
油性酵母菌における多不飽和脂肪酸生産のためのコドン最適化遺伝子
本発明は、リノール酸(LA)からγ−リノレン酸(GLA)、α−リノール酸(ALA)からステアリドン酸(STA)、GLAからジホモ−γ−リノール酸(DGLA)、STAからエイコサテトラエン酸(ETA)、DGLAからETA、エイコサペンタエン酸(EPA)からドコサペンタエン酸(DPA)、およびアラキドン酸(ARA)からEPAへの転換を触媒できる脂肪酸デサチュラーゼおよび鎖長延長酵素に関する。コドン最適化デサチュラーゼおよび鎖長延長酵素をコードする核酸配列、それにハイブリダイズする核酸配列、コドン最適化デサチュラーゼまたは鎖長延長酵素を含んでなるDNA構築物、および増大するレベルのデサチュラーゼまたは鎖長延長酵素を発現する組換え宿主微生物について記載される。

【発明の詳細な説明】
【関連出願との関係】
【0001】
本願は2003年5月7日に出願された米国仮出願第60/468677号、および2003年5月7日に出願された米国仮特許出願第60/468718号の優先権の利益を主張する。
【技術分野】
【0002】
本発明はバイオテクノロジー分野に関する。より具体的には本発明は、油性酵母菌における長鎖多不飽和脂肪酸(PUFA)の生産に有用な酵素をコードする核酸断片の合成に関する。
【背景技術】
【0003】
特定の多不飽和脂肪酸、すなわちPUFAが健康な細胞の重要な生物学的構成要素であることが、長期にわたり認識されている。例えばこのようなPUFAは、
哺乳類において新規に(de novo)合成できず、食餌中で得られなくてはならない、またはリノール酸(LA)またはα−リノレン酸(ALA)のさらなる不飽和化と延長によって誘導されなくてはならない「必須」脂肪酸、
リン脂質またはトリグリセリドなどの形態で見いだされてもよい細胞原形質膜の構成物、
特に成長中の幼児の脳において適切な発育、そして組織形成および修復に必要である、
プロスタサイクリン、エイコサノイド、ロイコトリエン、およびプロスタグランジンをはじめとする、哺乳類において重要ないくつかの生物学的に活性なエイコサノイドの前駆物質として認識されている。
【0004】
1970年代に、グリーンランドのエスキモーの観察から、心疾患の低発生率と長鎖ω−3PUFAの高摂取量とが結びつけられた(非特許文献1、非特許文献2)。より最近の研究はω−3PUFAの心臓血管保護効果を確証した(非特許文献3、非特許文献4)。さらに血管形成術後の再狭窄率、炎症および関節リウマチ、喘息、乾癬および湿疹の症状などのいくつかの障害はω−3脂肪酸による処置に反応することが見いだされている。γ−リノレン酸(GLA、ω−6PUFA)はストレスに関係した血圧上昇を低下させ、算術試験能力を改善することが示されている。GLAおよびジホモ−γ−リノレン酸(DGLA、もう1つのω−6PUFA)は、血小板凝集を阻害し、血管拡張を引き起こし、コレステロールレベルを低下させ、血管壁平滑筋および繊維組織の増殖を阻害することが示されている(非特許文献5)。GLAまたはDGLAの単独でのまたはエイコサペンタエン酸(EPA、ω−3PUFA)との組み合わせでの投与は、非ステロイド性抗炎症薬によって引き起こされる消化管出血およびその他の副作用を低下させ、または防止することが示されている(特許文献1)。さらにGLAおよびDGLAは、子宮内膜症および月経前症候群を防止または治療し(特許文献2)、筋痛性脳脊髄炎およびウィルス感染後の慢性疲労(特許文献3)を治療することが示されている。その他の証拠は、PUFAがカルシウム代謝調節に関与するかもしれないことを示唆し、骨粗鬆症および腎臓または尿道結石の治療または防止においてそれらが有用であるかもしれないことを示唆する。最後にPUFAは癌および糖尿病の治療において使用できる(特許文献4、非特許文献6)。
【0005】
PUFAは、必須脂肪酸であるLAおよびALAそれぞれの不飽和化および延長によって誘導される、2つの主要なクラス(ω−6およびω−3脂肪酸からなる)に概して分けられる(図1)。天然供給源からの多様なPUFAの商業的供給源(例えば月見草、ルリヂサ、およびクロフサスグリの種子や糸状菌(モルティエレラ(Mortierella))、チノリモ属(Porphyridium)(紅藻)、魚油および海洋性プランクト
ン(キクロテラ(Cyclotella)、ニッチア(Nitzschia)、クリプテコジニウム(Crypthecodinium))にもかかわらず、これらの生産方法と結びついたいくつかの不都合がある。第1に魚および植物などの天然供給源は、高度に不均一な油組成物を有しがちである。したがってこれらの供給源から得られた油は、所望のPUFAの1つもしくはそれ以上を分離または濃縮するために大規模な精製を必要とすることがある。天然供給源はまた、制御できない供給のばらつきを被りやすい(例えば天候、疾患、または魚資源の乱獲による)。PUFAを生産する作物は、食物生産のために開発されたハイブリッド作物と、経済的競争力がないことが多い。またPUFAを自然に生産するいくつかの生物体(例えばポルフィリディウム(Porphyridium)、モルティエレラ(Mortierella))の大規模発酵は、商業的規模で培養するのが高価および/または困難なことがある。
【0006】
上述の限界の結果として、以下に向けた大規模な研究が行われている。1.)商業的に容易に生産できるPUFAの組換え供給源の開発、および2.)所望のPUFA生産を可能にする脂肪酸生合成経路の修正。例えば過去数年間にわたり、様々な生物体からの脂肪酸デサチュラーゼおよび鎖長延長酵素(elongase)遺伝子の単離、クローン化、および操作において進歩があった。これらの遺伝子配列の知識は、PUFAを自然に生産しない新しい宿主生物体中で、所望の脂肪酸および/または脂肪酸組成物を生産する見込みを提供する。文献は、サッカロマイセス・セレヴィシエ(Saccharomyces
cerevisiae)における以下のようないくつかの例を報告する。
1.非特許文献7では、海洋珪藻フェオダクチルム・トリコルヌーツム(Phaeodactylum tricornutum)からの2個のデサチュラーゼがS.セレヴィシエ(S.cerevisiae)中にクローン化され、EPAの生産をもたらす。
2.非特許文献8では、線虫(Caenorhabditis elegans)からの遺伝子を使用して、ω−3およびω−6PUFA生合成経路がS.セレヴィシエ(S.cerevisiae)中に再構成される。
3.非特許文献9では、植物脂肪酸デサチュラーゼ(FAD2およびFAD3)がS.セレヴィシエ(S.cerevisiae)中に発現し、ALAの生産をもたらす。
4.アボット・ラボラトリーズ(Abbott Laboratories)のナットゾン(Knutzon)らに付与された特許文献5では西洋油菜(Brassica napus)からの1つのデサチュラーゼおよび菌・カビ類モルティエレラ・アルピナ(Mortierella alpina)からの2個のデサチュラーゼが、S.セレヴィシエ(S.cerevisiae)中にクローン化され、LA、GLA、ALA、およびSTAの生産をもたらす。
【0007】
しかしこれらのタイプの遺伝子を発現させて、商業的量の1つもしくはそれ以上のPUFAの経済的生産を提供できる適切な微生物のシステムに対する必要性がなおもある。さらに特にEPAおよびDHAである特定のPUFAが濃縮されている油に対する必要性が存在する。
【0008】
PUFAの生産プラットフォームとしてこれまで調査されていない1つのクラスの微生物が、油性酵母菌である。これらの生物体は、乾燥細胞重量の80%までの油を蓄積できる。高い油含量で油性酵母菌を生育させる技術は十分に開発されており(例えば特許文献6、非特許文献10を参照)、ω−3またはω−6PUFA生産のための商業的な微細藻類発酵と比べてコスト優位性を提供するかもしれない。そのままの酵母菌細胞はまた、機能食品および動物飼料サプリメントで使用するためのω−3またはω−6PUFA−濃縮油を封入する都合よい方法になるかもしれない。
【0009】
【特許文献1】米国特許第4,666,701号明細書
【特許文献2】米国特許第4,758,592号明細書
【特許文献3】米国特許第5,116,871号明細書
【特許文献4】米国特許第4,826,877号明細書
【特許文献5】米国特許第6,136,574号明細書
【特許文献6】欧州特許第0005277B1号明細書
【非特許文献1】ダイヤーバーグ(Dyerberg)J.ら、Amer.J.Clin Nutr.28:958〜966(1975)
【非特許文献2】ダイヤーバーグ(Dyerberg)J.ら、Lancet 2(8081):117〜119(1978年7月15日)
【非特許文献3】シモカワ(Shimokawa)H.、World Rev Nutr Diet、88:100〜108(2001)
【非特許文献4】フォン・シャッキー(von Schacky)C.およびダイヤーバーグ(Dyerberg)J.、World Rev Nutr Diet、88:90〜99(2001)
【非特許文献5】ブレナー(Brenner)ら、Adv.Exp.Med.Biol.83:85〜101(1976)
【非特許文献6】(Horrobin)ら、Am.J.Clin.Nutr.57(付録):732S〜737S(1993)
【非特許文献7】ドマーグ(Domergue)F.ら、Eur.J.Biochem.269:4105〜4113(2002)
【非特許文献8】ボードイン(Beaudoin)F.ら、Proc.Natl.Acad.Sci.U.S.A.97(12):6421〜6(2000)
【非特許文献9】ダイヤー(Dyer)J.M.ら、Appl.Eniv.Microbiol.、59:224〜230(2002)
【非特許文献10】ラトレッジ(Ratledge)C.、Prog.Ind.Microbiol.16:119〜206(1982)
【発明の開示】
【発明が解決しようとする課題】
【0010】
上述の利点にもかかわらず、油性酵母菌中の自然に生産されるPUFAは、18:2脂肪酸(そしてあまり一般的ではなく18:3脂肪酸)に限定されるので、これらの生物体は自然にはω−6およびω−3PUFAを欠いている。したがって解決すべき問題は、ω−3および/またはω−6脂肪酸が濃縮された油を蓄積する油性酵母菌を開発することである。このような目的で、油性酵母菌におけるω−3および/またはω−6脂肪酸の合成および蓄積を可能にする、飽和化酵素および鎖長延長酵素を導入することが必要である。多数の供給源から多様なデサチュラーゼおよび鎖長延長酵素遺伝子が入手できるにもかかわらず、これらの遺伝子は油性酵母菌などの別の宿主中では、遺伝子中のコドンが別の宿主生物体の典型的なコドン使用を反映しないので、最適効率で発現しない。したがって油性酵母菌におけるPUFA遺伝子の発現を最適化し、これらの特定の宿主生物体におけるω−3および/またはω−6脂肪酸の高レベルの生産および蓄積を可能にするために、コドン使用に関連した問題を克服しなくてはならない。
【0011】
出願人は、油性宿主、ヤロウィア・リポリティカ(Yarrowia lipolytica)における発現に適したデサチュラーゼおよび鎖長延長酵素遺伝子をコドン最適化する手段を開発することで既述の問題を解決した。ここで最適化される例示的な遺伝子は、Δ6デサチュラーゼ、Δ17デサチュラーゼ、および高親和力PUFA鎖長延長酵素をコードする遺伝子であり、コドン最適化は、Y.リポリティカ(Y.lipolytica)中でLAからGLAへの%基質変換(Δ6デサチュラーゼ)をおよそ40%改善し、ARAからEPAへの%基質変換(Δ17デサチュラーゼ)を約2倍にし、GLAからDGLAへの%基質変換(鎖長延長酵素)を約57%改善した。
【課題を解決するための手段】
【0012】
本発明は、ヤロウィア(Yarrowia)種における最適発現のためのω−3/ω−6脂肪酸生合成経路における様々な遺伝子の最適化に関する。したがって本発明は、
a)配列番号25に記載のΔ6デサチュラーゼをコードする単離された核酸分子、または
b)(a)と完全に相補的である単離された核酸分子
よりなる群から選択される単離された核酸分子を提供する。
【0013】
同様に本発明は、配列番号2に記載のΔ6デサチュラーゼをコードする単離された核酸分子を提供し、そこではヤロウィア(Yarrowia)種における発現のために少なくとも144個のコドンがコドン最適化される。
【0014】
本発明の別の実施態様では、
a)配列番号62に記載のΔ17デサチュラーゼをコードする単離された核酸分子、または
b)(a)と完全に相補的である単離された核酸分子
よりなる群から選択される、単離された核酸分子が提供される。
【0015】
本発明の別の実施態様では、配列番号4に記載のΔ17デサチュラーゼをコードする単離された核酸分子が提供され、そこではヤロウィア(Yarrowia)種における発現のために少なくとも117個のコドンがコドン最適化される。
【0016】
同様に本発明は、
a)配列番号91に記載の鎖長延長酵素をコードする単離された核酸分子、または
b)(a)と完全に相補的である単離された核酸分子よりなる群から選択される単離された核酸分子を提供する。
【0017】
代案としては本発明は、配列番号6に記載の鎖長延長酵素をコードする単離された核酸分子を提供し、そこではヤロウィア(Yarrowia)種における発現のために少なくとも85個のコドンがコドン最適化される。
【0018】
さらに本発明は、本発明の遺伝子の遺伝的キメラ、および該物質で形質転換された宿主細胞を提供する。
【0019】
本発明の特定の実施態様では、ヤロウィア(Yarrowia)種中で本発明のコドン最適化遺伝子を使用して、適切な前駆物質からの一段階酵素的反応によって、γ−リノレン酸(GLA)、ジホモ−γ−リノール酸(DGLA)、ステアリドン酸(STA)、エイコサテトラエン酸(ETA)、エイコサペンタエン酸(EPA)、およびドコサペンタエン酸(DPA)などの特定のω−3およびω−6脂肪酸生産が提供される。
【0020】
本発明の別の実施態様では、
a)油性酵母菌種についてヌクレオチドコード領域および対応するポリペプチドの配列を得て、コドンのデータベースを形成し、
b)コドンのデータベースを分析し、いずれのコドンが各アミノ酸を優先的にコードするのか判定し、
c)油性酵母菌種中で発現させる遺伝子の配列を得て、
d)ステップ(c)の配列中の好ましくないコドンをステップ(b)の好ましいコドンで置き換えて、遺伝子が油性酵母菌種における発現のためにコドン最適化すること
を含んでなる、油性酵母菌における発現のために遺伝子を最適化する方法が提供される。
【0021】
本発明の代案の実施態様では、配列番号122に記載のヤロウィア(Yarrowia)翻訳開始部位を含んでなる単離された核酸分子が提供される。さらに
a)ヤロウィア(Yarrowia)中で発現させる外来遺伝子を提供し、
b)ステップ(a)の遺伝子を配列番号122に記載のヤロウィア(Yarrowia)翻訳開始部位に作動可能に連結させ、外来遺伝子をヤロウィア(Yarrowia)における発現のために最適化すること
を含んでなる、ヤロウィア(Yarrowia)宿主における遺伝子の発現を最適化するための方法も提供される。
【0022】
配列説明
本願明細書の一部を形成する以下の詳細な説明および添付の配列説明によって、本発明をより完全に理解できるであろう。
【0023】
以下の配列は、37C.F.R.§1.821〜1.825(「ヌクレオチド配列および/またはアミノ酸配列開示を含む特許出願の要件−配列規則(Requirements for Patent Applications Containing Nucleotide Sequences and/or Amino Acid Sequence Disclosures − the Sequence Rules)」)を満たし、世界知的所有権機関(WIPO)標準ST.25(1998)およびEPOおよびPCTの配列表要件(規則5.2および49.5(aの2)、および実施細則第208号および附属書C)に一致する。ヌクレオチドおよびアミノ酸配列データのために使用される記号および型式は、37C.F.R.§1.822で述べられる規則に従う。
【0024】
配列番号1は、モルティエレラ・アルピナ(Mortierella alpina)Δ6デサチュラーゼ遺伝子のDNA配列を示し、他方配列番号2は、M.アルピナ(M.alpina)Δ6デサチュラーゼのアミノ酸配列を示す。
【0025】
配列番号3は、サプロレグニア・ディクリナ(Saprolegnia diclina)Δ17デサチュラーゼ遺伝子のDNA配列を示し、他方配列番号4は、S.ディクリナ(S.diclina)Δ17デサチュラーゼの対応するアミノ酸配列を示す。
【0026】
配列番号5は、モルティエレラ・アルピナ(Mortierella alpina)高親和力鎖長延長酵素遺伝子のDNA配列を示し、他方配列番号6はM.アルピナ(M.alpina)高親和力鎖長延長酵素のアミノ酸配列を示す。
【0027】
配列番号7および8は、TEFプロモーターを単離するのに使用されるプライマーTEF5’およびTEF3’にそれぞれ対応する。
【0028】
配列番号9および10は、XPR2転写ターミネーターを単離するのに使用される、プライマーXPR5’およびXPR3’にそれぞれ対応する。
【0029】
配列番号11〜24は、プラスミド構築のために使用されるプライマーYL1、YL2、YL3、YL4、YL23、YL24、YL5、YL6、YL9、YL10、YL7、YL8、YL61、およびYL62にそれぞれ対応する。
【0030】
配列番号25は、ヤロウィア・リポリティカ(Yarrowia lipolytica)における発現のためにコドン最適化された、合成Δ6デサチュラーゼ遺伝子のDNA配列を示す。
【0031】
配列番号26〜53は、M.アルピナ(M.alpina)Δ6デサチュラーゼ遺伝子
のコドン最適化コード領域全体を一緒に構成する、14対のオリゴヌクレオチドに対応する(例えばそれぞれD6−1A、D6−1B、D6−2A、D6−2B、D6−3A、D6−3B、D6−4A、D6−4B、D6−5A、D6−5B、D6−6A、D6−6B、D6−7A、D6−7B、D6−8A、D6−8B、D6−9A、D6−9B、D6−10A、D6−10B、D6−11A、D6−11B、D6−12A、D6−12B、D6−13A、D6−13B、D6−14A、およびD6−14B)。
【0032】
配列番号54〜61は、コドン最適化Δ6デサチュラーゼ遺伝子の合成中に、PCR増幅のために使用されるプライマーD6−1、D6−4R、D6−5、D6−7R、D6−8、D6−10R、D6−11、およびD6−14Rにそれぞれの対応する。
【0033】
配列番号62は、ヤロウィア・リポリティカ(Yarrowia lipolytica)における発現のためにコドン最適化された、合成Δ17デサチュラーゼ遺伝子のDNA配列を示す。
【0034】
配列番号63〜84は、S.ディクリナ(S.diclina)Δ17デサチュラーゼ遺伝子のコドン最適化コード領域全体を一緒に構成する、11対のオリゴヌクレオチドに対応する(例えばそれぞれD17−1A、D17−1B、D17−2A、D17−2B、D17−3A、D17−3B、D17−4A、D17−4B、D17−5A、D17−5B、D17−6A、D17−6B、D17−7A、D17−7B、D17−8A、D17−8B、D17−9A、D17−9B、D17−10A、D17−10B、D17−11A、およびD17−11B)。
【0035】
配列番号85〜90は、コドン最適化Δ17デサチュラーゼ遺伝子の合成中に、PCR増幅のために使用されるプライマーD17−1、D17−4R、D17−5、D17−8D、D17−8U、およびD17−11にそれぞれ対応する。
【0036】
配列番号91は、ヤロウィア・リポリティカ(Yarrowia lipolytica)における発現のためにコドン最適化された、合成高親和力鎖長延長酵素遺伝子のDNA配列を示す。
【0037】
配列番号92〜111は、M.アルピナ(M.alpina)高親和力鎖長延長酵素遺伝子のコドン最適化コード領域全体を一緒に構成する、10対のオリゴヌクレオチドに対応する(例えばそれぞれEL−1A、EL−1B、EL−2A、EL−2B、EL−3A、EL−3B、EL−4A、EL−4B、EL−5A、EL−5B、EL−6A、EL−6B、EL−7A、EL−7B、EL−8A、EL−8B、EL−9A、EL−9B、EL−10A、およびEL−10B)。
【0038】
配列番号112〜115は、コドン最適化鎖長延長酵素遺伝子の合成中に、PCR増幅のために使用されるプライマーEL−1、EL−5R、EL−6、およびEL−10Rにそれぞれ対応する。
【0039】
配列番号116および117は、部位特異的変異誘発のために使用されてpELSを生じる、プライマーEL−M1およびEL−M2に対応する。
【0040】
配列番号118および119は、プラスミドpRSP19からS.ディクリナ(S.diclina)の野生型Δ17デサチュラーゼ遺伝子を増幅するために使用される、プライマーYL21AおよびYL22に対応する。
【0041】
配列番号120および121は、部位特異的変異誘発のために使用されてpYSD17
Mを生じる、プライマーYL53およびYL54に対応する。
【0042】
配列番号122は、ヤロウィア(Yarrowia)種中で最適に発現する遺伝子のためのコドン最適化翻訳開始部位に対応する。
【発明を実施するための最良の形態】
【0043】
主題の発明にしたがって、出願人は油性酵母菌中、ヤロウィア・リポリティカ(Yarrowia lipolytica)における構造的な遺伝子のコドン使用を判定した。Δ6デサチュラーゼ(配列番号25)、Δ17デサチュラーゼ(配列番号62)および高親和力鎖長延長酵素(配列番号91)をコードするコドン最適化遺伝子、ならびにY.リポリティカ(Y.lipolytica)宿主細胞における前記遺伝子に発現のためのDNAカセットが提示される。さらにY.リポリティカ(Y.lipolytica)などの油性酵母菌の長鎖多不飽和脂肪酸(PUFA)含量の修正を可能にする、方法および組成物が提供される。
【0044】
主題発明には多くの用途がある。ここで開示される方法によって製造されるPUFA、またはその誘導体は、食餌代用品、またはサプリメント、特に乳児用調製粉乳として、静脈内栄養補給を受けている患者のために、または栄養不良を防止または処置するために使用できる。代案としては、精製されたPUFA(またはその誘導体)は、正常な使用で受領者が食餌栄養補給のための所望量を受容するように調合された、料理用油、脂肪またはマーガリンに組み込まれてもよい。PUFAはまた、乳児用調製粉乳、栄養サプリメントまたはその他の食物生産物に組み込まれてもよく、抗炎症薬またはコレステロール低下剤としての用途があるかもしれない。場合により組成物は、医薬品用途(ヒトまたは獣医学)のために使用されてもよい。この場合、PUFAは概して経口投与されるが、例えば非経口的(例えば皮下、筋肉内または静脈内)、経直腸、経腟または局所的(例えば皮膚用軟膏またはローションとして)など、それによって成功裏に吸収されるあらゆる経路で投与することができる。
【0045】
組換え手段によって製造されたPUFAによるヒトまたは動物の栄養補給は、追加的なPUFA、ならびにそれらの代謝子孫の増大したレベルをもたらすことができる。例えばアラキドン酸(ARA)による処置は、ARAの増大したレベルだけでなく、プロスタグランジンなどのARAの下流生産物をももたらすことができる。複雑な制御機序は、このような機序を防止、制御または克服して、個々の特定のPUFAの所望のレベルを達成するために、様々なPUFAを組み合わせ、または異なるPUFAコンジュゲートを追加することを望ましいものにできる。
【0046】
定義
本開示では、いくつかの用語および略語が使用される。以下の定義が提供される。
【0047】
「読み取り枠」はORFと略記する。
【0048】
「ポリメラーゼ連鎖反応」はPCRと略記する。
【0049】
「米国微生物系統保存機関」はATCCと略記する。
【0050】
「多不飽和脂肪酸」はPUFAと略記する。
【0051】
「脂肪酸」という用語は、(より長い、およびより短い鎖長の酸の双方も知られているが)約C12〜C22の様々な鎖長の長鎖脂肪族酸(アルカン酸)を指す。優勢な鎖長は、C16〜C22の間である。脂肪酸の構造は単純な表記法システム「X:Y」によって
表され、ここでXは特に脂肪酸の炭素(C)原子の総数であり、Yは二重結合の数である。
【0052】
概して脂肪酸は、飽和または不飽和として分類される。「飽和脂肪酸」という用語は、炭素主鎖間に「二重結合」を有さない脂肪酸を指す。対照的に「不飽和脂肪酸」は、それらの炭素主鎖に沿って「二重結合」を有するcis−異性体である。「一不飽和脂肪酸」は、(例えばパルミトレイン酸(16:1)およびオレイン酸(18:1)のように通常9および10番目の炭素原子の間に)炭素主鎖に沿って1つの「二重結合」のみを有し、他方「多不飽和脂肪酸」(または「PUFA」)は、(例えばリノール酸(18:2)のように9番目および10番目、および12番目および13番目の炭素原子間、およびα−リノレン酸(18:3)のように9番目および10番目、12番目および13番目目、および15番目および16番目の炭素原子間に)炭素主鎖に沿って少なくとも2個の二重結合を有する。
【0053】
「PUFA」は、(脂肪酸炭素鎖のメチル末端に最も近い第1の二重結合の位置(n)次第で)2つの主要ファミリーに分類できる。したがって「オメガ−6脂肪酸」(ω−6またはn−6)は、第1の不飽和二重結合を分子のω(メチル)末端から6個めの炭素原子に有し、さらに分子のカルボキシル末端に向かって3個めの追加的な炭素原子にそれぞれの続く不飽和がある、全部で2つ以上の二重結合を有する。対照的に「オメガ−3脂肪酸」(ω−3またはn−3)は、第1の不飽和二重結合を分子のω末端から3個離れた炭素原子に有し、さらに分子のカルボキシル末端に向かって3個目の追加的な炭素原子にそれぞれの続く不飽和がある、全部で3個以上の二重結合を有する。
【0054】
本開示の目的のために、ω参照システムを使用して、炭素数、二重結合数、およびω炭素(この目的では1番目とする)から数えたω炭素に最も近い二重結合の位置を示す。この命名法を下の表1で、「省略表記法」と題された欄に示す。表の残りは、ω−3およびω−6脂肪酸の共通の名称、明細書全体で使用される略語、および各化合物の化学名をまとめる。
【0055】
【表1】
【0056】
「必須脂肪酸」と言う用語は、固体が特定の必須脂肪酸を新規に(de novo)合成できないことから、生きるために摂取しなくてはならない特定のPUFAを指す。例えば、哺乳類は必須脂肪酸LA(18:2、ω−6)を合成できない。その他の必須脂肪酸としては、GLA(ω−6)、DGLA(ω−6)、ARA(ω−6)、EPA(ω−3)、およびDHA(ω−3)が挙げられる。
【0057】
「脂肪」と言う用語は、25℃で固体であり、通常、飽和である脂質物質を指す。
【0058】
「油」と言う用語は、25℃で液体であり、通常、多不飽和である脂質物質を指す。PUFAは、いくつかの藻類、油性酵母菌および糸状菌の油に見られる。「微生物油」または「単細胞油」は、微生物がそれらの寿命中に自然に生産する油である。このような油は長鎖PUFAを含有することができる。
【0059】
「PUFA生合成経路酵素」と言う用語は、PUFAの生合成に関連した以下の酵素(および前記酵素をコードする遺伝子)のいずれかを指す。Δ4デサチュラーゼ、Δ5デサチュラーゼ、Δ6デサチュラーゼ、Δ12デサチュラーゼ、Δ15デサチュラーゼ、Δ17デサチュラーゼ、Δ9デサチュラーゼ、および/または鎖長延長酵素。
【0060】
「ω−3/ω−6脂肪酸生合成経路」と言う用語は、適切な条件下で発現すると、ω−3およびω−6脂肪酸の片方または双方の生産を触媒する酵素をコードする一組の遺伝子を指す。典型的にω−3/ω−6脂肪酸生合成経路に関与する遺伝子は、以下の酵素のいくつかまたは全てをコードする。Δ12デサチュラーゼ、Δ6デサチュラーゼ、鎖長延長酵素、Δ5デサチュラーゼ、Δ17デサチュラーゼ、Δ15デサチュラーゼ、Δ9デサチュラーゼ、およびΔ4デサチュラーゼ。共通の供給源からいかにしてω−3およびω−6脂肪酸の双方が生産できるかを実証する代表的な経路は図1に示され、DHAへの様々な中間体を通じて、オレイン酸の変換を提供する。
【0061】
「デサチュラーゼ」と言う用語は、1つもしくはそれ以上の脂肪酸を不飽和化して関心のある一不飽和または多不飽和脂肪酸または前駆物質を生産できる、多酵素複合体のポリペプチド構成要素を指す。特定の脂肪酸に関して、本願明細書全体を通じてω参照システムを使用するのにもかかわらず、Δシステムを使用して基質のカルボキシル末端から数えることで、デサチュラーゼの活性を示す方が都合よい。ここで特に関心が高いのは、1.)分子のカルボキシル末端から数えて17番目および18番目の炭素原子間で脂肪酸を不飽和化し、例えばARAからEPAへのおよび/またはDGLAからETAへの転換を触媒するΔ17デサチュラーゼ、2.)LAからGLAへのおよび/またはALAからSTAへの転換を触媒するΔ6デサチュラーゼ、3.)DGLAからARAへのおよび/またはETAからEPAへの転換を触媒するΔ5デサチュラーゼ、4.)DPAからDHAへの転換を触媒するΔ4デサチュラーゼ、5.)オレイン酸からLAへの転換を触媒するΔ12デサチュラーゼ、6.)LAからALAへの転換を触媒するΔ15デサチュラーゼ、および7.)パルミチン酸からパルミトレイン酸(16:1)および/またはステアリン酸からオレイン酸(18:1)への転換を触媒するΔ9デサチュラーゼである。
【0062】
「鎖長延長酵素」と言う用語は、脂肪酸炭素鎖を伸長して、鎖長延長酵素が作用した脂肪酸基質よりも炭素2個分長い一不飽和または多不飽和脂肪酸が生産できる、多酵素複合体のポリペプチド構成要素を指す。この延長プロセスは、CoAがアシルキャリアである脂肪酸合成酵素と関連した多段階機序で起きる(ラスナー(Lassner)ら、The
Plant Cell 8:281〜292(1996))。手短に述べると、マロニル−CoAが長鎖アシル−CoAと縮合して、CO2およびβ−ケトアシル−CoA(アシル部分が炭素原子2個分伸長された)を生じる。引き続く反応には、β−ヒドロキシアシル−CoAへの還元、エノイル−CoAへの脱水、および伸長されたアシル−CoAを生じる第2の還元が含まれる。鎖長延長酵素によって触媒される反応の例は、GLAからDGLA、STAからETA、およびEPAからDPAへの転換である。したがって鎖長延長酵素は、異なる特異性を有することができる(例えばC16/18鎖長延長酵素はC16基質を好み、C18/20鎖長延長酵素はC18基質を好み、C20/22鎖長延長酵素はC20基質を好む)。
【0063】
「高親和力鎖長延長酵素」と言う用語は、その基質特異性が、好ましくはGLAに向けたものである(鎖長延長酵素反応の生産物としてのDGLAを伴う)鎖長延長酵素を指す。このような鎖長延長酵素の1つについて、国際公開第00/12720号パンフレットで述べられている。
【0064】
「変換効率」および「%基質変換」と言う用語は、それによって特定の酵素(例えばデサチュラーゼまたは鎖長延長酵素)が基質から生産物に変換できる効率を指す。変換効率は以下の式に従って測定される。([生産物]/[基質+生産物])×100(式中、「生産物」には即時の生産物およびそれから誘導される経路中の全生産物が含まれる)。
【0065】
「油性」と言う用語は、それらのエネルギー源を脂質の形態で保存する傾向がある生物
体を指す(ウィーテ(Weete)「真菌脂質生化学(Fungal Lipid Biochemistry)」第二版、プレナム(Plenum)、1980)。概してこれらの微生物の細胞PUFA含量はS字形曲線に従い、対数後期または初期定常増殖相において脂質濃度が最大に達するまで増大し、次に後期定常および死滅期中に徐々に減少する(ヨンマニットチャイ(Yongmanitchai)およびワード(Ward)、Appl.Environ.Microbiol.57:419〜25(1991))。
【0066】
「油性酵母菌」と言う用語は、少なくとも25%のそれらの乾燥細胞重量を油として蓄積できる酵母菌として分類される微生物を指す。油性酵母菌の例としては、ヤロウィア(Yarrowia)、カンジダ(Candida)、ロドトルラ(Rhodotorula)、ロドスポリジウム(Rhodosporidium)、クリプトコッカス(Cryptococcus)、トリコスポロン(Trichosporon)およびリポマイセス(Lipomyces)属が挙げられるが、決してこれに限定されるものではない。
【0067】
「発酵性炭素源」と言う用語は、微生物が代謝してエネルギーを引き出す炭素源を意味する。本発明の典型的な炭素源としては、単糖類、少糖類、多糖類、アルカン、脂肪酸、脂肪酸のエステル、モノグリセリド、ジグリセリド、トリグリセリド、二酸化炭素、メタノール、ホルムアルデヒド、ギ酸、および炭素含有アミンが挙げられるが、これに限定されるものではない。
【0068】
「コドン最適化された」と言う用語は、様々な宿主の形質転換のための遺伝子または核酸分子のコード領域を指し、DNAによってコードされるポリペプチドを改変することなく、宿主生物体の典型的なコドン使用を反映するための遺伝子または核酸分子のコード領域中のコドンの改変を指す。本発明の文脈で、遺伝子およびDNAコード領域は、表4で収集された情報を使用して、ヤロウィア(Yarrowia)種における最適発現のためにコドン最適化される。
【0069】
ここでの用法では、「単離された核酸断片」とは、場合により合成、非天然または改変ヌクレオチド塩基を含有する一本鎖または二本鎖のRNAまたはDNAポリマーである。DNAポリマーの形態の単離された核酸断片は、cDNA、ゲノムDNAまたは合成DNAの1つもしくはそれ以上のセグメントを含んでなってもよい。
【0070】
アミノ酸またはヌクレオチド配列の「かなりの部分」とは、当業者による配列の手動評価によって、あるいはBLAST(「基礎的局在性整列化検索ツール(Basic Local Alignment Search Tool)」アルトシュル(Altschul)S.F.ら、J.Mol.Biol.215:403〜410(1993))などのアルゴリズムを使用したコンピュータ支援配列比較および同定によって、遺伝子またはポリペプチドの推定上の同定を得るのに十分なポリペプチドのアミノ酸配列または遺伝子のヌクレオチド配列を含んでなる部分である。概してヌクレオチド配列の「かなりの部分」は、配列を含んでなる核酸断片を特異的に同定および/または単離できるようにする十分な配列(例えば20〜30個の連続(contiguous)ヌクレオチド)を含んでなる。本願明細書は、1つもしくはそれ以上の特定のタンパク質をコードする、部分的または完全なヌクレオチド配列を教示する。ここで報告される配列の利点を有した当業者は、当業者には既知の目的のために、開示される配列の全てまたはかなりの部分を使用することができる。したがって本発明は、添付の配列表で報告される完全な配列、ならびに上述の配列のかなりの部分を含んでなる。
【0071】
「相補的」と言う用語は、互いにハイブリッドできるヌクレオチド塩基間の関係について述べるために用いられる。例えばDNAに関して、アデノシンはチミンに相補的であり、シトシンはグアニンに相補的である。したがって本発明はまた、添付の配列表で報告さ
れる完全な配列に相補的な単離された核酸断片、ならびに実質的に同様の核酸配列を含む。
【0072】
「コドン縮重」とは、コードされるポリペプチドのアミノ酸配列に影響しないヌクレオチド配列の変動を可能にする遺伝子コードの性質を指す。当業者は、特定のアミノ酸を特定化するヌクレオチドコドンの使用における、特定の宿主細胞によって示される「コドン−バイアス」についてよく知っている。したがって宿主細胞中の改善された発現のために遺伝子を合成する場合、そのコドン使用頻度が、宿主細胞の好むコドン使用頻度に近くなるように遺伝子をデザインすることが望ましい。
【0073】
「化学的に合成された」とは、DNA配列に関連して構成要素ヌクレオチドが、生体外で(in vitro)構築されたことを意味する。DNAの手動化学合成は確立した手順を使用して達成されてもよく、あるいはいくつかの市販の機器の1つを使用して自動化学合成を実施できる。「合成遺伝子」は、当業者に知られる手順を使用して、化学的に合成されるオリゴヌクレオチド構成単位から構築できる。これらの構成単位をライゲートしアニールして、遺伝子断片を形成し、次にそれを酵素的に構築して所望の遺伝子全体を構成する。したがって遺伝子をヌクレオチド配列の最適化に基づいて、最適な遺伝子発現のために調整し、宿主細胞のコドンバイアスを反映させることができる。当業者は、コドン利用が宿主によって好まれるコドンに偏っている場合の遺伝子発現成功の見込みを理解する。好ましいコドンの判定は、配列情報が利用できる宿主細胞から誘導される遺伝子の調査に基づくことができる。
【0074】
「遺伝子」とは、コード配列に先行する(5’非コード配列)およびそれに続く(3’非コード配列)制御配列をはじめとする、特定のタンパク質を発現する核酸断片を指す。「天然遺伝子」とは、自然界にそれ自体の制御配列と共に見られる遺伝子を指す。「キメラ遺伝子」とは、自然界に共に見られない制御およびコード配列を含んでなる天然遺伝子でないあらゆる遺伝子を指す。したがってキメラ遺伝子は、異なる供給源から誘導される制御配列およびコード配列、あるいは同一供給源から誘導されるが、自然界に見られるのとは異なるやり方で配列される制御配列およびコード配列を含んでなってもよい。「内因性の遺伝子」とは、生物体ゲノムにおいてその天然位置にある天然遺伝子を指す。「外来性の」遺伝子とは、宿主生物体において常態では見られないが、遺伝子移入によって宿主生物体に導入される遺伝子を指す。外来性の遺伝子は、非天然生物体中に挿入された天然遺伝子、あるいはキメラ遺伝子を含んでなることができる。「導入遺伝子」とは、形質転換によってゲノム中に導入される遺伝子である。「コドン最適化遺伝子」とは、そのコドン使用頻度が宿主細胞の好むコドン使用頻度を模倣するようにデザインされる遺伝子である。
【0075】
「コード配列」とは、特定のアミノ酸配列をコードするDNA配列を指す。「適切な調節配列」とは、コード配列の上流(5’非コード配列)、配列内、または下流(3’非コード配列)に位置して、転写、RNAプロセシングまたは安定性、または関連コード配列の翻訳に影響を及ぼすヌクレオチド配列を指す。調節配列は、プロモーター、翻訳リーダー配列、イントロン、ポリアデニル化認識配列、RNAプロセッシング部位、エフェクター結合部位、およびステム−ループ構造を含んでもよい。
【0076】
「プロモーター」とは、コード配列または機能RNAの発現を調節できるDNA配列を指す。概してコード配列は、プロモーター配列に対して3’に位置する。プロモーターは、そっくりそのまま天然遺伝子から誘導されてもよく、あるいは自然界に見られる異なるプロモーターから誘導される異なる要素からなってもよく、あるいは合成DNAセグメントを含んでなってさえよい。異なるプロモーターは、異なる組織または細胞タイプ中で、あるいは異なる発達段階において、あるいは異なる環境または生理学的条件に呼応して、
遺伝子の発現を導いてもよいことが当業者には理解される。ほとんどの細胞タイプ中でほとんどの場合に遺伝子の発現を引き起こすプロモーターは、一般に「構成プロモーター」と称される。ほとんどの場合、制御配列のはっきりした境界は完全に限定されていないので、異なる長さのDNA断片が同一のプロモーター活性を有してもよいこともさらに認識される。
【0077】
「3’非翻訳配列」または「転写ターミネーター」と言う用語は、コード配列の下流に位置するDNA配列を指す。これには、mRNAプロセッシングまたは遺伝子発現に影響できる調節シグナルをコードするポリアデニル化認識配列およびその他の配列が含まれる。ポリアデニル化シグナルは、通常、mRNA前駆物質の3’末端へのポリアデニル酸トラクトの付加に影響することで特性決定される。3’領域は、転写、RNAプロセッシングまたは安定性、または関連コード配列の翻訳に影響できる。
【0078】
「RNA転写物」とは、DNA配列のRNAポリメラーゼが触媒する転写から得られる生産物を指す。RNA転写物がDNA配列の完全な相補的コピーである場合、それは一次転写物と称され、あるいはそれは一次転写物の転写後プロセッシングから誘導されるRNA配列であるかもしれず、成熟RNAと称される。「メッセンジャーRNA」または「mRNA」とはイントロンがなく、細胞によってタンパク質に翻訳されることができるRNAを指す。「cDNA」とは、mRNAに対して相補的であり、それから誘導される二重鎖DNAを指す。「センスRNA」とは、mRNAを含み、細胞によってタンパク質に翻訳されることができるRNA転写物を指す。「アンチセンスRNA」とは、標的一次転写物またはmRNAの全部または一部に相補的であり、標的遺伝子の発現をブロックするRNA転写物を指す(米国特許第5,170,065号明細書、国際公開第99/28508号パンフレット)。アンチセンスRNAの相補性は、特定の遺伝子転写物のあらゆる部分、すなわち5’非コード配列、3’非コード配列、またはコード配列にあっても良い。「機能RNA」とは、翻訳されないがそれでもなお細胞過程に影響するアンチセンスRNA、リボザイムRNA、またはその他のRNAを指す。
【0079】
「作動可能に連結した」と言う用語は、1つの機能が他方の機能によって影響される、単一核酸断片上の核酸配列のつながりを指す。例えばプロモーターはコード配列の発現に影響できる場合、そのコード配列と作動可能に連結する(すなわちコード配列がプロモーターの転写調節の下にある)。コード配列は、センスまたはアンチセンスオリエンテーションで制御配列に作動可能に連結できる。
【0080】
「発現」と言う用語は、ここでの用法では、本発明の核酸断片から誘導されるセンス(mRNA)またはアンチセンスRNAの転写および安定した蓄積を指す。発現はまた、mRNAのポリペプチドへの翻訳を指してもよい。
【0081】
「成熟した」タンパク質とは、翻訳後にプロセスされたポリペプチド、すなわちそれから一次翻訳産物中に存在するあらゆるプレ−またはプロペプチドが除去されたものを指す。「前駆」タンパク質とは、mRNAの一次翻訳産物、すなわちプレ−およびプロペプチドが未だに存在するものを指す。プレ−およびプロペプチドは細胞内局在化シグナルであってもよい(がそれに限定されない)。
【0082】
「形質転換」とは、遺伝的に安定した遺伝形質をもたらす、宿主生物体中への核酸分子の転移を指す。核酸分子は、例えば自律的に複製するプラスミドであってもよく、またはそれは宿主生物体のゲノム中に組み込まれてもよい。形質転換核酸断片を含有する宿主生物体は、「遺伝子組換え」または「組換え」または「形質転換」生物体と称される。
【0083】
「プラスミド」、「ベクター」、および「カセット」と言う用語は、細胞の中心的代謝
の一部ではない遺伝子を運ぶことが多く、通常環状二本鎖DNA断片の形態である染色体外因子を指す。このような因子は、あらゆる供給源から誘導される一本鎖または二本鎖DNAまたはRNAの配列、ゲノム一体化配列、直鎖または環状のファージまたはヌクレオチド配列を自律的に複製するかもしれず、そこではいくつかのヌクレオチド配列が独自の構成に連結または組み換えされ、それは選択された遺伝子産物のために、適切な3’非翻訳配列と共にプロモーター断片およびDNA配列を細胞中に導入することができる。
【0084】
「形質転換カセット」とは、外来性遺伝子を含有し、外来遺伝子に加えて特定の宿主細胞の形質転換を容易にする因子を有する特定のベクターを指す。「発現カセット」とは、外来性遺伝子を含有し、外来性遺伝子に加えて外来性宿主におけるその遺伝子の促進された発現を可能にする因子を有する特定のベクターを指す。
【0085】
「改変された生物学的活性」と言う用語は、アッセイ法によって測定できるヌクレオチド配列によってコードされるタンパク質に関連した活性を指し、そこでは活性は、未変性配列に関連した活性を超えるか、またはそれ未満である。「増強された生物学的活性」とは、未変性配列に結びついたものを超える改変された活性を指す。「低下した生物学的活性」とは、未変性配列に結びついたもの未満の改変された活性を指す。
【0086】
