
油脂分解菌及び油脂分解剤
【課題】 至適油脂分解条件(温度、pH等)がBOD分解菌の至適生育条件(温度、pH等)の範囲内に入らず、十分な増殖速度及び油脂分解速度を有する油脂分解菌を提供すること。
【解決手段】 本発明は、Pseudmonas属に属し、例えば、以下の菌学的性質を示す、油脂分解菌を提供する。
37℃での生育 +
45℃での生育 +
嫌気性条件下での生育 −
硝酸塩還元活性 +
インドール産生活性 −
アルギニンジヒドロラーゼ活性 +
ウレアーゼ活性 −
ゼラチン加水分解活性 +
β−ガラクトシダーゼ活性 −
チトクロームオキシダーゼ活性 +
ブドウ糖資化性 +
L−アラビノース資化性 −
D−マンノース資化性 −
D−マンニトール資化性 −
N−アセチル−D−グルコサミン資化性 +
マルトース資化性 −
グルコン酸カリウム資化性 +
なお、「+」は陽性であること、「−」は陰性であることを示す。
【解決手段】 本発明は、Pseudmonas属に属し、例えば、以下の菌学的性質を示す、油脂分解菌を提供する。
37℃での生育 +
45℃での生育 +
嫌気性条件下での生育 −
硝酸塩還元活性 +
インドール産生活性 −
アルギニンジヒドロラーゼ活性 +
ウレアーゼ活性 −
ゼラチン加水分解活性 +
β−ガラクトシダーゼ活性 −
チトクロームオキシダーゼ活性 +
ブドウ糖資化性 +
L−アラビノース資化性 −
D−マンノース資化性 −
D−マンニトール資化性 −
N−アセチル−D−グルコサミン資化性 +
マルトース資化性 −
グルコン酸カリウム資化性 +
なお、「+」は陽性であること、「−」は陰性であることを示す。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、油脂分解菌及び油脂分解剤に関する。
【背景技術】
【0002】
微生物を利用した排水処理方法として、一般に、好気性微生物による活性汚泥法(好気性処理法)及び嫌気性微生物によるメタン発酵法(嫌気性処理法)が知られている。好気性処理法では曝気するためのエネルギーを要するのに対し、嫌気性処理法は消費エネルギーが少なく、またメタンなどを主成分とするバイオガスを得ることもできる。このため、嫌気性処理法は、好気性処理法よりもエネルギー効率の点で有利であるといわれている。
【0003】
溶解性有機物を高濃度に含む高負荷排水をメタン発酵法で処理するための、種々のタイプの上向流式嫌気性処理槽が開発されている。例えば、UASB(Upflow Anaerobic Sludge Blanket)法やこれを改良したEGSB(Expanded Glanular Sludge Bed)法を利用した処理槽が知られている。これらの上向流式嫌気性処理槽は、グラニュール汚泥と呼ばれる粒状の汚泥を槽内に収容することができ、被処理水がグラニュール汚泥と接触しながら上方に流れることによって、有機物が効率的に分解される。
【0004】
ところで、製造工程で油脂を扱う食品工場の排水など、油脂を多く含む有機性排水をグラニュール汚泥で処理する場合、油脂がグラニュール汚泥に付着し、これによって排水とグラニュール汚泥との接触効率が低下して有機物の処理が不十分となりやすい。このような問題を解決する手段として、特許文献1には上向流式嫌気性処理槽に被処理水を導入するに先立ち、被処理水から油分を分離し、これをリパーゼ生成菌によって分解する油脂分解槽を備えた嫌気性処理装置が記載されている。
【0005】
また、油脂分解性を有するPseudomonas属菌が開示されている(特許文献2及び特許文献3)。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2005−270862号公報
【特許文献2】特開2008−220225号公報
【特許文献3】特開平6−153922号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、従来知られている油脂分解菌は、その至適油脂分解条件(温度、pH等)が、油脂分解菌よりも増殖速度の速い溶解性有機物分解菌(以下、BOD分解菌という。)の至適生育条件(温度、pH等)の範囲内に入ってしまう。そのため、BOD分解菌が油脂分解槽の汚泥中で優勢となり、油脂の分解効率低下に加えて溶解性有機物(以下、BOD成分という。)の分解により大量の余剰汚泥が生じる問題があった。また、BOD分解菌が油脂分解槽の汚泥中で優占的となり、油脂分解菌が存在しなくなるという問題もあった。
【0008】
そこで、本発明は、至適油脂分解条件(温度、pH等)がBOD分解菌の至適生育条件(温度、pH等)の範囲内に入らず、十分な増殖速度及び油脂分解速度を有する油脂分解菌の提供を目的とする。本発明はまた、上記油脂分解菌を含む油脂分解剤の提供も目的とする。
【課題を解決するための手段】
【0009】
本発明は、Pseudmonas属に属し、以下の菌学的性質を示す、油脂分解菌。
細胞形態 桿菌
細胞の直径 0.6〜0.7×1.0〜1.5μm
胞子の有無 −
グラム染色性 −
運動性 +
37℃での生育 +
45℃での生育 +
嫌気性条件下での生育 −
カタラーゼ +
オキシダーゼ +
グルコースからの酸/ガス産生 −/−
グルコースの酸化/発酵 +/−
硝酸塩還元活性 +
インドール産生活性 −
ブドウ糖酸性化活性 −
アルギニンジヒドロラーゼ活性 +
ウレアーゼ活性 −
エスクリン加水分解活性 −
ゼラチン加水分解活性 +
β−ガラクトシダーゼ活性 −
チトクロームオキシダーゼ活性 +
ブドウ糖資化性 +
L−アラビノース資化性 −
D−マンノース資化性 −
D−マンニトール資化性 −
N−アセチル−D−グルコサミン資化性 +
マルトース資化性 −
グルコン酸カリウム資化性 +
n−カプリン酸資化性 +
アジピン酸資化性 +
dl−リンゴ酸資化性 +
クエン酸ナトリウム資化性 +
酢酸フェニル資化性 −
なお、「+」は陽性であること、「−」は陰性であることを示す。
【0010】
上記油脂分解菌は、寒天培地上において、30℃、24時間培養で直径2〜3mmの周縁波状の円形、表面がスムーズ、扁平状で、不透明な緑色コロニーを形成することが好ましい。
【0011】
また、上記油脂分解菌は、配列番号1で示される16S rDNA塩基配列を有することが好ましい。
【0012】
上記油脂分解菌として、独立行政法人製品評価技術基盤機構特許微生物寄託センター(郵便番号292−0818、日本国千葉県木更津市かずさ鎌足2丁目5番地8号 NITEバイオテクノロジー本部)に受託番号NITE P−847で2009年11月25日に受託されているPseudmonas aeruginosa SS−219株とすることができる。
【0013】
本発明はまた、上述の油脂分解菌を含む油脂分解剤を提供する。
【0014】
上記油脂分解菌は、アルカリ条件及び高温条件下で効率よく増殖及び油脂分解が可能であるため、BOD分解菌の増殖を抑制しながら油脂を分解することができる。また、上記油脂分解菌は従来の油脂分解菌と比べて油脂分解速度が速いため、高濃度の油脂を含む排水の処理に好適に利用することができる。
【発明の効果】
【0015】
本発明の油脂分解菌は、油脂分解条件を高アルカリ及び高温とすることができるため、BOD分解菌の増殖を抑え、油脂分解菌を優勢又は優占的にすることができる。また、本発明の油脂分解菌は、増殖速度及び油脂分解速度が速いため、高濃度の油脂を含む排水の処理が可能である。さらに、油脂分解過程でBOD成分の分解が生じにくくなるため、高濃度の油脂分解生成物を得ることが可能となり、BOD成分濃度を高く維持した油脂分解排水を得ることができる。これは、油脂分解排水を後段で嫌気処理する場合に有利である。
