
環境ストレス応答性転写因子をコードする遺伝子
【課題】ストレス応答性転写因子をコードする遺伝子の提供。
【解決手段】以下の(a)、(b)又は(c)のDNAを含む遺伝子。 (a).特定の配列から選ばれるいずれかの塩基配列からなるDNA (b).特定の配列から選ばれるいずれかの塩基配列において1若しくは複数の塩基が欠失、置換若しくは付加された塩基配列からなり、環境ストレス応答性転写因子をコードするDNA (c).特定の配列から選ばれるいずれかの塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ環境ストレス応答性転写因子をコードするDNA
【解決手段】以下の(a)、(b)又は(c)のDNAを含む遺伝子。 (a).特定の配列から選ばれるいずれかの塩基配列からなるDNA (b).特定の配列から選ばれるいずれかの塩基配列において1若しくは複数の塩基が欠失、置換若しくは付加された塩基配列からなり、環境ストレス応答性転写因子をコードするDNA (c).特定の配列から選ばれるいずれかの塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ環境ストレス応答性転写因子をコードするDNA
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、環境ストレス応答性転写因子をコードする遺伝子に関する。
【背景技術】
【0002】
遺伝子の配列決定プロジェクトによって、数種の生物について大量のゲノム配列及びcDNA配列が決定されており、植物モデルであるシロイヌナズナ(Arabidopsis thaliana)では、2つの染色体の完全なゲノム配列が決定されている(Lin, X. et al., (1999) Nature 402, 761-768.; Mayer, K. et al., (1999) Nature 402, 769-777.)。
【0003】
EST(expressed sequence tag)プロジェクトも、発現遺伝子の発見に大いに貢献している(Hofte, H. et al., (1993) Plant J. 4, 1051-1061.; Newman, T. et al., (1994) Plant Physiol. 106, 1241-1255.; Cooke, R. et al., (1996) Plant J. 9, 1O1-124. Asamizu, E. et al., (2000) DNA Res. 7, 175-180.)。例えば、dbEST(National Center for Biotechnology Information(NCBI)のESTデータベース)には部分cDNA配列が含まれており、全遺伝子の半分以上(即ち、約28,000遺伝子)が再現されている(完全に配列決定されたシロイヌナズナの2番染色体の遺伝子含有量から推定[Lin, X. et al., (1999) Nature 402, 761-768.])。
【0004】
近年、ゲノムスケールの遺伝子発現を分析するのにマイクロアレイ(DNAチップ)技術が有用な手段となっている(Schena, M. et al., (1995) Science 270, 467-470.; Eisen, M. B. and Brown, P. O. (1999) Methods Enzymol. 303, 179-205.)。このDNAチップを用いる技術は、cDNA配列をスライドガラス上に1,000遺伝子/cm2以上の密度で配列させるものである。このように配列させたcDNA配列を、異なる細胞型又は組織型のRNAサンプルから調製した2色蛍光標識cDNAプローブ対に同時にハイブリダイズさせることで、遺伝子発現を直接かつ大量に比較分析することが可能となる。この技術は、最初、48個のシロイヌナズナ遺伝子を根及び苗条におけるディファレンシャル発現について分析することで実証された(Schena, M. et al., (1995) Science 270, 467-470.)。さらに、マイクロアレイは、熱ショック及びプロテインキナーゼC活性化に応答する新規な遺伝子を同定するため、ヒトcDNAライブラリーからランダムに採取した1,000個のクローンを調査するのに使用されている(Schena, M. et al., (1996) Proc. Natl. Acad. Sci. USA, 93, 10614-10619.)。
【0005】
一方、このDNAチップを用いる方法によって、各種の誘導条件下における炎症性疾患関連遺伝子の発現プロフィールの分析が行われている(Heller, R. A. et al., (1997) Proc. Natl. Acad. Sci. USA, 94, 2150-2155.)。さらに、マイクロアレイを用いて、6,000個を超えるコード配列からなる酵母ゲノムの動的発現についても分析が行われている(DeRisi, J.L. et al., (1997) Science 278, 680-686.; Wodicka, L. et al., (1997) Nature Biotechnol. 15, 1359-1367.)。
【0006】
しかしながら、植物の分野では、マイクロアレイ分析に対しては若干の報告がなされているに過ぎない(Schena, M. et al., (1995) Science 270, 467-470.; Ruan, Y. et al., (1998) Plant J. 15, 821-833.; Aharoni. A. et al., (2000) Plant Cell 12, 647-661.; Reymond, P. et al., (2000) Plant Cell 12, 707-719.)。
【0007】
植物の生育は、乾燥、高塩濃度及び低温等の環境ストレスの影響を顕著に受ける。これらのストレスのうち乾燥又は水分欠乏が、植物の生育及び作物の生産にとって最も厳しい制限因子となる。乾燥ストレスは、植物に様々な生化学的及び生理学的な応答を引き起こす。
【0008】
植物は、これらのストレス条件下で生き抜くために、ストレスに対する応答性及び順応性を獲得する。近年、転写レベルで乾燥に応答する数種の遺伝子が記載されている(Bohnert, H.J. et al., (1995) Plant Cell 7, 1099-1111.; Ingram, J., and Bartels, D. (1996) Plant Mol. Biol. 47, 377-403.; Bray, E. A. (1997) Trends Plant Sci. 2, 48-54.; Shinozaki. K., and Yamaguchi-Shinozaki, K. (1997) Plant Physiol. 115, 327-334. ; Shinozaki, K., and Yamaguchi-Shinozaki, K. (1999). Molecular responses to drought stress. Molecular responses to cold, drought, heat and salt stress in higher plants. Edited by Shinozaki, K. and Yamaguchi-Shinozaki, K. R. G. Landes Company.;Shinozaki, K., and Yamaguchi-Shinozaki, K. (2000) Curr. Opin. Plant Biol. 3, 217-223.)。
【0009】
一方、遺伝子導入によって植物のストレス耐性を向上させるために、ストレス誘導性遺伝子が使用されている(Holmberg, N., and Bulow, L. (1998) Trends Plant Sci. 3, 61-66.; Bajaj, S. et al., (1999) Mol. Breed. 5, 493-503.)。高等植物のストレス耐性とストレス応答の分子機構をさらに解明するためだけでなく、遺伝子操作によって作物のストレス耐性を向上させるためにも、ストレス誘導性遺伝子の機能を分析することが重要である。
【0010】
DRE/CRT(乾燥応答性エレメント/C-反復配列)は、乾燥、高塩分濃度及び低温ストレス応答性遺伝子のABA(アブシジン酸:植物ホルモンの一種で種子の休眠や環境ストレスのシグナル伝達因子として機能する。)に依存しない発現において重要なシス作動性エレメントとして同定されている(Yamaguchi-Shinozaki, K., and Shinozaki, K. (1994) Plant Cell 6, 251-264.; Thomashow, M.F. et al., (1999) Plant Mol. Biol. 50, 571-599.; Shinozaki, K., and Yamaguchi-Shinozaki, K. (2000) Curr. Opin. Plant Biol. 3, 217-223.)。また、DRE/CRT応答性遺伝子発現に関与する転写因子(DREB/CBF)がクローニングされている(Stockinger. E.J. et al., (1997) Proc. Natl. Acad. Sci. USA 94, 1035-1040.; Liu, Q. et al., (1998) Plant Cell 10, 1391-1406.; Shinwari, Z.K. et al., (1998) Biochem. Biophys. Res. Commun. 250, 161-170.; Gilmour, S.J.et al., (1998) Plant J. 16, 433-443.)。DREB1/CBFは低温応答性遺伝子発現において機能すると考えられ、DREB2は乾燥応答性遺伝子発現に関与している。カリフラワーモザイクウイルス(CaMV)35Sプロモーターの制御下でCBF1(DREB1B)cDNAを過剰発現するトランスジェニック・シロイヌナズナ植物では、凍結ストレスに対する強力な耐性が観察されている(Jaglo-Ottosen, K.R. et al., (1998) Science 280, 104-106.)。
【0011】
本発明者らは、CaMV 35Sプロモーター又はストレス誘導性rd29Aプロモーターの制御下におけるトランスジェニック植物でのDREB1A(CBF3)cDNAの過剰発現によって、ストレス誘導性DREB1A標的遺伝子の強力な構成的発現が引き起こされ、凍結ストレス、乾燥ストレス及び塩ストレスに対する耐性が向上することを報告している(Liu, Q. et al., (1998) Plant Cell 10, 1391-1406.; Kasuga, M. et al., (1999) Nature Biotechnol. 17, 287-291.)。また、既に本発明者らは、rd29A/lti78/cor78、kin1、kin2/cor6.6、cor15a、rd17/cor47及びerd10等の6個のDREB1A標的遺伝子を同定している(Kasuga, M. et al., (1999) Nature Biotechnol. 17, 287-291.)。しかしながら、トランスジェニック植物におけるDREB1A cDNAの過剰発現が凍結、乾燥及び塩分に対するストレス耐性をどのように高めているのかは、十分には解明されていない。乾燥及び凍結耐性の分子機構を研究するためには、DREB1Aによって制御される遺伝子をより多く同定・分析することが重要である。
【発明の開示】
【発明が解決しようとする課題】
【0012】
ところで、環境ストレスに対する応答性を示す転写因子としては、DREB1Aが知られている(特開2000-60558公報)。DREB1Aに関しては、植物体内でDREB1A遺伝子を過剰に発現させることによって、当該植物体の環境ストレス耐性を上昇させられることができる。しかしながら、植物体の環境ストレス耐性を上昇させられるような転写因子としては、あまり多く知られていないのが現状である。
【0013】
そこで、本発明は、環境ストレス応答性転写因子をコードする遺伝子を提供することを目的とする。
【課題を解決するための手段】
【0014】
本発明者は、上記課題を解決するため鋭意研究を行った結果、cDNAマイクロアレイ分析を応用して、低温ストレス、乾燥ストレス及び塩ストレス等の環境ストレスに特異的に応答性を示す、転写因子をコードする遺伝子を同定することに成功し、本発明を完成するに至った。
【0015】
すなわち、本発明は、以下を包含する。
(1)以下の(a)又は(b)のアミノ酸配列を含む環境ストレス応答性転写因子をコードする遺伝子。
(a) 配列番号2n(n=1〜36の整数)から選ばれるいずれかのアミノ酸配列
(b) 配列番号2n(n=1〜36の整数)から選ばれるいずれかのアミノ酸配列において1若しくは複数のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、環境ストレスに応答性を示し、かつ転写因子活性を有するアミノ酸配列
【0016】
(2)以下の(a)、(b)又は(c)のDNAを含む(1)記載の遺伝子。
【0017】
(a)配列番号2n-1(n=1〜36の整数)から選ばれるいずれかの塩基配列からなるDNA
(b)配列番号2n-1(n=1〜36の整数)から選ばれるいずれかの塩基配列において1若しくは複数の塩基が欠失、置換若しくは付加された塩基配列からなり、環境ストレス応答性転写因子をコードするDNA
(c)配列番号2n-1(n=1〜36の整数)から選ばれるいずれかの塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ環境ストレス応答性転写因子をコードするDNA
【0018】
(3)環境ストレスが低温ストレス、乾燥ストレス及び塩ストレスからなる群から選択される少なくとも1つである(1)又は(2)記載の遺伝子。
(4)(1)又は(2)記載の遺伝子を含む発現ベクター。
(5)(4)記載の発現ベクターを含む形質転換体。
(6)(4)記載の記載の発現ベクターを含むトランスジェニック植物。
【0019】
(7)植物が、植物体、植物器官、植物組織又は植物培養細胞である(6)記載のトランスジェニック植物。
【0020】
(8)(6)又は(7)記載のトランスジェニック植物を培養又は栽培することを特徴とするストレス耐性植物の製造方法。
【発明の効果】
【0021】
本発明により、ストレス応答性転写因子をコードする遺伝子が提供される。本発明の遺伝子は、環境ストレス耐性植物の分子育種に使用できる点で有用である。
【発明を実施するための最良の形態】
【0022】
以下、本発明を詳細に説明する。
本発明者らは、乾燥処理植物及び低温処理植物等の条件の異なる植物から、ビオチン化CAPトラッパー法(Carninci. P. et al., (1996) Genomics, 37, 327-336.)によってシロイヌナズナの完全長cDNAライブラリーを構築し (Seki. M. et al., (1998) Plant J. 15, 707-720.)、ストレス誘導性遺伝子を含む約7,000個の完全長cDNAを用いてシロイヌナズナの完全長cDNAマイクロアレイをそれぞれ調製した。また、これらの乾燥・低温誘導性の完全長cDNAに加えて、ストレス応答性遺伝子の発現をコントロールする転写制御因子であるDREB1Aの標的となる遺伝子を用いてcDNAマイクロアレイを作成した。そして、乾燥ストレス及び低温ストレス下における遺伝子の発現パターンをモニターし、ストレス応答性遺伝子を網羅的に解析した。その結果、約7,000個の完全長cDNAを含むcDNAマイクロアレイから、277個の乾燥誘導性遺伝子、53個の低温誘導性遺伝子及び194個の高塩濃度ストレス誘導性遺伝子を単離した。
【0023】
そして、これら環境ストレス応答性遺伝子の核酸塩基配列をアミノ酸配列に変換したものをGenBank Databaseに登録されているアミノ酸配列Dataに対してblast Xサーチする事により、転写因子をコードするもの(既知の転写因子をコードする遺伝子とE-value:e-10よりも高いスコアで相同性を示すもの)を同定することに成功したものである。以下の説明において、本発明で同定された転写因子を環境ストレス応答性転写因子と称する場合もある。
【0024】
以上のように、完全長cDNAマイクロアレイは、シロイヌナズナの乾燥・低温ストレス誘導性遺伝子の発現様式の解析やストレス関連転写制御因子の標的遺伝子の解析にとって有効なツールである。
1.転写因子の単離
