
治療用エピトープ及びそれらの使用
【課題】初期段階でセリアック病を診断するのに使用できる試験を開発する。
【解決手段】
個体から得られたサンプルにおいてセリアック病を検出するか又はセリアック病に感受性であることを検出する方法であって、i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び
ii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及び
iii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を、個体からのサンプルに接触させ;そして
サンプル中のT細胞が上記薬剤を認識するか否かをインビトロにて決定するが、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示すことを含む、方法を提供する。
【解決手段】
個体から得られたサンプルにおいてセリアック病を検出するか又はセリアック病に感受性であることを検出する方法であって、i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び
ii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及び
iii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を、個体からのサンプルに接触させ;そして
サンプル中のT細胞が上記薬剤を認識するか否かをインビトロにて決定するが、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示すことを含む、方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、セリアック病の診断及び治療に有用なエピトープに関し、診断剤、治療剤、キット、及び前記の使用方法を含む。
【背景技術】
【0002】
ダイエット中のグリアジン(グルテンの成分)に対する免疫反応がセリアック病を引き起こす。腸組織中の免疫応答は腸のトランスグルタミナーゼにより修飾されたグリアジンに優先的に応答する。セリアック病は抗−筋内膜抗体の検出により診断されるが、これは腸の生検内のリンパ球の炎症の発見による確認を必要とする。そのような生検の取得は患者にとって快適ではない。
【0003】
研究者は、腸のT細胞応答のみがグリアジンに対する免疫応答の正確な指標を提供すると、以前に仮定した。よって、彼らは、腸の組織内のT細胞の応答の研究に集中してきた1。トランスグルタミナーゼの修飾を必要とする(それらが免疫系により認識される前に)グリアジンエピトープは公知である2。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】1. MolbergO, etal. NatureMed.4, 713-717 (1998).
【非特許文献2】2. Quarsten H, et al. Eur. J. Immunol. 29, 2506-2514 (1999).
【非特許文献3】3. Greenberg CS et al. FASEB 5, 3071-3077 (1991).
【非特許文献4】4. Mantzaris G, Jewell D. Scand. J. Gastroenterol. 26, 392-398 (1991).
【非特許文献5】5.Mauri L, et al. Scand. J. Gastroenterol. 31, 247-253 (1996).
【非特許文献6】6. BuncoM, et al. Tissue Antigens 46, 355-367 (1995).
【非特許文献7】7. OlerupO, etal. Tissueantigens 41, 119-134 (1993).
【非特許文献8】8. Mullighan CG, et al. Tissue-Antigens. 50, 688-92 (1997).
【非特許文献9】9. PlebanskiM et al. Eur. J. Immunol. 28, 4345-4355 (1998).
【非特許文献10】10. Anderson DO, Greene FC. The alpha-gliadin gene family. II. DNA and protein sequence variation, subfamily structure, and origins of pseudogenes. Theor Appl Genet (1997) 95 : 59-65.
【非特許文献11】11. Arentz-HansenH, Korner R, Molberg O,Quarsten H, Van der Wal Y, Kooy YMC, Lundin KEA, Koning F, Roepstorff P, Sollid LM, McAdam SN. The intestinal T cell response to alpha-gliadin in adult celiac disease is focused on a single deamidated glutamine targeted by tissue transglutaminase. J Exp Med. 2000 ; 191 : 603-12.
【非特許文献12】12. Vader LW, de Ru A, van der Wal, Kooy YMC, Benckhuijsen W, Mearin ML, DrijfhoutJW, van Veelen P, Koning F. Specificity of tissue transglutaminase explains cereal toxicity in celiac disease. J Exp Med 2002 ; 195 : 643-649.
【非特許文献13】13. van der Wal Y, Kooy Y, van Veelan P, Pena S, Mearin L, Papadopoulos G, Koning F. Selective deamidation by tissue transglutaminase strongly enhances gliadin-specific T cell reactivity. J Immunol. 1998 ; 161 : 1585-8.
【非特許文献14】14. van der Wal Y, Kooy Y, van Veelan P, Pena S, Mearin L, Molberg O,Lundin KEA, Sollid L, Mutis T, BenckhuijsenWE, Drijfhout JW, Koning F. Proc Natl Acad Sci USA 1998 ; 95 : 10050-10054.
【非特許文献15】15. Vader W, Kooy Y, Van Veelen Petal. The gluten response in children with celiac disease is directed toward multiple gliadin and glutenin peptides. Gastroenterology2002, 122 : 1729-37 16. Arentz-Hansen H, McAdam SN, Molberg O, et al. Celiac lesion T cells recognize epitopes that cluster in regions of gliadin rich in proline residues. Gastroenterology 2002, 123 : 803-809.
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明者らは、セリアック病において免疫系により認識される免疫上優勢なT細胞A−グリアジンエピトープを発見し、そしてこれがセリアック病の個体の末梢血内でT細胞により認識されることを示した(WO 01/25793)。そのようなT細胞は、再刺激無しに検出可能なほど十分高い頻度で存在することを発見した(即ち、「フレッシュな応答」検出系が使用できた)。上記エピトープは非T細胞クローニングに基づく方法を用いて同定され、上記エピトープのより正確な反射(reflection)が認識された。上記の免疫上優勢なエピトープは免疫系が認識する前にトランスグルタミナーゼの修飾を必要とする(特定のグルタミンのグルタミン酸への置換を引き起こす)。
【0006】
この研究に基づいて、本発明者らは、初期段階でセリアック病を診断するのに使用できる試験を開発した。当該試験は、末梢血からのサンプル上で実施してよく、よって、腸の生検は必要ない。当該試験は現在使用されている抗体試験よりも感度が高い。
【課題を解決するための手段】
【0007】
発明は、即ち、個体においてセリアック病を診断するか又はセリアック病に対する感受性を診断する方法を提供し、
(a)宿主からのサンプルを、(i)配列番号:1(PQPELPY)又は配列番号:2(QLQPFPQPELPYPQPQS)である配列、又は配列番号:3により表されるグリアジンの天然に生じる相同体からの均等な配列を含むエピトープ、(ii)グリアジン蛋白質由来の単離されたオリゴペプチドである、配列番号:1を含むか又は配列番号:3により表されるグリアジンの天然に生じる相同体からの均等な配列(表1に示される)を含むエピトープ、(iii)(i)又は(ii)を認識するT細胞受容体により認識され得る(i)又は(ii)の類似体であって、ペプチドの場合は類似体が50アミノ酸の長さを超えない上記類似体、又は(iv)(i),(ii)又は(iii)において定義された2つ又はそれより多い薬剤(agent)を含む生成物からなる群から選択される薬剤と接触させ、そして(b)サンプル中のT細胞が薬剤を認識するか否かをインビトロにて決定して、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示す
ことを含む。
【0008】
コムギのグリアジンT細胞のエピトープの包括的なマッピングを通して(実施例13参照)、本発明者らは、類似のコア配列(例えば、配列番号:18−22)及び類似の完全長配列(例えば、配列番号:31−36)を有する他のコムギグリアジン中のセリアック病のHLA−DQ2+患者において、並びにライムギのセカリン及びオオムギのホールダイン(配列番号:39−41)の生物学上活性なエピトープをも発見した;表20及び21も参照。さらに、セリアック病においてHLA−DQ8+患者にて生物学上活性な幾つかのエピトープが同定された(配列番号42−44、46)。この包括的なマッピングは、即ち、セリアック病の患者においてT細胞により認識される優勢なエピトープを提供する。即ち、上記の方法及び本明細書に記載される他の方法は、これらの追加の同定されたエピトープ、それらの類似体及び均等物の何れかを使用して実施してよい;本明細書の(i)及び(ii)はこれらの追加のエピトープを含む。即ち、本発明の薬剤は新規なエピ
トープも含む。
【0009】
本発明は、個体において、セリアック病又はセリアック病に対する感受性を診断する方法における使用のための診断手段の製造のための薬剤の使用を提供し、当該方法は、当該個体のT細胞が薬剤を認識するか否かを決定することを含み、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示す。
【0010】
トランスグルタミナーゼにより修飾された免疫上優勢なエピトープ(並びに本明細書において定義された追加の他のエピトープ)の発見は、このエピトープに対する他の種類の免疫応答が存在するか否かを決定することに基づいたセリアック病の診断も可能にする。即ち、本発明は、個体において、セリアック病又はセリアック病に対する感受性を診断する方法も提供し、当該個体からのサンプル中のエピトープに結合する抗体の存在を決定することを含み、抗体の存在が、個体がセリアック病を有するか又はセリアック病に感受性であることを示す。
【0011】
本発明は、さらに、上記薬剤を認識するT細胞を寛容にするセリアック病を治療又は予防する方法における使用のための上記薬剤を、任意にキャリアーと共に提供する。また、T細胞と拮抗することによりセリアック病を治療又は予防する方法における使用のための、(i)又は(ii)を認識するT細胞受容体を有するT細胞のアンタゴニストを、任意にキャリアーと共に提供する。さらに、抗体の生産を阻害するために個体を寛容にすることにより個体においてセリアック病を治療又は予防する方法における使用のための、抗体(上記薬剤に結合する)を結合する薬剤又は類似体が提供される。
【0012】
本発明は、組成物がセリアック病を引き起こすか否かを決定する方法を提供し、トランスグルタミナーゼにより上で定義されたオリゴペプチド配列へ修飾される蛋白質が組成物中に存在するか否かを決定することを含み、蛋白質の存在が、組成物がセリアック病を引き起こし得ることを示す。
【0013】
本発明は、その野生型配列がトランスグルタミナーゼにより上で定義された配列を含むエピトープを含む配列に修飾され得る変異体グリアジン蛋白質も提供するが、しかし、当該変異体グリアジン蛋白質は、そのようなエピトープを含む配列を含む配列へトランスグルタミナーゼにより修飾され得る配列を含まない様式にて修飾されており;又は、本発明は、上記の様式により修飾されたそのような変異体グリアジン蛋白質の断片も提供する。
【0014】
本発明は、T細胞受容体へ結合することができる配列を含む蛋白質も提供し、但し、T細胞受容体は上記の薬剤を認識し、そして上記配列はそのようなT細胞受容体を有するT細胞の拮抗作用を引き起こすことができる。
【0015】
さらに、本発明は、上で定義された蛋白質を含む食品を提供する。
【図面の簡単な説明】
【0016】
【図1A】図1は、最初のうちはグルテンフリーダイエットのあとに緩解して、次に1日目から3日間200gのパン(bread)で毎日チャレンジされた、セリアック病被験者1における、A−グリアジン51−85にわたる5つのオーバーラップする15マーを含むトランスグルタミナーゼ(tTG)−処理されたものと処理されないペプチドプール3(各ペプチド10μg/ml)(表1参照)及びキモトリプシン−消化されたグリアジン(40μg/ml)に対する、新たに単離されたPBMC(末梢血単核細胞)IFNγELISPOT応答(垂直軸は106PBMCあたりのスポット形成細胞を示す)を示す(a)。
【図1B】10日間の間のパンチャレンジの間の全A−グリアジン蛋白質にわたるtTG−処理されたA−グリアジンペプチドプール1−10に対する被験者2によるPBMC IFNγELISPOT応答(b)。水平軸はパン開始後の日にちを示す。
【図2】図2は、最初のうちはグルテンフリーダイエットのあとに緩解して、次に3日間パンでチャレンジされた、セリアック病の7人の被験者における、tTG−処理されたペプチドプール3(A−グリアジン51−85にわたる)に対する、PBMC IFNγELISPOT応答(垂直軸は106PBMCあたりのスポット形成細胞を示す)を示す(1から3日)。垂直軸はパンを初めた後の日にちを示す。(a)。プール3中に含まれるtTG処理のオーバーラップ15マーに対する応答;バーは6日目又は7日目の6人のセリアック病の被験者の個々のペプチド(10μg/ml)に対する、平均(±SEM)の応答を表す(b)。(個々の被験者において、ペプチドに対するELISPOT応答はペプチド12により導かれた応答の%として計算された−水平軸により示されるとおり。)
【図3】図3は、A−グリアジン56−75のtTG−処理された末端削除体(0.1μM)に対するPBMC IFNγELISPOT応答を示す。バーは5人のセリアック病被験者の平均(±SEM)を表す。)。(個々の被験者において、応答は試験されたペプチドにより導かれた最大応答の%として計算された−水平軸により示されるとおり。)
【図4】図4は、優勢なA−グリアジンエピトープの最小構造が如何にしてtTG−処理された7−17マーのA−グリアジンペプチド(0.1μM)を用いてマップされたかを示し、配列PQPQLPY(配列番号:4)(A−グリアジン62−68)(a)、及びtTG処理しなかったがQ→E65の置換を伴う同じペプチドを含む(b)。各線はパンを1−3日目に摂取してから6又は7日目の3人のセリアック病被験者におけるPBMC IγELISPOT応答を表す。)。(個々の被験者において、ELISPOT応答は17マー、A−グリアジン57−73により導かれた応答の%として計算された。)
【図5】図5はtTGにより脱アミド化されたアミノ酸を示す。A−グリアジン56−75LQLQPFPQPQLPYPQPQSFP(配列番号:5)(0.1μM)をtTG(50μg/ml)と共に37℃において2時間インキュベートした。逆相HPLCにより単一の生成物が同定されて精製された。アミノ酸分析により、tTGに帰し得るA−グリアジン56−75内の各Gln残基の%脱アミド化(Q→E)の計算が可能になった(水平軸)。
【図6】図6は、パンを1−3日目に摂取してから6又は7日目の3人のセリアック病被験者における、17マー:QLQPFPQPELPYPQPES(配列番号:6)(E57,65),QLQPFPQPELPYPQPES(配列番号:7)(E65,72),ELQPFPQPELPYPQPES(配列番号:8)(E57,65,72),及びQLQPFPQPELPYPQPQS(配列番号:9)(E65)を用いた、Aグリアジン57−73中のQ65に加えた他の位置においてQ→Eを置換した効果を示す。水平軸はE65応答の%を示す。
【図7】図7は、tTG処理されたA−グリアジン56−75(0.1μM)が(a)CD4及びCD8の磁気ビーズ除去した(depleted)PBMCにおいてIFN−gELISPOT応答を導いたことを示す。(バーはCD4除去PBMC応答をCD8除去PBMC応答の%として表す;百万のCD8除去PBMCあたりのスポット形成細胞は:被験者4:29、及び被験者6:535であった。)(b)HLA−DQaI*0501,bI*0201に関して同型接合体の2人のセリアック病被験者における、tTG−処理56−75(0.1μM)の1時間前の、HLA−DR(L243),−DQ(L2)及び−DP(B7.21)(10μg/ml)に対するモノクローナル抗体とのインキュベーション後のPBMC IFNγELISPOT応答(スポット形成細胞/百万PBMC)。
【図8】図8は、免疫上優勢なエピトープ内の65位における他のアミノ酸のGluによる置換の効果を示す。水平軸は免疫上優勢なエピトープに関しての3人の被験者における%応答を示す。
【図9】図9は、トランスグルタミナーゼ処理を伴う場合(影)及び伴わない場合(透明)の配列PQLPY(配列番号:12)を含む、天然に存在するグリアジンペプチドの免疫反応性を示す(3人の被験者の応答を測定)。
【図10】図10は、グルテンチャレンジを開始して、次にインターフェロンガンマELISpotを行ってから6日後の2人のセリアック病被験者からの末梢血単核細胞のCD8,CD4,β7,及びαE−特異的免疫磁気ビーズ除去を示す。A−グリアジン57−73 QE65(25mcg/ml),tTG−処理されたキモトリプシン−消化グリアジン(100mcg/ml)又はPPD(10mcg/ml)を抗原として用いた。
【図11】図11は、最適なT細胞エピトープの長さを示す。
【図12A】図12aは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12B】図12bは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12C】図12cは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12D】図12dは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12E】図12eは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12F】図12fは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12G】図12gは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12H】図12Hは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図13A】図13aは、グリアジンとA−グリアジン57−73 QE65特異的応答の比較を示す。
【図13B】図13bは、グリアジンとA−グリアジン57−73 QE65特異的応答の比較を示す。
【図13C】図13cは、グリアジンとA−グリアジン57−73 QE65特異的応答の比較を示す。
【図14A】図14aは、セリアック病被験者におけるグリアジンの多型の生物活性を示す。
【図14B】図14bは、セリアック病被験者におけるグリアジンの多型の生物活性を示す。
【図14C】図14cは、セリアック病被験者におけるグリアジンの多型の生物活性を示す。
【図14D】図14dは、セリアック病被験者におけるグリアジンの多型の生物活性を示す。
【図14E】図14eは、セリアック病被験者におけるグリアジンの多型の生物活性を示す。
【図15】図15は、コアエピトープ配列の定義を示す。
【図16】図16は、コアエピトープ配列の定義を示す。
【図17】図17は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図18】図18は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図19】図19は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図20】図20は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図21】図21は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図22】図22は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図23】図23は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図24】図24は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図25】図25は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図26】図26は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図27】図27は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図28A】図28aは、異なる患者群における応答を示す。
【図28B】図28bは、異なる患者群における応答を示す。
【図28C】図28cは、異なる患者群における応答を示す。
【図28D】図28dは、異なる患者群における応答を示す。
【図28E】図28eは、異なる患者群における応答を示す。
【図28F】図28fは、異なる患者群における応答を示す。
【図28G】図28gは、異なる患者群における応答を示す。
【図29】図29は、A−グリアジン57−73のプロラミン相同体の生物活性を示す。
【図30】図30は、健康なHLA−DQ2被験者についての、tTG−脱アミド化されたグリアジンペプチドプールに対するELISpot応答の変化を示す。
【図31】図31は、セリアック病のHLA−DQ2被験者についての、tTG−脱アミド化されたグリアジンペプチドプールに対するELISpot応答の変化を示す。
【図32】図32は、「要約された」グリアジンペプチド応答に対する個々のペプチドの寄与を示す。
【図33】図33は、セリアック病のHLA−DQ2/8被験者C08に関しての、tTG−脱アミド化されたグリアジンペプチドプールに対する、グルテンチャレンジが導いたIFNγELISpot応答を示す。
【図34】図34は、セリアック病のHLA−DQ2/8被験者C07に関しての、tTG−脱アミド化されたグリアジンペプチドプールに対する、グルテンチャレンジが導いたIFNγELISpot応答を示す。
【図35】図35は、セリアック病のHLA−DQ8/7被験者C12に関しての、tTG−脱アミド化されたグリアジンペプチドプールに対する、グルテンチャレンジが導いたIFNγELISpot応答を示す。
【図36】図36は、セリアック病のHLA−DQ6/8被験者C11に関しての、tTG−脱アミド化されたグリアジンペプチドプールに対する、グルテンチャレンジが導いたIFNγELISpot応答を示す。
【発明を実施するための形態】
【0017】
本発明はセリアック病を治療又は予防する方法を提供し、a)配列番号:18−22、31−36、39−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びb)a)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、a)の類似体;及びc)配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を個体に投与するこ
とを含む。幾つかの態様において、上記薬剤は、HLA−DQ2−制限されるか、HLA−DQ8−制限されるか、又は一つの薬剤がHLA−DQ2−制限されており、そして第2の薬剤がHLA−DQ8−制限されている。幾つかの態様において、上記薬剤は、コムギのエピトープ、ライムギのエピトープ、オオムギのエピトープ又はそれらの組み合わせの何れかを、単一の薬剤として又は複数の薬剤として含む。
【0018】
本発明は、上記の薬剤及び薬学上受容可能なキャリアー又は希釈剤を含む薬学組成物を個体に投与することを含む、セリアック病を予防又は治療する方法も提供する。
【0019】
本発明は、上で定義されたT細胞受容体を有するT細胞のアンタゴニスト及び薬学上受容可能なキャリアー又は希釈剤を含む薬学組成物を個体に投与することを含む、セリアック病を予防又は治療する方法も提供する。
【0020】
本発明は、T細胞の生産又は上で定義された薬剤に対する抗体応答を抑圧するために個体をグリアジン蛋白質に対して寛容にするための組成物を個体に投与することを含む、セリアック病を予防又は治療する方法も提供する。
【0021】
本発明は、1)a)i)配列番号:18−22、31−36、39−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を、宿主からのサンプルに接触させ;そしてサンプル中のT細胞が上記薬剤を認識するか否かをインビトロにて決定するが、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示すか;又はb)上で定義された薬剤を投与し、そして個体中のT細胞が上記薬剤を認識するか否かをインビボにて決定するが、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示すかの何れかにより、個体においてセリアック病を診断し;そして2)セリアック病を有するか又はセリアック病に感受性であると診断された個体に、セリアック病を予防又は治療するための治療用薬剤を投与する
ことによりセリアック病を予防するか又は治療する方法も提供する。
【0022】
本発明は、上記薬剤を認識するT細胞を寛容にすることによりセリアック病を予防又は治療する方法における使用のための、上で定義された薬剤も、任意にキャリアーと共に、提供する。
【0023】
本発明は、T細胞と拮抗することによりセリアック病気を予防又は治療する方法における使用のための、上で定義されたT細胞受容体を認識するT細胞のアンタゴニストも、任意にキャリアーと共に、提供する。
【0024】
本発明は、T細胞受容体へ結合することができる配列を含む蛋白質も提供し、但し、T細胞受容体は上で定義された薬剤を認識し、そして上記配列はそのようなT細胞受容体を有するT細胞の拮抗作用を引き起こすことができる。
【0025】
本発明は、上で定義された薬剤又はアンタゴニスト及び薬学上受容可能なキャリアー又は希釈剤を含む薬学組成物も提供する。
【0026】
本発明は、T細胞の生産又は上で定義された薬剤に対する抗体応答を抑圧するために個体をグリアジン蛋白質に対して寛容にするための組成物も提供し、当該組成物は上で定義
された薬剤を含む。
【0027】
本発明は、上で定義された薬剤に対するT細胞応答に拮抗するための組成物も提供し、当該組成物は上で定義されたアンタゴニストを含む。
【0028】
本発明は、その野生型配列がトランスグルタミナーゼにより請求項1において定義された薬剤である配列に修飾され得る変異体グリアジン蛋白質も提供し、当該変異体グリアジン蛋白質は、上で定義された薬剤である配列へのトランスグルタミナーゼによるそれの修飾を阻害する変異を含み;又は、本発明は少なくとも15アミノ酸の長さであり、且つ上記変異を含む、そのような変異体グリアジン蛋白質の断片を提供する。
【0029】
本発明は、上で定義された蛋白質又は断片をコードするコーディング配列を含むポリヌクレオチドも提供する。
【0030】
本発明は、上で定義されたポリヌクレオチドを含む細胞又はそのようなポリヌクレオチドにより形質転換された細胞も提供する。
【0031】
本発明は、上で定義されたT細胞受容体を発現する哺乳類も提供する。
【0032】
本発明は、個体においてセリアック病を診断するか又はセリアック病に対する感受性を診断する方法も提供し、a)i)配列番号:18−22、31−36、39−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を宿主に投与し;そしてb)サンプル中のT細胞が薬剤を認識するか否かをインビトロにて決定して、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示すことを含む。
【0033】
本発明は、組成物がセリアック病を引き起こすことができるか否かを決定する方法も提供し、トランスグルタミナーゼによりオリゴペプチド配列に修飾され得る蛋白質が組成物中に存在するか否かを決定することを含み、蛋白質の存在が、組成物がセリアック病を引き起こし得ることを示す。
【0034】
本発明は、上で定義された薬剤を認識するT細胞のアンタゴニストを同定する方法も提供し、候補物質をT細胞と接触させ、そして当該物質がT細胞が抗原特異的応答を受ける能力の低下を引き起こすか否かを決定することを含み、上記能力のそのようなあらゆる低下の検出が、上記物質がアンタゴニストであることを示す。
【0035】
本発明は、上記の方法の何れかを実施するためのキットも提供し、上で定義された薬剤及びT細胞によるペプチドの認識を検出する手段を含む。
【0036】
本発明は、セリアック病の治療用の生成物を同定する方法も提供し、セリアック病を有するか又はセリアック病に感受性の候補物質を哺乳類に投与し、そして物質が哺乳類においてセリアック病を予防又は治療するか否かを決定することを含み、セリアック病の予防又は治療が、当該物質が治療用生成物であることを示す。
【0037】
本発明は、上で定義されたコーディング配列によりコードされる蛋白質の生産のためのプロセスも提供し、a)上記蛋白質の発現を可能にする条件下で上記細胞を培養し;そし
て、b)発現された蛋白質を回収することを含む。
【0038】
本発明は、トランスジェニック植物細胞を得るための方法も提供し、トランスジェニック植物細胞を供給するために上で定義されたベクターにより植物を形質転換することを含む。
【0039】
本発明は、初代トランスジェニック植物を得るための方法も提供し、トランスジェニック植物を提供するために上で定義されたベクターにより形質転換されたトランスジェニック植物細胞を再生することを含む。
【0040】
本発明は、トランスジェニック植物の種子を得るための方法も提供し、上で定義されたとおりに取得可能なトランスジェニック植物からトランスジェニック種子を得ることを含む。
【0041】
本発明は、トランスジェニック子孫植物を得るための方法も提供し、上で定義された方法により取得可能な初代トランスジェニック植物から第2世代のトランスジェニック子孫植物を得て、そして任意に、そうして得られた第2世代の子孫から一つ又はそれ以上の世代のトランスジェニック植物を得ることを含む。
【0042】
本発明は、上記の方法の何れかにより取得可能な、トランスジェニック植物細胞、植物、植物種子又は子孫植物も提供する。
【0043】
本発明は、上記の植物細胞を含むトランスジェニック植物又は植物種子も提供する。
【0044】
本発明は、上で定義されたトランスジェニック植物細胞、初代植物、植物種子又は子孫から取得可能な上記植物細胞を含む、トランスジェニック植物細胞カルスも提供する。
【0045】
本発明は、収穫生成物を得るための方法も提供し、上記の方法の何れかに従い植物から収穫生成物を回収し、そして任意にさらに回収された生成物を加工することを含む。
【0046】
本発明は、上で定義された蛋白質を含む食物も提供する。
発明の詳細な説明
用語「セリアック病」は、グルテンの感度を変更することにより引き起こされる症状の範囲を包含し、小腸の粘膜により特徴付けられる重度の形態(過形成絨毛萎縮)及び温和な兆候により特徴付けされる他の形態を含む。
【0047】
上で言及された個体(individual)(診断又は治療のコンテクストにおいて)はヒトである。彼らはセリアック病を有するか(兆候があるか又は無兆候)又はそれを有する疑いがある。彼らはグルテンダイエット中であってよい。彼らは急性の相の応答にあってよい(例えば、彼らはセリアック病を有してよいが、彼らがグルテンフリーダイエットを14から28日間行って最後の24時間はグルテンのみを摂取した)。
【0048】
個体は、セリアック病に感受性であってよく、例えば遺伝的に感受性であってよい(例えば、セリアック病の親戚を有するか又はセリアック病に対する疾病素因を引き起こす遺伝子を所有する個体により決定される)。
薬剤
薬剤は、典型的にはペプチドであり、例えば7から50アミノ酸の長さ、例えば10から40、又は15から30アミノ酸の長さである。
【0049】
配列番号:1はPQPELPYである。配列番号:2はQLQPFPQPELPYPQ
Sである。配列番号:3は表1に示され、そしてA−グリアジンの全配列である。配列番号:1の4位(配列番号:2の9位に均等)のグルタミン酸は、A−グリアジンのトランスグルタミナーゼ処理により生成される。
【0050】
薬剤は、配列番号:1又は2により表されるペプチド又はグリアジン蛋白質由来の単離されたオリゴペプチドである配列番号:1を含む配列を含むエピトープ;又は配列番号:3の相同体である天然のグリアジン蛋白質からのこれら配列の均等物であってよい。即ち、上記エピトープは配列番号:3により表される蛋白質の誘導体であってよい。そのような誘導体は、典型的には、グリアジンの断片、又は蛋白質全体又は断片の変異誘導体である。よって、発明のエピトープは、この天然のグリアジン蛋白質全体を含まず、そして他の天然のグリアジン全体を含まない。
【0051】
上記エピトープは、即ち、A−グリアジンの断片(例えば、配列番号:3)であり、配列番号:1の配列を含み、トランスグルタミナーゼにより処理すること(完全又は部分的)により得られるものであって、即ち、1、2、3又はそれ以上のグルタミンがグルタミン酸に置換されている(配列番号:1内での置換を含む)。
【0052】
そのような断片は、表1に示す配列番号:3の55から70、58から73、61から77位により表される配列であるか又はそれらの配列を含んでよい。典型的には、そのような断片は、配列番号:1又は2により表されるペプチドがセリアック病の患者からのサンプルを用いて本明細書にて記載されたアッセイの何れかにおいて認識されるのと少なくとも同じ程度にてT細胞により認識される。
【0053】
さらに、上記薬剤は、配列番号:18−22、31−36、39−44、及び46の何れかにより表されるペプチド、又は配列番号:18−22、31−36、39−44、及び46の何れかに相当する配列を含む蛋白質(例えば、グリアジンをトランスグルタミナーゼで処理した後に配列番号:18−22、31−36、39−44、及び46の何れかを含むグリアジンの断片)。そのような配列の生物学上活性な断片も発明の薬剤である。配列番号:18−22、31−36、39−44、及び46又はこれらの配列の類似体の何れかの配列均等物も、発明の薬剤である。
【0054】
上記エピトープが別のグリアジン(例えば、本明細書にて言及されたグリアジン蛋白質の何れか又はセリアック病を引き起こす何れかのグリアジン)由来の上記エピトープ(断片を含む)に均等な配列を含む場合、そのような均等配列は、トランスグルタミナーゼにより典型的に処理された(完全又は部分的)グリアジン蛋白質の断片に相当する。他のグリアジン蛋白質の配列を、オリジナルのエピトープが由来するグリアジン、例えば、配列番号:3と整列させる(aligning)ことにより(例えば、本明細書にて言及されたプログラムの何れかを用いて)、そのような均等ペプチドを決定することができる。トランスグルタミナーゼは市販されている(Sigma T−5398)。表4は、適当な均等配列の僅かな例を提供する。
【0055】
類似体である薬剤は、(i)又は(ii)を認識するTCRにより認識され得る。よって、一般には、上記類似体が(i)又は(ii)の存在下、典型的には抗原提示細胞(APC)(例えば、本明細書にて言及されたAPCsの何れか)の存在下でも、T細胞に添加された場合、当該類似体は(i)又は(ii)とそのような系において競合し得る。
【0056】
上記類似体は(i)又は(ii)を認識するTCRを結合することができるものであってよい。そのような結合は、標準の技術により試験することができる。そのようなTCRsは(例えば、本発明の方法を用いて)(i)又は(ii)を認識することが示されたT細胞から単離することができる。上記類似体のTCRsへの結合の証明は、類似体に結合
する物質、例えば類似体に対する抗体への類似体の結合をTCRsが阻害するか否かを決定することにより、のちに示すことができる。典型的には、上記類似体は、結合アッセイのそのような阻害においてクラスII MHC分子(例えば、HLA−DQ2)に結合する。
【0057】
典型的には、上記類似体は(i)又は(ii)のTCRへの結合を阻害する。この場合、類似体存在下でTCRを結合できる(i)又は(ii)の量は低下する。これは、類似体がTCRを結合し、よってTCRへの結合に関して(i)又は(ii)と競合するためである。
【0058】
上記の結合実験に使用するための細胞は、セリアック病の患者から、例えば本発明の方法を用いて単離することができる。
【0059】
類似体の他の結合特性は(i)又は(ii)のそれと同じであってよく、即ち典型的には、類似体はペプチドが結合するのと同じMHCクラスII分子に結合する(HLA−DQ2又は−DQ8)。類似体は、典型的には(i)又は(ii)に特異的な抗体に結合し、即ち、(i)又は(ii)のそのような抗体への結合を阻害する。
【0060】
類似体は、典型的にはペプチドである。それは、(i)又は(ii)と少なくとも70%の相同性、好ましくは少なくとも80%、90%、95%、97%又は99%の相同性を、例えば少なくとも15又はそれ以上(例えば類似体及び/又は(i)又は(ii)の完全長、又はTCRに接触するか又はMHC分子を結合する領域にわたり)連続するアミノ酸の領域にわたり有してよい。蛋白質の相同性を測定する方法は当業界公知であり、当業者には本コンテクストにおいて、相同性がアミノ酸同一性に基づいて計算されることが理解される(ときに、「ハードな相同性」と呼ばれる)。
【0061】
例えば、UWGCGパッケージは相同性を計算するために使用できるBESTFITプログラムを提供している(例えば、そのデフォルトセッティングに使用される)(Devereux et al(1984)Nucleic Acids Research 12,p387−395)。PILEUP及びBLASTアルゴリズムは相同性を計算するか又は配列をラインアップするのに使用することができ(典型的にはそれらのデフォルトセッティング上で)、Altschul S.F.(1993)J Mol Evol
36:290−300;Altschul,S.F et al(1990)J Mol 215:403−10に記載されるとおりである
BLAST分析を実施するためのソフトウエアは、例えば、「www.ncbi.nlm.nih.gov/」においてインターネットを通して全世界のウエブ上でNational Center for Biotechnology Informationを通して公的に利用可能である。このアルゴリズムは、データベース配列中で同じ長さのワードと並べられたときに幾つかの正の値の閾値スコアに適合するか又は当該値を満たす質問配列中の長さWの短いワードを同定することにより、高いスコアリングの配列対(HSPs)を最初に同定することを含む。Tは隣接ワードスコア閾値を意味する(Altschul et al,前出)。これらの最初の隣接ワードのヒットは、それらを含むHSPsを見つけるための検索を始めるための種として作用する。蓄積したアラインメントスコアがその最大に達成された値から量Xだけ低下する場合;一つ又は複数の負のスコアリング剰余アラインメントの蓄積のため、蓄積したスコアがゼロ又はそれ以下になった場合;又は何れかの配列の末端に到達した場合、各方向におけるワードヒットに関する延長が停止する。BLASTアルゴリズムのパラメーターW,T及びXは、アラインメントの感度と速度を決定する。BLASTプログラムはデフォルトとして11のワードの長さ(W)、50のBLOSUM62スコアリングマトリックス(Henikoff and Henikoff(1992)Proc.Natl.Acad.Sci.USA 89:
10915−1を参照)アラインメント(B)、10の期待値(E),M=5,N=4,及び両方の鎖の比較を用いる。
【0062】
BLASTアルゴリズムは2つの配列の間の類似性の統計分析を実施する;例えば、Larlin and Altschul(1993)Proc.Natl.Acad.Sci.USA 90:5873−5787を参照されたい。BLASTアルゴリズムにより提供される類似性の一つの測定値は、最小合計確率(P(N))であり、2つのヌクレオチド又はアミノ酸配列間で偶然に適合する確率の指標を提供する。例えば、第1の配列の第2の配列に対する比較において最小合計確率が約1より低く、好ましくは約0.1より低く、より好ましくは約0.01より低く、そしてもっと好ましくは約0.001より低いなら、配列が他方の配列に類似であると考えられる。
【0063】
相同なペプチド類似体は、(i)又は(ii)と1、2、3、4、5、6、7、8又はそれより多い変異だけ異なるのが典型である(置換、欠失又は挿入であってよい)。これらの変異は、相同性を計算することに関して上記の領域の何れかを横切って測定されてよい。置換は、好ましくは「保存的」である。これらは以下の表により定義される。第2列の同じブロックのアミノ酸、及び好ましくは第3の列の同じラインのアミノ酸を、互いに置換してよい。
【0064】
【表1】

【0065】
典型的には、MHC分子を結合するのに寄与するか又はTCRによる認識に必須の、(i)又は(ii)中のアミノ酸に均等な位置における類似体中のアミノ酸は同じか又は保存されている。
【0066】
典型的には、類似体ペプチドは一つ又は複数の修飾を含み、天然の翻訳後修飾又は人工修飾であってよい。修飾は科学的なモイエティ(典型的には、例えばC−H結合の水素の置換による)、例えばアミノ、アセチル、ヒドロキシ又はハロゲン(例えば、フッ素)基又は糖質基を提供してよい。典型的には、修飾はN又はC末端上に存在する。
【0067】
類似体は、一つ又は複数の天然アミノ酸、例えば天然アミノ酸とは側鎖が異なるアミノ酸を含んでよい。一般に、非天然アミノ酸はN末端及び/又はC末端を有する。非天然アミノ酸はL−又はD−アミノ酸であってよい。
【0068】
類似体は、典型的には、(i)又は(ii)と実質類似の形態、サイズ、柔軟性又は電子配置を有する。それは、典型的には、(i)又は(ii)の誘導体である。一つの態様において、類似体は、配列番号:1又は2の配列を含む融合蛋白質であるか又は本明細書にて言及された他のペプチドの何れか;及び非グリアジン配列である。
【0069】
一つの態様において、類似体は、MHCクラス分子に結合した(i)又は(ii)であるか、又は、を模倣する。2、3、4、又はそれ以上のそのような複合体を、例えば、ビオチン/ストレプトアビジンに基づく系を用いて互いに対合させるか又は結合させてよく、典型的には2、3、又は4つのビオチン標識MHC分子がストレプトアビジンモイエティに結合する。この類似体は、典型的には、(i)又は(ii)/MHCクラスII分子のTCR又は当該複合体に特異的な抗体への結合を阻害する。
【0070】
類似体は、典型的には、抗体又は抗体の断片、例えばFab又は(Fab)2断片である。類似体は固相上に固定化され、特に、MHC分子に結合したペプチドを模倣する類似体の場合である。
【0071】
類似体は、典型的には、コンピューター手段によりデザインされて、当業界公知の方法を用いて合成される。或いは、類似体は、化合物のライブラリーから選択できる。ライブラリーはコンビナトリアルライブラリー、ディスプレイライブラリー、例えばファージディスプレイライブラリーであってよい。化合物のライブラリーはMHCクラスII分子に結合した形態、例えばHLA−DQ2又は−DQ8においてディスプレイライブラリー中で発現されてよい。類似体は、一般に、結合特性(i)又は(ii)を模倣するそれらの能力に基づいてライブラリーから選択される。即ち、それらはTCR又は(i)又は(ii)を認識する抗体を結合する能力に基づいて選択してよい。
【0072】
典型的には、類似体は、薬剤(i)又は(ii)の何れかと少なくとも同じ程度、例えば均等なエピトープと少なくとも同じ程度まで、好ましくは配列番号:2により表されるペプチドと同じ程度までT細胞により認識され、本明細書にて記載されたアッセイの何れかにおいて認識され、典型的には、セリアック病の患者由来の細胞を用いる。類似体は、インビボにおいてこれらの程度まで認識されてよく、即ち、本明細書にて言及された薬剤の何れかと少なくとも同じ程度までセリアック病の兆候を誘導することができてよい(例えば、ヒト患者又は動物モデルにおいて)。
【0073】
類似体は、発明のエピトープを認識するT細胞受容体により候補の物質が認識されるか否かを決定することを含み、物質の認識が物質が類似体を認識することを示す方法において同定されてよい。そのようなTCRsは本明細書にて言及されたTCRsの何れかであってよく、T細胞上に存在してよい。本明細書にて言及された如何なる適切なアッセイも類似体を同定するのに使用することができる。一つの態様において、この方法はインビボにおいて実施される。上記のとおり、好ましい類似体はペプチド配列番号:2と少なくとも同じ程度まで認識され、よって、上記の方法はこの程度まで認識される類似体を同定するのに使用してよい。
【0074】
一つの態様において、上記方法は、発明のエピトープの認識を候補配列が阻害するか否かを決定することを含み、認識の阻害が、物質が類似体であることを示す。
【0075】
上記薬剤は、(i),(ii)又は(iii)により定義される少なくとも2、5、10又は20の薬剤を含む生成物であってよい。典型的には、上記組成物は、異なるグリアジン、例えば本明細書にて言及されたグリアジンの種又はバラエティ又は種類の何れかからの発明のエピトープ(又は均等な類似体)を含む。好ましい組成物は、本明細書にて言及された種又はバラエティの何れかの中に存在するグリアジンのすべて、又は2、3、4、又はそれ以上の本明細書にて言及された種からの(コムギ、ライムギ、オオムギ、エンバク、及びライコムギからなる群から選択される種のパネルから)、発明の少なくとも一つのエピトープ、又は均等類似体を含む。即ち、薬剤は単価又は多価であってよい。
診断
上記のとおり、発明の診断方法は、薬剤を結合するT細胞の検出又は薬剤を認識する抗体の検出に基づいてよい。
【0076】
上記の方法(上記の使用を含む)において薬剤を認識するT細胞は、一般に、インビボにおいて予めグリアジンに対して感作されたT細胞である。上記のとおり、そのような抗原−経験T細胞は、末梢血中に存在することがわかった。
【0077】
上記の方法において、T細胞はインビトロ又はインビボにおいて薬剤と接触させることができ、そしてT細胞が薬剤を認識するか否かを決定することをインビトロ又はインビボにて実施することができる。即ち、発明は、ヒトのからだで実施される診断方法においての使用のための薬剤を提供する。異なる薬剤が、そのような方法における、同時、別々、又は連続の使用のために提供される。
【0078】
インビトロの方法は、薬剤が添加された水性溶液内で実施されるのが典型である。当該溶液はT細胞も含む(及び特定の態様においては、以下に論じられるAPCsも)。本明細書にて使用される用語「接触させる」は、特定の物質を溶液に加えることを含む。
【0079】
T細胞が薬剤を認識するか否かを決定することは、一般には、薬剤の存在下でT細胞の状態の変化を検出するか、又はT細胞が薬剤を結合するか否かを決定することにより、達成される。状態の変化は、一般に、TCRが薬剤に結合した後に、T細胞の抗原特異的機能活性により引き起こされる。状態の変化は、T細胞の内部(例えば、蛋白質の細胞内発現の変化)又は外部(例えば、分泌された配列の検出)で測定してよい。
【0080】
T細胞の状態の変化は、T細胞からの物質、例えばサイトカイン、特にIFN−γ、IL−2又はTNF−αの分泌の開始又は増加であってよい。IFN−γの分泌の決定が特に好ましい。当該物質は、特異的結合薬剤へのそれの結合を可能にさせ、次に特異的結合薬剤/物質の複合体の存在を測定することにより検出されるのが典型である。特異的結合薬剤は、典型的には、抗体、例えばモノクローナル抗体である。サイトカインに対する抗体は、市販されており、或いは標準技術を用いて作成することができる。
【0081】
典型的には、特異的結合薬剤は、固相支持体上に固定化される。物質を固相支持体上に固定化させた後に、任意に洗浄することにより、薬剤に非特異的に結合した物質を除去することができる。薬剤/物質複合体は複合体を結合する第2の結合薬剤を用いることにより検出してよい。典型的には、第2薬剤は、第1薬剤を結合する部位とは異なる部位において物質を結合する。第2薬剤は、好ましくは抗体であり、そして、検出可能な標識により直接又は間接に標識される。
【0082】
即ち、第2薬剤は、検出可能な標識により直接又は間接に典型的には標識された第3薬剤により検出してよい。例えば、第2薬剤はビオチンモイエティを含んでよく、ストレプトアビジンモイエティ及び典型的にはアルカリホスファターゼを検出可能な標識として含む第3の薬剤による検出を可能にさせる。
【0083】
