
混在した細胞性組成物からのras介在性新生物細胞のレオウイルスクリアランス
【課題】新生物細胞を含む可能性のある自己移植片を清浄化するための、かなりの収率を有する非常に選択的な方法に対する必要性が存在する。
【解決手段】本発明は、ras介在性新生物細胞を選択的に溶解するレオウイルスで、混在した細胞性組成物を感染することによる、混在した細胞性組成物からras介在性新生物細胞を除去する方法に関する。多くの新生物細胞が活性化ras経路と関連しているため、本発明は広範囲の混在した細胞性組成物から、混在したまたは自然発生的な新生物細胞を除去するために使用できる。
【解決手段】本発明は、ras介在性新生物細胞を選択的に溶解するレオウイルスで、混在した細胞性組成物を感染することによる、混在した細胞性組成物からras介在性新生物細胞を除去する方法に関する。多くの新生物細胞が活性化ras経路と関連しているため、本発明は広範囲の混在した細胞性組成物から、混在したまたは自然発生的な新生物細胞を除去するために使用できる。
【発明の詳細な説明】
【技術分野】
【0001】
関連発明
本出願は、35 U.S.C. 119(e)の下で、2000年5月3日に出願された米国予備出願60/201,990、2000年5月19日に出願された60/205,389、2001年2月13日に出願された60/268,054の利益を享受する。前述の予備出願のそれぞれの全体の開示は、参考としてここに取り込まれる。
【0002】
本発明は、組成物中のras介在性新生物細胞を選択的に溶解するレオウイルスで、混在した細胞性組成物を感染することによる、混在した細胞性組成物からras介在性新生物細胞を除去する方法に関する。
【背景技術】
【0003】
通常細胞増殖は、増殖促進シグナルと増殖抑制シグナルの両者によって調節される。各細胞に対してこれらの2種類のシグナルは、特定の細胞に対する身体の必要性を反映する態様で、通常バランスを与えているであろう。もし細胞が増殖抑制シグナルに応答しない場合、または増殖促進シグナルに過剰応答する場合、その細胞は以上に迅速に増殖し(新生物細胞と称する)、悪性新生物であるガンに段階的に発展するであろう。
【0004】
ガンを治療するために現在行われている方法である化学療法は、ガン細胞の迅速な増殖特性に一般的に基づいている。ガン細胞は迅速に増殖するため、それらは細胞増殖を阻害する薬剤により感受性である。理論的には、化学療法薬の投与量を注意深く選択することによって、正常細胞を重大に損傷することなくガン細胞の増殖を阻害できる。しかしながら、造血幹細胞のようないくつかの正常細胞もまた迅速に増殖する。それ故、ガン細胞に有害なある投与量は、しばしば造血幹細胞にも有害である。他方で、もし投与量がガン細胞を殺傷するのに十分な程高くないと、化学療法が終結した後、ガンが短期間で再発する危険が存在する。
【0005】
選択的にガン細胞を殺傷する投与量を見出すことは困難であるため、高投与量の化学療法、引き続き異種造血始原幹細胞移植は、多くのガンにおける治療アプローチとして過度の適用が実施されている(例えば、Winter, 1999; Nieto及びShpall, 1999参照)。このアプローチでは、造血幹細胞の部分がガン患者から除去されて、次いで患者は、ガン細胞と造血幹細胞のような迅速に増殖する細胞に対して致死的な高投与量化学療法で治療される。その後患者は、同じ患者から以前に除去された自己造血幹細胞の移植を受け、造血系が再生する。
【0006】
この治療の重大な欠点は、造血始原幹細胞が患者から除去される場合、それらにしばしばガン細胞が混在している点である。これは特に、患者が造血系起源のガンを有する場合に問題であるが、特にもし固形腫瘍が転移したならば、固形腫瘍を有する患者でも、造血幹細胞の混在に苦しむ可能性がある。結果として、除去された細胞が戻されて移植され、造血系を再生する場合に、あるガン細胞がガン患者に戻されて配置され、その場合にそのガンが再び増殖して、ガンの再発に寄与する可能性がある。それ故、移植前の自己移植片を清浄化することが望ましい。
【0007】
自己移植片を清浄化するためにいくつかの方法が利用されている(Spyridonidis等, 1998;Bensinger, 1998)。自己移植片を化学療法で処理し、in vitroで混在した新生物細胞を殺傷できる。しかしながら、前述のように、新生物細胞またはガン細胞を選択的に殺傷するが、清浄な造血幹細胞を完全なままにする化学療法薬の投与量を見出すことは困難である。自己移植片はまた、新生物細胞に特異的である抗原を認識する抗体に接合された毒素で治療することもできるが、そのような腫瘍特異的抗原がいつも存在するとは限らない。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】U.S. Pat. No. 6,136,307
【特許文献2】WO 99/08692, published Feb. 25, 1999.
【非特許文献】
【0009】
【非特許文献1】Armstrong, G. D. et al., "Studies on reovirus receptors of L cells: virus binding characteristics and comparison with reovirus receptors of erythrocytes", Virology 138:37: 37-48 (1984)
【非特許文献2】Bensinger, W. I., "Should we purge?", Bone Marrow Tranplant. 21: 113-115 (1998)
【非特許文献3】Bos, J. L., "Ras Oncogenes in Human Cancer: A Review", Canc. Res. 49(17): 4682-4689 (1989)
【非特許文献4】Chandron and Nibert, "Protease cleavage of reovirus capsid protein mu1 and mu1C is blocked by alkyl sulfate detergents, yielding a new type of infectious subvirion particle", J. of Virology 72(1):467-75 (1998)
【非特許文献5】Coffey, M. C., et al., "Reovirus therapy of tumors with activated Ras pathway", Science 282: 1332-1334 (1998)
【非特許文献6】Duggan, P. R., et al., "Predictive factors for long-term engraftment of autologous blood stem cells", Bone Marrow Transplantation 26(12): 1299-1304 (2000)
【非特許文献7】Fields, B. N. et al., Fundamental Virology. 3rd Edition, Lippincott-Raven (1996)
【非特許文献8】Gao, J., B. Tombal and J. T. Isaacs, "Rapid in situ hybridization technique for detecting malignant mouse cell contamination in human xenograft tissue from nude mice and in vitro cultures from such xenografts", Prostate 39(1): 67-70, 1999
【非特許文献9】Gentsch, J. R. K. and Pacitti, A. F., "Effect of neuraminidase treatment of cells and effect of soluable glycoproteins of type 3 reovirus attachment to murine L cells", J. Virol. 56:356: 356-64 (1985)
【非特許文献10】Neito, Y. and E. J. Shpall, "Autologous stem-cell transplantation for solid tumors in adults", Hematol. Oncol. Clin. North Am. 13(5): 939-968 (1999)
【非特許文献11】Norman, K. and P. Lee, "Reovirus as a novel oncolytic agent", J. Clin. Invest. 105 (8): 1035-1038 (2000)
【非特許文献12】Paul R. W. et al., "The alpha-anameric form of sialic acid is the minimal receptor determinant recognized by reovirus", Virology 172: 382-385 (1989)
【非特許文献13】Sabin, A. B., Science 130: 966 (1959)
【非特許文献14】Spyridonidis, A. et al., "Minimal residual disease in autologous hematopoietic harvests from breast cancer patients", Annals of Oncology 9: 821-826 (1998)
【非特許文献15】Stewart, D. A., et al., "Superior autologous blood stem cell mobilization from dose-intensive cyclophosphamide, etoposide, cisplatin plus G-CSF than from less intensive chemotherapy regimens", Bone Marrow Transplant. 23(2): 111-117 (1999)
【非特許文献16】Strong, J. E., et al., "The molecular basis of viral oncolysis: usurpation of the Ras 40 signaling pathway by reovirus", EMBO J. 17: 3351-3362 (1998)
【非特許文献17】Strong, J. E. and P. W. Lee, "The v-erbV oncogene confers enhanced cellular susceptibility to reovirus infection", J. Virol. 70: 612-616 (1996)
【非特許文献18】Strong, J. E., et al., "Evidence that the Epidermal Growth Factor Receptor on Host Cells Confers Reovirus Infection Efficiency", Virology 197(1): 405-411 (1993)
【非特許文献19】Winter, J. N., "High-dose therapy with stem-cell transplantation in the malignant 5 lymphomas", Oncology (Huntingt) 13(12): 1635-1645 (1999)
【発明の概要】
【発明が解決しようとする課題】
【0010】
フローサイトメトリー、アフィニティーカラム、または磁性ビーズを使用することによって、幹細胞特異的表面マーカー(CD34)に基づいて、幹細胞を他の細胞から分離することも可能である。しかしながら、特定の造血細胞、例えばCD34+細胞のみを選択することによって、T細胞、B細胞、単球、及びナチュラルキラー細胞のような他の造血細胞もまた除去され、免疫回復が遅延する可能性がある(Bensinger, 1998)。この方法はまた、CD34+細胞の約半分の損失と、混在したガン細胞のある部分の維持を引き起こす(Spyridonidis, 1998)。それ故、新生物細胞を含む可能性のある自己移植片を清浄化するための、かなりの収率を有する非常に選択的な方法に対する必要性が存在する。
