
炭酸ガス吸収材料
【課題】本発明は、炭酸ガス吸収材料及びそれを用いる炭酸ガスの吸収方法の提供。
【解決手段】2−アルケナールリダクターゼを過剰発現する植物体を含む、炭酸ガス吸収材料。
【解決手段】2−アルケナールリダクターゼを過剰発現する植物体を含む、炭酸ガス吸収材料。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、炭酸ガス吸収材料及びそれを用いる炭酸ガスの吸収方法に関する。
【背景技術】
【0002】
世界の食糧とバイオ燃料の需要は増大しており、バイオマス生産性の向上は農学にとってますます重要な課題になっている。作物生産は乾燥、高・低温、強光、紫外線などの環境ストレスにより大きく阻害されている。環境ストレス耐性が向上した作物の開発が望まれている。
【0003】
また、炭酸ガスの増大による弊害とくに地球温暖化の問題が指摘されている。日本の炭酸ガス 排出量は全世界の5%を占めると言われ、毎年膨大な量の炭酸ガスが大気中に排出されている。したがって、炭酸ガスの効率的な吸収方法の開発が待たれている。
【発明の概要】
【発明が解決しようとする課題】
【0004】
本発明は、炭酸ガス吸収材料及びそれを用いる炭酸ガスの吸収方法を提供する。
【課題を解決するための手段】
【0005】
本発明者らは、鋭意研究の結果、酸化的ストレスに対する耐性が向上した2−アルケナールリダクターゼ(2-alkenal reductase: AER)過剰発現植物が、炭酸ガス吸収能力も向上していることを見いだし、本発明を完成させた。
【0006】
すなわち、本発明は、
[1]2−アルケナールリダクターゼを過剰発現する植物体を含む、炭酸ガス吸収材料。
[2]植物体が、タバコである、上記1に記載の炭酸ガス吸収材料。
[3]2−アルケナールリダクターゼがシロイヌナズナに由来する、上記2に記載の炭酸ガス吸収材料。
[4]植物体が、P1#11、P1#14又はP1#18である、上記3に記載の炭酸ガス吸収材料。
[5]2−アルケナールリダクターゼを過剰発現する植物体を用いることを含む、炭酸ガスの吸収方法。
[6]植物体が、タバコである、上記5に記載の炭酸ガスの吸収方法。
[7]2−アルケナールリダクターゼがシロイヌナズナに由来する、上記6に記載の炭酸ガスの吸収方法。
[8]植物体が、P1#11、P1#14又はP1#18である、上記7に記載の炭酸ガスの吸収方法。
【発明の効果】
【0007】
本発明の植物材料は、環境中の炭酸ガスを効率的に吸収することができる。
【図面の簡単な説明】
【0008】
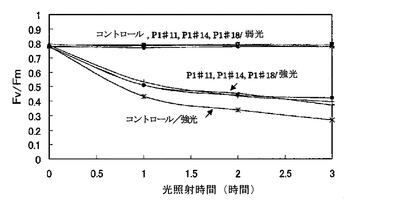
【図1−1】強光および弱光照射による光合成活性の変化である。強光は、2000μmol/m2/sであり、弱光は、10μmol/m2/sである。
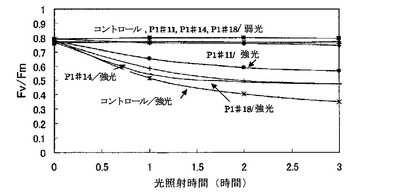
【図1−2】強光および弱光照射による光合成活性の変化である。強光は、1000μmol/m2/sであり、弱光は、10μmol/m2/sである。
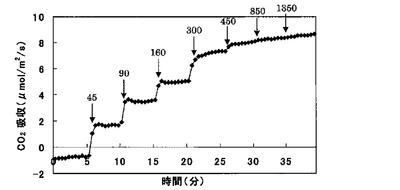
【図2】CO2吸収の時間変化である。
【発明を実施するための形態】
【0009】
本発明の2−アルケナールリダクターゼは、以下の特性を有する。国際生化学分子生物学連合の酵素委員会に、EC1.3.1.74として登録されている。また、NAD(P)H−依存アルケナール/オンオキシドリダクターゼ、NADPH:2−アルケナールα,β−ヒドロゲナーゼとも呼ばれる。触媒する反応は、n−アルカナール+NAD(P)+=アルクー2−エナール+NAD(P)H+H+である。4−ヒドロキシノン−2−エナール及びノン−2−エナールに極めて特異的である。短鎖の2−アルケナールには、親和性が低い。but−3−エン−2−オンやpent−3−エン−2−オンのような2−アルケノンについても高い活性を示す。α,β−不飽和アルデヒド及びケトンの解毒に関与する。
【0010】
シロイヌナズナの2−アルケナールリダクターゼ(AER)をコードする遺伝子は、シロイヌナズナArabidopsis thalianaエコタイプColumbia−0の遺伝子At5g16970である。この遺伝子の完全長cDNA(クローン番号RAFL−05−11−I02)は独立行政法人理化学研究所バイオリソースセンターで購入できる。
