
消化力増強飼料、臭気軽減飼料、整腸剤型飼料、カロリー吸収率調整型飼料、肉質改善飼料、免疫力増強飼料、ならびに受胎率改善飼料、飼料型水質浄化剤、多機能性食品
【課題】消化力の増強、整腸作用、ならびに肉質改善、および免疫力の増強と、水質を浄化することが可能となる多機能性飼料ならびに多機能性食品の提供。
【解決手段】好熱性微生物を用いて飼料中の有機物の一部を発酵させることにより、有機物の分解を促進して有用な代謝成分を豊富に持つ飼料を作製し動物の消化力の増強や整腸作用の向上、肉質改善を図る。さらには免疫力の増強や水質の浄化を併せて図ることのできる多機能性の飼料及び食品を作製する。
【解決手段】好熱性微生物を用いて飼料中の有機物の一部を発酵させることにより、有機物の分解を促進して有用な代謝成分を豊富に持つ飼料を作製し動物の消化力の増強や整腸作用の向上、肉質改善を図る。さらには免疫力の増強や水質の浄化を併せて図ることのできる多機能性の飼料及び食品を作製する。
【発明の詳細な説明】
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、消化力の増強、整腸作用、ならびに肉質改善、および免疫力の増強と、水質を浄化する機能をもつ機能性飼料ならびに機能性食品に関する。
【従来技術】
【0002】
従来より、飼料は栄養源として養殖、養豚、養鶏、などの畜水産分野で活用されている。
【0003】
また、近年、栄養的付加価値を向上させるために、アミノ酸やビタミン類を含有する飼料が開発され、また常温の微生物由来の酵素含有するものとして、フィターゼなどの酵素入りの飼料が普及している。
【0004】
しかし、これらは常温の酵素ならびに酵素群であり、必ずしも複数の分解反応を必要とする反応系で有益とはいえなかった。
【0005】
ヒトを含めた動物の腸内では真正細菌70門と古細菌13門が存在すると考えられているが、ヒトでは真正細菌ではBacteroidesとFirmicutesが、古細菌ではメタン産生古細菌のMethanobevibacter smithiiが大部分を占めている (EckburgPB,et.al.2005,Diversity of the human intestinal microbial flora.Science 308,1635)。さらに、これらの腸内細菌の有無が、栄養素の吸収、粘液バリアの強化、異物代謝、血管新生、生後の腸管の成熟を含む腸の重要な機能に関係する遺伝子の発現に影響を与えることが示されている(Xu J&Gordon JI,2003,Honor the symbionts Proc.Natl.Acad.Sci.USA 100,10452)。
【0006】
これらのデータのうち腸内細菌相と肥満との関係に関する細菌の研究報告では、ワシントン大学のGordon研究室のBackhedらが、腸内細菌相が宿主の体脂肪の増加に関与していることを示した(Backhed F,et.al.,2004The gutmicrobiota as an environmental factor that regulates fat storage.Proc.Natl.Acad.Sci.USA,101,15718)。さらに、同研究室のSamuelらは、人の腸内で最も一般的な真正細菌であるBacterodes thetaiotaomicronと腸内常在古細菌のMethanobevibacter smithiiとの共生生活が、宿主に及ぼす影響を与えた結果、これらいずれかの細菌が単独で存在するよりも、二つの細菌が共存することによって、100倍以上の多くの腸内細菌相が定着することが明らかになった。さらに、Bacterodes thetaiotaomicronの代謝に関わる遺伝子発現も増強した(Samuel BS&Gordon JI 2006 A humanized gnotobiotic mouse model of host−archaeal−bacterial mutualism.Proc Natl Acad Sci USA,103,10011)。
【0007】
このようなデータから腸内細菌が互いに連関し合い、その中でもこれまで注目されていなかった古細菌の腸内における存在意義が明らかにされつつあるが、いずれも常温の微生物の機能性を確認しているものである。
【0008】
常温菌ではなく、高温で生息する好熱性微生物の動物に与える影響に関する研究は、例えば代表的には有胞子性/耐熱性乳酸菌であるBacillus coagulans(建木節男.1978飼料と飼料工業 18:41−44)やStreptococcus thermophilus(Kimura N,et.al.1983 Bifidobacteria Microflora 2:41−55)等が知られている。また、好熱性微生物の共生系による食品の有効性を示した研究については、Streptococcus thermophilus と Lactobacillus bulgaricusの2菌株を用いて研究されている程度であり(Pette JW and H Lolkema.1950.Neth.Milk Dairy J.4:197)、その他の好熱菌を食品に応用した研究や3株以上の複合系の有効性に関する報告は見あたらず、その作用機序は不明な点が多い。
【0009】
一方、当特許申請に先立ち、宮本らはBacillus brevisならびにBacillusstearothermophilusなどを用いた好熱性微生物を用いた発酵飼料の特性を明らかにし、特許登録するとともに(特許第3314302号)、複数種の好熱性微生物からなるATCC国際登録済みの微生物群PTA−1773を用いた飼料の効能について特許公開している(特開2003−219864号)。これらの飼料は、その断片的な有効性について記載しているものの、消化力を増強する機能、臭気軽減機能、整腸機能、カロリー吸収率調整機能、肉質改善機能、免疫力増強機能、ならびに受胎率改善機能、飼料型水質浄化機能を複合的に有するものではなかった。
【本発明が解決しようとする課題】
【0010】
本発明の課題は、動物に対する飼料中の栄養成分を効率良く消化吸収し、さらに、飼料の栄養的価値以外に、肉質の改善や、免疫機能などの獲得を高効率可能とする多機能性の飼料としての活用が可能となることにある。
【問題を解決するための手段】
【0011】
前記目的を達成するために、好熱性微生物を用いて飼料中の有機物の一部を発酵させることによって、目的とする有機物の分解を促進する酵素、酵素活性を保持する分子シャペロン機能ならびにATPアーゼ活性を有する熱ショックプロテイン(HSP)ならびにその他の有用な代謝成分を豊富にもつ飼料を作製し、飼料としての栄養的な価値を向上させる。
【0012】
好熱性微生物の発酵物は、高温下の発酵により、常温下の発酵では得られない代謝成分を豊富に含ませる。例えば非酵素的な反応(メイラード反応)によって、メラノイジンのような抗酸化・抗がん物質が生成される可能性がある。
【0013】
次に、60〜90℃の高温の極限環境下で生息する好熱性微生物によって、その微生物が発現する耐熱性酵素は、常温においても酵素反応を示し、さらに耐久性に優れている。これらの酵素群は、動物体内の消化酵素の機能を補完し、濃厚飼料の消化を促進する。
【0014】
また、60〜90℃の高温の極限環境下で生息する好熱性微生物由来の熱ショックプロテイン(HSP)は、分子シャペロンとして働く分子もあるが、これらの分子は、耐熱性分子シャペロンであり、常温微生物由来の分子シャペロンよりも、構造上安定であるばかりでなく、動物体内の消化酵素の構造の劣化を防ぐ機能が高い。
【0015】
好熱性微生物由来の熱ショックプロテイン(HSP)は、サブユニットとしてもATPアーゼ(ATP合成酵素)活性を安定的に発揮することが知られているため、動物体内で働くATPアーゼの活性化に寄与することが想定される。
【0016】
さらに、腸管における熱ショックプロテインの透過性ならびに浸潤性が高まった場合、これらのタンパク質は、免疫担当細胞の表面認識部位に位置するMHC−クラスIと親和性があることから、抗原に対するアジュバンド活性があることが期待される。
【0017】
本発明は、このような複合機能をもつ成分を有機物残渣から大量生産するとともに、それらを経口投与することによって有効活用することを可能とすることにある。
【発明の実態】
【0018】
この実施形態に係る飼料は、キチン質を含む有機質や魚介類、あるいはおから、ビール粕、焼酎粕、海草などの植物性成分を50〜80℃で高温発酵させた発酵物を含み、発酵微生物としては好熱性微生物が望ましい。
【0019】
キチン質を含む有機質としては、カニ、エビなどの甲殻類、ハエ、コガネムシなどの昆虫などが挙げられる。
【0020】
発酵微生物としてはキチン質を分解するキチン質分解酵素を産生する微生物が望ましい。
【0021】