【図面の簡単な説明】
【0016】
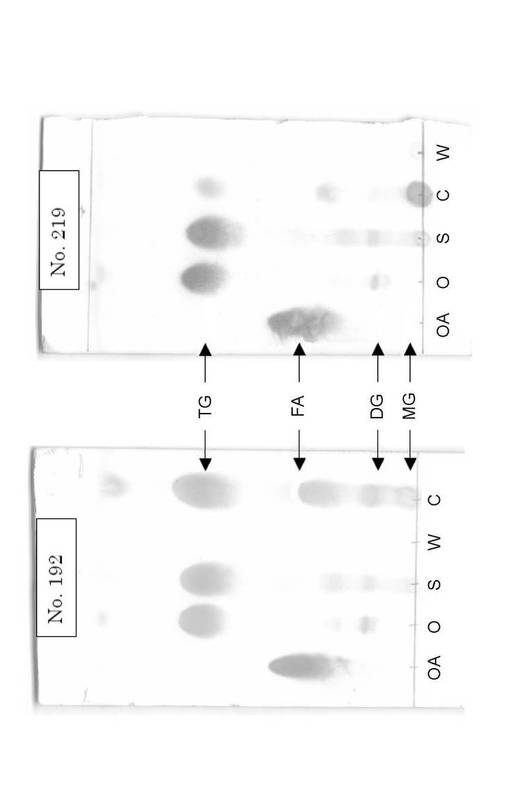
【図1】油脂分解生成物のTLC(薄層クロマトグラフィー)分析結果を示す写真である。図1中、TG、DG、MG、FAは、それぞれ、トリグリセリド、ジグリセリド、モノグリセリド、脂肪酸を表し、OA、O、S、W、Cは、それぞれ、オレイン酸、オリーブ油、混合油脂(サラダ油/ラード/牛脂、質量比1:1:1)、抽出後の水層、抽出後のクロロホルム層を表す。
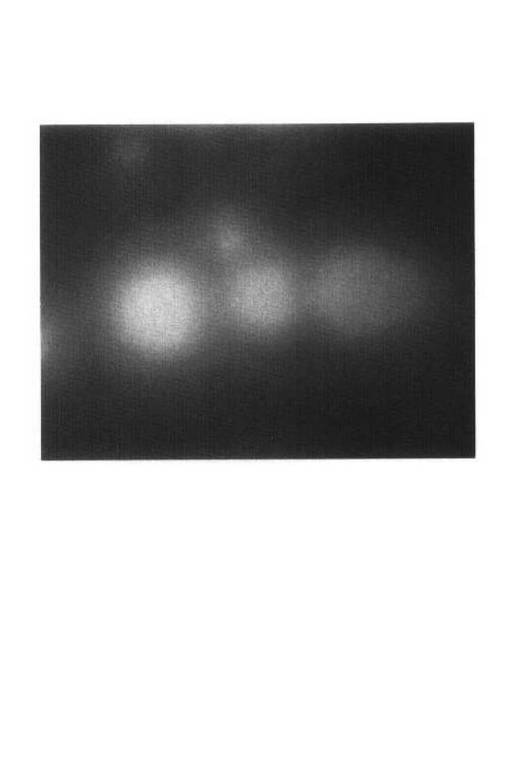
【図2】Pseudmonas aeruginosa SS−219株(NITE P−847)のコロニー形態を示す写真である。
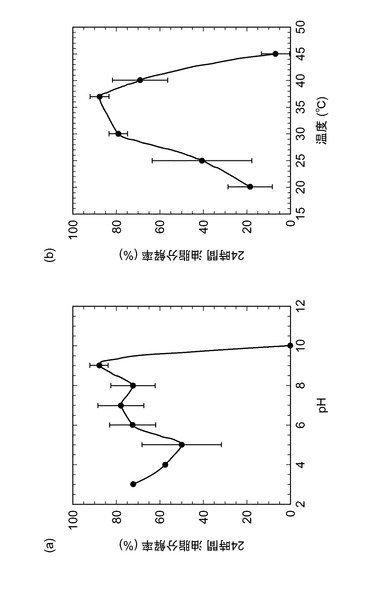
【図3】(a)培養液のpH変動又は(b)培養温度の変動に対する油脂分解率を示すグラフである。
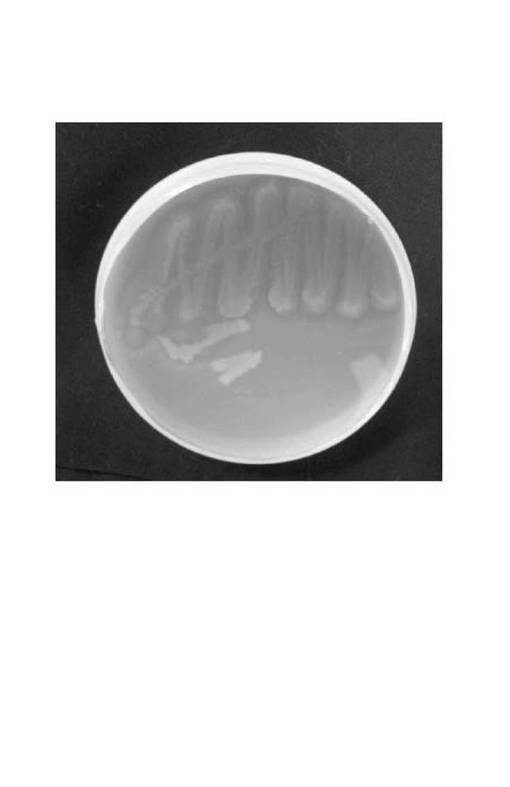
【図4】NITE P−847(Pseudmonas aeruginosa SS−219株)を油脂乳化プレートで生育させた様子を示す写真である。
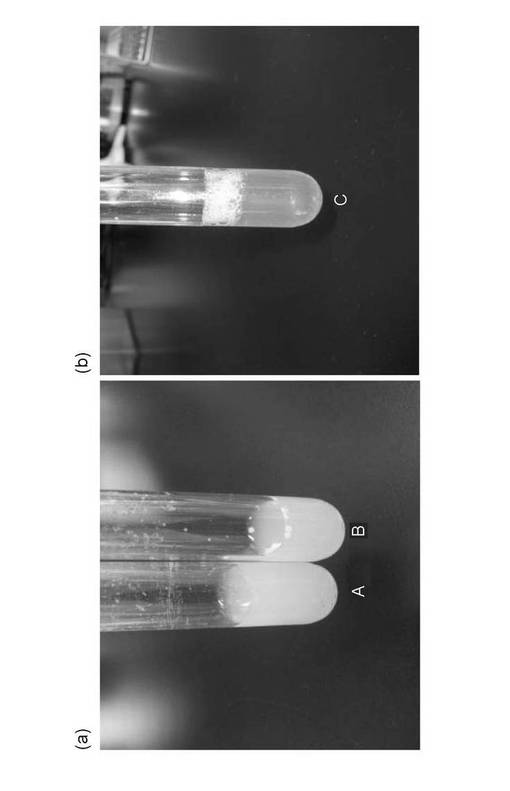
【図5】試験管培養したNITE P−847(Pseudmonas aeruginosa SS−219株)を示す写真である。(a)培養日数1日目(A)及び3日目(B)、(b)培養日数12日目(C)の写真である。
【発明を実施するための形態】
【0017】
以下、本発明を実施形態に基づいて詳細に説明する。
【0018】
本発明の油脂分解菌は、上述したとおり上記菌学的性質等を有するものである。上記油脂分解菌は、例えば、培養温度が30〜40℃、培養液のpHが8.5〜9.5という高アルカリ及び高温条件下で高い油脂分解活性を発揮することができる。また、増殖速度及び油脂分解速度が速く、サラダ油、ラード、牛脂等の食用油脂を24時間で82.8〜90.9%分解できる。従来の油脂分解菌によりこの程度の油脂分解率を達成するには48時間以上要するのが通常であり、本発明の油脂分解菌による油脂分解速度の速さがわかる。
【0019】
また、本実施形態に係る油脂分解菌としては、Pseudmonas aeruginosa SS−219株(NITE P−847)とすることが好ましい。Pseudmonas aeruginosa SS−219株は上述の菌学的性質等を全て備える油脂分解菌の具体例である。好アルカリ性菌として知られるAlcaligene属、Achromobacter属ではないものの、高アルカリ性条件下で増殖でき、油脂分解速度が速い。
【0020】
本発明に係る油脂分解菌が分解する対象となる油脂は、主として、動植物由来の油脂であるが、それらに限られるものではなく、種々の合成化学的手法を含む方法で得られた人工油脂、その誘導体をも含むものである。具体的には、排水に含まれる食用油脂、工業用油脂等とすることができ、特に、サラダ油、コーン油、大豆油、乳脂、バター、ラード、牛脂等の食用油脂とすることができる。
【0021】
本発明に係る油脂分解菌は、様々な環境から採取した細菌群を、油脂を含むアルカリ性の培地中で培養し、油脂分解能を指標としたスクリーニングにより単離することができる。
【0022】
具体的な方法は、例えば、以下のとおりである。油脂又は混合油脂を含むアルカリ性培地(例えば、pH8〜10)を試験管に添加し、さらに様々な環境(例えば、土壌、砂、泥、海水、湖沼など)から採取したサンプルを少量添加する。アルミホイル等で試験管にふたをし、震盪培養器中で適切な条件下(例えば、37℃、170回/分、24時間)で震盪培養した後、油脂分解が生じているサンプルを選定する。
【0023】
震盪培養した後に油脂分解が生じているサンプルの分析は、目視により残存油脂分が存在するかどうかの判定によることもできるが、例えば、JIS規格に基づくn−ヘキサン抽出による残存油脂分の定量又は参考文献(Journal of Bioscience and Bioengineering,2007年,103(4),325−330頁)に記載の方法によることもできる。
【0024】
油脂分解が生じていることを確認したサンプルから、純粋分離により、油脂分解菌を単離することができる。純粋分離は当業者によく知られた方法で実施することが可能であり、例えば、寒天培地上に接種して培養することにより、純粋分離されたコロニーを形成させることができ、そのコロニーをピックアップすることで目的とする油脂分解菌を単離することができる。
【0025】
また、油脂分解が生じているサンプルを選定した後、そのサンプルの一部を、新鮮な油脂又は混合油脂を含むアルカリ性培地(例えば、pH8〜10)を添加した試験管に再度接種して、再度震盪培養を行うこともできる。この震盪培養後に油脂分解を生じているサンプルを選定する。選定したサンプルに対し、上述の純粋分離を行うこともできるし、再びこの震盪培養サイクルを繰り返すこともできる。震盪培養サイクルを増やすことにより、目的とする油脂分解菌を集積させることができる。
【0026】
上述のようにして環境から採取したサンプルより単離した油脂分解菌の同定、性状及び性質等の解析は、公知の種々の同定試験方法又は市販されている同定キットを用いて行うことができる。また、16S rDNA塩基配列をシークエンスし、相同性検索、分子系統解析により、油脂分解菌の同定を行うこともできる。
【0027】
本発明の油脂分解剤は、本発明の油脂分解菌を製剤化して得ることができる。具体的には、例えば、「応用微生物学改訂版、村尾澤夫、荒井基夫共編、培風館、1993年」、「生物工学序論、佐田栄三ら著、講談社、1996年」、「産業用酵素、上島孝之著、丸善、1995年」などに記載の方法により、固定化製剤、液状化製剤、粉末状製剤とすることができる。
【0028】
本発明の油脂分解菌及び油脂分解剤は、これに限定されるものではないが製造工程で油脂を扱う食品工場の排水等、油脂を多く含む有機性排水の処理に用いることができる。排水の処理方法については特に制限はされず、従来公知である油脂分解微生物等を用いた排水処理において、油脂分解微生物等に代えて本発明の油脂分解菌又は油脂分解剤を用いることができる。また、本発明の油脂分解菌又は油脂分解剤を用いる場合、処理する排水の温度を30〜40℃に設定し、処理する排水のpHを8.5〜9.5に設定することが好ましい。これにより、本発明の油脂分解菌が高い油脂分解能を示す条件となるばかりでなく、BOD分解菌の増殖を抑えることができるため、汚泥発生量を少なくでき、かつ油脂分解効率を更に上昇させることができる。
【0029】
