
特定レクチン沈殿法を用いて糖タンパク質を同定する方法

本発明は、腫瘍の発生又は転移に関与する多様な糖タンパク質におけるグリコシル化の変化を測定する分析方法と関連したものである。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、腫瘍発生及び癌転移過程でグリコシル化の変化を示す糖タンパク質を検出する方法であって、より詳細には、癌の発生及び進行の間に修飾されるN−連結型糖タンパク質の質量分析方法に基づく同定に関するものである。
【背景技術】
【0002】
2次元電気泳動は、MALDI−TOF(Matrix Assisted Laser Desorption/Ionization Time−of−Flight)質量分析機及びアミノ酸配列分析機などの非常に複雑な分析機器と連係すれば、容易にかつ高い処理率でタンパク質を確認して特徴付けることができ、このような発展は、「ポストゲノム、プロテオミクス時代」を可能にした。しかし、プロテオミクス的接近は、生物学的力動性を示すより、プロテオームの停止状態を示すという限界があった。実際に、細胞内の信号伝達経路は、一般的にタンパク質の発現水準及び翻訳後修飾(post−translational modification)面で力動性を示す。
さらに、タンパク質が細胞内に少量しか存在していないので、信号伝達経路上のタンパク質を2次元ゲル上にディスプレイしてモニタすることはしばしば難しい。タンパク質のグリコシル化は、このような限界を克服し、細胞を力動性に基づいて観察することを可能にする。特定レクチンでグリコシル化の変化を追跡すれば、単純染色では識別不可能なタンパク質水準の変化が容易にモニタされる。
最近、このような「グリコミクス」と呼ばれる方法は、伝統的なプロテオミクスの限界を克服し、有望な分野として注目されている。グリコミクスは、主に翻訳後修飾の主要な種類であるタンパク質のグリコシル化での変化を追跡することに基づいている。
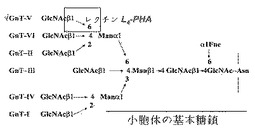
生物学的撹乱のうち一つがタンパク質の異常グリコシル化(aberrant glycosylation)であって、これは、特定の腫瘍遺伝子の信号によって誘導された後、細胞接着、細胞−細胞認識で機能障害を引き起こし、結果として、癌発生及び腫瘍悪化を引き起こす(Hakomori and Kannagi,1983,J.Natl.Cancer Inst.,71:231−251;Feizi,1985,Nature,314:53−571)。成熟されたmRNAは、小胞体上で翻訳され、小胞体でN−連結型グリコシル化の基本的な糖鎖部分が作られる。その後、糖タンパク質は、ゴルジ体に移動し、図1に示すような糖転移酵素によって追加的な糖鎖が付着される。各糖転移酵素は、ras、rafなどの特定の腫瘍遺伝子によって発現される信号によって活性化される。注目される糖転移酵素のうち一つがFUT8であるが、これは、N−糖鎖の基本N−アセチルグルコサミン(GlcNAc)にフコースを付加することを触媒する。アルファ−フェトプロテイン(アルファ−fetoprotein;AFP)とアンチトリプシン(antitrypsin)は、腫瘍悪性化でフコシル化の重要性を実証するものとして広く知られている(Miyoshi,E.,Ko,J.H.et al.(1999)Biochim.Biophys.Acta 1473:9−20)。フコシル化を担当するFUT8は、日本の谷口(N.Taniguchi)研究チームによって精製され、そのcDNAがクローン化された。また、fut8−細胞感染されたHep3B(Hep3B−FT)を免疫欠乏マウス(athymic nude mice)の脾臓に注射したとき、母体細胞に比べて肝での腫瘍発生が著しく抑制された(Miyoshi,E.,Ko,J.H.et al.(1999)Cancer Research 59:2237−2243)。レンズマメレクチン(LCA)は、N−糖鎖の最も内側のGlcNAcに付着されたフコースに結合され、ConAのように基本N−糖鎖にも結合される。
N−アセチルグルコサミン転移酵素V(GnT−V)によって誘導される異常グリコシル化は、腫瘍の侵潤と転移に関与するタンパク質修飾の代表的な例である(Dennis et al.1987,Science,236:582−5853)。小胞体上で基本糖鎖の付着後に付加される糖鎖は、主に6種類のN−アセチルグルコサミン転移酵素 (I−VI)によってゴルジ体で付加される。これらのうち、GnT−Vは、標的糖タンパク質を修飾し、腫瘍細胞の認識及び細胞付着特性を変化させることによって、腫瘍細胞の転移能力に関与するものと知られている。
GnT−Vは、ベータ1,6側鎖が腫瘍組織の成長又は転移時に著しく発生するというデニスなど(1987)の報告で初めて知られた(Dennis et al.1987,Science,236:582−5853)。細胞表面タンパク質gp130は、GnT−Vの一つの又は主要な標的タンパク質であって、ベータ1,6−N−アセチルグルコサミンを加えたときに高い腫瘍転移活性を示す。胚性幹細胞株においてGnT−Vが欠損したGnt−Vノックアウトマウスを構築し、癌を誘発させるために、ポリオーマウイルスの中型T抗原(以下、「PyMT」と称する)ウィルス性腫瘍遺伝子を導入した。結果として、PyMTによって誘導された腫瘍の成長及び転移は、PyMTのみが過剰発現された他の正常マウスグループと比べたとき、GnT−Vノックアウトマウスでは著しく抑制され(Granovsky et al.2000,Nature Med.,6:306−312)、乳房癌マウスでのベータ1,6側鎖の成長は、特に高い転移の原因となった。上述したように、GnT−Vは、多様な組織で腫瘍転移に関与し、高い腫瘍転移活性を示す。GnT−Vは、ヒトの肺腫瘍細胞及びマウスの腎臓で精製され、cDNAがクローン化され、プロモータ及び遺伝子構造が明らかになった(Gu et al.1993,J Biochem,113:614−619;Soreibah et al.,1993,J Biol.Chem.,268:15381−15385;Kang et al.,1996,J.Biol.Chem.,271:26706−26712)。本発明者は、転写酵素ets−1がGnT−Vの発現に関与すると報告した(Ko,et al.1999,J Biol.chem.,274(33):22941−22948)。薬剤を基本にする腫瘍治療の限界は、腫瘍に対する医学研究分野で治療可能な段階の早期発見へとパラダイムの変化を引き起こし、異常グリコシル化は、腫瘍早期発見マーカーとしての利用可能性が試験されている。
