
狂犬病ウイルスを中和することができる結合分子およびそれらの用途
本発明は狂犬病ウイルスに特異的に結合し、ウイルスを中和することのできる結合分子を提供する。さらに、本発明は、結合分子をコードする核酸分子、結合分子を含む組成物、および結合分子を同定または生成する方法を提供する。結合分子を、狂犬病ウイルスに起因する症状の診断、予防、および/または処置において使用することが可能である。好適には、結合分子は狂犬病の暴露後予防に使用することが可能である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は医学に関する。特に、本発明は、狂犬病ウイルスを中和する結合分子に関する。これらの結合分子は、狂犬病の暴露後予防に役立つ。
【背景技術】
【0002】
狂犬病はほぼ全世界に分布するウイルス感染症であって、主に野生動物や家畜に影響を及ぼすが、人間にも感染し、破壊的な、ほぼ不変の致命的な脳炎をひきおこす。年間70,000人以上の人間が死亡すると推定され、その他の何百万人もが暴露後の処置を必要とする。
【0003】
狂犬病ウイルスは弾丸型でエンベロープを有する一本鎖RNAウイルスで、ラブドウイルス科リッサウイルス属に分類される。狂犬病ウイルスのゲノムは以下の5つのウイルスタンパク質をコードする:RNA依存RNAポリメラーゼ(L);核タンパク質(N);リン酸化されたタンパク質(P);ウイルスタンパク質のエンベロープの内側にあるマトリクスタンパク質(M);および外表面糖蛋白質(G)。
【0004】
Gタンパク質(62〜67kDa)は、505個のアミノ酸から成るI型糖蛋白質であり、2〜4個の潜在的なN-グリコシル化部位を有するが、ウイルス株に依存して、そのうちの1個または2個のみがグリコシル化される。Gタンパク質は、ビリオンのエンベロープの外表面を覆う突起を形成し、ウイルス中和抗体を誘発することが知られている。
【0005】
狂犬病は、能動免疫法と受動免疫法のどちらによっても治療または予防が可能である。狂犬病の暴露後予防は、局部的な傷口の迅速な手当て、および受動免疫(抗狂犬病免疫グロブリン)および能動免疫(ワクチン)双方の実施から構成される。
【0006】
現在、抗狂犬病免疫グロブリン(RIG)は、ヒト抗狂犬病免疫グロブリン(HRIG)またはウマ抗狂犬病免疫グロブリン(ERIG)の血清試料から作られている。ERIGおよびHRIGの欠点は、それらを充分量において利用することができず、HRIGの場合には、高価すぎる点である。更に、ERIGの使用は、アナフィラキシー・ショックなどの副作用につながる可能性がある。既知あるいは未知の病原体による汚染の可能性は、HRIGに伴う更なる懸念である。これらの欠点を克服するために、暴露後予防において狂犬病ウイルスを中和することができるモノクローナル抗体を使用することが提案されてきた。狂犬病ウイルスを不活性化するマウス・モノクローナル抗体は、従来技術において周知である(Schumacher et al., 1989参照)。しかし、生体内でマウス抗体を使用することは、例えば、血中半減期が短い、特定のヒトエフェクター機能を誘発できない、およびヒト体内でマウス抗体に対する過剰な免疫反応(ヒト抗マウス抗体 "human anti-mouse antibody" (HAMA)による反応)が誘発されてしまうなどといった、マウス抗体をヒトに投与することに伴う問題があるため制限される。
【0007】
近年、狂犬病ウイルス中和ヒトモノクローナル抗体について述べられている (Dietzschold et al. , 1990, Champion et al., 2000、およびHanlon et al., 2001参照)。抗狂犬病ヒトモノクローナル抗体が暴露後予防においてHRIGと同等の効力をもつには、モノクローナル抗体の混合物を用いなければならない。そのような混合物においては、それぞれの抗体がウイルス上の別々のエピトープまたは部位に結合し、抵抗性のあるウイルス変異体の生存を防がなくてはならない。
【0008】
現在、暴露後予防の可能性を高める新規の狂犬病ウイルス中和ヒトモノクローナル抗体、特に異なるエピトープ認識特異性を持つ抗体の必要性が依然として高い。本発明は、幅広い狂犬病ウイルスおよびその中和抵抗性変異株に対する暴露後予防に役立つ、混合物として使用できる可能性を持つヒトモノクローナル抗体を提供する。
【0009】
(発明の説明)
以下、本発明で使用する用語の定義を記す。
【0010】
定義
結合分子
本明細書において用いる「結合分子」という用語は、モノクローナル抗体(キメラ、ヒト化、またはヒトモノクローナル抗体など)を含むインタクト型免疫グロブリンを指すか、または、インタクト型免疫グロブリンと競合して免疫グロブリンの結合パートナー(例えば狂犬病ウイルス、あるいはGタンパク質などの狂犬病ウイルスのフラグメント)と特異的結合を形成するような免疫グロブリンのフラグメントを備える、抗原結合領域および/または可変領域を指す。その構造に関係なく、抗原結合性フラグメントは、インタクト型免疫グロブリンによって認識される抗原と同一の抗原に結合する。抗原結合性フラグメントを、結合分子のアミノ酸配列のうちの、少なくとも2連続のアミノ酸残基、少なくとも5連続のアミノ酸残基、少なくとも10連続のアミノ酸残基、少なくとも15連続のアミノ酸残基、少なくとも20連続のアミノ酸残基、少なくとも25連続のアミノ酸残基、少なくとも30連続のアミノ酸残基、少なくとも35連続のアミノ酸残基、少なくとも40連続のアミノ酸残基、少なくとも50連続のアミノ酸残基、少なくとも60連続のアミノ酸残基、少なくとも70連続のアミノ酸残基、少なくとも80連続のアミノ酸残基、少なくとも90連続のアミノ酸残基、少なくとも100連続のアミノ酸残基、少なくとも125連続のアミノ酸残基、少なくとも150連続のアミノ酸残基、少なくとも175連続のアミノ酸残基、少なくとも200連続のアミノ酸残基、または少なくとも250連続のアミノ酸残基、のいずれかのアミノ酸配列を含むペプチドあるいはポリペプチドから構成することが可能である。
【0011】
本明細書において用いる「結合分子」という用語は、従来技術において知られている全てのクラスおよびサブクラスの免疫グロブリンを含む。重鎖の定常領域のアミノ酸配列によって、結合分子を、インタクト抗体の5つの大きなクラス、すなわちIgA, IgD, IgE, IgGおよびIgMに分類することができ、これらのいくつかを、例えばIgAl, IgA2, IgGl, IgG2, IgG3およびIgG4といったサブクラス(イソタイプ)にさらに分類することができる。
【0012】
抗原結合性フラグメントには、とりわけ、Fab、F (ab')、F(ab')2、Fv、dAb、Fd、相補性決定領域(CDR)フラグメント、単鎖抗体(scFv)、二価の単鎖抗体、単鎖ファージ抗体、二重特異性抗体(diabody)、三重特異性抗体(triabody)、四重特異性抗体(tetrabody)、(ポリ)ペプチドと特異的な抗原結合を形成するのに十分な、免疫グロブリンのフラグメントを少なくとも含む(ポリ)ペプチド、などが含まれる。上述のフラグメントは、合成によって作製するか、インタクト型免疫グロブリンを酵素または化学作用を用いて切断することによって作製するか、あるいは組み換えDNA技術を用いて遺伝子工学的に作製することができる。作製の方法は当技術分野では周知であり、例えば“Antibodies: A Laboratory Manual, Edited by:E. Harlow and D, Lane (1988), Cold Spring Harbor Laboratory, Cold Spring Harbor, New York”に記載されており、参照することにより本願明細書に組み込まれる。結合分子またはその抗原結合性フラグメントは、1つまたは複数の結合部位を有する可能性がある。2つ以上の結合部位を有する場合、結合部位は互いに同一である可能性も異なる可能性もある。
【0013】
結合分子は、むき出しであるか非コンジュゲート型の結合分子である可能性もあるが、免疫コンジュゲートを形成している可能性もある。むき出しであるか非コンジュゲート型の結合分子とは、エフェクター部分やタグ(とりわけ有害物質、放射性物質、リポソーム、酵素など)とコンジュゲートを形成していないか、作動可能に結合していないか、または他の物理的あるいは機能的な結合をしていない結合分子を指すよう意図されている。当然のことながら、むき出しであるか非コンジュゲート型の結合分子には、エフェクター部分やタグの結合以外であって、安定化、多量体化、ヒト化、あるいは任意の他の方法による操作を受けた結合分子を除外するものではない。従って、翻訳後修飾を受けた全てのむき出しであるか非コンジュゲート型の結合分子は「結合分子」に含まれるのであって、天然の結合分子産生細胞の環境において、組み換え型結合分子産生細胞によって、何処かに修飾がなされたもの、および、初めに結合分子を調製した後にヒトの手によって何処かに修飾が導入されたものも含まれる。当然、「むき出しであるか非コンジュゲート型の結合分子」という用語は、体内に投与された後にエフェクター細胞および/または分子と機能的な結合を形成する能力をも含むものであり、このような相互作用には生物学的作用を及ぼすために必要なものもあるためである。従って、結合しているエフェクターグループやタグが存在しないことは、試験管内では「むき出しであるか非コンジュゲート型の結合分子」の定義にあてはまるが、生体内においてはあてはまらない。
【0014】
相補性決定領域(CDR)
本明細書において用いる「相補性決定領域」という用語は、免疫グロブリンなどの結合分子の可変領域中に存在する塩基配列を意味し、通常、抗原上の認識されるエピトープに対して形状および電荷分布において相補的である抗原結合部位に大きな影響を及ぼす。CDR領域はタンパク質あるいはタンパク質断片の直線状エピトープ、不連続エピトープ、あるいは立体構造エピトープに特異的である可能性があり、これらのエピトープは天然の構造をとるタンパク質上に存在するか、または、ある場合には、例えばSDS中で溶解することによって変性したタンパク質上に存在する。また、エピトープはタンパク質の翻訳後修飾から構成される可能性もある。
【0015】
機能的変異体
本明細書において用いる「機能的変異体」という用語は、元の結合分子(親結合分子)のヌクレオチドおよび/またはアミノ酸配列と比較して1つ以上のヌクレオチドおよび/またはアミノ酸が変化したヌクレオチドおよび/またはアミノ酸配列を有しており、かつ、例えば狂犬病ウイルスまたはその断片などといった結合パートナーに対する結合を、依然として親結合分子と競合することができる結合分子を指す。つまり、親結合分子のヌクレオチドおよび/またはアミノ酸配列における修飾は、ヌクレオチド配列によってコードされる結合分子、あるいはアミノ酸配列を含む結合分子の結合能に重大な影響を与えたりこれを変化させたりはせず、すなわちその結合分子は依然としてターゲットを認識し結合することができる。機能的変異体は、ヌクレオチドおよびアミノ酸の置換、挿入、および欠失などの、保存的な配列修飾を有する可能性がある。これらの修飾は、例えば部位特異的突然変異導入法やPCRを用いたランダム突然変異導入法などの、当技術分野では周知の標準的な技術によって導入可能である。
【0016】
保存的なアミノ酸置換には、あるアミノ酸残基が、構造や化学的性質が似た別のアミノ酸残基によって置き換えられる置換が含まれる。類似したアミノ酸側鎖をもつアミノ酸残基のファミリーが当技術分野において定められている。このファミリーには、塩基性側鎖を有するアミノ酸(例:リジン、アルギニン、ヒスチジン)、酸性側鎖を有するアミノ酸(例:アスパラギン酸、グルタミン酸)、非荷電極性側鎖を有するアミノ酸(例:アスパラギン、グルタミン、セリン、トレオニン、チロシン、システイン、トリプトファン)、非極性側鎖を有するアミノ酸(例:グリシン、アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン)、β分岐側鎖を有するアミノ酸(例:トレオニン、バリン、イソロイシン)、芳香族側鎖を有するアミノ酸(例:チロシン、フェニルアラニン、トリプトファン)が含まれる。上で述べたのとは別のアミノ酸残基ファミリーの分類法も使用可能であることは、当業者に明らかであろう。更には、変異体は、例えばあるアミノ酸が、構造や化学的性質の異なるアミノ酸残基により置換されるといった、非保存的なアミノ酸置換を有する可能性もある。これに類似したマイナーな変異には、アミノ酸の欠失や挿入、或いはその双方が含まれる可能性がある。どのアミノ酸残基を免疫学的活性を損うことなく置換、挿入、あるいは欠失することが可能であるかについての指針は、当技術分野において周知であるコンピュータープログラムを用いて見出すことが可能である。
【0017】
ヌクレオチド配列における変異は、例えば転位変異または転換変異のように、遺伝子座における単一の変化(点変異)とすることも可能であるし、もしくは代わりに、単一の遺伝子座において複数のヌクレオチドを挿入、欠失あるいは置換することも可能である。さらに、1つのヌクレオチド配列中のかなり多数の遺伝子座において、1つまたは複数の変化を加えることが可能である。変異は、当技術分野において知られている適切な方法によって引き起こすことができる。
【0018】
ホスト
本明細書において用いる「ホスト」という用語は、クローニングベクターまたは発現ベクターのようなベクターが導入された生物あるいは細胞を指すことを意図されている。生物あるいは細胞は、原核細胞であっても真核細胞であってもよい。この用語は、対象となる特定の生物あるいは細胞だけでなく、その生物あるいは細胞の子孫にも当てはまるということが理解されなくてはならない。後の世代に突然変異あるいは環境の影響のいずれかによる特定の修飾が生じる可能性があるため、これらの子孫は実際には元の生物あるいは細胞と同一ではない可能性があるが、それでもなお、本明細書において用いる「ホスト」という用語の範囲内に含まれる。
【0019】
ヒト(Human)
「ヒト」という用語は、本願明細書において定義される結合分子に適用した際には、ヒトから直接得られた分子か、ヒトの塩基配列に基づいた分子かのいずれかを意味する。結合分子がヒトの塩基配列に由来するか又はヒトの塩基配列に基づいているのなら、その後修飾を加えたとしても、本明細書の全体を通して使用する場合には依然として「ヒト」結合分子であると見なす。言い換えると、「ヒト」という用語は、結合分子に適用した際には、可変領域または定常領域のいずれかに基づくヒト生殖細胞系列免疫グロブリン塩基配列に由来する可変領域および定常領域を有する結合分子、ヒトの体内またはヒトリンパ球内で生じたのではない結合分子、または修飾された形態の結合分子を含むことを意図されている。従って、ヒト結合分子は、ヒト生殖細胞系列免疫グロブリンの塩基配列によってコードされていないアミノ酸残基を含む可能性があり、置換および/または欠失(例:試験管内で、例えばランダム突然変異導入法または部位特異的突然変異導入法によって導入された変異、または生体内で体細胞変異によってもたらされた変異)を含むことがある。本明細書において用いる「〜に基づいた」という言葉は、核酸配列が鋳型から正確に複製される状況か、例えばエラーを起こしやすいPCR法によってマイナーな変異を伴う状況か、あるいは鋳型に正確に合致するかまたはマイナーな変更を伴って合成される状況を指す。ヒトの塩基配列に基づいた半合成分子もまた、本明細書において用いる「ヒト」結合分子あるとみなす。
【0020】
モノクローナル抗体
本明細書において用いる「モノクローナル抗体」という用語は、単一の分子構成(すなわち一次構造)を有する抗体分子調製物、すなわち単一のアミノ酸配列を有する抗体分子調製物を意味する。モノクローナル抗体は、特定のエピトープに対して単一の結合特異性および親和性を示す。従って、「ヒトモノクローナル抗体」という用語は、単一の結合特異性を示す抗体であって、ヒト生殖細胞系列免疫グロブリンの塩基配列に由来するか、基づいた可変領域および定常領域、または合成配列に完全に由来する可変領域および定常領域を有する該抗体を指す。モノクローナル抗体を作製する方法は問題とされない。
【0021】
核酸分子
本発明で用いる「核酸分子」という用語は、ヌクレオチドの多量体型を指し、RNA、cDNA、ゲノムDNA、およびこれらの合成型、および前記物質の混合ポリマーの、センス鎖およびアンチセンス鎖双方を含む。ヌクレオチドとは、リボヌクレオチド、デオキシヌクレオチド、またはどちらかの種類のヌクレオチドの修飾型を指す。この用語には、一本鎖および二本鎖のDNAも含まれる。加えて、ポリヌクレオチドには、天然ヌクレオチドおよび修飾ヌクレオチドのいずれかまたは両方が、天然のヌクレオチド結合および/または非天然のヌクレオチド結合によって結合したポリヌクレオチドが含まれる可能性もある。当業者ならば容易に理解できるように、核酸分子は化学的または生化的に修飾されうるし、もしくは非天然ヌクレオチド塩基又は誘導体化ヌクレオチド塩基を含みうる。このような修飾には、例えば、ラベル化、メチル化、1つまたは複数の天然ヌクレオチドのアナログによる置換、ヌクレオチド間の修飾(例えば、非荷電性の結合(例:メチルホスホネート、リン酸トリエステル、アミド亜リン酸エステル、カルバミン酸エステルなど)、荷電結合(例:ホスホロチオネート、ホスホロジチオエートなど)、懸垂部分(pendent moieties)(例:ポリペプチド)、インターカレータ(例:アクリジン、プソラレンなど)、キレート剤、アルキル化剤、および修飾された結合(例:α‐アノマー核酸など))が含まれる。上記の用語はまた、一本鎖、二本鎖、部分的二重らせん、三重らせん、ヘアピン構造、円形、および南京錠型の立体構造などの、任意のトポロジー構造を含むよう意図されている。また、ポリヌクレオチドの、水素結合やその他の化学的相互作用を介して目的の配列に結合する能力を模倣した合成分子も含まれる。このような分子は当技術分野においては周知であり、例えば、分子骨格においてリン酸結合をペプチド結合で置換した分子などが含まれる。核酸の塩基配列について言及した場合、特に明記しない限りは、その相補鎖も含まれる。従って、特定の配列を有する核酸分子について言及した場合、その相補鎖およびその相補的配列も含まれると理解すべきである。相補鎖もまた、例えばアンチセンス療法、ハイブリッド形成プローブ、およびPCRプライマーに有用である。
【0022】
薬学的に許容される賦形剤
「薬学的に許容される賦形剤」とは、好ましい剤形または便利な剤形を調製するために、薬物、薬剤、あるいは結合分子などの活性分子と配合するあらゆる不活性物質を意味する。この「薬学的に許容される賦形剤」は、非毒性であるか、または、少なくとも、使用する用量および濃度では、使用目的において、毒性が服用者にとって許容される程度であるかであって、そして薬物、薬剤または結合分子を含む製剤の他の成分と適合性を有する賦形剤である。
【0023】
暴露後予防
「暴露後予防(PEP)」は狂犬病にかかった動物に暴露したおそれがある人に行なう必要がある。起こり得る暴露には、動物咬傷などの咬傷暴露(例:歯による皮膚へのあらゆる侵入)および非咬傷暴露が含まれる。非咬傷暴露には、研究室や洞穴における大量のエアロゾル化狂犬病ウイルスへの暴露、および、狂犬病により死亡した患者から角膜移植手術を受けたレシピエントが含まれる。開放創、擦り傷、粘膜、あるいは理論的にはかき傷が、狂犬病にかかった動物の唾液または他の潜在的に感染性を有する物質(例えば神経組織)によって汚染した場合にも、非咬傷暴露が生じる。例えば狂犬病にかかった動物をなでたり、狂犬病にかかった動物の血液、尿、または糞便との接触したりするなどの他の接触は、それだけでは暴露とならず、暴露後予防の適応とはならない。PEPは暴露後できるだけ早く開始しなければならない。暴露が起こっていない場合は、暴露後予防は必要でない。全ての暴露後予防の処方計画において、過去に免疫化された人を除いて、能動免疫法および受動免疫法を並行して行なわなくてはならない。
【0024】
特異的に結合
結合分子の相互作用(例えば抗体およびその結合パートナー(例:抗原))に関して、本願明細書において用いる「特異的に結合」という用語は、この相互作用が、ある特定の構造(例えば結合パートナー上の抗原決定基またはエピトープ)の存在に依存していることを意味する。換言すれば、結合パートナーが他の分子や生物の混合物中に存在するときでも、抗体は選択的に結合パートナーに結合するか結合パートナーを認識する。結合は、共有結合性相互作用または非共有結合性相互作用、あるいは両者の組み合わせによって媒介される。さらに言い換えると、「特異的に結合」という用語は、免疫特異的に抗原またはその断片に結合し、他の抗原には免疫特異的に結合しないことを意味する。免疫特異的に抗原に結合する結合分子は、他のペプチドやポリペプチドに対してより低い親和性で結合することができ、これは例えばラジオイムノアッセイ(RIA)、酵素結合抗体免疫吸着アッセイ(ELISA)、BIACORE、または当技術分野において周知の他のアッセイによって測定することができる。免疫特異的に抗原に結合する結合分子またはそのフラグメントは、関連抗原と交差反応する可能性がある。免疫特異的に抗原に結合する結合分子またはそのフラグメントは、関連抗原と交差反応しないことが望ましい。
【0025】
治療に有効な量
「治療に有効な量」という用語は、狂犬病の暴露後予防に有効である、本明細書において定義される結合分子の量を意味する。
【0026】
ベクター
「ベクター」という用語は、第2の核酸分子をホストに導入するために挿入することができる核酸分子を意味し、ホスト内で複製され、場合によっては発現する。換言すると、ベクターは、自身に結合した核酸分子を輸送することができる。本願明細書において用いる「ベクター」という用語は、クローニングベクターおよび発現ベクターの双方を意図している。ベクターには、プラスミド、コスミド、細菌人工染色体(BAC)および酵母人工染色体(YAC)、そしてバクテリオファージまたは植物または動物(ヒトを含む)のウイルスに由来するベクターが含まれるが、これらに限定されない。ベクターは、予定されるホストによって認識される複製開始点を備え、そして発現ベクターの場合には、ホストによって認識されるプロモータおよび他の制御領域を備える。第2の核酸分子を含むベクターは、例えば形質転換、トランスフェクション、または細菌あるいはウイルスの侵入機構を利用して、細胞内に導入される。例えば、植物細胞および同類のものにおいてしばしば使用するエレクトロポレーションまたは微粒子銃などの、核酸を細胞内に導入する他の方法も周知である。核酸を細胞内に導入する方法は、とりわけ細胞の種類やその他いろいろに依存する。これは、本発明に決定的なものではない。ある種のベクターは、自身が導入されるホスト内で自律複製できる(例えば、細菌複製開始点を有するベクターは、細菌内で複製することができる)。他のベクターは、ホストへ導入された直後にホストのゲノムに組み込まれ、その結果、ホストのゲノムと共に複製される。
【発明の開示】
【0027】
(発明の概要)
本発明は、狂犬病ウイルスに特異的に結合して中和することができる結合分子を提供する。さらに、本発明は、これらの結合分子の少なくとも結合領域をコードする核酸分子に関する。さらに本発明は、狂犬病ウイルスに起因する状態を発現する危険のある被験者への、暴露後予防における本発明の結合分子の用途を提供する。
【発明を実施するための最良の形態】
【0028】
第1の態様において、本発明は、狂犬病ウイルスに特異的に結合することができる結合分子を備える。本発明の結合分子はまた、狂犬病ウイルスを中和する活性を有することが望ましい。好適には、本発明の結合分子はヒト結合分子である。あるいは、結合分子は他の動物の結合分子であっても良い。狂犬病ウイルスは、リッサウイルス属に含まれる。リッサウイルス属は、合計で11の遺伝子型を含む:すなわち、狂犬病ウイルス(遺伝子型1)、ラゴスコウモリウイルス(遺伝型2)、モコラウイルス(遺伝子型3)、ドゥベンヘイグウイルス(遺伝子型4)、ヨーロッパコウモリリッサウイルス1(遺伝子型5)、ヨーロッパコウモリリッサウイルス2(遺伝子型6)、オーストラリアコウモリリッサウイルス(遺伝子型7)、アラバンウイルス(遺伝子型8)、クージャンウイルス(遺伝子型9)、イルクーツウイルス(遺伝子型10)およびウェストコーカシアンウイルス(遺伝子型11)である。本発明の結合分子は、狂犬病ウイルスに結合するだけでなく、リッサウイルス属に含まれる他の遺伝子型にも結合する能力を持つ可能性がある。好適には、結合分子は、リッサウイルス属に含まれる他の遺伝子型をも中和する能力も持つ。更には、本発明の結合分子は、ラブドウイルス科のリッサウイルス属以外のウイルスを結合および/または中和する能力をも持ち得る。ラブドウイルス科には、サイトラブドウイルス、エファメロウイルス、リッサウイルス、ヌクレオラブドウイルス、ラブドウイルスおよびベシクロウイルスが含まれる。
【0029】
結合分子は、狂犬病ウイルスに、天然型であれ不活性型や弱毒化型であれ特異的に結合し得る。狂犬病ウイルスの失活は、とりわけ、β‐プロピオラクトン(BPL)での処理(White and Chappel, 1982)、56℃での30分間以上の加熱、ガンマ線照射、アセチルエチレンイミンまたはエチルイミンでの処理あるいはアスコルビン酸および硫酸銅での72時間の処理(Madhusudana et al., 2004)といった処理によって行なうことができる。例えば、とりわけパストリゼーション(湿性温熱処理)、乾熱処理、蒸気熱処理、低いpHでの処理、有機溶剤/界面活性剤での処理、ナノ濾過、紫外線照射などの、当業者にとって周知の一般的なウイルス失活法を使用することもできる。好適には、失活をβ‐プロピオラクトン(BPL)を用いた処理によって実行するのがよい。狂犬病ウイルスが依然感染力を持つか、あるいは部分的にまたは完全に不活化されているかどうか検査する方法は当業者にとって周知であり、とりわけ、” Laboratory techniques in rabies , Edited by: F. -X Meslin, M. M. Kaplan and H. Koprowski (1996), 4th edition, Chapter 36, World Health Organization, Geneva” に記載されている。
【0030】
結合分子は、狂犬病ウイルスの1つまたは複数の断片(例えば、とりわけ、一種類もしくは複数の種類のタンパク質からなる調製液、および/または狂犬病ウイルスに由来する(ポリ)ペプチドまたは狂犬病ウイルスタンパク質および/または(ポリ)ペプチドで形質転換された細胞から得られた(ポリ)ペプチド)にも特異的に結合することができる可能性がある。例えば狂犬病ウイルスの暴露後予防などの、治療法および/または予防法のためには、結合分子は、M(Ameyama et al. 2003参照)またはGタンパク質といった、狂犬病ウイルスの接触可能な表面タンパク質に特異的に結合できることが望ましい。診断の目的のためには、ヒト結合分子が、狂犬病ウイルスの表面には存在しないタンパク質に特異的に結合することができても良い。様々な既存の狂犬病ウイルス株の接触可能な表面タンパク質および内部タンパク質のアミノ酸配列は、EMBL-データベースおよび/または他のデータベースで検索することができる。
【0031】
望ましくは、この断片は、少なくとも、本発明のヒト結合分子によって認識される抗原決定基を備える。本願明細書において使用する「抗原決定基」とは、例えば狂犬病ウイルス(ポリ)ペプチド、(糖)タンパク質、またはその類似体または断片などの部分のことであり、本発明のヒト結合分子に十分に高い親和性で結合し、検出可能な抗原‐結合分子複合体を形成することのできる部分のことである。
【0032】
本発明による結合分子は、ポリクローナル抗体またはモノクローナル抗体などのインタクト型免疫グロブリン分子であってもよいし、あるいは結合分子は、Fab、F(ab')、F(ab')2、Fv、dAb、Fd、相補性決定領域(CDR)フラグメント、単鎖抗体(scFv)、二価の単鎖抗体、単鎖ファージ抗体、二重特異性抗体(diabody)、三重特異性抗体(triabody)、四重特異性抗体(tetrabody)、および狂犬病ウイルスまたはそのフラグメントに特異的な抗原結合を形成するのに十分な免疫グロブリンの断片を少なくとも含む(ポリ)ペプチド、などの抗原結合性断片であっても良いが、これらに限定されない。本発明の結合分子は、非単離型であっても単離型であっても使用することができる。更に、本発明の結合分子は、単独でも、または少なくとも1つのヒト結合分子(あるいはその変異体または断片)を含む混合物としても使用することができる。換言すれば、結合分子を、例えば2つ以上の結合分子あるいはそれらの変異体や断片から成る医薬品組成物として、組み合わせて使うことができる。例えば、目標とする予防、治療または診断の効果を達成するために、狂犬病ウイルス中和活性を有する複数の結合分子を1回の治療において組み合わせることができる。
【0033】
狂犬病ウイルスなどのRNAウイルスは、ウイルス複製の間、自身のRNAポリメラーゼを使用する。これらのRNAポリメラーゼは、エラーを起こしやすい傾向がある。このことによって、ウイルス感染の期間中にいわゆる擬似種が形成される結果となる。各々の擬似種は独自のRNAゲノムを有しており、その結果、ウイルスタンパク質のアミノ酸組成に相違が生じうる。もしもこのような変異がウイルスの構造タンパク質において生ずれば、ウイルスは、T細胞またはB細胞のエピトープの変化に起因して、潜在的にホストの免疫系から逃れることができる。こうしたことが起こる可能性は、ポリクローナル抗体の混合物(HRIG)によって個人を治療する場合よりも、例えばヒトモノクローナル抗体などの結合分子、2種類の混合物によって個人を治療する場合の方が高い。従って、狂犬病の治療のために2種類のヒトモノクローナル抗体を混合する際の必要条件は、2種類の抗体が、標的抗原、すなわち狂犬病ウイルス糖タンパク質上の互いに重複したりや競合したりしないエピトープを認識することである。この結果、狂犬病エスケープウイルスが出現する確率を最小化できる。その結果として、好適には、本発明の結合分子は、互いに異なる、互いに重複したり競合したりしない狂犬病ウイルスのエピトープ(例えば狂犬病ウイルスGタンパク質上のエピトープなど)と反応することができる。結合分子の混合物は、例えば狂犬病の暴露後予防に適する薬剤などの他の治療薬剤の少なくとも1つを、さらに含む可能性がある。
【0034】
通常、本発明による結合分子は、それぞれの結合パートナー(すなわち狂犬病ウイルスや、狂犬病ウイルスタンパク質などの狂犬病ウイルスのフラグメント)と、0.2×10-4 M、1.0×10-5M、1.0×10-6 M、1.0×10-7Mよりも小さく、好適には1.0×10-8 Mよりも小さく、より好適には1.0×10-9 Mよりも小さく、より好適には1.0×10-10 Mよりも小さく、更により好適には1.0×-11Mよりも小さく、特に1.0×10-12 Mよりも小さい親和定数(Kd値)で結合することができる。親和定数は、抗体アイソタイプによって変化する可能性がある。例えば、IgMアイソタイプの親和結合とは、少なくとも約1.0×10-7 Mの結合親和力を指す。親和定数は、例えば、表面プラズモン共鳴、すなわち、バイオセンサ・マトリックス内部のタンパク質濃度変化を検出することによって、生体特異的相互作用のリアルタイム分析を可能とする光学現象、を用いることによって、例えばBIACOREシステム(Pharmacia Biosensor AB, Uppsala, Sweden)を用いて測定することができる。
【0035】
本発明による結合分子は、精製された/単離された型の狂犬病ウイルスにも、精製されていない/単離されていない型の狂犬病ウイルスにも結合しうる。結合分子は、例えば試料中の可溶型の形態にある狂犬病ウイルスに結合しうるし、あるいは、例えばマイクロタイタープレート、膜、およびビーズなどの担体または基質に結合しているか付着している狂犬病ウイルスに結合しうる。担体または基質は、ガラス、プラスチック(例えばポリスチレン)、多糖類、ナイロン、ニトロセルロースまたはテフロン(登録商標)などでできていてよい。こうした支持材の表面は、固形でも多孔性でもよく、そして都合の良い任意の形状であってもよい。代わって、結合分子は、例えば狂犬病ウイルスのタンパク質または(ポリ)ペプチドなどの、狂犬病ウイルスのフラグメントにも結合しうる。一実施形態では、結合分子は狂犬病ウイルスGタンパク質またはそのフラグメントに特異的に結合することができる。狂犬病ウイルスタンパク質または(ポリ)ペプチドは、可溶性の形態である可能性もあるし、あるいは上記の通りに担体または基質に結合しているか付着している狂犬病ウイルスに結合する可能性もある。もう1つの実施例では、Gタンパク質で形質転換された細胞を、結合分子の結合パートナーとして用いることができる。
【0036】
本発明の好適な実施形態においては、本発明の結合分子は狂犬病ウイルスの感染性を中和する。これは、とりわけマウスニューロトロフィン受容体p75、神経細胞接着分子(CD56)、およびアセチルコリン受容体などの、宿主細胞上の受容体に狂犬病ウイルスが結合することを防ぐことによって、あるいは細胞の細胞質中へのRNA放出を阻害することによって、あるいはRNAの転写または翻訳を阻止することによって達成可能である。ある特定の実施形態においては、本発明の結合分子は、狂犬病ウイルスによる宿主細胞への感染を、前記結合分子の非存在下における狂犬病ウイルスによる宿主細胞への感染と比較して、少なくとも99%、少なくとも95%、少なくとも90%、少なくとも85%、少なくとも80%、少なくとも75%、少なくとも70%、少なくとも60%、少なくとも50%、少なくとも45%、少なくとも40%、少なくとも45%、少なくとも35%、少なくとも30%、少なくとも25%、少なくとも20%、または少なくとも10%阻止する。中和の測定は、例えば、“Laboratory techniques in rabies, Edited by: F. -X Meslin, M. M. Kaplan and H. Koprowski (1996), 4th edition, Chapters 15-17, World Health Organization, Geneva”に記載の通りに行うことができる。更に、本発明のヒト結合分子は、エンベロープを持つ狂犬病ウイルスの溶菌を補助することができる、補体結合性の結合分子である可能性がある。本発明のヒト結合分子はオプソニンとしても作用し、FcまたはC3b受容体を介した取り込みを促進するか、あるいは狂犬病ウイルスを凝集させて食作用を受けやすくするかのいずれかによって、狂犬病ウイルスの受ける食作用を増強させる可能性がある。
【0037】
好適な実施形態において、本発明による結合分子は少なくともCDR3領域を備えるが、該領域は配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、 配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23そして配列番号24からなる群から選択されたアミノ酸配列を含む。ある実施形態では、CDR3領域は重鎖のCDR3領域である。
【0038】
さらに別の実施形態では、本発明による結合分子は重鎖可変領域を備えるが、該領域は、基本的に、配列番号26、配列番号27、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番号44、配列番号45、配列番号46、配列番号47、配列番号48 および配列番号49からなる群から選択されたアミノ酸配列を含む。好適な実施形態では、本発明による結合分子は、基本的に配列番号335のアミノ酸1-119を含むアミノ酸配列から成る重鎖可変領域を備える。
【0039】
さらなる実施形態において、本発明による結合分子は、配列番号26のアミノ酸配列を含む重鎖可変領域および配列番号50のアミノ酸配列を含む軽鎖可変領域、配列番号27のアミノ酸配列を含む重鎖可変領域および配列番号51のアミノ酸配列を含む軽鎖可変領域、配列番号28のアミノ酸配列を含む重鎖可変領域および配列番号52のアミノ酸配列を含む軽鎖可変領域、配列番号29のアミノ酸配列を含む重鎖可変領域および配列番号53のアミノ酸配列を含む軽鎖可変領域、配列番号30のアミノ酸配列を含む重鎖可変領域および配列番号54のアミノ酸配列を含む軽鎖可変領域、配列番号31のアミノ酸配列を含む重鎖可変領域および配列番号55のアミノ酸配列を含む軽鎖可変領域、配列番号32のアミノ酸配列を含む重鎖可変領域および配列番号56のアミノ酸配列を含む軽鎖可変領域、配列番号33のアミノ酸配列を含む重鎖可変領域および配列番号57のアミノ酸配列を含む軽鎖可変領域、配列番号34のアミノ酸配列を含む重鎖可変領域および配列番号58のアミノ酸配列を含む軽鎖可変領域、配列番号35のアミノ酸配列を含む重鎖可変領域および配列番号59のアミノ酸配列を含む軽鎖可変領域、配列番号36のアミノ酸配列を含む重鎖可変領域および配列番号60のアミノ酸配列を含む軽鎖可変領域、配列番号37のアミノ酸配列を含む重鎖可変領域および配列番号61のアミノ酸配列を含む軽鎖可変領域、配列番号38のアミノ酸配列を含む重鎖可変領域および配列番号62のアミノ酸配列を含む軽鎖可変領域、配列番号39のアミノ酸配列を含む重鎖可変領域および配列番号63のアミノ酸配列を含む軽鎖可変領域、配列番号40のアミノ酸配列を含む重鎖可変領域および配列番号64のアミノ酸配列を含む軽鎖可変領域、配列番号41のアミノ酸配列を含む重鎖可変領域および配列番号65のアミノ酸配列を含む軽鎖可変領域、配列番号42のアミノ酸配列を含む重鎖可変領域および配列番号66のアミノ酸配列を含む軽鎖可変領域、配列番号43のアミノ酸配列を含む重鎖可変領域および配列番号67のアミノ酸配列を含む軽鎖可変領域、配列番号44のアミノ酸配列を含む重鎖可変領域および配列番号68のアミノ酸配列を含む軽鎖可変領域、配列番号45のアミノ酸配列を含む重鎖可変領域および配列番号69のアミノ酸配列を含む軽鎖可変領域、配列番号46のアミノ酸配列を含む重鎖可変領域および配列番号70のアミノ酸配列を含む軽鎖可変領域、配列番号47のアミノ酸配列を含む重鎖可変領域および配列番号71のアミノ酸配列を含む軽鎖可変領域、配列番号48のアミノ酸配列を含む重鎖可変領域および配列番号72のアミノ酸配列を含む軽鎖可変領域、配列番号49のアミノ酸配列を含む重鎖可変領域および配列番号73のアミノ酸配列を含む軽鎖可変領域、を備える。好適な実施形態において、本発明によるヒト結合分子は、配列番号335のアミノ酸1-119を含むアミノ酸配列を含む重鎖可変領域および配列番号337のアミノ酸1-107を含むアミノ酸配列を含む軽鎖可変領域を備える。
【0040】
好適な実施形態において、本発明の狂犬病ウイルス中和活性を有する結合分子は、IgG型で、好ましくはIgG1型で投与される。
【0041】
本発明の別の態様は、本願明細書で定義される結合分子の機能的変異体を含む。分子を本発明による結合分子の機能的変異体であるとみなすのは、該変異体が、狂犬病ウイルスまたはその断片に対して特異的に結合するために、本発明による親結合分子と競合することができる場合である。換言すれば、機能的変異体が、狂犬病ウイルスまたはその断片に結合する能力を依然として有する場合である。機能的変異体もまた、狂犬病ウイルスを中和する活性を有さなければならない。機能的変異体には、主要な構造配列において実質的に親結合分子に類似した誘導体であって、しかし親の結合分子には存在しない、化学的および/または生化学的な、試験管内あるいは生体内における修飾を有するような誘導体が含まれるが、これに限定されない。上記の修飾には、とりわけ、アセチル化、アシル化、ADPリボシル化、アミド化、フラビンの共有結合、ヘム部分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトールの共有結合、架橋、環化、ジスルフィド結合形成、脱メチル化、共有結合性架橋の形成、シスチンの形成、ピログルタミン酸の形成、ホルミル化、γ-カルボキシル化、グリコシル化、GPIアンカー形成、ヒドロキシル化、ヨウ化、メチル化、ミリストイル化、酸化、PEG化(pegylation)、タンパク質分解処理、リン酸化、プレニル化、ラセミ化、セレノ化、硫酸化、アルギニン化などのtRNAを介するタンパク質へのアミノ酸付加、ユビキチン化などが含まれる。
【0042】
あるいは、機能的変異体を、本発明において定義される結合分子であって、親の結合分子のアミノ酸配列と比較して1つ以上のアミノ酸に、置換、挿入、欠失またはそれらの組み合わせを含むアミノ酸配列を含むような結合分子とすることもできる。そのうえ、機能的変異体は、アミノ末端とカルボキシ末端の一方あるいは両方でアミノ酸配列の切断を受けている可能性がある。本発明による機能的変異体は、親結合分子と比較して、同一の結合親和性、あるいはより高いかより低いかの異なる結合親和性を有することが可能だが、依然として、狂犬病ウイルスまたはその断片に結合することができ、依然として狂犬病ウイルスを中和することができる。例えば、本発明による機能的変異体は、狂犬病ウイルスまたはその断片に対して、親結合分子と比較して、より高いかより低い結合親和性を有する可能性があり、あるいは、より高いかより低い狂犬病ウイルス中和活性を有する可能性がある。望ましくは、フレームワーク領域、超可変領域、とりわけCDR3領域を含む(ただし必ずしもこれらに限定されない)可変領域のアミノ酸配列に修飾が加えられる。概して、軽鎖および重鎖の可変領域は3つの超可変領域を備え、該可変領域は3つのCDRおよびより保存性の高い領域、いわゆるフレームワーク領域(FRs)、を備える。超可変領域は、CDR由来のアミノ酸残基および超可変ループ由来のアミノ酸残基を含む。本発明の範囲内にあることを意図された機能的変異体は、本願明細書で定義される親結合分子と、少なくとも約50%から約99%の、好適には少なくとも約60%から約99%の、より好適には少なくとも約70%から約99%の、更に好適には少なくとも約80%から約99%の、最も好適には少なくとも約90%から約99%の、とりわけ少なくとも約95%から約99%の、そしてとりわけ少なくとも約97%から約99%のアミノ酸配列の相同性を有する。とりわけ、ギャップまたはベストフィットなどの、当業者にとって周知のコンピュータ・アルゴリズムを、比較しようとするアミノ酸配列を最適にアライメントして、類似又は同一のアミノ酸残基を定義するために使用することができる。
【0043】
別の実施形態では、親結合分子が配列中にグリコシル化部位を含む場合に機能的変異体が生産されうるが、真核細胞で発現した場合には結合分子のグリコシル化が生じる結果となり、それ故に、抗原への結合が無効となる可能性がある。生産された機能的変異体はグリコシル化部位をもはや含まないが、狂犬病ウイルスへの結合能があり、依然として中和活性を有するであろう。
【0044】
機能的変異体は、親結合分子やその一部分を、エラープローンPCR法、オリゴヌクレオチド特異的突然変異導入法、および部位特異的突然変異導入法(ただしこれらに限らない)を含む、当技術分野では周知の一般的な分子生物学的手法によって変化させることによって得ることができる。そのうえ、機能的変異体は補体結合活性を有する可能性があり、エンベロープを持つ狂犬病ウイルスの溶菌を補助することができ、および/または、オプソニンとして作用し、そしてFcあるいはC3b受容体を介した取り込みを促進することによるか、または狂犬病ウイルスを凝集させてより食作用を受けやすくするかによって、狂犬病ウイルスの受ける食作用を増強させることができる。
【0045】
そのうえ更なる態様において、本発明は免疫コンジュゲートを含むが、この免疫コンジュゲートとは、本願明細書で定義される結合分子あるいはその機能的変異体を少なくとも1つ含み、そしてさらに少なくとも1つのタグ(とりわけ、検出可能な部分/薬剤など)を含む分子のことである。本発明においては、本発明による免疫コンジュゲートの混合液、あるいは本発明による少なくとも1つの免疫コンジュゲートと他の分子(例えば治療薬または他の結合分子または免疫コンジュゲート)との混合液も想定されている。更なる実施形態において、本発明の免疫コンジュゲートは、1つ以上のタグを備えることができる。これらのタグは、互いに同一でも異なっていても良く、結合分子に、非共有結合的に結合/共役させることができる。これらのタグを、ジスルフィド結合、水素結合、静電相互作用、組換え融合および立体配座結合(ただしこれらに限定されない)を含む共有結合によって、直接結合分子に結合/共役させることもできる。あるいは、これらのタグを、1つまたは複数の連結化合物(linking compound)によって結合分子に結合/共役させることもできる。結合分子にタグを共役させる技術は、当業者にとって周知である。
【0046】
本発明の免疫コンジュゲートのタグは治療薬であってもよいが、しかし、好適にはそれらは検出可能な部分/薬剤である。例えば、被験者が狂犬病ウイルスに感染しているかどうかを評価するために、または例えば、所定の治療計画の有効性を判断するための、臨床試験手順の一部として狂犬病ウイルス感染の発現や進行をモニターするために、検出可能な薬剤を含む免疫コンジュゲートを診断のために使用することができる。しかし、その他の検出および/または分析および/または診断の目的のためにこれらを使うこともできる。検出可能な部分/薬剤には、酵素、補欠分子族、蛍光物質、発光物質、生物発光物質、放射性物質、陽電子放出金属および非放射性の正磁性金属イオンが含まれるが、これらに限定されない。
【0047】
検出および/または分析および/または診断の目的のために結合分子をラベルするために使用されるタグは、使用する特定の検出/分析/診断技術、および/または方法に依存しており、例えば、とりわけ、(組織)サンプルの免疫組織化学的染色、フローサイトメトリー検出、レーザースキャンサイトメトリー検出、蛍光免疫法、酵素免疫測定吸着法(ELISA)、ラジオイムノアッセイ(RIA)、バイオアッセイ(例:中和アッセイ)、ウエスタンブロット法の利用などが挙げられる。組織サンプルを免疫組織化学的に染色するための好適なラベルは、検出可能な生成物の生成および局所堆積を触媒する酵素である。免疫組織化学的な視覚化を可能とするために結合分子に通常共役される酵素は周知であり、アセチルコリンエステラーゼ、アルカリ性ホスファターゼ、β‐ガラクトシダーゼ、グルコースオキシダーゼ、西洋わさびペルオキシダーゼおよびウレアーゼが含まれるが、これらに限定されない。視覚的に検出できる生成物を生成し、および堆積させる典型的な基質も、当技術分野において当業者には周知である。加えて、本発明の免疫コンジュゲートは、コロイド金を用いてラベルすることが可能であるか、または、例えば33P、32P、35S、3Hおよび125Iなどの放射性同位体)でラベルすることが可能である。本発明の結合分子を、直接的に、あるいは当技術分野において周知の方法によってキレート化剤を介して間接的に、放射性核種に付着させることが可能である。
【0048】
本発明の結合分子をフローサイトメトリー検出、レーザースキャンサイトメトリー検出、または蛍光免疫法に使用する場合は、蛍光体でラベルするのが有用である。本発明の結合分子に蛍光ラベルをつけるのに有用な多種多様な蛍光体は、当業者には周知である。本発明の結合分子を、ラベル付のアビジン、ストレプトアビジン、CaptAvidinまたはニュートラアビジンを使用した二次検出のために使用する際には、適切な補欠分子族の複合体を形成するために結合分子をビオチンでラベルすることが可能である。
【0049】
本発明の免疫コンジュゲートを生体内での診断上の目的で使用する際には、結合分子は、例えばガドリニウム・ジエチレントリアミンペンタ酢酸などの磁気共鳴画像(MRI)コントラスト剤、超音波コントラスト剤、あるいはX線コントラスト剤へ共役することによって、あるいはラジオアイソトープによりラベル化することによって、検出可能となる。
【0050】
さらにまた、本発明の結合分子、その機能的変異体、または免疫コンジュゲートを、固体の支持材に結合することも可能であり、特に狂犬病ウイルスまたはそのフラグメントの試験管内での免疫学的アッセイまたは精製に役立つ。このような固体の支持材は多孔性であっても非多孔性であってもよく、平面的であっても非平面的であってもよく、ガラス、セルロース、ポリアクリルアミド、ナイロン、ポリスチレン、ポリ塩化ビニルまたはポリプロピレンの支持材を含むが、これらに限定されない。ヒト結合分子を、例えば、免疫親和性クロマトグラフィーを目的として、例えばNHS活性化セファロースあるいはCNBr活性化セファロースなどのろ過媒体と有効に共役させることもできる。結合分子を、典型的にはビオチン-ストレプトアビジンの相互作用によって、常磁性のミクロスフィアに有効に結合することも可能である。ミクロスフィアは、狂犬病ウイルスまたはそのフラグメントを含むサンプルから、狂犬病ウイルスまたはそのフラグメントを単離するために使用することができる。別の例として、本発明のヒト結合分子はELISA用のミクロタイタープレートの表面に有効に結合することができる。
【0051】
本発明の結合分子またはその機能的変異体を、例えば精製を容易にするペプチドなどの、マーカー配列と融合することができる。具体例として、ヘキサヒスチジンタグ、ヘマグルチニン(HA)タグ、mycタグ、またはflagタグが挙げられるが、これらに限定されない。
【0052】
あるいは、抗体を第二の抗体と結合させて、抗体のヘテロコンジュゲートを形成させることができる。別の態様においては、本発明のヒト結合分子は、1つまたは複数の抗原に結合/付着されていてよい。好適には、これらの抗原は、結合分子-抗原の複合体を投与する対象の免疫系によって認識される抗原である。これらの抗原は互いに同一でもよいが、異なっていても良い。これらの抗原と結合分子を付着させる共役方法は、当技術分野では周知であって、架橋剤の使用を含むが、これに限定されるものではない。ヒト結合分子は狂犬病ウイルスに結合し、ヒト結合分子に付着した抗原は、複合体に対する強力なT細胞の攻撃を誘発し、それが最終的には狂犬病ウイルスの破壊につながる。
【0053】
免疫コンジュゲートを直接的に、あるいは例えばリンカーを介して間接的に結合させることによって、化学的に生成するのとは別に、免疫コンジュゲートを、本発明によるヒト結合分子および適切なタグが備わった融合タンパク質として生成することができる。融合タンパク質は、例えば、適切なタグをコードするヌクレオチド配列核酸分子を、ヒト結合分子をコードしているヌクレオチド配列とフレームを合わせて備える核酸分子を作製することによる、組み換え技術を用いるといった、当技術分野では周知の方法によって生産することができる。
【0054】
少なくとも本発明による結合分子またはその機能的変異体をコードする核酸分子を提供することは、本発明の別の態様である。そのような核酸分子は、例えば上記に述べたような親和性成熟の過程で、クローニングの目的のための中間体として用いられる可能性がある。好適な実施形態において、核酸分子は単離あるいは精製される。
【0055】
当業者であれば、これらの核酸分子の機能変異体もまた、本発明の一部に含まれることを意図されていることが理解できるであろう。機能的変異体は、親核酸分子から翻訳されるアミノ酸配列と同一のアミノ酸配列を提供するために、標準的な遺伝暗号を用いて直接翻訳されうる核酸配列である。
【0056】
好適には、核酸分子はCDR3領域を備える結合分子をコードするが、該領域は好適には重鎖CDR3領域であって、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、 配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23および配列番号24からなる群から選択されたアミノ酸配列を含む。
