
炭水化物結合分子
本発明は、炭水化物結合分子を含むかまたは用いる化合物、組成物、薬および方法を提供する。より詳しくは、本発明は、病原体、特に微生物の病原体が引き起こすかまたは一因となった疾病および/または疾患を治療する手段および炭水化物のスクリーニング、識別、検出、タギングおよび/またはラベリングする方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、炭水化物結合分子を備えるかまたは使用する化合物、組成物、薬剤および方法を提供する。特に、本発明は、病原体、特に微生物の病原体によって引き起こされるかまたは一因となる疾病および/または疾患を治療する手段、並びに炭水化物のスクリーニング、識別、検出タギングおよび/または標識付けの方法を提供する。
【背景技術】
【0002】
大部分の哺乳動物細胞の表面は複合糖質が豊富であり、多くの病原体が初期段階の発病中に細胞付着のためこれら複合糖質の末端炭水化物を利用している。例えば、インフルエンザおよびパラインフルエンザ両方のウイルス性病原体が哺乳類の細胞表面でシアル酸レセプターと結合する。
【0003】
シアル酸認識は、レクチンまたはレクチン様分子およびその対応するレセプターを介してもたらされる(3)。ほとんどの場合、いくつかのウイルス糖タンパク質および細菌毒素(2,4)並びにシグレック(5)およびセレクチン(6)のような哺乳類のレクチン上科も含むシアル酸結合レクチンは、これら分子の多価性により比較的高い親和性でレセプターと結合し、したがって大部分のタンパク質-炭水化物相互作用に関連する低い固有の親和性を緩和する(7,8)。一般に、かかるレクチンによる1価および2価のシアロシドの結合の会合定数(Ka)は、104M-1に達することができる。しかしなから、その多価性のため、いくつかのシアル酸結合レクチンが多価の細胞表面グリカンと相互作用して、結合活性効果によって109M-1に及ぶ親和性を達成することができる。これら増強された親和性は、好ましい結合エネルギー論に関連したリガンド結合によって促進されるタンパク質の改良された構造的パッキングによることが部分的に示された(9-11)。最もよく研究された多価のレクチン-シアル酸相互作用のうちの一つは、インフルエンザウイルス三量体血球凝集素であり、独立体の一つまたは両方が多価の状態でない場合に約4 x 102M-1と対照に最高108M-1親和性を達成することができる(12)。
【0004】
シアリダーゼ、すなわちノイラミニダーゼは、種々の複合糖質からシアル酸の加水分解を触媒し、大抵タンパク質の触媒コアに付着したアクセサリモジュールを含むモジュラー酵素である。これらモジュールのいくつかは、炭水化物結合モジュール(CBM)として識別される。CBMはグリコシドヒドロラーゼで広く見られ、親酵素をその効率的な加水分解用の基質にターゲットするために基質表面で酵素濃度を上げることによって主として存在する個々の非触媒性モジュールである(13)。該モジュールは、グリコシルヒドロラーゼの構造内でシングル、タンデムまたは複合物である。研究の結果、これらが親分子から分離された際に独立してその特異的なグリカンと結合することができ、またタンデムで分離された際に協同的に動くことができることを示した(14,15)。現在、CBMは一次配列類似性に基づき52の族に分類される(http://www.cazy.org/fam/acc_CBM.html)。CBMの構造における微妙な相違は多様なリガンド特異性をもたらし、CBMをタンパク質-炭水化物メカニズムを解明するための魅力的なシステムとすることができる。
【0005】
「Engineering Multivalent Sialic Acid Recognition using the 20 CBM40 module from Vibrio cholerae Sialidase」という題のポスター(2008年5月17日掲載:http://www.biochem.emory.edu/conferences/glycot/Images/GlycoTProgram-Posters.pdf参照)には、多価性によるシアル酸に対し増大した親和性を有する試薬の開発が開示されている。しかしながら、該ポスターは、かかる試薬が病原体によって引き起こされた疾病および/または疾患の治療に有用性を有することを記載していない。
【発明の概要】
【発明が解決しようとする課題】
【0006】
現在利用可能なインフルエンザ抗ウイルス剤(特に、ローチェのタミフル)に対する耐性が増えていることが十分文書化され、これはインフルエンザに対抗するための代替方法に目を向ける必要があることを強調した。以前の研究で、エルダーベリー由来のSNAレクチンのような非毒性レクチンをインフルエンザウイルス抑制剤として使用することが示されたが、これはシアル酸への弱い結合を示し、認識および効果的抑制のために2つまたは3つの異なる糖成分の存在を必要とした。組換え型シアリダーゼ-融合タンパク質を使用した最近の研究は、DAS181(NexBio社によって開発されたフルダーゼ)が現在これらの別の選択肢として調査されていることを示した。このタンパク質は、上皮細胞の表面からシアル酸を効果的に取り除き、ウイルスをレセプターと結合できなくする。しかしなから、シアリダーゼを用いてシアル酸を取り除くことによって、これはまた他の病原体用のレセプターとして役立ち得る隠れたレセプターを顕在化させることができる。
【0007】
本発明は、病原体によって引き起こされる疾病および/または疾患の治療に有用な化合物、組成物、薬剤および方法を提供し、かつ従来技術に伴う課題を回避することを目的とする。
【課題を解決するための手段】
【0008】
本発明は、炭水化物-結合分子(CBM)を活用してウイルスおよび/または細菌性病原体での感染の結果として生じる疾病または疾患と闘うおよび/または予防することができることを見出したことに基づく。
【0009】
細胞に結合/付着、コロニー形成または入り込むために、多くの病原体が炭水化物の存在を細胞表面上で利用することが既知である。例えば、オルトミクソウイルス科またはパラミキソウイルス属科に属するウイルスのような呼吸性の病原体は、種々の哺乳類の組織の特異的な細胞型に結合して入り込むために細胞表面炭水化物を利用する。同じように、連鎖球菌属に属するもののようなバクテリアは、特定の細胞に結合/付着するおよび/または入り込むための手段として細胞表面炭水化物を利用する。
【0010】
哺乳動物細胞の表面は、分子の多数の異なるタイプを備える。特に、哺乳動物細胞は、複合糖質が豊富で、その末端炭水化物は通常シアル酸である。従って、特定の病原体は、細胞表面でシアル酸の存在を活用してこれら細胞に結合/付着するおよび/または入り込むように進展する。
【0011】
CBM群のメンバーは、通常シアリダーゼまたはノイラミニダーゼのようなグリコシルヒドロラーゼ酵素をその効率的な加水分解用基質にターゲットまたは向かわせるように機能する。このように、CBMは例えば、シアル酸、ガラクトース、フコース、N-アセチルグルコサミンおよび血液型抗原などの細胞表面炭水化物に対し親和性を有する。当業者は、細胞表面炭水化物に対し親和性を呈する化合物を用いて、該炭水化物に結合または認識する病原体の結合能力を遮断し、したがって病原体が細胞のコロニー化および/または入り込みになることを防ぐことができることを認めるであろう。
【0012】
従って、本発明者らは、CBMの炭水化物への親和性を活用することによって、発病中に細胞表面炭水化物を結合/付着するかあるいはそれに伴う病原体が引き起こしたまたは一因となった様々な疾病および/または疾患の治療に有用な化合物、組成物、薬剤および/または方法を提供することか可能であることを発見した。
【0013】
従って、本発明の第1の態様は、一つ以上の病原体が引き起こしたまたは一因となった疾病および/または疾患を治療または予防するのに用いる炭水化物結合モジュール(CBM)を提供する。
【0014】
本発明の第2の態様は、一つ以上の病原体により引き起こされたまたは一因となった疾病および/または疾患の治療または予防用の薬剤製造への炭水化物結合モジュール(CBM)の使用を提供する。
【0015】
上記のものに加え、本発明の第3の態様は、一つ以上の病原体が引き起こしたまたは一因となった疾病および/または疾患の治療または予防用のCBMを含む製薬組成物を提供する。
【0016】
本発明によって提供される製薬組成物は、無菌の製薬組成物として処方されるのが好ましく、適当な賦形剤、キャリアまたは希釈剤としては、例えば、水、食塩水、リン酸塩緩衝食塩水、ブドウ糖、グリセロール、エタノール、イオン交換体、アルミナ、ステアリン酸アルミニウム、レシチン、血清アルブミンのような血清タンパク質、リン酸塩のような緩衝物質、グリシン、ソルビン酸、ソルビン酸カリウム、飽和植物性脂肪酸の部分的なグリセリド混合物、プロタミン硫酸エステルのような水塩類または電解質、リン酸水素二ナトリウム、リン酸水素カリウム、塩化ナトリウム、亜鉛塩、コロイダルシリカ、三ケイ酸マグネシウム、ポリビニルピロリドン、セルロース系物質、ポリエチレングリコン、カルボキシメチルセルロースナトリウム、ポリアクリラート、ワックス、ポリエチレン-ポリプロピレンブロックポリマー、ポリエチレングリコールおよび羊毛脂等またはその組合せを挙げることができる。
【0017】
前記製薬組成物は、例えば、経口、非経口または局所的投与に適した形態で処方することができる。
【0018】
当業者は、特異的な炭水化物を結合する個々のまたは単一CBMの使用に加えて、ここで提供される化合物、組成物および薬剤が代替の炭水化物に結合するかまたは親和性を示す、若しくは同じ炭水化物に異なる親和性を示す他のCBMを更に備えることができることを認識するであろう。
【0019】
前述のように、種々の疾病および/または疾患を治療または予防するのに用いる化合物、組成物および/または薬剤を提供することに加え、本発明はまた、病原体、特に細胞表面炭水化物を結合し得る病原体が引き起したまたは一因となった疾病および/または疾患に苦しむかまたは病にかかるリスクのある被検体、ヒト被検体の治療に有用な方法を提供する。
【0020】
従って、本発明の第4の態様は、病原体が引き起こしたまたは一因となった疾病を治療または予防する方法を提供するもので、該方法は炭水化物結合モジュール(CBM)の治療上有効量を投与するステップを備える。
【0021】
本発明は、病原体(例えばウイルス性および/または細菌性呼吸性病原体)が引き起こしたまたは一因となった疾病および/または疾患の既存の治療をこえて、例えば細胞表面炭水化物の酵素的除去を含む明白な利点がある。このタイプの戦略は、細胞を感染に対して脆弱にする他の病原体への隠れたレセプターを露出し得る。細胞表面から除去するよりもむしろ細胞表面炭水化物へのアクセスを遮断することによってこの課題が回避され、さらに、ここで記載したCBMが末端炭水化物残基(例えばシアル酸)と結合するので、CBMは免疫応答を顕在化しないかもしれない。
【0022】
ここで用いる用語「病原体」は、細胞表面炭水化物と結合、認識または逆に伴うことができるあらゆる病原体、特に細胞に結合/付着するかおよび/または入り込む手段として細胞表面炭水化物の存在を活用または利用するのに展開するものを含むと理解されたい。そのようなものとして、一実施形態における用語「病原体」は、微生物の病原菌を包含し、オルトミクソウイルス科またはパラミキソウイルス属科に属するウイルス、例えばインフルエンザおよびヒトパラインフルエンザウイルスのような呼吸性の病原菌、並びに肺炎連鎖球菌および/またはヘモフィルス属インフルエンザ、緑膿菌のような連鎖球菌属に属するバクテリアがあり、いずれもが哺乳動物細胞の表面上で炭水化物を結合することができる。当業者は、ここに記載するタイプの病原体によって最も頻繁にコロニー形成/感染された哺乳類の細胞は上皮細胞、特に粘膜路を裏打ちするもの(例えば呼吸性の上皮細胞)であると理解するであろう。
【0023】
上記を鑑みて、ここに提供される化合物、組成物、薬剤または方法を使用して治療可能または予防可能な疾病および/または疾患は多く、さまざまであり、ここで言及されるあらゆる病原体が引き起こしたまたは一因となったものとして考えるべきである。従って、例えばインフルエンザおよび/またはパラインフルエンザウイルスのようなウイルス性呼吸性病原体、および例えば肺炎連鎖球菌のような細菌性病原体が引き起こしたおよび/または一因となった疾病および/または疾患が、ここで提供される様々な化合物、組成物、薬剤および/または方法の使用により利益が得られる。この点に関しては、本発明は、インフルエンザのような疾病または疾患および偽膜性喉頭炎、肺炎および気管支炎のようなウイルス性および/または細菌性呼吸器病を治療および/または予防するのに用いる化合物、組成物、薬剤および/または方法を提供する。上記を考慮して、本発明の一実施形態は、インフルエンザウイルス、パラインフルエンザウイルスおよび肺炎連鎖球菌からなる群から選択した一つ以上の病原体が引き起こしたまたは一因となった疾病を治療するのに用いる化合物、組成物、薬剤および/または方法を提供する。
【0024】
用語「炭水化物」は、あらゆる炭水化物、特に細胞表面に存在するもの、とりわけ哺乳類の上皮細胞のような哺乳動物細胞の表面に存在するものを包含すると理解されたい。用語「炭水化物」は単糖単位を包含し得るが、また多糖、グリカン鎖および/または複合糖質の炭水化物部分を包含してもよいと理解されたい。このように、用語「炭水化物」としては例えば、シアル酸、細胞表面のグリカン鎖の末端(または遠位)にある9-炭素主鎖を有する炭水化物の系統を挙げることができる。シアル酸系統は、C4、C5、C7、C8およびC9でのアセチル化、グリコリル化、ラクトン化およびメチル化に由来し得る多数(およそ50)の誘導体を包含する。さらに、シアル酸はGalおよびGalNAcへのα(2,3)若しくはα(2,6)または別のシアル酸へのα(2,8)若しくはα(2,9)の結合で見られる。従って、用語「シアル酸」が本明細書の全体にわたって使用されるが、全ての誘導体、その類似物または変異体(自然に生じたまたは合成的に生成された)並びにこれを含むダイマー、トリマー、オリゴマー、ポリマーまたはコンカテマーを包含すると理解するのが重要である。他の細胞表面炭水化物は、N-アセチルグルコサミン、ガラクトース、N-アセチルガラクトーサミン、フコース、マンノースおよび血液型抗原としてかかるエピトープにおいて見いだされるそれらの複合体を含むことができる。
【0025】
CBMは、自然界で広く見いだされ、種々の異なる炭水化物に対し広範囲にわたる親和性を示す。これまで、CBMは一次配列類似性に基づいて52の属に分類されてきたが、本発明があらゆるCBMに関し得ることを理解されたい。勿論、特定の細胞表面炭水化物に結合する病原体によって生じることが既知の疾病または疾患を治療するために、該炭水化物に対して結合するかまたは親和性を示すCBMが、その疾病または疾患の治療または予防に特に有用である。
【0026】
本発明の一実施形態は、シアル酸に対し親和性を示すかまたは結合するCBM分子に関する。当業者はシアル酸に結合するかまたは親和性を示すCBMが、発病中にシアル酸に結合するかまたは逆に関連する病原体が引き起こしたまたは一因となった疾病および/または疾患を治療する化合物として、または組成物、薬剤または方法に有用であると認識するであろう。
【0027】
多くの異なるCBMが、特定の炭水化物に対して親和性を示し、その親和性の大きさが変化し得ることに留意されたい。しかし、シアル酸のような細胞表面炭水化物に結合するCBMの場合、シアル酸に対する親和性を示すあらゆるCBMが潜在的に有用であると考慮すべきであると理解されたい。
【0028】
これら潜在的に有用なCBMのうち、科40に属するものは“CBM40”のCBMとして既知である。コレラ菌およびウェルシュ菌に由来するCBMがCBM40科に含まれる。
【0029】
コレラ菌に由来するCBM40はシアル酸に対し比較的高い親和性(Kd〜30μM)を示すが、ウェルシュ菌に由来するCBM40はシアル酸に対して高いミリモル親和性を示す。
【0030】
上記に加えて、本発明者らは、単一のCBM分子が特定の炭水化物(例えばシアル酸)に対する親和性を示すことができるが、二つ以上のCBM分子を結合するかまたは一緒に組み合わせることにより結合活性を介して当該炭水化物に対しより高い親和性を有する分子を生成することが可能であることを発見した。
【0031】
したがって、本発明はまた、いくつかの手段によって一緒に接続、結合または共役された一つ以上のCBM(またはモノマー)を備える以後「CBMポリマー」と称する多価のCBM分子またはポリペプチドに関する。
【0032】
従って、本発明の第5の態様は、二つ以上のCBM(CBMモノマー)から成るCBMポリマーを提供する。
【0033】
用語「ポリマー」は、多くの結合単位から成る分子を含むことであると当業者によって容易に理解される。この点に関して、用語「CBM ポリマー」を用いて、CBM-ダイマー(すなわち2つのCBMモノマー)、CBM-トリマー(3つのCBMモノマー)、CBM-テトラマー(4つのCBMモノマー)およびより大きなCBM-オリゴマー(5つ以上のCBMモノマー)を説明することができる。
【0034】
当業者は、同一の反復CBMモノマー(例えば、コレラ菌由来のCBM40モノマーを反復する)を含むCBMポリマーを生成することが望ましいが、それぞれ異なる炭水化物を結合できるおよび/または特定の炭水化物に対し異なる親和性を示す多くの異なるCBMモノマーを含むCBMポリマーを作成することもまた可能であることを認識するであろう。同様に、異なった細胞表面炭水化物を結合するかまたは親和性を示すCBMモノマーを含むCBMポリマーもまた本発明の範囲内である。このように、本発明のCBMポリマーは、シアル酸に対し親和性を有するCBMモノマー、およびいくつかの他の炭水化物、例えばマンノース、ガラクトース、フコース、N-アセチルグルコサミンに対し親和性を有するかまたは結合するCBMモノマーを含むことができる。
【0035】
上述したように、CBMモノマーを一緒に接続、結合または共役することによって、特定の炭水化物または多くの炭水化物に対し結合活性を介してより大きな親和性を示すより大きな重合体分子をつくることが可能である。したがって、特定のCBMモノマー、例えば、コレラ菌に由来したCBM40がシアル酸に対し親和性を示し得るが、該CBM40モノマーの二つ以上を一緒に結合、接続または共役する場合、生成したポリマーは、シアル酸に対し増大した親和性を示し得る。