これによって、本発明者らは、GnT−Vが過剰発現される腫瘍細胞から2次元電気泳動及び質量分析を用いてGnT−Vによってベータ1,6−GlcNAcを有する候補タンパク質の目録を報告した(Kim et al.,2004,Proteomics,4(11):3353−3358;Kim et al.,2006,Proteomics,6(4):1187−1191)。しかし、従来のゲルに基づいた分析方法によれば、少量のタンパク質を同定するにおいて、本質的に偽陽性(false−positive)が起こる確率が高い。これを克服するために、本発明者らは、同定のためにL−PHAレクチンを用いてベータ1,6−GlcNAcを有する糖タンパク質を濃縮することによって本発明を完成した。
【発明の概要】
【発明が解決しようとする課題】
【0003】
本発明の目的は、発癌及び腫瘍転移に関与する異常糖タンパク質と呼ばれる変化した糖構造を獲得した糖タンパク質を高精度で同定する方法を提供することである。
本発明の他の目的は、発癌及び腫瘍転移に関与する異常糖タンパク質をより簡単な方法で探知して同定することである。
【課題を解決するための手段】
【0004】
前記目的を達成するために、本発明者らは、ベータ1,6−N−アセチルグルコサミン部分を有するN−連結型糖タンパク質を認識するレクチンであるL−PHAを用いて調査することによって、ベータ1,6−N−アセチルグルコサミンを有する異常糖タンパク質を探知する方法を開発した。
【発明の効果】
【0005】
本発明は、腫瘍発生及び転移に関与するベータ1,6−N−アセチルグルコサミンを有する異常糖タンパク質をレクチン沈殿で濃縮する方法を提供することによって、少量のタンパク質も探知することができる。そして、本発明は、全ての腫瘍に適用することができ、尿及び血液を含む生体材料に適用することができる。
本発明は、レクチン沈殿法を用いて癌の発生及び転移に関与する異常グリコシル化を探知する分析方法を提供し、腫瘍診断マーカー候補として異常糖タンパク質の目録を提供する
【図面の簡単な説明】
【0006】
【図1】多様な糖転移酵素によって形成されるN−連結型糖鎖連結構造及びL−PHAが結合されるベータ1,6−N−アセチルグルコサミン部位を示す図である。
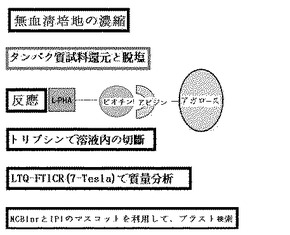
【図2】実施例1に記載された方法の概略図である。
【図3】実施例1の方法によって同定されたタンパク質を示すもので、非糖タンパク質が多く、陽性対照群TIMP−1が探知されていないことを示す図である。
【図4】実施例2に記載された方法の概略図である。
【図5】実施例2の方法によって同定されたタンパク質を示すもので、低いスコア値の糖タンパク質が比較的多く、TIMP−1が依然として探知されていないことを示す図である。
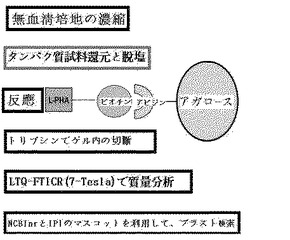
【図6】実施例3に記載された方法の概略図である。
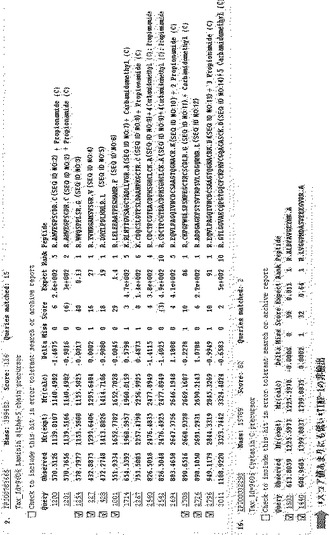
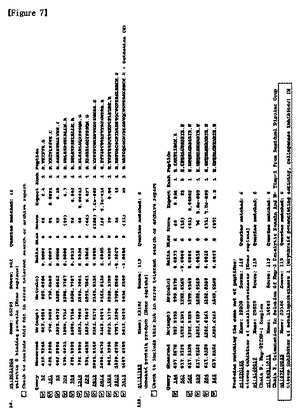
【図7】実施例3の方法によって同定されたタンパク質を示すもので、TIMP−1を含む多くの糖タンパク質が高いスコア値で現れたことを示す図である。
【図8】実施例3の方法と従来のレクチンブロット分析で共通的に確認されるタンパク質を示す図である。
【図9】実施例3の方法で確認され、従来のレクチンブロット分析では確認されていないタンパク質を示す図である。
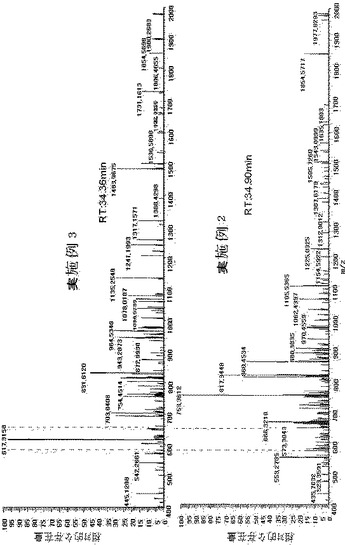
【図10】実施例2及び実施例3の結果を比較したもので、質量分析ピークが34.36分の滞留時間で実施例3の方法のみによって探知されることを示す図である。
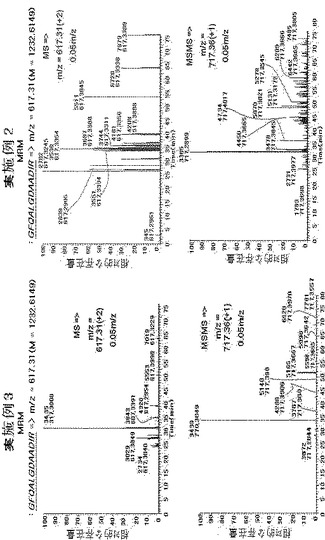
【図11】多重反応モニタリング(MRM)の結果として、TIMP−1ペプチドが実施例3のみで探知されることを示す図である。
【発明を実施するための形態】
【0007】
本発明は、癌の発生及び転移に関与する異常グリコシル化を有する糖タンパク質をレクチン沈殿法で高精度で分析する方法に関するものである。
前記癌の種類は、大腸癌、胃癌、肺癌、肝癌、子宮癌、乳房癌及び膵臓癌からなる群から選択されるが、上述した癌のみに限定されることはなく、癌の全ての種類に拡大することができる。
本発明は、癌細胞からベータ1,6−N−アセチルグルコサミン部分を獲得したN−連結型糖タンパク質を同定する高精度な分析方法を提供する。ベータ1,6−N−アセチルグルコサミンの形成を触媒するGnT−Vは、多様な腫瘍組織で幅広く発現され、その病理現象に関与する。ベータ1,6−N−アセチルグルコサミンがフィトヘマグルチニン−L4(以下、「L4−PHA」又は「L−PHA」と略称する)によって認識されるので、ビオチン標識されたL−PHAは、アビジン−アガロースビードに非共有結合で結合される。この複合体は、タンパク質の調合液からベータ1,6−N−アセチルグルコサミンを有する糖タンパク質を濃縮するのに用いられる。
本発明の一側面は、タンパク質をトリプシン処理し、7−テスラのフーリエ変換イオン共鳴型質量分析機(Fourier Transform Ion Cyclotron Resonance;FTICR)を用いたアミノ酸配列決定に適用し、ベータ1,6−N−アセチルグルコサミンを有する少量の糖タンパク質を同定する方法を提供する。
本発明は、リガンドが結合されたL4−PHAをリガンド受容体が付着されたグリカンビードに反応させ、L4−PHA−グリカンビード複合体を形成する段階と、糖タンパク質を得るために、試料を前記L4−PHA−グリカンビード複合体と反応させる段階と、前記糖タンパク質をゲル電気泳動で展開する段階と、前記糖タンパク質を含む結果物のゲルをペプチドにゲル内部で(in−gel)切断する段階と、前記切断されたペプチドを質量分析機で分析する段階とを含む糖タンパク質の分析方法を提供する。
本発明の他の側面は、前記グリカンビードがアガロースビード及びセファロースビードで構成されたグループから選択される前記方法を提供する。
本発明の他の側面は、前記リガンド及びリガンド受容体がビオチン及びアビジンである前記方法を提供する。
本発明の他の側面は、前記試料が自然状態又は変性形態である前記方法を提供する。
本発明の他の側面は、前記変性形態がベータ−メルカプトエタノール、ヨード酢酸及びヨードアセトアミドからなるグループから選択された少なくとも一つによって生成される前記方法を提供する。
