
無差別HER−2/NeuCD4T細胞エピトープ
本発明は異なるHLA−DR対立遺伝子少なくとも25個に対して無差別(promiscuous)であるヒトHER−2/Neu蛋白の新規なT細胞エピトープの発見に関する。本発明は又新規なエピトープの1つを含有する組成物、又はそのようなエピトープと非相同ポリペプチドの融合ペプチドに関する。更に、エピトープ又はその融合ペプチドの使用、及び、エピトープ又はその融合ペプチドを含有する組成物を本明細書において開示する。これらのエピトープの無差別性は、これらのエピトープをHLA型に関係なくHER−2/Neu特異的免疫応答を評価するための価値ある手段としている。
【発明の詳細な説明】
【技術分野】
【0001】
関連する出願
本願は、2007年8月11日に出願された米国特許出願第60/837,209号に対する優先権を主張するものであり、米国特許出願第60/837,209号の内容はその全体が本明細書中に参考として援用される。
【背景技術】
【0002】
HLAクラスII制限CD4+ T細胞は細胞性免疫において重要な役割を果たしており、そして抗腫瘍免疫応答の重要な成分である。CD4+ T細胞は腫瘍特異的CTLに対して必要な補助を提供し(非特許文献1)、そしてサイトカイン、例えばインターフェロンガンマ(IFNγ)を生産し、これは抗原提示細胞を活性化し、そして他の免疫学的作用を媒介することができる(非特許文献2)。数種の系における実験の結果は、CD4+ T細胞は有効な抗腫瘍免疫応答のためには必須であることを示している。強い免疫応答を発生させる場合にCD4+ T細胞が重要であることを鑑みれば、最適に設計された癌の免疫療法又は抗腫瘍ワクチンは最大の薬効のために腫瘍特異的CD4+ T細胞及びCD8+ T細胞の両方を誘導するはずである。
【0003】
癌の免疫療法及びワクチンの設計は腫瘍関連抗原の発見により大きく利益を得ている。1つのそのような抗原、HER−2/Neuは、それが乳癌及び卵巣癌を包含する種々の癌において増幅されることから、上記方策のための主要な標的となっている。HER−2/Neuオンコジーンは表皮成長因子受容体に対する相同性を有する膜貫通糖蛋白をコードしている(非特許文献3)。HER−2/Neuの過剰発現は乳腺癌の約30%で起こり、そして進行性疾患及び不良な予後を伴う。その結果、HER−2/Neu蛋白のT細胞認識を増大させるように設計された数種の免疫学的方策が臨床治験において試験されている。そのような試験において生じたHER−2/Neu特異的T細胞応答を特性化することは、HER−2/Neu蛋白内部の数種のHLAクラスI及びクラスII限定T細胞エピトープの発見をもたらしている(非特許文献4)。これらのエピトープの発見は、HER−2/Neu特異的T細胞応答を検出及び定量する本発明者等の能力を増強した。そのような情報は有効な免疫療法のための向上した設計をもたらし、HER−2/Neu発現腫瘍を根絶する際のHER−2/Neu特異的T細胞の役割のより良好な理解を可能にする。
【0004】
所定のT細胞エピトープの有用性はそのHLA制限により限定される。ペプチドエピトープは典型的にはHLA対立遺伝子の少数と生産的ペプチド−MHC複合体を形成し、これらの対立遺伝子を発現する個体においてのみT細胞応答を刺激する。このことは免疫学的試験及び臨床治験を特定のHLA型の個体に限定するものであり、それは一般的集団の20%未満である場合が多い。HLA対立遺伝子の多数により提示され得る、いわゆる無差別(promiscuous)T細胞エピトープが数種の腫瘍抗原に関して報告されている。無差別T細胞エピトープは複数のHLA対立遺伝子に結合して抗原特異的T細胞を刺激することができ、これにより異なるHLA型の個体におけるT細胞応答の誘導及び試験を可能にする。更に又、無差別エピトープは、これらのエピトープに基づいた免疫療法及びワクチンが癌の治療及び予防のために一般的集団に対して広範に適用できるため、価値あるものである。即ち、腫瘍抗原の以前は未知であった無差別エピトープに関連する新しい情報が必要とされていることは明らかである。
【0005】
本発明者等はHER−2/Neu蛋白配列における一連の新規無差別T細胞エピトープを発見した。HER−2/Neu蛋白の270〜284又は268〜286の領域内に位置するこれらのエピトープはHER−2/Neu過剰発現癌腫に対して自己活性細胞免疫療法を用いて治療された患者から作成されたCD4+ T細胞クローンにより認識される(非特許文献5)。T細胞クローンは少なくとも25種の異なるHLA−DRB1*対立遺伝子の関連において提示されるこれらのペプチドエピトープを認識する。抗体ブロッキング実験によって、認識はHLA−DR限定であることが確認される。更に又、これらのエピトープは外因性の蛋白抗原からも天然にプロセシングされて提示される。異なるHLA−DRB1*対立遺伝子に対するこれらのエピトープの無差別性は、これらのエピトープをHLA型に関係なくHER−2/Neu特異的免疫応答を評価するための価値ある手段としている。更に又、これらのエピトープはHER−2/Neu+癌の治療のためのペプチド系ワクチン又は免疫療法においてユニバーサルなCD4Tヘルパー細胞エピトープとして使用できる。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Topalian,1994.Curr Opin Immunol 6:741−745
【非特許文献2】Corthay等、2005.Immunity 22:371−383
【非特許文献3】Coussens等、1985、Science230:1132−1139
【非特許文献4】Sotiriadou等、2001、Br J Cancer85:1527−1534
【非特許文献5】Valone等、2001、Cancer J7 Suppl 2:S53−61
【発明の概要】
【課題を解決するための手段】
【0007】
本発明はHER−2/Neu特異的T細胞応答を誘導するための多くの異なる種類のHLA対立遺伝子の抗原提示細胞により提示することができる新しいHER−2/Neuエピトープについて記載する。第1の態様において、本発明はHER−2/Neu蛋白配列のセグメントから誘導された単離されたペプチドを提供する(残基268〜286、即ち配列番号1)。ペプチドは(a)配列番号1の残基3〜17の12〜15隣接アミノ酸;又は(b)配列番号1の16〜18隣接アミノ酸よりなる。同様に非相同ポリペプチドに融合したHER−2/Neu誘導ペプチドを含む融合産物も提供される。一部の場合においては、ペプチドはペプチド結合を介して非相同ポリペプチドに融合され、これにより融合産物は本質的に組み換え融合蛋白となる。
【0008】
好ましくは、HER−2/Neu配列から誘導された単離されたペプチド又はその融合産物は、少なくとも10種の異なるHLA−DR対立遺伝子、より好ましくは少なくとも12、13、14、15、16、17、18、19、20、21、22、23、24、25又はそれ以上の異なる種類のHLA−DR対立遺伝子の抗原提示細胞により提示された場合に、HER−2/Neu蛋白に特異的なT細胞免疫応答を誘導することができる。
【0009】
一部の実施形態においては、HLA−DR対立遺伝子は0101、0102、0103、1503、160201、0301、0302、0401、0402、040301、040501、1101、1102、1103、1104、110401、1201、1301、1302、1401、1402、0701、080101、080201及び0901よりなる群から選択される。
【0010】
一部の実施形態においては、HER−2/Neu誘導ペプチドは配列番号2のアミノ酸配列を有する。別の実施形態においては、非相同ポリペプチドは顆粒球マクロファージコロニー刺激因子(GM−CSF)である。
【0011】
第2の態様において、本発明は上記したHER−2/Neu誘導ペプチド又はHER−2/Neu誘導ペプチドと非相同ポリペプチドをペプチド結合により連結した融合蛋白をコードするポリヌクレオチド配列を含む単離された核酸、核酸を含む発現カセット、及び発現カセットを含む宿主細胞を提供する。
【0012】
一部の場合においては、ポリヌクレオチド配列は配列番号2のアミノ酸配列を有するペプチドをコードする。別の場合においては、ポリヌクレオチド配列は非相同ポリペプチドがGM−CSFである融合蛋白をコードする。
【0013】
一部の実施形態においては、発現カセットは組み換えウィルスベクターである。他の実施形態においては、発現カセットは配列番号2のアミノ酸配列を有するペプチド又は非相同ポリペプチドがGM−CSFである組み換え融合蛋白の発現を指向する。
【0014】
第3の態様において、本発明は上記したHER−2/Neu誘導ペプチド又は非相同ポリペプチドと融合させたペプチドの融合産物を、生理学的に許容される賦形剤とともに含む組成物を提供する。
【0015】
一部の実施形態においては、ペプチドは配列番号2のアミノ酸配列を有する。別の実施形態においては、非相同ポリペプチドは顆粒球マクロファージコロニー刺激因子(GM−CSF)である。更に別の実施形態においては、組成物は更に抗原提示細胞を含み、これは細胞表面上の主要組織適合性複合体(MHC)分子と複合体を形成するHER−2/Neu誘導ペプチドを有する。
【0016】
第4の態様において、本発明はHER−2/Neu蛋白に対して特異的なT細胞免疫応答を患者において誘導するための方法を提供する。この方法は上記したHER−2/Neu誘導ペプチド又は非相同ポリペプチドと融合させたペプチドの融合産物、並びに生理学的に許容される賦形剤を含む組成物の有効量を患者に投与する工程を含む。
【0017】
一部の実施形態においては、ペプチドは配列番号2のアミノ酸配列を有する。別の実施形態においては、非相同ポリペプチドは顆粒球マクロファージコロニー刺激因子(GM−CSF)である。
【0018】
第5の態様において、本発明はHER−2/Neu蛋白に対して特異的なT細胞免疫応答を患者において検出するための方法を提供する。この方法は以下の工程、即ち(a)患者から抗原提示細胞及びT細胞を得る工程;(b)抗原提示細胞及びT細胞をHER−2/Neu誘導ペプチド、又は該ペプチドおよび非相同ポリペプチドを含む融合産物に接触させる工程;及び(c)T細胞応答を検出し、ここでT細胞応答の検出は患者におけるHER−2/Neu蛋白に対して特異的なT細胞免疫応答の存在を示すものである工程、を含む。
【0019】
一部の実施形態においては、工程(c)はELISPOT、増殖試験、又はフローサイトメトリーにより実施される。別の実施形態においてはHER−2/Neu誘導ペプチドは配列番号2のアミノ酸配列を有する。
【図面の簡単な説明】
【0020】
【図1】T細胞クローンHER500.23c21はHER−2/Neuに対して特異的である。(A)HER500.23c21によるHER500(アミノ酸■及びBA7072(HER500/GM−CSF融合蛋白)◆特異的IFNγ生産を抗原の自己PBMC提示の後にELISAを用いて測定した;バックグラウンドも試験した▲。試験は10%FBS添加完全IMDM培地中96ウェル丸底プレートにおいて3回で設定した。自己PBMCは2×105細胞/ウェルで使用し、T細胞クローンHER500.23c21は1×105細胞/ウェルで添加した。抗原は図示する終濃度とした。試験は5%CO2下37℃において48時間インキュベートし、上澄みをウェルから取り出してサイトカイン生産を試験した。(A)IFNγ生産。示したデータポイントは各抗原濃度に関する計算されたpg/mlを示す。(B)同じ試験に関するIL−2生産をHT−2細胞増殖を用いて計測した。データポイントは各抗原濃度に関する平均のCPM値を示す。
【図2】T細胞クローンHER500.23.21はペプチド#63に対して特異的である(HERp270−284)。HER−2/neuエピトープをクローンHER500.23.21に関してマッピングし、自己EBV−LcL及び各個々のHER500ペプチド#1−125(15アミノ酸長)を用いて特異性を測定した。試験は自己EBV−LcLは2×105細胞/ウェルで、T細胞クローンHER500.23.21は1×105細胞/ウェルで添加しながら96ウェル丸底プレート中に設定した。各ペプチドは終濃度1ug/mlで使用した。試験は5%CO2下37℃において48時間インキュベートし、その時点で上澄みをウェルから取り出してサイトカイン生産を試験した。(A)クローンHER500.23.21によるIFNγ生産はELISAを用いて計測し、計算されたpg/mlは各HER500ペプチドに対して示した。(B)HT−2細胞増殖を用いてIL−2の生産を計測した。
【図3】HLA−DR制限を示す。ブロッキング抗MHCクラスII抗体の存在下および非存在下において2×105細胞/mLの自己EBV−LcL及びHERp270−284を使用してT細胞クローンHER500.23c21を刺激した(1×105細胞/ウェル)。抗HLA−DR◆及び抗HLA−DQ/抗HLA−DP■をcRPMI+10%FBS中において図示した濃度において滴定した。ウェルには又、EBV−LcL、HER500.23c21T細胞クローン及びペプチド(1μg/ml)を含有させた。試験は5%CO2下37℃において48時間インキュベートした。上澄みを採取し、−20℃で凍結し、そして後日、ELISAを用いてIFNγ生産に関して分析した。複数の実験を実施し、1つの代表的実験に対して平均の3連の値を示す。
【図4】HERp270−284は無差別性のMHCクラスII HER−2/Neuエピトープである。(A)種々のHLA−DR対立遺伝子に対してホモ接合であるEBV−LcL系統を、クローンHER500.23c21に対してHERp270−284を提示するその能力に関して試験した。HER500.23c21による抗原特異的IFNγの生産は全てのEBV−LcL系統においてIFNγ ELISAの検出上限(4000pg/ml)を超えていた。陰性対照には無関係のHER500ペプチド、無ペプチド、及び各EBV−LcL系統単独を使用した(全値が検出不可能であり、記載しなかった)。試験は2×105EBV−LcL/ウェル及び1×105HER500.23c21細胞/ウェルにおいてcRPMI+10%FBS中、96ウェル丸底プレート中に設定した。ペプチドは終濃度1μg/mlで使用した。試験は5%CO2下37℃において48時間インキュベートした。上澄みを採取し、−20℃で凍結し、そしてELISAによりIFNγに関して分析した。結果は1つの代表的実験に関して示した。(B)HERp270−284を各EBV−LcL系統で滴定することにより異なる対立遺伝子に対するHER500.23c21の感受性を測定した。試験はHERp270−284の濃度を10ng/ml、33ng/ml、66ng/ml、100ng/ml及び250ng/mlとした以外は(A)と同様に設定した。250ng/ml超の全てのHERp270−284の濃度はIFNγ ELISAの検出上限(4000pg/ml)より高値のHER500.23c21によるIFNγの生産をもたらした。結果は1つの代表的実験に関して示す。
【図5】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す。ペプチドはHER−2/Neuペプチド270−284内の9量体であった。
【図6】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す。ペプチドはHER−2/Neuペプチド270−284内の10又は11量体であった。
【図7】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す(492nmの吸光度により測定)。ペプチドはHER−2/Neuペプチド270−284内の12量体であった。
【図8】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す(492nmの吸光度により示す)。ペプチドはHER−2/Neuペプチド270−284内の13量体であった。
【図9】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す(492nmの吸光度により示す)。ペプチドはHER−2/Neuペプチド270−284内の14量体であった。
【図10】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す(492nmの吸光度により示す)。ペプチドはHER−2/Neuペプチド268−286内の16量体であった。
【図11】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す(492nmの吸光度により示す)。ペプチドはHER−2/Neuペプチド268−286内の17量体であった。
【発明を実施するための形態】
【0021】
(定義)
「単離された」という用語は、核酸又は蛋白に適用する場合、核酸又は蛋白が、それが天然の状態で会合している他の細胞成分を本質的に含有しないことを指す。それは乾燥又は水溶液の何れかであることもできるが、好ましくは均質な状態である。純度及び均質性は典型的には分析化学の手法、例えばポリアクリルアミドゲル電気泳動又は高速液体クロマトグラフィーを用いて測定する。調製品中に存在する主な物質種である蛋白は実質的に精製されている。特に、単離された遺伝子は該遺伝子の側面に位置し目的の遺伝子以外の蛋白をコードするオープンリーディングフレームから分離されている。「精製された」という用語は核酸又は蛋白が電気泳動ゲルにおいて本質的に1つのバンドを生じさせることを指す。特にこのことは核酸又は蛋白が少なくとも85%純粋、より好ましくは少なくとも95%純粋、そして最も好ましくは少なくとも99%純粋であることを意味する。
【0022】
本出願において、「アミノ酸」という用語は天然に存在する、及び合成のアミノ酸、並びに天然に存在するアミノ酸と同様の態様において機能するアミノ酸類縁体及びアミノ酸ミメティックを指す。天然に存在するアミノ酸は遺伝子コードによりコードされているもの、並びに、後に修飾されているアミノ酸、例えばヒドロキシプロリン、γ−カルボキシグルタミン酸、及びO−ホスホセリンである。アミノ酸類縁体とは天然に存在するアミノ酸と同じ基本的化学構造、即ち水素、カルボキシル基、アミノ基、及びR基に結合しているα炭素を有する化合物、例えばホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウムを指す。そのような類縁体は修飾されたR基(例えばノルロイシン)又は修飾されたペプチド骨格を有するが、天然に存在するアミノ酸と同じ基本的化学構造を保持している。アミノ酸ミメティックはアミノ酸の一般的化学構造とは異なる構造を有するが天然に存在するアミノ酸と同様の態様において機能することができる化合物を指す。
【0023】
「核酸」又は「ポリヌクレオチド」という用語は1本鎖又は2本鎖の形態の何れかにおけるデオキシリボヌクレオチド又はリボヌクレオチド及びそれらの重合体を指す。特段の制約が無い限り、用語は参照核酸と同様の結合特性を有し、そして天然に存在するヌクレオチドと同様の態様において代謝される天然のヌクレオチドの既知類縁体を含有する核酸を包含する。特段の記載が無い限り、特定の核酸配列は保存的に修飾されたその変異体(例えば縮重コドン置換)及び相補配列並びに明示された配列をすべて包含する。特に、縮重コドン置換は1つ以上の選択された(又は全ての)コドンの第3位が混合塩基及び/又はデオキシイノシン残基により置換されている配列を形成することにより達成してよい(Batzer等、Nucleic Acid Res.19:5081(1991);Ohtsuka等、J.Biol.Chem.260:2605−2608(1985);及びRossolini等、Mol.Cell.Probes 8:91−98(1994))。核酸という用語は遺伝子、cDNA、又は遺伝子によりコードされたmRNAと互換的に使用する。
【0024】
ポリヌクレオチド配列におけるエレメントの相対的位置が関わる場合、「下流の」位置とは参照点の3’側にあるものであり、「上流の」位置とは参照点の5’側にあるものである。
【0025】
「ポリペプチド」、「ペプチド」及び「蛋白」という用語はアミノ酸残基の重合体を指すために本明細書において互換的に使用する。用語はアミノ酸残基1つ以上が相当する天然に存在するアミノ酸の人工的化学的ミメティックであるアミノ酸重合体に対して、並びに、天然に存在するアミノ酸重合体及び天然に存在するものではないアミノ酸の重合体に対して適用する。本明細書において、用語はアミノ酸残基が共有結合によるペプチド結合により連結されている完全長の蛋白(即ち抗原)を包含する何れかの長さのアミノ酸鎖を包含する。本出願においては、ポリペプチドのアミノ酸配列はN末端からC末端に向けて示す。換言すれば、ペプチドのアミノ酸配列を記載する場合、N末端から最初のアミノ酸を「第1のアミノ酸」と称する。
【0026】
融合蛋白の相手を記載する際に使用する場合、「非相同」という用語は1つのペプチド融合相手のもう1つのペプチド融合相手に対する関連性を指し:融合相手が融合ペプチド内に存在する態様が天然に存在する蛋白に観察できるものではない。例えば、HER−2/Neuエピトープと融合して融合ペプチドを形成している「非相同ポリペプチド」は顆粒球マクロファージコロニー刺激因子(GM−CSF)のようなHER−2/Neu蛋白以外の蛋白を起源とするものであってよい。一方、「非相同ポリペプチド」がHER−2/Neuエピトープに直ぐ隣接しないHER−2/Neu蛋白の別の部分から誘導されたものであってよい。「非相同ポリペプチド」は天然に存在する蛋白配列又はその一部分の修飾、例えばアミノ酸残基1つ以上の欠失、付加、又は置換を含有してよい。「非相同ポリペプチド」の起源(即ちそれがHER−2/Neu蛋白又は別の蛋白からの何れから誘導されているか)には関わりなく、融合ペプチドは配列番号1のアミノ酸配列を包含するヒトHER−2/Neuのサブ配列を含有してはならず、それは18アミノ酸長より大きい。一部の例示される実施形態においては、本発明において使用する「非相同ポリペプチド」は15〜20アミノ酸長以下を有し;別の実施形態においては「非相同ポリペプチド」は少なくとも100アミノ酸長を有する。
【0027】
「融合」又は「融合した」という単語は非相同ポリペプチドに連結したHER−2/Neuエピトープを含む本発明のペプチドを説明する際に使用する場合は、ペプチド結合を包含する何れかの共有結合によるエピトープと非相同ポリペプチドの間の連結を指す。