転写因子は、遺伝子の上流に存在するシスエレメントと結合して、その下流の遺伝子の転写を活性化する機能を有するものである。本発明で単離された転写因子は、低温、乾燥、高塩濃度などの環境ストレスにより誘導される。
【0025】
環境ストレス応答性転写因子は、DREBファミリーに属するもの、ERFファミリーに属するもの、ジンクフィンガーファミリーに属するもの、WRKYファミリーに属するもの、MYBファミリーに属するもの、bHLHファミリーに属するもの、NACファミリーに属するもの、ホメオドメインファミリーに属するもの及びbZIPファミリーに属するものに大別される。
【0026】
転写因子を単離するにあたり、まず、マイクロアレイを用いてストレス応答性遺伝子を単離する。マイクロアレイとしては、シロイヌナズナ(Arabidopsis)の全長cDNAライブラリーから単離した遺伝子のほか、RD(Responsive to Dehydration)遺伝子、ERD(Early Responsive to Dehydration)遺伝子、内部標準としてλコントロール鋳型DNA断片(TX803、宝酒造株式会社製)から得られたPCR増幅断片、さらにネガティブコントロールとしてマウスのニコチン酸アセチルコリンレセプターのエプシロンサブユニット(nAChRE)遺伝子及びマウスのグルココルチコイドレセプターの相同性遺伝子からなる計約7000のcDNAを用いることができる。
【0027】
Kurabo製プラスミド調製装置を用いて抽出したプラスミドDNAをシーケンス解析に用いて、DNAシーケンサー(ABI PRISM 3700. PE Applied Biosystems, CA, USA)により配列を決定する。GenBank/EMBLデータベースをもとに、BLASTプログラムを用いて配列のホモロジー検索を行う。
【0028】
次に、ポリAセレクション後,逆転写反応をおこない2本鎖DNAを合成し、cDNAをベクターに挿入する。
【0029】
cDNAライブラリー作成用ベクターに挿入されたcDNAを、cDNAの両側のベクターの配列と相補的なプライマーを用いてPCR法により増幅する。ベクターとしては、λZAPII、λPS等が挙げられる。
【0030】
マイクロアレイは、通常の方法に従って作製することができ、特に限定されるものではない。例えば、gene tipマイクロアレイスタンプマシンGTMASS SYSTEM(Nippon Laser & Electronics Lab.製)を使って、上記PCR産物をマイクロタイタープレートからロードし、マイクロスライドガラスの上に所定間隔でスポットする。その後、非特異的なシグナルの発現を防ぐためにスライドをブロッキング・ソルーションに浸す。
【0031】
植物材料としては野生型のほか、特定の遺伝子の破壊株等が挙げられるが、DREB1AのcDNAが導入されたトランスジェニック植物を用いることができる。植物種は、シロイヌナズナ、タバコ,イネ等が挙げられるが、シロイヌナズナが好ましい。
【0032】
乾燥及び低温ストレス処理は公知方法の方法で行うことができる(Yamaguchi-Shinozaki, K., and Shinozaki, K. (1994) Plant Cell 6, 251-264.)。
【0033】
ストレス処理にさらした後は、植物体(野生型及びDREB1A過剰発現型形質転換体)をサンプリングし、液体窒素を用いて凍結保存する。野生型及びDREB1A過剰発現型形質転換体を、DREB1Aの標的遺伝子を同定するための実験に用いる。植物体から、公知方法又はキットを用いてmRNAを単離精製する。
【0034】
標識用Cy3 dUTP又はCy5 dUTP(Amersham Pharmacia)の存在下でそれぞれのmRNAサンプルの逆転写を行い、ハイブリダイゼーションに用いる。
【0035】
ハイブリダイゼーション後は、走査レーザー顕微鏡等を用いてマイクロアレイをスキャンする。マイクロアレイのデータ解析用プログラムとして、Imagene Ver 2.0(BioDiscovery)とQuantArray(GSI Lumonics)等を用いることができる。
【0036】
スキャン後は、目的とする遺伝子をもつプラスミドを調製することにより、遺伝子が単離される。
【0037】
転写因子の決定は、上記単離された遺伝子の塩基配列を解析し、データベース(GenBank/EMBL, ABRC)のゲノム情報をもとに、遺伝子解析用プログラムを用いて行われる。単離された遺伝子は、乾燥ストレス誘導性及び低温ストレス誘導性の両性質を有するもの、乾燥ストレス誘導性に特異的なもの、低温ストレス誘導性に特異的なものに分類することができる。遺伝子解析用プログラムによれば、上記遺伝子の中から36種の転写因子をコードする遺伝子が同定される。これら36種類の転写因子をコードする遺伝子の塩基配列を配列番号2n-1(nは1〜36の整数)、転写因子のアミノ酸配列を配列番号2n(nは1〜36の整数)に示す。配列番号と転写因子をコードする遺伝子の遺伝子名とを表1に示す
【0038】
【表1】

【0039】
但し、本発明の転写因子が環境ストレス応答性転写因子として機能する限り、配列番号2n-1(nは1〜36の整数)から選ばれるいずれかの塩基配列において1又は複数個、好ましくは1又は数個(例えば1〜10個、1〜5個)の塩基が欠失、置換又は付加された塩基配列を有するものでもよい。さらに、配列番号2n-1(nは1〜36の整数)から選ばれるいずれかの塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ環境ストレス応答性転写因子をコードするDNAも、本発明の転写因子に含まれる。ここで、ストリンジェントな条件とは、ナトリウム濃度が25〜500mM、好ましくは25〜300mMであり、温度が42〜68℃、好ましくは42〜65℃である。より具体的には、5×SSC(83mM NaCl、83mMクエン酸ナトリウム)、温度42℃である。
【0040】
本発明で単離した36種類の転写因子については以下のように分類することができる。
1)DREBファミリー:RAFL05-11-M11、RAFL06-11-K21、RAFL05-16-H23、RAFL08-16-D16
2)ERFファミリー:RAFL08-16-G17、RAFL06-08-H20
3)ジンクフィンガーファミリー:RAFL07-10-G04、RAFL04-17-D16、RAFL05-19-M20、RAFL08-11-M13、RAFL04-15-K19、RAFL05-11-L01、RAFL05-14-C11、RAFL05-19-G24、RAFL05-20-N02
4)WRKYファミリー:RAFL05-18-H12、RAFL05-19-E19、RAFL06-10-D22、RAFL06-12-M01
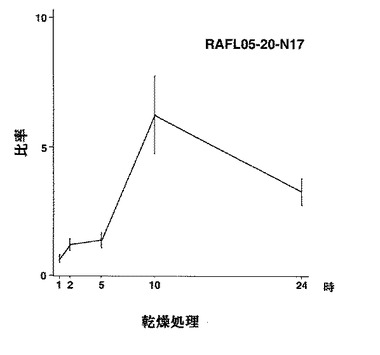
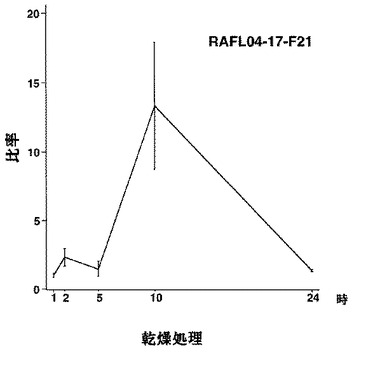
5)MYBファミリー:RAFL05-14-D24、RAFL05-20-N17、RAFL04-17-F21
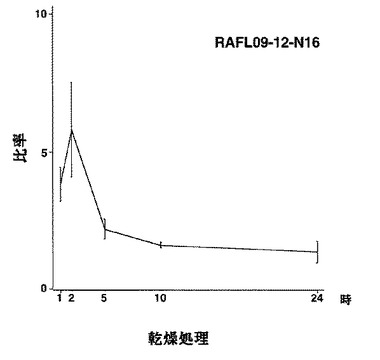
6)bHLHファミリー:RAFL09-12-N16
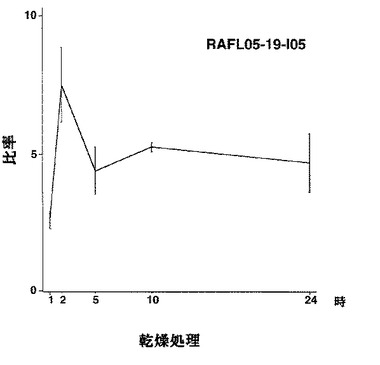
7)NACファミリー:RAFL05-19-I05、RAFL05-21-I22、RAFL08-11-H20、RAFL05-21-C17、RAFL05-08-D06
8)ホメオドメインファミリー:RAFL05-20-M16、RAFL11-01-J18、RAFL11-09-C20
9)bZIPファミリー:RAFL05-18-N16、RAFL11-10-D10、RAFL04-17-N22、RAFL05-09-G15
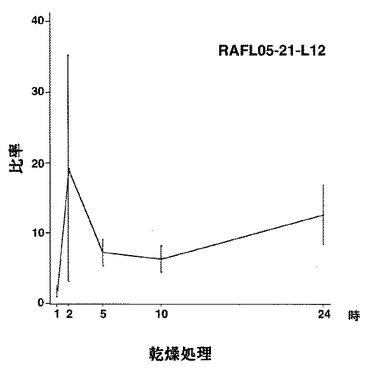
なお、RAFL05-21-L12については上記1)〜9)に分類できない。
【0041】
一旦本発明の転写因子をコードする遺伝子の塩基配列が確定されると、その後は化学合成によって、又はクローニングされたプローブを鋳型としたPCRによって、あるいは該塩基配列を有するDNA断片をプローブとしてハイブリダイズさせることによって、本発明の転写因子をコードする遺伝子を得ることができる。さらに、部位特異的突然変異誘発法等によって本発明の転写因子をコードする遺伝子の変異型であって変異前の転写因子をコードする遺伝子と同等の機能を有するものを合成することもできる。
【0042】
なお、転写因子をコードする遺伝子の塩基配列に変異を導入するには、Kunkel法、Gapped duplex法等の公知の手法又はこれに準ずる方法を採用することができる。例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-K(TAKARA社製)やMutant-G(TAKARA社製))などを用いて、あるいは、TAKARA社のLA PCR in vitro Mutagenesis シリーズキットを用いて変異の導入が行われる。
【0043】
ここで、「環境ストレス」とは、一般には非生物的ストレスを意味し、例えば乾燥ストレス、低温ストレス、高塩濃度ストレス、強光ストレス等をいう。「乾燥」とは水分が欠乏した状態を意味し、「低温」とはそれぞれの生物種の生活至適温度よりも低い温度にさらされた状態(例えばシロイヌナズナの場合-20〜+21℃の温度を継続的に1時間〜数週間さらすことをいう。また、「高塩濃度」とは、50mM〜600mMの濃度のNaClを継続的に0.5時間〜数週間処理したときの状態を意味する。「強光ストレス」とは、光合成能を超える強光が植物に照射された状態を意味し、例えば5,000〜10,000Lx以上の光が照射した場合が該当する。これらの環境ストレスは、1種類のものを負荷してもよく、複数種類のものを負荷してもよい。
【0044】
2.発現ベクターの構築
本発明の発現ベクターは、適当なベクターに本発明の転写因子をコードする遺伝子を連結(挿入)することにより得ることができる。本発明の転写因子をコードする遺伝子を挿入するためのベクターは、宿主中で複製可能なものであれば特に限定されず、例えばプラスミド、シャトルベクター、ヘルパープラスミドなどが挙げられる。
【0045】
プラスミド DNAとしては、大腸菌由来のプラスミド(例えばpBR322、pBR325、pUC118、pUC119、pUC18、pUC19、pBluescript等)、枯草菌由来のプラスミド(例えばpUB110、pTP5等)、酵母由来のプラスミド(例えばYEp13、YCp50等)などが挙げられ、ファージDNAとしてはλファージ(Charon4A、Charon21A、EMBL3、EMBL4、λgt10、λgt11、λZAP等)が挙げられる。さらに、レトロウイルス又はワクシニアウイルスなどの動物ウイルス、バキュロウイルスなどの昆虫ウイルスベクターを用いることもできる。
【0046】
ベクターに本発明の転写因子をコードする遺伝子を挿入するには、まず、精製されたDNAを適当な制限酵素で切断し、適当なベクター DNAの制限酵素部位又はマルチクローニングサイトに挿入してベクターに連結する方法などが採用される。
【0047】
本発明の転写因子をコードする遺伝子は、その3'末端にレポーター遺伝子、例えば、植物で広く用いられているGUS遺伝子を連結して用いれば、GUS活性を調べることで当該遺伝子の発現の強さを容易に評価することができる。なお、レポーター遺伝子としては、GUS遺伝子以外にも、ルシフェラーゼ、グリーンフルオレセイントプロテインなども用いることができる。
【0048】
3.形質転換体の作製
本発明の形質転換体は、本発明の発現ベクターを宿主中に導入することにより得ることができる。ここで、宿主としては、本発明の転写因子をコードする遺伝子を発現できるものであれば特に限定されるものではないが、植物が好ましい。宿主が植物である場合は、形質転換植物(トランスジェニック植物)は以下のようにして得ることができる。
【0049】
本発明において形質転換の対象となる植物は、植物体全体、植物器官(例えば葉、花弁、茎、根、種子等)、植物組織(例えば表皮、師部、柔組織、木部、維管束等)又は植物培養細胞のいずれをも意味するものである。形質転換に用いられる植物としては、アブラナ科、イネ科、ナス科、マメ科等に属する植物(下記参照)が挙げられるが、これらの植物に限定されるものではない。
アブラナ科:シロイヌナズナ(Arabidopsis thaliana)
ナス科:タバコ(Nicotiana tabacum)
イネ科:トウモロコシ(Zea mays) 、イネ(Oryza sativa)
マメ科:ダイズ(Glycine max)
【0050】
上記組換えベクターは、通常の形質転換方法、例えば電気穿孔法(エレクトロポレーション法)、アグロバクテリウム法、パーティクルガン法、PEG法等によって植物中に導入することができる。
【0051】
例えばエレクトロポレーション法を用いる場合は、パルスコントローラーを備えたエレクトロポレーション装置により、電圧500〜1600V、25〜1000μF、20〜30msecの条件で処理し、遺伝子を宿主に導入する。
【0052】
また、パーティクルガン法を用いる場合は、植物体、植物器官、植物組織自体をそのまま使用してもよく、切片を調製した後に使用してもよく、プロトプラストを調製して使用してもよい。このように調製した試料を遺伝子導入装置(例えばBio-Rad社のPDS-1000/He等)を用いて処理することができる。処理条件は植物又は試料により異なるが、通常は1000〜1800psi程度の圧力、5〜6cm程度の距離で行う。
【0053】
また、植物ウイルスをベクターとして利用することによって、目的遺伝子を植物体に導入することができる。利用可能な植物ウイルスとしては、例えば、カリフラワーモザイクウイルスが挙げられる。すなわち、まず、ウイルスゲノムを大腸菌由来のベクターなどに挿入して組換え体を調製した後、ウイルスのゲノム中に、これらの目的遺伝子を挿入する。このようにして修飾されたウイルスゲノムを制限酵素によって組換え体から切り出し、植物宿主に接種することによって、目的遺伝子を植物宿主に導入することができる。
【0054】
アグロバクテリウムのTiプラスミドを利用する方法においては、アグロバクテリウム(Agrobacterium)属に属する細菌が植物に感染すると、それが有するプラスミドDNAの一部を植物ゲノム中に移行させるという性質を利用して、目的遺伝子を植物宿主に導入する。アグロバクテリウム属に属する細菌のうちアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)は、植物に感染してクラウンゴールと呼ばれる腫瘍を形成し、また、アグロバクテリウム・リゾゲネス(Agrobacteriumu rhizogenes)は、植物に感染して毛状根を発生させる。これらは、感染の際にTiプラスミド又はRiプラスミドと呼ばれる各々の細菌中に存在するプラスミド上のT-DNA領域(Transferred DNA)と呼ばれる領域が植物中に移行し、植物のゲノム中に組み込まれることに起因するものである。
【0055】
Ti又はRiプラスミド上のT-DNA領域中に、植物ゲノム中に組み込みたいDNAを挿入しておけば、アグロバクテリウム属の細菌が植物宿主に感染する際に目的とするDNAを植物ゲノム中に組込むことができる。
【0056】
形質転換の結果得られる腫瘍組織やシュート、毛状根などは、そのまま細胞培養、組織培養又は器官培養に用いることが可能であり、また従来知られている植物組織培養法を用い、適当な濃度の植物ホルモン(オーキシン、サイトカイニン、ジベレリン、アブシジン酸、エチレン、ブラシノライド等)の投与などにより植物体に再生させることができる。
【0057】
本発明のベクターは、上記植物宿主に導入するのみならず、大腸菌(Escherichia coli)等のエッシェリヒア属、バチルス・ズブチリス(Bacillus subtilis)等のバチルス属、又はシュードモナス・プチダ(Pseudomonas putida)等のシュードモナス属に属する細菌、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)等の酵母、COS細胞、CHO細胞等の動物細胞、あるいはSf9等の昆虫細胞などに導入して形質転換体を得ることもできる。大腸菌、酵母等の細菌を宿主とする場合は、本発明の組換えベクターが該細菌中で自律複製可能であると同時に、本発明の転写因子をコードする遺伝子、当該遺伝子の上流に位置するリボソーム結合配列及びプロモーター配列、当該遺伝子の下流に位置する転写終結配列により構成されていることが好ましい。