また、本発明の油脂分解菌及び油脂分解剤は、溶解性有機物を高濃度に含む高負荷排水をメタン発酵法で処理するための、種々のタイプの上向流式嫌気性処理槽と組合わせて使用することがより好ましい。上向流式嫌気性処理槽としては、例えば、UASB(Upflow Anaerobic Sludge Blanket)法やこれを改良したEGSB(Expanded Glanular Sludge Bed)法を利用した処理槽とすることができる。油脂を多く含む有機性排水を前処理工程で本発明の油脂分解菌及び油脂分解剤により処理して油脂を除くことにより、上向流式嫌気性処理槽による後処理工程での排水とグラニュール汚泥との接触効率の低下を防ぐことができ、効率よく排水を処理できる。また、前処理工程でBOD成分の分解が生じにくいため、後処理工程へと供給する排水中には高濃度の油脂分解生成物がBOD成分として含まれる。したがって、後処理工程で嫌気性処理を行うことによって、高濃度のBOD成分が高いエネルギーに変換されるうえ、汚泥発生量も少ないため、高い処理効率を達成することができる。
【実施例】
【0030】
以下、実施例を用いて本発明を更に具体的に説明するが、本発明はこれに限定されるものではない。
【0031】
[アルカリ耐性油脂分解菌の探索]
神奈川県、福島県、愛媛県で採取した土壌を試料とし、滅菌した生理食塩水に懸濁した後、超音波洗浄器にて5分間超音波処理を行った。処理後5分間ほど静置し、上澄み液を3000ppmとなるよう混合油脂(サラダ油/ラード/牛脂、質量比1:1:1)を添加したアルカリ人工下水培地1(表1)又はアルカリ人工下水培地2(表2)各5mlが入った18φ×180mm試験管に1%(v/v)となるように接種した。
【0032】
【表1】

【0033】
【表2】
【0034】
上記試験管にアルミホイルでフタをし、震盪培養器にて37℃、170回/分の条件で震盪培養した。微生物の生育が認められた試料については、3000ppmとなるように上記混合油脂を添加した新鮮なアルカリ人工下水培地1又はアルカリ人工下水培地2に、1%(v/v)となるように接種し、同様に震盪培養した。これを繰り返し行うことで目的微生物を集積させた。3回以上繰り返した際に生育しており、かつ油分分解が観察された試料について、フラスコ試験を実施した。
【0035】
フラスコ試験は、以下の手順に従って行った。500mlバッフルフラスコに3000ppmとなるよう上記混合油脂を添加したアルカリ人工下水培地1又はアルカリ人工下水培地2を100ml入れ、試験管培養液を1%(v/v)となるように接種し、37℃、120rpmで24時間震盪培養した。これをオートクレーブ(121℃、20分)処理した後、JIS K0101に準拠したn−ヘキサン抽出法により、残存している油脂を定量し、菌を接種しないコントロールとの差から油脂分解率を算出した。
【0036】
アルカリ人工下水培地1を用いて25試料に対して、アルカリ人工下水培地2を用いて49試料に対してスクリーニングを実施した。アルカリ人工下水培地2を用いたスクリーニングの結果、78〜88%/dという高い油脂分解率を示した2種類のサンプル(サンプルNo.192及びNo.219)が得られた。サンプルNo.192は住友重機械工業株式会社横須賀製造所内レール付近の土壌から得た試料に由来し、サンプルNo.219は愛媛県内焚き火跡土壌(灰)から得た試料に由来するものであった。
【0037】
これらのサンプル中に含まれる微生物を純粋分離した結果、どちらのサンプルも集積培養によって単一菌の状態になっていた。分離したコロニーからマスタープレート、グリセロール保存を作成した。マスタープレートからピックアップした各菌をアガープレート上で培養し(30℃、24時間)、同定分析用プレートを各菌それぞれ1枚ずつ作成した。
【0038】
[油脂分解性の測定]
サンプルNo.192及びNo.219から純粋分離した菌株の油脂分解性について、再現性の確認を行った。n−ヘキサン抽出では抽出時に中間層及びエマルション形成により、残存している油脂の抽出精度が低いと考えられたため、クロロホルム/メタノール(3:1)40mlで抽出し、遠心分離(5,000rpm、30min)した後、静かに分液ロートに移し、クロロホルム(下)層を新しい遠心管に入れ、再び遠心分離(10,000rpm、10min)した。遠心分離後、静かに分液ロートに移し、クロロホルム(下)層をフラスコに取り、無水硫酸マグネシウムにて脱水後、ろ過、エバポレートすることにより残存油分の測定を行った。その結果、両菌株は再現性良く、高い油脂分解性を示すことがわかった(表3)。
【0039】
【表3】
【0040】
[油脂分解過程のTLC分析]
油脂分解過程を把握するために、上記抽出した残存油分のTLC分析を行った。その結果を図1に示す。No.192株(左パネル)は、左レーンから順に、オレイン酸(OA)、オリーブ油(トリオレイン;O)、混合油脂(サラダ油/ラード/牛脂、質量比1:1:1;S)、抽出後の水層(W)、抽出した残存油分が含まれるクロロホルム層(C)をロードした。同様に、No.219株(右パネル)は、左レーンから順に、オレイン酸(OA)、オリーブ油(トリオレイン;O)、混合油脂(サラダ油/ラード/牛脂、質量比1:1:1;S)、抽出した残存油分が含まれるクロロホルム層(C)、抽出後の水層(W)をロードした。No.192株及びNo.219株のいずれも水層(W)からは油脂が検出されず、残存油脂は完全にクロロホルム層(C)へ抽出されたことがわかった。No.219株に比べ、No.192株の方がジグリセリド(DG)、モノグリセリド(MG)、脂肪酸(FA)存在量が多いことがわかった。これは、No.192株の油脂分解過程ではリパーゼによるトリグリセリド(TG)の分解と脂肪酸の代謝(β酸化)が律速となっていることを示唆するものである。一方、No.219株では抽出油分中にDG、MGがほとんど観察されず、FA存在量も少ないことから、グリセリドの分解が迅速であると推察される。
【0041】
以下、サンプルNo.219から純粋分離した菌株(No.219株)について更に詳細な解析を実施した。
【0042】
[油脂分解菌の分類学上の同定]
16S rDNAの塩基配列解析により、No.219株の同定を行った。No.219株からのDNA抽出を、InstaGene Matrix(BioRad社製、CA、USA)を用いて行った。全長16S rDNAをPCR反応により増幅した。PCR反応には、PrimeSTAR HS DNA Polymerase(タカラバイオ社製)を用いた。得られたPCR産物を鋳型とし、BigDye Terminator v3.1 Cycle Sequencing Kit(アプライドバイオシステム社製、CA、USA)を用いてサイクルシークエンスを実施した。ABI PRISM 3130x1 Genetic Analyzer System(アプライドバイオシステム社製、CA、USA)によりシークエンスを解析し、ChromasPro1.4(Technelysium Pty Ltd.,Tewantin,AUS)ソフトウェアを用いて塩基配列を決定した。決定したNo.219株の16S rDNA塩基配列を、アポロン2.0(テクノスルガ・ラボ社製)ソフトウェア及びアポロンDB−BA ver4.0(テクノスルガ・ラボ社製)データベースによる相同性検索及び簡易分子系統解析に供した。その結果、No.219株の16S rDNA塩基配列は、Pseudmonas aeruginosa ATCC10145株の16S rDNA塩基配列と99.9%の高い相同率を示した(表4)。また、国際塩基配列データベース(GenBank/DDBJ/EMBL)に対しても相同性検索を実施した結果、No.219株の16S rDNA塩基配列は、P.aeruginosa MCCB123株、MML2212株、NK2.1B−1株等の16S rDNA塩基配列と100%の相同率を示したが、基準株に由来する16S rDNAは検索されなかった。
