
熱非対称インターレース(TAIL)PCRを用いたランダムホモ接合性遺伝子摂動(RHGP)
【課題】哺乳動物細胞においてインフルエンザ感染性を低下させる組成物の提供。
【解決手段】治療介入のための潜在的標的用の宿主遺伝子及びコードされるタンパク質を同定するための方法では、レンチウイルス又はMMLVベースのいずれかである遺伝子探索ベクターを用いており、ゲノム配列の事前の知識なしに細胞ゲノム全体を照合するために使用することができる。このランダムホモ接合性遺伝子摂動(RUGP)技術は、迅速に検証可能であり、インフルエンザ、HIV、及び他のウイルス感染についての介入のための潜在的な宿主標的を同定するために使用される。熱非対称インターレース(TAIL)PCRを使用し、有望な標的の同定のための期間を、数ヶ月から数週間又はそれ以下に低下させる。標的としてNedd4に対する抗体を医薬的に許容可能な担体中に含む組成物。
【解決手段】治療介入のための潜在的標的用の宿主遺伝子及びコードされるタンパク質を同定するための方法では、レンチウイルス又はMMLVベースのいずれかである遺伝子探索ベクターを用いており、ゲノム配列の事前の知識なしに細胞ゲノム全体を照合するために使用することができる。このランダムホモ接合性遺伝子摂動(RUGP)技術は、迅速に検証可能であり、インフルエンザ、HIV、及び他のウイルス感染についての介入のための潜在的な宿主標的を同定するために使用される。熱非対称インターレース(TAIL)PCRを使用し、有望な標的の同定のための期間を、数ヶ月から数週間又はそれ以下に低下させる。標的としてNedd4に対する抗体を医薬的に許容可能な担体中に含む組成物。
【発明の詳細な説明】
【技術分野】
【0001】
優先データ及び参照による組み入れ
本願は、以下に対する優先権の利益を主張する:米国特許仮出願第61/142,664号(2009年1月6日出願);米国特許仮出願第61/153,012号(2009年2月17日出願);米国特許仮出願第61/158,015号(2009年3月6日出願);米国特許仮出願第61/160,737号(2009年3月17日出願);米国特許仮出願第61/178,112号(2009年5月14日出願);及び米国特許仮出願第61/180,457号(2009年5月22日出願);それらの全てが参照によりその全体が組み入れられる。
【0002】
発明の背景
発明の分野
本発明は、熱非対称インターレースPCR、又はTAIL−PCRといわれるプロセスを通じてスポットされた標的の同定と合わせた、ランダムホモ接合性遺伝子摂動(RHGP)による治療用宿主標的を同定するための遺伝子探索ベクターシステム(GSV)の使用に関する。RHGPとTAIL PCRとの組み合わせは、本願を通して、RHGPといわれる。
【0003】
ヒトゲノムの配列決定は、即時に、かつて期待された処置のための治療標的のカスケードをもたらしてこなかった。遺伝子配列の存在を知ることによって、1人(それ以上はない)が、遺伝子又は遺伝子システムが関与する表現型、又は遺伝子が細胞の進行中の進化をどのように媒介するかを特徴付けることはできない。それにもかかわらず、宿主標的の同定は、多くの研究者にとって重要性の増加する分野となっている。ウイルス疾患は、特に、ウイルス侵入者によって攻撃された哺乳動物、及び特にヒトを予防及び処置する方法を開発する人の独創性に挑戦してきた。本発明は広い適用性を有するが、本願におけ焦点は、治療介入のための宿主標的を同定するためのデバイスとしてのRHGPの有用性である。
【0004】
ウイルスは、効果的な治療法を開発する能力を妨げてきた。部分的には、突然変異してウイルスを標的化する薬物の有効性から離れるそれらの能力のためである。ウイルスは、また、それらに対して利用可能な広いパレットの差動的な作用を有する。それらは迅速な侵入者(インフルエンザなど)である。又は、感染細胞の機構のウイルスによる乗っ取りは緩徐で、経時的に遅延されうる(HIVなど)。侵入者の標的化に対する代替物として、Functional Genetics, Inc.(本願の譲受人)は、ウイルスを標的化する治療法の設計から離れたパラダイムシフトに導いており、宿主タンパク質(例えばTSG101及び他の宿主タンパク質など)を標的化する抗体及び小分子治療の両方を開発している。宿主タンパク質を標的化することにより、ウイルスが突然変異して離れる圧力が低下する。ウイルスは所与の細胞に感染することができるが、しかし、そのライフサイクルのために必須の宿主タンパク質へのウイルスの接近を拒むことにより、細胞から細胞へのウイルスの伝播が予防される。
【0005】
潜在的な宿主の治療タンパク質及び遺伝子の同定を推進するために、ある種の先端サーチエンジンが要求される:
質問は基本的なものである。問題の遺伝子、及び/又はそれがコードする産物は、ウイルス侵入者に対してある種の耐性を付与するか。質問は、本質的に、イエス/ノーの質問でありうる。他の状況において(他の疾患を伴う)、質問は異なりうる。このように、RHGPを使用し、ヒト遺伝子(その発現がアルツハイマー患者において見出されるプラーク形成表現型と相関する)を同定してもよい。それらの結果は、米国特許出願シリアル番号12/566,951(2009年9月25日出願)において報告される。種々の技術が長年にわたり開発されており、種々の疾患に関与する宿主標的を同定してきた。例示的な技術は、競合RNA干渉(RNAi)である。例えば、Nature, Revealing the World of iRNA, 431, pp.338-342 (September, 2004)を参照のこと(参照により本明細書において組み入れられる)。低分子干渉(siRNA)は、標的発見において補助するためのゲノム規模のsiRNAライブラリーの生成のために多大な注目を受けてきた。Becket et al, Cancer Research, 69(2), (2009)。これらの方法は、同定される標的、及び関連するゲノムのある程度の事前の知識を要求する。過去十年間に、多くの生命を脅かす癌の診断及び処置における著しい進歩が目撃されてきた。進展の大部分は、癌細胞の挙動を制御する基本的機構の理解の上に築かれる。そのような知識によって、癌細胞により選択的に利用される分子変化及び調節機構に対する標的治療の合理的設計のための基礎が形成された。顕著な成功は、トラスツズマブ(Herceptin(登録商標))を用いた乳癌細胞上のHER2、ボルテゾミブ(Velcade(登録商標))を用いた多発性骨髄腫におけるプロテオソーム、及びゲフィチニブ(Iressa(登録商標))を用いた非小細胞肺癌細胞上のEGFRの標的化を含む(Ataergin et al., 2009; Campos, 2008; Madarnas et al., 2008; Pytel et al., 2009; Yang et al., 2009)。しかし、今日開発中の合理的に設計された治療の大半では、相対的に狭い一連の標的に焦点が合わされている。この強調は、論理的には、これらの標的についての知識の増加及び同じ又は類似の分子を標的化する早い模倣薬物を刺激する既存の薬物を用いた商業的成功に続く。それにもかかわらず、これらの傾向は、本来なら、過小評価される又は未知の標的もしくは機構から生じうる飛躍的進歩の邪魔をしうる。鍵は、従って、癌細胞の薬物感受性、転移能力、及び疾患の病態生理の他の特徴を決定的に制御する標的を同定する新規手段を開発することである。
【0006】
過去十年間に、多くの生命を脅かす癌の診断及び処置における著しい進歩が目撃されてきた。進展の大部分は、癌細胞の挙動を制御する基本的機構の理解の上に築かれる。そのような知識によって、癌細胞により選択的に利用される分子変化及び調節機構に対する標的治療の合理的設計のための基礎が形成された。顕著な成功は、トラスツズマブ(Herceptin(登録商標))を用いた乳癌細胞上のHER2、ボルテゾミブ(Velcade(登録商標))を用いた多発性骨髄腫におけるプロテオソーム、及びゲフィチニブ(Iressa(登録商標))を用いた非小細胞肺癌細胞上のEGFRの標的化を含む(Ataergin et al., 2009; Campos, 2008; Madarnas et al., 2008; Pytel et al., 2009; Yang et al., 2009)。しかし、今日開発中の合理的に設計された治療の大半では、相対的に狭い一連の標的に焦点が合わされている。この強調は、論理的には、これらの標的についての知識の増加及び同じ又は類似の分子を標的化する早い模倣薬物を刺激する既存の薬物を用いた商業的成功に続く。それにもかかわらず、これらの傾向は、本来なら、過小評価される又は未知の標的もしくは機構から生じうる飛躍的進歩の邪魔をしうる。鍵は、従って、癌細胞の薬物感受性、転移能力、及び疾患の病態生理の他の特徴を決定的に制御する標的を同定する新規手段を開発することである。
【0007】
相対的に単純な生物(例えば細菌など)のゲノムでの初期の調査において、単一コピーのDNAだけ、従って、標的遺伝子の単一対立遺伝子の存在が、標的遺伝子の迅速同定に役立った。標的組換え(相同組換え)により遺伝子を「ノックアウト」することによって、問題の遺伝子の機能を削除することができた。対照的に、二倍体生物(哺乳動物を含む)は、2コピーの各遺伝子を提示する。細胞中に存在する1本のDNA上の単一コピーだけを破壊することによって、遺伝子、又はそれに関連する表現型は除去されない。この問題に対する解決が、米国特許第5,679,523号(Li et al.)に提示されており、参照により本明細書において組み入れられる。それはランダムホモ接合性ノックアウト又は「RHKO」技術に向けられる。この方法において、遺伝子探索ベクターがゲノム中に挿入され、アンチセンス鎖の生成を起こすトランス活性化因子を保有する。このように、1本鎖中へのベクターの挿入によって1つの対立遺伝子が発現停止され、アンチセンス鎖の生成は、反対の対立遺伝子に結合し、それを不活性化又は発現停止する。
【0008】
RHKOに対するいくつかの限定がある。最初に、それはノックアウト効果に限定される。発現のアップレギュレーションの効果を決定し、アップレギュレーションの標的として貴重でありうる潜在的な標的を同定することができない。遺伝子(その増強された発現が重要な表現型を付与する)の発現を増強する方法については米国特許第5,272,071号を参照のこと。RHKO技術により提示される追加の課題は、改変細胞の複数ラウンドの培養についての必要性である。培養のために要求される時間と、変化した表現型(ここでベクターが挿入された)に関与する摂動の同定のために要求される時間の間に、多大な「ダウンタイム」がシステム中に導入される。
【0009】
米国特許出願シリアル番号11/928,393(Duan et al.)(参照により本明細書において組み入れられる)において、抗体を低レベルで発現する細胞において抗体の発現レベルを増強するための方法が開示され、ランダムホモ接合性遺伝子摂動、又はRHGPといわれる。この方法において、遺伝子探索ベクター、又はGSVは、アンチセンスRNAを生成し、それは挿入ベクターと組み合わされ、摂動されるいずれの遺伝子の発現も低下又は除去し、細胞集団にトランスフェクションするために使用される。本願の実施例は、特定の遺伝子の標的摂動を含み、再び、大部分がノックアウト技術に向けられ、増強された抗体産生(所望の表現型)という結果の検証のために特定の細胞の発現を抑制する。この技術は、また、摂動に起因する細胞を増大及び培養する必要性、及び結果として生じる遅延を含む。
【0010】
標的サーチエンジンは、任意の特定の疾患又は表現型の型に限定されるべきではない。このように、ウイルス疾患に加えて、種々の状況(抗体発現に加えて)を想像することができ、そこでは、個人、例えばヒトなど(宿主細胞だけではない)は、改善された表現型から利益を得うる。腫瘍学での薬物開発の歴史は、癌細胞の挙動の増加する理解と共に進化してきた。薬物開発の最初の期間において、化合物ライブラリーが、腫瘍細胞の増殖又は生存の阻害について分析された。これらのアプローチは、実験室での腫瘍細胞の培養における進歩の結果として可能になった。そのような改善によって、培養された腫瘍細胞の増殖を殺す又は予防する化合物を同定するための実験的スクリーニングを開発することが可能になった。これらの評価は機能第1(Function−First)アプローチを含み、そこでは、最終的な結果に最大の価値を、抗腫瘍活性の標的又は機構的基礎の同一性にはより少ない強調を置いた。これらのアプローチは、ハイスループットスクリーニング手順における改善を要求及び刺激し、化学構造のアレイを、新規天然産物の単離を介して、新規化学ライブラリー及び/又はその2つの組み合わせを作製することにより多様化させるための努力を促進した。結果として得られる細胞傷害性治療は、今日の本発明者らの抗腫瘍薬の武器庫の大半を含み、アルキル化剤(プラチナ化合物)、抗生物質(ドキソルビシン)、抗代謝剤(メトトレキセート、5−フルオロウラシル)、アルカロイド(ビンクリスチン)、及びタキサン(タキソル)を含む。しばしば、治療法のための標的の同一性は未知である、又は機構的基礎は後に同定された。効果的であるが、これらの細胞傷害性の抗腫瘍薬剤は、しばしば、不要な副作用を伴った。それによって長期間にわたる化合物の使用が限定されうる。これを複雑化することとして、P糖タンパク質の発現増加が、腫瘍細胞の迅速な突然変異速度と合わさって、薬物耐性腫瘍の選択を支持する傾向がある(Gottesman and Ling, 2006)。
【0011】
実験室において腫瘍細胞を培養する能力によって、また、研究者が良性細胞と悪性細胞を区別する変化を理解し始めることができるようになり、それは標的介入の時代を予告した。悪性形質転換に寄与するがん遺伝子の理解における基本的な進歩は、次に、癌細胞の挙動(増殖、生存、及び浸潤)を支配するシグナル伝達経路の同定を促進した。この理解の改善によって、公知の標的に基づく化学療法剤の新たな生成が促進された。結果として得られる化合物は、公知の標的、例えばトポイソメラーゼ(ポドフィロトキシン)及び増殖因子又はそれらの受容体などに影響を与えた。腫瘍細胞についての改善された選択性は、より効果的で選択的な抗腫瘍薬物をもたらした。標的薬物をスクリーニング又は操作し、特定の標的を選択的に阻害したため、新たな分子医薬は、しばしば、関連する診断用薬剤を伴ったが、それは患者の適格性又は処置結果を予測したであろう。我々は標的介入のこの時代の真っ最中に依然としているが、同じ歴史的な障害(不要な副作用及び薬物耐性の獲得を含む)の多くが残存する。これを複雑化することとして、結果として得られる薬物の多くが、相対的に狭い一連の標的(例、チューブリン、増殖因子、及びキナーゼ)に焦点を合わせている。この優先順位付けは、検証された標的を集中的に研究するための必要性の論理的結果であった。しかし、意図していない結果は、研究下にある標的の着実に狭まる型であった。標的同定への種々のアプローチの開発を図1に示す。
【0012】
疾患に対する遺伝的関連の理解の改善に伴い、癌の遺伝的基礎の増加する認識によって、腫瘍細胞のゲノム及び次にプロテオミクスサンプリングにおける興味が促された。癌細胞についての潜在的な標的を同定するための「−オミクス」の成長は、DNA配列決定における劇的な技術的進歩と同時期に起こり、多くの注目を集め、学界及び医薬業界を、その技術への相当なインフラの投資に導いた。ゲノミクス、プロテオミクス、及び関連する戦略によって、標的の数及び幅が増加した。このように、この分野では、それ自体が、幅広い標的にあふれていることが見出され、継続的な標的発見についての認知された必要性は底に達した。しかし、「−オミクス」を使用して同定された相関関連によって、しばしば、疾患病理を起こす又はそれに寄与する標的が、疾患プロセスの副作用として生じる標的から区別されないことがますます理解された。このあいまい性は、原因と結果の周辺の問題を解決するための相当な投資を要求し、このように、新規の腫瘍学標的についての一般的な欲求を減少させてきた。なぜなら、多くの機関が、「−オミクス」研究及びその結果として得られる標的を研究するために必要とされる技術に、相当なインフラを投資していたからである。
【0013】
したがって、潜在的な治療候補の迅速な検出のための方法を開発することが依然として当業者の目標であり、それは、同定された場合、それらの治療的な潜在力を開発する(発現低下(例、遺伝子治療)又はそれによりコードされるタンパク質のサイレンシング(リガンド結合治療)のいずれかを通じて)又は単に診断の対象とすべき遺伝子(それらの存在、又は複数コピーでの存在が、個人が、問題の遺伝子と相関する全ての表現型についての可能性の高い候補であることを示す)を同定するためのさらなる実験の対象となりうる。理想的には、この技術は、アップレギュレーションならびにダウンレギュレーションに起因する表現型を同定することが可能であるはずであり、ゲノム、又は標的遺伝子の知識は要求しない。不必要な多額のお金及び人的資源の出費を避けるために、この技術は、検証可能な潜在的標的(即ち、新たな表現型が、ゲノム自体の変化に起因し、それ自体の改変、又は攻撃状態に対する暴露のいずれかの結果ではない)を得る必要がある。それは、潜在的な候補のクローニング及び増大に起因する時間遅延を低下させる基礎に関する結果を得る必要がある。
【0014】
発明の概要
ランダムホモ接合性遺伝子摂動(RHGP)は、所与の細胞集団において遺伝子をランダムに破壊するための方法であり、任意の所望の表現型のスクリーニング及びその関連遺伝子の同定を可能にする。RHGPの能力に対して役立つのは、そのアンチセンス技術であり、それによって遺伝子の全コピーの同時不活性化が可能になる(所与の細胞において全てのコピーを無効にする反復標的化を要求する従来のノックアウトアプローチとは異なる)。さらに、RHGPは、競合RNA干渉(RNAi)技術よりも強力である。なぜなら、それは以下の利点を与えるからである:標的又は経路の事前の知識なしに、偏りのない方法で任意の遺伝子座に影響を与える能力;そのエピジェネティック状態(サイレンスされているか又はサイレンスされていないか)にかかわらず、任意の標的(マイクロRNAを含む)又はタンパク質ドメインを過剰発現又はノックダウンする能力;表現型の即時検証のための組み込み可逆性;及び同じベクターシステムを用いてインビボで表現型を試験する能力。また、RHGPは、RNAiのオフターゲットの影響を受けない。なぜなら、RHGPベクターがゲノム中に組込まれ、その活性のための基礎として相同性を使用しないからである。RHGPは、その現在の具体化において、しかし、RNAiと比較し、遺伝子標的を同定するために実質的により長い時間が必要である。なぜなら、それは、目的の全ての標的化された遺伝子座が細菌中にクローン化され、配列決定を通じて個々に同定されることを要求するからである。本発明において、本発明者らは、全てのRHGP挿入標的を、PCRを使用して迅速に同定するための方法の適応について記載しており、それによりRNAi方法論に沿った標的遺伝子発見のスピードをもたらす。この強力な技術は、ウイルス疾患又は細菌疾患と戦うための潜在的な宿主タンパク質の同定に限定されない。宿主タンパク質が関与する任意の症候群、例えば癌などを、RHGPの強力なプロセスに供することができる。
【0015】
RHKOの技術の上に構築し、そこでは最初のアプローチが開発され、真核細胞が一般的に2組の染色体を有するとの事実に起因するバリアを克服した(Li and Cohen, 1996)。RHGPでは同じ基本的アプローチを用いる。真核細胞の二倍体の性質によって、細菌経路の本発明者らの理解において非常に強力であることが証明されている標的遺伝子の単純なノックアウトベースの評価が妨げられた。siRNA(公知の標的のノックアウトに限定される)とは異なり、RHGPベースのアプローチは、任意の標的遺伝子の両方のコピーを同時にノックダウンする能力を含み、その遺伝子の任意の事前の知識又はアノテーションに非依存的である。強力な阻害が可能である。なぜなら、組込み事象自体が、遺伝子の第1コピーをノックアウトするために十分であり、標的遺伝子のアンチセンス発現が、第2の対立遺伝子をノックダウンするためである(図2)。RHGPが、また、遺伝子発現(全遺伝子又は個々のドメインを含む)をアップレギュレーションすることができる(図3A)。今日まで、今日までにRHGPを使用して同定された標的の約半分が、センス方向での組込みを示し、残りの半分がアンチセンス組込み事象を示している。結果的に、RHGPは、研究下にある表現型の原因となる任意の遺伝子変化についてゲノム全体を照合する手段を提供する。
【0016】
RHGPの中心となる特性は、固有のレンチウイルスベースの(又はサルマウス白血病ウイルスベースの)遺伝子エレメントであり、遺伝子探索ベクター(GSV)として公知であり、それは、ゲノム全体を照合し、目的の表現型を起こす標的遺伝子を同定するように設計された。GSVはゲノム内の単一部位に組込まれ、そこで、それは誘導プロモーターを介して標的遺伝子の発現を調節する(図4)。1個の細胞当たり単一の組込みが、相対的に低い感染効率(MOI)でGSVを用いた細胞の形質導入により制御される。ベクター自体が、自己不活性化レンチウイルスLTRをコードし、それは、GSVが形質導入細胞から再出現することを予防する。
【0017】
GSVは、ゲノム中に、センス又はアンチセンスのいずれかの方向で組込まれうる。アンチセンス配置において、組込み事象自体が1つの対立遺伝子を不活性化し、アンチセンスコンストラクトの発現を促進し、それによって他の対立遺伝子上でコードされる遺伝子がノックダウンされる(図3A)。反対(センス)方向に組込まれた場合、RHGPは、標的遺伝子の過剰発現又は非発現停止を促進することができる。この結果は、遺伝子全体の単純な過剰発現を超えて拡大しうる(例、開始部位の上流での挿入)、又は、開始部位の下流に組込まれた場合、特定のドメインの過剰発現を引き起こし、それは、野生型遺伝子の機能のドミナントネガティブな阻害剤を産生しうる。そのようなものとして、RHGPによって細胞ゲノム全体の照合ができるようになり、異なる型の標的事象を同定する。
【0018】
RHGPは、GSV形質導入細胞の「ライブラリー」の構築を要求する(図3B)。このライブラリーは、ゲノム全体をカバーするGSV組込み事象(過剰発現及びアンチセンス発現の両方に関する)を含む。RHGPは、柔軟で、分析される細胞型(リンパ系もしくは付着性)、種(ヒトもしくは非ヒト)、又は形質転換特徴(初代もしくは不死化もしくは形質転換)により偏らない。RHGP戦略では、内因的に有毒であるGSV形質導入事象が除かれる。例えば、重要遺伝子の発現を破壊するRHGP形質導入事象は、宿主細胞に対して有毒でありうる。このように、これらのクローンは、RHGPライブラリーから除去されうる。一旦形質導入されると、同じライブラリーが記録され、他のスクリーニングのために使用することができる。
【0019】
RHGP−TAILを使用し、疾患又は不全状態に応答性でありうる種々の現象及び表現型が試験されてきた。本明細書において考察されるのは、インフルエンザに対する耐性、ヒト免疫不全ウイルス(HIV)及び関連するAIDS、ならびに癌に対する耐性に関与する表現型を試験するためのRHGPの使用からの種々の結果である。これらのRHGP試験により生成された標的も追跡調査される。
【図面の簡単な説明】
【0020】
【図1】図1は、標的発見方法における最近の開発を反映するチャートである。
【図2】図2は、本願の本発明の遺伝子探索ベクター(GSV)の使用を実証する概略図である。
【図3A】図3Aは、本発明のGSVのセンス及びアンチセンス挿入を実証する概略図である。
【図3B】図3Bは、本発明のGSVのセンス及びアンチセンス挿入を実証する概略図である。
【図4】図4は、インフルエンザ関連標的についての完全なRHGP研究の実施に関与する段階を描写するフローチャートである。
【図5A】図5Aは、本発明において使用されるGSV pRHGP−12RSNについての遺伝子地図である。
【図5B】図5Bは、本発明において使用されるGSV pRHGP−12RSNについての遺伝子地図である。
【図6A】図6Aは、PTCH1の同定を、RHGP技術によるインフルエンザの治療的阻害のための可能な標的として描写する概略図である。
【図6B】図6Bは、PTCH1の同定を、RHGP技術によるインフルエンザの治療的阻害のための可能な標的として描写する概略図である。
【図6C】図6Cは、PTCH1の同定を、RHGP技術によるインフルエンザの治療的阻害のための可能な標的として描写する概略図である。
【図7】図7は、コントロールと比較した、PTCH1の阻害により起こされたウイルス産生における相対的な阻害を示すグラフである。
【図8A】図8Aは、異なるMOIでのPTCH1の阻害効果のグラフ同定である。
【図8B】図8Bは、異なるMOIでのPTCH1の阻害効果のグラフ同定である。
【図8C】図8Cは、異なるMOIでのPTCH1の阻害効果のグラフ同定である。
【図9A】図9Aは、生物学的プロセス及び分子機能によるRHGP同定されたインフルエンザ標的の分類を示すチャートである。
【図9B】図9Bは、生物学的プロセス及び分子機能によるRHGP同定されたインフルエンザ標的の分類を示すチャートである。
【図10】図10は、RHGPにより同定された治療標的の阻害により起こされたウイルス産生における低下を反映するグラフである。
【図11A】図11Aは、HIV感染を制御するための標的としてのRobo1の同定を例示する。
【図11B】図11Bは、HIV感染を制御するための標的としてのRobo1の同定を例示する。
【図11C】図11Cは、HIV感染を制御するための標的としてのRobo1の同定を例示する。
【図12A】図12Aは、HIV感染性に対する、RHGP同定されたRobo1を阻害する効果を反映する。
【図12B】図12Bは、HIV感染性に対する、RHGP同定されたRobo1を阻害する効果を反映する。
【図13】図13は、Nedd4(抗体が産生されるエピトープ)に対する種々の抗体の生成及び感染MDCK細胞の表面上でのNedd4の同定を描写する。
【図14】図14は、Nedd4抗体により起こされたウイルス複製の低下を実証するグラフである。
【図15】図15は、RHGPを使用して癌転移を阻害するための標的の同定において取るべき段階を例示するフローチャートである。
【図16A】図16Aは、転移癌の形成を阻害するために使用されうるRHGPにより同定された標的の重要な特徴を特定するチャートである。
【図16B】図16Bは、転移癌の形成を阻害するために使用されうるRHGPにより同定された標的の重要な特徴を特定するチャートである。
【0021】
発明の詳細な説明
遺伝子探索ベクター
RHGP遺伝子探索ベクターは、2つの特色で来る:モロニーマウス白血病ウイルス(MMLV)バージョン及びレンチウイルスバージョン。探索ベクターがどのようにコンストラクションされ、使用されるかを理解することが、本発明の詳細な実施を理解する際での重要な第一段階である。
【0022】
MMLVベクターは、それらが、遺伝子のプロモーター領域中への挿入を好むという利点を有し、それにより遺伝子全体がアップレギュレーション又はダウンレギュレーションされることを確実にする。さらに、MMLVは活性遺伝子に向かって偏っておらず、それらを相対的にランダムにゲノム全体に挿入することができる。レンチウイルスベクターは、MMLVと比較して感染の広い宿主範囲を有しており(それらが水疱性口内炎ウイルスGタンパク質、すなわちVSV−Gを用いてシュードタイピングされているとの理由で)、それらは分裂細胞及び非分裂細胞の両方に感染する能力を有する。しかし、それらは、それらが活発に転写された遺伝子を好む点で、それらが相対的に非ランダムに挿入されるという事実に悩まされる。レンチウイルスバージョンは、MMLVバージョン(プロモーター領域を好む傾向がある)と異なり、遺伝子自体の内に挿入の偏りを有さない。レンチウイルスバージョンは、未知の理由で、MMLVベクターと比較してより高い力価を一貫して産出している。
【0023】
MMLVベクターは、それらがいずれかの末端にMMLV 5’及び3’LTR(末端反復配列)を有する点で、同じ一般構造を有し、組込み後にウイルスの非重要エレメントを自動的に切り出すために使用されるCre遺伝子が隣接する(LoxPエレメントがあり、Cre及び不要なエレメントが自動的に除去される)。
【0024】
特定のベクターの特性
RHGP48:このベクターは、14コピーのテトラサイクリン応答エレメント(14×TRE)を伴うプロモーターを含み、転写ターミネーターなしにピューロマイシン耐性遺伝子の発現を促進させ、アンチセンス転写物を作製する(このアンチセンスは、RHGPベースの遺伝子不活性化の基礎である)。TREは、テトラサイクリンの非存在において、プロモーターが活性であり、アンチセンス発現を促す点でTet−Offプロモーターである。一旦テトラサイクリンが培地に加えられると、プロモーターが遮断され、アンチセンス転写物も遮断され、それはRHGPの可逆特性の基礎である。
【0025】
RHGP48−RSB:このベクターはRHGP48の構造と類似であるが、しかし、TREがアンチセンス発現を促す代わりに、それはNew England BioLabsのRheoSwitchプロモーターを使用し、それはRSL1リガンドにより活性化される。5コピーのRheoSwitchエレメントがあり、それらにRheoSwitchアクチベーターが結合する。このプロモーターは、ブラストサイジン耐性遺伝子の発現を促し、それも転写ターミネーターなしであり、アンチセンス産物を作ることができる。
【0026】
RHGP48−RSN:このMMLVベクターはRheoSwitch調節プロモーターを含み、アンチセンス転写物を促すが、しかし、アンチセンスから転写される耐性遺伝子はない。この「ネイキッド」プロモーターは、標的遺伝子のダウンレギュレーション及びアップレギュレーションの両方を促進すると考えられる。なぜなら、理論的に、プロモーターの下流に遺伝子を有することによって、下流の標的を翻訳する能力が損なわれうるからである(なぜなら、遺伝子、例えばブラストサイジン又はピューロマイシンなど翻訳終止コドンを有する)。挿入事象の選択を可能にするために、常時発現性のホスホグリセリン酸キナーゼ(PGK)プロモーターが存在し、ブラストサイジン耐性遺伝子を促す(これはセンス方向の正常遺伝子であり、アンチセンス転写物を産生しない)。
【0027】
レンチウイルスベクターは5’及び3’LTRの両方を含むが、しかし、3’LTRは自己不活性化し(3’SIN−LTRと呼ばれる)、ゲノム中への挿入後、ベクターはウイルスを複製し、産生することができない。これによって安全性プロファイルが劇的に増加する。これらのベクターはCre遺伝子を含まず、しかし、それらは全てがLoxP部位を有し、それらは、ゲノムにおいて、Creを外因的に加えることにより除去(逆転)することができる。これらのベクターは、また、RSVエンハンサーエレメントを5’LTR中に含み、レンチウイルスのより高い転写、故に、産生を可能にする。
【0028】
RHGP11:このベクターはEGFPを駆動する14×TREを有し、調節されたアンチセンス産物、ならびに緑色蛍光タンパク質を産生することを促す。これによって、細胞を蛍光顕微鏡下で見ることが可能になり、必要な場合、FACSにより選別される。また、このベクターはゼオシン耐性遺伝子を有し、PGKプロモーターを促し、薬物選択を可能にする。
【0029】
RHGP11−CMVE:このベクターはRHGP11と類似であるが、しかし、EGFPを伴うアンチセンス発現を促す調節されたTREの代わりに、アンチセンスは常時発現性である。なぜなら、CMVプロモーターがEGFPアンチセンスコンストラクトを駆動するからである。これは、表現型を簡単に逆転させることができない場合(例えばインビボ試験など)において重要でありうる。
【0030】
RHGP22:このベクターは、典型的なTREの代わりに、pTight TREを有する。5コピーのpTight応答エレメントがある。pTightは、また、アンチセンスのためのテトラサイクリン調節プロモーターであるが、しかし、本来のTREほどリーキーではない。このpTightは、ブラストサイジン耐性遺伝子を通じてアンチセンス発現を駆動している。
【0031】
RHGP12−RSN:このベクターはRheoSwitchプロモーターを使用し、「ネイキッド」アンチセンス転写物を促す。また、ベクターを、PGKプロモーターがブラストサイジン遺伝子の発現を駆動しているという事実により、薬物により選択することができる。また、EGFP遺伝子をブラストサイジン遺伝子の下流にIRES(配列内リボソーム進入部位)エレメントを介して置き、PGKプロモーターが両方のタンパク質の産生を駆動することができる。このベクターについての遺伝子地図を図5に示す。
【0032】
RHGP13−CMVN:このベクターもアンチセンス転写物を生成する「ネイキッド」プロモーターを有するが、しかし、このプロモーターは常時発現性であり(CMVプロモーター)、RheoSwitch又はTRE/pTightプロモーターように調節されない。これは、再び、インビボ試験のために有用であり、そこでは安定な非調節アンチセンス産生が意味をなす。このベクターは、RHGP12−RSNベクターと同じPGK−BS−IRES−EGFPコンストラクトを有する。
【0033】
RHGP−TAILプロトコール
RHGPの成功スクリーニングは、表現型をスクリーニングするために使用されるアッセイに依存的である。他のスクリーニング手順と同様に、所望の表現型は明確であり、頑健でなければならない。一旦それらの最小限の基準が満たされると、RHGPを実質的に任意の用途に適用することができ、それにおいて、所望の表現型によって細胞を物理的に単離することが可能になる。一例として、本発明者らの実験室及びその他ではRHGPを利用し、腫瘍細胞が、細胞傷害性薬剤を用いた化学療法攻撃(Lih, Wei, and Cohen, 2006; Reiske et al., 2009)又はホルモン療法に対して耐性となる標的を同定してきた。これらの条件下で、薬物感受性細胞が消滅し、RHGP摂動細胞のサブセットが生き残った。「オミクス」ベースのアプローチを典型的に表わす相関する知見とは異なり、薬物攻撃に直面した生存は、RHGP形質導入事象により直接的に起こされた。所望の表現型の選択は、細胞ベースのアッセイに限定されず、RHGPを同様にインビボでの適用のために使用することができる。例えば、動物モデルにおいて腫瘍形成能力又は転移能力を獲得する細胞を単離することができる。結果としての生存を超えて、RHGPは、また、他の特性(例えば特定のマーカーを発現する細胞の親和性ベースの単離など)について選択することができる。そのような結果は、特定のマーカー(所望の表現型として)の発現を使用し、フローサイトメトリー又はパニング手順を使用して細胞を物理的に単離することができる場合に可能である。
【0034】
相当な努力が投資され、RHGPの頑健性及び効率が最適化されてきた。GSVからの発現を促す可逆的プロモーターは、目的の標的を迅速に検証する効率的な手段を提供する。所望の表現型が得られた後、実験を、サイレンシングさせたGSVプロモーターを用いて繰り返し、表現型を逆転させることができる。表現型が、サイレンシングされたGSVを使用して持続する場合、そのクローンは優先順位から外されるであろう。なぜなら、表現型が、無関係のアーチファクトの結果として生じたかもしれないからである。そのようなものとして、一旦RHGPライブラリーが構築されると、目的の表現型について選択し、表現型の可逆性を検証することが可能になる(全て単一の試験において)。
【0035】
一旦表現型が、可逆的プロモーターを使用して検証されると、GSVは、標的遺伝子を同定し、GSV組込みの方向(例、センス又はアンチセンス)を決定するためのモチーフをコードする。RHGPの初期バージョンでは、宿主ゲノムDNAの物理的単離、それに続く従来の配列決定が要求された。この手順では、細菌形質転換のために十分なゲノムDNAを単離するための十分な数のクローンの増殖が要求された。個々の細菌クローンを次に単離し、プラスミドを、GSV組込み事象の配列決定のために単離した。このプロセスでは数週間の努力が要求され、限定されたスループットを伴った。標的同定は、熱非対称インターレース(TAIL)PCRの改変を使用して認められている。TAIL−PCRは、本来は、植物における遺伝子挿入を同定するために開発された(Liu et al., 1995)。より最近では、それを使用し、マウスゲノム中の遺伝子挿入が同定されている(Pillai et al., 2008)。本発明者らはTAIL−PCRを組み入れ、GSV挿入部位のハイスループット同定を実施した。なぜなら、少数の細胞が、ゲノムDNAを単離するために必要とされるからである。これらの改善によって、ハイスループットスクリーニングが促進され、GSV挿入部位についての情報が数週間ではなく数日以内に産生される。
【0036】
これは、トランスジーン挿入部位のハイスループットゲノム規模同定のためのTAIL−PCRの最初の適用、ならびにヒト細胞におけるトランスジーン挿入部位同定のためのTAIL−PCRの最初の適用である。
【0037】
本来のRHGPクローニング技術は、制限消化及び自己連結のための実質的な量のゲノムDNAを単離するために十分な数まで(例えば、6ウェルプレート密度まで)の哺乳動物細胞の増殖を含む。その後、自己連結プラスミドを細菌中に形質転換し、多くのクローンを個々に選び、一晩増殖させ、プラスミドをこれらの培養物から次の日に単離する。これらのプラスミドを、次に、トランスジーン挿入の部位を同定するための配列決定のために送る。
【0038】
TAIL−PCRを用いて、哺乳動物細胞を、96ウェルプレート中でのゲノム抽出のために直接的に処理することができ、それにより手順がハイスループットスクリーニングのために受け入れられるようになる。DNAを、次に、96ウェルプレートフォーマット中での、公知の及びランダムなプライマーの混合物を使用した3サイクルのTAIL−PCRに供し、次にプレートを、第3PCRラウンド後の配列決定のために直接提出し、挿入部位情報をわずか3日間で産出する。以下の表では種々の技術及びそれらの相対速度の推定を比較する(最適効率で):
【0039】
【表1】

【0040】
約50〜100ngのゲノムDNA/ウェルをPCR反応において使用する。より少ないDNAを、DNAが限定している場合に、確実に使用することができる。例えば、通常のゲノムDNA調製物は、0.2〜0.8μg/μlの範囲の産出量を有し、そのため、PCR反応のためのTE又はH2O中での1:10希釈で十分に作用する。各DNAサンプルを、1つのフォワード及び6つのリバースPCRプライマーの1つを有する6つの反応中に分けるため(全てが独立したウェル中)、1ゲノムDNAサンプル当たり8反応のプレミックスを、Immomix Red polymerase(Biolineから)を使用して調製する。フォワードプライマーの原液濃度は1μMであり、リバースプライマーは各々10μMである(それぞれ最終濃度0.15μM及び2μMについて)。
【0041】
【表2】
【0042】
一次TAIL−PCRサイクリング条件(約6時間):
【表3】
【0043】
レンチウイルスRHGPベクターについて、以下のプライマーを1DNAサンプル当たりに使用する(各々6反応):
一次TAIL−PCR:ISP1(フォワード)+AD1〜AD6(リバース)
二次TAIL−PCR:GSP3new(フォワード)+AD1〜AD6(リバース)
三次TAIL−PCR:ISP2(フォワード)+AD1〜AD6(リバース)
【0044】
フォワードプライマー:
【表4】
【0045】
リバースプライマー:
【表5】
【0046】
MMLV RHGPベクターについて、以下のプライマーを使用する:
【0047】
フォワードプライマー:
【表6】
【0048】
リバースプライマー(上と同じ)
【0049】
#1のサンプルを1%アガロースゲル上で流し、バンドを可視化することができる。その後、各サンプルの1:200希釈を行う。これは、最初に、各サンプルの水中への1:20希釈を行い、次に、この1μlを、二次TAIL−PCRのための続く10μlのPCR反応(1:200最終希釈比を達成するために)において使用することにより行う。時折、PCRサンプルにおける容積喪失があり、そのため、5μlの水を、各ウェルに(所望の場合)希釈前に加えることができる(同じことを三次TAIL−PCRの前に行うことができる)。二次TAIL−PCR用の1反応当たりのプレミックスは以下の通りである:
【0050】
【表7】
【0051】
リバースプライマー及び鋳型DNAを、マルチチャネルピペットを使用して分注することができる。二次TAIL−PCRサイクリング条件(約4時間):
【表8】
【0052】
#2のサンプルをゲル上で流し(所望の場合)、#2に示す通りに1:200希釈を行い、#2の同じプレミックスを使用し、三次TAIL−PCRを、以下のサイクリング条件を用いて行う(約2時間):
【表9】
【0053】
#3のサンプルを1%アガロースゲル上で流すことができる(所望の場合)。プレートを、次に、最終的な特異的プライマー(例えばレンチウイルス用のISP2、又はMMLV用の48RS3など)を使用した配列決定のために直接送ることができる。低容積についての懸念がある場合、15μlの水をより簡単な取り扱いのために各ウェルに加えることができる。
【0054】
この手順によって、クローンにおいてウイルス組込みの同じ部位が成功裏に同定されており、そこで、組込み部位が、本来のRHGPクローニングアプローチを使用し、RHGPレンチウイルス及びMMLVベクターの両方をヒト細胞株及び非ヒト細胞株において使用して同定されてきた。
【0055】
実施例1−インフルエンザ因子の研究及びPTCH1の同定