【課題を解決するための手段】
【0011】
本発明は、ras介在性新生物細胞を選択的に溶解するレオウイルスで、混在した細胞性組成物を感染することによる、混在した細胞性組成物からras介在性新生物細胞を除去する方法に関する。多くの新生物細胞が活性化ras経路と関連しているため、本発明は広範囲の混在した細胞性組成物から、混在したまたは自然発生的な新生物細胞を除去するために使用できる。従って、本発明の一つの特徴点は、ras介在性新生物細胞を含む疑いのある細胞性組成物からras介在性新生物細胞を除去する方法であって、ras介在性新生物細胞の腫瘍崩壊を引き起こす条件下で、前記細胞性組成物をレオウイルスと接触させることを含む方法を提供する。
【0012】
本発明の好ましい実施態様では、造血幹細胞を含む細胞性組成物が、混在したまたは自然発生的に存在する新生物細胞として細胞性組成物において存在するras介在性新生物細胞を除去するために処理される。レオウイルスは非常に選択的であり、ras介在性細胞のみに溶解的に感染するため、この方法は、造血系を分化し再構成する造血幹細胞の能力には影響しない。本発明の方法は、骨髄または血液由来の造血幹細胞を清浄化するために使用できる。造血幹細胞は、自己由来、同種由来、または異種由来であっても良い。
【0013】
本発明の別の実施態様では、移植の前に組織または器官移植物からras介在性新生物細胞を除去するために、レオウイルスが使用される。この方法は、移植物またはras介在性細胞のタイプや年齢と無関係に適用できるため、及びレオウイルスは正常細胞及び組織に必須に無害であるため、この方法は、移植の前にいずれかの移植物を「クリーンアップ」するための通常の実施として使用できる。
【0014】
本発明の別の実施態様では、ras経路の活性化のため自然発生的にトランスフォームされた細胞を除去するため、レオウイルスが培養細胞系を処理するために使用できる。この方法はさらに、人工授精または他の再生産関連方法の前に、精子またはドナーの卵を処理するためにも使用できる。
【0015】
本発明において有用なレオウイルスは、いずれのレオウイルスでも良い。好ましくはレオウイルスは、哺乳動物レオウイルス、より好ましくはヒトレオウイルスである。さらにより好ましくはレオウイルスは血清型3であり、最も好ましくはDearing株のレオウイルスである。
【0016】
本発明の好ましい実施態様では、本発明の方法は、DMSOを含む溶液中にレオウイルスで処理された組成物を凍結して貯蔵する工程をさらに含む。DMSOは動物細胞を凍結し貯蔵するために一般的に使用されているが、レオウイルスを変性する。それ故、この処理は、細胞性組成物から感染性レオウイルスを除去する一方で、長期間に亘って凍結状態にある組成物の活性を保存する。
【0017】
本発明の別の実施態様では、レオウイルスを溶解するために、抗レオウイルス抗体、または抗レオウイルス抗体と補体の組み合わせに混合物をかけることによって、レオウイルスで処理された細胞性組成物からレオウイルスが除去される。別法としてまたは加えて、レオウイルス粒子の表面にある分子を認識する抗ウイルス抗体が、この抗体を固定化し、固定化抗体に細胞性組成物を適用し、及び抗体に結合していない組成物の部分を回収することによって、レオウイルスを除去するために使用されても良い。
【0018】
同様に、抗レオウイルス抗体はin vivoでレオウイルスを除去するために移植受容者に投与でき、または受容者はこの目的を達成するために免疫刺激剤を与えられることができる。
【0019】
本発明の別の実施態様では、細胞からレオウイルスを分離できる勾配を使用することによって、レオウイルスで処理された細胞性組成物からレオウイルスが除去される。
【0020】
ras介在性新生物細胞を除去するためにレオウイルスで処理されている細胞性組成物もまた提供される。そのような組成物は、in vitro研究、または移植、受精、または他のin vivo方のために使用されても良い。移植は、自己由来、同種由来、または異種由来であっても良い。好ましくは移植は自己移植である。より好ましくは組成物は造血幹細胞を含む。
【0021】
本発明は、レオウイルスを使用することによる細胞性組成物からras介在性新生物細胞を除去する方法、並びにこれらの方法に従って処理されている組成物に関する。
【0022】
本発明をさらに詳細に記載する前に、この明細書において使用される用語は、他に記載がなければ以下のように定義される。
【図面の簡単な説明】
【0023】
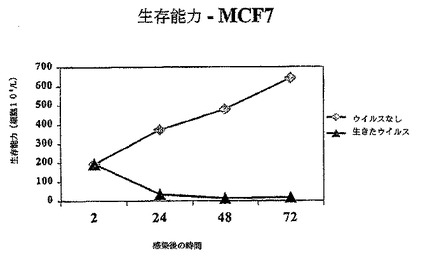
【図1A】図1Aは、示されているように生存レオウイルス、死んだウイルス、ウイルスなしで感染されたMCF7における生存細胞の数を示す図である。
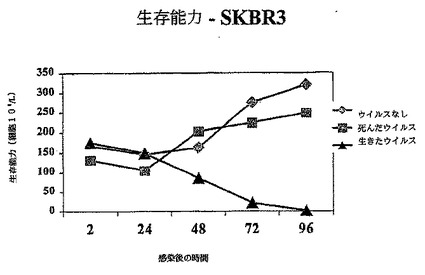
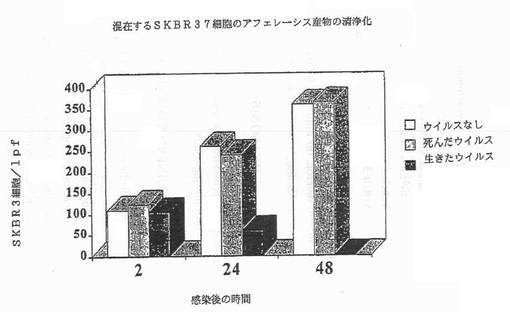
【図1B】図1Bは、示されているように生存レオウイルス、死んだウイルス、ウイルスなしで感染されたSKRB3における生存細胞の数を示す図である。
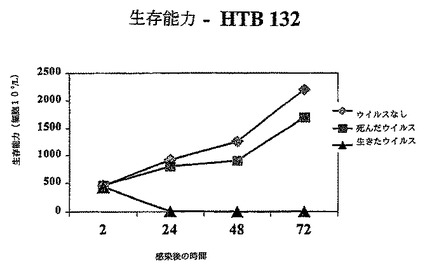
【図1C】図1Cは、示されているように生存レオウイルス、死んだウイルス、ウイルスなしで感染されたHTB132における生存細胞の数を示す図である。
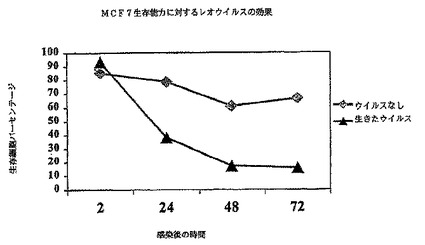
【図1D】図1Dは、レオウイルス感染の後の各種の時点での生存中のMCF7細胞のパーセンテージを示す図である。
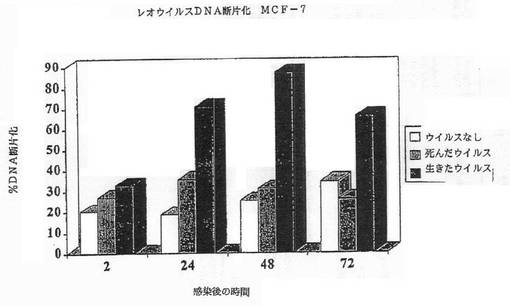
【図2A】図2Aは、MCF7細胞におけるレオウイルス感染によってアポトーシスが誘導されたことを示す図である。レオウイルス感染の後に断片化したDNAのパーセンテージが示されている。
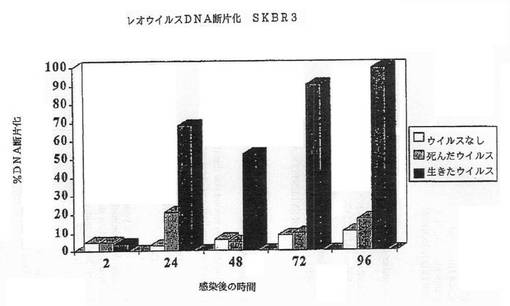
【図2B】図2Bは、SKRB3細胞におけるレオウイルス感染によってアポトーシスが誘導されたことを示す図である。レオウイルス感染の後に断片化したDNAのパーセンテージが示されている。
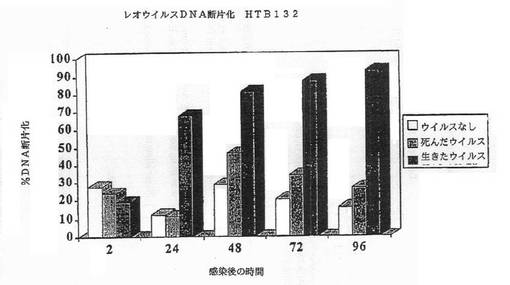
【図2C】図2Cは、HTB1327細胞におけるレオウイルス感染によってアポトーシスが誘導されたことを示す図である。レオウイルス感染の後に断片化したDNAのパーセンテージが示されている。
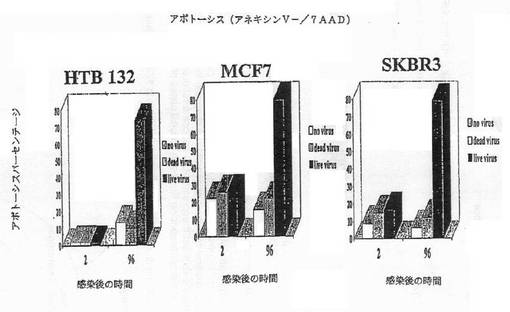
【図2D】図2Dは、レオウイルス感染の後のアポトーシスマーカーアネキシンV染色のパーセンテージを示す図である。
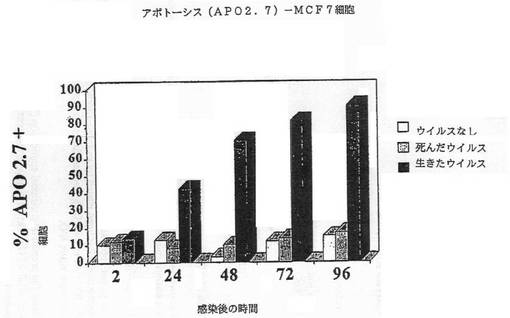
【図2E】図2Eは、MCF7細胞におけるAPO2.7+細胞のパーセンテージを示す図である。
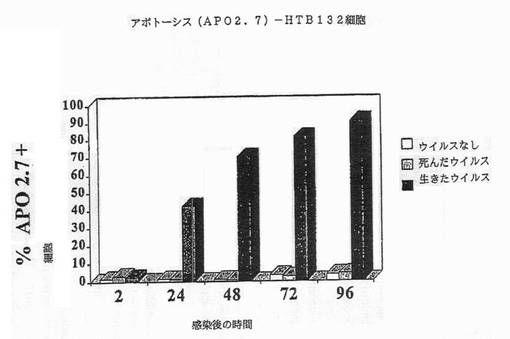
【図2F】図2Fは、HTB132細胞におけるAPO2.7+細胞のパーセンテージを示す図である。
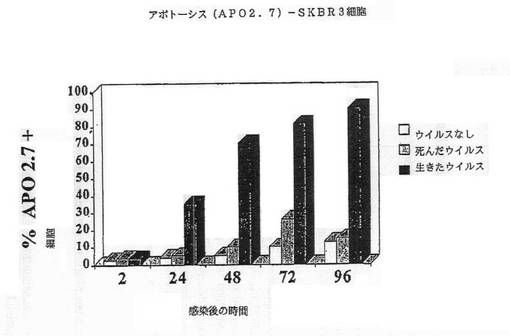
【図2G】図2Gは、SKRB3細胞におけるAPO2.7+細胞のパーセンテージを示す図である。
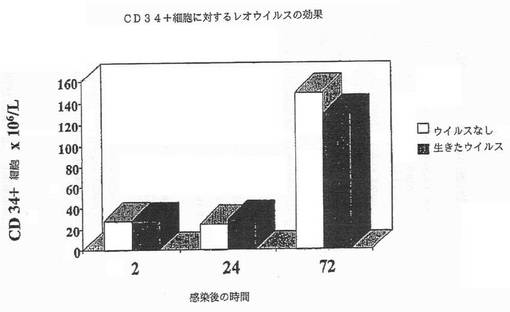
【図3A】図3Aは、CD34+幹細胞がレオウイルスで感染された後の各種の時点での生存細胞の数を示す図である。
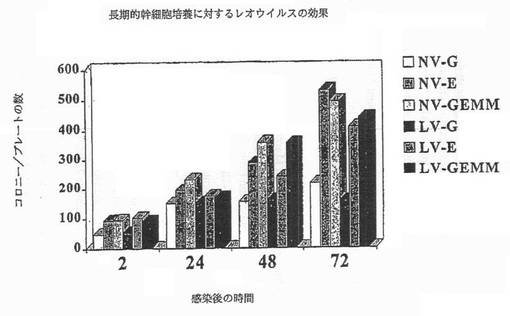
【図3B】図3Bは、長期的な幹細胞培養に対するレオウイルスの効果を示す図である。幹細胞をレオウイルスで感染し、それぞれ2,24,48または72時間インキュベートし、次いで細胞を希釈して細胞を14日間培養し、個々のコロニーを形成させた。顆粒球(G)、赤血球(E)、または顆粒球赤血球マクロファージ巨核球(GEMM)の各種のコロニーの数を、それぞれウイルスなし(NV)及び生存ウイルス(LV)で感染した細胞について測定した。例えば、NV-Gはウイルスなしで処理された細胞から由来する顆粒球コロニーを示し、LV-Gは生存ウイルスで処理された細胞から由来する顆粒球を示す。
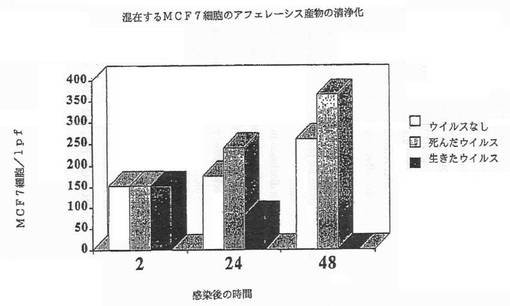
【図4A】図4Aは、MCF7細胞を有するアフェレーシス産物の混合物に対するレオウイルスの清浄化効果を示す図である。
【図4B】図4Bは、MDA MB 468細胞を有するアフェレーシス産物の混合物に対するレオウイルスの清浄化効果を示す図である。
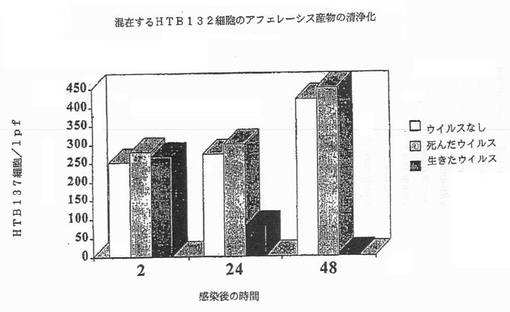
【図4C】図4Cは、SKRB3細胞を有するアフェレーシス産物の混合物に対するレオウイルスの清浄化効果を示す図である。
【発明を実施するための形態】
【0024】
定義
ここで使用される用語、「新生物細胞」、または「増殖性疾患を有する細胞」は、正常な増殖阻害特性を有さずに増殖する細胞を指す。新生物細胞を含む新たな増殖物は、新生物または腫瘍である。新生物は、一般的に別個の塊を形成する異常な組織増殖であり、正常な組織増殖より迅速な細胞増殖によって増殖する。新生物は、正常な組織との構造的組織化及び機能的強調の部分的または完全な欠如を示すであろう。ここで使用される新生物は、造血性新生物、並びに固形新生物を包含するように企図される。
新生物は、良性(良性腫瘍)または悪性(悪性腫瘍またはガン)であって良い。悪性腫瘍は、3種の主なタイプに広く分類できる。上皮構造から生ずる悪性新生物はカルシノーマと称され、筋肉、軟骨、脂肪、または骨のような結合組織から由来する悪性新生物はサルコーマと称され、免疫系の構成成分を含む造血構造(血液細胞の形成と関係する構造)に影響する悪性腫瘍は白血病及びリンパ腫と称される。他の新生物は、神経芽腫を制限することなく含む。
【0025】
ここで使用される用語、「ras活性化新生物細胞」または「ras介在性新生物細胞」は、少なくとも部分的にras経路の活性化のために異常に速い速度で増殖する細胞を指す。ras経路は、ras遺伝子の構造的ミューテーション、ras遺伝子発現の増大したレベル、ras遺伝子メッセージの増大した安定性、あるいはras、またはras経路におけるrasから下流または上流の因子の活性化を導くいずれかのミューテーションまたは他のメカニズムによって活性化され、それによってras経路活性を増大するであろう。例えば、EGFレセプター、PDGFレセプター、またはSosの活性化は、ras経路の活性化を導く。ras介在性新生物細胞は、ras介在性ガン細胞を制限することなく含み、それはras経路の活性のによる悪性の態様で増殖する細胞である。
【0026】
ここで使用される用語、「細胞性組成物」は、細胞を含む組成物を意味する。前記組成物は、非細胞物を含んでも良い。例えば全血は、赤血球及び白血球のような細胞に加えて血漿、血小板、ホルモン、及び他の非細胞物を含む細胞性組成物である。細胞性組成物は、各種のタイプの細胞、器官、または構築物を含んでも良い。例えば、所定の構造で整列した各種の細胞タイプを含む組織または器官は、細胞性組成物と考慮される。
【0027】
ここで使用される用語、「レオウイルス」は、各種の属に分類されるいずれのウイルスをも指す。レオウイルスの名称(Respiratory and enteric orphan virus)は、これらのウイルスが、ヒトにおけるいずれかの周知の疾患と関係していないが、呼吸器及び腸管の両者から単離できることを示唆する説明的な頭字語である(Sabin, 1959)。用語「レオウイルス」は、レオウイルス属に分類される全てのウイルスを指す。
【0028】
ヒトのレオウイルスは3種の血清型からなる:タイプ1(Lang株またはT1L)、タイプ2(Jones株、T2J)及びタイプ3(Dearing株またはAbney株、T3D)。この3種の血清型は、中和及びヘマグルチニン阻害アッセイを基礎に容易に同定可能である(例えばFields等, 1996参照)。
【0029】