好熱性微生物は、難分解性成分や有毒物質等の共存する場所をはじめとした劣悪な環境下でも、タンパク構造が変化しにくく酵素活性を保持できる耐熱性酵素を産生する。また、好熱性微生物由来の熱ショックプロテイン(HSP)は、常温微生物由来の熱ショックプロテインより構造的に安定であり、その機能として酵素の構造保持を保護する分子シャペロンを発現する。これによって、通常の常温微生物由来の成分では補うことのできない消化器官内における酵素活性を維持し、各種機能を有する代謝成分の生合成を可能にするという利点がある。好熱性微生物としては、例えば、好熱性種菌PTA−1773をはじめとした前期好熱菌群があげられる。尚、好熱性種菌PTA−1773は、発明者によって、2000年5月1日付けでATCC(American Type Culture Collection,10801 University Boulevard Manassas,Virginia 20110−2209 U.S.A.)に国際寄託されている(受託番号:PTA−1773)。これらの微生物は、高度な有機物分解能を持っており、70〜90℃の発酵熱エネルギーを発することができる微生物群である。好熱性種菌PTA−1773を培養するための栄養源としては、腐敗していない生のエビやカニの残渣等とともに、90℃程度の高温下でも分解されにくい多孔体である炭等を用いて微生物の付着部分を増やし、好気条件下で好熱性種菌PTA−1773を12時間以上培養する。この時、自然発酵熱は少なくとも60〜90℃に保たれる。
【0022】
さらに、Firmicutesに属するBacillus thermocloacae、Anoxybacillus toebii、Bacillus thermoalkalophilusなどの好熱性微生物群が挙げられる。
【0023】
60〜90℃の高温の極限環境下で生息する好熱性微生物によって、その微生物が発現する耐熱性酵素は、常温においても酵素反応を示し、さらに耐久性に優れている。これらの酵素群は、動物体内の消化酵素の機能を補完し、濃厚飼料の消化を促進する。
【0024】
さらに、60〜90℃の高温の極限環境下で生息する好熱性微生物由来の熱ショックプロテイン(HSP)は、分子シャペロンとして働く分子もある。これらの分子は、耐熱性分子シャペロンであり、常温微生物由来の分子シャペロンよりも、構造上安定であるばかりでなく、動物体内の消化酵素の構造の劣化を防ぐことが想定される。
【0025】
これらの相乗的な効果によって、動物体内の消化が促進されるため、成長促進ならびに糞便中の臭気成分の軽減などが図られると考えられる。
【0026】
さらに、好熱性微生物の発酵産物ならびにその含有成分が、腸内細菌相あるいは、腸管の細胞に作用することが想定される。
【0027】
例えば、好熱性微生物由来の熱ショックプロテイン(HSP)は、サブユニットとしてもATPアーゼ活性を安定的に発揮することが知られているため、腸内の消化吸収に働くATPアーゼの活性化に寄与すると考えられる。
【0027】
実際、これらの微生物ならびにその含有成分を豚の飲水に0.4%添加し、飼育すると、六ヶ月以内に糞中のリン酸イオンの値が減少する。この作用機序のメカニズムは明確ではないが、好熱性微生物由来の熱ショックプロテイン(HSP)は、サブユニットとしてもATPアーゼ活性を安定的に発揮することから、腸管内におけるATP合成活性が上がるために、消化吸収に寄与するトランスポーターが活性化すると考えると矛盾しない。
【0028】
さらに、有機性のリン化合物として腸内細菌に含有されるために、腸内細菌相が活性化する、あるいは体内に吸収されることによって動物の代謝エネルギーが補助されることも想定される。
【0029】
次に、栄養吸収が促進されることによって肝機能が改善することが期待されるが、実際に一般的な麻酔薬からの覚醒時間が短くなる効果が確認されている。
【0030】
また、腸管における熱ショックプロテインの透過性ならびに浸潤性が高まった場合、好熱性微生物の発酵産物を溶菌した液を豚や鶏に対して飲水添加する、あるいは魚の餌に添加することによって死亡率が減少することが判明している。
【0031】
この原因としては、当該発酵産物を溶菌することによって好熱性微生物由来の熱ショックプロテインが溶出し、それらが腸管を介して免疫担当細胞に認識されることが想定される。一般に、熱ショックプロテイン(HSP)は、MHCクラスI抗原提示経路において、抗原認識部位に位置するMHC−クラスIのタンパク分子の溝に認識されやすい。したがって、好熱性微生物由来の熱ショックプロテインは、経口投与を介して抗原のアジュバンド的な役割を果たし、抗原提示による免疫活性が上がることが考えられる。
【0032】
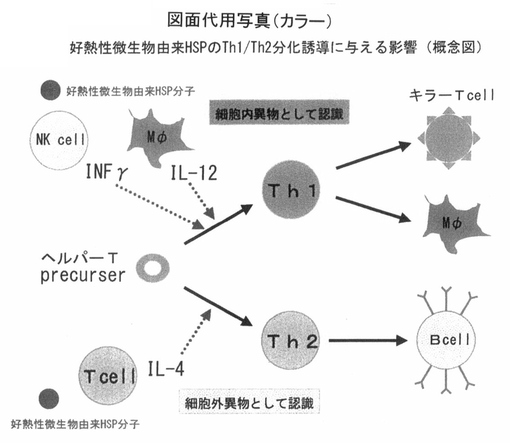
次に、腸内細菌相を改善することによって、腸管免疫系が活性化し、ベルパーT細胞のサブセット(Th1/Th2バランス)のそれぞれの活性化の程度に影響を与えることが一般的に知られている。細胞内の寄生体がNK細胞ならびにマクロファージを刺激することによって、インターフェロンガンマ(IFNγ)やインターロイキン12(IL−12)が産生され、ヘルパーT細胞の前駆細胞がTh1に変化するが、例えば、好熱性微生物由来の熱ショックプロテインについては、タンパク構造上において宿主細胞の熱ショックタンパクと類似しているため、細胞外の異物タンバクとして認識されてTh2を活性化するスキームとしてのみならず、細胞内寄生体として認識されてTh1が活性化することが考えられる。
一般的に、Th1が活性化されるとキラーT細胞ならびにマクロファージの働きが高まるため、免疫力が総体的に活性化する。
したがって、好熱性微生物群ならびにその含有成分を経口投与することによって、腸管免疫系が活性化する原因は、好熱性微生物由来の熱ショックプロテインが安定なタンパク質であり、構造的にも宿主と類似していることから、腸管内の透過性が高まる条件下にかぎり、図1のようなスキームで免疫系を活性化することが想定される。
【0033】
次に、好熱性微生物の発酵物は、高温下の発酵により、常温下の発酵では得られない代謝成分を豊富に含ませる。例えば非酵素的な反応(メイラード反応)によって、メラノイジンのような抗酸化・抗がん物質が生成される可能性があり、これらが腸内ならびに腸管を介して生体の体調機能に影響を与えることが想定される。
【0034】
以上の相乗効果として、図2に示したように、豚や鶏に対して、消化促進あるいは免疫力向上効果などが表れると考えられる。また、魚に対しても、図3で示したように消化力増強作用ならびに免疫力向上効果があると考えられる。次に実施例により、更に詳細に説明するが、この発明は係る実施例に限定されるものではない。
【実施例1】
【0035】
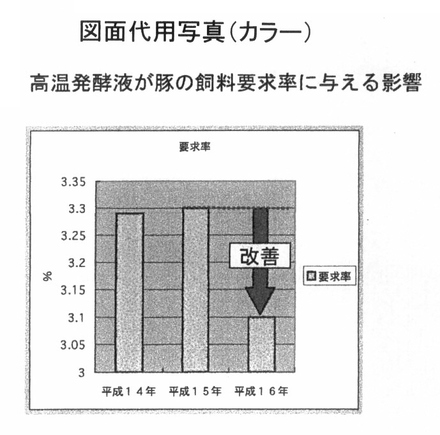
図2は、PTA−1773を含む高温発酵飼料600グラムを100リットルの水に溶解し、曝気条件下で60℃12時間処理した液を、豚の飲み水に対して0.4%添加して、春期の3月より6ヶ月間、継続飼育した場合の飼料要求率を示している。
【0036】
結果として、例年の飼料要求率が約3.3程度であるのに対して、3.1まで改善し、1キロの飼料当りの増体重が上昇する傾向が認められた。尚、試験対象の規模は、母豚150頭の一貫経営であり出荷頭数として、1年間当り約3000頭の養豚場であり、肥育豚として19.5m2当り27頭の飼育頭数で試験を実施した。
【実施例2】
【0037】
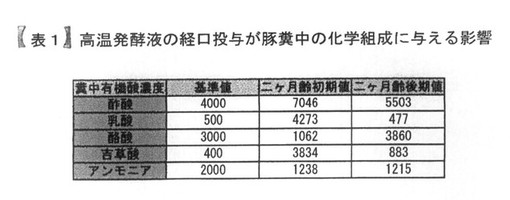
同じく、表1は別の養豚場において、前記PTA−1773を含む高温発酵飼料の処理液を、豚の飲み水に対して0.4%添加して6ヶ月間、継続飼育した場合の豚糞中の臭気関連成分の分析結果を示している。
【0038】
豚糞中の臭気関連成分としては、酢酸、乳酸、酪酸、吉草酸、アンモニアを対象として、キャピラリー電気泳動法にて分析した。
試料3gを50mlチューブに入れ、99.5%エタノール(和光純薬)10gを加え10分間振とう後、20gの超純水を入れてさらに10分間振とうさせてから一晩放置し、各成分の抽出を行った。遠心分離(3000rpm、10分間)により得られた上清を分画分子量3000Daの遠心限外ろ過フィルター(Microcon YM−3;MILLIPORE)を用いて除タンパクしたものを試料とした。
分析にはフォトダイオードアレイを搭載したキャピラリー電気泳動装置(G1600A;Agilent Technologies)を用いて測定した。Agilent Basic Anion Buffer(Agilent)を泳動バッファーとし、全長112.5cm(有効長104cm)・内径50μmのフューズドシリカキャピラリー(Agilent)に20℃で−30kVの電圧を印加し各成分を分離させた。試料の注入は50mbarの圧力で2秒間行った。検出は間接吸光度法により行い350nmと200nmの差を信号とした。各分析の間では、高圧で泳動バッファーを4分間流しキャピラリーの洗浄を行った。予めスタンダードとしてD.W.を加えて作成しておいた標準液を泳動に供し、これらスタンダードとサンプルとのピーク位置および面積の比較によって各成分の同定および濃度決定を行った。
【0039】
