
炭素数が7である脂肪酸を含むトリグリセリドを含有する栄養補助剤または医薬調製物
【課題】栄養的または食事療法的組成物または補助剤の提供。
【解決手段】炭素数が7である脂肪酸、好ましくはn-ヘプタン酸およびそのトリグリセライドを含む組成物。これらは、遺伝性代謝疾患または後天性代謝障害、特に長鎖脂肪酸代謝不全を有する患者のための優れたエネルギー源である。また、炭素数が7である脂肪酸(n−ヘプタン酸)は、脂肪酸代謝に由来するエネルギー生産の増加を必要とする患者のための栄養補助剤に加えることができる。
【解決手段】炭素数が7である脂肪酸、好ましくはn-ヘプタン酸およびそのトリグリセライドを含む組成物。これらは、遺伝性代謝疾患または後天性代謝障害、特に長鎖脂肪酸代謝不全を有する患者のための優れたエネルギー源である。また、炭素数が7である脂肪酸(n−ヘプタン酸)は、脂肪酸代謝に由来するエネルギー生産の増加を必要とする患者のための栄養補助剤に加えることができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、栄養的または食事療法的組成物または補助剤に関する。
【背景技術】
【0002】
脂肪酸はエネルギー産生に重要な役割を果たし、空腹時には必須である。骨格および/または心筋の弱りから乳幼児突然死症候群に似た致死性の代謝無呼吸異常症状における脂肪酸代謝の重篤な障害が起り得る。これらの障害は、重篤な心筋症、低血糖症、筋障害、罹患器官の微小血管脂肪堆積、および/または劇症肝炎と共に現れる。脂肪酸代謝の先天性遺伝欠陥をわずらう患者は、脂肪酸代謝経由でエネルギーを産生することができず、しばしば致命的なまたは反復性の重篤な衰弱異常症状を経験する。未熟児は高血糖レベルの維持が必要である。しばしば、患者の日常食事は十分な量の炭水化物エネルギー源を与えず、かつ誕生時の患者の脂肪代謝酵素は効率的でない。高齢患者も、食欲不振および非効率的な代謝により、血糖レベルの調節の困難なことがある。
【0003】
飽和脂肪酸は、次の構造により表される:
【化1】

