
活性化カルボキシペプチダーゼの製造方法
本発明は、Kex2部位がカルボキシペプチダーゼのプロ配列に導入されているカルボキシペプチダーゼのプロ型をコードするDNA配列を導入し、プロカルボキシペプチダーゼの発現に適した条件下で真菌細胞を培養し、細胞内でプロ配列を切断し、遊離の活性型のカルボキシペプチダーゼを放出させることを含む、真菌細胞において活性化したカルボキシペプチダーゼを製造する方法に関する。また本発明は、活性化したカルボキシペプチダーゼ酵素を使用することによる、成熟ヒトインスリン又はヒトインスリンアナログを製造する方法に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、活性化カルボキシペプチダーゼの製造方法及び真菌細胞での成熟ヒトインスリン又はヒトインスリンアナログの生産のための活性化カルボキシペプチダーゼの使用方法に関する。
【背景技術】
【0002】
インスリンは、ランゲルハンス島のベータ細胞で生産されるポリペプチドホルモンである。活性なインスリン分子は、2つのジスルフィド架橋により結合しているB及びA鎖からなる2本鎖分子である。インスリンは構造B-C-Aを有する前駆体分子プロインスリンとして合成され、ここでCペプチド鎖は、B鎖のC末端アミノ酸残基をA鎖のN末端アミノ酸残基に連結させている。成熟2本鎖インスリンは、A及びB鎖の接合部に位置する塩基性アミノ酸残基対にてCペプチドをインビボで切断することにより形成される。A及びB鎖は、それぞれA7とB7、及びA20とB19Cys残基の間の2つのジスルフィド架橋により互いに保持されている。さらに、生物学的に活性なインスリン分子は、A6及びA11位にあるCys残基間に内部ジスルフィド架橋を有している。
【0003】
組換えDNA技術の発展後、遺伝的に改変された宿主細胞中でインスリン及びその前駆体を生産するための多くの方法が記載されている。大腸菌は、発現されたポリペプチドを折り畳むための細胞機構を有しておらず、成熟インスリンにおいてA及びB鎖を連結するジスルフィド架橋を確立するために、この方法は、多くのインビトロプロセシング工程、例えば再折り畳み中でのジスルフィド架橋のインビトロでの確立と、続くCペプチドの切断を含む。
【0004】
大腸菌と比較して、真核生物は、折り畳み及びジスルフィド架橋の確立に必要な機構を有しており、よって、遺伝的に改変された生物での成熟インスリンの生産のための良好な候補であると思われる。Thimら, FEBS Letters, 第212巻, 2号, 307-312には、ヒトプロインスリンの発現、及びある種の修飾Cペプチドを有する多くのインスリン前駆体が開示されている。米国特許第4914026号には、酵母α−因子リーダー配列に結合したヒトプロインスリン遺伝子を酵母宿主細胞に挿入することにより、酵母において成熟インスリンを作製し、プロインスリンが発現し成熟型で分泌させる条件下で、形質転換された酵母細胞を栄養培地中で増殖させる方法が開示されている。
【0005】
国際公開第97/03089号には、式BZAを有するインスリン前駆体の発現が開示されており、ここでB及びAは、少なくとも一のジスルフィド結合により結合したヒトインスリンのA及びBペプチド鎖であり、Zは少なくとも一のタンパク分解的切断部位を有するポリペプチドである。国際公開第90/10075号には、発酵ブロスからの最初の回収に続いて、成熟インスリン又はインスリンアナログに酵素的に転換されるインスリン及びインスリンアナログを、酵母中でのインスリン又はインスリンアナログの前駆体の発現をベースにして製造する方法が開示されている。前駆体分子はある種の修飾Cペプチドを含んでおり、インスリンB鎖のN末端伸長部をさらに含んでいてもよい。修飾Cペプチド及びBペプチドの可能なN末端伸長部は、酵母細胞中で切断されないように設計され、よって前駆体は、A及びB鎖が修飾Cペプチドにまだ結合しているが、正確に位置付けされたジスルフィド架橋は有している単鎖ペプチドとして分泌される。ついで、成熟インスリン又はインスリンアナログ産物が、Cペプチドと場合によってはN末端伸長部を切断する複数の後続するインビトロ酵素的工程により得られる。これらの酵素的工程は、時間がかかり、多くの場合はコストが高く、高価なクロマトグラフィー工程等、さらなる下流プロセス工程で除去しなければならない付加的な汚染物質が入り込むおそれがある。
【0006】
天然には分泌顆粒を形成できない遺伝子操作動物細胞において成熟インスリンを製造する方法は米国特許第6348327号に開示されている。
【0007】
本発明の目的は、プロテイナーゼA及びBとは独立して細胞内的に活性化させるカルボキシペプチダーゼを生産可能な真菌株を開発すること、さらに、高価で時間のかかる下流精製プロセス工程が回避されるように別法で活性化されたカルボキシペプチダーゼにより、十分にプロセシングされた成熟インスリン又はインスリンアナログを製造する方法を開発することにある。
【発明の概要】
【0008】
一態様では、本発明は、挿入Kex2切断部位を含むカルボキシペプチダーゼの修飾されたプロ型(プロフォーム)をコードするDNA配列を真菌細胞中で発現させることを含む活性化したカルボキシペプチダーゼを製造する方法に関し、ここでカルボキシペプチダーゼのプロ型が細胞内で切断されて、遊離の活性型カルボキシペプチダーゼが放出される。
本発明の一実施態様では、本方法は、真菌細胞から活性型のカルボキシペプチダーゼを単離する工程をさらに含む。
本発明のさらなる実施態様では、真菌細胞は、非機能性PEP4遺伝子(プロテイナーゼAをコードする遺伝子)を有する。
本発明の一実施態様では、真菌細胞は欠失したPEP4遺伝子を有する。
Kex2部位は、カルボキシペプチダーゼのプロ型の、カルボキシペプチダーゼのプロ型が効果的に切断されてカルボキシペプチダーゼの活性化成熟型を形成させる位置に挿入される。
【0009】
本発明の一実施態様では、Kex2部位は、野生型カルボキシペプチダーゼの天然N末端アミノ酸残基に直接結合している。
他の実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基の比較的近接した位置に導入される。しかしながら、Kex2部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基に対して、好ましくは約30以下のアミノ酸残基上流又は下流に存在すべきである。
【0010】
本発明の一実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基に対して、1〜約30アミノ酸残基上流又は下流の距離に導入される。
本発明の一実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基に対して、1〜約30アミノ酸残基上流の距離に導入される。
本発明のさらなる実施態様では、Kex2切断部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基に対して、2-30アミノ酸残基上流の距離に導入される。
本発明のさらなる実施態様では、Kex2切断部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基に対して、5-30アミノ酸残基上流の距離に導入される。
本発明のさらなる実施態様では、Kex2切断部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基に対して、5-20アミノ酸残基上流の距離に導入される。
【0011】
他の実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の、2-30、2-29、2-28、2-27、2-26、2-25、2-24、2-23、2-22、2-21、又は2-20アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の、3-30、3-29、3-28、3-27、3-26、3-25、3-24、3-23、3-22、3-21、又は3-20アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の、4-30、4-29、4-28、4-27、4-26、4-25、4-24、4-23、4-22、4-21、又は4-20アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の、5-30、5-29、5-28、5-27、5-26、5-25、5-24、5-23、5-22、5-21、又は5-20アミノ酸残基上流又は下流の距離に導入される。
【0012】
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の1アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の2アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の3アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の4アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の5アミノ酸残基上流又は下流の距離に導入される。
【0013】
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の6アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の7アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の8アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の9アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の10アミノ酸残基上流又は下流の距離に導入される。
【0014】
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の12アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の13アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の14アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の15アミノ酸残基上流又は下流の距離に導入される。
【0015】
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の16アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の17アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の18アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の19アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の20アミノ酸残基上流又は下流の距離に導入される。
【0016】
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の21アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の22アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の23アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の24アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の25アミノ酸残基上流又は下流の距離に導入される。
【0017】
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の26アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の27アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の28アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の29アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の30アミノ酸残基上流又は下流の距離に導入される。
【0018】
さらなる実施態様では、本発明は、上述した実施態様のいずれかの方法により生成されたカルボキシペプチダーゼと、B鎖のC末端伸長部を有するヒトインスリン又はそのアナログの前躯体とを反応させることによる、成熟ヒトインスリン又はそのアナログを製造する方法であって、C末端伸長部が切断されて、成熟ヒトインスリン又はそのアナログが得られる方法に関する。
【0019】
一態様では、本発明は、成熟ヒトインスリン又はそのアナログを製造する方法であって、真菌細胞において、
i)B鎖のC末端伸長部を有するヒトインスリン又はそのアナログの前躯体をコードするDNA配列と、
ii)そのプロ型に挿入Kex2切断部位を有するカルボキシペプチダーゼのプロ型をコードするDNAと
を同時発現させることを含み、
ヒトインスリン前駆体分子におけるB鎖のC末端伸長部を、同時発現され活性化されたカルボキシペプチダーゼにより真菌細胞内で切断し、成熟ヒトインスリン又はそのアナログを培養培地から単離する方法に関する。
カルボキシペプチダーゼは、任意の適切なカルボキシペプチダーゼであってよい。しかしながら、本発明の一実施態様では、カルボキシペプチダーゼは宿主真菌細胞に内在している。さらなる実施態様では、カルボキシペプチダーゼはCPYである。
【0020】
本方法の一実施態様では、ヒトインスリン又はそのアナログの前躯体は、ヒトインスリン又はそのアナログのB鎖、ヒトインスリン又はそのアナログのA鎖、及びB鎖とA鎖を連結するCペプチドを含有し、ここでCペプチドは少なくとも一のKex2切断部位を含有し、B鎖は真菌細胞内におけるCペプチドのより効果的なKex2切断を促進させ、さらにカルボキシペプチダーゼを用いた切断が可能なC末端伸長部を含む。
【0021】
B鎖のC末端アミノ酸残基に結合するアミノ酸配列は比較的短く、典型的には、1-4又は1-3のアミノ酸残基、特に2のアミノ酸残基を有するであろう。この配列におけるアミノ酸残基は疎水性アミノ酸残基であり、典型的にはPhe、Leu、Ile、Tyr、Trp、Val、Met及びAlaからなる群から選択されるであろう。
B鎖のC末端伸長部の非限定的例は、Leu-Gly;Leu-Ala;Leu-Leu;Leu-Met及びLeu-Ileである。
【0022】
本発明の一実施態様では、インスリン前駆体におけるCペプチドは、A鎖においてN末端アミノ酸残基に直接結合した単一のKex2部位を含み、この位置で確実に切断される。
本発明のさらなる実施態様では、Cペプチドは、2つのKex2部位の間に挿入されてペプチド配列を有する、2つのKex2切断部位を含む。2つのKex2部位の間のペプチド配列の長さ及びアミノ酸組成は、発現した単鎖インスリン前駆体の折り畳みが可能であり、インスリン前駆体分子内に、正確に位置付けされたジスルフィド架橋が確立可能であるならば、変えてもよい。
天然Cペプチドの大きさは、35アミノ酸残基である。よって本発明の一態様では、2つのKex2部位の間のペプチド配列は、天然Cペプチドとほぼ同じ長さである。
【0023】
一実施態様では、2つのKex2部位の間のペプチド配列は、1-35、1-34、1-33、1-31、1-30、1-29、1-28、1-27、1-26、1-25、1-24、1-23、1-22、1-21、1-20、1-19、1-18、1-17、1-16、1-15、1-14、1-13、1-12、1-11、1-10、1-9、1-8、1-7、1-6、1-5、1-4、1-3又は1-2アミノ酸残基長である。
【0024】
本発明の一実施態様では、ヒトインスリン前駆体は、次のアミノ酸配列:
B(1-30)-X1-X2-Z-X3-X4-A(1-21)
[上式中、B(1-30)は、ヒトインスリンのB鎖又はそのアナログであり、A(1-21)はヒトインスリンのA鎖又はそのアナログであり、X1は、真菌細胞内でのさらに効果的なKex2切断を促進するであろう、1-5のアミノ酸残基のペプチド配列であり、X2はKex2切断部位であり、Zは、1〜約35のアミノ酸残基を有するペプチド配列又はペプチド結合であり、X3はKex2切断部位又はペプチド結合であり、X4はアミノペプチダーゼ切断部位又はペプチド結合である]
を有する。
【0025】
本発明の一実施態様では、X3はKex2切断部位であり、Zはペプチド配列であり、X4はペプチド結合である。
本発明の他の実施態様では、X3及びZはペプチド結合であり、X4はアミノペプチダーゼ切断部位である。
【0026】
X1は、典型的には、1-4又は1-3のアミノ酸残基長であり、一実施態様では、X1は、2つのアミノ酸残基のペプチド配列である。X1におけるアミノ酸残基は、好ましくは疎水性アミノ酸残基であり、典型的にはPhe、Leu、Ile、Tyr、Trp、Val、Met、Ala、Asp及びGlyからなる群から選択される。
他の実施態様では、X1におけるアミノ酸残基は、Phe、Leu、Ile、Tyr、Trp、Val、Met、Ala、Asp及びGlyからなる群から選択される。
本発明の一実施態様では、X1はLeu-Alaである。
本発明の他の実施態様では、X1はPhe-Leuである。
本発明の他の実施態様では、X1はLeu-Glyである。
本発明の他の実施態様では、X1はLeu-Leuである。
本発明の他の実施態様では、X1はLeu-Metである。
本発明の他の実施態様では、X1はLeu-Ileである。
【図面の簡単な説明】
【0027】
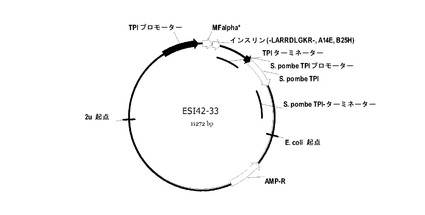
【図1】図1はプラスミドESI42-33を示す。
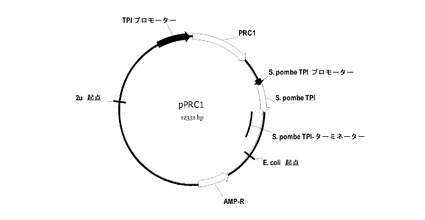
【図2】図2はプラスミドpRC1を示す。
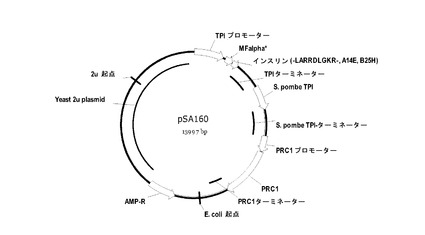
【図3】図3はプラスミドpSA160を示す。
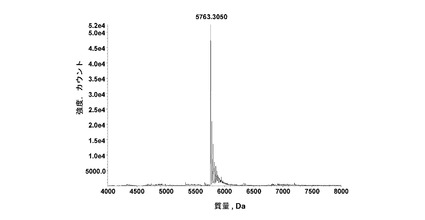
【図4】図4は、菌株包含pSA160を24時間培養した上清のデコンボリューション質量スペクトルを示す。質量5763は、十分にプロセシングされたA14E、B25Hインスリンに相当する。
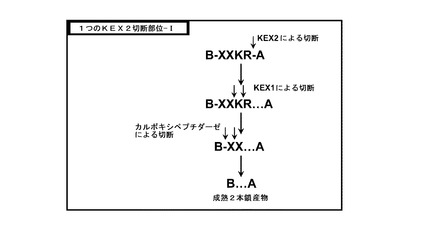