「配列分析ソフトウェア」と言う用語は、ヌクレオチドまたはアミノ酸配列の分析のために有用なあらゆるコンピュータアルゴリズムまたはソフトウェアプログラムを指す。「配列分析ソフトウェア」は、市販のものでも、あるいは独立して開発されても良い。典型的な配列分析ソフトウェアとしては、1.)ウィスコンシン州マディソンのジェネティック・コンピュータ・グループからのGCGパッケージプログラム(Wisconsin Package Version 9.0、Genetics Computer Group(GCG),Madison,WI)、2.)BLASTP、BLASTN、BLASTX(アルシュール(Altschul)ら著、J.Mol.Biol.215:403〜410(1990))、および3.)ウィスコンシン州マディソンのDNASTAR(DNASTAR,Inc.Madison、WI)、および4.スミス−ウォーターマン(Smith−Waterman)・アルゴリズムを組み入れたFASTAプログラム(W.R.ピアソン(Pearson)著、Comput.Methods Genome Res.[Proc.Int.Symp.](1994)、1992年会議、111〜20、編集者:スハイ,サンドール(Suhai,Sandor)、プレナム(Plenum)、ニューヨーク州ニューヨーク(New York、NY))が挙げられるが、これに限定されるものではない。本願明細書の文脈内では、配列分析ソフトウェアを分析のために使用する場合、分析結果は特に断りのない限り、言及されるプログラムの「デフォルト値」に基づくものと理解される。ここでの用法では、「デフォルト値」とは、最初に初期化されたときにソフトウェアに元々ロードされた、あらゆる値またはパラメータの組を意味する。
【0087】
ここで使用される標準組換えDNAおよび分子クローニング技術は技術分野で良く知られており、サムブルック(Sambrook)J.、フリッチュ(Fritsch)E.F.、およびマニアティス(Maniatis)T.「分子クローニング:実験室マニュアル(Molecular Cloning:A Laboratory Manual)」第二版、コールドスプリングハーバーラボラトリ(Cold Spring Harbor Laboratory)、ニューヨーク州コールドスプリングハーバー(Cold Spring Harbor、NY)(1989)(以下マニアティス(Maniatis));シルハビー(Silhavy)T.J.、ベンナン(Bennan)M.L.およびエンクイスト(Enquist)L.W.「遺伝子融合実験(Experiments with Gene Fusions)」コールドスプリングハーバーラボラトリ:ニューヨーク州コールドスプリングハーバー(Cold Spring Harbo
r、NY)(1984);およびオースベル(Ausubel)F.M.ら「分子生物学現代プロトコル(Current Protocols in Molecular Biology)」グリーンパブリッシングアッソシエーツ(Greene Publishing Assoc.)およびワイリーインターサイエンス(Wiley−Interscience)による出版(1987)で述べられている。
【0088】
脂肪酸の微生物生合成
一般に油性微生物中の脂質の蓄積は、増殖培地中に存在する全体的な炭素と窒素の比率に応答してトリガーされる。細胞が利用できる窒素供給を消耗した場合(例えば炭素と窒素の比率が約40を超える場合)、細胞アデノシン一リン酸(AMP)の枯渇は、ミトコンドリア中のAMP−依存イソクエン酸デヒドロゲナーゼ活性の休止、およびクエン酸の蓄積、クエン酸の細胞質ゾル内への輸送、そして引き続くATP−クエン酸リアーゼによるクエン酸の切断をもたらして、アセチル−CoAが生産する。アセチル−CoAは、脂肪酸の新規(de novo)生合成の主要(principle)構成ブロックである。効果的に代謝されてアセチル−CoAを生産できるあらゆる化合物が、脂肪酸前駆物質の役割を果たせるが、このタイプの反応ではグルコースが炭素の主な供給源である。グルコースは解糖を通じてピルビン酸に変換され、次にピルビン酸はミトコンドリア中に輸送され、そこでピルビン酸デヒドロゲナーゼによってアセチル−CoAに変換される。アセチル−CoAはミトコンドリア膜を越えて細胞質中に直接輸送できないので、アセチル−CoAからの2個の炭素がオキサロ酢酸と縮合して、クエン酸を生産する(クエン酸合成酵素によって触媒される)。クエン酸は細胞質内に直接輸送されて、そこでにATP−クエン酸リアーゼによって切断され、アセチル−CoAおよびオキサロ酢酸が再生する。オキサロ酢酸は、リンゴ酸への転換を通じて、トリカルボン酸サイクルに再び入る。
【0089】
マロニル−CoAの合成は、細胞質内で起きる脂肪酸生合成の第1の前駆ステップである。マロニル−CoAは、アセチル−CoAカルボキシラーゼによって、アセチル−CoAのカルボキシル化を通じて生産される。脂肪酸合成は、多酵素脂肪酸合成酵素複合体(「FAS」)によって触媒され、8個の二炭素断片(アセチル−CoAからのアセチル基)の縮合によって起き、炭素16個の飽和脂肪酸であるパルミチン酸が形成する。より具体的には、FASは以下が関与する一連の7つの反応を触媒する(スミス(Smith)S.、FASEB J.、8(15):1248〜59(1994))。
1.アセチル−CoAおよびマロニル−CoAがFASのアシルキャリアペプチド(ACP)に転移される。次にアセチル基がマロニル基に転移されてβ−ケトブチリル−ACPが形成し、CO2を放出する。
2.β−ケトブチリル−ACPが還元(β−ケトアシル還元酵素による)および脱水(β−ヒドロキシアシルデヒドラターゼによる)を被り、トランス−単不飽和脂肪アシル基が形成する。
3.二重結合がNADPHによって還元され、最初のものよりも炭素が2個分長い飽和脂肪−アシル基が生じる。次に新しいマロニル基と縮合して延長プロセスを繰り返すブチリル基の能力が再生する。
4.脂肪アシル基が炭素16個の長さになったら、チオエステラーゼ活性がそれを加水分解して遊離パルミチン酸を放出する。
【0090】
パルミチン酸(16:0)は、小胞体膜に存在する鎖長延長酵素およびデサチュラーゼの作用を通じた、より鎖長が長い飽和および不飽和脂肪酸(例えばステアリン酸(18:0)、パルミトレイン酸(16:1)、およびオレイン酸(18:1))の前駆物質である。パルミチン酸およびステアリン酸は、Δ9デサチュラーゼの作用によってそれらの不飽和(unsatuared)誘導体であるパルミトレイン酸(16:1)およびオレイン酸(18:1)酸にそれぞれ変換する。
【0091】
トリアシルグリセロール(脂肪酸の主な貯蔵単位)は、リン酸1,2−ジアシルグリセロール(一般にホスファチジン酸として同定される)を生じる、アシル−CoAの2個の分子からグリセロール−3−リン酸へのエステル化によって形成される。次にホスファチジン酸ホスファターゼによってリン酸が除去され、1,2−ジアシルグリセロールが生産する。第3の脂肪酸を添加すると、ジアシルグリセロール−アシルトランスフェラーゼの作用によってトリアシルグリセロールが形成する。
【0092】
Ω脂肪酸の生合成
非常に単純化すると、LAをGLA、DGLA、およびARA(ω−6経路)に、ALAをSTA、ETA、EPA、およびDHA(ω−3経路)に変換する代謝プロセスは、炭素原子の付加を通じた炭素鎖の延長と、二重結合の添加を通じた分子の不飽和化を伴う(図1)。これは小胞体膜に存在する一連の特別な不飽和化および延長酵素を必要とする。
【0093】
ω−6脂肪酸
オレイン酸は、Δ12デサチュラーゼの作用によって、最初のω−6脂肪酸であるLA(18:2)に変換される。引き続くω−6脂肪酸は、次のようにして生産される。1.)LAがΔ6デサチュラーゼ活性によってGLAに変換され、2.)GLAが鎖長延長酵素の作用によってDGLAに変換され、3.)DGLAがΔ5デサチュラーゼの作用によってARAに変換される。
【0094】
ω−3脂肪酸
リノール酸(LA)は、Δ15デサチュラーゼの作用によって、最初のω−3脂肪酸であるALAに変換される。引き続くω−3脂肪酸は、ω−6脂肪酸と類似した一連のステップで生産される。具体的には、1.)ALAがΔ6デサチュラーゼ活性によってSTAに変換され、2.)STAが鎖長延長酵素活性によってETAに変換され、3.)ETAがΔ5デサチュラーゼ活性によってEPAに変換される。代案としては、ETAおよびEPAは、Δ17デサチュラーゼ活性によって、DGLAおよびARAからそれぞれ生産できる。EPAは、鎖長延長酵素およびΔ4デサチュラーゼ活性によって、DHAにさらに変換できる。
【0095】
Ω脂肪酸生産に関与する遺伝子
藻類、細菌、カビおよび酵母菌をはじめとする多くの微生物は、通常の細胞代謝経路内でPUFAおよびω脂肪酸を合成できる。特によく適するのは、ヤブレツボカビ(Thraustochytrium)属の種であるスキゾキトリウム・アグレガトム(Schizochytrium aggregatm)、およびモルティエレラ・アルピナ(Morteriella alpina)をはじめとする菌・カビ類である。さらに多くの渦鞭毛藻類(渦鞭毛藻綱(Dinophyceaae))は、高濃度のPUFAを自然に生産する。したがって油生産に関与する多様な遺伝子が遺伝的手段を通じて同定されており、これらのいくつかの遺伝子のDNA配列は公的に入手できる(制限を意図しない例を下の表2に示す)。
【0096】
【表2】
【0097】
【表3】
【0098】
さらに特許文献は、PUFA生産に関与する多くの追加的な遺伝子のDNA配列(および/または上の遺伝子のいくつかに関する詳細およびそれらの単離方法)を提供する。例えば米国特許第5,968,809号明細書(Δ6デサチュラーゼ)、米国特許出願公開第2003/0196217 A1号明細書(Δ17デサチュラーゼ)、米国特許第5,972,664号明細書および米国特許第6,075,183号明細書(Δ5デサチュラーゼ)、国際公開第91/13972号パンフレットおよび米国特許第5,057,419号明細書(Δ9デサチュラーゼ)、国際公開第93/11245号パンフレット(Δ15デサチュラーゼ)、国際公開第94/11516号パンフレット、米国特許第5,443,974号明細書および国際公開第03/099216号パンフレット(Δ12デサチュラーゼ)、国際公開第02/090493号パンフレット(Δ4デサチュラーゼ)、および国際公開第00/12720号パンフレットおよび米国特許出願公開第2002/0139974 A1号明細書(鎖長延長酵素)を参照されたい。これらの各特許および出願は、その内容全体を参照によってここに援用したものとする。
【0099】
当業者には明らかなように、特定のPUFA最終生産物の生産のために宿主生物体に導入する必要がある特定機能は、宿主細胞(およびその未変性PUFAプロフィールおよび/またはデサチュラーゼプロフィール)、基質の入手可能性および所望の最終生産物に左右される。同時係属の米国仮特許出願第60/467677号(その内容全体を本願明細書に引用したものとする)で述べられ、図1に示すように、LA、GLA、DGLA、A
RA、ALA、STA、ETA、EPA、DPA、およびDHAは、以下のPUFA酵素機能の様々な組み合わせを導入することで、全て油性酵母菌中で生産されてもよい。Δ4デサチュラーゼ、Δ5デサチュラーゼ、Δ6デサチュラーゼ、Δ12デサチュラーゼ、Δ15デサチュラーゼ、Δ17デサチュラーゼ、Δ9デサチュラーゼ、および/または鎖長延長酵素。当業者は、公的に入手できる文献(例えばジェンバンク(GenBank))、特許文献、およびPUFAを製造する能力を有する微生物の実験的分析に従って、上の各酵素をコードする様々な候補遺伝子を同定できるであろう。配列は、例えば天然供給源などから(細菌、藻類、菌・カビ類、植物、動物などから)単離され、半合成または新規(de novo)合成経路を通じて生産される、あらゆる供給源に由来してもよい。しかし当業者には明らかなように、異種性遺伝子は別の宿主中では様々な効率で発現する。したがってω−3および/またはω−6PUFAの生産は、異種性宿主における発現レベルが、関心のある宿主生物体における別のデサチュラーゼまたは鎖長延長酵素の発現に比べて好ましい、特定のデサチュラーゼまたは鎖長延長酵素の選択によって最適化されてもよい。
【0100】
宿主に導入されるデサチュラーゼおよび鎖長延長酵素遺伝子の特定の供給源は重要でないが、デサチュラーゼまたは鎖長延長酵素活性を有する特定のポリペプチドを選択する上での考慮としては、次が挙げられる。1.)ポリペプチドの基質特異性、2.)ポリペプチドまたはその構成要素が律速酵素であるかどうか、3.)デサチュラーゼまたは鎖長延長酵素が所望のPUFAの合成のために必須であるかどうか、および/または4.)ポリペプチドによって必要とされる補助因子。発現するポリペプチドは、好ましくは宿主細胞内のその位置の生化学的環境と適合性のパラメーターを有する。例えばポリペプチドは、宿主細胞内で基質獲得のためにその他の酵素と競争しなくてはならないかもしれない。したがってポリペプチドのKMおよび特異的活性分析は、特定の宿主細胞内でPUFA生産を修正するために、特定のポリペプチドの適性を判定する上で考慮される。特定の宿主細胞で使用されるポリペプチドは、意図される宿主細胞中に存在する生化学的条件下で機能できるものであるが、その他の点では所望のPUFAを修正できる、デサチュラーゼまたは鎖長延長酵素活性を有するあらゆるポリペプチドであることができる。
【0101】
しかし究極の目的がω−3および/またはω−6脂肪酸が濃縮された油を蓄積する油性酵母菌の開発である、ここでの研究目的のためには、油性酵母菌中において比較的効率的に機能する、デサチュラーゼおよび鎖長延長酵素活性を有するポリペプチドを同定することが望ましかった。したがって、様々なデサチュラーゼおよび鎖長延長酵素が油性酵母菌中において発現され、基質供給試験においてω−3および/またはω−6脂肪酸を生産するそれらの能力についてスクリーンされた。ひとたび適切な酵素が同定されると(全体的%基質変換に基づいて)、次にこれらの遺伝子に後述するコドン最適化技術を施して、別の油性酵母菌宿主における各酵素の発現をさらに最適化した。これによってω−3および/またはω−6脂肪酸の最大の生産が可能になった。
【0102】
当業者は、ここでコドン最適化のために選択される特異的PUFA遺伝子が単に例示的であり、ここで本発明を制限することを意図しないことを理解するであろう。様々な供給源からの多数のその他の異種性デサチュラーゼ(Δ4、Δ5、Δ6、Δ9、Δ12、Δ15、および/またはΔ17デサチュラーゼ活性を有する)および鎖長延長酵素は、コドン最適化して油性酵母菌宿主におけるそれらの発現を改善できる。
【0103】
油性酵母菌における発現のためのΩ脂肪酸遺伝子コドンの最適化
当業者にはよく知られているように、宿主が好むコドンの使用は、ポリペプチドをコードする外来遺伝子の発現を実質的に増強できる。一般に宿主が好むコドンは、タンパク質中のコドン使用を調べてどのコドンが最高頻度で使用されるか判定することで、関心のある特定の宿主種(好ましくは最大量を発現するもの)中で判定できる。次に宿主種で好ま
しいコドンを使用して、デサチュラーゼまたは鎖長延長酵素活性を有する関心のあるポリペプチドのためのコード配列の全体または一部が合成できる。したがって関心のある特定の宿主生物体における発現のために遺伝子を最適化する方法は、概して
a)関心のある特定の宿主生物体についてヌクレオチドコード領域および対応するポリペプチドの配列を得て、コドンのデータベースを形成し、
b)コドンのデータベースを分析し、いずれのコドンが各アミノ酸を優先的にコードするのか判定し、
c)関心のある特定の宿主生物体中で発現させる遺伝子の配列(例えばデサチュラーゼまたは鎖長延長酵素)を得て、
d)ステップ(c)の配列中の好ましくないコドンをステップ(b)の好ましいコドンで置き換えて、遺伝子が関心のある特定の宿主生物体における発現のためにコドン最適化されることを必要とする。
【0104】
またDNAの全部(または一部)を合成して、転写mRNA中に存在する、二次構造のあらゆる不安定化配列または領域を除去できる。またDNAの全部(または一部)を合成して、塩基組成物を所望の宿主細胞中でより好ましいものに変更できる。
【0105】
本発明の目的のためには、修正されたポリペプチドが、別の宿主(すなわち油性酵母菌)によって好ましいコドンを使用するように、外来宿主中で発現させるPUFAデサチュラーゼおよび鎖長延長酵素活性を有する特定のポリペプチドをコードするコドンの一部を修正することが望ましかった。具体的にはΔ17デサチュラーゼ活性、Δ6デサチュラーゼ活性、および鎖長延長酵素活性を有するポリペプチドをコードするコドンの一部を修正して、ヤロウィア・リポリティカ(Yarrowia lipolytica)における遺伝子の発現を増強することが望ましかった。したがってY.リポリティカ(Y.lipolytica)コドン使用プロフィールは表4(実施例3)に示すように決定された。さらに翻訳開始コドン「ATG」を取り囲むヌクレオチド配列は、酵母菌細胞における発現に影響することが分かっている。酵母菌におけるポリペプチドの発現が不良であれば、外来性遺伝子のヌクレオチド配列を修正し、効率的な酵母菌翻訳開始配列を含めて、最適な遺伝子発現を得ることができる。したがってY.リポリティカ(Y.lipolytica)における遺伝子発現をさらに最適化するために、「ATG」開始コドン周囲の共通配列もまた決定された(図4、配列番号122)。
【0106】
Y.リポリティカ(Y.lipolytica)コドン使用プロフィールおよび「ATG」開始コドン周囲の共通配列に基づいて、Δ17デサチュラーゼ遺伝子、Δ6デサチュラーゼ遺伝子、および鎖長延長酵素遺伝子の核酸を配列を修正して、宿主が好むコドンを用いた。より具体的には、モルティエレラ・アルピナ(Mortierella alpina)(ATCC番号16266)Δ6デサチュラーゼ(ジェンバンク(GenBank)登録番号AF465281、米国特許第5,968,809号明細書)を選択して、LAをGLAに、および/またはALAをSTAに変換する酵素的能力を導入した。この野生型デサチュラーゼは、457個のアミノ酸(配列番号2)および51.8kDの予想分子量を有した。ここで作り出したコドン最適化遺伝子では、1374bpのコード領域(144個のコドンに対応する)の152bpがコドン最適化され、翻訳開始部位が修正された。同様にして野生型サプロレグニア・ディクリナ(Saprolegnia diclina)(ATCC番号56851)Δ17デサチュラーゼ(配列番号4、米国特許出願公開2003/0196217A1号明細書)を選択して、DGLAをETAに、および/またはARAをEPAに変換する酵素的能力を導入したが、翻訳開始部位は修正され、1077bpコード領域(117個のコドンを含んでなる)の127bpがコドン最適化された。最後にGLAからDGLA、STAからETA、および/またはEPAからDPAへの転換のため、油性酵母菌におけるその発現の最適化のために、318個のアミノ酸(配列番号6)および40.5kDの予想分子量を有する野生型M.アルピナ(M.
alpina)高親和力PUFA鎖長延長酵素(ジェンバンク(GenBank)登録番号AX464731、国際公開第00/12720号パンフレット)を選択した。具体的には957bpのコード領域(85個のコドンに対応する)の94bpをコドン最適化し、翻訳開始部位を修正した。したがって本発明は、添付の配列表で報告するような合成コドン最適化遺伝子の完全な配列、これらの完全な配列の相補的配列、およびこれらの配列のかなりの部分を含んでなる。
【0107】
当業者は、ここで述べる最適化方法がその他の遺伝子(例えばω−3/ω−6脂肪酸生合成経路中の遺伝子)に等しく適用でき、S.ディクリナ(S.diclina)Δ17デサチュラーゼ、およびM.アルピナ(M.alpina)Δ6デサチュラーゼ、およびM.アルピナ(M.alpina)鎖長延長酵素の調節が、単に例示的であることを理解するであろう。
【0108】
遺伝子変異を通じたコドン最適化遺伝子の最適化
コドン最適化は、油性酵母菌における機能のためのより望ましい物理的および動力学的パラメーターがあるデサチュラーゼまたは鎖長延長酵素活性をそれぞれ有する、ポリペプチドを生体内(in vivo)で生産する有用な手段であるが、宿主細胞におけるポリペプチドの活性をさらに増強する追加的手段が利用できる。具体的には配列を合成する方法および配列を一緒にする方法は、文献においてよく確立している。例えば生体外(in
vitro)変異誘発および選択、部位特異的変異誘発、誤りがちなPCR(メルニコフ(Melnikov)ら、Nucleic Acids Research、27(4):1056〜1062(1999年2月15日))、「遺伝子シャフリング」またはその他の手段を用いて、自然発生的またはコドン最適化デサチュラーゼまたは鎖長延長酵素遺伝子の変異を得ることができる。これによって例えば生体内(in vivo)で所望のPUFAの半減期がより長く、または生産速度がより早い、デサチュラーゼまたは鎖長延長酵素ポリペプチドそれぞれの生産が可能になる。
【0109】
必要に応じて、酵素活性に重要なコドン最適化デサチュラーゼまたは鎖長延長酵素ポリペプチドの領域は、ルーチンの変異誘発、得られる変異体ポリペプチドの発現、およびそれらの活性の判定を通じて判定できる。変異体は、欠失、挿入、および点変異、またはそれらの組み合わせを含んでもよい。典型的な機能分析は、欠失変異誘発に始まって機能に必要なタンパク質のN−およびC−末端限界を判定し、次に内部欠損、挿入または点変異体を作り出し、機能に必要な領域をさらに判定する。カセット変異誘発または完全合成などのその他の技術もまた使用できる。欠失変異誘発は例えば、エキソヌクレアーゼを使用して、5’または3’コード領域を逐次除去して達成される。このような技術のためのキットが利用できる。欠失後、開始または停止コドンを含有するオリゴヌクレオチドをそれぞれ5’または3’欠失後に、欠失コード領域にライゲーションして、コード領域が完成する。代案としては、部位特異的変異誘発、変異原性PCRをはじめとする多様な方法によって、または既存の制限部位において消化されたDNA上へのライゲーションによって、開始または停止コドンをコードするオリゴヌクレオチドをコード領域に挿入する。内部欠損は、部位特異的変異誘発または変異原性PCRを通じた変異原性プライマーの使用によって、DNA中の既存の制限部位の使用をはじめとする多様な方法を通じて同様に作り出せる。挿入は、リンカー−スキャニング変異誘発、部位特異的変異誘発または変異原性PCRなどの方法を通じて作り出される。点変異は、部位特異的変異誘発または変異原性PCRなどの技術を通じて作り出される。
【0110】
化学変異誘発もまた、活性に重要なデサチュラーゼまたは鎖長延長酵素ポリペプチドの領域を同定するために使用できる。変異したコンストラクトが発現し、得られる改変タンパク質がデサチュラーゼまたは鎖長延長酵素として機能する能力がアッセイされる。このような構造−機能分析は、どの領域が欠失してもよいか、どの領域が挿入を許容するか、
どの点変異によって変異体タンパク質が、未変性(またはコドン最適化)デサチュラーゼまたは未変性鎖長延長酵素と実質的に同じように機能できるようにするかを判定できる。
【0111】
ω−3および/またはω−6脂肪酸の微生物による生産
微生物によるω−3および/またはω−6脂肪酸の生産には、魚または植物などの天然供給源からの精製に比べて、例えば以下のようないくつかの利点がある。
1.)多くの微生物は、油組成が高等生物のものと比べてはるかに単純であることが知られており、所望の構成要素の精製を容易にする。
2.)微生物による生産には、天候および食物供給などの外部変数によって引き起こされるばらつきがない。
3.)微生物的に生産された油は、実質的に環境汚染物質による混入物がない。
4.)微生物は特定用途を有するかもしれない特定の形態で、PUFAを提供できる。
5.)微生物による油生産は、培養条件を制御することで、特に微生物的に発現される酵素のために特定の基質を提供することで、または化合物の添加または遺伝子工学アプローチによって望まれない生化学的経路を抑制することで操作できる。
【0112】
これらの利点に加えて、組換え微生物からのω−3および/またはω−6脂肪酸の生産は、宿主中に新しい生合成経路を提供することで、または望まれない経路を抑制することで、所望のPUFAまたはそれらのコンジュゲートされた形態のレベルを増大させて、望まれないPUFAのレベルを低下させ、自然発生的な微生物の脂肪酸プロフィールを変更する能力を提供する。例えばこのようにして生産したω−3とω−6脂肪酸の比率を修正する、他のω脂肪酸生産を排除しながらω−3またはω−6脂肪酸のどちらか一方だけを生産する、または他のPUFAの下流または上流生産物の顕著な蓄積なしに特異的PUFAの生産を操作することが可能である。
【0113】
発現システム、カセット、およびベクター
ここで述べるコドン最適化遺伝子および関心のある配列の遺伝子生産物は、異種性微生物の宿主細胞、特に油性酵母菌(例えばヤロウィア・リポリティカ(Yarrowia lipolytica))の細胞中で生産されてもよい。組換え微生物宿主における発現は、様々なPUFA経路中間体を生産するため、またはこれまで宿主を使用してできなかった新しい生産物を合成するため、宿主に既存のPUFA経路を調節するのに有用かもしれない。
【0114】
微生物の発現システム、および外来タンパク質の高レベル発現を導く調節配列を含有する発現ベクターは、当業者によく知られている。これらのいずれも本コドン最適化配列のあらゆる遺伝子生産物を生産するためのキメラ遺伝子を構築するのに使用できる。次に形質転換を通じてこれらのキメラ遺伝子を適切な微生物に導入して、コードされた酵素の高レベル発現を提供できる。
【0115】
したがって適切なプロモーター制御下における、PUFA生合成経路(例えばここで述べるコドン最適化Δ6デサチュラーゼ、Δ17デサチュラーゼ、および鎖長延長酵素)をコードするキメラ遺伝子の導入は、ω−3および/またはω−6脂肪酸の増大する生産をもたらすことが予期される。本コドン最適化遺伝子の様々な組み合わせを宿主微生物中で一緒に発現することが有用であることが考察される。
【0116】
さらにベクターがまた、ここで述べるコドン最適化遺伝子の1つもしくはそれ以上に加えて、その他の酵素をコードする1つもしくはそれ以上の遺伝子を含んでなってもよいことが考察される。例えばΔ4デサチュラーゼ、Δ5デサチュラーゼ、Δ6デサチュラーゼ、Δ12デサチュラーゼ、Δ15デサチュラーゼ、Δ17デサチュラーゼ、Δ9デサチュラーゼ、および/または鎖長延長酵素の1つもしくはそれ以上の酵素活性をコードする遺
伝子(それによってこれらのあらゆる遺伝子が、場合により特定の宿主生物体での増強された発現のためにコドン最適化される)を含んでなる発現カセットを構築することが望ましいかもしれない。当業者にはよく知られているように、特定の発現カセット内に含まれる特定の遺伝子は、宿主細胞(おそびそのPUFAプロフィールおよび/またはデサチュラーゼプロフィール)、基質の入手可能性、および所望の最終生産物に左右される。
【0117】
したがって本発明は、基質が所望の脂肪酸生産物に変換するように、脂肪酸基質をここで述べるPUFA酵素に暴露することを含んでなる、PUFAを生産する方法を包含する。したがってここで述べる各PUFA遺伝子および対応する酵素生産物は、PUFAの生産のために直接にまたは間接に使用できる。PUFAの直接的生産は、脂肪酸基質が中間ステップまたは経路中間体なしに直接に所望の脂肪酸生産物に変換する場合に起きる。例えばEPAの生産は、Δ17デサチュラーゼ活性を提供する発現カセットを細胞に添加または導入することで、ARAを生産する宿主細胞、またはARAが提供される宿主細胞中で起きる。
【0118】
対照的にPUFA生合成経路をコードする複数遺伝子を一連の反応が起きて所望のPUFAを生産するように組み合わせて使用してもよい。例えば鎖長延長酵素、Δ5デサチュラーゼ、Δ17デサチュラーゼ、およびΔ4デサチュラーゼ活性(そこでは各遺伝子が場合により宿主における発現のためにコドン最適化される)をコードする発現カセットは、自然にGLAを生産する宿主細胞が、代わりにDHAを生産できるようにする(GLAが鎖長延長酵素によってDGLAに変換し、次にDGLAはΔ5デサチュラーゼによってARAに変換し、次にARAはΔ17デサチュラーゼによってEPAに変換し、それは次には鎖長延長酵素によってDPAに変換してもよく、DPAはΔ4デサチュラーゼによってDHAに変換する)。宿主細胞が油性酵母菌である好ましい実施態様では、これらの生物体において自然に生産されるPUFAは18:2脂肪酸(すなわちLA)、もっとまれには18:3脂肪酸(すなわちALA)に限られるので、PUFA生合成に必要な各酵素をコードする発現カセットを生物体に導入しなくてはならない。代案としては基質供給が必要かもしれない。
【0119】
適切な宿主細胞の形質転換に有用なベクターまたはDNAカセットは、技術分野でよく知られている。コンストラクト中に存在する配列の特定の選択は、所望の発現生産物(上述)、宿主細胞の性質、および提案される形質転換細胞と非形質転換細胞とを分離する手段に左右される。しかし典型的に、ベクターまたはカセットは、関連遺伝子の転写および翻訳を導く配列、選択性マーカー、および自律性複製または染色体組み込みを可能にする配列を含有する。適切なベクターは、転写開始を制御する遺伝子の5’領域、および転写終結を制御するDNA断片の3’領域を含んでなる。双方の制御領域が、形質転換された宿主細胞の遺伝子に由来することが最も好ましいが、このような制御領域は、必ずしも生産宿主として選択された特定種に天然の遺伝子に由来しなくてよいものと理解される。
【0120】
所望の宿主細胞中で、デサチュラーゼおよび/または鎖長延長酵素ORFの発現を推進するのに有用な開始制御領域またはプロモーターは多数あり、当業者にはなじみが深い。選択された宿主細胞中でのこれらの遺伝子の発現を導くことができる、実質的にあらゆるプロモーターが本発明に適する。宿主細胞中での発現は、一過性または安定様式で達成できる。一過性発現は、関心のある遺伝子に作動可能に連結された、調節可能プロモーターの活性を誘導することで達成できる。安定発現は、関心のある遺伝子に作動可能に連結された構成プロモーターの使用によって達成できる。一例として宿主細胞が酵母菌の場合、酵母菌細胞中で機能する転写および翻訳領域が、特に宿主種から提供される。転写開始調節領域は、例えば、以下から得ることができる。1.)アルコールデヒドロゲナーゼ、グリセルアルデヒド3−リン酸−デヒドロゲナーゼ(米国特許出願第60/482263号明細書参照)、ホスホグリセリン酸ムターゼ(米国特許出願第60/482263号明細
書参照)、フルクトース−ビスリン染色体外因子アルドラーゼ(米国特許出願第60/519971号明細書参照)、ホスホグルコース−異性化酵素、ホスホグリセラートキナーゼなどの解糖経路中の遺伝子、または2.)酸ホスファターゼ、ラクターゼ、メタロチオネイン、グルコアミラーゼ、翻訳延長因子EF1−α(TEF)タンパク質(米国特許第6,265,185号明細書)、リボソームタンパク質S7(米国特許第6,265,185号明細書)などの調節可能遺伝子。構成または誘導転写が所望されるかどうか、関心のあるORFを発現する上でのプロモーター効率、構築の容易さなど次第で、いくつかの調節配列のいずれの1つでも使用できる。
【0121】
酵母菌での最適遺伝子発現は、それらが効率的な酵母菌翻訳開始列を含むように、外来性遺伝子の翻訳開始コドン「ATG」を取り囲むヌクレオチド配列を修正することで得ることができる。具体的には非効率的に発現する遺伝子の発現は部位特異的変異誘発によって増大でき、そこでは非効率的に発現する遺伝子が、好ましくは高度に発現する遺伝子である内因性酵母菌遺伝子にインフレーム融合する。代案としてはここでの発明でヤロウィア・リポリティカ(Yarrowia lipolytica)において実証されるように、宿主におけるコンセンサス翻訳開始配列を判定して、関心のある宿主でのそれらの最適発現のために、この配列を異種性遺伝子内に遺伝子操作できる。
【0122】
終結領域は、遺伝子の3’領域から誘導されることができ、開始領域はそれから、または異なる遺伝子から得られた。多数の終結領域が知られており、多様な宿主において満足に機能する(それらが由来する同一のおよび異なる、属および種の双方で使用した際に)。終結領域は、通常、特定の特性のためと言うよりも、便宜上で選択される。好ましくは、終結領域は、特にサッカロミセス(Saccharomyces)、分裂酵母(Schizosaccharomyces)、カンジダ(Candida)、ヤロウィア(Yarrowia)またはクリヴェロミセス(Kluyveromyces)である酵母菌遺伝子から誘導される。γ−インターフェロンおよびα−2インターフェロンをコードする哺乳類の遺伝子の3’−領域もまた、酵母菌中で機能することが知られている。終結制御領域もまた、好ましい宿主に天然の様々な遺伝子から誘導されてもよい。場合により終結部位は不必要かもしれないが、含まれることが最も好ましい。
【0123】
当業者は気づいているように、遺伝子をクローン化ベクターに単に挿入するだけでは、それが必要なレベルで成功裏に発現することを確証しない。高発現率の必要性に答えて、転写、翻訳、タンパク質安定性、酸素限界、および宿主細胞からの分泌の側面を制御するいくつかの異なる遺伝的要素を操作することで、多くの特殊化した発現ベクターが作り出されている。より具体的には、遺伝子発現を制御するように操作される分子の特徴のいくつかとして以下が挙げられる。1.)関連転写プロモーターおよびターミネーター配列の性質、2.)クローンされた遺伝子のコピー数、および遺伝子がプラスミド上にあるかまたは宿主細胞のゲノム中に組み込まれているかどうか、3.)合成された外来タンパク質の最終細胞内位置、4.)宿主生物体中の翻訳効率、および5.)宿主細胞内のクローン化遺伝子タンパク質の本質的な安定性。これらの各タイプの修正は、コドン最適化PUFA生合成経路酵素の発現をさらに最適化する手段として、本発明中に包含される。
【0124】
微生物宿主の形質転換
油性酵母菌中での発現に適したコドン最適化デサチュラーゼまたは鎖長延長酵素ポリペプチドをコードするDNAがひとたび得られると、それは宿主細胞中で自律複製できるプラスミドベクター中に入れられ、またはそれは宿主細胞のゲノム中に直接組み込まれる。発現カセットの組み込みは、宿主ゲノム内で無作為に起きることができ、または宿主遺伝子座内で遺伝子組み換えを標的とするのに十分な宿主ゲノムとの相同性領域を含有するコンストラクトの使用を通じて、標的を定めることができる。コンストラクトが内因性遺伝子座に標的を定めると、全てまたはいくつかの転写および翻訳調節領域が、内因性遺伝子
座によって提供できる。
【0125】
別々の複製ベクターから2つ以上の遺伝子が発現する場合、各ベクターが異なる選択手段を有することが望ましく、他のコンストラクトに対する相同性を欠いて、安定した発現を維持し、コンストラクト中における要素の再集合を防止すべきである。調節領域、選択手段、および導入コンストラクト増殖方法の思慮深い選択は、全ての導入された遺伝子が必要なレベルで発現して、所望の生産物の合成を提供するように実験的に判定できる。
【0126】
関心のある遺伝子を含んでなるコンストラクトは、あらゆる標準的技術によって宿主細胞に導入してもよい。これらの技術としては、形質転換(例えば酢酸リチウム形質転換[Methods in Enzymology、194:186〜187(1991)])、プロトプラスト融合、弾道衝撃(bolistic impact)、電気穿孔、マイクロインジェクション、または宿主細胞中に関心のある遺伝子を導入するその他のあらゆる方法が挙げられる。油性酵母菌(すなわち、ヤロウィア・リポリティカ(Yarrowia lipolytica))に適用できるより具体的な教示としては、米国特許第4,880,741号明細書および米国特許第5,071,764号明細書、およびチェン(Chen)D.C.ら(Appl Microbiol Biotechnol.48(2):232〜235(1997))が挙げられる。
【0127】
便宜上、DNA配列(例えば発現カセット)を取り込むように、あらゆる方法によって操作されている宿主細胞を「形質転換された」または「組換え」とここで称する。形質転換された宿主は、遺伝子がゲノム中に組み込まれるか、増幅されるか、または複数のコピー数を有する染色体外因子上に存在するかどうか次第で、発現コンストラクトの少なくとも1つのコピーを有し、2つ以上を有してもよい。形質転換された宿主細胞は、導入されたコンストラクト上に含有されるマーカーの選択によって同定できる。多くの形質転換技術は、多くのDNA分子を宿主細胞中に導入するので、代案としては、別々のマーカーコンストラクトを所望のコンストラクトと共に同時形質転換してもよい。典型的に、形質転換された宿主は、選択的培地上で生育するそれらの能力について選択される。選択培地には抗生物質が組み込まれていてもよく、または栄養素または成長因子などの非形質転換宿主の生育に必要な要素が欠如していてもよい。導入されたマーカー遺伝子は、形質転換された宿主中で発現すると、抗生物質抵抗性を与え、または必須成長因子または酵素をコードしてもよく、それによって選択培地上での生育を可能にしてもよい。また発現したマーカータンパク質が直接または間接に検出できる場合に、形質転換された宿主の選択ができる。マーカータンパク質は単独で、または別のタンパク質と融合して発現してもよい。マーカータンパク質は、次によって検出できる。1.)その酵素活性(例えばβ−ガラクトシダーゼは、基質X−gal[5−ブロモ−4−クロロ−3−インドリル−β−D−ガラクトピラノシド]を着色生産物に変換でき、ルシフェラーゼはルシフェリンを発光生産物に変換できる)、または2.)その光生産または修正特性(例えばオワンクラゲ(Aequorea victoria)の緑色蛍光タンパク質は青色光で照明されると蛍光を発する)。代案としては、抗体を使用して、マーカータンパク質、または例えば関心のあるタンパク質の上の分子タグを検出できる。マーカータンパク質またはタグを発現する細胞は、例えば視覚的に、またはFACS、または抗体を使用したパニングなどの技術によって選択できる。酵母菌形質転換体の選択のためには、酵母菌で機能するあらゆるマーカーを使用してもよい。望ましくは、重要であるカナマイシン、ハイグロマイシン、およびアミノグリコシドG418に対する抵抗性、ならびにウラシルまたはロイシン欠乏培地上で生育する能力である。
【0128】
形質転換に続いて、本コドン最適化配列の遺伝子産物(そして場合により、宿主細胞内で発現するその他のPUFA酵素)に適した基質が、宿主によって自然にまたは遺伝子組換え的に生産されてもよく、またはそれらは外来性に提供されてもよい。
【0129】
微生物中でのω−3および/またはω−6脂肪酸生合成の代謝エンジニアリング
本遺伝子のコドン最適化配列の知識、および油性酵母菌における発現のためにその他のPUFA遺伝子を最適化するのに必要な方法は、油性酵母菌、特にヤロウィア・リポリティカ(Yarrowia lipolytica)におけるω−3および/またはω−6脂肪酸生合成を操作するために有用であろう。これは直接PUFA生合成経路内での代謝エンジニアリング、または炭素をPUFA生合成経路に与える経路の追加的操作を必要とするかもしれない。生化学的経路を操作するのに有用な方法は、当業者にはよく知られている。
【0130】
望ましい生合成経路を上方制御する技術
デサチュラーゼおよび鎖長延長酵素遺伝子の追加的コピーを宿主に導入して、ω−3および/またはω−6脂肪酸生合成経路の産出量を増大させてもよい。デサチュラーゼまたは鎖長延長酵素遺伝子の発現はまた、mRNAまたはコードされたタンパク質のいずれかから不安定化配列を除去/消去することで、または安定化配列をmRNA(米国特許第4,910,141号明細書)に追加することで、(制御されたまたは構成的な)より強力なプロモーターの使用を通じて増大する発現を引き起こして、転写レベルで増大できる。本発明で実証されるように、デサチュラーゼまたは鎖長延長酵素遺伝子の発現を増大させるさらに別のアプローチは、選択された宿主微生物中での最適遺伝子発現のためのコドンで、未変性遺伝子中のコドンを置換することにより、コードされたmRNAの翻訳効率を増大させることである。
【0131】
望ましくない生合成経路を下方制御する技術
反対に、エネルギーまたは炭素についてω−3および/またはω−6脂肪酸生合成経路と競合する生化学的経路、または特定のPUFA最終生産物の生産を妨げる未変性PUFA生合成経路酵素を遺伝子混乱によって除去またはその他の手段(例えばアンチセンスmRNA)によって下方制御してもよい。遺伝子混乱では、そのコード配列を妨害し、それによって遺伝子を機能的に不活性化するために、混乱させたい構造的遺伝子中に外来DNA断片(典型的に選択可能マーカー遺伝子)を挿入し、混乱させる。混乱カセットの宿主細胞中への形質転換は、相同的組み換えによって、機能的未変性遺伝子の非機能的混乱遺伝子による置換をもたらす(例えばハミルトン(Hamilton)ら、J.Bacteriol.171:4617〜4622(1989);バルバス(Balbas)ら、Genes、136:211〜213(1993);ゲルデナー(Gueldener)ら、Nucleic Acid Res.24:2519〜2524(1996);およびスミス(Smith)ら、Methods Mol.Cell.Biol.5:270〜277(1996)参照)。
【0132】
アンチセンス技術は、標的遺伝子配列が知られている場合に遺伝子を下方制御する別の方法である。これを達成するために、所望の遺伝子からの核酸セグメントをクローン化して、RNAのアンチセンス鎖が転写されるようにプロモーターに作動可能に連結する。次にこのコンストラクトを宿主細胞に導入し、RNAのアンチセンス鎖を生産する。アンチセンスRNAは、関心のあるタンパク質をコードするmRNAの蓄積を防止することで、遺伝子発現を阻害する。当業者は、特定の遺伝子の発現を低下させるために、特別な考察がアンチセンス技術の使用に関連することを理解する。例えば、アンチセンス遺伝子の適切な発現レベルは、当業者には知られている異なる調節因子を使用した、異なるキメラ遺伝子の使用を必要とするかもしれない。
【0133】
配列が知られている場合、標的を定めた遺伝子混乱およびアンチセンス技術が遺伝子を下方制御する効果的手段を提供するが、配列ベースではないその他のより特異性が低い方法が開発されている。例えば細胞をUV放射線暴露して、次に所望の表現型についてスク
リーンしてもよい。変異体を作り出すために、化学薬品による変異誘発もまた効果的であり、一般に使用される物質としては、非複製DNAに影響する化学物質(例えばHNO2およびNH2OH)、ならびに複製DNAに影響する薬剤(例えばフレームシフト変異を引き起こすことで注目に値するアクリジン染料)が挙げられる。放射線または化学薬品を使用して変異体を作り出す特定の方法は、技術分野でよく文書化されている。例えばトーマスD.ブロック(Thomas D.Brock)著「バイオテクノロジー:工業微生物学テキストブック(Biotechnology:A Textbook of Industrial Microbiology)」第二版(1989)Sinauer Associates:マサチューセッツ州サンダーランド(Sunderland、MA)、またはデシュパンデ,ムカンド(Deshpande,Mukund)V.、Appl.Biochem.Biotechnol.、36:227(1992)を参照されたい。
【0134】
別の非特異的遺伝子混乱方法は、転移因子またはトランスポゾンの使用である。トランスポゾンは、DNAに無作為に挿入される遺伝的因子であるが、後で配列に基づいて検索して、挿入がどこで起きたのか判定できる。生体内(in vivo)および生体外(in vitro)転位法の双方が知られている。どちらの方法にもトランスポザーゼ酵素と組み合わされた転移因子の使用が関与する。転移因子またはトランスポゾンがトランスポザーゼ存在下で核酸断片に接触すると、転移因子が核酸断片中に無作為に挿入される。混乱された遺伝子は転移因子配列に基づいて同定されてもよいので、技術は、ランダム変異誘発のため、そして遺伝子単離のために有用である。生体外(in vitro)転位のためのキットは市販される。例えば1.)ニュージャージー州ブランチバーグのパーキン・エルマー・アプライド・バイオシステムズ(Perkin Elmer Applied Biosystems(Branchburg、NJ))から入手できる酵母菌Ty1因子に基づく、プライマー・アイランド転位キット、2.)マサチューセッツ州ビバリーのニュー・イングランド・バイオ・ラブズ(New England Biolabs、(Beverly、MA))から入手できる細菌性トランスポゾンTn7に基づくゲノム・プライミング・システム、および3.)ウィスコンシン州マディソンのエピセンター・テクノロジーズ(Epicentre Technologies(Madison、WI))から入手できる、Tn5細菌性転移因子に基づくEZ::TNトランスポゾン挿入システムを参照されたい。
