
特異的ヨウ素輸送体(NIS)をコードする組換えアデノウイルス
【課題】、腫瘍の遺伝子治療方法を提供する。
【解決手段】細胞中でのヨウ素の蓄積を促進するためのアデノウイルスベクターを使用した、腫瘍細胞中のヨウ素特異的キャリア(Na+/I−共輸送体)(NIS)をコードする遺伝子の挿入方法。また、nis遺伝子を含む複製用の欠陥組換えアデノウイルスならびに腫瘍細胞へのnis遺伝子の導入と代謝放射線治療法とを組み合わせた癌治療法におけるベクターの使用方法。
【解決手段】細胞中でのヨウ素の蓄積を促進するためのアデノウイルスベクターを使用した、腫瘍細胞中のヨウ素特異的キャリア(Na+/I−共輸送体)(NIS)をコードする遺伝子の挿入方法。また、nis遺伝子を含む複製用の欠陥組換えアデノウイルスならびに腫瘍細胞へのnis遺伝子の導入と代謝放射線治療法とを組み合わせた癌治療法におけるベクターの使用方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、腫瘍の遺伝子療法および治療の分野に関する。より詳細には、本発明は、細胞中へのヨウ素の蓄積を促進するためのアデノウイルスベクターによる腫瘍細胞への特異的ヨウ素輸送体(Na+/I−共輸送体)をコードする遺伝子の移入に関する。本発明はまた、nis遺伝子を含む複製欠陥組換えアデノウイルスおよび腫瘍細胞へのnis遺伝子の導入とヨウ素131での代謝放射線治療法とを組み合わせた癌治療法でのこれらのベクターの使用に関する。
【背景技術】
【0002】
ヨウ素131は、50年以上分化型甲状腺癌の治療に使用されてきた。その治療効果は、腫瘍組織に照射された放射線量に関連する。例えば、転移の場合、ヨウ素131での治療後の腫瘍反応は、腫瘍組織に送達した放射線量が80グレイを超えた場合に認められるが、35グレイ未満の線量では腫瘍反応レベルは低いかゼロである(Maxon、NEJM、309、937〜941、1983)。一般に、腫瘍体積の治療または減少に必要な全線量はその放射線感受性にしたがって40グレイと60グレイとの間で変化することを評価可能である。分化型甲状腺癌の固定化した転移の治療の間で行われるように、この全線量を、ヨウ素131を使用したいくつかの治療で送達させることができる。
【0003】
腫瘍組織に送達される放射線量は、2つの生物学的因子(腫瘍組織中のヨウ素131の有効半減期(Teff)および放射能濃度)に依存する。
【0004】
腫瘍組織中のヨウ素131の有効半減期(Teff)は、関連式I/Teff=I/Tbiol+I/Tphysによる生物学的半減期(Tbiol)および物理的半減期(Tphys)に依存する。固定細胞中のヨウ素131の生物学的半減期(Tbiol)は、ヨウ素の有機化能力に依存する。例えば非甲状腺細胞のように、有機化しない場合は、生物学的半減期は短く、数時間〜数十時間である。ヨウ素131の物理的半減期(Tphys)は、8.02日である。したがって、腫瘍組織中のヨウ素131の有効半減期は数時間〜数十時間であることを単に評価可能である。
【0005】
放射能濃度は、腫瘍組織によるヨウ素131の全固定とこの組織との間の比である。
【0006】
例として、1gの組織あたりに投与された0.1%の活性のヨウ素固定および1.5日の有効半減期では、3.7ギガベクレル(すなわち100mCi)の投与により、腫瘍組織への30グレイの線量が送達されると評価可能である。
【0007】
ヨウ素131の全固定は、一方では患者に投与されたヨウ素131の活性に依存し、一方ではヨウ素131を濃縮する腫瘍組織の能力に依存するパラメータである。
【0008】
患者に投与することができるヨウ素131の最大活性は、健常な組織、特に骨髄に送達する放射線量に起因するヨウ素131のヨウ素毒性によって制限される。したがって、甲状腺癌の場合、甲状腺機能低下症の患者に投与される治療活性量は、一般に3.7GBqと10GBq(100〜300mCi)との間である。ヨウ素の保持が甲状腺機能低下症で認められた活性量の半分であるので、甲状腺機能正常患者に投与することができる最大活性量はより多くなり、8.5と22.2GBqとの間の値(500〜600mCi)に達する。
【0009】
患者に投与された所与の活性量についての正常な組織のヨウ素131の濃縮能力は、ヨウ素共輸送体の発現レベルおよびその生物活性に依存する。これは、正常および罹患ヒト甲状腺組織の研究およびNIS活性のインビトロ研究によって証明された。ヨウ素131の比活性は高いが、投与されたヨウ素131の活性の増加に伴う飽和減少は認められなかった。
【0010】
患者に投与することができるヨウ素131の最大活性の制限のために、ヨウ素131の全固定を増加させるためにヨウ素131を濃縮する腫瘍組織の能力を増加させることができることが望ましい。
【0011】
腫瘍組織中でヨウ素固定が均一に増加することも重要である。実際、固定組織中のヨウ素の固定は、細胞間で、または腫瘍間で非常に不均一であり得る。腫瘍組織中の放射線量は、本質的にはヨウ素131のβ放射によって送達される。しかし、生体組織中のその経路は、せいぜい2〜3mmである。さらに、局在部周囲では、放射線量は距離ごとに指数関数的に減少していく。この元素は、固定腫瘍領域中の高い放射能濃度を達成する重要性を強化し、一方で、可能な限り均一にヨウ素が固定される必要がある。
【0012】
ヒト甲状腺によるヨウ素131の固定およびそれによる照射を、1μg/kg/日のL−トリヨードチロニンを1日3回にわけて3週間投与することによって容易に抑制する(ヨウ素131の測定不可能な固定)ことができることに留意すべきである。
【0013】
NIS輸送体は、甲状腺細胞による循環ヨウ素(ヨウ素I−の形態)の濃縮を担う(概説として、P.Thomopoulos、Medecine et Science、4、10、825〜828を参照のこと)。これらの細胞中のヨウ素濃縮は、以下の特徴を示す:ヨウ素は電気化学勾配に対して30〜40倍の因子によって濃縮されること;ナトリウム(Na+)が存在する必要があること;能動輸送に関連すること;一定の陰イオン(チオシアン酸塩、過塩素酸塩、過テクネチウム酸塩など)によって競合的に阻害されること。甲状腺細胞によるヨウ素の濃縮後、ヨウ素が有機化し、ヨードチロシンおよびチロイドホルモン(チロキシンT4およびトリヨードチロニンT3)が合成される。
【0014】
NISヨウ素のマウス輸送体(Daiら、Nature、379、458〜460、1996)およびヒト輸送体(Smanik P.A.ら、Biochem.Biophys.Res.Commun.、226、339〜345、1996)をコードする遺伝子が単離されており、これらは84%同一である。この遺伝子は、ヒト種では染色体19p上に存在する。これは14個のイントロンで分離された15個のエクソンを含む。その転写によって、選択的スプライシングによる2つの形態が得られるが、そのうち甲状腺では長い方の形態が主である。そのタンパク質の分子量は55kDaであり、グリコシル化後80kDaに達する。これは甲状腺細胞の外側の基底膜に存在する。そのアミノ末端およびカルボキシ末端は細胞内である一方で、ペプチド鎖の残部は、細胞内および細胞外ループによって連結された13個の膜貫通セグメントを含む(Levyら、1998、J.Biol.Chem.、273、22657〜22663)。NISをコードする遺伝子の非甲状腺細胞への移入により、非甲状腺細胞の甲状腺細胞と同一の性質であるヨウ捕獲能力(ナトリウム(Na+)の存在が必要であることおよび過塩素酸塩陰イオンによる阻害)が付与される。
【0015】
nis遺伝子の転写は、TSHによって甲状腺細胞中で活性化される。この効果は、サイクリックAMPによって媒介される。マウス甲状腺細胞中でのタンパク質の半減期は4日間である(Paire A.ら、J.Biol.Chem.、272、18245〜18249、1997)。甲状腺外組織では、遺伝子の活性は、基底状態での甲状腺のそれよりも低い。これは、おそらく甲状腺nis遺伝子のプロモーターへ結合することができるが、他の組織のプロモーターに結合しない特異的転写因子TTF−1(甲状腺転写因子1)による甲状腺細胞中のnis遺伝子の転写の刺激による(Endo T.ら、Mol.Endocrinol.、11、1747〜1755、1997)。甲状腺以外に、一定の組織(特に唾液腺および胃粘膜)がヨウ素を捕獲および濃縮することができる。
【0016】
最近の研究により、ヒトまたはマウス腫瘍細胞へのレトロウイルスによるnis遺伝子の導入が報告されている(Mandellら、Cancer Research、59、661〜668、1999)。これらの結果は、これらの細胞が非甲状腺細胞中でnis遺伝子を発現し、天然ではヨウ素を蓄積しない細胞中でのヨウ素の濃縮が認められるという点で興味深い。しかし、治療への適用を考慮するためのヨウ素の十分な固定レベルを達成するためには克服すべき多数の制限要因が残されている。
【0017】
第1の要因は、腫瘍細胞中でのnis遺伝子発現レベルの低さである。例えば、上記の研究では、インビトロで得られた結果は、非甲状腺細胞で達成されたヨウ素の濃縮が甲状腺細胞中で蓄積されたヨウ素濃度の1/2のままであることが示されている。
【0018】
さらに、腫瘍細胞が膜レベルで示される大量の破壊を考慮すると、非腫瘍性甲状腺細胞に匹敵する効率を有する輸送体の機能的取り込みを確保するのは困難なようであう。
【0019】
別の制限要因は、非甲状腺細胞中でヨウ素が有機化しないことである。しかし、このヨウ素の有機化は細胞中でのヨウ素の維持に必要な構成要素である。したがって、上記で報告の研究では、非甲状腺細胞におけるインビトロで蓄積されたヨウ素の非常に急速な流出(30分と60分との間)が認められる。
【0020】
最後に、腫瘍細胞の不均一性および膜レベルでのこれらの細胞の広範な改変を考慮すると、腫瘍細胞膜中のNIS輸送体の均一な分配を達成することは困難なようであう。
【0021】
したがって、現在記載されているアプローチでは、非甲状腺腫瘍治療用のヨウ素131の使用を考慮するのに十分な発現レベルで非甲状腺腫瘍中にヨウ素を蓄積させることは未だ不可能である。
【先行技術文献】
【非特許文献】
【0022】
【非特許文献1】Maxon、NEJM、309、937〜941、1983
【非特許文献2】P.Thomopoulos、Medecine et Science、4、10、825〜828
【非特許文献3】Daiら、Nature、379、458〜460、1996
【非特許文献4】Smanik P.A.ら、Biochem.Biophys.Res.Commun.、226、339〜345、1996
【非特許文献5】Levyら、1998、J.Biol.Chem.、273、22657〜22663
【非特許文献6】Paire A.ら、J.Biol.Chem.、272、18245〜18249、1997
【非特許文献7】Endo T.ら、Mol.Endocrinol.、11、1747〜1755、1997
【非特許文献8】Mandellら、Cancer Research、59、661〜668、1999
【発明の概要】
【発明が解決しようとする課題】
【0023】
本発明は、遺伝子治療とヨウ素131を使用した放射線治療とを組み合わせた腫瘍の改良された治療法を提供する。
【0024】
本発明は、特に、非甲状腺腫瘍レベルでのヨウ素の固定効率の増大を可能にし、それにより、甲状腺癌治療用に首尾よく開発された放射線治療の原理を非甲状腺腫瘍に適用可能な方法を記載する。
【課題を解決するための手段】
【0025】
本発明は、欠陥組換えアデノウイルスによる特異的ヨウ素輸送体(Na+/I−共輸送体)NISをコードする遺伝子の導入により、非甲状腺腫瘍細胞中のヨウ素の非常に高い蓄積が可能となるという証明に基づく。特定の実施形態では、nis遺伝子を発現する欠陥アデノウイルスの供給により甲状腺細胞で認められる濃度の約5倍の濃度でヨウ素を蓄積可能である。この驚くべき非甲状腺腫瘍細胞のヨウ素125の蓄積能力により、今まで一定の甲状腺腫瘍の治療に限定されていたヨウ素131を使用した代謝放射線治療に基づくこの型の腫瘍の新規の治療アプローチを考慮可能である。
【0026】
代謝放射線治療の主な利点は、周辺組織に有意に照射することなく固定した大放射線量の放射性同位元素を組織に送達させることである。
【0027】
通常はヨウ素を蓄積不可能な腫瘍細胞中に実質的にヨウ素を濃縮させることが有効に可能であるという発見により、非甲状腺起源の腫瘍治療用の多数の適応症においてこの方法の適用を考慮可能である。より詳細には、以下の2つの適応症の型が考慮される:その位置により外部からの放射線治療ではほとんどアクセス不可能な腫瘍(例として、手術不可能な前立腺癌または大脳内腫瘍)、既に照射されており、最大線量が既に投与されているので更なる外部からの放射線治療が不可能な腫瘍(これは、照射領域で発症するか再発する全ての癌に関する)。この方法はまた、ヨウ素捕獲能力を喪失し、代謝治療に反応しない甲状腺腫瘍の治療に適用可能である。
【0028】
分化型甲状腺癌の転移の場合に行われる代謝放射線治療と外部放射線治療との組み合わせも考慮可能である。この組み合わせにより、毒性を増加させることなく放射線治療の治療有効性を増大可能である。この組み合わせは、特に、放射線治療にあまり感受性を示さない腫瘍に有利なようである。さらに、過度に巨大なサイズのために代謝放射線治療でアクセス不可能な腫瘍へのこの技術の適用を考慮可能である。
【0029】
したがって、本発明の第1の目的は、特異的ヨウ素輸送体(Na+/I−共輸送体)NISまたはその誘導体をコードする少なくとも1つの異種DNA配列を含む欠陥組換えアデノウイルスにある。
【発明を実施するための形態】
【0030】
本発明の目的では、特異的ヨウ素輸送体(Na+/I−共輸送体)NISの表現「誘導体」は、NISポリペプチドに由来し、特異的ヨウ素輸送活性を保持する任意のアナログ、フラグメント、または変異形態を意味すると理解される。種々の誘導体は、天然の状態で存在し得る。これらの誘導体は、NISをコードする構造遺伝子のヌクレオチド配列の相違によって特徴づけられる対立遺伝子であるか特異的スプライシングまたは翻訳後修飾に起因し得る。これらの誘導体を、1つまたは複数のアミノ酸残基の置換、欠失、付加、および/または修飾によって得ることができる。これらの修飾を、当業者に公知の任意の技術によって行うことができる。これらの誘導体は、特に、その基質に高い親和性を有する分子、インビボでの改良された発現が可能な配列、プロテアーゼに対してより高い耐性を示す分子、生物学的有効性がより高いか副作用が低いか、おそらく新規の生物学的性質を有する分子である。本発明の文脈で使用することができる他の誘導体は、特に、1つまたは複数の残基が置換された分子、望ましくない活性が考慮されるか発現される結合部位との相互作用に関連しないかあまり関連しない領域の欠失によって得られた誘導体、および天然の配列に関する更なる残基(例えば、分泌シグナルおよび/または結合ペプチドなど)を含む誘導体である。
【0031】
本発明の文脈で産生される特異的ヨウ素輸送体(Na+/I−共輸送体)NISまたはその誘導体は、cDNA、ゲノムDNA(gDNA)、または例えば1つまたは複数のイントロンが挿入されたcDNAからなるハイブリッド構築物であり得る。これには、合成または半合成配列も含み得る。有利には、cDNAまたはgDNAが使用される。特に、gDNAの使用によりヒト細胞でのより良好な発現が可能である。
【0032】
第1の実施形態によれば、これは、マウス起源の特異的ヨウ素輸送体(Na+/I−共輸送体)NISをコードするcDNA配列である。本発明の好ましい実施形態によれば、これは、ヒト起源の特異的ヨウ素輸送体(Na+/I−共輸送体)NISをコードするcDNA配列である。
