
治療用ペプチドおよびワクチン
多様なHIV分岐群に対する抗体を誘導する広範なHIV治療剤およびワクチンを誘導し、HIVgp120によって誘導された短いペプチド配列免疫原を同定する能力に関する組成物、および、同定されたCCR5結合部位を構成するペプチドから作製される様々な治療用組成物を開示する。さらに、薬物候補となる可能性があるペプチド配列を選択する方法も開示し、これは、アルツハイマー病、乾癬、多発性硬化症およびその他のヒトの炎症性のカスケードに関連する病気、同様に、関連するレトロウイルス、例えばHTLV−I、熱帯性痙性不全対麻痺の原因のような領域において有効な治療を提供すると予想される。
【発明の詳細な説明】
【発明の開示】
【0001】
本願は、2005年6月23日付けで出願された仮出願番号60/693,087;2005年6月23日付けで出願された仮出願番号60/693,088、および、2005年6月23日付けで出願された仮出願番号60/693,089に基づく優先権を主張する。本願はまた、2006年6月23日付けで出願された出願番号(発明の名称「Stabilizing Alkylglycoside Compositions and Methods Thereof」)にも関する(これらの内容は、この参照により開示に含まれる)。
【0002】
発明の背景
一般的に、HIV−1感染やAIDSを防御することができるワクチンは、それらの地理的な由来、サブタイプまたは遺伝子型の特異性に関わりなく、それらウイルスの一次的な単離物に対する有力な中和活性を有する高レベルの抗体を誘導する免疫原を必要とすると考えられており、これが、現行のワクチン候補では実現できていない目的の一つである。それゆえに、合理的なワクチン設計および開発には広範な中和をもたらすエピトープの同定が重要である。具体的に重要なのは、優勢な北米の分岐群Bサブタイプに対して発展途上国に存在し、世界的に関連を有する発生株を含む分岐群Cサブタイプに対するワクチン候補の抗体の開発である。
【0003】
HIVに感染した個体において、中和抗体の優勢な標的はHIVエンベロープである。ある種の中和エピトープのクラスターは、一次および/または単球親和性の単離物のgp120envタンパク質に少なくとも部分的に晒されるようである。20年にわたり大規模な研究が行われているにもかかわらず、広範なワクチンの中和を惹起することができる免疫原は見出されていない。その理由として、HIV感染に対するワクチンは定義が難しいことがあった。
【0004】
ある発明は、多様なHIV株に有力な中和活性を有する抗体を導くことができるペプチド、および、HIV−1防御のための免疫化戦略の重要な成分であるペプチドを同定する補助受容体CCR5に結合するgp120の短い配列の同定に関与する。これらの結合配列に生産され、結合部位のペプチドの免疫原を含む抗体は、系統に関わりなくHIVウイルス感染からの防御が付与されると期待され、さらに治療にも役立つと予想されている。すなわち、それらの受動的な投与は、ウイルスレベルを減少させ、すでにHIVに感染した人間に臨床的な利点を提供すると予想される。
【0005】
中和抗体は、HIV−1感染に応答する。初感染後のウイルス血症から数ヶ月で、まずは抗ヒト免疫不全ウィルスタイプの中和抗体が出現する。初期に生じる中和抗体は、高度な型特異性を有する。感染中に、中和抗体反応はその後も広がる可能性があるが(16)、通常わずかであり、長期間感染した個体を含む患者の大半において散発的に発生する。
【0006】
中和抗体は、HIV−1エンベロープの糖タンパク質を認識し、これはgp120エクステリアとgp41膜貫通糖タンパク質とからなる。gp120糖タンパク質は、標的であるCD4、ならびにケモカイン受容体CCR5およびCXCR4に結合する。gp120がCD4との複合体を形成した後にのみ発現されるgp120のエピトープに対して抗体が発生する例はほとんどない。望ましいワクチン標的である実際の補助受容体結合部位は、CD4とgp120との会合の際にのみ形成されると考えられる。これらのCD4誘導性(CD4i)抗体は、gp120−sCD4複合体のCCR5およびCXCR4結合をブロックする。HIV−1に感染した人間から単離されたCD4i抗体の数は少ない。
【0007】
最も初期のHIV−1ワクチンを用いた研究は、中和抗体を誘発するワクチンを開発することに向けられていた。これらの中和抗体は、それらの作用が限定的であり、動物モデルにおいて接種されたウイルスのほぼ全ての系統に対して特異的である。それゆえにこれらの抗体は広範な特異性を有しておらず、優れたワクチン候補を形成しないだろう。
【0008】
最も成功したワクチンは、その受容体(HIVの場合、補助受容体CXCR4およびCC5である)に結合するウイルスをブロックする(1,2)。過去20年間にわたって懸命な調査の対象である補助受容体結合部位の同定は現在まで達成されておらず、従って、これらを同定することによって、望ましいワクチン用途に適した多様な系統の広範な中和を実現することができる免疫原が生産されることが期待される。
【0009】
本願の目的の一つは、生物学的に活性な補助受容体結合部位に対する抗体である。インビトロでの研究において、この抗体はCCR5受容体に結合するgp120−sCD4複合体の有力な阻害を示し、さらに、適切なペプチドコンフォメーションに対して生じた抗体は中和抗体を広範に誘導することから、それによってHIV−1感染に対する防御免疫、すなわちワクチンを提供することが期待される。
【0010】
10年以上にわたり、ほとんどのHIV−1単離物の広範かつ有力な中和を仲介するヒトモノクローナル抗体はほんのわずかしか単離されていない。これらのモノクローナル抗体のうち2種は、gp120(モノクローナル抗体b12および2G12)の表面に結合し(3,4)、1種は、gp41の膜貫通ドメインの直近位に存在するエピトープに結合する。モノクローナル抗体b12は、gp120のC3−V4領域と炭水化物とを含む複雑な不連続エピトープを認識する。モノクローナル抗体2F5は、gp41の細胞外ドメイン上にある直鎖状のエピトープに結合するが、このエピトープがシンプルかは疑わしく、恐らく6個の残基からなる直鎖状配列よりも複雑であり、モノクローナル抗体b12、2G12および2F5は、インビトロで、多種多様の一次単離物に対して中和活性を示した。これまで、広範な中和を仲介するが一次単離物に対して有意な中和抗体を発生させないことがわかっているエピトープも含めてあらゆる組換えエンベロープベースのワクチン候補が臨床試験で試験された。ペプチドベースのワクチン候補も、十分な免疫反応を引き起こすことができない。
【0011】
ペプチドDAPTAに対する相同配列(デルタ1−ペプチドT−アミド(5))は、多数のHIV−1単離物のV2内に存在する(我々は4,078種の配列を試験した)。ペプチドDAPTAに対するポリクローナルおよび精製IgG抗体を用いた我々の結果によれば、一次および実験用に適合させた系統に対する有意な中和活性が示され、このことから、架橋を形成するシート近傍にあるV2ドメインのカルボキシル末端から8個の配列が、新しい中和エピトープの一つであるという見込みが得られた。
【0012】
ポリクローナルペプチドDAPTA抗血清の生成
親和性によって精製した抗体を生成するために、以下を連続して皮下注射することによって6匹のウサギを免疫化した:1)フロイント完全アジュバントに懸濁した75μgのペプチドDAPTA(1日目)、2)フロイント不完全アジュバントに懸濁した75μgのDAPTA(20日目)、および、3)アジュバントなしの150μgのDAPTA(35日目)。セロコンバージョンに関して試験するために、1日目と週1回にウサギの耳の静脈から採血し、その後、精製したペプチドDAPTA、および、ホースラディッシュペルオキシダーゼに結合させたヤギ抗ウサギIgGで被覆したマイクロタイタープレートを用いた酸素結合免疫吸着検査法(ELISA)によって血清を試験した。その後、血清陽性のウサギから心臓穿刺によって採血した。抗血清中に存在するペプチドDAPTA抗体を、精製したペプチドDAPTAとカップリングしたアフィニティーセファロースカラムを用いたアフィニティークロマトグラフィーで精製した。
【0013】
発明の簡単な要約
以下はいくつかの好ましい実施態様の要約であって、本発明の範囲を限定する意図は全くない。多様なHIV分岐群、すなわち有効なワクチンの主要な成分に対する中和抗体を広範に誘導する組成物を開示する。
【0014】
いくつかの実施態様は、HIVgp120によって誘導された短いペプチド配列の免疫原を同定する能力、および、同定されたCCR5結合部位を構成するペプチドから製造された様々な治療用組成物に関する。このようなペプチドは、多様なHIV株由来のペプチド配列、関連ウイルス株由来の配列、その他のウイルスエンベロープ由来の配列、および、選択された神経ペプチド由来の配列を解析することによって選択される。
【0015】
さらに、その他の実施態様は、HIV治療剤として作用する組成物、または、ヒトに投与すると、広範に活性な抗HIVワクチン抗体を誘導する免疫原として作用する組成物を提供する。
【0016】
さらにその他の実施態様は、治療上有用な調合物において、安定化させ、凝集を減少させ、ペプチドの免疫原性を強化する能力に関する。より具体的には、本発明は、少なくとも1種の自己会合または自己凝集するペプチドまたはタンパク質薬物、および、少なくとも1種の界面活性剤を含む治療用組成物を提供し、ここで、界面活性剤はさらに、少なくとも1種のアルキルグリコシド、および/または、糖類とアルキルとのエステルで構成される。
【0017】
さらにその他の実施態様は、アルツハイマー病、乾癬、多発性硬化症、および、その他のヒトの炎症のカスケードに関連する疾患、同様に、関連するレトロウイルス、例えばHTLV−1、熱帯性痙性不全対麻痺の原因のような領域において有効な治療を提供すると予想される薬物候補になる可能性があるペプチド配列を選択する新規の方法に関する。
【0018】
図面の簡単な説明
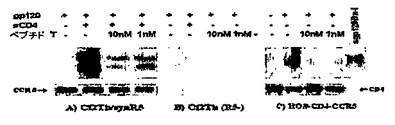
図1において、DAPTAは、CCR5受容体とのgp120−CD4複合体の共免疫沈降反応を防ぐ。
【0019】
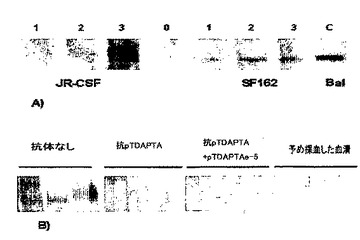
図2は、gp120のCCR5の結合分析である。
図3A〜Bにおいて、蛍光標識したgp120(FITC−gp120)は、sCD4が存在するときのみにCf2Th/synCCR5細胞に効率的に結合した。
【0020】
図4A〜Cにおいて、DAPTAは、gp120エンベロープタンパク質のCCR5への結合を阻害する。
図5A〜Bは、抗ペプチドDAPTAのgp120一量体への結合を示す。
【0021】
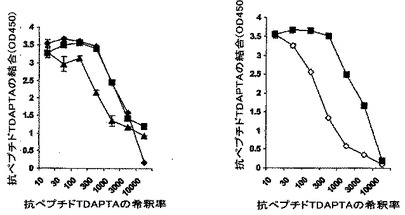
図6は、抗ペプチドTDAPTA血清のgp120に対する結合親和性を示す。
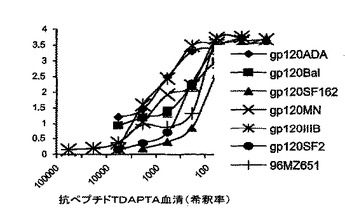
図7は、抗ペプチドTDAPTA IgGのgp120タンパク質への溶解性CD4依存性の結合を示す。
【0022】
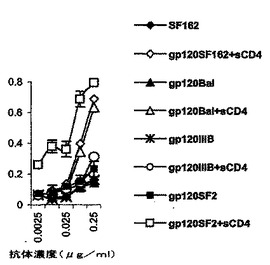
図8は、抗ペプチドTIgGおよびポリクローナル血清の、膜に固定されたウイルスおよび組換えgp120タンパク質への親和性結合を示す。
発明の詳細な説明
以前の研究の生成物および方法に対して、本明細書において開示された生成物および方法は、有用な抗HIV活性、およびその他の治療特性を示す予想外の特性を示す。以下の実施例において、様々な本発明の実施態様のいくつか(ただし全てではない)を説明する。
【0023】
本明細書において、補助受容体結合をブロックする可変性だが相同な配列を有するgp120のV2領域における生物活性を有するペンタペプチド群を説明する。以前の研究では、V2領域は、gp120の受容体への結合および感染力において重要な役割を有さないと考えられていた。V2が欠失した突然変異体は、感染力を有すると考えられていた(6,7)。V2ループに対して、V3ループは、gp120の受容体結合エピトープを含むことが一般的に提唱されてきた(7〜9)。なおこのような相同配列はさらに、それらが発生し得るV2以外のgp160の領域から得られる場合がある。
【0024】
本発明は、CCR5において受容体活性を有するペプチドを1種またはそれより多く含む調合物を説明する。ペプチドTに対する相同配列(SF−2単離物から誘導された)は、多数のHIV−1単離物のV2内の同様の位置に存在する。我々は、Entrezデータベースにおいて入手可能な4,078種のエンベロープ(gp120)V2配列を試験した。これらのうち、60%がxTxYxまたはxNxYxの形態を取り、23%がxSxYxの形態を取っているが、残りの17%は、普及率2〜7%の10種程度のその他のマイナーなモチーフに割り当てられる。これらの多様なエンベロープから誘導されたペンタペプチド配列は、受容体活性を有するモチーフ、例えば利用可能な配列の80%より多くを含む2種の主要なモチーフを定義するために重要であり、より十分に説明するためにいくつかの例を示す。このような構造の収集から、治療剤またはワクチン免疫原として外見的に多様なHIV株に対して広範な特異性を有するペプチドを、単独で、または、混合物として設計することができる。
【0025】
我々が研究してきたV2配列は、全部ではないが多数のHIV単離物においてほぼ同じ位置(幹部近傍のV2)に発生しており、ペプチドTDAPTAの末端の5個のアミノ酸にいくらか相同である。このようなペプチドは、VIP/PACAP/GHRHペプチドに属するペプチドと相同性を有する。これらのウイルスによって誘導された神経ペプチドの多く(例えば表1に記載されたもの)が合成され、試験されており、さらに、ペプチドTDAPTAオクタペプチドと同等か、またはそれよりも有効な生物活性を有することが示されている。薬理学的に、このようなペンタペプチドは、ケモカインの走化性の部分アンタゴニストである(10)。このようなDAPTAのアンタゴニストの作用は、そのgp120−sCD4のCCR5への結合をブロックする能力(11)、または、インビトロにおいてgp120によって誘導されるニューロンの死滅(12)をブロックする能力によるものである。
【0026】
gp120からのV2関連ペンタペプチドの免疫原ライブラリーにおける治療剤およびワクチンの同定
以下の表1に、19種の有用なペンタペプチドのライブラリーを示すが、これから治療用ペプチドおよびワクチン成分が選択されると予想される。
【0027】
【表1】

【0028】
以下の表で、V2ペンタペプチド、および、それらに対応するHIV単離物を列挙する。