【0057】
更に好適には、核酸分子は、重鎖可変領域を含むヒト結合分子をコードするが、該領域は、配列番号26、配列番号27、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、 配列番号41、配列番号42、配列番号43、配列番号44、配列番号45、配列番号46、配列番号47、配列番号48 および配列番号49からなる群から選択されたアミノ酸配列を原則的に含む。特に好適な実施形態において、核酸分子は、配列番号335のアミノ酸1-119を含んだアミノ酸配列を基本的に含む重鎖可変領域を備えた結合分子をコードする。
【0058】
さらに別の実施形態では、核酸分子は、配列番号26のアミノ酸配列を含む重鎖可変領域および配列番号50のアミノ酸配列を含む軽鎖可変領域を備える結合分子をコードするか、配列番号27のアミノ酸配列を含む重鎖可変領域および配列番号51のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号28のアミノ酸配列を含む重鎖可変領域および配列番号52のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号29のアミノ酸配列を含む重鎖可変領域および配列番号53のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号30のアミノ酸配列を含む重鎖可変領域および配列番号54のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号31のアミノ酸配列を含む重鎖可変領域および配列番号55のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号32のアミノ酸配列を含む重鎖可変領域および配列番号56のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号33のアミノ酸配列を含む重鎖可変領域および配列番号57のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号34のアミノ酸配列を含む重鎖可変領域および配列番号58のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号35のアミノ酸配列を含む重鎖可変領域および配列番号59のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号36のアミノ酸配列を含む重鎖可変領域および配列番号60のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号37のアミノ酸配列を含む重鎖可変領域および配列番号61のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号38のアミノ酸配列を含む重鎖可変領域および配列番号62のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号39のアミノ酸配列を含む重鎖可変領域および配列番号63のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号40のアミノ酸配列を含む重鎖可変領域および配列番号64のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号41のアミノ酸配列を含む重鎖可変領域および配列番号65のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号42のアミノ酸配列を含む重鎖可変領域および配列番号66のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号43のアミノ酸配列を含む重鎖可変領域および配列番号67のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号44のアミノ酸配列を含む重鎖可変領域および配列番号68のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号45のアミノ酸配列を含む重鎖可変領域および配列番号69のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号46のアミノ酸配列を含む重鎖可変領域および配列番号70のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号47のアミノ酸配列を含む重鎖可変領域および配列番号71のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号48のアミノ酸配列を含む重鎖可変領域および配列番号72のアミノ酸配列を含む軽鎖可変領域をコードするか、あるいは、配列番号49のアミノ酸配列を含む重鎖可変領域および配列番号73のアミノ酸配列を含む軽鎖可変領域をコードする。好適な実施例において、核酸分子は、配列番号335のアミノ酸1-119を含んだアミノ酸配列を含む重鎖可変領域および配列番号337のアミノ酸1-107を含んだアミノ酸配列を含む軽鎖可変領域を備えたヒト結合分子をコードする。
【0059】
本発明の特定の実施形態において、本発明の結合分子の重鎖可変領域をコードする核酸分子は、配列番号74、配列番号75、配列番号76、配列番号77、配列番号78、配列番号79、配列番号80、配列番号81、配列番号82、配列番号83、配列番号84、配列番号85、配列番号86、配列番号87、配列番号88、配列番号89、配列番号90、 配列番号91、配列番号92、配列番号93、配列番号94、配列番号95、配列番号96および配列番号97からなる群から選択されたヌクレオチド配列を基本的に含む。望ましくは、本発明の結合分子の重鎖可変領域をコードする核酸分子は、基本的に配列番号334のヌクレオチド1-357を含むヌクレオチド配列を備える。
【0060】
本発明の更なる特定の実施形態において、本発明の結合分子の軽鎖可変領域をコードする核酸分子は、配列番号98、配列番号99、配列番号100、配列番号101、配列番号102、配列番号103、配列番号104、配列番号105、配列番号106、配列番号107、配列番号108、配列番号109、配列番号110、配列番号111、配列番号112、配列番号113、配列番号114、配列番号115、配列番号116、配列番号117、配列番号118、配列番号119、配列番号120および配列番号121からなる群から選択されたヌクレオチド配列を基本的に含む。望ましくは、本発明のヒト結合分子の軽鎖可変領域をコードする核酸分子は、基本的に配列番号336のヌクレオチド1-321を含んだヌクレオチド配列を備える。
【0061】
本発明による核酸分子を1つまたは複数含むベクター、すなわち核酸コンストラクトを供給することは、本発明の別の態様である。ベクターは、プラスミド、例えばF、Rl、RP1、Col、pBR322、TOL、Ti、など;コスミド;ファージ、例えばラムダ、ラムダ様、M13、Mu、P1、P22、Qb、T-even、T-odd、T2、T4、T7、など;植物ウイルス、例えば、とりわけ、アルファルファモザイクウイルス、ブロモウイルス、カピロウイルス、カーラウイルス、カーモウイルス、caulivirus、クロステロウイルス、コモウイルス、クリプトウイルス、ククモウイルス、ダイアンソウイルス、ファバウイルス、フィジーウイルス、フロウイルス、ジェミニウイルス、ホルデイウイルス、イラルウイルス、ルテオウイルス、machlovirus、マラフィウイルス、ネクロウイルス、ネポウイルス、phytorepvirus、植物ラブドウイルス、ポテックスウイルス、ポチウイルス、ソベモウイルス、テヌイウイルス、タバモウイルス、トブラウイルス、トマト黄化えそウイルス、トンブスウイルス、チモウイルスなど;または動物ウイルス、例えば、とりわけ、アデノウイルス、アレナウイルス科、バキュロウイルス科 、ビルナウイルス科、ブニヤウイルス科、カルキウイルス科、カルディオウイルス、コロナウイルス科 、コルチコウイルス科 、シストウイルス科、Epstein-Barrウイルス、エンテロウイルス、フィロウイルス科 、フラビウイルス科、口蹄疫ウイルス、ヘパドナウイルス科、肝炎ウイルス、ヘルペスウイルス科、免疫不全症ウイルス、インフルエンザウイルス、イノウイルス科、イリドウイルス科、オルトミクソウイルス科、パポバウイルス、パラミクソウイルス科、パルボウイルス科、ピコルナウイルス科、ポリオウイルス、ポリドナウイルス科、ポックスウイルス科、レオウイルス科、レトロウイルス、ラブドウイルス科、ライノウイルス、セムリキ森林熱ウイルス、テトラウイルス科、トガウイルス科、トロウイルス科、ワクシニアウイルス、水疱性口内炎ウイルスなど、から得ることができる。ベクターは、クローニングするために、および/または本発明のヒト結合分子を発現させるために使用することができ、遺伝子治療目的のためにさえも使用することができる。1つまたは複数の発現制御核酸分子を作用可能なように結合された、本発明による核酸分子を1つまたは複数含むベクターも、本発明の範囲に含まれる。ベクターの選択は、その後の組換え手順および使用するホストに依存する。宿主細胞へのベクターの導入は、とりわけ、リン酸カルシウム・トランスフェクション法、ウイルス感染、DEAE-デキストラン・トランスフェクション法、リポフェクタミン・トランスフェクション法またはエレクトロポレーションによって行なうことができる。ベクターは自律的に複製してもよいし、自身が組み込まれた染色体とともに複製してもよい。好適には、ベクターは、1つまたは複数の選択マーカーを含む。マーカーの選択は、選択する宿主細胞に依存するが、これは当業者にとって周知のように本発明に決定的なものではない。選択マーカーは、カナマイシン、ネオマイシン、ピューロマイシン、ハイグロマイシン、ゼオシン、単純ヘルペスウイルス由来のチミジンキナーゼ遺伝子(HSV-TK)、マウス由来のジヒドロ葉酸還元酵素遺伝子(dhfr)を含むが、これらに限定されるものではない。上述のヒト結合分子をコードする核酸分子を1つまたは複数含むベクターで、結合分子の単離に用いることができるタンパク質あるいはペプチドをコードする1つまたは複数の核酸分子が作用可能な状態で結合されたベクターもまた、本発明の範囲に含まれる。これらのタンパク質またはペプチドには、グルタチオン-S-トランスフェラーゼ、マルトース結合タンパク質、金属結合性のポリヒスチジン、緑色蛍光タンパク質、ルシフェラーゼおよびβ‐ガラクトシダーゼが含まれるが、これらに限定されない。
【0062】
前述のベクターの1つまたは複数のコピーを含むホストは、本発明の更なる対象である。好適には、ホストは宿主細胞である。宿主細胞には、哺乳類、植物、昆虫、真菌類または細菌起源の細胞が含まれるが、これらに限定されない。細菌細胞には、バチルス属、放線菌およびブドウ球菌のいくつかの種などのグラム陽性菌の細胞、またはエシェリヒア属(例えば大腸菌および緑膿菌)のいくつかの種などのグラム陰性菌の細胞が含まれるが、これらに限定されるものではない。真菌細胞のグループのなかでは、酵母細胞を用いるのが好適である。酵母内での発現は、とりわけピキア・パストリス(Pichia pastoris)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)およびハンゼヌラ・ポリモルファ(Hansenula polymorpha)などの酵母株を用いて達成可能である。さらにまた、ショウジョウバエおよびSf9の細胞などの昆虫細胞も宿主細胞として用いることができる。その他に、宿主細胞は、植物細胞であっても良い。形質転換(組み換え)植物あるいは植物細胞は、例えば、アグロバクテリウム媒介性の遺伝子導入、葉片の形質転換、ポリエチレングリコール誘導のDNA導入によるプロトプラスト形質転換、エレクトロポレーション、音波処理、マイクロインジェクションまたは遺伝子銃などの周知の方法によって、作製することできる。また、適切な発現系をバキュロウイルス系とすることができる。チャイニーズハムスター卵巣(CHO)細胞、COS細胞、BHK細胞またはボーズメラノーマ細胞などの哺乳類細胞を用いた発現系が、本発明において好適である。哺乳類細胞は、哺乳類由来の天然の分子と最も類似した翻訳後修飾を伴うタンパク質を供給する。本発明はヒトに投与される可能性のある分子を取扱うので、ヒトの発現系と完全に一致する発現系はとりわけ好まれる。従って、さらにより好適には、宿主細胞をヒト細胞とする。ヒト細胞の具体例として、とりわけ、HeLa細胞、911細胞、AT1080細胞、A549細胞、293細胞およびHEK293T細胞が挙げられる。好適な哺乳類細胞は、911細胞、または、欧州細胞培養コレクション(ECACC), CAMR, Salisbury, Wiltshire SP4 OJG, Great Britainに、1996年2月29日にNo.96022940として寄託され、登録商標PER.C6(商標登録)(PER. C6は、Crucell Holland B.V.の登録商標である)として市場に出された細胞系などのヒト網膜細胞である。本出願の目的上、「PER.C6」とは、No.96022940として寄託された細胞あるいはその祖先細胞、上流あるいは下流の継代細胞、および寄託された細胞の祖先細胞の子孫細胞、および前述した任意の細胞の派生株を意味する。
【0063】
他の好ましい実施形態では、ヒトプロデューサー細胞は、少なくとも、アデノウイルスEl領域を発現可能な形態でコードする核酸配列の機能的な部分を含む。さらにより好ましい実施形態において、前記宿主細胞はヒト網膜由来で、アデノウイルスEl配列を含む核酸を不死化されており、例えば欧州細胞培養コレクション(ECACC), CAMR, Salisbury, Wiltshire SP4 OJG, Great BritainにNo.96022940として1996年2月29日に寄託されPER.C6(商標登録)として市場に出された細胞系が挙げられる。宿主細胞内での組み換えタンパク質の生産は、当技術分野において周知の方法に従って実行できる。登録商標PER.C6(商標登録)という名前で市場に出された細胞の、生産したいタンパク質を生産する場所としての用途はWO00/63403に記載されており、その明細書全体を参照することにより本明細書に組み込まれる。
【0064】
本発明による結合分子や機能的変異体を生成する方法は、本発明の付加的な部分である。この方法は、a)結合分子やその機能的変異体の発現を起こすような条件の下で本発明によるホストを培養する工程、およびb)必要に応じて、発現した結合分子やその機能的変異体を回収する工程から構成される。発現した結合分子またはその機能的変異体を、無細胞抽出液から回収しても良いが、好適には培養液から回収する。例えば結合分子などのタンパク質を無細胞抽出液または培養液から回収する方法は、当業者にとって周知である。上記の方法によって得ることができる結合分子またはその機能的変異体も、本発明の一部である。
【0065】
あるいは、宿主細胞などのホスト内での発現とは別に、本発明による結合分子またはその機能的変異体を、従来のペプチド・シンセサイザによって、または本発明によるDNA分子に由来するRNA核酸を使用した無細胞翻訳系において、合成的に生産することが可能である。上記した合成的な生産方法や無細胞翻訳系によって得られる結合分子またはその機能的変異体もまた、本発明の一部である。
【0066】
もう1つの実施形態では、本発明による結合分子またはその機能的変異体を、例えばトランスジェニック・マウスまたはウサギなどの、ヒト免疫グロブリン遺伝子を発現するヒト以外のトランスジェニック哺乳動物によって作り出すことができる。好適には、ヒト以外のトランスジェニック哺乳動物は、上記のヒト結合分子の全部分または一部をコードする、ヒト重鎖トランス遺伝子およびヒト軽鎖トランス遺伝子を含むゲノムを有する。ヒト以外のトランスジェニック哺乳動物は、狂犬病ウイルスまたはその断片を精製または濃縮した調製物を用いて免疫化することができる。ヒト以外の哺乳動物を免疫化するためのプロトコルは、当技術分野において確立されている。“Using Antibodies: A Laboratory Manual, Edited by: E. Harlow, D. Lane (1998), Cold Spring Harbor Laboratory, Cold Spring Harbor, New York”、および“Current Protocols in Immunology, Edited by: J. E. Coligan, A.M. Kruisbeek, D. H. Margulies, E.M. Shevach, W. Strober (2001), John Wiley & Sons Inc., New York”を参照されたく、その開示は参照することにより本明細書に組み込まれる。
【0067】
更なる態様において、本発明は、例えば本発明によるヒトモノクローナル抗体またはその断片、または本発明による核酸分子などの結合分子を同定する方法を提供するが、該方法は狂犬病ウイルスに特異的に結合することが可能であり、そしてa)結合を起こす条件の下で、複製可能な遺伝子パッケージ(genetic package)の表面に存在する結合分子の集合体と狂犬病ウイルスまたはその断片とを接触させる工程と、b)狂犬病ウイルスまたはその断片に結合する複製可能な遺伝子パッケージを少なくとも1回選択する工程と、およびc)狂犬病ウイルスまたはその断片に結合する複製可能な遺伝子パッケージを単離および回収する工程とから構成される。
【0068】
選択の工程は、狂犬病ウイルスの存在下で実行することができる。狂犬病ウイルスは、単離型であってもよいし、または、例えば感染した個体の血清中および/または血液中に存在するような、非単離型であってもよい。別の実施形態においては、狂犬病ウイルスは不活性化される。あるいは、選択の工程は、例えば狂犬病ウイルスの細胞外部分、Gタンパク質などの狂犬病ウイルスに由来する1つまたは複数の(ポリ)ペプチド、これらのタンパク質または(ポリ)ペプチドから成る融合タンパク質などの、狂犬病ウイルスの断片が存在する状態で実行することができる。もう1つの実施形態では、狂犬病ウイルスGタンパク質をトランスフェクションされた細胞を選択の手順に用いる。
【0069】
また更なる態様において、本発明は、本発明による結合分子または核酸分子を得る方法を提供するものであって、前記方法は、a)例えば本発明によるヒトモノクローナル抗体またはその断片、または本発明による核酸分子などの結合分子を同定するという、上述の方法を実行する工程、およびb)回収した複製可能な遺伝子パッケージから、結合分子および/または結合分子をコードする核酸を単離する工程から構成される。結合分子または結合分子をコードする核酸分子を同定する上述の方法によって、新規のモノクローナル抗体を一旦立証または同定した後は、scFvまたはFabをコードするDNAを細菌または複製可能な遺伝子パッケージから単離することができ、標準的な分子生物学的な技術と組み合わせることによって、2価のscFv、すなわち所望の特異性を有する完全なヒト免疫グロブリン(例えば、 IgG、IgA、またはIgM)をコードするコンストラクトを作製することができる。これらのコンストラクトは、適切な細胞系にトランスフェクションすることが可能であり、完全なヒトモノクローナル抗体を生産することが可能である(Huls et al., 1999; Boel et al., 2000参照)。
【0070】
本願明細書において用いる「複製可能な遺伝子パッケージ」は原核生物のものであっても真核生物のものであっても良く、細胞、胞子、細菌、ウイルス、(バクテリオ)ファージおよびポリゾームを含む。好適な複製可能な遺伝子パッケージは、ファージである。例えば単鎖Fvなどのヒト結合分子は、複製可能な遺伝子パッケージ上に露出しており、すなわち、複製可能な遺伝子パッケージの外表面に位置する官能基または分子に付着している。複製可能な遺伝子パッケージは、結合分子をコードする核酸分子に結合した、選別すべきヒト結合分子から構成される選別ユニットである。核酸分子は生体内でも(例:ベクターとして)、試験管内でも(例:PCR法による転写および翻訳)複製可能でなければならない。生体内での複製は、(細胞に関しては)自立的に、(ウイルスに関しては)ホストの因子の助けを借りて、あるいは(ファージミドに関しては)ホストおよびヘルパーウイルス両者の助けを借りて行われる。ヒト結合分子のコレクションを露出する複製可能な遺伝子パッケージは、ヒト結合分子外因性の結合分子をコードする核酸分子を複製可能な遺伝子パッケージのゲノム中に導入し、複製可能な遺伝子パッケージの外表面に通常発現される内在性のタンパク質との融合タンパク質を形成することによって、形成される。融合タンパク質の発現、外表面への輸送およびアセンブリの結果、複製可能な遺伝子パッケージの外表面に、外因性の結合分子が呈示される。更なる態様において、本発明は、狂犬病ウイルスまたはその断片に結合可能であって、上記の同定方法によって得ることのできるヒト結合分子に関する。
【0071】
また更なる態様において、本発明は、狂犬病ウイルスを中和する活性を潜在的に有する結合分子を同定する方法に関し、前記方法は、(a)結合を起こす条件の下で、複製可能な遺伝子パッケージの表面に存在する結合分子の集合体と狂犬病ウイルスとを接触させる工程と、(b)狂犬病ウイルスに結合する結合分子を、狂犬病ウイルスに結合しない結合分子から単離および回収する工程と、(c)少なくとも1つの回収した結合分子を単離する工程と、および(d)単離した結合分子が狂犬病ウイルスに対する中和活性を有するか否かを確認する工程とから構成され、集合体aにおける狂犬病ウイルスは不活性化されていることを特徴とする。不活性された狂犬病ウイルスは、不活性化される前の時点で精製することができる。精製は、例えばグリセロール・クッション(glycerol cushion)による遠心分離などの、ウイルスに適した周知の精製方法によって実行できる。集合体aで不活性化された狂犬病ウイルスを、使用する前に適切な材料に固定することが可能である。あるいは、集合体aでの狂犬病ウイルスは、依然として活性を持つ可能性がある。別の代わりの実施形態において、例えば、Gタンパク質などの狂犬病ウイルスのポリペプチドなどの、狂犬病ウイルスの断片が工程aで使用される。さらにもう1つの実施形態においては、狂犬病ウイルスGタンパク質をトランスフェクションされた細胞が、狂犬病ウイルスに対する中和活性を有する可能性のある結合分子を選択するために使用される。本願明細書において示されるように、選択された中和抗体の個数は、狂犬病ウイルスGタンパク質を発現する細胞が選択方法に含まれる場合に、精製された狂犬病ウイルスGタンパク質および/または不活性化された狂犬病ウイルスのみを使用することを特徴とする選択方法と比較して多かった。
【0072】
更なる実施形態において、上述した、狂犬病ウイルスに対する中和活性を有する可能性のある結合分子を同定する方法は、重鎖可変生殖系列遺伝子3−30を含むヒト結合分子を単離および回収し、必要に応じて単離する工程からさらに構成される。当業者は、例えばとりわけヌクレオチド配列決定法などの当業者にとって周知の技術によって、特定の生殖細胞系遺伝子を同定することができる。重鎖可変生殖系列遺伝子3−30を含む結合分子を単離および回収する工程は、工程cの前または後に実行することができる。下記の通り、本発明で発見された狂犬病ウイルスを中和するヒトモノクローナル抗体の大多数は、この特定のVH生殖細胞系遺伝子を含む。
【0073】
抗体などの、(中和活性をもつ)結合分子を同定および獲得するためのファージディスプレイ法は、今では当業者には周知の確立された手法である。それらは、例えば米国特許第5,696,108号、 Burton and Barbas, 1994, de Kruif et al., 1995b、および“Phage Display: A Laboratory Manual. Edited by: CF Barbas, DR Burton, JK Scott and GJ Silverman (2001), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York”に記載されている。これら全ては、その全体を参照することにより本明細書に組み込まれる。
【0074】
ファージ・ディスプレイ・ライブラリの構築のためには、ヒトモノクローナル抗体の重鎖および軽鎖の可変領域のコレクションが、バクテリオファージ粒子、好適には繊維状バクテリオファージ粒子の表面に、例えば単鎖Fv(scFv)またはFabの形態で発現される(de Kruif et al., 1995b参照)。抗体フラグメントを発現するファージの大きなライブラリは、概して1.0×109より大きな抗体特異性を有しており、免疫化された個体または免疫化されていない個体のBリンパ球内で発現する免疫グロブリンV領域からアセンブルすることが可能である。本発明の特定の実施形態において、ヒト結合分子のファージライブラリ、好適にはscFvファージライブラリは、狂犬病のワクチン接種を受けた被験者、あるいは狂犬病ウイルスに暴露された被験者から得た細胞より単離したRNAによって作られている。RNAは、とりわけ、骨髄または末梢血、好適には末梢血リンパ球から単離できる。被験者は、ワクチン接種を受けた動物または狂犬病ウイルスに暴露された動物とすれば良いが、好適には、ワクチン接種を受けたヒトまたは狂犬病ウイルスに暴露されたヒトを被験者とする。好適には、被験者となるヒトはワクチン接種を受けている。上記の通りの、例えばscFvファージライブラリなどの、複製可能な遺伝子パッケージの表面上に存在するヒト結合分子の集合体は、本発明の別の態様である。
【0075】
あるいは、ファージ・ディスプレイ・ライブラリは、ライブラリ内に付加的な抗体多様性をもたらすために試験管内で部分的にアセンブルした免疫グロブリン可変領域から構築することができる(半合成ライブラリ)。例えば、試験管内でアセンブルした可変領域は、合成によって生産された、ランダム化されたDNA領域または部分的にランダム化されたDNA領域を、例えばCDR領域などの抗体特異性にとって重要である分子内の領域に含む。狂犬病ウイルス特異性ファージ抗体は、ライブラリから選択することができるが、これは、例えば狂犬病ウイルスの抗原などの標的抗原を固相に固定化して、続いて標的抗原をファージライブラリにさらすことによって、固相に結合した抗原に特異的な抗体の断片を呈示するファージの結合を可能にすることによってなされる。結合を形成しなかったファージは洗浄によって取り除かれ、結合を形成したファージは、大腸菌(E.coli)へ感染させた後に引き続いて増殖させるために、固相から溶出される。ファージが標的抗原に対して特異的に結合するのを十分に強化するためには、複数回にわたって選択し増殖させることが通常は必要となる。必要に応じて、ファージライブラリを標的抗原にさらす前に、ファージライブラリを固相に結合した標的ではない抗原にさらすことによって、最初にファージライブラリの数を減らすことができる。また、例えば狂犬病ウイルスのタンパク質または(ポリ)ペプチドの複合混合物などの複合抗原に対する結合、1つまたは複数の狂犬病ウイルスのタンパク質または(ポリ)ペプチドを発現している宿主細胞に対する結合、または(不活性)狂犬病ウイルス自体に対する結合に関してファージを選択することも可能である。抗原特異的なファージ抗体は、不活性化された狂犬病ウイルスの調製物を取り付けられた固相をファージ抗体ライブラリと共にインキュベートし、例えばファージのscFv 部分またはFab部分を、狂犬病ウイルス調製物のタンパク質/ポリペプチドに結合させることによって、ライブラリから選択することができる。インキュベーションを行い、そして結合していないファージおよびゆるく結合したファージを取り除くために複数回の洗浄を行った後、調製物にscFv 部分またはFab部分を結合したファージを溶出して大腸菌に感染させることにより、新規の特異性を増幅することが可能となる。通常、過剰に存在する結合していないファージの中から、求めるファージを単離するためには、1回または複数回の選択の工程を必要とする。あるいは、狂犬病ウイルスの周知のタンパク質または(ポリ)ペプチドを宿主細胞内で発現させて、そしてこれらの細胞をタンパク質または(ボリ)ペプチドに対して特異的なファージ抗体を選択するために用いることができる。こうした宿主細胞を用いたファージディスプレイ法は、スクリーニングの操作中に、標的分子を含まない宿主細胞、または標的と似ているが同一ではない非標的分子を含む宿主細胞を過剰に加えることにより、特異的に結合しない結合分子を取り除くことで拡張および改善することができ、それによって適切な結合分子を見つけられる可能性が大いに増す(このプロセスは、MAbstract(登録商標)プロセスと呼ばれる。MAbstract(登録商標)はCrucell Holland B. V.の登録商標であり、 米国特許第6,265,150号も参照されたく、参照することにより本明細書に組み込まれる)。
【0076】
また更なる態様において、本発明は、少なくとも1つの結合分子、少なくとも1つの機能的変異体またはその断片、少なくとも1つの本発明による免疫コンジュゲート、またはそれらの組み合わせから構成される組成物を提供する。組成物は、アルブミンまたはポリエチレングリコールなどの安定化分子、または塩をさらに含む可能性がある。好適には、使用する塩は、ヒト結合分子の所望の生物学的活性を保持し、望ましくない有害な効果をもたらさない塩である。必要に応じて、本発明によるヒト結合分子を、酸による作用、または結合分子を不活性化する恐れのあるその他の天然/非天然の条件から保護するために、物質で覆うことが可能である。
【0077】
また更なる態様において、本発明は、本発明によって定義された核酸分子を少なくとも1つ含む組成物を提供する。組成物は、例えば、塩(例;NaClまたは上述の塩)を含む水溶液などの水溶液、界面活性剤(例;SDS)、および/または他の適切な構成要素を含むことが可能である。
【0078】
さらにまた本発明は、少なくとも1つの本発明による結合分子、少なくとも1つの機能的変異体またはその断片、少なくとも1つの本発明による免疫コンジュゲート、少なくとも1つの本発明による組成物、またはそれらの組み合わせから構成される医薬品組成物に関連する。本発明による医薬品組成物はさらに、薬学的に許容される賦形剤を少なくとも1つ含む。
【0079】
好適な実施形態において、本発明による医薬品組成物は、少なくとも1つの結合分子を追加的に含む。すなわち、医薬品組成物を結合分子のカクテル/混合物とすることが可能である。医薬品組成物は本発明による結合分子を少なくとも2つ含むか、あるいは、本発明による結合分子を少なくとも1つ含むのに加えて少なくとも1つの更なる抗狂犬病ウイルス結合分子を含みうる。前記の更なる結合分子は、好適には、配列番号25のアミノ酸配列を含むCDR3領域を備える。配列番号25のアミノ酸配列を含むCDR3領域を備える結合分子は、キメラモノクローナル抗体またはヒトモノクローナル抗体またはその機能的な断片であれば良いが、好適には、ヒトモノクローナル抗体またはその機能的な断片である。ある実施形態において、結合分子は、配列番号273のアミノ酸配列を含む重鎖可変領域を備える。別の実施形態において、結合分子は、配列番号275のアミノ酸配列を含む軽鎖可変領域を備える。さらにもう1つの実施形態において、結合分子は、配列番号123および配列番号125のアミノ酸配列をそれぞれ含む重鎖および軽鎖を備える。医薬品組成物に含まれる複数の結合分子は、狂犬病ウイルスの、互いに異なり互いに競合しないエピトープとそれぞれ反応できなくてはならない。エピトープは、狂犬病ウイルスのGタンパク質上に存在する可能性があり、互いに異なる重複しないエピトープである可能性がある。結合分子は、高い親和性を有さなければならず、広い特異性を有さなければならない。好適には、それらの結合分子は、できるだけ多くの狂犬病ウイルスの固定毒株および街上毒株(street strain)を中和する。さらにより好適には、それらの結合分子はリッサウイルス属の他の遺伝子型、またはラブドウイルス科の他のウイルスにさえも中和活性を示し、一方で、他のウイルスや通常の細胞タンパク質とは交差反応性を示さない。好適には、結合分子は、カクテル中の他の結合分子のエスケープ変異体を中和することができる。
【0080】
本発明の別の態様は、狂犬病ウイルス中和活性を有する結合分子を少なくとも2つ含む医薬品組成物に関し、好適には、前記結合分子は、狂犬病ウイルスの互いに異なり互いに競合しないエピトープに結合できることを特徴とする、本発明による(ヒト)結合分子である。ある実施形態において、医薬品組成物は、狂犬病ウイルスGタンパク質の抗原部位Iに位置するエピトープと結合することが可能な、狂犬病ウイルス中和活性を有する第一の結合分子と、狂犬病ウイルスGタンパク質の抗原部位IIIに位置するエピトープと結合することが可能な、狂犬病ウイルス中和活性を有する第二の結合分子とを備える。狂犬病糖タンパク質の抗原構造は、最初にLafon氏ら (1983)によって定義された。抗原部位は、マウスのmAbおよびそれらのそれぞれのmAb抵抗性ウイルス変異体のパネルを使用して同定された。それ以来、mAb抵抗性ウイルス変異体の糖タンパク質におけるアミノ酸変異を同定することによって、抗原部位は決定されてきた(Seif et al. , 1985、Prehaud et al. , 1988、およびBenmansour et al. , 1991を参照)。狂犬病ウイルス中和活性を有するmAbの大多数は抗原部位IIに対する抗体であり(Benmansour et al. , 1991参照)、抗原構造IIは、アミノ酸34-42およびアミノ酸198-200を含む、不連続な立体構造エピトープである(Prehaud et al. , 1988参照)。抗原部位IIIはアミノ酸330-338の位置に存在する連続な立体構造エピトープで、2つの荷電残基K330およびR333を擁し、これらの荷電残基はウイルスの病原性に影響を及ぼす(Seif et al., 1985、Coulon et al., 1998、および“Dietzschold et al., 1983参照)。立体的な抗原部位Iは、ただ1つのmAb、509-6によって定義され、アミノ酸231位置に存在する(Benmansour et al., 1991、Lafon et al., 1983参照)。抗原部位IVは、互いに重複する直線状エピトープを擁することが知られている(Tordo, 1996、Bunschoten et al., 1989、Luo et al., 1997、およびNi et al., 1995参照)。Benmansour et al. (1991) も、アミノ酸342-343に位置し、抗原部位IIIに近接しているにもかかわらず抗原部位IIIとは別のマイナーサイトaの存在について記載している。CR-57エピトープを、現在周知である狂犬病糖タンパク質上の直線状および立体構造の中和エピトープとアライメントすることにより(図10)、CR-57エピトープは、単一のmAb、509-6によって定義される抗原部位Iと同じ場所に位置することが明らかになった。CR04-098のエスケープウイルスの糖タンパク質のヌクレオチド配列およびアミノ酸配列に基づいて考えると、この抗体によって認識されるエピトープは連続的な立体構造をとる抗原部位IIIと同一の領域に位置するように思われる。
【0081】
好ましい実施形態において、医薬品組成物は、配列番号25のアミノ酸配列を含むCDR3領域(好適には重鎖CDR3領域)を少なくとも備える、狂犬病ウイルス中和活性を有する第一の結合分子と、配列番号4、配列番号10、配列番号14、配列番号15、配列番号16および配列番号22から構成される群から選択されたアミノ酸配列を含むCDR3領域(好適には重鎖CDR3領域)を少なくとも備える、狂犬病ウイルス中和活性を有する第二の結合分子とを備える。該領域は、より好適には、狂犬病ウイルス中和活性を有する第二の結合分子は、配列番号14のアミノ酸配列を含むCDR3領域(好適には重鎖CDR3領域)を少なくとも備える。好適には、狂犬病ウイルス中和活性を有する第一の結合分子は、配列番号123および配列番号125のアミノ酸配列をそれぞれ含む重鎖および軽鎖を備え、狂犬病ウイルス中和活性を有する第二の結合分子は、配列番号335および配列番号337のアミノ酸配列をそれぞれ含む重鎖および軽鎖を備える。好適には、狂犬病ウイルス中和活性を有する第一の結合分子の重鎖および軽鎖は、それぞれ配列番号122および配列番号124によってコードされ、狂犬病ウイルス中和活性を有する第二の結合分子の重鎖および軽鎖は、それぞれ配列番号334および配列番号336によってコードされる。
【0082】
2つの結合分子から成る医薬品組成物で、結合分子の等電点(pI)が互いに異なる場合、両方の結合分子を最適に安定化させる適切なバッファを選択するのが困難である可能性がある。一方の結合分子の安定性が増すように組成物のバッファのpHを調整すると、もう一方の結合分子の安定性が減少する可能性がある。結合分子の安定性が減少してしまうと、あるいは不安定性が減少してしまう場合でさえも、結合分子の沈殿または集合、あるいは結合分子の自発的な分解につながる可能性があり、その結果として結合分子の機能が失われる可能性がある。従って、別の態様においては、本発明は少なくとも2つの結合分子から成る医薬品組成物を提供するが、該結合分子は好適にはヒト結合分子であり、該結合分子は、互いの等電点(pI)の差が約1.5、1.4、1.3、1.2、1.1、1.0、0.9、0.8、0.7、0.6、0.5、0.4、0.3よりも小さく、好適には0.25pI単位以下であることを特徴とする。等電点は、例えば等電点電気泳動などを用いて実験的に測定可能であり、または結合分子のアミノ酸配列に基づいて計算することもできる。1つの実施形態において、結合分子は本発明による結合分子であり、医薬品組成物は本発明による医薬品組成物である。好適には、結合分子はモノクローナル抗体であり、その例えばIgGl 抗体などのヒトモノクローナル抗体がある。好適には、結合分子は、ウイルス、細菌、酵母、真菌または寄生虫などの感染性の病原体を結合および/または中和する能力がある。ある実施形態において、結合分子は、狂犬病ウイルスなどのリッサウイルスを結合および/または中和することができる。特定の実施形態において、双方の結合分子の算出等電点は、8.0〜9.5の範囲、好適には8.1〜9.2の範囲、より好適には8.2〜8.5間の範囲内に収まる。好適には、2つの結合分子は、それぞれ、配列番号14および配列番号25の重鎖CDR3領域を有する。
【0083】
別の実施形態では、本発明は、2つ以上のヒト結合分子または他の動物の結合分子(抗体を含むがこれに限定されない)のカクテルを提供するが、少なくとも1つの結合分子は抗体ファージに由来するか複製可能なパッケージを表示する他の技術に由来し、そして少なくとも1つの結合分子はハイブリドーマ技術によって得られる。異なる技術を使用する際には、それぞれの結合分子が保管、処理およびその後の使用の際に十分安定であるような組成物を得るために、適合する等電点を有する結合分子を選択することも非常に有益である。
【0084】
別の実施形態では、本発明の医薬品組成物中に存在する結合分子は、互いの中和活性を向上させる。すなわち、組み合わせて用いると結合分子は相乗的に作用する。換言すれば、医薬品組成物は、相乗効果的な、狂犬病ウイルス中和活性および更にはリッサウイルス中和活性を示しうる。本明細書において用いる「相乗効果的な」という用語は、複数の結合分子を組み合わせて用いた場合に得られる複合効果は、単独で用いた場合に得られる効果を足し合わせたものよりも大きいことを意味する。本発明の医薬品組成物の構成要素の範囲および比率は、構成要素の個々の効力に基づいて決定すべきであり、試験管内での中和アッセイや、ハムスターなどの動物モデルを用いて検証するべきである。
【0085】
さらにまた、本発明による医薬品組成物は、少なくとも1つの他の治療薬、予防薬および/または診断用薬を含むことができる。前記のさらなる治療薬および/または予防薬は、リバビリンやインターフェロンαなどの抗ウイルス薬でとしてよい。
【0086】
本発明の結合分子または医薬品組成物は、ヒトに使用する前に、適切な動物モデル系において検証することができる。そのような動物モデル系には、マウス、ラット、ハムスター、サルなどが含まれるが、これらに限定されない。
【0087】
一般的に、医薬品組成物は、製造および保管の条件の下で、無菌的かつ安定でなくてはならない。本発明のヒト結合分子、変異体やその断片、免疫コンジュゲート、核酸分子、または組成物を、投与する前や投与時に、薬学的に許容される適切な賦形剤に再構成するために、粉末形態とすることも可能である。無菌的な注射剤を調製するための無菌粉末の場合には、より好適な調製方法は真空乾燥およびフリーズドライ(凍結乾燥)であり、この調製方法によって、活性成分に所望の追加的な所望の成分が加わった粉末を、事前に除菌したそれらの溶液から得ることができる。
【0088】
あるいは、本発明のヒト結合分子、変異体やその断片、免疫コンジュゲート、核酸分子、または組成物は、溶液中に存在していてもよく、投与する前や投与時に、薬学的に許容される適切な賦形剤を添加および/または混合することによって、単位投与量の注射剤型を提供することが可能である。好適には、本発明において使用する薬学的に許容される賦形剤は、高い薬剤濃度に適しており、適当な流動性を維持することが可能で、必要に応じて吸収を遅らせることができる。
【0089】
医薬品組成物の最適な投与経路の選択はいくつかの要因に影響され、それには、組成物中の活性分子の物理化学的性質、臨床病状の緊急性、および所望の治療効果に対する血漿中の活性分子濃度の関係が含まれる。例えば、本発明のヒト結合分子は、必要に応じて、急速に放出されてしまうことからそれらを保護する担体と共に調製されることが可能であり、例えば、インプラント、経皮貼布、およびマイクロカプセル化したデリバリーシステムを含む、放出制御製剤が挙げられる。エチレン酢酸ビニル、ポリ無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル類およびポリ乳酸などの、生分解性の、生体適合性ポリマーがとりわけ使用されうる。更に、ヒト結合分子の不活性化を阻止するような材料または化合物によって、ヒト結合分子を覆うか、あるいはヒト結合分子と共に投与することが必要である可能性がある。例えば、ヒト結合分子は、例えばリポソームまたは希釈剤などの適切な担体の形で、対象者に投与されうる。
【0090】
投与経路は、一般的には、経口投与と非経口投与の2つの主要なカテゴリーに分類することができる。本発明のヒト結合分子および医薬品組成物の投与は、傷口の内部およびその周辺、そして臀部の筋肉内に行なうのが好適である。ヒト結合分子および医薬品組成物の調製は、投与経路に依存する。
【0091】
更なる態様において、結合分子、機能的変異体、免疫コンジュゲート、組成物、または医薬品組成物は、薬剤として使用することができる。ゆえに、本発明のヒト結合分子、機能的変異体、免疫コンジュゲート、組成物、または医薬品組成物を使用した、リッサウイルス感染症の治療法および/または予防法は本発明の他の一部分である。リッサウイルスは、既存の遺伝子型の任意のウイルスであり得るが、好適には狂犬病ウイルスである。上述の分子や組成物は、狂犬病の暴露後予防に使用することができる。
【0092】
上述の分子や組成物は、狂犬病ウイルスの診断、予防および/または治療に役立つ他の分子と併せて使用することが可能である。それらは、試験管内、生体外または生体内で使用することができる。例えば、本発明のヒト結合分子、機能的変異体、免疫コンジュゲート、または医薬品組成物を、狂犬病に対するワクチンと共に投与することができる。あるいは、ワクチンを、本発明の分子または組成物を投与する前または後に投与しても良い。本発明の分子または組成物をワクチンと共に投与することは、暴露後予防に適している。狂犬病ワクチンは、精製ニワトリ胚細胞(PCEC)ワクチン(RabAvert)、ヒト二倍体細胞ワクチン(HDCV; Imovaxワクチン)または、吸着狂犬病ワクチン(RVA; rabies vaccine adsorbed)を含むが、これらに限定されない。
【0093】
典型的には、これらの分子は、本発明の組成物および医薬品組成物の中に、治療上または診断上効果のある量として調製される。投与計画は、最適な所望の効果(例えば治療効果)をもたらすように調製することが可能である。適切な用量範囲は、例えば0.1〜100IU/体重(kg)、好適には1.0〜50IU/体重(kg)、より好適には10〜30IU/体重(kg)(例えば20IU/体重(kg))である可能性がある。
【0094】
好適には、本発明の結合分子または医薬品組成物の単回ボーラス投与が行われる。本発明による分子および医薬品組成物は好適には無菌である。これらの分子および組成物を無菌にする方法は当技術分野では周知である。暴露後予防の投薬計画は、狂犬病ウイルスに対する予防接種を以前に受けていない個人に対し、暴露後0日、3日、7日、14日および28日の計5回、狂犬病ワクチンを三角筋に筋肉内投与するというものである。本発明によるヒト結合分子または医薬品組成物は、暴露後0日に、そうでなければ暴露後出来る限り早く、傷口の内部および周辺に投与するべきであり、残りの量については、ワクチンから離れた場所に筋肉内投与する。予防接種を受けていない個体に対しては、抗狂犬病ウイルスヒト結合分子の投与が勧められるが、予防接種を受けた個体に対しても、このような治療が必要な際には抗狂犬病ウイルスヒト結合分子を投与して良いことは、当業者には明らかである。
【0095】
別の態様において、本発明は、リッサウイルスによる感染症の結果生じた状態の診断、予防、治療、またはそれらの組み合わせのために用いる薬剤を調製する際の、結合分子又はその機能的変異体、本発明による免疫コンジュゲート、本発明による核酸分子、本発明による組成物又は医薬品組成物の用途に関する。リッサウイルスは、既存の遺伝子型の任意のウイルスであり得るが、好適には狂犬病ウイルスである。好適には、上述の分子は、狂犬病の暴露後予防のための薬剤の調製に用いられる。
【0096】
次に、少なくとも1つの本発明による結合分子、少なくとも1つの本発明による結合分子の機能的変異体、少なくとも1つの本発明による免疫コンジュゲート、少なくとも1つの本発明による核酸分子、少なくとも1つの本発明による組成物、少なくとも1つの本発明による医薬品組成物、少なくとも1つの本発明によるベクター、少なくとも1つの本発明によるホスト、またはそれらの組み合わせを含むキットもまた、本発明の一部である。必要に応じて、上記の本発明のキットの構成要素は、適切な容器にパックされ、適応となる病気の診断、予防および/または治療用にラベルを貼られる。上述の構成要素は、好適には無菌の水溶液として、あるいは好適には無菌の凍結乾燥製剤として、1回量あるいは複数回分の用量を入れられる容器(例えば密封されたアンプル、バイアル、ビン、注射器および試験管など)の中に保管されうる。容器は、ガラスまたはプラスチックなどの種々の材料から形成することが可能で、無菌のアクセスポートを有することが可能である(例えば、この容器は、皮下注射針を刺し通すことが可能な栓を有する静脈内用溶液バッグまたはバイアルである可能性がある)。このキットは、例えばリン酸緩衝食塩水、リンガー溶液およびブドウ糖溶液などの、薬学的に許容されるバッファを含む、より多くの容器からさらに構成されうる。キットは、その他のバッファ、希釈剤、フィルタ、針、注射器、1つまたは複数の適切なホストのための培養液を含む、商業的およびユーザの見地から望ましい他の材料を更に含むことが可能である。キットには、治療、予防または診断用の製品の商業用パッケージには慣習的に含まれる取扱説明書をつけることができ、そのような治療、予防または診断用の製品を使用するに際しての、指示、用法、用量、製造、投与、禁忌および/または警告などの情報が含まれる。
【0097】
現在、HRIG製品が狂犬病の暴露後予防のために使われている。成人に対する1回投与量である1500 IU (体重75 kgの人, 20 IU/kg)は、10mlの体積としてのみ利用可能である。現在入手可能な10mlの1回投与量には、合計1〜1.5グラムのIgGが含まれるため、より濃縮されたHRIG製品を得ることは不可能である。その観点では、現在のHRIG製品には、2つの欠点がある。第一に、推奨される1回投与量を傷口の内部およびその周囲に投与するのは解剖学的に実行不可能であること、そして第二に、現在のHRIGの1回投与量であっても投与の際にかなりの痛みが伴うことである。本発明は、望まれる場合には、約2mlまたはそれ未満の体積中に成人の1回分の全投与量を含む医薬品組成物を提供することにより、これらの欠点に対する解決策を提供する。この種の医薬品組成物は、例えば、狂犬病ウイルスを中和する能力を有する2つの結合分子を含む可能性があり、好適にはCR57およびCR04-098である。医薬品組成物は、薬学的に許容される賦形剤を更に含み、そして約2mlの体積を有する。より大きな体積とすることも可能であるが、より大きな体積を注射した際に生じる痛みの観点からあまり望ましくはない。2ml未満の体積とすることも可能である。医薬品組成物は、暴露後予防の成功のために必要な成人の全投与量(IU単位)を含む。ある実施形態において、医薬品組成物は、例えば、10mlの栓付きのready-to-useバイアル(I型ガラス)などの、10mlバイアルの中に保存される。10mlのバイアルを提供することによって、個人が大きな傷口表面積を示した場合に、医薬品組成物をより大きな体積へと希釈するという選択肢が生ずる。本発明はまた、医薬品組成物を含む容器(例えばバイアル)を少なくとも備えるキットを提供する。キットは、より大きな体積へと医薬品組成物を希釈するのに適した希釈剤を入れることができる第二の容器をさらに備えることが可能である。適切な希釈剤は、医薬品組成物の薬学的に許容される賦形剤、および食塩水を含むが、これらに限定されない。さらにまた、キットは、医薬品組成物を希釈するための取扱説明書、および/または、希釈の有無にかかわらず、医薬品組成物を投与するための取扱説明書を備えることが可能である。
【0098】
本発明は更に、サンプル中の狂犬病ウイルスを検出する方法に関し、該方法は、a) 診断上効果のある量の、本発明による結合分子、機能的変異体、または免疫コンジュゲートをサンプルに接触させる工程、およびb) 結合分子、機能的変異体、または免疫コンジュゲートが、サンプルの分子と特異的に結合しているか否かを決定する工程から構成される。サンプルは生物学的サンプルとすることができ、(潜在的に)感染している対象由来の血液、血清、組織または他の生物学的材料を含むが、これらに限定されない。(潜在的に)感染している対象はヒトであっても良いが、狂犬病ウイルスのキャリアである疑いのある動物についても、本発明のヒト結合分子、機能的変異体、または免疫コンジュゲートを用いて、狂犬病ウイルスの存在を検査することができる。検出方法により適したものとするために、サンプルを最初に操作することができる。操作とは、とりわけ、狂犬病ウイルスがタンパク質、(ポリ)ペプチドまたは他の抗原断片などの抗原構成要素へ分解するような方法で、狂犬病ウイルスを含む疑いのあるサンプルおよび/または狂犬病ウイルスを含むサンプルを処理することを意味する。好適には、ヒト結合分子、およびサンプル中に存在する可能性のある狂犬病ウイルスまたはその抗原構成要素との間で免疫複合体が形成されるような条件の下で、本発明の結合分子、機能的変異体、または免疫コンジュゲートをサンプルと接触させる。サンプル中に狂犬病ウイルスが存在することを示唆する免疫複合体の形成が少しでも生じるとすれば、適切な手段によって検出および測定する。この種の方法には、とりわけ、例えばラジオイムノアッセイ(RIA)、ELISA、免疫蛍光検査、免疫組織化学、FACS、BIACOREおよびウエスタンブロット分析などの、均一系および不均一系に結合させるイムノアッセイが含まれる。
