
水痘帯状疱疹ウイルスに特異的な抗体
本発明は、水痘帯状ヘルペスウイルス(VZV)に結合し、そして、VZV感染を中和する新規抗体配列を提供する。新規配列は、VZV感染の医療管理のために、具体的には、ウイルスを検出するために、あるいは、VZV感染の予防的処置又は治療的処置に使用される医薬組成物を調製するために使用される。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ウイルスに特異的な生物学的活性を有する、ファージ・ディスプレイ・ライブラリーから単離された新規抗体配列に関する。
【背景技術】
【0002】
ファージ・ディスプレイ技術は、ポリペプチド(抗体フラグメント、生理活性ペプチド、酵素等)をクローニング、選択、及び設計するために、細菌細胞(例えば、エシェリキア・コリ(Escherichia coli)細胞)に感染する(M13などの)線状ファージのゲノムの小さな寸法と順応性を利用する。前記ポリペプチドは、線状ファージ表面上に発現され、そして、標的とそれらの相互作用により生じる生物学的機能を発揮する。
【0003】
論文(Bradbury A and Marks J,2004;Mancini et al,2004;Conrad U and Scheller J,2005;Hust M and Dubel S,2005)及び書籍(“Phage display:A Practical Approach”,vol.266,ed.Clackson and Lowman H,Oxford Univ.Press,2004;“Phage Display:A Laboratory Manual”,ed.Burton D et al,CSHL Press,2001)で概説されるように、いくつかのクローニング及び発現ストラテジー、ベクター、ライブラリー、ファージの伝播方法、及びスクリーニング・アッセイが、様々な適用のために開発された。
【0004】
ファージ・ディスプレイ・ライブラリーは、組み換えファージの集団で作られており、それぞれがタンパク質配列のレパートリーの1つの要素を提示している。特異的なタンパク質を発現するファージは、繰り返される親和性ベースの、及び/又は機能ベースの選択過程(「パンニング」)によってライブラリーから単離される。例えば、タンパク質は、精製抗原に対するそれら親和性、又は生物学的アッセイ法における活性に基づいて単離及び特徴づけされ得る、(一般的にFabsと名付けられる)可変重鎖/軽鎖ヘテロ二量体、又は一本鎖可変フラグメント(scFv)の形態の抗体フラグメントである。
【0005】
具体的には、前記結合特性に関係する関連生物学的活性を伴うこともある、病原菌及び生物学的標的に対する高い親和性と特異性を有する抗体フラグメントを同定するためのスクリーニング過程が開発された。実際には、(受動免疫療法又は受動血清療法と名付けられた)治療方法全体が、ヒト又はヒト以外の治療標的に向けられた抗体及び抗体フラグメントの抗原結合特性の上に成り立っている(Dunrnan P and Nesin M,2003;Keller M and Stiehm E,2000)。受動免疫療法は、病原性抗原(例えば、毒素、ヒト・タンパク質、ウイルス、又は寄生生物)に対して規定の結合特異性を有する治療抗体を含んで成る医薬組成物の個体への投与から成る。
【0006】
受動免疫療法は、臨床診療に取り入れられ、(感染症、免疫媒介疾患、及び癌を含めた)様々な疾患を治療する機会が急速に増大した。このアプローチは、免疫系が標的とされた分子を妨げる、及び/又は排除するのに必要とされる量、及び/又は特異性を有する抗体を産生することができない患者において特に有効である(Chatenoud L,2005;Laffly E and Sodoyer R,2005)。
【0007】
治療抗体を使用して標的とされ得る病原性抗原の中で、ヒト細胞に感染するウイルスは、特別な重要性を持つ。このような抗体の投与で、患者におけるウイルス増殖を妨げることができ、そして、集団におけるウイルス感染の大発生を阻む可能性がある。あるいは、前記抗体は、通常、免疫が保たれている人の健康に重大な、及び/又は永続的な影響を及ぼさないものを含めた感染症に対してより一層感受性が高くなる、程度の差はあれ長期間、免疫系が弱まっている患者(例えば、免疫抑制された人、高齢者、又は移植を受けた人)に投与され得る。
【0008】
水痘帯状疱疹ウイルス(VZV、水痘)は、免疫障害を持つ人にとってそのような命にかかわる可能性のある感染の原因となるウイルスの1種である。VZVは、少なくとも70個の異なったオープン・リーディング・フレーム(ORFs)をコードするアルファ・ヘルペスウイルスであって、その大部分が単純ヘルペスウイルスと近縁関係にある(Mori I and Nishiyama Y,2005)。VZVは、皮膚病変又は呼吸器分泌物中の感染性ウイルスとの直接接触によってヒト集団内で伝播され、皮膚とT細胞に対して特異的親和性を示す。第一次VZV感染は、リンパ球に向けられ、それに続いて、細胞関連ウイルス血症、臓器内でのウイルス複製、そして、第二次ウイルス血症に起因する広汎性皮膚疹が起こる。VZV感染は、ビリオンの放出、又は細胞間伝播によって拡散することが可能であり、知覚神経節内への潜伏を確立する(Arvin A,1996;Quinlivan M and Breuer J,2006)。
【0009】
通例、1回のVZV侵襲又はワクチン接種が、一生にわたる保護をもたらす。しかしながら、VZVには、ウイルスが先天的な免疫応答、及び後天的に得た免疫応答の両方を回避することを可能にする固有の特性がある。ワクチン接種後の検出可能な細胞性免疫にもかかわらず、VZV特異的抗体が失われる場合もあるので、VZVの再感染又は再活性化がさらに可能である(Ludwig B et al.,2006)。
【0010】
感染及びワクチン接種に対する免疫応答の機構と効力は、ヒト及び動物モデルにおいて(Maple P et al.,2006;Matsuo K et al.,2003;Kutinova L et al,2001;Massaer M et al.,1999;Haumont M et al.,1997;Hasnie F et al.,2007)又は試験管内培養系において(Finnen R et al.,2006;Andrei G et al.,2005a)研究された。VZV発症機序と免疫生物学は、トランスジェニック・マウス・モデルにおいて、生体外におけるヒト皮膚又は神経節モデルを使用して研究できる(Baiker A et al.,2004;Ku C et al.,2005;Taylor S and Moffat J,2005;Zerboni L et al.,2005)。このように、細胞型特異的VZVアポトーシス活性(Hood C et al.,2003)、及びVZV感染の細胞性/ウイルス性伝達物質(Berarducci B et al.,2006;Chen J et al.,2004;Li Q et al.,2007;Hambleton S et al.,2007)が同定された。
【0011】
弱毒化生水痘ワクチン(Oka/Merck菌株)が入手可能であり、その安全と有効性を鑑みて、上記ワクチンが日常的な幼児期予防接種と成人に推奨される(Oxman M et al.,2006;Arvin A,1996)。VZVワクチン接種が先進工業国において幅広く定着し、且つ、短期間/中期間、すべての形態の水痘(特に重症疾患)を低減するのに非常に効果的であるにもかかわらず、集団において潜在型又は進化した野生型ウイルスのプールが継続的な脅威になっている(Hambleton S and Gershon A,2005)。
【0012】
VZV菌株は、VZV転写産物の特異的な遺伝子変異(Grose C,2006)、及び3種類の大きく異なる遺伝子型(Norberg P et al.,2006)と関連する。遺伝的変異は、予防接種されたヒト宿主の中に検出された皮疹を形成するVZV遺伝子型に関連した(Quinlivan M et al.,2007)。そのうえ、VZV免疫のレベルは、異なったヒト集団において、例えば、欧州地域の中で、大きく異なる(Nardone A et al.,2007)。
【0013】
帯状ヘルペス(帯状疱疹)は、臨床診療において、特に加齢に伴って低下した細胞性免疫の結果として高齢者において、別の年齢分布においてはVZV再活性化の危険因子に起因して、そして、癌、免疫抑制治療、又はステロイド療法による随伴性免疫不全と共に、いまだに頻繁に観察されている。帯状ヘルペスの合併症は、神経学的合併症(ヘルペス後神経痛、脳血管炎、脳炎、無菌性髄膜炎、脳神経麻痺、又は髄膜脳炎)、目に関する合併症(眼帯状ヘルペス、ブドウ膜炎、網膜の壊死、視神経炎、又は角膜炎)、あるいは、内臓合併症(肝炎、肺炎、心筋炎)である。帯状ヘルペスの危険因子は、さらによく理解されるようになってきているが、それらの数の増加、並びに知覚神経節及び眼表面へのVZV持続性は、成人における、より広範囲のワクチン接種、及びより効果的な抗VZV治療の必要性を示唆している(Dworkin R et al.,2007;Weinberg J,2007;Liesegang T,2004)。
【0014】
VZV感染は、ワクチンの使用が妥当でない免疫抑制患者において非常に危険である。実際に、免疫抑制状態にある1人の人の単独のVZV再活性化又は再感染が、脊髄又は脳動脈を含めたいくつかの組織にVZVが侵入し得るような数人の患者に感染する可能性がある移植病棟において、VZVは重大な問題である(Chaves Tdo S.et al.,2005;Gilden D,2004)。妊娠中、VZV感染はまた、子宮内伝播又は新生児期感染によって胎児にも広まり、重大な皮膚病変及び四肢と臓器の発生の異常を伴った、子宮内胎児死亡又は先天性水痘症候群を引き起こす(Schleiss M,2003;Sauerbrei A and Wutzler P,2007)。
【0015】
いくつかの重要な禁忌(Arvin A,1996)があるワクチン接種は別として、抗ウイルス治療が利用可能である(アシクロビア、バラシクロビル、ファムシクロビル、及びブリブジンなど)。しかしながら、薬剤耐性VZV菌株の問題に加えて、これらの化合物はまた、例えば、ヒト酵素との相互作用のために、禁忌がある可能性がある(Andrei G et al.,2005b;Abdel−Haq N et al.2006)。コルチコステロイド又は抗痙攣薬などの他の治療が、追加的な鎮痛を与えるために使用されるが、それらの有害影響特性が、特にヘルペス後神経痛に対する、使用を制限する(Tyring S,2007)。これにより、予防のための現在の手段が適用不可能であるか、又はもはや有効性でない場合、代替のアプローチが必要である(Hambleton S and Gershon A,2005)。ワクチン不全とワクチン誘発免疫の低下を考慮して、現在、二次ワクチン投与が推奨されている(Chaves S et al.,2007)。
【0016】
VZV感染を早期に同定するために、いくつかの診断検査が、同様の患者集団において、並びにVZV感染の危険に晒された高齢者又は免疫不全の人において、VZV特異的抗体、抗原、又は転写産物を検出するために開発された(Hambleton S and Gershon A,2005;Smith J et al.,2001)。
【0017】
VZVに関連する眼感染(Kezuka,T. 2004)、血管障害(Burgoon M et al.,2003)、又は疫学的研究(Talukder Y et al.,2005)に関連して血清、脳脊髄液、又は口腔液中で、VZV特異的免疫活性が同定された。VZV暴露後の予防が提案され、そして、具体的には、感染を予防するために投与され得る有意な抗VZV力価を有するヒト抗体の調製物を使用して試験された(Keller M and Stiehm E,2000)。特異的な、抗VZV抗体の高い力価を有するヒト免疫グロブリンの静脈内注射用調製物が記載された(US4717564;CDC,2006)。単独での、又はVZV特異的な抗ウイルス剤としての他の化合物と組み合わせたその利用は、移植を受けた患者、妊婦、又は新生児で試験された(Huang Y et al.,001;Carby M et al.,2007;Koren G et al.,2002)。
【0018】
VZV−特異的抗原、VZV糖タンパク質の変異体、及びそれらの免疫原性エピトープが、異なる抗体調製物、例えば、ヒト血清(US6960442;Fowler W et al.,1995;Kjartansdottir A et al.,1996;WO96/01900)、ヒト以外のポリクローナル血清(US5306635;WO92/06989)など、を使用して説明された。VZVに対するモノクローナル抗体は、単離され、そして、ヒト起源(WO86/02092;WO91/16448;WO95/04080;EP148644;Nemeckova S et al.,1996;Foung S et al.,1985;Sugano T et al.,1987;Yokoyama T et al.,2001;Ito M et al.,1993)、又はマウス起源(EP321249;Vafai A and Yang W,1991;Montalvo E and Grose C 1986;Forghani B et al.,1994;Grose C et al.,1983;Lloyd−Evans P and Gilmour J,2000;Shankar V et al.,2005;Garcia−Valcarcel M et al.,1997)のトリオーマ又はハイブリドーマを使用して発現された。組み換え、VZV結合、及び/又は中和抗体フラグメントが、ファージ・ディスプレイ・ライブラリー中で同定され、そして、一本鎖可変フラグメント(Kausmally L et al.,2004;Drew P et al.,2001)又はFabs(Suzuki K et al.,2007;Williamson R et al.,1993)として発現された。抗VZVマウス・モノクローナル抗体は、ヒト化された(WO 95/31546)。
【0019】
ワクチンと全身的抗ウイルス剤の両方に大幅な改善がもたらされたにもかかわらず、疾患は存続している。療法は、予測不可能なパターンで発症するVZV合併症の多くを軽減するが、それを解消するわけではない。今もなお、集団におけるVZVの感染及び増殖をより効率的に検出し、そして、阻むことができる新規抗体及び抗体フラグメントの同定及び製造は、この感染症の療法、及び/又は予防のための改善された治療を確立するのに特に重要なことである。
【発明の概要】
【課題を解決するための手段】
【0020】
本発明は、VZVに結合し、中和する新規抗体配列、並びにVZV感染又はVZV関連疾患を検出する、治療する、抑制する、予防する、及び/又は改善するために使用できる新規抗体配列を提供する。
【0021】
ヒト抗体配列のパネルを組み換えファージ上に提示させ、そして、VZV特異的結合活性をファージ・ライブラリーにおいて検出した。2つの抗体フラグメントの重鎖及び軽鎖可変領域をコードし、且つ、VZV中和活性を有するDNA配列を同定し、DDF−VZV1とDDF−VZV2と命名した。対応するタンパク質配列、及びVZV特異的な生物学的活性に関与する相補性決定領域(CDRs)を決定した。
【0022】
本発明の配列は、組み換えタンパク質を製造するのに適当な技術を使用した、完全な抗体、抗体フラグメント、又はいずれか他の形式の機能タンパク質(具体的には融合タンパク質)の形態で、VZV特異的結合及び中和特性を有する組み換えタンパク質を製造するのに使用できる。
【0023】
VZV感染及びVZV関連疾患の管理において治療上、予防上、及び/又は診断上の有用性を有する組成物が、本発明のタンパク質を使用することで調製できる。このような組成物は、抗ウイルス化合物、及び/又は免疫グロブリン静注(IVIg)調製物に基づく現在のVZV治療を補うか、又はそれを置き換えるのに使用できる。
【0024】
単離されたDNA及びタンパク質配列、ベクター、組み換えファージ、及び宿主細胞、並びに医学方法や用途を含めた本発明のさらなる態様が、以下の説明の中で提供される。
【図面の簡単な説明】
【0025】
【図1】DDF−VZV1が(VZV−1)、DDF−VZV2(VZV−2)、又は陰性対照として使用された無関係なヒトFab(e−137)を発現する組み換えファージの調製物に対するVZV結合活性の特異性。結合活性は、(MRC−5細胞について)総タンパク質量抽出物、又は(ウシ血清アルブミン、BSAについて)精製タンパク質の形態でプレート・コーティングに使用される表示抗原(the indicated antigens)を使用したELISAにより計測された。
【図2】(A)DDF−VZV1の重鎖の可変領域のDNA(小文字)配列とタンパク質(大文字)配列の配列比較(DDF−VZV1 VH;配列番号1及び2)。予測されるCDRs(HCDR1、HCDR2、及びHCDR3;配列番号3、4、及び5)には下線が引かれている。(B)DDF−VZV1の軽鎖の可変領域のDNA(小文字)配列とタンパク質(大文字)配列の配列比較(DDF−VZV1 VL;配列番号6及び7)。予測されるCDRs(LCDR1、LCDR2、及びLCDR3)には下線が引かれている。
【図3】(A)DDF−VZV2の重鎖の可変領域のDNA(小文字)配列とタンパク質(大文字)配列の配列比較(DDF−VZV2 VH;配列番号8及び9)。予測されるCDRs(HCDR1、HCDR2、HCDR3;配列番号10、11、及び12)には下線が引かれている。(B)DDF−VZV2の軽鎖の可変領域のDNA(小文字)配列とタンパク質(大文字)配列の配列比較(DDF−VZV2 VL;配列番号13及び14)。予測されるCDRs(LCDR1、LCDR2、LCDR3)には下線が引かれている。
【図4】DDF−VZV1(A)及びDDF−VZV2(B)で染色した、VZV感染MRC−5細胞の免疫蛍光。細胞膜が白線で示されており、そして、核膜が白色点線で示されている。未感染細胞においてこれらのFabsを使用することで染色を得られなかった。
【図5】ファージ・コスト・タンパク質上に提示された、ある程度精製されたヒト組み換え型Fabsとして発現された場合のDDF−VZV1(A)及びDDF−VZV2(B)のVZV中和活性。プラーク減少の用量‐応答分析は、無関係なヒトFab(e137)の対応する濃度と平行して実施された。パーセンテージ値は、全くFabを含まずにプレインキュベートされたVZVを使用することで得られたデータと比較することによって計算された。
【図6】(A)対応する逆方向プライマー(pDD−FLAGhis逆方向:配列番号16)と対形成させ、SpeIとNheIで消化し、そして、cp3*配列(WO 07/007154)を置換するためにpDDベクター内にクローンニングした後に、このオリゴヌクレオチドによって連続してコードされるタンパク質タグ(FLAGHisタグ;配列番号17)の目印を伴ったFLAGhisタグ(pDD−FLAGhis順方向:配列番号15)のための順方向リンカー。(B)pDLacI−FLAGhisベクター内にクローンニングされた抗体フラグメントの重鎖(HC)及び軽鎖(LC)を含むDNAフラグメントの概要地図。HC配列は、PelBシグナル配列(PelB)とFLAGhisタグ配列(FLAGhis)の間のフレーム内にクローンニングされている。LC配列はPelBシグナル配列(PelB)と共にフレーム内にクローンニングされ、そして、その発現はLacZプロモーター(LacZ)によって駆動される。2つの発現ユニットの間で、マーカー遺伝子(ゼオシン(Zeocin);Zeo遺伝子)とLacZプロモーターを制御する遺伝子(Lac I遺伝子)は、それら自身のプロモーターと共にクローンニングされる。cp8*(cp8*)をコードする配列は、機能的プロモーターと開始点の不存在下では、転写及び翻訳されない。関連する制限部位、具体的には、HC(SpeIとXhoI)、LC(XbaIとSacI)、及びLacI遺伝子(StuI)をクローニングするために使用されるものを示す。pDLacシステムを使用してDDF−VZV1を発現するための対応するNheI−BglIフラグメントを、配列番号18として提供する。(C)pDLac−VZV1−FLAGhis精製を使用して発現されたDDF−VZV1−FLAGhisのクマシー染色。この図面は、アフィニティー・クロマトグラフィー・カラムから溶出された5つの連続した画分を示している。画分は、一緒に貯留され、濃縮され、そして、さらなる使用のために−20℃にて保存された。
【図7】(A)pDLac−VZV1−FLAGhis(DDF−VZV1 CHTag;配列番号20)を使用することで発現されるヒトFab DDF−VZV1の重鎖のタンパク質配列。対応するDNA配列は、配列番号19として提供される。pDDベクターを使用することで元々クローンニングされたこの重鎖の可変領域(配列番号2)には下線が引かれている。PelB配列は、第1アミノ酸と第26アミノ酸の間に包含されている。第155〜260アミノ酸は、ヒトIgガンマ−1鎖C領域の第1〜106アミノ酸に相当する(SwissProt Ace.No.:P01857)。第262〜275アミノ酸は、FLAGhis配列に相当する。(B)pDLac−VZV1−FLAGhis(DDF−VZV1 CL;配列番号22)を使用して発現されるヒトFab DDF−VZV1の軽鎖のタンパク質配列。対応するDNA配列は、配列番号19として提供される。pDDベクターを使用することで元々クローンニングされたこの軽鎖の可変領域(配列番号7)には下線が引かれている。PelB配列は、第1アミノ酸と第22アミノ酸の間に包含されている。第129〜234アミノ酸は、Igカッパ鎖C領域の第1〜106アミノ酸に相当する(SwissProt Ace.No.:P01834)。(C)pDLac−VZV2−FLAGhis(DDF−VZV2 CHTag;配列番号24)を使用して発現されるヒトFab DDF−VZV2の重鎖のタンパク質配列。対応するDNA配列は、配列番号23として提供される。pDDベクターを使用することで元々クローンニングされたこの重鎖の可変領域(配列番号9)には下線が引かれている。PelB配列は、第1アミノ酸と第26アミノ酸の間に包含されている。第152〜257アミノ酸は、ヒトIgガンマ−1鎖C領域の第1〜106アミノ酸に相当する(SwissProt Ace.No.:P01857)。第259〜272アミノ酸は、FLAGhis配列に相当している。(D)pDLac−VZV2−FLAGhis(DDF−VZV2 CL;配列番号26)を使用することで発現されるヒトFab DDF−VZV2の軽鎖のタンパク質配列。対応するDNA配列は、配列番号25として提供される。pDDベクターを使用することで元々クローンニングされたこの軽鎖の可変領域(配列番号14)には下線が引かれている。PelB配列は、第1アミノ酸と第22アミノ酸の間に包含されている。第135〜240アミノ酸は、Igカッパ鎖C領域の第1〜106アミノ酸に相当する(SwissProt Ace.No.:P01834)。
【図8】pDLacシステムを使用することで発現され、そして、精製されるヒトFabsのDDF−VZV1及びDDF−VZV2のVZV中和活性。(A)各Fabは示した濃度にて使用され、そして、プラーク減少が計算された。(B)各Fabは、単独、又は等量の各Fabを含んで成るFab調製物のいずれかで、示した濃度にて使用された。プラーク減少の用量‐応答分析が、同じ系(el37)を使用することで製造された無関係な対応する濃度のヒトFabと平行して実施された。パーセンテージ値は、全くFabを含まずにプレインキュベートされたVZVを使用することで得られたデータ(陰性対照)と比較することによって計算された。
【発明を実施するための形態】
【0026】
発明の詳細な説明
pDDファージミド及びWO07/007154に記載の関連方法は、2つの所定のファージ・コート・タンパク質の一方又はもう一方のいずれかに融合されたタンパク質配列のクローニング、発現、及び選別を可能にする。このアプローチは、表面上に又は組み換えファージにより示差的に発現又は提示されることができ、その結果、ファージ・ディスプレイ・ライブラリーと異なった効率で選別され得るタンパク質配列識別の選択を可能にする。