本発明の他の側面は、対照区及び実験区の培養細胞株を、リガンドが結合されたL4−PHAをリガンド受容体が付着されたグリカンビードに結合することによって形成されるL4−PHA−グリカンビード複合体にそれぞれ加え、ベータ1,6−N−アセチルグルコサミンに対するグリカン構造において差を示す糖タンパク質を探索する方法を提供する。
本発明の他の側面は、正常な血液及び実験区の血液を、リガンドが結合されたL4−PHAをリガンド受容体が付着されたグリカンビードに結合することによって形成されるL4−PHA−グリカンビード複合体にそれぞれ加え、ベータ1,6−N−アセチルグルコサミンに対するグリカン構造において差を示す糖タンパク質を探索する方法を提供する。
本発明の更に他の側面は、アルブミン、グロブリン及びトランスフェリンのような血液内に豊富に存在するタンパク質によって、最小化した阻害を有する従来に発見された腫瘍バイオマーカー候補を確定する同定方法を提供し、確認効率を最大化するためにSISCAPA(Stable Isotope Standards and Capture by Anti−peptide Antibodies:Pearson et al,2004,J.roteome Res.3,235−44)方法と組み合わせることができる。
腫瘍の転移は、細胞認識及び接着の機能障害から発生し、数多くの膜タンパク質又は分泌タンパク質がこの過程に関与している。これは、バイオマーカーが存在する血液及び尿などの体液をバイオマーカー探知用試料として利用できることを示す。大腸癌−特異的バイオマーカーを探すために、本発明者らはグリコミクス研究を行い、GnT−V遺伝子は、GnT−V発現水準の低い大腸癌細胞株であるWiDr内に安定的に浸透された。培養培地で得たタンパク質は、2次元電気泳動ゲル上で展開するが、これらは、クーマシーブリリアントブルー又はレクチンブロット分析で染色され、対照群及び浸透WiDr細胞間に異なるグリカン構造を示すタンパク質が選択された(表1参照)。しかし、レクチンブロット分析自体が非特異的結合又はゲル内の汚染タンパク質を含む可能性があるので、付加的な確認段階が必要である。さらに、2次元電気泳動ゲルは分解能に限界を有しており、結果として、少量のタンパク質は、ゲルに基づいたシステム下では確認が難しい。このような限界を克服するために、本発明者らは、ベータ1,6−N−アセチルグルコサミンが含まれた糖タンパク質を特定レクチンを用いて直接探知できる方法を発明した。
[表1−1]

[表1−2]
【0008】
a;配列範囲は、質量分析機で確認されたペプチドの百分率を意味する。
b;総値は、個別ペプチドから得られたスコア値の合計である。スコアは−10xLog(P)であって、Pは、観察された符合が偶然の事件である確率である。これは、MS/MSデータとしてMASCOTサーチプログラムを用いたNCBInrデータベースに基づく。
c;分子量Mr及び等電点pIは、グリカン残基を計算に考慮していないときの理論値である。
L−PHAは、ベータ1,6−N−アセチルグルコサミンが含まれた糖タンパク質に非常に特異的に結合されるが、利用可能な溶出方法がないので、結合されたタンパク質を剥がすことが難しかった。この問題を解決するために、本発明者らは、L−PHA−結合されたタンパク質をSDS−PAGEゲル上に最小に展開し、ゲルは、切断し、タンパク質同定のためにトリプシン処理した。
【0009】
以下、本発明の多様な側面と具体例を下記の実施例を通してより具体的に説明する。
【実施例1】
【0010】
レクチンを用いたベータ1,6−N−アセチルグルコサミンが含まれた糖タンパク質の濃縮
GnT−V発現の低いWiDr:mock細胞及びGnT−Vを浸透させたWiDr:GnT−Vをモデル細胞株として構築し、調節された無血清培地を細胞株から収去した。腫瘍の転移が細胞認識及び接着の機能障害によって起こること(Egeblad et al.,2002,Nature Rev.2:161−174;Kannagi et al.,2004,Cancer Sci.95(5):377−384)と、多くの膜タンパク質又は分泌タンパク質がこの過程に関与することが知られている。GnT−Vによって生成されるベータ1,6−N−アセチルグルコサミンが含まれた糖タンパク質を培地から精製した。収集された培地は濃縮し、1mgのタンパク質をアビジン−アガロースビードに結合させ非特異的に結合するタンパク質を予め除去した。非特異的に結合したタンパク質を予め除去したタンパク質は、L−PHA−ビオチン−アビジン−アガロースビード複合体と一晩中4℃で反応させた。マンノース−1,6−N−アセチルグルコサミンがL−PHA結合タンパク質を剥がすのに用いられるが、この二糖類は価格面や効率面で証明されていない。ある研究チームでは、酸性溶出方法で剥がすことを試みたが、この方法も証明されておらず、低いpHはタンパク質の完全性に損傷を与える。このような理由により、タンパク質は、複合体に結合された状態で切断した。このために、タンパク質複合体を過量のPBS(Phosphate buffered saline)緩衝液で洗浄し、10Uのトリプシン(Promega社)で一晩中37℃でトリプシン処理した。切断されたペプチドは、蒸留水とアセトニトリルで抽出し、真空乾燥し、7テスラのFTICR/LTQ質量分析機で質量分析した。タンパク質は、Mascotサーチプログラム(バージョン2.0)でペプチド配列を決定して確認した(図2及び図3)。
サーチデータベースは、NCBInr 20051212(taxonomy;human、entries;103、913 human sequence entries)であった。その結果、ペプチドは、効果的に配列が決定されておらず、完全なタンパク質がトリプシンによって充分に切断されていないと考えられる。TIMP−1(Tissue inhibitor of metalloproteinase 1)は、GnT−Vの標的タンパク質として既に確認されており、変更されたグリカン構造が質量分析で決定され、参考分子としてTIMP−1が用いられる。しかし、TIMP−1は、この方法では確認されておらず、その代わりに、糖タンパク質でない多くのタンパク質が確認された。
【実施例2】
【0011】
レクチンを用いたベータ1,6−N−アセチルグルコサミンが含まれた糖タンパク質の濃縮段階以前のタンパク質還元及び脱塩
実施例1に示された非特異的結合を減少させて敏感性を増大させるために、収集された培地から得たタンパク質を1%(v/v)ベータ−メルカプトエタノールに還元させ、未反応ベータ−メルカプトエタノール及び塩は、ゲルろ過カラムで除去した(図4)。1mgのタンパク質を沈殿させ、実施例1に記載したように確認した。結果は、より多くの糖タンパク質が確認されたことを示すが、スコア値が低く、TIMP−1も確認されなかった。