【0011】
AER完全長cDNAを制限酵素NotIで切断し、バイナリベクターpGSVE9のXbaI部位に挿入する。このバイナリベクターのT−DNA領域には、挿入遺伝子を駆動するため挿入部位上流にカリフラワーモザイクウイルス35Sプロモーター、および選抜マーカーとなるハイグロマイシン耐性遺伝子(ハイグロマイシンBホスホトランスフェラーゼCDS)とそれを駆動するノパリンシンターゼプロモーターを有する。AER遺伝子を挿入したバイナリベクターをAgrobacterium tumefaciens C51C1Rifr(pGV2260)株に導入し、A. tumefaciens C51C1Rifr(pGV2260;p35SP1S)株を得る。タバコNicotiana tabacum Petit Havana SR1株から得たリーフディスクとA. tumefaciens C51C1Rifr(pGV2260;p35SP1S)とを共培養し、T−DNAが導入されたタバコカルスを得る。このカルスから再分化させた個体のRNAゲルブロットハイブリダイーションによりAER遺伝子の発現レベルの高い個体を選抜する。自家受粉により得られた第二世代から種子を採取し、一部の種子をハイグロマイシンを含む無菌培地で発芽させる。種子がすべてハイグロマイシン耐性を示した第二世代はホモ接合体であり、P1#11、P1#14およびP1#18の3系統が得られた。これらの種子を採取し、使用する。
【0012】
植物体P1#18は、平成21年2月2日付けで、独立行政法人 産業技術総合研究所 特許生物寄託センターに寄託されており、その受託番号は、FERM P−21762である。
【実施例】
【0013】
シロイヌナズナの2−アルケナールリダクターゼ(AER)は、2−アルケナールのα,β不飽和結合を還元する。AER過剰発現タバコは、細胞質でAERを過剰発現しており、この植物はコントロール株に比べて、酸化的ストレス(強光ストレス、MVストレス、オゾンストレス、UV−Bストレス)の耐性が向上することが分かっている)。
【0014】
また、AER過剰発現タバコは、強光照射による光化学系II(PSII)の失活が軽微であることから、PSIIの失活または強光による細胞死に、2−アルケナールが関与していると考えられる。
【0015】
1.強光照射によるAER過剰発現タバコ株のPSII活性の変化
AER過剰発現タバコ
カリフラワーモザイクウイルスの強力な構成的プロモーター35sの制御下にあるバイナリーベクターpGSC1705のNot1−Not1フラグメントにシロイヌナズナに由来するAERのcDNAを導入した。このベクターは遺伝子マーカーとしてヒグロマイシンBホスホトランスフェラーゼ遺伝子を持つ。A.tumefaciens C51C1 RifR〔pGV2260〕とNicotiana tabacum cv. Petit Havana SR1の葉を用いた標準的なアグロバクテリウム−リーフディスクトランスフォーメーションを行った。トランスジェニック植物は30mg/LヒグロマイシンBを含むMSプレートにおけるカルスの形成、生育能力によって選別した。選別後、植物を土に移し生育させた。
【0016】
<材料>
タバコの植物体として、コントロール(ベクターのみ)、P1#11、P1#14、P1#18を用いた。
週齢は、13週齢であり、明期12時間及び暗期12時間の条件下で生育させた。
【0017】
タバコの栽培条件
バットにバーミキュライトを平らに敷き詰め、端から静かに水を注ぎ、湿った土に均一にタバコを播種した。上からラップをして光(100−150μmol/m2/s)の下で育て、播種後4週目に500倍希釈したハイポネックス(登録商標)を染み込ませた土を入れたプラスチックポットに植え替えた。
【0018】
<方法>
タバコの植物体に葉がついたままの状態で暗黒下に5分おき、葉の一部をMINI−PAMでPSHの量子収率(Fv/Fm)を測定し、その後、1時間、2000μmol/m2/sの光を照射した。その後暗黒下に5分間置き、MINI−PAMで光照射の中心部と弱光(10μmol/m2/s)照射部について、それぞれFv/Fmを測定し、再び1時間2000μmol/m2/sの光を照射した。これを繰り返し、3時間後までFv/Fmを測定した(図1−1)。また、強光を1000μmol/m2/sに変えて同様の実験を行った(図1−2)。