インフルエンザ感染は、感染性疾患媒介性の罹患及び死亡の主な原因のままである。蓄積する証拠は、季節性及び汎発性インフルエンザの大半のバリアントが、従来の治療に対する耐性を発生していることを示す。そのような情報は、インフルエンザを標的化するための新規アプローチの特定において新たな興味を生じている。本発明者らの実験室では、宿主志向性の治療法の新たな戦略を開発しており、それによって、ウイルスが疾患を起こすことを予防する安全で効果的な様式で宿主分子を標的化することを追求する。改善された発見技術(RHGP)を使用し、本発明者らは、PTCH1を、インフルエンザウイルス感染を決定的に制御する必須の宿主標的として同定する。本発明者らは、さらに、抗体又はsiRNAを使用したPTCH1に対する標的介入によってインフルエンザ感染が減少することを実証する。最終的に、本発明者らは、実験室の外部でのインフルエンザ感染におけるPTCH1の関与を、PTCH1の遺伝的変異が野外における減少した疾患の罹患率に関連することを示すことにより実証する。全体で、これらの知見は、インフルエンザ疾患の管理を改善するための新規の宿主指向の治療法の開発についての重要な意味を有する。
【0056】
インフルエンザは、感染性疾患媒介性の苦痛及び死亡の最も大きな原因の1つのままである。平均的な年において、インフルエンザは20〜30,000の生命を奪い、経済的な損害は何十億ドルにもなる。その広範囲の流行にもかかわらず、ワクチン接種は散発性のままである。最近の試験では、ワクチンが、処置された人々の半分未満において有効でありうることが示されている。これを複雑化することとして、わずか一握りの治療法が、インフルエンザ感染を処置するために開発されてきた。増加する証拠は、多くの季節性株及び汎発性(トリ)株がこれらの化合物に対して耐性になっており、このようにそれらの適用を取り除いていることを示す。薬物耐性株の不可避の増殖は、インフルエンザに固有ではない。インフルエンザ、HIV、ヘルペス、及び他のウイルスの耐性株の出現は、医学界が抗ウイルス治療のための戦略を再考することを強いてきた。
【0057】
従来の抗体ウイルス剤では、ウイルスによりコードされる経路が標的化されてきた。基本的な理論的根拠では、ウイルス経路の標的化によって正常宿主細胞に対する毒性が最小限になりうることが想定されてきた。しかし、結果として得られる化合物の多くが、実際に非常に有毒であり、依然として悪く、ウイルス標的への焦点は、病原体に対して選択圧力を置き、治療に耐性であるバリアントを引き出す。代替物についてのこの必要性は、本明細書において実施されるアプローチを促してきたが、それは、薬剤耐性に対して非感受性である新たな治療法を開発することを追求し、それは、広域スペクトルの異なるウイルス型に適用することができる。
【0058】
本発明者らの実験室では、感染性疾患のための宿主志向性の治療法の概念が開発されてきた。このアプローチでは、ウイルス感染細胞において誤発現される又は機能的に変化している宿主由来標的を同定することを追求する。そのように行う際、本発明者らは、変化した発現又は機能が、ウイルス疾患を処置する際に安全で効果的である新規治療法のための機会を提供しうると仮定してきた。異なる型のインフルエンザに必須である経路を同定、そして次に標的化することにより、本発明者らは、広域スペクトルの適用を伴う十分に必要とされる抗ウイルス剤を開発するであろう。知識における明らかなギャップは、安全で効果的な宿主標的及び、理想的には、それらの標的を遂行する手段を同定する必要性である。その目的のために、本発明者らは、本発明者らの中核技術(RHGP)を成功裏に利用した。本発明者らの実験戦略では、ゲノム中の単一部位でのGSVの組込みを中心とした。そこでは、それは標的遺伝子の過剰発現又は発現の喪失のいずれかを起こしうる。そのようなものとして、RHGPによって、本発明者らは、細胞ゲノム全体を照合し、宿主細胞がインフルエンザ感染に耐性となる又は生き残ることを可能にする異なる型の事象を同定することが可能になる。RHGPを使用し、PTCH1を、インフルエンザ感染における必須の宿主標的として同定した。PTCH1に対する標的介入によってインフルエンザ感染を遮断することができる。最終的に、SNP分析によって、PTCH1が、ブタ集団におけるインフルエンザの自然大流行の間での疾患転帰に関連することが実証される。
【0059】
材料及び方法
細胞培養。ヒトMDA−MB231乳癌細胞をAmerican Type Culture Collection(ATCC)から購入した。Phoenix A細胞株はNolan博士(Stanford University)からの寄贈であり、マウスN2a細胞株はXu博士(Rockefeller University)からの寄贈であった。MDA−MB231細胞をDMEM(10% FBSを含む)中で培養し、N2a細胞を50% DMEM培地及び50% Opti−MEM培地(5% FBSを含む)中で培養した。
【0060】
RHGPライブラリーの生成及びスクリーニング。RHGP遺伝子探索ベクターを適用し、MDCK細胞においてRHGPライブラリーを生成した。RHGPライブラリーを、インフルエンザウイルスA/Udorn/72を用いて、感染効率(MOI)0.1又は0.001で示された通りに感染させた。第2ラウンドの感染から2週間後、ゲノムDNAを、BIO−RAD AquaPure Genomic DNA Isolation Kitを使用して得た。自己連結ゲノムDNAを濃縮し、E.coli細胞中に電気穿孔し、クロラムフェニコールを使用して選択した。
【0061】
siRNAを用いた宿主標的遺伝子の検証。PTCH1についてのヒト二本鎖siRNAホモログを、製造者により推奨される通りに調製した。siRNA NP−1496配列(GGAUCUUAUUUCUUCGGAGUU)は、インフルエンザウイルスのヌクレオカプシド(NP)遺伝子を標的化し、ポジティブコントロールを提供した(Ge et al., 2003)。非標的siRNA(siCONTROL1)は、ネガティブコントロールを提供した。HEK293細胞を、24ウェルプレート中に、1×105個細胞/ウェルでそれぞれプレーティングした。24時間のインキュベーション後、細胞を、20nMのsiRNA及びTransIT−TKOを用いて、製造者の指示(Mirus)に従ってトランスフェクションした。第2ラウンドのトランスフェクションから24時間後、サンプルを、MEMを用いて洗浄し、インフルエンザウイルスA/Udorn/72(MOI 1)を用いた感染が続く。細胞を、1時間にわたり、15分毎に穏やかに揺らしながらインキュベーションした。各ウェルからの培養培地を感染後48時間目に回収し、培地中の子孫ウイルスを、標準的なプラークアッセイを使用して滴定した。
【0062】
フローサイトメトリー。フローサイトメトリー分析のために、MDCK細胞を、トリプシンを使用して浮遊させ、1×105個細胞を氷上で10mg/mL抗体を用いて30分間にわたりインキュベーションした。細胞をPBS(PBS中1% BSA(Sigma, St. Louis, MO)を含む)を用いて氷上で3回洗浄し、FITC抱合ヤギ抗ウサギ(Becton Dickinson, San Jose, CA)を用いて30分間にわたり氷上でインキュベーションした。洗浄後、細胞を、1%パラホルムアルデヒドを伴うPBS中で固定した。データは、EasyCyte Flow Cytometer(Guava Technologies, Hayward, CA)において取得され、FlowJo分析ソフトウェアを使用して分析した。
【0063】
SNP及びデータセット。PTCH1 SNPを選択し、2つの基準を満たした:(a)許容可能な対立遺伝子の頻度;及び(b)外部ジェノタイピングプロバイダー(Sequenom)による成功裏のフォーマッティング。
【0064】
インフルエンザデータセットを、商業用ブタ産生システムにおいて、PIC遺伝学を使用して収集した。試験期間の間に、農場は、ブタインフルエンザウイルスの大流行を経験した。4つの成長形質を測定した:(1)成長:これは、高及び低成長動物を比較した0/1形質である;(2)生命DG:生涯1日増体量(g/日);(3)LDG死体増体量:(グラム/日)及び(4)離乳:終点1日増体量(g/日)。
【0065】
LSMの好ましい遺伝子型−LSMの好ましくない遺伝子型/2
【0066】
結果
宿主細胞をインフルエンザに耐性にする標的を同定するために、RHGPライブラリーを、上に記載する通りにMDCK細胞において生成し、インフルエンザ攻撃を示した通りに実施した(図6A)。ライブラリーを、インフルエンザA/Udorn/72を用いた感染により攻撃し、インフルエンザ耐性細胞について選択した。本発明者らは、以前に、A/Udorn/72(MOI10−1)を用いた感染によって全てのMDCK細胞が48時間以内に再現性よく殺されることを確証していた(示さず)。コントロールとして、偽形質導入細胞の並行培養物を同じように処理し、生存体は48時間後に観察されなかった。
【0067】
複数ラウンドのインフルエンザ攻撃を生き残ったRHGP形質導入細胞についての選択後、本発明者らは、1つの特定の培養物(R26−7)に焦点を合わせ、それはインフルエンザ感染を生き残った。RHGP遺伝子探索ベクターによって、本発明者らは、標的遺伝子を効率的に位置付け、組込み事象の方向(センス又はアンチセンス)を決定することができた。ゲノムDNAをR26−7細胞から単離した。RHGPベクター挿入部位に隣接するゲノムDNA配列を配列決定し、イヌゲノムに対して、UCSC Genome Browserを使用してマッピングした(図6B)。R26−7における組込み事象が、イヌ染色体において、アンチセンス方向で、PTCH1遺伝子中で生じた(図6C)。そのようなものとして、組込みは、PTCH1の第1のコピーを物理的に破壊した。結果として得られるアンチセンスは、第2の対立遺伝子の発現をノックダウンした。
【0068】
天然細胞を使用したPTCH1の検証
本発明者らは、次に、PTCH1を、インフルエンザ媒介性の宿主細胞の殺傷を予防する候補標的として検証することを追求した。これらの試験は、RHGP又はMDCK細胞の使用から生じうるアーチファクトを除くことを意味した。PTCH1は、イヌ由来MDCK細胞から単離されていたため、本発明者らは、そのヒトホモログを同定し、ヒトHEK−293細胞においてこれらの標的について選択的なsiRNAを発現させた。二本鎖siRNAを生成し、HEK293細胞中にトランスフェクトした非標的siRNAは、トランスフェクションのための一致させたコントロール及び参照基準を提供した。NP−1496二本鎖siRNAは、決定的なインフルエンザ遺伝子を標的化し、インフルエンザ感染を阻害することが実証されており、ポジティブコントロールを提供した(Ge et al., 2003)。インフルエンザ感染はHEK293細胞を効率的に殺さないため、本発明者らは、本発明者らの実験プロトコールを改変し、ウイルス力価(宿主細胞の生存の代わりに)を、効率についての主要エンドポイントとして測定した。siRNA処理したHEK293細胞を、A/Udorn/72を用いて48時間にわたり感染させ、ウイルス力価をプラークアッセイにより測定した。PTCH1二本鎖siRNAは、インフルエンザウイルス産生を、ポジティブコントロール(NP−1496 siRNA)と同程度のレベルで減少させた。(図7)。
【0069】
追加の標的
同様の細胞において同じ手順を使用し、同様の攻撃を用いて、RHGP技術を使用し、追加の標的を同定した。これらの内で、SLC25A25(以前に、膜を横断するミトコンドリア転移及び恐らくはMg−ATP交換に関与するタンパク質として特徴付けられた)、Rgnef(細胞付着、細胞運動性、及びBリンパ球活性化、ならびに恐らくは細胞アポトーシスにおいて役割を有することが観察された)、Nedd4及びHerc6(両方ともユビキチンリガーゼ)(全てがインフルエンザのための潜在的な標的として)、ならびにRobo1(細胞運動性及び軸索ガイダンスに関与する受容体)(HIVのための潜在的な標的として)がある。これらの一部が以下で考察される。これらのタンパク質は、ウイルス感染様式で細胞表面上に現れるため、各々は、モノクローナル抗体、ペプチド、アンチセンス、siRNAなどを使用して標的化されうる。
【0070】
PTCH1の抗体標的化によってインフルエンザ感染が減少する。
PTCH1は、複数回貫通型膜タンパク質である。そのようなものとして、本発明者らは、それが、抗体ベースの介入のために標的を提供しうると考えた。この可能性を評価するために、親和性精製された抗血清を得て、インフルエンザ感染細胞の表面上のPTCH1を認識するそれらの能力についてテストした。MDCK細胞を、A/Udornインフルエンザを用いて、インタクトな細胞の染色の前に18時間にわたり感染させた。この実験計画を利用し、ウイルスを用いた長期のインキュベーション又は固定化(それらのいずれかによって膜の完全性が破壊されうる)の結果として生じうるアーチファクトについての潜在力を最小限にした。ウイルスによりコードされる赤血球凝集素及び一致させたコントロール(PTCH1を認識しない)について特異的な抗体は、ポジティブコントロール及びネガティブコントロールをそれぞれ提供した。PTCH1抗体はインフルエンザ感染細胞に結合し、かなり多くの染色が、非感染コントロールよりも、感染細胞を用いて観察された(図8A及び8B)。この結果は、膜の完全性の喪失を表わさなかった。なぜなら、細胞は、トリパンブルー及び他の生体染色色素を、利用された条件下で排除し続けたからである。
【0071】
PTCH1抗体がインフルエンザ感染細胞に結合する能力に基づき、本発明者らは、次に、これらの抗体を用いた処理がインフルエンザ感染性を変化させうるか否かを問うた。MDCK細胞を、インフルエンザを用いて、PTCH1抗体の存在又は非存在において感染させた。培養上清を次に収集し、感染性インフルエンザ粒子の数を、標準的なプラークアッセイを使用して定量化した。これらの試験によって、PTCH1抗体が、インフルエンザ増殖を、未処理細胞又は非特異的抗血清を用いて処理されていたコントロールと比較し、2〜3対数(100〜1000倍)だけ減少させることが実証された。本発明者らは、次に、この結果が、最初の感染又はウイルスライフサイクルの後の段階の抗体阻害を表わしうるか否かを問うた。この疑問は、より高い感染効率(MOI)0.1(0.001よりむしろ)で感染を繰り返すことにより対処した。本発明者らは、PTCH1抗体が、相対的に高いMOIでインフルエンザを比較的阻害する場合、それらは、最初の付着又は感染細胞中への侵入を遮断しているであろうと推論した。他方で、抗体がライフサイクルの後の段階(例、出芽又は放出)に影響を与えた場合、阻害活性はより高いMOIで減少しうる。後者の考え方と一致して、PTCH1抗体処理の抗ウイルス効果は、より高いMOIで減少し、インフルエンザにおける1対数(10倍)の減少を与えた。(図8C)これらの結果は、PTCH1が抗体治療のための適した標的を提供しうること、及び、抗体がウイルスライフサイクルの相対的に後の段階を遮断するように見えることを示唆する。
【0072】
PTCH1を、実験室から、野外におけるインフルエンザ感染に翻訳する。
細胞ベースのアッセイを用いた興味をそそる知見に基づき、本発明者らは、PTCH1が、実験室の外部でのインフルエンザ感受性に関連しうるか否かを問うた。このために、本発明者らは、ブタ集団における頑健形質の単一ヌクレオチド多型(SNP)評価を、ブタインフルエンザウイルス大流行の間に利用した。具体的には、本発明者らは、PTCH1 SNPがブタ体重増加における変化(ブタにおけるインフルエンザ感染の理解された結果である)に関連するか否かを問うた。
【0073】
PTCH1内又はその近くの一連の異なるSNP(619、661、662、及び663として公知)を、いくつかの基準(有意水準、対立遺伝子頻度、相加対優性効果、効果の大きさ(表現型の標準偏差))に基づいて選択した。本発明者らは、次に、SNPが野外においてインフルエンザ耐性に関連するか否かを問うた。このために、野外においてウイルス感染に暴露されたブタからのDNAを抽出し、各SNPについて遺伝子型を同定し、SNP遺伝子型と性能形質の間での統計的な関連性を決定した。試験では、データセット回収の期間中にブタインフルエンザ大流行を受けていた大農場からのデータセットを利用した。本発明者らの分析では、2つの異なるPTCH1 SNP(619及び660)を、離乳の時期以降での増加した平均1日体重増加及び体重増加と関連付けた(表I)。これらの結果は統計的に有意であり、PTCH1が体重増加(ブタ集団におけるインフルエンザ罹患の有意な指標である)と関連しうることを示唆する。
【0074】
このように、RHGP技術を使用し、主要な知見は、PTCH1細胞表面受容体がインフルエンザウイルスによる感染のために必須であること、及び、PTCH1に対する標的介入(siRNA又は抗体のいずれかを使用)によってインフルエンザ感染が減少しうることの実証である。本発明者らは、また、PTCH1が、多くの異なる種(ヒト、イヌ、及びブタを含む)からの細胞の感染の間にインフルエンザにより広く利用されることを示している。最終的に、本発明者らは、これらの基本的な知見の重要性を野外における状況に翻訳する(PTCH1が、ブタ集団の自然感染の間に疾患転帰に関連することを実証することによる)。
【0075】
本発明者らの試験の1つの新規局面は、PTCH1が宿主細胞のインフルエンザ感染を調節することの実証である。PTCH1は、ソニックヘッジホッグ(胚発生の間での細胞の組織化、増殖、及び生存を支配する分泌分子)のための細胞受容体のヒトホモログである。PTCH1は、また、腫瘍抑制因子としての役割に関与しており、種々の上皮性悪性腫瘍と関連付けられる。機構的な観点から、インフルエンザの基礎は未知であるが、しかし、異なるMOIでの抗体処理の差動的な効果は、ウイルスライフサイクルの相対的に後の段階におけるPTCH1についての役割を示唆しうる。
【0076】
本発明者らの本報告での別の重要な局面は、抗ウイルス疾患のための新規の宿主指向の標的を同定するためのRHGPの利用である。RHGPは、研究下にある表現型を調節する細胞中の任意の標的を同定するための機会を提供する(過剰発現又は発現の喪失を介して)。RHGPの中心となる特性は、疾患プロセスにおけるその関与を決定するための標的の事前の知識を有する必要がないことである。本発明者らの結果は、この概念をさらに支持する。なぜなら、PTCHは、以前に、インフルエンザに関連付けられていなかったからである。
【0077】
疾患管理の観点から、本発明者らの本知見についての興味深く重要な潜在的な意味がある。第1に、PTCH1は、PTCH1発現又は機能を標的化する抗体又は他のアプローチを使用して、インフルエンザによる感染に対して介入するための機会を提供しうる。比較可能な知見が、複数の種(ヒト、イヌ、ブタ)を使用して得られたとの事実は、PTCH1が、インフルエンザ感染のために使用される重要な中核機構を表わすことを示しうる。感染性疾患を標的化するための本発明者らの宿主指向のアプローチの中心となる教義は、基本的な経路の同定が、複数のウイルス株を標的化するための手段を提供しうるとの考え方に焦点を合わせている。今後の研究によって、異なる季節性及び汎発性形態のウイルス病原体におけるPTCH1の役割がさらに詳述されるはずである。宿主指向の治療から生じる第2の機会は、ウイルスが、重要な宿主機構に対する代替物を発生しなかったであろうという概念である。正しい場合、PTCH1の効果的な標的化は、インフルエンザの処置を長く悩ませてきた薬物耐性の獲得に関連する問題を克服する機会を提供しうる。
【0078】
表I。PTCH1多型は、ブタインフルエンザ大流行の状況におけるブタ集団の増加した体重増加と関連する−(好ましい対立遺伝子(主要対立遺伝子と比べて)の1コピーでの成長(グラム/日)に対するPTCH1多型の効果。全ての結果が、主要対立遺伝子と比べた、体重増加における相対的な増加として示される(p値)。生涯1日増体量=最終的な体重−出生時体重離乳終了時の1日増体量=最終的な体重−離乳時の体重)
【0079】
【表10】
【0080】
実施例2−他のインフルエンザ因子を同定するために使用されるRHGP
ウイルス病原体の治療的標的化のための従来のアプローチでは、耐性株の発生及び広域スペクトル用途の欠如から生じる障害に一貫して直面してきた。インフルエンザは特に問題となる治療課題を表わす。なぜなら、季節性抗原シフト及びドリフトは、高いウイルス突然変異率と合わせて、多くの従来の抗ウイルス剤を混乱させてきたからである。新たに出現する又は操作されたインフルエンザ株は、インフルエンザのトリ亜型における最近の興味により典型的に表わされる通り、さらにより大きな脅威を表わす。ウイルスによりコードされる分子の標的化に関連する限定に基づき、本発明者らは、ウイルス増殖を予防する又は宿主をウイルス媒介性の病原性から回避させる様式で、宿主経路を標的化する直交アプローチを取ってきた。この目的に対して、ランダムホモ接合性遺伝子摂動(RHGP)を使用し、正常細胞において十分に耐容されるが、しかし、インフルエンザを阻害する宿主志向性の標的が同定された。RHGPを用いたTAIL−PCRの使用によって、ゲノム全体の徹底的なスクリーニングが促進され(過剰発現又は発現の喪失の両方について)、宿主細胞をインフルエンザ感染に対して耐性にする標的が同定された。本発明者らは、インフルエンザが宿主細胞を殺すことを予防する一連の宿主志向性の標的を同定し、複数のアプローチを使用してこれらの標的を検証する。これらの試験では、ウイルス疾患と戦うための新たなパラダイムが支持され、新規の標的及び機構を同定するためのRHGPの能力が実証される。
【0081】
ウイルス感染に対処するための1つの新たなアプローチは、ウイルス疾患の病原性に必須である宿主因子を標的化することである。HIV/AIDSでの戦争からの顕著な例は、CCR5のアンタゴニストの開発である。これらのアンタゴニストは、結合及び内在化のために必要である表面暴露された共受容体の機能を予防することを追求する。初期の知見では、これらの薬剤が、CCR5栄養性ウイルスを用いて感染された患者のサブセットの処置のための十分に必要とされる選択肢を提供しうることを示唆する。
【0082】
実施例2において、哺乳動物(ヒトを含む)におけるインフルエンザに関する治療介入のための可能な標的の異なる研究を調べる。上で注目した通り、本発明は、改善された形態のRHGPの適用を含み、それは、また、細胞において任意の標的(全遺伝子又は個々のドメインを含む)のアップレギュレーションを起こすことができる。結果的に、RHGPは、研究下にある表現型の原因となる任意の遺伝子変化についてゲノム全体を照合する手段を提供する。本発明者らはRHGPを適用し、インフルエンザ感染について要求される一連の宿主志向性の標的を同定し、検証した。
【0083】
材料及び方法
化学物質及び細胞培養。MDCK細胞、ヒト293HEK細胞、及びMEM培地を、American Type Culture Collection(ATCC)から購入した。Fugene 6をRocheから購入した。Lipofectamine 2000、ドキシサイクリン(Dox)、T4 DNAリガーゼ、ElectroMax DH10Bコンピテント細胞、及びViraPower PackagingキットをInvitrogenから購入した。ベクターpIREShyg3及びpTRE−lucをClontechから得た。RNase A及びプロテイナーゼKをSigmaから購入した。インフルエンザウイルスA/Udorn/72はCharles River Labにより提供された。全ての制限酵素をNew England BioLabから購入した。細胞培養培地及び補助剤は、Invitrogen又はHycloneのいずれかからであった。全ての短鎖siRNAオリゴヌクレオチドをDharmacon Researchにより合成した。TransIT−TKO(siRNAトランスフェクション試薬)をMirusから購入した。MDCK細胞をMEM(10% FBSを含む)中で培養した。全ての細胞培養物を5% CO2を用いて37℃でインキュベーションした。Bright−Gloルシフェラーゼアッセイシステム(Dox誘導アッセイのために使用される)をPromegaから購入した。
【0084】
pTet−Offトランスアクチベーター−ベクターのコンストラクション及びMDCK/Tet−off安定細胞株。pTet−Offトランスアクチベーターベクターを、pIREShyg3ベクターを骨格として使用してコンストラクションした。PCR生成されたtTAフラグメントを、pIREShyg3ベクターのBamHI及びSofI部位中に挿入し、pTet−Offトランスアクチベーターベクター(pIREShyg3−tTA)を生成した。
【0085】
MDCK/Tet−Off細胞株を作製するために、10cmプレート中の1〜2×106個のMDCK細胞を、10μgのpIREShyg3−tTAベクター及び30μlのfugene 6を用いて、製造者の指示に従いトランスフェクトした。トランスフェクションから24時間後、MDCK細胞を1:10希釈により分割し、200μg/mlのハイグロマイシンを用いて培養した。培地を、3日毎に、非トランスフェクト細胞が殺され、トランスフェクトされた安定な細胞コロニーが形成されるまで交換した。目に見える個々のコロニーを選び、24ウェルプレート中で継代培養した。ハイグロマイシン耐性クローンを、Dox誘導活性について、pTRE−lucベクター及びBright−Gloルシフェラーゼアッセイキットを使用し、製造者の指示に従いスクリーニングした。40倍を上回る誘導性を伴う安定クローンを、RHGPライブラリーを作製するための標的細胞として使用した。
【0086】
RHGP遺伝子探索ベクター(pRHGP22)のコンストラクション。RHGP遺伝子探索ベクター(pRHGP22)を、pLESTベクターをCohen実験室(17, Lu, 2004)からの骨格として使用して構築した。pLEST中のNeoR−TRE−CMVのDNA配列を、最初に、pTRE−Tight−CMV−BlasticidinRの発現カセットを用いて置換した。3’UTR WPREエレメントを除去し、p15A ori−クロラムフェニコールアセチルトランスフェラーゼ(CAT)耐性遺伝子を同じ部位に挿入した。2つのLoxP部位を、BlasticidinR遺伝子の下流及びp15A ori−CATの上流にそれぞれ挿入した。Creレコンビナーゼを伴うLoxP部位によって、本発明者らは、TRE−CMVプロモーターを染色体から切り出すことにより表現型を可逆的に検証することができるようになるであろう。
【0087】
RHGPウイルスの産生及びRHGPライブラリーの構築。RHGPレンチウイルスを、InvitrogenのViroPower Lentiviral Expression Systemを使用して産生した。293FTの細胞を、10cmプレート中に、約1〜2×106個細胞/プレートでそれぞれプレーティングした。24時間のインキュベーション後、細胞を、3μgのRHGP22ベクター及び9μgのViraPower Packaging Mixを用いて、Lipofectimine 2000を使用してトランスフェクトした。培地を、新鮮なトランスフェクション培地を用いて、37℃での5時間のインキュベーション後に交換した。48時間のインキュベーション後、培地中のウイルスを回収し、0.45μmフィルターを通じてろ過した。レンチウイルスを、製造者の指示に従い滴定した。
【0088】
RHGPライブラリーを構築するために、MDCK/Tet−off細胞を、150mmプレート中に、2×106個細胞/プレートで、それぞれRHGPウイルスの収集の1日前にプレーティングした。次の日、細胞(4.6×106個細胞/プレート)を、10mlのPBSを用いて洗浄し、10mlの新鮮に収集されたRHGPウイルスを、6μg/mlのポリブレンを用いて形質導入した。細胞を37℃で2時間にわたりインキュベーションし、20分毎に揺らすことにより混合した。2時間のインキュベーション後、培地を、10mlの形質導入培地を用いて補充した。形質導入から24時間後、細胞を10mlのPBSを用いて洗浄し、インフルエンザウイルスA/Udorn/72(MOI0.1)を用いた2時間にわたる5mlの感染DMEM培地(0.2%のBSA、200μg/mlのハイグロマイシン、及び6μg/mlのブラストサイジンを含む)中での感染が続く。細胞を、穏やかに15分毎に揺らすことにより混合した。2時間の感染後、細胞を15mlの感染培地に加えた。24時間の感染後、細胞を、PBSを用いて洗浄し、インフルエンザウイルスA/Udorn/72を用いて再び同じMOIで感染させた。第2ラウンドの感染から2日後、細胞を、10mlのPBSを用いて洗浄し、20mlの50%条件培地(200μg/mlのハイグロマイシン及び6μg/mlのブラストサイジンを含む)に加えた。培地を、3日毎に、非形質導入細胞が殺され、ウイルス耐性コロニーが形成されるまで交換した。第2ラウンドの感染から2週間後、目に見える個々のコロニーを選び、24ウェルプレート中で継代培養した。
【0089】
インフルエンザウイルス耐性細胞クローンについての可逆性アッセイ。インフルエンザウイルス耐性クローンの可逆性を検証するために、表現型及び親MDCK/Tet−Off細胞を、Doxの非存在又は存在において3日間にわたり培養した。細胞を、100mmプレート中で、2×106個細胞/プレートでプレーティングし、Doxを用いて又は用いずに培養した。24時間後、細胞を、インフルエンザウイルスA/Udorn/72(それぞれMOIが1×10−2及び1×10−5)を用いて感染させた。3日間のインキュベーション後、細胞を、クリスタルバイオレット染色に供した。
【0090】
インフルエンザウイルス耐性クローンからの候補遺伝子の同定。典型的には、各クローンの1〜10×106個細胞を回収し、PBS溶液を用いて1回洗浄した。細胞ペレットを、10mlの溶解緩衝液(0.32Mスクロース、10mM Tris pH7.5、5mM MgCl2、及び1% Triton X−100を含む)中に再浮遊させた。ライセートを1500×gで15分間にわたり遠心分離した。ペレットを、0.5mlのプロテイナーゼK緩衝液(25mM EDTA、150mM NaCl、及び40mM Tris pH7.5を含む)中の100μgのプロテイナーゼKを用いて処理し、0.5% SDSの添加が続いた。混合物を一晩37℃でインキュベーションした。プロテイナーゼK処理されたライセートに50μgのRNase Aを加え、37℃で2〜4時間にわたりインキュベーションした。ゲノムDNAを、フェノール/イソアミルアルコール/クロロホルムを用いて2回抽出し、等容積のイソプロパノールを用いた沈殿が続いた。DNAペレットを、70%エタノールを用いて洗浄し、200μlのTE(pH7.5)緩衝液中に溶解した。
【0091】
250μl中のゲノムDNA(10μg)を、制限酵素BamHI又はXbaIを用いて2〜4時間にわたり消化した。消化されたDNAをフェノール/イソアミルアルコール/クロロホルムを用いて抽出し、エタノールを用いて沈殿し、30μlのTE緩衝液中に溶解した。250μl中の消化されたDNA(2.5μg)を、20単位のT4リガーゼを用いて16℃で一晩自己連結させた。連結されたDNAを、エタノールを用いて沈殿させ、70%エタノールを用いて洗浄した。DNAペレットを20μlのTE緩衝液中に溶解させ、DH10B ElectroMaxコンピテント細胞中に電気穿孔した。各々の連結DNAからのコロニー(8〜16)を、プラスミドDNA調製及びサイズ分析のための制限酵素を用いた消化のために選んだ。候補プラスミドDNAをさらに使用し、表現型に関与する標的遺伝子をDNA配列決定及びゲノムマッピングにより同定した。
【0092】
siRNAを用いた宿主標的遺伝子の検証。候補遺伝子MRPL42、COX5A、SLC25A25、及びTAPT1についてのヒト二本鎖siRNAホモログを、製造者によるsiGENOME SMART poolとして調製した。インフルエンザウイルスのヌクレオカプシド(NP)遺伝子に対するsiRNA NP−1496配列(GGAUCUUAUUUCUUCGGAGUU)を、ポジティブコントロールとして使用した(18, Ge, 2003)。非標的siRNA(siCONTROL 1)をネガティブコントロールとして使用した。HEK293細胞を24ウェルプレート中に1×105個細胞/ウェルでそれぞれプレーティングした。24時間のインキュベーション後、細胞を、20nMのsiRNA及び1.5μlのTransIT-TKOを用いて、製造者の指示に従いトランスフェクトした。細胞を、24時間のインキュベーション後、siRNAを用いて再トランスフェクトした。第2ラウンドのトランスフェクションから24時間後、細胞を0.5mlのMEMを用いて洗浄し、インフルエンザウイルスA/Udorn/72(MOI1.0)を用いた300μlのDMEM培地及び0.05% FBS中での感染が続く。細胞を、15分毎に穏やかに揺らしながら1時間にわたりインキュベーションした。細胞を、MEMを用いて2回洗浄し、ウイルス産生のために0.5mlの感染培地に加えた。各ウェルからの培地を、トランスフェクションから24又は48時間後に回収した。培地中の子孫ウイルスを、プラークアッセイにより滴定した。
【0093】
結果
RHGP細胞ライブラリーの構築
RHGPによって、細胞ゲノム全体を照合し、過剰発現及びノックアウト事象の両方を評価することを追求する。この課題では、宿主の全ゲノムのカバーを確実にするための十分な組込み事象を伴うRHGPライブラリーの作製が要求される。また、ライブラリーは、細胞当たり単一のRHGP組込み事象に限定され、同じ細胞での複数の摂動から生じる複雑さを防止することが重要である。ヒトゲノム中での平均遺伝子サイズが27kbと推定されるため(19)、本発明者らは、105の独立した組込み事象が、全ゲノムのランダムなカバーを確実にするために要求されると計算した(1遺伝子当たり1ベクター挿入)。RHKOを用いた先行する試験では、MMLVベースの組込みベクターに焦点を合わせていた(16)。しかし、これらのベクターの力価は、一般的に、103形質導入単位(TU)/mLに限定される。アプローチの効率を改善するために、レンチウイルスシステムを開発した。材料及び方法に概説されている通りに、pRHGP22を、ブラストサイジン耐性遺伝子を促すpTRE−Tight−CMVプロモーターからなる改善された発現カセットを使用して構築し、厳密な誘導制御下で選択可能マーカーを提供した。ori−クロラムフェニコールアセチルトランスフェラーゼ(CAT)耐性遺伝子は、独立したレポーターを提供し、宿主ゲノムDNAをレスキューし、標的遺伝子及びベクター組込みの部位を同定した。
【0094】
インフルエンザウイルス耐性表現型のスクリーニング
宿主細胞をインフルエンザに対して耐性にする標的を同定するために、RHGPライブラリーをMDCK細胞において生成した。この特定の細胞モデルを、インフルエンザ感染に対するその十分に確証された応答性に基づいて選択した。pRHGP22ベクターを用いた形質導入を介したライブラリー構築に続き、RHGP細胞ライブラリーを作製し、ヒトゲノムの100倍のカバーを確実にした。単一細胞内での複数の挿入についての潜在力を最小限にするために、低MOI(0.15)を、RHGPライブラリー調製のための標的細胞の形質導入の間に用いた。
【0095】
MDCK細胞ライブラリーを、本来なら致死濃度のブラストサイジン(感染から24時間後に投与)を使用して選択し、ベクター組込みを確実にした。ライブラリーを、また、インフルエンザA/Udorn/72を(相対的に高いMOI0.1で)用いた感染により攻撃し、インフルエンザ耐性細胞について選択した。インフルエンザ感受性細胞の徹底的な除去を確実にするために、培養物を複数ラウンドのインフルエンザ感染に供した。コントロールとして、一致させたコントロールを含めた。それは、RHGP形質導入を欠くが、しかし、その他の点では同様に処理された。一般的に、第2ラウンドのインフルエンザ攻撃後、生存細胞は、一致させたコントロールにおいて検出されなかった。その後、RHGP形質導入群中の生存細胞をクローン化し、増大させ、約3,000クローン/試験を産出した(表2)。これらから、合計194の個々のコロニーを、さらなる分析のために、単一ライブラリーから単離した。個々のクローンがインフルエンザ耐性表現型を保持したことを問うために、これらを、インフルエンザを用いて再攻撃した。再攻撃試験によって、本発明者らは、インフルエンザ攻撃に対していくらか感受性であるクローンを優先順位から外すことができた。14の高度に耐性のクローンのサブセットが、さらなる特徴付けのために選択された。
【0096】
ウイルス耐性クローンの可逆性アッセイ
RHGP技術の重要な特性は、誘導プロモーターによる調節を介して候補標的を検証する能力である。これによって、本発明者らは、自然突然変異又は感染アッセイでの他のアーチファクトの結果としてインフルエンザに対して耐性になったであろう任意の候補クローンを除去することができた。具体的には、インフルエンザ耐性クローン中に取り込まれたRHGPベクター用のプロモーターは、Tet−offシステムの制御下にあった。従って、外因的に加えられたテトラサイクリン(Tet)又はドキシサイクリン(Dox)の存在において、RHGPベクターを「オフにする」ことができる。この特性によって、本発明者らは、外因的に加えられたDoxの存在において攻撃された場合にインフルエンザに対して耐性のままであるそれらの候補を優先順位から外すことができた。これらの試験の経過において、本発明者らは、高い感染効率(MOI)でのインフルエンザ耐性が、インフルエンザライフサイクルの初期段階に関連する標的又は経路を好みうるのに対し、低MOIが、インフルエンザライフサイクルの後期段階のために要求される機構を好みうると考えた。このように、インフルエンザ攻撃を、2つの広く異なる感染効率(10−5又は10−2)で行い、アッセイシステムが、ウイルス耐性の初期又は後期のいずれかの段階の機構について偏りうる可能性を除いた。細胞生存を、クリスタルバイオレットを用いた感染から72時間後の染色により決定した。RHGP組込み事象を欠くコントロール細胞は、宿主細胞のウイルス媒介性の殺傷についてのポジティブコントロールを提供した。分析された14クローンの内、12(86%)がDoxの存在においてインフルエンザに対する感受性を再獲得し、残りの2つがさらなる研究のために優先順位から外された。
【0097】
RHGP遺伝子探索ベクターを設計し、標的遺伝子を効率的に位置付け、組込み事象の方向(センス又はアンチセンス)を決定した。具体的には、遺伝子探索ベクターはCATレポーター遺伝子をコードし、それは、制限酵素ベースのゲノムDNAクローニングによりレスキューすることができる。ゲノムDNAを、検証され、インフルエンザ耐性のDox媒介性の可逆性が実証されたそれらのクローンから単離した。ゲノムDNAを、適した制限酵素を用いて消化し、自己連結させ、環状ベクターゲノムDNAを得た。これらの構築物を使用し、E. coliを形質転換させ、クロラムフェニコール耐性サブクローンを単離し、配列決定した。ベクター組込みのゲノム部位を確認するために、少なくとも8〜16の異なるクロラムフェニコール耐性コロニーを単離し、各MDCKクローンから配列決定した。RHGPベクター挿入部位に隣接する結果として得られるゲノムDNA配列を、イヌゲノムに対する、標準的な探索アルゴリズムを使用したゲノムマッピングに供した。実験計画と一致して、大半の標的が、MDCK細胞当たり単一の組込み部位からなった。しかし、一部のクローンは複数の組込み事象を含み、それらは、複数のRHGP組込み部位の結果として又はその特定の候補が複数のクローンからなっていた場合に生じたであろう。また、本発明者らは、技術的な限定に起因して、他の候補の小さなサブセットから候補遺伝子を単離することができなかった。全体で、ゲノムDNAクローニングによって、インフルエンザ耐性に関連する合計70の異なる標的遺伝子が同定された。
【0098】
ナイーブな細胞を使用した標的遺伝子の検証
RHGPによって、インフルエンザ感染に対して耐性を付与する一連の標的が同定された。表現型の可逆性によって、実験システム内での標的の適用性が検証された。本発明者らは、また、独立した実験システムを使用してこれらの候補の検証を追求し、RHGP、宿主細胞システム(MDCK)の選択、又はインフルエンザバリアント(Udornウイルス)に固有でありうる予想外の結果についての潜在力を除いた。
【0099】
RHGPを使用して同定された標的の独立した確認として、これらの標的のサブセットを選択し、より従来のアプローチを使用してテストした。具体的には、本発明者らは標的を強調しており、そこでは減少した発現がインフルエンザ耐性に関連した。なぜなら、部分的には、ノックダウン標的が、将来の標的戦略に受け入れられ、小分子又は生物製剤ベースの治療を使用して発現又は機能を同程度にノックダウンしうるからである。重要なことには、MDCK細胞からの結果によってイヌ標的が同定され、siRNAを、標的遺伝子のヒトホモログに対して設計した。二本鎖siRNAを、4つのそのような標的(MRPL42、COX5A、TAPT1、及びSLC25A25)について生成し、HEK293細胞をトランスフェクトするために使用した。インフルエンザは一般的にHEK293に対して毒性がないため、ウイルス力価の測定は、効力を評価するためのエンドポイントとしての役割を果たした。本発明者らの最初の一連の試験のために、siRNA処理した293細胞を、RHGP試験のために使用した同じインフルエンザ株(A/Udorn/72)を用いて感染させた。Udornを用いた攻撃に続き、ウイルス力価を評価し、siRNAトランスフェクト細胞においてより低いレベルのウイルスを実証した。サイトカインの同化に対するsiRNAのアーチファクトベースの効果についての最近の証拠に照らして、並行試験を、無関係のsiRNAコンストラクトを用いて行い、このように、目的の標的についての抗ウイルス効果の特異性を検証した。
【0100】
Udornを使用した知見の検証後、本発明者らは、次に、結果を他のインフルエンザバリアントに拡大した。例えば、siRNAは、また、異なるインフルエンザ株(A/WSN/33)に対する耐性を付与した。比較可能な知見が、異なる感染効率で得られた。全体で、これらの知見は、新規の宿主ベースの標的を同定するためのRHGPの適用を検証し、異なる形態のインフルエンザに対する広域スペクトルの適用のための潜在力を示唆する。
【0101】
この実施例において、RHGPが、インフルエンザウイルス感染のために要求される任意の宿主因子(過剰発現又はノックダウンされている標的を含む)のゲノム規模のスクリーニングを行い、RHGPベクター内の内部エレメントを使用してこれらの標的を検証するための効率的な手段を提供することができることが示されている。このゲノム規模のスクリーニングでは、インフルエンザについての効果的な宿主志向性の標的としてMRPL42、COX5A、TAPT1、及びSLC25A25タンパク質に優先順位が付けられた。これらの標的の効率は、独立した試験において従来のsiRNAアプローチを使用してさらに検証された。