レオウイルスは天延に存在するものでも改変されていても良い。レオウイルスが天然のソースから単離でき、研究室でヒトによって意図的に改変されていない場合、レオウイルスは「天然に存在するもの」である。例えば、レオウイルスが「フィールドソース」由来であることができ、つまりレオウイルスで感染されたヒト由来であることができる。
【0030】
レオウイルスは、改変されても、活性なras経路を有する哺乳動物細胞を溶解的に感染可能であっても良い。レオウイルスは、増殖中の細胞に生化学的に投与される前に、化学的または生化学的に調製されても良い(例えばキモトリプシンまたはトリプシンのようなプロテアーゼでの処理によって)。プロテアーゼでの事前処理は、ウイルスの外側コートまたはキャプシドを除去し、ウイルスの感染能力を増大するであろう。レオウイルスは、レオウイルスに対する発達した免疫を有する哺乳動物から免疫応答を減少または妨げるために、リポソームまたはミセルにコートされても良い(Chandron及びNibert, 1998)。例えば、新規な感染性サブぶりオン粒子を生産するために、ビリオンがミセル形成濃度の硫酸アルキル変性剤の存在下でキモトリプシンで処理されても良い。
【0031】
レオウイルスが異なる抗原決定基を有し、それによってレオウイルス血清型に以前にさらされた哺乳動物による免疫応答を減少するまたは妨げるように、レオウイルスは、異なる病原性表現型を有する二つ以上のタイプのレオウイルスから得られた組換えレオウイルスであっても良い。そのような組換えビリオンは、生成したビリオンキャプシド内への、異なるサブタイプのコートタンパク質の生成したリソーティングと取り込みで、異なるサブタイプのレオウイルスを有する哺乳動物細胞の共感染によって生産できる。
【0032】
レオウイルス感染に対する細胞の「耐性」は、ウイルスを有する細胞の感染が、有意なウイルス生産または収率を生じないことを示す。「感受性」である細胞は、細胞変性効果の誘導、ウイルスタンパク質合成、及び/またはウイルス生産を示すものである。レオウイルス感染に対する耐性は、早期の転写よりもむしろ遺伝子翻訳のレベルで見出された:ウイルス転写物は生産される一方、ウイルスタンパク質は発現されなかった。一つの理論に制限されるものではないが、耐性細胞におけるウイルス遺伝子転写は、約65kDa細胞タンパク質のリン酸化と強調し、それは二本鎖RNA活性化プロテインキナーゼ(PKR)と決定され、それはトランスフォーム化細胞では観察されなかった。PKRのリン酸化は、転写の阻害を導く。リン酸化が周知のPKRのインヒビターである2−アミノプリンによって抑制された場合、レオウイルスタンパク質合成の促進がトランスフォーム化細胞で生じた。
【0033】
ここで使用される用語、「腫瘍崩壊」または「新生物細胞の生存能力の減少」は、標的新生物細胞の生存能力の少なくとも約20%の減少を意味する。生存能力は処理された細胞の生存可能な細胞のカウントによって測定でき、減少の度合いは、非処理細胞における生存可能な細胞の数と処理細胞における生存可能な細胞の数を比較することによって、またはウイルス処理の前後の生存可能な細胞のカウントを比較することによって測定できる。生存能力の減少は好ましくは、少なくとも約50%、より好ましくは少なくとも約70%、さらにより好ましくは少なくとも約80%、最も好ましくは少なくとも約90%である。
【0034】
ここで使用される用語、「移植受容者」は、細胞性組成物の移植を受ける哺乳動物である。好ましくは前記受容者はヒトであり、より好ましくは前記受容者は、ガンの治療において移植を受けるヒトである。
【0035】
ここで使用される用語、アポトーシスマーカーとしての「CPE」は、細胞膜気泡化、核の凝集、及びクロマチン凝集のような、顕微鏡の下で観察可能なアポトーシスと典型的に関連する細胞変性効果を指す。
【0036】
方法
本発明は、ras介在性新生物細胞を含む疑いのある細胞性組成物からras介在性新生物細胞を除去するレオウイルスの使用に関する。活性化ras経路は多くの腫瘍と関連しているため、本発明は広範囲の新生物細胞のいずれを有する疑いのある細胞性組成物にも適用できる。
【0037】
哺乳動物レオウイルスにとって、細胞表面認識シグナルはシアル酸である(Armstrong, 1984; Gentsch及びPacitti, 1985; Paul等, 1989)。シアル酸の独特な性質のため、レオウイルスは多数の細胞系に有効に結合し、そのため多くの各種の組織を標的化できるであろう;しかしながら、細胞系の間でレオウイルス感染に対する感受性の有意な差異が存在する。この感受性の矛盾の一つの理由は、各細胞系におけるras経路の活性であろう。
【0038】
レオウイルスが、in vitro、in vivo及びex vivoで活性化新生物細胞を選択的に溶解することを、我々は最近発見した(Coffey等, 1998; WO 99/08692)。普通では、細胞はレオウイルス感染に感受性ではない。しかしながら、ras経路が活性化されたならば、レオウイルスは細胞において成功して複製でき、最終的に宿主細胞の溶解を引き起こす。例えば、レオウイルス耐性NIH 3T3細胞が、rasを活性化するタンパク質である活性化rasまたはSosでトランスフォームされた場合、レオウイルス感染は増大された(Strong等, 1998)。同様に、レオウイルス感染に耐性であるマウス線維芽細胞は、EGFレセプター遺伝子またはv-erbB原ガン遺伝子でのトランスフェクションの後感受性となった(Strong等, 1993; Strong等, 1996)。
【0039】
一つの理論に制限されるものではないが、レオウイルス複製は翻訳レベルで調節されているようである(Strong等, 1998; Norman等, 2000)。非トランスフォーム化NIH 3T3細胞では、早期のウイルス転写物が二本鎖RNA活性化プロテインキナーゼ(PKR)を活性化し、それが翻訳を阻害し、それによってウイルス複製を阻害する。活性化ras(またはras経路の活性化エレメント)は、おそらくPKR活性化を阻害または不活性化するであろう。それ故、ウイルスタンパク質合成が進行し、ウイルス粒子が作製され、細胞は最終的に溶解する。
【0040】
ras原ガン遺伝子は、多数の腫瘍の原因である。ras遺伝子自体の活性化ミューテーションは、全てのヒト腫瘍の約30%で生じ(Bos, J.L., 1989)、主に膵臓(90%)、散発生大腸(50%)及び肺(40%)カルシノーマ、並びにミエロイド白血病(30%)で生じている。ras経路におけるrasの上流または下流因子の活性化も、腫瘍と関連している。例えば、HER2/Neu/ErbB2または上皮増殖因子(EGF)レセプターの過剰発現は、乳ガンにおいて一般的であり(25−30%)、血小板由来増殖因子(PDGF)レセプターまたはEGFレセプターの過剰発現は、グリオーマ及びグリオブラストーマ(40−50%)で一般的である。EGFレセプターとPDGFレセプターは、それらのそれぞれのリガンドに対する結合に際してrasを活性化することが知られており、v-erbBは細胞外ドメインを欠く構成的に活性化されたレセプターをコードする。
【0041】
我々は、レオウイルスが、感染細胞においてアポトーシスを誘導することによる、3種の乳ガンモデルシステム、MCF7、SKRB3及びHTB 132の腫瘍崩壊を有効に導くことを本発明において示した(実施例1)。かくしてレオウイルス処理は、MCF7、SKRB3及びHTB 132細胞の生存能力の顕著な減少を引き起こす一方、ウイルスなしまたは死んだウイルスで処理されたコントロールは普通に増殖した(図1A−1D)。生存能力の減少は、DNA断片化、アネキシンVまたはAPO2.7染色陽性(図2A−2G)、並びに顕微鏡で観察される細胞膜の気泡、核の凝集、及びクロマチン凝集のような細胞変性効果といった、アポトーシスと関連する特徴を伴った。
【0042】
レオウイルス感染は、正常細胞において翻訳レベルで通常ブロックされるが、ras介在性新生物細胞においてはブロックされないため、レオウイルス処理MCF7細胞及びCD34+幹細胞におけるタンパク質合成の度合いを調べた(図2)。実際、ウイルスタンパク質は、レオウイルス感染ガン細胞系で合成されるが、レオウイルスで処理されたCD34+幹細胞では合成されなかった(データ示さず)。この結果は、レオウイルスタンパク質がレオウイルス処理幹細胞では合成されず、細胞タンパク質合成が普通に進行しているため、レオウイルスでの造血幹細胞の処理が安全であることを示唆している。この点を確認するために、レオウイルス処理CD34+細胞の生存能力を、レオウイルス処理の各種の時点で測定した(実施例3)。生きたレオウイルスまたはウイルスなしで処理された集団における細胞数は各時点で同じであり、CD34+細胞がレオウイルス感染に感受性ではないことを示した。
【0043】
レオウイルスが、高投与量化学療法治療における造血幹細胞の清浄化に有用であるために、レオウイルス処理が、全部の造血系を再構成するそれぞれ及び毎回の造血系列へ分化する幹細胞の能力を変えないことが必須である。それ故、レオウイルス処理の長期的な効果が評価された(実施例3)。ウイルスなしまたは生きたウイルスのいずれかで処理されたCD34+細胞は、レオウイルス処理の72時間後でさえ、白血球、赤血球または白血球赤血球マクロファージ巨核球へ分化する能力に必須に差異を示さなかった(図3B)。これらの3種の系列の間の比は、この長期的な処理の後同じままであった。従って、レオウイルス処理は、CD34+細胞を殺傷せず、造血系を再構成するするそれらの可能性を変化しなかった。
【0044】
さらにレオウイルスは、ガン細胞の混合物中のMCF7、SKRB3またはHTB 132細胞の選択的な殺傷によって示されるように、混在した細胞性組成物、及びCD34+幹細胞を含むアフェレーシス産物を清浄化できる(実施例4)。CD34、並びにMCF7、SKRB3またはHTB 132のような上皮細胞に対する特異的なマーカーであるサイトケラチンを測定することによって、レオウイルスが混在した細胞性組成物からガン細胞を必須に除去する一方(図4A−4C)、幹細胞は完全なまま維持することが示された。それ故、レオウイルス処理は、造血幹細胞組成物から新生物細胞を清浄化する有効な方法である。
【0045】
従って、本発明の好ましい実施態様では、幹細胞含有自己移植片が、混在したまたは自然発生的なras活性化新生物細胞を除去するために、移植の前にレオウイルスで処理される。これは、高投与量化学療法/同種幹細胞移植治療の効力を増大する。高投与量化学療法及び同種幹細胞移植が、腫瘍を有する患者において有効に実施されているため、ホジキン病、多発性骨髄腫、非ホジキンリンパ腫、急性骨髄性白血病、生殖細胞(精巣)ガン、脳腫瘍、並びに乳ガンの治療は特に興味深いであろう。しかしながら、ras経路の活性化はいずれの細胞または組織タイプでも生じ得るため、本発明の方法は、他のガンにおいても有用であり、いずれのras活性化新生物細胞を除去するためにも有用であろう。
【0046】
造血始原幹細胞は、治療に先立って患者の骨髄から得ることができる。別法として、従来の非高投与量化学療法を受けているガン患者では、多くの幹細胞が典型的に末梢血で出現する。それ故、造血始原幹細胞は、アフェレーシス産物として血液から得ることができ、それは移植される前に長期に亘り貯蔵できる。本発明は、骨髄及び血液を含むいずれかの組織ソースから回収された幹細胞含有自己移植片に適用できる。
【0047】
レオウイルスは、いずれかの周知の疾患と関連するものではないが、化学療法のため免疫系が弱化されたガン患者にはより感染性であるかもしれない。従って、本発明の別の実施態様では、レオウイルスで処理された細胞性組成物は、DMSOを含む溶液で凍結され、移植の前に解凍される。DMSOは動物細胞を凍結して貯蔵するために一般的に使用される一方、それはレオウイルスを変性し、それによって幹細胞調製物から感染性レオウイルスを除去する。これはレオウイルスが幹細胞移植を介して移植受容者内に導入された場合の非所望の感染を引き起こす危険を減少する。
【0048】
別の実施態様では、レオウイルス処理された細胞組成物は、レオウイルスを不活性化または溶解するために、抗レオウイルス抗体、または抗レオウイルス抗体と補体の組み合わせで処理される。別法としてまたは加えて、レオウイルス粒子の表面の分子を認識する特異的な抗体が、レオウイルス処理された細胞性組成物からレオウイルス粒子を除去するために使用されても良い。かくして、前記抗体は、当該技術分野で周知のカラム、ビーズ、またはいずれかの他の物質に固定化され、細胞性組成物を固定化抗体に適用し、抗体に結合しない組成物の一部を、固定化の特定の方法に適した方法に従って回収する。