【0036】
従って、本発明および特に本発明の第1、第2、第3および第4の態様において用いる用語「CBM」は、ここに記載したCBMポリマーを含んでいるとして理解すべきである。
【0037】
当業者は、異なる炭水化物に結合または親和性を有するCBMモノマーを含むCBMポリマーそれ自体が同じ炭水化物に対し結合または親和性を示すことができることを認識するだろう。例えば、CBMポリマーが二つの異なる細胞表面炭水化物に対し結合または親和性を有するCBMモノマーを含む場合、生成したCBMポリマーはまた、同じ二つの細胞表面炭水化物に対し結合または親和性を示すことができる。
【0038】
多くの異なるCBMモノマーを含み、その結果多くの異なる炭水化物分子に対し結合または親和性を示すCBMポリマーは、複数の病原体が引き起こしたまたは一因となった疾病および/または疾患を治療または予防するための組成物、化合物、薬剤および/または方法に特に有用であり得る。
【0039】
例えば、それぞれ異なる細胞表面炭水化物に結合する(細胞のコロニー形成および/または入り込み手段として)二つの異なる病原体が引き起こしたまたは一因となった少なくとも二つの疾病または疾患に苦しむかまたはかかる傾向のある被検体にとって、これら炭水化物を認識、結合または逆に伴うCBMモノマーを含むCBMポリマーは前記疾病および/または疾患の治療または予防に有用であり得る。或いはまた、二つ以上の病原体によって生じる疾病および/または疾患は、関連したモノマーCBMを利用した化合物または組成物、薬剤若しくは方法を用いて治療することができる。
【0040】
当業者は、アミノ酸、タンパク質および/またはペプチドを一緒に結合し得る多くの方法があることを認識するであろう。一実施形態において、アミノ酸、ペプチドおよび/またはタンパク質を化学的手段によりおよび/またはクローニングおよび/またはPCR技術によって接続、結合または共役することができる。
【0041】
例えば、関連したCBMをエンコード化するヌクレオチド配列をPCRによって増幅し、その一つ以上のコピーを一緒にまたは他のCBMエンコード化ヌクレオチド配列に結合できるように修正することができる。加えて、CBMヌクレオチド配列を、一つ以上のアミノ酸残基を含むリンカー部分をエンコード化する配列をその3’および/または5’末端で含むように更に修飾させることができる。
【0042】
CBMポリマー(または多価のCBM)はまた、例えば、オリゴマー形成領域を含むように修飾したCBMモノマーを用いて発生させることができる。かかる領域を有する分子が自己集合してオリゴマー構造を形成することができる。一実施形態において、オリゴマー化領域を、例えばトリマー形成領域をコード化することが既知の緑膿菌のような細菌性種から誘導することができる。このように、PCRに基づく技術を用いて、CBM分子を修正することができる。
【0043】
次いで、増幅され、任意に修飾/結合されたCBM配列を、生成したCBMまたはCBMポリマーの発現および精製用の適当なベクターにクローン化することができる。組換え型CBMモノマー(例えばオリゴマー形成領域をエンコード化するもの)またはCBMポリマーを、例えば大腸菌において発現させることができる。
【0044】
上記を鑑みて、本発明の第6の態様は、CBMポリマーを生成する方法を提供するもので、該方法は
(a)CBMエンコード化ヌクレオチド配列を結合するステップ、および
(b)CBM ポリマーを生成するために結合したCBMヌクレオチド配列を発現するステップを備える。
【0045】
前述したように、本発明によって提供されるCBMポリマーは、一つ以上のアミノ酸の形態をとり得るリンカー部分のいくつかの形によって結合したCBMモノマーを含むことができる。リンカー部分を含むアミノ酸の正確な数は変り得るが、通常リンカー部分は、1からの約30アミノ酸までの任意の長さを含むであろう。
【0046】
CBMポリマーを生成する代替手段は、分子生物学的技術によりオリゴマー形成領域をCBMに結合することにある。例えば、緑膿菌のプシュードアミダーゼに見られるトリマー形成領域(残基335〜438、Xu, G., Ryan, C., Kiefel, M.J., Wilson, J.C. and Taylor, G.L.(2009) J. Mol.Biol. 386(3), 828 .840) は、リンカーペプチドによって一つ以上のCBMと結合することができた。次いで、単一構築物の発現は、結合活性効果により増大した親和性を有するトリマーを生成する。同様に、ヒト血管拡張を刺激したリンタンパク質(Kuhnel K., et al. PNAS 2004, 101, 17027-17032)からのテトラマー領域、すなわちテトラマーコイルドコイル構造を形成する45-残基ペプチドを、適当なリンカーを介して一つ以上のCBMに結合して、例えば、結合活性効果により増大した親和性を有するテトラマーオリゴマーを形成することができる。
【0047】
本発明の第7の態様は、疾病および/または疾患の治療に潜在的に有用なCBMをスクリーニングするかまたは識別する方法を提供するもので、該記方法は
(i) 細胞を結合または感染させるのに既知の病原体の存在下でCBMまたはCBMポリマーを前記細胞と接触させるステップ;
(ii) 病原体が結合したかまたは病原体が感染したこれら細胞を識別するステップ;および
(iii)かかる結果をCBMまたはCBMポリマーが加えられてない標準または対照アッセイと比較するステップとを備え、
ここで、対照アッセイに対し病原体が結合した細胞数の減少または病原体に感染した細胞数の減少が、該病原体が引き起こしたまたは一因となった疾病および/または疾患の治療に潜在的に有用なCBMまたはCBMポリマーを示す。
【0048】
CBM/CBMポリマーと接触した細胞は、組織培養条件下で培養した細胞単層若しくは組織生検または被検体から得た剥離物の形をとり得る。加えて、細胞が齧歯動物(マウス、ネズミ、モルモット、ウサギなど)のような動物に存在する場合がある。動物を本発明の第7の態様によって提供される方法に用いる場合、該動物に試験すべきCBMまたはCBMポリマーを投与し、また同時(またはその前後に)に病原体(例えば呼吸性の病原体)に感染させてもよい。さらに、標準または対照アッセイは、病原体に感染したがCBMまたはCBMポリマーを投与しなかった動物の形をとってもよい。
【0049】
動物を使用する場合、CBMまたはCBMポリマーが特定の病原体によって引き起こされたまたは一因となった疾病の治療または防止に潜在的に有用であるかどうかを確定するために、第7の態様によって提供された方法の完了後、細胞を動物から得、病原体が細胞に結合または感染させたか否かを決めるために検討することができる。加えてまたはあるいは、動物を疾病の徴候または症状のために観察してもよい。対照または標準のプロトコルを施した動物と比較する場合、症状の重症度の減少は、動物に投与したCBMまたはCBMポリマーが病原体によって引き起こされたまたは一因となった疾病および/または疾患の治療に有用であることを示すことができる。
【0050】
上記に加えて、本発明者らは、ここに記載した様々なCBMおよびCBMポリマーの異なる結合機能を用いて、炭水化物のスクリーニング、識別、検出、標識付けおよび/またはタギング手段を提供することができることを発見した。
【0051】
したがって、本発明の第8の態様は、サンプル中の炭水化物をスクリーニング、識別、標識付けおよび/またはタギングする方法を提供するもので、該方法は
(a) ここに記載したようなCBM/CBMポリマーと、それに結合するかまたは親和性を有する炭水化物との結合を可能にするに適した条件かで試料を該CBMまたはCBMポリマーと接触させるステップと、
(b) 未結合のCBMまたはCBMポリマーを除去するステップと、
(c)結合したCBMまたはCBMポリマーを検出するステップとを備える。
【0052】
当業者は、CBMが特定の炭水化物に対しある程度の特異性を示すので、ステップ(c)で検出したCBMまたはCBMポリマーがサンプルに存在する炭水化物の指標として役立つであろうことを認識するであろう。
【0053】
用語「サンプル」は、体液(尿、血液、血漿、血清、汗、唾液、精液など)のような生体試料、並びに例えば食品、飲料および水源(川、海)のような他の供給源に由来した試料を包含することを理解されたい。実際、炭水化物を含み、またCBMまたはCBMポリマーを添加し得ると考えられるほとんどすべてのサンプルを用いることができる。
【0054】
一実施態様において、サンプルは細胞、好ましくは組織生検、剥離物または分泌物に由来する哺乳類の細胞からなる。このように、本発明の第8の態様によって提供される方法を用いてサンプル中の特定の細胞の存在を検出することができる。当業者は、細胞がその表面でさまざまな炭水化物を発現するため、特定の細胞タイプで発現されるのが既知の炭水化物に特異的なCBMとサンプルを接触させ、CBMがサンプル中のあらゆる細胞に結合したか否かを検出することにより、特定の細胞の存在を不均質細胞集団内で識別することが可能と成り得ることを容易に理解するであろう。FACSのような技術を用いて、本発明に係るCBMまたはCBMポリマー(検出可能なタグを含むように任意に修飾された)で標識付けまたはタグ付けされた細胞を識別することができる。加えて、このタイプの技術は疾病または他の疾患を診断するのに有用である。例えば、癌細胞で発現されるのが既知の炭水化物に結合するかまたは親和性を有するCBMまたはCBMポリマーを用いてサンプル中の癌細胞の存在を識別または検出することができる。
【0055】
未結合のCBMまたはCBMポリマーを除去するために、上記ステップ(a)に由来するサンプル::CBM/CBMポリマー錯体に適切な緩衝材を用いる一つ以上の洗浄ステップを施してもよい。
【0056】
一実施形態において、サンプルを例えばプラスチック、ガラス、アガロース、ニトロセルロース、紙などの適切な基質に固定してもよい。
【0057】
当業者は、本発明によって提供されるCBMまたはCBMポリマーが一つ以上の検出可能なタグまたはラベルを含むように更に修飾され得ることを認識するであろう。このようにして、発色の化学発光反応を介してレベルを記録し得る酵素を含むようにCBMまたはCBMポリマーを修飾または共役させると、結合したCBMまたはCBMポリマーの検出を達成することができる。かかる酵素としては、西洋ワサビペルオキシダーゼ(HRP)およびアルカリフォスファターゼ(AlkP)を挙げることができるが、これに限定されるものではない。加えて、或いはまた、ここで提供するCBMまたはCBMポリマーは、例えばGFPのような蛍光分子およびFITC、ローダミンまたはテキサスレッドのような蛍光色素分子に共役させてもよい。ここに記載したCBMまたはCBMポリマーに共役し得る他のタイプの分子は、放射性標識化部分を含んでもよい。
【0058】
更なる実施形態において、本発明によって提供されるCBMポリマーまたはCBMは、細胞に送達すべき部分、例えば、抗体、小有機分子、核酸、薬または毒素を含むように更に修飾させることができる。細胞表面で発現した炭水化物を識別することによって、一つ以上の識別された炭水化物に結合するかまたは親和性を有するCBMまたはCBMポリマーを用いて、上述したタイプの部分を該細胞に送達するこができる。
【図面の簡単な説明】
【0059】
本発明は以下の図面を参照して詳細に説明する。
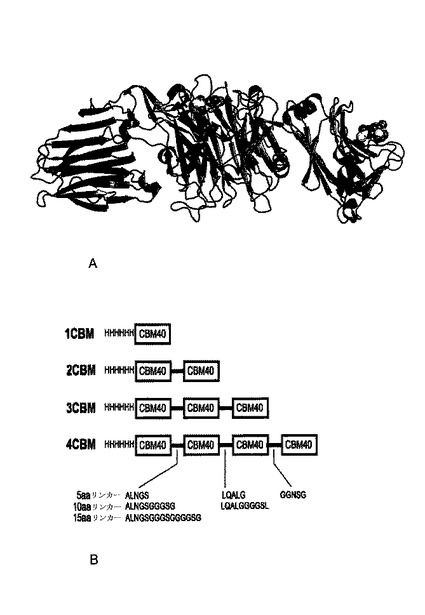
【図1】Aはレクチン様炭水化物-結合モジュール(CBM)が側面に位置した中央触媒領域を示すコレラ菌シアリダーゼの略図である。CBMを認識するシアル酸、すなわちCBM40が右手にある。Bは本研究で作成した構築物の略図である。
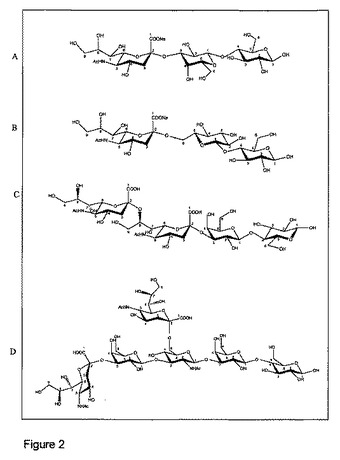
【図2】ITC研究に用いたシアロシドを示し、Aは3'シアリルラクトース、Bは6'シアリルラクトース、Cはジシアリルラクトース(DSL)、Dはジシアリル-ラクト-N-テトラオース(DSLNT)である。
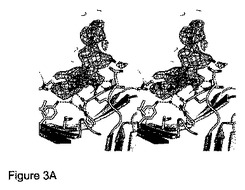
【図3】3'シアリルラクトースとの錯体におけるコレラ菌CBM40の結晶構造を示す。Aは点線として描画された水素結合を有するリガンドとCBMとの相互作用を示す。シグマ-a加重Fo-Fc電子密度マップを3σで描写する。Bは結晶の非対称ユニットにおける3つのCBM40モジュール間の関係を示す。
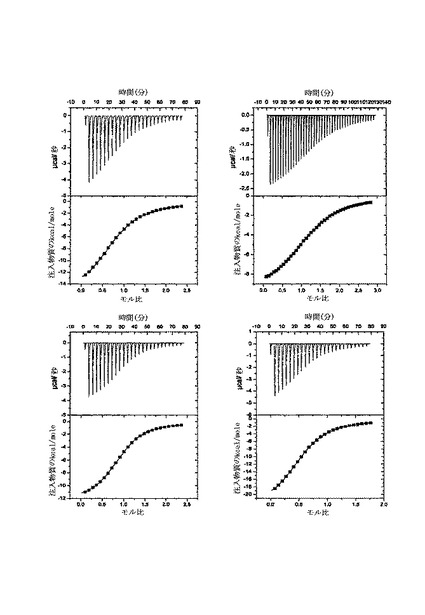
【図4】単離されたCBM40への様々なリガンドの結合を示す等温滴定熱量測定等温曲線であり、Aは3'SL、Bは6'SL、CはDSL、DはDSLNTである。
【図5】様々のCBM40構築物への3'SLの結合を示す等温滴定熱量測定等温曲線であり、Aは2CBM(5)、Bは2CBM(10)、Cは2CBM(15)、Dは3CBM(5)、Eは3CBM(10)、Fは4CBM(5)である。
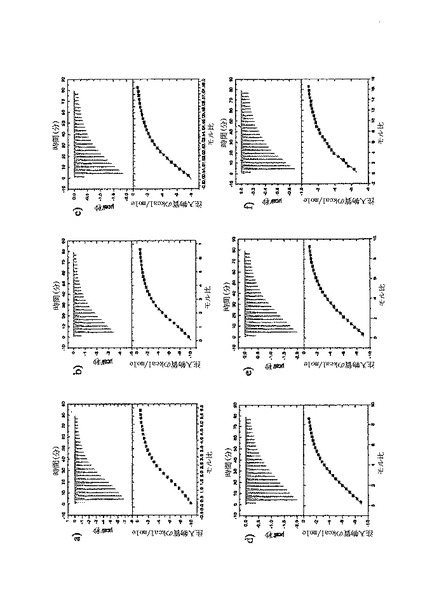
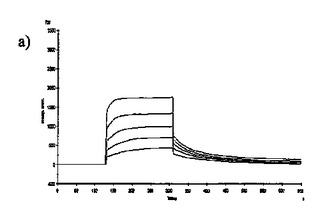
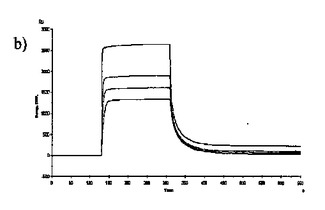
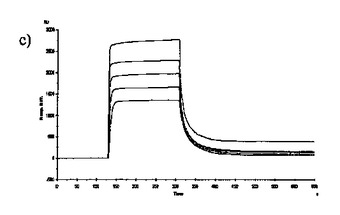
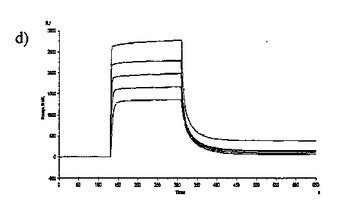
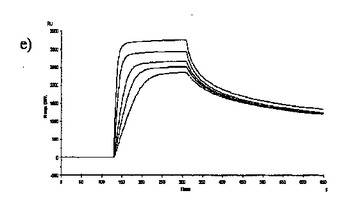
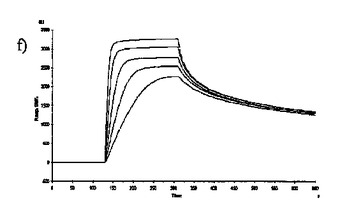
【図6】固定した3'シアリルラクトースレセプタへのCBM40モジュールの結合を示す表面プラスモン共鳴(SPR)センサーグラムであり、Aは1CBM, Bは2CBM(5), Cは2CBM(10), Dは2CBM(15), Eは3CBM(5), Fは3CBM(10), Gは4CBM(5)である。
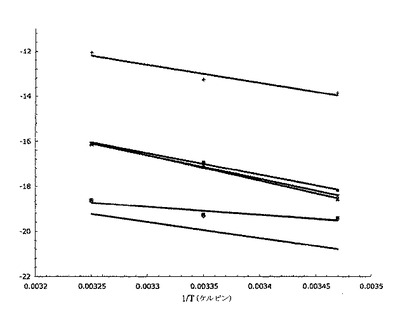
【図7】種々の温度でのSPR実験で得られたファント・ホッフの値をプロットしたものである。
【図8】多価CBM40ポリペプチドが生体外でインフルエンザウイルスの結合を低減し得ることを実証するプラークアッセイを示す。
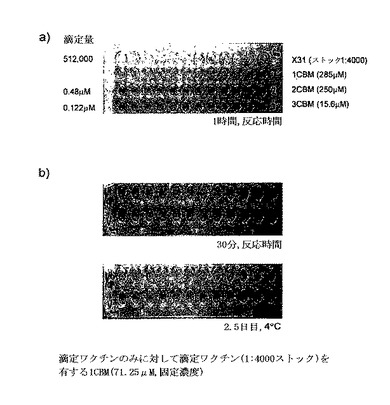
【図9】(a)はワクチンX31、単量体の多価CBM40を有するチキン赤血球の赤血球凝集反応アッセイ、(b)はワクチン株X31からのインフルエンザAウイルスの存在の有無でのチキン赤血球の赤血球凝集反応アッセイを示す。
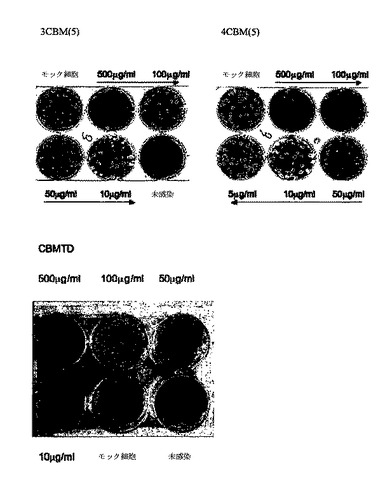
【図10】インフルエンザ株A/Udorn/72(H3N2)に対する3CBM(5)、4CBM(5)およびCBMTDポリペプチドによる哺乳類のMDCK細胞防御を示す。
【図11】(A)はMDCK細胞の細胞表面に結合するGFP-3CBM40(0.5mg/ml)を示し、(B)はUdornウイルスの72日目におけるGFP-3CBM40で培養したMDCK細胞の位相差顕微鏡写真である。パネル(a)はCBM(0.5mg/ml)のみ、(b)〜(f)はUdornウイルス存在での0.5、0.1、0.05、0.01および0.005mg/ml CBM、(g)はUdornウイルスのみ、および(h)は非感染細胞である。
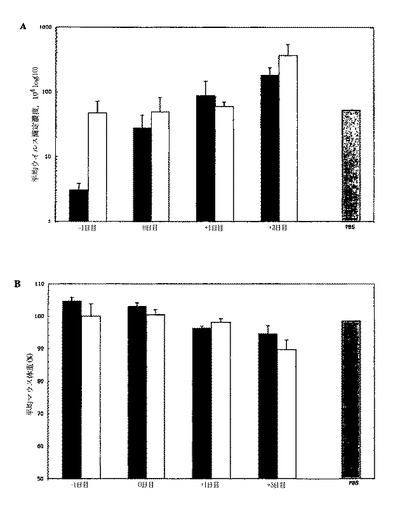
【図12】(A)は肺ウイルス滴定量に対する3CBM40の効果、および(B)はインフルエンザAウイルスに感染したBALB/cマウスのマウス重量に対する3CBM40の効果を示す。H1N1感染(103のウイルスpfu)の1日前、当日、または1または3日後に3CBM40(黒い棒)またはBSA(白い棒)をマウスに投与した。データを、6日p.i.で1グループにつき4匹のマウスから集めた。ウイルス対照グループ(0日)を棒グラフのPBSとして示す。
【0060】
(材料および方法)
(組換えDNA技術)
コレラ菌シアリダーゼ/ノイラミニダーゼ(VCNA)、残基25-216の科40 CBMをコード化するDNA断片を、表2の配列アウトラインに基づくオリゴヌクレオチドプライマペア1Fおよび1Rを用いてnanH遺伝子(16)を含む構築物pET30b+からPCRによって増幅した。573bpの増幅DNA断片をNco IおよびXho Iで消化し、pEHISTEVベクター(タバコエッチ病ウイルス(TEV)プロテアーゼによって切断されるN末端基6xHisタグを有するpET30の改変体)(Dr H. Liu、未発表の研究)にクローン化し、同じ酵素で消化し、大腸菌DH5aを形質転換するのに用いた。構築物p1CBMをDNAシーケンシングによって検証した(ダンディ大学シーケンシングサービス、英国)。
【0061】
コレラ菌CBM40をコード化するDNA断片を5’および3’末端で修飾して、異なるプライマペアの使用により異なる制限エンドヌクレアーゼ部位を組み込む。これは、DNA断片の個々のコピーの連結反応を許可する用に行って、タンデムに2、3および、4つのコピーを作製した(図1B)。さらに、プライマペアは、個々のモジュールを結合するアミノ酸の長さを表すために5、10または15のコドンの挿入を可能にした。これら全ての変更は、テンプレートとして表2に示した種々のプライマペアおよびp1CBMを用いるPCR増幅によって達成された。生成する断片を、モジュールの所望の数が得られるまで、適切に消化されたpEHISTEVベクターにクローン化した。これらを、それぞれ括弧内にリンカーのアミノ酸の数を有する2および3個の反復シアル酸結合ドメインを示すp2CBM(5)、p2CBM(10)、p2CBM(15)、p3CBM(5)およびp3CBM(10)と標識した。p4CBM(5)については、最終的な遺伝子をpEHISTEVに組立てる前に、HindIII-EcoRI修飾およびEcoRI-XhoI修飾DNA断片を最初に適切な酵素で消化したpET17bにクローン化した。すべての構築物を大腸菌DH5aで増殖した。タンパク質生産用の発現宿主大腸菌BL21(DE3)(1CBM、2CBM(5)、3CBM(5)、3CBM(10)、4CBM(5))および大腸菌BL21(DE3)Gold(2CBM(10)および2CBM(15)それぞれ)の形質転換前に、陽性クローンをDNAシーケンシングによって検証した。
【0062】
(タンパク質生産および精製)