【0027】
この場合、ファージ・ライブラリーは、ヒト重鎖及び軽鎖免疫グロブリンの可変領域をpDDファージミド内にクローンニングすることによって構築された。ライブラリーは、VZVタンパク質抽出物に対してパンニングしてより分けられ、それに続いて、選択されたクローンが、実施例に示されているように、VZV中和活性があるものを決定するために細胞ベースのアッセイ方法で試験された。
【0028】
DDF−VZV1及びDDF−VZV2と名付けた、2つの最も有望なクローンをコードするDNA配列が決定され、次に、細菌発現のために適切なベクター内にクローンニングされた。これらのFabsのVZV中和活性は、VZV感染の試験管内モデルを使用して、細菌細胞培養物から精製した組み換えファージの表面上の融合タンパク質としても、アフィニティー精製した組み換えヒトFabsとしても試験された。
【0029】
本発明は、VZVに結合し、中和できる新規タンパク質配列を提供し、それには、FabsのDDF−VZV1又はDDF−VZV2により識別される特異的なCDR(相補性決定領域)が含まれる。具体的には、本発明の各HCDR3s(重鎖可変領域のCDR3)(配列番号5及び配列番号12)は、それぞれ、DDF−VZV1及びDDF−VZV2の抗原結合部分を特徴とする。
【0030】
HCDR3の抗体は、抗原に結合し、その結果、生物学的活性(例えば、実施例に示されているように、VZVに結合し、そして、中和する)を発揮することができる、そのような抗体の抗原結合部分を特徴とすると考えられる。抗体の数個又はすべてのCDRが、完全な抗原結合表面を得るのに通常必要とされるにもかかわらず、HCDR3は、配列に関してだけではなく、長さに関しても抗体の間で最も高い相違を示すCDRである。実際、HCDR3の配列と長さの多様性は、大部分の抗体に対する特異性を測定するための基本となる(Xu J and Davies M,2000;Barrios Y et al.2004;Bond Get al.,2003)。
【0031】
前記HCDR3が同定されたのと同じFabからの他のCDRと組み合わせて又は組み合わせずに、VZV結合部分として本発明の特異的なHCDR3を含むタンパク質が、抗体タンパク質フレームワーク内に作り出される(Knappik A et al.,2000)。CDRの組み合わせは、抗体構造と無関係なタンパク質フレームワーク内であっても、元の結合活性を乱すことなく、元の結合特性を持ち続ける非常に短いタンパク質の状態に互いに連結され得る(Ladner R,2007;Kiss C et al.,2006)。
【0032】
1つの態様において、本発明は、Fab DDF−VZV1のHCDR3と少なくとも90%の同一性を有する配列を含んで成るタンパク質を提供する。HCDR1及びHCDR2(配列番号3及び配列番号4;図2A)と共に、このHCDR3は、DDF−VZV1 Fabの重鎖の可変領域(DDF−VZV1 VH;配列番号2)内に含まれる。このFabを形成する軽鎖の可変領域(DDF−VZV1 VL:配列番号7)、そのうえ、その特異的なLCDRs(軽鎖可変領域のCDRs)が決定された(図2B)。
【0033】
他の態様において、本発明は、Fab DDF−VZV2のHCDR3と少なくとも90%の同一性を有する配列を含んで成るタンパク質を提供する。HCDR1及びHCDR2(配列番号10及び配列番号11;図3A)と共に、このHCDR3は、DDF−VZV2の重鎖の可変領域(DDF−VZV2 VH;配列番号9)内に含まれる。このFab(DDF−VZV1 VL;配列番号14)を形成する軽鎖の可変領域、そのうえ、その特異的なLCDRsが決定された(Fig.3B)。
【0034】
本発明のタンパク質がDDF−VZV1の配列に基づいているなら、それらは、配列番号5に対して少なくとも90%の同一性を有する配列を含んで成るべきである。特に、それらは、配列番号2に対して少なくとも90%の同一性を有する配列を含んで成るべきである。より特に、前記タンパク質はまた、配列3及び配列番号4から成る群から選択される1つ以上の配列も含むべきである。
【0035】
あるいは、本発明のタンパク質が、DDF−VZV2の配列に基づいているなら、それらは、配列番号12に対して少なくとも90%の同一性を有する配列を含んで成るべきである。特に、それらは、配列番号9に対して少なくとも90%の同一性を有する配列を含んで成るべきである。より特に、前記タンパク質はまた、配列番号10及び配列番号11から成る群から選択される1つ以上の配列も含むべきである。
【0036】
本発明のさらなる態様は、両Fabsの重鎖及び軽鎖の可変領域をコードするDNA配列であり、具体的には、DDF−VZV1(重鎖については配列番号1;軽鎖については配列番号6)及びDDF−VZV2(重鎖については配列番号8;軽鎖については配列番号13)の可変領域についてクローンニングされ、そして、決定された本来のDNA配列と少なくとも90%の同一性を有するものである。これらのDNA配列(又は、図2及び3から容易に決定され得る単離されたHCDRs及びLCDRsをコードするものなどの選択された部分)は、それらがVZV結合及び中和特性をもたらす組み換え抗体(例えば、完全抗体、親和性成熟抗体、CDRグラフト抗体、又はフラグメント)又は融合タンパク質の公知の形式の1つでそれらを発現するために他のベクター内に移されてもよい。
【0037】
同一性のレベルがどこに示されるとしても、この同一性のレベルは、本発明の関連配列の完全長で測定されるべきである。
【0038】
DDF−VZV1又はDDF−VZV2(又は単離されたHCDRsやLCDRsなどの選択された部分)のいずれかを形成する重鎖及び軽鎖の可変領域は、特定のアイソタイプを持つ抗体の中に、特に、完全なヒト組み換え抗体の中に包含されることができる。この抗体は、2本の軽鎖及び2本の重鎖によって形成された四量複合体の天然の立体構造で、軽鎖及び重鎖可変領域としてDDF−VZV1又はDDF−VZV2のいずれかのVL及びVH配列を含んで成ることができる。完全なヒト抗体が望ましいとき、抗体は、ヒトIgG1、IgG2、IgG3、IgG4、IgM、IgA、及びIgE定常領域から成る群から選択される重鎖定常領域をさらに含んで成るべきである。例えば、IgGイソタイプは、ほとんどすべての承認された治療抗体の抗体形式である(Laffly E and Sodoyer R,2005)。しかしながら、ヒトIgG1から単離された抗原結合部分は、ヒトIgA配列に移されることができ、そして、得られた組み換え抗体が、HIV感染を抑制することができる抗体で最近示されたように、本来のIgG1の活性を維持した(Mantis N et al.,2007)。
【0039】
あるいは、DDF−VZV1又はDDF−VZV2(又は単離されたHCDRsやLCDRsなどの選択された部分)のいずれかを形成する重鎖及び軽鎖の可変領域は、Scfv(一本鎖可変フラグメント)、Fab(可変重鎖/軽鎖ヘテロ二量体)、二重特異性抗体、ペプタボディ(peptabody)、VHH(重鎖抗体の可変ドメイン)、単離された重鎖若しくは軽鎖、二重特異性抗体、及び非臨床/臨床適用のためのその他の遺伝子操作抗体変異体(Jain M et al.,2007;Laffly E and Sodoyer R,2005)などの異なった名称で文献に記載される機能的抗体フラグメントに関するあらゆる他のタンパク質形式に含まれるかもしれない。DDF−VZV1又はDDF−VZV2の組み換え変異体は、タグ付与したFabsの形態でpDLac−FLAGhisと呼ばれるpDD適合発現ベクターを使用することで製造された(図6Bは関連DNAフラグメントの概要地図に関し;配列番号18はDDF−VZV1を製造するために作り出されたこのようなフラグメントのDNA配列の例に関する)。DDF−VZV1及びDDF−VZV2のタグ付与バージョンを含んでいるpDLac−FLAGhisベースのベクターが作り出され、そして、DDF−VZV1 HCtagを製造するためにDDF−VZV1 LC(配列番号19〜22、図7A及びB)と共に、そして、DDF−VZV2 HCtagを製造するためにDDF−VZV2 LC(配列番号23〜26;図7C及びD)と共に使用された。
【0040】
追加抗体及び抗体フラグメントは、シャフリング軽鎖の過程を通してDDF−VZV1又はDDF−VZV2のいずれかの配列を使用することで作り出される。実際に、いくつかの異なった抗体が作り出され、そして、例えば、一般的なファージ・ディスプレイ技術、又はWO 07/007154に記載の技術を使用して、VL配列に関するライブラリーと組み合わせた1本重鎖可変ドメインVH(DDF−VZV1又はDDF−VZV2のいずれか一方など)を使用することで特異的な生物学的活性について試験され得る。このアプローチは、親和性、安定性、特異性、及び/又は組み換え体の製造に関して改善された特性を有するVH/VL組み合わせの決定を可能できる(Ohlin M et al.,1996;Rojas G et al.,2004;Suzuki K et al,2007)。
【0041】
そのうえ、特に(より良い薬物動態学的プロフィール、又は抗原に対するより高い親和性などの)臨床適用に関して、改善された特徴を有する抗体にするために特定の位置で抗体を修飾できることが知られている。これらの変更は、CDRs、及び/又はDDF−VZV1又はDDF−VZV2のいずれかのフレームワークに加えられる。配列は、親和性成熟及びその他の方法を利用して行われる抗体の合理的設計のための専用技術のいずれかを適用することによって決定できる(Kim S et al.,2005;Jain M et al.,2007)。
【0042】
新しい生理活性ペプチドを開発するための抗体ベースのストラテジーはまた、L−アミノ酸、及び/又はD−アミノ酸を含むCDR誘導ペプチド合成の実現可能性を示した。これらの分子は、より適切な薬理学的特性を伴った、本来の活性の比活性を維持する(Levi M et al.,2000;Wijkhuisen A et al.,2003)。そのため、本発明のHCDR3のそれぞれ、並びにそれらと高度に類似した配列、それらを含む融合タンパク質、そして、(例えば、D−アミノ酸を含むか、又はレトロ‐インベルソ型立体構造にある)それらに由来する合成ペプチドが、VZV結合タンパク質として使用及び試験され得る。
【0043】
本発明のタンパク質は、VZVに結合し、そして、中和する、抗体、抗体フラグメント、生理活性ペプチド、又は融合タンパク質として提供される。これらの代替タンパク質は、DDF−VZV1及びDDF−VZV2 Fabsについて測定されるそのような特性を強化しないにしても、維持するべきである。融合タンパク質の場合には、異種タンパク質配列は、その正しい発現と生物学的活性に影響することなく、VZV特異的部分(例えば、抗体フラグメントの特異的HCDR3又は可変領域)に対してN又はC末端位置に位置し得る。
【0044】
用語「異種タンパク質」は、VZV特異的部分(例えば、抗体フラグメント)に対してN又はC末端位置に、タンパク質配列が天然には存在していないことを示す。このタンパク質配列をコードするDNA配列は、一般的に組み換えDNA技術によって融合され、そして、少なくとも5つのアミノ酸をコードする配列を含んで成る。存在しているこの異種配列は、一般的にVZV特異的融合タンパク質に追加特性を与えるために選ばれている。このような追加特性の例には、検出若しくは精製のためのより優れた手段、追加の結合部分若しくは生物学的リガンド、又は融合タンパク質の翻訳後修飾(例えば、リン酸化、グリコシル化、ユビキチン化、SUMO化、若しくはタンパク質内部分解性開裂)が含まれる。
【0045】
あるいは(又は1つ以上の異種タンパク質配列への融合に加えて)、本発明のタンパク質の活性は、治療薬、安定化剤、又は診断薬などの異なった化合物との結合によって改善することができる。これらの作用物質の例は、化学リンカー又は重合体を使用して結合され得る検出可能な標識(例えば、放射性同位元素、蛍光化合物、毒素、金属原子、コロイド状金属、化学発光化合物、生物発光化合物、又は酵素)である。本発明のタンパク質のVZV特異的生物学的活性は、診断上の又は治療上の適用において代謝、及び/又は安定性を変える重合体のような化合物との融合によって改善される。治療活性は、別の抗ウイルス・タンパク質、あるいは、細胞の代謝、及び/又は活性を変えるタンパク質などの別の治療用タンパク質との融合によって改善される。
【0046】
タンパク質部分、リガンド、及び適切なリンカーの選択及び設計方法、並びに融合タンパク質の構築、精製、検出、及び使用のための方法及びストラテジーは、文献(ニルソンら、1997;「キメラ遺伝子と雑種タンパク質の適用」Methods Enzymol Vol.326−328、アカデミックプレス社、2000;WO01/77137)内に提供されており、且つ、臨床検査室及び研究室で一般的に利用可能である。例えば、融合タンパク質は、融合タンパク質の生体内、及び/又は試験管内における識別、又はその精製を容易にできる(ポリヒスチジン、FLAG、c−Myc、若しくはHAタグなどのタグを含めた)商業的な抗体によって認識される配列を含む。他のタンパク質配列は、(緑色蛍光タンパク質の場合のように)直接蛍光分析によって、又は(例えば、タンパク分解部位を使用した)特異的基質又は酵素によって容易に識別できる。
【0047】
VZV特異的抗体、抗体フラグメント、及び融合タンパク質の安定性は、ファージ・コート・タンパク質(単離されたか若しくは組み換えファージ内に含まれているcp3又はcp8)、マルトース結合タンパク質(MBP)、ウシ血清アルブミン(BSA)、又はグルタチオン−S−トランスフェラーゼ(GST)などの周知の担体タンパク質との融合により改善される。
【0048】
本発明のタンパク質はまた、VZVエンベロープに対する中和抗原を特徴づけするためにも使用できる。実際、DDF−VZV1及びDDF−VZV2は、最初、ELISAによる、VZV感染細胞株から得られた細胞抽出物へのそれらの特異的結合のためにクローンニングされ(図1)、そして、VZV感染を中和するそれらの能力はまた、VZV基準菌株を使用した試験管内における中和アッセイによって測定された(図5及び8)。その結果、本発明のタンパク質は、DDF−VZV1又はDDF−VZV2と競合する(例えば、完全な抗体、Fabs及び他の抗体フラグメント、生理活性ペプチド、又は融合タンパク質の形態の)他のVZV結合タンパク質を規定するのに使用できる。このような競争タンパク質は、任意に、本来のDDF−VZV1又はDDF−VZV2配列において確認されたものと一部又は完全に異なったHCDRs及びLCDRsと共に、先に規定されたHCDR3配列のいずれかを単に含む。
【0049】
(本発明の抗体、抗体フラグメント、生理活性ペプチド、又は融合タンパク質に関して)そのような競合タンパク質は、実施例又は文献に記載の関連アッセイによって測定される、本発明のタンパク質と競合するそれらの能力、それから、VZV感染を中和するそれらの能力を実証することによってスクリーニングされ、そして、単離され得る。本発明の背景は、類似活性を測定するための異なるアプローチに関するいくつかの基準を提供する。具体的には、本発明の抗体、抗体フラグメント、及び他のタンパク質は、抗体フラグメント(Kausmally L et al.,2004;Drew P et al.,2001;Suzuki K et al.,2007)、マウス若しくはヒト・モノクローナル抗体(Grose C et al.,1983;Montalvo E and Grose C,1986;Sugano T et al.,1987;Forghani B et al.,1994;Lloyd−Evans P and Gilmour J,2000)、並びにヒト以外/ヒト血清(Fowler W et al.,1995;Haumont M et al.,1997;Garcia−Valcarcel M et al.,1997)についてVZV特異的生物学的活性及びエピトープを特徴づけるために使用されるアッセイにより試験され得る。
【0050】
免疫蛍光研究(図4)は、他の抗体及び抗体フラグメントについてこれまでに示されたように、DDF−VZV1とDDF−VZV2が異なったVZV抗原を認識することを示した(Suzuki K et al.,2007;Wu L and Forghani B,1997;Grose C et al.,1983)。文献は、VZV抗原と各Fabによって認識される特異的エピトープが測定でき、且つ、過去に測定したものと比較できることを使用した技術に関するいくつかの例を提供する。例えば、VZVタンパク質、及び関連する切断変異体若しくは合成ペプチドを使用したELISA、免疫沈降反応、又はウエスタン・ブロットを、使用して、関連エピトープを同定した(Krah D,1996,Sauerbrei A and Wutzler P,2006)。具体的には、前記エピトープは、糖タンパク質E又は糖タンパク質B(Fowler W et al.,1995;Haumont M et al.,1997;Garcia−Valcarcel M et al.,1997;Kjartansdottir A et al.,1996)、及び糖タンパク質L:糖タンパク質H複合体(Forghani B et al.,1994;Yokoyama T et al.,2001;Suzuki K et al.,2007)内で同定された。
【0051】
本発明のタンパク質のVZV関連の予防上、診断上、及び治療学上の使用についてのより詳しい特徴づけとバリデーションは、その次に、本発明の背景にまとめられているとおり、VZV発症機序と免疫生物学を研究するための文献に開示されている試験管内又は生体内におけるアッセイ(組織若しくは細胞ベースのアッセイ、齧歯動物において確立された病態モデル)の1つ以上を使用することで実施できる(Forghani B et al.,1994: Vafai A et al.,1991;Wu and Forghani,1997;Fowler W et al,,1995;Maple P et al.,2006;Matsuo K et al.,2003;Kutinova L et al.,2001;Massaer M et al.,1999;Haumont M et al.,1997;Grose C,2006;Baiker 2004;Ku C et al.,2005;Taylor S and Moffat L,2005;Zerboni L et al.,2005)。
【0052】
本発明のさらなる目的は、先に規定された抗体、抗体フラグメント、融合タンパク質、又は単離CDRのいずれかをコードする核酸である。実施例は、具体的には、DDF−VZV1又はDDF2−VZV2重鎖及び軽鎖の完全な可変領域をコードする配列を提供する(配列番号1、6、8、及び13)。前記核酸は、配列番号1、配列番号6、配列番号8、及び/又は配列番号13と少なくとも90%の同一性を有していなければならない。前記配列、具体的には、特異的なCDRs(図2と3を参照のこと)に関連するそれらの中の配列は、例えば、発現ベクター内のプロモーターに作動できるように連結されるか、若しくはpDDベースのファージミド内にクローンニングされて、ベクター及びDNA発現カセット内に、並びにいずれか他のファージミド内に含まれ得る。これにより、(実施例に示されているように)本発明のタンパク質を発現するファージミド・ベクターを含んで成る組み換えファージは、VZV感染の検出手段、及び/又は中和手段として使用され得る。
【0053】
完全ヒト抗体が望ましい場合には、発現ベクターは、IgG1、IgG2、IgG3、IgG4、IgM、IgA、及びIgE定常領域から成る群から選択される重鎖の定常領域をさらに含んで成るはずである。着目の重鎖及び軽鎖の関連可変領域をコードする核酸配列は、それらが適切な調節配列(例えば、プロモーター、転写ターミネータ)に作動できるように連結される、1つのベクター又は異なったベクターの発現カセット内に適切にクローンニングされるべきである。発現カセットは、所望のタンパク質のための単シストロン性若しくは2シストロン性の転写産物の発現を駆動できるプロモーター、(必要であれば)リボソーム結合部位、開始/終止コドン、及びリーダー/分泌配列を含むはずである。
【0054】
VZVに結合し、そして、中和できると先に規定された本発明の抗体、抗体フラグメント、HCDRs、融合タンパク質、及びいずれか他の化合物は、適切な宿主細胞を形質転換するためのそのようなベクター、及び組み換えタンパク質としてのそれらの発現を可能にする十分に確立された技術を使用して製造され得る。これらの調製物は、VZV関連の予防上、診断上、及び治療上の使用に関するより詳細な特徴づけとバリデーションを実施するのに十分な量の組み換えタンパク質(マイクログラムからミリグラムの範囲)を提供するべきである。
【0055】
本発明の配列がクローンニングされ、そして、対応する組み換えファージの手段によって特徴づけされたpDDベースのファージミドは、本来のFabs、又はそれらから得られたタンパク質配列が(pDLac−FLAGhis系を用いて実施例で示されているように)宿主細胞内で組み換えタンパク質として適切に発現されるベクター内に(一部又は全体として)移され得るDNA配列を含む。ベクターは、構成的に活性であるか又は誘導性であるように選択された転写開始/終止調節配列の制御下、原核生物又は真核生物宿主細胞において組み換えタンパク質の発現を可能にするべきである。
【0056】
本発明の核酸を含んで成る宿主細胞は、原核生物又は真核生物宿主細胞であり、所望の組み換えタンパク質の分泌を可能にすべきである。そして、このような細胞が実質的に豊富になった細胞株は、単離されて、安定細胞株をもたらす。前記のタンパク質の製造方法には、タンパク質の発現に好適な条件下、それらのコード配列を含んで成る発現ベクターで形質転換された宿主細胞を培養し、そして、その宿主細胞培養物からタンパク質を回収するステップが含まれる。
【0057】
これらの核酸、組み換えファージ、及び宿主細胞は、一般的な組み換えDNA技術を適用することによって本発明のタンパク質を製造するのに使用できる。簡単に言えば、所望のDNA配列は、制限酵素でファージミドを消化することによって、又はポリメラーゼ連鎖反応(PCR法)の鋳型として本来のファージミドを使用し、及び重鎖及び軽鎖の完全な可変領域若しくは(HCDR3などの)それらの一部分だけを特異的に増幅するためのPCRプライマーを使用して増幅することによって抽出できる。
【0058】
次に、前記DNAフラグメントは、オックスフォード大学出版より刊行された「A Practical Approach」というシリーズの中のいくつかのタイトル(“DNA Cloning 2: Expression Systems”,1995;“DNA Cloning 4: Mammalian Systems”,1996;“Protein Expression”,1999;“Protein Purification Techniques”,2001)を含めた、組み換えタンパク質をどのようにクローンニングし、そして、製造するかに関する多くの書籍と総説に記載のとおり、原核生物又は真核生物宿主細胞内でのさらなる一時変異、及び/又は発現のためにより適切なベクターに移されてもよい。
【0059】