【0019】
2000μmol/m2/sの強光照射後のタバコ葉におけるPSH活性の時間変化は、いずれの株でも弱光(10μmol/m2/s)照射部の活性は低下しなかった。強光照射部では、コントロール株で最も活性が低下し、3時間までは急な低下は見られず、同じ割合で低下が見られた。コントロール株に比べるとAER過剰発現タバコのPSH活性の低下が軽微だった。また、1000μmol/m2/sの強光照射後のタバコ葉におけるPSII活性の時間変化は、弱光(10μmol/m2/s)照射部の活性は同様にいずれの株でも低下しなかった。強光照射部では、コントロール、P1#14、P1#18は、2000μmol/m2/sの強光照射とほぼ同様の結果を示したが、光が弱かったために全体的に低下は緩やかで、特に1時間後のPSII活性ではあまり差が見られなかった。
【0020】
またP1#11は、特に耐性が強くPSII活性の低下があまり見られなかった。今回の実験から、1000μmol/m2/sの強光照射でもコントロールとAER過剰発現タ
バコのPSl活性に差が出ることが分かったが、光が弱いために照射1時間の葉ではあまり差が見られない可能性が高いので2000μmol/m2/sの強光ストレスで実験を行った。
【0021】
光合成は植物やシアノバクテリアの持つ代謝系で、光エネルギーで水を光分解し酸素を発生する過程で作られるATPとNADPHを利用して、二酸化炭素をグルコースなどの有機化合物に固定する。光化学反応とカルビン回路に分けられる光合成過程のうち炭酸固定反応を持つのはカルビン回路である。光合成の炭酸固定の律速の原因となっているのは、リブロース−1,5−ビスホスフェートカルボキシラーゼオキシゲナーゼ(Rubisco)である。この酵素反応の効率の悪さ・およびこの酵素によるオキシゲナーゼ反応から始まる光呼吸(二酸化炭素の放出)が炭酸固定の効率を低下させる。光呼吸とは、Rubiscoのオキシゲナーゼ作用で生成するホスホグリコール酸の代謝によるグリコール酸回路でのO2吸収とCO2発生の現象でRubisco周辺にCO2が不足する条件、つまり強光・低CO2・高O2・高温下で顕著となる。CO2が不足したままではカルビン回路が回らず、光化学系で生じた還元力、プロトン勾配、ATPが消費されず、この条件下では光エネルギーは活性酸素を生成し光合成装置を破壊することになるので、O2を消費しCO2を発生する光呼吸は活性酸素の過剰な発生を抑制し、光阻害を回避する機能を持つといわれている。
【0022】
AER過剰発現タバコ株の強光耐性の仕組みを調べるために、PSIIの量子収率を測定した結果から、光の強度が強くなるほどコントロール株に比べてAER過剰発現株の電子伝達速度は大きくなった。相物に光が当たると、光エネルギーはPSIIで、熱あるいは電子伝達に変換される。前の実験で測定した電子伝達速度とはPSIIからPSIに流れる電子、つまり電子を受容するカルビン回路と光呼吸を足したものである。光呼吸は、Rubisco周辺にCO2が不足する条件で顕著となるので、最大電子伝達速度がコントロール株に比べてAER過剰発現株のほうが高いという結果が炭酸固定と光呼吸どちらかによるものかを区別するためにCO2濃度を変えて炭酸固定速度の測定を行った。
【0023】
CO2濃度350ppmは、ほぼ大気の濃度に等しく、炭酸固定のCO2発生と光呼吸のCO2吸収が合わさって実際よりも低い炭酸固定のCO2吸収速度が測定される。またCO2濃度2000ppmでは、光呼吸が抑制されるので炭酸固定のCO2吸収速度のみが測定される。
【0024】
本実験の目的は、AER過剰発現株において、強光での電子伝達活性が高いのは、カルビン回路での炭酸固定能力または光呼吸活性がコントロール株に比べて高いのではないかという仮説を実証するために、ストレスを与える前のAER過剰発現株とコントロール株で2つのCO2濃度条件で、炭酸固定で消費されるCO2吸収速度を比較した。
【0025】
〈材料〉
タバコの植物体(コントロール、P1#11、P1#18)
10週齢・光強度100μmol/m2/s・明期16時間、暗期8時間