【0058】
細菌への組換えベクターの導入方法は、細菌にDNAを導入する方法であれば特に限定されるものではない。例えばカルシウムイオンを用いる方法、エレクトロポレーション法等が挙げられる。
【0059】
酵母を宿主とする場合は、例えばサッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)などが用いられる。酵母への組換えベクターの導入方法は、酵母にDNAを導入する方法であれば特に限定されず、例えばエレクトロポレーション法、スフェロプラスト法、酢酸リチウム法等が挙げられる。
【0060】
動物細胞を宿主とする場合は、サル細胞COS-7、Vero、チャイニーズハムスター卵巣細胞(CHO細胞)、マウスL細胞などが用いられる。動物細胞への組換えベクターの導入方法としては、例えばエレクトロポレーション法、リン酸カルシウム法、リポフェクション法等が挙げられる。
【0061】
昆虫細胞を宿主とする場合は、Sf9細胞などが用いられる。昆虫細胞への組換えベクターの導入方法としては、例えばリン酸カルシウム法、リポフェクション法、エレクトロポレーション法などが挙げられる。
【0062】
遺伝子が宿主に組み込まれたか否かの確認は、PCR法、サザンハイブリダイゼーション法、ノーザンハイブリダイゼーション法等により行うことができる。例えば、形質転換体からDNAを調製し、DNA特異的プライマーを設計してPCRを行う。PCRは、前記プラスミドを調製するために使用した条件と同様の条件で行われる。その後は、増幅産物についてアガロースゲル電気泳動、ポリアクリルアミドゲル電気泳動又はキャピラリー電気泳動等を行い、臭化エチジウム、SYBR Green液等により染色し、そして増幅産物を1本のバンドとして検出することにより、形質転換されたことを確認する。また、予め蛍光色素等により標識したプライマーを用いてPCRを行い、増幅産物を検出することもできる。さらに、マイクロプレート等の固相に増幅産物を結合させ、蛍光又は酵素反応等により増幅産物を確認する方法も採用してもよい。
【0063】
4.植物の製造
本発明においては、上記形質転換植物細胞等から形質転換植物体に再生することができる。再生方法としては、カルス状の形質転換細胞をホルモンの種類、濃度を変えた培地へ移して培養し、不定胚を形成させ、完全な植物体を得る方法が採用される。使用する培地としては、LS培地、MS培地などが例示される。
【0064】
本発明の「植物体を製造する方法」は、上記環境ストレス応答性転写因子をコードする遺伝子を挿入した植物発現ベクターを宿主細胞に導入して形質転換植物細胞を得て、該形質転換植物細胞から形質転換植物体を再生し、得られた形質転換植物体から植物種子を得て、該植物種子から植物体を生産する工程を含む。
【0065】
形質転換植物体から植物種子を得るには、例えば、形質転換植物体を発根培地から採取し、水を含んだ土を入れたポットに移植し、一定温度下で生育させて、花を形成させ、最終的に種子を形成させる。また、種子から植物体を生産するには、例えば、形質転換植物体上で形成された種子が成熟したところで、単離して、水を含んだ土に播種し、一定温度、照度下で生育させることにより、植物体を生産する。このようにして育種された植物は、導入された環境ストレス応答性転写因子をコードする遺伝子のストレス応答性に応じた環境ストレス耐性植物となる。
【実施例】
【0066】
以下、実施例により本発明をさらに具体的に説明する。但し、本発明はこれら実施例にその技術的範囲が限定されるものではない。
【0067】
〔実施例1〕 環境ストレス応答性転写因子をコードする遺伝子の単離
1.材料と方法
(1) Arabidopsis cDNAクローン
Arabidopsisの全長cDNAライブラリーから単離した遺伝子に加えて、RD(Responsive to Dehydration)遺伝子、ERD(Early Responsive to Dehydration)遺伝子、kin1遺伝子、kin2遺伝子、cor15a遺伝子、また内部標準としてλコントロール鋳型DNAをPCRで増幅した断片、さらにネガティブコントロールとしてマウスのニコチン酸アセチルコリンレセプターのエプシロンサブユニット(nAChRE)遺伝子及びマウスのグルココルチコイドレセプターの相同性遺伝子の計約7000個のcDNAをマイクロアレイ作成に用いた。
陽性対照:乾燥誘導遺伝子(脱水応答性:rd、及び初期脱水応答遺伝子:erd)
内部標準:λコントロール鋳型DNAをPCRで増幅した断片(TX803、宝酒造社製、以下「コントロール断片」と称する)
陰性対照:非特異的ハイブリダイゼーションを評価するためにArabidopsisのデータベースにある任意の配列とは実質的にホモロジーを有さないニコチン性アセチルコリン受容体εサブユニット(nAChRE)遺伝子及びマウスグルココルチコイド受容体ホモログ遺伝子
【0068】
(2) Arabidopsis 全長cDNAマイクロアレイ
ビオチニル化CAPトラッパー法を用いて、本発明者はArabidopsisの植物体から、異なる条件(例えば、発芽から成熟種子までの種々の成長段階における乾燥処理、低温処理及び未処理)で全長cDNAライブラリーを構築した。全長cDNAライブラリーから、本発明者は、約7000個の独立したArabidopsis全長cDNAをそれぞれ単離した。公知の手法(Eisen and Brown, 1999)に従って、PCRで増幅したcDNA断片をスライドグラス上の整列させた。本発明者は、以下の遺伝子を含む約7000個のArabidopsisの全長cDNAを含有する全長cDNAマイクロアレイを調製した。
【0069】
(3) cDNAマイクロアレイを用いた乾燥誘導性遺伝子、低温誘導性遺伝子及び高塩濃度誘導性遺伝子
本例では、約7000個のArabidopsisの全長cDNAを含有する全長cDNAマイクロアレイを用いて、乾燥誘導性遺伝子、低温誘導性遺伝子及び高塩濃度誘導性遺伝子を単離した。
【0070】
上述した各種ストレスを受けた植物及びストレスを受けていない植物のCy3及びCy5蛍光標識プローブを混合し、約7000個のArabidopsisの全長cDNAを含有する全長cDNAマイクロアレイとハイブリダイズさせた。一方のmRNAサンプルをCy3-dUTPで標識し、他方のmRNAサンプルをCy5-dUTPで標識するcDNAプローブ対の二重標識によって、マイクロアレイ上のDNAエレメントへの同時ハイブリダイゼーションが可能となり、2種の異なる条件間(即ち、ストレス有り又はストレス無し)における遺伝子発現の直接的な定量測定が容易になる。ハイブリダイズさせたマイクロアレイを、各DNAエレメントからのCy3及びCy5発光について2つの別個のレーザーチャネルによって走査した。次いで、各DNAエレメントの2つの蛍光シグナルの強度比を相対値として測定し、マイクロアレイ上のcDNAスポットで表される遺伝子のディファレンシャル発現の変化を判定した。本実施例では、分析を行う2種の実験条件下で発現レベルがほぼ同等であるα-チューブリン遺伝子を内部対照遺伝子として使用した。
約7000個のArabidopsisの全長cDNAを含有する全長cDNAマイクロアレイにおける、乾燥誘導性遺伝子、低温誘導性遺伝子及び高塩濃度誘導性遺伝子の同定手順を示す。
【0071】
上述したストレスのうち1種類のストレスを負荷した植物由来のmRNA及びストレスを受けていない野生型植物由来のmRNAを、それぞれCy3-標識cDNAプローブ及びCy5-標識cDNAプローブの調製に使用した。これらのcDNAプローブを混合し、cDNAマイクロアレイとハイブリダイズさせた。本実施例では、2種の条件下で発現レベルがほぼ同等であるコントロール断片を内部対照遺伝子として使用した。発現比率(乾燥/ストレス無し、低温/ストレス無し、又は高塩濃度/ストレス無し)がコントロール断片の5倍を超える遺伝子を、負荷したストレスによって誘導される遺伝子とした。
【0072】
(4) 配列の解析
遺伝子配列のホモロジー検索をおこなうため、DNA抽出装置(model Biomek、ベックマンコールター社製)を用いて抽出しマルチスクリーン96穴フィルタープレート(ミリポア社製)を用いて精製したプラスミドDNAを配列解析に用いた。配列解析は、DNAシーケンサー(ABI PRISM 3700. PE Applied Biosystems, CA, USA)を用いてダイターミネーターサイクルシーケンス法により決定した。GenBank/EMBLデータベースをもとに、BLASTプログラムを用いて配列のホモロジー検索を行った。
【0073】
(5) cDNAの増幅
cDNAライブラリー作製用ベクターとして、λZAP及びλFLC-1を用いた。ライブラリー用のベクターに挿入されたcDNAを、cDNAの両側のベクターの配列と相補的なプライマーを用いてPCR法により増幅した。
プライマーの配列は以下の通りである。
FL forward 1224:5'-CGCCAGGGTTTTCCCAGTCACGA(配列番号73)
FL reverse 1233:5'-AGCGGATAACAATTTCACACAGGA(配列番号74)
【0074】
100 μl のPCR混合液(0.25 mM dNTP,0.2 μM PCRプライマー,1 X Ex Taq バッファー,1.25 U Ex Taqポリメラーゼ (宝酒造製))に、テンプレートとしてプラスミド(1-2 ng)を加えた。PCRは、最初に94℃で3分反応させた後、続いて95℃で1分、60℃で30秒及び72℃で3分のサイクルを35サイクル、最後に72℃で3分の条件で行った。PCR産物をエタノール沈澱させた後、25μlの3 X SSCに溶かした。0.7%アガロースゲルを用いた電気泳動により、得られたDNAの質とPCRの増幅効率を確認した。
【0075】
(6) cDNAマイクロアレイの作製
gene tipマイクロアレイスタンプマシンGTMASS SYSTEM(Nippon Laser & Electronics Lab.製)を使って、0.5μlのPCR産物(500-1000 ng/ml)を384穴のマイクロタイタープレートからロードし、48枚のマイクロスライドガラス(model Super Aldehyde substrate、Telechem International製)の上に300μmの間隔で0.5nlずつスポットした。スポット後のスライドを、相対湿度30%以下の雰囲気中で乾燥処理し、架橋反応のための紫外線照射に供した。
【0076】
その後、スライドを0.2%SDS中で2分間震盪する処理を3回行った後、蒸留水中で浸透する処理を2回行った。その後、スライドをスライドラックに置き、熱湯を入れたチャンバーにスライドラックを入れて2分間放置した。その後、ブロッキング溶液(1gのホウ化水素、300mlのPBS及び90mlの100%エタノールを含む)チャンバー注いだ。スライドラックを入れたガラスチャンバーを緩やかに振った後、0.2%SDSを含むチャンバーにスライドラックを移して1分間静かに震盪する処理を3回行った。その後、蒸留水を入れたガラスチャンバーにスライドラックを移して1分間緩やかに振った後、20分間遠心処理することによって乾燥させた。
【0077】
(7) 植物材料とRNAの単離
植物材料として、寒天培地に播種して3週間栽培した(Yamaguchi-ShinozakiとShinozaki,1994)野生型及びカリフラワーモザイクウイルスの35SプロモーターにDREB1AのcDNA(Kasugaなど,1999)をつないで導入したシロイヌナズナ(コロンビア種)の植物体を用いた。乾燥及び低温ストレス処理はYamaguchi-ShinozakiとShinozaki(1994)の方法で行った。すなわち、寒天培地から引き抜いた植物体をろ紙上に置き,22℃,相対湿度60%の条件で乾燥処理をおこなった。22℃で栽培した植物体を4℃に移すことにより低温処理をおこなった。また、高塩濃度ストレス処理は、250mMのNaClを含む水溶液内で水耕することによりおこなった。
【0078】
野生型の植物体を2時間又は10時間のストレス処理にさらした後サンプリングし、液体窒素を用いて凍結保存した。また、カナマイシンを加えない寒天培地で栽培した野生型及びDREB1A過剰発現型形質転換体を、DREB1Aの標的遺伝子を同定するための実験に用いた。DREB1A過剰発現型形質転換体に対しては、ストレス処理を行わなかった。植物体から、ISOGEN(Nippon gene, Tokyo, Japan)を用いてトータルRNAを単離し、Oligotex-dT30 mRNA精製キット(Takara,Tokyo,Japan)を用いてmRNAを単離精製した。
【0079】
(8) プローブの蛍光標識
Cy3 dUTP又はCy5 dUTP(Amersham Pharmacia)の存在下でそれぞれのmRNAサンプルの逆転写を行った。具体的に逆転写反応は、内部標準として1ngのλポリA+RNA-A(TX802、宝酒造社製)を含む1μgの変性ポリ(A)+、50ng/μlの12〜18merオリゴdTプライマー(ライフテクノロジー社製)、それぞれ0.5mMのdATP、dGDP及びdCTP、0.2mMのdTTP、0.1mMのCy3 dUTP若しくはCy5 dUTP、100UのRnase阻害剤、10mMのDTT及び200UのSuperscript II逆転写酵素を含む1×Superscript first-strand buffer(50mM Tris-HCl, pH8.3, 75mM KCl, 3mM MgCl2, 20mM DTTを含有。ライフテクノロジー社製)中で全量20μlとして行った。
【0080】
上記組成の反応溶液を42℃で35分間インキュベーションした後、200UのSuperscript II逆転写酵素を加え、更に42℃で35分間インキュベーションした。その後、反応溶液に0.5MのEDTAを5μl、1Nの水酸化ナトリウムを10μl及び20μLの蒸留水を添加することで、当該反応溶液における酵素反応を停止させるとともに鋳型を分解した。その後、反応溶液を65℃で1時間インキュベーションした。その後、反応溶液を1MのTris-HCl(pH7.5)で中和した。
【0081】
反応溶液をMicrocon 30 micro concentrator(アミコン社製)に移した。250μlのTEバッファーを加えてバッファー量が10μlになるまで遠心し、流出液を捨てる工程を4回繰り返した。反応溶液に含まれるプローブを遠心によって回収し数μlの蒸留水を加えた。得られたプローブに、5.1μlの20 X SSC、2μg/μlの酵母 tRNA、4.8μlの2%SDSを加えた。さらに、サンプルを100℃で2分間変成処理し、室温に5分間置いた後ハイブリダイゼーションに用いた。
【0082】
(9) マイクロアレイハイブリダイゼーション及びスキャニング
プローブをbenchtop micro centrifugeを用いて1分間の高速遠心にかけた。泡の発生を避けるために、プローブをアレイの中央に置きその上にカバースリップをかぶせた。スライドガラス上に5μlの3 X SSCを4滴落として、チェンバーを適度な湿度に保ち、ハイブリダイゼーション中のプローブの乾燥を防いだ。スライドガラスをハイブリダイゼーション用のカセット(THC-1,BM機器)に入れて密封した後、65℃で12〜16時間処理した。スライドガラスを取り出してスライドラックに置き、溶液1(2X SSC,0.03%SDS)中でカバースリップを慎重にはずした後ラックを振って洗浄し、ラックを溶液2(1X SSC)中に移して2分間洗浄した。さらにラックを溶液3(0.05X SSC)に移して2分間放置し、遠心(2500g, 1min)にかけて乾燥させた。
【0083】
走査レーザー顕微鏡(ScanArray4000; GSI Lumonics,Watertown,MA)を用いて1ピクセルあたり10μmの解像度でマイクロアレイをスキャンした。マイクロアレイのデータ解析用プログラムとして、QuantArrayバージョン2.0(GSI Lumonics)を用いた。バックグラウンドの蛍光は、陰性対照遺伝子(ニコチン性アセチルコリン受容体εサブユニット(nAChRE)遺伝子及びマウスグルココルチコイド受容体ホモログ遺伝子)の蛍光シグナルに基づいて算出した。蛍光シグナル値が1000未満(この値はバックグラウンドの蛍光シグナル値の2倍未満を意味する)のサンプルに関しては分析対象としなかった。遺伝子クラスタリング分析はGenespring(シリコンジェネティック社製)を用いて行った。