真核生物宿主(例えば、酵母、昆虫、又は哺乳動物細胞)に関して、宿主の性質によって、異なった転写及び翻訳調節配列を用いることができる。それらは、調節シグナルが、発現レベルが高い特定の遺伝子に関連するアデノウイルス、ウシ乳頭腫ウイルス、シミアンウイルス、又は同様のものなどのウイルス由来である。例は、ヘルペスウイルスのTKプロモーター、SV40初期プロモーター、酵母gal4遺伝子プロモーターなどである。一過性の(構成的)の抑制及び活性化を可能にし、その結果、修飾遺伝子発現を可能にする転写開始調節シグナルが選択できる。さらなるクローニング・ステップの間、抗体又は融合タンパク質をコードする配列は、例えば、特定のベクター及び宿主細胞を使用して組み換えタンパク質をクローンニングし、そして、発現するためのコドン使用及び制限部位が最も適当であるDNA配列を選択するためのソフトウェアを使用することで、決定され得るDNAレベルにおいてのみ、又はDNA及びタンパク質レベルの両方における特異的修飾のために、他のベクター内に適合させ、そして、再クローニングし得る(Rodi D et al,,2002;Grote A et al.,2005;Carton J et al.,2007)。タンパク質配列もまた、(Scfv、Fab、完全ヒト抗体などの)所望の抗体形式、又は1つ以上の内部アミノ酸の挿入、置換、又は除去と関連して追加され得る。
【0060】
これらの技術もまた、抗体の治療学的特性のさらなる構造的及び機能的な特徴づけ及び最適化のために(Kim S et al.,2005)、あるいは、それらの安定した生体内デリバリーを可能にするベクターを製造するために使用され得る(Fang J et al.,2005)。例えば、組み換え抗体もまた、可変領域に融合される特異的なFc領域を選択することによって(Furebring C et al.,2002;Logtenberg T,2007)、組み換え一本鎖抗体フラグメントを製造することによって(Gilliland L et al.,1996)、安定化ペプチド配列を融合することによって(WO01/49713)、又は化学的に修飾した残基に放射性化学物質又は重合体を追加することによって(Chapman A et al.,1999)、構造、及び/又は活性レベルで修飾され得る。
【0061】
いったん好適なエピソマー、又は非相同若しくは相同の組み込み型ベクター(integrating vector)内に挿入された、ディスプレイ及び選択されたタンパク質配列をコードするDNA配列は、(形質転換、トランスフェクション、接合、プロトプラスト融合、エレクトロポレーション、リン酸カルシウム沈殿、ダイレクト・マイクロインジェクションなどの)好適な手段のいずれかによって適切な宿主細胞内に導入されて、それらを形質転換することができる。特定のプラスミド又はウイルス・ベクターを選択する場合に考慮される重要な要因は:ベクターを含まない宿主細胞の中からベクターを含む宿主細胞を認識し、そして、選択することの容易さ;特定の宿主体内で所望されるベクターのコピー数;そして、異なる種の宿主細胞間のベクターである「シャトル」にそれが望ましいかどうかである。
【0062】
導入されたDNAによって安定に形質転換された細胞は、発現ベクターを含む宿主細胞の選別を可能にする1つ以上のマーカーを導入することによっても選択され得る。前記マーカーはまた、栄養要求性宿主に対して光合成栄養、殺生物剤抵抗性、例えば、抗生物質、若しくは銅などの重金属、又は同様のもの、に対する抵抗性も与え、そして、必要性であれば、切断可能であるか又は抑制される。選択マーカー遺伝子は、発現されるべきDNA遺伝子配列に直接連結され得るか、又は同時トランスフェクションによって同じ細胞内に挿入され得る。追加の転写制御要素もまた、最適な発現に必要である。
【0063】
宿主細胞は、原核生物又は真核生物のいずれかである。原核生物宿主細胞の中で、好ましいものは、B.ズブチリス(B.subtilis)とE.コリ(E.coli)である。真核生物宿主細胞の中で、好ましいものは、酵母、(バキュロウイルス・ベースの発現系を使用した)昆虫細胞、又は哺乳動物細胞、例えば、ヒト、サル、マウス、(バキュロウイルス・ベースの発現系を使用した)昆虫、及びチャイニーズハムスター卵巣(CHO)細胞などである。なぜならば、哺乳動物細胞が、正しい折りたたみ、又は正しい部位でのグリコシル化の特定の形態を含めた、タンパク質分子に対する翻訳後修飾を提供するからである。また、酵母細胞もグリコシル化を含めた翻訳後ペプチド修飾を行う。酵母において所望のタンパク質の製造に利用できる強力なプロモーター配列と高いプラスミド・コピー数を利用する、多くの組み換えDNAストラテジーが存在する。酵母は、クローンニングされた哺乳動物遺伝子産物、及びリーダー配列を担持している分泌ペプチド(すなわち、プレ‐ペプチド)の中のリーダー配列を認識する。
【0064】
発現のための宿主として利用可能な哺乳動物細胞株は、当該技術分野で公知であり、そして、それには、これだけに制限されることなく、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、ベビーハムスター腎臓(BHK)細胞、サル腎臓(COS)細胞、C127細胞、3T3細胞、BHK細胞、HEK 293細胞、Per.C6細胞、Bowes黒色腫細胞、及びヒト肝細胞癌(例えば、Hep G2)細胞、及びその他の多くの細胞株を含めたアメリカン・タイプ・カルチャー・コレクション(ATCC)から入手可能な多数の不死化細胞株が含まれる。バキュロウイルス系において、バキュロウイルス/昆虫細胞発現系の材料は、(例えば、Invitrogenによって商品化されている)キット形態で市販されている。
【0065】
長期間にわたる、高収率産生の組み換えポリペプチドの、安定的な発現が好まれる。例えば、着目のポリペプチドを安定して発現する細胞株は、同じか、又は別のベクター上にウイルス複製開始点、及び/又は内因性発現要素、並びに選択マーカー遺伝子を含む発現ベクターを使用することで形質転換される。ベクターの導入に続いて、細胞を、強化培地中で1〜2日間、増殖させ、その後、それらを選択培地に切り換える。選択マーカーの目的は、選択に対する耐性を与えることであり、その存在が導入された配列を首尾よく発現する細胞の増殖と回復を可能にする。安定に形質転換された細胞の耐性クローンは、細胞型に適切な組織培養技術を使用することで増殖する。そして、このような細胞が実質的に豊富になった細胞株は、単離されて、安定な細胞株を提供し得る。宿主細胞は、組み換えタンパク質の発現レベルに基づいてさらに選択される。
【0066】
ファージ・ディスプレイ技術を使用して単離される免疫グロブリン可変鎖(具体的には、ヒト免疫グロブリン可変鎖)の場合、重要な変更は、選択されたFab又はscFVの、好ましいアイソタイプ及び定常領域を持つ完全な免疫グロブリン・タンパク質への転換である。こういう種類の変更は、例えば、ファージ・ディスプレイ・ライブラリー由来の単鎖Fv又はFabsから構築されたすべてのアイソタイプの完全なヒト・モノクローナル抗体を製造し、そして、哺乳動物又は昆虫細胞内で発現することを可能にする。文献に広く記載のとおり(Persic L et al.,1997;Guttieri M et al.,2003)、ベクターは、所望のアイソタイプ(例えば、ヒトIgGガンマ)の定常(Fc)領域に対するこの配列の融合を可能にする発現抗体のために特異的に設計される。抗体又は融合タンパク質は、一過性に又は安定に形質転換された細胞として高レベルの発現を可能にする原核生物細胞(例えば、エシェリキア・コリ(Escherichia coli);Sorensen H and Mortensen K,2005;Venturi M et al.,2002)、植物細胞(Ma J et al.,2005)、又は真核生物細胞において組み換えタンパク質として発現される(Dinnis D and James D,2005)。これは、具体的には、抗体の特徴づけが、より困難なアッセイ、及び/又は生体内アッセイを使用して実施されなければならない場合に必要であるだろう。
【0067】
文献は、記事及び書籍の章で概説されるとおり、原核生物宿主細胞において、Fabとしてタンパク質を、又は抗体フラグメントと同様の形式を発現するための異なったストラテジーを提供する(“Phage display:A Practical Approach”,vol.266,ed.Clackson and Lowman H,Oxford Univ.Press,2004;“Phage Display:A Laboratory Manual”,ed.Burton D et al.,CSHL Press,2001;Corisdeo S and Wang B,2004;Benhar I,2001)。
【0068】
タンパク質、特に抗体が真核生物宿主細胞(具体的には、哺乳動物細胞株)において発現されるとき、異なったベクター及び発現系が、トランスフェクト細胞株の安定したプールを作り出すように設計される(Aldrich T et ah,2003;Bianchi A and McGrew J,2003)。組み換え抗体の高レベルの、最適化された、安定した発現は、細胞培養条件の最適化によっても(Gmnberg J et al.,2003;Yoon S et al.,2004)、そして、より高いレベルの抗体産生及び分泌を有するクローンを選択するか又は設計することによって(Bohm E et al.,2004;Butler M,2005)達成された(Schlatter S et al.,2005)。
【0069】
VZVに結合し、中和できると先に規定した抗体、抗体フラグメント、生理活性ペプチド、融合タンパク質、及びあらゆるその他タンパク質は、細胞培養物又は合成調製物からの非組み換え/組み換えタンパク質のいずれかの分離を可能にする十分に確立された技術、すなわち、抽出、沈殿、クロマトグラフィー、電気泳動、又は同様のものを含めた従来手順のいずれかを使用して精製され得る。抗体精製の方法は、カラム内に含まれる固定されたゲルマトリックス(Nisnevitch M and Firer M,2001;Huse K et al.,2002;Horenstein A et al.,2003)、具体的には、プロテインA若しくはプロテインGなどの基質、又は合成基質に対する抗体の一般的な親和性によるもの(Verdoliva A et al.,2002;Roque A et al.,2004)、並びに抗原又はエピトープ・ベースのアフィニティー・クロマトグラフィ(Murray A et al.,2002;Jensen L et al.,2004)を活用できる。洗浄の後に、タンパク質は、pH又はイオン強度の変化によってゲルから溶出される。あるいは、HPLC(高速液体クロマトグラフィー)が使用できる。溶出は、タンパク質精製のために一般的に用いられる水‐アセトニトリル・ベースの溶媒を使用することで行われ得る。
【0070】
VZVに結合し、中和できると先に規定された抗体、抗体フラグメント、生理活性ペプチド、融合タンパク質、及びあらゆるその他の化合物が、VZV感染を検出、治療、抑制、予防、及び/又は改善するために使用できる。この目的のために、前記化合物が、VZV感染及びVZV関連疾患の医療管理のための診断用、治療用、又は予防用の組成物を調製するのに使用できる。
【0071】
これらの組成物は、ヒトDDF−VZV1及びDDF−VZV2の配列と活性に基づいて先に規定された抗体、抗体フラグメント、融合タンパク質、CDR、及びあらゆるその他の化合物を含んで成る。前記組成物は、異なるVZV中和抗体若しくは抗体フラグメント、免疫グロブリン静注(IVIg)調製物、ステロイド、及び/又は抗ウイルス化合物をさらに含んで成る。異なるVZV中和抗体又は抗体フラグメントは、既に文献に記載されているものなどの異なるエピトープを特徴とするべきである。実際、文献には、ウイルス又はヒト標的に向けられた2種類以上の抗体が医薬組成物中で組み合わせられた場合に、得られた組成物が単なる相加効果ではなく、特定の相乗効果による改善された治療効果を有することができる多くの例が示されている(Logtenberg T,2007)。
【0072】
医薬組成物は、任意に、いずれかの医薬として許容されるビヒクル又は担体を含んで成る。これらの組成物は、ワクチン、免疫調節免疫グロブリン静注製剤、ステロイド、又は抗ウイルス化合物などのあらゆる追加の治療薬又は予防薬をさらに含んで成る(又はそれらと一緒に投与できる)。文献は、VZV複製に作用し、且つ、単独又は免疫グロブリン静注調製物と組み合わせて、ヒトで既に試験されている前記化合物に関するいくつかの例を提供する(Huang Y et al.,2001;Carby M et al.,2007;Koren G et al.,2002)。そのうえ、最近の文献は、前記医薬組成物の頻度、及び/又は投薬量を低減する機会を与える、ヒト・モノクローナル抗体が免疫グロブリン静注調製物、ステロイド、及び/又は抗ウイルス化合物などの現在の治療を補う(可能であれば、置き換える)ために使用され得ることを示唆している(Bayry J et al.,2007)。
【0073】
先に規定されたタンパク質(例えば、抗体、抗体フラグメント、融合タンパク質、生理活性ペプチド)及び核酸のいずれかを含んで成る組成物は、VZVに関連した診断上、治療上、又は予防上の目的がある人に投与され得る。これらの組成物は、VZVビリオンをターゲッティングすることによって、治療された患者のウイルスの増殖を抑制できる、及び集団内でのウイルス感染の大発生を妨げる可能性のある治療用化合物(具体的には、治療抗体若しくは治療抗体フラグメント)を提供するVZV特異的な受動免疫のための手段として投与され得る。
【0074】
特定の使用に依存して、組成物は、より長い又はより短い期間、(VZVに感染しているか、又は入院、免疫抑制薬若しくは化学療法剤治療、又はVZVに感染した人との接触のためVZVの危険性があると考えられるあらゆる幼児、妊婦、高齢者、又はその他の人である)ヒト対象に化合物を提供するべきである。この目的のために、組成物は、噴霧装置、吸入器によって、又は点眼薬として、非生分解性/生分解性基質物質中、あるいは、マイクロビーズなどの粒状薬物デリバリー・システムを使用して、様々な経路:筋肉内、静脈内、皮下、局所、粘膜を通じて、単回又は複数回の投薬量で、及び/又は適切なデバイスを使用して投与され得る。
【0075】
具体的には、組成物は、皮膚及び眼におけるVZVの存在を考慮すると、有用なアプローチである局所的投与及び眼内投与を可能にする(Arvin A,1996;Quinlivan M and Breuer J,2006;Liesegang T,2004)。そのうえ、抗体及び抗体フラグメントは、角膜に対して局所的に適用された場合に有効である(Brereton H et al.,2005;Nwanegbo E et al.,2007)。
【0076】
医薬組成物は、第二次ウイルス血症に関連する皮膚疹におけるウイルスの存在を考慮すると、化合物が、特に局所投与に関して、十分な期間その活性を発揮することを可能にする治療的又は予防的に有効量の化合物を対象に対して提供すべきである。望ましい効果は、VZV感染、再活性化、及び/又は再感染を制御することによって、並びにVZV感染の臨床徴候の少なくともいくつかを低減することによって、患者の体質を改善することである。例えば、前記組成物は、投与経路、投与される投薬量の数、及びその人の体質によって、約0.005〜約50mg/kg/体重の有効量にて投与されるべきである。
【0077】
診断用途のある組成物の場合、化合物は、生物学的サンプル中のウイルスを検出するための臨床検査室及び研究室において一般的に確立された技術(例えば、ELISA又はその他の血清学的なアッセイ)を使用して、又は生体内で対象に投与された場合には、投与後少なくとも1、2、5、10、24時間、又はそれより長い時間、検出されるべきである。VZVの検出は、免疫応答性宿主及び免疫不全宿主の両方から成る危険性のある集団における慢性又は急性VZV感染を観察するために確立された公知の手段及び手順と置換して又は合わせて、本発明のタンパク質を使用して実施され得る。
【0078】
本発明のタンパク質はまた、VZV感染、並びにVZV関連疾患を検出、治療、抑制、予防、及び/又は改善するための組成物の調製にも使用できる。これらの疾患は、VZV感染の合併症から生じる可能性がある(Dworkin R et al.,2007;Weinberg J,2007)。
【0079】
背景技術に示されているとおり、ヘルペス後神経痛の症例のように、劇的、且つ、衰弱させる効果を持つ可能性がある神経学的、眼性、又は臓器を冒す効果を持っている数多くの帯状ヘルペスの帯状ヘルペス合併症が存在する(Oxman M et al.,2006;Liesegang T,2004)。そのうえ、VZVの再活性化及び関連する合併症はまた、癌患者(Sandherr M et al.,2006)又は炎症性膠原病に罹患している患者、そして、通常、コルチコステロイドなどの免疫抑制治療、又は化学療法及びその他の抗体ベースの免疫抑制レジメン下にある患者においても見られた。
【0080】
VZV又はVZV関連疾患の治療、予防、又は診断方法は、先に規定されたタンパク質又は核酸の投与を含んで成る。前記方法は、異なるVZV中和抗体若しくは抗体フラグメント、免疫グロブリン静注(IVIg)調製物、ステロイド、及び/又は抗ウイルス化合物の投与をさらに含んで成る。
【0081】
臨床開発及び使用は、抗体の薬物動態学及び薬力学の特徴づけ(Lobo E et al.,2004)、前臨床的及び臨床的安全性データ(Tabrizi M and Riskos L,2007)、並びに治療学用組み換え抗体の商業的な製造規模の製剤と分析的特徴づけのための公的な要件の順守(Harris R et al.,2004)に基づくべきである。
【0082】
どのような形であっても本発明を制限すると解釈されるべきでない以下の実施例によって、本発明は説明されるだろう。
【実施例】
【0083】
実施例1:ELISAによるヒトFabs結合VZVタンパク質抽出物の発現と選別
材料と方法
ライブラリー構築
ヒトIgG1の重鎖及び軽鎖をコードするcDNAを、文献に従ってVZV血清陽性個体から得られたリンパ球から入手した(Burioni et al.,1998;“Phage Display:A laboratory Manual”,Burton DR et al.,CSHL Press,2001)。ファージ・ライブラリーを、PCT特許出願WO 07/007154に記載の技術に従ってpDDベクターと適合するクローニング・カセットを使用することで構築し、そして、Fabsを、ライブラリーの中の組み換えファージ表面上に発現させた。
【0084】
pDDベースのFabライブラリーのパンニング、そして、陽性クローンの配列決定によるヒトFabsの選別を、文献に記載されたとおり実施した(Burioni R et al.1998)。
【0085】
特異的なFabsのCDRsを、IMGT/V−QUEST(Giudicelli V et al.,2004)によって提供された予測及び配列比較と、ヨーロッパ生物情報科学研究所によって提供されるもの及びFASTA(http://www.ebi.ac.uk/fasta33/index.html)を使用して検索可能なものなどのヒト抗体のタンパク質配列を含む他のデータベースを比較することによって規定した。
【0086】
培養ヒト細胞からのタンパク質抽出物の調製
VZV分離及び増殖のために一般的に使用されるヒト胎児肺繊維芽細胞株である細胞株MRC−5(ATCC受入番号CCL−171)を、ファージ・ディスプレイ・ライブラリーのパンニングのため、及びELISAによりFabsを試験するために、VZVに特異的な物質の調製に使用した。MRC−5細胞を、10%の不活性化ウシ胎仔血清(FBS)、50μg/mlのペニシリン、100μg/mlのストレプトマイシン、及び2mMのL−グルタミンを含む変法イーグル培地中で維持する。
【0087】
細胞(VZV感染又は未感染MRC−5)を、掻き取り、そして、250mlの溶解バッファー(50mMのTris−HCl pH8.0;150mMのNaCl;0.02%のアジ化ナトリウム;0.5%のTritonX)中に再懸濁し、氷上で20分間インキューベートし、次に、4℃、12000rpmにて2分間遠心分離した。得られた上清のタンパク質濃度を、BCATM Protein Assay Kit(Pierce)を使用して、二重反復試験で測定した。未知試料のタンパク質濃度を、測定し、そして、既知濃度(0mg/ml=ブランク〜2.000mg/ml=最高値)のウシ血清アルブミン(BSA)の段階希釈を基準にして報告した。すべてのサンプルの吸収度を、540nmにセットした分光光度計を用いて計測した。
【0088】
タンパク質抽出物を使用したパンニングとELISA
タンパク質コーティングを、以下の抗原:VZV(ELLEN菌株;ATCC受入番号VR−586)を感染させたMRC−5ヒト線維芽細胞の細胞溶解物;インフルエンザ・ウイルス抗原(Virion Ltd.)の市販調製物;未感染MRC−5の細胞溶解物;ウシ血清アルブミン(BSA)を使用して、96ウェル・プレート上に実施した。各サンプルを、炭酸バッファーで希釈し(1ウェルあたり25□lの終量中に100ナノグラムの総タンパク質)、そして、そのプレートを4℃にて一晩インキューベートした。蒸留水で洗浄した後、そのプレートを、1%のBSAを含むPBS中、37℃にて1時間のインキュベーションによってブロッキングした。
【0089】
ELISAを、文献中に開示されているプロトコールを使用し、以下のFabs:DDF−VZV1、DDF−VZV2、及び調製された無関係なFabであるe137、を含む40□lの未希釈サンプルを使用して実施した(Bugli F et al.,2001)。前記Fabsを、二重反復試験のウェル内で試験した。各Fabとの37℃にて1時間のインキュベーション、そして、0.1%のTween−20、40μlのヤギ抗ヒトFabを含むPBSでの5回の洗浄の後に、ペルオキシダーゼ・コンジュゲート(Sigma;カタログ番号A0293)を、加え、そして、37℃にて1時間インキューベートした。前述のとおりプレート洗浄を繰り返し、そして、40μlの基質(TMB Substrate Kit;Pierce)を各ウェルに加えることによって、酵素反応を進めた。ELISA反応を、37℃にて15分間進めた。酵素活性を、停止溶液(H2SO4)を加えることによって停止させ、そして、吸収度を、450ナノメートルにセットした分光光度計を用いて計測した。
【0090】
ELISA及び中和アッセイのためのFab調製
プロトコールは、pDD、又はpGem変異体などの他のファージミドを使用した文献に記載のものと同様であった(“Molecular Cloning:A Laboratory Manual”,Sambrook et al.,Cold Spring Harbor Press,NY,1989;Burioni R et al.,1998;Burioni R et al.,2001;WO 07/007154)。
【0091】
簡単に言えば、ライブラリーの個々のE.コリ・クローンを、抗生物質を含む200mlのSuper Broth(SB;3.5%のバクト−トリプトン、2%の酵母抽出物、0.5%のNaCl)培地中で培養し、IPTGを補い、集菌し、そして、リン酸緩衝食塩水(PBS)で洗浄した。制御パルスを用いて4℃にて超音波処理することによって、溶解を細胞膜周辺腔に限定した。周辺質抽出物中のFabsを、JA−10ローターによる4℃、12,000rpmにて45分間の超遠心分離によってある程度精製した。生成物を、濾過し、そして、Centriconフィルタを用いて10回濃縮した。
【0092】
ある程度精製したFabsの濃度を、ImmunoPure Goat Anti−Human IgG[F(ab’)2](Pierce;カタログ番号31132)を使用したサンドイッチELISA法によって周辺質抽出物中で測定し、それを、96ウェル・プレート(Costar;カタログ番号3690)の表面上に結合させた。37℃にて1時間のインキュベーションの後に、プレートを、脱イオン水で6回洗浄し、そして、3%のBSAを含む170μl/ウェルのPBSを使用してブロッキングした。37°にて1時間のさらなるインキュベーションの後に、1%のBSAを含むPBS中、それぞれのFab、又は既知濃度の対照ヒトFab(Cappel;カタログ番号6001−0100)の一連の3倍希釈物50μlを、各ウェルに加え、そして、37℃にて1時間インキューベートした。次に、プレートを、TPBS(0.05%のTween−20を含むPBS)で6回洗浄した。次に、50μlのアルカリ性ホスファターゼ結合ヤギ抗ヒト抗体(Pierce;カタログ番号31312)を加え、そして、37℃にて1時間インキューベートすることによって、抗体結合を測定した。プレート洗浄を、前述のとおりTPBSを用いて繰り返し、そして、100μlのp−ニトロフェニル・リン酸二ナトリウム(Sigma)を各ウェルに加えた。ELISA反応を60分間進め、そして、結果を対照ヒトFabに対してプロットした。