炭酸固定の測定装置(GFS−3000ガス交換蛍光システム)
タバコの植物体(コントロール、P1#14)
8、9週齢・光強度100μmol/m2/s・明期16時間、暗期8時間
炭酸固定の測定装置(HCM−1000ガス交換蛍光システム)
【0026】
〈方法〉
タバコの葉の気孔を開かせるために、20%O2、1000ppmCO2、光強度560μmol/m2/sで30分置いた。350ppmCO2(大気の濃度にほぼ等しい)に下げ、光強度0にして値が安定するまで5分待ち、光強度0の値を5分間測定し、光強度を45、90、160、300、450、850、1350μmol/m2/sで5分ごとに上げ、CO2吸収量を測定した。これと同時に葉のFv/Fm値、ΔF/Fm’を計測した。同様の実験を、測定する時のCO2濃度を350ppmから2000ppmに変えて行った。
コントロール、P1#11、P1#18は、GFS−3000ガス交換蛍光システムで測定し、コントロール、P1#14は、HCM−1000ガス交換蛍光システムで測定した。
【0027】
その結果、図2に示すように、光強度を上げるとCO2吸収量は増加した。これは、すべての株でほぼ同様の結果が得られた。
【0028】
全ての株で最大の炭酸固定速度までは、光に依存して吸収速度は高くなり、最大に達すると一定になった。350ppmでは、AER過裂発現株のほうが光飽和時に高いCO2吸収速度を示した。2000ppmでは光呼吸のCO2発生分だけコントロール株でもAER過剰発現株でもCO2吸収速度が高くなった。しかし、その割合をみてみるとAER過剰発現株のほうが光飽和時に1.4〜2.2倍高いCO2吸収速度を示した。
【0029】
CO2吸収に対応する気孔伝導度(気孔の開閉度)の差は見られなかった。
【0030】
葉内CO2濃度を表すCiにも差がなかった。また、CO2吸収に対応する気孔伝導度に差がないことと、Rubiscoの周りのCO2濃度にほぼ等しいCiに差がないことから、葉緑体に対するCO2供給は同じであることが分かった。したがって、炭酸固定と光呼吸で観測されたコントロール株とAER過剰発現株の差は、葉内CO2濃度の差によるものではなく、カルビン回路、光呼吸におけるRubisco活性に差があると考えられた。
【0031】
コントロール株に比べて、AER過剰発現株では、PSIIの量子収率の低下が少なかった。PSIIの量子収率の低下はカルビン回路の上限を越えて電子を流すことで、PSIでのスーパーオキシド生成や、PSII反応中心での一重項酸素(1O2)生成を促進し、これらの活性酸素によって周囲の分子が損傷を受けることを防ぐための制御であると考えられている。つまりカルビン回路の上限がコントロール株に比べてAER過剰発現株のほうが高いと考えられる。
【産業上の利用可能性】
【0032】
本発明の植物材料は、環境中の炭酸ガス吸収のために有用である。
【技術分野】
【0001】
本発明は、炭酸ガス吸収材料及びそれを用いる炭酸ガスの吸収方法に関する。
【背景技術】
【0002】
世界の食糧とバイオ燃料の需要は増大しており、バイオマス生産性の向上は農学にとってますます重要な課題になっている。作物生産は乾燥、高・低温、強光、紫外線などの環境ストレスにより大きく阻害されている。環境ストレス耐性が向上した作物の開発が望まれている。
【0003】
また、炭酸ガスの増大による弊害とくに地球温暖化の問題が指摘されている。日本の炭酸ガス 排出量は全世界の5%を占めると言われ、毎年膨大な量の炭酸ガスが大気中に排出されている。したがって、炭酸ガスの効率的な吸収方法の開発が待たれている。
【発明の概要】
【発明が解決しようとする課題】
【0004】
本発明は、炭酸ガス吸収材料及びそれを用いる炭酸ガスの吸収方法を提供する。
【課題を解決するための手段】
【0005】
本発明者らは、鋭意研究の結果、酸化的ストレスに対する耐性が向上した2−アルケナールリダクターゼ(2-alkenal reductase: AER)過剰発現植物が、炭酸ガス吸収能力も向上していることを見いだし、本発明を完成させた。
【0006】
すなわち、本発明は、
[1]2−アルケナールリダクターゼを過剰発現する植物体を含む、炭酸ガス吸収材料。
[2]植物体が、タバコである、上記1に記載の炭酸ガス吸収材料。