【0033】
本発明の文脈で使用されるアデノウイルスは、組換えアデノウイルス(すなわち、異種DNA配列を含む)である。有利には、これらは、欠陥組換えアデノウイルス(すなわち、標的細胞中で自律複製不可能なアデノウイルス)である。
【0034】
本発明のアデノウイルスの構築には、種々の血清型を使用することができる。実際に多数の抗原型が存在し、その構造および性質はいくらか異なるが、類似の遺伝子構成を示す。より詳細には、組換えアデノウイルスは、ヒト起源であっても動物起源であってもよい。ヒト起源のアデノウイルスに関して、好ましくはC群(特に、アデノウイルス2型(Ad2)、5型(Ad5)、7型(Ad7)、または12型(Ad12))に分類され得る。動物起源の種々のアデノウイルスのうち、好ましくは、イヌ起源のアデノウイルス(特に、全てのCAV2アデノウイルス株(例えば、マンハッタン株またはA26/61株(ATCC VR−800)))であり得る。動物起源の他のアデノウイルスは、特に、WO 94/26914に引用されている。
【0035】
アデノウイルスゲノムは、特に、各末端の逆末端反復配列(ITR)、キャプシド形成配列(Psi)、初期遺伝子、および後期遺伝子を含む。主な初期遺伝子は、E1、E2、E3、およびE4領域中に含まれる。これらのうち、E1領域に含まれる遺伝子は、特に、ウイルス増殖に必要である。主な後期遺伝子は、L1〜L5領域に含まれる。Ad5アデノウイルスゲノムは完全に配列決定されており、データベースでアクセス可能である(特に、Genbank M73260を参照のこと)。同様に、他のアデノウイルスゲノムの一部またはさらに全部(Ad2、Ad7、Ad12など)も配列決定されている。
【0036】
上記のように、本発明のアデノウイルスは欠陥であるので、標的細胞中で自律複製不可能である。この効果により、アデノウイルス由来の種々の構築物が調製され、種々の治療遺伝子に組み込まれている。これらの各構築物では、感染細胞中で複製不可能なようにアデノウイルスを改変されていた。したがって、先行技術に記載の構築物は、ウイルス増殖に必須のE1領域が欠失され、異種DNA配列が挿入されたアデノウイルスである(Levreroら、Gene、101、1991、195、Gosh−Choudhuryら、Gene、50、1986、161)。さらに、ベクターの性質を改良するために、アデノウイルスゲノム中に他の欠失または改変を作製することが提案されている。したがって、熱感受性点変異をts125変異体に移入して、72kDaのDNA結合タンパク質(DBP)を不活化させることができる(Van der Vlietら、J.Virol.、1975、15、2、348〜354)。他のベクターは、ウイルスの複製および/または増殖に必須の別の領域(E4領域)の欠失を含む。E4領域は、実際に、後期遺伝子の発現の調製、後期核RNAの安定性、宿主細胞タンパク質の発現の消滅、およびウイルスDNAの複製効率に関連する。したがって、E1およびE4領域が欠失したアデノウイルスベクターは、転写バックグラウンドノイズを有し、且つウイルス遺伝子発現が非常に減少している。このようなベクターは、例えば、WO 94/28152に記載されている。さらに、IVa2遺伝子が改変されたベクターも記載されている(WO 96/1008)。
【0037】
本発明の好ましい実施形態では、組換えアデノウイルスは、C型ヒトアデノウイルスである。より詳細には、これはAd2またはAd5アデノウイルスである。
【0038】
有利には、本発明の文脈で使用される組換えアデノウイルスは、そのゲノムのE1領域が欠失している。さらにより詳細には、E1aおよびE1b領域が欠失している。例としては、影響を与えるヌクレオチド454〜3328、386〜3446、459〜3510、または357〜4020(Ad5のゲノムを基準とする)の欠失であり得る。
【0039】
別の変形形態では、本発明の文脈で使用される組換えアデノウイルスは、少なくともE1およびE3領域の全てまたは一部が欠失している。
【0040】
別の変形形態では、本発明の文脈で使用される組換えアデノウイルスは、E1領域の欠失に加えて、そのゲノム中のE4領域の影響を与える全てまたは一部が欠失している。より詳細には、E4領域の欠失は、全ての読み取り枠に影響を与える。詳細な例としては、出願WO 95/02697およびWO 96/22378に記載のE4領域の33466〜35535もしくは33093〜35535または一部のみの欠失であり得る。
【0041】
例えば、いわゆる第3世代組換えアデノウイルス(すなわち、E1およびE4領域が全体または部分的に欠失しており、任意選択的にE3領域が欠失している)であり得る。本発明の特定の変形形態は、以下の領域の全てまたは一部に影響を与える欠失を有するアデノウイルスの使用からなる:
−E1、E4、およびE3、
−E1、E4、およびE2、
−E1、E4、E2、およびE3、
−上記の領域ならびに後期アデノウイルス機能(L1〜L6)をコードする遺伝子の全部または一部、
−全てのコードウイルス領域。
【0042】
後期機能(「最小」ベクター)または全てのコード領域(「ガットレス」ベクター)をさらに欠くアデノウイルスに関して、その構築物は、例えば、Parksら、PNAS、93、1996、13565およびLieberら、J.Virol.、70、1996、8944に記載されている。
【0043】
これには、ハイブリッドアデノウイルスベクター(出願WO 99/60144に記載のものなど)も含んでもよく、このアデノウイルスは別のウイルス用のベクター(特に、レトロウイルスベクター)として使用される。
【0044】
ヨウ素輸送体(NIS)をコードする核酸を含む発現カセットを、組換えゲノムの別の部位に挿入することができる。これを、欠失配列の代わりまたは付加的にE1、E3、またはE4領域に挿入することができる。これを、cis中のウイルス産生に必要な配列(ITR配列およびキャプシド形成配列)の外側の任意の他の部位に挿入することもできる。
【0045】
本発明の目的のために、核酸の、表現「発現カセット」は、ベクターの特異的制限部位に挿入することができるDNAフラグメントを意味すると理解される。このDNAフラグメントは、目的のRNAまたはポリペプチドをコードするヌクレオチド配列に加えて、目的の配列の発現に必要な配列(エンハンサー、プロモーター、ポリアデニル化配列など)を含む。転写および/または翻訳用の適切な読み枠への前記フラグメントの挿入を確保するようにDNAフラグメントおよび制限部位を設計する。
【0046】
組換えアデノウイルスを、キャプシド形成株(すなわち、組換えアデノウイルスゲノム中の1つまたは複数の欠陥機能をトランスで補足することができる細胞株)産生させる。当業者に公知のキャプシド形成株のうち、例としては、アデノウイルスゲノムの一部が取り込まれた293株であり得る。より詳細には、293株は、左側のITR、キャプシド形成領域、E1領域(E1aおよびE1bを含む)、pIXタンパク質コード領域、およびpIVa2タンパク質コード領域の一部を含む血清型5アデノウイルス(Ad5)ゲノムの左側の末端(11〜12%)を含むヒト胚腎臓細胞株である。この株は、E1領域を欠く(すなわち、E1領域の全てまたは一部を欠く)組換えアデノウイルスをトランスで補足することができ、高力価のウイルスストックを産生することができる。この株はまた、許容温度(32℃)で熱感受性E2変異をさらに含むウイルスストックを産生することができる。特に、ヒト肺癌細胞A549(WO 94/28152)またはヒト網膜芽細胞(Hum.Gen.Ther.、1996、215)に基づいて、E1領域を補足することができる他の細胞株が記載されている。さらに、いくつかのアデノウイルス機能をトランスで補足することができる株が記載されている。特に、E1およびE4領域を補足する株(Yehら、J.Virol.、70、1996、559〜565、Cancer Gen.Ther.、2、1995、322、Krougliakら、Hum.Gen.Ther.、6、1995、1575)ならびにE1およびE2領域を補足する株(WO 94/28152、WO 95/02697、WO 95/27071)または特にこのようなウイルスの構築に関連する部位特異的リコンビナーゼ活性をさらに発現することにより最小アデノウイルスの産生に使用することができるそれら由来の株であり得る。
【0047】
腫瘍レベルでの感染効率を増加させるために、組換えアデノウイルスのキャプシド構造を改変することもできる。例えば、キャプシドは、腫瘍細胞へのアデノウイルスの標的化を可能にするuPARリガンドまたはRGDモチーフを含むことができ、このようなベクターおよび標的配列は、特に出願WO 00/12738に記載されている。標的配列を、ヘキソンタンパク質または繊維タンパク質に挿入することができる。好ましくは、繊維タンパク質またはヘキソンタンパク質の欠失レベルで標的配列を挿入する。有利には、Ad5のヘキソンのポリペプチド配列279〜292に対応するヘキソン13アミノ酸のポリペプチド配列から欠失する。有利には、Ad5の繊維(HIループ)のポリペプチド配列539〜547に対応する繊維11アミノ酸のポリペプチド配列から欠失する。
【0048】
組換えアデノウイルスは、通常、キャプシド形株へのウイルスDNAの移入から約2〜3日後(アデノウイルスのサイクル速度は24〜36時間である)に細胞を溶解することによって産生される。この方法を実行するために、移入されたウイルスDNAは、細胞に感染する、任意選択的に細菌(WO 96/25506)または酵母(WO 95/03400)中で構築された完全な組換えウイルスゲノムであり得る。これはまた、キャプシド形成株の感染に使用される組換えウイルスであり得る。ウイルスDNAを、それぞれ組換えウイルスゲノムの一部およびキャプシド形成細胞への移入後に種々のフラグメント間での相同組換えによって組換えウイルスを再構成可能な相同領域を有するフラグメント形態に挿入することもできる。
【0049】
細胞溶解後、組換えウイルス粒子を、塩化セシウム勾配遠心分離またはクロマトグラフィーなどの任意の公知の技術によって単離することができる。別の方法は、出願WO 98/00528に記載されている。
【0050】
発現調節配列。特異的ヨウ素キャリア(Na+/I−共輸送体、NIS)をコードする遺伝子を、機能的で宿主腫瘍細胞での発現を可能にする発現を調節する任意の配列(例えば、プロモーターまたはプロモーター/エンハンサーなど)の調節下に置くことができる。
【0051】
発現調節配列は、プロモーター領域に加えて、特異的ヨウ素輸送体をコードする遺伝子の3’に存在し、且つ転写終結シグナルおよびポリアデニル化部位が得られる領域を含む。これら全てのエレメントが発現カセットを構成する。
【0052】
プロモーターは、構成性プロモーターであっても調節(誘導)プロモーターであってもよい。これは、実際の遺伝子プロモーターであり得る。これはまた、異なる起源(他のタンパク質の発現を担うか合成プロモーター)の配列を含み得る。特に、これは、真核生物またはウイルス遺伝子のプロモーター配列を含み得る。例えば、これは、トランスフェクトが望まれる細胞のゲノム由来のプロモーター配列を含み得る。同様に、これは、ウイルスゲノム(使用されるウイルスを含む)由来のプロモーター配列を含み得る。これに関して、例として、E1A、MLP、CMV、およびRSV−LTR、MT−1、SV40などのプロモーターであり得る。
【0053】
さらに、これらの発現配列を、一定の組織中で組織特異的または主に発現可能な活性化配列または調節配列(エノラーゼプロモーターGFAPなど)の付加によって改変することができる。実際、ウイルスが腫瘍細胞に有効に感染した場合のみ、治療遺伝子がその効果を発現または産生するように腫瘍細胞中で特異的または主に活性である発現シグナルを使用することが特に有利であり得る。発現の特異的または主な特徴とは、プロモーター活性が腫瘍細胞中で有意により高いことを意味する。非特異的発現が他の細胞に存在し得るにもかかわらず、対応する活性レベルは、一般に腫瘍細胞中で認められるものよりも非常に低く(無視できるほど)、一般に少なくとも1/10ほどである。
【0054】
本発明の文脈で使用することができるプロモーターのうちでは、遍在性プロモーター(HPRT、ビメンチン、αアクチン、チューブリンなど)、中間径フィラメントのプロモーター(デスミン、神経フィラメント、ケラチン、GFAP)、治療目的の遺伝子のプロモーター(MDR、CFTR、第VIII因子など)、組織特異的プロモーター(デシン、ミオシン、クレアチンキナーゼ、ホスホグリセリン酸キナーゼ遺伝子のプロモーター)、増殖中の細胞でより特異的に活性なプロモーターまたは刺激に対応するプロモーター(天然のホルモン(ステロイドホルモンレセプター、レチノイン酸レセプターなど)に対応するプロモーターなど)、抗生物質(テトラサイクリン、ラパマイシンなど)によって調節されるプロモーター、天然または合成起源の他の分子に対応する他のプロモーター、またはウイルスゲノム由来のプロモーター配列(サイトメガロウイルスCMVエンハンサー/プロモーター、レトロウイルスLTRプロモーター、SV40プロモーター、E1A遺伝子プロモーター、MLPプロモーター)であり得る。テトラサイクリンによって調節することができるプロモーターおよびCMVプロモーターは、WO 96/01313、米国特許第5,168,062号、および同第5,385,839号に記載されている。
【0055】
本発明の実施に使用することができるプロモーターのうちでは、特に、サイトメガロウイルス(CMV)プロモーター、SV40初期プロモーター領域(Benoist and Chambon、1981、Nature、290、304〜310)、ラウス肉腫ウイルス3’LTR領域に含まれるプロモーター(Yamamotoら、1980、Cell、22、787〜797)、ヘルペスウイルスチミジンキナーゼプロモーター(Wagnerら、1981、Proc.Natl.Acad.Sci.U.S.A.、78、1441〜1445)、メタロチオネイン遺伝子の調節配列(Brinsterら、1982、Nature、296、39〜42)、原核生物起源のプロモーター(βラクタマーゼプロモーター(Villa−Kamaroffら、1978、Proc.Natl.Acad.Sci.U.S.A.、75、3727〜3731)、またはtacプロモーター(DeBoerら、1983、Proc.Natl.Acad.Sci.U.S.A.、80、21〜25)など(「組換え細菌由来の有用なタンパク質」、Scientific American、1980、242、74〜94もまた参照のこと))、酵母プロモーター(Gal4、ADC(アルコールデヒドロゲナーゼ)、PGK(ホスホグリセロールキナーゼ)、およびアルカリ性ホスファターゼなど)、および組織特異性を示し且つトランスジェニック動物に使用される以下の動物起源の転写緒節配列:膵臓の腺房細胞中で活性なエラスターゼI遺伝子の調節配列(Swift、1984、Cell、38、639〜646、Ornitzら、1986、Cold Spring Harbor Symp.Quant.Biol.、50、399〜409、MacDonald、1987、Hepatology、7、425〜515)、膵臓のβ細胞中で活性な遺伝子[lacuna]インスリン調節配列(Hanahan、1985、Nature、315、115〜122)、リンパ球様細胞中で活性な免疫グロブリンの発現を調節する配列(Grosschedlら、1984、Cell、38、647〜658、Adamesら、1985、Nature、318、533〜538、Alexxanderら、1987、Mol.Cell.Biol.、7、1436〜1444)、精巣細胞および乳房細胞、リンパ球および肥満細胞中で活性なマウス乳腺癌ウイルスの調節配列(Lederら、1986、Cell、45、485〜495)、前立腺癌中で活性なPSA遺伝子の調節配列、肝臓中で活性なアルブミン遺伝子の調節配列(Pinkertら、1987、Genes and Devel.、1、268〜276)、肝臓中で活性なαフェトプロテインの調節配列(Krumlaufら、1985、Mol.Cell.Biol.、5、1639〜1648、Hammerら、1987、Science、235、53〜58)、肝臓中で活性なα1抗トリプシン遺伝子の調節配列(Kelseyら、1987、Genes and Devel.、1、161〜171)、骨髄性細胞中で活性なβグロビン遺伝子の調節配列(Mogramら、1985、Nature、315、338〜340、Kolliasら、1986、Cell、46、89〜94)、脳の希突起膠細胞中で活性な塩基性ミエリン遺伝子の調節配列(Readheadら、1987、Cell、48、703〜712)、骨格筋中で活性なミオシン軽鎖2遺伝子の調節配列(Sani、1985、Nature、314、283〜286)、および視床下部で活性なゴナドトロピン放出ホルモン遺伝子の調節配列(Masonら、1986、Science、234、1372〜1378)であり得る。