【0093】
結果
組み換えファージに関するライブラリーを、pDD技術(WO 07/007154)によって作り出し、そして、VZVの臨床分離株を感染させたヒト線維芽細胞の細胞株から得られたタンパク質抽出物に対してパンニングした。5ラウンドのパンニングもまた、VZVに感染していない同じ細胞株からの対照タンパク質抽出物を使用することで、同じライブラリーを用いて平行して実施した。
【0094】
第3ラウンドまでに、対照タンパク質抽出物に対してパンニングしたサンプルのファージ力価は104未満であり、その一方で、第3ラウンドにおいてVZV抽出物に対してパンニングしたサンプルのファージ力価は104より高く、第5ラウンドのときに105に達した。この値は、Fabs結合VZV抗原をその表面上に発現する組み換えファージ内でのライブラリーの連続的な濃縮を実証している。
【0095】
第5ラウンドのパンニング後に得られた260個を超えるクローンを、ELISAにより個別に試験し、そして、それらの中の86個を陽性と確認した。選択されるクローンにおけるHCDR3のPCRと配列分析では、ヒトFabsを特徴とする2個の重鎖を同定し、ここで、DDF−VZV1及びDDF−VZV2と名付けた。DDF−VZV1又はDDF−VZV2を発現する組み換えファージの反応性を、VZV特異タンパク質抽出物に対して、並びに無関係な抗原(未感染細胞、ウシ血清アルブミン)に対して試験し、ELISAフォーマットによりそれらの強い結合活性を確認した(図1)。結合の特異性もまた、選択されたFabsが全く有意な親和性を示さなかったインフルエンザ・ウイルス抗原を使用して確認した。
【0096】
これらのFabsの完全な重鎖及び軽鎖可変領域のDNA配列を、DDF−VZV1及びDDF−VZV2について、対応するCDRsと共に測定した(図2及び3)。また、タンパク質発現用のE.コリ・ベースのシステムを使用して、さらなるアッセイに十分な量の組み換えタンパク質を得るために、これらのFabsを他のベクター内に再クローニングした。
【0097】
実施例2:VZV感染細胞培養において試験したDDF−VZV1とDDF−VZV2の特性
材料と方法
VZV特異的ヒトFabsの中和アッセイ
プラーク減少アッセイを、37℃にて72時間のインキュベーション後にコンフルエントな単層が通常形成される条件下でプレート内に植菌された104〜105個のMRC−5細胞を使用してCostarの24ウェル・プレート内で実施した。使用されるVZVウイルス株は、臨床分離株であった。
【0098】
Fabsを、実施例1に示したようにある程度精製し、そして、維持培地中に懸濁された等量のVZV無細胞ストック(ELLEN菌株;感染多重度0.01)と、様々な濃度(0.01、0.1、1、10、及び50μg/ml)で混合した。対照を、Fabsの不存在下(ブランク対照)、又はヒトC型肝炎ウイルスに特異的な無関係なFab(e137;Bugli F et al.,2001)の存在下、等量の維持培地とウイルスから構成した。
【0099】
37℃にて1時間のインキュベーション後に、250μlのウイルス・フラグメントFab混合物又は対照混合物を、培地を取り除いたウェル内に(二重反復試験の状態に)植菌した。プレートを、37℃にて2時間インキューベートして、未中和ウイルスを吸着させた。接種材料を取り除き、そして、1.5mlの維持培地を加えた。細胞培養条件下、1週間のインキュベーション後に、細胞を、PBSで洗浄し、エタノールを用いて室温にて10分間固定し、そして、1%のクリスタル・バイオレット溶液で10分間染色した。プレートを、蒸留水によって3回洗浄し、そして、溶解プラークをカウントした。それぞれのFabの中和能力を、単一溶解プラークをカウントし、対照サンプルと比較したウイルス・プラーク・カウントの減少のパーセンテージについて計算することによって測定した。
【0100】
それぞれのFabの中和能力を、蛍光顕微鏡(Olympus)によって単一蛍光発光細胞をカウントし、そして、対照サンプルと比較してVZV陽性細胞数の減少のパーセンテージを計算することによって測定した。
【0101】
免疫蛍光分析
MRC−5細胞を、無菌のガラス・カバーガラスを含むCostarの24ウェル・プレート内で培養した。細胞培養が(通常、37℃にて7日間のインキュベーション後に形成する)単層としてコンフルエントになったときに、以下のとおり、細胞にウイルス‐Fab混合物又は対照混合物を感染させた。培地を取り除き、そして、細胞単層に、37℃にて1時間のインキュベーションの間、指示濃度の半精製Fabsと共にプレインキュベートしたVZV Ellen基準菌株を使用して高い感染多重度にて感染させた(250μl/ウェル;感染多重度0.1〜1)。対照を、Fabsの不存在下(ブランク対照)、又はヒトC型肝炎ウイルス(e8)に特異的な無関係なFabの存在下、等量の維持培地とウイルスから構成した。前記プレートを37℃にて2時間インキューベートし、未中和ウイルスを吸着させた。接種材料は取り除き、そして、2%のFBSを含む1.5mlのMEMをそれぞれのウェルに入れた。
【0102】
免疫蛍光アッセイを、VZV感染の72時間後に実施した。培地を取り除いた後に、細胞単層を、PBSで1回洗浄し、そして、冷メタノール‐アセトン溶液(1:2の比率;−20℃にて保存)中、室温にて10分間固定した。固定細胞を、一次抗体としてのDDF−VZV1又はDDF−VZV2の調製物と共に湿潤大気中、37℃にて30分間インキューベートし、PBSで洗浄し、そして最後に、抗ヒトIgG Fab特異的FITC‐コンジュゲート(Sigma)と共に湿潤大気中、37℃にて30分間インキューベートした。対照として、細胞を、製造業者の取扱説明書に従って使用される市販の抗VZV抗体(Argene;カタログ番号11−017)と共に、二次FITC標識抗体だけを使用することで調製した。
【0103】
さらに、サイトメガロウイルスを感染させたMRC−5細胞を、一次抗体としてDDF−VZV1又はDDF−VZV2の同じ調製物を使用して試験した。スライドを、エバンスブルーによって対比染色し、グリセロール・バッファーで封入し、そして最後に、蛍光顕微鏡(Olympus)によって観察した。
【0104】
結果
DDF−VZV1とDDF−VZV2の両方に関する中和及び結合活性のさらなる分析を、ある程度精製したFabsの調製物を使用して実施した。
免疫蛍光では、感染細胞において核染色を示す市販のマウス・モノクローナル抗体と比較して、DDF−VZV1は、VZVビリオンが組み立てられる核(周辺)領域においてVZV感染細胞をより強く染色する。DDF−VZV2は、細胞質領域においてVZV感染細胞をより強く染色する(図4)。一次抗体の不存在下において、あるいは、感染させなかった、又はサイトメガロウイルスを感染させたMRC−5細胞に対するDDF−VZV1及びDDF−VZV2の使用において、染色は観察されなかった。これらのデータはまた、一時的に培養し、その後、免疫蛍光で観察した、VZVに感染している人の皮膚から得られた細胞に対して実施した免疫蛍光によっても確認された。
【0105】
プラーク減少に関するデータは、VZVとプレインキュベートした場合に、DDF−VZV1及びDDF−VZV2が強力な中和活性を与えられることを示している。実際、対照(Fabを伴わない、又は無関係なFabを伴ったVZV)と比較すると、これらのFabsの添加は、用量依存的様式でのプラーク形成の減少を引き起こす(図5)。
【0106】
実施例3:pDD適合発現ベクターを使用したDDF−VZV1及びDDF−VZV2の製造とバリデーション
材料と方法
pDLac−FLAGhisベクターの設計と構築
ゼオシン(Zeocine)遺伝子がマーカー遺伝子として使用されるDDbカセット(WO07/007154)を含むpDDベクターを、cp3*配列を含んでいるSpeI−NheIフラグメントを、2つのタグと終止コドンを含んでいるNheI−SpeI合成リンカーで置換することによって修飾した。前述のリンカーを、2つのオリゴヌクレオチド(配列番号15及び16)をアニーリングすることによって作り出した。得られた二本鎖DNA分子を、SpeIとNheIで消化し、そして、対応する線状化pDDベクター内へのクローニング・ステップに備えた。2つのタグ配列のインフレーム挿入を確認するために、最終的なベクターを、制限分析と配列分析によって特徴づけした。LacI遺伝子を、pET28(Invitrogen)という市販のプラスミドからPCR増幅し、そして次に、ゼオシン遺伝子と、FLAGhisタグに融合されるべきタンパク質の発現を駆動するLacZプロモーターの間のStuI部位に挿入した。
【0107】
PCR反応を、DNA鋳型(20〜30ng)、市販のPCRバッファー(1×;Invitrogen)、プライマー(0.2μM)、dNTP(0.25mM)、Taq DNAポリメラーゼ(0.25単位;Invitrogen)、及び水(50μlの終量まで)を含む混合物を使用して実施した。その反応を、サーマル・サイクラー(Perkin−Elmer)により、以下のとおり実施した:94℃にて5分間、そして、94℃にて30秒、50〜55℃にて30秒間、72℃にて1分間を30サイクル。部位特異的突然変異誘発PCR法を、Pfu融合Taqポリメラーゼ(Stratagene)を使用して、先に説明したのと同じ増幅条件で実施した。制限酵素を用いたDNAの消化を、製造業者の取扱説明書(New England Biolabs)に従って実施した。バクテリオファージT4 DNAリガーゼ(Boehringer)を、適切なベクター内への調製したDNAフラグメントの連結反応に使用した。ベクター:挿入DNAのモル比は1:3であり、そして、DNAの総濃度は約50ngであった。連結反応を、以下のとおり20μl中で実施した:DNA(50ng)、連結反応バッファー(1×)、10mMのATP(1μl)、T4 DNAリガーゼ(4単位)、及び水(20μlまで)。T4連結反応を、15℃にて一晩実施した。TG1又はXL−1 Blueエシェリキア・コリ・コンピテント細胞を、CaCl2プロトコールを使用して調製し、そして、pDLac−FLAGhisベクターを含む連結反応混合物で形質転換した。
【0108】
DDF−VZV1とDDF−VZV2の重鎖及び軽鎖をコードする配列を、適切な制限部位を使用することでpDLac−FLAGhis内にクローンニングした。最終的なベクター(pDLac−VZV1−FLAGhisとpDLac−VZV2−FLAGhis)を、2つの抗体配列のインフレーム挿入を確認するために、制限分析及び配列分析によって特徴づけした。
【0109】
FLAGhisタグ付与DDF−VZV1及びDDF−VZV2の発現、精製、及び検出
E.コリXL−1 Blueを、pDLac−VZV1−FLAGhis、pDLac−VZV2−FLAGhis、又はe509を発現するpDLac−FLAGhis変異体、対照として使用されるHCV特異的Fab(Bugli et al.,2001)で形質転換した。
【0110】
得られた菌株を、細菌培養によるFabsの製造に使用した。このために、形質転換から得られた単一コロニーを、テトラサイクリン(10μg/ml)、アンピシリン(100μg/ml)、及びゼオシン(zeocin)(50μg/ml)を含む10mlのSB培地中に植菌した。細菌を、37℃にて12時間増殖させた。インキュベーション時間の後に、500μlのそれらを、同じ抗生物質を補った100mLのSBで希釈し、0.6のODに達するまで6〜8時間増殖させた。イソプロピル−ベータ−D−チオ−ガラクトピラノシド(IPTG)を、1mMの終濃度まで加え、そして、細胞を、ロータリー振盪機により30℃にてさらに14時間インキューベートした。細菌を、5000gにて20分間の遠心分離によって集菌し、1mLのリン酸緩衝生理食塩水(PBS)中に再懸濁し、そして、凍結‐融解手順(−80℃及び37℃にて3ステップ)で溶解させた。細胞破片を、微量遠心管による15,000g、室温にて5分間の遠心分離によって取り除いた。
【0111】
Fab発現を確認するために、上清を、抗Fab及び抗His抗体検出を用いたウエスタン・ブロット分析に使用した。SDS−PAGEを、β−メルカプトエタノールを含む又は含まない(還元又は非還元条件)サンプルを添加して作製し、次に、ブロッティングを350mAにて2時間実施した;ニトロセルロース紙を、存在するブロッティングを確認するためにPonceaus Redで最初に染色し、次に、それを10%のミルク/PBS/Tween 0.1%(ブロッキング溶液)と共に4℃にて一晩撹拌した。PBS/Tween 0.1%で1回洗浄した後に、前記ニトロセルロースを、西洋ワサビ・ペルオキシダーゼ(HRP)コンジュゲート抗Fab抗体(Sigma)又はHRPコンジュゲート抗His−タグ抗体(Roche)と共に撹拌しながら室温で1時間インキューベートした。PBS/Tween 0.1%で3回洗浄した後に、SuperSignal West Pico Chemiluminescent基質(Pierce)を加え、そして、フィルムを、それぞれ30秒、1、及び5分間現像した。
【0112】
抗Fabを用いて検出したシグナルを、抗His HRPコンジュゲート検出と比較した。それらの両方が、(浄化した細胞抽出物を非還元条件を使用したゲルに泳動したとき)Fabフラグメントの天然型に対応する〜60kDaの主要なバンドを明らかにした。同じサンプルを還元条件を使用したSDS−PAGEに供したとき、抗His抗体を使用した場合に、重鎖(〜25kDa)だけが存在した。抗Fabを用いた検出では、タグを付与されていなかったので検出できなかった軽鎖の存在を明らかにした。
【0113】
FLAGhisタグ付与fabsのより大規模な製造において、テトラサイクリン(10μg/mL)、アンピシリン(100μg/mL)、及びゼオシン(50ug/ml)を含むスーパー・ブロス(super broth)の1リットル・アリコートに、特定のpDLac−FLAGhisベクターで形質転換した細菌コロニー1つを植菌し、ロータリー振盪機内で37℃にて7時間培養し、示したようにIPTGで誘導し、そして、30℃にて一晩培養した。細胞を、遠心分離によって集菌し、25mLのPBS中に再懸濁し、そして、超音波処理した。細胞破片を、遠心分離(15,000gにて50分間)によって除去し、そして、0.22μmで濾過した上清をIMACによって精製した。標準的なQiagen精製プロトコールに従った:NiNTA樹脂(Qiagen)を結合バッファー溶液(リン酸緩衝液、10mMのイミダゾール)で平衡化し、次に、濾過した上清をその樹脂と混合し(300μLの樹脂/100mLの培養物)、そして、4℃にて1時間撹拌した。この溶液を、400gにて5分間遠心分離して、樹脂に連結したFabフラグメントを回収し、そして、流れ出た物を収集した。その樹脂を、5倍量の結合バッファー10mMイミダゾールで洗浄して、非特異的結合を除去した。HisフラグFabフラグメントを、3倍量の溶出緩衝液(リン酸緩衝液、500mMのNaCl、500mMのイミダゾール)で溶出した。溶出したアリコートを、SDS/12.5%ポリアクリルアミドゲル電気泳動によって分析し、そして、クマシー・ブリリアントブルー染色及びウエスタン・ブロットで検出した。
【0114】
結果
DDF−VZV1及びDDF−VZV2の活性を、最初、ある程度精製したFabs及び組み換えファージとして試験した。さらなるバリデーションには、このような抗体フラグメントを適切な細菌発現系を使用した精製された組み換えタンパク質の形態で試験する必要がある。
【0115】
着目のFabを持つファージミドは、可溶性分子の産生のために好適な発現ベクター、及び/又は適切な菌株(例えば、アンバー非サプレッサー(amber non−suppressor))に通常移される。分子クローニングのこのような手間がかかったステップを避ける目的で、より効果的なアプローチは、前記Fabsを同定するのに使用した元々のpDDベクター内のものと適合する制限部位を持つ発現ベクターを製造することだろう。そのうえ、タグ配列の存在は、より簡単なFabの精製及び検出を可能にするだろう。
【0116】
pDLac−FLAGhisと名付けたこのような発現ベクターを、設計し、そして、構築した。それは、cp3*のコード配列を、終止コドンが後に続く2つのタンパク質タグ(8つのアミノ酸を含むFLAGペプチド、及び6−ヒスチジン・タグ)をコードする配列で置換したpDDベクターに基づいている(図6A)。ヒスチジン・タグは、例えば、IMAC(固定金属アフィニティー・クロマトグラフィー)精製システムを使用したアフィニティー・クロマトグラフィーによってFabを精製することを可能にする。FLAGタグは、(ELISAなどの)検出アッセイにおいて非常に有用であるかもしれない。これらの2つのタグがクローンニングされ、そして、組み換えタンパク質に融合されたベクター、おまけに、FLAGhisタグ付与タンパク質を検出又は精製する手段が、文献に記載されていた(Pagny S et al.,2000;Koivunen P et al.,2004;Komatsu M et al.,2004)。
【0117】
最初に同定され、そして、pDDファージミドを使用して特徴づけられたFab(又は、いずれかのその他タンパク質配列)は、その後、pDLac−FLAGhisベクター内に一方向性にクローンニングされるDDカセットを使用することで抽出できる。このように、元々コート・タンパク質に融合していた配列が、ここで、FLAGタグ及びポリヒスチジン・タグと共にC末端にてフレーム内にクローンニングされる。得られたベクターは、DDカセット内に本来のマーカー遺伝子(例えば、ゼオシン遺伝子)を含み、所望の挿入物を含むベクターの選択を可能にする。pDLac−FLAGhisベクター内のLacZプロモーターを使用して発現されるべきタンパク質の転写を制御するための手段として、LacI遺伝子を本来のDDカセット内に挿入した(図6B)。
【0118】
このクローニング・ストラテジーは、2つの利点をもたらした。第1の利点は、起こり得る相同組み換えを回避する、本来のDDカセット内の2つのLacZプロモーター領域の間の空間を増やすことである。第2に、オペレーター領域にそれが結合することによりLacZプロモーターを遮断して、発現系を下方制御できることが知られているLacI遺伝子からのリプレッサー・タンパク質である。そして、誘導は、適切な濃度のIPTG、及び/又はグルコースを使用して調整される。誘導により、LacZプロモーターの制御下にクローンニングされた配列/PelBシグナル配列の両方が、発現され、そして、pelB配列が酵素シグナル・ペプチダーゼによってそれに続いて切断されるところの周辺質に向けられる。周辺質内では、適切な酸化条件が、組み換えタンパク質内のジスルフィド結合の形成、及び正しいタンパク質の折り畳みを可能にする(例えば、ヘテロ二量体タンパク質に組み立てられるFabの軽鎖及び重鎖)。アフィニティー・クロマトグラフィーのカラムを溶出し、そして、試験することによって、FLAGhisタグ付与タンパク質を均質にまで精製できる(図6B)。次に、このタンパク質を、例えば、(Sigmaから入手可能な)市販の抗FLAGモノクローナル抗体を使用したウエスタン・ブロット又は免疫蛍光により同定できる。
【0119】
このアプローチを使用して、DDF−VZV1及びDDF−VZV2(図7)の、並びに対照fabのFLAGhisタグ付与バージョンを、製造し、そして、先に記載したアッセイにより試験した。選択したFabsのこれらの組み換え変異体で、2μg/ml前後のIC50を示す、VZV中和活性に関する本来の観察結果を確認した(図8A)。しかしながら、2つのFabsが1つの調製物に組み合わせられるとき、この調製物のVZV中和活性は、同量の単独のFabを含む調製物より優れた結果となった。実際、Fab濃度が1〜2μg/mlに達するときに、単独のFabを含む調製物がその活性を10〜15%増強する場合に、1μg/mlのDDF−VZV1と1μg/mlのDDF−VZV1を含む調製物は、2μg/ml未満のIC50値を有するはるかに高いVZV中和活性を有する(図8B)。この効果は、おそらく、2つのFabsによって認識される異なるVZV特異的エピトープのためであり(図4を参照のこと)、DDF−VZV1とDDF−VZV2(同様の特性を有する2つの抗体又は抗体フラグメント)を含む医薬組成物が、VZV感染及びVZV関連疾患に対してより良好な治療効果又は予防効果をもたらし得ることを示唆している。
【0120】
ここに提示した実験的証拠が、VZV感染及びVZV関連疾患に関係する診断上の、治療上の、又は予防上の適用のためのDDF−VZV1及びDDF−VZV2(又はそれらの特異的なHCDR3sに基づき、且つ、同様の特性を示す代替タンパク質配列)候補化合物をもたらす。
【0121】
参考文献
【0122】
【化1】
【0123】
【化2】
【0124】
【化3】
【0125】
【化4】
【0126】
【化5】
【技術分野】
【0001】
本発明は、ウイルスに特異的な生物学的活性を有する、ファージ・ディスプレイ・ライブラリーから単離された新規抗体配列に関する。
【背景技術】
【0002】
ファージ・ディスプレイ技術は、ポリペプチド(抗体フラグメント、生理活性ペプチド、酵素等)をクローニング、選択、及び設計するために、細菌細胞(例えば、エシェリキア・コリ(Escherichia coli)細胞)に感染する(M13などの)線状ファージのゲノムの小さな寸法と順応性を利用する。前記ポリペプチドは、線状ファージ表面上に発現され、そして、標的とそれらの相互作用により生じる生物学的機能を発揮する。
【0003】
論文(Bradbury A and Marks J,2004;Mancini et al,2004;Conrad U and Scheller J,2005;Hust M and Dubel S,2005)及び書籍(“Phage display:A Practical Approach”,vol.266,ed.Clackson and Lowman H,Oxford Univ.Press,2004;“Phage Display:A Laboratory Manual”,ed.Burton D et al,CSHL Press,2001)で概説されるように、いくつかのクローニング及び発現ストラテジー、ベクター、ライブラリー、ファージの伝播方法、及びスクリーニング・アッセイが、様々な適用のために開発された。
【0004】
ファージ・ディスプレイ・ライブラリーは、組み換えファージの集団で作られており、それぞれがタンパク質配列のレパートリーの1つの要素を提示している。特異的なタンパク質を発現するファージは、繰り返される親和性ベースの、及び/又は機能ベースの選択過程(「パンニング」)によってライブラリーから単離される。例えば、タンパク質は、精製抗原に対するそれら親和性、又は生物学的アッセイ法における活性に基づいて単離及び特徴づけされ得る、(一般的にFabsと名付けられる)可変重鎖/軽鎖ヘテロ二量体、又は一本鎖可変フラグメント(scFv)の形態の抗体フラグメントである。
【0005】
具体的には、前記結合特性に関係する関連生物学的活性を伴うこともある、病原菌及び生物学的標的に対する高い親和性と特異性を有する抗体フラグメントを同定するためのスクリーニング過程が開発された。実際には、(受動免疫療法又は受動血清療法と名付けられた)治療方法全体が、ヒト又はヒト以外の治療標的に向けられた抗体及び抗体フラグメントの抗原結合特性の上に成り立っている(Dunrnan P and Nesin M,2003;Keller M and Stiehm E,2000)。受動免疫療法は、病原性抗原(例えば、毒素、ヒト・タンパク質、ウイルス、又は寄生生物)に対して規定の結合特異性を有する治療抗体を含んで成る医薬組成物の個体への投与から成る。