【0043】
【表4】
【0044】
No.219株の16S rDNA配列とアポロンDB−BA ver4.0に対する相同性検索上位10株の16S rDNA配列を用いて簡易分子系統解析を実施した(表5)。なお、表5中、系統枝の分岐に位置する数字はブートストラップ値を示す。
【0045】
【表5】
【0046】
簡易分子系統解析の結果、No.219株の16S rDNAはP.aeruginosaの16S rDNAとクラスターを形成し、同一の分子系統学的位置を示した(表5)。以上の結果から、本菌株はP.aeruginosa種に帰属する菌株であると考えられる。そこで、本菌株は、Pseudmonas aeruginosa SS−219株と命名した。
【0047】
[油脂分解菌の菌学的特性の分析]
Pseudmonas aeruginosa SS−219株について、光学顕微鏡(BX50F4、オリンパス社製)による形態観察、Barrowらの方法(Cowan and Steel’s Manual for the Identification of Medical Bacteria. 3rd edition. 1993年,Cambridge University Press.)に基づいて、カタラーゼ反応、オキシダーゼ反応、グラム染色性、ブドウ糖からの酸/ガス産生、ブドウ糖の酸化/発酵(O/F)等の試験を実施した。グラム染色性解析にはフェイバーG「ニッスイ」(日水製薬社製)を用いた。結果を表6に示す。また、図2にPseudmonas aeruginosa SS−219株のコロニー形態を示す。
【0048】
【表6】
【0049】
さらに、API20NEキット(ビオメリュー社製、フランス)を用い、表7に記載した各種性状の試験を行った。判定はキットに添付のマニュアルに従った。加えて、表8に記載した事項についての試験を実施した。
【0050】
【表7】
【0051】
【表8】
【0052】
その結果、Pseudmonas aeruginosa SS−219株は、培養温度45℃での生育が可能であり(表6)、特許文献2及び特許文献3に記載されたPseudmonas族菌と相違していた。また、L−アラビノース、D−マンノースを資化しない点において、特許文献2に記載されたPseudmonas族菌と相違していた(表7)。
【0053】
[培養液pHと油脂分解率の関係]
Pseudmonas aeruginosa SS−219株の油脂分解率に対する培養液pHの影響を解析した。培地は、pH調整剤(1M Tris−HCl、pH9)以外はアルカリ人工下水培地2と同じ組成のものを用いた。この培地をベースとし、pH3〜8の培地はpH調整剤として塩酸又は水酸化ナトリウムを用い、pH9の培地はpH調整剤として1M Tris−HCl(pH9)を用い、pH10の培地はpH調整剤として1Mグリシン/水酸化ナトリウム緩衝液(pH10)を用いて、それぞれpHを調整した。これらの培地に3000ppmとなるように混合油脂(サラダ油/ラード/牛脂、質量比1:1:1)を添加したものを500mlバッフルフラスコに100ml入れ、24時間予備培養した試験管培養液を1%(v/v)となるように接種し、120rpm、37℃で24時間震盪培養した。培養後、オートクレーブ(121℃、20分)処理し、クロロホルム/メタノール(3:1)40mlで抽出し、遠心分離(5,000rpm、30min)した後、静かに分液ロートに移し、クロロホルム(下)層を新しい遠心管に入れ、再び遠心分離(10,000rpm、10min)した。遠心分離後、静かに分液ロートに移し、クロロホルム(下)層をフラスコに取り、無水硫酸マグネシウムにて脱水後、ろ過、エバポレートすることにより残存油分を定量した。菌を接種しないコントロールとの差から油脂分解率を算出した。
【0054】
結果を図3(a)に示す。培養液のpHが3、4、5、6、7、8、9、10のとき、Pseudmonas aeruginosa SS−219株の24時間培養後の油脂分解率は、それぞれ、72、58、63、80、85、80、83%であった。培地pH8.5〜9.5で高い油脂分解率を示し、最適培地pHは9付近であった。また、pH6〜8、pH3でも70%以上の油脂分解率を示した。
【0055】
[培養温度と油脂分解率の関係]
Pseudmonas aeruginosa SS−219株の油脂分解率に対する培養温度の影響を解析した。500mlバッフルフラスコに、3000ppmとなるように混合油脂(サラダ油/ラード/牛脂、質量比1:1:1)を添加したアルカリ人工下水培地2を100ml入れ、24時間予備培養した試験管培養液を1%(v/v)となるように接種した。培養温度を20、25、30、37、40、45℃にそれぞれ設定し、120rpmで24時間震盪培養した。その後、上記と同様にして残存油分を定量し、油脂分解率を算出した。
【0056】
結果を図3(b)に示す。培養温度が20、25、30、37、40、45℃のとき、Pseudmonas aeruginosa SS−219株の24時間培養後の油脂分解率は、それぞれ、17、65、75、83、70、13%であった。油脂分解率は30〜40℃で高い値を示し、最適温度は37℃付近であった。
【0057】
[その他の特徴]
抗生物質含有プレートを用いて、抗生物質感受性試験を行った。その結果、Pseudmonas aeruginosa SS−219株はアンピシリン、クロラムフェニコール抵抗性を示した。
【0058】
Pseudmonas aeruginosa SS−219株を油脂乳化プレートで生育させた様子を図4に示した。Pseudmonas aeruginosa SS−219株のコロニー周囲にはクリアハローが形成されており、菌体外にリパーゼを分泌していることが示唆された。Pseudmonas aeruginosa SS−219株の試験管培養の様子を図5(a)及び(b)に示した。Pseudmonas aeruginosa SS−219株は黄色の蛍光色素を産生しており、培養日数1日目(A)、3日目(B)と、培養日数の増加とともにその色調が濃くなった。さらに、培養12日目(C)には油脂分はほぼ完全に分解されていた。また、培養液の濁度も著しく低下していたことから、溶菌が起こっているものと考えられる。
【技術分野】
【0001】
本発明は、油脂分解菌及び油脂分解剤に関する。
【背景技術】
【0002】
微生物を利用した排水処理方法として、一般に、好気性微生物による活性汚泥法(好気性処理法)及び嫌気性微生物によるメタン発酵法(嫌気性処理法)が知られている。好気性処理法では曝気するためのエネルギーを要するのに対し、嫌気性処理法は消費エネルギーが少なく、またメタンなどを主成分とするバイオガスを得ることもできる。このため、嫌気性処理法は、好気性処理法よりもエネルギー効率の点で有利であるといわれている。
【0003】
溶解性有機物を高濃度に含む高負荷排水をメタン発酵法で処理するための、種々のタイプの上向流式嫌気性処理槽が開発されている。例えば、UASB(Upflow Anaerobic Sludge Blanket)法やこれを改良したEGSB(Expanded Glanular Sludge Bed)法を利用した処理槽が知られている。これらの上向流式嫌気性処理槽は、グラニュール汚泥と呼ばれる粒状の汚泥を槽内に収容することができ、被処理水がグラニュール汚泥と接触しながら上方に流れることによって、有機物が効率的に分解される。