[3]2−アルケナールリダクターゼがシロイヌナズナに由来する、上記2に記載の炭酸ガス吸収材料。
[4]植物体が、P1#11、P1#14又はP1#18である、上記3に記載の炭酸ガス吸収材料。
[5]2−アルケナールリダクターゼを過剰発現する植物体を用いることを含む、炭酸ガスの吸収方法。
[6]植物体が、タバコである、上記5に記載の炭酸ガスの吸収方法。
[7]2−アルケナールリダクターゼがシロイヌナズナに由来する、上記6に記載の炭酸ガスの吸収方法。
[8]植物体が、P1#11、P1#14又はP1#18である、上記7に記載の炭酸ガスの吸収方法。
【発明の効果】
【0007】
本発明の植物材料は、環境中の炭酸ガスを効率的に吸収することができる。
【図面の簡単な説明】
【0008】
【図1−1】強光および弱光照射による光合成活性の変化である。強光は、2000μmol/m2/sであり、弱光は、10μmol/m2/sである。
【図1−2】強光および弱光照射による光合成活性の変化である。強光は、1000μmol/m2/sであり、弱光は、10μmol/m2/sである。
【図2】CO2吸収の時間変化である。
【発明を実施するための形態】
【0009】
本発明の2−アルケナールリダクターゼは、以下の特性を有する。国際生化学分子生物学連合の酵素委員会に、EC1.3.1.74として登録されている。また、NAD(P)H−依存アルケナール/オンオキシドリダクターゼ、NADPH:2−アルケナールα,β−ヒドロゲナーゼとも呼ばれる。触媒する反応は、n−アルカナール+NAD(P)+=アルクー2−エナール+NAD(P)H+H+である。4−ヒドロキシノン−2−エナール及びノン−2−エナールに極めて特異的である。短鎖の2−アルケナールには、親和性が低い。but−3−エン−2−オンやpent−3−エン−2−オンのような2−アルケノンについても高い活性を示す。α,β−不飽和アルデヒド及びケトンの解毒に関与する。
【0010】
シロイヌナズナの2−アルケナールリダクターゼ(AER)をコードする遺伝子は、シロイヌナズナArabidopsis thalianaエコタイプColumbia−0の遺伝子At5g16970である。この遺伝子の完全長cDNA(クローン番号RAFL−05−11−I02)は独立行政法人理化学研究所バイオリソースセンターで購入できる。
【0011】
AER完全長cDNAを制限酵素NotIで切断し、バイナリベクターpGSVE9のXbaI部位に挿入する。このバイナリベクターのT−DNA領域には、挿入遺伝子を駆動するため挿入部位上流にカリフラワーモザイクウイルス35Sプロモーター、および選抜マーカーとなるハイグロマイシン耐性遺伝子(ハイグロマイシンBホスホトランスフェラーゼCDS)とそれを駆動するノパリンシンターゼプロモーターを有する。AER遺伝子を挿入したバイナリベクターをAgrobacterium tumefaciens C51C1Rifr(pGV2260)株に導入し、A. tumefaciens C51C1Rifr(pGV2260;p35SP1S)株を得る。タバコNicotiana tabacum Petit Havana SR1株から得たリーフディスクとA. tumefaciens C51C1Rifr(pGV2260;p35SP1S)とを共培養し、T−DNAが導入されたタバコカルスを得る。このカルスから再分化させた個体のRNAゲルブロットハイブリダイーションによりAER遺伝子の発現レベルの高い個体を選抜する。自家受粉により得られた第二世代から種子を採取し、一部の種子をハイグロマイシンを含む無菌培地で発芽させる。種子がすべてハイグロマイシン耐性を示した第二世代はホモ接合体であり、P1#11、P1#14およびP1#18の3系統が得られた。これらの種子を採取し、使用する。
【0012】
植物体P1#18は、平成21年2月2日付けで、独立行政法人 産業技術総合研究所 特許生物寄託センターに寄託されており、その受託番号は、FERM P−21762である。
【実施例】
【0013】
シロイヌナズナの2−アルケナールリダクターゼ(AER)は、2−アルケナールのα,β不飽和結合を還元する。AER過剰発現タバコは、細胞質でAERを過剰発現しており、この植物はコントロール株に比べて、酸化的ストレス(強光ストレス、MVストレス、オゾンストレス、UV−Bストレス)の耐性が向上することが分かっている)。