【実施例3】
【0012】
SDS−PAGEゲルで最大化されたタンパク質切断及びレクチン沈殿以後、ベータ1,6−N−アセチルグルコサミンが含まれた糖タンパク質確認
実施例1に記載されたように培養培地から収集されたタンパク質を、実施例2に記載されたように1%(v/v)ベータ−メルカプトエタノールに還元させ、未反応ベータ−メルカプトエタノール及び塩は、ゲルろ過カラムで除去した。1mgのタンパク質をアビジン−アガロースビードと1時間反応させて先に精製し、先に精製されたタンパク質は、L−PHA−ビオチン−アビジン−アガロースビード複合体と一晩中4℃で反応させた。PBSで洗浄した後、結合されたタンパク質に1×SDS−PAGE変性緩衝液を加えて少しの間沸騰させてビード複合体から分離した。タンパク質は、8%SDS−PAGEゲル上で分離され、ゲル内でトリプシンで切断された。切断されたペプチドは、ゲルで溶出され、アジレント1100ナノフローシステム(Agilent Technologies)を用いて分析され、7テスラのFT−LTQ質量分析機(Thermo Electron)と連結されたSurveyorシステムで配列が分析された。ペプチドは、10cmのReproSil−Pur C18−AQ樹脂(Ammerbuch−Entringen)で充填されたシリカカラム(75mm)上で結合されて分離された。ペプチド混合物は、100mm−C18カラム(5μm)上にオートサンプラ(Surveyor)で20ml/分の速度でローディングされた。移動相AとBは、それぞれ0%及び100%のACNで構成され、それぞれ0.1%のギ酸を含む。濃度勾配は、15分間5%Bから初めて、3分間20%B、47分間50%B、最終的に95%に上昇した。溶出されたペプチドは、直接質量分析機にエレクトロスプレーした。質量分析機は、Xcaliburソフトウェア(Thermo Electron Version 1.4SR1)で制御した。図7は、一つの結果を示す。この実施例によれば、1)高いスコア値(P<0.05)を有する糖タンパク質を確認することができ、2)図7のようにTIMP−1を探知することができ、3)従来のレクチンブロットに基づいた同定方法で確認されたタンパク質を再び確認することができ(図8)、4)高いスコア値を有しながら低く発現されるタンパク質を探知することができた(図9)。
【実施例4】
【0013】
レクチン沈殿法に基づいて同定されたタンパク質を質量分析方法で確認した。
実施例2と実施例3を比較するために、既に探知されたTIMP−1ペプチド、GFQALGDAADIRを質量分析機でモニタした。図10のように、GFQALGDAADIRは、実施例3では、分子量が約1232Daであって、2電荷を帯びた状態で617.31m/zであった。実施例3では、ペプチドは滞留時間34分で探知されたが、実施例2では探知されず、ペプチド配列はGFQALGDAADIRであった。また、図3によれば、実施例3では、m/z617.31で探知されるペプチドの親イオン及びm/z 717.36のフラグメントイオンが示されるが、実施例2ではこれが全く探知されておらず、これは、トリプシン切断と抽出がよく行われていないことに起因すると予想される。
したがって、本発明は、グリコシル化で変化を示す少量のタンパク質を確認するための2次元電気泳動に基づいたより敏感な方法を提供する。
【産業上の利用可能性】
【0014】
本発明によれば、正常対照群と試験試料との間に水準及び構造の差を示す糖タンパク質を区分することによって、早期に癌を容易に診断することができる。
(配列目録テキスト)
<110> Korea Research Institute of Bioscience and Biotechnology
<120> An identification method of glycoproteins using a specific lectin precipitation
<130> jhko-0711-lectinaffi2
<150> KR10-2007-100916
<151> 2007-10-08
<160> 1
<170> KopatentIn 1.71
<210> 1
<211> 12
<212> PRT
<213> Homo sapiens
<400> 1
Gly Phe Gln Ala Leu Gly Asp Ala Ala Asp Ile Arg
1 5 10
【技術分野】
【0001】
本発明は、腫瘍発生及び癌転移過程でグリコシル化の変化を示す糖タンパク質を検出する方法であって、より詳細には、癌の発生及び進行の間に修飾されるN−連結型糖タンパク質の質量分析方法に基づく同定に関するものである。
【背景技術】
【0002】
2次元電気泳動は、MALDI−TOF(Matrix Assisted Laser Desorption/Ionization Time−of−Flight)質量分析機及びアミノ酸配列分析機などの非常に複雑な分析機器と連係すれば、容易にかつ高い処理率でタンパク質を確認して特徴付けることができ、このような発展は、「ポストゲノム、プロテオミクス時代」を可能にした。しかし、プロテオミクス的接近は、生物学的力動性を示すより、プロテオームの停止状態を示すという限界があった。実際に、細胞内の信号伝達経路は、一般的にタンパク質の発現水準及び翻訳後修飾(post−translational modification)面で力動性を示す。
さらに、タンパク質が細胞内に少量しか存在していないので、信号伝達経路上のタンパク質を2次元ゲル上にディスプレイしてモニタすることはしばしば難しい。タンパク質のグリコシル化は、このような限界を克服し、細胞を力動性に基づいて観察することを可能にする。特定レクチンでグリコシル化の変化を追跡すれば、単純染色では識別不可能なタンパク質水準の変化が容易にモニタされる。
最近、このような「グリコミクス」と呼ばれる方法は、伝統的なプロテオミクスの限界を克服し、有望な分野として注目されている。グリコミクスは、主に翻訳後修飾の主要な種類であるタンパク質のグリコシル化での変化を追跡することに基づいている。