【0099】
さらにまた、本発明の結合分子は、Gタンパク質などの狂犬病ウイルスタンパク質のエピトープを同定するために用いることができる。エピトープは、直線状であってもよいが、構造的および/または立体配置的であってもよい。ある実施形態では、狂犬病ウイルス由来のタンパク質(例えば狂犬病ウイルスGタンパク質)の、一連の重複するペプチド(例えば15-merペプチドなど)に対する結合分子の結合は、PEPSCAN分析を用いて分析することができる(とりわけ、WO 84/03564、WO 93/09872、Slootstra et al. 1996参照)。それぞれのペプチドに対するヒト結合分子の結合は、PEPSCANに基づいた酵素免疫測定法(ELISA)によって試験することができる。他の実施形態では、本発明のヒト結合分子と結合することができるペプチドを同定するために、狂犬病ウイルスタンパク質由来のペプチドを含むランダムなペプチドライブラリをスクリーニングすることができる。上記のアッセイにおいて、狂犬病ウイルスを中和することができるヒト結合分子を使用することによって、1つまたは複数の中和エピトープが同定される可能性がある。発見されたペプチド/エピトープは、ワクチンとして使用することも可能であるし、狂犬病の診断のために使用することも可能である。
【0100】
更なる態様において、本発明は、本発明の結合分子または機能的変異体が結合するエピトープとは異なる、好適には互いに重複しない、狂犬病ウイルスのエピトープに対する特異的な結合を同定するために、結合分子または結合分子の機能的変異体のスクリーニングを行なう方法を提供するものであって、該方法は、a) スクリーニング対象の結合分子または機能的変異体と、本発明の結合分子または機能的変異体と、狂犬病ウイルスまたはその断片(例えば狂犬病ウイルスGタンパク質)とを接触させる工程、およびb) スクリーニング対象の結合分子または機能的変異体が、狂犬病ウイルスまたはその断片に対する特異的な結合をめぐって、本発明による結合分子または機能的変異体と競合する能力があるかを測定する工程から構成される。もしも競合が測定されないのならば、スクリーニング対象の結合分子または機能的変異体は、異なるエピトープに結合する。上記のスクリーニング方法の特定の実施形態において、配列番号25のアミノ酸配列を含むCD3領域を備えた結合分子によって認識されるエピトープとは異なるエピトープへの結合能を有する、ヒト結合分子または機能的変異体を同定するために、ヒト結合分子またはその機能的変異体のスクリーニングを行なうことが可能である。好適には、エピトープは互いに重複しないか、または互いに競合しない。上記のスクリーニング方法を、同一のエピトープへの結合能を有する結合分子またはその機能的変異体を同定するために用いることができることは、当業者にとって明らかである。更なる工程においては、狂犬病ウイルスまたはその断片に対する特異的な結合をめぐって競合する能力を有さない、スクリーニングされた結合分子が、中和活性を有するかどうかを決定することが可能である。また、狂犬病ウイルスまたはその断片に対する特異的な結合をめぐって競合する能力を有する、スクリーニングされた結合分子が、中和活性を有するかどうかを決定することも可能である。前記スクリーニング方法によって得られた、中和能力を有する抗狂犬病ウイルス結合分子またはその機能的変異体は、本発明の一部である。前記スクリーニング方法において、「同一のエピトープに対して特異的に結合する」とは、本発明のヒト結合分子が結合するエピトープと実質的にまたは本質的に同一のエピトープに対して特異的に結合することをも意図する。本発明のヒト結合分子が狂犬病ウイルスに結合するのを阻止したり、またはこのような結合と競合したりする能力があるということは、典型的には、スクリーニング対象の結合分子は、本発明の結合分子によって免疫特異的に認識される狂犬病ウイルス上の結合部位と構造的に重複する、狂犬病ウイルスのエピトープまたは結合部位と結合することを示唆する。あるいは、このことは、スクリーニング対象の結合分子は、本発明の結合分子によって免疫特異的に認識される結合部位と十分に近いエピトープまたは結合部位に結合するため、本発明による結合分子が狂犬病ウイルスまたはその断片に結合することを立体的にまたは別の方法によって阻止する、ということを示唆する可能性もある。
【0101】
一般的に、競合阻害はアッセイを用いて測定されるが、このアッセイにおいては、抗原組成物、すなわち、狂犬病ウイルスまたはその(Gタンパク質などの)断片から成る組成物を、対照結合分子およびスクリーニング対象の結合分子と混合する。ある実施形態においては、対照結合分子を本発明によるヒト結合分子の1つとしてよく、スクリーニング対象の結合分子を本発明の別のヒト結合分子としてよい。別の実施形態では、対照結合分子は配列番号25のアミノ酸配列を含むCDR3領域を備えた結合分子としてよく、スクリーニング対象の結合分子を本発明によるヒト結合分子の1つとしてよい。さらに別の実施形態においては、対照結合分子を本発明のヒト結合分子の1つとしてよく、スクリーニング対象の結合分子を配列番号25のアミノ酸配列を含むCDR3領域を備えた結合分子としてよい。通常、スクリーニング対象の結合分子は過剰に存在する。このような単純な競合を研究するためには、ELISAに基づいたプロトコルを用いるのが適当である。ある実施形態では、対照結合分子を、様々な量のスクリーニング対象の結合分子と予め混合し(例えば、1:10、1:20、1:30、1:40、1:50、1:60、1:70、1:80、1:90または1:100)、一定の時間をおいてから、それらを抗原組成物に加えることが可能である。他の実施形態では、対照結合分子およびスクリーニング対象の結合分子を、抗原組成物に暴露している間に単純に混合することが可能である。いずれにしても、動物種またはアイソタイプに対する二次抗体を使用することによって、結合した対照結合分子のみを検出することができるのだが、この結合力は、実質的に同一のエピトープを認識するスクリーニング対象の結合分子が存在する場合には減少する。対照結合分子およびスクリーニング対象の結合分子の間での、結合分子の競合についての研究を行なう際には(動物種またはアイソタイプにかかわらない)、まず、対照結合分子を、例えばビオチン、酵素標識、放射性標識またはその後の同定を可能とする別の標識などの、検出可能なラベルで標識することが可能である。こうした場合には、標識化された対照結合分子とスクリーニング対象の結合分子とを様々な比率(例えば、1:10、1:20、1:30、1:40、1:50、1:60、1:70、1:80、1:90または1:100)で予め混合するかインキュベートし、そして、(適当な時間が経過した後に、任意で、)標識化された対照結合分子の反応性を検定し、この値を、競合する可能性のある結合分子がインキュベーションの際に含まれていない場合のコントロール値と比較する。再度言うが、このアッセイは、抗体のハイブリダイゼーションに基づく免疫学的アッセイの範囲内のうちの任意の1つであればよく、対照結合分子は標識を検出することによって検出され、例えば、ビオチン化した対照結合分子の場合にはストレプトアビジンを使用することによって、または酵素標識に関連する発色性基質を使用することによって(3,3'5,5'-テトラメチルベンゼン(TMB)基質とペルオキシダーゼ)、または単純に放射性標識を検出することによって検出される。対照結合分子と同一のエピトープに結合するスクリーニング対象の結合分子は、結合をめぐって効果的に競合することができ、従って対照結合分子の結合は有意に減少するのだが、このことは結合度の減少によっても明らかである。互いに異なり、互いに競合しないエピトープに結合する結合分子の場合には、結合度は減少しない。全く関連性のない結合分子が存在しない時の(標識化された)対照結合分子の反応性を、コントロール・ハイバリュー(control high value)とする。コントロール・ローバリュー(control low value)は、標識化された対照結合分子を、標識化されていない正確に同じ種類の対照結合分子と共にインキュベートすることによって得ることができるが、この際には、競合が生じて、標識化された対照結合分子の結合度は減少する。試験アッセイでは、スクリーニング対象の結合分子の存在下で、標識化された結合分子の反応性に有意な減少がみられたことから、同一のエピトープを認識する結合分子(すなわち、標識化された対照結合分子)と「交差反応」する結合分子の存在が示唆される。減少がみられない場合、結合分子は、互いに異なる互いに競合しないエピトープに結合する可能性がある。
【0102】
これらの競合アッセイによって同定された結合分子(「競合結合分子」)には、対照結合分子が結合するエピトープまたは結合部位に結合する、抗体、抗体断片および他の結合薬剤も含まれるし、同時に、対照結合分子が結合するエピトープに十分近く、スクリーニング対象の結合分子および対照結合分子間での競合的結合が生じる位置に存在するようなエピトープまたは結合部位に結合する、抗体、抗体断片および他の結合薬剤も含まれるが、これらに限定されるものではない。好適には、本発明の競合結合分子は、過剰に存在する場合には、特定の標的に対する対照結合分子の特異的結合を、少なくとも10%、好適には少なくとも25%、より好適には少なくとも50%、そして最も好適には少なくとも75〜90%またはそれ以上阻害する。本発明の結合分子とほぼ同一のエピトープ、実質的に同一のエピトープ、本質的に同一のエピトープ、または全く同一のエピトープに結合する、1つまたは複数の競合結合分子を同定することは、単純に技術的な事項である。競合結合分分子の同定は対照結合分子との比較によってなされるため、対照結合分子と同一のエピトープまたは実質的に同一のエピトープに結合する競合結合分子を同定するために、対照結合分子および競合結合分子が結合するエピトープを実際に決定する必要性は全くないということが理解できるであろう。あるいは、これらの競合アッセイによって同定された、互いに異なる互いに競合しないエピトープに結合する結合分子にもまた、抗体、抗体断片、および他の結合薬剤が含まれるが、これらに限定されない。
【0103】
別の態様では、本発明は、生物に病気を引き起こす感染性物質を中和する活性を潜在的に有する結合分子、または生物に病気を引き起こす感染性物質を中和する活性を潜在的に有する結合分子をコードする核酸分子を同定する方法を提供するものであって、該方法は、a) 複製可能な遺伝子パッケージの表面に存在する結合分子の集合体に、少なくとも生物に病気を引き起こす感染性物質のタンパク質を表面に発現している細胞を、結合を起こす条件の下で接触させる工程と、b) 生物に病気を引き起こす感染性物質のタンパク質を表面に発現している細胞に結合する結合分子を、前記細胞に結合しない結合分子から分離および回収する工程と、c) 少なくとも1つの回収した結合分子を単離する工程と、およびd) 単離された結合分子が生物に病気を引き起こす感染性物質に対する中和活性を有するかどうかを確認する工程とから構成される。生物に病気を引き起こす感染性物質のタンパク質を表面に発現している細胞は、タンパク質をトランスフェクションされた細胞であってよい。当業者は、この方法では、タンパク質以外の感染性物質の抗原も、うまく利用できることを知っている。特定の実施形態においては、細胞はPER.C6(商標登録)細胞である。しかし、BHK細胞、CHO細胞、NS0細胞、HEK293細胞、または911細胞などの他の(E1‐不死化)細胞系列を、タンパク質を発現するために使用することも可能である。ある実施形態では、結合分子はヒト結合分子である。感染性物質は、ウイルス、バクテリア、酵母、真菌または寄生虫である可能性がある。ある実施形態では、タンパク質は、感染性物質の表面に通常発現するタンパク質であるか、あるいは、表面に存在するタンパク質の少なくとも一部を含む。ある特定の実施形態においては、複製可能な遺伝子パッケージの表面に存在する結合分子のコレクションは、感染性物質のタンパク質を発現するために使用した細胞によってサブストラクション/対抗選択され、すなわち、この細胞は、感染性物質のタンパク質を細胞表面に発現しないという点を除いては、ステップaで使用された細胞と同一である。サブストラクション/対抗選択に使用する細胞は、トランスフェクションされていない細胞であってよい。あるいは、細胞に、感染性物質の対応するタンパク質と配列や構造が類似する、および/または高度な相同性を示す、および/または同じ科又は同じ属の感染性物質由来の、タンパク質またはその(細胞外)部分をトランスフェクションすることも可能である。
【0104】
本発明の別の態様は、狂犬病ウイルス中和活性を有する、本発明において定義される結合分子に関するものであって、このヒト結合分子は配列番号25のアミノ酸配列を含む重鎖CD3領域を備えることを特徴とし、更に、このヒト結合分子は少なくとも2500 IU/mgタンパク質の狂犬病ウイルス中和活性を有することを特徴とする。より好適には、前記ヒト結合分子は狂犬病ウイルス中和活性を有するが、該中和活性は、少なくとも2800 IU/mgタンパク質、少なくとも3000 IU/mgタンパク質、少なくとも3200 IU/mgタンパク質、少なくとも3400 IU/mgタンパク質、少なくとも3600 IU/mgタンパク質、少なくとも3800 IU/mgタンパク質、少なくとも4000 IU/mgタンパク質、少なくとも4200 IU/mgタンパク質、少なくとも4400 IU/mgタンパク質、少なくとも4600 IU/mgタンパク質、少なくとも4800 IU/mgタンパク質、少なくとも5000 IU/mgタンパク質、少なくとも5200 IU/mgタンパク質、少なくとも5400 IU/mgタンパク質である。結合分子の中和活性は、試験管内での中和アッセイ(改良RFFIT (rapid fluorescent focus inhibition test))によって測定された。このアッセイについては、下の実施例の項で詳しく説明する。
【0105】
ある実施形態において、結合分子は、配列番号273のアミノ酸配列を含む重鎖可変領域を備える。別の実施形態において、結合分子は、配列番号123のアミノ酸配列を含む重鎖を備える。結合分子の軽鎖可変領域は配列番号275のアミノ酸配列を含む可能性がある。結合分子の軽鎖は、配列番号125のアミノ酸配列を含む可能性がある。
【0106】
上述の、結合分子をコードする核酸分子も、本発明の一部である。好適には、核酸分子は、配列番号122のヌクレオチド配列を含む。加えて、核酸分子は、配列番号124のヌクレオチド配列をも含む可能性がある。核酸分子を含むベクターおよび該ベクターを含む宿主細胞も、本願明細書において提供される。好適には、宿主細胞はヒトの細胞などの哺乳類細胞である。ヒト結合分子の生産に適した細胞の例は、とりわけ、HeLa細胞、911細胞、AT1080細胞、A549細胞、293細胞、およびHEK293T細胞である。好適な哺乳類細胞は911細胞、もしくは、欧州細胞培養コレクション(ECACC), CAMR, Salisbury, Wiltshire SP4 OJG, Great Britainに、No.96022940として1996年2月29日に寄託され、登録商標PER.C6(商標登録)(PER. C6は、Crucell Holland B.V.の登録商標である)として市場に出された細胞系などの、ヒト網膜細胞である。本出願の目的上、「PER.C6」とは、No.96022940として寄託された細胞あるいはその祖先細胞、上流あるいは下流の継代細胞、および寄託された細胞の祖先細胞の子孫細胞、および前述した任意の細胞の派生株を意味する。
【実施例】
【0107】
本発明を説明するために、以下の実施例を提供する。実施例は、いかなる形であれ本発明の範囲を限定することを目的としない。
【0108】
実施例1
ヒト抗狂犬病抗体CR-57およびCR-JBのエピトープの認識
CR-57およびCR-JBと呼ばれるヒト抗狂犬病抗体が互いに重複したり互いに競合したりしないエピトープを認識するかどうかという問題に取り組むために、CR-57およびCR-JBと呼ばれるヒト抗狂犬病抗体のエスケープウイルスを生成した。CR-57およびCR-JBは、対応する抗体遺伝子の重鎖および軽鎖可変領域をコードする領域をpcDNA3002(Neo)という名の単一のヒトIgG1発現ベクターへ導入する操作を経て、基本的には記載された通りに生成した(Jones et al., 2003参照)。その結果得られたベクターであるpgSO57C11およびpgSOJBC11は、欧州細胞培養コレクション(ECACC), CAMR, Salisbury, Wiltshire SP4 OJG, Great Britainに、1996年2月29日にNo.96022940として寄託され、登録商標PER.C6(登録商標)として市場に出された細胞系由来の細胞において、一時的発現をさせるために使用した。これらの抗体の重鎖および軽鎖のヌクレオチド配列およびアミノ酸配列を、それぞれ配列番号122 〜 129に示す。狂犬病ウイルス株CVS-11(0.5ml)の連続希釈液(10−1〜10−8の幅の希釈液)を一定量(〜4 IU/ml) のCR-57またはCR-JB抗体(0.5ml)と共に、37℃/5%CO2の条件の下で1時間インキュベートしたのち、マウス神経芽腫細胞(MNA細胞)またはBSR細胞(ベビーハムスター腎様細胞系列;Baby Hamster Kidney-like cell line)を含むウェルに加えた。ヒトモノクローナル抗体CR-57またはCR-JBのいずれかの存在下で3日間の選択(セレクション)の後、潜在的なエスケープウイルスを含む培地(1ml)を回収し、更に使用するまで4℃で保存した。続いて、それらの細胞を4℃で20分間アセトン固定し、抗狂犬病N-FITC抗体複合体(Centocor社)を用いて37℃/5%CO2の条件の下で、オーバーナイトで染色した。ウェル毎の細胞増殖巣の数を免疫蛍光測定法により記録し、1〜6個の細胞増殖巣を含むウェルの培地を、ウイルスを増幅させるために選択した。全てのE57エスケープウイルスは、E57B1という例外を除き、単一の細胞増殖巣から生成された(E57B1は3個の細胞増殖巣)。EJBエスケープウイルスは、1個の細胞増殖巣(EJB3F)、3個の細胞増殖巣(EJB2B)、4個の細胞増殖巣(EJB2C)、5個の細胞増殖巣(EJB2E, 2F)、または6個の細胞増殖巣(EJB2D)からそれぞれ単離された。各々のエスケープウイルスを、成長特性に基づき、BSRまたはMNA細胞を用いてスモールスケールでまず増幅した。その後、これらの小さなウイルスバッチを、BSRまたはMNA細胞においてウイルスをラージスケールで更に増幅するために用いた。続いて、それぞれのエスケープウイルスバッチのタイター、およびウイルス中和アッセイでの使用に最適なエスケープウイルス希釈物(24時間後に80〜100%を感染させる濃度)を決定するために、増幅されたウイルスをMNA細胞において漸増した。
【0109】
E57(CR-57のエスケープウイルス)およびEJB(CR-JBのエスケープウイルス)の、それぞれCR-JBおよびCR-57との干渉効果を調べるために、修正迅速蛍光フォーカス抑制試験(RFFIT)アッセイを行った。そのために、CR-57または CR-JBを、1:5希釈から始めて、3倍連続希釈した。それぞれの希釈物に、狂犬病ウイルス(CVS-11株)を80〜100%の感染率を与える濃度で加えた。ウイルス/IgGの混合物を、MNA細胞に添加する前に、37℃/5%CO2の条件で1時間インキュベートした。感染後24時間(34℃/5%CO2の条件)の時点で、細胞を4℃で20分間アセトン固定し、抗狂犬病ウイルスN-FITC抗体複合体(Centocor社)を用いて最低3時間染色した。その後、蛍光顕微鏡を用いて狂犬病ウイルス感染についてウェルを分析し、50%エンドポイントの希釈物を決定した。50%エンドポイントの希釈物とは、このアッセイで、ウイルス感染が50%阻害される希釈物を指す。効力を計算するために、国際標準物質(laboratory of Standards and Testing (DMPQ/CBER/FDA)による標準物質である狂犬病イムノグロブリン・ロットR3)を各修正迅速蛍光フォーカス抑制試験アッセイ中に含めた。この標準物質の50%エンドポイントの希釈物は、2U/mlの効能に対応する。ヒトモノクローナル抗体CR-57およびCR-JBの、単独での中和効力、およびこれらの抗体を組み合わせた場合の中和効力を試験した。
【0110】
EJBウイルスは、もはやCR-JB またはCR-57によって中和されず(表1参照)、これら2つの抗体は両方とも狂犬病ウイルス糖タンパク質の類似した部位に結合し、狂犬病ウイルス糖タンパク質の類似した部位のアミノ酸変化をもたらすことを示唆している。E57ウイルスはもはやCR-57によって中和されなかったが、6つのE57ウイルスのうち4つは、より弱い効力ではあるが、依然としてCR- JBによって中和された(表1参照)。抗体CR-57および抗体CR-JBの(1:1IU/mgの比率での)混合物は、単一の抗体による結果と同様の結果を示した(データは示さず)。
【0111】
狂犬病ウイルス糖タンパク質に起こりうる変異を同定するために、EJBおよびE57エスケープウイルスの、各々の糖タンパク質のオープン・リーディング・フレーム(ORF)のヌクレオチド配列を決定した。各々のエスケープウイルスおよびCVS-11のウイルスRNAを、ウイルス感染MNA細胞から単離し、標準的なRT-PCR法によりcDNAに変換した。続いて、変異を同定するために、cDNAを利用して狂犬病ウイルス糖タンパク質のORFのヌクレオチド配列を決定した。
【0112】
E57およびEJBエスケープウイルスは共に、糖タンパク質の同じ領域に変異を示した(それぞれ、図1および図2参照;図1および図2に記載された全配列については、配列番号130〜151を参照)。これは、2つの抗体が重複するエピトープを認識することを意味する。上記より、カクテル中のCR-57とCR-JBの組み合わせは、中和耐性を有する変異株のエスケープを妨げず、ゆえに、狂犬病暴露後予防において理想的な免疫グロブリン製剤ではないと結論づけることが可能である。
【0113】
実施例2
狂犬病ワクチンを接種されたドナーの末梢血リンパ球を用いたScFvファージ・ディスプレイ・ライブラリの構築
狂犬病ワクチンの接種を受けた4人のヒトの被験者の静脈から、最後のブーストの一週間後に、50mlの血液を採取した。Ficollの細胞密度分画を用いて、これらの血液サンプルから末梢血リンパ球(PBL)を単離した。血清は−20℃で冷凍保存した。狂犬病ウイルス糖タンパク質をトランスフェクションされた293T細胞のFACS染色により、血清中に抗狂犬病抗体が存在に関して、陽性が確認された。有機相分離(TRIZOL(登録商標))、続いてエタノール沈殿を用いて、PBLから全RNAを調整した。得られたRNAをDEPCで処理した超純水に溶かし、その濃度を260nmでの吸光度測定によって決定した。その後、RNAを、100 ng/μlの濃度に希釈した。次に、以下のように、1μgのRNAをcDNAに変換した:全10μlのRNAに、13μlのDEPC処理した超純水および1μlのランダムな6量体(DNA)(500 ng/μl)を加え、得られた混合物を65℃で5分間加熱し、氷浴上で急冷した。そこへ、8μlの5×First-Strand buffer、2μlのdNTP(それぞれ10mM)、2μlのDTT(0.1M)、2μlのRnase阻害剤(40 U/μl)、および2μlのSuperscript(登録商標)III MMLV 逆転写酵素(200 U/μl)を加え、室温で5分間インキュベートした後、50℃で1時間インキュベートした。 反応は、熱不活性化、すなわち、混合物を75℃で15分間インキュベートすることにより、停止させた。
【0114】
得られたcDNA産物を、DEPC処理した超純水を用いて200μlの最終容積に希釈した。この得られたcDNA産物の希釈物を(10mMトリス・バッファー中で)50倍希釈した溶液を260nmで吸光度測定した結果、0.1という値が得られた。
【0115】
それぞれのドナーにつき、5μlから10μlの希釈されたcDNA産物を、免疫グロブリンのガンマ重鎖ファミリー、およびカッパまたはラムダ軽鎖の塩基配列をPCR増幅するためのテンプレートとして、特定のオリゴヌクレオチドプライマーと共に用いた(表2〜7参照)。PCR反応溶液は、希釈されたcDNA産物の他に、25pmolのセンスプライマーおよび25pmolのアンチセンスプライマーを含み、20mM Tris-HCl (pH8.4)、50mM KCl、2.5mM MgCl2、250μM dNTPs、および1.25ユニットのTaqポリメラーゼから成る最終体積50μlの溶液であった。96℃の温度のヒートリッド・サーマルサイクラーにおいて、得られた混合物を2分間で急速に融解した後、96℃で30秒、60℃で30秒、72℃で60秒のサイクルを30回繰り返した。
【0116】
第1回目の増幅では、17個の軽鎖可変領域のセンスプライマー(ラムダ軽鎖の11個(表2参照)およびカッパ軽鎖の6個(表3参照))のそれぞれを、HuCκと呼ばれるC-カッパ 5'-ACACTCTCCCCTGTTGAAGCT CTT-3'(配列番号152参照)、またはC-ラムダ定常領域であるHuCλ2 5'-TGAACATTCTGTAGGGGCCACTG-3'(配列番号153参照)およびHuCλ7 5'-AGAGCATTCTGCAGGGGCCACTG-3' (配列番号154参照)を認識するアンチセンスプライマーと組み合わせて用いて(HuCλ2およびHuCλ7アンチセンスプライマーは使用前に等モルで混合した)、4×17個の600塩基対程の生成物を得た。これらの生成物を2%アガロースゲルで精製し、Qiagenゲル抽出カラムを用いて、ゲルから単離した。単離した生成物それぞれの1/10量を、同じ17個のセンスプライマーを用いた上記と同一のPCR反応に用い、ここにおいては、それぞれのラムダ軽鎖センスプライマーをJラムダ領域に特異的な3種類のアンチセンスプライマーのうちの1つと組み合わせて使用し、それぞれのカッパ軽鎖センスプライマーをJカッパ領域に特異的な5種類のアンチセンスプライマーのうちの1つと組み合わせて使用した。2ラウンド目の増幅のためのプライマーは、ファージ・ディスプレイ・ベクターPDV-C06(図3および配列番号155参照)において直接クローニングすることを可能にするため、制限酵素認識部位を付け加えたものを用いた(表4参照)。この結果、4×63個のおよそ350塩基対の生成物が得られ、それらを10の分画に貯留した。分画を10個という数にした理由は、ライブラリ内で、様々な軽鎖の系統の自然な分布を維持するためで、ある特定の系統を過剰にまたは過少に発現させないためである。1つのライブラリ内での発現量を判断するために、1つの系統中での対立遺伝子の数を用いた(表5参照)。次の工程では、貯留した分画のうち2.5μg、およびPDV-C06ベクター100μgを、SalIおよびNotIで制限酵素処理し、ゲルから精製した。その後、以下の様にオーバーナイトで16℃でライゲーションを行った。500ngのPDV-C06ベクターに70ngの貯留した分画を加え、50mMのTris-HCl(pH7.5)、10mMのMgCl2、10mMのDTT、1mMのATP、25 mg/mlのBSAおよび2.5μlのT4DNAリガーゼ(400 U/μl)を含む、全容積50μlのライゲーション混合溶液とした。貯留した分画それぞれについて、この手順を引き続き実行した。ライゲーション混合物を、フェノール/クロロホルムで精製した後、クロロホルムで抽出してエタノールで沈殿したが、これは当業者にとって周知の方法である。得られたDNAを50μlの超純水に溶解し、各ライゲーション混合物につき2.5μlを2度、製造者(Stratagene社)のプロトコルに従って、40μlのTG1コンピテント病原性大腸菌にエレクトロポレーションした。形質転換体を37℃でオーバーナイトで培養し、それには、50ig/mlのアンピシリンと4.5%グルコースを加えた2TYアガーを含む計30個のシャーレ(貯留された分画それぞれにつき3つのシャーレ;シャーレの寸法:240mm×240mm)を用いた。アガープレートから形質転換体をこすり取ることによって軽鎖可変領域の(サブ)ライブラリが得られた。この(サブ)ライブラリは、Qiagen(登録商標)のQIAFilter MAXI prep kitを用いたプラスミドDNAの調製に直接用いた。
【0117】
それぞれのドナーについて、重鎖免疫グロブリン配列の増幅を、上記に示した軽鎖領域の場合と同じcDNA調合液を用いて、同様の2ラウンドのPCRの手順を踏んで、表6および表7に示したプライマーを用いて行なった。増幅の最初のラウンドでは9つのセンス方向プライマー(表6参照;重鎖可変領域の全ての系統をカバーする)のセットを使用し、そのセンス方向プライマーそれぞれをHuCIgGと呼ばれるIgGに特異的な定常領域のアンチセンスプライマー、5'-GTC CAC CTT GGT GTT GCT GGG CTT-3' (配列番号156)と組み合わせて用い、4×9個の650塩基対程の生成物を得た。これらの生成物を2%アガロースゲル上で精製し、Qiagenゲル抽出カラムを用いて、ゲルから単離した。単離した生成物それぞれの1/10量を、同じ9個のセンスプライマーを用いた上記と同一のPCR反応に用い、ここにおいては、それぞれの重鎖センスプライマーを、JH領域に特異的な4種類のアンチセンスプライマーのうちの1つと組み合わせて使用した。2ラウンド目に用いたプライマーには、軽鎖(サブ)ライブラリベクターに直接クローニングすることを可能にするため、制限酵素認識部位を付け加えた(表7参照)。この結果、各ドナーにつき36種類の、350塩基対ほどの生成物が得られた。これらの生成物を、それぞれのドナーにつき、使用された(VH)センスプライマーに基づいて9つの分画に貯留した。得られた生成物をQiagenPCR精製カラムを用いて精製した。次に、それぞれの分画をSfiIおよびXhoIで制限酵素処理し、同じ制限酵素で切断した軽鎖(サブ)ライブラリベクター中に、上記の軽鎖(サブ)ライブラリにおいての場合と同じ方法および容積で、ライゲーションした。あるいは、それぞれの分画をNcoIおよびXhoIで制限酵素処理し、同じ制限酵素で切断した軽鎖(サブ)ライブラリベクター中に、上記の軽鎖(サブ)ライブラリにおいての場合と同じ方法および容積で、ライゲーションした。ライゲーション後の精製、およびそれに続く、生成した最終的なライブラリの形質転換を、上記の軽鎖(サブ)ライブラリにおける方法と同様に実行し、またこの時点で、各ドナーのライゲーション混合物を各VH貯留液毎に混合した。形質転換体を、50μg/mlのアンピシリンと4.5%グルコースを加えた2TYアガーを含む27個のシャーレ(貯留された分画それぞれにつき3つのシャーレ;シャーレの寸法:240mm×240mm)で培養した。全バクテリアを、50ig/mlのアンピシリンと4.5%グルコースを含む2TY培地から回収し、15%(v/v)までグリセロールを加えて混合し、1.5mlの一定量中で、−80℃で凍らせた。各ライブラリのレスキューおよび選択は以下のように行なった。
【0118】
実施例3
狂犬病ウイルス糖タンパク質を特異的に認識する単鎖Fvフラグメントを有するファージの選択
抗体フラグメントの選択は、抗体ファージ・ディスプレイ・ライブラリ、一般的なファージディスプレイ技術、およびMAbstract(登録商標)技術を用いて、基本的には米国特許番号第6,265,150号およびWO 98/15833(これらは共に参照することによって本明細書に組み込まれる)に記載の通りに行なった。使用した抗体ファージライブラリは、2つの異なる半合成scFvファージライブラリ(JK1994およびWT2000)と、上の実施例2に記載した通りに調製した免疫scFvファージライブラリ(RAB-03-G01およびRAB-04-G01)である。第一の半合成scFvファージライブラリ(JK1994)は“de Kruif et al. (1995b)”に記載されており、第二の半合成scFvファージライブラリ(WT2000)は、基本的に“de Kruif et al. (1995b)”に記載された通りに作製した。簡潔には、このライブラリは、CDR領域に変異を組み込む変性オリゴヌクレオチドを用いて、重鎖および軽鎖V遺伝子の内部に変異を組み込まれた半合成体の形式を有する。VH3重鎖遺伝子のみを、カッパおよびラムダ軽鎖と組み合わせて使用した。重鎖のCDR1およびCDR3、そして軽鎖のCDR3を、"de Kruif et al. (1995b)"の記載と同様のPCRをベースにした方法で合成的に再現した。このように作製したV領域遺伝子を順次scFvの形式としてファージミドベクターにクローン化し、ファージライブラリを生成するために前述の方法で増幅した。更に、WO 02/103012(参照することによって本明細書に組み込まれる)に記載されている方法およびヘルパーファージを本発明で使用した。狂犬病ウイルス糖タンパク質を認識するファージ抗体を同定するために、全狂犬病ウイルス(狂犬病ウイルスピットマン・ムーア(Pitman-Moore)株)をβ‐プロピオラクトンでの処理によって不活性化したもの、狂犬病ウイルス糖タンパク質(狂犬病ウイルスERA株)を精製したもの、および/または狂犬病ウイルスGタンパク質(狂犬病ウイルスERA株)を発現するトランスフェクション細胞を用いて、ファージ選択実験を行った。
【0119】
Gタンパク質は以下のように狂犬病ウイルスERA株から精製した。ウイルス溶液に1/10容積の10%オクチル−β−グルコピラノシドを加え、穏やかに混合した。4℃で30分間のインキュベーションし、すぐにサンプルをSW51ローターで遠心分離(36,000rpm、4℃)した。上澄みを回収し、0.1M Tris/EDTAに対して4℃でオーバーナイトで透析を行った。続いて、糖タンパク質を透析チャンバーから回収、分割し、更なる使用の時まで−80℃で保管した。タンパク質の濃度を280nmでの吸光度測定によって決定し、Gたんぱく質の完全性をSDS- PAGEによって分析した。
【0120】
不活性化された狂犬病ウイルス全体または狂犬病ウイルスGタンパク質をリン酸緩衝食塩水(PBS)に溶解し、2〜3mlをマキシソープNunc-イムノチューブ (Nunc社)に加え、回転装置上で4℃でオーバーナイトでインキュベーションした。ファージライブラリの一定量(500μl、約1013cfu、CTヘルパーファージを用いて増幅(WO 02/103012参照))のブロッキングを、ブロッキングバッファー(PBS溶液中に2%Protifarを含む)中で、室温で1〜2時間行なった。ブロッキングされたファージライブラリをイムノチューブ(CR-57によって認識されるエピトープをブロックするために、CR-57 scFvの存在下または非存在下のいずれかでプレインキュベートしたもの)に加え、室温で2時間インキュベーションを行い、結合していないファージを除去するために洗浄バッファ(PBS溶液中に0.1% Tween-20 (Serva社)を含む)で洗浄した。その後、50mM グリシン-HCl pH2.2を1ml用いて、10分間室温でインキュベーションすることにより、結合したファージを抗原から溶出した。続いて、溶出されたファージと1M Tris-HCl pH7.5の0.5mlとを混合することによりpHを中和した。この混合物を、600nmでの吸光度が約0.3となるまで37℃で培養された大腸菌XL1-Blue株培養液5mlを感染させるために使用した。37℃で30分間、ファージをXL1-Blueバクテリアに感染させた。その後、混合物を10分間3200×gで室温で遠心分離し、バクテリアのペレットを0.5mlの2−トリプトン酵母エキス(2TY)培地中に再懸濁した。得られたバクテリアの懸濁液を、テトラサイクリン、アンピシリンおよびグルコースが加えられた2枚の2TYアガープレート上に分割した。プレートを37℃でオーバーナイトでインキュベーションした後、コロニーをプレートから擦り取り、“De Kruif et al. (1995a)”およびWO 02/103012の記載されたのと基本的に同様の方法で、濃縮したファージライブラリを作製するために使用した。簡潔に述べると、擦り取ったバクテリアを、アンピシリン、テトラサイクリンおよびグルコースを含む2TY培地に植菌するために用い、600nmでの吸光度が約0.3となるまで37℃で培養した。CTヘルパーファージを加え、バクテリアを感染させることを可能とした後に、培地をアンピシリン、テトラサイクリンおよびカナマイシンを含む2TYへ変更した。インキュベーションを、引き続き30℃でオーバーナイトで行った。翌日、バクテリアを遠心分離によって2TY培地から除去した後、培地中のファージをポリエチレングリコール(PEG)6000/NaClを用いて沈殿させた。最終的に、ファージを1%のウシ血清アルブミン(BSA)を含む2mlのPBSに溶かし、フィルター滅菌し、選択の次のラウンドのために使用した。
【0121】
狂犬病ウイルス糖タンパク質をトランスフェクトした細胞を用いることによってもファージの選択を行った。使用した細胞は、欧州細胞培養コレクション(ECACC), CAMR, Salisbury, Wiltshire SP4 OJG, Great Britainに、1996年2月29日にNo.96022940として寄託され、登録商標PER.C6(登録商標)として市場に出された細胞系由来の細胞である。これらの細胞を、本願明細書において今後、PER. C6(商標登録)細胞と呼ぶ。ここではまず、ブロッキングされたファージライブラリ(2ml)を、(DMEM/10% FBS中の)1×107のサブトラクター細胞に加え、回転装置上で4℃で1時間インキュベートした。サブトラクター細胞は、狂犬病ウイルス膜貫通ドメインおよび細胞質ドメインと外表面上で融合した、水疱性口内炎ウイルス(VSV)糖タンパク質の細胞外ドメインを発現したPER. C6(商標登録)細胞であった。このサブトラクションステップによって、VSV糖タンパク質、またはPER. C6(商標登録)細胞に特異的な抗原のいずれかを認識するファージを、ファージライブラリから取り除いた。ファージ/細胞の混合物を遠心して(4℃、500xgで5分)細胞に結合したファージを取り除き、上澄みを1*107のサブトラクター細胞を3ml含む新しいチューブに加えた。このサブトラクションステップは、各回とも上澄みを取り出して2回繰り返した。続いて、サブトラクションを行ったファージを狂犬病ウイルス糖タンパク質を発現したトランスフェクション細胞と共に、回転装置上で4℃で1.5時間インキュベートした。その前に、トランスフェクション細胞を、CR-57によって認識されるエピトープをブロックするため、CR-57 scFvの存在下または非存在下のいずれかでプレインキュベートした。インキュベーション後、1mlのDMEM/10%FBSで細胞を5回洗浄し(それぞれの洗浄につき、細胞を再懸濁し、新品のチューブに移した)、上記の通りにファージを溶出、処理した。
【0122】
典型的には、各ファージ抗体の単離を行なう前に2ラウンドの選択を行った。2ラウンド目の選択の後、各大腸菌コロニーを使用してモノクローナルファージ抗体を作製した。基本的には、それぞれのコロニーを96ウェルプレート上で対数増殖期に入るまで増殖させ、VCSM13ヘルパーファージを感染させた後に、ファージ抗体の生産をオーバーナイトで行なうことを可能とする。産生されたファージ抗体を、PEG/NaCl沈殿およびフィルタ滅菌し、ELISAによって、不活性化した狂犬病ウイルス全体、精製した狂犬病ウイルスGタンパク質の両方への結合を試す。選択によって、不活性化した狂犬病ウイルス全体および狂犬病ウイルスGタンパク質の双方への結合を示すファージ抗体が大量に得られた(下の実施例を参照)。上記の免疫ライブラリでは、2つの選択のストラテジーをとった。1つ目のストラテジーでは、不活性化したウイルスまたは精製したGタンパク質を1回目および2回目の選択のラウンドで用いて、2回の選択のラウンドの後に736個のファージ抗体を得た。2つ目のストラテジーでは、1回目のラウンドでは細胞表面に発現させた組み替えGタンパク質を用い、2回目のラウンドで不活性化したウイルスまたは精製したGタンパク質を用いることで、2回の選択のラウンドの後に736個のファージ抗体を得た。1つ目のストラテジーを用いて得られたファージ抗体は97種類であった一方で、2つ目のストラテジーでは70種類の独自のファージ抗体が得られた。1つ目のストラテジーを用いて得た97種類の独自のファージ抗体は18個の中和抗体を生じ、2つ目のストラテジーを用いて得た70種類の独自のクローンは33個の中和抗体を生じた。このことは、狂犬病ウイルス糖タンパク質をトランスフェクトした細胞、すなわち細胞表面に発現した組み替えGタンパク質、を抗原として用いて選択を行った場合、精製したGタンパク質および/または不活性化されたウイルスのみを用いて選択を行った場合と比較して、より多くの中和抗体を生じるらしいということを明確に示す。
【0123】
実施例4
狂犬病ウイルス糖タンパク質に特異的な単鎖ファージ抗体の検証
上記のスクリーニングによって選択した単鎖ファージ抗体の特異性、すなわち狂犬病ウイルスGタンパク質への結合をELISAによって検証した。加えて、単鎖ファージ抗体の、5%FBSへの結合も検証した。この目的のために、狂犬病ウイルスGタンパク質または5%FBSの調合液をMaxisorp(登録商標)ELISAプレートにコートした。コーティングの後、プレートをPBS/1%Protifar中で、室温で1時間ブロッキングを行なった。選択された単鎖ファージ抗体を、等容積のPBS/1%Protifar中で15分間インキュベートすることによって、ブロッキングされたファージ抗体を得た。プレートを空にし、ブロッキングされたファージ抗体をウェルに加えた。続いて1時間のインキュベーションへと進むことが可能となり、プレートを0.1% Tween-20を含むPBS中で洗浄し、結合したファージ抗体をペルオキシダーゼに結合した抗M13抗体を用いて(492nmでの吸光度測定を用いて)検出した。同時にコントロールとして、この手順を単鎖ファージ抗体を使用しないか、ネガティブコントロールとしてCD8 (SC02-007)に対する単鎖ファージ抗体を使用するか、またはポジティブコントロールとして狂犬病ウイルス糖タンパク質(scFvSO57)に対する単鎖ファージ抗体を用いることによって行った。表8に示すように、選択されたファージ抗体で、SC04-001、SC04-004、SC04-008、SC04- 010、SC04-018、SC04-021、SC04-026、SC04-031、SC04-038、SC04- 040、SC04-060、SC04-073、SC04-097、SC04-098、SC04-103、SC04- 104、SC04-108、SC04-120、SC04-125、SC04-126、SC04-140、SC04-144、SC04-146およびSC04-164と呼ばれるファージ抗体が、固定された精製狂犬病ウイルスGタンパク質への有意な結合を示した一方、FBSへの結合は観測されなかった。上記の通りに作製した、不活性化された完全な狂犬病ウイルスを用いたELISA分析においても、同一の結果が得られた(データは図示せず)。
【0124】
実施例5
狂犬病ウイルスに特異的なscFvの特徴
選択された特異的な単鎖ファージ抗体(scFv)のクローンからプラスミドDNAを得て、標準的な技術によってヌクレオチド配列を決定した。SC04-001、SC04-004、SC04-008、SC04-010、SC04-018、SC04-021、SC04-026、SC04-031、SC04-038、SC04-040、SC04-060、SC04-073、SC04-097、SC04-098、SC04-103、SC04-104、SC04-108、SC04-120、SC04-125、SC04-126、SC04-140、SC04-144、SC04-146およびSC04-164と呼ばれるscFvのヌクレオチド配列(クローニングのための制限酵素認識部位を含む)は、それぞれ、配列番号157、配列番号159、配列番号161、配列番号163、配列番号165、配列番号167、配列番号169、配列番号171、配列番号173、配列番号175、配列番号177、配列番号179、配列番号181、配列番号183、配列番号185、配列番号187、配列番号189、配列番号191、配列番号193、配列番号195、配列番号197、配列番号199、配列番号201および配列番号203に示される。SC04-001、SC04-004、SC04-008、SC04-010、SC04-018、SC04-021、SC04-026、SC04-031、SC04-038、SC04-040、SC04-060、SC04-073、SC04-097、SC04-098、SC04-103、SC04-104、SC04-108、SC04-120、SC04-125、SC04-126、SC04-140、SC04-144、SC04-146およびSC04-164と呼ばれるscFvのアミノ酸配列は、それぞれ、配列番号158、配列番号160、配列番号162、配列番号164、配列番号166、配列番号168、配列番号170、配列番号172、配列番号174、配列番号176、配列番号178、配列番号180、配列番号182、配列番号184、配列番号186、配列番号188、配列番号190、配列番号192、配列番号194、配列番号196、配列番号198、配列番号200、配列番号202および配列番号204に示される。
【0125】
VH遺伝子およびVL遺伝子の特性(“Tomlinson IM, Williams SC, Ignatovitch O, Corbett SJ, Winter G. V-BASE Sequence Directory. Cambridge United Kingdom: MRC Centre for Protein Engineering (1997) ”参照)、および狂犬病ウイルスGタンパク質に特異的に結合するscFvの重鎖CDR3組成を表9に示す。
【0126】
実施例6
狂犬病ウイルスに特異的なscFvによる試験管内での狂犬病ウイルスの中和(修正RFFIT)
選択されたscFvが狂犬病ウイルス感染を阻止する能力があるかを判断するために、試験管内での中和アッセイ(修正RFFIT)を行った。scFvの調合液を1:5希釈から始めて、3倍連続希釈した。狂犬病ウイルス(CVS-11株)を、80〜100%の感染率を与える濃度で各希釈物に加えた。ウイルス/scFvの混合物を、MNA細胞に添加する前に37℃/5%CO2の条件で1時間インキュベートした。感染後24時間(34℃/5%CO2の条件)の時点で、細胞を4℃で20分間アセトン固定し、抗狂犬病N-FITC抗体複合体(Centocor社)を用いて最低3時間染色した。その後、蛍光顕微鏡を用いて狂犬病ウイルス感染があるかについて細胞を分析し、50%エンドポイントの希釈物を決定した。50%エンドポイントの希釈物とは、このアッセイ中でウイルス感染が50%阻害される希釈物を指す(実施例1参照)。複数のscFvが狂犬病ウイルスに対して中和活性を示すことが確認された(表10参照)。
【0127】
加えて、選択されたscFvが、実施例1で作製したE57エスケープウイルス(E57A2、E57A3、E57B1、E57B2、E57B3およびE57C3)を中和する能力を有するかどうかを、上述した試験管内での中和アッセイ(修正RFFIT)を用いて調査した。いくつかのscFvは、E57エスケープウイルスに対して中和活性を示すことが確認された(表11Aおよび11B参照)。
【0128】
実施例7
scFvを用いた狂犬病ウイルスGタンパク質競合ELISA
互いに重複したり競合したりしないエピトープに結合する抗体を同定するために、狂犬病糖タンパク質の競合ELISA分析を実施した。精製された狂犬病ウイルス糖タンパク質(1mg/ml;狂犬病ウイルスERA株)を PBS中で1:1000希釈した液(50μl)を用いて、Nunc-Immuno(登録商標) Maxisorp F96プレート (Nunc)を4℃でオーバーナイトでコートした。コートされなかったタンパク質を洗い流し、ウェルを100μl PBS/1%Protifarを用いて室温で1時間ブロッキングした。続いて、ブロッキング溶液を取り除き、PBS/1%Protifar溶液中の精製されていない抗狂犬病ウイルスscFv(2倍希釈液)を50μl加えた。ウェルを100μlのPBS/0.05% Tween-20で5回洗浄した。その後、ビオチン化した抗狂犬病ウイルス競合IgGであるCR-57bioを各ウェルに50μlずつ加え、5分間室温でインキュベートし、その後ウェルを100μlのPBS/0.05% Tween-20で5回洗浄した。CR-57bioの結合を検出するために、ストレプトアビジン-HRP抗体(Becton Dickinson)の1:2000希釈物をウェルに50μl加え、室温で1時間インキュベートした。ウェルを再び上記の通りに洗浄し、OPD reagens (Sigma)を100μl加えることによって、ELISA分析をさらに進めた。1M H2SO4を50μl加えることによって反応を停止させ、492nmでの吸光度測定を行った。
【0129】
CR-57bio単独によって得られるシグナルは、CR-57のscFv型であるscFv SO57(SO57のヌクレオチド配列およびアミノ酸配列についてはそれぞれ、配列番号205および206を参照されたい)または、CR-JBのscFv型であるscFv SOJB(SOJBのヌクレオチド配列およびアミノ酸配列についてはそれぞれ、配列番号312および313を参照されたい)と共にインキュベートした場合、バックグラウンドのレベルにまで下がりうる。これは、scFv のSO57およびSOJBが、CR-57bioと同じエピトープや重なり合うエピトープと結合することによって、狂犬病ウイルス糖タンパク質との相互作用を巡ってCR-57bioと競合することを示す。対照的に、CD8に結合するscFvである、SC02-007と呼ばれる関連のないscFvは、結合を巡って競合しなかった。SC04-004、SC04- 010、SC04-024、SC04-060、SC04-073、SC04-097、SC04-098、SC04- 103、SC04-104、SC04-120、SC04-125、SC04-127、SC04-140、SC04- 144およびSC04-146と呼ばれる抗狂犬病ウイルスscFvもまた、CR-57bioと競合せず、これらのscFvはCR-57の認識するエピトープとは異なったエピトープに結合することを示している(図4参照)。
【0130】
以下の実験によっても同様の結果が得られた。まず、狂犬病ウイルス抗体CR-57を、狂犬病ウイルスGタンパク質でコートしたウェルに加えた。次に、競合するscFvを加えた。この設定では、scFvのアミノ酸配列中にVSVタグが存在することを利用して、抗狂犬病ウイルスscFvを抗VSV-HRPを用いて検出した(図5参照)。
【0131】
実施例8
選択された抗狂犬病ウイルス単鎖Fvからの、完全なヒト免疫グロブリン分子(ヒトモノクローナル抗狂犬病ウイルス抗体)の作製