結果として、一般的に養豚場内の臭気が少ない場合の値(基準値)に対して、前記処理を実施していない数値は大きくずれていた。一方、前記処理6ヶ月後の数値は、比較的数値が基準値に近い値だった。また、この時の養豚場内の隣接境界領域の臭気は養豚場の存在の有無に気づかない程度の臭気まで軽減していた。
【実施例3】
【0040】
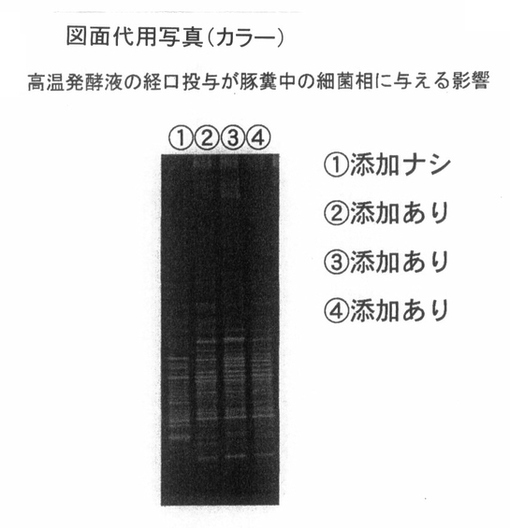
次に、図3は、2ヶ月齡の仔豚の糞中の細菌相と、高温発酵飼料を添加せずに飼育した仔豚の糞中の細菌相をPCR−DGGE法を用いて比較したデータである。尚、添加ありでは、前記PTA−1773を含む高温発酵飼料の処理液を、豚の飲み水に対して0.4%添加して6ヶ月間以上、継続飼育した。
【0041】
PCR−DGGE法は、微生物の16SrDNAに対するユニバーサルプライマーのforwardとしてGC−PRBA338fを用いて、reverseとしてPRUN518rを用いて、16SrDNA上の236bpの部位を増幅した。さらに、DGGEでは、尿素入りのgradient gelの濃度勾配を60〜30%の条件とした。具体的には、サンプル0.5gをDNA抽出装置(QuickGene−800:富士フィルム)に供し、サンプル中のDNAを抽出した。抽出したDNAを鋳型とし、PCR法により16SrRNAV3領域に対応するDNA配列中の236bpを増幅した。プライマーはGCクランプ(5’−CGC CCG CCG CGC GCG GCG GGC GGG GCG GGG GCA CGG GGG GAC TCC TAC GGG AGG CAG CAG−3’)を5’末端に付加したPRBA338f(5’−ACT CCT ACG GGA GGC AGC AG−3’)とPRUN518r(5’−ATT ACC GCG GCT GCT GG−3’)を使用した。DNAポリメラーゼにはpuReTaqReady−To−Go−PCR Beads、サーマルサイクラーにはTaKaRa PCR Thremal Cycler PERSONAL TP240を用いた。PCRの条件は92℃,2分、(92℃,1分;55℃,30秒;72℃,1分)×30サイクル、72℃,6分で行った。
PCRにより増幅されたDNAフラグメントを変性剤濃度勾配ゲル電気泳動装置(D−Coad:BioRad)に供し菌叢解析を行った。変性剤には尿素とホルムアミドを用い、泳動方向に30%〜60%の濃度勾配をかけて8%ポリアクリルアミドゲルを作成した。ゲルの作成に必要な試薬はすべてBioRad社製のDGGEシステムを使用した。泳動条件は、電圧170V、泳動時間6時間、温度60℃、泳動バッファー1×TAEとした。泳動後、SYBR Green II(タカラバイオ)で染色しトランスイルミネーターを用い310nmのUV照射下で検出した。
【0042】
図3の結果として、未添加群に対して、添加後6ヶ月では、豚糞中の細菌相のバンドバターンを比較すると、10%以下の一致率の差が認められた。また、添加群間の一致率は高く、未添加群間の一致率は低かった。
【0043】
これらのデータは、糞中の細菌相が、豚の腸内細菌相を反映していると想定できることから、添加群では、好熱性微生物、あるいはその含有成分の経口的な投与によって、常温微生物で構成されている腸内細菌相のバラツキが少なくなり、整腸作用が認められることを意味している。
【実施例4】
【0044】
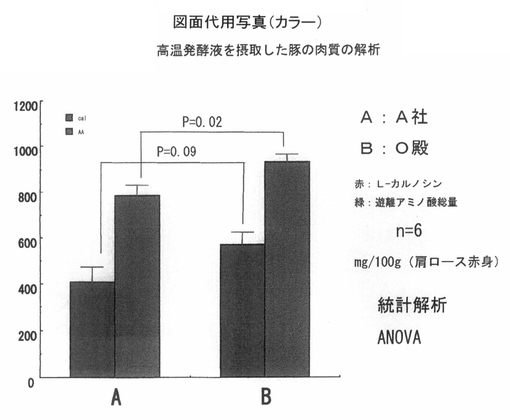
次に 図4は、豚肉肩ロース赤身部分の遊離アミノ酸濃度ならびにL−カルノシン濃度を、個体数n=6で解析して、その平均値を棒グラフで、標準誤差をエラーバーで示している。肉質中の遊離アミノ酸濃度ならびにL−カルノシン濃度を前記キャピラリー電気泳動法の諸条件にて測定したデータである。なお、分析結果は、分散分析(ANOVA)によって統計解析を実施した。
【0045】
養豚場A(対照群)ならびにB(比較群)は、双方とも同じ飼料で飼育されていたが、Bについては、前記PTA−1773を含む高温発酵飼料の処理液を、豚の飲み水に対して0.4%添加して12ヶ月間以上、継続使用されていた。分析対象としたのは、生後6ヶ月を経過した出荷用肥育豚とした。
【0046】
図に示して有るように、遊離アミノ酸濃度ならびにカルノシン濃度を比較すると、対照群Aに対して比較群Bはそれぞれ、p<0.02ならびにp<0.09で増加傾向にあった。
【0047】
肉質中の遊離アミノ酸濃度は味覚に関与している成分である。またカルノシンは、ラットを用いた遊泳実験において疲労回復性が示されている成分であり、今回の結果から、前記PFA−1773を含む高温発酵飼料の処理液を、豚の飲み水として使用することによって、豚肉の肉質の旨味ならびに健康機能性に影響を与えることと想定された。
【実施例5】
【0048】
表2は、陸上養殖ヒラメに対して、前記PTA−1773を含む高温発酵飼料の処理液を、ヒラメの餌に対して0.5%添加して継続給与して6ヶ月後、12ヶ月後のヒラメ肉質中のタウリン濃度を高速液体クロマトグラフィー(HPLC)で分析した結果を示している。
【0049】
生魚は直ちに背部を採取後包丁で細断し、飼料は、乳鉢、乳棒を用い摩砕後、分析に供するまで−30℃に保管した。各試料5gを50ml遠沈管内に精秤し、5%過塩素酸を30ml加え、ホモジナイザーで約1分間攪拌後、2,500rpm、10分間遠心分離を行った。上澄みを別の50ml遠沈管にデカンテーションし、2N水酸化カリウム溶液を適量添加しpHメーターを用いて確認しながら、pH2.0に調整した。シーロンフィルムで密栓後、氷冷中に30分放置し、2,500rpm、10分間遠心分離を行った。上澄み全てを50mlメスフラスコに入れ、0.01N HClで50mlに定容し、エキス成分とした。エキス成分溶液200μlを1.5mlマイクロチューブに入れ、100mMフェニルイソチオシアネート試薬を200μl、1Mトリエチルアミン試薬を200μl加え。撹拌後、37℃で60分間反応させた。ヘキサンを400μl加え、遠心分離機で2,000rpm、5min遠心分離後、上層を除き、下層10μlを高速液体クロマトグラフィー(HPLC)で分析した。
【0050】
表2に示したとおり、前記PTA−1773を含む高温発酵飼料の処理液を餌に添加して6ヶ月の継続給与時の肉質は、通常飼料使用時のデータに対して2倍程度まで上昇することが明らかになった。さらに、12ヶ月の継続給与時の肉質は、さらに上昇する傾向が認められた。これらのデータはn=5で、有意差p<0.05であった。
【0051】
次にエキス成分のDPPH法による抗酸化活性を測定した。各試料5gを50ml遠沈管内に精秤し、エタノールを30ml加え、ホモジナイザーで約1分間攪拌後、2,500rpm、10分間遠心分離を行った。上清をエバポレーターで濃縮後、エタノールを加え10mlに定容し、エキス成分(抗酸化活性分析用)とした。エキス成分1mlをねじ付試験管に入れた。これに、エタノール1ml、100μM DPPH溶液1mlを加え撹拌し、30℃、20分反応させた。反応後、エタノール1mlを加え、517nmの吸光度を測定し、ラジカルの消去能を測定した。消去能はIC50値で求めた。
【0051】
結果として、前記PTA−1773を含む高温発酵飼料の処理液を餌に添加して6ヶ月ならびに12ヶ月の継続給与時のヒラメの肉質中の抗酸化活性は上昇する傾向が認められた。
【実施例6】
【0052】
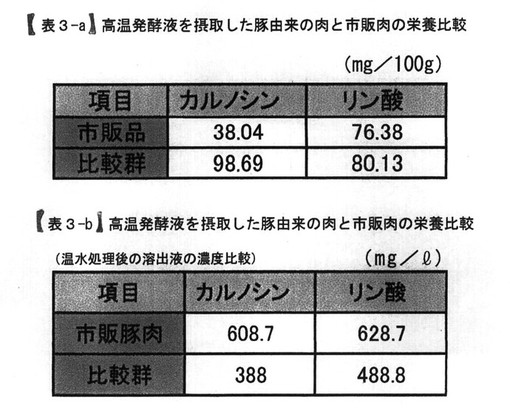
表3−aは、前記PTA−1773を含む高温発酵飼料の処理液を、飲水に0.5%添加している飼育環境において、24ヶ月経過した養豚場の出荷用肥育豚の豚肉を比較群として、市販品の豚肉Aとともに、肉質中のカルノシンならびにリン酸イオン濃度を測定した結果である。その結果、水溶性のリン酸イオンの濃度は、市販品と比較群ともに同程度であった。一方、カルノシンについては、比較群が、市販品の2倍以上に増量していた。
【0053】
次に、表3−bでは、これらの両群の肉の肉質中の細胞骨格の強度を確認するために、温水中での前記肉汁成分の漏出の度合を確認した。ファルコンチューブに80℃の温水50ccを投入し、その温度帯に保持したまま、10gのバラ肉を5秒間浸水し、その際に温水中に溶出したエキス成分を回収して分析した。
【0054】
その結果、溶液中のカルノシン濃度は、肉質中のカルノシン濃度が高かった比較群が、388mg/lの溶出量であるのに対して、市販品の方が約1.5倍を超える608.7mg/l溶出した。また、水溶性のリン酸イオンについても、比較群よりも市販品においいて溶出量が多かった。
【0055】
以上の結果は、通常の豚肉よりも、前記PTA−1773を含む高温発酵飼料の処理液を飲水に0.4%添加している飼育環境において、24ヶ月継続飼育した養豚場の出荷用肥育豚において、肉質中の細胞骨格の強度が強いことを意味している。
【実施例7】
【0056】
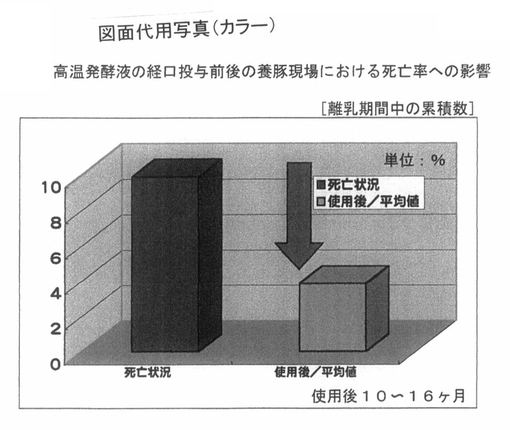