【図5】図5は、一つのKex2部位を有するインスリン前駆体の切断の概略図を示す。
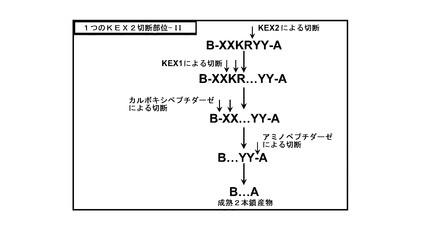
【図6】図6は、一つのKex2部位とアミノペプチダーゼ部位を有するインスリン前駆体の切断を示す。
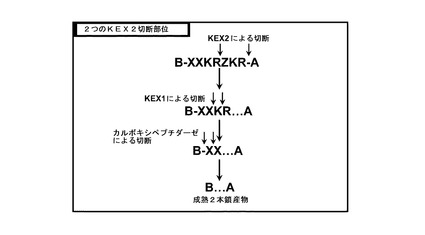
【図7】図7は、2つのKex2部位を有するインスリン前駆体の切断を示す。
【発明を実施するための形態】
【0028】
カルボキシペプチダーゼY(CPY)は、酵母サッカロミセス・セレビシエ(Saccharomyces cerevisiae)において、不活性なプロ型(プロCPY)として合成される。小胞体(ER)内におけるタンパク質の転座後、それは液胞に運搬される。到着時、それは、プロテイナーゼAにより開始されるタンパク質分解事象であるプロペプチドの除去により活性化される。2つのシナリオが、プロCPYのCPYへの効果的な転換を防止する:1)PEP4遺伝子(プロテイナーゼAをコードする遺伝子)が欠失しているか又は不活性であるならば、全てのCPYは不活性なプロ型のままであるだろう。2)PRC1(CPYをコードする遺伝子)が過剰発現しているならば、多量のプロCPYが発酵ブロスに分泌されるであろう。プロテイナーゼAが液胞に位置しており、よって分泌されたプロCPYから分離しているために、これはプロ型で残存しているであろう。これらの状況で、プロCPYを十分に活性化可能とするために、代替となる活性経路が必要とされている。
【0029】
プロペプチドと成熟したCPYとの間に界面に、プロテイナーゼAにより認識可能な配列モチーフが位置しており、よって特異的な切断に至る。切断時に、プロペプチドは成熟したCPYから分離する。本発明において、多くの変異体が作製され、ここでプロペプチドと成熟したCPYとの間の界面に近接した残基は、Kex2により認識される二塩基性切断部位が挿入される方法で変異される。これにより、Kex2pが位置するコルジ体において、プロCPYからCPYへの転換が確実になる。これで、CPYの活性化におけるプロテイナーゼAの必要性はなくなる。さらに、多量の活性化CPYを分泌可能とするために、変異した遺伝子は過剰発現している。
【0030】
以下、野生型酵素の天然N末端アミノ酸残基の31上流の位置から野生型CPYカルボキシペプチダーゼのプロ配列のアミノ酸配列、及び野生型酵素のN末端から最初の9のアミノ酸残基、及びKex2部位(KR)が、N末端アミノ酸残基に対して、種々の位置の上流又は下流に挿入されている変異体A-Gの対応する配列を示す。プロ配列が切断される切断位置を矢印で示す。
野生型:KPKFPEAIKTKKDWDFVVKNDAIENYQLRVN↓KIKDPKILG(配列番号:1)。
変異体A:KPKFPEAIKTKKDWDFVVKNDAIENYQLRVNKIKR↓DPKILG(配列番号:2)。
変異体B:KPKFPEAIKTKKDWDFVVKNDAIENYQLRVLGKR↓DPKILG(配列番号:3)。
変異体C:LGKR↓EFPEAIKTKKDWDFVVKNDAIENYQLRVNKIKDPKILG(配列番号:4)。
変異体D:KPKFPEAIKTKR↓DWDFVVKNDAIENYQLRVNKIKDPKILG(配列番号:5)。
変異体E:KPKFPEAIKTKKDWDFVKR↓NDAIENYQLRVNKIKDPKILG(配列番号:6)。
変異体F:KPKFPEAIKTKKDWDFVVKLDKR↓AIENYQLRVNKIKDPKILG(配列番号:7)。
変異体G:KPKFPEAIKTKKDWDFVVKNDAIENYQLRVNKIKDPKR↓GGILG(配列番号:8)。
【0031】
ここで使用される場合、用語「プロペプチド」又は「プロ配列」は、その機能が、折り畳み、及び発現したポリペプチドの、小胞体から、分泌経路内又は細胞外におけるその最終的な目的地までの運搬を促進させるものである、ポリペプチド配列を意味する。さらにプロペプチドは、カルボキシペプチダーゼの酵素活性を阻害する。
用語「プロ型」は、プロ配列と活性化ポリペプチド配列の融合産物を意味する。
【0032】
カルボキシペプチダーゼは、真菌細胞で発現可能であり、真菌細胞で発現した内在性Kex2p酵素により活性化可能な任意のカルボキシペプチダーゼであってよい。
カルボキシペプチダーゼの代表例は、カルボキシペプチダーゼA(EC3.4.17.1)、カルボキシペプチダーゼA2(EC3.4.17.15)、カルボキシペプチダーゼB(EC3.4.17.2)、カルボキシペプチダーゼE(EC3.4.17.10)、カルボキシペプチダーゼM(EC3.4.17.12)、カルボキシペプチダーゼT(EC3.4.17.18)、カルボキシペプチダーゼU(EC3.4.17.20)、及びカルボキシペプチダーゼY(EC3.4.16.5)である。
本発明の一実施態様では、カルボキシペプチダーゼはCPYである。
【0033】
導入されたKex2部位が、カルボキシペプチダーゼにおいてN末端アミノ酸残基に直接結合しているならば、活性型は、カルボキシペプチダーゼの天然形態であろう。
Kex2部位が、天然N末端アミノ酸に直接結合していないならば、放出されたカルボキシペプチダーゼは可変長のN末端伸長部を有するか、又はKex2部位の位置に応じて、N末端における一又は複数の天然アミノ酸残基が欠失しているであろう。
【0034】
活性化カルボキシペプチダーゼは、任意の種類の融合タンパク質のプロセシングに使用可能である。
カルボキシペプチダーゼは、それ自身のプロモーターを含む、任意の適切なプロモーターから発現してよい。しかしながら、カルボキシペプチダーゼレベルが、インスリン前駆体の異常なプロセシングに至しめる、あまりの高さになるおそれがあることがわかっている。インスリン前駆体と活性化カルボキシペプチダーゼとの間の適切な比率を発見するため、カルボキシペプチダーゼ変異体の発現レベルが、カルボキシペプチダーゼそれ自身のプロモーターを、代替プロモーターで置き換えることにより調節されてもよい。
カルボキシペプチダーゼがCPY酵素であるならば、適切な代替プロモーターは、CYC1、KEX2、MF(アルファ)1及びMPD1の遺伝子からのプロモーター領域である。
【0035】
また、カルボキシペプチダーゼのプロ型の量は、当該問題におけるプロモーターの転写因子の量を変えることにより、そのプロモーターからの発現性を調節することで調整されうる。
本発明の一実施態様では、カルボキシペプチダーゼのプロモーターはKex2プロモーターである。
【0036】
カルボキシペプチダーゼの修飾されたプロ型とヒトインスリン前駆体を、真菌細胞において同時発現させることで、さらなる高価な下流プロセシング工程を必要とすることなく、十分にプロセシングされたヒトインスリンを、発酵ブロスに分泌させることができるであろう。第1の工程として、発現したヒトインスリン前駆体を、ゴルジプロテアーゼKex1p及びKex2により、細胞内でプロセシングするであろう。ヒトインスリン前駆体は、インスリンのB鎖において、C末端アミノ酸残基に外部アミノ酸残基(X)を付加することにより修飾されているCペプチドを含む。これらの外部アミノ酸残基は、真菌細胞内において、インスリン分子のA及びB鎖からの、Cペプチドの効果的な切断を促進させるように設計されている。Kex1p及びKex2pによる完全なプロセシングにより、B鎖のC末端の伸長部Xを有する、2本鎖ヒトインスリン前駆体分子になる。前駆体は形態B-(X)n・・・Aを有し、ここでBはヒトインスリンのB鎖であり、AはヒトインスリンのA鎖であり、Xは外部アミノ酸残基であり、A及びB鎖はヒトインスリンのように、2つのジスルフィド架橋で結合している。
【0037】
ついで、このC末端で伸長している2本鎖ヒトインスリンは、B鎖C末端伸長部を切断するカルボキシプロテアーゼを同時発現及び活性化させることによる切断で、成熟ヒトインスリン又はそのアナログに転換され、成熟した2本鎖ヒトインスリン又はそのアナログが得られるであろう。
【0038】
B30位にあるアミノ酸残基に応じて、カルボキシペプチダーゼはB30アミノ酸残基を離し、desB30ヒトインスリンアナログになってもよい。さらにカルボキシペプチダーゼはB鎖のC末端から5までのアミノ酸残基が離れ、desB(29-30)-、desB28-30)-、desB(27-30)-、又はdesB(26-30)ヒトインスリンアナログになったもよい。
【0039】
インスリン前駆体の種々のプロセシングパターンを、図5-7において概略的に例証する。
図5において、インスリン前駆体コンストラクトは、単一のKex2部位、KR、及び結合ペプチドにおいて配列XXにより例証されるB鎖のC末端伸長部を有する。第1のプロセシング工程では、Kex2により、Kex2部位で切断されて、単鎖構造が、B鎖に結合した配列XXKRを有する2本鎖構造に転換される。続いて、酵素Kex1がKR配列を切断し、最終的に、カルボキシペプチダーゼがB鎖のXX伸長部を切断し、成熟した2本鎖インスリン生成物が得られる。
【0040】
図6には他の実施態様が例証されており、ここでインスリン前駆体は、単一のKex2部位及びアミノペプチダーゼ切断部位、YYを結合ペプチド中に有する。図5のように、Kex2p、Kex1p及びカルボキシペプチダーゼにより、XX-KR配列が除去されるであろう。しかしながら、この実施態様では、最後のアミノペプチダーゼ切断により、YY配列が除去されて、成熟した2本鎖インスリン生成物が得られる。
図7に例証した実施態様では、インスリン前駆体は、ペプチド鎖Zにより結合した2つのKex2切断部位を有する。Kex2を用いた第1の切断によりZ-KR配列が除去され、Kex1によりKR配列が除去され、最終的に、カルボキシペプチダーゼによりXX配列が除去される。
【0041】
真菌細胞から、成熟ヒトインスリン又はインスリンアナログが大量に生成されると、製薬目的のために、十分高純度のインスリン生成物を生成するのに必要な、下流精製工程の数をかなり減少させることができる。例えば、米国特許第4916212号に開示されている酵母においてインスリンを製造する方法では、インスリン前駆体は2つの工程、すなわち単鎖インスリン前駆体B(1-29)-Alal-Ala-Lys-A(1-21)をヒトインスリンのエステルへ転換させるペプチド転移、ついで、インスリンエステルのヒトインスリンへの加水分解において、ヒトインスリンに転換される。各転換工程は、最初の分離工程と、後続する少なくとも一の精製工程が必要である。よって、成熟インスリンの生成には、少なくとも一の酵素転換を含む、少なくとも6つの付加的な工程が必要である。
【0042】
欧州特許出願第0163529A号、PCT特許出願、国際公開第95/02059号及び国際公開第90/10075号には、酵母において、インスリン又はインスリンアナログの前駆体を発現させ、続いて、発酵ブロスからの最初の回収物を成熟インスリン又はインスリンアナログに酵素的に転換させることに基づいた、インスリン及びインスリンアナログを製造する方法が開示されている。前駆体分子は所定の修飾されたCペプチドを含有しており、インスリンB鎖のN末端伸長部をさらに含有していてもよい。修飾されたCペプチド、及び場合によってはBペプチドのN末端伸長部は、酵母細胞において切断されないように設計されており、よって前駆体は単鎖ペプチドとして分泌され、ここでA及びB鎖は修飾されたCペプチドにより結合しているが、正確に位置付けされたジスルフィド架橋は有している。ついで、成熟インスリン又はインスリンアナログ生成物は、Cペプチド、及び場合によってはN末端伸長部を切断する、複数の後続するインビトロ酵素的工程により得られる。これらの酵素的工程は、時間がかかり、多くの場合はコストが高く、高価なクロマトグラフィー工程等、さらなる下流プロセス工程で除去しなければならない付加的な汚染物質が入り込む。
【0043】
酵素的切断では、製薬用生成物の場合は効果的に除去されなければならない残存する未切断の不純物又は部分的に切断された不純物の100%切断には達しないことはよく知られている。よって、各切断工程には、少なくとも一の単離又は精製工程、典型的には交換クロマトグラフィー、ゲル濾過クロマトグラフィー、アフィニティークロマトグラフィー等によるクロマトグラフィー精製が続くであろう。
商業的規模で使用されているクロマトグラフィー用カラム材料は、非常に高価であり、よって、このようなクロマトグラフィー工程の数を減らすことは、生産経済において、かなりの影響を有するものである。下流の転換及び精製工程を減らすと、労働者の仕事量及びプロセスに費やされる時間がさらに減少し、よって生産経済がさらに改善される。
【0044】
本プロセスにおいて、成熟インスリン又はそのアナログは、培養ブロスから直接単離することができ、製薬用途に十分な純度の生成物を生成させるのに必要な下流のプロセス工程がさらに減少する。
【0045】
インスリン分子は、修飾が得られるインスリンアナログのインスリン活性に悪影響を及ぼさない限りは、A及び/又はB鎖でさらに修飾されていてもよい。
よって、ここで使用される場合、「インスリンアナログ」とは、天然インスリン中に生じる少なくとも一つのアミノ酸残基を欠失させ、及び/又は置換することにより、及び/又は少なくとも一つのアミノ酸残基を付加することによって、天然に生じるインスリン、例えばヒトインスリンの構造から形式的には誘導することができる分子構造を有するポリペプチドを意味する。付加された及び/又は置換されたアミノ酸残基は、コード可能なアミノ酸残基又は他の天然に生じるアミノ酸残基又は純粋に合成のアミノ酸残基の何れでもよい。
インスリンアナログは、ヒトインスリンと比較して、典型的には約7を越える変異、より典型的には5を越える変異、最も典型的には最大3の変異を含まない。
【0046】
長年にわたって、インスリンA及び/又はB鎖におけるかなり多数の変異が開示されている。例えば、B鎖の28位は、天然のPro残基から、Asp、Lys、又はIleに修飾されていてもよく、またB29位のLysはProに修飾されていてもよい。
また、A21位のAsnは、Ala、Gln、Glu、Gly、His、Ile、Leu、Met、Ser、Thr、Trp、Tyr又はVal、特にGly、Ala、Ser、又はThr、特にGlyに修飾されてもよい。さらに、B3位のAsnは、Lys又はAspに修飾されてもよい。インスリンアナログのさらなる具体例は、des(B30)ヒトインスリン、B1及びB2の一方又は双方が欠失したインスリンアナログ;A鎖及び/又はB鎖がN末端伸長を有するインスリンアナログ、及びA鎖及び/又はB鎖がC末端伸長を有するインスリンアナログである。またA18位にある天然アミノ酸残基はGln残基に変化していてもよく、またB26-B30位にある一又は複数のアミノ酸残基は欠失している。
【0047】
本方法で生成可能なインスリンアナログの具体例は、GlyA21ヒトインスリン、GlyA21des(B30)ヒトインスリン、desB1ヒトインスリン、des B30ヒトインスリン、AspB28ヒトインスリン、及びLysB28ProB29ヒトインスリンである。
インスリンアナログのさらなる具体例は、A21、B10、A8、A14、B25、B27及びB1の一又は複数の位置に変異を含む、ヒトインスリンアナログである。
【0048】
真菌細胞は、全ての真菌が、提供された種類のインスリン前駆体分子を切断し、結合ペプチドを切断し、2本鎖分子をライゲーションさせるのに必須のタンパク質分解活性を有するならば、任意の真菌細胞であってよい。しかしながら、長年にわたって、酵母が、インスリンサイズの小ペプチドの発現及び分泌に有効な細胞腫であることがわかっている。特に、酵母サッカロミセス・セレビシエが有用であることが判明している。
よって、本発明の一実施態様における真菌細胞は酵母細胞であり、さらなる実施態様では、酵母細胞はサッカロミセス・セレビシエである。
【0049】
本発明で使用されるDNA配列は、ゲノム又はcDNA由来のものでよく、例えばゲノム又はcDNAライブラリーを調製し、標準的な技術(例えば、Sambrook, J, Fritsch, EF及びManiatis, T, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, New York、1989)に従い、合成オリゴヌクレオチドプローブを使用するハイブリッド形成により、ポリペプチドの全体又は一部をコードするDNA配列をスクリーニングすることにより得られる。またインスリン前駆体をコードするDNA配列は、確立された標準的な方法、例えばBeaucage及びCaruthers, Tetrahedron Letters 22 (1981), 1859-1869に記載されたホスホアミジト(phosphoamidite)法、又はMatthesら, EMBO Journal 3 (1984), 801-805により記載された方法により合成的に調製されてもよい。さらに、DNA配列は、例えば米国特許第4683202号、又はSaikiら, Science 239 (1988), 487-491に記載されたようにして、特定のプライマーを使用するポリメラーゼ連鎖反応により調製されてもよい。
【0050】
DNA配列は、組換えDNA手順にかけられうる任意のベクターに挿入されてよく、多くの場合、ベクターの選択は、それが導入される宿主細胞に依存する。例えば、ベクターは自己複製可能なベクターであってよく、すなわちベクターは染色体外実体として存在しており、その複製は染色体複製、例えばプラスミドとは独立している。また、ベクターは、宿主細胞に導入される場合、宿主細胞ゲノムに組み込まれ、それが組み込まれる染色体(群)と共に複製される。
【0051】
ベクターは好ましくは発現ベクターであり、そこでインスリン前駆体をコードするDNA配列は、プロモーター等、DNAの転写に必要な付加的なセグメントに作用可能に結合する。プロモーターは宿主細胞の選択において転写活性を示す任意のDNA配列であってよく、宿主細胞と相同又は異種であるタンパク質をコードする遺伝子から誘導されてよい。
【0052】
酵母宿主細胞への使用に適したプロモーターの例には、酵母解糖遺伝子(Hitzemanら, J. Biol. Chem. 255(1980), 12073-12080; Alber及びKawasaki, J. Mol. Appl. Gen. 1(1982), 419-434、又はアルコールデヒドロゲナーゼ遺伝子(Youngら, Genetic Engineering of Microorganisms for Chemicals(Hollaenderら編), Plenum Press, New York, 1982)、又はTPI1(米国特許第4599311号)又はADH2-4c(Russellら, Nature 304(1983), 652-654)プロモーターが含まれる。
【0053】