【0049】
レオウイルス処理された混合物からウイルスを除去するために使用されても良い別の方法は、ウイルスから細胞を分離し、細胞のみを含む層を回収する勾配に混合物をかけることである。
【0050】
別の実施態様では、移植受容者は、レオウイルス感染の危険を減少するために免疫系を刺激する治療を受ける。この治療は、移植の前、同時、または後に実施されても良いが、移植の前に実施されるのが好ましい。別法の治療として、または免疫系刺激剤と組み合わせて、レオウイルス感染の危険を減少するために、受容者は抗レオウイルス抗体が与えられることができる。
【0051】
造血幹細胞に加えて、本発明は、多くの他の細胞性組成物からras活性化新生物細胞を除去するために広く適用できる。例えばレオウイルスは、いずれかの組織または器官移植物を「クリーンアップ」(それらからのras活性化新生物細胞の除去)のために通常の実施として使用できる。前述のように、レオウイルスに対するレセプターは豊富であり、レオウイルス複製を阻害する正常細胞の機構、PKRもまた豊富であるため、本発明に応用は細胞または組織タイプによって制限されない。それ故、いずれの細胞もras活性化新生物細胞となり、レオウイルス感染に感受性となる。後の輸血のための全血またはそのいずれかの一部をクリーンアップするための本発明の方法は特に興味深い。同様に、組織または器官移植はますます一般的となり、移植の前にras活性化新生物細胞を除去するように移植物が処理できれば有益であろう。肝臓、腎臓、心臓、角膜、皮膚移植片、膵島細胞、骨髄、またはそれらのいずれかの一部は、本発明を適用できる組織または器官の例である。
【0052】
組織または器官は、自己物、同種物、または異種物であることができる。組織または器官はまた、トランスジェニック動物から由来しても良く、幹細胞からin vitroで形成された物、またはex vivoで増大したものである組織/器官でも良い。レオウイルスで処理される組織または器官は、胚性または成人由来であることができる。例えば、胚性神経細胞は、アルツハイマー患者に移植される前に処理できる。同様に本発明は、ex vivoで精子またはドナーの卵を処理するために使用できる。
【0053】
本発明の応用は、移植物に制限されない。むしろいずれの細胞性組成物も、いずれかの目的でレオウイルスで「クリーンアップ」できる。かくして、前述の全ての例は、組織または器官が移植のためのものを意味しなくても応用可能である。
【0054】
細胞系はまた、自然発生的または混在したras活性化新生物細胞に対して保護するために通常処理されても良い。再現すると、もちろんras経路の活性化によってトランスフォームされた細胞系を除き、いずれの細胞系もこの方法のための適格な候補であろう。
【0055】
最近多くの研究室が、免疫抑制マウス中に接種されたヒト前立腺ガン組織の連続的に移植可能な異種移植片を確立することを試みている。しかしながら、マウスガン細胞の混在が、移植片の連続する継代培養の間でしばしば生じており、これらの細胞を最終的にヒト前立腺ガン細胞に成長させ得る(Gao等, 1999)。混在したガンがras介在性であり、移植片がそうでなければ、本発明のこの問題に対する簡単な解決策となるであろう。
【0056】
以下の実施例は、本発明を説明するために与えられ、いずれの態様でも本発明の範囲を制限するものとしては考慮されるべきではない。
【実施例】
【0057】
以下の実施例では、以下の略語は以下の意味を有する。定義されていない略語は、一般的に受け入れられている意味を有する
℃ = 摂氏の温度
hr = 時間
min = 分
μM = マイクロモーラー
mM = ミリモーラー
M = モーラー
ml = ミリリットル
μl = マイクロリットル
mg = ミリグラム
μg = マイクログラム
PAGE = ポリアクリルアミドゲル電気泳動
rpm = 回転/分
FBS = 胎児ウシ血清
DTT = ジチオスレイトール
SDS = ドデシル硫酸ナトリウム
PBS = リン酸緩衝生理食塩水
DMEM = ダルベッコ修飾イーグル培地
α−MEM = α−修飾イーゲル培地
β−ME = β−メルカプトエタノール
MOI = 感染の多様性
PFU = プラーク形成ユニット
PKR = 二本鎖RNA活性化プロテインキナーゼ
EGF = 上皮増殖因子
PDGF = 血小板由来増殖因子
DMSO = ジメチルスルホキシド
CPE = 細胞変性効果
GCSF = 顆粒球コロニー刺激因子
【0058】
実施例1:レオウイルスは乳ガン細胞におけるレオウイルス誘導腫瘍崩壊とアポトーシスを誘導した
新生物細胞の生存能力に対するレオウイルスの効果を測定するために、最初に3種の乳ガンモデル系、MCF7(ATCCナンバーHTB-22)、SKBR3(ATCCナンバーHTB-30)、及びMDA MB 468(ATCCナンバーHTB 132)を使用した。各細胞系の細胞を50−60%の密度に増殖し、40の感染の多様性でレオウイルス血清型3、Dearing株で感染した。レオウイルスは、米国特許第6,136,307号に記載されたように得て維持した。レオウイルス感染及び非感染細胞を、感染の0、24、48及び72時間後に回収し、生存能力を測定した。
【0059】
その結果は図1A−1Dに示されている。レオウイルス感染MCF7(図1A)、SKBR3(図1B)、またはMDA MB 468細胞(図1C)における生存細胞カウントは感染の後に有意に減少する一方、死んだウイルスまたはウイルスなしで感染された細胞は、予測されるように増殖した。レオウイルス処理は、MCF7(図1D)及びSKBR3の生存の能力を感染の後72時間までに93%から16%の減少を引き起こした。MDA MB 468細胞では、ウイルス処理された完全な細胞数は、感染の24、48及び72時間までに、それぞれ元の細胞数の12.7%、8.8%及び3.6%に減少した。かくしてレオウイルスは、全ての3種のガン細胞において有効に腫瘍崩壊を引き起こした。
【0060】
細胞はアポトーシスによって死んだ。CPE、アネキシンV及びDNA断片化のような典型的なアポトーシスマーカーが、生存能力の減少と平行にタイムコースで観察できた。図2A−2Gは、レオウイルス感染の後の各種の時点でのDNA断片化(図2A−2C)、アネキシンV染色(2D)、またはAPO2.7+細胞(2E−2G)のパーセンテージを示す。レオウイルスで処理された細胞は、ウイルスなしまたは死んだ細胞でのコントロールと比較して、劇的なレベルでアポトーシスの全ての兆候を示し、これら3種の細胞系の全てでレオウイルスがアポトーシスを誘導したことを示した。コントロールにおけるアポトーシスは、時間と共にゆっくりと増大するようであり、おそらく細胞があまりに高密度で増殖した場合に死に始めるためであろう。
【0061】
実施例2:レオウイルスはガン細胞においてタンパク質合成を阻害するが、CD34+幹細胞では阻害しなかった
幹細胞の選択的なウイルス感染のさらなる証拠のために、ウイルスタンパク質の35Sラベリング/SDS/PAGEを実施した。ウイルスタンパク質合成は、レオウイルスで感染されたMCF7細胞において1−2日後に明らかであった一方、細胞のタンパク質合成は同じ時点で減少し、レオウイルスが細胞マシネリーを乗っ取っていることが示された。感染の4日後では、タンパク質合成はさらには検出できず、全ての細胞が死んでいることを示唆した。細胞を死んだレオウイルスまたはウイルスなしで感染したコントロール実験では、ウイルスタンパク質合成は存在しない一方、細胞のタンパク質合成は正常レベルであった。対照的に、レオウイルスの存在下または不存在下でのCD34+幹細胞の35Sラベリングは、ウイルスの添加の後72時間までウイルスタンパク質合成を示さなかった。それ故、レオウイルスはMCF7細胞に選択的に感染するが、CD34+幹細胞には感染しない。
【0062】
実施例3:レオウイルス処理はCD34+細胞の細胞増殖を阻害せず、分化能力も改変しなかった
タンパク質合成の結果と一致して、生存細胞カウントは、レオウイルス処理が、ウイルスなしのコントロールと比較してCD34+細胞における生存細胞の数を減少しないことを示した(図3A)。
【0063】
CD34+細胞の数はレオウイルス感染によって影響されない一方で、適切な割合で全ての造血系列に分化するCD34+細胞の能力を、レオウイルスは変化しないかどうか疑問が残った。もしこの場合であれば、レオウイルスで処理された幹細胞は、全部の造血系の再構成のための適切な候補足り得ないであろう。この可能性を調べるために、CD34+細胞をレオウイルス細胞でそれぞれ2、24、48または72時間インキュベートした。次いでレオウイルスを除去し、細胞を希釈して、14日間新鮮な培地で培養し、コロニーを形成させた。各コロニーを、白血球、赤血球、または白血球赤血球マクロファージ巨核球に属するかどうか測定するために調べた。図3Bに召されるように、生存ウイルスで処理された幹細胞(LV)は、ウイルスなし(NV)のコントロールと同様な白血球(G)、赤血球(E)または白血球赤血球マクロファージ巨核球(GEMM)の数を生産した。それ故、レオウイルス処理は、CD34+細胞の分化能力を変えなかった。
【0064】
実施例4:レオウイルスは混在した細胞性組成物からガン細胞を選択的に除去した
新生物細胞をアフェレーシス産物と混合し、レオウイルス感染にかけ、レオウイルスが混在した細胞性組成物から新生物細胞を選択的に除去できるかを調べた。アフェレーシス産物は、以前に記載された方法に従って調製した(Stewart等, 1999; Duggan等, 2000)。アフェレーシス産物(90%)とMCF7(10%)の混合物をレオウイルスで処理し、細胞カウントを生存能力について毎日試験すると、サイトケラチンポジティブMCF7細胞の数の100倍の枯渇が存在する一方、CD34+細胞は完全で生存したままであった。図4A−4Cは、MCF7、SKBR3またはMDA MB 468細胞を有するアフェレーシス産物の混合物に対するレオウイルスの清浄化効果を示す。これらの結果は、レオウイルスが細胞混合物中の新生物細胞を選択的に殺傷し、幹細胞は完全なまま維持すること示す。
【技術分野】
【0001】
関連発明
本出願は、35 U.S.C. 119(e)の下で、2000年5月3日に出願された米国予備出願60/201,990、2000年5月19日に出願された60/205,389、2001年2月13日に出願された60/268,054の利益を享受する。前述の予備出願のそれぞれの全体の開示は、参考としてここに取り込まれる。
【0002】
本発明は、組成物中のras介在性新生物細胞を選択的に溶解するレオウイルスで、混在した細胞性組成物を感染することによる、混在した細胞性組成物からras介在性新生物細胞を除去する方法に関する。
【背景技術】
【0003】
通常細胞増殖は、増殖促進シグナルと増殖抑制シグナルの両者によって調節される。各細胞に対してこれらの2種類のシグナルは、特定の細胞に対する身体の必要性を反映する態様で、通常バランスを与えているであろう。もし細胞が増殖抑制シグナルに応答しない場合、または増殖促進シグナルに過剰応答する場合、その細胞は以上に迅速に増殖し(新生物細胞と称する)、悪性新生物であるガンに段階的に発展するであろう。
【0004】
ガンを治療するために現在行われている方法である化学療法は、ガン細胞の迅速な増殖特性に一般的に基づいている。ガン細胞は迅速に増殖するため、それらは細胞増殖を阻害する薬剤により感受性である。理論的には、化学療法薬の投与量を注意深く選択することによって、正常細胞を重大に損傷することなくガン細胞の増殖を阻害できる。しかしながら、造血幹細胞のようないくつかの正常細胞もまた迅速に増殖する。それ故、ガン細胞に有害なある投与量は、しばしば造血幹細胞にも有害である。他方で、もし投与量がガン細胞を殺傷するのに十分な程高くないと、化学療法が終結した後、ガンが短期間で再発する危険が存在する。
【0005】