SC04-001、SC04-008、SC04-018、SC04-040およびSC04-126と呼ばれるscFvの重鎖可変領域および軽鎖可変領域を、オリゴヌクレオチドを用いてPCRで増幅し、それぞれに制限酵素認識部位、および/またはIgG発現ベクターpSyn-C03-HCg1(配列番号277参照)およびpSyn-C04-Cl(配列番号278参照)における発現のための配列を付け加えた。VH遺伝子およびVL遺伝子を表12および表13に示したオリゴヌクレオチドを用いてそれぞれ増幅し、PCR産物をpSyn-C03-HCg1ベクターおよびpSyn-C04-Clベクターへそれぞれクローニングした。
【0132】
SC04-004、SC04-010、SC04-021、SC04-026、SC04-031、SC04-038、SC04-060、SC04-073、SC04-097、SC04-098、SC04-103、SC04-104、SC04-108、SC04-120、SC04-125、SC04-140、SC04-144、SC04-146およびSC04-164と呼ばれるscFvの重鎖可変領域および軽鎖可変領域をもまた、オリゴヌクレオチドを用いてPCRで増幅し、それぞれに制限酵素認識部位および/または、IgG発現ベクターpSyn-C03-HCg1およびpSyn-C05-Cκ(配列番号279参照)における発現のための配列を付け加えた。VH遺伝子およびVL遺伝子を表12および表13に示したオリゴヌクレオチドを用いてそれぞれ増幅し、PCR産物をpSyn-C03-HCa1ベクターおよびpSyn-C05-Cκベクターへそれぞれクローニングした。これらのオリゴヌクレオチドは、ライブラリの構築時に導入された生殖細胞系配列由来のいかなる変異をも修正するように設計されているのだが、これは、広いレパートリーの抗体遺伝子を増幅するために使用したオリゴヌクレオチドの数が限られていたために可能となった。全てのコンストラクトのヌクレオチド配列を熟練した当業者にとっては周知の標準的な技術によって検証した。
【0133】
結果として生じた、抗狂犬病ウイルスヒトIgG1重鎖をコードするpgG104-001C03、pgG104-008C03、pgG104-018C03、pgG104-040C03およびpgG104-126C03の発現コンストラクト、および対応する軽鎖をコードする該当するpSyn-C04-Vλ コンストラクトを組み合わせて293T細胞中で一時的に発現させ、IgG1抗体を含む上澄みを得た。抗狂犬病ウイルスヒトIgG1重鎖をコードするpgG104-004C03、pgG104-010C03、pgG104-021C03、pgG104-026C03、pgG104-031C03、pgG104-038C03、pgG104-060C03、pgG104-073C03、pgG104-097C03、pgG104-098C03、pgG104-103C03、pgG104-104C03、pgG104-108C03、pgG104-120C03、pgG104-125C03、pgG104-140C03、pgG104-144C03、pgG104-146C03およびpgG104-164C03の発現コンストラクト、および、対応する軽鎖をコードする該当するpSyn-C05-Vκコンストラクトを組み合わせて293T細胞中で一時的に発現させ、IgG1抗体を含む上澄みを得た。
【0134】
CR04-001、CR04-004、CR04-008、CR04-010、CR04-018、CR04-021、CR04-026、CR04-031、CR04-038、CR04-040、CR04-060、CR04-073、CR04-097、CR04-098、CR04-103、CR04-104、CR04-108、CR04-120、CR04-125、CR04-126、CR04-140、CR04-144、CR04-146およびCR04-164と呼ばれる抗体の、重鎖および軽鎖のヌクレオチド配列およびアミノ酸配列を標準的な技術によって決定した。続いて、通常免疫グロブリンで用いる標準的な精製方法(例えば、参照することにより本明細書に組み込まれるWO00/63403を参照)を用いて、組み換えヒトモノクローナル抗体をprotein-Aカラムで精製し、続いて脱塩カラム上でバッファ交換した。
【0135】
追加的に、CR04-098に関して、CR-57 およびCR-JBをそれぞれコードするpgSO57C11ベクターおよびpgSOJBC11ベクター(実施例1参照)に関して上述したのと同様にしてpgG104-098C10と呼ばれる単一のヒトIgG1発現ベクターを生成した。pgG104-098C10ベクターによってコードされる抗体CR04-098の重鎖および軽鎖のヌクレオチド配列およびアミノ酸配列を、それぞれ配列番号334〜337に示す。pgSO57C11ベクター(実施例1参照)およびpgG104-098C10ベクターを、CR-57およびCR04-098を、欧州細胞培養コレクション(ECACC), CAMR, Salisbury, Wiltshire SP4 OJG, Great Britainに1996年2月29日にNo.96022940として寄託され、登録商標PER.C6(登録商標)として市場に出された細胞系由来の細胞において安定的に発現させるためにそれぞれ使用した。安定的に生成したCR-57およびCR04-098の算出された等電点はそれぞれ、8.22および8.46である。実験的に観測された等電点は、CR-57については8.1から8.3の間で、CR04-098については9.0から9.2の間である。組み換えヒトモノクローナル抗体を、上述の通りに精製した。特に記していない限りは、CR04-001、CR04-004、CR04-008、CR04-010、CR04-018、CR04-021、CR04-026、CR04-031、CR04-038、CR04-040、CR04-060、CR04-073、CR04-097、CR04-098、CR04-103、CR04-104、CR04-108、CR04-120、CR04-125、CR04-126、CR04-140、CR04-144、CR04-146およびCR04-164については、上述の2つのベクターシステムによって一時的に発現された組み換えヒトモノクローナル抗体を利用し、CR57については、実施例1に記載の1つのベクターシステムによって一時的に発現された組み換えヒトモノクローナル抗体を利用した。
【0136】
実施例9
IgGによる狂犬病ウイルスGタンパク質競合ELISA
ヒトモノクローナル抗狂犬病ウイルスGタンパク質IgGが互いに重複したり競合したりしないエピトープに結合するか調べるため、競合実験を行った。狂犬病Gタンパク質でコートしたウェルを、複数の濃度(0〜50ig/ml)の非標識抗狂犬病ウイルスGタンパク質のIgGと共に、1時間室温でインキュベートした。その後、それぞれのウェルに、異なるビオチン化抗狂犬病ウイルスIgG(1ig/ml)を50μl加え、室温で5分間インキュベートし、直ちに100μlのPBS/0.05% Tween-20で5回洗浄した。続いて、ストレプトアビジン-HRP抗体(Becton Dickinson)の1:2000の希釈物を50μl加えて室温で1時間インキュベートし、上記の通りに洗浄し発色させた。非標識IgGの濃度の増加に伴うシグナルの減少は、2つの抗体が互いに競合していて、同じエピトープまたは重なり合うエピトープを認識することを示す。
【0137】
あるいは、狂犬病ウイルスGタンパク質(ERA株)によってコートしたウェルを、50ig/mlの非標識抗狂犬病ウイルスGタンパク質IgGと共に、1時間室温でインキュベートした。その後、ビオチン化CR57(0.5〜5ig/ml;飽和濃度以下)を各ウェルに加えた。さらなるステップは、上述の通りに行った。得られたシグナルを、ビオチン化CR57のみを使用した場合に得られるシグナルと比較した(図6;「競合物質なし」のグラフを参照)。図6から、ネガティブコントロールとして作用するCR02-428と呼ばれる抗体によってシグナルが減少してしまう可能性はないと推測できる。反対に、非標識CR57(ポジティブコントロール)またはCR-JBとの競合によって、シグナルはバックグランドのレベルにまで減少した。さらに、図6から、抗狂犬病ウイルスGタンパク質のIgGで、CR-57と有意に競合するものは存在しないことが推測でき、これは実施例7に記載したscFvの競合データと一致する。
【0138】
さらに、狂犬病ウイルスGタンパク質(ERA株)をトランスフェクトしたPER. C6細胞についてもフローサイトメトリー を用いて競合実験を行った。トランスフェクションされた細胞を20μlの非標識抗狂犬病ウイルスGタンパク質のIgG(50ig/ml)と共に20分間4℃でインキュベートした。細胞を1%BSAを含むPBSで洗浄した後、それぞれのウェルにビオチン化CR57(0.5〜5ig/ml;飽和濃度以下)を20μl加え、4℃で5分間インキュベートし、直ちに1%のBSAを含む100μlのPBSで2回洗浄した。続いて、ストレプトアビジン-PE(Caltag)の1:200の希釈物20μlと共に4℃で15分間インキュベートし、上記の通りに洗浄して発色させた。ビオチン化CR57によって得られるシグナルは、ネガティブコントロール抗体CR02-428によっては有意に減少しなかった(図7参照)。対照的に、非標識CR57(ポジティブコントロール)またはCR-JBとの競合によって、シグナルはバックグランドのレベルにまで減少した。抗狂犬病ウイルスGタンパク質のIgGで、CR-57と有意に競合するものは、シグナルを約30%まで減少させたCR04-126を除いては存在しなかった(図7参照)。CR04-126は、ELISA分析においては競合を示さなかった(図6参照)。これは、糖タンパク質が抗体に呈示される方法が、ELISA実験とFACS実験では異なることに起因する可能性がある。CR04-126の結合は糖タンパク質の立体構造により依存的である結果、FACSに基づく競合アッセイではCR04-126による競合効果が観察され、ELISAに基づく競合アッセイでは観察されなかった可能性がある。また、CR04-008およびCR04-010はFACSに基づく競合アッセイではシグナルを約50%まで減少させたが、これらの抗体がCR57と競合する可能性があることを示している。しかしCR04-010については、このことはscFv競合データによっても、ELISAに基づく競合アッセイによっても確認されなかった。他のIgGについては、FACSのデータは、それに対応するELISAのデータと、scFvとIgG双方について一致した。
【0139】
実施例10
狂犬病ウイルスの試験管内での中和の際の抗狂犬病IgGの加法的/相乗的効果(修正RFFIT)
抗狂犬病ウイルスGタンパク質のIgGが狂犬病ウイルスの中和において加法的または相乗的効果をもたらすかを決定するために、様々な組み合わせのIgGを試験した。まず、修正RFFITによって、それぞれの抗体単独での(IU/mgの単位での)効力を決定した(実施例1参照)。次に、IU/mgの量を等しくそろえて抗体の組み合わせを調製し、修正RFFITの試験を行った。抗体の組み合わせ各々の効力を決定し、期待される効力と比較することができる。もしも、抗体の組み合わせの効力が、組み合わせの中に存在する個々の抗体の効力の和と同じ値になるならば、これらの抗体は相加的な効果を有する。もしも抗体の組み合わせの効力の方が大きいのならば、抗体は狂犬病ウイルスの中和において相乗的な効果を有する。
【0140】
あるいは、相加的あるいは相乗的効果を以下の実験によって決定することも可能である。まず、検討を行なう抗体、例としてCR-57およびCR04-098、の効力を標準のRFFITによって決定する(“Laboratory techniques in rabies, Edited by: F. -X Meslin, M. M. Kaplan and H. Koprowski (1996), 4th edition, Chapter 15, World Health Organization, Geneva”参照)。次に、抗体をIU/mlに基づいて1:1で混合する。この抗体混合物について、同じ濃度の個々の抗体と共に、6回の独立したRFFIT実験を行なうことによって、中和の50%エンドポイントを決定した。続いて、Chou et al.(1984)の記載に従って、式、CI = (C1/Cx1) + (C2/Cx2) + (C1C2/Cx1Cx2) を用いて、抗体混合物のcombination index(CI)を決定した。C1およびC2は、組み合わせて使用したときに50%の中和をもたらすモノクローナル抗体1およびモノクローナル抗体2の(μg単位の)量であり、Cx1およびCx2は、単独で使用したときに50%の中和をもたらすモノクローナル抗体1およびモノクローナル抗体2の(μg単位の)量である。CI = 1はモノクローナル抗体の相加的効果を示し、CI < 1は相乗的効果、CI > 1は拮抗効果を示す。
【0141】
実施例11
組み換えヒト抗狂犬病ウイルス抗体によって認識されるエピトープの、PEPSCAN-ELISAによる同定
狂犬病ウイルスERA株のGタンパク質の細胞外ドメインより、15mer直線状ペプチドおよびループ状/環状ペプチドを合成し(狂犬病ウイルスERA株の糖タンパク質Gの完全なアミノ酸配列については配列番号207を参照されたい、細胞外ドメインはアミノ酸20-458からなる;EMBLデータベースにおける狂犬病ウイルスERA株の糖タンパク質のタンパクIDはJ02293である)、前述のように、クレジットカード状のミニPEPSCANカード(1枚のカードにつき455個のペプチドのフォーマット)を用いてスクリーニングを行った(Slootstra et al. , 1996; WO 93/09872)。全てのペプチドをアミノ酸末端でアセチル化した。全てのループ状のペプチドにおいて、位置2および位置14をシステインで置き換えた(acetyl-XCXXXXXXXXXXXCX-minicard)。準備したペプチド中で、位置2および位置14のシステインの近傍に他のシステインが存在した場合、それらをアラニンで置き換えた。ループ状のペプチドは標準的なFmoc法で合成し、スカベンジャー付きのトリフルオロ酢酸を用いて脱保護した。続いて、脱保護したペプチドを、1,3- ビス(ブロモメチル)ベンゼンを含む炭酸水素アンモニウム(20mM、pH7.9/アセトニトリル(1:1(v/v))溶液0.5mM中で、カードの上で反応させた。カードを、溶液に完全に浸った状態で30〜60分間穏やかに振盪した。最後に、過剰のH2Oを用いてカードを十分に洗浄し、PBS中に1%SDS/0.1%β‐メルカプトエタノールを含む破壊バッファ(pH7.2)中において70℃で30分間超音波処理を行い、続いてH2O中で45分間超音波処理を行った。ヒトモノクローナル抗体を上述の通りに作製した。これらの抗体の、各直線状ペプチドおよびループ状ペプチドへの結合を、PEPSCANに基づいた酵素免疫測定法(ELISA)を用いて検証した。共有結合したペプチドを含む455個のウェルを有するクレジットカード状のポリプロピレンカードを、抗体(10ig/ml;5%のウマ血清(v/v)および5%のオボアルブミン(w/v)を含むブロッキング液中に溶解)と共にインキュベートした(4℃、オーバーナイト)。洗浄後、ペプチドを抗ヒト抗体ペルオキシダーゼ(1/1000希釈物)と共にインキュベートし(25℃で1時間)、続いて、ペルオキシダーゼ基質を洗浄後、2,2'-アジノビス(3-エチルベンゾチアゾリン-スルホン酸) (ABTS)および2il/mlの3%H2O2を加えた。(直線状ペプチドおよびループ状ペプチドに対する)コントロールを、抗ヒト抗体ペルオキシダーゼのみと共にインキュベートした。1時間後、発色現象を測定した。ELISAの発色現象を、CCDカメラおよび画像処理システムを用いて定量化した。この装置は、CCDカメラおよび55mmレンズ(ソニー製CCDビデオカメラXC-77RR、ニコン製“micro-nikkor 55 mm f/2.8 レンズ”)、カメラアダプター(ソニー製カメラアダプターDC-77RR)、および画像処理ソフトウェアパッケージOptimas, version 6.5(Media Cybernetics, Silver Spring, MD 20910, U.S.A.)から構成される。Optimasは、pentium II コンピュータシステム上で作動した。
【0142】
上記の通り、合成された15merの直線状およびループ状/環状のペプチドへの、ヒト抗狂犬病ウイルスGタンパク質モノクローナル抗体の結合を検証した。ペプチドが抗体に適切に結合していると考えられるのは、OD値(光学濃度)が(抗体あたりの)全ペプチドのOD値の平均の2倍以上の場合である。CR57、CRJB、およびCR04-010と呼ばれるヒトモノクローナル抗体の、狂犬病ウイルスERA株の糖タンパク質Gの細胞外ドメイン直線状ペプチドへの結合の結果を表14に示す。それぞれの抗体と有意な結合を示す領域を灰色でハイライトしてある(表14参照)。
【0143】
抗体CR57は、SLKGACKLKLCGVLG (配列番号314)、LKGACKLKLCGVLGL (配列番号315)、KGACKLKLCGVLGLR (配列番号316)、GACKLKLCGVLGLRL (配列番号317)、ACKLKLCGVLGLRLM (配列番号318)、CKLKLCGVLGLRLMD (配列番号319)、KLKLCGVLGLRLMDG (配列番号320)、LKLCGVLGLRLMDGT (配列番号321)、およびKLCGVLGLRLMDGTW (配列番号322)からなる群から選択したアミノ酸配列を有する直線状ペプチドに結合した(表14参照)。GACKLKLCGVLGLRL (配列番号317)、ACKLKLCGVLGLRLM (配列番号318)のアミノ酸配列を有するペプチドは、平均値の2倍よりも小さな OD値を有する。それにもかかわらずこれらのペプチドを請求した理由は、これらが抗体CR57によって認識される抗原ペプチドの領域の近傍に存在するためである。アミノ酸配列KLCGVLGLRLMDGTW (配列番号322)を有するペプチドに対する結合が最も顕著であった。
【0144】
抗体CR04-010は、GFGKAYTIFNKTLME (配列番号323)、FGKAYTIFNKTLMEA (配列番号 324)、GKAYTIFNKTLMEAD (配列番号325)、KAYTIFNKTLMEADA (配列番号326)、AYTIFNKTLMEADAH (配列番号327)、YTIFNKTLMEADAHY (配列番号328)、TIFNKTLMEADAHYK (配列番号329)、IFNKTLMEADAHYKS (配列番号330)、およびFNKTLMEADAHYKSV (配列番号331)からなる群から選択したアミノ酸配列を有する直線状ペプチドに結合した。AYTIFNKTLMEADAH (配列番号327)、YTIFNKTLMEADAHY (配列番号328) のアミノ酸配列を有するペプチドは、平均値の2倍よりも小さな OD値を有する。それにもかかわらずこれらのペプチドを請求した理由は、これらは、抗体CR04-010の認識する抗原ペプチド領域の近傍に存在するからである。アミノ酸配列TIFNKTLMEADAHYK (配列番号329)、IFNKTLMEADAHYKS (配列番号330)、およびFNKTLMEADAHYKSV (配列番号331) を有するペプチドに対する結合が最も顕著であった。
【0145】
CRJB、およびCR04-040、CR04-098およびCR04-103と呼ばれる抗体(データは示さず)は、直線状の抗原ペプチドの領域を認識しなかった。
【0146】
上記のペプチドまたはその一部のどちらとも、狂犬病ウイルスの中和エピトープの優良な候補であり、狂犬病ウイルス感染の治療および/または予防のためのワクチンや中和抗体産生の基礎を形成しうる。
【0147】
SLKGACKLKLCGVLGLRLMDGTW (配列番号332)およびGFGKAYTIFNKTLMEADAHYKSV (配列番号333) は、PEPSCAN分析において高い反応性を示すことから、糖タンパク質の特に興味深い領域である。
【0148】
上記のPEPSCANデータから、CR57およびCR04-010と呼ばれるヒトモノクローナル抗体は、狂犬病ウイルスGタンパク質の異なる領域に結合するとさらに推測することができ、これらは互いに競合しないエピトープを認識することを示唆している。
【0149】
実施例12
試験管内中和アッセイを用いた、抗狂犬病ウイルスGタンパク質IgGの中和効力の決定(修正RFFIT)
作製したヒトモノクローナル抗体の中和効力を実施例1に記載の通り、修正RFFITによって決定した。16個のIgGが狂犬病CVS-11株を1000 IU/mgより大きな効力で中和し、2 IU/mgより低い効力を有するIgGは2つだけであった(表15参照)。16個の抗体のうちの8個の効力は、一時的に発現させて作製したCR-57の効力を上回り、これらが狂犬病ウイルスの暴露後予防においてCR-57よりも効果的であることを示唆した。一時的に発現させて生成したCR-57の効力は、約3800 IU/mgタンパク質(表1および表15参照)であった一方、安定的に生成されたCR-57は5400 IU/mgタンパク質(データは示さず)の効力を示した。興味深いことに、同定された中和ヒトモノクローナル抗体の大半は、重鎖可変生殖系列遺伝子3−30を有する(表9参照)。
【0150】
狂犬病ウイルスへの抗体の親和性(データは示さず)、および修正RFFITアッセイにおける抗体の100%エンドポイント希釈度(データは示さない)に基づき、6つの独自のIgG、すなわち、CR04-010、CR04-040、CR04-098、CR04-103、CR04-104およびCR04-144からなるパネルを、更なる発色のために選択した。このパネルのなかでは抗体CR04-098が最も高い効力、すなわち、約7300 IU/mgタンパク質を示したため、特に興味深かった(表15参照)。安定的に生成されたCR04-098も同様の効力を示した(データは示さず)。
【0151】
実施例13
抗狂犬病ウイルスIgGによるE57エスケープウイルスの試験管内での中和
新規のヒトモノクローナル抗狂犬病抗体の特徴を更に調べるため、IgGのE57エスケープウイルスに対する中和活性を、上記のように修正RFFITによって調べた。抗狂犬病ウイルスIgGの大半は、6種類のE57エスケープウイルスの全てに対して良好な中和活性を示した(表16参照)。一方、CR04-008、CR04-018およびCR04-126は、それぞれ、E57エスケープウイルスの6/6、2/6、および3/6を中和しなかった。「中和しない」とは、1:100の抗体希釈物を用いても50%エンドポイントに到達しないことを意味する。CR04-021、CR04-108、CR04-120、CR04-125、およびCR04-164の中和活性は、エスケープウイルスの個数を増やすに従って有意に減少した。このことは、これらの抗体のエピトープは、E57エスケープウイルス糖タンパク質の影響を直接的または間接的に受けることを意味する。上記に基づくと、暴露後予防の処置のための抗狂犬病カクテルにおいて、複数の抗狂犬病ウイルスIgGはCR-57と適合する。特に、上記で同定した6つの独自のIgGのパネル、すなわち、抗体CR04-010、CR04-040、CR04-098、CR04-103、CR04-104、およびCR04-144は、E57エスケープウイルスに対して良好な中和効力を示し、このことは、これらの抗体によって認識されるエピトープはCR-57によって導入されたアミノ酸変異の影響を受けなかったことを示唆する。抗体CR04-098は各エスケープウイルスに対して3000 IU/mgよりも大きな効力を示しため、最も有望である。
【0152】
実施例14
抗狂犬病抗体CR-57およびCR04-098のエピトープ認識
CR-57およびCR04-098と呼ばれるヒトモノクローナル抗体が互いに重複たり、互いに競合したりしないエピトープを認識することを確認するため、CR04-098と呼ばれるヒトモノクローナル抗体のエスケープウイルスを、基本的にはCR57のエスケープウイルスについて記載したのと同様の方法で作製した(実施例1参照)。手短に言うと、ウェル毎の細胞増殖巣の数を免疫蛍光測定法により記録し、好適には1個の細胞増殖巣を含むウェルの培地を、ウイルスを増幅させるために選択した。全てのE98エスケープウイルスは、E982(2個の細胞増殖巣)およびE98-4(4個の細胞増殖巣)という例外を除き、単一の細胞増殖巣から生成された。中和指数がlog2.5よりも小さいウイルスをエスケープ変異体と定義した。中和指数は、「ウイルスおよびモノクローナル抗体(約4 IU/ml)を感染させたBSR細胞培養液中に産生された、1mlあたりの感染性ウイルス粒子の個数」を「ウイルスのみが感染したBSRまたはMNA細胞培養液中に産生された、1mlあたりの感染性ウイルス粒子の個数」から引くことによって求めた(log(モノクローナル抗体非存在下でのウイルス1mlあたりの細胞増殖巣形成単位数−log(モノクローナル抗体存在下でのウイルス1mlあたりの細胞増殖巣形成単位数))。log2.5よりも小さな指数はエスケープウイルスが存在する証拠とした。
【0153】
CR04-098がCR-57とは異なり、互いに重複したり競合したりしないエピトープと結合するかを更に調べるために、CR-57のE98エスケープウイルスに対する修正RFFITアッセイを上記のように行った。表17に示す通り、CR-57は全5種類のE98エスケープウイルスに対して良好な中和活性を有した。加えて、抗体CR04-010および抗体CR04-144のE98エスケープウイルスに対する中和活性を測定した。どちらの抗体もE98エスケープウイルスを中和せず(データは示さず)、両抗体によって認識されるエピトープは、抗体CR04-098によって導入されたアミノ酸変異の影響を直接的にまたは間接的に受けていることを示唆した。抗体CR04-018および抗体CR04-126については、E98エスケープウイルスのうち1種類、すなわちE98-4に対する中和活性のみを測定した。CR04-018はエスケープウイルスを中和する能力を有した一方、CR04-126はエスケープウイルスに対して弱い中和効力しか持たなかった。このことから、CR04-018によって認識されるエピトープは、抗体CR04-098によって導入された変異の影響を受けていないことが示唆される。加えて、抗体CR04-010、抗体CR04-038、抗体CR04-040、抗体CR04-073、抗体CR04-103、抗体CR04-104、抗体CR04-108、抗体CR04-120、抗体CR04-125、抗体CR04-164はE98-4を中和せず、これらの抗体がCR04-098と同じエピトープを認識することが示唆した(データは示さず)。
【0154】
各E98エスケープウイルスの狂犬病糖タンパク質において生じうる変異を同定するために、E57およびEJBエスケープウイルスについて記載したのと同じ方法で、糖タンパク質のオープン・リーディング・フレーム(ORF)のヌクレオチド配列を決定した。全てのE98エスケープウイルスは、狂犬病糖タンパク質のアミノ酸部位336においてNがDに置き換わる変異が存在した(図8参照)。糖タンパク質のこの領域は抗原部位IIIと定義され、アミノ酸330-338から構成される(アミノ酸番号はシグナルペプチドを含まずにふっている)。一方、CR-57はアミノ酸226-231(アミノ番号はシグナルペプチドを含まずにふっている)の位置に存在するエピトープを認識し、この場所は抗原部位Iと重なる。N336D変異に加えて、E98-5と呼ばれるE98エスケープウイルスは、狂犬病糖タンパク質のアミノ酸部位354においてHがQに置き換わる変異(コドンはCATからCAGに変化)を示した(データは示さず)。
【0155】
更に、(E57エスケープウイルスに存在するような)変異CR57エピトープを有するペプチドに対するCR57の結合を観測したpepscan分析によって、CR57の相互作用が失われたことが示された。驚くべきことに、フローサイトメトリーの測定から、CR04-098は依然としてPER.C6(登録商標)細胞において発現された(N336D変異を含む)変異糖タンパク質への結合能を有することがわかったが、この変異を含むウイルスはもはや中和されなかった。
【0156】
更に、BIAcore3000(登録商標)分析装置を用いた表面プラズモン共鳴分析によって、エピトープマッピング研究およびアフィニティーランキング研究を行った。精製された狂犬病糖タンパク質(ERA株)をresearch grade CM5 4-flow channel (Fc) センサーチップ(Biacore AB, Sweden)上にリガンドとして、アミノカップリングを用いて固定した。ランキングは、HBS-EP(Biacore AB, Sweden)をランニングバッファーとして、25℃で行った。各抗体50μlを20μl/分の一定流速で注入した。次に、750秒間ランニングバッファーを流した後、5μlの2M NaOH、5μlの45mM HCl、および5μlの2mM NaOHを用いてCM5チップの再生を行った。共鳴単位(RU)で表される共鳴シグナルを時間の関数としてプロットし、それぞれ結合および解離の尺度として用いるRUの増加および減少を決定し、抗体のランキングのために使用した。表面プラズモン共鳴分析によって決定されたCR57およびCR04-098の実際の KD値はそれぞれ2.4nmおよび4.5nmであった。更にエピトープマッピング研究によって、CR57およびCR04-098は狂犬病糖タンパク質上の異なったエピトープに結合することが確認された。CR57の注入によって58RUの共鳴が得生じた(データは示さず)。CR04-098の注入後、反応レベルが更に増加した(24RU)ため、CR04-098の結合部位は占有されなかったことが示唆される(データは示さない)。逆の順序で行っても同様の結果が得られ、各抗体は注入の順序に関係なく同程度のRUレベルに達することが示された(データは示さず)。更にこれらの結果は、CR57およびCR04-098は、狂犬病ウイルス糖タンパク質上の互いに異なったエピトープに同時に結合し、認識できることを示している。
【0157】
概して、上記のデータは、抗体CR-57およびCR04-098が、互いに異なり互いに重なり合わないエピトープ、すなわち抗原認識部位IおよびIIIにあるエピトープをそれぞれ認識することをさらに裏付ける。このデータは、CR-57およびCR04-098がERA Gへの結合を巡って競合しないこと、および、全てのE57エスケープウイルスに対して抗体CR04-098が良好な中和活性を示すことを示している、ELISA/FACSによる競合データと十分に一致する。これらの結果、および、CR57およびCR04-098の組み合わせを狂犬病ウイルスへ試験管内で暴露することによってエスケープウイルスが得られなったこと(どちらかの抗体4 IU/mlの存在下で選択を行った)に基づくと、抗体CR-57およびCR04-098は互いに重なり合ったり競合したりしないエピトープを認識し、暴露後予防の処置に用いる抗狂犬病ウイルス抗体カクテルに用いるのに好都合であると結論できる。
【0158】
実施例15
CR57およびCR04-098によって認識されるエピトープの保存の評価
CR-57の最小結合領域(配列番号332中のアミノ酸KLCGVL;CR57によって認識される狂犬病ウイルス糖タンパク質の領域であることが、PEPSCANおよびアラニンスキャニング技術によって決定された領域)を229個の遺伝子型1の狂犬病ウイルス分離株のヌクレオチド配列と比較し、エピトープの保存を評価した(表18参照)。サンプル集合には、ヒト分離株、コウモリ分離株、およびイヌ分離株または狂犬病のイヌに噛まれたであろうと最も考えられる家畜からの分離株が含まれた。最小結合領域の各部位のアミノ酸の出現頻度分析から、エピトープを形成する必須のな残基が高度に保存されていることが判明した。位置1に存在するリジンは99.6%の分離株で保存され、229個の分離株のうち1つにおいてのみ、保存的なK>R変異が観測された。位置2および3(LおよびC)は完全に保存されていた。中央部のシステイン残基は糖タンパク質のフォールディングに構造的にかかわり、全リッサウイルスを通して保存されると考えられている(Badrane and Tordo, 2001を参照)。位置4のグリシンは98.7%の分離株で保存され、229個の分離株のうち3つにおいて荷電アミノ酸への変異がみられた(1/229の分離株でG>R変異、2/229の分離株でG>E変異)。5番目の位置もまた、1つの分離株で保存的なV>I変異が観測されたという以外を除き、保存されていた。アラニン置換スキャンによって必須の残基ではないと決定された6番目の位置においては、街上分離株(srreet isolates)に有意な多様性がみられた;すなわち、70.7%の株でL、26.7%の株でP、2.6%の株でSであった。総合すると、遭遇する可能性のある狂犬病ウイルスの約99%は、CR-57によって認識されると予測される。
【0159】
これら229個のウイルス分離株のうち123個について、CR-57およびCR04-098のエピトープ双方において変異が存在するかどうか分析した。これら123個の街上ウイルス(street viruses)はどちらのエピトープにおいても変異を含まなかった。E98エスケープウイルスにおいて観測されたN>D変異は、5つのウイルス分離株のみに存在した。これらのウイルスは地理的に異なり、アフリカの動物から分離されたものである(系統図は図9参照;5つのウイルス分離株、すなわちAF325483、AF325482、AF325481、AF325480、およびAF325485は太字で示した)。糖タンパク質配列の系統発生解析によって、変異したCR57エピトープを有する狂犬病ウイルスは、変異したCR04-098エピトープを有する狂犬病ウイルスとは遠縁にあたることがわかった。故に、CR-57およびCR04-098のカクテルによる中和に対して耐性を示す狂犬病ウイルスに遭遇する可能性は実質的に皆無である。
【0160】
【表1】

【0161】
【表2】
【0162】
【表3】
【0163】
【表4−1】
【0164】
【表4−2】
【0165】
【表4−3】
【0166】
【表5】
【0167】
【表6】
【0168】
【表7−1】
【0169】
【表7−2】
【0170】
【表8】
【0171】
【表9−1】
【0172】
【表9−2】
【0173】
【表10】
【0174】
【表11A】
【0175】
【表11B】
【0176】
【表12−1】
【0177】
【表12−2】
【0178】
【表12−3】
【0179】
【表13−1】
【0180】
【表13−2】
【0181】
【表13−3】
【0182】
【表14−1】
【0183】
【表14−2】
【0184】
【表14−3】
【0185】
【表14−4】
【0186】
【表14−5】
【0187】
【表14−6】
【0188】
【表14−7】
【0189】
【表14−8】
【0190】
【表14−9】
【0191】
【表14−10】
【0192】
【表15】
【0193】
【表16】
【0194】
【表17】
【0195】
【表18】
【0196】
参考文献
【図面の簡単な説明】
【0197】
【図1】図1は狂犬病ウイルス株CVS-11およびE57のエスケープウイルスのアミノ酸配列の比較を示す。ウイルス感染細胞を感染2日後に回収し、全RNAを単離した。cDNAを作製し、DNA塩基配列解読のために使用した。変異を含む領域を図示しており、変異を太字で示す。図1Aはヌクレオチド配列の比較を示す。アミノ酸の上の番号は、シグナルペプチドを含む、狂犬病ウイルス糖タンパク質のアミノ酸番号を示す。図1Bは、アミノ酸配列の比較を示す。狂犬病ウイルス糖タンパク質の略図を上部に示す。黒い四角形部分がシグナルペプチドを示し、灰色の四角形部分が膜貫通領域を示す。図1の配列は、配列番号130-141によっても表される。
【図2】図2は狂犬病ウイルス株CVS-11およびEJBのエスケープウイルスのアミノ酸配列の比較を示す。ウイルス感染細胞を感染2日後に回収し、全RNAを単離した。cDNAを作製し、DNA塩基配列解読のために使用した。変異を含む領域を図示しており、変異を太字で示す。図2Aはヌクレオチド配列の比較を示す。アミノ酸の上の番号は、シグナルペプチドを含む、狂犬病ウイルス糖タンパク質のアミノ酸番号を示す。図2Bは、アミノ酸配列の比較を示す。狂犬病ウイルス糖タンパク質の略図を上部に示す。黒い四角形部分がシグナルペプチドを示し、灰色の四角形部分が膜貫通領域を示す。図2の配列は、配列番号142-151 によっても表される。
【図3】PDV-C06ベクターを示す。
【図4】図4は抗狂犬病ウイルスscFvと、CR-57と呼ばれるビオチン化抗狂犬病ウイルス抗体との競合ELISAを示す。精製された狂犬病ウイルスGタンパク質によってコートされたELISAプレートを、各scFvと共にインキュベートした後、CR-57bio (0.5μg/ml)を加えた。続いて、scFvの存在下および非存在下においてCR-57bioの結合を観測した。
【図5】図5は抗狂犬病ウイルスscFvと、CR-57と呼ばれる抗狂犬病ウイルス抗体との競合ELISAを示す。精製された狂犬病ウイルスGタンパク質によってコートされたELISAプレートを、CR-57 (1μg/ml)と共にインキュベートした後、過剰量のscFvを加えた。続いて、CR-57の存在下および非存在下においてscFvの結合を観測した。
【図6】図6は抗狂犬病ウイルスGタンパク質IgGと、CR-57と呼ばれる抗狂犬病ウイルス抗体との競合ELIZA分析を示す。Gタンパク質(ERA株)を非標識IgGと共にインキュベートした(x軸上に示す)。ビオチン化CR57(CR57bio)を加えてGタンパクに結合させた後、ストレプトアビジン-HRPを加えることによって視覚化を行った。ELISAシグナルを、CR57bioの結合単独の場合のシグナル対するパーセントとして示す。
【図7】図7は抗狂犬病ウイルスGタンパク質IgGと、CR-57と呼ばれる抗狂犬病ウイルス抗体との競合FACS分析を示す。Gタンパク質(ERA株)を発現するPER.C6細胞を非標識IgGと共にインキュベートした(x軸上に示す)。ビオチン化したCR57(CR57bio)を加え、Gタンパク発現細胞に結合させた後、ストレプトアビジン-PEによる視覚化を行った。FACSシグナルを、CR57bioの結合単独の場合のシグナル対するパーセントとして示す。
【図8】図8はCVS-11およびE98のエスケープウイルスのアミノ酸配列の比較を示す。ウイルス感染細胞を感染2日後に回収し、全RNAを単離した。cDNAを作製し、DNA塩基配列解読のために使用した。点変異を含む領域を図示しており、変異を太字で示す。図8Aはヌクレオチド配列の比較を示す。ヌクレオチド上の番号は、シグナルペプチド配列を含まない、狂犬病ウイルス糖タンパク質のオープン・リーディング・フレームにおける、変異ヌクレオチド(太字で示す)を示す。図8Bはアミノ酸配列の比較を示す。アミノ酸上の番号は、シグナルペプチド配列を除いた狂犬病ウイルス糖タンパク質における、変異ヌクレオチド(太字で示す)を示す。
【図9】図9は123種類の狂犬病ウイルス街上毒株の系統樹を示す(123種類の狂犬病ウイルスG糖タンパク質の配列をもとに、近隣結合法、Kimuraの2−パラメータ法、および500回のブートストラップ法を用いた)。太字はE98のエスケープウイルスで観測されるのと同様のN>D変異を有するウイルスを示す。
【図10】図10は狂犬病糖タンパク質上の中和エピトープを示す。新規のCR57エピトープを含む抗原性部位を記載した、狂犬病ウイルス糖タンパク質の概略図を示す。シグナルペプチド(19アミノ酸)および膜貫通領域を、黒い四角形によって示す。ジスルフィド結合による架橋を示す。アミノ酸番号は、シグナルペプチド配列を除いた成熟タンパク質から付けられている。
【技術分野】
【0001】
本発明は医学に関する。特に、本発明は、狂犬病ウイルスを中和する結合分子に関する。これらの結合分子は、狂犬病の暴露後予防に役立つ。
【背景技術】
【0002】
狂犬病はほぼ全世界に分布するウイルス感染症であって、主に野生動物や家畜に影響を及ぼすが、人間にも感染し、破壊的な、ほぼ不変の致命的な脳炎をひきおこす。年間70,000人以上の人間が死亡すると推定され、その他の何百万人もが暴露後の処置を必要とする。
【0003】
狂犬病ウイルスは弾丸型でエンベロープを有する一本鎖RNAウイルスで、ラブドウイルス科リッサウイルス属に分類される。狂犬病ウイルスのゲノムは以下の5つのウイルスタンパク質をコードする:RNA依存RNAポリメラーゼ(L);核タンパク質(N);リン酸化されたタンパク質(P);ウイルスタンパク質のエンベロープの内側にあるマトリクスタンパク質(M);および外表面糖蛋白質(G)。
【0004】
Gタンパク質(62〜67kDa)は、505個のアミノ酸から成るI型糖蛋白質であり、2〜4個の潜在的なN-グリコシル化部位を有するが、ウイルス株に依存して、そのうちの1個または2個のみがグリコシル化される。Gタンパク質は、ビリオンのエンベロープの外表面を覆う突起を形成し、ウイルス中和抗体を誘発することが知られている。
【0005】
狂犬病は、能動免疫法と受動免疫法のどちらによっても治療または予防が可能である。狂犬病の暴露後予防は、局部的な傷口の迅速な手当て、および受動免疫(抗狂犬病免疫グロブリン)および能動免疫(ワクチン)双方の実施から構成される。
【0006】
現在、抗狂犬病免疫グロブリン(RIG)は、ヒト抗狂犬病免疫グロブリン(HRIG)またはウマ抗狂犬病免疫グロブリン(ERIG)の血清試料から作られている。ERIGおよびHRIGの欠点は、それらを充分量において利用することができず、HRIGの場合には、高価すぎる点である。更に、ERIGの使用は、アナフィラキシー・ショックなどの副作用につながる可能性がある。既知あるいは未知の病原体による汚染の可能性は、HRIGに伴う更なる懸念である。これらの欠点を克服するために、暴露後予防において狂犬病ウイルスを中和することができるモノクローナル抗体を使用することが提案されてきた。狂犬病ウイルスを不活性化するマウス・モノクローナル抗体は、従来技術において周知である(Schumacher et al., 1989参照)。しかし、生体内でマウス抗体を使用することは、例えば、血中半減期が短い、特定のヒトエフェクター機能を誘発できない、およびヒト体内でマウス抗体に対する過剰な免疫反応(ヒト抗マウス抗体 "human anti-mouse antibody" (HAMA)による反応)が誘発されてしまうなどといった、マウス抗体をヒトに投与することに伴う問題があるため制限される。
【0007】
近年、狂犬病ウイルス中和ヒトモノクローナル抗体について述べられている (Dietzschold et al. , 1990, Champion et al., 2000、およびHanlon et al., 2001参照)。抗狂犬病ヒトモノクローナル抗体が暴露後予防においてHRIGと同等の効力をもつには、モノクローナル抗体の混合物を用いなければならない。そのような混合物においては、それぞれの抗体がウイルス上の別々のエピトープまたは部位に結合し、抵抗性のあるウイルス変異体の生存を防がなくてはならない。
【0008】
現在、暴露後予防の可能性を高める新規の狂犬病ウイルス中和ヒトモノクローナル抗体、特に異なるエピトープ認識特異性を持つ抗体の必要性が依然として高い。本発明は、幅広い狂犬病ウイルスおよびその中和抵抗性変異株に対する暴露後予防に役立つ、混合物として使用できる可能性を持つヒトモノクローナル抗体を提供する。
【0009】
(発明の説明)
以下、本発明で使用する用語の定義を記す。
【0010】
定義
結合分子
本明細書において用いる「結合分子」という用語は、モノクローナル抗体(キメラ、ヒト化、またはヒトモノクローナル抗体など)を含むインタクト型免疫グロブリンを指すか、または、インタクト型免疫グロブリンと競合して免疫グロブリンの結合パートナー(例えば狂犬病ウイルス、あるいはGタンパク質などの狂犬病ウイルスのフラグメント)と特異的結合を形成するような免疫グロブリンのフラグメントを備える、抗原結合領域および/または可変領域を指す。その構造に関係なく、抗原結合性フラグメントは、インタクト型免疫グロブリンによって認識される抗原と同一の抗原に結合する。抗原結合性フラグメントを、結合分子のアミノ酸配列のうちの、少なくとも2連続のアミノ酸残基、少なくとも5連続のアミノ酸残基、少なくとも10連続のアミノ酸残基、少なくとも15連続のアミノ酸残基、少なくとも20連続のアミノ酸残基、少なくとも25連続のアミノ酸残基、少なくとも30連続のアミノ酸残基、少なくとも35連続のアミノ酸残基、少なくとも40連続のアミノ酸残基、少なくとも50連続のアミノ酸残基、少なくとも60連続のアミノ酸残基、少なくとも70連続のアミノ酸残基、少なくとも80連続のアミノ酸残基、少なくとも90連続のアミノ酸残基、少なくとも100連続のアミノ酸残基、少なくとも125連続のアミノ酸残基、少なくとも150連続のアミノ酸残基、少なくとも175連続のアミノ酸残基、少なくとも200連続のアミノ酸残基、または少なくとも250連続のアミノ酸残基、のいずれかのアミノ酸配列を含むペプチドあるいはポリペプチドから構成することが可能である。
【0011】
本明細書において用いる「結合分子」という用語は、従来技術において知られている全てのクラスおよびサブクラスの免疫グロブリンを含む。重鎖の定常領域のアミノ酸配列によって、結合分子を、インタクト抗体の5つの大きなクラス、すなわちIgA, IgD, IgE, IgGおよびIgMに分類することができ、これらのいくつかを、例えばIgAl, IgA2, IgGl, IgG2, IgG3およびIgG4といったサブクラス(イソタイプ)にさらに分類することができる。
【0012】
抗原結合性フラグメントには、とりわけ、Fab、F (ab')、F(ab')2、Fv、dAb、Fd、相補性決定領域(CDR)フラグメント、単鎖抗体(scFv)、二価の単鎖抗体、単鎖ファージ抗体、二重特異性抗体(diabody)、三重特異性抗体(triabody)、四重特異性抗体(tetrabody)、(ポリ)ペプチドと特異的な抗原結合を形成するのに十分な、免疫グロブリンのフラグメントを少なくとも含む(ポリ)ペプチド、などが含まれる。上述のフラグメントは、合成によって作製するか、インタクト型免疫グロブリンを酵素または化学作用を用いて切断することによって作製するか、あるいは組み換えDNA技術を用いて遺伝子工学的に作製することができる。作製の方法は当技術分野では周知であり、例えば“Antibodies: A Laboratory Manual, Edited by:E. Harlow and D, Lane (1988), Cold Spring Harbor Laboratory, Cold Spring Harbor, New York”に記載されており、参照することにより本願明細書に組み込まれる。結合分子またはその抗原結合性フラグメントは、1つまたは複数の結合部位を有する可能性がある。2つ以上の結合部位を有する場合、結合部位は互いに同一である可能性も異なる可能性もある。
【0013】
結合分子は、むき出しであるか非コンジュゲート型の結合分子である可能性もあるが、免疫コンジュゲートを形成している可能性もある。むき出しであるか非コンジュゲート型の結合分子とは、エフェクター部分やタグ(とりわけ有害物質、放射性物質、リポソーム、酵素など)とコンジュゲートを形成していないか、作動可能に結合していないか、または他の物理的あるいは機能的な結合をしていない結合分子を指すよう意図されている。当然のことながら、むき出しであるか非コンジュゲート型の結合分子には、エフェクター部分やタグの結合以外であって、安定化、多量体化、ヒト化、あるいは任意の他の方法による操作を受けた結合分子を除外するものではない。従って、翻訳後修飾を受けた全てのむき出しであるか非コンジュゲート型の結合分子は「結合分子」に含まれるのであって、天然の結合分子産生細胞の環境において、組み換え型結合分子産生細胞によって、何処かに修飾がなされたもの、および、初めに結合分子を調製した後にヒトの手によって何処かに修飾が導入されたものも含まれる。当然、「むき出しであるか非コンジュゲート型の結合分子」という用語は、体内に投与された後にエフェクター細胞および/または分子と機能的な結合を形成する能力をも含むものであり、このような相互作用には生物学的作用を及ぼすために必要なものもあるためである。従って、結合しているエフェクターグループやタグが存在しないことは、試験管内では「むき出しであるか非コンジュゲート型の結合分子」の定義にあてはまるが、生体内においてはあてはまらない。
【0014】
相補性決定領域(CDR)
本明細書において用いる「相補性決定領域」という用語は、免疫グロブリンなどの結合分子の可変領域中に存在する塩基配列を意味し、通常、抗原上の認識されるエピトープに対して形状および電荷分布において相補的である抗原結合部位に大きな影響を及ぼす。CDR領域はタンパク質あるいはタンパク質断片の直線状エピトープ、不連続エピトープ、あるいは立体構造エピトープに特異的である可能性があり、これらのエピトープは天然の構造をとるタンパク質上に存在するか、または、ある場合には、例えばSDS中で溶解することによって変性したタンパク質上に存在する。また、エピトープはタンパク質の翻訳後修飾から構成される可能性もある。
【0015】
機能的変異体
本明細書において用いる「機能的変異体」という用語は、元の結合分子(親結合分子)のヌクレオチドおよび/またはアミノ酸配列と比較して1つ以上のヌクレオチドおよび/またはアミノ酸が変化したヌクレオチドおよび/またはアミノ酸配列を有しており、かつ、例えば狂犬病ウイルスまたはその断片などといった結合パートナーに対する結合を、依然として親結合分子と競合することができる結合分子を指す。つまり、親結合分子のヌクレオチドおよび/またはアミノ酸配列における修飾は、ヌクレオチド配列によってコードされる結合分子、あるいはアミノ酸配列を含む結合分子の結合能に重大な影響を与えたりこれを変化させたりはせず、すなわちその結合分子は依然としてターゲットを認識し結合することができる。機能的変異体は、ヌクレオチドおよびアミノ酸の置換、挿入、および欠失などの、保存的な配列修飾を有する可能性がある。これらの修飾は、例えば部位特異的突然変異導入法やPCRを用いたランダム突然変異導入法などの、当技術分野では周知の標準的な技術によって導入可能である。
【0016】
保存的なアミノ酸置換には、あるアミノ酸残基が、構造や化学的性質が似た別のアミノ酸残基によって置き換えられる置換が含まれる。類似したアミノ酸側鎖をもつアミノ酸残基のファミリーが当技術分野において定められている。このファミリーには、塩基性側鎖を有するアミノ酸(例:リジン、アルギニン、ヒスチジン)、酸性側鎖を有するアミノ酸(例:アスパラギン酸、グルタミン酸)、非荷電極性側鎖を有するアミノ酸(例:アスパラギン、グルタミン、セリン、トレオニン、チロシン、システイン、トリプトファン)、非極性側鎖を有するアミノ酸(例:グリシン、アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン)、β分岐側鎖を有するアミノ酸(例:トレオニン、バリン、イソロイシン)、芳香族側鎖を有するアミノ酸(例:チロシン、フェニルアラニン、トリプトファン)が含まれる。上で述べたのとは別のアミノ酸残基ファミリーの分類法も使用可能であることは、当業者に明らかであろう。更には、変異体は、例えばあるアミノ酸が、構造や化学的性質の異なるアミノ酸残基により置換されるといった、非保存的なアミノ酸置換を有する可能性もある。これに類似したマイナーな変異には、アミノ酸の欠失や挿入、或いはその双方が含まれる可能性がある。どのアミノ酸残基を免疫学的活性を損うことなく置換、挿入、あるいは欠失することが可能であるかについての指針は、当技術分野において周知であるコンピュータープログラムを用いて見出すことが可能である。
