
熱ショックタンパク質とオリゴ糖または多糖とから形成されるコンジュゲート
【課題】従来使用されてきたキャリアで達成されたよりも良好な免疫原的特性をコンジュゲートに与える新規なタンパク質キャリアを同定し、オリゴ糖および多糖に対する免疫原性応答を高める、確実に高力価のワクチン接種を可能にすること。
【解決手段】少なくとも1つの熱ショックタンパク質または少なくとも1つの免疫刺激ドメインを合有する熱ショックタンパク質の一部と、病原菌の少なくとも1つの莢膜のオリゴ糖または多糖とを含有する、コンジュゲート化合物。
【解決手段】少なくとも1つの熱ショックタンパク質または少なくとも1つの免疫刺激ドメインを合有する熱ショックタンパク質の一部と、病原菌の少なくとも1つの莢膜のオリゴ糖または多糖とを含有する、コンジュゲート化合物。
【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、熱ショックタンパク質と、多糖またはオリゴ糖(特に、病原性微生物の莢膜由来の多糖またはオリゴ糖)とからなるコンジュゲート化合物に関する。本化合物は、抗多糖抗体の形成を誘導し得る。そのため、本化合物は、ヒトおよび動物に使用するワクチンとして有用である。
【背景技術】
【0002】
(技術状況)
細菌は、広範囲の疾患状態に対する病原物質である。
【0003】
このような疾患の例には、Neisseria meningitidisによって引き起こされる髄膜炎、およびHaemophilus influenzae b型(Hib)またはStreptococcus(Pneumococcusを含む)によって引き起こされる他の感染症、Salmonella typhiの感染によって引き起こされる腸チフス、非腸チフス原性のSalmonella菌またはShigella菌によって起こる腸疾患が含まれる。
【0004】
莢膜を有する細菌に対する防御免疫は、莢膜中の多糖に対する抗体により介在されることが知られている。免疫系を充分に刺激するには、莢膜の多糖を、キャリアタンパク質に結合させる必要があることも知られている(Robbinsら、J.Infect.Dis.1990,161,821−832)。
【0005】
特に、多糖(例えば、C群髄膜炎菌多糖(MenC)、HibおよびA群髄膜炎菌多糖(MenA))およびCRM−197(Corynebcterium diphtheriae由来のペプチド)のようなタンパク質、TD(ジフテリア毒素)またはTT(破傷風毒素−Peetersら、Inf.Immun.,(1991年10月)、3504−3510;Claessonら、J.Pediatrics St Louis,112(5)、695−702、(1988年5月)を参照のこと)からなるコンジュゲート化合物が文献中に記載されている。
【0006】
このようなワクチンのうち数種は、すでに臨床的に実用化されており、良好な結果が得られている。しかし、従来使用されてきたキャリアで達成されたよりも良好な免疫原的特性をコンジュゲートに与える新規なタンパク質キャリアを同定する必要がある。
【0007】
本発明は、オリゴ糖および多糖の免疫原性応答を高めるタンパク質キャリアとしての熱ショックタンパク質の使用に関する。
【0008】
熱ショックタンパク質は、非常に多数のTエピトープを含有し、そのため細胞の免疫系を刺激することが知られている。
【0009】
マラリア性エピトープのキャリアである、Mycobacterium bovisの熱ショックタンパク質のコンジュゲート化合物(65kDa)は、Bacillus Calmette−Guerin(BCG)で予め免疫化した動物に、アジュバントを必要とすることなく、著しい免疫化を誘導することが記載されている(Lussowら Eur.J.Immunol.,1991,21,2297−2302)。しかし、熱ショックタンパク質に結合した(T−細胞依存性であることが周知の)ペプチドによって示されるT細胞依存効果に関するLussowらに観察された効果は、注目に値する。
【0010】
より詳細には、熱ショックタンパク質は、株および型が異なる細菌間でもよく保存されるので、継続的プロセスである細菌感染により、確実に、免疫系が熱ショックタンパク質に対して感作された状態なる。そのため、最初のワクチン接種または追加免疫ワクチン接種における投与で、本発明のコンジュゲート化合物に対する良好な応答が保証される。
【0011】
本発明により、(アジュバントの使用も可能だが)アジュバントを用いずに、莢膜を有する細菌の多糖およびオリゴ糖の使用が可能となる。
【0012】
大勢の子供たちが、結核予防のためにBCGワクチン(細菌の熱ショックタンパク質を含む)を接種する。そのため、熱ショックタンパク質をキャリアとして含有する本発明のコンジュゲートは、このBCGワクチン接種の実施の結果、すでにキャリアで予備免疫化された多くの被検体を見出す。
【0013】
その上、熱ショックタンパク質は非常に保存されているので、BCGでワクチン接種を受けていない個体群でさえ(他の細菌との相互作用の結果として)容易に免疫を得られる。そのため、この個体群は、熱ショックタンパク質およびT細胞依存抗原(オリゴ糖または多糖)で構成されるコンジュゲートでのワクチン接種の後に良好な免疫応答を発現し得る状態にある。このように、本発明のキャリアは、確実に高力価のワクチン接種をするために、細菌間の熱ショックタンパク質の高い保存性およびT細胞記憶を、独自に利用するものである。
【発明の概要】
【課題を解決するための手段】
【0014】
(発明の要旨)
本発明は、少なくとも1つの熱ショックタンパク質または少なくとも1つの免疫刺激ドメインを含有する熱ショックタンパク質の一部と、少なくとも1つのオリゴ糖または多糖とを含有するコンジュゲート化合物を提供する。
【0015】
熱ショックタンパク質は、動物、好ましくはヒトにおいて、免疫刺激効果を示し得る如何なる熱ショックタンパク質でもよい。
【0016】
熱ショックタンパク質は、細菌、寄生生物および哺乳動物に高度に保存されている。重大な悪影響なしに意図する被免疫被検体にポジティブな免疫刺激効果を示す限り、あらゆる熱ショックタンパク質が本発明のコンジュゲート中に使用され得る。個々の例としては、Helicobacter pylori、P.aeruginosa、C.trachomatisおよびM.leprae由来の熱ショックタンパク質、特にhsp60群の熱ショックタンパク質が挙げられるが、これらに限定されない。
【0017】
より詳細には、3種の熱ショックタンパク質、すなわち、M.bovis BCG GroEL型の65kDaのhsp(hspR65)、組換えM.tuberculosis DnaK型の70kDaのhsp(hspR70)およびH.pylori由来の新規な熱ショックタンパク質の具体例を、本明細書中に示す。
【0018】
H.pylori熱ショックタンパク質(hsp)は、図3に示すヌクレオチド配列およびアミノ酸配列を有するタンパク質であり、その分子量は54〜62kDaの範囲にあり、好ましくは約58〜60kDaである。このhspは、グラム陰性菌熱ショックタンパク質の群であるhdp60に属する。一般的にhdpは全ての生物、つまり原核または真核の動物または植物に、最もよく保存されているタンパク質であり、そして、その保存は配列全体にわたっている。
【0019】
コンジュゲートは、1種またはそれ以上の熱ショックタンパク質またはその免疫刺激ドメインを含み得る。熱ショックタンパク質は、同一でも異なっていてもよい。しかし、1つの熱ショックタンパク質、あるいは1つまたはそれ以上の免疫刺激ドメインを含有する部分が存在することが好ましい。
【0020】
本明細書中で使用のように、用語「免疫刺激ドメイン」は、哺乳動物被検体のドメインを含有するコンジュゲート化合物の多糖またはオリゴ糖成分に対する、免疫を増強させ得る熱ショックタンパク質のアミノ酸配列の領域を指す。
【0021】
完全な熱ショックタンパク質由来の特定のドメインのみを使用することの利点は、ヒト熱ショックタンパク質に共通のドメインを、選択的に含まないようにすることが可能な点である。ヒトヘのワクチン投与に対して、これは有利なことである。それは、このような領域が「自己」と認識されるので、熱ショックタンパク質の免疫刺激効果に影響を与えないからである。さらに、このような「自己」の領域に対して刺激を受ける全ての免疫は、自己免疫になり得るからである。
【0022】
熱ショックタンパク質のhsp60ファミリーの好適なドメインを同定し、ヒト熱ショックタンパク質と相同性が減少した配列に下線を引いて図2に載せてある。図2に示したドメインのうち、機能的なサブドメインもまた、ドメインおよびサブドメインの組合せとして同様に使用され得る。
【0023】
当業者は、そのドメインまたはエピトープが免疫刺激反応を起こす所定の熱ショックタンパク質を容易に確認し得、そしてこれらのドメイン、またはそのサブセットのみを含む改変熱ショックタンパク質を、容易に調製し得る。
【0024】
コンジュゲート化合物のオリゴ糖成分または多糖成分は、ワクチン投与しようとする全ての病原性微生物の完全な莢膜の多糖またはオリゴ糖、または防御免疫を誘導し得るその一部であり得る。オリゴ糖または多糖は、単一の細菌由来、あるいは2種またはそれ以上の細菌由来であり得る。
【0025】
標的とされ得る細菌の個々の例には:Haemophilus influenzaeb型(Hib)、Streptococcus(pneumococcusを含む)、Salmonella特にSalmonella typhi、非タイポイダル(typoidal)Salmonella細菌またはShigella細菌によって引き起こされる腸疾患が挙げられるが、これらに限定されない。
【0026】
本発明の特定の実施態様によると、Meningococci C(MenC)群のオリゴ糖およびhspから成るコンジュゲートが調製された。
【0027】
本発明のさらに特定の実施態様によると、本発明の目的に使用されたhspは、hspR65およびhspR70である。
【0028】
本発明の第2の局面では、熱ショックタンパク質または少なくとも1つの免疫刺激ドメインを含むその一部を、少なくとも1つのオリゴ糖または多糖に共有結合させることを包含する、本発明のコンジュゲート化合物を生産するプロセスが提供される。
【0029】
オリゴ糖または多糖は、好ましくは標的とする細菌から単離されるが、合成によっても生成され得る。
【0030】
熱ショックタンパク質は、その自然発生源から単離され得、または合成的に生産される。好ましくは、熱ショックタンパク質は、本明細書の記載の一般的な中に記載された技法を用いる組換えDNA技術によって生産される。
【0031】
好ましくは、オリゴ糖または多糖は、結合に対する反応部位を提供するように、熱ショックタンパク質またはその部分と結合する前に改変される。好ましくは、この改変はオリゴ糖または多糖の末端基へのアミノ基のような活性官能基の導入を包含する。次いで、このように改変されたオリゴ糖または多糖を、次いでスクシンイミドのような連結基を用いて活性化し、熱ショックタンパク質またはそれらのタンパク質に結合させる。
【0032】
本発明の第3の局面では、製剤、好ましくはワクチンとして使用する、本発明の第1の局面に記載のコンジュゲート化合物が提供される。
【0033】
本発明の第4の局面では、細菌感染に対するワクチン接種用の薬剤の製造における、本発明の第1の局面に記載のコンジュゲート化合物の使用が提供される。
【0034】
本発明の第5の局面では、本発明の第1の局面に記載のコンジュゲート化合物の免疫学的に有効な量の投与する工程を包含するワクチン接種の方法が提供される。
【0035】
発明の第6の局面では、本発明の第1の局面に記載の1つ以上のコンジュゲート化合物と、薬学的に受容可能なキャリアとを含むワクチンまたは治療用組成物が提供される。
【0036】
好ましくは組成物は、ワクチン組成物であり、そして必要であればアジュバントのような他の賦形剤を含み得る(以下に記載の「ワクチン」と表題された節を参照せよ)。
【0037】
本発明の第7の局面では、1つ以上の本発明の第1の局面のコンジュゲート化合物を、薬学的に受容可能なキャリアおよび任意にアジュバントと組み合わせる工程を包含する、ワクチンの調製法が提供される。
【0038】
従って、本発明は、以下を提供する:
(項目1) 少なくとも1つの熱ショックタンパク質または少なくとも1つの免疫刺激ドメインを含有する熱ショックタンパク質の一部と、少なくとも1つのオリゴ糖または多糖とを含有する、コンジュゲート化合物。
【0039】
(項目2) Meningococci C(MenC)群のオリゴ糖、および熱ショックタンパク質を合有する、項目1に記載のコンジュゲート化合物。
【0040】
(項目3) 前記熱ショックタンパク質が、M.bovis BCG GroEL型65kDa hsp(hspR65)、組換えM.tuberculosis DnaK型70kDa hsp(hspR70)、およびH.pylori由来の熱ショックタンパク質から選択される、項目1または2に記載のコンジュゲート化合物。
【0041】
(項目4) 熱ショックタンパク質または少なくとも1つの免疫刺激ドメインを含むその一部を、少なくとも1つのオリゴ糖または多糖に共有結合させる工程を包含する、項目1から3のいずれかに記載のコンジュゲート化合物を生成する方法。
【0042】
(項目5) 製剤として使用する、項目1から3のいずれかに記載のコンジュゲート化合物。
【0043】
(項目6) 細菌感染に対するワクチン接種用の薬剤の製造における、項目1から3のいずれかに記載のコンジュゲート化合物の使用。
【0044】
(項目7) 免疫学的に有効な量の項目1から3のいずれかに記載のコンジュゲート化合物の投与する工程を包含する、ワクチン接種の方法。
【0045】
(項目8) 項目1から項目3のいずれかに記載の1つ以上のコンジュゲート化合物および薬学的に受容可能なキャリアを含有する、ワクチンまたは治療用組成物。
(項目9) 項目1から項目3のいずれかに記載の1つ以上のコンジュゲート化合物を、薬学的に受容可能なキャリアと結合させる工程を包含する、ワクチンの調製方法。
【図面の簡単な説明】
【0046】
【図1】図1は、hspR65/MenCコンジュゲートでマウスを免疫化した結果を示し、そして、血中の抗MenCに対するELISAの結果を含む。図1Aには、BCGで予め免疫化したマウスを記載し、図1BにはBCGで予め免疫化しなかったマウスを記載している。図は、PBS中のhspR65−MenC(白丸)コンジュゲートまたはhspR70−MenC(白菱形)コンジュゲートとでの結果を示す。コントロール群のマウスは、MenC多糖のみ(白三角)または水酸化アルミニウム中のCEM197−MenCコンジュゲートワクチン(白四角)で免疫化した。
【図2A】図2Aは、Helicobacter pyloriの熱ショックタンパク質のアミノ酸配列を示し、そして、それとP.aeruginos、C.trachomatis、M.lepraeおよびH.sapiens由来の関連する熱ショックタンパク質とを比較している。配列の下線は、配列1〜4とヒト熱ショックタンパク質との間で、相同性が減少したドメインを示す。
【図2B】図2Bは、図2Aの続きである。
【図2C】図2Cは、図2Bの続きである。
【図3A】図3Aは、Helicobacter pyloriの熱ショックタンパク質の、ヌクレオチド配列およびアミノ酸配列である。
【図3B】図3Bは、図2Aの続きである。
【図3C】図3Cは、図2Bの続きである。
【発明を実施するための形態】
【0047】
(実施態様の詳細な説明)
(I.一般的な方法論)
本発明の実施には、他に示されていなければ、当該分野の分子生物学、微生物学、組換えDNA、および免疫学の従来の方法を使用する。このような方法は、文献に十分に説明されている。例えば、Sambrookら、MOLECULAR CLONING;A LABORATORY MANUAL,第二版(1289):DNA CLONING,VOLUMES IおよびII(D.N Glover編 1985):OLIGONUCLEOTIDE SYNTHESIS(M.J.Gait編 1984):NUCLEIC ACID HYBRIDIZATION(B.D.HamesおよびS.J.Higgins編 1984);TRANSCRIPTION AND TRANSLATION(B.D.HamesおよびS.J.Higgins編 1984);ANIMAL
CELL CULTURE(R.I.Freshney編 1986);IMMOBILIZED CELLS AND ENZYMES(IRL Press,1986);B.Perbal,A PRACTICAL GUIDE TO MOLECULAR CLONIHG(1984);METHODS IN EHZYMOLOGYシリーズ(Academic Press,Inc.);GENE TRANSFER VECTORS FOR MAMMALlAN CELLS(J.H.MillerおよびM.P.Calos編 1987,Cold Spring Harbor Laboratory)、Methods in Enzymology Vol.154およびVol.155(それぞれに、WuおよびGrossman、およびWu編)、MayerおよびWalker編(1987),IMMHNOCHEMICAL METHODS IN CELL AND MOLECULAR BIOLOGY(Academic Press,London)、Scopes,(1987),PROTEIN PURIFICATION:PRINCIPLES AND PRACTICE,第二版(Springer−Verlag,N.Y.)、および、HANDBOOK OF EXPERIMENTAL IMMUNOLOGY,VOLUMES I−IV(D.M.WeirおよびC.C.Blackwell編 1986)を参照のこと。
【0048】
ヌクレオチドおよびアミノ酸の標準的な省略形が、本明細書中で使用されている。本明細書中に掲載されている、全ての刊行物、特許、および特許出願は、参考として援用されている。
【0049】
(2.定義)
本発明に使用し得る熱ショックタンパク質には、上記のポリペプチド、および、上記タンパク質の天然のアミノ酸配列から少数のアミノ酸が変異したポリペプチドがあげられ;特に保存的なアミノ酸置換が予期される。
【0050】
保存的置換は、関連する側鎖を有するアミノ酸ファミリー内で起こる置換である。遺伝子的にコードされたアミノ酸は、一般的に4つのファミリーに分類される:(1)酸性=アスパラギン酸、グルタミン酸;(2)塩基性=リジン、アルギニン、ヒスチジン;(3)非極性=アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン;および、(4)非荷電極性=グリシン、アスパラギン、グルタミン、システイン、セリン、トレオニン、チロシン。フェニルアラニン、トリプトファン、およびチロシンは、しばしば、芳香族アミノ酸として、一緒に分類される。例えば、ロイシンとイソロイシンまたはバリンの、アスパラギン酸とグルタミン酸の、トレオニンとセリンとの単独の置換、あるいは、あるアミノ酸と、構造が関連するアミノ酸との同様の保存的置換が、生物学的活性に大きな影響を与えないことは当然予想し得る。タンパク質として実質的に同じアミノ酸配列を有するが、機能面に実質的に影響しない少数のアミノ酸置換を有するポリペプチド分子は、本タンパク質の定義の範囲に含まれる。
【0051】
天然の供給源からタンパク質を単離精製するよりも、組換えDNA技術により熱ショックタンパク質を生産する大きな長所は、天然の供給源からタンパク質を単離するのに要求されるよりも少ない開始物質を用いて、同量のタンパク質が生産され得ることである。組換え技術によるタンパク質の生産は、細胞中に通常存在するいくつかの分子の非存在下でも、タンパク質を単離を可能にする。実際、ヒトタンパク質混入の痕跡が全くないタンパク質組成物を容易に生産し得る。なぜなら、組換え非ヒト宿主により生産されるヒトタンパク質は、問題の組換えタンパク質のみだからである。天然の供給源からの潜在的なウイルス性物質およびヒトに対して発病性のウイルス成分もまた、避けられる。
【0052】
本明細書中に使用されているように、用語「組換えポリヌクレオチド」は、ゲノム、cDNA、半合成または合成起源のポリヌクレチドを示し、その起源または操作により:(1)天然では結合しているポリヌクレオチドの全てまたは一部と結合しない、(2)天然ではそれに結合しているポリヌクレオチド以外のポリヌクレオチドに結合する、または(3)天然では存在しない。
【0053】
本明細書中に使用されているように、用語「ポリヌクレオチド」とは、任意の長さのヌクレオチド、好ましくはデオキシリボヌクレオチドのポリマー形態のことであり、本明細書中では、用語「オリゴヌクレオチド」および「オリゴマー」と相互変換的に使用される。この用語は、分子の一次構造のみを示す。従って、この用語には、二本鎖および一本鎖DNAおよびアンチセンスポリヌクレオチドが包含される。これにはさらに、既知の型の改変、例えば、当該分野で公知の標識の存在、メチル化、末端「caps」、1つ以上の天然に存在するヌクレオチドのアナログとの置換、ヌクレオチド間の改変(例えば、ある型の非荷電性結合(例えば、ホスホン酸メチル、ホスホトリエステル、ホスホアミデート、カルバミン酸エステルなど)、または、荷電性結合(例えば、ホスホロチオエート、ホスホロジチオエートなど)との置換)、ペンダント部分の導入(例えば、タンパク質(ヌクレアーゼ、毒素、抗体、シグナルペプチド、ポリ−L−リシンなど)、挿入剤(例えば、アクリジン、ソラレン(psoralen)など)、キレート剤(例えば、金属、放射性物質、ホウ素、酸化成分など)、アルキル化剤(例えば、αアノマー性核酸など))が、包含される。
【0054】
用語「ゲノムの」により、ベクターにクローン化された制限フラグメント由来のDNA分子のコレクションまたはライブラリーを意味する。これには、微生物の遺伝子物質の全てまたは一部が包含され得る。
【0055】
用語「cDNA」により、mRNAの相補鎖にハイブリダイズする相補mRNA配列を意味する。
【0056】
本明細書中で使用されているように、用語「オリゴマー」は、プライマーおよびプローブの両方を意味し、本明細書では用語「ポリヌクレオチド」と相互変換的に使用される。用語オリゴマーは、分子の大きさを意味しない。しかし、典型的にオリゴマーは、1000ヌクレオチド未満、より典型的には500ヌクレオチド未満、さらに典型的には250ヌクレオチド未満であり、それらは100ヌクレオチド未満であり得、75ヌクレオチド未満、そしてさらに50ヌクレオチド未満の長さであり得る。
【0057】
本明細書中で使用されているように、用語「プライマー」とは、適切な条件下で使用されるときに、ポリヌクレオチド鎖の合成開始点として作用し得るオリゴマーのことである。プライマーは、コピーされるべきポリヌクレオチド鎖の領域に、完全にまたは実質的に相補的である。従って、ハイブリダイゼーションを行う条件下で、プライマーは、分析物鎖の相補領域にアニールする。適切な反応物(例えば、ポリメラーゼ、ヌクレオチドトリホスフェートなど)の添加により、プライマーは、重合剤により伸長されて分析物鎖のコピーを形成する。プライマーは一本鎖であり得るか、または、部分的または全体に二本鎖であり得る。
【0058】
用語「分析物ポリヌクレオチド」および「分析物鎖」とは、標的領域を含有すると思われる、一本鎖または二本鎖核酸分子のことであり、生物学的試料中に存在し得る。
【0059】
本明細書中で使用されているように、用語「プローブ」とは、標的領域中の配列とプローブ中の少なくとも1つの配列との相補性により、標的配列とハイブリッド構造を形成するポリヌクレオチドを含有する構造のことである。プローブのポリヌクレオチド領域は、DNAおよび/またはRNA、および/または合成ヌクレオチドアナログから構成され得る。「捕獲プローブ」および「標識プローブ」は、プローブ内に包含される。
【0060】
本明細書中で使用されているように、用語「標的領域」とは、増幅および/または検出されるべき核酸領域のことである。用語「標的配列」とは、プローブまたはプライマーが、所望の条件下で安定なハイブリッドを形成し得る配列のことである。
【0061】
本明細書中で使用されているように、用語「捕獲プローブ」とは、結合パートナーに結合する一本鎖ポリヌクレオチドを有するポリヌクレオチドプローブのことである。この一本鎖ポリヌクレオチドは、標的するポリヌクレオチド配列に含有され、これは、分析物ポリヌクレオチド中の検出されるべき標的領域内の標的配列に相補的である。この相補領域は、二本鎖に、分析物ポリヌクレオチドを固体表面に固定する(結合パートナーを介して)のに十分に安定性を与えるために、十分な長さからなり、標的配列に対して相補的である。結合パートナーは、第二の結合パートナーに特異的であり、第二の結合パートナーは、固体支持体の表面に結合され得るか、または、その他の構造物または結合パートナーを介して間接的に固体支持体に結合され得る。
【0062】
本明細書中で使用されているように、用語「標的するポリヌクレオチド配列」とは、標的ヌクレオチド配列に相補的であるヌクレオチドが含有されるポリヌクレチド配列のことであり、その配列は、意図される目的に十分な安定性を有する二本鎖を形成するのに、十分な長さからなり、標的配列に相補的である。
【0063】
本明細書中で使用されているように、用語「結合パートナー」とは、例えば、抗原とそれに特異的な抗体のような、高度な特異性を有するリガンド分子を結合し得る分子のことである。一般的に、特異的結合パートナーは、単離条件下で、分析物コピー/相補鎖の二本鎖(捕獲プローブの場合)を固定するのに十分な親和性で結合しなければならない。特異的結合パートナーは、当該分野で公知であり、例えば、ビオチンとアビジンまたはストレプトアビジン、IgGとプロテインA、多くの既知のレセプター−リガンド対、および、相補的ポリヌクレオチド鎖を包含する。相補的ポリヌクレオチド結合パートナーの場合には、パートナーは通常、少なくとも約15塩基の長さであり、少なくとも40塩基であり得、さらに、少なくとも約40%から約60%のG+C含有量を有する。ポリヌクレオチドは、DNA、RNAまたは合成ヌクレオチドアナログを含み得る。
【0064】
本明細書中で使用されているように、用語「カップルした(coupled)」とは、共有結合によるまたは非共有結合の強い相互作用(例えば、疎水相互作用、水素結合など)による付着のことである。共有結合は、例えば、エステル、エーテル、リン酸エステル、アミド、ペプチド、イミド、炭素−イオウ結合、炭素−リン結合などであり得る。
【0065】
用語「支持体」とは、所望の結合パートナーが固定され得る、いずれもの固体または半固体の表面のことである。適切な支持体には、ガラス、プラスチック、金属、ポリマーゲルなどが含まれ、ビーズ、ウエル、ディップスティック、膜などの形態にされ得る。
【0066】
本明細書中で使用されているように、用語「標識」とは、検出し得る(好ましくは定量し得る)シグナルを提供するために使用され得、ポリヌクレオチドまたはポリペプチドに結合され得る、いずれもの元素または部分のことである。
【0067】
本明細書中で使用されているように、用語「標識プローブ」とは、分析物ポリヌクレオチド中の検出されるべき標的配列に相補的である、標的するポリヌクレオチド配列を含有するポリヌクレオチドプローブのことである。この相補領域は、「標識プローブ」および「標識配列」を含有する二本鎖が、標識により検出されるように、十分な長さからなり、標的配列に相補的である。標識プローブは、直接に、または、マルチマーを含む、互いに高度な特異性を有する1組のリガンド分子を介して間接に、標識とカップルする。
【0068】
本明細書中で使用されているように、用語「マルチマー」とは、同じ繰り返しの一本鎖ポリヌクレオチドユニットまたは異なる一本鎖ポリヌクレオチドユニットの、直鎖状または分枝状ポリマーのことである。ユニットの少なくとも1つは、目的の第一の一本鎖ヌクレオチド配列、典型的には分析物または分析物に結合したポリヌクレオチドプローブ(例えば、標識プローブ)に特異的にハイブリダイズし得る、配列、長さ、および組成を有する。このような特異性および安定性を得るために、このユニットは、通常は少なくとも約15ヌクレオチドの長さであり、典型的には約50ヌクレオチド以下の長さ、そして好ましくは約30ヌクレオチドの長さであり;さらに、G+C含有量は、通常は少なくとも約40%、ほとんどは約60%であるロこのようなユニットに加えて、マルチマーには、目的の第二の一本鎖ヌクレオチド、典型的には標識ポリヌクレオチドまたはその他のマルチマーに、特異的および安定にハイブリダイズし得る、多くのユニットが包含される。これらのユニットは、上記に考察されたマルチマーのように、一般にはほぼ同じ大きさおよび組成である。マルチマーがその他のマルチマーにハイブリダイズされるように設計されているときには、第一および第二のオリゴヌクレオチドユニットは異質(異なる)であり、選択されたアッセイの条件下では、互いにはハイブリダイズしない。従って、マルチマーは、標識プローブであり得るか、または標識をプローブにカップルするリガンドであり得る。