【0056】
本発明の特定の実施形態では、好ましくはRSV−LTRまたはCMV初期プロモーターから選択されるウイルスプロモーターの調節下で特異的ヨウ素輸送体(Na+/I−共輸送体)NISをコードする遺伝子を含む欠陥組換えアデノウイルスを使用する。
【0057】
本発明のベクターへのNIS遺伝子の移入とヨウ素131の生物学的半減期を増加可能な生物系への移入とを組み合わせることも可能である。したがって、NIS遺伝子およびヨウ素有機化機構に関連するポリペプチドをコードする1つの遺伝子またはいくつかの遺伝子を腫瘍細胞に同時または首尾よく導入可能である。例えば、グルコースオキシダーゼをコードする遺伝子などのペルオキシダーゼ系に関連するポリペプチドをコードする遺伝子であり得る。また、ヨウ素有機化過程に関連する甲状腺細胞中の酵素である甲状腺ペルオキシダーゼをコードする遺伝子(Nucleic Acids Res.、15、6735〜6735、1987、配列GenBank g37251)であり得る。甲状腺ペルオキシダーゼをコードする遺伝子の移入を、サイログロブリンの全てまたは一部(チロシン残基が豊富なサイログロブリン部分)をコードする核酸配列の移入と組み合わせることができる。
【0058】
いくつかの導入遺伝子を発現する1つのベクター、2つのベクターの同時感染、NIS遺伝子を発現するアデノウイルスベクターおよび別のベクター、ウイルスもしくはプラスミドベクター、ヨウ素有機化機構に関連するポリペプチドをコードする1つまたは複数の遺伝子を発現するウイルスまたはプラスミドベクターによる腫瘍細胞へのNIS遺伝子およびヨウ素有機化機構に関連するポリペプチドをコードする1つの遺伝子またはいくつかの遺伝子の送達も考慮可能である。
【0059】
さらに、本発明のベクターへのNIS遺伝子の移入と薬物(例えば、リチウム塩)によってヨウ素流出の抑制させる系の投与とを組み合わせ可能である。
【0060】
別の変形形態では、NIS遺伝子の移入と癌治療用の目的の治療遺伝子との組み合わせも考慮可能である。これには、ヘルペスチミジンキナーゼ遺伝子またはシトシンデアミナーゼ遺伝子などの自殺遺伝子を含み得る。これには、アポトーシスを誘導する遺伝子(p53、Bax、BclX−s、Bad、またはBc12およびBclX−1の任意の他のアンタゴニストなど)を含み得る。これには、改良された性質を示すこれらのタンパク質変異型(p53の変異型)(CTS−1、WO 97/04092)をコードする遺伝子を含み得る。これにはまた、抗血管形成または血管形成阻害因子(特に、Tie−1およびTie−2のリガンド、アンギオスタチン、エンドスタチン、ATF因子、プラスミノゲン誘導体、エンドセリン、トロンボスポンジン1および2、PF−4、インターフェロンαおよびβ、インターロイキン12、TNFα、ウロキナーゼレセプター、flt1、KDR、PAI1、PAI2、TIMP1、プロラクチンフラグメント)をコードする遺伝子を含み得る。これはまた、抗腫用免疫を誘導するか免疫応答を刺激することができるタンパク質(IL2、GM−CSF、IL12など)をコードする遺伝子を含み得る。癌治療目的のタンパク質をコードする遺伝子のうちで、抗体、抗体一本鎖抗体(ScFv)のフラグメント、または腫瘍治療の免疫療法用の認識能力を有する任意の他の抗体フラグメント(抗RAS抗体(WO 9429446))を強調することも重要である。
【0061】
有利には、これには、その発現により腫瘍細胞が放射線増感される遺伝子(p53遺伝子)を含み得る。
【0062】
NIS輸送体をコードする遺伝子を含むアデノウイルスベクターまたはウイルスまたは非ウイルス性の個別のベクターへの1つまたは複数のこれらの遺伝子移入によって、治療目的のこれらの遺伝子または上記の2つまたはそれ以上のこれらの遺伝子の組み合わせの投与を行うことができる。
【0063】
別の変形形態では、本発明のベクターを、抗癌剤(特に、タキソール、タキソテール、および他のタキソイドなど(特に、米国特許第4,857,653号、同第4,814,470号、同第4,924,011号、同第5,290,957号、同第5,292,921号、同第5,438,072号、同第5,587,493号、欧州特許第0253738号、およびWO 91・17976、WO 93/00928、WO 93/00929、およびWO 9601815に記載されている))または他の治療用抗癌剤(シスプラチンおよび白金誘導体、エトポシドおよびリン酸エトポシド、ブレオマイシン、マイトマイシンC、CCNU、ドキソルビシン、ダウノルビシン、イダルビシン、イホスファミドなど)と組み合わせて投与することができる。
【0064】
本発明はまた、上記のアデノウイルスベクターおよび生理学的に受容可能な賦形剤を含む薬学的組成物に関する。本発明の薬学的組成物は注射形態が好ましく、腫瘍内投与用または、経口、非経口、鼻腔内、動脈内、静脈内、または気管内経路などの投与用に処方することができる。
【0065】
好ましくは、薬学的組成物は、腫瘍内経路による投与を意図した処方用の薬学的に受容可能な賦形剤を含む。
【0066】
本発明の組成物は、適用の考慮および種々のパラメータ、特に使用した投与様式または所望の発現の持続時間による当業者が容易に調整することができる種々の用量の組換えアデノウイルスを含み得る。一般に、本発明の組換えウイルスを、104と1014pfuとの間、好ましくは106と1011pfuとの間の用量の形態で処方して投与する。用語「pfu(「プラーク形成単位」)」は、ウイルスの感染力に対応し、適切な細胞培養物への感染および感染細胞プラーク数の計数によって同定される。ウイルス溶液のpfu力価の同定技術は、文献に十分に記載されている。
【0067】
さらに、本発明の組成物はまた、化学導入剤または生物導入剤を含み得る。要路「化学導入剤または生物導入剤」は、細胞への核酸の浸透を容易にする任意の化合物(すなわち、組換えウイルス以外)を意味すると理解される。これには、陽イオン性非ウイルス薬(陽イオン性リピド、ペプチド、ポリマー(ポリエチレンイミン、ポリリジン)、ナノ粒子など)または非陽イオン性非ウイルス薬(非陽イオン性リポソーム、ポリマー、または非陽イオン性ナノ粒子など)を含み得る。
【0068】
好ましい実施形態では、本発明の組成物は、ヨウ素輸送体NISをコードする遺伝子を含む欠陥組換えベクターを含み、これは腫瘍内投与用に処方される。有利には、本発明の組成物は、104と1015pfuとの間、好ましくは107と1012pfuとの間を含む。
【0069】
本発明の主題はまた、ヨウ素輸送体(NIS)をコードする核酸を含む組換えベクターを1つまたは複数の適合性アジュバントおよび薬学的に受容可能なアジュバントと混合することを特徴とする、腫瘍の予防、改善、および/または治療に有用な薬物の調製法である。
【0070】
本発明はまた、ヨウ素輸送体(NIS)をコードする核酸を含む欠陥組換えアデノウイルスベクターの有効量を投与する工程を包含する、哺乳動物、特にヒトの治療法に関する。
【0071】
用語「有効量」は、ヨウ素輸送体(NIS)をコードする核酸を含むアデノウイルスの投与をヨウ素135での代謝放射線治療と組み合わせた場合、腫瘍体積の少なくとも約15%、好ましくは少なくとも50%、より好ましくは少なくとも90%を減少させるのみに十分な量またはさらに好ましくは腫瘍を消滅させるのに十分な量を示す。
【0072】
本発明は、腫瘍治療、より詳細には固形腫瘍の治療に関する。本発明の主題によって治療することができる固形腫瘍のうち、特に、肉腫および悪性腫瘍(例として、線維肉腫、骨原性肉腫、血管肉腫、内皮肉腫、リンパ管肉腫、ユーイング肉腫、大腸癌、膵臓癌、卵巣癌、前立腺癌、腺癌、腎臓、肝臓、または胆管の癌、ウィルムス腫瘍、子宮頸癌、精巣癌、肺癌、非小細胞肺癌、膀胱癌、上皮癌、神経膠腫、星状細胞腫、黒色腫、神経芽細胞腫、および網膜芽細胞腫であるが、これらに限定されない)であり得る。
【0073】
本発明はまた、上皮組織(子宮頚管、食道、および肺の上皮など)の増殖性障害(化生および異形成)の予防および/または治療に関する。これに関して、本発明は、特に、過形成、化生、より詳細には異形成などの非新生細胞の増殖が起こる状態での腫瘍形成または癌への進行に先立つことが公知であるか疑われる病態の治療に関する(これらの異常な増殖状態についての概説は、Robbins and Angell、1976、Basic Pathology、第2版、W.B.Sauders Co.、Philadelphia、68〜79を参照のこと)。過形成は、器官の顕著な構造的または機能的変化を伴わない器官中での細胞数の増加を含む制御された細胞増殖の形態である。例えば、子宮内膜の過形成は、子宮内膜癌に先駆けて起こる。化生は、成体または完全に分化された細胞型が別の細胞型に置換される制御された細胞増殖の形態である。化生は、上皮組織または結合組織で起こり得る。異形成は、しばしば癌の早期危険信号であり、主に上皮で見出される。これは、新生細胞増殖の最も頻繁な形態であり、各細胞の一様性の喪失および細胞の構造上の配向性の喪失を含む。異形成は、典型的には、慢性的刺激作用または炎症が存在する場合に起こり、しばしば子宮頚管、気道、声帯腔、および膀胱壁に認められる。概説として、Fishmanら、1985、Medicine、第2版、J.B.Lippincott Co.、Philadelphiaを参照のこと。
【0074】
本発明は、以下の実施例によってより詳細に記載されるが、これらは例示であり、本発明を制限しない。
【図面の簡単な説明】
【0075】
【図1】プラスミドpAB1、プラスミドpXL3048、およびプラスミドpAB2の線図である。プラスミドpXL3048は、アデノウイルス5型ゲノムの左側の末端(ヌクレオチド1〜382)、3つの固有のクローニング部位を含むポリリンカー、およびpIX遺伝子の一部(ヌクレオチド3446〜4296)を含む。プラスミドpAB2は、EcoRVで先に線状化したpXL3048へのpAB1のSspI−EcoRVフラグメントのクローニングに起因する。
【図2】Crouzetら、PNAS、94、1414、1997に記載の方法による、プラスミドpAB2およびpXL3215由来の二重組換えによって作製されたプラスミドpXL3215の産生を示す図である。プラスミドpXL3215は、E1およびE3領域が欠失されたアデノウイルス5型ゲノムを含み、且つNIS発現カセットを含む。
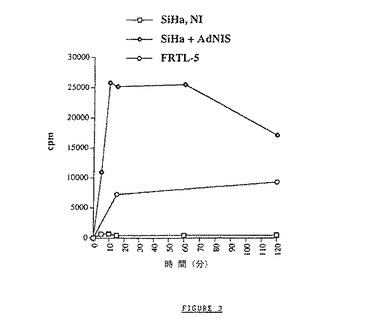
【図3】Ad−NISベクターで感染された(感染多重度10)FTRL−5およびSiHa細胞によるヨウ素125の蓄積速度を示す図である。速度の結果を、数/分/106細胞で示す。ヨウ素接触時のウェルあたりの細胞数の判定は、2つの測定の平均である。
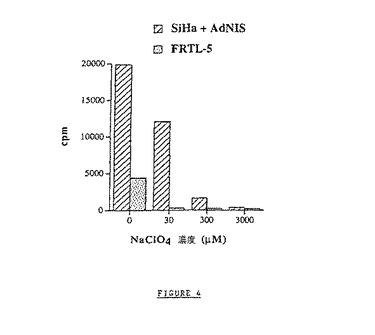
【図4】30、300、および3000μMの過塩素酸塩(NaClO4)の存在下でのNISによるヨウ素輸送の特異的阻害を示す図である。細胞とヨウ素125との間の接触時間は15分間である。
【図5A】Ad−NISベクターを感染させてない腫瘍によるインビボでのヨウ素蓄積を示す図である(コントロール)。MCF−7細胞(ヒト乳がんの腫瘍細胞)を、ヌードマウスに皮下注射した(5×106細胞)。15日後(腫瘍の出現)、チロキシンの腹腔内注射(2μg/動物/日)を毎日15日間行った。次いで、Ad−CMV−NISベクターを一定の動物に腫瘍内経路で注射した(2×109pfu/腫瘍、腫瘍サイズ3〜6mm)。注射から3日後、6μCiの125Iを、腹腔内経路でマウスに注射する。甲状腺上、腫瘍上、腫瘍のすぐ隣にプローブを置くことによって一定の時間間隔で10秒間計数した。結果を、時間の関数として検出された数(10秒間の測定)として示す。
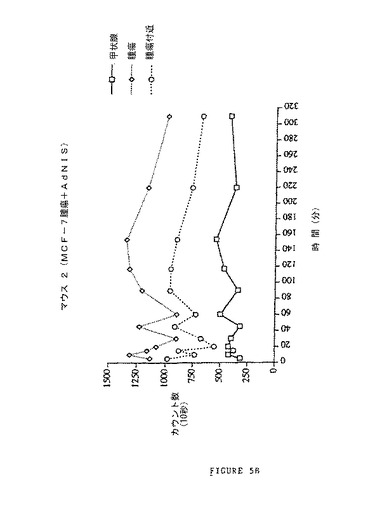
【図5B】Ad−NISベクターを感染させた腫瘍によるインビボでのヨウ素蓄積を示す図である。MCF−7細胞(ヒト乳がんの腫瘍細胞)を、ヌードマウスに皮下注射した(5×106細胞)。15日後(腫瘍の出現)、チロキシンの腹腔内注射(2μg/動物/日)を毎日15日間行った。次いで、Ad−CMV−NISベクターを一定の動物に腫瘍内経路で注射した(2×109pfu/腫瘍、腫瘍サイズ3〜6mm)。注射から3日後、6μCiの125Iを、腹腔内経路でマウスに注射する。甲状腺上、腫瘍上、腫瘍のすぐ隣にプローブを置くことによって一定の時間間隔で10秒間計数した。結果を、時間の関数として検出された数(10秒間の測定)として示す。
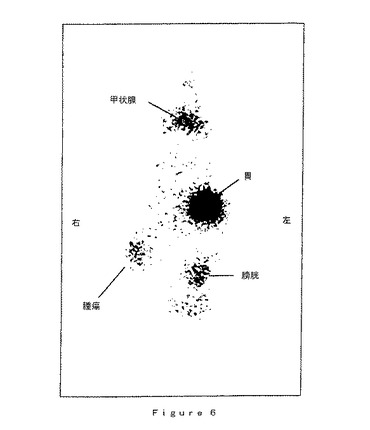
【図6】腫瘍をAd−NISベクターで感染させたマウスのシンチグラフィーである。MCF−7細胞(ヒト腫瘍細胞、乳癌)を、ヌードマウスに皮下注射した(5×106細胞)。15日後(腫瘍の出現)、チロキシンの腹腔内注射(2μg/動物/日)を毎日15日間行った。次いで、Ad−NISベクターを腫瘍内経路で注射した(2×109pfu/腫瘍、腫瘍サイズ3〜6mm)。注射から3日後、50μCiの125Iを、腹腔内経路でマウスに注射する。ヨウ素注射から1時間後に計数をおこなった。画像は、動物の縦断図である。
【0076】
材料と方法
一般的な分子生物学技術
分子生物学で慣習的に使用されている方法(プラスミドDNAの予備的抽出、塩化セシウム勾配によるプラスミドDNAの遠心分離、アガロースまたはアクリルアミドゲル電気泳動、電気溶出によるDNAフラグメントの精製、フェノールまたはフェノール−クロロホルムによるタンパク質抽出、エタノールまたはイソプロパノールによる生理食塩水培地中でのDNAの沈殿、Esxherichia coliの形質転換など)は、当業者に周知であり、文献中に多数記載されている(Maniatis T.ら、「分子クローニング、実験マニュアル」、Cold Spring Harbor Laboratory、Cold Spring Harbor,N.Y。、1982、Ausubel F.M.ら編、「現代の分子生物学プロトコール」、John Wiley & Sons、New York、1987)。
【0077】
ライゲーションでは、DNAフラグメントをアガロースまたはアクリルアミドゲル電気泳動によってそのサイズごとに分離し、フェノールまたはフェノール/クロロホルム混合物で抽出し、エタノールで沈殿させて、供給者の指示に従ってT4ファージDNAリガーゼ(Biolabs)の存在下でインキュベートすることができる。