【0102】
本発明者らの本試験の1つの固有の局面は、改善された形態のRHGPの使用である。以前の試験では、MMLVベースのベクターが用いられ、二倍体細胞において遺伝子の両方のコピーが阻害された。しかし、MMLVベクターは、内因的に限定された低力価の産生であった。本発明者らは、レンチウイルスシステムによって以前の限定が克服されることを実証する。結果として得られるシステムは、ヒトゲノム全体の100倍のカバーを確実とするために十分であった。それは、過剰発現された標的及びノックダウンされた標的の両方の徹底的な評価を行うために重要である。さらに、レンチウイルスベクターの使用は、活性な遺伝子転写の部位中への単一部位挿入を強く好む。それはスクリーニング手順の効率を増加させる。
【0103】
1つの異なる利点は、RHGPが標的の事前の知識により偏らず、同定された標的遺伝子がしばしばまだアノテートされていないことである。しかしながら、本発明者らは、PANTHER Classification System(Mi, 2005)を用いてRHGPインフルエンザ遺伝子のヒトホモログを分析し、推定上の生物学的経路を同定した。これらの標的は、生物学的プロセス及び分子機能のカテゴリーに広く分類された(図9)。70の標的の内、61(87%)を分類することができ、9(13%)はできなかった。それらの61の遺伝子によって、7つの主要な生物学的プロセス及び8つの主要な分子機能が同定された。生物学的プロセスの内で顕著なのは、核酸及びタンパク質代謝、シグナル伝達、及び細胞内タンパク質輸送に関連する経路であった。具体的には、標的機能は、核酸結合に関連する内因活性、酵素活性(キナーゼ、リガーゼ、ヒドロラーゼ、レドックス)、細胞シグナル伝達、及び細胞内トランスポートを含んだ。初期の指標は、これらの標的クラスがランダムではないことを示唆する。なぜなら、本明細書において同定されたキナーゼ、膜輸送タンパク質、リガーゼ、ヒドロラーゼ、及びトランスファー/キャリアタンパク質の頻度は、公知の標的クラスのランダム検索を行う場合に予測されうるものより少なくとも3倍高かったからである。
【0104】
増加する証拠は、ウイルス疾患のための宿主志向性の治療法の概念を支持してきた。特に、一連の最近のsiRNAベースの試験が行われ、インフルエンザ、HIV、又はウエストナイルウイルスによる感染と関連する宿主標的が同定されている。siRNAは、標的発見のためにますます利用されているが、しかし、siRNAの頑健で持続的な過剰発現の必要性により内因的に。一部のsiRNA知見の結果は、技術自体が、本来なら、細胞挙動をどのように変化させうるかについての懸念により曇らされてきた。特に、siRNAがサイトカイン発現を変化させうるとの最近の報告は、感染性疾患への適用について特に問題である。本発明者らは、RHGPが改善された代替物を提供しうると考える。なぜなら、それは、これらの同じ制約により限定されないからである。実際に、本明細書において同定された標的及び経路の大半(73%)が新規である。この結果は、RHGPが、また、まだアノテートされていない標的ならびに過剰発現されるタンパク質又はドメインを同定する能力を有することを反映する可能性が高い。対照的に、siRNAアプローチは、一般的に、公知の又はアノテートされた標的のノックダウンに限定される。興味深いことに、MDCK細胞のRHGP照合を使用して本明細書において同定された標的又は経路の27%が、ヒト細胞のHIV又はウエストナイル感染と関連する遺伝子とある程度の関係(配列、タンパク質ファミリー、又は機構的経路に関して)を有する(表3)。この観察は、異なるウイルス病原体が収束的に進化し、特定の決定的な宿主経路を共有することを示唆する。これらの経路は、しばしば、異なる哺乳動物種(例、イヌ及びヒト)の間で保存される。とりわけ、本明細書において同定された遺伝子は、最近のインフルエンザ試験とはあまり似てない。本発明者らは、この外見上の矛盾が、インフルエンザを用いた以前の試験でDrosophila細胞株が利用されたとの事実を表わしていると仮定する。それは重要である。なぜなら、ショウジョウバエは、哺乳動物細胞とは遺伝的に異なるからであり、なぜなら、Drosophilaゲノムは、約14,000遺伝子(ヒト又はイヌのゲノムより少ない)に限定されるからである。
【0105】
さらなる研究によって、RHGPを使用して同定された標的の一部をウイルス感染と関連付ける潜在的な機構情報が明らかになる。例えば、減少したMRPL42、COX5A、TAPT1、及びSLC25A25発現が、インフルエンザ感染に対する感受性を減少させる(図10)。MRPL42は、ミトコンドリアリボソームタンパク質(核遺伝子によりコードされる)であり、ミトコンドリアのタンパク質合成に関与する。最近の報告では、CD4+T細胞におけるSIV−Nefの導入が、関連する宿主ミトコンドリアタンパク質(MRPL1、MRPL14、及びMRPL19)の発現を刺激することが実証された。この報告では、これらの因子のアップレギュレーションが、ウイルスライフサイクルの後期段階で要求されるウイルスメッセージの産生を促進しうることが示唆された。正しい場合、本発明者らの結果は、この知見をMRPL42まで拡大させ、インフルエンザが同様の機構を利用しうることを示す。同様の知見によって、本明細書において同定された別の標的(COX5A)がウイルス感染と関連付けられる。COX5Aは、チトクロムCオキシダーゼ(COX)複合体の1成分である。異なるサブユニット(COX6A1)が、報告によると、インフルエンザウイルス複製のために要求される。興味深いことに、膜貫通タンパク質TAPT1(膜貫通前後形質転換1)は、脊椎動物胚における発生の間での軸骨格パターン形成のために要求され、遺伝子の一部が、HCMV gH受容体(gH受容体に対するモノクローナル抗体を用いたcDNA発現ライブラリーのスクリーニングにより同定された)をコードすることが明らかにされた。TAPT1が病原性に関与する機構は不明のままであるが、TAPT1が、また、インフルエンザウイルス感染に関与しうることも可能である。SLC25A25(可溶性キャリアファミリー25、メンバー25)タンパク質は、ミトコンドリア及びリン酸キャリアのスーパーファミリーのメンバーである。スーパーファミリーにおける多数の同様のタンパク質が、報告によると、HIV及びWNV感染に関与してきた。
【0106】
遺伝子不活性化に加えて、他の遺伝子のRHGP誘導によってインフルエンザ感受性が減少する。これらの内で顕著なのは、B細胞CLL/リンパ腫2(BCL2)及びプラスミノーゲンアクチベーターウロキナーゼ(urakinase)(PLAU)遺伝子である。プロトがん遺伝子BCL2が、細胞生存の十分に特徴付けられた調節因子である。最近の報告では、BCL2は、シンドビス、インフルエンザウイルス、又はHIV−1により起こされたアポトーシスに続く細胞生存の調節と関連付けられている。同様に、RHPGは、また、PLAUセリンプロテアーゼの過剰発現をインフルエンザと関連付けた。とりわけ、PLAUのアミノ末端フラグメントが、最近、感染細胞からのHIV−1の組み立て及び出芽を抑制することが示された(Wada, 2001)。ゲノム相関によって、また、PLAU及びその受容体(PLAUR)が、ヒト呼吸器合胞体ウイルスによる感染と関連付けられている。
【0107】
実施例3−RHGPを使用した潜在的なインフルエンザ標的のさらなる同定
上の実施例2に示すものと非常に類似の技術を使用し、インフルエンザウイルス感染を予防又は処置するための治療介入のための追加の潜在的な標的が同定された。
【0108】
材料及び方法
化学物質及び細胞培養。MDCK細胞、ヒト293HEK細胞、及びMEM培地をAmerican Type Culture Collection(ATCC)から購入した。インフルエンザウイルスA/Udorn/72をCharles River Labから購入した。全てのsiRNAオリゴヌクレオチドはDharmacon Researchにより合成された。
【0109】
pTet−Offトランスアクチベーターベクターの構築及びMDCK/Tet−off安定細胞株。pTet−Offトランスアクチベーターベクターを、pIREShyg3(Clontech)を骨格として使用して構築した。PCR生成されたtTAフラグメントを、pIREShyg3ベクターのBamHI及びSofI部位中に挿入し、pTet−Offトランスアクチベーターベクター(pIREShyg3−tTA)を生成した。
【0110】
MDCK/Tet−Off細胞株を作製するために、2×106個のMDCK細胞を10cmプレート中で培養し、10μgのpIREShyg3−tTAベクターを用いて、fugene 6(Roche)を使用してトランスフェクトした。トランスフェクションから24時間後、MDCK細胞の対数期の培養物を、200μg/mlハイグロマイシンを使用して選択し、培地を、3日毎に、全てのコントロール細胞が殺されるまで交換した。ハイグロマイシン耐性クローンを、pTRE−luc(Clontech)ベクター及びBright−Gloルシフェラーゼアッセイキット(Promega)を使用し、ドキシサイクリン(Doxycylcine)(Dox;Invitrogen)誘導活性についてスクリーニングした。少なくとも40倍の誘導を伴う安定クローンを使用し、RHGPライブラリーを作製した。
【0111】
RHGP遺伝子探索ベクター(pRHGP22)のコンストラクション。RHGP遺伝子探索ベクター(pRHGP22)を、pLESTベクター(Stanley Cohen博士(Stanford)により提供)(Lu et al., 2004)を骨格として使用してコンストラクションした。pTRE−Tight−CMV−BlasticidinRを、プロモーターとして置換した。3’UTR WPREエレメントを除去し、p15A ori−クロラムフェニコールアセチルトランスフェラーゼ(CAT)耐性遺伝子を同じ部位に挿入した。2つのLoxP部位を、BlasticidinR遺伝子から下流及びp15A ori−CATから上流にそれぞれ挿入した。
【0112】
RHGPウイルスの産生及びRHGPライブラリーの構築。RHGPレンチウイルスを、ViroPower Expression System(Invitrogen)を使用して産生した。HEK293FT細胞を、10cmプレート中に106個細胞/プレートでプレーティングした。24時間のインキュベーション後、細胞を、3μgのRHGP22及び9μgのViroPower Packaging Mixを用いて、Lipofectimine 2000(Invitrogen)を使用してトランスフェクトした。培地を、37℃で5時間のインキュベーション後に交換した。48時間後、培養培地中のウイルスを、0.45μmフィルターを通じてろ過し、製造者の指示に従い滴定した。
【0113】
RHGPライブラリーを構築するために、MDCK/Tet−off細胞を、15cmプレート中に、2×106個細胞/プレートでプレーティングした。次の日に、サンプルを、PBSを用いて洗浄し、6μg/mlポリブレン中のRHGPウイルスを用いて形質導入した。サンプルを、新鮮培地を用いた補充の前に混合しながら37℃で2時間にわたりインキュベーションした。形質導入から48時間後、細胞を、PBSを用いて洗浄し、インフルエンザウイルスA/Udorn/72を用いて、感染効率(MOI)0.1で2時間にわたり、DMEM培地(0.2% BSA、200μg/mlハイグロマイシン、2μg/mlウシトリプシン、及び6μg/mlブラストサイジンを含む)中で、15分毎に穏やかに混合しながら感染させた。培地を、次に、MEMを用いて補充した。2日後、細胞を、PBSを用いて洗浄し、上に示すとおりに、インフルエンザウイルスを用いて再び感染させた。第2ラウンドの感染後、サンプルを、PBSを用いて洗浄し、条件培地(200μg/mlのハイグロマイシン及び6μg/mlのブラストサイジンを含む)中でインキュベーションした。培地を、3日毎に、全ての非トランスフェクト細胞が殺され、ウイルス耐性コロニーが形成されるまで交換した。第2ラウンドの感染から2週間後、目に見える個々のコロニーを単離し、24ウェルプレート中で継代培養した。
【0114】
インフルエンザウイルス耐性細胞クローンについての可逆性アッセイ。RHGP事象の可逆性を検証するために、MDCK/Tet−Off細胞を、Doxの非存在又は存在において3日間にわたり培養した。細胞を、10cmプレート中で、2×106個細胞/プレートでプレーティングし、Doxを用いて又は用いずに培養した。24時間後、サンプルを、インフルエンザウイルスA/Udorn/72(それぞれMOIが1×10−2及び1×10−5)を用いて感染させた。3日後、生存細胞を、クリスタルバイオレットを用いて染色し、残りの細胞を可視化した。
【0115】
インフルエンザウイルス耐性クローンからの候補遺伝子の同定各クローンからの1×106個細胞を、溶解緩衝液(0.32Mスクロース、10mM Tris pH7.5、5mM MgCl2、及び1% Triton X−100を含む)中に浮遊させた。ライセートを1500×gで15分間にわたり遠心分離した。ペレットを、プロテイナーゼK緩衝液(25mM EDTA、150mM NaCl及び40mM Tris、0.5% SDS、pH7.5を含む)中の100μgのプロテイナーゼK(Sigma)を用いて処理した。混合物を一晩37℃でインキュベーションし、50μgのRNase A(Sigma)を用いて処理し、37℃で2時間にわたりインキュベーションした。ゲノムDNAを、フェノール/イソアミルアルコール/クロロホルムを用いて抽出し、イソプロパノールを用いた沈殿が続いた。DNAペレットを、70%エタノールを用いて洗浄し、TE緩衝液(pH7.5)中に溶解した。
【0116】
ゲノムDNA(10μg)を、制限酵素BamHI又はXbaIを用いて消化し、フェノール/イソアミルアルコール/クロロホルムを用いて抽出し、エタノールを用いて沈殿し、TE緩衝液中に溶解した。消化されたDNA(2.5μg)を、T4リガーゼ(Invitrogen)を使用して16℃で一晩自己連結させた。連結されたDNAを、エタノールを用いて沈殿させ、70%エタノールを用いて洗浄した。DNAペレットをTE緩衝液中に溶解させ、DH10B ElectroMaxコンピテント細胞(Invitrogen)中に電気穿孔した。複数のコロニーを、プラスミドDNA調製及び制限酵素消化のために単離した。プラスミドDNAをさらに使用し、標的遺伝子をDNA配列決定及びゲノムマッピングにより同定した。
【0117】
siRNAを用いた宿主標的遺伝子の検証。候補遺伝子MRPL42、COX5A、SLC25A25、及びTAPT1についてのヒト二本鎖siRNAホモログを、製造者により推奨される通りに調製した。siRNA NP−1496配列(GGAUCUUAUUUCUUCGGAGUU)は、インフルエンザウイルスのヌクレオカプシド(NP)遺伝子を標的化し、ポジティブコントロールを提供した(18, Ge, 2003)。非標的siRNA(siCONTROL1)がネガティブコントロールを提供した。HEK293細胞を24ウェルプレート中に1×105個細胞/ウェルでそれぞれプレーティングした。24時間のインキュベーション後、細胞を、20nMのsiRNA及びTransIT-TKOを用いて、製造者の指示(Mirus)に従いトランスフェクトした。細胞を、24時間のインキュベーション後、siRNAを用いて再トランスフェクトした。第2ラウンドのトランスフェクションから24時間後、サンプルを、MEMを用いて洗浄し、インフルエンザウイルスA/Udorn/72(MOI1)を用いた感染が続く。細胞を、15分毎に穏やかに揺らしながら1時間にわたりインキュベーションした。各ウェルからの培養培地を、トランスフェクションから48時間後に回収した。培地中の子孫ウイルスを、標準的なプラークアッセイを使用して滴定した。
【0118】
結果
RHGP細胞ライブラリーの構築
RHGPの中心となる特性は、固有のレンチウイルスベースの遺伝子エレメントであり、遺伝子探索ベクター(GSV)として公知であり、それは、ゲノム全体を照合し、宿主細胞がインフルエンザ感染に耐性となる又は生き残ることを可能にする標的を同定するように設計された。本発明者らの実験戦略では、ゲノム中の単一部位でのGSVの組込みを中心とした。そこで、それは誘導プロモーターを介して標的遺伝子の発現を調節した。ベクターは自己不活性化レンチウイルスLTRをコードし、それは、GSVが形質導入細胞から再出現することを予防した。RSV E/Pプロモーターは、GSVの産生のためだけに使用され、宿主ゲノム中へのGSVの組込みにより除去された。
【0119】
GSVは、センス又はアンチセンスのいずれかの方向で組込まれうる。アンチセンス配置において、組込み事象自体が1つの対立遺伝子を不活性化し、アンチセンス構築物の発現を促進し、それによって他の対立遺伝子上でコードされる遺伝子がノックダウンされた。この方法において、RHGPは、二倍体細胞における両方の遺伝子コピーのホモ接合性摂動を促進した。反対(センス)方向では、RHGPは、標的遺伝子の過剰発現を促進した。この結果は、遺伝子全体の単純な過剰発現を超えて拡大しうる(例、開始部位の上流での挿入)。なぜなら、開始部位の下流への組込みはドメインの過剰発現を引き起こし、野生型遺伝子の機能のドミナントネガティブな阻害剤を産生しうる。そのようなものとして、RHGPによって、本発明者らは、細胞ゲノム全体を照合し、宿主細胞がインフルエンザ感染に耐性となる又は生き残ることを可能にする異なる型の事象を同定することができた。
【0120】
RHGPを促進するために、MDCK細胞のライブラリーを、レンチウイルスベースのGSVを使用して形質導入する。ゲノム中での平均遺伝子サイズが27kbと推定されるため(Gupta and Varshney, 2004)、本発明者らは、105の独立した組込み事象が、ゲノム全体のランダムなカバーを確実にしうると計算した(Li and Cohen, 1996)。材料及び方法に概説されている通り、pRHGP22を、ブラストサイジン耐性遺伝子を促すpTRE−Tight−CMVプロモーターからなる発現カセットを使用して構築し、厳密な誘導制御下で選択可能マーカーを提供した。形質導入された宿主細胞のライブラリーを、次に、ブラストサイジンを使用して選択し、生存細胞を、以下で詳述する通りに、インフルエンザ耐性についてスクリーニングした。
【0121】
インフルエンザウイルス耐性表現型のスクリーニング
宿主細胞をインフルエンザに対して耐性にする標的を同定するために、RHGPライブラリーをMDCK細胞において生成し、インフルエンザ攻撃を実施した。この特定の細胞モデルを、インフルエンザ感染に対するその十分に確証された応答性に基づいて選択した。なぜなら、イヌゲノムがアノテートされており、ヒトホモログと比較することができるからである。本発明者らは予備試験を行い、MDCK細胞が、選択のために利用される条件下でA/Udorn/72により効率的かつ完全に殺されることを確実にした。この結果は、生存細胞が、RHGP摂動の結果として(インフルエンザに対する自然耐性のアーチファクトとしてではなく)生じることを確実にするために必須であった。pRHGP22 GSVを用いたライブラリー構築に続き、少なくとも107の独立したMDCKクローンからなるライブラリーを作製し、ゲノムの100倍のカバーを確実にした。低MOI(0.1)をライブラリー作製の間に用いて、1を上回る異なるGSVによる任意の細胞の形質導入を最小限にした。GSV組込みを確認する追加の手段として、MDCKライブラリーを、致死濃度のブラストサイジン(感染から48時間後に投与)を用いてインキュベーションした。
【0122】
ライブラリーを、インフルエンザA/Udorn/72を用いた感染により攻撃し、インフルエンザ耐性細胞について選択した。本発明者らは、以前に、A/Udorn/72(MOI10−1)を用いた感染によって全てのMDCK細胞が48時間以内に再現性よく殺されることを確証していた。コントロールとして、偽形質導入細胞の並行培養物を同じように処理し、生存体は48時間後に観察されなかった。本発明者らは、さらに、生存細胞を複数ラウンドの致死攻撃に供することによりアーチファクトを最小限にすることを追求した。このように、サンプルを少なくとも3ラウンドのインフルエンザ攻撃に供し、その時に、生存細胞は、一致させたコントロールのいずれにおいても検出されなかった。
【0123】
これらの試験の経過において、本発明者らは、高い相対的MOIのインフルエンザでの攻撃が特定の型の標的について偏りうると考えた。例えば、高いMOIでの生存が、意図的にではなく、感染の初期段階に関連する標的を好みうる、又は、後の段階での事象に関連する標的に対して偏りうる。そのような偏りを予防するために、並行した一連の試験を行い、低い力価でのウイルス攻撃に関連する生存を比較した。このために、クローンを、低い最初のMOI(MOI10−5)での感染に供した。培養物を、次に、経時的に、複数ラウンドの感染を使用して評価し、コントロールサンプル中の全ての細胞がインフルエンザにより殺されていたことを確実にする。
【0124】
インフルエンザを用いた攻撃を生き残ったRHGP形質導入細胞についての選択後、クローンを、単一細胞クローニングにより単離し、そして増大し、平均3,000クローン/実験を産出した。豊富なクローンを仮定し、303の異なるインフルエンザ耐性クローンのサブセット(全集団の10%)をランダムに選択し、第2ラウンドの単一細胞クローニングに供し、クローン性を確実にした(表2)。これらのサブクローンを、次に、一連の試験に供し、表現型を確認し、標的遺伝子を同定した。各サブクローンを、再び、インフルエンザを用いた致死攻撃を生き残る能力についてテストした。不活性Tet−off GSVを用いて形質導入されていた親MDCK宿主細胞は、一致させたネガティブコントロールを提供した。129の異なるサブクローン(最初の集団の43%)が、インフルエンザを用いた致死攻撃を生き残るそれらの能力に基づいて単離された。この段階での下方選択の大半が、単一細胞クローニングプロセスを生き残ることができなかったコロニーに起因した。
【0125】
ウイルス耐性細胞クローンの可逆性アッセイ
RHGP技術の重要な特性は、誘導プロモーターによる調節を介して候補標的を検証する能力である。これによって、本発明者らは、自然突然変異又はRHGPに関連しない他のアーチファクトの結果としてインフルエンザに対して耐性になったであろう候補を除去することができた。RHGPベクター用のプロモーターはTet−offシステムの制御下にあったため、本発明者らは、ドキシサイクリンの存在又は非存在において候補のインフルエンザ媒介性の殺傷を比較した。この試験によって、129の生存クローン中111(86%)でインフルエンザ攻撃に対する可逆性の耐性が実証された。残りの18クローン(14%)が優先順位から外され、RHGP、及び無関係ではないアーチファクトがインフルエンザ耐性表現型に関与することを確実にした。
【0126】
ゲノムDNAクローニングによる宿主遺伝子の同定
RHGP遺伝子探索ベクターを設計し、標的遺伝子を効率的に位置付け、組込み事象の方向(センス又はアンチセンス)を決定した。具体的には、遺伝子探索ベクターはOri−CATレポーター遺伝子をコードし、それは、制限酵素ベースのゲノムDNAクローニングによりレスキューすることができる。ゲノムDNAを、インフルエンザに対する実証された可逆的な耐性を有する111のクローンから単離し、合計110の標的遺伝子を産出した(表I)。RHGPベクター挿入部位に隣接する結果として得られるゲノムDNA配列を、イヌゲノムに対する、UCSC Genome Browserを使用したゲノムマッピングに供した。実験計画と一致して、大半の標的が、1クローン当たり単一の組込み部位からなった。本発明者らは、技術的な限定に起因して、候補の小さなサブセットから候補遺伝子を単離することができなかった。また、少数のサンプルが複数の組込み事象を含んだ。それらは、複数のRHGP組込み部位の結果として又はサブクローンがクローン性ではなかった場合に生じたであろう。このあいまい性に起因し、複数の組込み事象に関連する遺伝子を、さらなる検証のために優先順位から外した。
【0127】
RHGPにより提供される組込みの部位及び方向は、宿主細胞がインフルエンザを用いた攻撃を生き残ることを可能にする摂動の型に関する洞察を提供した(図4)。具体的には、RHGP摂動は広く3群に分けられた:(1)「アンチセンス」:1つの対立遺伝子の破壊及び他の対立遺伝子のアンチセンス阻害を促進させたアンチセンス組込み事象;(2)「センス下流」:センス方向での組込み(1つ又は複数のドメインの産生を促進すると予測されうる。全長遺伝子産物のドミナントネガティブ阻害剤として作用しうる);及び(3)「センス上流」:開始部位の上流のセンス方向での組込み(標的遺伝子全体の過剰発現を促進しうる)。RHGPを使用して同定された99の公知の標的の内、56の標的(全標的の56%)が標的発現の「アンチセンス」ノックダウンを表わした。標的の別の35%が「センス下流」事象を表わし、野生型遺伝子発現のドミナントネガティブ阻害剤の過剰発現を伝える可能性が高い。この解釈と一致して、本発明者らは、特定の標的PTBP1が2つの別々のクローン中の標的として独立して同定されることに注目した;1つのクローンは「アンチセンス」方向及び別は「センス下流」方向。残りの9%は「センス上流」挿入を表わした。11の標的遺伝子(本明細書において配列決定した110の標的の10%)の方向が確認されないであろう。なぜなら、遺伝子自体が、以前に、記載されておらず、従って、方向の割り当てが可能ではなかったからである。
【0128】
別々の評価において、本発明者らは、これらの110の標的遺伝子が以前にウイルス感染と関連付けられていたかを問うた。本明細書において同定される110の標的の内わずか4つ(MDN1、GRK6、AKT1、及びSTXBP1)がウイルス感染に直接的に関連付けられており(表II)、RHGPを使用して同定された106の標的(96%)についての新規情報を示唆している。本発明者らの分析を関連タンパク質に拡大することにより、本発明者らは、本明細書において同定される110の標的の内23(21%)が、以前にウイルス感染と関連付けられていた宿主標的と相同性を共有している又はそれらとスーパーファミリーの共通メンバーであることを見出した。これらの知見は、RHGPによって宿主標的の圧倒的多数(75%)についての新規機能が同定されたことを示唆する。本明細書において同定された2つの標的(C4orf32及びC21orf33)が、オープンリーディングフレーム(orf)として簡単に記載されており、帰する機能を伴わない。全体で、これらの知見は、宿主ゲノムのRHGPベースの照合によって両方の新規標的が同定され及び/又は新規機能が公知の遺伝子に帰されたことを示唆する。
【0129】
ナイーブな細胞を使用した標的遺伝子の検証
RHGPによってMDCK細胞中の一連の標的が同定され、それらはインフルエンザ感染に対して耐性を付与する。表現型の可逆性は、これらの標的についての最初の検証を提供した。本発明者らは、次に、独立した実験システムを使用してこれらの候補を検証することを追求し、MDCK細胞又はRHGP技術のアーチファクトとして生じうる結果を除いた。標的遺伝子がイヌ由来MDCK細胞から単離されていたため、本発明者らはそれらのヒトホモログを同定し、ヒトHEK−293細胞においてこれらの標的について選択的であるsiRNAを発現させた。
【0130】
RHGPによって、宿主細胞(過剰発現又はノックダウンされている標的を含む)のインフルエンザ媒介性の殺傷を予防する改変が同定された。本発明者らは、RHGPベクター内で誘導プロモーターを使用してこれらの標的を検証し、表現型を逆転させ、それにより、RHGP戦略に起因しない自然アーチファクトについての潜在力を最小限にできた。このゲノム規模のスクリーニングによって、本発明者らは、宿主細胞を、インフルエンザウイルスを用いた本来なら致死的な攻撃に対して耐性にする110の新規標的を同定することができた。これらの標的の内、大半(110中106)が、以前に、インフルエンザと関連付けられていた。また、本発明者らは、新規機能を、以前に未知である遺伝子及びorfに帰する。概念の証拠として、標的のサブセットを、従来のsiRNAアプローチを使用した独立した試験においてさらに検証した。
【0131】
最近の一連のsiRNAベースの試験によって、インフルエンザ、HIV、又はウエストナイルウイルスによる感染と関連する宿主標的が同定された(概要については表3を参照のこと)(Brass et al., 2008; Hao et al., 2008; Konig et al., 2008; Krishnan et al., 2008)。これらの初期の試験では、異なる種に由来する宿主細胞が利用された。それらは、本発明者らの本知見との比較の複雑さを増加させる。これを複雑化することとして、siRNAは、標的発見のためにますます利用されるが、しかし、siRNAの頑健で持続的な過剰発現の必要性により内因的に限定される。一部のsiRNAの知見での結果は、また、siRNAが宿主防御機構を非特異的に変化させるか否かの疑問により曇らされてきたが(Kleinman et al., 2008)、それは、抗ウイルス疾患へのsiRNA技術の適用について特に問題になりうる。RHGPは、また、潜在的な抗ウイルス宿主防御機構を誘導しうる。例えば、GSVのアンチセンス方向は、二本鎖RNAを誘導することが予測され、それ自体が宿主の抗ウイルス応答を引き起こすことができる。
【0132】
表
【表11】
【0133】
表2。示しているのは、宿主細胞を、本来なら致死的であるインフルエンザ感染に耐性にすることができる宿主標的のRHGP媒介性の同定と関連する事象の進行の概観である。RHGPを使用して同定された特定の標的(96%)及び標的ファミリー(75%)の大部分が固有であったことに注目して下さい。RHGPは、また、アノテートされていなかった標的の同定を促進した。
【0134】
【表12】
【0135】
表3。示しているのは、RHGP(本報告)又はsiRNAを使用して同定された標的及び標的ファミリーの評価である。この表は、本試験及び発表された知見(示す通り)の両方に共通である標的ファミリーに限定される。4つの標的(MDN1、GRK6、AKT1、及びSTXBP1、太字)が共有され、残りは固有であったが、一部の関連するスーパーファミリーメンバーは、異なるウイルスによる感染と関連付けられている。示されていないのは、以前に発表された情報と共通性を有さない、本明細書において同定された87の追加の標的である。
【0136】
実施例4−RHGPを使用し、HIV感染性を制御するための潜在的な標的を同定する。
ヒト免疫不全ウイルス(HIV−1)は、公衆衛生に対する世界的な脅威である。ウイルスを直接的に標的化する現在の医薬は、しばしば、薬物耐性ウイルスバリアントにおいて無効になりうる。薬物耐性と戦うための出現する概念は、ウイルスの機能、生存、及び増殖のために(しかし、宿主自体に対してではない)必須である宿主機構を標的化する考え方である。本明細書において、ヒトMT4 T細胞に対するランダムホモ接合摂動(RHGP)を使用し、本発明者らは、これらの細胞がHIV−1NL4−3を用いた致死感染に生き残るために必須である、安全で効果的な一連の宿主標的を同定した。RHGPは、細胞中の全ての標的をサンプリングすることができる新たな技術である。これは、遺伝子探索ベクター(GSV)の操作を通じて可能であり、それは、宿主染色体中にランダムに組込むことができる。GSVは、細胞において、誘導因子により調節される不可逆的な様式で、任意の標的をアップレギュレーション又はダウンレギュレーションすることができる。誘導因子の除去によるRHGP効果の逆転によって、操作された細胞は、HIV感染に再び感受性になる。siRNAアプローチを使用し、同定され、定義された遺伝子標的が検証され、CXCR4又はCCR5のいずれかに向性のウイルスを用いた感染のために必要であることが示された。これらの試験では、標的介入のための新規の遺伝子候補(機能の事前の知識を伴わない遺伝子配列(EST)を含む)を同定するためのRHGPの能力が実証される。これらの候補は、広域スペクトルの薬物開発のために直接的に適する。なぜなら、それらの発見の間での遺伝子破壊は、無視できる細胞傷害性を示すだけであるからである。
【0137】
三種混合薬物治療(すなわちHAART、高活性抗レトロウイルス治療)は、HIV−1複製を効果的に制御することができるが、HAART耐性HIV−1株が、エラープローンHIVウイルス逆転写酵素と頑健なウイルス複製及び不完全な患者のコンプライアンスとの組み合わせとして容赦なく出現している。一部の地域において、HARRTにおいて使用される三種薬物に対して耐性であるウイルスが、AIDS患者の20%近くから単離された(Grant, Hecht et al. 2002; Richman, Morton et al. 2004)。そのような知見によって、HIV−1疾患の処置のための新たなパラダイム(特に、耐性の発生に対して比較的非感受性である作用機構を伴うもの)を同定する緊急性が増加する。HIV−1感染(しかし、宿主自体ではない)に必須である宿主遺伝子産物を標的化する治療法は、処置のそのような新たなモダリティを提供しうる。なぜなら、十分に設計された宿主標的は、病原体に対して直接的な選択圧力を置かないかもしれないからである。
【0138】
ウイルスと宿主細胞の間での相互作用によってウイルス病原性の結果(ウイルスの除去から潜在的及び致死的な感染に及ぶ)が決定されることが十分に確証されている。HIV−1は、宿主細胞タンパク質と相互作用し、それらの複製を助け、免疫攻撃を回避することが公知である。この相互作用の例は、欠損した細胞表面受容体(CCR5)を保有し、従って、HIV−1感染に対して耐性である個人により実証される(Liu, Paxton et al. 1996)。同様の型の相互作用が、文献において報告されており、HIV−1ライフサイクルのほぼ全ての段階を包含する:細胞受容体CD4及び共受容体CXCR4又はCCR5へのEnv結合を要求するウイルス侵入から、Gag p6によりTSG101をハイジャックする必要性のあるウイルス放出まで(Garrus, von Schwedler et al. 2001)。HIV−1タンパク質と宿主細胞のものとの相互作用の完全な文書化は、このように、HIV−1複製及び病原性のプロセスを理解するために重大である。より重要なことには、そのような理解は、HIV/AIDSと戦うための治療法及び予防戦略を改善するための礎石を提供しうる。
【0139】
HIV−1感染における宿主因子の理解された重要性に照らして、増加する研究で、抗ウイルス治療のための宿主標的が考えられ始めている。宿主指向の治療法が、HIV/AIDS処置において、HIV結合及び内在化のために必要である受容体に影響を及ぼすための生物製剤及び小分子を用いてウイルス受容体CD4及び/共受容体CCR5(Reeves and Piefer 2005; Wheeler, McHale et al. 2007)を標的化することにより成功裏に使用されてきた。実験室での研究及び初期の臨床試験では、これらの薬剤がHIV/AIDSの処置のための選択肢を提供しうることが示唆される。
【0140】
発見された全ての宿主遺伝子が、治療標的として適するわけではない(特に、宿主自体に必須であるもの)。これらの標的の間で区別するために、本発明者らの実験室では新規技術であるランダムホモ接合性遺伝子摂動(RHGP)を用いて、HIV感染のために必須であるが、しかし、非感染細胞の増殖、生存、又は機能のためには必要ではない標的について選択している。RHGPは、真核細胞中の任意の遺伝子をアップレギュレーション又はダウンレギュレーションし、細胞挙動の客観的な測定を使用して結果を評価するように設計した。
【0141】
本明細書において、本発明者らは、RHGPを適用し、宿主細胞が致死的HIV感染に耐性になることを可能にする一連の宿主志向性の標的を同定した。これらの新規標的は、その機能が公知の生物学的経路に関連する遺伝子及び機能が割り当てられていないESTを含む。同定された遺伝子を、他の遺伝的アプローチ(siRNAを含む)を使用して検証した。従って、この研究では、HIV/AIDSに対する将来の薬物開発のための潜在的な新たな安全な標的が明らかになった。
【0142】
材料及び方法
細胞株及びウイルス
細胞(MT4、PM1細胞、及びTZM−blを含む)及びウイルス(薬物耐性突然変異体ウイルス及びR5向性バリアント)を、NIH AIDS Research及びReference Reagent Programから得た。HIV−1NL4−3を、HEK293から、プロウイルスDNA(NIH AIDSプログラム)のトランスフェクション後に作製し、MT4細胞中で増幅させた。
【0143】
HIV−1感染ならびに感染性及びp24産生の決定
MT4又はPM1細胞を、HIV−1(MOI0.001)を用いて、1時間にわたる低速遠心分離(1,200g)により感染させた。感染後に回収された上清を、次に、感染性の検査のために、TZM−bl指標細胞株(Wei, Decker et al. 2002)に移した。相対発光単位(RLU)を、TZM−bl細胞で、それらをBright−Glo Luciferase Assay System(Promega)を用いて3日後に処理した後に得た。
【0144】
回収された上清中のP24量を、また、HIV−1 p24 ELISAキット(Xpressbio, Thurmont, MD)を使用し、製造者の指示に従い測定した。
【0145】
MT−4 R1細胞株の構築
RheoSwitch(登録商標)Mammalian Inducible Expression SystemをNew England Biolabs(NEB)から購入した。トランスアクチベーターR1をコードするプラスミドpNEB−R1を、最初に、制限酵素ScaI(NEB)を使用して線状化した。MT4細胞を、次に、Eppendorf Multiporator(Eppendorf, AG 22331, Huammburg)を使用し、360v(電圧)及び100ms(時間)の条件下での電気穿孔によりトランスフェクションした。MT4細胞を、G−418(400μg/ml)を使用して選択し、G−418耐性細胞を連続限界希釈によりクローン化した。増大後、クローンを、R1応答性ルシフェラーゼレポーター遺伝子(pGluc)を用いたトランスフェクション後、Gaussia Luciferase Assay Kit(NEB)を使用して発光(相対発光単位(RLU))について少なくとも2回検討した。本発明者らは、これらの細胞クローンからの発光のRSL1誘導(倍)を以下の通りに決定した:誘導因子での処理なしのサンプルからのRLUにより除した、誘導因子の存在においてサンプルから得られたRLU。これらのクローンからの倍は2〜60倍に及んだ。最も高い誘導を伴う安定なクローン(#2−14)を使用し、RHGPライブラリーを作製した。
【0146】
RHGP遺伝子探索ベクターpRHGP12−RSNのコンストラクション
RHGP遺伝子探索ベクターpRHGP12−RSNは、企業において、レンチウイルスベースのpLESTベクター(寛大に、Stanley Cohen博士(Stanford)により提供された)を骨格として使用してコンストラクションされた一連のFGIのRHGPベクターの1つである(Lu, Wei et al. 2004)。このベクターは、RheoSwitch Mammalian Inducible System(NEB)を用いてコンストラクションされた。RheoSwitchシステムは、TATAボックスの上流に5コピーのGAL4応答エレメント(5×RE)を含み、RSL1リガンドの存在において低い基礎発現を伴う転写の高い誘導をもたらす。ベクターを構築するために、pLEST中のNeoR−TRE−CMVのDNA配列を、最初に、5’LTR部位の反対のOri−CAT−RSのRheoSwitch(RS)誘導性発現カセットを用いて置換した。選択マーカー及びブラストサイジン(BS)耐性遺伝子を含むレポーターカセットならびにPKGプロモーターにより制御されるEGFP遺伝子を、RS発現カセットの反対方向でNheI部位中に挿入した。2つのLoxP部位を、RSプロモーターの下流及びp15A ori−CATの上流にそれぞれ挿入した。
【0147】
GSVを保有するレンチウイルスの産生及びRHGPライブラリーの構築
RHGPレンチウイルスを、ViroPower Expression System(Invitrogen)を使用して産生した。HEK293FT細胞を、10cmプレート中に106個細胞/プレートでプレーティングした。24時間のインキュベーション後、細胞を、3μgのRHGP22−RSN及び9μgのViroPower Packaging Mixを用いて、Lipofectimine 2000(Invitrogen)を使用してトランスフェクトした。培地を、37℃で5時間のインキュベーション後に交換した。48時間後、培養培地中のウイルスを、0.45μmフィルターを通じてろ過し、製造者の指示に従い滴定した。
【0148】
RHGPライブラリーを構築するために、MT4−R1細胞を、RHGPウイルスを用いて、ポリブレン(6μg/ml)の存在において、1時間にわたる低速遠心分離(1,200g)により形質導入した。単一細胞内での複数の挿入についての潜在力を最小限にするために、低MOI(0.1)をライブラリー作製の間に用い、細胞が、1を上回る異なるGSVにより形質導入されうる可能性を最小限にする。GSV組込み細胞を、GBL培地(完全RPMI 1640培地、G418(400μg/ml)、ブラストサイジン(4μg/ml)、及びRSL1リガンド(0.5μM)を含む)を使用して選択した。
【0149】
HIV−1攻撃から生き残ったRHGP細胞クローンの選択
HIV−1NL4−3を用いた感染後、MT4−R1 RHGPライブラリーを、同じGBL培地中で培養した。個々の生存クローンを、連続限界希釈により確立し、GBL培地中で継続的に増大させた。細胞クローンを、さらに、HIV−1を用いて攻撃し、それらの耐性を確認した。