【0069】
「レプリコン」は、細胞内のポリヌクレオチド複製の自己複製ユニットとして挙動する、すなわち、自己制御下に複製し得るいずれもの遺伝子エレメント、例えば、プラスミド、染色体、ウイルス、コスミドなどである。これには選択マーカーが含まれ得る。
【0070】
「PCR」とは、Saikiら、Nature 524:163(1986);およびScharfら、Science(1986)233:1076−1078;および米国特許第4,683,195号;ならびに米国特許第4,683,202号に記載されているようなポリメラーゼ連鎖反応の技法である。
【0071】
本明細書中で使用されているように、本来xが、同じ様式でyに関連しない場合に、xはyに関して「異種」である。すなわち、天然ではxが全くyに関連しないか、または、xが天然に実在して認められるような同じ様式でyに関連しない。
【0072】
「相同性」とは、xとyとの間の類似性の程度のことである。1つの形態の配列と別の配列との対応は、当該分野で公知の方法により決定され得る。例えば、それらはポリヌクレオチドの配列情報の直接比較により決定され得る。または、相同性は、相同領域(例えば、S1消化の前に使用される)間で安定な二本鎖を形成する条件下でポリヌクレオチドをハイブイブリダイズし、その後、一本鎖特異ヌクレアーゼにより消化し、その後に消化されたフラグメントの大きさを決定することにより決定され得る。
【0073】
「ベクター」は、別のポリヌクレオチドセグメントが接続されて、この接続されたセグメントの複製および/または発現をもたらすレプリコンである。
【0074】
「制御配列」とは、それらが連結されるコーディング配列の発現をもたらすために必要であるポリヌクレオチド配列のことである。このような制御配列の性質は、宿主生物に依存して異なる;原核生物では、このような制御配列は、一般的にプロモーター、リボソーム結合部位、および転写終止配列を含み;真核生物では、一般的に、このような制御配列は、プロモーターおよび転写終止配列を含む。用語「制御配列」は、最少限、発現に必要とされる全成分を含み、さらに存在が有利である付加的な成分、例えば、リーダー配列および融合パートナー配列を含み得ると意図される。
【0075】
「作動可能に連結された」とは、記載されている成分が、それらの意図する様式で機能し得る関係にある並列のことである。コーディング配列に「作動可能に連結された」制御配列は、コーディング配列の発現が制御配列に適合した条件下で得られるように連結されている。
【0076】
「オープンリーディングフレーム」(ORF)は、ポリペプチドをコードするポリヌクレオチド配列の領域のことである;この領域は、コーディング配列の一部またはコーディング配列全てを示し得る。
【0077】
「コーディング配列」は、適切な調節配列の制御下におかれると、通常はmRNAを介してポリペプチドに翻訳されるポリヌクレオチド配列である。コーディング配列の境界は、5’末端の翻訳開始コドンおよびポ末端の翻訳終止コドンにより決定される。コーディング配列には、cDNAおよび組換えポリヌクレオチド配列が含まれる得るが、これらに限定されない。
【0078】
本明細書中で使用されているように、用語「ポリペプチド」は、アミノ酸のポリマーを指し、特定の長さの産物を指すわけではない;従って、ペプチド、オリゴペプチド、およびタンパク質が、ポリペプチドの定義内に包含される。この用語にはまた、ポリペプチドの発現後改変(例えば、グリコシル化、アセチル化、リン酸化など)を意味せず、あるいは除外しない。定義に包含されるのは、例えば、アミノ酸の1つ以上のアナログを含有するポリペプチド(例えば、非天然アミノ酸などを含む)、置換結合を有するポリペプチド、および当該分野で公知の天然に存在するおよび存在しない改変である。
【0079】
示された核酸配列「由来の」ポリペプチドまたはアミノ酸配列とは、配列にコードされたポリペプチドのアミノ酸配列と同一のアミノ酸配列を有するポリペプチド、またはそれらの部分、ここで、この部分は、少なくとも3−5アミノ酸、さらに好ましくは少なくとも8−10アミノ酸、そしてよりさらに好ましくは少なくとも11−15アミノ酸からなる、または配列にコードされたポリペプチドにより免疫学的に同定され得るポリペプチドのことである。この用語にはまた、示された核酸配列から発現されるポリペプチドも包含される。
【0080】
「免疫原性の」とは、アジュバントの存在または非存在下で、それのみで、またはキャリアに結合されて、体液性免疫応答および/または細胞性免疫応答を引き起こすポリペプチドの能力のことである。「中和」とは、感染因子の感染力を、部分的または全体的に抑制する免疫応答のことである。「エピトープ」とは、ペプチド、ポリペプチド、またはタンパク質の抗原決定基のことである;1つのエピトープは、そのエピトープに対して唯一の高次構造(spatial conformation)で、3つ以上のアミノ酸を含み得る。一般的には、エピトープは、少なくとも5つのこのようなアミノ酸からなり、さらに通常には少なくとも8−10のこのようなアミノ酸からなる。アミノ酸の高次構造の決定方法は当該分野では公知であり、例えば、これらには、x線結晶解析および二次元核磁気共鳴が含まれる。同じエピトープを認識する抗体は、1つの抗体が標的抗原に対するその他の抗原の結合を阻む能力を示す、簡単な免疫アッセイにおいて同定され得る。
【0081】
本明細書中で使用されているように、「処置(treatment)」とは、予防および/または治療(therapy)(すなわち、あらゆる疾患症状の調整)のことである。「個体(individual)」は、抗原性の莢膜多糖またはオリゴ糖構造を有する細菌の感染が疑われる動物を意味し、これにはヒトを含む霊長類が包含されるが、これらに限定されない。「ワクチン」は、免疫原性の、または、部分的または完全にこのような細菌に対する保護を誘起し得る、個体の処置に有用な組成物である。
【0082】
本発明のコンジュゲート化合物は、このタンパク質に特異的な、モノクローナルまたはポリクローナル抗体の産生に使用され得る。これらの抗体を産生する方法は、当該分野で公知である。
【0083】
「組換え宿主細胞」、「宿主細胞」、「細胞」、「細胞培養物」、およびその他のこのような用語は、例えば、組換えベクターまたは他の転移DNAの受容体として使用され得るかまたは使用されてきた、微生物、昆虫細胞、および哺乳類細胞を示し、これには、形質転換されたもとの細胞の子孫も包含される。1つの母細胞の子孫は、自然突然変異、偶発突然変異または意図的突然変異(deliberate mutation)のために、形態学的に、または、ゲノムDNAまたは全DNA補体が、必ずしも、もとの母細胞と完全に同一ではあり得ないことが理解される。哺乳類宿主細胞の例には、チャイニーズハムスター卵巣(CHO)細胞およびサル腎(COS)細胞が含まれる。
【0084】
詳細には、本明細書中で使用されているように、「細胞系」とは、インビトロにおいて継続してまたは長期間に増殖および分裂し得る細胞群のことである。しばしば、細胞系は1つの幹細胞(progenitor cell)からのクローン群である。さらに、このようなクローン群の貯蔵または転移の間に、核型に自発変化または誘導変化が起こり得ることが、当該分野では公知である。従って、いわゆる細胞系由来の細胞は、祖先細胞または培養物に正確には同一ではあり得ず、この細胞系には、このような変異体が包含される。用語「細胞系」はまた、不死化細胞を包含する。好ましくは、細胞系は、非ハイブリッド細胞系、または二細胞型のハイブリドーマを包含する。
【0085】
本明細書中で使用されているように、用語「微生物」には、細菌および真菌のような、原核微生物種および真核微生物種が包合され、後者には、酵母および糸状菌が含まれる。
【0086】
本明細書中で使用されているように、「形質転換」とは、挿入に用いられる方法(例えば、直接取り込み、形質導入、f−交配またはエレクトロポーレーション)に関係なく、外因性ポリヌクレオチドの宿主細胞への挿入のことである。外因性ポリヌクレオチドは、非組込みベクター(例えばプラスミド)として維持され得るか、または、宿主細胞ゲノムに組み込まれ得る。
【0087】
ポリペプチドまたはヌクレオチド配列に関するときには、「精製された」および「単離された」とは、示された分子が、同じ型のその他の生物学的高分子を実質的に含まずに存在することを意味する。本明細書中で使用されているように、用語「精製された」は、好ましくは、同じ型の生物学的高分子が、少なくとも75重量%、さらに好ましくは少なくとも85重量%、さらに好ましくは少なくとも95重量%、そして最も好ましくは少なくとも98重量%で存在することを意味する(しかし、水、緩衝液、およびその他の小分子、特に、1000未満の分子量を有する分子は存在し得る)。
【0088】
(3.発現系)
一旦、適切な熱ショックタンパク質のコーディング配列が単離されると、それは種々の異なる発現系、例えば、哺乳類細胞、バキュロウイルス、細菌、および酵母により用いられる発現系、で発現され得る。
【0089】
(3.1.哺乳類系)
哺乳類発現系は当該分野で公知である。哺乳類プロモーターは、哺乳類RNAポリメラーゼを結合し得、コーディング配列(例えば、構造遺伝子)のmRNAへの下流(3’)転写を開始し得る、いずれものDNA配列である。プロモーターは、通常はコーディング配列の5’末端近傍に位置する転写開始領域、および、通常は転写開始部位の上流25−30塩基対(bp)に位置するTATAボックスを有する。TATAボックスは、RNAポリメラーゼIIがRNA合成を適切な部位で開始させるのを導くと考えられている。哺乳類プロモーターはさらに、通常はTATAボックスの上流100から200bp内に位置する上流プロモーターエレメントを有する。上流プロモーターエレメントは、転写が開始される速度を決定し、いずれかの向きに作用し得る(Sambrookら、Molecular Cloning:A Laboratory Manual,第二版(1989))。
【0090】
哺乳類ウイルス遺伝子は、しばしば高度に発現され、広範囲の宿主を有する;従って、哺乳類ウイルス遺伝子をコードする配列は、特に有用なプロモーター配列を提供する。例には、SV40初期プロモーター、マウス乳癌ウイルスLTRプロモーター、アデノウイルス主要後期プロモーター(Ad MLP)、および単純ヘルペスウイルスプロモーターが含まれる。さらに、ネズミメタロチオネイン遺伝子のような非ウイルス遺伝子由来の配列もまた、有用なプロモーター配列を提供する。発現は、構成的または調節され得(誘導可能な)、プロモーターに依存して、ホルモン−応答細胞中でグルココルチコイドにより誘導され得る。
【0091】
上記のプロモーターエレメントに組み合わされたエンハンサーエレメント(エンハンサー)の存在は、通常は発現レベルを増大させる。エンハンサーは、相同または異種プロモーターに連結されたときに、通常のRNA開始部位で合成を開始して、最高1000倍まで転写を促進し得る調節DNA配列である。さらに、エンハンサーは、通常の向きまたは裏返しの向きで転写開始部位から上流または下流に、または、プロモーターから1000ヌクレオチドを超える位置に配置されるときに活性である(Maniatisら、Science 236:1237(1989);Albertsら、Molecular Biology of the Cell,第二版(1982))。ウイルス由来のエンハンサーエレメントは、通常は広範囲の宿主を有するので、特に有用であり得る。例には、SV40初期遺伝子エンハンサー(Dijkemaら、(1985)EMBO J.4:761)、および、ラウス肉腫ウイルスの長末端反復(LTR)由来(Gormanら、(1982)Proc.Natl.Acad.Sci.79:6777)およびヒトサイトメガロウイルス由来(Boshartら、(1985)Cell 41:5221)のエンハンサー/プロモーターが含まれる。さらに、いくつかのエンハンサーは、調節可能であり、ホルモンまたは金属イオンのような誘導物質が存在するときのみに活性になる(Sassone−Corsiら、(1986)Trends Genet.2:215;Maniatisら、(1987)Science 236:1237)。
【0092】
DNA分子は、哺乳類細胞において細胞内に発現され得る。プロモータ配列はDNA分子に直接連結され得るが、この場合にはいつも、組換えタンパク質のN末端の最初のアミノ酸がメチオニンであり、ATG開始コドンによりコードされる。所望であれば、N末端は、インビトロにおける臭化シアンとのインキュベーションにより、タンパク質から切断され得る。
【0093】
あるいは、外来タンパク質はまた、哺乳類細胞で外来タンパク質の分泌をなすリーダー配列フラグメントを含む、融合タンパク質をコードするキメラDNA分子をつくることにより、細胞から増殖培地内に分泌され得る。好ましくは、リーダーフラグメントと外来遺伝子との間でコードされるプロセッシング部位が存在し、これは、インビボまたはインビトロのいずれかにおいて切断され得る。リーダー配列フラグメントは通常、細胞からのタンパク質の分泌を導く、疎水性アミノ酸を含むシグナルペプチドをコードする。アデノウイルス3分節系リーダーは、哺乳類細胞内での外来タンパク質の分泌を提供するリーダー配列の例である。
【0094】
一般には、哺乳類細胞により認識される転写終止およびポリアデニル化配列は、翻訳終止コドンに対し3’側に位置する調節領域であり、従って、プロモーターエレメントとともに、コーディング配列に隣接する。成熟mRNAの3’末端は、部位特異的に翻訳後に切断され、ポリアデニル化されて形成される(Birnstielら、(1985)Cell 41:349;ProudfootおよびWhitelaw(1988)「真核RNAの終結および3’末端プロセッシング」Transcrition and splicing(B.D.HamesおよびD.M.Glover編);Proudfoot(1982)Trends Biochem.Sci.14:105)。これらの配列は、DNAによりコードされるポリペプチドに翻訳され得るmRNAの転写を導く。転写ターミネーター/ポリアデニル化シグナルの例には、SV40由来のものが含まれる(Sambrookら(1989),Molecular Cloning:A Laboratory Manual)。
【0095】
いくつかの遺伝子は、イントロン(介在配列とも呼ばれる)が存在するときに、より効率よく発現され得る。しかし、数個のcDNAは、スプライシングシグナル(スプライス供与部位および受容部位とも呼ばれる)が欠如しているベクターから効率よく発現された(例えば、GethingおよびSambrook(1981)Nature 293:620を参照のこと)。イントロンは、スプライス供与部位および受容部位を有するコーディング配列内の介在非コーディング配列である。それらは、一次転写物のポリアデニル化の後に、「スプライシング」と呼ばれるプロセスにより除去される(Nevins(1983)Annu、Rev.Biochem.52:441;Green(1986)Annu.Rev.Genet.20:671;Padgettら、(1986)Annu.Rev.Biochem.55:1119;KrainerおよびManiatis(1988)「RNAスプライシング」,Transcription and splicing(B.D.HamesおよびD.M.Glover編))。
【0096】
通常、プロモーター、ポリアデニル化シグナル、および転写終止配列を含有する上記の成分は、発現構築物中に組み立てられる。エンハンサー、機能性スプライス供与部位および受容部位を有するイントロン、およびリーダー配列もまた、所望であれば発現構築物中に含まれる。発現構築物は、しばしば、哺乳類細胞または細菌のような宿主中で安定に維持され得る染色体外エレメント(例えば、プラスミド)のような、レプリコン中に維持される。哺乳類複製系には、複製にトランス作用因子が必要とされる動物ウイルス由来のものが含まれる。例えば、SV40のようなパポーバウイルス(Gluzman(1981)Cell 23:175)またはポリオーマウイルスの複製系を含むプラスミドが、適切なウイルスT存在下で、非常に多数のコピーを複製する。哺乳類レプリコンのさらなる例には、ウシパピローマウイルスおよびエプスタイン・バーウイルス由来のものが含まれる。さらに、レプリコンは2つの複製系を有し得、このため、例えば、発現のために哺乳類細胞中に、およびクローニングおよび増幅のために原核生物宿主中に維持される。このような哺乳類−細菌シャトルベクターの例には、pMT2(Kaufmanら(1989)Mol,Cell Biol.9:946)、およびpHEBO(Shimizuら(1986)Mol.Cell,Biol.6:1074)が含まれる。
【0097】
使用される形質転換方法は、形質転換されるべき宿主に依存する。異種ポリヌクレオチドの哺乳類細胞への導入方法は、当該分野で公知であり、デキストラン介在トランスフェクション、リン酸カルシウム沈降法、ポリブレン介在トランスフェクション、プロトプラスト融合、エレクトロポーレーションポリヌクレオチドのリボソーム内へのカプセル化、および、DNAの核への直接マイクロインジェクションを包含する。
【0098】
発現用の宿主として入手可能な哺乳類細胞系は、当該分野で公知であり、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、ベビーハムスター腎(BHK)細胞、サル腎細胞(COS)、ヒト肝細胞癌腫細胞(例えば、HepG2)および多くのその他の細胞系を含む、アメリカンタイプカルチャーコレクション(ATCC)から入手可能な多くの不死化細胞系を包含するが、これらに限定されない。
【0099】
(ii.バキュロウイルス系)
タンパク質をコードするポリヌクレオチドもまた、適切な昆虫発現ベクターに挿入され得、ベクター内の制御エレメントに作動可能に連結される。ベクター構築には、当該分野で公知の方法が用いられる。
【0100】
一般的に、発現系の成分には転移ベクター、通常は細菌プラスミドが含まれ、これは、バキュロウイルスゲノムフラグメントと、発現されるべき異種の遺伝子または遺伝子群挿入のための便宜的な制限部位との両方;転移ベクター内のバキュロウイルス特異フラグメントに相同な配列を有する野生型バキュロウイルス(これは、バキュロウイルスゲノム内への異種遺伝子の相同的組換えを考慮する);および、適切な昆虫宿主細胞および増殖培地を含む。
【0101】
タンパク質をコードするDNA配列を転移ベクター内に挿入した後に、ベクターおよび野生型ウイルスゲノムが、昆虫宿主細胞内ヘトランスフェクトされ、そこでベクターとウイルスゲノムとの組換えが起こる。パッケージングされた組換えウイルスが発現され、組換えプラークが同定および精製される。バキュロウイルス/昆虫細胞発現系用の材料および方法は、キット形態で、特に、Invitrogen,San Diego CA(「MaxBac」キット)から市販されている。これらの方法は当業者に公知であり、SummersおよびSmith,Texas Agricultural Experiment Station Bulletin No,1555(1987)(以後、「SummersおよびSmith」)に十分に記載されている。
【0102】
タンパク質をコードするDNA配列をバキュロウイルスゲノムに挿入する前に、プロモーター、リーダー(所望であれば)、目的のコーディング配列、および転写終止配列を含む上記の成分が、通常は中間置換構築物(intermediate transplacement construct)(転移ベクター)に組み立てられる。この構築物は、1つの遺伝子および作動可能に連結された調節エレメント;各遺伝子がそれ自身の固有のセットの作動可能に連結された調節エレメントを有する複数の遺伝子;または、同じセットの調節エレメントに調節される複数の遺伝子を含み得る。中間置換構築物はしばしば、細菌のような宿主中で安定して維持され得る染色体外エレメント(例えば、プラスミド)のような、レプリコン中に維持される。レプリコンは1つの複製系を有し、このため、クローニングおよび増幅のために適切な宿主中に維持される。
【0103】
現在では、AcNPVへ外来遺伝子を導入するための最も一般的に使用される転移ベクターは、pAc373である。当業者に公知のその他の多くのベクターもまた設計されている。これらは、例えば、pVL985(ポリヘドリン開始コドンをATGからATTに変え、ATT下流32塩基対にBamHIクローニング部位を導入する;LuckowおよびSummers,Virology(1989)17:31を参照)。
【0104】
プラスミドは通常さらに、ポリヘドロンポリアデニル化シグナル(Millerら(1988)Ann.Rev.Microbiol.,42:177)、および、選択およびE.coli中での増殖のための原核アンピシリン耐性(amp)遺伝子および複製起点を含有する。
【0105】
バキュロウイルス転移ベクターは通常、バキュロウイルスプロモーターを含有する。バキュロウイルスプロモーターは、バキュロウイルスRNAポリメラーゼを結合し得、そしてコーディング配列(例えば、構造遺伝子)のmRNAへの下流(5’から3’)翻訳を開始し得る、いずれものDNA配列である。プロモーターは、コーディング配列の5’末端近傍に通常位置する転写開始領域を有する。この転写開始領域は通常、RNAポリメラーゼ結合部位および転写開始部位を含有する。バキュロウイルス転移ベクターもまた、エンハンサーと呼ばれる第二ドメインを有し得、これは存在する場合には、構造遺伝子の遠位に存在する。発現は調節され得るか、または構成的であり得る。
【0106】
ウイルス感染サイクルの後期に多量に転写された構造遺伝子は、特に有用なプロモーター配列を提供する。例には、ウイルスポリヘドロンタンパク質をコードする遺伝子由来の配列(Friesenら、(1986)「バキュロウイルス遺伝子発現の調節」,TheMolecular Biology of Baculoviruses(Walter Doerfler編);EPO公開No.127839およびNo.155476);および、p10タンパク質をコードする遺伝子(Vlakら、(1988),J.Gen.Virol.69:765)が含まれる。
【0107】
適切なシグナル配列をコードするDNAは、バキュロウイルスポリヘドリン遺伝子のような、昆虫またはバキュロウイルス分泌タンパク質の遺伝子由来であり得る(Carbonellら(1988)Gene,73:409)。あるいは、哺乳類細胞の翻訳後の改変(シグナルペプチド切断、タンパク質分解切断、およびリン酸化のような)のためのシグナルは、昆虫細胞により認識されると考えられており、そして分泌に必要なシグナルおよび核集積は、無脊椎動物細胞と脊椎動物細胞との間で保存されているとも考えられているので、ヒトα−インターフェロン(Maedaら、(1985),Nature 315:592);ヒトガストリン放出ペプチド(Lebacq−Verheydenら、(1988),Molec.Cell.Biol.8:3129);ヒトIL−2(Smithら、(1985)Proc.Nat’l Acad.Sci.USA,82:8404);マウスIL−3,(Miyajimaら、(1987)Gene 58:273;および、ヒトグルコセレブロシダーゼ(Martinら(1988)DNA 7:99)をコードする遺伝子由来のような、非昆虫起源のリーダーもまた、昆虫での分泌を提供するために使用され得る。
【0108】
組換えポリペプチドまたはポリタンパク質は、細胞内に発現され得るか、または、適切な調節配列で発現されれば分泌され得る。非融合外来タンパク質の良好な細胞内発現は通常、ATG開始シグナルの前に適切な翻訳開始シグナルを含有する短いリーダー配列を理想的に有する異種遺伝子を必要とする。所望であれば、N末端のメチオニンは、インビトロにおける臭化シアンとのインキュベーションにより、成熟タンパク質から切断され得る。
【0109】
あるいは、組換えポリタンパク質または天然では分泌されないタンパク質は、昆虫において外来タンパク質の分泌を提供するリーダー配列フラグメントを有する融合タンパク質をコードするキメラDNA分子をつくることにより、昆虫細胞から分泌され得る。リーダー配列フラグメントは通常、小胞体へのタンパク質の輸送(translocation)を導く、疎水性アミノ酸を有するシグナルペプチドをコードする。
【0110】
タンパク質の発現産物の前駆体をコードするDNA配列および/または遺伝子の挿入後に、昆虫細胞宿主を、転移ベクターの異種DNAと野生型バキュロウイルスのゲノムDNAとで共形質転換(通常は共トランスフェクション)する。構築物のプロモーターおよび転写終止配列は、通常バキュロウイルスゲノムの2−5kb断片を含有する。バキュロウイルスの所望の部位に異種DNAを導入する方法は、当該分野で公知である。(SummersおよびSmith:Juら(1987);Smithら、Mol.Cell.Biol.(1983)3:2156;および、LuckowおよびSummers(1989)を参照のこと)。例えば、挿入は、ポリヘドリン遺伝子のような遺伝子に、相同的二本鎖交差組換えによりなされ得る;挿入はまた、所望のバキュロウイルス遺伝子中に設計された制限酵素部位になされ得る。Millerら、(1989),Bioessays
4:91。
【0111】
DNA配列は、発現ベクターのポリヘドリン遺伝子の位置にクローニングされたときには、ポリヘドリン特異配列により、5’および3’両側に隣接され、ポリヘドリンプロモーターの下流に配置される。
【0112】
新たに形成されたバキュロウイルス発現ベクターは、次に、感染性組換えバキュロウイルス中にパッケージングされる。相同的組換えは低頻度(約1%と約5%との間)で生じるため、共トランスフェクション後に産生された主要なウイルスは、まだ野生型である。従って、組換えウイルスを同定する方法が必要である。発現系の利点は、視覚スクリーニングにより、組換えウイルスの区別が可能であることである。天然ウイルスにより産生されるポリヘドリンタンパク質は、ウイルス感染後、後期に感染細胞の核に非常に高レベルで産生される。集積されたポリヘドリンタンパク質は、埋め込まれた粒子をも含む封入体を形成する。これらの封入体は、15μmまでの大きさであり、非常に屈折性であるために、光学顕微鏡下で容易に視覚化される光沢のある外観となる。組換えウイルスに感染した細胞には、封入体がない。野生型ウイルスから組換えウイルスを区別するには、トランスフェクション上清を、当業者に公知の方法により、単層の昆虫細胞上でプラーク形成させる。すなわち、プラークを光学顕微鏡下で、封入体の存在(野生型ウイルスの指標)または非存在(組換えウイルスの指標)についてスクリーニングする。「Current Protocols in Microbiology」Vol.2(Ausubelら編)16.8(Supp.10,1990):SummersおよびSmith;Millerら(1899)。
【0113】
組換えバキュロウイルス発現ベクターは、数種の昆虫細胞への感染のために開発されている。例えば、組換えバキュロウイノレは、とりわけ、Aedes aegypti、Autographa californica、Bombyx mori、Drosophila melanogaster、Spodoptera frugiperda、およびTrichoplusa niのために開発されている(PCT公開No.WO89/046699;Carbonellら、(1985) J.Virol.56:153;Wright(1986)Nature 321:718;Smithら、(1983)Mol.Cell.Biol.3:2156:および一般的にはFraserら(1989)In Vitro Cell.Dev.Biol.25:225を参照のこと)。
【0114】
細胞および細胞培養培地は、バキュロウイルス/発現系での異種ポリペプチドの直接発現および融合発現市販されている;細胞培養法は、一般に当業者に公知である。例えば、SummersおよびSmithを参照のこと。