【0078】
供給者の指示に従って、E.coliのDNAポリメラーゼのクレノウフラグメント(Biolabs)を使用して、突出した5’末端の充填を行うことができる。供給者の指示に従って、T4ファージDNAポリメラーゼ(Biolabs)の存在下で、突出した3’末端の破壊を行う。S1ヌクレアーゼで調節された処理によって突出した5’末端の破壊を行う。
【0079】
Amershamから販売されているキットを使用し、Taylorら、Nucleic Acids Res.、13、1985、8749〜8764が開発した方法にしたがって、合成オリゴデオキシヌクレオチドによってインビトロで指向される変異誘発を行うことができる。
【0080】
いわゆるPCR技術(ポリメラーゼ触媒連鎖反応、Saiki R.K.ら、Science、230、1985、1350〜1354、Mullis K.B.and Faloona F.A.、Meth.Enzym.、155、1987、335〜350)によるDNAフラグメントの酵素増幅を、供給者の指示に従って、「DNAサーマルサイクラー」(Perkin Elmer Cetus)を使用して行うことができる。
【0081】
ヌクレオチド配列の評価を、Amershamから販売のキットを使用し、Sangerら、Proc.Natl.Acad.Sci.USA、74、1977、5463〜5467が開発した方法によって行うことができる。
【実施例】
【0082】
[実施例1]特異的ヨウ素輸送体(Na+/I−共輸送体)NISをコードする遺伝子を発現する欠陥組換えアデノウイルスの構築
本実施例は、CMVプロモーターに作動可能に連結された特異的ヨウ素輸送体(Na+/I−共輸送体)NIS(Genbank U60282を参照のこと)をコードする遺伝子を保有する欠陥アデノウイルスベクターの構築を記載する。
【0083】
AatII−HindIIIフラグメントの形態のラットnis遺伝子のヌクレオチド配列74〜2046を含むフラグメント(Daiら、1996、Nature、379、458〜460)を、プラスミドpCEP−4(Invitrogen)のPvuII−HindIII部位にクローン化した。AatII−HindIIIフラグメントのAtII部位を、Mung Beanヌクレアーゼ(Biolabs)での処理で予め平滑にし、プラスミドpCEP−4を挿入した。得られたプラスミドをpAB1と呼ぶ。
【0084】
プラスミドpAB1(図1)は、サイトメガロウイルスCMVエンハンサー/プロモーター(ヌクレオチド−522〜+72)の調節下で発現するnis遺伝子を含み(Boshartら、1985、Cell、41、521〜530)、その後ろにSV40ウイルスポリアデニル化部位(SV40ゲノムのヌクレオチド2538〜2759、Genbank遺伝子座SV4CG)を含む。
【0085】
(i)サイトメガロウイルスエンハンサー/プロモーターと、(ii)nis遺伝子のcDNAと、(iii)SV40ウイルスポリアデニル化部位との組み合わせを、以後NIS発現カセットと呼ぶ。
【0086】
次いで、上記で定義のNIS発現カセットを含むプラスミドpAB1のSspI−EcoRVフラグメント(3711bp)を、EcoRVで予め線状化したシャトルベクターpXL3048にクローン化する。得られたベクターを、pAB2と呼ぶ(図2)。プラスミドpXL3048は、5型アデノウイルスゲノムの左側の末端(ヌクレオチド1〜382)、3つの固有のクローニング部位を含むポリリンカー、およびpIX遺伝子の一部(ヌクレオチド3446〜4296)を含むプラスミドKan−SacB(Crouzetら、PNAS、94、1414〜1419、1997)由来のプラスミドである。
【0087】
アデノウイルスAd−NISを、Crouzetら、PNAS、94、1414〜1419、1997に記載の方法に従って、プラスミドpAB2とプラスミドpXL3215との間の相同組換えによって構築した。
【0088】
プラスミドpXL3215は、そのE1領域(ΔE1 386〜3446)にRSV−LacZカセットを含み、E3領域(ΔE3 28592〜30470)が欠失したアデノウイルスゲノムを含む。
【0089】
構築の原理を、図2に示す。この二重組換えによって作製されたプラスミドpXL3215は、E1およびE3領域が欠失され、CNV−NIS−ポリA SV40発現カセットを含む5型アデノウイルスゲノムを含む。この構築物を、NIS発現カセットの配列決定によって評価する。
【0090】
Ad−NISアデノウイルスを、293株(ATCC CRL−1573)へのPacIで消化したpXL3215のDNAの挿入によって作製する。次いで、得られたウイルス粒子を、この同一の株で増幅させ、産生されたウイルスストックを、二重CsCl勾配によって精製する。
【0091】
次いで、ウイルス粒子を使用して、種々の腫瘍株におけるNIS遺伝子の発現を研究する。
【0092】
[実施例2]Ad−NISベクター感染腫瘍細胞によるインビトロでのヨウ素の蓄積
Ad−NISベクター感染細胞におけるヨウ素の蓄積能力を以下の種々の株で試験した:上皮腫瘍細胞SiHa(ATCC HTB−35)、乳癌細胞MCF7(ATC HTB−22)およびT−47D(ATCC HTB−133)ならびに前立腺起源の腫瘍細胞DU−145(ATCCHTB−81)およびPC−3(ATCC CRL−1435)。
【0093】
細胞に感染多重度10でAd−NISベクターを感染させた。細胞のヨウ素捕獲能力を、Weiss らの方法(Endocrinology、1984、114、1090〜1098)にしたがって感染から28〜29時間後に試験する。感染細胞をHBSS緩衝液で洗浄し、0.1μCiのヨウ素125を含むHBSS緩衝液と共に図3に表示の時間接触させる。接触終了後、細胞を冷却HBSS緩衝液で1回洗浄し、冷却エタノールと20分間インキュベートする。細胞によって捕獲されたヨウ素125の量を、ガンマカウンターを使用したエタノール中の放射能のアッセイによって同定する。
【0094】
SiHa株を使用して得られた結果を図3に示し、これを、FRTL−5細胞(ヨウ素を天然に捕獲し、コントロールとして使用したラット甲状腺細胞)で得られた結果と比較することができる。
【0095】
過塩素酸塩の存在下でのNISによるヨウ素輸送体の特異的阻害を試験した。NaClO4をヨウ素125を含むHBSS緩衝液に添加して、30、300、および3000μMの最終濃度とする。細胞とヨウ素125との間の接触時間は15分間である。結果を図4に示す。結果を、同数の細胞についての数として示す。
【0096】
FRTL−5細胞(ポジティブコントロール)は、一定量のヨウ素を固定し、この現象は非常に急速である(接触から15分後にほぼ最大に達する)ことが認められる。得られた結果より、感染していないSiHa細胞(ヒト上皮腫瘍細胞)はヨウ素を捕獲しないことが示される。驚いたことに、Ad−NISベクターを感染させたSiHa細胞はFRTL−5細胞の5倍を超えるヨウ素を捕獲することが認められる。この現象の初速度はFRTL−5細胞で認められた速度に匹敵することにも注目すべきである。輸送特異性を、nis遺伝子を含まないコントロールアデノウイルスでの感染によって確認した(結果を示さず)。
【0097】
阻害実験の結果(図4)は、FRTL−5細胞のヨウ素固定が30μMのNaClO4濃度で90%を超えて阻害されることを示す(30μMでは93%、高濃度で95%)。一方、Ad−NISを感染SなせたSiHa細胞では、同レベルの阻害を得るために必要なインヒビター濃度は、更により高い(30μMのNaClO4ではわずか40%阻害、300μMのNaClO4では92%阻害、3mMのNaClO4では98%阻害)ことに注目することが興味深い。
【0098】
阻害実験により、NISによるヨウ素イオン輸送の競合的および特異的阻害であるこの現象はNaClO4によって阻害されるので、Ad−NIS感染SiHa細胞のヨウ素捕獲能力は確かにこれらの細胞中の発現NISによると結論付けることが可能である。他のヒト腫瘍細胞株(乳癌細胞:MCF−7およびT47−D、ならびに前立腺起源の腫瘍細胞:DU−145およびPC−3)についても類似の結果が得られた。
【0099】
NaClO4による阻害に関する結果により、腫瘍細胞のAd−NIS感染により特に高レベルでNISが発現することも確認されるが、これはこの現象の阻害に高濃度のNaClO4が必要であるからである(SiHa細胞およびFRTL−5細胞の関連細胞の場合、同レベルの阻害を得るために10倍を超える阻害濃度が必要である)。
【0100】
これら全ての結果により、アデノウイルスベクターは、nis遺伝子の導入に特に有利なベクターであり、このようなベクターの使用により非甲状腺腫瘍細胞においてヨウ素125を天然に捕獲する細胞で認められるレベルの5倍の蓄積レベルを獲得可能であることが示される。
【0101】
[実施例3]Ad−NISベクター感染腫瘍によるインビボでのヨウ素蓄積
MCF−7細胞(乳癌のヒト腫瘍細胞)を、ヌードマウスに皮下注射した(5×106細胞)。15日後(腫瘍の出現)、チロキシンの腹腔内注射(2μg/動物/日)を毎日15日間行い、動物の甲状腺を安静にした。次いで、Ad−NISベクターを一定の動物に腫瘍内経路で注射した(2×109pfu/腫瘍)。注射時の腫瘍の直径は3〜6mmであった。注射から3日後、6μCiの125Iを、腹腔内経路でマウスに注射し、動物の異なる部位に移動することができるプローブの支援によって10秒間にわたり計数した。甲状腺上、腫瘍上、腫瘍のすぐ隣にプローブを置くことによって一定の時間間隔で計数した。結果を図5に示す。結果を、時間の関数として検出された数(10秒間の測定)として示す。
【0102】
コントロール動物(Ad−NISを未投与)では、甲状腺は、少量のヨウ素を固定するが、チロキシンでの治療による甲状腺中のヨウ素の実質的な蓄積は長時間認められない。コントロール動物では、腫瘍中のヨウ素固定は認められない(図5Aを参照のこと)。NISが胃中に天然に存在するという事実およびヨウ素が尿から排泄されるという事実は、比較的高い値が認められることの説明となる(比較的高いバックグラウンドノイズ)。
【0103】
腫瘍内経路によって投与されたAd−NIS治療動物(図5B)では、甲状腺中の実質的なヨウ素蓄積が認められない(チロキシンの治療効果)。これらの動物では、腫瘍上にプローブを置くことによって検出されたヨウ素の量は、腫瘍付近で検出されたヨウ素量よりも系統的に高く、これはAd−NISベクターでの治療後に実際に腫瘍中でヨウ素が固定されたことを示す。
【0104】
腫瘍中でのヨウ素固定を確認するために、Ad−NISベクターで処理した動物におけるシンチグラフィーによる放射能測定を行った。得られた結果を図6に示す。50μ/Ciの123Iを腹腔内経路で注射する以外は上記のプロトコールにしたがって使用動物を調製し、ヨウ素注射から1時間後に画像を得た。
【0105】
画像は動物の縦断図である。以下の4つのヨウ素固定領域が明らかである:甲状腺、膀胱(ヨウ素排泄経路)、胃(天然でNISが発現される)、および腫瘍。これらのシンチグラフィーの結果により計数によって得られた先の結果が確認され、これは、本発明のベクターにより腫瘍中の実質的なヨウ素固定が可能であることを示す。
【0106】
[実施例4]Ad−NISベクター感染腫瘍によるヨウ素蓄積の定量的評価
SiHa細胞(5×106細胞)を、ヌードマウス(n=12)の両側腹に皮下注射した。15日後(腫瘍の出現)、チロキシンの腹腔内注射(2μg/動物/日)を毎日15日間行い、動物の甲状腺を安静にした。各動物について、Ad−NISベクターを腫瘍に腫瘍内経路で注射し(2×109pfu/腫瘍)、他の腫瘍をコントロールとして使用した。注射時の腫瘍の直径は5〜8mmであった。注射から3日後、6μCiの125Iを腹腔内経路でマウスに注射した。125I投与から90分後に動物を屠殺し、125Iの量を同定した。結果を表1に示す。結果を、1gの組織に投与した125Iの百分率として示す。
【0107】
【表1】

【0108】
得られた結果により、Ad−NIsベクターで治療した腫瘍はヨウ素125Iを非常に効率的に蓄積することができ、未処理腫瘍よりも25倍まで高いことが示される。これらの結果により先の結果が確認され、本発明のベクターにより腫瘍中のヨウ素が実質的に固定可能であることが示される。
【技術分野】
【0001】
本発明は、腫瘍の遺伝子療法および治療の分野に関する。より詳細には、本発明は、細胞中へのヨウ素の蓄積を促進するためのアデノウイルスベクターによる腫瘍細胞への特異的ヨウ素輸送体(Na+/I−共輸送体)をコードする遺伝子の移入に関する。本発明はまた、nis遺伝子を含む複製欠陥組換えアデノウイルスおよび腫瘍細胞へのnis遺伝子の導入とヨウ素131での代謝放射線治療法とを組み合わせた癌治療法でのこれらのベクターの使用に関する。
【背景技術】
【0002】
ヨウ素131は、50年以上分化型甲状腺癌の治療に使用されてきた。その治療効果は、腫瘍組織に照射された放射線量に関連する。例えば、転移の場合、ヨウ素131での治療後の腫瘍反応は、腫瘍組織に送達した放射線量が80グレイを超えた場合に認められるが、35グレイ未満の線量では腫瘍反応レベルは低いかゼロである(Maxon、NEJM、309、937〜941、1983)。一般に、腫瘍体積の治療または減少に必要な全線量はその放射線感受性にしたがって40グレイと60グレイとの間で変化することを評価可能である。分化型甲状腺癌の固定化した転移の治療の間で行われるように、この全線量を、ヨウ素131を使用したいくつかの治療で送達させることができる。
【0003】
腫瘍組織に送達される放射線量は、2つの生物学的因子(腫瘍組織中のヨウ素131の有効半減期(Teff)および放射能濃度)に依存する。
【0004】
腫瘍組織中のヨウ素131の有効半減期(Teff)は、関連式I/Teff=I/Tbiol+I/Tphysによる生物学的半減期(Tbiol)および物理的半減期(Tphys)に依存する。固定細胞中のヨウ素131の生物学的半減期(Tbiol)は、ヨウ素の有機化能力に依存する。例えば非甲状腺細胞のように、有機化しない場合は、生物学的半減期は短く、数時間〜数十時間である。ヨウ素131の物理的半減期(Tphys)は、8.02日である。したがって、腫瘍組織中のヨウ素131の有効半減期は数時間〜数十時間であることを単に評価可能である。
【0005】
放射能濃度は、腫瘍組織によるヨウ素131の全固定とこの組織との間の比である。
【0006】
例として、1gの組織あたりに投与された0.1%の活性のヨウ素固定および1.5日の有効半減期では、3.7ギガベクレル(すなわち100mCi)の投与により、腫瘍組織への30グレイの線量が送達されると評価可能である。
【0007】
ヨウ素131の全固定は、一方では患者に投与されたヨウ素131の活性に依存し、一方ではヨウ素131を濃縮する腫瘍組織の能力に依存するパラメータである。
【0008】
患者に投与することができるヨウ素131の最大活性は、健常な組織、特に骨髄に送達する放射線量に起因するヨウ素131のヨウ素毒性によって制限される。したがって、甲状腺癌の場合、甲状腺機能低下症の患者に投与される治療活性量は、一般に3.7GBqと10GBq(100〜300mCi)との間である。ヨウ素の保持が甲状腺機能低下症で認められた活性量の半分であるので、甲状腺機能正常患者に投与することができる最大活性量はより多くなり、8.5と22.2GBqとの間の値(500〜600mCi)に達する。
【0009】
患者に投与された所与の活性量についての正常な組織のヨウ素131の濃縮能力は、ヨウ素共輸送体の発現レベルおよびその生物活性に依存する。これは、正常および罹患ヒト甲状腺組織の研究およびNIS活性のインビトロ研究によって証明された。ヨウ素131の比活性は高いが、投与されたヨウ素131の活性の増加に伴う飽和減少は認められなかった。