【0006】
受動免疫療法は、臨床診療に取り入れられ、(感染症、免疫媒介疾患、及び癌を含めた)様々な疾患を治療する機会が急速に増大した。このアプローチは、免疫系が標的とされた分子を妨げる、及び/又は排除するのに必要とされる量、及び/又は特異性を有する抗体を産生することができない患者において特に有効である(Chatenoud L,2005;Laffly E and Sodoyer R,2005)。
【0007】
治療抗体を使用して標的とされ得る病原性抗原の中で、ヒト細胞に感染するウイルスは、特別な重要性を持つ。このような抗体の投与で、患者におけるウイルス増殖を妨げることができ、そして、集団におけるウイルス感染の大発生を阻む可能性がある。あるいは、前記抗体は、通常、免疫が保たれている人の健康に重大な、及び/又は永続的な影響を及ぼさないものを含めた感染症に対してより一層感受性が高くなる、程度の差はあれ長期間、免疫系が弱まっている患者(例えば、免疫抑制された人、高齢者、又は移植を受けた人)に投与され得る。
【0008】
水痘帯状疱疹ウイルス(VZV、水痘)は、免疫障害を持つ人にとってそのような命にかかわる可能性のある感染の原因となるウイルスの1種である。VZVは、少なくとも70個の異なったオープン・リーディング・フレーム(ORFs)をコードするアルファ・ヘルペスウイルスであって、その大部分が単純ヘルペスウイルスと近縁関係にある(Mori I and Nishiyama Y,2005)。VZVは、皮膚病変又は呼吸器分泌物中の感染性ウイルスとの直接接触によってヒト集団内で伝播され、皮膚とT細胞に対して特異的親和性を示す。第一次VZV感染は、リンパ球に向けられ、それに続いて、細胞関連ウイルス血症、臓器内でのウイルス複製、そして、第二次ウイルス血症に起因する広汎性皮膚疹が起こる。VZV感染は、ビリオンの放出、又は細胞間伝播によって拡散することが可能であり、知覚神経節内への潜伏を確立する(Arvin A,1996;Quinlivan M and Breuer J,2006)。
【0009】
通例、1回のVZV侵襲又はワクチン接種が、一生にわたる保護をもたらす。しかしながら、VZVには、ウイルスが先天的な免疫応答、及び後天的に得た免疫応答の両方を回避することを可能にする固有の特性がある。ワクチン接種後の検出可能な細胞性免疫にもかかわらず、VZV特異的抗体が失われる場合もあるので、VZVの再感染又は再活性化がさらに可能である(Ludwig B et al.,2006)。
【0010】
感染及びワクチン接種に対する免疫応答の機構と効力は、ヒト及び動物モデルにおいて(Maple P et al.,2006;Matsuo K et al.,2003;Kutinova L et al,2001;Massaer M et al.,1999;Haumont M et al.,1997;Hasnie F et al.,2007)又は試験管内培養系において(Finnen R et al.,2006;Andrei G et al.,2005a)研究された。VZV発症機序と免疫生物学は、トランスジェニック・マウス・モデルにおいて、生体外におけるヒト皮膚又は神経節モデルを使用して研究できる(Baiker A et al.,2004;Ku C et al.,2005;Taylor S and Moffat J,2005;Zerboni L et al.,2005)。このように、細胞型特異的VZVアポトーシス活性(Hood C et al.,2003)、及びVZV感染の細胞性/ウイルス性伝達物質(Berarducci B et al.,2006;Chen J et al.,2004;Li Q et al.,2007;Hambleton S et al.,2007)が同定された。
【0011】
弱毒化生水痘ワクチン(Oka/Merck菌株)が入手可能であり、その安全と有効性を鑑みて、上記ワクチンが日常的な幼児期予防接種と成人に推奨される(Oxman M et al.,2006;Arvin A,1996)。VZVワクチン接種が先進工業国において幅広く定着し、且つ、短期間/中期間、すべての形態の水痘(特に重症疾患)を低減するのに非常に効果的であるにもかかわらず、集団において潜在型又は進化した野生型ウイルスのプールが継続的な脅威になっている(Hambleton S and Gershon A,2005)。
【0012】
VZV菌株は、VZV転写産物の特異的な遺伝子変異(Grose C,2006)、及び3種類の大きく異なる遺伝子型(Norberg P et al.,2006)と関連する。遺伝的変異は、予防接種されたヒト宿主の中に検出された皮疹を形成するVZV遺伝子型に関連した(Quinlivan M et al.,2007)。そのうえ、VZV免疫のレベルは、異なったヒト集団において、例えば、欧州地域の中で、大きく異なる(Nardone A et al.,2007)。
【0013】
帯状ヘルペス(帯状疱疹)は、臨床診療において、特に加齢に伴って低下した細胞性免疫の結果として高齢者において、別の年齢分布においてはVZV再活性化の危険因子に起因して、そして、癌、免疫抑制治療、又はステロイド療法による随伴性免疫不全と共に、いまだに頻繁に観察されている。帯状ヘルペスの合併症は、神経学的合併症(ヘルペス後神経痛、脳血管炎、脳炎、無菌性髄膜炎、脳神経麻痺、又は髄膜脳炎)、目に関する合併症(眼帯状ヘルペス、ブドウ膜炎、網膜の壊死、視神経炎、又は角膜炎)、あるいは、内臓合併症(肝炎、肺炎、心筋炎)である。帯状ヘルペスの危険因子は、さらによく理解されるようになってきているが、それらの数の増加、並びに知覚神経節及び眼表面へのVZV持続性は、成人における、より広範囲のワクチン接種、及びより効果的な抗VZV治療の必要性を示唆している(Dworkin R et al.,2007;Weinberg J,2007;Liesegang T,2004)。
【0014】
VZV感染は、ワクチンの使用が妥当でない免疫抑制患者において非常に危険である。実際に、免疫抑制状態にある1人の人の単独のVZV再活性化又は再感染が、脊髄又は脳動脈を含めたいくつかの組織にVZVが侵入し得るような数人の患者に感染する可能性がある移植病棟において、VZVは重大な問題である(Chaves Tdo S.et al.,2005;Gilden D,2004)。妊娠中、VZV感染はまた、子宮内伝播又は新生児期感染によって胎児にも広まり、重大な皮膚病変及び四肢と臓器の発生の異常を伴った、子宮内胎児死亡又は先天性水痘症候群を引き起こす(Schleiss M,2003;Sauerbrei A and Wutzler P,2007)。
【0015】
いくつかの重要な禁忌(Arvin A,1996)があるワクチン接種は別として、抗ウイルス治療が利用可能である(アシクロビア、バラシクロビル、ファムシクロビル、及びブリブジンなど)。しかしながら、薬剤耐性VZV菌株の問題に加えて、これらの化合物はまた、例えば、ヒト酵素との相互作用のために、禁忌がある可能性がある(Andrei G et al.,2005b;Abdel−Haq N et al.2006)。コルチコステロイド又は抗痙攣薬などの他の治療が、追加的な鎮痛を与えるために使用されるが、それらの有害影響特性が、特にヘルペス後神経痛に対する、使用を制限する(Tyring S,2007)。これにより、予防のための現在の手段が適用不可能であるか、又はもはや有効性でない場合、代替のアプローチが必要である(Hambleton S and Gershon A,2005)。ワクチン不全とワクチン誘発免疫の低下を考慮して、現在、二次ワクチン投与が推奨されている(Chaves S et al.,2007)。
【0016】
VZV感染を早期に同定するために、いくつかの診断検査が、同様の患者集団において、並びにVZV感染の危険に晒された高齢者又は免疫不全の人において、VZV特異的抗体、抗原、又は転写産物を検出するために開発された(Hambleton S and Gershon A,2005;Smith J et al.,2001)。
【0017】
VZVに関連する眼感染(Kezuka,T. 2004)、血管障害(Burgoon M et al.,2003)、又は疫学的研究(Talukder Y et al.,2005)に関連して血清、脳脊髄液、又は口腔液中で、VZV特異的免疫活性が同定された。VZV暴露後の予防が提案され、そして、具体的には、感染を予防するために投与され得る有意な抗VZV力価を有するヒト抗体の調製物を使用して試験された(Keller M and Stiehm E,2000)。特異的な、抗VZV抗体の高い力価を有するヒト免疫グロブリンの静脈内注射用調製物が記載された(US4717564;CDC,2006)。単独での、又はVZV特異的な抗ウイルス剤としての他の化合物と組み合わせたその利用は、移植を受けた患者、妊婦、又は新生児で試験された(Huang Y et al.,001;Carby M et al.,2007;Koren G et al.,2002)。
【0018】
VZV−特異的抗原、VZV糖タンパク質の変異体、及びそれらの免疫原性エピトープが、異なる抗体調製物、例えば、ヒト血清(US6960442;Fowler W et al.,1995;Kjartansdottir A et al.,1996;WO96/01900)、ヒト以外のポリクローナル血清(US5306635;WO92/06989)など、を使用して説明された。VZVに対するモノクローナル抗体は、単離され、そして、ヒト起源(WO86/02092;WO91/16448;WO95/04080;EP148644;Nemeckova S et al.,1996;Foung S et al.,1985;Sugano T et al.,1987;Yokoyama T et al.,2001;Ito M et al.,1993)、又はマウス起源(EP321249;Vafai A and Yang W,1991;Montalvo E and Grose C 1986;Forghani B et al.,1994;Grose C et al.,1983;Lloyd−Evans P and Gilmour J,2000;Shankar V et al.,2005;Garcia−Valcarcel M et al.,1997)のトリオーマ又はハイブリドーマを使用して発現された。組み換え、VZV結合、及び/又は中和抗体フラグメントが、ファージ・ディスプレイ・ライブラリー中で同定され、そして、一本鎖可変フラグメント(Kausmally L et al.,2004;Drew P et al.,2001)又はFabs(Suzuki K et al.,2007;Williamson R et al.,1993)として発現された。抗VZVマウス・モノクローナル抗体は、ヒト化された(WO 95/31546)。
【0019】
ワクチンと全身的抗ウイルス剤の両方に大幅な改善がもたらされたにもかかわらず、疾患は存続している。療法は、予測不可能なパターンで発症するVZV合併症の多くを軽減するが、それを解消するわけではない。今もなお、集団におけるVZVの感染及び増殖をより効率的に検出し、そして、阻むことができる新規抗体及び抗体フラグメントの同定及び製造は、この感染症の療法、及び/又は予防のための改善された治療を確立するのに特に重要なことである。
【発明の概要】
【課題を解決するための手段】
【0020】
本発明は、VZVに結合し、中和する新規抗体配列、並びにVZV感染又はVZV関連疾患を検出する、治療する、抑制する、予防する、及び/又は改善するために使用できる新規抗体配列を提供する。
【0021】
ヒト抗体配列のパネルを組み換えファージ上に提示させ、そして、VZV特異的結合活性をファージ・ライブラリーにおいて検出した。2つの抗体フラグメントの重鎖及び軽鎖可変領域をコードし、且つ、VZV中和活性を有するDNA配列を同定し、DDF−VZV1とDDF−VZV2と命名した。対応するタンパク質配列、及びVZV特異的な生物学的活性に関与する相補性決定領域(CDRs)を決定した。
【0022】
本発明の配列は、組み換えタンパク質を製造するのに適当な技術を使用した、完全な抗体、抗体フラグメント、又はいずれか他の形式の機能タンパク質(具体的には融合タンパク質)の形態で、VZV特異的結合及び中和特性を有する組み換えタンパク質を製造するのに使用できる。
【0023】
VZV感染及びVZV関連疾患の管理において治療上、予防上、及び/又は診断上の有用性を有する組成物が、本発明のタンパク質を使用することで調製できる。このような組成物は、抗ウイルス化合物、及び/又は免疫グロブリン静注(IVIg)調製物に基づく現在のVZV治療を補うか、又はそれを置き換えるのに使用できる。
【0024】
単離されたDNA及びタンパク質配列、ベクター、組み換えファージ、及び宿主細胞、並びに医学方法や用途を含めた本発明のさらなる態様が、以下の説明の中で提供される。
【図面の簡単な説明】
【0025】
【図1】DDF−VZV1が(VZV−1)、DDF−VZV2(VZV−2)、又は陰性対照として使用された無関係なヒトFab(e−137)を発現する組み換えファージの調製物に対するVZV結合活性の特異性。結合活性は、(MRC−5細胞について)総タンパク質量抽出物、又は(ウシ血清アルブミン、BSAについて)精製タンパク質の形態でプレート・コーティングに使用される表示抗原(the indicated antigens)を使用したELISAにより計測された。
【図2】(A)DDF−VZV1の重鎖の可変領域のDNA(小文字)配列とタンパク質(大文字)配列の配列比較(DDF−VZV1 VH;配列番号1及び2)。予測されるCDRs(HCDR1、HCDR2、及びHCDR3;配列番号3、4、及び5)には下線が引かれている。(B)DDF−VZV1の軽鎖の可変領域のDNA(小文字)配列とタンパク質(大文字)配列の配列比較(DDF−VZV1 VL;配列番号6及び7)。予測されるCDRs(LCDR1、LCDR2、及びLCDR3)には下線が引かれている。
【図3】(A)DDF−VZV2の重鎖の可変領域のDNA(小文字)配列とタンパク質(大文字)配列の配列比較(DDF−VZV2 VH;配列番号8及び9)。予測されるCDRs(HCDR1、HCDR2、HCDR3;配列番号10、11、及び12)には下線が引かれている。(B)DDF−VZV2の軽鎖の可変領域のDNA(小文字)配列とタンパク質(大文字)配列の配列比較(DDF−VZV2 VL;配列番号13及び14)。予測されるCDRs(LCDR1、LCDR2、LCDR3)には下線が引かれている。
【図4】DDF−VZV1(A)及びDDF−VZV2(B)で染色した、VZV感染MRC−5細胞の免疫蛍光。細胞膜が白線で示されており、そして、核膜が白色点線で示されている。未感染細胞においてこれらのFabsを使用することで染色を得られなかった。
【図5】ファージ・コスト・タンパク質上に提示された、ある程度精製されたヒト組み換え型Fabsとして発現された場合のDDF−VZV1(A)及びDDF−VZV2(B)のVZV中和活性。プラーク減少の用量‐応答分析は、無関係なヒトFab(e137)の対応する濃度と平行して実施された。パーセンテージ値は、全くFabを含まずにプレインキュベートされたVZVを使用することで得られたデータと比較することによって計算された。
【図6】(A)対応する逆方向プライマー(pDD−FLAGhis逆方向:配列番号16)と対形成させ、SpeIとNheIで消化し、そして、cp3*配列(WO 07/007154)を置換するためにpDDベクター内にクローンニングした後に、このオリゴヌクレオチドによって連続してコードされるタンパク質タグ(FLAGHisタグ;配列番号17)の目印を伴ったFLAGhisタグ(pDD−FLAGhis順方向:配列番号15)のための順方向リンカー。(B)pDLacI−FLAGhisベクター内にクローンニングされた抗体フラグメントの重鎖(HC)及び軽鎖(LC)を含むDNAフラグメントの概要地図。HC配列は、PelBシグナル配列(PelB)とFLAGhisタグ配列(FLAGhis)の間のフレーム内にクローンニングされている。LC配列はPelBシグナル配列(PelB)と共にフレーム内にクローンニングされ、そして、その発現はLacZプロモーター(LacZ)によって駆動される。2つの発現ユニットの間で、マーカー遺伝子(ゼオシン(Zeocin);Zeo遺伝子)とLacZプロモーターを制御する遺伝子(Lac I遺伝子)は、それら自身のプロモーターと共にクローンニングされる。cp8*(cp8*)をコードする配列は、機能的プロモーターと開始点の不存在下では、転写及び翻訳されない。関連する制限部位、具体的には、HC(SpeIとXhoI)、LC(XbaIとSacI)、及びLacI遺伝子(StuI)をクローニングするために使用されるものを示す。pDLacシステムを使用してDDF−VZV1を発現するための対応するNheI−BglIフラグメントを、配列番号18として提供する。(C)pDLac−VZV1−FLAGhis精製を使用して発現されたDDF−VZV1−FLAGhisのクマシー染色。この図面は、アフィニティー・クロマトグラフィー・カラムから溶出された5つの連続した画分を示している。画分は、一緒に貯留され、濃縮され、そして、さらなる使用のために−20℃にて保存された。
【図7】(A)pDLac−VZV1−FLAGhis(DDF−VZV1 CHTag;配列番号20)を使用することで発現されるヒトFab DDF−VZV1の重鎖のタンパク質配列。対応するDNA配列は、配列番号19として提供される。pDDベクターを使用することで元々クローンニングされたこの重鎖の可変領域(配列番号2)には下線が引かれている。PelB配列は、第1アミノ酸と第26アミノ酸の間に包含されている。第155〜260アミノ酸は、ヒトIgガンマ−1鎖C領域の第1〜106アミノ酸に相当する(SwissProt Ace.No.:P01857)。第262〜275アミノ酸は、FLAGhis配列に相当する。(B)pDLac−VZV1−FLAGhis(DDF−VZV1 CL;配列番号22)を使用して発現されるヒトFab DDF−VZV1の軽鎖のタンパク質配列。対応するDNA配列は、配列番号19として提供される。pDDベクターを使用することで元々クローンニングされたこの軽鎖の可変領域(配列番号7)には下線が引かれている。PelB配列は、第1アミノ酸と第22アミノ酸の間に包含されている。第129〜234アミノ酸は、Igカッパ鎖C領域の第1〜106アミノ酸に相当する(SwissProt Ace.No.:P01834)。(C)pDLac−VZV2−FLAGhis(DDF−VZV2 CHTag;配列番号24)を使用して発現されるヒトFab DDF−VZV2の重鎖のタンパク質配列。対応するDNA配列は、配列番号23として提供される。pDDベクターを使用することで元々クローンニングされたこの重鎖の可変領域(配列番号9)には下線が引かれている。PelB配列は、第1アミノ酸と第26アミノ酸の間に包含されている。第152〜257アミノ酸は、ヒトIgガンマ−1鎖C領域の第1〜106アミノ酸に相当する(SwissProt Ace.No.:P01857)。第259〜272アミノ酸は、FLAGhis配列に相当している。(D)pDLac−VZV2−FLAGhis(DDF−VZV2 CL;配列番号26)を使用することで発現されるヒトFab DDF−VZV2の軽鎖のタンパク質配列。対応するDNA配列は、配列番号25として提供される。pDDベクターを使用することで元々クローンニングされたこの軽鎖の可変領域(配列番号14)には下線が引かれている。PelB配列は、第1アミノ酸と第22アミノ酸の間に包含されている。第135〜240アミノ酸は、Igカッパ鎖C領域の第1〜106アミノ酸に相当する(SwissProt Ace.No.:P01834)。
【図8】pDLacシステムを使用することで発現され、そして、精製されるヒトFabsのDDF−VZV1及びDDF−VZV2のVZV中和活性。(A)各Fabは示した濃度にて使用され、そして、プラーク減少が計算された。(B)各Fabは、単独、又は等量の各Fabを含んで成るFab調製物のいずれかで、示した濃度にて使用された。プラーク減少の用量‐応答分析が、同じ系(el37)を使用することで製造された無関係な対応する濃度のヒトFabと平行して実施された。パーセンテージ値は、全くFabを含まずにプレインキュベートされたVZVを使用することで得られたデータ(陰性対照)と比較することによって計算された。
【発明を実施するための形態】
【0026】
発明の詳細な説明
pDDファージミド及びWO07/007154に記載の関連方法は、2つの所定のファージ・コート・タンパク質の一方又はもう一方のいずれかに融合されたタンパク質配列のクローニング、発現、及び選別を可能にする。このアプローチは、表面上に又は組み換えファージにより示差的に発現又は提示されることができ、その結果、ファージ・ディスプレイ・ライブラリーと異なった効率で選別され得るタンパク質配列識別の選択を可能にする。
【0027】
この場合、ファージ・ライブラリーは、ヒト重鎖及び軽鎖免疫グロブリンの可変領域をpDDファージミド内にクローンニングすることによって構築された。ライブラリーは、VZVタンパク質抽出物に対してパンニングしてより分けられ、それに続いて、選択されたクローンが、実施例に示されているように、VZV中和活性があるものを決定するために細胞ベースのアッセイ方法で試験された。
【0028】
DDF−VZV1及びDDF−VZV2と名付けた、2つの最も有望なクローンをコードするDNA配列が決定され、次に、細菌発現のために適切なベクター内にクローンニングされた。これらのFabsのVZV中和活性は、VZV感染の試験管内モデルを使用して、細菌細胞培養物から精製した組み換えファージの表面上の融合タンパク質としても、アフィニティー精製した組み換えヒトFabsとしても試験された。
【0029】
本発明は、VZVに結合し、中和できる新規タンパク質配列を提供し、それには、FabsのDDF−VZV1又はDDF−VZV2により識別される特異的なCDR(相補性決定領域)が含まれる。具体的には、本発明の各HCDR3s(重鎖可変領域のCDR3)(配列番号5及び配列番号12)は、それぞれ、DDF−VZV1及びDDF−VZV2の抗原結合部分を特徴とする。
【0030】
HCDR3の抗体は、抗原に結合し、その結果、生物学的活性(例えば、実施例に示されているように、VZVに結合し、そして、中和する)を発揮することができる、そのような抗体の抗原結合部分を特徴とすると考えられる。抗体の数個又はすべてのCDRが、完全な抗原結合表面を得るのに通常必要とされるにもかかわらず、HCDR3は、配列に関してだけではなく、長さに関しても抗体の間で最も高い相違を示すCDRである。実際、HCDR3の配列と長さの多様性は、大部分の抗体に対する特異性を測定するための基本となる(Xu J and Davies M,2000;Barrios Y et al.2004;Bond Get al.,2003)。
【0031】
前記HCDR3が同定されたのと同じFabからの他のCDRと組み合わせて又は組み合わせずに、VZV結合部分として本発明の特異的なHCDR3を含むタンパク質が、抗体タンパク質フレームワーク内に作り出される(Knappik A et al.,2000)。CDRの組み合わせは、抗体構造と無関係なタンパク質フレームワーク内であっても、元の結合活性を乱すことなく、元の結合特性を持ち続ける非常に短いタンパク質の状態に互いに連結され得る(Ladner R,2007;Kiss C et al.,2006)。
【0032】
1つの態様において、本発明は、Fab DDF−VZV1のHCDR3と少なくとも90%の同一性を有する配列を含んで成るタンパク質を提供する。HCDR1及びHCDR2(配列番号3及び配列番号4;図2A)と共に、このHCDR3は、DDF−VZV1 Fabの重鎖の可変領域(DDF−VZV1 VH;配列番号2)内に含まれる。このFabを形成する軽鎖の可変領域(DDF−VZV1 VL:配列番号7)、そのうえ、その特異的なLCDRs(軽鎖可変領域のCDRs)が決定された(図2B)。
