
熱帯熱マラリア原虫ジャイレースB由来ポリペプチド
【課題】帯熱マラリア原虫DNAジャイレースのジャイレースB由来ポリペプチド、前記ポリペプチドをコードするポリヌクレオチド、前記ポリペプチドに対する抗体、当該抗体を含むマラリア原虫増殖抑制用組成物およびマラリア原虫検出用組成物の提供。
【解決手段】熱帯熱マラリア原虫DNAジャイレースのジャイレースBドメインポリペプチド、前記ポリペプチドをコードするポリヌクレオチド、および前記ポリペプチドに対するポリクローナル抗体、モノクローナル抗体、ヒト型化抗体、ヒトモノクローナル抗体抗体および、その抗イディオタイプ抗体。それらを含むマラリア原虫増殖抑制用組成物、および、マラリア原虫検出用組成物。
【解決手段】熱帯熱マラリア原虫DNAジャイレースのジャイレースBドメインポリペプチド、前記ポリペプチドをコードするポリヌクレオチド、および前記ポリペプチドに対するポリクローナル抗体、モノクローナル抗体、ヒト型化抗体、ヒトモノクローナル抗体抗体および、その抗イディオタイプ抗体。それらを含むマラリア原虫増殖抑制用組成物、および、マラリア原虫検出用組成物。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、熱帯熱マラリア原虫DNAジャイレースのジャイレースB由来ポリペプチド、前記ポリペプチドをコードするポリヌクレオチド、前記ポリペプチドに対する抗体、当該抗体を含むマラリア原虫増殖抑制用組成物およびマラリア原虫検出用組成物に関する。
【背景技術】
【0002】
熱帯熱マラリアはヒトを死に至らしめる原虫感染症の一つであり、年間300万人以上の人々が死亡している。近年、本原虫が既存のマラリア治療薬に対して薬剤耐性を獲得してきたことから、新規の治療薬の開発が望まれている。
【0003】
熱帯熱マラリア原虫の遺伝子の中にはヒトに対応遺伝子が存在しないもの、あるいはヒトの対応遺伝子には存在しないDNA挿入部分を有するものが存在する。かかる遺伝子がコードするタンパク質は、ヒトのタンパク質とは異なるアミノ酸配列を有し、立体構造も異なると考えられる。このようなタンパク質を標的とすることで、本原虫に特異的に作用しヒトには副作用がないマラリア治療薬の開発が可能になるものと考えられる。
【発明の概要】
【発明が解決しようとする課題】
【0004】
本発明者らは、熱帯熱マラリア治療薬の標的として、ヒトに対応遺伝子が存在しない熱帯熱マラリア原虫のDNAジャイレースに注目した。DNAジャイレースは、DNA複製に関与するDNAトポイソメラーゼII型の酵素であり、ジャイレースAおよびBの2種類のサブユニットからなっている。ジャイレースBにはいくつかのドメインが存在するが、そのうちジャイレースBドメインは、DNA複製時の二本鎖DNAの切断および再結合に関与すると考えられている。ジャイレースBの本ドメインに相当する部分ポリペプチドは、本ドメインを標的とするDNAジャイレースの阻害剤の探索に有用である。よって本発明は、熱帯熱マラリア原虫のジャイレースBドメインポリペプチドを提供することを目的とする。
【課題を解決するための手段】
【0005】
本発明は、配列番号2のアミノ酸配列からなるポリペプチド、前記ポリペプチドをコードするポリヌクレオチド、および前記ポリペプチドに対する抗体を提供する。
【発明の効果】
【0006】
本発明のポリペプチドは、ジャイレースBドメインの結晶化および構造解析に用いることができ、本ドメインに対する阻害剤の探索に有用である。
【図面の簡単な説明】
【0007】
【図1】ポリペプチドの大量発現時における発現量の培養温度依存性を示す。
【図2】Ni2+ アフィニティーカラムクロマトグラフィーの溶出結果を示す。
【図3】陰イオン交換カラムクロマトグラフィーの溶出結果を示す。
【図4】ゲル濾過カラムクロマトグラフィーの溶出結果を示す。
【図5】動的光散乱測定の結果を示す。
【図6】ウエスタンブロッティングの結果を示す。
【図7】熱帯熱マラリア原虫の蛍光抗体を用いた免疫染色の結果を示す。
【図8】抗ジャイレースBドメインモノクロナール抗体を処置して原虫を培養した時の赤血球(RBC)に感染したマラリア原虫の数(No. of protozoa)を示す。 なお、図中のGはgametocytes(生殖母体)、Dはdots forms(ドット状原虫、死亡虫体と考えられる)、Sはschizonts(分裂体)、Tはtrophozoites(栄養型原虫)、Rはring forms(輪状体)を示す。
【発明を実施するための形態】
【0008】
本発明は、配列番号2のアミノ酸配列からなるポリペプチドに関する。配列番号2のアミノ酸配列は、熱帯熱マラリア原虫DNAジャイレースのジャイレースBドメインに相当する。ジャイレースBの塩基配列およびアミノ酸配列は、DNAデータバンク(XM_001350753、GI:124806667)および論文(Gardner et al., Nature 419(6906), 498-511(2002))に開示されている(配列番号3および4)。配列番号2のアミノ酸配列は、配列番号4のアミノ酸配列の465-642位に相当する。
【0009】
本発明はまた、配列番号2のアミノ酸配列からなるポリペプチドの変異体に関し、具体的には、
(1)配列番号2のアミノ酸配列において1または数個のアミノ酸が欠失、置換または付加されたアミノ酸配列からなるポリペプチド、または、
(2)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列からなるポリペプチド
であって、配列番号2のアミノ酸配列からなるポリペプチドと同じ抗原性を有するポリペプチドに関する。
【0010】
アミノ酸の欠失、置換、または付加の数は、ポリペプチドの機能が維持されるかぎり限定されないが、好ましくは1〜20個、より好ましくは1〜10個、より一層好ましくは1〜5個、例えば1、2、3、4または5個である。かかる変異は、天然に存在する変異であってもよく、部位特異的突然変異誘発など公知の方法により人為的に導入したものであってもよい。アミノ酸配列の相同性は、Vector NTI、BLASTなど当業者に周知の配列比較プログラムにより決定することができ、90%以上、より好ましくは95%以上である。
アミノ酸の置換は、以下の(1)〜(6)のそれぞれの群内での置換である保存的アミノ酸置換であってもよい:(1)グリシン、アラニン、バリン、ロイシン、およびイソロイシン、(2)フェニルアラニン、チロシン、およびトリプトファン、(3)セリンおよびトレオニン、(4)アスパラギン酸およびグルタミン酸、(5)グルタミンおよびアスパラギン、および(6)リジン、アルギニン、およびヒスチジン。
【0011】
「配列番号2のアミノ酸配列からなるポリペプチドと同じ抗原性を有する」とは、配列番号2のアミノ酸配列からなるポリペプチドを特異的に認識する抗体により認識されることを意味する。かかる抗体は、配列番号2のアミノ酸配列からなるポリペプチドを免疫原として動物を免疫することにより得ることができる。抗体の作成方法は当業界にて周知である(Antibodies:A Laboratory Manual,Cold Spring Harber Laboratory Press,1988)。
【0012】
本発明はまた、タグが付加されたジャイレースBドメインポリペプチドに関する。本発明において「タグ」とは、ポリペプチドの精製、検出等のためポリペプチドに付加される部分を意味し、ヒスチジン(His)、グルタチオン-S-トランスフェラーゼ(GST)、マルトース結合タンパク質(MBP)、myc、FLAGタグなどが例示される。タグが付加された本発明のポリペプチドは、例えば、pET30a(Novagen社製)(Hisタグ用)、pGEX(GEヘルスケアバイオサイエンス株式会社製)(GSTタグ用)などの発現ベクターにジャイレースBドメインポリペプチドをコードするポリヌクレオチドを挿入し、適当な宿主細胞で発現させることで得られる。
【0013】
本発明はまた、本発明のポリペプチドをコードするポリヌクレオチドに関する。
【0014】
ある態様において、本発明のポリヌクレオチドは、配列番号2のアミノ酸配列からなるポリペプチドをコードし、具体的には、
(1)配列番号1の塩基配列、または
(2)配列番号1の塩基配列と縮重の関係にある塩基配列
を有するポリヌクレオチドである。「縮重の関係にある塩基配列」とは、もとの塩基配列中の1以上のコドンが同じアミノ酸をコードする別のコドンと置換されている塩基配列を意味する。
【0015】
本発明はまた、
(1)配列番号1の塩基配列からなるポリヌクレオチドにストリンジェントな条件下でハイブリダイズするポリヌクレオチド、または
(2)配列番号1の塩基配列と90%以上の相同性を有する塩基配列からなるポリヌクレオチド、
であって、配列番号2のアミノ酸配列からなるポリペプチドと同じ抗原性を有するポリペプチドをコードするポリヌクレオチド、に関する。
【0016】
「ストリンジェントな条件」とは、ポリヌクレオチド間の非特異的なハイブリダイセーションがおこらない条件を意味し、Molecular Cloning: A Laboratory Manual, 3rd edition (2001)などに基づき当業者が適宜決定しうるものであるが、例えば0.2xSSC、0.1%SDS、65℃などが挙げられる。
【0017】
塩基配列の相同性は、90%以上、好ましくは95%以上であり、Vector NTI、BLASTなど当業者に周知の配列比較プログラムにより決定することができる。
【0018】
本発明のポリペプチドおよびポリヌクレオチドは、Molecular Cloning: A Laboratory Manual, 3rd edition (2001)などに記載の公知の方法にて作製することができる。
【0019】
本発明はまた、本発明のポリヌクレオチドを含むベクターに関する。ある態様において、本発明のベクターは、適当な宿主細胞における本発明のポリペプチドの発現を可能とするものである。ベクターの種類は当業者が目的に応じて適宜決定しうるが、例えば、サブクローニング用ベクターとしてpBluescript(Takara-bio社製)、pGEM-T(Promega社製)など、発現用ベクターとしてpET30a(Novagen社製)、pGEX(GEヘルスケアバイオサイエンス株式会社製)などが挙げられる。
【0020】