【0004】
ところで、製造工程で油脂を扱う食品工場の排水など、油脂を多く含む有機性排水をグラニュール汚泥で処理する場合、油脂がグラニュール汚泥に付着し、これによって排水とグラニュール汚泥との接触効率が低下して有機物の処理が不十分となりやすい。このような問題を解決する手段として、特許文献1には上向流式嫌気性処理槽に被処理水を導入するに先立ち、被処理水から油分を分離し、これをリパーゼ生成菌によって分解する油脂分解槽を備えた嫌気性処理装置が記載されている。
【0005】
また、油脂分解性を有するPseudomonas属菌が開示されている(特許文献2及び特許文献3)。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2005−270862号公報
【特許文献2】特開2008−220225号公報
【特許文献3】特開平6−153922号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、従来知られている油脂分解菌は、その至適油脂分解条件(温度、pH等)が、油脂分解菌よりも増殖速度の速い溶解性有機物分解菌(以下、BOD分解菌という。)の至適生育条件(温度、pH等)の範囲内に入ってしまう。そのため、BOD分解菌が油脂分解槽の汚泥中で優勢となり、油脂の分解効率低下に加えて溶解性有機物(以下、BOD成分という。)の分解により大量の余剰汚泥が生じる問題があった。また、BOD分解菌が油脂分解槽の汚泥中で優占的となり、油脂分解菌が存在しなくなるという問題もあった。
【0008】
そこで、本発明は、至適油脂分解条件(温度、pH等)がBOD分解菌の至適生育条件(温度、pH等)の範囲内に入らず、十分な増殖速度及び油脂分解速度を有する油脂分解菌の提供を目的とする。本発明はまた、上記油脂分解菌を含む油脂分解剤の提供も目的とする。
【課題を解決するための手段】
【0009】
本発明は、Pseudmonas属に属し、以下の菌学的性質を示す、油脂分解菌。
細胞形態 桿菌
細胞の直径 0.6〜0.7×1.0〜1.5μm
胞子の有無 −
グラム染色性 −
運動性 +
37℃での生育 +
45℃での生育 +
嫌気性条件下での生育 −
カタラーゼ +
オキシダーゼ +
グルコースからの酸/ガス産生 −/−
グルコースの酸化/発酵 +/−
硝酸塩還元活性 +
インドール産生活性 −
ブドウ糖酸性化活性 −
アルギニンジヒドロラーゼ活性 +
ウレアーゼ活性 −
エスクリン加水分解活性 −
ゼラチン加水分解活性 +
β−ガラクトシダーゼ活性 −
チトクロームオキシダーゼ活性 +
ブドウ糖資化性 +
L−アラビノース資化性 −
D−マンノース資化性 −
D−マンニトール資化性 −
N−アセチル−D−グルコサミン資化性 +
マルトース資化性 −
グルコン酸カリウム資化性 +
n−カプリン酸資化性 +
アジピン酸資化性 +
dl−リンゴ酸資化性 +
クエン酸ナトリウム資化性 +
酢酸フェニル資化性 −
なお、「+」は陽性であること、「−」は陰性であることを示す。
【0010】
上記油脂分解菌は、寒天培地上において、30℃、24時間培養で直径2〜3mmの周縁波状の円形、表面がスムーズ、扁平状で、不透明な緑色コロニーを形成することが好ましい。
【0011】
また、上記油脂分解菌は、配列番号1で示される16S rDNA塩基配列を有することが好ましい。
【0012】
上記油脂分解菌として、独立行政法人製品評価技術基盤機構特許微生物寄託センター(郵便番号292−0818、日本国千葉県木更津市かずさ鎌足2丁目5番地8号 NITEバイオテクノロジー本部)に受託番号NITE P−847で2009年11月25日に受託されているPseudmonas aeruginosa SS−219株とすることができる。
【0013】
本発明はまた、上述の油脂分解菌を含む油脂分解剤を提供する。
【0014】
上記油脂分解菌は、アルカリ条件及び高温条件下で効率よく増殖及び油脂分解が可能であるため、BOD分解菌の増殖を抑制しながら油脂を分解することができる。また、上記油脂分解菌は従来の油脂分解菌と比べて油脂分解速度が速いため、高濃度の油脂を含む排水の処理に好適に利用することができる。
【発明の効果】
【0015】
本発明の油脂分解菌は、油脂分解条件を高アルカリ及び高温とすることができるため、BOD分解菌の増殖を抑え、油脂分解菌を優勢又は優占的にすることができる。また、本発明の油脂分解菌は、増殖速度及び油脂分解速度が速いため、高濃度の油脂を含む排水の処理が可能である。さらに、油脂分解過程でBOD成分の分解が生じにくくなるため、高濃度の油脂分解生成物を得ることが可能となり、BOD成分濃度を高く維持した油脂分解排水を得ることができる。これは、油脂分解排水を後段で嫌気処理する場合に有利である。
【図面の簡単な説明】
【0016】
【図1】油脂分解生成物のTLC(薄層クロマトグラフィー)分析結果を示す写真である。図1中、TG、DG、MG、FAは、それぞれ、トリグリセリド、ジグリセリド、モノグリセリド、脂肪酸を表し、OA、O、S、W、Cは、それぞれ、オレイン酸、オリーブ油、混合油脂(サラダ油/ラード/牛脂、質量比1:1:1)、抽出後の水層、抽出後のクロロホルム層を表す。
【図2】Pseudmonas aeruginosa SS−219株(NITE P−847)のコロニー形態を示す写真である。
【図3】(a)培養液のpH変動又は(b)培養温度の変動に対する油脂分解率を示すグラフである。
【図4】NITE P−847(Pseudmonas aeruginosa SS−219株)を油脂乳化プレートで生育させた様子を示す写真である。
【図5】試験管培養したNITE P−847(Pseudmonas aeruginosa SS−219株)を示す写真である。(a)培養日数1日目(A)及び3日目(B)、(b)培養日数12日目(C)の写真である。
【発明を実施するための形態】
【0017】
以下、本発明を実施形態に基づいて詳細に説明する。
【0018】
本発明の油脂分解菌は、上述したとおり上記菌学的性質等を有するものである。上記油脂分解菌は、例えば、培養温度が30〜40℃、培養液のpHが8.5〜9.5という高アルカリ及び高温条件下で高い油脂分解活性を発揮することができる。また、増殖速度及び油脂分解速度が速く、サラダ油、ラード、牛脂等の食用油脂を24時間で82.8〜90.9%分解できる。従来の油脂分解菌によりこの程度の油脂分解率を達成するには48時間以上要するのが通常であり、本発明の油脂分解菌による油脂分解速度の速さがわかる。
【0019】
また、本実施形態に係る油脂分解菌としては、Pseudmonas aeruginosa SS−219株(NITE P−847)とすることが好ましい。Pseudmonas aeruginosa SS−219株は上述の菌学的性質等を全て備える油脂分解菌の具体例である。好アルカリ性菌として知られるAlcaligene属、Achromobacter属ではないものの、高アルカリ性条件下で増殖でき、油脂分解速度が速い。
【0020】
本発明に係る油脂分解菌が分解する対象となる油脂は、主として、動植物由来の油脂であるが、それらに限られるものではなく、種々の合成化学的手法を含む方法で得られた人工油脂、その誘導体をも含むものである。具体的には、排水に含まれる食用油脂、工業用油脂等とすることができ、特に、サラダ油、コーン油、大豆油、乳脂、バター、ラード、牛脂等の食用油脂とすることができる。