【0014】
また、AER過剰発現タバコは、強光照射による光化学系II(PSII)の失活が軽微であることから、PSIIの失活または強光による細胞死に、2−アルケナールが関与していると考えられる。
【0015】
1.強光照射によるAER過剰発現タバコ株のPSII活性の変化
AER過剰発現タバコ
カリフラワーモザイクウイルスの強力な構成的プロモーター35sの制御下にあるバイナリーベクターpGSC1705のNot1−Not1フラグメントにシロイヌナズナに由来するAERのcDNAを導入した。このベクターは遺伝子マーカーとしてヒグロマイシンBホスホトランスフェラーゼ遺伝子を持つ。A.tumefaciens C51C1 RifR〔pGV2260〕とNicotiana tabacum cv. Petit Havana SR1の葉を用いた標準的なアグロバクテリウム−リーフディスクトランスフォーメーションを行った。トランスジェニック植物は30mg/LヒグロマイシンBを含むMSプレートにおけるカルスの形成、生育能力によって選別した。選別後、植物を土に移し生育させた。
【0016】
<材料>
タバコの植物体として、コントロール(ベクターのみ)、P1#11、P1#14、P1#18を用いた。
週齢は、13週齢であり、明期12時間及び暗期12時間の条件下で生育させた。
【0017】
タバコの栽培条件
バットにバーミキュライトを平らに敷き詰め、端から静かに水を注ぎ、湿った土に均一にタバコを播種した。上からラップをして光(100−150μmol/m2/s)の下で育て、播種後4週目に500倍希釈したハイポネックス(登録商標)を染み込ませた土を入れたプラスチックポットに植え替えた。
【0018】
<方法>
タバコの植物体に葉がついたままの状態で暗黒下に5分おき、葉の一部をMINI−PAMでPSHの量子収率(Fv/Fm)を測定し、その後、1時間、2000μmol/m2/sの光を照射した。その後暗黒下に5分間置き、MINI−PAMで光照射の中心部と弱光(10μmol/m2/s)照射部について、それぞれFv/Fmを測定し、再び1時間2000μmol/m2/sの光を照射した。これを繰り返し、3時間後までFv/Fmを測定した(図1−1)。また、強光を1000μmol/m2/sに変えて同様の実験を行った(図1−2)。
【0019】
2000μmol/m2/sの強光照射後のタバコ葉におけるPSH活性の時間変化は、いずれの株でも弱光(10μmol/m2/s)照射部の活性は低下しなかった。強光照射部では、コントロール株で最も活性が低下し、3時間までは急な低下は見られず、同じ割合で低下が見られた。コントロール株に比べるとAER過剰発現タバコのPSH活性の低下が軽微だった。また、1000μmol/m2/sの強光照射後のタバコ葉におけるPSII活性の時間変化は、弱光(10μmol/m2/s)照射部の活性は同様にいずれの株でも低下しなかった。強光照射部では、コントロール、P1#14、P1#18は、2000μmol/m2/sの強光照射とほぼ同様の結果を示したが、光が弱かったために全体的に低下は緩やかで、特に1時間後のPSII活性ではあまり差が見られなかった。
【0020】
またP1#11は、特に耐性が強くPSII活性の低下があまり見られなかった。今回の実験から、1000μmol/m2/sの強光照射でもコントロールとAER過剰発現タ
バコのPSl活性に差が出ることが分かったが、光が弱いために照射1時間の葉ではあまり差が見られない可能性が高いので2000μmol/m2/sの強光ストレスで実験を行った。
【0021】
光合成は植物やシアノバクテリアの持つ代謝系で、光エネルギーで水を光分解し酸素を発生する過程で作られるATPとNADPHを利用して、二酸化炭素をグルコースなどの有機化合物に固定する。光化学反応とカルビン回路に分けられる光合成過程のうち炭酸固定反応を持つのはカルビン回路である。光合成の炭酸固定の律速の原因となっているのは、リブロース−1,5−ビスホスフェートカルボキシラーゼオキシゲナーゼ(Rubisco)である。この酵素反応の効率の悪さ・およびこの酵素によるオキシゲナーゼ反応から始まる光呼吸(二酸化炭素の放出)が炭酸固定の効率を低下させる。光呼吸とは、Rubiscoのオキシゲナーゼ作用で生成するホスホグリコール酸の代謝によるグリコール酸回路でのO2吸収とCO2発生の現象でRubisco周辺にCO2が不足する条件、つまり強光・低CO2・高O2・高温下で顕著となる。CO2が不足したままではカルビン回路が回らず、光化学系で生じた還元力、プロトン勾配、ATPが消費されず、この条件下では光エネルギーは活性酸素を生成し光合成装置を破壊することになるので、O2を消費しCO2を発生する光呼吸は活性酸素の過剰な発生を抑制し、光阻害を回避する機能を持つといわれている。