【0028】
「〜をコードする核酸配列」という表現はrRNA、tRNA、又は特定の蛋白又はペプチドの第1アミノ酸配列又はトランス作用性の調節剤に関する結合部位のような構造RNAに関する配列情報を含有する核酸を指す。この表現は特に、ネイティブの配列又は特定の宿主細胞におけるコドンの優勢性に合致するように導入してよい配列の縮重コドン(即ち単一のアミノ酸をコードする異なるコドン)を包含する。
【0029】
「発現カセット」とは宿主細胞における特定のポリヌクレオチド配列の転写を可能にする、特定された核酸エレメントのシリーズを有する組み換え又は合成により形成された核酸構築物である。発現カセットはプラスミド、ウィルスゲノム、又は核酸フラグメントの部分であってよい。典型的には発現カセットはプロモーターに作動可能に連結した転写されるべきポリヌクレオチドを包含する。
【0030】
「組み換え」という用語は、例えば細胞又は核酸、蛋白、又はベクターに関して使用する場合は、細胞、核酸、蛋白又はベクターが、外部原料からの核酸又は蛋白の導入、又はネイティブの核酸又は蛋白の改変により修飾されていること、又は細胞がそのように修飾された細胞から誘導されることを表す。即ち、例えば組み換え細胞は細胞のネイティブ(非組み換え)型内部には存在しない遺伝子を発現するか、又は、別様には異常に発現されるか、過少発現されるか、又は全く発現されないネイティブの遺伝子を発現する。
【0031】
「投与」又は「投与する」という用語は哺乳類、特にヒトに物質を接触させる種々の方法を指す。投与の様式は例えば限定しないが、物質を静脈内、腹腔内、鼻内、経皮、局所、皮下、非経口、筋肉内、経口、又は全身投与により、そして任意の他の手段による注射、摂取、吸入、移植、又は吸収を介して接触させることを含む方法を包含する。本発明のHER−2/Neuペプチド又はHER−2/Neuペプチドと非相同ポリペプチドを含む融合ペプチドの投与の1つの例示的な手段は、静脈内送達を介するものであり、その場合ペプチド又は融合ペプチドは水溶液、懸濁液、又はエマルジョン等のような静脈内注射のために適当な形態の医薬組成物として製剤できる。本発明のHER−2/Neuペプチド又は融合ペプチドを送達するための別の手段は皮内注射、皮下注射、筋肉内注射、又はパッチ等を用いる経皮適用を包含する。
【0032】
特定の物質の「有効量」とは所望の結果をもたらすために十分である物質の量を指す。例えば、抗HER−2/Neu免疫を誘導することを意図される本発明のペプチドを含む組成物の有効量は、被験体に投与された場合に免疫を誘導するという目標を達成するために十分な量である。達成すべき作用は検出可能な程度までの、疾患/状態及び関連する合併症の症状の進行の防止、是正、又は抑制を包含する。「有効量」の厳密な量は投与の目的に依存し、周知の手法を用いながら当業者により確認できる(例えばLieberman,Pharmaceutical Dosage Forms(vols.1−3,1992);Lloyd,The Art,Science and Technology of Pharmaceutical Compounding(1999);及びPickar,Dosage Calculations(1999)を参照)。
【0033】
「生理学的に許容される賦形剤」とは本発明の組成物の製剤中に使用される不活性な成分であり、これはHER−2/Neuペプチド又はHER−2/Neuペプチドと非相同ポリペプチドを含む融合ペプチドの活性成分を含有し、そして、例えばそれを必要とする患者への注射による使用のために適している。この不活性成分は、本発明の組成物中に包含された場合に組成物の所望のpH、コンシステンシー、色、におい、又はフレーバーを与える物質であってよい。
【0034】
本明細書においては、「T細胞免疫応答」という用語は細胞表面上の分子の増殖、又は発現、又はサイトカインのような蛋白の分泌により測定した場合の抗原特異的T細胞の活性化を指す。
I.序論
本発明者等はHER−2/Neu蛋白に由来する一連の新規な無差別性のT細胞エピトープを発見した。これらのペプチドエピトープは、それらが少なくとも25種の異なるHLA−DRB1対立遺伝子の範囲内において提示され得ることから、顕著なHLA無差別性を呈している。そのような広範なHLA−DRB1対立遺伝子によるこれらのエピトープの提示のため、これらのエピトープは、一般的なヒトの集団におけるHER−2/Neuを過剰発現する癌の治療のためのワクチン製造又は免疫療法におけるユニバーサルCD4 ヘルパーT細胞エピトープとして極めて価値あるものとなっている。
II.ペプチドの化学合成
本発明のペプチド、特に比較的短鎖のもの(例えば50〜100アミノ酸以下)は従来のペプチド合成又は当該分野で良く知られている他のプロトコルを用いて化学合成してよい。
【0035】
ペプチドはMerrifield等、J.Am.Chem.Soc.85:2149−2156(1963);Barany and Merrifield,Solid−Phase Peptide Synthesis,in The Peptides:Analysis,Synthesis,Biology Gross and Meienhofer(eds.),Academic Press,N.Y.,vol.2,pp.3−284(1980);及びStewart等、Solid Phase Peptide Synthesis 2nd ed.,Pierce Chem.Co.,Rockford,III.(1984)により記載されているものと同様の操作法を用いながら固相ペプチド合成法により合成してよい。合成の間、保護された側鎖を有するN−α−保護アミノ酸を、自身のC末端により連結されている成長中のポリペプチド鎖に対し、そして、固体支持体、即ちポリスチレンビーズに対して段階的に添加する。ペプチドはジシクロヘキシルカルボジイミドのような試薬と反応させることにより活性化されているN−α−保護アミノ酸のα−カルボキシ基にN−α−脱保護アミノ酸のアミノ基を連結することにより合成される。活性化されたカルボキシルへの遊離のアミノ基の結合はペプチド結合の形成をもたらす。もっとも一般的に使用されているN−α−保護基は酸不安定性のBoc、及び塩基不安定性のFmocを包含する。
【0036】
固体支持体としての使用のために適する材料は当業者に良く知られており、例えば限定しないが、以下のもの、即ち:ハロメチル樹脂、例えばクロロメチル樹脂又はブロモメチル樹脂;ヒドロキシメチル樹脂;フェノール樹脂、例えば4−(α−[2,4−ジメトキシフェニル]−Fmoc−アミノメチル)フェノキシ樹脂;tert−アルキルオキシカルボニルヒドラジド化樹脂等を包含する。このような樹脂は市販されており、それらの製造方法は当業者に周知である。
【0037】
慨すれば、C末端N−α−保護アミノ酸をまず固体支持体に結合させる。次にN−α−保護基を除去する。脱保護されたα−アミノ基を次のN−α−保護アミノ酸の活性化されたα−カルボキシレート基にカップリングさせる。所望のペプチドが合成されるまでプロセスを反復する。次に得られたペプチドを不溶性重合体支持体から切断し、アミノ酸側鎖を脱保護する。より長いペプチドは保護されたペプチドフラグメントの縮合により誘導できる。適切な化学的手法、樹脂、保護基、保護されたアミノ酸及び試薬の詳細は当該分野で良く知られており、本明細書において詳細に考察しない(例えばAtherton等、Solid Phase Peptide Synthesis:A Practical Approach,IRL Press(1989)及びBodanszky,Peptide Chemistry,A Practical Textbook,2nd Ed.,Springer−Verlag(1993)を参照)。
III.ペプチドの組み換え生産
A.一般的な組み換え技術
組み換え遺伝子学の分野における一般的な方法及び手法を開示している基礎的なテキストはSambrook and Russell,Molecular Cloning,A Laboratory Manual(3rd ed.2001);Kriegler,Gene Transfer and Expression:A Laboratory Manual(1990);及びAusubel等編、Current Protocols in Molecular Biology(1994)を包含する。
【0038】
核酸に関しては、サイズはキロ塩基(kb)か塩基対(bp)の何れかで表示される。これらはアガロース又はアクリルアミドゲル電気泳動から、配列決定された核酸から、又は公開されたDNA配列から誘導される推定値である。蛋白に関しては、サイズはキロダルトン(kDa)又はアミノ酸残基数で表示される。蛋白サイズはゲル電気泳動から、配列決定された蛋白から、誘導されたアミノ酸配列から、又は公開された蛋白配列から推定される。
【0039】
市販されていないオリゴヌクレオチドは例えばVan Devanter等、Nucleic Acids Res.12:6159−6168(1984)に記載の通り自動合成装置を用いながら、Beaucage & Caruthers,Tetrahedron Lett.22:1859−1862(1981)において最初に報告された固相ホスホロアミダイトトリエステル法に従って化学合成することができる。オリゴヌクレオチドの精製は何れかの当該分野で認識されている方策、例えばネイティブのアクリルアミドゲル電気泳動又はアニオン交換HPLCを用いながら、Pearson & Reanier,J.Chrom.255:137−149(1983)に記載の通り実施する。
【0040】
組み換え生産は本発明のペプチド、特に比較的大きい分子量のもの、例えばHER−2/NeuエピトープとGM−CSFの融合ペプチドを得るための有効な手段である。本発明のペプチドをコードするポリヌクレオチド及び合成オリゴヌクレオチドの配列は、例えばWallace等、Gene16:21−26(1981)の2本鎖鋳型を配列決定するための鎖停止法を用いながらクローニング又はサブクローニングした後に確認できる。
B.発現カセットの構築
本発明のペプチドをコードするポリヌクレオチド配列の獲得
本発明のペプチドをコードするポリヌクレオチド配列は化学合成により得ることができ、あるいは、市販元より購入することができ、そしてこれを次に分子クローニングの標準的な手法を用いながらさらに操作してよい。
宿主生物における好ましいコドン使用のための核酸の修飾
本発明のペプチドをコードするポリヌクレオチド配列は場合により特定の宿主における好ましいコドンの使用に合致するように任意に改変することができる。例えば、細菌細胞の1つの系統における好ましいコドンの使用は、本発明のペプチドをコードし、この系統により好まれるコドンを包含するポリヌクレオチドを誘導するために使用できる。宿主細胞により示される好ましいコドンの使用の頻度は、宿主細胞により発現される遺伝子の大多数における好ましいコドンの使用の頻度を平均することにより計算できる(計算サービスはKazusa DNA Research Institute,Japanのウエブサイトから利用できる)。この分析は好ましくは宿主細胞により高度に発現される遺伝子に限定される。
【0041】
修飾終了時に、コーディング配列を配列決定により確認し、次に本発明のペプチドの組み換え製造のための適切な発現ベクターにサブクローニングする。
【0042】
コーディング配列の確認の後、組み換え遺伝子学の分野の定型的手法を用いて本発明のペプチドを製造できる。
C.発現系
本発明のペプチドをコードする核酸の高レベルの発現を実現するためには、典型的には、転写を指向する強力なプロモーター、転写/翻訳ターミネーター、及び翻訳開始のためのリボソーム結合部位を含有する発現ベクター内にペプチドをコードするポリヌクレオチドをサブクローニングする。適当な細菌プロモーターは当該分野で周知であり、上記したSambrook and Russell及び上記したAusubel等において説明されている。本発明のペプチドを発現させるための細菌の発現系は例えばE.coli、バチルス種、サルモネラ及びカウロバクターにおいて入手できる。そのような発現系のためのキットは市販されている。哺乳動物細胞、酵母及び昆虫細胞に対する真核生物の発現系は当該分野で周知であり、やはり市販されている。1つの実施形態において、真核生物発現ベクターはアデノウィルスベクター、アデノ関連ベクター、又はレトロウィルスベクターである。
【0043】
相同核酸の発現を指向するために使用されるプロモーターは、特定の用途に依存する。プロモーターは任意に、その天然の設定における転写開始部位からの距離と概ね同じ非相同転写開始部位からの距離に位置づけられる。しかしながら、当該分野で公知の通りこの距離におけるある程度の変動は、プロモーター機能を喪失することなく順応され得る。
【0044】
プロモーターの他に、発現ベクターは典型的には宿主細胞内の本発明のペプチドの発現のために必要な追加的エレメントの全てを含有する転写ユニット又は発現カセットを包含する。即ち典型的な発現カセットはペプチドをコードするポリヌクレオチド配列に作動可能に連結したプロモーター、及び、効率的な転写物のポリアデニル化、リボソーム結合部位、及び翻訳停止に必要なシグナルを含有する。ペプチドをコードする核酸配列は典型的には形質転換された細胞によるペプチドの分泌を促進するように切断可能なシグナルペプチド配列に連結される。そのようなシグナルペプチドは、特に、組織プラスミノーゲン活性化物質、インスリン、及びニューロン成長因子、及びニセアメリカタバコガ(Heliothis virescens)の幼若ホルモンエステラーゼ由来のシグナルペプチドを包含する。カセットの追加的エレメントはエンハンサーを、そして、ゲノムDNAを構造遺伝子(例えば非相同ポリペプチドをコードするもの)として使用する場合は機能的スプライスドナーを有するイントロン及びアクセプター部位を包含してよい。
【0045】
プロモーター配列に加えて、発現カセットは効率的な終止を可能とするために構造遺伝子の下流に転写終止領域を含有しなければならない。終止領域はプロモーター配列と同じ遺伝子から得てよく、或いは、異なる遺伝子から得てよい。
【0046】
細胞内に遺伝子情報を輸送するために使用される特定の発現ベクターは特に重要ではない。真核生物又は原核生物の細胞における発現のために使用されている従来のベクターの何れかを使用してよい。標準的な細菌の発現ベクターはプラスミド、例えばpBR322系のプラスミド、pSKF、pET23D、及び融合発現系、例えばGST及びLacZを包含する。エピトープタグも組み換え蛋白に添加することにより単離の好都合な方法としてよく、例えばc−mycが挙げられる。
【0047】
真核生物ウィルス由来の調節エレメントを含有する発現ベクターが典型的には真核生物発現ベクターにおいて使用され、例えばSV40ベクター、乳頭腫ウィルスベクター、及びエプスタイン・バーウィルス由来のベクターが挙げられる。他の例示的な真核生物ベクターはpMSG、pAV009/A+、pMTO10/A+、pMAMneo−5、バキュロウィルスpDSVE、及び、SV40初期プロモーター、SV40後期プロモーター、メタロチオネインプロモーター、ネズミ乳房腫瘍ウィルスプロモーター、ラウス肉腫ウィルスプロモーター、多核体プロモーター、又は真核生物細胞における発現のために効果的であることがわかっている他のプロモーターの指向下に蛋白の発現を可能とする何れかの他のベクターを包含する。
【0048】
一部の発現系はチミジンキナーゼ、ハイグロマイシンBホスホトランスフェラーゼ、及びジヒドロ葉酸還元酵素のような遺伝子増幅をもたらすマーカーを有する。或いは、遺伝子増幅をおこなわない高収率の発現系、例えば多核体プロモーター又は他の強力なバキュロウィルスプロモーターの指向下に本発明のペプチドをコードするポリヌクレオチド配列を有する昆虫細胞におけるバキュロウィルスベクターもまた適当である。
【0049】
発現ベクター内に典型的に包含されるエレメントは又、E.coli中で機能するレプリコン、組み換えプラスミドを保有している細菌の選択を可能にする抗生物質耐性をコードしている遺伝子、及び真核生物の配列の挿入を可能にするプラスミドの非必須領域のユニークな制限部位も包含する。選択される特定の抗生物質耐性遺伝子は重要ではなく当該分野で公知の多くの耐性遺伝子の何れも適当である。原核生物配列は任意に、必要に応じて真核生物細胞におけるDNAの複製を妨害しないようなものが選択される。抗生物質耐性選択マーカーと同様、既知の代謝経路に基づいた代謝選択マーカーも又形質転換された宿主細胞を選択するための手段として使用してよい。
【0050】
組み換え蛋白(例えば本発明のペプチド)のペリプラズムの発現が望まれる場合は、発現ベクターは更に、発現されるべき蛋白のコーディング配列の5’に直接連結されたE.coli OppA(ペリプラズムオリゴペプチド結合蛋白)分泌シグナル、又はその修飾された型のような分泌シグナルをコードする配列を含む。このシグナル配列は細胞質中に生産された組み換え蛋白を細胞膜通過によりペリプラズム空間内に指向させる。発現ベクターはさらにシグナルペプチダーゼ1に関するコーディング配列を含んでよく、これは組み換え蛋白のペリプラズム空間内への進入時にシグナル配列を酵素的に切断することができる。組み換え蛋白のペリプラズム生産に関するより詳細な説明は、例えばGray等、Gene39:247−254(1985)、米国特許第6,160,089号明細書及び同第6,436,674号明細書に記載されている。
C.トランスフェクション法
標準的なトランスフェクション法を用いて本発明のペプチドを大量に発現する細菌、哺乳類、酵母、昆虫、又は植物の細胞系統を作成し、それを次に標準的な手法で精製する(例えばColley等、J.Biol.Chem.264:17619−17622(1989);Guide to Protein Purification,in Methods in Enzymology,vol.182(Deutscher,ed.,1990)参照)。真核生物及び原核生物の細胞の形質転換は標準的手法に従って実施する(例えばMorrison,J.Bact.132:349−351(1977);Clark−Curtiss & Curtiss,Methods in Enzymology 101:347−362(Wu等編、1983)参照)。
【0051】
宿主細胞内に外来ヌクレオチド配列を導入するための周知の操作法のいずれかを使用してよい。これらにはリン酸カルシウムトランスフェクション、ポリブレン、プロトプラスト融合、エレクトロポレーション、リポソーム、マイクロインジェクション、プラズマベクター、ウィルスベクターの使用、及び宿主細胞内にクローニングされたゲノムDNA、cDNA、合成DNA、又は他の外来性の遺伝子材料を導入するための他の周知の方法が包含される(例えば、Sambrook and Russell,上出参照)。使用する特定の遺伝子操作法は、本発明のペプチドを発現できる宿主細胞内に少なくとも1つの遺伝子を良好に導入することができることのみが必要である。
D.宿主細胞内のペプチドの組み換え発現の検出
適切な宿主細胞内に発現ベクターを導入した後、トランスフェクトした細胞を本発明のペプチドの発現に好都合な条件下で培養する。次に組み換えペプチドの発現に関して細胞をスクリーニングし、これはその後、標準的方法を用いて培地から回収する(例えばScopes,Protein Purification:Principles and Practice(1982);米国特許第4,673,641号明細書;Ausubel等、上出;及びSambrook and Russell,上出参照)。
【0052】
遺伝子発現をスクリーニングするための数種の一般的方法は当業者の間で周知である。第1に、遺伝子発現は核酸レベルにおいて検出できる。核酸ハイブリダイゼーション手法を用いた特定のDNA及びRNAの測定の種々の方法が一般的に使用されている(例えばSambrook and Russell,上出)。一部の方法では電気泳動による分離を行うが(例えばDNAを検出するためのサザンブロット及びRNAを検出するためのノーザンブロット)、DNA又はRNAの検出は電気泳動を使用することなく実施することもできる(例えばドットブロットによる)。トランスフェクトされた細胞における本発明のペプチドをコードする核酸の存在は配列特異的プライマーを用いたPCR又はRT−PCRにより検出することもできる。
【0053】
第2に、遺伝子発現はポリペプチドレベルにおいて検出できる。特に、本発明のペプチド、特に十分大きい非相同ポリペプチドを含有するものと特異的に反応するポリクローナル又はモノクローナル抗体を用いながら遺伝子産物のレベルを計測するために、種々の免疫学的試験が当業者により類型的に使用されている(例えばHarlow and Lane,Antibodies,A Laboratory Manual,Chapter 14,Cold Spring Harbor,1988;Kohler and Milstein,Nature,256:495−497(1975))。このような手法はペプチド又はその抗原性部分に対して高い特異性を有する抗体を選択することによる抗体製造を必要とする。ポリクローナル及びモノクローナル抗体を形成する方法は十分確立されており、それらの説明は文献、例えばHarlow and Lane,上出;Kohler and Milstein,Eur.J.Immunol.,6:511−519(1976)に記載されている。
IV.ペプチドの精製
A.化学的に合成されたペプチドの精製
合成ペプチドの精製はクロマトグラフィーの種々の方法、例えば逆相HPLC、ゲル透過、イオン交換、サイズエクスクルージョン、アフィニティー、分配、又は向流分配を用いて実施される。適切なマトリックス及び緩衝物質の選択は当該分野で周知である。
B.組み換え生産されたペプチドの精製
1.細菌封入体からのペプチドの精製
発現は構成的なものとなるが、典型的にはプロモーター導入後に大量の形質転換された細菌から本発明のペプチドを組み換えにより製造する場合、ペプチドは不溶性の凝集塊を形成する場合がある。蛋白封入体の精製のために適当である数種のプロトコルが存在する。例えば凝集塊蛋白(以下、封入体と称する)の精製では典型的には例えば約100〜150μg/mlリゾチーム及び0.1% Nonidet P40(非イオン系界面活性剤)の緩衝液中のインキュベーションによる細菌細胞の破壊により、封入体の抽出、分離及び/又は精製を行う。細胞懸濁液はポリトロン粉砕機(Brinkman Instruments,Westbury,NY)を用いて粉砕できる。或いは、細胞は氷上で超音波処理できる。細菌を溶解する代替方法はともに上出のAusubel等、及びSambrook and Russellに記載されており、当業者に明らかである。
