をゼオシン(Invitrogen社製)100μg/mlを有するYPDSプレート上に展延し、コロニーが出現するまで30℃で数日間インキュベーションした。PIC5及びPIC3プライマーと2X・PCRマスターミックス(Fermentas社製)を用いるPCR法によりインサートの存在に関して幾つかの形質転換体を分析し、His6-sbTNFα発現に関して測定した。特定のコロニーにおいて、ゲノムDNA精製キット(Fermentas社製)を用いてゲノムDNAを単離し、そして、His6-sbTNFα含有DNA断片をEcoTaq・PULS・DNAポリメラーゼ(Ecogen社製)で増幅し、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学)によりその配列を決定した。
【0042】
His6-sbTNFαを有する組換アルコール資化酵母菌株をグリセロール含有最少培地25ml中で30℃、250rpmで、培養物がOD600〜6に達するまで、メタノール導入前にバイオマスを発生させるために成長させた。3000xgで5分間遠心分離することにより細胞を回収し、発現を誘発させるために、OD600〜1に達するまで、ペレットを0.5%メタノール含有最少培地200ml中に再懸濁させた。前記に述べたように培養物をインキュベーションし、発現誘導を維持するために24時間毎に0.5%メタノールを添加した。幾つかの時点でサンプルを回収し、タンパク質発現を分析した。タンパク質発現をチェックするために、全細胞抽出物を30秒間で10パルスの周期でBranson社製の超音波ホモジナイザーにより又は1分間で8パルスの周期でガラスビーズを用いるビードビーターにより粉砕した。粉砕処理完了後、細胞残留物を遠心分離により回収し、SDS-PAGE及び抗ポリヒスチジンmAb(Sigma社製)を用いるウエスタン・ブロット法による分析のためにSDS-ローディングバッファー液中で煮沸することにより上澄液のHis6-sbTNFαを測定した。
【実施例4】
【0043】
大腸菌におけるシーバスsbTNFαの発現
sbTNFαのpET15bへのクローニング
FE4及びRE5(図1参照)によるsbTNFαの増幅のためのPCRにおけるテンプレート(鋳型)として、シーバスの肝臓から得られたcDNAを使用した。両方のプライマーとも、プラスミドpET15b(Novagen社製)の同じ部位でPCR生成物を後クローニングするためのBamHI制限部位を有する。増幅はcDNAテンプレート、各dNTP50μM、プライマー0.2mM、MgCl2含有1X buffer PLUS及びEco Taq PULS DNAポリメラーゼ(Ecogen社製)1単位を含有するサンプル中で行った。サイクル反応はSmartサイクラー(Cepheid社製)内で、95℃・5分間を1サイクル、95℃・45秒間を30サイクル、55℃・45秒間を30サイクル及び72℃・90秒間を30サイクル続いて72℃・10分間を1サイクルの順で行った。0.8%アガロースゲル(Pronadisa社製)中で電気泳動することにより分離した後、PCR生成物をQIAquick・PCR精製キット(Qiagen社製)で精製した。精製断片及びプラスミドpET15bをBamHIを10単位使用し、37℃で2時間かけて消化し、PCR精製キットによる前記PCR精製物をT4DNAリガーゼ(Fermentas社製)1単位を用いて22℃でライゲーションさせた。ライゲーション混合物を形質転換大腸菌DH5αコンピテントセル(形質転換受容性細胞)に対して使用し、そして、アンピシリン含有LBプレート内に展延させた。このプレートを37℃でインキュベーションし、2X・PCRマスターミックス(fermentas社製)及びプライマーT7F及びT7R(図1参照)を用いるコロニーPCR法により組換体を識別した。幾つかの生成コロニーのプラスミドをQIAprep・Spin・Miniprepキット(Qiagen社製)で単離し、BamHI消化によるインサート遊離及びPvull消化によるインサート配向について試験した。組換プラスミドの配列はABI・Prism3130遺伝子分析器(SACE,Murcia大学製)で決定した。
【0044】
大腸菌の形質転換及び発現測定
シーバスHis6-sbTNFαの大腸菌の形質転換及び発現測定は実施例1に述べた方法と同様な方法で実施した。
【実施例5】
【0045】
アルコール資化酵母におけるシーバスsbTNFαの発現
His6-sbTNFαのpPICZAへのクローニング
シーバスのHis6-sbTNFαのpPICZAへのクローニングは実施例3に述べた方法と同様な同様な方法で行った。シーバスのHis6-sbTNFαを含有する生成プラスミドpPICZAの配列をABI・Prism3130遺伝子分析器(SACE,Murcia大学製)で決定した。実施例3で述べたように、sbTNFαのスターコドンATGは、酵母における外来遺伝子発現(Romanosらの1992年の論文参照)のために酵母のコンセンサス配列AAAAATGTCT(図4参照)中に存在する。このコンセンサス配列はプライマーTNF-ECOPP中に含まれる。