【0150】
可逆性
RHGP事象の可逆性を検証するために、ウイルス耐性MT4細胞クローンを、GBL培地又はRSL1を伴わないGBL培地中で少なくとも3日間にわたり培養した。HIV−1感染後、上清中でのウイルス産生(感染性及びp24)を上に記載する通りに検討した。
【0151】
ウイルス耐性クローンからの候補遺伝子の同定
RHGP遺伝子探索ベクターを設計し、標的遺伝子を効率的に発見し、組込み事象の方向(センス又はアンチセンス)を決定した。遺伝子探索ベクターはOri−CATレポーター遺伝子を含み(図1)、それは、制限酵素ベースのゲノムDNAクローニングによりレスキューすることができる。
【0152】
各クローンからの細胞(1×106)を、溶解緩衝液(0.32Mスクロース、10mM Tris pH7.5、5mM MgCl2、及び1% Triton X−100を含む)中に浮遊させた。遠心分離後、ペレットを、プロテイナーゼK緩衝液(25mM EDTA、150mM NaCl及び40mM Tris、0.5% SDS、pH7.5を含む)中のプロテイナーゼK(Sigma)及びRNase A(Sigma)を用いて処理した。ゲノムDNAを、フェノール/イソアミルアルコール/クロロホルムを用いて抽出し、イソプロパノールを用いた沈殿が続いた。DNAペレットを、70%エタノールを用いて洗浄し、TE緩衝液(pH7.5)中に溶解した。
【0153】
BamHI又はXbaIを用いた消化及びフェノール/イソアミルアルコール/クロロホルムを用いた抽出後、ゲノムDNA(10μg)を、エタノールを用いて沈殿し、TE緩衝液中に溶解した。消化されたDNA(2.5μg)を、T4リガーゼ(Invitrogen)を使用して16℃で一晩自己連結させた。連結されたDNAを、エタノールを用いて沈殿させ、70%エタノールを用いて洗浄した。DNAペレットをTE緩衝液中に溶解させ、DH10B ElectroMaxコンピテント細胞(Invitrogen)中に電気穿孔した。複数のコロニーを、プラスミドDNA調製及び制限酵素消化のために単離した。プラスミドDNAをさらに使用し、標的遺伝子をDNA配列決定及びゲノムマッピングにより同定した。RHGPベクター挿入部位に隣接する結果として得られるゲノムDNA配列を、ヒトゲノムに対する、UCSC Genome Browserを使用したゲノムマッピングに供した。
【0154】
siRNAを用いた宿主標的遺伝子の検証
RHGP同定遺伝子についてのヒト二本鎖siRNAホモログ(siGNOME SMART pool)を、製造者(Dharmacon)により推奨される通りに調製した。siRNA Rab 6A及びHIV−1 tatをポジティブコントロールとして用いた(Brass, Dykxhoorn et al. 2008)。非標的siRNA(siCONTROL1)をネガティブコントロールとして使用した。MT4又はPM1細胞を、完全RPMI 1640培地中で一晩培養した。対数期増殖中のMT4細胞を、1.2μMのsiRNAを用いて、電気穿孔プロトコールにより、製造者(Eppendorf)の指示に従いトランスフェクトした。PM1細胞を、同じプロトコール下で(パルス条件200v(電圧)及び200ms(時間)の使用を除く)、電気穿孔した。細胞を、HIV−1バリアントを用いて、トランスフェクションから24時間後に感染させた。培養培地を毎日新しくした。細胞生存をトリパンブルー色素排除アッセイにより毎日検討した。上清中でのウイルス産生(感染性及びp24)を上に記載する通りに検討した。
【0155】
結果
RHGP標的遺伝子の発見の概観
材料及び方法に概説されている通り、pRHGP12を、ブラストサイジン耐性遺伝子を促す常時発現性プロモーターからなる発現カセットを使用してコンストラクションした。GSVは、また、RheoSwitchプロモーターを含み、それはリガンドにより活性化することができる。リガンドの存在(プロモーターオン)は、このように、宿主転写物を産生し、RHGP効果の上方又は下方のいずれかの調節を誘発することができる。RHGPにより起こされる遺伝子摂動効果は、前に詳細に記載された(Sui, Bamba et al. 2009)。本発明者らはこの供給源を利用し、ヒトTリンパ球のゲノム全体を照合し、標的(アップレギュレーション又はダウンレギュレーションを問わず、これらの細胞を本来なら致死的なHIV−1感染に生き残ることを可能にする)を同定した。
【0156】
MT4 T細胞を操作し、トランスアクチベーターR1を安定的に発現させ、それはRHGP中の組込みプロモーターを活性化し、誘導因子RSL1の存在において転写物を産生することができる。遺伝子摂動の「RHGPライブラリー」を確立するためのRHGPベクターを用いた形質導入及び抗生物質選択に続き、宿主細胞を、HIV−1の致死感染を用いて攻撃した。
【0157】
生存細胞は、限界希釈によりクローン化された単一細胞であった。RHGPと無関係の事象の結果として生じたであろうそれらの生存体を除去するために、表現型を、RSL1の非存在においてHIV−1攻撃を繰り返すことにより確認した。この試験では、RHGP自体が(他のアーチファクト又は自然突然変異ではなく)、HIV−1感染に直面した細胞生存に直接的に関与することが確認された。生存クローンからのゲノムDNAを次に単離し、配列決定し、標的遺伝子ならびにGSV組込みの方向を特定した(結果が標的のアップレギュレーション又はダウンレギュレーションを表わすか否かを決定するために)。
【0158】
RHGP細胞ライブラリーの構築
105の組込み事象を伴うMT4−R1細胞のライブラリーを、レンチウイルスベースのpRHGP12 GSVを使用して形質導入し、宿主ゲノムのカバーを確実にした。ゲノム中での平均遺伝子サイズが27kbと推定されるため(Gupta and Varshney, 2004)、本発明者らは、これらの105の独立した組込み事象が、ゲノム全体のランダムなカバーを確実にしうると計算した。形質導入された宿主細胞のライブラリーを、次に、ブラストサイジンを使用して選択した。特定のブラストサイジン耐性細胞クローンから、本発明者らは、GSVが実際に宿主染色体中に組込まれ、アンチセンス転写物が誘導因子RSL1の処置時にだけ産生されることを見出した(データ示さず)。R1発現及びGSV組込みを確認及び維持する追加の手段として、MT4ライブラリーが、続く実験において、致死濃度のG418及びブラストサイジンと一緒に持続的にインキュベーションされた。
【0159】
HIV−1耐性細胞は、ウイルス誘導性細胞殺傷から生き残った。
MT4−R1細胞は、親MT4細胞と同様に、HIV−1NL4−3(MOI10−4)により、5日以内に効率的かつ完全に殺された。RSL1の添加はこの殺傷に影響を及ぼさなかった(図11A、左パネル)。これらの条件は、「プロモーターオン」を保つためのRSL1の存在において、生存細胞クローンについての続く選択のために厳密に利用された。追加のコントロールとして、偽形質導入細胞の並行培養物を同じように処理し、生存体は5日後に観察されなかった。この結果は、生存細胞が、RHGP摂動の結果として(HIV−1に対する自然耐性のアーチファクトとしてではなく)生じることを確実にするために必須であった。クローニング及び増大後、生存細胞を複数ラウンドの感染に供し、感受性細胞を除去した。最後に、本発明者らは、ウイルス誘導性殺傷に耐性である25のRHGP細胞クローンを得た。図11A(右パネル)は、HIV−1攻撃後でのそのような1つの耐性クローンの生存を示す。
【0160】
ゲノム配列決定と並行して、本発明者らは、これらの耐性細胞クローンが、HIV感染時に感染性ウイルス粒子を産生し続けるか否かを問うた。RSL1の存在におけるHIV−1NL4−3を用いたこれらの耐性細胞クローンの再攻撃後、上清を4dpiで回収し、TZM細胞に移し、それによって感染性の読み出しが提供される。図11Bは、得られたウイルス耐性RHGP細胞クローンが子孫ウイルスを産生及び放出しないとの代表的な実験を示す。これらの知見は、ELISAベースのアッセイにおけるp24の決定により独立して確認された。
【0161】
野生型HIV−1NL4−3を用いた結果が励みとなるが、本発明者らは、治療法についての大きな満たされない必要性は、従来の医薬に耐性であるウイルスバリアントへの新たな標的の適用であると考えた。従って、本発明者らは、プロテアーゼ阻害剤に対する確立された耐性を伴うHIV−1バリアントを用いた並行した一連の試験を実施した。野生型HIVを用いた攻撃に耐性であるRHGP形質導入クローンは、また、プロテアーゼ耐性バリアントに直面した攻撃に生き残った。この結果は、宿主細胞の生存に固有ではなかった。なぜなら、感染性アッセイならびにp24 ELISAによって、耐性細胞における突然変異体HIV−1による欠損した感染が確認された(図11C)。一緒に、これらの結果によって、本発明者らが得た細胞クローンが、WT及び薬物耐性HIV−1バリアントの両方による感染に耐性であることが確認された。
【0162】
ウイルス耐性細胞クローンの可逆性アッセイ
RHGP技術の重要な特性は、誘導プロモーターによる調節を介して候補標的を検証する能力である。本発明者らは、最初に、RHGPライブラリーを構築するために使用されているMT4−R1細胞においてトランスアクチベーターR1の誘導活性を検討した。MT4−R1細胞は、最初に、R1応答性ルシフェラーゼレポート遺伝子(NEB)を用いてトランスフェクトし、誘導因子RSL1の存在又は非存在において培養した。発光(RLU)を次に検討した。MT4−R1は、RSL1の存在において高く安定したレベルの発光を示したが、しかし、RSL1が存在しない場合には背景レベルだけであった(図12A)。この結果は、R1の活性化能力がRSL1により強固に制御されることを示した。
【0163】
これらのクローンでのHIV−1耐性の獲得が、本当にRHGPの組込みに起因することを確認するために、本発明者らは、次に、これらのクローンにおいてウイルス産生の可逆性を検討した。RHGPベクター用のプロモーターは、リガンド制御されたRheoSwitch誘導性トランスアクチベーターR1の制御下にあったため、本発明者らは、リガンドRSL1の存在又は非存在において候補のHIV媒介性殺傷を比較することができた。本発明者らの試験では、これらの生存クローンがHIV−1攻撃に対して可逆的な耐性を示すことが確認された。図12Bは、この一連の実験からの代表的な細胞クローンの結果を示す。リガンドが、HIV−1攻撃の間に、細胞培養中に存在しない場合、頑健なウイルス産生(親MT4細胞から産生されるものと同程度)がこの細胞クローンについて観察された。高レベルのウイルス放出が、細胞がウイルス媒介性殺傷で死ぬまでの多くの日数にわたり持続した。
【0164】
ゲノムDNAクローニングによる宿主遺伝子の同定
ゲノムDNAを、HIV−1に対して可逆的な耐性を示していたウイルス耐性クローンから単離した。GSV組込み部位を伴う25のHIV非感受性宿主細胞クローンが、21の異なる宿主細胞標的の同定をもたらした。これらのGSV組込みは、12の以前に記載された遺伝子及び2の非アノテートESTを含んだ。クローンの一部は、同じ親からの子孫に思われた。なぜなら、GSVは、同じ方向を伴う同じ遺伝子中に組込まれていたからである。3つのクローンが、遺伝子又はESTを伴わない領域中にRHGP挿入を有した。本発明者らは、Ori−CATレポーター遺伝子の部分的喪失に起因する4つの候補から候補遺伝子を単離することはできなかった。
【0165】
これらの遺伝子及びESTの特性を表4に列挙する。RHGPにより提供される組込みの部位及び方向は、宿主細胞がHIV−1攻撃を生き残ることを可能にする摂動の型に関する洞察を提供した。具体的には、RHGP摂動は広く3群に分けることができた:(1)「アンチセンス」:1つの対立遺伝子の破壊を促進させたアンチセンス組込み事象及び他の対立遺伝子のアンチセンス阻害;(2)「センス下流」:センス方向での組込み(内因性の遺伝子産物のドミナントネガティブ阻害剤の産生を促進すると予測されうる);及び(3)「センス上流」:開始部位の上流のセンス方向での組込み(標的遺伝子の過剰発現を促進すると予測されうる)。RHGPを使用して同定された12の遺伝子標的の内、8の標的が標的発現の「アンチセンス」ノックダウンを表わした。標的の他の6つが「センス下流」事象を表わし、野生型遺伝子発現のドミナントネガティブ阻害剤の過剰発現を表わす可能性が高い。「センス上流」挿入は、本試験において同定されなかった。これらの予測に基づき、候補遺伝子の全てがGSV組込み事象によりダウンレギュレーションされる可能性が高い。これによって、本発明者らは、ナイーブなMT4細胞でのsiRNAノックダウンアプローチを直接的に使用し、ウイルス耐性表現型を再現することができた。全体で、これらの知見は、宿主ゲノムのRHGPベースの照合によって両方の新規標的が同定され及び/又は新規機能が公知の遺伝子に帰されたことを示唆する。
【0166】
ナイーブな細胞を使用した標的遺伝子の検証
RHGPによってMT4細胞中の一連の標的が同定され、それらはHIV−1感染に対して耐性を付与する。表現型の可逆性は、これらの標的についての最初の検証を提供した。本発明者らは、次に、独立した実験システムを使用してこれらの候補を検証することを追求し、自然突然変異又は本発明者らのRHGP技術のアーチファクトとして生じうる結果を除いた。
【0167】
これらの耐性遺伝子に対する二本鎖siRNAをDharmaconから購入した(4つの個々のsiRNAを含むSmartpool)。非標的siRNAは、トランスフェクション及び参照基準のための一致されたコントロールを提供した。ウイルスtat及び細胞標的Rab6Aに特異的であるsiRNA構築物を、これらのsiRNAがHIV−1感染を効率的に阻害できるとの最近の報告に基づいて、ポジティブコントロールとして選択した(Brass, Dykxhoorn et al. 2008)。
【0168】
siRNAを、天然MT4細胞中に、電気穿孔を介して、HIV−1NL4−3感染の1日前にトランスフェクトした。上清を、感染から2日後に回収した。処理した細胞から産生された感染性ウイルス粒子を、TZM細胞ベースの読み出しを使用して検討した。図6Aに示す通り、12の選ばれた標的遺伝子に対する二本鎖siRNAによって、HIV−1ウイルス産生が、ポジティブコントロール(Tat及びRab6A)において観察されるものと同程度のレベルまで低下した。生存も検討し、siRNA処理からの可能な細胞傷害性効果を除いた。非常に同様のパーセンテージの生存細胞が、処理されたサンプルの全てについて観察された。これらの結果によって、RHGPにより同定されたこれらの遺伝子がウイルス複製において重要であるとの本発明者らの知見が確認され、新規の宿主ベースの標的を同定するためのRHGPの適用が検証された。
【0169】
HIV−1NL4−3(CXCR4向性ウイルス)誘導性殺傷から生き残った細胞から同定されたこれらの宿主遺伝子候補が、また、CCR5向性HIV−1ウイルスのウイルス複製に重要であるか否かを検討するために、同じsiRNAアプローチを、次に、CXCR4及びCCR5二重発現T細胞株PM1で使用し、R5向性HIV−1ME1を用いた感染が続いた。HIV−1ME1株のウイルス産生は、これらの12の遺伝子標的を標的化する特異的siRNAを用いて処理された細胞において有意に阻害された(図6B)。これらの結果によって、RHGPにより同定されたこれらの遺伝子がHIV−1ウイルス(X4及びR5向性バリアントの両方を含む)の複製において重要であるとの本発明者らの知見が確認され、新規の宿主ベースの標的を同定するためのRHGPの適用が検証された。
【0170】
本発明者らの本試験において、本発明者らはRHGP技術を適用し、HIV−1ウイルス感染のために要求される宿主因子についてのゲノム規模のスクリーニングを行った。本発明者らは、細胞を、HIV−1ウイルスを用いた本来なら致死的な攻撃に対して耐性にする新規の宿主ベースの標的を同定した。また、本発明者らは、新規の抗HIV−1機能を、以前に公知の遺伝子及びアノテートされていないESTに帰する。これらの標的を、最初に、表現型を逆転させるためのRHGPベクター内に組込まれた誘導プロモーターを使用し、次に、天然細胞において、従来のsiRNAアプローチを使用して検証した。本発明者らは、さらに、結果として得られる標的を、異なるHIVバリアント(薬物感受性及び耐性分離株ならびにCCR5及びCXCR4向性ウイルスを含む)に広く適用可能であることを見出した。
【0171】
本来なら致死的なインフルエンザ感染に直面した際の生存のために必須の宿主標的のパネルを同定した初期の試験において使用されたRHGPと同様に(Sui, Bamba et al. 2009)、本発明者らはレンチウイルスシステムを使用し、MMLVベースのRHKOにより起こされる低力価のGSV産生での先の限定を克服した(Li and Cohen 1996)。これらの改善によって、本発明者らは、宿主ゲノム全体をサンプリングし、改善されたGSVにより引き起こされる105の組込み事象から12の遺伝子標的を同定することができた。レンチウイルスベクターが、活性な遺伝子転写の部位中への単一部位挿入を支持するとの予測と一致して(Mitchell, Beitzel et al. 2004)、全ての組込みが活性な遺伝子領域において生じた。
【0172】
RHGPの1つの固有の特性は、標的発現が誘導可能なプロモーターの制御下にあることである。このように、表現型(HIV−1耐性)と摂動遺伝子の間での因果関係は、誘導因子の取り除きにより確認することができる。同じ実験設定内でこれらの標的を検証することにより、この特性は、治療候補の本発明者らの発見効率を増加させる。実際に、耐性クローンからの全てのRHGP標的が、天然細胞を用いた続くsiRNA試験において成功裏に確認された。候補遺伝子と比べたGSV挿入の位置及び方向に依存して、RHGPは、また、哺乳動物細胞において遺伝子の活性化、しばしば過剰発現を生成することができる。本発明者らの本試験において観察されなかったが、標的のサブセット(B細胞CLL/リンパ腫2(BCL2))の過剰発現によって、MDCK細胞が、インフルエンザウイルス感染に対する宿主遺伝子の発見の間にインフルエンザ感染に生き残ることができた(Sui, Bamba et al. 2009)。従来のアプローチ(例、siRNA)により生成される一過性のノックダウンの代わりに、RHGP表現型は高度に安定であり、それによって機構試験が促進され、HIV−1複製におけるこれらの摂動遺伝子の役割を特徴付けることができる。
【0173】
RHGPは、標的の事前の知識により偏らなかった。実際に、HIV−1耐性RHGP細胞クローンから、本発明者らは、2つのアノテートされていないESTがGSVにより標的化されることを発見した(表1)。さらに、本発明者らが、PANTHER Classification System(Mi, Lazareva-Ulitsky et al. 2005)を使用して遺伝子標的を分析した場合、それらの多くの推定分子機能が依然として分類されていなかった(CASD1、GDAP2、TNRC6A、及びTTC21B)、又は未知であった(CAMPSAP1L1及びGSTCD1)。それらの2つはユビキチン−タンパク質リガーゼに属する(HECW2及びDZIP3)。残りの標的において、各標的の1つが以下のファミリーにおいて見出される:免疫グロブリン受容体又は防御/免疫タンパク質(Robo1)、シグナル伝達分子(NLGN1)、mRNAポリアデニル化因子(CPSF1)、及びトランスポーター又はヒドロラーゼ(ATP8A1)。
【0174】
候補遺伝子の内、Robo1が非常に良好な治療標的としての役割を果たしうる。I型膜貫通タンパク質として、それは、本来は、神経発生の間に軸索ガイダンス受容体として同定された(Wong, Park et al. 2002)。Robo1は、また、CXCR4との相互作用(HIV感染との潜在的な連関)を通じてT細胞の化学走性において役割を果たすことが見出された(Wu, Feng et al. 2001; Prasad, Qamri et al. 2007)。それがCCR5にも結合するか否かを検討する価値がある。ウエスタンブロットを使用し、本発明者らは、Robo1発現がRHGP耐性クローン及びsiRNA処理細胞において低下されることを確認した(未発表データ)。さらなる研究が、Robo1欠損CD4+T細胞において、ウイルス複製が、恐らくは細胞膜のビリオン融合の間にウイルス侵入段階において遮断されることを明らかにするために必要とされる。Robo1は、HIV−1複製においてHIV−1共受容体CCXR4及びCCR5の活性を支持する潜在的な新たな役割を持ちうる。CCR5の治療標的と同様に、Robo1に対するアンタゴニスト及び/又はモノクローナル抗体が、従って、AIDSを処置するための新たな一連の治療計画を提供しうる。
【0175】
同定されたHIV−1耐性遺伝子のリストは、また、2つのEユビキチンリガーゼ(HECW2及びDZIP3)を含む。このファミリーからのタンパク質は、タンパク質分別経路及び小胞輸送プロセスに関与することが見出されており、それらは、また、ウイルス出芽及び放出の間にHIV−1及び他のウイルスにより使用される。1つの良好な例はTSG101(ウイルス脱出のためにHIV−1 Gag 6によりハイジャックされるユビキチン様リガーゼ)である(Garrus, von Schwedler et al. 2001)。HECW2は、別のE3ユビキチンリガーゼNEDD4と高度に相同である。HIV−1ウイルスがNEDD4を使用するとの直接的な証拠はないが、PPXYの後ドメインを伴う多くの他のエンベロープウイルスは、宿主細胞からのそれらの放出のためにNEDD4を要求しない。これら2つの遺伝子産物(HECW2及びbZIP3)は、また、HIV−1成熟及び脱出のために重要でありうることを推測することが興味をそそられる。
【0176】
【表13】
【0177】
実施例5−同定された潜在的な標的のさらなる研究
上で考察した試験によって、増殖性HIV感染のために必須である新規の宿主標的が同定された(Robo1)。Robo1は、可溶性リガンドのための膜貫通受容体であり、SLITとして公知である。Robo1は神経系の発生において主に発現され、軸索ガイダンスを調節する。Robo1は、また、リンパ系細胞で発現され、それは細胞間相互作用の調節において同様に役割を果たしうる。
【0178】
Robo1の細胞外構造は、それが5つの積み重なった免疫グロブリン(Ig)ドメインからなる点で顕著であり、それは3つのタンデムフィブロネクチンIII型(FN−III)ドメインにより支持される。そのようなものとして、Robo1は、細胞膜から比較的遠くに伸展すると予測される。そのような特性は、しばしば、細胞間接着を伴い観察される。なぜなら、グリコシル化タンパク質上の負の電荷を持つ糖(糖タンパク質及びプロテオグリカン)の静電反発力が、細胞を分離するバリアとしての役割を果たし、密接な相互作用(例えば膜融合など)を予防する。実際に、本発明者らは、この機構が進化し、部分的に、ウイルスが宿主細胞に侵入することを予防しうると推測する。
【0179】
本発明者らは、細胞膜からのRobo1の伸展突起が、ウイルス感染におけるその必須の役割についてのRHGPの証拠と組み合わせて、Robo1がウイルスの取り込みに関与し、恐らくはウイルス−宿主結合の初期部位又は共受容体として機能しうることを示唆すると仮定する。
【0180】
この考え方を支持し、Robo1が、文献において、ケモカイン受容体CCR5と相互作用することが報告されている。CCR5は、次に、HIV感染のための重要な共受容体として機能することが公知である。実際に、CCR5(ならびに別のケモカイン受容体CXCR4)は、HIV感染を処置するための一連の最近承認された又は治験中である新たな薬物の標的である。
【0181】
RJ Shattock and JP Moore, Nature Reviews Microbiology, 1, 25; 2003に示されている通り、CCR5は細胞膜から遠くに突出せず、HIVによりコードされるgp120へのCD4結合が安定性を提供し、十分に理解された静電反発力(本来であればHIVがT細胞と密接に接触することを予防しうる)を克服することが仮定される。この安定性は、次に、CCR5とウイルスタンパク質との続く相互作用を支持し、膜融合及び内在化を媒介しうる。
【0182】
このモデルは興味深いが、しかし、CD4細胞外ドメインが4つのIgドメインからなり、このように、CD4がRobo1よりもかなり短いことを認識することが重要である。本発明者らは、従って、既存のモデル上に拡大し、Robo1が追加の共受容体として機能しうると仮定し、それは、CD4により媒介される段階よりもウイルス相互作用のより初期段階を安定化するために必要である。
【0183】
この仮定は、ウイルス出芽を予防するための治療介入についての重要な潜在的な意味を保有する。例えば、Robo1又はそのウイルス結合パートナーの特異的阻害剤は、初期結合を予防する及び/又は感染の細胞間伝播を予防する役割を果たしうる。HIVウイルス産生をノックダウンするためのsiRNAトランスフェクションを含む実験において、Robo1がノックダウンし、又は、非常に低いウイルス産生値を与え、コントロール値の9%という低さを残した。このように、Robo1の阻害剤は、一旦、細胞の初期細胞感染が生じると、HIV−1の伝播を効果的に遮断しうる。これらの阻害剤は、必ずしも限定はされないが、小分子、アプタマー、及び生物製剤(抗体、ペプチド、及びそれらの派生物を含む)からなりうる。
【0184】
現在、Robo1についてのウイルス結合パートナーは公知ではないが、しかし、本発明者らは異なる可能性を仮定する。
【0185】
最初に、感染細胞から出芽する際にHIV粒子の外部エンベロープ中に取り込まれるSLITタンパク質(又はRobo1についての他の生理学的リガンド)及びこれらの残存タンパク質が、Robo1との最初の相互作用を促進するために十分でありうる。
【0186】
加えて又は代わりに、Robo1は、そのSLITリガンド上でコードされるロイシンリッチモチーフ(LRRモチーフ)を認識すると理解される。これを念頭に置き、HIVによりコードされるgp41が、比較的顕著にロイシン残基(123残基中16;13%)を含むことに注目すべきである;これらの残基中で特に豊富である局在化領域を伴う。gp41の親分子(gp160)は81のロイシンを含む(849の全残基中;9.5%)。gp160内でコードされる他の主要タンパク質はgp120であり、それは32のロイシン残基(合計455残基から;7.0%)を含むが、この分子中にはロイシンの顕著なクラスターもあり、その少なくとも1つがLRRモチーフと似うる。HIVによりコードされるnefは、また、ビリオンの表面上で暴露されうるが、17のロイシン残基(合計196アミノ酸から;8.8%)を含む。ロイシン(及びクラスター)についてのこの相対的な濃縮は、HIVの一般的な形質を表わさないように思われる。なぜなら、非膜暴露タンパク質(例えば逆転写酵素など)は、少数のロイシン(560中48;8.5%)を利用するためであり、それらは顕著にはクラスター形成しない。同様に、HIV−1 Gagは41のロイシン(508の全残基中;8.0%)を含む。
【0187】
ウイルス付着の直接的な阻害に加えて、フローサイトメトリーデータは、Robo1の特定のエピトープが、HIV感染細胞の表面上に優先的に暴露されることを示す。この知見は、宿主防御機構(ADCC、CDC)を介した除去のための感染細胞の抗体ベースの標的化についての可能性を生じる。また、これらの抗体は、毒素、放射性核種、又は感染細胞を標的化するための他の致死薬剤を用いて標識されうる。これらの型の活性は、必ずしも抗体に限定されないであろう。なぜなら、他の抗体、ペプチド、融合タンパク質、及びこれらのエピトープを標的化する生物製剤又は小分子も、感染細胞を標的化するための活性を有しうるからである。
【0188】
HIVを越えて、Fuctional Genetics, Inc.からの最近のデータは、Robo1上の特定のエピトープが、インフルエンザ感染細胞の表面上に優先的に暴露されることを示唆する。このデータは、Robo1ベースの治療法の広域適用についての意味を有しうる。Robo1が、HIVを越えたウイルスに関連する機構的基礎は公知ではなく、本研究の課題である。
【0189】
RHGPの実用的な能力を実証するために、上の実施例において同定された特定の標的が、ウイルス疾患の治療的な予防又は処置のための潜在的な宿主標的として、さらなる研究の対象であった。直ぐ上で考察したRobo1、及び上のインフルエンザに関連して考察されたPTCH1に加えて、Nedd4を、インフルエンザの処置のための可能な候補として上の通りに同定した。Nedd4は、また、共同所有される米国特許出願シリアル番号11/939,122(2007年11月13日出願)において考察され、その全体が参照により本明細書において組み入れられる。
【0190】
Nedd4は、ESCRTタンパク質と一般的にいわれるタンパク質のクラスのメンバーであり、典型的には細胞内で見出され、そこで、それは、ESCRTタンパク質ファミリーの他のメンバーと相互作用し、細胞内の1点から別の点にタンパク質を向ける又は運ぶ。Nedd4に対する抗体を、標準的な手段により調製した。ヒト身体の特定組織がNedd4について弱く染色されるが(例、結腸、小腸、肝臓の幹細胞)、一般的に、このタンパク質は表面で弱く発現される(仮にあっても)。産生された抗体の内、ヒトNedd4抗体Ab27979及びAb101APは、両方がFunctional Geneticsから利用可能であり、従来の方法を使用して調製されたが、しかし、任意の抗体産生方法を用いることができる。これら及び無関係の標的に対して産生されたコントロール抗体を用いて実施されたFACSによって、Nedd4は、感染されていないMDCK細胞の表面上で全く染色されず、インフルエンザ(A/Udorn/72)を用いて感染されたMDCK細胞は、表面上に多量のタンパク質を呈することが実証された。TSG101と同様に、ウイルス装置は感染細胞の細胞機能を乗っ取るため、Nedd4は、その通常の役割から「ハイジャック」され、使用されて、ビリオン、又は必須のビリオンタンパク質を細胞表面に輸送すると思われる。図13。
【0191】
Ab27979(Nedd4配列の中央、アミノ酸313−334から取られたエピトープに対して産生された)及びAb101AP(タンパク質のC末端、Nedd4アミノ酸860−900に対して産生された)(図13を参照のこと)は、インフルエンザ複製の強いADCC媒介性阻害を示した。濃度0.01μg/ml及びそれ以上で、これらの抗体は、反復インビトロ実験においてウイルス複製の80%より良好な阻害を示した。(図14)。
【0192】
上で考察された実験結果を、種々のインフルエンザ株に対して繰り返した。なぜなら、従来の治療からのインフルエンザについての「回避」の1つの主な手段が、処置での事前のワクチンによる中和を避けるための十分な違いである小さな突然変異、又は新たな株の導入であるからである。実験では、抗体結合が、比較されたインフルエンザ株、及び関連ウイルス(パラインフルエンザ、RSV)を通して持続的であることが実証された。抗体を使用し、MDCK細胞を使用したA/Udorn/72(H3N2)、MDCK細胞を使用したA/Hong Kong/8/68(H3N2)、HEK293細胞におけるA/Japan/305/57(H2N2)、HEK293細胞におけるA/Puerto Rico/8/34(H1N1)、HEK203細胞におけるA/WSN/33(H1N1)、MDCK細胞におけるB/Lee/40、HEp−2細胞におけるHPIC−3(ヒトパラインフルエンザウイルス3)、MRC−5細胞におけるCMV/AD−196、及びHEp−2細胞におけるRSV/A2に対するこれらの抗体による表面上での大量の染色が実証された。これらのウイルス全般での結果は、ヒト又は哺乳動物(他の哺乳動物は、ペット及び家畜、例えばイヌ、ネコ、ならびに関連する哺乳動物及び農業用動物、例えばウシ、ヒツジ、ブタ、ヤギなどを含む)を標的化することが、広く効果的でありうることを示唆しており、そこでは従来のウイルス標的処置は共通して株及び宿主に特異的である。明らかに、RHGP−TAIL技術は、種々の状況における介入のための標的としての開発のために遺伝子及びそれらのコードされるタンパク質を迅速に同定する能力を提供する。
【0193】
実施例6−転移関連遺伝子のRHGP同定
上で注目した通り、RHGPは、ウイルス感染性の阻害のための潜在的な標的の同定に限定されない。考えられている特定の条件について固有である表現型が特定できる限り、RHGPの使用は予測可能な結果を与え、そこでは同定された遺伝子はその表現型を促進又は阻害する。治療用薬剤が、遺伝子によりコードされるタンパク質に標的化される場合、遺伝子自体とは対照的に、薬剤は対応する効果を与えるはずである。
【0194】
このように、癌処置を改善させるための1つのアプローチは、化学療法に対する耐性を促進する薬剤を同定することである。多くの強力な化学療法剤が宿主に対して有毒であり、治癒の症例は疾患と同じくらい悪い。RHGPライブラリーを化学療法剤に暴露させる。薬物処置を生き残るものを同定し、摂動が変化した生存条件を起こした遺伝子を選択し、挿入されたRHGP挿入物の報告を決定し、上の通りに、過剰発現又は低下発現が所望の表現型に関与するか否かを決定する。
【0195】
生存は、しかし、RHGP探索において表現型として使用することができる唯一の特徴ではない。多くの癌において、それは、特に危険である転移状態の発生である。腫瘍の転移癌への形質転換を抑制しうる薬剤は、寿命を延長し、支持処置を助ける際に価値がありうる。腫瘍は、塞栓が解離し、血中に入ることにより転移する。効果的な転移が形成するために、腫瘍塞栓が、それらが移動している血管の内皮壁に結合しなければならない。図15におけるフローチャートにより示される通り、RHGPを使用して、付着を阻害又は促進するその能力について摂動細胞を照合することができる。付着を促進する候補標的の能力を予防する治療産物(例えば、候補に結合しうる抗体など)、又は付着の低下を増強する産物(例えば付着を阻害する候補の過剰発現又は投与など)は、同様に、転移の形成を遅延又は抑制しうる。
【0196】
図15のフローチャートに従った付着表現型のこのRHGP研究の適用は、研究のための複数の候補を産生している。主な候補を図16に同定しており、それらは、同定された候補だけでなく、付着、任意の同定された有用性又は機能などを促進又は低下させるか否かを反映する。これらの標的の各々での実行は、次に、効果的な癌処置薬剤である、追加の十分に耐容される薬剤を産生しうる。
【0197】
本特許出願を通して引用された参考文献、及び列挙された研究を、より良好な情報のために以下に列挙する。
【0198】
【表14】
【0199】
本願において開示する発明を、複数の具体的な実施例により例示し、ならびに、総称的に考察してきた。実施例(特定のベクターフラグメント、標的、及び手順を含む)は、限定することを意図しない。請求項において見出される具体的な列挙の非存在において、請求項は、任意の例示された実施態様に限定されるとは考えるべきではなく、開示の全ての局面と一致して読まれるべきである。
【技術分野】
【0001】
優先データ及び参照による組み入れ
本願は、以下に対する優先権の利益を主張する:米国特許仮出願第61/142,664号(2009年1月6日出願);米国特許仮出願第61/153,012号(2009年2月17日出願);米国特許仮出願第61/158,015号(2009年3月6日出願);米国特許仮出願第61/160,737号(2009年3月17日出願);米国特許仮出願第61/178,112号(2009年5月14日出願);及び米国特許仮出願第61/180,457号(2009年5月22日出願);それらの全てが参照によりその全体が組み入れられる。
【0002】
発明の背景
発明の分野
本発明は、熱非対称インターレースPCR、又はTAIL−PCRといわれるプロセスを通じてスポットされた標的の同定と合わせた、ランダムホモ接合性遺伝子摂動(RHGP)による治療用宿主標的を同定するための遺伝子探索ベクターシステム(GSV)の使用に関する。RHGPとTAIL PCRとの組み合わせは、本願を通して、RHGPといわれる。
【0003】
ヒトゲノムの配列決定は、即時に、かつて期待された処置のための治療標的のカスケードをもたらしてこなかった。遺伝子配列の存在を知ることによって、1人(それ以上はない)が、遺伝子又は遺伝子システムが関与する表現型、又は遺伝子が細胞の進行中の進化をどのように媒介するかを特徴付けることはできない。それにもかかわらず、宿主標的の同定は、多くの研究者にとって重要性の増加する分野となっている。ウイルス疾患は、特に、ウイルス侵入者によって攻撃された哺乳動物、及び特にヒトを予防及び処置する方法を開発する人の独創性に挑戦してきた。本発明は広い適用性を有するが、本願におけ焦点は、治療介入のための宿主標的を同定するためのデバイスとしてのRHGPの有用性である。
【0004】
ウイルスは、効果的な治療法を開発する能力を妨げてきた。部分的には、突然変異してウイルスを標的化する薬物の有効性から離れるそれらの能力のためである。ウイルスは、また、それらに対して利用可能な広いパレットの差動的な作用を有する。それらは迅速な侵入者(インフルエンザなど)である。又は、感染細胞の機構のウイルスによる乗っ取りは緩徐で、経時的に遅延されうる(HIVなど)。侵入者の標的化に対する代替物として、Functional Genetics, Inc.(本願の譲受人)は、ウイルスを標的化する治療法の設計から離れたパラダイムシフトに導いており、宿主タンパク質(例えばTSG101及び他の宿主タンパク質など)を標的化する抗体及び小分子治療の両方を開発している。宿主タンパク質を標的化することにより、ウイルスが突然変異して離れる圧力が低下する。ウイルスは所与の細胞に感染することができるが、しかし、そのライフサイクルのために必須の宿主タンパク質へのウイルスの接近を拒むことにより、細胞から細胞へのウイルスの伝播が予防される。
【0005】
潜在的な宿主の治療タンパク質及び遺伝子の同定を推進するために、ある種の先端サーチエンジンが要求される:
質問は基本的なものである。問題の遺伝子、及び/又はそれがコードする産物は、ウイルス侵入者に対してある種の耐性を付与するか。質問は、本質的に、イエス/ノーの質問でありうる。他の状況において(他の疾患を伴う)、質問は異なりうる。このように、RHGPを使用し、ヒト遺伝子(その発現がアルツハイマー患者において見出されるプラーク形成表現型と相関する)を同定してもよい。それらの結果は、米国特許出願シリアル番号12/566,951(2009年9月25日出願)において報告される。種々の技術が長年にわたり開発されており、種々の疾患に関与する宿主標的を同定してきた。例示的な技術は、競合RNA干渉(RNAi)である。例えば、Nature, Revealing the World of iRNA, 431, pp.338-342 (September, 2004)を参照のこと(参照により本明細書において組み入れられる)。低分子干渉(siRNA)は、標的発見において補助するためのゲノム規模のsiRNAライブラリーの生成のために多大な注目を受けてきた。Becket et al, Cancer Research, 69(2), (2009)。これらの方法は、同定される標的、及び関連するゲノムのある程度の事前の知識を要求する。過去十年間に、多くの生命を脅かす癌の診断及び処置における著しい進歩が目撃されてきた。進展の大部分は、癌細胞の挙動を制御する基本的機構の理解の上に築かれる。そのような知識によって、癌細胞により選択的に利用される分子変化及び調節機構に対する標的治療の合理的設計のための基礎が形成された。顕著な成功は、トラスツズマブ(Herceptin(登録商標))を用いた乳癌細胞上のHER2、ボルテゾミブ(Velcade(登録商標))を用いた多発性骨髄腫におけるプロテオソーム、及びゲフィチニブ(Iressa(登録商標))を用いた非小細胞肺癌細胞上のEGFRの標的化を含む(Ataergin et al., 2009; Campos, 2008; Madarnas et al., 2008; Pytel et al., 2009; Yang et al., 2009)。しかし、今日開発中の合理的に設計された治療の大半では、相対的に狭い一連の標的に焦点が合わされている。この強調は、論理的には、これらの標的についての知識の増加及び同じ又は類似の分子を標的化する早い模倣薬物を刺激する既存の薬物を用いた商業的成功に続く。それにもかかわらず、これらの傾向は、本来なら、過小評価される又は未知の標的もしくは機構から生じうる飛躍的進歩の邪魔をしうる。鍵は、従って、癌細胞の薬物感受性、転移能力、及び疾患の病態生理の他の特徴を決定的に制御する標的を同定する新規手段を開発することである。
【0006】
過去十年間に、多くの生命を脅かす癌の診断及び処置における著しい進歩が目撃されてきた。進展の大部分は、癌細胞の挙動を制御する基本的機構の理解の上に築かれる。そのような知識によって、癌細胞により選択的に利用される分子変化及び調節機構に対する標的治療の合理的設計のための基礎が形成された。顕著な成功は、トラスツズマブ(Herceptin(登録商標))を用いた乳癌細胞上のHER2、ボルテゾミブ(Velcade(登録商標))を用いた多発性骨髄腫におけるプロテオソーム、及びゲフィチニブ(Iressa(登録商標))を用いた非小細胞肺癌細胞上のEGFRの標的化を含む(Ataergin et al., 2009; Campos, 2008; Madarnas et al., 2008; Pytel et al., 2009; Yang et al., 2009)。しかし、今日開発中の合理的に設計された治療の大半では、相対的に狭い一連の標的に焦点が合わされている。この強調は、論理的には、これらの標的についての知識の増加及び同じ又は類似の分子を標的化する早い模倣薬物を刺激する既存の薬物を用いた商業的成功に続く。それにもかかわらず、これらの傾向は、本来なら、過小評価される又は未知の標的もしくは機構から生じうる飛躍的進歩の邪魔をしうる。鍵は、従って、癌細胞の薬物感受性、転移能力、及び疾患の病態生理の他の特徴を決定的に制御する標的を同定する新規手段を開発することである。