【0054】
細胞懸濁液は一般的に遠心分離し、そして封入体を含有するペレットを、封入体を溶解しないが洗浄する緩衝液、例えば20mM Tris−HCl(pH7.2)、1mM EDTA、150mM NaCl及び2% Triton−X100(非イオン系界面活性剤)中に再懸濁させる。可能な限り細胞破砕物を除去するために洗浄工程を反復することが必要な場合がある。封入体の残存ペレットを適切な緩衝液(例えば20mMリン酸ナトリウム、pH6.8,150mM NaCl)中に再懸濁してよい。他の適切な緩衝液は当業者に明らかである。
【0055】
洗浄工程の後、強力な水素受容体および強力な水素供与体の両方である溶媒(又はこのような特性の一方を各々が有する溶媒の組み合わせ)を添加することにより封入体を可溶化する。次に封入体を形成した蛋白を適合する緩衝液を用いた希釈又は透析により復元してよい。適当な溶媒は、限定しないが尿素(約4M〜約8M)、ホルムアミド(少なくとも約80%、体積/体積に基づく)、及び塩酸グアニジン(約4M〜約8M)を包含する。凝集塊形成蛋白を可溶化させることができる一部の溶媒、例えばSDS(ドデシル硫酸ナトリウム)及び70%ギ酸は、免疫原性及び/又は活性の欠如を伴った蛋白の不可逆的変性の可能性のために、この操作法における使用のためには不適切である場合がある。塩酸グアニジン及び同様の試薬は変性剤であるが、この変性は不可逆ではなく、そして変性剤の除去(例えば透析による)又は希釈時に復元がおこる場合があり、これにより目的の免疫学的及び/又は生物学的に活性な蛋白の再形成が可能となる。可溶化の後、蛋白は標準的分離手法により他の細菌蛋白から分離できる。細菌封入体から組み換えポリペプチドを精製することに関する説明は更に、例えばPatra等、Protein Expression and Purification 18:182−190(2000)に記載されている。
【0056】
或いは、組み換えポリペプチド、例えば本発明のペプチドを細菌のペリプラズムから精製することが可能である。細菌のペリプラズム内に組み換えポリペプチドをエクスポートする場合、細菌のペリプラズム画分は、当業者に周知の他の方法に加えて冷温浸透圧ショックにより単離できる(例えばAusubel等,上出参照)。ペリプラズムから組み換えペプチドを単離するためには、細菌細胞を遠心分離してペレットを生じさせる。ペレットを20%スクロース含有緩衝液に再懸濁する。細胞を溶解するために、細菌を遠心分離し、ペレットを氷冷5mM MgSO4中に再懸濁し、そして約10分間アイスバス中に保持する。細胞懸濁液を遠心分離し、上澄みを傾瀉して保存する。上澄み中に存在する組み換えペプチドは当業者に周知の標準的な分離手法により宿主蛋白から分離することができる。
2.精製のための標準的な蛋白分離手法
組み換えポリペプチド、例えば本発明のペプチドを可溶性形態で宿主細胞内に発現させる場合、その精製は以下に記載する標準的な蛋白精製操作法に従うことができる。この標準的精製操作法は又、化学合成により得られたペプチドを精製するためにも適している。
i.溶解度分画
多くの場合初期工程として、そして蛋白混合物が複合体である場合、初期の塩分画は目的の組み換え蛋白、例えば本発明のペプチドから望ましくない宿主細胞蛋白(又は細胞培養用の培地から誘導される蛋白)の多くを分離することができる。好ましい塩は硫酸アンモニウムである。硫酸アンモニウムは蛋白混合物中の水の量を効果的に低減することにより蛋白を沈殿させる。次に蛋白がその溶解度に基づいて沈殿する。蛋白が疎水性であるほど、より低い硫酸アンモニウム濃度において沈殿する可能性が高まる。典型的なプロトコルは結果として生じる硫酸アンモニウム濃度が20〜30%となるように蛋白溶液に飽和硫酸アンモニウムを添加することである。これにより大部分の疎水性蛋白が沈殿することになる。沈殿を廃棄(目的の蛋白が疎水性でない限り)し、目的蛋白を沈殿させることが分かっている濃度となるまで硫酸アンモニウムを上澄みに添加する。次に沈殿を緩衝液中に可溶化し、必要に応じて過剰な塩を透析又は透析濾過により除去する。蛋白の溶解度に依存している他の方法、例えば低温エタノール沈殿等は当業者に周知であり、複合体蛋白混合物を分画するために使用できる。
ii.サイズ示差濾過
計算された分子量に応じて、より大きい及び小さいサイズの蛋白を異なる孔径の膜(例えばAmicon又はMilliporeメンブレン)を通過させる限外濾過を用いて単離できる。第1の工程として、目的の蛋白、例えば本発明のペプチドの分子量より低値の分子量カットオフを有する孔径の膜を通過させて蛋白混合物を限外濾過する。次に限外濾過の保持物を、目的のペプチドの分子量より大きい分子カットオフを有する膜に対して限外濾過する。組み換え蛋白は膜を通過して濾液中に入ることになる。次に濾液を後述する通りクロマトグラフィーに付すことができる。
iii.カラムクロマトグラフィー
目的の蛋白(例えば本発明のペプチド)は又そのサイズ、実質的表面電荷、疎水性、又はリガンドに対する親和性に基づいて他の蛋白から分離できる。更に又、本発明のペプチドに対して作成された抗体をカラムマトリックスに結合し、ペプチドを免疫精製することもできる。これらの方法は全て当該分野で周知である。
【0057】
クロマトグラフィー手法は何れかの規模において、多くの異なる製造元から得た機材を用いながら実施できることは当業者に明らかである(例えばPharmacia Biotech)。
C.ペプチド配列の確認
本発明のペプチドのアミノ酸配列は多くの十分に確立された方法により確認できる。例えばEdman分解の従来法を用いてペプチドのアミノ酸配列を決定できる。Edman分解に基づく配列決定法の数種の変形例、例えばマイクロ配列決定、及び質量分析に基づく方法がこの目的のために頻繁に使用されている。
D.ペプチドの修飾
本発明のペプチドはより望ましい特性を達成するために修飾できる。化学修飾されたペプチド及び蛋白分解酵素による分解に対して抵抗性であるか向上した溶解性又は結合能力を有するペプチド模倣体の設計はよく知られている。
【0058】
本発明のHER−2/Neuペプチド又は融合ペプチドの修飾されたアミノ酸又は化学的誘導体は通常はHER−2/Neu蛋白の一部ではない修飾されたアミノ酸の追加的化学特徴部分を含有してよい。ペプチドの共有結合修飾は本発明の範囲に含まれる。そのような修飾は選択された側鎖又は末端残基と反応することができる有機誘導体形成剤にペプチドのターゲティングされたアミノ酸残基を反応させることによりペプチド内に導入してよい。化学誘導体の以下の例は例示として示すのみであり、限定するものではない。
【0059】
蛋白分解性酵素による分解に耐性であるペプチド模倣体の設計は当業者の知る通りである。例えばSawyer,Structure−Based Drug Design,P.Verapandia,Ed.,N.Y.(1997);米国特許第5,552,534号明細書及び同第5,550,251号明細書を参照。ペプチド骨格及び側鎖の修飾の両方を二次構造模倣の設計において使用してよい。可能な修飾はD−アミノ酸、Nα−Me−アミノ酸、Cα−Me−アミノ酸、及びデヒドロアミノ酸の置換を包含する。今日まで、種々の二次構造ミメティックが設計されており、そしてペプチド又はペプチドミメティックに組み込まれている。
【0060】
他の修飾には天然のアミノ酸の非天然のヒドロキシル化アミノ酸による置換、酸性アミノ酸のカルボキシ基のニトリル誘導体による置換、塩基性アミノ酸のヒドロキシル基のアルキル基による置換、又はメチオニンのメチオニンスルホキシドによる置換が包含される。更に又、本発明のHER−2/Neuペプチド又は融合ペプチドのアミノ酸は同じアミノ酸であるが逆のキラリティーのもので置き換えることができ、即ち、天然に存在するL−アミノ酸はそのD配置により置き換えられてよい。
V.非相同ポリペプチドとのHER−2/Neuエピトープの融合
本発明の1つの態様において、HER−2/Neu無差別性T細胞エピトープに相当するペプチドは、T細胞応答を誘導するHER−2/Neuエピトープの能力が増強されるように共有結合を介して非相同ポリペプチドに結合させることにより、融合ペプチドを形成する。頻繁にはこの共有結合はペプチド結合であり、そしてHER−2/Neuエピトープ及び非相同ポリペプチドは新しいポリペプチドを形成する。このペプチド結合はHER−2/Neuエピトープと非相同ポリペプチドの間の直接のペプチド結合であってよく、或いはそれはHER−2/Neuエピトープと非相同ポリペプチドの間のペプチドリンカーを用いて与えられる間接のペプチド結合であってよい。
【0061】
他の共有結合もまた非相同ポリペプチドにHER−2/Neuエピトープを融合させる目的のために適している。例えば、あるペプチドの官能基(例えば非末端アミン基、非末端カルボキシル酸基、ヒドロキシル基、及びスルフヒドリル基)は別のペプチドの官能基と容易に反応し、そして2つのペプチドを結合するペプチド結合以外の共有結合を確立する場合がある。HER−2/Neuエピトープのペプチドと非相同ポリペプチドの間の共有結合による連結は又、適当な官能基を有するリンカー分子を用いることにより得ることもできる。そのようなリンカー分子はペプチドリンカー又は非ペプチドリンカーであることができる。リンカーを結合の前に誘導体化することにより露出させるか、又は追加的な反応性官能基を結合させてよい。誘導体化においてはPierce Chemical Company,Rockford,Illinoisから入手可能なもののような多くの分子のいずれかの取り付けを行ってよい。
VI.機能的試験
本発明のHER−2/Neuエピトープ(又はHER−2/Neuペプチドと非相同ポリペプチドを含む融合ペプチド)は、少なくとも10種の異なるHLA−DR対立遺伝子、より好ましくは少なくとも15、20又は25種の異なるHLA−DR対立遺伝子の1つを有する場合がある抗原提示細胞によりエピトープが提示される場合に、HER−2/Neu蛋白に特異的なT細胞免疫応答を誘導するその能力のために有用である。種々の機能的試験を用いることにより、種々の異なるHLA−DR対立遺伝子の抗原提示細胞に関して無差別性の態様におけるそのようなHER−2/Neu特異的T細胞免疫応答を誘導するHER−2/Neuエピトープの能力を確認することができ、例えば増殖試験及びフローサイトメトリー試験によりT細胞受容体とペプチドエピトープの間の結合又はT細胞によるサイトカインの生産を検出する方法などが挙げられる。
【0062】
本出願の実施例において使用する機能的試験の系はこの目的のために特に適している。慨すれば異なるHLA−DR対立遺伝子に対して各々ホモ接合である少なくとも10種、好ましくは少なくとも15、20又は25種の抗原提示細胞系統のパネルを用いて、HER−2/Neu蛋白に対して特異的に応答(例えばIFNγ又はIL−2のようなサイトカインの生産による)するCD4+ T細胞のクローン(例えばクローンHER500.23c21)に対してHER−2/Neu誘導ペプチドを提示する。配列番号2のアミノ酸配列を有するエピトープ270〜284を陽性対照として使用し、そして非関連のHER−2/Neu誘導ペプチド、無ペプチド、及び各抗原提示細胞系統単独を試験の陰性対照として使用する。試験は各ウェル中抗原提示細胞及びCD4+ T細胞を含有する適切な培地中におけるマルチウェル細胞培養プレートとして設定する。ペプチドは適当な濃度まで希釈し、そして各ウェルに添加する。適切な時間インキュベートした後、上澄みをウェルから収集し、そして492nmにおける吸光度に基づいてELISAにより計測できるサイトカインの生産に関して分析する。典型的には、HER−2/Neu特異的CD4+ T細胞応答を誘導する場合の本発明のHER−2/NeuクラスII無差別エピトープの作用は同じ試験条件下において、例えば同じモル濃度において、HER−2/Neuエピトープ270〜284(配列番号2に示すアミノ酸配列を有する)の作用の少なくとも25%であり、同じ個別のHLA−DR対立遺伝子の抗原提示細胞により提示される。より好ましくはそのような作用は同じ条件下でHER−2/Neuエピトープ270〜284により示されるものよりも少なくとも30%、40%、50%、60%、70%、80%、又はそれより高値である。
VII.組成物及び投与
本発明は更に有効量の(1)HER−2/Neuペプチド;又は(2)HER−2/Neuペプチドと非相同ポリペプチドを含む融合ペプチド;又は(3)予防及び治療の両方の用途におけるHER−2/Neu蛋白に対して特異的なT細胞免疫応答を誘導するための細胞表面上のMHC分子との複合体を形成する(1)又は(2)のペプチドを伴った抗原提示細胞(APC)を含む組成物を提供する。本発明の医薬組成物は種々の薬物送達系における使用に適している。本発明で使用するための適当な製剤はRemington’s Pharmaceutical Sciences,Mack Publishing Company,Philadelphia,PA,17th ed.(1985)に記載されている。薬物送達方法に関する簡単な考察については、Langer,Science 249:1527−1533(1990)を参照されたい。
【0063】
抗原提示細胞(APC)は種々の方法によりペプチド負荷のために形成できる。出発原料は末梢血または、可動化をともなうかもしくはともなわない白血球除去物である。APCは多数の方法、例えば浮力密度遠心分離、水簸、磁気ビーズ及びプラスチック接着等を単独又は組み合わせて使用することにより単離できる。単離後、APCをサイトカイン、成長因子、活性化剤、及び成熟化剤の存在下又は非存在下に1〜14日間培養する。6〜48時間1μg〜1mg/mlの濃度において培地にペプチドを添加することによりAPCにペプチドを負荷させる。APCを採取し、洗浄し、そして注入のための適当な製剤中に再懸濁する。APCは新鮮時に送達することができ、或いは、後の時期における送達のために凍結保存することができる。
【0064】
本発明の医薬組成物は種々の経路で、例えば皮下、皮内、経皮、筋肉内、静脈内、又は腹腔内に投与できる。医薬組成物を投与する好ましい経路は、体重70kgの成人に対して本発明のペプチド約1μg〜10mg、好ましくは50μg〜1mgの2週毎用量において皮下又は皮内によるものである。適切な用量は1週毎、2週毎、又は1月毎の間隔において投与してよい。
【0065】
ペプチドパルスAPCは種々の経路で、例えば皮下、皮内、静脈内、又は腹腔内に投与できる。ペプチドパルスAPCは1週毎、2週毎、又は1月毎の間隔において細胞100万個〜100億個の用量で送達される。
【0066】
本発明のペプチドを含有する医薬組成物を製造するためには、不活性及び製薬上許容しうる賦形剤又は担体を使用する。液体の医薬組成物は例えば皮内、皮下、非経口、又は静脈内投与に適する溶液、懸濁液、及びエマルジョンを包含する。活性成分(例えばHER−2/Neuペプチド又は融合ペプチド)の滅菌された水溶液、又は水、緩衝された水、食塩水、PBS、エタノール、又はプロピレングリコールを含む溶媒中の活性成分の滅菌溶液が非経口投与に適する液体組成物の例である。組成物は生理学的状態に近似するために必要な製薬上許容しうる補助物質、例えばpH調節剤及び緩衝剤、張度調節剤、湿潤剤、界面活性剤などを含有してよい。
【0067】
滅菌溶液は所望の溶媒系中に活性成分(例えばHER−2/Neuペプチド又は融合ペプチド)を溶解し、そして次に得られた溶液をメンブレンフィルター通過により滅菌することにより、或いは、滅菌条件下で予め滅菌された溶媒に滅菌化合物を溶解することにより、製造できる。得られた水溶液はそのまま使用するためにパッケージングしてよく、又は、凍結乾燥し、凍結乾燥された製剤を投与前に滅菌水性担体と混合してよい。調製品のpHは典型的には3と11の間、より好ましくは5〜9、そして最も好ましくは7〜8となる。
【0068】
HER−2/Neuペプチド又は融合ペプチドを含有する医薬組成物は予防及び/又は治療の処置のために投与できる。治療用途においては、組成物は、状態及びその合併症の症状を防止、治癒、退行、又は少なくとも部分的に緩徐化又は停止させるために十分な量において、HER−2/Neu蛋白を過剰発現している腫瘍細胞の増殖により悪化する可能性がある状態にすでに罹患している患者に投与する。これを達成するために適当な量は「治療有効用量」と定義される。この使用のために有効な量は疾患又は状態の重症度及び患者の体重及び全身状態に依存するが、一般的には70kgの患者に対して2週毎にHER−2/Neuペプチド又は融合ペプチド約1μg〜約10mgの範囲となり、70kgの患者に対して2週毎にペプチド約50μg〜約1mgの用量がより一般的に使用される。適切な用量は1週毎、2週毎、又は1月毎の間隔において投与してよい。
【0069】
組成物の単回又は多数回の投与は担当医により選択される用量レベル及びパターンで実施できる。何れの場合においても、医薬組成物は治療目的のために患者においてHER−2/Neu過剰発現腫瘍細胞の増殖を効果的に抑制するために十分なHER−2/Neuペプチド又は融合ペプチドの量を備えなければならない。
VIII.HER−2/Neu蛋白に対して特異的なT細胞応答を検出するための方法
本発明は更にHER−2/Neu蛋白に対して特異的なT細胞免疫応答が患者に存在するかどうかを検出するための方法を提供する。この方法は以下の工程を含む:第1に、リンパ球、例えば少なくともT細胞、及び抗原提示細胞を患者から入手する。そのようなリンパ球が得られる適当な試料は血液、腫瘍浸潤液、及びリンパ節又はリンパ液を包含する。第2にT細胞に対する抗原提示細胞によってT細胞エピトープの適切な提示を可能とする条件下で、本発明のHER−2/Neuペプチド(又はHER−2/Neuペプチドと非相同ペプチドを含む融合ペプチド)にT細胞及び抗原提示細胞を曝露する。第3に、T細胞応答のサインを、当該分野で周知の手段、例えばELISPOT、増殖試験、又はフローサイトメトリーによりインビトロで計測する。T細胞応答がこれらの方法の何れかにより検出される場合、患者においてHER−2/Neu蛋白に対して特異的なT細胞免疫応答が存在すると結論付けることができる。
【実施例】
【0070】
以下の実施例は限定としてではなく説明としてのみ提示する。本質的に同様の結果を得るために変更又は改変することができる種々の非重要なパラメーターは当業者が容易に認識することになる。
(実施例1)
材料及び方法
組み換え蛋白及び合成ペプチド。BA7072はHER−2/neu+癌の治療のための治験中のワクチンAPC8024において使用するためにDendreon Corporation(Seattle,WA)により製造された専売品の組み換え融合蛋白である。BA7072はHER−2/neuの細胞外ドメイン(ECD)及び細胞内ドメイン(ICD)の両方に由来する蛋白配列を含有する。HER500はHER−2/neuのECDおよびICDの両方に由来する配列を含有するDendreon Corporationによりやはり製造されている組み換え蛋白であり、そして抗原BA7072のHER−2/neu部分である。インビトロにおけるHER500特異的免疫応答を定義するために、HER500蛋白配列から125ペプチドを作成した。これらのペプチドは15アミノ酸長であり、11量体分オーバーラップしている(Genemed Synthesis,South San Francisco,CA)。125のHER500ペプチドのHER500ペプチド#63(HERp270〜284、ALVTYNTDTFESMPN)は天然のHER−2/neu配列のアミノ酸270〜284に相当する。作成したHER500の15量体ペプチドに加えて、9−、10−、11−、12−、13−、14−、16−、及び17量体ペプチドをHER−2/Neu配列の残基268〜286から合成した。これらのペプチドはNH2−及びCOOH−末端のトランケーション、又はHERp270〜284への1〜2アミノ酸の付加の後に誘導され、記載したペプチドを得た。全てのHER500ペプチドを配列決定し、そして分析用HPLCおよび質量スペクトル分析(Genemed Synthesis)により>95%純度であることが測定された。
【0071】
被験体及び健常者ドナー試料収集。全ての被験体及び健常者ドナーの標本を適切なInvestigational Review Boardにより認可されている治験責任者依頼プロトコルに従って収集した。インフォームドコンセントを受領した後、ヘパリン処理したバキュテナチューブ又はシリンジ中に静脈穿刺により全血試料を収集し、輸送及び/又はプロセシングのために準備した。本発明者等の実験室で血液試料を受領した後、末梢血液単核細胞(PBMC)を密度勾配遠心分離により滅菌条件下で収集し、そして特定の試験における使用のために準備した。
【0072】
HER−2/neu特異的T細胞クローンのインビトロ作成。HER−2/neu陽性癌に対する治験中の治療薬であるAPC8024を投与された被験体のPBMCを2mM L−グルタミン、50U/mLペニシリン、50μg/mLストレプトマイシン、及び20mM HEPES緩衝液+10%ヒトAB血清を添加したRPMI1640(Gemini BioProducts,Calabasas,CA)(cRPMI+10%HS)中で、一晩BA7072を10μg/ml含有するT−25組織培養フラスコ中で刺激した。翌日、IFNγ分泌細胞をIFNγ分泌試験細胞富化検出キット(INFγ Secretion Assay Cell Enrichment and Detection kit)(Miltenyi Biotech,Auburn,CA)を用いてPBMC培養物から単離した。IFNγ富化集団は10U/mL組み換えヒトIL−2(Invitrogen)を用いて96ウェル丸底プレート内に限界希釈法により播種した。非IFNγ分泌細胞を放射線照射し(3000rad)、そして全ウェル内の最終容量が150μLとなるようにウェル当たり50μLで添加した。プレートを37℃5%CO2において7日間インキュベートした。EBV形質転換リンパ芽球様細胞(EBV−LcL)も又、増殖及び自己T細胞クローンの試験のために、自己PBMC及びB95−8細胞系統(ATCC、Manassas,VA)由来の上澄みを用いて本被験体から作成した。クローニング第7日に、IFNγ分泌細胞を前に記載したとおり96ウェルプレート中で非特異的に増殖させた(Yee等、2002、Proc.Natl.Acad.Sci.USA 99:16168−16173)。慨すれば、各ウェルに対し、25U/mL組み換えヒトIL−2及び10ng/mL抗ヒトCD3抗体を添加したcRPMI+10%HS培地(BD Pharmingen,San Diego,CA)100μLを、1×104/ウェルの照射自己EBV−LcL及び1×105/ウェルの照射同種異系PBMCとともに添加した。プレートを37℃で14日間インキュベートし、次にウェルを目視により検査することにより、陽性の増殖を調べた。増殖陽性のクローン、クローンHER.23c21及びその他を24ウェルプレートに移し、そしてrIL−2、抗−CD3、及びアクセサリー細胞を用いながら上記した通り増殖させた。各ウェルの最終容量は2.4mLであり、そしてアクセサリー細胞の数は増大してウェル当たり2×106/ウェルの照射同種異系PBMC及び1×105/ウェルの照射自己EBV−LcLとなった。