【0115】
次に、改変昆虫細胞は、適切な栄養培地で増殖され得る。この栄養培地により、改変昆虫宿主内に存在するプラスミドは安定に維持され得る。発現産物遺伝子が誘導制御下にある場合には、宿主は高密度に増殖され得、発現が誘導され得る。あるいは、発現が構成的である場合には、産物は培地に連続的に発現される。目的の産物を取り出し、枯渇した栄養分を補給しながら、栄養培地を連続的に取り替えねばならない。産物は、例えば、HPLC、アフィニティークロマトグラフィー、イオン交換クロマトグラフィーなどのクロマトグラフィー;電気泳動;密度勾配遠心分離;溶媒抽出などの方法で精製され得る。適切には、培地に分泌されるかまたは昆虫細胞溶解から生じるいずれもの昆虫タンパク質を実質的に除去するために、宿主の破片(例えば、タンパク質、脂質および多糖類)を少なくとも実質的に含まない産物を提供するために、必要であれば産物はさらに精製され得る。
【0116】
タンパク質発現を得るために、形質転換体由来の組換え宿主細胞は、配列をコードする組換えタンパク質の発現を可能にする条件下でインキュベートされる。これらの条件は、選択される宿主細胞に依存して変化する。しかし、条件は、当該分野で公知のものに基づいて、当業者に容易に確認され得る。
【0117】
(iii.細菌系)
細菌発現法は当該分野で公知である。細菌プロモーターは、細菌RNAポリメラーゼに結合し得、コーディング配列(例えば、構造遺伝子)のmRNAへの下流(3’)転写を開始し得るいずれものDNA配列である。プロモーターは、通常コーディング配列の5’末端近傍に位置する転写開始領域を有する。この転写開始領域は通常、RNAポリメラーゼ結合部位および転写開始部位を含む。細菌プロモーターはまた、オペレーターと呼ばれる第二ドメインを有し得、これは、RNA合成を開始する位置である隣接RNAポリメラーゼ結合部位に重複し得る。遺伝子リプレッサ一タンパク質はオペレターに結合し得、そのことにより特異遺伝子の転写を阻害し得るので、オペレーターは、負の調節(誘導可能な)転写を行い得る。構成的発現は、オペレーターのような負の調節エレメントの非存在下で起こり得る。さらに、正の調節は、遺伝子活性化タンパク質結合配列により達成され得、これは、存在する場合には、通常RNAポリメラーゼ結合配列(5’)の近くに存在する。遺伝子活性化タンパク質の例は、力タボライト活性化タンパク質(CAP)であり、これは、E.coliでのlacオペロンの転写開始を助ける(Raibaudら(1984)Annu.Rev.Genet.18:173)。従って、調節的発現は正または負のいずれかであり得、そのことにより転写を促進または減退する。
【0118】
代謝経路の酵素をコードする配列は、特に有用なプロモーター配列を提供する。例には、ガラクトース、ラクトース(lac)(Changら(1977)Nature 198:1056)、およびマルトースのような糖代謝酵素由来のプロモーター配列が含まれる。さらなる例には、トリプトファン(trp)のような生合成酵素由来のプロモーター配列が含まれる(Goeddelら(1980)Nuc.Acids Res.8:4057;Yelvertonら(1981)Nucl.Acids Res.9:731;米国特許第4,738,921号;EPO公開No.036776およびNo.121775)。g−ラオタマーゼ(g−laotamase)(bla)プロモーター系(Weissmann(1981)「インターフェロンのクローニングおよびその他の誤り(mistakes)」Interferon 3(I.Gresser編)、バクテリオファージλPL(Shimatakeら(1981)Nature 292:128)、およびT5(米国特許第4,689,406号)プロモーター系もまた、有用なプロモーター配列を提供する。
【0119】
さらに、天然に存在しない合成プロモーターもまた細菌プロモーターとして機能する。例えば、1つの細菌またはバクテリオファージプロモーターの転写活性化配列は、その他の細菌またはバクテリオファージプロモーターのオペロン配列に接続され得て、合成雑種プロモーターをつくる(米国特許第4,551,433号)。例えば、tacプロモーターは、lacリプレッサーにより調節される、trpプロモーターおよびlacオペロン両配列を含有する雑種trp−lacプロモーターである(Amannら(1983)Gene 25:167;de Boerら(1983)Proc.Natl.Acad.Sci.80:21)。さらに、細菌プロモーターは、細菌RNAポリメラーゼに結合し、転写を開始する能力を有する、非細菌起源の天然に存在するプロモーターを含み得る。非細菌起源の天然に存在するプロモーターはまた、適合するRNAポリメラーゼと結合して原核生物中である種の遺伝子を高レベルで発現し得る、バクテリオファージT7 RNAポリメラーゼ/プロモーター系は、結合されたプロモーター系の例である(Studierら(1986)J.Mol.Biol.189:113:Taborら(1985)Proc.Natl.Acad.Sci.82:1074)。さらに、雑種プロモーターもまた、バクテリオファージプロモーターおよびE.coliオペレーター領域を含有し得る(EPO公開No.267851)。
【0120】
機能プロモーター配列に加えて、能率のよいリボソーム結合部位もまた、原核生物での外来遺伝子の発現に有用である。E.coliでは、リボソーム結合部位はシャイン・ダルガーノ(SD)配列と呼ばれ、開始コドン(ATG)および開始コドンの3−11ヌクレオチド上流に位置する3−9ヌクレオチドの長さの配列を含む(Shineら(1975)Nature 254:34)。SD配列は、SD配列とE.coli 16S rRNAの3’末端(3’and)との間で塩基対合してmRNAのリボソームヘの結合を促進すると考えられている(Steitzら(1979)「メッセンジャーRNAにおける遺伝シグナルおよびヌクレオチド配列」,Biological Regulation and Develoment:Gene Expression(R.F.Goldberger編))。弱リボソーム結合部位を有する原核遺伝子および真核遺伝子を発現するためには、Sambrookら(1989),Molecular Clonin:A Laboratory Manualを参照のこと。
【0121】
DNA分子は細胞内に発現され得る。プロモーター配列は、直接DNA分子に連結され得、その場合、N末端の最初のアミノ酸はメチオニンであり、これはATG開始コドンによりコードされる。必要であれば、N末端のメチオニンは、インビトロにおける臭化シアンとのインキュベーション、または細菌メチオニンN末端ペプチダーゼとのインビボまたはインビトロインキュベーションにより、タンパク質から切断され得る(EPO公開No.219237)。
【0122】
融合タンパク質は、直接発現の代替を提供する。通常は、内因性細菌タンパク質またはその他の安定なタンパク質のN末端部分をコードするDNA配列が、異種コーディング配列の5’末端に融合される。発現に際し、この構築物は、2つのアミノ酸配列の融合を提供する。例えば、バクテリオファージλ細胞遺伝子が外来遺伝子の5’末端で連結され得、細菌内で発現され得る。得られた融合タンパク質は、好ましくは、外来遺伝子からバクテリオファージタンパク質を切断するためのプロセッシング酵素(因子Xa)に対する部位を保持している(Nagaiら(1984)Nature 309:810)。融合タンパク質はまたlacZ由来の配列を用いて生成され得る(Jiaら(1987)Gene 60:197,trpE,Allenら(1987)J.Biotechnol.5:93:Makoffら(189)J.Gen.Microbiol.135:11、およびEPO公開No.324647,genes)。2つのアミノ酸配列を接続したDNA配列は、切断部位をコードし得るか、またはコードし得ない。その他の例は、ユビキチン融合タンパク質である。このような融合タンパク質は、好ましくは、外来遺伝子からユビキチンを切断するためのプロセッシング酵素(例えば、ユビキチン特異的プロセッシングプロテアーゼ)に対する部位を保持しているユビキチン領域を用いて生成される。この方法によって、そのままの外来タンパク質が単離され得る(Millerら(1989)Bio/Technology 7:698)。
【0123】
あるいは、外来タンパク質はまた、細菌で外来タンパク質分泌を提供するシグナルペプチド配列フラグメントを含有する融合タンパク質をコードするキメラDNA分子をつくることにより、細胞から分泌され得る(米国特許第4,336,336号)。シグナル配列フラグメントは通常、細胞からのタンパク質分泌を導く疎水性アミノ酸を含有するシグナルペプチドをコードする。タンパク質は、増殖培地(グラム陽性細菌)、または、細胞の内膜と外膜との間に位置する細胞周辺腔(グラム陰性細菌)のいずれかに分泌される。好ましくは、シグナルペプチドフラグメントと外来遺伝子との間でコードされるプロセッシング部位が存在し、これはインビボまたはインビトロにおいて切断され得る。
【0124】
適切なシグナル配列をコードするDNAは、E.coli外膜タンパク質遺伝子(ompA)(Masuiら(1983),Experimental Manipulation of Gene Expression;Ghrayebら(1984)EMBO
J.3:2437)およびE.coliアルカリホスファターゼシグナル配列(phoA)(Okaら(1985)Proc.Natl Acad.Sci.82:7212)のような、分泌される細菌タンパク質の遺伝子由来であり得る。さらなる例として、種々のBacillus株由来のα−アミラーゼ遺伝子のシグナル配列は、B.subtilisから異種タンパク質を分泌するために使用され得る(Palvaら(1982)Proc.Natl.Acad.Sci.USA 79:5582:EPO公開No.244042)。
【0125】
通常、細菌により認識される転写終止配列は、翻訳終止コドンに対して3’側に位置する調節領域であり、従って、プロモーターとともにコーディング配列に隣接する。これらの配列は、DNAにコードされるポリペプチドに翻訳され得るmRNAの転写を導く。転写終止配列はしばしば、転写終止の助けとなるステムループ構造を形成し得る、約50ヌクレオチドのDNA配列を含む。例には、E.coliのtrp遺伝子およびその他の生合成遺伝子のような、強プロモーターを伴った遺伝子由来の転写終止配列が含まれる。
【0126】
通常、プロモーター、シグナル配列(所望であれば)、目的のコーディング配列、および転写終止配列を含む上記の成分は、発現構築物中に組み立てられる。発現構築物は、細菌のような宿主に安定して維持され得る染色体外エレメント(例えば、プラスミド)のような、レプリコン中に維持される。レプリコンは、1つの複製系を有し、このため、発現、またはクローニングおよび増幅のいずれかのために原核宿主中に維持される。さらに、レプリコンは、高コピー数または低コピー数のプラスミドであり得る。高コピー数のプラスミドは、一般的に、約5から約200、そして通常約10から約150、の範囲のコピー数を有する。高コピー数のプラスミドを含有する宿主は、好ましくは少なくとも約10、より好ましくは少なくとも約20のプラスミドを含有する。高コピー数または低コピー数のいずれかのベクターが、宿主でのベクターの効果および外来タンパク質に依存して選択され得る。
【0127】
あるいは、発現構築物は、組込みベクターを用いて細菌ゲノムに組み込まれ得る。組込みベクターは通常、ベクターの組込みを可能にする細菌染色体に相同な少なくとも1つの配列を含有する。組込みは、ベクター中の相同DNAと細菌染色体との間の組換えによると考えられる。例えば、種々のBacillus株由来のDNAで構築された組込みベクターは、Bacillus染色体に組み込む(EPO公開No.127328)。組込みベクターはまた、バクテリオファージまたはトランスポゾン配列を含有し得る。
【0128】
通常、染色体外および組込み発現構築物は、形質転換された細菌株の選択を可能にする選択マーカーを含有し得る。選択マーカーは、細菌宿主中で発現され得、アンピシリン、クロラムフェニコール、エリスロマイシン、カナマイシン(ネオマイシン)およびテトラサイクリンのような薬剤に対して細菌を耐性にする遺伝子を含有し得る。(Daviesら、(1978)Annu.Rev.Microbiol.82:469)。選択マーカーはまた、ヒスチジン、トリプトファン、およびロイシンの生合成経路でのような、生合成遺伝子を含み得る。
【0129】
あるいは、上記成分のいくつかは、形質転換ベクター中に組み立てられ得る。形質転換ベクターは通常、レプリコン中に維持されるかまたは組込みベクターに組み込まれる選択マーカーを含有する。
【0130】
染色体外レプリコンまたは組込みベクターである、発現および形質転換ベクターは、多くの細菌への形質転換のために開発されている。例えば、発現ベクターは、特に以下の細菌のために開発されている:Bacillus subtilis.Palvら(1982)Proc.Natl.Acad.Sci.USA 79:5582:EPO公開No.036259およびNo.063953;PCT公開No、Wo84/04541:E.coli,Shimatakeら(1981)Nature 292:128:Amannら(1985)Gene 40:183;Studierら(1986)J.Mol.Biol.189:113;EPO公開No.036776,No,136829およびNo.136907;Stretococcus cremoris.Powellら(1988)Appl.Environ.Microbiol.54:655;Streptococcus lividans,Powellら(1988)Appl.Environ.Microbiol.54:655;およびStretomyces lividans、米国特許第4,745,056号。
【0131】
外因性DNAの細菌宿主への導入方法は当該分野で周知であり、CaCl2、または、二価カチオンおよびDMSOのようなその他の試薬で処理された細菌の形質転換を包含する。DNAはまた、エレクトロポーレーションにより細菌細胞中に導入され得る。形質転換の方法は、通常形質転換されるべき細菌種により変化する。Massonら(1989)FEMS Microbiol.Lett.60:273;Palvaら(1982)Proc.Natl.Acad.Sci.USA 79:5582:EPO公開No.036259およびNo.063953:PCT公開No.WO84/04541、Bacillusに関する;Millerら(1988)Proc.Natl.Acad.Sci.85:856;Wangら(1990)J.Bacteriol.172:949、Campylobacterに関する;Cohenら(1973)Proc.Natl.Acad.Sci 69:2110:Dowerら(1988)Nucleic Acids Res 16:6127:Kushner(1978)「colE1由来プラスミドによるE.coli形質転換のための改良法」,Genetic Enineerings:Proceeding of the International Symposium on Genetic Engineering(H.W.BoyerおよびS.Nicosia編);Mandelら(1970)J.Mol.Biol.53:159;Taketo(1988)Biochim.Biophys.Acta 949:318、Escherichiaに関する;Chassyら(1987)FEMS Microbiol.Lett.44:173、Lactobacillusに関する;Fiedlerら(1988)Anal.Biochem 170:38、Pseudomonsに関する;Augustinら(1990)FEMS Microbiol.Lett.66:203、Staphylococcusに関する;Baranyら(1980)J.Bacteriol.144:698:Harlander(1987)「エレクトロポーレーションによるStretococcus lactisの形質転換」,Streptococcal Genetics(J.FerrettiおよびR.Curtiss III編);Perryら(1981)Infec.Immun.32:1295;Powellら(1988)Appl.Environ.Microbiol.54:655:Somkutiら(1987)Proc.4th Evr.Cong.Biotechnology 1:412、Stretococcusに関する、を参照のこと。
【0132】
(iv.酵母発現)
酵母発現系もまた当業者に公知である。酵母プロモーターは、酵母RNAポリメラーゼに結合し得、コーディング配列(例えば、構造遺伝子)のmRNAへの下流(3’)転写を開始し得る、いずれものDNA配列である。プロモーターは、コーディング配列の5’末端近傍に通常位置する転写開始領域を有する。この転写開始領域は通常、RNAポリメラーゼ結合部位(「TATAボックス」)および転写開始部位を含む。酵母プロモーターはさらに、上流活性化配列(UAS)と呼ばれる第二ドメインを有し得、これは、存在する場合には通常構造遺伝子の遠位に存在する。UASにより、調節(誘導可能な)発現が行われる。構成的発現は、UASの非存在下で起こる。調節的発現は正または負のいずれかであり得、そのことにより転写を促進または減退する。
【0133】
酵母は、活性代謝経路を有する発酵微生物であり、従って、代謝経路の酵素をコードする配列は特に有用なプロモーター配列を提供する。例には、アルコールデヒドロゲナーゼ(ADH)(EPO公開No.284044)、エノラーゼ、グルコキナーゼ、グルコース−6−リン酸イソメラーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(GAPまたはGAPDH)、ヘキソキナーゼ、ホスホフルクトキナーゼ、3−ホスホグリセリン酸ムターゼ、およびピルビン酸キナーゼ(PyK)(EPO公開No.329203)が包含される。酸性ホスファターゼをコードする酵母PHO5遺伝子もまた、有用なプロモーター配列を提供する(Myanoharaら(1983)Proc.Natl.Acad.Sci.USA 80:1)。
【0134】
さらに、天然に存在しない合成プロモーターもまた、酵母プロモーターとして機能する。例えば、1つの酵母プロモーターのUAS配列は、別の酵母プロモーターの転写活性化領域に接続され得、合成雑種プロモーターがつくられ得る。このような雑種プロモーターの例には、GAP転写活性化領域に連結されたADH調節配列が含まれる(米国特許第4,876,107号および米国特許第4,880,734号)。雑種プロモーターのその他の例には、GAPまたはPyKのような解糖酵素遺伝子の転写活性化領域に組み合わされた、ADH2、GAL4、GAL10、またはPHO5遺伝子のいずれかの調節配列からなるプロモーターが含まれる(EPO公開No.164556)。さらに、酵母プロモーターには、酵母RNAポリメラーゼに結合し、転写を開始する能力を有する、非酵母起源の天然に存在するプロモーターが含まれ得る。このようなプロモーターの例には、特に、Cohenら(1980)Proc.Natl.Acad.Sci.USA 77:1078;Henikoffら(1981)Nature 283:835:Hollenbergら(1981)Curr.Topics Microbiol.Immunol.96:119:Hollenbergら(1978)「酵母Saccharomyces cerevisiaeでの細菌抗生物質耐性遺伝子の発現」:Plasmids of Medical,Environmental and Commercial Imortance(K.N.TimmisおよびA.Puhler編);Mercerau−Puigalonら(1980)Gene 11:163;Panthierら(1980)Curr.Genet.2:109が、含まれる。
【0135】
DNA分子は酵母において細胞内に発現され得る。プロモーター配列は、DNA分子に直接に連結され得、その場合には、組換えタンパク質のN末端の最初のアミノ酸はいつもメチオニンであり、これはATG開始コドンによりコードされる。所望であれば、N末端メチオニンは、インビトロにおける臭化シアンとのインキュベーションにより、タンパク質から切断され得る。
【0136】
融合タンパク質は、哺乳類、バキュロウイルス、および細菌発現系と同様に、酵母発現系の代替を提供する。通常、内因性酵母タンパク質またはその他の安定なタンパク質のN末端部分をコードするDNA配列が、異種コーディング配列の5’に融合される。発現に際し、この構築物は、2つアミノ酸配列の融合を提供する。例えば、酵母またはヒトのスーパーオキシドジスムターゼ(SOD)遺伝子が、外来遺伝子の5’末端で連結され得、酵母で発現され得る。2つのアミノ酸配列の接続部位DNA配列は、切断部位をコードし得るか、またはコードし得ない。例えば、EPO公開No.196056を参照のこと。その他の例は、ユビキチン融合タンパク質である。このような融合タンパク質は、好ましくは、外来タンパク質からユビキチンを切断するためのプロセッシング酵素(例えば、ユビキチン特異的プロセッシングプロテアーゼ)に対する部位を保持しているユビキチン領域でつくられる。従って、この方法によってそのままの外来タンパク質が単離され得る(例えば、PCT公開No.WO88/024066を参照のこと)。
【0137】
あるいは、外来タンパク質はまた、酵母での外来タンパク質の分泌を提供するリーダー配列フラグメントを含有する融合タンパク質をコードする、キメラDNA分子をつくることにより、細胞から増殖培地に分泌され得る。好ましくは、リーダーフラグメントと外来遺伝子との間でコードされるプロセッシング部位が存在し、インビボまたはインビトロのいずれかで切断され得る。リーダー配列フラグメントは通常、細胞からのタンパク質の分泌を導く疎水性アミノ酸を含有するシグナルペプチドをコードする。
【0138】
適切なシグナル配列をコードするDNAは、酵母インベルターゼ遺伝子(EPO公開No.012873:JPO公開No.62,096,086)およびA因子遺伝子(米国特許第4,588,684号)のような、分泌酵母タンパク質の遺伝子に由来し得る。あるいは、インターフェロンリーダーのような、非酵母起源のリーダーが存在し、これもまた酵母での分泌を提供する(EPO公開No.060057)。
【0139】
分泌リーダーの好ましいクラスには、酵母α因子遺伝子のフラグメントを用いるリーダーがあり、これは「pre」シグナル配列および「pro」領域の両方を含有する。用いられ得るα因子フラグメントの型には、全長pre−pro α因子リーダー(約83アミノ酸残基)および切断型α因子リーダー(通常約25から約50アミノ酸残基)が含まれる(米国特許第4,546,083号および米国特許第4,870,008号;EPO公開No.324274)。分泌を提供するα因子リーダーフラグメントを用いる別のリーダーには、第一酵母のpre配列でつくられたハイブリッドα因子リーダーが含まれるが、第二酵母のα因子のpro領域は含まれない。(例えば、PCT公開No.WO89/02463を参照のこと)。
【0140】
通常、酵母により認識される転写終止配列は、翻訳終止コドンに対して3’側に位置する調節領域であり、従って、プロモーターとともにコーディング領域に隣接する。これらの配列は、DNAによりコードされるポリペプチドに翻訳され得るmRNAの転写を導く。転写終止配列およびその他の酵母認識終止配列の例は、解糖酵素をコードする配列などである。
【0141】
通常、プロモーター、リーダー(所望であれば)、目的のコーディング配列、および転写終止配列を含む上記の成分は、発現構築物中に組み立てられる。発現構築物はしばしば、酵母または細菌のような宿主中に安定して維持され得る染色体外エレメント(例えば、プラスミド)のような、レプリコン中に維持される。レプリコンは2つの複製系を有し得、このため、例えば、発現のために酵母中に、およびクローニングおよび増幅のために原核宿主中に維持される。このような酵母−細菌シャトルベクターの例には、YEp24(Botsteinら(1979)Gene 8:17−24);pCl/1(Brakeら(1984)Proc.Natl.Acad.Sci USA 81:4642−4646);およびYRp17(Stinchcombら(1982)J.Mol.Biol.158:157)が含まれる。さらに、レプリコンは、高コピー数または低コピー数のプラスミドであり得る。高コピー数のプラスミドは一般に、約5から約200の範囲のコピー数を有し、通常約10から約150のコピー数を有する。高コピー数のプラスミドを含有する宿主は、好ましくは少なくとも約10、より好ましくは少なくとも約20を有する。高コピー数または低コピー数のベクターは、宿主におけるベクターおよび外来タンパク質の効果に依存して選択され得る。
【0142】
あるいは、発現構築物は、組込みベクターを用いて酵母ゲノムに組み込まれ得る。組込みベクターは通常、ベクターの組込みを可能にする、酵母染色体に相同な少なくとも1つの配列を含有し、好ましくは、発現構築物に隣接する2つの相同な配列を含有する。組込みは、ベクター中の相同DNAと酵母染色体との間の組換えによると考えられる(Orr−Weaverら(1983)Methods in Enzymol.101:228−245)。組込みベクターは、ベクター中への封入に適切な相同配列を選択することにより、酵母の特異的な座に導かれ得る。1つ以上の発現構築物が組み込まれ得、おそらく産生される組換えタンパク質レベルに影響し得る(Rineら(1993)Proc.Natl.Acad.Sci.USA 80:6750)。ベクターに含まれる染色体配列は、ベクター中の1つのセグメントとして存在して、完全ベクターの組込みになり得るか、または、染色体中の隣接セグメントに相同で、ベクター中で発現構築物に隣接する2つのセグメントとして存在して、発現構築物のみの安定な組込みになり得る。
【0143】
通常は、染色体外および組込み発現構築物は、形質転換された酵母株の選択を可能にする選択マーカーを含有し得る。選択マーカーは、ADE2、HIS4、LEU2、TRP1、およびALG7のような、酵母宿主で発現され得る生合成遺伝子、および、ツニカマイシンおよびG418のそれぞれに対する酵母細胞において耐性を与えるG418耐性遺伝子を含み得る。さらに、適切な選択マーカーはまた、金属のような毒素化合物の存在下で増殖する能力を有する酵母を提供し得る。例えば、CUP1の存在は、銅イオンの存在下での酵母の増殖を可能にする(Buttら(1987)Microbiol.Rev.51:351)。
【0144】
あるいは、上記成分のいくつかは、形質転換ベクター中に組み立てれられ得る。形質転換ベクターは通常、レプリコン中に維持されるかまたは組込みベクターに組み込まれる選択マーカーを合有する。
【0145】
染色体外レプリコンまたは組込みベクターである、発現および形質転換ベクターは、多くの酵母の形質転換のために開発されている。例えば、発現ベクターは、特に以下の酵母のために開発されている:Candida albicans(Kurtzら(1986)Mol.Cell.Biol.6:142);Candida maltosa(Kunzeら(1985)J.Basic Microbiol.25:141):Hansenula polymorpha(Gleesonら(1986)J.Gen.Microbiol.132:3459;Roggenkampら(1986)Mol.Gen.Genet.202:302);Kluyveromyces fragilis(Dasら(1984)J.Bacteriol.158:1165);Kluyveromces lactis(De Louvencourtら(1983)J.Bacteriol.154:737;Van den Bergら(1990)Bio/Technology 8:135):Pichia guillerimondii(Kunzeら(1985)J.Basic Microbiol.25:141):Pichia pastoris(Creggら(1985)Mol.Cell.Biol.5:3376;米国特許第4,837,149号および米国特許第4,929,555号):Saccharomyces cerevisiae(Hinnenら(1978)Proc.Natl.Acad.Sci.USA 75:1929;Itoら(1983)J.Bacteriol.153:163):Schizosaccharomyces pombe(Beachら(1981)Nature 300:706);および、Yarrowia lipolytica(Davidowら(1985)Curr.Genet.10:380471 Gaillardinら(1985)Curr.Genet.10:49)。