VCNAについて述べられている(16)ように、全ての構築物を成長、発現させた。つまり、30μg/mlカナマイシンを含有するLB培地の1Lアリコートをシングルコロニーで接種し、培地の光学密度が600ナノメートルで0.4〜0.6に達するまで37℃で成長させた。細胞に42℃の水浴中で20分間熱ショックをかけた後、25℃まで冷却し、1mMのIPTGで誘導し、同じ温度で終夜振動した。細胞を遠心分離(8000xg、4℃)によって収集し、ペレットを要求されるまで-20℃で凍結した。
【0063】
全てのポリペプチドは、前述した(16)とおり、ニッケル親和性クロマトグラフィによって精製した。サンプルを、SDS PAGEを用いて分析し、部分的に精製したポリペプチドをTEVプロテアーゼ裂開緩衝材(PBS、0.3M NaCl、1mM DTT、0.5mM EDTA、20mM イミダゾール)で透析し、TEVプロテアーゼで終夜消化した。次いで、各ポリペプチドをPBS、0.3M NaCl、10mM イミダゾール緩衝液で更に透析した後、ニッケル充填カラムで二回目の精製を行って未消化のHis標識付けポリペプチドを除去した。未標識付けポリペプチドサンプルを10mMのHEPES緩衝材においてpH 7.4、0.15M NaClで広範囲に透析し、使用前に濃縮した。
【0064】
(結晶化および構造決定)
コレラ菌CBM40の単結晶を、シッテングドロップ法を用いて0.1M のMOPS、 pH 6.5、1.1Mの硫酸リチウム、沈殿剤としての0.6Mの硫酸アンモニュウム中で得た。結晶をまず10%(v/v)グリセロール-沈殿剤混合物へ移した後、20%(v/v)グリセロール-沈殿剤に放置することにより該結晶を凍結保護した。2.5Åに及ぶ回折データを、ESRFのビームライン BM-14で収集した。構造は分子交替プログラムPHASER(18)を用いて解析され、非対称ユニットに3つのCBM40モノマーを見いだした。微調整は、モデル構築(20)のために使用されるCOOTを有するREFMAC(19)を用いて実施された。データ収集および微調整統計を表3に示す。
【0065】
(等温滴定熱量計(ITC))
ITC実験を、1.4mlの細胞容積を有するMicrocal社(ノーサンプトン、MA)のVP-ITC微量熱量計で行った。明示しない限り、全てのITC滴定を0.15M NaClを含むpH7.4の10mMのHEPES緩衝材中で25℃で行った。単離されたCBM40(1CBM)の評価のために、以下のリガンド、すなわちDextra Labs(レディング、英国)から購入した3'シアリルラクトース(3'SL)、6'シアリルラクトース(6'SL)およびジシアリル-ラクト-N-テトラオース(DSLNT)、ならびに、およびシグマアルドリッチ(英国)から購入したジシアリルラクトース(DSL)を用いた(図2)。凍結乾燥されたシアロシドリガンドを、ペプチド構築物の透析のために用いた脱ガスのろ過したバッファに再懸濁させた。他の全てのポリペプチド構築物については、完全に3'シアリルラクトースをリガンドとして用いた。タンパク質濃度は、1CBM(38410M-1cm-1)、2CBM構築物(75860M-1cm-1)、3CBM構築物(113790M-1cm-1)および4CBM(5)(151720M-1cm-1)それぞれの算出モル吸光係数を使用してA280で決定された。CBM40ポリペプチドの濃度は0.007-0.084mMであり、シアロシドは0.45-2.14mMであった。シアロシドのアリコート(特記しない限り10μl)を各ポリペプチド溶液に滴定した。希釈熱は、MicroCal Origin softwareからの一結合部位モデルを用いる非線形回帰分析法によってデータをはめ込まれる前に、等温曲線データを結びつけることで差し引かれた。
【0066】
(表面プラスモン共鳴(SPR))
結合反応速度を、BIACORE 3000バイオセンサ機器(GE Biosystems)を用いるSPRによって求めた。使用の前に、ストレプトアビジン被覆(SA)バイオセンサチップを前記機器にドッキングし、50mM NaOH中に1M NaClの3回連続の1分噴射で予備調整した。ビオチニル化3'シアリルラクトース-PAA(Glycotech)は、HBS-P(10mM HEPES pH 7.4、0.15M NaClおよび0.005%サーファクタントP20)中で1μg/mlまで希釈した後、チップ中4つのうちの3つのフローセルに注入した。各細胞に対するリガンドの典型的固定化レペルは、およそ500RUであった。基準面をまたバルク効果の減算およびストレプトアビジンとの非特異的相互作用のために準備した。ランニングバッファは、100μl/分の流速で pH 7.4の10mMのHEPESからなる。
【0067】
各ペプチド構築物に対する固定化3'シアリルラクトースでの相互作用分析を、15、25および35℃での、ランニングバッファ中で行った。精製ペプチド構築物をHBS-Pに希釈して、1CBMについては0.625μM、1.25μM、 2.5μM、5μM、10μMの一連の濃度、2CBMについては20nM、125nM、250nM、500nMおよび1000nM、3CBMについては1nM、5nM、20nM、62.5nMおよび125nM、並びに4CBMについては0.18nM、0.5nM、1.6nM 5nMおよび15nMを得た。全ての検体を、30μl/分でフローセル表面上に注入した。表面からの検体の解離をランニングバッファにおいて3〜5分間同じ流速で達成した。表面は、2つの連続的な30μl/分の10mMのグリシン-HCl pH 2.5の30秒噴射によって再生された。平衡解離定数(KD)で記述される親和性は、BIA評価ソフトウェア(BIAcore)を用いてラングミュア(1:1)結合を仮定する動的同時ka/kdモデルに合わせることによって包括的に求められた。
【0068】
種々のCBMポリペプチド構築物のビオチニル化3'シアリルラクトース-PAAaとの相互作用の自由エネルギー変化は、各温度に対する動的化学反応速度定数の比率によって付与される平衡解離定数を用いて求められた。lnKD対1/Tのvan’t Hoffのプロットは、傾斜からΔH/Rおよびy切片からの-ΔS/Rに対する値をもたらした。
【0069】
(プラークアッセイ)
ウイルスプラークアッセイを用いて、インフルエンザAウイルスの存在下での結合多価CBM40ポリペプチドの有効性を実証した。6ウェルプレート中の集密的(106の細胞)なMDCK単層を無血清DMEMで2回洗浄し、2CBM40または3CBM40(10mg/ml、無血清DMEMで10倍に順次希釈)のどちらかの1mlの溶液によって37℃(+ 5%のCO2)で1時間培養した。無血清DMEMのみを平行して「モック細胞」培養に用いた。次いで、CBM40試薬を除去し、単層をインフルエンザウイルスA/Udorn/72[H3N2]の〜 80 PFUを用いて37℃(+ 5% CO2)で1時間培養した。ウイルス吸着後、接種材料を取り除き、細胞を2μg/mlトリプシンおよび1%(w/v)アガロースを加えたDMEM中で10mM HEPES(pH 7.4)で覆った。逆さにしたプレートを37℃(+ 5%のCO2)で2〜3日間培養した。プラークは、単層を5%のホルムアルデヒドで固定し、0.1%のクリスタルバイオレットで染色して明視化した。
【0070】
(血液凝集アッセイ)
アルセバー液のチキン赤血球を分離し、PBSで少なくとも4回洗浄した。該細胞を最終的に1%(v/v)PBSに再懸濁した。ワクチン株X31(香港/68 HAおよびNAの細胞表面を有する)を、PBSに1:4000ストックの2倍連続希釈を用いて96ウェルプレートで滴定し、これに等容積の赤血球細胞を添加した。該プレートを少なくとも30分間室温で培養した。終末点は、赤血球凝集反応が観測される最低ウイルス濃度として算出された。CBMをPBSで適切に希釈、滴定し、これに等容積の細胞を加えた。細胞に結合するCBMの終末点もまた算出した。次いで、滴定ウイルスを有する1CBMの固定濃度のチキンRBCで培養された際のみのウイルスに対する効果を求めるために実験を行った。
【0071】
(オリゴマー形成領域を用いるCBM.TDの開発)
1CBM40、p1CBMを含有するpEHISTEV構築物がC末端でオリゴマー形成領域を含むように修飾されて、大腸菌で発現された際にポリペプチドをオリゴマー構造に自己組織化させた。このために、緑膿菌プシュードミニダーゼ由来の三量体形成領域を用いた(Xu ら、2009年)。残基333〜440をコード化するDNA断片を、BamHIおよびXhoIを含むプライマーを用いるPCRによって増幅し、同じ酵素で予め消化されたp1CNMに結合した。次いで、これを、すべての他のCBM40構築物の生長および発現と類似した条件を用いて大腸菌BL21(DE3)Gold細胞に形質転換した。オリゴマーは、ニッケル親和性およびTEVプロテアーゼ消化を用いて精製し、ゲル濾過(データ示さず)後約100kDaの分子量を示した。
【0072】
(CBM40によりシアル酸を結合した細胞表面を調べるためのGFP-3CBM40の開発)
GFP融合3CBM40断片の構築については、3CBM40断片をp3CBM(5)由来のNcoI-XhoIで消化し、N末端基His標識下流の強化された緑色蛍光タンパク質(eGFP)をコード化する遺伝子を含む同様に消化されたpEHISTEVベクターに組み込んだ(Liu et al 2009; Connaris et al, 2009)。この構築物をしてCBM40が細胞表面でシアル酸に結合したかどうかを求めた。このために、96ウェルプレートにおけるMDCK細胞の集密的な単層を、種々の濃度のGFP-3CBMを用いて37℃で1時間培養し、その後インフルエンザ株A/Udorn/72(H3N2)ウイルス(moi = 0.002)を添加して同じ温度で更に1時間培養した。混合物を除去し、FCSのないDMEMを添加し、プレートを72時間培養した。
【0073】
(マウス研究:投薬量を求める実験:)
インフルエンザウイルスに対する抗ウイルス剤としてのCBMポリペプチドの予防/治療可能性を評価するために、初期の研究を行ってマウスに投与するのに必要なCBM40の投薬量を求めた。これら研究は、エジンバラの感染病センターのインフルエンザ調査のための動物実験設備で行われた。このために、5〜6週間目のBALB/cマウスを用いて、まず最初に生体外所見の投薬量に基づいた有害反応の検査をするための投薬量を求める実験を行った。3CBM40ポリペプチドを40μl PBS中50μg、また40μl PBS中500μgで用いてマウスの鼻腔内に投薬した。マウスは、臨床徴候を毎日調べられ、2日目以降から毎日計量された。マウスは、5日目に選別された(但し500μg投薬されたマウスのうちの2匹は選別する前14日間放置した)。
【0074】
(インフルエンザAウイルスに対するCBM40の有効性)
インフルエンザA/WSN/33(H1N1)に対するCBM40の予防/治療有効性を評価するために、BALB/cマウスを、3CBM40およびウイルスでの処理前にハロタン酸素ミックスで軽く麻酔した。実験用のCBM40投薬量は、鼻腔内に一度だけ送達するPBS(40μl)中に500μgの3CBM40 であった。BSAおよびPBSを対照物として用いた。マウスは、103のA/WSN/33 pfuで鼻腔内感染の日の1日前、若しくは当日、またはその後+1日目または+3日目に、CBM40またはプラセボで処理した。各グループのウイルス滴定濃度を、感染後マウスを選別する6日目に求め、肺をPBS中で均質化してプラークアッセイ用のウイルスを抽出した。選別したマウスの体重をまた測定して、体重減少を求めた。
【0075】
(結果)
(単離されたCBM40モジュールの構造)
コレラ菌シアリダーゼからのシアル酸結合モジュールCBM40をコード化する遺伝子断片をpEHISTEVにサブクローン化し、E.coliで発現させて1CBMを生成し、その後結合および構造的な研究のために精製した。まず最初に、1CBMを非常に高いレベルではあるが溶解せずに発現されたが、生長の対数期間の42℃での培養組織の熱ショックが1リットル当たり最高50〜70ミリグラムの量を絶えず生産するようにこのCBMの溶解性を改善した。コレラ菌CBM40の結晶を3'シアリルラクトースの存在下で成長させた。非対称ユニットは、3つのCBM40モノマーを含み、それぞれリガンドの3つ全ての糖残基に明白な電子密度を有した(図3A)。シアル酸だけがタンパク質との相互作用をなし、完全なコレラ菌シアリダーゼとの錯体におけるシアル酸のみで作られるのものと同じものである。結晶の非対称ユニットには、約750Å2(図3B)の界面を埋設するモノマーAおよびBを有する3つのCBM40モジュールがある。
【0076】
(シアロシドおよびコレラ菌CBM40の結合特異性の決定)
1CBMでの研究を行って種々のモノおよびジシアロシドに対する特異性を求めた。結合定数Ka並びにエンタルピー(DH)およびエントロピー(DS)の変化を各相互作用に対しITCを用いて直接測定し、化学量論的量(n、結合部位数)を上記の通りに一部位の結合モデルに対するデータの非線形最小二乗適合から求めた。希釈修正結合等温曲線の熱から得たデータは、α-シアル酸に対する1CBMの優先傾向を立証し(図4)、α(2,3)、α(2,6)およびα(2,8)-結合シアロシドに対する広い結合特異性を示す。試験したすべてのシアロシドが、10〜19μM(25℃で)の範囲の解離定数Kdを有する類似の親和性を示した(表4、図4)。ジシアロシド、DSLNTおよびDSLは、一価のリガンドと比較してシアル酸に対し1.5〜2倍より大きな親和性を示す。DSLNTは、DSL(-12.5kcal/モル)と比較して、-24kcal/モルのΔH値を有するエンタルピーの大きな変化を示した。DSLNTおよびDSLを結合するための化学量論的量を比較する場合、0.57および0.92の該当n値が観測され、DSLNTに対しては1CBMの2つの分子がDSLNTの1分子と結合するのに必要とされるが、一方DSLでは1:1相互作用があることを示す。この相違は、存在するシアル酸部分の数ではなく、そのシアロシド内の位置によるのである。構造的に、DSLNTは、DSLと異なり、分岐鎖の二価のシアロシドであり、直線型で一緒に結合した2つのシアル酸部分を有するので、末端シアル酸部分のみが認識される(図2)。この結果は、ほぼDSLNTのΔH値が一つの化学量論的量を有する3’シアリルラクトース、6’シアリルラクトースおよびDSLの個々の結合エンタルピーの総和であることを示す。
【0077】
(多価の、シアル酸特異性のCBM40ポリペプチドの作業)
1CBMの同一のコピーを一緒に結合して多価の種属を生成する場合にシアル酸結合が生じるかどうかを本発明者らは調査したかった。このために、CBM40をコード化する遺伝子のコピーを一緒にDNAリンカー(最高15のアミノ酸を示す)でつなげて、それぞれ2CBM(5)、2CBM(10)、2CBM(15)、3CBM(5),3CBM(10)および4CBM(5)(括弧の数字はリンカーアミノ酸の数を示す)として示された2、3および4モジュールのポリペプチドをタンデムに創生した(図1B)。これら遺伝子構築物の発現を大腸菌において行い、培養物に単離された1CBMのための熱ショックを施すまですべてのものが不溶性を示した。ニッケル親和性クロマトグラフィーでの精製後、全てのペプチド構築物のSDS PAGE分析は、それぞれ1CBM、2CBM、3CBMおよび4CBM構築物に対し〜21kDa、〜42kDa、〜63kDaおよび〜85kDaの単量体分子量を示した(データ示されず)。
【0078】
一価の3'シアリルラクトースを有する様々な設計されたCBM40ポリペプチドの結合等温曲線を図5に示す。ITCを用いると、設計されたCBM40ポリペプチドへのシアロシドの結合をエンタルピー的に駆動し、ΔH値が25℃で-12.3から-16.3 kcal/molの極めて類似の範囲にあることを明らかになった(表5)。各ポリペプチド相互作用について測定した他の熱力学パラメータのすべてにおいて相違もまたほとんどなかった。実際、シアル酸に対する多価のCBM40の結合親和性は1CBMのものと類似であると思われた。さらにまた、モジュール間のリンカーの長さがこの相互作用に著しく寄与することと熱力学的に示されなかった(表5)。一部位の結合モデルに基づいて、n値は、3’シアリルラクトースと相互作用する各々のCBM40ポリペプチドの部位の適切な数を示す。結合されたモジュールの数を上げると親和性の有意な増加がみられないという事実は、シアル酸−多価ポリペプチドの相互作用が、簡単な二分子会合を示す一価の単量体のものと類似していることを示唆する。
【0079】
(多価の3’シアリルラクトースに対する多価CBM40ポリペプチドの増強結合親和性)
シアル酸に対する結合力効果を多価のCBM40ポリペプチドで達成し得るかどうかを試験するために、表面プラスモン共鳴(SPR)をストレプトアビジンチップに固定したビオチン化3'シアリルラクトースを用いて行った。固定化3'シアリルラクトース上に注入した全てのCBM40ポリペプチドについてのセンサーグラムを図6に示す。ここに平衡解離定数Kdとして記述した各CBM40-3'SL相互作用の親和性は、ラングミュア1:1結合とみなす会合/解離速度定数(ka/kd)の比率に由来した包括的な適合モデルによって求めた(表6)。シアル酸に対する親和性の増加が、モジュール数を増加すると観察される。25℃での1CBM-3'SL相互作用について、ITCにより測定した対応する単量体の一価の相互作用のもの(Kd〜18μM)と比較して、結合の10倍の増加(Kd 〜1.8μM)がある。モジュールの数が2まで増加する場合に増強された親和性が観察され、ここでおよそ400〜500倍の結合の増加があり、25℃で38〜45nM間の親和性をもたらす(表6)。アミノ酸数を5からの15まで上げると、わずか1.2倍の親和性の増加が見られるので、リンカー長がこの場合わずかに結合活性に影響を与えると思われる。3および4つのCBM40モジュールを用いると、シアル酸に対する親和性が更に10〜20倍増強され、25℃で5aa-および10aa結合3CBMモジュールに対して約4nM、また4CBMモジュールに対して2.6nMの親和性に及ぶ。15℃で多価の3'SLを結合する場合、最も高い親和性は、Kd 〜861pMを有する4CBM(5)であった。したがって、相互作用の温度に依存して、一価の単量体の相互作用から多価のものへと進めることにより7000〜10000倍の親和性増加を達成し得ると思われる。15、25および35℃で測定した各CBM40ポリペプチド-シアル酸相互作用のファント・ホッフ値のプロットから得たデータ(図7、表7)は、1CBMに対し-7.8kcal/mol、2CBMに対し約-10kcal/mol、3CBMに対し-11.3kcal/molおよび4CBMに対し-12kcal/molのΔG値を示す。1CBMおよび2CBM間のΔG値の大きな相違はまた、相互作用のエンタルピーおよびエントロピーの変化に反映される。全てのCBM40ポリペプチドの中から、2CBMポリペプチドが、好適でない大きなエントロピー的寄与を補った約-20kcal/molのΔH値の最も大きなエンタルピー的変化を与えた(表7)。1CBMおよび2CBM相互作用間のエネルギー論におけるこの大きな相違はまた、1CBM、3CBMおよび4CBMポリペプチドと比較して、シアル酸に対する2CBMの増強された親和性に示され、リガンド-レセプター相互作用の共同性を示唆する。