本発明はまた、本発明のポリペプチドを特異的に認識する抗体、またはその抗原結合断片に関する。抗体には、ポリクローナル抗体およびモノクローナル抗体が含まれ、またキメラ抗体、ヒト化抗体、ヒト抗体も含まれる。抗原結合断片には、F(ab’)2断片、Fv断片、および一本鎖抗体が含まれる。本発明の抗体およびその抗原結合断片は、当業界にて周知の方法により調製可能である(例えば、Antibodies: A Laboratory Manual, Cold Spring Harber Laboratory Press, 1988、http://www.gene.mie-u.ac.jp/Protocol/Original/Antibody.html、米国特許第6331415号、米国特許第5693761号、米国特許第5225539号、米国特許第5981175号、米国特許第5612205号、米国特許第5814318号、米国特許第5545806号、米国特許第7145056号、米国特許第6492160号、米国特許第5871907号、米国特許第5733743号などを参照)。本発明のポリペプチドを特異的に認識する抗体は、本発明のポリペプチドおよび熱帯熱マラリア原虫由来タンパク質に特異的に結合する。特異的に結合するとは、もし当該抗体が、106M-1かそれより大きい、好ましくは107M-1かそれより大きい、さらに好ましくは108M-1かそれより大きい、および最も好ましくは109M-1かそれより大きい結合親和性(Ka)で本発明のポリペプチドまたはエピトープに結合すれば、特異的に結合するとみなされる。抗体の結合親和性は、たとえばスキャッチャード分析(Scatchard、Ann. NY Acad. Sci. 51: 660、1949)により、当業者によって容易に測定されることができる。
本発明のポリペプチドを特異的に認識する抗体は、本発明のポリペプチドを抗原として作成することができる。また、本発明のポリペプチドを特異的に認識する抗体は、配列番号2のアミノ酸配列からなるポリペプチドまたはその部分からなるエピトープペプチドを用いて作成してもよい。エピトープペプチドは、配列番号2に含まれている、少なくとも5個、8〜10個または15から30個までの間のアミノ酸配列からなる。
【0021】
本発明は、また、一態様として、本発明のポリペプチドを特異的に認識する抗体に、特異的に結合する抗イディオタイプ抗体を提供する。抗イディオタイプ抗体は、当業界にて周知の方法により調製可能である(例えば、米国特許第5,208,146号、米国特許第5,637,677号、およびVarthakavi & Minocha、J. Gen. Virol. 77 : 1875、1996参照)。
【0022】
本発明は、マウス細胞由来の抗体を提供するものであるが、さらに、各種動物由来抗体とのキメラ抗体、ヒト型化抗体などを提供する。
【0023】
本発明は、また、一態様として、本発明のポリペプチドを特異的に認識する抗体または抗原結合断片を含むマラリア原虫増殖抑制用組成物を提供する。
マラリア原虫増殖抑制は、輪状体の形成、栄養型原虫の形成、分裂体の形成の抑制を含んでもよい。よって、上記本発明のポリペプチドを特異的に認識する抗体または抗原結合断片を含むマラリア原虫増殖抑制用組成物は、マラリア原虫の増殖および/または分裂抑制用組成物であり得、または、マラリア原虫の輪状体の形成、栄養型原虫の形成および/または分裂体の形成の抑制用組成物であり得る。
上記組成物は、医薬品であり得るので、医薬品として許容できる担体(添加剤も含む)と共に製剤化することができる。医薬品として許容できる担体としては、例えば、賦形剤(例えば、デキストリン、ヒドロキシプロピルセルロース、ポリビニルピロリドン等)、崩壊剤(例えば、カルボキシメチルセルロース等)、滑沢剤(例えば、ステアリン酸マグネシウム等)、界面活性剤(例えば、ラウリル硫酸ナトリウム等)、溶剤(例えば、水、食塩水、大豆油等)、保存剤(例えば、p−ヒドロキシ安息香酸エステル等)などが挙げられるが、これらに限定されるものではない。
上記組成物の摂取または投与方法は、投与対象の年齢、体重、健康状態によって、当業者により適宜選択され得る。例えば、上記組成物は静脈内投与され得る。
上記組成物の投与対象は、マラリア原虫の増殖抑制を必要とする動物であれば特に限定されないが、例えば、ヒト、イヌ、ネコ、ニワトリ等が含まれる。よって、本発明は、上記組成物をヒトに投与するための医薬品の提供に加えて、イヌ、ネコ、ニワトリ等の動物の動物薬の提供も包含する。
【0024】
上記組成物に含まれる本発明のポリペプチドを特異的に認識する抗体または抗原結合断片の量は、本発明の効果を奏する限り、特に限定されるものではない。例えば、ヒト体重1kg当たり0.03〜300 mg、1〜100 mgまたは1〜10 mgを1回の投与量とし点滴静注とすることができる。上記組成物の投与方法および投与頻度も当業者が適宜設定できる。例えば、マウス由来細胞が産生したマウス型抗体をヒト化キメラ抗体とすることで、ヒトに何回でも投与することが可能になる。
製剤の用量単位(1製剤当たりの本発明のポリペプチドを特異的に認識する抗体の用量)は、当業者が適宜設定できる。
【0025】
本発明は、また、一態様として、本発明のポリペプチドを特異的に認識する抗体または抗原結合断片を含む、マラリア原虫検出用組成物を提供する。
本発明のマラリア原虫検出用組成物を用いて、動物(例えば、ヒト)由来の生物試料(例えば、血液)から、マラリア原虫を検出することができる。
マラリア原虫の検出には、本発明のポリペプチドを特異的に認識する抗体を用いた酵素抗体法(ELISA)、イムノブロッティング法、組織染色等を利用することができる。検出に際し必要な試薬(例えば、発色基質)を、本発明のマラリア原虫検出用組成物と別個に包装し、マラリア原虫検出用検出キットとして提供してもよい。本発明のマラリア原虫検出用組成物に含まれる本発明のポリペプチドを特異的に認識する抗体の量は、対象となる生物試料、測定条件等により適宜設定できるが、例えば、0.01〜100 μg/ml、0.1〜10 μg/ml、または1〜10 μg/mlであり得る。
【0026】
本発明は、また、一態様として、本発明のポリペプチドを特異的に認識する抗体または抗原結合断片を生物試料と接触させることを含む、生物試料においてマラリア原虫を検出する方法を提供する。
【0027】
以下、実施例により本発明をさらに説明するが、本発明はこれらに限定されるものではない。
【実施例】
【0028】
1)熱帯熱マラリア原虫の培養及びDNAの抽出
熱帯熱マラリア原虫(FVO株、FUP株(以上、大阪大学より分与)、SO株、NN株(以上、発明者が患者より分離))を、改良型RPMI1640培地(25 mM HEPES、23.8 mM 炭酸水素ナトリウム、2.1 mM グルタミン、0.267 mM ヒポキサンチンおよび10 % 不活化ウマ血清をそれぞれ添加したRPMI1640培地)で、5 % ヒト赤血球(O型)を用いて培養した。この原虫から、DNA抽出キット(QIAamp DNA Blood Mini Kit、Qiagen社製)を使用してDNAを抽出した。抽出したDNAをエタノール沈殿によって10倍に濃縮し、PCRに使用した。
【0029】
2)PCRプライマーの設計
DNAデータバンクおよび参考論文に開示される熱帯熱マラリア原虫のジャイレースBのDNA配列に基づき、以下の3種類のプライマーを設計した。
1:GyrB-pGEXBamHIFw
(38bp, 5’-atggattcggactaaatgaatatataacaaatataaca-3’ (配列番号5))
GyrB-pGEXEcRIRv
(41bp, 5’-gcgaattcttattttgatcttattaaatccctagctgcctt-3’ (配列番号6))
2:GyrB-ET30aNhisNdeIFw
(45bp, 5’-cgcatatgcatcatcatcatcatcatggactaaatgaatatataa-3’ (配列番号7))
GyrB-pET30aEcoRIRv
(41bp, 5’-atgaattcttattttgatcttattaaatccctagctgcctt-3’ (配列番号8))
3:GyrB-pET30aChisNdeIFw
(38bp, 5’-cgcatatgggactaaatgaatatataacaaatataaca-3’ (配列番号9))
GyrB-pET30aChisEcoRIRv
(45bp, 5’-atgaattcttaatgatgatgatgatgatgttttgatcttattaaa-3’ (配列番号10))
【0030】
3)PCRによるDNAの増幅および精製
TaKaRa ExTaq(Takara-bio社製)を用いて、以下の条件でPCRを行った。PCR産物を1.2%アガロース電気泳動で泳動し、約500 bpのバンドを得た。このバンドを切り出し、MiniElute Gel Extraction Kit (Qiagen社製)にて精製した。
[PCR条件]
プライマー:前述(配列番号5〜10)
反応条件:
10x ExTaq バッファー 5 μl
2.5 mM dNTP ミックス 2 μl
10 μM フォワード(FW)プライマー 2.5 μl
10 μM リバース(Rv)プライマー 2.5 μl
TaKaRa ExTaq 1 μl
DNA 1 μl
dH2O 36 μl
温度条件:
94℃ 3 分
94℃ 0.5 分
55℃ 1 分
72℃ 1 分
(94℃→55℃→72℃ 35サイクル)
72℃ 7 分
4℃ ∞
【0031】
4)TAクローニング
上記のDNA増幅産物をTAベクターであるpBluescript(Takara-bio社製)またはpGEM-T (Promega社製)にライゲーションし、クローニングを行った。出現した白色コロニーの内、それぞれ10個ずつ(合計30個)についてコロニーPCRを行い、DNA増幅産物を確認した4個のコロニーを増菌した(A1-3、A3-1、A3-3、A3-14)。増菌した大腸菌からプラスミドをQIAprep Spin Miniprep Kit(Qiagen社製)にて精製するとともに、インサートの確認のためPCRを行い、このPCR増幅産物を精製してシークエンスしたところ、A3-3、A3-14は点変異が見られた。よって、次の実験にはA1-3、A3-1のプラスミドを使用した。
[コロニーPCR条件]
プライマー:
pBluescript FW
5’-gtaatacgactcactatagggc-3’(配列番号11)(T7 フォワードプライマー)
pBluescript Rv
5’-ggaaacagctatgaccatg-3’ (配列番号12)(M13 リバースプライマー)
反応条件:
10x ExTaq バッファー 1.5μl
2.5 mM dNTP ミックス 1.5 μl
DMSO 0.75 μl
10 μM FW プライマー 1.5 μl
10 μM Rv プライマー 1.5 μl
TaKaRa ExTaq 0.15 μl
コロニー菌体 少々
dH2O 8.1 μl
温度条件:
94℃ 1 分
94℃ 0.5 分
55℃ 0.5 分
72℃ 1.5 分
(94℃→55℃→72℃ 35サイクル)
72℃ 2 分
4℃ ∞
[インサート確認用PCR条件]
プライマー:前述(配列番号5〜10)
反応条件:
10x ExTaq バッファー 5 μl
2.5 mM dNTP ミックス 2 μl
10 μM FW プライマー 2.5 μl
10 μM Rv プライマー 2.5 μl
TaKaRa ExTaq 1 μl
DNA 1 μl
dH2O 36 μl
温度条件:
94℃ 3分
94℃ 0.5 分
55℃ 1 分
72℃ 1 分
(94℃→55℃→72℃ 35サイクル)
72℃ 7 分
4℃ ∞
【0032】
5)発現ベクターの構築
候補のプラスミド(A1-3、A3-1)を制限酵素(A1-3用:BamHI(Toyobo社製)およびEcoRI(Toyobo社製)、あるいはA3-1用:NdeI(Takara-bio社製)およびEcoRI(Toyobo社製))で切断し、同時に同じ制限酵素で処理したpGEX6p-2ベクター(A1-3用、GEヘルスケアバイオサイエンス株式会社製)あるいはpET30aベクター(A3-1用、Novagen社製)にライゲーションした。ライゲーション後、DNA断片を挿入したプラスミドで大腸菌をトランスフォームしサブクローニングを行った。出現したコロニーについてコロニーPCR(前述)を行ったところ、A3-1由来DNA断片をライゲーションしたpET30aベクター(Gyrase B-NHis /pET-30a)で14個のコロニーのうち1個に目的のDNA配列が含まれることを確認した(A3-1-24)。この大腸菌を増菌後、プラスミドをQIAprep Spin Miniprep Kit(Qiagen社製)で精製し保存した。再度、インサート確認のためのPCR(前述)を行い、PCR増幅産物を精製し、シークエンスして、目的遺伝子と同一の配列であることを確認した(配列番号1)。
【0033】
6)ポリペプチドの発現および精製
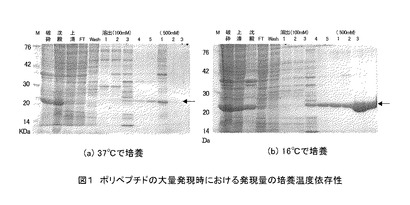
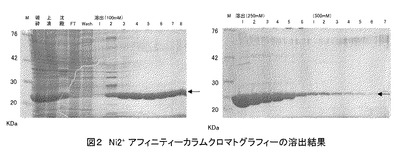
上記5)で得られた発現ベクターで大腸菌(BL21(DE3)、Takara-bio社製)をトランスフォームし、培養した。培養液を遠心分離して集菌し、沈査(菌体)を超音波で粉砕した後、さらに遠心分離し、上清をNi2+ アフィニティカラムクロマトグラフィー(HiTrap Chelating HP、GEヘルスケアバイオサイエンス社製)にかけた。培養温度条件として37℃および16℃、IPTG処理条件として0.3 mM、1 mM、および3 mMを検討したところ、温度16℃、IPTG 1 mMで36時間培養すると、効率よくポリペプチドが発現することがわかり、ポリペプチドを大量に作製することに成功した(図1)。また、Ni2+アフィニティカラムクロマトグラフィーでの精製条件を検討したところ、イミダゾール100〜250 mMで溶出されることが分かった(図2)。
【0034】
[培養条件]
発現ベクター(Gyrase B-NHis /pET-30a)を大腸菌(BL21(DE3)、Takara-bio社製)にトランスフオームして作製したグリセロールストックをLB-Kan(カナマイシン30 μg/ml)寒天プレートに植菌。
↓
37℃で一晩培養。
↓
コロニーをピックアップして、LB-Kan培地(カナマイシン30 μg/ml、5 ml)に植菌し、37℃で一晩培養。
↓
500 ml のTB-Kan培地(カナマイシン50 μg/ml)に前培養分5 mlを全量添加。
↓
37℃で培養(約3時間)。
↓
OD600〜0.8でシェイキングを中断し、16℃(または37℃)に設定後、30分間静置。
↓
IPTGを最終濃度で0.3 mM、1 mM、または3 mMとなるように添加。
↓
16℃(または37℃)でさらに36時間培養。
↓
培養を終え、集菌。
↓
菌体を-80℃に保存。
【0035】
[Ni2+ アフィニティカラムクロマトグラフィーによるポリペプチドの精製方法]
HiTrap Chelating HP(GEヘルスケアバイオサイエンス製、5 ml容量)を平衡化バッファーで平衡化(ペリスタポンプおよびフラクションコレクター(GEヘルスケアバイオサイエンス製)使用、流速 1 ml/分)。
平衡化バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、10 mM イミダゾール
↓
上清をカラムに通液し、フロースルーを回収(流速 1 ml/分)(FT画分とする)。
↓
平衡化バッファーで洗浄(25 ml、流速 1 ml/分)(Wash画分とする)。
↓
100 mM イミダゾールを含む溶出バッファーで溶出(3 ml X 10本、流速 1 ml/分)。
溶出バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、100 mM イミダゾール
↓
250 mM イミダゾールを含む溶出バッファ−で溶出(3 ml X 10本、流速 1 ml/分)。
溶出バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、250 mM イミダゾール
↓
500 mM イミダゾールを含む溶出バッファ−で溶出(3 ml X 10本、流速 1 ml/分)。
溶出バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、500 mM イミダゾール
↓
SDS-PAGE 後、溶出画分を回収。
↓
脱塩バッファーで透析(2Lのバッファーで、4 ℃、一晩)。
脱塩バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、1 mM DTT
【0036】
図1の各レーンのサンプルは以下のとおりである。
図1 ポリペプチドの大量発現時における発現量の培養温度依存性
(a)37℃での培養後の菌体を用いたポリアクリルアミド電気泳動像
レーンの番号
1: M (マーカー)
2: 破砕(超音波による菌体の破砕粗画分)
3: 沈殿(遠心後沈殿画分)
4: 上清(遠心後上清画分)
5: FT(Ni2+アフィニティカラムクロマトグラフィーに吸着しなかった画分)
6: Wash(Niカラムの洗浄画分)
7: 溶出(100 mM) 1(Niカラムの100 mMイミダゾール添加液での溶出画分1本目)
8: 溶出(100 mM) 2(Niカラムの100 mMイミダゾール添加液での溶出画分2本目)
9: 溶出(100 mM) 3(Niカラムの100 mMイミダゾール添加液での溶出画分3本目)
10: 溶出(100 mM) 4(Niカラムの100 mMイミダゾール添加液での溶出画分4本目)
11: 溶出(100 mM) 5(Niカラムの100 mMイミダゾール添加液での溶出画分5本目)
12: 溶出(500 mM) 1(Niカラムの500 mMイミダゾール添加液での溶出画分1本目)
13: 溶出(500 mM) 2(Niカラムの500 mMイミダゾール添加液での溶出画分2本目)
14: 溶出(500 mM) 3(Niカラムの500 mMイミダゾール添加液での溶出画分3本目)
(b)16℃での培養後の菌体を用いたポリアクリルアミド電気泳動像
レーンの番号
1: M (マーカー)
2: 破砕(超音波による菌体の破砕粗画分)
3: 沈殿(遠心後沈殿画分)
4: 上清(遠心後上清画分)
5: FT(Ni2+アフィニティカラムクロマトグラフィーに吸着しなかった画分)
6: Wash(Niカラムの洗浄画分)
7: 溶出(100 mM) 1(Niカラムの100 mMイミダゾール添加液での溶出画分1本目)
8: 溶出(100 mM) 2(Niカラムの100 mMイミダゾール添加液での溶出画分2本目)
9: 溶出(100 mM) 3(Niカラムの100 mMイミダゾール添加液での溶出画分3本目)
10: 溶出(100 mM) 4(Niカラムの100 mMイミダゾール添加液での溶出画分4本目)
11: 溶出(100 mM) 5(Niカラムの100 mMイミダゾール添加液での溶出画分5本目)
12: 溶出(500 mM) 1(Niカラムの500 mMイミダゾール添加液での溶出画分1本目)
13: 溶出(500 mM) 2(Niカラムの500 mMイミダゾール添加液での溶出画分2本目)
14: 溶出(500 mM) 3(Niカラムの500 mMイミダゾール添加液での溶出画分3本目)
【0037】
図2の各レーンのサンプルは以下のとおりである。
図2 Ni2+アフィニティカラムクロマトグラフィーの溶出結果
(左図)
レーンの番号
1: M (マーカー)
2: 破砕(超音波による菌体の破砕粗画分)
3: 上清(遠心後上清画分)
4: 沈殿(遠心後沈殿画分)
5: FT(Ni2+アフィニティカラムクロマトグラフィーに吸着しなかった画分)
6: Wash(Niカラムの洗浄画分)
7: 溶出(100 mM) 1(Niカラムの100 mMイミダゾール添加液での溶出画分1本目)
8: 溶出(100 mM) 2(Niカラムの100 mMイミダゾール添加液での溶出画分2本目)
9: 溶出(100 mM) 3(Niカラムの100 mMイミダゾール添加液での溶出画分3本目)
10: 溶出(100 mM) 4(Niカラムの100 mMイミダゾール添加液での溶出画分4本目)
11: 溶出(100 mM) 5(Niカラムの100 mMイミダゾール添加液での溶出画分5本目)
12: 溶出(100 mM) 6(Niカラムの100 mMイミダゾール添加液での溶出画分6本目)
13: 溶出(100 mM) 7(Niカラムの100 mMイミダゾール添加液での溶出画分7本目)
14: 溶出(100 mM) 8(Niカラムの100 mMイミダゾール添加液での溶出画分8本目)
(右図)
レーンの番号
1: M (マーカー)
2: 溶出(250 mM) 1(Niカラムの250 mMイミダゾール添加液での溶出画分1本目)
3: 溶出(250 mM) 2(Niカラムの250 mMイミダゾール添加液での溶出画分2本目)