【0146】
外因性DNAの酵母宿主への導入方法は、当該分野では公知であり、通常は、スフェロプラストまたは完全酵母細胞をアルカリカチオンで処理する形質転換を包含する。形質転換方法は通常、形質転換される酵母種により変化する。例えば、Kurtzら(1986)Mol.Cell.Biol.6:142;Kunzeら(1985)J.Basic
Microbiol.25:141、Candidaに関する;Gleesonら(1986)J.Gen.Microbioy.132:3459;Roggenkampら(1986)Mol.Gen.Genet.202:302、Hansenulaに関する;Dasら(1984)J.Bacteriol.158:1165;De Louvencourtら(1983)J.Bacteriol.154:1165:Van den Bergら(1990)Bio/Technology 8:135、Kluyveromycesに関する;Creggら(1985)Mol.Cell.Biol.5:3376;Kunzeら(1985)J.Basic Microbiol.25:141;米国特許第4,837,148号および米国特許第4,929,555号、Pichiaに関する;Hinnenら(1978)Proc.Natl.Acad.Sci.USA 75;1929;Itoら(1983)J.Bacteriol.153:163、Saccharomycesに関する;Beachら(1981)Nature 300:706、Schizosaccharomycesに関する;Davidowら(1985)Curr.Genet.10:39;Gaillardinら(1985)Curr.Genet.10:49、Yarrowiaに関する;参照のこと。
【0147】
(4.ワクチン)
本明細書中で考察されている各コンジュゲート化合物は、単一ワクチン候補物として、または他の病原由来の1つ以上のその他の抗原との組み合わせで使用され得る。これらのワクチンは、予防剤(感染を予防するため)または治療剤(感染後に疾患を治療するため)のいずれかであり得る。
【0148】
このようなワクチンは、コンジュゲート化合物を、通常は「薬学的に受容可能なキャリア」との組合せで含有し、このキャリアは、この組成物が与えられる個体に対して有害な抗体の産生をそれ自身が誘導しないいずれものキャリアを含む。適切なキャリアは、典型的には、タンパク質、多糖類、ポリ乳酸、ポリグリコール酸、アミノ酸ポリマー、アミノ酸コポリマー、脂質凝集体(例えば、油滴またはリボソーム)、および不活性ウイルス粒子のような、大きくて徐々に代謝される高分子である。このようなキャリアは、当業者に周知である。さらに、これらのキャリアは、免疫刺激剤(「アジュバント」)として機能し得る。さらに、抗原は、ジフテリア、破傷風などのような細菌トキソイドにコンジュゲートされ得る。
【0149】
組成物の効果を増強させる好ましいアジュバントには、以下が包含されるが、これらに限定されない:(1)アルミニウム塩(alum)(水酸化アルミニウム、リン酸アルミニウム、硫酸アルミニウムなど);(2)水中油型エマルジョン製剤〔ムラミルペプチド(下記を参照)のようなその他の特異的免疫刺激剤または細菌細胞壁成分を伴うまたは伴わない〕、例えば、(a)MF59(PCT公開No.WO90/14827):5%スクワレン、0.5% Tween 80、および0.5% Span85〔必須ではないが必要に応じて、種々の量のMTP−PE(下記を参照)を含有する〕を含有して、110Y型マイクロフルイダイザー(Microfluidics,Newton,MA)のようなマイクロフルイダイザーを使用して、サブミクロン粒子に処方される;(b)SAF:10%スクワラン、0.4% Tween 80、5%プルロニック(pluronic)−ブロックポリマーL121、およびthr−MDP(下記を参照)を含有し、マイクロフルイダイザーにかけてサブミクロン粒子エマルジョンにされるか、ボルテックスにかけてより大きな粒子サイズのエマルジョンを生成する;および、(c)RibiTMアジュバント系(RAS)(Ribi Immunochem,Hamilton,MT):2%スクワレン、0.2% Tween 80、および、1つ以上の細菌細胞壁成分(モノホスホリルリピド(monophosphorylipid)A(MPL)、トレハロースジミコレート(TDM)、および細胞壁骨格(CWS)よりなる群から、好ましくはMPL+CWS(DetoxTM))を含有する;(3)サポニンアジュバント(例えばStimulonTM(Cambridge Bioscience,Worcester,MA))が使用され得るか、または、ISCOM(免疫刺激複合体)のような、それらからつくられた粒子;(4)完全フロイントアジュバント(CFA)および不完全フロイントアジュバント(IFA);(5)サイトカイン(例えば、インターロイキン(IL−1、IL−2など)、マクロファージコロニー刺激因子(M−CSF)、腫瘍壊死因子(TNF)など);および、(6)組成物の効果を増強するための免疫刺激剤として作用するその他の物質。AlumおよびMF59が好ましい。
【0150】
上記のように、ムラミルペプチドには、N−アセチル−ムラミル−L−トレオニル−D−イソグルタミン(thr−MDP)、N−アセチル−ノルムラミル−L−アラニル−D−イソグルタミン(nor−MDP)、N−アセチルムラミル−L−アラニル−D−イソグルタミニル−L−アラニン−2−(1’−2’−ジパルミトイル−sn−グリセロ−3−ヒドロキシホスホリルオキシ(huydroxyphosphoryloxy))−エチルアミン(MTP−PE)などが含まれるが、これらに限定されない。
【0151】
免疫原性組成物(例えば、抗原、薬学的に受容可能なキャリア、およびアジュバント)は典型的に、水、生理食塩水、グリセロール、エタノールなどのような希釈剤を含有する。さらに、湿潤または乳化剤などのような補助物質、pH緩衝物質などが、このような賦形剤中に存在し得る。
【0152】
典型的には、免疫原性組成物は、溶液または懸濁液のいずれかとして注射用に調製される;注射の前に液体賦形剤に溶解または懸濁するのに適切な固形もまた、調製され得る。薬学的に受容可能なキャリアについて上記で考察したように、調製物はまた、アジュバント効果を増強するために、エマルジョンにされるか、またはリボソームにカプセル化され得る。
【0153】
ワクチンとして使用される免疫原性組成物は、免疫学的に有効な量の抗原ポリペプチド、および必要であれば、上記成分のいかなる他の成分をも含有する。「免疫学的に有効な量」とは、単回投与でまたは連続投与の一部として個体に投与される量が、治療または予防に有効であることを意味する。この量は、処置される個体の健康状態および身体条件、処置される個体の分類学上のグループ(例えば、ヒト以外の霊長類、霊長類など)、抗体を合成するための個体の免疫系の能力、所望の予防程度、ワクチン処方、医療状況での治療医の評価、およびその他の関連因子に依存して変化する。その量は比較的広範囲にあり、日常の試行により決定され得ると思われる。
【0154】
免疫原性組成物は従来より非経口的に、例えば、皮下または筋肉内注射により、投与される。その他の投与形態に適した処方には、経口および肺への処方、坐薬、および経皮投与が含まれる。投与処置は、単回投与スケジュールまたは複数回投与スケジュールであり得る。ワクチンは、その他の免疫調節剤と組み合わせて投与され得る。
【実施例】
【0155】
(5. 実施例1)
Meningococci C群の多糖類ならびに熱ショックタンパク質hspR70およびhsp65を含有するコンジュゲート化合物を構築し、ワクチンの効力について試験した。
【0156】
(5.1. Meningococci C(MenC)群の多糖の精製、およびMenCのオリゴ糖の生成)
C群髄膜炎菌の多糖を、Frasc C.E3.の「バイオテクノロジープロセスの進歩:バクテリアワクチン」(A.Mizrahi編),vol.13,pp.123−145,Wiley−Liss Inc.,New York(1990)に記載されているように精製した。精製多糖類(10mg/ml)を、pH5の0.01M酢酸緩衝液中で、100℃で8時間加水分解して解重合した。得られた生産物を、分析用クロマトグラフィー(Sephadex 6−50)で分析し、0.27のKd(分布係数)を示した。
【0157】
(5.2. 一級アミン基のオリゴ糖末端基への導入)
0.5Mの塩化アンモニウムおよび0.15Mのシアノボロヒドリドナトリウムを加水分解から得られた溶液に加えた。pHを7に上げて、得られた溶液を35℃で1週間置いた。次に、オリゴ糖をクロマトグラフィー(Sephadex 6−15)により、収集したアミン基およびカルボハイドレート基に関して化学活性を含有するボイド容量画分を精製した。一方、単糖および過剰の試薬を含有する画分は廃棄した。得られたMenCオリゴ糖は、アミン基、シアル酸基、およびO−アセチル基を測定することによって特徴付けられた。次のモル比が得られた:シアル酸/アミン基=20、O−アセチル/シアル酸=0.84。
【0158】
(5.3. 熱ショックタンパク質の調製)
M.bovis BCG GroEL−型 65kDa hsp(hspR65)をプラスミドpRIB1300を宿した組換えE.coli K12株から発現し(Tholeら、Infect.Immun.1985,50,800:Van Edenら、Nature,1989,331,171)、そして、Tholeら、Infect.Immune.,1987,55.1466に記述されたように精製した。
【0159】
組換えM.tuberculosis DnaK−型 70kDa hsp(hspR70)を得て、ATP−アガロースクロマトグラフィーで精製した(Mehlertら、Mol.Microbiol.,1989,3,125)。
【0160】
(5.4. MenC多糖とhspR65およびhspR70との糖コンジュゲートの調製)
MenCアミノ−オリゴ糖を、H2Oを10%含むジメチルスルホキシドに溶解し、それから(アミン基について)12倍過剰のアジピン酸のN−ヒドロキシスクシンイミドエステル[Hillら、「FEBS LETT.」102:282(1979)に従って調製した]と反応させた。ジオキサン(1〜4倍量)で沈澱させて精製した後、活性化オリゴ糖を真空中で乾燥し、N−ヒドロキシスクシンイミドエステルの含有量について分析した。次いで、pH7の0.1Mリン酸緩衝液中で5mg/mlの量のhspR65およびhspR70を、300倍モル過剰の活性化オリゴ糖と反応させた。それぞれ得られた糖コンジュゲートは、クロマトグラフィーにより未反応オリゴ糖から遊離し、濾過し、そして4℃で保存した。開始物質のMenC多糖中のシアル酸の割合に対する糖コンジュゲートのシアル酸含有量の比は、カップリングしたオリゴ糖の量を表す。調製したタンパク質の含有量は、Lowry「J.Biol.Chem.」193:265(1951)の方法により確認した。特に、hspR70とのコンジュゲートは、310μg/mlのタンパク質含有量および76μg/mlの糖含有量を有し、一方、hspR65とのコンジュゲートは、180μg/mlのタンパク質含有量および97μg/mlの糖含有量を有する。
【0161】
(5.5. マウスおよび免疫感作)
BALB/c(H−2d)、C567BL/6(H−2b)およびCBA/J(H−2k)の雌のマウス(8〜12週齢)を本発明者らの飼育施設で飼育した。
【0162】
最初のカップルはJackson Laboratory,Bar Harbor,MEから供給された。BALB/c nu/nu 胸腺欠失マウスをIffa Credo,L’Arbresle,Franceから得た。
【0163】
0日目、各々のマウスに106CFUのBCG(またはコントロール群の場合はPBS)を腹腔内投与し、続いて14日目と35日目に2用量のコンジュゲート(PBS中)を投与した。
【0164】
コントロール群は、MenCオリゴ糖のみ、または水酸化アルミニウム(1mg/用量、0.5ml中)に吸着したMenCオリゴ糖−CRM197コンジュゲートワクチンを接種した。各々の免疫感作において、マウスに2μgのMenCオリゴ糖を接種し、それは8.7μgのMenCオリゴ糖−CRM197コンジュゲートワクチン、8.4μgのMenCオリゴ糖−hspR70コンジュゲート、または3.7μgのMenCオリゴ糖−hspR65コンジュゲートに対応した。
【0165】
(5.6. ELISA法による抗体の測定)
毎週、マウスの後眼窩叢から血液を採取し、抗体はELISAで力価測定した。
【0166】
IgG抗MenC抗体を測定するために、96ウエル平底プレートをpH7.4のPBS中のMenC多糖(5μg/ml)で37℃で一晩のインキュベーションにより覆った(Nunc Immunoplate I,Nunc,Roskilde,Denmark)。0.054%のTween−20を含むPBS(PBS−T)で繰り返し洗浄し、そして5%のFCSを含有するPBS−Tの200μlで37℃、1時間インキュベーションした後、ウエルを5% FCSを含むPBS−Tで希釈したマウス血清100μlとともに4℃で一晩インキュベートした。洗浄を繰り返した後、プレートを再び、ペルオキシダーゼにコンジュゲートしたIgG抗マウス抗血清を適切に希釈したものの100μlとともに37℃で3時間インキュベートした。
【0167】
特異的抗体の存在を、2,2’−アジノ−ビス−(3−エチルベンゾチアゾリンスルホン酸)(ABTS;Kirkegaard and Pery Laboratories Inc.,Gaithersburg,MD)を基質として加えることにより明らかにした。結果は、414nmでの吸光度について測定した。試験した最初の希釈(1:50)で0.2未満の吸光度を有する血清サンプルは、陰性と考えた。
【0168】
(5.7. オリゴ糖ヘコンジュゲートしたhspR65およびhspR70のキャリア効果)
HspR65およびhspR70−MenCオリゴ糖コンジュゲートを、予めBCGで感作したまたは感作していないBALB/cおよびC57BL/6マウスの免疫に使用した。
【0169】
コントロールとして、数群のマウスにMenCのみまたは水酸化アルミニウム上のCRM197−MenCコンジュゲートワクチンを感作させた。
【0170】
図1は、抗MenC多糖IgG抗体が、水酸化アルミニウム上のCRM197−MenCコンジュゲートワクチンの場合に測定したのと同等またはそれ以上の量でhsp−MenCコンジュゲートの免疫の後産生されたことを示す(図1A中のC57BL/6)。この効果は、アジュバントが存在しない場合だけでなく、BCGで感作しなかったときのhspR70−MenCコンジュゲートの場合にも観察された(図1B)。これらの結果より、アジュバント非存在下またはBCG感作していないとき、ミコバクテリウムのhspを用いる免疫が、オリゴ糖抗原を用いても本当に達成し得ると述べ得る。
【0171】
従って、hsp分子(特にhspR70)は、BCGで感作していないマウスでも強力なキャリア効果を及ぼし、アジュバントの非存在下でさえ、移送した多糖に対して特異的なIgG抗体を誘導する能力のある分子の有効なキャリアとして作用することが確認された。
【0172】
アジュバントおよび前感作の非存在下で及ぼされる、このミコバクテリウムのhspの有効なキャリア効果により、記載されたコンジュゲートが、細菌感染に対する新規ワクチンの開発に特に有用になる。
【0173】
(6.実施例2)
新規のH.pylori熱ショックタンパク質が同定され、組換えDNA法を用いて産生された。
【0174】
(6.1. 材料および方法)
(6.1.1. H.pylori株および増殖条件)
使用したH.pylori株は:CCUG 17874、G39およびG33(Grosseto病院,Italy,で胃生検より単離)、Pylo 2U+およびPylo 2U−(F.Megraud,Pellegzin病院,Bordeaux,Franceにより供与)、BA96(Siena大学,Italy,で胃生検により単離)であった。Pylo 2U+株は非細胞毒性であり;Pylo 2U−株は非細胞毒性およびウレアーゼ陰性である。全ての株は、0.2%のシクロデキストリン、5μg/mlのセフスロジン、および5μg/mlのアムホテリシンBを含むコロンビアアガー上で、微好気性の条件下で、37^℃5〜6日間通常的に増殖した。細胞を採取し、PBSで洗浄した。ペレットをLaemmliサンプル緩衝液に再懸濁して、そして煮沸して細胞を溶解した。
【0175】
胃炎および潰瘍に冒されている患者の血清(A.Ponzetto,「Le Molinette」病院,Torino,Itaiyにより供与)および胃癌患者の血清(F.Roviello,Siena大学,Italyにより供与)を使用した。
【0176】
(6.1.2. ライブラリーの免疫スクリーニング)
λgt11 H.pylori DNA発現ライブラリーの50万個のプラークを、0.2%マルトースおよび10mM MgSO4を含むLB中で一晩増殖したE.coli
Y1090株の懸濁液5mlと混和し、そして0.5 O.D.になるように10mM
MgSO4に両懸濁した。37℃で10分インキュベーションした後、溶かした75mlのTopAgaroseを、細菌/ファージ混合物に注ぎ、そして全部をBBLプレート上で平板培養した(50,000プラーク/プレート)。42℃で平板培養したライブラリーを3.5時間インキュベーションした後、10mM IPTGで予め湿らせたニトロセルロース濾紙(SchleicherおよびSchuell,Dassel,Germany)をプレート上に載せ、そしてさらに37℃3.5時間、次いで4℃で一晩インキュベーションした。λタンパク質を有する濾紙を持ち上げてPBSで洗浄し、そしてTBST(10mM TRIS pH8、100mM NaCl、5M MgCl2)に溶解した5%脱脂乾燥乳で20分間飽和させた。最初のハイブリダイゼーション工程は、患者血清で行った;陽性プラークを呈色および可視化するために、製造者の指示に従って、AP緩衝液(100mM Tris pH9.5、100mM NaCl,5mM MgCl2)中でアルカリホスファターゼをコンジュゲートした抗ヒトIg抗体(Cappel,West Chester,PA)およびNBT/BCIPキット(Promega,Madison,WI)を用いた。
【0177】
(6.1.3. 組換えDNA手順)
使用した試薬および制限酵素は、Sigma(St.Louis,MO)およびBoehringer Mannheim(Germany)から得た。分子クローニング、一本鎖DNA精製、E、coliでの形質転換、プローブの放射標識化、H.pylori
DNA遺伝子ライブラリーのコロニースクリーニング、サザンブロット分析、PAGE、ウェスタンブロット分析については、標準的技法を用いた。
【0178】
(6.1.4. DNA配列分析)
DNAフラグメントをBluescriptSK+(Stratagene,San Diego,CA)でサブクローニングした。一本鎖DNA配列決定を、製造者の指示に従って、[33P]αdATP(New England Nuclear,Boston,MA)およびSequenaseキット(U.S.Biochemical Corp.,Cleveland,OH)を用いて行った。配列は両鎖について決定し、そして各鎖は平均2回配列決定した。コンピュータによる配列分析はGCGパッケージを用いて行った。
【0179】
(6.1.5. 組換えタンパク質)
MS2ポリメラーゼ融合タンパク質をpEX34A(pEX31の誘導体)ベクターを用いて生成した。挿入Hp67(図3のヌクレオチド445からヌクレオチド1402)およびEcoRIリンカーを、ベクターのEcoRI部位にフレーム内でクローニングした。停止コドンの位置を確認するために、HpG3’HindIIIフラグメントを、pEX34AのHindIII部位にフレーム内でクローニングした。組換えプラスミドをE.coli K12 H1△trpに形質転換した。誘導後の両方の場合で、予測した分子量の融合タンパク質を生成した。EcoRI/EcoRIフラグメントの場合は、誘導後に得られた融合タンパク質をウサギに免疫するために、標準プロトコルを用いて電気浴出した。
【0180】
(6.2. 結果)
(6.2.1. 発現ライブラリーのスクリーニングおよびH.pylori hspのクローニング)
H.pylori DNA発現ライブラリーのスクリーニングに適切な血清を見つけるために、H.pylori CCUG 17874株の超音波処理抽出物を、異種の胃炎によって冒された患者の血清に対するウェスタンブロット分析で試験した。異なった血清による抗原認識パターンは多岐にわたり、おそらく、個体の免疫応答の差異、および感染物に含まれる株により発現する抗原の差異のためであった。
【0181】
N°19の血清を、細菌の増殖の間にインビボで発現される、H.pylori特異的抗原を同定するためのλgt11 H.pylori DNA発現ライブラリーをスクリーニングするために選択した。この血清でライブラリーをスクリーニングした後、多くの陽性クローンを単離し、特徴付けた。これらの1つのヌクレオチド配列(いわゆるHp67)は、熱ショックタンパク質のhsp60ファミリーに高い相同性を有するタンパク質をコードする985塩基対のオープンリーディングフレームであることがわかった(Ellis,Nature 358:191−92(1992))。全コーディング領域を得るために、本発明者らは、フラグメントHp67を、異なる制限酵素で切断したH.pylori DNAのサザンブロット分析のプローブとして用いた。プローブHp67は、それぞれ、約800および1000塩基対の2つのHindIIIバンドを認識した。HindIII切断DNAのゲノムH.pyloriライブラリーをプローブHp67でスクリーニングし、そして予測した分子量の2つの陽性クローン(HpG5’およびHpG3’)を得た。プラスミドpHp60G2(約1から929のヌクレオチド)およびpHp60G5(約824から1838のヌクレオチド)を合むE.coliをアメリカンタイプカルチャーコレクション(ATCC)に寄託した。
【0182】
(6.3. 配列分析)
ヌクレオチド配列分析で、開始ATGの6塩基対上流の推定リボゾーム結合部位を有する1638塩基対のオープンリーディングフレームが明らかになった。図3に、H.pylori hspのヌクレオチド配列およびアミノ酸配列を示す。推定リボゾーム結合部位および中間HindIII部位を下線で示す。フラグメントHp67の445位のシトシンおよび1402位のグアニンは、それぞれ最初および最後のヌクレオチドである。チミジン1772を、独立因子ターミネーター領域の局在化に関する演算法を用いて、転写された最後の推定のヌクレオチドとして同定した。オープンリーディングフレームは、58.3KDaの予測分子量、および5.37の予測pIを有する546アミノ酸のタンパク質をコードしていた。この遺伝子のコドンの選択は、H.pyloriコドンの用法と一致する。
【0183】
疎水性のプロフィル分析では、予測されるリーダーペプチドまたは他の貫膜ドメインを除いて、ほとんど疎水性タンパク質であることがわかった。アミノ末端配列は、精製タンパク質について、Dunnら、Infect.Immun.60:1946−51(1992)により決定された30アミノ酸の配列に100%の相同性を示し、Evansら、Infect.Immun.60:2125−27(1992)によって公表された44アミノ酸の配列から1残基のみ(Lysに代わってSer42)異なっていた(Evansら、1992)。成熟hspタンパク質のN−末端配列は、開始メチオニンを含まず、このことは、翻訳後に除かれたことを示している。
【0184】
(6.4. hsp60ファミリーとの相同性)
アミノ酸配列分析で、全ての生物に存在するメンバーである熱ショックタンパク質hsp60ファミリーと非常に高い相同性が示された。異種のhsp60タンパク質問での相同の程度に基づいて、H.pylori hspは、グラム陰性細菌のhsp60タンパク質のサブグループに属する;しかし、hsp60ファミリーの他のタンパク質に対する相同の程度はかなり高い(少なくとも54%の一致)。
【0185】
他の熱ショックタンパク質とのH.pylori hspの相同性は図3に完全に例示した。H.pylori hspまたはその1つ以上の機能的な免疫刺激ドメインは、本発明に従ってコンジュゲート化合物を生成するために上記実施例1の手法を用いてオリゴ糖または多糖にコンジュゲートし得る。
【0186】
(6.5. 組換えタンパク質の発現およびポリクローナル抗血清の生成)
クローンHp67およびクローンHpG3’の挿入物を、MS2ポリメラーゼのアミノ末端に融合したこれらのオープンリーディングフレームを発現するために、発現ベクターpEX34Aにサブクローンした。クローンは予測したサイズの組換えタンパク質を生成し、そして最初のスクリーニングで用いたヒト血清により認識された。クローンHp67由来の融合タンパク質を電気浴出し、そして抗hsp特異的ポリクローナル抗血清を得るためにウサギの免疫に用いた。得られた抗血清は、融合タンパク質と、ウレアーゼ陰性株および非細胞毒性株を含有する、試験したいくつかのH.pylori株の全細胞抽出物の58KDaのタンパク質との両方を認識した。
【0187】
Hspは試験した全てのH.pylori株で発現されることが示され、そしてその発現はウレアーゼの存在または細胞毒性に関係ない。抗hsp抗血清により認識されるタンパク質は、H.pyloriの水溶性抽出物中に認められ、そしてウレアーゼサブユニットと同時に精製された。これは、このタンパク質が細菌外膜に弱い会合をしていることを示す。従って、hspはウレアーゼに会合し、そして表面が露出していると記載し得る細胞表面の局在化により、ほとんどのhspに相同なタンパク質が細胞質中またはミトコンドリアおよび色素体中に局在していることは驚くことである。hspにリーダーペプチドが欠けていることは、これが独特の輸送系により膜に輸送されるか、あるいはタンパク質が細胞質から放出され、細菌の死後に細菌の膜に受動的に吸着するかのいずれかであることを示唆する。
【0188】
(7. 生物学的材料の寄託)
以下の材料は、1992年12月15日および1993年1月22日に、本発明の譲渡人であるBiocine Sclavo,S.p.A.により、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約のもとに、アメリカンタイプカルチャーコレクション(ATCC)、メリーランド州,ロックビル,パークローンドライブ12301,電話(301)231−5519に寄託された。
ATCC No.69155 プラスミドpHp60G2を含むE.coli TG1
ATCC No.69156 プラスミドpHp605を含むE.coli TG1。
【0189】
これらの寄託物は、当業者の利便性のために提供され、寄託が、米国特許法112条、または本明細書の任意の指定国のいかなる相当規定のもとで要求されることを認めるものではない。これらの寄託物の核酸配列、およびそれにコードされるポリペプチドのアミノ酸配列は、本明細書中に参考として援用されており、寄託物の配列と比較して、本明細書に記載の配列中に任意の誤りがある場合には、参考にされるべきである。寄託物質を製造し、使用し、または売るためには許諾が必要であり得るが、ここでは、そのような許諾は承認されていない。
【産業上の利用可能性】
【0190】
このように、本発明のキャリアは、確実に高力価のワクチン接種をするために、細菌間の熱ショックタンパク質の高い保存性およびT細胞記憶を、独自に利用するものである。この化合物は、アジュバントを用いることなく、抗多糖抗体の形成を誘導し得る。そのため、本化合物は、ヒトおよび動物に使用するワクチン、特に細菌感染予防用ワクチンの調製に有用である。
【技術分野】
【0001】
(発明の分野)
本発明は、熱ショックタンパク質と、多糖またはオリゴ糖(特に、病原性微生物の莢膜由来の多糖またはオリゴ糖)とからなるコンジュゲート化合物に関する。本化合物は、抗多糖抗体の形成を誘導し得る。そのため、本化合物は、ヒトおよび動物に使用するワクチンとして有用である。
【背景技術】
【0002】
(技術状況)
細菌は、広範囲の疾患状態に対する病原物質である。