一つの態様において、使用される検出系は、WO98/23960に記載されたエクスビボELISPOTアッセイである。そのアッセイにおいて、T細胞から分泌されるIFN−γは、固相支持体に固定化された第1のIFN−γ特異的抗体により結合される。結合されたIFN−γは、次に、検出可能な標識により標識された第2のIFN−γ特異的抗体を用いて検出される。そのような標識された抗体はMABTECH(ストックホルム、スエーデン)から得ることができる。使用可能な他の検出可能な標識は以下に論じられる。
【0084】
測定され得るT細胞の状態の変化は、T細胞による物質の取り込み、例えばチミジンの取り込みの増加であってよい。状態の変化は、T細胞のサイズ、又はT細胞の増殖の増加、又はT細胞上の細胞表面マーカーの変化であってよい。
【0085】
一つの態様において、状態の変化は、蛋白質の細胞内発現の変化、例えば上記サイトカインの何れかの細胞内発現の増加を測定することにより、検出される。そのような細胞内変化は、特定の様式にて発現された蛋白質を結合するモイエティにT細胞内部を接触させることにより検出してよく、フローサイトメトリーによるT細胞の分離を可能にさせる。
【0086】
一つの態様において、TCRを結合するとき、薬剤はMHCクラスII分子(典型的にはHLA−DQ2又は−DQ8)に結合し、典型的には抗原提示細胞(APC)の表面上に存在する。しかしながら、本明細書にて言及されたとおり、他の薬剤はMHC分子にも結合する要求なしにTCRを結合することができる。
【0087】
一般に、上記の方法において接触させたT細胞は固体の血液から採取されるが、T細胞を含む他の種類のT細胞を使用することができる。サンプルは、直接アッセイに添加するか又は最初に加工してよい。典型的には、加工は、例えば、水又はバッファーによるサンプルの希釈を含んでよい。典型的には、サンプルを1.5から100倍、例えば2から50倍又は5から10倍希釈する。
【0088】
加工は、サンプルの成分の分離を含んでよい。典型的には、単核細胞(MCs)をサンプルから分離する。MCsはT細胞とAPCsを含むことになる。即ち、上記の方法において、分離されたMCs内に存在するAPCsは上記ペプチドをT細胞上に提示し得る。別の態様においては、T細胞のみ、例えばCD4 T細胞のみがサンプルから精製され得る。PBMCs,MCs、及びT細胞は、当業界公知の技術、例えばLalvani et al(1997)J.Exp.Med.186,p859−865に記載された技術を用いてサンプルから分離することができる。
【0089】
一つの態様において、上記のアッセイにおいて使用されるT細胞は、未加工の形態であるか、又は希釈されたサンプルであるか、又は新たに単離されたT細胞であり(例えば、新たに単離されたMCs又はPBMCsの形態)、エクスビボにて直接使用され、即ち、それらは上記方法において使用される前に培養されない。即ち、T細胞は抗原特異的様式にてインビトロにおいて再刺激されなかった。しかしながら、T細胞は、使用の前に、例えば一つ又は複数の薬剤及び一般には外来の成長促進サイトカインの存在下で培養することができる。培養の間、薬剤は典型的にはAPCs,例えば上記方法で使用されるAPCの表面上に存在する。T細胞の前培養は上記方法の感度の増加を導くかもしれない。即ち、T細胞は、細胞系、例えば短期間の細胞系に変換することができる(例えば、Ota et al(1990)Nature 346,p183−187に記載されたとおり)。
上記の方法に典型的に存在するAPCはT細胞と同じ個体から又は別の宿主からであってよい。APCは天然のAPC又は人工のAPCであってよい。APCは上記ペプチドを細胞に提示することができる細胞である。それは典型的にはB細胞、樹状細胞又はマクロファージである。それは典型的にはT細胞と同じサンプルから分離され、典型的にはT細胞と同時精製される。即ち、APCはMCs又はPBMCs上に存在してよい。APCは典型的には新たに単離されたエクスビボ細胞又は培養細胞である。それは細胞系の形態、例えば短期間細胞系又は不死の細胞系であってよい。APCはその表面上に空のMHCクラスII分子を発現してよい。
【0090】
上記の方法において、一つ又は複数の(異なる)薬剤を使用してよい。典型的には、サンプル由来のT細胞をアッセイ中において試験を意図する全ての薬剤と共に置くことがで
きるか、又は各々が一つ又は複数の薬剤を含む別々のアッセイ中に分配して置くことができる。
【0091】
本発明は、同時、分離又は連続用途のための(例えばインビボ用途)本明細書にて言及された薬剤の何れか2つ又はそれ以上のような薬剤も提供する(例えば、上で論じられた組成物中に存在する薬剤の組み合わせ)。
【0092】
一つの態様においては、薬剤それ自体を直接T細胞及びAPCsを含むアッセイに加える。上で論じられたとおり、そのようなアッセイにおいてT細胞とAPCsはMCsの形態であり得る。APCsによる提示に関する要求なしにT細胞により認識され得る薬剤を用いる場合、APCsは必要ない。MHC分子に結合させたオリジナルの(i)又は(ii)を模倣した類似体がそのような薬剤の例である。
【0093】
一つの態様において、上記薬剤はT細胞の不在下でAPCに提供される。APCは次に、典型的には薬剤をその表面に提示させた後に、T細胞に提供される。上記ペプチドはAPCの内部に拾い上げられて提示されるか、又はAPCの内部に入り込まずに単に表面上に拾い上げられる。
【0094】
薬剤がT細胞と接触する期間は、ペプチドの認識を決定するのに使用される方法しだいで変更される。典型的には105から107、好ましくは5x105から106のPBMCを各アッセイに添加する。薬剤を直接アッセイに添加する場合、その濃度は10−1から10−3μg/ml、好ましくは0.5から50μg/ml又は1から10μg/mlである。
【0095】
典型的には、T細胞を薬剤とインキュベートする時間の長さは、4から24時間、好ましくは6から16時間である。エクスビボにてPBMCを使用する場合、0.3x106のPBMCsを10μg/mlのペプチド中で12時間37℃においてインキュベートできることを発見した。
【0096】
T細胞による薬剤の認識の決定は、薬剤のT細胞への結合を測定することにより行ってよい(これは本明細書にて論じられる何れかの適当な結合アッセイを用いて実施できる)。典型的には、薬剤を結合するT細胞は、例えばFACSマシンを用いてこの結合に基づいて細胞を分離することができる。薬剤を認識するT細胞の存在は、薬剤を用いて分離される細胞の頻度が「対照」値を超える場合に生じるとみなされる。抗原を経験したT細胞の頻度は106中1から103中1であり、よって、分離された細胞が抗原経験T細胞であるか否かを決定することができる。
【0097】
T細胞による薬剤の認識の決定はインビボにおいて測定してよい。典型的には、薬剤を宿に投与して、次に、薬剤の認識を示す応答を測定してよい。薬剤は、典型的には、皮内又は表皮投与である。薬剤は、典型的には、皮膚の外部に接触させることにより投与され、そしてプラスター又は包帯の補助により当該部位に保持されてよい。或いは、薬剤は針により、例えば注射により投与してよいが、他の方法、例えば弾道(ballistics)(例えば、核酸をデリバリーするために使用される弾道技術)により投与することもできる。EP−A−0693119は、薬剤を投与するために典型的に使用できる技術を記載する。典型的には、0.001から1000μg、例えば0.01から100μg又は0.1から10μgの薬剤を投与する。
【0098】
一つの態様において、インビボにおいて薬剤を提供することができる生成物を投与できる。即ち、薬剤を発現できるポリヌクレオチドを投与することができ、典型的には、薬剤の投与のための上で記載された様式の何れかによる。ポリヌクレオチドは、典型的には、
以下に論じられる、発明により提供されるポリヌクレオチドの特性の何れかを有する。典型的には、0.001から1000μg、例えば0.01から100μg又は0.1から10μgのポリヌクレオチドを投与する。
【0099】
皮膚に投与される薬剤の認識は、典型的には、投与部位における炎症の発症により示唆される(例えば、硬化、紅斑又は浮腫)。これは、一般に、当該部位における目視試験により判断される。
【0100】
薬剤を結合する抗体の検出に基づく診断法は、典型的には、個体からのサンプル(本明細書にて言及されたサンプルの何れかで、任意に本明細書にて言及された様式の何れかにより加工された)を薬剤と接触させ、そしてサンプル中の抗体が薬剤を結合するか否かを決定することにより実施され、そのような結合が、個体がセリアック病を有するか又は感受性であるかを示す。結合アッセイのあらゆる適当なフォーマット、例えば本明細書にて言及されたあらゆるそのようなフォーマットを使用してよい。
治療
本明細書にて記載された免疫上優勢なエピトープ及び他のエピトープの同定は、このエピトープを認識するT細胞を標的とする治療用生成物が作成されるのを可能にさせる(そのようなT細胞はグリアジンに対する免疫応答に参加するものである)。これらの発見は、抗体又はエピトープに対するT細胞応答を抑圧(寛容)することによりセリアック病の予防又は治療を可能にもする。
【0101】
発明の特定の薬剤は、発明のエピトープを認識するTCRを結合し(上で論じられた結合アッセイの何れかを用いて測定されるとおり)、そしてTCRを有するT細胞の寛容を導く。そのような薬剤は、任意にキャリアーと共に、従ってセリアック病を予防又は治療するのに使用することができる。
【0102】
一般に、寛容は、(TCRによりされた後に)T細胞の抗原特異的機能活性を導き得る同じペプチドにより導かれ得る(例えば、本明細書にて言及されたそのような活性、例えばサイトカインの分泌)。そのような薬剤は「寛容」コンテクストにおいて免疫系に対して提示される時に寛容を導く。
【0103】
寛容は免疫系によるT細胞又は抗体エピトープの認識の低下を導く。T細胞エピトープの場合、これは、エピトープを認識するT細胞の欠失又はアネルギーにより導かれ得る。即ち、エピトープに応答するT細胞活性(例えば、本明細書にて言及された適当なアッセイにおいて測定されたとおり)が低下する。抗体応答の寛容は、エピトープが投与されたときに、エピトープに対する特異的抗体の量の低下が生じることを意味する。
【0104】
そのようなコンテクストにおいて免疫系に対して抗原を提示する方法は公知であり、例えば、Yoshida et al.Clin.Immunol.Immunopathol 82,207−215(1997),Thuau et al.Clin Exp.Immunol.109,370−6(1997)、及びWeiner et al.Res.Immunol.148,528−33(1997)に記載されている。特に、特別の投与経路が寛容を導き得て、例えば、経口、鼻内又は腹膜内である。寛容は樹状細胞及びテトラマー提示ペプチドにより達成してもよい。寛容を導く特定の生成物を個体に投与してよい(例えば、薬剤も含む組成物中で)。そのような生成物は、サイトカイン、例えばTh2応答を好むサイトカインを含む(例えば、IL−4,TGF−β又はIL−10)。生成物又は薬剤は、寛容を導く用量にて投与してよい。
【0105】
発明は、T細胞のアンタゴニストとして作用することができる配列を含む蛋白質を提供する(そのT細胞は薬剤を認識する)。そのような蛋白質及びそのようなアンタゴニスト
はセリアック病を予防又は治療するのに使用することもできる。アンタゴニストはT細胞応答の低下を導く。一つの態様において、アンタゴニストはT細胞のTCRに結合する(一般に、HLA−DQ2又は−DQ8との複合体の形態にて)が、正常な機能活性化を導く代わりに、T細胞内シグナリングカスケードを通して異常なシグナルを通過させることを導き、T細胞が低下した機能活性を持つように導く(例えば、エピトープの認識に応答して、典型的には本明細書にて言及された適当なアンタゴニストの何れかにより測定される)。
【0106】
一つの態様において、アンタゴニストはエピトープとMHCプロセシング及び提示の経路の成分への結合、例えばMHC分子(典型的には、HLA−DQ2又は−DQ8)に関して競合する。即ち、アンタゴニストは、HLA−DQ2又は−DQ8(即ち、このMHC分子により提示されるペプチドである)、例えばペプチドTP(表10)又はその相同体を結合してよい。
【0107】
拮抗作用を導く方法は、当業界公知である。一つの態様において、アンタゴニストは上で言及されたエピトープの相同体であり、そして配列の何れか、結合又は薬剤の他の特性を有してよい(特に、類似体)。アンタゴニストは、典型的には、上記のエピトープの何れかとは(T細胞内で正常な抗原特異的機能を導くことができる)、1、2、3、4又はそれ以上の変異だけ(各々は置換、挿入又は欠失であってよい)異なる。そのようなアンタゴニストは当業界において「変更されたペプチドリガンド」又は「APL」と呼ばれる。変異は、典型的には、TCRに接触するアミノ酸位置においてである。
【0108】
アンタゴニストはA−グリアジンのアミノ酸65から67により表される配列に均等な配列内の置換によりエピトープと異なってよい(表9に示すようなアンタゴニスト)。即ち、好ましくは、アンタゴニストは64、65又は67位の均等なものにおいて置換を有する。好ましくは、置換は、64W,67W,67M又は65Tである。
【0109】
個体における発明のエピトープに対するT細胞の免疫応答はポリクローナルであるから、1より多いアンタゴニストが投与されることにより、異なるTCRsを有する応答のT細胞の拮抗作用を導いてよい。よって、アンタゴニストは少なくとも2、4、6又はそれより多いアンタゴニストを含む組成物中で投与されてよく、各々が異なるT細胞と拮抗する。
【0110】
発明は、(上記薬剤を認識する)T細胞のアンタゴニストを同定する方法も提供し、候補物質をT細胞と接触させ、そしてT細胞が抗原特異的応答を経る能力の低下を当該物質が導くか否かを検出することを含み(例えば、本明細書にて言及された何れかの適当なアンタゴニストを用いて)、上記能力のあらゆるそのような低下の検出が物質がアンタゴニストであることを示す。
【0111】
一つの態様において、アンタゴニスト(特定のエピトープに対する複数のアンタゴニストの組み合わせを含む)又は寛容(T細胞及び抗体寛容)薬剤は、発明の異なるエピトープに対して、例えば1より多い物質を含む生成物である薬剤に関して上で論じられたエピトープの組み合わせに対して、拮抗するか又は寛容する少なくとも2、4、6又はそれより多いアンタゴニスト又は薬剤を含む組成物中に存在する。
組成物がセリアック病を引き起こすか否かを試験する
上で論じられたとおり、発明は、組成物がセリアック病を引き起こすことができるか否かを決定する方法を提供し、トランスグルタミナーゼにより発明の薬剤又はエピトープを含む配列に修飾され得る蛋白質配列の存在を検出することを含む(そのようなトランスグルタミナーゼ活性はヒトの腸のトランスグルタミナーゼ活性であってよい)。典型的には、これは、特定の様式にて配列に結合するモイエティを組成物に接触させる結合アッセイ
を用いて実施され、そして配列/モイエティ複合体の形成を検出して薬剤の存在を確認するのに使用する。そのようなモイエティは、本明細書にて言及された何れかの適当な物質(又は物質の種類)であってよく、そして典型的には特異的抗体である。結合アッセイのあらゆる適当なフォーマットを使用することができる(例えば、本明細書にて言及されたもの)。
【0112】
一つの態様において、上記組成物を、異なるグリアジン由来の発明のエピトープに特異的な少なくとも2、5、10又はそれより多い抗体、例えば、1よりも多い物質を含む生成物である発明の薬剤に関して上で論じられたエピトープの組み合わせを認識することができる抗体のパネルに接触させる。
【0113】
上記組成物は、典型的には、セリアック病を引き起こし得るグリアジンを発現する植物由来の材料を含む(例えば、本明細書にて言及されたグリアジン又は植物の何れも)。そのような材料は植物の一部、例えば収穫された作物(例えば、種子)であってよい。材料は植物材料の加工された生成物(例えば、本明細書にて言及されたあらゆるそのような生成物)、例えばグリアジンを含む穀粉又は食物であってよい。食物材料の加工及び適当な結合アッセイにおける試験は日常的であり、例えばKricka LJ,J.Biolumin.Chemilumin.13,189−93(1998)に言及されたとおりである。
結合アッセイ
本明細書にて言及された2つの物質間の結合の決定は、結合に際して変化する一方又は両方の物質の特性、例えば分光学上の変化を測定することにより実施してよい。
【0114】
結合アッセイフォーマットは「バンドシフト」系であってよい。これは、一つの物質(例えば、候補物質)の存在がゲル電気泳動の間に他の物質の進行を進めさせるか又は遅らせるかを決定することを含む。
【0115】
上記フォーマットは、一つの物質が他の物質、例えば特異的抗体に結合することが知られている薬剤に対する他の物質の結合を阻害できるか否かを決定する、競合結合法であってよい。
変異体グリアジン蛋白質
発明は、本発明の配列又はそのような配列を提供するためにトランスグルタミナーゼにより修飾され得る配列が変異することによりもはやエピトープを認識するT細胞応答を導かないか又はエピトープを認識するT細胞応答により認識されないような、グリアジン蛋白質を提供する。このコンテクストにおいて、用語認識はT細胞の正常な(非拮抗)抗原−特異的機能活性が生じるような様式におけるエピトープのTCR結合を意味する。
【0116】
他のグリアジンにおいて均等なエピトープを同定する方法は、上で論じられている。変異したグリアジンの野生型は、セリアック病を引き起こすものである。そのようなグリアジンは、例えば配列番号:3の全域又は配列番号:3の15、30、60、100又は200の連続するアミノ酸にわたり上記の程度(類似体に関して)、配列番号:3に相同性を有してよい。同様に、他の非−A−グリアジンに関しては、そのグリアジンの変異体と天然形態の間に相同性が存在することになる。他の天然グリアジン蛋白質の配列は当業界公知である。
【0117】
変異したグリアジンはセリアック病を引き起こさないか、又はセリアック病の兆候の低下を導く。典型的には、エピトープがT細胞応答を誘導する能力を変異が低下させる。変異したエピトープはHLA−DQ2又は−DQ8に対する低下した結合、APCにより提示される能力の低下、又は薬剤を認識するT細胞に結合するか又は当該T細胞により認識される能力(即ち、抗原−特異的機能活性)の低下を有してよい。変異したグリアジン又
はエピトープは、よって、発明の診断の側面に関して、本明細書にて言及されたアッセイの何れかにおいて認識を示さないか又は低下した認識を示すことになる。
【0118】
上記変異は、上記エピトープ中の1から3、4から6、6から10、11から15又はそれより長い長さの一つ又はそれより多い欠失、付加又は置換であってよく、例えば、配列番号:2の配列にわたるか、配列番号:18−22、31−36、39−44及び46の何れかにわたるか、又はそれらの均等物にわたる。好ましくは、上記変異グリアジンは、配列番号:1の配列内に少なくとも一つの変異を有する。好ましい変異は、A−グリアジン中の65位である(又は他のグリアジン中の均等な位置)。典型的には、この位置において天然に生じるグルタミンは、表3に示されるアミノ酸の何れか、好ましくはヒスチジン、チロシン、トリプトファン、リジン、プロリン又はアルギニンにより置換される。
【0119】
発明は、即ち、グリアジン蛋白質のエピトープ内の変異(本明細書にて論じられる何れかの配列中の変異のそのような変異)の用途も提供し、当該エピトープは本発明のエピトープであり、グリアジン蛋白質がセリアック病を引き起こす能力を低下させる。
【0120】
一つの態様において、変異した配列はアンタゴニストとして作用することができる。即ち、発明は、T細胞受容体に結合することができる配列を含む蛋白質を提供し、T細胞受容体は発明の薬剤を認識し、そして配列はそのようなT細胞受容体を認識するT細胞の拮抗作用を導くことができる。
【0121】
発明は、上記変異グリアジン蛋白質の断片である蛋白質も提供し、それらは少なくとも15アミノ酸の長さ(例えば、少なくとも30、60、100、150、200又は250アミノ酸長)であり、そしてグリアジンが認識される能力を低下させる上で論じられた変異を含む。上で言及された変異蛋白質の何れも(断片を含む)、例えば他のグリアジン又は非グリアジン蛋白質と融合蛋白質の形態で存在してもよい。
【0122】
変異グリアジン蛋白質に対する均等な野生型蛋白質は、典型的には、イネ科の単子葉植物、例えばコムギ(Triticum)属の植物、例えば、コムギ、ライムギ、オオムギ、エンバク又はライコムギに由来する。蛋白質は、典型的には、α、αβ、γ又はωグリアジンである。グリアジンはA−グリアジンであってよい。
キット
発明は、一つ又はそれより多い薬剤、及び任意にT細胞による薬剤の検出のための手段を含む、上記方法を実施するためのキットも提供する。典型的には、異なる薬剤を、同時、別々又は連続する使用のために提供する。提供、認識を検出する手段は、上で論じられた技術に基づいて検出を可能にするか又は補助する。
【0123】
即ち、上記手段は、認識後にT細胞により分泌された物質の検出を可能にしてよい。上記キットは、即ち、物質、例えば抗体のための特異的結合モイエティをさらに含んでよい。当該モイエティは、典型的には、IFN−γに特異的である。当該モイエティは典型的には固相支持体に固定化される。これは、モイエティを結合した後に、上記物質がそれを分泌するT細胞の付近に残ることを意味する。即ち、物質/モイエティ複合体の「スポット」が支持体上に形成され、各スポットが物質を分泌するT細胞を表す。スポットを定量し、そして典型的には対照に対して比較することにより、薬剤の認識の決定を可能にする。
【0124】
上記キットは、物質/モイエティ複合体を検出する手段も含む。検出可能な変化、例えば色の変化が、物質を結合後にモイエティ自身に生じる。或いは、第2のモイエティが直接又は間接に検出のために標識され、スポットの決定を可能にするように物質/モイエティ複合体を結合することを可能にしてよい。上で論じられたとおり、第2モイエティは物
質に対して特異的であってよいが、第1モイエティとは上記物質に対して異なる部位において結合する。
【0125】
固定化された支持体はウエルを有するプレート、例えばマイクロタイタープレートであってよい。各アッセイは、よって、プレート中の別々のウエル内で実施され得る。
【0126】
上記キットは、さらに、T細胞のための培地、検出モイエティ又は検出工程において使用される洗浄バッファーを含んでよい。上記キットは、さらに、サンプルからの分離に適した試薬、例えばサンプルからのPBMCs又はT細胞の分離に適した試薬を含んでよい。上記キットは、サンプルの成分の如何なる分離も無しに、サンプルからのT細胞の直接の同定を可能にするようにデザインしてよい。
【0127】
上記キットは、対照、例えば陽性対照又は陰性対照も含んでよい。陽性対照は検出系が試験されるのを可能にしてよい。即ち、陽性対照は、典型的には、上記方法の何れかにおいて薬剤の認識を模倣する。典型的には、インビトロにおいて認識を決定するためにデザインされたキットにおいては、陽性対照はサイトカインである。薬剤のインビボの認識を検出するためにデザインされたキットにおいては、陽性対照はほとんどの個体が認識すべき抗原であってよい。
【0128】
上記キットは、宿主からのT細胞を含むサンプル、例えば血液サンプルを採取するための手段も含んでよい。上記キットは、宿主からのサンプルから単核細胞又はT細胞を分離するための手段を含んでよい。
ポリヌクレオチド、細胞、トランスジェニック哺乳類及び抗体
発明は、薬剤又は変異グリアジン蛋白質を提供するための発現可能なポリヌクレオチドも提供する。典型的には、当該ポリヌクレオチドは、DNA又はRNAで、単鎖又は二重鎖である。当該ポリヌクレオチドは、好ましくは、少なくとも50塩基又は塩基対、例えば50から100、100から500、500から1000又は1000から2000又はそれより多い塩基又は塩基対を含むことになる。当該ポリヌクレオチドは、よって、配列番号:1又は2又は本明細書にて言及された他の薬剤の何れかの配列をコードする配列を含む。このコーディング配列の5’及び3’に、発明のポリヌクレオチドは対応するグリアジン遺伝子内のこれら配列に対して5’及び3’の配列又はコドンと異なる配列又はコドンを有する。
【0129】
上記ペプチドをコードする配列に対して5’及び/又は3’に、上記ポリヌクレオチドはコーディング配列又は非コーディング配列を有する。コーディング配列に対して5’及び/又は3’の配列は、上記薬剤をコードする配列の発現、例えば、転写及び/又は翻訳を助ける配列を含んでよい。上記ポリヌクレオチドは、原核又は真核細胞において上記薬剤を発現することができてよい。一つの態様において、上記ポリヌクレオチドは、哺乳類細胞、例えばヒト、霊長類又は齧歯類(例えば、マウス又はラット)細胞内で上記薬剤を発現することができる。
【0130】
発明のポリヌクレオチドは、バックグラウンドを顕著に超えるレベルで配列番号:3をコードするポリヌクレオチドに選択的にハイブリダイズしてよい。選択的ハイブリダイゼーションは、典型的には、中から高いストリンジェンシー(例えば、0.03M 塩化ナトリウム及び0.03Mクエン酸ナトリウム、約50℃から約60℃において)の条件下で達成される。しかしながら、そのようなハイブリダイゼーションは当業界公知のあらゆる適切な条件下で実施してよい(Sambrook et al(1989),Molecular Cloning:A Laboratory Manualを参照)。例えば、高いストリンジェンシーが必要なら、適切な条件は0.2xSSC、60℃を含む。低いストリンジェンシーが必要なら、適切な条件は2xSSC、60℃を含む。
【0131】
発明の薬剤又は蛋白質は、本明細書にて記載されるポリヌクレオチドによりコードされてよい。
【0132】
当該ポリヌクレオチドは、複製可能なベクターを形成するか又はベクターに取り込まれてよい。そのようなベクターは、適当な細胞中で複製することができる。当該ベクターは、発現ベクターであってよい。そのようなベクターにおいて、発明のポリヌクレオチドは、ポリヌクレオチドの発現を提供することができる制御配列に作動可能に連結される。当該ベクターは、選択可能なマーカー、例えばアンピシリン耐性遺伝子を含んでよい。
【0133】
上記ポリヌクレオチド又はベクターは細胞中に存在してよい。そのような細胞は上記ポリヌクレオチド又はベクターにより形質転換されていてよい。当該細胞は上記薬剤を発現してよい。当該細胞は上記ベクターに適合可能なように選択され、そして、例えば、原核(細菌)、酵母、昆虫又は哺乳類の細胞であってよい。当該ポリヌクレオチド又はベクターは、リン酸カルシウム沈殿法、DEAE−デキストラントランスフェクション、又はエレクトロポレーションを含む慣用技術を用いて宿主細胞に導入されてよい。
【0134】
発明は、組換え手段による発明の蛋白質の生産のためのプロセスを提供する。これは、(a)蛋白質の発現を可能にさせる条件下で上で定義された形質転換細胞を培養し;そして好ましくは(b)発現されたポリペプチドを回収することを含んでよい。任意に、上記ポリペプチドは、当業界公知の技術により単離され、及び/又は、精製されてよい。
【0135】
発明は、そのような認識(又は結合)が可能な薬剤又はその断片を認識する(又は結合する)TCRsも提供する。これらは、発明の蛋白質に関して本明細書にて論じられた本明細書にて言及された如何なる形態でも存在し得る(例えば、精製度)。発明は、発明の細胞に関して本明細書にて論じられた如何なる形態でも存在し得る(例えば、精製度)そのようなTCRsを発現するT細胞も提供する。
【0136】
発明は、薬剤(例えば、発明のエピトープ)を特異的に認識しそして発明の変異グリアジン蛋白質を認識する(そして典型的には均等な野生型グリアジンを認識しない)薬剤を特異的に認識するモノクローナル又はポリクローナル抗体、及びそのような抗体を作成する方法も提供する。発明の抗体は発明のこれらの物質に特異的に結合する。
【0137】
この発明のために、用語「抗体」は、抗体断片、例えば、Fv,F(ab)及びF(ab)2断片、並びに一本鎖抗体を含む。
【0138】
ポリクローナル抗体を生産する方法は、適当な宿主動物、例えば実験動物を免疫源で免疫し、そして血清からイムノグロブリンを単離することを含む。上記動物は、よって、免疫源を接種されてよく、次に血液を当該動物から採取し、そしてIgG分画を精製する。モノクローナル抗体を生産する方法は、所望の抗体を生産する細胞を不死にすることを含む。ハイブリドーマ細胞は、接種された実験動物からの脾臓細胞を腫瘍細胞と融合することにより生産してよい(Kohler and Milstein(1975)Nature 256,495−497)。
【0139】
所望の抗体を生産する不死の細胞を慣用の手法により選択してよい。ハイブリドーマは、培養液中で培養されるか、又は腹水液の形成のために腹膜内に注射されるか、又は同種の宿主又は免疫上無防備状態の宿主の血流に注射されてよい。ヒトの抗体は、ヒトリンパ球のインビトロ免疫、続くエプスタイン−バールウイルスによるリンパ球の形質転換により製造してよい。
【0140】
モノクローナル抗体は及びポリクローナル抗体はの両方の生産のためには、実験動物は、ヤギ、ウサギ、ラット又はマウスが適している。所望なら、免疫源は、例えばアミノ酸残基の一つの側鎖を通して免疫源が適当なキャリアーにカップリングされるようにコンジュゲートとして投与してよい。得られた抗体は、単離し、そして所望なら精製してよい。
【0141】
発明のポリヌクレオチド、薬剤、蛋白質、又は抗体は、検出可能な標識を有してよい。分泌された物質を可視検査により、任意に光学拡大手段の補助により、分泌された物質の検出を可能にする検出可能な手段は、好ましい。そのような系は、典型的には、基質中の色の変化を導く酵素標識に基づき、例えば、基質中の色の変化を導くアルカリホスファターゼである。そのような基質は市販されており、例えば、BioRadからである。他の適当な標識は、他の酵素、例えばペルオキシダーゼ、又は蛋白質標識、例えばビオチン;又は放射性標識、例えば32P又は35Sを含む。上記の標識は、公知の技術を用いて検出してよい。
【0142】
発明のポリヌクレオチド、薬剤、蛋白質、又は抗体は、実質上精製形態であってよい。それらは、製造物中のポリヌクレオチド、ペプチド抗体、細胞又は乾燥塊(dry mass)の少なくとも80%、例えば少なくとも90、95、97又は99%を一般に含む、実質上の単離された形態にあってよい。ポリヌクレオチド、薬剤、蛋白質又は抗体は、典型的には、他の細胞成分を実質上含まない。当該ポリヌクレオチド、薬剤、蛋白質又は抗体は、そのような実質上単離されたか、精製されたか、又は遊離の形態にて上記方法において使用されるか、又はそのような形態で容器キット中に存在してよい。
【0143】
発明は、発明のTCRを発現するトランスジェニック非ヒト哺乳類も提供する。これは、本明細書にて論じられた哺乳類の何れかであってよい(例えば、上記抗体の生産に関して)。好ましくは、当該哺乳類は、セリアック病を有するか又は感受性である。当該哺乳類はHLA−DQ2又は−DQ8又はHLA−DR3−DQ2を発現してもよく、及び/又は、セリアック病を引き起こすグリアジンを含むダイエットを与えられてよい(例えば、本明細書にて言及されたグリアジン蛋白質の何れか)。即ち、上記哺乳類はセリアック病の動物モデルとして機能してよい。
【0144】
発明は、セリアック病の治療用の生成物を同定する方法も提供し、セリアック病を有するか又は感受性の発明の動物に候補物質を投与し、そして当該哺乳類において物質がセリアック病を予防するか又は治療するか否かを決定することを含み、セリアック病の予防又は治療が、上記物質が治療用生成物であることを示す。そのような生成物はセリアック病を予防又は治療するのに使用してよい。
【0145】
発明は、治療用(予防用を含む)薬剤又は診断用(発明の薬剤、蛋白質及びポリヌクレオチド)薬剤を提供する。これらの物質は、薬学上受容可能なキャリアー又は希釈剤とそれらを混合することにより臨床投与のために製剤化される。例えば、それらは、局所、非経口、静脈内、筋肉内、皮下、眼球内、皮膚内、表皮内、経皮投与のために製剤化され得る。上記物質は、薬学上受容可能であって且つ所望の投与経路のために適切なあらゆる媒体と混合してよい。注射のための薬学上受容可能なキャリアー又は希釈剤は、例えば滅菌又は等張溶液、例えば注射用の水又は生理食塩水、又は弾道デリバリー用のキャリアー粒子であってよい。
【0146】
物質の用量は、様々なパラメーターに従い、特に使用される薬剤;処置される患者の年齢、体重及び状態;使用される投与様式;処置される症状の重度;及び要求される臨床上の養生法に従い調節してよい。規準として、注射により投与される物質の量は、0.01mg/kgから30mg/kg、好ましくは0.1mg/kgから10mg/kgが適している。
【0147】
記載された投与経路及び用量は規準としてのみ意図されるが、何故ならば熟練した開業医はあらゆる特定の患者及び症状に関して投与の最適な経路及び用量を決定することができるからである。
【0148】
発明の物質は、即ちヒト又は動物の身体の治療方法、又はヒト身体に実施される診断法において使用してよい。特に、それらは、セリアック病を治療又は予防する方法において使用してよい。発明は、セリアック病を治療又は予防するための医薬の製造法における使用のための薬剤も提供する。即ち、発明は、その要求のあるヒトへ発明の物質を投与することを含むセリアック病の予防又は治療法を提供する(典型的には、その非毒性有効量)。
【0149】
発明の薬剤は、標準の合成価額技術、例えば自動化された合成器の使用により作成することができる。上記薬剤は、より長いポリペプチド、例えば融合蛋白質から作成してよく、そのようなポリペプチドは上記ペプチドの配列を含む。当該ペプチドは、例えばポリペプチドを加水分解することにより、例えばプロテアーゼの使用により;又は物理的にポリペプチドを破壊することにより、ポリペプチドに由来し得る。発明のポリヌクレオチドは標準の技術、例えば合成器を使用することにより、作成することができる。
変異グリアジン蛋白質を発現するか又はアンタゴニストとして作用し得る配列を含む蛋白質を発現する植物細胞又は植物
発明の細胞は、植物細胞、例えばイネ科の単子葉植物種の細胞であってよい。当該種は、その野生型形態がグリアジン、本明細書にて言及されたグリアジン蛋白質の何れかを発現する種であってよい(配列番号:3に対して何れかの程度の相同性を有するグリアジンを含む)。そのようなグリアジンはヒトにおいてセリアック病を引き起こすかもしれない。上記細胞は、コムギ、トウモロコシ、エンバク、ライムギ、イネ、オオムギ、ライコムギ、モロコシ、又はサトウキビの細胞であってよい。典型的には、上記細胞は、コムギ(Triticum)属、例えばパンコムギ(aestivum)、スペルトコムギ(spelta)、ポーランドコムギ(polonicum)又はヒトツブコムギ(monococcum)の細胞である。
【0150】
発明の植物細胞は、典型的には、野生型グリアジンを発現しない(例えば、セリアック病を引き起こすかもしれない本明細書にて言及された何れかのグリアジン)か、又は薬剤を認識するT細胞により認識され得る配列を含むグリアジンを発現しないものである。即ち、野生型植物細胞がそのようなグリアジンを発現するなら、そのときは、そのようなグリアジンの発現を阻害するか低下させるか又はもはやセリアック病を引き起こさないようにグリアジンのアミノ酸配列を変化させるように操作してよい(典型的には、発明のエピトープをもはや発現させないことにより)。
【0151】
これは、変異を、例えば1、2、3又はそれより多いか又は全てのそのようなグリアジン遺伝子へ細胞内で導入すること、例えばコーディング又は非コーディング(例えば、プロモーター)へ導入することにより実施することができる。そのような変異は、本明細書にて論じられた変異の種類又は長さの何れかであり得る(例えば、相同蛋白質に関して)。上記変異は、直接様式(例えば、部位特異的変異導入又は相同組換え技術を用いて)又はランダムな様式(例えば、変異源を用い、次に典型的にはグリアジン(又はセリアック病を引き起こすグリアジン配列)をもはや発現しない変異導入された細胞を選択することにより)にて導入することができる。
【0152】
アンタゴニストとして作用し得る配列を含む蛋白質を発現する植物又は植物細胞の場合、そのような植物又は植物細胞は野生型グリアジン蛋白質を発現してよい(例えば、セリアック病を引き起こすもの)。好ましくは、アンタゴニスト配列の存在は、上記植物又は
植物細胞由来の蛋白質を含む食物を摂取した個体において低下した兆候(例えば兆候なし)のセリアック病を引き起こす。
【0153】
植物細胞内に存在する(か又はに形質転換された)ポリヌクレオチドは、一般に、植物細胞内で変異グリアジン蛋白質を発現できるプロモーターを含む。所望の発現のパターンに依存して、上記プロモーターは、構成的、組織特異的、ステージ特異的;及び/又は誘導可能性であってよい。例えば、植物における強い構成的発現は、CAMV35S,Rubisco ssu又はヒストンプロモーターにより得ることができる。また、組織特異的又はステージ特異的プロモーターは、トランスジェニック植物において特定の組織へ又はその発生において特定のステージへ、発明の蛋白質の発現を標的化するために使用してよい。即ち、例えば、種子特異的、根特異的、葉特異的、花特異的等のプロモーターを使用してよい。種子特異的プロモーターは、Dalta et al(Biotechnology Ann.Rev.(1997),3,pp.269−296)に記載されたものを含む。種子特異的プロモーターの特定の例は、napinプロモーター(EP−A−0 255,378)、ファセオリンプロモーター、グルテニンプロモーター、ヘリアンセニンプロモーター(WO98/45461)及びATS1及びATS3プロモーター(PCT/US98/06798)を含む。
【0154】
上記細胞はあらゆる形態であってよい。例えば、単離された細胞、例えばプロトプラストであってよいか、又は植物組織の一部、例えばカルス又は植物から切り出された組織であってよいか、又は植物全体の一部であってよい。上記細胞はあらゆる種類の細胞であってよい(例えば、植物の部分のあらゆる種類)。例えば、分化しない細胞、例えばカルス細胞;又は分化した細胞、胚、花粉、根、若枝(shoots)又は葉に見いだされる種類の細胞。植物の部分は、根;若枝;葉及び生殖に関与する部分、例えば、花粉、卵、おしべ、葯、花弁、萼片、及び他の花の部分を含む。
【0155】
発明は、トランスジェニック植物細胞を提供するように発明のポリヌクレオチド又はベクターにより植物細胞を形質転換することを含む、トランスジェニック植物細胞を得るための方法を提供する。あらゆる適切な形質転換方法を使用してよい(コムギの場合、Vasil V et al,Biotechnology 10,667−674(1992)に開示された技術を使用してよい)。好ましい形質転換技術は、植物プロトプラストのエレクトロポレーション及び粒子の砲撃を含む。形質転換は、即ち、幾つかの細胞がトランスジェニックで幾つかはそうでないようなキメラ組織又は植物を生じてよい。
【0156】
発明の細胞又はそうして得られた細胞は、当業界公知の技術によりトランスジェニック植物へ再生されてよい。これらは、トランスジェニック細胞の生育及び/又は分割を刺激するための、オーキシン、ジベレリン及び/又はサイトキニンのような植物成長物質の使用を含んでよい。同様に、体性胚形成及び分裂組織培養のような技術を使用してよい。再生技術は当業界にて公知であり、例えば、米国特許第4,459,355号、米国特許第4,536,475号、米国特許第5,464,763号、米国特許第5,177,010号、米国特許第5,187,073号、欧州特許267,159,欧州特許604,662,欧州特許672,752、米国特許第4,945,050号、米国特許第5,036,006号、米国特許第5,100,792号、米国特許第5,371,014号、米国特許第5,478,744号、米国特許第5,179,022号、米国特許第5,565,346号、米国特許第5,484,956号、米国特許第5,508,468号、米国特許第5,538,877号、米国特許第5,554,798号、米国特許第5,489,520号、米国特許第5,510,318号、米国特許第5,204,253号、米国特許第5,405,765号、欧州特許442,174,欧州特許486,233、欧州特許486,234、欧州特許539,563、欧州特許674,725、WO91/02071及びWO95/06128に見いだすことができる。
【0157】
そのような多くの技術において、一つの工程は、細胞を膨張させる及び/又は分割することを含む、カルス、即ち細胞組織の形成である。そのようなカルスは他の種類の植物細胞培養物及び植物の部分であるように、発明の別の側面である。即ち、例えば、発明は、トランスジェニック植物組織及び部分を提供するが、胚、分裂組織、種子、若枝、根、幹、葉及び花の部分を含む。これらは、それらの細胞の幾つかが発明の細胞であり幾つかはそうでない感覚において、キメラであってよい。発明のトランスジェニック植物部分及び組織、植物及び/又は種子は、本明細書にて言及された植物種の何れかのものであってよい。
【0158】
再生手法は、典型的には、マーカー遺伝子による、形質転換された細胞の選択を含む。
【0159】
再生工程は、第1世代のトランスジェニック植物を生じる。発明は、この第1世代植物からのさらなる再生のトランスジェニック植物を得るための方法も提供する。これらは子孫トランスジェニック植物として知られる。第2、第3、第4、第5、第6及びさらなる世代の子孫植物は、当業界公知のあらゆる手段により第1世代のトランスジェニック植物から得てよい。
【0160】
即ち、発明は、トランスジェニック子孫植物を得る方法を提供し、発明の第1世代のトランスジェニック植物から第2世代のトランスジェニック子孫植物を得て、そして任意に第2世代のトランスジェニック子孫植物から一つ又はそれより多い世代のトランスジェニック植物を得ることを含む。
【0161】
子孫植物はあらゆる公知の技術による前の世代のそれらの先祖から生じさせてよい。特に、子孫植物は:
前の世代に属する発明のトランスジェニック植物からトランスジェニック種子を得て、次にトランスジェニック種子を成長させることにより新世代の発明のトランスジェニック子孫植物を得て;そして/又は
前の世代に属する発明のトランスジェニック植物をクローンとして増殖させることにより新世代に属する発明のトランスジェニック子孫植物を提供し;そして/又は
前の世代に属する発明の第1世代のトランスジェニック植物を別の匹敵する植物と交配することにより、新世代に属する発明のトランスジェニック子孫植物を提供し;そして/任意に
そうして得られた子孫から一つ又はそれより多いさらなる世代のトランスジェニック子孫植物を得ること
により生産してよい。
【0162】
これらの技術はあらゆる組み合わせにおいて使用してよい。例えば、クローンとしての増殖及び有性増殖を、別の点において、培養のために適したトランスジェニック植物を生じるプロセスにおいて、使用してよい。特に、農業経済学上望まれる特性を有する植物分類群との繰り返しの戻し交配を企ててよい。植物及びそれから再生した新規な植物からの細胞の取り出しの別の工程も実施してよい。
【0163】
また、細胞、植物組織、植物又は種子をあらゆる適切なステージにおいて、さらに望まれる特性を導入することにより、発明のポリヌクレオチド以外の所望のコーディング配列を導入してよい。これは、発明のポリヌクレオチドの導入のために本明細書にて記載された技術により実施してよい。
【0164】
例えば、他の除草剤耐性特性、例えばグリフォセート(Glyphosate)に対する寛容(例えば、EPSPシンターゼ遺伝子(例えば、EP−A−0293,358)又
はグリフォセートオキシドレダクターゼ(WO92/000377)遺伝子)をコードするトランス遺伝子から;又は、フォサメチン;ジハロベンゾニトリル;グルフォシネートに対する寛容をコードするトランス遺伝子から、例えばホスフィノチリシナセチルトランスフェラーゼ(PAT)又はグルタミンシンターゼ遺伝子(EP−A−0242,236参照)を用いて;アスラム、例えば、ジヒドロプテロエートシンターゼ遺伝子(EP−A−0 369,367参照);又はスルフォニルウレア、例えば、ALS遺伝子を用いて;ジフェニルエーテル、例えば、アシフルオルフェン又はオキサジアゾン、例えば、プロトポルフィロジェンオキシダーゼ遺伝子;環状イミド、例えば、クロロフサリム;フェニルピラゾール、例えば、TNP、又はフェノピレート又はそのカルバメート類似体;さらなるトランス遺伝子を選択してよい。
【0165】
同様に、除草剤寛容以外の有益な特性に関する遺伝子を導入してよい。例えば、昆虫耐性遺伝子を導入してよく、注目に値するのはバチルスサリンジェネシス(Bt)の毒素をコードする遺伝子である。同様に、疾患耐性遺伝子を導入してよく、例えば、WO91/02701又はWO95/06128におけるとおりである。
【0166】
典型的には、発明の蛋白質を発明の植物内で発現させる。使用されるプロモーターにより、この発現は構成的又は誘導可能であってよい。同様に、組織特異的か又はステージ特異的であってよく、即ち、特定の植物の組織(例えば、本明細書にて言及された組織の何れか)又は植物の発生におけるステージに直接向けられる。
【0167】
発明は、作物を得るための方法も提供し、発明のトランスジェニック植物を収穫し、そして任意にさらに加工することを含む。作物は、作物植物から得ることができるあらゆる有用な生成物を意味する。
変異グリアジン蛋白質又はアンタゴニストとして作用することができる配列を含む蛋白質を含む生成物
発明は、変異グリアジン蛋白質又はアンタゴニストとして作用することができる配列を含む蛋白質を含む生成物を提供する。これは、典型的には、そのような蛋白質を発現する本明細書にて言及された植物に由来するか又は植物の部分を含む。そのような生成物は、発明の植物を直接収穫するか又は間接に収穫し、そしてさらに加工することにより得ることが可能であってよい。直接に得ることが可能な生成物は穀物を含む。あるいは、そのような生成物は、回収してさらに加工することにより間接に得ることが可能であってよい。さらに加工することにより得ることが可能な生成物の例は、小麦粉又は蒸留されたアルコール飲料;直接得ることができるか又はさらに加工した材料、例えば小麦粉から作られた焼いた製品(例えば、パン)から作られる食物製品である。典型的には、そのような食物製品はヒト個体により摂取可能であって消化可能である(即ち、非毒性であって栄養価値がある)。
【0168】
アンタゴニスト配列を含む蛋白質を含む食物製品の場合、食物製品は野生型グリアジンを含んでもよいが、好ましくは当該アンタゴニストはそのような食物を摂取した後にセリアック病の兆候の低下(又は完全に)を導くことができる。
【0169】
発明は、以下の非限定実施例により例示される。
【実施例】
【0170】
実施例1
我々は、完全に特性決定されたグリアジン、「A−グリアジン」の全配列にわたる51セットの合成15マーペプチドを使用することによりセリアック病のエピトープマッピングを実施した(表1参照)。A−グリアジンペプチドはtTGにより個々に処理されることにより、インビボで生成したものを模倣したかもしれない生成物を生成した3。我々は
、実験上の感染及び自己免疫疾患において記載されたとおり、エピトープの「拡散」又は「消費」が起きたかもしれない可能性を避けるために、疾患の消滅の開始点においてセリアック病患者を研究しようとも努めた。
3日及び10日のパンチャレンジによる臨床上及びA−グリアジン特異的なT細胞応答
パイロット研究において、セリアック病の2人の患者が緩解したが、血清の抗−筋内膜抗体(EMA)の不在により定義され、グルテンフリーダイエットは彼らの通常のグルテンフリーダイエットに加えて、標準のグルテン含有白パン4スライスを毎日給仕された。被験者1は3日後に腹痛、口の潰瘍及びゆるい下痢のためにパンを停止したが、被験者2は1週間目に軽い吐き気があっただけで10日間継続した。EMAは被験者2においてパンチャレンジの1週間後に陽性になったことから、使用したパンがセリアック病の再発を導いたことを示す。しかし、被験者1においては、EMAはパンチャレンジから2カ月まで陰性のままであった。両被験者において、パンチャレンジにより出現した兆候はグルテンフリーダイエットに戻って2日以内に解消した。
【0171】
IFNγELISPOTアッセイのA−グリアジンに対するPBMC応答は、パンチャレンジの前と間に見いだされなかった。しかし、被験者1においてはパンの消費の1日後に(4日目)、tTGにより処理したA−グリアジン51−85にわたる5つのオーバーラップするペプチドの単一のプール(プール3)が有力なIFNγ応答を示した(図1a参照)。被験者1においては、A−グリアジンペプチドに対するPBMC IFNγ応答はプール3単独に標的化されたままであり、8日目に最大であった。プール3に対する応答の動力学及び大きさは、α−キモトリプシン消化されたグリアジンにより導かれるのと同様であった。tTG処理したプール3に対するPBMC IFNγ応答はtTG処理されなかったプール3より5から12倍の大きさに一貫し、そしてα−キモトリプシン消化したグリアジンに対する応答はtTG処理した場合の3から10倍大きかった。被験者2においては、tTG処理されたプール3も、8日目にA−グリアジンペプチドの免疫源セットのみであったが、この応答は被験者1よりも弱く、4日目には観察されず、11日目にはプール3に対する応答は減少し、そしてA−グリアジンペプチドの他のtTG処理プールがより強いIFNγ応答を導いた(図1b参照)。
【0172】
パイロット研究は、これらのセリアック病被験者における初期T細胞応答が5つのペプチドの単一のtTG処理A−グリアジンプールに対してであったこと、及び末梢血において容易に測定されたことを示した。抗原暴露が3日ではなく10日間続いたなら、他のA−グリアジンペプチドに対するT細胞応答が、エピトープの拡散に一致して、出現する。tTG処理されたA−グリアジンペプチドによるセリアック病特異的IFN−g誘導
グルテンフリーダイエットの別の6人のセリアック病被験者の内の5人において(表1参照)、3日間のパンチャレンジはtTG処理されたペプチドをプール3に同定し、特に、PBMCからIFNγを導く唯一のA−グリアジン成分としての56−70(12)及び60−75(13)に相当するペプチドであった(図2参照)。IFNγ ELISPOTと平行して流したIL−10 ELISPOTは、tTG処理されたペプチド12又は13に対してIL−10の応答を示さなかった。一人の被験者においては、パンチャレンジの4日前、4日間又は4日目まで、あらゆるA−グリアジンペプチド又はα−キモトリプシン消化されたグリアジンに対してIFNγ応答はなかった。パンチャレンジの2カ月後まで測定した場合に、これらのセリアック病被験者の誰にも基底ラインからEMA状態が変化しなかった。
【0173】