所望の生成物をコードするDNA配列は、もし必要ならば、適切なターミネーター、ポリアデニル化シグナル、転写エンハンサー配列、及び翻訳エンハンサー配列に作用可能に結合していてもよい。さらに本発明の組換えベクターは、当該問題においては、ベクターが宿主細胞内で複製可能なDNA配列をさらに含んでよい。
【0054】
宿主細胞の分泌経路にインスリンを方向付けるために、分泌シグナル配列(リーダー配列、プレプロ配列、又はプレ配列としても公知)は、組換えベクター内に提供されてもよい。分泌シグナル配列は正確なリーディングフレームにおいて、インスリン前駆体をコードするDNA配列に結合している。分泌シグナル配列は、通常、ペプチドをコードするDNA配列に対し、5'に位置している。シグナルペプチドは、天然に生じるシグナルペプチド、又はその機能的部分であってよく、又は合成ペプチドであってもよい。
酵母における効果的な分泌のために、リーダーペプチドをコードする配列は、シグナル配列の下流と所望の生成物をコードするDNA配列の上流に挿入されてよい。
【0055】
異種DNAで酵母細胞を形質転換させ、異種ポリペプチドを生成させる方法は、例えば米国特許第4599311号、米国特許第4931373号、同第4870008号、同第5037743号、及び同第4845075号に記載されている。形質転換細胞は、選択可能マーカーにより決定されたフェノタイプ、一般的には薬剤耐性、又は特定の栄養素、例えばロイシンの不在下で成長する能力によって選択される。酵母に使用される好ましいベクターは、米国特許第4931373号に記載のPOT1ベクターである。
【0056】
発酵プロセスにおいて細胞培養に使用される培地は、宿主細胞の増殖に適した任意の従来からの培地、例えば適切なサプリメントを含有する最小又は複合培地であってよい。適切な培地は商業的供給者から入手可能であり、又は公開されているレシピ(例えば、American Type Culture Collectionのカタログ)に従い調製されてもよい。例えば、培地は、少なくとも一の炭素源、一又は複数の窒素源、カリウム、ナトリウム、マグネシウムの塩、リン酸塩、硝酸塩及び硫酸塩を含む必須の塩、微量金属、水溶性ビタミン類、限定されるものではないが、プロセス補助剤、例えばプロテアーゼインヒビター、安定剤、リガンド、消泡剤及び誘導因子を含有する。培地は、熱による滅菌を含む、いくつかの操作条件において、液体培地に部分的に沈殿又は分散する成分を含有していてもよい。培地は、いくつかの液体とガス状溶液とを混合することにより作製可能である。これらの溶液は発酵タンクに入れる前に混合することもできるし、所定の比率で添加された別々の液体流として、発酵タンクに供給される。培地成分の異なる液体溶液の比率は、発酵プロセスの様々な段階で変化し、これは培地の全ての成分が、発酵の経過によって変化し得ることを意味する。
【0057】
ついで、細胞により生成されるペプチドは、遠心分離又は濾過、硫酸アンモニウム等の塩により上清又は濾液のタンパク質様成分を沈殿させることによって、培地から宿主細胞を分離させ、当該分野のタンパク質の種類に応じて、イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー、アフィニティークロマトグラフィー等の多様なクロマトグラフィー手順により精製することを含む従来からの手順により、培養培地から回収されてよい。
【0058】
培養ブロスから単離させた後、成熟インスリン又はインスリンアナログを、特にB29Lys残基のε-アミノ基をアシル化することにより、例えばアシル化形態に転換させてもよい。インスリンをアシル化させる方法は当該技術でよく知られており、例えば欧州特許第792290号及び同第894095号、米国特許第5693609号、同第5646242号、同第5922675号、同第5750497号及び同第6011007号に開示されている。
【0059】
アシル化されたインスリンの例は、NεB29-テトラデカノイルdes(B30)ヒトインスリン、NεB29-リトコロリル-γ-グルタミルdes(B30)ヒトインスリン、NεB29-(Nα-(HOOC(CH2)14CO)-γ-Glu)des(B30)ヒトインスリン、又はNεB29-(Nα-(HOOC(CH2)16CO)-γ-Glu)des(B30)ヒトインスリンである。
【0060】
「desB30」又は「B(1-29)」とは、B30アミノ酸残基を欠く天然インスリンB鎖又はそのアナログを意味する。
B(1-30)はヒトインスリンの天然B鎖を意味し、「A(1-21)」は天然インスリンA鎖を意味する。A18Qヒトインスリンは、ヒトインスリンA鎖のA18位にGlnを有するインスリンアナログである。B10E、A8H、A14Eは、それぞれB10位にGlu、A8位にHis、及びA14位にGluを有するインスリンアナログである。
「B1」、「A1」等とは、それぞれインスリンのB鎖の1位(N末端から数える)のアミノ酸残基及びインスリンのA鎖の1位(N末端から数える)のアミノ酸残基を意味する。特定の位置にあるアミノ酸残基は、例えばB1位のアミノ酸残基がフェニルアラニン残基であることを意味するPheB1として表されてもよい。
【0061】
「Cペプチド」とは、インスリン分子のA及びBペプチド鎖を互いに結合させるペプチド配列を意味する。
「成熟インスリン」とは、正しいアミノ酸残基組成と、天然のヒトインスリン分子と同様の立体構造、すなわちCysA7とCysB7との間、及びCysA20とCysB19との間のジスルフィド架橋、及びCysA6とCysA11との間の内部ジスルフィド架橋、及びインスリン活性を有する2本鎖インスリンを意味する。よって、本発明の成熟インスリンはヒトインスリンである。成熟ヒトインスリンアナログは、B28Aspヒトインスリン、desB30ヒトインスリン、A14Glu、B25Hisヒトインスリン、及びB31Leu、B32Alaヒトインスリンであってよい。
ここで使用される場合、「インスリン誘導体」とは、天然に生じるインスリン、又は、例えばインスリン骨格の一又は複数の位置に側鎖が導入されることにより、もしくはインスリンのアミノ酸残基の基を酸化又は還元することにより、もしくは遊離のアミノ基又はヒドロキシ基をアシル化することにより化学的に修飾されたインスリンアナログを意味する。
【0062】
「Kex2」又は「Kex2p」とは、2つの塩基性残基(リジン又はアルギニン)の配列の後に、好ましくは切断を触媒するサブチリシン様エンドプロテアーゼを意味する(Rockwell, NC, Krysan、DJ、Komiyama, T & Fuller, RS 2002 Precursor Processing by Kex2/Furin Proteases. Chem. Rev. 102: 4525-4548)。
「Kex1」又は「Kex1p」とは、好ましくはC末端のリジル及び/又はアルギニル残基の除去を触媒するセリンカルボキシペプチダーゼを意味する(Shilton BH, Thomas DY, Cygler M 1997 Crystal structure of Kex1deltap, a prohormone-processing carboxypeptidase from Saccharomyces cerevisiae. Biochemistry 36: 9002-9012)。
CPYとは、好ましくは疎水性又は大きなC末端アミノ酸残基、例えばPhe及びLeuの除去を触媒するカルボキシペプチダーゼである、カルボキシペプチダーゼYを意味する(Remington, S.J. & Breddam, K. (1994) Carboxypeptidases C and D. Methods Enzymol. 244、231-248)。CPYのアミノ酸配列は、「Vallsら, 1987, Cell, 48(5):887-897」に開示されている。
【0063】
「正確にプロセシング」とは、正確なアミノ酸残基配列を有する所望の生成物が得られるように、所望の切断点で酵素的に切断することを意味する。
「効果的な切断」とは、少なくとも80%、好ましくは少なくとも85%、さらに好ましくは少なくとも95%の切断を意味することを意図している。
「POT」は、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)トリオースリン酸イソメラーゼ遺伝子であり、「TPI1」はサッカロミセス・セレビシエ・トリオースリン酸イソメラーゼ遺伝子である。
「リーダー」とは、プレペプチド(シグナルペプチド)及びプロペプチドからなるアミノ酸配列を意味する。
【0064】
「シグナルペプチド」なる用語は、タンパク質の前駆体形態において、N末端配列として存在するプレペプチドを意味すると理解される。シグナルペプチドの機能は、異種タンパク質の小胞体への転座を促進できることである。シグナルペプチドはこのプロセス中に通常は切断される。シグナルペプチドはタンパク質を生成する宿主生物体に対して異種又は同族であってもよい。
酵母宿主細胞に有用なシグナルペプチドは、サッカロミセス・セレビシエのα因子及びサッカロミセス・セレビシエのインベルターゼに対する遺伝子から得られる。本発明のDNAコンストラクトと共に使用され得る多くのシグナルペプチドには、酵母アスパラギン酸プロテアーゼ3(YAP3)シグナルペプチド、又は任意の機能的アナログ(Egel-Mitaniら、(1990) YEAST 6:127-137及び米国特許第5726038号)、MFα1遺伝子のα-因子シグナル(Thorner(1981), The Molecular Biology of the Yeast Saccharomyces cerevisiae, Strathern等編, pp 143-180, Cold Spring Harbor Laboratory, NY、及び米国特許第4870008号)、マウス唾液アミラーゼのシグナルペプチド(O. Hagenbuchleら, Nature 289、1981, pp. 643-646を参照)、修飾されたカルボキシペプチダーゼのシグナルペプチド(L.A. Vallsら, Cell 48,1987, pp. 887-897を参照)、及び酵母BAR1シグナルペプチド(国際公開第87/02670号を参照)が含まれる。
【0065】
本発明は、選択された微生物又は宿主細胞中で複製可能であり、本発明のインスリン生成物をコードするポリヌクレオチド配列を担持するベクターを包含する。組換えベクターは、自己複製ベクター、すなわち染色体外実体として存在するベクター、染色体複製とは独立した複製、例えばプラスミド、染色体外エレメント、ミニ染色体、又は人工染色体でありうる。ベクターは自己複製を保証する任意の手段を含みうる。あるいは、ベクターは、宿主細胞に導入されたときに、ゲノムに組み込まれ、それが組み込まれた染色体と共に複製されるものでありうる。さらに、単一のベクター又はプラスミド又は宿主細胞のゲノムに導入される全DNAを合わせて含む2以上のベクター又はプラスミド、あるいはトランスポゾンが使用されうる。ベクターは直鎖又は閉環プラスミドであってよく、宿主細胞のゲノムへのベクターの安定した組込、又はゲノムとは独立した細胞におけるベクターの自己複製を可能にするエレメント(類)を好ましくは含む。
一実施態様では、組換え発現ベクターは酵母中で複製可能である。ベクターを酵母中で複製させることができる配列の例は酵母プラスミド2μm複製遺伝子REP1-3及び複製起点である。
【0066】
またベクターは、選択可能なマーカー、例えば遺伝子、宿主細胞における欠失を補完する生成物、又は例えばアンピシリン、カナマイシン、テトラサイクリン、クロラムフェニコール、ネオマイシン、ハイグロマイシン、又はメトトレキサート等の薬剤に対する耐性を付与するものをさらに含んでもよい。
酵母宿主細胞のための適切なマーカーは、ADE2、HIS3、LEU2、LYS2、MET3、TRP1、及びURA3である。酵母とって好ましい選択可能なマーカーはシゾサッカロミセス・ポンベTPI遺伝子(Russell (1985) Gene 40:125-130)である。
酵母宿主において、有用なプロモーターは、サッカロミセス・セレビシエMFα1、TPI、ADH又はPGKプロモーターである。
本発明のポリヌクレオチドコンストラクトは典型的には適切なターミネーターに作用可能に結合しているであろう。酵母において、適切なターミネーターはTPIターミネーター(Alberら(1982) J. Mol. Appl. Genet. 1:419-434)である。
【0067】
インスリン前駆体をコードするDNA配列、プロモーター、及び場合によってはターミネーター、及び/又は分泌シグナル配列をそれぞれライゲーションし、複製に必要な情報を含む適切なベクターにそれらを挿入するのに使用される手順は、当業者によく知られている(例えば、Sambrookら, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, New York, 1989を参照)。
ベクターは、まず本発明のインスリン前駆体をコードする全DNA配列を含有するDNAコンストラクトを調製し、続いて適切な発現ベクターにこのフラグメントを挿入するか、又は個々のエレメントについての遺伝的情報を含むDNAフラグメントを挿入し、続いてライゲーションすることにより構築されると理解されるであろう。
【0068】
本発明で使用される宿主細胞は真菌細胞である。ここで使用される場合、「真菌」には、子嚢菌門、担子菌門、ツボカビ門、及び接合菌門(Hawksworthらによる定義, Ainsworth and Bisby's Dictionary of The Fungi, 8th edition, 1995, CAB International, University Press, Cambridge, UK)、並びに卵菌門(Hawksworth, 1995, 上掲, 171頁を引用)及び全ての栄養胞子形成菌(Hawksworthら, 1995, 上掲)が含まれる。
【0069】
一実施態様では、真菌宿主細胞は酵母細胞である。ここで使用される「酵母」には、有子嚢胞子(ascosporogenous)酵母(エンドマイセス属)、担子胞子形成(basidiosporogenous)酵母、及び不完全菌類に属する酵母(ブラストミセス属)が含まれる。有子嚢胞子酵母はスペルモフトラ科(Spermophthoraceae)とサッカロミセス科(Saccharomycetaceae)に分けられる。後者は、4つの亜科、すなわちスキゾサッカロミセス(Schizosaccharomycoideae)亜科(例えばスキゾサッカロミセス(Schizosaccharomyces) 属) 、ナドソニオイデ(Nadsonioideae)亜科 、リポミコイデー(Lipomycoideae) 亜科、及びサッカロミセス(Saccharomycoideae) 亜科(例えばピキア(Pichia)、クルイベロミセス(Kluyveromyces) 及びサッカロミセス(Saccharomyces) 属)からなる。担子胞子形成酵母には、ロイコスポリジウム(Leucosporidim)属、ロドスポリジウム(Rhodosporidium)属、スポリジオボラス(Sporidiovolus)属、フィロバシジウム(Filobasidium)属、及びフィロバシジエラ(Filobasidiella)属が含まれる。不完全菌類に属する酵母は、2つの科、つまりスポロボロミセス(Sporobolomycetaceae) 科(例えばスポロボロミセス(Sorobolomyces) 属及びブレラ(Bullera) 属) 及びクリプトコッカス(Cryptococcaceae) 科(例えばカンジダ(Candida) 属)に分けられる。酵母の分類は今後変更されることがあり得るので、本発明の目的のためには、酵母はBiology and Activities of Yeast (Skinner, F.A. 、Passmore, S.M.及びDavenport, R.R.編、Soc. App. Bacteriol. Symposium Series No.9, 1980)に記載されたように定義される。酵母の生物学及び酵母遺伝子の操作は、当該技術分野においてよく知られている(例えば、Biochemistry and Genetics of Yeast, Bacil, M., Horecker, B.J., 及びStopani, A.O.M.編, 第2版, 1987;The Yeasts, Rose, A.H., 及びHarrison, J.S.編, 第2版, 1987;及びThe Molecular Biology of the Yeast Saccharomyces, Strathernら編, 1981を参照)。
【0070】
酵母宿主細胞は、カンジダ、クルイベロミセス、サッカロミセス、スキゾサッカロミセス、ピキア、ハンゼヌラ又はヤロウィア(Yarrowia)属の種の細胞から選択されうる。一実施態様では、酵母宿主細胞は、サッカロミセス・カールスバージェンシス(Saccharomyces carlsbergensis)、サッカロミセス・セレビシエ、サッカロミセス・ディアスタティカス(Saccharomyces diastaticus)、サッカロミセス・ダグラシ(Saccharomyces douglasii)、サッカロミセス・クルイベリ(Saccharomyces kluyveri)、サッカロミセス・ノルベンシス(Saccharomyces norbensis)、サッカロミセス・オビフォルミス(Saccharomyces oviformis)、シゾサッカロミセス・ポンベ、 サッカロミセス・ウバルム(Sacchoromyces uvarum)、ピキア・クルイベリ(Pichia kluyveri)、ヤロイア・リポリチカ(Yarrowia lipolytica)、トルラ酵母(Candida utilis)、カンジダ・カカオイ(Candida cacaoi)、及びゲオトリクム・ファーメンタンス(Geotrichum fermentans)である。他の有用な酵母宿主細胞は、クルイベロミセス・ラクチス(Kluyveromyces lactis)、クルイベロミセス・フラジリス(Kluyveromyces fragilis)、ハンセヌラ・ポリモルファ(Hansenula polymorpha)、ピキア・パストリス(Pichia pastoris)、ヤロイア・リポリチカ(Yarrowia lipolytica)、シゾサッカロミセス・ポンベ、トウモロコシ黒穂病菌(Ustilgo maylis)、カンジダ・マルトース(Candida maltose)、ピキア・ギリエルモンデイ(Pichia guillermondii)及びピキア・メタノリオール(Pichia methanoliol)である(Gleesonら, J. Gen. Microbiol. 132, 1986, pp.3459-3465;米国特許第4882279号及び同第4879231号を参照)。
【0071】