【0029】
【表2】
【0030】
注釈:*は、ペプチドTの配列であり、末端ペンタペプチドが強調表示されており、#は、4位にD−チロシンを有する不活性な類似体である(12)。V2ペプチドに関するHIV単離物を列挙し、続いて実際のペプチド配列を列挙する。このようなペプチドは、チロシン(Y)が同じ位置に存在し、セリン(S)とスレオニン(T)とはメチル基が一つ異なる構造的に関連するアミノ酸であるため、類似体として認識される。改変型がわずかながら知られており、これらの構造的な詳細がこれまでに考察されている(18,57)。興味深いことに、ペプチドTは、実験用に適合させたウイルスSF−2(以前はARV,(13))から誘導されるが、CCR5に対して活性である。これは、これらのペプチドのうちいくつかは、関連するCXCR4侵入の受容体、および、数種のVIP受容体において固有の活性を有することを示す(14,15)。わずかな改変によって、HIV進行の後期におけるいわゆる「二重細胞指向性」ウイルス単離物の出現(これもCXCR4侵入の受容体を利用する)に伴う受容体の利用度、インビボで生じる作用が広がると予想される。
【0031】
脳のウイルス単離物は、低いレベルのCD4およびCCR5を発現する脳の小グリア細胞およびマクロファージにおける複製に適応した別個のエンベロープタンパク質の表現型を示すようである(16)。それゆえに、脳のウイルス単離物から誘導されたエンベロープタンパク質のペンタペプチドは、低いCD5または補助受容体の発現に適応したHIV株(例えば脳に感染するHIV株)をブロックすることに関して特定の治療上の利点を提供する可能性がある。低いCD4および/またはCCR5が細胞に感染する能力は、T細胞とマクロファージとの両方に関してより広範な親和性与えると予想され、それゆえに、脳に適応した株からのペンタペプチドは、これらに限定されないが、脳のマクロファージまたは小グリア細胞への、多様な組織および細胞における低い受容体の数に適応したHIV株感染を抑制する、または、ワクチンまたは治療抗体を生成させるための免疫原を作るのにも使用可能である。低い受容体数に適応したウイルス単離物の抑制は、HIV治療剤開発における次世代の領域である可能性が高く、高い効力を有するリガンドが最も有効であると予想される。天然のウイルスから生じた配列を精査することによって、生物学的関連性と実際の有用性が最適化されると予想される。候補の治療用ペプチドの受容体の効力は容易に試験することが可能であり、安定性および生物学的利用率に関してさらなる改変を施すことによって、最も高度な活性を有するペンタペプチドが治験薬として開発され得る。
【0032】
脳で同定されたV2受容体活性を有するペプチド治療剤および免疫原、ならびに、選択された神経ペプチドで同定されたワクチンのペンタペプチド
(16)で説明されている5種の脳単離物から典型的な配列を同定し、それを以下の表に示す。
【0033】
【表3】
【0034】
gp120のV2領域内のペンタペプチドに加えて、我々は、このペプチドのモチーフが、gp120のその他の部分、gp41(膜結合した)、および、VTP/PACAP/GHRHに関連する神経ペプチドに存在することを特筆する(17)。以下の表に、これらの例を列挙する。
【0035】
【表4】
【0036】
従って、V2に関連するペプチドは生物学的に活性であることが示され、最も簡単な説明は、これらは、特異的なケモカイン受容体において薬理学的に活性であることである。
HIVエンベロープタンパク質GP41において同定されたV2に関連するペンタペプチド
以下の表で、治療剤またはワクチン免疫原として使用可能なGP41のHIV配列を説明する。
【0037】
【表5】
【0038】
我々は、Entrezデータベースからの702種のgp41配列を試験した。記録された全てのペンタペプチド配列は、これらのペプチド類似体が有する共通の結合機能に相当する同じ位置、すなわちgp41のおよそ126位に位置していた。本発明は、高濃度か低濃度かに関わらず少なくとも1種のペプチドまたはタンパク質を含む調合物を説明する。
【0039】
CCR5に結合して、HIVgp120のCCR5への結合をブロックする多様なV2に関連するペンタペプチドの同定
上述した(11)で詳述された方法、すなわち、競合的な小さいgp120ペプチドを添加した場合、または添加しない場合のgp120/sCD4複合体のChfTh−CCR5発現細胞への結合を用いて、我々は、V2によって誘導されたDAPTAに相同なペプチドも、gp120のCCR5への結合をブロックする有力な能力を有する一般的なケースを示す。構造と機能活性を示すが、これは特異性の指標であり、ペプチドTTNYTにおいて、L型アミノ酸からD型アミノ酸への置換は効力を大きく減少させ、ペプチドINNYTにおいて、NからDへの置換によってペプチドIDNYTが形成されると、活性がより低くなる。驚くべきことに、ペプチドTTNYTのD型アミノ酸の形態はいずれも高い活性を有しており、これは、これらのペンタペプチドのD型アミノ酸の形態はいずれも有用であることを示しており、それらの予想されるペプチダーゼ分解に対する安定性のために好ましい場合がある。
【0040】
【表6】
【0041】
ペプチドTDAPTAは、gp120−sCD4複合体のCCR5受容体への結合を防ぐ
図1に示したように、DAPTAは、gp120−CD4複合体とCCR5受容体との共免疫沈降反応を防ぐ。パネルAで説明した研究において、hCCR5受容体を発現するCf2Th/synR5細胞が用いられた。可溶化したCCR5受容体を抗tag抗体ID4と共に免疫沈降させ、タンパク質A/Gアガロースで捕捉し、DAPTAの存在下または非存在下でgp120Bal−sCD4複合体とインキュベートした。ヒトHIVIgを用いたウェスタンブロッティングによって免疫沈降したgp120を検出した。同じ膜を裂いて、CCR5(NT)受容体に対するウサギポリクローナル抗血清を用いてハイブリダイズさせた(下部のパネルA)。パネルBは、Cf2Thの親の胸腺細胞系からの細胞溶解産物を示す。免疫沈降したgp120タンパク質は検出されなかった。パネルCは、gp120CM235タンパク質とインキュベートする前にDAPTAで処理したHOSCD4.CCR5細胞を示す。約2×1.107からの膜に結合したCCR5が、CCR5に対するモノクローナル抗体と免疫沈降した。ウェスタンブロットによって、共免疫沈降したgp120タンパク質、および、膜に結合したCD4受容体が検出された。
【0042】
DAPTAは、CD4依存性のgp120のCCR5への結合をブロックする
図2で説明したように、本発明者等は、gp120/sCD4複合体の、既知のCCR5リガンド、例えばMIP1−□またはCCR5「ブロッキング」抗体による阻害を受けやすいCCR5を発現する細胞への結合を分析した。抗ウイルス性の分析は従来技術で詳細に説明されており(Ruff等, 2001, Antiviral Res, 52, 63-75)、これは、参照により本発明に含め、本明細書においては簡単に説明するのみとする。
【0043】
我々は、蛍光タンパク質標識キット(ロシュ・ダイアグノスティックス社(Roche Diagnostics GmBh))を用いて、可溶性gp120タンパク質(25□g/ml)から新規のFITC標識トレーサーを製造した。結合分析は、50mMのHEPES(ギブコ(Gibco))、pH=7.4、5mMのMgCl2、1mMのCaCl2.6H2O(シグマ(Sigma))、5%BSA(シグマ)、および、0.1%NaN3を含み、最終容量100□1の結合緩衝液中で行われた。指定された通りの数種のCCR5を発現する系統を用いて、96ウェルのフィルタープレート(ミリポア(Millipore))中で37℃で1時間結合させた。未結合の標識タンパク質を、96−ウェルプレートマニホールドを用いた迅速な真空ろ過と洗浄によって除去した。蛍光プレートリーダー(ヒューレット・パッカード(Hewlett Packard))を495/530nmで用いてフィルターを計数した。sCD4の非存在下で非特異的な結合を決定し、それを100nMのsCD4(全ての結合)の存在下でのgp120−FITC結合から差し引いた。
【0044】
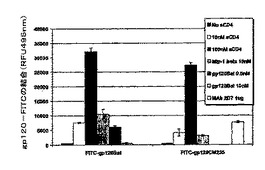
蛍光標識したgp120(FITC−gp120)は、sCD4の存在下のみでCf2Th/synCCR5細胞に効率的に結合した
分析中にsCD4が存在しない場合、結合はほぼ検出不可能であった。FITC−gp120の最大の結合は、それに続いて用いられた100nMのsCD4で得られた。
【0045】
図3で説明したように、gp120(Bal、および、CM235)結合の特異性は、標識されていないHIV−1Bal、または、MIP−1□、既知のCCR5リガンドとの競合により示された(18〜22)(図2,上記)。標識したFITC−gp120Bal(0.5nM)の結合は、等モル量(0.5nM)の標識されていないHIV−1Balの存在下で80%を超えて阻害されるか、または、20倍過量の(10nM)標識されていないHIV−1Balを用いた場合バックグラウンドレベルまで減少した。gp120タンパク質BalおよびCM235の両方の結合は、CCR5特異的なリガンドMIP−1□(10nM)を用いた場合に阻害され、これまでに述べられている通りである(23〜25)。また、FITC−gp120CM235の結合も、イヌの胸腺細胞を、gp120結合をブロックするCCR5に対する2D7抗体(図1)とインキュベートすることによって阻害された(26)。FITC−gp120タンパク質は、100nMのsCD4の存在下で、親のCCR5陰性のイヌ胸腺細胞Cf2Th細胞に感知できるほどに結合しなかった。これらの研究から、gp120−FITCのCCR5受容体への結合はCD4依存性であることが示される(26〜30)。
【0046】
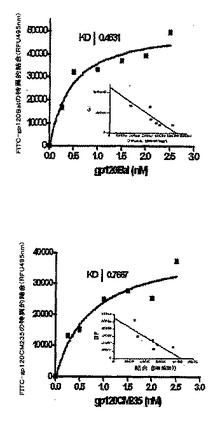
2種のCCR5結合gp120タンパク質(Bal、CM235)それぞれの飽和結合状態(図3)を、(Cf2Th/synR5)細胞にFITC−で標識されたされたgp120の量を高めて(0.25〜2.5nM)添加することによって研究した(図2,右)。sCD4の非存在下で非特異的な結合を決定し、それを100nMのsCD4存在下での結合から差し引いた。gp120Balの飽和可能な結合および高親和性の結合は、Kdが、0.46+0.17nM(P<0.05)で生じ、gp120CM2351に関してはKdが、0.77+0.35nM(P<0.05)で生じ、この結果は、その他にも一致する(27,31,31,32)。
【0047】
DAPTAは、gp120エンベロープタンパク質のCCR5への結合を阻害する
gp120のCCR5受容体への結合のペプチド阻害剤の効力を定義するために、濃度を増加させたCCR5選択的なケモカインリガンドMIP−□、および、DAPTAの存在下で固定濃度のsCD4/gp120複合体を用いて阻害の研究を行った。これらの研究は図4で説明される。特異的な結合の総量は、sCD4(100nM)を添加した場合、または、それらを添加しない場合での、FITIC−gp120の結合の差として定義した。
【0048】
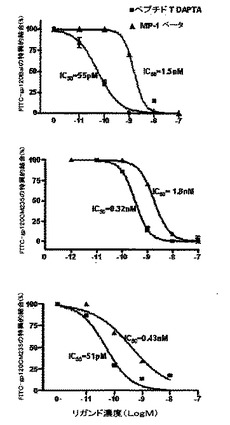
gp120BaL/sCD4のCCR5(Cf2Th/synR5)細胞への結合は、MIP−1□(IC50=1.5+0.002nM、P<0.05)、および、DAPTA(IC50=55+0.08pM、P<0.05)で完全に阻害された(図3A)。DAPTAに関するヒル(Hill)の傾きは−1.07であり、これは、1部位における競合結合モデルと一致した。同様に、我々はgp120CMCM235/sCD4の同じ細胞への結合阻害を研究したところ、再度、MIP−1□(IC50=1.8+0.006nM、P<0.05)、および、DAPTA(IC50=0.32+.03nM、P<0.05)による特異的結合の実質的な(>80%)阻害が示された(図3B)。我々はさらに、gp120CM235/sCD4の異なるCD4を発現する細胞系(GHOST CD4.CCR5)への結合も、MIP−1□(IC50=0.43+0.07nM、P<0.05)、および、DAPTA(IC50=51+0.09pM、P<0.05)で阻害されたことも示した(図3C)。示されたデータ(パネルA〜C)は、3回の実験(それぞれ3連での決定)の平均およびSEMである。PRISMで解析を行った。
【0049】
これらの臨床的および基礎的な調査の発見の関連性は、gp120のペプチドT領域が、ケモカイン補助受容体CCR5に結合するエピトープと同定されたことを証明するものである(11)。ほぼ25年にわたる努力によっても、CCR5の臨床的に有効な短いペプチド受容体の競合物は得られなかった。例えば、gp120分子全体を含むあらゆる包括的な20merを2セット製造し、抗ウイルス活性に関して試験した。(第二のセットは、第一のセットに比べて10個分の位置をフレームシフトさせた)。これらのサンプルはどれも、抗ウイルス活性をまったく示さなかった。加えて、10年にわたる完全な失敗、有効なHIVワクチンを同定しようとする300億ドルもの連邦または個人的な努力は、ワクチン研究センター(Vaccine Research Center)、NIHにおける臨床研究局長(Director of Clinical Studies)であるBarney Graham博士による講演、「有効なHIVは可能か(Is an Effective HIV Possible)」という表題の講演(2006年2月15日、メリーランド州ベセスダ)で認められているが、これは、広範な中和抗HIV抗体を作製する能力が導かれる本発明のアプローチが自明ではないことを示す。実際に、このような抗体を誘導することができないことが、過去および現在のワクチンに関するあらゆる試みの失敗の元となっている。
【0050】
CCR5受容体に結合するHIVエンベロープgp120のペプチドは、ウイルスの受容体への結合をブロックする免疫原として用いることができる。ほぼ全ての有効な抗体ワクチンは、ウイルスの受容体結合または付着をブロックすることによって作用する。
【0051】