【0021】
本発明に係る油脂分解菌は、様々な環境から採取した細菌群を、油脂を含むアルカリ性の培地中で培養し、油脂分解能を指標としたスクリーニングにより単離することができる。
【0022】
具体的な方法は、例えば、以下のとおりである。油脂又は混合油脂を含むアルカリ性培地(例えば、pH8〜10)を試験管に添加し、さらに様々な環境(例えば、土壌、砂、泥、海水、湖沼など)から採取したサンプルを少量添加する。アルミホイル等で試験管にふたをし、震盪培養器中で適切な条件下(例えば、37℃、170回/分、24時間)で震盪培養した後、油脂分解が生じているサンプルを選定する。
【0023】
震盪培養した後に油脂分解が生じているサンプルの分析は、目視により残存油脂分が存在するかどうかの判定によることもできるが、例えば、JIS規格に基づくn−ヘキサン抽出による残存油脂分の定量又は参考文献(Journal of Bioscience and Bioengineering,2007年,103(4),325−330頁)に記載の方法によることもできる。
【0024】
油脂分解が生じていることを確認したサンプルから、純粋分離により、油脂分解菌を単離することができる。純粋分離は当業者によく知られた方法で実施することが可能であり、例えば、寒天培地上に接種して培養することにより、純粋分離されたコロニーを形成させることができ、そのコロニーをピックアップすることで目的とする油脂分解菌を単離することができる。
【0025】
また、油脂分解が生じているサンプルを選定した後、そのサンプルの一部を、新鮮な油脂又は混合油脂を含むアルカリ性培地(例えば、pH8〜10)を添加した試験管に再度接種して、再度震盪培養を行うこともできる。この震盪培養後に油脂分解を生じているサンプルを選定する。選定したサンプルに対し、上述の純粋分離を行うこともできるし、再びこの震盪培養サイクルを繰り返すこともできる。震盪培養サイクルを増やすことにより、目的とする油脂分解菌を集積させることができる。
【0026】
上述のようにして環境から採取したサンプルより単離した油脂分解菌の同定、性状及び性質等の解析は、公知の種々の同定試験方法又は市販されている同定キットを用いて行うことができる。また、16S rDNA塩基配列をシークエンスし、相同性検索、分子系統解析により、油脂分解菌の同定を行うこともできる。
【0027】
本発明の油脂分解剤は、本発明の油脂分解菌を製剤化して得ることができる。具体的には、例えば、「応用微生物学改訂版、村尾澤夫、荒井基夫共編、培風館、1993年」、「生物工学序論、佐田栄三ら著、講談社、1996年」、「産業用酵素、上島孝之著、丸善、1995年」などに記載の方法により、固定化製剤、液状化製剤、粉末状製剤とすることができる。
【0028】
本発明の油脂分解菌及び油脂分解剤は、これに限定されるものではないが製造工程で油脂を扱う食品工場の排水等、油脂を多く含む有機性排水の処理に用いることができる。排水の処理方法については特に制限はされず、従来公知である油脂分解微生物等を用いた排水処理において、油脂分解微生物等に代えて本発明の油脂分解菌又は油脂分解剤を用いることができる。また、本発明の油脂分解菌又は油脂分解剤を用いる場合、処理する排水の温度を30〜40℃に設定し、処理する排水のpHを8.5〜9.5に設定することが好ましい。これにより、本発明の油脂分解菌が高い油脂分解能を示す条件となるばかりでなく、BOD分解菌の増殖を抑えることができるため、汚泥発生量を少なくでき、かつ油脂分解効率を更に上昇させることができる。
【0029】
また、本発明の油脂分解菌及び油脂分解剤は、溶解性有機物を高濃度に含む高負荷排水をメタン発酵法で処理するための、種々のタイプの上向流式嫌気性処理槽と組合わせて使用することがより好ましい。上向流式嫌気性処理槽としては、例えば、UASB(Upflow Anaerobic Sludge Blanket)法やこれを改良したEGSB(Expanded Glanular Sludge Bed)法を利用した処理槽とすることができる。油脂を多く含む有機性排水を前処理工程で本発明の油脂分解菌及び油脂分解剤により処理して油脂を除くことにより、上向流式嫌気性処理槽による後処理工程での排水とグラニュール汚泥との接触効率の低下を防ぐことができ、効率よく排水を処理できる。また、前処理工程でBOD成分の分解が生じにくいため、後処理工程へと供給する排水中には高濃度の油脂分解生成物がBOD成分として含まれる。したがって、後処理工程で嫌気性処理を行うことによって、高濃度のBOD成分が高いエネルギーに変換されるうえ、汚泥発生量も少ないため、高い処理効率を達成することができる。
【実施例】
【0030】
以下、実施例を用いて本発明を更に具体的に説明するが、本発明はこれに限定されるものではない。
【0031】
[アルカリ耐性油脂分解菌の探索]
神奈川県、福島県、愛媛県で採取した土壌を試料とし、滅菌した生理食塩水に懸濁した後、超音波洗浄器にて5分間超音波処理を行った。処理後5分間ほど静置し、上澄み液を3000ppmとなるよう混合油脂(サラダ油/ラード/牛脂、質量比1:1:1)を添加したアルカリ人工下水培地1(表1)又はアルカリ人工下水培地2(表2)各5mlが入った18φ×180mm試験管に1%(v/v)となるように接種した。
【0032】
【表1】
【0033】
【表2】
【0034】
上記試験管にアルミホイルでフタをし、震盪培養器にて37℃、170回/分の条件で震盪培養した。微生物の生育が認められた試料については、3000ppmとなるように上記混合油脂を添加した新鮮なアルカリ人工下水培地1又はアルカリ人工下水培地2に、1%(v/v)となるように接種し、同様に震盪培養した。これを繰り返し行うことで目的微生物を集積させた。3回以上繰り返した際に生育しており、かつ油分分解が観察された試料について、フラスコ試験を実施した。
【0035】
フラスコ試験は、以下の手順に従って行った。500mlバッフルフラスコに3000ppmとなるよう上記混合油脂を添加したアルカリ人工下水培地1又はアルカリ人工下水培地2を100ml入れ、試験管培養液を1%(v/v)となるように接種し、37℃、120rpmで24時間震盪培養した。これをオートクレーブ(121℃、20分)処理した後、JIS K0101に準拠したn−ヘキサン抽出法により、残存している油脂を定量し、菌を接種しないコントロールとの差から油脂分解率を算出した。
【0036】
アルカリ人工下水培地1を用いて25試料に対して、アルカリ人工下水培地2を用いて49試料に対してスクリーニングを実施した。アルカリ人工下水培地2を用いたスクリーニングの結果、78〜88%/dという高い油脂分解率を示した2種類のサンプル(サンプルNo.192及びNo.219)が得られた。サンプルNo.192は住友重機械工業株式会社横須賀製造所内レール付近の土壌から得た試料に由来し、サンプルNo.219は愛媛県内焚き火跡土壌(灰)から得た試料に由来するものであった。
【0037】
これらのサンプル中に含まれる微生物を純粋分離した結果、どちらのサンプルも集積培養によって単一菌の状態になっていた。分離したコロニーからマスタープレート、グリセロール保存を作成した。マスタープレートからピックアップした各菌をアガープレート上で培養し(30℃、24時間)、同定分析用プレートを各菌それぞれ1枚ずつ作成した。
【0038】
[油脂分解性の測定]