【0046】
アルコール資化酵母のエレクトロポレーション及び発現測定
エレクトロポレーションは実施例3に述べた方法と同様な方法で行った。しかし、プラスミドの線状化はPmel(Fermentas社製)で行った。選択されたコロニーにおいて、ゲノムDNA精製キット(Fermentas社製)でDNAを単離し、His6-sbTNFα含有DNA断片をEcoTaq・PULS・DNAポリメラーゼ(Ecogen社製)で増幅し、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学製)によりその配列を決定した。
【0047】
発現測定も実施例3に述べた方法と同様な方法で行った。
【実施例6】
【0048】
大腸菌におけるターボットsbTNFαの発現
His6-sbTNFαのpET15bへのクローニング
プライマーTNFRO-BAMF及びTNFRO-BAMR(図1参照)を用いるsbTNFα増幅のためのPCR法におけるテンプレートとしてターボットの肝臓から得られたcDNAを使用した。両方のプライマーとも、プラスミドpET15b(Novagen社製)の同じ部位でPCR生成物を後クローニングするためのBamHI制限部位を有する。増幅はcDNAテンプレート、各dNTP50μM、プライマー0.2mM、MgCl2含有1X buffer PLUS及びEco Taq PULS DNAポリメラーゼ(Ecogen社製)1単位を含有するサンプル中で行った。サイクル反応はSmartサイクラー(Cepheid社製)内で、95℃・5分間を1サイクル、95℃・45秒間を30サイクル、55℃・45秒間を30サイクル及び72℃・90秒間を30サイクル続いて72℃・10分間を1サイクルの順で行った。0.8%アガロースゲル(Pronadisa社製)で電気泳動することにより分離した後、PCR生成物をQIAquick・PCR精製キット(Qiagen社製)で精製した。精製断片及びプラスミドpET15bをBamHI10単位により37℃で2時間かけて消化し、PCR精製キットによる前記PCR精製物をT4DNAリガーゼ(Fermentas社製)1単位を用いて22℃でライゲーションさせた。ライゲーション混合物を形質転換大腸菌DH5αコンピテントセル(形質転換受容性細胞)に対して使用し、そして、アンピシリン含有LBプレート内に展延させた。このプレートを37℃でインキュベーションし、2X・PCRマスターミックス(fermentas社製)を用いるコロニーPCR法により組換体を識別した。幾つかの生成コロニーのプラスミドを単離し、BamHI消化によるインサート遊離及びPvull消化によるインサート配向について試験した。選択されたプラスミドの配列をABI・Prism377遺伝子分析器(CIB,CSIC社製)で決定した。
【0049】
大腸菌の形質転換及び発現測定
大腸菌の形質転換及びターボットHis6-sbTNFαの発現測定は実施例1に述べた方法と同様な方法で実施した。
【実施例7】
【0050】
アルコール資化酵母におけるターボットsbTNFαの発現
His6-sbTNFαのpPICZAへのクローニング
ターボットのHis6-sbTNFαのpPICZAへのクローニングは実施例3に述べた方法と同様な同様な方法で行った。ターボットのHis6-sbTNFαを含有する生成プラスミドpPICZAの配列をABI・Prism3130遺伝子分析器(SACE,Murcia大学製)で決定した。実施例3で述べたように、sbTNFαのスターコドンATGは、酵母における外来遺伝子発現(Romanosらの1992年の論文参照)のために酵母のコンセンサス配列AAAAATGTCT(図5参照)中に存在する。このコンセンサス配列はプライマーTNF-ECOPP中に含まれる。
【0051】
アルコール資化酵母のエレクトロポレーション及び発現測定
エレクトロポレーションは実施例3に述べた方法と同様な方法で行った。しかし、プラスミドの線状化はPmel(Fermentas社製)で行った。選択されたコロニーにおいて、ゲノムDNA精製キット(Fermentas社製)でDNAを単離し、His6-sbTNFα含有DNA断片をEcoTaq・PULS・DNAポリメラーゼ(Ecogen社製)で増幅し、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学製)によりその配列を決定した。
【0052】
発現測定も実施例3に述べた方法と同様な方法で行った。
【実施例8】
【0053】
クロマトグラフ法によるsbTNFαの精製