HLA−DQ対立遺伝子α1*0501,β1*0201(28−52歳、2人は女性)を有し、3日間パンチャレンジを受けて1カ月間グルテンフリーダイエットを受けた、4人の健康なEMA陰性被験者からのPBMCは、tTG処理をしてもしなくても、何れのA−グリアジンペプチドに対しても陰性対照を超えるIFNγ応答を示さなかった。即ち、PBMC中のIFNγのtTG処理プール3及びA−グリアジンペプチド56−70
(12)及び60−75(13)への誘導は、セリアック病特異的であった(7/8対0/4、p<0.01,カイ2乗分析による)。
最小のA−グリアジンT細胞エピトープのファインマッピング
A−グリアジン56−75の末端削除に相当するtTG処理したペプチドは、同じコアペプチド配列QPQLP(配列番号:9)が評価された5人のセリアック病被験者の全てにおいて抗原性に必須であったことを明らかにした(図3参照)。PBMC IFNγは、7マーのPQPQLPY(配列番号:4)で始まるこのコア配列にわたり、長さを伸ばしたtTG処理ペプチドに応答することから、tTG処理された17マーのQLQPFPQPQLPYPQPQS(配列番号:19)(A−グリアジン57−73)がIFNγ ELISPOTにおいて最大の活性を所有したことを示した(図4参照)。
tTGによるQ65の脱アミド化はA−グリアジン中の免疫上優勢なT細胞エピトープを生じる
HPLC分析は、A−グリアジンのtTG処理が、親ペプチドにおいてあとで隣接して溶出した単一の生成物を生じたことを証明した。アミノ酸配列決定は、A−グリアジン56−75中に含まれる6つのグルタミン(Q)残基の中から選択的にtTGにより脱アミド化されたことを示した(図5参照)。グルタミン酸(E)置換されたQ65のコアA−グリアジン62−68からの連続伸長物(serial expansions)に相当するペプチドの生物活性は、tTG処理後のQ65を有する同じペプチドと均等であった(図4a参照)。Q57及びQ72のEによる両方又は単独の置換にE65が伴うと、調査された3人のセリアック病被験者において、上記17マーの抗原性を増強しなかった(図6参照)。グリアジンペプチドにおいてグルタミン残基にプロリンが続くと、インビトロにおいてtTGにより脱アミド化されないため、Q57及びQ72を調査した(W.Vader et al,Proceedings 8th International
Symposium Coeliac Disease)。よって、免疫上優勢なT細胞エピトープをQLQPFPQPELPYPQPQS(配列番号:2)と定義した。
免疫上優勢なT細胞エピトープ応答はDQ−2制限され且つCD4依存性である
HLA−DQα1*0501,β1*0201に関して同型接合の(homozygous)2人のセリアック病被験者において、抗−DQモノクローナル抗体がtTG処理されたA−グリアジン56−75に対するELISPOT IFNγ応答をブロックしたが、抗−DP及び−DR抗体はブロックしなかった(図7参照)。2人のセリアック病被験者からのPBMCの抗−CD4及び−CD8磁気ビーズ除去(depletion)は、tTG処理されたA−グリアジン56−75に対するIFNγ応答がCD4 T細胞媒介性であることを示した。
考察
この研究において、我々は、配列:QLQPFPQPELPYPQPQS(配列番号:2)(残基57−73)のtTG処理されたA−グリアジン17マーに応答性のセリアック病被験者の末梢血においてCD4 T細胞の一過性集団を誘導するために、普通の白パンを用いたむしろ単純なダイエット抗原チャレンジを記載した。A−グリアジン(Q→E65)に対する免疫応答はセリアック病関連対立遺伝子DQα1*0501,β1*0201に限定される。インビトロにおける組織のトランスグルタミナーゼ作用はQ65を選択的に脱アミド化する。置換Q→E65を有する合成A−グリアジンペプチドに対する誘導された末梢血IFNgの応答は、tTG処理されたQ65 A−グリアジンペプチドに均等であり;両者は未修飾のQ65 A−グリアジンペプチドよりもIFNg ELISPOTにおいて10倍にまで、より多くのT細胞を刺激する。
【0174】
我々は、エピトープマッピングにおけるT細胞クローンの使用により生じるかもしれない方法論上の人工物の可能性を回避するため、インビボ抗原チャレンジ及び短期間エクスビボ免疫アッセイを用いて、このセリアック病特異的T細胞エピトープを慎重に定義した。我々の発見は、グルテンの摂取に対する末梢血のT細胞応答が迅速であるが短命であり、且つエピトープマッピングに利用可能であることを示す。インビボ抗原チャレンジは、
A−グリアジンペプチドに対する免疫応答の時間の経過順の階層(temporal hierarchy)が存在することを示し;tTGにより修飾されたA−グリアジン57−73はP中のもっとも強いIFNg応答を誘導するのみならず、第1のIFNg応答も出現する。
【0175】
我々はA−グリアジンにわたるペプチドしか評価しなかったので、セリアック病の病因において等しいか又はより重要な他のグリアジン内の別のエピトープが存在するかもしれない。事実、我々が同定したPQPQLPY(配列番号:4)であるA−グリアジン内のコアエピトープにおけるペプチド配列は幾つかのべつのグリアジンに共通する(SwissProt and Trembl受け入れ番号:P02863,Q41528,Q41531,Q41533,Q9ZP09,P04722,P04724,P18573)。しかしながら、生検チャレンジ及びインビボ研究において生物活性を所有することが以前に示されたA−グリアジンペプチド(例えば:31−43、44−55及び206−217)4,5は、セリアック病被験者において3日間のパンチャレンジ後にPBMCにおいてIFNg応答を誘導しなかった。これらのペプチドは、免疫応答の拡散を伴って現れる「二次」T細胞エピトープであるかもしれない。
実施例2
免疫上優勢なエピトープ内の置換に対するT細胞認識の効果
57−73A−グリアジンエピトープにおける65位のグルタミンの置換の効果を、合成ペプチドを用いてIFNγ ELISPOTアッセイにおいて置換されたエピトープに対する末梢血応答を測定することにより決定した(50μg/mlにおいて)。応答は3人のセリアック病患者においてグルテンチャレンジを始めて6日後に測定した(3日間4スライスのパンを毎日)。結果を表3及び図8に示す。グルタミン酸から、ヒスチジン、チロシン、トリプトファン、リジン、プロリン又はアルギニンへの置換において観察され得るように、免疫上優勢なエピトープに対する応答の大きさの10%より低い大きさの応答を刺激した。即ち、この位置におけるA−グリアジンの変異は、免疫反応性を低下させるか又は不在にする変異グリアジンを生成するのに使用することができた。
実施例3
他の天然に生じるグリアジンからの均等なペプチドの免疫反応性の試験
他の天然に生じるコムギのグリアジン由来の均等なペプチドの免疫反応性を、天然に生じる配列に相当する合成ペプチドをトランスグルタミナーゼにより処理して使用することにより、評価した。これらのペプチドは、ELISPOTにおいて、実施例2に記載されたのと同じ被験者からのPBMCを用いて同じ様式にて試験した。少なくとも5つのペプチドがA−グリアジン57−73 E65ペプチドに匹敵する免疫反応性を示す(トランスグルタミナーゼ処理後)ことから、コムギ中の他のグリアジン蛋白質も同ようにこのセリアック病特異的免疫応答を誘導することを示唆する(表4及び図9)。
方法
被験者:研究において使用した患者は、英国のオックスフォードにおいてセリアック病クリニックにかかっている。セリアック病は、特有の小腸の組織構造、及び兆候の標準化及びグルテンフリーダイエットの小腸組織構造に基づいて診断された。
組織型判定:組織型判定は、EDTA抗凝血末梢血から抽出されたDNAを用いて実施した。HLA−DQA及びDQB遺伝子型判定は、PCRにより配列特異的プライマーミックスを用いて実施した6−8。
抗筋内膜抗体アッセイ:EMAは、サルの食道(oesophagus)を用いて1:5に希釈された患者の血清を用い、続いてFITCコンジュゲートヤギ抗−ヒトIgAを用いた間接免疫蛍光により検出した。IgAをEMAの前に定量したが、どの被験者もIgA欠損ではなかった。
抗原チャレンジ:グルテンフリーダイエット後のセリアック病被験者に、4スライスのグルテン含有パンを毎日3又は10日間食べ尽くさせた(セインズベリーズの標準の「白サンドイッチパン」)。パンチャレンジを始める前の週及び2カ月後まで、EMAを査定し
た。グルテンフリーダイエットを4週間経験した健康な被験者は、3日間グルテン含有パン4スライスを含む彼らの通常のダイエットを食べつくし、そして次にさらに4日間グルテンフリーダイエットに戻った。
IFNγ及びIL−10 ELISPOT:Ficoll−Hypaque密度遠心分離により50−100mlの静脈血からPBMCを調製した。3回洗浄後に、PBMCを再度、10%熱不活性化ヒトAB血清を含む完全RPMIに懸濁した。IFNγ及びIL−10の単一細胞の分泌に関するELISPOTアッセイを、市販のキット(Mabtech;ストックホルム、スエーデン)を用いて96−ウエルプレート(MAIP−S−45;Millipore,ベッドフォード、MA)により、製造者の指示書(ほかで記載されたとおり9)に従い、2−5x105(IFNγ)又は二重のウエルにて査定し、そしてミコバクテリウムツベルクロシスの精製された蛋白質誘導体(PPD RT49)(Serum Institute;コペンハーゲン、デンマーク)(20μg/ml)を全てのアッセイにおいて陽性対照として含ませた。
ペプチド:合成ペプチドはResearch Genetics(ハンツビル、アラバマ)から購入し、質量分光分析及びHPLCにより確認されたペプチドの真正度は70%>精製度であった。グリアジン(Sigma;G−3375)(100mg/ml)のα−キモトリプシン(Sigma;C−3142)200:1(w/w)による消化は、0.1M NH4HCO3中で2M尿素と共に室温において実施し、98℃において10分間加熱することにより24時間後に中止した。遠心分離(13,000g,10分間)後に、グリアジン消化上清をフィルター滅菌した(0.2mm)。グルテンの消化をSDS−PAGEにより確認して、蛋白質濃度を査定した。α−キモトリプシン−消化されたグリアジン(640μg/ml)及び合成グリアジンペプチド(15マー:160μg/ml,他のペプチド:0.1mM)を個別にtTG(Sigma;T−5398)(50μg/ml)によりPBS中にてCaCl2を1mM加えて、2時間37℃において処理した。ペプチド及びペプチドプールを滅菌96−ウエルプレートに分配して、使用まで−20℃に凍結保存した。
ペプチドのアミノ酸配列決定:逆相HPLCを使用して、A−グリアジン56−75のtTG処理によるペプチドを精製した。単一生成物を同定して、アミノ酸配列決定(自動化配列決定機モデル494A,Applied Biosystems,フォスターシティー、カリフォルニア)に供した。未修飾のG56−75の配列はLQLQPFPQPQLPYPQPQSFP(配列番号:5)と確認され、そしてtTG処理されたG56−75はLQLQPFPQPELPYPQPQSFP(配列番号:11)と確認された。グルタミン残基の脱アミド化は、アミノ酸配列決定のサイクル2、4、8、10、15及び17において回収されたグルタミン及びグルタミン酸の合わせた量のパーセントとして表された、回収されたグルタミン酸の量(pmol)として定義された。tTGに帰する脱アミド化は、(tTG処理されたペプチド中のグルタミンの脱アミド化の%−未処理のペプチド中の脱アミド化の%)/(100−未処理のペプチド中の脱アミド化の%)として定義された。
CD4/CD8及びHLAクラスII制限:抗−CD4又は抗−CD8コートされた磁気ビーズ(Dynal,オスロ、ノルウエー)をRPMIにより4回洗浄し、次に10%熱不活性化ヒトAB血清を含む完全RPMI中のPBMCと共に、30分間氷の上でインキュベートした。ビーズを磁石により取り出して、残る細胞を計数した。tTG処理されたA−グリアジン56−75に対する免疫応答のインビボのHLA−クラスII制限は、PBMC(5x106細胞/ml)を抗−HLA−DR(L243),−DR(L2),及び−DP(B7.21)モノクローナル抗体(10μg/ml)と、室温において、ペプチドの添加前に1時間インキュベートすることにより、確立した。
実施例4
グリアジン−特異的末梢血リンパ球による粘膜インテグリンの発現
内皮とリンパ球のアドレシンの間の相互作用は、器官特異的なリンパ球のホーミングを促進させる。多数のアドレシンが公知である。ヘテロダイマーα4β7は粘膜固有層消化
管及び他の粘膜リンパ球に特異的であり、そしてαEβ7は消化管及び皮膚において内皮リンパ球内に特異的である。末梢血CD4 T細胞の約30%がα4β7を発現し、そして粘膜部位へ移動中であると推定され、末梢血CD4 T細胞の5%がαEβ7を発現する。αE又はβ7特異的な抗体をコートされた免疫磁気ビーズは、それぞれ、αEβ7又はαEβ7及びα4β7を発現する細胞のPBMCを除去する。ELISpotアッセイとの組み合わせにより、免疫磁気ビーズの除去は、これらの細胞を粘膜表面へのホーミングとして同定するかもしれないグリアジン特異的T細胞のアドレシン発現の決定を可能にする。おもしろいことに、インビボのグルテンチャレンジは小腸の粘膜固有層へのCD4
T細胞の迅速な流入に関連しており(内皮部位内ではなく)、90%を超えるリンパ球がα4β7を発現する。
【0176】
免疫磁気ビーズを製造して、3日間のグルテンチャレンジの後で6日目又は7日目にセリアック病被験者からのPBMCを除去するのに使用した。FACS分析は、αEビーズが約50%の陽性CD4 T細胞を除去し、一方β7ビーズは全てのβ7陽性CD4 T細胞を除去した。CD4−又はβ7−ビーズを用いたがCD8−又はαEビーズを用いないPBMC除去はインターフェロンガンマELISpotにおいて応答を完全に破壊した。tTGグリアジン及びPPD応答はCD4除去により完全に破壊されたが、インテグリン特異的ビーズ除去により一貫して影響された。
【0177】
即ち、セリアック病においてグルテンチャレンジ後に誘導されたA−グリアジン57−73 QE65特異的T細胞は、小腸中の粘膜固有層CD4 T細胞上に存在するインテグリンα4β7を発現する。
実施例5
最適なT細胞エピトープの長さ
A−グリアジン中の優勢なT細胞エピトープのコアにわたる7から17の長さのアミノ酸からのペプチドを試験する以前のデータは、3日間のグルテンチャレンジ開始から6日後にセリアック病のボランティアからの末梢血単核細胞(PBMC)を用いたインターフェロンガンマElispotにおいて17マーのA−グリアジン57−73 QE65(配列番号:2)が最大の応答を誘導したことを示した。
【0178】
A−グリアジン中の優勢なT細胞エピトープのコア配列からの伸長を示すペプチドを、3日間のグルテンチャレンジ開始から6日後にセリアック病のボランティアからの末梢血単核細胞(PBMC)を用いたIFNガンマELISPOTにおいて査定した(n=4)。ペプチド13:A−グリアジン59−71 QE65(13マー)、ペプチド15:58−72 QE65(15マー)、...、ペプチド27:52−78 SE65(27マー)。
【0179】
図11に示すとおり、A−グリアジン57−73 QE65の伸長はIFNガンマElispotにおいて応答を実質上は増強しない。次の実施例は、17マーペプチドを用いたA−グリアジン57−73 QE65のアゴニスト及びアンタゴニスト活性を特性決定する。
実施例6
A−グリアジン57−73 QE65とセリアック病における他のDQ2−制限されるT細胞エピトープの比較
セリアック病の腸の生検からのグルテン特異的T細胞クローン及び系列のT細胞エピトープに相当する未修飾及びトランスグルタミナーゼ処理ペプチドに相当するペプチドを用いて、用量応答研究を実施した。ペプチドに対する応答はA−グリアジン57−73 QE65に対する応答のパーセントとして表現した。全ての被験者はHLA−DQ2+であった(誰もDQ8+でなかった)。
【0180】
上記の研究は、A−グリアジン57−73 QE65が、グルテンチャレンジ後にセリアック病のPBMCを用いたELISpotアッセイにおいてインターフェロンガンマの誘導に関してもっとも有効性のあるグリアジンペプチドであることを示す(図12a−h,及び表5及び6を参照)。第2及び第3のエピトープは、より大きなペプチド、即ちA−グリアジン57−73 QE65及びGDA4 WHEAT P04724−84−100 QE92の準最適断片である。上記エピトープは適度に生物活性であるにすぎない(ブランクを差し引いた後ではA−グリアジン57−73 QE65より約1/20活性)。
【0181】
A−グリアジン57−73 QE65はセリアック病において他の公知のT細胞エピトープよりも有効性がある。配列決定されたグリアジン遺伝子の間でA−グリアジン57−73(配列PQLPY(配列番号:12)を含む)の16の多型が存在し、それらの生物活性を次で査定する。
実施例7
末梢血中でのグリアジン−及びA−グリアジン57−73 QE65の比較
セリアック病におけるグリアジンに対する全T細胞応答に対しての、優勢なエピトープA−グリアジン57−73 QE65の相対的な寄与は重大な問題である。ペプシン−トリプシン及びキモトリプシンにより消化されたグリアジンが、セリアック病におけるT細胞系列及びクローンの分化のための抗原として伝統的に使用されてきた。しかしながら、これらのプロテアーゼは特定のペプチドエピトープを分断するかもしれないと言える。事実、組換えα9−グリアジンのキモトリプシン消化はペプチドQLQPFPQPELPY(配列番号:13)を生じるが、最適エピトープ配列QLQPFPQPELPYPQPQS(配列番号:2)の末端削除体である(上を参照)。トランスグルタミナーゼ処理は、実質上、グリアジン特異的T細胞クローン及び系列の増殖アッセイにおいて、キモトリプシン消化されたグリアジンの能力を増加させる。よって、トランスグルタミナーゼ処理されたキモトリプシンで消化されたグリアジン(tTGグリアジン)は理想的な抗原ではないかもしれないが、この混合物に対する応答はグリアジンに特異的な末梢血リンパ球の「全」数に近似するかもしれない。ELISpotアッセイにおけるA−グリアジン57−73 QE65及びtTGグリアジンに対する応答の比較は、セリアック病におけるグリアジンに対する全免疫応答へのこの優勢なエピトープの寄与の指標を提供し、またエピトープ拡散の基準でもある。
【0182】
4人のセリアック病被験者においてグルテンチャレンジを開始して6日目又は7日目に回収されたPBMCを、キモトリプシン消化されたグリアジン+/−tTG処理を用いた用量応答研究において査定し、そして最適濃度のA−グリアジン57−73 QE65(25mcg/ml)に対するELISpot応答と比較した。グリアジンのtTG処理はELISpotにおいて約10倍PBMC応答を増強した(単独で査定した場合にtTGはブランクと同等であった)(図13a−c参照)。研究した4人のセリアック病被験者において、A−グリアジン57−73 QE65(25mcg/ml)はtTGグリアジン(500mcg/ml)の14から115%の間の応答を誘導し、そしてA−グリアジン57−73 QE65に対する応答が大きければ大きいほど、tTGグリアジン応答を表した比率が大きくなった。
【0183】
相対的に限られたデータから、A−グリアジン57−73 QE65応答が幾つかの被験者においてtTGグリアジンと同等であることが示唆される。より進化した抗−グリアジンT細胞応答に関連したエピトープ拡散は、幾つかの個体において末梢血の「全」応答へのA−グリアジン57−73 QE65の僅かな寄与の説明となるかもしれない。エピトープ拡散はさほど厳しくないグルテンフリーダイエットを受けた個体において保持されるかもしれない。
実施例8
セリアック病において生物活性なグリアジンペプチドの定義:A−グリアジン57−73の多型
A−グリアジンの全配列にわたるオーバーラップする15マーのペプチドを査定することにより、セリアック病の免疫上優勢な配列を同定した。A−グリアジンは最初に全配列を決定されたアルファグリアジン蛋白質及び遺伝子であったが、コムギにおいて約30−50の関連するアルファグリアジン蛋白質のひとつである。蛋白質データベースSwiss−Prot及びTREMBLを検索することにより、25の別々のアルファ−グリアジン遺伝子が同定され、さらに8つのアルファグリアジンを記述した。これら25のアルファグリアジンに含まれたのは、A−グリアジン57−73に相当する16の異なる多型である(表7参照)。
【0184】
トランスグルタミナーゼによるインビトロにおける処理、並びに10位において置換されたグルタミン酸による処理後、未修飾形態でのこれらの16の多型に相当する合成ペプチドを、通常のグルテンフリーダイエット後の、インターフェロンガンマELISpotアッセイにおけるグルテンチャレンジの6日又は7日後に、セリアック病の被験者からのPBMCを用いて査定した。グルタミン酸置換したペプチドを3つの濃度において(2.5、25及び250mcg/ml)査定し、未修飾のペプチド及びトランスグルタミナーゼ処理されたペプチドを25mcg/mlの濃度のみにて査定した。生物活性は、個々の被験者においてA−グリアジン57−73 QE65 25mcg/mlに付随した応答の%として表現した(n=4)(図14参照)。
「野生型」ペプチドの生物活性はトランスグルタミナーゼ処理により実質増加した(>5倍)。野生型ペプチドのトランスグルタミナーゼ処理は10位においてグルタミン酸により置換された同じペプチドの生物活性と同様な生物活性をもたらした。5つのグルタミン酸置換ペプチド(B,C,K,L,M)の生物活性はA−グリアジン57−73 QE65の>70%であったが、A−グリアジン57−73 QE65よりも顕著に高い生物活性のものはなかった。2.5及び250mcg/mlの濃度においてグルタミン酸置換されたペプチドに対するPBMC応答は25mcg/mlにおけるのと同等であった。6つのグルタミン酸置換グリアジンペプチド(H,I,J,N,O,P)は、A−グリアジン57−73 QE65の生物活性の<15%であった。他のペプチドは生物活性において中間であった。
【0185】
少なくとも6つのグリアジン由来のペプチドがトランスグルタミナーゼによる修飾語にA−グリアジン57−73 QE65に対する有効性が均等である。相対的に非生物活性のA−グリアジン57−73 QE65の多型も存在する。これらのデータは、トリチカムエステビウム、トリチカムウアルツ、トリチカムスペルタの幾つかのグリアジン由来のペプチドのトランスグルタミナーゼ修飾がセリアック病において免疫上優勢なT細胞エピトープを生成できるかもしれないことを示す。
【0186】
非−セリアック病−毒性コムギを生成するコムギの遺伝子修飾は、複数のグリアジン遺伝子の除去又は修飾を必要とするかもしれない。A−グリアジン57−73の変更されたペプチドリガンドアンタゴニストを定義する配列を取り込んだグリアジン又は他の蛋白質又はペプチドを含むコムギの生産は、セリアック病において「非毒性」よりむしろ治療用である遺伝学上修飾されたコムギを生成するための別の戦略である。
実施例9
コアエピトープ配列の定義:
N−末端又はC−末端からのA−グリアジン56−75の末端削除に相当するペプチドの比較は、T細胞エピトープのコア配列がPELPY(A−グリアジン64−68)であることを示した。非アゴニスト及びアンタゴニストを定義する試みは、その生物活性に実質貢献する残基において置換されたA−グリアジンのバリアントに焦点を注ぐことになる。
【0187】
残基57から73を置換したアラニン(図15)又はリジン(図16)を有するA−グリアジン57−73 QE65に相当するペプチドを、3日間のグルテンチャレンジ開始から6日後にセリアック病のボランティアからの末梢血単核細胞(PBMC)を用いたIFNガンマELISPOTにおいて比較した(n=8)。(BLはブランク、EはA−グリアジン57−73 QE65:QLQPFPQPELPYPQPQS(配列番号:2))。
【0188】
A−グリアジン60−70 QE65(PFPQPELPYPQ(配列番号:14)に相当する残基はA−グリアジン57−73 QE65において生物活性に実質貢献することを見いだした。60−70位において置換されたA−グリアジン57−73 QE65のバリアントは2段階の手法により査定する。最初に、対照的な特性を有する、10の異なるアミノ酸を用いて60−70位において置換されたA−グリアジン57−73 QE65を査定した。A−グリアジン57−73 QE65のバリアントの第2群(全ての他の天然に生じるアミノ酸により置換されているが、修飾に敏感であると証明された位置のシステインを除く)を第2ラウンドにおいて査定した。
実施例10
A−グリアジン57−73 QE65の置換されたバリアントのアゴニスト活性
A−グリアジン60−70 QE65は、A−グリアジンにおいて優勢なT細胞エピトープのコア配列である。このエピトープのアンタゴニスト及び非アゴニストペプチドバリアントは、このコア配列の修飾により生成されるらしい。最初に、対照的な特性を有する、10の異なるアミノ酸を用いて60−70位において置換されたA−グリアジン57−73 QE65を、3日間のグルテンチャレンジ開始から6日後にセリアック病の被験者からのPBMCを用いたIFNガンマELISPOTにおいて査定する。A−グリアジン57−73 QE65のバリアントの第2群(システイン以外、全ての他の天然に生じるアミノ酸により置換されている)も第2ラウンドにおいて査定した。両群のペプチド(全て50mcg/mlにおいて、二重)を8人の被験者からのPBMCを用いて査定して、未修飾のペプチドと比較した(アッセイあたり20反復)。様々な研究が、このアッセイにおけるA−グリアジン57−73 QE65の最適な濃度が10から100mcg/mlの間であることを示す。
【0189】
結果を、スポット形成細胞(95%コンフルエントの間隔)の平均応答として、個々の個体の%A−G 57−73 QE65平均応答として表現する。対になっていないt−試験を使用することにより、修飾されたペプチドのELISPOT応答をA−G 57−73 QE65と比較する。スーパー−アゴニストを、p<0.01の有意性のレベルにおいてA−G 57−73 QE65よりも高い応答を有するものとして定義し;部分アゴニストはp<0.01の有意性のレベルにおいてA−G 57−73 QE65よりも低い応答を有するものとして定義し、そして非アゴニストはブランク(ペプチド無しのバッファー)と顕著に異ならないものと定義した。A−G 57−73 QE65のアゴニスト活性の30%又はそれ未満のアゴニスト活性を有するペプチドは、アンタゴニスト活性を評価するための「適切な」部分アンタゴニスト又は非アンタゴニストと考えた(表8及び図17−27参照)。
【0190】
PBMCのA−グリアジン57−73 QE65に対するIFNガンマELISPOT応答は分子レベルにおいて特異性が高い。64位のプロリン(P64)、65位のグルタミン酸(E65)及び66位のロイシン(L66)、及び程度は劣るがQ63,P67,Y68及びP69が修飾に対して特に感受性である。Y61及びY70の置換は共に親ペプチドより30%高い生物活性のスーパー−アゴニストを生じ、おそらくは、HLA−DQ2への結合を増強するためであるが、何故ならば、このHLA分子のモチーフが1位及び9位の巨大な疎水性残基に関する選択性を示すからである。18の非アゴニストペプチ
ドを同定した。バリアントの生物活性(50mcg/ml):P65,K64,K65及びY65(生物活性7−8%)はブランク(7%)と同等であった。全部で、A−グリアジン57−73 QE65の57の変異バリアントがA−グリアジン57−73 QE65の30%又はそれ未満の生物活性であった。
【0191】
優勢なエピトープ、A−グリアジン57−73 QE65に対する末梢血リンパ球(PBL)T細胞応答の分子特異性は、HLA−DQ2+セリアック病被験者の間で一貫して再現性があり、そして上記7アミノ酸の中の限定された数のアミノ酸に特異性が高い。A−グリアジン57−73 QE65の特定の単一アミノ酸バリアントは、全てのHLA−DQ2+セリアック病被験者において一貫して非アゴニストである。
実施例11
置換されたバリアントのアンタゴニスト活性
HLA−DQ2+被験者におけるA−グリアジン57−73 QE65に対するPBL
T細胞応答の均質性は、PBL T細胞応答がポリクローナル又はオリゴクローナルであるらしいとしても、PBMCにおいてか又はエクスビボにおいて拮抗可能な変更されたペプチドリガンド(APL)が存在するかもしれないことを示唆する。APLアンタゴニストは一般に弱いアゴニストである。30%又はそれ未満のアゴニスト活性のA−グリアジン57−73 QE65の57の単一アミノ酸置換バリアントが同定されて、APLアンタゴニストとしての適切な候補である。さらに、A−グリアジン57−73 QE65の特定の弱い生物活性の天然に生じる多型も同定されて(以下参照)、「天然に生じる」APLアンタゴニストかもしれない。また、MHCへの結合が抗原特異的T細胞免疫とも拮抗するかもしれないことが示唆された。よって、グルテンチャレンジ後にセリアックPBMCにおいてIFNガンマ応答を誘導しないがHLA−DQ2に結合することが知られている非グリアジンペプチドは、A−グリアジン57−73 QE65により誘導されるT細胞応答を低下させることができるかもしれない。HLA−DQ2へ貪欲に結合する2つのペプチドはHLAクラス1α46−60(HLA 1a)(PRAPWIEQEGPEYW(配列番号:15))及びチロイドペルオキシダーゼ(tp)632−645Y(IDVWLGGLLAENFLPY(配列番号:16))である。
【0192】
3日間のグルテンチャレンジを開始後のセリアックボランティア6人からのPBMCを用いたIFNガンマELISPOTにおけるペプチド(50μg/ml)又はバッファー及びA−グリアジン57−73 QE65(10μg/ml)の同時添加(n=5)。結果は、ペプチドプラスA−G 57−73 QE65による応答として(二重の平均)、バッファープラスA−G 57−73 QE65による応答(20反復の平均)の%応答として表した(表9参照)。
【0193】
A−グリアジン57−73 QE65の4つの単一アミノ酸置換バリアントは、A−グリアジン57−73 QE65に対するインターフェロンガンマPBMC ELISPOT応答(p<0.01)を25%から28%低下させ、別の13のペプチドバリアントはELISPOT応答を18%から24%低下させた(p<0.06)。HLA−DQ2バインダー、チロイドペプチド(tp)632−645Yは、A−グリアジン57−73 QE65に対するPBMCインターフェロンガンマ応答を31%低下させる(p<0.0001)が、他のHLA−DQ2バインダー、HLAクラス1a46−60は応答を変化させない(表9及び10参照)。A−グリアジン57−73のトランスグルタミナーゼ修飾された多型に相当するペプチド、SwissProt受け入れ番号P04725 82−98 QE90(PQPQPFPPELPYPQPQS(配列番号:17))は、A−グリアジン57−73 QE65に対する応答を19%低下させる(p<0.009)(表11参照)。
【0194】
ELISPOTアッセイにおいてのA−グリアジン57−73 QE65に対するPB
MCのインターフェロンガンマ応答は、特定の単一アミノ酸A−グリアジン57−73 QE65バリアント、A−グリアジン57−73 QE65の多型、及び5倍過剰にHLA−DQ2を結合することが公知の非関連ペプチドの同時投与により低下する。これらの発見は、A−グリアジン57−73 QE65の変更されたペプチドリガンドアンタゴニストが存在することを示唆する。仮想上のAPLのみならず、HLA−DQ2を結合する特定のペプチドも、A−グリアジン57−73 QE65に対するPBL T細胞応答を有効に低下させる。
【0195】
これらの発見は、HLA−DQ2+セリアック病において優勢なA−グリアジンエピトープに対するT細胞応答を破壊する2つの戦略を支持する。
【0196】
1.「伝統的な」ペプチド薬剤としての使用のため又はコムギにおけるグリアジン遺伝子の特異的遺伝的修飾のための、一つの位置より多い位置において(64−67)アミノ酸を置換することによる、APLアンタゴニストの最適化。
【0197】
2.HLA−DQ2に付随してA−グリアジン57−73 QE65の提示を競合阻害するための、高親和性HLA−DQ2結合ペプチドの使用。
【0198】
これらの2つのアプローチは相互に両立可能かもしれない。F61及びQ70をチロシン残基で置き換えることにより、スーパー−アゴニストを生成した。これらのスーパー−アゴニストはT細胞受容体との増強された接触よりむしろHLA−DQ2への結合の改善によりもたらされたらしい。これらの修飾を適度に有効なAPLアンタゴニストを生成する他の置換と組み合わせることにより、置換されたA−グリアジン57−73 QE65バリアントの阻害効果を実質上増強するかもしれない。
実施例12
PBMC及びA−グリアジン57−73 QE65及びP04724 84−100 QE92をセリアック病の診断法として使用したインターフェロンガンマELISpotの開発:新たに診断されたセリアック病における免疫応答の定義
インターフェロン番号ELISpotにおいて測定されたPBMC中の優勢なA−グリアジンT細胞エピトープに対する応答性の誘導を、長期間の厳格なグルテンフリーダイエット(GFD)を受けたほとんど全てのDQ2+セリアック病被験者においてはグルテンチャレンジに続いて行ったが、厳格なGFDを受けて4週間後の健康なDQ2+被験者においては続いて行わなかった。A−グリアジン57−73 QE65応答はグルテンチャレンジ前のセリアック病被験者のPBMCにおいて測定不可能であり、そしてパイロットデータはこれらの応答が未処理の被験者のPBMCにおいて測定できなかったことを示唆した。これらのデータは、セリアック病において、A−グリアジン57−73 QE65に対する免疫応答性が抗原排除(GFD)後に復元することを示唆する。ELISpotアッセイ及びPBMCを用いて診断試験が開発されるとしたら、グルテンチャレンジが
血液中のA−グリアジン57−73 QE65及び他の免疫反応性グリアジンペプチドに対する応答を誘導できる前に必要とされるGFDの期間を定義することが望まれる。
【0199】
新たに診断されたDQ2+セリアック病被験者を、胃腸学の外来患者サービスから補充した。PBMCを調製して、被験者がGFDを始める前及びGFDを初めて1週間又は2週間後に、インターフェロンガンマELISpotアッセイにおいて試験した。さらに、グルテンチャレンジ(3日間は4スライスの標準の白パンを摂取、200g/日)を、GFD開始の1又2週間後に実施した。PBMCを調製し、そしてグルテンチャレンジを開始して6日後にアッセイした。A−グリアジン57−73 QE65(A),P04724 84−100 QE92(B)(単独及び化合して)及びA−グリアジン57−73
QP65(P65)(非生物活性バリアント、上記参照)(全て25mcg/ml)を査定した。
【0200】
1人の患者を除いて皆、新たに診断されたセリアック病患者はDQ2+であった(一人はDQ8+であった)(n=11)。未処置か又はGFDの1又は2週間後の新たに診断されたセリアック病患者からのPBMCは、A−グリアジン57−73 QE65及びP04724 84−100 QE92(単独及び化合して)に対する応答を示さずブランク又は及びA−グリアジン57−73 QP65と顕著な差異はなかった(n=9)(図28参照)。たった1週間しかGFDを受けなかったセリアック病患者におけるグルテンチャレンジは、A−グリアジン57−73 QE65及びP04724 84−100 QE92(単独及び化合して)に対する応答を実質上増強しなかった。しかし、GFDを開始して2週間後のグルテンチャレンジは、A−グリアジン57−73 QE65及びP04724 84−100 QE92(単独及び化合して)に対する応答を誘導し、非生物活性バリアントA−グリアジン57−73 QP65及びブランクより顕著に高かった。グルテンチャレンジ2週間後のこれらの応答は価値のあることであるが、GFD開始後>2カ月の被験者よりは低いらしい。A−グリアジン57−73 QE65単独に対する応答は、P04724 84−100 QE92単独に対する応答又はA−グリアジン57−73 QE65と混合した場合の応答と等しいか又はより大きかった。被験者のだれもがグルテンチャレンジの苦痛の兆候を経験しなかった。
【0201】
A−グリアジンに対する免疫応答性(グルテンチャレンジ後にPBMC中で測定)はGFDを開始して2週間後に部分的に回復したことから、この優勢なT細胞エピトープに対する「免疫非応答性」は、未処置のセリアック病患者において且つGFD開始後の少なくとも1週間は優勢であることを示唆する。グルテンチャレンジ及びELISpotアッセイにおけるA−グリアジン57−73 QE65に対する応答の測定を用いたセリアック病の診断試験のタイミングは、GFDを開始して少なくとも2週間後である。
【0202】
A−グリアジン57−73 QE65に特異的なインターフェロンガンマ分泌T細胞は未処置のセリアック病患者においては末梢血中にて測定できず、GFDの少なくとも2週間後のグルテンチャレンジにより誘導することができる(抗原排除)。よって、この方法論を用いた診断試験のタイミングは非常に重大であり、そしてその最適化にはさらなる研究が必要である。これらの発見は、抗原排除により逆転した優勢なエピトープ、A−グリアジン57−73 QE65に特異的なT細胞の機能アネルギーと一致する。この現象は、ヒト疾患において以前には証明されておらず、T細胞アネルギーがセリアック病においてはペプチド治療により誘導可能かもしれないという可能性を支持する。
実施例13
コムギグリアジンT細胞エピトープの包括的マッピング
抗原チャレンジは末梢血中で抗原特異的T細胞を誘導する。セリアック病においては、グルテンがこの免疫媒介性疾患を維持する抗原である。グルテンフリーダイエットで処置されたセリアック病のグルテンチャレンジは末梢血中のグルテン特異的T細胞の出現を導き、グルテンT細胞エピトープの分子特異性の決定を可能にさせる。上記のとおり、我々はモデルグルテン蛋白質中の単一の優勢なエピトープ、A−グリアジン(Q65において脱アミド化された57−73)を同定した。この実施例においては、セリアック病患者のグルテンチャレンジを用いて、GenBank中の111エントリーに由来する各公知のコムギグリアジン蛋白質中の全ての有力な12のアミノ酸配列を試験した。全部で、652の20マーペプチドをHLA−DQ2及びHLA−DQ8関連のセリアック病において試験した。セリアック病において90%を超えて存在する古典的なHLA−DQ2複合体(HLA−DQA1*05,HLA−DQB1*02)のセリアック病被験者9人のうちの7人は、末梢血中に誘導可能なA−グリアジン57−73 QE65−及びグリアジン−特異的T細胞応答を有した。A−グリアジン57−73は、HLA−DQ2関連セリアック病において、唯一の重要なα−グルテンT細胞エピトープであり、並びにもっとも有力なグリアジンT細胞エピトープであった。さらに、インターフェロン−γ ELISp
otアッセイにおいてA−グリアジン57−73ほど有効性のある10%から70%の構造上関連したペプチドの5ファミリーほど存在した。これらの新規なT細胞エピトープは、γ−及びω−グリアジンに由来しており、そしてA−グリアジン57−73のコア配列(コア配列:FPQPQLPYP(配列番号:18))、例えばFPQPQQPFP(配列番号:19)及びPQQPQQPFP(配列番号:20)と構造上極めて類似しているが同一ではない共通配列を含んだ。A−グリアジン57−73の相同体はライムギ又はオオムギにおいて見いだされなかったが、他の2つの穀類はセリアック病において毒性であり、γ−及びω−グリアジン中の新たに定義されたT細胞エピトープはライムギ及びオオムギの保存蛋白質において完全に適合した(それぞれ、セカリン(secalins)及びホルデイン(hordeins))。
【0203】
HLA−DQ2に関連しないセリアック病はほとんど常にHLA−DQ8と関連していた。7人のHLA−DQ8+セリアック病被験者のだれもが、彼らが完全なHLA−DQ2複合体も所有していないなら、誘導可能なA−グリアジン57−73−特異的T細胞応答をグルテンチャレンジ後に有さなかった。完全なHLA−DQ2複合体を有さない4人のHLA−DQ8+セリアック病被験者のうちの2人は、誘導可能なグリアジンペプチド特異的T細胞応答をグルテンチャレンジ後に有した。一人のHLA−DQ8被験者においては、新規な優勢T細胞エピトープが、コア配列LQPQNPSQQQPQ(配列番号:21)を用いて同定された。このペプチドのトランスグルタミナーゼによる脱アミド化バージョンは非脱アミド化ペプチドよりも有効性であった。以前の研究は、トランスグルタミナーゼにより脱アミド化したペプチドが配列LQPENPSQEQPE(配列番号:22)を有するはずであることを示唆したが;この配列を確認するためには、さらなる研究が必要である。1カ月の間グルテンフリーダイエットを受けた健康なHLA−DQ2(10)及びHLA−DQ8(1)の被験者の間で、グリアジンペプチド特異的T細胞応答は不変であり、グルテンチャレンジによりほとんど変わらず、そして、セリアック病の被験者においてグルテンチャレンジにより明らかにされた有効性T細胞エピトープでは決してなかった。結論として、HLA−DQ2関連セリアック病において6より多い重要なT細胞エピトープが存在するらしく、そのうち、A−グリアジン57−73がもっと有効性である。HLA−DQ2−及びHLA−DQ8−関連セリアック病は同じT細胞特異性を共有しない。
【0204】
我々は、グルテンフリーダイエット後のセリアック病の個体の短期間のグルテンチャレンジが末梢血中にてグリアジン特異的T細胞を誘導することを示した。これらのT細胞の頻度は末梢血中で6日目に最大であり、続いて、次の週を通して迅速に弱まる。末梢血のグリアジン特異的T細胞は消化管の粘膜固有層へのホーミングに関連するインテグリンα47を発現する。我々は、原型的なグルテンα−グリアジン蛋白質、A−グリアジンにおいてセリアック病に関連したT細胞エピトープをマップするためにこのヒト抗原−チャレンジデザインを開発した。トランスグルタミナーゼ(tTG)による脱アミド化が有る場合も無い場合も10アミノ酸がオーバーラップする15マーのペプチドを用いて、我々は、末梢血中で誘導されたT細胞が、65位のグルタミン酸が脱アミド化されたひとつのA−グリアジンペプチドの残基57−73を最初に標的化することを証明した。当該エピトープはHLA−DQ2−制限されており、HLA−DQ2のセリアック病との密接な関連と一致する。
【0205】
コムギ、ライムギ、及びオオムギの暴露によりセリアック病は再活性化される。コムギのグルテンのα/β−グリアジン画分はセリアック病においては一致して毒性であり、そしてほとんどの研究はこれらの蛋白質に焦点を合わせてきた。α/β−グリアジンをコードする遺伝子クラスターはコムギ染色体の6C上に位置する。ライムギ又はオオムギにはα/β−グリアジンの相同物がない。しかしながら、コムギのグリアジンサブタイプの3つ全て(α/β、γ、及びω)がセリアック病において毒性である。γ−及びω−グリア
ジン遺伝子はコムギでは染色体1APL上に位置しており、ライムギ及びオオムギのセカリン及びホルデインに相同である。
【0206】
パンコムギ(Triticum aestivum)においては61のα−グリアジンに関して同定された遺伝子が今存在する。α−グルテン配列は密接に相同であるが、A−グリアジンにおいて優勢なエピトープは、α−グリアジン配列においてほとんど多型性の領域に由来する。Andersonら(1997)は、パンコムギ(T.aestivum)中に全部で約150の異なるα−グリアジン遺伝子が存在するが、多くは偽遺伝子であると見積もった。よって、セリアック病に関連するT−細胞のエピトープが公知のα−グリアジン配列中に含まれないことは、ありそうもない。
【0207】
我々の研究は、セリアック病に特異的な有効性のT細胞エピトープとしてA−グリアジン57−73にほとんど同一の、脱アミド化されたα−グリアジンペプチドのグループを同定した。90%を超えるセリアック病患者はHLA−DQ2+であり、これまで、我々はグルテンチャレンジ後のHLA−DQ2+セリアック病被験者のみを査定してきた。しかしながら、HLA−DQ2を発現しないセリアック病患者がほとんど皆HLA−DQ8を有する。よって、他のコムギ、ライムギ及びオオムギグルテン蛋白質中のA−グリアジン57−73及びその相同体が、HLA−DQ2+及びHLA−DQ8+セリアック病の両方においてグルテンチャレンジにより誘導されるT細胞により認識される唯一のT細胞エピトープであるか否かを知ることは非常に不可欠である。これが事実なら、セリアック病のための治療用のペプチドのデザインは、たった一つのペプチドのみしか必要としないかもしれない。
A−グリアジン57−73のT細胞エピトープとしての相同体
A−グリアジン57−73(PQLPY<配列番号:12>)のコア配列をコードする穀物遺伝子に関するSwissProt及びTrembl遺伝子データベースの最初の検索は、α/β−グリアジンを明らかにしただけであった。しかしながら、A−グリアジン57−73 QE65エピトープの我々の初期のマッピング研究は、コア領域(PQLP)中の許容可能な点の置換が限定された数であることを明らかにした(Q65はエピトープ中で実際に脱アミド化されている)。よって、我々は、配列XXXXXXXPQ[ILMP][PST]XXXXXX(配列番号:23)を有するペプチドをコードするSwissProt及びTremblデータベースの中の遺伝子に我々の検索を拡大した。相同体は、γ−グリアジン、グルテニン、ホルデイン及びセカリンの間に同定された(表12参照)。さらなる相同体は、GenBankにおいて3つのω−グリアジンエントリーの可視検索によりω−グリアジン中に同定された。
【0208】
A−グリアジン57−73のこれらの相同体は、tTGによる脱アミド化(又は4つの密接な相同体中のグルタミン酸(QE)−置換されたバリアントの合成)の後に、IFNγELISpotアッセイを用いて、末梢血単核細胞を用いて、セリアック病の被験者においてグルテンチャレンジ後に査定した。ω−グリアジン配列(AAG17702 141−157)は唯一の生物活性ペプチドであり、約半分がA−グリアジン57−73として有効である(表12、及び図29参照)。よって、優勢なA−グリアジンエピトープの相同体の検索は、γ−グリアジン、セカリン、及びホルデインの毒性に関して考慮することを怠った。
方法
全ての可能性のあるコムギグリアジンT−細胞エピトープにわたるペプチドのセットのデザイン
公知のパンコムギ(Triticum aestivum)のグリアジン遺伝子又は遺伝子断片によりコードされた全ての可能なT細胞エピトープを同定するため(GenBank中の61α/β−,47γ−,及び3ω−グリアジンエントリー)、遺伝子由来の蛋白質配列を、CustalWソフトウエア(MegAlign)を用いてアラインメント
を行い、そして分類することにより系統発生グループ化した(表22参照)。多くのエントリーは、より長い配列の末端削除を表し、そして多くの遺伝子セグメントは、ポリグルタミン繰り返しの長さ又は稀な置換を除いて同一であった。よって、公知のコムギ遺伝子によりコードされる全ての有力な12アミノ酸配列を有理化して(rationalize)652の20マーペプチドのセットに含ませることができた。(シグナルペプチド配列は含まれなかった)。ペプチド配列を表23に掲載する。
包括的エピトープマッピング
4週間グルテンフリーダイエットを受けた健康な対照(HLA−DQ2+ n=10,及びHLA−DQ8+ n=1)、及び長期間のグルテンフリーダイエットを受けたセリアック病の被験者(6人のHLA−DQ2,4人の複合同型接合HLA−DQ2/8、及び3人のHLA−DQ8/X)(表13参照)を、グルテンチャレンジの前及び後の6及び7日目に調査した(標準の白パン50gスライス4切れ−セインズベリーズサンドイッチパン、毎日)。末梢血(7日間にわたり全部で300g)を採血して、末梢血単核細胞(PBMC)をLymphoprep密度勾配により分離した。PBMCを6又は8の20マーペプチド、又はtTGにより脱アミド化されたか又はされていない単一のペプチドのプールと、一晩、インターフェロンガンマ(IFNγ)ELISpotアッセイにおいてインキュベートした。
【0209】
ペプチドはPepsets(Mimotopes Inc.,メルボルン、オーストラリア)において96のバッチ中で合成した。約0.6マイクロモルの各651の20マーを提供した。2つのマーカー20マーペプチドを96の各セットに含ませ(VLQQHNIAHGSSQVLQESTY)−ペプチド161(配列番号:24)、そしてIKDFHVYFRESRDALWKGPG(配列番号:25)を逆相−HPLC及びアミノ酸配列分析により特性決定した。これらのマーカーペプチドの平均純度はそれぞれ50%及び19%であった。ペプチドを最初にアセトニトリル(10%)及びHepes 100mMから10mg/mlに溶解した。
【0210】
IFNγELISpotアッセイのためにPBMCとインキュベートされたプール(又は単独)中の個々のペプチドの最終濃度は20μg/mlであった。ペプチドの5倍濃縮した溶液及び1mMの塩化カルシウムを含むPBS中のプールを分配して、ELISpotアッセイにおいて後に使用されるテンプレートに従い96ウエルプレート中に保存した。モルモット組織tTG(Sigma T5398)との100:32の比における2時間37℃のインキュベーションにより、脱アミド化されたペプチド及びペプチドのプールを調製した。ペプチド溶液を−20℃において保存して、使用前に新たに解凍した。
【0211】
エンドトキシンフリーの水及び2M尿素中のグリアジン(Sigma G3375)(100mg/ml)を10分間ボイルし、室温に冷却し、そしてフィルター(0.2μm)−滅菌されたHCl 0.02M中のペプシン(Sigma P6887)(2mg/ml)又は重炭酸アンモニウム中のキモトリプシン(C3142)(4mg/ml)とインキュベートした。4時間のインキュベーション後に、ペプシン消化されたグリアジンを水酸化ナトリウムにより中和し、そして次にペプシン−消化グリアジン及びキモトリプシン−消化グリアジンの両方を15分間ボイルした。グリアジンを除いたプロテアーゼとの同一のインキュベーションも実施した。サンプルを15 000gにおいて遠心分離し、次に、蛋白質濃度を上清中でBCA法(Pierce,USA)により見積もった。IFNγELISpotアッセイにおける最終的な使用の前に、グリアジン−プロテアーゼのアリコートをtTGと共に比率2500:64μg/mlにてインキュベートした。
【0212】
IFNγELISpotアッセイ(Mabtech,スエーデン)を96−ウエルプレート(MAIP S−45,Millipore)中で、各ウエルが25μlのペプチド溶液及び100μlのPBMC(2−8105/ウエル)を、10%熱不活性化ヒトAB