【0033】
他の態様において、本発明は、Fab DDF−VZV2のHCDR3と少なくとも90%の同一性を有する配列を含んで成るタンパク質を提供する。HCDR1及びHCDR2(配列番号10及び配列番号11;図3A)と共に、このHCDR3は、DDF−VZV2の重鎖の可変領域(DDF−VZV2 VH;配列番号9)内に含まれる。このFab(DDF−VZV1 VL;配列番号14)を形成する軽鎖の可変領域、そのうえ、その特異的なLCDRsが決定された(Fig.3B)。
【0034】
本発明のタンパク質がDDF−VZV1の配列に基づいているなら、それらは、配列番号5に対して少なくとも90%の同一性を有する配列を含んで成るべきである。特に、それらは、配列番号2に対して少なくとも90%の同一性を有する配列を含んで成るべきである。より特に、前記タンパク質はまた、配列3及び配列番号4から成る群から選択される1つ以上の配列も含むべきである。
【0035】
あるいは、本発明のタンパク質が、DDF−VZV2の配列に基づいているなら、それらは、配列番号12に対して少なくとも90%の同一性を有する配列を含んで成るべきである。特に、それらは、配列番号9に対して少なくとも90%の同一性を有する配列を含んで成るべきである。より特に、前記タンパク質はまた、配列番号10及び配列番号11から成る群から選択される1つ以上の配列も含むべきである。
【0036】
本発明のさらなる態様は、両Fabsの重鎖及び軽鎖の可変領域をコードするDNA配列であり、具体的には、DDF−VZV1(重鎖については配列番号1;軽鎖については配列番号6)及びDDF−VZV2(重鎖については配列番号8;軽鎖については配列番号13)の可変領域についてクローンニングされ、そして、決定された本来のDNA配列と少なくとも90%の同一性を有するものである。これらのDNA配列(又は、図2及び3から容易に決定され得る単離されたHCDRs及びLCDRsをコードするものなどの選択された部分)は、それらがVZV結合及び中和特性をもたらす組み換え抗体(例えば、完全抗体、親和性成熟抗体、CDRグラフト抗体、又はフラグメント)又は融合タンパク質の公知の形式の1つでそれらを発現するために他のベクター内に移されてもよい。
【0037】
同一性のレベルがどこに示されるとしても、この同一性のレベルは、本発明の関連配列の完全長で測定されるべきである。
【0038】
DDF−VZV1又はDDF−VZV2(又は単離されたHCDRsやLCDRsなどの選択された部分)のいずれかを形成する重鎖及び軽鎖の可変領域は、特定のアイソタイプを持つ抗体の中に、特に、完全なヒト組み換え抗体の中に包含されることができる。この抗体は、2本の軽鎖及び2本の重鎖によって形成された四量複合体の天然の立体構造で、軽鎖及び重鎖可変領域としてDDF−VZV1又はDDF−VZV2のいずれかのVL及びVH配列を含んで成ることができる。完全なヒト抗体が望ましいとき、抗体は、ヒトIgG1、IgG2、IgG3、IgG4、IgM、IgA、及びIgE定常領域から成る群から選択される重鎖定常領域をさらに含んで成るべきである。例えば、IgGイソタイプは、ほとんどすべての承認された治療抗体の抗体形式である(Laffly E and Sodoyer R,2005)。しかしながら、ヒトIgG1から単離された抗原結合部分は、ヒトIgA配列に移されることができ、そして、得られた組み換え抗体が、HIV感染を抑制することができる抗体で最近示されたように、本来のIgG1の活性を維持した(Mantis N et al.,2007)。
【0039】
あるいは、DDF−VZV1又はDDF−VZV2(又は単離されたHCDRsやLCDRsなどの選択された部分)のいずれかを形成する重鎖及び軽鎖の可変領域は、Scfv(一本鎖可変フラグメント)、Fab(可変重鎖/軽鎖ヘテロ二量体)、二重特異性抗体、ペプタボディ(peptabody)、VHH(重鎖抗体の可変ドメイン)、単離された重鎖若しくは軽鎖、二重特異性抗体、及び非臨床/臨床適用のためのその他の遺伝子操作抗体変異体(Jain M et al.,2007;Laffly E and Sodoyer R,2005)などの異なった名称で文献に記載される機能的抗体フラグメントに関するあらゆる他のタンパク質形式に含まれるかもしれない。DDF−VZV1又はDDF−VZV2の組み換え変異体は、タグ付与したFabsの形態でpDLac−FLAGhisと呼ばれるpDD適合発現ベクターを使用することで製造された(図6Bは関連DNAフラグメントの概要地図に関し;配列番号18はDDF−VZV1を製造するために作り出されたこのようなフラグメントのDNA配列の例に関する)。DDF−VZV1及びDDF−VZV2のタグ付与バージョンを含んでいるpDLac−FLAGhisベースのベクターが作り出され、そして、DDF−VZV1 HCtagを製造するためにDDF−VZV1 LC(配列番号19〜22、図7A及びB)と共に、そして、DDF−VZV2 HCtagを製造するためにDDF−VZV2 LC(配列番号23〜26;図7C及びD)と共に使用された。
【0040】
追加抗体及び抗体フラグメントは、シャフリング軽鎖の過程を通してDDF−VZV1又はDDF−VZV2のいずれかの配列を使用することで作り出される。実際に、いくつかの異なった抗体が作り出され、そして、例えば、一般的なファージ・ディスプレイ技術、又はWO 07/007154に記載の技術を使用して、VL配列に関するライブラリーと組み合わせた1本重鎖可変ドメインVH(DDF−VZV1又はDDF−VZV2のいずれか一方など)を使用することで特異的な生物学的活性について試験され得る。このアプローチは、親和性、安定性、特異性、及び/又は組み換え体の製造に関して改善された特性を有するVH/VL組み合わせの決定を可能できる(Ohlin M et al.,1996;Rojas G et al.,2004;Suzuki K et al,2007)。
【0041】
そのうえ、特に(より良い薬物動態学的プロフィール、又は抗原に対するより高い親和性などの)臨床適用に関して、改善された特徴を有する抗体にするために特定の位置で抗体を修飾できることが知られている。これらの変更は、CDRs、及び/又はDDF−VZV1又はDDF−VZV2のいずれかのフレームワークに加えられる。配列は、親和性成熟及びその他の方法を利用して行われる抗体の合理的設計のための専用技術のいずれかを適用することによって決定できる(Kim S et al.,2005;Jain M et al.,2007)。
【0042】
新しい生理活性ペプチドを開発するための抗体ベースのストラテジーはまた、L−アミノ酸、及び/又はD−アミノ酸を含むCDR誘導ペプチド合成の実現可能性を示した。これらの分子は、より適切な薬理学的特性を伴った、本来の活性の比活性を維持する(Levi M et al.,2000;Wijkhuisen A et al.,2003)。そのため、本発明のHCDR3のそれぞれ、並びにそれらと高度に類似した配列、それらを含む融合タンパク質、そして、(例えば、D−アミノ酸を含むか、又はレトロ‐インベルソ型立体構造にある)それらに由来する合成ペプチドが、VZV結合タンパク質として使用及び試験され得る。
【0043】
本発明のタンパク質は、VZVに結合し、そして、中和する、抗体、抗体フラグメント、生理活性ペプチド、又は融合タンパク質として提供される。これらの代替タンパク質は、DDF−VZV1及びDDF−VZV2 Fabsについて測定されるそのような特性を強化しないにしても、維持するべきである。融合タンパク質の場合には、異種タンパク質配列は、その正しい発現と生物学的活性に影響することなく、VZV特異的部分(例えば、抗体フラグメントの特異的HCDR3又は可変領域)に対してN又はC末端位置に位置し得る。
【0044】
用語「異種タンパク質」は、VZV特異的部分(例えば、抗体フラグメント)に対してN又はC末端位置に、タンパク質配列が天然には存在していないことを示す。このタンパク質配列をコードするDNA配列は、一般的に組み換えDNA技術によって融合され、そして、少なくとも5つのアミノ酸をコードする配列を含んで成る。存在しているこの異種配列は、一般的にVZV特異的融合タンパク質に追加特性を与えるために選ばれている。このような追加特性の例には、検出若しくは精製のためのより優れた手段、追加の結合部分若しくは生物学的リガンド、又は融合タンパク質の翻訳後修飾(例えば、リン酸化、グリコシル化、ユビキチン化、SUMO化、若しくはタンパク質内部分解性開裂)が含まれる。
【0045】
あるいは(又は1つ以上の異種タンパク質配列への融合に加えて)、本発明のタンパク質の活性は、治療薬、安定化剤、又は診断薬などの異なった化合物との結合によって改善することができる。これらの作用物質の例は、化学リンカー又は重合体を使用して結合され得る検出可能な標識(例えば、放射性同位元素、蛍光化合物、毒素、金属原子、コロイド状金属、化学発光化合物、生物発光化合物、又は酵素)である。本発明のタンパク質のVZV特異的生物学的活性は、診断上の又は治療上の適用において代謝、及び/又は安定性を変える重合体のような化合物との融合によって改善される。治療活性は、別の抗ウイルス・タンパク質、あるいは、細胞の代謝、及び/又は活性を変えるタンパク質などの別の治療用タンパク質との融合によって改善される。
【0046】
タンパク質部分、リガンド、及び適切なリンカーの選択及び設計方法、並びに融合タンパク質の構築、精製、検出、及び使用のための方法及びストラテジーは、文献(ニルソンら、1997;「キメラ遺伝子と雑種タンパク質の適用」Methods Enzymol Vol.326−328、アカデミックプレス社、2000;WO01/77137)内に提供されており、且つ、臨床検査室及び研究室で一般的に利用可能である。例えば、融合タンパク質は、融合タンパク質の生体内、及び/又は試験管内における識別、又はその精製を容易にできる(ポリヒスチジン、FLAG、c−Myc、若しくはHAタグなどのタグを含めた)商業的な抗体によって認識される配列を含む。他のタンパク質配列は、(緑色蛍光タンパク質の場合のように)直接蛍光分析によって、又は(例えば、タンパク分解部位を使用した)特異的基質又は酵素によって容易に識別できる。
【0047】
VZV特異的抗体、抗体フラグメント、及び融合タンパク質の安定性は、ファージ・コート・タンパク質(単離されたか若しくは組み換えファージ内に含まれているcp3又はcp8)、マルトース結合タンパク質(MBP)、ウシ血清アルブミン(BSA)、又はグルタチオン−S−トランスフェラーゼ(GST)などの周知の担体タンパク質との融合により改善される。
【0048】
本発明のタンパク質はまた、VZVエンベロープに対する中和抗原を特徴づけするためにも使用できる。実際、DDF−VZV1及びDDF−VZV2は、最初、ELISAによる、VZV感染細胞株から得られた細胞抽出物へのそれらの特異的結合のためにクローンニングされ(図1)、そして、VZV感染を中和するそれらの能力はまた、VZV基準菌株を使用した試験管内における中和アッセイによって測定された(図5及び8)。その結果、本発明のタンパク質は、DDF−VZV1又はDDF−VZV2と競合する(例えば、完全な抗体、Fabs及び他の抗体フラグメント、生理活性ペプチド、又は融合タンパク質の形態の)他のVZV結合タンパク質を規定するのに使用できる。このような競争タンパク質は、任意に、本来のDDF−VZV1又はDDF−VZV2配列において確認されたものと一部又は完全に異なったHCDRs及びLCDRsと共に、先に規定されたHCDR3配列のいずれかを単に含む。
【0049】
(本発明の抗体、抗体フラグメント、生理活性ペプチド、又は融合タンパク質に関して)そのような競合タンパク質は、実施例又は文献に記載の関連アッセイによって測定される、本発明のタンパク質と競合するそれらの能力、それから、VZV感染を中和するそれらの能力を実証することによってスクリーニングされ、そして、単離され得る。本発明の背景は、類似活性を測定するための異なるアプローチに関するいくつかの基準を提供する。具体的には、本発明の抗体、抗体フラグメント、及び他のタンパク質は、抗体フラグメント(Kausmally L et al.,2004;Drew P et al.,2001;Suzuki K et al.,2007)、マウス若しくはヒト・モノクローナル抗体(Grose C et al.,1983;Montalvo E and Grose C,1986;Sugano T et al.,1987;Forghani B et al.,1994;Lloyd−Evans P and Gilmour J,2000)、並びにヒト以外/ヒト血清(Fowler W et al.,1995;Haumont M et al.,1997;Garcia−Valcarcel M et al.,1997)についてVZV特異的生物学的活性及びエピトープを特徴づけるために使用されるアッセイにより試験され得る。
【0050】
免疫蛍光研究(図4)は、他の抗体及び抗体フラグメントについてこれまでに示されたように、DDF−VZV1とDDF−VZV2が異なったVZV抗原を認識することを示した(Suzuki K et al.,2007;Wu L and Forghani B,1997;Grose C et al.,1983)。文献は、VZV抗原と各Fabによって認識される特異的エピトープが測定でき、且つ、過去に測定したものと比較できることを使用した技術に関するいくつかの例を提供する。例えば、VZVタンパク質、及び関連する切断変異体若しくは合成ペプチドを使用したELISA、免疫沈降反応、又はウエスタン・ブロットを、使用して、関連エピトープを同定した(Krah D,1996,Sauerbrei A and Wutzler P,2006)。具体的には、前記エピトープは、糖タンパク質E又は糖タンパク質B(Fowler W et al.,1995;Haumont M et al.,1997;Garcia−Valcarcel M et al.,1997;Kjartansdottir A et al.,1996)、及び糖タンパク質L:糖タンパク質H複合体(Forghani B et al.,1994;Yokoyama T et al.,2001;Suzuki K et al.,2007)内で同定された。
【0051】
本発明のタンパク質のVZV関連の予防上、診断上、及び治療学上の使用についてのより詳しい特徴づけとバリデーションは、その次に、本発明の背景にまとめられているとおり、VZV発症機序と免疫生物学を研究するための文献に開示されている試験管内又は生体内におけるアッセイ(組織若しくは細胞ベースのアッセイ、齧歯動物において確立された病態モデル)の1つ以上を使用することで実施できる(Forghani B et al.,1994: Vafai A et al.,1991;Wu and Forghani,1997;Fowler W et al,,1995;Maple P et al.,2006;Matsuo K et al.,2003;Kutinova L et al.,2001;Massaer M et al.,1999;Haumont M et al.,1997;Grose C,2006;Baiker 2004;Ku C et al.,2005;Taylor S and Moffat L,2005;Zerboni L et al.,2005)。
【0052】
本発明のさらなる目的は、先に規定された抗体、抗体フラグメント、融合タンパク質、又は単離CDRのいずれかをコードする核酸である。実施例は、具体的には、DDF−VZV1又はDDF2−VZV2重鎖及び軽鎖の完全な可変領域をコードする配列を提供する(配列番号1、6、8、及び13)。前記核酸は、配列番号1、配列番号6、配列番号8、及び/又は配列番号13と少なくとも90%の同一性を有していなければならない。前記配列、具体的には、特異的なCDRs(図2と3を参照のこと)に関連するそれらの中の配列は、例えば、発現ベクター内のプロモーターに作動できるように連結されるか、若しくはpDDベースのファージミド内にクローンニングされて、ベクター及びDNA発現カセット内に、並びにいずれか他のファージミド内に含まれ得る。これにより、(実施例に示されているように)本発明のタンパク質を発現するファージミド・ベクターを含んで成る組み換えファージは、VZV感染の検出手段、及び/又は中和手段として使用され得る。
【0053】
完全ヒト抗体が望ましい場合には、発現ベクターは、IgG1、IgG2、IgG3、IgG4、IgM、IgA、及びIgE定常領域から成る群から選択される重鎖の定常領域をさらに含んで成るはずである。着目の重鎖及び軽鎖の関連可変領域をコードする核酸配列は、それらが適切な調節配列(例えば、プロモーター、転写ターミネータ)に作動できるように連結される、1つのベクター又は異なったベクターの発現カセット内に適切にクローンニングされるべきである。発現カセットは、所望のタンパク質のための単シストロン性若しくは2シストロン性の転写産物の発現を駆動できるプロモーター、(必要であれば)リボソーム結合部位、開始/終止コドン、及びリーダー/分泌配列を含むはずである。
【0054】
VZVに結合し、そして、中和できると先に規定された本発明の抗体、抗体フラグメント、HCDRs、融合タンパク質、及びいずれか他の化合物は、適切な宿主細胞を形質転換するためのそのようなベクター、及び組み換えタンパク質としてのそれらの発現を可能にする十分に確立された技術を使用して製造され得る。これらの調製物は、VZV関連の予防上、診断上、及び治療上の使用に関するより詳細な特徴づけとバリデーションを実施するのに十分な量の組み換えタンパク質(マイクログラムからミリグラムの範囲)を提供するべきである。
【0055】
本発明の配列がクローンニングされ、そして、対応する組み換えファージの手段によって特徴づけされたpDDベースのファージミドは、本来のFabs、又はそれらから得られたタンパク質配列が(pDLac−FLAGhis系を用いて実施例で示されているように)宿主細胞内で組み換えタンパク質として適切に発現されるベクター内に(一部又は全体として)移され得るDNA配列を含む。ベクターは、構成的に活性であるか又は誘導性であるように選択された転写開始/終止調節配列の制御下、原核生物又は真核生物宿主細胞において組み換えタンパク質の発現を可能にするべきである。
【0056】
本発明の核酸を含んで成る宿主細胞は、原核生物又は真核生物宿主細胞であり、所望の組み換えタンパク質の分泌を可能にすべきである。そして、このような細胞が実質的に豊富になった細胞株は、単離されて、安定細胞株をもたらす。前記のタンパク質の製造方法には、タンパク質の発現に好適な条件下、それらのコード配列を含んで成る発現ベクターで形質転換された宿主細胞を培養し、そして、その宿主細胞培養物からタンパク質を回収するステップが含まれる。
【0057】
これらの核酸、組み換えファージ、及び宿主細胞は、一般的な組み換えDNA技術を適用することによって本発明のタンパク質を製造するのに使用できる。簡単に言えば、所望のDNA配列は、制限酵素でファージミドを消化することによって、又はポリメラーゼ連鎖反応(PCR法)の鋳型として本来のファージミドを使用し、及び重鎖及び軽鎖の完全な可変領域若しくは(HCDR3などの)それらの一部分だけを特異的に増幅するためのPCRプライマーを使用して増幅することによって抽出できる。
【0058】
次に、前記DNAフラグメントは、オックスフォード大学出版より刊行された「A Practical Approach」というシリーズの中のいくつかのタイトル(“DNA Cloning 2: Expression Systems”,1995;“DNA Cloning 4: Mammalian Systems”,1996;“Protein Expression”,1999;“Protein Purification Techniques”,2001)を含めた、組み換えタンパク質をどのようにクローンニングし、そして、製造するかに関する多くの書籍と総説に記載のとおり、原核生物又は真核生物宿主細胞内でのさらなる一時変異、及び/又は発現のためにより適切なベクターに移されてもよい。
【0059】
真核生物宿主(例えば、酵母、昆虫、又は哺乳動物細胞)に関して、宿主の性質によって、異なった転写及び翻訳調節配列を用いることができる。それらは、調節シグナルが、発現レベルが高い特定の遺伝子に関連するアデノウイルス、ウシ乳頭腫ウイルス、シミアンウイルス、又は同様のものなどのウイルス由来である。例は、ヘルペスウイルスのTKプロモーター、SV40初期プロモーター、酵母gal4遺伝子プロモーターなどである。一過性の(構成的)の抑制及び活性化を可能にし、その結果、修飾遺伝子発現を可能にする転写開始調節シグナルが選択できる。さらなるクローニング・ステップの間、抗体又は融合タンパク質をコードする配列は、例えば、特定のベクター及び宿主細胞を使用して組み換えタンパク質をクローンニングし、そして、発現するためのコドン使用及び制限部位が最も適当であるDNA配列を選択するためのソフトウェアを使用することで、決定され得るDNAレベルにおいてのみ、又はDNA及びタンパク質レベルの両方における特異的修飾のために、他のベクター内に適合させ、そして、再クローニングし得る(Rodi D et al,,2002;Grote A et al.,2005;Carton J et al.,2007)。タンパク質配列もまた、(Scfv、Fab、完全ヒト抗体などの)所望の抗体形式、又は1つ以上の内部アミノ酸の挿入、置換、又は除去と関連して追加され得る。
【0060】
これらの技術もまた、抗体の治療学的特性のさらなる構造的及び機能的な特徴づけ及び最適化のために(Kim S et al.,2005)、あるいは、それらの安定した生体内デリバリーを可能にするベクターを製造するために使用され得る(Fang J et al.,2005)。例えば、組み換え抗体もまた、可変領域に融合される特異的なFc領域を選択することによって(Furebring C et al.,2002;Logtenberg T,2007)、組み換え一本鎖抗体フラグメントを製造することによって(Gilliland L et al.,1996)、安定化ペプチド配列を融合することによって(WO01/49713)、又は化学的に修飾した残基に放射性化学物質又は重合体を追加することによって(Chapman A et al.,1999)、構造、及び/又は活性レベルで修飾され得る。
【0061】
いったん好適なエピソマー、又は非相同若しくは相同の組み込み型ベクター(integrating vector)内に挿入された、ディスプレイ及び選択されたタンパク質配列をコードするDNA配列は、(形質転換、トランスフェクション、接合、プロトプラスト融合、エレクトロポレーション、リン酸カルシウム沈殿、ダイレクト・マイクロインジェクションなどの)好適な手段のいずれかによって適切な宿主細胞内に導入されて、それらを形質転換することができる。特定のプラスミド又はウイルス・ベクターを選択する場合に考慮される重要な要因は:ベクターを含まない宿主細胞の中からベクターを含む宿主細胞を認識し、そして、選択することの容易さ;特定の宿主体内で所望されるベクターのコピー数;そして、異なる種の宿主細胞間のベクターである「シャトル」にそれが望ましいかどうかである。
【0062】
導入されたDNAによって安定に形質転換された細胞は、発現ベクターを含む宿主細胞の選別を可能にする1つ以上のマーカーを導入することによっても選択され得る。前記マーカーはまた、栄養要求性宿主に対して光合成栄養、殺生物剤抵抗性、例えば、抗生物質、若しくは銅などの重金属、又は同様のもの、に対する抵抗性も与え、そして、必要性であれば、切断可能であるか又は抑制される。選択マーカー遺伝子は、発現されるべきDNA遺伝子配列に直接連結され得るか、又は同時トランスフェクションによって同じ細胞内に挿入され得る。追加の転写制御要素もまた、最適な発現に必要である。
【0063】