大腸菌、出芽酵母又はアルコール資化酵母において発現されたHis6-sbTNFαは、AKTA・Explorer・FPLC(GE Healthcare社製)を用いてアフィニティクロマトグラフ法により精製した。精製はNi2+による固定金属イオンアフィニティクロマトグラフ法(ヒスチジン標識タンパク質の精製方法)によりHisTrap・FFカラム(GE Healthcare社製)を用いて行った。280nmにおける吸収をUNICORN5.10により測定した。容量1mlのカラムを、少なくとも5カラム容量のバインディングバッファー(20mM燐酸ナトリウム、0.5M塩化ナトリウム、5mMイミダゾール,pH7.4)を流速1ml/分で平衡化させた。次いで、成長培養物のサンプルを超音波又はビーズビーターで粉砕し、更に遠心分離してペレットを除去し、明澄な上澄液を得た。実施例2で述べたように、この直後に、この上澄液2mLをカラム内に装填した。吸収が安定したベースラインに達するまで(10〜15カラム容量)、カラムをバインディングバッファーで洗浄した。最後に、カラムに結合したタンパク質を溶出するために、溶出バッファー(20mM燐酸ナトリウム、0.5M塩化ナトリウム、5mMイミダゾール,pH7.4)の量を増大させる20〜25カラム容量の直線勾配を適用した。自動画分収集器を使用し、溶出液1mlの画分を収集した。His6-sbTNFαの存在について、SDS-PAGE及び抗ポリヒスチジンmAb(Sigma社製)を用いる免疫ブロット法によりタンパク質を含有する画分を試験した。選択された画分を合わせ、Bradfold(Sigma社製)によりHis6-sbTNFαの量を測定した。
【0054】
必要に応じて、トロンビン−アガロース(Recom-T,Sigma社製)を用いてHis6タグ(標識)を除去した。
【実施例9】
【0055】
SDS-PAGE及び免疫ブロット法によるsbTNFαの検出
12.5%アクリルアミド/ビサクリルアミドを用いるSDS-ポリアクリルミドゲル電気泳動法(SDS-PAGE)によりHis6-sbTNFαを有するサンプルを分析した。ゲルをcoomassie(登録商標)0.1%で30分間染色し、His6-sbTNFαに対応する21kDaのバンドを区別した。
【0056】
前記のようにSDS-PAGEの後に免疫ブロット法を実施することによりHis6-sbTNFαを検出するために、ゲルをTBS(20mM Tris-HCl,150mM NaCl,0.1% Tween20)中で100Vで1時間かけてニトロセルロース膜(Sigma社製)に移行させた。この膜を5%BSAを有するTTBS(10mM Tris-HCl,100mM NaCl,0.1% Tween20)と共にインキュベーションし、4℃でタンパク質をブロックした。膜をTTBSで洗浄後、これをTTBS+5%BSA+1:1000抗ポリヒスチジンmAb(Sigma社製)と共に2時間インキュベーションした。次いで、ECL(Amersham Biosciences社製)で染色する前に、この膜をTTBS及びTBSで再び洗浄した。His6-sbTNFαに対応する21kDaの単一のバンドが現れた(図6参照)。
【実施例10】
【0057】
発酵により酵母の培養
発酵による形質転換酵母の培養をバイオリアクターシステム(Applicon社製)で行った。
【0058】
一例として、アルコール資化酵母菌株をYPGly培地を有するフラスコ内で、培養物のOD600が30超に達するまで、培養した。次いで、成長培養物をMMG(グリセロール含有最少培地)含有バイオリアクターシステム内で接種した。培養条件は次の通りであった。温度30℃、pH5.00(NH3を添加することにより調節した)、撹拌速度500−1250rpm、酸素濃度30%、撹拌しながらのカスケード30%。これらのパラメータはBioXpert・V2により測定した。発酵は、培養物のOD600が50〜80に達するまで、バッチ法で行った。培養物のOD600が50〜80に達した直後に、発酵を半回分培養法に変更し、グリセロール含有飼料の給餌を10時間続けた(培養物のOD600は260〜280に達した)。最後に、メタノール含有飼料の給餌を40時間続けた。結局、培養物のOD600は380〜420に達した。
【0059】
組換TNF His6-sbTNFαの発現分析は免疫ブロット法(実施例9参照)により行った。
【実施例11】
【0060】
組換酵母のマイクロカプセル化
発酵が所望のバイオマスに達し、かつ、組換TNF・His6-sbTNFαの最適発現が得られた後、培養液を無菌的に採取し、20%マロデキストリン及び10%保護ポリマーKollicoat(登録商標)・MAE100P(BASF社製)を用いて製剤化した。
【0061】
次いで、製剤化された懸濁液を噴霧乾燥機(Buchi社製)から噴出させた。入口温度は120℃、出口温度は85℃に制御した。
【0062】
組換酵母の最終マイクロカプセルを分析し、細胞生存性の不存在及びTNF His6-sbTNFαの存在を免疫ブロット法(実施例10参照)により測定した。