血清を含むRPMI中に含むように、実施した。脱アミド化されたペプチドプールを一つの96−ウエルELISpotプレート中で査定し、そして第2プレート上で脱アミド化なしのペプチドプール(同一のレイアウト)を0日目及び6日目に査定した。脱アミド化されたペプチドを含むプレート中の全てのウエルがtTGを含んだ(64μg/ml)。各ELISpotプレート中に、ペプチドプールを含む83のウエルが存在し(各ウエル中唯一のプール)、そして「対照」ペプチドのウエルのシリーズ(ペプチドは全て>90%の純度、MS及びHPLCにより特性決定された、Research Genetics):P04722 77−93(QLQPFPQPQLPYPQPQP(配列番号:26)、P04722 77−93 QE85(二重)(QLQPFPQPELPYPQPQP(配列番号:27)、P02863 77−93(QLQPFPQPQLPYSQPQP(配列番号:28)、P02863 77−93 QE85(QLQPFPQPELPYSQPQP(配列番号:29)、及びキモトリプシン消化されたグリアジン(500μg/ml)、ペプシン消化されたグリアジン(500μg/ml)、キモトリプシン(20μg/ml)単独、ペプシン(10μg/ml)単独、及びブランク(PBS+/−tTG)(二重)。
【0213】
展開及び乾燥後に、IFNγELISpotプレートをMAIP自動化スポット形成ELISpotカウンターを用いて査定した。HLA−DQ2の健康被験者及びセリアック病被験者において、IFNγELISpotアッセイにおいてペプチドプールによりスポット形成細胞(sfc)の誘導を、100万PBMCあたりのスポット形成細胞(sfc)マイナスブランクに適用した、一方をつないだ(one−tailed)Wilcoxon Matched−Pairs Signed−Ranks試験(SPSSソフトウエア)を用いて、6日目体0日目(「正味の応答」)にて試験した。インビボグルテンチャレンジによるPBMC中のペプチドプールに対するIFNγ応答の顕著な誘導を、少なくとも10sfc/100万PBMC及び有意性レベルp<0.05の中央値の「正味の応答」として定義した。6日目のペプチドの特定のプールに対する顕著な応答に、同日又は7日目に採られたPBMCを使用して各プール内において個々のペプチドの評価を続けた。
【0214】
個々のペプチドのIFNγELISpotアッセイに関して、生物活性を同じELISpotプレート中で査定されたP04722 77−93 QE85に対する応答のパーセントとして表現した。36万(範囲:0.3−0.72)、PBMCの中央値を用いて、ブランクに対する中央値の応答(PBS単独)はウエルあたり0.2sfc(範囲0−5)であり、そしてウエルあたり陽性対照(P04722 77−93 QE85)76.5(範囲25−383)sfcであった。よって、P04722 77−93 QE65のパーセンテージとして表現されたブランクに対する中央値の応答は0.2%(範囲:0−6.7)であった。P04722 QE85のそれより10%高い平均生物活性を有する個々のペプチドを共通の構造上のモチーフに関して分析した。
結果
健康なHLA−DQ2被験者
1カ月グルテンフリーダイエットを受けた健康なHLA−DQ2+被験者の誰も、グルテンチャレンジの前又は後にA−グリアジン57−73の相同体に対してIFNγELISpot応答を有さなかった。しかしながら、10人の健康な被験者のうち9人において、グルテンチャレンジが、グリアジンのペプシン消化及びキモトリプシン消化の両方に対するIFNγ応答の顕著な増加を伴い、0日目の0−4sfcの中央値/百万から16−29sfcの中央値/百万であった(表14参照)。健康な被験者のグリアジン応答は脱アミド化により影響されなかった(表15参照)。健康な被験者のうち、特定のグリアジンペプチドプールに対するIFNγ応答の、グルテンチャレンジとの一致した誘導はなかった(図30、及び表16参照)。IFNγELISpot応答は場合によりみられたが、弱く、そして脱アミド化により影響されなかった。プールに対するもっとも強い応答の
多くも0日目には存在しなかった(表17、被験者H2,H8及びH9参照)。4人の健康な被験者はプール50に対して明確な応答を示し、6日目にもっとも強い応答をもった2人は0日目にも応答を有した。両被験者において、プール50に対するチャレンジ後の応答はペプチド390(QQTYPQRPQQPFPQTQQPQQ(配列番号:30))によった。
HLA−DQ2セリアック病被験者
HLA−DQ2+セリアック病被験者におけるグルテンチャレンジの後、P04722
77−93 QE85に対する中央値のIFNγELISPpot応答は0から133sfc/百万の中央値から上がった(表4参照)。6人のセリアック病被験者のうちの一人(C06)はP04722 77−93 QE85(2sfc/百万)に応答せず、そしてグリアジンペプチドプールに弱く応答しただけであった(最大:プール50+tTG
27sfc/百万)。前の研究に一致して、野生型P04722の生物活性はtTGによる脱アミド化により6.5倍上昇した(表15参照)。グリアジン消化に対するインターフェロンガンマ応答はベースラインに存在したが、キモトリプシン−グリアジンに関しては20から92sfcの中央値/百万まで、そしてペプシン−グリアジンに関しては44から176sfcの中央値/百万まで、グルテンチャレンジにより実質上は増加した。グリアジンの脱アミド化は、キモトリプシン−グリアジンに関して3.2倍の中央値、そしてペプシン−グリアジンに関して1.9倍の中央値だけ生物活性を増加させた(表15参照)。(ペプシンによる消化に要求される酸性度はグリアジンの部分脱アミド化をもたらすらしい。)
健康な被験者とは対照的に、グルテンチャレンジは、α−,γ−、及びω−グリアジンを含む83の処理されたプールのうちの22に対してIFNγELISpot応答を誘導した(図31及び表17参照)。プールの生物活性は被験者間で一致していた(表18参照)。ペプチドプールにより誘導されたIFNγELISpot応答は脱アミド化によりほとんどいつも増加した(表17参照)。だが、脱アミド化によるプールの生物活性の増強は、A−グリアジン57−73の相同体を含むプールに関してさえ、P04722 77−73 QE85ほど顕著ではなかった。これは、Pepsetペプチドが合成又は調製の間に部分的に脱アミド化されていたことを示唆し、例えばPepsetペプチドはトリフルオロ酢酸(TFA)の塩としてTFA溶液からの凍結乾燥後にデリバリーされる。
【0215】
21のもっとも生物活性なプールからの170の個々のtTG脱アミド化ペプチドを別々に査定した。72の脱アミド化ペプチドは均等な濃度においてP04722 77−93 QE85より10%生物活性が高かった(20μg/ml)(表19参照)。5つのもっとも有効なペプチド(P04722 QE85の85−94%の生物活性)は、以前にα−グリアジン相同A−グリアジン57−73として同定された。50の生物活性ペプチドはA−グリアジン57−73と相同でなかったが、構造上関連する配列の6ファミリーに分類できた(表20参照)。各々のペプチドファミリーのもっとも生物活性な配列は:PQQPQQPQQPFPQPQQPFPW(配列番号:31)(ペプチド626、P04722 QE85の中央値72%生物活性)、QQPQQPFPQPQQPOLPFPQQ(配列番号:32)(343、34%)、QAFPQPQQTFPHQPQQQFPQ(配列番号:33)(355、27%)、TQQPQQPFPQQPQQPFPQTQ(配列番号:34)(396、23%)、PIQPQQPFPQQPQQPQQPFP(配列番号:35)(625、22%)、PQQSFSYQQQPFPQQPYPQQ(配列番号:36)(618、18%)(コア配列に下線を引く)であった。これらの配列の全ては、トランスグルタミナーゼによる脱アミド化に感受性であると予測されたグルタミン残基を含む(例えば、QXP,QXPF(配列番号:37),QXX[FY](配列番号:38))(Vader et al 2002参照)。幾つかの生物活性ペプチドは異なるファミリーからの2つのコア配列を含む。
【0216】
異なるT細胞集団が別のコア配列を有するエピトープに応答するという可能性と一致し
て、異なるファミリーからのペプチドの生物活性は付加的らしい。例えば、tTG処理されたプール81の中央値生物活性はP04722 QE85の141%であったが、個々のペプチドの生物活性は:ペプチド631(A−グリアジン57−73の相同体)61%、636(626の相同体)51%、そして635 19%、629 16%、そして634 13%(全て396の相同体)の順位の序列であった。
【0217】
単純化しすぎたかもしれないが、グリアジンペプチドに対する合計のIFNγELISpot応答に対する各「ペプチドファミリー」の貢献をHLA−DQ2+セリアック病被験者において比較した(図32参照)。従って、グリアジンペプチドに対する合計の応答に対するP04722 77−73 E85の貢献は、1/5と2/3の間であった。
【0218】
インターネットのワールドワイドウエブ、例えば「cbrg.inf.ethz.ch/subsection3_1_5.html」を通してアクセス可能な、ペプチド相同性検索プログラム、WWW PepPepSearchを用いて、及びライムギのセカリンのGenBank配列との直接の比較により、厳密な適合が、オオムギのホルデイン(HOR8)及びライムギセカリン(A23277,CAA26499,AAG35598)中にコア配列:QQPFPQPQQPFP(配列番号:39)及びオオムギホルデイン(HOG1及びHOR8)中にコア配列:QQPFPQQPQQPFP(配列番号:40)に関して厳密な適合が発見され、そしてPIQPQQPFPQQP(配列番号:41)に関して、オオムギホルデイン(HOR8)中に発見された。
HLA−DQ8関連セリアック病
7人のHLA−DQ8+セリアック病被験者をグルテンチャレンジの前と後に研究した。これらのHLA−DQ8+(HLA−DQA0*0301−3,HLA−DQB0*0302)被験者のうちの5人も、セリアック病関連HLA−DQ2複合体(DQA0*05,DQB0*02)の一方又は両方を有した。両方のセリアック病関連HLA−DQ複合体を有する3人の被験者のうちの2人は、質的及び量的にHLA−DQ2セリアック病被験者と同一であったグリアジンペプチドプール(及びP04722 77−93 QE85を含む個々のペプチド)に対して有効な応答性を有した(図33及び34、及び表18を参照)。脱アミド化されたペプチドプール74は両HLA−DQ2/8被験者において生物活性であったが、6人のHLA−DQ2/X被験者の一人においてのみであった。プール74のtTGによる前処理は生物活性を3.8倍から22倍増強し、そして3人の応答者におけるtTG処理されたプール74の生物活性は、P04722 77−93 E85の生物活性の78%及び350%の間に等しい。現在、何れのペプチドが被験者C02,C07及びC08においてプール74において生物活性であるかわかっていない。
【0219】
セリアック病に関連したHLA−DQ2対立遺伝子の両方又は一方を欠く4人のHLA−DQ8セリアック病被験者のうちの2人はグリアジンペプチドプールに対して極めて弱いIFNγELISpot応答しか示さなかったが、残りの2人はプロテアーゼ−消化されたグリアジン及び特異的ペプチドプールの両方に応答した。被験者C12(HLA−DQ7/8)は、脱アミド化されたプール1−3に対して激しく応答した(図35参照)。これらのプールにおける個々のペプチドの評価は、コア配列LQPQNPSQQQPQ(配列番号:42)を含む一連の密接に関連した生物活性ペプチドを同定した(表20参照)。以前の研究(我々による)は、この配列中の3つのグルタミン残基がtTG媒介脱アミド化に感受性であることを証明した(下線)。WWW PepPepSearchを用いた相同性検索は、コムギα−グリアジンにおいてのみLQPQNPSQQQPQ(配列番号:43)に対して密接な適合を同定した。
【0220】
4人のHLA−DQ8被験者(C11)は、tTG処理されたプール33に対して誘導可能なIFNγELISpot応答を有した(図36参照)。プール32及び33はtTGによる脱アミド化後に活性な以前に定義されたHLA−DQ8制限されたグリアジンエ
ピトープ(QQYPSGQGSFQPSQQNPQ(配列番号:44))の多型を含む(下線を引かれたGlnは脱アミド化されて生物活性を運ぶ)(van der Wal et al 1998)。現在、何れのペプチドがプール33において被験者C11において生物活性であるか分かっていない。
【0221】
インビボグルテンチャレンジ及びコムギグリアジン中の全ての公知の12アミノ酸配列にわたる652ペプチドのセットを使用した、HLA−DQ2関連セリアック病における包括的T細胞エピトープマッピングは、即ち、公知のα−グリアジンエピトープ、A−グリアジン57−73 QE65の10%の生物活性の少なくとも72のペプチドを同定した。しかしながら、これらの生物活性ペプチドはほんの5つほどの異なるが密接に関連したペプチドファミリーのセットに減らされ得る。ほとんど全てのこれらペプチドは、プロリン、グルタミン、フェニルアラニン、及び/又はチロシンが豊富であり、そして配列PQ(QL)P(FY)P(配列番号:45)を含む。この配列はtTGによる2位のQの脱アミド化を促進させる。A−グリアジン57−68の脱アミド化による類推により(Arentz−Hansen 2000)、tTGによる脱アミド化により広く見いだされたこれらのペプチドの増強された生物活性は、HLA−DQ2に関する結合の親和性の増加よるかもしれない。
【0222】
これらの生物活性グリアジンペプチドの一つより多くをインビボにおいて認識するT細胞の間の交差反応性は起こり得る。しかしながら、関連するペプチドの各セットが別のT細胞集団をインビボにおいて活性化するなら、A−グリアジン57−73 E65に相当するエピトープはもっとも有効であり、そしてグルテンチャレンジの後のグリアジンに対する応答においてIFNγを分泌する末梢血T細胞の少なくとも40%により広く認識される。
【0223】
非グリアジン−ペプチド特異的応答が、HLA−DQ2/X関連のセリアック病におけるものと質的に異なったHLA−DQ2/8セリアック病において見いだされた。しかしながら、HLA−DQ2対立遺伝子の両方を持たないHLA−DQ8+のセリアック病被験者の末梢血T細胞は、A−グリアジン57−73 E65相同体を認識しなかった。2つの異なるエピトープが2つのHLA−DQ8+セリアック病において優勢であった。れらのHLA−DQ8+の個体における優勢なエピトープは以前に同定されていなかった(LQPQNPSQQQPQ(配列番号:46))。
【0224】
本明細書中の教示を仮定すれば、共通に認識されるT細胞エピトープの全てを利用してセリアック病のための免疫治療のデザインが実用的になり、6より少ない異なるペプチドを含むかもしれない。コムギのγ−及びω−グリアジン中のエピトープは、オオムギホルデイン及びライムギセカリン中にも存在する。
実施例14
幾つかのELISpotアッセイを前記のとおりに実施して、以下の結果及び/又は結論を生じた:
PQLPYによる複数のα−グリアジン多型の拡大
A−グリアジン57−73QEの有効なアゴニスト(G01)は、QLQPFPQPELPYPQPQS(G01),PQL−Y−−−−−−−−−−−−−−P(G10),及びPQPQPFL−−−−−−−−−−−−−(G12)を含む。有効性が劣るのは、−−−−−−−−−−−−L−−−−−−−−P(G04),−−−−−−−−−−−−−R−−−−−−−−−−−P(G05),及び−−−−−−−−−−−−S−−−−−P(G06)を含む。有効性がさらに劣るのは、−−−−−−L−−−−−−−−S−−−−−P(G07),−−−−−S−−−−−−−−−−S−−−−−−−−−P(G08)−−−−−−−−−−−−−−−S−−S−−−−−P(G09),及びPQPQPFP−−−−−−−−−−−−−−−−−(G13)を含む。ダッシュは特定の位置にお
けるG01配列との同一性を示す。
グルテンチャレンジは新たに診断されたセリアック病においてたった2週間のグルテンフリーダイエット後にA−グリアジン57−73 QE65 T細胞を含む
追加の分析は、tTG脱アミド化されたグリアジン応答が新たに診断されたセリアック病において2週間後に変化することを示した。他の分析は、脱アミド化されたグリアジン特異的T細胞がCD4+α4β7+HLA−DQ2制限されることを示した。
最適なエピトープ(クローン対グルテンチャレンジ)
「優勢な」エピトープがγIFN ELISpotによりグルテンチャレンジ後に定義される。QLQPFPQPELPYPQPQS(100%ELISpot応答)。腸のT細胞クローンにより定義されたエピトープ:QLQPFPQPELPY(27%),PQPELPYPQPELPY(52%),及びOOLPQPEQPQQSFPEQERPF(9%)。
個々のペプチド配列間の順位
順位はコムギ又はライムギに依存する。コムギに関して、優勢なペプチドは、ペプチド番号89、90及び91を含む(表23における番号を参照)。ライムギに関して、優勢なペプチドは、ペプチド番号368、369、370、371、及び372を含む(表23における番号を参照)。635及び636(表23における番号を参照)を含む幾つかのペプチドは、ライムギ及びコムギの両方において活性を示した。
インビボグルテンチャレンジはT細胞エピトープのヒエラルキーがセリアック病に関して定義されることを可能にする
エピトープのヒエラルキーはHLA−DQ2+セリアック病の間で一致しているが、HLA−DQ8+セリアック病に関しては異なる。ヒエラルキーはコムギ穀物が消費されることに依存する。脱アミド化はほとんど全てのグリアジンエピトープを生成する。HLA−DQ2,DQ8及びDR4が脱アミド化されたペプチドを提示する。HLA−DQ2/8−関連のセリアック病は優先的にDQ2−関連グリアジンエピトープを提示する。グリアジンエピトープは、エピトープに基づく治療剤の開発を正当であると理由付けするのに十分制限される。
【0225】
他の分析は以下を示した:HLA−DR3−DQ2(85−95%)及びHLA−DR4−DQ8(5−15%)。
【0226】
他の分析は以下を示した:
【0227】
【表2】
【0228】
被験者12のプール1−3において個々のtTG−脱アミド化されたペプチドの生物活性を測定するために、別の分析を実施した。結果は次のとおりである(配列の番号は表23に掲載されたペプチドを指す):配列8(100%),配列5(85%)、配列6(82%)、配列3(77%)、配列1(67%)、配列2(59%)、配列9(49%)、配列7(49%)、配列10(33%)、配列4(15%)、配列12(8%)、配列11(0%)、配列23(26%)、配列14(18%)、配列15(18%)、配列17(18%)、配列16(13%)、配列14(8%)、配列22(5%)、配列18(3%)、配列19(3%)、配列20(0%)、配列21(0%)。予測された脱アミド化配列はLQPENPSQEQPEである。
PBMCによる個々のELISpot応答(ELISpotリーダーにより測定されたスポット形成細胞)
【0229】
【表3】
【0230】
交差反応性
29人の被験者の652のペプチドからのデータを扱うため、又は特定の応答が真実の陽性のペプチド特異的T細胞応答であるかを決定するため、又はペプチドに対する応答が別の構造上関連するペプチドとの交差反応性によるかを決定するために、特定のペプチド応答の発現を「優勢な」ペプチド応答のパーセンテージとすることができる。或いは、当該発現はペプチド応答間の相関係数としての「遅延(retardness)」とし得るか、又はバイオインフォマティックスにより得る。
追加のエピトープ
代表的な結果は次の通りである:
ペプチドのP04722Eとの組み合わせ(全て20mcg/ml)(n=4)
単独 P04722E+
Pep626 60 135
P04722E 100 110
HLAa 0 85
(P04722Eのパーセントとして表された)
626+tT:PQQPQQPQQPFPQPQQPFPW
04722E:QLQPFPQPELPYPQPQL
ペプチド626のtTG脱アミド化(n=12)
tTGなし=100%
tTG=170%
特定の位置における置換
ペプチド626PQQP[Q1]QP[Q2]QPFPQP[Q3]QPFPVの置換(n=12)
Glu Arg
Q1 95 90
Q2 145 80
Q3 155 10
(野生型ペプチドのパーセントとして表された)
tTG処理された15マーに及ぶペプチド626/627(PQQPQQPQQPFPQPQQPFPWQP)(n=8)
P1−15 5
P2−16 4
P3−17 3
P4−18 38
P5−19 65
P6−20 95
P7−21 65
P8−22 90
(最大の15マー応答のパーセントとして表された)
複数エピトープ:
P04722E:QLQPFPQPQLPYPQPQL
626+tTG:PQQPQQPQQPFPQPQQPFPW
最小エピトープ:QPQQPFPQPQQPFPW
抗−CD4によりコードされたビーズ及び抗インテグリンβ7によるPBMCの免疫磁気除去はIFNγELISpot応答を除去させたが、抗−CD8によりコードされたビーズ又は抗アルファEインテグリンによるPBMCの免疫磁気除去はそうでなかった。即ち、IFNγを分泌するPBMCはCD4+及びα4β7+であり、消化管内の粘膜固有層へのホーミングに関連した。
【0231】
HLA−DQ2に関して異型接合及び同型接合においては、抗−DQ抗体によりブロックされたが抗−DR抗体によりブロックされなかった。これは一つの配列中の複数のエピトープを暗示するかもしれない。
セリアック病のT細胞エピトープ
他の研究者らは、特定の腸T細胞クローンエピトープを特性決定した。例えば、Vader et al.,Gastroenterology 2002,122:1729−37;Arentz−Hansen et al.,Gastroenterology 2002,123:803−809を参照。これらは、T細胞をクローン化するために使用されたインビトロ技術のためにその関係がいくらよく見ても不明であるエピトープの例である。
腸対末梢血クローン
腸:1)腸生検、2)グルテンのペプシン−トリプシン消化に対して生じさせたT細胞クローン、3)全てHLA−DQ2制限される、4)クローンはトランスグルタミナーゼにより脱アミド化されたグリアジンに応答する。
末梢血:グルテンに対して生じさせたT細胞クローンはHLA−DR,DQ及びDP制限される。結果:腸T細胞クローンはセリアック病関連エピトープをマップするのに限定的に使用することができる。
GDA_9コムギ307aa定義アルファ/ベータ−グリアジンMM1前駆体(プロラミ
ン)受け入れ番号P18573−−GenBank(その全体を引用により本明細書に編入する)
腸T細胞クローンエピトープ
腸T細胞クローンエピトープの定義は、例えば、Arentz−Hansen et al.,J Exp Med.2000,191:603−12に見いだすことができる。また、腸T細胞クローンに関するグルテンエピトープも開示されている。脱アミド化されたQLQPFPQPQLPYがエピトープであり、QLQPFPQPELPYの脱アミド化配列もある。HLA−DQ2制限がある。相同性検索は他の生物活性γアルファ−グリアジンがPQPQLPYを単独又は二重に含むことを示す。大多数のT細胞クローンはDQ2−αI:QLQPFPQPELPY DQ2−αII:PQPELPYPQPELPYの何れかに応答する。
優勢なグリアジンT細胞エピトープ
全てトランスグルタミナーゼにより脱アミド化される。
グルテンチャレンジ6日後の末梢血:A−グリアジン57−73:
QLQPFPQPELPYPQPQS
腸T細胞クローン:DQ2−αI:QLQPFPQPELPY DQ2−αII:PQPELPYPQPELPY
腸T細胞クローンエピトープマッピング
【0232】
【表4】
【0233】
末梢血におけるIFNγ−分泌A−グリアジン57−73 QE65−特異的T細胞のグルテン暴露及び誘導
未処理のセリアック病を1、2又は8週間グルテンフリーダイエットさせ、次にグルテン暴露し(3日 パン200g/日)、次にグルテンフリーダイエットをさせた。
【0234】
結果1:グルテンフリーダイエットの期間及びグルテンチャレンジの0日目及び6日目のIFNγELISpot応答:A−グリアジン57−73 QE65(IFNγ特異的スポット/百万PPBMCとして表された結果)
0日目:なし(0)、1週(1)、2週(2)、8週(1)
6日目:なし(0)、1週(4)、2週(28)、8週(48)
結果2:グルテンフリーダイエットの期間及びグルテンチャレンジの0日目及び6日目のIFNγELISpot応答:tTG−グリアジン57−73 QE65(IFNγ特
異的スポット/百万PPBMCとして表された結果)
0日目:なし(45)、1週(62)、2週(5)、8週(5)
6日目:なし(0)、1週(67)、2週(40)、8週(60)
結果3:グルテンフリーダイエットの期間及びグルテンチャレンジの0日目及び6日目のIFNγELISpot応答:A−グリアジン57−73 P65(IFNγ特異的スポット/百万PPBMCとして表された結果)
0日目:なし(1)、1週(2)、2週(1)、8週(1)
6日目:なし(0)、1週(0)、2週(0)、8週(0)
結果4:グルテンフリーダイエットの期間及びグルテンチャレンジの0日目及び6日目のIFNγELISpot応答:PPD(IFNγ特異的スポット/百万PPBMCとして表された結果)
0日目:なし(90)、1週(88)、2週(210)、8週(150)
6日目:なし(0)、1週(100)、2週(210)、8週(100)
結果5:グルテンフリーダイエットの期間及びグルテンチャレンジの0日目及び6日目のIFNγELISpot応答:tTG(IFNγ特異的スポット/百万PPBMCとして表された結果)
0日目:なし(5)、1週(4)、2週(3)、8週(2)
6日目:なし(0)、1週(4)、2週(1)、8週(2)
長期間のグルテンに対するHLA−DQ2セリアック病におけるグルテンチャレンジ
抗グリアジンT細胞応答の特性決定を、末梢血において、3日間のグルテンチャレンジの6−8日後に実施した。
結果1:PBMC 6日目長期間グルテンフリーダイエット(抗HLA−DR及び−DQ抗体と前インキュベーション)(%阻害として表現)
DR−:tTG−グリアジン100mcg/ml(105),A−グリアジン57−73 QE65 50mcg/ml(90),PPD 5mcg/ml(30)
DQ−:tTG−グリアジン 100mcg/ml(5),A−グリアジン57−73
QE65 50mcg/ml(22),PPD 5mcg/ml(78)。
【0235】
結果2:PBMC 6日目長期間グルテンフリーダイエット(%CD8−除去PBMC応答として表現)
B7除去:tTG−グリアジン n=6(7),A−グリアジン57−73 n=9(6),PPD n=8(62)
AE除去:tTG−グリアジン n=6(120),A−グリアジン57−73 ん=9(80)、PPD n=8(110)。
【0236】
CD4除去:tTG−グリアジン n=6(10),A−グリアジン57−73 n=9(9),PPD n=8(10)。
治療用ペプチドは限定ではないが以下を含む
【0237】
【表5】
【0238】
簡単には、経口の抗原チャレンジ後に、末梢血T細胞の特異性が腸のT細胞クローンの特異性を反映する。末梢血においては、腸T細胞クローンのエピトープは、最適なα−グリアジンエピトープであるA−グリアジン57−73 QE65に比較して準最適である。実施例15
以下のとおりのマッピング目的のためにもELISpotアッセイを実施した。
優勢なDQ−8関連エピトープのファインマッピング
【0239】
【表6】
【0240】
優勢なエピトープのファインマッピング(2)
【0241】
【表7】
【0242】
実施例16
IFNγ−ELISpotにおけるグリアジンエピトープの生物活性(25mcg/ml,n=6)(A−グリアジン57−73 QE65応答の%として表された)
DQ2−AII:野生型(WT)(4),WT+tTG(52),Glu−置換された(52)
DQ2−AI:野生型(WT)(2),WT+tTG(22),Glu−置換された(28)
GDA09:野生型(WT)(1),tTG(7),Glu−置換された(8)
A−G31−49:野生型(WT)(2),tTG(3),Glu−置換された(0)A−グリアジン57−73 QE65(G01E)の用量応答(n=8)(G01Eの最大応答の%として表された)
0.025mcg/ml(1),0.05mcg/ml(8),0.1mcg/ml(10),0.25mcg/ml(22),0.5mcg/ml(38)<1mcg/ml(43),2.5mcg/ml(52),5mcg/ml(70),10mcg/ml(81),25mcg/ml(95),50mcg/ml(90),100mcg/ml(85)。
【0243】
単独又はA−グリアジン57−75(G01E)と混合されたグリアジンエピトープに対するIFNγELISpot応答(全て50mcg/ml,tTG−グリアジン100mcg/ml,PPD 5mcg/ml;n=9)(G01E応答の%として表された)
単独:DQ2−A1(20),DQ2−A2(55),オメガG1(50),tTGグリアジン(80)、PPD(220),DQ2バインダー(0)
G01E+:DQ2−A1(90),DQ2−A2(95),オメガG1(100),tTGグリアジン(120)、PPD(280),DQ2バインダー(80)
個々のセリアック病被験者におけるIFNγELISpot応答に対するA−グリアジン57−73 QE65のアラニン及びリジン置換の効果(n=8)
エピトープ配列:QLQPFPQPELPYPQPQS
57−59位及び72−73位におけるアラニンの置換はA−グリアジン57−73 QE65応答の%において、ほとんど又は全く低下を示さなかった。60−62位及び68−71位におけるアラニンの置換はA−グリアジン57−73 QE65応答の%において穏やかな低下を示した。63−67位におけるアラニンの置換はA−グリアジン57−73 QE65応答の%において最大の低下を示した。
個々のセリアック病被験者におけるIFNγELISpot応答に対するA−グリアジン57−73 QE65のリジン置換の効果(n=8)
エピトープ配列:QLQPFPQPELPYPQPQS
57−59位及び71−73位におけるリジンの置換はA−グリアジン57−73 QE65応答の%において、ほとんど又は全く低下を示さなかった。60−61位及び69−70位におけるリジンの置換はA−グリアジン57−73 QE65応答の%において穏やかな低下を示した。62−68位におけるリジンの置換はA−グリアジン57−73
QE65応答の%において最大の低下を示した。
実施例17
表24は、コムギ又はライムギでチャレンジした数人の患者を用いた652のペプチドを試験する分析の結果を示す。
【0244】
【表8】
【0245】
本明細書において引用されたか又は言及されたPCTの公表、合衆国特許、他の特許、定期刊行物、及びあらゆるその他の刊行物はそれらの全体を引用により本明細書に編入する。
【0246】
【表9】
【0247】
【表10】
【0248】
【表11】
【0249】
【表12】
【0250】
【表13】
【0251】
【表14】
【0252】
【表15】
【0253】
【表16】
【0254】
【表17】
【0255】
【表18】
【0256】
【表19】
【0257】
【表20】
【0258】
【表21】
【0259】
【表22】
【0260】
【表23】
【0261】
【表24】
【0262】
【表25】
【0263】
【表26】
【0264】
【表27】
【0265】
【表28】
【0266】
【表29】
【0267】
【表30】
【0268】
【表31】
【0269】
【表32】
【0270】
【表33】
【0271】
【表34】
【0272】
【表35】
【0273】
【表36】
【0274】
【表37】
【0275】
【表38】
【0276】
【表39】
【0277】
【表40】
【0278】
【表41】
【0279】
【表42】
【0280】
【表43】
【0281】
【表44】
【0282】
【表45】
【0283】
【表46】
【0284】
【表47】
【0285】
【表48】
【技術分野】
【0001】
本発明は、セリアック病の診断及び治療に有用なエピトープに関し、診断剤、治療剤、キット、及び前記の使用方法を含む。
【背景技術】
【0002】
ダイエット中のグリアジン(グルテンの成分)に対する免疫反応がセリアック病を引き起こす。腸組織中の免疫応答は腸のトランスグルタミナーゼにより修飾されたグリアジンに優先的に応答する。セリアック病は抗−筋内膜抗体の検出により診断されるが、これは腸の生検内のリンパ球の炎症の発見による確認を必要とする。そのような生検の取得は患者にとって快適ではない。
【0003】
研究者は、腸のT細胞応答のみがグリアジンに対する免疫応答の正確な指標を提供すると、以前に仮定した。よって、彼らは、腸の組織内のT細胞の応答の研究に集中してきた1。トランスグルタミナーゼの修飾を必要とする(それらが免疫系により認識される前に)グリアジンエピトープは公知である2。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】1. MolbergO, etal. NatureMed.4, 713-717 (1998).
【非特許文献2】2. Quarsten H, et al. Eur. J. Immunol. 29, 2506-2514 (1999).
【非特許文献3】3. Greenberg CS et al. FASEB 5, 3071-3077 (1991).
【非特許文献4】4. Mantzaris G, Jewell D. Scand. J. Gastroenterol. 26, 392-398 (1991).
【非特許文献5】5.Mauri L, et al. Scand. J. Gastroenterol. 31, 247-253 (1996).
【非特許文献6】6. BuncoM, et al. Tissue Antigens 46, 355-367 (1995).
【非特許文献7】7. OlerupO, etal. Tissueantigens 41, 119-134 (1993).
【非特許文献8】8. Mullighan CG, et al. Tissue-Antigens. 50, 688-92 (1997).
【非特許文献9】9. PlebanskiM et al. Eur. J. Immunol. 28, 4345-4355 (1998).
【非特許文献10】10. Anderson DO, Greene FC. The alpha-gliadin gene family. II. DNA and protein sequence variation, subfamily structure, and origins of pseudogenes. Theor Appl Genet (1997) 95 : 59-65.
【非特許文献11】11. Arentz-HansenH, Korner R, Molberg O,Quarsten H, Van der Wal Y, Kooy YMC, Lundin KEA, Koning F, Roepstorff P, Sollid LM, McAdam SN. The intestinal T cell response to alpha-gliadin in adult celiac disease is focused on a single deamidated glutamine targeted by tissue transglutaminase. J Exp Med. 2000 ; 191 : 603-12.
【非特許文献12】12. Vader LW, de Ru A, van der Wal, Kooy YMC, Benckhuijsen W, Mearin ML, DrijfhoutJW, van Veelen P, Koning F. Specificity of tissue transglutaminase explains cereal toxicity in celiac disease. J Exp Med 2002 ; 195 : 643-649.
【非特許文献13】13. van der Wal Y, Kooy Y, van Veelan P, Pena S, Mearin L, Papadopoulos G, Koning F. Selective deamidation by tissue transglutaminase strongly enhances gliadin-specific T cell reactivity. J Immunol. 1998 ; 161 : 1585-8.
【非特許文献14】14. van der Wal Y, Kooy Y, van Veelan P, Pena S, Mearin L, Molberg O,Lundin KEA, Sollid L, Mutis T, BenckhuijsenWE, Drijfhout JW, Koning F. Proc Natl Acad Sci USA 1998 ; 95 : 10050-10054.
【非特許文献15】15. Vader W, Kooy Y, Van Veelen Petal. The gluten response in children with celiac disease is directed toward multiple gliadin and glutenin peptides. Gastroenterology2002, 122 : 1729-37 16. Arentz-Hansen H, McAdam SN, Molberg O, et al. Celiac lesion T cells recognize epitopes that cluster in regions of gliadin rich in proline residues. Gastroenterology 2002, 123 : 803-809.
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明者らは、セリアック病において免疫系により認識される免疫上優勢なT細胞A−グリアジンエピトープを発見し、そしてこれがセリアック病の個体の末梢血内でT細胞により認識されることを示した(WO 01/25793)。そのようなT細胞は、再刺激無しに検出可能なほど十分高い頻度で存在することを発見した(即ち、「フレッシュな応答」検出系が使用できた)。上記エピトープは非T細胞クローニングに基づく方法を用いて同定され、上記エピトープのより正確な反射(reflection)が認識された。上記の免疫上優勢なエピトープは免疫系が認識する前にトランスグルタミナーゼの修飾を必要とする(特定のグルタミンのグルタミン酸への置換を引き起こす)。
【0006】
この研究に基づいて、本発明者らは、初期段階でセリアック病を診断するのに使用できる試験を開発した。当該試験は、末梢血からのサンプル上で実施してよく、よって、腸の生検は必要ない。当該試験は現在使用されている抗体試験よりも感度が高い。
【課題を解決するための手段】
【0007】
発明は、即ち、個体においてセリアック病を診断するか又はセリアック病に対する感受性を診断する方法を提供し、
(a)宿主からのサンプルを、(i)配列番号:1(PQPELPY)又は配列番号:2(QLQPFPQPELPYPQPQS)である配列、又は配列番号:3により表されるグリアジンの天然に生じる相同体からの均等な配列を含むエピトープ、(ii)グリアジン蛋白質由来の単離されたオリゴペプチドである、配列番号:1を含むか又は配列番号:3により表されるグリアジンの天然に生じる相同体からの均等な配列(表1に示される)を含むエピトープ、(iii)(i)又は(ii)を認識するT細胞受容体により認識され得る(i)又は(ii)の類似体であって、ペプチドの場合は類似体が50アミノ酸の長さを超えない上記類似体、又は(iv)(i),(ii)又は(iii)において定義された2つ又はそれより多い薬剤(agent)を含む生成物からなる群から選択される薬剤と接触させ、そして(b)サンプル中のT細胞が薬剤を認識するか否かをインビトロにて決定して、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示す
ことを含む。
【0008】
コムギのグリアジンT細胞のエピトープの包括的なマッピングを通して(実施例13参照)、本発明者らは、類似のコア配列(例えば、配列番号:18−22)及び類似の完全長配列(例えば、配列番号:31−36)を有する他のコムギグリアジン中のセリアック病のHLA−DQ2+患者において、並びにライムギのセカリン及びオオムギのホールダイン(配列番号:39−41)の生物学上活性なエピトープをも発見した;表20及び21も参照。さらに、セリアック病においてHLA−DQ8+患者にて生物学上活性な幾つかのエピトープが同定された(配列番号42−44、46)。この包括的なマッピングは、即ち、セリアック病の患者においてT細胞により認識される優勢なエピトープを提供する。即ち、上記の方法及び本明細書に記載される他の方法は、これらの追加の同定されたエピトープ、それらの類似体及び均等物の何れかを使用して実施してよい;本明細書の(i)及び(ii)はこれらの追加のエピトープを含む。即ち、本発明の薬剤は新規なエピ
トープも含む。
【0009】
本発明は、個体において、セリアック病又はセリアック病に対する感受性を診断する方法における使用のための診断手段の製造のための薬剤の使用を提供し、当該方法は、当該個体のT細胞が薬剤を認識するか否かを決定することを含み、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示す。
【0010】
トランスグルタミナーゼにより修飾された免疫上優勢なエピトープ(並びに本明細書において定義された追加の他のエピトープ)の発見は、このエピトープに対する他の種類の免疫応答が存在するか否かを決定することに基づいたセリアック病の診断も可能にする。即ち、本発明は、個体において、セリアック病又はセリアック病に対する感受性を診断する方法も提供し、当該個体からのサンプル中のエピトープに結合する抗体の存在を決定することを含み、抗体の存在が、個体がセリアック病を有するか又はセリアック病に感受性であることを示す。
【0011】
本発明は、さらに、上記薬剤を認識するT細胞を寛容にするセリアック病を治療又は予防する方法における使用のための上記薬剤を、任意にキャリアーと共に提供する。また、T細胞と拮抗することによりセリアック病を治療又は予防する方法における使用のための、(i)又は(ii)を認識するT細胞受容体を有するT細胞のアンタゴニストを、任意にキャリアーと共に提供する。さらに、抗体の生産を阻害するために個体を寛容にすることにより個体においてセリアック病を治療又は予防する方法における使用のための、抗体(上記薬剤に結合する)を結合する薬剤又は類似体が提供される。
【0012】
本発明は、組成物がセリアック病を引き起こすか否かを決定する方法を提供し、トランスグルタミナーゼにより上で定義されたオリゴペプチド配列へ修飾される蛋白質が組成物中に存在するか否かを決定することを含み、蛋白質の存在が、組成物がセリアック病を引き起こし得ることを示す。
【0013】
本発明は、その野生型配列がトランスグルタミナーゼにより上で定義された配列を含むエピトープを含む配列に修飾され得る変異体グリアジン蛋白質も提供するが、しかし、当該変異体グリアジン蛋白質は、そのようなエピトープを含む配列を含む配列へトランスグルタミナーゼにより修飾され得る配列を含まない様式にて修飾されており;又は、本発明は、上記の様式により修飾されたそのような変異体グリアジン蛋白質の断片も提供する。
【0014】
本発明は、T細胞受容体へ結合することができる配列を含む蛋白質も提供し、但し、T細胞受容体は上記の薬剤を認識し、そして上記配列はそのようなT細胞受容体を有するT細胞の拮抗作用を引き起こすことができる。
【0015】
さらに、本発明は、上で定義された蛋白質を含む食品を提供する。
【図面の簡単な説明】
【0016】
【図1A】図1は、最初のうちはグルテンフリーダイエットのあとに緩解して、次に1日目から3日間200gのパン(bread)で毎日チャレンジされた、セリアック病被験者1における、A−グリアジン51−85にわたる5つのオーバーラップする15マーを含むトランスグルタミナーゼ(tTG)−処理されたものと処理されないペプチドプール3(各ペプチド10μg/ml)(表1参照)及びキモトリプシン−消化されたグリアジン(40μg/ml)に対する、新たに単離されたPBMC(末梢血単核細胞)IFNγELISPOT応答(垂直軸は106PBMCあたりのスポット形成細胞を示す)を示す(a)。
【図1B】10日間の間のパンチャレンジの間の全A−グリアジン蛋白質にわたるtTG−処理されたA−グリアジンペプチドプール1−10に対する被験者2によるPBMC IFNγELISPOT応答(b)。水平軸はパン開始後の日にちを示す。
【図2】図2は、最初のうちはグルテンフリーダイエットのあとに緩解して、次に3日間パンでチャレンジされた、セリアック病の7人の被験者における、tTG−処理されたペプチドプール3(A−グリアジン51−85にわたる)に対する、PBMC IFNγELISPOT応答(垂直軸は106PBMCあたりのスポット形成細胞を示す)を示す(1から3日)。垂直軸はパンを初めた後の日にちを示す。(a)。プール3中に含まれるtTG処理のオーバーラップ15マーに対する応答;バーは6日目又は7日目の6人のセリアック病の被験者の個々のペプチド(10μg/ml)に対する、平均(±SEM)の応答を表す(b)。(個々の被験者において、ペプチドに対するELISPOT応答はペプチド12により導かれた応答の%として計算された−水平軸により示されるとおり。)
【図3】図3は、A−グリアジン56−75のtTG−処理された末端削除体(0.1μM)に対するPBMC IFNγELISPOT応答を示す。バーは5人のセリアック病被験者の平均(±SEM)を表す。)。(個々の被験者において、応答は試験されたペプチドにより導かれた最大応答の%として計算された−水平軸により示されるとおり。)
【図4】図4は、優勢なA−グリアジンエピトープの最小構造が如何にしてtTG−処理された7−17マーのA−グリアジンペプチド(0.1μM)を用いてマップされたかを示し、配列PQPQLPY(配列番号:4)(A−グリアジン62−68)(a)、及びtTG処理しなかったがQ→E65の置換を伴う同じペプチドを含む(b)。各線はパンを1−3日目に摂取してから6又は7日目の3人のセリアック病被験者におけるPBMC IγELISPOT応答を表す。)。(個々の被験者において、ELISPOT応答は17マー、A−グリアジン57−73により導かれた応答の%として計算された。)
【図5】図5はtTGにより脱アミド化されたアミノ酸を示す。A−グリアジン56−75LQLQPFPQPQLPYPQPQSFP(配列番号:5)(0.1μM)をtTG(50μg/ml)と共に37℃において2時間インキュベートした。逆相HPLCにより単一の生成物が同定されて精製された。アミノ酸分析により、tTGに帰し得るA−グリアジン56−75内の各Gln残基の%脱アミド化(Q→E)の計算が可能になった(水平軸)。
【図6】図6は、パンを1−3日目に摂取してから6又は7日目の3人のセリアック病被験者における、17マー:QLQPFPQPELPYPQPES(配列番号:6)(E57,65),QLQPFPQPELPYPQPES(配列番号:7)(E65,72),ELQPFPQPELPYPQPES(配列番号:8)(E57,65,72),及びQLQPFPQPELPYPQPQS(配列番号:9)(E65)を用いた、Aグリアジン57−73中のQ65に加えた他の位置においてQ→Eを置換した効果を示す。水平軸はE65応答の%を示す。
【図7】図7は、tTG処理されたA−グリアジン56−75(0.1μM)が(a)CD4及びCD8の磁気ビーズ除去した(depleted)PBMCにおいてIFN−gELISPOT応答を導いたことを示す。(バーはCD4除去PBMC応答をCD8除去PBMC応答の%として表す;百万のCD8除去PBMCあたりのスポット形成細胞は:被験者4:29、及び被験者6:535であった。)(b)HLA−DQaI*0501,bI*0201に関して同型接合体の2人のセリアック病被験者における、tTG−処理56−75(0.1μM)の1時間前の、HLA−DR(L243),−DQ(L2)及び−DP(B7.21)(10μg/ml)に対するモノクローナル抗体とのインキュベーション後のPBMC IFNγELISPOT応答(スポット形成細胞/百万PBMC)。
【図8】図8は、免疫上優勢なエピトープ内の65位における他のアミノ酸のGluによる置換の効果を示す。水平軸は免疫上優勢なエピトープに関しての3人の被験者における%応答を示す。
【図9】図9は、トランスグルタミナーゼ処理を伴う場合(影)及び伴わない場合(透明)の配列PQLPY(配列番号:12)を含む、天然に存在するグリアジンペプチドの免疫反応性を示す(3人の被験者の応答を測定)。
【図10】図10は、グルテンチャレンジを開始して、次にインターフェロンガンマELISpotを行ってから6日後の2人のセリアック病被験者からの末梢血単核細胞のCD8,CD4,β7,及びαE−特異的免疫磁気ビーズ除去を示す。A−グリアジン57−73 QE65(25mcg/ml),tTG−処理されたキモトリプシン−消化グリアジン(100mcg/ml)又はPPD(10mcg/ml)を抗原として用いた。
【図11】図11は、最適なT細胞エピトープの長さを示す。
【図12A】図12aは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12B】図12bは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12C】図12cは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12D】図12dは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12E】図12eは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12F】図12fは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12G】図12gは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図12H】図12Hは、A−グリアジン57−73 QE65と他のペプチドとの用量応答研究における比較を示す。
【図13A】図13aは、グリアジンとA−グリアジン57−73 QE65特異的応答の比較を示す。
【図13B】図13bは、グリアジンとA−グリアジン57−73 QE65特異的応答の比較を示す。