近年、製剤化されたペプチドの特性の解析、および、詳細な構造的な研究(MacPhee,未発表)から、DAPTAは、水溶液に迅速に溶解させると、極めて強い凝集傾向を示し、それにより、生物学的利用率と抗ウイルス活性との両方の損失が起こることが解明されている。目下、このDAPTAの特性は、準最適な臨床結果しか得られない場合があり(33)、インビトロでの結果が不当に陰性になることさえあることが明らかである。例えば、ペプチドT溶液は、高濃度になり、「ゲル」になることが報告されており、それにより、粘膜(例えば鼻上皮)を介して輸送される活性および/または能力の見込みが失われると考えられている。調合物から塩化ナトリウムを除去し、濃度を5mg/mLに低くすることが、問題を解決するようである。しかしながら分光偏光解析によれば、わずか1mg/mLでも、室温で、それより低濃度の0.1mg/mlの205.4nmでの大きいピークから237.2nmでの大きいピークにシフトすることが明らかになり、これは、ゲル化が起こり得る凝集工程において、ペプチドTは、より高濃度の場合、それ自身と相互作用することを示す。電子顕微鏡法により、ペプチドTは細線維を形成することが確認され、我々の知る限りの知識によれば、ペプチドTは、すでに説明されているその他の小さいペプチドのどれよりも容易に細線維を形成する。
【0052】
細線維の形成の決定
ペプチドTまたはそれらの類似体の細線維の形成は、電子顕微鏡法を用いて測定することができる。DAPTA水溶液の2μlのアリコートを、フォームバー(formvar)/炭素で被覆されたニッケル製のEMグリッドに塗布した。このグリッドを10μlの蒸留水で3回リンスし、10μlの2%酢酸ウラニルで染色した。このサンプルをLaB6フィラメント(120kv)を備えたFEI TEM Tecnai顕微鏡で試験し、メガビューII(Megaview II)CCDカメラでイメージ化した。
【0053】
また、ペプチドTまたはそれらの類似体の細線維の形成は、色素結合を用いて決定することもできる。コンゴレッドを、PBS(5mMのリン酸カリウム、150mMのNaCl、pH7.4)に7μg/mLの濃度に溶解させた。この溶液を4℃に冷却し、DAPTAを10mg/mLのストック水溶液として添加し、0.48mg/mLの色素溶液中で最終的なペプチド濃度にした。粉末を溶解させた直後のペプチド溶液を、凝集したペプチドを含む時間を置いたストック溶液と比較した。4℃で400〜700nmでスペクトルを回収した。
【0054】
またペプチドTまたはそれらの類似体の細線維の形成は、円二色性(CD)分光分析を用いて決定することもできる。新たに製造されたペプチド、または、繊維性の凝集体を含むペプチドのいずれかの10mg/mLの水溶液を、4℃で蒸留水に50μg/mLの濃度まで添加した。ジャスコ(Jasco)のモデルJ−810スペクトロメーターで、0.1cmのパス長の石英キュベットを用いて、190〜250nmの範囲で、1分間のインターバル、および、2秒の応答時間を用いてCDスペクトルを得た。
【0055】
さらにその他のペプチドTまたはそれらの類似体の細線維の形成の決定方法は、フーリエ変換赤外(FTIR)分光分析を用いて行われる。DAPTAを重水に10mg/mlの濃度まで溶解させ、細線維の形成を促進する温度および時間条件下でインキュベートした。次に、25μlのサンプルを、6μmのスペーサーで仕切ったNaClウィンドウを備えた予め冷却した透過セル中に入れた。FTIRスペクトルを、DTGS検出器を用いて、バイオ・ラッド(BioRad)のFTS−175Cフーリエ変換スペクトロメーターで透過モードで回収した。2cm−1の解像度で2506枚の干渉写真を記録した。水蒸気を差し引いて、スペクトルの基線を修正した。
【0056】
免疫原性の特性
抗ペプチドDAPTA抗体のgp120単量体への結合
図5は、抗ペプチドDAPTA抗体のgp120単量体への結合を説明する。パネルAにおいて、IIIBgp120(●);gp120ADA()、および、gp120Bal(▲)は、マイクロプレートに直接被覆されている(被覆されたELISA)。そのウェルに25倍まで連続的に希釈した抗ペプチドDAPTAを添加した。結合した抗体を抗ウサギIgG−HPR−ロバ(KP&L)を1:1000にしたもので検出し、450nmにおける光学密度として測定した。バックグラウンドを一般的なウサギ血清コントロールによって推定した。試験された3種全てのgp120タンパク質への最大限の結合の半分に到達するのに必要なV2抗体の濃度は、8〜25ng/mlの間であった。これらのデータは、代表的な実験から得られたものだが、各データポイントは、3連の実験からの平均値である。パネルBは、sCD4(■)の存在下、および、sCD4なし(◇)でのgp120IIIBと抗ペプチドDAPTAとの反応性を示す。これらの抗体は、sCD4の存在下では高親和性で結合する。この結果は、2回繰り返した代表的な実験から得られたものである。各データポイントは、3連の実験からの平均値である。
【0057】
抗ペプチドTDAPTA抗体の可溶化したgp120sへの結合
抗ペプチドTDAPTA抗体、および、精製したIgG分子の、様々な実験用に適合させたウイルス株由来のgp120組換えタンパク質への親和性をELISA分析によって測定した。ポリクローナル抗ペプチドTDAPTA血清は、可溶化したgp120タンパク質に結合したが、gp120ADA、および、gp120IIIBには高親和性で結合し、SF162gp120には最も低い親和性で結合した。これは、図6で説明されている。捕捉ELISAからの結果は、中和活性との優れた相関を示し、従って、我々が本明細書において開発したような単純なELISA試験によって、ワクチン候補抗体を選別することができることが示される。
【0058】
gp120s(1μg/ml)を、マイクロプレート上のポリクローナルヤギ抗HIV−1SFgp120血清によって捕捉した(捕捉ELISA)。3倍まで連続的に希釈した抗ペプチドTDAPTA血清を加熱不活性化し、これをそのウェルに添加した。結合した抗体をロバ抗ウサギIgG−HRPで検出し、450nmにおける光学密度として測定した。バックグラウンドを、結合したペプチドTDAPTAに対して免疫化する前のウサギ血清の量によって推定し、それを差し引いた。このラインは、3連で行われた2回の実験からのデータのうち代表的なものである。
【0059】
抗ペプチドTDAPTA抗体の可溶化したgp120sへの結合は、CD4依存性である
溶解性CD4(sCD4)を添加した場合、および、それらを添加しない場合の精製した抗付着部位IgGのgp120sへの親和性結合を、ペプチドTをローディングした親和性ELISAから精製した抗ペプチドTDAPTAウサギ抗血清全体から誘導された免疫グロブリンのトラクションを用いて研究した。
【0060】
これは、図7で説明されている。可溶性gp120タンパク質を、抗HIV−1SF2血清、または、sCD4(1μg/ml)で被覆されたマイクロプレートウェル上で捕捉した。3倍まで連続的に希釈した抗ペプチドTDAPTA IgGを、各希釈率ごとに3連で添加した。結合した抗ペプチドTDAPTA IgGを、2%の一般的なヤギ血清を含むブロッキング緩衝液中のHPR−ロバ抗ウサギIgGによって検出した。その結果は、3連で行われた2回の実験の平均である。エラーバーは、SD(p<0.05)を示した。
【0061】
CD4分子に結合するとおおむねgp120を認識する抗体が特に興味深く、ワクチン関連性を有する。いわゆるCD4誘導性(CD4i)抗体は、CCR5、および、CXCR4のgp120−CD4複合体への結合を阻害することから(34〜36)、これら抗体は、gp120の補助受容体結合を許容するコンフォメーションに対して向けられていると認識される。しかしながら、ヒトから単離されたCD4i抗体の例は限定的であり、そのプロトタイプは、一次HIV−1単離物に対して弱い中和活性しか示さない。これらの抗体が一見希少であることの原因は、CD4iエピトープが比較的入手困難であることにある(6,37,38)。
【0062】
このようなgp120のCD4誘導性(CD4i)部位が見出された抗体は、極めて限られた数しか存在しない。CD4によって引き起こされるgp120のコンフォメーションを認識する抗体は、補助受容体結合エピトープに対して向けられている可能性が高い。抗ペプチドT抗体は、本明細書において、CD4誘導性エピトープを選択的に認識することが示されており、従って、補助受容体結合部位の近傍で、またはその場で結合すると推測される。
【0063】
抗付着部位抗体は、gp120/sCD4のCCR5侵入の補助受容体への結合をブロックする
抗ペプチドTDAPTA抗血清、および、精製した抗ペプチドTDAPTA IgGのgp120sへの親和性を、ウェスタンブロットでさらに研究した。2種のHIV−1ウイルス株JR−CSF、および、SF162からのウイルス粒子を濃縮し、NP40の存在下で溶解させ、4〜12%のポリアクリルアミドゲルを用いたタンパク質を電気泳動で分離した。p24抗原に関して3種の異なる濃度を用いられ、スロットあたり1、2および3ngの溶解産物で測定された。タンパク質をNEMPVDFメンブレンに移し、精製した抗ペプチドTDAPTA IgG(1:200)とハイブリダイズさせた。これらの抗体は、用量依存性の方式でウイルスのgp120タンパク質を認識した。
【0064】
これは、図8で説明されている。パネルAにおいて、ウイルス粒子を濃縮し、4〜12回のSDSPAGEで分離した。ELISAによって測定したところ1、2および3ngのp24タンパク質を含むウイルスの溶解産物をローディングした。このメンブレンを、ペプチドTDAPTA IgG(1:200)に対する親和性によって精製した抗ペプチドTDAPTAとハイブリダイズさせ、HRP−ロバ抗ウサギIgG(1:2000)で染色した。このフィルムをECL溶液(アマシャム(Amersham))で展開した。この実験を2回繰り返した。パネルBにおいて、IIIBHIV−1ウイルス株、および、可溶性gp120IIIB組換えタンパク質からの溶解産物全体(濃度21μg/ml)を10%SDSPAGEでの電気泳動で分離し、トランスファーさせた。このメンブレンを、ペプチドTDAPTA(10μM)の存在下での、抗ペプチドTDAPTAポリクローナル血清(#195)、予め採血した血清および抗ペプチドTDAPTA血清とインキュベートした。抗ペプチドTDAPTA抗体のHIV−1gp120IIIBエンベロープタンパク質に対する特異性が示された。ペプチドTDAPTAは、抗ペプチドTDAPTAのgp120タンパク質への結合をブロックした(図8A)。
【0065】
V2ペプチドベースの抗体の抗ペプチドDAPTAの中和特性
HELACD4+CXCR4/CCR5+細胞(P7)において融合細胞をカウントする分析で、HIV−1の中和を評価した。顕微鏡下で青色のフォーカスユニット(BFU/ウェル)をカウントした。以下の表1にまとめたこのデータは、抗体の固定濃度0.125μg/ウェルを用いて得られた。さらに精製したIgGの中和活性の滴定も行った。IgG分画は、より低い親和性を有するペプチドDAPTAに対するポリクローナル抗血清の滴定曲線に類似した滴定曲線に従っていた(データ示さず)。感染の阻害は3連の平均値であり、以下の式によって計算された:阻害(%):100−[(BFU(抗体を含む): BFU(抗体を含まない))×100]。
【0066】
【表7】
【0067】
数種の一般的に研究された分岐群B単離物に関するペプチドT/DAPTAに対する抗体の効力(50%[IC50]阻害を達成するのに必要な濃度)を比較するために、我々は、既定の補助受容体特異性を有する細胞系(HELA CD4−βガラクトシダーゼ、および、HELA CD4.CCR5)を用いたフォーカス形成分析において一回のウイルス感染を用いた(39)(表1)。
【0068】
上記の実験において、感染分析は以下のように行われた。HeLa細胞系(MAGI)が用いられ、この細胞系は、いずれも高レベルのCD4を発現し、トランケーションされたヒト免疫不全ウィルス1型(HIV−1)ロングターミナルリピート(LTR)の制御下にあるベータ−ガラクトシダーゼ遺伝子の単一の統合されたコピー(40)を含み、このCCR5遺伝子(MAGI−CCR5)を有する細胞系(Pirounaki等, 2000)は、NIAID AIDSレポジトリー(NIAID AIDS Repository)から入手した。ペプチドTのケモカイン受容体サブタイプの感受性を決定するために、MAGIおよびMAGI−CCR5を、以下に説明されているようにして用い、以前のプロトコール(40〜43)を改変させた。簡単に言えば、10,000個のMAGIまたはMAGI−CCR5細胞/ウェルを96ウェルプレートにシーディングした。1日後、培地を除去し、20□g/mlのDEAE−デキストリンを含むOpti−MEM培地(ギブコ・BRL(Gibco BRL),ライフ・テクノロジーズ(Life Technologies))でのペプチドTまたはMIP−1□の希釈液を添加した。このプレートを、60分間、37℃、5%CO2で培養し、次に、ウイルスの希釈液を添加し、さらに1.5時間培養した。接種されたウイルスを除去し、ウェルを0.2mLのOpti−MEM培地で2回洗浄し、新しい培地(150□l/ウェルのDMEM)を添加した。さらに46時間培養した後、細胞を、PBS中の1%ホルムアルデヒド、0.2%グルタルアルデヒドで室温で5分間固定し、PBSで2回洗浄し、PBS中の4mMのフェロシアン化カリウム、4mMのフェリシアン化カリウム、2mMのMgCl、および、0.4mg/mlの5−ブロモ−4−クロロ−3−インドリル−□−D−ガラクトピラノシド(X−gal染色、イナルコ・ファーマシューティカルズ(Inalco Pharmaceuticals),セントルイス・オビスポ,カリフォルニア州)で、37℃で、5%CO2で、50分間染色し、PBSで2回洗浄した。顕微鏡で青色のフォーカスを計数し、感染レベルを青色のフォーカスユニット(BFU/ウェル)として記録した。全ての分析において、BFU/ウェルのバックグラウンドレベルは概して3未満であった。
【0069】
上述した感染分析におけるCCR5株SF162、および、JR−CSFの感染力の阻害に関して、未精製の抗血清、および、免疫化する前のコントロール血清を試験した。中和力価(IC50)は、免疫血清において1:20〜1:40の希釈率の範囲であり、免疫化する前の血清における非特異的な中和はごくわずかであった(示さず)。
【0070】
特異性および効力を強化するために、親和性によって精製した抗血清を、ペプチドT/DAPTAカラムを用いたさらなる研究のために製造した。CCR5株SF162、および、JR−CSFに関する親和性によって精製した抗体分画の中和力価は、SF162は0.25〜125μg/mlの間であり、JR−CSFウイルス株は0.125および0.06μg/mlであった(示さず)。精製したIgG分画の中和活性の滴定は、gp120補助受容体の付着部位に対するポリクローナル血清の滴定曲線に類似した滴定曲線に従っており、SF162ワクチン株に関する親和性はより低かった。予め採血した血清を同じ解析で試験した。バックグラウンドの阻害は、1:20に希釈した血清サンプルで2〜4パーセントの範囲であった。続いて、gp120補助受容体の付着部位の親和性によって精製したIgの活性を、様々な分岐群のなかからそれらの多様性および外見に関して選択されたより大規模なHIV単離物サンプルで評価した。