【実施例12】
【0063】
マイクロカプセル化組換酵母によるアルテミア・ノープリウス幼生(Artemia nauplii)の富化
滅菌完全海水500mlが入ったフラスコ内でアルテミア・ノープリウス幼生を孵化させた。フラスコ内の海水は水槽用空気ポンプに接続された空気ホースにより曝気し、往復振盪水浴中で28℃に維持した。孵化してから24時間後に、ノープリウス幼生を富化システムに採取した。このシステムは、容量100mlの複数個のフラスコからなり、各フラスコには新鮮な海水が入れられており、各フラスコは全て同じ水浴中に配置され、そして、それぞれ別々に曝気されている。全ての実験について、海水0.5mlのサンプルを採取することにより、孵化率及びノープリウス幼生の個数を見積もった。
【0064】
マイクロカプセル化組換酵母による富化アルテミア・ノープリウス幼生の製造最適時間は、DTAFによる酵母の染色により評価した。次いで、アルテミア・ノープリウス幼生に給餌するために染色酵母を使用した。給餌後、0、30、45、60及び120分の各時点でアルテミア・ノープリウス幼生を殺した。富化アルテミア・ノープリウス幼生は蛍光鏡検法により観察した。
【0065】
結果を図7に示す。図示されているように、給餌開始後、初めのうちはマイクロカプセル化組換酵母の量の増加は口部領域で観察され、その後、腸管全体がマイクロカプセル化組換酵母で満たされ、個々の細胞は認められなかった。
【0066】
富化アルテミア・ノープリウス幼生の製造最適時間は60分であった。
【実施例13】
【0067】
アルコール資化酵母におけるヨーロッパヘダイsbTNFαの発現及び分泌
His6-sbTNFαのpPICZαAへのクローニング
615pbのHis6-sbTNFα含有断片の増幅のために、TNF-ECOF及びTNF-ECOR(図1参照)によるPCR法におけるテンプレートとしてsbTNFα含有プラスミドpET15bを使用した。両方のプライマーとも、プラスミドpPICZαAの同じ部位内でPCR生成物をクローニングするためのEcoRI制限部位を含有する。増幅は、テンプレートを5ng、MgCl2を2.5mM、各dNTPを50μM、プライマーを0.2mM、1X・High・Speec添加剤、1X buffer PLUS及びEco・Taq・PLUS・DNAポリメラーゼ(Ecogen社製)を2単位含有するサンプル中で行った。サイクル反応はSmartサイクラーII(Cepheid社製)中で、95℃・5分間を1サイクル、95℃・30秒間を30サイクル、55℃・45秒間を30サイクル及び72℃・2分間を30サイクル、続いて72℃・10分間を1サイクルの順で行った。臭化エチジウムを0.5μg/ml含有する0.8%アガロースゲル(Pronadisa社製)中で電気泳動することによりPCR生成物を分離し、QIAquickゲル抽出キット(Qiagen社製)で精製した。精製断片及びpPICZαA200ngをEcoRI(Fermentas社製)10単位により37℃で消化させ、その後、T4DNAリガーゼ(Fermentas社製)5単位を用いて22℃でライゲーションさせた。ライゲーション反応生成物を大腸菌TOP10F'コンピテントセルに形質転換させ、ゼオシン(Invitrogen社製)25μg/mlを有するLBプレート内でプラスミド含有コロニーを選別した。2X・PCR・Master・Mix(Fermentas社製)及びPICα及びPIC3プライマーを用いるコロニーPCRにより組換体を識別した。得られたHis6-sbTNFα含有プラスミドpPICZαAをQIAprep・Spin・Miniprepキット(Qiagen社製)で精製し、EcoRIによる消化後のインサート遊離及び消化Smal及びNcolによるインサート配向について試験した。また、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学)によりプラスミドの配列を決定した。
【0068】
アルコール資化酵母のエレクトロポレーション及び発現測定
アルコール資化酵母のエレクトロポレーションの前に、後のアルコール資化酵母のゲノムDNAにおける組換及び組込みのために、His6-sbTNFα含有プラスミドpPICZαAを37℃でSacl又はPmelによる消化を通じて線状化させた。線状プラスミドの全量5μgをMiniElute・Reaction・Cleanupキット(Qiagen社製)で精製し、滅菌水10μlで溶離した。エレクトロポレーションに使用したアルコール資化酵母菌株はX-33であった。エレクトロポレーション用のX-33を作製するために、OD600=1.5の培養物100mlの細胞を1500xgで4℃で5分間遠心分離した。滅菌水で2回洗浄後、細胞を1Mのソルビトール中に再懸濁させ、前記のように遠心分離し、そして最後に1Mのソルビトール100ml中に再懸濁させた。