【0084】
(10) ノーザン解析
トータルRNAを用いてノーザン解析を行った(Yamaguchi-ShinozakiとShinozaki,1994)。シロイヌナズナの全長cDNAライブラリーからPCR法によって単離したDNA断片をノーザンハイブリダイゼーションのプローブとして用いた。
【0085】
(11) 転写因子をコードする遺伝子の決定
データベース(GenBank/EMBL, ABRC)のシロイヌナズナのゲノム情報をもとに、遺伝子解析用プログラムBLASTを用いて転写因子をコードする遺伝子を解析した。
【0086】
2.結果
(1) ストレス誘導性遺伝子の同定
ストレスを受けていないシロイヌナズナ植物から単離したmRNAから、Cy5-dUTPの存在下で逆転写を行って蛍光標識cDNAを調製した。乾燥処理、低温処理(2時間)或いは高塩濃度処理を行った植物から、Cy3-dUTPで標識した第2のプローブを調製した。両プローブを約7000個のシロイヌナズナcDNAクローンを含むcDNAマイクロアレイへ同時にハイブリダイズさせた後、疑似カラー像を作製した。
【0087】
ストレスによって誘導された遺伝子及び抑制された遺伝子は、それぞれ赤色及び緑色のシグナルで表される。両方の処理においてほぼ同レベルで発現した遺伝子は、黄色のシグナルとなる。各スポットの強度は、各遺伝子の発現量の絶対値に相当する。低温誘導性遺伝子(rd29A)は赤色のシグナルでとなり、コントロール断片(内部対照)は黄色シグナルとなる。
【0088】
マイクロアレイをスキャンした結果、乾燥処理によって特異的に誘導された277個の遺伝子を同定し、低温処理によって特異的に誘導された53個の遺伝子を同定し、高塩濃度処理によって特異的に誘導された194個の遺伝子を同定した。なお、コントロール断片の発現比と比較して5倍以上の発現比で誘導された遺伝子を、各種ストレスによって誘導された遺伝子とした。
【0089】
データベースを用いた解析の結果、以下のファミリーに分類できる35種類の転写因子を同定することができた。なお、RAFL05-21-L12については以下のファミリーに分類することはできなかったが、核酸塩基配列をアミノ酸配列に変換したものをGenBank Databaseに登録されているアミノ酸配列Dataに対してblast Xサーチしたところ、既知の転写因子をコードするもの(heat shock transcription factor-like protein)とE-value:e-100のスコアで相同性を示したことから転写因子として同定することができた。すなわち、本実施例によって、36種類の転写因子を同定できた。
1)DREBファミリー:RAFL05-11-M11、RAFL06-11-K21、RAFL05-16-H23、RAFL08-16-D06
2)ERFファミリー:RAFL08-16-G17、RAFL06-08-H20
3)ジンクフィンガーファミリー:RAFL07-10-G04、RAFL04-17-D16、RAFL05-19-M20、RAFL08-11-M13、RAFL04-15-K19、RAFL05-11-L01、RAFL05-14-C11、RAFL05-19-G24、RAFL05-20-N02
4)WRKYファミリー:RAFL05-18-H12、RAFL05-19-E19、RAFL06-10-D22、RAFL06-12-M01
5)MYBファミリー:RAFL05-14-D24、RAFL05-20-N17、RAFL04-17-F21
6)bHLHファミリー:RAFL09-12-N16
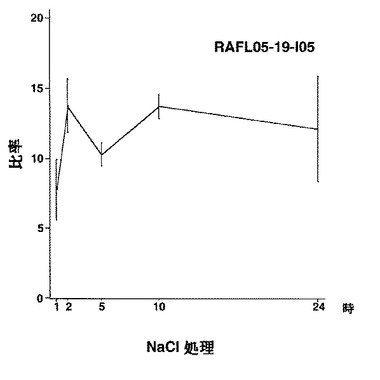
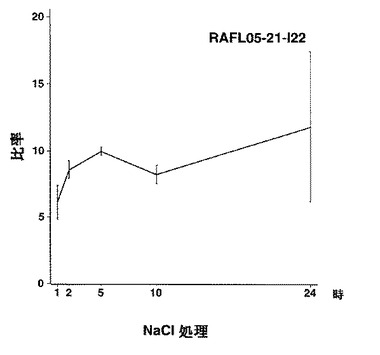
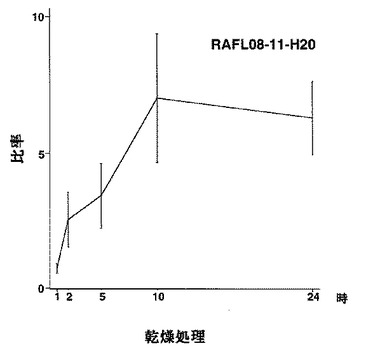
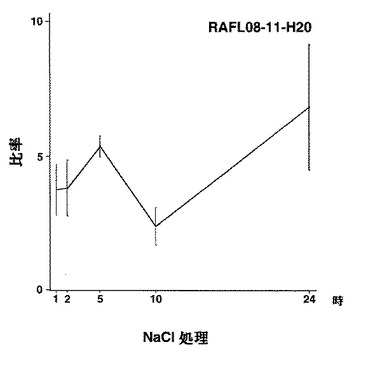
7)NACファミリー:RAFL05-19-I05、RAFL05-21-I22、RAFL08-11-H20、RAFL05-21-C17、RAFL05-08-D06
8)ホメオドメインファミリー:RAFL05-20-M16、RAFL11-01-J18、RAFL11-09-C20
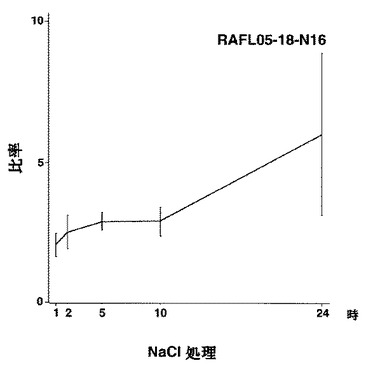
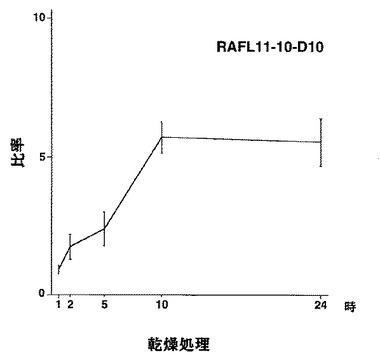
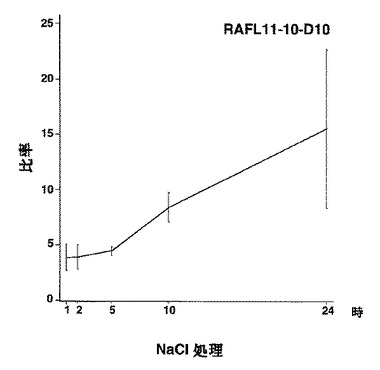
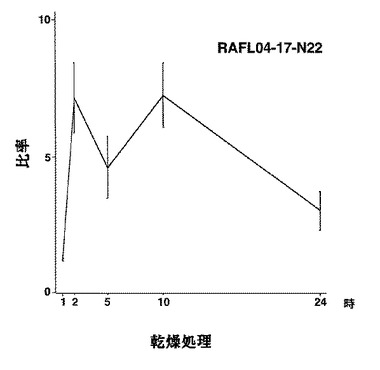
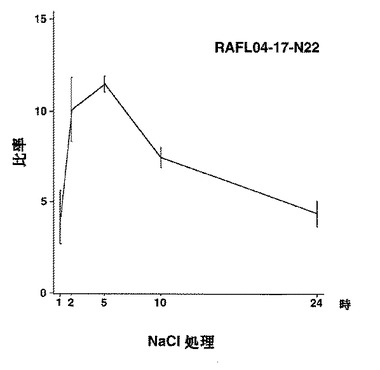
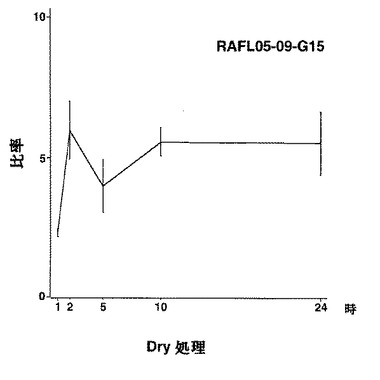
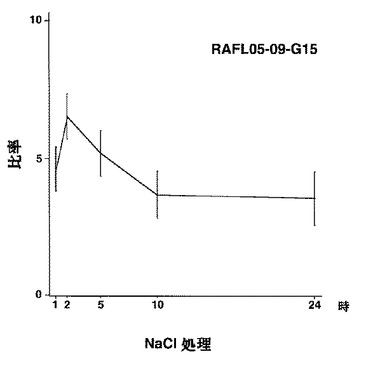
9)bZIPファミリー:RAFL05-18-N16、RAFL11-10-D10、RAFL04-17-N22、RAFL05-09-G15
【0090】
(2) 各種ストレス処理時間と発現比率との関係
上述したように単離された36種類の各種ストレス応答性転写因子をコードする遺伝子について、以下のように、各種ストレス処理時間と発現比率との関係を検討した結果を図1〜図57に示す。図1〜図57で示す遺伝子名とストレス処理との対応を表2に示した。
【0091】
【表2】
【0092】
図1〜図57において、縦軸は遺伝子の発現比率を示しており、以下のように算出される。なお、下記式においてFIは、蛍光強度を示す。
発現比率=[(ストレス負荷条件での各cDNAのFI)/(ストレスを負荷しない条件での各cDNAのFI)]÷[(ストレス負荷条件でのコントロール断片のFI)/(ストレスを負荷しない条件でのコントロール断片のFI)]
【0093】
これら図1〜図57に示すように、本方法により単離されたストレス応答性転写因子をコードする遺伝子は、それぞれ異なるプロファイルであるが、各種ストレスの付加により発現誘導されていることが判る。
【図面の簡単な説明】
【0094】
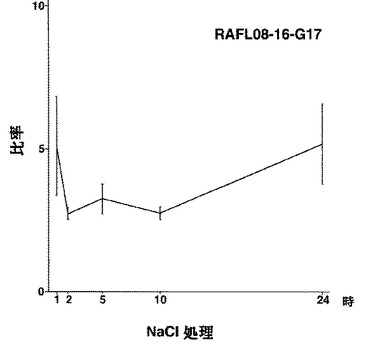
【図1】RAFL08-16-G17について高塩濃度ストレスと発現比率との関係を示す特性図である。
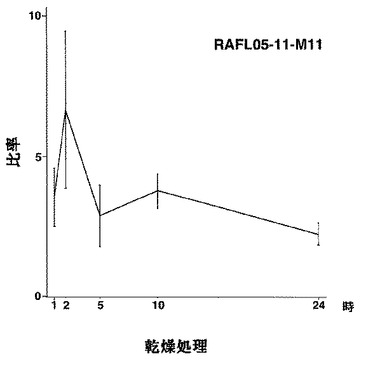
【図2】RAFL05-11-M11について乾燥ストレスと発現比率との関係を示す特性図である。
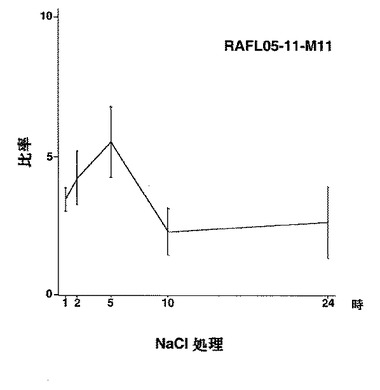
【図3】RAFL05-11-M11について高塩濃度ストレスと発現比率との関係を示す特性図である。
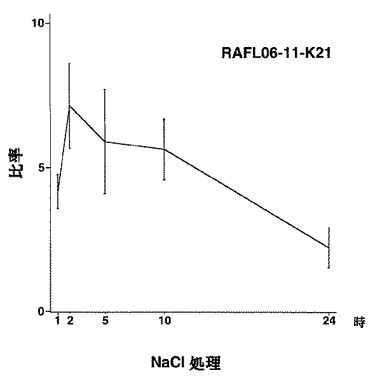
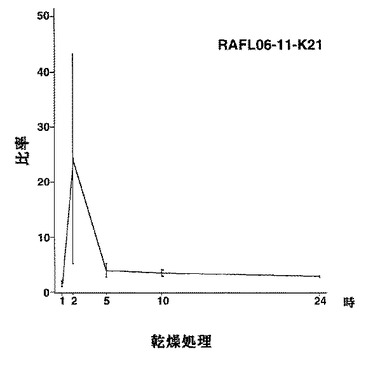
【図4】RAFL06-11-K21について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図5】RAFL06-11-K21について乾燥ストレスと発現比率との関係を示す特性図である。
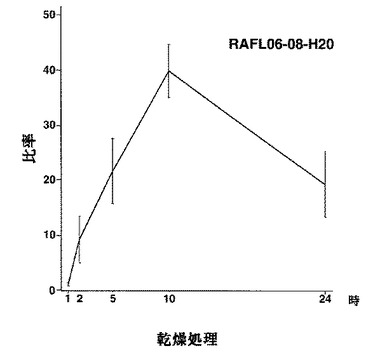
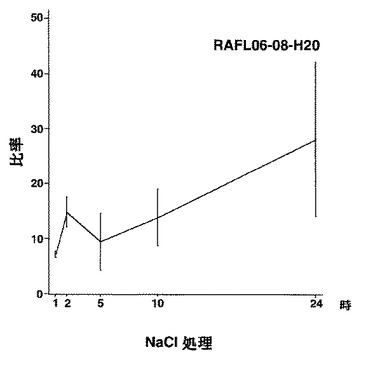
【図6】RAFL06-08-H20について乾燥ストレスと発現比率との関係を示す特性図である。
【図7】RAFL06-08-H20について高塩濃度ストレスと発現比率との関係を示す特性図である。
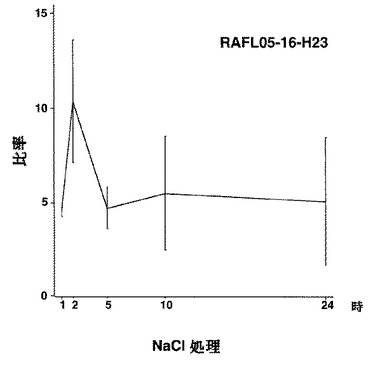
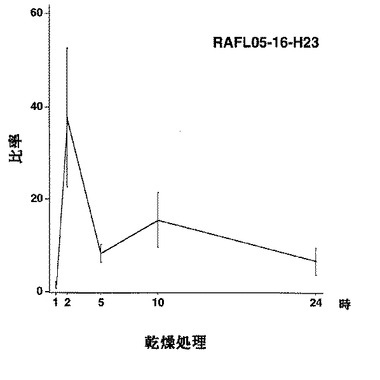
【図8】RAFL05-16-H23について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図9】RAFL05-16-H23について乾燥ストレスと発現比率との関係を示す特性図である。
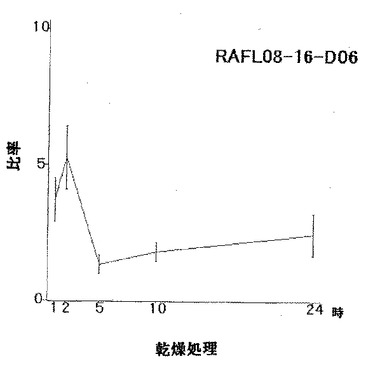
【図10】RAFL08-16-D06について乾燥ストレスと発現比率との関係を示す特性図である。
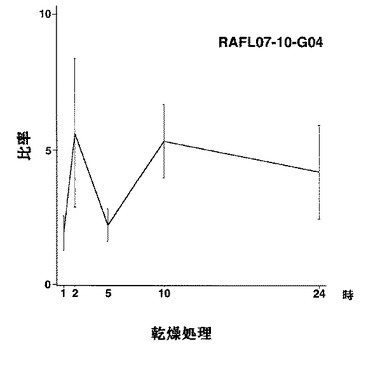
【図11】RAFL07-10-G04について乾燥ストレスと発現比率との関係を示す特性図である。
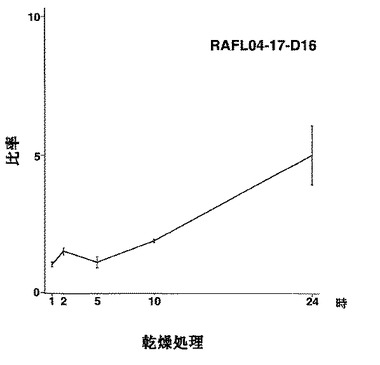
【図12】RAFL04-17-D16について乾燥ストレスと発現比率との関係を示す特性図である。
【図13】RAFL05-19-M20について乾燥ストレスと発現比率との関係を示す特性図である。
【図14】RAFL08-11-M13について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図15】RAFL04-15-K19について乾燥ストレスと発現比率との関係を示す特性図である。
【図16】RAFL04-15-K19について低温ストレスと発現比率との関係を示す特性図である。
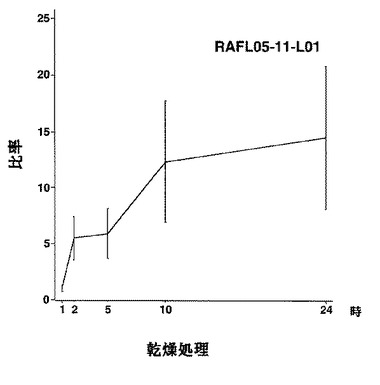
【図17】RAFL05-11-L01について乾燥ストレスと発現比率との関係を示す特性図である。
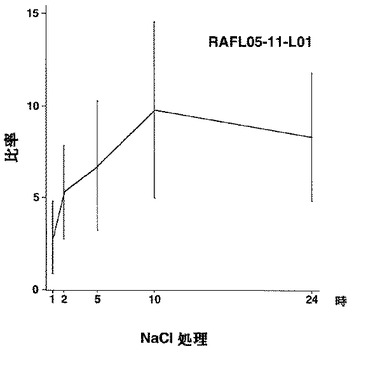
【図18】RAFL05-11-L01について高塩濃度ストレスと発現比率との関係を示す特性図である。
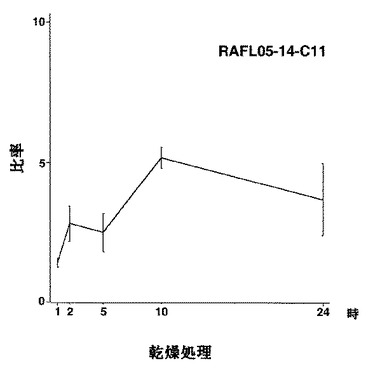
【図19】RAFL05-14-C11について乾燥ストレスと発現比率との関係を示す特性図である。
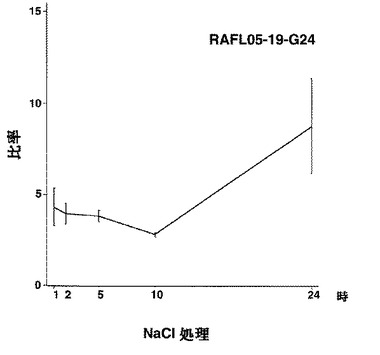
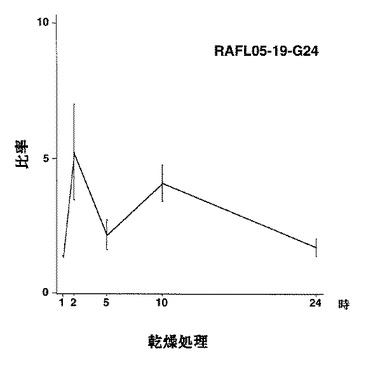
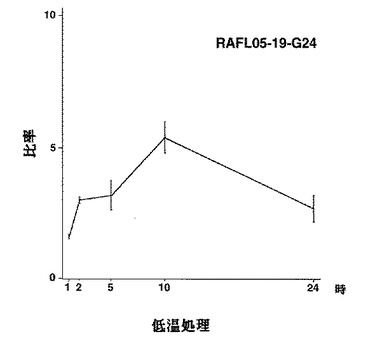
【図20】RAFL05-19-G24について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図21】RAFL05-19-G24について乾燥ストレスと発現比率との関係を示す特性図である。
【図22】RAFL05-19-G24について低温ストレスと発現比率との関係を示す特性図である。
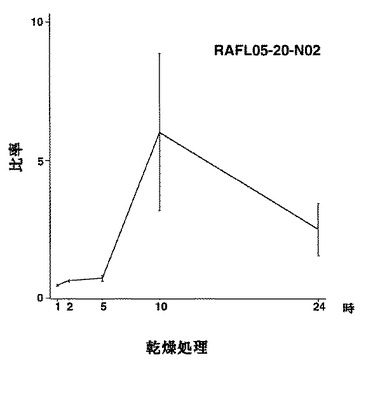
【図23】RAFL05-20-N02について乾燥ストレスと発現比率との関係を示す特性図である。
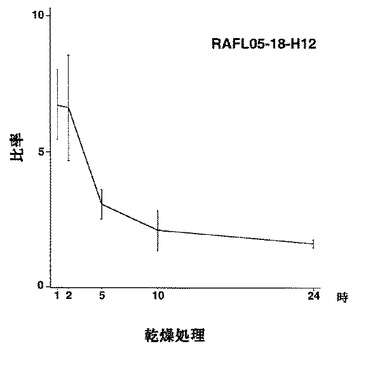
【図24】RAFL05-18-H12について乾燥ストレスと発現比率との関係を示す特性図である。
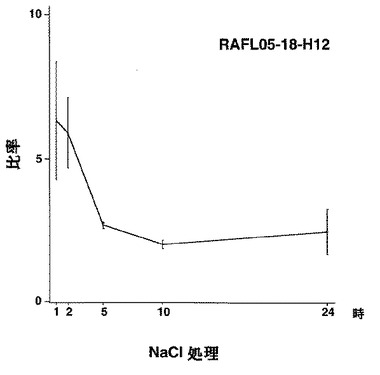
【図25】RAFL05-18-H12について高塩濃度ストレスと発現比率との関係を示す特性図である。
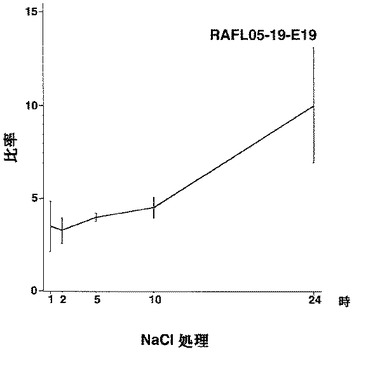
【図26】RAFL05-19-E19について高塩濃度ストレスと発現比率との関係を示す特性図である。
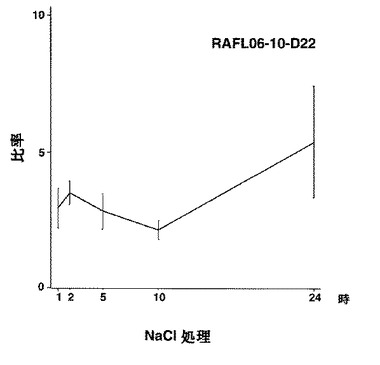
【図27】RAFL06-10-D22について高塩濃度ストレスと発現比率との関係を示す特性図である。
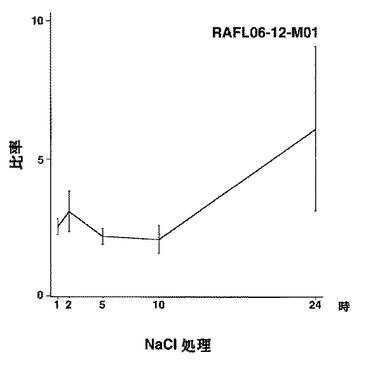
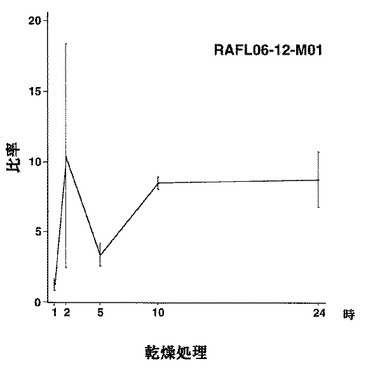
【図28】RAFL06-12-M01について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図29】RAFL06-12-M01について乾燥ストレスと発現比率との関係を示す特性図である。
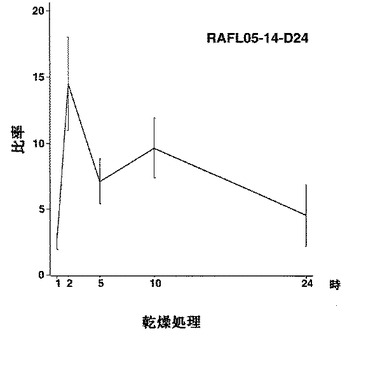
【図30】RAFL05-14-D24について乾燥ストレスと発現比率との関係を示す特性図である。
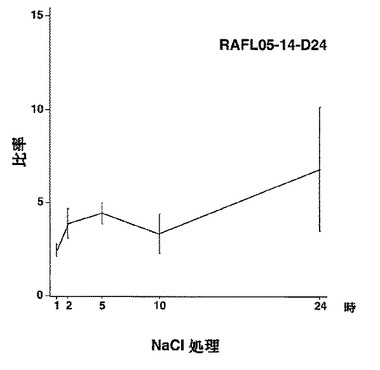
【図31】RAFL05-14-D24について高塩濃度ストレスと発現比率との関係を示す特性図である。
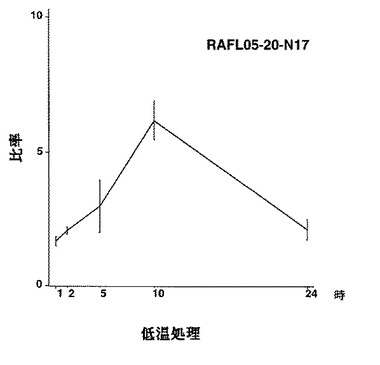
【図32】RAFL05-20-N17について低温ストレスと発現比率との関係を示す特性図である。
【図33】RAFL05-20-N17について乾燥ストレスと発現比率との関係を示す特性図である。
【図34】RAFL04-17-F21について乾燥ストレスと発現比率との関係を示す特性図である。
【図35】RAFL09-12-N16について乾燥ストレスと発現比率との関係を示す特性図である。
【図36】AFL05-19-I05について乾燥ストレスと発現比率との関係を示す特性図である。
【図37】RAFL05-19-I05について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図38】RAFL05-21-I22について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図39】RAFL08-11-H20について乾燥ストレスと発現比率との関係を示す特性図である。
【図40】RAFL08-11-H20について高塩濃度ストレスと発現比率との関係を示す特性図である。
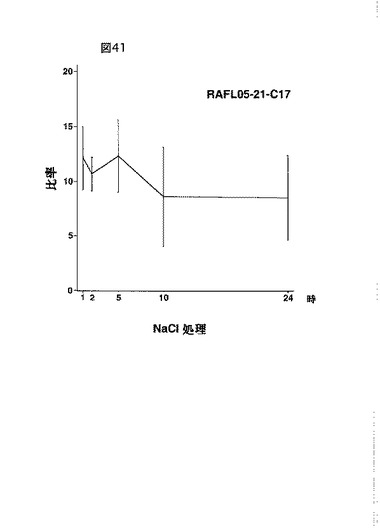
【図41】RAFL05-21-C17について高塩濃度ストレスと発現比率との関係を示す特性図である。
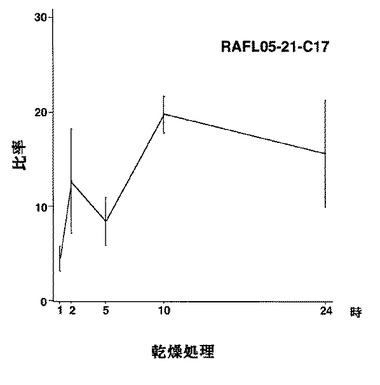
【図42】RAFL05-21-C17について乾燥ストレスと発現比率との関係を示す特性図である。
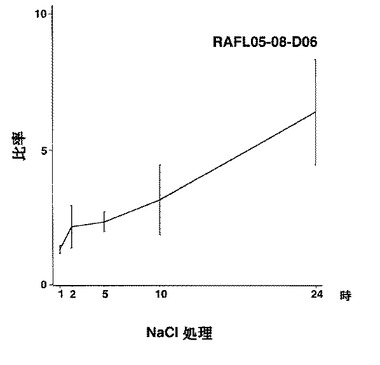
【図43】RAFL05-08-D06について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図44】RAFL05-20-M16について乾燥ストレスと発現比率との関係を示す特性図である。
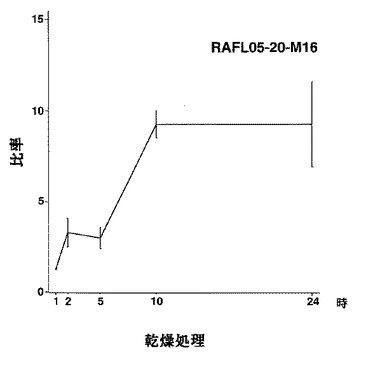
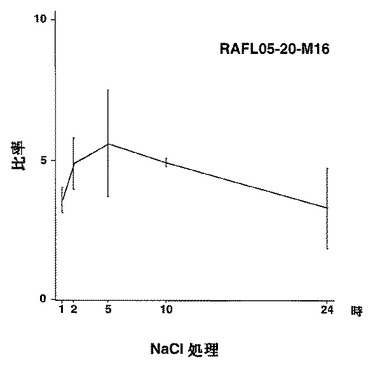
【図45】RAFL05-20-M16について高塩濃度ストレスと発現比率との関係を示す特性図である。
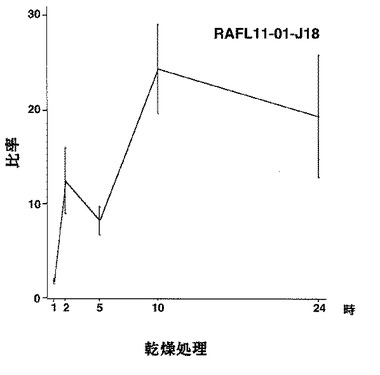
【図46】RAFL11-01-J18について乾燥ストレスと発現比率との関係を示す特性図である。
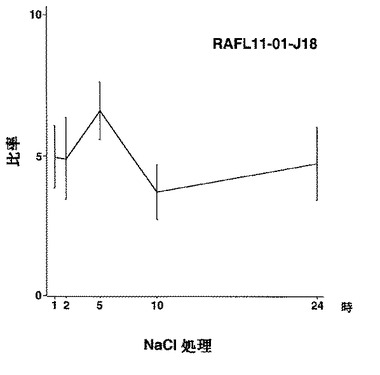
【図47】RAFL11-01-J18について高塩濃度ストレスと発現比率との関係を示す特性図である。
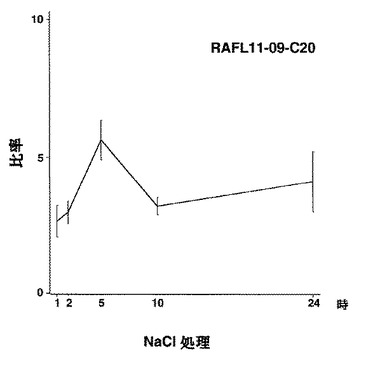
【図48】RAFL11-09-C20について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図49】RAFL05-18-N16について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図50】RAFL11-10-D10について乾燥ストレスと発現比率との関係を示す特性図である。
【図51】RAFL11-10-D10について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図52】RAFL04-17-N22について乾燥ストレスと発現比率との関係を示す特性図である。
【図53】RAFL04-17-N22について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図54】RAFL05-09-G15について乾燥ストレスと発現比率との関係を示す特性図である。
【図55】RAFL05-09-G15について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図56】RAFL05-21-L12について乾燥ストレスと発現比率との関係を示す特性図である。
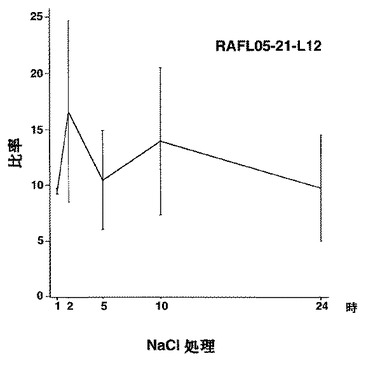
【図57】RAFL05-21-L12について高塩濃度ストレスと発現比率との関係を示す特性図である。