生物学的撹乱のうち一つがタンパク質の異常グリコシル化(aberrant glycosylation)であって、これは、特定の腫瘍遺伝子の信号によって誘導された後、細胞接着、細胞−細胞認識で機能障害を引き起こし、結果として、癌発生及び腫瘍悪化を引き起こす(Hakomori and Kannagi,1983,J.Natl.Cancer Inst.,71:231−251;Feizi,1985,Nature,314:53−571)。成熟されたmRNAは、小胞体上で翻訳され、小胞体でN−連結型グリコシル化の基本的な糖鎖部分が作られる。その後、糖タンパク質は、ゴルジ体に移動し、図1に示すような糖転移酵素によって追加的な糖鎖が付着される。各糖転移酵素は、ras、rafなどの特定の腫瘍遺伝子によって発現される信号によって活性化される。注目される糖転移酵素のうち一つがFUT8であるが、これは、N−糖鎖の基本N−アセチルグルコサミン(GlcNAc)にフコースを付加することを触媒する。アルファ−フェトプロテイン(アルファ−fetoprotein;AFP)とアンチトリプシン(antitrypsin)は、腫瘍悪性化でフコシル化の重要性を実証するものとして広く知られている(Miyoshi,E.,Ko,J.H.et al.(1999)Biochim.Biophys.Acta 1473:9−20)。フコシル化を担当するFUT8は、日本の谷口(N.Taniguchi)研究チームによって精製され、そのcDNAがクローン化された。また、fut8−細胞感染されたHep3B(Hep3B−FT)を免疫欠乏マウス(athymic nude mice)の脾臓に注射したとき、母体細胞に比べて肝での腫瘍発生が著しく抑制された(Miyoshi,E.,Ko,J.H.et al.(1999)Cancer Research 59:2237−2243)。レンズマメレクチン(LCA)は、N−糖鎖の最も内側のGlcNAcに付着されたフコースに結合され、ConAのように基本N−糖鎖にも結合される。
N−アセチルグルコサミン転移酵素V(GnT−V)によって誘導される異常グリコシル化は、腫瘍の侵潤と転移に関与するタンパク質修飾の代表的な例である(Dennis et al.1987,Science,236:582−5853)。小胞体上で基本糖鎖の付着後に付加される糖鎖は、主に6種類のN−アセチルグルコサミン転移酵素 (I−VI)によってゴルジ体で付加される。これらのうち、GnT−Vは、標的糖タンパク質を修飾し、腫瘍細胞の認識及び細胞付着特性を変化させることによって、腫瘍細胞の転移能力に関与するものと知られている。
GnT−Vは、ベータ1,6側鎖が腫瘍組織の成長又は転移時に著しく発生するというデニスなど(1987)の報告で初めて知られた(Dennis et al.1987,Science,236:582−5853)。細胞表面タンパク質gp130は、GnT−Vの一つの又は主要な標的タンパク質であって、ベータ1,6−N−アセチルグルコサミンを加えたときに高い腫瘍転移活性を示す。胚性幹細胞株においてGnT−Vが欠損したGnt−Vノックアウトマウスを構築し、癌を誘発させるために、ポリオーマウイルスの中型T抗原(以下、「PyMT」と称する)ウィルス性腫瘍遺伝子を導入した。結果として、PyMTによって誘導された腫瘍の成長及び転移は、PyMTのみが過剰発現された他の正常マウスグループと比べたとき、GnT−Vノックアウトマウスでは著しく抑制され(Granovsky et al.2000,Nature Med.,6:306−312)、乳房癌マウスでのベータ1,6側鎖の成長は、特に高い転移の原因となった。上述したように、GnT−Vは、多様な組織で腫瘍転移に関与し、高い腫瘍転移活性を示す。GnT−Vは、ヒトの肺腫瘍細胞及びマウスの腎臓で精製され、cDNAがクローン化され、プロモータ及び遺伝子構造が明らかになった(Gu et al.1993,J Biochem,113:614−619;Soreibah et al.,1993,J Biol.Chem.,268:15381−15385;Kang et al.,1996,J.Biol.Chem.,271:26706−26712)。本発明者は、転写酵素ets−1がGnT−Vの発現に関与すると報告した(Ko,et al.1999,J Biol.chem.,274(33):22941−22948)。薬剤を基本にする腫瘍治療の限界は、腫瘍に対する医学研究分野で治療可能な段階の早期発見へとパラダイムの変化を引き起こし、異常グリコシル化は、腫瘍早期発見マーカーとしての利用可能性が試験されている。
これによって、本発明者らは、GnT−Vが過剰発現される腫瘍細胞から2次元電気泳動及び質量分析を用いてGnT−Vによってベータ1,6−GlcNAcを有する候補タンパク質の目録を報告した(Kim et al.,2004,Proteomics,4(11):3353−3358;Kim et al.,2006,Proteomics,6(4):1187−1191)。しかし、従来のゲルに基づいた分析方法によれば、少量のタンパク質を同定するにおいて、本質的に偽陽性(false−positive)が起こる確率が高い。これを克服するために、本発明者らは、同定のためにL−PHAレクチンを用いてベータ1,6−GlcNAcを有する糖タンパク質を濃縮することによって本発明を完成した。
【発明の概要】
【発明が解決しようとする課題】
【0003】
本発明の目的は、発癌及び腫瘍転移に関与する異常糖タンパク質と呼ばれる変化した糖構造を獲得した糖タンパク質を高精度で同定する方法を提供することである。
本発明の他の目的は、発癌及び腫瘍転移に関与する異常糖タンパク質をより簡単な方法で探知して同定することである。
【課題を解決するための手段】
【0004】
前記目的を達成するために、本発明者らは、ベータ1,6−N−アセチルグルコサミン部分を有するN−連結型糖タンパク質を認識するレクチンであるL−PHAを用いて調査することによって、ベータ1,6−N−アセチルグルコサミンを有する異常糖タンパク質を探知する方法を開発した。
【発明の効果】
【0005】
本発明は、腫瘍発生及び転移に関与するベータ1,6−N−アセチルグルコサミンを有する異常糖タンパク質をレクチン沈殿で濃縮する方法を提供することによって、少量のタンパク質も探知することができる。そして、本発明は、全ての腫瘍に適用することができ、尿及び血液を含む生体材料に適用することができる。
本発明は、レクチン沈殿法を用いて癌の発生及び転移に関与する異常グリコシル化を探知する分析方法を提供し、腫瘍診断マーカー候補として異常糖タンパク質の目録を提供する
【図面の簡単な説明】
【0006】
【図1】多様な糖転移酵素によって形成されるN−連結型糖鎖連結構造及びL−PHAが結合されるベータ1,6−N−アセチルグルコサミン部位を示す図である。
【図2】実施例1に記載された方法の概略図である。
【図3】実施例1の方法によって同定されたタンパク質を示すもので、非糖タンパク質が多く、陽性対照群TIMP−1が探知されていないことを示す図である。
【図4】実施例2に記載された方法の概略図である。
【図5】実施例2の方法によって同定されたタンパク質を示すもので、低いスコア値の糖タンパク質が比較的多く、TIMP−1が依然として探知されていないことを示す図である。