【0007】
相対的に単純な生物(例えば細菌など)のゲノムでの初期の調査において、単一コピーのDNAだけ、従って、標的遺伝子の単一対立遺伝子の存在が、標的遺伝子の迅速同定に役立った。標的組換え(相同組換え)により遺伝子を「ノックアウト」することによって、問題の遺伝子の機能を削除することができた。対照的に、二倍体生物(哺乳動物を含む)は、2コピーの各遺伝子を提示する。細胞中に存在する1本のDNA上の単一コピーだけを破壊することによって、遺伝子、又はそれに関連する表現型は除去されない。この問題に対する解決が、米国特許第5,679,523号(Li et al.)に提示されており、参照により本明細書において組み入れられる。それはランダムホモ接合性ノックアウト又は「RHKO」技術に向けられる。この方法において、遺伝子探索ベクターがゲノム中に挿入され、アンチセンス鎖の生成を起こすトランス活性化因子を保有する。このように、1本鎖中へのベクターの挿入によって1つの対立遺伝子が発現停止され、アンチセンス鎖の生成は、反対の対立遺伝子に結合し、それを不活性化又は発現停止する。
【0008】
RHKOに対するいくつかの限定がある。最初に、それはノックアウト効果に限定される。発現のアップレギュレーションの効果を決定し、アップレギュレーションの標的として貴重でありうる潜在的な標的を同定することができない。遺伝子(その増強された発現が重要な表現型を付与する)の発現を増強する方法については米国特許第5,272,071号を参照のこと。RHKO技術により提示される追加の課題は、改変細胞の複数ラウンドの培養についての必要性である。培養のために要求される時間と、変化した表現型(ここでベクターが挿入された)に関与する摂動の同定のために要求される時間の間に、多大な「ダウンタイム」がシステム中に導入される。
【0009】
米国特許出願シリアル番号11/928,393(Duan et al.)(参照により本明細書において組み入れられる)において、抗体を低レベルで発現する細胞において抗体の発現レベルを増強するための方法が開示され、ランダムホモ接合性遺伝子摂動、又はRHGPといわれる。この方法において、遺伝子探索ベクター、又はGSVは、アンチセンスRNAを生成し、それは挿入ベクターと組み合わされ、摂動されるいずれの遺伝子の発現も低下又は除去し、細胞集団にトランスフェクションするために使用される。本願の実施例は、特定の遺伝子の標的摂動を含み、再び、大部分がノックアウト技術に向けられ、増強された抗体産生(所望の表現型)という結果の検証のために特定の細胞の発現を抑制する。この技術は、また、摂動に起因する細胞を増大及び培養する必要性、及び結果として生じる遅延を含む。
【0010】
標的サーチエンジンは、任意の特定の疾患又は表現型の型に限定されるべきではない。このように、ウイルス疾患に加えて、種々の状況(抗体発現に加えて)を想像することができ、そこでは、個人、例えばヒトなど(宿主細胞だけではない)は、改善された表現型から利益を得うる。腫瘍学での薬物開発の歴史は、癌細胞の挙動の増加する理解と共に進化してきた。薬物開発の最初の期間において、化合物ライブラリーが、腫瘍細胞の増殖又は生存の阻害について分析された。これらのアプローチは、実験室での腫瘍細胞の培養における進歩の結果として可能になった。そのような改善によって、培養された腫瘍細胞の増殖を殺す又は予防する化合物を同定するための実験的スクリーニングを開発することが可能になった。これらの評価は機能第1(Function−First)アプローチを含み、そこでは、最終的な結果に最大の価値を、抗腫瘍活性の標的又は機構的基礎の同一性にはより少ない強調を置いた。これらのアプローチは、ハイスループットスクリーニング手順における改善を要求及び刺激し、化学構造のアレイを、新規天然産物の単離を介して、新規化学ライブラリー及び/又はその2つの組み合わせを作製することにより多様化させるための努力を促進した。結果として得られる細胞傷害性治療は、今日の本発明者らの抗腫瘍薬の武器庫の大半を含み、アルキル化剤(プラチナ化合物)、抗生物質(ドキソルビシン)、抗代謝剤(メトトレキセート、5−フルオロウラシル)、アルカロイド(ビンクリスチン)、及びタキサン(タキソル)を含む。しばしば、治療法のための標的の同一性は未知である、又は機構的基礎は後に同定された。効果的であるが、これらの細胞傷害性の抗腫瘍薬剤は、しばしば、不要な副作用を伴った。それによって長期間にわたる化合物の使用が限定されうる。これを複雑化することとして、P糖タンパク質の発現増加が、腫瘍細胞の迅速な突然変異速度と合わさって、薬物耐性腫瘍の選択を支持する傾向がある(Gottesman and Ling, 2006)。
【0011】
実験室において腫瘍細胞を培養する能力によって、また、研究者が良性細胞と悪性細胞を区別する変化を理解し始めることができるようになり、それは標的介入の時代を予告した。悪性形質転換に寄与するがん遺伝子の理解における基本的な進歩は、次に、癌細胞の挙動(増殖、生存、及び浸潤)を支配するシグナル伝達経路の同定を促進した。この理解の改善によって、公知の標的に基づく化学療法剤の新たな生成が促進された。結果として得られる化合物は、公知の標的、例えばトポイソメラーゼ(ポドフィロトキシン)及び増殖因子又はそれらの受容体などに影響を与えた。腫瘍細胞についての改善された選択性は、より効果的で選択的な抗腫瘍薬物をもたらした。標的薬物をスクリーニング又は操作し、特定の標的を選択的に阻害したため、新たな分子医薬は、しばしば、関連する診断用薬剤を伴ったが、それは患者の適格性又は処置結果を予測したであろう。我々は標的介入のこの時代の真っ最中に依然としているが、同じ歴史的な障害(不要な副作用及び薬物耐性の獲得を含む)の多くが残存する。これを複雑化することとして、結果として得られる薬物の多くが、相対的に狭い一連の標的(例、チューブリン、増殖因子、及びキナーゼ)に焦点を合わせている。この優先順位付けは、検証された標的を集中的に研究するための必要性の論理的結果であった。しかし、意図していない結果は、研究下にある標的の着実に狭まる型であった。標的同定への種々のアプローチの開発を図1に示す。
【0012】
疾患に対する遺伝的関連の理解の改善に伴い、癌の遺伝的基礎の増加する認識によって、腫瘍細胞のゲノム及び次にプロテオミクスサンプリングにおける興味が促された。癌細胞についての潜在的な標的を同定するための「−オミクス」の成長は、DNA配列決定における劇的な技術的進歩と同時期に起こり、多くの注目を集め、学界及び医薬業界を、その技術への相当なインフラの投資に導いた。ゲノミクス、プロテオミクス、及び関連する戦略によって、標的の数及び幅が増加した。このように、この分野では、それ自体が、幅広い標的にあふれていることが見出され、継続的な標的発見についての認知された必要性は底に達した。しかし、「−オミクス」を使用して同定された相関関連によって、しばしば、疾患病理を起こす又はそれに寄与する標的が、疾患プロセスの副作用として生じる標的から区別されないことがますます理解された。このあいまい性は、原因と結果の周辺の問題を解決するための相当な投資を要求し、このように、新規の腫瘍学標的についての一般的な欲求を減少させてきた。なぜなら、多くの機関が、「−オミクス」研究及びその結果として得られる標的を研究するために必要とされる技術に、相当なインフラを投資していたからである。
【0013】
したがって、潜在的な治療候補の迅速な検出のための方法を開発することが依然として当業者の目標であり、それは、同定された場合、それらの治療的な潜在力を開発する(発現低下(例、遺伝子治療)又はそれによりコードされるタンパク質のサイレンシング(リガンド結合治療)のいずれかを通じて)又は単に診断の対象とすべき遺伝子(それらの存在、又は複数コピーでの存在が、個人が、問題の遺伝子と相関する全ての表現型についての可能性の高い候補であることを示す)を同定するためのさらなる実験の対象となりうる。理想的には、この技術は、アップレギュレーションならびにダウンレギュレーションに起因する表現型を同定することが可能であるはずであり、ゲノム、又は標的遺伝子の知識は要求しない。不必要な多額のお金及び人的資源の出費を避けるために、この技術は、検証可能な潜在的標的(即ち、新たな表現型が、ゲノム自体の変化に起因し、それ自体の改変、又は攻撃状態に対する暴露のいずれかの結果ではない)を得る必要がある。それは、潜在的な候補のクローニング及び増大に起因する時間遅延を低下させる基礎に関する結果を得る必要がある。
【0014】
発明の概要
ランダムホモ接合性遺伝子摂動(RHGP)は、所与の細胞集団において遺伝子をランダムに破壊するための方法であり、任意の所望の表現型のスクリーニング及びその関連遺伝子の同定を可能にする。RHGPの能力に対して役立つのは、そのアンチセンス技術であり、それによって遺伝子の全コピーの同時不活性化が可能になる(所与の細胞において全てのコピーを無効にする反復標的化を要求する従来のノックアウトアプローチとは異なる)。さらに、RHGPは、競合RNA干渉(RNAi)技術よりも強力である。なぜなら、それは以下の利点を与えるからである:標的又は経路の事前の知識なしに、偏りのない方法で任意の遺伝子座に影響を与える能力;そのエピジェネティック状態(サイレンスされているか又はサイレンスされていないか)にかかわらず、任意の標的(マイクロRNAを含む)又はタンパク質ドメインを過剰発現又はノックダウンする能力;表現型の即時検証のための組み込み可逆性;及び同じベクターシステムを用いてインビボで表現型を試験する能力。また、RHGPは、RNAiのオフターゲットの影響を受けない。なぜなら、RHGPベクターがゲノム中に組込まれ、その活性のための基礎として相同性を使用しないからである。RHGPは、その現在の具体化において、しかし、RNAiと比較し、遺伝子標的を同定するために実質的により長い時間が必要である。なぜなら、それは、目的の全ての標的化された遺伝子座が細菌中にクローン化され、配列決定を通じて個々に同定されることを要求するからである。本発明において、本発明者らは、全てのRHGP挿入標的を、PCRを使用して迅速に同定するための方法の適応について記載しており、それによりRNAi方法論に沿った標的遺伝子発見のスピードをもたらす。この強力な技術は、ウイルス疾患又は細菌疾患と戦うための潜在的な宿主タンパク質の同定に限定されない。宿主タンパク質が関与する任意の症候群、例えば癌などを、RHGPの強力なプロセスに供することができる。
【0015】
RHKOの技術の上に構築し、そこでは最初のアプローチが開発され、真核細胞が一般的に2組の染色体を有するとの事実に起因するバリアを克服した(Li and Cohen, 1996)。RHGPでは同じ基本的アプローチを用いる。真核細胞の二倍体の性質によって、細菌経路の本発明者らの理解において非常に強力であることが証明されている標的遺伝子の単純なノックアウトベースの評価が妨げられた。siRNA(公知の標的のノックアウトに限定される)とは異なり、RHGPベースのアプローチは、任意の標的遺伝子の両方のコピーを同時にノックダウンする能力を含み、その遺伝子の任意の事前の知識又はアノテーションに非依存的である。強力な阻害が可能である。なぜなら、組込み事象自体が、遺伝子の第1コピーをノックアウトするために十分であり、標的遺伝子のアンチセンス発現が、第2の対立遺伝子をノックダウンするためである(図2)。RHGPが、また、遺伝子発現(全遺伝子又は個々のドメインを含む)をアップレギュレーションすることができる(図3A)。今日まで、今日までにRHGPを使用して同定された標的の約半分が、センス方向での組込みを示し、残りの半分がアンチセンス組込み事象を示している。結果的に、RHGPは、研究下にある表現型の原因となる任意の遺伝子変化についてゲノム全体を照合する手段を提供する。
【0016】
RHGPの中心となる特性は、固有のレンチウイルスベースの(又はサルマウス白血病ウイルスベースの)遺伝子エレメントであり、遺伝子探索ベクター(GSV)として公知であり、それは、ゲノム全体を照合し、目的の表現型を起こす標的遺伝子を同定するように設計された。GSVはゲノム内の単一部位に組込まれ、そこで、それは誘導プロモーターを介して標的遺伝子の発現を調節する(図4)。1個の細胞当たり単一の組込みが、相対的に低い感染効率(MOI)でGSVを用いた細胞の形質導入により制御される。ベクター自体が、自己不活性化レンチウイルスLTRをコードし、それは、GSVが形質導入細胞から再出現することを予防する。
【0017】
GSVは、ゲノム中に、センス又はアンチセンスのいずれかの方向で組込まれうる。アンチセンス配置において、組込み事象自体が1つの対立遺伝子を不活性化し、アンチセンスコンストラクトの発現を促進し、それによって他の対立遺伝子上でコードされる遺伝子がノックダウンされる(図3A)。反対(センス)方向に組込まれた場合、RHGPは、標的遺伝子の過剰発現又は非発現停止を促進することができる。この結果は、遺伝子全体の単純な過剰発現を超えて拡大しうる(例、開始部位の上流での挿入)、又は、開始部位の下流に組込まれた場合、特定のドメインの過剰発現を引き起こし、それは、野生型遺伝子の機能のドミナントネガティブな阻害剤を産生しうる。そのようなものとして、RHGPによって細胞ゲノム全体の照合ができるようになり、異なる型の標的事象を同定する。
【0018】
RHGPは、GSV形質導入細胞の「ライブラリー」の構築を要求する(図3B)。このライブラリーは、ゲノム全体をカバーするGSV組込み事象(過剰発現及びアンチセンス発現の両方に関する)を含む。RHGPは、柔軟で、分析される細胞型(リンパ系もしくは付着性)、種(ヒトもしくは非ヒト)、又は形質転換特徴(初代もしくは不死化もしくは形質転換)により偏らない。RHGP戦略では、内因的に有毒であるGSV形質導入事象が除かれる。例えば、重要遺伝子の発現を破壊するRHGP形質導入事象は、宿主細胞に対して有毒でありうる。このように、これらのクローンは、RHGPライブラリーから除去されうる。一旦形質導入されると、同じライブラリーが記録され、他のスクリーニングのために使用することができる。
【0019】
RHGP−TAILを使用し、疾患又は不全状態に応答性でありうる種々の現象及び表現型が試験されてきた。本明細書において考察されるのは、インフルエンザに対する耐性、ヒト免疫不全ウイルス(HIV)及び関連するAIDS、ならびに癌に対する耐性に関与する表現型を試験するためのRHGPの使用からの種々の結果である。これらのRHGP試験により生成された標的も追跡調査される。
【図面の簡単な説明】
【0020】
【図1】図1は、標的発見方法における最近の開発を反映するチャートである。
【図2】図2は、本願の本発明の遺伝子探索ベクター(GSV)の使用を実証する概略図である。
【図3A】図3Aは、本発明のGSVのセンス及びアンチセンス挿入を実証する概略図である。
【図3B】図3Bは、本発明のGSVのセンス及びアンチセンス挿入を実証する概略図である。
【図4】図4は、インフルエンザ関連標的についての完全なRHGP研究の実施に関与する段階を描写するフローチャートである。
【図5A】図5Aは、本発明において使用されるGSV pRHGP−12RSNについての遺伝子地図である。
【図5B】図5Bは、本発明において使用されるGSV pRHGP−12RSNについての遺伝子地図である。
【図6A】図6Aは、PTCH1の同定を、RHGP技術によるインフルエンザの治療的阻害のための可能な標的として描写する概略図である。
【図6B】図6Bは、PTCH1の同定を、RHGP技術によるインフルエンザの治療的阻害のための可能な標的として描写する概略図である。
【図6C】図6Cは、PTCH1の同定を、RHGP技術によるインフルエンザの治療的阻害のための可能な標的として描写する概略図である。
【図7】図7は、コントロールと比較した、PTCH1の阻害により起こされたウイルス産生における相対的な阻害を示すグラフである。
【図8A】図8Aは、異なるMOIでのPTCH1の阻害効果のグラフ同定である。
【図8B】図8Bは、異なるMOIでのPTCH1の阻害効果のグラフ同定である。
【図8C】図8Cは、異なるMOIでのPTCH1の阻害効果のグラフ同定である。
【図9A】図9Aは、生物学的プロセス及び分子機能によるRHGP同定されたインフルエンザ標的の分類を示すチャートである。
【図9B】図9Bは、生物学的プロセス及び分子機能によるRHGP同定されたインフルエンザ標的の分類を示すチャートである。
【図10】図10は、RHGPにより同定された治療標的の阻害により起こされたウイルス産生における低下を反映するグラフである。
【図11A】図11Aは、HIV感染を制御するための標的としてのRobo1の同定を例示する。
【図11B】図11Bは、HIV感染を制御するための標的としてのRobo1の同定を例示する。
【図11C】図11Cは、HIV感染を制御するための標的としてのRobo1の同定を例示する。
【図12A】図12Aは、HIV感染性に対する、RHGP同定されたRobo1を阻害する効果を反映する。
【図12B】図12Bは、HIV感染性に対する、RHGP同定されたRobo1を阻害する効果を反映する。
【図13】図13は、Nedd4(抗体が産生されるエピトープ)に対する種々の抗体の生成及び感染MDCK細胞の表面上でのNedd4の同定を描写する。
【図14】図14は、Nedd4抗体により起こされたウイルス複製の低下を実証するグラフである。
【図15】図15は、RHGPを使用して癌転移を阻害するための標的の同定において取るべき段階を例示するフローチャートである。
【図16A】図16Aは、転移癌の形成を阻害するために使用されうるRHGPにより同定された標的の重要な特徴を特定するチャートである。
【図16B】図16Bは、転移癌の形成を阻害するために使用されうるRHGPにより同定された標的の重要な特徴を特定するチャートである。
【0021】
発明の詳細な説明
遺伝子探索ベクター
RHGP遺伝子探索ベクターは、2つの特色で来る:モロニーマウス白血病ウイルス(MMLV)バージョン及びレンチウイルスバージョン。探索ベクターがどのようにコンストラクションされ、使用されるかを理解することが、本発明の詳細な実施を理解する際での重要な第一段階である。
【0022】
MMLVベクターは、それらが、遺伝子のプロモーター領域中への挿入を好むという利点を有し、それにより遺伝子全体がアップレギュレーション又はダウンレギュレーションされることを確実にする。さらに、MMLVは活性遺伝子に向かって偏っておらず、それらを相対的にランダムにゲノム全体に挿入することができる。レンチウイルスベクターは、MMLVと比較して感染の広い宿主範囲を有しており(それらが水疱性口内炎ウイルスGタンパク質、すなわちVSV−Gを用いてシュードタイピングされているとの理由で)、それらは分裂細胞及び非分裂細胞の両方に感染する能力を有する。しかし、それらは、それらが活発に転写された遺伝子を好む点で、それらが相対的に非ランダムに挿入されるという事実に悩まされる。レンチウイルスバージョンは、MMLVバージョン(プロモーター領域を好む傾向がある)と異なり、遺伝子自体の内に挿入の偏りを有さない。レンチウイルスバージョンは、未知の理由で、MMLVベクターと比較してより高い力価を一貫して産出している。
【0023】
MMLVベクターは、それらがいずれかの末端にMMLV 5’及び3’LTR(末端反復配列)を有する点で、同じ一般構造を有し、組込み後にウイルスの非重要エレメントを自動的に切り出すために使用されるCre遺伝子が隣接する(LoxPエレメントがあり、Cre及び不要なエレメントが自動的に除去される)。
【0024】
特定のベクターの特性
RHGP48:このベクターは、14コピーのテトラサイクリン応答エレメント(14×TRE)を伴うプロモーターを含み、転写ターミネーターなしにピューロマイシン耐性遺伝子の発現を促進させ、アンチセンス転写物を作製する(このアンチセンスは、RHGPベースの遺伝子不活性化の基礎である)。TREは、テトラサイクリンの非存在において、プロモーターが活性であり、アンチセンス発現を促す点でTet−Offプロモーターである。一旦テトラサイクリンが培地に加えられると、プロモーターが遮断され、アンチセンス転写物も遮断され、それはRHGPの可逆特性の基礎である。
【0025】
RHGP48−RSB:このベクターはRHGP48の構造と類似であるが、しかし、TREがアンチセンス発現を促す代わりに、それはNew England BioLabsのRheoSwitchプロモーターを使用し、それはRSL1リガンドにより活性化される。5コピーのRheoSwitchエレメントがあり、それらにRheoSwitchアクチベーターが結合する。このプロモーターは、ブラストサイジン耐性遺伝子の発現を促し、それも転写ターミネーターなしであり、アンチセンス産物を作ることができる。
【0026】
RHGP48−RSN:このMMLVベクターはRheoSwitch調節プロモーターを含み、アンチセンス転写物を促すが、しかし、アンチセンスから転写される耐性遺伝子はない。この「ネイキッド」プロモーターは、標的遺伝子のダウンレギュレーション及びアップレギュレーションの両方を促進すると考えられる。なぜなら、理論的に、プロモーターの下流に遺伝子を有することによって、下流の標的を翻訳する能力が損なわれうるからである(なぜなら、遺伝子、例えばブラストサイジン又はピューロマイシンなど翻訳終止コドンを有する)。挿入事象の選択を可能にするために、常時発現性のホスホグリセリン酸キナーゼ(PGK)プロモーターが存在し、ブラストサイジン耐性遺伝子を促す(これはセンス方向の正常遺伝子であり、アンチセンス転写物を産生しない)。
【0027】
レンチウイルスベクターは5’及び3’LTRの両方を含むが、しかし、3’LTRは自己不活性化し(3’SIN−LTRと呼ばれる)、ゲノム中への挿入後、ベクターはウイルスを複製し、産生することができない。これによって安全性プロファイルが劇的に増加する。これらのベクターはCre遺伝子を含まず、しかし、それらは全てがLoxP部位を有し、それらは、ゲノムにおいて、Creを外因的に加えることにより除去(逆転)することができる。これらのベクターは、また、RSVエンハンサーエレメントを5’LTR中に含み、レンチウイルスのより高い転写、故に、産生を可能にする。
【0028】
RHGP11:このベクターはEGFPを駆動する14×TREを有し、調節されたアンチセンス産物、ならびに緑色蛍光タンパク質を産生することを促す。これによって、細胞を蛍光顕微鏡下で見ることが可能になり、必要な場合、FACSにより選別される。また、このベクターはゼオシン耐性遺伝子を有し、PGKプロモーターを促し、薬物選択を可能にする。
【0029】
RHGP11−CMVE:このベクターはRHGP11と類似であるが、しかし、EGFPを伴うアンチセンス発現を促す調節されたTREの代わりに、アンチセンスは常時発現性である。なぜなら、CMVプロモーターがEGFPアンチセンスコンストラクトを駆動するからである。これは、表現型を簡単に逆転させることができない場合(例えばインビボ試験など)において重要でありうる。
【0030】
RHGP22:このベクターは、典型的なTREの代わりに、pTight TREを有する。5コピーのpTight応答エレメントがある。pTightは、また、アンチセンスのためのテトラサイクリン調節プロモーターであるが、しかし、本来のTREほどリーキーではない。このpTightは、ブラストサイジン耐性遺伝子を通じてアンチセンス発現を駆動している。
【0031】
RHGP12−RSN:このベクターはRheoSwitchプロモーターを使用し、「ネイキッド」アンチセンス転写物を促す。また、ベクターを、PGKプロモーターがブラストサイジン遺伝子の発現を駆動しているという事実により、薬物により選択することができる。また、EGFP遺伝子をブラストサイジン遺伝子の下流にIRES(配列内リボソーム進入部位)エレメントを介して置き、PGKプロモーターが両方のタンパク質の産生を駆動することができる。このベクターについての遺伝子地図を図5に示す。
【0032】
RHGP13−CMVN:このベクターもアンチセンス転写物を生成する「ネイキッド」プロモーターを有するが、しかし、このプロモーターは常時発現性であり(CMVプロモーター)、RheoSwitch又はTRE/pTightプロモーターように調節されない。これは、再び、インビボ試験のために有用であり、そこでは安定な非調節アンチセンス産生が意味をなす。このベクターは、RHGP12−RSNベクターと同じPGK−BS−IRES−EGFPコンストラクトを有する。
【0033】
RHGP−TAILプロトコール
RHGPの成功スクリーニングは、表現型をスクリーニングするために使用されるアッセイに依存的である。他のスクリーニング手順と同様に、所望の表現型は明確であり、頑健でなければならない。一旦それらの最小限の基準が満たされると、RHGPを実質的に任意の用途に適用することができ、それにおいて、所望の表現型によって細胞を物理的に単離することが可能になる。一例として、本発明者らの実験室及びその他ではRHGPを利用し、腫瘍細胞が、細胞傷害性薬剤を用いた化学療法攻撃(Lih, Wei, and Cohen, 2006; Reiske et al., 2009)又はホルモン療法に対して耐性となる標的を同定してきた。これらの条件下で、薬物感受性細胞が消滅し、RHGP摂動細胞のサブセットが生き残った。「オミクス」ベースのアプローチを典型的に表わす相関する知見とは異なり、薬物攻撃に直面した生存は、RHGP形質導入事象により直接的に起こされた。所望の表現型の選択は、細胞ベースのアッセイに限定されず、RHGPを同様にインビボでの適用のために使用することができる。例えば、動物モデルにおいて腫瘍形成能力又は転移能力を獲得する細胞を単離することができる。結果としての生存を超えて、RHGPは、また、他の特性(例えば特定のマーカーを発現する細胞の親和性ベースの単離など)について選択することができる。そのような結果は、特定のマーカー(所望の表現型として)の発現を使用し、フローサイトメトリー又はパニング手順を使用して細胞を物理的に単離することができる場合に可能である。
【0034】
相当な努力が投資され、RHGPの頑健性及び効率が最適化されてきた。GSVからの発現を促す可逆的プロモーターは、目的の標的を迅速に検証する効率的な手段を提供する。所望の表現型が得られた後、実験を、サイレンシングさせたGSVプロモーターを用いて繰り返し、表現型を逆転させることができる。表現型が、サイレンシングされたGSVを使用して持続する場合、そのクローンは優先順位から外されるであろう。なぜなら、表現型が、無関係のアーチファクトの結果として生じたかもしれないからである。そのようなものとして、一旦RHGPライブラリーが構築されると、目的の表現型について選択し、表現型の可逆性を検証することが可能になる(全て単一の試験において)。
【0035】
一旦表現型が、可逆的プロモーターを使用して検証されると、GSVは、標的遺伝子を同定し、GSV組込みの方向(例、センス又はアンチセンス)を決定するためのモチーフをコードする。RHGPの初期バージョンでは、宿主ゲノムDNAの物理的単離、それに続く従来の配列決定が要求された。この手順では、細菌形質転換のために十分なゲノムDNAを単離するための十分な数のクローンの増殖が要求された。個々の細菌クローンを次に単離し、プラスミドを、GSV組込み事象の配列決定のために単離した。このプロセスでは数週間の努力が要求され、限定されたスループットを伴った。標的同定は、熱非対称インターレース(TAIL)PCRの改変を使用して認められている。TAIL−PCRは、本来は、植物における遺伝子挿入を同定するために開発された(Liu et al., 1995)。より最近では、それを使用し、マウスゲノム中の遺伝子挿入が同定されている(Pillai et al., 2008)。本発明者らはTAIL−PCRを組み入れ、GSV挿入部位のハイスループット同定を実施した。なぜなら、少数の細胞が、ゲノムDNAを単離するために必要とされるからである。これらの改善によって、ハイスループットスクリーニングが促進され、GSV挿入部位についての情報が数週間ではなく数日以内に産生される。
【0036】
これは、トランスジーン挿入部位のハイスループットゲノム規模同定のためのTAIL−PCRの最初の適用、ならびにヒト細胞におけるトランスジーン挿入部位同定のためのTAIL−PCRの最初の適用である。
【0037】
本来のRHGPクローニング技術は、制限消化及び自己連結のための実質的な量のゲノムDNAを単離するために十分な数まで(例えば、6ウェルプレート密度まで)の哺乳動物細胞の増殖を含む。その後、自己連結プラスミドを細菌中に形質転換し、多くのクローンを個々に選び、一晩増殖させ、プラスミドをこれらの培養物から次の日に単離する。これらのプラスミドを、次に、トランスジーン挿入の部位を同定するための配列決定のために送る。
【0038】
TAIL−PCRを用いて、哺乳動物細胞を、96ウェルプレート中でのゲノム抽出のために直接的に処理することができ、それにより手順がハイスループットスクリーニングのために受け入れられるようになる。DNAを、次に、96ウェルプレートフォーマット中での、公知の及びランダムなプライマーの混合物を使用した3サイクルのTAIL−PCRに供し、次にプレートを、第3PCRラウンド後の配列決定のために直接提出し、挿入部位情報をわずか3日間で産出する。以下の表では種々の技術及びそれらの相対速度の推定を比較する(最適効率で):
【0039】
【表1】
【0040】
約50〜100ngのゲノムDNA/ウェルをPCR反応において使用する。より少ないDNAを、DNAが限定している場合に、確実に使用することができる。例えば、通常のゲノムDNA調製物は、0.2〜0.8μg/μlの範囲の産出量を有し、そのため、PCR反応のためのTE又はH2O中での1:10希釈で十分に作用する。各DNAサンプルを、1つのフォワード及び6つのリバースPCRプライマーの1つを有する6つの反応中に分けるため(全てが独立したウェル中)、1ゲノムDNAサンプル当たり8反応のプレミックスを、Immomix Red polymerase(Biolineから)を使用して調製する。フォワードプライマーの原液濃度は1μMであり、リバースプライマーは各々10μMである(それぞれ最終濃度0.15μM及び2μMについて)。
【0041】
【表2】
【0042】
一次TAIL−PCRサイクリング条件(約6時間):
【表3】
【0043】
レンチウイルスRHGPベクターについて、以下のプライマーを1DNAサンプル当たりに使用する(各々6反応):
一次TAIL−PCR:ISP1(フォワード)+AD1〜AD6(リバース)
二次TAIL−PCR:GSP3new(フォワード)+AD1〜AD6(リバース)
三次TAIL−PCR:ISP2(フォワード)+AD1〜AD6(リバース)
【0044】
フォワードプライマー:
【表4】
【0045】
リバースプライマー:
【表5】
【0046】
MMLV RHGPベクターについて、以下のプライマーを使用する:
【0047】
フォワードプライマー:
【表6】
【0048】
リバースプライマー(上と同じ)
【0049】
#1のサンプルを1%アガロースゲル上で流し、バンドを可視化することができる。その後、各サンプルの1:200希釈を行う。これは、最初に、各サンプルの水中への1:20希釈を行い、次に、この1μlを、二次TAIL−PCRのための続く10μlのPCR反応(1:200最終希釈比を達成するために)において使用することにより行う。時折、PCRサンプルにおける容積喪失があり、そのため、5μlの水を、各ウェルに(所望の場合)希釈前に加えることができる(同じことを三次TAIL−PCRの前に行うことができる)。二次TAIL−PCR用の1反応当たりのプレミックスは以下の通りである:
【0050】
【表7】
【0051】
リバースプライマー及び鋳型DNAを、マルチチャネルピペットを使用して分注することができる。二次TAIL−PCRサイクリング条件(約4時間):
【表8】
【0052】
#2のサンプルをゲル上で流し(所望の場合)、#2に示す通りに1:200希釈を行い、#2の同じプレミックスを使用し、三次TAIL−PCRを、以下のサイクリング条件を用いて行う(約2時間):
【表9】
【0053】
#3のサンプルを1%アガロースゲル上で流すことができる(所望の場合)。プレートを、次に、最終的な特異的プライマー(例えばレンチウイルス用のISP2、又はMMLV用の48RS3など)を使用した配列決定のために直接送ることができる。低容積についての懸念がある場合、15μlの水をより簡単な取り扱いのために各ウェルに加えることができる。
【0054】
この手順によって、クローンにおいてウイルス組込みの同じ部位が成功裏に同定されており、そこで、組込み部位が、本来のRHGPクローニングアプローチを使用し、RHGPレンチウイルス及びMMLVベクターの両方をヒト細胞株及び非ヒト細胞株において使用して同定されてきた。
【0055】
実施例1−インフルエンザ因子の研究及びPTCH1の同定
インフルエンザ感染は、感染性疾患媒介性の罹患及び死亡の主な原因のままである。蓄積する証拠は、季節性及び汎発性インフルエンザの大半のバリアントが、従来の治療に対する耐性を発生していることを示す。そのような情報は、インフルエンザを標的化するための新規アプローチの特定において新たな興味を生じている。本発明者らの実験室では、宿主志向性の治療法の新たな戦略を開発しており、それによって、ウイルスが疾患を起こすことを予防する安全で効果的な様式で宿主分子を標的化することを追求する。改善された発見技術(RHGP)を使用し、本発明者らは、PTCH1を、インフルエンザウイルス感染を決定的に制御する必須の宿主標的として同定する。本発明者らは、さらに、抗体又はsiRNAを使用したPTCH1に対する標的介入によってインフルエンザ感染が減少することを実証する。最終的に、本発明者らは、実験室の外部でのインフルエンザ感染におけるPTCH1の関与を、PTCH1の遺伝的変異が野外における減少した疾患の罹患率に関連することを示すことにより実証する。全体で、これらの知見は、インフルエンザ疾患の管理を改善するための新規の宿主指向の治療法の開発についての重要な意味を有する。
【0056】
インフルエンザは、感染性疾患媒介性の苦痛及び死亡の最も大きな原因の1つのままである。平均的な年において、インフルエンザは20〜30,000の生命を奪い、経済的な損害は何十億ドルにもなる。その広範囲の流行にもかかわらず、ワクチン接種は散発性のままである。最近の試験では、ワクチンが、処置された人々の半分未満において有効でありうることが示されている。これを複雑化することとして、わずか一握りの治療法が、インフルエンザ感染を処置するために開発されてきた。増加する証拠は、多くの季節性株及び汎発性(トリ)株がこれらの化合物に対して耐性になっており、このようにそれらの適用を取り除いていることを示す。薬物耐性株の不可避の増殖は、インフルエンザに固有ではない。インフルエンザ、HIV、ヘルペス、及び他のウイルスの耐性株の出現は、医学界が抗ウイルス治療のための戦略を再考することを強いてきた。
【0057】
従来の抗体ウイルス剤では、ウイルスによりコードされる経路が標的化されてきた。基本的な理論的根拠では、ウイルス経路の標的化によって正常宿主細胞に対する毒性が最小限になりうることが想定されてきた。しかし、結果として得られる化合物の多くが、実際に非常に有毒であり、依然として悪く、ウイルス標的への焦点は、病原体に対して選択圧力を置き、治療に耐性であるバリアントを引き出す。代替物についてのこの必要性は、本明細書において実施されるアプローチを促してきたが、それは、薬剤耐性に対して非感受性である新たな治療法を開発することを追求し、それは、広域スペクトルの異なるウイルス型に適用することができる。
【0058】
本発明者らの実験室では、感染性疾患のための宿主志向性の治療法の概念が開発されてきた。このアプローチでは、ウイルス感染細胞において誤発現される又は機能的に変化している宿主由来標的を同定することを追求する。そのように行う際、本発明者らは、変化した発現又は機能が、ウイルス疾患を処置する際に安全で効果的である新規治療法のための機会を提供しうると仮定してきた。異なる型のインフルエンザに必須である経路を同定、そして次に標的化することにより、本発明者らは、広域スペクトルの適用を伴う十分に必要とされる抗ウイルス剤を開発するであろう。知識における明らかなギャップは、安全で効果的な宿主標的及び、理想的には、それらの標的を遂行する手段を同定する必要性である。その目的のために、本発明者らは、本発明者らの中核技術(RHGP)を成功裏に利用した。本発明者らの実験戦略では、ゲノム中の単一部位でのGSVの組込みを中心とした。そこでは、それは標的遺伝子の過剰発現又は発現の喪失のいずれかを起こしうる。そのようなものとして、RHGPによって、本発明者らは、細胞ゲノム全体を照合し、宿主細胞がインフルエンザ感染に耐性となる又は生き残ることを可能にする異なる型の事象を同定することが可能になる。RHGPを使用し、PTCH1を、インフルエンザ感染における必須の宿主標的として同定した。PTCH1に対する標的介入によってインフルエンザ感染を遮断することができる。最終的に、SNP分析によって、PTCH1が、ブタ集団におけるインフルエンザの自然大流行の間での疾患転帰に関連することが実証される。
【0059】
材料及び方法
細胞培養。ヒトMDA−MB231乳癌細胞をAmerican Type Culture Collection(ATCC)から購入した。Phoenix A細胞株はNolan博士(Stanford University)からの寄贈であり、マウスN2a細胞株はXu博士(Rockefeller University)からの寄贈であった。MDA−MB231細胞をDMEM(10% FBSを含む)中で培養し、N2a細胞を50% DMEM培地及び50% Opti−MEM培地(5% FBSを含む)中で培養した。
【0060】
RHGPライブラリーの生成及びスクリーニング。RHGP遺伝子探索ベクターを適用し、MDCK細胞においてRHGPライブラリーを生成した。RHGPライブラリーを、インフルエンザウイルスA/Udorn/72を用いて、感染効率(MOI)0.1又は0.001で示された通りに感染させた。第2ラウンドの感染から2週間後、ゲノムDNAを、BIO−RAD AquaPure Genomic DNA Isolation Kitを使用して得た。自己連結ゲノムDNAを濃縮し、E.coli細胞中に電気穿孔し、クロラムフェニコールを使用して選択した。
【0061】
siRNAを用いた宿主標的遺伝子の検証。PTCH1についてのヒト二本鎖siRNAホモログを、製造者により推奨される通りに調製した。siRNA NP−1496配列(GGAUCUUAUUUCUUCGGAGUU)は、インフルエンザウイルスのヌクレオカプシド(NP)遺伝子を標的化し、ポジティブコントロールを提供した(Ge et al., 2003)。非標的siRNA(siCONTROL1)は、ネガティブコントロールを提供した。HEK293細胞を、24ウェルプレート中に、1×105個細胞/ウェルでそれぞれプレーティングした。24時間のインキュベーション後、細胞を、20nMのsiRNA及びTransIT−TKOを用いて、製造者の指示(Mirus)に従ってトランスフェクションした。第2ラウンドのトランスフェクションから24時間後、サンプルを、MEMを用いて洗浄し、インフルエンザウイルスA/Udorn/72(MOI 1)を用いた感染が続く。細胞を、1時間にわたり、15分毎に穏やかに揺らしながらインキュベーションした。各ウェルからの培養培地を感染後48時間目に回収し、培地中の子孫ウイルスを、標準的なプラークアッセイを使用して滴定した。
【0062】
フローサイトメトリー。フローサイトメトリー分析のために、MDCK細胞を、トリプシンを使用して浮遊させ、1×105個細胞を氷上で10mg/mL抗体を用いて30分間にわたりインキュベーションした。細胞をPBS(PBS中1% BSA(Sigma, St. Louis, MO)を含む)を用いて氷上で3回洗浄し、FITC抱合ヤギ抗ウサギ(Becton Dickinson, San Jose, CA)を用いて30分間にわたり氷上でインキュベーションした。洗浄後、細胞を、1%パラホルムアルデヒドを伴うPBS中で固定した。データは、EasyCyte Flow Cytometer(Guava Technologies, Hayward, CA)において取得され、FlowJo分析ソフトウェアを使用して分析した。
【0063】
SNP及びデータセット。PTCH1 SNPを選択し、2つの基準を満たした:(a)許容可能な対立遺伝子の頻度;及び(b)外部ジェノタイピングプロバイダー(Sequenom)による成功裏のフォーマッティング。
【0064】
インフルエンザデータセットを、商業用ブタ産生システムにおいて、PIC遺伝学を使用して収集した。試験期間の間に、農場は、ブタインフルエンザウイルスの大流行を経験した。4つの成長形質を測定した:(1)成長:これは、高及び低成長動物を比較した0/1形質である;(2)生命DG:生涯1日増体量(g/日);(3)LDG死体増体量:(グラム/日)及び(4)離乳:終点1日増体量(g/日)。
【0065】
LSMの好ましい遺伝子型−LSMの好ましくない遺伝子型/2
【0066】
結果
宿主細胞をインフルエンザに耐性にする標的を同定するために、RHGPライブラリーを、上に記載する通りにMDCK細胞において生成し、インフルエンザ攻撃を示した通りに実施した(図6A)。ライブラリーを、インフルエンザA/Udorn/72を用いた感染により攻撃し、インフルエンザ耐性細胞について選択した。本発明者らは、以前に、A/Udorn/72(MOI10−1)を用いた感染によって全てのMDCK細胞が48時間以内に再現性よく殺されることを確証していた(示さず)。コントロールとして、偽形質導入細胞の並行培養物を同じように処理し、生存体は48時間後に観察されなかった。
【0067】