【0073】
HER−2/neu特異的T細胞クローンの発見及び特性化。クローンを自己PBMC及び抗原又は自己EBV−LcL及びHer500ペプチドを用いて抗原特異性に関してスクリーニングした。刺激はcRPMI+10%FBS培地中において96ウェル丸底プレート中に設定し、5%CO2下37℃において48時間インキュベートした。更に又、クローンはフローサイトメトリーによりCD4及びCD8表面発現に関して染色した。
【0074】
サイトカイン生産。抗原特異的刺激試験におけるサイトカイン生産を測定するために、48時間後、上澄み200μLを培養物から採取し、IL−2及びIFNγの両方の生産について試験した。IL−2の生産はIL−2依存性系統HT−2(ATCC)を用いて計測した。HT−2細胞を10%FBS及び100uM NEAA、1mMピルビン酸ナトリウム、2mM L−グルタミン、50mMペニシリン、50U/mLストレプトマイシン、20mM HEPES及び20uM 2−メルカプトエタノールを添加したIMDM(cIMDM+10%FBS)中で生育させ、そして20ng/mLのrIL−2を週2回供給した。試験のために、HT−2細胞を継代した4日後に細胞をIMDM+10%FBSで洗浄することによりすべてのrIL−2を除去し、細胞をcIMDM+10%FBS中1×104細胞/ウェルで96ウェル丸底プレートに添加した。抗原特異的刺激に由来する上澄みをウェルに添加し、プレートを37℃で24〜30時間インキュベートした。翌日1μCiの3[H]TdRを試験の最終6時間に添加し、プレートをガラス繊維フィルターマットに回収し、そして放射性同位体のDNA取り込み、又は増殖応答を液体シンチレーションカウンター(PerkinElmer Life and Analytical Sciences,Inc.Boston,MA)を用いて分当たりの計数により求めた。INFγ生産は、ELISA用の抗ヒトIFNγ抗体対(BD Pharmingen,San Diego,CA)を用いて計測した。慨すれば、イムロン4プレート(Thermo Labsystems/VWR,Brisbane,CA)を3μg/mlの精製した抗ヒトIFNγ抗体(NIB42)で一晩コーティングした。翌日コーティング抗体を廃棄し、そしてPBS(Invitrogen)中4%ウシ血清アルブミン(BSA)(Sigma,St.Louis,MO)をウェルに添加し、そしてプレートを37℃で2時間インキュベートした。プレートをPBS+0.05%Tween20で洗浄し、抗原特異的刺激に由来する上澄み試料100μLをウェルに添加し、そして1.25〜2時間室温でインキュベートした。プレートを洗浄し、ビオチニル化抗ヒトIFNγ抗体(4S.B3)をPBS中1%BSA中に希釈(1μg/ml)し、そして室温で1時間プレートに添加した。プレートを洗浄した後、ストレプ−アビジンHRP(BD Pharmingen)をPBST中1:1000に希釈し、室温で30分間ウェルに添加した。最後にプレートを洗浄し、Sigma(登録商標) Fast OPDを暗所で15分間添加した。2M HClを添加することにより反応を停止させ、分光光度計においてプレートの492nmの吸光度を読み取った。
【0075】
HLA−DR制約性及び無差別性。T細胞エピトープHERp270〜284のHLA−DR制約性を測定するために、抗HLA−DRmAbL243、HLA−DQmAb1a3又はHLA−DPmAbB7/21(20〜1.25μg/ml)をT細胞クローンHER.23c21、ペプチドHERp270〜284及び自己EBV−LcLと共にcRPMI+10%FBS培地中で培養した。上澄みを48時間後に採取し、IL−2及びIFNγの生産に関して試験した。HERp270〜284も又、フランスのストラスブールで開催された第12回国際組織適合性ワークショップ(IHW)を起源とするEuropian Collection of Cell Culturesより購入したEBV−LcL系統を用いてHLA−DR無差別性に関して試験した。表Iに列挙したIHW系統は種々のHLA−DRβ1対立遺伝子に対してホモ接合であり、10%ウシ胎児血清、20mM HEPES、2mM L−グルタミン、50mMペニシリン及び50U/mLストレプトマイシンを添加したRPMI1640培地(cRPMI+10%FBS)(Invitrogen,Carlsbad,CA)中で増殖させた。MHCクラスII無差別性を試験するために、HERp270〜284を1μg/mlにおいて各々別個のEBV−LcL系統(2×105細胞/ウェル)及びT細胞クローンHER.23c21(1×105細胞/ウェル)と共に37℃のcRPMI+10%FBS培地中96ウェル丸底プレート中に添加した。上澄みを48時間後に採取し、IFNγの生産に関して試験した。
結果
APC8024はHer−2/neu発現乳癌に対する治験中の自己細胞免疫療法である。この免疫療法により誘導される免疫応答を特性化するために、本発明者等はAPC8024を投与した臨床治験被験体からT細胞クローンを単離した。作成したCD4+ T細胞クローンの1つであるHER500.23c21は自己PBMCにより提示される外因性のHER−2/Neu蛋白配列に対して特異的な応答を呈した(図1)。この実験において、HER500.23c21をHER−2/Neuの細胞内及び細胞外のドメインを含有する組み換え蛋白であるHER500及びhGM−CSFとの融合蛋白として発現されるHER500より成るBA7072の漸増用量で刺激した。HER500.23c21は両方の抗原に応答してIFNγ及びIL−2を特異的に生産したが、抗原の非存在下においてはサイトカインの生産は観察されなかった。HER500.23c21は用量依存的な態様においてこれらの蛋白の各々に応答し、外因性蛋白抗原より天然にプロセシングされて提示されるHER−2/NeuエピトープをHER500.23c21が認識することを示している。
【0076】
HER−2/Neu蛋白配列に対するHER500.23c21の明確な応答のために、HER500.23c21により認識される特異的エピトープをペプチドを用いてマッピングした。HER500内にHER−2/Neu蛋白配列を包含している125の個別のオーバーラップした15量体のペプチドのパネルに対してHER500.23.c21を試験した。各ペプチドは抗原提示細胞としての自己EBV LCL細胞と共に1ug/mlで使用した。これらの実験において、HER500.23c21は、IFNγ又はIL2生産の何れかにより計測した場合、125全ての個別のペプチドのプールに対して強力に応答した(図2A及びB,上棒グラフ)。更に又、HER500.23c21は個別のペプチドの一つ、ペプチド63のみに応答し、このペプチドがHER500.23c21により認識されるエピトープを含有していることを示していた(図2)。ペプチド63はHER−2/Neu蛋白配列のアミノ酸270〜284に相当し、そして隣接ペプチドと11アミノ酸分オーバーラップしている。即ち、ペプチド62又は64への応答が欠如していることは、HER500.23c21 T細胞エピトープはHERp270〜284配列に完全に含有されており、そして側面に位置するペプチドに共通のアミノ酸はこのT細胞クローンに対する完全なエピトープを含有しないことを示唆している。HER500.23c21により認識される最小のエピトープを定義するために、本発明者等はHERp270〜284のNH2−及びCOOH−末端のトランケーションを有する9量体、10量体又は11量体のペプチドを設計した。これらのペプチドによるHER500.23c21の刺激は、HLA−DRB1対立遺伝子*0301、*0302、*1301、及び*1402を与えるEBV LcL系統により提示される場合に、11量体配列VTYNTDTFESM及びTYNTDTFESMPのみにおいて良好であった。
【0077】
どのHLA分子がHER500.23c21に対してHERp270〜284を提示することを担っているかを決定するために、HLAブロッキング抗体を用いて提示及びT細胞活性化を抑制した。HER500.23c21をHLA−DR又はHLA−DP/−DQに対して特異的な抗体の漸増濃度の非存在下又は存在下において1ug/mlのHERp270〜284及び自己EBV−LcLで刺激した。T細胞刺激はIFNγ生産により測定した。HLA−DRに対する抗体は用量依存的な態様においてHERp270〜284によるHER500.23c21の刺激を抑制したのに対し、HLA−DP/−DQブロッキング抗体は20ug/mlの高濃度においてもT細胞活性化に対して作用を示さなかった(図3)。これらの結果はHERp270〜284のHER500.23c21認識がHLA−DR制約性であることを明らかにしている。
【0078】
HLA制約性を更に定義するために、本発明者等は13のDR血清学的ファミリーを与える異なるHLA−DRB1*対立遺伝子に対してホモ接合である25のEBV LcL系統のパネルを使用してHER500.23c21を活性化させるHERp270〜284の能力を試験した(表1)。試験した各EBV−LcL系統は抗原特異的な態様においてHER500.23c21を効率的に刺激することができ、HERp270〜284エピトープは少なくとも25のHLA−DRB1*対立遺伝子に対して無差別であることが示された(図4A)。これらの実験において、1ug/mlのHERp270〜284ペプチドは使用したEBV−LcL系統とは無関係に最大のT細胞応答を誘導した。しかしながら、HER500.23c21を刺激する種々の対立遺伝子の能力における相違は、より低い濃度のペプチドにおいて顕著となり(図4B)、一部の対立遺伝子、例えばDRB1*0401は、10ng/mLもの低濃度で刺激することができ、そして他の対立遺伝子は250ng/mL未満では刺激することができなかった(DRB1*1103)。これらの相違はおそらくは異なるDRB1*対立遺伝子に対するHERp270〜284の結合親和性の範囲を反映していると考えられる。しかしながらこれらの実験において観察された高度な無差別性は、HERp270〜284がHER−2/Neuに対するユニバーサルなヘルパーT細胞エピトープを含有している場合があることを示唆している。
考察
抗腫瘍免疫応答の試験は試薬が限られているため特定のHLA型により提示される抗原少数に制限される場合が多い。無差別T細胞エピトープの発見は多様なHLA型の個体における抗腫瘍免疫応答の分析を可能にすることにより、これらの制限の緩和に寄与すると考えられる。研究用ツール及び潜在的な治療薬の両方としてのそれらの価値のため、多くの研究は無差別性のCD4及びCD8 T細胞エピトープの発見及び特性化に着目してきた。自身のHLA結合において多様な程度の無差別性を有するペプチドエピトープが感染性疾患の抗原であるHIV(van der Burg等、1999、J Immunol162:152−160)、マイコバクテリア(Valle等、2001、Clin Exp Immunol 123:226−232)、及びp.falciparum(Contreras等、1998、Infect Immun 66:3579−3590)、ならびに腫瘍抗原、例えばNYESO(Zarour等、2002、Cancer Res 62:213−218)、MAGE(Consogno等、2003、Blood 101:1038−1044)、Tert(Schroers等、2003.Clin Cancer Res 9:4743−4755)、及びHer2/neu(Kobayashi等、2000、Cancer Res 60:5228−5236)において発見されている。TEPITOPE(Bian and Hammer,2004,Methods 34:468−475)のようなコンピュータープログラムは蛋白配列に基づいて無差別エピトープを予測するために既知の共通の結合モチーフを使用しており、種々の原料に由来する多くの潜在的な新規T細胞エピトープを発見している。インシリコで発見されたエピトープの生物学的関連性は多くの系において着目されている(Ruiz等、2004、Clin Cancer Res 10:2860−2867;及びAl−Attiyah and Mustafa 2004.Scand J Immunol 59:16−24)。
【0079】
本試験において、新規な無差別T細胞エピトープが腫瘍関連抗原HER−2/Neuから発見された。このT細胞エピトープはHER−2/Neu+癌に対する自己細胞免疫療法で治療された患者から単離されたCD4+ T細胞クローンと共に発見された。エピトープはHER−2/Neu配列のアミノ酸270〜284内に含有されており、外因性の蛋白抗原より天然にプロセシングされて提示される。このエピトープに対して特異的なT細胞クローンがHER−2/Neu特異的免疫療法で治療された個体から単離されたという事実は、このエピトープが臨床的に関連性のある抗腫瘍免疫応答の部分としてインビボで役割を果たしている可能性があることを示唆している。生物学的に関連する範囲において発見されたことのほかに、このペプチドエピトープはそれが極めて広範なHLA−DR無差別性を有し、13の血清学的なDRファミリーを与える少なくとも25の異なるHLA−DRB1*対立遺伝子によりT細胞クローンHER500.23c21に提示されることができることから、興味深いものである。無差別T細胞エピトープとして発見された他のペプチドの大部分はわずか数種の異なるHLA対立遺伝子により提示されるのみであるが、本発明者等はなお、T細胞クローンHER500.23c21に対してHERp270〜284を提示することができないHLA−DRB1*対立遺伝子を発見しなければならない。このエピトープの提示に関するMHC制約性が相対的に乏しいことは、これをユニバーサルなHER−2/Neu CD4 T細胞エピトープの理想的な候補としている。
【0080】
腫瘍免疫学の主要な目標は腫瘍関連抗原に対抗して効果的な癌免疫療法及びワクチンを開発することである。そのような治療は抗腫瘍免疫応答を刺激して腫瘍を根絶するように設計される。特定の腫瘍関連抗原に着目することは、抗腫瘍免疫応答を良好に生じさせ、一部の腫瘍抗原内の特定のT細胞エピトープの発見をもたらした。初期の研究の大部分が腫瘍特異的CD8 T細胞応答を生じさせることにのみ着目していたが、効果的な抗腫瘍免疫を生じさせる際のCD4 T細胞の重要性の認識も高まっていた。このため、抗腫瘍CD4 T細胞応答及び腫瘍関連抗原からのクラスII制約型のT細胞エピトープの発見の研究が進展した。そのようなエピトープの利用は、そのようなエピトープが1つより多いHLA型により提示されることができれば大きく増大する。ペプチド系癌ワクチンの方策はワクチン内のペプチドエピトープのHLA制約性により妨害されている。本明細書に記載したような無差別T細胞エピトープを包含させることは一般的集団内のそのようなワクチンの有用性を広げる。無差別エピトープによる腫瘍抗原に対する免疫応答の誘導は一般的集団のより大きいパーセントに到達するための効率的な方法である。そのような無差別エピトープはまた応答を分析するための有用な手段を与える。
【0081】
【表1A】

【0082】
【表1B】
細胞系統はECACC European Collection of Cell Culturesより入手し、それらはIMGT/HLA細胞要覧に記載されている(ウエブサイト:ebi.ac.uk/imgt/hla/cell query.html)。
(実施例2)
この一連の実験においては、HER−2/Neu蛋白から誘導された別のペプチドを無差別MHC クラスII HER−2/neuエピトープとしてのその潜在性に関して試験した。エピトープHERp270〜284(配列番号2)を実験の陽性対照として使用した。種々のHLA−DR対立遺伝子に対してホモ接合であるEBV−LcL系統のパネルを用いて、HERp270〜284並びにHER−2/Neu268〜286(配列番号1)の15アミノ酸配列内の種々の長さのペプチド又は19アミノ酸配列内のより長いペプチドをクローンHER500.23c21に対して提示した。試験の陰性対照には非関連HER500ペプチド、無ペプチド、及び各EBV−LcL系統単独を使用した。自己EBV−LcLも又、クローンHER500.23c21特異性に関する対照として各条件で使用した。試験は2×105 EBV−LcL/ウェル及び1×105 HER500.23c21細胞/ウェルでcRPMI+10%FBS中、96ウェル丸底プレート中に設定した。ペプチドを終濃度1μg/mlに希釈し、100μL/ウェルで添加した。試験は5% CO2下37℃において48時間インキュベートした。上澄みを採取し、ELISAを用いてIFNγ生産に関して分析した。結果は各ペプチドシリーズに対して1つの代表的な実験について示した(図5〜11)。ペプチド及び各HLA−DR対立遺伝子に応答してクローンHER500.23c21により生産されたIFNγの結果はIFNγのpg/mL量において、又は492nmにおける吸光度として報告する。
【0083】
本出願において引用した全ての特許、特許出願、及び他の公開物は、公開されたアミノ酸又はポリヌクレオチドの配列も含めて、あらゆる目的のために、参照としてその全体が援用される。
【0084】
配列表
配列番号1 (HER−2/Neu蛋白の268−286)
CPALVTYNTDTFESMPNPE
配列番号2 (HER−2/Neu蛋白の270−284、または配列番号1の残基3−17)
ALVTYNTDTFESMPN
【技術分野】
【0001】
関連する出願
本願は、2007年8月11日に出願された米国特許出願第60/837,209号に対する優先権を主張するものであり、米国特許出願第60/837,209号の内容はその全体が本明細書中に参考として援用される。
【背景技術】
【0002】
HLAクラスII制限CD4+ T細胞は細胞性免疫において重要な役割を果たしており、そして抗腫瘍免疫応答の重要な成分である。CD4+ T細胞は腫瘍特異的CTLに対して必要な補助を提供し(非特許文献1)、そしてサイトカイン、例えばインターフェロンガンマ(IFNγ)を生産し、これは抗原提示細胞を活性化し、そして他の免疫学的作用を媒介することができる(非特許文献2)。数種の系における実験の結果は、CD4+ T細胞は有効な抗腫瘍免疫応答のためには必須であることを示している。強い免疫応答を発生させる場合にCD4+ T細胞が重要であることを鑑みれば、最適に設計された癌の免疫療法又は抗腫瘍ワクチンは最大の薬効のために腫瘍特異的CD4+ T細胞及びCD8+ T細胞の両方を誘導するはずである。
【0003】
癌の免疫療法及びワクチンの設計は腫瘍関連抗原の発見により大きく利益を得ている。1つのそのような抗原、HER−2/Neuは、それが乳癌及び卵巣癌を包含する種々の癌において増幅されることから、上記方策のための主要な標的となっている。HER−2/Neuオンコジーンは表皮成長因子受容体に対する相同性を有する膜貫通糖蛋白をコードしている(非特許文献3)。HER−2/Neuの過剰発現は乳腺癌の約30%で起こり、そして進行性疾患及び不良な予後を伴う。その結果、HER−2/Neu蛋白のT細胞認識を増大させるように設計された数種の免疫学的方策が臨床治験において試験されている。そのような試験において生じたHER−2/Neu特異的T細胞応答を特性化することは、HER−2/Neu蛋白内部の数種のHLAクラスI及びクラスII限定T細胞エピトープの発見をもたらしている(非特許文献4)。これらのエピトープの発見は、HER−2/Neu特異的T細胞応答を検出及び定量する本発明者等の能力を増強した。そのような情報は有効な免疫療法のための向上した設計をもたらし、HER−2/Neu発現腫瘍を根絶する際のHER−2/Neu特異的T細胞の役割のより良好な理解を可能にする。
【0004】
所定のT細胞エピトープの有用性はそのHLA制限により限定される。ペプチドエピトープは典型的にはHLA対立遺伝子の少数と生産的ペプチド−MHC複合体を形成し、これらの対立遺伝子を発現する個体においてのみT細胞応答を刺激する。このことは免疫学的試験及び臨床治験を特定のHLA型の個体に限定するものであり、それは一般的集団の20%未満である場合が多い。HLA対立遺伝子の多数により提示され得る、いわゆる無差別(promiscuous)T細胞エピトープが数種の腫瘍抗原に関して報告されている。無差別T細胞エピトープは複数のHLA対立遺伝子に結合して抗原特異的T細胞を刺激することができ、これにより異なるHLA型の個体におけるT細胞応答の誘導及び試験を可能にする。更に又、無差別エピトープは、これらのエピトープに基づいた免疫療法及びワクチンが癌の治療及び予防のために一般的集団に対して広範に適用できるため、価値あるものである。即ち、腫瘍抗原の以前は未知であった無差別エピトープに関連する新しい情報が必要とされていることは明らかである。
【0005】
本発明者等はHER−2/Neu蛋白配列における一連の新規無差別T細胞エピトープを発見した。HER−2/Neu蛋白の270〜284又は268〜286の領域内に位置するこれらのエピトープはHER−2/Neu過剰発現癌腫に対して自己活性細胞免疫療法を用いて治療された患者から作成されたCD4+ T細胞クローンにより認識される(非特許文献5)。T細胞クローンは少なくとも25種の異なるHLA−DRB1*対立遺伝子の関連において提示されるこれらのペプチドエピトープを認識する。抗体ブロッキング実験によって、認識はHLA−DR限定であることが確認される。更に又、これらのエピトープは外因性の蛋白抗原からも天然にプロセシングされて提示される。異なるHLA−DRB1*対立遺伝子に対するこれらのエピトープの無差別性は、これらのエピトープをHLA型に関係なくHER−2/Neu特異的免疫応答を評価するための価値ある手段としている。更に又、これらのエピトープはHER−2/Neu+癌の治療のためのペプチド系ワクチン又は免疫療法においてユニバーサルなCD4Tヘルパー細胞エピトープとして使用できる。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Topalian,1994.Curr Opin Immunol 6:741−745