「コード可能なアミノ酸」又は「コード可能なアミノ酸残基」なる表現は、ヌクレオチドのトリプレット(「コドン」) によりコード可能なアミノ酸又はアミノ酸残基を示すために使用される。
本記載において、アミノ酸の3文字又は1文字表記は、それらの従来からの意味で使用される。明確に示されない限りは、ここに挙げたアミノ酸はLアミノ酸である。さらにペプチドのアミノ酸配列の左及び右端は、特に記載しない限りは、それぞれN及びC末端である。
【0072】
ここに引用された刊行物、特許出願及び特許を含む全ての文献は、各文献が、出典明示により個々にかつ特に援用され、その全内容がここに説明されるかの如く、その全体が出典明示によりここに援用される(法律により許容される最大範囲)。
全ての表題及び副題は、ここでは便宜的に使用され、決して本発明を限定するものと解してはならない。
ここに提供される任意かつ全ての例、又は例示的言語(例えば「等」)の使用は、単に本発明をより明らかにすることを意図しており、特に請求項に記載がない限り、本発明の範囲に限定をもたらすものではない。明細書中の如何なる語句も請求項に記載していない要素が本発明の実施に必須であることを示しているものと解してはならない。
ここでの特許文献の引用及び援用は単に便宜上なされているもので、そのような特許文献の有効性、特許性、及び/又は権利行使性についての見解を反映させるものではない。 この発明は、適用される法律に容認される場合、ここに付加される請求項に列挙された主題事項の全ての修正点及び等価物を含む。
【実施例】
【0073】
一般的手順
全ての発現プラスミドは、欧州特許第171,142号に記載されているものと同様の、C-POT型のものである。これらは、プラスミドの選択、及びサッカロミセス・セレビシエの安定化の目的のために、シゾサッカロミセス・ポンベ・トリオースリン酸イソメラーゼ遺伝子(POT)を含有することにより特徴付けられる2μベースの発現ベクターである。また、プラスミドは、サッカロミセス・セレビシエ・トリオースリン酸イソメラーゼプロモーター及びターミネーター(図1)を含む。これらの配列は、次の:1)インスリン生成物とリーダーの融合タンパク質をコードするEcoRI-Xbalフラグメントの配列、及び2)発現ベクターの2μ領域におけるNcoI部位の除去に帰するサイレント変異の導入、を除き、全ての配列である場合、プラスミドpKFN1003(国際公開第9010075号に記載)における対応配列に類似している。種々の融合タンパク質のクローニングを促進させるために、MFα1プレプロリーダーをコードするDNA配列を変化させてNcoI部位を導入し(図2を参照)、MFα1*プレプロリーダーと称する。このようにNcoI-Xbalフラグメントは、関心あるインスリンコンストラクトをコードするNcoI-Xbalフラグメントで、簡単に置き換えられる。このようなNcoI-Xbalフラグメントは、標準的な技術に従い、合成オリゴヌクレオチド及びPCRを使用して合成されてよい。アルファ-リーダーに加えて、他のリーダーを使用することもできる。
【0074】
酵母形質転換を、宿主菌株であるサッカロミセス・セレビシエ株MT663又はME1719の形質転換により調製した。酵母菌株MT663(MATa/MATα pep4-3/pep4-3 HIS4/his4 Δtpi::LEU2/Δtpi::LEU2 Cir’)は、国際公開第92l11378号に関連して、Deutsche Sammlung von Mikroorganismen und Zellkulturenに寄託されており、寄託番号DSM6278が付与されている。サッカロミセス・セレビシエ菌株ME1719(MATa/α leu2/leu2 pep4-3/pep4-3 Δtpi::LEU2/Δtpi::LEU2 Δura3/Δura3 Δyps1::URA3/Δyps1::ura3 Cir+)は国際公開第98/01535号に記載されている。
【0075】
MT663又はME1719を、600nmでの光学密度が0.6になるまで、YPGaL(1%のBacto酵母抽出物、2%のBactoペプトン、2%のガラクトース、1%のラクタート)において増殖させた。遠心分離により100mlの培養体を収集し、10mlの水で洗浄し、再度遠心分離にかけ、1.2Mのソルビトール、25mMのNa2EDTA、pH=8.0及び6.7mg/mlのジチオトレイトールを含有する10mlの溶液に再懸濁させた。懸濁液を30℃で15分インキュベートし、遠心分離にかけ、細胞を、1.2Mのソルビトール、10mMのNa2EDTA、0.1Mのクエン酸ナトリウム、pH05.8、及び2mgのNovozymC3234を含有する10mlの溶液に再懸濁させた。懸濁液を30℃で30分インキュベートし、遠心分離により細胞を収集し、10mlの1.2Mソルビトールと10mlのCAS(1.2Mのソルビトール、10mMのCaCl2、10mMのトリスHCI(トリス=トリス(ヒドロキシメチル1)-アミノメタン)pH=7.5)で洗浄し、2mlのCASに再懸濁させた。形質転換のために、1mlのCAS懸濁細胞を、約0.1mgのプラスミドDNAと混合し、室温で15分放置した。1mlの(20%のポリエチレングリコール4000、10mMのCaCl2、10mMのトリスHCI、pH=7.5)を添加し、混合物を室温でさらに30分放置した。混合物を遠心分離にかけ、ペレットを0.1mlのSOS(1.2Mのソルビトール、33%vlvのYPD、6.7mMのCaCl2)に再懸濁させ、30℃で2時間インキュベートした。ついで、懸濁液を遠心分離にかけ、ペレットを0.5mlの1.2Mソルビトールに再懸濁させた。ついで、6mlの上層寒天(1.2Mのソルビトールと2.5%の寒天を含有する、Shermanら (1982) Methods in Yeast Genetics, Cold Spring Harbor LaboratoryのSC培地)を52℃で添加し、同様の寒天固形化されたソルビトール含有培地を含むプレートの頂部に懸濁液を注いだ。
【0076】
実施例1
ヒトインスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリン用の酵母発現システムの構築
図1には、pESI42-33と称される酵母プラスミドが示されている。プラスミドは、サッカロミセス・セレビシエTPI遺伝子の転写プロモーターと転写ターミネーターとの間のプラスミドに挿入されたEcoRI-XbaIフラグメントを含有する発現カセットを含む。プラスミドpESI42-33において、EcoRI-XbaIフラグメントは、MFα1*プレプロリーダー、二塩基プロセシングエンドペプチダーゼKex2に対するLys-Arg切断部位、及びインスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリンからなる融合産物をコードする。
【0077】
インスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリンをコードする配列を含有するDNAフラグメントを、合成オリゴヌクレオチド及び標準的なPCR増幅を使用して構築した。得られたPCRフラグメントを精製し、NcoI及びXbaIで消化し、修飾されたcPOT型の発現ベクターのNcoI-XbaIベクターフラグメントにライゲーションさせた(図1)。
発現プラスミドを、アンピシリンの存在下で成長させた大腸菌にて増殖させ、標準的な技術を使用し単離させた(Sambrookら, 1989)。プラスミドDNAを適切な制限ヌクレアーゼにより挿入についてチェックし(例えば、EcoRI、NcoI、XbaI)、配列分析により、インスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリンの適切な配列を含有することが示された。
プラスミドをサッカロミセス・セレビシエ菌株MT663において形質転換させた。プラスミドを包含する酵母形質転換体を、YPD(1%の酵母抽出物、2%のペプトン、2%のグルコース)寒天(2%)プレートにおける炭素源としてのグルコース利用によって選択した。
【0078】
実施例2
CPY活性変異体用の酵母発現システムの構築
図2には、pPRC1と称される酵素プラスミドが示されている。プラスミドは、サッカロミセス・セレビシエTPI遺伝子の転写プロモーターと転写ターミネーターとの間のプラスミドに挿入されたClaI-NheIフラグメントを含有する発現カセットを含む。プラスミドpPRC1、ClaI-NheIフラグメントは、野生型プレプロCPYをコードする。全てのCPY変異プラスミドは、関連点変異が、二塩基性Kex2切断モチーフの導入用に作製されていることを除けば、pPRC1と同一である。
Kex2切断部位の挿入により変異したPRC1をコードする配列を含有するDNAフラグメントを、合成オリゴヌクレオチド、テンプレートとしてゲノム酵母DNA、及び標準的なPCR増幅を使用して構築した。得られたPCRフラグメントを精製し、ClaI及びNheIで消化し、修飾されたcPOT型の発現ベクターのClaI-NheIベクターフラグメントにライゲーションさせた。
【0079】
発現プラスミドを、アンピシリンの存在下で成長させた大腸菌にて増殖させ、標準的な技術を使用し単離させた(Sambrookら, 1989)。プラスミドDNAを適切な制限ヌクレアーゼにより挿入についてチェックし、配列分析により、PRC1の適切な配列を含有することが示された。
プラスミドをサッカロミセス・セレビシエ菌株MT663において形質転換させた。プラスミドを包含する酵母形質転換体を、YPD(1%の酵母抽出物、2%のペプトン、2%のグルコース)寒天(2%)プレートにおける炭素源としてのグルコース利用によって選択した。
【0080】
実施例3
インスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリン及びCPY活性変異体の同時発現用の酵母発現システムの構築
図3には、pSA160と称される酵母プラスミドが示されている。プラスミドは、サッカロミセス・セレビシエTPI遺伝子の転写プロモーターと転写ターミネーターとの間のプラスミドに挿入されたEcoRI-XbaIフラグメントを含有する発現カセットを含む。プラスミドpSA160において、EcoRI-XbaIフラグメントは、MFα1*プレプロリーダー、二塩基プロセシングエンドペプチダーゼKex2に対するLys-Arg切断部位、及びインスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリンからなる融合産物をコードする。またプラスミドは、サッカロミセス・セレビシエPRC1遺伝子の転写プロモーターの後のプラスミド中に挿入されるKpnI-SacIフラグメントを含むCPY変異体「F」をコードする発現カセットも含む。
インスリンカセットを、適切なcPOT型の発現ベクターにおいてサブクローニングした。続いて、PRC1プロモーターを、テンプレートとしてゲノム酵母DNAを使用するPRCにより増幅し、SalI-KpnIフラグメントとして、上述したプラスミドにおいてクローニングした。最終的に、ターミネータープラスCPY変異体をコードするDNAを、PCRを使用して増幅させ、プラスミドにおいてクローニングした。
【0081】
発現プラスミドを、アンピシリンの存在下で成長させた大腸菌にて増殖させ、標準的な技術を使用し単離させた(Sambrookら, 1989)。プラスミドDNAを適切な制限ヌクレアーゼにより挿入についてチェックし、配列分析により、PRC1オープンリーディングフレームの適切な配列を含有することが示された。
プラスミドをサッカロミセス・セレビシエ菌株MT663において形質転換させた。プラスミドを包含する酵母形質転換体を、YPD(1%の酵母抽出物、2%のペプトン、2%のグルコース)寒天(2%)プレートにおける炭素源としてのグルコース利用によって選択した。
【0082】
実施例4
複数のCPY変異体を構築した。プラスミドを酵母において形質転換させ、研究室規模のバッチ発酵を、30℃で72時間、定められた培地における培養により実施した。合成基質N-(3-[2-フリル]アクリロイル)-Phe-Pheを使用し、CPY活性について発酵ブロスをアッセイした。この基質を使用すると、337nmでの吸光度における変化として、CPY活性を分光学的に追跡することができる。既知の濃度のCPY標準体の活性度に対して、活性度を相互比較することにより、上清における濃度を推定することができる。これらのアッセイに基づき、9つの変異体が、0-2.3mg/Lの濃度で存在することが推定された。
【0083】
プロセシングの度合いを調査するために、発酵上清をウエスタンブロット分析にかけ、CPY抗体で探索した。プロセシングされていないCPY(プロCPY)とプロセシングされたCPYとの間の比率を、ブロットから推定した。これにより、ほとんどの変異体において、成熟したCPYへの〜80%の転換率が示された。
【0084】
実施例5
ヒトインスリン前駆体(B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリンとCPY活性変異体「F」との同時発現用のプラスミドpSA160を、酵母において形質転換させ、研究室規模のバッチ発酵を、30℃で定められた培地における培養により実施した。インスリン種の培養24時間後、発酵ブロスをLC-MSにより分析した。これにより、分泌されたインスリン種の>90%が、十分にプロセシングされたA14E,B25Hヒトインスリンアナログであったことが示された。図4には、分泌された活性CPYにより、インスリンB鎖のC末端から、LA伸長部が成功裏に除去されたことが示されている。
【0085】
実施例6
図3に記載された同時発現プラスミドを包含する酵素菌株の連続発酵により、CPYレベルがあまりに高まり、ヒトインスリン前駆体の異常なプロセシングに至ることが示された。ヒトインスリン前駆体と活性CPYとの間の適切な比率を見出すために、PRC1プロモーターを代替プロモーターに置き換えることにより、CPY変異体の発現レベルを調節した。
CYC1、KEX2、MF(アルファ)1、及びMPD1の遺伝子からのプロモーター領域をPCRにより増幅させ、PRC1プロモーター配列を、それぞれ代替CYC1、KEX2、MF(アルファ)1、及びMPD1のプロモーター配列と交換することで得られた、実施例3に記載の同時発現プラスミドのSalI−KpnI部位においてクローニングした。得られたプラスミドをサッカロミセス・セレビシエ菌株MT663において形質転換させた。プラスミドを包含する酵母形質転換体を、YPD(1%の酵母抽出物、2%のペプトン、2%のグルコース)寒天(2%)プレートにおける炭素源としてのグルコース利用によって選択した。
【0086】
実施例7
研究室規模のバッチ発酵を、実施例6に記載したプラスミドを包含する新規の酵母菌株を用いて実施した。菌株を定められた培地において、30℃で培養した。72時間後、発酵ブロス中の活性CPYの濃度を、発色基質FA-Phe-Pheを使用し、活性度を測定することにより決定した。結果は広範囲の発現レベルを示した。
【0087】
実施例8
連続発酵を、実施例6に記載のCPY活性変異体と、ヒトインスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリンとを同時発現させるための、同時発現プラスミドを包含する酵母菌株を用いて実施した。KEX2プロモーターにより、異常にプロセシングされたA14,B25ヒトインスリンのパーセンテージがかなり低下する結果となるCPYの発現レベルとなった。
【技術分野】
【0001】
本発明は、活性化カルボキシペプチダーゼの製造方法及び真菌細胞での成熟ヒトインスリン又はヒトインスリンアナログの生産のための活性化カルボキシペプチダーゼの使用方法に関する。
【背景技術】
【0002】
インスリンは、ランゲルハンス島のベータ細胞で生産されるポリペプチドホルモンである。活性なインスリン分子は、2つのジスルフィド架橋により結合しているB及びA鎖からなる2本鎖分子である。インスリンは構造B-C-Aを有する前駆体分子プロインスリンとして合成され、ここでCペプチド鎖は、B鎖のC末端アミノ酸残基をA鎖のN末端アミノ酸残基に連結させている。成熟2本鎖インスリンは、A及びB鎖の接合部に位置する塩基性アミノ酸残基対にてCペプチドをインビボで切断することにより形成される。A及びB鎖は、それぞれA7とB7、及びA20とB19Cys残基の間の2つのジスルフィド架橋により互いに保持されている。さらに、生物学的に活性なインスリン分子は、A6及びA11位にあるCys残基間に内部ジスルフィド架橋を有している。
【0003】
組換えDNA技術の発展後、遺伝的に改変された宿主細胞中でインスリン及びその前駆体を生産するための多くの方法が記載されている。大腸菌は、発現されたポリペプチドを折り畳むための細胞機構を有しておらず、成熟インスリンにおいてA及びB鎖を連結するジスルフィド架橋を確立するために、この方法は、多くのインビトロプロセシング工程、例えば再折り畳み中でのジスルフィド架橋のインビトロでの確立と、続くCペプチドの切断を含む。
【0004】
大腸菌と比較して、真核生物は、折り畳み及びジスルフィド架橋の確立に必要な機構を有しており、よって、遺伝的に改変された生物での成熟インスリンの生産のための良好な候補であると思われる。Thimら, FEBS Letters, 第212巻, 2号, 307-312には、ヒトプロインスリンの発現、及びある種の修飾Cペプチドを有する多くのインスリン前駆体が開示されている。米国特許第4914026号には、酵母α−因子リーダー配列に結合したヒトプロインスリン遺伝子を酵母宿主細胞に挿入することにより、酵母において成熟インスリンを作製し、プロインスリンが発現し成熟型で分泌させる条件下で、形質転換された酵母細胞を栄養培地中で増殖させる方法が開示されている。
【0005】
国際公開第97/03089号には、式BZAを有するインスリン前駆体の発現が開示されており、ここでB及びAは、少なくとも一のジスルフィド結合により結合したヒトインスリンのA及びBペプチド鎖であり、Zは少なくとも一のタンパク分解的切断部位を有するポリペプチドである。国際公開第90/10075号には、発酵ブロスからの最初の回収に続いて、成熟インスリン又はインスリンアナログに酵素的に転換されるインスリン及びインスリンアナログを、酵母中でのインスリン又はインスリンアナログの前駆体の発現をベースにして製造する方法が開示されている。前駆体分子はある種の修飾Cペプチドを含んでおり、インスリンB鎖のN末端伸長部をさらに含んでいてもよい。修飾Cペプチド及びBペプチドの可能なN末端伸長部は、酵母細胞中で切断されないように設計され、よって前駆体は、A及びB鎖が修飾Cペプチドにまだ結合しているが、正確に位置付けされたジスルフィド架橋は有している単鎖ペプチドとして分泌される。ついで、成熟インスリン又はインスリンアナログ産物が、Cペプチドと場合によってはN末端伸長部を切断する複数の後続するインビトロ酵素的工程により得られる。これらの酵素的工程は、時間がかかり、多くの場合はコストが高く、高価なクロマトグラフィー工程等、さらなる下流プロセス工程で除去しなければならない付加的な汚染物質が入り込むおそれがある。