エレクトロポレーションを行うために、DNA10μlを細胞80μlと混合し、氷冷された0.2cmのエレクトロポレーション・キュベット(BioRad社製)に移送した。5分間インキュベーションした後、MicroPulserエレクトロポレーター(BioRad社製)により25μF、1.5kV及び400Ωの条件でパルスを印加した。その直後に、氷冷された1Mソルビトール溶液1mlをキュベットに添加し、得られた混合物を30℃で1時間インキュベーションした。細胞をゼオシン(Invitrogen社製)100μg/mlを有するYPDSプレート上に展延し、コロニーが出現するまで30℃で数日間インキュベーションした。PIC5及びPIC3プライマーと2X・PCRマスターミックス(Fermentas社製)を用いるPCR法によりインサートの存在に関して幾つかの形質転換体を分析し、His6-sbTNFα発現に関して測定した。特定のコロニーにおいて、ゲノムDNA精製キット(Fermentas社製)を用いてゲノムDNAを単離し、そして、His6-sbTNFα含有DNA断片をEcoTaq・PULS・DNAポリメラーゼ(Ecogen社製)で増幅し、ABI・Prism3130遺伝子アナライザー(SACE,Murcia大学)によりその配列を決定した。
【0069】
His6-sbTNFαを有する組換アルコール資化酵母菌株をグリセロール含有最少培地25ml中で30℃、250rpmで、培養物がOD600〜6に達するまで、メタノール導入前にバイオマスを発生させるために成長させた。3000xgで5分間遠心分離することにより細胞を回収し、発現を誘発させるために、OD600〜1に達するまで、ペレットを0.5%メタノール含有最少培地200ml中に再懸濁させた。前記に述べたように培養物をインキュベーションし、発現誘導を維持するために24時間毎に0.5%メタノールを添加した。幾つかの時点でサンプルを回収し、タンパク質発現を分析した。タンパク質発現をチェックするために、SDS-PAGE及び抗ポリヒスチジンmAb(Sigma社製)を用いるウエスタン・ブロット法による分析のためにSDS-ローディングバッファー液中で煮沸することにより上澄液のHis6-sbTNFαを測定した。
【実施例14】
【0070】
マイクロカプセル化組換酵母の効力
体重120gの健康なヨーロッパヘダイ(Sparus aurata L.)(硬骨魚類,スズキ目,タイ科)のサンプルを容量2m3の遊泳海水水槽内で飼育した。海水には20%水槽容量/時の流量で酸素6ppmを溶解させた。水槽は常温及び日長に維持され、市販のペレット飼料(スペインのブルゴスに所在するTrouvit社製)を1日に2回給餌した。試験群には、基礎飼料に対照酵母又は成熟(活性)sbTNFα過剰発現マイクロカプセル化酵母を0.1、1又は10%追加した餌を2日毎に給餌した。全てのサンプリングタイム(処置から2日、4日、6日及び10日後)時点で、標本魚の体重を測定し、頭腎及び腸管を採取し、光学顕微鏡で検査し、かつ、遺伝子発現を調査した。ここに述べた試験法は、実験動物の使用に関する欧州連合理事会(European Union Council)(86/609/EU)のガイドライン及びスペインのMurcia大学の生命倫理委員会のガイドラインに従っている。呼吸性バースト活性は、異なる刺激時間後の頭腎ロイコサイト懸濁液により生成されたルミノール依存性化学ルミネッセンス(Sepulcreらの2007年の論文参照)として測定した。全ての試験は5匹の魚と3重のサンプルを用いて行った。結果の有効性を確保するために、同じ試験を2回繰り返した。SPSS・13.0.1統計ソフトウエアパッケージを全ての統計分析に使用した。試験群間の差を決定するために、データは平方偏差(ANOVA)及びテューキー(Tukey)多重範囲検定により分析した。
【0071】
魚の食欲、成長速度、遊泳行動及び外的形態は処置により影響を受けなかった。また、試験中の魚の死亡は認められなかった。更に、10日間にわたってsbTNFα含有酵母を給餌された魚の腸管内には、食細胞(すなわち、好酸性顆粒又はマクロファージ)の浸潤ではない病理組織学的病変は認められなかった。酵母単独の投与は、頭腎ロイコサイトの呼吸性バーストとして測定した場合、魚の免疫状態に殆ど作用を及ぼさなかった。これと際立って対称的に、過剰発現sbTNFα含有マイクロカプセル化酵母を0.1及び1%含有する魚類飼料は、治療開始から4日後には、投与量に依存して増大する呼吸性バーストを示した(図8参照)。驚くべきことに、TLR9の例外を除いて、酵母の投与は頭腎及び腸管における幾つかの炎症性遺伝子のmRNAレベルに対して殆ど影響力を有していなかった。TLR9の発現は顕著に上方制御された(図9参照)。
【0072】