サンプルNo.192及びNo.219から純粋分離した菌株の油脂分解性について、再現性の確認を行った。n−ヘキサン抽出では抽出時に中間層及びエマルション形成により、残存している油脂の抽出精度が低いと考えられたため、クロロホルム/メタノール(3:1)40mlで抽出し、遠心分離(5,000rpm、30min)した後、静かに分液ロートに移し、クロロホルム(下)層を新しい遠心管に入れ、再び遠心分離(10,000rpm、10min)した。遠心分離後、静かに分液ロートに移し、クロロホルム(下)層をフラスコに取り、無水硫酸マグネシウムにて脱水後、ろ過、エバポレートすることにより残存油分の測定を行った。その結果、両菌株は再現性良く、高い油脂分解性を示すことがわかった(表3)。
【0039】
【表3】
【0040】
[油脂分解過程のTLC分析]
油脂分解過程を把握するために、上記抽出した残存油分のTLC分析を行った。その結果を図1に示す。No.192株(左パネル)は、左レーンから順に、オレイン酸(OA)、オリーブ油(トリオレイン;O)、混合油脂(サラダ油/ラード/牛脂、質量比1:1:1;S)、抽出後の水層(W)、抽出した残存油分が含まれるクロロホルム層(C)をロードした。同様に、No.219株(右パネル)は、左レーンから順に、オレイン酸(OA)、オリーブ油(トリオレイン;O)、混合油脂(サラダ油/ラード/牛脂、質量比1:1:1;S)、抽出した残存油分が含まれるクロロホルム層(C)、抽出後の水層(W)をロードした。No.192株及びNo.219株のいずれも水層(W)からは油脂が検出されず、残存油脂は完全にクロロホルム層(C)へ抽出されたことがわかった。No.219株に比べ、No.192株の方がジグリセリド(DG)、モノグリセリド(MG)、脂肪酸(FA)存在量が多いことがわかった。これは、No.192株の油脂分解過程ではリパーゼによるトリグリセリド(TG)の分解と脂肪酸の代謝(β酸化)が律速となっていることを示唆するものである。一方、No.219株では抽出油分中にDG、MGがほとんど観察されず、FA存在量も少ないことから、グリセリドの分解が迅速であると推察される。
【0041】
以下、サンプルNo.219から純粋分離した菌株(No.219株)について更に詳細な解析を実施した。
【0042】
[油脂分解菌の分類学上の同定]
16S rDNAの塩基配列解析により、No.219株の同定を行った。No.219株からのDNA抽出を、InstaGene Matrix(BioRad社製、CA、USA)を用いて行った。全長16S rDNAをPCR反応により増幅した。PCR反応には、PrimeSTAR HS DNA Polymerase(タカラバイオ社製)を用いた。得られたPCR産物を鋳型とし、BigDye Terminator v3.1 Cycle Sequencing Kit(アプライドバイオシステム社製、CA、USA)を用いてサイクルシークエンスを実施した。ABI PRISM 3130x1 Genetic Analyzer System(アプライドバイオシステム社製、CA、USA)によりシークエンスを解析し、ChromasPro1.4(Technelysium Pty Ltd.,Tewantin,AUS)ソフトウェアを用いて塩基配列を決定した。決定したNo.219株の16S rDNA塩基配列を、アポロン2.0(テクノスルガ・ラボ社製)ソフトウェア及びアポロンDB−BA ver4.0(テクノスルガ・ラボ社製)データベースによる相同性検索及び簡易分子系統解析に供した。その結果、No.219株の16S rDNA塩基配列は、Pseudmonas aeruginosa ATCC10145株の16S rDNA塩基配列と99.9%の高い相同率を示した(表4)。また、国際塩基配列データベース(GenBank/DDBJ/EMBL)に対しても相同性検索を実施した結果、No.219株の16S rDNA塩基配列は、P.aeruginosa MCCB123株、MML2212株、NK2.1B−1株等の16S rDNA塩基配列と100%の相同率を示したが、基準株に由来する16S rDNAは検索されなかった。
【0043】
【表4】
【0044】
No.219株の16S rDNA配列とアポロンDB−BA ver4.0に対する相同性検索上位10株の16S rDNA配列を用いて簡易分子系統解析を実施した(表5)。なお、表5中、系統枝の分岐に位置する数字はブートストラップ値を示す。
【0045】
【表5】
【0046】
簡易分子系統解析の結果、No.219株の16S rDNAはP.aeruginosaの16S rDNAとクラスターを形成し、同一の分子系統学的位置を示した(表5)。以上の結果から、本菌株はP.aeruginosa種に帰属する菌株であると考えられる。そこで、本菌株は、Pseudmonas aeruginosa SS−219株と命名した。
【0047】
[油脂分解菌の菌学的特性の分析]
Pseudmonas aeruginosa SS−219株について、光学顕微鏡(BX50F4、オリンパス社製)による形態観察、Barrowらの方法(Cowan and Steel’s Manual for the Identification of Medical Bacteria. 3rd edition. 1993年,Cambridge University Press.)に基づいて、カタラーゼ反応、オキシダーゼ反応、グラム染色性、ブドウ糖からの酸/ガス産生、ブドウ糖の酸化/発酵(O/F)等の試験を実施した。グラム染色性解析にはフェイバーG「ニッスイ」(日水製薬社製)を用いた。結果を表6に示す。また、図2にPseudmonas aeruginosa SS−219株のコロニー形態を示す。
【0048】
【表6】
【0049】
さらに、API20NEキット(ビオメリュー社製、フランス)を用い、表7に記載した各種性状の試験を行った。判定はキットに添付のマニュアルに従った。加えて、表8に記載した事項についての試験を実施した。
【0050】
【表7】
【0051】
【表8】
【0052】
その結果、Pseudmonas aeruginosa SS−219株は、培養温度45℃での生育が可能であり(表6)、特許文献2及び特許文献3に記載されたPseudmonas族菌と相違していた。また、L−アラビノース、D−マンノースを資化しない点において、特許文献2に記載されたPseudmonas族菌と相違していた(表7)。
【0053】
[培養液pHと油脂分解率の関係]
Pseudmonas aeruginosa SS−219株の油脂分解率に対する培養液pHの影響を解析した。培地は、pH調整剤(1M Tris−HCl、pH9)以外はアルカリ人工下水培地2と同じ組成のものを用いた。この培地をベースとし、pH3〜8の培地はpH調整剤として塩酸又は水酸化ナトリウムを用い、pH9の培地はpH調整剤として1M Tris−HCl(pH9)を用い、pH10の培地はpH調整剤として1Mグリシン/水酸化ナトリウム緩衝液(pH10)を用いて、それぞれpHを調整した。これらの培地に3000ppmとなるように混合油脂(サラダ油/ラード/牛脂、質量比1:1:1)を添加したものを500mlバッフルフラスコに100ml入れ、24時間予備培養した試験管培養液を1%(v/v)となるように接種し、120rpm、37℃で24時間震盪培養した。培養後、オートクレーブ(121℃、20分)処理し、クロロホルム/メタノール(3:1)40mlで抽出し、遠心分離(5,000rpm、30min)した後、静かに分液ロートに移し、クロロホルム(下)層を新しい遠心管に入れ、再び遠心分離(10,000rpm、10min)した。遠心分離後、静かに分液ロートに移し、クロロホルム(下)層をフラスコに取り、無水硫酸マグネシウムにて脱水後、ろ過、エバポレートすることにより残存油分を定量した。菌を接種しないコントロールとの差から油脂分解率を算出した。