複数ラウンドのインフルエンザ攻撃を生き残ったRHGP形質導入細胞についての選択後、本発明者らは、1つの特定の培養物(R26−7)に焦点を合わせ、それはインフルエンザ感染を生き残った。RHGP遺伝子探索ベクターによって、本発明者らは、標的遺伝子を効率的に位置付け、組込み事象の方向(センス又はアンチセンス)を決定することができた。ゲノムDNAをR26−7細胞から単離した。RHGPベクター挿入部位に隣接するゲノムDNA配列を配列決定し、イヌゲノムに対して、UCSC Genome Browserを使用してマッピングした(図6B)。R26−7における組込み事象が、イヌ染色体において、アンチセンス方向で、PTCH1遺伝子中で生じた(図6C)。そのようなものとして、組込みは、PTCH1の第1のコピーを物理的に破壊した。結果として得られるアンチセンスは、第2の対立遺伝子の発現をノックダウンした。
【0068】
天然細胞を使用したPTCH1の検証
本発明者らは、次に、PTCH1を、インフルエンザ媒介性の宿主細胞の殺傷を予防する候補標的として検証することを追求した。これらの試験は、RHGP又はMDCK細胞の使用から生じうるアーチファクトを除くことを意味した。PTCH1は、イヌ由来MDCK細胞から単離されていたため、本発明者らは、そのヒトホモログを同定し、ヒトHEK−293細胞においてこれらの標的について選択的なsiRNAを発現させた。二本鎖siRNAを生成し、HEK293細胞中にトランスフェクトした非標的siRNAは、トランスフェクションのための一致させたコントロール及び参照基準を提供した。NP−1496二本鎖siRNAは、決定的なインフルエンザ遺伝子を標的化し、インフルエンザ感染を阻害することが実証されており、ポジティブコントロールを提供した(Ge et al., 2003)。インフルエンザ感染はHEK293細胞を効率的に殺さないため、本発明者らは、本発明者らの実験プロトコールを改変し、ウイルス力価(宿主細胞の生存の代わりに)を、効率についての主要エンドポイントとして測定した。siRNA処理したHEK293細胞を、A/Udorn/72を用いて48時間にわたり感染させ、ウイルス力価をプラークアッセイにより測定した。PTCH1二本鎖siRNAは、インフルエンザウイルス産生を、ポジティブコントロール(NP−1496 siRNA)と同程度のレベルで減少させた。(図7)。
【0069】
追加の標的
同様の細胞において同じ手順を使用し、同様の攻撃を用いて、RHGP技術を使用し、追加の標的を同定した。これらの内で、SLC25A25(以前に、膜を横断するミトコンドリア転移及び恐らくはMg−ATP交換に関与するタンパク質として特徴付けられた)、Rgnef(細胞付着、細胞運動性、及びBリンパ球活性化、ならびに恐らくは細胞アポトーシスにおいて役割を有することが観察された)、Nedd4及びHerc6(両方ともユビキチンリガーゼ)(全てがインフルエンザのための潜在的な標的として)、ならびにRobo1(細胞運動性及び軸索ガイダンスに関与する受容体)(HIVのための潜在的な標的として)がある。これらの一部が以下で考察される。これらのタンパク質は、ウイルス感染様式で細胞表面上に現れるため、各々は、モノクローナル抗体、ペプチド、アンチセンス、siRNAなどを使用して標的化されうる。
【0070】
PTCH1の抗体標的化によってインフルエンザ感染が減少する。
PTCH1は、複数回貫通型膜タンパク質である。そのようなものとして、本発明者らは、それが、抗体ベースの介入のために標的を提供しうると考えた。この可能性を評価するために、親和性精製された抗血清を得て、インフルエンザ感染細胞の表面上のPTCH1を認識するそれらの能力についてテストした。MDCK細胞を、A/Udornインフルエンザを用いて、インタクトな細胞の染色の前に18時間にわたり感染させた。この実験計画を利用し、ウイルスを用いた長期のインキュベーション又は固定化(それらのいずれかによって膜の完全性が破壊されうる)の結果として生じうるアーチファクトについての潜在力を最小限にした。ウイルスによりコードされる赤血球凝集素及び一致させたコントロール(PTCH1を認識しない)について特異的な抗体は、ポジティブコントロール及びネガティブコントロールをそれぞれ提供した。PTCH1抗体はインフルエンザ感染細胞に結合し、かなり多くの染色が、非感染コントロールよりも、感染細胞を用いて観察された(図8A及び8B)。この結果は、膜の完全性の喪失を表わさなかった。なぜなら、細胞は、トリパンブルー及び他の生体染色色素を、利用された条件下で排除し続けたからである。
【0071】
PTCH1抗体がインフルエンザ感染細胞に結合する能力に基づき、本発明者らは、次に、これらの抗体を用いた処理がインフルエンザ感染性を変化させうるか否かを問うた。MDCK細胞を、インフルエンザを用いて、PTCH1抗体の存在又は非存在において感染させた。培養上清を次に収集し、感染性インフルエンザ粒子の数を、標準的なプラークアッセイを使用して定量化した。これらの試験によって、PTCH1抗体が、インフルエンザ増殖を、未処理細胞又は非特異的抗血清を用いて処理されていたコントロールと比較し、2〜3対数(100〜1000倍)だけ減少させることが実証された。本発明者らは、次に、この結果が、最初の感染又はウイルスライフサイクルの後の段階の抗体阻害を表わしうるか否かを問うた。この疑問は、より高い感染効率(MOI)0.1(0.001よりむしろ)で感染を繰り返すことにより対処した。本発明者らは、PTCH1抗体が、相対的に高いMOIでインフルエンザを比較的阻害する場合、それらは、最初の付着又は感染細胞中への侵入を遮断しているであろうと推論した。他方で、抗体がライフサイクルの後の段階(例、出芽又は放出)に影響を与えた場合、阻害活性はより高いMOIで減少しうる。後者の考え方と一致して、PTCH1抗体処理の抗ウイルス効果は、より高いMOIで減少し、インフルエンザにおける1対数(10倍)の減少を与えた。(図8C)これらの結果は、PTCH1が抗体治療のための適した標的を提供しうること、及び、抗体がウイルスライフサイクルの相対的に後の段階を遮断するように見えることを示唆する。
【0072】
PTCH1を、実験室から、野外におけるインフルエンザ感染に翻訳する。
細胞ベースのアッセイを用いた興味をそそる知見に基づき、本発明者らは、PTCH1が、実験室の外部でのインフルエンザ感受性に関連しうるか否かを問うた。このために、本発明者らは、ブタ集団における頑健形質の単一ヌクレオチド多型(SNP)評価を、ブタインフルエンザウイルス大流行の間に利用した。具体的には、本発明者らは、PTCH1 SNPがブタ体重増加における変化(ブタにおけるインフルエンザ感染の理解された結果である)に関連するか否かを問うた。
【0073】
PTCH1内又はその近くの一連の異なるSNP(619、661、662、及び663として公知)を、いくつかの基準(有意水準、対立遺伝子頻度、相加対優性効果、効果の大きさ(表現型の標準偏差))に基づいて選択した。本発明者らは、次に、SNPが野外においてインフルエンザ耐性に関連するか否かを問うた。このために、野外においてウイルス感染に暴露されたブタからのDNAを抽出し、各SNPについて遺伝子型を同定し、SNP遺伝子型と性能形質の間での統計的な関連性を決定した。試験では、データセット回収の期間中にブタインフルエンザ大流行を受けていた大農場からのデータセットを利用した。本発明者らの分析では、2つの異なるPTCH1 SNP(619及び660)を、離乳の時期以降での増加した平均1日体重増加及び体重増加と関連付けた(表I)。これらの結果は統計的に有意であり、PTCH1が体重増加(ブタ集団におけるインフルエンザ罹患の有意な指標である)と関連しうることを示唆する。
【0074】
このように、RHGP技術を使用し、主要な知見は、PTCH1細胞表面受容体がインフルエンザウイルスによる感染のために必須であること、及び、PTCH1に対する標的介入(siRNA又は抗体のいずれかを使用)によってインフルエンザ感染が減少しうることの実証である。本発明者らは、また、PTCH1が、多くの異なる種(ヒト、イヌ、及びブタを含む)からの細胞の感染の間にインフルエンザにより広く利用されることを示している。最終的に、本発明者らは、これらの基本的な知見の重要性を野外における状況に翻訳する(PTCH1が、ブタ集団の自然感染の間に疾患転帰に関連することを実証することによる)。
【0075】
本発明者らの試験の1つの新規局面は、PTCH1が宿主細胞のインフルエンザ感染を調節することの実証である。PTCH1は、ソニックヘッジホッグ(胚発生の間での細胞の組織化、増殖、及び生存を支配する分泌分子)のための細胞受容体のヒトホモログである。PTCH1は、また、腫瘍抑制因子としての役割に関与しており、種々の上皮性悪性腫瘍と関連付けられる。機構的な観点から、インフルエンザの基礎は未知であるが、しかし、異なるMOIでの抗体処理の差動的な効果は、ウイルスライフサイクルの相対的に後の段階におけるPTCH1についての役割を示唆しうる。
【0076】
本発明者らの本報告での別の重要な局面は、抗ウイルス疾患のための新規の宿主指向の標的を同定するためのRHGPの利用である。RHGPは、研究下にある表現型を調節する細胞中の任意の標的を同定するための機会を提供する(過剰発現又は発現の喪失を介して)。RHGPの中心となる特性は、疾患プロセスにおけるその関与を決定するための標的の事前の知識を有する必要がないことである。本発明者らの結果は、この概念をさらに支持する。なぜなら、PTCHは、以前に、インフルエンザに関連付けられていなかったからである。
【0077】
疾患管理の観点から、本発明者らの本知見についての興味深く重要な潜在的な意味がある。第1に、PTCH1は、PTCH1発現又は機能を標的化する抗体又は他のアプローチを使用して、インフルエンザによる感染に対して介入するための機会を提供しうる。比較可能な知見が、複数の種(ヒト、イヌ、ブタ)を使用して得られたとの事実は、PTCH1が、インフルエンザ感染のために使用される重要な中核機構を表わすことを示しうる。感染性疾患を標的化するための本発明者らの宿主指向のアプローチの中心となる教義は、基本的な経路の同定が、複数のウイルス株を標的化するための手段を提供しうるとの考え方に焦点を合わせている。今後の研究によって、異なる季節性及び汎発性形態のウイルス病原体におけるPTCH1の役割がさらに詳述されるはずである。宿主指向の治療から生じる第2の機会は、ウイルスが、重要な宿主機構に対する代替物を発生しなかったであろうという概念である。正しい場合、PTCH1の効果的な標的化は、インフルエンザの処置を長く悩ませてきた薬物耐性の獲得に関連する問題を克服する機会を提供しうる。
【0078】
表I。PTCH1多型は、ブタインフルエンザ大流行の状況におけるブタ集団の増加した体重増加と関連する−(好ましい対立遺伝子(主要対立遺伝子と比べて)の1コピーでの成長(グラム/日)に対するPTCH1多型の効果。全ての結果が、主要対立遺伝子と比べた、体重増加における相対的な増加として示される(p値)。生涯1日増体量=最終的な体重−出生時体重離乳終了時の1日増体量=最終的な体重−離乳時の体重)
【0079】
【表10】
【0080】
実施例2−他のインフルエンザ因子を同定するために使用されるRHGP
ウイルス病原体の治療的標的化のための従来のアプローチでは、耐性株の発生及び広域スペクトル用途の欠如から生じる障害に一貫して直面してきた。インフルエンザは特に問題となる治療課題を表わす。なぜなら、季節性抗原シフト及びドリフトは、高いウイルス突然変異率と合わせて、多くの従来の抗ウイルス剤を混乱させてきたからである。新たに出現する又は操作されたインフルエンザ株は、インフルエンザのトリ亜型における最近の興味により典型的に表わされる通り、さらにより大きな脅威を表わす。ウイルスによりコードされる分子の標的化に関連する限定に基づき、本発明者らは、ウイルス増殖を予防する又は宿主をウイルス媒介性の病原性から回避させる様式で、宿主経路を標的化する直交アプローチを取ってきた。この目的に対して、ランダムホモ接合性遺伝子摂動(RHGP)を使用し、正常細胞において十分に耐容されるが、しかし、インフルエンザを阻害する宿主志向性の標的が同定された。RHGPを用いたTAIL−PCRの使用によって、ゲノム全体の徹底的なスクリーニングが促進され(過剰発現又は発現の喪失の両方について)、宿主細胞をインフルエンザ感染に対して耐性にする標的が同定された。本発明者らは、インフルエンザが宿主細胞を殺すことを予防する一連の宿主志向性の標的を同定し、複数のアプローチを使用してこれらの標的を検証する。これらの試験では、ウイルス疾患と戦うための新たなパラダイムが支持され、新規の標的及び機構を同定するためのRHGPの能力が実証される。
【0081】
ウイルス感染に対処するための1つの新たなアプローチは、ウイルス疾患の病原性に必須である宿主因子を標的化することである。HIV/AIDSでの戦争からの顕著な例は、CCR5のアンタゴニストの開発である。これらのアンタゴニストは、結合及び内在化のために必要である表面暴露された共受容体の機能を予防することを追求する。初期の知見では、これらの薬剤が、CCR5栄養性ウイルスを用いて感染された患者のサブセットの処置のための十分に必要とされる選択肢を提供しうることを示唆する。
【0082】
実施例2において、哺乳動物(ヒトを含む)におけるインフルエンザに関する治療介入のための可能な標的の異なる研究を調べる。上で注目した通り、本発明は、改善された形態のRHGPの適用を含み、それは、また、細胞において任意の標的(全遺伝子又は個々のドメインを含む)のアップレギュレーションを起こすことができる。結果的に、RHGPは、研究下にある表現型の原因となる任意の遺伝子変化についてゲノム全体を照合する手段を提供する。本発明者らはRHGPを適用し、インフルエンザ感染について要求される一連の宿主志向性の標的を同定し、検証した。
【0083】
材料及び方法
化学物質及び細胞培養。MDCK細胞、ヒト293HEK細胞、及びMEM培地を、American Type Culture Collection(ATCC)から購入した。Fugene 6をRocheから購入した。Lipofectamine 2000、ドキシサイクリン(Dox)、T4 DNAリガーゼ、ElectroMax DH10Bコンピテント細胞、及びViraPower PackagingキットをInvitrogenから購入した。ベクターpIREShyg3及びpTRE−lucをClontechから得た。RNase A及びプロテイナーゼKをSigmaから購入した。インフルエンザウイルスA/Udorn/72はCharles River Labにより提供された。全ての制限酵素をNew England BioLabから購入した。細胞培養培地及び補助剤は、Invitrogen又はHycloneのいずれかからであった。全ての短鎖siRNAオリゴヌクレオチドをDharmacon Researchにより合成した。TransIT−TKO(siRNAトランスフェクション試薬)をMirusから購入した。MDCK細胞をMEM(10% FBSを含む)中で培養した。全ての細胞培養物を5% CO2を用いて37℃でインキュベーションした。Bright−Gloルシフェラーゼアッセイシステム(Dox誘導アッセイのために使用される)をPromegaから購入した。
【0084】
pTet−Offトランスアクチベーター−ベクターのコンストラクション及びMDCK/Tet−off安定細胞株。pTet−Offトランスアクチベーターベクターを、pIREShyg3ベクターを骨格として使用してコンストラクションした。PCR生成されたtTAフラグメントを、pIREShyg3ベクターのBamHI及びSofI部位中に挿入し、pTet−Offトランスアクチベーターベクター(pIREShyg3−tTA)を生成した。
【0085】
MDCK/Tet−Off細胞株を作製するために、10cmプレート中の1〜2×106個のMDCK細胞を、10μgのpIREShyg3−tTAベクター及び30μlのfugene 6を用いて、製造者の指示に従いトランスフェクトした。トランスフェクションから24時間後、MDCK細胞を1:10希釈により分割し、200μg/mlのハイグロマイシンを用いて培養した。培地を、3日毎に、非トランスフェクト細胞が殺され、トランスフェクトされた安定な細胞コロニーが形成されるまで交換した。目に見える個々のコロニーを選び、24ウェルプレート中で継代培養した。ハイグロマイシン耐性クローンを、Dox誘導活性について、pTRE−lucベクター及びBright−Gloルシフェラーゼアッセイキットを使用し、製造者の指示に従いスクリーニングした。40倍を上回る誘導性を伴う安定クローンを、RHGPライブラリーを作製するための標的細胞として使用した。
【0086】
RHGP遺伝子探索ベクター(pRHGP22)のコンストラクション。RHGP遺伝子探索ベクター(pRHGP22)を、pLESTベクターをCohen実験室(17, Lu, 2004)からの骨格として使用して構築した。pLEST中のNeoR−TRE−CMVのDNA配列を、最初に、pTRE−Tight−CMV−BlasticidinRの発現カセットを用いて置換した。3’UTR WPREエレメントを除去し、p15A ori−クロラムフェニコールアセチルトランスフェラーゼ(CAT)耐性遺伝子を同じ部位に挿入した。2つのLoxP部位を、BlasticidinR遺伝子の下流及びp15A ori−CATの上流にそれぞれ挿入した。Creレコンビナーゼを伴うLoxP部位によって、本発明者らは、TRE−CMVプロモーターを染色体から切り出すことにより表現型を可逆的に検証することができるようになるであろう。
【0087】
RHGPウイルスの産生及びRHGPライブラリーの構築。RHGPレンチウイルスを、InvitrogenのViroPower Lentiviral Expression Systemを使用して産生した。293FTの細胞を、10cmプレート中に、約1〜2×106個細胞/プレートでそれぞれプレーティングした。24時間のインキュベーション後、細胞を、3μgのRHGP22ベクター及び9μgのViraPower Packaging Mixを用いて、Lipofectimine 2000を使用してトランスフェクトした。培地を、新鮮なトランスフェクション培地を用いて、37℃での5時間のインキュベーション後に交換した。48時間のインキュベーション後、培地中のウイルスを回収し、0.45μmフィルターを通じてろ過した。レンチウイルスを、製造者の指示に従い滴定した。
【0088】
RHGPライブラリーを構築するために、MDCK/Tet−off細胞を、150mmプレート中に、2×106個細胞/プレートで、それぞれRHGPウイルスの収集の1日前にプレーティングした。次の日、細胞(4.6×106個細胞/プレート)を、10mlのPBSを用いて洗浄し、10mlの新鮮に収集されたRHGPウイルスを、6μg/mlのポリブレンを用いて形質導入した。細胞を37℃で2時間にわたりインキュベーションし、20分毎に揺らすことにより混合した。2時間のインキュベーション後、培地を、10mlの形質導入培地を用いて補充した。形質導入から24時間後、細胞を10mlのPBSを用いて洗浄し、インフルエンザウイルスA/Udorn/72(MOI0.1)を用いた2時間にわたる5mlの感染DMEM培地(0.2%のBSA、200μg/mlのハイグロマイシン、及び6μg/mlのブラストサイジンを含む)中での感染が続く。細胞を、穏やかに15分毎に揺らすことにより混合した。2時間の感染後、細胞を15mlの感染培地に加えた。24時間の感染後、細胞を、PBSを用いて洗浄し、インフルエンザウイルスA/Udorn/72を用いて再び同じMOIで感染させた。第2ラウンドの感染から2日後、細胞を、10mlのPBSを用いて洗浄し、20mlの50%条件培地(200μg/mlのハイグロマイシン及び6μg/mlのブラストサイジンを含む)に加えた。培地を、3日毎に、非形質導入細胞が殺され、ウイルス耐性コロニーが形成されるまで交換した。第2ラウンドの感染から2週間後、目に見える個々のコロニーを選び、24ウェルプレート中で継代培養した。
【0089】
インフルエンザウイルス耐性細胞クローンについての可逆性アッセイ。インフルエンザウイルス耐性クローンの可逆性を検証するために、表現型及び親MDCK/Tet−Off細胞を、Doxの非存在又は存在において3日間にわたり培養した。細胞を、100mmプレート中で、2×106個細胞/プレートでプレーティングし、Doxを用いて又は用いずに培養した。24時間後、細胞を、インフルエンザウイルスA/Udorn/72(それぞれMOIが1×10−2及び1×10−5)を用いて感染させた。3日間のインキュベーション後、細胞を、クリスタルバイオレット染色に供した。
【0090】
インフルエンザウイルス耐性クローンからの候補遺伝子の同定。典型的には、各クローンの1〜10×106個細胞を回収し、PBS溶液を用いて1回洗浄した。細胞ペレットを、10mlの溶解緩衝液(0.32Mスクロース、10mM Tris pH7.5、5mM MgCl2、及び1% Triton X−100を含む)中に再浮遊させた。ライセートを1500×gで15分間にわたり遠心分離した。ペレットを、0.5mlのプロテイナーゼK緩衝液(25mM EDTA、150mM NaCl、及び40mM Tris pH7.5を含む)中の100μgのプロテイナーゼKを用いて処理し、0.5% SDSの添加が続いた。混合物を一晩37℃でインキュベーションした。プロテイナーゼK処理されたライセートに50μgのRNase Aを加え、37℃で2〜4時間にわたりインキュベーションした。ゲノムDNAを、フェノール/イソアミルアルコール/クロロホルムを用いて2回抽出し、等容積のイソプロパノールを用いた沈殿が続いた。DNAペレットを、70%エタノールを用いて洗浄し、200μlのTE(pH7.5)緩衝液中に溶解した。
【0091】
250μl中のゲノムDNA(10μg)を、制限酵素BamHI又はXbaIを用いて2〜4時間にわたり消化した。消化されたDNAをフェノール/イソアミルアルコール/クロロホルムを用いて抽出し、エタノールを用いて沈殿し、30μlのTE緩衝液中に溶解した。250μl中の消化されたDNA(2.5μg)を、20単位のT4リガーゼを用いて16℃で一晩自己連結させた。連結されたDNAを、エタノールを用いて沈殿させ、70%エタノールを用いて洗浄した。DNAペレットを20μlのTE緩衝液中に溶解させ、DH10B ElectroMaxコンピテント細胞中に電気穿孔した。各々の連結DNAからのコロニー(8〜16)を、プラスミドDNA調製及びサイズ分析のための制限酵素を用いた消化のために選んだ。候補プラスミドDNAをさらに使用し、表現型に関与する標的遺伝子をDNA配列決定及びゲノムマッピングにより同定した。
【0092】
siRNAを用いた宿主標的遺伝子の検証。候補遺伝子MRPL42、COX5A、SLC25A25、及びTAPT1についてのヒト二本鎖siRNAホモログを、製造者によるsiGENOME SMART poolとして調製した。インフルエンザウイルスのヌクレオカプシド(NP)遺伝子に対するsiRNA NP−1496配列(GGAUCUUAUUUCUUCGGAGUU)を、ポジティブコントロールとして使用した(18, Ge, 2003)。非標的siRNA(siCONTROL 1)をネガティブコントロールとして使用した。HEK293細胞を24ウェルプレート中に1×105個細胞/ウェルでそれぞれプレーティングした。24時間のインキュベーション後、細胞を、20nMのsiRNA及び1.5μlのTransIT-TKOを用いて、製造者の指示に従いトランスフェクトした。細胞を、24時間のインキュベーション後、siRNAを用いて再トランスフェクトした。第2ラウンドのトランスフェクションから24時間後、細胞を0.5mlのMEMを用いて洗浄し、インフルエンザウイルスA/Udorn/72(MOI1.0)を用いた300μlのDMEM培地及び0.05% FBS中での感染が続く。細胞を、15分毎に穏やかに揺らしながら1時間にわたりインキュベーションした。細胞を、MEMを用いて2回洗浄し、ウイルス産生のために0.5mlの感染培地に加えた。各ウェルからの培地を、トランスフェクションから24又は48時間後に回収した。培地中の子孫ウイルスを、プラークアッセイにより滴定した。
【0093】
結果
RHGP細胞ライブラリーの構築
RHGPによって、細胞ゲノム全体を照合し、過剰発現及びノックアウト事象の両方を評価することを追求する。この課題では、宿主の全ゲノムのカバーを確実にするための十分な組込み事象を伴うRHGPライブラリーの作製が要求される。また、ライブラリーは、細胞当たり単一のRHGP組込み事象に限定され、同じ細胞での複数の摂動から生じる複雑さを防止することが重要である。ヒトゲノム中での平均遺伝子サイズが27kbと推定されるため(19)、本発明者らは、105の独立した組込み事象が、全ゲノムのランダムなカバーを確実にするために要求されると計算した(1遺伝子当たり1ベクター挿入)。RHKOを用いた先行する試験では、MMLVベースの組込みベクターに焦点を合わせていた(16)。しかし、これらのベクターの力価は、一般的に、103形質導入単位(TU)/mLに限定される。アプローチの効率を改善するために、レンチウイルスシステムを開発した。材料及び方法に概説されている通りに、pRHGP22を、ブラストサイジン耐性遺伝子を促すpTRE−Tight−CMVプロモーターからなる改善された発現カセットを使用して構築し、厳密な誘導制御下で選択可能マーカーを提供した。ori−クロラムフェニコールアセチルトランスフェラーゼ(CAT)耐性遺伝子は、独立したレポーターを提供し、宿主ゲノムDNAをレスキューし、標的遺伝子及びベクター組込みの部位を同定した。
【0094】
インフルエンザウイルス耐性表現型のスクリーニング
宿主細胞をインフルエンザに対して耐性にする標的を同定するために、RHGPライブラリーをMDCK細胞において生成した。この特定の細胞モデルを、インフルエンザ感染に対するその十分に確証された応答性に基づいて選択した。pRHGP22ベクターを用いた形質導入を介したライブラリー構築に続き、RHGP細胞ライブラリーを作製し、ヒトゲノムの100倍のカバーを確実にした。単一細胞内での複数の挿入についての潜在力を最小限にするために、低MOI(0.15)を、RHGPライブラリー調製のための標的細胞の形質導入の間に用いた。
【0095】
MDCK細胞ライブラリーを、本来なら致死濃度のブラストサイジン(感染から24時間後に投与)を使用して選択し、ベクター組込みを確実にした。ライブラリーを、また、インフルエンザA/Udorn/72を(相対的に高いMOI0.1で)用いた感染により攻撃し、インフルエンザ耐性細胞について選択した。インフルエンザ感受性細胞の徹底的な除去を確実にするために、培養物を複数ラウンドのインフルエンザ感染に供した。コントロールとして、一致させたコントロールを含めた。それは、RHGP形質導入を欠くが、しかし、その他の点では同様に処理された。一般的に、第2ラウンドのインフルエンザ攻撃後、生存細胞は、一致させたコントロールにおいて検出されなかった。その後、RHGP形質導入群中の生存細胞をクローン化し、増大させ、約3,000クローン/試験を産出した(表2)。これらから、合計194の個々のコロニーを、さらなる分析のために、単一ライブラリーから単離した。個々のクローンがインフルエンザ耐性表現型を保持したことを問うために、これらを、インフルエンザを用いて再攻撃した。再攻撃試験によって、本発明者らは、インフルエンザ攻撃に対していくらか感受性であるクローンを優先順位から外すことができた。14の高度に耐性のクローンのサブセットが、さらなる特徴付けのために選択された。
【0096】
ウイルス耐性クローンの可逆性アッセイ
RHGP技術の重要な特性は、誘導プロモーターによる調節を介して候補標的を検証する能力である。これによって、本発明者らは、自然突然変異又は感染アッセイでの他のアーチファクトの結果としてインフルエンザに対して耐性になったであろう任意の候補クローンを除去することができた。具体的には、インフルエンザ耐性クローン中に取り込まれたRHGPベクター用のプロモーターは、Tet−offシステムの制御下にあった。従って、外因的に加えられたテトラサイクリン(Tet)又はドキシサイクリン(Dox)の存在において、RHGPベクターを「オフにする」ことができる。この特性によって、本発明者らは、外因的に加えられたDoxの存在において攻撃された場合にインフルエンザに対して耐性のままであるそれらの候補を優先順位から外すことができた。これらの試験の経過において、本発明者らは、高い感染効率(MOI)でのインフルエンザ耐性が、インフルエンザライフサイクルの初期段階に関連する標的又は経路を好みうるのに対し、低MOIが、インフルエンザライフサイクルの後期段階のために要求される機構を好みうると考えた。このように、インフルエンザ攻撃を、2つの広く異なる感染効率(10−5又は10−2)で行い、アッセイシステムが、ウイルス耐性の初期又は後期のいずれかの段階の機構について偏りうる可能性を除いた。細胞生存を、クリスタルバイオレットを用いた感染から72時間後の染色により決定した。RHGP組込み事象を欠くコントロール細胞は、宿主細胞のウイルス媒介性の殺傷についてのポジティブコントロールを提供した。分析された14クローンの内、12(86%)がDoxの存在においてインフルエンザに対する感受性を再獲得し、残りの2つがさらなる研究のために優先順位から外された。
【0097】
RHGP遺伝子探索ベクターを設計し、標的遺伝子を効率的に位置付け、組込み事象の方向(センス又はアンチセンス)を決定した。具体的には、遺伝子探索ベクターはCATレポーター遺伝子をコードし、それは、制限酵素ベースのゲノムDNAクローニングによりレスキューすることができる。ゲノムDNAを、検証され、インフルエンザ耐性のDox媒介性の可逆性が実証されたそれらのクローンから単離した。ゲノムDNAを、適した制限酵素を用いて消化し、自己連結させ、環状ベクターゲノムDNAを得た。これらの構築物を使用し、E. coliを形質転換させ、クロラムフェニコール耐性サブクローンを単離し、配列決定した。ベクター組込みのゲノム部位を確認するために、少なくとも8〜16の異なるクロラムフェニコール耐性コロニーを単離し、各MDCKクローンから配列決定した。RHGPベクター挿入部位に隣接する結果として得られるゲノムDNA配列を、イヌゲノムに対する、標準的な探索アルゴリズムを使用したゲノムマッピングに供した。実験計画と一致して、大半の標的が、MDCK細胞当たり単一の組込み部位からなった。しかし、一部のクローンは複数の組込み事象を含み、それらは、複数のRHGP組込み部位の結果として又はその特定の候補が複数のクローンからなっていた場合に生じたであろう。また、本発明者らは、技術的な限定に起因して、他の候補の小さなサブセットから候補遺伝子を単離することができなかった。全体で、ゲノムDNAクローニングによって、インフルエンザ耐性に関連する合計70の異なる標的遺伝子が同定された。
【0098】
ナイーブな細胞を使用した標的遺伝子の検証
RHGPによって、インフルエンザ感染に対して耐性を付与する一連の標的が同定された。表現型の可逆性によって、実験システム内での標的の適用性が検証された。本発明者らは、また、独立した実験システムを使用してこれらの候補の検証を追求し、RHGP、宿主細胞システム(MDCK)の選択、又はインフルエンザバリアント(Udornウイルス)に固有でありうる予想外の結果についての潜在力を除いた。
【0099】
RHGPを使用して同定された標的の独立した確認として、これらの標的のサブセットを選択し、より従来のアプローチを使用してテストした。具体的には、本発明者らは標的を強調しており、そこでは減少した発現がインフルエンザ耐性に関連した。なぜなら、部分的には、ノックダウン標的が、将来の標的戦略に受け入れられ、小分子又は生物製剤ベースの治療を使用して発現又は機能を同程度にノックダウンしうるからである。重要なことには、MDCK細胞からの結果によってイヌ標的が同定され、siRNAを、標的遺伝子のヒトホモログに対して設計した。二本鎖siRNAを、4つのそのような標的(MRPL42、COX5A、TAPT1、及びSLC25A25)について生成し、HEK293細胞をトランスフェクトするために使用した。インフルエンザは一般的にHEK293に対して毒性がないため、ウイルス力価の測定は、効力を評価するためのエンドポイントとしての役割を果たした。本発明者らの最初の一連の試験のために、siRNA処理した293細胞を、RHGP試験のために使用した同じインフルエンザ株(A/Udorn/72)を用いて感染させた。Udornを用いた攻撃に続き、ウイルス力価を評価し、siRNAトランスフェクト細胞においてより低いレベルのウイルスを実証した。サイトカインの同化に対するsiRNAのアーチファクトベースの効果についての最近の証拠に照らして、並行試験を、無関係のsiRNAコンストラクトを用いて行い、このように、目的の標的についての抗ウイルス効果の特異性を検証した。
【0100】
Udornを使用した知見の検証後、本発明者らは、次に、結果を他のインフルエンザバリアントに拡大した。例えば、siRNAは、また、異なるインフルエンザ株(A/WSN/33)に対する耐性を付与した。比較可能な知見が、異なる感染効率で得られた。全体で、これらの知見は、新規の宿主ベースの標的を同定するためのRHGPの適用を検証し、異なる形態のインフルエンザに対する広域スペクトルの適用のための潜在力を示唆する。
【0101】
この実施例において、RHGPが、インフルエンザウイルス感染のために要求される任意の宿主因子(過剰発現又はノックダウンされている標的を含む)のゲノム規模のスクリーニングを行い、RHGPベクター内の内部エレメントを使用してこれらの標的を検証するための効率的な手段を提供することができることが示されている。このゲノム規模のスクリーニングでは、インフルエンザについての効果的な宿主志向性の標的としてMRPL42、COX5A、TAPT1、及びSLC25A25タンパク質に優先順位が付けられた。これらの標的の効率は、独立した試験において従来のsiRNAアプローチを使用してさらに検証された。
【0102】
本発明者らの本試験の1つの固有の局面は、改善された形態のRHGPの使用である。以前の試験では、MMLVベースのベクターが用いられ、二倍体細胞において遺伝子の両方のコピーが阻害された。しかし、MMLVベクターは、内因的に限定された低力価の産生であった。本発明者らは、レンチウイルスシステムによって以前の限定が克服されることを実証する。結果として得られるシステムは、ヒトゲノム全体の100倍のカバーを確実とするために十分であった。それは、過剰発現された標的及びノックダウンされた標的の両方の徹底的な評価を行うために重要である。さらに、レンチウイルスベクターの使用は、活性な遺伝子転写の部位中への単一部位挿入を強く好む。それはスクリーニング手順の効率を増加させる。
【0103】
1つの異なる利点は、RHGPが標的の事前の知識により偏らず、同定された標的遺伝子がしばしばまだアノテートされていないことである。しかしながら、本発明者らは、PANTHER Classification System(Mi, 2005)を用いてRHGPインフルエンザ遺伝子のヒトホモログを分析し、推定上の生物学的経路を同定した。これらの標的は、生物学的プロセス及び分子機能のカテゴリーに広く分類された(図9)。70の標的の内、61(87%)を分類することができ、9(13%)はできなかった。それらの61の遺伝子によって、7つの主要な生物学的プロセス及び8つの主要な分子機能が同定された。生物学的プロセスの内で顕著なのは、核酸及びタンパク質代謝、シグナル伝達、及び細胞内タンパク質輸送に関連する経路であった。具体的には、標的機能は、核酸結合に関連する内因活性、酵素活性(キナーゼ、リガーゼ、ヒドロラーゼ、レドックス)、細胞シグナル伝達、及び細胞内トランスポートを含んだ。初期の指標は、これらの標的クラスがランダムではないことを示唆する。なぜなら、本明細書において同定されたキナーゼ、膜輸送タンパク質、リガーゼ、ヒドロラーゼ、及びトランスファー/キャリアタンパク質の頻度は、公知の標的クラスのランダム検索を行う場合に予測されうるものより少なくとも3倍高かったからである。
【0104】
増加する証拠は、ウイルス疾患のための宿主志向性の治療法の概念を支持してきた。特に、一連の最近のsiRNAベースの試験が行われ、インフルエンザ、HIV、又はウエストナイルウイルスによる感染と関連する宿主標的が同定されている。siRNAは、標的発見のためにますます利用されているが、しかし、siRNAの頑健で持続的な過剰発現の必要性により内因的に。一部のsiRNA知見の結果は、技術自体が、本来なら、細胞挙動をどのように変化させうるかについての懸念により曇らされてきた。特に、siRNAがサイトカイン発現を変化させうるとの最近の報告は、感染性疾患への適用について特に問題である。本発明者らは、RHGPが改善された代替物を提供しうると考える。なぜなら、それは、これらの同じ制約により限定されないからである。実際に、本明細書において同定された標的及び経路の大半(73%)が新規である。この結果は、RHGPが、また、まだアノテートされていない標的ならびに過剰発現されるタンパク質又はドメインを同定する能力を有することを反映する可能性が高い。対照的に、siRNAアプローチは、一般的に、公知の又はアノテートされた標的のノックダウンに限定される。興味深いことに、MDCK細胞のRHGP照合を使用して本明細書において同定された標的又は経路の27%が、ヒト細胞のHIV又はウエストナイル感染と関連する遺伝子とある程度の関係(配列、タンパク質ファミリー、又は機構的経路に関して)を有する(表3)。この観察は、異なるウイルス病原体が収束的に進化し、特定の決定的な宿主経路を共有することを示唆する。これらの経路は、しばしば、異なる哺乳動物種(例、イヌ及びヒト)の間で保存される。とりわけ、本明細書において同定された遺伝子は、最近のインフルエンザ試験とはあまり似てない。本発明者らは、この外見上の矛盾が、インフルエンザを用いた以前の試験でDrosophila細胞株が利用されたとの事実を表わしていると仮定する。それは重要である。なぜなら、ショウジョウバエは、哺乳動物細胞とは遺伝的に異なるからであり、なぜなら、Drosophilaゲノムは、約14,000遺伝子(ヒト又はイヌのゲノムより少ない)に限定されるからである。
【0105】
さらなる研究によって、RHGPを使用して同定された標的の一部をウイルス感染と関連付ける潜在的な機構情報が明らかになる。例えば、減少したMRPL42、COX5A、TAPT1、及びSLC25A25発現が、インフルエンザ感染に対する感受性を減少させる(図10)。MRPL42は、ミトコンドリアリボソームタンパク質(核遺伝子によりコードされる)であり、ミトコンドリアのタンパク質合成に関与する。最近の報告では、CD4+T細胞におけるSIV−Nefの導入が、関連する宿主ミトコンドリアタンパク質(MRPL1、MRPL14、及びMRPL19)の発現を刺激することが実証された。この報告では、これらの因子のアップレギュレーションが、ウイルスライフサイクルの後期段階で要求されるウイルスメッセージの産生を促進しうることが示唆された。正しい場合、本発明者らの結果は、この知見をMRPL42まで拡大させ、インフルエンザが同様の機構を利用しうることを示す。同様の知見によって、本明細書において同定された別の標的(COX5A)がウイルス感染と関連付けられる。COX5Aは、チトクロムCオキシダーゼ(COX)複合体の1成分である。異なるサブユニット(COX6A1)が、報告によると、インフルエンザウイルス複製のために要求される。興味深いことに、膜貫通タンパク質TAPT1(膜貫通前後形質転換1)は、脊椎動物胚における発生の間での軸骨格パターン形成のために要求され、遺伝子の一部が、HCMV gH受容体(gH受容体に対するモノクローナル抗体を用いたcDNA発現ライブラリーのスクリーニングにより同定された)をコードすることが明らかにされた。TAPT1が病原性に関与する機構は不明のままであるが、TAPT1が、また、インフルエンザウイルス感染に関与しうることも可能である。SLC25A25(可溶性キャリアファミリー25、メンバー25)タンパク質は、ミトコンドリア及びリン酸キャリアのスーパーファミリーのメンバーである。スーパーファミリーにおける多数の同様のタンパク質が、報告によると、HIV及びWNV感染に関与してきた。
【0106】
遺伝子不活性化に加えて、他の遺伝子のRHGP誘導によってインフルエンザ感受性が減少する。これらの内で顕著なのは、B細胞CLL/リンパ腫2(BCL2)及びプラスミノーゲンアクチベーターウロキナーゼ(urakinase)(PLAU)遺伝子である。プロトがん遺伝子BCL2が、細胞生存の十分に特徴付けられた調節因子である。最近の報告では、BCL2は、シンドビス、インフルエンザウイルス、又はHIV−1により起こされたアポトーシスに続く細胞生存の調節と関連付けられている。同様に、RHPGは、また、PLAUセリンプロテアーゼの過剰発現をインフルエンザと関連付けた。とりわけ、PLAUのアミノ末端フラグメントが、最近、感染細胞からのHIV−1の組み立て及び出芽を抑制することが示された(Wada, 2001)。ゲノム相関によって、また、PLAU及びその受容体(PLAUR)が、ヒト呼吸器合胞体ウイルスによる感染と関連付けられている。
【0107】
実施例3−RHGPを使用した潜在的なインフルエンザ標的のさらなる同定
上の実施例2に示すものと非常に類似の技術を使用し、インフルエンザウイルス感染を予防又は処置するための治療介入のための追加の潜在的な標的が同定された。
【0108】
材料及び方法
化学物質及び細胞培養。MDCK細胞、ヒト293HEK細胞、及びMEM培地をAmerican Type Culture Collection(ATCC)から購入した。インフルエンザウイルスA/Udorn/72をCharles River Labから購入した。全てのsiRNAオリゴヌクレオチドはDharmacon Researchにより合成された。
【0109】
pTet−Offトランスアクチベーターベクターの構築及びMDCK/Tet−off安定細胞株。pTet−Offトランスアクチベーターベクターを、pIREShyg3(Clontech)を骨格として使用して構築した。PCR生成されたtTAフラグメントを、pIREShyg3ベクターのBamHI及びSofI部位中に挿入し、pTet−Offトランスアクチベーターベクター(pIREShyg3−tTA)を生成した。
【0110】