【0010】
患者に投与することができるヨウ素131の最大活性の制限のために、ヨウ素131の全固定を増加させるためにヨウ素131を濃縮する腫瘍組織の能力を増加させることができることが望ましい。
【0011】
腫瘍組織中でヨウ素固定が均一に増加することも重要である。実際、固定組織中のヨウ素の固定は、細胞間で、または腫瘍間で非常に不均一であり得る。腫瘍組織中の放射線量は、本質的にはヨウ素131のβ放射によって送達される。しかし、生体組織中のその経路は、せいぜい2〜3mmである。さらに、局在部周囲では、放射線量は距離ごとに指数関数的に減少していく。この元素は、固定腫瘍領域中の高い放射能濃度を達成する重要性を強化し、一方で、可能な限り均一にヨウ素が固定される必要がある。
【0012】
ヒト甲状腺によるヨウ素131の固定およびそれによる照射を、1μg/kg/日のL−トリヨードチロニンを1日3回にわけて3週間投与することによって容易に抑制する(ヨウ素131の測定不可能な固定)ことができることに留意すべきである。
【0013】
NIS輸送体は、甲状腺細胞による循環ヨウ素(ヨウ素I−の形態)の濃縮を担う(概説として、P.Thomopoulos、Medecine et Science、4、10、825〜828を参照のこと)。これらの細胞中のヨウ素濃縮は、以下の特徴を示す:ヨウ素は電気化学勾配に対して30〜40倍の因子によって濃縮されること;ナトリウム(Na+)が存在する必要があること;能動輸送に関連すること;一定の陰イオン(チオシアン酸塩、過塩素酸塩、過テクネチウム酸塩など)によって競合的に阻害されること。甲状腺細胞によるヨウ素の濃縮後、ヨウ素が有機化し、ヨードチロシンおよびチロイドホルモン(チロキシンT4およびトリヨードチロニンT3)が合成される。
【0014】
NISヨウ素のマウス輸送体(Daiら、Nature、379、458〜460、1996)およびヒト輸送体(Smanik P.A.ら、Biochem.Biophys.Res.Commun.、226、339〜345、1996)をコードする遺伝子が単離されており、これらは84%同一である。この遺伝子は、ヒト種では染色体19p上に存在する。これは14個のイントロンで分離された15個のエクソンを含む。その転写によって、選択的スプライシングによる2つの形態が得られるが、そのうち甲状腺では長い方の形態が主である。そのタンパク質の分子量は55kDaであり、グリコシル化後80kDaに達する。これは甲状腺細胞の外側の基底膜に存在する。そのアミノ末端およびカルボキシ末端は細胞内である一方で、ペプチド鎖の残部は、細胞内および細胞外ループによって連結された13個の膜貫通セグメントを含む(Levyら、1998、J.Biol.Chem.、273、22657〜22663)。NISをコードする遺伝子の非甲状腺細胞への移入により、非甲状腺細胞の甲状腺細胞と同一の性質であるヨウ捕獲能力(ナトリウム(Na+)の存在が必要であることおよび過塩素酸塩陰イオンによる阻害)が付与される。
【0015】
nis遺伝子の転写は、TSHによって甲状腺細胞中で活性化される。この効果は、サイクリックAMPによって媒介される。マウス甲状腺細胞中でのタンパク質の半減期は4日間である(Paire A.ら、J.Biol.Chem.、272、18245〜18249、1997)。甲状腺外組織では、遺伝子の活性は、基底状態での甲状腺のそれよりも低い。これは、おそらく甲状腺nis遺伝子のプロモーターへ結合することができるが、他の組織のプロモーターに結合しない特異的転写因子TTF−1(甲状腺転写因子1)による甲状腺細胞中のnis遺伝子の転写の刺激による(Endo T.ら、Mol.Endocrinol.、11、1747〜1755、1997)。甲状腺以外に、一定の組織(特に唾液腺および胃粘膜)がヨウ素を捕獲および濃縮することができる。
【0016】
最近の研究により、ヒトまたはマウス腫瘍細胞へのレトロウイルスによるnis遺伝子の導入が報告されている(Mandellら、Cancer Research、59、661〜668、1999)。これらの結果は、これらの細胞が非甲状腺細胞中でnis遺伝子を発現し、天然ではヨウ素を蓄積しない細胞中でのヨウ素の濃縮が認められるという点で興味深い。しかし、治療への適用を考慮するためのヨウ素の十分な固定レベルを達成するためには克服すべき多数の制限要因が残されている。
【0017】
第1の要因は、腫瘍細胞中でのnis遺伝子発現レベルの低さである。例えば、上記の研究では、インビトロで得られた結果は、非甲状腺細胞で達成されたヨウ素の濃縮が甲状腺細胞中で蓄積されたヨウ素濃度の1/2のままであることが示されている。
【0018】
さらに、腫瘍細胞が膜レベルで示される大量の破壊を考慮すると、非腫瘍性甲状腺細胞に匹敵する効率を有する輸送体の機能的取り込みを確保するのは困難なようであう。
【0019】
別の制限要因は、非甲状腺細胞中でヨウ素が有機化しないことである。しかし、このヨウ素の有機化は細胞中でのヨウ素の維持に必要な構成要素である。したがって、上記で報告の研究では、非甲状腺細胞におけるインビトロで蓄積されたヨウ素の非常に急速な流出(30分と60分との間)が認められる。
【0020】
最後に、腫瘍細胞の不均一性および膜レベルでのこれらの細胞の広範な改変を考慮すると、腫瘍細胞膜中のNIS輸送体の均一な分配を達成することは困難なようであう。
【0021】
したがって、現在記載されているアプローチでは、非甲状腺腫瘍治療用のヨウ素131の使用を考慮するのに十分な発現レベルで非甲状腺腫瘍中にヨウ素を蓄積させることは未だ不可能である。
【先行技術文献】
【非特許文献】
【0022】
【非特許文献1】Maxon、NEJM、309、937〜941、1983
【非特許文献2】P.Thomopoulos、Medecine et Science、4、10、825〜828
【非特許文献3】Daiら、Nature、379、458〜460、1996
【非特許文献4】Smanik P.A.ら、Biochem.Biophys.Res.Commun.、226、339〜345、1996
【非特許文献5】Levyら、1998、J.Biol.Chem.、273、22657〜22663
【非特許文献6】Paire A.ら、J.Biol.Chem.、272、18245〜18249、1997
【非特許文献7】Endo T.ら、Mol.Endocrinol.、11、1747〜1755、1997
【非特許文献8】Mandellら、Cancer Research、59、661〜668、1999
【発明の概要】
【発明が解決しようとする課題】
【0023】
本発明は、遺伝子治療とヨウ素131を使用した放射線治療とを組み合わせた腫瘍の改良された治療法を提供する。
【0024】
本発明は、特に、非甲状腺腫瘍レベルでのヨウ素の固定効率の増大を可能にし、それにより、甲状腺癌治療用に首尾よく開発された放射線治療の原理を非甲状腺腫瘍に適用可能な方法を記載する。
【課題を解決するための手段】
【0025】
本発明は、欠陥組換えアデノウイルスによる特異的ヨウ素輸送体(Na+/I−共輸送体)NISをコードする遺伝子の導入により、非甲状腺腫瘍細胞中のヨウ素の非常に高い蓄積が可能となるという証明に基づく。特定の実施形態では、nis遺伝子を発現する欠陥アデノウイルスの供給により甲状腺細胞で認められる濃度の約5倍の濃度でヨウ素を蓄積可能である。この驚くべき非甲状腺腫瘍細胞のヨウ素125の蓄積能力により、今まで一定の甲状腺腫瘍の治療に限定されていたヨウ素131を使用した代謝放射線治療に基づくこの型の腫瘍の新規の治療アプローチを考慮可能である。
【0026】
代謝放射線治療の主な利点は、周辺組織に有意に照射することなく固定した大放射線量の放射性同位元素を組織に送達させることである。
【0027】
通常はヨウ素を蓄積不可能な腫瘍細胞中に実質的にヨウ素を濃縮させることが有効に可能であるという発見により、非甲状腺起源の腫瘍治療用の多数の適応症においてこの方法の適用を考慮可能である。より詳細には、以下の2つの適応症の型が考慮される:その位置により外部からの放射線治療ではほとんどアクセス不可能な腫瘍(例として、手術不可能な前立腺癌または大脳内腫瘍)、既に照射されており、最大線量が既に投与されているので更なる外部からの放射線治療が不可能な腫瘍(これは、照射領域で発症するか再発する全ての癌に関する)。この方法はまた、ヨウ素捕獲能力を喪失し、代謝治療に反応しない甲状腺腫瘍の治療に適用可能である。
【0028】
分化型甲状腺癌の転移の場合に行われる代謝放射線治療と外部放射線治療との組み合わせも考慮可能である。この組み合わせにより、毒性を増加させることなく放射線治療の治療有効性を増大可能である。この組み合わせは、特に、放射線治療にあまり感受性を示さない腫瘍に有利なようである。さらに、過度に巨大なサイズのために代謝放射線治療でアクセス不可能な腫瘍へのこの技術の適用を考慮可能である。
【0029】
したがって、本発明の第1の目的は、特異的ヨウ素輸送体(Na+/I−共輸送体)NISまたはその誘導体をコードする少なくとも1つの異種DNA配列を含む欠陥組換えアデノウイルスにある。
【発明を実施するための形態】
【0030】
本発明の目的では、特異的ヨウ素輸送体(Na+/I−共輸送体)NISの表現「誘導体」は、NISポリペプチドに由来し、特異的ヨウ素輸送活性を保持する任意のアナログ、フラグメント、または変異形態を意味すると理解される。種々の誘導体は、天然の状態で存在し得る。これらの誘導体は、NISをコードする構造遺伝子のヌクレオチド配列の相違によって特徴づけられる対立遺伝子であるか特異的スプライシングまたは翻訳後修飾に起因し得る。これらの誘導体を、1つまたは複数のアミノ酸残基の置換、欠失、付加、および/または修飾によって得ることができる。これらの修飾を、当業者に公知の任意の技術によって行うことができる。これらの誘導体は、特に、その基質に高い親和性を有する分子、インビボでの改良された発現が可能な配列、プロテアーゼに対してより高い耐性を示す分子、生物学的有効性がより高いか副作用が低いか、おそらく新規の生物学的性質を有する分子である。本発明の文脈で使用することができる他の誘導体は、特に、1つまたは複数の残基が置換された分子、望ましくない活性が考慮されるか発現される結合部位との相互作用に関連しないかあまり関連しない領域の欠失によって得られた誘導体、および天然の配列に関する更なる残基(例えば、分泌シグナルおよび/または結合ペプチドなど)を含む誘導体である。
【0031】
本発明の文脈で産生される特異的ヨウ素輸送体(Na+/I−共輸送体)NISまたはその誘導体は、cDNA、ゲノムDNA(gDNA)、または例えば1つまたは複数のイントロンが挿入されたcDNAからなるハイブリッド構築物であり得る。これには、合成または半合成配列も含み得る。有利には、cDNAまたはgDNAが使用される。特に、gDNAの使用によりヒト細胞でのより良好な発現が可能である。
【0032】
第1の実施形態によれば、これは、マウス起源の特異的ヨウ素輸送体(Na+/I−共輸送体)NISをコードするcDNA配列である。本発明の好ましい実施形態によれば、これは、ヒト起源の特異的ヨウ素輸送体(Na+/I−共輸送体)NISをコードするcDNA配列である。
【0033】
本発明の文脈で使用されるアデノウイルスは、組換えアデノウイルス(すなわち、異種DNA配列を含む)である。有利には、これらは、欠陥組換えアデノウイルス(すなわち、標的細胞中で自律複製不可能なアデノウイルス)である。
【0034】
本発明のアデノウイルスの構築には、種々の血清型を使用することができる。実際に多数の抗原型が存在し、その構造および性質はいくらか異なるが、類似の遺伝子構成を示す。より詳細には、組換えアデノウイルスは、ヒト起源であっても動物起源であってもよい。ヒト起源のアデノウイルスに関して、好ましくはC群(特に、アデノウイルス2型(Ad2)、5型(Ad5)、7型(Ad7)、または12型(Ad12))に分類され得る。動物起源の種々のアデノウイルスのうち、好ましくは、イヌ起源のアデノウイルス(特に、全てのCAV2アデノウイルス株(例えば、マンハッタン株またはA26/61株(ATCC VR−800)))であり得る。動物起源の他のアデノウイルスは、特に、WO 94/26914に引用されている。
【0035】
アデノウイルスゲノムは、特に、各末端の逆末端反復配列(ITR)、キャプシド形成配列(Psi)、初期遺伝子、および後期遺伝子を含む。主な初期遺伝子は、E1、E2、E3、およびE4領域中に含まれる。これらのうち、E1領域に含まれる遺伝子は、特に、ウイルス増殖に必要である。主な後期遺伝子は、L1〜L5領域に含まれる。Ad5アデノウイルスゲノムは完全に配列決定されており、データベースでアクセス可能である(特に、Genbank M73260を参照のこと)。同様に、他のアデノウイルスゲノムの一部またはさらに全部(Ad2、Ad7、Ad12など)も配列決定されている。
【0036】
上記のように、本発明のアデノウイルスは欠陥であるので、標的細胞中で自律複製不可能である。この効果により、アデノウイルス由来の種々の構築物が調製され、種々の治療遺伝子に組み込まれている。これらの各構築物では、感染細胞中で複製不可能なようにアデノウイルスを改変されていた。したがって、先行技術に記載の構築物は、ウイルス増殖に必須のE1領域が欠失され、異種DNA配列が挿入されたアデノウイルスである(Levreroら、Gene、101、1991、195、Gosh−Choudhuryら、Gene、50、1986、161)。さらに、ベクターの性質を改良するために、アデノウイルスゲノム中に他の欠失または改変を作製することが提案されている。したがって、熱感受性点変異をts125変異体に移入して、72kDaのDNA結合タンパク質(DBP)を不活化させることができる(Van der Vlietら、J.Virol.、1975、15、2、348〜354)。他のベクターは、ウイルスの複製および/または増殖に必須の別の領域(E4領域)の欠失を含む。E4領域は、実際に、後期遺伝子の発現の調製、後期核RNAの安定性、宿主細胞タンパク質の発現の消滅、およびウイルスDNAの複製効率に関連する。したがって、E1およびE4領域が欠失したアデノウイルスベクターは、転写バックグラウンドノイズを有し、且つウイルス遺伝子発現が非常に減少している。このようなベクターは、例えば、WO 94/28152に記載されている。さらに、IVa2遺伝子が改変されたベクターも記載されている(WO 96/1008)。
【0037】
本発明の好ましい実施形態では、組換えアデノウイルスは、C型ヒトアデノウイルスである。より詳細には、これはAd2またはAd5アデノウイルスである。
【0038】
有利には、本発明の文脈で使用される組換えアデノウイルスは、そのゲノムのE1領域が欠失している。さらにより詳細には、E1aおよびE1b領域が欠失している。例としては、影響を与えるヌクレオチド454〜3328、386〜3446、459〜3510、または357〜4020(Ad5のゲノムを基準とする)の欠失であり得る。
【0039】
別の変形形態では、本発明の文脈で使用される組換えアデノウイルスは、少なくともE1およびE3領域の全てまたは一部が欠失している。
【0040】
別の変形形態では、本発明の文脈で使用される組換えアデノウイルスは、E1領域の欠失に加えて、そのゲノム中のE4領域の影響を与える全てまたは一部が欠失している。より詳細には、E4領域の欠失は、全ての読み取り枠に影響を与える。詳細な例としては、出願WO 95/02697およびWO 96/22378に記載のE4領域の33466〜35535もしくは33093〜35535または一部のみの欠失であり得る。