【0080】
対照的に、3CBMポリペプチドは小さい有利なエントロピー的利得を示すが、相互作用の自由エネルギーの変化はより望ましい有利なエンタルピー的ペナルティによりまだ強くネガティブだった(表7)。面白いことに、3CBMおよび4CBMポリペプチドの両方は、相互作用の自由エネルギーが多分親和性の利得に寄与するモジュールの数の増加と共に増加したという事実にもかかわらず、1CBMおよび2CBMポリペプチドと比較して、リガンド結合に対するΔHおよびTΔSの両方へのネガティブな寄与の少ないことを示した。この見解は、多くの因子に基づくことができ、例えば、モジュールの立体配座配置、リガンド結合部位のアクセスしやすさ、分子内部および分子間結合のような結合のモード並びにモジュールの内部構造的なパッキングにより相互作用がリンカー柔軟性に敏感である。しかし、3CBMポリペプチド間のリンカー長の影響は、異なる2CBMポリペプチドと同様に結合エネルギーおよび親和性に関して無視し得るものであった。親和性の関連利得が25℃で4CBM(5)によって得られなかったので、ポリペプチドの更なる設計を行わないことが決定された。
【0081】
(赤血球凝集反応アッセイ)
単量体のおよび多価のCBM40が赤血球細胞に結合し、インフルエンザウイルスの結合を予防したかどうかを試験するために、初期実験を赤血球細胞に対し滴定されたCBMおよび滴定されたウイルス、ワクチン株X31で行った。予備結果は、1CBM40の場合凝集がみられないことを示し、またこのCBMが単量体であるので、細胞表面シアル酸に一価の型、すなわち 1:1会合で結合すると思われる(図9a)。多価CBM40に関しては、2および3CBMの両方が、ワクチンのX31のように赤血球細胞を凝集した(図9a)。加えて、細胞を凝集するのに必要な多価CBMの終点濃度は、3CBM40が2CBMよりも低く(それぞれ0.122μMおよび0.48μM)、これは繰り返しモジュールの数の増加がシアル酸に対する結合親和性を増大させることを示唆する。しかしながら、多価CBMが赤血球細胞を凝集するので、1CBM(71.25μM)の固定濃度を有する滴定されたワクチン混合物をワクチンのみに対して試験して凝集を観察することを決定した。図9bに示すように、ワクチンでテストした際の1CBMの存在は、モック細胞(ウイルスのみ)と比較すると、ウイルスの結合を防止する。プレートを4℃で2.5日間放置した際に、凝集が1CBMおよびウイルスの存在下で認められず、競争分子の結合をブロックするCBM40の有効性を示唆する。
【0082】
(プラークアッセイ)
プラークアッセイの予備結果は、10mg/mlの 3CBM(5)の1/100希釈液が同一希釈濃度の2CBM(10)よりも細胞へのウイルス性侵入をブロックするのにより効果的なことを示唆する強い証拠を示す(図8参照)。特に、モック細胞と比較した際に、異なる繰り返しCBMでMDCK細胞を培養後のプラーク数に相違があり、また3CBM(5)を用いる場合、H3N2の存在下でが極めて少ないプラークが現れることが分る。
【0083】
3CBM(5)(1/100希釈)では、H3N2ウイルス・プラーク形成単位の80-85%の減少がある。2CBM(10)の結果では、ウイルス遮断効果があるが、同じ濃度の同一希釈は3CBM(5)と同じ効果を有さなかった。このように、CBMモジュール数の増加は細胞表面でのシアル酸に対するウイルス結合を効果的に遮断するのに重要であったことがわかる。
【0084】
(プラーク研究(CBMTDの使用))
ウイルス感染MDCK細胞:インフルエンザ株A/Udorn/72(H3N2)ウイルスをCBM40ポリペプチドによりブロックした。図10は、Udornウイルスに対するMDCK細胞上のポリペプチド3CBM40、4CBM40およびCBMTDの細胞防護効果を示す。結合したCBM40の数が3から4CBM40まで増加すると、インフルエンザ株に対するCBM阻害作用強度の増加がわかる。また、ウイルスの添加前のオリゴマーCBM.TDでのMDCK細胞の培養は、ウイルス対照物と比較して0.5mg/mlで有効な抑制と、0.1mg/ml CBMでプラーク数の顕著な減少を示した(図10)。これらのデータは、最高4ドメイン反複のコレラ菌シアリダーゼCBM40がタンデム結合したかまたは自己集合したオリゴマーとしてウイルス感染からMDCK細胞を効果的に保護することができることを示す。
【0085】
(CBM40によりシアル酸を結合した細胞表面の監視)
図11aは、ウイルス添加1時間後のMDCK細胞の細胞表面へのGFP-3CBM40の結合を示す。図11bは、ウイルスのみのウェルと比較した際に、感染72時間後でもGFP-3CBM40がウイルス感染から相当な保護を付与することを示す。パネル(a)はまた、ウイルスの不在下でMDCK細胞に加えたCBM(0.5mg/ml)が72時間後毒作用を示さず、未感染対照物[パネル(h)]に対し類似の形態を有する細胞が生存能力を有すると認められたことを示した。
【0086】
(マウス研究(投薬量を求める実験))
表1は、研究中のマウスの最初と最終体重の結果を示す。臨床見解から、マウスすべてが非常に健康であり、5日および14日の指定された期間中それら全てが3CBM40に副作用なしで重くなったことがわかった。抗体反応および肺組織の組織学に関する更なる研究は、まだ実施されていない。
【0087】
(インフルエンザAウイルスに対するCBM40の有効性)
予備結果は、WSNウイルスチャレンジの1日前の3CBM40の投与がマウスのウイルス感染の効果を著しく減らすことを示し、CBM40がインフルエンザAウイルスに対する予防薬として効果的であることを実証したことを示唆した。これは、103のウイルスpfuの初回量から1.5ログ滴定濃度の増加のみが測定されたBSAおよびPBS対照と比較して、感染6日後に選別されたマウスのウイルス滴定濃度の減少で明白だった(図12A)。このグループのマウスはまた、体重が増え、ウイルスチャレンジからの副作用を示さなかった(図12B)。3CBM40をウイルスと同時に投与した場合(0日目)、ウイルスの2.4ログ増加が感染6日後に100%を超える体重増加とともに認められ、ウイルス滴定濃度の増加にもかかわらずマウスがウイルス感染に耐えたことを示す。しかしながら、CBM40をウイルス感染後の1および3日目に投与する場合、マウスは軽くなり、ウイルス滴定濃度が初期の負荷と比較して3〜3.25ログだけ増加したことが分り、CBM40は治療薬として付与した際に少なくとも本方法で記述する条件下で効果的ではないことを示唆した。1回の投与ではなく感染中毎日の投薬および投与手段の最適化を含む更なる研究は、CBM40が特にインフルエンザ感染の初期段階の治療の抗ウイルス薬として考えることができたかどうかを決定することができる。予防法として、このタンパク質生物製剤を鼻腔内に送達してウイルス攻撃から呼吸性上皮粘膜を保護することができるであろう。
【0088】
(考察)
本発明者らは、コレラ菌シアリダーゼからのシアル酸特異性CBMをその親酵素からうまく単離し、シアル酸を結合する多価のポリペプチドを高い親和性で作り出すのに活用することができることを示した。ITCおよびSPRの組合せを使用し、本発明者らはまた、これまで多くのCBM−炭水化物相互作用である(Boraston et al 2004, etc)ようにエンタルピー的に駆動されるが、多価の表面に導入される際に結合モジュール数が2を超える場合にエントロピー的に駆動される一価および二価の様態であるシアル酸へのコレラ菌CBM40の一価および多価の様態の結合の熱力学を実証した。
【0089】
リガンドおよび結合特異性に関して、単離されたCBM40はマイクロモル濃度の親和性でα(2,3)、α(2,6)およびα(2,8)-結合シアル酸を優先して結合する。この結合親和性は、非加水分解性チオシアロシドとのコレラ菌シアリダーゼ相互作用について報告されたものに類似している(16)。異なる結合シアロシドの間でほとんど区別がないという事実は、3'シアリルラクトースで錯化したCBM40の結晶構造においてみられる。該構造は、シアル酸との分子間相互作用の大規模なネットワークを明らかにする。ガラクトースおよびグルコース部分はタンパク質ドメインと相互作用せず、これは異なる結合シアロシドについてここで見られる類似のKd値および結合の自由エネルギーに反映する。
【0090】
シアル酸に対するコレラ菌CBM40の比較的高い一価の親和性を活用することによって、本発明者らは、多価の表面と相互作用する際にサブナノモーラー結合活性を得るためにn=4までのタンデム結合反複ポリペプチドをうまく設計した。これは、異なる長さのリンカーペプチドを使用して同一のコピーを結合させることによって操作されたCBMのおそらく第1の報告である。植物細胞壁ポリマーの分析用の分子プローブとして単離されたCBMの使用の報告(21)、並びに別のモジュールまたはそれらの機能を決定するためにその隣接したモジュールと組合せのいずれかとして単離されたマルチモジュラーグリコシドヒドロラーゼで天然に見られるCBMの研究が以前にあった(14,17,22)。
【0091】
配座の制約がこれら設計されたポリペプチドの結合を防止するかどうかを決定するために、本発明者らはレセプターとの多価の相互作用の熱力学を調査したかった。タンパク質サブユニット間のリンカー長が相互作用の熱力学に影響を及ぼすことができ、また多くの場合、共有結合サブユニットが解離のエントロピー的駆動力を減らすことによって相互作用のオリゴマー安定性を強化することができることは周知である(23)。本発明者らは、リンカー長が設計されたCBMポリペプチドのシアル酸に対する親和性を著しく強化するかどうか決めるために、5つのアミノ酸から15のアミノ酸までの範囲にあるCBMモジュール間の柔軟なペプチドリンカーを設計した。2タンデム結合CBMの場合、ペプチドリンカーを10アミノ酸まで増加すると、異なる2CBM構築物間のシアル酸に対する親和性に1.5〜2倍の増加を認めることにもかかわらず、配座のエントロピーの有意な減少を与えた。これは、モジュールのある程度の柔軟性が2つのCBMを一緒に結合する際に生起したことを示唆する。シアル酸に対する2CBMの結合相互作用はまた、大きな好適でないエントロピー的寄与を補う結合エンタルピーの増加を示し、結合自由エネルギーの増加および1CBM―多価3SL相互作用と比較した際シアル酸に対する親和性の強化をもたらした。これは、シアル酸に対する2CBMの結合が非常に安定した分子内相互作用であり、1CBMに対してエンタルピー的に駆動されることを示唆する。しかしながら、固定された多価3SLと相互作用する3CBMに関し、異なる長さのペプチドの間では有意差がなかった。実際、異なる3CBM-シアル酸の相互作用のvan’t Hoffプロットから得たエントロピーデータはすべて、エントロピーの増加を示す。また、結合エンタルピーの有意な減少および親和性の10倍の強化は、2から3CBMになるときに認められた。シアル酸へ結合する3CBMの場合に認められる親和性の強化は、SPRチップの表面で生じる安定した集合体の有望な形成に起因し、また大きなエントロピー変化が密着したタンパク質-タンパク質パッキングによって影響される水和殻から解放される構造的に秩序ある水分子によるものである。これは、3つの1CBMモノマーを一緒に密接にパックされた1CBM-3SL錯体の構造から明らかである(図3B)。エントロピー駆動相互作用は、特に分子間結合が生じる他の多価のタンパク質−炭水化物システムでみられ、集合体の形成にいたる(24)(25)、(26)。面白いことに、これはわずかに固定された3’SLとの1CBM相互作用のものより少ないが、結合モジュール数を4まで増加すると、3CBMsのものへの好適なエントロピー寄与が少なく、結合エンタルピーが好ましいことを示した。結合の自由エネルギーは、3から4CBMまでの親和性の僅かな増大に対応する全ての他のCBMより僅かに大きかった。設計されたタンデム結合ポリペプチドの結合価は、おそらくオリゴマーの安定化およびそれらの相互作用に重要な役割を果たし得る。プーン(20)は、タンデム結合された単一ポリペプチド鎖の結合価を増大すると、熱力学的に言えば、より安定になる結合オリゴマーの分子数の対応する減少をもたらし得ることを記載する。実際に、プーンは、オリゴマーの分子数より高い結合価のタンデムは、または分割し得ないタンデムが交差結合したオリゴマーをもたらすことを述べた。したがって、4CBMの場合のような4量体オリゴマーに関しては、タンデムダイマーまたはテトラマーが好ましいが、三量体または五量体オリゴマーは、それぞれ同じ結合価のタンデムを必要とする。
【0092】
これらの研究は、同一コピーを結合してそのシアル酸に対する親和性を結合活性により増強することによってCBM40を単離し、操作することができることを示した。さらにまた、本発明者らは、モノマーおよびコレラ菌シアリダーゼから設計した多量体CBM40をインフルエンザAウイルスに対する潜在的抗ウイルス薬として用い得ることを示した。従って、この種の技術を使用すると、他のCBMをグリカンスクリーニングおよびプロファイリング用の高親和性ツールとしての使用のために探利子、設計することができるであろう。加えて、そのグリカンとの錯体におけるCBM構造の決定は、グライコミクスの分野での使用への選択的試薬の発達に役立ちえる。
【0093】
(参考文献)

【0094】
【表1】
【0095】
【表2】
【0096】
【表3】
【0097】
【表4】
【0098】
【表5】
【0099】
【表6】
【0100】
【表7】
【技術分野】
【0001】
本発明は、炭水化物結合分子を備えるかまたは使用する化合物、組成物、薬剤および方法を提供する。特に、本発明は、病原体、特に微生物の病原体によって引き起こされるかまたは一因となる疾病および/または疾患を治療する手段、並びに炭水化物のスクリーニング、識別、検出タギングおよび/または標識付けの方法を提供する。
【背景技術】
【0002】
大部分の哺乳動物細胞の表面は複合糖質が豊富であり、多くの病原体が初期段階の発病中に細胞付着のためこれら複合糖質の末端炭水化物を利用している。例えば、インフルエンザおよびパラインフルエンザ両方のウイルス性病原体が哺乳類の細胞表面でシアル酸レセプターと結合する。
【0003】
シアル酸認識は、レクチンまたはレクチン様分子およびその対応するレセプターを介してもたらされる(3)。ほとんどの場合、いくつかのウイルス糖タンパク質および細菌毒素(2,4)並びにシグレック(5)およびセレクチン(6)のような哺乳類のレクチン上科も含むシアル酸結合レクチンは、これら分子の多価性により比較的高い親和性でレセプターと結合し、したがって大部分のタンパク質-炭水化物相互作用に関連する低い固有の親和性を緩和する(7,8)。一般に、かかるレクチンによる1価および2価のシアロシドの結合の会合定数(Ka)は、104M-1に達することができる。しかしなから、その多価性のため、いくつかのシアル酸結合レクチンが多価の細胞表面グリカンと相互作用して、結合活性効果によって109M-1に及ぶ親和性を達成することができる。これら増強された親和性は、好ましい結合エネルギー論に関連したリガンド結合によって促進されるタンパク質の改良された構造的パッキングによることが部分的に示された(9-11)。最もよく研究された多価のレクチン-シアル酸相互作用のうちの一つは、インフルエンザウイルス三量体血球凝集素であり、独立体の一つまたは両方が多価の状態でない場合に約4 x 102M-1と対照に最高108M-1親和性を達成することができる(12)。
【0004】
シアリダーゼ、すなわちノイラミニダーゼは、種々の複合糖質からシアル酸の加水分解を触媒し、大抵タンパク質の触媒コアに付着したアクセサリモジュールを含むモジュラー酵素である。これらモジュールのいくつかは、炭水化物結合モジュール(CBM)として識別される。CBMはグリコシドヒドロラーゼで広く見られ、親酵素をその効率的な加水分解用の基質にターゲットするために基質表面で酵素濃度を上げることによって主として存在する個々の非触媒性モジュールである(13)。該モジュールは、グリコシルヒドロラーゼの構造内でシングル、タンデムまたは複合物である。研究の結果、これらが親分子から分離された際に独立してその特異的なグリカンと結合することができ、またタンデムで分離された際に協同的に動くことができることを示した(14,15)。現在、CBMは一次配列類似性に基づき52の族に分類される(http://www.cazy.org/fam/acc_CBM.html)。CBMの構造における微妙な相違は多様なリガンド特異性をもたらし、CBMをタンパク質-炭水化物メカニズムを解明するための魅力的なシステムとすることができる。
【0005】
「Engineering Multivalent Sialic Acid Recognition using the 20 CBM40 module from Vibrio cholerae Sialidase」という題のポスター(2008年5月17日掲載:http://www.biochem.emory.edu/conferences/glycot/Images/GlycoTProgram-Posters.pdf参照)には、多価性によるシアル酸に対し増大した親和性を有する試薬の開発が開示されている。しかしながら、該ポスターは、かかる試薬が病原体によって引き起こされた疾病および/または疾患の治療に有用性を有することを記載していない。
【発明の概要】
【発明が解決しようとする課題】
【0006】