選択的にガン細胞を殺傷する投与量を見出すことは困難であるため、高投与量の化学療法、引き続き異種造血始原幹細胞移植は、多くのガンにおける治療アプローチとして過度の適用が実施されている(例えば、Winter, 1999; Nieto及びShpall, 1999参照)。このアプローチでは、造血幹細胞の部分がガン患者から除去されて、次いで患者は、ガン細胞と造血幹細胞のような迅速に増殖する細胞に対して致死的な高投与量化学療法で治療される。その後患者は、同じ患者から以前に除去された自己造血幹細胞の移植を受け、造血系が再生する。
【0006】
この治療の重大な欠点は、造血始原幹細胞が患者から除去される場合、それらにしばしばガン細胞が混在している点である。これは特に、患者が造血系起源のガンを有する場合に問題であるが、特にもし固形腫瘍が転移したならば、固形腫瘍を有する患者でも、造血幹細胞の混在に苦しむ可能性がある。結果として、除去された細胞が戻されて移植され、造血系を再生する場合に、あるガン細胞がガン患者に戻されて配置され、その場合にそのガンが再び増殖して、ガンの再発に寄与する可能性がある。それ故、移植前の自己移植片を清浄化することが望ましい。
【0007】
自己移植片を清浄化するためにいくつかの方法が利用されている(Spyridonidis等, 1998;Bensinger, 1998)。自己移植片を化学療法で処理し、in vitroで混在した新生物細胞を殺傷できる。しかしながら、前述のように、新生物細胞またはガン細胞を選択的に殺傷するが、清浄な造血幹細胞を完全なままにする化学療法薬の投与量を見出すことは困難である。自己移植片はまた、新生物細胞に特異的である抗原を認識する抗体に接合された毒素で治療することもできるが、そのような腫瘍特異的抗原がいつも存在するとは限らない。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】U.S. Pat. No. 6,136,307
【特許文献2】WO 99/08692, published Feb. 25, 1999.
【非特許文献】
【0009】
【非特許文献1】Armstrong, G. D. et al., "Studies on reovirus receptors of L cells: virus binding characteristics and comparison with reovirus receptors of erythrocytes", Virology 138:37: 37-48 (1984)
【非特許文献2】Bensinger, W. I., "Should we purge?", Bone Marrow Tranplant. 21: 113-115 (1998)
【非特許文献3】Bos, J. L., "Ras Oncogenes in Human Cancer: A Review", Canc. Res. 49(17): 4682-4689 (1989)
【非特許文献4】Chandron and Nibert, "Protease cleavage of reovirus capsid protein mu1 and mu1C is blocked by alkyl sulfate detergents, yielding a new type of infectious subvirion particle", J. of Virology 72(1):467-75 (1998)
【非特許文献5】Coffey, M. C., et al., "Reovirus therapy of tumors with activated Ras pathway", Science 282: 1332-1334 (1998)
【非特許文献6】Duggan, P. R., et al., "Predictive factors for long-term engraftment of autologous blood stem cells", Bone Marrow Transplantation 26(12): 1299-1304 (2000)
【非特許文献7】Fields, B. N. et al., Fundamental Virology. 3rd Edition, Lippincott-Raven (1996)
【非特許文献8】Gao, J., B. Tombal and J. T. Isaacs, "Rapid in situ hybridization technique for detecting malignant mouse cell contamination in human xenograft tissue from nude mice and in vitro cultures from such xenografts", Prostate 39(1): 67-70, 1999
【非特許文献9】Gentsch, J. R. K. and Pacitti, A. F., "Effect of neuraminidase treatment of cells and effect of soluable glycoproteins of type 3 reovirus attachment to murine L cells", J. Virol. 56:356: 356-64 (1985)
【非特許文献10】Neito, Y. and E. J. Shpall, "Autologous stem-cell transplantation for solid tumors in adults", Hematol. Oncol. Clin. North Am. 13(5): 939-968 (1999)
【非特許文献11】Norman, K. and P. Lee, "Reovirus as a novel oncolytic agent", J. Clin. Invest. 105 (8): 1035-1038 (2000)
【非特許文献12】Paul R. W. et al., "The alpha-anameric form of sialic acid is the minimal receptor determinant recognized by reovirus", Virology 172: 382-385 (1989)
【非特許文献13】Sabin, A. B., Science 130: 966 (1959)
【非特許文献14】Spyridonidis, A. et al., "Minimal residual disease in autologous hematopoietic harvests from breast cancer patients", Annals of Oncology 9: 821-826 (1998)
【非特許文献15】Stewart, D. A., et al., "Superior autologous blood stem cell mobilization from dose-intensive cyclophosphamide, etoposide, cisplatin plus G-CSF than from less intensive chemotherapy regimens", Bone Marrow Transplant. 23(2): 111-117 (1999)
【非特許文献16】Strong, J. E., et al., "The molecular basis of viral oncolysis: usurpation of the Ras 40 signaling pathway by reovirus", EMBO J. 17: 3351-3362 (1998)
【非特許文献17】Strong, J. E. and P. W. Lee, "The v-erbV oncogene confers enhanced cellular susceptibility to reovirus infection", J. Virol. 70: 612-616 (1996)
【非特許文献18】Strong, J. E., et al., "Evidence that the Epidermal Growth Factor Receptor on Host Cells Confers Reovirus Infection Efficiency", Virology 197(1): 405-411 (1993)
【非特許文献19】Winter, J. N., "High-dose therapy with stem-cell transplantation in the malignant 5 lymphomas", Oncology (Huntingt) 13(12): 1635-1645 (1999)
【発明の概要】
【発明が解決しようとする課題】
【0010】
フローサイトメトリー、アフィニティーカラム、または磁性ビーズを使用することによって、幹細胞特異的表面マーカー(CD34)に基づいて、幹細胞を他の細胞から分離することも可能である。しかしながら、特定の造血細胞、例えばCD34+細胞のみを選択することによって、T細胞、B細胞、単球、及びナチュラルキラー細胞のような他の造血細胞もまた除去され、免疫回復が遅延する可能性がある(Bensinger, 1998)。この方法はまた、CD34+細胞の約半分の損失と、混在したガン細胞のある部分の維持を引き起こす(Spyridonidis, 1998)。それ故、新生物細胞を含む可能性のある自己移植片を清浄化するための、かなりの収率を有する非常に選択的な方法に対する必要性が存在する。
【課題を解決するための手段】
【0011】
本発明は、ras介在性新生物細胞を選択的に溶解するレオウイルスで、混在した細胞性組成物を感染することによる、混在した細胞性組成物からras介在性新生物細胞を除去する方法に関する。多くの新生物細胞が活性化ras経路と関連しているため、本発明は広範囲の混在した細胞性組成物から、混在したまたは自然発生的な新生物細胞を除去するために使用できる。従って、本発明の一つの特徴点は、ras介在性新生物細胞を含む疑いのある細胞性組成物からras介在性新生物細胞を除去する方法であって、ras介在性新生物細胞の腫瘍崩壊を引き起こす条件下で、前記細胞性組成物をレオウイルスと接触させることを含む方法を提供する。
【0012】
本発明の好ましい実施態様では、造血幹細胞を含む細胞性組成物が、混在したまたは自然発生的に存在する新生物細胞として細胞性組成物において存在するras介在性新生物細胞を除去するために処理される。レオウイルスは非常に選択的であり、ras介在性細胞のみに溶解的に感染するため、この方法は、造血系を分化し再構成する造血幹細胞の能力には影響しない。本発明の方法は、骨髄または血液由来の造血幹細胞を清浄化するために使用できる。造血幹細胞は、自己由来、同種由来、または異種由来であっても良い。
【0013】
本発明の別の実施態様では、移植の前に組織または器官移植物からras介在性新生物細胞を除去するために、レオウイルスが使用される。この方法は、移植物またはras介在性細胞のタイプや年齢と無関係に適用できるため、及びレオウイルスは正常細胞及び組織に必須に無害であるため、この方法は、移植の前にいずれかの移植物を「クリーンアップ」するための通常の実施として使用できる。
【0014】
本発明の別の実施態様では、ras経路の活性化のため自然発生的にトランスフォームされた細胞を除去するため、レオウイルスが培養細胞系を処理するために使用できる。この方法はさらに、人工授精または他の再生産関連方法の前に、精子またはドナーの卵を処理するためにも使用できる。
【0015】
本発明において有用なレオウイルスは、いずれのレオウイルスでも良い。好ましくはレオウイルスは、哺乳動物レオウイルス、より好ましくはヒトレオウイルスである。さらにより好ましくはレオウイルスは血清型3であり、最も好ましくはDearing株のレオウイルスである。
【0016】
本発明の好ましい実施態様では、本発明の方法は、DMSOを含む溶液中にレオウイルスで処理された組成物を凍結して貯蔵する工程をさらに含む。DMSOは動物細胞を凍結し貯蔵するために一般的に使用されているが、レオウイルスを変性する。それ故、この処理は、細胞性組成物から感染性レオウイルスを除去する一方で、長期間に亘って凍結状態にある組成物の活性を保存する。
【0017】