【0071】
19%のカットオフ値は、1:5の希釈率で試験された一般的なウサギ血清を用いて得られた95%の信頼レベルに基づく(Laboratory of Antiviral Drug Mechanisms, Science Applications International Corporation at Frederick National Cancer Institute-Frederick Cancer Research and Development Center)。表Xに示した結果は、少なくとも2回の独立した実験(ウイルス株ごとに3連で行われた)から得られたものである。
【0072】
gp120補助受容体の付着部位の親和性によって精製した免疫グロブリンは、2種の一次単離物、分岐群B/F由来の93BR019、および、93IN101分岐群Cのウイルス株に対して極めて高い中和活性、および、2株を除く全てに対する有意な中和活性を示した。X4ウイルス単離物に対する活性は予想外であり、これは、これらのペプチドは、R5、X4、および、二重細胞指向性HIV株に対する免疫原または治療剤を作製するのに有用であり得ることを示す。
【0073】
親和性によって純粋にした抗ペプチドTによるHIVの中和
IC50が0.045μg/mlの抗付着部位IgGの、HIV−1分岐群C単離物(Dul23、および、Dul56)に対する中和活性を、デューク大学(Duke University)のAIDSワクチン研究所(AIDS Vaccine Lab)(David Montefiori、個人的な人脈)による一回のルシフェラーゼレポーター遺伝子分析によって確認した。そのデータは、以下の通りである:
【0074】
【表8】
【0075】
値は、相対的な発光単位(RLU)が、TZM−bl細胞においてウイルスコントロールウェル(試験サンプルなし)と比較して50%減少した時点の血清の希釈濃度である。活性を有するサンプルは、太字で示す。
【0076】
反復配列
有用なペンタペプチド配列は、直線状のアレイで2〜5回の頻度で生じるそれらの反復モチーフによっても同定が可能であり、このようなモチーフは、分離されていないか、または、どこかで1〜数百個のアミノ酸で分離されているかのいずれかであり、すなわちエンベロープタンパク質のそれ以外のいずれか一部分を除きV2には存在しない。これらのペプチドの望ましくない凝集特性は、エンベロープ内の「ステープル様の」付着部位として役立つこれらのペンタペプチド配列を含むウイルスエンベロープの三次構造を構築することに有用な場合がある。どのような場合においても、いずれの所定のウイルス内におけるペンタペプチド配列の反復は、規則に従っていなくても、追加の有用な配列を示す可能性がある。例えば、EYIYT反復配列は5回反復される。この反復は、コンセンサス配列の近傍に続いて存在しない。これらは反復しているが、活性な化合物のライブラリーに加えられる。
【0077】
CCR5受容体活性の存在
有用な候補は、CCR5受容体への保存的な親和性を有すると予想される。従って、CCR5受容体への相対的な親和性に関して、様々なペンタペプチド組成物(すなわち、それらの様々なリンカーおよび免疫原が結合している)の活性を分析すること(結合分析、抗ウイルス性またはその他のCCR5受容体の相対的な生物活性の分析)は、さらなる有用なスクリーニングであると予想される。CCR5受容体への親和性が高ければ高いほど、最終産物が治療活性を有する可能性は高くなる。
【0078】
有用な候補は、概して、免疫原として用いられる前に広範な抗ウイルス活性を示すと予想される。従って、これらのワクチンはそれ自身、治療剤であると期待される。
本発明の特に重要な形態は、これらのペプチドの混合型のアンタゴニストとしての薬理学的作用に関する。Nobelist Arvid Carlsson(Medicine/Physiology, 2000)によって述べられたように、薬物の混合型の(部分)アゴニストの形態は、受容体相互作用の臨床的に好ましいタイプである。混合型/部分アゴニスト薬物の場合、完全アンタゴニストの除感作、耐性および副作用を回避することが一般的であり、このタイプの成功した薬物が製薬会社によって作製されている。部分アゴニスト薬物は、受容体活性のバランスをとるように作用し(DAPTA作用の混合型アゴニストの形態について示したRedwine, 1999を参照)、従ってこれらは好ましい治療剤である。我々は、本明細書で説明するような、これらのペプチドの固有の生物活性を決定する方法を開発し、予想通りにヒトへ投与してから6ヶ月後に受容体における耐性の誘導を示さない混合型アゴニスト(例えば表6に記載のペプチド)が得られることを見出した。
【0079】
HIV−1gp120は、最も広範囲にわたりグリコシル化されたタンパク質の1種である。これらは、23または24個のN結合型糖付加部位、および、タンパク質の総質量の約半分を占めるこれらの部位に結合したグリカンを含む。糖付加、および、グリコシダーゼ阻害剤を用いた多数の研究により、HIV−1エンベロープ糖タンパク質のコンフォメーションを決定することにおける炭水化物部分の重要性が解明されている。部位特異的変異誘発から、糖付加部位の多くはいずれも非必須であることが示された。24部位のうち、いずれもgp120のアミノ末端側の半分に位置する5部位のみ(アミノ酸88、141、197、262および276)が、ウイルス感染力に影響を与えた。197部位は、V2の近傍に存在するか、または、我々が定義した結合部位のペプチドの多くの位置に存在する。例えば、ここで留意すべきは、本発明のペプチドに、糖付加モチーフT/SxNが存在することが多いことである。我々の結果によれば、N結合型糖付加部位が、本明細書に記載のペプチドの近傍、その場、またはその範囲内に存在することが多く、さらに、補助受容体様CCR5へのウイルス結合エピトープを定義することが示される。糖付加が存在する場合、糖付加は、構造的な特異性に寄与するか、または、この重要なエピトープの免疫監視を回避する手段として天然の抗体による中和をマスクする可能性がある。
【0080】
本発明の一実施態様は、免疫原性特性を有する組成物である。この組成物は、一般的に、A−L−P−C部分の組み合わせとして説明され、ここで、Aは、あらゆるアジュバントであり、Lは、あらゆる結合部分であり、Pは、ペプチド部分であり、Cは、保護基である。
【0081】
開示された化合物において、あらゆるアジュバントが使用できる。ペプチドとリンカー領域とは、アジュバントとは独立して作用すると考えられるので、アジュバントの選択はまったく制限されない。いくつかの有用なアジュバントの例は、これらに限定されないが、ヒトにおいて許容できるあらゆるアジュバント、肝炎のコア抗原、ポリリジンなどである。この開示の後に開発されたアジュバントも作用すると考えられ、その前に開発されたアジュバントも同様である。
【0082】
あるいは、アジュバントは、所定の環境に応じて調合してもよい。例えばペプチドを凝集するようにする場合である。
その他の本発明の実施態様は、治療特性を有する組成物である。この組成物は、一般的に、L−P−C部分の組み合わせとして説明され、ここで、Lは、あらゆる部分であり、ただし好ましくは短いペプチド部分であり、Pは、その他のペプチド部分であり、Cは、保護基である。
【0083】
連結させたLおよびP領域の長さに関して、いずれも特定の長さ(約7〜約16個)のアミノ酸が機能すると考えられる。好ましい実施態様において、このような連結は、7〜11個の範囲である。さらに好ましい実施態様において、このような連結は、7〜9個の範囲である。よりさらに好ましい実施態様において、8である。あるいは、天然の神経性のペプチドは28個のアミノ酸からなることを示すため、開示された組成物のペプチド領域の長さは、約28個のアミノ酸であると考えられる。(表4を参照)。
【0084】
本明細書において開示された様々な化合物は、単独で、または、カクテルとして組み合わせて作用するように設計される。多種多様な化合物が使用でき、限定されない。最も有用なカクテルは、2〜7種の異なる化合物からなると考えられる。
【0085】
C部分は、あらゆる保護基が可能である。好ましい実施態様は、アミドである。
本発明の開示はまた、以下の活性:HIVウイルスに感染したヒトにおける、単球によって誘導されたマクロファージ(MDM)、小グリア細胞、または、T細胞におけるR5または二重細胞指向性(R5/X4)株におけるHIVの抑制を示す化合物、および、その化合物に関してスクリーニングする方法も包含する。その他の活性としては、CCR5受容体を調節することによって治療可能なあらゆるヒトの病気が挙げられ、例えば、これらに限定されないが、アルツハイマーを引き起こす神経炎症、多発性硬化症、乾癬、および、関節炎である。これらの病気は、抑制されるか、または、解消されると予想され、病気の経過は止まるか、または、回復すると予想され、および、臨床的な利点が生じると予想される。
【0086】
最後に、本発明の開示は、組成物の安定性に関してスクリーニングする方法を包含する。これは、ウイルスの感染力、走化性、マップキナーゼ活性化、gp120結合、CCR5活性化もしくは阻害、または、Gタンパク質シグナル伝達からなる群より選択される生物学的に特徴付ける方法によって決定される。
【0087】
引用文献
【0088】
【表9】
【0089】
【表10】
【0090】
【表11】
【0091】
【表12】
【図面の簡単な説明】
【0092】
【図1】DAPTAは、CCR5受容体とのgp120−CD4複合体の共免疫沈降反応を防ぐ。
【図2】gp120のCCR5の結合分析である。
【図3】蛍光標識したgp120(FITC−gp120)は、sCD4が存在するときのみにCf2Th/synCCR5細胞に効率的に結合した。
【図4】DAPTAは、gp120エンベロープタンパク質のCCR5への結合を阻害する。
【図5】抗ペプチドDAPTAのgp120一量体への結合を示す。
【図6】抗ペプチドTDAPTA血清のgp120に対する結合親和性を示す。
【図7】抗ペプチドTDAPTA IgGのgp120タンパク質への溶解性CD4依存性の結合を示す。
【図8】抗ペプチドTIgGおよびポリクローナル血清の、膜に固定されたウイルスおよび組換えgp120タンパク質への親和性結合を示す。
【発明の開示】
【0001】
本願は、2005年6月23日付けで出願された仮出願番号60/693,087;2005年6月23日付けで出願された仮出願番号60/693,088、および、2005年6月23日付けで出願された仮出願番号60/693,089に基づく優先権を主張する。本願はまた、2006年6月23日付けで出願された出願番号(発明の名称「Stabilizing Alkylglycoside Compositions and Methods Thereof」)にも関する(これらの内容は、この参照により開示に含まれる)。
【0002】
発明の背景
一般的に、HIV−1感染やAIDSを防御することができるワクチンは、それらの地理的な由来、サブタイプまたは遺伝子型の特異性に関わりなく、それらウイルスの一次的な単離物に対する有力な中和活性を有する高レベルの抗体を誘導する免疫原を必要とすると考えられており、これが、現行のワクチン候補では実現できていない目的の一つである。それゆえに、合理的なワクチン設計および開発には広範な中和をもたらすエピトープの同定が重要である。具体的に重要なのは、優勢な北米の分岐群Bサブタイプに対して発展途上国に存在し、世界的に関連を有する発生株を含む分岐群Cサブタイプに対するワクチン候補の抗体の開発である。
【0003】
HIVに感染した個体において、中和抗体の優勢な標的はHIVエンベロープである。ある種の中和エピトープのクラスターは、一次および/または単球親和性の単離物のgp120envタンパク質に少なくとも部分的に晒されるようである。20年にわたり大規模な研究が行われているにもかかわらず、広範なワクチンの中和を惹起することができる免疫原は見出されていない。その理由として、HIV感染に対するワクチンは定義が難しいことがあった。
【0004】
ある発明は、多様なHIV株に有力な中和活性を有する抗体を導くことができるペプチド、および、HIV−1防御のための免疫化戦略の重要な成分であるペプチドを同定する補助受容体CCR5に結合するgp120の短い配列の同定に関与する。これらの結合配列に生産され、結合部位のペプチドの免疫原を含む抗体は、系統に関わりなくHIVウイルス感染からの防御が付与されると期待され、さらに治療にも役立つと予想されている。すなわち、それらの受動的な投与は、ウイルスレベルを減少させ、すでにHIVに感染した人間に臨床的な利点を提供すると予想される。
【0005】
中和抗体は、HIV−1感染に応答する。初感染後のウイルス血症から数ヶ月で、まずは抗ヒト免疫不全ウィルスタイプの中和抗体が出現する。初期に生じる中和抗体は、高度な型特異性を有する。感染中に、中和抗体反応はその後も広がる可能性があるが(16)、通常わずかであり、長期間感染した個体を含む患者の大半において散発的に発生する。
【0006】
中和抗体は、HIV−1エンベロープの糖タンパク質を認識し、これはgp120エクステリアとgp41膜貫通糖タンパク質とからなる。gp120糖タンパク質は、標的であるCD4、ならびにケモカイン受容体CCR5およびCXCR4に結合する。gp120がCD4との複合体を形成した後にのみ発現されるgp120のエピトープに対して抗体が発生する例はほとんどない。望ましいワクチン標的である実際の補助受容体結合部位は、CD4とgp120との会合の際にのみ形成されると考えられる。これらのCD4誘導性(CD4i)抗体は、gp120−sCD4複合体のCCR5およびCXCR4結合をブロックする。HIV−1に感染した人間から単離されたCD4i抗体の数は少ない。
【0007】
最も初期のHIV−1ワクチンを用いた研究は、中和抗体を誘発するワクチンを開発することに向けられていた。これらの中和抗体は、それらの作用が限定的であり、動物モデルにおいて接種されたウイルスのほぼ全ての系統に対して特異的である。それゆえにこれらの抗体は広範な特異性を有しておらず、優れたワクチン候補を形成しないだろう。
【0008】
最も成功したワクチンは、その受容体(HIVの場合、補助受容体CXCR4およびCC5である)に結合するウイルスをブロックする(1,2)。過去20年間にわたって懸命な調査の対象である補助受容体結合部位の同定は現在まで達成されておらず、従って、これらを同定することによって、望ましいワクチン用途に適した多様な系統の広範な中和を実現することができる免疫原が生産されることが期待される。