【0135】
本発明の文脈では、上述のいずれか1つの方法によって脂肪酸生合成経路の発現を調節することが有用であるかもしれない。例えば本発明は、ω−3および/またはω−6脂肪酸の生産をもたらす生合成経路中の重要な酵素をコードするコドン最適化遺伝子を提供する。これらのコドン最適化遺伝子には、Δ6デサチュラーゼ、Δ17デサチュラーゼ、およびPUFA鎖長延長酵素が含まれる。ω−3および/またはω−6脂肪酸生合成経路を自然に有さない油性酵母菌中でこれらの遺伝子を発現させ、これらおよびその他のPUFA生合成遺伝子の遺伝子の発現を調節して、宿主生物体の代謝工学のための様々な手段を使用して好ましいPUFA生産物の生産を最大化することが特に有用であろう。同様にこれらの遺伝子によってPUFA生産を最大化するためには、PUFA生合成に向けて導かれる炭素の流れと競合する経路を混乱させることが必要かもしれない。
【0136】
コドン最適化遺伝子の組換え発現のための好ましい微生物宿主
本コドン最適化遺伝子および核酸断片の発現のための宿主細胞は、広範な温度およびpHで、単純または複合炭水化物、有機酸およびアルコール、および/または炭化水素をはじめとする多様な原材料上で生育する微生物の宿主を含んでもよい。本発明で述べられる遺伝子は、油性酵母菌、特にヤロウィア・リポリティカ(Yarrowia lipolytica)における発現のためにコドン最適化されているが、転写、翻訳、およびタンパク質生合成器官は高度に保存されているので、あらゆる細菌、酵母菌および/または糸
状菌が、本核酸断片の発現に適した宿主になることが考察される。
【0137】
好ましい微生物の宿主は、油性酵母菌である。これらの生物体は自然に油の合成および蓄積ができ、油は、細胞乾燥重量の約25%を超え、より好ましくは細胞乾燥重量の約30%を超え、最も好ましくは細胞乾燥重量の約40%を超える量を構成できる。油性酵母菌として典型的に同定されている属としては、ヤロウィア(Yarrowia)、カンジダ(Candida)、ロドトルラ(Rhodotorula)、ロドスポリジウム(Rhodosporidium)、クリプトコッカス(Cryptococcus)、トリコスポロン(Trichosporon)、およびリポマイセス(Lipomyces)が挙げられるが、これに限定されるものではない。より具体的には、例証的な油合成酵母菌としては、ロドスポリジウム・トルイデス(Rhodosporidium toruloides)、リポマイセス・スターケイ(Lipomyces starkeyii)、L.リポフェラス(L.lipoferus)、カンジダ・レブカウフィ(Candida revkaufi)、C.プルケリマ(C.pulcherrima)、C.トロピカリス(C.tropicalis)、C.ユチリス(C.utilis)、トリコスポロン・プランス(Trichosporon pullans)、T.クタネウム(T.cutaneum)、ロドトルラ・グルティナス(Rhodotorula glutinus)、R.グラミニス(R.graminis)、およびヤロウィア・リポリティカ(Yarrowia lipolytica)(以前はカンジダ・リポリティカ(Candida lipolytica)として分類された)が挙げられる。
【0138】
最も好ましいのは油性酵母菌ヤロウィア・リポリティカ(Yarrowia lipolytica)であり、さらなる実施態様で最も好ましいのは、ATCC番号20362、ATCC番号8862、ATCC番号18944、ATCC番号76982および/またはLGAMS(7)1と称されるY.リポリティカ(Y.lipolytica)株である(パパニコラオウ(Papanikolaou)S.、およびアゲリス(Aggelis)G.、Bioresour.Technol.82(1):43〜9(2002))。
【0139】
PUFA生産のための発酵プロセス
形質転換された微生物宿主細胞をデサチュラーゼおよび鎖長延長酵素活性を最適化させる条件下で生育させ、好ましいPUFAの最大かつ最も経済的な収率を得る。一般に最適化されてもよい培地条件としては、炭素源のタイプおよび量、窒素源のタイプおよび量、炭素−対−窒素比率、酸素レベル、生育温度、pH、バイオマス生産相の長さ、油蓄積相の長さ、および細胞収穫時間が挙げられる。油性酵母菌などの関心のある微生物を複合培地(例えば酵母菌抽出物−ペプトン−デキストロース液体培地(YPD))上で生育させ、または生育に必要な構成要素が欠如する合成(defined)最少培地(例えばミシガン州デトロイトのディフコ・ラボラトリーズ(DIFCO Laboratories(Detroit、MI))からの酵母菌窒素塩基)上で生育させて、所望の発現カセットの選択を強要する。
【0140】
本発明における発酵培地は、適切な炭素源を含有しなくてはならない。適切な炭素源としては、単糖類(例えばグルコース、フルクトース)、二糖類(例えばラクトース、スクロース)、少糖類、多糖類(例えばデンプン、セルロースまたはそれらの混合物)、糖アルコール(例えばグリセロール)または再生可能原材料からの混合物(例えば乳清透過液、コーンスティープリーカー、甜菜モラセス、大麦の麦芽)が挙げられるが、これに限定されるものではない。さらに炭素源としては、アルカン、脂肪酸、脂肪酸エステル、モノグリセリド、ジグリセリド、トリグリセリド、リン脂質、および植物油(例えばダイズ油)および動物脂肪をはじめとする脂肪酸の様々な商業的供給源が挙げられる。さらに炭素源としては、重要な生化学的中間体への代謝転換が実証されている一炭素源(例えば二酸
化炭素、メタノール、ホルムアルデヒド、ギ酸および炭素−含有アミン)が挙げられる。したがって本発明で使用される炭素源は多種多様な炭素含有源を包含し、および宿主生物体の選択によってのみ制限されることが考察された。上述の全ての炭素源およびそれらの混合物が本発明で適切であることが期待されるが、好ましい炭素源は糖および/または脂肪酸である。最も好ましいのは、10〜22個の炭素含有グルコースおよび/または脂肪酸である。
【0141】
窒素は、無機(例えば(NH4)2SO4)または有機源(例えば尿素またはグルタミン酸)から供給されてもよい。適切な炭素および窒素源に加えて、発酵培地はまた、適切なミネラル、塩、補助因子、緩衝液、ビタミン、および当業者には知られている微生物の生育とPUFA生産に必要な酵素的経路の促進に適したその他の構成要素を含有しなくてはならない。脂質およびPUFAの合成を促進するいくつかの金属イオン(例えばMn+2、Co+2、Zn+2、Mg+2)が注目されている(ナカハラ(Nakahara)T.ら、「単細胞油の工業的応用(Ind.Appl.of Single Cell Oils)」、D.J.カイル(Kyle)およびR.コリン(Colin)編、p61〜97(1992))。
【0142】
本発明における好ましい増殖培地は、ミシガン州デトロイトのディフコ・ラボラトリーズ(DIFCO Laboratories(Detroit、MI))からの酵母菌窒素塩基などの一般的な商業的調製培地である。またその他の合成(defined)または合成(synthetic)増殖培地が使用されてもよく、特定の微生物の生育に適した培地は、微生物学または発酵科学の当業者に知られている。発酵に適したpH範囲は、典型的に約pH4.0〜pH8.0の間であり、pH5.5〜pH7.0が最初の生育条件の範囲として好ましい。発酵は好気性または好気性条件下で実施されてもよく、マイクロ好気性条件が好ましい。
【0143】
典型的に、油性酵母菌細胞におけるPUFAの高レベルの蓄積は、代謝状態が生育と脂肪合成/貯蔵間で「バランスが取れて」いなくてはならないので、二段階プロセスを必要とする。したがって最も好ましくは、油性酵母菌におけるPUFA生産には、二段階発酵プロセスが必要である。このアプローチでは、第1の発酵状態が細胞集団の生産および蓄積に供され、迅速な細胞生育および細胞分割によって特徴づけられる。発酵の第2段階では、培養内の窒素欠乏条件を確立して、高レベルの脂質蓄積を促進することが好ましい。この窒素欠乏の効果は、細胞内のAMPの効果的濃度を低下させることにより、ミトコンドリアのNAD−依存イソクエン酸デヒドロゲナーゼ活性を低下させることである。これが起きるとクエン酸が蓄積するので、細胞質中にアセチル−CoAの豊富なプールが形成し、脂肪酸合成の下準備をする。したがってこの相は、細胞分割休止と、それに続く脂肪酸合成および油蓄積によって特徴づけられる。
【0144】
細胞は典型的に約30℃で生育させるが、いくつかの研究は、より低い温度における不飽和脂肪酸の増大した合成を示している(ヨンマニットチャイ(Yongmanitchai)およびワード(Ward)、Appl.Environ.Microbiol.57:419〜25(1991))。プロセスの経済に基づけば、この温度シフトは、大部分の生物体の生育が起きる二段階発酵の第1相後に起きるはずである。
【0145】
このコドン最適化遺伝子を使用したω脂肪酸の商業生産が所望される場合、多様な発酵プロセスデザインを適用してもよいことが考察される。例えば組換え微生物宿主からのPUFAの商業生産は、バッチ、供給バッチまたは連続発酵プロセスによって生産されてもよい。
【0146】
バッチ発酵プロセスは閉鎖システムであり、培地組成物がプロセス開始時に定められ、
プロセス中にpHおよび酸素レベル維持のために必要とされるもの以外は、さらなる追加を受けない。したがって培養プロセスの始めに所望の生物体を培地に接種し、培地への追加的供給源(すなわち炭素および窒素源)の添加なしに、生育または代謝活性を生じさせる。バッチプロセスでは、システムの代謝産物およびバイオマス組成物は、培養が終結するまで絶えず変化する。典型的なバッチプロセスでは、細胞は静止対数相から高生育対数相を通過して、最終的に発育速度が減少または停止する定常相に進む。処置されない場合、定常相の細胞は次第に死滅する。標準バッチプロセスのバリエーションが供給バッチプロセスであり、炭素源は発酵プロセス経過中に発酵槽に連続的に添加される。供給バッチプロセスもまた、本発明に適している。供給バッチプロセスは分解産物抑制が、細胞代謝を阻害する傾向がある場合に、またはあらゆる時点で培地中に限定量の源を有することが望ましい場合に有用である。供給バッチシステム中の炭素源濃度の測定は困難であるので、pH、溶解酸素、および排ガス(例えばCO2)などの測定可能な要素の変化に基づいて推定してもよい。バッチおよび供給バッチ培養方法は一般的で技術分野でよく知られており、実例が以下にある。トーマス(Thomas)D.ブロック(Brock)著、「バイオテクノロジー:工業的微生物学テキストブック(Biotechnology:A
Textbook of Industrial Microbiology)」第二版、Sinauer Associates:マサチューセッツ州サンダーランド(Sunderland、MA)(1989)、またはデシュパンデ,ムカンド(Deshpande,Mukund)V.、Appl.Biochem.Biotechnol.、36:227(1992)。これらは参照によってここに援用される。
【0147】
このコドン最適化遺伝子を使用したω脂肪酸の商業生産はまた、連続発酵プロセスによって達成されてもよく、そこでは生産物の回収のため等量の培養物を除去するのと同時に、合成培地をバイオリアクター内に連続的に添加する。連続培養は、概して細胞を一定細胞密度の対数増殖期に保つ。連続または半連続培養法は、細胞生育または最終生産物濃度に影響する1つの因子またはあらゆるいくつかの因子の調節を可能にする。例えば1つのアプローチでは炭素源を制限して、あらゆるその他のパラメータが代謝を調節できるようにしてもよい。その他のシステムでは、培地濁度によって測定される細胞濃度を一定に保ちながら、生育に影響するいくつかの因子を連続的に変化させてもよい。連続システムは定常状態生育を維持することを目指すので、細胞生育率は、培養から培地が抜き取られることによる細胞損失に対してバランスが取れていなくてはならない。連続的培養プロセスのために栄養素および成長因子を調節する方法、ならびに生産物形成速度を最大化する技術は工業微生物学の技術分野でよく知られており、多様な方法が上記のブロック(Brock)で詳述される。
【0148】
PUFAの精製
PUFAは、遊離脂肪酸として、またはアシルグリセロール、リン脂質、スルホリピドまたは糖脂質などのエステル化形態で宿主微生物中に見ることができ、技術分野でよく知られている多様な手段を通じて宿主細胞から抽出されてもよい。酵母菌脂質の抽出技術、品質分析、および許容性基準に関する1つのレビューは、Z.ジェーコブス(Jacobs)(Critical Reviews in Biotechnology、12(5/6):463〜491(1992))である。下流プロセスに関する簡潔なレビューは、A.シン(Singh)およびO.ワード(Ward)(Adv.Appl.Microbiol.45:271〜312(1997))にもある。
【0149】
一般にPUFA精製手段は、有機溶剤、超音波処理、超臨界流体抽出(例えば二酸化炭素を使用して)による抽出と、鹸化と、圧搾などの物理的手段またはそれらの組み合わせを含んでもよい。特に興味深いのは、水存在下でのメタノールおよびクロロホルムによる抽出である(E.G.ブライ(Bligh)およびW.J.ダイヤー(Dyer)、Can.J.Biochem.Physiol.37:911〜917(1959))。望ま
しい場合、水性層を酸性化して負の電荷を帯びた部分をプロトン化することで、有機層中への所望の生産物の分配を増大できる。抽出後、有機溶剤は窒素流の下で蒸発によって除去できる。コンジュゲートされた形態で単離されると、生産物は酵素的にまたは化学的に切断されて、関心のある遊離脂肪酸またはより単純なコンジュゲートを放出してもよく、次にさらに操作されて所望の最終生産物を生産してもよい。望ましくはコンジュゲートされた形態の脂肪酸は、水酸化カリウムによって切断される。
【0150】
さらに精製が必要ならば、標準法を用いることができる。このような方法としては、抽出、尿素処理、分別結晶化、HPLC、分留、シリカゲルクロマトグラフィー、高速遠心分離または蒸留、またはこれらの技術の組み合わせが挙げられる。酸またはアルケニル基などの反応性基の保護は、既知の技術(例えばアルキル化、ヨウ素化)を通じて、あらゆるステップで実施してもよい。使用される方法としては、メチルエステルを生産するための脂肪酸のメチル化が挙げられる。同様に、保護基はあらゆるステップで除去されてもよい。望ましくはGLA、STA、ARA、DHA、およびEPAを含有する画分の精製は、尿素および/または分留による処置によって達成されてもよい。
【0151】
好ましい実施態様の説明
ここで述べる研究の究極の目的は、ω−3および/またはω−6脂肪酸が濃縮された油を蓄積する油性酵母菌の開発である。このような目的で、油性酵母菌中において効率的に機能するω−3および/またはω−6脂肪酸生合成経路のデサチュラーゼおよび鎖長延長酵素を同定し、これらの宿主におけるω脂肪酸の合成および蓄積を可能にしなくてはならない。
【0152】
本発明では、出願人は、デサチュラーゼおよび鎖長延長酵素遺伝子のコドン最適化に適する技術を実証し、そこでは得られた合成コドン最適化遺伝子が、ヤロウィア・リポリティカ(Yarrowia lipolytica)などの油性酵母菌において増大する変換効率で機能する。これらの方法は、ここでの教示において具現化され、具体的にはコドン最適化Δ6デサチュラーゼ(LAをGLAに、および/またはALAをSTAに変換する酵素的能力を有する)、コドン最適化Δ17デサチュラーゼ(DGLAをETAに、および/またはARAをEPAに変換する役割を果たす)、およびコドン最適化高親和力PUFA鎖長延長酵素(GLAをDGLAに、STAをETAに、および/またはEPAをDPAに変換できる)の合成に標的を定める。当業者は、容易にここで述べる技術を適用して、ω−3および/またはω−6生合成脂肪酸経路中のその他の遺伝子を最適化させ、ヤロウィア(Yarrowia)中で発現すると増大する変換効率を有することが期待される合成遺伝子を作り出せるであろう。したがってここでの教示は、ω−3および/またはω−6脂肪酸が濃縮された油を蓄積する油性酵母菌の開発において大きな有用性を持つ。
【0153】
出願人は、モデル油性酵母菌、ヤロウィア・リポリティカ(Yarrowia lipolytica)におけるコドン最適化のために3つの例示的な野生型遺伝子を選択した。これらの各野生型遺伝子は、下の表3で述べるようにオハイオ州コロンバスのロス・プロダクツ(Ross Products(Columbus、OH))からのプラスミド中で得られた。
【0154】
【表4】
【0155】
微生物宿主における各野生型酵素活性の確証(基質供給試験による)に続いて、各遺伝子をコドン最適化した(少なくとも9%の未変性コドンが、宿主の好むコドンを用いるように修正された)。
【0156】
Δ6デサチュラーゼ、Δ17デサチュラーゼ、および高親和力PUFA鎖長延長酵素遺伝子のコドン最適化は、最初にヤロウィア・リポリティカ(Yarrowia lipolytica)におけるコドン使用および構造遺伝子のシグネチャの判定を必要とした。次にコドン最適化遺伝子をデザインし、遺伝子配列全長を合成する所要時間を実質的に短縮するプロトコルを使用して、生体外(in vitro)で合成した。このプロトコルは、ここで合成される特定の遺伝子に限定されないので、多様な一般用途に広く適用できる。生体外(in vitro)遺伝子合成に伴う基本的ステップは、以下からなった。
1.複数の対のオリゴヌクレオチド(各約100bp長さ)を合成する。各オリゴヌクレオチド配列は、合成される遺伝子配列全長の一部に対応し、各センス鎖オリゴヌクレオチド配列は、配列の5’末端で対応するアンチセンス−鎖オリゴヌクレオチドと約4bpの重なりを有する。
2.全オリゴヌクレオチドの5’および3’末端をキナーゼ反応でリン酸化し、次にセンスおよびアンチセンスオリゴヌクレオチド対に個々のアニール反応を施して、短い(約100bp)二本鎖DNA断片を生産する。
3.次にオリゴヌクレオチド合成時にデザインされた約4bpの重なりを使用して、短い(約100bp)二本鎖DNA断片のプールを一緒にライゲーションし、より長い(約300〜400bp)DNA断片を生産する。
4.ライゲーションに続いて、より長い断片をPCRによって増幅し、その生産物を適切なベクター中にクローン化し、宿主細胞中に形質転換する。
5.各PCR生産物を含有するプラスミドDNAを宿主細胞から精製し、次に合成する遺伝子の各約300〜400bpの断片に対応するPCR生産物単離のための制限酵素消化を施す。
6.合成する約300〜400bpの遺伝子断片を一緒にライゲーションし、次にPCR増幅を施して合成配列全長を生産する。
【0157】
コドン最適化Δ6デサチュラーゼ、Δ17デサチュラーゼ、および高親和力PUFA鎖長延長酵素遺伝子が合成されると、それぞれ個々にヤロウィア・リポリティカ(Yarrowia lipolytica)中に形質転換した。供給実験(適切な基質を使用した)は、Y.リポリティカ(Y.lipolytica)においてコドン最適化遺伝子が、対応する未変性野生型遺伝子よりも効率的に発現すると判定した。具体的にはY.リポリティカ(Y.lipolytica)において発現すると、同一生物学的条件下で、コド
ン最適化Δ6デサチュラーゼは野生型対応酵素よりもおよそ40%多くのLAをGLAに変換し、コドン最適化Δ17デサチュラーゼは野生型酵素の約2倍のARAをEPAにを変換し、コドン最適化高親和力PUFA鎖長延長酵素はおよそ57%より多くのGLAをDGLAに変換した。
【実施例】
【0158】
以下の実施例で本発明をさらに定義する。これらの実施例は、本発明の好ましい実施態様を示しながら、例証のみの目的で提供されるものとする。上の考察およびこれらの実施例から、当業者は本発明の必須特性を把握でき、その精神と範囲を逸脱することなく、本発明の様々な変更および修正を行って、それを様々な使用法および条件に適合できる。
【0159】
一般方法
実施例で使用する標準組換えDNAおよび分子クローニング技術は、技術分野でよく知られており、1.)サムブルック(Sambrook)、J.、フリッチュ(Fritsch)、E.F.およびマニアティス(Maniatis)T.「分子クローニング:実験室マニュアル(Molecular Cloning:A Laboratory Manual)」、コールドスプリングハーバーラボラトリ:ニューヨーク州コールドスプリングハーバー(Cold Spring Harbor、NY)(1989)(マニアティス(Maniatis))、2.)T.J.シルハビー(Silhavy)、M.L.ベンナン(Bennan)およびL.W.エンクイスト(Enquist)、「遺伝子融合実験(Experiments with Gene Fusions)」(コールドスプリングハーバーラボラトリ:Cold Spring Harbor,NY,1984)、および3.)オースベル(Ausubel)F.M.ら「分子生物学現代プロトコル(Current Protocols in Molecular Biology)」(グリーンパブリッシングおよびワイリーインターサイエンスによる出版;1987)で述べられる。
【0160】
微生物培養の維持および生育に適した材料および方法は、技術分野でよく知られている。以下の実施例で使用するのに適した技術は、次で述べられる。「一般微生物学方法マニュアル(Manual of Methods for General Bacteriology)」、フィリップス・ゲアハルト(Phillipp Gerhardt)、R.G.E.マレー(R.G.E.Murray)、ラルフN.コスティロウ(Ralph N.Costilow)、ユージーンW.ネスター(Eugene W.Nester)、ウィリスA.ウッド(Willis A.Wood)、ノエルR.クリーグ(Noel R.Krieg)、およびG.ブリッグス・フィリップス(G.Briggs Phillips)編、米国微生物学会(American Society for Microbiology)、ワシントンD.C.(Washington,D.C.)(1994)またはトーマス(Thomas)D.ブロック(Brock)著、バイオテクノロジー:工業的微生物学テキストブック(Biotechnology:A Textbook of Industrial Microbiology)第二版、Sinauer Associates:(Sunderland、MA)(1989)。微生物細胞の生育および維持のために使用される全ての試薬、制限酵素および材料は、特に断りのない限り、ウィスコンシン州ミルウォーキーのアルドリッチ・ケミカルズ(Aldrich Chemicals(Milwaukee、WI))、ミシガン州デトロイトのディフコ・ラボラトリーズ(DIFCO Laboratories(Detroit、MI))、メリーランド州ゲーサーズバーグのギブコ/BRL(GIBCO/BRL(Gaithersburg、MD))、またはミズーリ州セントルイスのシグマケミカル(Sigma Chemical Company(St.Louis、MO))から得た。
【0161】
カリフォルニア州カールスバッドのインビトロゲン(Invitrogen(Carlsbad、CA))から大腸菌(E.coli)TOP10細胞および大腸菌(E.coli)エレクトロマックス(electromax)DH10B細胞を得た。メリーランド州ゲイサーズバーグのギブコ/BRL(GIBCO/BRL)(Gaithersburg、MD))から大腸菌(E.coli)DH5αの最大効率コンピテント細胞を得た。カリフォルニア州サンディエゴのストラタジーン社(Stratagene Company(San Diego、CA))から大腸菌(E.coli)(XL1−Blue)コンピテント細胞を購入した。大腸菌(E. coli)株は典型的にルリア・ベルターニ(Luria Bertani)(LB)プレート上で37℃で生育させた。
【0162】
ヤロウィア・リポリティカ(Yarrowia lipolytica)ロイシン独立栄養株(ATCC番号76982)をメリーランド州ロックビル(Rockville、MD)の米国微生物系統保存機関(American Type Culture Collection)から購入し、機能性アッセイで使用した。Y.リポリティカ(Y.lipolytica)株は通常、YPD寒天(1%酵母菌抽出物、2%バクトペプトン、2%グルコース、2%寒天)上で28℃で生育させた。形質転換体の選択のために、ミシガン州デトロイトのジフコ・ラボラトリーズ(DIFCO Laboratories(Detroit、MI))からの最少培地(硫酸アンモニウムまたはアミノ酸を含まない0.17%酵母菌窒素ベース、2%グルコース、0.1%プロリン、pH6.1)を使用した。アデニン、ロイシン、リジン、および/またはウラシルサプリメントを最終濃度0.01%で添加した。
【0163】
一般分子クローニングは、標準法に従って実施された(サムブルック(Sambrook)ら、同上)。オリゴヌクレオチドはシグマ−ジェノシス(Sigma−Genosys、テキサス州スプリング(Spring、TX))によって合成した。部位特異的変異誘発は、ストラタジーン(Stratagene)のクイックチェンジ(QuickChange)TM部位特異的変異誘発キットを製造業者の説明書に従って使用して実施した。ポリメラーゼ連鎖反応(PCR)または部位特異的変異誘発がサブクローニングに関与する場合、コンストラクトは、配列中に確実にいかなる誤りも導入されないように配列された。PCR生産物は、ウィスコンシン州マディソンのプロメガ(Promega(Madison、WI))からのpGEM−T−イージーベクター中にクローン化した。
【0164】
ウィスコンシン州マディソンのジーンズ・コンピューター・グループ(GCG)社(Genes Computer Group Inc.(Madison、WI))から入手できる一組のプログラム、ウィスコンシン・パッケージ(Wisconsin Package Version9.0)を使用して、遺伝的配列の操作が達成された。ギャップ作成デフォルト値12、およびギャップ延長デフォルト値4でGCGプログラム「パイルアップ(Pileup)」を使用した。デフォルトギャップ作成ペナルティ50、およびデフォルトギャップ延長ペナルティ3で、GCG「ギャップ(ギャップ)」または「ベストフィット(Bestfit)」プログラムを使用した。特に断りのない限り、全てのその他の場合において、GCGプログラムのデフォルトパラメータを使用した。
【0165】
略語の意味は以下の通り。「sec」は秒を意味し、「min」は分を意味し、「h」は時間を意味し、「d」は日を意味し、「μL」はマイクロリットルを意味し、「mL」はミリリットルを意味し、「L」はリットルを意味し、「μM」はマイクロモル(micromolar)を意味し、「mM」はミリモル(millimolar)を意味し、「M」はモル(molar)を意味し、「mmol」はミリモル(millimole)を意味し、「μmole」マイクロモル(micromole)を意味し、「g」はグラムを意味し、「μg」はマイクログラムを意味し、「ng」はナノグラムを意味し、「U」は単位を意味し、「bp」は塩基対を意味し、「kB」はキロベースを意味する。
【0166】
実施例1
ヤロウィア・リポリティカ(Yarrowia lipolytica)中の異種性遺伝子発現に適したプラスミドの構造
図2に図解するように、pINA532誘導体であるプラスミドpY5(Insitut National Agronomics、Centre de biotechnologie Agro−Industrielle、laboratoire de Genetique Moleculaire et Cellularie INRA−CNRS、F−78850、Thiverval−Grignon、Franceのクロード・ガィヤルダン(Claude Gaillardin)博士からの贈与)をヤロウィア・リポリティカ(Yarrowia lipolytica)における異種性遺伝子発現のために構築した。
【0167】
最初に、pINA532のARS18配列およびLEU2遺伝子を含有する、部分的に消化された3598bpのEcoRI断片をカリフォルニア州サンディエゴのストラタジーン社(Stratagene Company(San Diego、CA))からのpBluescriptのEcoRI部位中にサブクローン化してpY2を生産した。
【0168】
TEF5’(配列番号7)およびTEF3’(配列番号8)をプライマーとして使用して、PCRによりヤロウィア・リポリティカ(Yarrowia lipolytica)ゲノムのDNAからTEFプロモーターを増幅した(ミュラー(Muller)S.ら「酵母菌(Yeast)」14:1267〜1283(1998))。100ngのヤロウィア(Yarrowia)ゲノムDNAと、10mM KCl、10mM(NH4)2SO4、20mMトリス−HCl(pH8.75)、2mM MgSO4、0.1%TritonX−100、100μg/mL BSA(最終濃度)を含有するPCR緩衝液と、各200ミュートμMのデオキシリボヌクレオチド三リン酸と、10pmoleの各プライマーと、カリフォルニア州サンディエゴのストラタジーン(Stratagene、San Diego、CA)からの1μLのPfuTurbo DNAポリメラーゼとを含有する、50μLの総容積でPCR増幅を実施した。増幅は以下のように実施した。95℃で3minの初期変性、それに続いて95℃で1min、56℃で30sec、72℃で1minを35サイクル。72℃で10minの最終延長サイクル実施し、それに続いて4℃での反応終結。418bpのPCR生産物をpCR−BluntにライゲーションしてpIP−tefを生産した。pIP−tefのBamHI/EcoRV断片をpY2のBamHI/SmaI部位にサブクローン化して、pY4を生産した。
【0169】
テンプレートとしてpINA532、プライマーとしてXPR5’(配列番号9)およびXPR3’(配列番号10)を使用して、PCRによってXPR2転写ターミネーターを増幅した。上述の構成要素および条件を使用して、PCR増幅を50μLの総容積中で実施した。179bpのPCR生産物をSacIIで消化し、pY4のSacII部位にライゲーションしてpY5を生産した。したがってpY5(図2および3に示す)は、以下を含有するヤロウィア(Yarrowia)−大腸菌(E. coli)シャトルプラスミドとして有用である。
1)ヤロウィア(Yarrowia)自律性複製配列(ARS18)、
2)ColE1プラスミド複製起点、
3)大腸菌(E. coli)の選択のためのアンピシリン−抵抗性遺伝子(AmpR)、
4)ヤロウィア(Yarrowia)の選択のためのヤロウィア(Yarrowia)LEU2遺伝子、
5)ヤロウィア(Yarrowia)における異種性遺伝子発現のための翻訳延長プロモーター(TEFP)、
6)ヤロウィア(Yarrowia)における異種性遺伝子発現の転写終結のための細胞外プロテアーゼ遺伝子ターミネーター(XPR2)。
【0170】
pY5−4およびpY5−13(図3)をpY5の誘導体として構築して、ヤロウィア・リポリティカ(Yarrowia lipolytica)中のサブクローン化および異種性遺伝子発現を容易にした。
【0171】
具体的にはpY5をテンプレートとして使用して、3ラウンドの部位特異的変異誘発によってpY5−4を構築した。オリゴヌクレオチドYL1およびYL2(配列番号11および12)を使用して、LEU2レポーター遺伝子内に位置するNcoI部位をpY5から除去し、pY5−1を生産した。オリゴヌクレオチドYL3およびYL4(配列番号13および14)を使用して、特異的変異誘発によって、NcoI部位をTEFプロモーターとXPR2転写ターミネーター部位の間でpY5−1内に導入し、pY5−2を生産した。次にオリゴヌクレオチドYL23およびYL24(配列番号15および16)を使用して、PacI部位をTEFプロモーターとXPR2転写ターミネーターの間でY5−2に導入し、pY5−4を生産した。
【0172】
pY5をテンプレートとして使用して、6ラウンドの部位特異的変異誘発によってpY5−13構築した。オリゴヌクレオチドYL5およびYL6(配列番号17および18)を使用して、部位特異的変異誘発によって、SalIおよびClaI部位の双方をpY5から除去して、pY5−5を生産した。オリゴヌクレオチドYL9およびYL10(配列番号19および20)を使用して、部位特異的変異誘発によって、SalI部位をLEU2遺伝子とTEFプロモーターの間でpY5−5中に導入し、pY5−6を生産した。オリゴヌクレオチドYL7およびYL8(配列番号21および22)を使用して、PacI部位をLEU2遺伝子とARS18の間でpY5−6中に導入し、pY5−8を生産した。オリゴヌクレオチドYL3およびYL4(配列番号13および14)を使用して、NcoI部位をTEFプロモーターの翻訳開始コドン周囲のpY5−8中に導入しpY5−9を生産した。YL1およびYL2オリゴヌクレオチド(配列番号11および12)を使用して、pY5−9のLEU2遺伝子中のNcoI部位を除去し、pY5−12を生産した。最後にオリゴヌクレオチドYL61およびYL62(配列番号23および24)を使用して、BsiWI部位をColEIとXPR2領域の間でpY5−12中に導入し、pY5−13を生産した。
【0173】
実施例2
ヤロウィア・リポリティカ(Yarrowia lipolytica)における野生型Δ6およびΔ17デサチュラーゼおよび高親和力PUFA鎖長延長酵素の変換効率分析
ヤロウィア・リポリティカ(Yarrowia lipolytica)における野生型Δ6デサチュラーゼ、鎖長延長酵素、およびΔ17デサチュラーゼの機能性を(コドン最適化に先だって)確証するために、各野生型タンパク質の変換効率を測定した。具体的にはモルティエレラ・アルピナ(Mortierella alpina)Δ6デサチュラーゼ、サプロレグニア・ディクリナ(Saprolegnia diclina)Δ17デサチュラーゼ、およびM.アルピナ(M.alpina)高親和力PUFA鎖長延長酵素を別の宿主において発現させ、基質供給試験で活性についてスクリーニングした。各酵素は基質の少なくとも23%を生産物に変換できることが分かった。
【0174】
野生型モルティエレラ・アルピナ(Mortierella alpina)(登録番号AF465281)Δ6デサチュラーゼ
M.アルピナ(M.alpina)Δ6デサチュラーゼ遺伝子(配列番号1)を含有するpCGR5(米国特許第5,968,809号明細書)の1384bpのNcoI/NotI断片をpY5−2(実施例1)のNcoI/NotI部位に挿入し、pY54を生
産した。
【0175】
野生型サプロレグニア・ディクリナ(Saprolegnia diclina)(ATCC番号56851)Δ17デサチュラーゼ
プライマーとしてオリゴヌクレオチドYL21A(配列番号118)およびYL22(配列番号119)を使用して、PCRによって、プラスミドpRSP19(米国特許出願公開第2003/0196217 A1号明細書)からS.ディクリナ(S.diclina)の野生型Δ17デサチュラーゼ遺伝子を増幅した。PCR増幅は、10ngのテンプレート、10mM KCl、10mM(NH4)2SO4、20mMトリス−HCl(pH8.75)、2mM MgSO4、0.1%TritonX−100、および100μg/mL BSA(最終濃度)を含有するPCR緩衝液と、200μMの各デオキシリボヌクレオチド三リン酸と、10pmoleの各プライマーと、カリフォルニア州サンディエゴのストラタジーン(Stratagene、San Diego、CA)からの1μLのPfuTurbo DNAポリメラーゼを含んでなる50μLの総容積で実施した。増幅を以下のようにして実施した。95℃で3minの初期変性、それに続いて95℃で1min、56℃で30sec、72℃で1minを35サイクル。72℃で10minの最終延長サイクルの実施と、それに続く4℃での反応終結。PCR生産物をNcoI/PacIで消化し、次にNcoI/PacIで消化したpY5−4(図3、実施例1で述べられる)にライゲーションしてpYSD17を生産した。
【0176】
野生型モルティエレラ・アルピナ(Mortierella alpina)(登録番号AX464731)高親和力鎖長延長酵素
M.アルピナ(M.alpina)高親和力PUFA鎖長延長酵素遺伝子(配列番号5)のコード領域を含有するpRPB2(国際公開第00/12720号パンフレット)の973bpのNotI断片をpY5のNotI部位(実施例1、図2および3)に挿入し、pY58を生産した(図13B)。
【0177】
ヤロウィア・リポリティカ(Yarrowia lipolytica)の形質転換
チェン(Chen)D.C.ら(Appl Microbiol Biotechnol.48(2):232〜235(1997))の方法に従って、プラスミドpY54、pYSD17、およびpY58を別々にY.リポリティカ(Y.lipolytica)ATCC番号76982中に形質転換した。
【0178】
簡単に述べると、ヤロウィア(Yarrowia)のロイシン栄養要求株をYPDプレート上に画線し、30℃でおよそ18時間生育させた。いくつかの大型白金耳を満たす細胞をプレートからこすり取り、以下を含有する1mLの形質転換緩衝液に再懸濁した。
2.25mLの50%PEG、平均分子量3350、
0.125mLの2M酢酸Li、pH6.0、
0.125mLの2MDTT、および
50μgの剪断サケ精子DNA。
【0179】
約500ngのプラスミドDNAを100μLの再懸濁細胞内で培養し、15min間隔でボルテックス混合しながら39℃に1hr維持した。細胞をロイシン欠乏最少培地プレートに蒔いて、30℃で2〜3日間維持した。
【0180】
%基質変換の判定
pY54、pYSD17またはpY58を含有する形質転換体Y.リポリティカ(Y.lipolytica)の単一コロニーをそれぞれ3mLの最少培地(20g/Lグルコース、1.7g/Lのアミノ酸なしの酵母窒素ベース、1g/LのL−プロリン、0.1g/LのL−アデニン、0.1g/LのL−リジン、pH6.1)中で、30℃でOD6
00が約1.0になるまで生育させた。基質供給のために、次に10μgの基質を含有する3mLの最少培地内で100μLの細胞を30℃で約24hr継代培養した。引き続いてブライ(Bligh)E.G.およびダイヤー(Dyer)W.J.(Can.J.Biochem.Physiol.37:911〜917(1959))で述べられるように、細胞を遠心分離し収集して脂質を抽出した。ナトリウムメトキシド(ローガン(Roughan)G.およびニシダ(Nishida)I.、Arch Biochem Biophys.276(1):38〜46(1990))による脂質抽出物のエステル交換反応によって、脂肪酸メチルエステルを調製し、30m×0.25mm(内径)HP−INNOWAXヒューレットパッカード(Hewlett−Packard)カラムを装着したヒューレットパッカード(Hewlett−Packard)6890 GCで引き続き分析した。オーブン温度は170℃(25分間保持)から185℃に3.5℃/minで上昇させた。%基質変換を以下のように計算した([生産物]/[基質+生産物])×100)。
【0181】
野生型M.アルピナ(M.alpina)Δ6デサチュラーゼによる%基質変換
M.アルピナ(M.alpina)Δ6デサチュラーゼは、LAをGLAに、および/またはALAをSTAに変換する。pY54を含有するY.リポリティカ(Y.lipolytica)株を上述のように(基質供給は必要でない)生育させ、脂質を分析した。結果は、pY54のあるヤロウィア(Yarrowia)株が、約30%のLAをGLAに変換することを示した。
【0182】
野生型S.ディクリナ(S.diclina)Δ17デサチュラーゼの%基質変換
S.ディクリナ(S.diclina)Δ17デサチュラーゼは、ARAをEPAに、および/またはDGLAをETAに変換する。pYSD17を含有するY.リポリティカ(Y.lipolytica)株を単一コロニーから生育させ、10μgのARAを含有する最少培地で継代培養し、上述のように脂質分析にかけた。ARA供給実験の結果は、pYSD17を有するヤロウィア(Yarrowia)株が約23%の細胞内ARAをEPAに変換したことを示した。
【0183】
野生型M.アルピナ(M.alpina)高親和力鎖長延長酵素の%基質変換
M.アルピナ(M.alpina)高親和力PUFA鎖長延長酵素は、GLAをDGLAに、STAをETAに、および/またはEPAをDPAに変換する。pY58を含有するY.リポリティカ(Y.lipolytica)株を単一コロニーから生育させ、10μgのGLAを含有する最少培地で継代培養し、上述のように脂質分析にかけた。GLA供給実験の結果は、pY58を有するヤロウィア(Yarrowia)株が、約30%の細胞内GLAをDGLAに変換したことを示した。
【0184】
ヤロウィア・リポリティカ(Yarrowia lipolytica)において好まれるコドン使用の判定
Y.リポリティカ(Y.lipolytica)のおよそ100個の遺伝子が、アメリカ国立バイオテクノロジー情報センター公共データベースにある。121,167bpを含んでなるこれらの遺伝子のコード領域をDNAStarのEditseqプログラムによって対応する40,389個のアミノ酸に翻訳し、表4に示すY.リポリティカ(Y.lipolytica)のコドン使用プロフィールを判定するために表にした。「No.」と題された欄は、特定のコドンが、40,389個のアミノ酸のサンプル中で特定のアミノ酸をコードする回数を指す。「%」と題された欄は、特定のコドンが特定のアミノ酸をコードする頻度を指す。太字で示されるエントリはY・リポリティカ(Yarrowia lipolytica)中で好まれるコドンを表す。
【0185】
【表5】
【0186】
Y.リポリティカ(Y.lipolytica)中でのさらなる遺伝子発現の最適化の
ために、79個の遺伝子の「ATG」開始コドン周囲の共通配列を調査した。図4では、下線を引いたATG翻訳開始コドンの第1の「A」は、+1と見なされる。分析された遺伝子の77%が、−3の位置に「A」を有し、この位置における「A」に対する強い優先度が示された。また−4、−2および−1の位置で「A」または「C」に、+5の位置で「A」、「C」または「T」に、+6の位置で「G」または「C」に対する優先度があった。したがってY.リポリティカ(Y.lipolytica)中における遺伝子の最適発現のためのコドン最適化翻訳開始部位の好ましい共通配列は、「MAMMATGNHS」(配列番号122)であり、そこで使用された核酸退縮コードは、以下のようである。M=A/C、S=C/G、H=A/C/T、およびN=A/C/G/T。
【0187】
実施例4
コドン最適化Δ6デサチュラーゼ遺伝子の合成
モルティエレラ・アルピナ(Mortierella alpina)(配列番号1)からのΔ6デサチュラーゼ遺伝子は長さが1374bpであった(米国特許第5,968,809号明細書、ジェンバンク(GenBank)#AF465281)。ヤロウィア(Yarrowia)コドン使用パターン、ATG翻訳開始コドン周囲の共通配列、およびRNA安定性の一般規則(グハニヨギ(Guhaniyogi)G.およびJ.ブルーアー(Brewer)、Gene 265(1〜2):11〜23(2001))に従って、M.アルピナ(M.alpina)DNA配列に基づいて、コドン最適化Δ6デサチュラーゼ遺伝子をデザインした。翻訳開始部位を修正するのに加えて、1347bpのコード領域(144のコドンを含んでなる)の152bpもまたコドン最適化した。このコドン最適化遺伝子(配列番号25)とM.アルピナ(M.alpina)からの野生型配列全長(配列番号1)との比較を図5に示し、そこでは太字のヌクレオチドが、コドン最適化遺伝子において修正されたヌクレオチドに対応する。コドン最適化遺伝子中の修正のいずれも、コードされたタンパク質(配列番号2)のアミノ酸配列を変化させなかった。
【0188】