[式中、Rはアルキル基を表す]。高等植物および動物の脂質に由来する天然に存在する脂肪酸は、飽和および不飽和の両方の偶数炭素鎖を含むものである。最も豊富な天然に存在する飽和脂肪酸は、パルミチン酸(16個の炭素;C16)およびステアリン酸(18個の炭素;C18)である。それより短鎖の脂肪酸(12-14個の炭素;C12-C14)およびそれより長鎖の脂肪酸(28個の炭素;C28までの)が小量、天然に存在する。10個未満の炭素をもつ脂肪酸はミルク脂肪を除く動物脂質には稀にしか存在しないが、ミルク脂肪はオレイン酸(不飽和C18)約32%、パルミチン酸(C16)約15%、ミリスチン酸(C14)約20%、ステアリン酸(C18)約15%、ラウリン酸(C12)約6%、および4〜10個の炭素をもつ脂肪酸(C4-C10)約10%を含有する。
【0004】
脂肪酸は一般的に、カルボキシル基に結合する炭素鎖長により分類される:4〜6個の炭素(C4-C6)の短鎖脂肪酸、8〜14個の炭素(C8-C14)の中鎖脂肪酸、16〜18個の炭素(C16-C18)の長鎖脂肪酸、および20〜28個の炭素(C20-C28)の極長鎖脂肪酸である。
【0005】
脂肪酸が代謝されるプロセスは、細胞のミトコンドリア内のミトコンドリアβ酸化を含む。図1に図解するように、パルミチン酸のような長鎖脂肪酸の脂肪酸酸化は、形質膜性カルニチントランスポーターを介して形質膜を通過する脂肪酸の輸送に始まる。脂肪酸がミトコンドリア外膜を通過すると、脂肪酸は、補酵素A(CoASII)およびアシルCoAシンセターゼの存在下で、ATPを消費して補酵素Aの脂肪酸エステル(脂肪族アシル-CoA)に転化される。脂肪族アシル-CoAは、カルニチンおよびカルニチンパルミトイルトランスフェラーゼI(CPT I)の存在下で脂肪族アシルカルニチンに転化される。次に、脂肪族アシルカルニチンはミトコンドリアの内膜を通過し、この工程はカルニチン/アシルカルニチントランスロカーゼ(carnitine/acylcarnitine translocase)の触媒作用を受ける。ミトコンドリアの内側に入ると、脂肪族アシルカルニチンは、次に、カルニチンパルミトイルトランスフェラーゼII(CPT II)の存在下で転化され、脂肪族アシルCoAに戻される。ミトコンドリア内の酸化サイクルで、脂肪族アシルCoAは、鎖特異的アシルCoAデヒドロゲナーゼを介して、αおよびβ炭素原子から1対の水素原子の除去による脱水素を受け、α、β-不飽和アシルCoA、または2-trans-エノイルCoAを生じる。適当なアシルCoAデヒドロゲナーゼは、脂肪族アシルCoAの炭素鎖長により決定され、すなわち、長鎖アシルCoAデヒドロゲナーゼ(LCAD;C12-C18)、中鎖アシルCoAデヒドロゲナーゼ(MCAD;C4-C12)、短鎖アシルCoAデヒドロゲナーゼ(SCAD;C4-C6)、または極長鎖アシルCoAデヒドロゲナーゼ(VLCAD;C14-C20)である。次に、α,β-不飽和アシルCoAは、2-エノイルCoAヒドラターゼを介して酵素作用により水和され、L-3-ヒドロキシアシルCoAを生成し、これは、次に、鎖特異的L-3-ヒドロキシアシルCoAデヒドロゲナーゼの触媒作用によりNAD連結反応で脱水素され、β-ケトアシルCoAを生成する。適当なL-3-ヒドロキシアシルCoAデヒドロゲナーゼは、L-3-ヒドロキシアシルCoAの炭素鎖長により決定され、すなわち、長鎖L-3-ヒドロキシアシルCoAデヒドロゲナーゼ(LCHAD;C12-C18)、または、短鎖L-3-ヒドロキシアシルCoAデヒドロゲナーゼ(SCHAD;C4-C16、鎖長の増加とともに活性は低下する)である。β-ケトアシルCoAエステルは、3-ケトアシルCoAチオラーゼの存在下で第2のCoA分子のチオール基の攻撃により酵素分解を受け、脂肪族アシルCoAおよび元来の脂肪酸のαカルボキシルおよびβ炭素原子に由来するアセチルCoAを生成する。他の産物、すなわち原料脂肪酸より2個少ない炭素原子を有する長鎖飽和脂肪族アシルCoAは、今度は次回の反応の基質となって、最初の脱水素工程に始まり、最後に2回目の2個の炭素をもつ断片はアセチルCoAとして除去される。この循環型プロセスを通過する毎に、脂肪酸鎖はアセチルCoAとして2個の炭素断片および2対の水素原子を特定の受容体に向けて失う。
【0006】
脂肪酸酸化プロセスの各工程は、重複する炭素鎖長特異性をもつ酵素による触媒作用を受ける。これらの酵素による触媒作用の消失に関連する脂肪酸酸化の遺伝性障害が確認されている。これらの障害には、形質膜カルニチン輸送;CPT IおよびII;カルニチン/アシルカルニチントランスロカーゼ;極長鎖、中鎖、および短鎖アシルCoAデヒドロゲナーゼ(すなわち、それぞれVLCAD、MCAD、およびSCAD);2,4-ジエノイルCoAレダクターゼ;長鎖3-ヒドロキシアシルCoAデヒドロゲナーゼアシルCoA(LCHAD)の欠損症、およびミトコンドリア三機能性(trifunctional)タンパク質(MTP)不全が含まれる。今までに、中鎖デヒドロゲナーゼ(MCAD)不全に対する治療は見出されている。しかし、他の不全症は、出生1年以内の患者にはしばしば致命的であり、利用できる有効な治療は知られていない。特に、重篤なカルニチン/アシルカルニチントランスロカーゼ不全をわずらう患者は通常死亡し、生存者は知られてないし、そして治療は見出されてない。
【0007】
これらの障害を治療する試みは、欠陥のある酵素による触媒作用の欠失を迂回する食物源を提供することに重点が置かれている。例えば、欠陥のあるカルニチン/アシルカルチニントランスロカーゼにより引き起される長鎖脂肪酸代謝不全(以後、「トランスロカーゼ不全」と呼ぶ)は、しばしば新生児期の死をもたらす。あるトランスロカーゼ不全の患者に、カルチニン、高炭水化物食、および中鎖トリグリセリドを与えたが、脂肪酸代謝不全を克服することはできなかった。中鎖脂肪酸は自由にミトコンドリアに入ると予想されていたので、中鎖脂肪酸の代謝はカルニチン/アシルカルチニントランスロカーゼを必要としないだろうと考えられた。従って、トランスロカーゼ欠陥をバイパスすると予想される偶数個の炭素数をもつ中鎖トリグリセリド(MCT)(例えば、84% C8、8% C6および8% C10)を含有する乳幼児処方が開発された。これらの処方による治療の試みに関わらず、志望者の発生は続いている。
【0008】
ペラゴン酸(9個の炭素;C9をもつ飽和脂肪酸)は例外として、奇数炭素数脂肪酸は高等植物および動物脂質中に稀である。ある特定の奇数炭素数をもつ合成トリグリセリドが、潜在的な脂肪酸源としての食品、および食品の製造に利用するために試験されている。C7およびC9トリグリセリド由来の奇数鎖脂肪酸の酸化速度が単離した子ブタ肝細胞中でin vitroで試験されている(Odleら, 1991. 「新生仔の子ブタによる中鎖トリグリセリドの利用;偶数および奇数炭素数脂肪酸の鎖長と見かけの消化/吸収および肝代謝」, J Nutr 121:605-614;Lin, X,ら, 1996. 「酢酸エステルは、新生仔の子ブタから単離した肝細胞におけるヘプタン酸エステルおよびオクタン酸エステルβ酸化の主要産物である」, Biochem J 318:235-240;およびOdle, J. 1997.「新生仔による中鎖トリグリセリドの利用についての新しい洞察:子ブタモデルからの観察」, J Nutr 127:1061-1067)。奇数鎖脂肪酸、プロピオン酸エステル(C3)、吉草酸エステル(C5)、およびノナン酸エステル(C9)の糖新生前駆体としての重要性が、飢餓ラット由来の肝細胞において評価された(Sugden,ら, 1984.「飢餓ラット由来の肝細胞中の奇数炭素脂肪酸代謝」, Biochem Int'l 8:61-67)。放射標識したマルガル酸(C17)エステルの酸化がラット肝切片において試験された(Boyer,ら, 1970.「1-14Cオクタン酸および1-14Cマルガル酸の肝代謝」, Lipids 4:615-617)。
【0009】
C3、C5、C7、C9、C11、およびC17を利用するin vivo研究も、モルモット、ウサギ、およびラットについてin vivoで実施されている。体系的に注入されたC7およびC9トリグリセリド由来の中鎖脂肪酸およびC7/C9トリグリセリド混合物のin vivo酸化速度が新生仔のブタで試験されている(Odel,ら, 1992.「連続注入放射性トレーサーキネティクス法を使った、新生仔の子ブタによる[1-14C]-中鎖脂肪酸酸化の評価」, J Nutr 122:2183-2189;およびOdle,ら, 1989.「新生仔の子ブタによる中鎖トリグリセリドの利用:II.出生の最初2日にわたる偶数および奇数鎖トリグリセリド消費が血液代謝物および尿窒素排泄に与える影響」, J Anima Sci 67:3340-3351)。トリウンデカノイン(飽和C11)を与えたラットは、非飢餓血糖レベルを延長された飢餓期間にわたり維持することが観察された(Anderson,ら, 1975.「飢餓ラットにおけるラード、サフラワー油、およびトリウンデカノインのグルコース形成およびケト体生成能」, J Nutr 105:185-189)。トリノナノイン(C9)および長鎖トリグリセリドの乳剤がウサギに注入され、長期総非経口性栄養(long-term total parenteral nutrition)として評価された(Linseisen,ら, 1993.「総非経口性栄養の奇数中鎖トリグリセリド(トリノナノイン):ウサギの脂肪代謝パラメーターに与える影響」, J Parenteral and Enteral Nutr 17:522-528)。7個の炭素をもつ飽和脂肪族n-ヘプタン酸(C7)を含有するトリグリセリドトリヘプタノインは、欧州において、農業飼料、バター製造のトレーサ分子として、およびチョコレートおよび他の菓子の製造における遊離剤(reasing agent)として使われたことが報じられている。しかし、今まで、7炭素脂肪酸がヒトによる摂取に安全であるとかまたはヒトに対するいずれか特定の栄養上の利点を有するという指摘はなかった。
【0010】
このほど、本発明者らは、後天的代謝撹乱および遺伝性代謝障害、特に脂肪酸代謝不全症は、n-ヘプタン酸のような炭素数が7である脂肪酸を含有する栄養組成物を使って克服できることを見出した。欠陥のあるまたは低下のある脂肪酸代謝を有している患者を、非常に効率的なエネルギー源としてn-ヘプタン酸および/またはそのトリグリセリドであるトリヘプタノインのような炭素数が7である脂肪酸を含有する栄養組成物を用いて、治療することができる。また迅速なエネルギーを必要とする患者も、炭素数が7である脂肪酸またはそのトリグリセリドを消費することにより利益を得ることができる。
【発明の概要】
【0011】
1つの態様において、本発明は、飽和炭素数が7である脂肪酸を含む栄養補助剤である。本発明に有用な炭素数が7である脂肪酸は、n-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノイン(triheptanoin)が、本発明のこの態様に最も有用である。好ましくは、炭素数が7である脂肪酸は、総食事カロリーの少なくとも約25%を供給する濃度である。
【0012】
別の態様において、本発明は炭素数が7である脂肪酸を含む製剤である。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。この製剤は、経口または非経口投与することができる。
【0013】
さらに別の態様において、本発明は、炭素数が7である脂肪酸を含む、少なくとも1つの代謝障害を有する患者に治療上の効果を与えるための投与に適合した一回量剤形とした製剤である。上記代謝障害は、たとえば、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼII、カルニチン/アシルカルニチントランスロカーゼ、心臓型極長鎖アシルCoAデヒドロゲナーゼ、低血糖型極長鎖アシルCoAデヒドロゲナーゼ、筋型極長鎖アシルCoAデヒドロゲナーゼ、ミトコンドリア三機能性タンパク質、長鎖L-3-ヒドロキシアシルCoAデヒドロゲナーゼ、および短鎖アシルCoAデヒドロゲナーゼなどの脂肪酸代謝不全の結果として生じ得る。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。この製剤は経口または非経口投与することができる。
【0014】
さらに別の態様において、本発明は炭素数が7である脂肪酸および製薬上許容される担体を含む一回量医薬剤形であって、この剤形は、少なくとも1つの代謝障害を有する患者の脂肪酸代謝を増大させるのに有効な量で提供される。上記代謝障害は、たとえば、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼII、カルニチン/アシルカルニチントランスロカーゼ、心臓型極長鎖アシルCoAデヒドロゲナーゼ、低血糖型極長鎖アシルCoAデヒドロゲナーゼ、筋型極長鎖アシルCoAデヒドロゲナーゼ、ミトコンドリア三機能性タンパク質、長鎖L-3-ヒドロキシアシルCoAデヒドロゲナーゼ、および短鎖アシルCoAデヒドロゲナーゼなどの、脂肪酸代謝不全の結果として生じ得る。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。この一回量医薬剤形は経口または非経口投与することができる。
【0015】
別の態様において、本発明は、炭素数が7である脂肪酸を含む栄養補助剤の、ヒトにおける脂肪酸代謝を増大させるための使用である。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。好ましくは、炭素数が7である脂肪酸は、上記補助剤の総カロリーの少なくとも約25%を供給する。
【0016】
別の態様において、本発明は、炭素数が7である脂肪酸を含む栄養補助剤の、グルコースの栄養源としての使用である。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。好ましくは、炭素数が7である脂肪酸は、上記補助剤の総カロリーの少なくとも約25%を供給する。
【0017】
別の態様において、本発明は、炭素数が7である脂肪酸を含む一回量剤形とした製剤を、少なくとも1つの代謝障害を有する患者に治療効果を与えるために使用することである。上記代謝障害は、たとえば、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼII、カルニチン/アシルカルニチントランスロカーゼ、心臓型極長鎖アシルCoAデヒドロゲナーゼ、低血糖型極長鎖アシルCoAデヒドロゲナーゼ、筋型極長鎖アシルCoAデヒドロゲナーゼ、ミトコンドリア三機能性タンパク質、長鎖L-3-ヒドロキシアシルCoAデヒドロゲナーゼ、および短鎖アシルCoAデヒドロゲナーゼなどの脂肪酸代謝不全の結果として生じ得る。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。好ましくは、炭素数が7である脂肪酸は、補助剤の総カロリーの少なくとも約25%を供給する。
【0018】
さらに別の態様において、本発明は、食物に炭素数が7である脂肪酸を加えることを含む、食物1グラムあたりのエネルギーポテンシャルを増大させる方法である。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。好ましくは、該食物は経口または非経口投与される。
【0019】
別の態様において、本発明は、炭素数が7である脂肪酸を含む栄養補助剤を用いて、少なくとも1つの代謝障害を有する患者を治療する方法である。上記代謝障害は、たとえば、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼII、カルニチン/アシルカルニチントランスロカーゼ、心臓型極長鎖アシルCoAデヒドロゲナーゼ、低血糖型極長鎖アシルCoAデヒドロゲナーゼ、筋型極長鎖アシルCoAデヒドロゲナーゼ、ミトコンドリア三機能性タンパク質、長鎖L-3-ヒドロキシアシルCoAデヒドロゲナーゼ、および短鎖アシルCoAデヒドロゲナーゼなどの脂肪酸代謝不全の結果として生じ得る。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。治療効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。好ましくは、炭素数が7である脂肪酸を、総食事カロリーの少なくとも約25%を供給するような濃度で添加する。
【0020】
さらに別の態様においては、本発明は、炭素数が7である脂肪酸を含む栄養補助剤を患者に投与することにより、脂肪酸代謝の効率を増大させる方法である。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。治療効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。好ましくは、炭素数が7である脂肪酸を、総食事カロリーの少なくとも約25%を供給するような濃度で添加する。
【0021】
詳細な説明 この度、炭素数が7(C7)である脂肪酸またはそれらのトリグリセリドは、エネルギー産生のため長鎖脂肪酸をミトコンドリア中に輸送するために必要な通常の酵素、すなわち、カルニチン/アシルカルニチントランスフェラーゼ、カルニチンパルミトイルトランスロカーゼ(「CPT」)IおよびCPT IIを必要としないことを確認した。従って、炭素数が7である脂肪酸から構成されるトリグリセリドは、このような酵素を必要とする脂肪酸代謝不全を克服するために有用である。炭素数が7である脂肪酸を含有する栄養補助剤または医薬調製物は、遺伝性代謝障害ならびに後天的代謝撹乱の治療に有用である。
【0022】
好ましい炭素数が7である脂肪酸はn-ヘプタン酸である。n-ヘプタン酸は、次の構造:
【化2】
による炭素数7をもつ飽和直鎖脂肪酸である。トリヘプタノインは、3つのn-ヘプタン酸分子とグリセロールとのエステル化により調製されるトリグリセリドである。治療に関して、用語「ヘプタン酸」、「ヘプタン酸エステル」、および「トリヘプタノイン」は、以下の説明において互換的に使うことができる。また、当業者であれば、ヘプタン酸、ヘプタン酸エステル、およびトリヘプタノインは以下の説明全体を通じて本発明の炭素数7をもつ脂肪酸源の例として使われ、本発明の説明を意図するが、いかなる方法でも本発明の範囲を限定するものでないと解釈されることを理解するであろう。置換された、不飽和、または分枝ヘプタン酸エステル、ならびに他の改変された炭素数7をもつ脂肪酸を、本発明の範囲から逸脱することなく、使うことができる。
【0023】
トリヘプタノインは、最初に、ヘプタン酸の3つの分子とグリセロールに分解される。図2に説明したように、次いで、第1サイクルにおいて、ヘプタン酸は、通常のβ酸化過程を通してn-バレリルCoA(C5)およびアセチルCoA(C2)に分解される。次いで、第2サイクルにおいて、このn-バレリルCoA(C5)はn-プロピオニルCoA(C3)およびアセチルCoA(C2)に分解されるが、これらは両方ともクレブス回路(Kreb's cycle)およびエネルギー産生の燃料として重要な前駆体である。従って、トリヘプタノインは、エネルギー産生の燃料の効率的な供給源として有用である。さらに、プロピオニルCoAはグルコース産生の直接前駆体である。従って、トリヘプタノインは低血糖症に罹患しやすい患者、特に未熟児および高齢者に対する食事補助剤として有用である。トリヘプタノインはまた、未熟児の成長速度刺激剤として使うこともでき、入院期間を短縮し、それによってこれらの幼児に対する医療費を削減する。さらに、脂肪酸は心組織の主要燃料であり、かつトリヘプタノインは、糖新生物性(gluconeogenic)を有するので、心臓またはその他のリスクの高い外科手術から回復しつつある成人の心組織の直接燃料補給に使うことができる。
【0024】
ヘプタン酸は様々なフーゼル油にかなりの量が見出されており、当業界で周知の方法により抽出することができる。またヘプタン酸は、ヘプトアルデヒド(heptaldehyd)を希硫酸中で過マンガン酸カリウムにより酸化して合成することもできる(Ruhoff, Org Syn Coll. vol II, 315 (1943))。ヘプタン酸はまた、Sigma Chemical Co.(St. Louis, MO)から購入することもできる。
【0025】
トリヘプタノインは、当業界で周知の方法により、ヘプタン酸とグリセロールとのエステル化によって得ることができる。トリヘプタノインはCondea Chemie GmbH(Witten, ドイツ)からSpecial Oil 107として購入することもできる。
【0026】
不飽和ヘプタン酸エステルも脂肪酸代謝不全を克服するための栄養補助剤として利用することができる。さらに、特別な輸送酵素なしにミトコンドリアに容易に入り込む置換した不飽和および/または分枝の炭素数が7である脂肪酸も、本発明に利用することができる。例えば、4-メチルヘキサン酸エステル、4-メチルヘキセン酸エステル、および3-ヒドロキシ-4-メチルヘキサン酸エステルは、通常のβ酸化により2-メチル酪酸まで分解され、最終分解はイソロイシン経路で達成される。同様に、5-メチルヘキサン酸エステル、5-メチルヘキセン酸エステル、および3-ヒドロキシ-5-メチルヘキサン酸エステルは、通常のβ酸化によりイソ吉草酸まで分解され、最終分解はロイシン経路で達成される。
【0027】
本発明の炭素数が7であるトリグリセリドは、経口的に、非経口的に、または腹腔内に投与することができる。好ましくは、上記トリグリセリドは、トリヘプタノインのような炭素数が7である脂肪酸源を含有する食物の摂取を経由して、治療レベルを達成するために有効な濃度にて投与することができる。あるいは、カプセルとしてまたはリポソームに包んで、溶液または懸濁液で、単独または他の栄養素、追加の甘味剤および/または香料と一緒に、投与することができる。カプセルおよび錠剤は、公知のように、糖、シェラックおよび他の腸溶剤によりコーティングすることができる。
【0028】
投与方法は患者の年齢および脂肪酸代謝不全の程度により決定する。脂肪酸代謝欠陥、特にトランスロカーゼ不全をもつ乳幼児の治療に対して、トリヘプタノインは、栄養補助剤として低脂肪および/または削減(reduced)した長鎖脂肪酸を含有する食事乳幼児処方に加えることが好ましい。トリヘプタノインと共に使う市販の乳幼児処方の例として、Tolerex(Novartis Nutritionals, Minneapolis, MN)、Vivonex(Ross Laboratories, Columbus, OH)、およびPortagen and Pregestamil(Mead Johnson, Evansville, IN)が挙げられる。トリヘプタノインは、治療効果を達成するために有効な濃度にて処方に加える。栄養補充を必要とする子供または成人患者、例えば、化学療法を受けている外科または腫瘍患者に対して、トリヘプタノインを栄養ドリンクまたは全非経口栄養投与の一部として補充することが好ましい。
【0029】
代謝の先天性過誤により脂肪酸代謝経路が完全に破壊されている患者に対しては、トリヘプタノインを、24時間当たりの全カロリーのほぼ15%〜40%、好ましくは20%〜35%、そして最も好ましくはほぼ25%を与える濃度にて利用する。
【0030】
脂肪酸代謝経路機能の効率が低下している患者(例えば、未熟児、高齢者、心臓病患者)に対しては、トリヘプタノインを、24時間当たりの全カロリーのほぼ15%〜40%、好ましくは20%〜35%、そして最も好ましくはほぼ25%を与える濃度にて用いる。
【0031】
プロピオニルCoAはトリヘプタノイン酸化の副生物であるので、プロピオン酸の血液濃度が増加しうる。さらに、プロピオニルCoAは他の酵素反応に入り、クレブス回路および尿素回路に影響を与える毒性化合物を生じ得る。従って、特に血清プロピオン酸のビルドアップを示す患者にn-ヘプタン酸および/またはトリヘプタノイン補助剤のような炭素数が7である脂肪酸を投与するには、カルニチン補助剤および/またはビオチンとビタミンB12の組合わせを投与することが必要である。過剰のL-カルニチンおよびミトコンドリア酵素カルニチンアシルトランスフェラーゼの存在のもとで、プロピオニルCoAは、無毒物質であるプロピオニルカルニチンに転化されて尿中に排泄される。ビオチンは、プロピオニルCoAのメチルマロニルCoAへの転化の触媒となる酵素プロピオニルCoAカルボキシラーゼが必要とするビタミン補因子である。シアノコバラミンはビタミンB12の1つの形態であり、メチルマロニルCoAのスクシニルCoAへの転化の触媒となる酵素メチルマロニルCoAムターゼの補因子として働く。スクシニルCoAはクレブス回路に容易に引き抜かれる。従って、患者血液中の過剰のプロピニルCoAはスクシニルCoAへの転化により除去される。
【実施例1】
【0032】
細胞系への補充