【0022】
AER過剰発現タバコ株の強光耐性の仕組みを調べるために、PSIIの量子収率を測定した結果から、光の強度が強くなるほどコントロール株に比べてAER過剰発現株の電子伝達速度は大きくなった。相物に光が当たると、光エネルギーはPSIIで、熱あるいは電子伝達に変換される。前の実験で測定した電子伝達速度とはPSIIからPSIに流れる電子、つまり電子を受容するカルビン回路と光呼吸を足したものである。光呼吸は、Rubisco周辺にCO2が不足する条件で顕著となるので、最大電子伝達速度がコントロール株に比べてAER過剰発現株のほうが高いという結果が炭酸固定と光呼吸どちらかによるものかを区別するためにCO2濃度を変えて炭酸固定速度の測定を行った。
【0023】
CO2濃度350ppmは、ほぼ大気の濃度に等しく、炭酸固定のCO2発生と光呼吸のCO2吸収が合わさって実際よりも低い炭酸固定のCO2吸収速度が測定される。またCO2濃度2000ppmでは、光呼吸が抑制されるので炭酸固定のCO2吸収速度のみが測定される。
【0024】
本実験の目的は、AER過剰発現株において、強光での電子伝達活性が高いのは、カルビン回路での炭酸固定能力または光呼吸活性がコントロール株に比べて高いのではないかという仮説を実証するために、ストレスを与える前のAER過剰発現株とコントロール株で2つのCO2濃度条件で、炭酸固定で消費されるCO2吸収速度を比較した。
【0025】
〈材料〉
タバコの植物体(コントロール、P1#11、P1#18)
10週齢・光強度100μmol/m2/s・明期16時間、暗期8時間
炭酸固定の測定装置(GFS−3000ガス交換蛍光システム)
タバコの植物体(コントロール、P1#14)
8、9週齢・光強度100μmol/m2/s・明期16時間、暗期8時間
炭酸固定の測定装置(HCM−1000ガス交換蛍光システム)
【0026】
〈方法〉
タバコの葉の気孔を開かせるために、20%O2、1000ppmCO2、光強度560μmol/m2/sで30分置いた。350ppmCO2(大気の濃度にほぼ等しい)に下げ、光強度0にして値が安定するまで5分待ち、光強度0の値を5分間測定し、光強度を45、90、160、300、450、850、1350μmol/m2/sで5分ごとに上げ、CO2吸収量を測定した。これと同時に葉のFv/Fm値、ΔF/Fm’を計測した。同様の実験を、測定する時のCO2濃度を350ppmから2000ppmに変えて行った。
コントロール、P1#11、P1#18は、GFS−3000ガス交換蛍光システムで測定し、コントロール、P1#14は、HCM−1000ガス交換蛍光システムで測定した。
【0027】
その結果、図2に示すように、光強度を上げるとCO2吸収量は増加した。これは、すべての株でほぼ同様の結果が得られた。
【0028】
全ての株で最大の炭酸固定速度までは、光に依存して吸収速度は高くなり、最大に達すると一定になった。350ppmでは、AER過裂発現株のほうが光飽和時に高いCO2吸収速度を示した。2000ppmでは光呼吸のCO2発生分だけコントロール株でもAER過剰発現株でもCO2吸収速度が高くなった。しかし、その割合をみてみるとAER過剰発現株のほうが光飽和時に1.4〜2.2倍高いCO2吸収速度を示した。
【0029】
CO2吸収に対応する気孔伝導度(気孔の開閉度)の差は見られなかった。
【0030】
葉内CO2濃度を表すCiにも差がなかった。また、CO2吸収に対応する気孔伝導度に差がないことと、Rubiscoの周りのCO2濃度にほぼ等しいCiに差がないことから、葉緑体に対するCO2供給は同じであることが分かった。したがって、炭酸固定と光呼吸で観測されたコントロール株とAER過剰発現株の差は、葉内CO2濃度の差によるものではなく、カルビン回路、光呼吸におけるRubisco活性に差があると考えられた。