コドン最適化Δ6デサチュラーゼ遺伝子を合成するのに使用される方法は、図6で図示される。最初に14対のオリゴヌクレオチドがデザインされて、M.アルピナ(M.alpina)Δ6デサチュラーゼ遺伝子(例えば配列番号26〜53に対応するD6−1A、D6−1B、D6−2A、D6−2B、D6−3A、D6−3B、D6−4A、D6−4B、D6−5A、D6−5B、D6−6A、D6−6B、D6−7A、D6−7B、D6−8A、D6−8B、D6−9A、D6−9B、D6−10A、D6−10B、D6−11A、D6−11B、D6−12A、D6−12B、D6−13A、D6−13B、D6−14AおよびD6−14B)のコドン最適化コード領域の全長(すなわち1374bp)が延長される。各5’末端の4bpのオーバーハングを例外として、センス(A)およびアンチセンス(B)オリゴヌクレオチドの各対は相補的である。引き続くサブクローン化のために、プライマーD6−1Aはその5’末端にNcoI部位を含有し、プライマーD6−4BおよびD6−5AはStuI部位を含有し、プライマーD6−7B、D6−8A、およびD6−10BはそれぞれBamHI部位を含有した。50mMトリス−HCl(pH7.5)、10mM MgCl2、10mM DTT、0.5mMスペルミジン、0.5mM ATPおよび10単位のT4ポリヌクレオチドキナーゼを含有する20μLの容積中で、100ngの各オリゴヌクレオチドを37℃で1hrリン酸化した。センスおよびアンチセンスオリゴヌクレオチドの各対を混合し、以下のパラメーターを使用して、サーモサイクラー中でアニールした。95℃(2min)、85℃(2min)、65℃(15min)、37℃(15min)、24℃(15min)および4℃(15min)。このようにしてD6−1A(配列番号26)をD6−1B(配列番号27)にアニールし、二本鎖生産物「D6−1AB」を生産した。同様に、D6−2A(配列番号28)をD6−2B(配列番号29)にアニールし、二本鎖生産物「D6−2AB」を生産した。
【0189】
次に以下に示すように4個の別個のアニールされた二本鎖オリゴヌクレオチドのプールを一緒にライゲーションした。
・プール1:D6−1AB、D6−2AB、D6−3AB、およびD6−4ABを含んでなる、
・プール2:D6−5AB、D6−6AB、およびD6−7ABを含んでなる、
・プール3:D6−8AB、D6−9AB、およびD6−10ABを含んでなる、
・プール4:D6−11AB、D6−12AB、D6−13AB、およびD6−14ABを含んでなる。
【0190】
アニールされたオリゴヌクレオチドの各プールを20μLの容積中で10単位のT4DNAリガーゼと共に混合し、ライゲーション反応を一晩16℃で培養した。
【0191】
次に各ライゲーション反応の生産物をPCRによって増幅した。具体的には、テンプレートとしてライゲーションされた「プール1」混合物(すなわちD6−1AB、D6−2AB、D6−3AB、およびD6−4AB)、プライマーとしてオリゴヌクレオチドD6−1(配列番号54)およびD6−4R(配列番号55)を使用して、コドン最適化Δ6デサチュラーゼ遺伝子の第1の部分をPCRによって増幅した。PCR増幅を10mM KCl、10mM (NH4)2SO4、20mMトリス−HCl(pH8.75)、2mM MgSO4、0.1%TritonX−100、100μg/mL BSA(最終濃度)を含有するPCR緩衝液と、各200μMのデオキシリボヌクレオチド三リン酸と、10pmoleの各プライマーと、カリフォルニア州サンディエゴのストラタジーン(Stratagene、San Diego、CA)からの1μLのPfuTurbo DNAポリメラーゼを含んでなる50μLの総容積中で実施した。増幅を以下のようにして実施した。95℃で3minの初期変性、それに続いて95℃で1min、56℃で30sec、72℃で40secの35サイクル。72℃で10minの最終延長サイクルの実施と、それに続く4℃での反応終結。380bpのPCR断片をpGEM−Tイージーベクター(プロメガ(Promega))中にサブクローン化して、pT6(1−4)を生産した。
【0192】
テンプレートとしてライゲーションされた「プール2」混合物(すなわちD6−5AB、D6−6AB、およびD6−7AB)、プライマーとしてオリゴヌクレオチドD6−5(配列番号56)およびD6−7R(配列番号57)を使用して、コドン最適化Δ6デサチュラーゼ遺伝子の第2の部分を同様にPCRによって増幅し、pGEM−T−イージーベクター中にクローン化してpT6(5−7)を生産した。テンプレートとして「プール3」ライゲーション混合物(すなわち、D6−8AB、D6−9AB,およびD6−10AB)、プライマーとしてオリゴヌクレオチドD6−8(配列番号58)およびD6−10R(配列番号59)を使用して、同様にPCRによってコドン最適化Δ6デサチュラーゼ遺伝子の第3の部分を増幅し、pGEM−T−イージーベクター中にクローン化してpT6(8−10)を生産した。最後にテンプレートとして「プール4」ライゲーション混合物(すなわちD6−11AB、D6−12AB、D6−13AB、およびD6−14AB)、プライマーとしてオリゴヌクレオチドD6−11(配列番号60)およびD6−14R(配列番号61)を使用して、同様にPCRによってコドン最適化Δ6デサチュラーゼ遺伝子の第4の部分を増幅し、pGEM−T−イージーベクター中にクローン化してpT6(11−14)を生産した。
【0193】
大腸菌(E.coli)をpT6(1−4)、pT6(5−7)、pT6(8−10)、およびpT6(11−14)で別々に形質転換し、アンピシリン−抵抗性形質転換体から単離されたプラスミドDNAを精製して適切な制限エンドヌクレアーゼで消化し、380bpのpT6(1〜4)のNcoI/StuI断片、310bpのpT6(5−7)のStuI/BamHI断片、320bpのpT6(8−10)のBamHI断片、および
410bpのpT6(11―14)のBamHI/NotI断片を遊離した。次にこれらの断片を組み合わせて正しい方向に一緒にライゲーションし、pY5−13のNcoI/NotI部位に挿入してpYD6Sを生産した(図7)。
【0194】
実施例5
コドン最適化Δ17デサチュラーゼ遺伝子の合成
サプロレグニア・ディクリナ(Saprolegnia diclina)(配列番号3)からのΔ17デサチュラーゼ遺伝子は長さが1077bpであった。ヤロウィア(Yarrowia)コドン使用パターン、ATG翻訳開始コドン周囲の共通配列、およびRNA安定性の一般規則(グハニヨギ(Guhaniyogi)およびブルーアー(Brewer)前出)に従って、S.ディクリナ(S.diclina)DNA配列に基づいて、コドン最適化Δ17デサチュラーゼ遺伝子をデザインした。翻訳開始部位の修正に加えて、1077bpのコード領域(117のコドンを含んでなる)の127bpをコドン最適化した。このコドン最適化DNA配列(配列番号62)とS.ディクリナ(S.diclina)Δ17デサチュラーゼ遺伝子DNA配列(配列番号3)との比較を図8に示し、そこでは太字のヌクレオチドが、コドン最適化遺伝子において修正されたヌクレオチドに対応する。コドン最適化遺伝子中の修正のいずれも、コードされたタンパク質(配列番号4)のアミノ酸配列を変化させなかった。
【0195】
コドン最適化Δ17デサチュラーゼ遺伝子を合成するのに使用される方法は、図9で図示される。最初に11対のオリゴヌクレオチドがデザインされて、S.ディクリナ(S.diclina)Δ17デサチュラーゼ遺伝子(例えば配列番号63〜84に対応するD17−1A、D17−1B、D17−2A、D17−2B、D17−3A、D17−3B、D17−4A、D17−4B、D17−5A、D17−5B、D17−6A、D17−6B、D17−7A、D17−7B、D17−8A、D17−8B、D17−9A、D17−9B、D17−10A、D17−10B、D17−11AおよびD17−11B、対応する)のコドン最適化コード領域の全長が延長される。各5’末端の4bpのオーバーハングを例外として、センス(A)およびアンチセンス(B)オリゴヌクレオチドの各対は相補的である。さらにプライマーD17−1A、D17−4B、D17−5A、D17−8A、およびD17−8Bもまた、引き続くサブクローン化のためにNcoI、BglIIおよびSalI制限部位にそれぞれ導入される。
【0196】
実施例4で使用した方法に続いて、100ngの各オリゴヌクレオチドをT4ポリヌクレオチドキナーゼでリン酸化した。次に各センスおよびアンチセンスオリゴヌクレオチド対を混合し、アニールした。したがって、D17−1A(配列番号63)をD17−1B(配列番号64)にアニールして二本鎖生産物「D17−1AB」を生産した。同様にD17−2A(配列番号65)をD17−2B(配列番号66)にアニールして二本鎖生産物「D17−2AB」などを生産した。
【0197】
次に以下に示すように3個の別個のアニールされた二本鎖オリゴヌクレオチドのプールを一緒にライゲーションした。
・プール1:D17−1AB、D17−2AB、D17−3AB、およびD17−4ABを含んでなる、
・プール2:D17−5AB、D17−6AB、D17−7AB、およびD17−8ABを含んでなる、そして
・プール3:D17−9AB、D17−10AB、およびD17−11ABを含んでなる。
【0198】
アニールされたオリゴヌクレオチドの各プールを20μLの容積中で一晩16℃でライゲーションした。
【0199】
次に各ライゲーション反応の生産物をPCRによって増幅した。具体的には、テンプレートとしてライゲーションされた「プール1」混合物(すなわちD17−1AB、D17−2AB、D17−3AB、およびD17−4AB)、プライマーとしてオリゴヌクレオチドD17−1(配列番号85)およびD17−4R(配列番号86)を使用して、コドン最適化Δ17デサチュラーゼ遺伝子の第1の部分をPCRによって増幅した。実施例4で述べられるPCR条件およびサーモサイクリングプログラムを使用して、PCR増幅を50μLの総容積中で実施した。430bpのPCR断片をpGEM−Tイージーベクター(プロメガ(Promega))中にサブクローン化して、pT17(1−4)を生産した。
【0200】
テンプレートとしてライゲーションされた「プール2」混合物(すなわちD17−5AB、D17−6AB、D17−7AB、およびD17−8AB)、プライマーとしてオリゴヌクレオチドD17−5(配列番号87)およびD17−8D(配列番号88)を使用して、コドン最適化Δ17デサチュラーゼ遺伝子の第2の部分を同様にPCRによって増幅し、pGEM−T−イージーベクター中にクローン化してpT17(5−8)を生産した。最後に、テンプレートとして「プール3」ライゲーション混合物(すなわちD17−9AB、D17−10AB、およびD17−11AB)、プライマーとしてオリゴヌクレオチドD17−8U(配列番号89)、D17−11(配列番号90)を使用して、同様にPCRによってコドン最適化Δ17デサチュラーゼ遺伝子の第3の部分を増幅し、pGEM−T−イージーベクター中にクローン化してpT17(9−11)を生産した。
【0201】
大腸菌(E. coli)をpT17(1−4)、pT17(5−8)、およびpT17(9−11)で別々に形質転換し、プラスミドDNAをアンピシリン−抵抗性形質転換体から単離した。プラスミドDNAを適切な制限エンドヌクレアーゼで精製し消化して、pT17(1―4)の420bp NcoI/BglII断片、pT17(5−8)の400bp BglII/SalI断片、およびpT17(9−11)の300bp SalI/NotI断片を遊離した。次にこれらの断片を組み合わせて一緒にライゲーションして、合成Δ17デサチュラーゼ遺伝子全体の増幅のためのテンプレートとして使用し、D17−1(配列番号85)およびD17−11(配列番号90)をプライマーとして使用した。上述のΔ17デサチュラーゼ遺伝子の各部分のための条件、および以下のサーモサイクリングを使用して、50μLの総容積中でPCR増幅を実施した。95℃で3minの初期変性、それに続いて95℃で1min、56℃で30sec、72℃で1.1minの35サイクル。72℃で10minの最終延長サイクルの実施と、それに続く4℃での反応終結。1.1kBのPCR生産物をNcoI/NotIで消化し、NcoI/NotI消化したpY5−13中にサブクローン化して、pYSD17Sを生産した(図10A)。
【0202】
野生型およびコドン最適化Δ17デサチュラーゼと共に比較基質供給試験で使用するための追加的な「対照」として、プライマーとしてYL53(配列番号120)およびYL54(配列番号121)を使用して、部位特異的変異誘発によってpYSD17中のATに富んだPacI部位(実施例2で述べられる)を除去し、pYSD17Mを生産した(図10B)。
【0203】
実施例6
コドン最適化高親和力PUFA鎖長延長酵素遺伝子の合成
M.アルピナ(M.alpina)(配列番号5)からの高親和力PUFA鎖長延長酵素遺伝子は長さが957bpであった(ジェンバンク(GenBank)#AX464731、国際公開第00/12720号パンフレット)。ヤロウィア(Yarrowia)コドン使用パターン、「ATG」翻訳開始コドン周囲の共通配列、およびRNA安定性の
一般規則(グハニヨギ(Guhaniyogi)およびブルーアー(Brewer)前出)に従って、M.アルピナ(M.alpina)DNA配列に基づいて、コドン最適化高親和力PUFA鎖長延長酵素遺伝子をデザインした。翻訳開始部位を修正するのに加えて、957bpのコード領域(85のコドンを含んでなる)の94bpもまたコドン最適化した。このコドン最適化遺伝子(配列番号91)とM.アルピナ(M.alpina)からの野生型配列全長(配列番号5)との比較を図11に示し、そこでは太字のヌクレオチドが、コドン最適化遺伝子において修正されたヌクレオチドに対応する。コドン最適化遺伝子中の修正のいずれも、コードされたタンパク質(配列番号6)のアミノ酸配列を変化させなかった。
【0204】
高親和力鎖長延長酵素遺伝子を合成するのに使用される方法は、図12で図示される。具体的には10対のオリゴヌクレオチドがデザインされて、M.アルピナ(M.alpina)高親和力鎖長延長酵素のコード領域(すなわち配列番号92−111に対応するEL−1A、EL−1B、EL−2A、EL−2B、EL−3A、EL−3B、EL−4A、EL−4B、EL−5A、EL−5B、EL−6A、EL−6B、EL−7A、EL−7B、EL−8A、EL−8B、EL−9A、EL−9B、EL−10A、およびEL−10B)の長さに沿って延長される。5’末端の4bpのオーバーハングを例外として、センス(A)およびアンチセンス(B)オリゴヌクレオチドの各対は相補的である。
【0205】
実施例4の方法に続いて、100ngの各オリゴヌクレオチドをT4ポリヌクレオチドキナーゼでリン酸化した。次に各センスおよびアンチセンスオリゴヌクレオチド対を混合し、アニールした。したがってEL−1A(配列番号92)をEL1−1B(配列番号93)にアニールして、二本鎖生産物「EL−1AB」を生産した。同様にEL−2A(配列番号95)をEL−2B(配列番号95)にアニールして、二本鎖生産物「EL−2AB」などを生産した。
【0206】
次に以下に示すように2個の別個のアニールされた二本鎖オリゴヌクレオチドのプールを一緒にライゲーションした。
・プール1:EL−1AB、EL−2AB、EL−3AB、EL−4ABおよびEL−5ABを含んでなる、そして
・プール2:EL−6AB、EL−7AB、EL−8AB、EL−9ABおよびEL−10ABを含んでなる。
【0207】
アニールされたオリゴヌクレオチドの各プールを10UのT4DNAリガーゼと共に20μLの容積中で一晩16℃でライゲーションした。
【0208】
次に各ライゲーション反応の生産物をPCRによって増幅した。具体的にはテンプレートとしてライゲーションされた「プール1」混合物(すなわち、EL−1AB、EL−2AB、EL−3AB、EL−4AB、およびEL−5AB)、プライマーとしてオリゴヌクレオチドEL−1(配列番号112)およびEL−5R(配列番号113)を使用して、コドン最適化鎖長延長酵素の第1の部分をPCRによって増幅した。実施例4述べられるようにしてPCR増幅を50μLの反応混合物中で実施した。増幅を以下のようにして実施した。95℃で3minの初期変性、それに続いて95℃で1min、56℃で30sec、72℃で1minの35サイクル。72℃で10minの最終延長サイクルの実施と、それに続く4℃での反応終結。500bpのPCR断片をpGEM−Tイージーベクター(プロメガ(Promega))中にサブクローン化して、pTEL(1−5)を生産した。
【0209】
テンプレートとしてライゲーションされた「プール2」混合物(すなわち、EL−6AB、EL−7AB、EL−8AB、EL−9ABおよびEL−10AB)、プライマーと
してオリゴヌクレオチドEL−6(配列番号114)およびEL−10R(配列番号115)を使用して、コドン最適化鎖長延長酵素遺伝子の第2の部分を同様にPCRによって増幅し、pGEM−T−イージーベクター中にサブクローン化してpTEL(6−10)を生産した。
【0210】
大腸菌(E. coli)細胞をpTEL(1−5)およびpTEL(6−10)で別々に形質転換し、アンピシリン抵抗性形質転換体からのプラスミドDNAを精製し、適切な制限エンドヌクレアーゼで消化して、500bpのpTEL(1−5)のNcoI/SalI断片および470bpのpTEL(6−10)のSalI/NotI断片を遊離した。次にこれらの断片を混合し、NcoI/NotI消化したpY5−13にライゲーションしてpELS−1を生産した。
【0211】
pELS−1インサートのDNA配列分析は、Thr(ACC)からSer(AGC)へのアミノ酸変化をもたらす、+65の位置における1つの「C」から「T」への塩基置換の存在を同定した(「ATG」翻訳コドンの「A」を+1とする)。引き続きプライマーとしてオリゴヌクレオチドEL−M1(配列番号116)およびEL−M2(配列番号117)を使用して、部位特異的変異誘発によってこの変異を修正して、pELS(図13A)を生産した。
【0212】
実施例7
コドン最適化Δ6デサチュラーゼ、Δ17デサチュラーゼ、および高親和力PUFA鎖長延長酵素遺伝子によるヤロウィア・リポリティカ(Yarrowia lipolytica)の形質転換
実施例2で述べた方法に従って、野生型およびコドン最適化Δ6デサチュラーゼ、Δ17デサチュラーゼ、および高親和力PUFA鎖長延長酵素遺伝子を含有するプラスミドをY.リポリティカ(Y.lipolytica)ATCC番号76982中で別々に形質転換した。この技術を使用して、以下のプラスミドを含有する形質転換体を得た。
【0213】
【表6】
【0214】
実施例8
ヤロウィア・リポリティカ(Yarrowia lipolytica)におけるコドン最適化Δ6およびΔ17デサチュラーゼおよび高親和力鎖長延長酵素の変換効率の分析
以下の野生型およびコドン最適化Δ6デサチュラーゼ、Δ17デサチュラーゼ、および高親和力鎖長延長酵素それぞれの変換効率の比較では、コドン最適化がY.リポリティカ(Y.lipolytica)において、LAからGLA(Δ6デサチュラーゼ)への%基質変換をおよそ40%、ARAからEPA(Δ17デサチュラーゼ)への%基質変換を
約2倍に、そしてGLAからDGLA(鎖長延長酵素)への%基質変換を約57%改善したと判定された。
【0215】
コドン最適化Δ6デサチュラーゼによる%基質変換
Δ6デサチュラーゼは、LAをGLAに、および/またはALAをSTAに変換する(図1参照)。野生型とコドン最適化Δ6デサチュラーゼ遺伝子の変換効率を比較するために、それぞれ別のプラスミドコンストラクト(すなわちpYSD6またはpYD6S)を含有するヤロウィア・リポリティカ(Yarrowia lipolytica)において、%基質変換([生産物]/[基質+生産物])×100)を求めた。具体的には3mLの最少培地中でpYSD6またはpYD6Sのいずれかを含有するヤロウィア・リポリティカ(Yarrowia lipolytica)を単一コロニーから生育させ、10μgのLAを含有する最少培地中で継代培養して、実施例2で述べるように脂質分析を施した。
【0216】
実験の結果は、pYSD6を含有するヤロウィア(Yarrowia)株が、約30%の基質LAをGLAに変換し(図14A)、他方pYD6Sを含有するものは約42%のLAをGLAに変換する(図14B)ことを示唆した。これに基づき、コドン最適化Δ6デサチュラーゼ遺伝子を含有するヤロウィア(Yarrowia)は、Y.リポリティカ(Y.lipolytica)中の野生型M.アルピナ(M.alpina)Δ6デサチュラーゼ遺伝子よりもおよそ40%多くのLAを変換した。
【0217】
コドン最適化Δ17デサチュラーゼによる%基質転換
Δ17デサチュラーゼはDGLAをETAに、および/またはARAをEPAに変換する(図1参照)。野生型とコドン最適化Δ17デサチュラーゼ遺伝子の変換効率を比較するために、pYSD17、pYSD17M、およびpYSD17Sを含有するヤロウィア・リポリティカ(Yarrowia lipolytica)における%基質変換を求めた。3mLの最小培地中で各形質転換体を単一コロニーから生育させ、10μgのARAを含有する最少培地中で継代培養して、実施例2で述べるように脂質分析を施した。
【0218】
ARA供給実験の結果は、対照プラスミドpYSD17またはpYSD17Mを有するヤロウィア(Yarrowia)株が約23%の細胞内ARAをEPAに変換し(図15A)、他方pYSD17S上にコドン最適化Δ17デサチュラーゼ遺伝子を含有するものが約45%の細胞内ARAをEPAに変換する(図15B)ことを示した。したがってコドン最適化Δ17デサチュラーゼ遺伝子を含有するヤロウィア(Yarrowia)は、野生型S.ディクリナ(S.diclina)遺伝子を含有する株よりも約2倍多いARAを変換した。
【0219】
コドン最適化鎖長延長酵素による%基質転換
M.アルピナ(M.alpina)の高親和力PUFA鎖長延長酵素は、主にGLAからDGLAへの転換を触媒する役目を果たす(図1参照、国際公開第00/12720号パンフレット)。野生型とコドン最適化鎖長延長酵素遺伝子の変換効率を比較するために、pY58およびpELSを含有するヤロウィア・リポリティカ(Yarrowia lipolytica)中で%基質変換を求めた。形質転換体を3mLの最少培地中で単一コロニーから生育させ、10μgのGLAを含有する最少培地中で継代培養し、実施例2で述べるように脂質分析を施した。
【0220】
GLA供給実験の結果は、pY58を含有するヤロウィア・リポリティカ(Yarrowia lipolytica)が約30%の細胞内基質GLAをDGLAに変換し(図16A)、他方pELSを含有するものが約47%のGLAをDGLAに変換する(図16B)ことを示した。これに基づいて、コドン最適化鎖長延長酵素遺伝子は、Y.リポリ
ティカ(Y.lipolytica)中の野生型M.アルピナ(M.alpina)鎖長延長酵素遺伝子よりもおよそ57%多いGLAを変換した。
【図面の簡単な説明】
【0221】
【図1】ω−3およびω−6脂肪酸生合成経路を図示する。
【図2】ヤロウィア・リポリティカ(Yarrowia lipolytica)中で使用するためのプラスミドベクターpY5の構築を図示する。
【図3】Y.リポリティカ(Y.lipolytica)における遺伝子発現のためのプラスミドベクターpY5−13およびpY5−4の構築を図示する。
【図4】Y.リポリティカ(Y.lipolytica)中の翻訳開始コドン「ATG」周囲の好ましい共通配列を図示する。
【図5】モルティエレラ・アルピナ(Mortierella alpina)Δ6デサチュラーゼ遺伝子と、Y.リポリティカ(Y.lipolytica)における発現のためにコドン最適化された合成遺伝子とのDNA配列の比較を示す。
【図6】コドン最適化Δ6デサチュラーゼ遺伝子の生体外(in vitro)合成のために用いられるストラテジーを図示する。
【図7】Y.リポリティカ(Y.lipolytica)におけるコドン最適化Δ6デサチュラーゼ遺伝子の発現のためのプラスミドを示す。
【図8】サプロレグニア・ディクリナ(Saprolegnia diclina)Δ17デサチュラーゼ遺伝子と、Y.リポリティカ(Y.lipolytica)における発現のためにコドン最適化された合成遺伝子とのDNA配列の比較を示す。
【図9】コドン最適化Δ17デサチュラーゼ遺伝子の生体外(in vitro)合成のために用いられるストラテジーを図示する。
【図10】Y.リポリティカ(Y.lipolytica)におけるコドン最適化および野生型Δ17デサチュラーゼ遺伝子発現のためのプラスミドを示す。
【図11】モルティエレラ・アルピナ(Mortierella alpina)鎖長延長酵素遺伝子と、Y.リポリティカ(Y.lipolytica)における発現のためにコドン最適化された合成遺伝子とのDNA配列の比較を示す。
【図12】コドン最適化鎖長延長酵素遺伝子の生体外(in vitro)合成のために用いられるストラテジーを図示する。
【図13】Y.リポリティカ(Y.lipolytica)におけるコドン最適化および野生型鎖長延長酵素遺伝子発現のためのプラスミドを示す。
【図14A】約30%の基質LAからGLAへの変換を示す、コドン最適化および野生型Δ6デサチュラーゼ遺伝子で形質転換されたY.リポリティカ(Y.lipolytica)中で生産された脂肪酸のガスクロマトグラフィー分析の結果を示す。
【図14B】約42%の基質LAからGLAへの変換を示す、コドン最適化および野生型Δ6デサチュラーゼ遺伝子で形質転換されたY.リポリティカ(Y.lipolytica)中で生産された脂肪酸のガスクロマトグラフィー分析の結果を示す。
【図15】約23%の細胞内ARAからEPAへの変換を示す、コドン最適化および野生型Δ17デサチュラーゼ遺伝子で形質転換されたY.リポリティカ(Y.lipolytica)中で生産された脂肪酸のガスクロマトグラフィー分析の結果を示す。
【図15B】約45%の細胞内ARAからEPAへの変換を示す、コドン最適化および野生型Δ17デサチュラーゼ遺伝子で形質転換されたY.リポリティカ(Y.lipolytica)中で生産された脂肪酸のガスクロマトグラフィー分析の結果を示す。
【図16】約30%の細胞内GLAからDGLAへの変換を示す、コドン最適化および野生型鎖長延長酵素遺伝子で形質転換されたY.リポリティカ(Y.lipolytica)中で生産された脂肪酸のガスクロマトグラフィー分析の結果を示す。
【図16B】約47%の細胞内GLAからDGLAへの変換を示す、コドン最適化および野生型鎖長延長酵素遺伝子で形質転換されたY.リポリティカ(Y.lipolytica)中で生産された脂肪酸のガスクロマトグラフィー分析の結果を示す。
【関連出願との関係】
【0001】
本願は2003年5月7日に出願された米国仮出願第60/468677号、および2003年5月7日に出願された米国仮特許出願第60/468718号の優先権の利益を主張する。
【技術分野】
【0002】
本発明はバイオテクノロジー分野に関する。より具体的には本発明は、油性酵母菌における長鎖多不飽和脂肪酸(PUFA)の生産に有用な酵素をコードする核酸断片の合成に関する。
【背景技術】
【0003】
特定の多不飽和脂肪酸、すなわちPUFAが健康な細胞の重要な生物学的構成要素であることが、長期にわたり認識されている。例えばこのようなPUFAは、
哺乳類において新規に(de novo)合成できず、食餌中で得られなくてはならない、またはリノール酸(LA)またはα−リノレン酸(ALA)のさらなる不飽和化と延長によって誘導されなくてはならない「必須」脂肪酸、
リン脂質またはトリグリセリドなどの形態で見いだされてもよい細胞原形質膜の構成物、
特に成長中の幼児の脳において適切な発育、そして組織形成および修復に必要である、
プロスタサイクリン、エイコサノイド、ロイコトリエン、およびプロスタグランジンをはじめとする、哺乳類において重要ないくつかの生物学的に活性なエイコサノイドの前駆物質として認識されている。
【0004】
1970年代に、グリーンランドのエスキモーの観察から、心疾患の低発生率と長鎖ω−3PUFAの高摂取量とが結びつけられた(非特許文献1、非特許文献2)。より最近の研究はω−3PUFAの心臓血管保護効果を確証した(非特許文献3、非特許文献4)。さらに血管形成術後の再狭窄率、炎症および関節リウマチ、喘息、乾癬および湿疹の症状などのいくつかの障害はω−3脂肪酸による処置に反応することが見いだされている。γ−リノレン酸(GLA、ω−6PUFA)はストレスに関係した血圧上昇を低下させ、算術試験能力を改善することが示されている。GLAおよびジホモ−γ−リノレン酸(DGLA、もう1つのω−6PUFA)は、血小板凝集を阻害し、血管拡張を引き起こし、コレステロールレベルを低下させ、血管壁平滑筋および繊維組織の増殖を阻害することが示されている(非特許文献5)。GLAまたはDGLAの単独でのまたはエイコサペンタエン酸(EPA、ω−3PUFA)との組み合わせでの投与は、非ステロイド性抗炎症薬によって引き起こされる消化管出血およびその他の副作用を低下させ、または防止することが示されている(特許文献1)。さらにGLAおよびDGLAは、子宮内膜症および月経前症候群を防止または治療し(特許文献2)、筋痛性脳脊髄炎およびウィルス感染後の慢性疲労(特許文献3)を治療することが示されている。その他の証拠は、PUFAがカルシウム代謝調節に関与するかもしれないことを示唆し、骨粗鬆症および腎臓または尿道結石の治療または防止においてそれらが有用であるかもしれないことを示唆する。最後にPUFAは癌および糖尿病の治療において使用できる(特許文献4、非特許文献6)。
【0005】
PUFAは、必須脂肪酸であるLAおよびALAそれぞれの不飽和化および延長によって誘導される、2つの主要なクラス(ω−6およびω−3脂肪酸からなる)に概して分けられる(図1)。天然供給源からの多様なPUFAの商業的供給源(例えば月見草、ルリヂサ、およびクロフサスグリの種子や糸状菌(モルティエレラ(Mortierella))、チノリモ属(Porphyridium)(紅藻)、魚油および海洋性プランクト
ン(キクロテラ(Cyclotella)、ニッチア(Nitzschia)、クリプテコジニウム(Crypthecodinium))にもかかわらず、これらの生産方法と結びついたいくつかの不都合がある。第1に魚および植物などの天然供給源は、高度に不均一な油組成物を有しがちである。したがってこれらの供給源から得られた油は、所望のPUFAの1つもしくはそれ以上を分離または濃縮するために大規模な精製を必要とすることがある。天然供給源はまた、制御できない供給のばらつきを被りやすい(例えば天候、疾患、または魚資源の乱獲による)。PUFAを生産する作物は、食物生産のために開発されたハイブリッド作物と、経済的競争力がないことが多い。またPUFAを自然に生産するいくつかの生物体(例えばポルフィリディウム(Porphyridium)、モルティエレラ(Mortierella))の大規模発酵は、商業的規模で培養するのが高価および/または困難なことがある。
【0006】
上述の限界の結果として、以下に向けた大規模な研究が行われている。1.)商業的に容易に生産できるPUFAの組換え供給源の開発、および2.)所望のPUFA生産を可能にする脂肪酸生合成経路の修正。例えば過去数年間にわたり、様々な生物体からの脂肪酸デサチュラーゼおよび鎖長延長酵素(elongase)遺伝子の単離、クローン化、および操作において進歩があった。これらの遺伝子配列の知識は、PUFAを自然に生産しない新しい宿主生物体中で、所望の脂肪酸および/または脂肪酸組成物を生産する見込みを提供する。文献は、サッカロマイセス・セレヴィシエ(Saccharomyces
cerevisiae)における以下のようないくつかの例を報告する。
1.非特許文献7では、海洋珪藻フェオダクチルム・トリコルヌーツム(Phaeodactylum tricornutum)からの2個のデサチュラーゼがS.セレヴィシエ(S.cerevisiae)中にクローン化され、EPAの生産をもたらす。
2.非特許文献8では、線虫(Caenorhabditis elegans)からの遺伝子を使用して、ω−3およびω−6PUFA生合成経路がS.セレヴィシエ(S.cerevisiae)中に再構成される。
3.非特許文献9では、植物脂肪酸デサチュラーゼ(FAD2およびFAD3)がS.セレヴィシエ(S.cerevisiae)中に発現し、ALAの生産をもたらす。
4.アボット・ラボラトリーズ(Abbott Laboratories)のナットゾン(Knutzon)らに付与された特許文献5では西洋油菜(Brassica napus)からの1つのデサチュラーゼおよび菌・カビ類モルティエレラ・アルピナ(Mortierella alpina)からの2個のデサチュラーゼが、S.セレヴィシエ(S.cerevisiae)中にクローン化され、LA、GLA、ALA、およびSTAの生産をもたらす。
【0007】
しかしこれらのタイプの遺伝子を発現させて、商業的量の1つもしくはそれ以上のPUFAの経済的生産を提供できる適切な微生物のシステムに対する必要性がなおもある。さらに特にEPAおよびDHAである特定のPUFAが濃縮されている油に対する必要性が存在する。
【0008】
PUFAの生産プラットフォームとしてこれまで調査されていない1つのクラスの微生物が、油性酵母菌である。これらの生物体は、乾燥細胞重量の80%までの油を蓄積できる。高い油含量で油性酵母菌を生育させる技術は十分に開発されており(例えば特許文献6、非特許文献10を参照)、ω−3またはω−6PUFA生産のための商業的な微細藻類発酵と比べてコスト優位性を提供するかもしれない。そのままの酵母菌細胞はまた、機能食品および動物飼料サプリメントで使用するためのω−3またはω−6PUFA−濃縮油を封入する都合よい方法になるかもしれない。
【0009】
【特許文献1】米国特許第4,666,701号明細書
【特許文献2】米国特許第4,758,592号明細書
【特許文献3】米国特許第5,116,871号明細書
【特許文献4】米国特許第4,826,877号明細書
【特許文献5】米国特許第6,136,574号明細書
【特許文献6】欧州特許第0005277B1号明細書
【非特許文献1】ダイヤーバーグ(Dyerberg)J.ら、Amer.J.Clin Nutr.28:958〜966(1975)
【非特許文献2】ダイヤーバーグ(Dyerberg)J.ら、Lancet 2(8081):117〜119(1978年7月15日)
【非特許文献3】シモカワ(Shimokawa)H.、World Rev Nutr Diet、88:100〜108(2001)
【非特許文献4】フォン・シャッキー(von Schacky)C.およびダイヤーバーグ(Dyerberg)J.、World Rev Nutr Diet、88:90〜99(2001)
【非特許文献5】ブレナー(Brenner)ら、Adv.Exp.Med.Biol.83:85〜101(1976)
【非特許文献6】(Horrobin)ら、Am.J.Clin.Nutr.57(付録):732S〜737S(1993)
【非特許文献7】ドマーグ(Domergue)F.ら、Eur.J.Biochem.269:4105〜4113(2002)
【非特許文献8】ボードイン(Beaudoin)F.ら、Proc.Natl.Acad.Sci.U.S.A.97(12):6421〜6(2000)
【非特許文献9】ダイヤー(Dyer)J.M.ら、Appl.Eniv.Microbiol.、59:224〜230(2002)
【非特許文献10】ラトレッジ(Ratledge)C.、Prog.Ind.Microbiol.16:119〜206(1982)
【発明の開示】
【発明が解決しようとする課題】
【0010】
上述の利点にもかかわらず、油性酵母菌中の自然に生産されるPUFAは、18:2脂肪酸(そしてあまり一般的ではなく18:3脂肪酸)に限定されるので、これらの生物体は自然にはω−6およびω−3PUFAを欠いている。したがって解決すべき問題は、ω−3および/またはω−6脂肪酸が濃縮された油を蓄積する油性酵母菌を開発することである。このような目的で、油性酵母菌におけるω−3および/またはω−6脂肪酸の合成および蓄積を可能にする、飽和化酵素および鎖長延長酵素を導入することが必要である。多数の供給源から多様なデサチュラーゼおよび鎖長延長酵素遺伝子が入手できるにもかかわらず、これらの遺伝子は油性酵母菌などの別の宿主中では、遺伝子中のコドンが別の宿主生物体の典型的なコドン使用を反映しないので、最適効率で発現しない。したがって油性酵母菌におけるPUFA遺伝子の発現を最適化し、これらの特定の宿主生物体におけるω−3および/またはω−6脂肪酸の高レベルの生産および蓄積を可能にするために、コドン使用に関連した問題を克服しなくてはならない。
【0011】
出願人は、油性宿主、ヤロウィア・リポリティカ(Yarrowia lipolytica)における発現に適したデサチュラーゼおよび鎖長延長酵素遺伝子をコドン最適化する手段を開発することで既述の問題を解決した。ここで最適化される例示的な遺伝子は、Δ6デサチュラーゼ、Δ17デサチュラーゼ、および高親和力PUFA鎖長延長酵素をコードする遺伝子であり、コドン最適化は、Y.リポリティカ(Y.lipolytica)中でLAからGLAへの%基質変換(Δ6デサチュラーゼ)をおよそ40%改善し、ARAからEPAへの%基質変換(Δ17デサチュラーゼ)を約2倍にし、GLAからDGLAへの%基質変換(鎖長延長酵素)を約57%改善した。
【課題を解決するための手段】
【0012】
本発明は、ヤロウィア(Yarrowia)種における最適発現のためのω−3/ω−6脂肪酸生合成経路における様々な遺伝子の最適化に関する。したがって本発明は、
a)配列番号25に記載のΔ6デサチュラーゼをコードする単離された核酸分子、または
b)(a)と完全に相補的である単離された核酸分子
よりなる群から選択される単離された核酸分子を提供する。
【0013】
同様に本発明は、配列番号2に記載のΔ6デサチュラーゼをコードする単離された核酸分子を提供し、そこではヤロウィア(Yarrowia)種における発現のために少なくとも144個のコドンがコドン最適化される。
【0014】
本発明の別の実施態様では、
a)配列番号62に記載のΔ17デサチュラーゼをコードする単離された核酸分子、または
b)(a)と完全に相補的である単離された核酸分子
よりなる群から選択される、単離された核酸分子が提供される。
【0015】
本発明の別の実施態様では、配列番号4に記載のΔ17デサチュラーゼをコードする単離された核酸分子が提供され、そこではヤロウィア(Yarrowia)種における発現のために少なくとも117個のコドンがコドン最適化される。
【0016】
同様に本発明は、
a)配列番号91に記載の鎖長延長酵素をコードする単離された核酸分子、または
b)(a)と完全に相補的である単離された核酸分子よりなる群から選択される単離された核酸分子を提供する。
【0017】
代案としては本発明は、配列番号6に記載の鎖長延長酵素をコードする単離された核酸分子を提供し、そこではヤロウィア(Yarrowia)種における発現のために少なくとも85個のコドンがコドン最適化される。
【0018】
さらに本発明は、本発明の遺伝子の遺伝的キメラ、および該物質で形質転換された宿主細胞を提供する。
【0019】
本発明の特定の実施態様では、ヤロウィア(Yarrowia)種中で本発明のコドン最適化遺伝子を使用して、適切な前駆物質からの一段階酵素的反応によって、γ−リノレン酸(GLA)、ジホモ−γ−リノール酸(DGLA)、ステアリドン酸(STA)、エイコサテトラエン酸(ETA)、エイコサペンタエン酸(EPA)、およびドコサペンタエン酸(DPA)などの特定のω−3およびω−6脂肪酸生産が提供される。
【0020】
本発明の別の実施態様では、
a)油性酵母菌種についてヌクレオチドコード領域および対応するポリペプチドの配列を得て、コドンのデータベースを形成し、
b)コドンのデータベースを分析し、いずれのコドンが各アミノ酸を優先的にコードするのか判定し、
c)油性酵母菌種中で発現させる遺伝子の配列を得て、
d)ステップ(c)の配列中の好ましくないコドンをステップ(b)の好ましいコドンで置き換えて、遺伝子が油性酵母菌種における発現のためにコドン最適化すること
を含んでなる、油性酵母菌における発現のために遺伝子を最適化する方法が提供される。
【0021】
本発明の代案の実施態様では、配列番号122に記載のヤロウィア(Yarrowia)翻訳開始部位を含んでなる単離された核酸分子が提供される。さらに
a)ヤロウィア(Yarrowia)中で発現させる外来遺伝子を提供し、
b)ステップ(a)の遺伝子を配列番号122に記載のヤロウィア(Yarrowia)翻訳開始部位に作動可能に連結させ、外来遺伝子をヤロウィア(Yarrowia)における発現のために最適化すること
を含んでなる、ヤロウィア(Yarrowia)宿主における遺伝子の発現を最適化するための方法も提供される。
【0022】
配列説明
本願明細書の一部を形成する以下の詳細な説明および添付の配列説明によって、本発明をより完全に理解できるであろう。
【0023】
以下の配列は、37C.F.R.§1.821〜1.825(「ヌクレオチド配列および/またはアミノ酸配列開示を含む特許出願の要件−配列規則(Requirements for Patent Applications Containing Nucleotide Sequences and/or Amino Acid Sequence Disclosures − the Sequence Rules)」)を満たし、世界知的所有権機関(WIPO)標準ST.25(1998)およびEPOおよびPCTの配列表要件(規則5.2および49.5(aの2)、および実施細則第208号および附属書C)に一致する。ヌクレオチドおよびアミノ酸配列データのために使用される記号および型式は、37C.F.R.§1.822で述べられる規則に従う。
【0024】
配列番号1は、モルティエレラ・アルピナ(Mortierella alpina)Δ6デサチュラーゼ遺伝子のDNA配列を示し、他方配列番号2は、M.アルピナ(M.alpina)Δ6デサチュラーゼのアミノ酸配列を示す。
【0025】
配列番号3は、サプロレグニア・ディクリナ(Saprolegnia diclina)Δ17デサチュラーゼ遺伝子のDNA配列を示し、他方配列番号4は、S.ディクリナ(S.diclina)Δ17デサチュラーゼの対応するアミノ酸配列を示す。
【0026】
配列番号5は、モルティエレラ・アルピナ(Mortierella alpina)高親和力鎖長延長酵素遺伝子のDNA配列を示し、他方配列番号6はM.アルピナ(M.alpina)高親和力鎖長延長酵素のアミノ酸配列を示す。
【0027】
配列番号7および8は、TEFプロモーターを単離するのに使用されるプライマーTEF5’およびTEF3’にそれぞれ対応する。
【0028】
配列番号9および10は、XPR2転写ターミネーターを単離するのに使用される、プライマーXPR5’およびXPR3’にそれぞれ対応する。
【0029】
配列番号11〜24は、プラスミド構築のために使用されるプライマーYL1、YL2、YL3、YL4、YL23、YL24、YL5、YL6、YL9、YL10、YL7、YL8、YL61、およびYL62にそれぞれ対応する。
【0030】
配列番号25は、ヤロウィア・リポリティカ(Yarrowia lipolytica)における発現のためにコドン最適化された、合成Δ6デサチュラーゼ遺伝子のDNA配列を示す。
【0031】
配列番号26〜53は、M.アルピナ(M.alpina)Δ6デサチュラーゼ遺伝子
のコドン最適化コード領域全体を一緒に構成する、14対のオリゴヌクレオチドに対応する(例えばそれぞれD6−1A、D6−1B、D6−2A、D6−2B、D6−3A、D6−3B、D6−4A、D6−4B、D6−5A、D6−5B、D6−6A、D6−6B、D6−7A、D6−7B、D6−8A、D6−8B、D6−9A、D6−9B、D6−10A、D6−10B、D6−11A、D6−11B、D6−12A、D6−12B、D6−13A、D6−13B、D6−14A、およびD6−14B)。
【0032】
配列番号54〜61は、コドン最適化Δ6デサチュラーゼ遺伝子の合成中に、PCR増幅のために使用されるプライマーD6−1、D6−4R、D6−5、D6−7R、D6−8、D6−10R、D6−11、およびD6−14Rにそれぞれの対応する。
【0033】
配列番号62は、ヤロウィア・リポリティカ(Yarrowia lipolytica)における発現のためにコドン最適化された、合成Δ17デサチュラーゼ遺伝子のDNA配列を示す。