致死型トランスロカーゼ不全をもつ患者から採取した培養細胞(織維芽細胞)にN-ヘプタン酸を加えると酸化に成功した。
【0033】
同胞は4日齢で重篤なトランスロカーゼ不全により死亡していたので、胎児から得た羊膜細胞を脂肪酸代謝の能力について試験した。試験結果は、胎児も重篤なトランスロカーゼ不全を有することを示した。
【0034】
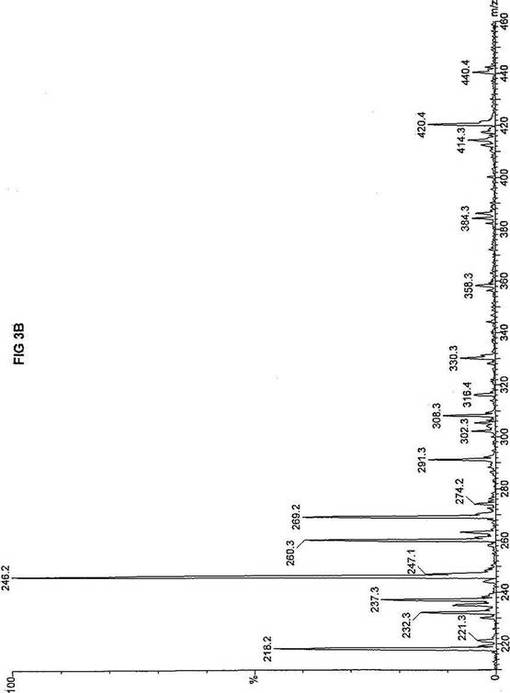
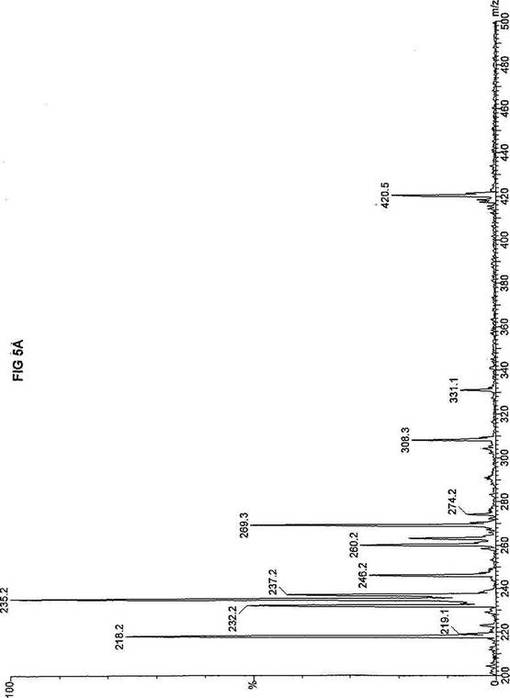
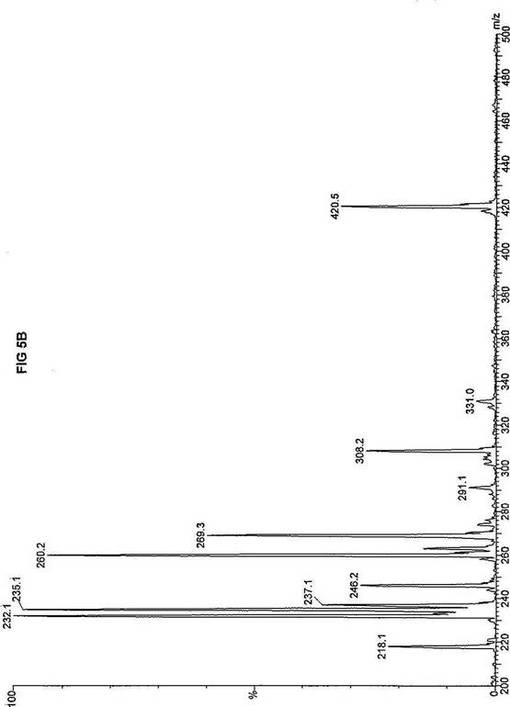
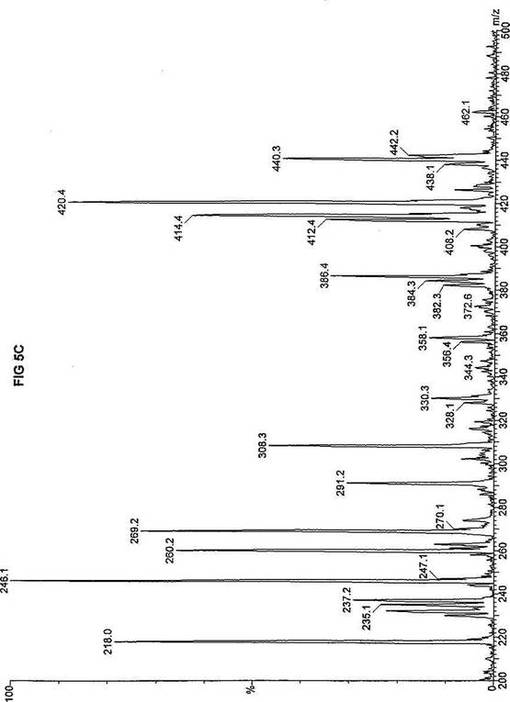
死亡した同胞から採取した織維芽細胞および羊膜細胞の両方を、先に報じられたタンデム質量分析アッセイを使い、n-ヘプタン酸(C7)の脂肪酸代謝について評価した(Yang,ら, 1998.「カルニチンパルミトイルトランスフェラーゼII(CPT II)不全をもつ患者における4つの新規突然変異の同定(Identification of four novel mutations in patients with carnitine palmitoyltransferase II (CPT II) deficiency)」Mol Genet Metab 64:229-236)。質量分析結果を、死亡した同胞から採取した織維芽細胞に対しては、パルミチン酸エステルについて図3Aにおよびトリヘプタノインについて図3Bに、ならびに、胎児から採取した羊膜細胞に対しては、パルミチン酸エステルについて図4Aにおよびトリヘプタノインについて図4Bに報じた。研究結果は、両方の細胞系のトランスロカーゼ不全にも関わらず、n-ヘプタン酸(図3Bおよび4B)はカルニチン/アシルカルニチントランスロカーゼに依存せず、容易にプロピオニルCoAに酸化されることを示した。重篤なトランスロカーゼ不全を有する2つの細胞系によるn-ヘプタン酸代謝の成功に基づいて、正常患者からおよび他の協力研究室で直接酵素アッセイにより立証された次の脂肪酸化の遺伝性欠損症に罹っている患者から採取した織維芽細胞系について、タンデム質量分析アッセイを実施した:カルニチンパルミトイルトランスフェラーゼI(CPT I);重篤なカルニチン/アシルカルニチントランスロカーゼ(トランスロカーゼ(TRANSLOCASE));カルニチンパルミトイルトランスフェラーゼII(CPT II);極長鎖アシルCoAデヒドロゲナーゼ「心(cardiac)」型(VLCAD-C);極長鎖アシルCoAデヒドロゲナーゼ「低血糖」型(VLCAD-H);ミトコンドリア三機能タンパク質(トリファンクショナル(TRIFUNCTIONAL));長鎖L-3-ヒドロキシ-アシルCoAデヒドロゲナーゼ(LCHAD);中鎖アシルCoAデヒドロゲナーゼ(MCAD);短鎖アシルCoAデヒドロゲナーゼ(SCAD);電子伝達フラビンタンパク質QOデヒドロゲナーゼ-マイルド(ETF-DHマイルド);および電子伝達フラビンタンパク質QOデヒドロゲナーゼ-シビア(ETF-DHシビア)。各細胞系を別々に、7-2H3-ヘプタン酸エステル(D3-C7)、8-2H3-オクタン酸エステル(D3-C8)、9-2H3-ノナン酸エステル(D3-C9)、12-2H3-ドデカン酸エステル(D3-C12)、および16-2H3-パルミチン酸エステル(D3-C16)と共に培養した。タンデム質量分析の結果を、D3-C7について図5A-Lに;D3−C8について図6A-Lに;D3-C9について図7A-Lに;D3-C12について図8A-Lに;およびD3-C16について図9A-Lに示した。
【0035】
正常細胞系および11種の異常細胞系は3グループについて分析した。定量性をもたせるため、標識した内部標準を各分析に含ませ、各グループの最初のプロフィールに「IS」と記した。内部標準の質量数は、m/z420(2H6-パルミチン酸エステル-C16)、m/z308(2H6-オクタン酸エステル-C8)、m/z269(2H9-イソ吉草酸-C5)、およびm/z237.0(2H5-プロピオン酸エステル-C3)であり、ここにm/zは質量:電荷比である。
【0036】
図9Aに示すように、正常細胞をD3-C16と共に培養すると、標識したアシルカルニチン中間体がC16から下方C4(C4を含む)まで観察することができる。これらの2H3標識したアシルカルニチンの質量数はメチルエステルとして、m/z417(C16)、m/z389(C14)、m/z361(C12)、m/z333(C10)、m/z305(C8)、m/z277(C6)、およびm/z249(C4)である。
【0037】
16-2H3-パルミチン酸エステルD3-C16を用いて培養した各種細胞系(図9A-L)を観察すると、CPT I細胞(図9B)においては実質的に酸化が起らず、そして予期したとおり、パルミチン酸エステルがミトコンドリア中への輸送のためのパルミトイルカルニチンに容易に転化されないので、D3-C16(m/z417(C16))から最小量のパルミトイルカルニチンしか観察されない。トランスロカーゼ(図9C)およびCPT II(図9D)不全細胞系は両方とも、酸化は起らないが、CPT Iが存在する結果として、D3-C16(m/z417(C16))から大量の標識したパルミトイルカルニチンが蓄積する。VLCAD-C(図9E)、VLCAD-H(図9F)、トリファンクショナル(図9G)、LCHAD(図9H)、ETE-DH-マイルド(図9K)、およびETE-DH-シビア(図9L)中の標識したアシルカルニチンの異常なプロフィールは、失われた酵素活性の炭素鎖長特異性に対応する蓄積を反映する。MCAD(図9I)においては、酸化は、下方C8(m/z305.3)のレベルまで明らかに進行し、この点で顕著な蓄積があって失われたMCAD酵素活性の基質特異性を反映する。同様に、SCAD(図9J)においては、酸化はm/z249(2H3-ブチルカルニチン-C4)で停止する。これらの結果は、CPT I、トランスロカーゼ、CPT II、VLCAD、トリファンクショナル、LCHAD、ETE-DHは全て、パルミチン酸エステルの完全な酸化に必要であることを示す。D3-C12プロフィール(図8A-L)は、D3-C16プロフィールと同様である。
【0038】
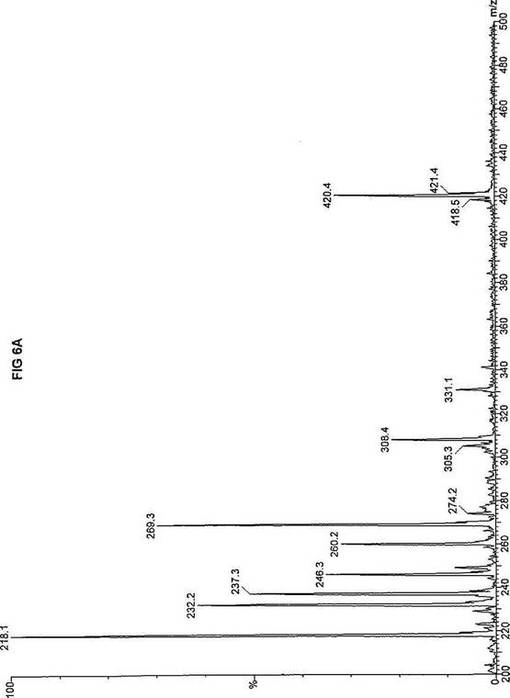
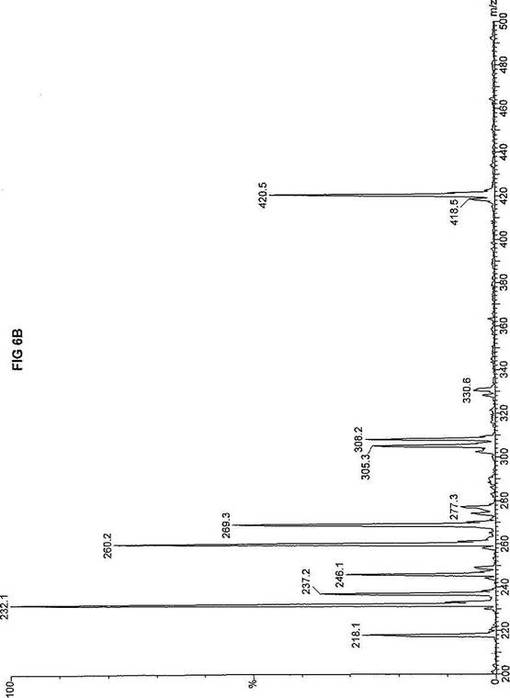
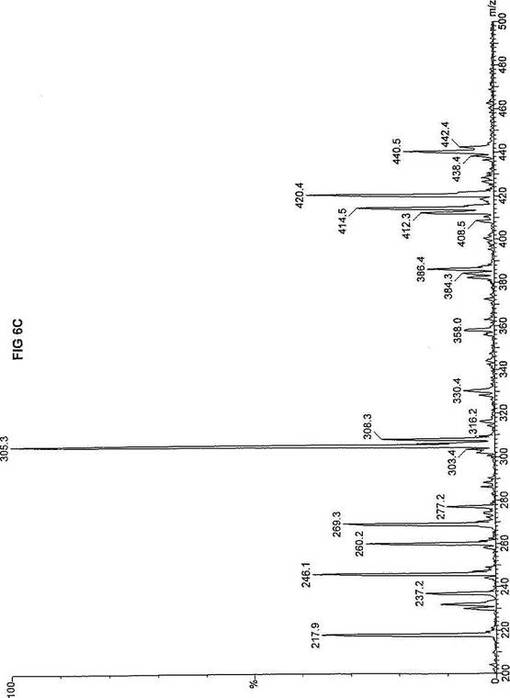
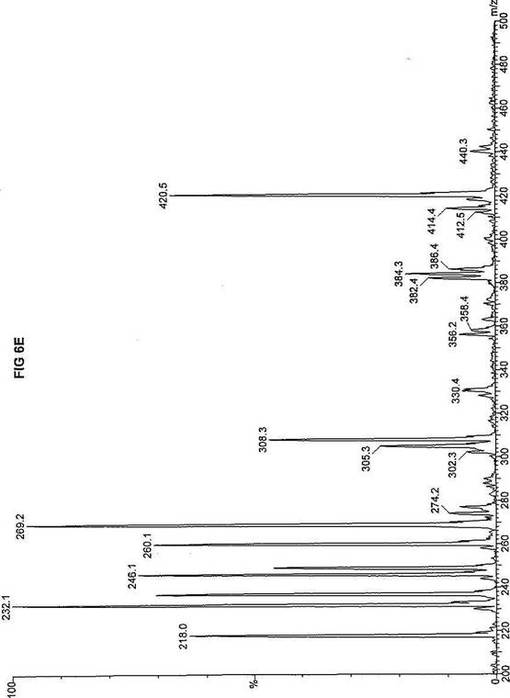
D3-C8(図6A-L)の場合、m/z305(2H3-オクタン酸エステル-C8)の相対的蓄積が起り、完全酸化のためにトランスロカーゼ(図6C)とMCAD(図6I)の両方が明らかに必要であることを示す。主要成分がオクタン酸エステルである市販中鎖トリグリセリド(MCT)はCPT I、トランスロカーゼ、およびCPT IIに依存しないと考えられているが、2H3-オクタン酸-C8に対するこのデータは、MCTが重篤なトランスロカーゼ不全に対する有効な治療でないことを示す。さらに、このデータは、MCTがMCAD不全のための適当な治療でないことを説明する。
【0039】
奇数炭素基質D3-C7(図5A-L図)およびD3-C9(図7A-L)を用いて処置した細胞系についての有利な効果は次に基づく:(1)培地で無標識内因性脂質の酸化からある程度産生されうる診断プロフィールが不在であること;および(2)正常な対照細胞(D3-C7は図5A、またはD3-C9は図7A)に見られるのと比較した、標識した奇数炭素分解最終産物であるm/z235(2H3-プロピオン酸エステル-C3)の相対量。このm/z235(2H3-プロピオン酸エステル-C3)の相対量は、m/z269(2H9-イソ吉草酸-C5)およびm/z237.0(2H5-プロピオン酸エステル-C3)の内部標準のレベルと比較される。D3-C9については、トランスロカーゼ、CPT II、およびLCHAD細胞系ではm/z319(9-2H3-ノナン酸エステル)にて増加が観察された。これらの結果は、ノナン酸エステルの完全分解にトランスロカーゼ、CPT II、およびLCHADが全て必要であることを示す。
【0040】
D3-C7については、正常細胞と、CPT I、トランスロカーゼ、CPT II、VLCAD、トリファンクショナル、LCHAD、およびSCAD異常細胞系(図5A-HおよびJ)とにおいて産生される2H3-プロピオン酸エステル-C3(m/z235)の相対量は、比較し得るかまたは正常細胞に見られる量を超過するので、前駆体の有利な酸化が起ったことを示す。観察された例外の1つは、MCAD不全(図5I)であり、D3-C7は酸化のためにMCADを必要とすると予想され、MCADが不在であるとm/z291(2H3-ヘプタノイルカルニチン-C7)が顕著に増加する。ETF-DHについては、標識した7-2H3-ヘプタン酸エステルの酸化は観察されなかった。これらの結果は、n-ヘプタン酸補充組成物は、MCADおよびETFデヒドロゲナーゼを除いて、以下に挙げる脂肪酸欠損症の治療に使用できるを示す:トランスロカーゼ不全;カルニチンパルミトイルトランスフェラーゼIおよびII不全;L-3-ヒドロキシアシル-CoAデヒドロゲナーゼ(LCHAD)不全;極長鎖アシルCoAデヒドロゲナーゼ(VLCAD)不全、および短鎖アシル-CoAデヒドロゲナーゼ(SCAD)不全。
【実施例2】
【0041】
重篤なトランスロカーゼ不全患者におけるトリヘプタノインの補充のin vivoでの利用
実施例1において確認した重篤な新生児トランスロカーゼ不全をもつ乳幼児の治療に、トリヘプタノイン補充した低脂肪処方を使って成功した。さらに、トリヘプタノイン療法に対する臨床結果と乳幼児の羊膜細胞のin vitro質量分析との間の相関を支持する確証がある。
【0042】
妊娠38週に、実施例1に記載した重篤なトランスロカーゼ不全の試験結果が陽性であった羊膜をもつ乳幼児の分娩を実施した。臍帯血を、全および遊離カルニチンレベルならびに個々のアシルカルニチンのレベルについて、タンデム質量分析により分析した(Yang,ら, 1998.「カルニチンパルミトイルトランスフェラーゼ II(CPT II)不全をもつ患者における4つの新規突然変異の同定(Identification of four novel mutations in patients with carnitine palmitoyltransferase II (CPT II) deficiency)」, Mol Genet Metab 64:229-236)。母性血も分娩時に、これらの上記レベルを検定した。結果は、乳幼児が重篤なトランスロカーゼ不全を患っていることを立証した。
【0043】
分娩後の最初の12時間内に、乳幼児に、経鼻胃管(nasogastric tube)を経由してトリヘプタノインを補充した低脂肪処方を与えた。続いて、トリヘプタノイン補充した処方による給食を、任意の正期産児と同じ頻度で与えた。カルニチン、ビオチン、およびシアノコバラミンの補充は必要でなかった。
【0044】
動脈血ガス(ABG)、電解質、血清尿素窒素(BUN)、クレアチニン、アンモニア、グルコース、血清クレアチンホスホキナーゼ(CPK)、ALT、AST、ヘモグロビン(Hgb)、およびヘマトクリット(Hct)を、標準の新生児集中看護手順に従ってモニターした。アシルカルニチンは、1日2回タンデム質量分析により定量分析した。尿有機酸の定量分析を実施すると共に、尿中に存在するジカルボン酸の量をモニターした。
【0045】
トリヘプタノインを補充する処方の介入は、トランスロカーゼ不全の影響を抑制するのに全体として成功した。乳幼児の入院中、上記の様々な生理学的パラメーターは正常範囲内にある結果が得られた。乳幼児は7-8週齢で退院し、トリヘプタノイン補充処方を用いる食事管理が完全であることを示した。平均処方を与えた乳幼児の1日当たり20-25gの平均重量取得と比較すると、トリヘプタノイン補充処方に基づく継続看護期間中、該乳幼児は1日当たり35gの1日当たり平均重量取得を維持した。4ヶ月半齢において、乳幼児は、トリヘプタノイン補充処方に基づいてよく育ち続けて、カルニチン、ビオチン、またはビタミンB12の補充を必要としなかった。
【図面の簡単な説明】
【0046】
【図1】図1は、長鎖脂肪酸のミトコンドリアβ酸化経路を描く図であり、所要の輸送体および酵素をイタリック字体で記しかつ3つの指摘した膜を二重線で表現している。
【図2】図2は、n-ヘプタン酸のミトコンドリアβ酸化経路を描く図であり、所要の輸送体および酵素をイタリック字体で記しかつ指摘した内側ミトコンドリア膜を二重線で表現している。
【図3】図3Aは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、重篤なトランスロカーゼ不全をわずらう死亡した子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図3Bは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、図3Aに報じた重篤なトランスロカーゼ不全をわずらう死亡した子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。
【図4】図4Aは、D3-C16(2H3-パルミチン酸エステル-C16)を用いて処理した羊膜細胞に対する、タンデム型質量分析プロフィールを描くグラフである。羊膜細胞は、重篤なトランスロカーゼ不全と診断された胎児から取得し、その同胞(sibling)は図3Aおよび3Bに報じた死亡した子供であった。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図4Bは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した羊膜細胞に対する、タンデム型質量分析プロフィールを描くグラフである。羊膜細胞は、図4Aに報じた重篤なトランスロカーゼ不全と診断された胎児から取得し、その同胞は図3Aおよび3Bに報じた死亡した子供であった。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。
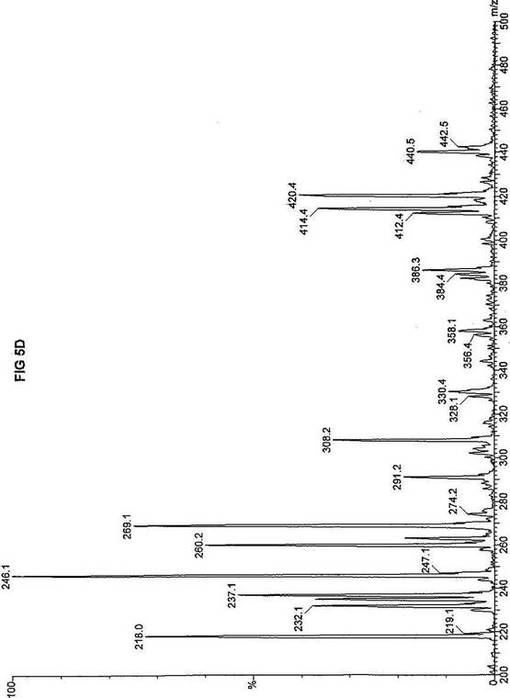
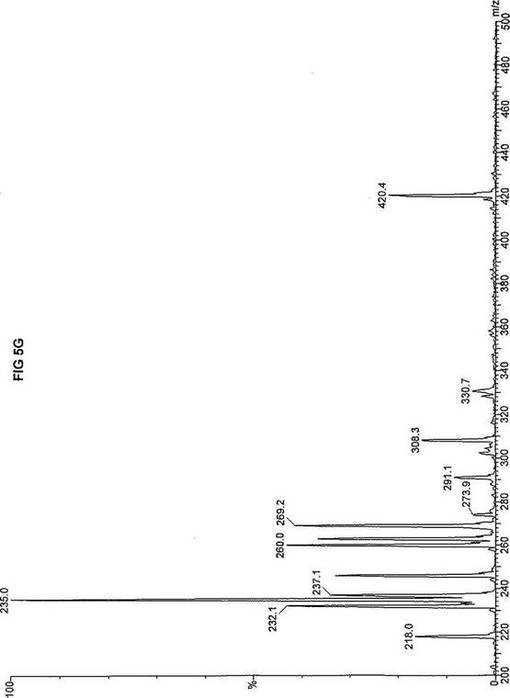
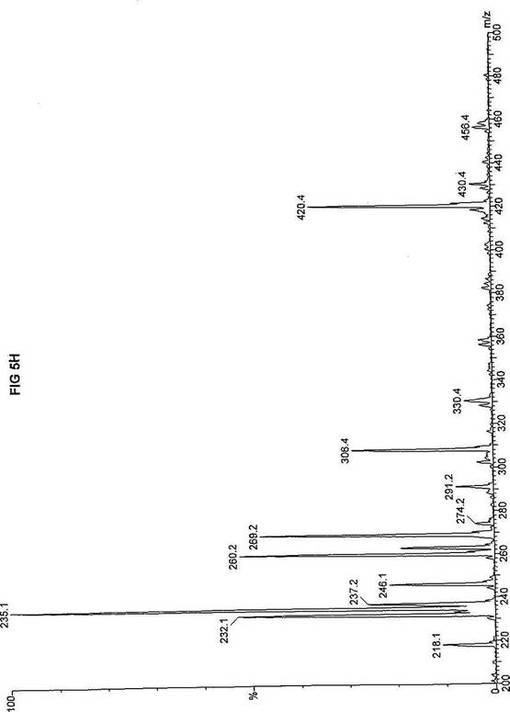
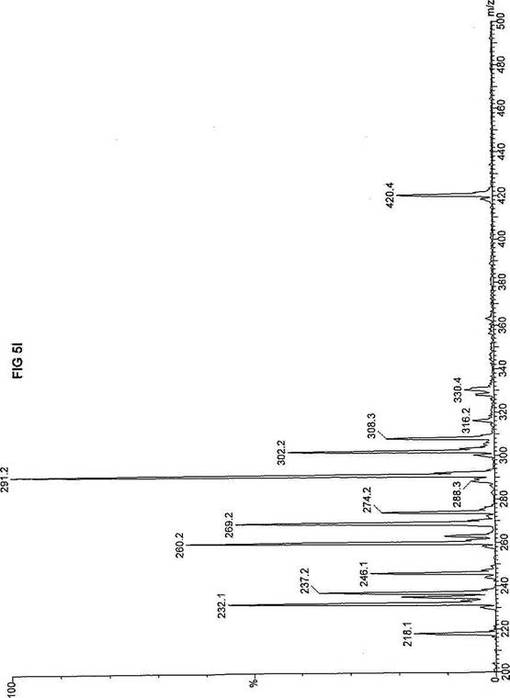
【図5】図5Aは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した正常な織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図5A-5Cのプロフィールに対する内部標準は、m/z420.3(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.1(2H9-イソ吉草酸-C5)、およびm/z237.0(2H5-プロピオン酸エステル-C3)に位置し、ここにおいてm/zとは質量:電荷比である。ピークm/z291はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Bは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼI(CPT I)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235.0は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Cは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、トランスロカーゼ不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291.3はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Dは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼII(CPT II)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図5D-5Fのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.1(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z291.1はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Eは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「心臓」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-C)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235.1は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Fは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「低血糖」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-H)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291.4はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235.1は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Gは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、ミトコンドリア三機能タンパク質(トリファンクショナル)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図5G-5Iのプロフィールに対する内部標準は、m/z420.3(2H6-パルミチン酸エステル-C16)、m/z308.1(2H6-オクタン酸エステル-C8)、m/z269.0(2H9-イソ吉草酸-C5)、およびm/z237.2(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z291はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235.1は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Hは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、長鎖L-3-ヒドロキシ-アシルCoAデヒドロゲナーゼ(LCHAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291.1はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235.1は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Iは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、中鎖アシルCoAデヒドロゲナーゼ(MCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291.2はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Jは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、短鎖アシルCoAデヒドロゲナーゼ(SCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図5J-5Lのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.0(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z291.1はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235.1は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Kは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-マイルド(ETF-DHマイルド)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291.3はD3-C7(7-2H3-ヘプタン酸エステル)を表す。図5Lは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-シビア(ETF-DHシビア)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291はD3-C7(7-2H3-ヘプタン酸エステル)を表す。
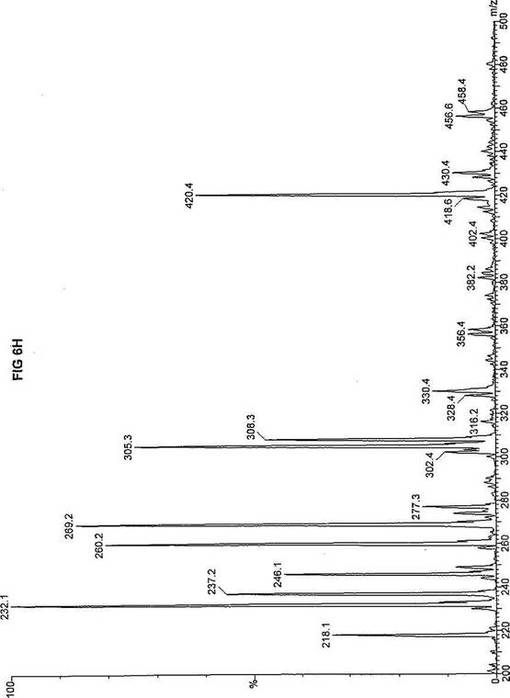
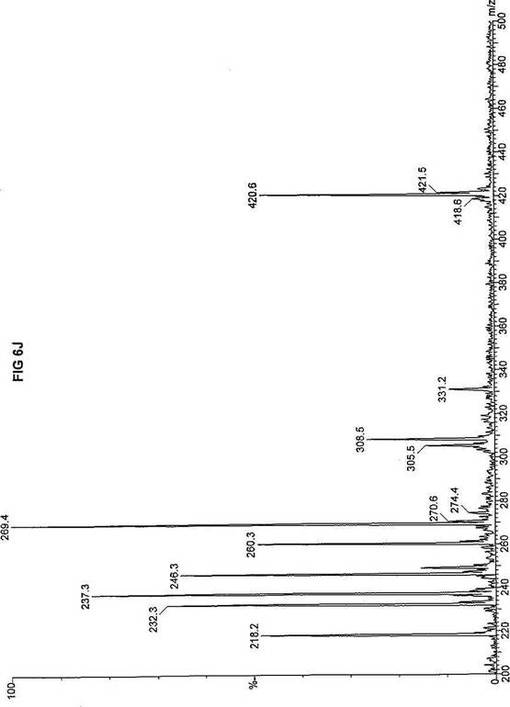
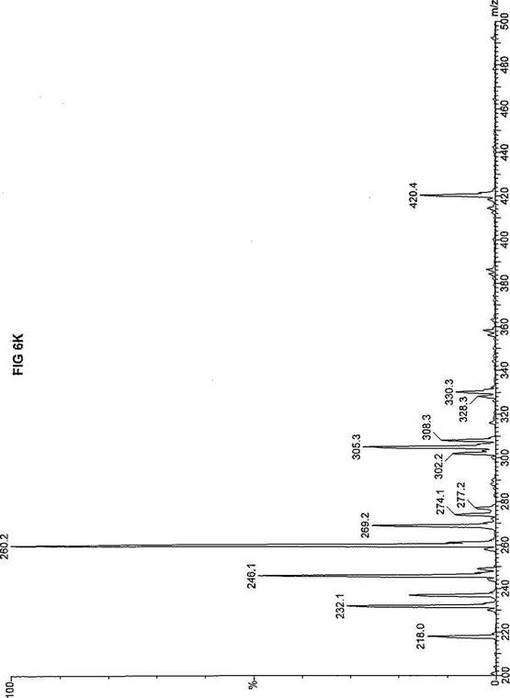
【図6】図6Aは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した正常な織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図6A-6Cのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.1(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z305.3はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Bは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼI(CPT I)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.0はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Cは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、トランスロカーゼ不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.3はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Dは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼII(CPT II)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図6D-6Fのプロフィールに対する内部標準は、m/z420.3(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.2(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z305.3はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Eは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「心臓」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-C)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.3はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Fは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「低血糖」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-H)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.2はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Gは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、ミトコンドリア三機能タンパク質(トリファンクショナル)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図6G-6Iのプロフィールに対する内部標準は、m/z420.5(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.2(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z305.3はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Hは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、長鎖L-3-ヒドロキシ-アシルCoAデヒドロゲナーゼ(LCHAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Iは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、中鎖アシルCoAデヒドロゲナーゼ(MCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.2はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Jは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、短鎖アシルCoAデヒドロゲナーゼ(SCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図6J-6Lのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.1(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z305.0はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Kは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-マイルド(ETF-DHマイルド)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.2はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Lは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-シビア(ETF-DHシビア)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.3はD3-C8(8-2H3-オクタン酸エステル)を表す。