【0017】
ヌクレオチド配列における変異は、例えば転位変異または転換変異のように、遺伝子座における単一の変化(点変異)とすることも可能であるし、もしくは代わりに、単一の遺伝子座において複数のヌクレオチドを挿入、欠失あるいは置換することも可能である。さらに、1つのヌクレオチド配列中のかなり多数の遺伝子座において、1つまたは複数の変化を加えることが可能である。変異は、当技術分野において知られている適切な方法によって引き起こすことができる。
【0018】
ホスト
本明細書において用いる「ホスト」という用語は、クローニングベクターまたは発現ベクターのようなベクターが導入された生物あるいは細胞を指すことを意図されている。生物あるいは細胞は、原核細胞であっても真核細胞であってもよい。この用語は、対象となる特定の生物あるいは細胞だけでなく、その生物あるいは細胞の子孫にも当てはまるということが理解されなくてはならない。後の世代に突然変異あるいは環境の影響のいずれかによる特定の修飾が生じる可能性があるため、これらの子孫は実際には元の生物あるいは細胞と同一ではない可能性があるが、それでもなお、本明細書において用いる「ホスト」という用語の範囲内に含まれる。
【0019】
ヒト(Human)
「ヒト」という用語は、本願明細書において定義される結合分子に適用した際には、ヒトから直接得られた分子か、ヒトの塩基配列に基づいた分子かのいずれかを意味する。結合分子がヒトの塩基配列に由来するか又はヒトの塩基配列に基づいているのなら、その後修飾を加えたとしても、本明細書の全体を通して使用する場合には依然として「ヒト」結合分子であると見なす。言い換えると、「ヒト」という用語は、結合分子に適用した際には、可変領域または定常領域のいずれかに基づくヒト生殖細胞系列免疫グロブリン塩基配列に由来する可変領域および定常領域を有する結合分子、ヒトの体内またはヒトリンパ球内で生じたのではない結合分子、または修飾された形態の結合分子を含むことを意図されている。従って、ヒト結合分子は、ヒト生殖細胞系列免疫グロブリンの塩基配列によってコードされていないアミノ酸残基を含む可能性があり、置換および/または欠失(例:試験管内で、例えばランダム突然変異導入法または部位特異的突然変異導入法によって導入された変異、または生体内で体細胞変異によってもたらされた変異)を含むことがある。本明細書において用いる「〜に基づいた」という言葉は、核酸配列が鋳型から正確に複製される状況か、例えばエラーを起こしやすいPCR法によってマイナーな変異を伴う状況か、あるいは鋳型に正確に合致するかまたはマイナーな変更を伴って合成される状況を指す。ヒトの塩基配列に基づいた半合成分子もまた、本明細書において用いる「ヒト」結合分子あるとみなす。
【0020】
モノクローナル抗体
本明細書において用いる「モノクローナル抗体」という用語は、単一の分子構成(すなわち一次構造)を有する抗体分子調製物、すなわち単一のアミノ酸配列を有する抗体分子調製物を意味する。モノクローナル抗体は、特定のエピトープに対して単一の結合特異性および親和性を示す。従って、「ヒトモノクローナル抗体」という用語は、単一の結合特異性を示す抗体であって、ヒト生殖細胞系列免疫グロブリンの塩基配列に由来するか、基づいた可変領域および定常領域、または合成配列に完全に由来する可変領域および定常領域を有する該抗体を指す。モノクローナル抗体を作製する方法は問題とされない。
【0021】
核酸分子
本発明で用いる「核酸分子」という用語は、ヌクレオチドの多量体型を指し、RNA、cDNA、ゲノムDNA、およびこれらの合成型、および前記物質の混合ポリマーの、センス鎖およびアンチセンス鎖双方を含む。ヌクレオチドとは、リボヌクレオチド、デオキシヌクレオチド、またはどちらかの種類のヌクレオチドの修飾型を指す。この用語には、一本鎖および二本鎖のDNAも含まれる。加えて、ポリヌクレオチドには、天然ヌクレオチドおよび修飾ヌクレオチドのいずれかまたは両方が、天然のヌクレオチド結合および/または非天然のヌクレオチド結合によって結合したポリヌクレオチドが含まれる可能性もある。当業者ならば容易に理解できるように、核酸分子は化学的または生化的に修飾されうるし、もしくは非天然ヌクレオチド塩基又は誘導体化ヌクレオチド塩基を含みうる。このような修飾には、例えば、ラベル化、メチル化、1つまたは複数の天然ヌクレオチドのアナログによる置換、ヌクレオチド間の修飾(例えば、非荷電性の結合(例:メチルホスホネート、リン酸トリエステル、アミド亜リン酸エステル、カルバミン酸エステルなど)、荷電結合(例:ホスホロチオネート、ホスホロジチオエートなど)、懸垂部分(pendent moieties)(例:ポリペプチド)、インターカレータ(例:アクリジン、プソラレンなど)、キレート剤、アルキル化剤、および修飾された結合(例:α‐アノマー核酸など))が含まれる。上記の用語はまた、一本鎖、二本鎖、部分的二重らせん、三重らせん、ヘアピン構造、円形、および南京錠型の立体構造などの、任意のトポロジー構造を含むよう意図されている。また、ポリヌクレオチドの、水素結合やその他の化学的相互作用を介して目的の配列に結合する能力を模倣した合成分子も含まれる。このような分子は当技術分野においては周知であり、例えば、分子骨格においてリン酸結合をペプチド結合で置換した分子などが含まれる。核酸の塩基配列について言及した場合、特に明記しない限りは、その相補鎖も含まれる。従って、特定の配列を有する核酸分子について言及した場合、その相補鎖およびその相補的配列も含まれると理解すべきである。相補鎖もまた、例えばアンチセンス療法、ハイブリッド形成プローブ、およびPCRプライマーに有用である。
【0022】
薬学的に許容される賦形剤
「薬学的に許容される賦形剤」とは、好ましい剤形または便利な剤形を調製するために、薬物、薬剤、あるいは結合分子などの活性分子と配合するあらゆる不活性物質を意味する。この「薬学的に許容される賦形剤」は、非毒性であるか、または、少なくとも、使用する用量および濃度では、使用目的において、毒性が服用者にとって許容される程度であるかであって、そして薬物、薬剤または結合分子を含む製剤の他の成分と適合性を有する賦形剤である。
【0023】
暴露後予防
「暴露後予防(PEP)」は狂犬病にかかった動物に暴露したおそれがある人に行なう必要がある。起こり得る暴露には、動物咬傷などの咬傷暴露(例:歯による皮膚へのあらゆる侵入)および非咬傷暴露が含まれる。非咬傷暴露には、研究室や洞穴における大量のエアロゾル化狂犬病ウイルスへの暴露、および、狂犬病により死亡した患者から角膜移植手術を受けたレシピエントが含まれる。開放創、擦り傷、粘膜、あるいは理論的にはかき傷が、狂犬病にかかった動物の唾液または他の潜在的に感染性を有する物質(例えば神経組織)によって汚染した場合にも、非咬傷暴露が生じる。例えば狂犬病にかかった動物をなでたり、狂犬病にかかった動物の血液、尿、または糞便との接触したりするなどの他の接触は、それだけでは暴露とならず、暴露後予防の適応とはならない。PEPは暴露後できるだけ早く開始しなければならない。暴露が起こっていない場合は、暴露後予防は必要でない。全ての暴露後予防の処方計画において、過去に免疫化された人を除いて、能動免疫法および受動免疫法を並行して行なわなくてはならない。
【0024】
特異的に結合
結合分子の相互作用(例えば抗体およびその結合パートナー(例:抗原))に関して、本願明細書において用いる「特異的に結合」という用語は、この相互作用が、ある特定の構造(例えば結合パートナー上の抗原決定基またはエピトープ)の存在に依存していることを意味する。換言すれば、結合パートナーが他の分子や生物の混合物中に存在するときでも、抗体は選択的に結合パートナーに結合するか結合パートナーを認識する。結合は、共有結合性相互作用または非共有結合性相互作用、あるいは両者の組み合わせによって媒介される。さらに言い換えると、「特異的に結合」という用語は、免疫特異的に抗原またはその断片に結合し、他の抗原には免疫特異的に結合しないことを意味する。免疫特異的に抗原に結合する結合分子は、他のペプチドやポリペプチドに対してより低い親和性で結合することができ、これは例えばラジオイムノアッセイ(RIA)、酵素結合抗体免疫吸着アッセイ(ELISA)、BIACORE、または当技術分野において周知の他のアッセイによって測定することができる。免疫特異的に抗原に結合する結合分子またはそのフラグメントは、関連抗原と交差反応する可能性がある。免疫特異的に抗原に結合する結合分子またはそのフラグメントは、関連抗原と交差反応しないことが望ましい。
【0025】
治療に有効な量
「治療に有効な量」という用語は、狂犬病の暴露後予防に有効である、本明細書において定義される結合分子の量を意味する。
【0026】
ベクター
「ベクター」という用語は、第2の核酸分子をホストに導入するために挿入することができる核酸分子を意味し、ホスト内で複製され、場合によっては発現する。換言すると、ベクターは、自身に結合した核酸分子を輸送することができる。本願明細書において用いる「ベクター」という用語は、クローニングベクターおよび発現ベクターの双方を意図している。ベクターには、プラスミド、コスミド、細菌人工染色体(BAC)および酵母人工染色体(YAC)、そしてバクテリオファージまたは植物または動物(ヒトを含む)のウイルスに由来するベクターが含まれるが、これらに限定されない。ベクターは、予定されるホストによって認識される複製開始点を備え、そして発現ベクターの場合には、ホストによって認識されるプロモータおよび他の制御領域を備える。第2の核酸分子を含むベクターは、例えば形質転換、トランスフェクション、または細菌あるいはウイルスの侵入機構を利用して、細胞内に導入される。例えば、植物細胞および同類のものにおいてしばしば使用するエレクトロポレーションまたは微粒子銃などの、核酸を細胞内に導入する他の方法も周知である。核酸を細胞内に導入する方法は、とりわけ細胞の種類やその他いろいろに依存する。これは、本発明に決定的なものではない。ある種のベクターは、自身が導入されるホスト内で自律複製できる(例えば、細菌複製開始点を有するベクターは、細菌内で複製することができる)。他のベクターは、ホストへ導入された直後にホストのゲノムに組み込まれ、その結果、ホストのゲノムと共に複製される。
【発明の開示】
【0027】
(発明の概要)
本発明は、狂犬病ウイルスに特異的に結合して中和することができる結合分子を提供する。さらに、本発明は、これらの結合分子の少なくとも結合領域をコードする核酸分子に関する。さらに本発明は、狂犬病ウイルスに起因する状態を発現する危険のある被験者への、暴露後予防における本発明の結合分子の用途を提供する。
【発明を実施するための最良の形態】
【0028】
第1の態様において、本発明は、狂犬病ウイルスに特異的に結合することができる結合分子を備える。本発明の結合分子はまた、狂犬病ウイルスを中和する活性を有することが望ましい。好適には、本発明の結合分子はヒト結合分子である。あるいは、結合分子は他の動物の結合分子であっても良い。狂犬病ウイルスは、リッサウイルス属に含まれる。リッサウイルス属は、合計で11の遺伝子型を含む:すなわち、狂犬病ウイルス(遺伝子型1)、ラゴスコウモリウイルス(遺伝型2)、モコラウイルス(遺伝子型3)、ドゥベンヘイグウイルス(遺伝子型4)、ヨーロッパコウモリリッサウイルス1(遺伝子型5)、ヨーロッパコウモリリッサウイルス2(遺伝子型6)、オーストラリアコウモリリッサウイルス(遺伝子型7)、アラバンウイルス(遺伝子型8)、クージャンウイルス(遺伝子型9)、イルクーツウイルス(遺伝子型10)およびウェストコーカシアンウイルス(遺伝子型11)である。本発明の結合分子は、狂犬病ウイルスに結合するだけでなく、リッサウイルス属に含まれる他の遺伝子型にも結合する能力を持つ可能性がある。好適には、結合分子は、リッサウイルス属に含まれる他の遺伝子型をも中和する能力も持つ。更には、本発明の結合分子は、ラブドウイルス科のリッサウイルス属以外のウイルスを結合および/または中和する能力をも持ち得る。ラブドウイルス科には、サイトラブドウイルス、エファメロウイルス、リッサウイルス、ヌクレオラブドウイルス、ラブドウイルスおよびベシクロウイルスが含まれる。
【0029】
結合分子は、狂犬病ウイルスに、天然型であれ不活性型や弱毒化型であれ特異的に結合し得る。狂犬病ウイルスの失活は、とりわけ、β‐プロピオラクトン(BPL)での処理(White and Chappel, 1982)、56℃での30分間以上の加熱、ガンマ線照射、アセチルエチレンイミンまたはエチルイミンでの処理あるいはアスコルビン酸および硫酸銅での72時間の処理(Madhusudana et al., 2004)といった処理によって行なうことができる。例えば、とりわけパストリゼーション(湿性温熱処理)、乾熱処理、蒸気熱処理、低いpHでの処理、有機溶剤/界面活性剤での処理、ナノ濾過、紫外線照射などの、当業者にとって周知の一般的なウイルス失活法を使用することもできる。好適には、失活をβ‐プロピオラクトン(BPL)を用いた処理によって実行するのがよい。狂犬病ウイルスが依然感染力を持つか、あるいは部分的にまたは完全に不活化されているかどうか検査する方法は当業者にとって周知であり、とりわけ、” Laboratory techniques in rabies , Edited by: F. -X Meslin, M. M. Kaplan and H. Koprowski (1996), 4th edition, Chapter 36, World Health Organization, Geneva” に記載されている。
【0030】
結合分子は、狂犬病ウイルスの1つまたは複数の断片(例えば、とりわけ、一種類もしくは複数の種類のタンパク質からなる調製液、および/または狂犬病ウイルスに由来する(ポリ)ペプチドまたは狂犬病ウイルスタンパク質および/または(ポリ)ペプチドで形質転換された細胞から得られた(ポリ)ペプチド)にも特異的に結合することができる可能性がある。例えば狂犬病ウイルスの暴露後予防などの、治療法および/または予防法のためには、結合分子は、M(Ameyama et al. 2003参照)またはGタンパク質といった、狂犬病ウイルスの接触可能な表面タンパク質に特異的に結合できることが望ましい。診断の目的のためには、ヒト結合分子が、狂犬病ウイルスの表面には存在しないタンパク質に特異的に結合することができても良い。様々な既存の狂犬病ウイルス株の接触可能な表面タンパク質および内部タンパク質のアミノ酸配列は、EMBL-データベースおよび/または他のデータベースで検索することができる。
【0031】
望ましくは、この断片は、少なくとも、本発明のヒト結合分子によって認識される抗原決定基を備える。本願明細書において使用する「抗原決定基」とは、例えば狂犬病ウイルス(ポリ)ペプチド、(糖)タンパク質、またはその類似体または断片などの部分のことであり、本発明のヒト結合分子に十分に高い親和性で結合し、検出可能な抗原‐結合分子複合体を形成することのできる部分のことである。
【0032】
本発明による結合分子は、ポリクローナル抗体またはモノクローナル抗体などのインタクト型免疫グロブリン分子であってもよいし、あるいは結合分子は、Fab、F(ab')、F(ab')2、Fv、dAb、Fd、相補性決定領域(CDR)フラグメント、単鎖抗体(scFv)、二価の単鎖抗体、単鎖ファージ抗体、二重特異性抗体(diabody)、三重特異性抗体(triabody)、四重特異性抗体(tetrabody)、および狂犬病ウイルスまたはそのフラグメントに特異的な抗原結合を形成するのに十分な免疫グロブリンの断片を少なくとも含む(ポリ)ペプチド、などの抗原結合性断片であっても良いが、これらに限定されない。本発明の結合分子は、非単離型であっても単離型であっても使用することができる。更に、本発明の結合分子は、単独でも、または少なくとも1つのヒト結合分子(あるいはその変異体または断片)を含む混合物としても使用することができる。換言すれば、結合分子を、例えば2つ以上の結合分子あるいはそれらの変異体や断片から成る医薬品組成物として、組み合わせて使うことができる。例えば、目標とする予防、治療または診断の効果を達成するために、狂犬病ウイルス中和活性を有する複数の結合分子を1回の治療において組み合わせることができる。
【0033】
狂犬病ウイルスなどのRNAウイルスは、ウイルス複製の間、自身のRNAポリメラーゼを使用する。これらのRNAポリメラーゼは、エラーを起こしやすい傾向がある。このことによって、ウイルス感染の期間中にいわゆる擬似種が形成される結果となる。各々の擬似種は独自のRNAゲノムを有しており、その結果、ウイルスタンパク質のアミノ酸組成に相違が生じうる。もしもこのような変異がウイルスの構造タンパク質において生ずれば、ウイルスは、T細胞またはB細胞のエピトープの変化に起因して、潜在的にホストの免疫系から逃れることができる。こうしたことが起こる可能性は、ポリクローナル抗体の混合物(HRIG)によって個人を治療する場合よりも、例えばヒトモノクローナル抗体などの結合分子、2種類の混合物によって個人を治療する場合の方が高い。従って、狂犬病の治療のために2種類のヒトモノクローナル抗体を混合する際の必要条件は、2種類の抗体が、標的抗原、すなわち狂犬病ウイルス糖タンパク質上の互いに重複したりや競合したりしないエピトープを認識することである。この結果、狂犬病エスケープウイルスが出現する確率を最小化できる。その結果として、好適には、本発明の結合分子は、互いに異なる、互いに重複したり競合したりしない狂犬病ウイルスのエピトープ(例えば狂犬病ウイルスGタンパク質上のエピトープなど)と反応することができる。結合分子の混合物は、例えば狂犬病の暴露後予防に適する薬剤などの他の治療薬剤の少なくとも1つを、さらに含む可能性がある。
【0034】
通常、本発明による結合分子は、それぞれの結合パートナー(すなわち狂犬病ウイルスや、狂犬病ウイルスタンパク質などの狂犬病ウイルスのフラグメント)と、0.2×10-4 M、1.0×10-5M、1.0×10-6 M、1.0×10-7Mよりも小さく、好適には1.0×10-8 Mよりも小さく、より好適には1.0×10-9 Mよりも小さく、より好適には1.0×10-10 Mよりも小さく、更により好適には1.0×-11Mよりも小さく、特に1.0×10-12 Mよりも小さい親和定数(Kd値)で結合することができる。親和定数は、抗体アイソタイプによって変化する可能性がある。例えば、IgMアイソタイプの親和結合とは、少なくとも約1.0×10-7 Mの結合親和力を指す。親和定数は、例えば、表面プラズモン共鳴、すなわち、バイオセンサ・マトリックス内部のタンパク質濃度変化を検出することによって、生体特異的相互作用のリアルタイム分析を可能とする光学現象、を用いることによって、例えばBIACOREシステム(Pharmacia Biosensor AB, Uppsala, Sweden)を用いて測定することができる。
【0035】
本発明による結合分子は、精製された/単離された型の狂犬病ウイルスにも、精製されていない/単離されていない型の狂犬病ウイルスにも結合しうる。結合分子は、例えば試料中の可溶型の形態にある狂犬病ウイルスに結合しうるし、あるいは、例えばマイクロタイタープレート、膜、およびビーズなどの担体または基質に結合しているか付着している狂犬病ウイルスに結合しうる。担体または基質は、ガラス、プラスチック(例えばポリスチレン)、多糖類、ナイロン、ニトロセルロースまたはテフロン(登録商標)などでできていてよい。こうした支持材の表面は、固形でも多孔性でもよく、そして都合の良い任意の形状であってもよい。代わって、結合分子は、例えば狂犬病ウイルスのタンパク質または(ポリ)ペプチドなどの、狂犬病ウイルスのフラグメントにも結合しうる。一実施形態では、結合分子は狂犬病ウイルスGタンパク質またはそのフラグメントに特異的に結合することができる。狂犬病ウイルスタンパク質または(ポリ)ペプチドは、可溶性の形態である可能性もあるし、あるいは上記の通りに担体または基質に結合しているか付着している狂犬病ウイルスに結合する可能性もある。もう1つの実施例では、Gタンパク質で形質転換された細胞を、結合分子の結合パートナーとして用いることができる。
【0036】
本発明の好適な実施形態においては、本発明の結合分子は狂犬病ウイルスの感染性を中和する。これは、とりわけマウスニューロトロフィン受容体p75、神経細胞接着分子(CD56)、およびアセチルコリン受容体などの、宿主細胞上の受容体に狂犬病ウイルスが結合することを防ぐことによって、あるいは細胞の細胞質中へのRNA放出を阻害することによって、あるいはRNAの転写または翻訳を阻止することによって達成可能である。ある特定の実施形態においては、本発明の結合分子は、狂犬病ウイルスによる宿主細胞への感染を、前記結合分子の非存在下における狂犬病ウイルスによる宿主細胞への感染と比較して、少なくとも99%、少なくとも95%、少なくとも90%、少なくとも85%、少なくとも80%、少なくとも75%、少なくとも70%、少なくとも60%、少なくとも50%、少なくとも45%、少なくとも40%、少なくとも45%、少なくとも35%、少なくとも30%、少なくとも25%、少なくとも20%、または少なくとも10%阻止する。中和の測定は、例えば、“Laboratory techniques in rabies, Edited by: F. -X Meslin, M. M. Kaplan and H. Koprowski (1996), 4th edition, Chapters 15-17, World Health Organization, Geneva”に記載の通りに行うことができる。更に、本発明のヒト結合分子は、エンベロープを持つ狂犬病ウイルスの溶菌を補助することができる、補体結合性の結合分子である可能性がある。本発明のヒト結合分子はオプソニンとしても作用し、FcまたはC3b受容体を介した取り込みを促進するか、あるいは狂犬病ウイルスを凝集させて食作用を受けやすくするかのいずれかによって、狂犬病ウイルスの受ける食作用を増強させる可能性がある。
【0037】
好適な実施形態において、本発明による結合分子は少なくともCDR3領域を備えるが、該領域は配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、 配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23そして配列番号24からなる群から選択されたアミノ酸配列を含む。ある実施形態では、CDR3領域は重鎖のCDR3領域である。
【0038】
さらに別の実施形態では、本発明による結合分子は重鎖可変領域を備えるが、該領域は、基本的に、配列番号26、配列番号27、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番号44、配列番号45、配列番号46、配列番号47、配列番号48 および配列番号49からなる群から選択されたアミノ酸配列を含む。好適な実施形態では、本発明による結合分子は、基本的に配列番号335のアミノ酸1-119を含むアミノ酸配列から成る重鎖可変領域を備える。
【0039】
さらなる実施形態において、本発明による結合分子は、配列番号26のアミノ酸配列を含む重鎖可変領域および配列番号50のアミノ酸配列を含む軽鎖可変領域、配列番号27のアミノ酸配列を含む重鎖可変領域および配列番号51のアミノ酸配列を含む軽鎖可変領域、配列番号28のアミノ酸配列を含む重鎖可変領域および配列番号52のアミノ酸配列を含む軽鎖可変領域、配列番号29のアミノ酸配列を含む重鎖可変領域および配列番号53のアミノ酸配列を含む軽鎖可変領域、配列番号30のアミノ酸配列を含む重鎖可変領域および配列番号54のアミノ酸配列を含む軽鎖可変領域、配列番号31のアミノ酸配列を含む重鎖可変領域および配列番号55のアミノ酸配列を含む軽鎖可変領域、配列番号32のアミノ酸配列を含む重鎖可変領域および配列番号56のアミノ酸配列を含む軽鎖可変領域、配列番号33のアミノ酸配列を含む重鎖可変領域および配列番号57のアミノ酸配列を含む軽鎖可変領域、配列番号34のアミノ酸配列を含む重鎖可変領域および配列番号58のアミノ酸配列を含む軽鎖可変領域、配列番号35のアミノ酸配列を含む重鎖可変領域および配列番号59のアミノ酸配列を含む軽鎖可変領域、配列番号36のアミノ酸配列を含む重鎖可変領域および配列番号60のアミノ酸配列を含む軽鎖可変領域、配列番号37のアミノ酸配列を含む重鎖可変領域および配列番号61のアミノ酸配列を含む軽鎖可変領域、配列番号38のアミノ酸配列を含む重鎖可変領域および配列番号62のアミノ酸配列を含む軽鎖可変領域、配列番号39のアミノ酸配列を含む重鎖可変領域および配列番号63のアミノ酸配列を含む軽鎖可変領域、配列番号40のアミノ酸配列を含む重鎖可変領域および配列番号64のアミノ酸配列を含む軽鎖可変領域、配列番号41のアミノ酸配列を含む重鎖可変領域および配列番号65のアミノ酸配列を含む軽鎖可変領域、配列番号42のアミノ酸配列を含む重鎖可変領域および配列番号66のアミノ酸配列を含む軽鎖可変領域、配列番号43のアミノ酸配列を含む重鎖可変領域および配列番号67のアミノ酸配列を含む軽鎖可変領域、配列番号44のアミノ酸配列を含む重鎖可変領域および配列番号68のアミノ酸配列を含む軽鎖可変領域、配列番号45のアミノ酸配列を含む重鎖可変領域および配列番号69のアミノ酸配列を含む軽鎖可変領域、配列番号46のアミノ酸配列を含む重鎖可変領域および配列番号70のアミノ酸配列を含む軽鎖可変領域、配列番号47のアミノ酸配列を含む重鎖可変領域および配列番号71のアミノ酸配列を含む軽鎖可変領域、配列番号48のアミノ酸配列を含む重鎖可変領域および配列番号72のアミノ酸配列を含む軽鎖可変領域、配列番号49のアミノ酸配列を含む重鎖可変領域および配列番号73のアミノ酸配列を含む軽鎖可変領域、を備える。好適な実施形態において、本発明によるヒト結合分子は、配列番号335のアミノ酸1-119を含むアミノ酸配列を含む重鎖可変領域および配列番号337のアミノ酸1-107を含むアミノ酸配列を含む軽鎖可変領域を備える。
【0040】
好適な実施形態において、本発明の狂犬病ウイルス中和活性を有する結合分子は、IgG型で、好ましくはIgG1型で投与される。
【0041】
本発明の別の態様は、本願明細書で定義される結合分子の機能的変異体を含む。分子を本発明による結合分子の機能的変異体であるとみなすのは、該変異体が、狂犬病ウイルスまたはその断片に対して特異的に結合するために、本発明による親結合分子と競合することができる場合である。換言すれば、機能的変異体が、狂犬病ウイルスまたはその断片に結合する能力を依然として有する場合である。機能的変異体もまた、狂犬病ウイルスを中和する活性を有さなければならない。機能的変異体には、主要な構造配列において実質的に親結合分子に類似した誘導体であって、しかし親の結合分子には存在しない、化学的および/または生化学的な、試験管内あるいは生体内における修飾を有するような誘導体が含まれるが、これに限定されない。上記の修飾には、とりわけ、アセチル化、アシル化、ADPリボシル化、アミド化、フラビンの共有結合、ヘム部分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトールの共有結合、架橋、環化、ジスルフィド結合形成、脱メチル化、共有結合性架橋の形成、シスチンの形成、ピログルタミン酸の形成、ホルミル化、γ-カルボキシル化、グリコシル化、GPIアンカー形成、ヒドロキシル化、ヨウ化、メチル化、ミリストイル化、酸化、PEG化(pegylation)、タンパク質分解処理、リン酸化、プレニル化、ラセミ化、セレノ化、硫酸化、アルギニン化などのtRNAを介するタンパク質へのアミノ酸付加、ユビキチン化などが含まれる。
【0042】
あるいは、機能的変異体を、本発明において定義される結合分子であって、親の結合分子のアミノ酸配列と比較して1つ以上のアミノ酸に、置換、挿入、欠失またはそれらの組み合わせを含むアミノ酸配列を含むような結合分子とすることもできる。そのうえ、機能的変異体は、アミノ末端とカルボキシ末端の一方あるいは両方でアミノ酸配列の切断を受けている可能性がある。本発明による機能的変異体は、親結合分子と比較して、同一の結合親和性、あるいはより高いかより低いかの異なる結合親和性を有することが可能だが、依然として、狂犬病ウイルスまたはその断片に結合することができ、依然として狂犬病ウイルスを中和することができる。例えば、本発明による機能的変異体は、狂犬病ウイルスまたはその断片に対して、親結合分子と比較して、より高いかより低い結合親和性を有する可能性があり、あるいは、より高いかより低い狂犬病ウイルス中和活性を有する可能性がある。望ましくは、フレームワーク領域、超可変領域、とりわけCDR3領域を含む(ただし必ずしもこれらに限定されない)可変領域のアミノ酸配列に修飾が加えられる。概して、軽鎖および重鎖の可変領域は3つの超可変領域を備え、該可変領域は3つのCDRおよびより保存性の高い領域、いわゆるフレームワーク領域(FRs)、を備える。超可変領域は、CDR由来のアミノ酸残基および超可変ループ由来のアミノ酸残基を含む。本発明の範囲内にあることを意図された機能的変異体は、本願明細書で定義される親結合分子と、少なくとも約50%から約99%の、好適には少なくとも約60%から約99%の、より好適には少なくとも約70%から約99%の、更に好適には少なくとも約80%から約99%の、最も好適には少なくとも約90%から約99%の、とりわけ少なくとも約95%から約99%の、そしてとりわけ少なくとも約97%から約99%のアミノ酸配列の相同性を有する。とりわけ、ギャップまたはベストフィットなどの、当業者にとって周知のコンピュータ・アルゴリズムを、比較しようとするアミノ酸配列を最適にアライメントして、類似又は同一のアミノ酸残基を定義するために使用することができる。
【0043】
別の実施形態では、親結合分子が配列中にグリコシル化部位を含む場合に機能的変異体が生産されうるが、真核細胞で発現した場合には結合分子のグリコシル化が生じる結果となり、それ故に、抗原への結合が無効となる可能性がある。生産された機能的変異体はグリコシル化部位をもはや含まないが、狂犬病ウイルスへの結合能があり、依然として中和活性を有するであろう。
【0044】
機能的変異体は、親結合分子やその一部分を、エラープローンPCR法、オリゴヌクレオチド特異的突然変異導入法、および部位特異的突然変異導入法(ただしこれらに限らない)を含む、当技術分野では周知の一般的な分子生物学的手法によって変化させることによって得ることができる。そのうえ、機能的変異体は補体結合活性を有する可能性があり、エンベロープを持つ狂犬病ウイルスの溶菌を補助することができ、および/または、オプソニンとして作用し、そしてFcあるいはC3b受容体を介した取り込みを促進することによるか、または狂犬病ウイルスを凝集させてより食作用を受けやすくするかによって、狂犬病ウイルスの受ける食作用を増強させることができる。
【0045】
そのうえ更なる態様において、本発明は免疫コンジュゲートを含むが、この免疫コンジュゲートとは、本願明細書で定義される結合分子あるいはその機能的変異体を少なくとも1つ含み、そしてさらに少なくとも1つのタグ(とりわけ、検出可能な部分/薬剤など)を含む分子のことである。本発明においては、本発明による免疫コンジュゲートの混合液、あるいは本発明による少なくとも1つの免疫コンジュゲートと他の分子(例えば治療薬または他の結合分子または免疫コンジュゲート)との混合液も想定されている。更なる実施形態において、本発明の免疫コンジュゲートは、1つ以上のタグを備えることができる。これらのタグは、互いに同一でも異なっていても良く、結合分子に、非共有結合的に結合/共役させることができる。これらのタグを、ジスルフィド結合、水素結合、静電相互作用、組換え融合および立体配座結合(ただしこれらに限定されない)を含む共有結合によって、直接結合分子に結合/共役させることもできる。あるいは、これらのタグを、1つまたは複数の連結化合物(linking compound)によって結合分子に結合/共役させることもできる。結合分子にタグを共役させる技術は、当業者にとって周知である。
【0046】
本発明の免疫コンジュゲートのタグは治療薬であってもよいが、しかし、好適にはそれらは検出可能な部分/薬剤である。例えば、被験者が狂犬病ウイルスに感染しているかどうかを評価するために、または例えば、所定の治療計画の有効性を判断するための、臨床試験手順の一部として狂犬病ウイルス感染の発現や進行をモニターするために、検出可能な薬剤を含む免疫コンジュゲートを診断のために使用することができる。しかし、その他の検出および/または分析および/または診断の目的のためにこれらを使うこともできる。検出可能な部分/薬剤には、酵素、補欠分子族、蛍光物質、発光物質、生物発光物質、放射性物質、陽電子放出金属および非放射性の正磁性金属イオンが含まれるが、これらに限定されない。
【0047】
検出および/または分析および/または診断の目的のために結合分子をラベルするために使用されるタグは、使用する特定の検出/分析/診断技術、および/または方法に依存しており、例えば、とりわけ、(組織)サンプルの免疫組織化学的染色、フローサイトメトリー検出、レーザースキャンサイトメトリー検出、蛍光免疫法、酵素免疫測定吸着法(ELISA)、ラジオイムノアッセイ(RIA)、バイオアッセイ(例:中和アッセイ)、ウエスタンブロット法の利用などが挙げられる。組織サンプルを免疫組織化学的に染色するための好適なラベルは、検出可能な生成物の生成および局所堆積を触媒する酵素である。免疫組織化学的な視覚化を可能とするために結合分子に通常共役される酵素は周知であり、アセチルコリンエステラーゼ、アルカリ性ホスファターゼ、β‐ガラクトシダーゼ、グルコースオキシダーゼ、西洋わさびペルオキシダーゼおよびウレアーゼが含まれるが、これらに限定されない。視覚的に検出できる生成物を生成し、および堆積させる典型的な基質も、当技術分野において当業者には周知である。加えて、本発明の免疫コンジュゲートは、コロイド金を用いてラベルすることが可能であるか、または、例えば33P、32P、35S、3Hおよび125Iなどの放射性同位体)でラベルすることが可能である。本発明の結合分子を、直接的に、あるいは当技術分野において周知の方法によってキレート化剤を介して間接的に、放射性核種に付着させることが可能である。
【0048】
本発明の結合分子をフローサイトメトリー検出、レーザースキャンサイトメトリー検出、または蛍光免疫法に使用する場合は、蛍光体でラベルするのが有用である。本発明の結合分子に蛍光ラベルをつけるのに有用な多種多様な蛍光体は、当業者には周知である。本発明の結合分子を、ラベル付のアビジン、ストレプトアビジン、CaptAvidinまたはニュートラアビジンを使用した二次検出のために使用する際には、適切な補欠分子族の複合体を形成するために結合分子をビオチンでラベルすることが可能である。
【0049】
本発明の免疫コンジュゲートを生体内での診断上の目的で使用する際には、結合分子は、例えばガドリニウム・ジエチレントリアミンペンタ酢酸などの磁気共鳴画像(MRI)コントラスト剤、超音波コントラスト剤、あるいはX線コントラスト剤へ共役することによって、あるいはラジオアイソトープによりラベル化することによって、検出可能となる。
【0050】
さらにまた、本発明の結合分子、その機能的変異体、または免疫コンジュゲートを、固体の支持材に結合することも可能であり、特に狂犬病ウイルスまたはそのフラグメントの試験管内での免疫学的アッセイまたは精製に役立つ。このような固体の支持材は多孔性であっても非多孔性であってもよく、平面的であっても非平面的であってもよく、ガラス、セルロース、ポリアクリルアミド、ナイロン、ポリスチレン、ポリ塩化ビニルまたはポリプロピレンの支持材を含むが、これらに限定されない。ヒト結合分子を、例えば、免疫親和性クロマトグラフィーを目的として、例えばNHS活性化セファロースあるいはCNBr活性化セファロースなどのろ過媒体と有効に共役させることもできる。結合分子を、典型的にはビオチン-ストレプトアビジンの相互作用によって、常磁性のミクロスフィアに有効に結合することも可能である。ミクロスフィアは、狂犬病ウイルスまたはそのフラグメントを含むサンプルから、狂犬病ウイルスまたはそのフラグメントを単離するために使用することができる。別の例として、本発明のヒト結合分子はELISA用のミクロタイタープレートの表面に有効に結合することができる。
【0051】
本発明の結合分子またはその機能的変異体を、例えば精製を容易にするペプチドなどの、マーカー配列と融合することができる。具体例として、ヘキサヒスチジンタグ、ヘマグルチニン(HA)タグ、mycタグ、またはflagタグが挙げられるが、これらに限定されない。
【0052】
あるいは、抗体を第二の抗体と結合させて、抗体のヘテロコンジュゲートを形成させることができる。別の態様においては、本発明のヒト結合分子は、1つまたは複数の抗原に結合/付着されていてよい。好適には、これらの抗原は、結合分子-抗原の複合体を投与する対象の免疫系によって認識される抗原である。これらの抗原は互いに同一でもよいが、異なっていても良い。これらの抗原と結合分子を付着させる共役方法は、当技術分野では周知であって、架橋剤の使用を含むが、これに限定されるものではない。ヒト結合分子は狂犬病ウイルスに結合し、ヒト結合分子に付着した抗原は、複合体に対する強力なT細胞の攻撃を誘発し、それが最終的には狂犬病ウイルスの破壊につながる。
【0053】
免疫コンジュゲートを直接的に、あるいは例えばリンカーを介して間接的に結合させることによって、化学的に生成するのとは別に、免疫コンジュゲートを、本発明によるヒト結合分子および適切なタグが備わった融合タンパク質として生成することができる。融合タンパク質は、例えば、適切なタグをコードするヌクレオチド配列核酸分子を、ヒト結合分子をコードしているヌクレオチド配列とフレームを合わせて備える核酸分子を作製することによる、組み換え技術を用いるといった、当技術分野では周知の方法によって生産することができる。
【0054】
少なくとも本発明による結合分子またはその機能的変異体をコードする核酸分子を提供することは、本発明の別の態様である。そのような核酸分子は、例えば上記に述べたような親和性成熟の過程で、クローニングの目的のための中間体として用いられる可能性がある。好適な実施形態において、核酸分子は単離あるいは精製される。
【0055】
当業者であれば、これらの核酸分子の機能変異体もまた、本発明の一部に含まれることを意図されていることが理解できるであろう。機能的変異体は、親核酸分子から翻訳されるアミノ酸配列と同一のアミノ酸配列を提供するために、標準的な遺伝暗号を用いて直接翻訳されうる核酸配列である。
【0056】
好適には、核酸分子はCDR3領域を備える結合分子をコードするが、該領域は好適には重鎖CDR3領域であって、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、 配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23および配列番号24からなる群から選択されたアミノ酸配列を含む。
【0057】
更に好適には、核酸分子は、重鎖可変領域を含むヒト結合分子をコードするが、該領域は、配列番号26、配列番号27、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、 配列番号41、配列番号42、配列番号43、配列番号44、配列番号45、配列番号46、配列番号47、配列番号48 および配列番号49からなる群から選択されたアミノ酸配列を原則的に含む。特に好適な実施形態において、核酸分子は、配列番号335のアミノ酸1-119を含んだアミノ酸配列を基本的に含む重鎖可変領域を備えた結合分子をコードする。
【0058】
さらに別の実施形態では、核酸分子は、配列番号26のアミノ酸配列を含む重鎖可変領域および配列番号50のアミノ酸配列を含む軽鎖可変領域を備える結合分子をコードするか、配列番号27のアミノ酸配列を含む重鎖可変領域および配列番号51のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号28のアミノ酸配列を含む重鎖可変領域および配列番号52のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号29のアミノ酸配列を含む重鎖可変領域および配列番号53のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号30のアミノ酸配列を含む重鎖可変領域および配列番号54のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号31のアミノ酸配列を含む重鎖可変領域および配列番号55のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号32のアミノ酸配列を含む重鎖可変領域および配列番号56のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号33のアミノ酸配列を含む重鎖可変領域および配列番号57のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号34のアミノ酸配列を含む重鎖可変領域および配列番号58のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号35のアミノ酸配列を含む重鎖可変領域および配列番号59のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号36のアミノ酸配列を含む重鎖可変領域および配列番号60のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号37のアミノ酸配列を含む重鎖可変領域および配列番号61のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号38のアミノ酸配列を含む重鎖可変領域および配列番号62のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号39のアミノ酸配列を含む重鎖可変領域および配列番号63のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号40のアミノ酸配列を含む重鎖可変領域および配列番号64のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号41のアミノ酸配列を含む重鎖可変領域および配列番号65のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号42のアミノ酸配列を含む重鎖可変領域および配列番号66のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号43のアミノ酸配列を含む重鎖可変領域および配列番号67のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号44のアミノ酸配列を含む重鎖可変領域および配列番号68のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号45のアミノ酸配列を含む重鎖可変領域および配列番号69のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号46のアミノ酸配列を含む重鎖可変領域および配列番号70のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号47のアミノ酸配列を含む重鎖可変領域および配列番号71のアミノ酸配列を含む軽鎖可変領域をコードするか、配列番号48のアミノ酸配列を含む重鎖可変領域および配列番号72のアミノ酸配列を含む軽鎖可変領域をコードするか、あるいは、配列番号49のアミノ酸配列を含む重鎖可変領域および配列番号73のアミノ酸配列を含む軽鎖可変領域をコードする。好適な実施例において、核酸分子は、配列番号335のアミノ酸1-119を含んだアミノ酸配列を含む重鎖可変領域および配列番号337のアミノ酸1-107を含んだアミノ酸配列を含む軽鎖可変領域を備えたヒト結合分子をコードする。
【0059】
本発明の特定の実施形態において、本発明の結合分子の重鎖可変領域をコードする核酸分子は、配列番号74、配列番号75、配列番号76、配列番号77、配列番号78、配列番号79、配列番号80、配列番号81、配列番号82、配列番号83、配列番号84、配列番号85、配列番号86、配列番号87、配列番号88、配列番号89、配列番号90、 配列番号91、配列番号92、配列番号93、配列番号94、配列番号95、配列番号96および配列番号97からなる群から選択されたヌクレオチド配列を基本的に含む。望ましくは、本発明の結合分子の重鎖可変領域をコードする核酸分子は、基本的に配列番号334のヌクレオチド1-357を含むヌクレオチド配列を備える。
【0060】
本発明の更なる特定の実施形態において、本発明の結合分子の軽鎖可変領域をコードする核酸分子は、配列番号98、配列番号99、配列番号100、配列番号101、配列番号102、配列番号103、配列番号104、配列番号105、配列番号106、配列番号107、配列番号108、配列番号109、配列番号110、配列番号111、配列番号112、配列番号113、配列番号114、配列番号115、配列番号116、配列番号117、配列番号118、配列番号119、配列番号120および配列番号121からなる群から選択されたヌクレオチド配列を基本的に含む。望ましくは、本発明のヒト結合分子の軽鎖可変領域をコードする核酸分子は、基本的に配列番号336のヌクレオチド1-321を含んだヌクレオチド配列を備える。
【0061】
本発明による核酸分子を1つまたは複数含むベクター、すなわち核酸コンストラクトを供給することは、本発明の別の態様である。ベクターは、プラスミド、例えばF、Rl、RP1、Col、pBR322、TOL、Ti、など;コスミド;ファージ、例えばラムダ、ラムダ様、M13、Mu、P1、P22、Qb、T-even、T-odd、T2、T4、T7、など;植物ウイルス、例えば、とりわけ、アルファルファモザイクウイルス、ブロモウイルス、カピロウイルス、カーラウイルス、カーモウイルス、caulivirus、クロステロウイルス、コモウイルス、クリプトウイルス、ククモウイルス、ダイアンソウイルス、ファバウイルス、フィジーウイルス、フロウイルス、ジェミニウイルス、ホルデイウイルス、イラルウイルス、ルテオウイルス、machlovirus、マラフィウイルス、ネクロウイルス、ネポウイルス、phytorepvirus、植物ラブドウイルス、ポテックスウイルス、ポチウイルス、ソベモウイルス、テヌイウイルス、タバモウイルス、トブラウイルス、トマト黄化えそウイルス、トンブスウイルス、チモウイルスなど;または動物ウイルス、例えば、とりわけ、アデノウイルス、アレナウイルス科、バキュロウイルス科 、ビルナウイルス科、ブニヤウイルス科、カルキウイルス科、カルディオウイルス、コロナウイルス科 、コルチコウイルス科 、シストウイルス科、Epstein-Barrウイルス、エンテロウイルス、フィロウイルス科 、フラビウイルス科、口蹄疫ウイルス、ヘパドナウイルス科、肝炎ウイルス、ヘルペスウイルス科、免疫不全症ウイルス、インフルエンザウイルス、イノウイルス科、イリドウイルス科、オルトミクソウイルス科、パポバウイルス、パラミクソウイルス科、パルボウイルス科、ピコルナウイルス科、ポリオウイルス、ポリドナウイルス科、ポックスウイルス科、レオウイルス科、レトロウイルス、ラブドウイルス科、ライノウイルス、セムリキ森林熱ウイルス、テトラウイルス科、トガウイルス科、トロウイルス科、ワクシニアウイルス、水疱性口内炎ウイルスなど、から得ることができる。ベクターは、クローニングするために、および/または本発明のヒト結合分子を発現させるために使用することができ、遺伝子治療目的のためにさえも使用することができる。1つまたは複数の発現制御核酸分子を作用可能なように結合された、本発明による核酸分子を1つまたは複数含むベクターも、本発明の範囲に含まれる。ベクターの選択は、その後の組換え手順および使用するホストに依存する。宿主細胞へのベクターの導入は、とりわけ、リン酸カルシウム・トランスフェクション法、ウイルス感染、DEAE-デキストラン・トランスフェクション法、リポフェクタミン・トランスフェクション法またはエレクトロポレーションによって行なうことができる。ベクターは自律的に複製してもよいし、自身が組み込まれた染色体とともに複製してもよい。好適には、ベクターは、1つまたは複数の選択マーカーを含む。マーカーの選択は、選択する宿主細胞に依存するが、これは当業者にとって周知のように本発明に決定的なものではない。選択マーカーは、カナマイシン、ネオマイシン、ピューロマイシン、ハイグロマイシン、ゼオシン、単純ヘルペスウイルス由来のチミジンキナーゼ遺伝子(HSV-TK)、マウス由来のジヒドロ葉酸還元酵素遺伝子(dhfr)を含むが、これらに限定されるものではない。上述のヒト結合分子をコードする核酸分子を1つまたは複数含むベクターで、結合分子の単離に用いることができるタンパク質あるいはペプチドをコードする1つまたは複数の核酸分子が作用可能な状態で結合されたベクターもまた、本発明の範囲に含まれる。これらのタンパク質またはペプチドには、グルタチオン-S-トランスフェラーゼ、マルトース結合タンパク質、金属結合性のポリヒスチジン、緑色蛍光タンパク質、ルシフェラーゼおよびβ‐ガラクトシダーゼが含まれるが、これらに限定されない。
【0062】
前述のベクターの1つまたは複数のコピーを含むホストは、本発明の更なる対象である。好適には、ホストは宿主細胞である。宿主細胞には、哺乳類、植物、昆虫、真菌類または細菌起源の細胞が含まれるが、これらに限定されない。細菌細胞には、バチルス属、放線菌およびブドウ球菌のいくつかの種などのグラム陽性菌の細胞、またはエシェリヒア属(例えば大腸菌および緑膿菌)のいくつかの種などのグラム陰性菌の細胞が含まれるが、これらに限定されるものではない。真菌細胞のグループのなかでは、酵母細胞を用いるのが好適である。酵母内での発現は、とりわけピキア・パストリス(Pichia pastoris)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)およびハンゼヌラ・ポリモルファ(Hansenula polymorpha)などの酵母株を用いて達成可能である。さらにまた、ショウジョウバエおよびSf9の細胞などの昆虫細胞も宿主細胞として用いることができる。その他に、宿主細胞は、植物細胞であっても良い。形質転換(組み換え)植物あるいは植物細胞は、例えば、アグロバクテリウム媒介性の遺伝子導入、葉片の形質転換、ポリエチレングリコール誘導のDNA導入によるプロトプラスト形質転換、エレクトロポレーション、音波処理、マイクロインジェクションまたは遺伝子銃などの周知の方法によって、作製することできる。また、適切な発現系をバキュロウイルス系とすることができる。チャイニーズハムスター卵巣(CHO)細胞、COS細胞、BHK細胞またはボーズメラノーマ細胞などの哺乳類細胞を用いた発現系が、本発明において好適である。哺乳類細胞は、哺乳類由来の天然の分子と最も類似した翻訳後修飾を伴うタンパク質を供給する。本発明はヒトに投与される可能性のある分子を取扱うので、ヒトの発現系と完全に一致する発現系はとりわけ好まれる。従って、さらにより好適には、宿主細胞をヒト細胞とする。ヒト細胞の具体例として、とりわけ、HeLa細胞、911細胞、AT1080細胞、A549細胞、293細胞およびHEK293T細胞が挙げられる。好適な哺乳類細胞は、911細胞、または、欧州細胞培養コレクション(ECACC), CAMR, Salisbury, Wiltshire SP4 OJG, Great Britainに、1996年2月29日にNo.96022940として寄託され、登録商標PER.C6(商標登録)(PER. C6は、Crucell Holland B.V.の登録商標である)として市場に出された細胞系などのヒト網膜細胞である。本出願の目的上、「PER.C6」とは、No.96022940として寄託された細胞あるいはその祖先細胞、上流あるいは下流の継代細胞、および寄託された細胞の祖先細胞の子孫細胞、および前述した任意の細胞の派生株を意味する。