4: 溶出(250 mM) 3(Niカラムの250 mMイミダゾール添加液での溶出画分3本目)
5: 溶出(250 mM) 4(Niカラムの250 mMイミダゾール添加液での溶出画分4本目)
6: 溶出(250 mM) 5(Niカラムの250 mMイミダゾール添加液での溶出画分5本目)
7: 溶出(250 mM) 6(Niカラムの250 mMイミダゾール添加液での溶出画分6本目)
8: 溶出(500 mM) 1(Niカラムの500 mMイミダゾール添加液での溶出画分1本目)
9: 溶出(600 mM) 2(Niカラムの500 mMイミダゾール添加液での溶出画分2本目)
10: 溶出(500 mM) 3(Niカラムの500 mMイミダゾール添加液での溶出画分3本目)
11: 溶出(500 mM) 4(Niカラムの500 mMイミダゾール添加液での溶出画分4本目)
12: 溶出(500 mM) 5(Niカラムの500 mMイミダゾール添加液での溶出画分5本目)
13: 溶出(500 mM) 6(Niカラムの500 mMイミダゾール添加液での溶出画分6本目)
14: 溶出(500 mM) 7(Niカラムの500 mMイミダゾール添加液での溶出画分7本目)
【0038】
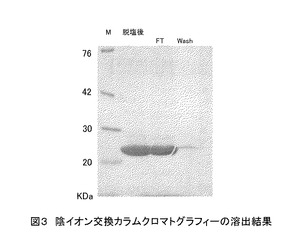
また、陰イオンカラムクロマトグラフィー(GEヘルスケアバイオサイエンス製、5 ml容量)での溶出条件の検討をしたところ、目的のポリペプチドがフロースルー (FT)画分に溶出され、不純物を除去できることがわかった(図3)。
[陰イオン交換カラムクロマトグラフィーによるポリペプチドの精製方法]
HiTrapQ FFカラム(GEヘルスケアバイオサイエンス製、5 ml容量)を平衡化バッファーで平衡化(ペリスタポンプおよびフラクションコレクター(GEヘルスケアバイオサイエンス製)使用、流速 1 ml/分)。
平衡化バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、5 mM DTT
↓
上清をカラムに通液;フロースルーを回収(流速 1 ml/分)(FT画分とする)。
↓
平衡化バッファーで洗浄(25 ml、流速 1 ml/分)(Wash画分とする)。
↓
溶出バッファーで溶出(流速 1 ml/分)。
溶出バッファーの組成:20 mM Tris-HCl pH8.0、1000 mM NaCl、5 mM DTT
↓
SDS-PAGE後、FTおよびWash画分を回収。
↓
YM-10(Amicon製)で約2 mlまで濃縮後、フイルタリング(PALL製 NANOCEP 0.2/m)。
【0039】
図3の各レーンのサンプルは以下のとおりである。
図3 陰イオン交換カラムクロマトグラフィーの溶出結果
レーンの番号
1: M (マーカー)
2: 脱塩後(超音波による菌体の破砕粗画分)
3: FT(陰イオン交換カラムクロマトグラフィーに吸着しなかった画分)
4: Wash(陰イオン交換カラムクロマトグラフィーの洗浄画分)
【0040】
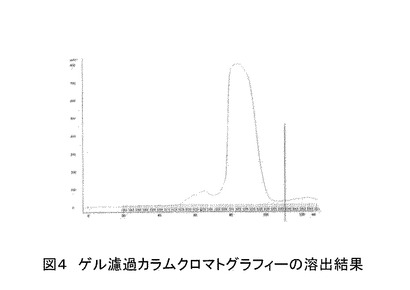
さらに、ゲル濾過カラムクロマトグラフィー(GEヘルスケアバイオサイエンス製、120 ml カラム容量)を用いることで、より純度の高いポリペプチドを得ることが出来た(図4)。[ゲルろ過カラムクロマトグラフィーによるポリペプチドの精製方法]
HiLoad 16/60 Superdex200カラム(GEヘルスケアバイオサイエンス製、120 mlカラム容量)をゲルろ過バッファーで平衡化。
ゲルろ過バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、5 mM DTT
↓
菌体破砕し、遠心分離後の上清画分をゲルろ過(流速 0.2 ml/分、1 ml/画分)。
↓
ゲルろ過終了後、SDS-PAGEで各画分を確認し、目的の画分を回収。
【0041】
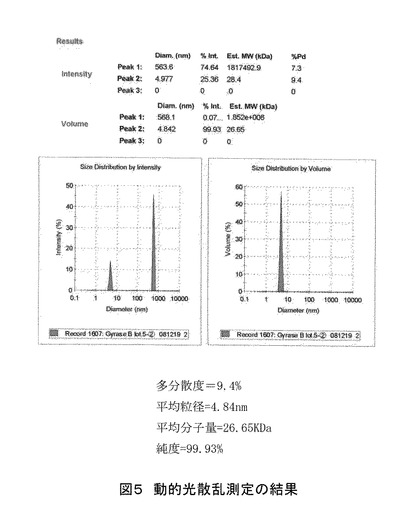
ゲルろ過後の精製サンプルについて、動的光散乱(DLS)の測定を行った(図5)。測定の結果は、多分散度9.4%、純度99.93%であり、高純度のポリペプチドが得られた。計算された平均分子量は26.65KDa であり、理論分子量とほぼ一致していた。精製後のポリペプチドの収量は、大腸菌の培養培地1リットルあたり約40 mgであった。
[動的光散乱(DLS)測定の実験条件]
動的光散乱装置:MALVERN社製 Zeta Sizer Nano-S
測定温度:4℃
測定に使用したポリペプチド濃度:2 mg/ml
【0042】
7)モノクローナル抗体の作成
a)免疫
ジャイレースBドメインを抗原としてIncomplete Freund’s adjuvantとともにマウスに投与した。抗原の1回の投与量は、マウスの場合1匹当たり50〜100 μgであった。4回目の免疫後、脾臓細胞を採集した。
【0043】
b)細胞融合
脾臓細胞とマウス由来のミエローマ細胞との細胞融合を、血清を含まない動物細胞培養用培地中で脾臓細胞とミエローマ細胞とを混合し、ポリエチレングリコールの存在下で行った。
【0044】
c)ハイブリドーマの選別
融合細胞をヒポキサンチン(100μM)、アミノプテリン(0.4μM)及びチミジン(16μM)を含む培地を用いて培養し、生育する細胞をハイブリドーマとして得た。
次に、増殖したハイブリドーマの培養上清中に、目的とする抗体が存在するか否かについて抗原を用いてELISAでスクリーニングした。
【0045】
d)クローニング
融合細胞のクローニングは、限界希釈法等により行い、最終的にモノクローナル抗体産生細胞であるハイブリドーマを樹立した。モノクローナル抗体産生細胞を培養し、その培養上清から抗体を取得した。
【0046】
e)精製
プロテインA(またはプロテインG)セファロースクロマトグラフィーを用いて抗体を精製できる。
【0047】
f)ジャイレースBドメインのウエスタンブロッティングによる検出
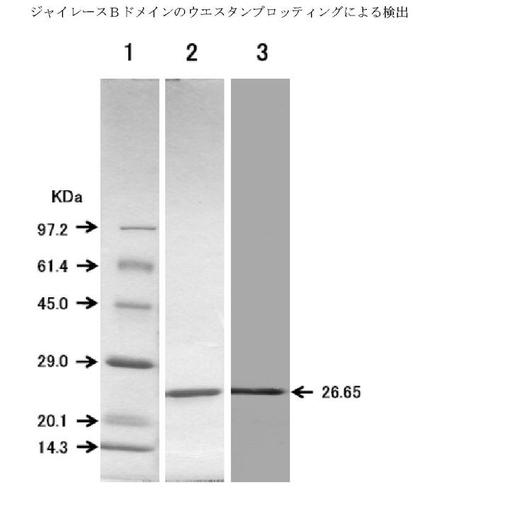
ジャイレースBドメインのポリアクリルアミドゲル電気泳動を行った。ゲルを切り取り、ニトロセルロース膜に転写し、上記で得られたモノクローナル抗体(02C12など)を用いウエスタンブロッティングを行った。その結果、モノクローナル抗体(02C12など)がジャイレースBドメインに結合し、ジャイレースBドメインを検出することが確認された(図6)。
Lane 1: Marker
Lane 2: ジャイレースBドメイント精製物(5 μg)のCBB染色像
Lane 3: ジャイレースBドメイント精製物(5 μg)のモノクローナル抗体による免疫染色像
1次抗体:抗ジャイレースBドメインモノクローナル抗体(02C12)
2次抗体:HRP標識抗マウスIgG(H+L)抗体
発色液:ジアミノベンジジン(DBB)
【0048】
g)熱帯熱マラリア原虫の蛍光抗体を用いた免疫染色
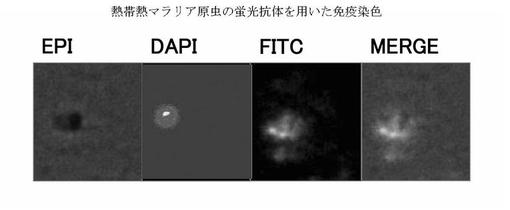
熱帯熱マラリア原虫を、上記で得られたモノクローナル抗体(02C12など)を1次抗体、FITC標識抗マウスIgG(H+L)抗体を2次抗体として用いて免疫染色した。その結果、モノクローナル抗体(02C12など)はマラリア原虫の栄養型原虫を染色し、マラリア原虫内(EPI)で、モノクローナル抗体(02C12など)によるジャイレースBドメインの染色像(FITC)とDNAの染色像(DAPI)が重なった(MERGE)。これは、ジャイレースBドメインがDNA複製時の二本鎖DNAレベルの切断および再結合に関与するという知見と一致する結果である(図7)。
EPI:透過光
DAPI:原虫の核のDNAのDAPIによる染色
FITC:抗ジャイレースBドメインモノクローナル抗体との反応
熱帯熱マラリア原虫栄養型原虫
+抗ジャイレースBドメインモノクローナル抗体(02C12)(1次抗体)
+FITC標識抗マウスIgG(H+L)抗体(2次抗体)
MERGE:EPI+DAPI+FITCの重ね合わせ
【0049】
8)モノクローナル抗体による熱帯熱マラリア原虫の増殖抑制
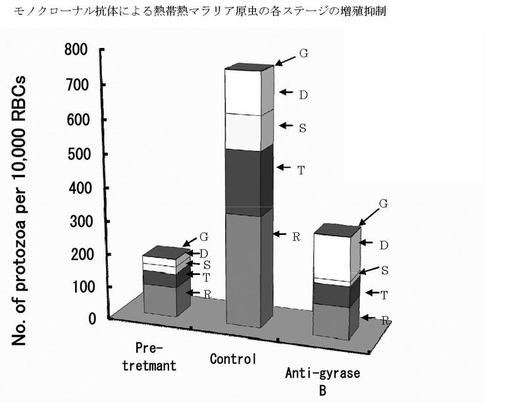
熱帯熱マラリア原虫を培養し、モノクローナル抗体(02C12など)が感染赤血球率に及ぼす影響を検討した。モノクローナル抗体(02C12など)産生細胞の培養上清(02C12など)を原虫培地の1/10量添加して2日間培養した。その結果、モノクローナル抗体(02C12など) 産生細胞の培養上清により熱帯熱マラリア原虫の感染赤血球率が減少し、原虫の増殖および分裂が抑制された(表1および図8)。すなわち、未処置群(control)では、2日間の培養で原虫が増殖し分裂し、輪状体(Ring forms, R)、栄養型(Trophozoites, T)及び分裂体(Schizonts, S)が増加したのに対して、モノクローナル抗体を含む培養上清を用いた処置群(anti-gyraseB)では原虫数の増加が認められなかった。
【表1】

【技術分野】
【0001】
本発明は、熱帯熱マラリア原虫DNAジャイレースのジャイレースB由来ポリペプチド、前記ポリペプチドをコードするポリヌクレオチド、前記ポリペプチドに対する抗体、当該抗体を含むマラリア原虫増殖抑制用組成物およびマラリア原虫検出用組成物に関する。
【背景技術】
【0002】
熱帯熱マラリアはヒトを死に至らしめる原虫感染症の一つであり、年間300万人以上の人々が死亡している。近年、本原虫が既存のマラリア治療薬に対して薬剤耐性を獲得してきたことから、新規の治療薬の開発が望まれている。
【0003】
熱帯熱マラリア原虫の遺伝子の中にはヒトに対応遺伝子が存在しないもの、あるいはヒトの対応遺伝子には存在しないDNA挿入部分を有するものが存在する。かかる遺伝子がコードするタンパク質は、ヒトのタンパク質とは異なるアミノ酸配列を有し、立体構造も異なると考えられる。このようなタンパク質を標的とすることで、本原虫に特異的に作用しヒトには副作用がないマラリア治療薬の開発が可能になるものと考えられる。
【発明の概要】
【発明が解決しようとする課題】
【0004】
本発明者らは、熱帯熱マラリア治療薬の標的として、ヒトに対応遺伝子が存在しない熱帯熱マラリア原虫のDNAジャイレースに注目した。DNAジャイレースは、DNA複製に関与するDNAトポイソメラーゼII型の酵素であり、ジャイレースAおよびBの2種類のサブユニットからなっている。ジャイレースBにはいくつかのドメインが存在するが、そのうちジャイレースBドメインは、DNA複製時の二本鎖DNAの切断および再結合に関与すると考えられている。ジャイレースBの本ドメインに相当する部分ポリペプチドは、本ドメインを標的とするDNAジャイレースの阻害剤の探索に有用である。よって本発明は、熱帯熱マラリア原虫のジャイレースBドメインポリペプチドを提供することを目的とする。