【0041】
例えば、いわゆる第3世代組換えアデノウイルス(すなわち、E1およびE4領域が全体または部分的に欠失しており、任意選択的にE3領域が欠失している)であり得る。本発明の特定の変形形態は、以下の領域の全てまたは一部に影響を与える欠失を有するアデノウイルスの使用からなる:
−E1、E4、およびE3、
−E1、E4、およびE2、
−E1、E4、E2、およびE3、
−上記の領域ならびに後期アデノウイルス機能(L1〜L6)をコードする遺伝子の全部または一部、
−全てのコードウイルス領域。
【0042】
後期機能(「最小」ベクター)または全てのコード領域(「ガットレス」ベクター)をさらに欠くアデノウイルスに関して、その構築物は、例えば、Parksら、PNAS、93、1996、13565およびLieberら、J.Virol.、70、1996、8944に記載されている。
【0043】
これには、ハイブリッドアデノウイルスベクター(出願WO 99/60144に記載のものなど)も含んでもよく、このアデノウイルスは別のウイルス用のベクター(特に、レトロウイルスベクター)として使用される。
【0044】
ヨウ素輸送体(NIS)をコードする核酸を含む発現カセットを、組換えゲノムの別の部位に挿入することができる。これを、欠失配列の代わりまたは付加的にE1、E3、またはE4領域に挿入することができる。これを、cis中のウイルス産生に必要な配列(ITR配列およびキャプシド形成配列)の外側の任意の他の部位に挿入することもできる。
【0045】
本発明の目的のために、核酸の、表現「発現カセット」は、ベクターの特異的制限部位に挿入することができるDNAフラグメントを意味すると理解される。このDNAフラグメントは、目的のRNAまたはポリペプチドをコードするヌクレオチド配列に加えて、目的の配列の発現に必要な配列(エンハンサー、プロモーター、ポリアデニル化配列など)を含む。転写および/または翻訳用の適切な読み枠への前記フラグメントの挿入を確保するようにDNAフラグメントおよび制限部位を設計する。
【0046】
組換えアデノウイルスを、キャプシド形成株(すなわち、組換えアデノウイルスゲノム中の1つまたは複数の欠陥機能をトランスで補足することができる細胞株)産生させる。当業者に公知のキャプシド形成株のうち、例としては、アデノウイルスゲノムの一部が取り込まれた293株であり得る。より詳細には、293株は、左側のITR、キャプシド形成領域、E1領域(E1aおよびE1bを含む)、pIXタンパク質コード領域、およびpIVa2タンパク質コード領域の一部を含む血清型5アデノウイルス(Ad5)ゲノムの左側の末端(11〜12%)を含むヒト胚腎臓細胞株である。この株は、E1領域を欠く(すなわち、E1領域の全てまたは一部を欠く)組換えアデノウイルスをトランスで補足することができ、高力価のウイルスストックを産生することができる。この株はまた、許容温度(32℃)で熱感受性E2変異をさらに含むウイルスストックを産生することができる。特に、ヒト肺癌細胞A549(WO 94/28152)またはヒト網膜芽細胞(Hum.Gen.Ther.、1996、215)に基づいて、E1領域を補足することができる他の細胞株が記載されている。さらに、いくつかのアデノウイルス機能をトランスで補足することができる株が記載されている。特に、E1およびE4領域を補足する株(Yehら、J.Virol.、70、1996、559〜565、Cancer Gen.Ther.、2、1995、322、Krougliakら、Hum.Gen.Ther.、6、1995、1575)ならびにE1およびE2領域を補足する株(WO 94/28152、WO 95/02697、WO 95/27071)または特にこのようなウイルスの構築に関連する部位特異的リコンビナーゼ活性をさらに発現することにより最小アデノウイルスの産生に使用することができるそれら由来の株であり得る。
【0047】
腫瘍レベルでの感染効率を増加させるために、組換えアデノウイルスのキャプシド構造を改変することもできる。例えば、キャプシドは、腫瘍細胞へのアデノウイルスの標的化を可能にするuPARリガンドまたはRGDモチーフを含むことができ、このようなベクターおよび標的配列は、特に出願WO 00/12738に記載されている。標的配列を、ヘキソンタンパク質または繊維タンパク質に挿入することができる。好ましくは、繊維タンパク質またはヘキソンタンパク質の欠失レベルで標的配列を挿入する。有利には、Ad5のヘキソンのポリペプチド配列279〜292に対応するヘキソン13アミノ酸のポリペプチド配列から欠失する。有利には、Ad5の繊維(HIループ)のポリペプチド配列539〜547に対応する繊維11アミノ酸のポリペプチド配列から欠失する。
【0048】
組換えアデノウイルスは、通常、キャプシド形株へのウイルスDNAの移入から約2〜3日後(アデノウイルスのサイクル速度は24〜36時間である)に細胞を溶解することによって産生される。この方法を実行するために、移入されたウイルスDNAは、細胞に感染する、任意選択的に細菌(WO 96/25506)または酵母(WO 95/03400)中で構築された完全な組換えウイルスゲノムであり得る。これはまた、キャプシド形成株の感染に使用される組換えウイルスであり得る。ウイルスDNAを、それぞれ組換えウイルスゲノムの一部およびキャプシド形成細胞への移入後に種々のフラグメント間での相同組換えによって組換えウイルスを再構成可能な相同領域を有するフラグメント形態に挿入することもできる。
【0049】
細胞溶解後、組換えウイルス粒子を、塩化セシウム勾配遠心分離またはクロマトグラフィーなどの任意の公知の技術によって単離することができる。別の方法は、出願WO 98/00528に記載されている。
【0050】
発現調節配列。特異的ヨウ素キャリア(Na+/I−共輸送体、NIS)をコードする遺伝子を、機能的で宿主腫瘍細胞での発現を可能にする発現を調節する任意の配列(例えば、プロモーターまたはプロモーター/エンハンサーなど)の調節下に置くことができる。
【0051】
発現調節配列は、プロモーター領域に加えて、特異的ヨウ素輸送体をコードする遺伝子の3’に存在し、且つ転写終結シグナルおよびポリアデニル化部位が得られる領域を含む。これら全てのエレメントが発現カセットを構成する。
【0052】
プロモーターは、構成性プロモーターであっても調節(誘導)プロモーターであってもよい。これは、実際の遺伝子プロモーターであり得る。これはまた、異なる起源(他のタンパク質の発現を担うか合成プロモーター)の配列を含み得る。特に、これは、真核生物またはウイルス遺伝子のプロモーター配列を含み得る。例えば、これは、トランスフェクトが望まれる細胞のゲノム由来のプロモーター配列を含み得る。同様に、これは、ウイルスゲノム(使用されるウイルスを含む)由来のプロモーター配列を含み得る。これに関して、例として、E1A、MLP、CMV、およびRSV−LTR、MT−1、SV40などのプロモーターであり得る。
【0053】
さらに、これらの発現配列を、一定の組織中で組織特異的または主に発現可能な活性化配列または調節配列(エノラーゼプロモーターGFAPなど)の付加によって改変することができる。実際、ウイルスが腫瘍細胞に有効に感染した場合のみ、治療遺伝子がその効果を発現または産生するように腫瘍細胞中で特異的または主に活性である発現シグナルを使用することが特に有利であり得る。発現の特異的または主な特徴とは、プロモーター活性が腫瘍細胞中で有意により高いことを意味する。非特異的発現が他の細胞に存在し得るにもかかわらず、対応する活性レベルは、一般に腫瘍細胞中で認められるものよりも非常に低く(無視できるほど)、一般に少なくとも1/10ほどである。
【0054】
本発明の文脈で使用することができるプロモーターのうちでは、遍在性プロモーター(HPRT、ビメンチン、αアクチン、チューブリンなど)、中間径フィラメントのプロモーター(デスミン、神経フィラメント、ケラチン、GFAP)、治療目的の遺伝子のプロモーター(MDR、CFTR、第VIII因子など)、組織特異的プロモーター(デシン、ミオシン、クレアチンキナーゼ、ホスホグリセリン酸キナーゼ遺伝子のプロモーター)、増殖中の細胞でより特異的に活性なプロモーターまたは刺激に対応するプロモーター(天然のホルモン(ステロイドホルモンレセプター、レチノイン酸レセプターなど)に対応するプロモーターなど)、抗生物質(テトラサイクリン、ラパマイシンなど)によって調節されるプロモーター、天然または合成起源の他の分子に対応する他のプロモーター、またはウイルスゲノム由来のプロモーター配列(サイトメガロウイルスCMVエンハンサー/プロモーター、レトロウイルスLTRプロモーター、SV40プロモーター、E1A遺伝子プロモーター、MLPプロモーター)であり得る。テトラサイクリンによって調節することができるプロモーターおよびCMVプロモーターは、WO 96/01313、米国特許第5,168,062号、および同第5,385,839号に記載されている。
【0055】
本発明の実施に使用することができるプロモーターのうちでは、特に、サイトメガロウイルス(CMV)プロモーター、SV40初期プロモーター領域(Benoist and Chambon、1981、Nature、290、304〜310)、ラウス肉腫ウイルス3’LTR領域に含まれるプロモーター(Yamamotoら、1980、Cell、22、787〜797)、ヘルペスウイルスチミジンキナーゼプロモーター(Wagnerら、1981、Proc.Natl.Acad.Sci.U.S.A.、78、1441〜1445)、メタロチオネイン遺伝子の調節配列(Brinsterら、1982、Nature、296、39〜42)、原核生物起源のプロモーター(βラクタマーゼプロモーター(Villa−Kamaroffら、1978、Proc.Natl.Acad.Sci.U.S.A.、75、3727〜3731)、またはtacプロモーター(DeBoerら、1983、Proc.Natl.Acad.Sci.U.S.A.、80、21〜25)など(「組換え細菌由来の有用なタンパク質」、Scientific American、1980、242、74〜94もまた参照のこと))、酵母プロモーター(Gal4、ADC(アルコールデヒドロゲナーゼ)、PGK(ホスホグリセロールキナーゼ)、およびアルカリ性ホスファターゼなど)、および組織特異性を示し且つトランスジェニック動物に使用される以下の動物起源の転写緒節配列:膵臓の腺房細胞中で活性なエラスターゼI遺伝子の調節配列(Swift、1984、Cell、38、639〜646、Ornitzら、1986、Cold Spring Harbor Symp.Quant.Biol.、50、399〜409、MacDonald、1987、Hepatology、7、425〜515)、膵臓のβ細胞中で活性な遺伝子[lacuna]インスリン調節配列(Hanahan、1985、Nature、315、115〜122)、リンパ球様細胞中で活性な免疫グロブリンの発現を調節する配列(Grosschedlら、1984、Cell、38、647〜658、Adamesら、1985、Nature、318、533〜538、Alexxanderら、1987、Mol.Cell.Biol.、7、1436〜1444)、精巣細胞および乳房細胞、リンパ球および肥満細胞中で活性なマウス乳腺癌ウイルスの調節配列(Lederら、1986、Cell、45、485〜495)、前立腺癌中で活性なPSA遺伝子の調節配列、肝臓中で活性なアルブミン遺伝子の調節配列(Pinkertら、1987、Genes and Devel.、1、268〜276)、肝臓中で活性なαフェトプロテインの調節配列(Krumlaufら、1985、Mol.Cell.Biol.、5、1639〜1648、Hammerら、1987、Science、235、53〜58)、肝臓中で活性なα1抗トリプシン遺伝子の調節配列(Kelseyら、1987、Genes and Devel.、1、161〜171)、骨髄性細胞中で活性なβグロビン遺伝子の調節配列(Mogramら、1985、Nature、315、338〜340、Kolliasら、1986、Cell、46、89〜94)、脳の希突起膠細胞中で活性な塩基性ミエリン遺伝子の調節配列(Readheadら、1987、Cell、48、703〜712)、骨格筋中で活性なミオシン軽鎖2遺伝子の調節配列(Sani、1985、Nature、314、283〜286)、および視床下部で活性なゴナドトロピン放出ホルモン遺伝子の調節配列(Masonら、1986、Science、234、1372〜1378)であり得る。
【0056】
本発明の特定の実施形態では、好ましくはRSV−LTRまたはCMV初期プロモーターから選択されるウイルスプロモーターの調節下で特異的ヨウ素輸送体(Na+/I−共輸送体)NISをコードする遺伝子を含む欠陥組換えアデノウイルスを使用する。
【0057】
本発明のベクターへのNIS遺伝子の移入とヨウ素131の生物学的半減期を増加可能な生物系への移入とを組み合わせることも可能である。したがって、NIS遺伝子およびヨウ素有機化機構に関連するポリペプチドをコードする1つの遺伝子またはいくつかの遺伝子を腫瘍細胞に同時または首尾よく導入可能である。例えば、グルコースオキシダーゼをコードする遺伝子などのペルオキシダーゼ系に関連するポリペプチドをコードする遺伝子であり得る。また、ヨウ素有機化過程に関連する甲状腺細胞中の酵素である甲状腺ペルオキシダーゼをコードする遺伝子(Nucleic Acids Res.、15、6735〜6735、1987、配列GenBank g37251)であり得る。甲状腺ペルオキシダーゼをコードする遺伝子の移入を、サイログロブリンの全てまたは一部(チロシン残基が豊富なサイログロブリン部分)をコードする核酸配列の移入と組み合わせることができる。
【0058】
いくつかの導入遺伝子を発現する1つのベクター、2つのベクターの同時感染、NIS遺伝子を発現するアデノウイルスベクターおよび別のベクター、ウイルスもしくはプラスミドベクター、ヨウ素有機化機構に関連するポリペプチドをコードする1つまたは複数の遺伝子を発現するウイルスまたはプラスミドベクターによる腫瘍細胞へのNIS遺伝子およびヨウ素有機化機構に関連するポリペプチドをコードする1つの遺伝子またはいくつかの遺伝子の送達も考慮可能である。
【0059】
さらに、本発明のベクターへのNIS遺伝子の移入と薬物(例えば、リチウム塩)によってヨウ素流出の抑制させる系の投与とを組み合わせ可能である。
【0060】
別の変形形態では、NIS遺伝子の移入と癌治療用の目的の治療遺伝子との組み合わせも考慮可能である。これには、ヘルペスチミジンキナーゼ遺伝子またはシトシンデアミナーゼ遺伝子などの自殺遺伝子を含み得る。これには、アポトーシスを誘導する遺伝子(p53、Bax、BclX−s、Bad、またはBc12およびBclX−1の任意の他のアンタゴニストなど)を含み得る。これには、改良された性質を示すこれらのタンパク質変異型(p53の変異型)(CTS−1、WO 97/04092)をコードする遺伝子を含み得る。これにはまた、抗血管形成または血管形成阻害因子(特に、Tie−1およびTie−2のリガンド、アンギオスタチン、エンドスタチン、ATF因子、プラスミノゲン誘導体、エンドセリン、トロンボスポンジン1および2、PF−4、インターフェロンαおよびβ、インターロイキン12、TNFα、ウロキナーゼレセプター、flt1、KDR、PAI1、PAI2、TIMP1、プロラクチンフラグメント)をコードする遺伝子を含み得る。これはまた、抗腫用免疫を誘導するか免疫応答を刺激することができるタンパク質(IL2、GM−CSF、IL12など)をコードする遺伝子を含み得る。癌治療目的のタンパク質をコードする遺伝子のうちで、抗体、抗体一本鎖抗体(ScFv)のフラグメント、または腫瘍治療の免疫療法用の認識能力を有する任意の他の抗体フラグメント(抗RAS抗体(WO 9429446))を強調することも重要である。