【0063】
他の好ましい実施形態では、ヒトプロデューサー細胞は、少なくとも、アデノウイルスEl領域を発現可能な形態でコードする核酸配列の機能的な部分を含む。さらにより好ましい実施形態において、前記宿主細胞はヒト網膜由来で、アデノウイルスEl配列を含む核酸を不死化されており、例えば欧州細胞培養コレクション(ECACC), CAMR, Salisbury, Wiltshire SP4 OJG, Great BritainにNo.96022940として1996年2月29日に寄託されPER.C6(商標登録)として市場に出された細胞系が挙げられる。宿主細胞内での組み換えタンパク質の生産は、当技術分野において周知の方法に従って実行できる。登録商標PER.C6(商標登録)という名前で市場に出された細胞の、生産したいタンパク質を生産する場所としての用途はWO00/63403に記載されており、その明細書全体を参照することにより本明細書に組み込まれる。
【0064】
本発明による結合分子や機能的変異体を生成する方法は、本発明の付加的な部分である。この方法は、a)結合分子やその機能的変異体の発現を起こすような条件の下で本発明によるホストを培養する工程、およびb)必要に応じて、発現した結合分子やその機能的変異体を回収する工程から構成される。発現した結合分子またはその機能的変異体を、無細胞抽出液から回収しても良いが、好適には培養液から回収する。例えば結合分子などのタンパク質を無細胞抽出液または培養液から回収する方法は、当業者にとって周知である。上記の方法によって得ることができる結合分子またはその機能的変異体も、本発明の一部である。
【0065】
あるいは、宿主細胞などのホスト内での発現とは別に、本発明による結合分子またはその機能的変異体を、従来のペプチド・シンセサイザによって、または本発明によるDNA分子に由来するRNA核酸を使用した無細胞翻訳系において、合成的に生産することが可能である。上記した合成的な生産方法や無細胞翻訳系によって得られる結合分子またはその機能的変異体もまた、本発明の一部である。
【0066】
もう1つの実施形態では、本発明による結合分子またはその機能的変異体を、例えばトランスジェニック・マウスまたはウサギなどの、ヒト免疫グロブリン遺伝子を発現するヒト以外のトランスジェニック哺乳動物によって作り出すことができる。好適には、ヒト以外のトランスジェニック哺乳動物は、上記のヒト結合分子の全部分または一部をコードする、ヒト重鎖トランス遺伝子およびヒト軽鎖トランス遺伝子を含むゲノムを有する。ヒト以外のトランスジェニック哺乳動物は、狂犬病ウイルスまたはその断片を精製または濃縮した調製物を用いて免疫化することができる。ヒト以外の哺乳動物を免疫化するためのプロトコルは、当技術分野において確立されている。“Using Antibodies: A Laboratory Manual, Edited by: E. Harlow, D. Lane (1998), Cold Spring Harbor Laboratory, Cold Spring Harbor, New York”、および“Current Protocols in Immunology, Edited by: J. E. Coligan, A.M. Kruisbeek, D. H. Margulies, E.M. Shevach, W. Strober (2001), John Wiley & Sons Inc., New York”を参照されたく、その開示は参照することにより本明細書に組み込まれる。
【0067】
更なる態様において、本発明は、例えば本発明によるヒトモノクローナル抗体またはその断片、または本発明による核酸分子などの結合分子を同定する方法を提供するが、該方法は狂犬病ウイルスに特異的に結合することが可能であり、そしてa)結合を起こす条件の下で、複製可能な遺伝子パッケージ(genetic package)の表面に存在する結合分子の集合体と狂犬病ウイルスまたはその断片とを接触させる工程と、b)狂犬病ウイルスまたはその断片に結合する複製可能な遺伝子パッケージを少なくとも1回選択する工程と、およびc)狂犬病ウイルスまたはその断片に結合する複製可能な遺伝子パッケージを単離および回収する工程とから構成される。
【0068】
選択の工程は、狂犬病ウイルスの存在下で実行することができる。狂犬病ウイルスは、単離型であってもよいし、または、例えば感染した個体の血清中および/または血液中に存在するような、非単離型であってもよい。別の実施形態においては、狂犬病ウイルスは不活性化される。あるいは、選択の工程は、例えば狂犬病ウイルスの細胞外部分、Gタンパク質などの狂犬病ウイルスに由来する1つまたは複数の(ポリ)ペプチド、これらのタンパク質または(ポリ)ペプチドから成る融合タンパク質などの、狂犬病ウイルスの断片が存在する状態で実行することができる。もう1つの実施形態では、狂犬病ウイルスGタンパク質をトランスフェクションされた細胞を選択の手順に用いる。
【0069】
また更なる態様において、本発明は、本発明による結合分子または核酸分子を得る方法を提供するものであって、前記方法は、a)例えば本発明によるヒトモノクローナル抗体またはその断片、または本発明による核酸分子などの結合分子を同定するという、上述の方法を実行する工程、およびb)回収した複製可能な遺伝子パッケージから、結合分子および/または結合分子をコードする核酸を単離する工程から構成される。結合分子または結合分子をコードする核酸分子を同定する上述の方法によって、新規のモノクローナル抗体を一旦立証または同定した後は、scFvまたはFabをコードするDNAを細菌または複製可能な遺伝子パッケージから単離することができ、標準的な分子生物学的な技術と組み合わせることによって、2価のscFv、すなわち所望の特異性を有する完全なヒト免疫グロブリン(例えば、 IgG、IgA、またはIgM)をコードするコンストラクトを作製することができる。これらのコンストラクトは、適切な細胞系にトランスフェクションすることが可能であり、完全なヒトモノクローナル抗体を生産することが可能である(Huls et al., 1999; Boel et al., 2000参照)。
【0070】
本願明細書において用いる「複製可能な遺伝子パッケージ」は原核生物のものであっても真核生物のものであっても良く、細胞、胞子、細菌、ウイルス、(バクテリオ)ファージおよびポリゾームを含む。好適な複製可能な遺伝子パッケージは、ファージである。例えば単鎖Fvなどのヒト結合分子は、複製可能な遺伝子パッケージ上に露出しており、すなわち、複製可能な遺伝子パッケージの外表面に位置する官能基または分子に付着している。複製可能な遺伝子パッケージは、結合分子をコードする核酸分子に結合した、選別すべきヒト結合分子から構成される選別ユニットである。核酸分子は生体内でも(例:ベクターとして)、試験管内でも(例:PCR法による転写および翻訳)複製可能でなければならない。生体内での複製は、(細胞に関しては)自立的に、(ウイルスに関しては)ホストの因子の助けを借りて、あるいは(ファージミドに関しては)ホストおよびヘルパーウイルス両者の助けを借りて行われる。ヒト結合分子のコレクションを露出する複製可能な遺伝子パッケージは、ヒト結合分子外因性の結合分子をコードする核酸分子を複製可能な遺伝子パッケージのゲノム中に導入し、複製可能な遺伝子パッケージの外表面に通常発現される内在性のタンパク質との融合タンパク質を形成することによって、形成される。融合タンパク質の発現、外表面への輸送およびアセンブリの結果、複製可能な遺伝子パッケージの外表面に、外因性の結合分子が呈示される。更なる態様において、本発明は、狂犬病ウイルスまたはその断片に結合可能であって、上記の同定方法によって得ることのできるヒト結合分子に関する。
【0071】
また更なる態様において、本発明は、狂犬病ウイルスを中和する活性を潜在的に有する結合分子を同定する方法に関し、前記方法は、(a)結合を起こす条件の下で、複製可能な遺伝子パッケージの表面に存在する結合分子の集合体と狂犬病ウイルスとを接触させる工程と、(b)狂犬病ウイルスに結合する結合分子を、狂犬病ウイルスに結合しない結合分子から単離および回収する工程と、(c)少なくとも1つの回収した結合分子を単離する工程と、および(d)単離した結合分子が狂犬病ウイルスに対する中和活性を有するか否かを確認する工程とから構成され、集合体aにおける狂犬病ウイルスは不活性化されていることを特徴とする。不活性された狂犬病ウイルスは、不活性化される前の時点で精製することができる。精製は、例えばグリセロール・クッション(glycerol cushion)による遠心分離などの、ウイルスに適した周知の精製方法によって実行できる。集合体aで不活性化された狂犬病ウイルスを、使用する前に適切な材料に固定することが可能である。あるいは、集合体aでの狂犬病ウイルスは、依然として活性を持つ可能性がある。別の代わりの実施形態において、例えば、Gタンパク質などの狂犬病ウイルスのポリペプチドなどの、狂犬病ウイルスの断片が工程aで使用される。さらにもう1つの実施形態においては、狂犬病ウイルスGタンパク質をトランスフェクションされた細胞が、狂犬病ウイルスに対する中和活性を有する可能性のある結合分子を選択するために使用される。本願明細書において示されるように、選択された中和抗体の個数は、狂犬病ウイルスGタンパク質を発現する細胞が選択方法に含まれる場合に、精製された狂犬病ウイルスGタンパク質および/または不活性化された狂犬病ウイルスのみを使用することを特徴とする選択方法と比較して多かった。
【0072】
更なる実施形態において、上述した、狂犬病ウイルスに対する中和活性を有する可能性のある結合分子を同定する方法は、重鎖可変生殖系列遺伝子3−30を含むヒト結合分子を単離および回収し、必要に応じて単離する工程からさらに構成される。当業者は、例えばとりわけヌクレオチド配列決定法などの当業者にとって周知の技術によって、特定の生殖細胞系遺伝子を同定することができる。重鎖可変生殖系列遺伝子3−30を含む結合分子を単離および回収する工程は、工程cの前または後に実行することができる。下記の通り、本発明で発見された狂犬病ウイルスを中和するヒトモノクローナル抗体の大多数は、この特定のVH生殖細胞系遺伝子を含む。
【0073】
抗体などの、(中和活性をもつ)結合分子を同定および獲得するためのファージディスプレイ法は、今では当業者には周知の確立された手法である。それらは、例えば米国特許第5,696,108号、 Burton and Barbas, 1994, de Kruif et al., 1995b、および“Phage Display: A Laboratory Manual. Edited by: CF Barbas, DR Burton, JK Scott and GJ Silverman (2001), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York”に記載されている。これら全ては、その全体を参照することにより本明細書に組み込まれる。
【0074】
ファージ・ディスプレイ・ライブラリの構築のためには、ヒトモノクローナル抗体の重鎖および軽鎖の可変領域のコレクションが、バクテリオファージ粒子、好適には繊維状バクテリオファージ粒子の表面に、例えば単鎖Fv(scFv)またはFabの形態で発現される(de Kruif et al., 1995b参照)。抗体フラグメントを発現するファージの大きなライブラリは、概して1.0×109より大きな抗体特異性を有しており、免疫化された個体または免疫化されていない個体のBリンパ球内で発現する免疫グロブリンV領域からアセンブルすることが可能である。本発明の特定の実施形態において、ヒト結合分子のファージライブラリ、好適にはscFvファージライブラリは、狂犬病のワクチン接種を受けた被験者、あるいは狂犬病ウイルスに暴露された被験者から得た細胞より単離したRNAによって作られている。RNAは、とりわけ、骨髄または末梢血、好適には末梢血リンパ球から単離できる。被験者は、ワクチン接種を受けた動物または狂犬病ウイルスに暴露された動物とすれば良いが、好適には、ワクチン接種を受けたヒトまたは狂犬病ウイルスに暴露されたヒトを被験者とする。好適には、被験者となるヒトはワクチン接種を受けている。上記の通りの、例えばscFvファージライブラリなどの、複製可能な遺伝子パッケージの表面上に存在するヒト結合分子の集合体は、本発明の別の態様である。
【0075】
あるいは、ファージ・ディスプレイ・ライブラリは、ライブラリ内に付加的な抗体多様性をもたらすために試験管内で部分的にアセンブルした免疫グロブリン可変領域から構築することができる(半合成ライブラリ)。例えば、試験管内でアセンブルした可変領域は、合成によって生産された、ランダム化されたDNA領域または部分的にランダム化されたDNA領域を、例えばCDR領域などの抗体特異性にとって重要である分子内の領域に含む。狂犬病ウイルス特異性ファージ抗体は、ライブラリから選択することができるが、これは、例えば狂犬病ウイルスの抗原などの標的抗原を固相に固定化して、続いて標的抗原をファージライブラリにさらすことによって、固相に結合した抗原に特異的な抗体の断片を呈示するファージの結合を可能にすることによってなされる。結合を形成しなかったファージは洗浄によって取り除かれ、結合を形成したファージは、大腸菌(E.coli)へ感染させた後に引き続いて増殖させるために、固相から溶出される。ファージが標的抗原に対して特異的に結合するのを十分に強化するためには、複数回にわたって選択し増殖させることが通常は必要となる。必要に応じて、ファージライブラリを標的抗原にさらす前に、ファージライブラリを固相に結合した標的ではない抗原にさらすことによって、最初にファージライブラリの数を減らすことができる。また、例えば狂犬病ウイルスのタンパク質または(ポリ)ペプチドの複合混合物などの複合抗原に対する結合、1つまたは複数の狂犬病ウイルスのタンパク質または(ポリ)ペプチドを発現している宿主細胞に対する結合、または(不活性)狂犬病ウイルス自体に対する結合に関してファージを選択することも可能である。抗原特異的なファージ抗体は、不活性化された狂犬病ウイルスの調製物を取り付けられた固相をファージ抗体ライブラリと共にインキュベートし、例えばファージのscFv 部分またはFab部分を、狂犬病ウイルス調製物のタンパク質/ポリペプチドに結合させることによって、ライブラリから選択することができる。インキュベーションを行い、そして結合していないファージおよびゆるく結合したファージを取り除くために複数回の洗浄を行った後、調製物にscFv 部分またはFab部分を結合したファージを溶出して大腸菌に感染させることにより、新規の特異性を増幅することが可能となる。通常、過剰に存在する結合していないファージの中から、求めるファージを単離するためには、1回または複数回の選択の工程を必要とする。あるいは、狂犬病ウイルスの周知のタンパク質または(ポリ)ペプチドを宿主細胞内で発現させて、そしてこれらの細胞をタンパク質または(ボリ)ペプチドに対して特異的なファージ抗体を選択するために用いることができる。こうした宿主細胞を用いたファージディスプレイ法は、スクリーニングの操作中に、標的分子を含まない宿主細胞、または標的と似ているが同一ではない非標的分子を含む宿主細胞を過剰に加えることにより、特異的に結合しない結合分子を取り除くことで拡張および改善することができ、それによって適切な結合分子を見つけられる可能性が大いに増す(このプロセスは、MAbstract(登録商標)プロセスと呼ばれる。MAbstract(登録商標)はCrucell Holland B. V.の登録商標であり、 米国特許第6,265,150号も参照されたく、参照することにより本明細書に組み込まれる)。
【0076】
また更なる態様において、本発明は、少なくとも1つの結合分子、少なくとも1つの機能的変異体またはその断片、少なくとも1つの本発明による免疫コンジュゲート、またはそれらの組み合わせから構成される組成物を提供する。組成物は、アルブミンまたはポリエチレングリコールなどの安定化分子、または塩をさらに含む可能性がある。好適には、使用する塩は、ヒト結合分子の所望の生物学的活性を保持し、望ましくない有害な効果をもたらさない塩である。必要に応じて、本発明によるヒト結合分子を、酸による作用、または結合分子を不活性化する恐れのあるその他の天然/非天然の条件から保護するために、物質で覆うことが可能である。
【0077】
また更なる態様において、本発明は、本発明によって定義された核酸分子を少なくとも1つ含む組成物を提供する。組成物は、例えば、塩(例;NaClまたは上述の塩)を含む水溶液などの水溶液、界面活性剤(例;SDS)、および/または他の適切な構成要素を含むことが可能である。
【0078】
さらにまた本発明は、少なくとも1つの本発明による結合分子、少なくとも1つの機能的変異体またはその断片、少なくとも1つの本発明による免疫コンジュゲート、少なくとも1つの本発明による組成物、またはそれらの組み合わせから構成される医薬品組成物に関連する。本発明による医薬品組成物はさらに、薬学的に許容される賦形剤を少なくとも1つ含む。
【0079】
好適な実施形態において、本発明による医薬品組成物は、少なくとも1つの結合分子を追加的に含む。すなわち、医薬品組成物を結合分子のカクテル/混合物とすることが可能である。医薬品組成物は本発明による結合分子を少なくとも2つ含むか、あるいは、本発明による結合分子を少なくとも1つ含むのに加えて少なくとも1つの更なる抗狂犬病ウイルス結合分子を含みうる。前記の更なる結合分子は、好適には、配列番号25のアミノ酸配列を含むCDR3領域を備える。配列番号25のアミノ酸配列を含むCDR3領域を備える結合分子は、キメラモノクローナル抗体またはヒトモノクローナル抗体またはその機能的な断片であれば良いが、好適には、ヒトモノクローナル抗体またはその機能的な断片である。ある実施形態において、結合分子は、配列番号273のアミノ酸配列を含む重鎖可変領域を備える。別の実施形態において、結合分子は、配列番号275のアミノ酸配列を含む軽鎖可変領域を備える。さらにもう1つの実施形態において、結合分子は、配列番号123および配列番号125のアミノ酸配列をそれぞれ含む重鎖および軽鎖を備える。医薬品組成物に含まれる複数の結合分子は、狂犬病ウイルスの、互いに異なり互いに競合しないエピトープとそれぞれ反応できなくてはならない。エピトープは、狂犬病ウイルスのGタンパク質上に存在する可能性があり、互いに異なる重複しないエピトープである可能性がある。結合分子は、高い親和性を有さなければならず、広い特異性を有さなければならない。好適には、それらの結合分子は、できるだけ多くの狂犬病ウイルスの固定毒株および街上毒株(street strain)を中和する。さらにより好適には、それらの結合分子はリッサウイルス属の他の遺伝子型、またはラブドウイルス科の他のウイルスにさえも中和活性を示し、一方で、他のウイルスや通常の細胞タンパク質とは交差反応性を示さない。好適には、結合分子は、カクテル中の他の結合分子のエスケープ変異体を中和することができる。
【0080】
本発明の別の態様は、狂犬病ウイルス中和活性を有する結合分子を少なくとも2つ含む医薬品組成物に関し、好適には、前記結合分子は、狂犬病ウイルスの互いに異なり互いに競合しないエピトープに結合できることを特徴とする、本発明による(ヒト)結合分子である。ある実施形態において、医薬品組成物は、狂犬病ウイルスGタンパク質の抗原部位Iに位置するエピトープと結合することが可能な、狂犬病ウイルス中和活性を有する第一の結合分子と、狂犬病ウイルスGタンパク質の抗原部位IIIに位置するエピトープと結合することが可能な、狂犬病ウイルス中和活性を有する第二の結合分子とを備える。狂犬病糖タンパク質の抗原構造は、最初にLafon氏ら (1983)によって定義された。抗原部位は、マウスのmAbおよびそれらのそれぞれのmAb抵抗性ウイルス変異体のパネルを使用して同定された。それ以来、mAb抵抗性ウイルス変異体の糖タンパク質におけるアミノ酸変異を同定することによって、抗原部位は決定されてきた(Seif et al. , 1985、Prehaud et al. , 1988、およびBenmansour et al. , 1991を参照)。狂犬病ウイルス中和活性を有するmAbの大多数は抗原部位IIに対する抗体であり(Benmansour et al. , 1991参照)、抗原構造IIは、アミノ酸34-42およびアミノ酸198-200を含む、不連続な立体構造エピトープである(Prehaud et al. , 1988参照)。抗原部位IIIはアミノ酸330-338の位置に存在する連続な立体構造エピトープで、2つの荷電残基K330およびR333を擁し、これらの荷電残基はウイルスの病原性に影響を及ぼす(Seif et al., 1985、Coulon et al., 1998、および“Dietzschold et al., 1983参照)。立体的な抗原部位Iは、ただ1つのmAb、509-6によって定義され、アミノ酸231位置に存在する(Benmansour et al., 1991、Lafon et al., 1983参照)。抗原部位IVは、互いに重複する直線状エピトープを擁することが知られている(Tordo, 1996、Bunschoten et al., 1989、Luo et al., 1997、およびNi et al., 1995参照)。Benmansour et al. (1991) も、アミノ酸342-343に位置し、抗原部位IIIに近接しているにもかかわらず抗原部位IIIとは別のマイナーサイトaの存在について記載している。CR-57エピトープを、現在周知である狂犬病糖タンパク質上の直線状および立体構造の中和エピトープとアライメントすることにより(図10)、CR-57エピトープは、単一のmAb、509-6によって定義される抗原部位Iと同じ場所に位置することが明らかになった。CR04-098のエスケープウイルスの糖タンパク質のヌクレオチド配列およびアミノ酸配列に基づいて考えると、この抗体によって認識されるエピトープは連続的な立体構造をとる抗原部位IIIと同一の領域に位置するように思われる。
【0081】
好ましい実施形態において、医薬品組成物は、配列番号25のアミノ酸配列を含むCDR3領域(好適には重鎖CDR3領域)を少なくとも備える、狂犬病ウイルス中和活性を有する第一の結合分子と、配列番号4、配列番号10、配列番号14、配列番号15、配列番号16および配列番号22から構成される群から選択されたアミノ酸配列を含むCDR3領域(好適には重鎖CDR3領域)を少なくとも備える、狂犬病ウイルス中和活性を有する第二の結合分子とを備える。該領域は、より好適には、狂犬病ウイルス中和活性を有する第二の結合分子は、配列番号14のアミノ酸配列を含むCDR3領域(好適には重鎖CDR3領域)を少なくとも備える。好適には、狂犬病ウイルス中和活性を有する第一の結合分子は、配列番号123および配列番号125のアミノ酸配列をそれぞれ含む重鎖および軽鎖を備え、狂犬病ウイルス中和活性を有する第二の結合分子は、配列番号335および配列番号337のアミノ酸配列をそれぞれ含む重鎖および軽鎖を備える。好適には、狂犬病ウイルス中和活性を有する第一の結合分子の重鎖および軽鎖は、それぞれ配列番号122および配列番号124によってコードされ、狂犬病ウイルス中和活性を有する第二の結合分子の重鎖および軽鎖は、それぞれ配列番号334および配列番号336によってコードされる。
【0082】
2つの結合分子から成る医薬品組成物で、結合分子の等電点(pI)が互いに異なる場合、両方の結合分子を最適に安定化させる適切なバッファを選択するのが困難である可能性がある。一方の結合分子の安定性が増すように組成物のバッファのpHを調整すると、もう一方の結合分子の安定性が減少する可能性がある。結合分子の安定性が減少してしまうと、あるいは不安定性が減少してしまう場合でさえも、結合分子の沈殿または集合、あるいは結合分子の自発的な分解につながる可能性があり、その結果として結合分子の機能が失われる可能性がある。従って、別の態様においては、本発明は少なくとも2つの結合分子から成る医薬品組成物を提供するが、該結合分子は好適にはヒト結合分子であり、該結合分子は、互いの等電点(pI)の差が約1.5、1.4、1.3、1.2、1.1、1.0、0.9、0.8、0.7、0.6、0.5、0.4、0.3よりも小さく、好適には0.25pI単位以下であることを特徴とする。等電点は、例えば等電点電気泳動などを用いて実験的に測定可能であり、または結合分子のアミノ酸配列に基づいて計算することもできる。1つの実施形態において、結合分子は本発明による結合分子であり、医薬品組成物は本発明による医薬品組成物である。好適には、結合分子はモノクローナル抗体であり、その例えばIgGl 抗体などのヒトモノクローナル抗体がある。好適には、結合分子は、ウイルス、細菌、酵母、真菌または寄生虫などの感染性の病原体を結合および/または中和する能力がある。ある実施形態において、結合分子は、狂犬病ウイルスなどのリッサウイルスを結合および/または中和することができる。特定の実施形態において、双方の結合分子の算出等電点は、8.0〜9.5の範囲、好適には8.1〜9.2の範囲、より好適には8.2〜8.5間の範囲内に収まる。好適には、2つの結合分子は、それぞれ、配列番号14および配列番号25の重鎖CDR3領域を有する。
【0083】
別の実施形態では、本発明は、2つ以上のヒト結合分子または他の動物の結合分子(抗体を含むがこれに限定されない)のカクテルを提供するが、少なくとも1つの結合分子は抗体ファージに由来するか複製可能なパッケージを表示する他の技術に由来し、そして少なくとも1つの結合分子はハイブリドーマ技術によって得られる。異なる技術を使用する際には、それぞれの結合分子が保管、処理およびその後の使用の際に十分安定であるような組成物を得るために、適合する等電点を有する結合分子を選択することも非常に有益である。
【0084】
別の実施形態では、本発明の医薬品組成物中に存在する結合分子は、互いの中和活性を向上させる。すなわち、組み合わせて用いると結合分子は相乗的に作用する。換言すれば、医薬品組成物は、相乗効果的な、狂犬病ウイルス中和活性および更にはリッサウイルス中和活性を示しうる。本明細書において用いる「相乗効果的な」という用語は、複数の結合分子を組み合わせて用いた場合に得られる複合効果は、単独で用いた場合に得られる効果を足し合わせたものよりも大きいことを意味する。本発明の医薬品組成物の構成要素の範囲および比率は、構成要素の個々の効力に基づいて決定すべきであり、試験管内での中和アッセイや、ハムスターなどの動物モデルを用いて検証するべきである。
【0085】
さらにまた、本発明による医薬品組成物は、少なくとも1つの他の治療薬、予防薬および/または診断用薬を含むことができる。前記のさらなる治療薬および/または予防薬は、リバビリンやインターフェロンαなどの抗ウイルス薬でとしてよい。
【0086】
本発明の結合分子または医薬品組成物は、ヒトに使用する前に、適切な動物モデル系において検証することができる。そのような動物モデル系には、マウス、ラット、ハムスター、サルなどが含まれるが、これらに限定されない。
【0087】
一般的に、医薬品組成物は、製造および保管の条件の下で、無菌的かつ安定でなくてはならない。本発明のヒト結合分子、変異体やその断片、免疫コンジュゲート、核酸分子、または組成物を、投与する前や投与時に、薬学的に許容される適切な賦形剤に再構成するために、粉末形態とすることも可能である。無菌的な注射剤を調製するための無菌粉末の場合には、より好適な調製方法は真空乾燥およびフリーズドライ(凍結乾燥)であり、この調製方法によって、活性成分に所望の追加的な所望の成分が加わった粉末を、事前に除菌したそれらの溶液から得ることができる。
【0088】
あるいは、本発明のヒト結合分子、変異体やその断片、免疫コンジュゲート、核酸分子、または組成物は、溶液中に存在していてもよく、投与する前や投与時に、薬学的に許容される適切な賦形剤を添加および/または混合することによって、単位投与量の注射剤型を提供することが可能である。好適には、本発明において使用する薬学的に許容される賦形剤は、高い薬剤濃度に適しており、適当な流動性を維持することが可能で、必要に応じて吸収を遅らせることができる。
【0089】
医薬品組成物の最適な投与経路の選択はいくつかの要因に影響され、それには、組成物中の活性分子の物理化学的性質、臨床病状の緊急性、および所望の治療効果に対する血漿中の活性分子濃度の関係が含まれる。例えば、本発明のヒト結合分子は、必要に応じて、急速に放出されてしまうことからそれらを保護する担体と共に調製されることが可能であり、例えば、インプラント、経皮貼布、およびマイクロカプセル化したデリバリーシステムを含む、放出制御製剤が挙げられる。エチレン酢酸ビニル、ポリ無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル類およびポリ乳酸などの、生分解性の、生体適合性ポリマーがとりわけ使用されうる。更に、ヒト結合分子の不活性化を阻止するような材料または化合物によって、ヒト結合分子を覆うか、あるいはヒト結合分子と共に投与することが必要である可能性がある。例えば、ヒト結合分子は、例えばリポソームまたは希釈剤などの適切な担体の形で、対象者に投与されうる。
【0090】
投与経路は、一般的には、経口投与と非経口投与の2つの主要なカテゴリーに分類することができる。本発明のヒト結合分子および医薬品組成物の投与は、傷口の内部およびその周辺、そして臀部の筋肉内に行なうのが好適である。ヒト結合分子および医薬品組成物の調製は、投与経路に依存する。
【0091】
更なる態様において、結合分子、機能的変異体、免疫コンジュゲート、組成物、または医薬品組成物は、薬剤として使用することができる。ゆえに、本発明のヒト結合分子、機能的変異体、免疫コンジュゲート、組成物、または医薬品組成物を使用した、リッサウイルス感染症の治療法および/または予防法は本発明の他の一部分である。リッサウイルスは、既存の遺伝子型の任意のウイルスであり得るが、好適には狂犬病ウイルスである。上述の分子や組成物は、狂犬病の暴露後予防に使用することができる。
【0092】
上述の分子や組成物は、狂犬病ウイルスの診断、予防および/または治療に役立つ他の分子と併せて使用することが可能である。それらは、試験管内、生体外または生体内で使用することができる。例えば、本発明のヒト結合分子、機能的変異体、免疫コンジュゲート、または医薬品組成物を、狂犬病に対するワクチンと共に投与することができる。あるいは、ワクチンを、本発明の分子または組成物を投与する前または後に投与しても良い。本発明の分子または組成物をワクチンと共に投与することは、暴露後予防に適している。狂犬病ワクチンは、精製ニワトリ胚細胞(PCEC)ワクチン(RabAvert)、ヒト二倍体細胞ワクチン(HDCV; Imovaxワクチン)または、吸着狂犬病ワクチン(RVA; rabies vaccine adsorbed)を含むが、これらに限定されない。
【0093】
典型的には、これらの分子は、本発明の組成物および医薬品組成物の中に、治療上または診断上効果のある量として調製される。投与計画は、最適な所望の効果(例えば治療効果)をもたらすように調製することが可能である。適切な用量範囲は、例えば0.1〜100IU/体重(kg)、好適には1.0〜50IU/体重(kg)、より好適には10〜30IU/体重(kg)(例えば20IU/体重(kg))である可能性がある。
【0094】
好適には、本発明の結合分子または医薬品組成物の単回ボーラス投与が行われる。本発明による分子および医薬品組成物は好適には無菌である。これらの分子および組成物を無菌にする方法は当技術分野では周知である。暴露後予防の投薬計画は、狂犬病ウイルスに対する予防接種を以前に受けていない個人に対し、暴露後0日、3日、7日、14日および28日の計5回、狂犬病ワクチンを三角筋に筋肉内投与するというものである。本発明によるヒト結合分子または医薬品組成物は、暴露後0日に、そうでなければ暴露後出来る限り早く、傷口の内部および周辺に投与するべきであり、残りの量については、ワクチンから離れた場所に筋肉内投与する。予防接種を受けていない個体に対しては、抗狂犬病ウイルスヒト結合分子の投与が勧められるが、予防接種を受けた個体に対しても、このような治療が必要な際には抗狂犬病ウイルスヒト結合分子を投与して良いことは、当業者には明らかである。
【0095】
別の態様において、本発明は、リッサウイルスによる感染症の結果生じた状態の診断、予防、治療、またはそれらの組み合わせのために用いる薬剤を調製する際の、結合分子又はその機能的変異体、本発明による免疫コンジュゲート、本発明による核酸分子、本発明による組成物又は医薬品組成物の用途に関する。リッサウイルスは、既存の遺伝子型の任意のウイルスであり得るが、好適には狂犬病ウイルスである。好適には、上述の分子は、狂犬病の暴露後予防のための薬剤の調製に用いられる。
【0096】
次に、少なくとも1つの本発明による結合分子、少なくとも1つの本発明による結合分子の機能的変異体、少なくとも1つの本発明による免疫コンジュゲート、少なくとも1つの本発明による核酸分子、少なくとも1つの本発明による組成物、少なくとも1つの本発明による医薬品組成物、少なくとも1つの本発明によるベクター、少なくとも1つの本発明によるホスト、またはそれらの組み合わせを含むキットもまた、本発明の一部である。必要に応じて、上記の本発明のキットの構成要素は、適切な容器にパックされ、適応となる病気の診断、予防および/または治療用にラベルを貼られる。上述の構成要素は、好適には無菌の水溶液として、あるいは好適には無菌の凍結乾燥製剤として、1回量あるいは複数回分の用量を入れられる容器(例えば密封されたアンプル、バイアル、ビン、注射器および試験管など)の中に保管されうる。容器は、ガラスまたはプラスチックなどの種々の材料から形成することが可能で、無菌のアクセスポートを有することが可能である(例えば、この容器は、皮下注射針を刺し通すことが可能な栓を有する静脈内用溶液バッグまたはバイアルである可能性がある)。このキットは、例えばリン酸緩衝食塩水、リンガー溶液およびブドウ糖溶液などの、薬学的に許容されるバッファを含む、より多くの容器からさらに構成されうる。キットは、その他のバッファ、希釈剤、フィルタ、針、注射器、1つまたは複数の適切なホストのための培養液を含む、商業的およびユーザの見地から望ましい他の材料を更に含むことが可能である。キットには、治療、予防または診断用の製品の商業用パッケージには慣習的に含まれる取扱説明書をつけることができ、そのような治療、予防または診断用の製品を使用するに際しての、指示、用法、用量、製造、投与、禁忌および/または警告などの情報が含まれる。
【0097】
現在、HRIG製品が狂犬病の暴露後予防のために使われている。成人に対する1回投与量である1500 IU (体重75 kgの人, 20 IU/kg)は、10mlの体積としてのみ利用可能である。現在入手可能な10mlの1回投与量には、合計1〜1.5グラムのIgGが含まれるため、より濃縮されたHRIG製品を得ることは不可能である。その観点では、現在のHRIG製品には、2つの欠点がある。第一に、推奨される1回投与量を傷口の内部およびその周囲に投与するのは解剖学的に実行不可能であること、そして第二に、現在のHRIGの1回投与量であっても投与の際にかなりの痛みが伴うことである。本発明は、望まれる場合には、約2mlまたはそれ未満の体積中に成人の1回分の全投与量を含む医薬品組成物を提供することにより、これらの欠点に対する解決策を提供する。この種の医薬品組成物は、例えば、狂犬病ウイルスを中和する能力を有する2つの結合分子を含む可能性があり、好適にはCR57およびCR04-098である。医薬品組成物は、薬学的に許容される賦形剤を更に含み、そして約2mlの体積を有する。より大きな体積とすることも可能であるが、より大きな体積を注射した際に生じる痛みの観点からあまり望ましくはない。2ml未満の体積とすることも可能である。医薬品組成物は、暴露後予防の成功のために必要な成人の全投与量(IU単位)を含む。ある実施形態において、医薬品組成物は、例えば、10mlの栓付きのready-to-useバイアル(I型ガラス)などの、10mlバイアルの中に保存される。10mlのバイアルを提供することによって、個人が大きな傷口表面積を示した場合に、医薬品組成物をより大きな体積へと希釈するという選択肢が生ずる。本発明はまた、医薬品組成物を含む容器(例えばバイアル)を少なくとも備えるキットを提供する。キットは、より大きな体積へと医薬品組成物を希釈するのに適した希釈剤を入れることができる第二の容器をさらに備えることが可能である。適切な希釈剤は、医薬品組成物の薬学的に許容される賦形剤、および食塩水を含むが、これらに限定されない。さらにまた、キットは、医薬品組成物を希釈するための取扱説明書、および/または、希釈の有無にかかわらず、医薬品組成物を投与するための取扱説明書を備えることが可能である。
【0098】
本発明は更に、サンプル中の狂犬病ウイルスを検出する方法に関し、該方法は、a) 診断上効果のある量の、本発明による結合分子、機能的変異体、または免疫コンジュゲートをサンプルに接触させる工程、およびb) 結合分子、機能的変異体、または免疫コンジュゲートが、サンプルの分子と特異的に結合しているか否かを決定する工程から構成される。サンプルは生物学的サンプルとすることができ、(潜在的に)感染している対象由来の血液、血清、組織または他の生物学的材料を含むが、これらに限定されない。(潜在的に)感染している対象はヒトであっても良いが、狂犬病ウイルスのキャリアである疑いのある動物についても、本発明のヒト結合分子、機能的変異体、または免疫コンジュゲートを用いて、狂犬病ウイルスの存在を検査することができる。検出方法により適したものとするために、サンプルを最初に操作することができる。操作とは、とりわけ、狂犬病ウイルスがタンパク質、(ポリ)ペプチドまたは他の抗原断片などの抗原構成要素へ分解するような方法で、狂犬病ウイルスを含む疑いのあるサンプルおよび/または狂犬病ウイルスを含むサンプルを処理することを意味する。好適には、ヒト結合分子、およびサンプル中に存在する可能性のある狂犬病ウイルスまたはその抗原構成要素との間で免疫複合体が形成されるような条件の下で、本発明の結合分子、機能的変異体、または免疫コンジュゲートをサンプルと接触させる。サンプル中に狂犬病ウイルスが存在することを示唆する免疫複合体の形成が少しでも生じるとすれば、適切な手段によって検出および測定する。この種の方法には、とりわけ、例えばラジオイムノアッセイ(RIA)、ELISA、免疫蛍光検査、免疫組織化学、FACS、BIACOREおよびウエスタンブロット分析などの、均一系および不均一系に結合させるイムノアッセイが含まれる。
【0099】
さらにまた、本発明の結合分子は、Gタンパク質などの狂犬病ウイルスタンパク質のエピトープを同定するために用いることができる。エピトープは、直線状であってもよいが、構造的および/または立体配置的であってもよい。ある実施形態では、狂犬病ウイルス由来のタンパク質(例えば狂犬病ウイルスGタンパク質)の、一連の重複するペプチド(例えば15-merペプチドなど)に対する結合分子の結合は、PEPSCAN分析を用いて分析することができる(とりわけ、WO 84/03564、WO 93/09872、Slootstra et al. 1996参照)。それぞれのペプチドに対するヒト結合分子の結合は、PEPSCANに基づいた酵素免疫測定法(ELISA)によって試験することができる。他の実施形態では、本発明のヒト結合分子と結合することができるペプチドを同定するために、狂犬病ウイルスタンパク質由来のペプチドを含むランダムなペプチドライブラリをスクリーニングすることができる。上記のアッセイにおいて、狂犬病ウイルスを中和することができるヒト結合分子を使用することによって、1つまたは複数の中和エピトープが同定される可能性がある。発見されたペプチド/エピトープは、ワクチンとして使用することも可能であるし、狂犬病の診断のために使用することも可能である。
【0100】
更なる態様において、本発明は、本発明の結合分子または機能的変異体が結合するエピトープとは異なる、好適には互いに重複しない、狂犬病ウイルスのエピトープに対する特異的な結合を同定するために、結合分子または結合分子の機能的変異体のスクリーニングを行なう方法を提供するものであって、該方法は、a) スクリーニング対象の結合分子または機能的変異体と、本発明の結合分子または機能的変異体と、狂犬病ウイルスまたはその断片(例えば狂犬病ウイルスGタンパク質)とを接触させる工程、およびb) スクリーニング対象の結合分子または機能的変異体が、狂犬病ウイルスまたはその断片に対する特異的な結合をめぐって、本発明による結合分子または機能的変異体と競合する能力があるかを測定する工程から構成される。もしも競合が測定されないのならば、スクリーニング対象の結合分子または機能的変異体は、異なるエピトープに結合する。上記のスクリーニング方法の特定の実施形態において、配列番号25のアミノ酸配列を含むCD3領域を備えた結合分子によって認識されるエピトープとは異なるエピトープへの結合能を有する、ヒト結合分子または機能的変異体を同定するために、ヒト結合分子またはその機能的変異体のスクリーニングを行なうことが可能である。好適には、エピトープは互いに重複しないか、または互いに競合しない。上記のスクリーニング方法を、同一のエピトープへの結合能を有する結合分子またはその機能的変異体を同定するために用いることができることは、当業者にとって明らかである。更なる工程においては、狂犬病ウイルスまたはその断片に対する特異的な結合をめぐって競合する能力を有さない、スクリーニングされた結合分子が、中和活性を有するかどうかを決定することが可能である。また、狂犬病ウイルスまたはその断片に対する特異的な結合をめぐって競合する能力を有する、スクリーニングされた結合分子が、中和活性を有するかどうかを決定することも可能である。前記スクリーニング方法によって得られた、中和能力を有する抗狂犬病ウイルス結合分子またはその機能的変異体は、本発明の一部である。前記スクリーニング方法において、「同一のエピトープに対して特異的に結合する」とは、本発明のヒト結合分子が結合するエピトープと実質的にまたは本質的に同一のエピトープに対して特異的に結合することをも意図する。本発明のヒト結合分子が狂犬病ウイルスに結合するのを阻止したり、またはこのような結合と競合したりする能力があるということは、典型的には、スクリーニング対象の結合分子は、本発明の結合分子によって免疫特異的に認識される狂犬病ウイルス上の結合部位と構造的に重複する、狂犬病ウイルスのエピトープまたは結合部位と結合することを示唆する。あるいは、このことは、スクリーニング対象の結合分子は、本発明の結合分子によって免疫特異的に認識される結合部位と十分に近いエピトープまたは結合部位に結合するため、本発明による結合分子が狂犬病ウイルスまたはその断片に結合することを立体的にまたは別の方法によって阻止する、ということを示唆する可能性もある。
【0101】
一般的に、競合阻害はアッセイを用いて測定されるが、このアッセイにおいては、抗原組成物、すなわち、狂犬病ウイルスまたはその(Gタンパク質などの)断片から成る組成物を、対照結合分子およびスクリーニング対象の結合分子と混合する。ある実施形態においては、対照結合分子を本発明によるヒト結合分子の1つとしてよく、スクリーニング対象の結合分子を本発明の別のヒト結合分子としてよい。別の実施形態では、対照結合分子は配列番号25のアミノ酸配列を含むCDR3領域を備えた結合分子としてよく、スクリーニング対象の結合分子を本発明によるヒト結合分子の1つとしてよい。さらに別の実施形態においては、対照結合分子を本発明のヒト結合分子の1つとしてよく、スクリーニング対象の結合分子を配列番号25のアミノ酸配列を含むCDR3領域を備えた結合分子としてよい。通常、スクリーニング対象の結合分子は過剰に存在する。このような単純な競合を研究するためには、ELISAに基づいたプロトコルを用いるのが適当である。ある実施形態では、対照結合分子を、様々な量のスクリーニング対象の結合分子と予め混合し(例えば、1:10、1:20、1:30、1:40、1:50、1:60、1:70、1:80、1:90または1:100)、一定の時間をおいてから、それらを抗原組成物に加えることが可能である。他の実施形態では、対照結合分子およびスクリーニング対象の結合分子を、抗原組成物に暴露している間に単純に混合することが可能である。いずれにしても、動物種またはアイソタイプに対する二次抗体を使用することによって、結合した対照結合分子のみを検出することができるのだが、この結合力は、実質的に同一のエピトープを認識するスクリーニング対象の結合分子が存在する場合には減少する。対照結合分子およびスクリーニング対象の結合分子の間での、結合分子の競合についての研究を行なう際には(動物種またはアイソタイプにかかわらない)、まず、対照結合分子を、例えばビオチン、酵素標識、放射性標識またはその後の同定を可能とする別の標識などの、検出可能なラベルで標識することが可能である。こうした場合には、標識化された対照結合分子とスクリーニング対象の結合分子とを様々な比率(例えば、1:10、1:20、1:30、1:40、1:50、1:60、1:70、1:80、1:90または1:100)で予め混合するかインキュベートし、そして、(適当な時間が経過した後に、任意で、)標識化された対照結合分子の反応性を検定し、この値を、競合する可能性のある結合分子がインキュベーションの際に含まれていない場合のコントロール値と比較する。再度言うが、このアッセイは、抗体のハイブリダイゼーションに基づく免疫学的アッセイの範囲内のうちの任意の1つであればよく、対照結合分子は標識を検出することによって検出され、例えば、ビオチン化した対照結合分子の場合にはストレプトアビジンを使用することによって、または酵素標識に関連する発色性基質を使用することによって(3,3'5,5'-テトラメチルベンゼン(TMB)基質とペルオキシダーゼ)、または単純に放射性標識を検出することによって検出される。対照結合分子と同一のエピトープに結合するスクリーニング対象の結合分子は、結合をめぐって効果的に競合することができ、従って対照結合分子の結合は有意に減少するのだが、このことは結合度の減少によっても明らかである。互いに異なり、互いに競合しないエピトープに結合する結合分子の場合には、結合度は減少しない。全く関連性のない結合分子が存在しない時の(標識化された)対照結合分子の反応性を、コントロール・ハイバリュー(control high value)とする。コントロール・ローバリュー(control low value)は、標識化された対照結合分子を、標識化されていない正確に同じ種類の対照結合分子と共にインキュベートすることによって得ることができるが、この際には、競合が生じて、標識化された対照結合分子の結合度は減少する。試験アッセイでは、スクリーニング対象の結合分子の存在下で、標識化された結合分子の反応性に有意な減少がみられたことから、同一のエピトープを認識する結合分子(すなわち、標識化された対照結合分子)と「交差反応」する結合分子の存在が示唆される。減少がみられない場合、結合分子は、互いに異なる互いに競合しないエピトープに結合する可能性がある。
【0102】
これらの競合アッセイによって同定された結合分子(「競合結合分子」)には、対照結合分子が結合するエピトープまたは結合部位に結合する、抗体、抗体断片および他の結合薬剤も含まれるし、同時に、対照結合分子が結合するエピトープに十分近く、スクリーニング対象の結合分子および対照結合分子間での競合的結合が生じる位置に存在するようなエピトープまたは結合部位に結合する、抗体、抗体断片および他の結合薬剤も含まれるが、これらに限定されるものではない。好適には、本発明の競合結合分子は、過剰に存在する場合には、特定の標的に対する対照結合分子の特異的結合を、少なくとも10%、好適には少なくとも25%、より好適には少なくとも50%、そして最も好適には少なくとも75〜90%またはそれ以上阻害する。本発明の結合分子とほぼ同一のエピトープ、実質的に同一のエピトープ、本質的に同一のエピトープ、または全く同一のエピトープに結合する、1つまたは複数の競合結合分子を同定することは、単純に技術的な事項である。競合結合分分子の同定は対照結合分子との比較によってなされるため、対照結合分子と同一のエピトープまたは実質的に同一のエピトープに結合する競合結合分子を同定するために、対照結合分子および競合結合分子が結合するエピトープを実際に決定する必要性は全くないということが理解できるであろう。あるいは、これらの競合アッセイによって同定された、互いに異なる互いに競合しないエピトープに結合する結合分子にもまた、抗体、抗体断片、および他の結合薬剤が含まれるが、これらに限定されない。
【0103】
別の態様では、本発明は、生物に病気を引き起こす感染性物質を中和する活性を潜在的に有する結合分子、または生物に病気を引き起こす感染性物質を中和する活性を潜在的に有する結合分子をコードする核酸分子を同定する方法を提供するものであって、該方法は、a) 複製可能な遺伝子パッケージの表面に存在する結合分子の集合体に、少なくとも生物に病気を引き起こす感染性物質のタンパク質を表面に発現している細胞を、結合を起こす条件の下で接触させる工程と、b) 生物に病気を引き起こす感染性物質のタンパク質を表面に発現している細胞に結合する結合分子を、前記細胞に結合しない結合分子から分離および回収する工程と、c) 少なくとも1つの回収した結合分子を単離する工程と、およびd) 単離された結合分子が生物に病気を引き起こす感染性物質に対する中和活性を有するかどうかを確認する工程とから構成される。生物に病気を引き起こす感染性物質のタンパク質を表面に発現している細胞は、タンパク質をトランスフェクションされた細胞であってよい。当業者は、この方法では、タンパク質以外の感染性物質の抗原も、うまく利用できることを知っている。特定の実施形態においては、細胞はPER.C6(商標登録)細胞である。しかし、BHK細胞、CHO細胞、NS0細胞、HEK293細胞、または911細胞などの他の(E1‐不死化)細胞系列を、タンパク質を発現するために使用することも可能である。ある実施形態では、結合分子はヒト結合分子である。感染性物質は、ウイルス、バクテリア、酵母、真菌または寄生虫である可能性がある。ある実施形態では、タンパク質は、感染性物質の表面に通常発現するタンパク質であるか、あるいは、表面に存在するタンパク質の少なくとも一部を含む。ある特定の実施形態においては、複製可能な遺伝子パッケージの表面に存在する結合分子のコレクションは、感染性物質のタンパク質を発現するために使用した細胞によってサブストラクション/対抗選択され、すなわち、この細胞は、感染性物質のタンパク質を細胞表面に発現しないという点を除いては、ステップaで使用された細胞と同一である。サブストラクション/対抗選択に使用する細胞は、トランスフェクションされていない細胞であってよい。あるいは、細胞に、感染性物質の対応するタンパク質と配列や構造が類似する、および/または高度な相同性を示す、および/または同じ科又は同じ属の感染性物質由来の、タンパク質またはその(細胞外)部分をトランスフェクションすることも可能である。