【0006】
天然には分泌顆粒を形成できない遺伝子操作動物細胞において成熟インスリンを製造する方法は米国特許第6348327号に開示されている。
【0007】
本発明の目的は、プロテイナーゼA及びBとは独立して細胞内的に活性化させるカルボキシペプチダーゼを生産可能な真菌株を開発すること、さらに、高価で時間のかかる下流精製プロセス工程が回避されるように別法で活性化されたカルボキシペプチダーゼにより、十分にプロセシングされた成熟インスリン又はインスリンアナログを製造する方法を開発することにある。
【発明の概要】
【0008】
一態様では、本発明は、挿入Kex2切断部位を含むカルボキシペプチダーゼの修飾されたプロ型(プロフォーム)をコードするDNA配列を真菌細胞中で発現させることを含む活性化したカルボキシペプチダーゼを製造する方法に関し、ここでカルボキシペプチダーゼのプロ型が細胞内で切断されて、遊離の活性型カルボキシペプチダーゼが放出される。
本発明の一実施態様では、本方法は、真菌細胞から活性型のカルボキシペプチダーゼを単離する工程をさらに含む。
本発明のさらなる実施態様では、真菌細胞は、非機能性PEP4遺伝子(プロテイナーゼAをコードする遺伝子)を有する。
本発明の一実施態様では、真菌細胞は欠失したPEP4遺伝子を有する。
Kex2部位は、カルボキシペプチダーゼのプロ型の、カルボキシペプチダーゼのプロ型が効果的に切断されてカルボキシペプチダーゼの活性化成熟型を形成させる位置に挿入される。
【0009】
本発明の一実施態様では、Kex2部位は、野生型カルボキシペプチダーゼの天然N末端アミノ酸残基に直接結合している。
他の実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基の比較的近接した位置に導入される。しかしながら、Kex2部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基に対して、好ましくは約30以下のアミノ酸残基上流又は下流に存在すべきである。
【0010】
本発明の一実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基に対して、1〜約30アミノ酸残基上流又は下流の距離に導入される。
本発明の一実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基に対して、1〜約30アミノ酸残基上流の距離に導入される。
本発明のさらなる実施態様では、Kex2切断部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基に対して、2-30アミノ酸残基上流の距離に導入される。
本発明のさらなる実施態様では、Kex2切断部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基に対して、5-30アミノ酸残基上流の距離に導入される。
本発明のさらなる実施態様では、Kex2切断部位は、カルボキシペプチダーゼの天然N末端アミノ酸残基に対して、5-20アミノ酸残基上流の距離に導入される。
【0011】
他の実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の、2-30、2-29、2-28、2-27、2-26、2-25、2-24、2-23、2-22、2-21、又は2-20アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の、3-30、3-29、3-28、3-27、3-26、3-25、3-24、3-23、3-22、3-21、又は3-20アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の、4-30、4-29、4-28、4-27、4-26、4-25、4-24、4-23、4-22、4-21、又は4-20アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の、5-30、5-29、5-28、5-27、5-26、5-25、5-24、5-23、5-22、5-21、又は5-20アミノ酸残基上流又は下流の距離に導入される。
【0012】
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の1アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の2アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の3アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の4アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の5アミノ酸残基上流又は下流の距離に導入される。
【0013】
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の6アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の7アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の8アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の9アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の10アミノ酸残基上流又は下流の距離に導入される。
【0014】
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の12アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の13アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の14アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の15アミノ酸残基上流又は下流の距離に導入される。
【0015】
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の16アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の17アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の18アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の19アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の20アミノ酸残基上流又は下流の距離に導入される。
【0016】
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の21アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の22アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の23アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の24アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の25アミノ酸残基上流又は下流の距離に導入される。
【0017】
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の26アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の27アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の28アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の29アミノ酸残基上流又は下流の距離に導入される。
さらなる実施態様では、Kex2部位は、カルボキシペプチダーゼの天然N末端酸残基の30アミノ酸残基上流又は下流の距離に導入される。
【0018】
さらなる実施態様では、本発明は、上述した実施態様のいずれかの方法により生成されたカルボキシペプチダーゼと、B鎖のC末端伸長部を有するヒトインスリン又はそのアナログの前躯体とを反応させることによる、成熟ヒトインスリン又はそのアナログを製造する方法であって、C末端伸長部が切断されて、成熟ヒトインスリン又はそのアナログが得られる方法に関する。
【0019】
一態様では、本発明は、成熟ヒトインスリン又はそのアナログを製造する方法であって、真菌細胞において、
i)B鎖のC末端伸長部を有するヒトインスリン又はそのアナログの前躯体をコードするDNA配列と、
ii)そのプロ型に挿入Kex2切断部位を有するカルボキシペプチダーゼのプロ型をコードするDNAと
を同時発現させることを含み、
ヒトインスリン前駆体分子におけるB鎖のC末端伸長部を、同時発現され活性化されたカルボキシペプチダーゼにより真菌細胞内で切断し、成熟ヒトインスリン又はそのアナログを培養培地から単離する方法に関する。
カルボキシペプチダーゼは、任意の適切なカルボキシペプチダーゼであってよい。しかしながら、本発明の一実施態様では、カルボキシペプチダーゼは宿主真菌細胞に内在している。さらなる実施態様では、カルボキシペプチダーゼはCPYである。
【0020】
本方法の一実施態様では、ヒトインスリン又はそのアナログの前躯体は、ヒトインスリン又はそのアナログのB鎖、ヒトインスリン又はそのアナログのA鎖、及びB鎖とA鎖を連結するCペプチドを含有し、ここでCペプチドは少なくとも一のKex2切断部位を含有し、B鎖は真菌細胞内におけるCペプチドのより効果的なKex2切断を促進させ、さらにカルボキシペプチダーゼを用いた切断が可能なC末端伸長部を含む。
【0021】
B鎖のC末端アミノ酸残基に結合するアミノ酸配列は比較的短く、典型的には、1-4又は1-3のアミノ酸残基、特に2のアミノ酸残基を有するであろう。この配列におけるアミノ酸残基は疎水性アミノ酸残基であり、典型的にはPhe、Leu、Ile、Tyr、Trp、Val、Met及びAlaからなる群から選択されるであろう。
B鎖のC末端伸長部の非限定的例は、Leu-Gly;Leu-Ala;Leu-Leu;Leu-Met及びLeu-Ileである。
【0022】
本発明の一実施態様では、インスリン前駆体におけるCペプチドは、A鎖においてN末端アミノ酸残基に直接結合した単一のKex2部位を含み、この位置で確実に切断される。
本発明のさらなる実施態様では、Cペプチドは、2つのKex2部位の間に挿入されてペプチド配列を有する、2つのKex2切断部位を含む。2つのKex2部位の間のペプチド配列の長さ及びアミノ酸組成は、発現した単鎖インスリン前駆体の折り畳みが可能であり、インスリン前駆体分子内に、正確に位置付けされたジスルフィド架橋が確立可能であるならば、変えてもよい。
天然Cペプチドの大きさは、35アミノ酸残基である。よって本発明の一態様では、2つのKex2部位の間のペプチド配列は、天然Cペプチドとほぼ同じ長さである。
【0023】
一実施態様では、2つのKex2部位の間のペプチド配列は、1-35、1-34、1-33、1-31、1-30、1-29、1-28、1-27、1-26、1-25、1-24、1-23、1-22、1-21、1-20、1-19、1-18、1-17、1-16、1-15、1-14、1-13、1-12、1-11、1-10、1-9、1-8、1-7、1-6、1-5、1-4、1-3又は1-2アミノ酸残基長である。
【0024】
本発明の一実施態様では、ヒトインスリン前駆体は、次のアミノ酸配列:
B(1-30)-X1-X2-Z-X3-X4-A(1-21)
[上式中、B(1-30)は、ヒトインスリンのB鎖又はそのアナログであり、A(1-21)はヒトインスリンのA鎖又はそのアナログであり、X1は、真菌細胞内でのさらに効果的なKex2切断を促進するであろう、1-5のアミノ酸残基のペプチド配列であり、X2はKex2切断部位であり、Zは、1〜約35のアミノ酸残基を有するペプチド配列又はペプチド結合であり、X3はKex2切断部位又はペプチド結合であり、X4はアミノペプチダーゼ切断部位又はペプチド結合である]
を有する。
【0025】
本発明の一実施態様では、X3はKex2切断部位であり、Zはペプチド配列であり、X4はペプチド結合である。
本発明の他の実施態様では、X3及びZはペプチド結合であり、X4はアミノペプチダーゼ切断部位である。
【0026】
X1は、典型的には、1-4又は1-3のアミノ酸残基長であり、一実施態様では、X1は、2つのアミノ酸残基のペプチド配列である。X1におけるアミノ酸残基は、好ましくは疎水性アミノ酸残基であり、典型的にはPhe、Leu、Ile、Tyr、Trp、Val、Met、Ala、Asp及びGlyからなる群から選択される。
他の実施態様では、X1におけるアミノ酸残基は、Phe、Leu、Ile、Tyr、Trp、Val、Met、Ala、Asp及びGlyからなる群から選択される。
本発明の一実施態様では、X1はLeu-Alaである。
本発明の他の実施態様では、X1はPhe-Leuである。
本発明の他の実施態様では、X1はLeu-Glyである。
本発明の他の実施態様では、X1はLeu-Leuである。
本発明の他の実施態様では、X1はLeu-Metである。
本発明の他の実施態様では、X1はLeu-Ileである。
【図面の簡単な説明】
【0027】
【図1】図1はプラスミドESI42-33を示す。
【図2】図2はプラスミドpRC1を示す。
【図3】図3はプラスミドpSA160を示す。
【図4】図4は、菌株包含pSA160を24時間培養した上清のデコンボリューション質量スペクトルを示す。質量5763は、十分にプロセシングされたA14E、B25Hインスリンに相当する。
【図5】図5は、一つのKex2部位を有するインスリン前駆体の切断の概略図を示す。
【図6】図6は、一つのKex2部位とアミノペプチダーゼ部位を有するインスリン前駆体の切断を示す。
【図7】図7は、2つのKex2部位を有するインスリン前駆体の切断を示す。
【発明を実施するための形態】
【0028】
カルボキシペプチダーゼY(CPY)は、酵母サッカロミセス・セレビシエ(Saccharomyces cerevisiae)において、不活性なプロ型(プロCPY)として合成される。小胞体(ER)内におけるタンパク質の転座後、それは液胞に運搬される。到着時、それは、プロテイナーゼAにより開始されるタンパク質分解事象であるプロペプチドの除去により活性化される。2つのシナリオが、プロCPYのCPYへの効果的な転換を防止する:1)PEP4遺伝子(プロテイナーゼAをコードする遺伝子)が欠失しているか又は不活性であるならば、全てのCPYは不活性なプロ型のままであるだろう。2)PRC1(CPYをコードする遺伝子)が過剰発現しているならば、多量のプロCPYが発酵ブロスに分泌されるであろう。プロテイナーゼAが液胞に位置しており、よって分泌されたプロCPYから分離しているために、これはプロ型で残存しているであろう。これらの状況で、プロCPYを十分に活性化可能とするために、代替となる活性経路が必要とされている。
【0029】
プロペプチドと成熟したCPYとの間に界面に、プロテイナーゼAにより認識可能な配列モチーフが位置しており、よって特異的な切断に至る。切断時に、プロペプチドは成熟したCPYから分離する。本発明において、多くの変異体が作製され、ここでプロペプチドと成熟したCPYとの間の界面に近接した残基は、Kex2により認識される二塩基性切断部位が挿入される方法で変異される。これにより、Kex2pが位置するコルジ体において、プロCPYからCPYへの転換が確実になる。これで、CPYの活性化におけるプロテイナーゼAの必要性はなくなる。さらに、多量の活性化CPYを分泌可能とするために、変異した遺伝子は過剰発現している。
【0030】
以下、野生型酵素の天然N末端アミノ酸残基の31上流の位置から野生型CPYカルボキシペプチダーゼのプロ配列のアミノ酸配列、及び野生型酵素のN末端から最初の9のアミノ酸残基、及びKex2部位(KR)が、N末端アミノ酸残基に対して、種々の位置の上流又は下流に挿入されている変異体A-Gの対応する配列を示す。プロ配列が切断される切断位置を矢印で示す。
野生型:KPKFPEAIKTKKDWDFVVKNDAIENYQLRVN↓KIKDPKILG(配列番号:1)。
変異体A:KPKFPEAIKTKKDWDFVVKNDAIENYQLRVNKIKR↓DPKILG(配列番号:2)。
変異体B:KPKFPEAIKTKKDWDFVVKNDAIENYQLRVLGKR↓DPKILG(配列番号:3)。
変異体C:LGKR↓EFPEAIKTKKDWDFVVKNDAIENYQLRVNKIKDPKILG(配列番号:4)。
変異体D:KPKFPEAIKTKR↓DWDFVVKNDAIENYQLRVNKIKDPKILG(配列番号:5)。
変異体E:KPKFPEAIKTKKDWDFVKR↓NDAIENYQLRVNKIKDPKILG(配列番号:6)。
変異体F:KPKFPEAIKTKKDWDFVVKLDKR↓AIENYQLRVNKIKDPKILG(配列番号:7)。
変異体G:KPKFPEAIKTKKDWDFVVKNDAIENYQLRVNKIKDPKR↓GGILG(配列番号:8)。
【0031】
ここで使用される場合、用語「プロペプチド」又は「プロ配列」は、その機能が、折り畳み、及び発現したポリペプチドの、小胞体から、分泌経路内又は細胞外におけるその最終的な目的地までの運搬を促進させるものである、ポリペプチド配列を意味する。さらにプロペプチドは、カルボキシペプチダーゼの酵素活性を阻害する。
用語「プロ型」は、プロ配列と活性化ポリペプチド配列の融合産物を意味する。
【0032】
カルボキシペプチダーゼは、真菌細胞で発現可能であり、真菌細胞で発現した内在性Kex2p酵素により活性化可能な任意のカルボキシペプチダーゼであってよい。
カルボキシペプチダーゼの代表例は、カルボキシペプチダーゼA(EC3.4.17.1)、カルボキシペプチダーゼA2(EC3.4.17.15)、カルボキシペプチダーゼB(EC3.4.17.2)、カルボキシペプチダーゼE(EC3.4.17.10)、カルボキシペプチダーゼM(EC3.4.17.12)、カルボキシペプチダーゼT(EC3.4.17.18)、カルボキシペプチダーゼU(EC3.4.17.20)、及びカルボキシペプチダーゼY(EC3.4.16.5)である。