現在利用可能なインフルエンザ抗ウイルス剤(特に、ローチェのタミフル)に対する耐性が増えていることが十分文書化され、これはインフルエンザに対抗するための代替方法に目を向ける必要があることを強調した。以前の研究で、エルダーベリー由来のSNAレクチンのような非毒性レクチンをインフルエンザウイルス抑制剤として使用することが示されたが、これはシアル酸への弱い結合を示し、認識および効果的抑制のために2つまたは3つの異なる糖成分の存在を必要とした。組換え型シアリダーゼ-融合タンパク質を使用した最近の研究は、DAS181(NexBio社によって開発されたフルダーゼ)が現在これらの別の選択肢として調査されていることを示した。このタンパク質は、上皮細胞の表面からシアル酸を効果的に取り除き、ウイルスをレセプターと結合できなくする。しかしなから、シアリダーゼを用いてシアル酸を取り除くことによって、これはまた他の病原体用のレセプターとして役立ち得る隠れたレセプターを顕在化させることができる。
【0007】
本発明は、病原体によって引き起こされる疾病および/または疾患の治療に有用な化合物、組成物、薬剤および方法を提供し、かつ従来技術に伴う課題を回避することを目的とする。
【課題を解決するための手段】
【0008】
本発明は、炭水化物-結合分子(CBM)を活用してウイルスおよび/または細菌性病原体での感染の結果として生じる疾病または疾患と闘うおよび/または予防することができることを見出したことに基づく。
【0009】
細胞に結合/付着、コロニー形成または入り込むために、多くの病原体が炭水化物の存在を細胞表面上で利用することが既知である。例えば、オルトミクソウイルス科またはパラミキソウイルス属科に属するウイルスのような呼吸性の病原体は、種々の哺乳類の組織の特異的な細胞型に結合して入り込むために細胞表面炭水化物を利用する。同じように、連鎖球菌属に属するもののようなバクテリアは、特定の細胞に結合/付着するおよび/または入り込むための手段として細胞表面炭水化物を利用する。
【0010】
哺乳動物細胞の表面は、分子の多数の異なるタイプを備える。特に、哺乳動物細胞は、複合糖質が豊富で、その末端炭水化物は通常シアル酸である。従って、特定の病原体は、細胞表面でシアル酸の存在を活用してこれら細胞に結合/付着するおよび/または入り込むように進展する。
【0011】
CBM群のメンバーは、通常シアリダーゼまたはノイラミニダーゼのようなグリコシルヒドロラーゼ酵素をその効率的な加水分解用基質にターゲットまたは向かわせるように機能する。このように、CBMは例えば、シアル酸、ガラクトース、フコース、N-アセチルグルコサミンおよび血液型抗原などの細胞表面炭水化物に対し親和性を有する。当業者は、細胞表面炭水化物に対し親和性を呈する化合物を用いて、該炭水化物に結合または認識する病原体の結合能力を遮断し、したがって病原体が細胞のコロニー化および/または入り込みになることを防ぐことができることを認めるであろう。
【0012】
従って、本発明者らは、CBMの炭水化物への親和性を活用することによって、発病中に細胞表面炭水化物を結合/付着するかあるいはそれに伴う病原体が引き起こしたまたは一因となった様々な疾病および/または疾患の治療に有用な化合物、組成物、薬剤および/または方法を提供することか可能であることを発見した。
【0013】
従って、本発明の第1の態様は、一つ以上の病原体が引き起こしたまたは一因となった疾病および/または疾患を治療または予防するのに用いる炭水化物結合モジュール(CBM)を提供する。
【0014】
本発明の第2の態様は、一つ以上の病原体により引き起こされたまたは一因となった疾病および/または疾患の治療または予防用の薬剤製造への炭水化物結合モジュール(CBM)の使用を提供する。
【0015】
上記のものに加え、本発明の第3の態様は、一つ以上の病原体が引き起こしたまたは一因となった疾病および/または疾患の治療または予防用のCBMを含む製薬組成物を提供する。
【0016】
本発明によって提供される製薬組成物は、無菌の製薬組成物として処方されるのが好ましく、適当な賦形剤、キャリアまたは希釈剤としては、例えば、水、食塩水、リン酸塩緩衝食塩水、ブドウ糖、グリセロール、エタノール、イオン交換体、アルミナ、ステアリン酸アルミニウム、レシチン、血清アルブミンのような血清タンパク質、リン酸塩のような緩衝物質、グリシン、ソルビン酸、ソルビン酸カリウム、飽和植物性脂肪酸の部分的なグリセリド混合物、プロタミン硫酸エステルのような水塩類または電解質、リン酸水素二ナトリウム、リン酸水素カリウム、塩化ナトリウム、亜鉛塩、コロイダルシリカ、三ケイ酸マグネシウム、ポリビニルピロリドン、セルロース系物質、ポリエチレングリコン、カルボキシメチルセルロースナトリウム、ポリアクリラート、ワックス、ポリエチレン-ポリプロピレンブロックポリマー、ポリエチレングリコールおよび羊毛脂等またはその組合せを挙げることができる。
【0017】
前記製薬組成物は、例えば、経口、非経口または局所的投与に適した形態で処方することができる。
【0018】
当業者は、特異的な炭水化物を結合する個々のまたは単一CBMの使用に加えて、ここで提供される化合物、組成物および薬剤が代替の炭水化物に結合するかまたは親和性を示す、若しくは同じ炭水化物に異なる親和性を示す他のCBMを更に備えることができることを認識するであろう。
【0019】
前述のように、種々の疾病および/または疾患を治療または予防するのに用いる化合物、組成物および/または薬剤を提供することに加え、本発明はまた、病原体、特に細胞表面炭水化物を結合し得る病原体が引き起したまたは一因となった疾病および/または疾患に苦しむかまたは病にかかるリスクのある被検体、ヒト被検体の治療に有用な方法を提供する。
【0020】
従って、本発明の第4の態様は、病原体が引き起こしたまたは一因となった疾病を治療または予防する方法を提供するもので、該方法は炭水化物結合モジュール(CBM)の治療上有効量を投与するステップを備える。
【0021】
本発明は、病原体(例えばウイルス性および/または細菌性呼吸性病原体)が引き起こしたまたは一因となった疾病および/または疾患の既存の治療をこえて、例えば細胞表面炭水化物の酵素的除去を含む明白な利点がある。このタイプの戦略は、細胞を感染に対して脆弱にする他の病原体への隠れたレセプターを露出し得る。細胞表面から除去するよりもむしろ細胞表面炭水化物へのアクセスを遮断することによってこの課題が回避され、さらに、ここで記載したCBMが末端炭水化物残基(例えばシアル酸)と結合するので、CBMは免疫応答を顕在化しないかもしれない。
【0022】
ここで用いる用語「病原体」は、細胞表面炭水化物と結合、認識または逆に伴うことができるあらゆる病原体、特に細胞に結合/付着するかおよび/または入り込む手段として細胞表面炭水化物の存在を活用または利用するのに展開するものを含むと理解されたい。そのようなものとして、一実施形態における用語「病原体」は、微生物の病原菌を包含し、オルトミクソウイルス科またはパラミキソウイルス属科に属するウイルス、例えばインフルエンザおよびヒトパラインフルエンザウイルスのような呼吸性の病原菌、並びに肺炎連鎖球菌および/またはヘモフィルス属インフルエンザ、緑膿菌のような連鎖球菌属に属するバクテリアがあり、いずれもが哺乳動物細胞の表面上で炭水化物を結合することができる。当業者は、ここに記載するタイプの病原体によって最も頻繁にコロニー形成/感染された哺乳類の細胞は上皮細胞、特に粘膜路を裏打ちするもの(例えば呼吸性の上皮細胞)であると理解するであろう。
【0023】
上記を鑑みて、ここに提供される化合物、組成物、薬剤または方法を使用して治療可能または予防可能な疾病および/または疾患は多く、さまざまであり、ここで言及されるあらゆる病原体が引き起こしたまたは一因となったものとして考えるべきである。従って、例えばインフルエンザおよび/またはパラインフルエンザウイルスのようなウイルス性呼吸性病原体、および例えば肺炎連鎖球菌のような細菌性病原体が引き起こしたおよび/または一因となった疾病および/または疾患が、ここで提供される様々な化合物、組成物、薬剤および/または方法の使用により利益が得られる。この点に関しては、本発明は、インフルエンザのような疾病または疾患および偽膜性喉頭炎、肺炎および気管支炎のようなウイルス性および/または細菌性呼吸器病を治療および/または予防するのに用いる化合物、組成物、薬剤および/または方法を提供する。上記を考慮して、本発明の一実施形態は、インフルエンザウイルス、パラインフルエンザウイルスおよび肺炎連鎖球菌からなる群から選択した一つ以上の病原体が引き起こしたまたは一因となった疾病を治療するのに用いる化合物、組成物、薬剤および/または方法を提供する。
【0024】
用語「炭水化物」は、あらゆる炭水化物、特に細胞表面に存在するもの、とりわけ哺乳類の上皮細胞のような哺乳動物細胞の表面に存在するものを包含すると理解されたい。用語「炭水化物」は単糖単位を包含し得るが、また多糖、グリカン鎖および/または複合糖質の炭水化物部分を包含してもよいと理解されたい。このように、用語「炭水化物」としては例えば、シアル酸、細胞表面のグリカン鎖の末端(または遠位)にある9-炭素主鎖を有する炭水化物の系統を挙げることができる。シアル酸系統は、C4、C5、C7、C8およびC9でのアセチル化、グリコリル化、ラクトン化およびメチル化に由来し得る多数(およそ50)の誘導体を包含する。さらに、シアル酸はGalおよびGalNAcへのα(2,3)若しくはα(2,6)または別のシアル酸へのα(2,8)若しくはα(2,9)の結合で見られる。従って、用語「シアル酸」が本明細書の全体にわたって使用されるが、全ての誘導体、その類似物または変異体(自然に生じたまたは合成的に生成された)並びにこれを含むダイマー、トリマー、オリゴマー、ポリマーまたはコンカテマーを包含すると理解するのが重要である。他の細胞表面炭水化物は、N-アセチルグルコサミン、ガラクトース、N-アセチルガラクトーサミン、フコース、マンノースおよび血液型抗原としてかかるエピトープにおいて見いだされるそれらの複合体を含むことができる。
【0025】
CBMは、自然界で広く見いだされ、種々の異なる炭水化物に対し広範囲にわたる親和性を示す。これまで、CBMは一次配列類似性に基づいて52の属に分類されてきたが、本発明があらゆるCBMに関し得ることを理解されたい。勿論、特定の細胞表面炭水化物に結合する病原体によって生じることが既知の疾病または疾患を治療するために、該炭水化物に対して結合するかまたは親和性を示すCBMが、その疾病または疾患の治療または予防に特に有用である。
【0026】
本発明の一実施形態は、シアル酸に対し親和性を示すかまたは結合するCBM分子に関する。当業者はシアル酸に結合するかまたは親和性を示すCBMが、発病中にシアル酸に結合するかまたは逆に関連する病原体が引き起こしたまたは一因となった疾病および/または疾患を治療する化合物として、または組成物、薬剤または方法に有用であると認識するであろう。
【0027】
多くの異なるCBMが、特定の炭水化物に対して親和性を示し、その親和性の大きさが変化し得ることに留意されたい。しかし、シアル酸のような細胞表面炭水化物に結合するCBMの場合、シアル酸に対する親和性を示すあらゆるCBMが潜在的に有用であると考慮すべきであると理解されたい。
【0028】
これら潜在的に有用なCBMのうち、科40に属するものは“CBM40”のCBMとして既知である。コレラ菌およびウェルシュ菌に由来するCBMがCBM40科に含まれる。
【0029】
コレラ菌に由来するCBM40はシアル酸に対し比較的高い親和性(Kd〜30μM)を示すが、ウェルシュ菌に由来するCBM40はシアル酸に対して高いミリモル親和性を示す。
【0030】
上記に加えて、本発明者らは、単一のCBM分子が特定の炭水化物(例えばシアル酸)に対する親和性を示すことができるが、二つ以上のCBM分子を結合するかまたは一緒に組み合わせることにより結合活性を介して当該炭水化物に対しより高い親和性を有する分子を生成することが可能であることを発見した。
【0031】
したがって、本発明はまた、いくつかの手段によって一緒に接続、結合または共役された一つ以上のCBM(またはモノマー)を備える以後「CBMポリマー」と称する多価のCBM分子またはポリペプチドに関する。
【0032】
従って、本発明の第5の態様は、二つ以上のCBM(CBMモノマー)から成るCBMポリマーを提供する。
【0033】
用語「ポリマー」は、多くの結合単位から成る分子を含むことであると当業者によって容易に理解される。この点に関して、用語「CBM ポリマー」を用いて、CBM-ダイマー(すなわち2つのCBMモノマー)、CBM-トリマー(3つのCBMモノマー)、CBM-テトラマー(4つのCBMモノマー)およびより大きなCBM-オリゴマー(5つ以上のCBMモノマー)を説明することができる。
【0034】
当業者は、同一の反復CBMモノマー(例えば、コレラ菌由来のCBM40モノマーを反復する)を含むCBMポリマーを生成することが望ましいが、それぞれ異なる炭水化物を結合できるおよび/または特定の炭水化物に対し異なる親和性を示す多くの異なるCBMモノマーを含むCBMポリマーを作成することもまた可能であることを認識するであろう。同様に、異なった細胞表面炭水化物を結合するかまたは親和性を示すCBMモノマーを含むCBMポリマーもまた本発明の範囲内である。このように、本発明のCBMポリマーは、シアル酸に対し親和性を有するCBMモノマー、およびいくつかの他の炭水化物、例えばマンノース、ガラクトース、フコース、N-アセチルグルコサミンに対し親和性を有するかまたは結合するCBMモノマーを含むことができる。
【0035】
上述したように、CBMモノマーを一緒に接続、結合または共役することによって、特定の炭水化物または多くの炭水化物に対し結合活性を介してより大きな親和性を示すより大きな重合体分子をつくることが可能である。したがって、特定のCBMモノマー、例えば、コレラ菌に由来したCBM40がシアル酸に対し親和性を示し得るが、該CBM40モノマーの二つ以上を一緒に結合、接続または共役する場合、生成したポリマーは、シアル酸に対し増大した親和性を示し得る。
【0036】
従って、本発明および特に本発明の第1、第2、第3および第4の態様において用いる用語「CBM」は、ここに記載したCBMポリマーを含んでいるとして理解すべきである。
【0037】
当業者は、異なる炭水化物に結合または親和性を有するCBMモノマーを含むCBMポリマーそれ自体が同じ炭水化物に対し結合または親和性を示すことができることを認識するだろう。例えば、CBMポリマーが二つの異なる細胞表面炭水化物に対し結合または親和性を有するCBMモノマーを含む場合、生成したCBMポリマーはまた、同じ二つの細胞表面炭水化物に対し結合または親和性を示すことができる。
【0038】
多くの異なるCBMモノマーを含み、その結果多くの異なる炭水化物分子に対し結合または親和性を示すCBMポリマーは、複数の病原体が引き起こしたまたは一因となった疾病および/または疾患を治療または予防するための組成物、化合物、薬剤および/または方法に特に有用であり得る。
【0039】
例えば、それぞれ異なる細胞表面炭水化物に結合する(細胞のコロニー形成および/または入り込み手段として)二つの異なる病原体が引き起こしたまたは一因となった少なくとも二つの疾病または疾患に苦しむかまたはかかる傾向のある被検体にとって、これら炭水化物を認識、結合または逆に伴うCBMモノマーを含むCBMポリマーは前記疾病および/または疾患の治療または予防に有用であり得る。或いはまた、二つ以上の病原体によって生じる疾病および/または疾患は、関連したモノマーCBMを利用した化合物または組成物、薬剤若しくは方法を用いて治療することができる。
【0040】
当業者は、アミノ酸、タンパク質および/またはペプチドを一緒に結合し得る多くの方法があることを認識するであろう。一実施形態において、アミノ酸、ペプチドおよび/またはタンパク質を化学的手段によりおよび/またはクローニングおよび/またはPCR技術によって接続、結合または共役することができる。
【0041】
例えば、関連したCBMをエンコード化するヌクレオチド配列をPCRによって増幅し、その一つ以上のコピーを一緒にまたは他のCBMエンコード化ヌクレオチド配列に結合できるように修正することができる。加えて、CBMヌクレオチド配列を、一つ以上のアミノ酸残基を含むリンカー部分をエンコード化する配列をその3’および/または5’末端で含むように更に修飾させることができる。
【0042】
CBMポリマー(または多価のCBM)はまた、例えば、オリゴマー形成領域を含むように修飾したCBMモノマーを用いて発生させることができる。かかる領域を有する分子が自己集合してオリゴマー構造を形成することができる。一実施形態において、オリゴマー化領域を、例えばトリマー形成領域をコード化することが既知の緑膿菌のような細菌性種から誘導することができる。このように、PCRに基づく技術を用いて、CBM分子を修正することができる。
【0043】
次いで、増幅され、任意に修飾/結合されたCBM配列を、生成したCBMまたはCBMポリマーの発現および精製用の適当なベクターにクローン化することができる。組換え型CBMモノマー(例えばオリゴマー形成領域をエンコード化するもの)またはCBMポリマーを、例えば大腸菌において発現させることができる。
【0044】
上記を鑑みて、本発明の第6の態様は、CBMポリマーを生成する方法を提供するもので、該方法は
(a)CBMエンコード化ヌクレオチド配列を結合するステップ、および
(b)CBM ポリマーを生成するために結合したCBMヌクレオチド配列を発現するステップを備える。
【0045】
前述したように、本発明によって提供されるCBMポリマーは、一つ以上のアミノ酸の形態をとり得るリンカー部分のいくつかの形によって結合したCBMモノマーを含むことができる。リンカー部分を含むアミノ酸の正確な数は変り得るが、通常リンカー部分は、1からの約30アミノ酸までの任意の長さを含むであろう。
【0046】
CBMポリマーを生成する代替手段は、分子生物学的技術によりオリゴマー形成領域をCBMに結合することにある。例えば、緑膿菌のプシュードアミダーゼに見られるトリマー形成領域(残基335〜438、Xu, G., Ryan, C., Kiefel, M.J., Wilson, J.C. and Taylor, G.L.(2009) J. Mol.Biol. 386(3), 828 .840) は、リンカーペプチドによって一つ以上のCBMと結合することができた。次いで、単一構築物の発現は、結合活性効果により増大した親和性を有するトリマーを生成する。同様に、ヒト血管拡張を刺激したリンタンパク質(Kuhnel K., et al. PNAS 2004, 101, 17027-17032)からのテトラマー領域、すなわちテトラマーコイルドコイル構造を形成する45-残基ペプチドを、適当なリンカーを介して一つ以上のCBMに結合して、例えば、結合活性効果により増大した親和性を有するテトラマーオリゴマーを形成することができる。