【図6】実施例3に記載された方法の概略図である。
【図7】実施例3の方法によって同定されたタンパク質を示すもので、TIMP−1を含む多くの糖タンパク質が高いスコア値で現れたことを示す図である。
【図8】実施例3の方法と従来のレクチンブロット分析で共通的に確認されるタンパク質を示す図である。
【図9】実施例3の方法で確認され、従来のレクチンブロット分析では確認されていないタンパク質を示す図である。
【図10】実施例2及び実施例3の結果を比較したもので、質量分析ピークが34.36分の滞留時間で実施例3の方法のみによって探知されることを示す図である。
【図11】多重反応モニタリング(MRM)の結果として、TIMP−1ペプチドが実施例3のみで探知されることを示す図である。
【発明を実施するための形態】
【0007】
本発明は、癌の発生及び転移に関与する異常グリコシル化を有する糖タンパク質をレクチン沈殿法で高精度で分析する方法に関するものである。
前記癌の種類は、大腸癌、胃癌、肺癌、肝癌、子宮癌、乳房癌及び膵臓癌からなる群から選択されるが、上述した癌のみに限定されることはなく、癌の全ての種類に拡大することができる。
本発明は、癌細胞からベータ1,6−N−アセチルグルコサミン部分を獲得したN−連結型糖タンパク質を同定する高精度な分析方法を提供する。ベータ1,6−N−アセチルグルコサミンの形成を触媒するGnT−Vは、多様な腫瘍組織で幅広く発現され、その病理現象に関与する。ベータ1,6−N−アセチルグルコサミンがフィトヘマグルチニン−L4(以下、「L4−PHA」又は「L−PHA」と略称する)によって認識されるので、ビオチン標識されたL−PHAは、アビジン−アガロースビードに非共有結合で結合される。この複合体は、タンパク質の調合液からベータ1,6−N−アセチルグルコサミンを有する糖タンパク質を濃縮するのに用いられる。
本発明の一側面は、タンパク質をトリプシン処理し、7−テスラのフーリエ変換イオン共鳴型質量分析機(Fourier Transform Ion Cyclotron Resonance;FTICR)を用いたアミノ酸配列決定に適用し、ベータ1,6−N−アセチルグルコサミンを有する少量の糖タンパク質を同定する方法を提供する。
本発明は、リガンドが結合されたL4−PHAをリガンド受容体が付着されたグリカンビードに反応させ、L4−PHA−グリカンビード複合体を形成する段階と、糖タンパク質を得るために、試料を前記L4−PHA−グリカンビード複合体と反応させる段階と、前記糖タンパク質をゲル電気泳動で展開する段階と、前記糖タンパク質を含む結果物のゲルをペプチドにゲル内部で(in−gel)切断する段階と、前記切断されたペプチドを質量分析機で分析する段階とを含む糖タンパク質の分析方法を提供する。
本発明の他の側面は、前記グリカンビードがアガロースビード及びセファロースビードで構成されたグループから選択される前記方法を提供する。
本発明の他の側面は、前記リガンド及びリガンド受容体がビオチン及びアビジンである前記方法を提供する。
本発明の他の側面は、前記試料が自然状態又は変性形態である前記方法を提供する。
本発明の他の側面は、前記変性形態がベータ−メルカプトエタノール、ヨード酢酸及びヨードアセトアミドからなるグループから選択された少なくとも一つによって生成される前記方法を提供する。
本発明の他の側面は、対照区及び実験区の培養細胞株を、リガンドが結合されたL4−PHAをリガンド受容体が付着されたグリカンビードに結合することによって形成されるL4−PHA−グリカンビード複合体にそれぞれ加え、ベータ1,6−N−アセチルグルコサミンに対するグリカン構造において差を示す糖タンパク質を探索する方法を提供する。
本発明の他の側面は、正常な血液及び実験区の血液を、リガンドが結合されたL4−PHAをリガンド受容体が付着されたグリカンビードに結合することによって形成されるL4−PHA−グリカンビード複合体にそれぞれ加え、ベータ1,6−N−アセチルグルコサミンに対するグリカン構造において差を示す糖タンパク質を探索する方法を提供する。
本発明の更に他の側面は、アルブミン、グロブリン及びトランスフェリンのような血液内に豊富に存在するタンパク質によって、最小化した阻害を有する従来に発見された腫瘍バイオマーカー候補を確定する同定方法を提供し、確認効率を最大化するためにSISCAPA(Stable Isotope Standards and Capture by Anti−peptide Antibodies:Pearson et al,2004,J.roteome Res.3,235−44)方法と組み合わせることができる。
腫瘍の転移は、細胞認識及び接着の機能障害から発生し、数多くの膜タンパク質又は分泌タンパク質がこの過程に関与している。これは、バイオマーカーが存在する血液及び尿などの体液をバイオマーカー探知用試料として利用できることを示す。大腸癌−特異的バイオマーカーを探すために、本発明者らはグリコミクス研究を行い、GnT−V遺伝子は、GnT−V発現水準の低い大腸癌細胞株であるWiDr内に安定的に浸透された。培養培地で得たタンパク質は、2次元電気泳動ゲル上で展開するが、これらは、クーマシーブリリアントブルー又はレクチンブロット分析で染色され、対照群及び浸透WiDr細胞間に異なるグリカン構造を示すタンパク質が選択された(表1参照)。しかし、レクチンブロット分析自体が非特異的結合又はゲル内の汚染タンパク質を含む可能性があるので、付加的な確認段階が必要である。さらに、2次元電気泳動ゲルは分解能に限界を有しており、結果として、少量のタンパク質は、ゲルに基づいたシステム下では確認が難しい。このような限界を克服するために、本発明者らは、ベータ1,6−N−アセチルグルコサミンが含まれた糖タンパク質を特定レクチンを用いて直接探知できる方法を発明した。
[表1−1]
[表1−2]
【0008】
a;配列範囲は、質量分析機で確認されたペプチドの百分率を意味する。
b;総値は、個別ペプチドから得られたスコア値の合計である。スコアは−10xLog(P)であって、Pは、観察された符合が偶然の事件である確率である。これは、MS/MSデータとしてMASCOTサーチプログラムを用いたNCBInrデータベースに基づく。
c;分子量Mr及び等電点pIは、グリカン残基を計算に考慮していないときの理論値である。
L−PHAは、ベータ1,6−N−アセチルグルコサミンが含まれた糖タンパク質に非常に特異的に結合されるが、利用可能な溶出方法がないので、結合されたタンパク質を剥がすことが難しかった。この問題を解決するために、本発明者らは、L−PHA−結合されたタンパク質をSDS−PAGEゲル上に最小に展開し、ゲルは、切断し、タンパク質同定のためにトリプシン処理した。