【0003】
このような疾患の例には、Neisseria meningitidisによって引き起こされる髄膜炎、およびHaemophilus influenzae b型(Hib)またはStreptococcus(Pneumococcusを含む)によって引き起こされる他の感染症、Salmonella typhiの感染によって引き起こされる腸チフス、非腸チフス原性のSalmonella菌またはShigella菌によって起こる腸疾患が含まれる。
【0004】
莢膜を有する細菌に対する防御免疫は、莢膜中の多糖に対する抗体により介在されることが知られている。免疫系を充分に刺激するには、莢膜の多糖を、キャリアタンパク質に結合させる必要があることも知られている(Robbinsら、J.Infect.Dis.1990,161,821−832)。
【0005】
特に、多糖(例えば、C群髄膜炎菌多糖(MenC)、HibおよびA群髄膜炎菌多糖(MenA))およびCRM−197(Corynebcterium diphtheriae由来のペプチド)のようなタンパク質、TD(ジフテリア毒素)またはTT(破傷風毒素−Peetersら、Inf.Immun.,(1991年10月)、3504−3510;Claessonら、J.Pediatrics St Louis,112(5)、695−702、(1988年5月)を参照のこと)からなるコンジュゲート化合物が文献中に記載されている。
【0006】
このようなワクチンのうち数種は、すでに臨床的に実用化されており、良好な結果が得られている。しかし、従来使用されてきたキャリアで達成されたよりも良好な免疫原的特性をコンジュゲートに与える新規なタンパク質キャリアを同定する必要がある。
【0007】
本発明は、オリゴ糖および多糖の免疫原性応答を高めるタンパク質キャリアとしての熱ショックタンパク質の使用に関する。
【0008】
熱ショックタンパク質は、非常に多数のTエピトープを含有し、そのため細胞の免疫系を刺激することが知られている。
【0009】
マラリア性エピトープのキャリアである、Mycobacterium bovisの熱ショックタンパク質のコンジュゲート化合物(65kDa)は、Bacillus Calmette−Guerin(BCG)で予め免疫化した動物に、アジュバントを必要とすることなく、著しい免疫化を誘導することが記載されている(Lussowら Eur.J.Immunol.,1991,21,2297−2302)。しかし、熱ショックタンパク質に結合した(T−細胞依存性であることが周知の)ペプチドによって示されるT細胞依存効果に関するLussowらに観察された効果は、注目に値する。
【0010】
より詳細には、熱ショックタンパク質は、株および型が異なる細菌間でもよく保存されるので、継続的プロセスである細菌感染により、確実に、免疫系が熱ショックタンパク質に対して感作された状態なる。そのため、最初のワクチン接種または追加免疫ワクチン接種における投与で、本発明のコンジュゲート化合物に対する良好な応答が保証される。
【0011】
本発明により、(アジュバントの使用も可能だが)アジュバントを用いずに、莢膜を有する細菌の多糖およびオリゴ糖の使用が可能となる。
【0012】
大勢の子供たちが、結核予防のためにBCGワクチン(細菌の熱ショックタンパク質を含む)を接種する。そのため、熱ショックタンパク質をキャリアとして含有する本発明のコンジュゲートは、このBCGワクチン接種の実施の結果、すでにキャリアで予備免疫化された多くの被検体を見出す。
【0013】
その上、熱ショックタンパク質は非常に保存されているので、BCGでワクチン接種を受けていない個体群でさえ(他の細菌との相互作用の結果として)容易に免疫を得られる。そのため、この個体群は、熱ショックタンパク質およびT細胞依存抗原(オリゴ糖または多糖)で構成されるコンジュゲートでのワクチン接種の後に良好な免疫応答を発現し得る状態にある。このように、本発明のキャリアは、確実に高力価のワクチン接種をするために、細菌間の熱ショックタンパク質の高い保存性およびT細胞記憶を、独自に利用するものである。
【発明の概要】
【課題を解決するための手段】
【0014】
(発明の要旨)
本発明は、少なくとも1つの熱ショックタンパク質または少なくとも1つの免疫刺激ドメインを含有する熱ショックタンパク質の一部と、少なくとも1つのオリゴ糖または多糖とを含有するコンジュゲート化合物を提供する。
【0015】
熱ショックタンパク質は、動物、好ましくはヒトにおいて、免疫刺激効果を示し得る如何なる熱ショックタンパク質でもよい。
【0016】
熱ショックタンパク質は、細菌、寄生生物および哺乳動物に高度に保存されている。重大な悪影響なしに意図する被免疫被検体にポジティブな免疫刺激効果を示す限り、あらゆる熱ショックタンパク質が本発明のコンジュゲート中に使用され得る。個々の例としては、Helicobacter pylori、P.aeruginosa、C.trachomatisおよびM.leprae由来の熱ショックタンパク質、特にhsp60群の熱ショックタンパク質が挙げられるが、これらに限定されない。
【0017】
より詳細には、3種の熱ショックタンパク質、すなわち、M.bovis BCG GroEL型の65kDaのhsp(hspR65)、組換えM.tuberculosis DnaK型の70kDaのhsp(hspR70)およびH.pylori由来の新規な熱ショックタンパク質の具体例を、本明細書中に示す。
【0018】
H.pylori熱ショックタンパク質(hsp)は、図3に示すヌクレオチド配列およびアミノ酸配列を有するタンパク質であり、その分子量は54〜62kDaの範囲にあり、好ましくは約58〜60kDaである。このhspは、グラム陰性菌熱ショックタンパク質の群であるhdp60に属する。一般的にhdpは全ての生物、つまり原核または真核の動物または植物に、最もよく保存されているタンパク質であり、そして、その保存は配列全体にわたっている。
【0019】
コンジュゲートは、1種またはそれ以上の熱ショックタンパク質またはその免疫刺激ドメインを含み得る。熱ショックタンパク質は、同一でも異なっていてもよい。しかし、1つの熱ショックタンパク質、あるいは1つまたはそれ以上の免疫刺激ドメインを含有する部分が存在することが好ましい。
【0020】
本明細書中で使用のように、用語「免疫刺激ドメイン」は、哺乳動物被検体のドメインを含有するコンジュゲート化合物の多糖またはオリゴ糖成分に対する、免疫を増強させ得る熱ショックタンパク質のアミノ酸配列の領域を指す。
【0021】
完全な熱ショックタンパク質由来の特定のドメインのみを使用することの利点は、ヒト熱ショックタンパク質に共通のドメインを、選択的に含まないようにすることが可能な点である。ヒトヘのワクチン投与に対して、これは有利なことである。それは、このような領域が「自己」と認識されるので、熱ショックタンパク質の免疫刺激効果に影響を与えないからである。さらに、このような「自己」の領域に対して刺激を受ける全ての免疫は、自己免疫になり得るからである。
【0022】
熱ショックタンパク質のhsp60ファミリーの好適なドメインを同定し、ヒト熱ショックタンパク質と相同性が減少した配列に下線を引いて図2に載せてある。図2に示したドメインのうち、機能的なサブドメインもまた、ドメインおよびサブドメインの組合せとして同様に使用され得る。
【0023】
当業者は、そのドメインまたはエピトープが免疫刺激反応を起こす所定の熱ショックタンパク質を容易に確認し得、そしてこれらのドメイン、またはそのサブセットのみを含む改変熱ショックタンパク質を、容易に調製し得る。
【0024】
コンジュゲート化合物のオリゴ糖成分または多糖成分は、ワクチン投与しようとする全ての病原性微生物の完全な莢膜の多糖またはオリゴ糖、または防御免疫を誘導し得るその一部であり得る。オリゴ糖または多糖は、単一の細菌由来、あるいは2種またはそれ以上の細菌由来であり得る。
【0025】
標的とされ得る細菌の個々の例には:Haemophilus influenzaeb型(Hib)、Streptococcus(pneumococcusを含む)、Salmonella特にSalmonella typhi、非タイポイダル(typoidal)Salmonella細菌またはShigella細菌によって引き起こされる腸疾患が挙げられるが、これらに限定されない。
【0026】
本発明の特定の実施態様によると、Meningococci C(MenC)群のオリゴ糖およびhspから成るコンジュゲートが調製された。
【0027】
本発明のさらに特定の実施態様によると、本発明の目的に使用されたhspは、hspR65およびhspR70である。
【0028】
本発明の第2の局面では、熱ショックタンパク質または少なくとも1つの免疫刺激ドメインを含むその一部を、少なくとも1つのオリゴ糖または多糖に共有結合させることを包含する、本発明のコンジュゲート化合物を生産するプロセスが提供される。
【0029】
オリゴ糖または多糖は、好ましくは標的とする細菌から単離されるが、合成によっても生成され得る。
【0030】
熱ショックタンパク質は、その自然発生源から単離され得、または合成的に生産される。好ましくは、熱ショックタンパク質は、本明細書の記載の一般的な中に記載された技法を用いる組換えDNA技術によって生産される。
【0031】
好ましくは、オリゴ糖または多糖は、結合に対する反応部位を提供するように、熱ショックタンパク質またはその部分と結合する前に改変される。好ましくは、この改変はオリゴ糖または多糖の末端基へのアミノ基のような活性官能基の導入を包含する。次いで、このように改変されたオリゴ糖または多糖を、次いでスクシンイミドのような連結基を用いて活性化し、熱ショックタンパク質またはそれらのタンパク質に結合させる。
【0032】
本発明の第3の局面では、製剤、好ましくはワクチンとして使用する、本発明の第1の局面に記載のコンジュゲート化合物が提供される。
【0033】
本発明の第4の局面では、細菌感染に対するワクチン接種用の薬剤の製造における、本発明の第1の局面に記載のコンジュゲート化合物の使用が提供される。
【0034】
本発明の第5の局面では、本発明の第1の局面に記載のコンジュゲート化合物の免疫学的に有効な量の投与する工程を包含するワクチン接種の方法が提供される。
【0035】
発明の第6の局面では、本発明の第1の局面に記載の1つ以上のコンジュゲート化合物と、薬学的に受容可能なキャリアとを含むワクチンまたは治療用組成物が提供される。
【0036】
好ましくは組成物は、ワクチン組成物であり、そして必要であればアジュバントのような他の賦形剤を含み得る(以下に記載の「ワクチン」と表題された節を参照せよ)。
【0037】
本発明の第7の局面では、1つ以上の本発明の第1の局面のコンジュゲート化合物を、薬学的に受容可能なキャリアおよび任意にアジュバントと組み合わせる工程を包含する、ワクチンの調製法が提供される。
【0038】
従って、本発明は、以下を提供する:
(項目1) 少なくとも1つの熱ショックタンパク質または少なくとも1つの免疫刺激ドメインを含有する熱ショックタンパク質の一部と、少なくとも1つのオリゴ糖または多糖とを含有する、コンジュゲート化合物。
【0039】
(項目2) Meningococci C(MenC)群のオリゴ糖、および熱ショックタンパク質を合有する、項目1に記載のコンジュゲート化合物。
【0040】
(項目3) 前記熱ショックタンパク質が、M.bovis BCG GroEL型65kDa hsp(hspR65)、組換えM.tuberculosis DnaK型70kDa hsp(hspR70)、およびH.pylori由来の熱ショックタンパク質から選択される、項目1または2に記載のコンジュゲート化合物。
【0041】
(項目4) 熱ショックタンパク質または少なくとも1つの免疫刺激ドメインを含むその一部を、少なくとも1つのオリゴ糖または多糖に共有結合させる工程を包含する、項目1から3のいずれかに記載のコンジュゲート化合物を生成する方法。
【0042】
(項目5) 製剤として使用する、項目1から3のいずれかに記載のコンジュゲート化合物。
【0043】
(項目6) 細菌感染に対するワクチン接種用の薬剤の製造における、項目1から3のいずれかに記載のコンジュゲート化合物の使用。
【0044】
(項目7) 免疫学的に有効な量の項目1から3のいずれかに記載のコンジュゲート化合物の投与する工程を包含する、ワクチン接種の方法。
【0045】
(項目8) 項目1から項目3のいずれかに記載の1つ以上のコンジュゲート化合物および薬学的に受容可能なキャリアを含有する、ワクチンまたは治療用組成物。
(項目9) 項目1から項目3のいずれかに記載の1つ以上のコンジュゲート化合物を、薬学的に受容可能なキャリアと結合させる工程を包含する、ワクチンの調製方法。
【図面の簡単な説明】
【0046】
【図1】図1は、hspR65/MenCコンジュゲートでマウスを免疫化した結果を示し、そして、血中の抗MenCに対するELISAの結果を含む。図1Aには、BCGで予め免疫化したマウスを記載し、図1BにはBCGで予め免疫化しなかったマウスを記載している。図は、PBS中のhspR65−MenC(白丸)コンジュゲートまたはhspR70−MenC(白菱形)コンジュゲートとでの結果を示す。コントロール群のマウスは、MenC多糖のみ(白三角)または水酸化アルミニウム中のCEM197−MenCコンジュゲートワクチン(白四角)で免疫化した。
【図2A】図2Aは、Helicobacter pyloriの熱ショックタンパク質のアミノ酸配列を示し、そして、それとP.aeruginos、C.trachomatis、M.lepraeおよびH.sapiens由来の関連する熱ショックタンパク質とを比較している。配列の下線は、配列1〜4とヒト熱ショックタンパク質との間で、相同性が減少したドメインを示す。
【図2B】図2Bは、図2Aの続きである。
【図2C】図2Cは、図2Bの続きである。
【図3A】図3Aは、Helicobacter pyloriの熱ショックタンパク質の、ヌクレオチド配列およびアミノ酸配列である。
【図3B】図3Bは、図2Aの続きである。
【図3C】図3Cは、図2Bの続きである。
【発明を実施するための形態】
【0047】
(実施態様の詳細な説明)
(I.一般的な方法論)
本発明の実施には、他に示されていなければ、当該分野の分子生物学、微生物学、組換えDNA、および免疫学の従来の方法を使用する。このような方法は、文献に十分に説明されている。例えば、Sambrookら、MOLECULAR CLONING;A LABORATORY MANUAL,第二版(1289):DNA CLONING,VOLUMES IおよびII(D.N Glover編 1985):OLIGONUCLEOTIDE SYNTHESIS(M.J.Gait編 1984):NUCLEIC ACID HYBRIDIZATION(B.D.HamesおよびS.J.Higgins編 1984);TRANSCRIPTION AND TRANSLATION(B.D.HamesおよびS.J.Higgins編 1984);ANIMAL
CELL CULTURE(R.I.Freshney編 1986);IMMOBILIZED CELLS AND ENZYMES(IRL Press,1986);B.Perbal,A PRACTICAL GUIDE TO MOLECULAR CLONIHG(1984);METHODS IN EHZYMOLOGYシリーズ(Academic Press,Inc.);GENE TRANSFER VECTORS FOR MAMMALlAN CELLS(J.H.MillerおよびM.P.Calos編 1987,Cold Spring Harbor Laboratory)、Methods in Enzymology Vol.154およびVol.155(それぞれに、WuおよびGrossman、およびWu編)、MayerおよびWalker編(1987),IMMHNOCHEMICAL METHODS IN CELL AND MOLECULAR BIOLOGY(Academic Press,London)、Scopes,(1987),PROTEIN PURIFICATION:PRINCIPLES AND PRACTICE,第二版(Springer−Verlag,N.Y.)、および、HANDBOOK OF EXPERIMENTAL IMMUNOLOGY,VOLUMES I−IV(D.M.WeirおよびC.C.Blackwell編 1986)を参照のこと。
【0048】
ヌクレオチドおよびアミノ酸の標準的な省略形が、本明細書中で使用されている。本明細書中に掲載されている、全ての刊行物、特許、および特許出願は、参考として援用されている。
【0049】
(2.定義)
本発明に使用し得る熱ショックタンパク質には、上記のポリペプチド、および、上記タンパク質の天然のアミノ酸配列から少数のアミノ酸が変異したポリペプチドがあげられ;特に保存的なアミノ酸置換が予期される。
【0050】
保存的置換は、関連する側鎖を有するアミノ酸ファミリー内で起こる置換である。遺伝子的にコードされたアミノ酸は、一般的に4つのファミリーに分類される:(1)酸性=アスパラギン酸、グルタミン酸;(2)塩基性=リジン、アルギニン、ヒスチジン;(3)非極性=アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン;および、(4)非荷電極性=グリシン、アスパラギン、グルタミン、システイン、セリン、トレオニン、チロシン。フェニルアラニン、トリプトファン、およびチロシンは、しばしば、芳香族アミノ酸として、一緒に分類される。例えば、ロイシンとイソロイシンまたはバリンの、アスパラギン酸とグルタミン酸の、トレオニンとセリンとの単独の置換、あるいは、あるアミノ酸と、構造が関連するアミノ酸との同様の保存的置換が、生物学的活性に大きな影響を与えないことは当然予想し得る。タンパク質として実質的に同じアミノ酸配列を有するが、機能面に実質的に影響しない少数のアミノ酸置換を有するポリペプチド分子は、本タンパク質の定義の範囲に含まれる。
【0051】
天然の供給源からタンパク質を単離精製するよりも、組換えDNA技術により熱ショックタンパク質を生産する大きな長所は、天然の供給源からタンパク質を単離するのに要求されるよりも少ない開始物質を用いて、同量のタンパク質が生産され得ることである。組換え技術によるタンパク質の生産は、細胞中に通常存在するいくつかの分子の非存在下でも、タンパク質を単離を可能にする。実際、ヒトタンパク質混入の痕跡が全くないタンパク質組成物を容易に生産し得る。なぜなら、組換え非ヒト宿主により生産されるヒトタンパク質は、問題の組換えタンパク質のみだからである。天然の供給源からの潜在的なウイルス性物質およびヒトに対して発病性のウイルス成分もまた、避けられる。
【0052】
本明細書中に使用されているように、用語「組換えポリヌクレオチド」は、ゲノム、cDNA、半合成または合成起源のポリヌクレチドを示し、その起源または操作により:(1)天然では結合しているポリヌクレオチドの全てまたは一部と結合しない、(2)天然ではそれに結合しているポリヌクレオチド以外のポリヌクレオチドに結合する、または(3)天然では存在しない。
【0053】
本明細書中に使用されているように、用語「ポリヌクレオチド」とは、任意の長さのヌクレオチド、好ましくはデオキシリボヌクレオチドのポリマー形態のことであり、本明細書中では、用語「オリゴヌクレオチド」および「オリゴマー」と相互変換的に使用される。この用語は、分子の一次構造のみを示す。従って、この用語には、二本鎖および一本鎖DNAおよびアンチセンスポリヌクレオチドが包含される。これにはさらに、既知の型の改変、例えば、当該分野で公知の標識の存在、メチル化、末端「caps」、1つ以上の天然に存在するヌクレオチドのアナログとの置換、ヌクレオチド間の改変(例えば、ある型の非荷電性結合(例えば、ホスホン酸メチル、ホスホトリエステル、ホスホアミデート、カルバミン酸エステルなど)、または、荷電性結合(例えば、ホスホロチオエート、ホスホロジチオエートなど)との置換)、ペンダント部分の導入(例えば、タンパク質(ヌクレアーゼ、毒素、抗体、シグナルペプチド、ポリ−L−リシンなど)、挿入剤(例えば、アクリジン、ソラレン(psoralen)など)、キレート剤(例えば、金属、放射性物質、ホウ素、酸化成分など)、アルキル化剤(例えば、αアノマー性核酸など))が、包含される。
【0054】
用語「ゲノムの」により、ベクターにクローン化された制限フラグメント由来のDNA分子のコレクションまたはライブラリーを意味する。これには、微生物の遺伝子物質の全てまたは一部が包含され得る。
【0055】
用語「cDNA」により、mRNAの相補鎖にハイブリダイズする相補mRNA配列を意味する。
【0056】
本明細書中で使用されているように、用語「オリゴマー」は、プライマーおよびプローブの両方を意味し、本明細書では用語「ポリヌクレオチド」と相互変換的に使用される。用語オリゴマーは、分子の大きさを意味しない。しかし、典型的にオリゴマーは、1000ヌクレオチド未満、より典型的には500ヌクレオチド未満、さらに典型的には250ヌクレオチド未満であり、それらは100ヌクレオチド未満であり得、75ヌクレオチド未満、そしてさらに50ヌクレオチド未満の長さであり得る。
【0057】
本明細書中で使用されているように、用語「プライマー」とは、適切な条件下で使用されるときに、ポリヌクレオチド鎖の合成開始点として作用し得るオリゴマーのことである。プライマーは、コピーされるべきポリヌクレオチド鎖の領域に、完全にまたは実質的に相補的である。従って、ハイブリダイゼーションを行う条件下で、プライマーは、分析物鎖の相補領域にアニールする。適切な反応物(例えば、ポリメラーゼ、ヌクレオチドトリホスフェートなど)の添加により、プライマーは、重合剤により伸長されて分析物鎖のコピーを形成する。プライマーは一本鎖であり得るか、または、部分的または全体に二本鎖であり得る。
【0058】
用語「分析物ポリヌクレオチド」および「分析物鎖」とは、標的領域を含有すると思われる、一本鎖または二本鎖核酸分子のことであり、生物学的試料中に存在し得る。
【0059】
本明細書中で使用されているように、用語「プローブ」とは、標的領域中の配列とプローブ中の少なくとも1つの配列との相補性により、標的配列とハイブリッド構造を形成するポリヌクレオチドを含有する構造のことである。プローブのポリヌクレオチド領域は、DNAおよび/またはRNA、および/または合成ヌクレオチドアナログから構成され得る。「捕獲プローブ」および「標識プローブ」は、プローブ内に包含される。
【0060】
本明細書中で使用されているように、用語「標的領域」とは、増幅および/または検出されるべき核酸領域のことである。用語「標的配列」とは、プローブまたはプライマーが、所望の条件下で安定なハイブリッドを形成し得る配列のことである。
【0061】
本明細書中で使用されているように、用語「捕獲プローブ」とは、結合パートナーに結合する一本鎖ポリヌクレオチドを有するポリヌクレオチドプローブのことである。この一本鎖ポリヌクレオチドは、標的するポリヌクレオチド配列に含有され、これは、分析物ポリヌクレオチド中の検出されるべき標的領域内の標的配列に相補的である。この相補領域は、二本鎖に、分析物ポリヌクレオチドを固体表面に固定する(結合パートナーを介して)のに十分に安定性を与えるために、十分な長さからなり、標的配列に対して相補的である。結合パートナーは、第二の結合パートナーに特異的であり、第二の結合パートナーは、固体支持体の表面に結合され得るか、または、その他の構造物または結合パートナーを介して間接的に固体支持体に結合され得る。
【0062】
本明細書中で使用されているように、用語「標的するポリヌクレオチド配列」とは、標的ヌクレオチド配列に相補的であるヌクレオチドが含有されるポリヌクレチド配列のことであり、その配列は、意図される目的に十分な安定性を有する二本鎖を形成するのに、十分な長さからなり、標的配列に相補的である。
【0063】
本明細書中で使用されているように、用語「結合パートナー」とは、例えば、抗原とそれに特異的な抗体のような、高度な特異性を有するリガンド分子を結合し得る分子のことである。一般的に、特異的結合パートナーは、単離条件下で、分析物コピー/相補鎖の二本鎖(捕獲プローブの場合)を固定するのに十分な親和性で結合しなければならない。特異的結合パートナーは、当該分野で公知であり、例えば、ビオチンとアビジンまたはストレプトアビジン、IgGとプロテインA、多くの既知のレセプター−リガンド対、および、相補的ポリヌクレオチド鎖を包含する。相補的ポリヌクレオチド結合パートナーの場合には、パートナーは通常、少なくとも約15塩基の長さであり、少なくとも40塩基であり得、さらに、少なくとも約40%から約60%のG+C含有量を有する。ポリヌクレオチドは、DNA、RNAまたは合成ヌクレオチドアナログを含み得る。
【0064】
本明細書中で使用されているように、用語「カップルした(coupled)」とは、共有結合によるまたは非共有結合の強い相互作用(例えば、疎水相互作用、水素結合など)による付着のことである。共有結合は、例えば、エステル、エーテル、リン酸エステル、アミド、ペプチド、イミド、炭素−イオウ結合、炭素−リン結合などであり得る。
【0065】
用語「支持体」とは、所望の結合パートナーが固定され得る、いずれもの固体または半固体の表面のことである。適切な支持体には、ガラス、プラスチック、金属、ポリマーゲルなどが含まれ、ビーズ、ウエル、ディップスティック、膜などの形態にされ得る。
【0066】
本明細書中で使用されているように、用語「標識」とは、検出し得る(好ましくは定量し得る)シグナルを提供するために使用され得、ポリヌクレオチドまたはポリペプチドに結合され得る、いずれもの元素または部分のことである。
【0067】
本明細書中で使用されているように、用語「標識プローブ」とは、分析物ポリヌクレオチド中の検出されるべき標的配列に相補的である、標的するポリヌクレオチド配列を含有するポリヌクレオチドプローブのことである。この相補領域は、「標識プローブ」および「標識配列」を含有する二本鎖が、標識により検出されるように、十分な長さからなり、標的配列に相補的である。標識プローブは、直接に、または、マルチマーを含む、互いに高度な特異性を有する1組のリガンド分子を介して間接に、標識とカップルする。
【0068】
本明細書中で使用されているように、用語「マルチマー」とは、同じ繰り返しの一本鎖ポリヌクレオチドユニットまたは異なる一本鎖ポリヌクレオチドユニットの、直鎖状または分枝状ポリマーのことである。ユニットの少なくとも1つは、目的の第一の一本鎖ヌクレオチド配列、典型的には分析物または分析物に結合したポリヌクレオチドプローブ(例えば、標識プローブ)に特異的にハイブリダイズし得る、配列、長さ、および組成を有する。このような特異性および安定性を得るために、このユニットは、通常は少なくとも約15ヌクレオチドの長さであり、典型的には約50ヌクレオチド以下の長さ、そして好ましくは約30ヌクレオチドの長さであり;さらに、G+C含有量は、通常は少なくとも約40%、ほとんどは約60%であるロこのようなユニットに加えて、マルチマーには、目的の第二の一本鎖ヌクレオチド、典型的には標識ポリヌクレオチドまたはその他のマルチマーに、特異的および安定にハイブリダイズし得る、多くのユニットが包含される。これらのユニットは、上記に考察されたマルチマーのように、一般にはほぼ同じ大きさおよび組成である。マルチマーがその他のマルチマーにハイブリダイズされるように設計されているときには、第一および第二のオリゴヌクレオチドユニットは異質(異なる)であり、選択されたアッセイの条件下では、互いにはハイブリダイズしない。従って、マルチマーは、標識プローブであり得るか、または標識をプローブにカップルするリガンドであり得る。
【0069】
「レプリコン」は、細胞内のポリヌクレオチド複製の自己複製ユニットとして挙動する、すなわち、自己制御下に複製し得るいずれもの遺伝子エレメント、例えば、プラスミド、染色体、ウイルス、コスミドなどである。これには選択マーカーが含まれ得る。
【0070】
「PCR」とは、Saikiら、Nature 524:163(1986);およびScharfら、Science(1986)233:1076−1078;および米国特許第4,683,195号;ならびに米国特許第4,683,202号に記載されているようなポリメラーゼ連鎖反応の技法である。
【0071】