【0034】
配列番号63〜84は、S.ディクリナ(S.diclina)Δ17デサチュラーゼ遺伝子のコドン最適化コード領域全体を一緒に構成する、11対のオリゴヌクレオチドに対応する(例えばそれぞれD17−1A、D17−1B、D17−2A、D17−2B、D17−3A、D17−3B、D17−4A、D17−4B、D17−5A、D17−5B、D17−6A、D17−6B、D17−7A、D17−7B、D17−8A、D17−8B、D17−9A、D17−9B、D17−10A、D17−10B、D17−11A、およびD17−11B)。
【0035】
配列番号85〜90は、コドン最適化Δ17デサチュラーゼ遺伝子の合成中に、PCR増幅のために使用されるプライマーD17−1、D17−4R、D17−5、D17−8D、D17−8U、およびD17−11にそれぞれ対応する。
【0036】
配列番号91は、ヤロウィア・リポリティカ(Yarrowia lipolytica)における発現のためにコドン最適化された、合成高親和力鎖長延長酵素遺伝子のDNA配列を示す。
【0037】
配列番号92〜111は、M.アルピナ(M.alpina)高親和力鎖長延長酵素遺伝子のコドン最適化コード領域全体を一緒に構成する、10対のオリゴヌクレオチドに対応する(例えばそれぞれEL−1A、EL−1B、EL−2A、EL−2B、EL−3A、EL−3B、EL−4A、EL−4B、EL−5A、EL−5B、EL−6A、EL−6B、EL−7A、EL−7B、EL−8A、EL−8B、EL−9A、EL−9B、EL−10A、およびEL−10B)。
【0038】
配列番号112〜115は、コドン最適化鎖長延長酵素遺伝子の合成中に、PCR増幅のために使用されるプライマーEL−1、EL−5R、EL−6、およびEL−10Rにそれぞれ対応する。
【0039】
配列番号116および117は、部位特異的変異誘発のために使用されてpELSを生じる、プライマーEL−M1およびEL−M2に対応する。
【0040】
配列番号118および119は、プラスミドpRSP19からS.ディクリナ(S.diclina)の野生型Δ17デサチュラーゼ遺伝子を増幅するために使用される、プライマーYL21AおよびYL22に対応する。
【0041】
配列番号120および121は、部位特異的変異誘発のために使用されてpYSD17
Mを生じる、プライマーYL53およびYL54に対応する。
【0042】
配列番号122は、ヤロウィア(Yarrowia)種中で最適に発現する遺伝子のためのコドン最適化翻訳開始部位に対応する。
【発明を実施するための最良の形態】
【0043】
主題の発明にしたがって、出願人は油性酵母菌中、ヤロウィア・リポリティカ(Yarrowia lipolytica)における構造的な遺伝子のコドン使用を判定した。Δ6デサチュラーゼ(配列番号25)、Δ17デサチュラーゼ(配列番号62)および高親和力鎖長延長酵素(配列番号91)をコードするコドン最適化遺伝子、ならびにY.リポリティカ(Y.lipolytica)宿主細胞における前記遺伝子に発現のためのDNAカセットが提示される。さらにY.リポリティカ(Y.lipolytica)などの油性酵母菌の長鎖多不飽和脂肪酸(PUFA)含量の修正を可能にする、方法および組成物が提供される。
【0044】
主題発明には多くの用途がある。ここで開示される方法によって製造されるPUFA、またはその誘導体は、食餌代用品、またはサプリメント、特に乳児用調製粉乳として、静脈内栄養補給を受けている患者のために、または栄養不良を防止または処置するために使用できる。代案としては、精製されたPUFA(またはその誘導体)は、正常な使用で受領者が食餌栄養補給のための所望量を受容するように調合された、料理用油、脂肪またはマーガリンに組み込まれてもよい。PUFAはまた、乳児用調製粉乳、栄養サプリメントまたはその他の食物生産物に組み込まれてもよく、抗炎症薬またはコレステロール低下剤としての用途があるかもしれない。場合により組成物は、医薬品用途(ヒトまたは獣医学)のために使用されてもよい。この場合、PUFAは概して経口投与されるが、例えば非経口的(例えば皮下、筋肉内または静脈内)、経直腸、経腟または局所的(例えば皮膚用軟膏またはローションとして)など、それによって成功裏に吸収されるあらゆる経路で投与することができる。
【0045】
組換え手段によって製造されたPUFAによるヒトまたは動物の栄養補給は、追加的なPUFA、ならびにそれらの代謝子孫の増大したレベルをもたらすことができる。例えばアラキドン酸(ARA)による処置は、ARAの増大したレベルだけでなく、プロスタグランジンなどのARAの下流生産物をももたらすことができる。複雑な制御機序は、このような機序を防止、制御または克服して、個々の特定のPUFAの所望のレベルを達成するために、様々なPUFAを組み合わせ、または異なるPUFAコンジュゲートを追加することを望ましいものにできる。
【0046】
定義
本開示では、いくつかの用語および略語が使用される。以下の定義が提供される。
【0047】
「読み取り枠」はORFと略記する。
【0048】
「ポリメラーゼ連鎖反応」はPCRと略記する。
【0049】
「米国微生物系統保存機関」はATCCと略記する。
【0050】
「多不飽和脂肪酸」はPUFAと略記する。
【0051】
「脂肪酸」という用語は、(より長い、およびより短い鎖長の酸の双方も知られているが)約C12〜C22の様々な鎖長の長鎖脂肪族酸(アルカン酸)を指す。優勢な鎖長は、C16〜C22の間である。脂肪酸の構造は単純な表記法システム「X:Y」によって
表され、ここでXは特に脂肪酸の炭素(C)原子の総数であり、Yは二重結合の数である。
【0052】
概して脂肪酸は、飽和または不飽和として分類される。「飽和脂肪酸」という用語は、炭素主鎖間に「二重結合」を有さない脂肪酸を指す。対照的に「不飽和脂肪酸」は、それらの炭素主鎖に沿って「二重結合」を有するcis−異性体である。「一不飽和脂肪酸」は、(例えばパルミトレイン酸(16:1)およびオレイン酸(18:1)のように通常9および10番目の炭素原子の間に)炭素主鎖に沿って1つの「二重結合」のみを有し、他方「多不飽和脂肪酸」(または「PUFA」)は、(例えばリノール酸(18:2)のように9番目および10番目、および12番目および13番目の炭素原子間、およびα−リノレン酸(18:3)のように9番目および10番目、12番目および13番目目、および15番目および16番目の炭素原子間に)炭素主鎖に沿って少なくとも2個の二重結合を有する。
【0053】
「PUFA」は、(脂肪酸炭素鎖のメチル末端に最も近い第1の二重結合の位置(n)次第で)2つの主要ファミリーに分類できる。したがって「オメガ−6脂肪酸」(ω−6またはn−6)は、第1の不飽和二重結合を分子のω(メチル)末端から6個めの炭素原子に有し、さらに分子のカルボキシル末端に向かって3個めの追加的な炭素原子にそれぞれの続く不飽和がある、全部で2つ以上の二重結合を有する。対照的に「オメガ−3脂肪酸」(ω−3またはn−3)は、第1の不飽和二重結合を分子のω末端から3個離れた炭素原子に有し、さらに分子のカルボキシル末端に向かって3個目の追加的な炭素原子にそれぞれの続く不飽和がある、全部で3個以上の二重結合を有する。
【0054】
本開示の目的のために、ω参照システムを使用して、炭素数、二重結合数、およびω炭素(この目的では1番目とする)から数えたω炭素に最も近い二重結合の位置を示す。この命名法を下の表1で、「省略表記法」と題された欄に示す。表の残りは、ω−3およびω−6脂肪酸の共通の名称、明細書全体で使用される略語、および各化合物の化学名をまとめる。
【0055】
【表1】
【0056】
「必須脂肪酸」と言う用語は、固体が特定の必須脂肪酸を新規に(de novo)合成できないことから、生きるために摂取しなくてはならない特定のPUFAを指す。例えば、哺乳類は必須脂肪酸LA(18:2、ω−6)を合成できない。その他の必須脂肪酸としては、GLA(ω−6)、DGLA(ω−6)、ARA(ω−6)、EPA(ω−3)、およびDHA(ω−3)が挙げられる。
【0057】
「脂肪」と言う用語は、25℃で固体であり、通常、飽和である脂質物質を指す。
【0058】
「油」と言う用語は、25℃で液体であり、通常、多不飽和である脂質物質を指す。PUFAは、いくつかの藻類、油性酵母菌および糸状菌の油に見られる。「微生物油」または「単細胞油」は、微生物がそれらの寿命中に自然に生産する油である。このような油は長鎖PUFAを含有することができる。
【0059】
「PUFA生合成経路酵素」と言う用語は、PUFAの生合成に関連した以下の酵素(および前記酵素をコードする遺伝子)のいずれかを指す。Δ4デサチュラーゼ、Δ5デサチュラーゼ、Δ6デサチュラーゼ、Δ12デサチュラーゼ、Δ15デサチュラーゼ、Δ17デサチュラーゼ、Δ9デサチュラーゼ、および/または鎖長延長酵素。
【0060】
「ω−3/ω−6脂肪酸生合成経路」と言う用語は、適切な条件下で発現すると、ω−3およびω−6脂肪酸の片方または双方の生産を触媒する酵素をコードする一組の遺伝子を指す。典型的にω−3/ω−6脂肪酸生合成経路に関与する遺伝子は、以下の酵素のいくつかまたは全てをコードする。Δ12デサチュラーゼ、Δ6デサチュラーゼ、鎖長延長酵素、Δ5デサチュラーゼ、Δ17デサチュラーゼ、Δ15デサチュラーゼ、Δ9デサチュラーゼ、およびΔ4デサチュラーゼ。共通の供給源からいかにしてω−3およびω−6脂肪酸の双方が生産できるかを実証する代表的な経路は図1に示され、DHAへの様々な中間体を通じて、オレイン酸の変換を提供する。
【0061】
「デサチュラーゼ」と言う用語は、1つもしくはそれ以上の脂肪酸を不飽和化して関心のある一不飽和または多不飽和脂肪酸または前駆物質を生産できる、多酵素複合体のポリペプチド構成要素を指す。特定の脂肪酸に関して、本願明細書全体を通じてω参照システムを使用するのにもかかわらず、Δシステムを使用して基質のカルボキシル末端から数えることで、デサチュラーゼの活性を示す方が都合よい。ここで特に関心が高いのは、1.)分子のカルボキシル末端から数えて17番目および18番目の炭素原子間で脂肪酸を不飽和化し、例えばARAからEPAへのおよび/またはDGLAからETAへの転換を触媒するΔ17デサチュラーゼ、2.)LAからGLAへのおよび/またはALAからSTAへの転換を触媒するΔ6デサチュラーゼ、3.)DGLAからARAへのおよび/またはETAからEPAへの転換を触媒するΔ5デサチュラーゼ、4.)DPAからDHAへの転換を触媒するΔ4デサチュラーゼ、5.)オレイン酸からLAへの転換を触媒するΔ12デサチュラーゼ、6.)LAからALAへの転換を触媒するΔ15デサチュラーゼ、および7.)パルミチン酸からパルミトレイン酸(16:1)および/またはステアリン酸からオレイン酸(18:1)への転換を触媒するΔ9デサチュラーゼである。
【0062】
「鎖長延長酵素」と言う用語は、脂肪酸炭素鎖を伸長して、鎖長延長酵素が作用した脂肪酸基質よりも炭素2個分長い一不飽和または多不飽和脂肪酸が生産できる、多酵素複合体のポリペプチド構成要素を指す。この延長プロセスは、CoAがアシルキャリアである脂肪酸合成酵素と関連した多段階機序で起きる(ラスナー(Lassner)ら、The
Plant Cell 8:281〜292(1996))。手短に述べると、マロニル−CoAが長鎖アシル−CoAと縮合して、CO2およびβ−ケトアシル−CoA(アシル部分が炭素原子2個分伸長された)を生じる。引き続く反応には、β−ヒドロキシアシル−CoAへの還元、エノイル−CoAへの脱水、および伸長されたアシル−CoAを生じる第2の還元が含まれる。鎖長延長酵素によって触媒される反応の例は、GLAからDGLA、STAからETA、およびEPAからDPAへの転換である。したがって鎖長延長酵素は、異なる特異性を有することができる(例えばC16/18鎖長延長酵素はC16基質を好み、C18/20鎖長延長酵素はC18基質を好み、C20/22鎖長延長酵素はC20基質を好む)。
【0063】
「高親和力鎖長延長酵素」と言う用語は、その基質特異性が、好ましくはGLAに向けたものである(鎖長延長酵素反応の生産物としてのDGLAを伴う)鎖長延長酵素を指す。このような鎖長延長酵素の1つについて、国際公開第00/12720号パンフレットで述べられている。
【0064】
「変換効率」および「%基質変換」と言う用語は、それによって特定の酵素(例えばデサチュラーゼまたは鎖長延長酵素)が基質から生産物に変換できる効率を指す。変換効率は以下の式に従って測定される。([生産物]/[基質+生産物])×100(式中、「生産物」には即時の生産物およびそれから誘導される経路中の全生産物が含まれる)。
【0065】
「油性」と言う用語は、それらのエネルギー源を脂質の形態で保存する傾向がある生物
体を指す(ウィーテ(Weete)「真菌脂質生化学(Fungal Lipid Biochemistry)」第二版、プレナム(Plenum)、1980)。概してこれらの微生物の細胞PUFA含量はS字形曲線に従い、対数後期または初期定常増殖相において脂質濃度が最大に達するまで増大し、次に後期定常および死滅期中に徐々に減少する(ヨンマニットチャイ(Yongmanitchai)およびワード(Ward)、Appl.Environ.Microbiol.57:419〜25(1991))。
【0066】
「油性酵母菌」と言う用語は、少なくとも25%のそれらの乾燥細胞重量を油として蓄積できる酵母菌として分類される微生物を指す。油性酵母菌の例としては、ヤロウィア(Yarrowia)、カンジダ(Candida)、ロドトルラ(Rhodotorula)、ロドスポリジウム(Rhodosporidium)、クリプトコッカス(Cryptococcus)、トリコスポロン(Trichosporon)およびリポマイセス(Lipomyces)属が挙げられるが、決してこれに限定されるものではない。
【0067】
「発酵性炭素源」と言う用語は、微生物が代謝してエネルギーを引き出す炭素源を意味する。本発明の典型的な炭素源としては、単糖類、少糖類、多糖類、アルカン、脂肪酸、脂肪酸のエステル、モノグリセリド、ジグリセリド、トリグリセリド、二酸化炭素、メタノール、ホルムアルデヒド、ギ酸、および炭素含有アミンが挙げられるが、これに限定されるものではない。
【0068】
「コドン最適化された」と言う用語は、様々な宿主の形質転換のための遺伝子または核酸分子のコード領域を指し、DNAによってコードされるポリペプチドを改変することなく、宿主生物体の典型的なコドン使用を反映するための遺伝子または核酸分子のコード領域中のコドンの改変を指す。本発明の文脈で、遺伝子およびDNAコード領域は、表4で収集された情報を使用して、ヤロウィア(Yarrowia)種における最適発現のためにコドン最適化される。
【0069】
ここでの用法では、「単離された核酸断片」とは、場合により合成、非天然または改変ヌクレオチド塩基を含有する一本鎖または二本鎖のRNAまたはDNAポリマーである。DNAポリマーの形態の単離された核酸断片は、cDNA、ゲノムDNAまたは合成DNAの1つもしくはそれ以上のセグメントを含んでなってもよい。
【0070】
アミノ酸またはヌクレオチド配列の「かなりの部分」とは、当業者による配列の手動評価によって、あるいはBLAST(「基礎的局在性整列化検索ツール(Basic Local Alignment Search Tool)」アルトシュル(Altschul)S.F.ら、J.Mol.Biol.215:403〜410(1993))などのアルゴリズムを使用したコンピュータ支援配列比較および同定によって、遺伝子またはポリペプチドの推定上の同定を得るのに十分なポリペプチドのアミノ酸配列または遺伝子のヌクレオチド配列を含んでなる部分である。概してヌクレオチド配列の「かなりの部分」は、配列を含んでなる核酸断片を特異的に同定および/または単離できるようにする十分な配列(例えば20〜30個の連続(contiguous)ヌクレオチド)を含んでなる。本願明細書は、1つもしくはそれ以上の特定のタンパク質をコードする、部分的または完全なヌクレオチド配列を教示する。ここで報告される配列の利点を有した当業者は、当業者には既知の目的のために、開示される配列の全てまたはかなりの部分を使用することができる。したがって本発明は、添付の配列表で報告される完全な配列、ならびに上述の配列のかなりの部分を含んでなる。
【0071】
「相補的」と言う用語は、互いにハイブリッドできるヌクレオチド塩基間の関係について述べるために用いられる。例えばDNAに関して、アデノシンはチミンに相補的であり、シトシンはグアニンに相補的である。したがって本発明はまた、添付の配列表で報告さ
れる完全な配列に相補的な単離された核酸断片、ならびに実質的に同様の核酸配列を含む。
【0072】
「コドン縮重」とは、コードされるポリペプチドのアミノ酸配列に影響しないヌクレオチド配列の変動を可能にする遺伝子コードの性質を指す。当業者は、特定のアミノ酸を特定化するヌクレオチドコドンの使用における、特定の宿主細胞によって示される「コドン−バイアス」についてよく知っている。したがって宿主細胞中の改善された発現のために遺伝子を合成する場合、そのコドン使用頻度が、宿主細胞の好むコドン使用頻度に近くなるように遺伝子をデザインすることが望ましい。
【0073】
「化学的に合成された」とは、DNA配列に関連して構成要素ヌクレオチドが、生体外で(in vitro)構築されたことを意味する。DNAの手動化学合成は確立した手順を使用して達成されてもよく、あるいはいくつかの市販の機器の1つを使用して自動化学合成を実施できる。「合成遺伝子」は、当業者に知られる手順を使用して、化学的に合成されるオリゴヌクレオチド構成単位から構築できる。これらの構成単位をライゲートしアニールして、遺伝子断片を形成し、次にそれを酵素的に構築して所望の遺伝子全体を構成する。したがって遺伝子をヌクレオチド配列の最適化に基づいて、最適な遺伝子発現のために調整し、宿主細胞のコドンバイアスを反映させることができる。当業者は、コドン利用が宿主によって好まれるコドンに偏っている場合の遺伝子発現成功の見込みを理解する。好ましいコドンの判定は、配列情報が利用できる宿主細胞から誘導される遺伝子の調査に基づくことができる。
【0074】
「遺伝子」とは、コード配列に先行する(5’非コード配列)およびそれに続く(3’非コード配列)制御配列をはじめとする、特定のタンパク質を発現する核酸断片を指す。「天然遺伝子」とは、自然界にそれ自体の制御配列と共に見られる遺伝子を指す。「キメラ遺伝子」とは、自然界に共に見られない制御およびコード配列を含んでなる天然遺伝子でないあらゆる遺伝子を指す。したがってキメラ遺伝子は、異なる供給源から誘導される制御配列およびコード配列、あるいは同一供給源から誘導されるが、自然界に見られるのとは異なるやり方で配列される制御配列およびコード配列を含んでなってもよい。「内因性の遺伝子」とは、生物体ゲノムにおいてその天然位置にある天然遺伝子を指す。「外来性の」遺伝子とは、宿主生物体において常態では見られないが、遺伝子移入によって宿主生物体に導入される遺伝子を指す。外来性の遺伝子は、非天然生物体中に挿入された天然遺伝子、あるいはキメラ遺伝子を含んでなることができる。「導入遺伝子」とは、形質転換によってゲノム中に導入される遺伝子である。「コドン最適化遺伝子」とは、そのコドン使用頻度が宿主細胞の好むコドン使用頻度を模倣するようにデザインされる遺伝子である。
【0075】
「コード配列」とは、特定のアミノ酸配列をコードするDNA配列を指す。「適切な調節配列」とは、コード配列の上流(5’非コード配列)、配列内、または下流(3’非コード配列)に位置して、転写、RNAプロセシングまたは安定性、または関連コード配列の翻訳に影響を及ぼすヌクレオチド配列を指す。調節配列は、プロモーター、翻訳リーダー配列、イントロン、ポリアデニル化認識配列、RNAプロセッシング部位、エフェクター結合部位、およびステム−ループ構造を含んでもよい。
【0076】
「プロモーター」とは、コード配列または機能RNAの発現を調節できるDNA配列を指す。概してコード配列は、プロモーター配列に対して3’に位置する。プロモーターは、そっくりそのまま天然遺伝子から誘導されてもよく、あるいは自然界に見られる異なるプロモーターから誘導される異なる要素からなってもよく、あるいは合成DNAセグメントを含んでなってさえよい。異なるプロモーターは、異なる組織または細胞タイプ中で、あるいは異なる発達段階において、あるいは異なる環境または生理学的条件に呼応して、
遺伝子の発現を導いてもよいことが当業者には理解される。ほとんどの細胞タイプ中でほとんどの場合に遺伝子の発現を引き起こすプロモーターは、一般に「構成プロモーター」と称される。ほとんどの場合、制御配列のはっきりした境界は完全に限定されていないので、異なる長さのDNA断片が同一のプロモーター活性を有してもよいこともさらに認識される。
【0077】
「3’非翻訳配列」または「転写ターミネーター」と言う用語は、コード配列の下流に位置するDNA配列を指す。これには、mRNAプロセッシングまたは遺伝子発現に影響できる調節シグナルをコードするポリアデニル化認識配列およびその他の配列が含まれる。ポリアデニル化シグナルは、通常、mRNA前駆物質の3’末端へのポリアデニル酸トラクトの付加に影響することで特性決定される。3’領域は、転写、RNAプロセッシングまたは安定性、または関連コード配列の翻訳に影響できる。
【0078】
「RNA転写物」とは、DNA配列のRNAポリメラーゼが触媒する転写から得られる生産物を指す。RNA転写物がDNA配列の完全な相補的コピーである場合、それは一次転写物と称され、あるいはそれは一次転写物の転写後プロセッシングから誘導されるRNA配列であるかもしれず、成熟RNAと称される。「メッセンジャーRNA」または「mRNA」とはイントロンがなく、細胞によってタンパク質に翻訳されることができるRNAを指す。「cDNA」とは、mRNAに対して相補的であり、それから誘導される二重鎖DNAを指す。「センスRNA」とは、mRNAを含み、細胞によってタンパク質に翻訳されることができるRNA転写物を指す。「アンチセンスRNA」とは、標的一次転写物またはmRNAの全部または一部に相補的であり、標的遺伝子の発現をブロックするRNA転写物を指す(米国特許第5,170,065号明細書、国際公開第99/28508号パンフレット)。アンチセンスRNAの相補性は、特定の遺伝子転写物のあらゆる部分、すなわち5’非コード配列、3’非コード配列、またはコード配列にあっても良い。「機能RNA」とは、翻訳されないがそれでもなお細胞過程に影響するアンチセンスRNA、リボザイムRNA、またはその他のRNAを指す。
【0079】
「作動可能に連結した」と言う用語は、1つの機能が他方の機能によって影響される、単一核酸断片上の核酸配列のつながりを指す。例えばプロモーターはコード配列の発現に影響できる場合、そのコード配列と作動可能に連結する(すなわちコード配列がプロモーターの転写調節の下にある)。コード配列は、センスまたはアンチセンスオリエンテーションで制御配列に作動可能に連結できる。
【0080】
「発現」と言う用語は、ここでの用法では、本発明の核酸断片から誘導されるセンス(mRNA)またはアンチセンスRNAの転写および安定した蓄積を指す。発現はまた、mRNAのポリペプチドへの翻訳を指してもよい。
【0081】
「成熟した」タンパク質とは、翻訳後にプロセスされたポリペプチド、すなわちそれから一次翻訳産物中に存在するあらゆるプレ−またはプロペプチドが除去されたものを指す。「前駆」タンパク質とは、mRNAの一次翻訳産物、すなわちプレ−およびプロペプチドが未だに存在するものを指す。プレ−およびプロペプチドは細胞内局在化シグナルであってもよい(がそれに限定されない)。
【0082】
「形質転換」とは、遺伝的に安定した遺伝形質をもたらす、宿主生物体中への核酸分子の転移を指す。核酸分子は、例えば自律的に複製するプラスミドであってもよく、またはそれは宿主生物体のゲノム中に組み込まれてもよい。形質転換核酸断片を含有する宿主生物体は、「遺伝子組換え」または「組換え」または「形質転換」生物体と称される。
【0083】
「プラスミド」、「ベクター」、および「カセット」と言う用語は、細胞の中心的代謝
の一部ではない遺伝子を運ぶことが多く、通常環状二本鎖DNA断片の形態である染色体外因子を指す。このような因子は、あらゆる供給源から誘導される一本鎖または二本鎖DNAまたはRNAの配列、ゲノム一体化配列、直鎖または環状のファージまたはヌクレオチド配列を自律的に複製するかもしれず、そこではいくつかのヌクレオチド配列が独自の構成に連結または組み換えされ、それは選択された遺伝子産物のために、適切な3’非翻訳配列と共にプロモーター断片およびDNA配列を細胞中に導入することができる。
【0084】
「形質転換カセット」とは、外来性遺伝子を含有し、外来遺伝子に加えて特定の宿主細胞の形質転換を容易にする因子を有する特定のベクターを指す。「発現カセット」とは、外来性遺伝子を含有し、外来性遺伝子に加えて外来性宿主におけるその遺伝子の促進された発現を可能にする因子を有する特定のベクターを指す。
【0085】
「改変された生物学的活性」と言う用語は、アッセイ法によって測定できるヌクレオチド配列によってコードされるタンパク質に関連した活性を指し、そこでは活性は、未変性配列に関連した活性を超えるか、またはそれ未満である。「増強された生物学的活性」とは、未変性配列に結びついたものを超える改変された活性を指す。「低下した生物学的活性」とは、未変性配列に結びついたもの未満の改変された活性を指す。
【0086】
「配列分析ソフトウェア」と言う用語は、ヌクレオチドまたはアミノ酸配列の分析のために有用なあらゆるコンピュータアルゴリズムまたはソフトウェアプログラムを指す。「配列分析ソフトウェア」は、市販のものでも、あるいは独立して開発されても良い。典型的な配列分析ソフトウェアとしては、1.)ウィスコンシン州マディソンのジェネティック・コンピュータ・グループからのGCGパッケージプログラム(Wisconsin Package Version 9.0、Genetics Computer Group(GCG),Madison,WI)、2.)BLASTP、BLASTN、BLASTX(アルシュール(Altschul)ら著、J.Mol.Biol.215:403〜410(1990))、および3.)ウィスコンシン州マディソンのDNASTAR(DNASTAR,Inc.Madison、WI)、および4.スミス−ウォーターマン(Smith−Waterman)・アルゴリズムを組み入れたFASTAプログラム(W.R.ピアソン(Pearson)著、Comput.Methods Genome Res.[Proc.Int.Symp.](1994)、1992年会議、111〜20、編集者:スハイ,サンドール(Suhai,Sandor)、プレナム(Plenum)、ニューヨーク州ニューヨーク(New York、NY))が挙げられるが、これに限定されるものではない。本願明細書の文脈内では、配列分析ソフトウェアを分析のために使用する場合、分析結果は特に断りのない限り、言及されるプログラムの「デフォルト値」に基づくものと理解される。ここでの用法では、「デフォルト値」とは、最初に初期化されたときにソフトウェアに元々ロードされた、あらゆる値またはパラメータの組を意味する。
【0087】
ここで使用される標準組換えDNAおよび分子クローニング技術は技術分野で良く知られており、サムブルック(Sambrook)J.、フリッチュ(Fritsch)E.F.、およびマニアティス(Maniatis)T.「分子クローニング:実験室マニュアル(Molecular Cloning:A Laboratory Manual)」第二版、コールドスプリングハーバーラボラトリ(Cold Spring Harbor Laboratory)、ニューヨーク州コールドスプリングハーバー(Cold Spring Harbor、NY)(1989)(以下マニアティス(Maniatis));シルハビー(Silhavy)T.J.、ベンナン(Bennan)M.L.およびエンクイスト(Enquist)L.W.「遺伝子融合実験(Experiments with Gene Fusions)」コールドスプリングハーバーラボラトリ:ニューヨーク州コールドスプリングハーバー(Cold Spring Harbo
r、NY)(1984);およびオースベル(Ausubel)F.M.ら「分子生物学現代プロトコル(Current Protocols in Molecular Biology)」グリーンパブリッシングアッソシエーツ(Greene Publishing Assoc.)およびワイリーインターサイエンス(Wiley−Interscience)による出版(1987)で述べられている。
【0088】
脂肪酸の微生物生合成
一般に油性微生物中の脂質の蓄積は、増殖培地中に存在する全体的な炭素と窒素の比率に応答してトリガーされる。細胞が利用できる窒素供給を消耗した場合(例えば炭素と窒素の比率が約40を超える場合)、細胞アデノシン一リン酸(AMP)の枯渇は、ミトコンドリア中のAMP−依存イソクエン酸デヒドロゲナーゼ活性の休止、およびクエン酸の蓄積、クエン酸の細胞質ゾル内への輸送、そして引き続くATP−クエン酸リアーゼによるクエン酸の切断をもたらして、アセチル−CoAが生産する。アセチル−CoAは、脂肪酸の新規(de novo)生合成の主要(principle)構成ブロックである。効果的に代謝されてアセチル−CoAを生産できるあらゆる化合物が、脂肪酸前駆物質の役割を果たせるが、このタイプの反応ではグルコースが炭素の主な供給源である。グルコースは解糖を通じてピルビン酸に変換され、次にピルビン酸はミトコンドリア中に輸送され、そこでピルビン酸デヒドロゲナーゼによってアセチル−CoAに変換される。アセチル−CoAはミトコンドリア膜を越えて細胞質中に直接輸送できないので、アセチル−CoAからの2個の炭素がオキサロ酢酸と縮合して、クエン酸を生産する(クエン酸合成酵素によって触媒される)。クエン酸は細胞質内に直接輸送されて、そこでにATP−クエン酸リアーゼによって切断され、アセチル−CoAおよびオキサロ酢酸が再生する。オキサロ酢酸は、リンゴ酸への転換を通じて、トリカルボン酸サイクルに再び入る。
【0089】
マロニル−CoAの合成は、細胞質内で起きる脂肪酸生合成の第1の前駆ステップである。マロニル−CoAは、アセチル−CoAカルボキシラーゼによって、アセチル−CoAのカルボキシル化を通じて生産される。脂肪酸合成は、多酵素脂肪酸合成酵素複合体(「FAS」)によって触媒され、8個の二炭素断片(アセチル−CoAからのアセチル基)の縮合によって起き、炭素16個の飽和脂肪酸であるパルミチン酸が形成する。より具体的には、FASは以下が関与する一連の7つの反応を触媒する(スミス(Smith)S.、FASEB J.、8(15):1248〜59(1994))。
1.アセチル−CoAおよびマロニル−CoAがFASのアシルキャリアペプチド(ACP)に転移される。次にアセチル基がマロニル基に転移されてβ−ケトブチリル−ACPが形成し、CO2を放出する。
2.β−ケトブチリル−ACPが還元(β−ケトアシル還元酵素による)および脱水(β−ヒドロキシアシルデヒドラターゼによる)を被り、トランス−単不飽和脂肪アシル基が形成する。
3.二重結合がNADPHによって還元され、最初のものよりも炭素が2個分長い飽和脂肪−アシル基が生じる。次に新しいマロニル基と縮合して延長プロセスを繰り返すブチリル基の能力が再生する。
4.脂肪アシル基が炭素16個の長さになったら、チオエステラーゼ活性がそれを加水分解して遊離パルミチン酸を放出する。
【0090】
パルミチン酸(16:0)は、小胞体膜に存在する鎖長延長酵素およびデサチュラーゼの作用を通じた、より鎖長が長い飽和および不飽和脂肪酸(例えばステアリン酸(18:0)、パルミトレイン酸(16:1)、およびオレイン酸(18:1))の前駆物質である。パルミチン酸およびステアリン酸は、Δ9デサチュラーゼの作用によってそれらの不飽和(unsatuared)誘導体であるパルミトレイン酸(16:1)およびオレイン酸(18:1)酸にそれぞれ変換する。
【0091】
トリアシルグリセロール(脂肪酸の主な貯蔵単位)は、リン酸1,2−ジアシルグリセロール(一般にホスファチジン酸として同定される)を生じる、アシル−CoAの2個の分子からグリセロール−3−リン酸へのエステル化によって形成される。次にホスファチジン酸ホスファターゼによってリン酸が除去され、1,2−ジアシルグリセロールが生産する。第3の脂肪酸を添加すると、ジアシルグリセロール−アシルトランスフェラーゼの作用によってトリアシルグリセロールが形成する。
【0092】
Ω脂肪酸の生合成
非常に単純化すると、LAをGLA、DGLA、およびARA(ω−6経路)に、ALAをSTA、ETA、EPA、およびDHA(ω−3経路)に変換する代謝プロセスは、炭素原子の付加を通じた炭素鎖の延長と、二重結合の添加を通じた分子の不飽和化を伴う(図1)。これは小胞体膜に存在する一連の特別な不飽和化および延長酵素を必要とする。
【0093】
ω−6脂肪酸
オレイン酸は、Δ12デサチュラーゼの作用によって、最初のω−6脂肪酸であるLA(18:2)に変換される。引き続くω−6脂肪酸は、次のようにして生産される。1.)LAがΔ6デサチュラーゼ活性によってGLAに変換され、2.)GLAが鎖長延長酵素の作用によってDGLAに変換され、3.)DGLAがΔ5デサチュラーゼの作用によってARAに変換される。
【0094】
ω−3脂肪酸
リノール酸(LA)は、Δ15デサチュラーゼの作用によって、最初のω−3脂肪酸であるALAに変換される。引き続くω−3脂肪酸は、ω−6脂肪酸と類似した一連のステップで生産される。具体的には、1.)ALAがΔ6デサチュラーゼ活性によってSTAに変換され、2.)STAが鎖長延長酵素活性によってETAに変換され、3.)ETAがΔ5デサチュラーゼ活性によってEPAに変換される。代案としては、ETAおよびEPAは、Δ17デサチュラーゼ活性によって、DGLAおよびARAからそれぞれ生産できる。EPAは、鎖長延長酵素およびΔ4デサチュラーゼ活性によって、DHAにさらに変換できる。
【0095】
Ω脂肪酸生産に関与する遺伝子
藻類、細菌、カビおよび酵母菌をはじめとする多くの微生物は、通常の細胞代謝経路内でPUFAおよびω脂肪酸を合成できる。特によく適するのは、ヤブレツボカビ(Thraustochytrium)属の種であるスキゾキトリウム・アグレガトム(Schizochytrium aggregatm)、およびモルティエレラ・アルピナ(Morteriella alpina)をはじめとする菌・カビ類である。さらに多くの渦鞭毛藻類(渦鞭毛藻綱(Dinophyceaae))は、高濃度のPUFAを自然に生産する。したがって油生産に関与する多様な遺伝子が遺伝的手段を通じて同定されており、これらのいくつかの遺伝子のDNA配列は公的に入手できる(制限を意図しない例を下の表2に示す)。
【0096】
【表2】
【0097】
【表3】
【0098】
さらに特許文献は、PUFA生産に関与する多くの追加的な遺伝子のDNA配列(および/または上の遺伝子のいくつかに関する詳細およびそれらの単離方法)を提供する。例えば米国特許第5,968,809号明細書(Δ6デサチュラーゼ)、米国特許出願公開第2003/0196217 A1号明細書(Δ17デサチュラーゼ)、米国特許第5,972,664号明細書および米国特許第6,075,183号明細書(Δ5デサチュラーゼ)、国際公開第91/13972号パンフレットおよび米国特許第5,057,419号明細書(Δ9デサチュラーゼ)、国際公開第93/11245号パンフレット(Δ15デサチュラーゼ)、国際公開第94/11516号パンフレット、米国特許第5,443,974号明細書および国際公開第03/099216号パンフレット(Δ12デサチュラーゼ)、国際公開第02/090493号パンフレット(Δ4デサチュラーゼ)、および国際公開第00/12720号パンフレットおよび米国特許出願公開第2002/0139974 A1号明細書(鎖長延長酵素)を参照されたい。これらの各特許および出願は、その内容全体を参照によってここに援用したものとする。
【0099】
当業者には明らかなように、特定のPUFA最終生産物の生産のために宿主生物体に導入する必要がある特定機能は、宿主細胞(およびその未変性PUFAプロフィールおよび/またはデサチュラーゼプロフィール)、基質の入手可能性および所望の最終生産物に左右される。同時係属の米国仮特許出願第60/467677号(その内容全体を本願明細書に引用したものとする)で述べられ、図1に示すように、LA、GLA、DGLA、A
RA、ALA、STA、ETA、EPA、DPA、およびDHAは、以下のPUFA酵素機能の様々な組み合わせを導入することで、全て油性酵母菌中で生産されてもよい。Δ4デサチュラーゼ、Δ5デサチュラーゼ、Δ6デサチュラーゼ、Δ12デサチュラーゼ、Δ15デサチュラーゼ、Δ17デサチュラーゼ、Δ9デサチュラーゼ、および/または鎖長延長酵素。当業者は、公的に入手できる文献(例えばジェンバンク(GenBank))、特許文献、およびPUFAを製造する能力を有する微生物の実験的分析に従って、上の各酵素をコードする様々な候補遺伝子を同定できるであろう。配列は、例えば天然供給源などから(細菌、藻類、菌・カビ類、植物、動物などから)単離され、半合成または新規(de novo)合成経路を通じて生産される、あらゆる供給源に由来してもよい。しかし当業者には明らかなように、異種性遺伝子は別の宿主中では様々な効率で発現する。したがってω−3および/またはω−6PUFAの生産は、異種性宿主における発現レベルが、関心のある宿主生物体における別のデサチュラーゼまたは鎖長延長酵素の発現に比べて好ましい、特定のデサチュラーゼまたは鎖長延長酵素の選択によって最適化されてもよい。
【0100】
宿主に導入されるデサチュラーゼおよび鎖長延長酵素遺伝子の特定の供給源は重要でないが、デサチュラーゼまたは鎖長延長酵素活性を有する特定のポリペプチドを選択する上での考慮としては、次が挙げられる。1.)ポリペプチドの基質特異性、2.)ポリペプチドまたはその構成要素が律速酵素であるかどうか、3.)デサチュラーゼまたは鎖長延長酵素が所望のPUFAの合成のために必須であるかどうか、および/または4.)ポリペプチドによって必要とされる補助因子。発現するポリペプチドは、好ましくは宿主細胞内のその位置の生化学的環境と適合性のパラメーターを有する。例えばポリペプチドは、宿主細胞内で基質獲得のためにその他の酵素と競争しなくてはならないかもしれない。したがってポリペプチドのKMおよび特異的活性分析は、特定の宿主細胞内でPUFA生産を修正するために、特定のポリペプチドの適性を判定する上で考慮される。特定の宿主細胞で使用されるポリペプチドは、意図される宿主細胞中に存在する生化学的条件下で機能できるものであるが、その他の点では所望のPUFAを修正できる、デサチュラーゼまたは鎖長延長酵素活性を有するあらゆるポリペプチドであることができる。
【0101】
しかし究極の目的がω−3および/またはω−6脂肪酸が濃縮された油を蓄積する油性酵母菌の開発である、ここでの研究目的のためには、油性酵母菌中において比較的効率的に機能する、デサチュラーゼおよび鎖長延長酵素活性を有するポリペプチドを同定することが望ましかった。したがって、様々なデサチュラーゼおよび鎖長延長酵素が油性酵母菌中において発現され、基質供給試験においてω−3および/またはω−6脂肪酸を生産するそれらの能力についてスクリーンされた。ひとたび適切な酵素が同定されると(全体的%基質変換に基づいて)、次にこれらの遺伝子に後述するコドン最適化技術を施して、別の油性酵母菌宿主における各酵素の発現をさらに最適化した。これによってω−3および/またはω−6脂肪酸の最大の生産が可能になった。
【0102】
当業者は、ここでコドン最適化のために選択される特異的PUFA遺伝子が単に例示的であり、ここで本発明を制限することを意図しないことを理解するであろう。様々な供給源からの多数のその他の異種性デサチュラーゼ(Δ4、Δ5、Δ6、Δ9、Δ12、Δ15、および/またはΔ17デサチュラーゼ活性を有する)および鎖長延長酵素は、コドン最適化して油性酵母菌宿主におけるそれらの発現を改善できる。
【0103】
油性酵母菌における発現のためのΩ脂肪酸遺伝子コドンの最適化
当業者にはよく知られているように、宿主が好むコドンの使用は、ポリペプチドをコードする外来遺伝子の発現を実質的に増強できる。一般に宿主が好むコドンは、タンパク質中のコドン使用を調べてどのコドンが最高頻度で使用されるか判定することで、関心のある特定の宿主種(好ましくは最大量を発現するもの)中で判定できる。次に宿主種で好ま
しいコドンを使用して、デサチュラーゼまたは鎖長延長酵素活性を有する関心のあるポリペプチドのためのコード配列の全体または一部が合成できる。したがって関心のある特定の宿主生物体における発現のために遺伝子を最適化する方法は、概して
a)関心のある特定の宿主生物体についてヌクレオチドコード領域および対応するポリペプチドの配列を得て、コドンのデータベースを形成し、
b)コドンのデータベースを分析し、いずれのコドンが各アミノ酸を優先的にコードするのか判定し、
c)関心のある特定の宿主生物体中で発現させる遺伝子の配列(例えばデサチュラーゼまたは鎖長延長酵素)を得て、
d)ステップ(c)の配列中の好ましくないコドンをステップ(b)の好ましいコドンで置き換えて、遺伝子が関心のある特定の宿主生物体における発現のためにコドン最適化されることを必要とする。
【0104】
またDNAの全部(または一部)を合成して、転写mRNA中に存在する、二次構造のあらゆる不安定化配列または領域を除去できる。またDNAの全部(または一部)を合成して、塩基組成物を所望の宿主細胞中でより好ましいものに変更できる。
【0105】
本発明の目的のためには、修正されたポリペプチドが、別の宿主(すなわち油性酵母菌)によって好ましいコドンを使用するように、外来宿主中で発現させるPUFAデサチュラーゼおよび鎖長延長酵素活性を有する特定のポリペプチドをコードするコドンの一部を修正することが望ましかった。具体的にはΔ17デサチュラーゼ活性、Δ6デサチュラーゼ活性、および鎖長延長酵素活性を有するポリペプチドをコードするコドンの一部を修正して、ヤロウィア・リポリティカ(Yarrowia lipolytica)における遺伝子の発現を増強することが望ましかった。したがってY.リポリティカ(Y.lipolytica)コドン使用プロフィールは表4(実施例3)に示すように決定された。さらに翻訳開始コドン「ATG」を取り囲むヌクレオチド配列は、酵母菌細胞における発現に影響することが分かっている。酵母菌におけるポリペプチドの発現が不良であれば、外来性遺伝子のヌクレオチド配列を修正し、効率的な酵母菌翻訳開始配列を含めて、最適な遺伝子発現を得ることができる。したがってY.リポリティカ(Y.lipolytica)における遺伝子発現をさらに最適化するために、「ATG」開始コドン周囲の共通配列もまた決定された(図4、配列番号122)。
【0106】
Y.リポリティカ(Y.lipolytica)コドン使用プロフィールおよび「ATG」開始コドン周囲の共通配列に基づいて、Δ17デサチュラーゼ遺伝子、Δ6デサチュラーゼ遺伝子、および鎖長延長酵素遺伝子の核酸を配列を修正して、宿主が好むコドンを用いた。より具体的には、モルティエレラ・アルピナ(Mortierella alpina)(ATCC番号16266)Δ6デサチュラーゼ(ジェンバンク(GenBank)登録番号AF465281、米国特許第5,968,809号明細書)を選択して、LAをGLAに、および/またはALAをSTAに変換する酵素的能力を導入した。この野生型デサチュラーゼは、457個のアミノ酸(配列番号2)および51.8kDの予想分子量を有した。ここで作り出したコドン最適化遺伝子では、1374bpのコード領域(144個のコドンに対応する)の152bpがコドン最適化され、翻訳開始部位が修正された。同様にして野生型サプロレグニア・ディクリナ(Saprolegnia diclina)(ATCC番号56851)Δ17デサチュラーゼ(配列番号4、米国特許出願公開2003/0196217A1号明細書)を選択して、DGLAをETAに、および/またはARAをEPAに変換する酵素的能力を導入したが、翻訳開始部位は修正され、1077bpコード領域(117個のコドンを含んでなる)の127bpがコドン最適化された。最後にGLAからDGLA、STAからETA、および/またはEPAからDPAへの転換のため、油性酵母菌におけるその発現の最適化のために、318個のアミノ酸(配列番号6)および40.5kDの予想分子量を有する野生型M.アルピナ(M.