【0061】
有利には、これには、その発現により腫瘍細胞が放射線増感される遺伝子(p53遺伝子)を含み得る。
【0062】
NIS輸送体をコードする遺伝子を含むアデノウイルスベクターまたはウイルスまたは非ウイルス性の個別のベクターへの1つまたは複数のこれらの遺伝子移入によって、治療目的のこれらの遺伝子または上記の2つまたはそれ以上のこれらの遺伝子の組み合わせの投与を行うことができる。
【0063】
別の変形形態では、本発明のベクターを、抗癌剤(特に、タキソール、タキソテール、および他のタキソイドなど(特に、米国特許第4,857,653号、同第4,814,470号、同第4,924,011号、同第5,290,957号、同第5,292,921号、同第5,438,072号、同第5,587,493号、欧州特許第0253738号、およびWO 91・17976、WO 93/00928、WO 93/00929、およびWO 9601815に記載されている))または他の治療用抗癌剤(シスプラチンおよび白金誘導体、エトポシドおよびリン酸エトポシド、ブレオマイシン、マイトマイシンC、CCNU、ドキソルビシン、ダウノルビシン、イダルビシン、イホスファミドなど)と組み合わせて投与することができる。
【0064】
本発明はまた、上記のアデノウイルスベクターおよび生理学的に受容可能な賦形剤を含む薬学的組成物に関する。本発明の薬学的組成物は注射形態が好ましく、腫瘍内投与用または、経口、非経口、鼻腔内、動脈内、静脈内、または気管内経路などの投与用に処方することができる。
【0065】
好ましくは、薬学的組成物は、腫瘍内経路による投与を意図した処方用の薬学的に受容可能な賦形剤を含む。
【0066】
本発明の組成物は、適用の考慮および種々のパラメータ、特に使用した投与様式または所望の発現の持続時間による当業者が容易に調整することができる種々の用量の組換えアデノウイルスを含み得る。一般に、本発明の組換えウイルスを、104と1014pfuとの間、好ましくは106と1011pfuとの間の用量の形態で処方して投与する。用語「pfu(「プラーク形成単位」)」は、ウイルスの感染力に対応し、適切な細胞培養物への感染および感染細胞プラーク数の計数によって同定される。ウイルス溶液のpfu力価の同定技術は、文献に十分に記載されている。
【0067】
さらに、本発明の組成物はまた、化学導入剤または生物導入剤を含み得る。要路「化学導入剤または生物導入剤」は、細胞への核酸の浸透を容易にする任意の化合物(すなわち、組換えウイルス以外)を意味すると理解される。これには、陽イオン性非ウイルス薬(陽イオン性リピド、ペプチド、ポリマー(ポリエチレンイミン、ポリリジン)、ナノ粒子など)または非陽イオン性非ウイルス薬(非陽イオン性リポソーム、ポリマー、または非陽イオン性ナノ粒子など)を含み得る。
【0068】
好ましい実施形態では、本発明の組成物は、ヨウ素輸送体NISをコードする遺伝子を含む欠陥組換えベクターを含み、これは腫瘍内投与用に処方される。有利には、本発明の組成物は、104と1015pfuとの間、好ましくは107と1012pfuとの間を含む。
【0069】
本発明の主題はまた、ヨウ素輸送体(NIS)をコードする核酸を含む組換えベクターを1つまたは複数の適合性アジュバントおよび薬学的に受容可能なアジュバントと混合することを特徴とする、腫瘍の予防、改善、および/または治療に有用な薬物の調製法である。
【0070】
本発明はまた、ヨウ素輸送体(NIS)をコードする核酸を含む欠陥組換えアデノウイルスベクターの有効量を投与する工程を包含する、哺乳動物、特にヒトの治療法に関する。
【0071】
用語「有効量」は、ヨウ素輸送体(NIS)をコードする核酸を含むアデノウイルスの投与をヨウ素135での代謝放射線治療と組み合わせた場合、腫瘍体積の少なくとも約15%、好ましくは少なくとも50%、より好ましくは少なくとも90%を減少させるのみに十分な量またはさらに好ましくは腫瘍を消滅させるのに十分な量を示す。
【0072】
本発明は、腫瘍治療、より詳細には固形腫瘍の治療に関する。本発明の主題によって治療することができる固形腫瘍のうち、特に、肉腫および悪性腫瘍(例として、線維肉腫、骨原性肉腫、血管肉腫、内皮肉腫、リンパ管肉腫、ユーイング肉腫、大腸癌、膵臓癌、卵巣癌、前立腺癌、腺癌、腎臓、肝臓、または胆管の癌、ウィルムス腫瘍、子宮頸癌、精巣癌、肺癌、非小細胞肺癌、膀胱癌、上皮癌、神経膠腫、星状細胞腫、黒色腫、神経芽細胞腫、および網膜芽細胞腫であるが、これらに限定されない)であり得る。
【0073】
本発明はまた、上皮組織(子宮頚管、食道、および肺の上皮など)の増殖性障害(化生および異形成)の予防および/または治療に関する。これに関して、本発明は、特に、過形成、化生、より詳細には異形成などの非新生細胞の増殖が起こる状態での腫瘍形成または癌への進行に先立つことが公知であるか疑われる病態の治療に関する(これらの異常な増殖状態についての概説は、Robbins and Angell、1976、Basic Pathology、第2版、W.B.Sauders Co.、Philadelphia、68〜79を参照のこと)。過形成は、器官の顕著な構造的または機能的変化を伴わない器官中での細胞数の増加を含む制御された細胞増殖の形態である。例えば、子宮内膜の過形成は、子宮内膜癌に先駆けて起こる。化生は、成体または完全に分化された細胞型が別の細胞型に置換される制御された細胞増殖の形態である。化生は、上皮組織または結合組織で起こり得る。異形成は、しばしば癌の早期危険信号であり、主に上皮で見出される。これは、新生細胞増殖の最も頻繁な形態であり、各細胞の一様性の喪失および細胞の構造上の配向性の喪失を含む。異形成は、典型的には、慢性的刺激作用または炎症が存在する場合に起こり、しばしば子宮頚管、気道、声帯腔、および膀胱壁に認められる。概説として、Fishmanら、1985、Medicine、第2版、J.B.Lippincott Co.、Philadelphiaを参照のこと。
【0074】
本発明は、以下の実施例によってより詳細に記載されるが、これらは例示であり、本発明を制限しない。
【図面の簡単な説明】
【0075】
【図1】プラスミドpAB1、プラスミドpXL3048、およびプラスミドpAB2の線図である。プラスミドpXL3048は、アデノウイルス5型ゲノムの左側の末端(ヌクレオチド1〜382)、3つの固有のクローニング部位を含むポリリンカー、およびpIX遺伝子の一部(ヌクレオチド3446〜4296)を含む。プラスミドpAB2は、EcoRVで先に線状化したpXL3048へのpAB1のSspI−EcoRVフラグメントのクローニングに起因する。
【図2】Crouzetら、PNAS、94、1414、1997に記載の方法による、プラスミドpAB2およびpXL3215由来の二重組換えによって作製されたプラスミドpXL3215の産生を示す図である。プラスミドpXL3215は、E1およびE3領域が欠失されたアデノウイルス5型ゲノムを含み、且つNIS発現カセットを含む。
【図3】Ad−NISベクターで感染された(感染多重度10)FTRL−5およびSiHa細胞によるヨウ素125の蓄積速度を示す図である。速度の結果を、数/分/106細胞で示す。ヨウ素接触時のウェルあたりの細胞数の判定は、2つの測定の平均である。
【図4】30、300、および3000μMの過塩素酸塩(NaClO4)の存在下でのNISによるヨウ素輸送の特異的阻害を示す図である。細胞とヨウ素125との間の接触時間は15分間である。
【図5A】Ad−NISベクターを感染させてない腫瘍によるインビボでのヨウ素蓄積を示す図である(コントロール)。MCF−7細胞(ヒト乳がんの腫瘍細胞)を、ヌードマウスに皮下注射した(5×106細胞)。15日後(腫瘍の出現)、チロキシンの腹腔内注射(2μg/動物/日)を毎日15日間行った。次いで、Ad−CMV−NISベクターを一定の動物に腫瘍内経路で注射した(2×109pfu/腫瘍、腫瘍サイズ3〜6mm)。注射から3日後、6μCiの125Iを、腹腔内経路でマウスに注射する。甲状腺上、腫瘍上、腫瘍のすぐ隣にプローブを置くことによって一定の時間間隔で10秒間計数した。結果を、時間の関数として検出された数(10秒間の測定)として示す。
【図5B】Ad−NISベクターを感染させた腫瘍によるインビボでのヨウ素蓄積を示す図である。MCF−7細胞(ヒト乳がんの腫瘍細胞)を、ヌードマウスに皮下注射した(5×106細胞)。15日後(腫瘍の出現)、チロキシンの腹腔内注射(2μg/動物/日)を毎日15日間行った。次いで、Ad−CMV−NISベクターを一定の動物に腫瘍内経路で注射した(2×109pfu/腫瘍、腫瘍サイズ3〜6mm)。注射から3日後、6μCiの125Iを、腹腔内経路でマウスに注射する。甲状腺上、腫瘍上、腫瘍のすぐ隣にプローブを置くことによって一定の時間間隔で10秒間計数した。結果を、時間の関数として検出された数(10秒間の測定)として示す。
【図6】腫瘍をAd−NISベクターで感染させたマウスのシンチグラフィーである。MCF−7細胞(ヒト腫瘍細胞、乳癌)を、ヌードマウスに皮下注射した(5×106細胞)。15日後(腫瘍の出現)、チロキシンの腹腔内注射(2μg/動物/日)を毎日15日間行った。次いで、Ad−NISベクターを腫瘍内経路で注射した(2×109pfu/腫瘍、腫瘍サイズ3〜6mm)。注射から3日後、50μCiの125Iを、腹腔内経路でマウスに注射する。ヨウ素注射から1時間後に計数をおこなった。画像は、動物の縦断図である。
【0076】
材料と方法
一般的な分子生物学技術
分子生物学で慣習的に使用されている方法(プラスミドDNAの予備的抽出、塩化セシウム勾配によるプラスミドDNAの遠心分離、アガロースまたはアクリルアミドゲル電気泳動、電気溶出によるDNAフラグメントの精製、フェノールまたはフェノール−クロロホルムによるタンパク質抽出、エタノールまたはイソプロパノールによる生理食塩水培地中でのDNAの沈殿、Esxherichia coliの形質転換など)は、当業者に周知であり、文献中に多数記載されている(Maniatis T.ら、「分子クローニング、実験マニュアル」、Cold Spring Harbor Laboratory、Cold Spring Harbor,N.Y。、1982、Ausubel F.M.ら編、「現代の分子生物学プロトコール」、John Wiley & Sons、New York、1987)。
【0077】
ライゲーションでは、DNAフラグメントをアガロースまたはアクリルアミドゲル電気泳動によってそのサイズごとに分離し、フェノールまたはフェノール/クロロホルム混合物で抽出し、エタノールで沈殿させて、供給者の指示に従ってT4ファージDNAリガーゼ(Biolabs)の存在下でインキュベートすることができる。
【0078】
供給者の指示に従って、E.coliのDNAポリメラーゼのクレノウフラグメント(Biolabs)を使用して、突出した5’末端の充填を行うことができる。供給者の指示に従って、T4ファージDNAポリメラーゼ(Biolabs)の存在下で、突出した3’末端の破壊を行う。S1ヌクレアーゼで調節された処理によって突出した5’末端の破壊を行う。
【0079】
Amershamから販売されているキットを使用し、Taylorら、Nucleic Acids Res.、13、1985、8749〜8764が開発した方法にしたがって、合成オリゴデオキシヌクレオチドによってインビトロで指向される変異誘発を行うことができる。
【0080】
いわゆるPCR技術(ポリメラーゼ触媒連鎖反応、Saiki R.K.ら、Science、230、1985、1350〜1354、Mullis K.B.and Faloona F.A.、Meth.Enzym.、155、1987、335〜350)によるDNAフラグメントの酵素増幅を、供給者の指示に従って、「DNAサーマルサイクラー」(Perkin Elmer Cetus)を使用して行うことができる。
【0081】
ヌクレオチド配列の評価を、Amershamから販売のキットを使用し、Sangerら、Proc.Natl.Acad.Sci.USA、74、1977、5463〜5467が開発した方法によって行うことができる。
【実施例】
【0082】
[実施例1]特異的ヨウ素輸送体(Na+/I−共輸送体)NISをコードする遺伝子を発現する欠陥組換えアデノウイルスの構築
本実施例は、CMVプロモーターに作動可能に連結された特異的ヨウ素輸送体(Na+/I−共輸送体)NIS(Genbank U60282を参照のこと)をコードする遺伝子を保有する欠陥アデノウイルスベクターの構築を記載する。
【0083】
AatII−HindIIIフラグメントの形態のラットnis遺伝子のヌクレオチド配列74〜2046を含むフラグメント(Daiら、1996、Nature、379、458〜460)を、プラスミドpCEP−4(Invitrogen)のPvuII−HindIII部位にクローン化した。AatII−HindIIIフラグメントのAtII部位を、Mung Beanヌクレアーゼ(Biolabs)での処理で予め平滑にし、プラスミドpCEP−4を挿入した。得られたプラスミドをpAB1と呼ぶ。
【0084】
プラスミドpAB1(図1)は、サイトメガロウイルスCMVエンハンサー/プロモーター(ヌクレオチド−522〜+72)の調節下で発現するnis遺伝子を含み(Boshartら、1985、Cell、41、521〜530)、その後ろにSV40ウイルスポリアデニル化部位(SV40ゲノムのヌクレオチド2538〜2759、Genbank遺伝子座SV4CG)を含む。
【0085】
(i)サイトメガロウイルスエンハンサー/プロモーターと、(ii)nis遺伝子のcDNAと、(iii)SV40ウイルスポリアデニル化部位との組み合わせを、以後NIS発現カセットと呼ぶ。
【0086】
次いで、上記で定義のNIS発現カセットを含むプラスミドpAB1のSspI−EcoRVフラグメント(3711bp)を、EcoRVで予め線状化したシャトルベクターpXL3048にクローン化する。得られたベクターを、pAB2と呼ぶ(図2)。プラスミドpXL3048は、5型アデノウイルスゲノムの左側の末端(ヌクレオチド1〜382)、3つの固有のクローニング部位を含むポリリンカー、およびpIX遺伝子の一部(ヌクレオチド3446〜4296)を含むプラスミドKan−SacB(Crouzetら、PNAS、94、1414〜1419、1997)由来のプラスミドである。
【0087】
アデノウイルスAd−NISを、Crouzetら、PNAS、94、1414〜1419、1997に記載の方法に従って、プラスミドpAB2とプラスミドpXL3215との間の相同組換えによって構築した。
【0088】
プラスミドpXL3215は、そのE1領域(ΔE1 386〜3446)にRSV−LacZカセットを含み、E3領域(ΔE3 28592〜30470)が欠失したアデノウイルスゲノムを含む。
【0089】
構築の原理を、図2に示す。この二重組換えによって作製されたプラスミドpXL3215は、E1およびE3領域が欠失され、CNV−NIS−ポリA SV40発現カセットを含む5型アデノウイルスゲノムを含む。この構築物を、NIS発現カセットの配列決定によって評価する。
【0090】
Ad−NISアデノウイルスを、293株(ATCC CRL−1573)へのPacIで消化したpXL3215のDNAの挿入によって作製する。次いで、得られたウイルス粒子を、この同一の株で増幅させ、産生されたウイルスストックを、二重CsCl勾配によって精製する。