MDCK/Tet−Off細胞株を作製するために、2×106個のMDCK細胞を10cmプレート中で培養し、10μgのpIREShyg3−tTAベクターを用いて、fugene 6(Roche)を使用してトランスフェクトした。トランスフェクションから24時間後、MDCK細胞の対数期の培養物を、200μg/mlハイグロマイシンを使用して選択し、培地を、3日毎に、全てのコントロール細胞が殺されるまで交換した。ハイグロマイシン耐性クローンを、pTRE−luc(Clontech)ベクター及びBright−Gloルシフェラーゼアッセイキット(Promega)を使用し、ドキシサイクリン(Doxycylcine)(Dox;Invitrogen)誘導活性についてスクリーニングした。少なくとも40倍の誘導を伴う安定クローンを使用し、RHGPライブラリーを作製した。
【0111】
RHGP遺伝子探索ベクター(pRHGP22)のコンストラクション。RHGP遺伝子探索ベクター(pRHGP22)を、pLESTベクター(Stanley Cohen博士(Stanford)により提供)(Lu et al., 2004)を骨格として使用してコンストラクションした。pTRE−Tight−CMV−BlasticidinRを、プロモーターとして置換した。3’UTR WPREエレメントを除去し、p15A ori−クロラムフェニコールアセチルトランスフェラーゼ(CAT)耐性遺伝子を同じ部位に挿入した。2つのLoxP部位を、BlasticidinR遺伝子から下流及びp15A ori−CATから上流にそれぞれ挿入した。
【0112】
RHGPウイルスの産生及びRHGPライブラリーの構築。RHGPレンチウイルスを、ViroPower Expression System(Invitrogen)を使用して産生した。HEK293FT細胞を、10cmプレート中に106個細胞/プレートでプレーティングした。24時間のインキュベーション後、細胞を、3μgのRHGP22及び9μgのViroPower Packaging Mixを用いて、Lipofectimine 2000(Invitrogen)を使用してトランスフェクトした。培地を、37℃で5時間のインキュベーション後に交換した。48時間後、培養培地中のウイルスを、0.45μmフィルターを通じてろ過し、製造者の指示に従い滴定した。
【0113】
RHGPライブラリーを構築するために、MDCK/Tet−off細胞を、15cmプレート中に、2×106個細胞/プレートでプレーティングした。次の日に、サンプルを、PBSを用いて洗浄し、6μg/mlポリブレン中のRHGPウイルスを用いて形質導入した。サンプルを、新鮮培地を用いた補充の前に混合しながら37℃で2時間にわたりインキュベーションした。形質導入から48時間後、細胞を、PBSを用いて洗浄し、インフルエンザウイルスA/Udorn/72を用いて、感染効率(MOI)0.1で2時間にわたり、DMEM培地(0.2% BSA、200μg/mlハイグロマイシン、2μg/mlウシトリプシン、及び6μg/mlブラストサイジンを含む)中で、15分毎に穏やかに混合しながら感染させた。培地を、次に、MEMを用いて補充した。2日後、細胞を、PBSを用いて洗浄し、上に示すとおりに、インフルエンザウイルスを用いて再び感染させた。第2ラウンドの感染後、サンプルを、PBSを用いて洗浄し、条件培地(200μg/mlのハイグロマイシン及び6μg/mlのブラストサイジンを含む)中でインキュベーションした。培地を、3日毎に、全ての非トランスフェクト細胞が殺され、ウイルス耐性コロニーが形成されるまで交換した。第2ラウンドの感染から2週間後、目に見える個々のコロニーを単離し、24ウェルプレート中で継代培養した。
【0114】
インフルエンザウイルス耐性細胞クローンについての可逆性アッセイ。RHGP事象の可逆性を検証するために、MDCK/Tet−Off細胞を、Doxの非存在又は存在において3日間にわたり培養した。細胞を、10cmプレート中で、2×106個細胞/プレートでプレーティングし、Doxを用いて又は用いずに培養した。24時間後、サンプルを、インフルエンザウイルスA/Udorn/72(それぞれMOIが1×10−2及び1×10−5)を用いて感染させた。3日後、生存細胞を、クリスタルバイオレットを用いて染色し、残りの細胞を可視化した。
【0115】
インフルエンザウイルス耐性クローンからの候補遺伝子の同定各クローンからの1×106個細胞を、溶解緩衝液(0.32Mスクロース、10mM Tris pH7.5、5mM MgCl2、及び1% Triton X−100を含む)中に浮遊させた。ライセートを1500×gで15分間にわたり遠心分離した。ペレットを、プロテイナーゼK緩衝液(25mM EDTA、150mM NaCl及び40mM Tris、0.5% SDS、pH7.5を含む)中の100μgのプロテイナーゼK(Sigma)を用いて処理した。混合物を一晩37℃でインキュベーションし、50μgのRNase A(Sigma)を用いて処理し、37℃で2時間にわたりインキュベーションした。ゲノムDNAを、フェノール/イソアミルアルコール/クロロホルムを用いて抽出し、イソプロパノールを用いた沈殿が続いた。DNAペレットを、70%エタノールを用いて洗浄し、TE緩衝液(pH7.5)中に溶解した。
【0116】
ゲノムDNA(10μg)を、制限酵素BamHI又はXbaIを用いて消化し、フェノール/イソアミルアルコール/クロロホルムを用いて抽出し、エタノールを用いて沈殿し、TE緩衝液中に溶解した。消化されたDNA(2.5μg)を、T4リガーゼ(Invitrogen)を使用して16℃で一晩自己連結させた。連結されたDNAを、エタノールを用いて沈殿させ、70%エタノールを用いて洗浄した。DNAペレットをTE緩衝液中に溶解させ、DH10B ElectroMaxコンピテント細胞(Invitrogen)中に電気穿孔した。複数のコロニーを、プラスミドDNA調製及び制限酵素消化のために単離した。プラスミドDNAをさらに使用し、標的遺伝子をDNA配列決定及びゲノムマッピングにより同定した。
【0117】
siRNAを用いた宿主標的遺伝子の検証。候補遺伝子MRPL42、COX5A、SLC25A25、及びTAPT1についてのヒト二本鎖siRNAホモログを、製造者により推奨される通りに調製した。siRNA NP−1496配列(GGAUCUUAUUUCUUCGGAGUU)は、インフルエンザウイルスのヌクレオカプシド(NP)遺伝子を標的化し、ポジティブコントロールを提供した(18, Ge, 2003)。非標的siRNA(siCONTROL1)がネガティブコントロールを提供した。HEK293細胞を24ウェルプレート中に1×105個細胞/ウェルでそれぞれプレーティングした。24時間のインキュベーション後、細胞を、20nMのsiRNA及びTransIT-TKOを用いて、製造者の指示(Mirus)に従いトランスフェクトした。細胞を、24時間のインキュベーション後、siRNAを用いて再トランスフェクトした。第2ラウンドのトランスフェクションから24時間後、サンプルを、MEMを用いて洗浄し、インフルエンザウイルスA/Udorn/72(MOI1)を用いた感染が続く。細胞を、15分毎に穏やかに揺らしながら1時間にわたりインキュベーションした。各ウェルからの培養培地を、トランスフェクションから48時間後に回収した。培地中の子孫ウイルスを、標準的なプラークアッセイを使用して滴定した。
【0118】
結果
RHGP細胞ライブラリーの構築
RHGPの中心となる特性は、固有のレンチウイルスベースの遺伝子エレメントであり、遺伝子探索ベクター(GSV)として公知であり、それは、ゲノム全体を照合し、宿主細胞がインフルエンザ感染に耐性となる又は生き残ることを可能にする標的を同定するように設計された。本発明者らの実験戦略では、ゲノム中の単一部位でのGSVの組込みを中心とした。そこで、それは誘導プロモーターを介して標的遺伝子の発現を調節した。ベクターは自己不活性化レンチウイルスLTRをコードし、それは、GSVが形質導入細胞から再出現することを予防した。RSV E/Pプロモーターは、GSVの産生のためだけに使用され、宿主ゲノム中へのGSVの組込みにより除去された。
【0119】
GSVは、センス又はアンチセンスのいずれかの方向で組込まれうる。アンチセンス配置において、組込み事象自体が1つの対立遺伝子を不活性化し、アンチセンス構築物の発現を促進し、それによって他の対立遺伝子上でコードされる遺伝子がノックダウンされた。この方法において、RHGPは、二倍体細胞における両方の遺伝子コピーのホモ接合性摂動を促進した。反対(センス)方向では、RHGPは、標的遺伝子の過剰発現を促進した。この結果は、遺伝子全体の単純な過剰発現を超えて拡大しうる(例、開始部位の上流での挿入)。なぜなら、開始部位の下流への組込みはドメインの過剰発現を引き起こし、野生型遺伝子の機能のドミナントネガティブな阻害剤を産生しうる。そのようなものとして、RHGPによって、本発明者らは、細胞ゲノム全体を照合し、宿主細胞がインフルエンザ感染に耐性となる又は生き残ることを可能にする異なる型の事象を同定することができた。
【0120】
RHGPを促進するために、MDCK細胞のライブラリーを、レンチウイルスベースのGSVを使用して形質導入する。ゲノム中での平均遺伝子サイズが27kbと推定されるため(Gupta and Varshney, 2004)、本発明者らは、105の独立した組込み事象が、ゲノム全体のランダムなカバーを確実にしうると計算した(Li and Cohen, 1996)。材料及び方法に概説されている通り、pRHGP22を、ブラストサイジン耐性遺伝子を促すpTRE−Tight−CMVプロモーターからなる発現カセットを使用して構築し、厳密な誘導制御下で選択可能マーカーを提供した。形質導入された宿主細胞のライブラリーを、次に、ブラストサイジンを使用して選択し、生存細胞を、以下で詳述する通りに、インフルエンザ耐性についてスクリーニングした。
【0121】
インフルエンザウイルス耐性表現型のスクリーニング
宿主細胞をインフルエンザに対して耐性にする標的を同定するために、RHGPライブラリーをMDCK細胞において生成し、インフルエンザ攻撃を実施した。この特定の細胞モデルを、インフルエンザ感染に対するその十分に確証された応答性に基づいて選択した。なぜなら、イヌゲノムがアノテートされており、ヒトホモログと比較することができるからである。本発明者らは予備試験を行い、MDCK細胞が、選択のために利用される条件下でA/Udorn/72により効率的かつ完全に殺されることを確実にした。この結果は、生存細胞が、RHGP摂動の結果として(インフルエンザに対する自然耐性のアーチファクトとしてではなく)生じることを確実にするために必須であった。pRHGP22 GSVを用いたライブラリー構築に続き、少なくとも107の独立したMDCKクローンからなるライブラリーを作製し、ゲノムの100倍のカバーを確実にした。低MOI(0.1)をライブラリー作製の間に用いて、1を上回る異なるGSVによる任意の細胞の形質導入を最小限にした。GSV組込みを確認する追加の手段として、MDCKライブラリーを、致死濃度のブラストサイジン(感染から48時間後に投与)を用いてインキュベーションした。
【0122】
ライブラリーを、インフルエンザA/Udorn/72を用いた感染により攻撃し、インフルエンザ耐性細胞について選択した。本発明者らは、以前に、A/Udorn/72(MOI10−1)を用いた感染によって全てのMDCK細胞が48時間以内に再現性よく殺されることを確証していた。コントロールとして、偽形質導入細胞の並行培養物を同じように処理し、生存体は48時間後に観察されなかった。本発明者らは、さらに、生存細胞を複数ラウンドの致死攻撃に供することによりアーチファクトを最小限にすることを追求した。このように、サンプルを少なくとも3ラウンドのインフルエンザ攻撃に供し、その時に、生存細胞は、一致させたコントロールのいずれにおいても検出されなかった。
【0123】
これらの試験の経過において、本発明者らは、高い相対的MOIのインフルエンザでの攻撃が特定の型の標的について偏りうると考えた。例えば、高いMOIでの生存が、意図的にではなく、感染の初期段階に関連する標的を好みうる、又は、後の段階での事象に関連する標的に対して偏りうる。そのような偏りを予防するために、並行した一連の試験を行い、低い力価でのウイルス攻撃に関連する生存を比較した。このために、クローンを、低い最初のMOI(MOI10−5)での感染に供した。培養物を、次に、経時的に、複数ラウンドの感染を使用して評価し、コントロールサンプル中の全ての細胞がインフルエンザにより殺されていたことを確実にする。
【0124】
インフルエンザを用いた攻撃を生き残ったRHGP形質導入細胞についての選択後、クローンを、単一細胞クローニングにより単離し、そして増大し、平均3,000クローン/実験を産出した。豊富なクローンを仮定し、303の異なるインフルエンザ耐性クローンのサブセット(全集団の10%)をランダムに選択し、第2ラウンドの単一細胞クローニングに供し、クローン性を確実にした(表2)。これらのサブクローンを、次に、一連の試験に供し、表現型を確認し、標的遺伝子を同定した。各サブクローンを、再び、インフルエンザを用いた致死攻撃を生き残る能力についてテストした。不活性Tet−off GSVを用いて形質導入されていた親MDCK宿主細胞は、一致させたネガティブコントロールを提供した。129の異なるサブクローン(最初の集団の43%)が、インフルエンザを用いた致死攻撃を生き残るそれらの能力に基づいて単離された。この段階での下方選択の大半が、単一細胞クローニングプロセスを生き残ることができなかったコロニーに起因した。
【0125】
ウイルス耐性細胞クローンの可逆性アッセイ
RHGP技術の重要な特性は、誘導プロモーターによる調節を介して候補標的を検証する能力である。これによって、本発明者らは、自然突然変異又はRHGPに関連しない他のアーチファクトの結果としてインフルエンザに対して耐性になったであろう候補を除去することができた。RHGPベクター用のプロモーターはTet−offシステムの制御下にあったため、本発明者らは、ドキシサイクリンの存在又は非存在において候補のインフルエンザ媒介性の殺傷を比較した。この試験によって、129の生存クローン中111(86%)でインフルエンザ攻撃に対する可逆性の耐性が実証された。残りの18クローン(14%)が優先順位から外され、RHGP、及び無関係ではないアーチファクトがインフルエンザ耐性表現型に関与することを確実にした。
【0126】
ゲノムDNAクローニングによる宿主遺伝子の同定
RHGP遺伝子探索ベクターを設計し、標的遺伝子を効率的に位置付け、組込み事象の方向(センス又はアンチセンス)を決定した。具体的には、遺伝子探索ベクターはOri−CATレポーター遺伝子をコードし、それは、制限酵素ベースのゲノムDNAクローニングによりレスキューすることができる。ゲノムDNAを、インフルエンザに対する実証された可逆的な耐性を有する111のクローンから単離し、合計110の標的遺伝子を産出した(表I)。RHGPベクター挿入部位に隣接する結果として得られるゲノムDNA配列を、イヌゲノムに対する、UCSC Genome Browserを使用したゲノムマッピングに供した。実験計画と一致して、大半の標的が、1クローン当たり単一の組込み部位からなった。本発明者らは、技術的な限定に起因して、候補の小さなサブセットから候補遺伝子を単離することができなかった。また、少数のサンプルが複数の組込み事象を含んだ。それらは、複数のRHGP組込み部位の結果として又はサブクローンがクローン性ではなかった場合に生じたであろう。このあいまい性に起因し、複数の組込み事象に関連する遺伝子を、さらなる検証のために優先順位から外した。
【0127】
RHGPにより提供される組込みの部位及び方向は、宿主細胞がインフルエンザを用いた攻撃を生き残ることを可能にする摂動の型に関する洞察を提供した(図4)。具体的には、RHGP摂動は広く3群に分けられた:(1)「アンチセンス」:1つの対立遺伝子の破壊及び他の対立遺伝子のアンチセンス阻害を促進させたアンチセンス組込み事象;(2)「センス下流」:センス方向での組込み(1つ又は複数のドメインの産生を促進すると予測されうる。全長遺伝子産物のドミナントネガティブ阻害剤として作用しうる);及び(3)「センス上流」:開始部位の上流のセンス方向での組込み(標的遺伝子全体の過剰発現を促進しうる)。RHGPを使用して同定された99の公知の標的の内、56の標的(全標的の56%)が標的発現の「アンチセンス」ノックダウンを表わした。標的の別の35%が「センス下流」事象を表わし、野生型遺伝子発現のドミナントネガティブ阻害剤の過剰発現を伝える可能性が高い。この解釈と一致して、本発明者らは、特定の標的PTBP1が2つの別々のクローン中の標的として独立して同定されることに注目した;1つのクローンは「アンチセンス」方向及び別は「センス下流」方向。残りの9%は「センス上流」挿入を表わした。11の標的遺伝子(本明細書において配列決定した110の標的の10%)の方向が確認されないであろう。なぜなら、遺伝子自体が、以前に、記載されておらず、従って、方向の割り当てが可能ではなかったからである。
【0128】
別々の評価において、本発明者らは、これらの110の標的遺伝子が以前にウイルス感染と関連付けられていたかを問うた。本明細書において同定される110の標的の内わずか4つ(MDN1、GRK6、AKT1、及びSTXBP1)がウイルス感染に直接的に関連付けられており(表II)、RHGPを使用して同定された106の標的(96%)についての新規情報を示唆している。本発明者らの分析を関連タンパク質に拡大することにより、本発明者らは、本明細書において同定される110の標的の内23(21%)が、以前にウイルス感染と関連付けられていた宿主標的と相同性を共有している又はそれらとスーパーファミリーの共通メンバーであることを見出した。これらの知見は、RHGPによって宿主標的の圧倒的多数(75%)についての新規機能が同定されたことを示唆する。本明細書において同定された2つの標的(C4orf32及びC21orf33)が、オープンリーディングフレーム(orf)として簡単に記載されており、帰する機能を伴わない。全体で、これらの知見は、宿主ゲノムのRHGPベースの照合によって両方の新規標的が同定され及び/又は新規機能が公知の遺伝子に帰されたことを示唆する。
【0129】
ナイーブな細胞を使用した標的遺伝子の検証
RHGPによってMDCK細胞中の一連の標的が同定され、それらはインフルエンザ感染に対して耐性を付与する。表現型の可逆性は、これらの標的についての最初の検証を提供した。本発明者らは、次に、独立した実験システムを使用してこれらの候補を検証することを追求し、MDCK細胞又はRHGP技術のアーチファクトとして生じうる結果を除いた。標的遺伝子がイヌ由来MDCK細胞から単離されていたため、本発明者らはそれらのヒトホモログを同定し、ヒトHEK−293細胞においてこれらの標的について選択的であるsiRNAを発現させた。
【0130】
RHGPによって、宿主細胞(過剰発現又はノックダウンされている標的を含む)のインフルエンザ媒介性の殺傷を予防する改変が同定された。本発明者らは、RHGPベクター内で誘導プロモーターを使用してこれらの標的を検証し、表現型を逆転させ、それにより、RHGP戦略に起因しない自然アーチファクトについての潜在力を最小限にできた。このゲノム規模のスクリーニングによって、本発明者らは、宿主細胞を、インフルエンザウイルスを用いた本来なら致死的な攻撃に対して耐性にする110の新規標的を同定することができた。これらの標的の内、大半(110中106)が、以前に、インフルエンザと関連付けられていた。また、本発明者らは、新規機能を、以前に未知である遺伝子及びorfに帰する。概念の証拠として、標的のサブセットを、従来のsiRNAアプローチを使用した独立した試験においてさらに検証した。
【0131】
最近の一連のsiRNAベースの試験によって、インフルエンザ、HIV、又はウエストナイルウイルスによる感染と関連する宿主標的が同定された(概要については表3を参照のこと)(Brass et al., 2008; Hao et al., 2008; Konig et al., 2008; Krishnan et al., 2008)。これらの初期の試験では、異なる種に由来する宿主細胞が利用された。それらは、本発明者らの本知見との比較の複雑さを増加させる。これを複雑化することとして、siRNAは、標的発見のためにますます利用されるが、しかし、siRNAの頑健で持続的な過剰発現の必要性により内因的に限定される。一部のsiRNAの知見での結果は、また、siRNAが宿主防御機構を非特異的に変化させるか否かの疑問により曇らされてきたが(Kleinman et al., 2008)、それは、抗ウイルス疾患へのsiRNA技術の適用について特に問題になりうる。RHGPは、また、潜在的な抗ウイルス宿主防御機構を誘導しうる。例えば、GSVのアンチセンス方向は、二本鎖RNAを誘導することが予測され、それ自体が宿主の抗ウイルス応答を引き起こすことができる。
【0132】
表
【表11】
【0133】
表2。示しているのは、宿主細胞を、本来なら致死的であるインフルエンザ感染に耐性にすることができる宿主標的のRHGP媒介性の同定と関連する事象の進行の概観である。RHGPを使用して同定された特定の標的(96%)及び標的ファミリー(75%)の大部分が固有であったことに注目して下さい。RHGPは、また、アノテートされていなかった標的の同定を促進した。
【0134】
【表12】
【0135】
表3。示しているのは、RHGP(本報告)又はsiRNAを使用して同定された標的及び標的ファミリーの評価である。この表は、本試験及び発表された知見(示す通り)の両方に共通である標的ファミリーに限定される。4つの標的(MDN1、GRK6、AKT1、及びSTXBP1、太字)が共有され、残りは固有であったが、一部の関連するスーパーファミリーメンバーは、異なるウイルスによる感染と関連付けられている。示されていないのは、以前に発表された情報と共通性を有さない、本明細書において同定された87の追加の標的である。
【0136】
実施例4−RHGPを使用し、HIV感染性を制御するための潜在的な標的を同定する。
ヒト免疫不全ウイルス(HIV−1)は、公衆衛生に対する世界的な脅威である。ウイルスを直接的に標的化する現在の医薬は、しばしば、薬物耐性ウイルスバリアントにおいて無効になりうる。薬物耐性と戦うための出現する概念は、ウイルスの機能、生存、及び増殖のために(しかし、宿主自体に対してではない)必須である宿主機構を標的化する考え方である。本明細書において、ヒトMT4 T細胞に対するランダムホモ接合摂動(RHGP)を使用し、本発明者らは、これらの細胞がHIV−1NL4−3を用いた致死感染に生き残るために必須である、安全で効果的な一連の宿主標的を同定した。RHGPは、細胞中の全ての標的をサンプリングすることができる新たな技術である。これは、遺伝子探索ベクター(GSV)の操作を通じて可能であり、それは、宿主染色体中にランダムに組込むことができる。GSVは、細胞において、誘導因子により調節される不可逆的な様式で、任意の標的をアップレギュレーション又はダウンレギュレーションすることができる。誘導因子の除去によるRHGP効果の逆転によって、操作された細胞は、HIV感染に再び感受性になる。siRNAアプローチを使用し、同定され、定義された遺伝子標的が検証され、CXCR4又はCCR5のいずれかに向性のウイルスを用いた感染のために必要であることが示された。これらの試験では、標的介入のための新規の遺伝子候補(機能の事前の知識を伴わない遺伝子配列(EST)を含む)を同定するためのRHGPの能力が実証される。これらの候補は、広域スペクトルの薬物開発のために直接的に適する。なぜなら、それらの発見の間での遺伝子破壊は、無視できる細胞傷害性を示すだけであるからである。
【0137】
三種混合薬物治療(すなわちHAART、高活性抗レトロウイルス治療)は、HIV−1複製を効果的に制御することができるが、HAART耐性HIV−1株が、エラープローンHIVウイルス逆転写酵素と頑健なウイルス複製及び不完全な患者のコンプライアンスとの組み合わせとして容赦なく出現している。一部の地域において、HARRTにおいて使用される三種薬物に対して耐性であるウイルスが、AIDS患者の20%近くから単離された(Grant, Hecht et al. 2002; Richman, Morton et al. 2004)。そのような知見によって、HIV−1疾患の処置のための新たなパラダイム(特に、耐性の発生に対して比較的非感受性である作用機構を伴うもの)を同定する緊急性が増加する。HIV−1感染(しかし、宿主自体ではない)に必須である宿主遺伝子産物を標的化する治療法は、処置のそのような新たなモダリティを提供しうる。なぜなら、十分に設計された宿主標的は、病原体に対して直接的な選択圧力を置かないかもしれないからである。
【0138】
ウイルスと宿主細胞の間での相互作用によってウイルス病原性の結果(ウイルスの除去から潜在的及び致死的な感染に及ぶ)が決定されることが十分に確証されている。HIV−1は、宿主細胞タンパク質と相互作用し、それらの複製を助け、免疫攻撃を回避することが公知である。この相互作用の例は、欠損した細胞表面受容体(CCR5)を保有し、従って、HIV−1感染に対して耐性である個人により実証される(Liu, Paxton et al. 1996)。同様の型の相互作用が、文献において報告されており、HIV−1ライフサイクルのほぼ全ての段階を包含する:細胞受容体CD4及び共受容体CXCR4又はCCR5へのEnv結合を要求するウイルス侵入から、Gag p6によりTSG101をハイジャックする必要性のあるウイルス放出まで(Garrus, von Schwedler et al. 2001)。HIV−1タンパク質と宿主細胞のものとの相互作用の完全な文書化は、このように、HIV−1複製及び病原性のプロセスを理解するために重大である。より重要なことには、そのような理解は、HIV/AIDSと戦うための治療法及び予防戦略を改善するための礎石を提供しうる。
【0139】
HIV−1感染における宿主因子の理解された重要性に照らして、増加する研究で、抗ウイルス治療のための宿主標的が考えられ始めている。宿主指向の治療法が、HIV/AIDS処置において、HIV結合及び内在化のために必要である受容体に影響を及ぼすための生物製剤及び小分子を用いてウイルス受容体CD4及び/共受容体CCR5(Reeves and Piefer 2005; Wheeler, McHale et al. 2007)を標的化することにより成功裏に使用されてきた。実験室での研究及び初期の臨床試験では、これらの薬剤がHIV/AIDSの処置のための選択肢を提供しうることが示唆される。
【0140】
発見された全ての宿主遺伝子が、治療標的として適するわけではない(特に、宿主自体に必須であるもの)。これらの標的の間で区別するために、本発明者らの実験室では新規技術であるランダムホモ接合性遺伝子摂動(RHGP)を用いて、HIV感染のために必須であるが、しかし、非感染細胞の増殖、生存、又は機能のためには必要ではない標的について選択している。RHGPは、真核細胞中の任意の遺伝子をアップレギュレーション又はダウンレギュレーションし、細胞挙動の客観的な測定を使用して結果を評価するように設計した。
【0141】
本明細書において、本発明者らは、RHGPを適用し、宿主細胞が致死的HIV感染に耐性になることを可能にする一連の宿主志向性の標的を同定した。これらの新規標的は、その機能が公知の生物学的経路に関連する遺伝子及び機能が割り当てられていないESTを含む。同定された遺伝子を、他の遺伝的アプローチ(siRNAを含む)を使用して検証した。従って、この研究では、HIV/AIDSに対する将来の薬物開発のための潜在的な新たな安全な標的が明らかになった。
【0142】
材料及び方法
細胞株及びウイルス
細胞(MT4、PM1細胞、及びTZM−blを含む)及びウイルス(薬物耐性突然変異体ウイルス及びR5向性バリアント)を、NIH AIDS Research及びReference Reagent Programから得た。HIV−1NL4−3を、HEK293から、プロウイルスDNA(NIH AIDSプログラム)のトランスフェクション後に作製し、MT4細胞中で増幅させた。
【0143】
HIV−1感染ならびに感染性及びp24産生の決定
MT4又はPM1細胞を、HIV−1(MOI0.001)を用いて、1時間にわたる低速遠心分離(1,200g)により感染させた。感染後に回収された上清を、次に、感染性の検査のために、TZM−bl指標細胞株(Wei, Decker et al. 2002)に移した。相対発光単位(RLU)を、TZM−bl細胞で、それらをBright−Glo Luciferase Assay System(Promega)を用いて3日後に処理した後に得た。
【0144】
回収された上清中のP24量を、また、HIV−1 p24 ELISAキット(Xpressbio, Thurmont, MD)を使用し、製造者の指示に従い測定した。
【0145】
MT−4 R1細胞株の構築
RheoSwitch(登録商標)Mammalian Inducible Expression SystemをNew England Biolabs(NEB)から購入した。トランスアクチベーターR1をコードするプラスミドpNEB−R1を、最初に、制限酵素ScaI(NEB)を使用して線状化した。MT4細胞を、次に、Eppendorf Multiporator(Eppendorf, AG 22331, Huammburg)を使用し、360v(電圧)及び100ms(時間)の条件下での電気穿孔によりトランスフェクションした。MT4細胞を、G−418(400μg/ml)を使用して選択し、G−418耐性細胞を連続限界希釈によりクローン化した。増大後、クローンを、R1応答性ルシフェラーゼレポーター遺伝子(pGluc)を用いたトランスフェクション後、Gaussia Luciferase Assay Kit(NEB)を使用して発光(相対発光単位(RLU))について少なくとも2回検討した。本発明者らは、これらの細胞クローンからの発光のRSL1誘導(倍)を以下の通りに決定した:誘導因子での処理なしのサンプルからのRLUにより除した、誘導因子の存在においてサンプルから得られたRLU。これらのクローンからの倍は2〜60倍に及んだ。最も高い誘導を伴う安定なクローン(#2−14)を使用し、RHGPライブラリーを作製した。
【0146】
RHGP遺伝子探索ベクターpRHGP12−RSNのコンストラクション
RHGP遺伝子探索ベクターpRHGP12−RSNは、企業において、レンチウイルスベースのpLESTベクター(寛大に、Stanley Cohen博士(Stanford)により提供された)を骨格として使用してコンストラクションされた一連のFGIのRHGPベクターの1つである(Lu, Wei et al. 2004)。このベクターは、RheoSwitch Mammalian Inducible System(NEB)を用いてコンストラクションされた。RheoSwitchシステムは、TATAボックスの上流に5コピーのGAL4応答エレメント(5×RE)を含み、RSL1リガンドの存在において低い基礎発現を伴う転写の高い誘導をもたらす。ベクターを構築するために、pLEST中のNeoR−TRE−CMVのDNA配列を、最初に、5’LTR部位の反対のOri−CAT−RSのRheoSwitch(RS)誘導性発現カセットを用いて置換した。選択マーカー及びブラストサイジン(BS)耐性遺伝子を含むレポーターカセットならびにPKGプロモーターにより制御されるEGFP遺伝子を、RS発現カセットの反対方向でNheI部位中に挿入した。2つのLoxP部位を、RSプロモーターの下流及びp15A ori−CATの上流にそれぞれ挿入した。
【0147】
GSVを保有するレンチウイルスの産生及びRHGPライブラリーの構築
RHGPレンチウイルスを、ViroPower Expression System(Invitrogen)を使用して産生した。HEK293FT細胞を、10cmプレート中に106個細胞/プレートでプレーティングした。24時間のインキュベーション後、細胞を、3μgのRHGP22−RSN及び9μgのViroPower Packaging Mixを用いて、Lipofectimine 2000(Invitrogen)を使用してトランスフェクトした。培地を、37℃で5時間のインキュベーション後に交換した。48時間後、培養培地中のウイルスを、0.45μmフィルターを通じてろ過し、製造者の指示に従い滴定した。
【0148】
RHGPライブラリーを構築するために、MT4−R1細胞を、RHGPウイルスを用いて、ポリブレン(6μg/ml)の存在において、1時間にわたる低速遠心分離(1,200g)により形質導入した。単一細胞内での複数の挿入についての潜在力を最小限にするために、低MOI(0.1)をライブラリー作製の間に用い、細胞が、1を上回る異なるGSVにより形質導入されうる可能性を最小限にする。GSV組込み細胞を、GBL培地(完全RPMI 1640培地、G418(400μg/ml)、ブラストサイジン(4μg/ml)、及びRSL1リガンド(0.5μM)を含む)を使用して選択した。
【0149】
HIV−1攻撃から生き残ったRHGP細胞クローンの選択
HIV−1NL4−3を用いた感染後、MT4−R1 RHGPライブラリーを、同じGBL培地中で培養した。個々の生存クローンを、連続限界希釈により確立し、GBL培地中で継続的に増大させた。細胞クローンを、さらに、HIV−1を用いて攻撃し、それらの耐性を確認した。
【0150】
可逆性
RHGP事象の可逆性を検証するために、ウイルス耐性MT4細胞クローンを、GBL培地又はRSL1を伴わないGBL培地中で少なくとも3日間にわたり培養した。HIV−1感染後、上清中でのウイルス産生(感染性及びp24)を上に記載する通りに検討した。
【0151】
ウイルス耐性クローンからの候補遺伝子の同定
RHGP遺伝子探索ベクターを設計し、標的遺伝子を効率的に発見し、組込み事象の方向(センス又はアンチセンス)を決定した。遺伝子探索ベクターはOri−CATレポーター遺伝子を含み(図1)、それは、制限酵素ベースのゲノムDNAクローニングによりレスキューすることができる。
【0152】
各クローンからの細胞(1×106)を、溶解緩衝液(0.32Mスクロース、10mM Tris pH7.5、5mM MgCl2、及び1% Triton X−100を含む)中に浮遊させた。遠心分離後、ペレットを、プロテイナーゼK緩衝液(25mM EDTA、150mM NaCl及び40mM Tris、0.5% SDS、pH7.5を含む)中のプロテイナーゼK(Sigma)及びRNase A(Sigma)を用いて処理した。ゲノムDNAを、フェノール/イソアミルアルコール/クロロホルムを用いて抽出し、イソプロパノールを用いた沈殿が続いた。DNAペレットを、70%エタノールを用いて洗浄し、TE緩衝液(pH7.5)中に溶解した。
【0153】
BamHI又はXbaIを用いた消化及びフェノール/イソアミルアルコール/クロロホルムを用いた抽出後、ゲノムDNA(10μg)を、エタノールを用いて沈殿し、TE緩衝液中に溶解した。消化されたDNA(2.5μg)を、T4リガーゼ(Invitrogen)を使用して16℃で一晩自己連結させた。連結されたDNAを、エタノールを用いて沈殿させ、70%エタノールを用いて洗浄した。DNAペレットをTE緩衝液中に溶解させ、DH10B ElectroMaxコンピテント細胞(Invitrogen)中に電気穿孔した。複数のコロニーを、プラスミドDNA調製及び制限酵素消化のために単離した。プラスミドDNAをさらに使用し、標的遺伝子をDNA配列決定及びゲノムマッピングにより同定した。RHGPベクター挿入部位に隣接する結果として得られるゲノムDNA配列を、ヒトゲノムに対する、UCSC Genome Browserを使用したゲノムマッピングに供した。
【0154】
siRNAを用いた宿主標的遺伝子の検証
RHGP同定遺伝子についてのヒト二本鎖siRNAホモログ(siGNOME SMART pool)を、製造者(Dharmacon)により推奨される通りに調製した。siRNA Rab 6A及びHIV−1 tatをポジティブコントロールとして用いた(Brass, Dykxhoorn et al. 2008)。非標的siRNA(siCONTROL1)をネガティブコントロールとして使用した。MT4又はPM1細胞を、完全RPMI 1640培地中で一晩培養した。対数期増殖中のMT4細胞を、1.2μMのsiRNAを用いて、電気穿孔プロトコールにより、製造者(Eppendorf)の指示に従いトランスフェクトした。PM1細胞を、同じプロトコール下で(パルス条件200v(電圧)及び200ms(時間)の使用を除く)、電気穿孔した。細胞を、HIV−1バリアントを用いて、トランスフェクションから24時間後に感染させた。培養培地を毎日新しくした。細胞生存をトリパンブルー色素排除アッセイにより毎日検討した。上清中でのウイルス産生(感染性及びp24)を上に記載する通りに検討した。
【0155】
結果
RHGP標的遺伝子の発見の概観
材料及び方法に概説されている通り、pRHGP12を、ブラストサイジン耐性遺伝子を促す常時発現性プロモーターからなる発現カセットを使用してコンストラクションした。GSVは、また、RheoSwitchプロモーターを含み、それはリガンドにより活性化することができる。リガンドの存在(プロモーターオン)は、このように、宿主転写物を産生し、RHGP効果の上方又は下方のいずれかの調節を誘発することができる。RHGPにより起こされる遺伝子摂動効果は、前に詳細に記載された(Sui, Bamba et al. 2009)。本発明者らはこの供給源を利用し、ヒトTリンパ球のゲノム全体を照合し、標的(アップレギュレーション又はダウンレギュレーションを問わず、これらの細胞を本来なら致死的なHIV−1感染に生き残ることを可能にする)を同定した。
【0156】
MT4 T細胞を操作し、トランスアクチベーターR1を安定的に発現させ、それはRHGP中の組込みプロモーターを活性化し、誘導因子RSL1の存在において転写物を産生することができる。遺伝子摂動の「RHGPライブラリー」を確立するためのRHGPベクターを用いた形質導入及び抗生物質選択に続き、宿主細胞を、HIV−1の致死感染を用いて攻撃した。
【0157】
生存細胞は、限界希釈によりクローン化された単一細胞であった。RHGPと無関係の事象の結果として生じたであろうそれらの生存体を除去するために、表現型を、RSL1の非存在においてHIV−1攻撃を繰り返すことにより確認した。この試験では、RHGP自体が(他のアーチファクト又は自然突然変異ではなく)、HIV−1感染に直面した細胞生存に直接的に関与することが確認された。生存クローンからのゲノムDNAを次に単離し、配列決定し、標的遺伝子ならびにGSV組込みの方向を特定した(結果が標的のアップレギュレーション又はダウンレギュレーションを表わすか否かを決定するために)。
【0158】
RHGP細胞ライブラリーの構築
105の組込み事象を伴うMT4−R1細胞のライブラリーを、レンチウイルスベースのpRHGP12 GSVを使用して形質導入し、宿主ゲノムのカバーを確実にした。ゲノム中での平均遺伝子サイズが27kbと推定されるため(Gupta and Varshney, 2004)、本発明者らは、これらの105の独立した組込み事象が、ゲノム全体のランダムなカバーを確実にしうると計算した。形質導入された宿主細胞のライブラリーを、次に、ブラストサイジンを使用して選択した。特定のブラストサイジン耐性細胞クローンから、本発明者らは、GSVが実際に宿主染色体中に組込まれ、アンチセンス転写物が誘導因子RSL1の処置時にだけ産生されることを見出した(データ示さず)。R1発現及びGSV組込みを確認及び維持する追加の手段として、MT4ライブラリーが、続く実験において、致死濃度のG418及びブラストサイジンと一緒に持続的にインキュベーションされた。
【0159】
HIV−1耐性細胞は、ウイルス誘導性細胞殺傷から生き残った。
MT4−R1細胞は、親MT4細胞と同様に、HIV−1NL4−3(MOI10−4)により、5日以内に効率的かつ完全に殺された。RSL1の添加はこの殺傷に影響を及ぼさなかった(図11A、左パネル)。これらの条件は、「プロモーターオン」を保つためのRSL1の存在において、生存細胞クローンについての続く選択のために厳密に利用された。追加のコントロールとして、偽形質導入細胞の並行培養物を同じように処理し、生存体は5日後に観察されなかった。この結果は、生存細胞が、RHGP摂動の結果として(HIV−1に対する自然耐性のアーチファクトとしてではなく)生じることを確実にするために必須であった。クローニング及び増大後、生存細胞を複数ラウンドの感染に供し、感受性細胞を除去した。最後に、本発明者らは、ウイルス誘導性殺傷に耐性である25のRHGP細胞クローンを得た。図11A(右パネル)は、HIV−1攻撃後でのそのような1つの耐性クローンの生存を示す。
【0160】
ゲノム配列決定と並行して、本発明者らは、これらの耐性細胞クローンが、HIV感染時に感染性ウイルス粒子を産生し続けるか否かを問うた。RSL1の存在におけるHIV−1NL4−3を用いたこれらの耐性細胞クローンの再攻撃後、上清を4dpiで回収し、TZM細胞に移し、それによって感染性の読み出しが提供される。図11Bは、得られたウイルス耐性RHGP細胞クローンが子孫ウイルスを産生及び放出しないとの代表的な実験を示す。これらの知見は、ELISAベースのアッセイにおけるp24の決定により独立して確認された。
【0161】
野生型HIV−1NL4−3を用いた結果が励みとなるが、本発明者らは、治療法についての大きな満たされない必要性は、従来の医薬に耐性であるウイルスバリアントへの新たな標的の適用であると考えた。従って、本発明者らは、プロテアーゼ阻害剤に対する確立された耐性を伴うHIV−1バリアントを用いた並行した一連の試験を実施した。野生型HIVを用いた攻撃に耐性であるRHGP形質導入クローンは、また、プロテアーゼ耐性バリアントに直面した攻撃に生き残った。この結果は、宿主細胞の生存に固有ではなかった。なぜなら、感染性アッセイならびにp24 ELISAによって、耐性細胞における突然変異体HIV−1による欠損した感染が確認された(図11C)。一緒に、これらの結果によって、本発明者らが得た細胞クローンが、WT及び薬物耐性HIV−1バリアントの両方による感染に耐性であることが確認された。