【0047】
本発明の第7の態様は、疾病および/または疾患の治療に潜在的に有用なCBMをスクリーニングするかまたは識別する方法を提供するもので、該記方法は
(i) 細胞を結合または感染させるのに既知の病原体の存在下でCBMまたはCBMポリマーを前記細胞と接触させるステップ;
(ii) 病原体が結合したかまたは病原体が感染したこれら細胞を識別するステップ;および
(iii)かかる結果をCBMまたはCBMポリマーが加えられてない標準または対照アッセイと比較するステップとを備え、
ここで、対照アッセイに対し病原体が結合した細胞数の減少または病原体に感染した細胞数の減少が、該病原体が引き起こしたまたは一因となった疾病および/または疾患の治療に潜在的に有用なCBMまたはCBMポリマーを示す。
【0048】
CBM/CBMポリマーと接触した細胞は、組織培養条件下で培養した細胞単層若しくは組織生検または被検体から得た剥離物の形をとり得る。加えて、細胞が齧歯動物(マウス、ネズミ、モルモット、ウサギなど)のような動物に存在する場合がある。動物を本発明の第7の態様によって提供される方法に用いる場合、該動物に試験すべきCBMまたはCBMポリマーを投与し、また同時(またはその前後に)に病原体(例えば呼吸性の病原体)に感染させてもよい。さらに、標準または対照アッセイは、病原体に感染したがCBMまたはCBMポリマーを投与しなかった動物の形をとってもよい。
【0049】
動物を使用する場合、CBMまたはCBMポリマーが特定の病原体によって引き起こされたまたは一因となった疾病の治療または防止に潜在的に有用であるかどうかを確定するために、第7の態様によって提供された方法の完了後、細胞を動物から得、病原体が細胞に結合または感染させたか否かを決めるために検討することができる。加えてまたはあるいは、動物を疾病の徴候または症状のために観察してもよい。対照または標準のプロトコルを施した動物と比較する場合、症状の重症度の減少は、動物に投与したCBMまたはCBMポリマーが病原体によって引き起こされたまたは一因となった疾病および/または疾患の治療に有用であることを示すことができる。
【0050】
上記に加えて、本発明者らは、ここに記載した様々なCBMおよびCBMポリマーの異なる結合機能を用いて、炭水化物のスクリーニング、識別、検出、標識付けおよび/またはタギング手段を提供することができることを発見した。
【0051】
したがって、本発明の第8の態様は、サンプル中の炭水化物をスクリーニング、識別、標識付けおよび/またはタギングする方法を提供するもので、該方法は
(a) ここに記載したようなCBM/CBMポリマーと、それに結合するかまたは親和性を有する炭水化物との結合を可能にするに適した条件かで試料を該CBMまたはCBMポリマーと接触させるステップと、
(b) 未結合のCBMまたはCBMポリマーを除去するステップと、
(c)結合したCBMまたはCBMポリマーを検出するステップとを備える。
【0052】
当業者は、CBMが特定の炭水化物に対しある程度の特異性を示すので、ステップ(c)で検出したCBMまたはCBMポリマーがサンプルに存在する炭水化物の指標として役立つであろうことを認識するであろう。
【0053】
用語「サンプル」は、体液(尿、血液、血漿、血清、汗、唾液、精液など)のような生体試料、並びに例えば食品、飲料および水源(川、海)のような他の供給源に由来した試料を包含することを理解されたい。実際、炭水化物を含み、またCBMまたはCBMポリマーを添加し得ると考えられるほとんどすべてのサンプルを用いることができる。
【0054】
一実施態様において、サンプルは細胞、好ましくは組織生検、剥離物または分泌物に由来する哺乳類の細胞からなる。このように、本発明の第8の態様によって提供される方法を用いてサンプル中の特定の細胞の存在を検出することができる。当業者は、細胞がその表面でさまざまな炭水化物を発現するため、特定の細胞タイプで発現されるのが既知の炭水化物に特異的なCBMとサンプルを接触させ、CBMがサンプル中のあらゆる細胞に結合したか否かを検出することにより、特定の細胞の存在を不均質細胞集団内で識別することが可能と成り得ることを容易に理解するであろう。FACSのような技術を用いて、本発明に係るCBMまたはCBMポリマー(検出可能なタグを含むように任意に修飾された)で標識付けまたはタグ付けされた細胞を識別することができる。加えて、このタイプの技術は疾病または他の疾患を診断するのに有用である。例えば、癌細胞で発現されるのが既知の炭水化物に結合するかまたは親和性を有するCBMまたはCBMポリマーを用いてサンプル中の癌細胞の存在を識別または検出することができる。
【0055】
未結合のCBMまたはCBMポリマーを除去するために、上記ステップ(a)に由来するサンプル::CBM/CBMポリマー錯体に適切な緩衝材を用いる一つ以上の洗浄ステップを施してもよい。
【0056】
一実施形態において、サンプルを例えばプラスチック、ガラス、アガロース、ニトロセルロース、紙などの適切な基質に固定してもよい。
【0057】
当業者は、本発明によって提供されるCBMまたはCBMポリマーが一つ以上の検出可能なタグまたはラベルを含むように更に修飾され得ることを認識するであろう。このようにして、発色の化学発光反応を介してレベルを記録し得る酵素を含むようにCBMまたはCBMポリマーを修飾または共役させると、結合したCBMまたはCBMポリマーの検出を達成することができる。かかる酵素としては、西洋ワサビペルオキシダーゼ(HRP)およびアルカリフォスファターゼ(AlkP)を挙げることができるが、これに限定されるものではない。加えて、或いはまた、ここで提供するCBMまたはCBMポリマーは、例えばGFPのような蛍光分子およびFITC、ローダミンまたはテキサスレッドのような蛍光色素分子に共役させてもよい。ここに記載したCBMまたはCBMポリマーに共役し得る他のタイプの分子は、放射性標識化部分を含んでもよい。
【0058】
更なる実施形態において、本発明によって提供されるCBMポリマーまたはCBMは、細胞に送達すべき部分、例えば、抗体、小有機分子、核酸、薬または毒素を含むように更に修飾させることができる。細胞表面で発現した炭水化物を識別することによって、一つ以上の識別された炭水化物に結合するかまたは親和性を有するCBMまたはCBMポリマーを用いて、上述したタイプの部分を該細胞に送達するこができる。
【図面の簡単な説明】
【0059】
本発明は以下の図面を参照して詳細に説明する。
【図1】Aはレクチン様炭水化物-結合モジュール(CBM)が側面に位置した中央触媒領域を示すコレラ菌シアリダーゼの略図である。CBMを認識するシアル酸、すなわちCBM40が右手にある。Bは本研究で作成した構築物の略図である。
【図2】ITC研究に用いたシアロシドを示し、Aは3'シアリルラクトース、Bは6'シアリルラクトース、Cはジシアリルラクトース(DSL)、Dはジシアリル-ラクト-N-テトラオース(DSLNT)である。
【図3】3'シアリルラクトースとの錯体におけるコレラ菌CBM40の結晶構造を示す。Aは点線として描画された水素結合を有するリガンドとCBMとの相互作用を示す。シグマ-a加重Fo-Fc電子密度マップを3σで描写する。Bは結晶の非対称ユニットにおける3つのCBM40モジュール間の関係を示す。
【図4】単離されたCBM40への様々なリガンドの結合を示す等温滴定熱量測定等温曲線であり、Aは3'SL、Bは6'SL、CはDSL、DはDSLNTである。
【図5】様々のCBM40構築物への3'SLの結合を示す等温滴定熱量測定等温曲線であり、Aは2CBM(5)、Bは2CBM(10)、Cは2CBM(15)、Dは3CBM(5)、Eは3CBM(10)、Fは4CBM(5)である。
【図6】固定した3'シアリルラクトースレセプタへのCBM40モジュールの結合を示す表面プラスモン共鳴(SPR)センサーグラムであり、Aは1CBM, Bは2CBM(5), Cは2CBM(10), Dは2CBM(15), Eは3CBM(5), Fは3CBM(10), Gは4CBM(5)である。
【図7】種々の温度でのSPR実験で得られたファント・ホッフの値をプロットしたものである。
【図8】多価CBM40ポリペプチドが生体外でインフルエンザウイルスの結合を低減し得ることを実証するプラークアッセイを示す。
【図9】(a)はワクチンX31、単量体の多価CBM40を有するチキン赤血球の赤血球凝集反応アッセイ、(b)はワクチン株X31からのインフルエンザAウイルスの存在の有無でのチキン赤血球の赤血球凝集反応アッセイを示す。
【図10】インフルエンザ株A/Udorn/72(H3N2)に対する3CBM(5)、4CBM(5)およびCBMTDポリペプチドによる哺乳類のMDCK細胞防御を示す。
【図11】(A)はMDCK細胞の細胞表面に結合するGFP-3CBM40(0.5mg/ml)を示し、(B)はUdornウイルスの72日目におけるGFP-3CBM40で培養したMDCK細胞の位相差顕微鏡写真である。パネル(a)はCBM(0.5mg/ml)のみ、(b)〜(f)はUdornウイルス存在での0.5、0.1、0.05、0.01および0.005mg/ml CBM、(g)はUdornウイルスのみ、および(h)は非感染細胞である。
【図12】(A)は肺ウイルス滴定量に対する3CBM40の効果、および(B)はインフルエンザAウイルスに感染したBALB/cマウスのマウス重量に対する3CBM40の効果を示す。H1N1感染(103のウイルスpfu)の1日前、当日、または1または3日後に3CBM40(黒い棒)またはBSA(白い棒)をマウスに投与した。データを、6日p.i.で1グループにつき4匹のマウスから集めた。ウイルス対照グループ(0日)を棒グラフのPBSとして示す。
【0060】
(材料および方法)
(組換えDNA技術)
コレラ菌シアリダーゼ/ノイラミニダーゼ(VCNA)、残基25-216の科40 CBMをコード化するDNA断片を、表2の配列アウトラインに基づくオリゴヌクレオチドプライマペア1Fおよび1Rを用いてnanH遺伝子(16)を含む構築物pET30b+からPCRによって増幅した。573bpの増幅DNA断片をNco IおよびXho Iで消化し、pEHISTEVベクター(タバコエッチ病ウイルス(TEV)プロテアーゼによって切断されるN末端基6xHisタグを有するpET30の改変体)(Dr H. Liu、未発表の研究)にクローン化し、同じ酵素で消化し、大腸菌DH5aを形質転換するのに用いた。構築物p1CBMをDNAシーケンシングによって検証した(ダンディ大学シーケンシングサービス、英国)。
【0061】
コレラ菌CBM40をコード化するDNA断片を5’および3’末端で修飾して、異なるプライマペアの使用により異なる制限エンドヌクレアーゼ部位を組み込む。これは、DNA断片の個々のコピーの連結反応を許可する用に行って、タンデムに2、3および、4つのコピーを作製した(図1B)。さらに、プライマペアは、個々のモジュールを結合するアミノ酸の長さを表すために5、10または15のコドンの挿入を可能にした。これら全ての変更は、テンプレートとして表2に示した種々のプライマペアおよびp1CBMを用いるPCR増幅によって達成された。生成する断片を、モジュールの所望の数が得られるまで、適切に消化されたpEHISTEVベクターにクローン化した。これらを、それぞれ括弧内にリンカーのアミノ酸の数を有する2および3個の反復シアル酸結合ドメインを示すp2CBM(5)、p2CBM(10)、p2CBM(15)、p3CBM(5)およびp3CBM(10)と標識した。p4CBM(5)については、最終的な遺伝子をpEHISTEVに組立てる前に、HindIII-EcoRI修飾およびEcoRI-XhoI修飾DNA断片を最初に適切な酵素で消化したpET17bにクローン化した。すべての構築物を大腸菌DH5aで増殖した。タンパク質生産用の発現宿主大腸菌BL21(DE3)(1CBM、2CBM(5)、3CBM(5)、3CBM(10)、4CBM(5))および大腸菌BL21(DE3)Gold(2CBM(10)および2CBM(15)それぞれ)の形質転換前に、陽性クローンをDNAシーケンシングによって検証した。
【0062】
(タンパク質生産および精製)
VCNAについて述べられている(16)ように、全ての構築物を成長、発現させた。つまり、30μg/mlカナマイシンを含有するLB培地の1Lアリコートをシングルコロニーで接種し、培地の光学密度が600ナノメートルで0.4〜0.6に達するまで37℃で成長させた。細胞に42℃の水浴中で20分間熱ショックをかけた後、25℃まで冷却し、1mMのIPTGで誘導し、同じ温度で終夜振動した。細胞を遠心分離(8000xg、4℃)によって収集し、ペレットを要求されるまで-20℃で凍結した。
【0063】
全てのポリペプチドは、前述した(16)とおり、ニッケル親和性クロマトグラフィによって精製した。サンプルを、SDS PAGEを用いて分析し、部分的に精製したポリペプチドをTEVプロテアーゼ裂開緩衝材(PBS、0.3M NaCl、1mM DTT、0.5mM EDTA、20mM イミダゾール)で透析し、TEVプロテアーゼで終夜消化した。次いで、各ポリペプチドをPBS、0.3M NaCl、10mM イミダゾール緩衝液で更に透析した後、ニッケル充填カラムで二回目の精製を行って未消化のHis標識付けポリペプチドを除去した。未標識付けポリペプチドサンプルを10mMのHEPES緩衝材においてpH 7.4、0.15M NaClで広範囲に透析し、使用前に濃縮した。
【0064】
(結晶化および構造決定)
コレラ菌CBM40の単結晶を、シッテングドロップ法を用いて0.1M のMOPS、 pH 6.5、1.1Mの硫酸リチウム、沈殿剤としての0.6Mの硫酸アンモニュウム中で得た。結晶をまず10%(v/v)グリセロール-沈殿剤混合物へ移した後、20%(v/v)グリセロール-沈殿剤に放置することにより該結晶を凍結保護した。2.5Åに及ぶ回折データを、ESRFのビームライン BM-14で収集した。構造は分子交替プログラムPHASER(18)を用いて解析され、非対称ユニットに3つのCBM40モノマーを見いだした。微調整は、モデル構築(20)のために使用されるCOOTを有するREFMAC(19)を用いて実施された。データ収集および微調整統計を表3に示す。
【0065】
(等温滴定熱量計(ITC))
ITC実験を、1.4mlの細胞容積を有するMicrocal社(ノーサンプトン、MA)のVP-ITC微量熱量計で行った。明示しない限り、全てのITC滴定を0.15M NaClを含むpH7.4の10mMのHEPES緩衝材中で25℃で行った。単離されたCBM40(1CBM)の評価のために、以下のリガンド、すなわちDextra Labs(レディング、英国)から購入した3'シアリルラクトース(3'SL)、6'シアリルラクトース(6'SL)およびジシアリル-ラクト-N-テトラオース(DSLNT)、ならびに、およびシグマアルドリッチ(英国)から購入したジシアリルラクトース(DSL)を用いた(図2)。凍結乾燥されたシアロシドリガンドを、ペプチド構築物の透析のために用いた脱ガスのろ過したバッファに再懸濁させた。他の全てのポリペプチド構築物については、完全に3'シアリルラクトースをリガンドとして用いた。タンパク質濃度は、1CBM(38410M-1cm-1)、2CBM構築物(75860M-1cm-1)、3CBM構築物(113790M-1cm-1)および4CBM(5)(151720M-1cm-1)それぞれの算出モル吸光係数を使用してA280で決定された。CBM40ポリペプチドの濃度は0.007-0.084mMであり、シアロシドは0.45-2.14mMであった。シアロシドのアリコート(特記しない限り10μl)を各ポリペプチド溶液に滴定した。希釈熱は、MicroCal Origin softwareからの一結合部位モデルを用いる非線形回帰分析法によってデータをはめ込まれる前に、等温曲線データを結びつけることで差し引かれた。
【0066】
(表面プラスモン共鳴(SPR))
結合反応速度を、BIACORE 3000バイオセンサ機器(GE Biosystems)を用いるSPRによって求めた。使用の前に、ストレプトアビジン被覆(SA)バイオセンサチップを前記機器にドッキングし、50mM NaOH中に1M NaClの3回連続の1分噴射で予備調整した。ビオチニル化3'シアリルラクトース-PAA(Glycotech)は、HBS-P(10mM HEPES pH 7.4、0.15M NaClおよび0.005%サーファクタントP20)中で1μg/mlまで希釈した後、チップ中4つのうちの3つのフローセルに注入した。各細胞に対するリガンドの典型的固定化レペルは、およそ500RUであった。基準面をまたバルク効果の減算およびストレプトアビジンとの非特異的相互作用のために準備した。ランニングバッファは、100μl/分の流速で pH 7.4の10mMのHEPESからなる。
【0067】
各ペプチド構築物に対する固定化3'シアリルラクトースでの相互作用分析を、15、25および35℃での、ランニングバッファ中で行った。精製ペプチド構築物をHBS-Pに希釈して、1CBMについては0.625μM、1.25μM、 2.5μM、5μM、10μMの一連の濃度、2CBMについては20nM、125nM、250nM、500nMおよび1000nM、3CBMについては1nM、5nM、20nM、62.5nMおよび125nM、並びに4CBMについては0.18nM、0.5nM、1.6nM 5nMおよび15nMを得た。全ての検体を、30μl/分でフローセル表面上に注入した。表面からの検体の解離をランニングバッファにおいて3〜5分間同じ流速で達成した。表面は、2つの連続的な30μl/分の10mMのグリシン-HCl pH 2.5の30秒噴射によって再生された。平衡解離定数(KD)で記述される親和性は、BIA評価ソフトウェア(BIAcore)を用いてラングミュア(1:1)結合を仮定する動的同時ka/kdモデルに合わせることによって包括的に求められた。
【0068】