【0091】
次いで、ウイルス粒子を使用して、種々の腫瘍株におけるNIS遺伝子の発現を研究する。
【0092】
[実施例2]Ad−NISベクター感染腫瘍細胞によるインビトロでのヨウ素の蓄積
Ad−NISベクター感染細胞におけるヨウ素の蓄積能力を以下の種々の株で試験した:上皮腫瘍細胞SiHa(ATCC HTB−35)、乳癌細胞MCF7(ATC HTB−22)およびT−47D(ATCC HTB−133)ならびに前立腺起源の腫瘍細胞DU−145(ATCCHTB−81)およびPC−3(ATCC CRL−1435)。
【0093】
細胞に感染多重度10でAd−NISベクターを感染させた。細胞のヨウ素捕獲能力を、Weiss らの方法(Endocrinology、1984、114、1090〜1098)にしたがって感染から28〜29時間後に試験する。感染細胞をHBSS緩衝液で洗浄し、0.1μCiのヨウ素125を含むHBSS緩衝液と共に図3に表示の時間接触させる。接触終了後、細胞を冷却HBSS緩衝液で1回洗浄し、冷却エタノールと20分間インキュベートする。細胞によって捕獲されたヨウ素125の量を、ガンマカウンターを使用したエタノール中の放射能のアッセイによって同定する。
【0094】
SiHa株を使用して得られた結果を図3に示し、これを、FRTL−5細胞(ヨウ素を天然に捕獲し、コントロールとして使用したラット甲状腺細胞)で得られた結果と比較することができる。
【0095】
過塩素酸塩の存在下でのNISによるヨウ素輸送体の特異的阻害を試験した。NaClO4をヨウ素125を含むHBSS緩衝液に添加して、30、300、および3000μMの最終濃度とする。細胞とヨウ素125との間の接触時間は15分間である。結果を図4に示す。結果を、同数の細胞についての数として示す。
【0096】
FRTL−5細胞(ポジティブコントロール)は、一定量のヨウ素を固定し、この現象は非常に急速である(接触から15分後にほぼ最大に達する)ことが認められる。得られた結果より、感染していないSiHa細胞(ヒト上皮腫瘍細胞)はヨウ素を捕獲しないことが示される。驚いたことに、Ad−NISベクターを感染させたSiHa細胞はFRTL−5細胞の5倍を超えるヨウ素を捕獲することが認められる。この現象の初速度はFRTL−5細胞で認められた速度に匹敵することにも注目すべきである。輸送特異性を、nis遺伝子を含まないコントロールアデノウイルスでの感染によって確認した(結果を示さず)。
【0097】
阻害実験の結果(図4)は、FRTL−5細胞のヨウ素固定が30μMのNaClO4濃度で90%を超えて阻害されることを示す(30μMでは93%、高濃度で95%)。一方、Ad−NISを感染SなせたSiHa細胞では、同レベルの阻害を得るために必要なインヒビター濃度は、更により高い(30μMのNaClO4ではわずか40%阻害、300μMのNaClO4では92%阻害、3mMのNaClO4では98%阻害)ことに注目することが興味深い。
【0098】
阻害実験により、NISによるヨウ素イオン輸送の競合的および特異的阻害であるこの現象はNaClO4によって阻害されるので、Ad−NIS感染SiHa細胞のヨウ素捕獲能力は確かにこれらの細胞中の発現NISによると結論付けることが可能である。他のヒト腫瘍細胞株(乳癌細胞:MCF−7およびT47−D、ならびに前立腺起源の腫瘍細胞:DU−145およびPC−3)についても類似の結果が得られた。
【0099】
NaClO4による阻害に関する結果により、腫瘍細胞のAd−NIS感染により特に高レベルでNISが発現することも確認されるが、これはこの現象の阻害に高濃度のNaClO4が必要であるからである(SiHa細胞およびFRTL−5細胞の関連細胞の場合、同レベルの阻害を得るために10倍を超える阻害濃度が必要である)。
【0100】
これら全ての結果により、アデノウイルスベクターは、nis遺伝子の導入に特に有利なベクターであり、このようなベクターの使用により非甲状腺腫瘍細胞においてヨウ素125を天然に捕獲する細胞で認められるレベルの5倍の蓄積レベルを獲得可能であることが示される。
【0101】
[実施例3]Ad−NISベクター感染腫瘍によるインビボでのヨウ素蓄積
MCF−7細胞(乳癌のヒト腫瘍細胞)を、ヌードマウスに皮下注射した(5×106細胞)。15日後(腫瘍の出現)、チロキシンの腹腔内注射(2μg/動物/日)を毎日15日間行い、動物の甲状腺を安静にした。次いで、Ad−NISベクターを一定の動物に腫瘍内経路で注射した(2×109pfu/腫瘍)。注射時の腫瘍の直径は3〜6mmであった。注射から3日後、6μCiの125Iを、腹腔内経路でマウスに注射し、動物の異なる部位に移動することができるプローブの支援によって10秒間にわたり計数した。甲状腺上、腫瘍上、腫瘍のすぐ隣にプローブを置くことによって一定の時間間隔で計数した。結果を図5に示す。結果を、時間の関数として検出された数(10秒間の測定)として示す。
【0102】
コントロール動物(Ad−NISを未投与)では、甲状腺は、少量のヨウ素を固定するが、チロキシンでの治療による甲状腺中のヨウ素の実質的な蓄積は長時間認められない。コントロール動物では、腫瘍中のヨウ素固定は認められない(図5Aを参照のこと)。NISが胃中に天然に存在するという事実およびヨウ素が尿から排泄されるという事実は、比較的高い値が認められることの説明となる(比較的高いバックグラウンドノイズ)。
【0103】
腫瘍内経路によって投与されたAd−NIS治療動物(図5B)では、甲状腺中の実質的なヨウ素蓄積が認められない(チロキシンの治療効果)。これらの動物では、腫瘍上にプローブを置くことによって検出されたヨウ素の量は、腫瘍付近で検出されたヨウ素量よりも系統的に高く、これはAd−NISベクターでの治療後に実際に腫瘍中でヨウ素が固定されたことを示す。
【0104】
腫瘍中でのヨウ素固定を確認するために、Ad−NISベクターで処理した動物におけるシンチグラフィーによる放射能測定を行った。得られた結果を図6に示す。50μ/Ciの123Iを腹腔内経路で注射する以外は上記のプロトコールにしたがって使用動物を調製し、ヨウ素注射から1時間後に画像を得た。
【0105】
画像は動物の縦断図である。以下の4つのヨウ素固定領域が明らかである:甲状腺、膀胱(ヨウ素排泄経路)、胃(天然でNISが発現される)、および腫瘍。これらのシンチグラフィーの結果により計数によって得られた先の結果が確認され、これは、本発明のベクターにより腫瘍中の実質的なヨウ素固定が可能であることを示す。
【0106】
[実施例4]Ad−NISベクター感染腫瘍によるヨウ素蓄積の定量的評価
SiHa細胞(5×106細胞)を、ヌードマウス(n=12)の両側腹に皮下注射した。15日後(腫瘍の出現)、チロキシンの腹腔内注射(2μg/動物/日)を毎日15日間行い、動物の甲状腺を安静にした。各動物について、Ad−NISベクターを腫瘍に腫瘍内経路で注射し(2×109pfu/腫瘍)、他の腫瘍をコントロールとして使用した。注射時の腫瘍の直径は5〜8mmであった。注射から3日後、6μCiの125Iを腹腔内経路でマウスに注射した。125I投与から90分後に動物を屠殺し、125Iの量を同定した。結果を表1に示す。結果を、1gの組織に投与した125Iの百分率として示す。
【0107】
【表1】
【0108】
得られた結果により、Ad−NIsベクターで治療した腫瘍はヨウ素125Iを非常に効率的に蓄積することができ、未処理腫瘍よりも25倍まで高いことが示される。これらの結果により先の結果が確認され、本発明のベクターにより腫瘍中のヨウ素が実質的に固定可能であることが示される。
【特許請求の範囲】
【請求項1】
特異的ヨウ素輸送体(Na+/I−共輸送体)NISまたはその誘導体をコードする少なくとも1つのDNA配列を含むこと特徴とする、欠陥組換えアデノウイルス。
【請求項2】
前記DNA配列がcDNA配列であることを特徴とする、請求項1に記載のアデノウイルス。
【請求項3】
前記DNA配列がgDNA配列であることを特徴とする、請求項1に記載のアデノウイルス。
【請求項4】
前記DNA配列が特異的マウスヨウ素輸送体(Na+/I−共輸送体)NISをコードすることを特徴とする、請求項1〜3のいずれか1項に記載のアデノウイルス。
【請求項5】
前記DNA配列が特異的ヒトヨウ素輸送体(Na+/I−共輸送体)NISをコードすることを特徴とする、請求項1〜3のいずれか1項に記載のアデノウイルス
【請求項6】
前記DNA配列が腫瘍細胞中でのその発現を可能にする転写プロモーターの調節下に存在することを特徴とする、請求項1〜5のいずれか1項に記載のアデノウイルス。
【請求項7】
前記転写プロモーターがウイルスプロモーター、好ましくはE1A、MLP、CMV、およびRSV−LTR、MT−1、SV40のプロモーターから選択されることを特徴とする、請求項6に記載のアデノウイルス。
【請求項8】
CMVプロモーターの調節下でヒトヨウ素輸送体NISをコードするcDNA配列を含む、欠陥組換えアデノウイルス。
【請求項9】
腫瘍細胞中での主な発現を可能にするプロモーターの調節下でヨウ素輸送体NISまたはその誘導体をコードするDNA配列を含む、欠陥組換えアデノウイルス。
【請求項10】
前記プロモーターが、エラスターゼI遺伝子、インスリン遺伝子、免疫グロブリンの遺伝子、マウス乳腺癌ウイルス、PSA遺伝子、αフェトプロテイン遺伝子、α1抗トリプシン遺伝子、βグロビン遺伝子、塩基性ミエリン遺伝子、ミオシン軽鎖2の遺伝子、およびゴナドトロピン放出ホルモン遺伝子の調節配列から選択されることを特徴とする、請求項9に記載の欠陥組換えアデノウイルス。
【請求項11】
少なくともE1領域の全てまたは一部が欠失されており、且つE4領域の全部または一部が欠失されていることを特徴とする、請求項1〜10のいずれか1項に記載のアデノウイルス。
【請求項12】
E4領域の全てまたは一部がさらに欠失されていることを特徴とする、請求項11に記載のアデノウイルス。
【請求項13】
ヒトアデノウイルスAd2型またはAd5型またはイヌアデノウイルスCAV−2型であることを特徴とする、請求項1〜12のいずれか1項に記載のアデノウイルス。
【請求項14】
グルコースオキシダーゼまたは甲状腺ペルオキシダーゼの遺伝子などのペルオキシダーゼ系に関連するポリペプチドをコードする少なくとも1つの遺伝子をさらに含むことを特徴とする、請求項1〜13のいずれか1項に記載のアデノウイルス。
【請求項15】
腫瘍成長の治療および/または阻害を意図する薬学的組成物の調製用の、請求項1〜14のいずれか1項に記載のアデノウイルスの使用。
【請求項16】
請求項1〜14のいずれか1項に記載の1つまたは複数の欠陥組換えアデノウイルスを含む薬学的組成物。
【請求項17】
注射可能形態であることを特徴とする、請求項16に記載の薬学的組成物。
【請求項18】
104と1014pfu/mlとの間、好ましくは106と1011pfu/mlとの間の欠陥組換えアデノウイルスを含むことを特徴とする、請求項16または17に記載の薬学的組成物。
【請求項1】
特異的ヨウ素輸送体(Na+/I−共輸送体)NISまたはその誘導体をコードする少なくとも1つのDNA配列を含むこと特徴とする、欠陥組換えアデノウイルス。
【請求項2】
前記DNA配列がcDNA配列であることを特徴とする、請求項1に記載のアデノウイルス。
【請求項3】
前記DNA配列がgDNA配列であることを特徴とする、請求項1に記載のアデノウイルス。
【請求項4】
前記DNA配列が特異的マウスヨウ素輸送体(Na+/I−共輸送体)NISをコードすることを特徴とする、請求項1〜3のいずれか1項に記載のアデノウイルス。
【請求項5】
前記DNA配列が特異的ヒトヨウ素輸送体(Na+/I−共輸送体)NISをコードすることを特徴とする、請求項1〜3のいずれか1項に記載のアデノウイルス
【請求項6】
前記DNA配列が腫瘍細胞中でのその発現を可能にする転写プロモーターの調節下に存在することを特徴とする、請求項1〜5のいずれか1項に記載のアデノウイルス。
【請求項7】
前記転写プロモーターがウイルスプロモーター、好ましくはE1A、MLP、CMV、およびRSV−LTR、MT−1、SV40のプロモーターから選択されることを特徴とする、請求項6に記載のアデノウイルス。
【請求項8】
CMVプロモーターの調節下でヒトヨウ素輸送体NISをコードするcDNA配列を含む、欠陥組換えアデノウイルス。
【請求項9】
腫瘍細胞中での主な発現を可能にするプロモーターの調節下でヨウ素輸送体NISまたはその誘導体をコードするDNA配列を含む、欠陥組換えアデノウイルス。
【請求項10】
前記プロモーターが、エラスターゼI遺伝子、インスリン遺伝子、免疫グロブリンの遺伝子、マウス乳腺癌ウイルス、PSA遺伝子、αフェトプロテイン遺伝子、α1抗トリプシン遺伝子、βグロビン遺伝子、塩基性ミエリン遺伝子、ミオシン軽鎖2の遺伝子、およびゴナドトロピン放出ホルモン遺伝子の調節配列から選択されることを特徴とする、請求項9に記載の欠陥組換えアデノウイルス。
【請求項11】
少なくともE1領域の全てまたは一部が欠失されており、且つE4領域の全部または一部が欠失されていることを特徴とする、請求項1〜10のいずれか1項に記載のアデノウイルス。
【請求項12】
E4領域の全てまたは一部がさらに欠失されていることを特徴とする、請求項11に記載のアデノウイルス。
【請求項13】
ヒトアデノウイルスAd2型またはAd5型またはイヌアデノウイルスCAV−2型であることを特徴とする、請求項1〜12のいずれか1項に記載のアデノウイルス。
【請求項14】
グルコースオキシダーゼまたは甲状腺ペルオキシダーゼの遺伝子などのペルオキシダーゼ系に関連するポリペプチドをコードする少なくとも1つの遺伝子をさらに含むことを特徴とする、請求項1〜13のいずれか1項に記載のアデノウイルス。
【請求項15】
腫瘍成長の治療および/または阻害を意図する薬学的組成物の調製用の、請求項1〜14のいずれか1項に記載のアデノウイルスの使用。
【請求項16】
請求項1〜14のいずれか1項に記載の1つまたは複数の欠陥組換えアデノウイルスを含む薬学的組成物。
【請求項17】
注射可能形態であることを特徴とする、請求項16に記載の薬学的組成物。
【請求項18】
104と1014pfu/mlとの間、好ましくは106と1011pfu/mlとの間の欠陥組換えアデノウイルスを含むことを特徴とする、請求項16または17に記載の薬学的組成物。
【図1】


【図2】

【図3】
【図4】
【図5A】

【図5B】
【図6】
【図2】
【図3】
【図4】
【図5A】
【図5B】
【図6】
【公開番号】特開2013−48626(P2013−48626A)
【公開日】平成25年3月14日(2013.3.14)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−225794(P2012−225794)
【出願日】平成24年10月11日(2012.10.11)
【分割の表示】特願2001−502790(P2001−502790)の分割
【原出願日】平成12年6月8日(2000.6.8)
【出願人】(500152119)アバンテイス・フアルマ・エス・アー (65)
【Fターム(参考)】
【公開日】平成25年3月14日(2013.3.14)
【国際特許分類】
【出願番号】特願2012−225794(P2012−225794)
【出願日】平成24年10月11日(2012.10.11)
【分割の表示】特願2001−502790(P2001−502790)の分割
【原出願日】平成12年6月8日(2000.6.8)
【出願人】(500152119)アバンテイス・フアルマ・エス・アー (65)
【Fターム(参考)】
[ Back to top ]