【0009】
本願の目的の一つは、生物学的に活性な補助受容体結合部位に対する抗体である。インビトロでの研究において、この抗体はCCR5受容体に結合するgp120−sCD4複合体の有力な阻害を示し、さらに、適切なペプチドコンフォメーションに対して生じた抗体は中和抗体を広範に誘導することから、それによってHIV−1感染に対する防御免疫、すなわちワクチンを提供することが期待される。
【0010】
10年以上にわたり、ほとんどのHIV−1単離物の広範かつ有力な中和を仲介するヒトモノクローナル抗体はほんのわずかしか単離されていない。これらのモノクローナル抗体のうち2種は、gp120(モノクローナル抗体b12および2G12)の表面に結合し(3,4)、1種は、gp41の膜貫通ドメインの直近位に存在するエピトープに結合する。モノクローナル抗体b12は、gp120のC3−V4領域と炭水化物とを含む複雑な不連続エピトープを認識する。モノクローナル抗体2F5は、gp41の細胞外ドメイン上にある直鎖状のエピトープに結合するが、このエピトープがシンプルかは疑わしく、恐らく6個の残基からなる直鎖状配列よりも複雑であり、モノクローナル抗体b12、2G12および2F5は、インビトロで、多種多様の一次単離物に対して中和活性を示した。これまで、広範な中和を仲介するが一次単離物に対して有意な中和抗体を発生させないことがわかっているエピトープも含めてあらゆる組換えエンベロープベースのワクチン候補が臨床試験で試験された。ペプチドベースのワクチン候補も、十分な免疫反応を引き起こすことができない。
【0011】
ペプチドDAPTAに対する相同配列(デルタ1−ペプチドT−アミド(5))は、多数のHIV−1単離物のV2内に存在する(我々は4,078種の配列を試験した)。ペプチドDAPTAに対するポリクローナルおよび精製IgG抗体を用いた我々の結果によれば、一次および実験用に適合させた系統に対する有意な中和活性が示され、このことから、架橋を形成するシート近傍にあるV2ドメインのカルボキシル末端から8個の配列が、新しい中和エピトープの一つであるという見込みが得られた。
【0012】
ポリクローナルペプチドDAPTA抗血清の生成
親和性によって精製した抗体を生成するために、以下を連続して皮下注射することによって6匹のウサギを免疫化した:1)フロイント完全アジュバントに懸濁した75μgのペプチドDAPTA(1日目)、2)フロイント不完全アジュバントに懸濁した75μgのDAPTA(20日目)、および、3)アジュバントなしの150μgのDAPTA(35日目)。セロコンバージョンに関して試験するために、1日目と週1回にウサギの耳の静脈から採血し、その後、精製したペプチドDAPTA、および、ホースラディッシュペルオキシダーゼに結合させたヤギ抗ウサギIgGで被覆したマイクロタイタープレートを用いた酸素結合免疫吸着検査法(ELISA)によって血清を試験した。その後、血清陽性のウサギから心臓穿刺によって採血した。抗血清中に存在するペプチドDAPTA抗体を、精製したペプチドDAPTAとカップリングしたアフィニティーセファロースカラムを用いたアフィニティークロマトグラフィーで精製した。
【0013】
発明の簡単な要約
以下はいくつかの好ましい実施態様の要約であって、本発明の範囲を限定する意図は全くない。多様なHIV分岐群、すなわち有効なワクチンの主要な成分に対する中和抗体を広範に誘導する組成物を開示する。
【0014】
いくつかの実施態様は、HIVgp120によって誘導された短いペプチド配列の免疫原を同定する能力、および、同定されたCCR5結合部位を構成するペプチドから製造された様々な治療用組成物に関する。このようなペプチドは、多様なHIV株由来のペプチド配列、関連ウイルス株由来の配列、その他のウイルスエンベロープ由来の配列、および、選択された神経ペプチド由来の配列を解析することによって選択される。
【0015】
さらに、その他の実施態様は、HIV治療剤として作用する組成物、または、ヒトに投与すると、広範に活性な抗HIVワクチン抗体を誘導する免疫原として作用する組成物を提供する。
【0016】
さらにその他の実施態様は、治療上有用な調合物において、安定化させ、凝集を減少させ、ペプチドの免疫原性を強化する能力に関する。より具体的には、本発明は、少なくとも1種の自己会合または自己凝集するペプチドまたはタンパク質薬物、および、少なくとも1種の界面活性剤を含む治療用組成物を提供し、ここで、界面活性剤はさらに、少なくとも1種のアルキルグリコシド、および/または、糖類とアルキルとのエステルで構成される。
【0017】
さらにその他の実施態様は、アルツハイマー病、乾癬、多発性硬化症、および、その他のヒトの炎症のカスケードに関連する疾患、同様に、関連するレトロウイルス、例えばHTLV−1、熱帯性痙性不全対麻痺の原因のような領域において有効な治療を提供すると予想される薬物候補になる可能性があるペプチド配列を選択する新規の方法に関する。
【0018】
図面の簡単な説明
図1において、DAPTAは、CCR5受容体とのgp120−CD4複合体の共免疫沈降反応を防ぐ。
【0019】
図2は、gp120のCCR5の結合分析である。
図3A〜Bにおいて、蛍光標識したgp120(FITC−gp120)は、sCD4が存在するときのみにCf2Th/synCCR5細胞に効率的に結合した。
【0020】
図4A〜Cにおいて、DAPTAは、gp120エンベロープタンパク質のCCR5への結合を阻害する。
図5A〜Bは、抗ペプチドDAPTAのgp120一量体への結合を示す。
【0021】
図6は、抗ペプチドTDAPTA血清のgp120に対する結合親和性を示す。
図7は、抗ペプチドTDAPTA IgGのgp120タンパク質への溶解性CD4依存性の結合を示す。
【0022】
図8は、抗ペプチドTIgGおよびポリクローナル血清の、膜に固定されたウイルスおよび組換えgp120タンパク質への親和性結合を示す。
発明の詳細な説明
以前の研究の生成物および方法に対して、本明細書において開示された生成物および方法は、有用な抗HIV活性、およびその他の治療特性を示す予想外の特性を示す。以下の実施例において、様々な本発明の実施態様のいくつか(ただし全てではない)を説明する。
【0023】
本明細書において、補助受容体結合をブロックする可変性だが相同な配列を有するgp120のV2領域における生物活性を有するペンタペプチド群を説明する。以前の研究では、V2領域は、gp120の受容体への結合および感染力において重要な役割を有さないと考えられていた。V2が欠失した突然変異体は、感染力を有すると考えられていた(6,7)。V2ループに対して、V3ループは、gp120の受容体結合エピトープを含むことが一般的に提唱されてきた(7〜9)。なおこのような相同配列はさらに、それらが発生し得るV2以外のgp160の領域から得られる場合がある。
【0024】
本発明は、CCR5において受容体活性を有するペプチドを1種またはそれより多く含む調合物を説明する。ペプチドTに対する相同配列(SF−2単離物から誘導された)は、多数のHIV−1単離物のV2内の同様の位置に存在する。我々は、Entrezデータベースにおいて入手可能な4,078種のエンベロープ(gp120)V2配列を試験した。これらのうち、60%がxTxYxまたはxNxYxの形態を取り、23%がxSxYxの形態を取っているが、残りの17%は、普及率2〜7%の10種程度のその他のマイナーなモチーフに割り当てられる。これらの多様なエンベロープから誘導されたペンタペプチド配列は、受容体活性を有するモチーフ、例えば利用可能な配列の80%より多くを含む2種の主要なモチーフを定義するために重要であり、より十分に説明するためにいくつかの例を示す。このような構造の収集から、治療剤またはワクチン免疫原として外見的に多様なHIV株に対して広範な特異性を有するペプチドを、単独で、または、混合物として設計することができる。
【0025】
我々が研究してきたV2配列は、全部ではないが多数のHIV単離物においてほぼ同じ位置(幹部近傍のV2)に発生しており、ペプチドTDAPTAの末端の5個のアミノ酸にいくらか相同である。このようなペプチドは、VIP/PACAP/GHRHペプチドに属するペプチドと相同性を有する。これらのウイルスによって誘導された神経ペプチドの多く(例えば表1に記載されたもの)が合成され、試験されており、さらに、ペプチドTDAPTAオクタペプチドと同等か、またはそれよりも有効な生物活性を有することが示されている。薬理学的に、このようなペンタペプチドは、ケモカインの走化性の部分アンタゴニストである(10)。このようなDAPTAのアンタゴニストの作用は、そのgp120−sCD4のCCR5への結合をブロックする能力(11)、または、インビトロにおいてgp120によって誘導されるニューロンの死滅(12)をブロックする能力によるものである。
【0026】
gp120からのV2関連ペンタペプチドの免疫原ライブラリーにおける治療剤およびワクチンの同定
以下の表1に、19種の有用なペンタペプチドのライブラリーを示すが、これから治療用ペプチドおよびワクチン成分が選択されると予想される。
【0027】
【表1】
【0028】
以下の表で、V2ペンタペプチド、および、それらに対応するHIV単離物を列挙する。
【0029】
【表2】
【0030】
注釈:*は、ペプチドTの配列であり、末端ペンタペプチドが強調表示されており、#は、4位にD−チロシンを有する不活性な類似体である(12)。V2ペプチドに関するHIV単離物を列挙し、続いて実際のペプチド配列を列挙する。このようなペプチドは、チロシン(Y)が同じ位置に存在し、セリン(S)とスレオニン(T)とはメチル基が一つ異なる構造的に関連するアミノ酸であるため、類似体として認識される。改変型がわずかながら知られており、これらの構造的な詳細がこれまでに考察されている(18,57)。興味深いことに、ペプチドTは、実験用に適合させたウイルスSF−2(以前はARV,(13))から誘導されるが、CCR5に対して活性である。これは、これらのペプチドのうちいくつかは、関連するCXCR4侵入の受容体、および、数種のVIP受容体において固有の活性を有することを示す(14,15)。わずかな改変によって、HIV進行の後期におけるいわゆる「二重細胞指向性」ウイルス単離物の出現(これもCXCR4侵入の受容体を利用する)に伴う受容体の利用度、インビボで生じる作用が広がると予想される。
【0031】
脳のウイルス単離物は、低いレベルのCD4およびCCR5を発現する脳の小グリア細胞およびマクロファージにおける複製に適応した別個のエンベロープタンパク質の表現型を示すようである(16)。それゆえに、脳のウイルス単離物から誘導されたエンベロープタンパク質のペンタペプチドは、低いCD5または補助受容体の発現に適応したHIV株(例えば脳に感染するHIV株)をブロックすることに関して特定の治療上の利点を提供する可能性がある。低いCD4および/またはCCR5が細胞に感染する能力は、T細胞とマクロファージとの両方に関してより広範な親和性与えると予想され、それゆえに、脳に適応した株からのペンタペプチドは、これらに限定されないが、脳のマクロファージまたは小グリア細胞への、多様な組織および細胞における低い受容体の数に適応したHIV株感染を抑制する、または、ワクチンまたは治療抗体を生成させるための免疫原を作るのにも使用可能である。低い受容体数に適応したウイルス単離物の抑制は、HIV治療剤開発における次世代の領域である可能性が高く、高い効力を有するリガンドが最も有効であると予想される。天然のウイルスから生じた配列を精査することによって、生物学的関連性と実際の有用性が最適化されると予想される。候補の治療用ペプチドの受容体の効力は容易に試験することが可能であり、安定性および生物学的利用率に関してさらなる改変を施すことによって、最も高度な活性を有するペンタペプチドが治験薬として開発され得る。
【0032】
脳で同定されたV2受容体活性を有するペプチド治療剤および免疫原、ならびに、選択された神経ペプチドで同定されたワクチンのペンタペプチド
(16)で説明されている5種の脳単離物から典型的な配列を同定し、それを以下の表に示す。
【0033】
【表3】
【0034】
gp120のV2領域内のペンタペプチドに加えて、我々は、このペプチドのモチーフが、gp120のその他の部分、gp41(膜結合した)、および、VTP/PACAP/GHRHに関連する神経ペプチドに存在することを特筆する(17)。以下の表に、これらの例を列挙する。
【0035】
【表4】
【0036】
従って、V2に関連するペプチドは生物学的に活性であることが示され、最も簡単な説明は、これらは、特異的なケモカイン受容体において薬理学的に活性であることである。
HIVエンベロープタンパク質GP41において同定されたV2に関連するペンタペプチド
以下の表で、治療剤またはワクチン免疫原として使用可能なGP41のHIV配列を説明する。
【0037】
【表5】
【0038】
我々は、Entrezデータベースからの702種のgp41配列を試験した。記録された全てのペンタペプチド配列は、これらのペプチド類似体が有する共通の結合機能に相当する同じ位置、すなわちgp41のおよそ126位に位置していた。本発明は、高濃度か低濃度かに関わらず少なくとも1種のペプチドまたはタンパク質を含む調合物を説明する。
【0039】
CCR5に結合して、HIVgp120のCCR5への結合をブロックする多様なV2に関連するペンタペプチドの同定
上述した(11)で詳述された方法、すなわち、競合的な小さいgp120ペプチドを添加した場合、または添加しない場合のgp120/sCD4複合体のChfTh−CCR5発現細胞への結合を用いて、我々は、V2によって誘導されたDAPTAに相同なペプチドも、gp120のCCR5への結合をブロックする有力な能力を有する一般的なケースを示す。構造と機能活性を示すが、これは特異性の指標であり、ペプチドTTNYTにおいて、L型アミノ酸からD型アミノ酸への置換は効力を大きく減少させ、ペプチドINNYTにおいて、NからDへの置換によってペプチドIDNYTが形成されると、活性がより低くなる。驚くべきことに、ペプチドTTNYTのD型アミノ酸の形態はいずれも高い活性を有しており、これは、これらのペンタペプチドのD型アミノ酸の形態はいずれも有用であることを示しており、それらの予想されるペプチダーゼ分解に対する安定性のために好ましい場合がある。
【0040】
【表6】
【0041】
ペプチドTDAPTAは、gp120−sCD4複合体のCCR5受容体への結合を防ぐ