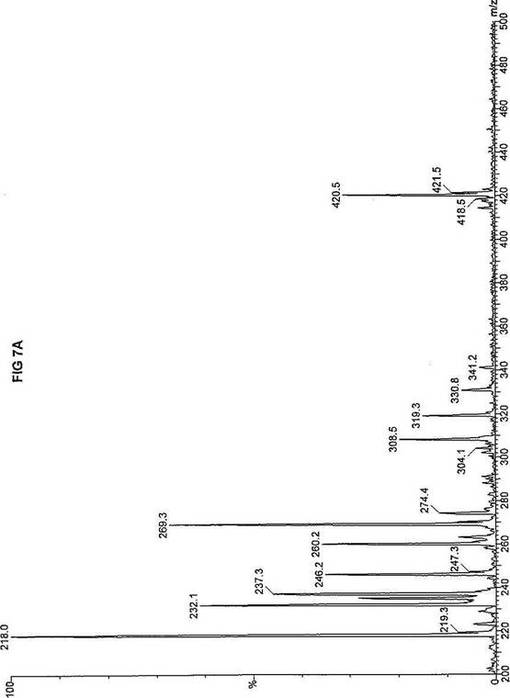
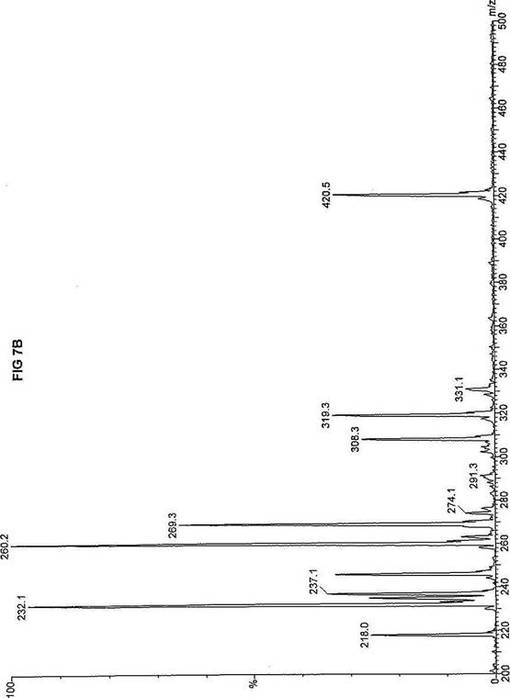
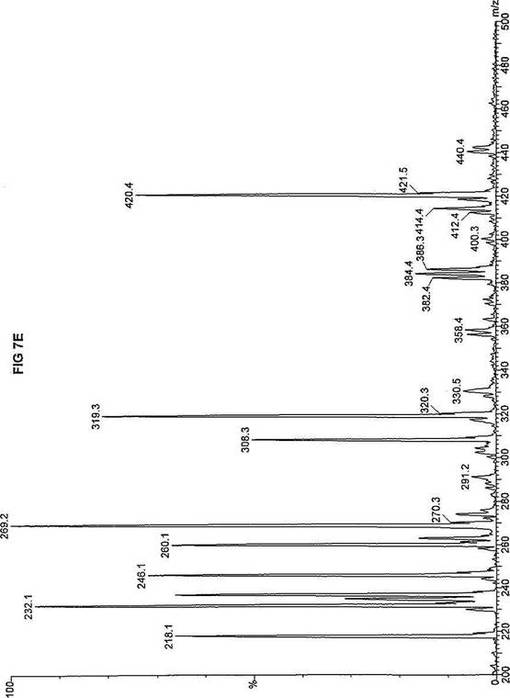
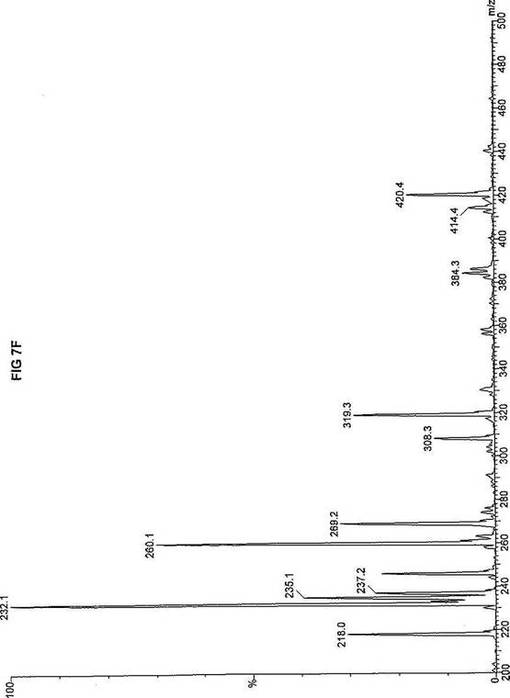
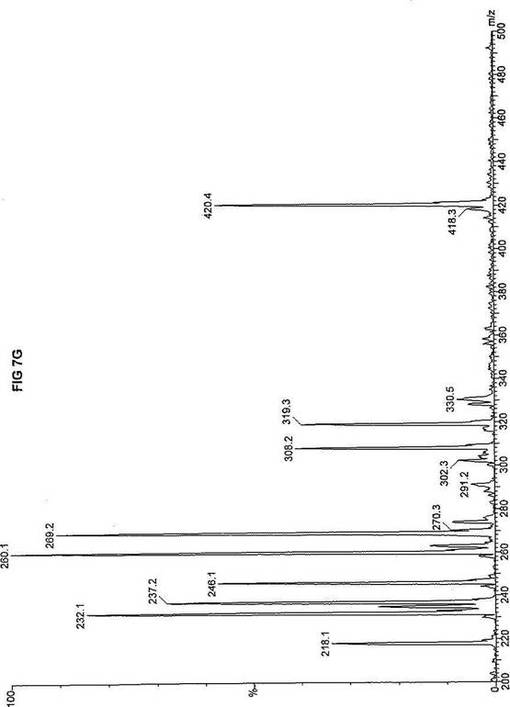
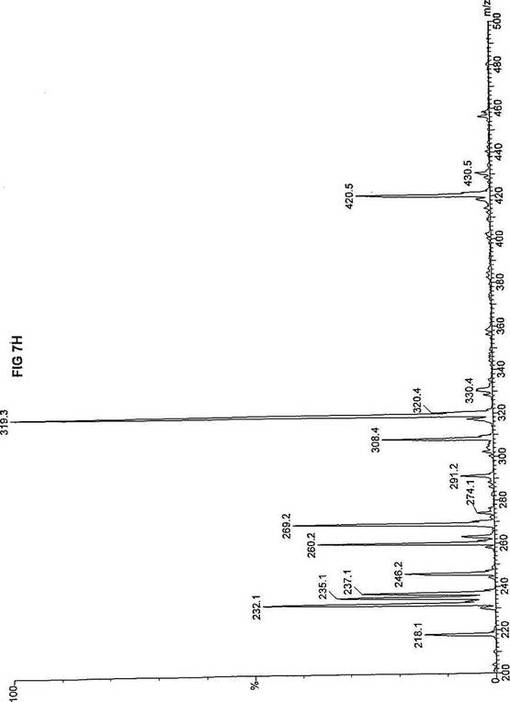
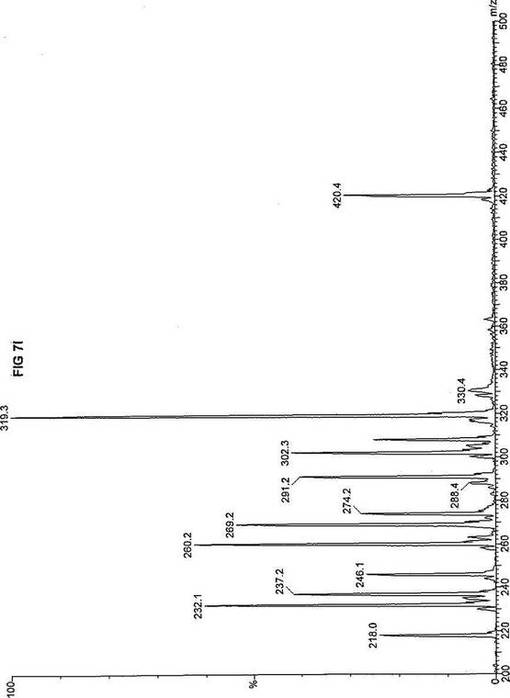
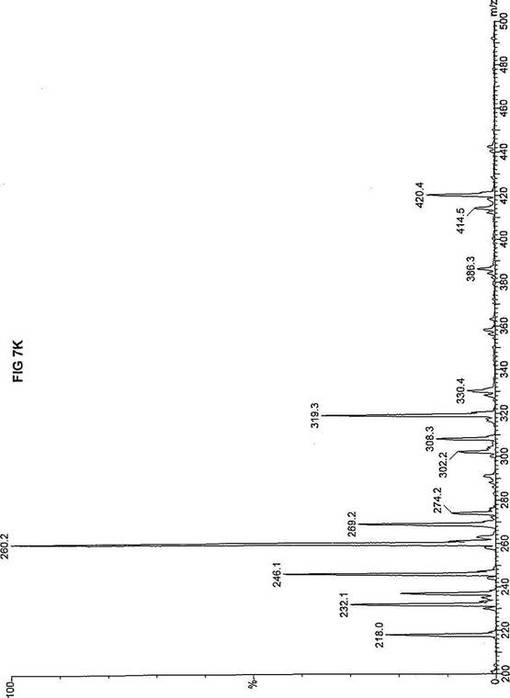
【図7】図7Aは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した正常な織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図7A-7Cのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.3(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Bは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼI(CPT I)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Cは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、トランスロカーゼ不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Dは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼII(CPT II)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図7D-7Fのプロフィールに対する内部標準は、m/z420.5(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.3(2H9-イソ吉草酸-C5)、およびm/z237.1(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Eは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「心臓」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-C)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Fは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「低血糖」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-H)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Gは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、ミトコンドリア三機能タンパク質(トリファンクショナル)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図7G-7Iのプロフィールに対する内部標準は、m/z420.5(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.3(2H9-イソ吉草酸-C5)、およびm/z237.2(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Hは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、長鎖L-3-ヒドロキシ-アシルCoAデヒドロゲナーゼ(LCHAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.2はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Iは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、中鎖アシルCoAデヒドロゲナーゼ(MCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.0はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Jは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、短鎖アシルCoAデヒドロゲナーゼ(SCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図7J-7Lのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.3(2H9-イソ吉草酸-C5)、およびm/z237.0(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Kは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-マイルド(ETF-DHマイルド)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Lは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-シビア(ETF-DHシビア)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。
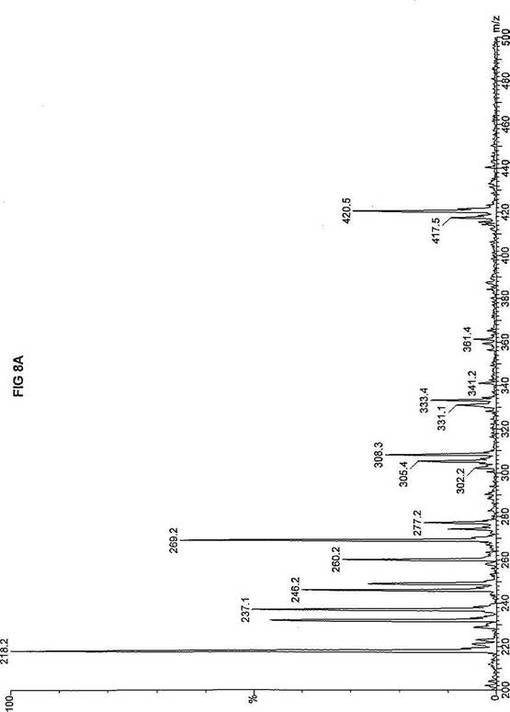
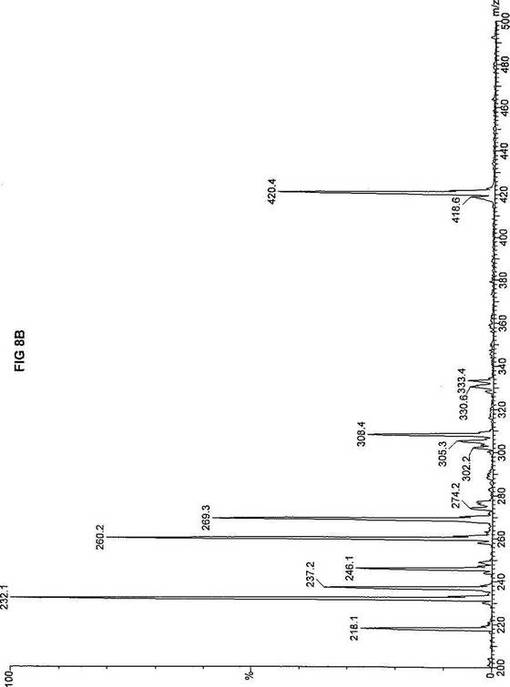
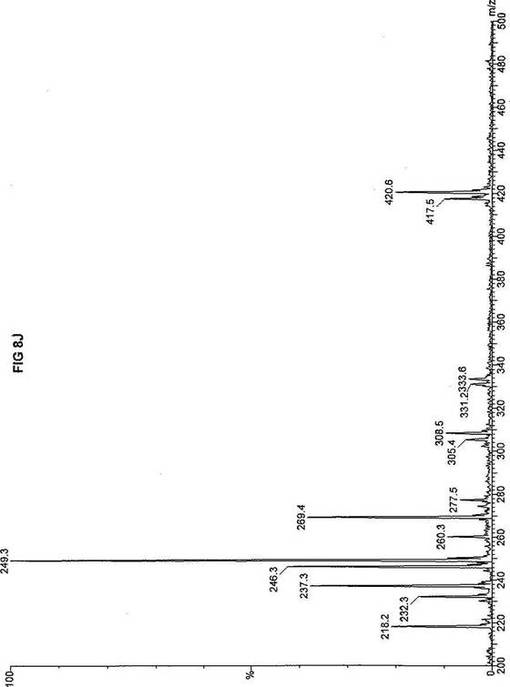
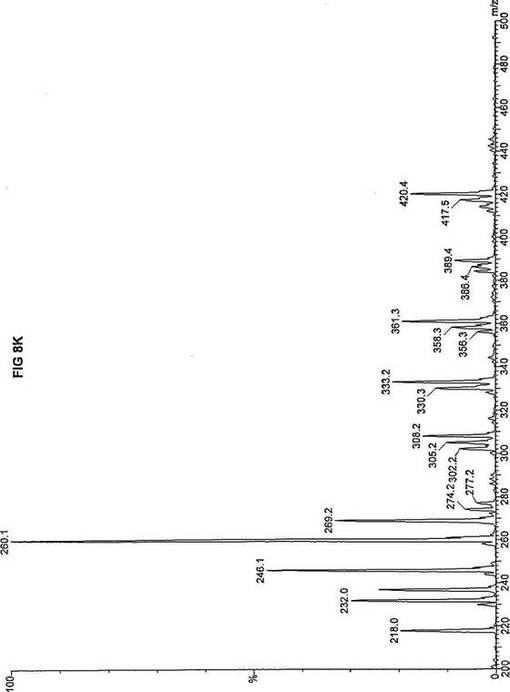
【図8】図8Aは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した正常な織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図8A-8Cのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.1(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.0(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z361.3はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Bは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼI(CPT I)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.3はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Cは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、トランスロカーゼ不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.3はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Dは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼII(CPT II)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図8D-8Fのプロフィールに対する内部標準は、m/z420.5(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.0(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z361.3はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Eは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「心臓」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-C)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.2はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Fは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「低血糖」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-H)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.3はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Gは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、ミトコンドリア三機能タンパク質(トリファンクショナル)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図8G-8Iのプロフィールに対する内部標準は、m/z420.5(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.1(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z361.1はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Hは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、長鎖L-3-ヒドロキシ-アシルCoAデヒドロゲナーゼ(LCHAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.4はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Iは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、中鎖アシルCoAデヒドロゲナーゼ(MCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.4はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Jは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、短鎖アシルCoAデヒドロゲナーゼ(SCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図8J-8Lのプロフィールに対する内部標準は、m/z420.2(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z361.4はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Kは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-マイルド(ETF-DHマイルド)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.4はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Lは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-シビア(ETF-DHシビア)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.3はD3-C12(12-2H3-ドデカン酸エステル)を表す。
【図9】図9Aは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した正常な織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図9A-9Cのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.1(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z417.0はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Bは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼI(CPT I)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.6はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Cは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、トランスロカーゼ不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.4はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Dは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼII(CPT II)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図9D-9Fのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.2(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z417.4はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Eは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「心臓」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-C)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.5はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Fは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「低血糖」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-H)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.5はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Gは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、ミトコンドリア三機能タンパク質(トリファンクショナル)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図9G-9Iのプロフィールに対する内部標準は、m/z420.5(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.0(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z417.4はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Hは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、長鎖L-3-ヒドロキシ-アシルCoAデヒドロゲナーゼ(LCHAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.4はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Iは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、中鎖アシルCoAデヒドロゲナーゼ(MCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Jは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、短鎖アシルCoAデヒドロゲナーゼ(SCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図9J-9Lのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308(2H6-オクタン酸エステル-C8)、m/z269.1(2H9-イソ吉草酸-C5)、およびm/z237(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z417はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Kは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-マイルド(ETF-DHマイルド)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.3はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Lは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-シビア(ETF-DHシビア)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.3はD3-C16(16-2H3-パルミチン酸エステル)を表す。形態に従ったマイクロレンズを示す断面図である。
【技術分野】
【0001】
本発明は、栄養的または食事療法的組成物または補助剤に関する。
【背景技術】
【0002】
脂肪酸はエネルギー産生に重要な役割を果たし、空腹時には必須である。骨格および/または心筋の弱りから乳幼児突然死症候群に似た致死性の代謝無呼吸異常症状における脂肪酸代謝の重篤な障害が起り得る。これらの障害は、重篤な心筋症、低血糖症、筋障害、罹患器官の微小血管脂肪堆積、および/または劇症肝炎と共に現れる。脂肪酸代謝の先天性遺伝欠陥をわずらう患者は、脂肪酸代謝経由でエネルギーを産生することができず、しばしば致命的なまたは反復性の重篤な衰弱異常症状を経験する。未熟児は高血糖レベルの維持が必要である。しばしば、患者の日常食事は十分な量の炭水化物エネルギー源を与えず、かつ誕生時の患者の脂肪代謝酵素は効率的でない。高齢患者も、食欲不振および非効率的な代謝により、血糖レベルの調節の困難なことがある。
【0003】
飽和脂肪酸は、次の構造により表される:
【化1】
[式中、Rはアルキル基を表す]。高等植物および動物の脂質に由来する天然に存在する脂肪酸は、飽和および不飽和の両方の偶数炭素鎖を含むものである。最も豊富な天然に存在する飽和脂肪酸は、パルミチン酸(16個の炭素;C16)およびステアリン酸(18個の炭素;C18)である。それより短鎖の脂肪酸(12-14個の炭素;C12-C14)およびそれより長鎖の脂肪酸(28個の炭素;C28までの)が小量、天然に存在する。10個未満の炭素をもつ脂肪酸はミルク脂肪を除く動物脂質には稀にしか存在しないが、ミルク脂肪はオレイン酸(不飽和C18)約32%、パルミチン酸(C16)約15%、ミリスチン酸(C14)約20%、ステアリン酸(C18)約15%、ラウリン酸(C12)約6%、および4〜10個の炭素をもつ脂肪酸(C4-C10)約10%を含有する。
【0004】
脂肪酸は一般的に、カルボキシル基に結合する炭素鎖長により分類される:4〜6個の炭素(C4-C6)の短鎖脂肪酸、8〜14個の炭素(C8-C14)の中鎖脂肪酸、16〜18個の炭素(C16-C18)の長鎖脂肪酸、および20〜28個の炭素(C20-C28)の極長鎖脂肪酸である。
【0005】