【図13C】図13cは、グリアジンとA−グリアジン57−73 QE65特異的応答の比較を示す。
【図14A】図14aは、セリアック病被験者におけるグリアジンの多型の生物活性を示す。
【図14B】図14bは、セリアック病被験者におけるグリアジンの多型の生物活性を示す。
【図14C】図14cは、セリアック病被験者におけるグリアジンの多型の生物活性を示す。
【図14D】図14dは、セリアック病被験者におけるグリアジンの多型の生物活性を示す。
【図14E】図14eは、セリアック病被験者におけるグリアジンの多型の生物活性を示す。
【図15】図15は、コアエピトープ配列の定義を示す。
【図16】図16は、コアエピトープ配列の定義を示す。
【図17】図17は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図18】図18は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図19】図19は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図20】図20は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図21】図21は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図22】図22は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図23】図23は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図24】図24は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図25】図25は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図26】図26は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図27】図27は、A−グリアジン57−73 QE65バリアントのアゴニスト活性を示す。
【図28A】図28aは、異なる患者群における応答を示す。
【図28B】図28bは、異なる患者群における応答を示す。
【図28C】図28cは、異なる患者群における応答を示す。
【図28D】図28dは、異なる患者群における応答を示す。
【図28E】図28eは、異なる患者群における応答を示す。
【図28F】図28fは、異なる患者群における応答を示す。
【図28G】図28gは、異なる患者群における応答を示す。
【図29】図29は、A−グリアジン57−73のプロラミン相同体の生物活性を示す。
【図30】図30は、健康なHLA−DQ2被験者についての、tTG−脱アミド化されたグリアジンペプチドプールに対するELISpot応答の変化を示す。
【図31】図31は、セリアック病のHLA−DQ2被験者についての、tTG−脱アミド化されたグリアジンペプチドプールに対するELISpot応答の変化を示す。
【図32】図32は、「要約された」グリアジンペプチド応答に対する個々のペプチドの寄与を示す。
【図33】図33は、セリアック病のHLA−DQ2/8被験者C08に関しての、tTG−脱アミド化されたグリアジンペプチドプールに対する、グルテンチャレンジが導いたIFNγELISpot応答を示す。
【図34】図34は、セリアック病のHLA−DQ2/8被験者C07に関しての、tTG−脱アミド化されたグリアジンペプチドプールに対する、グルテンチャレンジが導いたIFNγELISpot応答を示す。
【図35】図35は、セリアック病のHLA−DQ8/7被験者C12に関しての、tTG−脱アミド化されたグリアジンペプチドプールに対する、グルテンチャレンジが導いたIFNγELISpot応答を示す。
【図36】図36は、セリアック病のHLA−DQ6/8被験者C11に関しての、tTG−脱アミド化されたグリアジンペプチドプールに対する、グルテンチャレンジが導いたIFNγELISpot応答を示す。
【発明を実施するための形態】
【0017】
本発明はセリアック病を治療又は予防する方法を提供し、a)配列番号:18−22、31−36、39−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びb)a)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、a)の類似体;及びc)配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を個体に投与するこ
とを含む。幾つかの態様において、上記薬剤は、HLA−DQ2−制限されるか、HLA−DQ8−制限されるか、又は一つの薬剤がHLA−DQ2−制限されており、そして第2の薬剤がHLA−DQ8−制限されている。幾つかの態様において、上記薬剤は、コムギのエピトープ、ライムギのエピトープ、オオムギのエピトープ又はそれらの組み合わせの何れかを、単一の薬剤として又は複数の薬剤として含む。
【0018】
本発明は、上記の薬剤及び薬学上受容可能なキャリアー又は希釈剤を含む薬学組成物を個体に投与することを含む、セリアック病を予防又は治療する方法も提供する。
【0019】
本発明は、上で定義されたT細胞受容体を有するT細胞のアンタゴニスト及び薬学上受容可能なキャリアー又は希釈剤を含む薬学組成物を個体に投与することを含む、セリアック病を予防又は治療する方法も提供する。
【0020】
本発明は、T細胞の生産又は上で定義された薬剤に対する抗体応答を抑圧するために個体をグリアジン蛋白質に対して寛容にするための組成物を個体に投与することを含む、セリアック病を予防又は治療する方法も提供する。
【0021】
本発明は、1)a)i)配列番号:18−22、31−36、39−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を、宿主からのサンプルに接触させ;そしてサンプル中のT細胞が上記薬剤を認識するか否かをインビトロにて決定するが、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示すか;又はb)上で定義された薬剤を投与し、そして個体中のT細胞が上記薬剤を認識するか否かをインビボにて決定するが、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示すかの何れかにより、個体においてセリアック病を診断し;そして2)セリアック病を有するか又はセリアック病に感受性であると診断された個体に、セリアック病を予防又は治療するための治療用薬剤を投与する
ことによりセリアック病を予防するか又は治療する方法も提供する。
【0022】
本発明は、上記薬剤を認識するT細胞を寛容にすることによりセリアック病を予防又は治療する方法における使用のための、上で定義された薬剤も、任意にキャリアーと共に、提供する。
【0023】
本発明は、T細胞と拮抗することによりセリアック病気を予防又は治療する方法における使用のための、上で定義されたT細胞受容体を認識するT細胞のアンタゴニストも、任意にキャリアーと共に、提供する。
【0024】
本発明は、T細胞受容体へ結合することができる配列を含む蛋白質も提供し、但し、T細胞受容体は上で定義された薬剤を認識し、そして上記配列はそのようなT細胞受容体を有するT細胞の拮抗作用を引き起こすことができる。
【0025】
本発明は、上で定義された薬剤又はアンタゴニスト及び薬学上受容可能なキャリアー又は希釈剤を含む薬学組成物も提供する。
【0026】
本発明は、T細胞の生産又は上で定義された薬剤に対する抗体応答を抑圧するために個体をグリアジン蛋白質に対して寛容にするための組成物も提供し、当該組成物は上で定義
された薬剤を含む。
【0027】
本発明は、上で定義された薬剤に対するT細胞応答に拮抗するための組成物も提供し、当該組成物は上で定義されたアンタゴニストを含む。
【0028】
本発明は、その野生型配列がトランスグルタミナーゼにより請求項1において定義された薬剤である配列に修飾され得る変異体グリアジン蛋白質も提供し、当該変異体グリアジン蛋白質は、上で定義された薬剤である配列へのトランスグルタミナーゼによるそれの修飾を阻害する変異を含み;又は、本発明は少なくとも15アミノ酸の長さであり、且つ上記変異を含む、そのような変異体グリアジン蛋白質の断片を提供する。
【0029】
本発明は、上で定義された蛋白質又は断片をコードするコーディング配列を含むポリヌクレオチドも提供する。
【0030】
本発明は、上で定義されたポリヌクレオチドを含む細胞又はそのようなポリヌクレオチドにより形質転換された細胞も提供する。
【0031】
本発明は、上で定義されたT細胞受容体を発現する哺乳類も提供する。
【0032】
本発明は、個体においてセリアック病を診断するか又はセリアック病に対する感受性を診断する方法も提供し、a)i)配列番号:18−22、31−36、39−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を宿主に投与し;そしてb)サンプル中のT細胞が薬剤を認識するか否かをインビトロにて決定して、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示すことを含む。
【0033】
本発明は、組成物がセリアック病を引き起こすことができるか否かを決定する方法も提供し、トランスグルタミナーゼによりオリゴペプチド配列に修飾され得る蛋白質が組成物中に存在するか否かを決定することを含み、蛋白質の存在が、組成物がセリアック病を引き起こし得ることを示す。
【0034】
本発明は、上で定義された薬剤を認識するT細胞のアンタゴニストを同定する方法も提供し、候補物質をT細胞と接触させ、そして当該物質がT細胞が抗原特異的応答を受ける能力の低下を引き起こすか否かを決定することを含み、上記能力のそのようなあらゆる低下の検出が、上記物質がアンタゴニストであることを示す。
【0035】
本発明は、上記の方法の何れかを実施するためのキットも提供し、上で定義された薬剤及びT細胞によるペプチドの認識を検出する手段を含む。
【0036】
本発明は、セリアック病の治療用の生成物を同定する方法も提供し、セリアック病を有するか又はセリアック病に感受性の候補物質を哺乳類に投与し、そして物質が哺乳類においてセリアック病を予防又は治療するか否かを決定することを含み、セリアック病の予防又は治療が、当該物質が治療用生成物であることを示す。
【0037】
本発明は、上で定義されたコーディング配列によりコードされる蛋白質の生産のためのプロセスも提供し、a)上記蛋白質の発現を可能にする条件下で上記細胞を培養し;そし
て、b)発現された蛋白質を回収することを含む。
【0038】
本発明は、トランスジェニック植物細胞を得るための方法も提供し、トランスジェニック植物細胞を供給するために上で定義されたベクターにより植物を形質転換することを含む。
【0039】
本発明は、初代トランスジェニック植物を得るための方法も提供し、トランスジェニック植物を提供するために上で定義されたベクターにより形質転換されたトランスジェニック植物細胞を再生することを含む。
【0040】
本発明は、トランスジェニック植物の種子を得るための方法も提供し、上で定義されたとおりに取得可能なトランスジェニック植物からトランスジェニック種子を得ることを含む。
【0041】
本発明は、トランスジェニック子孫植物を得るための方法も提供し、上で定義された方法により取得可能な初代トランスジェニック植物から第2世代のトランスジェニック子孫植物を得て、そして任意に、そうして得られた第2世代の子孫から一つ又はそれ以上の世代のトランスジェニック植物を得ることを含む。
【0042】
本発明は、上記の方法の何れかにより取得可能な、トランスジェニック植物細胞、植物、植物種子又は子孫植物も提供する。
【0043】
本発明は、上記の植物細胞を含むトランスジェニック植物又は植物種子も提供する。
【0044】
本発明は、上で定義されたトランスジェニック植物細胞、初代植物、植物種子又は子孫から取得可能な上記植物細胞を含む、トランスジェニック植物細胞カルスも提供する。
【0045】
本発明は、収穫生成物を得るための方法も提供し、上記の方法の何れかに従い植物から収穫生成物を回収し、そして任意にさらに回収された生成物を加工することを含む。
【0046】
本発明は、上で定義された蛋白質を含む食物も提供する。
発明の詳細な説明
用語「セリアック病」は、グルテンの感度を変更することにより引き起こされる症状の範囲を包含し、小腸の粘膜により特徴付けられる重度の形態(過形成絨毛萎縮)及び温和な兆候により特徴付けされる他の形態を含む。
【0047】
上で言及された個体(individual)(診断又は治療のコンテクストにおいて)はヒトである。彼らはセリアック病を有するか(兆候があるか又は無兆候)又はそれを有する疑いがある。彼らはグルテンダイエット中であってよい。彼らは急性の相の応答にあってよい(例えば、彼らはセリアック病を有してよいが、彼らがグルテンフリーダイエットを14から28日間行って最後の24時間はグルテンのみを摂取した)。
【0048】
個体は、セリアック病に感受性であってよく、例えば遺伝的に感受性であってよい(例えば、セリアック病の親戚を有するか又はセリアック病に対する疾病素因を引き起こす遺伝子を所有する個体により決定される)。
薬剤
薬剤は、典型的にはペプチドであり、例えば7から50アミノ酸の長さ、例えば10から40、又は15から30アミノ酸の長さである。
【0049】
配列番号:1はPQPELPYである。配列番号:2はQLQPFPQPELPYPQ
Sである。配列番号:3は表1に示され、そしてA−グリアジンの全配列である。配列番号:1の4位(配列番号:2の9位に均等)のグルタミン酸は、A−グリアジンのトランスグルタミナーゼ処理により生成される。
【0050】
薬剤は、配列番号:1又は2により表されるペプチド又はグリアジン蛋白質由来の単離されたオリゴペプチドである配列番号:1を含む配列を含むエピトープ;又は配列番号:3の相同体である天然のグリアジン蛋白質からのこれら配列の均等物であってよい。即ち、上記エピトープは配列番号:3により表される蛋白質の誘導体であってよい。そのような誘導体は、典型的には、グリアジンの断片、又は蛋白質全体又は断片の変異誘導体である。よって、発明のエピトープは、この天然のグリアジン蛋白質全体を含まず、そして他の天然のグリアジン全体を含まない。
【0051】
上記エピトープは、即ち、A−グリアジンの断片(例えば、配列番号:3)であり、配列番号:1の配列を含み、トランスグルタミナーゼにより処理すること(完全又は部分的)により得られるものであって、即ち、1、2、3又はそれ以上のグルタミンがグルタミン酸に置換されている(配列番号:1内での置換を含む)。
【0052】
そのような断片は、表1に示す配列番号:3の55から70、58から73、61から77位により表される配列であるか又はそれらの配列を含んでよい。典型的には、そのような断片は、配列番号:1又は2により表されるペプチドがセリアック病の患者からのサンプルを用いて本明細書にて記載されたアッセイの何れかにおいて認識されるのと少なくとも同じ程度にてT細胞により認識される。
【0053】
さらに、上記薬剤は、配列番号:18−22、31−36、39−44、及び46の何れかにより表されるペプチド、又は配列番号:18−22、31−36、39−44、及び46の何れかに相当する配列を含む蛋白質(例えば、グリアジンをトランスグルタミナーゼで処理した後に配列番号:18−22、31−36、39−44、及び46の何れかを含むグリアジンの断片)。そのような配列の生物学上活性な断片も発明の薬剤である。配列番号:18−22、31−36、39−44、及び46又はこれらの配列の類似体の何れかの配列均等物も、発明の薬剤である。
【0054】
上記エピトープが別のグリアジン(例えば、本明細書にて言及されたグリアジン蛋白質の何れか又はセリアック病を引き起こす何れかのグリアジン)由来の上記エピトープ(断片を含む)に均等な配列を含む場合、そのような均等配列は、トランスグルタミナーゼにより典型的に処理された(完全又は部分的)グリアジン蛋白質の断片に相当する。他のグリアジン蛋白質の配列を、オリジナルのエピトープが由来するグリアジン、例えば、配列番号:3と整列させる(aligning)ことにより(例えば、本明細書にて言及されたプログラムの何れかを用いて)、そのような均等ペプチドを決定することができる。トランスグルタミナーゼは市販されている(Sigma T−5398)。表4は、適当な均等配列の僅かな例を提供する。
【0055】
類似体である薬剤は、(i)又は(ii)を認識するTCRにより認識され得る。よって、一般には、上記類似体が(i)又は(ii)の存在下、典型的には抗原提示細胞(APC)(例えば、本明細書にて言及されたAPCsの何れか)の存在下でも、T細胞に添加された場合、当該類似体は(i)又は(ii)とそのような系において競合し得る。
【0056】
上記類似体は(i)又は(ii)を認識するTCRを結合することができるものであってよい。そのような結合は、標準の技術により試験することができる。そのようなTCRsは(例えば、本発明の方法を用いて)(i)又は(ii)を認識することが示されたT細胞から単離することができる。上記類似体のTCRsへの結合の証明は、類似体に結合
する物質、例えば類似体に対する抗体への類似体の結合をTCRsが阻害するか否かを決定することにより、のちに示すことができる。典型的には、上記類似体は、結合アッセイのそのような阻害においてクラスII MHC分子(例えば、HLA−DQ2)に結合する。
【0057】
典型的には、上記類似体は(i)又は(ii)のTCRへの結合を阻害する。この場合、類似体存在下でTCRを結合できる(i)又は(ii)の量は低下する。これは、類似体がTCRを結合し、よってTCRへの結合に関して(i)又は(ii)と競合するためである。
【0058】
上記の結合実験に使用するための細胞は、セリアック病の患者から、例えば本発明の方法を用いて単離することができる。
【0059】
類似体の他の結合特性は(i)又は(ii)のそれと同じであってよく、即ち典型的には、類似体はペプチドが結合するのと同じMHCクラスII分子に結合する(HLA−DQ2又は−DQ8)。類似体は、典型的には(i)又は(ii)に特異的な抗体に結合し、即ち、(i)又は(ii)のそのような抗体への結合を阻害する。
【0060】
類似体は、典型的にはペプチドである。それは、(i)又は(ii)と少なくとも70%の相同性、好ましくは少なくとも80%、90%、95%、97%又は99%の相同性を、例えば少なくとも15又はそれ以上(例えば類似体及び/又は(i)又は(ii)の完全長、又はTCRに接触するか又はMHC分子を結合する領域にわたり)連続するアミノ酸の領域にわたり有してよい。蛋白質の相同性を測定する方法は当業界公知であり、当業者には本コンテクストにおいて、相同性がアミノ酸同一性に基づいて計算されることが理解される(ときに、「ハードな相同性」と呼ばれる)。
【0061】
例えば、UWGCGパッケージは相同性を計算するために使用できるBESTFITプログラムを提供している(例えば、そのデフォルトセッティングに使用される)(Devereux et al(1984)Nucleic Acids Research 12,p387−395)。PILEUP及びBLASTアルゴリズムは相同性を計算するか又は配列をラインアップするのに使用することができ(典型的にはそれらのデフォルトセッティング上で)、Altschul S.F.(1993)J Mol Evol
36:290−300;Altschul,S.F et al(1990)J Mol 215:403−10に記載されるとおりである
BLAST分析を実施するためのソフトウエアは、例えば、「www.ncbi.nlm.nih.gov/」においてインターネットを通して全世界のウエブ上でNational Center for Biotechnology Informationを通して公的に利用可能である。このアルゴリズムは、データベース配列中で同じ長さのワードと並べられたときに幾つかの正の値の閾値スコアに適合するか又は当該値を満たす質問配列中の長さWの短いワードを同定することにより、高いスコアリングの配列対(HSPs)を最初に同定することを含む。Tは隣接ワードスコア閾値を意味する(Altschul et al,前出)。これらの最初の隣接ワードのヒットは、それらを含むHSPsを見つけるための検索を始めるための種として作用する。蓄積したアラインメントスコアがその最大に達成された値から量Xだけ低下する場合;一つ又は複数の負のスコアリング剰余アラインメントの蓄積のため、蓄積したスコアがゼロ又はそれ以下になった場合;又は何れかの配列の末端に到達した場合、各方向におけるワードヒットに関する延長が停止する。BLASTアルゴリズムのパラメーターW,T及びXは、アラインメントの感度と速度を決定する。BLASTプログラムはデフォルトとして11のワードの長さ(W)、50のBLOSUM62スコアリングマトリックス(Henikoff and Henikoff(1992)Proc.Natl.Acad.Sci.USA 89:
10915−1を参照)アラインメント(B)、10の期待値(E),M=5,N=4,及び両方の鎖の比較を用いる。
【0062】
BLASTアルゴリズムは2つの配列の間の類似性の統計分析を実施する;例えば、Larlin and Altschul(1993)Proc.Natl.Acad.Sci.USA 90:5873−5787を参照されたい。BLASTアルゴリズムにより提供される類似性の一つの測定値は、最小合計確率(P(N))であり、2つのヌクレオチド又はアミノ酸配列間で偶然に適合する確率の指標を提供する。例えば、第1の配列の第2の配列に対する比較において最小合計確率が約1より低く、好ましくは約0.1より低く、より好ましくは約0.01より低く、そしてもっと好ましくは約0.001より低いなら、配列が他方の配列に類似であると考えられる。
【0063】
相同なペプチド類似体は、(i)又は(ii)と1、2、3、4、5、6、7、8又はそれより多い変異だけ異なるのが典型である(置換、欠失又は挿入であってよい)。これらの変異は、相同性を計算することに関して上記の領域の何れかを横切って測定されてよい。置換は、好ましくは「保存的」である。これらは以下の表により定義される。第2列の同じブロックのアミノ酸、及び好ましくは第3の列の同じラインのアミノ酸を、互いに置換してよい。
【0064】
【表1】
【0065】
典型的には、MHC分子を結合するのに寄与するか又はTCRによる認識に必須の、(i)又は(ii)中のアミノ酸に均等な位置における類似体中のアミノ酸は同じか又は保存されている。
【0066】
典型的には、類似体ペプチドは一つ又は複数の修飾を含み、天然の翻訳後修飾又は人工修飾であってよい。修飾は科学的なモイエティ(典型的には、例えばC−H結合の水素の置換による)、例えばアミノ、アセチル、ヒドロキシ又はハロゲン(例えば、フッ素)基又は糖質基を提供してよい。典型的には、修飾はN又はC末端上に存在する。
【0067】
類似体は、一つ又は複数の天然アミノ酸、例えば天然アミノ酸とは側鎖が異なるアミノ酸を含んでよい。一般に、非天然アミノ酸はN末端及び/又はC末端を有する。非天然アミノ酸はL−又はD−アミノ酸であってよい。
【0068】
類似体は、典型的には、(i)又は(ii)と実質類似の形態、サイズ、柔軟性又は電子配置を有する。それは、典型的には、(i)又は(ii)の誘導体である。一つの態様において、類似体は、配列番号:1又は2の配列を含む融合蛋白質であるか又は本明細書にて言及された他のペプチドの何れか;及び非グリアジン配列である。
【0069】
一つの態様において、類似体は、MHCクラス分子に結合した(i)又は(ii)であるか、又は、を模倣する。2、3、4、又はそれ以上のそのような複合体を、例えば、ビオチン/ストレプトアビジンに基づく系を用いて互いに対合させるか又は結合させてよく、典型的には2、3、又は4つのビオチン標識MHC分子がストレプトアビジンモイエティに結合する。この類似体は、典型的には、(i)又は(ii)/MHCクラスII分子のTCR又は当該複合体に特異的な抗体への結合を阻害する。
【0070】
類似体は、典型的には、抗体又は抗体の断片、例えばFab又は(Fab)2断片である。類似体は固相上に固定化され、特に、MHC分子に結合したペプチドを模倣する類似体の場合である。
【0071】
類似体は、典型的には、コンピューター手段によりデザインされて、当業界公知の方法を用いて合成される。或いは、類似体は、化合物のライブラリーから選択できる。ライブラリーはコンビナトリアルライブラリー、ディスプレイライブラリー、例えばファージディスプレイライブラリーであってよい。化合物のライブラリーはMHCクラスII分子に結合した形態、例えばHLA−DQ2又は−DQ8においてディスプレイライブラリー中で発現されてよい。類似体は、一般に、結合特性(i)又は(ii)を模倣するそれらの能力に基づいてライブラリーから選択される。即ち、それらはTCR又は(i)又は(ii)を認識する抗体を結合する能力に基づいて選択してよい。
【0072】
典型的には、類似体は、薬剤(i)又は(ii)の何れかと少なくとも同じ程度、例えば均等なエピトープと少なくとも同じ程度まで、好ましくは配列番号:2により表されるペプチドと同じ程度までT細胞により認識され、本明細書にて記載されたアッセイの何れかにおいて認識され、典型的には、セリアック病の患者由来の細胞を用いる。類似体は、インビボにおいてこれらの程度まで認識されてよく、即ち、本明細書にて言及された薬剤の何れかと少なくとも同じ程度までセリアック病の兆候を誘導することができてよい(例えば、ヒト患者又は動物モデルにおいて)。
【0073】
類似体は、発明のエピトープを認識するT細胞受容体により候補の物質が認識されるか否かを決定することを含み、物質の認識が物質が類似体を認識することを示す方法において同定されてよい。そのようなTCRsは本明細書にて言及されたTCRsの何れかであってよく、T細胞上に存在してよい。本明細書にて言及された如何なる適切なアッセイも類似体を同定するのに使用することができる。一つの態様において、この方法はインビボにおいて実施される。上記のとおり、好ましい類似体はペプチド配列番号:2と少なくとも同じ程度まで認識され、よって、上記の方法はこの程度まで認識される類似体を同定するのに使用してよい。
【0074】
一つの態様において、上記方法は、発明のエピトープの認識を候補配列が阻害するか否かを決定することを含み、認識の阻害が、物質が類似体であることを示す。
【0075】
上記薬剤は、(i),(ii)又は(iii)により定義される少なくとも2、5、10又は20の薬剤を含む生成物であってよい。典型的には、上記組成物は、異なるグリアジン、例えば本明細書にて言及されたグリアジンの種又はバラエティ又は種類の何れかからの発明のエピトープ(又は均等な類似体)を含む。好ましい組成物は、本明細書にて言及された種又はバラエティの何れかの中に存在するグリアジンのすべて、又は2、3、4、又はそれ以上の本明細書にて言及された種からの(コムギ、ライムギ、オオムギ、エンバク、及びライコムギからなる群から選択される種のパネルから)、発明の少なくとも一つのエピトープ、又は均等類似体を含む。即ち、薬剤は単価又は多価であってよい。
診断
上記のとおり、発明の診断方法は、薬剤を結合するT細胞の検出又は薬剤を認識する抗体の検出に基づいてよい。
【0076】
上記の方法(上記の使用を含む)において薬剤を認識するT細胞は、一般に、インビボにおいて予めグリアジンに対して感作されたT細胞である。上記のとおり、そのような抗原−経験T細胞は、末梢血中に存在することがわかった。
【0077】
上記の方法において、T細胞はインビトロ又はインビボにおいて薬剤と接触させることができ、そしてT細胞が薬剤を認識するか否かを決定することをインビトロ又はインビボにて実施することができる。即ち、発明は、ヒトのからだで実施される診断方法においての使用のための薬剤を提供する。異なる薬剤が、そのような方法における、同時、別々、又は連続の使用のために提供される。
【0078】
インビトロの方法は、薬剤が添加された水性溶液内で実施されるのが典型である。当該溶液はT細胞も含む(及び特定の態様においては、以下に論じられるAPCsも)。本明細書にて使用される用語「接触させる」は、特定の物質を溶液に加えることを含む。
【0079】
T細胞が薬剤を認識するか否かを決定することは、一般には、薬剤の存在下でT細胞の状態の変化を検出するか、又はT細胞が薬剤を結合するか否かを決定することにより、達成される。状態の変化は、一般に、TCRが薬剤に結合した後に、T細胞の抗原特異的機能活性により引き起こされる。状態の変化は、T細胞の内部(例えば、蛋白質の細胞内発現の変化)又は外部(例えば、分泌された配列の検出)で測定してよい。
【0080】
T細胞の状態の変化は、T細胞からの物質、例えばサイトカイン、特にIFN−γ、IL−2又はTNF−αの分泌の開始又は増加であってよい。IFN−γの分泌の決定が特に好ましい。当該物質は、特異的結合薬剤へのそれの結合を可能にさせ、次に特異的結合薬剤/物質の複合体の存在を測定することにより検出されるのが典型である。特異的結合薬剤は、典型的には、抗体、例えばモノクローナル抗体である。サイトカインに対する抗体は、市販されており、或いは標準技術を用いて作成することができる。
【0081】
典型的には、特異的結合薬剤は、固相支持体上に固定化される。物質を固相支持体上に固定化させた後に、任意に洗浄することにより、薬剤に非特異的に結合した物質を除去することができる。薬剤/物質複合体は複合体を結合する第2の結合薬剤を用いることにより検出してよい。典型的には、第2薬剤は、第1薬剤を結合する部位とは異なる部位において物質を結合する。第2薬剤は、好ましくは抗体であり、そして、検出可能な標識により直接又は間接に標識される。
【0082】
即ち、第2薬剤は、検出可能な標識により直接又は間接に典型的には標識された第3薬剤により検出してよい。例えば、第2薬剤はビオチンモイエティを含んでよく、ストレプトアビジンモイエティ及び典型的にはアルカリホスファターゼを検出可能な標識として含む第3の薬剤による検出を可能にさせる。
【0083】
一つの態様において、使用される検出系は、WO98/23960に記載されたエクスビボELISPOTアッセイである。そのアッセイにおいて、T細胞から分泌されるIFN−γは、固相支持体に固定化された第1のIFN−γ特異的抗体により結合される。結合されたIFN−γは、次に、検出可能な標識により標識された第2のIFN−γ特異的抗体を用いて検出される。そのような標識された抗体はMABTECH(ストックホルム、スエーデン)から得ることができる。使用可能な他の検出可能な標識は以下に論じられる。
【0084】
測定され得るT細胞の状態の変化は、T細胞による物質の取り込み、例えばチミジンの取り込みの増加であってよい。状態の変化は、T細胞のサイズ、又はT細胞の増殖の増加、又はT細胞上の細胞表面マーカーの変化であってよい。
【0085】
一つの態様において、状態の変化は、蛋白質の細胞内発現の変化、例えば上記サイトカインの何れかの細胞内発現の増加を測定することにより、検出される。そのような細胞内変化は、特定の様式にて発現された蛋白質を結合するモイエティにT細胞内部を接触させることにより検出してよく、フローサイトメトリーによるT細胞の分離を可能にさせる。
【0086】
一つの態様において、TCRを結合するとき、薬剤はMHCクラスII分子(典型的にはHLA−DQ2又は−DQ8)に結合し、典型的には抗原提示細胞(APC)の表面上に存在する。しかしながら、本明細書にて言及されたとおり、他の薬剤はMHC分子にも結合する要求なしにTCRを結合することができる。
【0087】
一般に、上記の方法において接触させたT細胞は固体の血液から採取されるが、T細胞を含む他の種類のT細胞を使用することができる。サンプルは、直接アッセイに添加するか又は最初に加工してよい。典型的には、加工は、例えば、水又はバッファーによるサンプルの希釈を含んでよい。典型的には、サンプルを1.5から100倍、例えば2から50倍又は5から10倍希釈する。
【0088】
加工は、サンプルの成分の分離を含んでよい。典型的には、単核細胞(MCs)をサンプルから分離する。MCsはT細胞とAPCsを含むことになる。即ち、上記の方法において、分離されたMCs内に存在するAPCsは上記ペプチドをT細胞上に提示し得る。別の態様においては、T細胞のみ、例えばCD4 T細胞のみがサンプルから精製され得る。PBMCs,MCs、及びT細胞は、当業界公知の技術、例えばLalvani et al(1997)J.Exp.Med.186,p859−865に記載された技術を用いてサンプルから分離することができる。
【0089】
一つの態様において、上記のアッセイにおいて使用されるT細胞は、未加工の形態であるか、又は希釈されたサンプルであるか、又は新たに単離されたT細胞であり(例えば、新たに単離されたMCs又はPBMCsの形態)、エクスビボにて直接使用され、即ち、それらは上記方法において使用される前に培養されない。即ち、T細胞は抗原特異的様式にてインビトロにおいて再刺激されなかった。しかしながら、T細胞は、使用の前に、例えば一つ又は複数の薬剤及び一般には外来の成長促進サイトカインの存在下で培養することができる。培養の間、薬剤は典型的にはAPCs,例えば上記方法で使用されるAPCの表面上に存在する。T細胞の前培養は上記方法の感度の増加を導くかもしれない。即ち、T細胞は、細胞系、例えば短期間の細胞系に変換することができる(例えば、Ota et al(1990)Nature 346,p183−187に記載されたとおり)。
上記の方法に典型的に存在するAPCはT細胞と同じ個体から又は別の宿主からであってよい。APCは天然のAPC又は人工のAPCであってよい。APCは上記ペプチドを細胞に提示することができる細胞である。それは典型的にはB細胞、樹状細胞又はマクロファージである。それは典型的にはT細胞と同じサンプルから分離され、典型的にはT細胞と同時精製される。即ち、APCはMCs又はPBMCs上に存在してよい。APCは典型的には新たに単離されたエクスビボ細胞又は培養細胞である。それは細胞系の形態、例えば短期間細胞系又は不死の細胞系であってよい。APCはその表面上に空のMHCクラスII分子を発現してよい。
【0090】
上記の方法において、一つ又は複数の(異なる)薬剤を使用してよい。典型的には、サンプル由来のT細胞をアッセイ中において試験を意図する全ての薬剤と共に置くことがで
きるか、又は各々が一つ又は複数の薬剤を含む別々のアッセイ中に分配して置くことができる。
【0091】
本発明は、同時、分離又は連続用途のための(例えばインビボ用途)本明細書にて言及された薬剤の何れか2つ又はそれ以上のような薬剤も提供する(例えば、上で論じられた組成物中に存在する薬剤の組み合わせ)。
【0092】
一つの態様においては、薬剤それ自体を直接T細胞及びAPCsを含むアッセイに加える。上で論じられたとおり、そのようなアッセイにおいてT細胞とAPCsはMCsの形態であり得る。APCsによる提示に関する要求なしにT細胞により認識され得る薬剤を用いる場合、APCsは必要ない。MHC分子に結合させたオリジナルの(i)又は(ii)を模倣した類似体がそのような薬剤の例である。
【0093】
一つの態様において、上記薬剤はT細胞の不在下でAPCに提供される。APCは次に、典型的には薬剤をその表面に提示させた後に、T細胞に提供される。上記ペプチドはAPCの内部に拾い上げられて提示されるか、又はAPCの内部に入り込まずに単に表面上に拾い上げられる。
【0094】
薬剤がT細胞と接触する期間は、ペプチドの認識を決定するのに使用される方法しだいで変更される。典型的には105から107、好ましくは5x105から106のPBMCを各アッセイに添加する。薬剤を直接アッセイに添加する場合、その濃度は10−1から10−3μg/ml、好ましくは0.5から50μg/ml又は1から10μg/mlである。
【0095】
典型的には、T細胞を薬剤とインキュベートする時間の長さは、4から24時間、好ましくは6から16時間である。エクスビボにてPBMCを使用する場合、0.3x106のPBMCsを10μg/mlのペプチド中で12時間37℃においてインキュベートできることを発見した。
【0096】
T細胞による薬剤の認識の決定は、薬剤のT細胞への結合を測定することにより行ってよい(これは本明細書にて論じられる何れかの適当な結合アッセイを用いて実施できる)。典型的には、薬剤を結合するT細胞は、例えばFACSマシンを用いてこの結合に基づいて細胞を分離することができる。薬剤を認識するT細胞の存在は、薬剤を用いて分離される細胞の頻度が「対照」値を超える場合に生じるとみなされる。抗原を経験したT細胞の頻度は106中1から103中1であり、よって、分離された細胞が抗原経験T細胞であるか否かを決定することができる。
【0097】
T細胞による薬剤の認識の決定はインビボにおいて測定してよい。典型的には、薬剤を宿に投与して、次に、薬剤の認識を示す応答を測定してよい。薬剤は、典型的には、皮内又は表皮投与である。薬剤は、典型的には、皮膚の外部に接触させることにより投与され、そしてプラスター又は包帯の補助により当該部位に保持されてよい。或いは、薬剤は針により、例えば注射により投与してよいが、他の方法、例えば弾道(ballistics)(例えば、核酸をデリバリーするために使用される弾道技術)により投与することもできる。EP−A−0693119は、薬剤を投与するために典型的に使用できる技術を記載する。典型的には、0.001から1000μg、例えば0.01から100μg又は0.1から10μgの薬剤を投与する。
【0098】
一つの態様において、インビボにおいて薬剤を提供することができる生成物を投与できる。即ち、薬剤を発現できるポリヌクレオチドを投与することができ、典型的には、薬剤の投与のための上で記載された様式の何れかによる。ポリヌクレオチドは、典型的には、
以下に論じられる、発明により提供されるポリヌクレオチドの特性の何れかを有する。典型的には、0.001から1000μg、例えば0.01から100μg又は0.1から10μgのポリヌクレオチドを投与する。
【0099】
皮膚に投与される薬剤の認識は、典型的には、投与部位における炎症の発症により示唆される(例えば、硬化、紅斑又は浮腫)。これは、一般に、当該部位における目視試験により判断される。
【0100】
薬剤を結合する抗体の検出に基づく診断法は、典型的には、個体からのサンプル(本明細書にて言及されたサンプルの何れかで、任意に本明細書にて言及された様式の何れかにより加工された)を薬剤と接触させ、そしてサンプル中の抗体が薬剤を結合するか否かを決定することにより実施され、そのような結合が、個体がセリアック病を有するか又は感受性であるかを示す。結合アッセイのあらゆる適当なフォーマット、例えば本明細書にて言及されたあらゆるそのようなフォーマットを使用してよい。
治療
本明細書にて記載された免疫上優勢なエピトープ及び他のエピトープの同定は、このエピトープを認識するT細胞を標的とする治療用生成物が作成されるのを可能にさせる(そのようなT細胞はグリアジンに対する免疫応答に参加するものである)。これらの発見は、抗体又はエピトープに対するT細胞応答を抑圧(寛容)することによりセリアック病の予防又は治療を可能にもする。
【0101】
発明の特定の薬剤は、発明のエピトープを認識するTCRを結合し(上で論じられた結合アッセイの何れかを用いて測定されるとおり)、そしてTCRを有するT細胞の寛容を導く。そのような薬剤は、任意にキャリアーと共に、従ってセリアック病を予防又は治療するのに使用することができる。
【0102】
一般に、寛容は、(TCRによりされた後に)T細胞の抗原特異的機能活性を導き得る同じペプチドにより導かれ得る(例えば、本明細書にて言及されたそのような活性、例えばサイトカインの分泌)。そのような薬剤は「寛容」コンテクストにおいて免疫系に対して提示される時に寛容を導く。
【0103】
寛容は免疫系によるT細胞又は抗体エピトープの認識の低下を導く。T細胞エピトープの場合、これは、エピトープを認識するT細胞の欠失又はアネルギーにより導かれ得る。即ち、エピトープに応答するT細胞活性(例えば、本明細書にて言及された適当なアッセイにおいて測定されたとおり)が低下する。抗体応答の寛容は、エピトープが投与されたときに、エピトープに対する特異的抗体の量の低下が生じることを意味する。
【0104】
そのようなコンテクストにおいて免疫系に対して抗原を提示する方法は公知であり、例えば、Yoshida et al.Clin.Immunol.Immunopathol 82,207−215(1997),Thuau et al.Clin Exp.Immunol.109,370−6(1997)、及びWeiner et al.Res.Immunol.148,528−33(1997)に記載されている。特に、特別の投与経路が寛容を導き得て、例えば、経口、鼻内又は腹膜内である。寛容は樹状細胞及びテトラマー提示ペプチドにより達成してもよい。寛容を導く特定の生成物を個体に投与してよい(例えば、薬剤も含む組成物中で)。そのような生成物は、サイトカイン、例えばTh2応答を好むサイトカインを含む(例えば、IL−4,TGF−β又はIL−10)。生成物又は薬剤は、寛容を導く用量にて投与してよい。
【0105】
発明は、T細胞のアンタゴニストとして作用することができる配列を含む蛋白質を提供する(そのT細胞は薬剤を認識する)。そのような蛋白質及びそのようなアンタゴニスト
はセリアック病を予防又は治療するのに使用することもできる。アンタゴニストはT細胞応答の低下を導く。一つの態様において、アンタゴニストはT細胞のTCRに結合する(一般に、HLA−DQ2又は−DQ8との複合体の形態にて)が、正常な機能活性化を導く代わりに、T細胞内シグナリングカスケードを通して異常なシグナルを通過させることを導き、T細胞が低下した機能活性を持つように導く(例えば、エピトープの認識に応答して、典型的には本明細書にて言及された適当なアンタゴニストの何れかにより測定される)。
【0106】
一つの態様において、アンタゴニストはエピトープとMHCプロセシング及び提示の経路の成分への結合、例えばMHC分子(典型的には、HLA−DQ2又は−DQ8)に関して競合する。即ち、アンタゴニストは、HLA−DQ2又は−DQ8(即ち、このMHC分子により提示されるペプチドである)、例えばペプチドTP(表10)又はその相同体を結合してよい。
【0107】
拮抗作用を導く方法は、当業界公知である。一つの態様において、アンタゴニストは上で言及されたエピトープの相同体であり、そして配列の何れか、結合又は薬剤の他の特性を有してよい(特に、類似体)。アンタゴニストは、典型的には、上記のエピトープの何れかとは(T細胞内で正常な抗原特異的機能を導くことができる)、1、2、3、4又はそれ以上の変異だけ(各々は置換、挿入又は欠失であってよい)異なる。そのようなアンタゴニストは当業界において「変更されたペプチドリガンド」又は「APL」と呼ばれる。変異は、典型的には、TCRに接触するアミノ酸位置においてである。
【0108】
アンタゴニストはA−グリアジンのアミノ酸65から67により表される配列に均等な配列内の置換によりエピトープと異なってよい(表9に示すようなアンタゴニスト)。即ち、好ましくは、アンタゴニストは64、65又は67位の均等なものにおいて置換を有する。好ましくは、置換は、64W,67W,67M又は65Tである。
【0109】
個体における発明のエピトープに対するT細胞の免疫応答はポリクローナルであるから、1より多いアンタゴニストが投与されることにより、異なるTCRsを有する応答のT細胞の拮抗作用を導いてよい。よって、アンタゴニストは少なくとも2、4、6又はそれより多いアンタゴニストを含む組成物中で投与されてよく、各々が異なるT細胞と拮抗する。
【0110】
発明は、(上記薬剤を認識する)T細胞のアンタゴニストを同定する方法も提供し、候補物質をT細胞と接触させ、そしてT細胞が抗原特異的応答を経る能力の低下を当該物質が導くか否かを検出することを含み(例えば、本明細書にて言及された何れかの適当なアンタゴニストを用いて)、上記能力のあらゆるそのような低下の検出が物質がアンタゴニストであることを示す。
【0111】
一つの態様において、アンタゴニスト(特定のエピトープに対する複数のアンタゴニストの組み合わせを含む)又は寛容(T細胞及び抗体寛容)薬剤は、発明の異なるエピトープに対して、例えば1より多い物質を含む生成物である薬剤に関して上で論じられたエピトープの組み合わせに対して、拮抗するか又は寛容する少なくとも2、4、6又はそれより多いアンタゴニスト又は薬剤を含む組成物中に存在する。
組成物がセリアック病を引き起こすか否かを試験する
上で論じられたとおり、発明は、組成物がセリアック病を引き起こすことができるか否かを決定する方法を提供し、トランスグルタミナーゼにより発明の薬剤又はエピトープを含む配列に修飾され得る蛋白質配列の存在を検出することを含む(そのようなトランスグルタミナーゼ活性はヒトの腸のトランスグルタミナーゼ活性であってよい)。典型的には、これは、特定の様式にて配列に結合するモイエティを組成物に接触させる結合アッセイ
を用いて実施され、そして配列/モイエティ複合体の形成を検出して薬剤の存在を確認するのに使用する。そのようなモイエティは、本明細書にて言及された何れかの適当な物質(又は物質の種類)であってよく、そして典型的には特異的抗体である。結合アッセイのあらゆる適当なフォーマットを使用することができる(例えば、本明細書にて言及されたもの)。
【0112】
一つの態様において、上記組成物を、異なるグリアジン由来の発明のエピトープに特異的な少なくとも2、5、10又はそれより多い抗体、例えば、1よりも多い物質を含む生成物である発明の薬剤に関して上で論じられたエピトープの組み合わせを認識することができる抗体のパネルに接触させる。
【0113】
上記組成物は、典型的には、セリアック病を引き起こし得るグリアジンを発現する植物由来の材料を含む(例えば、本明細書にて言及されたグリアジン又は植物の何れも)。そのような材料は植物の一部、例えば収穫された作物(例えば、種子)であってよい。材料は植物材料の加工された生成物(例えば、本明細書にて言及されたあらゆるそのような生成物)、例えばグリアジンを含む穀粉又は食物であってよい。食物材料の加工及び適当な結合アッセイにおける試験は日常的であり、例えばKricka LJ,J.Biolumin.Chemilumin.13,189−93(1998)に言及されたとおりである。
結合アッセイ
本明細書にて言及された2つの物質間の結合の決定は、結合に際して変化する一方又は両方の物質の特性、例えば分光学上の変化を測定することにより実施してよい。
【0114】
結合アッセイフォーマットは「バンドシフト」系であってよい。これは、一つの物質(例えば、候補物質)の存在がゲル電気泳動の間に他の物質の進行を進めさせるか又は遅らせるかを決定することを含む。
【0115】
上記フォーマットは、一つの物質が他の物質、例えば特異的抗体に結合することが知られている薬剤に対する他の物質の結合を阻害できるか否かを決定する、競合結合法であってよい。
変異体グリアジン蛋白質
発明は、本発明の配列又はそのような配列を提供するためにトランスグルタミナーゼにより修飾され得る配列が変異することによりもはやエピトープを認識するT細胞応答を導かないか又はエピトープを認識するT細胞応答により認識されないような、グリアジン蛋白質を提供する。このコンテクストにおいて、用語認識はT細胞の正常な(非拮抗)抗原−特異的機能活性が生じるような様式におけるエピトープのTCR結合を意味する。
【0116】
他のグリアジンにおいて均等なエピトープを同定する方法は、上で論じられている。変異したグリアジンの野生型は、セリアック病を引き起こすものである。そのようなグリアジンは、例えば配列番号:3の全域又は配列番号:3の15、30、60、100又は200の連続するアミノ酸にわたり上記の程度(類似体に関して)、配列番号:3に相同性を有してよい。同様に、他の非−A−グリアジンに関しては、そのグリアジンの変異体と天然形態の間に相同性が存在することになる。他の天然グリアジン蛋白質の配列は当業界公知である。
【0117】
変異したグリアジンはセリアック病を引き起こさないか、又はセリアック病の兆候の低下を導く。典型的には、エピトープがT細胞応答を誘導する能力を変異が低下させる。変異したエピトープはHLA−DQ2又は−DQ8に対する低下した結合、APCにより提示される能力の低下、又は薬剤を認識するT細胞に結合するか又は当該T細胞により認識される能力(即ち、抗原−特異的機能活性)の低下を有してよい。変異したグリアジン又
はエピトープは、よって、発明の診断の側面に関して、本明細書にて言及されたアッセイの何れかにおいて認識を示さないか又は低下した認識を示すことになる。
【0118】
上記変異は、上記エピトープ中の1から3、4から6、6から10、11から15又はそれより長い長さの一つ又はそれより多い欠失、付加又は置換であってよく、例えば、配列番号:2の配列にわたるか、配列番号:18−22、31−36、39−44及び46の何れかにわたるか、又はそれらの均等物にわたる。好ましくは、上記変異グリアジンは、配列番号:1の配列内に少なくとも一つの変異を有する。好ましい変異は、A−グリアジン中の65位である(又は他のグリアジン中の均等な位置)。典型的には、この位置において天然に生じるグルタミンは、表3に示されるアミノ酸の何れか、好ましくはヒスチジン、チロシン、トリプトファン、リジン、プロリン又はアルギニンにより置換される。
【0119】
発明は、即ち、グリアジン蛋白質のエピトープ内の変異(本明細書にて論じられる何れかの配列中の変異のそのような変異)の用途も提供し、当該エピトープは本発明のエピトープであり、グリアジン蛋白質がセリアック病を引き起こす能力を低下させる。
【0120】
一つの態様において、変異した配列はアンタゴニストとして作用することができる。即ち、発明は、T細胞受容体に結合することができる配列を含む蛋白質を提供し、T細胞受容体は発明の薬剤を認識し、そして配列はそのようなT細胞受容体を認識するT細胞の拮抗作用を導くことができる。
【0121】
発明は、上記変異グリアジン蛋白質の断片である蛋白質も提供し、それらは少なくとも15アミノ酸の長さ(例えば、少なくとも30、60、100、150、200又は250アミノ酸長)であり、そしてグリアジンが認識される能力を低下させる上で論じられた変異を含む。上で言及された変異蛋白質の何れも(断片を含む)、例えば他のグリアジン又は非グリアジン蛋白質と融合蛋白質の形態で存在してもよい。
【0122】
変異グリアジン蛋白質に対する均等な野生型蛋白質は、典型的には、イネ科の単子葉植物、例えばコムギ(Triticum)属の植物、例えば、コムギ、ライムギ、オオムギ、エンバク又はライコムギに由来する。蛋白質は、典型的には、α、αβ、γ又はωグリアジンである。グリアジンはA−グリアジンであってよい。
キット