図1に示したように、DAPTAは、gp120−CD4複合体とCCR5受容体との共免疫沈降反応を防ぐ。パネルAで説明した研究において、hCCR5受容体を発現するCf2Th/synR5細胞が用いられた。可溶化したCCR5受容体を抗tag抗体ID4と共に免疫沈降させ、タンパク質A/Gアガロースで捕捉し、DAPTAの存在下または非存在下でgp120Bal−sCD4複合体とインキュベートした。ヒトHIVIgを用いたウェスタンブロッティングによって免疫沈降したgp120を検出した。同じ膜を裂いて、CCR5(NT)受容体に対するウサギポリクローナル抗血清を用いてハイブリダイズさせた(下部のパネルA)。パネルBは、Cf2Thの親の胸腺細胞系からの細胞溶解産物を示す。免疫沈降したgp120タンパク質は検出されなかった。パネルCは、gp120CM235タンパク質とインキュベートする前にDAPTAで処理したHOSCD4.CCR5細胞を示す。約2×1.107からの膜に結合したCCR5が、CCR5に対するモノクローナル抗体と免疫沈降した。ウェスタンブロットによって、共免疫沈降したgp120タンパク質、および、膜に結合したCD4受容体が検出された。
【0042】
DAPTAは、CD4依存性のgp120のCCR5への結合をブロックする
図2で説明したように、本発明者等は、gp120/sCD4複合体の、既知のCCR5リガンド、例えばMIP1−□またはCCR5「ブロッキング」抗体による阻害を受けやすいCCR5を発現する細胞への結合を分析した。抗ウイルス性の分析は従来技術で詳細に説明されており(Ruff等, 2001, Antiviral Res, 52, 63-75)、これは、参照により本発明に含め、本明細書においては簡単に説明するのみとする。
【0043】
我々は、蛍光タンパク質標識キット(ロシュ・ダイアグノスティックス社(Roche Diagnostics GmBh))を用いて、可溶性gp120タンパク質(25□g/ml)から新規のFITC標識トレーサーを製造した。結合分析は、50mMのHEPES(ギブコ(Gibco))、pH=7.4、5mMのMgCl2、1mMのCaCl2.6H2O(シグマ(Sigma))、5%BSA(シグマ)、および、0.1%NaN3を含み、最終容量100□1の結合緩衝液中で行われた。指定された通りの数種のCCR5を発現する系統を用いて、96ウェルのフィルタープレート(ミリポア(Millipore))中で37℃で1時間結合させた。未結合の標識タンパク質を、96−ウェルプレートマニホールドを用いた迅速な真空ろ過と洗浄によって除去した。蛍光プレートリーダー(ヒューレット・パッカード(Hewlett Packard))を495/530nmで用いてフィルターを計数した。sCD4の非存在下で非特異的な結合を決定し、それを100nMのsCD4(全ての結合)の存在下でのgp120−FITC結合から差し引いた。
【0044】
蛍光標識したgp120(FITC−gp120)は、sCD4の存在下のみでCf2Th/synCCR5細胞に効率的に結合した
分析中にsCD4が存在しない場合、結合はほぼ検出不可能であった。FITC−gp120の最大の結合は、それに続いて用いられた100nMのsCD4で得られた。
【0045】
図3で説明したように、gp120(Bal、および、CM235)結合の特異性は、標識されていないHIV−1Bal、または、MIP−1□、既知のCCR5リガンドとの競合により示された(18〜22)(図2,上記)。標識したFITC−gp120Bal(0.5nM)の結合は、等モル量(0.5nM)の標識されていないHIV−1Balの存在下で80%を超えて阻害されるか、または、20倍過量の(10nM)標識されていないHIV−1Balを用いた場合バックグラウンドレベルまで減少した。gp120タンパク質BalおよびCM235の両方の結合は、CCR5特異的なリガンドMIP−1□(10nM)を用いた場合に阻害され、これまでに述べられている通りである(23〜25)。また、FITC−gp120CM235の結合も、イヌの胸腺細胞を、gp120結合をブロックするCCR5に対する2D7抗体(図1)とインキュベートすることによって阻害された(26)。FITC−gp120タンパク質は、100nMのsCD4の存在下で、親のCCR5陰性のイヌ胸腺細胞Cf2Th細胞に感知できるほどに結合しなかった。これらの研究から、gp120−FITCのCCR5受容体への結合はCD4依存性であることが示される(26〜30)。
【0046】
2種のCCR5結合gp120タンパク質(Bal、CM235)それぞれの飽和結合状態(図3)を、(Cf2Th/synR5)細胞にFITC−で標識されたされたgp120の量を高めて(0.25〜2.5nM)添加することによって研究した(図2,右)。sCD4の非存在下で非特異的な結合を決定し、それを100nMのsCD4存在下での結合から差し引いた。gp120Balの飽和可能な結合および高親和性の結合は、Kdが、0.46+0.17nM(P<0.05)で生じ、gp120CM2351に関してはKdが、0.77+0.35nM(P<0.05)で生じ、この結果は、その他にも一致する(27,31,31,32)。
【0047】
DAPTAは、gp120エンベロープタンパク質のCCR5への結合を阻害する
gp120のCCR5受容体への結合のペプチド阻害剤の効力を定義するために、濃度を増加させたCCR5選択的なケモカインリガンドMIP−□、および、DAPTAの存在下で固定濃度のsCD4/gp120複合体を用いて阻害の研究を行った。これらの研究は図4で説明される。特異的な結合の総量は、sCD4(100nM)を添加した場合、または、それらを添加しない場合での、FITIC−gp120の結合の差として定義した。
【0048】
gp120BaL/sCD4のCCR5(Cf2Th/synR5)細胞への結合は、MIP−1□(IC50=1.5+0.002nM、P<0.05)、および、DAPTA(IC50=55+0.08pM、P<0.05)で完全に阻害された(図3A)。DAPTAに関するヒル(Hill)の傾きは−1.07であり、これは、1部位における競合結合モデルと一致した。同様に、我々はgp120CMCM235/sCD4の同じ細胞への結合阻害を研究したところ、再度、MIP−1□(IC50=1.8+0.006nM、P<0.05)、および、DAPTA(IC50=0.32+.03nM、P<0.05)による特異的結合の実質的な(>80%)阻害が示された(図3B)。我々はさらに、gp120CM235/sCD4の異なるCD4を発現する細胞系(GHOST CD4.CCR5)への結合も、MIP−1□(IC50=0.43+0.07nM、P<0.05)、および、DAPTA(IC50=51+0.09pM、P<0.05)で阻害されたことも示した(図3C)。示されたデータ(パネルA〜C)は、3回の実験(それぞれ3連での決定)の平均およびSEMである。PRISMで解析を行った。
【0049】
これらの臨床的および基礎的な調査の発見の関連性は、gp120のペプチドT領域が、ケモカイン補助受容体CCR5に結合するエピトープと同定されたことを証明するものである(11)。ほぼ25年にわたる努力によっても、CCR5の臨床的に有効な短いペプチド受容体の競合物は得られなかった。例えば、gp120分子全体を含むあらゆる包括的な20merを2セット製造し、抗ウイルス活性に関して試験した。(第二のセットは、第一のセットに比べて10個分の位置をフレームシフトさせた)。これらのサンプルはどれも、抗ウイルス活性をまったく示さなかった。加えて、10年にわたる完全な失敗、有効なHIVワクチンを同定しようとする300億ドルもの連邦または個人的な努力は、ワクチン研究センター(Vaccine Research Center)、NIHにおける臨床研究局長(Director of Clinical Studies)であるBarney Graham博士による講演、「有効なHIVは可能か(Is an Effective HIV Possible)」という表題の講演(2006年2月15日、メリーランド州ベセスダ)で認められているが、これは、広範な中和抗HIV抗体を作製する能力が導かれる本発明のアプローチが自明ではないことを示す。実際に、このような抗体を誘導することができないことが、過去および現在のワクチンに関するあらゆる試みの失敗の元となっている。
【0050】
CCR5受容体に結合するHIVエンベロープgp120のペプチドは、ウイルスの受容体への結合をブロックする免疫原として用いることができる。ほぼ全ての有効な抗体ワクチンは、ウイルスの受容体結合または付着をブロックすることによって作用する。
【0051】
近年、製剤化されたペプチドの特性の解析、および、詳細な構造的な研究(MacPhee,未発表)から、DAPTAは、水溶液に迅速に溶解させると、極めて強い凝集傾向を示し、それにより、生物学的利用率と抗ウイルス活性との両方の損失が起こることが解明されている。目下、このDAPTAの特性は、準最適な臨床結果しか得られない場合があり(33)、インビトロでの結果が不当に陰性になることさえあることが明らかである。例えば、ペプチドT溶液は、高濃度になり、「ゲル」になることが報告されており、それにより、粘膜(例えば鼻上皮)を介して輸送される活性および/または能力の見込みが失われると考えられている。調合物から塩化ナトリウムを除去し、濃度を5mg/mLに低くすることが、問題を解決するようである。しかしながら分光偏光解析によれば、わずか1mg/mLでも、室温で、それより低濃度の0.1mg/mlの205.4nmでの大きいピークから237.2nmでの大きいピークにシフトすることが明らかになり、これは、ゲル化が起こり得る凝集工程において、ペプチドTは、より高濃度の場合、それ自身と相互作用することを示す。電子顕微鏡法により、ペプチドTは細線維を形成することが確認され、我々の知る限りの知識によれば、ペプチドTは、すでに説明されているその他の小さいペプチドのどれよりも容易に細線維を形成する。
【0052】
細線維の形成の決定
ペプチドTまたはそれらの類似体の細線維の形成は、電子顕微鏡法を用いて測定することができる。DAPTA水溶液の2μlのアリコートを、フォームバー(formvar)/炭素で被覆されたニッケル製のEMグリッドに塗布した。このグリッドを10μlの蒸留水で3回リンスし、10μlの2%酢酸ウラニルで染色した。このサンプルをLaB6フィラメント(120kv)を備えたFEI TEM Tecnai顕微鏡で試験し、メガビューII(Megaview II)CCDカメラでイメージ化した。
【0053】
また、ペプチドTまたはそれらの類似体の細線維の形成は、色素結合を用いて決定することもできる。コンゴレッドを、PBS(5mMのリン酸カリウム、150mMのNaCl、pH7.4)に7μg/mLの濃度に溶解させた。この溶液を4℃に冷却し、DAPTAを10mg/mLのストック水溶液として添加し、0.48mg/mLの色素溶液中で最終的なペプチド濃度にした。粉末を溶解させた直後のペプチド溶液を、凝集したペプチドを含む時間を置いたストック溶液と比較した。4℃で400〜700nmでスペクトルを回収した。
【0054】
またペプチドTまたはそれらの類似体の細線維の形成は、円二色性(CD)分光分析を用いて決定することもできる。新たに製造されたペプチド、または、繊維性の凝集体を含むペプチドのいずれかの10mg/mLの水溶液を、4℃で蒸留水に50μg/mLの濃度まで添加した。ジャスコ(Jasco)のモデルJ−810スペクトロメーターで、0.1cmのパス長の石英キュベットを用いて、190〜250nmの範囲で、1分間のインターバル、および、2秒の応答時間を用いてCDスペクトルを得た。
【0055】
さらにその他のペプチドTまたはそれらの類似体の細線維の形成の決定方法は、フーリエ変換赤外(FTIR)分光分析を用いて行われる。DAPTAを重水に10mg/mlの濃度まで溶解させ、細線維の形成を促進する温度および時間条件下でインキュベートした。次に、25μlのサンプルを、6μmのスペーサーで仕切ったNaClウィンドウを備えた予め冷却した透過セル中に入れた。FTIRスペクトルを、DTGS検出器を用いて、バイオ・ラッド(BioRad)のFTS−175Cフーリエ変換スペクトロメーターで透過モードで回収した。2cm−1の解像度で2506枚の干渉写真を記録した。水蒸気を差し引いて、スペクトルの基線を修正した。
【0056】
免疫原性の特性
抗ペプチドDAPTA抗体のgp120単量体への結合
図5は、抗ペプチドDAPTA抗体のgp120単量体への結合を説明する。パネルAにおいて、IIIBgp120(●);gp120ADA()、および、gp120Bal(▲)は、マイクロプレートに直接被覆されている(被覆されたELISA)。そのウェルに25倍まで連続的に希釈した抗ペプチドDAPTAを添加した。結合した抗体を抗ウサギIgG−HPR−ロバ(KP&L)を1:1000にしたもので検出し、450nmにおける光学密度として測定した。バックグラウンドを一般的なウサギ血清コントロールによって推定した。試験された3種全てのgp120タンパク質への最大限の結合の半分に到達するのに必要なV2抗体の濃度は、8〜25ng/mlの間であった。これらのデータは、代表的な実験から得られたものだが、各データポイントは、3連の実験からの平均値である。パネルBは、sCD4(■)の存在下、および、sCD4なし(◇)でのgp120IIIBと抗ペプチドDAPTAとの反応性を示す。これらの抗体は、sCD4の存在下では高親和性で結合する。この結果は、2回繰り返した代表的な実験から得られたものである。各データポイントは、3連の実験からの平均値である。
【0057】
抗ペプチドTDAPTA抗体の可溶化したgp120sへの結合
抗ペプチドTDAPTA抗体、および、精製したIgG分子の、様々な実験用に適合させたウイルス株由来のgp120組換えタンパク質への親和性をELISA分析によって測定した。ポリクローナル抗ペプチドTDAPTA血清は、可溶化したgp120タンパク質に結合したが、gp120ADA、および、gp120IIIBには高親和性で結合し、SF162gp120には最も低い親和性で結合した。これは、図6で説明されている。捕捉ELISAからの結果は、中和活性との優れた相関を示し、従って、我々が本明細書において開発したような単純なELISA試験によって、ワクチン候補抗体を選別することができることが示される。
【0058】
gp120s(1μg/ml)を、マイクロプレート上のポリクローナルヤギ抗HIV−1SFgp120血清によって捕捉した(捕捉ELISA)。3倍まで連続的に希釈した抗ペプチドTDAPTA血清を加熱不活性化し、これをそのウェルに添加した。結合した抗体をロバ抗ウサギIgG−HRPで検出し、450nmにおける光学密度として測定した。バックグラウンドを、結合したペプチドTDAPTAに対して免疫化する前のウサギ血清の量によって推定し、それを差し引いた。このラインは、3連で行われた2回の実験からのデータのうち代表的なものである。