宿主細胞は、原核生物又は真核生物のいずれかである。原核生物宿主細胞の中で、好ましいものは、B.ズブチリス(B.subtilis)とE.コリ(E.coli)である。真核生物宿主細胞の中で、好ましいものは、酵母、(バキュロウイルス・ベースの発現系を使用した)昆虫細胞、又は哺乳動物細胞、例えば、ヒト、サル、マウス、(バキュロウイルス・ベースの発現系を使用した)昆虫、及びチャイニーズハムスター卵巣(CHO)細胞などである。なぜならば、哺乳動物細胞が、正しい折りたたみ、又は正しい部位でのグリコシル化の特定の形態を含めた、タンパク質分子に対する翻訳後修飾を提供するからである。また、酵母細胞もグリコシル化を含めた翻訳後ペプチド修飾を行う。酵母において所望のタンパク質の製造に利用できる強力なプロモーター配列と高いプラスミド・コピー数を利用する、多くの組み換えDNAストラテジーが存在する。酵母は、クローンニングされた哺乳動物遺伝子産物、及びリーダー配列を担持している分泌ペプチド(すなわち、プレ‐ペプチド)の中のリーダー配列を認識する。
【0064】
発現のための宿主として利用可能な哺乳動物細胞株は、当該技術分野で公知であり、そして、それには、これだけに制限されることなく、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、ベビーハムスター腎臓(BHK)細胞、サル腎臓(COS)細胞、C127細胞、3T3細胞、BHK細胞、HEK 293細胞、Per.C6細胞、Bowes黒色腫細胞、及びヒト肝細胞癌(例えば、Hep G2)細胞、及びその他の多くの細胞株を含めたアメリカン・タイプ・カルチャー・コレクション(ATCC)から入手可能な多数の不死化細胞株が含まれる。バキュロウイルス系において、バキュロウイルス/昆虫細胞発現系の材料は、(例えば、Invitrogenによって商品化されている)キット形態で市販されている。
【0065】
長期間にわたる、高収率産生の組み換えポリペプチドの、安定的な発現が好まれる。例えば、着目のポリペプチドを安定して発現する細胞株は、同じか、又は別のベクター上にウイルス複製開始点、及び/又は内因性発現要素、並びに選択マーカー遺伝子を含む発現ベクターを使用することで形質転換される。ベクターの導入に続いて、細胞を、強化培地中で1〜2日間、増殖させ、その後、それらを選択培地に切り換える。選択マーカーの目的は、選択に対する耐性を与えることであり、その存在が導入された配列を首尾よく発現する細胞の増殖と回復を可能にする。安定に形質転換された細胞の耐性クローンは、細胞型に適切な組織培養技術を使用することで増殖する。そして、このような細胞が実質的に豊富になった細胞株は、単離されて、安定な細胞株を提供し得る。宿主細胞は、組み換えタンパク質の発現レベルに基づいてさらに選択される。
【0066】
ファージ・ディスプレイ技術を使用して単離される免疫グロブリン可変鎖(具体的には、ヒト免疫グロブリン可変鎖)の場合、重要な変更は、選択されたFab又はscFVの、好ましいアイソタイプ及び定常領域を持つ完全な免疫グロブリン・タンパク質への転換である。こういう種類の変更は、例えば、ファージ・ディスプレイ・ライブラリー由来の単鎖Fv又はFabsから構築されたすべてのアイソタイプの完全なヒト・モノクローナル抗体を製造し、そして、哺乳動物又は昆虫細胞内で発現することを可能にする。文献に広く記載のとおり(Persic L et al.,1997;Guttieri M et al.,2003)、ベクターは、所望のアイソタイプ(例えば、ヒトIgGガンマ)の定常(Fc)領域に対するこの配列の融合を可能にする発現抗体のために特異的に設計される。抗体又は融合タンパク質は、一過性に又は安定に形質転換された細胞として高レベルの発現を可能にする原核生物細胞(例えば、エシェリキア・コリ(Escherichia coli);Sorensen H and Mortensen K,2005;Venturi M et al.,2002)、植物細胞(Ma J et al.,2005)、又は真核生物細胞において組み換えタンパク質として発現される(Dinnis D and James D,2005)。これは、具体的には、抗体の特徴づけが、より困難なアッセイ、及び/又は生体内アッセイを使用して実施されなければならない場合に必要であるだろう。
【0067】
文献は、記事及び書籍の章で概説されるとおり、原核生物宿主細胞において、Fabとしてタンパク質を、又は抗体フラグメントと同様の形式を発現するための異なったストラテジーを提供する(“Phage display:A Practical Approach”,vol.266,ed.Clackson and Lowman H,Oxford Univ.Press,2004;“Phage Display:A Laboratory Manual”,ed.Burton D et al.,CSHL Press,2001;Corisdeo S and Wang B,2004;Benhar I,2001)。
【0068】
タンパク質、特に抗体が真核生物宿主細胞(具体的には、哺乳動物細胞株)において発現されるとき、異なったベクター及び発現系が、トランスフェクト細胞株の安定したプールを作り出すように設計される(Aldrich T et ah,2003;Bianchi A and McGrew J,2003)。組み換え抗体の高レベルの、最適化された、安定した発現は、細胞培養条件の最適化によっても(Gmnberg J et al.,2003;Yoon S et al.,2004)、そして、より高いレベルの抗体産生及び分泌を有するクローンを選択するか又は設計することによって(Bohm E et al.,2004;Butler M,2005)達成された(Schlatter S et al.,2005)。
【0069】
VZVに結合し、中和できると先に規定した抗体、抗体フラグメント、生理活性ペプチド、融合タンパク質、及びあらゆるその他タンパク質は、細胞培養物又は合成調製物からの非組み換え/組み換えタンパク質のいずれかの分離を可能にする十分に確立された技術、すなわち、抽出、沈殿、クロマトグラフィー、電気泳動、又は同様のものを含めた従来手順のいずれかを使用して精製され得る。抗体精製の方法は、カラム内に含まれる固定されたゲルマトリックス(Nisnevitch M and Firer M,2001;Huse K et al.,2002;Horenstein A et al.,2003)、具体的には、プロテインA若しくはプロテインGなどの基質、又は合成基質に対する抗体の一般的な親和性によるもの(Verdoliva A et al.,2002;Roque A et al.,2004)、並びに抗原又はエピトープ・ベースのアフィニティー・クロマトグラフィ(Murray A et al.,2002;Jensen L et al.,2004)を活用できる。洗浄の後に、タンパク質は、pH又はイオン強度の変化によってゲルから溶出される。あるいは、HPLC(高速液体クロマトグラフィー)が使用できる。溶出は、タンパク質精製のために一般的に用いられる水‐アセトニトリル・ベースの溶媒を使用することで行われ得る。
【0070】
VZVに結合し、中和できると先に規定された抗体、抗体フラグメント、生理活性ペプチド、融合タンパク質、及びあらゆるその他の化合物が、VZV感染を検出、治療、抑制、予防、及び/又は改善するために使用できる。この目的のために、前記化合物が、VZV感染及びVZV関連疾患の医療管理のための診断用、治療用、又は予防用の組成物を調製するのに使用できる。
【0071】
これらの組成物は、ヒトDDF−VZV1及びDDF−VZV2の配列と活性に基づいて先に規定された抗体、抗体フラグメント、融合タンパク質、CDR、及びあらゆるその他の化合物を含んで成る。前記組成物は、異なるVZV中和抗体若しくは抗体フラグメント、免疫グロブリン静注(IVIg)調製物、ステロイド、及び/又は抗ウイルス化合物をさらに含んで成る。異なるVZV中和抗体又は抗体フラグメントは、既に文献に記載されているものなどの異なるエピトープを特徴とするべきである。実際、文献には、ウイルス又はヒト標的に向けられた2種類以上の抗体が医薬組成物中で組み合わせられた場合に、得られた組成物が単なる相加効果ではなく、特定の相乗効果による改善された治療効果を有することができる多くの例が示されている(Logtenberg T,2007)。
【0072】
医薬組成物は、任意に、いずれかの医薬として許容されるビヒクル又は担体を含んで成る。これらの組成物は、ワクチン、免疫調節免疫グロブリン静注製剤、ステロイド、又は抗ウイルス化合物などのあらゆる追加の治療薬又は予防薬をさらに含んで成る(又はそれらと一緒に投与できる)。文献は、VZV複製に作用し、且つ、単独又は免疫グロブリン静注調製物と組み合わせて、ヒトで既に試験されている前記化合物に関するいくつかの例を提供する(Huang Y et al.,2001;Carby M et al.,2007;Koren G et al.,2002)。そのうえ、最近の文献は、前記医薬組成物の頻度、及び/又は投薬量を低減する機会を与える、ヒト・モノクローナル抗体が免疫グロブリン静注調製物、ステロイド、及び/又は抗ウイルス化合物などの現在の治療を補う(可能であれば、置き換える)ために使用され得ることを示唆している(Bayry J et al.,2007)。
【0073】
先に規定されたタンパク質(例えば、抗体、抗体フラグメント、融合タンパク質、生理活性ペプチド)及び核酸のいずれかを含んで成る組成物は、VZVに関連した診断上、治療上、又は予防上の目的がある人に投与され得る。これらの組成物は、VZVビリオンをターゲッティングすることによって、治療された患者のウイルスの増殖を抑制できる、及び集団内でのウイルス感染の大発生を妨げる可能性のある治療用化合物(具体的には、治療抗体若しくは治療抗体フラグメント)を提供するVZV特異的な受動免疫のための手段として投与され得る。
【0074】
特定の使用に依存して、組成物は、より長い又はより短い期間、(VZVに感染しているか、又は入院、免疫抑制薬若しくは化学療法剤治療、又はVZVに感染した人との接触のためVZVの危険性があると考えられるあらゆる幼児、妊婦、高齢者、又はその他の人である)ヒト対象に化合物を提供するべきである。この目的のために、組成物は、噴霧装置、吸入器によって、又は点眼薬として、非生分解性/生分解性基質物質中、あるいは、マイクロビーズなどの粒状薬物デリバリー・システムを使用して、様々な経路:筋肉内、静脈内、皮下、局所、粘膜を通じて、単回又は複数回の投薬量で、及び/又は適切なデバイスを使用して投与され得る。
【0075】
具体的には、組成物は、皮膚及び眼におけるVZVの存在を考慮すると、有用なアプローチである局所的投与及び眼内投与を可能にする(Arvin A,1996;Quinlivan M and Breuer J,2006;Liesegang T,2004)。そのうえ、抗体及び抗体フラグメントは、角膜に対して局所的に適用された場合に有効である(Brereton H et al.,2005;Nwanegbo E et al.,2007)。
【0076】
医薬組成物は、第二次ウイルス血症に関連する皮膚疹におけるウイルスの存在を考慮すると、化合物が、特に局所投与に関して、十分な期間その活性を発揮することを可能にする治療的又は予防的に有効量の化合物を対象に対して提供すべきである。望ましい効果は、VZV感染、再活性化、及び/又は再感染を制御することによって、並びにVZV感染の臨床徴候の少なくともいくつかを低減することによって、患者の体質を改善することである。例えば、前記組成物は、投与経路、投与される投薬量の数、及びその人の体質によって、約0.005〜約50mg/kg/体重の有効量にて投与されるべきである。
【0077】
診断用途のある組成物の場合、化合物は、生物学的サンプル中のウイルスを検出するための臨床検査室及び研究室において一般的に確立された技術(例えば、ELISA又はその他の血清学的なアッセイ)を使用して、又は生体内で対象に投与された場合には、投与後少なくとも1、2、5、10、24時間、又はそれより長い時間、検出されるべきである。VZVの検出は、免疫応答性宿主及び免疫不全宿主の両方から成る危険性のある集団における慢性又は急性VZV感染を観察するために確立された公知の手段及び手順と置換して又は合わせて、本発明のタンパク質を使用して実施され得る。
【0078】
本発明のタンパク質はまた、VZV感染、並びにVZV関連疾患を検出、治療、抑制、予防、及び/又は改善するための組成物の調製にも使用できる。これらの疾患は、VZV感染の合併症から生じる可能性がある(Dworkin R et al.,2007;Weinberg J,2007)。
【0079】
背景技術に示されているとおり、ヘルペス後神経痛の症例のように、劇的、且つ、衰弱させる効果を持つ可能性がある神経学的、眼性、又は臓器を冒す効果を持っている数多くの帯状ヘルペスの帯状ヘルペス合併症が存在する(Oxman M et al.,2006;Liesegang T,2004)。そのうえ、VZVの再活性化及び関連する合併症はまた、癌患者(Sandherr M et al.,2006)又は炎症性膠原病に罹患している患者、そして、通常、コルチコステロイドなどの免疫抑制治療、又は化学療法及びその他の抗体ベースの免疫抑制レジメン下にある患者においても見られた。
【0080】
VZV又はVZV関連疾患の治療、予防、又は診断方法は、先に規定されたタンパク質又は核酸の投与を含んで成る。前記方法は、異なるVZV中和抗体若しくは抗体フラグメント、免疫グロブリン静注(IVIg)調製物、ステロイド、及び/又は抗ウイルス化合物の投与をさらに含んで成る。
【0081】
臨床開発及び使用は、抗体の薬物動態学及び薬力学の特徴づけ(Lobo E et al.,2004)、前臨床的及び臨床的安全性データ(Tabrizi M and Riskos L,2007)、並びに治療学用組み換え抗体の商業的な製造規模の製剤と分析的特徴づけのための公的な要件の順守(Harris R et al.,2004)に基づくべきである。
【0082】
どのような形であっても本発明を制限すると解釈されるべきでない以下の実施例によって、本発明は説明されるだろう。
【実施例】
【0083】
実施例1:ELISAによるヒトFabs結合VZVタンパク質抽出物の発現と選別
材料と方法
ライブラリー構築
ヒトIgG1の重鎖及び軽鎖をコードするcDNAを、文献に従ってVZV血清陽性個体から得られたリンパ球から入手した(Burioni et al.,1998;“Phage Display:A laboratory Manual”,Burton DR et al.,CSHL Press,2001)。ファージ・ライブラリーを、PCT特許出願WO 07/007154に記載の技術に従ってpDDベクターと適合するクローニング・カセットを使用することで構築し、そして、Fabsを、ライブラリーの中の組み換えファージ表面上に発現させた。
【0084】
pDDベースのFabライブラリーのパンニング、そして、陽性クローンの配列決定によるヒトFabsの選別を、文献に記載されたとおり実施した(Burioni R et al.1998)。
【0085】
特異的なFabsのCDRsを、IMGT/V−QUEST(Giudicelli V et al.,2004)によって提供された予測及び配列比較と、ヨーロッパ生物情報科学研究所によって提供されるもの及びFASTA(http://www.ebi.ac.uk/fasta33/index.html)を使用して検索可能なものなどのヒト抗体のタンパク質配列を含む他のデータベースを比較することによって規定した。
【0086】
培養ヒト細胞からのタンパク質抽出物の調製
VZV分離及び増殖のために一般的に使用されるヒト胎児肺繊維芽細胞株である細胞株MRC−5(ATCC受入番号CCL−171)を、ファージ・ディスプレイ・ライブラリーのパンニングのため、及びELISAによりFabsを試験するために、VZVに特異的な物質の調製に使用した。MRC−5細胞を、10%の不活性化ウシ胎仔血清(FBS)、50μg/mlのペニシリン、100μg/mlのストレプトマイシン、及び2mMのL−グルタミンを含む変法イーグル培地中で維持する。
【0087】
細胞(VZV感染又は未感染MRC−5)を、掻き取り、そして、250mlの溶解バッファー(50mMのTris−HCl pH8.0;150mMのNaCl;0.02%のアジ化ナトリウム;0.5%のTritonX)中に再懸濁し、氷上で20分間インキューベートし、次に、4℃、12000rpmにて2分間遠心分離した。得られた上清のタンパク質濃度を、BCATM Protein Assay Kit(Pierce)を使用して、二重反復試験で測定した。未知試料のタンパク質濃度を、測定し、そして、既知濃度(0mg/ml=ブランク〜2.000mg/ml=最高値)のウシ血清アルブミン(BSA)の段階希釈を基準にして報告した。すべてのサンプルの吸収度を、540nmにセットした分光光度計を用いて計測した。
【0088】
タンパク質抽出物を使用したパンニングとELISA
タンパク質コーティングを、以下の抗原:VZV(ELLEN菌株;ATCC受入番号VR−586)を感染させたMRC−5ヒト線維芽細胞の細胞溶解物;インフルエンザ・ウイルス抗原(Virion Ltd.)の市販調製物;未感染MRC−5の細胞溶解物;ウシ血清アルブミン(BSA)を使用して、96ウェル・プレート上に実施した。各サンプルを、炭酸バッファーで希釈し(1ウェルあたり25□lの終量中に100ナノグラムの総タンパク質)、そして、そのプレートを4℃にて一晩インキューベートした。蒸留水で洗浄した後、そのプレートを、1%のBSAを含むPBS中、37℃にて1時間のインキュベーションによってブロッキングした。
【0089】
ELISAを、文献中に開示されているプロトコールを使用し、以下のFabs:DDF−VZV1、DDF−VZV2、及び調製された無関係なFabであるe137、を含む40□lの未希釈サンプルを使用して実施した(Bugli F et al.,2001)。前記Fabsを、二重反復試験のウェル内で試験した。各Fabとの37℃にて1時間のインキュベーション、そして、0.1%のTween−20、40μlのヤギ抗ヒトFabを含むPBSでの5回の洗浄の後に、ペルオキシダーゼ・コンジュゲート(Sigma;カタログ番号A0293)を、加え、そして、37℃にて1時間インキューベートした。前述のとおりプレート洗浄を繰り返し、そして、40μlの基質(TMB Substrate Kit;Pierce)を各ウェルに加えることによって、酵素反応を進めた。ELISA反応を、37℃にて15分間進めた。酵素活性を、停止溶液(H2SO4)を加えることによって停止させ、そして、吸収度を、450ナノメートルにセットした分光光度計を用いて計測した。
【0090】
ELISA及び中和アッセイのためのFab調製
プロトコールは、pDD、又はpGem変異体などの他のファージミドを使用した文献に記載のものと同様であった(“Molecular Cloning:A Laboratory Manual”,Sambrook et al.,Cold Spring Harbor Press,NY,1989;Burioni R et al.,1998;Burioni R et al.,2001;WO 07/007154)。
【0091】
簡単に言えば、ライブラリーの個々のE.コリ・クローンを、抗生物質を含む200mlのSuper Broth(SB;3.5%のバクト−トリプトン、2%の酵母抽出物、0.5%のNaCl)培地中で培養し、IPTGを補い、集菌し、そして、リン酸緩衝食塩水(PBS)で洗浄した。制御パルスを用いて4℃にて超音波処理することによって、溶解を細胞膜周辺腔に限定した。周辺質抽出物中のFabsを、JA−10ローターによる4℃、12,000rpmにて45分間の超遠心分離によってある程度精製した。生成物を、濾過し、そして、Centriconフィルタを用いて10回濃縮した。
【0092】
ある程度精製したFabsの濃度を、ImmunoPure Goat Anti−Human IgG[F(ab’)2](Pierce;カタログ番号31132)を使用したサンドイッチELISA法によって周辺質抽出物中で測定し、それを、96ウェル・プレート(Costar;カタログ番号3690)の表面上に結合させた。37℃にて1時間のインキュベーションの後に、プレートを、脱イオン水で6回洗浄し、そして、3%のBSAを含む170μl/ウェルのPBSを使用してブロッキングした。37°にて1時間のさらなるインキュベーションの後に、1%のBSAを含むPBS中、それぞれのFab、又は既知濃度の対照ヒトFab(Cappel;カタログ番号6001−0100)の一連の3倍希釈物50μlを、各ウェルに加え、そして、37℃にて1時間インキューベートした。次に、プレートを、TPBS(0.05%のTween−20を含むPBS)で6回洗浄した。次に、50μlのアルカリ性ホスファターゼ結合ヤギ抗ヒト抗体(Pierce;カタログ番号31312)を加え、そして、37℃にて1時間インキューベートすることによって、抗体結合を測定した。プレート洗浄を、前述のとおりTPBSを用いて繰り返し、そして、100μlのp−ニトロフェニル・リン酸二ナトリウム(Sigma)を各ウェルに加えた。ELISA反応を60分間進め、そして、結果を対照ヒトFabに対してプロットした。
【0093】
結果
組み換えファージに関するライブラリーを、pDD技術(WO 07/007154)によって作り出し、そして、VZVの臨床分離株を感染させたヒト線維芽細胞の細胞株から得られたタンパク質抽出物に対してパンニングした。5ラウンドのパンニングもまた、VZVに感染していない同じ細胞株からの対照タンパク質抽出物を使用することで、同じライブラリーを用いて平行して実施した。
【0094】
第3ラウンドまでに、対照タンパク質抽出物に対してパンニングしたサンプルのファージ力価は104未満であり、その一方で、第3ラウンドにおいてVZV抽出物に対してパンニングしたサンプルのファージ力価は104より高く、第5ラウンドのときに105に達した。この値は、Fabs結合VZV抗原をその表面上に発現する組み換えファージ内でのライブラリーの連続的な濃縮を実証している。
【0095】
第5ラウンドのパンニング後に得られた260個を超えるクローンを、ELISAにより個別に試験し、そして、それらの中の86個を陽性と確認した。選択されるクローンにおけるHCDR3のPCRと配列分析では、ヒトFabsを特徴とする2個の重鎖を同定し、ここで、DDF−VZV1及びDDF−VZV2と名付けた。DDF−VZV1又はDDF−VZV2を発現する組み換えファージの反応性を、VZV特異タンパク質抽出物に対して、並びに無関係な抗原(未感染細胞、ウシ血清アルブミン)に対して試験し、ELISAフォーマットによりそれらの強い結合活性を確認した(図1)。結合の特異性もまた、選択されたFabsが全く有意な親和性を示さなかったインフルエンザ・ウイルス抗原を使用して確認した。
【0096】
これらのFabsの完全な重鎖及び軽鎖可変領域のDNA配列を、DDF−VZV1及びDDF−VZV2について、対応するCDRsと共に測定した(図2及び3)。また、タンパク質発現用のE.コリ・ベースのシステムを使用して、さらなるアッセイに十分な量の組み換えタンパク質を得るために、これらのFabsを他のベクター内に再クローニングした。
【0097】
実施例2:VZV感染細胞培養において試験したDDF−VZV1とDDF−VZV2の特性
材料と方法
VZV特異的ヒトFabsの中和アッセイ
プラーク減少アッセイを、37℃にて72時間のインキュベーション後にコンフルエントな単層が通常形成される条件下でプレート内に植菌された104〜105個のMRC−5細胞を使用してCostarの24ウェル・プレート内で実施した。使用されるVZVウイルス株は、臨床分離株であった。
【0098】
Fabsを、実施例1に示したようにある程度精製し、そして、維持培地中に懸濁された等量のVZV無細胞ストック(ELLEN菌株;感染多重度0.01)と、様々な濃度(0.01、0.1、1、10、及び50μg/ml)で混合した。対照を、Fabsの不存在下(ブランク対照)、又はヒトC型肝炎ウイルスに特異的な無関係なFab(e137;Bugli F et al.,2001)の存在下、等量の維持培地とウイルスから構成した。