本明細書中で使用されているように、本来xが、同じ様式でyに関連しない場合に、xはyに関して「異種」である。すなわち、天然ではxが全くyに関連しないか、または、xが天然に実在して認められるような同じ様式でyに関連しない。
【0072】
「相同性」とは、xとyとの間の類似性の程度のことである。1つの形態の配列と別の配列との対応は、当該分野で公知の方法により決定され得る。例えば、それらはポリヌクレオチドの配列情報の直接比較により決定され得る。または、相同性は、相同領域(例えば、S1消化の前に使用される)間で安定な二本鎖を形成する条件下でポリヌクレオチドをハイブイブリダイズし、その後、一本鎖特異ヌクレアーゼにより消化し、その後に消化されたフラグメントの大きさを決定することにより決定され得る。
【0073】
「ベクター」は、別のポリヌクレオチドセグメントが接続されて、この接続されたセグメントの複製および/または発現をもたらすレプリコンである。
【0074】
「制御配列」とは、それらが連結されるコーディング配列の発現をもたらすために必要であるポリヌクレオチド配列のことである。このような制御配列の性質は、宿主生物に依存して異なる;原核生物では、このような制御配列は、一般的にプロモーター、リボソーム結合部位、および転写終止配列を含み;真核生物では、一般的に、このような制御配列は、プロモーターおよび転写終止配列を含む。用語「制御配列」は、最少限、発現に必要とされる全成分を含み、さらに存在が有利である付加的な成分、例えば、リーダー配列および融合パートナー配列を含み得ると意図される。
【0075】
「作動可能に連結された」とは、記載されている成分が、それらの意図する様式で機能し得る関係にある並列のことである。コーディング配列に「作動可能に連結された」制御配列は、コーディング配列の発現が制御配列に適合した条件下で得られるように連結されている。
【0076】
「オープンリーディングフレーム」(ORF)は、ポリペプチドをコードするポリヌクレオチド配列の領域のことである;この領域は、コーディング配列の一部またはコーディング配列全てを示し得る。
【0077】
「コーディング配列」は、適切な調節配列の制御下におかれると、通常はmRNAを介してポリペプチドに翻訳されるポリヌクレオチド配列である。コーディング配列の境界は、5’末端の翻訳開始コドンおよびポ末端の翻訳終止コドンにより決定される。コーディング配列には、cDNAおよび組換えポリヌクレオチド配列が含まれる得るが、これらに限定されない。
【0078】
本明細書中で使用されているように、用語「ポリペプチド」は、アミノ酸のポリマーを指し、特定の長さの産物を指すわけではない;従って、ペプチド、オリゴペプチド、およびタンパク質が、ポリペプチドの定義内に包含される。この用語にはまた、ポリペプチドの発現後改変(例えば、グリコシル化、アセチル化、リン酸化など)を意味せず、あるいは除外しない。定義に包含されるのは、例えば、アミノ酸の1つ以上のアナログを含有するポリペプチド(例えば、非天然アミノ酸などを含む)、置換結合を有するポリペプチド、および当該分野で公知の天然に存在するおよび存在しない改変である。
【0079】
示された核酸配列「由来の」ポリペプチドまたはアミノ酸配列とは、配列にコードされたポリペプチドのアミノ酸配列と同一のアミノ酸配列を有するポリペプチド、またはそれらの部分、ここで、この部分は、少なくとも3−5アミノ酸、さらに好ましくは少なくとも8−10アミノ酸、そしてよりさらに好ましくは少なくとも11−15アミノ酸からなる、または配列にコードされたポリペプチドにより免疫学的に同定され得るポリペプチドのことである。この用語にはまた、示された核酸配列から発現されるポリペプチドも包含される。
【0080】
「免疫原性の」とは、アジュバントの存在または非存在下で、それのみで、またはキャリアに結合されて、体液性免疫応答および/または細胞性免疫応答を引き起こすポリペプチドの能力のことである。「中和」とは、感染因子の感染力を、部分的または全体的に抑制する免疫応答のことである。「エピトープ」とは、ペプチド、ポリペプチド、またはタンパク質の抗原決定基のことである;1つのエピトープは、そのエピトープに対して唯一の高次構造(spatial conformation)で、3つ以上のアミノ酸を含み得る。一般的には、エピトープは、少なくとも5つのこのようなアミノ酸からなり、さらに通常には少なくとも8−10のこのようなアミノ酸からなる。アミノ酸の高次構造の決定方法は当該分野では公知であり、例えば、これらには、x線結晶解析および二次元核磁気共鳴が含まれる。同じエピトープを認識する抗体は、1つの抗体が標的抗原に対するその他の抗原の結合を阻む能力を示す、簡単な免疫アッセイにおいて同定され得る。
【0081】
本明細書中で使用されているように、「処置(treatment)」とは、予防および/または治療(therapy)(すなわち、あらゆる疾患症状の調整)のことである。「個体(individual)」は、抗原性の莢膜多糖またはオリゴ糖構造を有する細菌の感染が疑われる動物を意味し、これにはヒトを含む霊長類が包含されるが、これらに限定されない。「ワクチン」は、免疫原性の、または、部分的または完全にこのような細菌に対する保護を誘起し得る、個体の処置に有用な組成物である。
【0082】
本発明のコンジュゲート化合物は、このタンパク質に特異的な、モノクローナルまたはポリクローナル抗体の産生に使用され得る。これらの抗体を産生する方法は、当該分野で公知である。
【0083】
「組換え宿主細胞」、「宿主細胞」、「細胞」、「細胞培養物」、およびその他のこのような用語は、例えば、組換えベクターまたは他の転移DNAの受容体として使用され得るかまたは使用されてきた、微生物、昆虫細胞、および哺乳類細胞を示し、これには、形質転換されたもとの細胞の子孫も包含される。1つの母細胞の子孫は、自然突然変異、偶発突然変異または意図的突然変異(deliberate mutation)のために、形態学的に、または、ゲノムDNAまたは全DNA補体が、必ずしも、もとの母細胞と完全に同一ではあり得ないことが理解される。哺乳類宿主細胞の例には、チャイニーズハムスター卵巣(CHO)細胞およびサル腎(COS)細胞が含まれる。
【0084】
詳細には、本明細書中で使用されているように、「細胞系」とは、インビトロにおいて継続してまたは長期間に増殖および分裂し得る細胞群のことである。しばしば、細胞系は1つの幹細胞(progenitor cell)からのクローン群である。さらに、このようなクローン群の貯蔵または転移の間に、核型に自発変化または誘導変化が起こり得ることが、当該分野では公知である。従って、いわゆる細胞系由来の細胞は、祖先細胞または培養物に正確には同一ではあり得ず、この細胞系には、このような変異体が包含される。用語「細胞系」はまた、不死化細胞を包含する。好ましくは、細胞系は、非ハイブリッド細胞系、または二細胞型のハイブリドーマを包含する。
【0085】
本明細書中で使用されているように、用語「微生物」には、細菌および真菌のような、原核微生物種および真核微生物種が包合され、後者には、酵母および糸状菌が含まれる。
【0086】
本明細書中で使用されているように、「形質転換」とは、挿入に用いられる方法(例えば、直接取り込み、形質導入、f−交配またはエレクトロポーレーション)に関係なく、外因性ポリヌクレオチドの宿主細胞への挿入のことである。外因性ポリヌクレオチドは、非組込みベクター(例えばプラスミド)として維持され得るか、または、宿主細胞ゲノムに組み込まれ得る。
【0087】
ポリペプチドまたはヌクレオチド配列に関するときには、「精製された」および「単離された」とは、示された分子が、同じ型のその他の生物学的高分子を実質的に含まずに存在することを意味する。本明細書中で使用されているように、用語「精製された」は、好ましくは、同じ型の生物学的高分子が、少なくとも75重量%、さらに好ましくは少なくとも85重量%、さらに好ましくは少なくとも95重量%、そして最も好ましくは少なくとも98重量%で存在することを意味する(しかし、水、緩衝液、およびその他の小分子、特に、1000未満の分子量を有する分子は存在し得る)。
【0088】
(3.発現系)
一旦、適切な熱ショックタンパク質のコーディング配列が単離されると、それは種々の異なる発現系、例えば、哺乳類細胞、バキュロウイルス、細菌、および酵母により用いられる発現系、で発現され得る。
【0089】
(3.1.哺乳類系)
哺乳類発現系は当該分野で公知である。哺乳類プロモーターは、哺乳類RNAポリメラーゼを結合し得、コーディング配列(例えば、構造遺伝子)のmRNAへの下流(3’)転写を開始し得る、いずれものDNA配列である。プロモーターは、通常はコーディング配列の5’末端近傍に位置する転写開始領域、および、通常は転写開始部位の上流25−30塩基対(bp)に位置するTATAボックスを有する。TATAボックスは、RNAポリメラーゼIIがRNA合成を適切な部位で開始させるのを導くと考えられている。哺乳類プロモーターはさらに、通常はTATAボックスの上流100から200bp内に位置する上流プロモーターエレメントを有する。上流プロモーターエレメントは、転写が開始される速度を決定し、いずれかの向きに作用し得る(Sambrookら、Molecular Cloning:A Laboratory Manual,第二版(1989))。
【0090】
哺乳類ウイルス遺伝子は、しばしば高度に発現され、広範囲の宿主を有する;従って、哺乳類ウイルス遺伝子をコードする配列は、特に有用なプロモーター配列を提供する。例には、SV40初期プロモーター、マウス乳癌ウイルスLTRプロモーター、アデノウイルス主要後期プロモーター(Ad MLP)、および単純ヘルペスウイルスプロモーターが含まれる。さらに、ネズミメタロチオネイン遺伝子のような非ウイルス遺伝子由来の配列もまた、有用なプロモーター配列を提供する。発現は、構成的または調節され得(誘導可能な)、プロモーターに依存して、ホルモン−応答細胞中でグルココルチコイドにより誘導され得る。
【0091】
上記のプロモーターエレメントに組み合わされたエンハンサーエレメント(エンハンサー)の存在は、通常は発現レベルを増大させる。エンハンサーは、相同または異種プロモーターに連結されたときに、通常のRNA開始部位で合成を開始して、最高1000倍まで転写を促進し得る調節DNA配列である。さらに、エンハンサーは、通常の向きまたは裏返しの向きで転写開始部位から上流または下流に、または、プロモーターから1000ヌクレオチドを超える位置に配置されるときに活性である(Maniatisら、Science 236:1237(1989);Albertsら、Molecular Biology of the Cell,第二版(1982))。ウイルス由来のエンハンサーエレメントは、通常は広範囲の宿主を有するので、特に有用であり得る。例には、SV40初期遺伝子エンハンサー(Dijkemaら、(1985)EMBO J.4:761)、および、ラウス肉腫ウイルスの長末端反復(LTR)由来(Gormanら、(1982)Proc.Natl.Acad.Sci.79:6777)およびヒトサイトメガロウイルス由来(Boshartら、(1985)Cell 41:5221)のエンハンサー/プロモーターが含まれる。さらに、いくつかのエンハンサーは、調節可能であり、ホルモンまたは金属イオンのような誘導物質が存在するときのみに活性になる(Sassone−Corsiら、(1986)Trends Genet.2:215;Maniatisら、(1987)Science 236:1237)。
【0092】
DNA分子は、哺乳類細胞において細胞内に発現され得る。プロモータ配列はDNA分子に直接連結され得るが、この場合にはいつも、組換えタンパク質のN末端の最初のアミノ酸がメチオニンであり、ATG開始コドンによりコードされる。所望であれば、N末端は、インビトロにおける臭化シアンとのインキュベーションにより、タンパク質から切断され得る。
【0093】
あるいは、外来タンパク質はまた、哺乳類細胞で外来タンパク質の分泌をなすリーダー配列フラグメントを含む、融合タンパク質をコードするキメラDNA分子をつくることにより、細胞から増殖培地内に分泌され得る。好ましくは、リーダーフラグメントと外来遺伝子との間でコードされるプロセッシング部位が存在し、これは、インビボまたはインビトロのいずれかにおいて切断され得る。リーダー配列フラグメントは通常、細胞からのタンパク質の分泌を導く、疎水性アミノ酸を含むシグナルペプチドをコードする。アデノウイルス3分節系リーダーは、哺乳類細胞内での外来タンパク質の分泌を提供するリーダー配列の例である。
【0094】
一般には、哺乳類細胞により認識される転写終止およびポリアデニル化配列は、翻訳終止コドンに対し3’側に位置する調節領域であり、従って、プロモーターエレメントとともに、コーディング配列に隣接する。成熟mRNAの3’末端は、部位特異的に翻訳後に切断され、ポリアデニル化されて形成される(Birnstielら、(1985)Cell 41:349;ProudfootおよびWhitelaw(1988)「真核RNAの終結および3’末端プロセッシング」Transcrition and splicing(B.D.HamesおよびD.M.Glover編);Proudfoot(1982)Trends Biochem.Sci.14:105)。これらの配列は、DNAによりコードされるポリペプチドに翻訳され得るmRNAの転写を導く。転写ターミネーター/ポリアデニル化シグナルの例には、SV40由来のものが含まれる(Sambrookら(1989),Molecular Cloning:A Laboratory Manual)。
【0095】
いくつかの遺伝子は、イントロン(介在配列とも呼ばれる)が存在するときに、より効率よく発現され得る。しかし、数個のcDNAは、スプライシングシグナル(スプライス供与部位および受容部位とも呼ばれる)が欠如しているベクターから効率よく発現された(例えば、GethingおよびSambrook(1981)Nature 293:620を参照のこと)。イントロンは、スプライス供与部位および受容部位を有するコーディング配列内の介在非コーディング配列である。それらは、一次転写物のポリアデニル化の後に、「スプライシング」と呼ばれるプロセスにより除去される(Nevins(1983)Annu、Rev.Biochem.52:441;Green(1986)Annu.Rev.Genet.20:671;Padgettら、(1986)Annu.Rev.Biochem.55:1119;KrainerおよびManiatis(1988)「RNAスプライシング」,Transcription and splicing(B.D.HamesおよびD.M.Glover編))。
【0096】
通常、プロモーター、ポリアデニル化シグナル、および転写終止配列を含有する上記の成分は、発現構築物中に組み立てられる。エンハンサー、機能性スプライス供与部位および受容部位を有するイントロン、およびリーダー配列もまた、所望であれば発現構築物中に含まれる。発現構築物は、しばしば、哺乳類細胞または細菌のような宿主中で安定に維持され得る染色体外エレメント(例えば、プラスミド)のような、レプリコン中に維持される。哺乳類複製系には、複製にトランス作用因子が必要とされる動物ウイルス由来のものが含まれる。例えば、SV40のようなパポーバウイルス(Gluzman(1981)Cell 23:175)またはポリオーマウイルスの複製系を含むプラスミドが、適切なウイルスT存在下で、非常に多数のコピーを複製する。哺乳類レプリコンのさらなる例には、ウシパピローマウイルスおよびエプスタイン・バーウイルス由来のものが含まれる。さらに、レプリコンは2つの複製系を有し得、このため、例えば、発現のために哺乳類細胞中に、およびクローニングおよび増幅のために原核生物宿主中に維持される。このような哺乳類−細菌シャトルベクターの例には、pMT2(Kaufmanら(1989)Mol,Cell Biol.9:946)、およびpHEBO(Shimizuら(1986)Mol.Cell,Biol.6:1074)が含まれる。
【0097】
使用される形質転換方法は、形質転換されるべき宿主に依存する。異種ポリヌクレオチドの哺乳類細胞への導入方法は、当該分野で公知であり、デキストラン介在トランスフェクション、リン酸カルシウム沈降法、ポリブレン介在トランスフェクション、プロトプラスト融合、エレクトロポーレーションポリヌクレオチドのリボソーム内へのカプセル化、および、DNAの核への直接マイクロインジェクションを包含する。
【0098】
発現用の宿主として入手可能な哺乳類細胞系は、当該分野で公知であり、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、ベビーハムスター腎(BHK)細胞、サル腎細胞(COS)、ヒト肝細胞癌腫細胞(例えば、HepG2)および多くのその他の細胞系を含む、アメリカンタイプカルチャーコレクション(ATCC)から入手可能な多くの不死化細胞系を包含するが、これらに限定されない。
【0099】
(ii.バキュロウイルス系)
タンパク質をコードするポリヌクレオチドもまた、適切な昆虫発現ベクターに挿入され得、ベクター内の制御エレメントに作動可能に連結される。ベクター構築には、当該分野で公知の方法が用いられる。
【0100】
一般的に、発現系の成分には転移ベクター、通常は細菌プラスミドが含まれ、これは、バキュロウイルスゲノムフラグメントと、発現されるべき異種の遺伝子または遺伝子群挿入のための便宜的な制限部位との両方;転移ベクター内のバキュロウイルス特異フラグメントに相同な配列を有する野生型バキュロウイルス(これは、バキュロウイルスゲノム内への異種遺伝子の相同的組換えを考慮する);および、適切な昆虫宿主細胞および増殖培地を含む。
【0101】
タンパク質をコードするDNA配列を転移ベクター内に挿入した後に、ベクターおよび野生型ウイルスゲノムが、昆虫宿主細胞内ヘトランスフェクトされ、そこでベクターとウイルスゲノムとの組換えが起こる。パッケージングされた組換えウイルスが発現され、組換えプラークが同定および精製される。バキュロウイルス/昆虫細胞発現系用の材料および方法は、キット形態で、特に、Invitrogen,San Diego CA(「MaxBac」キット)から市販されている。これらの方法は当業者に公知であり、SummersおよびSmith,Texas Agricultural Experiment Station Bulletin No,1555(1987)(以後、「SummersおよびSmith」)に十分に記載されている。
【0102】
タンパク質をコードするDNA配列をバキュロウイルスゲノムに挿入する前に、プロモーター、リーダー(所望であれば)、目的のコーディング配列、および転写終止配列を含む上記の成分が、通常は中間置換構築物(intermediate transplacement construct)(転移ベクター)に組み立てられる。この構築物は、1つの遺伝子および作動可能に連結された調節エレメント;各遺伝子がそれ自身の固有のセットの作動可能に連結された調節エレメントを有する複数の遺伝子;または、同じセットの調節エレメントに調節される複数の遺伝子を含み得る。中間置換構築物はしばしば、細菌のような宿主中で安定して維持され得る染色体外エレメント(例えば、プラスミド)のような、レプリコン中に維持される。レプリコンは1つの複製系を有し、このため、クローニングおよび増幅のために適切な宿主中に維持される。
【0103】
現在では、AcNPVへ外来遺伝子を導入するための最も一般的に使用される転移ベクターは、pAc373である。当業者に公知のその他の多くのベクターもまた設計されている。これらは、例えば、pVL985(ポリヘドリン開始コドンをATGからATTに変え、ATT下流32塩基対にBamHIクローニング部位を導入する;LuckowおよびSummers,Virology(1989)17:31を参照)。
【0104】
プラスミドは通常さらに、ポリヘドロンポリアデニル化シグナル(Millerら(1988)Ann.Rev.Microbiol.,42:177)、および、選択およびE.coli中での増殖のための原核アンピシリン耐性(amp)遺伝子および複製起点を含有する。
【0105】
バキュロウイルス転移ベクターは通常、バキュロウイルスプロモーターを含有する。バキュロウイルスプロモーターは、バキュロウイルスRNAポリメラーゼを結合し得、そしてコーディング配列(例えば、構造遺伝子)のmRNAへの下流(5’から3’)翻訳を開始し得る、いずれものDNA配列である。プロモーターは、コーディング配列の5’末端近傍に通常位置する転写開始領域を有する。この転写開始領域は通常、RNAポリメラーゼ結合部位および転写開始部位を含有する。バキュロウイルス転移ベクターもまた、エンハンサーと呼ばれる第二ドメインを有し得、これは存在する場合には、構造遺伝子の遠位に存在する。発現は調節され得るか、または構成的であり得る。
【0106】
ウイルス感染サイクルの後期に多量に転写された構造遺伝子は、特に有用なプロモーター配列を提供する。例には、ウイルスポリヘドロンタンパク質をコードする遺伝子由来の配列(Friesenら、(1986)「バキュロウイルス遺伝子発現の調節」,TheMolecular Biology of Baculoviruses(Walter Doerfler編);EPO公開No.127839およびNo.155476);および、p10タンパク質をコードする遺伝子(Vlakら、(1988),J.Gen.Virol.69:765)が含まれる。
【0107】
適切なシグナル配列をコードするDNAは、バキュロウイルスポリヘドリン遺伝子のような、昆虫またはバキュロウイルス分泌タンパク質の遺伝子由来であり得る(Carbonellら(1988)Gene,73:409)。あるいは、哺乳類細胞の翻訳後の改変(シグナルペプチド切断、タンパク質分解切断、およびリン酸化のような)のためのシグナルは、昆虫細胞により認識されると考えられており、そして分泌に必要なシグナルおよび核集積は、無脊椎動物細胞と脊椎動物細胞との間で保存されているとも考えられているので、ヒトα−インターフェロン(Maedaら、(1985),Nature 315:592);ヒトガストリン放出ペプチド(Lebacq−Verheydenら、(1988),Molec.Cell.Biol.8:3129);ヒトIL−2(Smithら、(1985)Proc.Nat’l Acad.Sci.USA,82:8404);マウスIL−3,(Miyajimaら、(1987)Gene 58:273;および、ヒトグルコセレブロシダーゼ(Martinら(1988)DNA 7:99)をコードする遺伝子由来のような、非昆虫起源のリーダーもまた、昆虫での分泌を提供するために使用され得る。
【0108】
組換えポリペプチドまたはポリタンパク質は、細胞内に発現され得るか、または、適切な調節配列で発現されれば分泌され得る。非融合外来タンパク質の良好な細胞内発現は通常、ATG開始シグナルの前に適切な翻訳開始シグナルを含有する短いリーダー配列を理想的に有する異種遺伝子を必要とする。所望であれば、N末端のメチオニンは、インビトロにおける臭化シアンとのインキュベーションにより、成熟タンパク質から切断され得る。
【0109】
あるいは、組換えポリタンパク質または天然では分泌されないタンパク質は、昆虫において外来タンパク質の分泌を提供するリーダー配列フラグメントを有する融合タンパク質をコードするキメラDNA分子をつくることにより、昆虫細胞から分泌され得る。リーダー配列フラグメントは通常、小胞体へのタンパク質の輸送(translocation)を導く、疎水性アミノ酸を有するシグナルペプチドをコードする。
【0110】
タンパク質の発現産物の前駆体をコードするDNA配列および/または遺伝子の挿入後に、昆虫細胞宿主を、転移ベクターの異種DNAと野生型バキュロウイルスのゲノムDNAとで共形質転換(通常は共トランスフェクション)する。構築物のプロモーターおよび転写終止配列は、通常バキュロウイルスゲノムの2−5kb断片を含有する。バキュロウイルスの所望の部位に異種DNAを導入する方法は、当該分野で公知である。(SummersおよびSmith:Juら(1987);Smithら、Mol.Cell.Biol.(1983)3:2156;および、LuckowおよびSummers(1989)を参照のこと)。例えば、挿入は、ポリヘドリン遺伝子のような遺伝子に、相同的二本鎖交差組換えによりなされ得る;挿入はまた、所望のバキュロウイルス遺伝子中に設計された制限酵素部位になされ得る。Millerら、(1989),Bioessays
4:91。
【0111】
DNA配列は、発現ベクターのポリヘドリン遺伝子の位置にクローニングされたときには、ポリヘドリン特異配列により、5’および3’両側に隣接され、ポリヘドリンプロモーターの下流に配置される。
【0112】
新たに形成されたバキュロウイルス発現ベクターは、次に、感染性組換えバキュロウイルス中にパッケージングされる。相同的組換えは低頻度(約1%と約5%との間)で生じるため、共トランスフェクション後に産生された主要なウイルスは、まだ野生型である。従って、組換えウイルスを同定する方法が必要である。発現系の利点は、視覚スクリーニングにより、組換えウイルスの区別が可能であることである。天然ウイルスにより産生されるポリヘドリンタンパク質は、ウイルス感染後、後期に感染細胞の核に非常に高レベルで産生される。集積されたポリヘドリンタンパク質は、埋め込まれた粒子をも含む封入体を形成する。これらの封入体は、15μmまでの大きさであり、非常に屈折性であるために、光学顕微鏡下で容易に視覚化される光沢のある外観となる。組換えウイルスに感染した細胞には、封入体がない。野生型ウイルスから組換えウイルスを区別するには、トランスフェクション上清を、当業者に公知の方法により、単層の昆虫細胞上でプラーク形成させる。すなわち、プラークを光学顕微鏡下で、封入体の存在(野生型ウイルスの指標)または非存在(組換えウイルスの指標)についてスクリーニングする。「Current Protocols in Microbiology」Vol.2(Ausubelら編)16.8(Supp.10,1990):SummersおよびSmith;Millerら(1899)。
【0113】
組換えバキュロウイルス発現ベクターは、数種の昆虫細胞への感染のために開発されている。例えば、組換えバキュロウイノレは、とりわけ、Aedes aegypti、Autographa californica、Bombyx mori、Drosophila melanogaster、Spodoptera frugiperda、およびTrichoplusa niのために開発されている(PCT公開No.WO89/046699;Carbonellら、(1985) J.Virol.56:153;Wright(1986)Nature 321:718;Smithら、(1983)Mol.Cell.Biol.3:2156:および一般的にはFraserら(1989)In Vitro Cell.Dev.Biol.25:225を参照のこと)。
【0114】
細胞および細胞培養培地は、バキュロウイルス/発現系での異種ポリペプチドの直接発現および融合発現市販されている;細胞培養法は、一般に当業者に公知である。例えば、SummersおよびSmithを参照のこと。
【0115】
次に、改変昆虫細胞は、適切な栄養培地で増殖され得る。この栄養培地により、改変昆虫宿主内に存在するプラスミドは安定に維持され得る。発現産物遺伝子が誘導制御下にある場合には、宿主は高密度に増殖され得、発現が誘導され得る。あるいは、発現が構成的である場合には、産物は培地に連続的に発現される。目的の産物を取り出し、枯渇した栄養分を補給しながら、栄養培地を連続的に取り替えねばならない。産物は、例えば、HPLC、アフィニティークロマトグラフィー、イオン交換クロマトグラフィーなどのクロマトグラフィー;電気泳動;密度勾配遠心分離;溶媒抽出などの方法で精製され得る。適切には、培地に分泌されるかまたは昆虫細胞溶解から生じるいずれもの昆虫タンパク質を実質的に除去するために、宿主の破片(例えば、タンパク質、脂質および多糖類)を少なくとも実質的に含まない産物を提供するために、必要であれば産物はさらに精製され得る。
【0116】
タンパク質発現を得るために、形質転換体由来の組換え宿主細胞は、配列をコードする組換えタンパク質の発現を可能にする条件下でインキュベートされる。これらの条件は、選択される宿主細胞に依存して変化する。しかし、条件は、当該分野で公知のものに基づいて、当業者に容易に確認され得る。