【課題を解決するための手段】
【0005】
本発明は、配列番号2のアミノ酸配列からなるポリペプチド、前記ポリペプチドをコードするポリヌクレオチド、および前記ポリペプチドに対する抗体を提供する。
【発明の効果】
【0006】
本発明のポリペプチドは、ジャイレースBドメインの結晶化および構造解析に用いることができ、本ドメインに対する阻害剤の探索に有用である。
【図面の簡単な説明】
【0007】
【図1】ポリペプチドの大量発現時における発現量の培養温度依存性を示す。
【図2】Ni2+ アフィニティーカラムクロマトグラフィーの溶出結果を示す。
【図3】陰イオン交換カラムクロマトグラフィーの溶出結果を示す。
【図4】ゲル濾過カラムクロマトグラフィーの溶出結果を示す。
【図5】動的光散乱測定の結果を示す。
【図6】ウエスタンブロッティングの結果を示す。
【図7】熱帯熱マラリア原虫の蛍光抗体を用いた免疫染色の結果を示す。
【図8】抗ジャイレースBドメインモノクロナール抗体を処置して原虫を培養した時の赤血球(RBC)に感染したマラリア原虫の数(No. of protozoa)を示す。 なお、図中のGはgametocytes(生殖母体)、Dはdots forms(ドット状原虫、死亡虫体と考えられる)、Sはschizonts(分裂体)、Tはtrophozoites(栄養型原虫)、Rはring forms(輪状体)を示す。
【発明を実施するための形態】
【0008】
本発明は、配列番号2のアミノ酸配列からなるポリペプチドに関する。配列番号2のアミノ酸配列は、熱帯熱マラリア原虫DNAジャイレースのジャイレースBドメインに相当する。ジャイレースBの塩基配列およびアミノ酸配列は、DNAデータバンク(XM_001350753、GI:124806667)および論文(Gardner et al., Nature 419(6906), 498-511(2002))に開示されている(配列番号3および4)。配列番号2のアミノ酸配列は、配列番号4のアミノ酸配列の465-642位に相当する。
【0009】
本発明はまた、配列番号2のアミノ酸配列からなるポリペプチドの変異体に関し、具体的には、
(1)配列番号2のアミノ酸配列において1または数個のアミノ酸が欠失、置換または付加されたアミノ酸配列からなるポリペプチド、または、
(2)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列からなるポリペプチド
であって、配列番号2のアミノ酸配列からなるポリペプチドと同じ抗原性を有するポリペプチドに関する。
【0010】
アミノ酸の欠失、置換、または付加の数は、ポリペプチドの機能が維持されるかぎり限定されないが、好ましくは1〜20個、より好ましくは1〜10個、より一層好ましくは1〜5個、例えば1、2、3、4または5個である。かかる変異は、天然に存在する変異であってもよく、部位特異的突然変異誘発など公知の方法により人為的に導入したものであってもよい。アミノ酸配列の相同性は、Vector NTI、BLASTなど当業者に周知の配列比較プログラムにより決定することができ、90%以上、より好ましくは95%以上である。
アミノ酸の置換は、以下の(1)〜(6)のそれぞれの群内での置換である保存的アミノ酸置換であってもよい:(1)グリシン、アラニン、バリン、ロイシン、およびイソロイシン、(2)フェニルアラニン、チロシン、およびトリプトファン、(3)セリンおよびトレオニン、(4)アスパラギン酸およびグルタミン酸、(5)グルタミンおよびアスパラギン、および(6)リジン、アルギニン、およびヒスチジン。
【0011】
「配列番号2のアミノ酸配列からなるポリペプチドと同じ抗原性を有する」とは、配列番号2のアミノ酸配列からなるポリペプチドを特異的に認識する抗体により認識されることを意味する。かかる抗体は、配列番号2のアミノ酸配列からなるポリペプチドを免疫原として動物を免疫することにより得ることができる。抗体の作成方法は当業界にて周知である(Antibodies:A Laboratory Manual,Cold Spring Harber Laboratory Press,1988)。
【0012】
本発明はまた、タグが付加されたジャイレースBドメインポリペプチドに関する。本発明において「タグ」とは、ポリペプチドの精製、検出等のためポリペプチドに付加される部分を意味し、ヒスチジン(His)、グルタチオン-S-トランスフェラーゼ(GST)、マルトース結合タンパク質(MBP)、myc、FLAGタグなどが例示される。タグが付加された本発明のポリペプチドは、例えば、pET30a(Novagen社製)(Hisタグ用)、pGEX(GEヘルスケアバイオサイエンス株式会社製)(GSTタグ用)などの発現ベクターにジャイレースBドメインポリペプチドをコードするポリヌクレオチドを挿入し、適当な宿主細胞で発現させることで得られる。
【0013】
本発明はまた、本発明のポリペプチドをコードするポリヌクレオチドに関する。
【0014】
ある態様において、本発明のポリヌクレオチドは、配列番号2のアミノ酸配列からなるポリペプチドをコードし、具体的には、
(1)配列番号1の塩基配列、または
(2)配列番号1の塩基配列と縮重の関係にある塩基配列
を有するポリヌクレオチドである。「縮重の関係にある塩基配列」とは、もとの塩基配列中の1以上のコドンが同じアミノ酸をコードする別のコドンと置換されている塩基配列を意味する。
【0015】
本発明はまた、
(1)配列番号1の塩基配列からなるポリヌクレオチドにストリンジェントな条件下でハイブリダイズするポリヌクレオチド、または
(2)配列番号1の塩基配列と90%以上の相同性を有する塩基配列からなるポリヌクレオチド、
であって、配列番号2のアミノ酸配列からなるポリペプチドと同じ抗原性を有するポリペプチドをコードするポリヌクレオチド、に関する。
【0016】
「ストリンジェントな条件」とは、ポリヌクレオチド間の非特異的なハイブリダイセーションがおこらない条件を意味し、Molecular Cloning: A Laboratory Manual, 3rd edition (2001)などに基づき当業者が適宜決定しうるものであるが、例えば0.2xSSC、0.1%SDS、65℃などが挙げられる。
【0017】
塩基配列の相同性は、90%以上、好ましくは95%以上であり、Vector NTI、BLASTなど当業者に周知の配列比較プログラムにより決定することができる。
【0018】
本発明のポリペプチドおよびポリヌクレオチドは、Molecular Cloning: A Laboratory Manual, 3rd edition (2001)などに記載の公知の方法にて作製することができる。
【0019】
本発明はまた、本発明のポリヌクレオチドを含むベクターに関する。ある態様において、本発明のベクターは、適当な宿主細胞における本発明のポリペプチドの発現を可能とするものである。ベクターの種類は当業者が目的に応じて適宜決定しうるが、例えば、サブクローニング用ベクターとしてpBluescript(Takara-bio社製)、pGEM-T(Promega社製)など、発現用ベクターとしてpET30a(Novagen社製)、pGEX(GEヘルスケアバイオサイエンス株式会社製)などが挙げられる。
【0020】
本発明はまた、本発明のポリペプチドを特異的に認識する抗体、またはその抗原結合断片に関する。抗体には、ポリクローナル抗体およびモノクローナル抗体が含まれ、またキメラ抗体、ヒト化抗体、ヒト抗体も含まれる。抗原結合断片には、F(ab’)2断片、Fv断片、および一本鎖抗体が含まれる。本発明の抗体およびその抗原結合断片は、当業界にて周知の方法により調製可能である(例えば、Antibodies: A Laboratory Manual, Cold Spring Harber Laboratory Press, 1988、http://www.gene.mie-u.ac.jp/Protocol/Original/Antibody.html、米国特許第6331415号、米国特許第5693761号、米国特許第5225539号、米国特許第5981175号、米国特許第5612205号、米国特許第5814318号、米国特許第5545806号、米国特許第7145056号、米国特許第6492160号、米国特許第5871907号、米国特許第5733743号などを参照)。本発明のポリペプチドを特異的に認識する抗体は、本発明のポリペプチドおよび熱帯熱マラリア原虫由来タンパク質に特異的に結合する。特異的に結合するとは、もし当該抗体が、106M-1かそれより大きい、好ましくは107M-1かそれより大きい、さらに好ましくは108M-1かそれより大きい、および最も好ましくは109M-1かそれより大きい結合親和性(Ka)で本発明のポリペプチドまたはエピトープに結合すれば、特異的に結合するとみなされる。抗体の結合親和性は、たとえばスキャッチャード分析(Scatchard、Ann. NY Acad. Sci. 51: 660、1949)により、当業者によって容易に測定されることができる。
本発明のポリペプチドを特異的に認識する抗体は、本発明のポリペプチドを抗原として作成することができる。また、本発明のポリペプチドを特異的に認識する抗体は、配列番号2のアミノ酸配列からなるポリペプチドまたはその部分からなるエピトープペプチドを用いて作成してもよい。エピトープペプチドは、配列番号2に含まれている、少なくとも5個、8〜10個または15から30個までの間のアミノ酸配列からなる。
【0021】
本発明は、また、一態様として、本発明のポリペプチドを特異的に認識する抗体に、特異的に結合する抗イディオタイプ抗体を提供する。抗イディオタイプ抗体は、当業界にて周知の方法により調製可能である(例えば、米国特許第5,208,146号、米国特許第5,637,677号、およびVarthakavi & Minocha、J. Gen. Virol. 77 : 1875、1996参照)。
【0022】
本発明は、マウス細胞由来の抗体を提供するものであるが、さらに、各種動物由来抗体とのキメラ抗体、ヒト型化抗体などを提供する。
【0023】