次に、図5は、養豚場における離乳期経過後の死亡率に与える影響を解析した結果である。前記PTA−1773を含む高温発酵飼料の処理液を、飲水に0.4%添加している飼育環境において、10〜16ヶ月経過した養豚場における死亡率の平均値は、未処理時が10%であるのに対して、4%以下まで低下しており、統計的に有意差(p<0.05)があり、仔豚の死亡率の軽減効果は結果として60%程度であった。
【実施例8】
【0057】
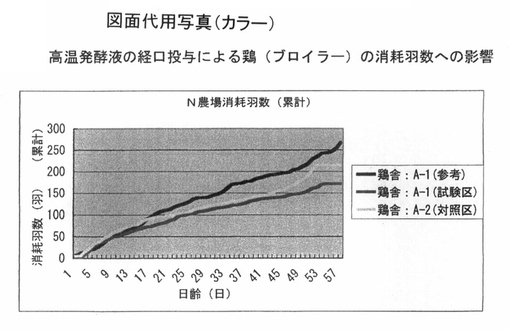
図6は、ブロイラーの消耗率に与える影響を解析した結果である。前記PTA−1773を含む高温発酵飼料の処理液を、飲水に0.4%添加している飼育環境において、未処理群と比較して、出荷までの累積消耗羽数は、未処理群が一万羽の飼育規模あたり210羽以上であるのに対して、試験区では、170羽程度であり、全体として減少する傾向が認められた。
【実施例9】
【0058】
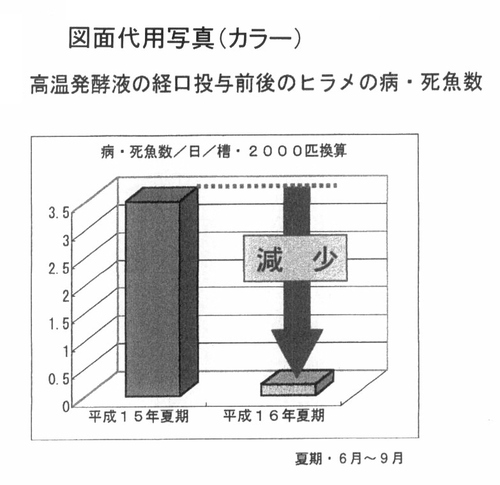
図7は、ヒラメの夏期の病・死魚率に与える影響を解析した結果である。前記PTA−1773を含む高温発酵飼料の処理液を餌に0.5%添加している飼育環境の養殖池の病・死魚数は、0.5以下であり、当該添加をしていなかった15年度の3.3匹/日と比較して、毎日の病・死魚数は、3分の1以下であった。この経済効果は、一池当り45万円程度の損失回避であった。
【0059】
さらに、病原菌のエドワジエラによる腹水発症の稚魚数の減少が確認されたことから、魚体において免疫力が強化されていることが示唆された。
【実施例10】
【0060】
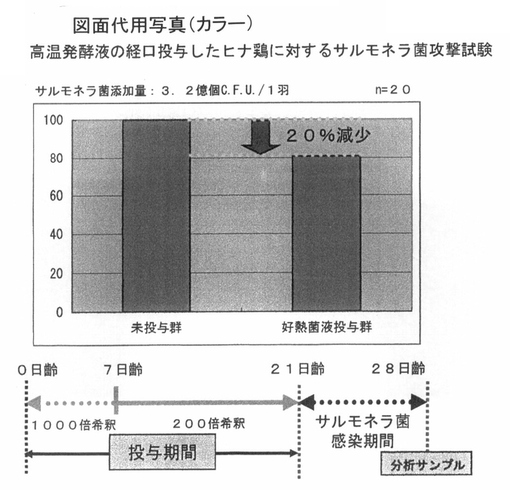
図8は、採卵鶏のヒナ鶏に対して、前記PTA−1773を含む高温発酵飼料の処理液を飲水に対して、0.5%添加している飼育環境にある好熱菌液投与群と、未処理群とで、サルモネラに対する攻撃試験の結果を示したものである。
【0061】
20日齡まで、図の棒グラフで示したように、未投与群ならびに好熱菌液投与群を飼育し、21日に1羽当りの3.2億個のサルモネラ菌を添加し、1週間後に、採取したサルモネラ菌数を測定した
【0062】
その結果、好熱菌液投与群において、20%程度の減少傾向が認められた。この結果は、強いサルモネラ菌に対する抑制効果は認められないものの、内在性の腸内細菌相を活性化している可能性が示唆された。これらの結果から、サルモネラ菌の増殖抑制細菌の作用を増給する可能性があると考えられた。したがって、このような増殖抑制細菌とともに利用することが好ましいと考えられる。
【実施例11】
【0063】
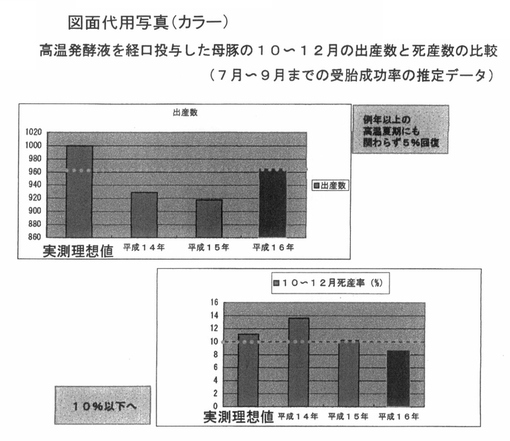
次に、母豚の出産数と死産数に与える影響を解析した。図9は、前記PTA−1773を含む高温発酵飼料の処理液を飲水に対して、0.4%添加している飼育環境にある養豚場において、使用以前の出産率ならびに死産率との比較をしたデータである。
【0064】
平成16年の夏は、例年よりも高温期が継続したため、出産数は減少する傾向にあることが想定されたが、図9に示したように、10〜12月の出産数ならびに死産数ともに改善した。すなわち、出産数は例年よりも多く、5%回復し、死産数は例年よりも減少し、10%以下になっていた。
【0065】
以上の結果から、前記PTA−1773を含む高温発酵飼料の成分が、7〜9月までの受胎の成功率を向上させている可能性が想定された。
【実施例12】
【0066】
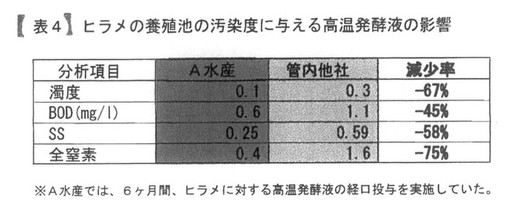
表4は、前記PTA−1773を含む高温発酵飼料の処理液を餌に0.5%添加している飼育環境でヒラメ養殖をしているA水産の養殖池の汚染度を、管内の他の池と比較した結果を示している。
【0067】
餌を投入する直前の同時刻の水を採取し分析項目として濁度、BOD、SS、全窒素を確認した。結果として、管内他社に比べて、45〜75%低い値であった。すなわち、前記PTA−1773を含む高温発酵飼料の処理液を餌に含浸することによって、水質浄化をもたらすことが明らかになった。
【0068】
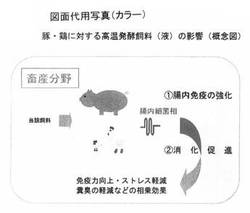
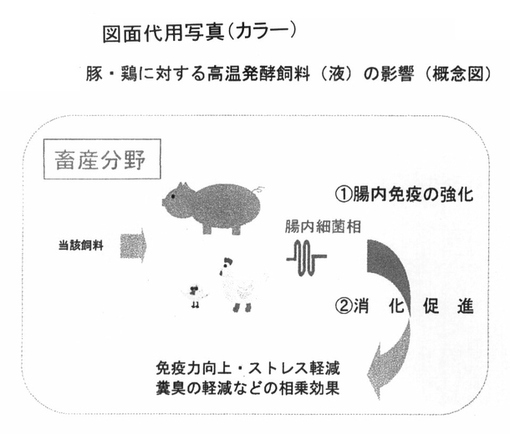
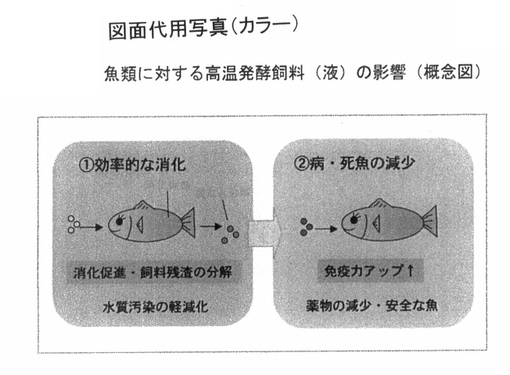
以上の結果は、豚ならびに鶏に対して、図10のような概念図として示すことができるまた魚類については、図11のような概念図で示すことができる。
【発明の効果】
【0069】
当該発明は、消化力の増強、整腸作用、ならびに肉質改善、および免疫力の増強と、水質を浄化することが可能となる飼料ならびに機能性食品であり、これによって、幅広い健康維持を可能とする高品質の食品を消費者に供給することが実現する。
【図面の簡単な説明】
【0070】
【図1】 図1は、好熱性微生物由来の熱ショックプロテインが経口投与を介して、腸管免疫系を活性化し、Th1/Th2の分化誘導に与える影響についての概念図を示している。
【図2】 図2は、高温発酵飼料を液状化し、飲水添加することによって、豚の飼料要求率に与える影響を示している。
【図3】 図3は、高温発酵飼料を液状化し、飲水添加することによって、豚の腸内細菌相に与える影響を示している。
【図4】 図4は、高温発酵飼料を液状化し、飲水添加することによって、豚の肉質に与える影響に与える影響を示している
【図5】 図5は、高温発酵飼料を液状化し、飲水添加することによって、豚の離乳期間中における死亡率に与える影響を示している。
【図6】 図6は、高温発酵飼料を液状化し、飲水添加することによって、ブロイラーの消耗羽数に与える影響を示している。
【図7】 図7は、高温発酵飼料を液状化し、飲水添加することによって、ヒラメの病・死魚数に与える影響を示している。
【図8】 図8は、高温発酵飼料を液状化し、飲水添加したヒナ鶏に対するサルモネラ菌の攻撃試験の結果を示している。
【図9】 図9は、高温発酵飼料を液状化し、飲水添加した母豚の10月〜12月における出産数と死産数の比較データを示している。
【図10】 図10は、高温発酵飼料を鶏ならびに豚に投与する場合の効能に関しての概念図を示している。
【図11】 図11は、高温発酵飼料を魚類に投与する場合の効能に関しての概念図を示している。
【表1】
表1は、高温発酵飼料を液状化し、飲水添加することによって、豚の糞中の有機酸組成に与える影響を示している。
【表2】
表2は、高温発酵飼料を飼料添加することによって、ヒラメの肉質のタウリン濃度に与える影響を示している。
【表3】
表3−aは、高温発酵飼料を液状化し、飲水添加して飼育した豚の肉質のカルノシン濃度ならびにリン酸濃度と、市販の豚肉の栄養成分を比較している。表3−bは、それらの肉を80℃の温水に浸した後の溶液中の化学組成を分析したデータである。
【表4】
表4は、ヒラメの養殖池の汚染度に与える影響を分析したデータである。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、消化力の増強、整腸作用、ならびに肉質改善、および免疫力の増強と、水質を浄化する機能をもつ機能性飼料ならびに機能性食品に関する。
【従来技術】
【0002】
従来より、飼料は栄養源として養殖、養豚、養鶏、などの畜水産分野で活用されている。
【0003】
また、近年、栄養的付加価値を向上させるために、アミノ酸やビタミン類を含有する飼料が開発され、また常温の微生物由来の酵素含有するものとして、フィターゼなどの酵素入りの飼料が普及している。
【0004】
しかし、これらは常温の酵素ならびに酵素群であり、必ずしも複数の分解反応を必要とする反応系で有益とはいえなかった。
【0005】
ヒトを含めた動物の腸内では真正細菌70門と古細菌13門が存在すると考えられているが、ヒトでは真正細菌ではBacteroidesとFirmicutesが、古細菌ではメタン産生古細菌のMethanobevibacter smithiiが大部分を占めている (EckburgPB,et.al.2005,Diversity of the human intestinal microbial flora.Science 308,1635)。さらに、これらの腸内細菌の有無が、栄養素の吸収、粘液バリアの強化、異物代謝、血管新生、生後の腸管の成熟を含む腸の重要な機能に関係する遺伝子の発現に影響を与えることが示されている(Xu J&Gordon JI,2003,Honor the symbionts Proc.Natl.Acad.Sci.USA 100,10452)。
【0006】