【0104】
本発明の別の態様は、狂犬病ウイルス中和活性を有する、本発明において定義される結合分子に関するものであって、このヒト結合分子は配列番号25のアミノ酸配列を含む重鎖CD3領域を備えることを特徴とし、更に、このヒト結合分子は少なくとも2500 IU/mgタンパク質の狂犬病ウイルス中和活性を有することを特徴とする。より好適には、前記ヒト結合分子は狂犬病ウイルス中和活性を有するが、該中和活性は、少なくとも2800 IU/mgタンパク質、少なくとも3000 IU/mgタンパク質、少なくとも3200 IU/mgタンパク質、少なくとも3400 IU/mgタンパク質、少なくとも3600 IU/mgタンパク質、少なくとも3800 IU/mgタンパク質、少なくとも4000 IU/mgタンパク質、少なくとも4200 IU/mgタンパク質、少なくとも4400 IU/mgタンパク質、少なくとも4600 IU/mgタンパク質、少なくとも4800 IU/mgタンパク質、少なくとも5000 IU/mgタンパク質、少なくとも5200 IU/mgタンパク質、少なくとも5400 IU/mgタンパク質である。結合分子の中和活性は、試験管内での中和アッセイ(改良RFFIT (rapid fluorescent focus inhibition test))によって測定された。このアッセイについては、下の実施例の項で詳しく説明する。
【0105】
ある実施形態において、結合分子は、配列番号273のアミノ酸配列を含む重鎖可変領域を備える。別の実施形態において、結合分子は、配列番号123のアミノ酸配列を含む重鎖を備える。結合分子の軽鎖可変領域は配列番号275のアミノ酸配列を含む可能性がある。結合分子の軽鎖は、配列番号125のアミノ酸配列を含む可能性がある。
【0106】
上述の、結合分子をコードする核酸分子も、本発明の一部である。好適には、核酸分子は、配列番号122のヌクレオチド配列を含む。加えて、核酸分子は、配列番号124のヌクレオチド配列をも含む可能性がある。核酸分子を含むベクターおよび該ベクターを含む宿主細胞も、本願明細書において提供される。好適には、宿主細胞はヒトの細胞などの哺乳類細胞である。ヒト結合分子の生産に適した細胞の例は、とりわけ、HeLa細胞、911細胞、AT1080細胞、A549細胞、293細胞、およびHEK293T細胞である。好適な哺乳類細胞は911細胞、もしくは、欧州細胞培養コレクション(ECACC), CAMR, Salisbury, Wiltshire SP4 OJG, Great Britainに、No.96022940として1996年2月29日に寄託され、登録商標PER.C6(商標登録)(PER. C6は、Crucell Holland B.V.の登録商標である)として市場に出された細胞系などの、ヒト網膜細胞である。本出願の目的上、「PER.C6」とは、No.96022940として寄託された細胞あるいはその祖先細胞、上流あるいは下流の継代細胞、および寄託された細胞の祖先細胞の子孫細胞、および前述した任意の細胞の派生株を意味する。
【実施例】
【0107】
本発明を説明するために、以下の実施例を提供する。実施例は、いかなる形であれ本発明の範囲を限定することを目的としない。
【0108】
実施例1
ヒト抗狂犬病抗体CR-57およびCR-JBのエピトープの認識
CR-57およびCR-JBと呼ばれるヒト抗狂犬病抗体が互いに重複したり互いに競合したりしないエピトープを認識するかどうかという問題に取り組むために、CR-57およびCR-JBと呼ばれるヒト抗狂犬病抗体のエスケープウイルスを生成した。CR-57およびCR-JBは、対応する抗体遺伝子の重鎖および軽鎖可変領域をコードする領域をpcDNA3002(Neo)という名の単一のヒトIgG1発現ベクターへ導入する操作を経て、基本的には記載された通りに生成した(Jones et al., 2003参照)。その結果得られたベクターであるpgSO57C11およびpgSOJBC11は、欧州細胞培養コレクション(ECACC), CAMR, Salisbury, Wiltshire SP4 OJG, Great Britainに、1996年2月29日にNo.96022940として寄託され、登録商標PER.C6(登録商標)として市場に出された細胞系由来の細胞において、一時的発現をさせるために使用した。これらの抗体の重鎖および軽鎖のヌクレオチド配列およびアミノ酸配列を、それぞれ配列番号122 〜 129に示す。狂犬病ウイルス株CVS-11(0.5ml)の連続希釈液(10−1〜10−8の幅の希釈液)を一定量(〜4 IU/ml) のCR-57またはCR-JB抗体(0.5ml)と共に、37℃/5%CO2の条件の下で1時間インキュベートしたのち、マウス神経芽腫細胞(MNA細胞)またはBSR細胞(ベビーハムスター腎様細胞系列;Baby Hamster Kidney-like cell line)を含むウェルに加えた。ヒトモノクローナル抗体CR-57またはCR-JBのいずれかの存在下で3日間の選択(セレクション)の後、潜在的なエスケープウイルスを含む培地(1ml)を回収し、更に使用するまで4℃で保存した。続いて、それらの細胞を4℃で20分間アセトン固定し、抗狂犬病N-FITC抗体複合体(Centocor社)を用いて37℃/5%CO2の条件の下で、オーバーナイトで染色した。ウェル毎の細胞増殖巣の数を免疫蛍光測定法により記録し、1〜6個の細胞増殖巣を含むウェルの培地を、ウイルスを増幅させるために選択した。全てのE57エスケープウイルスは、E57B1という例外を除き、単一の細胞増殖巣から生成された(E57B1は3個の細胞増殖巣)。EJBエスケープウイルスは、1個の細胞増殖巣(EJB3F)、3個の細胞増殖巣(EJB2B)、4個の細胞増殖巣(EJB2C)、5個の細胞増殖巣(EJB2E, 2F)、または6個の細胞増殖巣(EJB2D)からそれぞれ単離された。各々のエスケープウイルスを、成長特性に基づき、BSRまたはMNA細胞を用いてスモールスケールでまず増幅した。その後、これらの小さなウイルスバッチを、BSRまたはMNA細胞においてウイルスをラージスケールで更に増幅するために用いた。続いて、それぞれのエスケープウイルスバッチのタイター、およびウイルス中和アッセイでの使用に最適なエスケープウイルス希釈物(24時間後に80〜100%を感染させる濃度)を決定するために、増幅されたウイルスをMNA細胞において漸増した。
【0109】
E57(CR-57のエスケープウイルス)およびEJB(CR-JBのエスケープウイルス)の、それぞれCR-JBおよびCR-57との干渉効果を調べるために、修正迅速蛍光フォーカス抑制試験(RFFIT)アッセイを行った。そのために、CR-57または CR-JBを、1:5希釈から始めて、3倍連続希釈した。それぞれの希釈物に、狂犬病ウイルス(CVS-11株)を80〜100%の感染率を与える濃度で加えた。ウイルス/IgGの混合物を、MNA細胞に添加する前に、37℃/5%CO2の条件で1時間インキュベートした。感染後24時間(34℃/5%CO2の条件)の時点で、細胞を4℃で20分間アセトン固定し、抗狂犬病ウイルスN-FITC抗体複合体(Centocor社)を用いて最低3時間染色した。その後、蛍光顕微鏡を用いて狂犬病ウイルス感染についてウェルを分析し、50%エンドポイントの希釈物を決定した。50%エンドポイントの希釈物とは、このアッセイで、ウイルス感染が50%阻害される希釈物を指す。効力を計算するために、国際標準物質(laboratory of Standards and Testing (DMPQ/CBER/FDA)による標準物質である狂犬病イムノグロブリン・ロットR3)を各修正迅速蛍光フォーカス抑制試験アッセイ中に含めた。この標準物質の50%エンドポイントの希釈物は、2U/mlの効能に対応する。ヒトモノクローナル抗体CR-57およびCR-JBの、単独での中和効力、およびこれらの抗体を組み合わせた場合の中和効力を試験した。
【0110】
EJBウイルスは、もはやCR-JB またはCR-57によって中和されず(表1参照)、これら2つの抗体は両方とも狂犬病ウイルス糖タンパク質の類似した部位に結合し、狂犬病ウイルス糖タンパク質の類似した部位のアミノ酸変化をもたらすことを示唆している。E57ウイルスはもはやCR-57によって中和されなかったが、6つのE57ウイルスのうち4つは、より弱い効力ではあるが、依然としてCR- JBによって中和された(表1参照)。抗体CR-57および抗体CR-JBの(1:1IU/mgの比率での)混合物は、単一の抗体による結果と同様の結果を示した(データは示さず)。
【0111】
狂犬病ウイルス糖タンパク質に起こりうる変異を同定するために、EJBおよびE57エスケープウイルスの、各々の糖タンパク質のオープン・リーディング・フレーム(ORF)のヌクレオチド配列を決定した。各々のエスケープウイルスおよびCVS-11のウイルスRNAを、ウイルス感染MNA細胞から単離し、標準的なRT-PCR法によりcDNAに変換した。続いて、変異を同定するために、cDNAを利用して狂犬病ウイルス糖タンパク質のORFのヌクレオチド配列を決定した。
【0112】
E57およびEJBエスケープウイルスは共に、糖タンパク質の同じ領域に変異を示した(それぞれ、図1および図2参照;図1および図2に記載された全配列については、配列番号130〜151を参照)。これは、2つの抗体が重複するエピトープを認識することを意味する。上記より、カクテル中のCR-57とCR-JBの組み合わせは、中和耐性を有する変異株のエスケープを妨げず、ゆえに、狂犬病暴露後予防において理想的な免疫グロブリン製剤ではないと結論づけることが可能である。
【0113】
実施例2
狂犬病ワクチンを接種されたドナーの末梢血リンパ球を用いたScFvファージ・ディスプレイ・ライブラリの構築
狂犬病ワクチンの接種を受けた4人のヒトの被験者の静脈から、最後のブーストの一週間後に、50mlの血液を採取した。Ficollの細胞密度分画を用いて、これらの血液サンプルから末梢血リンパ球(PBL)を単離した。血清は−20℃で冷凍保存した。狂犬病ウイルス糖タンパク質をトランスフェクションされた293T細胞のFACS染色により、血清中に抗狂犬病抗体が存在に関して、陽性が確認された。有機相分離(TRIZOL(登録商標))、続いてエタノール沈殿を用いて、PBLから全RNAを調整した。得られたRNAをDEPCで処理した超純水に溶かし、その濃度を260nmでの吸光度測定によって決定した。その後、RNAを、100 ng/μlの濃度に希釈した。次に、以下のように、1μgのRNAをcDNAに変換した:全10μlのRNAに、13μlのDEPC処理した超純水および1μlのランダムな6量体(DNA)(500 ng/μl)を加え、得られた混合物を65℃で5分間加熱し、氷浴上で急冷した。そこへ、8μlの5×First-Strand buffer、2μlのdNTP(それぞれ10mM)、2μlのDTT(0.1M)、2μlのRnase阻害剤(40 U/μl)、および2μlのSuperscript(登録商標)III MMLV 逆転写酵素(200 U/μl)を加え、室温で5分間インキュベートした後、50℃で1時間インキュベートした。 反応は、熱不活性化、すなわち、混合物を75℃で15分間インキュベートすることにより、停止させた。
【0114】
得られたcDNA産物を、DEPC処理した超純水を用いて200μlの最終容積に希釈した。この得られたcDNA産物の希釈物を(10mMトリス・バッファー中で)50倍希釈した溶液を260nmで吸光度測定した結果、0.1という値が得られた。
【0115】
それぞれのドナーにつき、5μlから10μlの希釈されたcDNA産物を、免疫グロブリンのガンマ重鎖ファミリー、およびカッパまたはラムダ軽鎖の塩基配列をPCR増幅するためのテンプレートとして、特定のオリゴヌクレオチドプライマーと共に用いた(表2〜7参照)。PCR反応溶液は、希釈されたcDNA産物の他に、25pmolのセンスプライマーおよび25pmolのアンチセンスプライマーを含み、20mM Tris-HCl (pH8.4)、50mM KCl、2.5mM MgCl2、250μM dNTPs、および1.25ユニットのTaqポリメラーゼから成る最終体積50μlの溶液であった。96℃の温度のヒートリッド・サーマルサイクラーにおいて、得られた混合物を2分間で急速に融解した後、96℃で30秒、60℃で30秒、72℃で60秒のサイクルを30回繰り返した。
【0116】
第1回目の増幅では、17個の軽鎖可変領域のセンスプライマー(ラムダ軽鎖の11個(表2参照)およびカッパ軽鎖の6個(表3参照))のそれぞれを、HuCκと呼ばれるC-カッパ 5'-ACACTCTCCCCTGTTGAAGCT CTT-3'(配列番号152参照)、またはC-ラムダ定常領域であるHuCλ2 5'-TGAACATTCTGTAGGGGCCACTG-3'(配列番号153参照)およびHuCλ7 5'-AGAGCATTCTGCAGGGGCCACTG-3' (配列番号154参照)を認識するアンチセンスプライマーと組み合わせて用いて(HuCλ2およびHuCλ7アンチセンスプライマーは使用前に等モルで混合した)、4×17個の600塩基対程の生成物を得た。これらの生成物を2%アガロースゲルで精製し、Qiagenゲル抽出カラムを用いて、ゲルから単離した。単離した生成物それぞれの1/10量を、同じ17個のセンスプライマーを用いた上記と同一のPCR反応に用い、ここにおいては、それぞれのラムダ軽鎖センスプライマーをJラムダ領域に特異的な3種類のアンチセンスプライマーのうちの1つと組み合わせて使用し、それぞれのカッパ軽鎖センスプライマーをJカッパ領域に特異的な5種類のアンチセンスプライマーのうちの1つと組み合わせて使用した。2ラウンド目の増幅のためのプライマーは、ファージ・ディスプレイ・ベクターPDV-C06(図3および配列番号155参照)において直接クローニングすることを可能にするため、制限酵素認識部位を付け加えたものを用いた(表4参照)。この結果、4×63個のおよそ350塩基対の生成物が得られ、それらを10の分画に貯留した。分画を10個という数にした理由は、ライブラリ内で、様々な軽鎖の系統の自然な分布を維持するためで、ある特定の系統を過剰にまたは過少に発現させないためである。1つのライブラリ内での発現量を判断するために、1つの系統中での対立遺伝子の数を用いた(表5参照)。次の工程では、貯留した分画のうち2.5μg、およびPDV-C06ベクター100μgを、SalIおよびNotIで制限酵素処理し、ゲルから精製した。その後、以下の様にオーバーナイトで16℃でライゲーションを行った。500ngのPDV-C06ベクターに70ngの貯留した分画を加え、50mMのTris-HCl(pH7.5)、10mMのMgCl2、10mMのDTT、1mMのATP、25 mg/mlのBSAおよび2.5μlのT4DNAリガーゼ(400 U/μl)を含む、全容積50μlのライゲーション混合溶液とした。貯留した分画それぞれについて、この手順を引き続き実行した。ライゲーション混合物を、フェノール/クロロホルムで精製した後、クロロホルムで抽出してエタノールで沈殿したが、これは当業者にとって周知の方法である。得られたDNAを50μlの超純水に溶解し、各ライゲーション混合物につき2.5μlを2度、製造者(Stratagene社)のプロトコルに従って、40μlのTG1コンピテント病原性大腸菌にエレクトロポレーションした。形質転換体を37℃でオーバーナイトで培養し、それには、50ig/mlのアンピシリンと4.5%グルコースを加えた2TYアガーを含む計30個のシャーレ(貯留された分画それぞれにつき3つのシャーレ;シャーレの寸法:240mm×240mm)を用いた。アガープレートから形質転換体をこすり取ることによって軽鎖可変領域の(サブ)ライブラリが得られた。この(サブ)ライブラリは、Qiagen(登録商標)のQIAFilter MAXI prep kitを用いたプラスミドDNAの調製に直接用いた。
【0117】
それぞれのドナーについて、重鎖免疫グロブリン配列の増幅を、上記に示した軽鎖領域の場合と同じcDNA調合液を用いて、同様の2ラウンドのPCRの手順を踏んで、表6および表7に示したプライマーを用いて行なった。増幅の最初のラウンドでは9つのセンス方向プライマー(表6参照;重鎖可変領域の全ての系統をカバーする)のセットを使用し、そのセンス方向プライマーそれぞれをHuCIgGと呼ばれるIgGに特異的な定常領域のアンチセンスプライマー、5'-GTC CAC CTT GGT GTT GCT GGG CTT-3' (配列番号156)と組み合わせて用い、4×9個の650塩基対程の生成物を得た。これらの生成物を2%アガロースゲル上で精製し、Qiagenゲル抽出カラムを用いて、ゲルから単離した。単離した生成物それぞれの1/10量を、同じ9個のセンスプライマーを用いた上記と同一のPCR反応に用い、ここにおいては、それぞれの重鎖センスプライマーを、JH領域に特異的な4種類のアンチセンスプライマーのうちの1つと組み合わせて使用した。2ラウンド目に用いたプライマーには、軽鎖(サブ)ライブラリベクターに直接クローニングすることを可能にするため、制限酵素認識部位を付け加えた(表7参照)。この結果、各ドナーにつき36種類の、350塩基対ほどの生成物が得られた。これらの生成物を、それぞれのドナーにつき、使用された(VH)センスプライマーに基づいて9つの分画に貯留した。得られた生成物をQiagenPCR精製カラムを用いて精製した。次に、それぞれの分画をSfiIおよびXhoIで制限酵素処理し、同じ制限酵素で切断した軽鎖(サブ)ライブラリベクター中に、上記の軽鎖(サブ)ライブラリにおいての場合と同じ方法および容積で、ライゲーションした。あるいは、それぞれの分画をNcoIおよびXhoIで制限酵素処理し、同じ制限酵素で切断した軽鎖(サブ)ライブラリベクター中に、上記の軽鎖(サブ)ライブラリにおいての場合と同じ方法および容積で、ライゲーションした。ライゲーション後の精製、およびそれに続く、生成した最終的なライブラリの形質転換を、上記の軽鎖(サブ)ライブラリにおける方法と同様に実行し、またこの時点で、各ドナーのライゲーション混合物を各VH貯留液毎に混合した。形質転換体を、50μg/mlのアンピシリンと4.5%グルコースを加えた2TYアガーを含む27個のシャーレ(貯留された分画それぞれにつき3つのシャーレ;シャーレの寸法:240mm×240mm)で培養した。全バクテリアを、50ig/mlのアンピシリンと4.5%グルコースを含む2TY培地から回収し、15%(v/v)までグリセロールを加えて混合し、1.5mlの一定量中で、−80℃で凍らせた。各ライブラリのレスキューおよび選択は以下のように行なった。
【0118】
実施例3
狂犬病ウイルス糖タンパク質を特異的に認識する単鎖Fvフラグメントを有するファージの選択
抗体フラグメントの選択は、抗体ファージ・ディスプレイ・ライブラリ、一般的なファージディスプレイ技術、およびMAbstract(登録商標)技術を用いて、基本的には米国特許番号第6,265,150号およびWO 98/15833(これらは共に参照することによって本明細書に組み込まれる)に記載の通りに行なった。使用した抗体ファージライブラリは、2つの異なる半合成scFvファージライブラリ(JK1994およびWT2000)と、上の実施例2に記載した通りに調製した免疫scFvファージライブラリ(RAB-03-G01およびRAB-04-G01)である。第一の半合成scFvファージライブラリ(JK1994)は“de Kruif et al. (1995b)”に記載されており、第二の半合成scFvファージライブラリ(WT2000)は、基本的に“de Kruif et al. (1995b)”に記載された通りに作製した。簡潔には、このライブラリは、CDR領域に変異を組み込む変性オリゴヌクレオチドを用いて、重鎖および軽鎖V遺伝子の内部に変異を組み込まれた半合成体の形式を有する。VH3重鎖遺伝子のみを、カッパおよびラムダ軽鎖と組み合わせて使用した。重鎖のCDR1およびCDR3、そして軽鎖のCDR3を、"de Kruif et al. (1995b)"の記載と同様のPCRをベースにした方法で合成的に再現した。このように作製したV領域遺伝子を順次scFvの形式としてファージミドベクターにクローン化し、ファージライブラリを生成するために前述の方法で増幅した。更に、WO 02/103012(参照することによって本明細書に組み込まれる)に記載されている方法およびヘルパーファージを本発明で使用した。狂犬病ウイルス糖タンパク質を認識するファージ抗体を同定するために、全狂犬病ウイルス(狂犬病ウイルスピットマン・ムーア(Pitman-Moore)株)をβ‐プロピオラクトンでの処理によって不活性化したもの、狂犬病ウイルス糖タンパク質(狂犬病ウイルスERA株)を精製したもの、および/または狂犬病ウイルスGタンパク質(狂犬病ウイルスERA株)を発現するトランスフェクション細胞を用いて、ファージ選択実験を行った。
【0119】
Gタンパク質は以下のように狂犬病ウイルスERA株から精製した。ウイルス溶液に1/10容積の10%オクチル−β−グルコピラノシドを加え、穏やかに混合した。4℃で30分間のインキュベーションし、すぐにサンプルをSW51ローターで遠心分離(36,000rpm、4℃)した。上澄みを回収し、0.1M Tris/EDTAに対して4℃でオーバーナイトで透析を行った。続いて、糖タンパク質を透析チャンバーから回収、分割し、更なる使用の時まで−80℃で保管した。タンパク質の濃度を280nmでの吸光度測定によって決定し、Gたんぱく質の完全性をSDS- PAGEによって分析した。
【0120】
不活性化された狂犬病ウイルス全体または狂犬病ウイルスGタンパク質をリン酸緩衝食塩水(PBS)に溶解し、2〜3mlをマキシソープNunc-イムノチューブ (Nunc社)に加え、回転装置上で4℃でオーバーナイトでインキュベーションした。ファージライブラリの一定量(500μl、約1013cfu、CTヘルパーファージを用いて増幅(WO 02/103012参照))のブロッキングを、ブロッキングバッファー(PBS溶液中に2%Protifarを含む)中で、室温で1〜2時間行なった。ブロッキングされたファージライブラリをイムノチューブ(CR-57によって認識されるエピトープをブロックするために、CR-57 scFvの存在下または非存在下のいずれかでプレインキュベートしたもの)に加え、室温で2時間インキュベーションを行い、結合していないファージを除去するために洗浄バッファ(PBS溶液中に0.1% Tween-20 (Serva社)を含む)で洗浄した。その後、50mM グリシン-HCl pH2.2を1ml用いて、10分間室温でインキュベーションすることにより、結合したファージを抗原から溶出した。続いて、溶出されたファージと1M Tris-HCl pH7.5の0.5mlとを混合することによりpHを中和した。この混合物を、600nmでの吸光度が約0.3となるまで37℃で培養された大腸菌XL1-Blue株培養液5mlを感染させるために使用した。37℃で30分間、ファージをXL1-Blueバクテリアに感染させた。その後、混合物を10分間3200×gで室温で遠心分離し、バクテリアのペレットを0.5mlの2−トリプトン酵母エキス(2TY)培地中に再懸濁した。得られたバクテリアの懸濁液を、テトラサイクリン、アンピシリンおよびグルコースが加えられた2枚の2TYアガープレート上に分割した。プレートを37℃でオーバーナイトでインキュベーションした後、コロニーをプレートから擦り取り、“De Kruif et al. (1995a)”およびWO 02/103012の記載されたのと基本的に同様の方法で、濃縮したファージライブラリを作製するために使用した。簡潔に述べると、擦り取ったバクテリアを、アンピシリン、テトラサイクリンおよびグルコースを含む2TY培地に植菌するために用い、600nmでの吸光度が約0.3となるまで37℃で培養した。CTヘルパーファージを加え、バクテリアを感染させることを可能とした後に、培地をアンピシリン、テトラサイクリンおよびカナマイシンを含む2TYへ変更した。インキュベーションを、引き続き30℃でオーバーナイトで行った。翌日、バクテリアを遠心分離によって2TY培地から除去した後、培地中のファージをポリエチレングリコール(PEG)6000/NaClを用いて沈殿させた。最終的に、ファージを1%のウシ血清アルブミン(BSA)を含む2mlのPBSに溶かし、フィルター滅菌し、選択の次のラウンドのために使用した。
【0121】
狂犬病ウイルス糖タンパク質をトランスフェクトした細胞を用いることによってもファージの選択を行った。使用した細胞は、欧州細胞培養コレクション(ECACC), CAMR, Salisbury, Wiltshire SP4 OJG, Great Britainに、1996年2月29日にNo.96022940として寄託され、登録商標PER.C6(登録商標)として市場に出された細胞系由来の細胞である。これらの細胞を、本願明細書において今後、PER. C6(商標登録)細胞と呼ぶ。ここではまず、ブロッキングされたファージライブラリ(2ml)を、(DMEM/10% FBS中の)1×107のサブトラクター細胞に加え、回転装置上で4℃で1時間インキュベートした。サブトラクター細胞は、狂犬病ウイルス膜貫通ドメインおよび細胞質ドメインと外表面上で融合した、水疱性口内炎ウイルス(VSV)糖タンパク質の細胞外ドメインを発現したPER. C6(商標登録)細胞であった。このサブトラクションステップによって、VSV糖タンパク質、またはPER. C6(商標登録)細胞に特異的な抗原のいずれかを認識するファージを、ファージライブラリから取り除いた。ファージ/細胞の混合物を遠心して(4℃、500xgで5分)細胞に結合したファージを取り除き、上澄みを1*107のサブトラクター細胞を3ml含む新しいチューブに加えた。このサブトラクションステップは、各回とも上澄みを取り出して2回繰り返した。続いて、サブトラクションを行ったファージを狂犬病ウイルス糖タンパク質を発現したトランスフェクション細胞と共に、回転装置上で4℃で1.5時間インキュベートした。その前に、トランスフェクション細胞を、CR-57によって認識されるエピトープをブロックするため、CR-57 scFvの存在下または非存在下のいずれかでプレインキュベートした。インキュベーション後、1mlのDMEM/10%FBSで細胞を5回洗浄し(それぞれの洗浄につき、細胞を再懸濁し、新品のチューブに移した)、上記の通りにファージを溶出、処理した。
【0122】
典型的には、各ファージ抗体の単離を行なう前に2ラウンドの選択を行った。2ラウンド目の選択の後、各大腸菌コロニーを使用してモノクローナルファージ抗体を作製した。基本的には、それぞれのコロニーを96ウェルプレート上で対数増殖期に入るまで増殖させ、VCSM13ヘルパーファージを感染させた後に、ファージ抗体の生産をオーバーナイトで行なうことを可能とする。産生されたファージ抗体を、PEG/NaCl沈殿およびフィルタ滅菌し、ELISAによって、不活性化した狂犬病ウイルス全体、精製した狂犬病ウイルスGタンパク質の両方への結合を試す。選択によって、不活性化した狂犬病ウイルス全体および狂犬病ウイルスGタンパク質の双方への結合を示すファージ抗体が大量に得られた(下の実施例を参照)。上記の免疫ライブラリでは、2つの選択のストラテジーをとった。1つ目のストラテジーでは、不活性化したウイルスまたは精製したGタンパク質を1回目および2回目の選択のラウンドで用いて、2回の選択のラウンドの後に736個のファージ抗体を得た。2つ目のストラテジーでは、1回目のラウンドでは細胞表面に発現させた組み替えGタンパク質を用い、2回目のラウンドで不活性化したウイルスまたは精製したGタンパク質を用いることで、2回の選択のラウンドの後に736個のファージ抗体を得た。1つ目のストラテジーを用いて得られたファージ抗体は97種類であった一方で、2つ目のストラテジーでは70種類の独自のファージ抗体が得られた。1つ目のストラテジーを用いて得た97種類の独自のファージ抗体は18個の中和抗体を生じ、2つ目のストラテジーを用いて得た70種類の独自のクローンは33個の中和抗体を生じた。このことは、狂犬病ウイルス糖タンパク質をトランスフェクトした細胞、すなわち細胞表面に発現した組み替えGタンパク質、を抗原として用いて選択を行った場合、精製したGタンパク質および/または不活性化されたウイルスのみを用いて選択を行った場合と比較して、より多くの中和抗体を生じるらしいということを明確に示す。
【0123】
実施例4
狂犬病ウイルス糖タンパク質に特異的な単鎖ファージ抗体の検証
上記のスクリーニングによって選択した単鎖ファージ抗体の特異性、すなわち狂犬病ウイルスGタンパク質への結合をELISAによって検証した。加えて、単鎖ファージ抗体の、5%FBSへの結合も検証した。この目的のために、狂犬病ウイルスGタンパク質または5%FBSの調合液をMaxisorp(登録商標)ELISAプレートにコートした。コーティングの後、プレートをPBS/1%Protifar中で、室温で1時間ブロッキングを行なった。選択された単鎖ファージ抗体を、等容積のPBS/1%Protifar中で15分間インキュベートすることによって、ブロッキングされたファージ抗体を得た。プレートを空にし、ブロッキングされたファージ抗体をウェルに加えた。続いて1時間のインキュベーションへと進むことが可能となり、プレートを0.1% Tween-20を含むPBS中で洗浄し、結合したファージ抗体をペルオキシダーゼに結合した抗M13抗体を用いて(492nmでの吸光度測定を用いて)検出した。同時にコントロールとして、この手順を単鎖ファージ抗体を使用しないか、ネガティブコントロールとしてCD8 (SC02-007)に対する単鎖ファージ抗体を使用するか、またはポジティブコントロールとして狂犬病ウイルス糖タンパク質(scFvSO57)に対する単鎖ファージ抗体を用いることによって行った。表8に示すように、選択されたファージ抗体で、SC04-001、SC04-004、SC04-008、SC04- 010、SC04-018、SC04-021、SC04-026、SC04-031、SC04-038、SC04- 040、SC04-060、SC04-073、SC04-097、SC04-098、SC04-103、SC04- 104、SC04-108、SC04-120、SC04-125、SC04-126、SC04-140、SC04-144、SC04-146およびSC04-164と呼ばれるファージ抗体が、固定された精製狂犬病ウイルスGタンパク質への有意な結合を示した一方、FBSへの結合は観測されなかった。上記の通りに作製した、不活性化された完全な狂犬病ウイルスを用いたELISA分析においても、同一の結果が得られた(データは図示せず)。
【0124】
実施例5
狂犬病ウイルスに特異的なscFvの特徴
選択された特異的な単鎖ファージ抗体(scFv)のクローンからプラスミドDNAを得て、標準的な技術によってヌクレオチド配列を決定した。SC04-001、SC04-004、SC04-008、SC04-010、SC04-018、SC04-021、SC04-026、SC04-031、SC04-038、SC04-040、SC04-060、SC04-073、SC04-097、SC04-098、SC04-103、SC04-104、SC04-108、SC04-120、SC04-125、SC04-126、SC04-140、SC04-144、SC04-146およびSC04-164と呼ばれるscFvのヌクレオチド配列(クローニングのための制限酵素認識部位を含む)は、それぞれ、配列番号157、配列番号159、配列番号161、配列番号163、配列番号165、配列番号167、配列番号169、配列番号171、配列番号173、配列番号175、配列番号177、配列番号179、配列番号181、配列番号183、配列番号185、配列番号187、配列番号189、配列番号191、配列番号193、配列番号195、配列番号197、配列番号199、配列番号201および配列番号203に示される。SC04-001、SC04-004、SC04-008、SC04-010、SC04-018、SC04-021、SC04-026、SC04-031、SC04-038、SC04-040、SC04-060、SC04-073、SC04-097、SC04-098、SC04-103、SC04-104、SC04-108、SC04-120、SC04-125、SC04-126、SC04-140、SC04-144、SC04-146およびSC04-164と呼ばれるscFvのアミノ酸配列は、それぞれ、配列番号158、配列番号160、配列番号162、配列番号164、配列番号166、配列番号168、配列番号170、配列番号172、配列番号174、配列番号176、配列番号178、配列番号180、配列番号182、配列番号184、配列番号186、配列番号188、配列番号190、配列番号192、配列番号194、配列番号196、配列番号198、配列番号200、配列番号202および配列番号204に示される。
【0125】
VH遺伝子およびVL遺伝子の特性(“Tomlinson IM, Williams SC, Ignatovitch O, Corbett SJ, Winter G. V-BASE Sequence Directory. Cambridge United Kingdom: MRC Centre for Protein Engineering (1997) ”参照)、および狂犬病ウイルスGタンパク質に特異的に結合するscFvの重鎖CDR3組成を表9に示す。
【0126】
実施例6
狂犬病ウイルスに特異的なscFvによる試験管内での狂犬病ウイルスの中和(修正RFFIT)
選択されたscFvが狂犬病ウイルス感染を阻止する能力があるかを判断するために、試験管内での中和アッセイ(修正RFFIT)を行った。scFvの調合液を1:5希釈から始めて、3倍連続希釈した。狂犬病ウイルス(CVS-11株)を、80〜100%の感染率を与える濃度で各希釈物に加えた。ウイルス/scFvの混合物を、MNA細胞に添加する前に37℃/5%CO2の条件で1時間インキュベートした。感染後24時間(34℃/5%CO2の条件)の時点で、細胞を4℃で20分間アセトン固定し、抗狂犬病N-FITC抗体複合体(Centocor社)を用いて最低3時間染色した。その後、蛍光顕微鏡を用いて狂犬病ウイルス感染があるかについて細胞を分析し、50%エンドポイントの希釈物を決定した。50%エンドポイントの希釈物とは、このアッセイ中でウイルス感染が50%阻害される希釈物を指す(実施例1参照)。複数のscFvが狂犬病ウイルスに対して中和活性を示すことが確認された(表10参照)。
【0127】
加えて、選択されたscFvが、実施例1で作製したE57エスケープウイルス(E57A2、E57A3、E57B1、E57B2、E57B3およびE57C3)を中和する能力を有するかどうかを、上述した試験管内での中和アッセイ(修正RFFIT)を用いて調査した。いくつかのscFvは、E57エスケープウイルスに対して中和活性を示すことが確認された(表11Aおよび11B参照)。
【0128】
実施例7
scFvを用いた狂犬病ウイルスGタンパク質競合ELISA
互いに重複したり競合したりしないエピトープに結合する抗体を同定するために、狂犬病糖タンパク質の競合ELISA分析を実施した。精製された狂犬病ウイルス糖タンパク質(1mg/ml;狂犬病ウイルスERA株)を PBS中で1:1000希釈した液(50μl)を用いて、Nunc-Immuno(登録商標) Maxisorp F96プレート (Nunc)を4℃でオーバーナイトでコートした。コートされなかったタンパク質を洗い流し、ウェルを100μl PBS/1%Protifarを用いて室温で1時間ブロッキングした。続いて、ブロッキング溶液を取り除き、PBS/1%Protifar溶液中の精製されていない抗狂犬病ウイルスscFv(2倍希釈液)を50μl加えた。ウェルを100μlのPBS/0.05% Tween-20で5回洗浄した。その後、ビオチン化した抗狂犬病ウイルス競合IgGであるCR-57bioを各ウェルに50μlずつ加え、5分間室温でインキュベートし、その後ウェルを100μlのPBS/0.05% Tween-20で5回洗浄した。CR-57bioの結合を検出するために、ストレプトアビジン-HRP抗体(Becton Dickinson)の1:2000希釈物をウェルに50μl加え、室温で1時間インキュベートした。ウェルを再び上記の通りに洗浄し、OPD reagens (Sigma)を100μl加えることによって、ELISA分析をさらに進めた。1M H2SO4を50μl加えることによって反応を停止させ、492nmでの吸光度測定を行った。
【0129】
CR-57bio単独によって得られるシグナルは、CR-57のscFv型であるscFv SO57(SO57のヌクレオチド配列およびアミノ酸配列についてはそれぞれ、配列番号205および206を参照されたい)または、CR-JBのscFv型であるscFv SOJB(SOJBのヌクレオチド配列およびアミノ酸配列についてはそれぞれ、配列番号312および313を参照されたい)と共にインキュベートした場合、バックグラウンドのレベルにまで下がりうる。これは、scFv のSO57およびSOJBが、CR-57bioと同じエピトープや重なり合うエピトープと結合することによって、狂犬病ウイルス糖タンパク質との相互作用を巡ってCR-57bioと競合することを示す。対照的に、CD8に結合するscFvである、SC02-007と呼ばれる関連のないscFvは、結合を巡って競合しなかった。SC04-004、SC04- 010、SC04-024、SC04-060、SC04-073、SC04-097、SC04-098、SC04- 103、SC04-104、SC04-120、SC04-125、SC04-127、SC04-140、SC04- 144およびSC04-146と呼ばれる抗狂犬病ウイルスscFvもまた、CR-57bioと競合せず、これらのscFvはCR-57の認識するエピトープとは異なったエピトープに結合することを示している(図4参照)。
【0130】
以下の実験によっても同様の結果が得られた。まず、狂犬病ウイルス抗体CR-57を、狂犬病ウイルスGタンパク質でコートしたウェルに加えた。次に、競合するscFvを加えた。この設定では、scFvのアミノ酸配列中にVSVタグが存在することを利用して、抗狂犬病ウイルスscFvを抗VSV-HRPを用いて検出した(図5参照)。
【0131】
実施例8
選択された抗狂犬病ウイルス単鎖Fvからの、完全なヒト免疫グロブリン分子(ヒトモノクローナル抗狂犬病ウイルス抗体)の作製
SC04-001、SC04-008、SC04-018、SC04-040およびSC04-126と呼ばれるscFvの重鎖可変領域および軽鎖可変領域を、オリゴヌクレオチドを用いてPCRで増幅し、それぞれに制限酵素認識部位、および/またはIgG発現ベクターpSyn-C03-HCg1(配列番号277参照)およびpSyn-C04-Cl(配列番号278参照)における発現のための配列を付け加えた。VH遺伝子およびVL遺伝子を表12および表13に示したオリゴヌクレオチドを用いてそれぞれ増幅し、PCR産物をpSyn-C03-HCg1ベクターおよびpSyn-C04-Clベクターへそれぞれクローニングした。
【0132】
SC04-004、SC04-010、SC04-021、SC04-026、SC04-031、SC04-038、SC04-060、SC04-073、SC04-097、SC04-098、SC04-103、SC04-104、SC04-108、SC04-120、SC04-125、SC04-140、SC04-144、SC04-146およびSC04-164と呼ばれるscFvの重鎖可変領域および軽鎖可変領域をもまた、オリゴヌクレオチドを用いてPCRで増幅し、それぞれに制限酵素認識部位および/または、IgG発現ベクターpSyn-C03-HCg1およびpSyn-C05-Cκ(配列番号279参照)における発現のための配列を付け加えた。VH遺伝子およびVL遺伝子を表12および表13に示したオリゴヌクレオチドを用いてそれぞれ増幅し、PCR産物をpSyn-C03-HCa1ベクターおよびpSyn-C05-Cκベクターへそれぞれクローニングした。これらのオリゴヌクレオチドは、ライブラリの構築時に導入された生殖細胞系配列由来のいかなる変異をも修正するように設計されているのだが、これは、広いレパートリーの抗体遺伝子を増幅するために使用したオリゴヌクレオチドの数が限られていたために可能となった。全てのコンストラクトのヌクレオチド配列を熟練した当業者にとっては周知の標準的な技術によって検証した。
【0133】
結果として生じた、抗狂犬病ウイルスヒトIgG1重鎖をコードするpgG104-001C03、pgG104-008C03、pgG104-018C03、pgG104-040C03およびpgG104-126C03の発現コンストラクト、および対応する軽鎖をコードする該当するpSyn-C04-Vλ コンストラクトを組み合わせて293T細胞中で一時的に発現させ、IgG1抗体を含む上澄みを得た。抗狂犬病ウイルスヒトIgG1重鎖をコードするpgG104-004C03、pgG104-010C03、pgG104-021C03、pgG104-026C03、pgG104-031C03、pgG104-038C03、pgG104-060C03、pgG104-073C03、pgG104-097C03、pgG104-098C03、pgG104-103C03、pgG104-104C03、pgG104-108C03、pgG104-120C03、pgG104-125C03、pgG104-140C03、pgG104-144C03、pgG104-146C03およびpgG104-164C03の発現コンストラクト、および、対応する軽鎖をコードする該当するpSyn-C05-Vκコンストラクトを組み合わせて293T細胞中で一時的に発現させ、IgG1抗体を含む上澄みを得た。
【0134】
CR04-001、CR04-004、CR04-008、CR04-010、CR04-018、CR04-021、CR04-026、CR04-031、CR04-038、CR04-040、CR04-060、CR04-073、CR04-097、CR04-098、CR04-103、CR04-104、CR04-108、CR04-120、CR04-125、CR04-126、CR04-140、CR04-144、CR04-146およびCR04-164と呼ばれる抗体の、重鎖および軽鎖のヌクレオチド配列およびアミノ酸配列を標準的な技術によって決定した。続いて、通常免疫グロブリンで用いる標準的な精製方法(例えば、参照することにより本明細書に組み込まれるWO00/63403を参照)を用いて、組み換えヒトモノクローナル抗体をprotein-Aカラムで精製し、続いて脱塩カラム上でバッファ交換した。
【0135】
追加的に、CR04-098に関して、CR-57 およびCR-JBをそれぞれコードするpgSO57C11ベクターおよびpgSOJBC11ベクター(実施例1参照)に関して上述したのと同様にしてpgG104-098C10と呼ばれる単一のヒトIgG1発現ベクターを生成した。pgG104-098C10ベクターによってコードされる抗体CR04-098の重鎖および軽鎖のヌクレオチド配列およびアミノ酸配列を、それぞれ配列番号334〜337に示す。pgSO57C11ベクター(実施例1参照)およびpgG104-098C10ベクターを、CR-57およびCR04-098を、欧州細胞培養コレクション(ECACC), CAMR, Salisbury, Wiltshire SP4 OJG, Great Britainに1996年2月29日にNo.96022940として寄託され、登録商標PER.C6(登録商標)として市場に出された細胞系由来の細胞において安定的に発現させるためにそれぞれ使用した。安定的に生成したCR-57およびCR04-098の算出された等電点はそれぞれ、8.22および8.46である。実験的に観測された等電点は、CR-57については8.1から8.3の間で、CR04-098については9.0から9.2の間である。組み換えヒトモノクローナル抗体を、上述の通りに精製した。特に記していない限りは、CR04-001、CR04-004、CR04-008、CR04-010、CR04-018、CR04-021、CR04-026、CR04-031、CR04-038、CR04-040、CR04-060、CR04-073、CR04-097、CR04-098、CR04-103、CR04-104、CR04-108、CR04-120、CR04-125、CR04-126、CR04-140、CR04-144、CR04-146およびCR04-164については、上述の2つのベクターシステムによって一時的に発現された組み換えヒトモノクローナル抗体を利用し、CR57については、実施例1に記載の1つのベクターシステムによって一時的に発現された組み換えヒトモノクローナル抗体を利用した。
【0136】
実施例9
IgGによる狂犬病ウイルスGタンパク質競合ELISA
ヒトモノクローナル抗狂犬病ウイルスGタンパク質IgGが互いに重複したり競合したりしないエピトープに結合するか調べるため、競合実験を行った。狂犬病Gタンパク質でコートしたウェルを、複数の濃度(0〜50ig/ml)の非標識抗狂犬病ウイルスGタンパク質のIgGと共に、1時間室温でインキュベートした。その後、それぞれのウェルに、異なるビオチン化抗狂犬病ウイルスIgG(1ig/ml)を50μl加え、室温で5分間インキュベートし、直ちに100μlのPBS/0.05% Tween-20で5回洗浄した。続いて、ストレプトアビジン-HRP抗体(Becton Dickinson)の1:2000の希釈物を50μl加えて室温で1時間インキュベートし、上記の通りに洗浄し発色させた。非標識IgGの濃度の増加に伴うシグナルの減少は、2つの抗体が互いに競合していて、同じエピトープまたは重なり合うエピトープを認識することを示す。
【0137】
あるいは、狂犬病ウイルスGタンパク質(ERA株)によってコートしたウェルを、50ig/mlの非標識抗狂犬病ウイルスGタンパク質IgGと共に、1時間室温でインキュベートした。その後、ビオチン化CR57(0.5〜5ig/ml;飽和濃度以下)を各ウェルに加えた。さらなるステップは、上述の通りに行った。得られたシグナルを、ビオチン化CR57のみを使用した場合に得られるシグナルと比較した(図6;「競合物質なし」のグラフを参照)。図6から、ネガティブコントロールとして作用するCR02-428と呼ばれる抗体によってシグナルが減少してしまう可能性はないと推測できる。反対に、非標識CR57(ポジティブコントロール)またはCR-JBとの競合によって、シグナルはバックグランドのレベルにまで減少した。さらに、図6から、抗狂犬病ウイルスGタンパク質のIgGで、CR-57と有意に競合するものは存在しないことが推測でき、これは実施例7に記載したscFvの競合データと一致する。
【0138】
さらに、狂犬病ウイルスGタンパク質(ERA株)をトランスフェクトしたPER. C6細胞についてもフローサイトメトリー を用いて競合実験を行った。トランスフェクションされた細胞を20μlの非標識抗狂犬病ウイルスGタンパク質のIgG(50ig/ml)と共に20分間4℃でインキュベートした。細胞を1%BSAを含むPBSで洗浄した後、それぞれのウェルにビオチン化CR57(0.5〜5ig/ml;飽和濃度以下)を20μl加え、4℃で5分間インキュベートし、直ちに1%のBSAを含む100μlのPBSで2回洗浄した。続いて、ストレプトアビジン-PE(Caltag)の1:200の希釈物20μlと共に4℃で15分間インキュベートし、上記の通りに洗浄して発色させた。ビオチン化CR57によって得られるシグナルは、ネガティブコントロール抗体CR02-428によっては有意に減少しなかった(図7参照)。対照的に、非標識CR57(ポジティブコントロール)またはCR-JBとの競合によって、シグナルはバックグランドのレベルにまで減少した。抗狂犬病ウイルスGタンパク質のIgGで、CR-57と有意に競合するものは、シグナルを約30%まで減少させたCR04-126を除いては存在しなかった(図7参照)。CR04-126は、ELISA分析においては競合を示さなかった(図6参照)。これは、糖タンパク質が抗体に呈示される方法が、ELISA実験とFACS実験では異なることに起因する可能性がある。CR04-126の結合は糖タンパク質の立体構造により依存的である結果、FACSに基づく競合アッセイではCR04-126による競合効果が観察され、ELISAに基づく競合アッセイでは観察されなかった可能性がある。また、CR04-008およびCR04-010はFACSに基づく競合アッセイではシグナルを約50%まで減少させたが、これらの抗体がCR57と競合する可能性があることを示している。しかしCR04-010については、このことはscFv競合データによっても、ELISAに基づく競合アッセイによっても確認されなかった。他のIgGについては、FACSのデータは、それに対応するELISAのデータと、scFvとIgG双方について一致した。
【0139】
実施例10
狂犬病ウイルスの試験管内での中和の際の抗狂犬病IgGの加法的/相乗的効果(修正RFFIT)
抗狂犬病ウイルスGタンパク質のIgGが狂犬病ウイルスの中和において加法的または相乗的効果をもたらすかを決定するために、様々な組み合わせのIgGを試験した。まず、修正RFFITによって、それぞれの抗体単独での(IU/mgの単位での)効力を決定した(実施例1参照)。次に、IU/mgの量を等しくそろえて抗体の組み合わせを調製し、修正RFFITの試験を行った。抗体の組み合わせ各々の効力を決定し、期待される効力と比較することができる。もしも、抗体の組み合わせの効力が、組み合わせの中に存在する個々の抗体の効力の和と同じ値になるならば、これらの抗体は相加的な効果を有する。もしも抗体の組み合わせの効力の方が大きいのならば、抗体は狂犬病ウイルスの中和において相乗的な効果を有する。
【0140】
あるいは、相加的あるいは相乗的効果を以下の実験によって決定することも可能である。まず、検討を行なう抗体、例としてCR-57およびCR04-098、の効力を標準のRFFITによって決定する(“Laboratory techniques in rabies, Edited by: F. -X Meslin, M. M. Kaplan and H. Koprowski (1996), 4th edition, Chapter 15, World Health Organization, Geneva”参照)。次に、抗体をIU/mlに基づいて1:1で混合する。この抗体混合物について、同じ濃度の個々の抗体と共に、6回の独立したRFFIT実験を行なうことによって、中和の50%エンドポイントを決定した。続いて、Chou et al.(1984)の記載に従って、式、CI = (C1/Cx1) + (C2/Cx2) + (C1C2/Cx1Cx2) を用いて、抗体混合物のcombination index(CI)を決定した。C1およびC2は、組み合わせて使用したときに50%の中和をもたらすモノクローナル抗体1およびモノクローナル抗体2の(μg単位の)量であり、Cx1およびCx2は、単独で使用したときに50%の中和をもたらすモノクローナル抗体1およびモノクローナル抗体2の(μg単位の)量である。CI = 1はモノクローナル抗体の相加的効果を示し、CI < 1は相乗的効果、CI > 1は拮抗効果を示す。