脂肪酸が代謝されるプロセスは、細胞のミトコンドリア内のミトコンドリアβ酸化を含む。図1に図解するように、パルミチン酸のような長鎖脂肪酸の脂肪酸酸化は、形質膜性カルニチントランスポーターを介して形質膜を通過する脂肪酸の輸送に始まる。脂肪酸がミトコンドリア外膜を通過すると、脂肪酸は、補酵素A(CoASII)およびアシルCoAシンセターゼの存在下で、ATPを消費して補酵素Aの脂肪酸エステル(脂肪族アシル-CoA)に転化される。脂肪族アシル-CoAは、カルニチンおよびカルニチンパルミトイルトランスフェラーゼI(CPT I)の存在下で脂肪族アシルカルニチンに転化される。次に、脂肪族アシルカルニチンはミトコンドリアの内膜を通過し、この工程はカルニチン/アシルカルニチントランスロカーゼ(carnitine/acylcarnitine translocase)の触媒作用を受ける。ミトコンドリアの内側に入ると、脂肪族アシルカルニチンは、次に、カルニチンパルミトイルトランスフェラーゼII(CPT II)の存在下で転化され、脂肪族アシルCoAに戻される。ミトコンドリア内の酸化サイクルで、脂肪族アシルCoAは、鎖特異的アシルCoAデヒドロゲナーゼを介して、αおよびβ炭素原子から1対の水素原子の除去による脱水素を受け、α、β-不飽和アシルCoA、または2-trans-エノイルCoAを生じる。適当なアシルCoAデヒドロゲナーゼは、脂肪族アシルCoAの炭素鎖長により決定され、すなわち、長鎖アシルCoAデヒドロゲナーゼ(LCAD;C12-C18)、中鎖アシルCoAデヒドロゲナーゼ(MCAD;C4-C12)、短鎖アシルCoAデヒドロゲナーゼ(SCAD;C4-C6)、または極長鎖アシルCoAデヒドロゲナーゼ(VLCAD;C14-C20)である。次に、α,β-不飽和アシルCoAは、2-エノイルCoAヒドラターゼを介して酵素作用により水和され、L-3-ヒドロキシアシルCoAを生成し、これは、次に、鎖特異的L-3-ヒドロキシアシルCoAデヒドロゲナーゼの触媒作用によりNAD連結反応で脱水素され、β-ケトアシルCoAを生成する。適当なL-3-ヒドロキシアシルCoAデヒドロゲナーゼは、L-3-ヒドロキシアシルCoAの炭素鎖長により決定され、すなわち、長鎖L-3-ヒドロキシアシルCoAデヒドロゲナーゼ(LCHAD;C12-C18)、または、短鎖L-3-ヒドロキシアシルCoAデヒドロゲナーゼ(SCHAD;C4-C16、鎖長の増加とともに活性は低下する)である。β-ケトアシルCoAエステルは、3-ケトアシルCoAチオラーゼの存在下で第2のCoA分子のチオール基の攻撃により酵素分解を受け、脂肪族アシルCoAおよび元来の脂肪酸のαカルボキシルおよびβ炭素原子に由来するアセチルCoAを生成する。他の産物、すなわち原料脂肪酸より2個少ない炭素原子を有する長鎖飽和脂肪族アシルCoAは、今度は次回の反応の基質となって、最初の脱水素工程に始まり、最後に2回目の2個の炭素をもつ断片はアセチルCoAとして除去される。この循環型プロセスを通過する毎に、脂肪酸鎖はアセチルCoAとして2個の炭素断片および2対の水素原子を特定の受容体に向けて失う。
【0006】
脂肪酸酸化プロセスの各工程は、重複する炭素鎖長特異性をもつ酵素による触媒作用を受ける。これらの酵素による触媒作用の消失に関連する脂肪酸酸化の遺伝性障害が確認されている。これらの障害には、形質膜カルニチン輸送;CPT IおよびII;カルニチン/アシルカルニチントランスロカーゼ;極長鎖、中鎖、および短鎖アシルCoAデヒドロゲナーゼ(すなわち、それぞれVLCAD、MCAD、およびSCAD);2,4-ジエノイルCoAレダクターゼ;長鎖3-ヒドロキシアシルCoAデヒドロゲナーゼアシルCoA(LCHAD)の欠損症、およびミトコンドリア三機能性(trifunctional)タンパク質(MTP)不全が含まれる。今までに、中鎖デヒドロゲナーゼ(MCAD)不全に対する治療は見出されている。しかし、他の不全症は、出生1年以内の患者にはしばしば致命的であり、利用できる有効な治療は知られていない。特に、重篤なカルニチン/アシルカルニチントランスロカーゼ不全をわずらう患者は通常死亡し、生存者は知られてないし、そして治療は見出されてない。
【0007】
これらの障害を治療する試みは、欠陥のある酵素による触媒作用の欠失を迂回する食物源を提供することに重点が置かれている。例えば、欠陥のあるカルニチン/アシルカルチニントランスロカーゼにより引き起される長鎖脂肪酸代謝不全(以後、「トランスロカーゼ不全」と呼ぶ)は、しばしば新生児期の死をもたらす。あるトランスロカーゼ不全の患者に、カルチニン、高炭水化物食、および中鎖トリグリセリドを与えたが、脂肪酸代謝不全を克服することはできなかった。中鎖脂肪酸は自由にミトコンドリアに入ると予想されていたので、中鎖脂肪酸の代謝はカルニチン/アシルカルチニントランスロカーゼを必要としないだろうと考えられた。従って、トランスロカーゼ欠陥をバイパスすると予想される偶数個の炭素数をもつ中鎖トリグリセリド(MCT)(例えば、84% C8、8% C6および8% C10)を含有する乳幼児処方が開発された。これらの処方による治療の試みに関わらず、志望者の発生は続いている。
【0008】
ペラゴン酸(9個の炭素;C9をもつ飽和脂肪酸)は例外として、奇数炭素数脂肪酸は高等植物および動物脂質中に稀である。ある特定の奇数炭素数をもつ合成トリグリセリドが、潜在的な脂肪酸源としての食品、および食品の製造に利用するために試験されている。C7およびC9トリグリセリド由来の奇数鎖脂肪酸の酸化速度が単離した子ブタ肝細胞中でin vitroで試験されている(Odleら, 1991. 「新生仔の子ブタによる中鎖トリグリセリドの利用;偶数および奇数炭素数脂肪酸の鎖長と見かけの消化/吸収および肝代謝」, J Nutr 121:605-614;Lin, X,ら, 1996. 「酢酸エステルは、新生仔の子ブタから単離した肝細胞におけるヘプタン酸エステルおよびオクタン酸エステルβ酸化の主要産物である」, Biochem J 318:235-240;およびOdle, J. 1997.「新生仔による中鎖トリグリセリドの利用についての新しい洞察:子ブタモデルからの観察」, J Nutr 127:1061-1067)。奇数鎖脂肪酸、プロピオン酸エステル(C3)、吉草酸エステル(C5)、およびノナン酸エステル(C9)の糖新生前駆体としての重要性が、飢餓ラット由来の肝細胞において評価された(Sugden,ら, 1984.「飢餓ラット由来の肝細胞中の奇数炭素脂肪酸代謝」, Biochem Int'l 8:61-67)。放射標識したマルガル酸(C17)エステルの酸化がラット肝切片において試験された(Boyer,ら, 1970.「1-14Cオクタン酸および1-14Cマルガル酸の肝代謝」, Lipids 4:615-617)。
【0009】
C3、C5、C7、C9、C11、およびC17を利用するin vivo研究も、モルモット、ウサギ、およびラットについてin vivoで実施されている。体系的に注入されたC7およびC9トリグリセリド由来の中鎖脂肪酸およびC7/C9トリグリセリド混合物のin vivo酸化速度が新生仔のブタで試験されている(Odel,ら, 1992.「連続注入放射性トレーサーキネティクス法を使った、新生仔の子ブタによる[1-14C]-中鎖脂肪酸酸化の評価」, J Nutr 122:2183-2189;およびOdle,ら, 1989.「新生仔の子ブタによる中鎖トリグリセリドの利用:II.出生の最初2日にわたる偶数および奇数鎖トリグリセリド消費が血液代謝物および尿窒素排泄に与える影響」, J Anima Sci 67:3340-3351)。トリウンデカノイン(飽和C11)を与えたラットは、非飢餓血糖レベルを延長された飢餓期間にわたり維持することが観察された(Anderson,ら, 1975.「飢餓ラットにおけるラード、サフラワー油、およびトリウンデカノインのグルコース形成およびケト体生成能」, J Nutr 105:185-189)。トリノナノイン(C9)および長鎖トリグリセリドの乳剤がウサギに注入され、長期総非経口性栄養(long-term total parenteral nutrition)として評価された(Linseisen,ら, 1993.「総非経口性栄養の奇数中鎖トリグリセリド(トリノナノイン):ウサギの脂肪代謝パラメーターに与える影響」, J Parenteral and Enteral Nutr 17:522-528)。7個の炭素をもつ飽和脂肪族n-ヘプタン酸(C7)を含有するトリグリセリドトリヘプタノインは、欧州において、農業飼料、バター製造のトレーサ分子として、およびチョコレートおよび他の菓子の製造における遊離剤(reasing agent)として使われたことが報じられている。しかし、今まで、7炭素脂肪酸がヒトによる摂取に安全であるとかまたはヒトに対するいずれか特定の栄養上の利点を有するという指摘はなかった。
【0010】
このほど、本発明者らは、後天的代謝撹乱および遺伝性代謝障害、特に脂肪酸代謝不全症は、n-ヘプタン酸のような炭素数が7である脂肪酸を含有する栄養組成物を使って克服できることを見出した。欠陥のあるまたは低下のある脂肪酸代謝を有している患者を、非常に効率的なエネルギー源としてn-ヘプタン酸および/またはそのトリグリセリドであるトリヘプタノインのような炭素数が7である脂肪酸を含有する栄養組成物を用いて、治療することができる。また迅速なエネルギーを必要とする患者も、炭素数が7である脂肪酸またはそのトリグリセリドを消費することにより利益を得ることができる。
【発明の概要】
【0011】
1つの態様において、本発明は、飽和炭素数が7である脂肪酸を含む栄養補助剤である。本発明に有用な炭素数が7である脂肪酸は、n-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノイン(triheptanoin)が、本発明のこの態様に最も有用である。好ましくは、炭素数が7である脂肪酸は、総食事カロリーの少なくとも約25%を供給する濃度である。
【0012】
別の態様において、本発明は炭素数が7である脂肪酸を含む製剤である。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。この製剤は、経口または非経口投与することができる。
【0013】
さらに別の態様において、本発明は、炭素数が7である脂肪酸を含む、少なくとも1つの代謝障害を有する患者に治療上の効果を与えるための投与に適合した一回量剤形とした製剤である。上記代謝障害は、たとえば、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼII、カルニチン/アシルカルニチントランスロカーゼ、心臓型極長鎖アシルCoAデヒドロゲナーゼ、低血糖型極長鎖アシルCoAデヒドロゲナーゼ、筋型極長鎖アシルCoAデヒドロゲナーゼ、ミトコンドリア三機能性タンパク質、長鎖L-3-ヒドロキシアシルCoAデヒドロゲナーゼ、および短鎖アシルCoAデヒドロゲナーゼなどの脂肪酸代謝不全の結果として生じ得る。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。この製剤は経口または非経口投与することができる。
【0014】
さらに別の態様において、本発明は炭素数が7である脂肪酸および製薬上許容される担体を含む一回量医薬剤形であって、この剤形は、少なくとも1つの代謝障害を有する患者の脂肪酸代謝を増大させるのに有効な量で提供される。上記代謝障害は、たとえば、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼII、カルニチン/アシルカルニチントランスロカーゼ、心臓型極長鎖アシルCoAデヒドロゲナーゼ、低血糖型極長鎖アシルCoAデヒドロゲナーゼ、筋型極長鎖アシルCoAデヒドロゲナーゼ、ミトコンドリア三機能性タンパク質、長鎖L-3-ヒドロキシアシルCoAデヒドロゲナーゼ、および短鎖アシルCoAデヒドロゲナーゼなどの、脂肪酸代謝不全の結果として生じ得る。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。この一回量医薬剤形は経口または非経口投与することができる。
【0015】
別の態様において、本発明は、炭素数が7である脂肪酸を含む栄養補助剤の、ヒトにおける脂肪酸代謝を増大させるための使用である。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。好ましくは、炭素数が7である脂肪酸は、上記補助剤の総カロリーの少なくとも約25%を供給する。
【0016】
別の態様において、本発明は、炭素数が7である脂肪酸を含む栄養補助剤の、グルコースの栄養源としての使用である。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。好ましくは、炭素数が7である脂肪酸は、上記補助剤の総カロリーの少なくとも約25%を供給する。
【0017】
別の態様において、本発明は、炭素数が7である脂肪酸を含む一回量剤形とした製剤を、少なくとも1つの代謝障害を有する患者に治療効果を与えるために使用することである。上記代謝障害は、たとえば、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼII、カルニチン/アシルカルニチントランスロカーゼ、心臓型極長鎖アシルCoAデヒドロゲナーゼ、低血糖型極長鎖アシルCoAデヒドロゲナーゼ、筋型極長鎖アシルCoAデヒドロゲナーゼ、ミトコンドリア三機能性タンパク質、長鎖L-3-ヒドロキシアシルCoAデヒドロゲナーゼ、および短鎖アシルCoAデヒドロゲナーゼなどの脂肪酸代謝不全の結果として生じ得る。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。好ましくは、炭素数が7である脂肪酸は、補助剤の総カロリーの少なくとも約25%を供給する。
【0018】
さらに別の態様において、本発明は、食物に炭素数が7である脂肪酸を加えることを含む、食物1グラムあたりのエネルギーポテンシャルを増大させる方法である。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。有益な効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。好ましくは、該食物は経口または非経口投与される。
【0019】
別の態様において、本発明は、炭素数が7である脂肪酸を含む栄養補助剤を用いて、少なくとも1つの代謝障害を有する患者を治療する方法である。上記代謝障害は、たとえば、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼII、カルニチン/アシルカルニチントランスロカーゼ、心臓型極長鎖アシルCoAデヒドロゲナーゼ、低血糖型極長鎖アシルCoAデヒドロゲナーゼ、筋型極長鎖アシルCoAデヒドロゲナーゼ、ミトコンドリア三機能性タンパク質、長鎖L-3-ヒドロキシアシルCoAデヒドロゲナーゼ、および短鎖アシルCoAデヒドロゲナーゼなどの脂肪酸代謝不全の結果として生じ得る。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。治療効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。好ましくは、炭素数が7である脂肪酸を、総食事カロリーの少なくとも約25%を供給するような濃度で添加する。
【0020】
さらに別の態様においては、本発明は、炭素数が7である脂肪酸を含む栄養補助剤を患者に投与することにより、脂肪酸代謝の効率を増大させる方法である。本発明に有用な炭素数が7である脂肪酸はn-ヘプタン酸である。炭素数が7である脂肪酸は、好ましくはn-ヘプタン酸を含む、トリグリセリドの形で提供することができる。治療効果を提供するのに十分な濃度のトリグリセリドトリヘプタノインが、本発明のこの態様に最も有用である。好ましくは、炭素数が7である脂肪酸を、総食事カロリーの少なくとも約25%を供給するような濃度で添加する。
【0021】
詳細な説明 この度、炭素数が7(C7)である脂肪酸またはそれらのトリグリセリドは、エネルギー産生のため長鎖脂肪酸をミトコンドリア中に輸送するために必要な通常の酵素、すなわち、カルニチン/アシルカルニチントランスフェラーゼ、カルニチンパルミトイルトランスロカーゼ(「CPT」)IおよびCPT IIを必要としないことを確認した。従って、炭素数が7である脂肪酸から構成されるトリグリセリドは、このような酵素を必要とする脂肪酸代謝不全を克服するために有用である。炭素数が7である脂肪酸を含有する栄養補助剤または医薬調製物は、遺伝性代謝障害ならびに後天的代謝撹乱の治療に有用である。
【0022】
好ましい炭素数が7である脂肪酸はn-ヘプタン酸である。n-ヘプタン酸は、次の構造:
【化2】
による炭素数7をもつ飽和直鎖脂肪酸である。トリヘプタノインは、3つのn-ヘプタン酸分子とグリセロールとのエステル化により調製されるトリグリセリドである。治療に関して、用語「ヘプタン酸」、「ヘプタン酸エステル」、および「トリヘプタノイン」は、以下の説明において互換的に使うことができる。また、当業者であれば、ヘプタン酸、ヘプタン酸エステル、およびトリヘプタノインは以下の説明全体を通じて本発明の炭素数7をもつ脂肪酸源の例として使われ、本発明の説明を意図するが、いかなる方法でも本発明の範囲を限定するものでないと解釈されることを理解するであろう。置換された、不飽和、または分枝ヘプタン酸エステル、ならびに他の改変された炭素数7をもつ脂肪酸を、本発明の範囲から逸脱することなく、使うことができる。
【0023】
トリヘプタノインは、最初に、ヘプタン酸の3つの分子とグリセロールに分解される。図2に説明したように、次いで、第1サイクルにおいて、ヘプタン酸は、通常のβ酸化過程を通してn-バレリルCoA(C5)およびアセチルCoA(C2)に分解される。次いで、第2サイクルにおいて、このn-バレリルCoA(C5)はn-プロピオニルCoA(C3)およびアセチルCoA(C2)に分解されるが、これらは両方ともクレブス回路(Kreb's cycle)およびエネルギー産生の燃料として重要な前駆体である。従って、トリヘプタノインは、エネルギー産生の燃料の効率的な供給源として有用である。さらに、プロピオニルCoAはグルコース産生の直接前駆体である。従って、トリヘプタノインは低血糖症に罹患しやすい患者、特に未熟児および高齢者に対する食事補助剤として有用である。トリヘプタノインはまた、未熟児の成長速度刺激剤として使うこともでき、入院期間を短縮し、それによってこれらの幼児に対する医療費を削減する。さらに、脂肪酸は心組織の主要燃料であり、かつトリヘプタノインは、糖新生物性(gluconeogenic)を有するので、心臓またはその他のリスクの高い外科手術から回復しつつある成人の心組織の直接燃料補給に使うことができる。
【0024】
ヘプタン酸は様々なフーゼル油にかなりの量が見出されており、当業界で周知の方法により抽出することができる。またヘプタン酸は、ヘプトアルデヒド(heptaldehyd)を希硫酸中で過マンガン酸カリウムにより酸化して合成することもできる(Ruhoff, Org Syn Coll. vol II, 315 (1943))。ヘプタン酸はまた、Sigma Chemical Co.(St. Louis, MO)から購入することもできる。
【0025】
トリヘプタノインは、当業界で周知の方法により、ヘプタン酸とグリセロールとのエステル化によって得ることができる。トリヘプタノインはCondea Chemie GmbH(Witten, ドイツ)からSpecial Oil 107として購入することもできる。
【0026】
不飽和ヘプタン酸エステルも脂肪酸代謝不全を克服するための栄養補助剤として利用することができる。さらに、特別な輸送酵素なしにミトコンドリアに容易に入り込む置換した不飽和および/または分枝の炭素数が7である脂肪酸も、本発明に利用することができる。例えば、4-メチルヘキサン酸エステル、4-メチルヘキセン酸エステル、および3-ヒドロキシ-4-メチルヘキサン酸エステルは、通常のβ酸化により2-メチル酪酸まで分解され、最終分解はイソロイシン経路で達成される。同様に、5-メチルヘキサン酸エステル、5-メチルヘキセン酸エステル、および3-ヒドロキシ-5-メチルヘキサン酸エステルは、通常のβ酸化によりイソ吉草酸まで分解され、最終分解はロイシン経路で達成される。
【0027】
本発明の炭素数が7であるトリグリセリドは、経口的に、非経口的に、または腹腔内に投与することができる。好ましくは、上記トリグリセリドは、トリヘプタノインのような炭素数が7である脂肪酸源を含有する食物の摂取を経由して、治療レベルを達成するために有効な濃度にて投与することができる。あるいは、カプセルとしてまたはリポソームに包んで、溶液または懸濁液で、単独または他の栄養素、追加の甘味剤および/または香料と一緒に、投与することができる。カプセルおよび錠剤は、公知のように、糖、シェラックおよび他の腸溶剤によりコーティングすることができる。
【0028】
投与方法は患者の年齢および脂肪酸代謝不全の程度により決定する。脂肪酸代謝欠陥、特にトランスロカーゼ不全をもつ乳幼児の治療に対して、トリヘプタノインは、栄養補助剤として低脂肪および/または削減(reduced)した長鎖脂肪酸を含有する食事乳幼児処方に加えることが好ましい。トリヘプタノインと共に使う市販の乳幼児処方の例として、Tolerex(Novartis Nutritionals, Minneapolis, MN)、Vivonex(Ross Laboratories, Columbus, OH)、およびPortagen and Pregestamil(Mead Johnson, Evansville, IN)が挙げられる。トリヘプタノインは、治療効果を達成するために有効な濃度にて処方に加える。栄養補充を必要とする子供または成人患者、例えば、化学療法を受けている外科または腫瘍患者に対して、トリヘプタノインを栄養ドリンクまたは全非経口栄養投与の一部として補充することが好ましい。
【0029】
代謝の先天性過誤により脂肪酸代謝経路が完全に破壊されている患者に対しては、トリヘプタノインを、24時間当たりの全カロリーのほぼ15%〜40%、好ましくは20%〜35%、そして最も好ましくはほぼ25%を与える濃度にて利用する。
【0030】
脂肪酸代謝経路機能の効率が低下している患者(例えば、未熟児、高齢者、心臓病患者)に対しては、トリヘプタノインを、24時間当たりの全カロリーのほぼ15%〜40%、好ましくは20%〜35%、そして最も好ましくはほぼ25%を与える濃度にて用いる。
【0031】
プロピオニルCoAはトリヘプタノイン酸化の副生物であるので、プロピオン酸の血液濃度が増加しうる。さらに、プロピオニルCoAは他の酵素反応に入り、クレブス回路および尿素回路に影響を与える毒性化合物を生じ得る。従って、特に血清プロピオン酸のビルドアップを示す患者にn-ヘプタン酸および/またはトリヘプタノイン補助剤のような炭素数が7である脂肪酸を投与するには、カルニチン補助剤および/またはビオチンとビタミンB12の組合わせを投与することが必要である。過剰のL-カルニチンおよびミトコンドリア酵素カルニチンアシルトランスフェラーゼの存在のもとで、プロピオニルCoAは、無毒物質であるプロピオニルカルニチンに転化されて尿中に排泄される。ビオチンは、プロピオニルCoAのメチルマロニルCoAへの転化の触媒となる酵素プロピオニルCoAカルボキシラーゼが必要とするビタミン補因子である。シアノコバラミンはビタミンB12の1つの形態であり、メチルマロニルCoAのスクシニルCoAへの転化の触媒となる酵素メチルマロニルCoAムターゼの補因子として働く。スクシニルCoAはクレブス回路に容易に引き抜かれる。従って、患者血液中の過剰のプロピニルCoAはスクシニルCoAへの転化により除去される。
【実施例1】
【0032】
細胞系への補充
致死型トランスロカーゼ不全をもつ患者から採取した培養細胞(織維芽細胞)にN-ヘプタン酸を加えると酸化に成功した。
【0033】
同胞は4日齢で重篤なトランスロカーゼ不全により死亡していたので、胎児から得た羊膜細胞を脂肪酸代謝の能力について試験した。試験結果は、胎児も重篤なトランスロカーゼ不全を有することを示した。
【0034】