【配列表フリーテキスト】
【0095】
配列番号73及び74は合成プライマーである。
【技術分野】
【0001】
本発明は、環境ストレス応答性転写因子をコードする遺伝子に関する。
【背景技術】
【0002】
遺伝子の配列決定プロジェクトによって、数種の生物について大量のゲノム配列及びcDNA配列が決定されており、植物モデルであるシロイヌナズナ(Arabidopsis thaliana)では、2つの染色体の完全なゲノム配列が決定されている(Lin, X. et al., (1999) Nature 402, 761-768.; Mayer, K. et al., (1999) Nature 402, 769-777.)。
【0003】
EST(expressed sequence tag)プロジェクトも、発現遺伝子の発見に大いに貢献している(Hofte, H. et al., (1993) Plant J. 4, 1051-1061.; Newman, T. et al., (1994) Plant Physiol. 106, 1241-1255.; Cooke, R. et al., (1996) Plant J. 9, 1O1-124. Asamizu, E. et al., (2000) DNA Res. 7, 175-180.)。例えば、dbEST(National Center for Biotechnology Information(NCBI)のESTデータベース)には部分cDNA配列が含まれており、全遺伝子の半分以上(即ち、約28,000遺伝子)が再現されている(完全に配列決定されたシロイヌナズナの2番染色体の遺伝子含有量から推定[Lin, X. et al., (1999) Nature 402, 761-768.])。
【0004】
近年、ゲノムスケールの遺伝子発現を分析するのにマイクロアレイ(DNAチップ)技術が有用な手段となっている(Schena, M. et al., (1995) Science 270, 467-470.; Eisen, M. B. and Brown, P. O. (1999) Methods Enzymol. 303, 179-205.)。このDNAチップを用いる技術は、cDNA配列をスライドガラス上に1,000遺伝子/cm2以上の密度で配列させるものである。このように配列させたcDNA配列を、異なる細胞型又は組織型のRNAサンプルから調製した2色蛍光標識cDNAプローブ対に同時にハイブリダイズさせることで、遺伝子発現を直接かつ大量に比較分析することが可能となる。この技術は、最初、48個のシロイヌナズナ遺伝子を根及び苗条におけるディファレンシャル発現について分析することで実証された(Schena, M. et al., (1995) Science 270, 467-470.)。さらに、マイクロアレイは、熱ショック及びプロテインキナーゼC活性化に応答する新規な遺伝子を同定するため、ヒトcDNAライブラリーからランダムに採取した1,000個のクローンを調査するのに使用されている(Schena, M. et al., (1996) Proc. Natl. Acad. Sci. USA, 93, 10614-10619.)。
【0005】
一方、このDNAチップを用いる方法によって、各種の誘導条件下における炎症性疾患関連遺伝子の発現プロフィールの分析が行われている(Heller, R. A. et al., (1997) Proc. Natl. Acad. Sci. USA, 94, 2150-2155.)。さらに、マイクロアレイを用いて、6,000個を超えるコード配列からなる酵母ゲノムの動的発現についても分析が行われている(DeRisi, J.L. et al., (1997) Science 278, 680-686.; Wodicka, L. et al., (1997) Nature Biotechnol. 15, 1359-1367.)。
【0006】
しかしながら、植物の分野では、マイクロアレイ分析に対しては若干の報告がなされているに過ぎない(Schena, M. et al., (1995) Science 270, 467-470.; Ruan, Y. et al., (1998) Plant J. 15, 821-833.; Aharoni. A. et al., (2000) Plant Cell 12, 647-661.; Reymond, P. et al., (2000) Plant Cell 12, 707-719.)。
【0007】
植物の生育は、乾燥、高塩濃度及び低温等の環境ストレスの影響を顕著に受ける。これらのストレスのうち乾燥又は水分欠乏が、植物の生育及び作物の生産にとって最も厳しい制限因子となる。乾燥ストレスは、植物に様々な生化学的及び生理学的な応答を引き起こす。
【0008】
植物は、これらのストレス条件下で生き抜くために、ストレスに対する応答性及び順応性を獲得する。近年、転写レベルで乾燥に応答する数種の遺伝子が記載されている(Bohnert, H.J. et al., (1995) Plant Cell 7, 1099-1111.; Ingram, J., and Bartels, D. (1996) Plant Mol. Biol. 47, 377-403.; Bray, E. A. (1997) Trends Plant Sci. 2, 48-54.; Shinozaki. K., and Yamaguchi-Shinozaki, K. (1997) Plant Physiol. 115, 327-334. ; Shinozaki, K., and Yamaguchi-Shinozaki, K. (1999). Molecular responses to drought stress. Molecular responses to cold, drought, heat and salt stress in higher plants. Edited by Shinozaki, K. and Yamaguchi-Shinozaki, K. R. G. Landes Company.;Shinozaki, K., and Yamaguchi-Shinozaki, K. (2000) Curr. Opin. Plant Biol. 3, 217-223.)。
【0009】
一方、遺伝子導入によって植物のストレス耐性を向上させるために、ストレス誘導性遺伝子が使用されている(Holmberg, N., and Bulow, L. (1998) Trends Plant Sci. 3, 61-66.; Bajaj, S. et al., (1999) Mol. Breed. 5, 493-503.)。高等植物のストレス耐性とストレス応答の分子機構をさらに解明するためだけでなく、遺伝子操作によって作物のストレス耐性を向上させるためにも、ストレス誘導性遺伝子の機能を分析することが重要である。
【0010】
DRE/CRT(乾燥応答性エレメント/C-反復配列)は、乾燥、高塩分濃度及び低温ストレス応答性遺伝子のABA(アブシジン酸:植物ホルモンの一種で種子の休眠や環境ストレスのシグナル伝達因子として機能する。)に依存しない発現において重要なシス作動性エレメントとして同定されている(Yamaguchi-Shinozaki, K., and Shinozaki, K. (1994) Plant Cell 6, 251-264.; Thomashow, M.F. et al., (1999) Plant Mol. Biol. 50, 571-599.; Shinozaki, K., and Yamaguchi-Shinozaki, K. (2000) Curr. Opin. Plant Biol. 3, 217-223.)。また、DRE/CRT応答性遺伝子発現に関与する転写因子(DREB/CBF)がクローニングされている(Stockinger. E.J. et al., (1997) Proc. Natl. Acad. Sci. USA 94, 1035-1040.; Liu, Q. et al., (1998) Plant Cell 10, 1391-1406.; Shinwari, Z.K. et al., (1998) Biochem. Biophys. Res. Commun. 250, 161-170.; Gilmour, S.J.et al., (1998) Plant J. 16, 433-443.)。DREB1/CBFは低温応答性遺伝子発現において機能すると考えられ、DREB2は乾燥応答性遺伝子発現に関与している。カリフラワーモザイクウイルス(CaMV)35Sプロモーターの制御下でCBF1(DREB1B)cDNAを過剰発現するトランスジェニック・シロイヌナズナ植物では、凍結ストレスに対する強力な耐性が観察されている(Jaglo-Ottosen, K.R. et al., (1998) Science 280, 104-106.)。
【0011】
本発明者らは、CaMV 35Sプロモーター又はストレス誘導性rd29Aプロモーターの制御下におけるトランスジェニック植物でのDREB1A(CBF3)cDNAの過剰発現によって、ストレス誘導性DREB1A標的遺伝子の強力な構成的発現が引き起こされ、凍結ストレス、乾燥ストレス及び塩ストレスに対する耐性が向上することを報告している(Liu, Q. et al., (1998) Plant Cell 10, 1391-1406.; Kasuga, M. et al., (1999) Nature Biotechnol. 17, 287-291.)。また、既に本発明者らは、rd29A/lti78/cor78、kin1、kin2/cor6.6、cor15a、rd17/cor47及びerd10等の6個のDREB1A標的遺伝子を同定している(Kasuga, M. et al., (1999) Nature Biotechnol. 17, 287-291.)。しかしながら、トランスジェニック植物におけるDREB1A cDNAの過剰発現が凍結、乾燥及び塩分に対するストレス耐性をどのように高めているのかは、十分には解明されていない。乾燥及び凍結耐性の分子機構を研究するためには、DREB1Aによって制御される遺伝子をより多く同定・分析することが重要である。
【発明の開示】
【発明が解決しようとする課題】
【0012】
ところで、環境ストレスに対する応答性を示す転写因子としては、DREB1Aが知られている(特開2000-60558公報)。DREB1Aに関しては、植物体内でDREB1A遺伝子を過剰に発現させることによって、当該植物体の環境ストレス耐性を上昇させられることができる。しかしながら、植物体の環境ストレス耐性を上昇させられるような転写因子としては、あまり多く知られていないのが現状である。
【0013】
そこで、本発明は、環境ストレス応答性転写因子をコードする遺伝子を提供することを目的とする。
【課題を解決するための手段】
【0014】
本発明者は、上記課題を解決するため鋭意研究を行った結果、cDNAマイクロアレイ分析を応用して、低温ストレス、乾燥ストレス及び塩ストレス等の環境ストレスに特異的に応答性を示す、転写因子をコードする遺伝子を同定することに成功し、本発明を完成するに至った。
【0015】
すなわち、本発明は、以下を包含する。
(1)以下の(a)又は(b)のアミノ酸配列を含む環境ストレス応答性転写因子をコードする遺伝子。
(a) 配列番号2n(n=1〜36の整数)から選ばれるいずれかのアミノ酸配列
(b) 配列番号2n(n=1〜36の整数)から選ばれるいずれかのアミノ酸配列において1若しくは複数のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、環境ストレスに応答性を示し、かつ転写因子活性を有するアミノ酸配列
【0016】
(2)以下の(a)、(b)又は(c)のDNAを含む(1)記載の遺伝子。
【0017】
(a)配列番号2n-1(n=1〜36の整数)から選ばれるいずれかの塩基配列からなるDNA
(b)配列番号2n-1(n=1〜36の整数)から選ばれるいずれかの塩基配列において1若しくは複数の塩基が欠失、置換若しくは付加された塩基配列からなり、環境ストレス応答性転写因子をコードするDNA
(c)配列番号2n-1(n=1〜36の整数)から選ばれるいずれかの塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ環境ストレス応答性転写因子をコードするDNA
【0018】
(3)環境ストレスが低温ストレス、乾燥ストレス及び塩ストレスからなる群から選択される少なくとも1つである(1)又は(2)記載の遺伝子。
(4)(1)又は(2)記載の遺伝子を含む発現ベクター。
(5)(4)記載の発現ベクターを含む形質転換体。
(6)(4)記載の記載の発現ベクターを含むトランスジェニック植物。
【0019】
(7)植物が、植物体、植物器官、植物組織又は植物培養細胞である(6)記載のトランスジェニック植物。
【0020】
(8)(6)又は(7)記載のトランスジェニック植物を培養又は栽培することを特徴とするストレス耐性植物の製造方法。
【発明の効果】
【0021】
本発明により、ストレス応答性転写因子をコードする遺伝子が提供される。本発明の遺伝子は、環境ストレス耐性植物の分子育種に使用できる点で有用である。
【発明を実施するための最良の形態】
【0022】
以下、本発明を詳細に説明する。
本発明者らは、乾燥処理植物及び低温処理植物等の条件の異なる植物から、ビオチン化CAPトラッパー法(Carninci. P. et al., (1996) Genomics, 37, 327-336.)によってシロイヌナズナの完全長cDNAライブラリーを構築し (Seki. M. et al., (1998) Plant J. 15, 707-720.)、ストレス誘導性遺伝子を含む約7,000個の完全長cDNAを用いてシロイヌナズナの完全長cDNAマイクロアレイをそれぞれ調製した。また、これらの乾燥・低温誘導性の完全長cDNAに加えて、ストレス応答性遺伝子の発現をコントロールする転写制御因子であるDREB1Aの標的となる遺伝子を用いてcDNAマイクロアレイを作成した。そして、乾燥ストレス及び低温ストレス下における遺伝子の発現パターンをモニターし、ストレス応答性遺伝子を網羅的に解析した。その結果、約7,000個の完全長cDNAを含むcDNAマイクロアレイから、277個の乾燥誘導性遺伝子、53個の低温誘導性遺伝子及び194個の高塩濃度ストレス誘導性遺伝子を単離した。
【0023】
そして、これら環境ストレス応答性遺伝子の核酸塩基配列をアミノ酸配列に変換したものをGenBank Databaseに登録されているアミノ酸配列Dataに対してblast Xサーチする事により、転写因子をコードするもの(既知の転写因子をコードする遺伝子とE-value:e-10よりも高いスコアで相同性を示すもの)を同定することに成功したものである。以下の説明において、本発明で同定された転写因子を環境ストレス応答性転写因子と称する場合もある。
【0024】
以上のように、完全長cDNAマイクロアレイは、シロイヌナズナの乾燥・低温ストレス誘導性遺伝子の発現様式の解析やストレス関連転写制御因子の標的遺伝子の解析にとって有効なツールである。
1.転写因子の単離
転写因子は、遺伝子の上流に存在するシスエレメントと結合して、その下流の遺伝子の転写を活性化する機能を有するものである。本発明で単離された転写因子は、低温、乾燥、高塩濃度などの環境ストレスにより誘導される。
【0025】
環境ストレス応答性転写因子は、DREBファミリーに属するもの、ERFファミリーに属するもの、ジンクフィンガーファミリーに属するもの、WRKYファミリーに属するもの、MYBファミリーに属するもの、bHLHファミリーに属するもの、NACファミリーに属するもの、ホメオドメインファミリーに属するもの及びbZIPファミリーに属するものに大別される。
【0026】
転写因子を単離するにあたり、まず、マイクロアレイを用いてストレス応答性遺伝子を単離する。マイクロアレイとしては、シロイヌナズナ(Arabidopsis)の全長cDNAライブラリーから単離した遺伝子のほか、RD(Responsive to Dehydration)遺伝子、ERD(Early Responsive to Dehydration)遺伝子、内部標準としてλコントロール鋳型DNA断片(TX803、宝酒造株式会社製)から得られたPCR増幅断片、さらにネガティブコントロールとしてマウスのニコチン酸アセチルコリンレセプターのエプシロンサブユニット(nAChRE)遺伝子及びマウスのグルココルチコイドレセプターの相同性遺伝子からなる計約7000のcDNAを用いることができる。