【0141】
実施例11
組み換えヒト抗狂犬病ウイルス抗体によって認識されるエピトープの、PEPSCAN-ELISAによる同定
狂犬病ウイルスERA株のGタンパク質の細胞外ドメインより、15mer直線状ペプチドおよびループ状/環状ペプチドを合成し(狂犬病ウイルスERA株の糖タンパク質Gの完全なアミノ酸配列については配列番号207を参照されたい、細胞外ドメインはアミノ酸20-458からなる;EMBLデータベースにおける狂犬病ウイルスERA株の糖タンパク質のタンパクIDはJ02293である)、前述のように、クレジットカード状のミニPEPSCANカード(1枚のカードにつき455個のペプチドのフォーマット)を用いてスクリーニングを行った(Slootstra et al. , 1996; WO 93/09872)。全てのペプチドをアミノ酸末端でアセチル化した。全てのループ状のペプチドにおいて、位置2および位置14をシステインで置き換えた(acetyl-XCXXXXXXXXXXXCX-minicard)。準備したペプチド中で、位置2および位置14のシステインの近傍に他のシステインが存在した場合、それらをアラニンで置き換えた。ループ状のペプチドは標準的なFmoc法で合成し、スカベンジャー付きのトリフルオロ酢酸を用いて脱保護した。続いて、脱保護したペプチドを、1,3- ビス(ブロモメチル)ベンゼンを含む炭酸水素アンモニウム(20mM、pH7.9/アセトニトリル(1:1(v/v))溶液0.5mM中で、カードの上で反応させた。カードを、溶液に完全に浸った状態で30〜60分間穏やかに振盪した。最後に、過剰のH2Oを用いてカードを十分に洗浄し、PBS中に1%SDS/0.1%β‐メルカプトエタノールを含む破壊バッファ(pH7.2)中において70℃で30分間超音波処理を行い、続いてH2O中で45分間超音波処理を行った。ヒトモノクローナル抗体を上述の通りに作製した。これらの抗体の、各直線状ペプチドおよびループ状ペプチドへの結合を、PEPSCANに基づいた酵素免疫測定法(ELISA)を用いて検証した。共有結合したペプチドを含む455個のウェルを有するクレジットカード状のポリプロピレンカードを、抗体(10ig/ml;5%のウマ血清(v/v)および5%のオボアルブミン(w/v)を含むブロッキング液中に溶解)と共にインキュベートした(4℃、オーバーナイト)。洗浄後、ペプチドを抗ヒト抗体ペルオキシダーゼ(1/1000希釈物)と共にインキュベートし(25℃で1時間)、続いて、ペルオキシダーゼ基質を洗浄後、2,2'-アジノビス(3-エチルベンゾチアゾリン-スルホン酸) (ABTS)および2il/mlの3%H2O2を加えた。(直線状ペプチドおよびループ状ペプチドに対する)コントロールを、抗ヒト抗体ペルオキシダーゼのみと共にインキュベートした。1時間後、発色現象を測定した。ELISAの発色現象を、CCDカメラおよび画像処理システムを用いて定量化した。この装置は、CCDカメラおよび55mmレンズ(ソニー製CCDビデオカメラXC-77RR、ニコン製“micro-nikkor 55 mm f/2.8 レンズ”)、カメラアダプター(ソニー製カメラアダプターDC-77RR)、および画像処理ソフトウェアパッケージOptimas, version 6.5(Media Cybernetics, Silver Spring, MD 20910, U.S.A.)から構成される。Optimasは、pentium II コンピュータシステム上で作動した。
【0142】
上記の通り、合成された15merの直線状およびループ状/環状のペプチドへの、ヒト抗狂犬病ウイルスGタンパク質モノクローナル抗体の結合を検証した。ペプチドが抗体に適切に結合していると考えられるのは、OD値(光学濃度)が(抗体あたりの)全ペプチドのOD値の平均の2倍以上の場合である。CR57、CRJB、およびCR04-010と呼ばれるヒトモノクローナル抗体の、狂犬病ウイルスERA株の糖タンパク質Gの細胞外ドメイン直線状ペプチドへの結合の結果を表14に示す。それぞれの抗体と有意な結合を示す領域を灰色でハイライトしてある(表14参照)。
【0143】
抗体CR57は、SLKGACKLKLCGVLG (配列番号314)、LKGACKLKLCGVLGL (配列番号315)、KGACKLKLCGVLGLR (配列番号316)、GACKLKLCGVLGLRL (配列番号317)、ACKLKLCGVLGLRLM (配列番号318)、CKLKLCGVLGLRLMD (配列番号319)、KLKLCGVLGLRLMDG (配列番号320)、LKLCGVLGLRLMDGT (配列番号321)、およびKLCGVLGLRLMDGTW (配列番号322)からなる群から選択したアミノ酸配列を有する直線状ペプチドに結合した(表14参照)。GACKLKLCGVLGLRL (配列番号317)、ACKLKLCGVLGLRLM (配列番号318)のアミノ酸配列を有するペプチドは、平均値の2倍よりも小さな OD値を有する。それにもかかわらずこれらのペプチドを請求した理由は、これらが抗体CR57によって認識される抗原ペプチドの領域の近傍に存在するためである。アミノ酸配列KLCGVLGLRLMDGTW (配列番号322)を有するペプチドに対する結合が最も顕著であった。
【0144】
抗体CR04-010は、GFGKAYTIFNKTLME (配列番号323)、FGKAYTIFNKTLMEA (配列番号 324)、GKAYTIFNKTLMEAD (配列番号325)、KAYTIFNKTLMEADA (配列番号326)、AYTIFNKTLMEADAH (配列番号327)、YTIFNKTLMEADAHY (配列番号328)、TIFNKTLMEADAHYK (配列番号329)、IFNKTLMEADAHYKS (配列番号330)、およびFNKTLMEADAHYKSV (配列番号331)からなる群から選択したアミノ酸配列を有する直線状ペプチドに結合した。AYTIFNKTLMEADAH (配列番号327)、YTIFNKTLMEADAHY (配列番号328) のアミノ酸配列を有するペプチドは、平均値の2倍よりも小さな OD値を有する。それにもかかわらずこれらのペプチドを請求した理由は、これらは、抗体CR04-010の認識する抗原ペプチド領域の近傍に存在するからである。アミノ酸配列TIFNKTLMEADAHYK (配列番号329)、IFNKTLMEADAHYKS (配列番号330)、およびFNKTLMEADAHYKSV (配列番号331) を有するペプチドに対する結合が最も顕著であった。
【0145】
CRJB、およびCR04-040、CR04-098およびCR04-103と呼ばれる抗体(データは示さず)は、直線状の抗原ペプチドの領域を認識しなかった。
【0146】
上記のペプチドまたはその一部のどちらとも、狂犬病ウイルスの中和エピトープの優良な候補であり、狂犬病ウイルス感染の治療および/または予防のためのワクチンや中和抗体産生の基礎を形成しうる。
【0147】
SLKGACKLKLCGVLGLRLMDGTW (配列番号332)およびGFGKAYTIFNKTLMEADAHYKSV (配列番号333) は、PEPSCAN分析において高い反応性を示すことから、糖タンパク質の特に興味深い領域である。
【0148】
上記のPEPSCANデータから、CR57およびCR04-010と呼ばれるヒトモノクローナル抗体は、狂犬病ウイルスGタンパク質の異なる領域に結合するとさらに推測することができ、これらは互いに競合しないエピトープを認識することを示唆している。
【0149】
実施例12
試験管内中和アッセイを用いた、抗狂犬病ウイルスGタンパク質IgGの中和効力の決定(修正RFFIT)
作製したヒトモノクローナル抗体の中和効力を実施例1に記載の通り、修正RFFITによって決定した。16個のIgGが狂犬病CVS-11株を1000 IU/mgより大きな効力で中和し、2 IU/mgより低い効力を有するIgGは2つだけであった(表15参照)。16個の抗体のうちの8個の効力は、一時的に発現させて作製したCR-57の効力を上回り、これらが狂犬病ウイルスの暴露後予防においてCR-57よりも効果的であることを示唆した。一時的に発現させて生成したCR-57の効力は、約3800 IU/mgタンパク質(表1および表15参照)であった一方、安定的に生成されたCR-57は5400 IU/mgタンパク質(データは示さず)の効力を示した。興味深いことに、同定された中和ヒトモノクローナル抗体の大半は、重鎖可変生殖系列遺伝子3−30を有する(表9参照)。
【0150】
狂犬病ウイルスへの抗体の親和性(データは示さず)、および修正RFFITアッセイにおける抗体の100%エンドポイント希釈度(データは示さない)に基づき、6つの独自のIgG、すなわち、CR04-010、CR04-040、CR04-098、CR04-103、CR04-104およびCR04-144からなるパネルを、更なる発色のために選択した。このパネルのなかでは抗体CR04-098が最も高い効力、すなわち、約7300 IU/mgタンパク質を示したため、特に興味深かった(表15参照)。安定的に生成されたCR04-098も同様の効力を示した(データは示さず)。
【0151】
実施例13
抗狂犬病ウイルスIgGによるE57エスケープウイルスの試験管内での中和
新規のヒトモノクローナル抗狂犬病抗体の特徴を更に調べるため、IgGのE57エスケープウイルスに対する中和活性を、上記のように修正RFFITによって調べた。抗狂犬病ウイルスIgGの大半は、6種類のE57エスケープウイルスの全てに対して良好な中和活性を示した(表16参照)。一方、CR04-008、CR04-018およびCR04-126は、それぞれ、E57エスケープウイルスの6/6、2/6、および3/6を中和しなかった。「中和しない」とは、1:100の抗体希釈物を用いても50%エンドポイントに到達しないことを意味する。CR04-021、CR04-108、CR04-120、CR04-125、およびCR04-164の中和活性は、エスケープウイルスの個数を増やすに従って有意に減少した。このことは、これらの抗体のエピトープは、E57エスケープウイルス糖タンパク質の影響を直接的または間接的に受けることを意味する。上記に基づくと、暴露後予防の処置のための抗狂犬病カクテルにおいて、複数の抗狂犬病ウイルスIgGはCR-57と適合する。特に、上記で同定した6つの独自のIgGのパネル、すなわち、抗体CR04-010、CR04-040、CR04-098、CR04-103、CR04-104、およびCR04-144は、E57エスケープウイルスに対して良好な中和効力を示し、このことは、これらの抗体によって認識されるエピトープはCR-57によって導入されたアミノ酸変異の影響を受けなかったことを示唆する。抗体CR04-098は各エスケープウイルスに対して3000 IU/mgよりも大きな効力を示しため、最も有望である。
【0152】
実施例14
抗狂犬病抗体CR-57およびCR04-098のエピトープ認識
CR-57およびCR04-098と呼ばれるヒトモノクローナル抗体が互いに重複たり、互いに競合したりしないエピトープを認識することを確認するため、CR04-098と呼ばれるヒトモノクローナル抗体のエスケープウイルスを、基本的にはCR57のエスケープウイルスについて記載したのと同様の方法で作製した(実施例1参照)。手短に言うと、ウェル毎の細胞増殖巣の数を免疫蛍光測定法により記録し、好適には1個の細胞増殖巣を含むウェルの培地を、ウイルスを増幅させるために選択した。全てのE98エスケープウイルスは、E982(2個の細胞増殖巣)およびE98-4(4個の細胞増殖巣)という例外を除き、単一の細胞増殖巣から生成された。中和指数がlog2.5よりも小さいウイルスをエスケープ変異体と定義した。中和指数は、「ウイルスおよびモノクローナル抗体(約4 IU/ml)を感染させたBSR細胞培養液中に産生された、1mlあたりの感染性ウイルス粒子の個数」を「ウイルスのみが感染したBSRまたはMNA細胞培養液中に産生された、1mlあたりの感染性ウイルス粒子の個数」から引くことによって求めた(log(モノクローナル抗体非存在下でのウイルス1mlあたりの細胞増殖巣形成単位数−log(モノクローナル抗体存在下でのウイルス1mlあたりの細胞増殖巣形成単位数))。log2.5よりも小さな指数はエスケープウイルスが存在する証拠とした。
【0153】
CR04-098がCR-57とは異なり、互いに重複したり競合したりしないエピトープと結合するかを更に調べるために、CR-57のE98エスケープウイルスに対する修正RFFITアッセイを上記のように行った。表17に示す通り、CR-57は全5種類のE98エスケープウイルスに対して良好な中和活性を有した。加えて、抗体CR04-010および抗体CR04-144のE98エスケープウイルスに対する中和活性を測定した。どちらの抗体もE98エスケープウイルスを中和せず(データは示さず)、両抗体によって認識されるエピトープは、抗体CR04-098によって導入されたアミノ酸変異の影響を直接的にまたは間接的に受けていることを示唆した。抗体CR04-018および抗体CR04-126については、E98エスケープウイルスのうち1種類、すなわちE98-4に対する中和活性のみを測定した。CR04-018はエスケープウイルスを中和する能力を有した一方、CR04-126はエスケープウイルスに対して弱い中和効力しか持たなかった。このことから、CR04-018によって認識されるエピトープは、抗体CR04-098によって導入された変異の影響を受けていないことが示唆される。加えて、抗体CR04-010、抗体CR04-038、抗体CR04-040、抗体CR04-073、抗体CR04-103、抗体CR04-104、抗体CR04-108、抗体CR04-120、抗体CR04-125、抗体CR04-164はE98-4を中和せず、これらの抗体がCR04-098と同じエピトープを認識することが示唆した(データは示さず)。
【0154】
各E98エスケープウイルスの狂犬病糖タンパク質において生じうる変異を同定するために、E57およびEJBエスケープウイルスについて記載したのと同じ方法で、糖タンパク質のオープン・リーディング・フレーム(ORF)のヌクレオチド配列を決定した。全てのE98エスケープウイルスは、狂犬病糖タンパク質のアミノ酸部位336においてNがDに置き換わる変異が存在した(図8参照)。糖タンパク質のこの領域は抗原部位IIIと定義され、アミノ酸330-338から構成される(アミノ酸番号はシグナルペプチドを含まずにふっている)。一方、CR-57はアミノ酸226-231(アミノ番号はシグナルペプチドを含まずにふっている)の位置に存在するエピトープを認識し、この場所は抗原部位Iと重なる。N336D変異に加えて、E98-5と呼ばれるE98エスケープウイルスは、狂犬病糖タンパク質のアミノ酸部位354においてHがQに置き換わる変異(コドンはCATからCAGに変化)を示した(データは示さず)。
【0155】
更に、(E57エスケープウイルスに存在するような)変異CR57エピトープを有するペプチドに対するCR57の結合を観測したpepscan分析によって、CR57の相互作用が失われたことが示された。驚くべきことに、フローサイトメトリーの測定から、CR04-098は依然としてPER.C6(登録商標)細胞において発現された(N336D変異を含む)変異糖タンパク質への結合能を有することがわかったが、この変異を含むウイルスはもはや中和されなかった。
【0156】
更に、BIAcore3000(登録商標)分析装置を用いた表面プラズモン共鳴分析によって、エピトープマッピング研究およびアフィニティーランキング研究を行った。精製された狂犬病糖タンパク質(ERA株)をresearch grade CM5 4-flow channel (Fc) センサーチップ(Biacore AB, Sweden)上にリガンドとして、アミノカップリングを用いて固定した。ランキングは、HBS-EP(Biacore AB, Sweden)をランニングバッファーとして、25℃で行った。各抗体50μlを20μl/分の一定流速で注入した。次に、750秒間ランニングバッファーを流した後、5μlの2M NaOH、5μlの45mM HCl、および5μlの2mM NaOHを用いてCM5チップの再生を行った。共鳴単位(RU)で表される共鳴シグナルを時間の関数としてプロットし、それぞれ結合および解離の尺度として用いるRUの増加および減少を決定し、抗体のランキングのために使用した。表面プラズモン共鳴分析によって決定されたCR57およびCR04-098の実際の KD値はそれぞれ2.4nmおよび4.5nmであった。更にエピトープマッピング研究によって、CR57およびCR04-098は狂犬病糖タンパク質上の異なったエピトープに結合することが確認された。CR57の注入によって58RUの共鳴が得生じた(データは示さず)。CR04-098の注入後、反応レベルが更に増加した(24RU)ため、CR04-098の結合部位は占有されなかったことが示唆される(データは示さない)。逆の順序で行っても同様の結果が得られ、各抗体は注入の順序に関係なく同程度のRUレベルに達することが示された(データは示さず)。更にこれらの結果は、CR57およびCR04-098は、狂犬病ウイルス糖タンパク質上の互いに異なったエピトープに同時に結合し、認識できることを示している。
【0157】
概して、上記のデータは、抗体CR-57およびCR04-098が、互いに異なり互いに重なり合わないエピトープ、すなわち抗原認識部位IおよびIIIにあるエピトープをそれぞれ認識することをさらに裏付ける。このデータは、CR-57およびCR04-098がERA Gへの結合を巡って競合しないこと、および、全てのE57エスケープウイルスに対して抗体CR04-098が良好な中和活性を示すことを示している、ELISA/FACSによる競合データと十分に一致する。これらの結果、および、CR57およびCR04-098の組み合わせを狂犬病ウイルスへ試験管内で暴露することによってエスケープウイルスが得られなったこと(どちらかの抗体4 IU/mlの存在下で選択を行った)に基づくと、抗体CR-57およびCR04-098は互いに重なり合ったり競合したりしないエピトープを認識し、暴露後予防の処置に用いる抗狂犬病ウイルス抗体カクテルに用いるのに好都合であると結論できる。
【0158】
実施例15
CR57およびCR04-098によって認識されるエピトープの保存の評価
CR-57の最小結合領域(配列番号332中のアミノ酸KLCGVL;CR57によって認識される狂犬病ウイルス糖タンパク質の領域であることが、PEPSCANおよびアラニンスキャニング技術によって決定された領域)を229個の遺伝子型1の狂犬病ウイルス分離株のヌクレオチド配列と比較し、エピトープの保存を評価した(表18参照)。サンプル集合には、ヒト分離株、コウモリ分離株、およびイヌ分離株または狂犬病のイヌに噛まれたであろうと最も考えられる家畜からの分離株が含まれた。最小結合領域の各部位のアミノ酸の出現頻度分析から、エピトープを形成する必須のな残基が高度に保存されていることが判明した。位置1に存在するリジンは99.6%の分離株で保存され、229個の分離株のうち1つにおいてのみ、保存的なK>R変異が観測された。位置2および3(LおよびC)は完全に保存されていた。中央部のシステイン残基は糖タンパク質のフォールディングに構造的にかかわり、全リッサウイルスを通して保存されると考えられている(Badrane and Tordo, 2001を参照)。位置4のグリシンは98.7%の分離株で保存され、229個の分離株のうち3つにおいて荷電アミノ酸への変異がみられた(1/229の分離株でG>R変異、2/229の分離株でG>E変異)。5番目の位置もまた、1つの分離株で保存的なV>I変異が観測されたという以外を除き、保存されていた。アラニン置換スキャンによって必須の残基ではないと決定された6番目の位置においては、街上分離株(srreet isolates)に有意な多様性がみられた;すなわち、70.7%の株でL、26.7%の株でP、2.6%の株でSであった。総合すると、遭遇する可能性のある狂犬病ウイルスの約99%は、CR-57によって認識されると予測される。
【0159】
これら229個のウイルス分離株のうち123個について、CR-57およびCR04-098のエピトープ双方において変異が存在するかどうか分析した。これら123個の街上ウイルス(street viruses)はどちらのエピトープにおいても変異を含まなかった。E98エスケープウイルスにおいて観測されたN>D変異は、5つのウイルス分離株のみに存在した。これらのウイルスは地理的に異なり、アフリカの動物から分離されたものである(系統図は図9参照;5つのウイルス分離株、すなわちAF325483、AF325482、AF325481、AF325480、およびAF325485は太字で示した)。糖タンパク質配列の系統発生解析によって、変異したCR57エピトープを有する狂犬病ウイルスは、変異したCR04-098エピトープを有する狂犬病ウイルスとは遠縁にあたることがわかった。故に、CR-57およびCR04-098のカクテルによる中和に対して耐性を示す狂犬病ウイルスに遭遇する可能性は実質的に皆無である。
【0160】
【表1】
【0161】
【表2】
【0162】
【表3】
【0163】
【表4−1】
【0164】
【表4−2】
【0165】
【表4−3】
【0166】
【表5】
【0167】
【表6】
【0168】
【表7−1】
【0169】
【表7−2】
【0170】
【表8】
【0171】
【表9−1】
【0172】
【表9−2】
【0173】
【表10】
【0174】
【表11A】
【0175】
【表11B】
【0176】
【表12−1】
【0177】
【表12−2】
【0178】
【表12−3】
【0179】
【表13−1】
【0180】
【表13−2】
【0181】
【表13−3】
【0182】
【表14−1】
【0183】
【表14−2】
【0184】
【表14−3】
【0185】
【表14−4】
【0186】
【表14−5】
【0187】
【表14−6】
【0188】
【表14−7】
【0189】
【表14−8】
【0190】
【表14−9】
【0191】
【表14−10】
【0192】
【表15】
【0193】
【表16】
【0194】
【表17】
【0195】
【表18】
【0196】
参考文献
【図面の簡単な説明】
【0197】
【図1】図1は狂犬病ウイルス株CVS-11およびE57のエスケープウイルスのアミノ酸配列の比較を示す。ウイルス感染細胞を感染2日後に回収し、全RNAを単離した。cDNAを作製し、DNA塩基配列解読のために使用した。変異を含む領域を図示しており、変異を太字で示す。図1Aはヌクレオチド配列の比較を示す。アミノ酸の上の番号は、シグナルペプチドを含む、狂犬病ウイルス糖タンパク質のアミノ酸番号を示す。図1Bは、アミノ酸配列の比較を示す。狂犬病ウイルス糖タンパク質の略図を上部に示す。黒い四角形部分がシグナルペプチドを示し、灰色の四角形部分が膜貫通領域を示す。図1の配列は、配列番号130-141によっても表される。
【図2】図2は狂犬病ウイルス株CVS-11およびEJBのエスケープウイルスのアミノ酸配列の比較を示す。ウイルス感染細胞を感染2日後に回収し、全RNAを単離した。cDNAを作製し、DNA塩基配列解読のために使用した。変異を含む領域を図示しており、変異を太字で示す。図2Aはヌクレオチド配列の比較を示す。アミノ酸の上の番号は、シグナルペプチドを含む、狂犬病ウイルス糖タンパク質のアミノ酸番号を示す。図2Bは、アミノ酸配列の比較を示す。狂犬病ウイルス糖タンパク質の略図を上部に示す。黒い四角形部分がシグナルペプチドを示し、灰色の四角形部分が膜貫通領域を示す。図2の配列は、配列番号142-151 によっても表される。
【図3】PDV-C06ベクターを示す。
【図4】図4は抗狂犬病ウイルスscFvと、CR-57と呼ばれるビオチン化抗狂犬病ウイルス抗体との競合ELISAを示す。精製された狂犬病ウイルスGタンパク質によってコートされたELISAプレートを、各scFvと共にインキュベートした後、CR-57bio (0.5μg/ml)を加えた。続いて、scFvの存在下および非存在下においてCR-57bioの結合を観測した。
【図5】図5は抗狂犬病ウイルスscFvと、CR-57と呼ばれる抗狂犬病ウイルス抗体との競合ELISAを示す。精製された狂犬病ウイルスGタンパク質によってコートされたELISAプレートを、CR-57 (1μg/ml)と共にインキュベートした後、過剰量のscFvを加えた。続いて、CR-57の存在下および非存在下においてscFvの結合を観測した。
【図6】図6は抗狂犬病ウイルスGタンパク質IgGと、CR-57と呼ばれる抗狂犬病ウイルス抗体との競合ELIZA分析を示す。Gタンパク質(ERA株)を非標識IgGと共にインキュベートした(x軸上に示す)。ビオチン化CR57(CR57bio)を加えてGタンパクに結合させた後、ストレプトアビジン-HRPを加えることによって視覚化を行った。ELISAシグナルを、CR57bioの結合単独の場合のシグナル対するパーセントとして示す。
【図7】図7は抗狂犬病ウイルスGタンパク質IgGと、CR-57と呼ばれる抗狂犬病ウイルス抗体との競合FACS分析を示す。Gタンパク質(ERA株)を発現するPER.C6細胞を非標識IgGと共にインキュベートした(x軸上に示す)。ビオチン化したCR57(CR57bio)を加え、Gタンパク発現細胞に結合させた後、ストレプトアビジン-PEによる視覚化を行った。FACSシグナルを、CR57bioの結合単独の場合のシグナル対するパーセントとして示す。
【図8】図8はCVS-11およびE98のエスケープウイルスのアミノ酸配列の比較を示す。ウイルス感染細胞を感染2日後に回収し、全RNAを単離した。cDNAを作製し、DNA塩基配列解読のために使用した。点変異を含む領域を図示しており、変異を太字で示す。図8Aはヌクレオチド配列の比較を示す。ヌクレオチド上の番号は、シグナルペプチド配列を含まない、狂犬病ウイルス糖タンパク質のオープン・リーディング・フレームにおける、変異ヌクレオチド(太字で示す)を示す。図8Bはアミノ酸配列の比較を示す。アミノ酸上の番号は、シグナルペプチド配列を除いた狂犬病ウイルス糖タンパク質における、変異ヌクレオチド(太字で示す)を示す。
【図9】図9は123種類の狂犬病ウイルス街上毒株の系統樹を示す(123種類の狂犬病ウイルスG糖タンパク質の配列をもとに、近隣結合法、Kimuraの2−パラメータ法、および500回のブートストラップ法を用いた)。太字はE98のエスケープウイルスで観測されるのと同様のN>D変異を有するウイルスを示す。
【図10】図10は狂犬病糖タンパク質上の中和エピトープを示す。新規のCR57エピトープを含む抗原性部位を記載した、狂犬病ウイルス糖タンパク質の概略図を示す。シグナルペプチド(19アミノ酸)および膜貫通領域を、黒い四角形によって示す。ジスルフィド結合による架橋を示す。アミノ酸番号は、シグナルペプチド配列を除いた成熟タンパク質から付けられている。
【特許請求の範囲】
【請求項1】
狂犬病ウイルス中和活性を有する結合分子であって、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23および配列番号24からなる群から選択されたアミノ酸配列を含むCDR3領域を少なくとも備えることを特徴とする結合分子。
【請求項2】
請求項1に記載の結合分子であって、配列番号26、配列番号27、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番号44、配列番号45、配列番号46、配列番号47、配列番号48、配列番号49、および配列番号335のアミノ酸1−119からなる群から選択されたアミノ酸配列を含む重鎖可変領域を備えることを特徴とすることを特徴とする結合分子。
【請求項3】
狂犬病ウイルス中和活性を有することを特徴とする請求項1または2に記載の結合分子の機能的変異体であって機能的変異体。
【請求項4】
請求項1または2に記載の結合分子または請求項3に記載の機能的変異体を備えた免疫コンジュゲートであって、更に少なくとも1つのタグを備えた免疫コンジュゲート。
【請求項5】
請求項1または2に記載の結合分子または請求項3に記載の機能的変異体をコードする核酸分子。
【請求項6】
請求項5に記載の核酸分子を少なくとも1つ備えたベクター。
【請求項7】
請求項6に記載のベクターを少なくとも1つ備えたホスト。
【請求項8】
ホストがヒト細胞由来の細胞である、請求項7に記載のホスト。
【請求項9】
請求項1または2に記載の結合分子または請求項3に記載の機能的変異体を生産する方法であって、
a)請求項7または8に記載のホストを結合分子または機能的変異体の発現を起こすような条件の下で培養する工程、および必要に応じて、
b)発現した結合分子または機能的変異体を回収する工程、
から構成される方法。
【請求項10】
請求項1または2に記載の結合分子を備えた医薬品組成物であって、さらに薬学的に許容される賦形剤を少なくとも1つ備えた医薬品組成物。
【請求項11】
少なくとも1つの追加的な結合分子を含む請求項10に記載の医薬品組成物であって、該追加的な結合分子は、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24および配列番号25からなる群から選択されたアミノ酸配列を含むCDR3領域を少なくとも備え、該結合分子は狂犬病ウイルスの互いに異なった、競合しないエピトープと反応できることを特徴とする組成物。
【請求項12】
請求項11に記載の医薬品組成物であって、追加的な結合分子は、配列番号26、配列番号27、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番44、配列番号45、配列番号46、配列番号47、配列番号48、配列番号49、配列番号273および配列番号335のアミノ酸1−119からなる群から選択されたアミノ酸配列を含む重鎖可変領域を備えることを特徴とする組成物。
【請求項13】
狂犬病ウイルス中和能力を有する結合分子を少なくとも2つ含む医薬品組成物であって、該結合分子は狂犬病ウイルスの互いに異なった、競合しないエピトープと反応できることを特徴とする組成物。
【請求項14】
請求項13に記載の医薬品組成物であって、第一の狂犬病ウイルス中和能力を有する結合分子は狂犬病ウイルスGタンパク質の抗原部位Iに位置するエピトープと反応することができ、第二の狂犬病ウイルス中和能力を有する結合分子は狂犬病ウイルスGタンパク質の抗原部位IIIに位置するエピトープと反応できることを特徴とする組成物。
【請求項15】
請求項13に記載の医薬品組成物であって、第一の狂犬病ウイルス中和能力を有する結合分子は配列番号25のアミノ酸配列を含むCDR3領域を少なくとも備え、第二の狂犬病ウイルス中和能力を有する結合分子は配列番号4、配列番号10、配列番号14、配列番号15、配列番号16および配列番号22からなる群から選択されたアミノ酸配列を含むCDR3領域を少なくとも備えることを特徴とする組成物。
【請求項16】
医薬品として使用される、請求項1または2に記載の結合分子、請求項3に記載の機能的変異体、請求項4に記載の免疫コンジュゲート、または請求項10〜15の何れか一つに記載の医薬品組成物。
【請求項17】
リッサウイルスに起因する症状の診断、予防、処置、またはそれらの組み合わせにおいて使用される、請求項1または2に記載の結合分子、請求項3に記載の機能的変異体、請求項4に記載の免疫コンジュゲート、または請求項10〜15の何れか一つに記載の医薬品組成物。
【請求項18】
リッサウイルスに起因する症状の診断、予防、処置、またはそれらの組み合わせのために使用する薬剤調剤において、請求項1または2に記載の結合分子、請求項3に記載の機能的変異体、請求項4に記載の免疫コンジュゲート、または請求項10〜15の何れか一つに記載の医薬品組成物を使用する方法。
【請求項19】
少なくとも1つの請求項1または2に記載の結合分子、少なくとも1つの請求項3に記載の機能的変異体、少なくとも1つの請求項4に記載の免疫コンジュゲート、または少なくとも1つの請求項10〜15の何れか一つに記載の医薬品組成物、またはこれらの組み合わせを備えたキット。
【請求項20】
狂犬病ウイルスに特異的に結合する結合分子、または狂犬病ウイルスに特異的に結合する結合分子をコードする核酸分子を同定する方法であって、
a)複製可能な遺伝子パッケージの表面に存在する結合分子の集合体に、狂犬病ウイルスまたはその断片を、結合を起こす条件の下で接触させる工程、
b)狂犬病ウイルスまたはその断片に結合する結合分子の選択を少なくとも1回行なう工程、および
c)狂犬病ウイルスまたはその断片に結合した結合分子を分離および回収する工程、
から構成される方法。
【請求項21】
狂犬病ウイルスに対する中和活性を潜在的に有する結合分子、または狂犬病ウイルスに対する中和活性を潜在的に有する結合分子をコードする核酸分子を同定する方法であって、
a)複製可能な遺伝子パッケージの表面に存在する結合分子の集合体に、狂犬病ウイルスまたはその断片を、結合を起こす条件の下で接触させる工程、
b)狂犬病ウイルスまたはその断片に結合する結合分子を、結合しない結合分子から分離および回収する工程、
c)少なくとも1つの回収された結合分子を単離する工程、
d)単離された結合分子が狂犬ウイルスに対する中和活性を有するかを確認する工程、
から構成される方法。
【請求項22】
請求項21に記載の方法であって、重鎖可変生殖系列遺伝子3−30を含む結合分子を分離および回収する工程をさらに含む方法。
【請求項23】
請求項20〜22の何れか一つに記載の方法であって、複製可能な遺伝子パッケージの表面に存在する結合分子の集合体が、狂犬病のワクチン接種を受けた被験者、あるいは狂犬病ウイルスに暴露された被験者から得られた細胞より単離されたRNAから調製されている方法。
【請求項24】
請求項20〜23の何れか一つに記載の方法であって、複製可能な遺伝子パッケージの表面に存在する結合分子の集合体が、単鎖Fvライブラリである方法。
【請求項25】
請求項23および24の何れかに記載の方法であって、該被験者が狂犬病のワクチン接種を受けたヒト個体である方法。
【請求項26】
請求項20〜25の何れか一つに記載の方法であって、複製可能な遺伝子パッケージは、ファージ粒子、細菌、酵母、真菌、微生物の胞子およびリボソームからなる群から選択されることを特徴とする方法。
【請求項27】
複製可能な遺伝子パッケージの表面に存在するヒト結合分子の集合体であって、狂犬病のワクチン接種を受けた被験者、あるいは狂犬病ウイルスに暴露された被験者から得られた細胞より単離されたRNAから調製されたことを特徴とする集合体。
【請求項28】
請求項27に記載の集合体であって、単鎖Fvライブラリであることを特徴とする集合体。
【請求項29】
請求項27または28に記載の集合体であって、該被験者が狂犬病のワクチン接種を受けたヒト個体であることを特徴とする集合体。
【請求項30】
生体内で疾患を引き起こす感染性物質に対する中和活性を潜在的に有する結合分子、または生物内で疾患を引き起こす感染性物質に対する中和活性を潜在的に有する結合分子をコードする核酸分子を同定する方法であって、
a)複製可能な遺伝子パッケージの表面に存在する結合分子の集合体に、少なくとも生体内で疾患を引き起こす感染性物質のタンパク質を表面に発現した細胞を、結合を起こす条件の下で接触させる工程、
b)生体内で疾患を引き起こす感染性物質のタンパク質を表面に発現した細胞に結合する結合分子を、前記細胞に結合しない結合分子から分離および回収する工程、
c)少なくとも1つの回収された結合分子を単離する工程、
d)単離された結合分子が生物内で疾患を引き起こす感染性物質に対する中和活性を有するか確認する工程、
から構成される方法。
【請求項31】
請求項30に記載の方法であって、該結合分子がヒト由来であることを特徴とする方法。
【請求項32】
請求項30または31に記載の方法であって、該感染性物質がウイルス、細菌、酵母、真菌または寄生虫であることを特徴とする方法。
【請求項33】
請求項30〜32の何れかに記載の方法であって、タンパク質が、感性性物質の表面に正常に発現したタンパク質であることを特徴とする方法。
【請求項34】
請求項30〜33の何れかに記載の方法であって、複製可能な遺伝子パッケージの表面に存在する結合分子の集合体は、生体内で疾患を引き起こす感染性物質のタンパク質をそれらの表面に発現していない細胞が取り除かれたものであることを特徴とする方法。
【請求項35】
配列番号25を含むアミノ酸配列を含む重鎖CDR3領域を少なくとも備えることを特徴とする狂犬病ウイルス中和活性を有する結合分子であって、少なくとも2500IU/mgタンパク質の中和活性を有することを特徴とする前記結合分子。
【請求項36】
請求項35に記載の結合分子であって、配列番号273を含むアミノ酸配列を含む重鎖可変領域を備えることを特徴とする結合分子。
【請求項37】
請求項35または36に記載の結合分子であって、配列番号123を含むアミノ酸配列を含む重鎖を備えることを特徴とする結合分子。
【請求項38】
請求項35〜37の何れか一つに記載の結合分子をコードする核酸分子。
【請求項39】
請求項38に記載の核酸分子であって、配列番号122を含むヌクレオチド配列を備えることを特徴とする核酸分子。
【請求項40】
少なくとも2つの結合分子を含む医薬品組成物であって、該結合分子の等電点の差は1.5pI単位未満であることを特徴とする医薬品組成物。
【請求項41】
請求項40に記載の医薬品組成物であって、該結合分子がヒトモノクローナル抗体であることを特徴とする医薬品組成物。
【請求項42】
請求項40または41に記載の医薬品組成物であって、両者の結合分子が感染性物質を中和する能力を有することを特徴とする医薬品組成物。
【請求項43】
請求項40〜42の何れかに記載の医薬品組成物であって、両者の結合分子が狂犬病ウイルスを中和する能力を有することを特徴とする医薬品組成物。
【請求項44】
請求項1、2または35〜37に記載の結合分子であって、ヒト由来であることを特徴とする結合分子。
【請求項1】
狂犬病ウイルス中和活性を有する結合分子であって、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23および配列番号24からなる群から選択されたアミノ酸配列を含むCDR3領域を少なくとも備えることを特徴とする結合分子。
【請求項2】
請求項1に記載の結合分子であって、配列番号26、配列番号27、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番号44、配列番号45、配列番号46、配列番号47、配列番号48、配列番号49、および配列番号335のアミノ酸1−119からなる群から選択されたアミノ酸配列を含む重鎖可変領域を備えることを特徴とすることを特徴とする結合分子。
【請求項3】
狂犬病ウイルス中和活性を有することを特徴とする請求項1または2に記載の結合分子の機能的変異体であって機能的変異体。
【請求項4】
請求項1または2に記載の結合分子または請求項3に記載の機能的変異体を備えた免疫コンジュゲートであって、更に少なくとも1つのタグを備えた免疫コンジュゲート。
【請求項5】
請求項1または2に記載の結合分子または請求項3に記載の機能的変異体をコードする核酸分子。
【請求項6】
請求項5に記載の核酸分子を少なくとも1つ備えたベクター。
【請求項7】
請求項6に記載のベクターを少なくとも1つ備えたホスト。
【請求項8】
ホストがヒト細胞由来の細胞である、請求項7に記載のホスト。
【請求項9】
請求項1または2に記載の結合分子または請求項3に記載の機能的変異体を生産する方法であって、
a)請求項7または8に記載のホストを結合分子または機能的変異体の発現を起こすような条件の下で培養する工程、および必要に応じて、
b)発現した結合分子または機能的変異体を回収する工程、
から構成される方法。
【請求項10】
請求項1または2に記載の結合分子を備えた医薬品組成物であって、さらに薬学的に許容される賦形剤を少なくとも1つ備えた医薬品組成物。
【請求項11】
少なくとも1つの追加的な結合分子を含む請求項10に記載の医薬品組成物であって、該追加的な結合分子は、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24および配列番号25からなる群から選択されたアミノ酸配列を含むCDR3領域を少なくとも備え、該結合分子は狂犬病ウイルスの互いに異なった、競合しないエピトープと反応できることを特徴とする組成物。
【請求項12】
請求項11に記載の医薬品組成物であって、追加的な結合分子は、配列番号26、配列番号27、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番44、配列番号45、配列番号46、配列番号47、配列番号48、配列番号49、配列番号273および配列番号335のアミノ酸1−119からなる群から選択されたアミノ酸配列を含む重鎖可変領域を備えることを特徴とする組成物。
【請求項13】
狂犬病ウイルス中和能力を有する結合分子を少なくとも2つ含む医薬品組成物であって、該結合分子は狂犬病ウイルスの互いに異なった、競合しないエピトープと反応できることを特徴とする組成物。
【請求項14】
請求項13に記載の医薬品組成物であって、第一の狂犬病ウイルス中和能力を有する結合分子は狂犬病ウイルスGタンパク質の抗原部位Iに位置するエピトープと反応することができ、第二の狂犬病ウイルス中和能力を有する結合分子は狂犬病ウイルスGタンパク質の抗原部位IIIに位置するエピトープと反応できることを特徴とする組成物。
【請求項15】
請求項13に記載の医薬品組成物であって、第一の狂犬病ウイルス中和能力を有する結合分子は配列番号25のアミノ酸配列を含むCDR3領域を少なくとも備え、第二の狂犬病ウイルス中和能力を有する結合分子は配列番号4、配列番号10、配列番号14、配列番号15、配列番号16および配列番号22からなる群から選択されたアミノ酸配列を含むCDR3領域を少なくとも備えることを特徴とする組成物。
【請求項16】
医薬品として使用される、請求項1または2に記載の結合分子、請求項3に記載の機能的変異体、請求項4に記載の免疫コンジュゲート、または請求項10〜15の何れか一つに記載の医薬品組成物。
【請求項17】
リッサウイルスに起因する症状の診断、予防、処置、またはそれらの組み合わせにおいて使用される、請求項1または2に記載の結合分子、請求項3に記載の機能的変異体、請求項4に記載の免疫コンジュゲート、または請求項10〜15の何れか一つに記載の医薬品組成物。
【請求項18】
リッサウイルスに起因する症状の診断、予防、処置、またはそれらの組み合わせのために使用する薬剤調剤において、請求項1または2に記載の結合分子、請求項3に記載の機能的変異体、請求項4に記載の免疫コンジュゲート、または請求項10〜15の何れか一つに記載の医薬品組成物を使用する方法。
【請求項19】
少なくとも1つの請求項1または2に記載の結合分子、少なくとも1つの請求項3に記載の機能的変異体、少なくとも1つの請求項4に記載の免疫コンジュゲート、または少なくとも1つの請求項10〜15の何れか一つに記載の医薬品組成物、またはこれらの組み合わせを備えたキット。
【請求項20】
狂犬病ウイルスに特異的に結合する結合分子、または狂犬病ウイルスに特異的に結合する結合分子をコードする核酸分子を同定する方法であって、
a)複製可能な遺伝子パッケージの表面に存在する結合分子の集合体に、狂犬病ウイルスまたはその断片を、結合を起こす条件の下で接触させる工程、
b)狂犬病ウイルスまたはその断片に結合する結合分子の選択を少なくとも1回行なう工程、および
c)狂犬病ウイルスまたはその断片に結合した結合分子を分離および回収する工程、
から構成される方法。
【請求項21】
狂犬病ウイルスに対する中和活性を潜在的に有する結合分子、または狂犬病ウイルスに対する中和活性を潜在的に有する結合分子をコードする核酸分子を同定する方法であって、
a)複製可能な遺伝子パッケージの表面に存在する結合分子の集合体に、狂犬病ウイルスまたはその断片を、結合を起こす条件の下で接触させる工程、
b)狂犬病ウイルスまたはその断片に結合する結合分子を、結合しない結合分子から分離および回収する工程、
c)少なくとも1つの回収された結合分子を単離する工程、
d)単離された結合分子が狂犬ウイルスに対する中和活性を有するかを確認する工程、
から構成される方法。
【請求項22】
請求項21に記載の方法であって、重鎖可変生殖系列遺伝子3−30を含む結合分子を分離および回収する工程をさらに含む方法。
【請求項23】
請求項20〜22の何れか一つに記載の方法であって、複製可能な遺伝子パッケージの表面に存在する結合分子の集合体が、狂犬病のワクチン接種を受けた被験者、あるいは狂犬病ウイルスに暴露された被験者から得られた細胞より単離されたRNAから調製されている方法。
【請求項24】
請求項20〜23の何れか一つに記載の方法であって、複製可能な遺伝子パッケージの表面に存在する結合分子の集合体が、単鎖Fvライブラリである方法。
【請求項25】
請求項23および24の何れかに記載の方法であって、該被験者が狂犬病のワクチン接種を受けたヒト個体である方法。
【請求項26】
請求項20〜25の何れか一つに記載の方法であって、複製可能な遺伝子パッケージは、ファージ粒子、細菌、酵母、真菌、微生物の胞子およびリボソームからなる群から選択されることを特徴とする方法。
【請求項27】
複製可能な遺伝子パッケージの表面に存在するヒト結合分子の集合体であって、狂犬病のワクチン接種を受けた被験者、あるいは狂犬病ウイルスに暴露された被験者から得られた細胞より単離されたRNAから調製されたことを特徴とする集合体。
【請求項28】
請求項27に記載の集合体であって、単鎖Fvライブラリであることを特徴とする集合体。
【請求項29】
請求項27または28に記載の集合体であって、該被験者が狂犬病のワクチン接種を受けたヒト個体であることを特徴とする集合体。
【請求項30】
生体内で疾患を引き起こす感染性物質に対する中和活性を潜在的に有する結合分子、または生物内で疾患を引き起こす感染性物質に対する中和活性を潜在的に有する結合分子をコードする核酸分子を同定する方法であって、
a)複製可能な遺伝子パッケージの表面に存在する結合分子の集合体に、少なくとも生体内で疾患を引き起こす感染性物質のタンパク質を表面に発現した細胞を、結合を起こす条件の下で接触させる工程、
b)生体内で疾患を引き起こす感染性物質のタンパク質を表面に発現した細胞に結合する結合分子を、前記細胞に結合しない結合分子から分離および回収する工程、
c)少なくとも1つの回収された結合分子を単離する工程、
d)単離された結合分子が生物内で疾患を引き起こす感染性物質に対する中和活性を有するか確認する工程、
から構成される方法。
【請求項31】
請求項30に記載の方法であって、該結合分子がヒト由来であることを特徴とする方法。
【請求項32】
請求項30または31に記載の方法であって、該感染性物質がウイルス、細菌、酵母、真菌または寄生虫であることを特徴とする方法。
【請求項33】
請求項30〜32の何れかに記載の方法であって、タンパク質が、感性性物質の表面に正常に発現したタンパク質であることを特徴とする方法。
【請求項34】
請求項30〜33の何れかに記載の方法であって、複製可能な遺伝子パッケージの表面に存在する結合分子の集合体は、生体内で疾患を引き起こす感染性物質のタンパク質をそれらの表面に発現していない細胞が取り除かれたものであることを特徴とする方法。
【請求項35】
配列番号25を含むアミノ酸配列を含む重鎖CDR3領域を少なくとも備えることを特徴とする狂犬病ウイルス中和活性を有する結合分子であって、少なくとも2500IU/mgタンパク質の中和活性を有することを特徴とする前記結合分子。
【請求項36】
請求項35に記載の結合分子であって、配列番号273を含むアミノ酸配列を含む重鎖可変領域を備えることを特徴とする結合分子。
【請求項37】
請求項35または36に記載の結合分子であって、配列番号123を含むアミノ酸配列を含む重鎖を備えることを特徴とする結合分子。
【請求項38】
請求項35〜37の何れか一つに記載の結合分子をコードする核酸分子。
【請求項39】
請求項38に記載の核酸分子であって、配列番号122を含むヌクレオチド配列を備えることを特徴とする核酸分子。
【請求項40】
少なくとも2つの結合分子を含む医薬品組成物であって、該結合分子の等電点の差は1.5pI単位未満であることを特徴とする医薬品組成物。
【請求項41】
請求項40に記載の医薬品組成物であって、該結合分子がヒトモノクローナル抗体であることを特徴とする医薬品組成物。
【請求項42】
請求項40または41に記載の医薬品組成物であって、両者の結合分子が感染性物質を中和する能力を有することを特徴とする医薬品組成物。
【請求項43】
請求項40〜42の何れかに記載の医薬品組成物であって、両者の結合分子が狂犬病ウイルスを中和する能力を有することを特徴とする医薬品組成物。
【請求項44】
請求項1、2または35〜37に記載の結合分子であって、ヒト由来であることを特徴とする結合分子。
【図1】

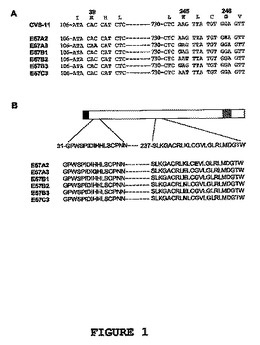
【図2】

【図3】
【図4】
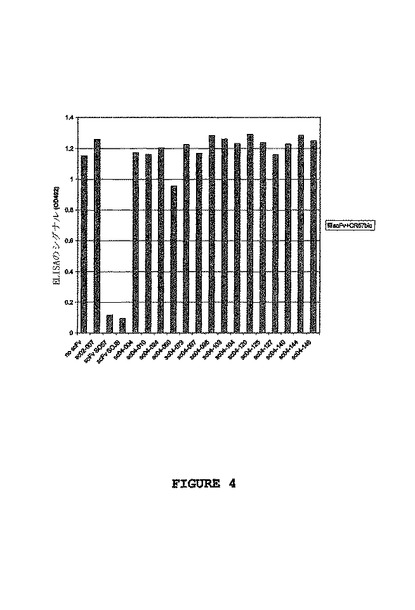
【図5】
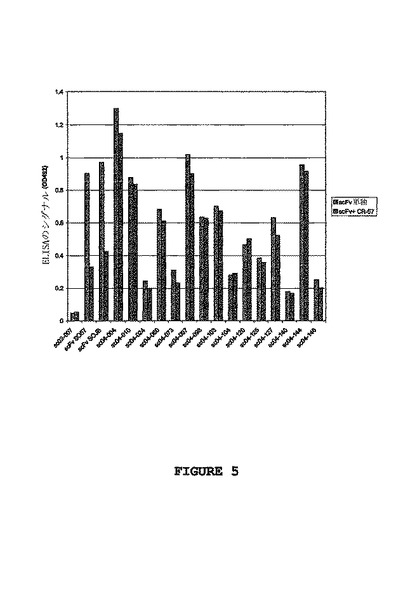
【図8】
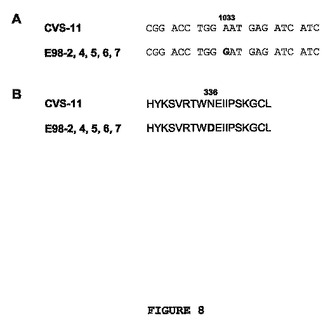
【図9】
【図10】
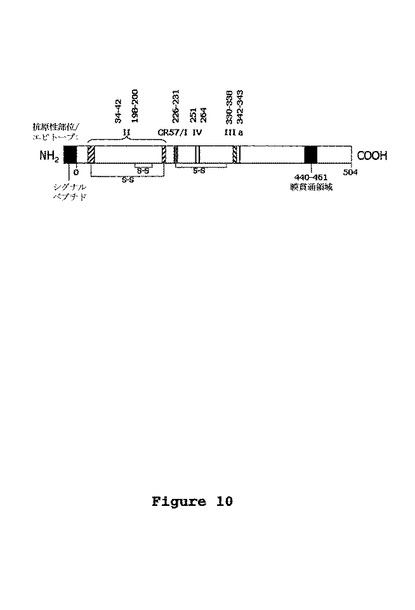
【図6】

【図7】

【図2】
【図3】
【図4】
【図5】
【図8】
【図9】
【図10】
【図6】
【図7】
【公表番号】特表2008−508859(P2008−508859A)
【公表日】平成20年3月27日(2008.3.27)
【国際特許分類】
【出願番号】特願2007−513935(P2007−513935)
【出願日】平成17年5月26日(2005.5.26)
【国際出願番号】PCT/EP2005/052410
【国際公開番号】WO2005/118644
【国際公開日】平成17年12月15日(2005.12.15)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.PENTIUM
【出願人】(301055549)クルセル ホランド ベー ヴェー (27)
【Fターム(参考)】
【公表日】平成20年3月27日(2008.3.27)
【国際特許分類】
【出願日】平成17年5月26日(2005.5.26)
【国際出願番号】PCT/EP2005/052410
【国際公開番号】WO2005/118644
【国際公開日】平成17年12月15日(2005.12.15)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.PENTIUM
【出願人】(301055549)クルセル ホランド ベー ヴェー (27)
【Fターム(参考)】
[ Back to top ]