本発明は、また、一態様として、本発明のポリペプチドを特異的に認識する抗体または抗原結合断片を含むマラリア原虫増殖抑制用組成物を提供する。
マラリア原虫増殖抑制は、輪状体の形成、栄養型原虫の形成、分裂体の形成の抑制を含んでもよい。よって、上記本発明のポリペプチドを特異的に認識する抗体または抗原結合断片を含むマラリア原虫増殖抑制用組成物は、マラリア原虫の増殖および/または分裂抑制用組成物であり得、または、マラリア原虫の輪状体の形成、栄養型原虫の形成および/または分裂体の形成の抑制用組成物であり得る。
上記組成物は、医薬品であり得るので、医薬品として許容できる担体(添加剤も含む)と共に製剤化することができる。医薬品として許容できる担体としては、例えば、賦形剤(例えば、デキストリン、ヒドロキシプロピルセルロース、ポリビニルピロリドン等)、崩壊剤(例えば、カルボキシメチルセルロース等)、滑沢剤(例えば、ステアリン酸マグネシウム等)、界面活性剤(例えば、ラウリル硫酸ナトリウム等)、溶剤(例えば、水、食塩水、大豆油等)、保存剤(例えば、p−ヒドロキシ安息香酸エステル等)などが挙げられるが、これらに限定されるものではない。
上記組成物の摂取または投与方法は、投与対象の年齢、体重、健康状態によって、当業者により適宜選択され得る。例えば、上記組成物は静脈内投与され得る。
上記組成物の投与対象は、マラリア原虫の増殖抑制を必要とする動物であれば特に限定されないが、例えば、ヒト、イヌ、ネコ、ニワトリ等が含まれる。よって、本発明は、上記組成物をヒトに投与するための医薬品の提供に加えて、イヌ、ネコ、ニワトリ等の動物の動物薬の提供も包含する。
【0024】
上記組成物に含まれる本発明のポリペプチドを特異的に認識する抗体または抗原結合断片の量は、本発明の効果を奏する限り、特に限定されるものではない。例えば、ヒト体重1kg当たり0.03〜300 mg、1〜100 mgまたは1〜10 mgを1回の投与量とし点滴静注とすることができる。上記組成物の投与方法および投与頻度も当業者が適宜設定できる。例えば、マウス由来細胞が産生したマウス型抗体をヒト化キメラ抗体とすることで、ヒトに何回でも投与することが可能になる。
製剤の用量単位(1製剤当たりの本発明のポリペプチドを特異的に認識する抗体の用量)は、当業者が適宜設定できる。
【0025】
本発明は、また、一態様として、本発明のポリペプチドを特異的に認識する抗体または抗原結合断片を含む、マラリア原虫検出用組成物を提供する。
本発明のマラリア原虫検出用組成物を用いて、動物(例えば、ヒト)由来の生物試料(例えば、血液)から、マラリア原虫を検出することができる。
マラリア原虫の検出には、本発明のポリペプチドを特異的に認識する抗体を用いた酵素抗体法(ELISA)、イムノブロッティング法、組織染色等を利用することができる。検出に際し必要な試薬(例えば、発色基質)を、本発明のマラリア原虫検出用組成物と別個に包装し、マラリア原虫検出用検出キットとして提供してもよい。本発明のマラリア原虫検出用組成物に含まれる本発明のポリペプチドを特異的に認識する抗体の量は、対象となる生物試料、測定条件等により適宜設定できるが、例えば、0.01〜100 μg/ml、0.1〜10 μg/ml、または1〜10 μg/mlであり得る。
【0026】
本発明は、また、一態様として、本発明のポリペプチドを特異的に認識する抗体または抗原結合断片を生物試料と接触させることを含む、生物試料においてマラリア原虫を検出する方法を提供する。
【0027】
以下、実施例により本発明をさらに説明するが、本発明はこれらに限定されるものではない。
【実施例】
【0028】
1)熱帯熱マラリア原虫の培養及びDNAの抽出
熱帯熱マラリア原虫(FVO株、FUP株(以上、大阪大学より分与)、SO株、NN株(以上、発明者が患者より分離))を、改良型RPMI1640培地(25 mM HEPES、23.8 mM 炭酸水素ナトリウム、2.1 mM グルタミン、0.267 mM ヒポキサンチンおよび10 % 不活化ウマ血清をそれぞれ添加したRPMI1640培地)で、5 % ヒト赤血球(O型)を用いて培養した。この原虫から、DNA抽出キット(QIAamp DNA Blood Mini Kit、Qiagen社製)を使用してDNAを抽出した。抽出したDNAをエタノール沈殿によって10倍に濃縮し、PCRに使用した。
【0029】
2)PCRプライマーの設計
DNAデータバンクおよび参考論文に開示される熱帯熱マラリア原虫のジャイレースBのDNA配列に基づき、以下の3種類のプライマーを設計した。
1:GyrB-pGEXBamHIFw
(38bp, 5’-atggattcggactaaatgaatatataacaaatataaca-3’ (配列番号5))
GyrB-pGEXEcRIRv
(41bp, 5’-gcgaattcttattttgatcttattaaatccctagctgcctt-3’ (配列番号6))
2:GyrB-ET30aNhisNdeIFw
(45bp, 5’-cgcatatgcatcatcatcatcatcatggactaaatgaatatataa-3’ (配列番号7))
GyrB-pET30aEcoRIRv
(41bp, 5’-atgaattcttattttgatcttattaaatccctagctgcctt-3’ (配列番号8))
3:GyrB-pET30aChisNdeIFw
(38bp, 5’-cgcatatgggactaaatgaatatataacaaatataaca-3’ (配列番号9))
GyrB-pET30aChisEcoRIRv
(45bp, 5’-atgaattcttaatgatgatgatgatgatgttttgatcttattaaa-3’ (配列番号10))
【0030】
3)PCRによるDNAの増幅および精製
TaKaRa ExTaq(Takara-bio社製)を用いて、以下の条件でPCRを行った。PCR産物を1.2%アガロース電気泳動で泳動し、約500 bpのバンドを得た。このバンドを切り出し、MiniElute Gel Extraction Kit (Qiagen社製)にて精製した。
[PCR条件]
プライマー:前述(配列番号5〜10)
反応条件:
10x ExTaq バッファー 5 μl
2.5 mM dNTP ミックス 2 μl
10 μM フォワード(FW)プライマー 2.5 μl
10 μM リバース(Rv)プライマー 2.5 μl
TaKaRa ExTaq 1 μl
DNA 1 μl
dH2O 36 μl
温度条件:
94℃ 3 分
94℃ 0.5 分
55℃ 1 分
72℃ 1 分
(94℃→55℃→72℃ 35サイクル)
72℃ 7 分
4℃ ∞
【0031】
4)TAクローニング
上記のDNA増幅産物をTAベクターであるpBluescript(Takara-bio社製)またはpGEM-T (Promega社製)にライゲーションし、クローニングを行った。出現した白色コロニーの内、それぞれ10個ずつ(合計30個)についてコロニーPCRを行い、DNA増幅産物を確認した4個のコロニーを増菌した(A1-3、A3-1、A3-3、A3-14)。増菌した大腸菌からプラスミドをQIAprep Spin Miniprep Kit(Qiagen社製)にて精製するとともに、インサートの確認のためPCRを行い、このPCR増幅産物を精製してシークエンスしたところ、A3-3、A3-14は点変異が見られた。よって、次の実験にはA1-3、A3-1のプラスミドを使用した。
[コロニーPCR条件]
プライマー:
pBluescript FW
5’-gtaatacgactcactatagggc-3’(配列番号11)(T7 フォワードプライマー)
pBluescript Rv
5’-ggaaacagctatgaccatg-3’ (配列番号12)(M13 リバースプライマー)
反応条件:
10x ExTaq バッファー 1.5μl
2.5 mM dNTP ミックス 1.5 μl
DMSO 0.75 μl
10 μM FW プライマー 1.5 μl
10 μM Rv プライマー 1.5 μl
TaKaRa ExTaq 0.15 μl
コロニー菌体 少々
dH2O 8.1 μl
温度条件:
94℃ 1 分
94℃ 0.5 分
55℃ 0.5 分
72℃ 1.5 分
(94℃→55℃→72℃ 35サイクル)
72℃ 2 分
4℃ ∞
[インサート確認用PCR条件]
プライマー:前述(配列番号5〜10)
反応条件:
10x ExTaq バッファー 5 μl
2.5 mM dNTP ミックス 2 μl
10 μM FW プライマー 2.5 μl
10 μM Rv プライマー 2.5 μl
TaKaRa ExTaq 1 μl
DNA 1 μl
dH2O 36 μl
温度条件:
94℃ 3分
94℃ 0.5 分
55℃ 1 分
72℃ 1 分
(94℃→55℃→72℃ 35サイクル)
72℃ 7 分
4℃ ∞
【0032】
5)発現ベクターの構築
候補のプラスミド(A1-3、A3-1)を制限酵素(A1-3用:BamHI(Toyobo社製)およびEcoRI(Toyobo社製)、あるいはA3-1用:NdeI(Takara-bio社製)およびEcoRI(Toyobo社製))で切断し、同時に同じ制限酵素で処理したpGEX6p-2ベクター(A1-3用、GEヘルスケアバイオサイエンス株式会社製)あるいはpET30aベクター(A3-1用、Novagen社製)にライゲーションした。ライゲーション後、DNA断片を挿入したプラスミドで大腸菌をトランスフォームしサブクローニングを行った。出現したコロニーについてコロニーPCR(前述)を行ったところ、A3-1由来DNA断片をライゲーションしたpET30aベクター(Gyrase B-NHis /pET-30a)で14個のコロニーのうち1個に目的のDNA配列が含まれることを確認した(A3-1-24)。この大腸菌を増菌後、プラスミドをQIAprep Spin Miniprep Kit(Qiagen社製)で精製し保存した。再度、インサート確認のためのPCR(前述)を行い、PCR増幅産物を精製し、シークエンスして、目的遺伝子と同一の配列であることを確認した(配列番号1)。
【0033】
6)ポリペプチドの発現および精製