前記の結果は、10日間の免疫賦活剤の投与は魚類に何らの副作用も及ぼさないことを示す。呼吸性バーストは魚類の免疫状態の指標として広く使用されているので(Muleroらの1988年の論文、Garcia-Castilloらの2004年の論文、Chaves-Pozoらの2005年の論文及びSepulcreらの2007年の論文参照)、TNFα含有酵母を有する魚類飼料におけるこの免疫応答の活性化は、本発明の免疫賦活剤が非常に有効であり、その結果、非生物的ストレス(すなわち、魚類操作により惹起されるストレス)及び生物的ストレス(すなわち、病原体により惹起されるストレス)に対して魚類を保護することを示している。更に、本発明の免疫賦活剤の投与による腸管内のTLR9の発現の上方制御(すなわち、過剰発現)も、魚類の腸管がウイルス及び細菌感染に対して良好に保護されていることを示唆している。なぜなら、この受容体(レセプター)はこれら病原体の認識の主要成分だからである(Ishii及びAkiraの2006年の論文参照)。総括的に、前記の結果も全て、組換sbTNFαは、生来の免疫細胞の系統的活性化及びTLR9の発現量の増大により、経口ワクチンの優れたアジュバントであることを示している。
【0073】
以上の説明は、本発明の一実施例に関するもので、この技術分野の当業者であれば、本発明の種々の変形例を考え得るが、それらはいずれも本発明の技術的範囲に包含される。
用語「又は」に関して、例えば「A又はB」は、「Aのみ」、「Bのみ」ならず、「AとBの両方」を選択することも含む。特に記載のない限り、装置又は手段の数は、単数か複数かを問わない。
【特許請求の範囲】
【請求項1】
少なくともマイクロカプセル化サイトカインと腸溶保護ポリマーからなり、前記サイトカインは魚類サイトカイン、軟体動物類サイトカイン又は甲殻類サイトカインであることを特徴とする経口投与可能な免疫賦活剤。
【請求項2】
前記サイトカインは組換サイトカインであることを特徴とする請求項1に記載の免疫賦活剤。
【請求項3】
前記マイクロカプセル化組換サイトカインは腫瘍壊死因子α(TNFα)であることを特徴とする請求項2に記載の免疫賦活剤。
【請求項4】
前記マイクロカプセル化組換サイトカインは酵母である宿主微生物中に過剰発現されることを特徴とする請求項2に記載の免疫賦活剤。
【請求項5】
前記酵母はアルコール資化酵母であることを特徴とする請求項4に記載の免疫賦活剤。
【請求項6】
前記サイトカインは、ヨーロッパヘダイ(Sparus aurata)、ヨーロピアンシーバス(Dicentrarchus labrax)、ニジマス(Oncorhynchus mykiss)、イシビラメ(Psetta maxima)、シマアジ(Dentex dentex)、シャープスナウト・シーブリーム(Diplodus puntazzo)、スペインダイ(Pagellus bogaraveo)、コルビナ(Argyrosomus regius)、ヨーロッパウナギ(Anguilla anguilla)、ミミックオクトパス(Octopus sp.)などのような水産養殖に適した海洋生物から誘導されることを特徴とする請求項1〜5の何れかに記載の免疫賦活剤。
【請求項7】
前記免疫賦活剤は多細胞生物に給餌されることを特徴とする請求項1〜6の何れかに記載の免疫賦活剤。
【請求項8】
前記多細胞生物はアルテミア(Artemia sp.)又はワムシ(phylum Rotifera)であることを特徴とする請求項7に記載の免疫賦活剤。
【請求項9】
前記サイトカイン及び/又は組換サイトカイン含有宿主微生物は腸溶保護ポリマーでマイクロカプセル化されていることを特徴とする請求項1〜8の何れかに記載の免疫賦活剤。
【請求項10】
(a)選択されたサイトカインをコーディング(解読)するcDNAの組織を、魚類、軟体動物類又は甲殻類から選別し、そして単離するステップと、
(b)少なくとも適当な宿主微生物におけるサイトカイン発現のための少なくとも発現ベクターに前記cDNAをクローニングするステップと、
(c)宿主微生物の宿主細胞を培養し、組換サイトカイン又は当該組換サイトカイン含有宿主微生物の培養物を得るステップと、
(d)組換サイトカイン又は当該組換サイトカイン含有宿主微生物をマイクロカプセル化し、マイクロカプセル化された組換サイトカイン又は当該組換サイトカイン含有宿主微生物を得るステップとからなることを特徴とする請求項2に記載の免疫賦活剤の製造方法。
【請求項11】
前記宿主微生物はアルコール資化酵母、出芽酵母及びクルイベロミセス・ラクティス(Kluyveromyces lactis)からなる群から選択される酵母であることを特徴とする請求項10に記載の製造方法。
【請求項12】
前記(d)マイクロカプセル化ステップは、腸溶保護ポリマーの存在下で噴霧して噴霧乾燥させることにより行われることを特徴とする請求項10又は11に記載の製造方法。
【請求項13】