本発明の別の実施態様では、レオウイルスを溶解するために、抗レオウイルス抗体、または抗レオウイルス抗体と補体の組み合わせに混合物をかけることによって、レオウイルスで処理された細胞性組成物からレオウイルスが除去される。別法としてまたは加えて、レオウイルス粒子の表面にある分子を認識する抗ウイルス抗体が、この抗体を固定化し、固定化抗体に細胞性組成物を適用し、及び抗体に結合していない組成物の部分を回収することによって、レオウイルスを除去するために使用されても良い。
【0018】
同様に、抗レオウイルス抗体はin vivoでレオウイルスを除去するために移植受容者に投与でき、または受容者はこの目的を達成するために免疫刺激剤を与えられることができる。
【0019】
本発明の別の実施態様では、細胞からレオウイルスを分離できる勾配を使用することによって、レオウイルスで処理された細胞性組成物からレオウイルスが除去される。
【0020】
ras介在性新生物細胞を除去するためにレオウイルスで処理されている細胞性組成物もまた提供される。そのような組成物は、in vitro研究、または移植、受精、または他のin vivo方のために使用されても良い。移植は、自己由来、同種由来、または異種由来であっても良い。好ましくは移植は自己移植である。より好ましくは組成物は造血幹細胞を含む。
【0021】
本発明は、レオウイルスを使用することによる細胞性組成物からras介在性新生物細胞を除去する方法、並びにこれらの方法に従って処理されている組成物に関する。
【0022】
本発明をさらに詳細に記載する前に、この明細書において使用される用語は、他に記載がなければ以下のように定義される。
【図面の簡単な説明】
【0023】
【図1A】図1Aは、示されているように生存レオウイルス、死んだウイルス、ウイルスなしで感染されたMCF7における生存細胞の数を示す図である。
【図1B】図1Bは、示されているように生存レオウイルス、死んだウイルス、ウイルスなしで感染されたSKRB3における生存細胞の数を示す図である。
【図1C】図1Cは、示されているように生存レオウイルス、死んだウイルス、ウイルスなしで感染されたHTB132における生存細胞の数を示す図である。
【図1D】図1Dは、レオウイルス感染の後の各種の時点での生存中のMCF7細胞のパーセンテージを示す図である。
【図2A】図2Aは、MCF7細胞におけるレオウイルス感染によってアポトーシスが誘導されたことを示す図である。レオウイルス感染の後に断片化したDNAのパーセンテージが示されている。
【図2B】図2Bは、SKRB3細胞におけるレオウイルス感染によってアポトーシスが誘導されたことを示す図である。レオウイルス感染の後に断片化したDNAのパーセンテージが示されている。
【図2C】図2Cは、HTB1327細胞におけるレオウイルス感染によってアポトーシスが誘導されたことを示す図である。レオウイルス感染の後に断片化したDNAのパーセンテージが示されている。
【図2D】図2Dは、レオウイルス感染の後のアポトーシスマーカーアネキシンV染色のパーセンテージを示す図である。
【図2E】図2Eは、MCF7細胞におけるAPO2.7+細胞のパーセンテージを示す図である。
【図2F】図2Fは、HTB132細胞におけるAPO2.7+細胞のパーセンテージを示す図である。
【図2G】図2Gは、SKRB3細胞におけるAPO2.7+細胞のパーセンテージを示す図である。
【図3A】図3Aは、CD34+幹細胞がレオウイルスで感染された後の各種の時点での生存細胞の数を示す図である。
【図3B】図3Bは、長期的な幹細胞培養に対するレオウイルスの効果を示す図である。幹細胞をレオウイルスで感染し、それぞれ2,24,48または72時間インキュベートし、次いで細胞を希釈して細胞を14日間培養し、個々のコロニーを形成させた。顆粒球(G)、赤血球(E)、または顆粒球赤血球マクロファージ巨核球(GEMM)の各種のコロニーの数を、それぞれウイルスなし(NV)及び生存ウイルス(LV)で感染した細胞について測定した。例えば、NV-Gはウイルスなしで処理された細胞から由来する顆粒球コロニーを示し、LV-Gは生存ウイルスで処理された細胞から由来する顆粒球を示す。
【図4A】図4Aは、MCF7細胞を有するアフェレーシス産物の混合物に対するレオウイルスの清浄化効果を示す図である。
【図4B】図4Bは、MDA MB 468細胞を有するアフェレーシス産物の混合物に対するレオウイルスの清浄化効果を示す図である。
【図4C】図4Cは、SKRB3細胞を有するアフェレーシス産物の混合物に対するレオウイルスの清浄化効果を示す図である。
【発明を実施するための形態】
【0024】
定義
ここで使用される用語、「新生物細胞」、または「増殖性疾患を有する細胞」は、正常な増殖阻害特性を有さずに増殖する細胞を指す。新生物細胞を含む新たな増殖物は、新生物または腫瘍である。新生物は、一般的に別個の塊を形成する異常な組織増殖であり、正常な組織増殖より迅速な細胞増殖によって増殖する。新生物は、正常な組織との構造的組織化及び機能的強調の部分的または完全な欠如を示すであろう。ここで使用される新生物は、造血性新生物、並びに固形新生物を包含するように企図される。
新生物は、良性(良性腫瘍)または悪性(悪性腫瘍またはガン)であって良い。悪性腫瘍は、3種の主なタイプに広く分類できる。上皮構造から生ずる悪性新生物はカルシノーマと称され、筋肉、軟骨、脂肪、または骨のような結合組織から由来する悪性新生物はサルコーマと称され、免疫系の構成成分を含む造血構造(血液細胞の形成と関係する構造)に影響する悪性腫瘍は白血病及びリンパ腫と称される。他の新生物は、神経芽腫を制限することなく含む。
【0025】
ここで使用される用語、「ras活性化新生物細胞」または「ras介在性新生物細胞」は、少なくとも部分的にras経路の活性化のために異常に速い速度で増殖する細胞を指す。ras経路は、ras遺伝子の構造的ミューテーション、ras遺伝子発現の増大したレベル、ras遺伝子メッセージの増大した安定性、あるいはras、またはras経路におけるrasから下流または上流の因子の活性化を導くいずれかのミューテーションまたは他のメカニズムによって活性化され、それによってras経路活性を増大するであろう。例えば、EGFレセプター、PDGFレセプター、またはSosの活性化は、ras経路の活性化を導く。ras介在性新生物細胞は、ras介在性ガン細胞を制限することなく含み、それはras経路の活性のによる悪性の態様で増殖する細胞である。
【0026】
ここで使用される用語、「細胞性組成物」は、細胞を含む組成物を意味する。前記組成物は、非細胞物を含んでも良い。例えば全血は、赤血球及び白血球のような細胞に加えて血漿、血小板、ホルモン、及び他の非細胞物を含む細胞性組成物である。細胞性組成物は、各種のタイプの細胞、器官、または構築物を含んでも良い。例えば、所定の構造で整列した各種の細胞タイプを含む組織または器官は、細胞性組成物と考慮される。
【0027】
ここで使用される用語、「レオウイルス」は、各種の属に分類されるいずれのウイルスをも指す。レオウイルスの名称(Respiratory and enteric orphan virus)は、これらのウイルスが、ヒトにおけるいずれかの周知の疾患と関係していないが、呼吸器及び腸管の両者から単離できることを示唆する説明的な頭字語である(Sabin, 1959)。用語「レオウイルス」は、レオウイルス属に分類される全てのウイルスを指す。
【0028】
ヒトのレオウイルスは3種の血清型からなる:タイプ1(Lang株またはT1L)、タイプ2(Jones株、T2J)及びタイプ3(Dearing株またはAbney株、T3D)。この3種の血清型は、中和及びヘマグルチニン阻害アッセイを基礎に容易に同定可能である(例えばFields等, 1996参照)。
【0029】
レオウイルスは天延に存在するものでも改変されていても良い。レオウイルスが天然のソースから単離でき、研究室でヒトによって意図的に改変されていない場合、レオウイルスは「天然に存在するもの」である。例えば、レオウイルスが「フィールドソース」由来であることができ、つまりレオウイルスで感染されたヒト由来であることができる。
【0030】
レオウイルスは、改変されても、活性なras経路を有する哺乳動物細胞を溶解的に感染可能であっても良い。レオウイルスは、増殖中の細胞に生化学的に投与される前に、化学的または生化学的に調製されても良い(例えばキモトリプシンまたはトリプシンのようなプロテアーゼでの処理によって)。プロテアーゼでの事前処理は、ウイルスの外側コートまたはキャプシドを除去し、ウイルスの感染能力を増大するであろう。レオウイルスは、レオウイルスに対する発達した免疫を有する哺乳動物から免疫応答を減少または妨げるために、リポソームまたはミセルにコートされても良い(Chandron及びNibert, 1998)。例えば、新規な感染性サブぶりオン粒子を生産するために、ビリオンがミセル形成濃度の硫酸アルキル変性剤の存在下でキモトリプシンで処理されても良い。
【0031】
レオウイルスが異なる抗原決定基を有し、それによってレオウイルス血清型に以前にさらされた哺乳動物による免疫応答を減少するまたは妨げるように、レオウイルスは、異なる病原性表現型を有する二つ以上のタイプのレオウイルスから得られた組換えレオウイルスであっても良い。そのような組換えビリオンは、生成したビリオンキャプシド内への、異なるサブタイプのコートタンパク質の生成したリソーティングと取り込みで、異なるサブタイプのレオウイルスを有する哺乳動物細胞の共感染によって生産できる。
【0032】
レオウイルス感染に対する細胞の「耐性」は、ウイルスを有する細胞の感染が、有意なウイルス生産または収率を生じないことを示す。「感受性」である細胞は、細胞変性効果の誘導、ウイルスタンパク質合成、及び/またはウイルス生産を示すものである。レオウイルス感染に対する耐性は、早期の転写よりもむしろ遺伝子翻訳のレベルで見出された:ウイルス転写物は生産される一方、ウイルスタンパク質は発現されなかった。一つの理論に制限されるものではないが、耐性細胞におけるウイルス遺伝子転写は、約65kDa細胞タンパク質のリン酸化と強調し、それは二本鎖RNA活性化プロテインキナーゼ(PKR)と決定され、それはトランスフォーム化細胞では観察されなかった。PKRのリン酸化は、転写の阻害を導く。リン酸化が周知のPKRのインヒビターである2−アミノプリンによって抑制された場合、レオウイルスタンパク質合成の促進がトランスフォーム化細胞で生じた。
【0033】
ここで使用される用語、「腫瘍崩壊」または「新生物細胞の生存能力の減少」は、標的新生物細胞の生存能力の少なくとも約20%の減少を意味する。生存能力は処理された細胞の生存可能な細胞のカウントによって測定でき、減少の度合いは、非処理細胞における生存可能な細胞の数と処理細胞における生存可能な細胞の数を比較することによって、またはウイルス処理の前後の生存可能な細胞のカウントを比較することによって測定できる。生存能力の減少は好ましくは、少なくとも約50%、より好ましくは少なくとも約70%、さらにより好ましくは少なくとも約80%、最も好ましくは少なくとも約90%である。
【0034】
ここで使用される用語、「移植受容者」は、細胞性組成物の移植を受ける哺乳動物である。好ましくは前記受容者はヒトであり、より好ましくは前記受容者は、ガンの治療において移植を受けるヒトである。
【0035】
ここで使用される用語、アポトーシスマーカーとしての「CPE」は、細胞膜気泡化、核の凝集、及びクロマチン凝集のような、顕微鏡の下で観察可能なアポトーシスと典型的に関連する細胞変性効果を指す。
【0036】
方法
本発明は、ras介在性新生物細胞を含む疑いのある細胞性組成物からras介在性新生物細胞を除去するレオウイルスの使用に関する。活性化ras経路は多くの腫瘍と関連しているため、本発明は広範囲の新生物細胞のいずれを有する疑いのある細胞性組成物にも適用できる。
【0037】
哺乳動物レオウイルスにとって、細胞表面認識シグナルはシアル酸である(Armstrong, 1984; Gentsch及びPacitti, 1985; Paul等, 1989)。シアル酸の独特な性質のため、レオウイルスは多数の細胞系に有効に結合し、そのため多くの各種の組織を標的化できるであろう;しかしながら、細胞系の間でレオウイルス感染に対する感受性の有意な差異が存在する。この感受性の矛盾の一つの理由は、各細胞系におけるras経路の活性であろう。