上記5)で得られた発現ベクターで大腸菌(BL21(DE3)、Takara-bio社製)をトランスフォームし、培養した。培養液を遠心分離して集菌し、沈査(菌体)を超音波で粉砕した後、さらに遠心分離し、上清をNi2+ アフィニティカラムクロマトグラフィー(HiTrap Chelating HP、GEヘルスケアバイオサイエンス社製)にかけた。培養温度条件として37℃および16℃、IPTG処理条件として0.3 mM、1 mM、および3 mMを検討したところ、温度16℃、IPTG 1 mMで36時間培養すると、効率よくポリペプチドが発現することがわかり、ポリペプチドを大量に作製することに成功した(図1)。また、Ni2+アフィニティカラムクロマトグラフィーでの精製条件を検討したところ、イミダゾール100〜250 mMで溶出されることが分かった(図2)。
【0034】
[培養条件]
発現ベクター(Gyrase B-NHis /pET-30a)を大腸菌(BL21(DE3)、Takara-bio社製)にトランスフオームして作製したグリセロールストックをLB-Kan(カナマイシン30 μg/ml)寒天プレートに植菌。
↓
37℃で一晩培養。
↓
コロニーをピックアップして、LB-Kan培地(カナマイシン30 μg/ml、5 ml)に植菌し、37℃で一晩培養。
↓
500 ml のTB-Kan培地(カナマイシン50 μg/ml)に前培養分5 mlを全量添加。
↓
37℃で培養(約3時間)。
↓
OD600〜0.8でシェイキングを中断し、16℃(または37℃)に設定後、30分間静置。
↓
IPTGを最終濃度で0.3 mM、1 mM、または3 mMとなるように添加。
↓
16℃(または37℃)でさらに36時間培養。
↓
培養を終え、集菌。
↓
菌体を-80℃に保存。
【0035】
[Ni2+ アフィニティカラムクロマトグラフィーによるポリペプチドの精製方法]
HiTrap Chelating HP(GEヘルスケアバイオサイエンス製、5 ml容量)を平衡化バッファーで平衡化(ペリスタポンプおよびフラクションコレクター(GEヘルスケアバイオサイエンス製)使用、流速 1 ml/分)。
平衡化バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、10 mM イミダゾール
↓
上清をカラムに通液し、フロースルーを回収(流速 1 ml/分)(FT画分とする)。
↓
平衡化バッファーで洗浄(25 ml、流速 1 ml/分)(Wash画分とする)。
↓
100 mM イミダゾールを含む溶出バッファーで溶出(3 ml X 10本、流速 1 ml/分)。
溶出バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、100 mM イミダゾール
↓
250 mM イミダゾールを含む溶出バッファ−で溶出(3 ml X 10本、流速 1 ml/分)。
溶出バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、250 mM イミダゾール
↓
500 mM イミダゾールを含む溶出バッファ−で溶出(3 ml X 10本、流速 1 ml/分)。
溶出バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、500 mM イミダゾール
↓
SDS-PAGE 後、溶出画分を回収。
↓
脱塩バッファーで透析(2Lのバッファーで、4 ℃、一晩)。
脱塩バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、1 mM DTT
【0036】
図1の各レーンのサンプルは以下のとおりである。
図1 ポリペプチドの大量発現時における発現量の培養温度依存性
(a)37℃での培養後の菌体を用いたポリアクリルアミド電気泳動像
レーンの番号
1: M (マーカー)
2: 破砕(超音波による菌体の破砕粗画分)
3: 沈殿(遠心後沈殿画分)
4: 上清(遠心後上清画分)
5: FT(Ni2+アフィニティカラムクロマトグラフィーに吸着しなかった画分)
6: Wash(Niカラムの洗浄画分)
7: 溶出(100 mM) 1(Niカラムの100 mMイミダゾール添加液での溶出画分1本目)
8: 溶出(100 mM) 2(Niカラムの100 mMイミダゾール添加液での溶出画分2本目)
9: 溶出(100 mM) 3(Niカラムの100 mMイミダゾール添加液での溶出画分3本目)
10: 溶出(100 mM) 4(Niカラムの100 mMイミダゾール添加液での溶出画分4本目)
11: 溶出(100 mM) 5(Niカラムの100 mMイミダゾール添加液での溶出画分5本目)
12: 溶出(500 mM) 1(Niカラムの500 mMイミダゾール添加液での溶出画分1本目)
13: 溶出(500 mM) 2(Niカラムの500 mMイミダゾール添加液での溶出画分2本目)
14: 溶出(500 mM) 3(Niカラムの500 mMイミダゾール添加液での溶出画分3本目)
(b)16℃での培養後の菌体を用いたポリアクリルアミド電気泳動像
レーンの番号
1: M (マーカー)
2: 破砕(超音波による菌体の破砕粗画分)
3: 沈殿(遠心後沈殿画分)
4: 上清(遠心後上清画分)
5: FT(Ni2+アフィニティカラムクロマトグラフィーに吸着しなかった画分)
6: Wash(Niカラムの洗浄画分)
7: 溶出(100 mM) 1(Niカラムの100 mMイミダゾール添加液での溶出画分1本目)
8: 溶出(100 mM) 2(Niカラムの100 mMイミダゾール添加液での溶出画分2本目)
9: 溶出(100 mM) 3(Niカラムの100 mMイミダゾール添加液での溶出画分3本目)
10: 溶出(100 mM) 4(Niカラムの100 mMイミダゾール添加液での溶出画分4本目)
11: 溶出(100 mM) 5(Niカラムの100 mMイミダゾール添加液での溶出画分5本目)
12: 溶出(500 mM) 1(Niカラムの500 mMイミダゾール添加液での溶出画分1本目)
13: 溶出(500 mM) 2(Niカラムの500 mMイミダゾール添加液での溶出画分2本目)
14: 溶出(500 mM) 3(Niカラムの500 mMイミダゾール添加液での溶出画分3本目)
【0037】
図2の各レーンのサンプルは以下のとおりである。
図2 Ni2+アフィニティカラムクロマトグラフィーの溶出結果
(左図)
レーンの番号
1: M (マーカー)
2: 破砕(超音波による菌体の破砕粗画分)
3: 上清(遠心後上清画分)
4: 沈殿(遠心後沈殿画分)
5: FT(Ni2+アフィニティカラムクロマトグラフィーに吸着しなかった画分)
6: Wash(Niカラムの洗浄画分)
7: 溶出(100 mM) 1(Niカラムの100 mMイミダゾール添加液での溶出画分1本目)
8: 溶出(100 mM) 2(Niカラムの100 mMイミダゾール添加液での溶出画分2本目)
9: 溶出(100 mM) 3(Niカラムの100 mMイミダゾール添加液での溶出画分3本目)
10: 溶出(100 mM) 4(Niカラムの100 mMイミダゾール添加液での溶出画分4本目)
11: 溶出(100 mM) 5(Niカラムの100 mMイミダゾール添加液での溶出画分5本目)
12: 溶出(100 mM) 6(Niカラムの100 mMイミダゾール添加液での溶出画分6本目)
13: 溶出(100 mM) 7(Niカラムの100 mMイミダゾール添加液での溶出画分7本目)
14: 溶出(100 mM) 8(Niカラムの100 mMイミダゾール添加液での溶出画分8本目)
(右図)
レーンの番号
1: M (マーカー)
2: 溶出(250 mM) 1(Niカラムの250 mMイミダゾール添加液での溶出画分1本目)
3: 溶出(250 mM) 2(Niカラムの250 mMイミダゾール添加液での溶出画分2本目)
4: 溶出(250 mM) 3(Niカラムの250 mMイミダゾール添加液での溶出画分3本目)
5: 溶出(250 mM) 4(Niカラムの250 mMイミダゾール添加液での溶出画分4本目)
6: 溶出(250 mM) 5(Niカラムの250 mMイミダゾール添加液での溶出画分5本目)
7: 溶出(250 mM) 6(Niカラムの250 mMイミダゾール添加液での溶出画分6本目)
8: 溶出(500 mM) 1(Niカラムの500 mMイミダゾール添加液での溶出画分1本目)
9: 溶出(600 mM) 2(Niカラムの500 mMイミダゾール添加液での溶出画分2本目)
10: 溶出(500 mM) 3(Niカラムの500 mMイミダゾール添加液での溶出画分3本目)
11: 溶出(500 mM) 4(Niカラムの500 mMイミダゾール添加液での溶出画分4本目)
12: 溶出(500 mM) 5(Niカラムの500 mMイミダゾール添加液での溶出画分5本目)
13: 溶出(500 mM) 6(Niカラムの500 mMイミダゾール添加液での溶出画分6本目)
14: 溶出(500 mM) 7(Niカラムの500 mMイミダゾール添加液での溶出画分7本目)
【0038】
また、陰イオンカラムクロマトグラフィー(GEヘルスケアバイオサイエンス製、5 ml容量)での溶出条件の検討をしたところ、目的のポリペプチドがフロースルー (FT)画分に溶出され、不純物を除去できることがわかった(図3)。
[陰イオン交換カラムクロマトグラフィーによるポリペプチドの精製方法]
HiTrapQ FFカラム(GEヘルスケアバイオサイエンス製、5 ml容量)を平衡化バッファーで平衡化(ペリスタポンプおよびフラクションコレクター(GEヘルスケアバイオサイエンス製)使用、流速 1 ml/分)。
平衡化バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、5 mM DTT
↓
上清をカラムに通液;フロースルーを回収(流速 1 ml/分)(FT画分とする)。
↓
平衡化バッファーで洗浄(25 ml、流速 1 ml/分)(Wash画分とする)。
↓
溶出バッファーで溶出(流速 1 ml/分)。
溶出バッファーの組成:20 mM Tris-HCl pH8.0、1000 mM NaCl、5 mM DTT
↓
SDS-PAGE後、FTおよびWash画分を回収。
↓
YM-10(Amicon製)で約2 mlまで濃縮後、フイルタリング(PALL製 NANOCEP 0.2/m)。
【0039】
図3の各レーンのサンプルは以下のとおりである。
図3 陰イオン交換カラムクロマトグラフィーの溶出結果
レーンの番号
1: M (マーカー)
2: 脱塩後(超音波による菌体の破砕粗画分)
3: FT(陰イオン交換カラムクロマトグラフィーに吸着しなかった画分)
4: Wash(陰イオン交換カラムクロマトグラフィーの洗浄画分)
【0040】