これらのデータのうち腸内細菌相と肥満との関係に関する細菌の研究報告では、ワシントン大学のGordon研究室のBackhedらが、腸内細菌相が宿主の体脂肪の増加に関与していることを示した(Backhed F,et.al.,2004The gutmicrobiota as an environmental factor that regulates fat storage.Proc.Natl.Acad.Sci.USA,101,15718)。さらに、同研究室のSamuelらは、人の腸内で最も一般的な真正細菌であるBacterodes thetaiotaomicronと腸内常在古細菌のMethanobevibacter smithiiとの共生生活が、宿主に及ぼす影響を与えた結果、これらいずれかの細菌が単独で存在するよりも、二つの細菌が共存することによって、100倍以上の多くの腸内細菌相が定着することが明らかになった。さらに、Bacterodes thetaiotaomicronの代謝に関わる遺伝子発現も増強した(Samuel BS&Gordon JI 2006 A humanized gnotobiotic mouse model of host−archaeal−bacterial mutualism.Proc Natl Acad Sci USA,103,10011)。
【0007】
このようなデータから腸内細菌が互いに連関し合い、その中でもこれまで注目されていなかった古細菌の腸内における存在意義が明らかにされつつあるが、いずれも常温の微生物の機能性を確認しているものである。
【0008】
常温菌ではなく、高温で生息する好熱性微生物の動物に与える影響に関する研究は、例えば代表的には有胞子性/耐熱性乳酸菌であるBacillus coagulans(建木節男.1978飼料と飼料工業 18:41−44)やStreptococcus thermophilus(Kimura N,et.al.1983 Bifidobacteria Microflora 2:41−55)等が知られている。また、好熱性微生物の共生系による食品の有効性を示した研究については、Streptococcus thermophilus と Lactobacillus bulgaricusの2菌株を用いて研究されている程度であり(Pette JW and H Lolkema.1950.Neth.Milk Dairy J.4:197)、その他の好熱菌を食品に応用した研究や3株以上の複合系の有効性に関する報告は見あたらず、その作用機序は不明な点が多い。
【0009】
一方、当特許申請に先立ち、宮本らはBacillus brevisならびにBacillusstearothermophilusなどを用いた好熱性微生物を用いた発酵飼料の特性を明らかにし、特許登録するとともに(特許第3314302号)、複数種の好熱性微生物からなるATCC国際登録済みの微生物群PTA−1773を用いた飼料の効能について特許公開している(特開2003−219864号)。これらの飼料は、その断片的な有効性について記載しているものの、消化力を増強する機能、臭気軽減機能、整腸機能、カロリー吸収率調整機能、肉質改善機能、免疫力増強機能、ならびに受胎率改善機能、飼料型水質浄化機能を複合的に有するものではなかった。
【本発明が解決しようとする課題】
【0010】
本発明の課題は、動物に対する飼料中の栄養成分を効率良く消化吸収し、さらに、飼料の栄養的価値以外に、肉質の改善や、免疫機能などの獲得を高効率可能とする多機能性の飼料としての活用が可能となることにある。
【問題を解決するための手段】
【0011】
前記目的を達成するために、好熱性微生物を用いて飼料中の有機物の一部を発酵させることによって、目的とする有機物の分解を促進する酵素、酵素活性を保持する分子シャペロン機能ならびにATPアーゼ活性を有する熱ショックプロテイン(HSP)ならびにその他の有用な代謝成分を豊富にもつ飼料を作製し、飼料としての栄養的な価値を向上させる。
【0012】
好熱性微生物の発酵物は、高温下の発酵により、常温下の発酵では得られない代謝成分を豊富に含ませる。例えば非酵素的な反応(メイラード反応)によって、メラノイジンのような抗酸化・抗がん物質が生成される可能性がある。
【0013】
次に、60〜90℃の高温の極限環境下で生息する好熱性微生物によって、その微生物が発現する耐熱性酵素は、常温においても酵素反応を示し、さらに耐久性に優れている。これらの酵素群は、動物体内の消化酵素の機能を補完し、濃厚飼料の消化を促進する。
【0014】
また、60〜90℃の高温の極限環境下で生息する好熱性微生物由来の熱ショックプロテイン(HSP)は、分子シャペロンとして働く分子もあるが、これらの分子は、耐熱性分子シャペロンであり、常温微生物由来の分子シャペロンよりも、構造上安定であるばかりでなく、動物体内の消化酵素の構造の劣化を防ぐ機能が高い。
【0015】
好熱性微生物由来の熱ショックプロテイン(HSP)は、サブユニットとしてもATPアーゼ(ATP合成酵素)活性を安定的に発揮することが知られているため、動物体内で働くATPアーゼの活性化に寄与することが想定される。
【0016】
さらに、腸管における熱ショックプロテインの透過性ならびに浸潤性が高まった場合、これらのタンパク質は、免疫担当細胞の表面認識部位に位置するMHC−クラスIと親和性があることから、抗原に対するアジュバンド活性があることが期待される。
【0017】
本発明は、このような複合機能をもつ成分を有機物残渣から大量生産するとともに、それらを経口投与することによって有効活用することを可能とすることにある。
【発明の実態】
【0018】
この実施形態に係る飼料は、キチン質を含む有機質や魚介類、あるいはおから、ビール粕、焼酎粕、海草などの植物性成分を50〜80℃で高温発酵させた発酵物を含み、発酵微生物としては好熱性微生物が望ましい。
【0019】
キチン質を含む有機質としては、カニ、エビなどの甲殻類、ハエ、コガネムシなどの昆虫などが挙げられる。
【0020】
発酵微生物としてはキチン質を分解するキチン質分解酵素を産生する微生物が望ましい。
【0021】
好熱性微生物は、難分解性成分や有毒物質等の共存する場所をはじめとした劣悪な環境下でも、タンパク構造が変化しにくく酵素活性を保持できる耐熱性酵素を産生する。また、好熱性微生物由来の熱ショックプロテイン(HSP)は、常温微生物由来の熱ショックプロテインより構造的に安定であり、その機能として酵素の構造保持を保護する分子シャペロンを発現する。これによって、通常の常温微生物由来の成分では補うことのできない消化器官内における酵素活性を維持し、各種機能を有する代謝成分の生合成を可能にするという利点がある。好熱性微生物としては、例えば、好熱性種菌PTA−1773をはじめとした前期好熱菌群があげられる。尚、好熱性種菌PTA−1773は、発明者によって、2000年5月1日付けでATCC(American Type Culture Collection,10801 University Boulevard Manassas,Virginia 20110−2209 U.S.A.)に国際寄託されている(受託番号:PTA−1773)。これらの微生物は、高度な有機物分解能を持っており、70〜90℃の発酵熱エネルギーを発することができる微生物群である。好熱性種菌PTA−1773を培養するための栄養源としては、腐敗していない生のエビやカニの残渣等とともに、90℃程度の高温下でも分解されにくい多孔体である炭等を用いて微生物の付着部分を増やし、好気条件下で好熱性種菌PTA−1773を12時間以上培養する。この時、自然発酵熱は少なくとも60〜90℃に保たれる。
【0022】
さらに、Firmicutesに属するBacillus thermocloacae、Anoxybacillus toebii、Bacillus thermoalkalophilusなどの好熱性微生物群が挙げられる。
【0023】
60〜90℃の高温の極限環境下で生息する好熱性微生物によって、その微生物が発現する耐熱性酵素は、常温においても酵素反応を示し、さらに耐久性に優れている。これらの酵素群は、動物体内の消化酵素の機能を補完し、濃厚飼料の消化を促進する。
【0024】
さらに、60〜90℃の高温の極限環境下で生息する好熱性微生物由来の熱ショックプロテイン(HSP)は、分子シャペロンとして働く分子もある。これらの分子は、耐熱性分子シャペロンであり、常温微生物由来の分子シャペロンよりも、構造上安定であるばかりでなく、動物体内の消化酵素の構造の劣化を防ぐことが想定される。
【0025】
これらの相乗的な効果によって、動物体内の消化が促進されるため、成長促進ならびに糞便中の臭気成分の軽減などが図られると考えられる。
【0026】
さらに、好熱性微生物の発酵産物ならびにその含有成分が、腸内細菌相あるいは、腸管の細胞に作用することが想定される。
【0027】
例えば、好熱性微生物由来の熱ショックプロテイン(HSP)は、サブユニットとしてもATPアーゼ活性を安定的に発揮することが知られているため、腸内の消化吸収に働くATPアーゼの活性化に寄与すると考えられる。
【0027】
実際、これらの微生物ならびにその含有成分を豚の飲水に0.4%添加し、飼育すると、六ヶ月以内に糞中のリン酸イオンの値が減少する。この作用機序のメカニズムは明確ではないが、好熱性微生物由来の熱ショックプロテイン(HSP)は、サブユニットとしてもATPアーゼ活性を安定的に発揮することから、腸管内におけるATP合成活性が上がるために、消化吸収に寄与するトランスポーターが活性化すると考えると矛盾しない。