【0038】
レオウイルスが、in vitro、in vivo及びex vivoで活性化新生物細胞を選択的に溶解することを、我々は最近発見した(Coffey等, 1998; WO 99/08692)。普通では、細胞はレオウイルス感染に感受性ではない。しかしながら、ras経路が活性化されたならば、レオウイルスは細胞において成功して複製でき、最終的に宿主細胞の溶解を引き起こす。例えば、レオウイルス耐性NIH 3T3細胞が、rasを活性化するタンパク質である活性化rasまたはSosでトランスフォームされた場合、レオウイルス感染は増大された(Strong等, 1998)。同様に、レオウイルス感染に耐性であるマウス線維芽細胞は、EGFレセプター遺伝子またはv-erbB原ガン遺伝子でのトランスフェクションの後感受性となった(Strong等, 1993; Strong等, 1996)。
【0039】
一つの理論に制限されるものではないが、レオウイルス複製は翻訳レベルで調節されているようである(Strong等, 1998; Norman等, 2000)。非トランスフォーム化NIH 3T3細胞では、早期のウイルス転写物が二本鎖RNA活性化プロテインキナーゼ(PKR)を活性化し、それが翻訳を阻害し、それによってウイルス複製を阻害する。活性化ras(またはras経路の活性化エレメント)は、おそらくPKR活性化を阻害または不活性化するであろう。それ故、ウイルスタンパク質合成が進行し、ウイルス粒子が作製され、細胞は最終的に溶解する。
【0040】
ras原ガン遺伝子は、多数の腫瘍の原因である。ras遺伝子自体の活性化ミューテーションは、全てのヒト腫瘍の約30%で生じ(Bos, J.L., 1989)、主に膵臓(90%)、散発生大腸(50%)及び肺(40%)カルシノーマ、並びにミエロイド白血病(30%)で生じている。ras経路におけるrasの上流または下流因子の活性化も、腫瘍と関連している。例えば、HER2/Neu/ErbB2または上皮増殖因子(EGF)レセプターの過剰発現は、乳ガンにおいて一般的であり(25−30%)、血小板由来増殖因子(PDGF)レセプターまたはEGFレセプターの過剰発現は、グリオーマ及びグリオブラストーマ(40−50%)で一般的である。EGFレセプターとPDGFレセプターは、それらのそれぞれのリガンドに対する結合に際してrasを活性化することが知られており、v-erbBは細胞外ドメインを欠く構成的に活性化されたレセプターをコードする。
【0041】
我々は、レオウイルスが、感染細胞においてアポトーシスを誘導することによる、3種の乳ガンモデルシステム、MCF7、SKRB3及びHTB 132の腫瘍崩壊を有効に導くことを本発明において示した(実施例1)。かくしてレオウイルス処理は、MCF7、SKRB3及びHTB 132細胞の生存能力の顕著な減少を引き起こす一方、ウイルスなしまたは死んだウイルスで処理されたコントロールは普通に増殖した(図1A−1D)。生存能力の減少は、DNA断片化、アネキシンVまたはAPO2.7染色陽性(図2A−2G)、並びに顕微鏡で観察される細胞膜の気泡、核の凝集、及びクロマチン凝集のような細胞変性効果といった、アポトーシスと関連する特徴を伴った。
【0042】
レオウイルス感染は、正常細胞において翻訳レベルで通常ブロックされるが、ras介在性新生物細胞においてはブロックされないため、レオウイルス処理MCF7細胞及びCD34+幹細胞におけるタンパク質合成の度合いを調べた(図2)。実際、ウイルスタンパク質は、レオウイルス感染ガン細胞系で合成されるが、レオウイルスで処理されたCD34+幹細胞では合成されなかった(データ示さず)。この結果は、レオウイルスタンパク質がレオウイルス処理幹細胞では合成されず、細胞タンパク質合成が普通に進行しているため、レオウイルスでの造血幹細胞の処理が安全であることを示唆している。この点を確認するために、レオウイルス処理CD34+細胞の生存能力を、レオウイルス処理の各種の時点で測定した(実施例3)。生きたレオウイルスまたはウイルスなしで処理された集団における細胞数は各時点で同じであり、CD34+細胞がレオウイルス感染に感受性ではないことを示した。
【0043】
レオウイルスが、高投与量化学療法治療における造血幹細胞の清浄化に有用であるために、レオウイルス処理が、全部の造血系を再構成するそれぞれ及び毎回の造血系列へ分化する幹細胞の能力を変えないことが必須である。それ故、レオウイルス処理の長期的な効果が評価された(実施例3)。ウイルスなしまたは生きたウイルスのいずれかで処理されたCD34+細胞は、レオウイルス処理の72時間後でさえ、白血球、赤血球または白血球赤血球マクロファージ巨核球へ分化する能力に必須に差異を示さなかった(図3B)。これらの3種の系列の間の比は、この長期的な処理の後同じままであった。従って、レオウイルス処理は、CD34+細胞を殺傷せず、造血系を再構成するするそれらの可能性を変化しなかった。
【0044】
さらにレオウイルスは、ガン細胞の混合物中のMCF7、SKRB3またはHTB 132細胞の選択的な殺傷によって示されるように、混在した細胞性組成物、及びCD34+幹細胞を含むアフェレーシス産物を清浄化できる(実施例4)。CD34、並びにMCF7、SKRB3またはHTB 132のような上皮細胞に対する特異的なマーカーであるサイトケラチンを測定することによって、レオウイルスが混在した細胞性組成物からガン細胞を必須に除去する一方(図4A−4C)、幹細胞は完全なまま維持することが示された。それ故、レオウイルス処理は、造血幹細胞組成物から新生物細胞を清浄化する有効な方法である。
【0045】
従って、本発明の好ましい実施態様では、幹細胞含有自己移植片が、混在したまたは自然発生的なras活性化新生物細胞を除去するために、移植の前にレオウイルスで処理される。これは、高投与量化学療法/同種幹細胞移植治療の効力を増大する。高投与量化学療法及び同種幹細胞移植が、腫瘍を有する患者において有効に実施されているため、ホジキン病、多発性骨髄腫、非ホジキンリンパ腫、急性骨髄性白血病、生殖細胞(精巣)ガン、脳腫瘍、並びに乳ガンの治療は特に興味深いであろう。しかしながら、ras経路の活性化はいずれの細胞または組織タイプでも生じ得るため、本発明の方法は、他のガンにおいても有用であり、いずれのras活性化新生物細胞を除去するためにも有用であろう。
【0046】
造血始原幹細胞は、治療に先立って患者の骨髄から得ることができる。別法として、従来の非高投与量化学療法を受けているガン患者では、多くの幹細胞が典型的に末梢血で出現する。それ故、造血始原幹細胞は、アフェレーシス産物として血液から得ることができ、それは移植される前に長期に亘り貯蔵できる。本発明は、骨髄及び血液を含むいずれかの組織ソースから回収された幹細胞含有自己移植片に適用できる。
【0047】
レオウイルスは、いずれかの周知の疾患と関連するものではないが、化学療法のため免疫系が弱化されたガン患者にはより感染性であるかもしれない。従って、本発明の別の実施態様では、レオウイルスで処理された細胞性組成物は、DMSOを含む溶液で凍結され、移植の前に解凍される。DMSOは動物細胞を凍結して貯蔵するために一般的に使用される一方、それはレオウイルスを変性し、それによって幹細胞調製物から感染性レオウイルスを除去する。これはレオウイルスが幹細胞移植を介して移植受容者内に導入された場合の非所望の感染を引き起こす危険を減少する。
【0048】
別の実施態様では、レオウイルス処理された細胞組成物は、レオウイルスを不活性化または溶解するために、抗レオウイルス抗体、または抗レオウイルス抗体と補体の組み合わせで処理される。別法としてまたは加えて、レオウイルス粒子の表面の分子を認識する特異的な抗体が、レオウイルス処理された細胞性組成物からレオウイルス粒子を除去するために使用されても良い。かくして、前記抗体は、当該技術分野で周知のカラム、ビーズ、またはいずれかの他の物質に固定化され、細胞性組成物を固定化抗体に適用し、抗体に結合しない組成物の一部を、固定化の特定の方法に適した方法に従って回収する。
【0049】
レオウイルス処理された混合物からウイルスを除去するために使用されても良い別の方法は、ウイルスから細胞を分離し、細胞のみを含む層を回収する勾配に混合物をかけることである。
【0050】
別の実施態様では、移植受容者は、レオウイルス感染の危険を減少するために免疫系を刺激する治療を受ける。この治療は、移植の前、同時、または後に実施されても良いが、移植の前に実施されるのが好ましい。別法の治療として、または免疫系刺激剤と組み合わせて、レオウイルス感染の危険を減少するために、受容者は抗レオウイルス抗体が与えられることができる。
【0051】
造血幹細胞に加えて、本発明は、多くの他の細胞性組成物からras活性化新生物細胞を除去するために広く適用できる。例えばレオウイルスは、いずれかの組織または器官移植物を「クリーンアップ」(それらからのras活性化新生物細胞の除去)のために通常の実施として使用できる。前述のように、レオウイルスに対するレセプターは豊富であり、レオウイルス複製を阻害する正常細胞の機構、PKRもまた豊富であるため、本発明に応用は細胞または組織タイプによって制限されない。それ故、いずれの細胞もras活性化新生物細胞となり、レオウイルス感染に感受性となる。後の輸血のための全血またはそのいずれかの一部をクリーンアップするための本発明の方法は特に興味深い。同様に、組織または器官移植はますます一般的となり、移植の前にras活性化新生物細胞を除去するように移植物が処理できれば有益であろう。肝臓、腎臓、心臓、角膜、皮膚移植片、膵島細胞、骨髄、またはそれらのいずれかの一部は、本発明を適用できる組織または器官の例である。
【0052】
組織または器官は、自己物、同種物、または異種物であることができる。組織または器官はまた、トランスジェニック動物から由来しても良く、幹細胞からin vitroで形成された物、またはex vivoで増大したものである組織/器官でも良い。レオウイルスで処理される組織または器官は、胚性または成人由来であることができる。例えば、胚性神経細胞は、アルツハイマー患者に移植される前に処理できる。同様に本発明は、ex vivoで精子またはドナーの卵を処理するために使用できる。
【0053】
本発明の応用は、移植物に制限されない。むしろいずれの細胞性組成物も、いずれかの目的でレオウイルスで「クリーンアップ」できる。かくして、前述の全ての例は、組織または器官が移植のためのものを意味しなくても応用可能である。
【0054】
細胞系はまた、自然発生的または混在したras活性化新生物細胞に対して保護するために通常処理されても良い。再現すると、もちろんras経路の活性化によってトランスフォームされた細胞系を除き、いずれの細胞系もこの方法のための適格な候補であろう。
【0055】
最近多くの研究室が、免疫抑制マウス中に接種されたヒト前立腺ガン組織の連続的に移植可能な異種移植片を確立することを試みている。しかしながら、マウスガン細胞の混在が、移植片の連続する継代培養の間でしばしば生じており、これらの細胞を最終的にヒト前立腺ガン細胞に成長させ得る(Gao等, 1999)。混在したガンがras介在性であり、移植片がそうでなければ、本発明のこの問題に対する簡単な解決策となるであろう。
【0056】
以下の実施例は、本発明を説明するために与えられ、いずれの態様でも本発明の範囲を制限するものとしては考慮されるべきではない。
【実施例】
【0057】
以下の実施例では、以下の略語は以下の意味を有する。定義されていない略語は、一般的に受け入れられている意味を有する
℃ = 摂氏の温度
hr = 時間
min = 分
μM = マイクロモーラー
mM = ミリモーラー
M = モーラー
ml = ミリリットル
μl = マイクロリットル
mg = ミリグラム
μg = マイクログラム
PAGE = ポリアクリルアミドゲル電気泳動
rpm = 回転/分
FBS = 胎児ウシ血清
DTT = ジチオスレイトール
SDS = ドデシル硫酸ナトリウム
PBS = リン酸緩衝生理食塩水
DMEM = ダルベッコ修飾イーグル培地
α−MEM = α−修飾イーゲル培地
β−ME = β−メルカプトエタノール
MOI = 感染の多様性
PFU = プラーク形成ユニット
PKR = 二本鎖RNA活性化プロテインキナーゼ
EGF = 上皮増殖因子