【0059】
抗ペプチドTDAPTA抗体の可溶化したgp120sへの結合は、CD4依存性である
溶解性CD4(sCD4)を添加した場合、および、それらを添加しない場合の精製した抗付着部位IgGのgp120sへの親和性結合を、ペプチドTをローディングした親和性ELISAから精製した抗ペプチドTDAPTAウサギ抗血清全体から誘導された免疫グロブリンのトラクションを用いて研究した。
【0060】
これは、図7で説明されている。可溶性gp120タンパク質を、抗HIV−1SF2血清、または、sCD4(1μg/ml)で被覆されたマイクロプレートウェル上で捕捉した。3倍まで連続的に希釈した抗ペプチドTDAPTA IgGを、各希釈率ごとに3連で添加した。結合した抗ペプチドTDAPTA IgGを、2%の一般的なヤギ血清を含むブロッキング緩衝液中のHPR−ロバ抗ウサギIgGによって検出した。その結果は、3連で行われた2回の実験の平均である。エラーバーは、SD(p<0.05)を示した。
【0061】
CD4分子に結合するとおおむねgp120を認識する抗体が特に興味深く、ワクチン関連性を有する。いわゆるCD4誘導性(CD4i)抗体は、CCR5、および、CXCR4のgp120−CD4複合体への結合を阻害することから(34〜36)、これら抗体は、gp120の補助受容体結合を許容するコンフォメーションに対して向けられていると認識される。しかしながら、ヒトから単離されたCD4i抗体の例は限定的であり、そのプロトタイプは、一次HIV−1単離物に対して弱い中和活性しか示さない。これらの抗体が一見希少であることの原因は、CD4iエピトープが比較的入手困難であることにある(6,37,38)。
【0062】
このようなgp120のCD4誘導性(CD4i)部位が見出された抗体は、極めて限られた数しか存在しない。CD4によって引き起こされるgp120のコンフォメーションを認識する抗体は、補助受容体結合エピトープに対して向けられている可能性が高い。抗ペプチドT抗体は、本明細書において、CD4誘導性エピトープを選択的に認識することが示されており、従って、補助受容体結合部位の近傍で、またはその場で結合すると推測される。
【0063】
抗付着部位抗体は、gp120/sCD4のCCR5侵入の補助受容体への結合をブロックする
抗ペプチドTDAPTA抗血清、および、精製した抗ペプチドTDAPTA IgGのgp120sへの親和性を、ウェスタンブロットでさらに研究した。2種のHIV−1ウイルス株JR−CSF、および、SF162からのウイルス粒子を濃縮し、NP40の存在下で溶解させ、4〜12%のポリアクリルアミドゲルを用いたタンパク質を電気泳動で分離した。p24抗原に関して3種の異なる濃度を用いられ、スロットあたり1、2および3ngの溶解産物で測定された。タンパク質をNEMPVDFメンブレンに移し、精製した抗ペプチドTDAPTA IgG(1:200)とハイブリダイズさせた。これらの抗体は、用量依存性の方式でウイルスのgp120タンパク質を認識した。
【0064】
これは、図8で説明されている。パネルAにおいて、ウイルス粒子を濃縮し、4〜12回のSDSPAGEで分離した。ELISAによって測定したところ1、2および3ngのp24タンパク質を含むウイルスの溶解産物をローディングした。このメンブレンを、ペプチドTDAPTA IgG(1:200)に対する親和性によって精製した抗ペプチドTDAPTAとハイブリダイズさせ、HRP−ロバ抗ウサギIgG(1:2000)で染色した。このフィルムをECL溶液(アマシャム(Amersham))で展開した。この実験を2回繰り返した。パネルBにおいて、IIIBHIV−1ウイルス株、および、可溶性gp120IIIB組換えタンパク質からの溶解産物全体(濃度21μg/ml)を10%SDSPAGEでの電気泳動で分離し、トランスファーさせた。このメンブレンを、ペプチドTDAPTA(10μM)の存在下での、抗ペプチドTDAPTAポリクローナル血清(#195)、予め採血した血清および抗ペプチドTDAPTA血清とインキュベートした。抗ペプチドTDAPTA抗体のHIV−1gp120IIIBエンベロープタンパク質に対する特異性が示された。ペプチドTDAPTAは、抗ペプチドTDAPTAのgp120タンパク質への結合をブロックした(図8A)。
【0065】
V2ペプチドベースの抗体の抗ペプチドDAPTAの中和特性
HELACD4+CXCR4/CCR5+細胞(P7)において融合細胞をカウントする分析で、HIV−1の中和を評価した。顕微鏡下で青色のフォーカスユニット(BFU/ウェル)をカウントした。以下の表1にまとめたこのデータは、抗体の固定濃度0.125μg/ウェルを用いて得られた。さらに精製したIgGの中和活性の滴定も行った。IgG分画は、より低い親和性を有するペプチドDAPTAに対するポリクローナル抗血清の滴定曲線に類似した滴定曲線に従っていた(データ示さず)。感染の阻害は3連の平均値であり、以下の式によって計算された:阻害(%):100−[(BFU(抗体を含む): BFU(抗体を含まない))×100]。
【0066】
【表7】
【0067】
数種の一般的に研究された分岐群B単離物に関するペプチドT/DAPTAに対する抗体の効力(50%[IC50]阻害を達成するのに必要な濃度)を比較するために、我々は、既定の補助受容体特異性を有する細胞系(HELA CD4−βガラクトシダーゼ、および、HELA CD4.CCR5)を用いたフォーカス形成分析において一回のウイルス感染を用いた(39)(表1)。
【0068】
上記の実験において、感染分析は以下のように行われた。HeLa細胞系(MAGI)が用いられ、この細胞系は、いずれも高レベルのCD4を発現し、トランケーションされたヒト免疫不全ウィルス1型(HIV−1)ロングターミナルリピート(LTR)の制御下にあるベータ−ガラクトシダーゼ遺伝子の単一の統合されたコピー(40)を含み、このCCR5遺伝子(MAGI−CCR5)を有する細胞系(Pirounaki等, 2000)は、NIAID AIDSレポジトリー(NIAID AIDS Repository)から入手した。ペプチドTのケモカイン受容体サブタイプの感受性を決定するために、MAGIおよびMAGI−CCR5を、以下に説明されているようにして用い、以前のプロトコール(40〜43)を改変させた。簡単に言えば、10,000個のMAGIまたはMAGI−CCR5細胞/ウェルを96ウェルプレートにシーディングした。1日後、培地を除去し、20□g/mlのDEAE−デキストリンを含むOpti−MEM培地(ギブコ・BRL(Gibco BRL),ライフ・テクノロジーズ(Life Technologies))でのペプチドTまたはMIP−1□の希釈液を添加した。このプレートを、60分間、37℃、5%CO2で培養し、次に、ウイルスの希釈液を添加し、さらに1.5時間培養した。接種されたウイルスを除去し、ウェルを0.2mLのOpti−MEM培地で2回洗浄し、新しい培地(150□l/ウェルのDMEM)を添加した。さらに46時間培養した後、細胞を、PBS中の1%ホルムアルデヒド、0.2%グルタルアルデヒドで室温で5分間固定し、PBSで2回洗浄し、PBS中の4mMのフェロシアン化カリウム、4mMのフェリシアン化カリウム、2mMのMgCl、および、0.4mg/mlの5−ブロモ−4−クロロ−3−インドリル−□−D−ガラクトピラノシド(X−gal染色、イナルコ・ファーマシューティカルズ(Inalco Pharmaceuticals),セントルイス・オビスポ,カリフォルニア州)で、37℃で、5%CO2で、50分間染色し、PBSで2回洗浄した。顕微鏡で青色のフォーカスを計数し、感染レベルを青色のフォーカスユニット(BFU/ウェル)として記録した。全ての分析において、BFU/ウェルのバックグラウンドレベルは概して3未満であった。
【0069】
上述した感染分析におけるCCR5株SF162、および、JR−CSFの感染力の阻害に関して、未精製の抗血清、および、免疫化する前のコントロール血清を試験した。中和力価(IC50)は、免疫血清において1:20〜1:40の希釈率の範囲であり、免疫化する前の血清における非特異的な中和はごくわずかであった(示さず)。
【0070】
特異性および効力を強化するために、親和性によって精製した抗血清を、ペプチドT/DAPTAカラムを用いたさらなる研究のために製造した。CCR5株SF162、および、JR−CSFに関する親和性によって精製した抗体分画の中和力価は、SF162は0.25〜125μg/mlの間であり、JR−CSFウイルス株は0.125および0.06μg/mlであった(示さず)。精製したIgG分画の中和活性の滴定は、gp120補助受容体の付着部位に対するポリクローナル血清の滴定曲線に類似した滴定曲線に従っており、SF162ワクチン株に関する親和性はより低かった。予め採血した血清を同じ解析で試験した。バックグラウンドの阻害は、1:20に希釈した血清サンプルで2〜4パーセントの範囲であった。続いて、gp120補助受容体の付着部位の親和性によって精製したIgの活性を、様々な分岐群のなかからそれらの多様性および外見に関して選択されたより大規模なHIV単離物サンプルで評価した。
【0071】
19%のカットオフ値は、1:5の希釈率で試験された一般的なウサギ血清を用いて得られた95%の信頼レベルに基づく(Laboratory of Antiviral Drug Mechanisms, Science Applications International Corporation at Frederick National Cancer Institute-Frederick Cancer Research and Development Center)。表Xに示した結果は、少なくとも2回の独立した実験(ウイルス株ごとに3連で行われた)から得られたものである。
【0072】
gp120補助受容体の付着部位の親和性によって精製した免疫グロブリンは、2種の一次単離物、分岐群B/F由来の93BR019、および、93IN101分岐群Cのウイルス株に対して極めて高い中和活性、および、2株を除く全てに対する有意な中和活性を示した。X4ウイルス単離物に対する活性は予想外であり、これは、これらのペプチドは、R5、X4、および、二重細胞指向性HIV株に対する免疫原または治療剤を作製するのに有用であり得ることを示す。
【0073】
親和性によって純粋にした抗ペプチドTによるHIVの中和
IC50が0.045μg/mlの抗付着部位IgGの、HIV−1分岐群C単離物(Dul23、および、Dul56)に対する中和活性を、デューク大学(Duke University)のAIDSワクチン研究所(AIDS Vaccine Lab)(David Montefiori、個人的な人脈)による一回のルシフェラーゼレポーター遺伝子分析によって確認した。そのデータは、以下の通りである:
【0074】
【表8】
【0075】
値は、相対的な発光単位(RLU)が、TZM−bl細胞においてウイルスコントロールウェル(試験サンプルなし)と比較して50%減少した時点の血清の希釈濃度である。活性を有するサンプルは、太字で示す。
【0076】
反復配列
有用なペンタペプチド配列は、直線状のアレイで2〜5回の頻度で生じるそれらの反復モチーフによっても同定が可能であり、このようなモチーフは、分離されていないか、または、どこかで1〜数百個のアミノ酸で分離されているかのいずれかであり、すなわちエンベロープタンパク質のそれ以外のいずれか一部分を除きV2には存在しない。これらのペプチドの望ましくない凝集特性は、エンベロープ内の「ステープル様の」付着部位として役立つこれらのペンタペプチド配列を含むウイルスエンベロープの三次構造を構築することに有用な場合がある。どのような場合においても、いずれの所定のウイルス内におけるペンタペプチド配列の反復は、規則に従っていなくても、追加の有用な配列を示す可能性がある。例えば、EYIYT反復配列は5回反復される。この反復は、コンセンサス配列の近傍に続いて存在しない。これらは反復しているが、活性な化合物のライブラリーに加えられる。
【0077】
CCR5受容体活性の存在
有用な候補は、CCR5受容体への保存的な親和性を有すると予想される。従って、CCR5受容体への相対的な親和性に関して、様々なペンタペプチド組成物(すなわち、それらの様々なリンカーおよび免疫原が結合している)の活性を分析すること(結合分析、抗ウイルス性またはその他のCCR5受容体の相対的な生物活性の分析)は、さらなる有用なスクリーニングであると予想される。CCR5受容体への親和性が高ければ高いほど、最終産物が治療活性を有する可能性は高くなる。
【0078】
有用な候補は、概して、免疫原として用いられる前に広範な抗ウイルス活性を示すと予想される。従って、これらのワクチンはそれ自身、治療剤であると期待される。
本発明の特に重要な形態は、これらのペプチドの混合型のアンタゴニストとしての薬理学的作用に関する。Nobelist Arvid Carlsson(Medicine/Physiology, 2000)によって述べられたように、薬物の混合型の(部分)アゴニストの形態は、受容体相互作用の臨床的に好ましいタイプである。混合型/部分アゴニスト薬物の場合、完全アンタゴニストの除感作、耐性および副作用を回避することが一般的であり、このタイプの成功した薬物が製薬会社によって作製されている。部分アゴニスト薬物は、受容体活性のバランスをとるように作用し(DAPTA作用の混合型アゴニストの形態について示したRedwine, 1999を参照)、従ってこれらは好ましい治療剤である。我々は、本明細書で説明するような、これらのペプチドの固有の生物活性を決定する方法を開発し、予想通りにヒトへ投与してから6ヶ月後に受容体における耐性の誘導を示さない混合型アゴニスト(例えば表6に記載のペプチド)が得られることを見出した。
【0079】