【0027】
Kurabo製プラスミド調製装置を用いて抽出したプラスミドDNAをシーケンス解析に用いて、DNAシーケンサー(ABI PRISM 3700. PE Applied Biosystems, CA, USA)により配列を決定する。GenBank/EMBLデータベースをもとに、BLASTプログラムを用いて配列のホモロジー検索を行う。
【0028】
次に、ポリAセレクション後,逆転写反応をおこない2本鎖DNAを合成し、cDNAをベクターに挿入する。
【0029】
cDNAライブラリー作成用ベクターに挿入されたcDNAを、cDNAの両側のベクターの配列と相補的なプライマーを用いてPCR法により増幅する。ベクターとしては、λZAPII、λPS等が挙げられる。
【0030】
マイクロアレイは、通常の方法に従って作製することができ、特に限定されるものではない。例えば、gene tipマイクロアレイスタンプマシンGTMASS SYSTEM(Nippon Laser & Electronics Lab.製)を使って、上記PCR産物をマイクロタイタープレートからロードし、マイクロスライドガラスの上に所定間隔でスポットする。その後、非特異的なシグナルの発現を防ぐためにスライドをブロッキング・ソルーションに浸す。
【0031】
植物材料としては野生型のほか、特定の遺伝子の破壊株等が挙げられるが、DREB1AのcDNAが導入されたトランスジェニック植物を用いることができる。植物種は、シロイヌナズナ、タバコ,イネ等が挙げられるが、シロイヌナズナが好ましい。
【0032】
乾燥及び低温ストレス処理は公知方法の方法で行うことができる(Yamaguchi-Shinozaki, K., and Shinozaki, K. (1994) Plant Cell 6, 251-264.)。
【0033】
ストレス処理にさらした後は、植物体(野生型及びDREB1A過剰発現型形質転換体)をサンプリングし、液体窒素を用いて凍結保存する。野生型及びDREB1A過剰発現型形質転換体を、DREB1Aの標的遺伝子を同定するための実験に用いる。植物体から、公知方法又はキットを用いてmRNAを単離精製する。
【0034】
標識用Cy3 dUTP又はCy5 dUTP(Amersham Pharmacia)の存在下でそれぞれのmRNAサンプルの逆転写を行い、ハイブリダイゼーションに用いる。
【0035】
ハイブリダイゼーション後は、走査レーザー顕微鏡等を用いてマイクロアレイをスキャンする。マイクロアレイのデータ解析用プログラムとして、Imagene Ver 2.0(BioDiscovery)とQuantArray(GSI Lumonics)等を用いることができる。
【0036】
スキャン後は、目的とする遺伝子をもつプラスミドを調製することにより、遺伝子が単離される。
【0037】
転写因子の決定は、上記単離された遺伝子の塩基配列を解析し、データベース(GenBank/EMBL, ABRC)のゲノム情報をもとに、遺伝子解析用プログラムを用いて行われる。単離された遺伝子は、乾燥ストレス誘導性及び低温ストレス誘導性の両性質を有するもの、乾燥ストレス誘導性に特異的なもの、低温ストレス誘導性に特異的なものに分類することができる。遺伝子解析用プログラムによれば、上記遺伝子の中から36種の転写因子をコードする遺伝子が同定される。これら36種類の転写因子をコードする遺伝子の塩基配列を配列番号2n-1(nは1〜36の整数)、転写因子のアミノ酸配列を配列番号2n(nは1〜36の整数)に示す。配列番号と転写因子をコードする遺伝子の遺伝子名とを表1に示す
【0038】
【表1】
【0039】
但し、本発明の転写因子が環境ストレス応答性転写因子として機能する限り、配列番号2n-1(nは1〜36の整数)から選ばれるいずれかの塩基配列において1又は複数個、好ましくは1又は数個(例えば1〜10個、1〜5個)の塩基が欠失、置換又は付加された塩基配列を有するものでもよい。さらに、配列番号2n-1(nは1〜36の整数)から選ばれるいずれかの塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ環境ストレス応答性転写因子をコードするDNAも、本発明の転写因子に含まれる。ここで、ストリンジェントな条件とは、ナトリウム濃度が25〜500mM、好ましくは25〜300mMであり、温度が42〜68℃、好ましくは42〜65℃である。より具体的には、5×SSC(83mM NaCl、83mMクエン酸ナトリウム)、温度42℃である。
【0040】
本発明で単離した36種類の転写因子については以下のように分類することができる。
1)DREBファミリー:RAFL05-11-M11、RAFL06-11-K21、RAFL05-16-H23、RAFL08-16-D16
2)ERFファミリー:RAFL08-16-G17、RAFL06-08-H20
3)ジンクフィンガーファミリー:RAFL07-10-G04、RAFL04-17-D16、RAFL05-19-M20、RAFL08-11-M13、RAFL04-15-K19、RAFL05-11-L01、RAFL05-14-C11、RAFL05-19-G24、RAFL05-20-N02
4)WRKYファミリー:RAFL05-18-H12、RAFL05-19-E19、RAFL06-10-D22、RAFL06-12-M01
5)MYBファミリー:RAFL05-14-D24、RAFL05-20-N17、RAFL04-17-F21
6)bHLHファミリー:RAFL09-12-N16
7)NACファミリー:RAFL05-19-I05、RAFL05-21-I22、RAFL08-11-H20、RAFL05-21-C17、RAFL05-08-D06
8)ホメオドメインファミリー:RAFL05-20-M16、RAFL11-01-J18、RAFL11-09-C20
9)bZIPファミリー:RAFL05-18-N16、RAFL11-10-D10、RAFL04-17-N22、RAFL05-09-G15
なお、RAFL05-21-L12については上記1)〜9)に分類できない。
【0041】
一旦本発明の転写因子をコードする遺伝子の塩基配列が確定されると、その後は化学合成によって、又はクローニングされたプローブを鋳型としたPCRによって、あるいは該塩基配列を有するDNA断片をプローブとしてハイブリダイズさせることによって、本発明の転写因子をコードする遺伝子を得ることができる。さらに、部位特異的突然変異誘発法等によって本発明の転写因子をコードする遺伝子の変異型であって変異前の転写因子をコードする遺伝子と同等の機能を有するものを合成することもできる。
【0042】
なお、転写因子をコードする遺伝子の塩基配列に変異を導入するには、Kunkel法、Gapped duplex法等の公知の手法又はこれに準ずる方法を採用することができる。例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-K(TAKARA社製)やMutant-G(TAKARA社製))などを用いて、あるいは、TAKARA社のLA PCR in vitro Mutagenesis シリーズキットを用いて変異の導入が行われる。
【0043】
ここで、「環境ストレス」とは、一般には非生物的ストレスを意味し、例えば乾燥ストレス、低温ストレス、高塩濃度ストレス、強光ストレス等をいう。「乾燥」とは水分が欠乏した状態を意味し、「低温」とはそれぞれの生物種の生活至適温度よりも低い温度にさらされた状態(例えばシロイヌナズナの場合-20〜+21℃の温度を継続的に1時間〜数週間さらすことをいう。また、「高塩濃度」とは、50mM〜600mMの濃度のNaClを継続的に0.5時間〜数週間処理したときの状態を意味する。「強光ストレス」とは、光合成能を超える強光が植物に照射された状態を意味し、例えば5,000〜10,000Lx以上の光が照射した場合が該当する。これらの環境ストレスは、1種類のものを負荷してもよく、複数種類のものを負荷してもよい。
【0044】
2.発現ベクターの構築
本発明の発現ベクターは、適当なベクターに本発明の転写因子をコードする遺伝子を連結(挿入)することにより得ることができる。本発明の転写因子をコードする遺伝子を挿入するためのベクターは、宿主中で複製可能なものであれば特に限定されず、例えばプラスミド、シャトルベクター、ヘルパープラスミドなどが挙げられる。
【0045】
プラスミド DNAとしては、大腸菌由来のプラスミド(例えばpBR322、pBR325、pUC118、pUC119、pUC18、pUC19、pBluescript等)、枯草菌由来のプラスミド(例えばpUB110、pTP5等)、酵母由来のプラスミド(例えばYEp13、YCp50等)などが挙げられ、ファージDNAとしてはλファージ(Charon4A、Charon21A、EMBL3、EMBL4、λgt10、λgt11、λZAP等)が挙げられる。さらに、レトロウイルス又はワクシニアウイルスなどの動物ウイルス、バキュロウイルスなどの昆虫ウイルスベクターを用いることもできる。
【0046】
ベクターに本発明の転写因子をコードする遺伝子を挿入するには、まず、精製されたDNAを適当な制限酵素で切断し、適当なベクター DNAの制限酵素部位又はマルチクローニングサイトに挿入してベクターに連結する方法などが採用される。
【0047】
本発明の転写因子をコードする遺伝子は、その3'末端にレポーター遺伝子、例えば、植物で広く用いられているGUS遺伝子を連結して用いれば、GUS活性を調べることで当該遺伝子の発現の強さを容易に評価することができる。なお、レポーター遺伝子としては、GUS遺伝子以外にも、ルシフェラーゼ、グリーンフルオレセイントプロテインなども用いることができる。
【0048】
3.形質転換体の作製
本発明の形質転換体は、本発明の発現ベクターを宿主中に導入することにより得ることができる。ここで、宿主としては、本発明の転写因子をコードする遺伝子を発現できるものであれば特に限定されるものではないが、植物が好ましい。宿主が植物である場合は、形質転換植物(トランスジェニック植物)は以下のようにして得ることができる。
【0049】
本発明において形質転換の対象となる植物は、植物体全体、植物器官(例えば葉、花弁、茎、根、種子等)、植物組織(例えば表皮、師部、柔組織、木部、維管束等)又は植物培養細胞のいずれをも意味するものである。形質転換に用いられる植物としては、アブラナ科、イネ科、ナス科、マメ科等に属する植物(下記参照)が挙げられるが、これらの植物に限定されるものではない。
アブラナ科:シロイヌナズナ(Arabidopsis thaliana)
ナス科:タバコ(Nicotiana tabacum)
イネ科:トウモロコシ(Zea mays) 、イネ(Oryza sativa)
マメ科:ダイズ(Glycine max)
【0050】
上記組換えベクターは、通常の形質転換方法、例えば電気穿孔法(エレクトロポレーション法)、アグロバクテリウム法、パーティクルガン法、PEG法等によって植物中に導入することができる。
【0051】
例えばエレクトロポレーション法を用いる場合は、パルスコントローラーを備えたエレクトロポレーション装置により、電圧500〜1600V、25〜1000μF、20〜30msecの条件で処理し、遺伝子を宿主に導入する。
【0052】
また、パーティクルガン法を用いる場合は、植物体、植物器官、植物組織自体をそのまま使用してもよく、切片を調製した後に使用してもよく、プロトプラストを調製して使用してもよい。このように調製した試料を遺伝子導入装置(例えばBio-Rad社のPDS-1000/He等)を用いて処理することができる。処理条件は植物又は試料により異なるが、通常は1000〜1800psi程度の圧力、5〜6cm程度の距離で行う。
【0053】
また、植物ウイルスをベクターとして利用することによって、目的遺伝子を植物体に導入することができる。利用可能な植物ウイルスとしては、例えば、カリフラワーモザイクウイルスが挙げられる。すなわち、まず、ウイルスゲノムを大腸菌由来のベクターなどに挿入して組換え体を調製した後、ウイルスのゲノム中に、これらの目的遺伝子を挿入する。このようにして修飾されたウイルスゲノムを制限酵素によって組換え体から切り出し、植物宿主に接種することによって、目的遺伝子を植物宿主に導入することができる。
【0054】
アグロバクテリウムのTiプラスミドを利用する方法においては、アグロバクテリウム(Agrobacterium)属に属する細菌が植物に感染すると、それが有するプラスミドDNAの一部を植物ゲノム中に移行させるという性質を利用して、目的遺伝子を植物宿主に導入する。アグロバクテリウム属に属する細菌のうちアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)は、植物に感染してクラウンゴールと呼ばれる腫瘍を形成し、また、アグロバクテリウム・リゾゲネス(Agrobacteriumu rhizogenes)は、植物に感染して毛状根を発生させる。これらは、感染の際にTiプラスミド又はRiプラスミドと呼ばれる各々の細菌中に存在するプラスミド上のT-DNA領域(Transferred DNA)と呼ばれる領域が植物中に移行し、植物のゲノム中に組み込まれることに起因するものである。
【0055】
Ti又はRiプラスミド上のT-DNA領域中に、植物ゲノム中に組み込みたいDNAを挿入しておけば、アグロバクテリウム属の細菌が植物宿主に感染する際に目的とするDNAを植物ゲノム中に組込むことができる。
【0056】
形質転換の結果得られる腫瘍組織やシュート、毛状根などは、そのまま細胞培養、組織培養又は器官培養に用いることが可能であり、また従来知られている植物組織培養法を用い、適当な濃度の植物ホルモン(オーキシン、サイトカイニン、ジベレリン、アブシジン酸、エチレン、ブラシノライド等)の投与などにより植物体に再生させることができる。
【0057】
本発明のベクターは、上記植物宿主に導入するのみならず、大腸菌(Escherichia coli)等のエッシェリヒア属、バチルス・ズブチリス(Bacillus subtilis)等のバチルス属、又はシュードモナス・プチダ(Pseudomonas putida)等のシュードモナス属に属する細菌、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)等の酵母、COS細胞、CHO細胞等の動物細胞、あるいはSf9等の昆虫細胞などに導入して形質転換体を得ることもできる。大腸菌、酵母等の細菌を宿主とする場合は、本発明の組換えベクターが該細菌中で自律複製可能であると同時に、本発明の転写因子をコードする遺伝子、当該遺伝子の上流に位置するリボソーム結合配列及びプロモーター配列、当該遺伝子の下流に位置する転写終結配列により構成されていることが好ましい。
【0058】
細菌への組換えベクターの導入方法は、細菌にDNAを導入する方法であれば特に限定されるものではない。例えばカルシウムイオンを用いる方法、エレクトロポレーション法等が挙げられる。
【0059】
酵母を宿主とする場合は、例えばサッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)などが用いられる。酵母への組換えベクターの導入方法は、酵母にDNAを導入する方法であれば特に限定されず、例えばエレクトロポレーション法、スフェロプラスト法、酢酸リチウム法等が挙げられる。
【0060】
動物細胞を宿主とする場合は、サル細胞COS-7、Vero、チャイニーズハムスター卵巣細胞(CHO細胞)、マウスL細胞などが用いられる。動物細胞への組換えベクターの導入方法としては、例えばエレクトロポレーション法、リン酸カルシウム法、リポフェクション法等が挙げられる。
【0061】
昆虫細胞を宿主とする場合は、Sf9細胞などが用いられる。昆虫細胞への組換えベクターの導入方法としては、例えばリン酸カルシウム法、リポフェクション法、エレクトロポレーション法などが挙げられる。
【0062】