発明は、一つ又はそれより多い薬剤、及び任意にT細胞による薬剤の検出のための手段を含む、上記方法を実施するためのキットも提供する。典型的には、異なる薬剤を、同時、別々又は連続する使用のために提供する。提供、認識を検出する手段は、上で論じられた技術に基づいて検出を可能にするか又は補助する。
【0123】
即ち、上記手段は、認識後にT細胞により分泌された物質の検出を可能にしてよい。上記キットは、即ち、物質、例えば抗体のための特異的結合モイエティをさらに含んでよい。当該モイエティは、典型的には、IFN−γに特異的である。当該モイエティは典型的には固相支持体に固定化される。これは、モイエティを結合した後に、上記物質がそれを分泌するT細胞の付近に残ることを意味する。即ち、物質/モイエティ複合体の「スポット」が支持体上に形成され、各スポットが物質を分泌するT細胞を表す。スポットを定量し、そして典型的には対照に対して比較することにより、薬剤の認識の決定を可能にする。
【0124】
上記キットは、物質/モイエティ複合体を検出する手段も含む。検出可能な変化、例えば色の変化が、物質を結合後にモイエティ自身に生じる。或いは、第2のモイエティが直接又は間接に検出のために標識され、スポットの決定を可能にするように物質/モイエティ複合体を結合することを可能にしてよい。上で論じられたとおり、第2モイエティは物
質に対して特異的であってよいが、第1モイエティとは上記物質に対して異なる部位において結合する。
【0125】
固定化された支持体はウエルを有するプレート、例えばマイクロタイタープレートであってよい。各アッセイは、よって、プレート中の別々のウエル内で実施され得る。
【0126】
上記キットは、さらに、T細胞のための培地、検出モイエティ又は検出工程において使用される洗浄バッファーを含んでよい。上記キットは、さらに、サンプルからの分離に適した試薬、例えばサンプルからのPBMCs又はT細胞の分離に適した試薬を含んでよい。上記キットは、サンプルの成分の如何なる分離も無しに、サンプルからのT細胞の直接の同定を可能にするようにデザインしてよい。
【0127】
上記キットは、対照、例えば陽性対照又は陰性対照も含んでよい。陽性対照は検出系が試験されるのを可能にしてよい。即ち、陽性対照は、典型的には、上記方法の何れかにおいて薬剤の認識を模倣する。典型的には、インビトロにおいて認識を決定するためにデザインされたキットにおいては、陽性対照はサイトカインである。薬剤のインビボの認識を検出するためにデザインされたキットにおいては、陽性対照はほとんどの個体が認識すべき抗原であってよい。
【0128】
上記キットは、宿主からのT細胞を含むサンプル、例えば血液サンプルを採取するための手段も含んでよい。上記キットは、宿主からのサンプルから単核細胞又はT細胞を分離するための手段を含んでよい。
ポリヌクレオチド、細胞、トランスジェニック哺乳類及び抗体
発明は、薬剤又は変異グリアジン蛋白質を提供するための発現可能なポリヌクレオチドも提供する。典型的には、当該ポリヌクレオチドは、DNA又はRNAで、単鎖又は二重鎖である。当該ポリヌクレオチドは、好ましくは、少なくとも50塩基又は塩基対、例えば50から100、100から500、500から1000又は1000から2000又はそれより多い塩基又は塩基対を含むことになる。当該ポリヌクレオチドは、よって、配列番号:1又は2又は本明細書にて言及された他の薬剤の何れかの配列をコードする配列を含む。このコーディング配列の5’及び3’に、発明のポリヌクレオチドは対応するグリアジン遺伝子内のこれら配列に対して5’及び3’の配列又はコドンと異なる配列又はコドンを有する。
【0129】
上記ペプチドをコードする配列に対して5’及び/又は3’に、上記ポリヌクレオチドはコーディング配列又は非コーディング配列を有する。コーディング配列に対して5’及び/又は3’の配列は、上記薬剤をコードする配列の発現、例えば、転写及び/又は翻訳を助ける配列を含んでよい。上記ポリヌクレオチドは、原核又は真核細胞において上記薬剤を発現することができてよい。一つの態様において、上記ポリヌクレオチドは、哺乳類細胞、例えばヒト、霊長類又は齧歯類(例えば、マウス又はラット)細胞内で上記薬剤を発現することができる。
【0130】
発明のポリヌクレオチドは、バックグラウンドを顕著に超えるレベルで配列番号:3をコードするポリヌクレオチドに選択的にハイブリダイズしてよい。選択的ハイブリダイゼーションは、典型的には、中から高いストリンジェンシー(例えば、0.03M 塩化ナトリウム及び0.03Mクエン酸ナトリウム、約50℃から約60℃において)の条件下で達成される。しかしながら、そのようなハイブリダイゼーションは当業界公知のあらゆる適切な条件下で実施してよい(Sambrook et al(1989),Molecular Cloning:A Laboratory Manualを参照)。例えば、高いストリンジェンシーが必要なら、適切な条件は0.2xSSC、60℃を含む。低いストリンジェンシーが必要なら、適切な条件は2xSSC、60℃を含む。
【0131】
発明の薬剤又は蛋白質は、本明細書にて記載されるポリヌクレオチドによりコードされてよい。
【0132】
当該ポリヌクレオチドは、複製可能なベクターを形成するか又はベクターに取り込まれてよい。そのようなベクターは、適当な細胞中で複製することができる。当該ベクターは、発現ベクターであってよい。そのようなベクターにおいて、発明のポリヌクレオチドは、ポリヌクレオチドの発現を提供することができる制御配列に作動可能に連結される。当該ベクターは、選択可能なマーカー、例えばアンピシリン耐性遺伝子を含んでよい。
【0133】
上記ポリヌクレオチド又はベクターは細胞中に存在してよい。そのような細胞は上記ポリヌクレオチド又はベクターにより形質転換されていてよい。当該細胞は上記薬剤を発現してよい。当該細胞は上記ベクターに適合可能なように選択され、そして、例えば、原核(細菌)、酵母、昆虫又は哺乳類の細胞であってよい。当該ポリヌクレオチド又はベクターは、リン酸カルシウム沈殿法、DEAE−デキストラントランスフェクション、又はエレクトロポレーションを含む慣用技術を用いて宿主細胞に導入されてよい。
【0134】
発明は、組換え手段による発明の蛋白質の生産のためのプロセスを提供する。これは、(a)蛋白質の発現を可能にさせる条件下で上で定義された形質転換細胞を培養し;そして好ましくは(b)発現されたポリペプチドを回収することを含んでよい。任意に、上記ポリペプチドは、当業界公知の技術により単離され、及び/又は、精製されてよい。
【0135】
発明は、そのような認識(又は結合)が可能な薬剤又はその断片を認識する(又は結合する)TCRsも提供する。これらは、発明の蛋白質に関して本明細書にて論じられた本明細書にて言及された如何なる形態でも存在し得る(例えば、精製度)。発明は、発明の細胞に関して本明細書にて論じられた如何なる形態でも存在し得る(例えば、精製度)そのようなTCRsを発現するT細胞も提供する。
【0136】
発明は、薬剤(例えば、発明のエピトープ)を特異的に認識しそして発明の変異グリアジン蛋白質を認識する(そして典型的には均等な野生型グリアジンを認識しない)薬剤を特異的に認識するモノクローナル又はポリクローナル抗体、及びそのような抗体を作成する方法も提供する。発明の抗体は発明のこれらの物質に特異的に結合する。
【0137】
この発明のために、用語「抗体」は、抗体断片、例えば、Fv,F(ab)及びF(ab)2断片、並びに一本鎖抗体を含む。
【0138】
ポリクローナル抗体を生産する方法は、適当な宿主動物、例えば実験動物を免疫源で免疫し、そして血清からイムノグロブリンを単離することを含む。上記動物は、よって、免疫源を接種されてよく、次に血液を当該動物から採取し、そしてIgG分画を精製する。モノクローナル抗体を生産する方法は、所望の抗体を生産する細胞を不死にすることを含む。ハイブリドーマ細胞は、接種された実験動物からの脾臓細胞を腫瘍細胞と融合することにより生産してよい(Kohler and Milstein(1975)Nature 256,495−497)。
【0139】
所望の抗体を生産する不死の細胞を慣用の手法により選択してよい。ハイブリドーマは、培養液中で培養されるか、又は腹水液の形成のために腹膜内に注射されるか、又は同種の宿主又は免疫上無防備状態の宿主の血流に注射されてよい。ヒトの抗体は、ヒトリンパ球のインビトロ免疫、続くエプスタイン−バールウイルスによるリンパ球の形質転換により製造してよい。
【0140】
モノクローナル抗体は及びポリクローナル抗体はの両方の生産のためには、実験動物は、ヤギ、ウサギ、ラット又はマウスが適している。所望なら、免疫源は、例えばアミノ酸残基の一つの側鎖を通して免疫源が適当なキャリアーにカップリングされるようにコンジュゲートとして投与してよい。得られた抗体は、単離し、そして所望なら精製してよい。
【0141】
発明のポリヌクレオチド、薬剤、蛋白質、又は抗体は、検出可能な標識を有してよい。分泌された物質を可視検査により、任意に光学拡大手段の補助により、分泌された物質の検出を可能にする検出可能な手段は、好ましい。そのような系は、典型的には、基質中の色の変化を導く酵素標識に基づき、例えば、基質中の色の変化を導くアルカリホスファターゼである。そのような基質は市販されており、例えば、BioRadからである。他の適当な標識は、他の酵素、例えばペルオキシダーゼ、又は蛋白質標識、例えばビオチン;又は放射性標識、例えば32P又は35Sを含む。上記の標識は、公知の技術を用いて検出してよい。
【0142】
発明のポリヌクレオチド、薬剤、蛋白質、又は抗体は、実質上精製形態であってよい。それらは、製造物中のポリヌクレオチド、ペプチド抗体、細胞又は乾燥塊(dry mass)の少なくとも80%、例えば少なくとも90、95、97又は99%を一般に含む、実質上の単離された形態にあってよい。ポリヌクレオチド、薬剤、蛋白質又は抗体は、典型的には、他の細胞成分を実質上含まない。当該ポリヌクレオチド、薬剤、蛋白質又は抗体は、そのような実質上単離されたか、精製されたか、又は遊離の形態にて上記方法において使用されるか、又はそのような形態で容器キット中に存在してよい。
【0143】
発明は、発明のTCRを発現するトランスジェニック非ヒト哺乳類も提供する。これは、本明細書にて論じられた哺乳類の何れかであってよい(例えば、上記抗体の生産に関して)。好ましくは、当該哺乳類は、セリアック病を有するか又は感受性である。当該哺乳類はHLA−DQ2又は−DQ8又はHLA−DR3−DQ2を発現してもよく、及び/又は、セリアック病を引き起こすグリアジンを含むダイエットを与えられてよい(例えば、本明細書にて言及されたグリアジン蛋白質の何れか)。即ち、上記哺乳類はセリアック病の動物モデルとして機能してよい。
【0144】
発明は、セリアック病の治療用の生成物を同定する方法も提供し、セリアック病を有するか又は感受性の発明の動物に候補物質を投与し、そして当該哺乳類において物質がセリアック病を予防するか又は治療するか否かを決定することを含み、セリアック病の予防又は治療が、上記物質が治療用生成物であることを示す。そのような生成物はセリアック病を予防又は治療するのに使用してよい。
【0145】
発明は、治療用(予防用を含む)薬剤又は診断用(発明の薬剤、蛋白質及びポリヌクレオチド)薬剤を提供する。これらの物質は、薬学上受容可能なキャリアー又は希釈剤とそれらを混合することにより臨床投与のために製剤化される。例えば、それらは、局所、非経口、静脈内、筋肉内、皮下、眼球内、皮膚内、表皮内、経皮投与のために製剤化され得る。上記物質は、薬学上受容可能であって且つ所望の投与経路のために適切なあらゆる媒体と混合してよい。注射のための薬学上受容可能なキャリアー又は希釈剤は、例えば滅菌又は等張溶液、例えば注射用の水又は生理食塩水、又は弾道デリバリー用のキャリアー粒子であってよい。
【0146】
物質の用量は、様々なパラメーターに従い、特に使用される薬剤;処置される患者の年齢、体重及び状態;使用される投与様式;処置される症状の重度;及び要求される臨床上の養生法に従い調節してよい。規準として、注射により投与される物質の量は、0.01mg/kgから30mg/kg、好ましくは0.1mg/kgから10mg/kgが適している。
【0147】
記載された投与経路及び用量は規準としてのみ意図されるが、何故ならば熟練した開業医はあらゆる特定の患者及び症状に関して投与の最適な経路及び用量を決定することができるからである。
【0148】
発明の物質は、即ちヒト又は動物の身体の治療方法、又はヒト身体に実施される診断法において使用してよい。特に、それらは、セリアック病を治療又は予防する方法において使用してよい。発明は、セリアック病を治療又は予防するための医薬の製造法における使用のための薬剤も提供する。即ち、発明は、その要求のあるヒトへ発明の物質を投与することを含むセリアック病の予防又は治療法を提供する(典型的には、その非毒性有効量)。
【0149】
発明の薬剤は、標準の合成価額技術、例えば自動化された合成器の使用により作成することができる。上記薬剤は、より長いポリペプチド、例えば融合蛋白質から作成してよく、そのようなポリペプチドは上記ペプチドの配列を含む。当該ペプチドは、例えばポリペプチドを加水分解することにより、例えばプロテアーゼの使用により;又は物理的にポリペプチドを破壊することにより、ポリペプチドに由来し得る。発明のポリヌクレオチドは標準の技術、例えば合成器を使用することにより、作成することができる。
変異グリアジン蛋白質を発現するか又はアンタゴニストとして作用し得る配列を含む蛋白質を発現する植物細胞又は植物
発明の細胞は、植物細胞、例えばイネ科の単子葉植物種の細胞であってよい。当該種は、その野生型形態がグリアジン、本明細書にて言及されたグリアジン蛋白質の何れかを発現する種であってよい(配列番号:3に対して何れかの程度の相同性を有するグリアジンを含む)。そのようなグリアジンはヒトにおいてセリアック病を引き起こすかもしれない。上記細胞は、コムギ、トウモロコシ、エンバク、ライムギ、イネ、オオムギ、ライコムギ、モロコシ、又はサトウキビの細胞であってよい。典型的には、上記細胞は、コムギ(Triticum)属、例えばパンコムギ(aestivum)、スペルトコムギ(spelta)、ポーランドコムギ(polonicum)又はヒトツブコムギ(monococcum)の細胞である。
【0150】
発明の植物細胞は、典型的には、野生型グリアジンを発現しない(例えば、セリアック病を引き起こすかもしれない本明細書にて言及された何れかのグリアジン)か、又は薬剤を認識するT細胞により認識され得る配列を含むグリアジンを発現しないものである。即ち、野生型植物細胞がそのようなグリアジンを発現するなら、そのときは、そのようなグリアジンの発現を阻害するか低下させるか又はもはやセリアック病を引き起こさないようにグリアジンのアミノ酸配列を変化させるように操作してよい(典型的には、発明のエピトープをもはや発現させないことにより)。
【0151】
これは、変異を、例えば1、2、3又はそれより多いか又は全てのそのようなグリアジン遺伝子へ細胞内で導入すること、例えばコーディング又は非コーディング(例えば、プロモーター)へ導入することにより実施することができる。そのような変異は、本明細書にて論じられた変異の種類又は長さの何れかであり得る(例えば、相同蛋白質に関して)。上記変異は、直接様式(例えば、部位特異的変異導入又は相同組換え技術を用いて)又はランダムな様式(例えば、変異源を用い、次に典型的にはグリアジン(又はセリアック病を引き起こすグリアジン配列)をもはや発現しない変異導入された細胞を選択することにより)にて導入することができる。
【0152】
アンタゴニストとして作用し得る配列を含む蛋白質を発現する植物又は植物細胞の場合、そのような植物又は植物細胞は野生型グリアジン蛋白質を発現してよい(例えば、セリアック病を引き起こすもの)。好ましくは、アンタゴニスト配列の存在は、上記植物又は
植物細胞由来の蛋白質を含む食物を摂取した個体において低下した兆候(例えば兆候なし)のセリアック病を引き起こす。
【0153】
植物細胞内に存在する(か又はに形質転換された)ポリヌクレオチドは、一般に、植物細胞内で変異グリアジン蛋白質を発現できるプロモーターを含む。所望の発現のパターンに依存して、上記プロモーターは、構成的、組織特異的、ステージ特異的;及び/又は誘導可能性であってよい。例えば、植物における強い構成的発現は、CAMV35S,Rubisco ssu又はヒストンプロモーターにより得ることができる。また、組織特異的又はステージ特異的プロモーターは、トランスジェニック植物において特定の組織へ又はその発生において特定のステージへ、発明の蛋白質の発現を標的化するために使用してよい。即ち、例えば、種子特異的、根特異的、葉特異的、花特異的等のプロモーターを使用してよい。種子特異的プロモーターは、Dalta et al(Biotechnology Ann.Rev.(1997),3,pp.269−296)に記載されたものを含む。種子特異的プロモーターの特定の例は、napinプロモーター(EP−A−0 255,378)、ファセオリンプロモーター、グルテニンプロモーター、ヘリアンセニンプロモーター(WO98/45461)及びATS1及びATS3プロモーター(PCT/US98/06798)を含む。
【0154】
上記細胞はあらゆる形態であってよい。例えば、単離された細胞、例えばプロトプラストであってよいか、又は植物組織の一部、例えばカルス又は植物から切り出された組織であってよいか、又は植物全体の一部であってよい。上記細胞はあらゆる種類の細胞であってよい(例えば、植物の部分のあらゆる種類)。例えば、分化しない細胞、例えばカルス細胞;又は分化した細胞、胚、花粉、根、若枝(shoots)又は葉に見いだされる種類の細胞。植物の部分は、根;若枝;葉及び生殖に関与する部分、例えば、花粉、卵、おしべ、葯、花弁、萼片、及び他の花の部分を含む。
【0155】
発明は、トランスジェニック植物細胞を提供するように発明のポリヌクレオチド又はベクターにより植物細胞を形質転換することを含む、トランスジェニック植物細胞を得るための方法を提供する。あらゆる適切な形質転換方法を使用してよい(コムギの場合、Vasil V et al,Biotechnology 10,667−674(1992)に開示された技術を使用してよい)。好ましい形質転換技術は、植物プロトプラストのエレクトロポレーション及び粒子の砲撃を含む。形質転換は、即ち、幾つかの細胞がトランスジェニックで幾つかはそうでないようなキメラ組織又は植物を生じてよい。
【0156】
発明の細胞又はそうして得られた細胞は、当業界公知の技術によりトランスジェニック植物へ再生されてよい。これらは、トランスジェニック細胞の生育及び/又は分割を刺激するための、オーキシン、ジベレリン及び/又はサイトキニンのような植物成長物質の使用を含んでよい。同様に、体性胚形成及び分裂組織培養のような技術を使用してよい。再生技術は当業界にて公知であり、例えば、米国特許第4,459,355号、米国特許第4,536,475号、米国特許第5,464,763号、米国特許第5,177,010号、米国特許第5,187,073号、欧州特許267,159,欧州特許604,662,欧州特許672,752、米国特許第4,945,050号、米国特許第5,036,006号、米国特許第5,100,792号、米国特許第5,371,014号、米国特許第5,478,744号、米国特許第5,179,022号、米国特許第5,565,346号、米国特許第5,484,956号、米国特許第5,508,468号、米国特許第5,538,877号、米国特許第5,554,798号、米国特許第5,489,520号、米国特許第5,510,318号、米国特許第5,204,253号、米国特許第5,405,765号、欧州特許442,174,欧州特許486,233、欧州特許486,234、欧州特許539,563、欧州特許674,725、WO91/02071及びWO95/06128に見いだすことができる。
【0157】
そのような多くの技術において、一つの工程は、細胞を膨張させる及び/又は分割することを含む、カルス、即ち細胞組織の形成である。そのようなカルスは他の種類の植物細胞培養物及び植物の部分であるように、発明の別の側面である。即ち、例えば、発明は、トランスジェニック植物組織及び部分を提供するが、胚、分裂組織、種子、若枝、根、幹、葉及び花の部分を含む。これらは、それらの細胞の幾つかが発明の細胞であり幾つかはそうでない感覚において、キメラであってよい。発明のトランスジェニック植物部分及び組織、植物及び/又は種子は、本明細書にて言及された植物種の何れかのものであってよい。
【0158】
再生手法は、典型的には、マーカー遺伝子による、形質転換された細胞の選択を含む。
【0159】
再生工程は、第1世代のトランスジェニック植物を生じる。発明は、この第1世代植物からのさらなる再生のトランスジェニック植物を得るための方法も提供する。これらは子孫トランスジェニック植物として知られる。第2、第3、第4、第5、第6及びさらなる世代の子孫植物は、当業界公知のあらゆる手段により第1世代のトランスジェニック植物から得てよい。
【0160】
即ち、発明は、トランスジェニック子孫植物を得る方法を提供し、発明の第1世代のトランスジェニック植物から第2世代のトランスジェニック子孫植物を得て、そして任意に第2世代のトランスジェニック子孫植物から一つ又はそれより多い世代のトランスジェニック植物を得ることを含む。
【0161】
子孫植物はあらゆる公知の技術による前の世代のそれらの先祖から生じさせてよい。特に、子孫植物は:
前の世代に属する発明のトランスジェニック植物からトランスジェニック種子を得て、次にトランスジェニック種子を成長させることにより新世代の発明のトランスジェニック子孫植物を得て;そして/又は
前の世代に属する発明のトランスジェニック植物をクローンとして増殖させることにより新世代に属する発明のトランスジェニック子孫植物を提供し;そして/又は
前の世代に属する発明の第1世代のトランスジェニック植物を別の匹敵する植物と交配することにより、新世代に属する発明のトランスジェニック子孫植物を提供し;そして/任意に
そうして得られた子孫から一つ又はそれより多いさらなる世代のトランスジェニック子孫植物を得ること
により生産してよい。
【0162】
これらの技術はあらゆる組み合わせにおいて使用してよい。例えば、クローンとしての増殖及び有性増殖を、別の点において、培養のために適したトランスジェニック植物を生じるプロセスにおいて、使用してよい。特に、農業経済学上望まれる特性を有する植物分類群との繰り返しの戻し交配を企ててよい。植物及びそれから再生した新規な植物からの細胞の取り出しの別の工程も実施してよい。
【0163】
また、細胞、植物組織、植物又は種子をあらゆる適切なステージにおいて、さらに望まれる特性を導入することにより、発明のポリヌクレオチド以外の所望のコーディング配列を導入してよい。これは、発明のポリヌクレオチドの導入のために本明細書にて記載された技術により実施してよい。
【0164】
例えば、他の除草剤耐性特性、例えばグリフォセート(Glyphosate)に対する寛容(例えば、EPSPシンターゼ遺伝子(例えば、EP−A−0293,358)又
はグリフォセートオキシドレダクターゼ(WO92/000377)遺伝子)をコードするトランス遺伝子から;又は、フォサメチン;ジハロベンゾニトリル;グルフォシネートに対する寛容をコードするトランス遺伝子から、例えばホスフィノチリシナセチルトランスフェラーゼ(PAT)又はグルタミンシンターゼ遺伝子(EP−A−0242,236参照)を用いて;アスラム、例えば、ジヒドロプテロエートシンターゼ遺伝子(EP−A−0 369,367参照);又はスルフォニルウレア、例えば、ALS遺伝子を用いて;ジフェニルエーテル、例えば、アシフルオルフェン又はオキサジアゾン、例えば、プロトポルフィロジェンオキシダーゼ遺伝子;環状イミド、例えば、クロロフサリム;フェニルピラゾール、例えば、TNP、又はフェノピレート又はそのカルバメート類似体;さらなるトランス遺伝子を選択してよい。
【0165】
同様に、除草剤寛容以外の有益な特性に関する遺伝子を導入してよい。例えば、昆虫耐性遺伝子を導入してよく、注目に値するのはバチルスサリンジェネシス(Bt)の毒素をコードする遺伝子である。同様に、疾患耐性遺伝子を導入してよく、例えば、WO91/02701又はWO95/06128におけるとおりである。
【0166】
典型的には、発明の蛋白質を発明の植物内で発現させる。使用されるプロモーターにより、この発現は構成的又は誘導可能であってよい。同様に、組織特異的か又はステージ特異的であってよく、即ち、特定の植物の組織(例えば、本明細書にて言及された組織の何れか)又は植物の発生におけるステージに直接向けられる。
【0167】
発明は、作物を得るための方法も提供し、発明のトランスジェニック植物を収穫し、そして任意にさらに加工することを含む。作物は、作物植物から得ることができるあらゆる有用な生成物を意味する。
変異グリアジン蛋白質又はアンタゴニストとして作用することができる配列を含む蛋白質を含む生成物
発明は、変異グリアジン蛋白質又はアンタゴニストとして作用することができる配列を含む蛋白質を含む生成物を提供する。これは、典型的には、そのような蛋白質を発現する本明細書にて言及された植物に由来するか又は植物の部分を含む。そのような生成物は、発明の植物を直接収穫するか又は間接に収穫し、そしてさらに加工することにより得ることが可能であってよい。直接に得ることが可能な生成物は穀物を含む。あるいは、そのような生成物は、回収してさらに加工することにより間接に得ることが可能であってよい。さらに加工することにより得ることが可能な生成物の例は、小麦粉又は蒸留されたアルコール飲料;直接得ることができるか又はさらに加工した材料、例えば小麦粉から作られた焼いた製品(例えば、パン)から作られる食物製品である。典型的には、そのような食物製品はヒト個体により摂取可能であって消化可能である(即ち、非毒性であって栄養価値がある)。
【0168】
アンタゴニスト配列を含む蛋白質を含む食物製品の場合、食物製品は野生型グリアジンを含んでもよいが、好ましくは当該アンタゴニストはそのような食物を摂取した後にセリアック病の兆候の低下(又は完全に)を導くことができる。
【0169】
発明は、以下の非限定実施例により例示される。
【実施例】
【0170】
実施例1
我々は、完全に特性決定されたグリアジン、「A−グリアジン」の全配列にわたる51セットの合成15マーペプチドを使用することによりセリアック病のエピトープマッピングを実施した(表1参照)。A−グリアジンペプチドはtTGにより個々に処理されることにより、インビボで生成したものを模倣したかもしれない生成物を生成した3。我々は
、実験上の感染及び自己免疫疾患において記載されたとおり、エピトープの「拡散」又は「消費」が起きたかもしれない可能性を避けるために、疾患の消滅の開始点においてセリアック病患者を研究しようとも努めた。
3日及び10日のパンチャレンジによる臨床上及びA−グリアジン特異的なT細胞応答
パイロット研究において、セリアック病の2人の患者が緩解したが、血清の抗−筋内膜抗体(EMA)の不在により定義され、グルテンフリーダイエットは彼らの通常のグルテンフリーダイエットに加えて、標準のグルテン含有白パン4スライスを毎日給仕された。被験者1は3日後に腹痛、口の潰瘍及びゆるい下痢のためにパンを停止したが、被験者2は1週間目に軽い吐き気があっただけで10日間継続した。EMAは被験者2においてパンチャレンジの1週間後に陽性になったことから、使用したパンがセリアック病の再発を導いたことを示す。しかし、被験者1においては、EMAはパンチャレンジから2カ月まで陰性のままであった。両被験者において、パンチャレンジにより出現した兆候はグルテンフリーダイエットに戻って2日以内に解消した。
【0171】
IFNγELISPOTアッセイのA−グリアジンに対するPBMC応答は、パンチャレンジの前と間に見いだされなかった。しかし、被験者1においてはパンの消費の1日後に(4日目)、tTGにより処理したA−グリアジン51−85にわたる5つのオーバーラップするペプチドの単一のプール(プール3)が有力なIFNγ応答を示した(図1a参照)。被験者1においては、A−グリアジンペプチドに対するPBMC IFNγ応答はプール3単独に標的化されたままであり、8日目に最大であった。プール3に対する応答の動力学及び大きさは、α−キモトリプシン消化されたグリアジンにより導かれるのと同様であった。tTG処理したプール3に対するPBMC IFNγ応答はtTG処理されなかったプール3より5から12倍の大きさに一貫し、そしてα−キモトリプシン消化したグリアジンに対する応答はtTG処理した場合の3から10倍大きかった。被験者2においては、tTG処理されたプール3も、8日目にA−グリアジンペプチドの免疫源セットのみであったが、この応答は被験者1よりも弱く、4日目には観察されず、11日目にはプール3に対する応答は減少し、そしてA−グリアジンペプチドの他のtTG処理プールがより強いIFNγ応答を導いた(図1b参照)。
【0172】
パイロット研究は、これらのセリアック病被験者における初期T細胞応答が5つのペプチドの単一のtTG処理A−グリアジンプールに対してであったこと、及び末梢血において容易に測定されたことを示した。抗原暴露が3日ではなく10日間続いたなら、他のA−グリアジンペプチドに対するT細胞応答が、エピトープの拡散に一致して、出現する。tTG処理されたA−グリアジンペプチドによるセリアック病特異的IFN−g誘導
グルテンフリーダイエットの別の6人のセリアック病被験者の内の5人において(表1参照)、3日間のパンチャレンジはtTG処理されたペプチドをプール3に同定し、特に、PBMCからIFNγを導く唯一のA−グリアジン成分としての56−70(12)及び60−75(13)に相当するペプチドであった(図2参照)。IFNγ ELISPOTと平行して流したIL−10 ELISPOTは、tTG処理されたペプチド12又は13に対してIL−10の応答を示さなかった。一人の被験者においては、パンチャレンジの4日前、4日間又は4日目まで、あらゆるA−グリアジンペプチド又はα−キモトリプシン消化されたグリアジンに対してIFNγ応答はなかった。パンチャレンジの2カ月後まで測定した場合に、これらのセリアック病被験者の誰にも基底ラインからEMA状態が変化しなかった。
【0173】
HLA−DQ対立遺伝子α1*0501,β1*0201(28−52歳、2人は女性)を有し、3日間パンチャレンジを受けて1カ月間グルテンフリーダイエットを受けた、4人の健康なEMA陰性被験者からのPBMCは、tTG処理をしてもしなくても、何れのA−グリアジンペプチドに対しても陰性対照を超えるIFNγ応答を示さなかった。即ち、PBMC中のIFNγのtTG処理プール3及びA−グリアジンペプチド56−70
(12)及び60−75(13)への誘導は、セリアック病特異的であった(7/8対0/4、p<0.01,カイ2乗分析による)。
最小のA−グリアジンT細胞エピトープのファインマッピング
A−グリアジン56−75の末端削除に相当するtTG処理したペプチドは、同じコアペプチド配列QPQLP(配列番号:9)が評価された5人のセリアック病被験者の全てにおいて抗原性に必須であったことを明らかにした(図3参照)。PBMC IFNγは、7マーのPQPQLPY(配列番号:4)で始まるこのコア配列にわたり、長さを伸ばしたtTG処理ペプチドに応答することから、tTG処理された17マーのQLQPFPQPQLPYPQPQS(配列番号:19)(A−グリアジン57−73)がIFNγ ELISPOTにおいて最大の活性を所有したことを示した(図4参照)。
tTGによるQ65の脱アミド化はA−グリアジン中の免疫上優勢なT細胞エピトープを生じる
HPLC分析は、A−グリアジンのtTG処理が、親ペプチドにおいてあとで隣接して溶出した単一の生成物を生じたことを証明した。アミノ酸配列決定は、A−グリアジン56−75中に含まれる6つのグルタミン(Q)残基の中から選択的にtTGにより脱アミド化されたことを示した(図5参照)。グルタミン酸(E)置換されたQ65のコアA−グリアジン62−68からの連続伸長物(serial expansions)に相当するペプチドの生物活性は、tTG処理後のQ65を有する同じペプチドと均等であった(図4a参照)。Q57及びQ72のEによる両方又は単独の置換にE65が伴うと、調査された3人のセリアック病被験者において、上記17マーの抗原性を増強しなかった(図6参照)。グリアジンペプチドにおいてグルタミン残基にプロリンが続くと、インビトロにおいてtTGにより脱アミド化されないため、Q57及びQ72を調査した(W.Vader et al,Proceedings 8th International
Symposium Coeliac Disease)。よって、免疫上優勢なT細胞エピトープをQLQPFPQPELPYPQPQS(配列番号:2)と定義した。
免疫上優勢なT細胞エピトープ応答はDQ−2制限され且つCD4依存性である
HLA−DQα1*0501,β1*0201に関して同型接合の(homozygous)2人のセリアック病被験者において、抗−DQモノクローナル抗体がtTG処理されたA−グリアジン56−75に対するELISPOT IFNγ応答をブロックしたが、抗−DP及び−DR抗体はブロックしなかった(図7参照)。2人のセリアック病被験者からのPBMCの抗−CD4及び−CD8磁気ビーズ除去(depletion)は、tTG処理されたA−グリアジン56−75に対するIFNγ応答がCD4 T細胞媒介性であることを示した。
考察
この研究において、我々は、配列:QLQPFPQPELPYPQPQS(配列番号:2)(残基57−73)のtTG処理されたA−グリアジン17マーに応答性のセリアック病被験者の末梢血においてCD4 T細胞の一過性集団を誘導するために、普通の白パンを用いたむしろ単純なダイエット抗原チャレンジを記載した。A−グリアジン(Q→E65)に対する免疫応答はセリアック病関連対立遺伝子DQα1*0501,β1*0201に限定される。インビトロにおける組織のトランスグルタミナーゼ作用はQ65を選択的に脱アミド化する。置換Q→E65を有する合成A−グリアジンペプチドに対する誘導された末梢血IFNgの応答は、tTG処理されたQ65 A−グリアジンペプチドに均等であり;両者は未修飾のQ65 A−グリアジンペプチドよりもIFNg ELISPOTにおいて10倍にまで、より多くのT細胞を刺激する。
【0174】
我々は、エピトープマッピングにおけるT細胞クローンの使用により生じるかもしれない方法論上の人工物の可能性を回避するため、インビボ抗原チャレンジ及び短期間エクスビボ免疫アッセイを用いて、このセリアック病特異的T細胞エピトープを慎重に定義した。我々の発見は、グルテンの摂取に対する末梢血のT細胞応答が迅速であるが短命であり、且つエピトープマッピングに利用可能であることを示す。インビボ抗原チャレンジは、
A−グリアジンペプチドに対する免疫応答の時間の経過順の階層(temporal hierarchy)が存在することを示し;tTGにより修飾されたA−グリアジン57−73はP中のもっとも強いIFNg応答を誘導するのみならず、第1のIFNg応答も出現する。
【0175】
我々はA−グリアジンにわたるペプチドしか評価しなかったので、セリアック病の病因において等しいか又はより重要な他のグリアジン内の別のエピトープが存在するかもしれない。事実、我々が同定したPQPQLPY(配列番号:4)であるA−グリアジン内のコアエピトープにおけるペプチド配列は幾つかのべつのグリアジンに共通する(SwissProt and Trembl受け入れ番号:P02863,Q41528,Q41531,Q41533,Q9ZP09,P04722,P04724,P18573)。しかしながら、生検チャレンジ及びインビボ研究において生物活性を所有することが以前に示されたA−グリアジンペプチド(例えば:31−43、44−55及び206−217)4,5は、セリアック病被験者において3日間のパンチャレンジ後にPBMCにおいてIFNg応答を誘導しなかった。これらのペプチドは、免疫応答の拡散を伴って現れる「二次」T細胞エピトープであるかもしれない。
実施例2
免疫上優勢なエピトープ内の置換に対するT細胞認識の効果
57−73A−グリアジンエピトープにおける65位のグルタミンの置換の効果を、合成ペプチドを用いてIFNγ ELISPOTアッセイにおいて置換されたエピトープに対する末梢血応答を測定することにより決定した(50μg/mlにおいて)。応答は3人のセリアック病患者においてグルテンチャレンジを始めて6日後に測定した(3日間4スライスのパンを毎日)。結果を表3及び図8に示す。グルタミン酸から、ヒスチジン、チロシン、トリプトファン、リジン、プロリン又はアルギニンへの置換において観察され得るように、免疫上優勢なエピトープに対する応答の大きさの10%より低い大きさの応答を刺激した。即ち、この位置におけるA−グリアジンの変異は、免疫反応性を低下させるか又は不在にする変異グリアジンを生成するのに使用することができた。
実施例3
他の天然に生じるグリアジンからの均等なペプチドの免疫反応性の試験
他の天然に生じるコムギのグリアジン由来の均等なペプチドの免疫反応性を、天然に生じる配列に相当する合成ペプチドをトランスグルタミナーゼにより処理して使用することにより、評価した。これらのペプチドは、ELISPOTにおいて、実施例2に記載されたのと同じ被験者からのPBMCを用いて同じ様式にて試験した。少なくとも5つのペプチドがA−グリアジン57−73 E65ペプチドに匹敵する免疫反応性を示す(トランスグルタミナーゼ処理後)ことから、コムギ中の他のグリアジン蛋白質も同ようにこのセリアック病特異的免疫応答を誘導することを示唆する(表4及び図9)。
方法
被験者:研究において使用した患者は、英国のオックスフォードにおいてセリアック病クリニックにかかっている。セリアック病は、特有の小腸の組織構造、及び兆候の標準化及びグルテンフリーダイエットの小腸組織構造に基づいて診断された。
組織型判定:組織型判定は、EDTA抗凝血末梢血から抽出されたDNAを用いて実施した。HLA−DQA及びDQB遺伝子型判定は、PCRにより配列特異的プライマーミックスを用いて実施した6−8。
抗筋内膜抗体アッセイ:EMAは、サルの食道(oesophagus)を用いて1:5に希釈された患者の血清を用い、続いてFITCコンジュゲートヤギ抗−ヒトIgAを用いた間接免疫蛍光により検出した。IgAをEMAの前に定量したが、どの被験者もIgA欠損ではなかった。
抗原チャレンジ:グルテンフリーダイエット後のセリアック病被験者に、4スライスのグルテン含有パンを毎日3又は10日間食べ尽くさせた(セインズベリーズの標準の「白サンドイッチパン」)。パンチャレンジを始める前の週及び2カ月後まで、EMAを査定し
た。グルテンフリーダイエットを4週間経験した健康な被験者は、3日間グルテン含有パン4スライスを含む彼らの通常のダイエットを食べつくし、そして次にさらに4日間グルテンフリーダイエットに戻った。
IFNγ及びIL−10 ELISPOT:Ficoll−Hypaque密度遠心分離により50−100mlの静脈血からPBMCを調製した。3回洗浄後に、PBMCを再度、10%熱不活性化ヒトAB血清を含む完全RPMIに懸濁した。IFNγ及びIL−10の単一細胞の分泌に関するELISPOTアッセイを、市販のキット(Mabtech;ストックホルム、スエーデン)を用いて96−ウエルプレート(MAIP−S−45;Millipore,ベッドフォード、MA)により、製造者の指示書(ほかで記載されたとおり9)に従い、2−5x105(IFNγ)又は二重のウエルにて査定し、そしてミコバクテリウムツベルクロシスの精製された蛋白質誘導体(PPD RT49)(Serum Institute;コペンハーゲン、デンマーク)(20μg/ml)を全てのアッセイにおいて陽性対照として含ませた。
ペプチド:合成ペプチドはResearch Genetics(ハンツビル、アラバマ)から購入し、質量分光分析及びHPLCにより確認されたペプチドの真正度は70%>精製度であった。グリアジン(Sigma;G−3375)(100mg/ml)のα−キモトリプシン(Sigma;C−3142)200:1(w/w)による消化は、0.1M NH4HCO3中で2M尿素と共に室温において実施し、98℃において10分間加熱することにより24時間後に中止した。遠心分離(13,000g,10分間)後に、グリアジン消化上清をフィルター滅菌した(0.2mm)。グルテンの消化をSDS−PAGEにより確認して、蛋白質濃度を査定した。α−キモトリプシン−消化されたグリアジン(640μg/ml)及び合成グリアジンペプチド(15マー:160μg/ml,他のペプチド:0.1mM)を個別にtTG(Sigma;T−5398)(50μg/ml)によりPBS中にてCaCl2を1mM加えて、2時間37℃において処理した。ペプチド及びペプチドプールを滅菌96−ウエルプレートに分配して、使用まで−20℃に凍結保存した。
ペプチドのアミノ酸配列決定:逆相HPLCを使用して、A−グリアジン56−75のtTG処理によるペプチドを精製した。単一生成物を同定して、アミノ酸配列決定(自動化配列決定機モデル494A,Applied Biosystems,フォスターシティー、カリフォルニア)に供した。未修飾のG56−75の配列はLQLQPFPQPQLPYPQPQSFP(配列番号:5)と確認され、そしてtTG処理されたG56−75はLQLQPFPQPELPYPQPQSFP(配列番号:11)と確認された。グルタミン残基の脱アミド化は、アミノ酸配列決定のサイクル2、4、8、10、15及び17において回収されたグルタミン及びグルタミン酸の合わせた量のパーセントとして表された、回収されたグルタミン酸の量(pmol)として定義された。tTGに帰する脱アミド化は、(tTG処理されたペプチド中のグルタミンの脱アミド化の%−未処理のペプチド中の脱アミド化の%)/(100−未処理のペプチド中の脱アミド化の%)として定義された。
CD4/CD8及びHLAクラスII制限:抗−CD4又は抗−CD8コートされた磁気ビーズ(Dynal,オスロ、ノルウエー)をRPMIにより4回洗浄し、次に10%熱不活性化ヒトAB血清を含む完全RPMI中のPBMCと共に、30分間氷の上でインキュベートした。ビーズを磁石により取り出して、残る細胞を計数した。tTG処理されたA−グリアジン56−75に対する免疫応答のインビボのHLA−クラスII制限は、PBMC(5x106細胞/ml)を抗−HLA−DR(L243),−DR(L2),及び−DP(B7.21)モノクローナル抗体(10μg/ml)と、室温において、ペプチドの添加前に1時間インキュベートすることにより、確立した。
実施例4
グリアジン−特異的末梢血リンパ球による粘膜インテグリンの発現
内皮とリンパ球のアドレシンの間の相互作用は、器官特異的なリンパ球のホーミングを促進させる。多数のアドレシンが公知である。ヘテロダイマーα4β7は粘膜固有層消化
管及び他の粘膜リンパ球に特異的であり、そしてαEβ7は消化管及び皮膚において内皮リンパ球内に特異的である。末梢血CD4 T細胞の約30%がα4β7を発現し、そして粘膜部位へ移動中であると推定され、末梢血CD4 T細胞の5%がαEβ7を発現する。αE又はβ7特異的な抗体をコートされた免疫磁気ビーズは、それぞれ、αEβ7又はαEβ7及びα4β7を発現する細胞のPBMCを除去する。ELISpotアッセイとの組み合わせにより、免疫磁気ビーズの除去は、これらの細胞を粘膜表面へのホーミングとして同定するかもしれないグリアジン特異的T細胞のアドレシン発現の決定を可能にする。おもしろいことに、インビボのグルテンチャレンジは小腸の粘膜固有層へのCD4
T細胞の迅速な流入に関連しており(内皮部位内ではなく)、90%を超えるリンパ球がα4β7を発現する。
【0176】
免疫磁気ビーズを製造して、3日間のグルテンチャレンジの後で6日目又は7日目にセリアック病被験者からのPBMCを除去するのに使用した。FACS分析は、αEビーズが約50%の陽性CD4 T細胞を除去し、一方β7ビーズは全てのβ7陽性CD4 T細胞を除去した。CD4−又はβ7−ビーズを用いたがCD8−又はαEビーズを用いないPBMC除去はインターフェロンガンマELISpotにおいて応答を完全に破壊した。tTGグリアジン及びPPD応答はCD4除去により完全に破壊されたが、インテグリン特異的ビーズ除去により一貫して影響された。
【0177】
即ち、セリアック病においてグルテンチャレンジ後に誘導されたA−グリアジン57−73 QE65特異的T細胞は、小腸中の粘膜固有層CD4 T細胞上に存在するインテグリンα4β7を発現する。
実施例5
最適なT細胞エピトープの長さ
A−グリアジン中の優勢なT細胞エピトープのコアにわたる7から17の長さのアミノ酸からのペプチドを試験する以前のデータは、3日間のグルテンチャレンジ開始から6日後にセリアック病のボランティアからの末梢血単核細胞(PBMC)を用いたインターフェロンガンマElispotにおいて17マーのA−グリアジン57−73 QE65(配列番号:2)が最大の応答を誘導したことを示した。
【0178】
A−グリアジン中の優勢なT細胞エピトープのコア配列からの伸長を示すペプチドを、3日間のグルテンチャレンジ開始から6日後にセリアック病のボランティアからの末梢血単核細胞(PBMC)を用いたIFNガンマELISPOTにおいて査定した(n=4)。ペプチド13:A−グリアジン59−71 QE65(13マー)、ペプチド15:58−72 QE65(15マー)、...、ペプチド27:52−78 SE65(27マー)。
【0179】
図11に示すとおり、A−グリアジン57−73 QE65の伸長はIFNガンマElispotにおいて応答を実質上は増強しない。次の実施例は、17マーペプチドを用いたA−グリアジン57−73 QE65のアゴニスト及びアンタゴニスト活性を特性決定する。
実施例6
A−グリアジン57−73 QE65とセリアック病における他のDQ2−制限されるT細胞エピトープの比較
セリアック病の腸の生検からのグルテン特異的T細胞クローン及び系列のT細胞エピトープに相当する未修飾及びトランスグルタミナーゼ処理ペプチドに相当するペプチドを用いて、用量応答研究を実施した。ペプチドに対する応答はA−グリアジン57−73 QE65に対する応答のパーセントとして表現した。全ての被験者はHLA−DQ2+であった(誰もDQ8+でなかった)。
【0180】
上記の研究は、A−グリアジン57−73 QE65が、グルテンチャレンジ後にセリアック病のPBMCを用いたELISpotアッセイにおいてインターフェロンガンマの誘導に関してもっとも有効性のあるグリアジンペプチドであることを示す(図12a−h,及び表5及び6を参照)。第2及び第3のエピトープは、より大きなペプチド、即ちA−グリアジン57−73 QE65及びGDA4 WHEAT P04724−84−100 QE92の準最適断片である。上記エピトープは適度に生物活性であるにすぎない(ブランクを差し引いた後ではA−グリアジン57−73 QE65より約1/20活性)。
【0181】
A−グリアジン57−73 QE65はセリアック病において他の公知のT細胞エピトープよりも有効性がある。配列決定されたグリアジン遺伝子の間でA−グリアジン57−73(配列PQLPY(配列番号:12)を含む)の16の多型が存在し、それらの生物活性を次で査定する。
実施例7
末梢血中でのグリアジン−及びA−グリアジン57−73 QE65の比較
セリアック病におけるグリアジンに対する全T細胞応答に対しての、優勢なエピトープA−グリアジン57−73 QE65の相対的な寄与は重大な問題である。ペプシン−トリプシン及びキモトリプシンにより消化されたグリアジンが、セリアック病におけるT細胞系列及びクローンの分化のための抗原として伝統的に使用されてきた。しかしながら、これらのプロテアーゼは特定のペプチドエピトープを分断するかもしれないと言える。事実、組換えα9−グリアジンのキモトリプシン消化はペプチドQLQPFPQPELPY(配列番号:13)を生じるが、最適エピトープ配列QLQPFPQPELPYPQPQS(配列番号:2)の末端削除体である(上を参照)。トランスグルタミナーゼ処理は、実質上、グリアジン特異的T細胞クローン及び系列の増殖アッセイにおいて、キモトリプシン消化されたグリアジンの能力を増加させる。よって、トランスグルタミナーゼ処理されたキモトリプシンで消化されたグリアジン(tTGグリアジン)は理想的な抗原ではないかもしれないが、この混合物に対する応答はグリアジンに特異的な末梢血リンパ球の「全」数に近似するかもしれない。ELISpotアッセイにおけるA−グリアジン57−73 QE65及びtTGグリアジンに対する応答の比較は、セリアック病におけるグリアジンに対する全免疫応答へのこの優勢なエピトープの寄与の指標を提供し、またエピトープ拡散の基準でもある。
【0182】
4人のセリアック病被験者においてグルテンチャレンジを開始して6日目又は7日目に回収されたPBMCを、キモトリプシン消化されたグリアジン+/−tTG処理を用いた用量応答研究において査定し、そして最適濃度のA−グリアジン57−73 QE65(25mcg/ml)に対するELISpot応答と比較した。グリアジンのtTG処理はELISpotにおいて約10倍PBMC応答を増強した(単独で査定した場合にtTGはブランクと同等であった)(図13a−c参照)。研究した4人のセリアック病被験者において、A−グリアジン57−73 QE65(25mcg/ml)はtTGグリアジン(500mcg/ml)の14から115%の間の応答を誘導し、そしてA−グリアジン57−73 QE65に対する応答が大きければ大きいほど、tTGグリアジン応答を表した比率が大きくなった。
【0183】
相対的に限られたデータから、A−グリアジン57−73 QE65応答が幾つかの被験者においてtTGグリアジンと同等であることが示唆される。より進化した抗−グリアジンT細胞応答に関連したエピトープ拡散は、幾つかの個体において末梢血の「全」応答へのA−グリアジン57−73 QE65の僅かな寄与の説明となるかもしれない。エピトープ拡散はさほど厳しくないグルテンフリーダイエットを受けた個体において保持されるかもしれない。
実施例8
セリアック病において生物活性なグリアジンペプチドの定義:A−グリアジン57−73の多型
A−グリアジンの全配列にわたるオーバーラップする15マーのペプチドを査定することにより、セリアック病の免疫上優勢な配列を同定した。A−グリアジンは最初に全配列を決定されたアルファグリアジン蛋白質及び遺伝子であったが、コムギにおいて約30−50の関連するアルファグリアジン蛋白質のひとつである。蛋白質データベースSwiss−Prot及びTREMBLを検索することにより、25の別々のアルファ−グリアジン遺伝子が同定され、さらに8つのアルファグリアジンを記述した。これら25のアルファグリアジンに含まれたのは、A−グリアジン57−73に相当する16の異なる多型である(表7参照)。
【0184】