さらに、ゲル濾過カラムクロマトグラフィー(GEヘルスケアバイオサイエンス製、120 ml カラム容量)を用いることで、より純度の高いポリペプチドを得ることが出来た(図4)。[ゲルろ過カラムクロマトグラフィーによるポリペプチドの精製方法]
HiLoad 16/60 Superdex200カラム(GEヘルスケアバイオサイエンス製、120 mlカラム容量)をゲルろ過バッファーで平衡化。
ゲルろ過バッファーの組成:20 mM Tris-HCl pH8.0、500 mM NaCl、5 mM DTT
↓
菌体破砕し、遠心分離後の上清画分をゲルろ過(流速 0.2 ml/分、1 ml/画分)。
↓
ゲルろ過終了後、SDS-PAGEで各画分を確認し、目的の画分を回収。
【0041】
ゲルろ過後の精製サンプルについて、動的光散乱(DLS)の測定を行った(図5)。測定の結果は、多分散度9.4%、純度99.93%であり、高純度のポリペプチドが得られた。計算された平均分子量は26.65KDa であり、理論分子量とほぼ一致していた。精製後のポリペプチドの収量は、大腸菌の培養培地1リットルあたり約40 mgであった。
[動的光散乱(DLS)測定の実験条件]
動的光散乱装置:MALVERN社製 Zeta Sizer Nano-S
測定温度:4℃
測定に使用したポリペプチド濃度:2 mg/ml
【0042】
7)モノクローナル抗体の作成
a)免疫
ジャイレースBドメインを抗原としてIncomplete Freund’s adjuvantとともにマウスに投与した。抗原の1回の投与量は、マウスの場合1匹当たり50〜100 μgであった。4回目の免疫後、脾臓細胞を採集した。
【0043】
b)細胞融合
脾臓細胞とマウス由来のミエローマ細胞との細胞融合を、血清を含まない動物細胞培養用培地中で脾臓細胞とミエローマ細胞とを混合し、ポリエチレングリコールの存在下で行った。
【0044】
c)ハイブリドーマの選別
融合細胞をヒポキサンチン(100μM)、アミノプテリン(0.4μM)及びチミジン(16μM)を含む培地を用いて培養し、生育する細胞をハイブリドーマとして得た。
次に、増殖したハイブリドーマの培養上清中に、目的とする抗体が存在するか否かについて抗原を用いてELISAでスクリーニングした。
【0045】
d)クローニング
融合細胞のクローニングは、限界希釈法等により行い、最終的にモノクローナル抗体産生細胞であるハイブリドーマを樹立した。モノクローナル抗体産生細胞を培養し、その培養上清から抗体を取得した。
【0046】
e)精製
プロテインA(またはプロテインG)セファロースクロマトグラフィーを用いて抗体を精製できる。
【0047】
f)ジャイレースBドメインのウエスタンブロッティングによる検出
ジャイレースBドメインのポリアクリルアミドゲル電気泳動を行った。ゲルを切り取り、ニトロセルロース膜に転写し、上記で得られたモノクローナル抗体(02C12など)を用いウエスタンブロッティングを行った。その結果、モノクローナル抗体(02C12など)がジャイレースBドメインに結合し、ジャイレースBドメインを検出することが確認された(図6)。
Lane 1: Marker
Lane 2: ジャイレースBドメイント精製物(5 μg)のCBB染色像
Lane 3: ジャイレースBドメイント精製物(5 μg)のモノクローナル抗体による免疫染色像
1次抗体:抗ジャイレースBドメインモノクローナル抗体(02C12)
2次抗体:HRP標識抗マウスIgG(H+L)抗体
発色液:ジアミノベンジジン(DBB)
【0048】
g)熱帯熱マラリア原虫の蛍光抗体を用いた免疫染色
熱帯熱マラリア原虫を、上記で得られたモノクローナル抗体(02C12など)を1次抗体、FITC標識抗マウスIgG(H+L)抗体を2次抗体として用いて免疫染色した。その結果、モノクローナル抗体(02C12など)はマラリア原虫の栄養型原虫を染色し、マラリア原虫内(EPI)で、モノクローナル抗体(02C12など)によるジャイレースBドメインの染色像(FITC)とDNAの染色像(DAPI)が重なった(MERGE)。これは、ジャイレースBドメインがDNA複製時の二本鎖DNAレベルの切断および再結合に関与するという知見と一致する結果である(図7)。
EPI:透過光
DAPI:原虫の核のDNAのDAPIによる染色
FITC:抗ジャイレースBドメインモノクローナル抗体との反応
熱帯熱マラリア原虫栄養型原虫
+抗ジャイレースBドメインモノクローナル抗体(02C12)(1次抗体)
+FITC標識抗マウスIgG(H+L)抗体(2次抗体)
MERGE:EPI+DAPI+FITCの重ね合わせ
【0049】
8)モノクローナル抗体による熱帯熱マラリア原虫の増殖抑制
熱帯熱マラリア原虫を培養し、モノクローナル抗体(02C12など)が感染赤血球率に及ぼす影響を検討した。モノクローナル抗体(02C12など)産生細胞の培養上清(02C12など)を原虫培地の1/10量添加して2日間培養した。その結果、モノクローナル抗体(02C12など) 産生細胞の培養上清により熱帯熱マラリア原虫の感染赤血球率が減少し、原虫の増殖および分裂が抑制された(表1および図8)。すなわち、未処置群(control)では、2日間の培養で原虫が増殖し分裂し、輪状体(Ring forms, R)、栄養型(Trophozoites, T)及び分裂体(Schizonts, S)が増加したのに対して、モノクローナル抗体を含む培養上清を用いた処置群(anti-gyraseB)では原虫数の増加が認められなかった。
【表1】
【特許請求の範囲】
【請求項1】
配列番号2のアミノ酸配列からなるポリペプチド。
【請求項2】
(1)配列番号2のアミノ酸配列において1または数個のアミノ酸が欠失、置換または付加されたアミノ酸配列からなるポリペプチド、または、
(2)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列からなるポリペプチド であって、配列番号2のアミノ酸配列からなるポリペプチドと同じ抗原性を有するポリペプチド。
【請求項3】
タグが付加された、請求項1または2のポリペプチド
【請求項4】
請求項1〜3のいずれかのポリペプチドをコードするポリヌクレオチド。
【請求項5】
(1)配列番号1の塩基配列、または
(2)配列番号1の塩基配列と縮重の関係にある塩基配列
からなる、請求項4のポリヌクレオチド。
【請求項6】
(1)配列番号1の塩基配列からなるポリヌクレオチドにストリンジェントな条件下でハイブリダイズするポリヌクレオチド、または
(2)配列番号1の塩基配列と90%以上の相同性を有する塩基配列からなるポリヌクレオチド、
であって、配列番号2のアミノ酸配列からなるポリペプチドと同じ抗原性を有するポリペプチドをコードするポリヌクレオチド。
【請求項7】
請求項4〜6のいずれかのポリヌクレオチドを含むベクター。
【請求項8】
請求項1または2のポリペプチドを特異的に認識する抗体またはその抗原結合断片。
【請求項9】
抗体が、
(a)ポリクローナル抗体;
(b)モノクローナル抗体;
(c)ヒト型化抗体;および
(d)ヒトモノクローナル抗体
からなる群から選択される請求項8に記載の抗体または抗原結合断片。
【請求項10】
請求項8または請求項9に記載の抗体または抗原結合断片に特異的に結合する抗イディオタイプ抗体。
【請求項11】
マラリア原虫に特異的に結合する、請求項8または9に記載の抗体または抗原結合断片。
【請求項12】
請求項8、9または11に記載の抗体または抗原結合断片を含む、マラリア原虫増殖抑制用組成物。
【請求項13】
請求項8、9または11に記載の抗体または抗原結合断片を含む、マラリア原虫検出用組成物。
【請求項1】
配列番号2のアミノ酸配列からなるポリペプチド。
【請求項2】
(1)配列番号2のアミノ酸配列において1または数個のアミノ酸が欠失、置換または付加されたアミノ酸配列からなるポリペプチド、または、
(2)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列からなるポリペプチド であって、配列番号2のアミノ酸配列からなるポリペプチドと同じ抗原性を有するポリペプチド。
【請求項3】
タグが付加された、請求項1または2のポリペプチド
【請求項4】
請求項1〜3のいずれかのポリペプチドをコードするポリヌクレオチド。
【請求項5】
(1)配列番号1の塩基配列、または
(2)配列番号1の塩基配列と縮重の関係にある塩基配列
からなる、請求項4のポリヌクレオチド。
【請求項6】
(1)配列番号1の塩基配列からなるポリヌクレオチドにストリンジェントな条件下でハイブリダイズするポリヌクレオチド、または
(2)配列番号1の塩基配列と90%以上の相同性を有する塩基配列からなるポリヌクレオチド、
であって、配列番号2のアミノ酸配列からなるポリペプチドと同じ抗原性を有するポリペプチドをコードするポリヌクレオチド。
【請求項7】
請求項4〜6のいずれかのポリヌクレオチドを含むベクター。
【請求項8】
請求項1または2のポリペプチドを特異的に認識する抗体またはその抗原結合断片。
【請求項9】
抗体が、
(a)ポリクローナル抗体;
(b)モノクローナル抗体;
(c)ヒト型化抗体;および
(d)ヒトモノクローナル抗体
からなる群から選択される請求項8に記載の抗体または抗原結合断片。
【請求項10】
請求項8または請求項9に記載の抗体または抗原結合断片に特異的に結合する抗イディオタイプ抗体。
【請求項11】
マラリア原虫に特異的に結合する、請求項8または9に記載の抗体または抗原結合断片。
【請求項12】
請求項8、9または11に記載の抗体または抗原結合断片を含む、マラリア原虫増殖抑制用組成物。
【請求項13】
請求項8、9または11に記載の抗体または抗原結合断片を含む、マラリア原虫検出用組成物。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2012−147703(P2012−147703A)
【公開日】平成24年8月9日(2012.8.9)
【国際特許分類】
【出願番号】特願2011−7714(P2011−7714)
【出願日】平成23年1月18日(2011.1.18)
【出願人】(505127721)公立大学法人大阪府立大学 (688)
【Fターム(参考)】
【公開日】平成24年8月9日(2012.8.9)
【国際特許分類】
【出願日】平成23年1月18日(2011.1.18)
【出願人】(505127721)公立大学法人大阪府立大学 (688)
【Fターム(参考)】
[ Back to top ]