本発明の一実施態様では、カルボキシペプチダーゼはCPYである。
【0033】
導入されたKex2部位が、カルボキシペプチダーゼにおいてN末端アミノ酸残基に直接結合しているならば、活性型は、カルボキシペプチダーゼの天然形態であろう。
Kex2部位が、天然N末端アミノ酸に直接結合していないならば、放出されたカルボキシペプチダーゼは可変長のN末端伸長部を有するか、又はKex2部位の位置に応じて、N末端における一又は複数の天然アミノ酸残基が欠失しているであろう。
【0034】
活性化カルボキシペプチダーゼは、任意の種類の融合タンパク質のプロセシングに使用可能である。
カルボキシペプチダーゼは、それ自身のプロモーターを含む、任意の適切なプロモーターから発現してよい。しかしながら、カルボキシペプチダーゼレベルが、インスリン前駆体の異常なプロセシングに至しめる、あまりの高さになるおそれがあることがわかっている。インスリン前駆体と活性化カルボキシペプチダーゼとの間の適切な比率を発見するため、カルボキシペプチダーゼ変異体の発現レベルが、カルボキシペプチダーゼそれ自身のプロモーターを、代替プロモーターで置き換えることにより調節されてもよい。
カルボキシペプチダーゼがCPY酵素であるならば、適切な代替プロモーターは、CYC1、KEX2、MF(アルファ)1及びMPD1の遺伝子からのプロモーター領域である。
【0035】
また、カルボキシペプチダーゼのプロ型の量は、当該問題におけるプロモーターの転写因子の量を変えることにより、そのプロモーターからの発現性を調節することで調整されうる。
本発明の一実施態様では、カルボキシペプチダーゼのプロモーターはKex2プロモーターである。
【0036】
カルボキシペプチダーゼの修飾されたプロ型とヒトインスリン前駆体を、真菌細胞において同時発現させることで、さらなる高価な下流プロセシング工程を必要とすることなく、十分にプロセシングされたヒトインスリンを、発酵ブロスに分泌させることができるであろう。第1の工程として、発現したヒトインスリン前駆体を、ゴルジプロテアーゼKex1p及びKex2により、細胞内でプロセシングするであろう。ヒトインスリン前駆体は、インスリンのB鎖において、C末端アミノ酸残基に外部アミノ酸残基(X)を付加することにより修飾されているCペプチドを含む。これらの外部アミノ酸残基は、真菌細胞内において、インスリン分子のA及びB鎖からの、Cペプチドの効果的な切断を促進させるように設計されている。Kex1p及びKex2pによる完全なプロセシングにより、B鎖のC末端の伸長部Xを有する、2本鎖ヒトインスリン前駆体分子になる。前駆体は形態B-(X)n・・・Aを有し、ここでBはヒトインスリンのB鎖であり、AはヒトインスリンのA鎖であり、Xは外部アミノ酸残基であり、A及びB鎖はヒトインスリンのように、2つのジスルフィド架橋で結合している。
【0037】
ついで、このC末端で伸長している2本鎖ヒトインスリンは、B鎖C末端伸長部を切断するカルボキシプロテアーゼを同時発現及び活性化させることによる切断で、成熟ヒトインスリン又はそのアナログに転換され、成熟した2本鎖ヒトインスリン又はそのアナログが得られるであろう。
【0038】
B30位にあるアミノ酸残基に応じて、カルボキシペプチダーゼはB30アミノ酸残基を離し、desB30ヒトインスリンアナログになってもよい。さらにカルボキシペプチダーゼはB鎖のC末端から5までのアミノ酸残基が離れ、desB(29-30)-、desB28-30)-、desB(27-30)-、又はdesB(26-30)ヒトインスリンアナログになったもよい。
【0039】
インスリン前駆体の種々のプロセシングパターンを、図5-7において概略的に例証する。
図5において、インスリン前駆体コンストラクトは、単一のKex2部位、KR、及び結合ペプチドにおいて配列XXにより例証されるB鎖のC末端伸長部を有する。第1のプロセシング工程では、Kex2により、Kex2部位で切断されて、単鎖構造が、B鎖に結合した配列XXKRを有する2本鎖構造に転換される。続いて、酵素Kex1がKR配列を切断し、最終的に、カルボキシペプチダーゼがB鎖のXX伸長部を切断し、成熟した2本鎖インスリン生成物が得られる。
【0040】
図6には他の実施態様が例証されており、ここでインスリン前駆体は、単一のKex2部位及びアミノペプチダーゼ切断部位、YYを結合ペプチド中に有する。図5のように、Kex2p、Kex1p及びカルボキシペプチダーゼにより、XX-KR配列が除去されるであろう。しかしながら、この実施態様では、最後のアミノペプチダーゼ切断により、YY配列が除去されて、成熟した2本鎖インスリン生成物が得られる。
図7に例証した実施態様では、インスリン前駆体は、ペプチド鎖Zにより結合した2つのKex2切断部位を有する。Kex2を用いた第1の切断によりZ-KR配列が除去され、Kex1によりKR配列が除去され、最終的に、カルボキシペプチダーゼによりXX配列が除去される。
【0041】
真菌細胞から、成熟ヒトインスリン又はインスリンアナログが大量に生成されると、製薬目的のために、十分高純度のインスリン生成物を生成するのに必要な、下流精製工程の数をかなり減少させることができる。例えば、米国特許第4916212号に開示されている酵母においてインスリンを製造する方法では、インスリン前駆体は2つの工程、すなわち単鎖インスリン前駆体B(1-29)-Alal-Ala-Lys-A(1-21)をヒトインスリンのエステルへ転換させるペプチド転移、ついで、インスリンエステルのヒトインスリンへの加水分解において、ヒトインスリンに転換される。各転換工程は、最初の分離工程と、後続する少なくとも一の精製工程が必要である。よって、成熟インスリンの生成には、少なくとも一の酵素転換を含む、少なくとも6つの付加的な工程が必要である。
【0042】
欧州特許出願第0163529A号、PCT特許出願、国際公開第95/02059号及び国際公開第90/10075号には、酵母において、インスリン又はインスリンアナログの前駆体を発現させ、続いて、発酵ブロスからの最初の回収物を成熟インスリン又はインスリンアナログに酵素的に転換させることに基づいた、インスリン及びインスリンアナログを製造する方法が開示されている。前駆体分子は所定の修飾されたCペプチドを含有しており、インスリンB鎖のN末端伸長部をさらに含有していてもよい。修飾されたCペプチド、及び場合によってはBペプチドのN末端伸長部は、酵母細胞において切断されないように設計されており、よって前駆体は単鎖ペプチドとして分泌され、ここでA及びB鎖は修飾されたCペプチドにより結合しているが、正確に位置付けされたジスルフィド架橋は有している。ついで、成熟インスリン又はインスリンアナログ生成物は、Cペプチド、及び場合によってはN末端伸長部を切断する、複数の後続するインビトロ酵素的工程により得られる。これらの酵素的工程は、時間がかかり、多くの場合はコストが高く、高価なクロマトグラフィー工程等、さらなる下流プロセス工程で除去しなければならない付加的な汚染物質が入り込む。
【0043】
酵素的切断では、製薬用生成物の場合は効果的に除去されなければならない残存する未切断の不純物又は部分的に切断された不純物の100%切断には達しないことはよく知られている。よって、各切断工程には、少なくとも一の単離又は精製工程、典型的には交換クロマトグラフィー、ゲル濾過クロマトグラフィー、アフィニティークロマトグラフィー等によるクロマトグラフィー精製が続くであろう。
商業的規模で使用されているクロマトグラフィー用カラム材料は、非常に高価であり、よって、このようなクロマトグラフィー工程の数を減らすことは、生産経済において、かなりの影響を有するものである。下流の転換及び精製工程を減らすと、労働者の仕事量及びプロセスに費やされる時間がさらに減少し、よって生産経済がさらに改善される。
【0044】
本プロセスにおいて、成熟インスリン又はそのアナログは、培養ブロスから直接単離することができ、製薬用途に十分な純度の生成物を生成させるのに必要な下流のプロセス工程がさらに減少する。
【0045】
インスリン分子は、修飾が得られるインスリンアナログのインスリン活性に悪影響を及ぼさない限りは、A及び/又はB鎖でさらに修飾されていてもよい。
よって、ここで使用される場合、「インスリンアナログ」とは、天然インスリン中に生じる少なくとも一つのアミノ酸残基を欠失させ、及び/又は置換することにより、及び/又は少なくとも一つのアミノ酸残基を付加することによって、天然に生じるインスリン、例えばヒトインスリンの構造から形式的には誘導することができる分子構造を有するポリペプチドを意味する。付加された及び/又は置換されたアミノ酸残基は、コード可能なアミノ酸残基又は他の天然に生じるアミノ酸残基又は純粋に合成のアミノ酸残基の何れでもよい。
インスリンアナログは、ヒトインスリンと比較して、典型的には約7を越える変異、より典型的には5を越える変異、最も典型的には最大3の変異を含まない。
【0046】
長年にわたって、インスリンA及び/又はB鎖におけるかなり多数の変異が開示されている。例えば、B鎖の28位は、天然のPro残基から、Asp、Lys、又はIleに修飾されていてもよく、またB29位のLysはProに修飾されていてもよい。
また、A21位のAsnは、Ala、Gln、Glu、Gly、His、Ile、Leu、Met、Ser、Thr、Trp、Tyr又はVal、特にGly、Ala、Ser、又はThr、特にGlyに修飾されてもよい。さらに、B3位のAsnは、Lys又はAspに修飾されてもよい。インスリンアナログのさらなる具体例は、des(B30)ヒトインスリン、B1及びB2の一方又は双方が欠失したインスリンアナログ;A鎖及び/又はB鎖がN末端伸長を有するインスリンアナログ、及びA鎖及び/又はB鎖がC末端伸長を有するインスリンアナログである。またA18位にある天然アミノ酸残基はGln残基に変化していてもよく、またB26-B30位にある一又は複数のアミノ酸残基は欠失している。
【0047】
本方法で生成可能なインスリンアナログの具体例は、GlyA21ヒトインスリン、GlyA21des(B30)ヒトインスリン、desB1ヒトインスリン、des B30ヒトインスリン、AspB28ヒトインスリン、及びLysB28ProB29ヒトインスリンである。
インスリンアナログのさらなる具体例は、A21、B10、A8、A14、B25、B27及びB1の一又は複数の位置に変異を含む、ヒトインスリンアナログである。
【0048】
真菌細胞は、全ての真菌が、提供された種類のインスリン前駆体分子を切断し、結合ペプチドを切断し、2本鎖分子をライゲーションさせるのに必須のタンパク質分解活性を有するならば、任意の真菌細胞であってよい。しかしながら、長年にわたって、酵母が、インスリンサイズの小ペプチドの発現及び分泌に有効な細胞腫であることがわかっている。特に、酵母サッカロミセス・セレビシエが有用であることが判明している。
よって、本発明の一実施態様における真菌細胞は酵母細胞であり、さらなる実施態様では、酵母細胞はサッカロミセス・セレビシエである。
【0049】
本発明で使用されるDNA配列は、ゲノム又はcDNA由来のものでよく、例えばゲノム又はcDNAライブラリーを調製し、標準的な技術(例えば、Sambrook, J, Fritsch, EF及びManiatis, T, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, New York、1989)に従い、合成オリゴヌクレオチドプローブを使用するハイブリッド形成により、ポリペプチドの全体又は一部をコードするDNA配列をスクリーニングすることにより得られる。またインスリン前駆体をコードするDNA配列は、確立された標準的な方法、例えばBeaucage及びCaruthers, Tetrahedron Letters 22 (1981), 1859-1869に記載されたホスホアミジト(phosphoamidite)法、又はMatthesら, EMBO Journal 3 (1984), 801-805により記載された方法により合成的に調製されてもよい。さらに、DNA配列は、例えば米国特許第4683202号、又はSaikiら, Science 239 (1988), 487-491に記載されたようにして、特定のプライマーを使用するポリメラーゼ連鎖反応により調製されてもよい。
【0050】
DNA配列は、組換えDNA手順にかけられうる任意のベクターに挿入されてよく、多くの場合、ベクターの選択は、それが導入される宿主細胞に依存する。例えば、ベクターは自己複製可能なベクターであってよく、すなわちベクターは染色体外実体として存在しており、その複製は染色体複製、例えばプラスミドとは独立している。また、ベクターは、宿主細胞に導入される場合、宿主細胞ゲノムに組み込まれ、それが組み込まれる染色体(群)と共に複製される。
【0051】
ベクターは好ましくは発現ベクターであり、そこでインスリン前駆体をコードするDNA配列は、プロモーター等、DNAの転写に必要な付加的なセグメントに作用可能に結合する。プロモーターは宿主細胞の選択において転写活性を示す任意のDNA配列であってよく、宿主細胞と相同又は異種であるタンパク質をコードする遺伝子から誘導されてよい。
【0052】
酵母宿主細胞への使用に適したプロモーターの例には、酵母解糖遺伝子(Hitzemanら, J. Biol. Chem. 255(1980), 12073-12080; Alber及びKawasaki, J. Mol. Appl. Gen. 1(1982), 419-434、又はアルコールデヒドロゲナーゼ遺伝子(Youngら, Genetic Engineering of Microorganisms for Chemicals(Hollaenderら編), Plenum Press, New York, 1982)、又はTPI1(米国特許第4599311号)又はADH2-4c(Russellら, Nature 304(1983), 652-654)プロモーターが含まれる。
【0053】
所望の生成物をコードするDNA配列は、もし必要ならば、適切なターミネーター、ポリアデニル化シグナル、転写エンハンサー配列、及び翻訳エンハンサー配列に作用可能に結合していてもよい。さらに本発明の組換えベクターは、当該問題においては、ベクターが宿主細胞内で複製可能なDNA配列をさらに含んでよい。
【0054】
宿主細胞の分泌経路にインスリンを方向付けるために、分泌シグナル配列(リーダー配列、プレプロ配列、又はプレ配列としても公知)は、組換えベクター内に提供されてもよい。分泌シグナル配列は正確なリーディングフレームにおいて、インスリン前駆体をコードするDNA配列に結合している。分泌シグナル配列は、通常、ペプチドをコードするDNA配列に対し、5'に位置している。シグナルペプチドは、天然に生じるシグナルペプチド、又はその機能的部分であってよく、又は合成ペプチドであってもよい。
酵母における効果的な分泌のために、リーダーペプチドをコードする配列は、シグナル配列の下流と所望の生成物をコードするDNA配列の上流に挿入されてよい。
【0055】
異種DNAで酵母細胞を形質転換させ、異種ポリペプチドを生成させる方法は、例えば米国特許第4599311号、米国特許第4931373号、同第4870008号、同第5037743号、及び同第4845075号に記載されている。形質転換細胞は、選択可能マーカーにより決定されたフェノタイプ、一般的には薬剤耐性、又は特定の栄養素、例えばロイシンの不在下で成長する能力によって選択される。酵母に使用される好ましいベクターは、米国特許第4931373号に記載のPOT1ベクターである。
【0056】
発酵プロセスにおいて細胞培養に使用される培地は、宿主細胞の増殖に適した任意の従来からの培地、例えば適切なサプリメントを含有する最小又は複合培地であってよい。適切な培地は商業的供給者から入手可能であり、又は公開されているレシピ(例えば、American Type Culture Collectionのカタログ)に従い調製されてもよい。例えば、培地は、少なくとも一の炭素源、一又は複数の窒素源、カリウム、ナトリウム、マグネシウムの塩、リン酸塩、硝酸塩及び硫酸塩を含む必須の塩、微量金属、水溶性ビタミン類、限定されるものではないが、プロセス補助剤、例えばプロテアーゼインヒビター、安定剤、リガンド、消泡剤及び誘導因子を含有する。培地は、熱による滅菌を含む、いくつかの操作条件において、液体培地に部分的に沈殿又は分散する成分を含有していてもよい。培地は、いくつかの液体とガス状溶液とを混合することにより作製可能である。これらの溶液は発酵タンクに入れる前に混合することもできるし、所定の比率で添加された別々の液体流として、発酵タンクに供給される。培地成分の異なる液体溶液の比率は、発酵プロセスの様々な段階で変化し、これは培地の全ての成分が、発酵の経過によって変化し得ることを意味する。
【0057】
ついで、細胞により生成されるペプチドは、遠心分離又は濾過、硫酸アンモニウム等の塩により上清又は濾液のタンパク質様成分を沈殿させることによって、培地から宿主細胞を分離させ、当該分野のタンパク質の種類に応じて、イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー、アフィニティークロマトグラフィー等の多様なクロマトグラフィー手順により精製することを含む従来からの手順により、培養培地から回収されてよい。
【0058】
培養ブロスから単離させた後、成熟インスリン又はインスリンアナログを、特にB29Lys残基のε-アミノ基をアシル化することにより、例えばアシル化形態に転換させてもよい。インスリンをアシル化させる方法は当該技術でよく知られており、例えば欧州特許第792290号及び同第894095号、米国特許第5693609号、同第5646242号、同第5922675号、同第5750497号及び同第6011007号に開示されている。
【0059】
アシル化されたインスリンの例は、NεB29-テトラデカノイルdes(B30)ヒトインスリン、NεB29-リトコロリル-γ-グルタミルdes(B30)ヒトインスリン、NεB29-(Nα-(HOOC(CH2)14CO)-γ-Glu)des(B30)ヒトインスリン、又はNεB29-(Nα-(HOOC(CH2)16CO)-γ-Glu)des(B30)ヒトインスリンである。
【0060】
「desB30」又は「B(1-29)」とは、B30アミノ酸残基を欠く天然インスリンB鎖又はそのアナログを意味する。
B(1-30)はヒトインスリンの天然B鎖を意味し、「A(1-21)」は天然インスリンA鎖を意味する。A18Qヒトインスリンは、ヒトインスリンA鎖のA18位にGlnを有するインスリンアナログである。B10E、A8H、A14Eは、それぞれB10位にGlu、A8位にHis、及びA14位にGluを有するインスリンアナログである。