【0099】
37℃にて1時間のインキュベーション後に、250μlのウイルス・フラグメントFab混合物又は対照混合物を、培地を取り除いたウェル内に(二重反復試験の状態に)植菌した。プレートを、37℃にて2時間インキューベートして、未中和ウイルスを吸着させた。接種材料を取り除き、そして、1.5mlの維持培地を加えた。細胞培養条件下、1週間のインキュベーション後に、細胞を、PBSで洗浄し、エタノールを用いて室温にて10分間固定し、そして、1%のクリスタル・バイオレット溶液で10分間染色した。プレートを、蒸留水によって3回洗浄し、そして、溶解プラークをカウントした。それぞれのFabの中和能力を、単一溶解プラークをカウントし、対照サンプルと比較したウイルス・プラーク・カウントの減少のパーセンテージについて計算することによって測定した。
【0100】
それぞれのFabの中和能力を、蛍光顕微鏡(Olympus)によって単一蛍光発光細胞をカウントし、そして、対照サンプルと比較してVZV陽性細胞数の減少のパーセンテージを計算することによって測定した。
【0101】
免疫蛍光分析
MRC−5細胞を、無菌のガラス・カバーガラスを含むCostarの24ウェル・プレート内で培養した。細胞培養が(通常、37℃にて7日間のインキュベーション後に形成する)単層としてコンフルエントになったときに、以下のとおり、細胞にウイルス‐Fab混合物又は対照混合物を感染させた。培地を取り除き、そして、細胞単層に、37℃にて1時間のインキュベーションの間、指示濃度の半精製Fabsと共にプレインキュベートしたVZV Ellen基準菌株を使用して高い感染多重度にて感染させた(250μl/ウェル;感染多重度0.1〜1)。対照を、Fabsの不存在下(ブランク対照)、又はヒトC型肝炎ウイルス(e8)に特異的な無関係なFabの存在下、等量の維持培地とウイルスから構成した。前記プレートを37℃にて2時間インキューベートし、未中和ウイルスを吸着させた。接種材料は取り除き、そして、2%のFBSを含む1.5mlのMEMをそれぞれのウェルに入れた。
【0102】
免疫蛍光アッセイを、VZV感染の72時間後に実施した。培地を取り除いた後に、細胞単層を、PBSで1回洗浄し、そして、冷メタノール‐アセトン溶液(1:2の比率;−20℃にて保存)中、室温にて10分間固定した。固定細胞を、一次抗体としてのDDF−VZV1又はDDF−VZV2の調製物と共に湿潤大気中、37℃にて30分間インキューベートし、PBSで洗浄し、そして最後に、抗ヒトIgG Fab特異的FITC‐コンジュゲート(Sigma)と共に湿潤大気中、37℃にて30分間インキューベートした。対照として、細胞を、製造業者の取扱説明書に従って使用される市販の抗VZV抗体(Argene;カタログ番号11−017)と共に、二次FITC標識抗体だけを使用することで調製した。
【0103】
さらに、サイトメガロウイルスを感染させたMRC−5細胞を、一次抗体としてDDF−VZV1又はDDF−VZV2の同じ調製物を使用して試験した。スライドを、エバンスブルーによって対比染色し、グリセロール・バッファーで封入し、そして最後に、蛍光顕微鏡(Olympus)によって観察した。
【0104】
結果
DDF−VZV1とDDF−VZV2の両方に関する中和及び結合活性のさらなる分析を、ある程度精製したFabsの調製物を使用して実施した。
免疫蛍光では、感染細胞において核染色を示す市販のマウス・モノクローナル抗体と比較して、DDF−VZV1は、VZVビリオンが組み立てられる核(周辺)領域においてVZV感染細胞をより強く染色する。DDF−VZV2は、細胞質領域においてVZV感染細胞をより強く染色する(図4)。一次抗体の不存在下において、あるいは、感染させなかった、又はサイトメガロウイルスを感染させたMRC−5細胞に対するDDF−VZV1及びDDF−VZV2の使用において、染色は観察されなかった。これらのデータはまた、一時的に培養し、その後、免疫蛍光で観察した、VZVに感染している人の皮膚から得られた細胞に対して実施した免疫蛍光によっても確認された。
【0105】
プラーク減少に関するデータは、VZVとプレインキュベートした場合に、DDF−VZV1及びDDF−VZV2が強力な中和活性を与えられることを示している。実際、対照(Fabを伴わない、又は無関係なFabを伴ったVZV)と比較すると、これらのFabsの添加は、用量依存的様式でのプラーク形成の減少を引き起こす(図5)。
【0106】
実施例3:pDD適合発現ベクターを使用したDDF−VZV1及びDDF−VZV2の製造とバリデーション
材料と方法
pDLac−FLAGhisベクターの設計と構築
ゼオシン(Zeocine)遺伝子がマーカー遺伝子として使用されるDDbカセット(WO07/007154)を含むpDDベクターを、cp3*配列を含んでいるSpeI−NheIフラグメントを、2つのタグと終止コドンを含んでいるNheI−SpeI合成リンカーで置換することによって修飾した。前述のリンカーを、2つのオリゴヌクレオチド(配列番号15及び16)をアニーリングすることによって作り出した。得られた二本鎖DNA分子を、SpeIとNheIで消化し、そして、対応する線状化pDDベクター内へのクローニング・ステップに備えた。2つのタグ配列のインフレーム挿入を確認するために、最終的なベクターを、制限分析と配列分析によって特徴づけした。LacI遺伝子を、pET28(Invitrogen)という市販のプラスミドからPCR増幅し、そして次に、ゼオシン遺伝子と、FLAGhisタグに融合されるべきタンパク質の発現を駆動するLacZプロモーターの間のStuI部位に挿入した。
【0107】
PCR反応を、DNA鋳型(20〜30ng)、市販のPCRバッファー(1×;Invitrogen)、プライマー(0.2μM)、dNTP(0.25mM)、Taq DNAポリメラーゼ(0.25単位;Invitrogen)、及び水(50μlの終量まで)を含む混合物を使用して実施した。その反応を、サーマル・サイクラー(Perkin−Elmer)により、以下のとおり実施した:94℃にて5分間、そして、94℃にて30秒、50〜55℃にて30秒間、72℃にて1分間を30サイクル。部位特異的突然変異誘発PCR法を、Pfu融合Taqポリメラーゼ(Stratagene)を使用して、先に説明したのと同じ増幅条件で実施した。制限酵素を用いたDNAの消化を、製造業者の取扱説明書(New England Biolabs)に従って実施した。バクテリオファージT4 DNAリガーゼ(Boehringer)を、適切なベクター内への調製したDNAフラグメントの連結反応に使用した。ベクター:挿入DNAのモル比は1:3であり、そして、DNAの総濃度は約50ngであった。連結反応を、以下のとおり20μl中で実施した:DNA(50ng)、連結反応バッファー(1×)、10mMのATP(1μl)、T4 DNAリガーゼ(4単位)、及び水(20μlまで)。T4連結反応を、15℃にて一晩実施した。TG1又はXL−1 Blueエシェリキア・コリ・コンピテント細胞を、CaCl2プロトコールを使用して調製し、そして、pDLac−FLAGhisベクターを含む連結反応混合物で形質転換した。
【0108】
DDF−VZV1とDDF−VZV2の重鎖及び軽鎖をコードする配列を、適切な制限部位を使用することでpDLac−FLAGhis内にクローンニングした。最終的なベクター(pDLac−VZV1−FLAGhisとpDLac−VZV2−FLAGhis)を、2つの抗体配列のインフレーム挿入を確認するために、制限分析及び配列分析によって特徴づけした。
【0109】
FLAGhisタグ付与DDF−VZV1及びDDF−VZV2の発現、精製、及び検出
E.コリXL−1 Blueを、pDLac−VZV1−FLAGhis、pDLac−VZV2−FLAGhis、又はe509を発現するpDLac−FLAGhis変異体、対照として使用されるHCV特異的Fab(Bugli et al.,2001)で形質転換した。
【0110】
得られた菌株を、細菌培養によるFabsの製造に使用した。このために、形質転換から得られた単一コロニーを、テトラサイクリン(10μg/ml)、アンピシリン(100μg/ml)、及びゼオシン(zeocin)(50μg/ml)を含む10mlのSB培地中に植菌した。細菌を、37℃にて12時間増殖させた。インキュベーション時間の後に、500μlのそれらを、同じ抗生物質を補った100mLのSBで希釈し、0.6のODに達するまで6〜8時間増殖させた。イソプロピル−ベータ−D−チオ−ガラクトピラノシド(IPTG)を、1mMの終濃度まで加え、そして、細胞を、ロータリー振盪機により30℃にてさらに14時間インキューベートした。細菌を、5000gにて20分間の遠心分離によって集菌し、1mLのリン酸緩衝生理食塩水(PBS)中に再懸濁し、そして、凍結‐融解手順(−80℃及び37℃にて3ステップ)で溶解させた。細胞破片を、微量遠心管による15,000g、室温にて5分間の遠心分離によって取り除いた。
【0111】
Fab発現を確認するために、上清を、抗Fab及び抗His抗体検出を用いたウエスタン・ブロット分析に使用した。SDS−PAGEを、β−メルカプトエタノールを含む又は含まない(還元又は非還元条件)サンプルを添加して作製し、次に、ブロッティングを350mAにて2時間実施した;ニトロセルロース紙を、存在するブロッティングを確認するためにPonceaus Redで最初に染色し、次に、それを10%のミルク/PBS/Tween 0.1%(ブロッキング溶液)と共に4℃にて一晩撹拌した。PBS/Tween 0.1%で1回洗浄した後に、前記ニトロセルロースを、西洋ワサビ・ペルオキシダーゼ(HRP)コンジュゲート抗Fab抗体(Sigma)又はHRPコンジュゲート抗His−タグ抗体(Roche)と共に撹拌しながら室温で1時間インキューベートした。PBS/Tween 0.1%で3回洗浄した後に、SuperSignal West Pico Chemiluminescent基質(Pierce)を加え、そして、フィルムを、それぞれ30秒、1、及び5分間現像した。
【0112】
抗Fabを用いて検出したシグナルを、抗His HRPコンジュゲート検出と比較した。それらの両方が、(浄化した細胞抽出物を非還元条件を使用したゲルに泳動したとき)Fabフラグメントの天然型に対応する〜60kDaの主要なバンドを明らかにした。同じサンプルを還元条件を使用したSDS−PAGEに供したとき、抗His抗体を使用した場合に、重鎖(〜25kDa)だけが存在した。抗Fabを用いた検出では、タグを付与されていなかったので検出できなかった軽鎖の存在を明らかにした。
【0113】
FLAGhisタグ付与fabsのより大規模な製造において、テトラサイクリン(10μg/mL)、アンピシリン(100μg/mL)、及びゼオシン(50ug/ml)を含むスーパー・ブロス(super broth)の1リットル・アリコートに、特定のpDLac−FLAGhisベクターで形質転換した細菌コロニー1つを植菌し、ロータリー振盪機内で37℃にて7時間培養し、示したようにIPTGで誘導し、そして、30℃にて一晩培養した。細胞を、遠心分離によって集菌し、25mLのPBS中に再懸濁し、そして、超音波処理した。細胞破片を、遠心分離(15,000gにて50分間)によって除去し、そして、0.22μmで濾過した上清をIMACによって精製した。標準的なQiagen精製プロトコールに従った:NiNTA樹脂(Qiagen)を結合バッファー溶液(リン酸緩衝液、10mMのイミダゾール)で平衡化し、次に、濾過した上清をその樹脂と混合し(300μLの樹脂/100mLの培養物)、そして、4℃にて1時間撹拌した。この溶液を、400gにて5分間遠心分離して、樹脂に連結したFabフラグメントを回収し、そして、流れ出た物を収集した。その樹脂を、5倍量の結合バッファー10mMイミダゾールで洗浄して、非特異的結合を除去した。HisフラグFabフラグメントを、3倍量の溶出緩衝液(リン酸緩衝液、500mMのNaCl、500mMのイミダゾール)で溶出した。溶出したアリコートを、SDS/12.5%ポリアクリルアミドゲル電気泳動によって分析し、そして、クマシー・ブリリアントブルー染色及びウエスタン・ブロットで検出した。
【0114】
結果
DDF−VZV1及びDDF−VZV2の活性を、最初、ある程度精製したFabs及び組み換えファージとして試験した。さらなるバリデーションには、このような抗体フラグメントを適切な細菌発現系を使用した精製された組み換えタンパク質の形態で試験する必要がある。
【0115】
着目のFabを持つファージミドは、可溶性分子の産生のために好適な発現ベクター、及び/又は適切な菌株(例えば、アンバー非サプレッサー(amber non−suppressor))に通常移される。分子クローニングのこのような手間がかかったステップを避ける目的で、より効果的なアプローチは、前記Fabsを同定するのに使用した元々のpDDベクター内のものと適合する制限部位を持つ発現ベクターを製造することだろう。そのうえ、タグ配列の存在は、より簡単なFabの精製及び検出を可能にするだろう。
【0116】
pDLac−FLAGhisと名付けたこのような発現ベクターを、設計し、そして、構築した。それは、cp3*のコード配列を、終止コドンが後に続く2つのタンパク質タグ(8つのアミノ酸を含むFLAGペプチド、及び6−ヒスチジン・タグ)をコードする配列で置換したpDDベクターに基づいている(図6A)。ヒスチジン・タグは、例えば、IMAC(固定金属アフィニティー・クロマトグラフィー)精製システムを使用したアフィニティー・クロマトグラフィーによってFabを精製することを可能にする。FLAGタグは、(ELISAなどの)検出アッセイにおいて非常に有用であるかもしれない。これらの2つのタグがクローンニングされ、そして、組み換えタンパク質に融合されたベクター、おまけに、FLAGhisタグ付与タンパク質を検出又は精製する手段が、文献に記載されていた(Pagny S et al.,2000;Koivunen P et al.,2004;Komatsu M et al.,2004)。
【0117】
最初に同定され、そして、pDDファージミドを使用して特徴づけられたFab(又は、いずれかのその他タンパク質配列)は、その後、pDLac−FLAGhisベクター内に一方向性にクローンニングされるDDカセットを使用することで抽出できる。このように、元々コート・タンパク質に融合していた配列が、ここで、FLAGタグ及びポリヒスチジン・タグと共にC末端にてフレーム内にクローンニングされる。得られたベクターは、DDカセット内に本来のマーカー遺伝子(例えば、ゼオシン遺伝子)を含み、所望の挿入物を含むベクターの選択を可能にする。pDLac−FLAGhisベクター内のLacZプロモーターを使用して発現されるべきタンパク質の転写を制御するための手段として、LacI遺伝子を本来のDDカセット内に挿入した(図6B)。
【0118】
このクローニング・ストラテジーは、2つの利点をもたらした。第1の利点は、起こり得る相同組み換えを回避する、本来のDDカセット内の2つのLacZプロモーター領域の間の空間を増やすことである。第2に、オペレーター領域にそれが結合することによりLacZプロモーターを遮断して、発現系を下方制御できることが知られているLacI遺伝子からのリプレッサー・タンパク質である。そして、誘導は、適切な濃度のIPTG、及び/又はグルコースを使用して調整される。誘導により、LacZプロモーターの制御下にクローンニングされた配列/PelBシグナル配列の両方が、発現され、そして、pelB配列が酵素シグナル・ペプチダーゼによってそれに続いて切断されるところの周辺質に向けられる。周辺質内では、適切な酸化条件が、組み換えタンパク質内のジスルフィド結合の形成、及び正しいタンパク質の折り畳みを可能にする(例えば、ヘテロ二量体タンパク質に組み立てられるFabの軽鎖及び重鎖)。アフィニティー・クロマトグラフィーのカラムを溶出し、そして、試験することによって、FLAGhisタグ付与タンパク質を均質にまで精製できる(図6B)。次に、このタンパク質を、例えば、(Sigmaから入手可能な)市販の抗FLAGモノクローナル抗体を使用したウエスタン・ブロット又は免疫蛍光により同定できる。
【0119】
このアプローチを使用して、DDF−VZV1及びDDF−VZV2(図7)の、並びに対照fabのFLAGhisタグ付与バージョンを、製造し、そして、先に記載したアッセイにより試験した。選択したFabsのこれらの組み換え変異体で、2μg/ml前後のIC50を示す、VZV中和活性に関する本来の観察結果を確認した(図8A)。しかしながら、2つのFabsが1つの調製物に組み合わせられるとき、この調製物のVZV中和活性は、同量の単独のFabを含む調製物より優れた結果となった。実際、Fab濃度が1〜2μg/mlに達するときに、単独のFabを含む調製物がその活性を10〜15%増強する場合に、1μg/mlのDDF−VZV1と1μg/mlのDDF−VZV1を含む調製物は、2μg/ml未満のIC50値を有するはるかに高いVZV中和活性を有する(図8B)。この効果は、おそらく、2つのFabsによって認識される異なるVZV特異的エピトープのためであり(図4を参照のこと)、DDF−VZV1とDDF−VZV2(同様の特性を有する2つの抗体又は抗体フラグメント)を含む医薬組成物が、VZV感染及びVZV関連疾患に対してより良好な治療効果又は予防効果をもたらし得ることを示唆している。
【0120】
ここに提示した実験的証拠が、VZV感染及びVZV関連疾患に関係する診断上の、治療上の、又は予防上の適用のためのDDF−VZV1及びDDF−VZV2(又はそれらの特異的なHCDR3sに基づき、且つ、同様の特性を示す代替タンパク質配列)候補化合物をもたらす。
【0121】
参考文献
【0122】
【化1】
【0123】
【化2】
【0124】
【化3】
【0125】
【化4】
【0126】
【化5】
【特許請求の範囲】
【請求項1】
配列番号5と少なくとも90%の同一性を有する配列を含んで成るタンパク質。
【請求項2】
前記タンパク質が、配列番号2と少なくとも90%の同一性を有する配列を含んで成る、請求項1に記載のタンパク質。
【請求項3】
前記タンパク質が、配列番号7と少なくとも90%の同一性を有する配列をさらに含んで成る、請求項1又は2に記載のタンパク質。
【請求項4】
前記タンパク質が、抗体、抗体フラグメント、生理活性ペプチド、又は融合タンパク質である、請求項1〜3のいずれか1項に記載のタンパク質。
【請求項5】
前記抗体フラグメントが、可変重鎖/軽鎖ヘテロ二量体であるか、又は一本鎖可変フラグメントである、請求項4に記載のタンパク質。
【請求項6】
前記タンパク質が、水痘帯状ヘルペスウイルス(VZV)に結合し、そして、中和する、請求項1〜5のいずれか1項に記載のタンパク質。
【請求項7】
請求項1〜6のいずれか1項に記載のタンパク質をコードする核酸分子。
【請求項8】
前記核酸が、配列番号1と少なくとも90%の同一性を有する、請求項7に記載の核酸分子。
【請求項9】
請求項7又は8に記載の核酸を含んで成るベクター。
【請求項10】
請求項7又は8に記載の核酸を含んで成る組み換えファージ、原核生物宿主細胞、又は真核生物宿主細胞。
【請求項11】
請求項1〜6のいずれか1項に記載のタンパク質を製造するための、請求項7〜9のいずれか1項に記載の核酸、あるいは、請求項10に記載の組み換えファージ又は宿主細胞の使用方法。
【請求項12】
VZV感染又はVZV関連疾患を検出、治療、抑制、予防、及び/又は改善するための組成物の調製のための、請求項1〜6のいずれか1項に記載のタンパク質の使用方法。
【請求項13】
請求項1〜6のいずれか1項に記載のタンパク質を含んで成る治療用組成物、予防用組成物、又は診断用組成物。
【請求項14】
前記組成物が、眼又は局所に投与される、請求項13に記載の組成物。
【請求項15】
異なるVZV中和抗体、VZV中和抗体フラグメント、免疫グロブリン静注調製物、ステロイド、及び/又は抗ウイルス化合物をさらに含んで成る、請求項13又は14に記載の組成物。
【請求項1】
配列番号5と少なくとも90%の同一性を有する配列を含んで成るタンパク質。
【請求項2】
前記タンパク質が、配列番号2と少なくとも90%の同一性を有する配列を含んで成る、請求項1に記載のタンパク質。
【請求項3】
前記タンパク質が、配列番号7と少なくとも90%の同一性を有する配列をさらに含んで成る、請求項1又は2に記載のタンパク質。
【請求項4】
前記タンパク質が、抗体、抗体フラグメント、生理活性ペプチド、又は融合タンパク質である、請求項1〜3のいずれか1項に記載のタンパク質。
【請求項5】
前記抗体フラグメントが、可変重鎖/軽鎖ヘテロ二量体であるか、又は一本鎖可変フラグメントである、請求項4に記載のタンパク質。
【請求項6】
前記タンパク質が、水痘帯状ヘルペスウイルス(VZV)に結合し、そして、中和する、請求項1〜5のいずれか1項に記載のタンパク質。
【請求項7】
請求項1〜6のいずれか1項に記載のタンパク質をコードする核酸分子。
【請求項8】
前記核酸が、配列番号1と少なくとも90%の同一性を有する、請求項7に記載の核酸分子。
【請求項9】
請求項7又は8に記載の核酸を含んで成るベクター。
【請求項10】
請求項7又は8に記載の核酸を含んで成る組み換えファージ、原核生物宿主細胞、又は真核生物宿主細胞。
【請求項11】
請求項1〜6のいずれか1項に記載のタンパク質を製造するための、請求項7〜9のいずれか1項に記載の核酸、あるいは、請求項10に記載の組み換えファージ又は宿主細胞の使用方法。
【請求項12】
VZV感染又はVZV関連疾患を検出、治療、抑制、予防、及び/又は改善するための組成物の調製のための、請求項1〜6のいずれか1項に記載のタンパク質の使用方法。
【請求項13】
請求項1〜6のいずれか1項に記載のタンパク質を含んで成る治療用組成物、予防用組成物、又は診断用組成物。
【請求項14】
前記組成物が、眼又は局所に投与される、請求項13に記載の組成物。
【請求項15】
異なるVZV中和抗体、VZV中和抗体フラグメント、免疫グロブリン静注調製物、ステロイド、及び/又は抗ウイルス化合物をさらに含んで成る、請求項13又は14に記載の組成物。
【図1】

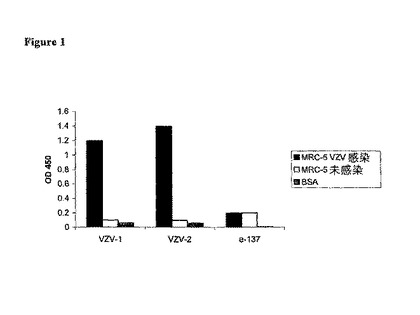
【図2A)】

【図2B)】

【図3A)】

【図3B)】

【図4A)】
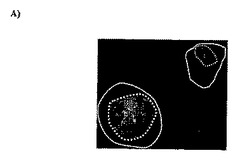
【図4B)】

【図5】
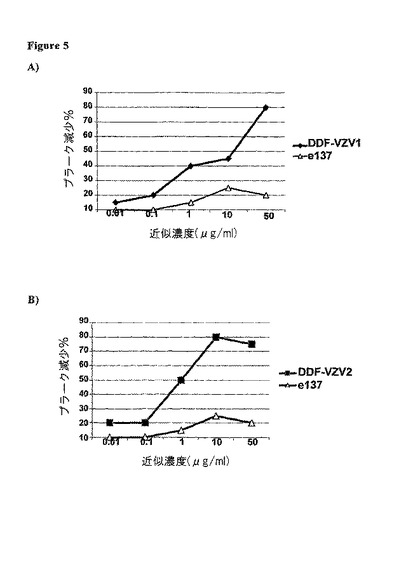
【図6】
【図7A)】

【図7B)】

【図7C)】

【図7D)】

【図8】
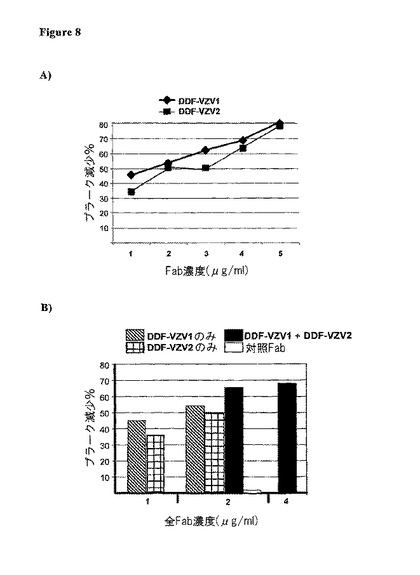
【図2A)】
【図2B)】
【図3A)】
【図3B)】
【図4A)】
【図4B)】
【図5】
【図6】
【図7A)】
【図7B)】
【図7C)】
【図7D)】
【図8】
【公表番号】特表2010−517557(P2010−517557A)
【公表日】平成22年5月27日(2010.5.27)
【国際特許分類】
【出願番号】特願2009−548760(P2009−548760)
【出願日】平成20年2月6日(2008.2.6)
【国際出願番号】PCT/IB2008/000266
【国際公開番号】WO2008/096242
【国際公開日】平成20年8月14日(2008.8.14)
【出願人】(508178478)リボバックス バイオテクノロジーズ ソシエテ アノニム (6)
【Fターム(参考)】
【公表日】平成22年5月27日(2010.5.27)
【国際特許分類】
【出願日】平成20年2月6日(2008.2.6)
【国際出願番号】PCT/IB2008/000266
【国際公開番号】WO2008/096242
【国際公開日】平成20年8月14日(2008.8.14)
【出願人】(508178478)リボバックス バイオテクノロジーズ ソシエテ アノニム (6)
【Fターム(参考)】
[ Back to top ]