トランスグルタミナーゼによるインビトロにおける処理、並びに10位において置換されたグルタミン酸による処理後、未修飾形態でのこれらの16の多型に相当する合成ペプチドを、通常のグルテンフリーダイエット後の、インターフェロンガンマELISpotアッセイにおけるグルテンチャレンジの6日又は7日後に、セリアック病の被験者からのPBMCを用いて査定した。グルタミン酸置換したペプチドを3つの濃度において(2.5、25及び250mcg/ml)査定し、未修飾のペプチド及びトランスグルタミナーゼ処理されたペプチドを25mcg/mlの濃度のみにて査定した。生物活性は、個々の被験者においてA−グリアジン57−73 QE65 25mcg/mlに付随した応答の%として表現した(n=4)(図14参照)。
「野生型」ペプチドの生物活性はトランスグルタミナーゼ処理により実質増加した(>5倍)。野生型ペプチドのトランスグルタミナーゼ処理は10位においてグルタミン酸により置換された同じペプチドの生物活性と同様な生物活性をもたらした。5つのグルタミン酸置換ペプチド(B,C,K,L,M)の生物活性はA−グリアジン57−73 QE65の>70%であったが、A−グリアジン57−73 QE65よりも顕著に高い生物活性のものはなかった。2.5及び250mcg/mlの濃度においてグルタミン酸置換されたペプチドに対するPBMC応答は25mcg/mlにおけるのと同等であった。6つのグルタミン酸置換グリアジンペプチド(H,I,J,N,O,P)は、A−グリアジン57−73 QE65の生物活性の<15%であった。他のペプチドは生物活性において中間であった。
【0185】
少なくとも6つのグリアジン由来のペプチドがトランスグルタミナーゼによる修飾語にA−グリアジン57−73 QE65に対する有効性が均等である。相対的に非生物活性のA−グリアジン57−73 QE65の多型も存在する。これらのデータは、トリチカムエステビウム、トリチカムウアルツ、トリチカムスペルタの幾つかのグリアジン由来のペプチドのトランスグルタミナーゼ修飾がセリアック病において免疫上優勢なT細胞エピトープを生成できるかもしれないことを示す。
【0186】
非−セリアック病−毒性コムギを生成するコムギの遺伝子修飾は、複数のグリアジン遺伝子の除去又は修飾を必要とするかもしれない。A−グリアジン57−73の変更されたペプチドリガンドアンタゴニストを定義する配列を取り込んだグリアジン又は他の蛋白質又はペプチドを含むコムギの生産は、セリアック病において「非毒性」よりむしろ治療用である遺伝学上修飾されたコムギを生成するための別の戦略である。
実施例9
コアエピトープ配列の定義:
N−末端又はC−末端からのA−グリアジン56−75の末端削除に相当するペプチドの比較は、T細胞エピトープのコア配列がPELPY(A−グリアジン64−68)であることを示した。非アゴニスト及びアンタゴニストを定義する試みは、その生物活性に実質貢献する残基において置換されたA−グリアジンのバリアントに焦点を注ぐことになる。
【0187】
残基57から73を置換したアラニン(図15)又はリジン(図16)を有するA−グリアジン57−73 QE65に相当するペプチドを、3日間のグルテンチャレンジ開始から6日後にセリアック病のボランティアからの末梢血単核細胞(PBMC)を用いたIFNガンマELISPOTにおいて比較した(n=8)。(BLはブランク、EはA−グリアジン57−73 QE65:QLQPFPQPELPYPQPQS(配列番号:2))。
【0188】
A−グリアジン60−70 QE65(PFPQPELPYPQ(配列番号:14)に相当する残基はA−グリアジン57−73 QE65において生物活性に実質貢献することを見いだした。60−70位において置換されたA−グリアジン57−73 QE65のバリアントは2段階の手法により査定する。最初に、対照的な特性を有する、10の異なるアミノ酸を用いて60−70位において置換されたA−グリアジン57−73 QE65を査定した。A−グリアジン57−73 QE65のバリアントの第2群(全ての他の天然に生じるアミノ酸により置換されているが、修飾に敏感であると証明された位置のシステインを除く)を第2ラウンドにおいて査定した。
実施例10
A−グリアジン57−73 QE65の置換されたバリアントのアゴニスト活性
A−グリアジン60−70 QE65は、A−グリアジンにおいて優勢なT細胞エピトープのコア配列である。このエピトープのアンタゴニスト及び非アゴニストペプチドバリアントは、このコア配列の修飾により生成されるらしい。最初に、対照的な特性を有する、10の異なるアミノ酸を用いて60−70位において置換されたA−グリアジン57−73 QE65を、3日間のグルテンチャレンジ開始から6日後にセリアック病の被験者からのPBMCを用いたIFNガンマELISPOTにおいて査定する。A−グリアジン57−73 QE65のバリアントの第2群(システイン以外、全ての他の天然に生じるアミノ酸により置換されている)も第2ラウンドにおいて査定した。両群のペプチド(全て50mcg/mlにおいて、二重)を8人の被験者からのPBMCを用いて査定して、未修飾のペプチドと比較した(アッセイあたり20反復)。様々な研究が、このアッセイにおけるA−グリアジン57−73 QE65の最適な濃度が10から100mcg/mlの間であることを示す。
【0189】
結果を、スポット形成細胞(95%コンフルエントの間隔)の平均応答として、個々の個体の%A−G 57−73 QE65平均応答として表現する。対になっていないt−試験を使用することにより、修飾されたペプチドのELISPOT応答をA−G 57−73 QE65と比較する。スーパー−アゴニストを、p<0.01の有意性のレベルにおいてA−G 57−73 QE65よりも高い応答を有するものとして定義し;部分アゴニストはp<0.01の有意性のレベルにおいてA−G 57−73 QE65よりも低い応答を有するものとして定義し、そして非アゴニストはブランク(ペプチド無しのバッファー)と顕著に異ならないものと定義した。A−G 57−73 QE65のアゴニスト活性の30%又はそれ未満のアゴニスト活性を有するペプチドは、アンタゴニスト活性を評価するための「適切な」部分アンタゴニスト又は非アンタゴニストと考えた(表8及び図17−27参照)。
【0190】
PBMCのA−グリアジン57−73 QE65に対するIFNガンマELISPOT応答は分子レベルにおいて特異性が高い。64位のプロリン(P64)、65位のグルタミン酸(E65)及び66位のロイシン(L66)、及び程度は劣るがQ63,P67,Y68及びP69が修飾に対して特に感受性である。Y61及びY70の置換は共に親ペプチドより30%高い生物活性のスーパー−アゴニストを生じ、おそらくは、HLA−DQ2への結合を増強するためであるが、何故ならば、このHLA分子のモチーフが1位及び9位の巨大な疎水性残基に関する選択性を示すからである。18の非アゴニストペプチ
ドを同定した。バリアントの生物活性(50mcg/ml):P65,K64,K65及びY65(生物活性7−8%)はブランク(7%)と同等であった。全部で、A−グリアジン57−73 QE65の57の変異バリアントがA−グリアジン57−73 QE65の30%又はそれ未満の生物活性であった。
【0191】
優勢なエピトープ、A−グリアジン57−73 QE65に対する末梢血リンパ球(PBL)T細胞応答の分子特異性は、HLA−DQ2+セリアック病被験者の間で一貫して再現性があり、そして上記7アミノ酸の中の限定された数のアミノ酸に特異性が高い。A−グリアジン57−73 QE65の特定の単一アミノ酸バリアントは、全てのHLA−DQ2+セリアック病被験者において一貫して非アゴニストである。
実施例11
置換されたバリアントのアンタゴニスト活性
HLA−DQ2+被験者におけるA−グリアジン57−73 QE65に対するPBL
T細胞応答の均質性は、PBL T細胞応答がポリクローナル又はオリゴクローナルであるらしいとしても、PBMCにおいてか又はエクスビボにおいて拮抗可能な変更されたペプチドリガンド(APL)が存在するかもしれないことを示唆する。APLアンタゴニストは一般に弱いアゴニストである。30%又はそれ未満のアゴニスト活性のA−グリアジン57−73 QE65の57の単一アミノ酸置換バリアントが同定されて、APLアンタゴニストとしての適切な候補である。さらに、A−グリアジン57−73 QE65の特定の弱い生物活性の天然に生じる多型も同定されて(以下参照)、「天然に生じる」APLアンタゴニストかもしれない。また、MHCへの結合が抗原特異的T細胞免疫とも拮抗するかもしれないことが示唆された。よって、グルテンチャレンジ後にセリアックPBMCにおいてIFNガンマ応答を誘導しないがHLA−DQ2に結合することが知られている非グリアジンペプチドは、A−グリアジン57−73 QE65により誘導されるT細胞応答を低下させることができるかもしれない。HLA−DQ2へ貪欲に結合する2つのペプチドはHLAクラス1α46−60(HLA 1a)(PRAPWIEQEGPEYW(配列番号:15))及びチロイドペルオキシダーゼ(tp)632−645Y(IDVWLGGLLAENFLPY(配列番号:16))である。
【0192】
3日間のグルテンチャレンジを開始後のセリアックボランティア6人からのPBMCを用いたIFNガンマELISPOTにおけるペプチド(50μg/ml)又はバッファー及びA−グリアジン57−73 QE65(10μg/ml)の同時添加(n=5)。結果は、ペプチドプラスA−G 57−73 QE65による応答として(二重の平均)、バッファープラスA−G 57−73 QE65による応答(20反復の平均)の%応答として表した(表9参照)。
【0193】
A−グリアジン57−73 QE65の4つの単一アミノ酸置換バリアントは、A−グリアジン57−73 QE65に対するインターフェロンガンマPBMC ELISPOT応答(p<0.01)を25%から28%低下させ、別の13のペプチドバリアントはELISPOT応答を18%から24%低下させた(p<0.06)。HLA−DQ2バインダー、チロイドペプチド(tp)632−645Yは、A−グリアジン57−73 QE65に対するPBMCインターフェロンガンマ応答を31%低下させる(p<0.0001)が、他のHLA−DQ2バインダー、HLAクラス1a46−60は応答を変化させない(表9及び10参照)。A−グリアジン57−73のトランスグルタミナーゼ修飾された多型に相当するペプチド、SwissProt受け入れ番号P04725 82−98 QE90(PQPQPFPPELPYPQPQS(配列番号:17))は、A−グリアジン57−73 QE65に対する応答を19%低下させる(p<0.009)(表11参照)。
【0194】
ELISPOTアッセイにおいてのA−グリアジン57−73 QE65に対するPB
MCのインターフェロンガンマ応答は、特定の単一アミノ酸A−グリアジン57−73 QE65バリアント、A−グリアジン57−73 QE65の多型、及び5倍過剰にHLA−DQ2を結合することが公知の非関連ペプチドの同時投与により低下する。これらの発見は、A−グリアジン57−73 QE65の変更されたペプチドリガンドアンタゴニストが存在することを示唆する。仮想上のAPLのみならず、HLA−DQ2を結合する特定のペプチドも、A−グリアジン57−73 QE65に対するPBL T細胞応答を有効に低下させる。
【0195】
これらの発見は、HLA−DQ2+セリアック病において優勢なA−グリアジンエピトープに対するT細胞応答を破壊する2つの戦略を支持する。
【0196】
1.「伝統的な」ペプチド薬剤としての使用のため又はコムギにおけるグリアジン遺伝子の特異的遺伝的修飾のための、一つの位置より多い位置において(64−67)アミノ酸を置換することによる、APLアンタゴニストの最適化。
【0197】
2.HLA−DQ2に付随してA−グリアジン57−73 QE65の提示を競合阻害するための、高親和性HLA−DQ2結合ペプチドの使用。
【0198】
これらの2つのアプローチは相互に両立可能かもしれない。F61及びQ70をチロシン残基で置き換えることにより、スーパー−アゴニストを生成した。これらのスーパー−アゴニストはT細胞受容体との増強された接触よりむしろHLA−DQ2への結合の改善によりもたらされたらしい。これらの修飾を適度に有効なAPLアンタゴニストを生成する他の置換と組み合わせることにより、置換されたA−グリアジン57−73 QE65バリアントの阻害効果を実質上増強するかもしれない。
実施例12
PBMC及びA−グリアジン57−73 QE65及びP04724 84−100 QE92をセリアック病の診断法として使用したインターフェロンガンマELISpotの開発:新たに診断されたセリアック病における免疫応答の定義
インターフェロン番号ELISpotにおいて測定されたPBMC中の優勢なA−グリアジンT細胞エピトープに対する応答性の誘導を、長期間の厳格なグルテンフリーダイエット(GFD)を受けたほとんど全てのDQ2+セリアック病被験者においてはグルテンチャレンジに続いて行ったが、厳格なGFDを受けて4週間後の健康なDQ2+被験者においては続いて行わなかった。A−グリアジン57−73 QE65応答はグルテンチャレンジ前のセリアック病被験者のPBMCにおいて測定不可能であり、そしてパイロットデータはこれらの応答が未処理の被験者のPBMCにおいて測定できなかったことを示唆した。これらのデータは、セリアック病において、A−グリアジン57−73 QE65に対する免疫応答性が抗原排除(GFD)後に復元することを示唆する。ELISpotアッセイ及びPBMCを用いて診断試験が開発されるとしたら、グルテンチャレンジが
血液中のA−グリアジン57−73 QE65及び他の免疫反応性グリアジンペプチドに対する応答を誘導できる前に必要とされるGFDの期間を定義することが望まれる。
【0199】
新たに診断されたDQ2+セリアック病被験者を、胃腸学の外来患者サービスから補充した。PBMCを調製して、被験者がGFDを始める前及びGFDを初めて1週間又は2週間後に、インターフェロンガンマELISpotアッセイにおいて試験した。さらに、グルテンチャレンジ(3日間は4スライスの標準の白パンを摂取、200g/日)を、GFD開始の1又2週間後に実施した。PBMCを調製し、そしてグルテンチャレンジを開始して6日後にアッセイした。A−グリアジン57−73 QE65(A),P04724 84−100 QE92(B)(単独及び化合して)及びA−グリアジン57−73
QP65(P65)(非生物活性バリアント、上記参照)(全て25mcg/ml)を査定した。
【0200】
1人の患者を除いて皆、新たに診断されたセリアック病患者はDQ2+であった(一人はDQ8+であった)(n=11)。未処置か又はGFDの1又は2週間後の新たに診断されたセリアック病患者からのPBMCは、A−グリアジン57−73 QE65及びP04724 84−100 QE92(単独及び化合して)に対する応答を示さずブランク又は及びA−グリアジン57−73 QP65と顕著な差異はなかった(n=9)(図28参照)。たった1週間しかGFDを受けなかったセリアック病患者におけるグルテンチャレンジは、A−グリアジン57−73 QE65及びP04724 84−100 QE92(単独及び化合して)に対する応答を実質上増強しなかった。しかし、GFDを開始して2週間後のグルテンチャレンジは、A−グリアジン57−73 QE65及びP04724 84−100 QE92(単独及び化合して)に対する応答を誘導し、非生物活性バリアントA−グリアジン57−73 QP65及びブランクより顕著に高かった。グルテンチャレンジ2週間後のこれらの応答は価値のあることであるが、GFD開始後>2カ月の被験者よりは低いらしい。A−グリアジン57−73 QE65単独に対する応答は、P04724 84−100 QE92単独に対する応答又はA−グリアジン57−73 QE65と混合した場合の応答と等しいか又はより大きかった。被験者のだれもがグルテンチャレンジの苦痛の兆候を経験しなかった。
【0201】
A−グリアジンに対する免疫応答性(グルテンチャレンジ後にPBMC中で測定)はGFDを開始して2週間後に部分的に回復したことから、この優勢なT細胞エピトープに対する「免疫非応答性」は、未処置のセリアック病患者において且つGFD開始後の少なくとも1週間は優勢であることを示唆する。グルテンチャレンジ及びELISpotアッセイにおけるA−グリアジン57−73 QE65に対する応答の測定を用いたセリアック病の診断試験のタイミングは、GFDを開始して少なくとも2週間後である。
【0202】
A−グリアジン57−73 QE65に特異的なインターフェロンガンマ分泌T細胞は未処置のセリアック病患者においては末梢血中にて測定できず、GFDの少なくとも2週間後のグルテンチャレンジにより誘導することができる(抗原排除)。よって、この方法論を用いた診断試験のタイミングは非常に重大であり、そしてその最適化にはさらなる研究が必要である。これらの発見は、抗原排除により逆転した優勢なエピトープ、A−グリアジン57−73 QE65に特異的なT細胞の機能アネルギーと一致する。この現象は、ヒト疾患において以前には証明されておらず、T細胞アネルギーがセリアック病においてはペプチド治療により誘導可能かもしれないという可能性を支持する。
実施例13
コムギグリアジンT細胞エピトープの包括的マッピング
抗原チャレンジは末梢血中で抗原特異的T細胞を誘導する。セリアック病においては、グルテンがこの免疫媒介性疾患を維持する抗原である。グルテンフリーダイエットで処置されたセリアック病のグルテンチャレンジは末梢血中のグルテン特異的T細胞の出現を導き、グルテンT細胞エピトープの分子特異性の決定を可能にさせる。上記のとおり、我々はモデルグルテン蛋白質中の単一の優勢なエピトープ、A−グリアジン(Q65において脱アミド化された57−73)を同定した。この実施例においては、セリアック病患者のグルテンチャレンジを用いて、GenBank中の111エントリーに由来する各公知のコムギグリアジン蛋白質中の全ての有力な12のアミノ酸配列を試験した。全部で、652の20マーペプチドをHLA−DQ2及びHLA−DQ8関連のセリアック病において試験した。セリアック病において90%を超えて存在する古典的なHLA−DQ2複合体(HLA−DQA1*05,HLA−DQB1*02)のセリアック病被験者9人のうちの7人は、末梢血中に誘導可能なA−グリアジン57−73 QE65−及びグリアジン−特異的T細胞応答を有した。A−グリアジン57−73は、HLA−DQ2関連セリアック病において、唯一の重要なα−グルテンT細胞エピトープであり、並びにもっとも有力なグリアジンT細胞エピトープであった。さらに、インターフェロン−γ ELISp
otアッセイにおいてA−グリアジン57−73ほど有効性のある10%から70%の構造上関連したペプチドの5ファミリーほど存在した。これらの新規なT細胞エピトープは、γ−及びω−グリアジンに由来しており、そしてA−グリアジン57−73のコア配列(コア配列:FPQPQLPYP(配列番号:18))、例えばFPQPQQPFP(配列番号:19)及びPQQPQQPFP(配列番号:20)と構造上極めて類似しているが同一ではない共通配列を含んだ。A−グリアジン57−73の相同体はライムギ又はオオムギにおいて見いだされなかったが、他の2つの穀類はセリアック病において毒性であり、γ−及びω−グリアジン中の新たに定義されたT細胞エピトープはライムギ及びオオムギの保存蛋白質において完全に適合した(それぞれ、セカリン(secalins)及びホルデイン(hordeins))。
【0203】
HLA−DQ2に関連しないセリアック病はほとんど常にHLA−DQ8と関連していた。7人のHLA−DQ8+セリアック病被験者のだれもが、彼らが完全なHLA−DQ2複合体も所有していないなら、誘導可能なA−グリアジン57−73−特異的T細胞応答をグルテンチャレンジ後に有さなかった。完全なHLA−DQ2複合体を有さない4人のHLA−DQ8+セリアック病被験者のうちの2人は、誘導可能なグリアジンペプチド特異的T細胞応答をグルテンチャレンジ後に有した。一人のHLA−DQ8被験者においては、新規な優勢T細胞エピトープが、コア配列LQPQNPSQQQPQ(配列番号:21)を用いて同定された。このペプチドのトランスグルタミナーゼによる脱アミド化バージョンは非脱アミド化ペプチドよりも有効性であった。以前の研究は、トランスグルタミナーゼにより脱アミド化したペプチドが配列LQPENPSQEQPE(配列番号:22)を有するはずであることを示唆したが;この配列を確認するためには、さらなる研究が必要である。1カ月の間グルテンフリーダイエットを受けた健康なHLA−DQ2(10)及びHLA−DQ8(1)の被験者の間で、グリアジンペプチド特異的T細胞応答は不変であり、グルテンチャレンジによりほとんど変わらず、そして、セリアック病の被験者においてグルテンチャレンジにより明らかにされた有効性T細胞エピトープでは決してなかった。結論として、HLA−DQ2関連セリアック病において6より多い重要なT細胞エピトープが存在するらしく、そのうち、A−グリアジン57−73がもっと有効性である。HLA−DQ2−及びHLA−DQ8−関連セリアック病は同じT細胞特異性を共有しない。
【0204】
我々は、グルテンフリーダイエット後のセリアック病の個体の短期間のグルテンチャレンジが末梢血中にてグリアジン特異的T細胞を誘導することを示した。これらのT細胞の頻度は末梢血中で6日目に最大であり、続いて、次の週を通して迅速に弱まる。末梢血のグリアジン特異的T細胞は消化管の粘膜固有層へのホーミングに関連するインテグリンα47を発現する。我々は、原型的なグルテンα−グリアジン蛋白質、A−グリアジンにおいてセリアック病に関連したT細胞エピトープをマップするためにこのヒト抗原−チャレンジデザインを開発した。トランスグルタミナーゼ(tTG)による脱アミド化が有る場合も無い場合も10アミノ酸がオーバーラップする15マーのペプチドを用いて、我々は、末梢血中で誘導されたT細胞が、65位のグルタミン酸が脱アミド化されたひとつのA−グリアジンペプチドの残基57−73を最初に標的化することを証明した。当該エピトープはHLA−DQ2−制限されており、HLA−DQ2のセリアック病との密接な関連と一致する。
【0205】
コムギ、ライムギ、及びオオムギの暴露によりセリアック病は再活性化される。コムギのグルテンのα/β−グリアジン画分はセリアック病においては一致して毒性であり、そしてほとんどの研究はこれらの蛋白質に焦点を合わせてきた。α/β−グリアジンをコードする遺伝子クラスターはコムギ染色体の6C上に位置する。ライムギ又はオオムギにはα/β−グリアジンの相同物がない。しかしながら、コムギのグリアジンサブタイプの3つ全て(α/β、γ、及びω)がセリアック病において毒性である。γ−及びω−グリア
ジン遺伝子はコムギでは染色体1APL上に位置しており、ライムギ及びオオムギのセカリン及びホルデインに相同である。
【0206】
パンコムギ(Triticum aestivum)においては61のα−グリアジンに関して同定された遺伝子が今存在する。α−グルテン配列は密接に相同であるが、A−グリアジンにおいて優勢なエピトープは、α−グリアジン配列においてほとんど多型性の領域に由来する。Andersonら(1997)は、パンコムギ(T.aestivum)中に全部で約150の異なるα−グリアジン遺伝子が存在するが、多くは偽遺伝子であると見積もった。よって、セリアック病に関連するT−細胞のエピトープが公知のα−グリアジン配列中に含まれないことは、ありそうもない。
【0207】
我々の研究は、セリアック病に特異的な有効性のT細胞エピトープとしてA−グリアジン57−73にほとんど同一の、脱アミド化されたα−グリアジンペプチドのグループを同定した。90%を超えるセリアック病患者はHLA−DQ2+であり、これまで、我々はグルテンチャレンジ後のHLA−DQ2+セリアック病被験者のみを査定してきた。しかしながら、HLA−DQ2を発現しないセリアック病患者がほとんど皆HLA−DQ8を有する。よって、他のコムギ、ライムギ及びオオムギグルテン蛋白質中のA−グリアジン57−73及びその相同体が、HLA−DQ2+及びHLA−DQ8+セリアック病の両方においてグルテンチャレンジにより誘導されるT細胞により認識される唯一のT細胞エピトープであるか否かを知ることは非常に不可欠である。これが事実なら、セリアック病のための治療用のペプチドのデザインは、たった一つのペプチドのみしか必要としないかもしれない。
A−グリアジン57−73のT細胞エピトープとしての相同体
A−グリアジン57−73(PQLPY<配列番号:12>)のコア配列をコードする穀物遺伝子に関するSwissProt及びTrembl遺伝子データベースの最初の検索は、α/β−グリアジンを明らかにしただけであった。しかしながら、A−グリアジン57−73 QE65エピトープの我々の初期のマッピング研究は、コア領域(PQLP)中の許容可能な点の置換が限定された数であることを明らかにした(Q65はエピトープ中で実際に脱アミド化されている)。よって、我々は、配列XXXXXXXPQ[ILMP][PST]XXXXXX(配列番号:23)を有するペプチドをコードするSwissProt及びTremblデータベースの中の遺伝子に我々の検索を拡大した。相同体は、γ−グリアジン、グルテニン、ホルデイン及びセカリンの間に同定された(表12参照)。さらなる相同体は、GenBankにおいて3つのω−グリアジンエントリーの可視検索によりω−グリアジン中に同定された。
【0208】
A−グリアジン57−73のこれらの相同体は、tTGによる脱アミド化(又は4つの密接な相同体中のグルタミン酸(QE)−置換されたバリアントの合成)の後に、IFNγELISpotアッセイを用いて、末梢血単核細胞を用いて、セリアック病の被験者においてグルテンチャレンジ後に査定した。ω−グリアジン配列(AAG17702 141−157)は唯一の生物活性ペプチドであり、約半分がA−グリアジン57−73として有効である(表12、及び図29参照)。よって、優勢なA−グリアジンエピトープの相同体の検索は、γ−グリアジン、セカリン、及びホルデインの毒性に関して考慮することを怠った。
方法
全ての可能性のあるコムギグリアジンT−細胞エピトープにわたるペプチドのセットのデザイン
公知のパンコムギ(Triticum aestivum)のグリアジン遺伝子又は遺伝子断片によりコードされた全ての可能なT細胞エピトープを同定するため(GenBank中の61α/β−,47γ−,及び3ω−グリアジンエントリー)、遺伝子由来の蛋白質配列を、CustalWソフトウエア(MegAlign)を用いてアラインメント
を行い、そして分類することにより系統発生グループ化した(表22参照)。多くのエントリーは、より長い配列の末端削除を表し、そして多くの遺伝子セグメントは、ポリグルタミン繰り返しの長さ又は稀な置換を除いて同一であった。よって、公知のコムギ遺伝子によりコードされる全ての有力な12アミノ酸配列を有理化して(rationalize)652の20マーペプチドのセットに含ませることができた。(シグナルペプチド配列は含まれなかった)。ペプチド配列を表23に掲載する。
包括的エピトープマッピング
4週間グルテンフリーダイエットを受けた健康な対照(HLA−DQ2+ n=10,及びHLA−DQ8+ n=1)、及び長期間のグルテンフリーダイエットを受けたセリアック病の被験者(6人のHLA−DQ2,4人の複合同型接合HLA−DQ2/8、及び3人のHLA−DQ8/X)(表13参照)を、グルテンチャレンジの前及び後の6及び7日目に調査した(標準の白パン50gスライス4切れ−セインズベリーズサンドイッチパン、毎日)。末梢血(7日間にわたり全部で300g)を採血して、末梢血単核細胞(PBMC)をLymphoprep密度勾配により分離した。PBMCを6又は8の20マーペプチド、又はtTGにより脱アミド化されたか又はされていない単一のペプチドのプールと、一晩、インターフェロンガンマ(IFNγ)ELISpotアッセイにおいてインキュベートした。
【0209】
ペプチドはPepsets(Mimotopes Inc.,メルボルン、オーストラリア)において96のバッチ中で合成した。約0.6マイクロモルの各651の20マーを提供した。2つのマーカー20マーペプチドを96の各セットに含ませ(VLQQHNIAHGSSQVLQESTY)−ペプチド161(配列番号:24)、そしてIKDFHVYFRESRDALWKGPG(配列番号:25)を逆相−HPLC及びアミノ酸配列分析により特性決定した。これらのマーカーペプチドの平均純度はそれぞれ50%及び19%であった。ペプチドを最初にアセトニトリル(10%)及びHepes 100mMから10mg/mlに溶解した。
【0210】
IFNγELISpotアッセイのためにPBMCとインキュベートされたプール(又は単独)中の個々のペプチドの最終濃度は20μg/mlであった。ペプチドの5倍濃縮した溶液及び1mMの塩化カルシウムを含むPBS中のプールを分配して、ELISpotアッセイにおいて後に使用されるテンプレートに従い96ウエルプレート中に保存した。モルモット組織tTG(Sigma T5398)との100:32の比における2時間37℃のインキュベーションにより、脱アミド化されたペプチド及びペプチドのプールを調製した。ペプチド溶液を−20℃において保存して、使用前に新たに解凍した。
【0211】
エンドトキシンフリーの水及び2M尿素中のグリアジン(Sigma G3375)(100mg/ml)を10分間ボイルし、室温に冷却し、そしてフィルター(0.2μm)−滅菌されたHCl 0.02M中のペプシン(Sigma P6887)(2mg/ml)又は重炭酸アンモニウム中のキモトリプシン(C3142)(4mg/ml)とインキュベートした。4時間のインキュベーション後に、ペプシン消化されたグリアジンを水酸化ナトリウムにより中和し、そして次にペプシン−消化グリアジン及びキモトリプシン−消化グリアジンの両方を15分間ボイルした。グリアジンを除いたプロテアーゼとの同一のインキュベーションも実施した。サンプルを15 000gにおいて遠心分離し、次に、蛋白質濃度を上清中でBCA法(Pierce,USA)により見積もった。IFNγELISpotアッセイにおける最終的な使用の前に、グリアジン−プロテアーゼのアリコートをtTGと共に比率2500:64μg/mlにてインキュベートした。
【0212】
IFNγELISpotアッセイ(Mabtech,スエーデン)を96−ウエルプレート(MAIP S−45,Millipore)中で、各ウエルが25μlのペプチド溶液及び100μlのPBMC(2−8105/ウエル)を、10%熱不活性化ヒトAB
血清を含むRPMI中に含むように、実施した。脱アミド化されたペプチドプールを一つの96−ウエルELISpotプレート中で査定し、そして第2プレート上で脱アミド化なしのペプチドプール(同一のレイアウト)を0日目及び6日目に査定した。脱アミド化されたペプチドを含むプレート中の全てのウエルがtTGを含んだ(64μg/ml)。各ELISpotプレート中に、ペプチドプールを含む83のウエルが存在し(各ウエル中唯一のプール)、そして「対照」ペプチドのウエルのシリーズ(ペプチドは全て>90%の純度、MS及びHPLCにより特性決定された、Research Genetics):P04722 77−93(QLQPFPQPQLPYPQPQP(配列番号:26)、P04722 77−93 QE85(二重)(QLQPFPQPELPYPQPQP(配列番号:27)、P02863 77−93(QLQPFPQPQLPYSQPQP(配列番号:28)、P02863 77−93 QE85(QLQPFPQPELPYSQPQP(配列番号:29)、及びキモトリプシン消化されたグリアジン(500μg/ml)、ペプシン消化されたグリアジン(500μg/ml)、キモトリプシン(20μg/ml)単独、ペプシン(10μg/ml)単独、及びブランク(PBS+/−tTG)(二重)。
【0213】
展開及び乾燥後に、IFNγELISpotプレートをMAIP自動化スポット形成ELISpotカウンターを用いて査定した。HLA−DQ2の健康被験者及びセリアック病被験者において、IFNγELISpotアッセイにおいてペプチドプールによりスポット形成細胞(sfc)の誘導を、100万PBMCあたりのスポット形成細胞(sfc)マイナスブランクに適用した、一方をつないだ(one−tailed)Wilcoxon Matched−Pairs Signed−Ranks試験(SPSSソフトウエア)を用いて、6日目体0日目(「正味の応答」)にて試験した。インビボグルテンチャレンジによるPBMC中のペプチドプールに対するIFNγ応答の顕著な誘導を、少なくとも10sfc/100万PBMC及び有意性レベルp<0.05の中央値の「正味の応答」として定義した。6日目のペプチドの特定のプールに対する顕著な応答に、同日又は7日目に採られたPBMCを使用して各プール内において個々のペプチドの評価を続けた。
【0214】
個々のペプチドのIFNγELISpotアッセイに関して、生物活性を同じELISpotプレート中で査定されたP04722 77−93 QE85に対する応答のパーセントとして表現した。36万(範囲:0.3−0.72)、PBMCの中央値を用いて、ブランクに対する中央値の応答(PBS単独)はウエルあたり0.2sfc(範囲0−5)であり、そしてウエルあたり陽性対照(P04722 77−93 QE85)76.5(範囲25−383)sfcであった。よって、P04722 77−93 QE65のパーセンテージとして表現されたブランクに対する中央値の応答は0.2%(範囲:0−6.7)であった。P04722 QE85のそれより10%高い平均生物活性を有する個々のペプチドを共通の構造上のモチーフに関して分析した。
結果
健康なHLA−DQ2被験者
1カ月グルテンフリーダイエットを受けた健康なHLA−DQ2+被験者の誰も、グルテンチャレンジの前又は後にA−グリアジン57−73の相同体に対してIFNγELISpot応答を有さなかった。しかしながら、10人の健康な被験者のうち9人において、グルテンチャレンジが、グリアジンのペプシン消化及びキモトリプシン消化の両方に対するIFNγ応答の顕著な増加を伴い、0日目の0−4sfcの中央値/百万から16−29sfcの中央値/百万であった(表14参照)。健康な被験者のグリアジン応答は脱アミド化により影響されなかった(表15参照)。健康な被験者のうち、特定のグリアジンペプチドプールに対するIFNγ応答の、グルテンチャレンジとの一致した誘導はなかった(図30、及び表16参照)。IFNγELISpot応答は場合によりみられたが、弱く、そして脱アミド化により影響されなかった。プールに対するもっとも強い応答の
多くも0日目には存在しなかった(表17、被験者H2,H8及びH9参照)。4人の健康な被験者はプール50に対して明確な応答を示し、6日目にもっとも強い応答をもった2人は0日目にも応答を有した。両被験者において、プール50に対するチャレンジ後の応答はペプチド390(QQTYPQRPQQPFPQTQQPQQ(配列番号:30))によった。
HLA−DQ2セリアック病被験者
HLA−DQ2+セリアック病被験者におけるグルテンチャレンジの後、P04722
77−93 QE85に対する中央値のIFNγELISPpot応答は0から133sfc/百万の中央値から上がった(表4参照)。6人のセリアック病被験者のうちの一人(C06)はP04722 77−93 QE85(2sfc/百万)に応答せず、そしてグリアジンペプチドプールに弱く応答しただけであった(最大:プール50+tTG
27sfc/百万)。前の研究に一致して、野生型P04722の生物活性はtTGによる脱アミド化により6.5倍上昇した(表15参照)。グリアジン消化に対するインターフェロンガンマ応答はベースラインに存在したが、キモトリプシン−グリアジンに関しては20から92sfcの中央値/百万まで、そしてペプシン−グリアジンに関しては44から176sfcの中央値/百万まで、グルテンチャレンジにより実質上は増加した。グリアジンの脱アミド化は、キモトリプシン−グリアジンに関して3.2倍の中央値、そしてペプシン−グリアジンに関して1.9倍の中央値だけ生物活性を増加させた(表15参照)。(ペプシンによる消化に要求される酸性度はグリアジンの部分脱アミド化をもたらすらしい。)
健康な被験者とは対照的に、グルテンチャレンジは、α−,γ−、及びω−グリアジンを含む83の処理されたプールのうちの22に対してIFNγELISpot応答を誘導した(図31及び表17参照)。プールの生物活性は被験者間で一致していた(表18参照)。ペプチドプールにより誘導されたIFNγELISpot応答は脱アミド化によりほとんどいつも増加した(表17参照)。だが、脱アミド化によるプールの生物活性の増強は、A−グリアジン57−73の相同体を含むプールに関してさえ、P04722 77−73 QE85ほど顕著ではなかった。これは、Pepsetペプチドが合成又は調製の間に部分的に脱アミド化されていたことを示唆し、例えばPepsetペプチドはトリフルオロ酢酸(TFA)の塩としてTFA溶液からの凍結乾燥後にデリバリーされる。
【0215】
21のもっとも生物活性なプールからの170の個々のtTG脱アミド化ペプチドを別々に査定した。72の脱アミド化ペプチドは均等な濃度においてP04722 77−93 QE85より10%生物活性が高かった(20μg/ml)(表19参照)。5つのもっとも有効なペプチド(P04722 QE85の85−94%の生物活性)は、以前にα−グリアジン相同A−グリアジン57−73として同定された。50の生物活性ペプチドはA−グリアジン57−73と相同でなかったが、構造上関連する配列の6ファミリーに分類できた(表20参照)。各々のペプチドファミリーのもっとも生物活性な配列は:PQQPQQPQQPFPQPQQPFPW(配列番号:31)(ペプチド626、P04722 QE85の中央値72%生物活性)、QQPQQPFPQPQQPOLPFPQQ(配列番号:32)(343、34%)、QAFPQPQQTFPHQPQQQFPQ(配列番号:33)(355、27%)、TQQPQQPFPQQPQQPFPQTQ(配列番号:34)(396、23%)、PIQPQQPFPQQPQQPQQPFP(配列番号:35)(625、22%)、PQQSFSYQQQPFPQQPYPQQ(配列番号:36)(618、18%)(コア配列に下線を引く)であった。これらの配列の全ては、トランスグルタミナーゼによる脱アミド化に感受性であると予測されたグルタミン残基を含む(例えば、QXP,QXPF(配列番号:37),QXX[FY](配列番号:38))(Vader et al 2002参照)。幾つかの生物活性ペプチドは異なるファミリーからの2つのコア配列を含む。
【0216】
異なるT細胞集団が別のコア配列を有するエピトープに応答するという可能性と一致し
て、異なるファミリーからのペプチドの生物活性は付加的らしい。例えば、tTG処理されたプール81の中央値生物活性はP04722 QE85の141%であったが、個々のペプチドの生物活性は:ペプチド631(A−グリアジン57−73の相同体)61%、636(626の相同体)51%、そして635 19%、629 16%、そして634 13%(全て396の相同体)の順位の序列であった。
【0217】
単純化しすぎたかもしれないが、グリアジンペプチドに対する合計のIFNγELISpot応答に対する各「ペプチドファミリー」の貢献をHLA−DQ2+セリアック病被験者において比較した(図32参照)。従って、グリアジンペプチドに対する合計の応答に対するP04722 77−73 E85の貢献は、1/5と2/3の間であった。
【0218】
インターネットのワールドワイドウエブ、例えば「cbrg.inf.ethz.ch/subsection3_1_5.html」を通してアクセス可能な、ペプチド相同性検索プログラム、WWW PepPepSearchを用いて、及びライムギのセカリンのGenBank配列との直接の比較により、厳密な適合が、オオムギのホルデイン(HOR8)及びライムギセカリン(A23277,CAA26499,AAG35598)中にコア配列:QQPFPQPQQPFP(配列番号:39)及びオオムギホルデイン(HOG1及びHOR8)中にコア配列:QQPFPQQPQQPFP(配列番号:40)に関して厳密な適合が発見され、そしてPIQPQQPFPQQP(配列番号:41)に関して、オオムギホルデイン(HOR8)中に発見された。
HLA−DQ8関連セリアック病
7人のHLA−DQ8+セリアック病被験者をグルテンチャレンジの前と後に研究した。これらのHLA−DQ8+(HLA−DQA0*0301−3,HLA−DQB0*0302)被験者のうちの5人も、セリアック病関連HLA−DQ2複合体(DQA0*05,DQB0*02)の一方又は両方を有した。両方のセリアック病関連HLA−DQ複合体を有する3人の被験者のうちの2人は、質的及び量的にHLA−DQ2セリアック病被験者と同一であったグリアジンペプチドプール(及びP04722 77−93 QE85を含む個々のペプチド)に対して有効な応答性を有した(図33及び34、及び表18を参照)。脱アミド化されたペプチドプール74は両HLA−DQ2/8被験者において生物活性であったが、6人のHLA−DQ2/X被験者の一人においてのみであった。プール74のtTGによる前処理は生物活性を3.8倍から22倍増強し、そして3人の応答者におけるtTG処理されたプール74の生物活性は、P04722 77−93 E85の生物活性の78%及び350%の間に等しい。現在、何れのペプチドが被験者C02,C07及びC08においてプール74において生物活性であるかわかっていない。
【0219】
セリアック病に関連したHLA−DQ2対立遺伝子の両方又は一方を欠く4人のHLA−DQ8セリアック病被験者のうちの2人はグリアジンペプチドプールに対して極めて弱いIFNγELISpot応答しか示さなかったが、残りの2人はプロテアーゼ−消化されたグリアジン及び特異的ペプチドプールの両方に応答した。被験者C12(HLA−DQ7/8)は、脱アミド化されたプール1−3に対して激しく応答した(図35参照)。これらのプールにおける個々のペプチドの評価は、コア配列LQPQNPSQQQPQ(配列番号:42)を含む一連の密接に関連した生物活性ペプチドを同定した(表20参照)。以前の研究(我々による)は、この配列中の3つのグルタミン残基がtTG媒介脱アミド化に感受性であることを証明した(下線)。WWW PepPepSearchを用いた相同性検索は、コムギα−グリアジンにおいてのみLQPQNPSQQQPQ(配列番号:43)に対して密接な適合を同定した。
【0220】
4人のHLA−DQ8被験者(C11)は、tTG処理されたプール33に対して誘導可能なIFNγELISpot応答を有した(図36参照)。プール32及び33はtTGによる脱アミド化後に活性な以前に定義されたHLA−DQ8制限されたグリアジンエ
ピトープ(QQYPSGQGSFQPSQQNPQ(配列番号:44))の多型を含む(下線を引かれたGlnは脱アミド化されて生物活性を運ぶ)(van der Wal et al 1998)。現在、何れのペプチドがプール33において被験者C11において生物活性であるか分かっていない。
【0221】
インビボグルテンチャレンジ及びコムギグリアジン中の全ての公知の12アミノ酸配列にわたる652ペプチドのセットを使用した、HLA−DQ2関連セリアック病における包括的T細胞エピトープマッピングは、即ち、公知のα−グリアジンエピトープ、A−グリアジン57−73 QE65の10%の生物活性の少なくとも72のペプチドを同定した。しかしながら、これらの生物活性ペプチドはほんの5つほどの異なるが密接に関連したペプチドファミリーのセットに減らされ得る。ほとんど全てのこれらペプチドは、プロリン、グルタミン、フェニルアラニン、及び/又はチロシンが豊富であり、そして配列PQ(QL)P(FY)P(配列番号:45)を含む。この配列はtTGによる2位のQの脱アミド化を促進させる。A−グリアジン57−68の脱アミド化による類推により(Arentz−Hansen 2000)、tTGによる脱アミド化により広く見いだされたこれらのペプチドの増強された生物活性は、HLA−DQ2に関する結合の親和性の増加よるかもしれない。
【0222】
これらの生物活性グリアジンペプチドの一つより多くをインビボにおいて認識するT細胞の間の交差反応性は起こり得る。しかしながら、関連するペプチドの各セットが別のT細胞集団をインビボにおいて活性化するなら、A−グリアジン57−73 E65に相当するエピトープはもっとも有効であり、そしてグルテンチャレンジの後のグリアジンに対する応答においてIFNγを分泌する末梢血T細胞の少なくとも40%により広く認識される。
【0223】
非グリアジン−ペプチド特異的応答が、HLA−DQ2/X関連のセリアック病におけるものと質的に異なったHLA−DQ2/8セリアック病において見いだされた。しかしながら、HLA−DQ2対立遺伝子の両方を持たないHLA−DQ8+のセリアック病被験者の末梢血T細胞は、A−グリアジン57−73 E65相同体を認識しなかった。2つの異なるエピトープが2つのHLA−DQ8+セリアック病において優勢であった。れらのHLA−DQ8+の個体における優勢なエピトープは以前に同定されていなかった(LQPQNPSQQQPQ(配列番号:46))。
【0224】
本明細書中の教示を仮定すれば、共通に認識されるT細胞エピトープの全てを利用してセリアック病のための免疫治療のデザインが実用的になり、6より少ない異なるペプチドを含むかもしれない。コムギのγ−及びω−グリアジン中のエピトープは、オオムギホルデイン及びライムギセカリン中にも存在する。
実施例14
幾つかのELISpotアッセイを前記のとおりに実施して、以下の結果及び/又は結論を生じた:
PQLPYによる複数のα−グリアジン多型の拡大
A−グリアジン57−73QEの有効なアゴニスト(G01)は、QLQPFPQPELPYPQPQS(G01),PQL−Y−−−−−−−−−−−−−−P(G10),及びPQPQPFL−−−−−−−−−−−−−(G12)を含む。有効性が劣るのは、−−−−−−−−−−−−L−−−−−−−−P(G04),−−−−−−−−−−−−−R−−−−−−−−−−−P(G05),及び−−−−−−−−−−−−S−−−−−P(G06)を含む。有効性がさらに劣るのは、−−−−−−L−−−−−−−−S−−−−−P(G07),−−−−−S−−−−−−−−−−S−−−−−−−−−P(G08)−−−−−−−−−−−−−−−S−−S−−−−−P(G09),及びPQPQPFP−−−−−−−−−−−−−−−−−(G13)を含む。ダッシュは特定の位置にお
けるG01配列との同一性を示す。
グルテンチャレンジは新たに診断されたセリアック病においてたった2週間のグルテンフリーダイエット後にA−グリアジン57−73 QE65 T細胞を含む
追加の分析は、tTG脱アミド化されたグリアジン応答が新たに診断されたセリアック病において2週間後に変化することを示した。他の分析は、脱アミド化されたグリアジン特異的T細胞がCD4+α4β7+HLA−DQ2制限されることを示した。
最適なエピトープ(クローン対グルテンチャレンジ)
「優勢な」エピトープがγIFN ELISpotによりグルテンチャレンジ後に定義される。QLQPFPQPELPYPQPQS(100%ELISpot応答)。腸のT細胞クローンにより定義されたエピトープ:QLQPFPQPELPY(27%),PQPELPYPQPELPY(52%),及びOOLPQPEQPQQSFPEQERPF(9%)。
個々のペプチド配列間の順位
順位はコムギ又はライムギに依存する。コムギに関して、優勢なペプチドは、ペプチド番号89、90及び91を含む(表23における番号を参照)。ライムギに関して、優勢なペプチドは、ペプチド番号368、369、370、371、及び372を含む(表23における番号を参照)。635及び636(表23における番号を参照)を含む幾つかのペプチドは、ライムギ及びコムギの両方において活性を示した。
インビボグルテンチャレンジはT細胞エピトープのヒエラルキーがセリアック病に関して定義されることを可能にする
エピトープのヒエラルキーはHLA−DQ2+セリアック病の間で一致しているが、HLA−DQ8+セリアック病に関しては異なる。ヒエラルキーはコムギ穀物が消費されることに依存する。脱アミド化はほとんど全てのグリアジンエピトープを生成する。HLA−DQ2,DQ8及びDR4が脱アミド化されたペプチドを提示する。HLA−DQ2/8−関連のセリアック病は優先的にDQ2−関連グリアジンエピトープを提示する。グリアジンエピトープは、エピトープに基づく治療剤の開発を正当であると理由付けするのに十分制限される。
【0225】
他の分析は以下を示した:HLA−DR3−DQ2(85−95%)及びHLA−DR4−DQ8(5−15%)。
【0226】
他の分析は以下を示した:
【0227】
【表2】
【0228】
被験者12のプール1−3において個々のtTG−脱アミド化されたペプチドの生物活性を測定するために、別の分析を実施した。結果は次のとおりである(配列の番号は表23に掲載されたペプチドを指す):配列8(100%),配列5(85%)、配列6(82%)、配列3(77%)、配列1(67%)、配列2(59%)、配列9(49%)、配列7(49%)、配列10(33%)、配列4(15%)、配列12(8%)、配列11(0%)、配列23(26%)、配列14(18%)、配列15(18%)、配列17(18%)、配列16(13%)、配列14(8%)、配列22(5%)、配列18(3%)、配列19(3%)、配列20(0%)、配列21(0%)。予測された脱アミド化配列はLQPENPSQEQPEである。
PBMCによる個々のELISpot応答(ELISpotリーダーにより測定されたスポット形成細胞)
【0229】
【表3】
【0230】
交差反応性
29人の被験者の652のペプチドからのデータを扱うため、又は特定の応答が真実の陽性のペプチド特異的T細胞応答であるかを決定するため、又はペプチドに対する応答が別の構造上関連するペプチドとの交差反応性によるかを決定するために、特定のペプチド応答の発現を「優勢な」ペプチド応答のパーセンテージとすることができる。或いは、当該発現はペプチド応答間の相関係数としての「遅延(retardness)」とし得るか、又はバイオインフォマティックスにより得る。
追加のエピトープ
代表的な結果は次の通りである:
ペプチドのP04722Eとの組み合わせ(全て20mcg/ml)(n=4)
単独 P04722E+
Pep626 60 135
P04722E 100 110
HLAa 0 85
(P04722Eのパーセントとして表された)
626+tT:PQQPQQPQQPFPQPQQPFPW
04722E:QLQPFPQPELPYPQPQL
ペプチド626のtTG脱アミド化(n=12)
tTGなし=100%
tTG=170%
特定の位置における置換
ペプチド626PQQP[Q1]QP[Q2]QPFPQP[Q3]QPFPVの置換(n=12)
Glu Arg
Q1 95 90
Q2 145 80
Q3 155 10
(野生型ペプチドのパーセントとして表された)
tTG処理された15マーに及ぶペプチド626/627(PQQPQQPQQPFPQPQQPFPWQP)(n=8)
P1−15 5
P2−16 4
P3−17 3
P4−18 38
P5−19 65
P6−20 95
P7−21 65