alpina)高親和力PUFA鎖長延長酵素(ジェンバンク(GenBank)登録番号AX464731、国際公開第00/12720号パンフレット)を選択した。具体的には957bpのコード領域(85個のコドンに対応する)の94bpをコドン最適化し、翻訳開始部位を修正した。したがって本発明は、添付の配列表で報告するような合成コドン最適化遺伝子の完全な配列、これらの完全な配列の相補的配列、およびこれらの配列のかなりの部分を含んでなる。
【0107】
当業者は、ここで述べる最適化方法がその他の遺伝子(例えばω−3/ω−6脂肪酸生合成経路中の遺伝子)に等しく適用でき、S.ディクリナ(S.diclina)Δ17デサチュラーゼ、およびM.アルピナ(M.alpina)Δ6デサチュラーゼ、およびM.アルピナ(M.alpina)鎖長延長酵素の調節が、単に例示的であることを理解するであろう。
【0108】
遺伝子変異を通じたコドン最適化遺伝子の最適化
コドン最適化は、油性酵母菌における機能のためのより望ましい物理的および動力学的パラメーターがあるデサチュラーゼまたは鎖長延長酵素活性をそれぞれ有する、ポリペプチドを生体内(in vivo)で生産する有用な手段であるが、宿主細胞におけるポリペプチドの活性をさらに増強する追加的手段が利用できる。具体的には配列を合成する方法および配列を一緒にする方法は、文献においてよく確立している。例えば生体外(in
vitro)変異誘発および選択、部位特異的変異誘発、誤りがちなPCR(メルニコフ(Melnikov)ら、Nucleic Acids Research、27(4):1056〜1062(1999年2月15日))、「遺伝子シャフリング」またはその他の手段を用いて、自然発生的またはコドン最適化デサチュラーゼまたは鎖長延長酵素遺伝子の変異を得ることができる。これによって例えば生体内(in vivo)で所望のPUFAの半減期がより長く、または生産速度がより早い、デサチュラーゼまたは鎖長延長酵素ポリペプチドそれぞれの生産が可能になる。
【0109】
必要に応じて、酵素活性に重要なコドン最適化デサチュラーゼまたは鎖長延長酵素ポリペプチドの領域は、ルーチンの変異誘発、得られる変異体ポリペプチドの発現、およびそれらの活性の判定を通じて判定できる。変異体は、欠失、挿入、および点変異、またはそれらの組み合わせを含んでもよい。典型的な機能分析は、欠失変異誘発に始まって機能に必要なタンパク質のN−およびC−末端限界を判定し、次に内部欠損、挿入または点変異体を作り出し、機能に必要な領域をさらに判定する。カセット変異誘発または完全合成などのその他の技術もまた使用できる。欠失変異誘発は例えば、エキソヌクレアーゼを使用して、5’または3’コード領域を逐次除去して達成される。このような技術のためのキットが利用できる。欠失後、開始または停止コドンを含有するオリゴヌクレオチドをそれぞれ5’または3’欠失後に、欠失コード領域にライゲーションして、コード領域が完成する。代案としては、部位特異的変異誘発、変異原性PCRをはじめとする多様な方法によって、または既存の制限部位において消化されたDNA上へのライゲーションによって、開始または停止コドンをコードするオリゴヌクレオチドをコード領域に挿入する。内部欠損は、部位特異的変異誘発または変異原性PCRを通じた変異原性プライマーの使用によって、DNA中の既存の制限部位の使用をはじめとする多様な方法を通じて同様に作り出せる。挿入は、リンカー−スキャニング変異誘発、部位特異的変異誘発または変異原性PCRなどの方法を通じて作り出される。点変異は、部位特異的変異誘発または変異原性PCRなどの技術を通じて作り出される。
【0110】
化学変異誘発もまた、活性に重要なデサチュラーゼまたは鎖長延長酵素ポリペプチドの領域を同定するために使用できる。変異したコンストラクトが発現し、得られる改変タンパク質がデサチュラーゼまたは鎖長延長酵素として機能する能力がアッセイされる。このような構造−機能分析は、どの領域が欠失してもよいか、どの領域が挿入を許容するか、
どの点変異によって変異体タンパク質が、未変性(またはコドン最適化)デサチュラーゼまたは未変性鎖長延長酵素と実質的に同じように機能できるようにするかを判定できる。
【0111】
ω−3および/またはω−6脂肪酸の微生物による生産
微生物によるω−3および/またはω−6脂肪酸の生産には、魚または植物などの天然供給源からの精製に比べて、例えば以下のようないくつかの利点がある。
1.)多くの微生物は、油組成が高等生物のものと比べてはるかに単純であることが知られており、所望の構成要素の精製を容易にする。
2.)微生物による生産には、天候および食物供給などの外部変数によって引き起こされるばらつきがない。
3.)微生物的に生産された油は、実質的に環境汚染物質による混入物がない。
4.)微生物は特定用途を有するかもしれない特定の形態で、PUFAを提供できる。
5.)微生物による油生産は、培養条件を制御することで、特に微生物的に発現される酵素のために特定の基質を提供することで、または化合物の添加または遺伝子工学アプローチによって望まれない生化学的経路を抑制することで操作できる。
【0112】
これらの利点に加えて、組換え微生物からのω−3および/またはω−6脂肪酸の生産は、宿主中に新しい生合成経路を提供することで、または望まれない経路を抑制することで、所望のPUFAまたはそれらのコンジュゲートされた形態のレベルを増大させて、望まれないPUFAのレベルを低下させ、自然発生的な微生物の脂肪酸プロフィールを変更する能力を提供する。例えばこのようにして生産したω−3とω−6脂肪酸の比率を修正する、他のω脂肪酸生産を排除しながらω−3またはω−6脂肪酸のどちらか一方だけを生産する、または他のPUFAの下流または上流生産物の顕著な蓄積なしに特異的PUFAの生産を操作することが可能である。
【0113】
発現システム、カセット、およびベクター
ここで述べるコドン最適化遺伝子および関心のある配列の遺伝子生産物は、異種性微生物の宿主細胞、特に油性酵母菌(例えばヤロウィア・リポリティカ(Yarrowia lipolytica))の細胞中で生産されてもよい。組換え微生物宿主における発現は、様々なPUFA経路中間体を生産するため、またはこれまで宿主を使用してできなかった新しい生産物を合成するため、宿主に既存のPUFA経路を調節するのに有用かもしれない。
【0114】
微生物の発現システム、および外来タンパク質の高レベル発現を導く調節配列を含有する発現ベクターは、当業者によく知られている。これらのいずれも本コドン最適化配列のあらゆる遺伝子生産物を生産するためのキメラ遺伝子を構築するのに使用できる。次に形質転換を通じてこれらのキメラ遺伝子を適切な微生物に導入して、コードされた酵素の高レベル発現を提供できる。
【0115】
したがって適切なプロモーター制御下における、PUFA生合成経路(例えばここで述べるコドン最適化Δ6デサチュラーゼ、Δ17デサチュラーゼ、および鎖長延長酵素)をコードするキメラ遺伝子の導入は、ω−3および/またはω−6脂肪酸の増大する生産をもたらすことが予期される。本コドン最適化遺伝子の様々な組み合わせを宿主微生物中で一緒に発現することが有用であることが考察される。
【0116】
さらにベクターがまた、ここで述べるコドン最適化遺伝子の1つもしくはそれ以上に加えて、その他の酵素をコードする1つもしくはそれ以上の遺伝子を含んでなってもよいことが考察される。例えばΔ4デサチュラーゼ、Δ5デサチュラーゼ、Δ6デサチュラーゼ、Δ12デサチュラーゼ、Δ15デサチュラーゼ、Δ17デサチュラーゼ、Δ9デサチュラーゼ、および/または鎖長延長酵素の1つもしくはそれ以上の酵素活性をコードする遺
伝子(それによってこれらのあらゆる遺伝子が、場合により特定の宿主生物体での増強された発現のためにコドン最適化される)を含んでなる発現カセットを構築することが望ましいかもしれない。当業者にはよく知られているように、特定の発現カセット内に含まれる特定の遺伝子は、宿主細胞(おそびそのPUFAプロフィールおよび/またはデサチュラーゼプロフィール)、基質の入手可能性、および所望の最終生産物に左右される。
【0117】
したがって本発明は、基質が所望の脂肪酸生産物に変換するように、脂肪酸基質をここで述べるPUFA酵素に暴露することを含んでなる、PUFAを生産する方法を包含する。したがってここで述べる各PUFA遺伝子および対応する酵素生産物は、PUFAの生産のために直接にまたは間接に使用できる。PUFAの直接的生産は、脂肪酸基質が中間ステップまたは経路中間体なしに直接に所望の脂肪酸生産物に変換する場合に起きる。例えばEPAの生産は、Δ17デサチュラーゼ活性を提供する発現カセットを細胞に添加または導入することで、ARAを生産する宿主細胞、またはARAが提供される宿主細胞中で起きる。
【0118】
対照的にPUFA生合成経路をコードする複数遺伝子を一連の反応が起きて所望のPUFAを生産するように組み合わせて使用してもよい。例えば鎖長延長酵素、Δ5デサチュラーゼ、Δ17デサチュラーゼ、およびΔ4デサチュラーゼ活性(そこでは各遺伝子が場合により宿主における発現のためにコドン最適化される)をコードする発現カセットは、自然にGLAを生産する宿主細胞が、代わりにDHAを生産できるようにする(GLAが鎖長延長酵素によってDGLAに変換し、次にDGLAはΔ5デサチュラーゼによってARAに変換し、次にARAはΔ17デサチュラーゼによってEPAに変換し、それは次には鎖長延長酵素によってDPAに変換してもよく、DPAはΔ4デサチュラーゼによってDHAに変換する)。宿主細胞が油性酵母菌である好ましい実施態様では、これらの生物体において自然に生産されるPUFAは18:2脂肪酸(すなわちLA)、もっとまれには18:3脂肪酸(すなわちALA)に限られるので、PUFA生合成に必要な各酵素をコードする発現カセットを生物体に導入しなくてはならない。代案としては基質供給が必要かもしれない。
【0119】
適切な宿主細胞の形質転換に有用なベクターまたはDNAカセットは、技術分野でよく知られている。コンストラクト中に存在する配列の特定の選択は、所望の発現生産物(上述)、宿主細胞の性質、および提案される形質転換細胞と非形質転換細胞とを分離する手段に左右される。しかし典型的に、ベクターまたはカセットは、関連遺伝子の転写および翻訳を導く配列、選択性マーカー、および自律性複製または染色体組み込みを可能にする配列を含有する。適切なベクターは、転写開始を制御する遺伝子の5’領域、および転写終結を制御するDNA断片の3’領域を含んでなる。双方の制御領域が、形質転換された宿主細胞の遺伝子に由来することが最も好ましいが、このような制御領域は、必ずしも生産宿主として選択された特定種に天然の遺伝子に由来しなくてよいものと理解される。
【0120】
所望の宿主細胞中で、デサチュラーゼおよび/または鎖長延長酵素ORFの発現を推進するのに有用な開始制御領域またはプロモーターは多数あり、当業者にはなじみが深い。選択された宿主細胞中でのこれらの遺伝子の発現を導くことができる、実質的にあらゆるプロモーターが本発明に適する。宿主細胞中での発現は、一過性または安定様式で達成できる。一過性発現は、関心のある遺伝子に作動可能に連結された、調節可能プロモーターの活性を誘導することで達成できる。安定発現は、関心のある遺伝子に作動可能に連結された構成プロモーターの使用によって達成できる。一例として宿主細胞が酵母菌の場合、酵母菌細胞中で機能する転写および翻訳領域が、特に宿主種から提供される。転写開始調節領域は、例えば、以下から得ることができる。1.)アルコールデヒドロゲナーゼ、グリセルアルデヒド3−リン酸−デヒドロゲナーゼ(米国特許出願第60/482263号明細書参照)、ホスホグリセリン酸ムターゼ(米国特許出願第60/482263号明細
書参照)、フルクトース−ビスリン染色体外因子アルドラーゼ(米国特許出願第60/519971号明細書参照)、ホスホグルコース−異性化酵素、ホスホグリセラートキナーゼなどの解糖経路中の遺伝子、または2.)酸ホスファターゼ、ラクターゼ、メタロチオネイン、グルコアミラーゼ、翻訳延長因子EF1−α(TEF)タンパク質(米国特許第6,265,185号明細書)、リボソームタンパク質S7(米国特許第6,265,185号明細書)などの調節可能遺伝子。構成または誘導転写が所望されるかどうか、関心のあるORFを発現する上でのプロモーター効率、構築の容易さなど次第で、いくつかの調節配列のいずれの1つでも使用できる。
【0121】
酵母菌での最適遺伝子発現は、それらが効率的な酵母菌翻訳開始列を含むように、外来性遺伝子の翻訳開始コドン「ATG」を取り囲むヌクレオチド配列を修正することで得ることができる。具体的には非効率的に発現する遺伝子の発現は部位特異的変異誘発によって増大でき、そこでは非効率的に発現する遺伝子が、好ましくは高度に発現する遺伝子である内因性酵母菌遺伝子にインフレーム融合する。代案としてはここでの発明でヤロウィア・リポリティカ(Yarrowia lipolytica)において実証されるように、宿主におけるコンセンサス翻訳開始配列を判定して、関心のある宿主でのそれらの最適発現のために、この配列を異種性遺伝子内に遺伝子操作できる。
【0122】
終結領域は、遺伝子の3’領域から誘導されることができ、開始領域はそれから、または異なる遺伝子から得られた。多数の終結領域が知られており、多様な宿主において満足に機能する(それらが由来する同一のおよび異なる、属および種の双方で使用した際に)。終結領域は、通常、特定の特性のためと言うよりも、便宜上で選択される。好ましくは、終結領域は、特にサッカロミセス(Saccharomyces)、分裂酵母(Schizosaccharomyces)、カンジダ(Candida)、ヤロウィア(Yarrowia)またはクリヴェロミセス(Kluyveromyces)である酵母菌遺伝子から誘導される。γ−インターフェロンおよびα−2インターフェロンをコードする哺乳類の遺伝子の3’−領域もまた、酵母菌中で機能することが知られている。終結制御領域もまた、好ましい宿主に天然の様々な遺伝子から誘導されてもよい。場合により終結部位は不必要かもしれないが、含まれることが最も好ましい。
【0123】
当業者は気づいているように、遺伝子をクローン化ベクターに単に挿入するだけでは、それが必要なレベルで成功裏に発現することを確証しない。高発現率の必要性に答えて、転写、翻訳、タンパク質安定性、酸素限界、および宿主細胞からの分泌の側面を制御するいくつかの異なる遺伝的要素を操作することで、多くの特殊化した発現ベクターが作り出されている。より具体的には、遺伝子発現を制御するように操作される分子の特徴のいくつかとして以下が挙げられる。1.)関連転写プロモーターおよびターミネーター配列の性質、2.)クローンされた遺伝子のコピー数、および遺伝子がプラスミド上にあるかまたは宿主細胞のゲノム中に組み込まれているかどうか、3.)合成された外来タンパク質の最終細胞内位置、4.)宿主生物体中の翻訳効率、および5.)宿主細胞内のクローン化遺伝子タンパク質の本質的な安定性。これらの各タイプの修正は、コドン最適化PUFA生合成経路酵素の発現をさらに最適化する手段として、本発明中に包含される。
【0124】
微生物宿主の形質転換
油性酵母菌中での発現に適したコドン最適化デサチュラーゼまたは鎖長延長酵素ポリペプチドをコードするDNAがひとたび得られると、それは宿主細胞中で自律複製できるプラスミドベクター中に入れられ、またはそれは宿主細胞のゲノム中に直接組み込まれる。発現カセットの組み込みは、宿主ゲノム内で無作為に起きることができ、または宿主遺伝子座内で遺伝子組み換えを標的とするのに十分な宿主ゲノムとの相同性領域を含有するコンストラクトの使用を通じて、標的を定めることができる。コンストラクトが内因性遺伝子座に標的を定めると、全てまたはいくつかの転写および翻訳調節領域が、内因性遺伝子
座によって提供できる。
【0125】
別々の複製ベクターから2つ以上の遺伝子が発現する場合、各ベクターが異なる選択手段を有することが望ましく、他のコンストラクトに対する相同性を欠いて、安定した発現を維持し、コンストラクト中における要素の再集合を防止すべきである。調節領域、選択手段、および導入コンストラクト増殖方法の思慮深い選択は、全ての導入された遺伝子が必要なレベルで発現して、所望の生産物の合成を提供するように実験的に判定できる。
【0126】
関心のある遺伝子を含んでなるコンストラクトは、あらゆる標準的技術によって宿主細胞に導入してもよい。これらの技術としては、形質転換(例えば酢酸リチウム形質転換[Methods in Enzymology、194:186〜187(1991)])、プロトプラスト融合、弾道衝撃(bolistic impact)、電気穿孔、マイクロインジェクション、または宿主細胞中に関心のある遺伝子を導入するその他のあらゆる方法が挙げられる。油性酵母菌(すなわち、ヤロウィア・リポリティカ(Yarrowia lipolytica))に適用できるより具体的な教示としては、米国特許第4,880,741号明細書および米国特許第5,071,764号明細書、およびチェン(Chen)D.C.ら(Appl Microbiol Biotechnol.48(2):232〜235(1997))が挙げられる。
【0127】
便宜上、DNA配列(例えば発現カセット)を取り込むように、あらゆる方法によって操作されている宿主細胞を「形質転換された」または「組換え」とここで称する。形質転換された宿主は、遺伝子がゲノム中に組み込まれるか、増幅されるか、または複数のコピー数を有する染色体外因子上に存在するかどうか次第で、発現コンストラクトの少なくとも1つのコピーを有し、2つ以上を有してもよい。形質転換された宿主細胞は、導入されたコンストラクト上に含有されるマーカーの選択によって同定できる。多くの形質転換技術は、多くのDNA分子を宿主細胞中に導入するので、代案としては、別々のマーカーコンストラクトを所望のコンストラクトと共に同時形質転換してもよい。典型的に、形質転換された宿主は、選択的培地上で生育するそれらの能力について選択される。選択培地には抗生物質が組み込まれていてもよく、または栄養素または成長因子などの非形質転換宿主の生育に必要な要素が欠如していてもよい。導入されたマーカー遺伝子は、形質転換された宿主中で発現すると、抗生物質抵抗性を与え、または必須成長因子または酵素をコードしてもよく、それによって選択培地上での生育を可能にしてもよい。また発現したマーカータンパク質が直接または間接に検出できる場合に、形質転換された宿主の選択ができる。マーカータンパク質は単独で、または別のタンパク質と融合して発現してもよい。マーカータンパク質は、次によって検出できる。1.)その酵素活性(例えばβ−ガラクトシダーゼは、基質X−gal[5−ブロモ−4−クロロ−3−インドリル−β−D−ガラクトピラノシド]を着色生産物に変換でき、ルシフェラーゼはルシフェリンを発光生産物に変換できる)、または2.)その光生産または修正特性(例えばオワンクラゲ(Aequorea victoria)の緑色蛍光タンパク質は青色光で照明されると蛍光を発する)。代案としては、抗体を使用して、マーカータンパク質、または例えば関心のあるタンパク質の上の分子タグを検出できる。マーカータンパク質またはタグを発現する細胞は、例えば視覚的に、またはFACS、または抗体を使用したパニングなどの技術によって選択できる。酵母菌形質転換体の選択のためには、酵母菌で機能するあらゆるマーカーを使用してもよい。望ましくは、重要であるカナマイシン、ハイグロマイシン、およびアミノグリコシドG418に対する抵抗性、ならびにウラシルまたはロイシン欠乏培地上で生育する能力である。
【0128】
形質転換に続いて、本コドン最適化配列の遺伝子産物(そして場合により、宿主細胞内で発現するその他のPUFA酵素)に適した基質が、宿主によって自然にまたは遺伝子組換え的に生産されてもよく、またはそれらは外来性に提供されてもよい。
【0129】
微生物中でのω−3および/またはω−6脂肪酸生合成の代謝エンジニアリング
本遺伝子のコドン最適化配列の知識、および油性酵母菌における発現のためにその他のPUFA遺伝子を最適化するのに必要な方法は、油性酵母菌、特にヤロウィア・リポリティカ(Yarrowia lipolytica)におけるω−3および/またはω−6脂肪酸生合成を操作するために有用であろう。これは直接PUFA生合成経路内での代謝エンジニアリング、または炭素をPUFA生合成経路に与える経路の追加的操作を必要とするかもしれない。生化学的経路を操作するのに有用な方法は、当業者にはよく知られている。
【0130】
望ましい生合成経路を上方制御する技術
デサチュラーゼおよび鎖長延長酵素遺伝子の追加的コピーを宿主に導入して、ω−3および/またはω−6脂肪酸生合成経路の産出量を増大させてもよい。デサチュラーゼまたは鎖長延長酵素遺伝子の発現はまた、mRNAまたはコードされたタンパク質のいずれかから不安定化配列を除去/消去することで、または安定化配列をmRNA(米国特許第4,910,141号明細書)に追加することで、(制御されたまたは構成的な)より強力なプロモーターの使用を通じて増大する発現を引き起こして、転写レベルで増大できる。本発明で実証されるように、デサチュラーゼまたは鎖長延長酵素遺伝子の発現を増大させるさらに別のアプローチは、選択された宿主微生物中での最適遺伝子発現のためのコドンで、未変性遺伝子中のコドンを置換することにより、コードされたmRNAの翻訳効率を増大させることである。
【0131】
望ましくない生合成経路を下方制御する技術
反対に、エネルギーまたは炭素についてω−3および/またはω−6脂肪酸生合成経路と競合する生化学的経路、または特定のPUFA最終生産物の生産を妨げる未変性PUFA生合成経路酵素を遺伝子混乱によって除去またはその他の手段(例えばアンチセンスmRNA)によって下方制御してもよい。遺伝子混乱では、そのコード配列を妨害し、それによって遺伝子を機能的に不活性化するために、混乱させたい構造的遺伝子中に外来DNA断片(典型的に選択可能マーカー遺伝子)を挿入し、混乱させる。混乱カセットの宿主細胞中への形質転換は、相同的組み換えによって、機能的未変性遺伝子の非機能的混乱遺伝子による置換をもたらす(例えばハミルトン(Hamilton)ら、J.Bacteriol.171:4617〜4622(1989);バルバス(Balbas)ら、Genes、136:211〜213(1993);ゲルデナー(Gueldener)ら、Nucleic Acid Res.24:2519〜2524(1996);およびスミス(Smith)ら、Methods Mol.Cell.Biol.5:270〜277(1996)参照)。
【0132】
アンチセンス技術は、標的遺伝子配列が知られている場合に遺伝子を下方制御する別の方法である。これを達成するために、所望の遺伝子からの核酸セグメントをクローン化して、RNAのアンチセンス鎖が転写されるようにプロモーターに作動可能に連結する。次にこのコンストラクトを宿主細胞に導入し、RNAのアンチセンス鎖を生産する。アンチセンスRNAは、関心のあるタンパク質をコードするmRNAの蓄積を防止することで、遺伝子発現を阻害する。当業者は、特定の遺伝子の発現を低下させるために、特別な考察がアンチセンス技術の使用に関連することを理解する。例えば、アンチセンス遺伝子の適切な発現レベルは、当業者には知られている異なる調節因子を使用した、異なるキメラ遺伝子の使用を必要とするかもしれない。
【0133】
配列が知られている場合、標的を定めた遺伝子混乱およびアンチセンス技術が遺伝子を下方制御する効果的手段を提供するが、配列ベースではないその他のより特異性が低い方法が開発されている。例えば細胞をUV放射線暴露して、次に所望の表現型についてスク
リーンしてもよい。変異体を作り出すために、化学薬品による変異誘発もまた効果的であり、一般に使用される物質としては、非複製DNAに影響する化学物質(例えばHNO2およびNH2OH)、ならびに複製DNAに影響する薬剤(例えばフレームシフト変異を引き起こすことで注目に値するアクリジン染料)が挙げられる。放射線または化学薬品を使用して変異体を作り出す特定の方法は、技術分野でよく文書化されている。例えばトーマスD.ブロック(Thomas D.Brock)著「バイオテクノロジー:工業微生物学テキストブック(Biotechnology:A Textbook of Industrial Microbiology)」第二版(1989)Sinauer Associates:マサチューセッツ州サンダーランド(Sunderland、MA)、またはデシュパンデ,ムカンド(Deshpande,Mukund)V.、Appl.Biochem.Biotechnol.、36:227(1992)を参照されたい。
【0134】
別の非特異的遺伝子混乱方法は、転移因子またはトランスポゾンの使用である。トランスポゾンは、DNAに無作為に挿入される遺伝的因子であるが、後で配列に基づいて検索して、挿入がどこで起きたのか判定できる。生体内(in vivo)および生体外(in vitro)転位法の双方が知られている。どちらの方法にもトランスポザーゼ酵素と組み合わされた転移因子の使用が関与する。転移因子またはトランスポゾンがトランスポザーゼ存在下で核酸断片に接触すると、転移因子が核酸断片中に無作為に挿入される。混乱された遺伝子は転移因子配列に基づいて同定されてもよいので、技術は、ランダム変異誘発のため、そして遺伝子単離のために有用である。生体外(in vitro)転位のためのキットは市販される。例えば1.)ニュージャージー州ブランチバーグのパーキン・エルマー・アプライド・バイオシステムズ(Perkin Elmer Applied Biosystems(Branchburg、NJ))から入手できる酵母菌Ty1因子に基づく、プライマー・アイランド転位キット、2.)マサチューセッツ州ビバリーのニュー・イングランド・バイオ・ラブズ(New England Biolabs、(Beverly、MA))から入手できる細菌性トランスポゾンTn7に基づくゲノム・プライミング・システム、および3.)ウィスコンシン州マディソンのエピセンター・テクノロジーズ(Epicentre Technologies(Madison、WI))から入手できる、Tn5細菌性転移因子に基づくEZ::TNトランスポゾン挿入システムを参照されたい。
【0135】
本発明の文脈では、上述のいずれか1つの方法によって脂肪酸生合成経路の発現を調節することが有用であるかもしれない。例えば本発明は、ω−3および/またはω−6脂肪酸の生産をもたらす生合成経路中の重要な酵素をコードするコドン最適化遺伝子を提供する。これらのコドン最適化遺伝子には、Δ6デサチュラーゼ、Δ17デサチュラーゼ、およびPUFA鎖長延長酵素が含まれる。ω−3および/またはω−6脂肪酸生合成経路を自然に有さない油性酵母菌中でこれらの遺伝子を発現させ、これらおよびその他のPUFA生合成遺伝子の遺伝子の発現を調節して、宿主生物体の代謝工学のための様々な手段を使用して好ましいPUFA生産物の生産を最大化することが特に有用であろう。同様にこれらの遺伝子によってPUFA生産を最大化するためには、PUFA生合成に向けて導かれる炭素の流れと競合する経路を混乱させることが必要かもしれない。
【0136】
コドン最適化遺伝子の組換え発現のための好ましい微生物宿主
本コドン最適化遺伝子および核酸断片の発現のための宿主細胞は、広範な温度およびpHで、単純または複合炭水化物、有機酸およびアルコール、および/または炭化水素をはじめとする多様な原材料上で生育する微生物の宿主を含んでもよい。本発明で述べられる遺伝子は、油性酵母菌、特にヤロウィア・リポリティカ(Yarrowia lipolytica)における発現のためにコドン最適化されているが、転写、翻訳、およびタンパク質生合成器官は高度に保存されているので、あらゆる細菌、酵母菌および/または糸
状菌が、本核酸断片の発現に適した宿主になることが考察される。
【0137】
好ましい微生物の宿主は、油性酵母菌である。これらの生物体は自然に油の合成および蓄積ができ、油は、細胞乾燥重量の約25%を超え、より好ましくは細胞乾燥重量の約30%を超え、最も好ましくは細胞乾燥重量の約40%を超える量を構成できる。油性酵母菌として典型的に同定されている属としては、ヤロウィア(Yarrowia)、カンジダ(Candida)、ロドトルラ(Rhodotorula)、ロドスポリジウム(Rhodosporidium)、クリプトコッカス(Cryptococcus)、トリコスポロン(Trichosporon)、およびリポマイセス(Lipomyces)が挙げられるが、これに限定されるものではない。より具体的には、例証的な油合成酵母菌としては、ロドスポリジウム・トルイデス(Rhodosporidium toruloides)、リポマイセス・スターケイ(Lipomyces starkeyii)、L.リポフェラス(L.lipoferus)、カンジダ・レブカウフィ(Candida revkaufi)、C.プルケリマ(C.pulcherrima)、C.トロピカリス(C.tropicalis)、C.ユチリス(C.utilis)、トリコスポロン・プランス(Trichosporon pullans)、T.クタネウム(T.cutaneum)、ロドトルラ・グルティナス(Rhodotorula glutinus)、R.グラミニス(R.graminis)、およびヤロウィア・リポリティカ(Yarrowia lipolytica)(以前はカンジダ・リポリティカ(Candida lipolytica)として分類された)が挙げられる。
【0138】
最も好ましいのは油性酵母菌ヤロウィア・リポリティカ(Yarrowia lipolytica)であり、さらなる実施態様で最も好ましいのは、ATCC番号20362、ATCC番号8862、ATCC番号18944、ATCC番号76982および/またはLGAMS(7)1と称されるY.リポリティカ(Y.lipolytica)株である(パパニコラオウ(Papanikolaou)S.、およびアゲリス(Aggelis)G.、Bioresour.Technol.82(1):43〜9(2002))。
【0139】
PUFA生産のための発酵プロセス
形質転換された微生物宿主細胞をデサチュラーゼおよび鎖長延長酵素活性を最適化させる条件下で生育させ、好ましいPUFAの最大かつ最も経済的な収率を得る。一般に最適化されてもよい培地条件としては、炭素源のタイプおよび量、窒素源のタイプおよび量、炭素−対−窒素比率、酸素レベル、生育温度、pH、バイオマス生産相の長さ、油蓄積相の長さ、および細胞収穫時間が挙げられる。油性酵母菌などの関心のある微生物を複合培地(例えば酵母菌抽出物−ペプトン−デキストロース液体培地(YPD))上で生育させ、または生育に必要な構成要素が欠如する合成(defined)最少培地(例えばミシガン州デトロイトのディフコ・ラボラトリーズ(DIFCO Laboratories(Detroit、MI))からの酵母菌窒素塩基)上で生育させて、所望の発現カセットの選択を強要する。
【0140】
本発明における発酵培地は、適切な炭素源を含有しなくてはならない。適切な炭素源としては、単糖類(例えばグルコース、フルクトース)、二糖類(例えばラクトース、スクロース)、少糖類、多糖類(例えばデンプン、セルロースまたはそれらの混合物)、糖アルコール(例えばグリセロール)または再生可能原材料からの混合物(例えば乳清透過液、コーンスティープリーカー、甜菜モラセス、大麦の麦芽)が挙げられるが、これに限定されるものではない。さらに炭素源としては、アルカン、脂肪酸、脂肪酸エステル、モノグリセリド、ジグリセリド、トリグリセリド、リン脂質、および植物油(例えばダイズ油)および動物脂肪をはじめとする脂肪酸の様々な商業的供給源が挙げられる。さらに炭素源としては、重要な生化学的中間体への代謝転換が実証されている一炭素源(例えば二酸
化炭素、メタノール、ホルムアルデヒド、ギ酸および炭素−含有アミン)が挙げられる。したがって本発明で使用される炭素源は多種多様な炭素含有源を包含し、および宿主生物体の選択によってのみ制限されることが考察された。上述の全ての炭素源およびそれらの混合物が本発明で適切であることが期待されるが、好ましい炭素源は糖および/または脂肪酸である。最も好ましいのは、10〜22個の炭素含有グルコースおよび/または脂肪酸である。
【0141】
窒素は、無機(例えば(NH4)2SO4)または有機源(例えば尿素またはグルタミン酸)から供給されてもよい。適切な炭素および窒素源に加えて、発酵培地はまた、適切なミネラル、塩、補助因子、緩衝液、ビタミン、および当業者には知られている微生物の生育とPUFA生産に必要な酵素的経路の促進に適したその他の構成要素を含有しなくてはならない。脂質およびPUFAの合成を促進するいくつかの金属イオン(例えばMn+2、Co+2、Zn+2、Mg+2)が注目されている(ナカハラ(Nakahara)T.ら、「単細胞油の工業的応用(Ind.Appl.of Single Cell Oils)」、D.J.カイル(Kyle)およびR.コリン(Colin)編、p61〜97(1992))。
【0142】
本発明における好ましい増殖培地は、ミシガン州デトロイトのディフコ・ラボラトリーズ(DIFCO Laboratories(Detroit、MI))からの酵母菌窒素塩基などの一般的な商業的調製培地である。またその他の合成(defined)または合成(synthetic)増殖培地が使用されてもよく、特定の微生物の生育に適した培地は、微生物学または発酵科学の当業者に知られている。発酵に適したpH範囲は、典型的に約pH4.0〜pH8.0の間であり、pH5.5〜pH7.0が最初の生育条件の範囲として好ましい。発酵は好気性または好気性条件下で実施されてもよく、マイクロ好気性条件が好ましい。
【0143】
典型的に、油性酵母菌細胞におけるPUFAの高レベルの蓄積は、代謝状態が生育と脂肪合成/貯蔵間で「バランスが取れて」いなくてはならないので、二段階プロセスを必要とする。したがって最も好ましくは、油性酵母菌におけるPUFA生産には、二段階発酵プロセスが必要である。このアプローチでは、第1の発酵状態が細胞集団の生産および蓄積に供され、迅速な細胞生育および細胞分割によって特徴づけられる。発酵の第2段階では、培養内の窒素欠乏条件を確立して、高レベルの脂質蓄積を促進することが好ましい。この窒素欠乏の効果は、細胞内のAMPの効果的濃度を低下させることにより、ミトコンドリアのNAD−依存イソクエン酸デヒドロゲナーゼ活性を低下させることである。これが起きるとクエン酸が蓄積するので、細胞質中にアセチル−CoAの豊富なプールが形成し、脂肪酸合成の下準備をする。したがってこの相は、細胞分割休止と、それに続く脂肪酸合成および油蓄積によって特徴づけられる。
【0144】
細胞は典型的に約30℃で生育させるが、いくつかの研究は、より低い温度における不飽和脂肪酸の増大した合成を示している(ヨンマニットチャイ(Yongmanitchai)およびワード(Ward)、Appl.Environ.Microbiol.57:419〜25(1991))。プロセスの経済に基づけば、この温度シフトは、大部分の生物体の生育が起きる二段階発酵の第1相後に起きるはずである。
【0145】
このコドン最適化遺伝子を使用したω脂肪酸の商業生産が所望される場合、多様な発酵プロセスデザインを適用してもよいことが考察される。例えば組換え微生物宿主からのPUFAの商業生産は、バッチ、供給バッチまたは連続発酵プロセスによって生産されてもよい。
【0146】
バッチ発酵プロセスは閉鎖システムであり、培地組成物がプロセス開始時に定められ、
プロセス中にpHおよび酸素レベル維持のために必要とされるもの以外は、さらなる追加を受けない。したがって培養プロセスの始めに所望の生物体を培地に接種し、培地への追加的供給源(すなわち炭素および窒素源)の添加なしに、生育または代謝活性を生じさせる。バッチプロセスでは、システムの代謝産物およびバイオマス組成物は、培養が終結するまで絶えず変化する。典型的なバッチプロセスでは、細胞は静止対数相から高生育対数相を通過して、最終的に発育速度が減少または停止する定常相に進む。処置されない場合、定常相の細胞は次第に死滅する。標準バッチプロセスのバリエーションが供給バッチプロセスであり、炭素源は発酵プロセス経過中に発酵槽に連続的に添加される。供給バッチプロセスもまた、本発明に適している。供給バッチプロセスは分解産物抑制が、細胞代謝を阻害する傾向がある場合に、またはあらゆる時点で培地中に限定量の源を有することが望ましい場合に有用である。供給バッチシステム中の炭素源濃度の測定は困難であるので、pH、溶解酸素、および排ガス(例えばCO2)などの測定可能な要素の変化に基づいて推定してもよい。バッチおよび供給バッチ培養方法は一般的で技術分野でよく知られており、実例が以下にある。トーマス(Thomas)D.ブロック(Brock)著、「バイオテクノロジー:工業的微生物学テキストブック(Biotechnology:A
Textbook of Industrial Microbiology)」第二版、Sinauer Associates:マサチューセッツ州サンダーランド(Sunderland、MA)(1989)、またはデシュパンデ,ムカンド(Deshpande,Mukund)V.、Appl.Biochem.Biotechnol.、36:227(1992)。これらは参照によってここに援用される。
【0147】
このコドン最適化遺伝子を使用したω脂肪酸の商業生産はまた、連続発酵プロセスによって達成されてもよく、そこでは生産物の回収のため等量の培養物を除去するのと同時に、合成培地をバイオリアクター内に連続的に添加する。連続培養は、概して細胞を一定細胞密度の対数増殖期に保つ。連続または半連続培養法は、細胞生育または最終生産物濃度に影響する1つの因子またはあらゆるいくつかの因子の調節を可能にする。例えば1つのアプローチでは炭素源を制限して、あらゆるその他のパラメータが代謝を調節できるようにしてもよい。その他のシステムでは、培地濁度によって測定される細胞濃度を一定に保ちながら、生育に影響するいくつかの因子を連続的に変化させてもよい。連続システムは定常状態生育を維持することを目指すので、細胞生育率は、培養から培地が抜き取られることによる細胞損失に対してバランスが取れていなくてはならない。連続的培養プロセスのために栄養素および成長因子を調節する方法、ならびに生産物形成速度を最大化する技術は工業微生物学の技術分野でよく知られており、多様な方法が上記のブロック(Brock)で詳述される。
【0148】
PUFAの精製
PUFAは、遊離脂肪酸として、またはアシルグリセロール、リン脂質、スルホリピドまたは糖脂質などのエステル化形態で宿主微生物中に見ることができ、技術分野でよく知られている多様な手段を通じて宿主細胞から抽出されてもよい。酵母菌脂質の抽出技術、品質分析、および許容性基準に関する1つのレビューは、Z.ジェーコブス(Jacobs)(Critical Reviews in Biotechnology、12(5/6):463〜491(1992))である。下流プロセスに関する簡潔なレビューは、A.シン(Singh)およびO.ワード(Ward)(Adv.Appl.Microbiol.45:271〜312(1997))にもある。
【0149】
一般にPUFA精製手段は、有機溶剤、超音波処理、超臨界流体抽出(例えば二酸化炭素を使用して)による抽出と、鹸化と、圧搾などの物理的手段またはそれらの組み合わせを含んでもよい。特に興味深いのは、水存在下でのメタノールおよびクロロホルムによる抽出である(E.G.ブライ(Bligh)およびW.J.ダイヤー(Dyer)、Can.J.Biochem.Physiol.37:911〜917(1959))。望ま
しい場合、水性層を酸性化して負の電荷を帯びた部分をプロトン化することで、有機層中への所望の生産物の分配を増大できる。抽出後、有機溶剤は窒素流の下で蒸発によって除去できる。コンジュゲートされた形態で単離されると、生産物は酵素的にまたは化学的に切断されて、関心のある遊離脂肪酸またはより単純なコンジュゲートを放出してもよく、次にさらに操作されて所望の最終生産物を生産してもよい。望ましくはコンジュゲートされた形態の脂肪酸は、水酸化カリウムによって切断される。
【0150】
さらに精製が必要ならば、標準法を用いることができる。このような方法としては、抽出、尿素処理、分別結晶化、HPLC、分留、シリカゲルクロマトグラフィー、高速遠心分離または蒸留、またはこれらの技術の組み合わせが挙げられる。酸またはアルケニル基などの反応性基の保護は、既知の技術(例えばアルキル化、ヨウ素化)を通じて、あらゆるステップで実施してもよい。使用される方法としては、メチルエステルを生産するための脂肪酸のメチル化が挙げられる。同様に、保護基はあらゆるステップで除去されてもよい。望ましくはGLA、STA、ARA、DHA、およびEPAを含有する画分の精製は、尿素および/または分留による処置によって達成されてもよい。
【0151】
好ましい実施態様の説明