死亡した同胞から採取した織維芽細胞および羊膜細胞の両方を、先に報じられたタンデム質量分析アッセイを使い、n-ヘプタン酸(C7)の脂肪酸代謝について評価した(Yang,ら, 1998.「カルニチンパルミトイルトランスフェラーゼII(CPT II)不全をもつ患者における4つの新規突然変異の同定(Identification of four novel mutations in patients with carnitine palmitoyltransferase II (CPT II) deficiency)」Mol Genet Metab 64:229-236)。質量分析結果を、死亡した同胞から採取した織維芽細胞に対しては、パルミチン酸エステルについて図3Aにおよびトリヘプタノインについて図3Bに、ならびに、胎児から採取した羊膜細胞に対しては、パルミチン酸エステルについて図4Aにおよびトリヘプタノインについて図4Bに報じた。研究結果は、両方の細胞系のトランスロカーゼ不全にも関わらず、n-ヘプタン酸(図3Bおよび4B)はカルニチン/アシルカルニチントランスロカーゼに依存せず、容易にプロピオニルCoAに酸化されることを示した。重篤なトランスロカーゼ不全を有する2つの細胞系によるn-ヘプタン酸代謝の成功に基づいて、正常患者からおよび他の協力研究室で直接酵素アッセイにより立証された次の脂肪酸化の遺伝性欠損症に罹っている患者から採取した織維芽細胞系について、タンデム質量分析アッセイを実施した:カルニチンパルミトイルトランスフェラーゼI(CPT I);重篤なカルニチン/アシルカルニチントランスロカーゼ(トランスロカーゼ(TRANSLOCASE));カルニチンパルミトイルトランスフェラーゼII(CPT II);極長鎖アシルCoAデヒドロゲナーゼ「心(cardiac)」型(VLCAD-C);極長鎖アシルCoAデヒドロゲナーゼ「低血糖」型(VLCAD-H);ミトコンドリア三機能タンパク質(トリファンクショナル(TRIFUNCTIONAL));長鎖L-3-ヒドロキシ-アシルCoAデヒドロゲナーゼ(LCHAD);中鎖アシルCoAデヒドロゲナーゼ(MCAD);短鎖アシルCoAデヒドロゲナーゼ(SCAD);電子伝達フラビンタンパク質QOデヒドロゲナーゼ-マイルド(ETF-DHマイルド);および電子伝達フラビンタンパク質QOデヒドロゲナーゼ-シビア(ETF-DHシビア)。各細胞系を別々に、7-2H3-ヘプタン酸エステル(D3-C7)、8-2H3-オクタン酸エステル(D3-C8)、9-2H3-ノナン酸エステル(D3-C9)、12-2H3-ドデカン酸エステル(D3-C12)、および16-2H3-パルミチン酸エステル(D3-C16)と共に培養した。タンデム質量分析の結果を、D3-C7について図5A-Lに;D3−C8について図6A-Lに;D3-C9について図7A-Lに;D3-C12について図8A-Lに;およびD3-C16について図9A-Lに示した。
【0035】
正常細胞系および11種の異常細胞系は3グループについて分析した。定量性をもたせるため、標識した内部標準を各分析に含ませ、各グループの最初のプロフィールに「IS」と記した。内部標準の質量数は、m/z420(2H6-パルミチン酸エステル-C16)、m/z308(2H6-オクタン酸エステル-C8)、m/z269(2H9-イソ吉草酸-C5)、およびm/z237.0(2H5-プロピオン酸エステル-C3)であり、ここにm/zは質量:電荷比である。
【0036】
図9Aに示すように、正常細胞をD3-C16と共に培養すると、標識したアシルカルニチン中間体がC16から下方C4(C4を含む)まで観察することができる。これらの2H3標識したアシルカルニチンの質量数はメチルエステルとして、m/z417(C16)、m/z389(C14)、m/z361(C12)、m/z333(C10)、m/z305(C8)、m/z277(C6)、およびm/z249(C4)である。
【0037】
16-2H3-パルミチン酸エステルD3-C16を用いて培養した各種細胞系(図9A-L)を観察すると、CPT I細胞(図9B)においては実質的に酸化が起らず、そして予期したとおり、パルミチン酸エステルがミトコンドリア中への輸送のためのパルミトイルカルニチンに容易に転化されないので、D3-C16(m/z417(C16))から最小量のパルミトイルカルニチンしか観察されない。トランスロカーゼ(図9C)およびCPT II(図9D)不全細胞系は両方とも、酸化は起らないが、CPT Iが存在する結果として、D3-C16(m/z417(C16))から大量の標識したパルミトイルカルニチンが蓄積する。VLCAD-C(図9E)、VLCAD-H(図9F)、トリファンクショナル(図9G)、LCHAD(図9H)、ETE-DH-マイルド(図9K)、およびETE-DH-シビア(図9L)中の標識したアシルカルニチンの異常なプロフィールは、失われた酵素活性の炭素鎖長特異性に対応する蓄積を反映する。MCAD(図9I)においては、酸化は、下方C8(m/z305.3)のレベルまで明らかに進行し、この点で顕著な蓄積があって失われたMCAD酵素活性の基質特異性を反映する。同様に、SCAD(図9J)においては、酸化はm/z249(2H3-ブチルカルニチン-C4)で停止する。これらの結果は、CPT I、トランスロカーゼ、CPT II、VLCAD、トリファンクショナル、LCHAD、ETE-DHは全て、パルミチン酸エステルの完全な酸化に必要であることを示す。D3-C12プロフィール(図8A-L)は、D3-C16プロフィールと同様である。
【0038】
D3-C8(図6A-L)の場合、m/z305(2H3-オクタン酸エステル-C8)の相対的蓄積が起り、完全酸化のためにトランスロカーゼ(図6C)とMCAD(図6I)の両方が明らかに必要であることを示す。主要成分がオクタン酸エステルである市販中鎖トリグリセリド(MCT)はCPT I、トランスロカーゼ、およびCPT IIに依存しないと考えられているが、2H3-オクタン酸-C8に対するこのデータは、MCTが重篤なトランスロカーゼ不全に対する有効な治療でないことを示す。さらに、このデータは、MCTがMCAD不全のための適当な治療でないことを説明する。
【0039】
奇数炭素基質D3-C7(図5A-L図)およびD3-C9(図7A-L)を用いて処置した細胞系についての有利な効果は次に基づく:(1)培地で無標識内因性脂質の酸化からある程度産生されうる診断プロフィールが不在であること;および(2)正常な対照細胞(D3-C7は図5A、またはD3-C9は図7A)に見られるのと比較した、標識した奇数炭素分解最終産物であるm/z235(2H3-プロピオン酸エステル-C3)の相対量。このm/z235(2H3-プロピオン酸エステル-C3)の相対量は、m/z269(2H9-イソ吉草酸-C5)およびm/z237.0(2H5-プロピオン酸エステル-C3)の内部標準のレベルと比較される。D3-C9については、トランスロカーゼ、CPT II、およびLCHAD細胞系ではm/z319(9-2H3-ノナン酸エステル)にて増加が観察された。これらの結果は、ノナン酸エステルの完全分解にトランスロカーゼ、CPT II、およびLCHADが全て必要であることを示す。
【0040】
D3-C7については、正常細胞と、CPT I、トランスロカーゼ、CPT II、VLCAD、トリファンクショナル、LCHAD、およびSCAD異常細胞系(図5A-HおよびJ)とにおいて産生される2H3-プロピオン酸エステル-C3(m/z235)の相対量は、比較し得るかまたは正常細胞に見られる量を超過するので、前駆体の有利な酸化が起ったことを示す。観察された例外の1つは、MCAD不全(図5I)であり、D3-C7は酸化のためにMCADを必要とすると予想され、MCADが不在であるとm/z291(2H3-ヘプタノイルカルニチン-C7)が顕著に増加する。ETF-DHについては、標識した7-2H3-ヘプタン酸エステルの酸化は観察されなかった。これらの結果は、n-ヘプタン酸補充組成物は、MCADおよびETFデヒドロゲナーゼを除いて、以下に挙げる脂肪酸欠損症の治療に使用できるを示す:トランスロカーゼ不全;カルニチンパルミトイルトランスフェラーゼIおよびII不全;L-3-ヒドロキシアシル-CoAデヒドロゲナーゼ(LCHAD)不全;極長鎖アシルCoAデヒドロゲナーゼ(VLCAD)不全、および短鎖アシル-CoAデヒドロゲナーゼ(SCAD)不全。
【実施例2】
【0041】
重篤なトランスロカーゼ不全患者におけるトリヘプタノインの補充のin vivoでの利用
実施例1において確認した重篤な新生児トランスロカーゼ不全をもつ乳幼児の治療に、トリヘプタノイン補充した低脂肪処方を使って成功した。さらに、トリヘプタノイン療法に対する臨床結果と乳幼児の羊膜細胞のin vitro質量分析との間の相関を支持する確証がある。
【0042】
妊娠38週に、実施例1に記載した重篤なトランスロカーゼ不全の試験結果が陽性であった羊膜をもつ乳幼児の分娩を実施した。臍帯血を、全および遊離カルニチンレベルならびに個々のアシルカルニチンのレベルについて、タンデム質量分析により分析した(Yang,ら, 1998.「カルニチンパルミトイルトランスフェラーゼ II(CPT II)不全をもつ患者における4つの新規突然変異の同定(Identification of four novel mutations in patients with carnitine palmitoyltransferase II (CPT II) deficiency)」, Mol Genet Metab 64:229-236)。母性血も分娩時に、これらの上記レベルを検定した。結果は、乳幼児が重篤なトランスロカーゼ不全を患っていることを立証した。
【0043】
分娩後の最初の12時間内に、乳幼児に、経鼻胃管(nasogastric tube)を経由してトリヘプタノインを補充した低脂肪処方を与えた。続いて、トリヘプタノイン補充した処方による給食を、任意の正期産児と同じ頻度で与えた。カルニチン、ビオチン、およびシアノコバラミンの補充は必要でなかった。
【0044】
動脈血ガス(ABG)、電解質、血清尿素窒素(BUN)、クレアチニン、アンモニア、グルコース、血清クレアチンホスホキナーゼ(CPK)、ALT、AST、ヘモグロビン(Hgb)、およびヘマトクリット(Hct)を、標準の新生児集中看護手順に従ってモニターした。アシルカルニチンは、1日2回タンデム質量分析により定量分析した。尿有機酸の定量分析を実施すると共に、尿中に存在するジカルボン酸の量をモニターした。
【0045】
トリヘプタノインを補充する処方の介入は、トランスロカーゼ不全の影響を抑制するのに全体として成功した。乳幼児の入院中、上記の様々な生理学的パラメーターは正常範囲内にある結果が得られた。乳幼児は7-8週齢で退院し、トリヘプタノイン補充処方を用いる食事管理が完全であることを示した。平均処方を与えた乳幼児の1日当たり20-25gの平均重量取得と比較すると、トリヘプタノイン補充処方に基づく継続看護期間中、該乳幼児は1日当たり35gの1日当たり平均重量取得を維持した。4ヶ月半齢において、乳幼児は、トリヘプタノイン補充処方に基づいてよく育ち続けて、カルニチン、ビオチン、またはビタミンB12の補充を必要としなかった。
【図面の簡単な説明】
【0046】
【図1】図1は、長鎖脂肪酸のミトコンドリアβ酸化経路を描く図であり、所要の輸送体および酵素をイタリック字体で記しかつ3つの指摘した膜を二重線で表現している。
【図2】図2は、n-ヘプタン酸のミトコンドリアβ酸化経路を描く図であり、所要の輸送体および酵素をイタリック字体で記しかつ指摘した内側ミトコンドリア膜を二重線で表現している。
【図3】図3Aは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、重篤なトランスロカーゼ不全をわずらう死亡した子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図3Bは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、図3Aに報じた重篤なトランスロカーゼ不全をわずらう死亡した子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。
【図4】図4Aは、D3-C16(2H3-パルミチン酸エステル-C16)を用いて処理した羊膜細胞に対する、タンデム型質量分析プロフィールを描くグラフである。羊膜細胞は、重篤なトランスロカーゼ不全と診断された胎児から取得し、その同胞(sibling)は図3Aおよび3Bに報じた死亡した子供であった。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図4Bは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した羊膜細胞に対する、タンデム型質量分析プロフィールを描くグラフである。羊膜細胞は、図4Aに報じた重篤なトランスロカーゼ不全と診断された胎児から取得し、その同胞は図3Aおよび3Bに報じた死亡した子供であった。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。
【図5】図5Aは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した正常な織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図5A-5Cのプロフィールに対する内部標準は、m/z420.3(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.1(2H9-イソ吉草酸-C5)、およびm/z237.0(2H5-プロピオン酸エステル-C3)に位置し、ここにおいてm/zとは質量:電荷比である。ピークm/z291はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Bは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼI(CPT I)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235.0は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Cは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、トランスロカーゼ不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291.3はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Dは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼII(CPT II)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図5D-5Fのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.1(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z291.1はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Eは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「心臓」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-C)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235.1は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Fは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「低血糖」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-H)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291.4はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235.1は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Gは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、ミトコンドリア三機能タンパク質(トリファンクショナル)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図5G-5Iのプロフィールに対する内部標準は、m/z420.3(2H6-パルミチン酸エステル-C16)、m/z308.1(2H6-オクタン酸エステル-C8)、m/z269.0(2H9-イソ吉草酸-C5)、およびm/z237.2(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z291はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235.1は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Hは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、長鎖L-3-ヒドロキシ-アシルCoAデヒドロゲナーゼ(LCHAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291.1はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235.1は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Iは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、中鎖アシルCoAデヒドロゲナーゼ(MCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291.2はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Jは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、短鎖アシルCoAデヒドロゲナーゼ(SCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図5J-5Lのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.0(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z291.1はD3-C7(7-2H3-ヘプタン酸エステル)を表す。ピークm/z235.1は奇数炭素分解の終点であるD3-C3(3-2H3-プロピオン酸エステル)を表す。図5Kは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-マイルド(ETF-DHマイルド)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291.3はD3-C7(7-2H3-ヘプタン酸エステル)を表す。図5Lは、D3-C7(7-2H3-ヘプタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-シビア(ETF-DHシビア)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z291はD3-C7(7-2H3-ヘプタン酸エステル)を表す。
【図6】図6Aは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した正常な織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図6A-6Cのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.1(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z305.3はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Bは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼI(CPT I)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.0はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Cは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、トランスロカーゼ不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.3はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Dは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼII(CPT II)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図6D-6Fのプロフィールに対する内部標準は、m/z420.3(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.2(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z305.3はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Eは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「心臓」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-C)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.3はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Fは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「低血糖」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-H)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.2はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Gは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、ミトコンドリア三機能タンパク質(トリファンクショナル)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図6G-6Iのプロフィールに対する内部標準は、m/z420.5(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.2(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z305.3はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Hは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、長鎖L-3-ヒドロキシ-アシルCoAデヒドロゲナーゼ(LCHAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Iは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、中鎖アシルCoAデヒドロゲナーゼ(MCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.2はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Jは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、短鎖アシルCoAデヒドロゲナーゼ(SCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図6J-6Lのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.1(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z305.0はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Kは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-マイルド(ETF-DHマイルド)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.2はD3-C8(8-2H3-オクタン酸エステル)を表す。図6Lは、D3-C8(8-2H3-オクタン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-シビア(ETF-DHシビア)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z305.3はD3-C8(8-2H3-オクタン酸エステル)を表す。