P8−22 90
(最大の15マー応答のパーセントとして表された)
複数エピトープ:
P04722E:QLQPFPQPQLPYPQPQL
626+tTG:PQQPQQPQQPFPQPQQPFPW
最小エピトープ:QPQQPFPQPQQPFPW
抗−CD4によりコードされたビーズ及び抗インテグリンβ7によるPBMCの免疫磁気除去はIFNγELISpot応答を除去させたが、抗−CD8によりコードされたビーズ又は抗アルファEインテグリンによるPBMCの免疫磁気除去はそうでなかった。即ち、IFNγを分泌するPBMCはCD4+及びα4β7+であり、消化管内の粘膜固有層へのホーミングに関連した。
【0231】
HLA−DQ2に関して異型接合及び同型接合においては、抗−DQ抗体によりブロックされたが抗−DR抗体によりブロックされなかった。これは一つの配列中の複数のエピトープを暗示するかもしれない。
セリアック病のT細胞エピトープ
他の研究者らは、特定の腸T細胞クローンエピトープを特性決定した。例えば、Vader et al.,Gastroenterology 2002,122:1729−37;Arentz−Hansen et al.,Gastroenterology 2002,123:803−809を参照。これらは、T細胞をクローン化するために使用されたインビトロ技術のためにその関係がいくらよく見ても不明であるエピトープの例である。
腸対末梢血クローン
腸:1)腸生検、2)グルテンのペプシン−トリプシン消化に対して生じさせたT細胞クローン、3)全てHLA−DQ2制限される、4)クローンはトランスグルタミナーゼにより脱アミド化されたグリアジンに応答する。
末梢血:グルテンに対して生じさせたT細胞クローンはHLA−DR,DQ及びDP制限される。結果:腸T細胞クローンはセリアック病関連エピトープをマップするのに限定的に使用することができる。
GDA_9コムギ307aa定義アルファ/ベータ−グリアジンMM1前駆体(プロラミ
ン)受け入れ番号P18573−−GenBank(その全体を引用により本明細書に編入する)
腸T細胞クローンエピトープ
腸T細胞クローンエピトープの定義は、例えば、Arentz−Hansen et al.,J Exp Med.2000,191:603−12に見いだすことができる。また、腸T細胞クローンに関するグルテンエピトープも開示されている。脱アミド化されたQLQPFPQPQLPYがエピトープであり、QLQPFPQPELPYの脱アミド化配列もある。HLA−DQ2制限がある。相同性検索は他の生物活性γアルファ−グリアジンがPQPQLPYを単独又は二重に含むことを示す。大多数のT細胞クローンはDQ2−αI:QLQPFPQPELPY DQ2−αII:PQPELPYPQPELPYの何れかに応答する。
優勢なグリアジンT細胞エピトープ
全てトランスグルタミナーゼにより脱アミド化される。
グルテンチャレンジ6日後の末梢血:A−グリアジン57−73:
QLQPFPQPELPYPQPQS
腸T細胞クローン:DQ2−αI:QLQPFPQPELPY DQ2−αII:PQPELPYPQPELPY
腸T細胞クローンエピトープマッピング
【0232】
【表4】
【0233】
末梢血におけるIFNγ−分泌A−グリアジン57−73 QE65−特異的T細胞のグルテン暴露及び誘導
未処理のセリアック病を1、2又は8週間グルテンフリーダイエットさせ、次にグルテン暴露し(3日 パン200g/日)、次にグルテンフリーダイエットをさせた。
【0234】
結果1:グルテンフリーダイエットの期間及びグルテンチャレンジの0日目及び6日目のIFNγELISpot応答:A−グリアジン57−73 QE65(IFNγ特異的スポット/百万PPBMCとして表された結果)
0日目:なし(0)、1週(1)、2週(2)、8週(1)
6日目:なし(0)、1週(4)、2週(28)、8週(48)
結果2:グルテンフリーダイエットの期間及びグルテンチャレンジの0日目及び6日目のIFNγELISpot応答:tTG−グリアジン57−73 QE65(IFNγ特
異的スポット/百万PPBMCとして表された結果)
0日目:なし(45)、1週(62)、2週(5)、8週(5)
6日目:なし(0)、1週(67)、2週(40)、8週(60)
結果3:グルテンフリーダイエットの期間及びグルテンチャレンジの0日目及び6日目のIFNγELISpot応答:A−グリアジン57−73 P65(IFNγ特異的スポット/百万PPBMCとして表された結果)
0日目:なし(1)、1週(2)、2週(1)、8週(1)
6日目:なし(0)、1週(0)、2週(0)、8週(0)
結果4:グルテンフリーダイエットの期間及びグルテンチャレンジの0日目及び6日目のIFNγELISpot応答:PPD(IFNγ特異的スポット/百万PPBMCとして表された結果)
0日目:なし(90)、1週(88)、2週(210)、8週(150)
6日目:なし(0)、1週(100)、2週(210)、8週(100)
結果5:グルテンフリーダイエットの期間及びグルテンチャレンジの0日目及び6日目のIFNγELISpot応答:tTG(IFNγ特異的スポット/百万PPBMCとして表された結果)
0日目:なし(5)、1週(4)、2週(3)、8週(2)
6日目:なし(0)、1週(4)、2週(1)、8週(2)
長期間のグルテンに対するHLA−DQ2セリアック病におけるグルテンチャレンジ
抗グリアジンT細胞応答の特性決定を、末梢血において、3日間のグルテンチャレンジの6−8日後に実施した。
結果1:PBMC 6日目長期間グルテンフリーダイエット(抗HLA−DR及び−DQ抗体と前インキュベーション)(%阻害として表現)
DR−:tTG−グリアジン100mcg/ml(105),A−グリアジン57−73 QE65 50mcg/ml(90),PPD 5mcg/ml(30)
DQ−:tTG−グリアジン 100mcg/ml(5),A−グリアジン57−73
QE65 50mcg/ml(22),PPD 5mcg/ml(78)。
【0235】
結果2:PBMC 6日目長期間グルテンフリーダイエット(%CD8−除去PBMC応答として表現)
B7除去:tTG−グリアジン n=6(7),A−グリアジン57−73 n=9(6),PPD n=8(62)
AE除去:tTG−グリアジン n=6(120),A−グリアジン57−73 ん=9(80)、PPD n=8(110)。
【0236】
CD4除去:tTG−グリアジン n=6(10),A−グリアジン57−73 n=9(9),PPD n=8(10)。
治療用ペプチドは限定ではないが以下を含む
【0237】
【表5】
【0238】
簡単には、経口の抗原チャレンジ後に、末梢血T細胞の特異性が腸のT細胞クローンの特異性を反映する。末梢血においては、腸T細胞クローンのエピトープは、最適なα−グリアジンエピトープであるA−グリアジン57−73 QE65に比較して準最適である。実施例15
以下のとおりのマッピング目的のためにもELISpotアッセイを実施した。
優勢なDQ−8関連エピトープのファインマッピング
【0239】
【表6】
【0240】
優勢なエピトープのファインマッピング(2)
【0241】
【表7】
【0242】
実施例16
IFNγ−ELISpotにおけるグリアジンエピトープの生物活性(25mcg/ml,n=6)(A−グリアジン57−73 QE65応答の%として表された)
DQ2−AII:野生型(WT)(4),WT+tTG(52),Glu−置換された(52)
DQ2−AI:野生型(WT)(2),WT+tTG(22),Glu−置換された(28)
GDA09:野生型(WT)(1),tTG(7),Glu−置換された(8)
A−G31−49:野生型(WT)(2),tTG(3),Glu−置換された(0)A−グリアジン57−73 QE65(G01E)の用量応答(n=8)(G01Eの最大応答の%として表された)
0.025mcg/ml(1),0.05mcg/ml(8),0.1mcg/ml(10),0.25mcg/ml(22),0.5mcg/ml(38)<1mcg/ml(43),2.5mcg/ml(52),5mcg/ml(70),10mcg/ml(81),25mcg/ml(95),50mcg/ml(90),100mcg/ml(85)。
【0243】
単独又はA−グリアジン57−75(G01E)と混合されたグリアジンエピトープに対するIFNγELISpot応答(全て50mcg/ml,tTG−グリアジン100mcg/ml,PPD 5mcg/ml;n=9)(G01E応答の%として表された)
単独:DQ2−A1(20),DQ2−A2(55),オメガG1(50),tTGグリアジン(80)、PPD(220),DQ2バインダー(0)
G01E+:DQ2−A1(90),DQ2−A2(95),オメガG1(100),tTGグリアジン(120)、PPD(280),DQ2バインダー(80)
個々のセリアック病被験者におけるIFNγELISpot応答に対するA−グリアジン57−73 QE65のアラニン及びリジン置換の効果(n=8)
エピトープ配列:QLQPFPQPELPYPQPQS
57−59位及び72−73位におけるアラニンの置換はA−グリアジン57−73 QE65応答の%において、ほとんど又は全く低下を示さなかった。60−62位及び68−71位におけるアラニンの置換はA−グリアジン57−73 QE65応答の%において穏やかな低下を示した。63−67位におけるアラニンの置換はA−グリアジン57−73 QE65応答の%において最大の低下を示した。
個々のセリアック病被験者におけるIFNγELISpot応答に対するA−グリアジン57−73 QE65のリジン置換の効果(n=8)
エピトープ配列:QLQPFPQPELPYPQPQS
57−59位及び71−73位におけるリジンの置換はA−グリアジン57−73 QE65応答の%において、ほとんど又は全く低下を示さなかった。60−61位及び69−70位におけるリジンの置換はA−グリアジン57−73 QE65応答の%において穏やかな低下を示した。62−68位におけるリジンの置換はA−グリアジン57−73
QE65応答の%において最大の低下を示した。
実施例17
表24は、コムギ又はライムギでチャレンジした数人の患者を用いた652のペプチドを試験する分析の結果を示す。
【0244】
【表8】
【0245】
本明細書において引用されたか又は言及されたPCTの公表、合衆国特許、他の特許、定期刊行物、及びあらゆるその他の刊行物はそれらの全体を引用により本明細書に編入する。
【0246】
【表9】
【0247】
【表10】
【0248】
【表11】
【0249】
【表12】
【0250】
【表13】
【0251】
【表14】
【0252】
【表15】
【0253】
【表16】
【0254】
【表17】
【0255】
【表18】
【0256】
【表19】
【0257】
【表20】
【0258】
【表21】
【0259】
【表22】
【0260】
【表23】
【0261】
【表24】
【0262】
【表25】
【0263】
【表26】
【0264】
【表27】
【0265】
【表28】
【0266】
【表29】
【0267】
【表30】
【0268】
【表31】
【0269】
【表32】
【0270】
【表33】
【0271】
【表34】
【0272】
【表35】
【0273】
【表36】
【0274】
【表37】
【0275】
【表38】
【0276】
【表39】
【0277】
【表40】
【0278】
【表41】
【0279】
【表42】
【0280】
【表43】
【0281】
【表44】
【0282】
【表45】
【0283】
【表46】
【0284】
【表47】
【0285】
【表48】
【特許請求の範囲】
【請求項1】
個体から得られたサンプルにおいてセリアック病を検出するか又はセリアック病に感受性であることを検出する方法であって、i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び
ii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及び
iii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を、個体からのサンプルに接触させ;そして
サンプル中のT細胞が上記薬剤を認識するか否かをインビトロにて決定するが、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示すことを含む、方法。
【請求項2】
セリアック病を治療又は予防するための医薬の製造のための薬剤の使用であって、薬剤は、
(a)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び
(b)(a)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、(a)の類似体;及び
(c)任意に、(a)及び(b)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチド
を含む、使用。
【請求項3】
薬剤がHLA−DQ2−制限される、請求項2記載の使用。
【請求項4】
薬剤がHLA−DQ8−制限される、請求項2記載の使用。
【請求項5】
一つの薬剤がHLA−DQ2−制限され、そして第2の薬剤がHLA−DQ8制限される、請求項2記載の使用。
【請求項6】
薬剤がコムギのエピトープを含む、請求項2記載の使用。
【請求項7】
一つの薬剤がコムギのエピトープを含み、そして一つの薬剤がライムギのエピトープを含む、請求項2記載の使用。
【請求項8】
一つの薬剤がコムギのエピトープを含み、そして一つの薬剤がオオムギのエピトープを含む、請求項2記載の使用。
【請求項9】
一つの薬剤がライムギのエピトープを含み、そして一つの薬剤がオオムギのエピトープを含む、請求項2記載の使用。
【請求項10】
一つの薬剤がコムギのエピトープを含み、一つの薬剤がオオムギのエピトープを含み、そして一つの薬剤がライムギのエピトープを含む、請求項2記載の使用。
【請求項11】
単一の薬剤がコムギのエピトープ、オオムギのエピトープ、及びライムギのエピトープを含む、請求項2記載の使用。
【請求項12】
薬剤が、薬学上受容可能なキャリアー又は希釈剤を含む薬学組成物中に存在する、請求項2記載の使用。
【請求項13】
薬剤が、請求項2記載のT細胞受容体を有するT細胞のアンタゴニスト、及び薬学上受容可能なキャリアー又は希釈剤を含む薬学組成物中に存在する、請求項2記載の使用。
【請求項14】
薬剤が、請求項2記載の薬剤に応答性のT細胞又は抗体の生産を抑圧するために個体を寛容化するための組成物中に含まれる、請求項2記載の使用。
【請求項15】
任意にキャリアーと共に、薬剤を認識するT細胞を寛容化することによりセリアック病疾患を治療又は予防する方法における、請求項2記載の薬剤の使用。
【請求項16】
任意にキャリアーを伴う、配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチドを認識するT細胞受容体を有するT細胞のアンタゴニストであって、そのようなT細胞と拮抗することによりセリアック病を予防又は治療する方法における使用のためのアンタゴニスト。
【請求項17】
個体を寛容にすることにより抗体の生産を阻害して個体においてセリアック病を治療又は予防する方法において使用するための、配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープに結合する抗体を結合する類似体。
【請求項18】
配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む蛋白質であって、但し、T細胞受容体は上で定義された薬剤を認識し、そして上記配列はそのようなT細胞受容体を有するT細胞の拮抗作用を引き起こすことができる、蛋白質。
【請求項19】
配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチドを認識するT細胞受容体を有するT細胞のアンタゴニスト。
【請求項20】
i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤又は
請求項19記載のアンタゴニスト及び
薬学上受容可能なキャリアー又は希釈剤
を含む薬学組成物。
【請求項21】
T細胞の生産又は
i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤に対する抗体応答
を抑圧するために個体をグリアジン蛋白質に対して寛容にするための組成物であって、
i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を含む組成物。
【請求項22】
i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤に対するT細胞応答に拮抗するための組成物であって、請求項19記載のアンタゴニストを含む組成物。
【請求項23】
その野生型配列がトランスグルタミナーゼにより配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列に修飾され得る変異体グリアジン蛋白質であって、変異体グリアジン蛋白質はトランスグルタミナーゼによるその上記配列への修飾を阻害する変異を含み;又は、本発明は少なくとも15アミノ酸の長さであり、且つ上記変異を含む、そのような変異体グリアジン蛋白質の断片。
【請求項24】
請求項23記載の断片又は18記載の蛋白質をコードするコーディング配列を含むポリヌクレオチド。
【請求項25】
コーディング配列に作動可能に連結された一つ又はそれより多い制御配列を追加して含むが、但し、制御配列は細胞中でコーディング配列の発現を保証することができる、請求項24記載のポリヌクレオチド。
【請求項26】
制御配列が原核又は哺乳類の細胞におけるコーディング配列の発現を可能にする、請求項25記載のポリヌクレオチド。
【請求項27】
ベクターであるか、又はベクターの形態である、請求項24乃至26の何れか1項記載のポリヌクレオチド。
【請求項28】
請求項24乃至26の何れか1項に記載のポリヌクレオチドを含むか、又はそのようなポリヌクレオチドにより形質転換された、単離された細胞。
【請求項29】
原核細胞又は哺乳類細胞である、請求項28記載の細胞。
【請求項30】
配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチドを認識するT細胞受容体を発現するヒト以外の哺乳類。
【請求項31】
個体から得られたサンプルにおいてセリアック病を検出するか又はセリアック病に感受性であることを検出する方法であって、
(a)(i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び
(ii)(i)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、(i)の類似体;及び
(iii)任意に、(i)及び(ii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤をサンプルに接触させ;そして
(b)サンプル中のT細胞が薬剤を認識するか否かをインビトロにて決定して、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示すこと
を含む方法。
【請求項32】
個体から得られたサンプルにおいてセリアック病を検出するか又はセリアック病に感受性であることを検出する方法における使用のための診断手段の製造のための請求項31記載の薬剤の使用であって、上記方法は、個体のT細胞が薬剤を認識するか否かを決定することを含み、T細胞による認識が個体がセリアック病を有するか又はセリアック病に感受性である、請求項31記載の薬剤の使用。
【請求項33】
薬剤が、(a)HLA分子又は(b)(i)又は(ii)を結合できるHLA分子の断片に結合した(i)又は(ii)を含む類似体(iii)である、請求項31記載の方法。
【請求項34】
薬剤が、(a)HLA分子又は(b)(i)又は(ii)を結合できるHLA分子の断片に結合した(i)又は(ii)を含む類似体(iii)である、請求項32記載の使用。
【請求項35】
HLA分子又は断片が、4つのHLA分子又はHLA分子の断片を含む複合体中にある、請求項33の方法。
【請求項36】
HLA分子又は断片が、4つのHLA分子又はHLA分子の断片を含む複合体中にある、請求項34の使用。
【請求項37】
方法が、薬剤を個体の皮膚に投与し、そして投与の部位において炎症の存在を検出することを含み、炎症の検出が、個体のT細胞が薬剤を認識することを示す、請求項32、34又は36記載の使用。
【請求項38】
サンプルが血液サンプルである、請求項31、33又は35記載の方法。
【請求項39】
決定の前に抗原特異的様式にてインビボにおいてT細胞が再刺激されない、請求項31、33、35又は38記載の方法。
【請求項40】
T細胞による薬剤の認識がT細胞によるサイトカインの分泌を検出することにより決定される、請求項31、33、35又は38記載の方法。
【請求項41】
T細胞による薬剤の認識がT細胞によるサイトカインの分泌を検出することにより決定される、請求項32、34、36又は37記載の使用。
【請求項42】
サイトカインがIFN−γである、請求項40記載の方法。
【請求項43】
サイトカインがIFN−γである、請求項41記載の使用。
【請求項44】
サイトカインに特異的な固定化された抗体へのサイトカインの結合を可能にし、そして次に抗体/サイトカイン複合体の存在を検出することによりサイトカインを検出する、請求項40又は請求項42記載の方法。
【請求項45】
サイトカインに特異的な固定化された抗体へのサイトカインの結合を可能にし、そして次に抗体/サイトカイン複合体の存在を検出することによりサイトカインを検出する、請求項41又は請求項43記載の使用。
【請求項46】
薬剤がT細胞受容体に結合するか否かを測定することにより上記決定を行う、請求項31、33、35又は38記載の方法。
【請求項47】
薬剤がT細胞受容体に結合するか否かを測定することにより上記決定を行う、請求項32、34、36又は37記載の使用。
【請求項48】
配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチドを認識するT細胞受容体により認識されることができる類似体であって、且つ50アミノ酸の長さを超えない上記類似体を同定する方法であって、配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含むエピトープを認識するT細胞受容体により候補物質が認識されるか否かを決定することを含み、物質の認識が、物質が類似体であることを示す、方法。
【請求項49】
個体から得られたサンプルにおいてセリアック病又はセリアック病に対する感受性を検出する方法であって、配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含むエピトープのエピトープに結合する抗体の、個体からのサンプル中の存在を決定することを含み、抗体の存在が、個体がセリアック病を有するか又はセリアック病に感受性であることを示す、方法。
【請求項50】
組成物がセリアック病を引き起こすか否かを決定する方法であって、配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列へトランスグルタミナーゼにより修飾され得る蛋白質が組成物中に存在するか否かを決定することを含み、蛋白質の存在が、組成物がセリアック病を引き起こすことを示す、方法。
【請求項51】
上記配列に修飾され得る配列に特異的な抗体に組成物を接触させることにより上記決定を行い、組成物中の蛋白質への抗体の結合が、組成物がセリアック病を引き起こすことを示す、請求項50記載の方法。
【請求項52】
i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を認識するT細胞のアンタゴニストを同定する方法であって、候補物質をT細胞と接触させ、そして当該物質がT細胞が抗原特異的応答を受ける能力の低下を引き起こすか否かを決定することを含み、上記能力のそのようなあらゆる低下の検出が、上記物質がアンタゴニストであることを示す、方法。
【請求項53】
請求項31乃至44の何れか1項記載の方法又は使用を実施するためのキットであって、
(i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び(ii)(i)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、(i)の類似体;及び(iii)任意に、(i)及び(ii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤及び
T細胞によるペプチドの認識を検出する手段
を含む、キット。
【請求項54】
認識を検出する手段がIFN−γに対する抗体を含む、請求項53記載のキット。
【請求項55】
抗体が固相支持体上に固定されており、そして任意にキットが抗体/IFN−γ複合体を検出する手段も含む、請求項54記載のキット。
【請求項56】
(i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び(ii)(i)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、(i)の類似体;及び(iii)任意に、(i)及び(ii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤又は
請求項19記載のアンタゴニスト又は
トランスグルタミナーゼにより配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列に修飾され得る変異体グリアジン蛋白質の野生型配列
の、上記薬剤、上記アンタゴニスト又は上記野生型配列に特異的な抗体を生産するための使用。
【請求項57】
セリアック病を引き起こすグリアジン蛋白質の能力を低下させるための、配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープ内の変異の使用。
【請求項58】
セリアック病の治療用の生成物を同定する方法であって、セリアック病を有するか又はセリアック病に感受性の候補物質を請求項30記載のヒト以外の哺乳類に投与し、そして物質が哺乳類においてセリアック病を予防又は治療するか否かを決定することを含み、セリアック病の予防又は治療が、当該物質が治療用生成物であることを示す、方法。
【請求項59】
セリアック病を予防又は治療するための方法において同定された治療用生成物。
【請求項60】
個体から得られたサンプルにおいてセリアック病又はセリアック病に対する感受性を検出する方法であって、(i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び(ii)(i)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、(i)の類似体;及び(iii)任意に、(i)及び(ii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤をサンプルに接触させ、そしてインビトロにおいてサンプル中のT細胞が薬剤を認識するか否かを決定することを含み、薬剤の認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示す、方法。
【請求項1】
個体から得られたサンプルにおいてセリアック病を検出するか又はセリアック病に感受性であることを検出する方法であって、i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び
ii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及び
iii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を、個体からのサンプルに接触させ;そして
サンプル中のT細胞が上記薬剤を認識するか否かをインビトロにて決定するが、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示すことを含む、方法。
【請求項2】
セリアック病を治療又は予防するための医薬の製造のための薬剤の使用であって、薬剤は、
(a)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び
(b)(a)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、(a)の類似体;及び
(c)任意に、(a)及び(b)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチド
を含む、使用。
【請求項3】
薬剤がHLA−DQ2−制限される、請求項2記載の使用。
【請求項4】
薬剤がHLA−DQ8−制限される、請求項2記載の使用。
【請求項5】
一つの薬剤がHLA−DQ2−制限され、そして第2の薬剤がHLA−DQ8制限される、請求項2記載の使用。
【請求項6】
薬剤がコムギのエピトープを含む、請求項2記載の使用。
【請求項7】
一つの薬剤がコムギのエピトープを含み、そして一つの薬剤がライムギのエピトープを含む、請求項2記載の使用。
【請求項8】
一つの薬剤がコムギのエピトープを含み、そして一つの薬剤がオオムギのエピトープを含む、請求項2記載の使用。
【請求項9】
一つの薬剤がライムギのエピトープを含み、そして一つの薬剤がオオムギのエピトープを含む、請求項2記載の使用。
【請求項10】
一つの薬剤がコムギのエピトープを含み、一つの薬剤がオオムギのエピトープを含み、そして一つの薬剤がライムギのエピトープを含む、請求項2記載の使用。
【請求項11】
単一の薬剤がコムギのエピトープ、オオムギのエピトープ、及びライムギのエピトープを含む、請求項2記載の使用。
【請求項12】
薬剤が、薬学上受容可能なキャリアー又は希釈剤を含む薬学組成物中に存在する、請求項2記載の使用。
【請求項13】
薬剤が、請求項2記載のT細胞受容体を有するT細胞のアンタゴニスト、及び薬学上受容可能なキャリアー又は希釈剤を含む薬学組成物中に存在する、請求項2記載の使用。
【請求項14】
薬剤が、請求項2記載の薬剤に応答性のT細胞又は抗体の生産を抑圧するために個体を寛容化するための組成物中に含まれる、請求項2記載の使用。
【請求項15】
任意にキャリアーと共に、薬剤を認識するT細胞を寛容化することによりセリアック病疾患を治療又は予防する方法における、請求項2記載の薬剤の使用。
【請求項16】
任意にキャリアーを伴う、配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチドを認識するT細胞受容体を有するT細胞のアンタゴニストであって、そのようなT細胞と拮抗することによりセリアック病を予防又は治療する方法における使用のためのアンタゴニスト。
【請求項17】
個体を寛容にすることにより抗体の生産を阻害して個体においてセリアック病を治療又は予防する方法において使用するための、配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープに結合する抗体を結合する類似体。
【請求項18】
配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む蛋白質であって、但し、T細胞受容体は上で定義された薬剤を認識し、そして上記配列はそのようなT細胞受容体を有するT細胞の拮抗作用を引き起こすことができる、蛋白質。
【請求項19】
配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチドを認識するT細胞受容体を有するT細胞のアンタゴニスト。
【請求項20】
i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤又は
請求項19記載のアンタゴニスト及び
薬学上受容可能なキャリアー又は希釈剤
を含む薬学組成物。
【請求項21】
T細胞の生産又は
i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤に対する抗体応答
を抑圧するために個体をグリアジン蛋白質に対して寛容にするための組成物であって、
i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を含む組成物。
【請求項22】
i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤に対するT細胞応答に拮抗するための組成物であって、請求項19記載のアンタゴニストを含む組成物。
【請求項23】
その野生型配列がトランスグルタミナーゼにより配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列に修飾され得る変異体グリアジン蛋白質であって、変異体グリアジン蛋白質はトランスグルタミナーゼによるその上記配列への修飾を阻害する変異を含み;又は、本発明は少なくとも15アミノ酸の長さであり、且つ上記変異を含む、そのような変異体グリアジン蛋白質の断片。
【請求項24】
請求項23記載の断片又は18記載の蛋白質をコードするコーディング配列を含むポリヌクレオチド。
【請求項25】
コーディング配列に作動可能に連結された一つ又はそれより多い制御配列を追加して含むが、但し、制御配列は細胞中でコーディング配列の発現を保証することができる、請求項24記載のポリヌクレオチド。
【請求項26】
制御配列が原核又は哺乳類の細胞におけるコーディング配列の発現を可能にする、請求項25記載のポリヌクレオチド。
【請求項27】
ベクターであるか、又はベクターの形態である、請求項24乃至26の何れか1項記載のポリヌクレオチド。
【請求項28】
請求項24乃至26の何れか1項に記載のポリヌクレオチドを含むか、又はそのようなポリヌクレオチドにより形質転換された、単離された細胞。
【請求項29】
原核細胞又は哺乳類細胞である、請求項28記載の細胞。
【請求項30】
配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチドを認識するT細胞受容体を発現するヒト以外の哺乳類。
【請求項31】
個体から得られたサンプルにおいてセリアック病を検出するか又はセリアック病に感受性であることを検出する方法であって、
(a)(i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び
(ii)(i)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、(i)の類似体;及び
(iii)任意に、(i)及び(ii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤をサンプルに接触させ;そして
(b)サンプル中のT細胞が薬剤を認識するか否かをインビトロにて決定して、T細胞による認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示すこと
を含む方法。
【請求項32】
個体から得られたサンプルにおいてセリアック病を検出するか又はセリアック病に感受性であることを検出する方法における使用のための診断手段の製造のための請求項31記載の薬剤の使用であって、上記方法は、個体のT細胞が薬剤を認識するか否かを決定することを含み、T細胞による認識が個体がセリアック病を有するか又はセリアック病に感受性である、請求項31記載の薬剤の使用。
【請求項33】
薬剤が、(a)HLA分子又は(b)(i)又は(ii)を結合できるHLA分子の断片に結合した(i)又は(ii)を含む類似体(iii)である、請求項31記載の方法。
【請求項34】
薬剤が、(a)HLA分子又は(b)(i)又は(ii)を結合できるHLA分子の断片に結合した(i)又は(ii)を含む類似体(iii)である、請求項32記載の使用。
【請求項35】
HLA分子又は断片が、4つのHLA分子又はHLA分子の断片を含む複合体中にある、請求項33の方法。
【請求項36】
HLA分子又は断片が、4つのHLA分子又はHLA分子の断片を含む複合体中にある、請求項34の使用。
【請求項37】
方法が、薬剤を個体の皮膚に投与し、そして投与の部位において炎症の存在を検出することを含み、炎症の検出が、個体のT細胞が薬剤を認識することを示す、請求項32、34又は36記載の使用。
【請求項38】
サンプルが血液サンプルである、請求項31、33又は35記載の方法。
【請求項39】
決定の前に抗原特異的様式にてインビボにおいてT細胞が再刺激されない、請求項31、33、35又は38記載の方法。
【請求項40】
T細胞による薬剤の認識がT細胞によるサイトカインの分泌を検出することにより決定される、請求項31、33、35又は38記載の方法。
【請求項41】
T細胞による薬剤の認識がT細胞によるサイトカインの分泌を検出することにより決定される、請求項32、34、36又は37記載の使用。
【請求項42】
サイトカインがIFN−γである、請求項40記載の方法。
【請求項43】
サイトカインがIFN−γである、請求項41記載の使用。
【請求項44】
サイトカインに特異的な固定化された抗体へのサイトカインの結合を可能にし、そして次に抗体/サイトカイン複合体の存在を検出することによりサイトカインを検出する、請求項40又は請求項42記載の方法。
【請求項45】
サイトカインに特異的な固定化された抗体へのサイトカインの結合を可能にし、そして次に抗体/サイトカイン複合体の存在を検出することによりサイトカインを検出する、請求項41又は請求項43記載の使用。
【請求項46】
薬剤がT細胞受容体に結合するか否かを測定することにより上記決定を行う、請求項31、33、35又は38記載の方法。
【請求項47】
薬剤がT細胞受容体に結合するか否かを測定することにより上記決定を行う、請求項32、34、36又は37記載の使用。
【請求項48】
配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチドを認識するT細胞受容体により認識されることができる類似体であって、且つ50アミノ酸の長さを超えない上記類似体を同定する方法であって、配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含むエピトープを認識するT細胞受容体により候補物質が認識されるか否かを決定することを含み、物質の認識が、物質が類似体であることを示す、方法。
【請求項49】
個体から得られたサンプルにおいてセリアック病又はセリアック病に対する感受性を検出する方法であって、配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含むエピトープのエピトープに結合する抗体の、個体からのサンプル中の存在を決定することを含み、抗体の存在が、個体がセリアック病を有するか又はセリアック病に感受性であることを示す、方法。
【請求項50】
組成物がセリアック病を引き起こすか否かを決定する方法であって、配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列へトランスグルタミナーゼにより修飾され得る蛋白質が組成物中に存在するか否かを決定することを含み、蛋白質の存在が、組成物がセリアック病を引き起こすことを示す、方法。
【請求項51】
上記配列に修飾され得る配列に特異的な抗体に組成物を接触させることにより上記決定を行い、組成物中の蛋白質への抗体の結合が、組成物がセリアック病を引き起こすことを示す、請求項50記載の方法。
【請求項52】
i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及びii)i)を認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、i)の類似体;及びiii)任意に、i)及びii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤を認識するT細胞のアンタゴニストを同定する方法であって、候補物質をT細胞と接触させ、そして当該物質がT細胞が抗原特異的応答を受ける能力の低下を引き起こすか否かを決定することを含み、上記能力のそのようなあらゆる低下の検出が、上記物質がアンタゴニストであることを示す、方法。
【請求項53】
請求項31乃至44の何れか1項記載の方法又は使用を実施するためのキットであって、
(i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び(ii)(i)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、(i)の類似体;及び(iii)任意に、(i)及び(ii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤及び
T細胞によるペプチドの認識を検出する手段
を含む、キット。
【請求項54】
認識を検出する手段がIFN−γに対する抗体を含む、請求項53記載のキット。
【請求項55】
抗体が固相支持体上に固定されており、そして任意にキットが抗体/IFN−γ複合体を検出する手段も含む、請求項54記載のキット。
【請求項56】
(i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び(ii)(i)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、(i)の類似体;及び(iii)任意に、(i)及び(ii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤又は
請求項19記載のアンタゴニスト又は
トランスグルタミナーゼにより配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列に修飾され得る変異体グリアジン蛋白質の野生型配列
の、上記薬剤、上記アンタゴニスト又は上記野生型配列に特異的な抗体を生産するための使用。
【請求項57】
セリアック病を引き起こすグリアジン蛋白質の能力を低下させるための、配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープ内の変異の使用。
【請求項58】
セリアック病の治療用の生成物を同定する方法であって、セリアック病を有するか又はセリアック病に感受性の候補物質を請求項30記載のヒト以外の哺乳類に投与し、そして物質が哺乳類においてセリアック病を予防又は治療するか否かを決定することを含み、セリアック病の予防又は治療が、当該物質が治療用生成物であることを示す、方法。
【請求項59】
セリアック病を予防又は治療するための方法において同定された治療用生成物。
【請求項60】
個体から得られたサンプルにおいてセリアック病又はセリアック病に対する感受性を検出する方法であって、(i)配列番号:18、20−22、31−36、40−44、及び46、及びそれらの均等物からなる群から選択される配列を含む少なくとも一つのエピトープを含むペプチド;及び(ii)(i)のペプチドを認識するT細胞受容体により認識されることができ、且つ50アミノ酸の長さを超えない、(i)の類似体;及び(iii)任意に、(i)及び(ii)から選択される薬剤に加えて、配列番号:1及び配列番号:2から選択される配列を含む少なくとも一つのエピトープを含むペプチドから選択される少なくとも一つの薬剤をサンプルに接触させ、そしてインビトロにおいてサンプル中のT細胞が薬剤を認識するか否かを決定することを含み、薬剤の認識が、個体がセリアック病を有するか又はセリアック病に感受性であることを示す、方法。
【図1A】

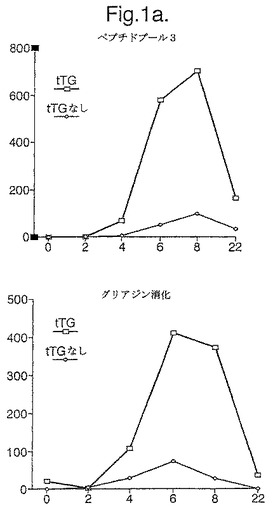
【図2】
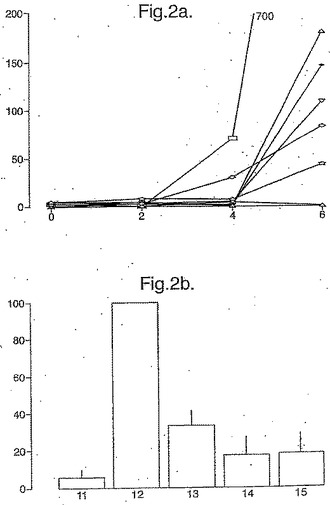
【図3】
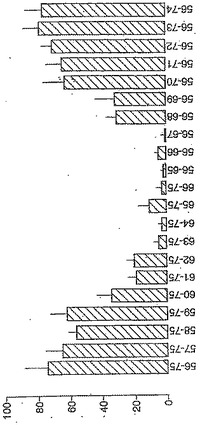
【図4】
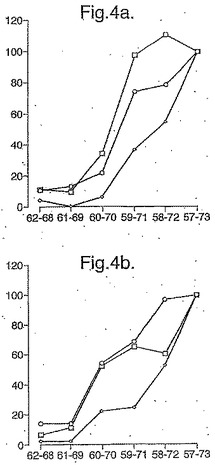
【図5】
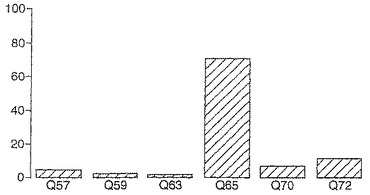
【図8】

【図9】
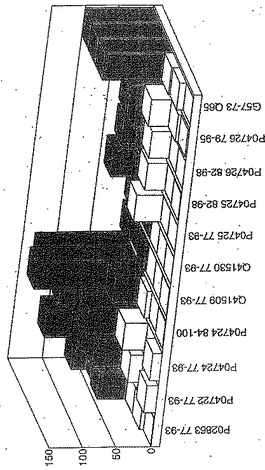
【図11】
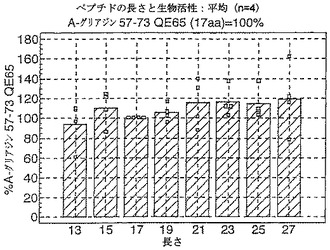
【図12A】
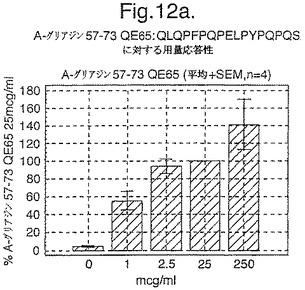
【図12B】
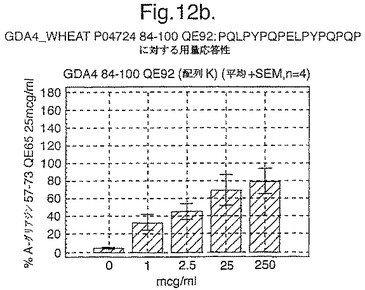
【図12C】
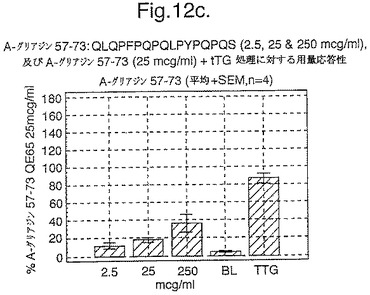
【図12D】
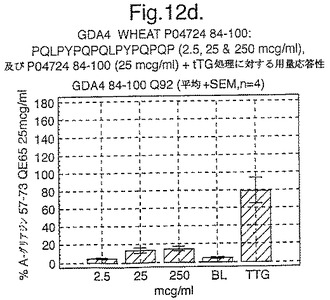
【図12E】
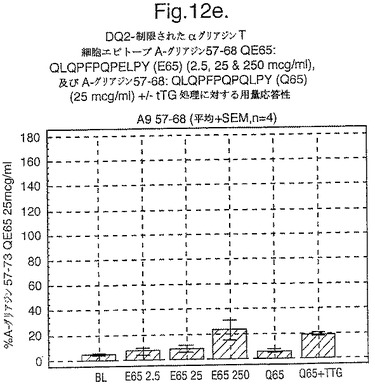
【図12F】
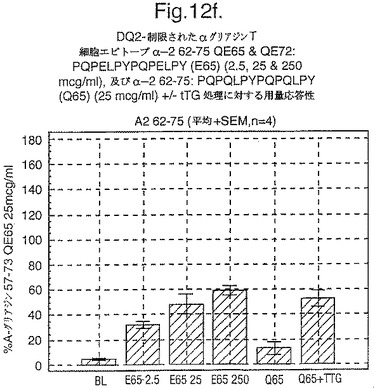
【図12G】
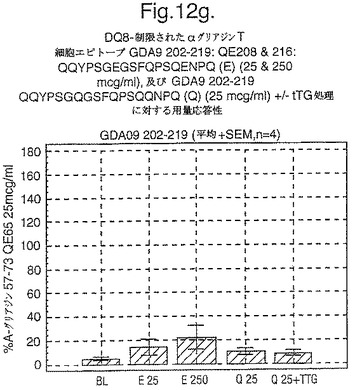
【図12H】
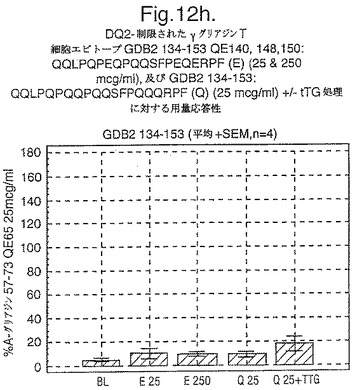
【図13A】
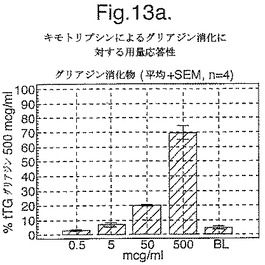
【図13B】
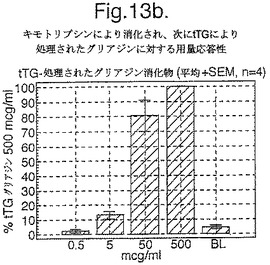
【図13C】
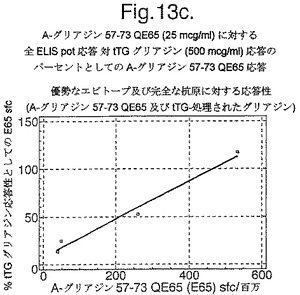
【図14A】
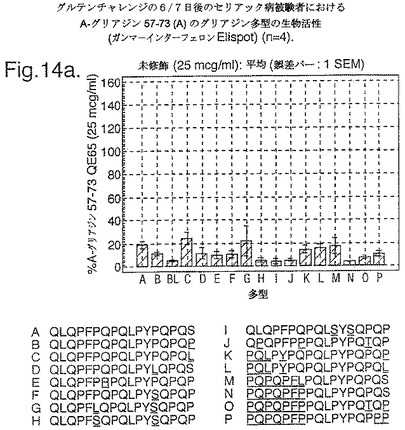
【図14B】
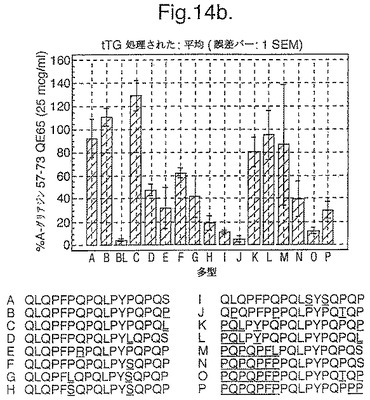
【図14C】
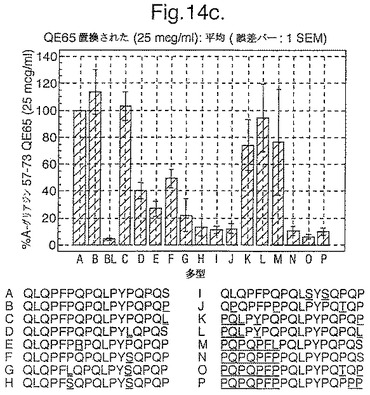
【図14D】
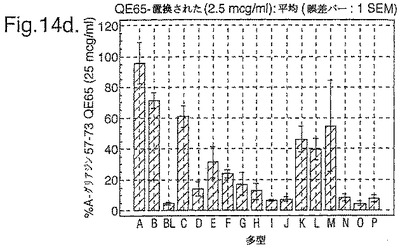
【図14E】
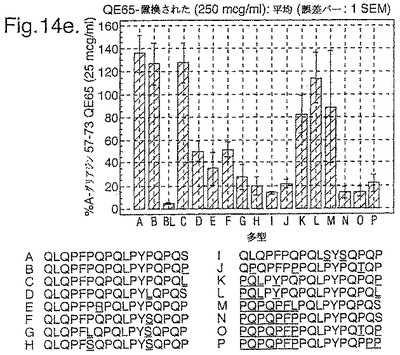
【図15】
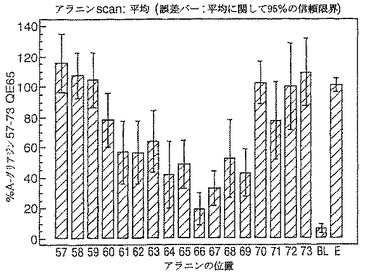
【図16】
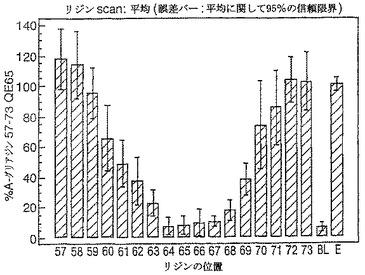
【図17】
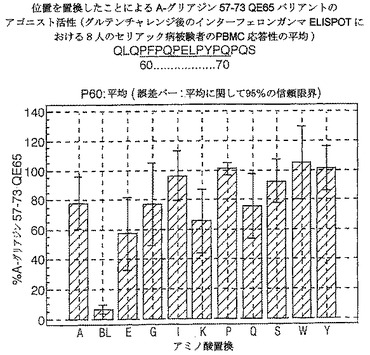
【図18】
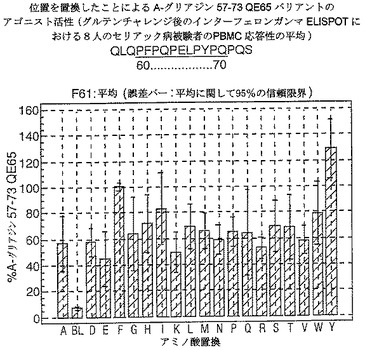
【図19】
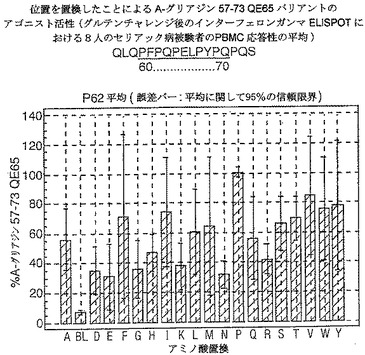
【図20】
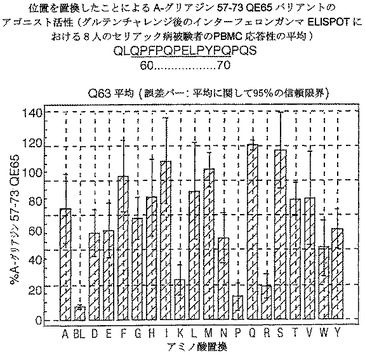
【図21】
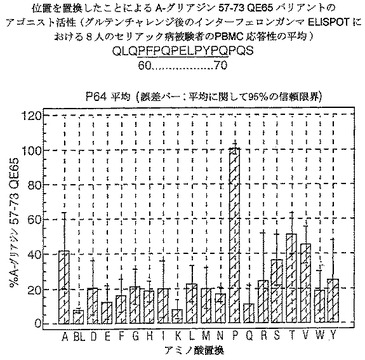
【図22】
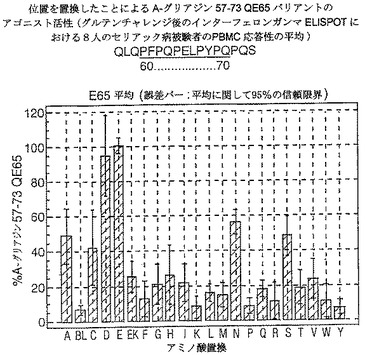
【図23】
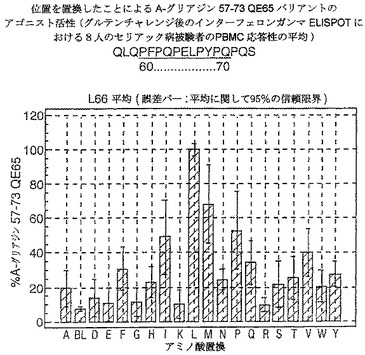
【図24】
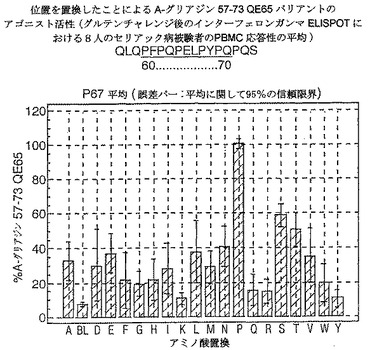
【図25】
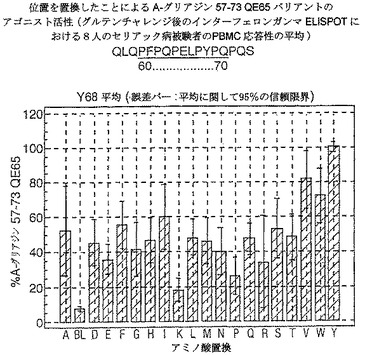
【図26】
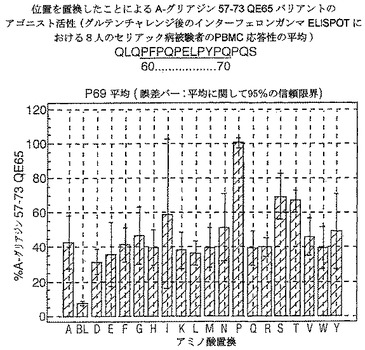
【図27】
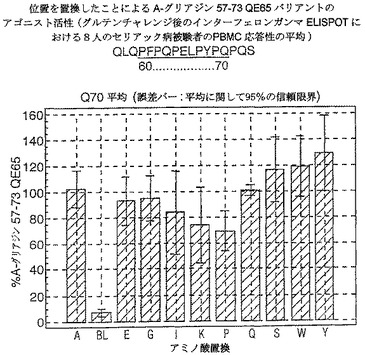
【図28A】
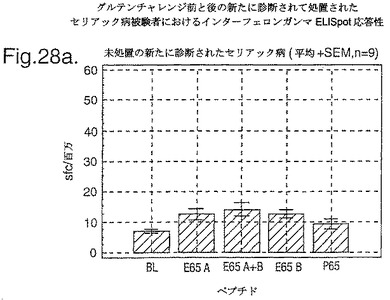
【図28B】
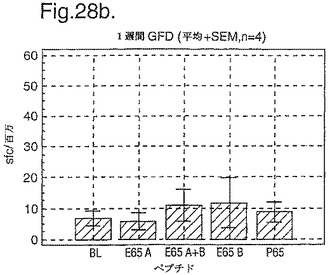
【図28C】
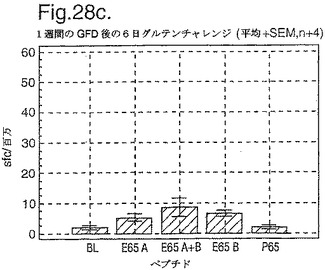
【図28D】
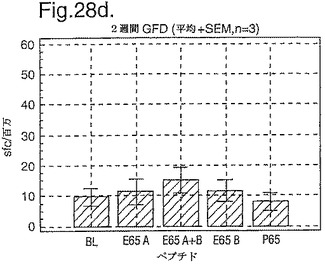
【図28E】
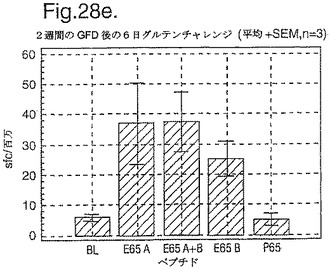
【図28F】
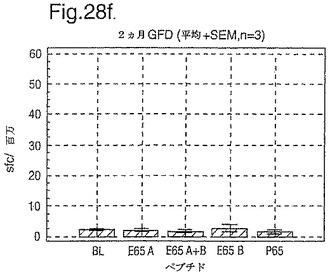
【図28G】
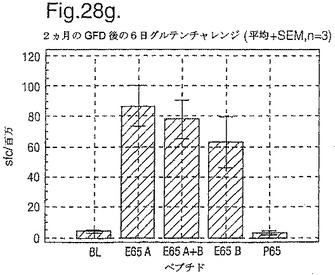
【図30】

【図31】

【図33】

【図34】
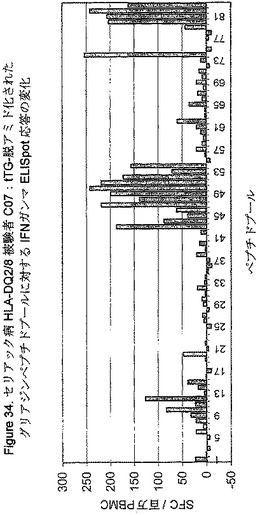
【図35】

【図36】

【図1B】
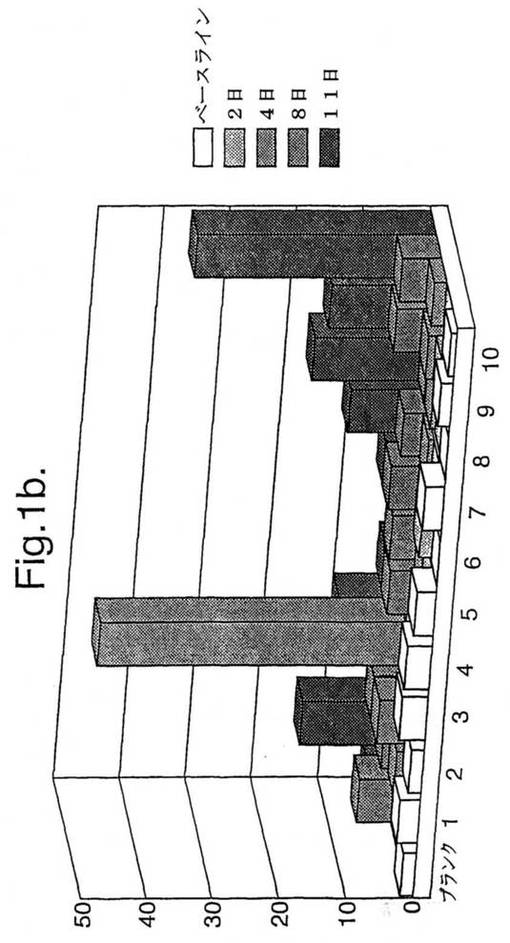
【図6】
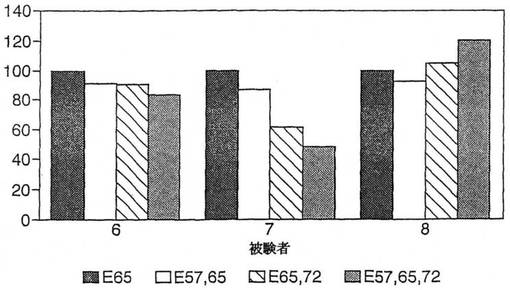
【図7】
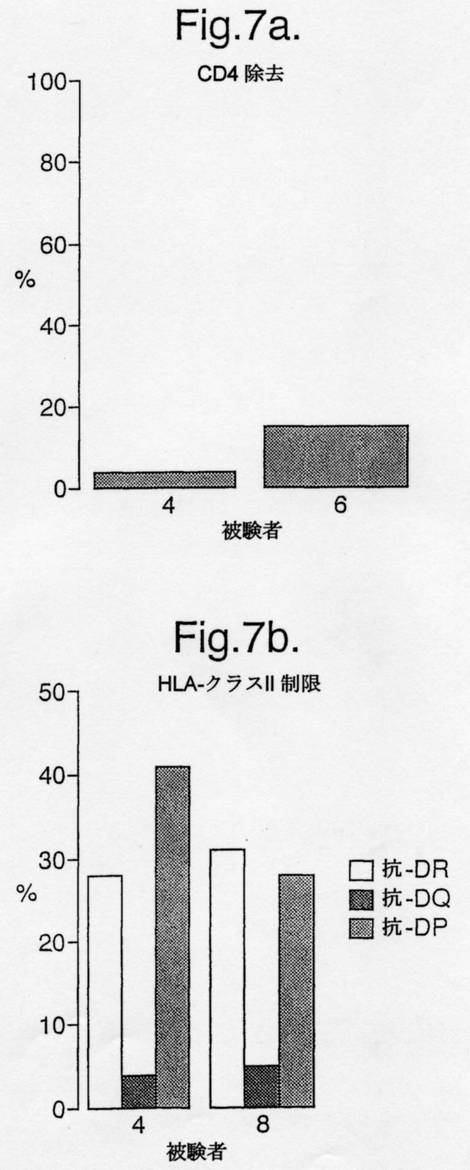
【図10】
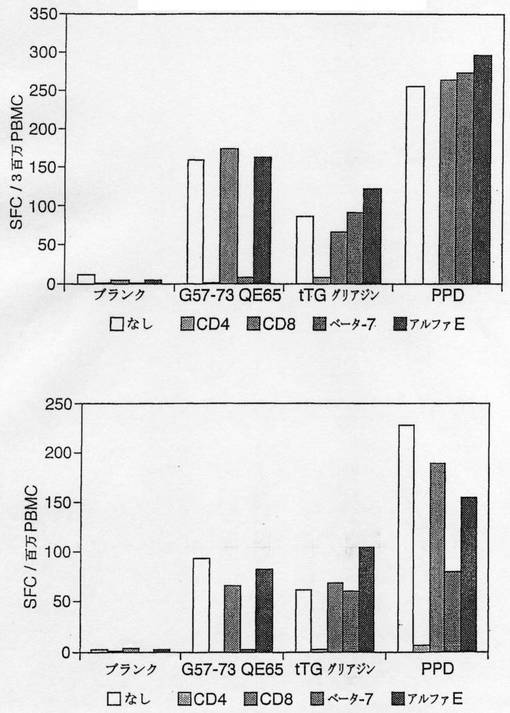
【図29】
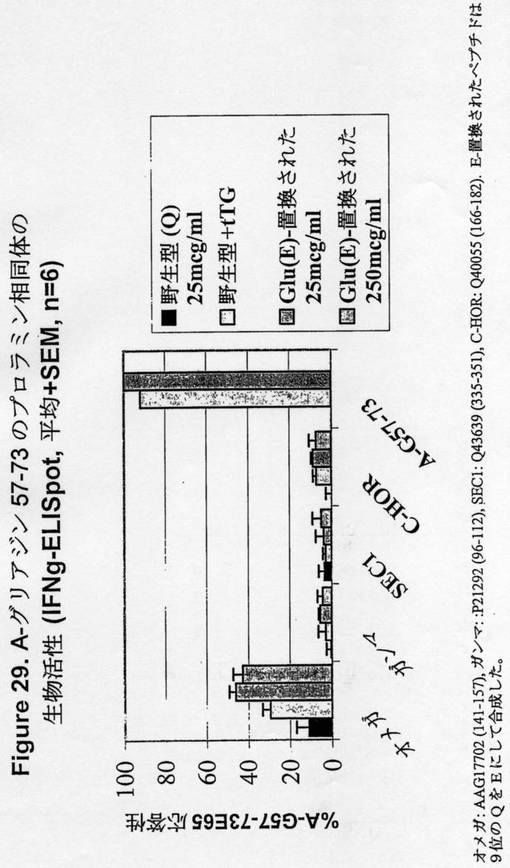
【図32】
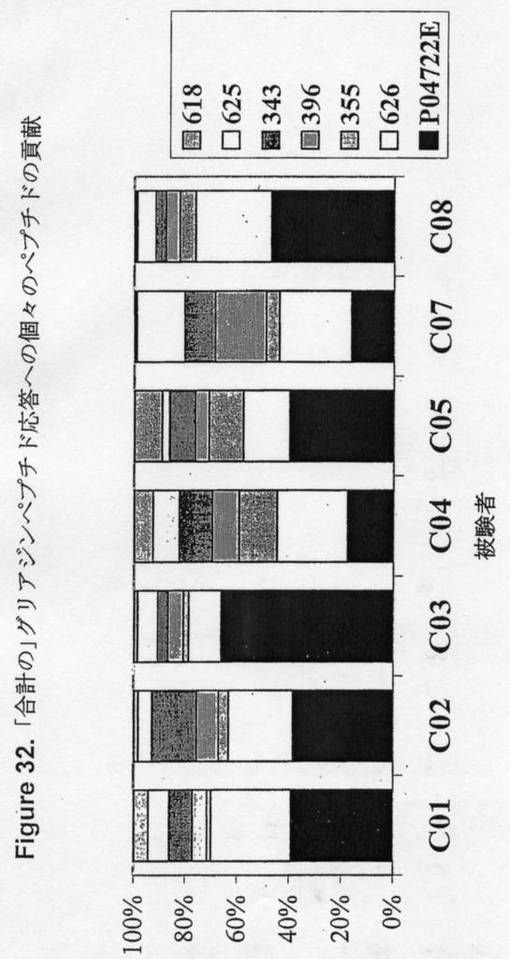
【図2】
【図3】
【図4】
【図5】
【図8】
【図9】
【図11】
【図12A】
【図12B】
【図12C】
【図12D】
【図12E】
【図12F】
【図12G】
【図12H】
【図13A】
【図13B】
【図13C】
【図14A】
【図14B】
【図14C】
【図14D】
【図14E】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28A】
【図28B】
【図28C】
【図28D】
【図28E】
【図28F】
【図28G】
【図30】
【図31】
【図33】
【図34】
【図35】
【図36】
【図1B】
【図6】
【図7】
【図10】
【図29】
【図32】
【公開番号】特開2013−101146(P2013−101146A)
【公開日】平成25年5月23日(2013.5.23)
【国際特許分類】
【出願番号】特願2013−21737(P2013−21737)
【出願日】平成25年2月6日(2013.2.6)
【分割の表示】特願2010−110683(P2010−110683)の分割
【原出願日】平成15年6月5日(2003.6.5)
【出願人】(500056231)アイシス・イノベーション・リミテッド (17)
【氏名又は名称原語表記】ISIS INNOVATION LIMITED
【Fターム(参考)】
【公開日】平成25年5月23日(2013.5.23)
【国際特許分類】
【出願日】平成25年2月6日(2013.2.6)
【分割の表示】特願2010−110683(P2010−110683)の分割
【原出願日】平成15年6月5日(2003.6.5)
【出願人】(500056231)アイシス・イノベーション・リミテッド (17)
【氏名又は名称原語表記】ISIS INNOVATION LIMITED
【Fターム(参考)】
[ Back to top ]