【非特許文献2】Corthay等、2005.Immunity 22:371−383
【非特許文献3】Coussens等、1985、Science230:1132−1139
【非特許文献4】Sotiriadou等、2001、Br J Cancer85:1527−1534
【非特許文献5】Valone等、2001、Cancer J7 Suppl 2:S53−61
【発明の概要】
【課題を解決するための手段】
【0007】
本発明はHER−2/Neu特異的T細胞応答を誘導するための多くの異なる種類のHLA対立遺伝子の抗原提示細胞により提示することができる新しいHER−2/Neuエピトープについて記載する。第1の態様において、本発明はHER−2/Neu蛋白配列のセグメントから誘導された単離されたペプチドを提供する(残基268〜286、即ち配列番号1)。ペプチドは(a)配列番号1の残基3〜17の12〜15隣接アミノ酸;又は(b)配列番号1の16〜18隣接アミノ酸よりなる。同様に非相同ポリペプチドに融合したHER−2/Neu誘導ペプチドを含む融合産物も提供される。一部の場合においては、ペプチドはペプチド結合を介して非相同ポリペプチドに融合され、これにより融合産物は本質的に組み換え融合蛋白となる。
【0008】
好ましくは、HER−2/Neu配列から誘導された単離されたペプチド又はその融合産物は、少なくとも10種の異なるHLA−DR対立遺伝子、より好ましくは少なくとも12、13、14、15、16、17、18、19、20、21、22、23、24、25又はそれ以上の異なる種類のHLA−DR対立遺伝子の抗原提示細胞により提示された場合に、HER−2/Neu蛋白に特異的なT細胞免疫応答を誘導することができる。
【0009】
一部の実施形態においては、HLA−DR対立遺伝子は0101、0102、0103、1503、160201、0301、0302、0401、0402、040301、040501、1101、1102、1103、1104、110401、1201、1301、1302、1401、1402、0701、080101、080201及び0901よりなる群から選択される。
【0010】
一部の実施形態においては、HER−2/Neu誘導ペプチドは配列番号2のアミノ酸配列を有する。別の実施形態においては、非相同ポリペプチドは顆粒球マクロファージコロニー刺激因子(GM−CSF)である。
【0011】
第2の態様において、本発明は上記したHER−2/Neu誘導ペプチド又はHER−2/Neu誘導ペプチドと非相同ポリペプチドをペプチド結合により連結した融合蛋白をコードするポリヌクレオチド配列を含む単離された核酸、核酸を含む発現カセット、及び発現カセットを含む宿主細胞を提供する。
【0012】
一部の場合においては、ポリヌクレオチド配列は配列番号2のアミノ酸配列を有するペプチドをコードする。別の場合においては、ポリヌクレオチド配列は非相同ポリペプチドがGM−CSFである融合蛋白をコードする。
【0013】
一部の実施形態においては、発現カセットは組み換えウィルスベクターである。他の実施形態においては、発現カセットは配列番号2のアミノ酸配列を有するペプチド又は非相同ポリペプチドがGM−CSFである組み換え融合蛋白の発現を指向する。
【0014】
第3の態様において、本発明は上記したHER−2/Neu誘導ペプチド又は非相同ポリペプチドと融合させたペプチドの融合産物を、生理学的に許容される賦形剤とともに含む組成物を提供する。
【0015】
一部の実施形態においては、ペプチドは配列番号2のアミノ酸配列を有する。別の実施形態においては、非相同ポリペプチドは顆粒球マクロファージコロニー刺激因子(GM−CSF)である。更に別の実施形態においては、組成物は更に抗原提示細胞を含み、これは細胞表面上の主要組織適合性複合体(MHC)分子と複合体を形成するHER−2/Neu誘導ペプチドを有する。
【0016】
第4の態様において、本発明はHER−2/Neu蛋白に対して特異的なT細胞免疫応答を患者において誘導するための方法を提供する。この方法は上記したHER−2/Neu誘導ペプチド又は非相同ポリペプチドと融合させたペプチドの融合産物、並びに生理学的に許容される賦形剤を含む組成物の有効量を患者に投与する工程を含む。
【0017】
一部の実施形態においては、ペプチドは配列番号2のアミノ酸配列を有する。別の実施形態においては、非相同ポリペプチドは顆粒球マクロファージコロニー刺激因子(GM−CSF)である。
【0018】
第5の態様において、本発明はHER−2/Neu蛋白に対して特異的なT細胞免疫応答を患者において検出するための方法を提供する。この方法は以下の工程、即ち(a)患者から抗原提示細胞及びT細胞を得る工程;(b)抗原提示細胞及びT細胞をHER−2/Neu誘導ペプチド、又は該ペプチドおよび非相同ポリペプチドを含む融合産物に接触させる工程;及び(c)T細胞応答を検出し、ここでT細胞応答の検出は患者におけるHER−2/Neu蛋白に対して特異的なT細胞免疫応答の存在を示すものである工程、を含む。
【0019】
一部の実施形態においては、工程(c)はELISPOT、増殖試験、又はフローサイトメトリーにより実施される。別の実施形態においてはHER−2/Neu誘導ペプチドは配列番号2のアミノ酸配列を有する。
【図面の簡単な説明】
【0020】
【図1】T細胞クローンHER500.23c21はHER−2/Neuに対して特異的である。(A)HER500.23c21によるHER500(アミノ酸■及びBA7072(HER500/GM−CSF融合蛋白)◆特異的IFNγ生産を抗原の自己PBMC提示の後にELISAを用いて測定した;バックグラウンドも試験した▲。試験は10%FBS添加完全IMDM培地中96ウェル丸底プレートにおいて3回で設定した。自己PBMCは2×105細胞/ウェルで使用し、T細胞クローンHER500.23c21は1×105細胞/ウェルで添加した。抗原は図示する終濃度とした。試験は5%CO2下37℃において48時間インキュベートし、上澄みをウェルから取り出してサイトカイン生産を試験した。(A)IFNγ生産。示したデータポイントは各抗原濃度に関する計算されたpg/mlを示す。(B)同じ試験に関するIL−2生産をHT−2細胞増殖を用いて計測した。データポイントは各抗原濃度に関する平均のCPM値を示す。
【図2】T細胞クローンHER500.23.21はペプチド#63に対して特異的である(HERp270−284)。HER−2/neuエピトープをクローンHER500.23.21に関してマッピングし、自己EBV−LcL及び各個々のHER500ペプチド#1−125(15アミノ酸長)を用いて特異性を測定した。試験は自己EBV−LcLは2×105細胞/ウェルで、T細胞クローンHER500.23.21は1×105細胞/ウェルで添加しながら96ウェル丸底プレート中に設定した。各ペプチドは終濃度1ug/mlで使用した。試験は5%CO2下37℃において48時間インキュベートし、その時点で上澄みをウェルから取り出してサイトカイン生産を試験した。(A)クローンHER500.23.21によるIFNγ生産はELISAを用いて計測し、計算されたpg/mlは各HER500ペプチドに対して示した。(B)HT−2細胞増殖を用いてIL−2の生産を計測した。
【図3】HLA−DR制限を示す。ブロッキング抗MHCクラスII抗体の存在下および非存在下において2×105細胞/mLの自己EBV−LcL及びHERp270−284を使用してT細胞クローンHER500.23c21を刺激した(1×105細胞/ウェル)。抗HLA−DR◆及び抗HLA−DQ/抗HLA−DP■をcRPMI+10%FBS中において図示した濃度において滴定した。ウェルには又、EBV−LcL、HER500.23c21T細胞クローン及びペプチド(1μg/ml)を含有させた。試験は5%CO2下37℃において48時間インキュベートした。上澄みを採取し、−20℃で凍結し、そして後日、ELISAを用いてIFNγ生産に関して分析した。複数の実験を実施し、1つの代表的実験に対して平均の3連の値を示す。
【図4】HERp270−284は無差別性のMHCクラスII HER−2/Neuエピトープである。(A)種々のHLA−DR対立遺伝子に対してホモ接合であるEBV−LcL系統を、クローンHER500.23c21に対してHERp270−284を提示するその能力に関して試験した。HER500.23c21による抗原特異的IFNγの生産は全てのEBV−LcL系統においてIFNγ ELISAの検出上限(4000pg/ml)を超えていた。陰性対照には無関係のHER500ペプチド、無ペプチド、及び各EBV−LcL系統単独を使用した(全値が検出不可能であり、記載しなかった)。試験は2×105EBV−LcL/ウェル及び1×105HER500.23c21細胞/ウェルにおいてcRPMI+10%FBS中、96ウェル丸底プレート中に設定した。ペプチドは終濃度1μg/mlで使用した。試験は5%CO2下37℃において48時間インキュベートした。上澄みを採取し、−20℃で凍結し、そしてELISAによりIFNγに関して分析した。結果は1つの代表的実験に関して示した。(B)HERp270−284を各EBV−LcL系統で滴定することにより異なる対立遺伝子に対するHER500.23c21の感受性を測定した。試験はHERp270−284の濃度を10ng/ml、33ng/ml、66ng/ml、100ng/ml及び250ng/mlとした以外は(A)と同様に設定した。250ng/ml超の全てのHERp270−284の濃度はIFNγ ELISAの検出上限(4000pg/ml)より高値のHER500.23c21によるIFNγの生産をもたらした。結果は1つの代表的実験に関して示す。
【図5】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す。ペプチドはHER−2/Neuペプチド270−284内の9量体であった。
【図6】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す。ペプチドはHER−2/Neuペプチド270−284内の10又は11量体であった。
【図7】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す(492nmの吸光度により測定)。ペプチドはHER−2/Neuペプチド270−284内の12量体であった。
【図8】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す(492nmの吸光度により示す)。ペプチドはHER−2/Neuペプチド270−284内の13量体であった。
【図9】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す(492nmの吸光度により示す)。ペプチドはHER−2/Neuペプチド270−284内の14量体であった。
【図10】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す(492nmの吸光度により示す)。ペプチドはHER−2/Neuペプチド268−286内の16量体であった。
【図11】多様なHLA−DRb1対立遺伝子のEBV LcL系統により提示されるペプチドへの曝露後のHER500.23c21によるIFNγの生産をグラフで示す(492nmの吸光度により示す)。ペプチドはHER−2/Neuペプチド268−286内の17量体であった。
【発明を実施するための形態】
【0021】
(定義)
「単離された」という用語は、核酸又は蛋白に適用する場合、核酸又は蛋白が、それが天然の状態で会合している他の細胞成分を本質的に含有しないことを指す。それは乾燥又は水溶液の何れかであることもできるが、好ましくは均質な状態である。純度及び均質性は典型的には分析化学の手法、例えばポリアクリルアミドゲル電気泳動又は高速液体クロマトグラフィーを用いて測定する。調製品中に存在する主な物質種である蛋白は実質的に精製されている。特に、単離された遺伝子は該遺伝子の側面に位置し目的の遺伝子以外の蛋白をコードするオープンリーディングフレームから分離されている。「精製された」という用語は核酸又は蛋白が電気泳動ゲルにおいて本質的に1つのバンドを生じさせることを指す。特にこのことは核酸又は蛋白が少なくとも85%純粋、より好ましくは少なくとも95%純粋、そして最も好ましくは少なくとも99%純粋であることを意味する。
【0022】
本出願において、「アミノ酸」という用語は天然に存在する、及び合成のアミノ酸、並びに天然に存在するアミノ酸と同様の態様において機能するアミノ酸類縁体及びアミノ酸ミメティックを指す。天然に存在するアミノ酸は遺伝子コードによりコードされているもの、並びに、後に修飾されているアミノ酸、例えばヒドロキシプロリン、γ−カルボキシグルタミン酸、及びO−ホスホセリンである。アミノ酸類縁体とは天然に存在するアミノ酸と同じ基本的化学構造、即ち水素、カルボキシル基、アミノ基、及びR基に結合しているα炭素を有する化合物、例えばホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウムを指す。そのような類縁体は修飾されたR基(例えばノルロイシン)又は修飾されたペプチド骨格を有するが、天然に存在するアミノ酸と同じ基本的化学構造を保持している。アミノ酸ミメティックはアミノ酸の一般的化学構造とは異なる構造を有するが天然に存在するアミノ酸と同様の態様において機能することができる化合物を指す。
【0023】
「核酸」又は「ポリヌクレオチド」という用語は1本鎖又は2本鎖の形態の何れかにおけるデオキシリボヌクレオチド又はリボヌクレオチド及びそれらの重合体を指す。特段の制約が無い限り、用語は参照核酸と同様の結合特性を有し、そして天然に存在するヌクレオチドと同様の態様において代謝される天然のヌクレオチドの既知類縁体を含有する核酸を包含する。特段の記載が無い限り、特定の核酸配列は保存的に修飾されたその変異体(例えば縮重コドン置換)及び相補配列並びに明示された配列をすべて包含する。特に、縮重コドン置換は1つ以上の選択された(又は全ての)コドンの第3位が混合塩基及び/又はデオキシイノシン残基により置換されている配列を形成することにより達成してよい(Batzer等、Nucleic Acid Res.19:5081(1991);Ohtsuka等、J.Biol.Chem.260:2605−2608(1985);及びRossolini等、Mol.Cell.Probes 8:91−98(1994))。核酸という用語は遺伝子、cDNA、又は遺伝子によりコードされたmRNAと互換的に使用する。
【0024】
ポリヌクレオチド配列におけるエレメントの相対的位置が関わる場合、「下流の」位置とは参照点の3’側にあるものであり、「上流の」位置とは参照点の5’側にあるものである。
【0025】
「ポリペプチド」、「ペプチド」及び「蛋白」という用語はアミノ酸残基の重合体を指すために本明細書において互換的に使用する。用語はアミノ酸残基1つ以上が相当する天然に存在するアミノ酸の人工的化学的ミメティックであるアミノ酸重合体に対して、並びに、天然に存在するアミノ酸重合体及び天然に存在するものではないアミノ酸の重合体に対して適用する。本明細書において、用語はアミノ酸残基が共有結合によるペプチド結合により連結されている完全長の蛋白(即ち抗原)を包含する何れかの長さのアミノ酸鎖を包含する。本出願においては、ポリペプチドのアミノ酸配列はN末端からC末端に向けて示す。換言すれば、ペプチドのアミノ酸配列を記載する場合、N末端から最初のアミノ酸を「第1のアミノ酸」と称する。
【0026】
融合蛋白の相手を記載する際に使用する場合、「非相同」という用語は1つのペプチド融合相手のもう1つのペプチド融合相手に対する関連性を指し:融合相手が融合ペプチド内に存在する態様が天然に存在する蛋白に観察できるものではない。例えば、HER−2/Neuエピトープと融合して融合ペプチドを形成している「非相同ポリペプチド」は顆粒球マクロファージコロニー刺激因子(GM−CSF)のようなHER−2/Neu蛋白以外の蛋白を起源とするものであってよい。一方、「非相同ポリペプチド」がHER−2/Neuエピトープに直ぐ隣接しないHER−2/Neu蛋白の別の部分から誘導されたものであってよい。「非相同ポリペプチド」は天然に存在する蛋白配列又はその一部分の修飾、例えばアミノ酸残基1つ以上の欠失、付加、又は置換を含有してよい。「非相同ポリペプチド」の起源(即ちそれがHER−2/Neu蛋白又は別の蛋白からの何れから誘導されているか)には関わりなく、融合ペプチドは配列番号1のアミノ酸配列を包含するヒトHER−2/Neuのサブ配列を含有してはならず、それは18アミノ酸長より大きい。一部の例示される実施形態においては、本発明において使用する「非相同ポリペプチド」は15〜20アミノ酸長以下を有し;別の実施形態においては「非相同ポリペプチド」は少なくとも100アミノ酸長を有する。
【0027】
「融合」又は「融合した」という単語は非相同ポリペプチドに連結したHER−2/Neuエピトープを含む本発明のペプチドを説明する際に使用する場合は、ペプチド結合を包含する何れかの共有結合によるエピトープと非相同ポリペプチドの間の連結を指す。
【0028】
「〜をコードする核酸配列」という表現はrRNA、tRNA、又は特定の蛋白又はペプチドの第1アミノ酸配列又はトランス作用性の調節剤に関する結合部位のような構造RNAに関する配列情報を含有する核酸を指す。この表現は特に、ネイティブの配列又は特定の宿主細胞におけるコドンの優勢性に合致するように導入してよい配列の縮重コドン(即ち単一のアミノ酸をコードする異なるコドン)を包含する。
【0029】
「発現カセット」とは宿主細胞における特定のポリヌクレオチド配列の転写を可能にする、特定された核酸エレメントのシリーズを有する組み換え又は合成により形成された核酸構築物である。発現カセットはプラスミド、ウィルスゲノム、又は核酸フラグメントの部分であってよい。典型的には発現カセットはプロモーターに作動可能に連結した転写されるべきポリヌクレオチドを包含する。
【0030】
「組み換え」という用語は、例えば細胞又は核酸、蛋白、又はベクターに関して使用する場合は、細胞、核酸、蛋白又はベクターが、外部原料からの核酸又は蛋白の導入、又はネイティブの核酸又は蛋白の改変により修飾されていること、又は細胞がそのように修飾された細胞から誘導されることを表す。即ち、例えば組み換え細胞は細胞のネイティブ(非組み換え)型内部には存在しない遺伝子を発現するか、又は、別様には異常に発現されるか、過少発現されるか、又は全く発現されないネイティブの遺伝子を発現する。
【0031】
「投与」又は「投与する」という用語は哺乳類、特にヒトに物質を接触させる種々の方法を指す。投与の様式は例えば限定しないが、物質を静脈内、腹腔内、鼻内、経皮、局所、皮下、非経口、筋肉内、経口、又は全身投与により、そして任意の他の手段による注射、摂取、吸入、移植、又は吸収を介して接触させることを含む方法を包含する。本発明のHER−2/Neuペプチド又はHER−2/Neuペプチドと非相同ポリペプチドを含む融合ペプチドの投与の1つの例示的な手段は、静脈内送達を介するものであり、その場合ペプチド又は融合ペプチドは水溶液、懸濁液、又はエマルジョン等のような静脈内注射のために適当な形態の医薬組成物として製剤できる。本発明のHER−2/Neuペプチド又は融合ペプチドを送達するための別の手段は皮内注射、皮下注射、筋肉内注射、又はパッチ等を用いる経皮適用を包含する。
【0032】
特定の物質の「有効量」とは所望の結果をもたらすために十分である物質の量を指す。例えば、抗HER−2/Neu免疫を誘導することを意図される本発明のペプチドを含む組成物の有効量は、被験体に投与された場合に免疫を誘導するという目標を達成するために十分な量である。達成すべき作用は検出可能な程度までの、疾患/状態及び関連する合併症の症状の進行の防止、是正、又は抑制を包含する。「有効量」の厳密な量は投与の目的に依存し、周知の手法を用いながら当業者により確認できる(例えばLieberman,Pharmaceutical Dosage Forms(vols.1−3,1992);Lloyd,The Art,Science and Technology of Pharmaceutical Compounding(1999);及びPickar,Dosage Calculations(1999)を参照)。
【0033】
「生理学的に許容される賦形剤」とは本発明の組成物の製剤中に使用される不活性な成分であり、これはHER−2/Neuペプチド又はHER−2/Neuペプチドと非相同ポリペプチドを含む融合ペプチドの活性成分を含有し、そして、例えばそれを必要とする患者への注射による使用のために適している。この不活性成分は、本発明の組成物中に包含された場合に組成物の所望のpH、コンシステンシー、色、におい、又はフレーバーを与える物質であってよい。
【0034】
本明細書においては、「T細胞免疫応答」という用語は細胞表面上の分子の増殖、又は発現、又はサイトカインのような蛋白の分泌により測定した場合の抗原特異的T細胞の活性化を指す。
I.序論
本発明者等はHER−2/Neu蛋白に由来する一連の新規な無差別性のT細胞エピトープを発見した。これらのペプチドエピトープは、それらが少なくとも25種の異なるHLA−DRB1対立遺伝子の範囲内において提示され得ることから、顕著なHLA無差別性を呈している。そのような広範なHLA−DRB1対立遺伝子によるこれらのエピトープの提示のため、これらのエピトープは、一般的なヒトの集団におけるHER−2/Neuを過剰発現する癌の治療のためのワクチン製造又は免疫療法におけるユニバーサルCD4 ヘルパーT細胞エピトープとして極めて価値あるものとなっている。
II.ペプチドの化学合成
本発明のペプチド、特に比較的短鎖のもの(例えば50〜100アミノ酸以下)は従来のペプチド合成又は当該分野で良く知られている他のプロトコルを用いて化学合成してよい。
【0035】