「B1」、「A1」等とは、それぞれインスリンのB鎖の1位(N末端から数える)のアミノ酸残基及びインスリンのA鎖の1位(N末端から数える)のアミノ酸残基を意味する。特定の位置にあるアミノ酸残基は、例えばB1位のアミノ酸残基がフェニルアラニン残基であることを意味するPheB1として表されてもよい。
【0061】
「Cペプチド」とは、インスリン分子のA及びBペプチド鎖を互いに結合させるペプチド配列を意味する。
「成熟インスリン」とは、正しいアミノ酸残基組成と、天然のヒトインスリン分子と同様の立体構造、すなわちCysA7とCysB7との間、及びCysA20とCysB19との間のジスルフィド架橋、及びCysA6とCysA11との間の内部ジスルフィド架橋、及びインスリン活性を有する2本鎖インスリンを意味する。よって、本発明の成熟インスリンはヒトインスリンである。成熟ヒトインスリンアナログは、B28Aspヒトインスリン、desB30ヒトインスリン、A14Glu、B25Hisヒトインスリン、及びB31Leu、B32Alaヒトインスリンであってよい。
ここで使用される場合、「インスリン誘導体」とは、天然に生じるインスリン、又は、例えばインスリン骨格の一又は複数の位置に側鎖が導入されることにより、もしくはインスリンのアミノ酸残基の基を酸化又は還元することにより、もしくは遊離のアミノ基又はヒドロキシ基をアシル化することにより化学的に修飾されたインスリンアナログを意味する。
【0062】
「Kex2」又は「Kex2p」とは、2つの塩基性残基(リジン又はアルギニン)の配列の後に、好ましくは切断を触媒するサブチリシン様エンドプロテアーゼを意味する(Rockwell, NC, Krysan、DJ、Komiyama, T & Fuller, RS 2002 Precursor Processing by Kex2/Furin Proteases. Chem. Rev. 102: 4525-4548)。
「Kex1」又は「Kex1p」とは、好ましくはC末端のリジル及び/又はアルギニル残基の除去を触媒するセリンカルボキシペプチダーゼを意味する(Shilton BH, Thomas DY, Cygler M 1997 Crystal structure of Kex1deltap, a prohormone-processing carboxypeptidase from Saccharomyces cerevisiae. Biochemistry 36: 9002-9012)。
CPYとは、好ましくは疎水性又は大きなC末端アミノ酸残基、例えばPhe及びLeuの除去を触媒するカルボキシペプチダーゼである、カルボキシペプチダーゼYを意味する(Remington, S.J. & Breddam, K. (1994) Carboxypeptidases C and D. Methods Enzymol. 244、231-248)。CPYのアミノ酸配列は、「Vallsら, 1987, Cell, 48(5):887-897」に開示されている。
【0063】
「正確にプロセシング」とは、正確なアミノ酸残基配列を有する所望の生成物が得られるように、所望の切断点で酵素的に切断することを意味する。
「効果的な切断」とは、少なくとも80%、好ましくは少なくとも85%、さらに好ましくは少なくとも95%の切断を意味することを意図している。
「POT」は、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)トリオースリン酸イソメラーゼ遺伝子であり、「TPI1」はサッカロミセス・セレビシエ・トリオースリン酸イソメラーゼ遺伝子である。
「リーダー」とは、プレペプチド(シグナルペプチド)及びプロペプチドからなるアミノ酸配列を意味する。
【0064】
「シグナルペプチド」なる用語は、タンパク質の前駆体形態において、N末端配列として存在するプレペプチドを意味すると理解される。シグナルペプチドの機能は、異種タンパク質の小胞体への転座を促進できることである。シグナルペプチドはこのプロセス中に通常は切断される。シグナルペプチドはタンパク質を生成する宿主生物体に対して異種又は同族であってもよい。
酵母宿主細胞に有用なシグナルペプチドは、サッカロミセス・セレビシエのα因子及びサッカロミセス・セレビシエのインベルターゼに対する遺伝子から得られる。本発明のDNAコンストラクトと共に使用され得る多くのシグナルペプチドには、酵母アスパラギン酸プロテアーゼ3(YAP3)シグナルペプチド、又は任意の機能的アナログ(Egel-Mitaniら、(1990) YEAST 6:127-137及び米国特許第5726038号)、MFα1遺伝子のα-因子シグナル(Thorner(1981), The Molecular Biology of the Yeast Saccharomyces cerevisiae, Strathern等編, pp 143-180, Cold Spring Harbor Laboratory, NY、及び米国特許第4870008号)、マウス唾液アミラーゼのシグナルペプチド(O. Hagenbuchleら, Nature 289、1981, pp. 643-646を参照)、修飾されたカルボキシペプチダーゼのシグナルペプチド(L.A. Vallsら, Cell 48,1987, pp. 887-897を参照)、及び酵母BAR1シグナルペプチド(国際公開第87/02670号を参照)が含まれる。
【0065】
本発明は、選択された微生物又は宿主細胞中で複製可能であり、本発明のインスリン生成物をコードするポリヌクレオチド配列を担持するベクターを包含する。組換えベクターは、自己複製ベクター、すなわち染色体外実体として存在するベクター、染色体複製とは独立した複製、例えばプラスミド、染色体外エレメント、ミニ染色体、又は人工染色体でありうる。ベクターは自己複製を保証する任意の手段を含みうる。あるいは、ベクターは、宿主細胞に導入されたときに、ゲノムに組み込まれ、それが組み込まれた染色体と共に複製されるものでありうる。さらに、単一のベクター又はプラスミド又は宿主細胞のゲノムに導入される全DNAを合わせて含む2以上のベクター又はプラスミド、あるいはトランスポゾンが使用されうる。ベクターは直鎖又は閉環プラスミドであってよく、宿主細胞のゲノムへのベクターの安定した組込、又はゲノムとは独立した細胞におけるベクターの自己複製を可能にするエレメント(類)を好ましくは含む。
一実施態様では、組換え発現ベクターは酵母中で複製可能である。ベクターを酵母中で複製させることができる配列の例は酵母プラスミド2μm複製遺伝子REP1-3及び複製起点である。
【0066】
またベクターは、選択可能なマーカー、例えば遺伝子、宿主細胞における欠失を補完する生成物、又は例えばアンピシリン、カナマイシン、テトラサイクリン、クロラムフェニコール、ネオマイシン、ハイグロマイシン、又はメトトレキサート等の薬剤に対する耐性を付与するものをさらに含んでもよい。
酵母宿主細胞のための適切なマーカーは、ADE2、HIS3、LEU2、LYS2、MET3、TRP1、及びURA3である。酵母とって好ましい選択可能なマーカーはシゾサッカロミセス・ポンベTPI遺伝子(Russell (1985) Gene 40:125-130)である。
酵母宿主において、有用なプロモーターは、サッカロミセス・セレビシエMFα1、TPI、ADH又はPGKプロモーターである。
本発明のポリヌクレオチドコンストラクトは典型的には適切なターミネーターに作用可能に結合しているであろう。酵母において、適切なターミネーターはTPIターミネーター(Alberら(1982) J. Mol. Appl. Genet. 1:419-434)である。
【0067】
インスリン前駆体をコードするDNA配列、プロモーター、及び場合によってはターミネーター、及び/又は分泌シグナル配列をそれぞれライゲーションし、複製に必要な情報を含む適切なベクターにそれらを挿入するのに使用される手順は、当業者によく知られている(例えば、Sambrookら, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, New York, 1989を参照)。
ベクターは、まず本発明のインスリン前駆体をコードする全DNA配列を含有するDNAコンストラクトを調製し、続いて適切な発現ベクターにこのフラグメントを挿入するか、又は個々のエレメントについての遺伝的情報を含むDNAフラグメントを挿入し、続いてライゲーションすることにより構築されると理解されるであろう。
【0068】
本発明で使用される宿主細胞は真菌細胞である。ここで使用される場合、「真菌」には、子嚢菌門、担子菌門、ツボカビ門、及び接合菌門(Hawksworthらによる定義, Ainsworth and Bisby's Dictionary of The Fungi, 8th edition, 1995, CAB International, University Press, Cambridge, UK)、並びに卵菌門(Hawksworth, 1995, 上掲, 171頁を引用)及び全ての栄養胞子形成菌(Hawksworthら, 1995, 上掲)が含まれる。
【0069】
一実施態様では、真菌宿主細胞は酵母細胞である。ここで使用される「酵母」には、有子嚢胞子(ascosporogenous)酵母(エンドマイセス属)、担子胞子形成(basidiosporogenous)酵母、及び不完全菌類に属する酵母(ブラストミセス属)が含まれる。有子嚢胞子酵母はスペルモフトラ科(Spermophthoraceae)とサッカロミセス科(Saccharomycetaceae)に分けられる。後者は、4つの亜科、すなわちスキゾサッカロミセス(Schizosaccharomycoideae)亜科(例えばスキゾサッカロミセス(Schizosaccharomyces) 属) 、ナドソニオイデ(Nadsonioideae)亜科 、リポミコイデー(Lipomycoideae) 亜科、及びサッカロミセス(Saccharomycoideae) 亜科(例えばピキア(Pichia)、クルイベロミセス(Kluyveromyces) 及びサッカロミセス(Saccharomyces) 属)からなる。担子胞子形成酵母には、ロイコスポリジウム(Leucosporidim)属、ロドスポリジウム(Rhodosporidium)属、スポリジオボラス(Sporidiovolus)属、フィロバシジウム(Filobasidium)属、及びフィロバシジエラ(Filobasidiella)属が含まれる。不完全菌類に属する酵母は、2つの科、つまりスポロボロミセス(Sporobolomycetaceae) 科(例えばスポロボロミセス(Sorobolomyces) 属及びブレラ(Bullera) 属) 及びクリプトコッカス(Cryptococcaceae) 科(例えばカンジダ(Candida) 属)に分けられる。酵母の分類は今後変更されることがあり得るので、本発明の目的のためには、酵母はBiology and Activities of Yeast (Skinner, F.A. 、Passmore, S.M.及びDavenport, R.R.編、Soc. App. Bacteriol. Symposium Series No.9, 1980)に記載されたように定義される。酵母の生物学及び酵母遺伝子の操作は、当該技術分野においてよく知られている(例えば、Biochemistry and Genetics of Yeast, Bacil, M., Horecker, B.J., 及びStopani, A.O.M.編, 第2版, 1987;The Yeasts, Rose, A.H., 及びHarrison, J.S.編, 第2版, 1987;及びThe Molecular Biology of the Yeast Saccharomyces, Strathernら編, 1981を参照)。
【0070】
酵母宿主細胞は、カンジダ、クルイベロミセス、サッカロミセス、スキゾサッカロミセス、ピキア、ハンゼヌラ又はヤロウィア(Yarrowia)属の種の細胞から選択されうる。一実施態様では、酵母宿主細胞は、サッカロミセス・カールスバージェンシス(Saccharomyces carlsbergensis)、サッカロミセス・セレビシエ、サッカロミセス・ディアスタティカス(Saccharomyces diastaticus)、サッカロミセス・ダグラシ(Saccharomyces douglasii)、サッカロミセス・クルイベリ(Saccharomyces kluyveri)、サッカロミセス・ノルベンシス(Saccharomyces norbensis)、サッカロミセス・オビフォルミス(Saccharomyces oviformis)、シゾサッカロミセス・ポンベ、 サッカロミセス・ウバルム(Sacchoromyces uvarum)、ピキア・クルイベリ(Pichia kluyveri)、ヤロイア・リポリチカ(Yarrowia lipolytica)、トルラ酵母(Candida utilis)、カンジダ・カカオイ(Candida cacaoi)、及びゲオトリクム・ファーメンタンス(Geotrichum fermentans)である。他の有用な酵母宿主細胞は、クルイベロミセス・ラクチス(Kluyveromyces lactis)、クルイベロミセス・フラジリス(Kluyveromyces fragilis)、ハンセヌラ・ポリモルファ(Hansenula polymorpha)、ピキア・パストリス(Pichia pastoris)、ヤロイア・リポリチカ(Yarrowia lipolytica)、シゾサッカロミセス・ポンベ、トウモロコシ黒穂病菌(Ustilgo maylis)、カンジダ・マルトース(Candida maltose)、ピキア・ギリエルモンデイ(Pichia guillermondii)及びピキア・メタノリオール(Pichia methanoliol)である(Gleesonら, J. Gen. Microbiol. 132, 1986, pp.3459-3465;米国特許第4882279号及び同第4879231号を参照)。
【0071】
「コード可能なアミノ酸」又は「コード可能なアミノ酸残基」なる表現は、ヌクレオチドのトリプレット(「コドン」) によりコード可能なアミノ酸又はアミノ酸残基を示すために使用される。
本記載において、アミノ酸の3文字又は1文字表記は、それらの従来からの意味で使用される。明確に示されない限りは、ここに挙げたアミノ酸はLアミノ酸である。さらにペプチドのアミノ酸配列の左及び右端は、特に記載しない限りは、それぞれN及びC末端である。
【0072】
ここに引用された刊行物、特許出願及び特許を含む全ての文献は、各文献が、出典明示により個々にかつ特に援用され、その全内容がここに説明されるかの如く、その全体が出典明示によりここに援用される(法律により許容される最大範囲)。
全ての表題及び副題は、ここでは便宜的に使用され、決して本発明を限定するものと解してはならない。
ここに提供される任意かつ全ての例、又は例示的言語(例えば「等」)の使用は、単に本発明をより明らかにすることを意図しており、特に請求項に記載がない限り、本発明の範囲に限定をもたらすものではない。明細書中の如何なる語句も請求項に記載していない要素が本発明の実施に必須であることを示しているものと解してはならない。
ここでの特許文献の引用及び援用は単に便宜上なされているもので、そのような特許文献の有効性、特許性、及び/又は権利行使性についての見解を反映させるものではない。 この発明は、適用される法律に容認される場合、ここに付加される請求項に列挙された主題事項の全ての修正点及び等価物を含む。
【実施例】
【0073】
一般的手順
全ての発現プラスミドは、欧州特許第171,142号に記載されているものと同様の、C-POT型のものである。これらは、プラスミドの選択、及びサッカロミセス・セレビシエの安定化の目的のために、シゾサッカロミセス・ポンベ・トリオースリン酸イソメラーゼ遺伝子(POT)を含有することにより特徴付けられる2μベースの発現ベクターである。また、プラスミドは、サッカロミセス・セレビシエ・トリオースリン酸イソメラーゼプロモーター及びターミネーター(図1)を含む。これらの配列は、次の:1)インスリン生成物とリーダーの融合タンパク質をコードするEcoRI-Xbalフラグメントの配列、及び2)発現ベクターの2μ領域におけるNcoI部位の除去に帰するサイレント変異の導入、を除き、全ての配列である場合、プラスミドpKFN1003(国際公開第9010075号に記載)における対応配列に類似している。種々の融合タンパク質のクローニングを促進させるために、MFα1プレプロリーダーをコードするDNA配列を変化させてNcoI部位を導入し(図2を参照)、MFα1*プレプロリーダーと称する。このようにNcoI-Xbalフラグメントは、関心あるインスリンコンストラクトをコードするNcoI-Xbalフラグメントで、簡単に置き換えられる。このようなNcoI-Xbalフラグメントは、標準的な技術に従い、合成オリゴヌクレオチド及びPCRを使用して合成されてよい。アルファ-リーダーに加えて、他のリーダーを使用することもできる。
【0074】
酵母形質転換を、宿主菌株であるサッカロミセス・セレビシエ株MT663又はME1719の形質転換により調製した。酵母菌株MT663(MATa/MATα pep4-3/pep4-3 HIS4/his4 Δtpi::LEU2/Δtpi::LEU2 Cir’)は、国際公開第92l11378号に関連して、Deutsche Sammlung von Mikroorganismen und Zellkulturenに寄託されており、寄託番号DSM6278が付与されている。サッカロミセス・セレビシエ菌株ME1719(MATa/α leu2/leu2 pep4-3/pep4-3 Δtpi::LEU2/Δtpi::LEU2 Δura3/Δura3 Δyps1::URA3/Δyps1::ura3 Cir+)は国際公開第98/01535号に記載されている。
【0075】