【図7】図7Aは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した正常な織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図7A-7Cのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.3(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Bは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼI(CPT I)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Cは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、トランスロカーゼ不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Dは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼII(CPT II)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図7D-7Fのプロフィールに対する内部標準は、m/z420.5(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.3(2H9-イソ吉草酸-C5)、およびm/z237.1(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Eは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「心臓」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-C)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Fは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「低血糖」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-H)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Gは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、ミトコンドリア三機能タンパク質(トリファンクショナル)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図7G-7Iのプロフィールに対する内部標準は、m/z420.5(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.3(2H9-イソ吉草酸-C5)、およびm/z237.2(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Hは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、長鎖L-3-ヒドロキシ-アシルCoAデヒドロゲナーゼ(LCHAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.2はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Iは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、中鎖アシルCoAデヒドロゲナーゼ(MCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.0はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Jは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、短鎖アシルCoAデヒドロゲナーゼ(SCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図7J-7Lのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.3(2H9-イソ吉草酸-C5)、およびm/z237.0(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Kは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-マイルド(ETF-DHマイルド)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。図7Lは、D3-C9(9-2H3-ノナン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-シビア(ETF-DHシビア)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z319.3はD3-C9(9-2H3-ノナン酸エステル)を表す。
【図8】図8Aは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した正常な織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図8A-8Cのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.1(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.0(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z361.3はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Bは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼI(CPT I)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.3はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Cは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、トランスロカーゼ不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.3はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Dは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼII(CPT II)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図8D-8Fのプロフィールに対する内部標準は、m/z420.5(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.0(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z361.3はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Eは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「心臓」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-C)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.2はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Fは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「低血糖」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-H)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.3はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Gは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、ミトコンドリア三機能タンパク質(トリファンクショナル)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図8G-8Iのプロフィールに対する内部標準は、m/z420.5(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.1(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z361.1はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Hは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、長鎖L-3-ヒドロキシ-アシルCoAデヒドロゲナーゼ(LCHAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.4はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Iは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、中鎖アシルCoAデヒドロゲナーゼ(MCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.4はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Jは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、短鎖アシルCoAデヒドロゲナーゼ(SCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図8J-8Lのプロフィールに対する内部標準は、m/z420.2(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z361.4はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Kは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-マイルド(ETF-DHマイルド)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.4はD3-C12(12-2H3-ドデカン酸エステル)を表す。図8Lは、D3-C12(12-2H3-ドデカン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-シビア(ETF-DHシビア)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z361.3はD3-C12(12-2H3-ドデカン酸エステル)を表す。
【図9】図9Aは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した正常な織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図9A-9Cのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.1(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z417.0はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Bは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼI(CPT I)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.6はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Cは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、トランスロカーゼ不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.4はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Dは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、カルニチンパルミトイルトランスフェラーゼII(CPT II)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図9D-9Fのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308.2(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.2(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z417.4はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Eは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「心臓」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-C)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.5はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Fは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、「低血糖」型極長鎖アシルCoAデヒドロゲナーゼ(VLCAD-H)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.5はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Gは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、ミトコンドリア三機能タンパク質(トリファンクショナル)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図9G-9Iのプロフィールに対する内部標準は、m/z420.5(2H6-パルミチン酸エステル-C16)、m/z308.3(2H6-オクタン酸エステル-C8)、m/z269.2(2H9-イソ吉草酸-C5)、およびm/z237.0(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z417.4はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Hは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、長鎖L-3-ヒドロキシ-アシルCoAデヒドロゲナーゼ(LCHAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.4はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Iは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、中鎖アシルCoAデヒドロゲナーゼ(MCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Jは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、短鎖アシルCoAデヒドロゲナーゼ(SCAD)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。図9J-9Lのプロフィールに対する内部標準は、m/z420.4(2H6-パルミチン酸エステル-C16)、m/z308(2H6-オクタン酸エステル-C8)、m/z269.1(2H9-イソ吉草酸-C5)、およびm/z237(2H5-プロピオン酸エステル-C3)に位置する。ピークm/z417はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Kは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-マイルド(ETF-DHマイルド)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.3はD3-C16(16-2H3-パルミチン酸エステル)を表す。図9Lは、D3-C16(16-2H3-パルミチン酸エステル)を用いて処理した織維芽細胞に対する、タンデム型質量分析プロフィールを描くグラフである。織維芽細胞は、電子伝達フラビンタンパク質QOデヒドロゲナーゼ-シビア(ETF-DHシビア)不全をわずらう子供から取得した。試験パラメーターは、99FB(高速原子衝撃)のペアレントおよびMCA取得であった。ピークm/z417.3はD3-C16(16-2H3-パルミチン酸エステル)を表す。形態に従ったマイクロレンズを示す断面図である。
【特許請求の範囲】
【請求項1】
低血糖症を予防するための、炭素数が7である脂肪酸化合物を含んで成る製剤であって、ここで前記の炭素数が7である脂肪酸化合物は、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼIIまたはカルニチン/アシルカルニチントランスロカーゼを必要としないトランスポートメカニズムによりミトコンドリアの内膜を通過する能力と、ミトコンドリアによるβ−酸化を受ける能力とによって特徴付けられるものであり、そして前記の炭素数が7である脂肪酸化合物が、n−ヘプタン酸である、上記製剤。
【請求項2】
心組織のための、炭素数が7である脂肪酸化合物を含んで成る製剤であって、ここで前記の炭素数が7である脂肪酸化合物は、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼIIまたはカルニチン/アシルカルニチントランスロカーゼを必要としないトランスポートメカニズムによりミトコンドリアの内膜を通過する能力と、ミトコンドリアによるβ−酸化を受ける能力とによって特徴付けられるものであり、そして前記の炭素数が7である脂肪酸化合物が、n−ヘプタン酸である、上記製剤。
【請求項3】
未熟児の成長速度を加速するための、炭素数が7である脂肪酸化合物を含んで成る製剤であって、ここで前記の炭素数が7である脂肪酸化合物は、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼIIまたはカルニチン/アシルカルニチントランスロカーゼを必要としないトランスポートメカニズムによりミトコンドリアの内膜を通過する能力と、ミトコンドリアによるβ−酸化を受ける能力とによって特徴付けられるものであり、そして前記の炭素数が7である脂肪酸化合物が、n−ヘプタン酸である、上記製剤。
【請求項4】
迅速なエネルギー源を提供するための、炭素数が7である脂肪酸化合物を含んで成る製剤であって、ここで前記の炭素数が7である脂肪酸化合物が、n−ヘプタン酸である、上記製剤。
【請求項5】
脂肪酸代謝不全である患者のための、炭素数が7である脂肪酸化合物の有効量を含んで成る栄養補助剤であって、ここで前記の炭素数が7である脂肪酸化合物は、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼIIまたはカルニチン/アシルカルニチントランスロカーゼを必要としないトランスポートメカニズムによりミトコンドリアの内膜を通過する能力と、ミトコンドリアによるβ−酸化を受ける能力とによって特徴付けられるものであり、そして前記の炭素数が7である脂肪酸化合物が、n−ヘプタン酸である、上記栄養補助剤。
【請求項6】
前記酵素がカルニチンパルミトイルトランスフェラーゼIである、請求項5記載の栄養補助剤。
【請求項7】
前記酵素がカルニチンパルミトイルトランスフェラーゼIIである、請求項5記載の栄養補助剤。
【請求項8】
前記酵素がカルニチン/アシルカルニチントランスロカーゼである、請求項5記載の栄養補助剤。
【請求項9】
前記酵素が心臓型極長鎖アシルCoAデヒドロゲナーゼである、請求項5記載の栄養補助剤。
【請求項10】
前記酵素が低血糖型極長鎖アシルCoAデヒドロゲナーゼである、請求項5記載の栄養補助剤。
【請求項11】
前記酵素が長鎖L−3−ヒドロキシアシルCoAデヒドロゲナーゼである、請求項5記載の栄養補助剤。
【請求項12】
前記酵素が短鎖アシルCoAデヒドロゲナーゼである、請求項5記載の栄養補助剤。
【請求項1】
低血糖症を予防するための、炭素数が7である脂肪酸化合物を含んで成る製剤であって、ここで前記の炭素数が7である脂肪酸化合物は、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼIIまたはカルニチン/アシルカルニチントランスロカーゼを必要としないトランスポートメカニズムによりミトコンドリアの内膜を通過する能力と、ミトコンドリアによるβ−酸化を受ける能力とによって特徴付けられるものであり、そして前記の炭素数が7である脂肪酸化合物が、n−ヘプタン酸である、上記製剤。
【請求項2】
心組織のための、炭素数が7である脂肪酸化合物を含んで成る製剤であって、ここで前記の炭素数が7である脂肪酸化合物は、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼIIまたはカルニチン/アシルカルニチントランスロカーゼを必要としないトランスポートメカニズムによりミトコンドリアの内膜を通過する能力と、ミトコンドリアによるβ−酸化を受ける能力とによって特徴付けられるものであり、そして前記の炭素数が7である脂肪酸化合物が、n−ヘプタン酸である、上記製剤。
【請求項3】
未熟児の成長速度を加速するための、炭素数が7である脂肪酸化合物を含んで成る製剤であって、ここで前記の炭素数が7である脂肪酸化合物は、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼIIまたはカルニチン/アシルカルニチントランスロカーゼを必要としないトランスポートメカニズムによりミトコンドリアの内膜を通過する能力と、ミトコンドリアによるβ−酸化を受ける能力とによって特徴付けられるものであり、そして前記の炭素数が7である脂肪酸化合物が、n−ヘプタン酸である、上記製剤。
【請求項4】
迅速なエネルギー源を提供するための、炭素数が7である脂肪酸化合物を含んで成る製剤であって、ここで前記の炭素数が7である脂肪酸化合物が、n−ヘプタン酸である、上記製剤。
【請求項5】
脂肪酸代謝不全である患者のための、炭素数が7である脂肪酸化合物の有効量を含んで成る栄養補助剤であって、ここで前記の炭素数が7である脂肪酸化合物は、カルニチンパルミトイルトランスフェラーゼI、カルニチンパルミトイルトランスフェラーゼIIまたはカルニチン/アシルカルニチントランスロカーゼを必要としないトランスポートメカニズムによりミトコンドリアの内膜を通過する能力と、ミトコンドリアによるβ−酸化を受ける能力とによって特徴付けられるものであり、そして前記の炭素数が7である脂肪酸化合物が、n−ヘプタン酸である、上記栄養補助剤。
【請求項6】
前記酵素がカルニチンパルミトイルトランスフェラーゼIである、請求項5記載の栄養補助剤。
【請求項7】
前記酵素がカルニチンパルミトイルトランスフェラーゼIIである、請求項5記載の栄養補助剤。
【請求項8】
前記酵素がカルニチン/アシルカルニチントランスロカーゼである、請求項5記載の栄養補助剤。
【請求項9】
前記酵素が心臓型極長鎖アシルCoAデヒドロゲナーゼである、請求項5記載の栄養補助剤。
【請求項10】
前記酵素が低血糖型極長鎖アシルCoAデヒドロゲナーゼである、請求項5記載の栄養補助剤。
【請求項11】
前記酵素が長鎖L−3−ヒドロキシアシルCoAデヒドロゲナーゼである、請求項5記載の栄養補助剤。
【請求項12】
前記酵素が短鎖アシルCoAデヒドロゲナーゼである、請求項5記載の栄養補助剤。
【図1】