【0117】
(iii.細菌系)
細菌発現法は当該分野で公知である。細菌プロモーターは、細菌RNAポリメラーゼに結合し得、コーディング配列(例えば、構造遺伝子)のmRNAへの下流(3’)転写を開始し得るいずれものDNA配列である。プロモーターは、通常コーディング配列の5’末端近傍に位置する転写開始領域を有する。この転写開始領域は通常、RNAポリメラーゼ結合部位および転写開始部位を含む。細菌プロモーターはまた、オペレーターと呼ばれる第二ドメインを有し得、これは、RNA合成を開始する位置である隣接RNAポリメラーゼ結合部位に重複し得る。遺伝子リプレッサ一タンパク質はオペレターに結合し得、そのことにより特異遺伝子の転写を阻害し得るので、オペレーターは、負の調節(誘導可能な)転写を行い得る。構成的発現は、オペレーターのような負の調節エレメントの非存在下で起こり得る。さらに、正の調節は、遺伝子活性化タンパク質結合配列により達成され得、これは、存在する場合には、通常RNAポリメラーゼ結合配列(5’)の近くに存在する。遺伝子活性化タンパク質の例は、力タボライト活性化タンパク質(CAP)であり、これは、E.coliでのlacオペロンの転写開始を助ける(Raibaudら(1984)Annu.Rev.Genet.18:173)。従って、調節的発現は正または負のいずれかであり得、そのことにより転写を促進または減退する。
【0118】
代謝経路の酵素をコードする配列は、特に有用なプロモーター配列を提供する。例には、ガラクトース、ラクトース(lac)(Changら(1977)Nature 198:1056)、およびマルトースのような糖代謝酵素由来のプロモーター配列が含まれる。さらなる例には、トリプトファン(trp)のような生合成酵素由来のプロモーター配列が含まれる(Goeddelら(1980)Nuc.Acids Res.8:4057;Yelvertonら(1981)Nucl.Acids Res.9:731;米国特許第4,738,921号;EPO公開No.036776およびNo.121775)。g−ラオタマーゼ(g−laotamase)(bla)プロモーター系(Weissmann(1981)「インターフェロンのクローニングおよびその他の誤り(mistakes)」Interferon 3(I.Gresser編)、バクテリオファージλPL(Shimatakeら(1981)Nature 292:128)、およびT5(米国特許第4,689,406号)プロモーター系もまた、有用なプロモーター配列を提供する。
【0119】
さらに、天然に存在しない合成プロモーターもまた細菌プロモーターとして機能する。例えば、1つの細菌またはバクテリオファージプロモーターの転写活性化配列は、その他の細菌またはバクテリオファージプロモーターのオペロン配列に接続され得て、合成雑種プロモーターをつくる(米国特許第4,551,433号)。例えば、tacプロモーターは、lacリプレッサーにより調節される、trpプロモーターおよびlacオペロン両配列を含有する雑種trp−lacプロモーターである(Amannら(1983)Gene 25:167;de Boerら(1983)Proc.Natl.Acad.Sci.80:21)。さらに、細菌プロモーターは、細菌RNAポリメラーゼに結合し、転写を開始する能力を有する、非細菌起源の天然に存在するプロモーターを含み得る。非細菌起源の天然に存在するプロモーターはまた、適合するRNAポリメラーゼと結合して原核生物中である種の遺伝子を高レベルで発現し得る、バクテリオファージT7 RNAポリメラーゼ/プロモーター系は、結合されたプロモーター系の例である(Studierら(1986)J.Mol.Biol.189:113:Taborら(1985)Proc.Natl.Acad.Sci.82:1074)。さらに、雑種プロモーターもまた、バクテリオファージプロモーターおよびE.coliオペレーター領域を含有し得る(EPO公開No.267851)。
【0120】
機能プロモーター配列に加えて、能率のよいリボソーム結合部位もまた、原核生物での外来遺伝子の発現に有用である。E.coliでは、リボソーム結合部位はシャイン・ダルガーノ(SD)配列と呼ばれ、開始コドン(ATG)および開始コドンの3−11ヌクレオチド上流に位置する3−9ヌクレオチドの長さの配列を含む(Shineら(1975)Nature 254:34)。SD配列は、SD配列とE.coli 16S rRNAの3’末端(3’and)との間で塩基対合してmRNAのリボソームヘの結合を促進すると考えられている(Steitzら(1979)「メッセンジャーRNAにおける遺伝シグナルおよびヌクレオチド配列」,Biological Regulation and Develoment:Gene Expression(R.F.Goldberger編))。弱リボソーム結合部位を有する原核遺伝子および真核遺伝子を発現するためには、Sambrookら(1989),Molecular Clonin:A Laboratory Manualを参照のこと。
【0121】
DNA分子は細胞内に発現され得る。プロモーター配列は、直接DNA分子に連結され得、その場合、N末端の最初のアミノ酸はメチオニンであり、これはATG開始コドンによりコードされる。必要であれば、N末端のメチオニンは、インビトロにおける臭化シアンとのインキュベーション、または細菌メチオニンN末端ペプチダーゼとのインビボまたはインビトロインキュベーションにより、タンパク質から切断され得る(EPO公開No.219237)。
【0122】
融合タンパク質は、直接発現の代替を提供する。通常は、内因性細菌タンパク質またはその他の安定なタンパク質のN末端部分をコードするDNA配列が、異種コーディング配列の5’末端に融合される。発現に際し、この構築物は、2つのアミノ酸配列の融合を提供する。例えば、バクテリオファージλ細胞遺伝子が外来遺伝子の5’末端で連結され得、細菌内で発現され得る。得られた融合タンパク質は、好ましくは、外来遺伝子からバクテリオファージタンパク質を切断するためのプロセッシング酵素(因子Xa)に対する部位を保持している(Nagaiら(1984)Nature 309:810)。融合タンパク質はまたlacZ由来の配列を用いて生成され得る(Jiaら(1987)Gene 60:197,trpE,Allenら(1987)J.Biotechnol.5:93:Makoffら(189)J.Gen.Microbiol.135:11、およびEPO公開No.324647,genes)。2つのアミノ酸配列を接続したDNA配列は、切断部位をコードし得るか、またはコードし得ない。その他の例は、ユビキチン融合タンパク質である。このような融合タンパク質は、好ましくは、外来遺伝子からユビキチンを切断するためのプロセッシング酵素(例えば、ユビキチン特異的プロセッシングプロテアーゼ)に対する部位を保持しているユビキチン領域を用いて生成される。この方法によって、そのままの外来タンパク質が単離され得る(Millerら(1989)Bio/Technology 7:698)。
【0123】
あるいは、外来タンパク質はまた、細菌で外来タンパク質分泌を提供するシグナルペプチド配列フラグメントを含有する融合タンパク質をコードするキメラDNA分子をつくることにより、細胞から分泌され得る(米国特許第4,336,336号)。シグナル配列フラグメントは通常、細胞からのタンパク質分泌を導く疎水性アミノ酸を含有するシグナルペプチドをコードする。タンパク質は、増殖培地(グラム陽性細菌)、または、細胞の内膜と外膜との間に位置する細胞周辺腔(グラム陰性細菌)のいずれかに分泌される。好ましくは、シグナルペプチドフラグメントと外来遺伝子との間でコードされるプロセッシング部位が存在し、これはインビボまたはインビトロにおいて切断され得る。
【0124】
適切なシグナル配列をコードするDNAは、E.coli外膜タンパク質遺伝子(ompA)(Masuiら(1983),Experimental Manipulation of Gene Expression;Ghrayebら(1984)EMBO
J.3:2437)およびE.coliアルカリホスファターゼシグナル配列(phoA)(Okaら(1985)Proc.Natl Acad.Sci.82:7212)のような、分泌される細菌タンパク質の遺伝子由来であり得る。さらなる例として、種々のBacillus株由来のα−アミラーゼ遺伝子のシグナル配列は、B.subtilisから異種タンパク質を分泌するために使用され得る(Palvaら(1982)Proc.Natl.Acad.Sci.USA 79:5582:EPO公開No.244042)。
【0125】
通常、細菌により認識される転写終止配列は、翻訳終止コドンに対して3’側に位置する調節領域であり、従って、プロモーターとともにコーディング配列に隣接する。これらの配列は、DNAにコードされるポリペプチドに翻訳され得るmRNAの転写を導く。転写終止配列はしばしば、転写終止の助けとなるステムループ構造を形成し得る、約50ヌクレオチドのDNA配列を含む。例には、E.coliのtrp遺伝子およびその他の生合成遺伝子のような、強プロモーターを伴った遺伝子由来の転写終止配列が含まれる。
【0126】
通常、プロモーター、シグナル配列(所望であれば)、目的のコーディング配列、および転写終止配列を含む上記の成分は、発現構築物中に組み立てられる。発現構築物は、細菌のような宿主に安定して維持され得る染色体外エレメント(例えば、プラスミド)のような、レプリコン中に維持される。レプリコンは、1つの複製系を有し、このため、発現、またはクローニングおよび増幅のいずれかのために原核宿主中に維持される。さらに、レプリコンは、高コピー数または低コピー数のプラスミドであり得る。高コピー数のプラスミドは、一般的に、約5から約200、そして通常約10から約150、の範囲のコピー数を有する。高コピー数のプラスミドを含有する宿主は、好ましくは少なくとも約10、より好ましくは少なくとも約20のプラスミドを含有する。高コピー数または低コピー数のいずれかのベクターが、宿主でのベクターの効果および外来タンパク質に依存して選択され得る。
【0127】
あるいは、発現構築物は、組込みベクターを用いて細菌ゲノムに組み込まれ得る。組込みベクターは通常、ベクターの組込みを可能にする細菌染色体に相同な少なくとも1つの配列を含有する。組込みは、ベクター中の相同DNAと細菌染色体との間の組換えによると考えられる。例えば、種々のBacillus株由来のDNAで構築された組込みベクターは、Bacillus染色体に組み込む(EPO公開No.127328)。組込みベクターはまた、バクテリオファージまたはトランスポゾン配列を含有し得る。
【0128】
通常、染色体外および組込み発現構築物は、形質転換された細菌株の選択を可能にする選択マーカーを含有し得る。選択マーカーは、細菌宿主中で発現され得、アンピシリン、クロラムフェニコール、エリスロマイシン、カナマイシン(ネオマイシン)およびテトラサイクリンのような薬剤に対して細菌を耐性にする遺伝子を含有し得る。(Daviesら、(1978)Annu.Rev.Microbiol.82:469)。選択マーカーはまた、ヒスチジン、トリプトファン、およびロイシンの生合成経路でのような、生合成遺伝子を含み得る。
【0129】
あるいは、上記成分のいくつかは、形質転換ベクター中に組み立てられ得る。形質転換ベクターは通常、レプリコン中に維持されるかまたは組込みベクターに組み込まれる選択マーカーを含有する。
【0130】
染色体外レプリコンまたは組込みベクターである、発現および形質転換ベクターは、多くの細菌への形質転換のために開発されている。例えば、発現ベクターは、特に以下の細菌のために開発されている:Bacillus subtilis.Palvら(1982)Proc.Natl.Acad.Sci.USA 79:5582:EPO公開No.036259およびNo.063953;PCT公開No、Wo84/04541:E.coli,Shimatakeら(1981)Nature 292:128:Amannら(1985)Gene 40:183;Studierら(1986)J.Mol.Biol.189:113;EPO公開No.036776,No,136829およびNo.136907;Stretococcus cremoris.Powellら(1988)Appl.Environ.Microbiol.54:655;Streptococcus lividans,Powellら(1988)Appl.Environ.Microbiol.54:655;およびStretomyces lividans、米国特許第4,745,056号。
【0131】
外因性DNAの細菌宿主への導入方法は当該分野で周知であり、CaCl2、または、二価カチオンおよびDMSOのようなその他の試薬で処理された細菌の形質転換を包含する。DNAはまた、エレクトロポーレーションにより細菌細胞中に導入され得る。形質転換の方法は、通常形質転換されるべき細菌種により変化する。Massonら(1989)FEMS Microbiol.Lett.60:273;Palvaら(1982)Proc.Natl.Acad.Sci.USA 79:5582:EPO公開No.036259およびNo.063953:PCT公開No.WO84/04541、Bacillusに関する;Millerら(1988)Proc.Natl.Acad.Sci.85:856;Wangら(1990)J.Bacteriol.172:949、Campylobacterに関する;Cohenら(1973)Proc.Natl.Acad.Sci 69:2110:Dowerら(1988)Nucleic Acids Res 16:6127:Kushner(1978)「colE1由来プラスミドによるE.coli形質転換のための改良法」,Genetic Enineerings:Proceeding of the International Symposium on Genetic Engineering(H.W.BoyerおよびS.Nicosia編);Mandelら(1970)J.Mol.Biol.53:159;Taketo(1988)Biochim.Biophys.Acta 949:318、Escherichiaに関する;Chassyら(1987)FEMS Microbiol.Lett.44:173、Lactobacillusに関する;Fiedlerら(1988)Anal.Biochem 170:38、Pseudomonsに関する;Augustinら(1990)FEMS Microbiol.Lett.66:203、Staphylococcusに関する;Baranyら(1980)J.Bacteriol.144:698:Harlander(1987)「エレクトロポーレーションによるStretococcus lactisの形質転換」,Streptococcal Genetics(J.FerrettiおよびR.Curtiss III編);Perryら(1981)Infec.Immun.32:1295;Powellら(1988)Appl.Environ.Microbiol.54:655:Somkutiら(1987)Proc.4th Evr.Cong.Biotechnology 1:412、Stretococcusに関する、を参照のこと。
【0132】
(iv.酵母発現)
酵母発現系もまた当業者に公知である。酵母プロモーターは、酵母RNAポリメラーゼに結合し得、コーディング配列(例えば、構造遺伝子)のmRNAへの下流(3’)転写を開始し得る、いずれものDNA配列である。プロモーターは、コーディング配列の5’末端近傍に通常位置する転写開始領域を有する。この転写開始領域は通常、RNAポリメラーゼ結合部位(「TATAボックス」)および転写開始部位を含む。酵母プロモーターはさらに、上流活性化配列(UAS)と呼ばれる第二ドメインを有し得、これは、存在する場合には通常構造遺伝子の遠位に存在する。UASにより、調節(誘導可能な)発現が行われる。構成的発現は、UASの非存在下で起こる。調節的発現は正または負のいずれかであり得、そのことにより転写を促進または減退する。
【0133】
酵母は、活性代謝経路を有する発酵微生物であり、従って、代謝経路の酵素をコードする配列は特に有用なプロモーター配列を提供する。例には、アルコールデヒドロゲナーゼ(ADH)(EPO公開No.284044)、エノラーゼ、グルコキナーゼ、グルコース−6−リン酸イソメラーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(GAPまたはGAPDH)、ヘキソキナーゼ、ホスホフルクトキナーゼ、3−ホスホグリセリン酸ムターゼ、およびピルビン酸キナーゼ(PyK)(EPO公開No.329203)が包含される。酸性ホスファターゼをコードする酵母PHO5遺伝子もまた、有用なプロモーター配列を提供する(Myanoharaら(1983)Proc.Natl.Acad.Sci.USA 80:1)。
【0134】
さらに、天然に存在しない合成プロモーターもまた、酵母プロモーターとして機能する。例えば、1つの酵母プロモーターのUAS配列は、別の酵母プロモーターの転写活性化領域に接続され得、合成雑種プロモーターがつくられ得る。このような雑種プロモーターの例には、GAP転写活性化領域に連結されたADH調節配列が含まれる(米国特許第4,876,107号および米国特許第4,880,734号)。雑種プロモーターのその他の例には、GAPまたはPyKのような解糖酵素遺伝子の転写活性化領域に組み合わされた、ADH2、GAL4、GAL10、またはPHO5遺伝子のいずれかの調節配列からなるプロモーターが含まれる(EPO公開No.164556)。さらに、酵母プロモーターには、酵母RNAポリメラーゼに結合し、転写を開始する能力を有する、非酵母起源の天然に存在するプロモーターが含まれ得る。このようなプロモーターの例には、特に、Cohenら(1980)Proc.Natl.Acad.Sci.USA 77:1078;Henikoffら(1981)Nature 283:835:Hollenbergら(1981)Curr.Topics Microbiol.Immunol.96:119:Hollenbergら(1978)「酵母Saccharomyces cerevisiaeでの細菌抗生物質耐性遺伝子の発現」:Plasmids of Medical,Environmental and Commercial Imortance(K.N.TimmisおよびA.Puhler編);Mercerau−Puigalonら(1980)Gene 11:163;Panthierら(1980)Curr.Genet.2:109が、含まれる。
【0135】
DNA分子は酵母において細胞内に発現され得る。プロモーター配列は、DNA分子に直接に連結され得、その場合には、組換えタンパク質のN末端の最初のアミノ酸はいつもメチオニンであり、これはATG開始コドンによりコードされる。所望であれば、N末端メチオニンは、インビトロにおける臭化シアンとのインキュベーションにより、タンパク質から切断され得る。
【0136】
融合タンパク質は、哺乳類、バキュロウイルス、および細菌発現系と同様に、酵母発現系の代替を提供する。通常、内因性酵母タンパク質またはその他の安定なタンパク質のN末端部分をコードするDNA配列が、異種コーディング配列の5’に融合される。発現に際し、この構築物は、2つアミノ酸配列の融合を提供する。例えば、酵母またはヒトのスーパーオキシドジスムターゼ(SOD)遺伝子が、外来遺伝子の5’末端で連結され得、酵母で発現され得る。2つのアミノ酸配列の接続部位DNA配列は、切断部位をコードし得るか、またはコードし得ない。例えば、EPO公開No.196056を参照のこと。その他の例は、ユビキチン融合タンパク質である。このような融合タンパク質は、好ましくは、外来タンパク質からユビキチンを切断するためのプロセッシング酵素(例えば、ユビキチン特異的プロセッシングプロテアーゼ)に対する部位を保持しているユビキチン領域でつくられる。従って、この方法によってそのままの外来タンパク質が単離され得る(例えば、PCT公開No.WO88/024066を参照のこと)。
【0137】
あるいは、外来タンパク質はまた、酵母での外来タンパク質の分泌を提供するリーダー配列フラグメントを含有する融合タンパク質をコードする、キメラDNA分子をつくることにより、細胞から増殖培地に分泌され得る。好ましくは、リーダーフラグメントと外来遺伝子との間でコードされるプロセッシング部位が存在し、インビボまたはインビトロのいずれかで切断され得る。リーダー配列フラグメントは通常、細胞からのタンパク質の分泌を導く疎水性アミノ酸を含有するシグナルペプチドをコードする。
【0138】
適切なシグナル配列をコードするDNAは、酵母インベルターゼ遺伝子(EPO公開No.012873:JPO公開No.62,096,086)およびA因子遺伝子(米国特許第4,588,684号)のような、分泌酵母タンパク質の遺伝子に由来し得る。あるいは、インターフェロンリーダーのような、非酵母起源のリーダーが存在し、これもまた酵母での分泌を提供する(EPO公開No.060057)。
【0139】
分泌リーダーの好ましいクラスには、酵母α因子遺伝子のフラグメントを用いるリーダーがあり、これは「pre」シグナル配列および「pro」領域の両方を含有する。用いられ得るα因子フラグメントの型には、全長pre−pro α因子リーダー(約83アミノ酸残基)および切断型α因子リーダー(通常約25から約50アミノ酸残基)が含まれる(米国特許第4,546,083号および米国特許第4,870,008号;EPO公開No.324274)。分泌を提供するα因子リーダーフラグメントを用いる別のリーダーには、第一酵母のpre配列でつくられたハイブリッドα因子リーダーが含まれるが、第二酵母のα因子のpro領域は含まれない。(例えば、PCT公開No.WO89/02463を参照のこと)。
【0140】
通常、酵母により認識される転写終止配列は、翻訳終止コドンに対して3’側に位置する調節領域であり、従って、プロモーターとともにコーディング領域に隣接する。これらの配列は、DNAによりコードされるポリペプチドに翻訳され得るmRNAの転写を導く。転写終止配列およびその他の酵母認識終止配列の例は、解糖酵素をコードする配列などである。
【0141】
通常、プロモーター、リーダー(所望であれば)、目的のコーディング配列、および転写終止配列を含む上記の成分は、発現構築物中に組み立てられる。発現構築物はしばしば、酵母または細菌のような宿主中に安定して維持され得る染色体外エレメント(例えば、プラスミド)のような、レプリコン中に維持される。レプリコンは2つの複製系を有し得、このため、例えば、発現のために酵母中に、およびクローニングおよび増幅のために原核宿主中に維持される。このような酵母−細菌シャトルベクターの例には、YEp24(Botsteinら(1979)Gene 8:17−24);pCl/1(Brakeら(1984)Proc.Natl.Acad.Sci USA 81:4642−4646);およびYRp17(Stinchcombら(1982)J.Mol.Biol.158:157)が含まれる。さらに、レプリコンは、高コピー数または低コピー数のプラスミドであり得る。高コピー数のプラスミドは一般に、約5から約200の範囲のコピー数を有し、通常約10から約150のコピー数を有する。高コピー数のプラスミドを含有する宿主は、好ましくは少なくとも約10、より好ましくは少なくとも約20を有する。高コピー数または低コピー数のベクターは、宿主におけるベクターおよび外来タンパク質の効果に依存して選択され得る。
【0142】
あるいは、発現構築物は、組込みベクターを用いて酵母ゲノムに組み込まれ得る。組込みベクターは通常、ベクターの組込みを可能にする、酵母染色体に相同な少なくとも1つの配列を含有し、好ましくは、発現構築物に隣接する2つの相同な配列を含有する。組込みは、ベクター中の相同DNAと酵母染色体との間の組換えによると考えられる(Orr−Weaverら(1983)Methods in Enzymol.101:228−245)。組込みベクターは、ベクター中への封入に適切な相同配列を選択することにより、酵母の特異的な座に導かれ得る。1つ以上の発現構築物が組み込まれ得、おそらく産生される組換えタンパク質レベルに影響し得る(Rineら(1993)Proc.Natl.Acad.Sci.USA 80:6750)。ベクターに含まれる染色体配列は、ベクター中の1つのセグメントとして存在して、完全ベクターの組込みになり得るか、または、染色体中の隣接セグメントに相同で、ベクター中で発現構築物に隣接する2つのセグメントとして存在して、発現構築物のみの安定な組込みになり得る。
【0143】
通常は、染色体外および組込み発現構築物は、形質転換された酵母株の選択を可能にする選択マーカーを含有し得る。選択マーカーは、ADE2、HIS4、LEU2、TRP1、およびALG7のような、酵母宿主で発現され得る生合成遺伝子、および、ツニカマイシンおよびG418のそれぞれに対する酵母細胞において耐性を与えるG418耐性遺伝子を含み得る。さらに、適切な選択マーカーはまた、金属のような毒素化合物の存在下で増殖する能力を有する酵母を提供し得る。例えば、CUP1の存在は、銅イオンの存在下での酵母の増殖を可能にする(Buttら(1987)Microbiol.Rev.51:351)。
【0144】
あるいは、上記成分のいくつかは、形質転換ベクター中に組み立てれられ得る。形質転換ベクターは通常、レプリコン中に維持されるかまたは組込みベクターに組み込まれる選択マーカーを合有する。
【0145】
染色体外レプリコンまたは組込みベクターである、発現および形質転換ベクターは、多くの酵母の形質転換のために開発されている。例えば、発現ベクターは、特に以下の酵母のために開発されている:Candida albicans(Kurtzら(1986)Mol.Cell.Biol.6:142);Candida maltosa(Kunzeら(1985)J.Basic Microbiol.25:141):Hansenula polymorpha(Gleesonら(1986)J.Gen.Microbiol.132:3459;Roggenkampら(1986)Mol.Gen.Genet.202:302);Kluyveromyces fragilis(Dasら(1984)J.Bacteriol.158:1165);Kluyveromces lactis(De Louvencourtら(1983)J.Bacteriol.154:737;Van den Bergら(1990)Bio/Technology 8:135):Pichia guillerimondii(Kunzeら(1985)J.Basic Microbiol.25:141):Pichia pastoris(Creggら(1985)Mol.Cell.Biol.5:3376;米国特許第4,837,149号および米国特許第4,929,555号):Saccharomyces cerevisiae(Hinnenら(1978)Proc.Natl.Acad.Sci.USA 75:1929;Itoら(1983)J.Bacteriol.153:163):Schizosaccharomyces pombe(Beachら(1981)Nature 300:706);および、Yarrowia lipolytica(Davidowら(1985)Curr.Genet.10:380471 Gaillardinら(1985)Curr.Genet.10:49)。
【0146】
外因性DNAの酵母宿主への導入方法は、当該分野では公知であり、通常は、スフェロプラストまたは完全酵母細胞をアルカリカチオンで処理する形質転換を包含する。形質転換方法は通常、形質転換される酵母種により変化する。例えば、Kurtzら(1986)Mol.Cell.Biol.6:142;Kunzeら(1985)J.Basic