MT663又はME1719を、600nmでの光学密度が0.6になるまで、YPGaL(1%のBacto酵母抽出物、2%のBactoペプトン、2%のガラクトース、1%のラクタート)において増殖させた。遠心分離により100mlの培養体を収集し、10mlの水で洗浄し、再度遠心分離にかけ、1.2Mのソルビトール、25mMのNa2EDTA、pH=8.0及び6.7mg/mlのジチオトレイトールを含有する10mlの溶液に再懸濁させた。懸濁液を30℃で15分インキュベートし、遠心分離にかけ、細胞を、1.2Mのソルビトール、10mMのNa2EDTA、0.1Mのクエン酸ナトリウム、pH05.8、及び2mgのNovozymC3234を含有する10mlの溶液に再懸濁させた。懸濁液を30℃で30分インキュベートし、遠心分離により細胞を収集し、10mlの1.2Mソルビトールと10mlのCAS(1.2Mのソルビトール、10mMのCaCl2、10mMのトリスHCI(トリス=トリス(ヒドロキシメチル1)-アミノメタン)pH=7.5)で洗浄し、2mlのCASに再懸濁させた。形質転換のために、1mlのCAS懸濁細胞を、約0.1mgのプラスミドDNAと混合し、室温で15分放置した。1mlの(20%のポリエチレングリコール4000、10mMのCaCl2、10mMのトリスHCI、pH=7.5)を添加し、混合物を室温でさらに30分放置した。混合物を遠心分離にかけ、ペレットを0.1mlのSOS(1.2Mのソルビトール、33%vlvのYPD、6.7mMのCaCl2)に再懸濁させ、30℃で2時間インキュベートした。ついで、懸濁液を遠心分離にかけ、ペレットを0.5mlの1.2Mソルビトールに再懸濁させた。ついで、6mlの上層寒天(1.2Mのソルビトールと2.5%の寒天を含有する、Shermanら (1982) Methods in Yeast Genetics, Cold Spring Harbor LaboratoryのSC培地)を52℃で添加し、同様の寒天固形化されたソルビトール含有培地を含むプレートの頂部に懸濁液を注いだ。
【0076】
実施例1
ヒトインスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリン用の酵母発現システムの構築
図1には、pESI42-33と称される酵母プラスミドが示されている。プラスミドは、サッカロミセス・セレビシエTPI遺伝子の転写プロモーターと転写ターミネーターとの間のプラスミドに挿入されたEcoRI-XbaIフラグメントを含有する発現カセットを含む。プラスミドpESI42-33において、EcoRI-XbaIフラグメントは、MFα1*プレプロリーダー、二塩基プロセシングエンドペプチダーゼKex2に対するLys-Arg切断部位、及びインスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリンからなる融合産物をコードする。
【0077】
インスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリンをコードする配列を含有するDNAフラグメントを、合成オリゴヌクレオチド及び標準的なPCR増幅を使用して構築した。得られたPCRフラグメントを精製し、NcoI及びXbaIで消化し、修飾されたcPOT型の発現ベクターのNcoI-XbaIベクターフラグメントにライゲーションさせた(図1)。
発現プラスミドを、アンピシリンの存在下で成長させた大腸菌にて増殖させ、標準的な技術を使用し単離させた(Sambrookら, 1989)。プラスミドDNAを適切な制限ヌクレアーゼにより挿入についてチェックし(例えば、EcoRI、NcoI、XbaI)、配列分析により、インスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリンの適切な配列を含有することが示された。
プラスミドをサッカロミセス・セレビシエ菌株MT663において形質転換させた。プラスミドを包含する酵母形質転換体を、YPD(1%の酵母抽出物、2%のペプトン、2%のグルコース)寒天(2%)プレートにおける炭素源としてのグルコース利用によって選択した。
【0078】
実施例2
CPY活性変異体用の酵母発現システムの構築
図2には、pPRC1と称される酵素プラスミドが示されている。プラスミドは、サッカロミセス・セレビシエTPI遺伝子の転写プロモーターと転写ターミネーターとの間のプラスミドに挿入されたClaI-NheIフラグメントを含有する発現カセットを含む。プラスミドpPRC1、ClaI-NheIフラグメントは、野生型プレプロCPYをコードする。全てのCPY変異プラスミドは、関連点変異が、二塩基性Kex2切断モチーフの導入用に作製されていることを除けば、pPRC1と同一である。
Kex2切断部位の挿入により変異したPRC1をコードする配列を含有するDNAフラグメントを、合成オリゴヌクレオチド、テンプレートとしてゲノム酵母DNA、及び標準的なPCR増幅を使用して構築した。得られたPCRフラグメントを精製し、ClaI及びNheIで消化し、修飾されたcPOT型の発現ベクターのClaI-NheIベクターフラグメントにライゲーションさせた。
【0079】
発現プラスミドを、アンピシリンの存在下で成長させた大腸菌にて増殖させ、標準的な技術を使用し単離させた(Sambrookら, 1989)。プラスミドDNAを適切な制限ヌクレアーゼにより挿入についてチェックし、配列分析により、PRC1の適切な配列を含有することが示された。
プラスミドをサッカロミセス・セレビシエ菌株MT663において形質転換させた。プラスミドを包含する酵母形質転換体を、YPD(1%の酵母抽出物、2%のペプトン、2%のグルコース)寒天(2%)プレートにおける炭素源としてのグルコース利用によって選択した。
【0080】
実施例3
インスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリン及びCPY活性変異体の同時発現用の酵母発現システムの構築
図3には、pSA160と称される酵母プラスミドが示されている。プラスミドは、サッカロミセス・セレビシエTPI遺伝子の転写プロモーターと転写ターミネーターとの間のプラスミドに挿入されたEcoRI-XbaIフラグメントを含有する発現カセットを含む。プラスミドpSA160において、EcoRI-XbaIフラグメントは、MFα1*プレプロリーダー、二塩基プロセシングエンドペプチダーゼKex2に対するLys-Arg切断部位、及びインスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリンからなる融合産物をコードする。またプラスミドは、サッカロミセス・セレビシエPRC1遺伝子の転写プロモーターの後のプラスミド中に挿入されるKpnI-SacIフラグメントを含むCPY変異体「F」をコードする発現カセットも含む。
インスリンカセットを、適切なcPOT型の発現ベクターにおいてサブクローニングした。続いて、PRC1プロモーターを、テンプレートとしてゲノム酵母DNAを使用するPRCにより増幅し、SalI-KpnIフラグメントとして、上述したプラスミドにおいてクローニングした。最終的に、ターミネータープラスCPY変異体をコードするDNAを、PCRを使用して増幅させ、プラスミドにおいてクローニングした。
【0081】
発現プラスミドを、アンピシリンの存在下で成長させた大腸菌にて増殖させ、標準的な技術を使用し単離させた(Sambrookら, 1989)。プラスミドDNAを適切な制限ヌクレアーゼにより挿入についてチェックし、配列分析により、PRC1オープンリーディングフレームの適切な配列を含有することが示された。
プラスミドをサッカロミセス・セレビシエ菌株MT663において形質転換させた。プラスミドを包含する酵母形質転換体を、YPD(1%の酵母抽出物、2%のペプトン、2%のグルコース)寒天(2%)プレートにおける炭素源としてのグルコース利用によって選択した。
【0082】
実施例4
複数のCPY変異体を構築した。プラスミドを酵母において形質転換させ、研究室規模のバッチ発酵を、30℃で72時間、定められた培地における培養により実施した。合成基質N-(3-[2-フリル]アクリロイル)-Phe-Pheを使用し、CPY活性について発酵ブロスをアッセイした。この基質を使用すると、337nmでの吸光度における変化として、CPY活性を分光学的に追跡することができる。既知の濃度のCPY標準体の活性度に対して、活性度を相互比較することにより、上清における濃度を推定することができる。これらのアッセイに基づき、9つの変異体が、0-2.3mg/Lの濃度で存在することが推定された。
【0083】
プロセシングの度合いを調査するために、発酵上清をウエスタンブロット分析にかけ、CPY抗体で探索した。プロセシングされていないCPY(プロCPY)とプロセシングされたCPYとの間の比率を、ブロットから推定した。これにより、ほとんどの変異体において、成熟したCPYへの〜80%の転換率が示された。
【0084】
実施例5
ヒトインスリン前駆体(B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリンとCPY活性変異体「F」との同時発現用のプラスミドpSA160を、酵母において形質転換させ、研究室規模のバッチ発酵を、30℃で定められた培地における培養により実施した。インスリン種の培養24時間後、発酵ブロスをLC-MSにより分析した。これにより、分泌されたインスリン種の>90%が、十分にプロセシングされたA14E,B25Hヒトインスリンアナログであったことが示された。図4には、分泌された活性CPYにより、インスリンB鎖のC末端から、LA伸長部が成功裏に除去されたことが示されている。
【0085】
実施例6
図3に記載された同時発現プラスミドを包含する酵素菌株の連続発酵により、CPYレベルがあまりに高まり、ヒトインスリン前駆体の異常なプロセシングに至ることが示された。ヒトインスリン前駆体と活性CPYとの間の適切な比率を見出すために、PRC1プロモーターを代替プロモーターに置き換えることにより、CPY変異体の発現レベルを調節した。
CYC1、KEX2、MF(アルファ)1、及びMPD1の遺伝子からのプロモーター領域をPCRにより増幅させ、PRC1プロモーター配列を、それぞれ代替CYC1、KEX2、MF(アルファ)1、及びMPD1のプロモーター配列と交換することで得られた、実施例3に記載の同時発現プラスミドのSalI−KpnI部位においてクローニングした。得られたプラスミドをサッカロミセス・セレビシエ菌株MT663において形質転換させた。プラスミドを包含する酵母形質転換体を、YPD(1%の酵母抽出物、2%のペプトン、2%のグルコース)寒天(2%)プレートにおける炭素源としてのグルコース利用によって選択した。
【0086】
実施例7
研究室規模のバッチ発酵を、実施例6に記載したプラスミドを包含する新規の酵母菌株を用いて実施した。菌株を定められた培地において、30℃で培養した。72時間後、発酵ブロス中の活性CPYの濃度を、発色基質FA-Phe-Pheを使用し、活性度を測定することにより決定した。結果は広範囲の発現レベルを示した。
【0087】
実施例8
連続発酵を、実施例6に記載のCPY活性変異体と、ヒトインスリン前駆体B(1-30)-LARRDLGKR(配列番号:9)-(A1-21)、(A14E,B25H)ヒトインスリンとを同時発現させるための、同時発現プラスミドを包含する酵母菌株を用いて実施した。KEX2プロモーターにより、異常にプロセシングされたA14,B25ヒトインスリンのパーセンテージがかなり低下する結果となるCPYの発現レベルとなった。
【特許請求の範囲】
【請求項1】
挿入Kex2切断部位を含むカルボキシペプチダーゼの修飾されたプロ型をコードするDNA配列を、カルボキシペプチダーゼの修飾されたプロ型の発現に適した条件下で発現させ、プロ配列を細胞内で切断し、遊離の活性型のカルボキシペプチダーゼを放出させる、真菌細胞における活性化カルボキシペプチダーゼの製造方法。
【請求項2】
真菌細胞から活性型のカルボキシペプチダーゼを単離する工程をさらに含む請求項1に記載の方法。
【請求項3】
真菌細胞が、非機能性PEP4遺伝子(プロテイナーゼAをコードする遺伝子)を有する請求項1又は2に記載の方法。
【請求項4】
カルボキシペプチダーゼが宿主真菌細胞に内在している請求項1ないし3のいずれか一項に記載の方法。
【請求項5】
Kex2部位が、カルボキシペプチダーゼのプロ型の、野生型カルボキシペプチダーゼの天然N末端アミノ酸残基に対して0〜約30のアミノ酸残基上流又は下流の位置に、挿入されている請求項1ないし4のいずれか一項に記載の方法。
【請求項6】
Kex2切断部位が、プロ配列の、カルボキシペプチダーゼの野生型形態の天然N末端アミノ酸残基に対して5-20、5-15又は5-10アミノ酸残基上流の距離に、導入される請求項5に記載の方法。
【請求項7】
Kex2切断部位が、野生型カルボキシペプチダーゼ酵素のN末端アミノ酸残基に対して2-20、2-15又は2-10アミノ酸残基上流の位置に挿入される請求項5に記載の方法。
【請求項8】
カルボキシペプチダーゼがCPYである請求項1ないし7のいずれか一項に記載の方法。
【請求項9】
B鎖のC末端伸長部を有するヒトインスリン又はそのアナログの前躯体を、請求項1ないし8のいずれか一項に記載の方法により製造されたカルボキシペプチダーゼと反応させることによる、成熟ヒトインスリン又はそのアナログを製造する方法であって、B鎖のC末端伸長部が切断されて、成熟ヒトインスリン又はそのアナログが得られる方法。
【請求項10】
成熟ヒトインスリン又はそのアナログを製造する方法であって、真菌細胞において、
i)B鎖のC末端伸長部を有するヒトインスリン又はそのアナログの前躯体をコードするDNA配列と、
ii)そのプロ型に挿入Kex2切断部位を有するカルボキシペプチダーゼのプロ型をコードするDNAと
を同時発現させることを含み、
ヒトインスリン前駆体分子におけるB鎖のC末端伸長部を、同時発現され活性化されたカルボキシペプチダーゼにより真菌細胞内で切断し、成熟ヒトインスリン又はそのアナログを培養培地から単離する方法。
【請求項11】
ヒトインスリン前駆体分子が、ヒトインスリン又はそのアナログのB鎖、ヒトインスリン又はそのアナログのA鎖、及びB鎖とA鎖を連結するCペプチドを含み、Cペプチドが少なくとも一のKex2切断部位を含み、B鎖が、真菌細胞内におけるCペプチドのより効果的なKex2切断を促進させるC末端伸長部を含む請求項10に記載の方法。
【請求項12】
B鎖のC末端伸長部が4までのアミノ酸残基を有する請求項9ないし11のいずれか一項に記載の方法。
【請求項13】
B鎖のC末端伸長部が、Leu-Ala、Phe-Leu、Leu-Gly、Leu-Leu、Leu-Met及びLeu-Ileからなる群から選択される請求項12に記載の方法。
【請求項14】
カルボキシペプチダーゼのプロ型が、それ自身のプロモーターとは異なるプロモーターの調節下で発現される請求項10ないし13のいずれか一項に記載の方法。
【請求項15】
カルボキシペプチダーゼが酵母CPY酵素であり、プロモーターがKex2プロモーターである請求項14に記載の方法。
【請求項1】
挿入Kex2切断部位を含むカルボキシペプチダーゼの修飾されたプロ型をコードするDNA配列を、カルボキシペプチダーゼの修飾されたプロ型の発現に適した条件下で発現させ、プロ配列を細胞内で切断し、遊離の活性型のカルボキシペプチダーゼを放出させる、真菌細胞における活性化カルボキシペプチダーゼの製造方法。
【請求項2】
真菌細胞から活性型のカルボキシペプチダーゼを単離する工程をさらに含む請求項1に記載の方法。
【請求項3】
真菌細胞が、非機能性PEP4遺伝子(プロテイナーゼAをコードする遺伝子)を有する請求項1又は2に記載の方法。
【請求項4】
カルボキシペプチダーゼが宿主真菌細胞に内在している請求項1ないし3のいずれか一項に記載の方法。
【請求項5】
Kex2部位が、カルボキシペプチダーゼのプロ型の、野生型カルボキシペプチダーゼの天然N末端アミノ酸残基に対して0〜約30のアミノ酸残基上流又は下流の位置に、挿入されている請求項1ないし4のいずれか一項に記載の方法。
【請求項6】
Kex2切断部位が、プロ配列の、カルボキシペプチダーゼの野生型形態の天然N末端アミノ酸残基に対して5-20、5-15又は5-10アミノ酸残基上流の距離に、導入される請求項5に記載の方法。
【請求項7】
Kex2切断部位が、野生型カルボキシペプチダーゼ酵素のN末端アミノ酸残基に対して2-20、2-15又は2-10アミノ酸残基上流の位置に挿入される請求項5に記載の方法。
【請求項8】
カルボキシペプチダーゼがCPYである請求項1ないし7のいずれか一項に記載の方法。
【請求項9】
B鎖のC末端伸長部を有するヒトインスリン又はそのアナログの前躯体を、請求項1ないし8のいずれか一項に記載の方法により製造されたカルボキシペプチダーゼと反応させることによる、成熟ヒトインスリン又はそのアナログを製造する方法であって、B鎖のC末端伸長部が切断されて、成熟ヒトインスリン又はそのアナログが得られる方法。
【請求項10】
成熟ヒトインスリン又はそのアナログを製造する方法であって、真菌細胞において、
i)B鎖のC末端伸長部を有するヒトインスリン又はそのアナログの前躯体をコードするDNA配列と、
ii)そのプロ型に挿入Kex2切断部位を有するカルボキシペプチダーゼのプロ型をコードするDNAと
を同時発現させることを含み、
ヒトインスリン前駆体分子におけるB鎖のC末端伸長部を、同時発現され活性化されたカルボキシペプチダーゼにより真菌細胞内で切断し、成熟ヒトインスリン又はそのアナログを培養培地から単離する方法。
【請求項11】
ヒトインスリン前駆体分子が、ヒトインスリン又はそのアナログのB鎖、ヒトインスリン又はそのアナログのA鎖、及びB鎖とA鎖を連結するCペプチドを含み、Cペプチドが少なくとも一のKex2切断部位を含み、B鎖が、真菌細胞内におけるCペプチドのより効果的なKex2切断を促進させるC末端伸長部を含む請求項10に記載の方法。
【請求項12】
B鎖のC末端伸長部が4までのアミノ酸残基を有する請求項9ないし11のいずれか一項に記載の方法。
【請求項13】
B鎖のC末端伸長部が、Leu-Ala、Phe-Leu、Leu-Gly、Leu-Leu、Leu-Met及びLeu-Ileからなる群から選択される請求項12に記載の方法。
【請求項14】
カルボキシペプチダーゼのプロ型が、それ自身のプロモーターとは異なるプロモーターの調節下で発現される請求項10ないし13のいずれか一項に記載の方法。
【請求項15】
カルボキシペプチダーゼが酵母CPY酵素であり、プロモーターがKex2プロモーターである請求項14に記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2010−509929(P2010−509929A)
【公表日】平成22年4月2日(2010.4.2)
【国際特許分類】
【出願番号】特願2009−537629(P2009−537629)
【出願日】平成19年11月21日(2007.11.21)
【国際出願番号】PCT/EP2007/062629
【国際公開番号】WO2008/062010
【国際公開日】平成20年5月29日(2008.5.29)
【出願人】(596113096)ノボ・ノルデイスク・エー/エス (241)
【Fターム(参考)】
【公表日】平成22年4月2日(2010.4.2)
【国際特許分類】
【出願日】平成19年11月21日(2007.11.21)
【国際出願番号】PCT/EP2007/062629
【国際公開番号】WO2008/062010
【国際公開日】平成20年5月29日(2008.5.29)
【出願人】(596113096)ノボ・ノルデイスク・エー/エス (241)
【Fターム(参考)】
[ Back to top ]