種々のCBMポリペプチド構築物のビオチニル化3'シアリルラクトース-PAAaとの相互作用の自由エネルギー変化は、各温度に対する動的化学反応速度定数の比率によって付与される平衡解離定数を用いて求められた。lnKD対1/Tのvan’t Hoffのプロットは、傾斜からΔH/Rおよびy切片からの-ΔS/Rに対する値をもたらした。
【0069】
(プラークアッセイ)
ウイルスプラークアッセイを用いて、インフルエンザAウイルスの存在下での結合多価CBM40ポリペプチドの有効性を実証した。6ウェルプレート中の集密的(106の細胞)なMDCK単層を無血清DMEMで2回洗浄し、2CBM40または3CBM40(10mg/ml、無血清DMEMで10倍に順次希釈)のどちらかの1mlの溶液によって37℃(+ 5%のCO2)で1時間培養した。無血清DMEMのみを平行して「モック細胞」培養に用いた。次いで、CBM40試薬を除去し、単層をインフルエンザウイルスA/Udorn/72[H3N2]の〜 80 PFUを用いて37℃(+ 5% CO2)で1時間培養した。ウイルス吸着後、接種材料を取り除き、細胞を2μg/mlトリプシンおよび1%(w/v)アガロースを加えたDMEM中で10mM HEPES(pH 7.4)で覆った。逆さにしたプレートを37℃(+ 5%のCO2)で2〜3日間培養した。プラークは、単層を5%のホルムアルデヒドで固定し、0.1%のクリスタルバイオレットで染色して明視化した。
【0070】
(血液凝集アッセイ)
アルセバー液のチキン赤血球を分離し、PBSで少なくとも4回洗浄した。該細胞を最終的に1%(v/v)PBSに再懸濁した。ワクチン株X31(香港/68 HAおよびNAの細胞表面を有する)を、PBSに1:4000ストックの2倍連続希釈を用いて96ウェルプレートで滴定し、これに等容積の赤血球細胞を添加した。該プレートを少なくとも30分間室温で培養した。終末点は、赤血球凝集反応が観測される最低ウイルス濃度として算出された。CBMをPBSで適切に希釈、滴定し、これに等容積の細胞を加えた。細胞に結合するCBMの終末点もまた算出した。次いで、滴定ウイルスを有する1CBMの固定濃度のチキンRBCで培養された際のみのウイルスに対する効果を求めるために実験を行った。
【0071】
(オリゴマー形成領域を用いるCBM.TDの開発)
1CBM40、p1CBMを含有するpEHISTEV構築物がC末端でオリゴマー形成領域を含むように修飾されて、大腸菌で発現された際にポリペプチドをオリゴマー構造に自己組織化させた。このために、緑膿菌プシュードミニダーゼ由来の三量体形成領域を用いた(Xu ら、2009年)。残基333〜440をコード化するDNA断片を、BamHIおよびXhoIを含むプライマーを用いるPCRによって増幅し、同じ酵素で予め消化されたp1CNMに結合した。次いで、これを、すべての他のCBM40構築物の生長および発現と類似した条件を用いて大腸菌BL21(DE3)Gold細胞に形質転換した。オリゴマーは、ニッケル親和性およびTEVプロテアーゼ消化を用いて精製し、ゲル濾過(データ示さず)後約100kDaの分子量を示した。
【0072】
(CBM40によりシアル酸を結合した細胞表面を調べるためのGFP-3CBM40の開発)
GFP融合3CBM40断片の構築については、3CBM40断片をp3CBM(5)由来のNcoI-XhoIで消化し、N末端基His標識下流の強化された緑色蛍光タンパク質(eGFP)をコード化する遺伝子を含む同様に消化されたpEHISTEVベクターに組み込んだ(Liu et al 2009; Connaris et al, 2009)。この構築物をしてCBM40が細胞表面でシアル酸に結合したかどうかを求めた。このために、96ウェルプレートにおけるMDCK細胞の集密的な単層を、種々の濃度のGFP-3CBMを用いて37℃で1時間培養し、その後インフルエンザ株A/Udorn/72(H3N2)ウイルス(moi = 0.002)を添加して同じ温度で更に1時間培養した。混合物を除去し、FCSのないDMEMを添加し、プレートを72時間培養した。
【0073】
(マウス研究:投薬量を求める実験:)
インフルエンザウイルスに対する抗ウイルス剤としてのCBMポリペプチドの予防/治療可能性を評価するために、初期の研究を行ってマウスに投与するのに必要なCBM40の投薬量を求めた。これら研究は、エジンバラの感染病センターのインフルエンザ調査のための動物実験設備で行われた。このために、5〜6週間目のBALB/cマウスを用いて、まず最初に生体外所見の投薬量に基づいた有害反応の検査をするための投薬量を求める実験を行った。3CBM40ポリペプチドを40μl PBS中50μg、また40μl PBS中500μgで用いてマウスの鼻腔内に投薬した。マウスは、臨床徴候を毎日調べられ、2日目以降から毎日計量された。マウスは、5日目に選別された(但し500μg投薬されたマウスのうちの2匹は選別する前14日間放置した)。
【0074】
(インフルエンザAウイルスに対するCBM40の有効性)
インフルエンザA/WSN/33(H1N1)に対するCBM40の予防/治療有効性を評価するために、BALB/cマウスを、3CBM40およびウイルスでの処理前にハロタン酸素ミックスで軽く麻酔した。実験用のCBM40投薬量は、鼻腔内に一度だけ送達するPBS(40μl)中に500μgの3CBM40 であった。BSAおよびPBSを対照物として用いた。マウスは、103のA/WSN/33 pfuで鼻腔内感染の日の1日前、若しくは当日、またはその後+1日目または+3日目に、CBM40またはプラセボで処理した。各グループのウイルス滴定濃度を、感染後マウスを選別する6日目に求め、肺をPBS中で均質化してプラークアッセイ用のウイルスを抽出した。選別したマウスの体重をまた測定して、体重減少を求めた。
【0075】
(結果)
(単離されたCBM40モジュールの構造)
コレラ菌シアリダーゼからのシアル酸結合モジュールCBM40をコード化する遺伝子断片をpEHISTEVにサブクローン化し、E.coliで発現させて1CBMを生成し、その後結合および構造的な研究のために精製した。まず最初に、1CBMを非常に高いレベルではあるが溶解せずに発現されたが、生長の対数期間の42℃での培養組織の熱ショックが1リットル当たり最高50〜70ミリグラムの量を絶えず生産するようにこのCBMの溶解性を改善した。コレラ菌CBM40の結晶を3'シアリルラクトースの存在下で成長させた。非対称ユニットは、3つのCBM40モノマーを含み、それぞれリガンドの3つ全ての糖残基に明白な電子密度を有した(図3A)。シアル酸だけがタンパク質との相互作用をなし、完全なコレラ菌シアリダーゼとの錯体におけるシアル酸のみで作られるのものと同じものである。結晶の非対称ユニットには、約750Å2(図3B)の界面を埋設するモノマーAおよびBを有する3つのCBM40モジュールがある。
【0076】
(シアロシドおよびコレラ菌CBM40の結合特異性の決定)
1CBMでの研究を行って種々のモノおよびジシアロシドに対する特異性を求めた。結合定数Ka並びにエンタルピー(DH)およびエントロピー(DS)の変化を各相互作用に対しITCを用いて直接測定し、化学量論的量(n、結合部位数)を上記の通りに一部位の結合モデルに対するデータの非線形最小二乗適合から求めた。希釈修正結合等温曲線の熱から得たデータは、α-シアル酸に対する1CBMの優先傾向を立証し(図4)、α(2,3)、α(2,6)およびα(2,8)-結合シアロシドに対する広い結合特異性を示す。試験したすべてのシアロシドが、10〜19μM(25℃で)の範囲の解離定数Kdを有する類似の親和性を示した(表4、図4)。ジシアロシド、DSLNTおよびDSLは、一価のリガンドと比較してシアル酸に対し1.5〜2倍より大きな親和性を示す。DSLNTは、DSL(-12.5kcal/モル)と比較して、-24kcal/モルのΔH値を有するエンタルピーの大きな変化を示した。DSLNTおよびDSLを結合するための化学量論的量を比較する場合、0.57および0.92の該当n値が観測され、DSLNTに対しては1CBMの2つの分子がDSLNTの1分子と結合するのに必要とされるが、一方DSLでは1:1相互作用があることを示す。この相違は、存在するシアル酸部分の数ではなく、そのシアロシド内の位置によるのである。構造的に、DSLNTは、DSLと異なり、分岐鎖の二価のシアロシドであり、直線型で一緒に結合した2つのシアル酸部分を有するので、末端シアル酸部分のみが認識される(図2)。この結果は、ほぼDSLNTのΔH値が一つの化学量論的量を有する3’シアリルラクトース、6’シアリルラクトースおよびDSLの個々の結合エンタルピーの総和であることを示す。
【0077】
(多価の、シアル酸特異性のCBM40ポリペプチドの作業)
1CBMの同一のコピーを一緒に結合して多価の種属を生成する場合にシアル酸結合が生じるかどうかを本発明者らは調査したかった。このために、CBM40をコード化する遺伝子のコピーを一緒にDNAリンカー(最高15のアミノ酸を示す)でつなげて、それぞれ2CBM(5)、2CBM(10)、2CBM(15)、3CBM(5),3CBM(10)および4CBM(5)(括弧の数字はリンカーアミノ酸の数を示す)として示された2、3および4モジュールのポリペプチドをタンデムに創生した(図1B)。これら遺伝子構築物の発現を大腸菌において行い、培養物に単離された1CBMのための熱ショックを施すまですべてのものが不溶性を示した。ニッケル親和性クロマトグラフィーでの精製後、全てのペプチド構築物のSDS PAGE分析は、それぞれ1CBM、2CBM、3CBMおよび4CBM構築物に対し〜21kDa、〜42kDa、〜63kDaおよび〜85kDaの単量体分子量を示した(データ示されず)。
【0078】
一価の3'シアリルラクトースを有する様々な設計されたCBM40ポリペプチドの結合等温曲線を図5に示す。ITCを用いると、設計されたCBM40ポリペプチドへのシアロシドの結合をエンタルピー的に駆動し、ΔH値が25℃で-12.3から-16.3 kcal/molの極めて類似の範囲にあることを明らかになった(表5)。各ポリペプチド相互作用について測定した他の熱力学パラメータのすべてにおいて相違もまたほとんどなかった。実際、シアル酸に対する多価のCBM40の結合親和性は1CBMのものと類似であると思われた。さらにまた、モジュール間のリンカーの長さがこの相互作用に著しく寄与することと熱力学的に示されなかった(表5)。一部位の結合モデルに基づいて、n値は、3’シアリルラクトースと相互作用する各々のCBM40ポリペプチドの部位の適切な数を示す。結合されたモジュールの数を上げると親和性の有意な増加がみられないという事実は、シアル酸−多価ポリペプチドの相互作用が、簡単な二分子会合を示す一価の単量体のものと類似していることを示唆する。
【0079】
(多価の3’シアリルラクトースに対する多価CBM40ポリペプチドの増強結合親和性)
シアル酸に対する結合力効果を多価のCBM40ポリペプチドで達成し得るかどうかを試験するために、表面プラスモン共鳴(SPR)をストレプトアビジンチップに固定したビオチン化3'シアリルラクトースを用いて行った。固定化3'シアリルラクトース上に注入した全てのCBM40ポリペプチドについてのセンサーグラムを図6に示す。ここに平衡解離定数Kdとして記述した各CBM40-3'SL相互作用の親和性は、ラングミュア1:1結合とみなす会合/解離速度定数(ka/kd)の比率に由来した包括的な適合モデルによって求めた(表6)。シアル酸に対する親和性の増加が、モジュール数を増加すると観察される。25℃での1CBM-3'SL相互作用について、ITCにより測定した対応する単量体の一価の相互作用のもの(Kd〜18μM)と比較して、結合の10倍の増加(Kd 〜1.8μM)がある。モジュールの数が2まで増加する場合に増強された親和性が観察され、ここでおよそ400〜500倍の結合の増加があり、25℃で38〜45nM間の親和性をもたらす(表6)。アミノ酸数を5からの15まで上げると、わずか1.2倍の親和性の増加が見られるので、リンカー長がこの場合わずかに結合活性に影響を与えると思われる。3および4つのCBM40モジュールを用いると、シアル酸に対する親和性が更に10〜20倍増強され、25℃で5aa-および10aa結合3CBMモジュールに対して約4nM、また4CBMモジュールに対して2.6nMの親和性に及ぶ。15℃で多価の3'SLを結合する場合、最も高い親和性は、Kd 〜861pMを有する4CBM(5)であった。したがって、相互作用の温度に依存して、一価の単量体の相互作用から多価のものへと進めることにより7000〜10000倍の親和性増加を達成し得ると思われる。15、25および35℃で測定した各CBM40ポリペプチド-シアル酸相互作用のファント・ホッフ値のプロットから得たデータ(図7、表7)は、1CBMに対し-7.8kcal/mol、2CBMに対し約-10kcal/mol、3CBMに対し-11.3kcal/molおよび4CBMに対し-12kcal/molのΔG値を示す。1CBMおよび2CBM間のΔG値の大きな相違はまた、相互作用のエンタルピーおよびエントロピーの変化に反映される。全てのCBM40ポリペプチドの中から、2CBMポリペプチドが、好適でない大きなエントロピー的寄与を補った約-20kcal/molのΔH値の最も大きなエンタルピー的変化を与えた(表7)。1CBMおよび2CBM相互作用間のエネルギー論におけるこの大きな相違はまた、1CBM、3CBMおよび4CBMポリペプチドと比較して、シアル酸に対する2CBMの増強された親和性に示され、リガンド-レセプター相互作用の共同性を示唆する。
【0080】
対照的に、3CBMポリペプチドは小さい有利なエントロピー的利得を示すが、相互作用の自由エネルギーの変化はより望ましい有利なエンタルピー的ペナルティによりまだ強くネガティブだった(表7)。面白いことに、3CBMおよび4CBMポリペプチドの両方は、相互作用の自由エネルギーが多分親和性の利得に寄与するモジュールの数の増加と共に増加したという事実にもかかわらず、1CBMおよび2CBMポリペプチドと比較して、リガンド結合に対するΔHおよびTΔSの両方へのネガティブな寄与の少ないことを示した。この見解は、多くの因子に基づくことができ、例えば、モジュールの立体配座配置、リガンド結合部位のアクセスしやすさ、分子内部および分子間結合のような結合のモード並びにモジュールの内部構造的なパッキングにより相互作用がリンカー柔軟性に敏感である。しかし、3CBMポリペプチド間のリンカー長の影響は、異なる2CBMポリペプチドと同様に結合エネルギーおよび親和性に関して無視し得るものであった。親和性の関連利得が25℃で4CBM(5)によって得られなかったので、ポリペプチドの更なる設計を行わないことが決定された。
【0081】
(赤血球凝集反応アッセイ)
単量体のおよび多価のCBM40が赤血球細胞に結合し、インフルエンザウイルスの結合を予防したかどうかを試験するために、初期実験を赤血球細胞に対し滴定されたCBMおよび滴定されたウイルス、ワクチン株X31で行った。予備結果は、1CBM40の場合凝集がみられないことを示し、またこのCBMが単量体であるので、細胞表面シアル酸に一価の型、すなわち 1:1会合で結合すると思われる(図9a)。多価CBM40に関しては、2および3CBMの両方が、ワクチンのX31のように赤血球細胞を凝集した(図9a)。加えて、細胞を凝集するのに必要な多価CBMの終点濃度は、3CBM40が2CBMよりも低く(それぞれ0.122μMおよび0.48μM)、これは繰り返しモジュールの数の増加がシアル酸に対する結合親和性を増大させることを示唆する。しかしながら、多価CBMが赤血球細胞を凝集するので、1CBM(71.25μM)の固定濃度を有する滴定されたワクチン混合物をワクチンのみに対して試験して凝集を観察することを決定した。図9bに示すように、ワクチンでテストした際の1CBMの存在は、モック細胞(ウイルスのみ)と比較すると、ウイルスの結合を防止する。プレートを4℃で2.5日間放置した際に、凝集が1CBMおよびウイルスの存在下で認められず、競争分子の結合をブロックするCBM40の有効性を示唆する。
【0082】
(プラークアッセイ)
プラークアッセイの予備結果は、10mg/mlの 3CBM(5)の1/100希釈液が同一希釈濃度の2CBM(10)よりも細胞へのウイルス性侵入をブロックするのにより効果的なことを示唆する強い証拠を示す(図8参照)。特に、モック細胞と比較した際に、異なる繰り返しCBMでMDCK細胞を培養後のプラーク数に相違があり、また3CBM(5)を用いる場合、H3N2の存在下でが極めて少ないプラークが現れることが分る。
【0083】
3CBM(5)(1/100希釈)では、H3N2ウイルス・プラーク形成単位の80-85%の減少がある。2CBM(10)の結果では、ウイルス遮断効果があるが、同じ濃度の同一希釈は3CBM(5)と同じ効果を有さなかった。このように、CBMモジュール数の増加は細胞表面でのシアル酸に対するウイルス結合を効果的に遮断するのに重要であったことがわかる。
【0084】
(プラーク研究(CBMTDの使用))
ウイルス感染MDCK細胞:インフルエンザ株A/Udorn/72(H3N2)ウイルスをCBM40ポリペプチドによりブロックした。図10は、Udornウイルスに対するMDCK細胞上のポリペプチド3CBM40、4CBM40およびCBMTDの細胞防護効果を示す。結合したCBM40の数が3から4CBM40まで増加すると、インフルエンザ株に対するCBM阻害作用強度の増加がわかる。また、ウイルスの添加前のオリゴマーCBM.TDでのMDCK細胞の培養は、ウイルス対照物と比較して0.5mg/mlで有効な抑制と、0.1mg/ml CBMでプラーク数の顕著な減少を示した(図10)。これらのデータは、最高4ドメイン反複のコレラ菌シアリダーゼCBM40がタンデム結合したかまたは自己集合したオリゴマーとしてウイルス感染からMDCK細胞を効果的に保護することができることを示す。
【0085】
(CBM40によりシアル酸を結合した細胞表面の監視)
図11aは、ウイルス添加1時間後のMDCK細胞の細胞表面へのGFP-3CBM40の結合を示す。図11bは、ウイルスのみのウェルと比較した際に、感染72時間後でもGFP-3CBM40がウイルス感染から相当な保護を付与することを示す。パネル(a)はまた、ウイルスの不在下でMDCK細胞に加えたCBM(0.5mg/ml)が72時間後毒作用を示さず、未感染対照物[パネル(h)]に対し類似の形態を有する細胞が生存能力を有すると認められたことを示した。
【0086】
(マウス研究(投薬量を求める実験))