【0028】
さらに、有機性のリン化合物として腸内細菌に含有されるために、腸内細菌相が活性化する、あるいは体内に吸収されることによって動物の代謝エネルギーが補助されることも想定される。
【0029】
次に、栄養吸収が促進されることによって肝機能が改善することが期待されるが、実際に一般的な麻酔薬からの覚醒時間が短くなる効果が確認されている。
【0030】
また、腸管における熱ショックプロテインの透過性ならびに浸潤性が高まった場合、好熱性微生物の発酵産物を溶菌した液を豚や鶏に対して飲水添加する、あるいは魚の餌に添加することによって死亡率が減少することが判明している。
【0031】
この原因としては、当該発酵産物を溶菌することによって好熱性微生物由来の熱ショックプロテインが溶出し、それらが腸管を介して免疫担当細胞に認識されることが想定される。一般に、熱ショックプロテイン(HSP)は、MHCクラスI抗原提示経路において、抗原認識部位に位置するMHC−クラスIのタンパク分子の溝に認識されやすい。したがって、好熱性微生物由来の熱ショックプロテインは、経口投与を介して抗原のアジュバンド的な役割を果たし、抗原提示による免疫活性が上がることが考えられる。
【0032】
次に、腸内細菌相を改善することによって、腸管免疫系が活性化し、ベルパーT細胞のサブセット(Th1/Th2バランス)のそれぞれの活性化の程度に影響を与えることが一般的に知られている。細胞内の寄生体がNK細胞ならびにマクロファージを刺激することによって、インターフェロンガンマ(IFNγ)やインターロイキン12(IL−12)が産生され、ヘルパーT細胞の前駆細胞がTh1に変化するが、例えば、好熱性微生物由来の熱ショックプロテインについては、タンパク構造上において宿主細胞の熱ショックタンパクと類似しているため、細胞外の異物タンバクとして認識されてTh2を活性化するスキームとしてのみならず、細胞内寄生体として認識されてTh1が活性化することが考えられる。
一般的に、Th1が活性化されるとキラーT細胞ならびにマクロファージの働きが高まるため、免疫力が総体的に活性化する。
したがって、好熱性微生物群ならびにその含有成分を経口投与することによって、腸管免疫系が活性化する原因は、好熱性微生物由来の熱ショックプロテインが安定なタンパク質であり、構造的にも宿主と類似していることから、腸管内の透過性が高まる条件下にかぎり、図1のようなスキームで免疫系を活性化することが想定される。
【0033】
次に、好熱性微生物の発酵物は、高温下の発酵により、常温下の発酵では得られない代謝成分を豊富に含ませる。例えば非酵素的な反応(メイラード反応)によって、メラノイジンのような抗酸化・抗がん物質が生成される可能性があり、これらが腸内ならびに腸管を介して生体の体調機能に影響を与えることが想定される。
【0034】
以上の相乗効果として、図2に示したように、豚や鶏に対して、消化促進あるいは免疫力向上効果などが表れると考えられる。また、魚に対しても、図3で示したように消化力増強作用ならびに免疫力向上効果があると考えられる。次に実施例により、更に詳細に説明するが、この発明は係る実施例に限定されるものではない。
【実施例1】
【0035】
図2は、PTA−1773を含む高温発酵飼料600グラムを100リットルの水に溶解し、曝気条件下で60℃12時間処理した液を、豚の飲み水に対して0.4%添加して、春期の3月より6ヶ月間、継続飼育した場合の飼料要求率を示している。
【0036】
結果として、例年の飼料要求率が約3.3程度であるのに対して、3.1まで改善し、1キロの飼料当りの増体重が上昇する傾向が認められた。尚、試験対象の規模は、母豚150頭の一貫経営であり出荷頭数として、1年間当り約3000頭の養豚場であり、肥育豚として19.5m2当り27頭の飼育頭数で試験を実施した。
【実施例2】
【0037】
同じく、表1は別の養豚場において、前記PTA−1773を含む高温発酵飼料の処理液を、豚の飲み水に対して0.4%添加して6ヶ月間、継続飼育した場合の豚糞中の臭気関連成分の分析結果を示している。
【0038】
豚糞中の臭気関連成分としては、酢酸、乳酸、酪酸、吉草酸、アンモニアを対象として、キャピラリー電気泳動法にて分析した。
試料3gを50mlチューブに入れ、99.5%エタノール(和光純薬)10gを加え10分間振とう後、20gの超純水を入れてさらに10分間振とうさせてから一晩放置し、各成分の抽出を行った。遠心分離(3000rpm、10分間)により得られた上清を分画分子量3000Daの遠心限外ろ過フィルター(Microcon YM−3;MILLIPORE)を用いて除タンパクしたものを試料とした。
分析にはフォトダイオードアレイを搭載したキャピラリー電気泳動装置(G1600A;Agilent Technologies)を用いて測定した。Agilent Basic Anion Buffer(Agilent)を泳動バッファーとし、全長112.5cm(有効長104cm)・内径50μmのフューズドシリカキャピラリー(Agilent)に20℃で−30kVの電圧を印加し各成分を分離させた。試料の注入は50mbarの圧力で2秒間行った。検出は間接吸光度法により行い350nmと200nmの差を信号とした。各分析の間では、高圧で泳動バッファーを4分間流しキャピラリーの洗浄を行った。予めスタンダードとしてD.W.を加えて作成しておいた標準液を泳動に供し、これらスタンダードとサンプルとのピーク位置および面積の比較によって各成分の同定および濃度決定を行った。
【0039】
結果として、一般的に養豚場内の臭気が少ない場合の値(基準値)に対して、前記処理を実施していない数値は大きくずれていた。一方、前記処理6ヶ月後の数値は、比較的数値が基準値に近い値だった。また、この時の養豚場内の隣接境界領域の臭気は養豚場の存在の有無に気づかない程度の臭気まで軽減していた。
【実施例3】
【0040】
次に、図3は、2ヶ月齡の仔豚の糞中の細菌相と、高温発酵飼料を添加せずに飼育した仔豚の糞中の細菌相をPCR−DGGE法を用いて比較したデータである。尚、添加ありでは、前記PTA−1773を含む高温発酵飼料の処理液を、豚の飲み水に対して0.4%添加して6ヶ月間以上、継続飼育した。
【0041】
PCR−DGGE法は、微生物の16SrDNAに対するユニバーサルプライマーのforwardとしてGC−PRBA338fを用いて、reverseとしてPRUN518rを用いて、16SrDNA上の236bpの部位を増幅した。さらに、DGGEでは、尿素入りのgradient gelの濃度勾配を60〜30%の条件とした。具体的には、サンプル0.5gをDNA抽出装置(QuickGene−800:富士フィルム)に供し、サンプル中のDNAを抽出した。抽出したDNAを鋳型とし、PCR法により16SrRNAV3領域に対応するDNA配列中の236bpを増幅した。プライマーはGCクランプ(5’−CGC CCG CCG CGC GCG GCG GGC GGG GCG GGG GCA CGG GGG GAC TCC TAC GGG AGG CAG CAG−3’)を5’末端に付加したPRBA338f(5’−ACT CCT ACG GGA GGC AGC AG−3’)とPRUN518r(5’−ATT ACC GCG GCT GCT GG−3’)を使用した。DNAポリメラーゼにはpuReTaqReady−To−Go−PCR Beads、サーマルサイクラーにはTaKaRa PCR Thremal Cycler PERSONAL TP240を用いた。PCRの条件は92℃,2分、(92℃,1分;55℃,30秒;72℃,1分)×30サイクル、72℃,6分で行った。
PCRにより増幅されたDNAフラグメントを変性剤濃度勾配ゲル電気泳動装置(D−Coad:BioRad)に供し菌叢解析を行った。変性剤には尿素とホルムアミドを用い、泳動方向に30%〜60%の濃度勾配をかけて8%ポリアクリルアミドゲルを作成した。ゲルの作成に必要な試薬はすべてBioRad社製のDGGEシステムを使用した。泳動条件は、電圧170V、泳動時間6時間、温度60℃、泳動バッファー1×TAEとした。泳動後、SYBR Green II(タカラバイオ)で染色しトランスイルミネーターを用い310nmのUV照射下で検出した。
【0042】
図3の結果として、未添加群に対して、添加後6ヶ月では、豚糞中の細菌相のバンドバターンを比較すると、10%以下の一致率の差が認められた。また、添加群間の一致率は高く、未添加群間の一致率は低かった。
【0043】
これらのデータは、糞中の細菌相が、豚の腸内細菌相を反映していると想定できることから、添加群では、好熱性微生物、あるいはその含有成分の経口的な投与によって、常温微生物で構成されている腸内細菌相のバラツキが少なくなり、整腸作用が認められることを意味している。
【実施例4】
【0044】