【0054】
結果を図3(a)に示す。培養液のpHが3、4、5、6、7、8、9、10のとき、Pseudmonas aeruginosa SS−219株の24時間培養後の油脂分解率は、それぞれ、72、58、63、80、85、80、83%であった。培地pH8.5〜9.5で高い油脂分解率を示し、最適培地pHは9付近であった。また、pH6〜8、pH3でも70%以上の油脂分解率を示した。
【0055】
[培養温度と油脂分解率の関係]
Pseudmonas aeruginosa SS−219株の油脂分解率に対する培養温度の影響を解析した。500mlバッフルフラスコに、3000ppmとなるように混合油脂(サラダ油/ラード/牛脂、質量比1:1:1)を添加したアルカリ人工下水培地2を100ml入れ、24時間予備培養した試験管培養液を1%(v/v)となるように接種した。培養温度を20、25、30、37、40、45℃にそれぞれ設定し、120rpmで24時間震盪培養した。その後、上記と同様にして残存油分を定量し、油脂分解率を算出した。
【0056】
結果を図3(b)に示す。培養温度が20、25、30、37、40、45℃のとき、Pseudmonas aeruginosa SS−219株の24時間培養後の油脂分解率は、それぞれ、17、65、75、83、70、13%であった。油脂分解率は30〜40℃で高い値を示し、最適温度は37℃付近であった。
【0057】
[その他の特徴]
抗生物質含有プレートを用いて、抗生物質感受性試験を行った。その結果、Pseudmonas aeruginosa SS−219株はアンピシリン、クロラムフェニコール抵抗性を示した。
【0058】
Pseudmonas aeruginosa SS−219株を油脂乳化プレートで生育させた様子を図4に示した。Pseudmonas aeruginosa SS−219株のコロニー周囲にはクリアハローが形成されており、菌体外にリパーゼを分泌していることが示唆された。Pseudmonas aeruginosa SS−219株の試験管培養の様子を図5(a)及び(b)に示した。Pseudmonas aeruginosa SS−219株は黄色の蛍光色素を産生しており、培養日数1日目(A)、3日目(B)と、培養日数の増加とともにその色調が濃くなった。さらに、培養12日目(C)には油脂分はほぼ完全に分解されていた。また、培養液の濁度も著しく低下していたことから、溶菌が起こっているものと考えられる。
【特許請求の範囲】
【請求項1】
Pseudmonas属に属し、以下の菌学的性質を示す、油脂分解菌。
細胞形態 桿菌
細胞の直径 0.6〜0.7×1.0〜1.5μm
胞子の有無 −
グラム染色性 −
運動性 +
37℃での生育 +
45℃での生育 +
嫌気性条件下での生育 −
カタラーゼ +
オキシダーゼ +
グルコースからの酸/ガス産生 −/−
グルコースの酸化/発酵 +/−
硝酸塩還元活性 +
インドール産生活性 −
ブドウ糖酸性化活性 −
アルギニンジヒドロラーゼ活性 +
ウレアーゼ活性 −
エスクリン加水分解活性 −
ゼラチン加水分解活性 +
β−ガラクトシダーゼ活性 −
チトクロームオキシダーゼ活性 +
ブドウ糖資化性 +
L−アラビノース資化性 −
D−マンノース資化性 −
D−マンニトール資化性 −
N−アセチル−D−グルコサミン資化性 +
マルトース資化性 −
グルコン酸カリウム資化性 +
n−カプリン酸資化性 +
アジピン酸資化性 +
dl−リンゴ酸資化性 +
クエン酸ナトリウム資化性 +
酢酸フェニル資化性 −
なお、「+」は陽性であること、「−」は陰性であることを示す。
【請求項2】
寒天培地上において、30℃、24時間培養で直径2〜3mmの周縁波状の円形、表面がスムーズ、扁平状で、不透明な緑色コロニーを形成する、請求項1に記載の油脂分解菌。
【請求項3】
配列番号1で示される16S rDNA塩基配列を有する、請求項1又は2に記載の油脂分解菌。
【請求項4】
独立行政法人製品評価技術基盤機構特許微生物寄託センター 受託番号NITE P−847(Pseudmonas aeruginosa SS−219株)で特定される油脂分解菌。
【請求項5】
請求項1〜4のいずれか一項に記載の油脂分解菌を含む油脂分解剤。
【請求項1】
Pseudmonas属に属し、以下の菌学的性質を示す、油脂分解菌。
細胞形態 桿菌
細胞の直径 0.6〜0.7×1.0〜1.5μm
胞子の有無 −
グラム染色性 −
運動性 +
37℃での生育 +
45℃での生育 +
嫌気性条件下での生育 −
カタラーゼ +
オキシダーゼ +
グルコースからの酸/ガス産生 −/−
グルコースの酸化/発酵 +/−
硝酸塩還元活性 +
インドール産生活性 −
ブドウ糖酸性化活性 −
アルギニンジヒドロラーゼ活性 +
ウレアーゼ活性 −
エスクリン加水分解活性 −
ゼラチン加水分解活性 +
β−ガラクトシダーゼ活性 −
チトクロームオキシダーゼ活性 +
ブドウ糖資化性 +
L−アラビノース資化性 −
D−マンノース資化性 −
D−マンニトール資化性 −
N−アセチル−D−グルコサミン資化性 +
マルトース資化性 −
グルコン酸カリウム資化性 +
n−カプリン酸資化性 +
アジピン酸資化性 +
dl−リンゴ酸資化性 +
クエン酸ナトリウム資化性 +
酢酸フェニル資化性 −
なお、「+」は陽性であること、「−」は陰性であることを示す。
【請求項2】
寒天培地上において、30℃、24時間培養で直径2〜3mmの周縁波状の円形、表面がスムーズ、扁平状で、不透明な緑色コロニーを形成する、請求項1に記載の油脂分解菌。
【請求項3】
配列番号1で示される16S rDNA塩基配列を有する、請求項1又は2に記載の油脂分解菌。
【請求項4】
独立行政法人製品評価技術基盤機構特許微生物寄託センター 受託番号NITE P−847(Pseudmonas aeruginosa SS−219株)で特定される油脂分解菌。
【請求項5】
請求項1〜4のいずれか一項に記載の油脂分解菌を含む油脂分解剤。
【図3】

【図1】
【図2】
【図4】
【図5】
【図1】
【図2】
【図4】
【図5】
【公開番号】特開2011−125825(P2011−125825A)
【公開日】平成23年6月30日(2011.6.30)
【国際特許分類】
【出願番号】特願2009−289107(P2009−289107)
【出願日】平成21年12月21日(2009.12.21)
【出願人】(000002107)住友重機械工業株式会社 (2,241)
【出願人】(505089614)国立大学法人福島大学 (34)
【Fターム(参考)】
【公開日】平成23年6月30日(2011.6.30)
【国際特許分類】
【出願日】平成21年12月21日(2009.12.21)
【出願人】(000002107)住友重機械工業株式会社 (2,241)
【出願人】(505089614)国立大学法人福島大学 (34)
【Fターム(参考)】
[ Back to top ]