【0009】
以下、本発明の多様な側面と具体例を下記の実施例を通してより具体的に説明する。
【実施例1】
【0010】
レクチンを用いたベータ1,6−N−アセチルグルコサミンが含まれた糖タンパク質の濃縮
GnT−V発現の低いWiDr:mock細胞及びGnT−Vを浸透させたWiDr:GnT−Vをモデル細胞株として構築し、調節された無血清培地を細胞株から収去した。腫瘍の転移が細胞認識及び接着の機能障害によって起こること(Egeblad et al.,2002,Nature Rev.2:161−174;Kannagi et al.,2004,Cancer Sci.95(5):377−384)と、多くの膜タンパク質又は分泌タンパク質がこの過程に関与することが知られている。GnT−Vによって生成されるベータ1,6−N−アセチルグルコサミンが含まれた糖タンパク質を培地から精製した。収集された培地は濃縮し、1mgのタンパク質をアビジン−アガロースビードに結合させ非特異的に結合するタンパク質を予め除去した。非特異的に結合したタンパク質を予め除去したタンパク質は、L−PHA−ビオチン−アビジン−アガロースビード複合体と一晩中4℃で反応させた。マンノース−1,6−N−アセチルグルコサミンがL−PHA結合タンパク質を剥がすのに用いられるが、この二糖類は価格面や効率面で証明されていない。ある研究チームでは、酸性溶出方法で剥がすことを試みたが、この方法も証明されておらず、低いpHはタンパク質の完全性に損傷を与える。このような理由により、タンパク質は、複合体に結合された状態で切断した。このために、タンパク質複合体を過量のPBS(Phosphate buffered saline)緩衝液で洗浄し、10Uのトリプシン(Promega社)で一晩中37℃でトリプシン処理した。切断されたペプチドは、蒸留水とアセトニトリルで抽出し、真空乾燥し、7テスラのFTICR/LTQ質量分析機で質量分析した。タンパク質は、Mascotサーチプログラム(バージョン2.0)でペプチド配列を決定して確認した(図2及び図3)。
サーチデータベースは、NCBInr 20051212(taxonomy;human、entries;103、913 human sequence entries)であった。その結果、ペプチドは、効果的に配列が決定されておらず、完全なタンパク質がトリプシンによって充分に切断されていないと考えられる。TIMP−1(Tissue inhibitor of metalloproteinase 1)は、GnT−Vの標的タンパク質として既に確認されており、変更されたグリカン構造が質量分析で決定され、参考分子としてTIMP−1が用いられる。しかし、TIMP−1は、この方法では確認されておらず、その代わりに、糖タンパク質でない多くのタンパク質が確認された。
【実施例2】
【0011】
レクチンを用いたベータ1,6−N−アセチルグルコサミンが含まれた糖タンパク質の濃縮段階以前のタンパク質還元及び脱塩
実施例1に示された非特異的結合を減少させて敏感性を増大させるために、収集された培地から得たタンパク質を1%(v/v)ベータ−メルカプトエタノールに還元させ、未反応ベータ−メルカプトエタノール及び塩は、ゲルろ過カラムで除去した(図4)。1mgのタンパク質を沈殿させ、実施例1に記載したように確認した。結果は、より多くの糖タンパク質が確認されたことを示すが、スコア値が低く、TIMP−1も確認されなかった。
【実施例3】
【0012】
SDS−PAGEゲルで最大化されたタンパク質切断及びレクチン沈殿以後、ベータ1,6−N−アセチルグルコサミンが含まれた糖タンパク質確認
実施例1に記載されたように培養培地から収集されたタンパク質を、実施例2に記載されたように1%(v/v)ベータ−メルカプトエタノールに還元させ、未反応ベータ−メルカプトエタノール及び塩は、ゲルろ過カラムで除去した。1mgのタンパク質をアビジン−アガロースビードと1時間反応させて先に精製し、先に精製されたタンパク質は、L−PHA−ビオチン−アビジン−アガロースビード複合体と一晩中4℃で反応させた。PBSで洗浄した後、結合されたタンパク質に1×SDS−PAGE変性緩衝液を加えて少しの間沸騰させてビード複合体から分離した。タンパク質は、8%SDS−PAGEゲル上で分離され、ゲル内でトリプシンで切断された。切断されたペプチドは、ゲルで溶出され、アジレント1100ナノフローシステム(Agilent Technologies)を用いて分析され、7テスラのFT−LTQ質量分析機(Thermo Electron)と連結されたSurveyorシステムで配列が分析された。ペプチドは、10cmのReproSil−Pur C18−AQ樹脂(Ammerbuch−Entringen)で充填されたシリカカラム(75mm)上で結合されて分離された。ペプチド混合物は、100mm−C18カラム(5μm)上にオートサンプラ(Surveyor)で20ml/分の速度でローディングされた。移動相AとBは、それぞれ0%及び100%のACNで構成され、それぞれ0.1%のギ酸を含む。濃度勾配は、15分間5%Bから初めて、3分間20%B、47分間50%B、最終的に95%に上昇した。溶出されたペプチドは、直接質量分析機にエレクトロスプレーした。質量分析機は、Xcaliburソフトウェア(Thermo Electron Version 1.4SR1)で制御した。図7は、一つの結果を示す。この実施例によれば、1)高いスコア値(P<0.05)を有する糖タンパク質を確認することができ、2)図7のようにTIMP−1を探知することができ、3)従来のレクチンブロットに基づいた同定方法で確認されたタンパク質を再び確認することができ(図8)、4)高いスコア値を有しながら低く発現されるタンパク質を探知することができた(図9)。
【実施例4】
【0013】
レクチン沈殿法に基づいて同定されたタンパク質を質量分析方法で確認した。
実施例2と実施例3を比較するために、既に探知されたTIMP−1ペプチド、GFQALGDAADIRを質量分析機でモニタした。図10のように、GFQALGDAADIRは、実施例3では、分子量が約1232Daであって、2電荷を帯びた状態で617.31m/zであった。実施例3では、ペプチドは滞留時間34分で探知されたが、実施例2では探知されず、ペプチド配列はGFQALGDAADIRであった。また、図3によれば、実施例3では、m/z617.31で探知されるペプチドの親イオン及びm/z 717.36のフラグメントイオンが示されるが、実施例2ではこれが全く探知されておらず、これは、トリプシン切断と抽出がよく行われていないことに起因すると予想される。