【0031】
コントロール株に比べて、AER過剰発現株では、PSIIの量子収率の低下が少なかった。PSIIの量子収率の低下はカルビン回路の上限を越えて電子を流すことで、PSIでのスーパーオキシド生成や、PSII反応中心での一重項酸素(1O2)生成を促進し、これらの活性酸素によって周囲の分子が損傷を受けることを防ぐための制御であると考えられている。つまりカルビン回路の上限がコントロール株に比べてAER過剰発現株のほうが高いと考えられる。
【産業上の利用可能性】
【0032】
本発明の植物材料は、環境中の炭酸ガス吸収のために有用である。
【特許請求の範囲】
【請求項1】
2−アルケナールリダクターゼを過剰発現する植物体を含む、炭酸ガス吸収材料。
【請求項2】
植物体が、タバコである、請求項1に記載の炭酸ガス吸収材料。
【請求項3】
2−アルケナールリダクターゼがシロイヌナズナに由来する、請求項2に記載の炭酸ガス吸収材料。
【請求項4】
植物体が、P1#11、P1#14又はP1#18である、請求項3に記載の炭酸ガス吸収材料。
【請求項5】
2−アルケナールリダクターゼを過剰発現する植物体を用いることを含む、炭酸ガスの吸収方法。
【請求項6】
植物体が、タバコである、請求項5に記載の炭酸ガスの吸収方法。
【請求項7】
2−アルケナールリダクターゼがシロイヌナズナに由来する、請求項6に記載の炭酸ガスの吸収方法。
【請求項8】
植物体が、P1#11、P1#14又はP1#18である、請求項7に記載の炭酸ガスの吸収方法。
【請求項1】
2−アルケナールリダクターゼを過剰発現する植物体を含む、炭酸ガス吸収材料。
【請求項2】
植物体が、タバコである、請求項1に記載の炭酸ガス吸収材料。
【請求項3】
2−アルケナールリダクターゼがシロイヌナズナに由来する、請求項2に記載の炭酸ガス吸収材料。
【請求項4】
植物体が、P1#11、P1#14又はP1#18である、請求項3に記載の炭酸ガス吸収材料。
【請求項5】
2−アルケナールリダクターゼを過剰発現する植物体を用いることを含む、炭酸ガスの吸収方法。
【請求項6】
植物体が、タバコである、請求項5に記載の炭酸ガスの吸収方法。
【請求項7】
2−アルケナールリダクターゼがシロイヌナズナに由来する、請求項6に記載の炭酸ガスの吸収方法。
【請求項8】
植物体が、P1#11、P1#14又はP1#18である、請求項7に記載の炭酸ガスの吸収方法。
【図1−1】


【図1−2】
【図2】
【図1−2】
【図2】
【公開番号】特開2010−239871(P2010−239871A)
【公開日】平成22年10月28日(2010.10.28)
【国際特許分類】
【出願番号】特願2009−89336(P2009−89336)
【出願日】平成21年4月1日(2009.4.1)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2009年3月16日 第50回日本植物生理学会年会委員会発行の「第50回日本植物生理学会年会要旨集」に発表 2009年3月16日 http://www.jspp.org/13member/2009suppl/Suppl_50.htmlを通じて発表 を通じて発表
【出願人】(304020177)国立大学法人山口大学 (579)
【出願人】(000000206)宇部興産株式会社 (2,022)
【Fターム(参考)】
【公開日】平成22年10月28日(2010.10.28)
【国際特許分類】
【出願日】平成21年4月1日(2009.4.1)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2009年3月16日 第50回日本植物生理学会年会委員会発行の「第50回日本植物生理学会年会要旨集」に発表 2009年3月16日 http://www.jspp.org/13member/2009suppl/Suppl_50.htmlを通じて発表 を通じて発表
【出願人】(304020177)国立大学法人山口大学 (579)
【出願人】(000000206)宇部興産株式会社 (2,022)
【Fターム(参考)】
[ Back to top ]