表1は、研究中のマウスの最初と最終体重の結果を示す。臨床見解から、マウスすべてが非常に健康であり、5日および14日の指定された期間中それら全てが3CBM40に副作用なしで重くなったことがわかった。抗体反応および肺組織の組織学に関する更なる研究は、まだ実施されていない。
【0087】
(インフルエンザAウイルスに対するCBM40の有効性)
予備結果は、WSNウイルスチャレンジの1日前の3CBM40の投与がマウスのウイルス感染の効果を著しく減らすことを示し、CBM40がインフルエンザAウイルスに対する予防薬として効果的であることを実証したことを示唆した。これは、103のウイルスpfuの初回量から1.5ログ滴定濃度の増加のみが測定されたBSAおよびPBS対照と比較して、感染6日後に選別されたマウスのウイルス滴定濃度の減少で明白だった(図12A)。このグループのマウスはまた、体重が増え、ウイルスチャレンジからの副作用を示さなかった(図12B)。3CBM40をウイルスと同時に投与した場合(0日目)、ウイルスの2.4ログ増加が感染6日後に100%を超える体重増加とともに認められ、ウイルス滴定濃度の増加にもかかわらずマウスがウイルス感染に耐えたことを示す。しかしながら、CBM40をウイルス感染後の1および3日目に投与する場合、マウスは軽くなり、ウイルス滴定濃度が初期の負荷と比較して3〜3.25ログだけ増加したことが分り、CBM40は治療薬として付与した際に少なくとも本方法で記述する条件下で効果的ではないことを示唆した。1回の投与ではなく感染中毎日の投薬および投与手段の最適化を含む更なる研究は、CBM40が特にインフルエンザ感染の初期段階の治療の抗ウイルス薬として考えることができたかどうかを決定することができる。予防法として、このタンパク質生物製剤を鼻腔内に送達してウイルス攻撃から呼吸性上皮粘膜を保護することができるであろう。
【0088】
(考察)
本発明者らは、コレラ菌シアリダーゼからのシアル酸特異性CBMをその親酵素からうまく単離し、シアル酸を結合する多価のポリペプチドを高い親和性で作り出すのに活用することができることを示した。ITCおよびSPRの組合せを使用し、本発明者らはまた、これまで多くのCBM−炭水化物相互作用である(Boraston et al 2004, etc)ようにエンタルピー的に駆動されるが、多価の表面に導入される際に結合モジュール数が2を超える場合にエントロピー的に駆動される一価および二価の様態であるシアル酸へのコレラ菌CBM40の一価および多価の様態の結合の熱力学を実証した。
【0089】
リガンドおよび結合特異性に関して、単離されたCBM40はマイクロモル濃度の親和性でα(2,3)、α(2,6)およびα(2,8)-結合シアル酸を優先して結合する。この結合親和性は、非加水分解性チオシアロシドとのコレラ菌シアリダーゼ相互作用について報告されたものに類似している(16)。異なる結合シアロシドの間でほとんど区別がないという事実は、3'シアリルラクトースで錯化したCBM40の結晶構造においてみられる。該構造は、シアル酸との分子間相互作用の大規模なネットワークを明らかにする。ガラクトースおよびグルコース部分はタンパク質ドメインと相互作用せず、これは異なる結合シアロシドについてここで見られる類似のKd値および結合の自由エネルギーに反映する。
【0090】
シアル酸に対するコレラ菌CBM40の比較的高い一価の親和性を活用することによって、本発明者らは、多価の表面と相互作用する際にサブナノモーラー結合活性を得るためにn=4までのタンデム結合反複ポリペプチドをうまく設計した。これは、異なる長さのリンカーペプチドを使用して同一のコピーを結合させることによって操作されたCBMのおそらく第1の報告である。植物細胞壁ポリマーの分析用の分子プローブとして単離されたCBMの使用の報告(21)、並びに別のモジュールまたはそれらの機能を決定するためにその隣接したモジュールと組合せのいずれかとして単離されたマルチモジュラーグリコシドヒドロラーゼで天然に見られるCBMの研究が以前にあった(14,17,22)。
【0091】
配座の制約がこれら設計されたポリペプチドの結合を防止するかどうかを決定するために、本発明者らはレセプターとの多価の相互作用の熱力学を調査したかった。タンパク質サブユニット間のリンカー長が相互作用の熱力学に影響を及ぼすことができ、また多くの場合、共有結合サブユニットが解離のエントロピー的駆動力を減らすことによって相互作用のオリゴマー安定性を強化することができることは周知である(23)。本発明者らは、リンカー長が設計されたCBMポリペプチドのシアル酸に対する親和性を著しく強化するかどうか決めるために、5つのアミノ酸から15のアミノ酸までの範囲にあるCBMモジュール間の柔軟なペプチドリンカーを設計した。2タンデム結合CBMの場合、ペプチドリンカーを10アミノ酸まで増加すると、異なる2CBM構築物間のシアル酸に対する親和性に1.5〜2倍の増加を認めることにもかかわらず、配座のエントロピーの有意な減少を与えた。これは、モジュールのある程度の柔軟性が2つのCBMを一緒に結合する際に生起したことを示唆する。シアル酸に対する2CBMの結合相互作用はまた、大きな好適でないエントロピー的寄与を補う結合エンタルピーの増加を示し、結合自由エネルギーの増加および1CBM―多価3SL相互作用と比較した際シアル酸に対する親和性の強化をもたらした。これは、シアル酸に対する2CBMの結合が非常に安定した分子内相互作用であり、1CBMに対してエンタルピー的に駆動されることを示唆する。しかしながら、固定された多価3SLと相互作用する3CBMに関し、異なる長さのペプチドの間では有意差がなかった。実際、異なる3CBM-シアル酸の相互作用のvan’t Hoffプロットから得たエントロピーデータはすべて、エントロピーの増加を示す。また、結合エンタルピーの有意な減少および親和性の10倍の強化は、2から3CBMになるときに認められた。シアル酸へ結合する3CBMの場合に認められる親和性の強化は、SPRチップの表面で生じる安定した集合体の有望な形成に起因し、また大きなエントロピー変化が密着したタンパク質-タンパク質パッキングによって影響される水和殻から解放される構造的に秩序ある水分子によるものである。これは、3つの1CBMモノマーを一緒に密接にパックされた1CBM-3SL錯体の構造から明らかである(図3B)。エントロピー駆動相互作用は、特に分子間結合が生じる他の多価のタンパク質−炭水化物システムでみられ、集合体の形成にいたる(24)(25)、(26)。面白いことに、これはわずかに固定された3’SLとの1CBM相互作用のものより少ないが、結合モジュール数を4まで増加すると、3CBMsのものへの好適なエントロピー寄与が少なく、結合エンタルピーが好ましいことを示した。結合の自由エネルギーは、3から4CBMまでの親和性の僅かな増大に対応する全ての他のCBMより僅かに大きかった。設計されたタンデム結合ポリペプチドの結合価は、おそらくオリゴマーの安定化およびそれらの相互作用に重要な役割を果たし得る。プーン(20)は、タンデム結合された単一ポリペプチド鎖の結合価を増大すると、熱力学的に言えば、より安定になる結合オリゴマーの分子数の対応する減少をもたらし得ることを記載する。実際に、プーンは、オリゴマーの分子数より高い結合価のタンデムは、または分割し得ないタンデムが交差結合したオリゴマーをもたらすことを述べた。したがって、4CBMの場合のような4量体オリゴマーに関しては、タンデムダイマーまたはテトラマーが好ましいが、三量体または五量体オリゴマーは、それぞれ同じ結合価のタンデムを必要とする。
【0092】
これらの研究は、同一コピーを結合してそのシアル酸に対する親和性を結合活性により増強することによってCBM40を単離し、操作することができることを示した。さらにまた、本発明者らは、モノマーおよびコレラ菌シアリダーゼから設計した多量体CBM40をインフルエンザAウイルスに対する潜在的抗ウイルス薬として用い得ることを示した。従って、この種の技術を使用すると、他のCBMをグリカンスクリーニングおよびプロファイリング用の高親和性ツールとしての使用のために探利子、設計することができるであろう。加えて、そのグリカンとの錯体におけるCBM構造の決定は、グライコミクスの分野での使用への選択的試薬の発達に役立ちえる。
【0093】
(参考文献)
【0094】
【表1】
【0095】
【表2】
【0096】
【表3】
【0097】
【表4】
【0098】
【表5】
【0099】
【表6】
【0100】
【表7】
【特許請求の範囲】
【請求項1】
一つ以上の病原体が引き起こすかまたは一因となった疾病および/または疾患の治療または予防に用いる炭水化物結合分子(CBM)。
【請求項2】
一つ以上の病原体が引き起こすかまたは一因となった疾病および/または疾患の治療または予防用の薬剤の製造への炭水化物結合分子(CBM)の使用。
【請求項3】
一つ以上の病原体が引き起こすかまたは一因となった疾病および/または疾患を治療または予防するのに用いるCBMおよび薬学的に許容し得る賦形剤を含む医薬品組成物。
【請求項4】
経口、非経口または局所的な投与用に処方した請求項3に記載の医薬品組成物。
【請求項5】
炭水化物結合モジュール(CBM)の治療上有効量を投与するステップを備える病原体によって生じるかまたは一因となった疾病を治療または予防する方法。
【請求項6】
前記疾病または疾患がウイルス性の呼吸性病原体および/または細菌性病原体によって生じる前記請求項のいずれかに記載のCBM、使用、組成物または方法。
【請求項7】
前記ウイルス性の呼吸性病原体がオルトミクソウイルスまたはパラミキソウイルス科に属する請求項6に記載のCBM、使用、組成物または方法。
【請求項8】
前記細菌性病原体がストレプトコッカス属、インフルエンザ菌またはシュードモーナス・エールジノーサである請求項6に記載のCBM、使用、組成物または方法。
【請求項9】
インフルエンザ、偽膜性喉頭炎、肺炎および/または気管支炎を治療および/または予防するための請求項1〜6に記載のCBM、組成物、使用および/または方法。
【請求項10】
前記CBMがシアル酸に親和性を示すかまたは結合する前記請求項のいずれかに記載のCBM、組成物、使用または方法。
【請求項11】
前記CBMが40CBM科に属する前記請求項のいずれかに記載のCBM、組成物、使用または方法。
【請求項12】
前記CBMをコレラ菌またはウェルシュ菌から得る前記請求項のいずれかに記載のCBM、組成物、使用または方法。
【請求項13】
前記CBMが2つ以上のCBMモノマーを含む多価のCBMである前記請求項のいずれかに記載のCBM、使用、組成物または方法。
【請求項14】
2つ以上のCBM分子を含むCBMポリマー。
【請求項15】
1つ以上の細胞表面炭水化物に結合するかまたは親和性を有する請求項14に記載のCBMポリマー。
【請求項16】
一つ以上の病原体が引き起こすかまたは一因となった疾病および/または疾患を治療または予防するための請求項15に記載のCBMポリマー。
【請求項17】
(a) CBMをコード化するヌクレオチド配列を結合するステップ;および
(b)結合CBMヌクレオチド配列を発現させてCBMポリマーを作り出すステップを備えるCBMポリマーの製造方法。
【請求項18】
(i) CBMまたはCBMポリマーを細胞と該細胞を結合または感染させるのに既知な病原体の存在下で接触させるステップ;
(ii) 病原体が結合したかまたは病原体が感染した細胞を識別するステップ;および
(iii) 結果をCBMまたはCBMポリマーを加えない標準または対照アッセイと比較するステップを備え、
対照アッセイと比べて病原体が結合した細胞数の減少または病原体に感染した細胞数の減少がその病原体が引き起こすかまたは一因となった疾病および/または疾患の治療に潜在的に有用なCBMまたはCBMポリマーを示すことを特徴とする疾病および/または疾患の治療に潜在的に有用なCBMをスクリーニングするかまたは識別するための方法。
【請求項19】
(a) サンプルを請求項14のCBMまたはCBMポリマーと該CBM/CBMポリマーおよびこれを結合するかまたは親和性を有する炭水化物との間で結合し得るのに適した条件下で接触させるステップ;
(b) 未結合のCBMまたはCBMポリマーを取り除くステップ;および
(c)結合CBMまたはCBMポリマーを検出するステップを備えるサンプル中の炭水化物をスクリーニング、識別、検出、ラベリングおよび/またはタギングする方法。
【請求項20】
前記サンプルが組織生検、剥離物または分泌物から得た細胞からなる請求項19に記載の方法。
【請求項21】
前記CBMまたはCBMポリマーを更に修飾して細胞に送達すべき部分を含める前記請求項のいずれかに記載のCBM、使用、組成物または方法。
【請求項1】
一つ以上の病原体が引き起こすかまたは一因となった疾病および/または疾患の治療または予防に用いる炭水化物結合分子(CBM)。
【請求項2】
一つ以上の病原体が引き起こすかまたは一因となった疾病および/または疾患の治療または予防用の薬剤の製造への炭水化物結合分子(CBM)の使用。
【請求項3】
一つ以上の病原体が引き起こすかまたは一因となった疾病および/または疾患を治療または予防するのに用いるCBMおよび薬学的に許容し得る賦形剤を含む医薬品組成物。
【請求項4】
経口、非経口または局所的な投与用に処方した請求項3に記載の医薬品組成物。
【請求項5】
炭水化物結合モジュール(CBM)の治療上有効量を投与するステップを備える病原体によって生じるかまたは一因となった疾病を治療または予防する方法。
【請求項6】
前記疾病または疾患がウイルス性の呼吸性病原体および/または細菌性病原体によって生じる前記請求項のいずれかに記載のCBM、使用、組成物または方法。
【請求項7】
前記ウイルス性の呼吸性病原体がオルトミクソウイルスまたはパラミキソウイルス科に属する請求項6に記載のCBM、使用、組成物または方法。
【請求項8】
前記細菌性病原体がストレプトコッカス属、インフルエンザ菌またはシュードモーナス・エールジノーサである請求項6に記載のCBM、使用、組成物または方法。
【請求項9】
インフルエンザ、偽膜性喉頭炎、肺炎および/または気管支炎を治療および/または予防するための請求項1〜6に記載のCBM、組成物、使用および/または方法。
【請求項10】
前記CBMがシアル酸に親和性を示すかまたは結合する前記請求項のいずれかに記載のCBM、組成物、使用または方法。
【請求項11】
前記CBMが40CBM科に属する前記請求項のいずれかに記載のCBM、組成物、使用または方法。
【請求項12】
前記CBMをコレラ菌またはウェルシュ菌から得る前記請求項のいずれかに記載のCBM、組成物、使用または方法。
【請求項13】
前記CBMが2つ以上のCBMモノマーを含む多価のCBMである前記請求項のいずれかに記載のCBM、使用、組成物または方法。
【請求項14】
2つ以上のCBM分子を含むCBMポリマー。
【請求項15】
1つ以上の細胞表面炭水化物に結合するかまたは親和性を有する請求項14に記載のCBMポリマー。
【請求項16】
一つ以上の病原体が引き起こすかまたは一因となった疾病および/または疾患を治療または予防するための請求項15に記載のCBMポリマー。
【請求項17】
(a) CBMをコード化するヌクレオチド配列を結合するステップ;および
(b)結合CBMヌクレオチド配列を発現させてCBMポリマーを作り出すステップを備えるCBMポリマーの製造方法。
【請求項18】
(i) CBMまたはCBMポリマーを細胞と該細胞を結合または感染させるのに既知な病原体の存在下で接触させるステップ;
(ii) 病原体が結合したかまたは病原体が感染した細胞を識別するステップ;および
(iii) 結果をCBMまたはCBMポリマーを加えない標準または対照アッセイと比較するステップを備え、
対照アッセイと比べて病原体が結合した細胞数の減少または病原体に感染した細胞数の減少がその病原体が引き起こすかまたは一因となった疾病および/または疾患の治療に潜在的に有用なCBMまたはCBMポリマーを示すことを特徴とする疾病および/または疾患の治療に潜在的に有用なCBMをスクリーニングするかまたは識別するための方法。
【請求項19】
(a) サンプルを請求項14のCBMまたはCBMポリマーと該CBM/CBMポリマーおよびこれを結合するかまたは親和性を有する炭水化物との間で結合し得るのに適した条件下で接触させるステップ;
(b) 未結合のCBMまたはCBMポリマーを取り除くステップ;および
(c)結合CBMまたはCBMポリマーを検出するステップを備えるサンプル中の炭水化物をスクリーニング、識別、検出、ラベリングおよび/またはタギングする方法。
【請求項20】
前記サンプルが組織生検、剥離物または分泌物から得た細胞からなる請求項19に記載の方法。
【請求項21】
前記CBMまたはCBMポリマーを更に修飾して細胞に送達すべき部分を含める前記請求項のいずれかに記載のCBM、使用、組成物または方法。
【図1】

【図2】
【図3A】
【図3B】

【図4】
【図5】
【図6a)】
【図6b)】
【図6c)】
【図6d)】
【図6e)】
【図6f)】
【図6g)】

【図7】
【図8】

【図9】
【図10】
【図11A】

【図11B】

【図12】
【図2】
【図3A】
【図3B】
【図4】
【図5】
【図6a)】
【図6b)】
【図6c)】
【図6d)】
【図6e)】
【図6f)】
【図6g)】
【図7】
【図8】
【図9】
【図10】
【図11A】
【図11B】
【図12】
【公表番号】特表2012−502086(P2012−502086A)
【公表日】平成24年1月26日(2012.1.26)
【国際特許分類】
【出願番号】特願2011−526555(P2011−526555)
【出願日】平成21年9月11日(2009.9.11)
【国際出願番号】PCT/GB2009/002189
【国際公開番号】WO2010/029312
【国際公開日】平成22年3月18日(2010.3.18)
【出願人】(507178453)ユニバーシティー コート オブ ザ ユニバーシティー オブ セント アンドリューズ (3)
【Fターム(参考)】
【公表日】平成24年1月26日(2012.1.26)
【国際特許分類】
【出願日】平成21年9月11日(2009.9.11)
【国際出願番号】PCT/GB2009/002189
【国際公開番号】WO2010/029312
【国際公開日】平成22年3月18日(2010.3.18)
【出願人】(507178453)ユニバーシティー コート オブ ザ ユニバーシティー オブ セント アンドリューズ (3)
【Fターム(参考)】
[ Back to top ]