次に 図4は、豚肉肩ロース赤身部分の遊離アミノ酸濃度ならびにL−カルノシン濃度を、個体数n=6で解析して、その平均値を棒グラフで、標準誤差をエラーバーで示している。肉質中の遊離アミノ酸濃度ならびにL−カルノシン濃度を前記キャピラリー電気泳動法の諸条件にて測定したデータである。なお、分析結果は、分散分析(ANOVA)によって統計解析を実施した。
【0045】
養豚場A(対照群)ならびにB(比較群)は、双方とも同じ飼料で飼育されていたが、Bについては、前記PTA−1773を含む高温発酵飼料の処理液を、豚の飲み水に対して0.4%添加して12ヶ月間以上、継続使用されていた。分析対象としたのは、生後6ヶ月を経過した出荷用肥育豚とした。
【0046】
図に示して有るように、遊離アミノ酸濃度ならびにカルノシン濃度を比較すると、対照群Aに対して比較群Bはそれぞれ、p<0.02ならびにp<0.09で増加傾向にあった。
【0047】
肉質中の遊離アミノ酸濃度は味覚に関与している成分である。またカルノシンは、ラットを用いた遊泳実験において疲労回復性が示されている成分であり、今回の結果から、前記PFA−1773を含む高温発酵飼料の処理液を、豚の飲み水として使用することによって、豚肉の肉質の旨味ならびに健康機能性に影響を与えることと想定された。
【実施例5】
【0048】
表2は、陸上養殖ヒラメに対して、前記PTA−1773を含む高温発酵飼料の処理液を、ヒラメの餌に対して0.5%添加して継続給与して6ヶ月後、12ヶ月後のヒラメ肉質中のタウリン濃度を高速液体クロマトグラフィー(HPLC)で分析した結果を示している。
【0049】
生魚は直ちに背部を採取後包丁で細断し、飼料は、乳鉢、乳棒を用い摩砕後、分析に供するまで−30℃に保管した。各試料5gを50ml遠沈管内に精秤し、5%過塩素酸を30ml加え、ホモジナイザーで約1分間攪拌後、2,500rpm、10分間遠心分離を行った。上澄みを別の50ml遠沈管にデカンテーションし、2N水酸化カリウム溶液を適量添加しpHメーターを用いて確認しながら、pH2.0に調整した。シーロンフィルムで密栓後、氷冷中に30分放置し、2,500rpm、10分間遠心分離を行った。上澄み全てを50mlメスフラスコに入れ、0.01N HClで50mlに定容し、エキス成分とした。エキス成分溶液200μlを1.5mlマイクロチューブに入れ、100mMフェニルイソチオシアネート試薬を200μl、1Mトリエチルアミン試薬を200μl加え。撹拌後、37℃で60分間反応させた。ヘキサンを400μl加え、遠心分離機で2,000rpm、5min遠心分離後、上層を除き、下層10μlを高速液体クロマトグラフィー(HPLC)で分析した。
【0050】
表2に示したとおり、前記PTA−1773を含む高温発酵飼料の処理液を餌に添加して6ヶ月の継続給与時の肉質は、通常飼料使用時のデータに対して2倍程度まで上昇することが明らかになった。さらに、12ヶ月の継続給与時の肉質は、さらに上昇する傾向が認められた。これらのデータはn=5で、有意差p<0.05であった。
【0051】
次にエキス成分のDPPH法による抗酸化活性を測定した。各試料5gを50ml遠沈管内に精秤し、エタノールを30ml加え、ホモジナイザーで約1分間攪拌後、2,500rpm、10分間遠心分離を行った。上清をエバポレーターで濃縮後、エタノールを加え10mlに定容し、エキス成分(抗酸化活性分析用)とした。エキス成分1mlをねじ付試験管に入れた。これに、エタノール1ml、100μM DPPH溶液1mlを加え撹拌し、30℃、20分反応させた。反応後、エタノール1mlを加え、517nmの吸光度を測定し、ラジカルの消去能を測定した。消去能はIC50値で求めた。
【0051】
結果として、前記PTA−1773を含む高温発酵飼料の処理液を餌に添加して6ヶ月ならびに12ヶ月の継続給与時のヒラメの肉質中の抗酸化活性は上昇する傾向が認められた。
【実施例6】
【0052】
表3−aは、前記PTA−1773を含む高温発酵飼料の処理液を、飲水に0.5%添加している飼育環境において、24ヶ月経過した養豚場の出荷用肥育豚の豚肉を比較群として、市販品の豚肉Aとともに、肉質中のカルノシンならびにリン酸イオン濃度を測定した結果である。その結果、水溶性のリン酸イオンの濃度は、市販品と比較群ともに同程度であった。一方、カルノシンについては、比較群が、市販品の2倍以上に増量していた。
【0053】
次に、表3−bでは、これらの両群の肉の肉質中の細胞骨格の強度を確認するために、温水中での前記肉汁成分の漏出の度合を確認した。ファルコンチューブに80℃の温水50ccを投入し、その温度帯に保持したまま、10gのバラ肉を5秒間浸水し、その際に温水中に溶出したエキス成分を回収して分析した。
【0054】
その結果、溶液中のカルノシン濃度は、肉質中のカルノシン濃度が高かった比較群が、388mg/lの溶出量であるのに対して、市販品の方が約1.5倍を超える608.7mg/l溶出した。また、水溶性のリン酸イオンについても、比較群よりも市販品においいて溶出量が多かった。
【0055】
以上の結果は、通常の豚肉よりも、前記PTA−1773を含む高温発酵飼料の処理液を飲水に0.4%添加している飼育環境において、24ヶ月継続飼育した養豚場の出荷用肥育豚において、肉質中の細胞骨格の強度が強いことを意味している。
【実施例7】
【0056】
次に、図5は、養豚場における離乳期経過後の死亡率に与える影響を解析した結果である。前記PTA−1773を含む高温発酵飼料の処理液を、飲水に0.4%添加している飼育環境において、10〜16ヶ月経過した養豚場における死亡率の平均値は、未処理時が10%であるのに対して、4%以下まで低下しており、統計的に有意差(p<0.05)があり、仔豚の死亡率の軽減効果は結果として60%程度であった。
【実施例8】
【0057】
図6は、ブロイラーの消耗率に与える影響を解析した結果である。前記PTA−1773を含む高温発酵飼料の処理液を、飲水に0.4%添加している飼育環境において、未処理群と比較して、出荷までの累積消耗羽数は、未処理群が一万羽の飼育規模あたり210羽以上であるのに対して、試験区では、170羽程度であり、全体として減少する傾向が認められた。
【実施例9】
【0058】
図7は、ヒラメの夏期の病・死魚率に与える影響を解析した結果である。前記PTA−1773を含む高温発酵飼料の処理液を餌に0.5%添加している飼育環境の養殖池の病・死魚数は、0.5以下であり、当該添加をしていなかった15年度の3.3匹/日と比較して、毎日の病・死魚数は、3分の1以下であった。この経済効果は、一池当り45万円程度の損失回避であった。
【0059】
さらに、病原菌のエドワジエラによる腹水発症の稚魚数の減少が確認されたことから、魚体において免疫力が強化されていることが示唆された。
【実施例10】
【0060】
図8は、採卵鶏のヒナ鶏に対して、前記PTA−1773を含む高温発酵飼料の処理液を飲水に対して、0.5%添加している飼育環境にある好熱菌液投与群と、未処理群とで、サルモネラに対する攻撃試験の結果を示したものである。
【0061】
20日齡まで、図の棒グラフで示したように、未投与群ならびに好熱菌液投与群を飼育し、21日に1羽当りの3.2億個のサルモネラ菌を添加し、1週間後に、採取したサルモネラ菌数を測定した
【0062】
その結果、好熱菌液投与群において、20%程度の減少傾向が認められた。この結果は、強いサルモネラ菌に対する抑制効果は認められないものの、内在性の腸内細菌相を活性化している可能性が示唆された。これらの結果から、サルモネラ菌の増殖抑制細菌の作用を増給する可能性があると考えられた。したがって、このような増殖抑制細菌とともに利用することが好ましいと考えられる。
【実施例11】
【0063】
次に、母豚の出産数と死産数に与える影響を解析した。図9は、前記PTA−1773を含む高温発酵飼料の処理液を飲水に対して、0.4%添加している飼育環境にある養豚場において、使用以前の出産率ならびに死産率との比較をしたデータである。
【0064】
平成16年の夏は、例年よりも高温期が継続したため、出産数は減少する傾向にあることが想定されたが、図9に示したように、10〜12月の出産数ならびに死産数ともに改善した。すなわち、出産数は例年よりも多く、5%回復し、死産数は例年よりも減少し、10%以下になっていた。
【0065】
以上の結果から、前記PTA−1773を含む高温発酵飼料の成分が、7〜9月までの受胎の成功率を向上させている可能性が想定された。
【実施例12】
【0066】
表4は、前記PTA−1773を含む高温発酵飼料の処理液を餌に0.5%添加している飼育環境でヒラメ養殖をしているA水産の養殖池の汚染度を、管内の他の池と比較した結果を示している。
【0067】
餌を投入する直前の同時刻の水を採取し分析項目として濁度、BOD、SS、全窒素を確認した。結果として、管内他社に比べて、45〜75%低い値であった。すなわち、前記PTA−1773を含む高温発酵飼料の処理液を餌に含浸することによって、水質浄化をもたらすことが明らかになった。
【0068】
以上の結果は、豚ならびに鶏に対して、図10のような概念図として示すことができるまた魚類については、図11のような概念図で示すことができる。