前記腸溶保護ポリマーは、酢酸フタル酸セルロース、フタル酸ヒドロキシプロピルセルロース、カルボキシメチルセルロース、メタクリル酸コポリマー、マルトデキストリン、キトサン、ゼラチン、スターチ及びアラビアゴムからなる群から選択されることを特徴とする請求項12に記載の製造方法。
【請求項14】
水産養殖される魚類、軟体動物類又は甲殻類の免疫刺激用としての請求項1〜9の何れかに記載の経口投与可能な免疫賦活剤の使用方法。
【請求項15】
前記サイトカインは、免疫刺激されるべき同じ魚類、軟体動物類又は甲殻類の種から由来することを特徴とする請求項14に記載の使用方法。
【請求項16】
前記魚類、軟体動物類又は甲殻類は、ヨーロッパヘダイ(Sparus aurata)、ヨーロピアンシーバス(Dicentrarchus labrax)、ニジマス(Oncorhynchus mykiss)、イシビラメ(Psetta maxima)、シマアジ(Dentex dentex)、シャープスナウト・シーブリーム(Diplodus puntazzo)、スペインダイ(Pagellus bogaraveo)、コルビナ(Argyrosomus regius)、ヨーロッパウナギ(Anguilla anguilla)、ミミックオクトパス(Octopus sp.)からなる群から選択されることを特徴とする請求項14又は15に記載の使用方法。
【請求項17】
前記免疫刺激は、給餌中に飼料と共に前記免疫賦活剤を投与することにより行われることを特徴とする請求項14〜16の何れかに記載の使用方法。
【請求項18】
前記免疫刺激は、
(a)免疫賦活剤を多細胞生物に投与するステップと、
(b)富化された多細胞生物を得るステップと、
(c)引き続いて、前記種に前記富化多細胞生物を給餌するステップと、
により行われることを特徴とする請求項14〜16の何れかに記載の使用方法。
【請求項19】
前記多細胞生物はアルテミア(Artemia sp.)又はワムシ(phylum Rotifera)であることを特徴とする請求項18に記載の使用方法。
【請求項20】
少なくともマイクロカプセル化された魚類、軟体動物類又は甲殻類サイトカインからなる経口投与可能な免疫賦活剤で富化された多細胞生物。
【請求項21】
ワクチンアジュバントとしての請求項1〜9の何れかに記載の経口投与可能な免疫賦活剤の使用方法。
【請求項22】
魚類、軟体動物類又は甲殻類用のワクチンを更に含有することを特徴とする請求項1〜9の何れかに記載の免疫賦活剤。
【請求項23】
前記ワクチンは前記サイトカインでマイクロカプセル化されていることを特徴とする請求項22に記載の免疫賦活剤。
【請求項1】
少なくともマイクロカプセル化サイトカインと腸溶保護ポリマーからなり、前記サイトカインは魚類サイトカイン、軟体動物類サイトカイン又は甲殻類サイトカインであることを特徴とする経口投与可能な免疫賦活剤。
【請求項2】
前記サイトカインは組換サイトカインであることを特徴とする請求項1に記載の免疫賦活剤。
【請求項3】
前記マイクロカプセル化組換サイトカインは腫瘍壊死因子α(TNFα)であることを特徴とする請求項2に記載の免疫賦活剤。
【請求項4】
前記マイクロカプセル化組換サイトカインは酵母である宿主微生物中に過剰発現されることを特徴とする請求項2に記載の免疫賦活剤。
【請求項5】
前記酵母はアルコール資化酵母であることを特徴とする請求項4に記載の免疫賦活剤。
【請求項6】
前記サイトカインは、ヨーロッパヘダイ(Sparus aurata)、ヨーロピアンシーバス(Dicentrarchus labrax)、ニジマス(Oncorhynchus mykiss)、イシビラメ(Psetta maxima)、シマアジ(Dentex dentex)、シャープスナウト・シーブリーム(Diplodus puntazzo)、スペインダイ(Pagellus bogaraveo)、コルビナ(Argyrosomus regius)、ヨーロッパウナギ(Anguilla anguilla)、ミミックオクトパス(Octopus sp.)などのような水産養殖に適した海洋生物から誘導されることを特徴とする請求項1〜5の何れかに記載の免疫賦活剤。
【請求項7】
前記免疫賦活剤は多細胞生物に給餌されることを特徴とする請求項1〜6の何れかに記載の免疫賦活剤。
【請求項8】
前記多細胞生物はアルテミア(Artemia sp.)又はワムシ(phylum Rotifera)であることを特徴とする請求項7に記載の免疫賦活剤。
【請求項9】
前記サイトカイン及び/又は組換サイトカイン含有宿主微生物は腸溶保護ポリマーでマイクロカプセル化されていることを特徴とする請求項1〜8の何れかに記載の免疫賦活剤。
【請求項10】
(a)選択されたサイトカインをコーディング(解読)するcDNAの組織を、魚類、軟体動物類又は甲殻類から選別し、そして単離するステップと、
(b)少なくとも適当な宿主微生物におけるサイトカイン発現のための少なくとも発現ベクターに前記cDNAをクローニングするステップと、