Microbiol.25:141、Candidaに関する;Gleesonら(1986)J.Gen.Microbioy.132:3459;Roggenkampら(1986)Mol.Gen.Genet.202:302、Hansenulaに関する;Dasら(1984)J.Bacteriol.158:1165;De Louvencourtら(1983)J.Bacteriol.154:1165:Van den Bergら(1990)Bio/Technology 8:135、Kluyveromycesに関する;Creggら(1985)Mol.Cell.Biol.5:3376;Kunzeら(1985)J.Basic Microbiol.25:141;米国特許第4,837,148号および米国特許第4,929,555号、Pichiaに関する;Hinnenら(1978)Proc.Natl.Acad.Sci.USA 75;1929;Itoら(1983)J.Bacteriol.153:163、Saccharomycesに関する;Beachら(1981)Nature 300:706、Schizosaccharomycesに関する;Davidowら(1985)Curr.Genet.10:39;Gaillardinら(1985)Curr.Genet.10:49、Yarrowiaに関する;参照のこと。
【0147】
(4.ワクチン)
本明細書中で考察されている各コンジュゲート化合物は、単一ワクチン候補物として、または他の病原由来の1つ以上のその他の抗原との組み合わせで使用され得る。これらのワクチンは、予防剤(感染を予防するため)または治療剤(感染後に疾患を治療するため)のいずれかであり得る。
【0148】
このようなワクチンは、コンジュゲート化合物を、通常は「薬学的に受容可能なキャリア」との組合せで含有し、このキャリアは、この組成物が与えられる個体に対して有害な抗体の産生をそれ自身が誘導しないいずれものキャリアを含む。適切なキャリアは、典型的には、タンパク質、多糖類、ポリ乳酸、ポリグリコール酸、アミノ酸ポリマー、アミノ酸コポリマー、脂質凝集体(例えば、油滴またはリボソーム)、および不活性ウイルス粒子のような、大きくて徐々に代謝される高分子である。このようなキャリアは、当業者に周知である。さらに、これらのキャリアは、免疫刺激剤(「アジュバント」)として機能し得る。さらに、抗原は、ジフテリア、破傷風などのような細菌トキソイドにコンジュゲートされ得る。
【0149】
組成物の効果を増強させる好ましいアジュバントには、以下が包含されるが、これらに限定されない:(1)アルミニウム塩(alum)(水酸化アルミニウム、リン酸アルミニウム、硫酸アルミニウムなど);(2)水中油型エマルジョン製剤〔ムラミルペプチド(下記を参照)のようなその他の特異的免疫刺激剤または細菌細胞壁成分を伴うまたは伴わない〕、例えば、(a)MF59(PCT公開No.WO90/14827):5%スクワレン、0.5% Tween 80、および0.5% Span85〔必須ではないが必要に応じて、種々の量のMTP−PE(下記を参照)を含有する〕を含有して、110Y型マイクロフルイダイザー(Microfluidics,Newton,MA)のようなマイクロフルイダイザーを使用して、サブミクロン粒子に処方される;(b)SAF:10%スクワラン、0.4% Tween 80、5%プルロニック(pluronic)−ブロックポリマーL121、およびthr−MDP(下記を参照)を含有し、マイクロフルイダイザーにかけてサブミクロン粒子エマルジョンにされるか、ボルテックスにかけてより大きな粒子サイズのエマルジョンを生成する;および、(c)RibiTMアジュバント系(RAS)(Ribi Immunochem,Hamilton,MT):2%スクワレン、0.2% Tween 80、および、1つ以上の細菌細胞壁成分(モノホスホリルリピド(monophosphorylipid)A(MPL)、トレハロースジミコレート(TDM)、および細胞壁骨格(CWS)よりなる群から、好ましくはMPL+CWS(DetoxTM))を含有する;(3)サポニンアジュバント(例えばStimulonTM(Cambridge Bioscience,Worcester,MA))が使用され得るか、または、ISCOM(免疫刺激複合体)のような、それらからつくられた粒子;(4)完全フロイントアジュバント(CFA)および不完全フロイントアジュバント(IFA);(5)サイトカイン(例えば、インターロイキン(IL−1、IL−2など)、マクロファージコロニー刺激因子(M−CSF)、腫瘍壊死因子(TNF)など);および、(6)組成物の効果を増強するための免疫刺激剤として作用するその他の物質。AlumおよびMF59が好ましい。
【0150】
上記のように、ムラミルペプチドには、N−アセチル−ムラミル−L−トレオニル−D−イソグルタミン(thr−MDP)、N−アセチル−ノルムラミル−L−アラニル−D−イソグルタミン(nor−MDP)、N−アセチルムラミル−L−アラニル−D−イソグルタミニル−L−アラニン−2−(1’−2’−ジパルミトイル−sn−グリセロ−3−ヒドロキシホスホリルオキシ(huydroxyphosphoryloxy))−エチルアミン(MTP−PE)などが含まれるが、これらに限定されない。
【0151】
免疫原性組成物(例えば、抗原、薬学的に受容可能なキャリア、およびアジュバント)は典型的に、水、生理食塩水、グリセロール、エタノールなどのような希釈剤を含有する。さらに、湿潤または乳化剤などのような補助物質、pH緩衝物質などが、このような賦形剤中に存在し得る。
【0152】
典型的には、免疫原性組成物は、溶液または懸濁液のいずれかとして注射用に調製される;注射の前に液体賦形剤に溶解または懸濁するのに適切な固形もまた、調製され得る。薬学的に受容可能なキャリアについて上記で考察したように、調製物はまた、アジュバント効果を増強するために、エマルジョンにされるか、またはリボソームにカプセル化され得る。
【0153】
ワクチンとして使用される免疫原性組成物は、免疫学的に有効な量の抗原ポリペプチド、および必要であれば、上記成分のいかなる他の成分をも含有する。「免疫学的に有効な量」とは、単回投与でまたは連続投与の一部として個体に投与される量が、治療または予防に有効であることを意味する。この量は、処置される個体の健康状態および身体条件、処置される個体の分類学上のグループ(例えば、ヒト以外の霊長類、霊長類など)、抗体を合成するための個体の免疫系の能力、所望の予防程度、ワクチン処方、医療状況での治療医の評価、およびその他の関連因子に依存して変化する。その量は比較的広範囲にあり、日常の試行により決定され得ると思われる。
【0154】
免疫原性組成物は従来より非経口的に、例えば、皮下または筋肉内注射により、投与される。その他の投与形態に適した処方には、経口および肺への処方、坐薬、および経皮投与が含まれる。投与処置は、単回投与スケジュールまたは複数回投与スケジュールであり得る。ワクチンは、その他の免疫調節剤と組み合わせて投与され得る。
【実施例】
【0155】
(5. 実施例1)
Meningococci C群の多糖類ならびに熱ショックタンパク質hspR70およびhsp65を含有するコンジュゲート化合物を構築し、ワクチンの効力について試験した。
【0156】
(5.1. Meningococci C(MenC)群の多糖の精製、およびMenCのオリゴ糖の生成)
C群髄膜炎菌の多糖を、Frasc C.E3.の「バイオテクノロジープロセスの進歩:バクテリアワクチン」(A.Mizrahi編),vol.13,pp.123−145,Wiley−Liss Inc.,New York(1990)に記載されているように精製した。精製多糖類(10mg/ml)を、pH5の0.01M酢酸緩衝液中で、100℃で8時間加水分解して解重合した。得られた生産物を、分析用クロマトグラフィー(Sephadex 6−50)で分析し、0.27のKd(分布係数)を示した。
【0157】
(5.2. 一級アミン基のオリゴ糖末端基への導入)
0.5Mの塩化アンモニウムおよび0.15Mのシアノボロヒドリドナトリウムを加水分解から得られた溶液に加えた。pHを7に上げて、得られた溶液を35℃で1週間置いた。次に、オリゴ糖をクロマトグラフィー(Sephadex 6−15)により、収集したアミン基およびカルボハイドレート基に関して化学活性を含有するボイド容量画分を精製した。一方、単糖および過剰の試薬を含有する画分は廃棄した。得られたMenCオリゴ糖は、アミン基、シアル酸基、およびO−アセチル基を測定することによって特徴付けられた。次のモル比が得られた:シアル酸/アミン基=20、O−アセチル/シアル酸=0.84。
【0158】
(5.3. 熱ショックタンパク質の調製)
M.bovis BCG GroEL−型 65kDa hsp(hspR65)をプラスミドpRIB1300を宿した組換えE.coli K12株から発現し(Tholeら、Infect.Immun.1985,50,800:Van Edenら、Nature,1989,331,171)、そして、Tholeら、Infect.Immune.,1987,55.1466に記述されたように精製した。
【0159】
組換えM.tuberculosis DnaK−型 70kDa hsp(hspR70)を得て、ATP−アガロースクロマトグラフィーで精製した(Mehlertら、Mol.Microbiol.,1989,3,125)。
【0160】
(5.4. MenC多糖とhspR65およびhspR70との糖コンジュゲートの調製)
MenCアミノ−オリゴ糖を、H2Oを10%含むジメチルスルホキシドに溶解し、それから(アミン基について)12倍過剰のアジピン酸のN−ヒドロキシスクシンイミドエステル[Hillら、「FEBS LETT.」102:282(1979)に従って調製した]と反応させた。ジオキサン(1〜4倍量)で沈澱させて精製した後、活性化オリゴ糖を真空中で乾燥し、N−ヒドロキシスクシンイミドエステルの含有量について分析した。次いで、pH7の0.1Mリン酸緩衝液中で5mg/mlの量のhspR65およびhspR70を、300倍モル過剰の活性化オリゴ糖と反応させた。それぞれ得られた糖コンジュゲートは、クロマトグラフィーにより未反応オリゴ糖から遊離し、濾過し、そして4℃で保存した。開始物質のMenC多糖中のシアル酸の割合に対する糖コンジュゲートのシアル酸含有量の比は、カップリングしたオリゴ糖の量を表す。調製したタンパク質の含有量は、Lowry「J.Biol.Chem.」193:265(1951)の方法により確認した。特に、hspR70とのコンジュゲートは、310μg/mlのタンパク質含有量および76μg/mlの糖含有量を有し、一方、hspR65とのコンジュゲートは、180μg/mlのタンパク質含有量および97μg/mlの糖含有量を有する。
【0161】
(5.5. マウスおよび免疫感作)
BALB/c(H−2d)、C567BL/6(H−2b)およびCBA/J(H−2k)の雌のマウス(8〜12週齢)を本発明者らの飼育施設で飼育した。
【0162】
最初のカップルはJackson Laboratory,Bar Harbor,MEから供給された。BALB/c nu/nu 胸腺欠失マウスをIffa Credo,L’Arbresle,Franceから得た。
【0163】
0日目、各々のマウスに106CFUのBCG(またはコントロール群の場合はPBS)を腹腔内投与し、続いて14日目と35日目に2用量のコンジュゲート(PBS中)を投与した。
【0164】
コントロール群は、MenCオリゴ糖のみ、または水酸化アルミニウム(1mg/用量、0.5ml中)に吸着したMenCオリゴ糖−CRM197コンジュゲートワクチンを接種した。各々の免疫感作において、マウスに2μgのMenCオリゴ糖を接種し、それは8.7μgのMenCオリゴ糖−CRM197コンジュゲートワクチン、8.4μgのMenCオリゴ糖−hspR70コンジュゲート、または3.7μgのMenCオリゴ糖−hspR65コンジュゲートに対応した。
【0165】
(5.6. ELISA法による抗体の測定)
毎週、マウスの後眼窩叢から血液を採取し、抗体はELISAで力価測定した。
【0166】
IgG抗MenC抗体を測定するために、96ウエル平底プレートをpH7.4のPBS中のMenC多糖(5μg/ml)で37℃で一晩のインキュベーションにより覆った(Nunc Immunoplate I,Nunc,Roskilde,Denmark)。0.054%のTween−20を含むPBS(PBS−T)で繰り返し洗浄し、そして5%のFCSを含有するPBS−Tの200μlで37℃、1時間インキュベーションした後、ウエルを5% FCSを含むPBS−Tで希釈したマウス血清100μlとともに4℃で一晩インキュベートした。洗浄を繰り返した後、プレートを再び、ペルオキシダーゼにコンジュゲートしたIgG抗マウス抗血清を適切に希釈したものの100μlとともに37℃で3時間インキュベートした。
【0167】
特異的抗体の存在を、2,2’−アジノ−ビス−(3−エチルベンゾチアゾリンスルホン酸)(ABTS;Kirkegaard and Pery Laboratories Inc.,Gaithersburg,MD)を基質として加えることにより明らかにした。結果は、414nmでの吸光度について測定した。試験した最初の希釈(1:50)で0.2未満の吸光度を有する血清サンプルは、陰性と考えた。
【0168】
(5.7. オリゴ糖ヘコンジュゲートしたhspR65およびhspR70のキャリア効果)
HspR65およびhspR70−MenCオリゴ糖コンジュゲートを、予めBCGで感作したまたは感作していないBALB/cおよびC57BL/6マウスの免疫に使用した。
【0169】
コントロールとして、数群のマウスにMenCのみまたは水酸化アルミニウム上のCRM197−MenCコンジュゲートワクチンを感作させた。
【0170】
図1は、抗MenC多糖IgG抗体が、水酸化アルミニウム上のCRM197−MenCコンジュゲートワクチンの場合に測定したのと同等またはそれ以上の量でhsp−MenCコンジュゲートの免疫の後産生されたことを示す(図1A中のC57BL/6)。この効果は、アジュバントが存在しない場合だけでなく、BCGで感作しなかったときのhspR70−MenCコンジュゲートの場合にも観察された(図1B)。これらの結果より、アジュバント非存在下またはBCG感作していないとき、ミコバクテリウムのhspを用いる免疫が、オリゴ糖抗原を用いても本当に達成し得ると述べ得る。
【0171】
従って、hsp分子(特にhspR70)は、BCGで感作していないマウスでも強力なキャリア効果を及ぼし、アジュバントの非存在下でさえ、移送した多糖に対して特異的なIgG抗体を誘導する能力のある分子の有効なキャリアとして作用することが確認された。
【0172】
アジュバントおよび前感作の非存在下で及ぼされる、このミコバクテリウムのhspの有効なキャリア効果により、記載されたコンジュゲートが、細菌感染に対する新規ワクチンの開発に特に有用になる。
【0173】
(6.実施例2)
新規のH.pylori熱ショックタンパク質が同定され、組換えDNA法を用いて産生された。
【0174】
(6.1. 材料および方法)
(6.1.1. H.pylori株および増殖条件)
使用したH.pylori株は:CCUG 17874、G39およびG33(Grosseto病院,Italy,で胃生検より単離)、Pylo 2U+およびPylo 2U−(F.Megraud,Pellegzin病院,Bordeaux,Franceにより供与)、BA96(Siena大学,Italy,で胃生検により単離)であった。Pylo 2U+株は非細胞毒性であり;Pylo 2U−株は非細胞毒性およびウレアーゼ陰性である。全ての株は、0.2%のシクロデキストリン、5μg/mlのセフスロジン、および5μg/mlのアムホテリシンBを含むコロンビアアガー上で、微好気性の条件下で、37^℃5〜6日間通常的に増殖した。細胞を採取し、PBSで洗浄した。ペレットをLaemmliサンプル緩衝液に再懸濁して、そして煮沸して細胞を溶解した。
【0175】
胃炎および潰瘍に冒されている患者の血清(A.Ponzetto,「Le Molinette」病院,Torino,Itaiyにより供与)および胃癌患者の血清(F.Roviello,Siena大学,Italyにより供与)を使用した。
【0176】
(6.1.2. ライブラリーの免疫スクリーニング)
λgt11 H.pylori DNA発現ライブラリーの50万個のプラークを、0.2%マルトースおよび10mM MgSO4を含むLB中で一晩増殖したE.coli
Y1090株の懸濁液5mlと混和し、そして0.5 O.D.になるように10mM
MgSO4に両懸濁した。37℃で10分インキュベーションした後、溶かした75mlのTopAgaroseを、細菌/ファージ混合物に注ぎ、そして全部をBBLプレート上で平板培養した(50,000プラーク/プレート)。42℃で平板培養したライブラリーを3.5時間インキュベーションした後、10mM IPTGで予め湿らせたニトロセルロース濾紙(SchleicherおよびSchuell,Dassel,Germany)をプレート上に載せ、そしてさらに37℃3.5時間、次いで4℃で一晩インキュベーションした。λタンパク質を有する濾紙を持ち上げてPBSで洗浄し、そしてTBST(10mM TRIS pH8、100mM NaCl、5M MgCl2)に溶解した5%脱脂乾燥乳で20分間飽和させた。最初のハイブリダイゼーション工程は、患者血清で行った;陽性プラークを呈色および可視化するために、製造者の指示に従って、AP緩衝液(100mM Tris pH9.5、100mM NaCl,5mM MgCl2)中でアルカリホスファターゼをコンジュゲートした抗ヒトIg抗体(Cappel,West Chester,PA)およびNBT/BCIPキット(Promega,Madison,WI)を用いた。
【0177】
(6.1.3. 組換えDNA手順)
使用した試薬および制限酵素は、Sigma(St.Louis,MO)およびBoehringer Mannheim(Germany)から得た。分子クローニング、一本鎖DNA精製、E、coliでの形質転換、プローブの放射標識化、H.pylori
DNA遺伝子ライブラリーのコロニースクリーニング、サザンブロット分析、PAGE、ウェスタンブロット分析については、標準的技法を用いた。
【0178】
(6.1.4. DNA配列分析)
DNAフラグメントをBluescriptSK+(Stratagene,San Diego,CA)でサブクローニングした。一本鎖DNA配列決定を、製造者の指示に従って、[33P]αdATP(New England Nuclear,Boston,MA)およびSequenaseキット(U.S.Biochemical Corp.,Cleveland,OH)を用いて行った。配列は両鎖について決定し、そして各鎖は平均2回配列決定した。コンピュータによる配列分析はGCGパッケージを用いて行った。
【0179】
(6.1.5. 組換えタンパク質)
MS2ポリメラーゼ融合タンパク質をpEX34A(pEX31の誘導体)ベクターを用いて生成した。挿入Hp67(図3のヌクレオチド445からヌクレオチド1402)およびEcoRIリンカーを、ベクターのEcoRI部位にフレーム内でクローニングした。停止コドンの位置を確認するために、HpG3’HindIIIフラグメントを、pEX34AのHindIII部位にフレーム内でクローニングした。組換えプラスミドをE.coli K12 H1△trpに形質転換した。誘導後の両方の場合で、予測した分子量の融合タンパク質を生成した。EcoRI/EcoRIフラグメントの場合は、誘導後に得られた融合タンパク質をウサギに免疫するために、標準プロトコルを用いて電気浴出した。
【0180】
(6.2. 結果)
(6.2.1. 発現ライブラリーのスクリーニングおよびH.pylori hspのクローニング)
H.pylori DNA発現ライブラリーのスクリーニングに適切な血清を見つけるために、H.pylori CCUG 17874株の超音波処理抽出物を、異種の胃炎によって冒された患者の血清に対するウェスタンブロット分析で試験した。異なった血清による抗原認識パターンは多岐にわたり、おそらく、個体の免疫応答の差異、および感染物に含まれる株により発現する抗原の差異のためであった。
【0181】
N°19の血清を、細菌の増殖の間にインビボで発現される、H.pylori特異的抗原を同定するためのλgt11 H.pylori DNA発現ライブラリーをスクリーニングするために選択した。この血清でライブラリーをスクリーニングした後、多くの陽性クローンを単離し、特徴付けた。これらの1つのヌクレオチド配列(いわゆるHp67)は、熱ショックタンパク質のhsp60ファミリーに高い相同性を有するタンパク質をコードする985塩基対のオープンリーディングフレームであることがわかった(Ellis,Nature 358:191−92(1992))。全コーディング領域を得るために、本発明者らは、フラグメントHp67を、異なる制限酵素で切断したH.pylori DNAのサザンブロット分析のプローブとして用いた。プローブHp67は、それぞれ、約800および1000塩基対の2つのHindIIIバンドを認識した。HindIII切断DNAのゲノムH.pyloriライブラリーをプローブHp67でスクリーニングし、そして予測した分子量の2つの陽性クローン(HpG5’およびHpG3’)を得た。プラスミドpHp60G2(約1から929のヌクレオチド)およびpHp60G5(約824から1838のヌクレオチド)を合むE.coliをアメリカンタイプカルチャーコレクション(ATCC)に寄託した。
【0182】
(6.3. 配列分析)
ヌクレオチド配列分析で、開始ATGの6塩基対上流の推定リボゾーム結合部位を有する1638塩基対のオープンリーディングフレームが明らかになった。図3に、H.pylori hspのヌクレオチド配列およびアミノ酸配列を示す。推定リボゾーム結合部位および中間HindIII部位を下線で示す。フラグメントHp67の445位のシトシンおよび1402位のグアニンは、それぞれ最初および最後のヌクレオチドである。チミジン1772を、独立因子ターミネーター領域の局在化に関する演算法を用いて、転写された最後の推定のヌクレオチドとして同定した。オープンリーディングフレームは、58.3KDaの予測分子量、および5.37の予測pIを有する546アミノ酸のタンパク質をコードしていた。この遺伝子のコドンの選択は、H.pyloriコドンの用法と一致する。
【0183】
疎水性のプロフィル分析では、予測されるリーダーペプチドまたは他の貫膜ドメインを除いて、ほとんど疎水性タンパク質であることがわかった。アミノ末端配列は、精製タンパク質について、Dunnら、Infect.Immun.60:1946−51(1992)により決定された30アミノ酸の配列に100%の相同性を示し、Evansら、Infect.Immun.60:2125−27(1992)によって公表された44アミノ酸の配列から1残基のみ(Lysに代わってSer42)異なっていた(Evansら、1992)。成熟hspタンパク質のN−末端配列は、開始メチオニンを含まず、このことは、翻訳後に除かれたことを示している。
【0184】
(6.4. hsp60ファミリーとの相同性)
アミノ酸配列分析で、全ての生物に存在するメンバーである熱ショックタンパク質hsp60ファミリーと非常に高い相同性が示された。異種のhsp60タンパク質問での相同の程度に基づいて、H.pylori hspは、グラム陰性細菌のhsp60タンパク質のサブグループに属する;しかし、hsp60ファミリーの他のタンパク質に対する相同の程度はかなり高い(少なくとも54%の一致)。
【0185】
他の熱ショックタンパク質とのH.pylori hspの相同性は図3に完全に例示した。H.pylori hspまたはその1つ以上の機能的な免疫刺激ドメインは、本発明に従ってコンジュゲート化合物を生成するために上記実施例1の手法を用いてオリゴ糖または多糖にコンジュゲートし得る。
【0186】
(6.5. 組換えタンパク質の発現およびポリクローナル抗血清の生成)
クローンHp67およびクローンHpG3’の挿入物を、MS2ポリメラーゼのアミノ末端に融合したこれらのオープンリーディングフレームを発現するために、発現ベクターpEX34Aにサブクローンした。クローンは予測したサイズの組換えタンパク質を生成し、そして最初のスクリーニングで用いたヒト血清により認識された。クローンHp67由来の融合タンパク質を電気浴出し、そして抗hsp特異的ポリクローナル抗血清を得るためにウサギの免疫に用いた。得られた抗血清は、融合タンパク質と、ウレアーゼ陰性株および非細胞毒性株を含有する、試験したいくつかのH.pylori株の全細胞抽出物の58KDaのタンパク質との両方を認識した。
【0187】
Hspは試験した全てのH.pylori株で発現されることが示され、そしてその発現はウレアーゼの存在または細胞毒性に関係ない。抗hsp抗血清により認識されるタンパク質は、H.pyloriの水溶性抽出物中に認められ、そしてウレアーゼサブユニットと同時に精製された。これは、このタンパク質が細菌外膜に弱い会合をしていることを示す。従って、hspはウレアーゼに会合し、そして表面が露出していると記載し得る細胞表面の局在化により、ほとんどのhspに相同なタンパク質が細胞質中またはミトコンドリアおよび色素体中に局在していることは驚くことである。hspにリーダーペプチドが欠けていることは、これが独特の輸送系により膜に輸送されるか、あるいはタンパク質が細胞質から放出され、細菌の死後に細菌の膜に受動的に吸着するかのいずれかであることを示唆する。
【0188】
(7. 生物学的材料の寄託)
以下の材料は、1992年12月15日および1993年1月22日に、本発明の譲渡人であるBiocine Sclavo,S.p.A.により、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約のもとに、アメリカンタイプカルチャーコレクション(ATCC)、メリーランド州,ロックビル,パークローンドライブ12301,電話(301)231−5519に寄託された。
ATCC No.69155 プラスミドpHp60G2を含むE.coli TG1
ATCC No.69156 プラスミドpHp605を含むE.coli TG1。
【0189】
これらの寄託物は、当業者の利便性のために提供され、寄託が、米国特許法112条、または本明細書の任意の指定国のいかなる相当規定のもとで要求されることを認めるものではない。これらの寄託物の核酸配列、およびそれにコードされるポリペプチドのアミノ酸配列は、本明細書中に参考として援用されており、寄託物の配列と比較して、本明細書に記載の配列中に任意の誤りがある場合には、参考にされるべきである。寄託物質を製造し、使用し、または売るためには許諾が必要であり得るが、ここでは、そのような許諾は承認されていない。
【産業上の利用可能性】
【0190】
このように、本発明のキャリアは、確実に高力価のワクチン接種をするために、細菌間の熱ショックタンパク質の高い保存性およびT細胞記憶を、独自に利用するものである。この化合物は、アジュバントを用いることなく、抗多糖抗体の形成を誘導し得る。そのため、本化合物は、ヒトおよび動物に使用するワクチン、特に細菌感染予防用ワクチンの調製に有用である。
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1】


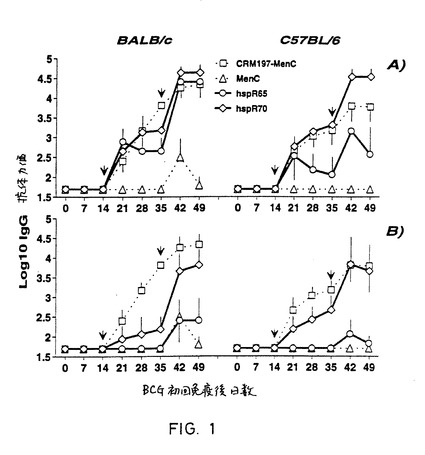
【図2A】
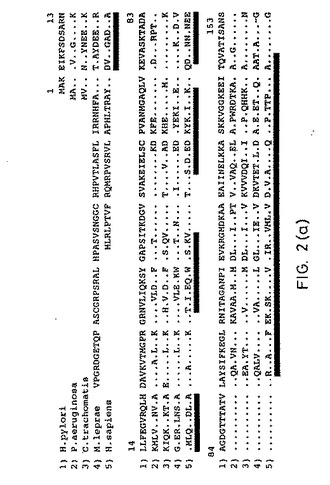
【図2B】
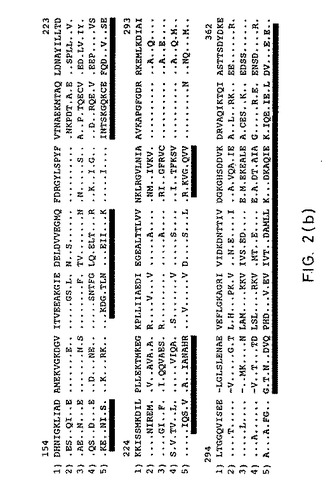
【図2C】
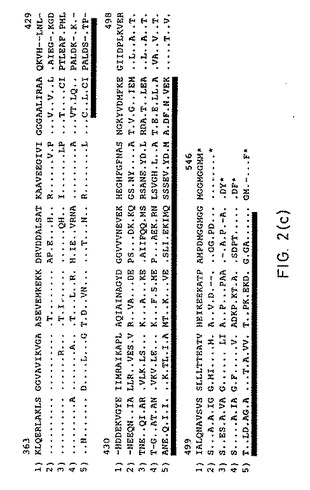
【図3A】

【図3B】

【図3C】

【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図3C】
【公開番号】特開2011−52000(P2011−52000A)
【公開日】平成23年3月17日(2011.3.17)
【国際特許分類】
【出願番号】特願2010−239145(P2010−239145)
【出願日】平成22年10月25日(2010.10.25)
【分割の表示】特願2008−312770(P2008−312770)の分割
【原出願日】平成5年3月8日(1993.3.8)
【出願人】(592243793)カイロン ソチエタ ア レスポンサビリタ リミタータ (107)
【Fターム(参考)】
【公開日】平成23年3月17日(2011.3.17)
【国際特許分類】
【出願日】平成22年10月25日(2010.10.25)
【分割の表示】特願2008−312770(P2008−312770)の分割
【原出願日】平成5年3月8日(1993.3.8)
【出願人】(592243793)カイロン ソチエタ ア レスポンサビリタ リミタータ (107)
【Fターム(参考)】
[ Back to top ]