【発明の効果】
【0069】
当該発明は、消化力の増強、整腸作用、ならびに肉質改善、および免疫力の増強と、水質を浄化することが可能となる飼料ならびに機能性食品であり、これによって、幅広い健康維持を可能とする高品質の食品を消費者に供給することが実現する。
【図面の簡単な説明】
【0070】
【図1】 図1は、好熱性微生物由来の熱ショックプロテインが経口投与を介して、腸管免疫系を活性化し、Th1/Th2の分化誘導に与える影響についての概念図を示している。
【図2】 図2は、高温発酵飼料を液状化し、飲水添加することによって、豚の飼料要求率に与える影響を示している。
【図3】 図3は、高温発酵飼料を液状化し、飲水添加することによって、豚の腸内細菌相に与える影響を示している。
【図4】 図4は、高温発酵飼料を液状化し、飲水添加することによって、豚の肉質に与える影響に与える影響を示している
【図5】 図5は、高温発酵飼料を液状化し、飲水添加することによって、豚の離乳期間中における死亡率に与える影響を示している。
【図6】 図6は、高温発酵飼料を液状化し、飲水添加することによって、ブロイラーの消耗羽数に与える影響を示している。
【図7】 図7は、高温発酵飼料を液状化し、飲水添加することによって、ヒラメの病・死魚数に与える影響を示している。
【図8】 図8は、高温発酵飼料を液状化し、飲水添加したヒナ鶏に対するサルモネラ菌の攻撃試験の結果を示している。
【図9】 図9は、高温発酵飼料を液状化し、飲水添加した母豚の10月〜12月における出産数と死産数の比較データを示している。
【図10】 図10は、高温発酵飼料を鶏ならびに豚に投与する場合の効能に関しての概念図を示している。
【図11】 図11は、高温発酵飼料を魚類に投与する場合の効能に関しての概念図を示している。
【表1】
表1は、高温発酵飼料を液状化し、飲水添加することによって、豚の糞中の有機酸組成に与える影響を示している。
【表2】
表2は、高温発酵飼料を飼料添加することによって、ヒラメの肉質のタウリン濃度に与える影響を示している。
【表3】
表3−aは、高温発酵飼料を液状化し、飲水添加して飼育した豚の肉質のカルノシン濃度ならびにリン酸濃度と、市販の豚肉の栄養成分を比較している。表3−bは、それらの肉を80℃の温水に浸した後の溶液中の化学組成を分析したデータである。
【表4】
表4は、ヒラメの養殖池の汚染度に与える影響を分析したデータである。
【特許請求の範囲】
【請求項1】
昆虫ならびに甲殻類のキチン質を50〜70℃で発酵した産物を含むことを特徴とする飼料。
【請求項2】
魚貝類を50〜70℃で発酵した産物を含むことを特徴とする飼料。
【請求項3】
好熱性微生物によって有機物を50〜70℃で発酵して生じた耐熱性酵素を含むことを特徴とする飼料。
【請求項4】
好熱性微生物由来の耐熱性分子シャペロンを含むことを特徴とする飼料。
【請求項4】
Firmicutesに属するBacillus thermocloacae、Anoxybacillus toebii、Bacillus thermoalkalophilusの全て、あるいはいずれかが有機物を発酵した産物を含むことを特徴とする飼料。
【請求項5】
複数種の好熱性微生物群で構成されるPTA−1773、あるいはその複合微生物群のいずれか一つが有機物を発酵した成分を含むことを特徴とする飼料。
【請求項6】
前記1乃至5記載の飼料であって、消化力を増強する機能を有する飼料。
【請求項7】
前記1乃至5記載の飼料であって、腸内の有機酸組成を変化することによって、動物糞の臭気を軽減することを特徴とする飼料。
【請求項8】
前記1乃至5記載の飼料であって、腸内の有機酸組成を変化することによって、増体あるいは減量させることを特徴とするカロリー吸収率調整型の飼料。
【請求項9】
前記1乃至5記載の飼料であって、動物腸内の微生物相を変化させることを腸内の微生物相を整えることを特徴とする整腸剤型の飼料。
【請求項10】
前記1乃至5記載の飼料であって、肉質中の遊離アミノ酸濃度を増加させることを特徴とする飼料。
【請求項11】
前記1乃至5記載の飼料であって、肉質中の遊離脂肪酸濃度を変化させることを特徴とする飼料。
【請求項12】
前記1乃至5記載の飼料であって、肉質中の細胞膜の細胞骨格を増強することを特徴とする飼料。
【請求項13】
前記1乃至5記載の飼料であって、肉質中の遊離カルノシン濃度を増加させることを特徴とする飼料。
【請求項14】
前記1乃至5記載の飼料であって、肉質中の遊離タウリン濃度を増加させることを特徴とする飼料。
【請求項15】
前記1乃至5記載の飼料であって、腸内細菌相の変化に伴い、動物の血液中の免疫賦活体を活性化し、動物の免疫力を増強することを特徴とする飼料。
【請求項16】
前記1乃至5記載の飼料であって、受胎効率を改善することを特徴とする飼料。
【請求項17】
前記1乃至5記載の飼料であって、当該飼料を給与された動物の消化促進に伴い、当該動物の排泄後の糞の浄化が促進し、水圏の有機物分解を促進させることを特徴とする飼料型の水質浄化剤。
【請求項18】
前記1乃至17記載の成分と機能を有する、多機能性飼料。
【請求項19】
前記1乃至17記載の成分と機能を有する、ヒトに対する多機能性食品。
【請求項1】
昆虫ならびに甲殻類のキチン質を50〜70℃で発酵した産物を含むことを特徴とする飼料。
【請求項2】
魚貝類を50〜70℃で発酵した産物を含むことを特徴とする飼料。
【請求項3】
好熱性微生物によって有機物を50〜70℃で発酵して生じた耐熱性酵素を含むことを特徴とする飼料。
【請求項4】
好熱性微生物由来の耐熱性分子シャペロンを含むことを特徴とする飼料。
【請求項4】
Firmicutesに属するBacillus thermocloacae、Anoxybacillus toebii、Bacillus thermoalkalophilusの全て、あるいはいずれかが有機物を発酵した産物を含むことを特徴とする飼料。
【請求項5】
複数種の好熱性微生物群で構成されるPTA−1773、あるいはその複合微生物群のいずれか一つが有機物を発酵した成分を含むことを特徴とする飼料。
【請求項6】
前記1乃至5記載の飼料であって、消化力を増強する機能を有する飼料。
【請求項7】
前記1乃至5記載の飼料であって、腸内の有機酸組成を変化することによって、動物糞の臭気を軽減することを特徴とする飼料。
【請求項8】
前記1乃至5記載の飼料であって、腸内の有機酸組成を変化することによって、増体あるいは減量させることを特徴とするカロリー吸収率調整型の飼料。
【請求項9】
前記1乃至5記載の飼料であって、動物腸内の微生物相を変化させることを腸内の微生物相を整えることを特徴とする整腸剤型の飼料。
【請求項10】
前記1乃至5記載の飼料であって、肉質中の遊離アミノ酸濃度を増加させることを特徴とする飼料。
【請求項11】
前記1乃至5記載の飼料であって、肉質中の遊離脂肪酸濃度を変化させることを特徴とする飼料。
【請求項12】
前記1乃至5記載の飼料であって、肉質中の細胞膜の細胞骨格を増強することを特徴とする飼料。
【請求項13】
前記1乃至5記載の飼料であって、肉質中の遊離カルノシン濃度を増加させることを特徴とする飼料。
【請求項14】
前記1乃至5記載の飼料であって、肉質中の遊離タウリン濃度を増加させることを特徴とする飼料。
【請求項15】
前記1乃至5記載の飼料であって、腸内細菌相の変化に伴い、動物の血液中の免疫賦活体を活性化し、動物の免疫力を増強することを特徴とする飼料。
【請求項16】
前記1乃至5記載の飼料であって、受胎効率を改善することを特徴とする飼料。
【請求項17】
前記1乃至5記載の飼料であって、当該飼料を給与された動物の消化促進に伴い、当該動物の排泄後の糞の浄化が促進し、水圏の有機物分解を促進させることを特徴とする飼料型の水質浄化剤。
【請求項18】
前記1乃至17記載の成分と機能を有する、多機能性飼料。
【請求項19】
前記1乃至17記載の成分と機能を有する、ヒトに対する多機能性食品。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2009−100728(P2009−100728A)
【公開日】平成21年5月14日(2009.5.14)
【国際特許分類】
【出願番号】特願2007−297257(P2007−297257)
【出願日】平成19年10月19日(2007.10.19)
【出願人】(501176303)日環科学株式会社 (10)
【出願人】(597149951)株式会社三六九 (6)
【出願人】(598092993)京葉プラントエンジニアリング株式会社 (8)
【Fターム(参考)】
【公開日】平成21年5月14日(2009.5.14)
【国際特許分類】
【出願日】平成19年10月19日(2007.10.19)
【出願人】(501176303)日環科学株式会社 (10)
【出願人】(597149951)株式会社三六九 (6)
【出願人】(598092993)京葉プラントエンジニアリング株式会社 (8)
【Fターム(参考)】
[ Back to top ]