ペプチドはMerrifield等、J.Am.Chem.Soc.85:2149−2156(1963);Barany and Merrifield,Solid−Phase Peptide Synthesis,in The Peptides:Analysis,Synthesis,Biology Gross and Meienhofer(eds.),Academic Press,N.Y.,vol.2,pp.3−284(1980);及びStewart等、Solid Phase Peptide Synthesis 2nd ed.,Pierce Chem.Co.,Rockford,III.(1984)により記載されているものと同様の操作法を用いながら固相ペプチド合成法により合成してよい。合成の間、保護された側鎖を有するN−α−保護アミノ酸を、自身のC末端により連結されている成長中のポリペプチド鎖に対し、そして、固体支持体、即ちポリスチレンビーズに対して段階的に添加する。ペプチドはジシクロヘキシルカルボジイミドのような試薬と反応させることにより活性化されているN−α−保護アミノ酸のα−カルボキシ基にN−α−脱保護アミノ酸のアミノ基を連結することにより合成される。活性化されたカルボキシルへの遊離のアミノ基の結合はペプチド結合の形成をもたらす。もっとも一般的に使用されているN−α−保護基は酸不安定性のBoc、及び塩基不安定性のFmocを包含する。
【0036】
固体支持体としての使用のために適する材料は当業者に良く知られており、例えば限定しないが、以下のもの、即ち:ハロメチル樹脂、例えばクロロメチル樹脂又はブロモメチル樹脂;ヒドロキシメチル樹脂;フェノール樹脂、例えば4−(α−[2,4−ジメトキシフェニル]−Fmoc−アミノメチル)フェノキシ樹脂;tert−アルキルオキシカルボニルヒドラジド化樹脂等を包含する。このような樹脂は市販されており、それらの製造方法は当業者に周知である。
【0037】
慨すれば、C末端N−α−保護アミノ酸をまず固体支持体に結合させる。次にN−α−保護基を除去する。脱保護されたα−アミノ基を次のN−α−保護アミノ酸の活性化されたα−カルボキシレート基にカップリングさせる。所望のペプチドが合成されるまでプロセスを反復する。次に得られたペプチドを不溶性重合体支持体から切断し、アミノ酸側鎖を脱保護する。より長いペプチドは保護されたペプチドフラグメントの縮合により誘導できる。適切な化学的手法、樹脂、保護基、保護されたアミノ酸及び試薬の詳細は当該分野で良く知られており、本明細書において詳細に考察しない(例えばAtherton等、Solid Phase Peptide Synthesis:A Practical Approach,IRL Press(1989)及びBodanszky,Peptide Chemistry,A Practical Textbook,2nd Ed.,Springer−Verlag(1993)を参照)。
III.ペプチドの組み換え生産
A.一般的な組み換え技術
組み換え遺伝子学の分野における一般的な方法及び手法を開示している基礎的なテキストはSambrook and Russell,Molecular Cloning,A Laboratory Manual(3rd ed.2001);Kriegler,Gene Transfer and Expression:A Laboratory Manual(1990);及びAusubel等編、Current Protocols in Molecular Biology(1994)を包含する。
【0038】
核酸に関しては、サイズはキロ塩基(kb)か塩基対(bp)の何れかで表示される。これらはアガロース又はアクリルアミドゲル電気泳動から、配列決定された核酸から、又は公開されたDNA配列から誘導される推定値である。蛋白に関しては、サイズはキロダルトン(kDa)又はアミノ酸残基数で表示される。蛋白サイズはゲル電気泳動から、配列決定された蛋白から、誘導されたアミノ酸配列から、又は公開された蛋白配列から推定される。
【0039】
市販されていないオリゴヌクレオチドは例えばVan Devanter等、Nucleic Acids Res.12:6159−6168(1984)に記載の通り自動合成装置を用いながら、Beaucage & Caruthers,Tetrahedron Lett.22:1859−1862(1981)において最初に報告された固相ホスホロアミダイトトリエステル法に従って化学合成することができる。オリゴヌクレオチドの精製は何れかの当該分野で認識されている方策、例えばネイティブのアクリルアミドゲル電気泳動又はアニオン交換HPLCを用いながら、Pearson & Reanier,J.Chrom.255:137−149(1983)に記載の通り実施する。
【0040】
組み換え生産は本発明のペプチド、特に比較的大きい分子量のもの、例えばHER−2/NeuエピトープとGM−CSFの融合ペプチドを得るための有効な手段である。本発明のペプチドをコードするポリヌクレオチド及び合成オリゴヌクレオチドの配列は、例えばWallace等、Gene16:21−26(1981)の2本鎖鋳型を配列決定するための鎖停止法を用いながらクローニング又はサブクローニングした後に確認できる。
B.発現カセットの構築
本発明のペプチドをコードするポリヌクレオチド配列の獲得
本発明のペプチドをコードするポリヌクレオチド配列は化学合成により得ることができ、あるいは、市販元より購入することができ、そしてこれを次に分子クローニングの標準的な手法を用いながらさらに操作してよい。
宿主生物における好ましいコドン使用のための核酸の修飾
本発明のペプチドをコードするポリヌクレオチド配列は場合により特定の宿主における好ましいコドンの使用に合致するように任意に改変することができる。例えば、細菌細胞の1つの系統における好ましいコドンの使用は、本発明のペプチドをコードし、この系統により好まれるコドンを包含するポリヌクレオチドを誘導するために使用できる。宿主細胞により示される好ましいコドンの使用の頻度は、宿主細胞により発現される遺伝子の大多数における好ましいコドンの使用の頻度を平均することにより計算できる(計算サービスはKazusa DNA Research Institute,Japanのウエブサイトから利用できる)。この分析は好ましくは宿主細胞により高度に発現される遺伝子に限定される。
【0041】
修飾終了時に、コーディング配列を配列決定により確認し、次に本発明のペプチドの組み換え製造のための適切な発現ベクターにサブクローニングする。
【0042】
コーディング配列の確認の後、組み換え遺伝子学の分野の定型的手法を用いて本発明のペプチドを製造できる。
C.発現系
本発明のペプチドをコードする核酸の高レベルの発現を実現するためには、典型的には、転写を指向する強力なプロモーター、転写/翻訳ターミネーター、及び翻訳開始のためのリボソーム結合部位を含有する発現ベクター内にペプチドをコードするポリヌクレオチドをサブクローニングする。適当な細菌プロモーターは当該分野で周知であり、上記したSambrook and Russell及び上記したAusubel等において説明されている。本発明のペプチドを発現させるための細菌の発現系は例えばE.coli、バチルス種、サルモネラ及びカウロバクターにおいて入手できる。そのような発現系のためのキットは市販されている。哺乳動物細胞、酵母及び昆虫細胞に対する真核生物の発現系は当該分野で周知であり、やはり市販されている。1つの実施形態において、真核生物発現ベクターはアデノウィルスベクター、アデノ関連ベクター、又はレトロウィルスベクターである。
【0043】
相同核酸の発現を指向するために使用されるプロモーターは、特定の用途に依存する。プロモーターは任意に、その天然の設定における転写開始部位からの距離と概ね同じ非相同転写開始部位からの距離に位置づけられる。しかしながら、当該分野で公知の通りこの距離におけるある程度の変動は、プロモーター機能を喪失することなく順応され得る。
【0044】
プロモーターの他に、発現ベクターは典型的には宿主細胞内の本発明のペプチドの発現のために必要な追加的エレメントの全てを含有する転写ユニット又は発現カセットを包含する。即ち典型的な発現カセットはペプチドをコードするポリヌクレオチド配列に作動可能に連結したプロモーター、及び、効率的な転写物のポリアデニル化、リボソーム結合部位、及び翻訳停止に必要なシグナルを含有する。ペプチドをコードする核酸配列は典型的には形質転換された細胞によるペプチドの分泌を促進するように切断可能なシグナルペプチド配列に連結される。そのようなシグナルペプチドは、特に、組織プラスミノーゲン活性化物質、インスリン、及びニューロン成長因子、及びニセアメリカタバコガ(Heliothis virescens)の幼若ホルモンエステラーゼ由来のシグナルペプチドを包含する。カセットの追加的エレメントはエンハンサーを、そして、ゲノムDNAを構造遺伝子(例えば非相同ポリペプチドをコードするもの)として使用する場合は機能的スプライスドナーを有するイントロン及びアクセプター部位を包含してよい。
【0045】
プロモーター配列に加えて、発現カセットは効率的な終止を可能とするために構造遺伝子の下流に転写終止領域を含有しなければならない。終止領域はプロモーター配列と同じ遺伝子から得てよく、或いは、異なる遺伝子から得てよい。
【0046】
細胞内に遺伝子情報を輸送するために使用される特定の発現ベクターは特に重要ではない。真核生物又は原核生物の細胞における発現のために使用されている従来のベクターの何れかを使用してよい。標準的な細菌の発現ベクターはプラスミド、例えばpBR322系のプラスミド、pSKF、pET23D、及び融合発現系、例えばGST及びLacZを包含する。エピトープタグも組み換え蛋白に添加することにより単離の好都合な方法としてよく、例えばc−mycが挙げられる。
【0047】
真核生物ウィルス由来の調節エレメントを含有する発現ベクターが典型的には真核生物発現ベクターにおいて使用され、例えばSV40ベクター、乳頭腫ウィルスベクター、及びエプスタイン・バーウィルス由来のベクターが挙げられる。他の例示的な真核生物ベクターはpMSG、pAV009/A+、pMTO10/A+、pMAMneo−5、バキュロウィルスpDSVE、及び、SV40初期プロモーター、SV40後期プロモーター、メタロチオネインプロモーター、ネズミ乳房腫瘍ウィルスプロモーター、ラウス肉腫ウィルスプロモーター、多核体プロモーター、又は真核生物細胞における発現のために効果的であることがわかっている他のプロモーターの指向下に蛋白の発現を可能とする何れかの他のベクターを包含する。
【0048】
一部の発現系はチミジンキナーゼ、ハイグロマイシンBホスホトランスフェラーゼ、及びジヒドロ葉酸還元酵素のような遺伝子増幅をもたらすマーカーを有する。或いは、遺伝子増幅をおこなわない高収率の発現系、例えば多核体プロモーター又は他の強力なバキュロウィルスプロモーターの指向下に本発明のペプチドをコードするポリヌクレオチド配列を有する昆虫細胞におけるバキュロウィルスベクターもまた適当である。
【0049】
発現ベクター内に典型的に包含されるエレメントは又、E.coli中で機能するレプリコン、組み換えプラスミドを保有している細菌の選択を可能にする抗生物質耐性をコードしている遺伝子、及び真核生物の配列の挿入を可能にするプラスミドの非必須領域のユニークな制限部位も包含する。選択される特定の抗生物質耐性遺伝子は重要ではなく当該分野で公知の多くの耐性遺伝子の何れも適当である。原核生物配列は任意に、必要に応じて真核生物細胞におけるDNAの複製を妨害しないようなものが選択される。抗生物質耐性選択マーカーと同様、既知の代謝経路に基づいた代謝選択マーカーも又形質転換された宿主細胞を選択するための手段として使用してよい。
【0050】
組み換え蛋白(例えば本発明のペプチド)のペリプラズムの発現が望まれる場合は、発現ベクターは更に、発現されるべき蛋白のコーディング配列の5’に直接連結されたE.coli OppA(ペリプラズムオリゴペプチド結合蛋白)分泌シグナル、又はその修飾された型のような分泌シグナルをコードする配列を含む。このシグナル配列は細胞質中に生産された組み換え蛋白を細胞膜通過によりペリプラズム空間内に指向させる。発現ベクターはさらにシグナルペプチダーゼ1に関するコーディング配列を含んでよく、これは組み換え蛋白のペリプラズム空間内への進入時にシグナル配列を酵素的に切断することができる。組み換え蛋白のペリプラズム生産に関するより詳細な説明は、例えばGray等、Gene39:247−254(1985)、米国特許第6,160,089号明細書及び同第6,436,674号明細書に記載されている。
C.トランスフェクション法
標準的なトランスフェクション法を用いて本発明のペプチドを大量に発現する細菌、哺乳類、酵母、昆虫、又は植物の細胞系統を作成し、それを次に標準的な手法で精製する(例えばColley等、J.Biol.Chem.264:17619−17622(1989);Guide to Protein Purification,in Methods in Enzymology,vol.182(Deutscher,ed.,1990)参照)。真核生物及び原核生物の細胞の形質転換は標準的手法に従って実施する(例えばMorrison,J.Bact.132:349−351(1977);Clark−Curtiss & Curtiss,Methods in Enzymology 101:347−362(Wu等編、1983)参照)。
【0051】
宿主細胞内に外来ヌクレオチド配列を導入するための周知の操作法のいずれかを使用してよい。これらにはリン酸カルシウムトランスフェクション、ポリブレン、プロトプラスト融合、エレクトロポレーション、リポソーム、マイクロインジェクション、プラズマベクター、ウィルスベクターの使用、及び宿主細胞内にクローニングされたゲノムDNA、cDNA、合成DNA、又は他の外来性の遺伝子材料を導入するための他の周知の方法が包含される(例えば、Sambrook and Russell,上出参照)。使用する特定の遺伝子操作法は、本発明のペプチドを発現できる宿主細胞内に少なくとも1つの遺伝子を良好に導入することができることのみが必要である。
D.宿主細胞内のペプチドの組み換え発現の検出
適切な宿主細胞内に発現ベクターを導入した後、トランスフェクトした細胞を本発明のペプチドの発現に好都合な条件下で培養する。次に組み換えペプチドの発現に関して細胞をスクリーニングし、これはその後、標準的方法を用いて培地から回収する(例えばScopes,Protein Purification:Principles and Practice(1982);米国特許第4,673,641号明細書;Ausubel等、上出;及びSambrook and Russell,上出参照)。
【0052】
遺伝子発現をスクリーニングするための数種の一般的方法は当業者の間で周知である。第1に、遺伝子発現は核酸レベルにおいて検出できる。核酸ハイブリダイゼーション手法を用いた特定のDNA及びRNAの測定の種々の方法が一般的に使用されている(例えばSambrook and Russell,上出)。一部の方法では電気泳動による分離を行うが(例えばDNAを検出するためのサザンブロット及びRNAを検出するためのノーザンブロット)、DNA又はRNAの検出は電気泳動を使用することなく実施することもできる(例えばドットブロットによる)。トランスフェクトされた細胞における本発明のペプチドをコードする核酸の存在は配列特異的プライマーを用いたPCR又はRT−PCRにより検出することもできる。
【0053】
第2に、遺伝子発現はポリペプチドレベルにおいて検出できる。特に、本発明のペプチド、特に十分大きい非相同ポリペプチドを含有するものと特異的に反応するポリクローナル又はモノクローナル抗体を用いながら遺伝子産物のレベルを計測するために、種々の免疫学的試験が当業者により類型的に使用されている(例えばHarlow and Lane,Antibodies,A Laboratory Manual,Chapter 14,Cold Spring Harbor,1988;Kohler and Milstein,Nature,256:495−497(1975))。このような手法はペプチド又はその抗原性部分に対して高い特異性を有する抗体を選択することによる抗体製造を必要とする。ポリクローナル及びモノクローナル抗体を形成する方法は十分確立されており、それらの説明は文献、例えばHarlow and Lane,上出;Kohler and Milstein,Eur.J.Immunol.,6:511−519(1976)に記載されている。
IV.ペプチドの精製
A.化学的に合成されたペプチドの精製
合成ペプチドの精製はクロマトグラフィーの種々の方法、例えば逆相HPLC、ゲル透過、イオン交換、サイズエクスクルージョン、アフィニティー、分配、又は向流分配を用いて実施される。適切なマトリックス及び緩衝物質の選択は当該分野で周知である。
B.組み換え生産されたペプチドの精製
1.細菌封入体からのペプチドの精製
発現は構成的なものとなるが、典型的にはプロモーター導入後に大量の形質転換された細菌から本発明のペプチドを組み換えにより製造する場合、ペプチドは不溶性の凝集塊を形成する場合がある。蛋白封入体の精製のために適当である数種のプロトコルが存在する。例えば凝集塊蛋白(以下、封入体と称する)の精製では典型的には例えば約100〜150μg/mlリゾチーム及び0.1% Nonidet P40(非イオン系界面活性剤)の緩衝液中のインキュベーションによる細菌細胞の破壊により、封入体の抽出、分離及び/又は精製を行う。細胞懸濁液はポリトロン粉砕機(Brinkman Instruments,Westbury,NY)を用いて粉砕できる。或いは、細胞は氷上で超音波処理できる。細菌を溶解する代替方法はともに上出のAusubel等、及びSambrook and Russellに記載されており、当業者に明らかである。
【0054】
細胞懸濁液は一般的に遠心分離し、そして封入体を含有するペレットを、封入体を溶解しないが洗浄する緩衝液、例えば20mM Tris−HCl(pH7.2)、1mM EDTA、150mM NaCl及び2% Triton−X100(非イオン系界面活性剤)中に再懸濁させる。可能な限り細胞破砕物を除去するために洗浄工程を反復することが必要な場合がある。封入体の残存ペレットを適切な緩衝液(例えば20mMリン酸ナトリウム、pH6.8,150mM NaCl)中に再懸濁してよい。他の適切な緩衝液は当業者に明らかである。
【0055】
洗浄工程の後、強力な水素受容体および強力な水素供与体の両方である溶媒(又はこのような特性の一方を各々が有する溶媒の組み合わせ)を添加することにより封入体を可溶化する。次に封入体を形成した蛋白を適合する緩衝液を用いた希釈又は透析により復元してよい。適当な溶媒は、限定しないが尿素(約4M〜約8M)、ホルムアミド(少なくとも約80%、体積/体積に基づく)、及び塩酸グアニジン(約4M〜約8M)を包含する。凝集塊形成蛋白を可溶化させることができる一部の溶媒、例えばSDS(ドデシル硫酸ナトリウム)及び70%ギ酸は、免疫原性及び/又は活性の欠如を伴った蛋白の不可逆的変性の可能性のために、この操作法における使用のためには不適切である場合がある。塩酸グアニジン及び同様の試薬は変性剤であるが、この変性は不可逆ではなく、そして変性剤の除去(例えば透析による)又は希釈時に復元がおこる場合があり、これにより目的の免疫学的及び/又は生物学的に活性な蛋白の再形成が可能となる。可溶化の後、蛋白は標準的分離手法により他の細菌蛋白から分離できる。細菌封入体から組み換えポリペプチドを精製することに関する説明は更に、例えばPatra等、Protein Expression and Purification 18:182−190(2000)に記載されている。
【0056】
或いは、組み換えポリペプチド、例えば本発明のペプチドを細菌のペリプラズムから精製することが可能である。細菌のペリプラズム内に組み換えポリペプチドをエクスポートする場合、細菌のペリプラズム画分は、当業者に周知の他の方法に加えて冷温浸透圧ショックにより単離できる(例えばAusubel等,上出参照)。ペリプラズムから組み換えペプチドを単離するためには、細菌細胞を遠心分離してペレットを生じさせる。ペレットを20%スクロース含有緩衝液に再懸濁する。細胞を溶解するために、細菌を遠心分離し、ペレットを氷冷5mM MgSO4中に再懸濁し、そして約10分間アイスバス中に保持する。細胞懸濁液を遠心分離し、上澄みを傾瀉して保存する。上澄み中に存在する組み換えペプチドは当業者に周知の標準的な分離手法により宿主蛋白から分離することができる。
2.精製のための標準的な蛋白分離手法
組み換えポリペプチド、例えば本発明のペプチドを可溶性形態で宿主細胞内に発現させる場合、その精製は以下に記載する標準的な蛋白精製操作法に従うことができる。この標準的精製操作法は又、化学合成により得られたペプチドを精製するためにも適している。
i.溶解度分画
多くの場合初期工程として、そして蛋白混合物が複合体である場合、初期の塩分画は目的の組み換え蛋白、例えば本発明のペプチドから望ましくない宿主細胞蛋白(又は細胞培養用の培地から誘導される蛋白)の多くを分離することができる。好ましい塩は硫酸アンモニウムである。硫酸アンモニウムは蛋白混合物中の水の量を効果的に低減することにより蛋白を沈殿させる。次に蛋白がその溶解度に基づいて沈殿する。蛋白が疎水性であるほど、より低い硫酸アンモニウム濃度において沈殿する可能性が高まる。典型的なプロトコルは結果として生じる硫酸アンモニウム濃度が20〜30%となるように蛋白溶液に飽和硫酸アンモニウムを添加することである。これにより大部分の疎水性蛋白が沈殿することになる。沈殿を廃棄(目的の蛋白が疎水性でない限り)し、目的蛋白を沈殿させることが分かっている濃度となるまで硫酸アンモニウムを上澄みに添加する。次に沈殿を緩衝液中に可溶化し、必要に応じて過剰な塩を透析又は透析濾過により除去する。蛋白の溶解度に依存している他の方法、例えば低温エタノール沈殿等は当業者に周知であり、複合体蛋白混合物を分画するために使用できる。
ii.サイズ示差濾過
計算された分子量に応じて、より大きい及び小さいサイズの蛋白を異なる孔径の膜(例えばAmicon又はMilliporeメンブレン)を通過させる限外濾過を用いて単離できる。第1の工程として、目的の蛋白、例えば本発明のペプチドの分子量より低値の分子量カットオフを有する孔径の膜を通過させて蛋白混合物を限外濾過する。次に限外濾過の保持物を、目的のペプチドの分子量より大きい分子カットオフを有する膜に対して限外濾過する。組み換え蛋白は膜を通過して濾液中に入ることになる。次に濾液を後述する通りクロマトグラフィーに付すことができる。