【0162】
ウイルス耐性細胞クローンの可逆性アッセイ
RHGP技術の重要な特性は、誘導プロモーターによる調節を介して候補標的を検証する能力である。本発明者らは、最初に、RHGPライブラリーを構築するために使用されているMT4−R1細胞においてトランスアクチベーターR1の誘導活性を検討した。MT4−R1細胞は、最初に、R1応答性ルシフェラーゼレポート遺伝子(NEB)を用いてトランスフェクトし、誘導因子RSL1の存在又は非存在において培養した。発光(RLU)を次に検討した。MT4−R1は、RSL1の存在において高く安定したレベルの発光を示したが、しかし、RSL1が存在しない場合には背景レベルだけであった(図12A)。この結果は、R1の活性化能力がRSL1により強固に制御されることを示した。
【0163】
これらのクローンでのHIV−1耐性の獲得が、本当にRHGPの組込みに起因することを確認するために、本発明者らは、次に、これらのクローンにおいてウイルス産生の可逆性を検討した。RHGPベクター用のプロモーターは、リガンド制御されたRheoSwitch誘導性トランスアクチベーターR1の制御下にあったため、本発明者らは、リガンドRSL1の存在又は非存在において候補のHIV媒介性殺傷を比較することができた。本発明者らの試験では、これらの生存クローンがHIV−1攻撃に対して可逆的な耐性を示すことが確認された。図12Bは、この一連の実験からの代表的な細胞クローンの結果を示す。リガンドが、HIV−1攻撃の間に、細胞培養中に存在しない場合、頑健なウイルス産生(親MT4細胞から産生されるものと同程度)がこの細胞クローンについて観察された。高レベルのウイルス放出が、細胞がウイルス媒介性殺傷で死ぬまでの多くの日数にわたり持続した。
【0164】
ゲノムDNAクローニングによる宿主遺伝子の同定
ゲノムDNAを、HIV−1に対して可逆的な耐性を示していたウイルス耐性クローンから単離した。GSV組込み部位を伴う25のHIV非感受性宿主細胞クローンが、21の異なる宿主細胞標的の同定をもたらした。これらのGSV組込みは、12の以前に記載された遺伝子及び2の非アノテートESTを含んだ。クローンの一部は、同じ親からの子孫に思われた。なぜなら、GSVは、同じ方向を伴う同じ遺伝子中に組込まれていたからである。3つのクローンが、遺伝子又はESTを伴わない領域中にRHGP挿入を有した。本発明者らは、Ori−CATレポーター遺伝子の部分的喪失に起因する4つの候補から候補遺伝子を単離することはできなかった。
【0165】
これらの遺伝子及びESTの特性を表4に列挙する。RHGPにより提供される組込みの部位及び方向は、宿主細胞がHIV−1攻撃を生き残ることを可能にする摂動の型に関する洞察を提供した。具体的には、RHGP摂動は広く3群に分けることができた:(1)「アンチセンス」:1つの対立遺伝子の破壊を促進させたアンチセンス組込み事象及び他の対立遺伝子のアンチセンス阻害;(2)「センス下流」:センス方向での組込み(内因性の遺伝子産物のドミナントネガティブ阻害剤の産生を促進すると予測されうる);及び(3)「センス上流」:開始部位の上流のセンス方向での組込み(標的遺伝子の過剰発現を促進すると予測されうる)。RHGPを使用して同定された12の遺伝子標的の内、8の標的が標的発現の「アンチセンス」ノックダウンを表わした。標的の他の6つが「センス下流」事象を表わし、野生型遺伝子発現のドミナントネガティブ阻害剤の過剰発現を表わす可能性が高い。「センス上流」挿入は、本試験において同定されなかった。これらの予測に基づき、候補遺伝子の全てがGSV組込み事象によりダウンレギュレーションされる可能性が高い。これによって、本発明者らは、ナイーブなMT4細胞でのsiRNAノックダウンアプローチを直接的に使用し、ウイルス耐性表現型を再現することができた。全体で、これらの知見は、宿主ゲノムのRHGPベースの照合によって両方の新規標的が同定され及び/又は新規機能が公知の遺伝子に帰されたことを示唆する。
【0166】
ナイーブな細胞を使用した標的遺伝子の検証
RHGPによってMT4細胞中の一連の標的が同定され、それらはHIV−1感染に対して耐性を付与する。表現型の可逆性は、これらの標的についての最初の検証を提供した。本発明者らは、次に、独立した実験システムを使用してこれらの候補を検証することを追求し、自然突然変異又は本発明者らのRHGP技術のアーチファクトとして生じうる結果を除いた。
【0167】
これらの耐性遺伝子に対する二本鎖siRNAをDharmaconから購入した(4つの個々のsiRNAを含むSmartpool)。非標的siRNAは、トランスフェクション及び参照基準のための一致されたコントロールを提供した。ウイルスtat及び細胞標的Rab6Aに特異的であるsiRNA構築物を、これらのsiRNAがHIV−1感染を効率的に阻害できるとの最近の報告に基づいて、ポジティブコントロールとして選択した(Brass, Dykxhoorn et al. 2008)。
【0168】
siRNAを、天然MT4細胞中に、電気穿孔を介して、HIV−1NL4−3感染の1日前にトランスフェクトした。上清を、感染から2日後に回収した。処理した細胞から産生された感染性ウイルス粒子を、TZM細胞ベースの読み出しを使用して検討した。図6Aに示す通り、12の選ばれた標的遺伝子に対する二本鎖siRNAによって、HIV−1ウイルス産生が、ポジティブコントロール(Tat及びRab6A)において観察されるものと同程度のレベルまで低下した。生存も検討し、siRNA処理からの可能な細胞傷害性効果を除いた。非常に同様のパーセンテージの生存細胞が、処理されたサンプルの全てについて観察された。これらの結果によって、RHGPにより同定されたこれらの遺伝子がウイルス複製において重要であるとの本発明者らの知見が確認され、新規の宿主ベースの標的を同定するためのRHGPの適用が検証された。
【0169】
HIV−1NL4−3(CXCR4向性ウイルス)誘導性殺傷から生き残った細胞から同定されたこれらの宿主遺伝子候補が、また、CCR5向性HIV−1ウイルスのウイルス複製に重要であるか否かを検討するために、同じsiRNAアプローチを、次に、CXCR4及びCCR5二重発現T細胞株PM1で使用し、R5向性HIV−1ME1を用いた感染が続いた。HIV−1ME1株のウイルス産生は、これらの12の遺伝子標的を標的化する特異的siRNAを用いて処理された細胞において有意に阻害された(図6B)。これらの結果によって、RHGPにより同定されたこれらの遺伝子がHIV−1ウイルス(X4及びR5向性バリアントの両方を含む)の複製において重要であるとの本発明者らの知見が確認され、新規の宿主ベースの標的を同定するためのRHGPの適用が検証された。
【0170】
本発明者らの本試験において、本発明者らはRHGP技術を適用し、HIV−1ウイルス感染のために要求される宿主因子についてのゲノム規模のスクリーニングを行った。本発明者らは、細胞を、HIV−1ウイルスを用いた本来なら致死的な攻撃に対して耐性にする新規の宿主ベースの標的を同定した。また、本発明者らは、新規の抗HIV−1機能を、以前に公知の遺伝子及びアノテートされていないESTに帰する。これらの標的を、最初に、表現型を逆転させるためのRHGPベクター内に組込まれた誘導プロモーターを使用し、次に、天然細胞において、従来のsiRNAアプローチを使用して検証した。本発明者らは、さらに、結果として得られる標的を、異なるHIVバリアント(薬物感受性及び耐性分離株ならびにCCR5及びCXCR4向性ウイルスを含む)に広く適用可能であることを見出した。
【0171】
本来なら致死的なインフルエンザ感染に直面した際の生存のために必須の宿主標的のパネルを同定した初期の試験において使用されたRHGPと同様に(Sui, Bamba et al. 2009)、本発明者らはレンチウイルスシステムを使用し、MMLVベースのRHKOにより起こされる低力価のGSV産生での先の限定を克服した(Li and Cohen 1996)。これらの改善によって、本発明者らは、宿主ゲノム全体をサンプリングし、改善されたGSVにより引き起こされる105の組込み事象から12の遺伝子標的を同定することができた。レンチウイルスベクターが、活性な遺伝子転写の部位中への単一部位挿入を支持するとの予測と一致して(Mitchell, Beitzel et al. 2004)、全ての組込みが活性な遺伝子領域において生じた。
【0172】
RHGPの1つの固有の特性は、標的発現が誘導可能なプロモーターの制御下にあることである。このように、表現型(HIV−1耐性)と摂動遺伝子の間での因果関係は、誘導因子の取り除きにより確認することができる。同じ実験設定内でこれらの標的を検証することにより、この特性は、治療候補の本発明者らの発見効率を増加させる。実際に、耐性クローンからの全てのRHGP標的が、天然細胞を用いた続くsiRNA試験において成功裏に確認された。候補遺伝子と比べたGSV挿入の位置及び方向に依存して、RHGPは、また、哺乳動物細胞において遺伝子の活性化、しばしば過剰発現を生成することができる。本発明者らの本試験において観察されなかったが、標的のサブセット(B細胞CLL/リンパ腫2(BCL2))の過剰発現によって、MDCK細胞が、インフルエンザウイルス感染に対する宿主遺伝子の発見の間にインフルエンザ感染に生き残ることができた(Sui, Bamba et al. 2009)。従来のアプローチ(例、siRNA)により生成される一過性のノックダウンの代わりに、RHGP表現型は高度に安定であり、それによって機構試験が促進され、HIV−1複製におけるこれらの摂動遺伝子の役割を特徴付けることができる。
【0173】
RHGPは、標的の事前の知識により偏らなかった。実際に、HIV−1耐性RHGP細胞クローンから、本発明者らは、2つのアノテートされていないESTがGSVにより標的化されることを発見した(表1)。さらに、本発明者らが、PANTHER Classification System(Mi, Lazareva-Ulitsky et al. 2005)を使用して遺伝子標的を分析した場合、それらの多くの推定分子機能が依然として分類されていなかった(CASD1、GDAP2、TNRC6A、及びTTC21B)、又は未知であった(CAMPSAP1L1及びGSTCD1)。それらの2つはユビキチン−タンパク質リガーゼに属する(HECW2及びDZIP3)。残りの標的において、各標的の1つが以下のファミリーにおいて見出される:免疫グロブリン受容体又は防御/免疫タンパク質(Robo1)、シグナル伝達分子(NLGN1)、mRNAポリアデニル化因子(CPSF1)、及びトランスポーター又はヒドロラーゼ(ATP8A1)。
【0174】
候補遺伝子の内、Robo1が非常に良好な治療標的としての役割を果たしうる。I型膜貫通タンパク質として、それは、本来は、神経発生の間に軸索ガイダンス受容体として同定された(Wong, Park et al. 2002)。Robo1は、また、CXCR4との相互作用(HIV感染との潜在的な連関)を通じてT細胞の化学走性において役割を果たすことが見出された(Wu, Feng et al. 2001; Prasad, Qamri et al. 2007)。それがCCR5にも結合するか否かを検討する価値がある。ウエスタンブロットを使用し、本発明者らは、Robo1発現がRHGP耐性クローン及びsiRNA処理細胞において低下されることを確認した(未発表データ)。さらなる研究が、Robo1欠損CD4+T細胞において、ウイルス複製が、恐らくは細胞膜のビリオン融合の間にウイルス侵入段階において遮断されることを明らかにするために必要とされる。Robo1は、HIV−1複製においてHIV−1共受容体CCXR4及びCCR5の活性を支持する潜在的な新たな役割を持ちうる。CCR5の治療標的と同様に、Robo1に対するアンタゴニスト及び/又はモノクローナル抗体が、従って、AIDSを処置するための新たな一連の治療計画を提供しうる。
【0175】
同定されたHIV−1耐性遺伝子のリストは、また、2つのEユビキチンリガーゼ(HECW2及びDZIP3)を含む。このファミリーからのタンパク質は、タンパク質分別経路及び小胞輸送プロセスに関与することが見出されており、それらは、また、ウイルス出芽及び放出の間にHIV−1及び他のウイルスにより使用される。1つの良好な例はTSG101(ウイルス脱出のためにHIV−1 Gag 6によりハイジャックされるユビキチン様リガーゼ)である(Garrus, von Schwedler et al. 2001)。HECW2は、別のE3ユビキチンリガーゼNEDD4と高度に相同である。HIV−1ウイルスがNEDD4を使用するとの直接的な証拠はないが、PPXYの後ドメインを伴う多くの他のエンベロープウイルスは、宿主細胞からのそれらの放出のためにNEDD4を要求しない。これら2つの遺伝子産物(HECW2及びbZIP3)は、また、HIV−1成熟及び脱出のために重要でありうることを推測することが興味をそそられる。
【0176】
【表13】
【0177】
実施例5−同定された潜在的な標的のさらなる研究
上で考察した試験によって、増殖性HIV感染のために必須である新規の宿主標的が同定された(Robo1)。Robo1は、可溶性リガンドのための膜貫通受容体であり、SLITとして公知である。Robo1は神経系の発生において主に発現され、軸索ガイダンスを調節する。Robo1は、また、リンパ系細胞で発現され、それは細胞間相互作用の調節において同様に役割を果たしうる。
【0178】
Robo1の細胞外構造は、それが5つの積み重なった免疫グロブリン(Ig)ドメインからなる点で顕著であり、それは3つのタンデムフィブロネクチンIII型(FN−III)ドメインにより支持される。そのようなものとして、Robo1は、細胞膜から比較的遠くに伸展すると予測される。そのような特性は、しばしば、細胞間接着を伴い観察される。なぜなら、グリコシル化タンパク質上の負の電荷を持つ糖(糖タンパク質及びプロテオグリカン)の静電反発力が、細胞を分離するバリアとしての役割を果たし、密接な相互作用(例えば膜融合など)を予防する。実際に、本発明者らは、この機構が進化し、部分的に、ウイルスが宿主細胞に侵入することを予防しうると推測する。
【0179】
本発明者らは、細胞膜からのRobo1の伸展突起が、ウイルス感染におけるその必須の役割についてのRHGPの証拠と組み合わせて、Robo1がウイルスの取り込みに関与し、恐らくはウイルス−宿主結合の初期部位又は共受容体として機能しうることを示唆すると仮定する。
【0180】
この考え方を支持し、Robo1が、文献において、ケモカイン受容体CCR5と相互作用することが報告されている。CCR5は、次に、HIV感染のための重要な共受容体として機能することが公知である。実際に、CCR5(ならびに別のケモカイン受容体CXCR4)は、HIV感染を処置するための一連の最近承認された又は治験中である新たな薬物の標的である。
【0181】
RJ Shattock and JP Moore, Nature Reviews Microbiology, 1, 25; 2003に示されている通り、CCR5は細胞膜から遠くに突出せず、HIVによりコードされるgp120へのCD4結合が安定性を提供し、十分に理解された静電反発力(本来であればHIVがT細胞と密接に接触することを予防しうる)を克服することが仮定される。この安定性は、次に、CCR5とウイルスタンパク質との続く相互作用を支持し、膜融合及び内在化を媒介しうる。
【0182】
このモデルは興味深いが、しかし、CD4細胞外ドメインが4つのIgドメインからなり、このように、CD4がRobo1よりもかなり短いことを認識することが重要である。本発明者らは、従って、既存のモデル上に拡大し、Robo1が追加の共受容体として機能しうると仮定し、それは、CD4により媒介される段階よりもウイルス相互作用のより初期段階を安定化するために必要である。
【0183】
この仮定は、ウイルス出芽を予防するための治療介入についての重要な潜在的な意味を保有する。例えば、Robo1又はそのウイルス結合パートナーの特異的阻害剤は、初期結合を予防する及び/又は感染の細胞間伝播を予防する役割を果たしうる。HIVウイルス産生をノックダウンするためのsiRNAトランスフェクションを含む実験において、Robo1がノックダウンし、又は、非常に低いウイルス産生値を与え、コントロール値の9%という低さを残した。このように、Robo1の阻害剤は、一旦、細胞の初期細胞感染が生じると、HIV−1の伝播を効果的に遮断しうる。これらの阻害剤は、必ずしも限定はされないが、小分子、アプタマー、及び生物製剤(抗体、ペプチド、及びそれらの派生物を含む)からなりうる。
【0184】
現在、Robo1についてのウイルス結合パートナーは公知ではないが、しかし、本発明者らは異なる可能性を仮定する。
【0185】
最初に、感染細胞から出芽する際にHIV粒子の外部エンベロープ中に取り込まれるSLITタンパク質(又はRobo1についての他の生理学的リガンド)及びこれらの残存タンパク質が、Robo1との最初の相互作用を促進するために十分でありうる。
【0186】
加えて又は代わりに、Robo1は、そのSLITリガンド上でコードされるロイシンリッチモチーフ(LRRモチーフ)を認識すると理解される。これを念頭に置き、HIVによりコードされるgp41が、比較的顕著にロイシン残基(123残基中16;13%)を含むことに注目すべきである;これらの残基中で特に豊富である局在化領域を伴う。gp41の親分子(gp160)は81のロイシンを含む(849の全残基中;9.5%)。gp160内でコードされる他の主要タンパク質はgp120であり、それは32のロイシン残基(合計455残基から;7.0%)を含むが、この分子中にはロイシンの顕著なクラスターもあり、その少なくとも1つがLRRモチーフと似うる。HIVによりコードされるnefは、また、ビリオンの表面上で暴露されうるが、17のロイシン残基(合計196アミノ酸から;8.8%)を含む。ロイシン(及びクラスター)についてのこの相対的な濃縮は、HIVの一般的な形質を表わさないように思われる。なぜなら、非膜暴露タンパク質(例えば逆転写酵素など)は、少数のロイシン(560中48;8.5%)を利用するためであり、それらは顕著にはクラスター形成しない。同様に、HIV−1 Gagは41のロイシン(508の全残基中;8.0%)を含む。
【0187】
ウイルス付着の直接的な阻害に加えて、フローサイトメトリーデータは、Robo1の特定のエピトープが、HIV感染細胞の表面上に優先的に暴露されることを示す。この知見は、宿主防御機構(ADCC、CDC)を介した除去のための感染細胞の抗体ベースの標的化についての可能性を生じる。また、これらの抗体は、毒素、放射性核種、又は感染細胞を標的化するための他の致死薬剤を用いて標識されうる。これらの型の活性は、必ずしも抗体に限定されないであろう。なぜなら、他の抗体、ペプチド、融合タンパク質、及びこれらのエピトープを標的化する生物製剤又は小分子も、感染細胞を標的化するための活性を有しうるからである。
【0188】
HIVを越えて、Fuctional Genetics, Inc.からの最近のデータは、Robo1上の特定のエピトープが、インフルエンザ感染細胞の表面上に優先的に暴露されることを示唆する。このデータは、Robo1ベースの治療法の広域適用についての意味を有しうる。Robo1が、HIVを越えたウイルスに関連する機構的基礎は公知ではなく、本研究の課題である。
【0189】
RHGPの実用的な能力を実証するために、上の実施例において同定された特定の標的が、ウイルス疾患の治療的な予防又は処置のための潜在的な宿主標的として、さらなる研究の対象であった。直ぐ上で考察したRobo1、及び上のインフルエンザに関連して考察されたPTCH1に加えて、Nedd4を、インフルエンザの処置のための可能な候補として上の通りに同定した。Nedd4は、また、共同所有される米国特許出願シリアル番号11/939,122(2007年11月13日出願)において考察され、その全体が参照により本明細書において組み入れられる。
【0190】
Nedd4は、ESCRTタンパク質と一般的にいわれるタンパク質のクラスのメンバーであり、典型的には細胞内で見出され、そこで、それは、ESCRTタンパク質ファミリーの他のメンバーと相互作用し、細胞内の1点から別の点にタンパク質を向ける又は運ぶ。Nedd4に対する抗体を、標準的な手段により調製した。ヒト身体の特定組織がNedd4について弱く染色されるが(例、結腸、小腸、肝臓の幹細胞)、一般的に、このタンパク質は表面で弱く発現される(仮にあっても)。産生された抗体の内、ヒトNedd4抗体Ab27979及びAb101APは、両方がFunctional Geneticsから利用可能であり、従来の方法を使用して調製されたが、しかし、任意の抗体産生方法を用いることができる。これら及び無関係の標的に対して産生されたコントロール抗体を用いて実施されたFACSによって、Nedd4は、感染されていないMDCK細胞の表面上で全く染色されず、インフルエンザ(A/Udorn/72)を用いて感染されたMDCK細胞は、表面上に多量のタンパク質を呈することが実証された。TSG101と同様に、ウイルス装置は感染細胞の細胞機能を乗っ取るため、Nedd4は、その通常の役割から「ハイジャック」され、使用されて、ビリオン、又は必須のビリオンタンパク質を細胞表面に輸送すると思われる。図13。
【0191】
Ab27979(Nedd4配列の中央、アミノ酸313−334から取られたエピトープに対して産生された)及びAb101AP(タンパク質のC末端、Nedd4アミノ酸860−900に対して産生された)(図13を参照のこと)は、インフルエンザ複製の強いADCC媒介性阻害を示した。濃度0.01μg/ml及びそれ以上で、これらの抗体は、反復インビトロ実験においてウイルス複製の80%より良好な阻害を示した。(図14)。
【0192】
上で考察された実験結果を、種々のインフルエンザ株に対して繰り返した。なぜなら、従来の治療からのインフルエンザについての「回避」の1つの主な手段が、処置での事前のワクチンによる中和を避けるための十分な違いである小さな突然変異、又は新たな株の導入であるからである。実験では、抗体結合が、比較されたインフルエンザ株、及び関連ウイルス(パラインフルエンザ、RSV)を通して持続的であることが実証された。抗体を使用し、MDCK細胞を使用したA/Udorn/72(H3N2)、MDCK細胞を使用したA/Hong Kong/8/68(H3N2)、HEK293細胞におけるA/Japan/305/57(H2N2)、HEK293細胞におけるA/Puerto Rico/8/34(H1N1)、HEK203細胞におけるA/WSN/33(H1N1)、MDCK細胞におけるB/Lee/40、HEp−2細胞におけるHPIC−3(ヒトパラインフルエンザウイルス3)、MRC−5細胞におけるCMV/AD−196、及びHEp−2細胞におけるRSV/A2に対するこれらの抗体による表面上での大量の染色が実証された。これらのウイルス全般での結果は、ヒト又は哺乳動物(他の哺乳動物は、ペット及び家畜、例えばイヌ、ネコ、ならびに関連する哺乳動物及び農業用動物、例えばウシ、ヒツジ、ブタ、ヤギなどを含む)を標的化することが、広く効果的でありうることを示唆しており、そこでは従来のウイルス標的処置は共通して株及び宿主に特異的である。明らかに、RHGP−TAIL技術は、種々の状況における介入のための標的としての開発のために遺伝子及びそれらのコードされるタンパク質を迅速に同定する能力を提供する。
【0193】
実施例6−転移関連遺伝子のRHGP同定
上で注目した通り、RHGPは、ウイルス感染性の阻害のための潜在的な標的の同定に限定されない。考えられている特定の条件について固有である表現型が特定できる限り、RHGPの使用は予測可能な結果を与え、そこでは同定された遺伝子はその表現型を促進又は阻害する。治療用薬剤が、遺伝子によりコードされるタンパク質に標的化される場合、遺伝子自体とは対照的に、薬剤は対応する効果を与えるはずである。
【0194】
このように、癌処置を改善させるための1つのアプローチは、化学療法に対する耐性を促進する薬剤を同定することである。多くの強力な化学療法剤が宿主に対して有毒であり、治癒の症例は疾患と同じくらい悪い。RHGPライブラリーを化学療法剤に暴露させる。薬物処置を生き残るものを同定し、摂動が変化した生存条件を起こした遺伝子を選択し、挿入されたRHGP挿入物の報告を決定し、上の通りに、過剰発現又は低下発現が所望の表現型に関与するか否かを決定する。
【0195】
生存は、しかし、RHGP探索において表現型として使用することができる唯一の特徴ではない。多くの癌において、それは、特に危険である転移状態の発生である。腫瘍の転移癌への形質転換を抑制しうる薬剤は、寿命を延長し、支持処置を助ける際に価値がありうる。腫瘍は、塞栓が解離し、血中に入ることにより転移する。効果的な転移が形成するために、腫瘍塞栓が、それらが移動している血管の内皮壁に結合しなければならない。図15におけるフローチャートにより示される通り、RHGPを使用して、付着を阻害又は促進するその能力について摂動細胞を照合することができる。付着を促進する候補標的の能力を予防する治療産物(例えば、候補に結合しうる抗体など)、又は付着の低下を増強する産物(例えば付着を阻害する候補の過剰発現又は投与など)は、同様に、転移の形成を遅延又は抑制しうる。
【0196】
図15のフローチャートに従った付着表現型のこのRHGP研究の適用は、研究のための複数の候補を産生している。主な候補を図16に同定しており、それらは、同定された候補だけでなく、付着、任意の同定された有用性又は機能などを促進又は低下させるか否かを反映する。これらの標的の各々での実行は、次に、効果的な癌処置薬剤である、追加の十分に耐容される薬剤を産生しうる。
【0197】
本特許出願を通して引用された参考文献、及び列挙された研究を、より良好な情報のために以下に列挙する。
【0198】
【表14】
【0199】
本願において開示する発明を、複数の具体的な実施例により例示し、ならびに、総称的に考察してきた。実施例(特定のベクターフラグメント、標的、及び手順を含む)は、限定することを意図しない。請求項において見出される具体的な列挙の非存在において、請求項は、任意の例示された実施態様に限定されるとは考えるべきではなく、開示の全ての局面と一致して読まれるべきである。
【特許請求の範囲】
【請求項1】
疾患状態に対処するための治療用薬物の開発のための標的を同定する方法であって、以下:
活性化因子を発現する哺乳動物細胞の集団(該細胞が宿主ゲノムを含む)を、誘導薬剤の非存在において不活性化される誘導可能プロモーターを含む遺伝子探索ベクターを含むウイルス粒子を用いて形質導入し、それにおいて該遺伝子探索ベクターが、該宿主ゲノム中の宿主遺伝子に、宿主遺伝子に対してセンス又は反対の方向で挿入され、該誘導薬剤の存在においてセンス又はアンチセンスRNAを生成し、それにより、該宿主遺伝子の発現が両方の対立遺伝子で抑制される又は該挿入により増強され、それにおいて十分な数の挿入事象が該集団において生じ、該ゲノムの実質的に全ての遺伝子座における該遺伝子探索ベクターの挿入を確実にすること;
哺乳動物細胞の形質導入集団を検証し、該疾患状態に固有の表現型を示す細胞を同定すること;
該遺伝子探索ベクター(GSV)の挿入点での各遺伝子配列を、該疾患状態に固有の表現型を示す細胞において同定すること;及び
該GSVの挿入点での各々の該遺伝子配列を特徴付け、該挿入により該ゲノムの摂動が、該遺伝子配列の過剰発現又は低下発現を起こしているか否かを決定し、それにおいて該配列が、該疾患状態に対処するための治療用薬物の開発のための標的として同定されること
を含む、方法。
【請求項2】
GSVの挿入点での遺伝子配列が、熱非対称インターレースポリメラーゼ連鎖反応(TAIL−PCR)の使用により特徴付けられる、請求項1記載の方法。
【請求項3】
疾患状態に固有の表現型を示す摂動細胞の同定に続き、該細胞を誘導薬剤の非存在において増殖させ、該誘導薬剤の非存在において該疾患状態に固有の該表現型を示さない細胞だけを、標的を同定する目的のためにさらに考慮する、請求項1記載の方法。
【請求項4】
ウイルス粒子が、GSVを用いてトランスフェクションされたレンチウイルス又はGSVを用いてトランスフェクションされたサルマウス白血病ウイルス(MMLV)のいずれかに基づく、請求項1記載の方法。
【請求項5】
ウイルスが、自己不活性化しており、GSVが、その挿入点から脱することができない、請求項4記載の方法。
【請求項6】
疾患状態に固有の表現型が、ウイルスにより攻撃された場合での生存であり、細胞を誘導薬剤の非存在において増殖させ、該ウイルスにより攻撃された場合、それらが生き残らない、請求項3記載の方法。
【請求項7】
ウイルスが、インフルエンザ、ヒト免疫不全ウイルス(HIV)、呼吸器合胞体ウイルス(RSV)、又はパラインフルエンザウイルスである、請求項6記載の方法。
【請求項8】
疾患状態に固有の表現型が、腫瘍細胞による転移特徴の獲得である、請求項1記載の方法。
【請求項9】
疾患状態に固有の表現型が、有毒化学療法薬物に対する耐性である、請求項1記載の方法。
【請求項10】
疾患状態に固有の表現型を示す摂動細胞の同定に続き、該細胞を誘導薬剤の非存在において及び有毒化学療法薬物の存在において増殖させ、該誘導薬剤の非存在において該有毒化学療法薬物に対して耐性である細胞を、治療用薬物の開発のための標的を反映しないとしてさらに特徴付けない、請求項3記載の方法。
【請求項11】
形質導入が、低い感染効率(MOI)を使用して行われ、形質導入された実質的に全ての細胞が、単一のGSV挿入事象を用いて形質導入されることを確実にする、請求項1記載の方法。
【請求項12】
MOIが0.2未満である、請求項11記載の方法。
【請求項13】
GSVが、少なくとも1つの遺伝子モチーフ又は検出可能な標識を含み、形質導入に続くGSVの挿入遺伝子座の同定を容易にする、請求項1記載の方法。
【請求項14】
細胞に、PTCH1遺伝子の発現を阻害する薬剤、又はPTCH1によりコードされるタンパク質に結合する薬剤を投与することを含む、哺乳動物細胞においてインフルエンザの感染性を阻害する方法。
【請求項15】
薬剤が、PTCH1タンパク質に対する抗体又はPTCH1タンパク質に結合する小分子を含む、請求項14記載の方法。
【請求項16】
医薬的に許容可能な担体中に懸濁されたPTCH1タンパク質に結合する抗体を含む、哺乳動物細胞においてインフルエンザの感染性を阻害するための組成物。
【請求項17】
哺乳動物細胞において、MRPL42、COX5A、TAPT1、又はSLCA25の少なくとも1つの発現レベルを低下させることを含む、哺乳動物細胞においてインフルエンザの感染性を阻害する方法。
【請求項18】
発現が、MRPL42、COX5A、TAPT1、又はSCLA25遺伝子の発現を阻害することにより低下される、請求項17記載の方法。
【請求項19】
方法が、細胞に、MRPL42タンパク質、COX5Aタンパク質、TAPT1タンパク質、又はSCLA25タンパク質に結合する薬剤を投与することを含む、請求項17記載の方法。
【請求項20】
薬剤が、抗体である、請求項19記載の方法。
【請求項21】
哺乳動物細胞においてRobo1の発現レベルを低下させることを含む、哺乳動物細胞においてHIVの感染性を阻害する方法。
【請求項22】
発現が、Robo1遺伝子の発現を阻害することにより低下される、請求項21記載の方法。
【請求項23】
方法が、細胞に、Robo1タンパク質に結合する薬剤を投与することを含む、請求項21記載の方法。
【請求項24】
薬剤が、抗体である、請求項23記載の方法。
【請求項25】
医薬的に許容可能な担体中に懸濁されたRobo1に対する抗体を含む、哺乳動物細胞においてHIVの感染性を低下させる際に効果的な組成物。
【請求項26】
哺乳動物細胞においてNedd4の発現レベルを低下させることを含む、哺乳動物細胞においてインフルエンザの感染性を阻害する方法。
【請求項27】
発現が、Nedd4をコードする遺伝子の発現を阻害することにより低下される、請求項26記載の方法。
【請求項28】
方法が、細胞に、Nedd4タンパク質に結合する薬剤を投与することを含む、請求項26記載の方法。
【請求項29】
薬剤が抗体である、請求項28記載の方法。
【請求項30】
Nedd4に対する抗体を医薬的に許容可能な担体中に含む、哺乳動物細胞においてインフルエンザ感染性を低下させる際に効果的な組成物。
【請求項1】
疾患状態に対処するための治療用薬物の開発のための標的を同定する方法であって、以下:
活性化因子を発現する哺乳動物細胞の集団(該細胞が宿主ゲノムを含む)を、誘導薬剤の非存在において不活性化される誘導可能プロモーターを含む遺伝子探索ベクターを含むウイルス粒子を用いて形質導入し、それにおいて該遺伝子探索ベクターが、該宿主ゲノム中の宿主遺伝子に、宿主遺伝子に対してセンス又は反対の方向で挿入され、該誘導薬剤の存在においてセンス又はアンチセンスRNAを生成し、それにより、該宿主遺伝子の発現が両方の対立遺伝子で抑制される又は該挿入により増強され、それにおいて十分な数の挿入事象が該集団において生じ、該ゲノムの実質的に全ての遺伝子座における該遺伝子探索ベクターの挿入を確実にすること;
哺乳動物細胞の形質導入集団を検証し、該疾患状態に固有の表現型を示す細胞を同定すること;
該遺伝子探索ベクター(GSV)の挿入点での各遺伝子配列を、該疾患状態に固有の表現型を示す細胞において同定すること;及び
該GSVの挿入点での各々の該遺伝子配列を特徴付け、該挿入により該ゲノムの摂動が、該遺伝子配列の過剰発現又は低下発現を起こしているか否かを決定し、それにおいて該配列が、該疾患状態に対処するための治療用薬物の開発のための標的として同定されること
を含む、方法。
【請求項2】
GSVの挿入点での遺伝子配列が、熱非対称インターレースポリメラーゼ連鎖反応(TAIL−PCR)の使用により特徴付けられる、請求項1記載の方法。
【請求項3】
疾患状態に固有の表現型を示す摂動細胞の同定に続き、該細胞を誘導薬剤の非存在において増殖させ、該誘導薬剤の非存在において該疾患状態に固有の該表現型を示さない細胞だけを、標的を同定する目的のためにさらに考慮する、請求項1記載の方法。
【請求項4】
ウイルス粒子が、GSVを用いてトランスフェクションされたレンチウイルス又はGSVを用いてトランスフェクションされたサルマウス白血病ウイルス(MMLV)のいずれかに基づく、請求項1記載の方法。
【請求項5】
ウイルスが、自己不活性化しており、GSVが、その挿入点から脱することができない、請求項4記載の方法。
【請求項6】
疾患状態に固有の表現型が、ウイルスにより攻撃された場合での生存であり、細胞を誘導薬剤の非存在において増殖させ、該ウイルスにより攻撃された場合、それらが生き残らない、請求項3記載の方法。
【請求項7】
ウイルスが、インフルエンザ、ヒト免疫不全ウイルス(HIV)、呼吸器合胞体ウイルス(RSV)、又はパラインフルエンザウイルスである、請求項6記載の方法。
【請求項8】
疾患状態に固有の表現型が、腫瘍細胞による転移特徴の獲得である、請求項1記載の方法。
【請求項9】
疾患状態に固有の表現型が、有毒化学療法薬物に対する耐性である、請求項1記載の方法。
【請求項10】
疾患状態に固有の表現型を示す摂動細胞の同定に続き、該細胞を誘導薬剤の非存在において及び有毒化学療法薬物の存在において増殖させ、該誘導薬剤の非存在において該有毒化学療法薬物に対して耐性である細胞を、治療用薬物の開発のための標的を反映しないとしてさらに特徴付けない、請求項3記載の方法。
【請求項11】
形質導入が、低い感染効率(MOI)を使用して行われ、形質導入された実質的に全ての細胞が、単一のGSV挿入事象を用いて形質導入されることを確実にする、請求項1記載の方法。
【請求項12】
MOIが0.2未満である、請求項11記載の方法。
【請求項13】
GSVが、少なくとも1つの遺伝子モチーフ又は検出可能な標識を含み、形質導入に続くGSVの挿入遺伝子座の同定を容易にする、請求項1記載の方法。
【請求項14】
細胞に、PTCH1遺伝子の発現を阻害する薬剤、又はPTCH1によりコードされるタンパク質に結合する薬剤を投与することを含む、哺乳動物細胞においてインフルエンザの感染性を阻害する方法。
【請求項15】
薬剤が、PTCH1タンパク質に対する抗体又はPTCH1タンパク質に結合する小分子を含む、請求項14記載の方法。
【請求項16】
医薬的に許容可能な担体中に懸濁されたPTCH1タンパク質に結合する抗体を含む、哺乳動物細胞においてインフルエンザの感染性を阻害するための組成物。
【請求項17】
哺乳動物細胞において、MRPL42、COX5A、TAPT1、又はSLCA25の少なくとも1つの発現レベルを低下させることを含む、哺乳動物細胞においてインフルエンザの感染性を阻害する方法。
【請求項18】
発現が、MRPL42、COX5A、TAPT1、又はSCLA25遺伝子の発現を阻害することにより低下される、請求項17記載の方法。
【請求項19】
方法が、細胞に、MRPL42タンパク質、COX5Aタンパク質、TAPT1タンパク質、又はSCLA25タンパク質に結合する薬剤を投与することを含む、請求項17記載の方法。
【請求項20】
薬剤が、抗体である、請求項19記載の方法。
【請求項21】
哺乳動物細胞においてRobo1の発現レベルを低下させることを含む、哺乳動物細胞においてHIVの感染性を阻害する方法。
【請求項22】
発現が、Robo1遺伝子の発現を阻害することにより低下される、請求項21記載の方法。
【請求項23】
方法が、細胞に、Robo1タンパク質に結合する薬剤を投与することを含む、請求項21記載の方法。
【請求項24】
薬剤が、抗体である、請求項23記載の方法。
【請求項25】
医薬的に許容可能な担体中に懸濁されたRobo1に対する抗体を含む、哺乳動物細胞においてHIVの感染性を低下させる際に効果的な組成物。
【請求項26】
哺乳動物細胞においてNedd4の発現レベルを低下させることを含む、哺乳動物細胞においてインフルエンザの感染性を阻害する方法。
【請求項27】
発現が、Nedd4をコードする遺伝子の発現を阻害することにより低下される、請求項26記載の方法。
【請求項28】
方法が、細胞に、Nedd4タンパク質に結合する薬剤を投与することを含む、請求項26記載の方法。
【請求項29】
薬剤が抗体である、請求項28記載の方法。
【請求項30】
Nedd4に対する抗体を医薬的に許容可能な担体中に含む、哺乳動物細胞においてインフルエンザ感染性を低下させる際に効果的な組成物。
【図1】


【図2】
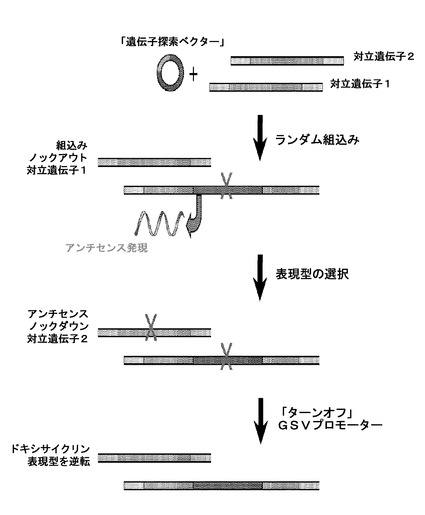
【図3A】

【図3B】
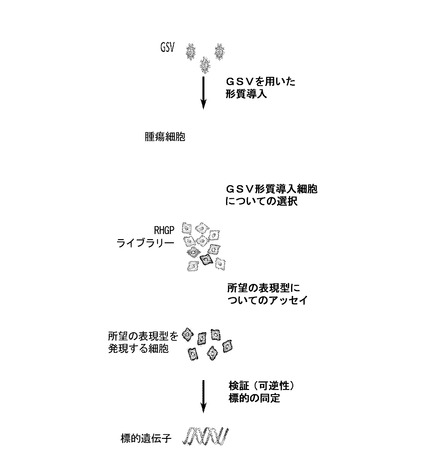
【図4】
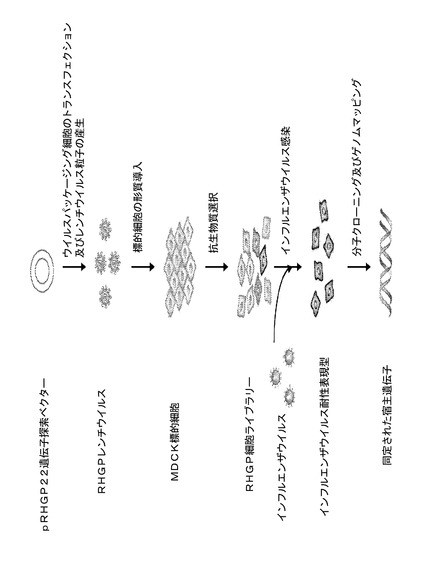
【図5A】
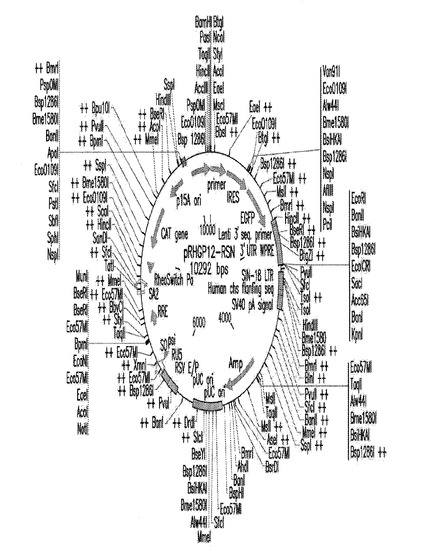
【図5B】
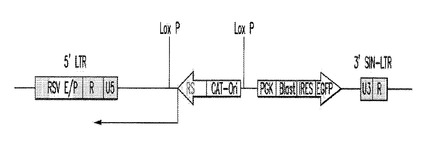
【図6A】
【図6B】

【図6C】
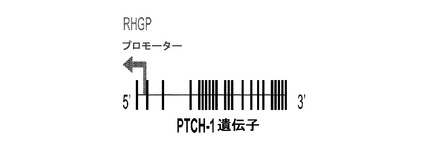
【図7】

【図8A】
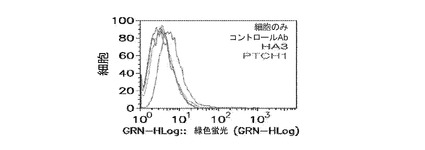
【図8B】
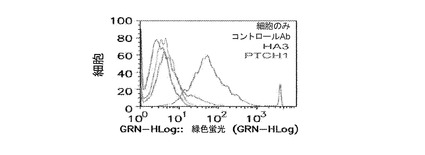
【図8C】
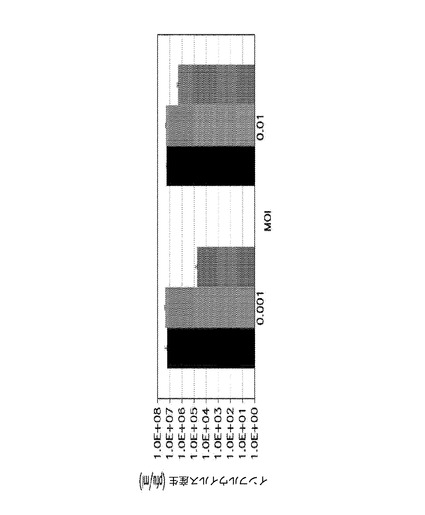
【図9A】
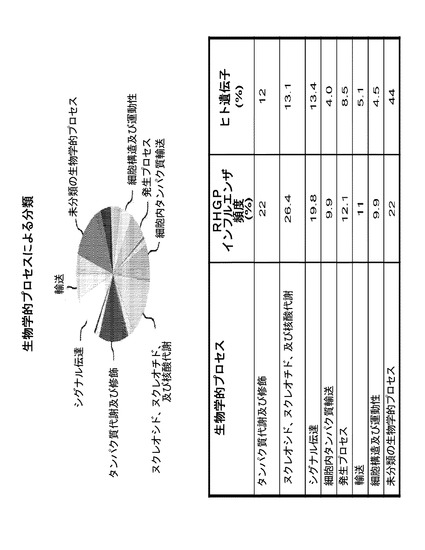
【図9B】
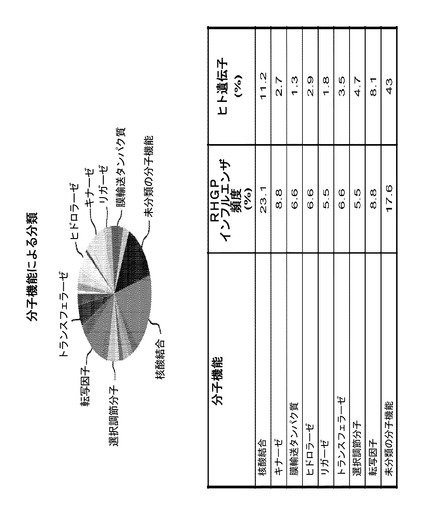
【図10】
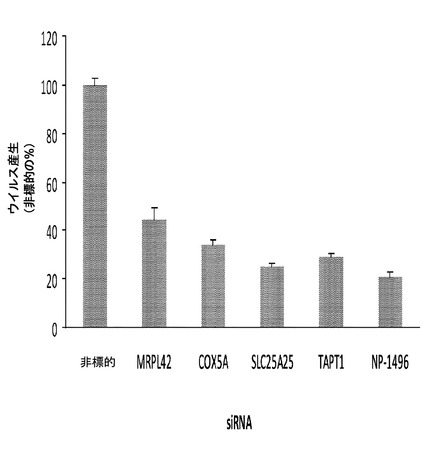
【図11A】
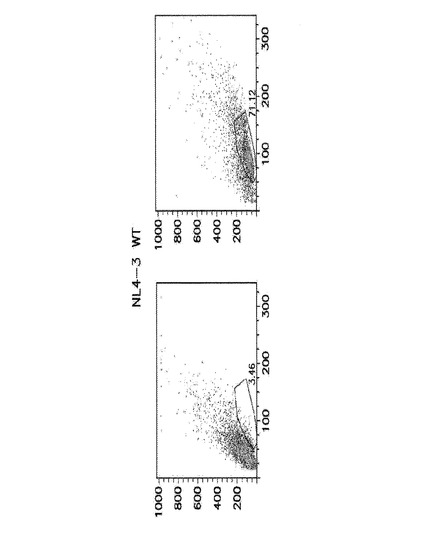
【図11B】

【図11C】
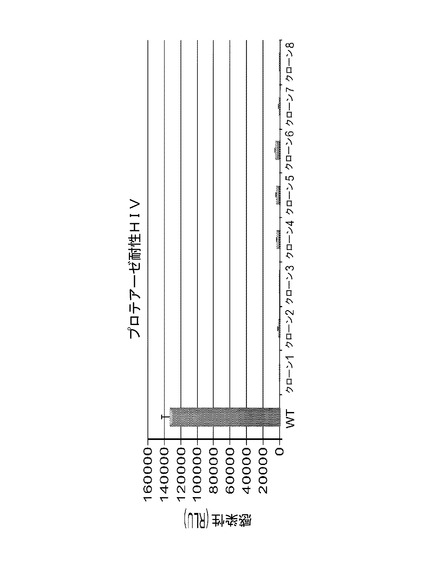
【図12A】
【図12B】
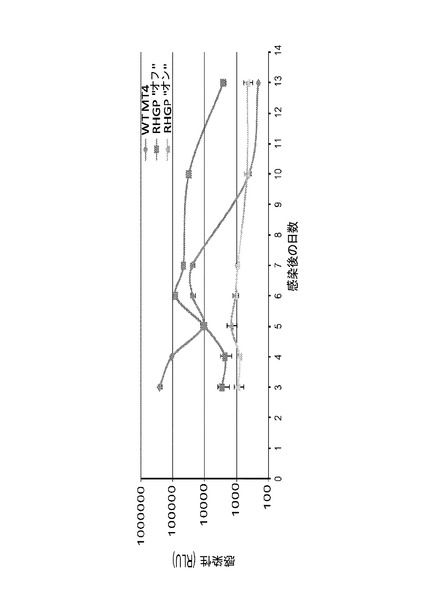
【図13】

【図14】
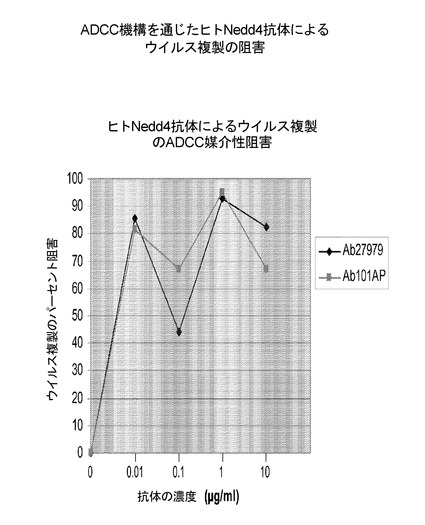
【図15】
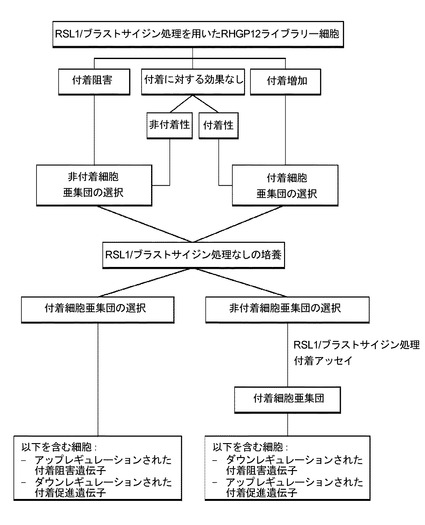
【図16A】
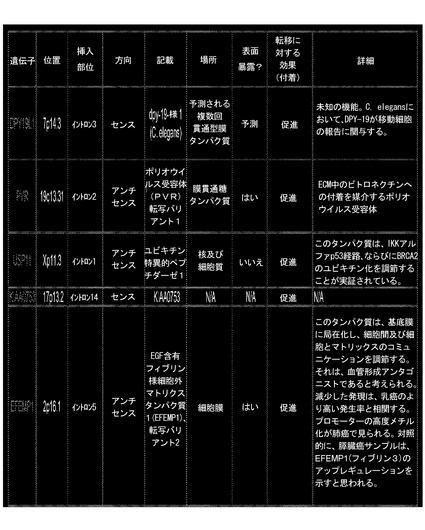
【図16B】

【図2】
【図3A】
【図3B】
【図4】
【図5A】
【図5B】
【図6A】
【図6B】
【図6C】
【図7】
【図8A】
【図8B】
【図8C】
【図9A】
【図9B】
【図10】
【図11A】
【図11B】
【図11C】
【図12A】
【図12B】
【図13】
【図14】
【図15】
【図16A】
【図16B】
【公開番号】特開2012−167093(P2012−167093A)
【公開日】平成24年9月6日(2012.9.6)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−71170(P2012−71170)
【出願日】平成24年3月27日(2012.3.27)
【分割の表示】特願2011−544669(P2011−544669)の分割
【原出願日】平成22年1月6日(2010.1.6)
【出願人】(509133377)ファンクショナル・ジェネティクス・インコーポレーテッド (7)
【氏名又は名称原語表記】FUNCTIONAL GENETICS,INC.
【Fターム(参考)】
【公開日】平成24年9月6日(2012.9.6)
【国際特許分類】
【出願番号】特願2012−71170(P2012−71170)
【出願日】平成24年3月27日(2012.3.27)
【分割の表示】特願2011−544669(P2011−544669)の分割
【原出願日】平成22年1月6日(2010.1.6)
【出願人】(509133377)ファンクショナル・ジェネティクス・インコーポレーテッド (7)
【氏名又は名称原語表記】FUNCTIONAL GENETICS,INC.
【Fターム(参考)】
[ Back to top ]