【図2】

【図3A】

【図3B】
【図4A】

【図4B】

【図5A】
【図5B】
【図5C】
【図5D】
【図5E】

【図5F】

【図5G】
【図5H】
【図5I】
【図5J】

【図5K】

【図5L】

【図6A】
【図6B】
【図6C】
【図6D】

【図6E】
【図6F】

【図6G】

【図6H】
【図6I】

【図6J】
【図6K】
【図6L】

【図7A】
【図7B】
【図7C】

【図7D】

【図7E】
【図7F】
【図7G】
【図7H】
【図7I】
【図7J】

【図7K】
【図7L】

【図8A】
【図8B】
【図8C】

【図8D】

【図8E】

【図8F】

【図8G】

【図8H】

【図8I】

【図8J】
【図8K】
【図8L】

【図2】
【図3A】
【図3B】
【図4A】
【図4B】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図5F】
【図5G】
【図5H】
【図5I】
【図5J】
【図5K】
【図5L】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図6F】
【図6G】
【図6H】
【図6I】
【図6J】
【図6K】
【図6L】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図7F】
【図7G】
【図7H】
【図7I】
【図7J】
【図7K】
【図7L】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図8F】
【図8G】
【図8H】
【図8I】
【図8J】
【図8K】
【図8L】
【公開番号】特開2012−82202(P2012−82202A)
【公開日】平成24年4月26日(2012.4.26)
【国際特許分類】
【出願番号】特願2011−249219(P2011−249219)
【出願日】平成23年11月15日(2011.11.15)
【分割の表示】特願2000−596781(P2000−596781)の分割
【原出願日】平成12年2月3日(2000.2.3)
【出願人】(509004712)ベイラー リサーチ インスティテュート (38)
【氏名又は名称原語表記】BAYLOR RESEARCH INSTITUTE
【Fターム(参考)】
【公開日】平成24年4月26日(2012.4.26)
【国際特許分類】
【出願日】平成23年11月15日(2011.11.15)
【分割の表示】特願2000−596781(P2000−596781)の分割
【原出願日】平成12年2月3日(2000.2.3)
【出願人】(509004712)ベイラー リサーチ インスティテュート (38)
【氏名又は名称原語表記】BAYLOR RESEARCH INSTITUTE
【Fターム(参考)】
[ Back to top ]