HIV−1gp120は、最も広範囲にわたりグリコシル化されたタンパク質の1種である。これらは、23または24個のN結合型糖付加部位、および、タンパク質の総質量の約半分を占めるこれらの部位に結合したグリカンを含む。糖付加、および、グリコシダーゼ阻害剤を用いた多数の研究により、HIV−1エンベロープ糖タンパク質のコンフォメーションを決定することにおける炭水化物部分の重要性が解明されている。部位特異的変異誘発から、糖付加部位の多くはいずれも非必須であることが示された。24部位のうち、いずれもgp120のアミノ末端側の半分に位置する5部位のみ(アミノ酸88、141、197、262および276)が、ウイルス感染力に影響を与えた。197部位は、V2の近傍に存在するか、または、我々が定義した結合部位のペプチドの多くの位置に存在する。例えば、ここで留意すべきは、本発明のペプチドに、糖付加モチーフT/SxNが存在することが多いことである。我々の結果によれば、N結合型糖付加部位が、本明細書に記載のペプチドの近傍、その場、またはその範囲内に存在することが多く、さらに、補助受容体様CCR5へのウイルス結合エピトープを定義することが示される。糖付加が存在する場合、糖付加は、構造的な特異性に寄与するか、または、この重要なエピトープの免疫監視を回避する手段として天然の抗体による中和をマスクする可能性がある。
【0080】
本発明の一実施態様は、免疫原性特性を有する組成物である。この組成物は、一般的に、A−L−P−C部分の組み合わせとして説明され、ここで、Aは、あらゆるアジュバントであり、Lは、あらゆる結合部分であり、Pは、ペプチド部分であり、Cは、保護基である。
【0081】
開示された化合物において、あらゆるアジュバントが使用できる。ペプチドとリンカー領域とは、アジュバントとは独立して作用すると考えられるので、アジュバントの選択はまったく制限されない。いくつかの有用なアジュバントの例は、これらに限定されないが、ヒトにおいて許容できるあらゆるアジュバント、肝炎のコア抗原、ポリリジンなどである。この開示の後に開発されたアジュバントも作用すると考えられ、その前に開発されたアジュバントも同様である。
【0082】
あるいは、アジュバントは、所定の環境に応じて調合してもよい。例えばペプチドを凝集するようにする場合である。
その他の本発明の実施態様は、治療特性を有する組成物である。この組成物は、一般的に、L−P−C部分の組み合わせとして説明され、ここで、Lは、あらゆる部分であり、ただし好ましくは短いペプチド部分であり、Pは、その他のペプチド部分であり、Cは、保護基である。
【0083】
連結させたLおよびP領域の長さに関して、いずれも特定の長さ(約7〜約16個)のアミノ酸が機能すると考えられる。好ましい実施態様において、このような連結は、7〜11個の範囲である。さらに好ましい実施態様において、このような連結は、7〜9個の範囲である。よりさらに好ましい実施態様において、8である。あるいは、天然の神経性のペプチドは28個のアミノ酸からなることを示すため、開示された組成物のペプチド領域の長さは、約28個のアミノ酸であると考えられる。(表4を参照)。
【0084】
本明細書において開示された様々な化合物は、単独で、または、カクテルとして組み合わせて作用するように設計される。多種多様な化合物が使用でき、限定されない。最も有用なカクテルは、2〜7種の異なる化合物からなると考えられる。
【0085】
C部分は、あらゆる保護基が可能である。好ましい実施態様は、アミドである。
本発明の開示はまた、以下の活性:HIVウイルスに感染したヒトにおける、単球によって誘導されたマクロファージ(MDM)、小グリア細胞、または、T細胞におけるR5または二重細胞指向性(R5/X4)株におけるHIVの抑制を示す化合物、および、その化合物に関してスクリーニングする方法も包含する。その他の活性としては、CCR5受容体を調節することによって治療可能なあらゆるヒトの病気が挙げられ、例えば、これらに限定されないが、アルツハイマーを引き起こす神経炎症、多発性硬化症、乾癬、および、関節炎である。これらの病気は、抑制されるか、または、解消されると予想され、病気の経過は止まるか、または、回復すると予想され、および、臨床的な利点が生じると予想される。
【0086】
最後に、本発明の開示は、組成物の安定性に関してスクリーニングする方法を包含する。これは、ウイルスの感染力、走化性、マップキナーゼ活性化、gp120結合、CCR5活性化もしくは阻害、または、Gタンパク質シグナル伝達からなる群より選択される生物学的に特徴付ける方法によって決定される。
【0087】
引用文献
【0088】
【表9】
【0089】
【表10】
【0090】
【表11】
【0091】
【表12】
【図面の簡単な説明】
【0092】
【図1】DAPTAは、CCR5受容体とのgp120−CD4複合体の共免疫沈降反応を防ぐ。
【図2】gp120のCCR5の結合分析である。
【図3】蛍光標識したgp120(FITC−gp120)は、sCD4が存在するときのみにCf2Th/synCCR5細胞に効率的に結合した。
【図4】DAPTAは、gp120エンベロープタンパク質のCCR5への結合を阻害する。
【図5】抗ペプチドDAPTAのgp120一量体への結合を示す。
【図6】抗ペプチドTDAPTA血清のgp120に対する結合親和性を示す。
【図7】抗ペプチドTDAPTA IgGのgp120タンパク質への溶解性CD4依存性の結合を示す。
【図8】抗ペプチドTIgGおよびポリクローナル血清の、膜に固定されたウイルスおよび組換えgp120タンパク質への親和性結合を示す。
【特許請求の範囲】
【請求項1】
ヒト免疫不全ウィルスに対して有効な治療用組成物およびワクチンの免疫原であって、
構造:A−L−P−C
[式中、
Aは、あらゆるアジュバントである、
Lは、あらゆる結合部分であり、
Pは、構造R1R2R3R4R5を有するペプチドであり、ここで:
R1は、N、S、T、D、R、K、A、または、Eからなる群より選択され、
R2は、N、S、T、D、G、I、または、Eからなる群より選択され、
R3は、N、S、T、D、G、R、K、I、Q、H、または、Eからなる群より選択され、
R4は、Yであり、
R5は、N、S、T、R、K、A、M、WもしくはG、またはそれらのアミド;および、その他のあらゆるアミノ酸からなる群より選択され、
Cは、あらゆる保護基である]
を含む、上記治療用組成物およびワクチンの免疫原。
【請求項2】
Lが、3個のアミノ酸部分からなり、Pが、5個のアミノ酸部分からなる、請求項1に記載の組成物。
【請求項3】
Lが、トリペプチド部分A−S−Tである、請求項1に記載の組成物。
【請求項4】
R2が、T、S、および、Nからなる群より選択される、請求項1に記載の組成物。
【請求項5】
Pが、EYIYT、INNYT、ISNYS、ISNYT、ISYRL、NNRYR、NNSYR、NSNYS、NSSYR、NTIYT、NTSYG、NTSYM、NTSYR、NTSYS、SSEYR、SSRYR、SSTYR、STNYT、STSYR、TISYR、TSNYS、TTNFT、TTNYT、全てD型のTTNYT、TTSYR、TTSYT、および、YTSYRからなる群より選択される、請求項1に記載の組成物。
【請求項6】
Aが、ヒトにおいて許容できるあらゆるアジュバントである、請求項1に記載の組成物。
【請求項7】
Cが、C末端のアミド基である、請求項1に記載の組成物。
【請求項8】
前記組成物が、2〜5種の成分のカクテルを含み、ここで、これらの成分は、P領域におけるアミノ酸部分の配列が異なる、請求項1に記載の組成物。
【請求項9】
L−P領域が、約7〜約16個のアミノ酸残基の長さを有するペプチドを含む、請求項1に記載の組成物。
【請求項10】
前記アミノ酸が、Dステレオ異性体である、請求項1に記載の組成物。
【請求項11】
構造:L−P−C
[式中、
Lは、あらゆるトリ−ペプチド配列を含み、
Pは、構造R1R2R3R4R5を有するペプチドを含み、ここで:
R1は、N、S、T、D、R、K、A、または、Eからなる群より選択され、
R2は、N、S、T、D、G、I、または、Eからなる群より選択され、
R3は、N、S、T、D、G、R、K、I、Q、H、または、Eからなる群より選択され、
R4は、Yであり、
R5は、N、S、T、R、K、A、M、WもしくはG、またはそれらのアミド;および、その他のあらゆるアミノ酸からなる群より選択され、
Cは、あらゆる保護基である]
を有する治療用組成物。
【請求項12】
Lが、3個のアミノ酸部分からなり、Pが、5個のアミノ酸部分からなる、請求項11に記載の組成物。
【請求項13】
Lが、トリペプチド部分A−S−Tである、請求項11に記載の組成物。
【請求項14】
R2が、T、S、および、Nからなる群より選択される、請求項11に記載の組成物。
【請求項15】
Pが、EYIYT、INNYT、ISNYS、ISNYT、ISYRL、NNRYR、NNSYR、NSNYS、NSSYR、NTIYT、NTSYG、NTSYM、NTSYR、NTSYS、SSEYR、SSRYR、SSTYR、STNYT、STSYR、TISYR、TSNYS、TTNFT、TTNYT、全てD型のTTNYT、TTSYR、TTSYT、および、YTSYRからなる群より選択される、請求項11に記載の組成物。
【請求項16】
Aが、ヒトにおいて許容できるあらゆるアジュバントである、請求項11に記載の組成物。
【請求項17】
Cが、C末端のアミド基である、請求項11に記載の組成物。
【請求項18】
前記組成物が、2〜5種の成分のカクテルを含み、ここで、これらの成分は、P領域におけるアミノ酸部分の配列が異なる、請求項11に記載の組成物。
【請求項19】
L−P領域が、約7〜約16個のアミノ酸残基の長さを有するペプチドを含む、請求項11に記載の組成物。
【請求項20】
前記アミノ酸が、Dステレオ異性体である、請求項11に記載の組成物。
【請求項1】
ヒト免疫不全ウィルスに対して有効な治療用組成物およびワクチンの免疫原であって、
構造:A−L−P−C
[式中、
Aは、あらゆるアジュバントである、
Lは、あらゆる結合部分であり、
Pは、構造R1R2R3R4R5を有するペプチドであり、ここで:
R1は、N、S、T、D、R、K、A、または、Eからなる群より選択され、
R2は、N、S、T、D、G、I、または、Eからなる群より選択され、
R3は、N、S、T、D、G、R、K、I、Q、H、または、Eからなる群より選択され、
R4は、Yであり、
R5は、N、S、T、R、K、A、M、WもしくはG、またはそれらのアミド;および、その他のあらゆるアミノ酸からなる群より選択され、
Cは、あらゆる保護基である]
を含む、上記治療用組成物およびワクチンの免疫原。
【請求項2】
Lが、3個のアミノ酸部分からなり、Pが、5個のアミノ酸部分からなる、請求項1に記載の組成物。
【請求項3】
Lが、トリペプチド部分A−S−Tである、請求項1に記載の組成物。
【請求項4】
R2が、T、S、および、Nからなる群より選択される、請求項1に記載の組成物。
【請求項5】
Pが、EYIYT、INNYT、ISNYS、ISNYT、ISYRL、NNRYR、NNSYR、NSNYS、NSSYR、NTIYT、NTSYG、NTSYM、NTSYR、NTSYS、SSEYR、SSRYR、SSTYR、STNYT、STSYR、TISYR、TSNYS、TTNFT、TTNYT、全てD型のTTNYT、TTSYR、TTSYT、および、YTSYRからなる群より選択される、請求項1に記載の組成物。
【請求項6】
Aが、ヒトにおいて許容できるあらゆるアジュバントである、請求項1に記載の組成物。
【請求項7】
Cが、C末端のアミド基である、請求項1に記載の組成物。
【請求項8】
前記組成物が、2〜5種の成分のカクテルを含み、ここで、これらの成分は、P領域におけるアミノ酸部分の配列が異なる、請求項1に記載の組成物。
【請求項9】
L−P領域が、約7〜約16個のアミノ酸残基の長さを有するペプチドを含む、請求項1に記載の組成物。
【請求項10】
前記アミノ酸が、Dステレオ異性体である、請求項1に記載の組成物。
【請求項11】
構造:L−P−C
[式中、
Lは、あらゆるトリ−ペプチド配列を含み、
Pは、構造R1R2R3R4R5を有するペプチドを含み、ここで:
R1は、N、S、T、D、R、K、A、または、Eからなる群より選択され、
R2は、N、S、T、D、G、I、または、Eからなる群より選択され、
R3は、N、S、T、D、G、R、K、I、Q、H、または、Eからなる群より選択され、
R4は、Yであり、
R5は、N、S、T、R、K、A、M、WもしくはG、またはそれらのアミド;および、その他のあらゆるアミノ酸からなる群より選択され、
Cは、あらゆる保護基である]
を有する治療用組成物。
【請求項12】
Lが、3個のアミノ酸部分からなり、Pが、5個のアミノ酸部分からなる、請求項11に記載の組成物。
【請求項13】
Lが、トリペプチド部分A−S−Tである、請求項11に記載の組成物。
【請求項14】
R2が、T、S、および、Nからなる群より選択される、請求項11に記載の組成物。
【請求項15】
Pが、EYIYT、INNYT、ISNYS、ISNYT、ISYRL、NNRYR、NNSYR、NSNYS、NSSYR、NTIYT、NTSYG、NTSYM、NTSYR、NTSYS、SSEYR、SSRYR、SSTYR、STNYT、STSYR、TISYR、TSNYS、TTNFT、TTNYT、全てD型のTTNYT、TTSYR、TTSYT、および、YTSYRからなる群より選択される、請求項11に記載の組成物。
【請求項16】
Aが、ヒトにおいて許容できるあらゆるアジュバントである、請求項11に記載の組成物。
【請求項17】
Cが、C末端のアミド基である、請求項11に記載の組成物。
【請求項18】
前記組成物が、2〜5種の成分のカクテルを含み、ここで、これらの成分は、P領域におけるアミノ酸部分の配列が異なる、請求項11に記載の組成物。
【請求項19】
L−P領域が、約7〜約16個のアミノ酸残基の長さを有するペプチドを含む、請求項11に記載の組成物。
【請求項20】
前記アミノ酸が、Dステレオ異性体である、請求項11に記載の組成物。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公表番号】特表2008−546803(P2008−546803A)
【公表日】平成20年12月25日(2008.12.25)
【国際特許分類】
【出願番号】特願2008−518460(P2008−518460)
【出願日】平成18年6月23日(2006.6.23)
【国際出願番号】PCT/US2006/024580
【国際公開番号】WO2006/138745
【国際公開日】平成18年12月28日(2006.12.28)
【出願人】(507421016)ラピッド・ファーマシューティカルズ・インコーポレーテッド (1)
【Fターム(参考)】
【公表日】平成20年12月25日(2008.12.25)
【国際特許分類】
【出願日】平成18年6月23日(2006.6.23)
【国際出願番号】PCT/US2006/024580
【国際公開番号】WO2006/138745
【国際公開日】平成18年12月28日(2006.12.28)
【出願人】(507421016)ラピッド・ファーマシューティカルズ・インコーポレーテッド (1)
【Fターム(参考)】
[ Back to top ]