iii.カラムクロマトグラフィー
目的の蛋白(例えば本発明のペプチド)は又そのサイズ、実質的表面電荷、疎水性、又はリガンドに対する親和性に基づいて他の蛋白から分離できる。更に又、本発明のペプチドに対して作成された抗体をカラムマトリックスに結合し、ペプチドを免疫精製することもできる。これらの方法は全て当該分野で周知である。
【0057】
クロマトグラフィー手法は何れかの規模において、多くの異なる製造元から得た機材を用いながら実施できることは当業者に明らかである(例えばPharmacia Biotech)。
C.ペプチド配列の確認
本発明のペプチドのアミノ酸配列は多くの十分に確立された方法により確認できる。例えばEdman分解の従来法を用いてペプチドのアミノ酸配列を決定できる。Edman分解に基づく配列決定法の数種の変形例、例えばマイクロ配列決定、及び質量分析に基づく方法がこの目的のために頻繁に使用されている。
D.ペプチドの修飾
本発明のペプチドはより望ましい特性を達成するために修飾できる。化学修飾されたペプチド及び蛋白分解酵素による分解に対して抵抗性であるか向上した溶解性又は結合能力を有するペプチド模倣体の設計はよく知られている。
【0058】
本発明のHER−2/Neuペプチド又は融合ペプチドの修飾されたアミノ酸又は化学的誘導体は通常はHER−2/Neu蛋白の一部ではない修飾されたアミノ酸の追加的化学特徴部分を含有してよい。ペプチドの共有結合修飾は本発明の範囲に含まれる。そのような修飾は選択された側鎖又は末端残基と反応することができる有機誘導体形成剤にペプチドのターゲティングされたアミノ酸残基を反応させることによりペプチド内に導入してよい。化学誘導体の以下の例は例示として示すのみであり、限定するものではない。
【0059】
蛋白分解性酵素による分解に耐性であるペプチド模倣体の設計は当業者の知る通りである。例えばSawyer,Structure−Based Drug Design,P.Verapandia,Ed.,N.Y.(1997);米国特許第5,552,534号明細書及び同第5,550,251号明細書を参照。ペプチド骨格及び側鎖の修飾の両方を二次構造模倣の設計において使用してよい。可能な修飾はD−アミノ酸、Nα−Me−アミノ酸、Cα−Me−アミノ酸、及びデヒドロアミノ酸の置換を包含する。今日まで、種々の二次構造ミメティックが設計されており、そしてペプチド又はペプチドミメティックに組み込まれている。
【0060】
他の修飾には天然のアミノ酸の非天然のヒドロキシル化アミノ酸による置換、酸性アミノ酸のカルボキシ基のニトリル誘導体による置換、塩基性アミノ酸のヒドロキシル基のアルキル基による置換、又はメチオニンのメチオニンスルホキシドによる置換が包含される。更に又、本発明のHER−2/Neuペプチド又は融合ペプチドのアミノ酸は同じアミノ酸であるが逆のキラリティーのもので置き換えることができ、即ち、天然に存在するL−アミノ酸はそのD配置により置き換えられてよい。
V.非相同ポリペプチドとのHER−2/Neuエピトープの融合
本発明の1つの態様において、HER−2/Neu無差別性T細胞エピトープに相当するペプチドは、T細胞応答を誘導するHER−2/Neuエピトープの能力が増強されるように共有結合を介して非相同ポリペプチドに結合させることにより、融合ペプチドを形成する。頻繁にはこの共有結合はペプチド結合であり、そしてHER−2/Neuエピトープ及び非相同ポリペプチドは新しいポリペプチドを形成する。このペプチド結合はHER−2/Neuエピトープと非相同ポリペプチドの間の直接のペプチド結合であってよく、或いはそれはHER−2/Neuエピトープと非相同ポリペプチドの間のペプチドリンカーを用いて与えられる間接のペプチド結合であってよい。
【0061】
他の共有結合もまた非相同ポリペプチドにHER−2/Neuエピトープを融合させる目的のために適している。例えば、あるペプチドの官能基(例えば非末端アミン基、非末端カルボキシル酸基、ヒドロキシル基、及びスルフヒドリル基)は別のペプチドの官能基と容易に反応し、そして2つのペプチドを結合するペプチド結合以外の共有結合を確立する場合がある。HER−2/Neuエピトープのペプチドと非相同ポリペプチドの間の共有結合による連結は又、適当な官能基を有するリンカー分子を用いることにより得ることもできる。そのようなリンカー分子はペプチドリンカー又は非ペプチドリンカーであることができる。リンカーを結合の前に誘導体化することにより露出させるか、又は追加的な反応性官能基を結合させてよい。誘導体化においてはPierce Chemical Company,Rockford,Illinoisから入手可能なもののような多くの分子のいずれかの取り付けを行ってよい。
VI.機能的試験
本発明のHER−2/Neuエピトープ(又はHER−2/Neuペプチドと非相同ポリペプチドを含む融合ペプチド)は、少なくとも10種の異なるHLA−DR対立遺伝子、より好ましくは少なくとも15、20又は25種の異なるHLA−DR対立遺伝子の1つを有する場合がある抗原提示細胞によりエピトープが提示される場合に、HER−2/Neu蛋白に特異的なT細胞免疫応答を誘導するその能力のために有用である。種々の機能的試験を用いることにより、種々の異なるHLA−DR対立遺伝子の抗原提示細胞に関して無差別性の態様におけるそのようなHER−2/Neu特異的T細胞免疫応答を誘導するHER−2/Neuエピトープの能力を確認することができ、例えば増殖試験及びフローサイトメトリー試験によりT細胞受容体とペプチドエピトープの間の結合又はT細胞によるサイトカインの生産を検出する方法などが挙げられる。
【0062】
本出願の実施例において使用する機能的試験の系はこの目的のために特に適している。慨すれば異なるHLA−DR対立遺伝子に対して各々ホモ接合である少なくとも10種、好ましくは少なくとも15、20又は25種の抗原提示細胞系統のパネルを用いて、HER−2/Neu蛋白に対して特異的に応答(例えばIFNγ又はIL−2のようなサイトカインの生産による)するCD4+ T細胞のクローン(例えばクローンHER500.23c21)に対してHER−2/Neu誘導ペプチドを提示する。配列番号2のアミノ酸配列を有するエピトープ270〜284を陽性対照として使用し、そして非関連のHER−2/Neu誘導ペプチド、無ペプチド、及び各抗原提示細胞系統単独を試験の陰性対照として使用する。試験は各ウェル中抗原提示細胞及びCD4+ T細胞を含有する適切な培地中におけるマルチウェル細胞培養プレートとして設定する。ペプチドは適当な濃度まで希釈し、そして各ウェルに添加する。適切な時間インキュベートした後、上澄みをウェルから収集し、そして492nmにおける吸光度に基づいてELISAにより計測できるサイトカインの生産に関して分析する。典型的には、HER−2/Neu特異的CD4+ T細胞応答を誘導する場合の本発明のHER−2/NeuクラスII無差別エピトープの作用は同じ試験条件下において、例えば同じモル濃度において、HER−2/Neuエピトープ270〜284(配列番号2に示すアミノ酸配列を有する)の作用の少なくとも25%であり、同じ個別のHLA−DR対立遺伝子の抗原提示細胞により提示される。より好ましくはそのような作用は同じ条件下でHER−2/Neuエピトープ270〜284により示されるものよりも少なくとも30%、40%、50%、60%、70%、80%、又はそれより高値である。
VII.組成物及び投与
本発明は更に有効量の(1)HER−2/Neuペプチド;又は(2)HER−2/Neuペプチドと非相同ポリペプチドを含む融合ペプチド;又は(3)予防及び治療の両方の用途におけるHER−2/Neu蛋白に対して特異的なT細胞免疫応答を誘導するための細胞表面上のMHC分子との複合体を形成する(1)又は(2)のペプチドを伴った抗原提示細胞(APC)を含む組成物を提供する。本発明の医薬組成物は種々の薬物送達系における使用に適している。本発明で使用するための適当な製剤はRemington’s Pharmaceutical Sciences,Mack Publishing Company,Philadelphia,PA,17th ed.(1985)に記載されている。薬物送達方法に関する簡単な考察については、Langer,Science 249:1527−1533(1990)を参照されたい。
【0063】
抗原提示細胞(APC)は種々の方法によりペプチド負荷のために形成できる。出発原料は末梢血または、可動化をともなうかもしくはともなわない白血球除去物である。APCは多数の方法、例えば浮力密度遠心分離、水簸、磁気ビーズ及びプラスチック接着等を単独又は組み合わせて使用することにより単離できる。単離後、APCをサイトカイン、成長因子、活性化剤、及び成熟化剤の存在下又は非存在下に1〜14日間培養する。6〜48時間1μg〜1mg/mlの濃度において培地にペプチドを添加することによりAPCにペプチドを負荷させる。APCを採取し、洗浄し、そして注入のための適当な製剤中に再懸濁する。APCは新鮮時に送達することができ、或いは、後の時期における送達のために凍結保存することができる。
【0064】
本発明の医薬組成物は種々の経路で、例えば皮下、皮内、経皮、筋肉内、静脈内、又は腹腔内に投与できる。医薬組成物を投与する好ましい経路は、体重70kgの成人に対して本発明のペプチド約1μg〜10mg、好ましくは50μg〜1mgの2週毎用量において皮下又は皮内によるものである。適切な用量は1週毎、2週毎、又は1月毎の間隔において投与してよい。
【0065】
ペプチドパルスAPCは種々の経路で、例えば皮下、皮内、静脈内、又は腹腔内に投与できる。ペプチドパルスAPCは1週毎、2週毎、又は1月毎の間隔において細胞100万個〜100億個の用量で送達される。
【0066】
本発明のペプチドを含有する医薬組成物を製造するためには、不活性及び製薬上許容しうる賦形剤又は担体を使用する。液体の医薬組成物は例えば皮内、皮下、非経口、又は静脈内投与に適する溶液、懸濁液、及びエマルジョンを包含する。活性成分(例えばHER−2/Neuペプチド又は融合ペプチド)の滅菌された水溶液、又は水、緩衝された水、食塩水、PBS、エタノール、又はプロピレングリコールを含む溶媒中の活性成分の滅菌溶液が非経口投与に適する液体組成物の例である。組成物は生理学的状態に近似するために必要な製薬上許容しうる補助物質、例えばpH調節剤及び緩衝剤、張度調節剤、湿潤剤、界面活性剤などを含有してよい。
【0067】
滅菌溶液は所望の溶媒系中に活性成分(例えばHER−2/Neuペプチド又は融合ペプチド)を溶解し、そして次に得られた溶液をメンブレンフィルター通過により滅菌することにより、或いは、滅菌条件下で予め滅菌された溶媒に滅菌化合物を溶解することにより、製造できる。得られた水溶液はそのまま使用するためにパッケージングしてよく、又は、凍結乾燥し、凍結乾燥された製剤を投与前に滅菌水性担体と混合してよい。調製品のpHは典型的には3と11の間、より好ましくは5〜9、そして最も好ましくは7〜8となる。
【0068】
HER−2/Neuペプチド又は融合ペプチドを含有する医薬組成物は予防及び/又は治療の処置のために投与できる。治療用途においては、組成物は、状態及びその合併症の症状を防止、治癒、退行、又は少なくとも部分的に緩徐化又は停止させるために十分な量において、HER−2/Neu蛋白を過剰発現している腫瘍細胞の増殖により悪化する可能性がある状態にすでに罹患している患者に投与する。これを達成するために適当な量は「治療有効用量」と定義される。この使用のために有効な量は疾患又は状態の重症度及び患者の体重及び全身状態に依存するが、一般的には70kgの患者に対して2週毎にHER−2/Neuペプチド又は融合ペプチド約1μg〜約10mgの範囲となり、70kgの患者に対して2週毎にペプチド約50μg〜約1mgの用量がより一般的に使用される。適切な用量は1週毎、2週毎、又は1月毎の間隔において投与してよい。
【0069】
組成物の単回又は多数回の投与は担当医により選択される用量レベル及びパターンで実施できる。何れの場合においても、医薬組成物は治療目的のために患者においてHER−2/Neu過剰発現腫瘍細胞の増殖を効果的に抑制するために十分なHER−2/Neuペプチド又は融合ペプチドの量を備えなければならない。
VIII.HER−2/Neu蛋白に対して特異的なT細胞応答を検出するための方法
本発明は更にHER−2/Neu蛋白に対して特異的なT細胞免疫応答が患者に存在するかどうかを検出するための方法を提供する。この方法は以下の工程を含む:第1に、リンパ球、例えば少なくともT細胞、及び抗原提示細胞を患者から入手する。そのようなリンパ球が得られる適当な試料は血液、腫瘍浸潤液、及びリンパ節又はリンパ液を包含する。第2にT細胞に対する抗原提示細胞によってT細胞エピトープの適切な提示を可能とする条件下で、本発明のHER−2/Neuペプチド(又はHER−2/Neuペプチドと非相同ペプチドを含む融合ペプチド)にT細胞及び抗原提示細胞を曝露する。第3に、T細胞応答のサインを、当該分野で周知の手段、例えばELISPOT、増殖試験、又はフローサイトメトリーによりインビトロで計測する。T細胞応答がこれらの方法の何れかにより検出される場合、患者においてHER−2/Neu蛋白に対して特異的なT細胞免疫応答が存在すると結論付けることができる。
【実施例】
【0070】
以下の実施例は限定としてではなく説明としてのみ提示する。本質的に同様の結果を得るために変更又は改変することができる種々の非重要なパラメーターは当業者が容易に認識することになる。
(実施例1)
材料及び方法
組み換え蛋白及び合成ペプチド。BA7072はHER−2/neu+癌の治療のための治験中のワクチンAPC8024において使用するためにDendreon Corporation(Seattle,WA)により製造された専売品の組み換え融合蛋白である。BA7072はHER−2/neuの細胞外ドメイン(ECD)及び細胞内ドメイン(ICD)の両方に由来する蛋白配列を含有する。HER500はHER−2/neuのECDおよびICDの両方に由来する配列を含有するDendreon Corporationによりやはり製造されている組み換え蛋白であり、そして抗原BA7072のHER−2/neu部分である。インビトロにおけるHER500特異的免疫応答を定義するために、HER500蛋白配列から125ペプチドを作成した。これらのペプチドは15アミノ酸長であり、11量体分オーバーラップしている(Genemed Synthesis,South San Francisco,CA)。125のHER500ペプチドのHER500ペプチド#63(HERp270〜284、ALVTYNTDTFESMPN)は天然のHER−2/neu配列のアミノ酸270〜284に相当する。作成したHER500の15量体ペプチドに加えて、9−、10−、11−、12−、13−、14−、16−、及び17量体ペプチドをHER−2/Neu配列の残基268〜286から合成した。これらのペプチドはNH2−及びCOOH−末端のトランケーション、又はHERp270〜284への1〜2アミノ酸の付加の後に誘導され、記載したペプチドを得た。全てのHER500ペプチドを配列決定し、そして分析用HPLCおよび質量スペクトル分析(Genemed Synthesis)により>95%純度であることが測定された。
【0071】
被験体及び健常者ドナー試料収集。全ての被験体及び健常者ドナーの標本を適切なInvestigational Review Boardにより認可されている治験責任者依頼プロトコルに従って収集した。インフォームドコンセントを受領した後、ヘパリン処理したバキュテナチューブ又はシリンジ中に静脈穿刺により全血試料を収集し、輸送及び/又はプロセシングのために準備した。本発明者等の実験室で血液試料を受領した後、末梢血液単核細胞(PBMC)を密度勾配遠心分離により滅菌条件下で収集し、そして特定の試験における使用のために準備した。
【0072】
HER−2/neu特異的T細胞クローンのインビトロ作成。HER−2/neu陽性癌に対する治験中の治療薬であるAPC8024を投与された被験体のPBMCを2mM L−グルタミン、50U/mLペニシリン、50μg/mLストレプトマイシン、及び20mM HEPES緩衝液+10%ヒトAB血清を添加したRPMI1640(Gemini BioProducts,Calabasas,CA)(cRPMI+10%HS)中で、一晩BA7072を10μg/ml含有するT−25組織培養フラスコ中で刺激した。翌日、IFNγ分泌細胞をIFNγ分泌試験細胞富化検出キット(INFγ Secretion Assay Cell Enrichment and Detection kit)(Miltenyi Biotech,Auburn,CA)を用いてPBMC培養物から単離した。IFNγ富化集団は10U/mL組み換えヒトIL−2(Invitrogen)を用いて96ウェル丸底プレート内に限界希釈法により播種した。非IFNγ分泌細胞を放射線照射し(3000rad)、そして全ウェル内の最終容量が150μLとなるようにウェル当たり50μLで添加した。プレートを37℃5%CO2において7日間インキュベートした。EBV形質転換リンパ芽球様細胞(EBV−LcL)も又、増殖及び自己T細胞クローンの試験のために、自己PBMC及びB95−8細胞系統(ATCC、Manassas,VA)由来の上澄みを用いて本被験体から作成した。クローニング第7日に、IFNγ分泌細胞を前に記載したとおり96ウェルプレート中で非特異的に増殖させた(Yee等、2002、Proc.Natl.Acad.Sci.USA 99:16168−16173)。慨すれば、各ウェルに対し、25U/mL組み換えヒトIL−2及び10ng/mL抗ヒトCD3抗体を添加したcRPMI+10%HS培地(BD Pharmingen,San Diego,CA)100μLを、1×104/ウェルの照射自己EBV−LcL及び1×105/ウェルの照射同種異系PBMCとともに添加した。プレートを37℃で14日間インキュベートし、次にウェルを目視により検査することにより、陽性の増殖を調べた。増殖陽性のクローン、クローンHER.23c21及びその他を24ウェルプレートに移し、そしてrIL−2、抗−CD3、及びアクセサリー細胞を用いながら上記した通り増殖させた。各ウェルの最終容量は2.4mLであり、そしてアクセサリー細胞の数は増大してウェル当たり2×106/ウェルの照射同種異系PBMC及び1×105/ウェルの照射自己EBV−LcLとなった。
【0073】
HER−2/neu特異的T細胞クローンの発見及び特性化。クローンを自己PBMC及び抗原又は自己EBV−LcL及びHer500ペプチドを用いて抗原特異性に関してスクリーニングした。刺激はcRPMI+10%FBS培地中において96ウェル丸底プレート中に設定し、5%CO2下37℃において48時間インキュベートした。更に又、クローンはフローサイトメトリーによりCD4及びCD8表面発現に関して染色した。
【0074】
サイトカイン生産。抗原特異的刺激試験におけるサイトカイン生産を測定するために、48時間後、上澄み200μLを培養物から採取し、IL−2及びIFNγの両方の生産について試験した。IL−2の生産はIL−2依存性系統HT−2(ATCC)を用いて計測した。HT−2細胞を10%FBS及び100uM NEAA、1mMピルビン酸ナトリウム、2mM L−グルタミン、50mMペニシリン、50U/mLストレプトマイシン、20mM HEPES及び20uM 2−メルカプトエタノールを添加したIMDM(cIMDM+10%FBS)中で生育させ、そして20ng/mLのrIL−2を週2回供給した。試験のために、HT−2細胞を継代した4日後に細胞をIMDM+10%FBSで洗浄することによりすべてのrIL−2を除去し、細胞をcIMDM+10%FBS中1×104細胞/ウェルで96ウェル丸底プレートに添加した。抗原特異的刺激に由来する上澄みをウェルに添加し、プレートを37℃で24〜30時間インキュベートした。翌日1μCiの3[H]TdRを試験の最終6時間に添加し、プレートをガラス繊維フィルターマットに回収し、そして放射性同位体のDNA取り込み、又は増殖応答を液体シンチレーションカウンター(PerkinElmer Life and Analytical Sciences,Inc.Boston,MA)を用いて分当たりの計数により求めた。INFγ生産は、ELISA用の抗ヒトIFNγ抗体対(BD Pharmingen,San Diego,CA)を用いて計測した。慨すれば、イムロン4プレート(Thermo Labsystems/VWR,Brisbane,CA)を3μg/mlの精製した抗ヒトIFNγ抗体(NIB42)で一晩コーティングした。翌日コーティング抗体を廃棄し、そしてPBS(Invitrogen)中4%ウシ血清アルブミン(BSA)(Sigma,St.Louis,MO)をウェルに添加し、そしてプレートを37℃で2時間インキュベートした。プレートをPBS+0.05%Tween20で洗浄し、抗原特異的刺激に由来する上澄み試料100μLをウェルに添加し、そして1.25〜2時間室温でインキュベートした。プレートを洗浄し、ビオチニル化抗ヒトIFNγ抗体(4S.B3)をPBS中1%BSA中に希釈(1μg/ml)し、そして室温で1時間プレートに添加した。プレートを洗浄した後、ストレプ−アビジンHRP(BD Pharmingen)をPBST中1:1000に希釈し、室温で30分間ウェルに添加した。最後にプレートを洗浄し、Sigma(登録商標) Fast OPDを暗所で15分間添加した。2M HClを添加することにより反応を停止させ、分光光度計においてプレートの492nmの吸光度を読み取った。
【0075】
HLA−DR制約性及び無差別性。T細胞エピトープHERp270〜284のHLA−DR制約性を測定するために、抗HLA−DRmAbL243、HLA−DQmAb1a3又はHLA−DPmAbB7/21(20〜1.25μg/ml)をT細胞クローンHER.23c21、ペプチドHERp270〜284及び自己EBV−LcLと共にcRPMI+10%FBS培地中で培養した。上澄みを48時間後に採取し、IL−2及びIFNγの生産に関して試験した。HERp270〜284も又、フランスのストラスブールで開催された第12回国際組織適合性ワークショップ(IHW)を起源とするEuropian Collection of Cell Culturesより購入したEBV−LcL系統を用いてHLA−DR無差別性に関して試験した。表Iに列挙したIHW系統は種々のHLA−DRβ1対立遺伝子に対してホモ接合であり、10%ウシ胎児血清、20mM HEPES、2mM L−グルタミン、50mMペニシリン及び50U/mLストレプトマイシンを添加したRPMI1640培地(cRPMI+10%FBS)(Invitrogen,Carlsbad,CA)中で増殖させた。MHCクラスII無差別性を試験するために、HERp270〜284を1μg/mlにおいて各々別個のEBV−LcL系統(2×105細胞/ウェル)及びT細胞クローンHER.23c21(1×105細胞/ウェル)と共に37℃のcRPMI+10%FBS培地中96ウェル丸底プレート中に添加した。上澄みを48時間後に採取し、IFNγの生産に関して試験した。