PDGF = 血小板由来増殖因子
DMSO = ジメチルスルホキシド
CPE = 細胞変性効果
GCSF = 顆粒球コロニー刺激因子
【0058】
実施例1:レオウイルスは乳ガン細胞におけるレオウイルス誘導腫瘍崩壊とアポトーシスを誘導した
新生物細胞の生存能力に対するレオウイルスの効果を測定するために、最初に3種の乳ガンモデル系、MCF7(ATCCナンバーHTB-22)、SKBR3(ATCCナンバーHTB-30)、及びMDA MB 468(ATCCナンバーHTB 132)を使用した。各細胞系の細胞を50−60%の密度に増殖し、40の感染の多様性でレオウイルス血清型3、Dearing株で感染した。レオウイルスは、米国特許第6,136,307号に記載されたように得て維持した。レオウイルス感染及び非感染細胞を、感染の0、24、48及び72時間後に回収し、生存能力を測定した。
【0059】
その結果は図1A−1Dに示されている。レオウイルス感染MCF7(図1A)、SKBR3(図1B)、またはMDA MB 468細胞(図1C)における生存細胞カウントは感染の後に有意に減少する一方、死んだウイルスまたはウイルスなしで感染された細胞は、予測されるように増殖した。レオウイルス処理は、MCF7(図1D)及びSKBR3の生存の能力を感染の後72時間までに93%から16%の減少を引き起こした。MDA MB 468細胞では、ウイルス処理された完全な細胞数は、感染の24、48及び72時間までに、それぞれ元の細胞数の12.7%、8.8%及び3.6%に減少した。かくしてレオウイルスは、全ての3種のガン細胞において有効に腫瘍崩壊を引き起こした。
【0060】
細胞はアポトーシスによって死んだ。CPE、アネキシンV及びDNA断片化のような典型的なアポトーシスマーカーが、生存能力の減少と平行にタイムコースで観察できた。図2A−2Gは、レオウイルス感染の後の各種の時点でのDNA断片化(図2A−2C)、アネキシンV染色(2D)、またはAPO2.7+細胞(2E−2G)のパーセンテージを示す。レオウイルスで処理された細胞は、ウイルスなしまたは死んだ細胞でのコントロールと比較して、劇的なレベルでアポトーシスの全ての兆候を示し、これら3種の細胞系の全てでレオウイルスがアポトーシスを誘導したことを示した。コントロールにおけるアポトーシスは、時間と共にゆっくりと増大するようであり、おそらく細胞があまりに高密度で増殖した場合に死に始めるためであろう。
【0061】
実施例2:レオウイルスはガン細胞においてタンパク質合成を阻害するが、CD34+幹細胞では阻害しなかった
幹細胞の選択的なウイルス感染のさらなる証拠のために、ウイルスタンパク質の35Sラベリング/SDS/PAGEを実施した。ウイルスタンパク質合成は、レオウイルスで感染されたMCF7細胞において1−2日後に明らかであった一方、細胞のタンパク質合成は同じ時点で減少し、レオウイルスが細胞マシネリーを乗っ取っていることが示された。感染の4日後では、タンパク質合成はさらには検出できず、全ての細胞が死んでいることを示唆した。細胞を死んだレオウイルスまたはウイルスなしで感染したコントロール実験では、ウイルスタンパク質合成は存在しない一方、細胞のタンパク質合成は正常レベルであった。対照的に、レオウイルスの存在下または不存在下でのCD34+幹細胞の35Sラベリングは、ウイルスの添加の後72時間までウイルスタンパク質合成を示さなかった。それ故、レオウイルスはMCF7細胞に選択的に感染するが、CD34+幹細胞には感染しない。
【0062】
実施例3:レオウイルス処理はCD34+細胞の細胞増殖を阻害せず、分化能力も改変しなかった
タンパク質合成の結果と一致して、生存細胞カウントは、レオウイルス処理が、ウイルスなしのコントロールと比較してCD34+細胞における生存細胞の数を減少しないことを示した(図3A)。
【0063】
CD34+細胞の数はレオウイルス感染によって影響されない一方で、適切な割合で全ての造血系列に分化するCD34+細胞の能力を、レオウイルスは変化しないかどうか疑問が残った。もしこの場合であれば、レオウイルスで処理された幹細胞は、全部の造血系の再構成のための適切な候補足り得ないであろう。この可能性を調べるために、CD34+細胞をレオウイルス細胞でそれぞれ2、24、48または72時間インキュベートした。次いでレオウイルスを除去し、細胞を希釈して、14日間新鮮な培地で培養し、コロニーを形成させた。各コロニーを、白血球、赤血球、または白血球赤血球マクロファージ巨核球に属するかどうか測定するために調べた。図3Bに召されるように、生存ウイルスで処理された幹細胞(LV)は、ウイルスなし(NV)のコントロールと同様な白血球(G)、赤血球(E)または白血球赤血球マクロファージ巨核球(GEMM)の数を生産した。それ故、レオウイルス処理は、CD34+細胞の分化能力を変えなかった。
【0064】
実施例4:レオウイルスは混在した細胞性組成物からガン細胞を選択的に除去した
新生物細胞をアフェレーシス産物と混合し、レオウイルス感染にかけ、レオウイルスが混在した細胞性組成物から新生物細胞を選択的に除去できるかを調べた。アフェレーシス産物は、以前に記載された方法に従って調製した(Stewart等, 1999; Duggan等, 2000)。アフェレーシス産物(90%)とMCF7(10%)の混合物をレオウイルスで処理し、細胞カウントを生存能力について毎日試験すると、サイトケラチンポジティブMCF7細胞の数の100倍の枯渇が存在する一方、CD34+細胞は完全で生存したままであった。図4A−4Cは、MCF7、SKBR3またはMDA MB 468細胞を有するアフェレーシス産物の混合物に対するレオウイルスの清浄化効果を示す。これらの結果は、レオウイルスが細胞混合物中の新生物細胞を選択的に殺傷し、幹細胞は完全なまま維持すること示す。
【特許請求の範囲】
【請求項1】
哺乳動物に細胞性組成物を移植する方法のための医薬の製造のためのレオウイルス及び細胞性組成物の使用であって、哺乳動物におけるレオウイルスに対する免疫応答を誘導するのに十分量で、レオウイルスを哺乳動物に投与し、細胞性組成物はレオウイルスで清浄化されており、レオウイルスの投与後に、前記哺乳動物に清浄化された細胞性組成物を移植する、使用。
【請求項2】
細胞性組成物が造血幹細胞を含む、請求項1記載の使用。
【請求項3】
造血幹細胞が前記哺乳動物の血液から収集されている、請求項2記載の使用。
【請求項4】
造血幹細胞が前記哺乳動物の骨髄から収集されている、請求項2記載の使用。
【請求項5】
細胞性組成物が、組織、器官、または組織若しくは器官のいずれかの一部を含む、請求項1記載の使用。
【請求項6】
組織または器官が、肝臓、腎臓、心臓、大腸、皮膚、肺、膵島細胞、及び全血からなる群から選択される、請求項5記載の使用。
【請求項7】
哺乳動物が新生物に罹患している、請求項1に記載の使用。
【請求項8】
新生物がras介在性新生物である、請求項7に記載の使用。
【請求項9】
新生物が肺ガン、前立腺ガン、大腸ガン、甲状腺ガン、腎臓ガン、副腎ガン、肝臓ガン、膵臓ガン、乳ガン、並びに中枢神経系及び末梢神経系のガンからなる群から選択される、請求項7に記載の使用。
【請求項10】
新生物がホジキン病、多発性骨髄腫、非ホジキンリンパ腫、急性骨髄性白血病、生殖細胞(精巣)ガン、脳腫瘍、並びに乳ガンからなる群から選択される、請求項7に記載の使用。
【請求項11】
哺乳動物が化学療法薬または放射線治療も受けている、請求項1に記載の使用。
【請求項12】
清浄化された細胞性組成物が、移植の前にDMSOの存在下で貯蔵されている、請求項1記載の使用。
【請求項13】
レオウイルスが血清型1レオウイルス、血清型2レオウイルス、及び血清型3レオウイルスからなる群から選択される、請求項1に記載の使用。
【請求項14】
レオウイルスが組換えレオウイルスである、請求項1に記載の使用。
【請求項15】
レオウイルスが変性レオウイルスである、請求項1に記載の使用。
【請求項16】
レオウイルスで処理された組成物を、抗レオウイルス抗体で処理した、請求項1記載の使用。
【請求項17】
レオウイルスで処理された細胞性組成物を、レオウイルスから前記細胞性組成物を分離する勾配に供した、請求項1記載の使用。
【請求項18】
細胞性組成物の細胞を含む層が、勾配分離の後に回収された、請求項17に記載の使用。
【請求項1】
哺乳動物に細胞性組成物を移植する方法のための医薬の製造のためのレオウイルス及び細胞性組成物の使用であって、哺乳動物におけるレオウイルスに対する免疫応答を誘導するのに十分量で、レオウイルスを哺乳動物に投与し、細胞性組成物はレオウイルスで清浄化されており、レオウイルスの投与後に、前記哺乳動物に清浄化された細胞性組成物を移植する、使用。
【請求項2】
細胞性組成物が造血幹細胞を含む、請求項1記載の使用。
【請求項3】
造血幹細胞が前記哺乳動物の血液から収集されている、請求項2記載の使用。
【請求項4】
造血幹細胞が前記哺乳動物の骨髄から収集されている、請求項2記載の使用。
【請求項5】
細胞性組成物が、組織、器官、または組織若しくは器官のいずれかの一部を含む、請求項1記載の使用。
【請求項6】
組織または器官が、肝臓、腎臓、心臓、大腸、皮膚、肺、膵島細胞、及び全血からなる群から選択される、請求項5記載の使用。
【請求項7】
哺乳動物が新生物に罹患している、請求項1に記載の使用。
【請求項8】
新生物がras介在性新生物である、請求項7に記載の使用。
【請求項9】
新生物が肺ガン、前立腺ガン、大腸ガン、甲状腺ガン、腎臓ガン、副腎ガン、肝臓ガン、膵臓ガン、乳ガン、並びに中枢神経系及び末梢神経系のガンからなる群から選択される、請求項7に記載の使用。
【請求項10】
新生物がホジキン病、多発性骨髄腫、非ホジキンリンパ腫、急性骨髄性白血病、生殖細胞(精巣)ガン、脳腫瘍、並びに乳ガンからなる群から選択される、請求項7に記載の使用。
【請求項11】
哺乳動物が化学療法薬または放射線治療も受けている、請求項1に記載の使用。
【請求項12】
清浄化された細胞性組成物が、移植の前にDMSOの存在下で貯蔵されている、請求項1記載の使用。
【請求項13】
レオウイルスが血清型1レオウイルス、血清型2レオウイルス、及び血清型3レオウイルスからなる群から選択される、請求項1に記載の使用。
【請求項14】
レオウイルスが組換えレオウイルスである、請求項1に記載の使用。
【請求項15】
レオウイルスが変性レオウイルスである、請求項1に記載の使用。
【請求項16】
レオウイルスで処理された組成物を、抗レオウイルス抗体で処理した、請求項1記載の使用。
【請求項17】
レオウイルスで処理された細胞性組成物を、レオウイルスから前記細胞性組成物を分離する勾配に供した、請求項1記載の使用。
【請求項18】
細胞性組成物の細胞を含む層が、勾配分離の後に回収された、請求項17に記載の使用。
【図1A】


【図1B】
【図1C】
【図1D】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図2G】
【図3A】
【図3B】
【図4A】
【図4B】
【図4C】
【図1B】
【図1C】
【図1D】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図2G】
【図3A】
【図3B】
【図4A】
【図4B】
【図4C】
【公開番号】特開2010−260875(P2010−260875A)
【公開日】平成22年11月18日(2010.11.18)
【国際特許分類】
【出願番号】特願2010−181396(P2010−181396)
【出願日】平成22年8月13日(2010.8.13)
【分割の表示】特願2008−190310(P2008−190310)の分割
【原出願日】平成13年5月2日(2001.5.2)
【出願人】(501332264)オンコリティクス バイオテク,インコーポレーテッド (16)
【Fターム(参考)】
【公開日】平成22年11月18日(2010.11.18)
【国際特許分類】
【出願日】平成22年8月13日(2010.8.13)
【分割の表示】特願2008−190310(P2008−190310)の分割
【原出願日】平成13年5月2日(2001.5.2)
【出願人】(501332264)オンコリティクス バイオテク,インコーポレーテッド (16)
【Fターム(参考)】
[ Back to top ]