ここで述べる研究の究極の目的は、ω−3および/またはω−6脂肪酸が濃縮された油を蓄積する油性酵母菌の開発である。このような目的で、油性酵母菌中において効率的に機能するω−3および/またはω−6脂肪酸生合成経路のデサチュラーゼおよび鎖長延長酵素を同定し、これらの宿主におけるω脂肪酸の合成および蓄積を可能にしなくてはならない。
【0152】
本発明では、出願人は、デサチュラーゼおよび鎖長延長酵素遺伝子のコドン最適化に適する技術を実証し、そこでは得られた合成コドン最適化遺伝子が、ヤロウィア・リポリティカ(Yarrowia lipolytica)などの油性酵母菌において増大する変換効率で機能する。これらの方法は、ここでの教示において具現化され、具体的にはコドン最適化Δ6デサチュラーゼ(LAをGLAに、および/またはALAをSTAに変換する酵素的能力を有する)、コドン最適化Δ17デサチュラーゼ(DGLAをETAに、および/またはARAをEPAに変換する役割を果たす)、およびコドン最適化高親和力PUFA鎖長延長酵素(GLAをDGLAに、STAをETAに、および/またはEPAをDPAに変換できる)の合成に標的を定める。当業者は、容易にここで述べる技術を適用して、ω−3および/またはω−6生合成脂肪酸経路中のその他の遺伝子を最適化させ、ヤロウィア(Yarrowia)中で発現すると増大する変換効率を有することが期待される合成遺伝子を作り出せるであろう。したがってここでの教示は、ω−3および/またはω−6脂肪酸が濃縮された油を蓄積する油性酵母菌の開発において大きな有用性を持つ。
【0153】
出願人は、モデル油性酵母菌、ヤロウィア・リポリティカ(Yarrowia lipolytica)におけるコドン最適化のために3つの例示的な野生型遺伝子を選択した。これらの各野生型遺伝子は、下の表3で述べるようにオハイオ州コロンバスのロス・プロダクツ(Ross Products(Columbus、OH))からのプラスミド中で得られた。
【0154】
【表4】
【0155】
微生物宿主における各野生型酵素活性の確証(基質供給試験による)に続いて、各遺伝子をコドン最適化した(少なくとも9%の未変性コドンが、宿主の好むコドンを用いるように修正された)。
【0156】
Δ6デサチュラーゼ、Δ17デサチュラーゼ、および高親和力PUFA鎖長延長酵素遺伝子のコドン最適化は、最初にヤロウィア・リポリティカ(Yarrowia lipolytica)におけるコドン使用および構造遺伝子のシグネチャの判定を必要とした。次にコドン最適化遺伝子をデザインし、遺伝子配列全長を合成する所要時間を実質的に短縮するプロトコルを使用して、生体外(in vitro)で合成した。このプロトコルは、ここで合成される特定の遺伝子に限定されないので、多様な一般用途に広く適用できる。生体外(in vitro)遺伝子合成に伴う基本的ステップは、以下からなった。
1.複数の対のオリゴヌクレオチド(各約100bp長さ)を合成する。各オリゴヌクレオチド配列は、合成される遺伝子配列全長の一部に対応し、各センス鎖オリゴヌクレオチド配列は、配列の5’末端で対応するアンチセンス−鎖オリゴヌクレオチドと約4bpの重なりを有する。
2.全オリゴヌクレオチドの5’および3’末端をキナーゼ反応でリン酸化し、次にセンスおよびアンチセンスオリゴヌクレオチド対に個々のアニール反応を施して、短い(約100bp)二本鎖DNA断片を生産する。
3.次にオリゴヌクレオチド合成時にデザインされた約4bpの重なりを使用して、短い(約100bp)二本鎖DNA断片のプールを一緒にライゲーションし、より長い(約300〜400bp)DNA断片を生産する。
4.ライゲーションに続いて、より長い断片をPCRによって増幅し、その生産物を適切なベクター中にクローン化し、宿主細胞中に形質転換する。
5.各PCR生産物を含有するプラスミドDNAを宿主細胞から精製し、次に合成する遺伝子の各約300〜400bpの断片に対応するPCR生産物単離のための制限酵素消化を施す。
6.合成する約300〜400bpの遺伝子断片を一緒にライゲーションし、次にPCR増幅を施して合成配列全長を生産する。
【0157】
コドン最適化Δ6デサチュラーゼ、Δ17デサチュラーゼ、および高親和力PUFA鎖長延長酵素遺伝子が合成されると、それぞれ個々にヤロウィア・リポリティカ(Yarrowia lipolytica)中に形質転換した。供給実験(適切な基質を使用した)は、Y.リポリティカ(Y.lipolytica)においてコドン最適化遺伝子が、対応する未変性野生型遺伝子よりも効率的に発現すると判定した。具体的にはY.リポリティカ(Y.lipolytica)において発現すると、同一生物学的条件下で、コド
ン最適化Δ6デサチュラーゼは野生型対応酵素よりもおよそ40%多くのLAをGLAに変換し、コドン最適化Δ17デサチュラーゼは野生型酵素の約2倍のARAをEPAにを変換し、コドン最適化高親和力PUFA鎖長延長酵素はおよそ57%より多くのGLAをDGLAに変換した。
【実施例】
【0158】
以下の実施例で本発明をさらに定義する。これらの実施例は、本発明の好ましい実施態様を示しながら、例証のみの目的で提供されるものとする。上の考察およびこれらの実施例から、当業者は本発明の必須特性を把握でき、その精神と範囲を逸脱することなく、本発明の様々な変更および修正を行って、それを様々な使用法および条件に適合できる。
【0159】
一般方法
実施例で使用する標準組換えDNAおよび分子クローニング技術は、技術分野でよく知られており、1.)サムブルック(Sambrook)、J.、フリッチュ(Fritsch)、E.F.およびマニアティス(Maniatis)T.「分子クローニング:実験室マニュアル(Molecular Cloning:A Laboratory Manual)」、コールドスプリングハーバーラボラトリ:ニューヨーク州コールドスプリングハーバー(Cold Spring Harbor、NY)(1989)(マニアティス(Maniatis))、2.)T.J.シルハビー(Silhavy)、M.L.ベンナン(Bennan)およびL.W.エンクイスト(Enquist)、「遺伝子融合実験(Experiments with Gene Fusions)」(コールドスプリングハーバーラボラトリ:Cold Spring Harbor,NY,1984)、および3.)オースベル(Ausubel)F.M.ら「分子生物学現代プロトコル(Current Protocols in Molecular Biology)」(グリーンパブリッシングおよびワイリーインターサイエンスによる出版;1987)で述べられる。
【0160】
微生物培養の維持および生育に適した材料および方法は、技術分野でよく知られている。以下の実施例で使用するのに適した技術は、次で述べられる。「一般微生物学方法マニュアル(Manual of Methods for General Bacteriology)」、フィリップス・ゲアハルト(Phillipp Gerhardt)、R.G.E.マレー(R.G.E.Murray)、ラルフN.コスティロウ(Ralph N.Costilow)、ユージーンW.ネスター(Eugene W.Nester)、ウィリスA.ウッド(Willis A.Wood)、ノエルR.クリーグ(Noel R.Krieg)、およびG.ブリッグス・フィリップス(G.Briggs Phillips)編、米国微生物学会(American Society for Microbiology)、ワシントンD.C.(Washington,D.C.)(1994)またはトーマス(Thomas)D.ブロック(Brock)著、バイオテクノロジー:工業的微生物学テキストブック(Biotechnology:A Textbook of Industrial Microbiology)第二版、Sinauer Associates:(Sunderland、MA)(1989)。微生物細胞の生育および維持のために使用される全ての試薬、制限酵素および材料は、特に断りのない限り、ウィスコンシン州ミルウォーキーのアルドリッチ・ケミカルズ(Aldrich Chemicals(Milwaukee、WI))、ミシガン州デトロイトのディフコ・ラボラトリーズ(DIFCO Laboratories(Detroit、MI))、メリーランド州ゲーサーズバーグのギブコ/BRL(GIBCO/BRL(Gaithersburg、MD))、またはミズーリ州セントルイスのシグマケミカル(Sigma Chemical Company(St.Louis、MO))から得た。
【0161】
カリフォルニア州カールスバッドのインビトロゲン(Invitrogen(Carlsbad、CA))から大腸菌(E.coli)TOP10細胞および大腸菌(E.coli)エレクトロマックス(electromax)DH10B細胞を得た。メリーランド州ゲイサーズバーグのギブコ/BRL(GIBCO/BRL)(Gaithersburg、MD))から大腸菌(E.coli)DH5αの最大効率コンピテント細胞を得た。カリフォルニア州サンディエゴのストラタジーン社(Stratagene Company(San Diego、CA))から大腸菌(E.coli)(XL1−Blue)コンピテント細胞を購入した。大腸菌(E. coli)株は典型的にルリア・ベルターニ(Luria Bertani)(LB)プレート上で37℃で生育させた。
【0162】
ヤロウィア・リポリティカ(Yarrowia lipolytica)ロイシン独立栄養株(ATCC番号76982)をメリーランド州ロックビル(Rockville、MD)の米国微生物系統保存機関(American Type Culture Collection)から購入し、機能性アッセイで使用した。Y.リポリティカ(Y.lipolytica)株は通常、YPD寒天(1%酵母菌抽出物、2%バクトペプトン、2%グルコース、2%寒天)上で28℃で生育させた。形質転換体の選択のために、ミシガン州デトロイトのジフコ・ラボラトリーズ(DIFCO Laboratories(Detroit、MI))からの最少培地(硫酸アンモニウムまたはアミノ酸を含まない0.17%酵母菌窒素ベース、2%グルコース、0.1%プロリン、pH6.1)を使用した。アデニン、ロイシン、リジン、および/またはウラシルサプリメントを最終濃度0.01%で添加した。
【0163】
一般分子クローニングは、標準法に従って実施された(サムブルック(Sambrook)ら、同上)。オリゴヌクレオチドはシグマ−ジェノシス(Sigma−Genosys、テキサス州スプリング(Spring、TX))によって合成した。部位特異的変異誘発は、ストラタジーン(Stratagene)のクイックチェンジ(QuickChange)TM部位特異的変異誘発キットを製造業者の説明書に従って使用して実施した。ポリメラーゼ連鎖反応(PCR)または部位特異的変異誘発がサブクローニングに関与する場合、コンストラクトは、配列中に確実にいかなる誤りも導入されないように配列された。PCR生産物は、ウィスコンシン州マディソンのプロメガ(Promega(Madison、WI))からのpGEM−T−イージーベクター中にクローン化した。
【0164】
ウィスコンシン州マディソンのジーンズ・コンピューター・グループ(GCG)社(Genes Computer Group Inc.(Madison、WI))から入手できる一組のプログラム、ウィスコンシン・パッケージ(Wisconsin Package Version9.0)を使用して、遺伝的配列の操作が達成された。ギャップ作成デフォルト値12、およびギャップ延長デフォルト値4でGCGプログラム「パイルアップ(Pileup)」を使用した。デフォルトギャップ作成ペナルティ50、およびデフォルトギャップ延長ペナルティ3で、GCG「ギャップ(ギャップ)」または「ベストフィット(Bestfit)」プログラムを使用した。特に断りのない限り、全てのその他の場合において、GCGプログラムのデフォルトパラメータを使用した。
【0165】
略語の意味は以下の通り。「sec」は秒を意味し、「min」は分を意味し、「h」は時間を意味し、「d」は日を意味し、「μL」はマイクロリットルを意味し、「mL」はミリリットルを意味し、「L」はリットルを意味し、「μM」はマイクロモル(micromolar)を意味し、「mM」はミリモル(millimolar)を意味し、「M」はモル(molar)を意味し、「mmol」はミリモル(millimole)を意味し、「μmole」マイクロモル(micromole)を意味し、「g」はグラムを意味し、「μg」はマイクログラムを意味し、「ng」はナノグラムを意味し、「U」は単位を意味し、「bp」は塩基対を意味し、「kB」はキロベースを意味する。
【0166】
実施例1
ヤロウィア・リポリティカ(Yarrowia lipolytica)中の異種性遺伝子発現に適したプラスミドの構造
図2に図解するように、pINA532誘導体であるプラスミドpY5(Insitut National Agronomics、Centre de biotechnologie Agro−Industrielle、laboratoire de Genetique Moleculaire et Cellularie INRA−CNRS、F−78850、Thiverval−Grignon、Franceのクロード・ガィヤルダン(Claude Gaillardin)博士からの贈与)をヤロウィア・リポリティカ(Yarrowia lipolytica)における異種性遺伝子発現のために構築した。
【0167】
最初に、pINA532のARS18配列およびLEU2遺伝子を含有する、部分的に消化された3598bpのEcoRI断片をカリフォルニア州サンディエゴのストラタジーン社(Stratagene Company(San Diego、CA))からのpBluescriptのEcoRI部位中にサブクローン化してpY2を生産した。
【0168】
TEF5’(配列番号7)およびTEF3’(配列番号8)をプライマーとして使用して、PCRによりヤロウィア・リポリティカ(Yarrowia lipolytica)ゲノムのDNAからTEFプロモーターを増幅した(ミュラー(Muller)S.ら「酵母菌(Yeast)」14:1267〜1283(1998))。100ngのヤロウィア(Yarrowia)ゲノムDNAと、10mM KCl、10mM(NH4)2SO4、20mMトリス−HCl(pH8.75)、2mM MgSO4、0.1%TritonX−100、100μg/mL BSA(最終濃度)を含有するPCR緩衝液と、各200ミュートμMのデオキシリボヌクレオチド三リン酸と、10pmoleの各プライマーと、カリフォルニア州サンディエゴのストラタジーン(Stratagene、San Diego、CA)からの1μLのPfuTurbo DNAポリメラーゼとを含有する、50μLの総容積でPCR増幅を実施した。増幅は以下のように実施した。95℃で3minの初期変性、それに続いて95℃で1min、56℃で30sec、72℃で1minを35サイクル。72℃で10minの最終延長サイクル実施し、それに続いて4℃での反応終結。418bpのPCR生産物をpCR−BluntにライゲーションしてpIP−tefを生産した。pIP−tefのBamHI/EcoRV断片をpY2のBamHI/SmaI部位にサブクローン化して、pY4を生産した。
【0169】
テンプレートとしてpINA532、プライマーとしてXPR5’(配列番号9)およびXPR3’(配列番号10)を使用して、PCRによってXPR2転写ターミネーターを増幅した。上述の構成要素および条件を使用して、PCR増幅を50μLの総容積中で実施した。179bpのPCR生産物をSacIIで消化し、pY4のSacII部位にライゲーションしてpY5を生産した。したがってpY5(図2および3に示す)は、以下を含有するヤロウィア(Yarrowia)−大腸菌(E. coli)シャトルプラスミドとして有用である。
1)ヤロウィア(Yarrowia)自律性複製配列(ARS18)、
2)ColE1プラスミド複製起点、
3)大腸菌(E. coli)の選択のためのアンピシリン−抵抗性遺伝子(AmpR)、
4)ヤロウィア(Yarrowia)の選択のためのヤロウィア(Yarrowia)LEU2遺伝子、
5)ヤロウィア(Yarrowia)における異種性遺伝子発現のための翻訳延長プロモーター(TEFP)、
6)ヤロウィア(Yarrowia)における異種性遺伝子発現の転写終結のための細胞外プロテアーゼ遺伝子ターミネーター(XPR2)。
【0170】
pY5−4およびpY5−13(図3)をpY5の誘導体として構築して、ヤロウィア・リポリティカ(Yarrowia lipolytica)中のサブクローン化および異種性遺伝子発現を容易にした。
【0171】
具体的にはpY5をテンプレートとして使用して、3ラウンドの部位特異的変異誘発によってpY5−4を構築した。オリゴヌクレオチドYL1およびYL2(配列番号11および12)を使用して、LEU2レポーター遺伝子内に位置するNcoI部位をpY5から除去し、pY5−1を生産した。オリゴヌクレオチドYL3およびYL4(配列番号13および14)を使用して、特異的変異誘発によって、NcoI部位をTEFプロモーターとXPR2転写ターミネーター部位の間でpY5−1内に導入し、pY5−2を生産した。次にオリゴヌクレオチドYL23およびYL24(配列番号15および16)を使用して、PacI部位をTEFプロモーターとXPR2転写ターミネーターの間でY5−2に導入し、pY5−4を生産した。
【0172】
pY5をテンプレートとして使用して、6ラウンドの部位特異的変異誘発によってpY5−13構築した。オリゴヌクレオチドYL5およびYL6(配列番号17および18)を使用して、部位特異的変異誘発によって、SalIおよびClaI部位の双方をpY5から除去して、pY5−5を生産した。オリゴヌクレオチドYL9およびYL10(配列番号19および20)を使用して、部位特異的変異誘発によって、SalI部位をLEU2遺伝子とTEFプロモーターの間でpY5−5中に導入し、pY5−6を生産した。オリゴヌクレオチドYL7およびYL8(配列番号21および22)を使用して、PacI部位をLEU2遺伝子とARS18の間でpY5−6中に導入し、pY5−8を生産した。オリゴヌクレオチドYL3およびYL4(配列番号13および14)を使用して、NcoI部位をTEFプロモーターの翻訳開始コドン周囲のpY5−8中に導入しpY5−9を生産した。YL1およびYL2オリゴヌクレオチド(配列番号11および12)を使用して、pY5−9のLEU2遺伝子中のNcoI部位を除去し、pY5−12を生産した。最後にオリゴヌクレオチドYL61およびYL62(配列番号23および24)を使用して、BsiWI部位をColEIとXPR2領域の間でpY5−12中に導入し、pY5−13を生産した。
【0173】
実施例2
ヤロウィア・リポリティカ(Yarrowia lipolytica)における野生型Δ6およびΔ17デサチュラーゼおよび高親和力PUFA鎖長延長酵素の変換効率分析
ヤロウィア・リポリティカ(Yarrowia lipolytica)における野生型Δ6デサチュラーゼ、鎖長延長酵素、およびΔ17デサチュラーゼの機能性を(コドン最適化に先だって)確証するために、各野生型タンパク質の変換効率を測定した。具体的にはモルティエレラ・アルピナ(Mortierella alpina)Δ6デサチュラーゼ、サプロレグニア・ディクリナ(Saprolegnia diclina)Δ17デサチュラーゼ、およびM.アルピナ(M.alpina)高親和力PUFA鎖長延長酵素を別の宿主において発現させ、基質供給試験で活性についてスクリーニングした。各酵素は基質の少なくとも23%を生産物に変換できることが分かった。
【0174】
野生型モルティエレラ・アルピナ(Mortierella alpina)(登録番号AF465281)Δ6デサチュラーゼ
M.アルピナ(M.alpina)Δ6デサチュラーゼ遺伝子(配列番号1)を含有するpCGR5(米国特許第5,968,809号明細書)の1384bpのNcoI/NotI断片をpY5−2(実施例1)のNcoI/NotI部位に挿入し、pY54を生
産した。
【0175】
野生型サプロレグニア・ディクリナ(Saprolegnia diclina)(ATCC番号56851)Δ17デサチュラーゼ
プライマーとしてオリゴヌクレオチドYL21A(配列番号118)およびYL22(配列番号119)を使用して、PCRによって、プラスミドpRSP19(米国特許出願公開第2003/0196217 A1号明細書)からS.ディクリナ(S.diclina)の野生型Δ17デサチュラーゼ遺伝子を増幅した。PCR増幅は、10ngのテンプレート、10mM KCl、10mM(NH4)2SO4、20mMトリス−HCl(pH8.75)、2mM MgSO4、0.1%TritonX−100、および100μg/mL BSA(最終濃度)を含有するPCR緩衝液と、200μMの各デオキシリボヌクレオチド三リン酸と、10pmoleの各プライマーと、カリフォルニア州サンディエゴのストラタジーン(Stratagene、San Diego、CA)からの1μLのPfuTurbo DNAポリメラーゼを含んでなる50μLの総容積で実施した。増幅を以下のようにして実施した。95℃で3minの初期変性、それに続いて95℃で1min、56℃で30sec、72℃で1minを35サイクル。72℃で10minの最終延長サイクルの実施と、それに続く4℃での反応終結。PCR生産物をNcoI/PacIで消化し、次にNcoI/PacIで消化したpY5−4(図3、実施例1で述べられる)にライゲーションしてpYSD17を生産した。
【0176】
野生型モルティエレラ・アルピナ(Mortierella alpina)(登録番号AX464731)高親和力鎖長延長酵素
M.アルピナ(M.alpina)高親和力PUFA鎖長延長酵素遺伝子(配列番号5)のコード領域を含有するpRPB2(国際公開第00/12720号パンフレット)の973bpのNotI断片をpY5のNotI部位(実施例1、図2および3)に挿入し、pY58を生産した(図13B)。
【0177】
ヤロウィア・リポリティカ(Yarrowia lipolytica)の形質転換
チェン(Chen)D.C.ら(Appl Microbiol Biotechnol.48(2):232〜235(1997))の方法に従って、プラスミドpY54、pYSD17、およびpY58を別々にY.リポリティカ(Y.lipolytica)ATCC番号76982中に形質転換した。
【0178】
簡単に述べると、ヤロウィア(Yarrowia)のロイシン栄養要求株をYPDプレート上に画線し、30℃でおよそ18時間生育させた。いくつかの大型白金耳を満たす細胞をプレートからこすり取り、以下を含有する1mLの形質転換緩衝液に再懸濁した。
2.25mLの50%PEG、平均分子量3350、
0.125mLの2M酢酸Li、pH6.0、
0.125mLの2MDTT、および
50μgの剪断サケ精子DNA。
【0179】
約500ngのプラスミドDNAを100μLの再懸濁細胞内で培養し、15min間隔でボルテックス混合しながら39℃に1hr維持した。細胞をロイシン欠乏最少培地プレートに蒔いて、30℃で2〜3日間維持した。
【0180】
%基質変換の判定
pY54、pYSD17またはpY58を含有する形質転換体Y.リポリティカ(Y.lipolytica)の単一コロニーをそれぞれ3mLの最少培地(20g/Lグルコース、1.7g/Lのアミノ酸なしの酵母窒素ベース、1g/LのL−プロリン、0.1g/LのL−アデニン、0.1g/LのL−リジン、pH6.1)中で、30℃でOD6
00が約1.0になるまで生育させた。基質供給のために、次に10μgの基質を含有する3mLの最少培地内で100μLの細胞を30℃で約24hr継代培養した。引き続いてブライ(Bligh)E.G.およびダイヤー(Dyer)W.J.(Can.J.Biochem.Physiol.37:911〜917(1959))で述べられるように、細胞を遠心分離し収集して脂質を抽出した。ナトリウムメトキシド(ローガン(Roughan)G.およびニシダ(Nishida)I.、Arch Biochem Biophys.276(1):38〜46(1990))による脂質抽出物のエステル交換反応によって、脂肪酸メチルエステルを調製し、30m×0.25mm(内径)HP−INNOWAXヒューレットパッカード(Hewlett−Packard)カラムを装着したヒューレットパッカード(Hewlett−Packard)6890 GCで引き続き分析した。オーブン温度は170℃(25分間保持)から185℃に3.5℃/minで上昇させた。%基質変換を以下のように計算した([生産物]/[基質+生産物])×100)。
【0181】
野生型M.アルピナ(M.alpina)Δ6デサチュラーゼによる%基質変換
M.アルピナ(M.alpina)Δ6デサチュラーゼは、LAをGLAに、および/またはALAをSTAに変換する。pY54を含有するY.リポリティカ(Y.lipolytica)株を上述のように(基質供給は必要でない)生育させ、脂質を分析した。結果は、pY54のあるヤロウィア(Yarrowia)株が、約30%のLAをGLAに変換することを示した。
【0182】
野生型S.ディクリナ(S.diclina)Δ17デサチュラーゼの%基質変換
S.ディクリナ(S.diclina)Δ17デサチュラーゼは、ARAをEPAに、および/またはDGLAをETAに変換する。pYSD17を含有するY.リポリティカ(Y.lipolytica)株を単一コロニーから生育させ、10μgのARAを含有する最少培地で継代培養し、上述のように脂質分析にかけた。ARA供給実験の結果は、pYSD17を有するヤロウィア(Yarrowia)株が約23%の細胞内ARAをEPAに変換したことを示した。
【0183】
野生型M.アルピナ(M.alpina)高親和力鎖長延長酵素の%基質変換
M.アルピナ(M.alpina)高親和力PUFA鎖長延長酵素は、GLAをDGLAに、STAをETAに、および/またはEPAをDPAに変換する。pY58を含有するY.リポリティカ(Y.lipolytica)株を単一コロニーから生育させ、10μgのGLAを含有する最少培地で継代培養し、上述のように脂質分析にかけた。GLA供給実験の結果は、pY58を有するヤロウィア(Yarrowia)株が、約30%の細胞内GLAをDGLAに変換したことを示した。
【0184】
ヤロウィア・リポリティカ(Yarrowia lipolytica)において好まれるコドン使用の判定
Y.リポリティカ(Y.lipolytica)のおよそ100個の遺伝子が、アメリカ国立バイオテクノロジー情報センター公共データベースにある。121,167bpを含んでなるこれらの遺伝子のコード領域をDNAStarのEditseqプログラムによって対応する40,389個のアミノ酸に翻訳し、表4に示すY.リポリティカ(Y.lipolytica)のコドン使用プロフィールを判定するために表にした。「No.」と題された欄は、特定のコドンが、40,389個のアミノ酸のサンプル中で特定のアミノ酸をコードする回数を指す。「%」と題された欄は、特定のコドンが特定のアミノ酸をコードする頻度を指す。太字で示されるエントリはY・リポリティカ(Yarrowia lipolytica)中で好まれるコドンを表す。
【0185】
【表5】
【0186】
Y.リポリティカ(Y.lipolytica)中でのさらなる遺伝子発現の最適化の
ために、79個の遺伝子の「ATG」開始コドン周囲の共通配列を調査した。図4では、下線を引いたATG翻訳開始コドンの第1の「A」は、+1と見なされる。分析された遺伝子の77%が、−3の位置に「A」を有し、この位置における「A」に対する強い優先度が示された。また−4、−2および−1の位置で「A」または「C」に、+5の位置で「A」、「C」または「T」に、+6の位置で「G」または「C」に対する優先度があった。したがってY.リポリティカ(Y.lipolytica)中における遺伝子の最適発現のためのコドン最適化翻訳開始部位の好ましい共通配列は、「MAMMATGNHS」(配列番号122)であり、そこで使用された核酸退縮コードは、以下のようである。M=A/C、S=C/G、H=A/C/T、およびN=A/C/G/T。
【0187】
実施例4
コドン最適化Δ6デサチュラーゼ遺伝子の合成
モルティエレラ・アルピナ(Mortierella alpina)(配列番号1)からのΔ6デサチュラーゼ遺伝子は長さが1374bpであった(米国特許第5,968,809号明細書、ジェンバンク(GenBank)#AF465281)。ヤロウィア(Yarrowia)コドン使用パターン、ATG翻訳開始コドン周囲の共通配列、およびRNA安定性の一般規則(グハニヨギ(Guhaniyogi)G.およびJ.ブルーアー(Brewer)、Gene 265(1〜2):11〜23(2001))に従って、M.アルピナ(M.alpina)DNA配列に基づいて、コドン最適化Δ6デサチュラーゼ遺伝子をデザインした。翻訳開始部位を修正するのに加えて、1347bpのコード領域(144のコドンを含んでなる)の152bpもまたコドン最適化した。このコドン最適化遺伝子(配列番号25)とM.アルピナ(M.alpina)からの野生型配列全長(配列番号1)との比較を図5に示し、そこでは太字のヌクレオチドが、コドン最適化遺伝子において修正されたヌクレオチドに対応する。コドン最適化遺伝子中の修正のいずれも、コードされたタンパク質(配列番号2)のアミノ酸配列を変化させなかった。
【0188】
コドン最適化Δ6デサチュラーゼ遺伝子を合成するのに使用される方法は、図6で図示される。最初に14対のオリゴヌクレオチドがデザインされて、M.アルピナ(M.alpina)Δ6デサチュラーゼ遺伝子(例えば配列番号26〜53に対応するD6−1A、D6−1B、D6−2A、D6−2B、D6−3A、D6−3B、D6−4A、D6−4B、D6−5A、D6−5B、D6−6A、D6−6B、D6−7A、D6−7B、D6−8A、D6−8B、D6−9A、D6−9B、D6−10A、D6−10B、D6−11A、D6−11B、D6−12A、D6−12B、D6−13A、D6−13B、D6−14AおよびD6−14B)のコドン最適化コード領域の全長(すなわち1374bp)が延長される。各5’末端の4bpのオーバーハングを例外として、センス(A)およびアンチセンス(B)オリゴヌクレオチドの各対は相補的である。引き続くサブクローン化のために、プライマーD6−1Aはその5’末端にNcoI部位を含有し、プライマーD6−4BおよびD6−5AはStuI部位を含有し、プライマーD6−7B、D6−8A、およびD6−10BはそれぞれBamHI部位を含有した。50mMトリス−HCl(pH7.5)、10mM MgCl2、10mM DTT、0.5mMスペルミジン、0.5mM ATPおよび10単位のT4ポリヌクレオチドキナーゼを含有する20μLの容積中で、100ngの各オリゴヌクレオチドを37℃で1hrリン酸化した。センスおよびアンチセンスオリゴヌクレオチドの各対を混合し、以下のパラメーターを使用して、サーモサイクラー中でアニールした。95℃(2min)、85℃(2min)、65℃(15min)、37℃(15min)、24℃(15min)および4℃(15min)。このようにしてD6−1A(配列番号26)をD6−1B(配列番号27)にアニールし、二本鎖生産物「D6−1AB」を生産した。同様に、D6−2A(配列番号28)をD6−2B(配列番号29)にアニールし、二本鎖生産物「D6−2AB」を生産した。
【0189】
次に以下に示すように4個の別個のアニールされた二本鎖オリゴヌクレオチドのプールを一緒にライゲーションした。
・プール1:D6−1AB、D6−2AB、D6−3AB、およびD6−4ABを含んでなる、
・プール2:D6−5AB、D6−6AB、およびD6−7ABを含んでなる、
・プール3:D6−8AB、D6−9AB、およびD6−10ABを含んでなる、
・プール4:D6−11AB、D6−12AB、D6−13AB、およびD6−14ABを含んでなる。
【0190】
アニールされたオリゴヌクレオチドの各プールを20μLの容積中で10単位のT4DNAリガーゼと共に混合し、ライゲーション反応を一晩16℃で培養した。
【0191】
次に各ライゲーション反応の生産物をPCRによって増幅した。具体的には、テンプレートとしてライゲーションされた「プール1」混合物(すなわちD6−1AB、D6−2AB、D6−3AB、およびD6−4AB)、プライマーとしてオリゴヌクレオチドD6−1(配列番号54)およびD6−4R(配列番号55)を使用して、コドン最適化Δ6デサチュラーゼ遺伝子の第1の部分をPCRによって増幅した。PCR増幅を10mM KCl、10mM (NH4)2SO4、20mMトリス−HCl(pH8.75)、2mM MgSO4、0.1%TritonX−100、100μg/mL BSA(最終濃度)を含有するPCR緩衝液と、各200μMのデオキシリボヌクレオチド三リン酸と、10pmoleの各プライマーと、カリフォルニア州サンディエゴのストラタジーン(Stratagene、San Diego、CA)からの1μLのPfuTurbo DNAポリメラーゼを含んでなる50μLの総容積中で実施した。増幅を以下のようにして実施した。95℃で3minの初期変性、それに続いて95℃で1min、56℃で30sec、72℃で40secの35サイクル。72℃で10minの最終延長サイクルの実施と、それに続く4℃での反応終結。380bpのPCR断片をpGEM−Tイージーベクター(プロメガ(Promega))中にサブクローン化して、pT6(1−4)を生産した。
【0192】
テンプレートとしてライゲーションされた「プール2」混合物(すなわちD6−5AB、D6−6AB、およびD6−7AB)、プライマーとしてオリゴヌクレオチドD6−5(配列番号56)およびD6−7R(配列番号57)を使用して、コドン最適化Δ6デサチュラーゼ遺伝子の第2の部分を同様にPCRによって増幅し、pGEM−T−イージーベクター中にクローン化してpT6(5−7)を生産した。テンプレートとして「プール3」ライゲーション混合物(すなわち、D6−8AB、D6−9AB,およびD6−10AB)、プライマーとしてオリゴヌクレオチドD6−8(配列番号58)およびD6−10R(配列番号59)を使用して、同様にPCRによってコドン最適化Δ6デサチュラーゼ遺伝子の第3の部分を増幅し、pGEM−T−イージーベクター中にクローン化してpT6(8−10)を生産した。最後にテンプレートとして「プール4」ライゲーション混合物(すなわちD6−11AB、D6−12AB、D6−13AB、およびD6−14AB)、プライマーとしてオリゴヌクレオチドD6−11(配列番号60)およびD6−14R(配列番号61)を使用して、同様にPCRによってコドン最適化Δ6デサチュラーゼ遺伝子の第4の部分を増幅し、pGEM−T−イージーベクター中にクローン化してpT6(11−14)を生産した。
【0193】
大腸菌(E.coli)をpT6(1−4)、pT6(5−7)、pT6(8−10)、およびpT6(11−14)で別々に形質転換し、アンピシリン−抵抗性形質転換体から単離されたプラスミドDNAを精製して適切な制限エンドヌクレアーゼで消化し、380bpのpT6(1〜4)のNcoI/StuI断片、310bpのpT6(5−7)のStuI/BamHI断片、320bpのpT6(8−10)のBamHI断片、および
410bpのpT6(11―14)のBamHI/NotI断片を遊離した。次にこれらの断片を組み合わせて正しい方向に一緒にライゲーションし、pY5−13のNcoI/NotI部位に挿入してpYD6Sを生産した(図7)。
【0194】
実施例5
コドン最適化Δ17デサチュラーゼ遺伝子の合成
サプロレグニア・ディクリナ(Saprolegnia diclina)(配列番号3)からのΔ17デサチュラーゼ遺伝子は長さが1077bpであった。ヤロウィア(Yarrowia)コドン使用パターン、ATG翻訳開始コドン周囲の共通配列、およびRNA安定性の一般規則(グハニヨギ(Guhaniyogi)およびブルーアー(Brewer)前出)に従って、S.ディクリナ(S.diclina)DNA配列に基づいて、コドン最適化Δ17デサチュラーゼ遺伝子をデザインした。翻訳開始部位の修正に加えて、1077bpのコード領域(117のコドンを含んでなる)の127bpをコドン最適化した。このコドン最適化DNA配列(配列番号62)とS.ディクリナ(S.diclina)Δ17デサチュラーゼ遺伝子DNA配列(配列番号3)との比較を図8に示し、そこでは太字のヌクレオチドが、コドン最適化遺伝子において修正されたヌクレオチドに対応する。コドン最適化遺伝子中の修正のいずれも、コードされたタンパク質(配列番号4)のアミノ酸配列を変化させなかった。
【0195】
コドン最適化Δ17デサチュラーゼ遺伝子を合成するのに使用される方法は、図9で図示される。最初に11対のオリゴヌクレオチドがデザインされて、S.ディクリナ(S.diclina)Δ17デサチュラーゼ遺伝子(例えば配列番号63〜84に対応するD17−1A、D17−1B、D17−2A、D17−2B、D17−3A、D17−3B、D17−4A、D17−4B、D17−5A、D17−5B、D17−6A、D17−6B、D17−7A、D17−7B、D17−8A、D17−8B、D17−9A、D17−9B、D17−10A、D17−10B、D17−11AおよびD17−11B、対応する)のコドン最適化コード領域の全長が延長される。各5’末端の4bpのオーバーハングを例外として、センス(A)およびアンチセンス(B)オリゴヌクレオチドの各対は相補的である。さらにプライマーD17−1A、D17−4B、D17−5A、D17−8A、およびD17−8Bもまた、引き続くサブクローン化のためにNcoI、BglIIおよびSalI制限部位にそれぞれ導入される。
【0196】
実施例4で使用した方法に続いて、100ngの各オリゴヌクレオチドをT4ポリヌクレオチドキナーゼでリン酸化した。次に各センスおよびアンチセンスオリゴヌクレオチド対を混合し、アニールした。したがって、D17−1A(配列番号63)をD17−1B(配列番号64)にアニールして二本鎖生産物「D17−1AB」を生産した。同様にD17−2A(配列番号65)をD17−2B(配列番号66)にアニールして二本鎖生産物「D17−2AB」などを生産した。
【0197】
次に以下に示すように3個の別個のアニールされた二本鎖オリゴヌクレオチドのプールを一緒にライゲーションした。
・プール1:D17−1AB、D17−2AB、D17−3AB、およびD17−4ABを含んでなる、
・プール2:D17−5AB、D17−6AB、D17−7AB、およびD17−8ABを含んでなる、そして
・プール3:D17−9AB、D17−10AB、およびD17−11ABを含んでなる。
【0198】
アニールされたオリゴヌクレオチドの各プールを20μLの容積中で一晩16℃でライゲーションした。
【0199】
次に各ライゲーション反応の生産物をPCRによって増幅した。具体的には、テンプレートとしてライゲーションされた「プール1」混合物(すなわちD17−1AB、D17−2AB、D17−3AB、およびD17−4AB)、プライマーとしてオリゴヌクレオチドD17−1(配列番号85)およびD17−4R(配列番号86)を使用して、コドン最適化Δ17デサチュラーゼ遺伝子の第1の部分をPCRによって増幅した。実施例4で述べられるPCR条件およびサーモサイクリングプログラムを使用して、PCR増幅を50μLの総容積中で実施した。430bpのPCR断片をpGEM−Tイージーベクター(プロメガ(Promega))中にサブクローン化して、pT17(1−4)を生産した。
【0200】
テンプレートとしてライゲーションされた「プール2」混合物(すなわちD17−5AB、D17−6AB、D17−7AB、およびD17−8AB)、プライマーとしてオリゴヌクレオチドD17−5(配列番号87)およびD17−8D(配列番号88)を使用して、コドン最適化Δ17デサチュラーゼ遺伝子の第2の部分を同様にPCRによって増幅し、pGEM−T−イージーベクター中にクローン化してpT17(5−8)を生産した。最後に、テンプレートとして「プール3」ライゲーション混合物(すなわちD17−9AB、D17−10AB、およびD17−11AB)、プライマーとしてオリゴヌクレオチドD17−8U(配列番号89)、D17−11(配列番号90)を使用して、同様にPCRによってコドン最適化Δ17デサチュラーゼ遺伝子の第3の部分を増幅し、pGEM−T−イージーベクター中にクローン化してpT17(9−11)を生産した。
【0201】