結果
APC8024はHer−2/neu発現乳癌に対する治験中の自己細胞免疫療法である。この免疫療法により誘導される免疫応答を特性化するために、本発明者等はAPC8024を投与した臨床治験被験体からT細胞クローンを単離した。作成したCD4+ T細胞クローンの1つであるHER500.23c21は自己PBMCにより提示される外因性のHER−2/Neu蛋白配列に対して特異的な応答を呈した(図1)。この実験において、HER500.23c21をHER−2/Neuの細胞内及び細胞外のドメインを含有する組み換え蛋白であるHER500及びhGM−CSFとの融合蛋白として発現されるHER500より成るBA7072の漸増用量で刺激した。HER500.23c21は両方の抗原に応答してIFNγ及びIL−2を特異的に生産したが、抗原の非存在下においてはサイトカインの生産は観察されなかった。HER500.23c21は用量依存的な態様においてこれらの蛋白の各々に応答し、外因性蛋白抗原より天然にプロセシングされて提示されるHER−2/NeuエピトープをHER500.23c21が認識することを示している。
【0076】
HER−2/Neu蛋白配列に対するHER500.23c21の明確な応答のために、HER500.23c21により認識される特異的エピトープをペプチドを用いてマッピングした。HER500内にHER−2/Neu蛋白配列を包含している125の個別のオーバーラップした15量体のペプチドのパネルに対してHER500.23.c21を試験した。各ペプチドは抗原提示細胞としての自己EBV LCL細胞と共に1ug/mlで使用した。これらの実験において、HER500.23c21は、IFNγ又はIL2生産の何れかにより計測した場合、125全ての個別のペプチドのプールに対して強力に応答した(図2A及びB,上棒グラフ)。更に又、HER500.23c21は個別のペプチドの一つ、ペプチド63のみに応答し、このペプチドがHER500.23c21により認識されるエピトープを含有していることを示していた(図2)。ペプチド63はHER−2/Neu蛋白配列のアミノ酸270〜284に相当し、そして隣接ペプチドと11アミノ酸分オーバーラップしている。即ち、ペプチド62又は64への応答が欠如していることは、HER500.23c21 T細胞エピトープはHERp270〜284配列に完全に含有されており、そして側面に位置するペプチドに共通のアミノ酸はこのT細胞クローンに対する完全なエピトープを含有しないことを示唆している。HER500.23c21により認識される最小のエピトープを定義するために、本発明者等はHERp270〜284のNH2−及びCOOH−末端のトランケーションを有する9量体、10量体又は11量体のペプチドを設計した。これらのペプチドによるHER500.23c21の刺激は、HLA−DRB1対立遺伝子*0301、*0302、*1301、及び*1402を与えるEBV LcL系統により提示される場合に、11量体配列VTYNTDTFESM及びTYNTDTFESMPのみにおいて良好であった。
【0077】
どのHLA分子がHER500.23c21に対してHERp270〜284を提示することを担っているかを決定するために、HLAブロッキング抗体を用いて提示及びT細胞活性化を抑制した。HER500.23c21をHLA−DR又はHLA−DP/−DQに対して特異的な抗体の漸増濃度の非存在下又は存在下において1ug/mlのHERp270〜284及び自己EBV−LcLで刺激した。T細胞刺激はIFNγ生産により測定した。HLA−DRに対する抗体は用量依存的な態様においてHERp270〜284によるHER500.23c21の刺激を抑制したのに対し、HLA−DP/−DQブロッキング抗体は20ug/mlの高濃度においてもT細胞活性化に対して作用を示さなかった(図3)。これらの結果はHERp270〜284のHER500.23c21認識がHLA−DR制約性であることを明らかにしている。
【0078】
HLA制約性を更に定義するために、本発明者等は13のDR血清学的ファミリーを与える異なるHLA−DRB1*対立遺伝子に対してホモ接合である25のEBV LcL系統のパネルを使用してHER500.23c21を活性化させるHERp270〜284の能力を試験した(表1)。試験した各EBV−LcL系統は抗原特異的な態様においてHER500.23c21を効率的に刺激することができ、HERp270〜284エピトープは少なくとも25のHLA−DRB1*対立遺伝子に対して無差別であることが示された(図4A)。これらの実験において、1ug/mlのHERp270〜284ペプチドは使用したEBV−LcL系統とは無関係に最大のT細胞応答を誘導した。しかしながら、HER500.23c21を刺激する種々の対立遺伝子の能力における相違は、より低い濃度のペプチドにおいて顕著となり(図4B)、一部の対立遺伝子、例えばDRB1*0401は、10ng/mLもの低濃度で刺激することができ、そして他の対立遺伝子は250ng/mL未満では刺激することができなかった(DRB1*1103)。これらの相違はおそらくは異なるDRB1*対立遺伝子に対するHERp270〜284の結合親和性の範囲を反映していると考えられる。しかしながらこれらの実験において観察された高度な無差別性は、HERp270〜284がHER−2/Neuに対するユニバーサルなヘルパーT細胞エピトープを含有している場合があることを示唆している。
考察
抗腫瘍免疫応答の試験は試薬が限られているため特定のHLA型により提示される抗原少数に制限される場合が多い。無差別T細胞エピトープの発見は多様なHLA型の個体における抗腫瘍免疫応答の分析を可能にすることにより、これらの制限の緩和に寄与すると考えられる。研究用ツール及び潜在的な治療薬の両方としてのそれらの価値のため、多くの研究は無差別性のCD4及びCD8 T細胞エピトープの発見及び特性化に着目してきた。自身のHLA結合において多様な程度の無差別性を有するペプチドエピトープが感染性疾患の抗原であるHIV(van der Burg等、1999、J Immunol162:152−160)、マイコバクテリア(Valle等、2001、Clin Exp Immunol 123:226−232)、及びp.falciparum(Contreras等、1998、Infect Immun 66:3579−3590)、ならびに腫瘍抗原、例えばNYESO(Zarour等、2002、Cancer Res 62:213−218)、MAGE(Consogno等、2003、Blood 101:1038−1044)、Tert(Schroers等、2003.Clin Cancer Res 9:4743−4755)、及びHer2/neu(Kobayashi等、2000、Cancer Res 60:5228−5236)において発見されている。TEPITOPE(Bian and Hammer,2004,Methods 34:468−475)のようなコンピュータープログラムは蛋白配列に基づいて無差別エピトープを予測するために既知の共通の結合モチーフを使用しており、種々の原料に由来する多くの潜在的な新規T細胞エピトープを発見している。インシリコで発見されたエピトープの生物学的関連性は多くの系において着目されている(Ruiz等、2004、Clin Cancer Res 10:2860−2867;及びAl−Attiyah and Mustafa 2004.Scand J Immunol 59:16−24)。
【0079】
本試験において、新規な無差別T細胞エピトープが腫瘍関連抗原HER−2/Neuから発見された。このT細胞エピトープはHER−2/Neu+癌に対する自己細胞免疫療法で治療された患者から単離されたCD4+ T細胞クローンと共に発見された。エピトープはHER−2/Neu配列のアミノ酸270〜284内に含有されており、外因性の蛋白抗原より天然にプロセシングされて提示される。このエピトープに対して特異的なT細胞クローンがHER−2/Neu特異的免疫療法で治療された個体から単離されたという事実は、このエピトープが臨床的に関連性のある抗腫瘍免疫応答の部分としてインビボで役割を果たしている可能性があることを示唆している。生物学的に関連する範囲において発見されたことのほかに、このペプチドエピトープはそれが極めて広範なHLA−DR無差別性を有し、13の血清学的なDRファミリーを与える少なくとも25の異なるHLA−DRB1*対立遺伝子によりT細胞クローンHER500.23c21に提示されることができることから、興味深いものである。無差別T細胞エピトープとして発見された他のペプチドの大部分はわずか数種の異なるHLA対立遺伝子により提示されるのみであるが、本発明者等はなお、T細胞クローンHER500.23c21に対してHERp270〜284を提示することができないHLA−DRB1*対立遺伝子を発見しなければならない。このエピトープの提示に関するMHC制約性が相対的に乏しいことは、これをユニバーサルなHER−2/Neu CD4 T細胞エピトープの理想的な候補としている。
【0080】
腫瘍免疫学の主要な目標は腫瘍関連抗原に対抗して効果的な癌免疫療法及びワクチンを開発することである。そのような治療は抗腫瘍免疫応答を刺激して腫瘍を根絶するように設計される。特定の腫瘍関連抗原に着目することは、抗腫瘍免疫応答を良好に生じさせ、一部の腫瘍抗原内の特定のT細胞エピトープの発見をもたらした。初期の研究の大部分が腫瘍特異的CD8 T細胞応答を生じさせることにのみ着目していたが、効果的な抗腫瘍免疫を生じさせる際のCD4 T細胞の重要性の認識も高まっていた。このため、抗腫瘍CD4 T細胞応答及び腫瘍関連抗原からのクラスII制約型のT細胞エピトープの発見の研究が進展した。そのようなエピトープの利用は、そのようなエピトープが1つより多いHLA型により提示されることができれば大きく増大する。ペプチド系癌ワクチンの方策はワクチン内のペプチドエピトープのHLA制約性により妨害されている。本明細書に記載したような無差別T細胞エピトープを包含させることは一般的集団内のそのようなワクチンの有用性を広げる。無差別エピトープによる腫瘍抗原に対する免疫応答の誘導は一般的集団のより大きいパーセントに到達するための効率的な方法である。そのような無差別エピトープはまた応答を分析するための有用な手段を与える。
【0081】
【表1A】
【0082】
【表1B】
細胞系統はECACC European Collection of Cell Culturesより入手し、それらはIMGT/HLA細胞要覧に記載されている(ウエブサイト:ebi.ac.uk/imgt/hla/cell query.html)。
(実施例2)
この一連の実験においては、HER−2/Neu蛋白から誘導された別のペプチドを無差別MHC クラスII HER−2/neuエピトープとしてのその潜在性に関して試験した。エピトープHERp270〜284(配列番号2)を実験の陽性対照として使用した。種々のHLA−DR対立遺伝子に対してホモ接合であるEBV−LcL系統のパネルを用いて、HERp270〜284並びにHER−2/Neu268〜286(配列番号1)の15アミノ酸配列内の種々の長さのペプチド又は19アミノ酸配列内のより長いペプチドをクローンHER500.23c21に対して提示した。試験の陰性対照には非関連HER500ペプチド、無ペプチド、及び各EBV−LcL系統単独を使用した。自己EBV−LcLも又、クローンHER500.23c21特異性に関する対照として各条件で使用した。試験は2×105 EBV−LcL/ウェル及び1×105 HER500.23c21細胞/ウェルでcRPMI+10%FBS中、96ウェル丸底プレート中に設定した。ペプチドを終濃度1μg/mlに希釈し、100μL/ウェルで添加した。試験は5% CO2下37℃において48時間インキュベートした。上澄みを採取し、ELISAを用いてIFNγ生産に関して分析した。結果は各ペプチドシリーズに対して1つの代表的な実験について示した(図5〜11)。ペプチド及び各HLA−DR対立遺伝子に応答してクローンHER500.23c21により生産されたIFNγの結果はIFNγのpg/mL量において、又は492nmにおける吸光度として報告する。
【0083】
本出願において引用した全ての特許、特許出願、及び他の公開物は、公開されたアミノ酸又はポリヌクレオチドの配列も含めて、あらゆる目的のために、参照としてその全体が援用される。
【0084】
配列表
配列番号1 (HER−2/Neu蛋白の268−286)
CPALVTYNTDTFESMPNPE
配列番号2 (HER−2/Neu蛋白の270−284、または配列番号1の残基3−17)
ALVTYNTDTFESMPN
【特許請求の範囲】
【請求項1】
(a)配列番号1の残基3〜17の12〜15隣接アミノ酸;又は(b)配列番号1の16〜18隣接アミノ酸よりなる単離されたペプチド。
【請求項2】
非相同ポリペプチドに融合した請求項1記載のペプチドを含む融合ペプチド。
【請求項3】
請求項1のペプチドがペプチド結合を介して非相同ポリペプチドに融合されている請求項2記載の融合ペプチド。
【請求項4】
異なるHLA−DR対立遺伝子少なくとも15個の抗原提示細胞により提示された場合にHER−2/Neu蛋白に対して特異的なT細胞免疫応答を誘導する請求項1又は2記載のペプチド。
【請求項5】
異なるHLA−DR対立遺伝子少なくとも25個の抗原提示細胞により提示された場合にHER−2/Neu蛋白に対して特異的なT細胞免疫応答を誘導する請求項4記載のペプチド。
【請求項6】
HLA−DR対立遺伝子が0101、0102、0103、1503、160201、0301、0302、0401、0402、040301、040501、1101、1102、1103、1104、110401、1201、1301、1302、1401、1402、0701、080101、080201及び0901よりなる群から選択される請求項4記載のペプチド。
【請求項7】
配列番号2のアミノ酸配列を有する請求項1記載のペプチド。
【請求項8】
非相同ポリペプチドが顆粒球マクロファージコロニー刺激因子(GM−CSF)である請求項2記載のペプチド。
【請求項9】
請求項1又は3のペプチドをコードするポリヌクレオチド配列を含む単離された核酸。
【請求項10】
ポリヌクレオチド配列が請求項7のペプチドをコードする請求項9記載の核酸。
【請求項11】
ポリヌクレオチド配列が請求項8のペプチドをコードする請求項9記載の核酸。
【請求項12】
請求項9の核酸を含む発現カセット。
【請求項13】
組み換えウィルスベクターである請求項12記載の発現カセット。
【請求項14】
請求項7又は8のペプチドの発現を指向する請求項12記載の発現カセット。
【請求項15】
請求項12又は13の発現カセットを含む宿主細胞。
【請求項16】
請求項1又は2のペプチド及び生理学的に許容しうる賦形剤を含む組成物。
【請求項17】
ペプチドが配列番号2のアミノ酸配列を有する請求項16記載の組成物。
【請求項18】
非相同ポリペプチドが顆粒球マクロファージコロニー刺激因子(GM−CSF)である請求項16記載の組成物。
【請求項19】
細胞の表面上の主要組織適合性複合体(MHC)分子と複合体を形成する請求項1記載のペプチドを有する抗原提示細胞を更に含む請求項16記載の組成物。
【請求項20】
HER−2/Neu蛋白に対して特異的なT細胞免疫応答を患者において誘導するための方法であって、該方法が患者に請求項16の組成物の有効量を投与する工程を含む方法。
【請求項21】
非相同ポリペプチドが顆粒球マクロファージコロニー刺激因子(GM−CSF)である請求項20記載の方法。
【請求項22】
HER−2/Neu蛋白に対して特異的なT細胞免疫応答を患者において検出するための方法であって、該方法は、(a)患者から抗原提示細胞及びT細胞を得る工程;(b)抗原提示細胞及びT細胞を請求項1又は2記載のペプチドに接触させる工程;及び(c)T細胞応答を検出する工程であって、ここでT細胞応答の検出は患者におけるHER−2/Neu蛋白に対して特異的なT細胞免疫応答の存在を示すものである工程、を含む方法。
【請求項23】
工程(c)がELISPOT、増殖試験、又はフローサイトメトリーにより実施される請求項22記載の方法。
【請求項24】
ペプチドが配列番号2のアミノ酸配列を有する請求項22記載の方法。
【請求項1】
(a)配列番号1の残基3〜17の12〜15隣接アミノ酸;又は(b)配列番号1の16〜18隣接アミノ酸よりなる単離されたペプチド。
【請求項2】
非相同ポリペプチドに融合した請求項1記載のペプチドを含む融合ペプチド。
【請求項3】
請求項1のペプチドがペプチド結合を介して非相同ポリペプチドに融合されている請求項2記載の融合ペプチド。
【請求項4】
異なるHLA−DR対立遺伝子少なくとも15個の抗原提示細胞により提示された場合にHER−2/Neu蛋白に対して特異的なT細胞免疫応答を誘導する請求項1又は2記載のペプチド。
【請求項5】
異なるHLA−DR対立遺伝子少なくとも25個の抗原提示細胞により提示された場合にHER−2/Neu蛋白に対して特異的なT細胞免疫応答を誘導する請求項4記載のペプチド。
【請求項6】
HLA−DR対立遺伝子が0101、0102、0103、1503、160201、0301、0302、0401、0402、040301、040501、1101、1102、1103、1104、110401、1201、1301、1302、1401、1402、0701、080101、080201及び0901よりなる群から選択される請求項4記載のペプチド。
【請求項7】
配列番号2のアミノ酸配列を有する請求項1記載のペプチド。
【請求項8】
非相同ポリペプチドが顆粒球マクロファージコロニー刺激因子(GM−CSF)である請求項2記載のペプチド。
【請求項9】
請求項1又は3のペプチドをコードするポリヌクレオチド配列を含む単離された核酸。
【請求項10】
ポリヌクレオチド配列が請求項7のペプチドをコードする請求項9記載の核酸。
【請求項11】
ポリヌクレオチド配列が請求項8のペプチドをコードする請求項9記載の核酸。
【請求項12】
請求項9の核酸を含む発現カセット。
【請求項13】
組み換えウィルスベクターである請求項12記載の発現カセット。
【請求項14】
請求項7又は8のペプチドの発現を指向する請求項12記載の発現カセット。
【請求項15】
請求項12又は13の発現カセットを含む宿主細胞。
【請求項16】
請求項1又は2のペプチド及び生理学的に許容しうる賦形剤を含む組成物。
【請求項17】
ペプチドが配列番号2のアミノ酸配列を有する請求項16記載の組成物。
【請求項18】
非相同ポリペプチドが顆粒球マクロファージコロニー刺激因子(GM−CSF)である請求項16記載の組成物。
【請求項19】
細胞の表面上の主要組織適合性複合体(MHC)分子と複合体を形成する請求項1記載のペプチドを有する抗原提示細胞を更に含む請求項16記載の組成物。
【請求項20】
HER−2/Neu蛋白に対して特異的なT細胞免疫応答を患者において誘導するための方法であって、該方法が患者に請求項16の組成物の有効量を投与する工程を含む方法。
【請求項21】
非相同ポリペプチドが顆粒球マクロファージコロニー刺激因子(GM−CSF)である請求項20記載の方法。
【請求項22】
HER−2/Neu蛋白に対して特異的なT細胞免疫応答を患者において検出するための方法であって、該方法は、(a)患者から抗原提示細胞及びT細胞を得る工程;(b)抗原提示細胞及びT細胞を請求項1又は2記載のペプチドに接触させる工程;及び(c)T細胞応答を検出する工程であって、ここでT細胞応答の検出は患者におけるHER−2/Neu蛋白に対して特異的なT細胞免疫応答の存在を示すものである工程、を含む方法。
【請求項23】
工程(c)がELISPOT、増殖試験、又はフローサイトメトリーにより実施される請求項22記載の方法。
【請求項24】
ペプチドが配列番号2のアミノ酸配列を有する請求項22記載の方法。
【図1】

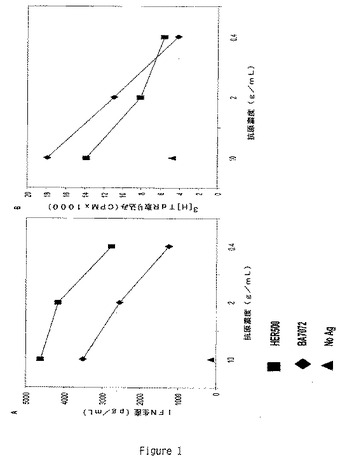
【図2】
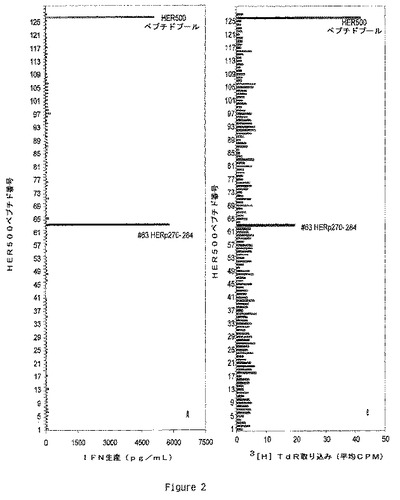
【図3】
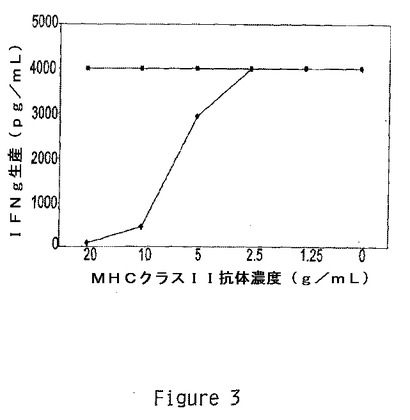
【図4】
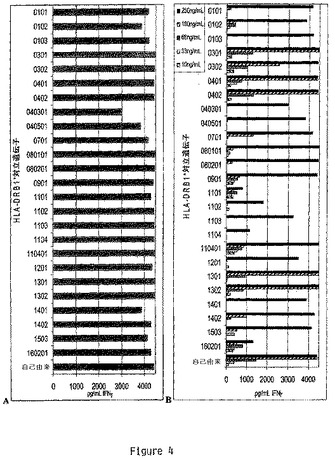
【図5】
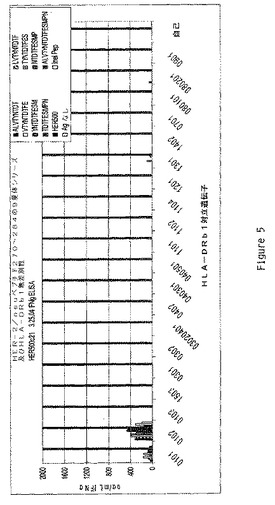
【図6】
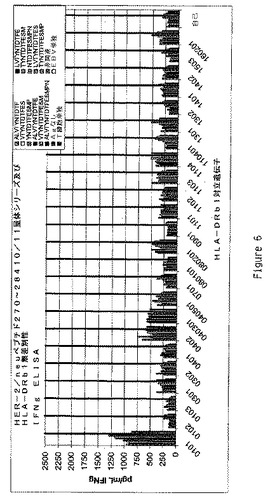
【図7】

【図8】

【図9】
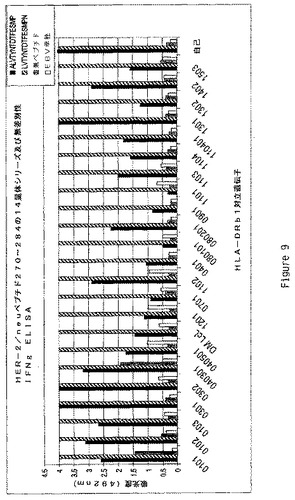
【図10】

【図11】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2010−501160(P2010−501160A)
【公表日】平成22年1月21日(2010.1.21)
【国際特許分類】
【出願番号】特願2009−524009(P2009−524009)
【出願日】平成19年8月10日(2007.8.10)
【国際出願番号】PCT/US2007/075681
【国際公開番号】WO2008/022019
【国際公開日】平成20年2月21日(2008.2.21)
【出願人】(500072714)デンドレオン コーポレイション (11)
【Fターム(参考)】
【公表日】平成22年1月21日(2010.1.21)
【国際特許分類】
【出願日】平成19年8月10日(2007.8.10)
【国際出願番号】PCT/US2007/075681
【国際公開番号】WO2008/022019
【国際公開日】平成20年2月21日(2008.2.21)
【出願人】(500072714)デンドレオン コーポレイション (11)
【Fターム(参考)】
[ Back to top ]