(c)宿主微生物の宿主細胞を培養し、組換サイトカイン又は当該組換サイトカイン含有宿主微生物の培養物を得るステップと、
(d)組換サイトカイン又は当該組換サイトカイン含有宿主微生物をマイクロカプセル化し、マイクロカプセル化された組換サイトカイン又は当該組換サイトカイン含有宿主微生物を得るステップとからなることを特徴とする請求項2に記載の免疫賦活剤の製造方法。
【請求項11】
前記宿主微生物はアルコール資化酵母、出芽酵母及びクルイベロミセス・ラクティス(Kluyveromyces lactis)からなる群から選択される酵母であることを特徴とする請求項10に記載の製造方法。
【請求項12】
前記(d)マイクロカプセル化ステップは、腸溶保護ポリマーの存在下で噴霧して噴霧乾燥させることにより行われることを特徴とする請求項10又は11に記載の製造方法。
【請求項13】
前記腸溶保護ポリマーは、酢酸フタル酸セルロース、フタル酸ヒドロキシプロピルセルロース、カルボキシメチルセルロース、メタクリル酸コポリマー、マルトデキストリン、キトサン、ゼラチン、スターチ及びアラビアゴムからなる群から選択されることを特徴とする請求項12に記載の製造方法。
【請求項14】
水産養殖される魚類、軟体動物類又は甲殻類の免疫刺激用としての請求項1〜9の何れかに記載の経口投与可能な免疫賦活剤の使用方法。
【請求項15】
前記サイトカインは、免疫刺激されるべき同じ魚類、軟体動物類又は甲殻類の種から由来することを特徴とする請求項14に記載の使用方法。
【請求項16】
前記魚類、軟体動物類又は甲殻類は、ヨーロッパヘダイ(Sparus aurata)、ヨーロピアンシーバス(Dicentrarchus labrax)、ニジマス(Oncorhynchus mykiss)、イシビラメ(Psetta maxima)、シマアジ(Dentex dentex)、シャープスナウト・シーブリーム(Diplodus puntazzo)、スペインダイ(Pagellus bogaraveo)、コルビナ(Argyrosomus regius)、ヨーロッパウナギ(Anguilla anguilla)、ミミックオクトパス(Octopus sp.)からなる群から選択されることを特徴とする請求項14又は15に記載の使用方法。
【請求項17】
前記免疫刺激は、給餌中に飼料と共に前記免疫賦活剤を投与することにより行われることを特徴とする請求項14〜16の何れかに記載の使用方法。
【請求項18】
前記免疫刺激は、
(a)免疫賦活剤を多細胞生物に投与するステップと、
(b)富化された多細胞生物を得るステップと、
(c)引き続いて、前記種に前記富化多細胞生物を給餌するステップと、
により行われることを特徴とする請求項14〜16の何れかに記載の使用方法。
【請求項19】
前記多細胞生物はアルテミア(Artemia sp.)又はワムシ(phylum Rotifera)であることを特徴とする請求項18に記載の使用方法。
【請求項20】
少なくともマイクロカプセル化された魚類、軟体動物類又は甲殻類サイトカインからなる経口投与可能な免疫賦活剤で富化された多細胞生物。
【請求項21】
ワクチンアジュバントとしての請求項1〜9の何れかに記載の経口投与可能な免疫賦活剤の使用方法。
【請求項22】
魚類、軟体動物類又は甲殻類用のワクチンを更に含有することを特徴とする請求項1〜9の何れかに記載の免疫賦活剤。
【請求項23】
前記ワクチンは前記サイトカインでマイクロカプセル化されていることを特徴とする請求項22に記載の免疫賦活剤。
【図1】



【図2】
【図3】

【図4】

【図5】

【図6A】
【図6B】

【図7】
【図8】

【図9】
【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図7】
【図8】
【図9】
【公表番号】特表2012−505193(P2012−505193A)
【公表日】平成24年3月1日(2012.3.1)
【国際特許分類】
【出願番号】特願2011−530581(P2011−530581)
【出願日】平成20年10月10日(2008.10.10)
【国際出願番号】PCT/IB2008/002697
【国際公開番号】WO2010/041096
【国際公開日】平成22年4月15日(2010.4.15)
【出願人】(509211022)プロベルテ ファーマ,エス.エー. (5)
【Fターム(参考)】
【公表日】平成24年3月1日(2012.3.1)
【国際特許分類】
【出願日】平成20年10月10日(2008.10.10)
【国際出願番号】PCT/IB2008/002697
【国際公開番号】WO2010/041096
【国際公開日】平成22年4月15日(2010.4.15)
【出願人】(509211022)プロベルテ ファーマ,エス.エー. (5)
【Fターム(参考)】
[ Back to top ]