したがって、本発明は、グリコシル化で変化を示す少量のタンパク質を確認するための2次元電気泳動に基づいたより敏感な方法を提供する。
【産業上の利用可能性】
【0014】
本発明によれば、正常対照群と試験試料との間に水準及び構造の差を示す糖タンパク質を区分することによって、早期に癌を容易に診断することができる。
(配列目録テキスト)
<110> Korea Research Institute of Bioscience and Biotechnology
<120> An identification method of glycoproteins using a specific lectin precipitation
<130> jhko-0711-lectinaffi2
<150> KR10-2007-100916
<151> 2007-10-08
<160> 1
<170> KopatentIn 1.71
<210> 1
<211> 12
<212> PRT
<213> Homo sapiens
<400> 1
Gly Phe Gln Ala Leu Gly Asp Ala Ala Asp Ile Arg
1 5 10
【特許請求の範囲】
【請求項1】
リガンドが結合されたL4−PHA(Phytohemagglutinin−L4)とリガンド受容体が付着されたグリカンビードを反応させてL4−PHA−グリカンビード複合体を形成し、
糖タンパク質を得るために、試料を前記L4−PHA−グリカンビード複合体と反応させ、
ゲル電気泳動で前記糖タンパク質を展開し、
前記糖タンパク質を含むゲルをゲル内部で切断(in−gel digesting)してペプチドを得て、
前記切断されたペプチドを質量分析機で分析することを含む糖タンパク質の分析方法。
【請求項2】
前記グリカンビードは、アガロースビード及びセファロースビードからなる群から選択されることを特徴とする、請求項1に記載の糖タンパク質の分析方法。
【請求項3】
前記リガンド及び前記リガンド受容体は、ビオチン及びアビジンであることを特徴とする、請求項1に記載の糖タンパク質の分析方法。
【請求項4】
前記試料は、自然状態又は変性形態であることを特徴とする、請求項1に記載の糖タンパク質の分析方法。
【請求項5】
前記変性状態は、ベータ−メルカプトエタノール、ヨード酢酸及びヨードアセトアミドからなる群から選択された少なくとも1つによって生成されることを特徴とする、請求項4に記載の糖タンパク質の分析方法。
【請求項6】
リガンドが結合されたL4−PHA(Phytohemagglutinin−L4)とリガンド受容体が付着されたグリカンビードを反応させて製造されるL4−PHA−グリカンビード複合体に対照群と試験用培養細胞株をそれぞれ加え、ベータ1,6−N−アセチルグルコサミンのグリカン構造に差を示す糖タンパク質を探知する方法。
【請求項7】
リガンドが結合されたL4−PHA(Phytohemagglutinin−L4)とリガンド受容体が付着されたグリカンビードを反応させて製造されるL4−PHA−グリカンビード複合体に正常血清と試験用血清をそれぞれ加え、ベータ1,6−N−アセチルグルコサミンのグリカン構造に差を示す糖タンパク質を探知する方法。
【請求項1】
リガンドが結合されたL4−PHA(Phytohemagglutinin−L4)とリガンド受容体が付着されたグリカンビードを反応させてL4−PHA−グリカンビード複合体を形成し、
糖タンパク質を得るために、試料を前記L4−PHA−グリカンビード複合体と反応させ、
ゲル電気泳動で前記糖タンパク質を展開し、
前記糖タンパク質を含むゲルをゲル内部で切断(in−gel digesting)してペプチドを得て、
前記切断されたペプチドを質量分析機で分析することを含む糖タンパク質の分析方法。
【請求項2】
前記グリカンビードは、アガロースビード及びセファロースビードからなる群から選択されることを特徴とする、請求項1に記載の糖タンパク質の分析方法。
【請求項3】
前記リガンド及び前記リガンド受容体は、ビオチン及びアビジンであることを特徴とする、請求項1に記載の糖タンパク質の分析方法。
【請求項4】
前記試料は、自然状態又は変性形態であることを特徴とする、請求項1に記載の糖タンパク質の分析方法。
【請求項5】
前記変性状態は、ベータ−メルカプトエタノール、ヨード酢酸及びヨードアセトアミドからなる群から選択された少なくとも1つによって生成されることを特徴とする、請求項4に記載の糖タンパク質の分析方法。
【請求項6】
リガンドが結合されたL4−PHA(Phytohemagglutinin−L4)とリガンド受容体が付着されたグリカンビードを反応させて製造されるL4−PHA−グリカンビード複合体に対照群と試験用培養細胞株をそれぞれ加え、ベータ1,6−N−アセチルグルコサミンのグリカン構造に差を示す糖タンパク質を探知する方法。
【請求項7】
リガンドが結合されたL4−PHA(Phytohemagglutinin−L4)とリガンド受容体が付着されたグリカンビードを反応させて製造されるL4−PHA−グリカンビード複合体に正常血清と試験用血清をそれぞれ加え、ベータ1,6−N−アセチルグルコサミンのグリカン構造に差を示す糖タンパク質を探知する方法。
【図1】

【図2】

【図3】

【図4】
【図5】
【図6】
【図7】
【図8】

【図9】

【図10】
【図11】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2010−540956(P2010−540956A)
【公表日】平成22年12月24日(2010.12.24)
【国際特許分類】
【出願番号】特願2010−527866(P2010−527866)
【出願日】平成19年11月30日(2007.11.30)
【国際出願番号】PCT/KR2007/006124
【国際公開番号】WO2009/048196
【国際公開日】平成21年4月16日(2009.4.16)
【出願人】(500197682)コリア リサーチ インスティチュート オブ バイオサイエンス アンド バイオテクノロジー (4)
【Fターム(参考)】
【公表日】平成22年12月24日(2010.12.24)
【国際特許分類】
【出願日】平成19年11月30日(2007.11.30)
【国際出願番号】PCT/KR2007/006124
【国際公開番号】WO2009/048196
【国際公開日】平成21年4月16日(2009.4.16)
【出願人】(500197682)コリア リサーチ インスティチュート オブ バイオサイエンス アンド バイオテクノロジー (4)
【Fターム(参考)】
[ Back to top ]