遺伝子が宿主に組み込まれたか否かの確認は、PCR法、サザンハイブリダイゼーション法、ノーザンハイブリダイゼーション法等により行うことができる。例えば、形質転換体からDNAを調製し、DNA特異的プライマーを設計してPCRを行う。PCRは、前記プラスミドを調製するために使用した条件と同様の条件で行われる。その後は、増幅産物についてアガロースゲル電気泳動、ポリアクリルアミドゲル電気泳動又はキャピラリー電気泳動等を行い、臭化エチジウム、SYBR Green液等により染色し、そして増幅産物を1本のバンドとして検出することにより、形質転換されたことを確認する。また、予め蛍光色素等により標識したプライマーを用いてPCRを行い、増幅産物を検出することもできる。さらに、マイクロプレート等の固相に増幅産物を結合させ、蛍光又は酵素反応等により増幅産物を確認する方法も採用してもよい。
【0063】
4.植物の製造
本発明においては、上記形質転換植物細胞等から形質転換植物体に再生することができる。再生方法としては、カルス状の形質転換細胞をホルモンの種類、濃度を変えた培地へ移して培養し、不定胚を形成させ、完全な植物体を得る方法が採用される。使用する培地としては、LS培地、MS培地などが例示される。
【0064】
本発明の「植物体を製造する方法」は、上記環境ストレス応答性転写因子をコードする遺伝子を挿入した植物発現ベクターを宿主細胞に導入して形質転換植物細胞を得て、該形質転換植物細胞から形質転換植物体を再生し、得られた形質転換植物体から植物種子を得て、該植物種子から植物体を生産する工程を含む。
【0065】
形質転換植物体から植物種子を得るには、例えば、形質転換植物体を発根培地から採取し、水を含んだ土を入れたポットに移植し、一定温度下で生育させて、花を形成させ、最終的に種子を形成させる。また、種子から植物体を生産するには、例えば、形質転換植物体上で形成された種子が成熟したところで、単離して、水を含んだ土に播種し、一定温度、照度下で生育させることにより、植物体を生産する。このようにして育種された植物は、導入された環境ストレス応答性転写因子をコードする遺伝子のストレス応答性に応じた環境ストレス耐性植物となる。
【実施例】
【0066】
以下、実施例により本発明をさらに具体的に説明する。但し、本発明はこれら実施例にその技術的範囲が限定されるものではない。
【0067】
〔実施例1〕 環境ストレス応答性転写因子をコードする遺伝子の単離
1.材料と方法
(1) Arabidopsis cDNAクローン
Arabidopsisの全長cDNAライブラリーから単離した遺伝子に加えて、RD(Responsive to Dehydration)遺伝子、ERD(Early Responsive to Dehydration)遺伝子、kin1遺伝子、kin2遺伝子、cor15a遺伝子、また内部標準としてλコントロール鋳型DNAをPCRで増幅した断片、さらにネガティブコントロールとしてマウスのニコチン酸アセチルコリンレセプターのエプシロンサブユニット(nAChRE)遺伝子及びマウスのグルココルチコイドレセプターの相同性遺伝子の計約7000個のcDNAをマイクロアレイ作成に用いた。
陽性対照:乾燥誘導遺伝子(脱水応答性:rd、及び初期脱水応答遺伝子:erd)
内部標準:λコントロール鋳型DNAをPCRで増幅した断片(TX803、宝酒造社製、以下「コントロール断片」と称する)
陰性対照:非特異的ハイブリダイゼーションを評価するためにArabidopsisのデータベースにある任意の配列とは実質的にホモロジーを有さないニコチン性アセチルコリン受容体εサブユニット(nAChRE)遺伝子及びマウスグルココルチコイド受容体ホモログ遺伝子
【0068】
(2) Arabidopsis 全長cDNAマイクロアレイ
ビオチニル化CAPトラッパー法を用いて、本発明者はArabidopsisの植物体から、異なる条件(例えば、発芽から成熟種子までの種々の成長段階における乾燥処理、低温処理及び未処理)で全長cDNAライブラリーを構築した。全長cDNAライブラリーから、本発明者は、約7000個の独立したArabidopsis全長cDNAをそれぞれ単離した。公知の手法(Eisen and Brown, 1999)に従って、PCRで増幅したcDNA断片をスライドグラス上の整列させた。本発明者は、以下の遺伝子を含む約7000個のArabidopsisの全長cDNAを含有する全長cDNAマイクロアレイを調製した。
【0069】
(3) cDNAマイクロアレイを用いた乾燥誘導性遺伝子、低温誘導性遺伝子及び高塩濃度誘導性遺伝子
本例では、約7000個のArabidopsisの全長cDNAを含有する全長cDNAマイクロアレイを用いて、乾燥誘導性遺伝子、低温誘導性遺伝子及び高塩濃度誘導性遺伝子を単離した。
【0070】
上述した各種ストレスを受けた植物及びストレスを受けていない植物のCy3及びCy5蛍光標識プローブを混合し、約7000個のArabidopsisの全長cDNAを含有する全長cDNAマイクロアレイとハイブリダイズさせた。一方のmRNAサンプルをCy3-dUTPで標識し、他方のmRNAサンプルをCy5-dUTPで標識するcDNAプローブ対の二重標識によって、マイクロアレイ上のDNAエレメントへの同時ハイブリダイゼーションが可能となり、2種の異なる条件間(即ち、ストレス有り又はストレス無し)における遺伝子発現の直接的な定量測定が容易になる。ハイブリダイズさせたマイクロアレイを、各DNAエレメントからのCy3及びCy5発光について2つの別個のレーザーチャネルによって走査した。次いで、各DNAエレメントの2つの蛍光シグナルの強度比を相対値として測定し、マイクロアレイ上のcDNAスポットで表される遺伝子のディファレンシャル発現の変化を判定した。本実施例では、分析を行う2種の実験条件下で発現レベルがほぼ同等であるα-チューブリン遺伝子を内部対照遺伝子として使用した。
約7000個のArabidopsisの全長cDNAを含有する全長cDNAマイクロアレイにおける、乾燥誘導性遺伝子、低温誘導性遺伝子及び高塩濃度誘導性遺伝子の同定手順を示す。
【0071】
上述したストレスのうち1種類のストレスを負荷した植物由来のmRNA及びストレスを受けていない野生型植物由来のmRNAを、それぞれCy3-標識cDNAプローブ及びCy5-標識cDNAプローブの調製に使用した。これらのcDNAプローブを混合し、cDNAマイクロアレイとハイブリダイズさせた。本実施例では、2種の条件下で発現レベルがほぼ同等であるコントロール断片を内部対照遺伝子として使用した。発現比率(乾燥/ストレス無し、低温/ストレス無し、又は高塩濃度/ストレス無し)がコントロール断片の5倍を超える遺伝子を、負荷したストレスによって誘導される遺伝子とした。
【0072】
(4) 配列の解析
遺伝子配列のホモロジー検索をおこなうため、DNA抽出装置(model Biomek、ベックマンコールター社製)を用いて抽出しマルチスクリーン96穴フィルタープレート(ミリポア社製)を用いて精製したプラスミドDNAを配列解析に用いた。配列解析は、DNAシーケンサー(ABI PRISM 3700. PE Applied Biosystems, CA, USA)を用いてダイターミネーターサイクルシーケンス法により決定した。GenBank/EMBLデータベースをもとに、BLASTプログラムを用いて配列のホモロジー検索を行った。
【0073】
(5) cDNAの増幅
cDNAライブラリー作製用ベクターとして、λZAP及びλFLC-1を用いた。ライブラリー用のベクターに挿入されたcDNAを、cDNAの両側のベクターの配列と相補的なプライマーを用いてPCR法により増幅した。
プライマーの配列は以下の通りである。
FL forward 1224:5'-CGCCAGGGTTTTCCCAGTCACGA(配列番号73)
FL reverse 1233:5'-AGCGGATAACAATTTCACACAGGA(配列番号74)
【0074】
100 μl のPCR混合液(0.25 mM dNTP,0.2 μM PCRプライマー,1 X Ex Taq バッファー,1.25 U Ex Taqポリメラーゼ (宝酒造製))に、テンプレートとしてプラスミド(1-2 ng)を加えた。PCRは、最初に94℃で3分反応させた後、続いて95℃で1分、60℃で30秒及び72℃で3分のサイクルを35サイクル、最後に72℃で3分の条件で行った。PCR産物をエタノール沈澱させた後、25μlの3 X SSCに溶かした。0.7%アガロースゲルを用いた電気泳動により、得られたDNAの質とPCRの増幅効率を確認した。
【0075】
(6) cDNAマイクロアレイの作製
gene tipマイクロアレイスタンプマシンGTMASS SYSTEM(Nippon Laser & Electronics Lab.製)を使って、0.5μlのPCR産物(500-1000 ng/ml)を384穴のマイクロタイタープレートからロードし、48枚のマイクロスライドガラス(model Super Aldehyde substrate、Telechem International製)の上に300μmの間隔で0.5nlずつスポットした。スポット後のスライドを、相対湿度30%以下の雰囲気中で乾燥処理し、架橋反応のための紫外線照射に供した。
【0076】
その後、スライドを0.2%SDS中で2分間震盪する処理を3回行った後、蒸留水中で浸透する処理を2回行った。その後、スライドをスライドラックに置き、熱湯を入れたチャンバーにスライドラックを入れて2分間放置した。その後、ブロッキング溶液(1gのホウ化水素、300mlのPBS及び90mlの100%エタノールを含む)チャンバー注いだ。スライドラックを入れたガラスチャンバーを緩やかに振った後、0.2%SDSを含むチャンバーにスライドラックを移して1分間静かに震盪する処理を3回行った。その後、蒸留水を入れたガラスチャンバーにスライドラックを移して1分間緩やかに振った後、20分間遠心処理することによって乾燥させた。
【0077】
(7) 植物材料とRNAの単離
植物材料として、寒天培地に播種して3週間栽培した(Yamaguchi-ShinozakiとShinozaki,1994)野生型及びカリフラワーモザイクウイルスの35SプロモーターにDREB1AのcDNA(Kasugaなど,1999)をつないで導入したシロイヌナズナ(コロンビア種)の植物体を用いた。乾燥及び低温ストレス処理はYamaguchi-ShinozakiとShinozaki(1994)の方法で行った。すなわち、寒天培地から引き抜いた植物体をろ紙上に置き,22℃,相対湿度60%の条件で乾燥処理をおこなった。22℃で栽培した植物体を4℃に移すことにより低温処理をおこなった。また、高塩濃度ストレス処理は、250mMのNaClを含む水溶液内で水耕することによりおこなった。
【0078】
野生型の植物体を2時間又は10時間のストレス処理にさらした後サンプリングし、液体窒素を用いて凍結保存した。また、カナマイシンを加えない寒天培地で栽培した野生型及びDREB1A過剰発現型形質転換体を、DREB1Aの標的遺伝子を同定するための実験に用いた。DREB1A過剰発現型形質転換体に対しては、ストレス処理を行わなかった。植物体から、ISOGEN(Nippon gene, Tokyo, Japan)を用いてトータルRNAを単離し、Oligotex-dT30 mRNA精製キット(Takara,Tokyo,Japan)を用いてmRNAを単離精製した。
【0079】
(8) プローブの蛍光標識
Cy3 dUTP又はCy5 dUTP(Amersham Pharmacia)の存在下でそれぞれのmRNAサンプルの逆転写を行った。具体的に逆転写反応は、内部標準として1ngのλポリA+RNA-A(TX802、宝酒造社製)を含む1μgの変性ポリ(A)+、50ng/μlの12〜18merオリゴdTプライマー(ライフテクノロジー社製)、それぞれ0.5mMのdATP、dGDP及びdCTP、0.2mMのdTTP、0.1mMのCy3 dUTP若しくはCy5 dUTP、100UのRnase阻害剤、10mMのDTT及び200UのSuperscript II逆転写酵素を含む1×Superscript first-strand buffer(50mM Tris-HCl, pH8.3, 75mM KCl, 3mM MgCl2, 20mM DTTを含有。ライフテクノロジー社製)中で全量20μlとして行った。
【0080】
上記組成の反応溶液を42℃で35分間インキュベーションした後、200UのSuperscript II逆転写酵素を加え、更に42℃で35分間インキュベーションした。その後、反応溶液に0.5MのEDTAを5μl、1Nの水酸化ナトリウムを10μl及び20μLの蒸留水を添加することで、当該反応溶液における酵素反応を停止させるとともに鋳型を分解した。その後、反応溶液を65℃で1時間インキュベーションした。その後、反応溶液を1MのTris-HCl(pH7.5)で中和した。
【0081】
反応溶液をMicrocon 30 micro concentrator(アミコン社製)に移した。250μlのTEバッファーを加えてバッファー量が10μlになるまで遠心し、流出液を捨てる工程を4回繰り返した。反応溶液に含まれるプローブを遠心によって回収し数μlの蒸留水を加えた。得られたプローブに、5.1μlの20 X SSC、2μg/μlの酵母 tRNA、4.8μlの2%SDSを加えた。さらに、サンプルを100℃で2分間変成処理し、室温に5分間置いた後ハイブリダイゼーションに用いた。
【0082】
(9) マイクロアレイハイブリダイゼーション及びスキャニング
プローブをbenchtop micro centrifugeを用いて1分間の高速遠心にかけた。泡の発生を避けるために、プローブをアレイの中央に置きその上にカバースリップをかぶせた。スライドガラス上に5μlの3 X SSCを4滴落として、チェンバーを適度な湿度に保ち、ハイブリダイゼーション中のプローブの乾燥を防いだ。スライドガラスをハイブリダイゼーション用のカセット(THC-1,BM機器)に入れて密封した後、65℃で12〜16時間処理した。スライドガラスを取り出してスライドラックに置き、溶液1(2X SSC,0.03%SDS)中でカバースリップを慎重にはずした後ラックを振って洗浄し、ラックを溶液2(1X SSC)中に移して2分間洗浄した。さらにラックを溶液3(0.05X SSC)に移して2分間放置し、遠心(2500g, 1min)にかけて乾燥させた。
【0083】
走査レーザー顕微鏡(ScanArray4000; GSI Lumonics,Watertown,MA)を用いて1ピクセルあたり10μmの解像度でマイクロアレイをスキャンした。マイクロアレイのデータ解析用プログラムとして、QuantArrayバージョン2.0(GSI Lumonics)を用いた。バックグラウンドの蛍光は、陰性対照遺伝子(ニコチン性アセチルコリン受容体εサブユニット(nAChRE)遺伝子及びマウスグルココルチコイド受容体ホモログ遺伝子)の蛍光シグナルに基づいて算出した。蛍光シグナル値が1000未満(この値はバックグラウンドの蛍光シグナル値の2倍未満を意味する)のサンプルに関しては分析対象としなかった。遺伝子クラスタリング分析はGenespring(シリコンジェネティック社製)を用いて行った。
【0084】
(10) ノーザン解析
トータルRNAを用いてノーザン解析を行った(Yamaguchi-ShinozakiとShinozaki,1994)。シロイヌナズナの全長cDNAライブラリーからPCR法によって単離したDNA断片をノーザンハイブリダイゼーションのプローブとして用いた。
【0085】
(11) 転写因子をコードする遺伝子の決定
データベース(GenBank/EMBL, ABRC)のシロイヌナズナのゲノム情報をもとに、遺伝子解析用プログラムBLASTを用いて転写因子をコードする遺伝子を解析した。
【0086】
2.結果
(1) ストレス誘導性遺伝子の同定
ストレスを受けていないシロイヌナズナ植物から単離したmRNAから、Cy5-dUTPの存在下で逆転写を行って蛍光標識cDNAを調製した。乾燥処理、低温処理(2時間)或いは高塩濃度処理を行った植物から、Cy3-dUTPで標識した第2のプローブを調製した。両プローブを約7000個のシロイヌナズナcDNAクローンを含むcDNAマイクロアレイへ同時にハイブリダイズさせた後、疑似カラー像を作製した。
【0087】
ストレスによって誘導された遺伝子及び抑制された遺伝子は、それぞれ赤色及び緑色のシグナルで表される。両方の処理においてほぼ同レベルで発現した遺伝子は、黄色のシグナルとなる。各スポットの強度は、各遺伝子の発現量の絶対値に相当する。低温誘導性遺伝子(rd29A)は赤色のシグナルでとなり、コントロール断片(内部対照)は黄色シグナルとなる。
【0088】
マイクロアレイをスキャンした結果、乾燥処理によって特異的に誘導された277個の遺伝子を同定し、低温処理によって特異的に誘導された53個の遺伝子を同定し、高塩濃度処理によって特異的に誘導された194個の遺伝子を同定した。なお、コントロール断片の発現比と比較して5倍以上の発現比で誘導された遺伝子を、各種ストレスによって誘導された遺伝子とした。