大腸菌(E. coli)をpT17(1−4)、pT17(5−8)、およびpT17(9−11)で別々に形質転換し、プラスミドDNAをアンピシリン−抵抗性形質転換体から単離した。プラスミドDNAを適切な制限エンドヌクレアーゼで精製し消化して、pT17(1―4)の420bp NcoI/BglII断片、pT17(5−8)の400bp BglII/SalI断片、およびpT17(9−11)の300bp SalI/NotI断片を遊離した。次にこれらの断片を組み合わせて一緒にライゲーションして、合成Δ17デサチュラーゼ遺伝子全体の増幅のためのテンプレートとして使用し、D17−1(配列番号85)およびD17−11(配列番号90)をプライマーとして使用した。上述のΔ17デサチュラーゼ遺伝子の各部分のための条件、および以下のサーモサイクリングを使用して、50μLの総容積中でPCR増幅を実施した。95℃で3minの初期変性、それに続いて95℃で1min、56℃で30sec、72℃で1.1minの35サイクル。72℃で10minの最終延長サイクルの実施と、それに続く4℃での反応終結。1.1kBのPCR生産物をNcoI/NotIで消化し、NcoI/NotI消化したpY5−13中にサブクローン化して、pYSD17Sを生産した(図10A)。
【0202】
野生型およびコドン最適化Δ17デサチュラーゼと共に比較基質供給試験で使用するための追加的な「対照」として、プライマーとしてYL53(配列番号120)およびYL54(配列番号121)を使用して、部位特異的変異誘発によってpYSD17中のATに富んだPacI部位(実施例2で述べられる)を除去し、pYSD17Mを生産した(図10B)。
【0203】
実施例6
コドン最適化高親和力PUFA鎖長延長酵素遺伝子の合成
M.アルピナ(M.alpina)(配列番号5)からの高親和力PUFA鎖長延長酵素遺伝子は長さが957bpであった(ジェンバンク(GenBank)#AX464731、国際公開第00/12720号パンフレット)。ヤロウィア(Yarrowia)コドン使用パターン、「ATG」翻訳開始コドン周囲の共通配列、およびRNA安定性の
一般規則(グハニヨギ(Guhaniyogi)およびブルーアー(Brewer)前出)に従って、M.アルピナ(M.alpina)DNA配列に基づいて、コドン最適化高親和力PUFA鎖長延長酵素遺伝子をデザインした。翻訳開始部位を修正するのに加えて、957bpのコード領域(85のコドンを含んでなる)の94bpもまたコドン最適化した。このコドン最適化遺伝子(配列番号91)とM.アルピナ(M.alpina)からの野生型配列全長(配列番号5)との比較を図11に示し、そこでは太字のヌクレオチドが、コドン最適化遺伝子において修正されたヌクレオチドに対応する。コドン最適化遺伝子中の修正のいずれも、コードされたタンパク質(配列番号6)のアミノ酸配列を変化させなかった。
【0204】
高親和力鎖長延長酵素遺伝子を合成するのに使用される方法は、図12で図示される。具体的には10対のオリゴヌクレオチドがデザインされて、M.アルピナ(M.alpina)高親和力鎖長延長酵素のコード領域(すなわち配列番号92−111に対応するEL−1A、EL−1B、EL−2A、EL−2B、EL−3A、EL−3B、EL−4A、EL−4B、EL−5A、EL−5B、EL−6A、EL−6B、EL−7A、EL−7B、EL−8A、EL−8B、EL−9A、EL−9B、EL−10A、およびEL−10B)の長さに沿って延長される。5’末端の4bpのオーバーハングを例外として、センス(A)およびアンチセンス(B)オリゴヌクレオチドの各対は相補的である。
【0205】
実施例4の方法に続いて、100ngの各オリゴヌクレオチドをT4ポリヌクレオチドキナーゼでリン酸化した。次に各センスおよびアンチセンスオリゴヌクレオチド対を混合し、アニールした。したがってEL−1A(配列番号92)をEL1−1B(配列番号93)にアニールして、二本鎖生産物「EL−1AB」を生産した。同様にEL−2A(配列番号95)をEL−2B(配列番号95)にアニールして、二本鎖生産物「EL−2AB」などを生産した。
【0206】
次に以下に示すように2個の別個のアニールされた二本鎖オリゴヌクレオチドのプールを一緒にライゲーションした。
・プール1:EL−1AB、EL−2AB、EL−3AB、EL−4ABおよびEL−5ABを含んでなる、そして
・プール2:EL−6AB、EL−7AB、EL−8AB、EL−9ABおよびEL−10ABを含んでなる。
【0207】
アニールされたオリゴヌクレオチドの各プールを10UのT4DNAリガーゼと共に20μLの容積中で一晩16℃でライゲーションした。
【0208】
次に各ライゲーション反応の生産物をPCRによって増幅した。具体的にはテンプレートとしてライゲーションされた「プール1」混合物(すなわち、EL−1AB、EL−2AB、EL−3AB、EL−4AB、およびEL−5AB)、プライマーとしてオリゴヌクレオチドEL−1(配列番号112)およびEL−5R(配列番号113)を使用して、コドン最適化鎖長延長酵素の第1の部分をPCRによって増幅した。実施例4述べられるようにしてPCR増幅を50μLの反応混合物中で実施した。増幅を以下のようにして実施した。95℃で3minの初期変性、それに続いて95℃で1min、56℃で30sec、72℃で1minの35サイクル。72℃で10minの最終延長サイクルの実施と、それに続く4℃での反応終結。500bpのPCR断片をpGEM−Tイージーベクター(プロメガ(Promega))中にサブクローン化して、pTEL(1−5)を生産した。
【0209】
テンプレートとしてライゲーションされた「プール2」混合物(すなわち、EL−6AB、EL−7AB、EL−8AB、EL−9ABおよびEL−10AB)、プライマーと
してオリゴヌクレオチドEL−6(配列番号114)およびEL−10R(配列番号115)を使用して、コドン最適化鎖長延長酵素遺伝子の第2の部分を同様にPCRによって増幅し、pGEM−T−イージーベクター中にサブクローン化してpTEL(6−10)を生産した。
【0210】
大腸菌(E. coli)細胞をpTEL(1−5)およびpTEL(6−10)で別々に形質転換し、アンピシリン抵抗性形質転換体からのプラスミドDNAを精製し、適切な制限エンドヌクレアーゼで消化して、500bpのpTEL(1−5)のNcoI/SalI断片および470bpのpTEL(6−10)のSalI/NotI断片を遊離した。次にこれらの断片を混合し、NcoI/NotI消化したpY5−13にライゲーションしてpELS−1を生産した。
【0211】
pELS−1インサートのDNA配列分析は、Thr(ACC)からSer(AGC)へのアミノ酸変化をもたらす、+65の位置における1つの「C」から「T」への塩基置換の存在を同定した(「ATG」翻訳コドンの「A」を+1とする)。引き続きプライマーとしてオリゴヌクレオチドEL−M1(配列番号116)およびEL−M2(配列番号117)を使用して、部位特異的変異誘発によってこの変異を修正して、pELS(図13A)を生産した。
【0212】
実施例7
コドン最適化Δ6デサチュラーゼ、Δ17デサチュラーゼ、および高親和力PUFA鎖長延長酵素遺伝子によるヤロウィア・リポリティカ(Yarrowia lipolytica)の形質転換
実施例2で述べた方法に従って、野生型およびコドン最適化Δ6デサチュラーゼ、Δ17デサチュラーゼ、および高親和力PUFA鎖長延長酵素遺伝子を含有するプラスミドをY.リポリティカ(Y.lipolytica)ATCC番号76982中で別々に形質転換した。この技術を使用して、以下のプラスミドを含有する形質転換体を得た。
【0213】
【表6】
【0214】
実施例8
ヤロウィア・リポリティカ(Yarrowia lipolytica)におけるコドン最適化Δ6およびΔ17デサチュラーゼおよび高親和力鎖長延長酵素の変換効率の分析
以下の野生型およびコドン最適化Δ6デサチュラーゼ、Δ17デサチュラーゼ、および高親和力鎖長延長酵素それぞれの変換効率の比較では、コドン最適化がY.リポリティカ(Y.lipolytica)において、LAからGLA(Δ6デサチュラーゼ)への%基質変換をおよそ40%、ARAからEPA(Δ17デサチュラーゼ)への%基質変換を
約2倍に、そしてGLAからDGLA(鎖長延長酵素)への%基質変換を約57%改善したと判定された。
【0215】
コドン最適化Δ6デサチュラーゼによる%基質変換
Δ6デサチュラーゼは、LAをGLAに、および/またはALAをSTAに変換する(図1参照)。野生型とコドン最適化Δ6デサチュラーゼ遺伝子の変換効率を比較するために、それぞれ別のプラスミドコンストラクト(すなわちpYSD6またはpYD6S)を含有するヤロウィア・リポリティカ(Yarrowia lipolytica)において、%基質変換([生産物]/[基質+生産物])×100)を求めた。具体的には3mLの最少培地中でpYSD6またはpYD6Sのいずれかを含有するヤロウィア・リポリティカ(Yarrowia lipolytica)を単一コロニーから生育させ、10μgのLAを含有する最少培地中で継代培養して、実施例2で述べるように脂質分析を施した。
【0216】
実験の結果は、pYSD6を含有するヤロウィア(Yarrowia)株が、約30%の基質LAをGLAに変換し(図14A)、他方pYD6Sを含有するものは約42%のLAをGLAに変換する(図14B)ことを示唆した。これに基づき、コドン最適化Δ6デサチュラーゼ遺伝子を含有するヤロウィア(Yarrowia)は、Y.リポリティカ(Y.lipolytica)中の野生型M.アルピナ(M.alpina)Δ6デサチュラーゼ遺伝子よりもおよそ40%多くのLAを変換した。
【0217】
コドン最適化Δ17デサチュラーゼによる%基質転換
Δ17デサチュラーゼはDGLAをETAに、および/またはARAをEPAに変換する(図1参照)。野生型とコドン最適化Δ17デサチュラーゼ遺伝子の変換効率を比較するために、pYSD17、pYSD17M、およびpYSD17Sを含有するヤロウィア・リポリティカ(Yarrowia lipolytica)における%基質変換を求めた。3mLの最小培地中で各形質転換体を単一コロニーから生育させ、10μgのARAを含有する最少培地中で継代培養して、実施例2で述べるように脂質分析を施した。
【0218】
ARA供給実験の結果は、対照プラスミドpYSD17またはpYSD17Mを有するヤロウィア(Yarrowia)株が約23%の細胞内ARAをEPAに変換し(図15A)、他方pYSD17S上にコドン最適化Δ17デサチュラーゼ遺伝子を含有するものが約45%の細胞内ARAをEPAに変換する(図15B)ことを示した。したがってコドン最適化Δ17デサチュラーゼ遺伝子を含有するヤロウィア(Yarrowia)は、野生型S.ディクリナ(S.diclina)遺伝子を含有する株よりも約2倍多いARAを変換した。
【0219】
コドン最適化鎖長延長酵素による%基質転換
M.アルピナ(M.alpina)の高親和力PUFA鎖長延長酵素は、主にGLAからDGLAへの転換を触媒する役目を果たす(図1参照、国際公開第00/12720号パンフレット)。野生型とコドン最適化鎖長延長酵素遺伝子の変換効率を比較するために、pY58およびpELSを含有するヤロウィア・リポリティカ(Yarrowia lipolytica)中で%基質変換を求めた。形質転換体を3mLの最少培地中で単一コロニーから生育させ、10μgのGLAを含有する最少培地中で継代培養し、実施例2で述べるように脂質分析を施した。
【0220】
GLA供給実験の結果は、pY58を含有するヤロウィア・リポリティカ(Yarrowia lipolytica)が約30%の細胞内基質GLAをDGLAに変換し(図16A)、他方pELSを含有するものが約47%のGLAをDGLAに変換する(図16B)ことを示した。これに基づいて、コドン最適化鎖長延長酵素遺伝子は、Y.リポリ
ティカ(Y.lipolytica)中の野生型M.アルピナ(M.alpina)鎖長延長酵素遺伝子よりもおよそ57%多いGLAを変換した。
【図面の簡単な説明】
【0221】
【図1】ω−3およびω−6脂肪酸生合成経路を図示する。
【図2】ヤロウィア・リポリティカ(Yarrowia lipolytica)中で使用するためのプラスミドベクターpY5の構築を図示する。
【図3】Y.リポリティカ(Y.lipolytica)における遺伝子発現のためのプラスミドベクターpY5−13およびpY5−4の構築を図示する。
【図4】Y.リポリティカ(Y.lipolytica)中の翻訳開始コドン「ATG」周囲の好ましい共通配列を図示する。
【図5】モルティエレラ・アルピナ(Mortierella alpina)Δ6デサチュラーゼ遺伝子と、Y.リポリティカ(Y.lipolytica)における発現のためにコドン最適化された合成遺伝子とのDNA配列の比較を示す。
【図6】コドン最適化Δ6デサチュラーゼ遺伝子の生体外(in vitro)合成のために用いられるストラテジーを図示する。
【図7】Y.リポリティカ(Y.lipolytica)におけるコドン最適化Δ6デサチュラーゼ遺伝子の発現のためのプラスミドを示す。
【図8】サプロレグニア・ディクリナ(Saprolegnia diclina)Δ17デサチュラーゼ遺伝子と、Y.リポリティカ(Y.lipolytica)における発現のためにコドン最適化された合成遺伝子とのDNA配列の比較を示す。
【図9】コドン最適化Δ17デサチュラーゼ遺伝子の生体外(in vitro)合成のために用いられるストラテジーを図示する。
【図10】Y.リポリティカ(Y.lipolytica)におけるコドン最適化および野生型Δ17デサチュラーゼ遺伝子発現のためのプラスミドを示す。
【図11】モルティエレラ・アルピナ(Mortierella alpina)鎖長延長酵素遺伝子と、Y.リポリティカ(Y.lipolytica)における発現のためにコドン最適化された合成遺伝子とのDNA配列の比較を示す。
【図12】コドン最適化鎖長延長酵素遺伝子の生体外(in vitro)合成のために用いられるストラテジーを図示する。
【図13】Y.リポリティカ(Y.lipolytica)におけるコドン最適化および野生型鎖長延長酵素遺伝子発現のためのプラスミドを示す。
【図14A】約30%の基質LAからGLAへの変換を示す、コドン最適化および野生型Δ6デサチュラーゼ遺伝子で形質転換されたY.リポリティカ(Y.lipolytica)中で生産された脂肪酸のガスクロマトグラフィー分析の結果を示す。
【図14B】約42%の基質LAからGLAへの変換を示す、コドン最適化および野生型Δ6デサチュラーゼ遺伝子で形質転換されたY.リポリティカ(Y.lipolytica)中で生産された脂肪酸のガスクロマトグラフィー分析の結果を示す。
【図15】約23%の細胞内ARAからEPAへの変換を示す、コドン最適化および野生型Δ17デサチュラーゼ遺伝子で形質転換されたY.リポリティカ(Y.lipolytica)中で生産された脂肪酸のガスクロマトグラフィー分析の結果を示す。
【図15B】約45%の細胞内ARAからEPAへの変換を示す、コドン最適化および野生型Δ17デサチュラーゼ遺伝子で形質転換されたY.リポリティカ(Y.lipolytica)中で生産された脂肪酸のガスクロマトグラフィー分析の結果を示す。
【図16】約30%の細胞内GLAからDGLAへの変換を示す、コドン最適化および野生型鎖長延長酵素遺伝子で形質転換されたY.リポリティカ(Y.lipolytica)中で生産された脂肪酸のガスクロマトグラフィー分析の結果を示す。
【図16B】約47%の細胞内GLAからDGLAへの変換を示す、コドン最適化および野生型鎖長延長酵素遺伝子で形質転換されたY.リポリティカ(Y.lipolytica)中で生産された脂肪酸のガスクロマトグラフィー分析の結果を示す。
【特許請求の範囲】
【請求項1】
a)配列番号25に記載のΔ6デサチュラーゼをコードする単離された核酸分子、または
b)(a)と完全に相補的である単離された核酸分子
よりなる群から選択される単離された核酸分子。
【請求項2】
少なくとも144個のコドンがヤロウィア(Yarrowia)における発現のためにコドン最適化される、配列番号2に記載のΔ6デサチュラーゼをコードする単離された核酸分子。
【請求項3】
a)配列番号62に記載のΔ17デサチュラーゼをコードする単離された核酸分子、または
b)(a)と完全に相補的である単離された核酸分子
よりなる群から選択される単離された核酸分子。
【請求項4】
少なくとも117個のコドンがヤロウィア(Yarrowia)における発現のためにコドン最適化されている、配列番号4に記載のΔ17デサチュラーゼをコードする単離された核酸分子。
【請求項5】
a)配列番号91に記載の鎖長延長酵素をコードする単離された核酸分子、または
b)(a)と完全に相補的である単離された核酸分子
よりなる群から選択される単離された核酸分子。
【請求項6】
少なくとも85個のコドンがヤロウィア(Yarrowia)における発現のためにコドン最適化されている、配列番号6に記載の鎖長延長酵素をコードする単離された核酸分子。
【請求項7】
適切な調節配列に作動可能に連結された、請求項1〜6のいずれかに記載の単離された核酸分子を含んでなる、キメラ遺伝子。
【請求項8】
請求項7に記載のキメラ遺伝子を含んでなる、形質転換されたヤロウィア(Yarrowia)種。
【請求項9】
ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号20362、ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号8862、ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号18944、ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号76982、およびヤロウィア・リポリティカ(Yarrowia lipolytica)LGAMS(7)1よりなる群から選択される、請求項8に記載の形質転換されたヤロウィア(Yarrowia)種。
【請求項10】
a)(i)適切な調節配列の制御下にあり、Δ6デサチュラーゼポリペプチドをコードする、請求項1または2に記載の核酸分子、および
(ii)リノール酸からなるデサチュラーゼ基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)のヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項1または2に記載の核酸分子が発現し、前記リノール酸がγ−リノレン酸に変換し、
c)場合により、ステップ(b)のγ−リノレン酸を回収すること
を含んでなるγ−リノレン酸の生産方法。
【請求項11】
デサチュラーゼ基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項10に記載の方法。
【請求項12】
a)(i)適切な調節配列の制御下にあり、Δ6デサチュラーゼポリペプチドをコードする、請求項1または2に記載の核酸分子、および
(ii)α−リノール酸からなるデサチュラーゼ基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)のヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項1または2に記載の核酸分子が発現し、前記α−リノール酸がステアリドン酸に変換し、
c)場合により、ステップ(b)のステアリドン酸を回収すること
を含んでなるステアリドン酸の生産方法。
【請求項13】
デサチュラーゼ基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項12に記載の方法。
【請求項14】
a)(i)適切な調節配列の制御下にあり、Δ17デサチュラーゼポリペプチドをコードする、請求項3または4に記載の核酸分子、および
(ii)ジホモ−γ−リノール酸からなるデサチュラーゼ基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)のヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項3または4に記載の核酸分子が発現し、ジホモ−γ−リノール酸がエイコサテトラエン酸に変換し、
c)場合により、ステップ(b)のエイコサテトラエン酸を回収すること
を含んでなるエイコサテトラエン酸の生産方法。
【請求項15】
デサチュラーゼ基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項14に記載の方法。
【請求項16】
a)(i)適切な調節配列の制御下にあり、Δ17デサチュラーゼポリペプチドをコードする、請求項3または4に記載の核酸分子、および
(ii)アラキドン酸からなるデサチュラーゼ基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)のヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項3または4に記載の核酸分子が発現し、前記アラキドン酸がエイコサペンタエン酸に変換し、
c)場合により、ステップ(b)のエイコサペンタエン酸を回収すること
を含んでなるエイコサペンタエン酸の生産方法。
【請求項17】
デサチュラーゼ基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項16に記載の方法。
【請求項18】
a)(i)適切な調節配列の制御下にあり、鎖長延長酵素ポリペプチドをコードする、請求項5または6に記載の核酸分子、および
(ii)γ−リノレン酸からなる鎖長延長酵素基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)のヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項5または6に記載の核酸分子が発現し、前記γ−リノレン酸がジホモ
−γ−リノール酸に変換し、
c)場合により、ステップ(b)の前記ジホモ−γ−リノール酸を回収すること
を含んでなるジホモ−γ−リノール酸の生産方法。
【請求項19】
鎖長延長酵素基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項18に記載の方法。
【請求項20】
a)(i)適切な調節配列の制御下にあり、鎖長延長酵素ポリペプチドをコードする、請求項5または6に記載の核酸分子、および
(ii)ステアリドン酸からなる鎖長延長酵素基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)の前記ヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項5または6に記載の核酸分子が発現し、前記ステアリドン酸がエイコサテトラエン酸に変換し、
c)場合により、ステップ(b)の前記エイコサテトラエン酸を回収すること
を含んでなるエイコサテトラエン酸の生産方法。
【請求項21】
鎖長延長酵素基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項20に記載の方法。
【請求項22】
a)(i)適切な調節配列の制御下で鎖長延長酵素ポリペプチドをコードする、請求項5または6に記載の核酸分子、および
(ii)エイコサペンタエン酸からなる鎖長延長酵素基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)のヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項5または6に記載の核酸分子が発現し、前記エイコサペンタエン酸がドコサペンタエン酸に変換し、
c)場合により、ステップ(b)のドコサペンタエン酸を回収すること
を含んでなるドコサペンタエン酸の生産方法。
【請求項23】
鎖長延長酵素基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項22に記載の方法。
【請求項24】
a)油性酵母菌種について、ヌクレオチドコード領域配列および対応するポリペプチド配列を得て、コドンのデータベースを形成し、
b)前記コドンのデータベースを分析し、いずれのコドンが各アミノ酸を優先的にコードするのか判定し、
c)油性酵母菌種において発現させる遺伝子の配列を得て、
d)ステップ(c)の配列において好ましくないコドンをステップ(b)の好ましいコドンで置き換えことで、前記遺伝子が油性酵母菌種における発現のためにコドン最適化されること
を含んでなる、油性酵母菌における発現のために遺伝子を最適化する方法。
【請求項25】
油性酵母菌が、ヤロウィア(Yarrowia)、カンジダ(Candida)、ロドトルラ(Rhodotorula)、ロドスポリジウム(Rhodosporidium)、クリプトコッカス(Cryptococcus)、トリコスポロン(Trichosporon)、およびリポマイセス(Lipomyces)よりなる群から選択される、請求項24に記載の方法。
【請求項26】
宿主細胞が、ヤロウィア・リポリティカ(Yarrowia lipolytica)
ATCC番号20362、ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号8862、ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号18944、ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号76982、およびヤロウィア・リポリティカ(Yarrowia lipolytica)LGAMS(7)1よりなる群から選択される、請求項25に記載の方法。
【請求項27】
最適化させる遺伝子が、Δ12デサチュラーゼ、Δ6デサチュラーゼ、Δ9デサチュラーゼ、鎖長延長酵素、Δ5デサチュラーゼ、Δ4デサチュラーゼ、Δ17デサチュラーゼ、およびΔ15デサチュラーゼよりなる群から選択される酵素をコードする、請求項24に記載の方法。
【請求項28】
ヤロウィア(Yarrowia)翻訳開始部位を含んでなる、配列番号122に記載の単離された核酸分子。
【請求項29】
a)ヤロウィア(Yarrowia)において発現させる外来遺伝子を提供し、
b)ステップ(a)の遺伝子を請求項28の単離された核酸分子と作動可能に連結して、前記外来遺伝子がヤロウィア(Yarrowia)における発現のために最適化されること
を含んでなる、ヤロウィア(Yarrowia)宿主における遺伝子の発現を最適化する方法。
【請求項30】
外来遺伝子が、Δ12デサチュラーゼ、Δ6デサチュラーゼ、鎖長延長酵素、Δ5デサチュラーゼ、Δ17デサチュラーゼ、Δ15デサチュラーゼ、Δ9デサチュラーゼ、およびΔ4デサチュラーゼよりなる群から選択される酵素をコードする、請求項29に記載の方法。
【請求項31】
請求項10〜27のいずれか一項の方法によって生産される微生物油。
【請求項1】
a)配列番号25に記載のΔ6デサチュラーゼをコードする単離された核酸分子、または
b)(a)と完全に相補的である単離された核酸分子
よりなる群から選択される単離された核酸分子。
【請求項2】
少なくとも144個のコドンがヤロウィア(Yarrowia)における発現のためにコドン最適化される、配列番号2に記載のΔ6デサチュラーゼをコードする単離された核酸分子。
【請求項3】
a)配列番号62に記載のΔ17デサチュラーゼをコードする単離された核酸分子、または
b)(a)と完全に相補的である単離された核酸分子
よりなる群から選択される単離された核酸分子。
【請求項4】
少なくとも117個のコドンがヤロウィア(Yarrowia)における発現のためにコドン最適化されている、配列番号4に記載のΔ17デサチュラーゼをコードする単離された核酸分子。
【請求項5】
a)配列番号91に記載の鎖長延長酵素をコードする単離された核酸分子、または
b)(a)と完全に相補的である単離された核酸分子
よりなる群から選択される単離された核酸分子。
【請求項6】
少なくとも85個のコドンがヤロウィア(Yarrowia)における発現のためにコドン最適化されている、配列番号6に記載の鎖長延長酵素をコードする単離された核酸分子。
【請求項7】
適切な調節配列に作動可能に連結された、請求項1〜6のいずれかに記載の単離された核酸分子を含んでなる、キメラ遺伝子。
【請求項8】
請求項7に記載のキメラ遺伝子を含んでなる、形質転換されたヤロウィア(Yarrowia)種。
【請求項9】
ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号20362、ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号8862、ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号18944、ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号76982、およびヤロウィア・リポリティカ(Yarrowia lipolytica)LGAMS(7)1よりなる群から選択される、請求項8に記載の形質転換されたヤロウィア(Yarrowia)種。
【請求項10】
a)(i)適切な調節配列の制御下にあり、Δ6デサチュラーゼポリペプチドをコードする、請求項1または2に記載の核酸分子、および
(ii)リノール酸からなるデサチュラーゼ基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)のヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項1または2に記載の核酸分子が発現し、前記リノール酸がγ−リノレン酸に変換し、
c)場合により、ステップ(b)のγ−リノレン酸を回収すること
を含んでなるγ−リノレン酸の生産方法。
【請求項11】
デサチュラーゼ基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項10に記載の方法。
【請求項12】
a)(i)適切な調節配列の制御下にあり、Δ6デサチュラーゼポリペプチドをコードする、請求項1または2に記載の核酸分子、および
(ii)α−リノール酸からなるデサチュラーゼ基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)のヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項1または2に記載の核酸分子が発現し、前記α−リノール酸がステアリドン酸に変換し、
c)場合により、ステップ(b)のステアリドン酸を回収すること
を含んでなるステアリドン酸の生産方法。
【請求項13】
デサチュラーゼ基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項12に記載の方法。
【請求項14】
a)(i)適切な調節配列の制御下にあり、Δ17デサチュラーゼポリペプチドをコードする、請求項3または4に記載の核酸分子、および
(ii)ジホモ−γ−リノール酸からなるデサチュラーゼ基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)のヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項3または4に記載の核酸分子が発現し、ジホモ−γ−リノール酸がエイコサテトラエン酸に変換し、
c)場合により、ステップ(b)のエイコサテトラエン酸を回収すること
を含んでなるエイコサテトラエン酸の生産方法。
【請求項15】
デサチュラーゼ基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項14に記載の方法。
【請求項16】
a)(i)適切な調節配列の制御下にあり、Δ17デサチュラーゼポリペプチドをコードする、請求項3または4に記載の核酸分子、および
(ii)アラキドン酸からなるデサチュラーゼ基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)のヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項3または4に記載の核酸分子が発現し、前記アラキドン酸がエイコサペンタエン酸に変換し、
c)場合により、ステップ(b)のエイコサペンタエン酸を回収すること
を含んでなるエイコサペンタエン酸の生産方法。
【請求項17】
デサチュラーゼ基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項16に記載の方法。
【請求項18】
a)(i)適切な調節配列の制御下にあり、鎖長延長酵素ポリペプチドをコードする、請求項5または6に記載の核酸分子、および
(ii)γ−リノレン酸からなる鎖長延長酵素基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)のヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項5または6に記載の核酸分子が発現し、前記γ−リノレン酸がジホモ
−γ−リノール酸に変換し、
c)場合により、ステップ(b)の前記ジホモ−γ−リノール酸を回収すること
を含んでなるジホモ−γ−リノール酸の生産方法。
【請求項19】
鎖長延長酵素基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項18に記載の方法。
【請求項20】
a)(i)適切な調節配列の制御下にあり、鎖長延長酵素ポリペプチドをコードする、請求項5または6に記載の核酸分子、および
(ii)ステアリドン酸からなる鎖長延長酵素基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)の前記ヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項5または6に記載の核酸分子が発現し、前記ステアリドン酸がエイコサテトラエン酸に変換し、
c)場合により、ステップ(b)の前記エイコサテトラエン酸を回収すること
を含んでなるエイコサテトラエン酸の生産方法。
【請求項21】
鎖長延長酵素基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項20に記載の方法。
【請求項22】
a)(i)適切な調節配列の制御下で鎖長延長酵素ポリペプチドをコードする、請求項5または6に記載の核酸分子、および
(ii)エイコサペンタエン酸からなる鎖長延長酵素基質の供給源
を含んでなるヤロウィア(Yarrowia)種を提供し、
b)ステップ(a)のヤロウィア(Yarrowia)種を適切な発酵性炭素源存在下で生育させ、請求項5または6に記載の核酸分子が発現し、前記エイコサペンタエン酸がドコサペンタエン酸に変換し、
c)場合により、ステップ(b)のドコサペンタエン酸を回収すること
を含んでなるドコサペンタエン酸の生産方法。
【請求項23】
鎖長延長酵素基質の供給源が前記ヤロウィア(Yarrowia)種に内因性である、請求項22に記載の方法。
【請求項24】
a)油性酵母菌種について、ヌクレオチドコード領域配列および対応するポリペプチド配列を得て、コドンのデータベースを形成し、
b)前記コドンのデータベースを分析し、いずれのコドンが各アミノ酸を優先的にコードするのか判定し、
c)油性酵母菌種において発現させる遺伝子の配列を得て、
d)ステップ(c)の配列において好ましくないコドンをステップ(b)の好ましいコドンで置き換えことで、前記遺伝子が油性酵母菌種における発現のためにコドン最適化されること
を含んでなる、油性酵母菌における発現のために遺伝子を最適化する方法。
【請求項25】
油性酵母菌が、ヤロウィア(Yarrowia)、カンジダ(Candida)、ロドトルラ(Rhodotorula)、ロドスポリジウム(Rhodosporidium)、クリプトコッカス(Cryptococcus)、トリコスポロン(Trichosporon)、およびリポマイセス(Lipomyces)よりなる群から選択される、請求項24に記載の方法。
【請求項26】
宿主細胞が、ヤロウィア・リポリティカ(Yarrowia lipolytica)
ATCC番号20362、ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号8862、ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号18944、ヤロウィア・リポリティカ(Yarrowia lipolytica)ATCC番号76982、およびヤロウィア・リポリティカ(Yarrowia lipolytica)LGAMS(7)1よりなる群から選択される、請求項25に記載の方法。
【請求項27】
最適化させる遺伝子が、Δ12デサチュラーゼ、Δ6デサチュラーゼ、Δ9デサチュラーゼ、鎖長延長酵素、Δ5デサチュラーゼ、Δ4デサチュラーゼ、Δ17デサチュラーゼ、およびΔ15デサチュラーゼよりなる群から選択される酵素をコードする、請求項24に記載の方法。
【請求項28】
ヤロウィア(Yarrowia)翻訳開始部位を含んでなる、配列番号122に記載の単離された核酸分子。
【請求項29】
a)ヤロウィア(Yarrowia)において発現させる外来遺伝子を提供し、
b)ステップ(a)の遺伝子を請求項28の単離された核酸分子と作動可能に連結して、前記外来遺伝子がヤロウィア(Yarrowia)における発現のために最適化されること
を含んでなる、ヤロウィア(Yarrowia)宿主における遺伝子の発現を最適化する方法。
【請求項30】
外来遺伝子が、Δ12デサチュラーゼ、Δ6デサチュラーゼ、鎖長延長酵素、Δ5デサチュラーゼ、Δ17デサチュラーゼ、Δ15デサチュラーゼ、Δ9デサチュラーゼ、およびΔ4デサチュラーゼよりなる群から選択される酵素をコードする、請求項29に記載の方法。
【請求項31】
請求項10〜27のいずれか一項の方法によって生産される微生物油。
【図1】

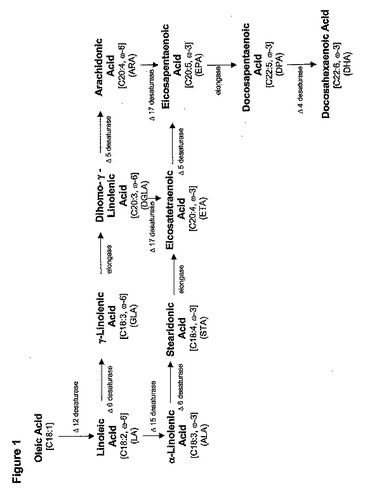
【図2】
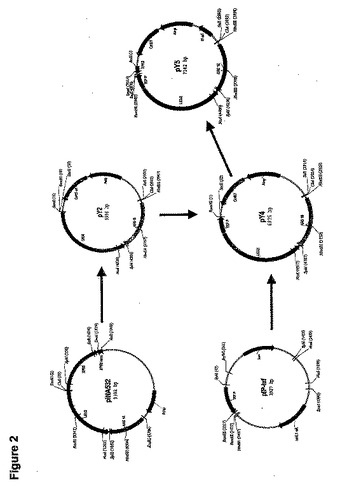
【図3】
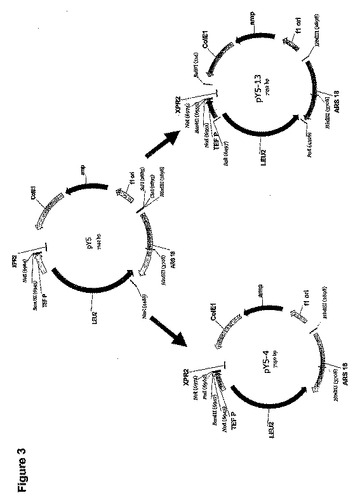
【図4】
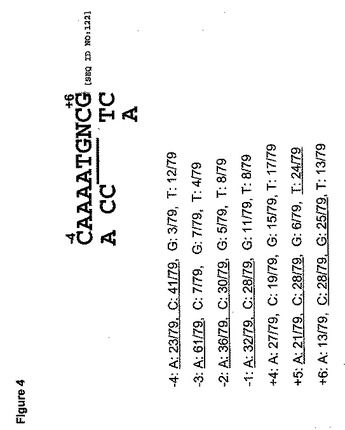
【図5A】

【図5A】

【図6】
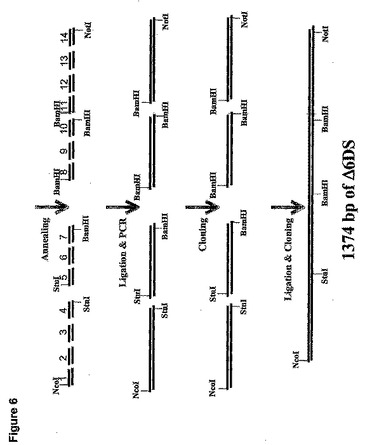
【図7】
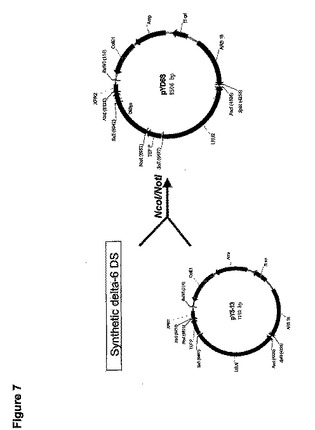
【図8A】

【図8A】

【図9】
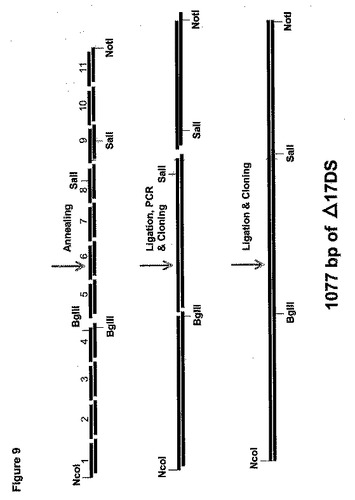
【図10】
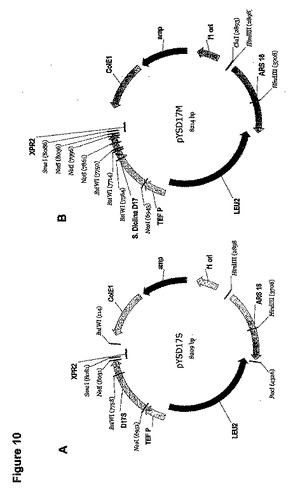
【図11】

【図12】
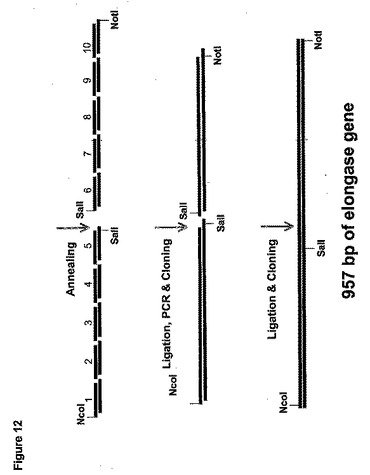
【図13】
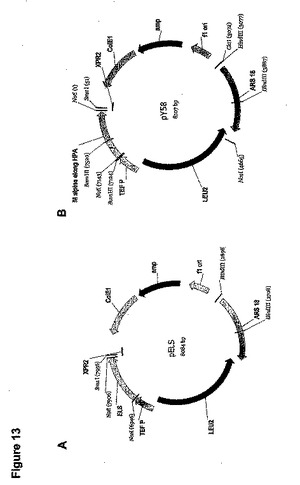
【図14】
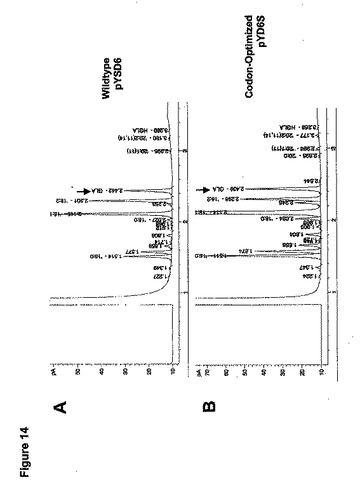
【図15】
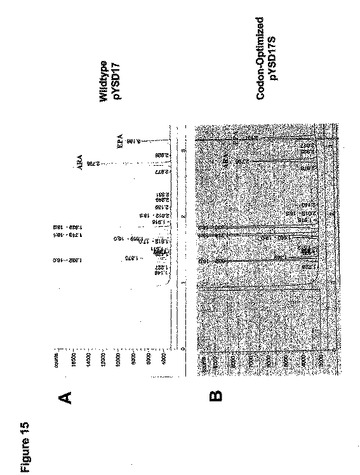
【図16】
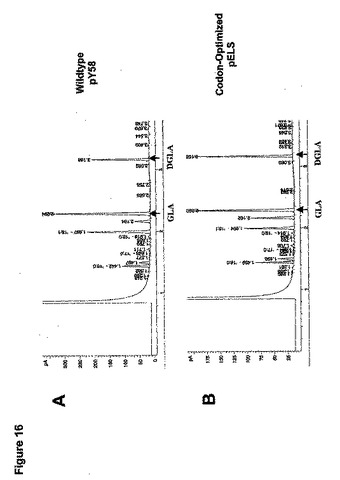
【図2】
【図3】
【図4】
【図5A】
【図5A】
【図6】
【図7】
【図8A】
【図8A】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【公表番号】特表2007−504838(P2007−504838A)
【公表日】平成19年3月8日(2007.3.8)
【国際特許分類】
【出願番号】特願2006−532884(P2006−532884)
【出願日】平成16年5月7日(2004.5.7)
【国際出願番号】PCT/US2004/014442
【国際公開番号】WO2004/101753
【国際公開日】平成16年11月25日(2004.11.25)
【出願人】(390023674)イー・アイ・デュポン・ドウ・ヌムール・アンド・カンパニー (2,692)
【氏名又は名称原語表記】E.I.DU PONT DE NEMOURS AND COMPANY
【Fターム(参考)】
【公表日】平成19年3月8日(2007.3.8)
【国際特許分類】
【出願日】平成16年5月7日(2004.5.7)
【国際出願番号】PCT/US2004/014442
【国際公開番号】WO2004/101753
【国際公開日】平成16年11月25日(2004.11.25)
【出願人】(390023674)イー・アイ・デュポン・ドウ・ヌムール・アンド・カンパニー (2,692)
【氏名又は名称原語表記】E.I.DU PONT DE NEMOURS AND COMPANY
【Fターム(参考)】
[ Back to top ]