【0089】
データベースを用いた解析の結果、以下のファミリーに分類できる35種類の転写因子を同定することができた。なお、RAFL05-21-L12については以下のファミリーに分類することはできなかったが、核酸塩基配列をアミノ酸配列に変換したものをGenBank Databaseに登録されているアミノ酸配列Dataに対してblast Xサーチしたところ、既知の転写因子をコードするもの(heat shock transcription factor-like protein)とE-value:e-100のスコアで相同性を示したことから転写因子として同定することができた。すなわち、本実施例によって、36種類の転写因子を同定できた。
1)DREBファミリー:RAFL05-11-M11、RAFL06-11-K21、RAFL05-16-H23、RAFL08-16-D06
2)ERFファミリー:RAFL08-16-G17、RAFL06-08-H20
3)ジンクフィンガーファミリー:RAFL07-10-G04、RAFL04-17-D16、RAFL05-19-M20、RAFL08-11-M13、RAFL04-15-K19、RAFL05-11-L01、RAFL05-14-C11、RAFL05-19-G24、RAFL05-20-N02
4)WRKYファミリー:RAFL05-18-H12、RAFL05-19-E19、RAFL06-10-D22、RAFL06-12-M01
5)MYBファミリー:RAFL05-14-D24、RAFL05-20-N17、RAFL04-17-F21
6)bHLHファミリー:RAFL09-12-N16
7)NACファミリー:RAFL05-19-I05、RAFL05-21-I22、RAFL08-11-H20、RAFL05-21-C17、RAFL05-08-D06
8)ホメオドメインファミリー:RAFL05-20-M16、RAFL11-01-J18、RAFL11-09-C20
9)bZIPファミリー:RAFL05-18-N16、RAFL11-10-D10、RAFL04-17-N22、RAFL05-09-G15
【0090】
(2) 各種ストレス処理時間と発現比率との関係
上述したように単離された36種類の各種ストレス応答性転写因子をコードする遺伝子について、以下のように、各種ストレス処理時間と発現比率との関係を検討した結果を図1〜図57に示す。図1〜図57で示す遺伝子名とストレス処理との対応を表2に示した。
【0091】
【表2】
【0092】
図1〜図57において、縦軸は遺伝子の発現比率を示しており、以下のように算出される。なお、下記式においてFIは、蛍光強度を示す。
発現比率=[(ストレス負荷条件での各cDNAのFI)/(ストレスを負荷しない条件での各cDNAのFI)]÷[(ストレス負荷条件でのコントロール断片のFI)/(ストレスを負荷しない条件でのコントロール断片のFI)]
【0093】
これら図1〜図57に示すように、本方法により単離されたストレス応答性転写因子をコードする遺伝子は、それぞれ異なるプロファイルであるが、各種ストレスの付加により発現誘導されていることが判る。
【図面の簡単な説明】
【0094】
【図1】RAFL08-16-G17について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図2】RAFL05-11-M11について乾燥ストレスと発現比率との関係を示す特性図である。
【図3】RAFL05-11-M11について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図4】RAFL06-11-K21について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図5】RAFL06-11-K21について乾燥ストレスと発現比率との関係を示す特性図である。
【図6】RAFL06-08-H20について乾燥ストレスと発現比率との関係を示す特性図である。
【図7】RAFL06-08-H20について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図8】RAFL05-16-H23について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図9】RAFL05-16-H23について乾燥ストレスと発現比率との関係を示す特性図である。
【図10】RAFL08-16-D06について乾燥ストレスと発現比率との関係を示す特性図である。
【図11】RAFL07-10-G04について乾燥ストレスと発現比率との関係を示す特性図である。
【図12】RAFL04-17-D16について乾燥ストレスと発現比率との関係を示す特性図である。
【図13】RAFL05-19-M20について乾燥ストレスと発現比率との関係を示す特性図である。
【図14】RAFL08-11-M13について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図15】RAFL04-15-K19について乾燥ストレスと発現比率との関係を示す特性図である。
【図16】RAFL04-15-K19について低温ストレスと発現比率との関係を示す特性図である。
【図17】RAFL05-11-L01について乾燥ストレスと発現比率との関係を示す特性図である。
【図18】RAFL05-11-L01について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図19】RAFL05-14-C11について乾燥ストレスと発現比率との関係を示す特性図である。
【図20】RAFL05-19-G24について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図21】RAFL05-19-G24について乾燥ストレスと発現比率との関係を示す特性図である。
【図22】RAFL05-19-G24について低温ストレスと発現比率との関係を示す特性図である。
【図23】RAFL05-20-N02について乾燥ストレスと発現比率との関係を示す特性図である。
【図24】RAFL05-18-H12について乾燥ストレスと発現比率との関係を示す特性図である。
【図25】RAFL05-18-H12について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図26】RAFL05-19-E19について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図27】RAFL06-10-D22について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図28】RAFL06-12-M01について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図29】RAFL06-12-M01について乾燥ストレスと発現比率との関係を示す特性図である。
【図30】RAFL05-14-D24について乾燥ストレスと発現比率との関係を示す特性図である。
【図31】RAFL05-14-D24について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図32】RAFL05-20-N17について低温ストレスと発現比率との関係を示す特性図である。
【図33】RAFL05-20-N17について乾燥ストレスと発現比率との関係を示す特性図である。
【図34】RAFL04-17-F21について乾燥ストレスと発現比率との関係を示す特性図である。
【図35】RAFL09-12-N16について乾燥ストレスと発現比率との関係を示す特性図である。
【図36】AFL05-19-I05について乾燥ストレスと発現比率との関係を示す特性図である。
【図37】RAFL05-19-I05について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図38】RAFL05-21-I22について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図39】RAFL08-11-H20について乾燥ストレスと発現比率との関係を示す特性図である。
【図40】RAFL08-11-H20について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図41】RAFL05-21-C17について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図42】RAFL05-21-C17について乾燥ストレスと発現比率との関係を示す特性図である。
【図43】RAFL05-08-D06について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図44】RAFL05-20-M16について乾燥ストレスと発現比率との関係を示す特性図である。
【図45】RAFL05-20-M16について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図46】RAFL11-01-J18について乾燥ストレスと発現比率との関係を示す特性図である。
【図47】RAFL11-01-J18について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図48】RAFL11-09-C20について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図49】RAFL05-18-N16について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図50】RAFL11-10-D10について乾燥ストレスと発現比率との関係を示す特性図である。
【図51】RAFL11-10-D10について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図52】RAFL04-17-N22について乾燥ストレスと発現比率との関係を示す特性図である。
【図53】RAFL04-17-N22について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図54】RAFL05-09-G15について乾燥ストレスと発現比率との関係を示す特性図である。
【図55】RAFL05-09-G15について高塩濃度ストレスと発現比率との関係を示す特性図である。
【図56】RAFL05-21-L12について乾燥ストレスと発現比率との関係を示す特性図である。
【図57】RAFL05-21-L12について高塩濃度ストレスと発現比率との関係を示す特性図である。
【配列表フリーテキスト】
【0095】
配列番号73及び74は合成プライマーである。
【特許請求の範囲】
【請求項1】
以下の(a)又は(b)のアミノ酸配列を含む乾燥ストレス応答性転写因子をコードする遺伝子。
(a)配列番号8のアミノ酸配列
(b)配列番号8のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、乾燥ストレスに応答性を示し、かつ転写因子活性を有するアミノ酸配列
【請求項2】
以下の(a)、(b)又は(c)のDNAを含む、請求項1記載の遺伝子。
(a)配列番号7の塩基配列からなるDNA
(b)配列番号7の塩基配列において1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列からなり、乾燥ストレス応答性転写因子をコードするDNA
(c)配列番号7の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ乾燥ストレス応答性転写因子をコードするDNA
【請求項3】
請求項1又は2記載の遺伝子を含む発現ベクター。
【請求項4】
請求項3記載の発現ベクターを含む形質転換体。
【請求項5】
請求項3記載の発現ベクターを含むトランスジェニック植物。
【請求項6】
植物が、植物体、植物器官、植物組織又は植物培養細胞である請求項5記載のトランスジェニック植物。
【請求項7】
請求項5又は6記載のトランスジェニック植物を培養又は栽培することを特徴とする乾燥ストレス耐性植物の製造方法。
【請求項8】
以下の(a)又は(b)のアミノ酸配列を含む乾燥ストレス応答性転写因子をコードする遺伝子。
(a)配列番号10のアミノ酸配列
(b)配列番号10のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、乾燥ストレスに応答性を示し、かつ転写因子活性を有するアミノ酸配列
【請求項1】
以下の(a)又は(b)のアミノ酸配列を含む乾燥ストレス応答性転写因子をコードする遺伝子。
(a)配列番号8のアミノ酸配列
(b)配列番号8のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、乾燥ストレスに応答性を示し、かつ転写因子活性を有するアミノ酸配列
【請求項2】
以下の(a)、(b)又は(c)のDNAを含む、請求項1記載の遺伝子。
(a)配列番号7の塩基配列からなるDNA
(b)配列番号7の塩基配列において1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列からなり、乾燥ストレス応答性転写因子をコードするDNA
(c)配列番号7の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ乾燥ストレス応答性転写因子をコードするDNA
【請求項3】
請求項1又は2記載の遺伝子を含む発現ベクター。
【請求項4】
請求項3記載の発現ベクターを含む形質転換体。
【請求項5】
請求項3記載の発現ベクターを含むトランスジェニック植物。
【請求項6】
植物が、植物体、植物器官、植物組織又は植物培養細胞である請求項5記載のトランスジェニック植物。
【請求項7】
請求項5又は6記載のトランスジェニック植物を培養又は栽培することを特徴とする乾燥ストレス耐性植物の製造方法。
【請求項8】
以下の(a)又は(b)のアミノ酸配列を含む乾燥ストレス応答性転写因子をコードする遺伝子。
(a)配列番号10のアミノ酸配列
(b)配列番号10のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、乾燥ストレスに応答性を示し、かつ転写因子活性を有するアミノ酸配列
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
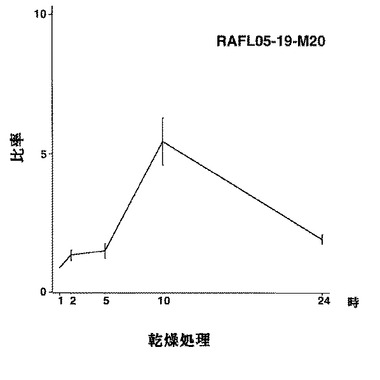
【図14】
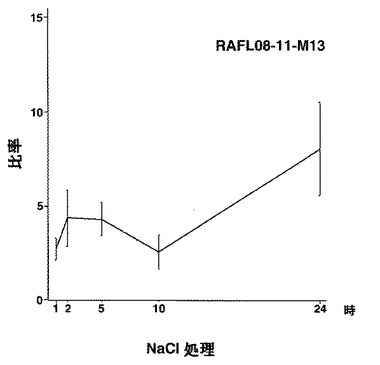
【図15】
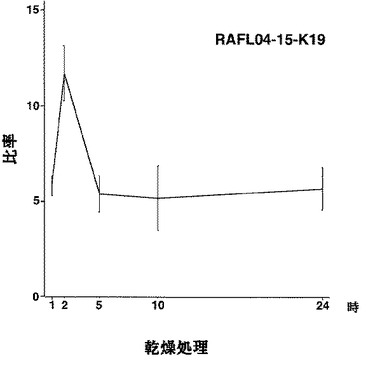
【図16】
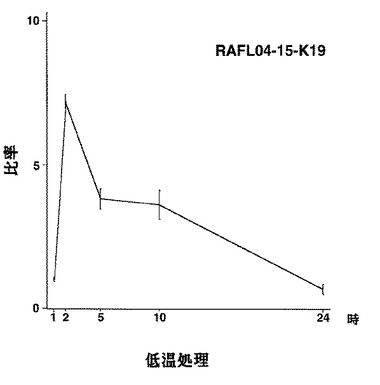
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【図42】
【図43】
【図44】
【図45】
【図46】
【図47】
【図48】
【図49】
【図50】
【図51】
【図52】
【図53】
【図54】
【図55】
【図56】
【図57】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【図42】
【図43】
【図44】
【図45】
【図46】
【図47】
【図48】
【図49】
【図50】
【図51】
【図52】
【図53】
【図54】
【図55】
【図56】
【図57】
【公開番号】特開2007−82557(P2007−82557A)
【公開日】平成19年4月5日(2007.4.5)
【国際特許分類】
【出願番号】特願2006−318803(P2006−318803)
【出願日】平成18年11月27日(2006.11.27)
【分割の表示】特願2002−20329(P2002−20329)の分割
【原出願日】平成14年1月29日(2002.1.29)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【Fターム(参考)】
【公開日】平成19年4月5日(2007.4.5)
【国際特許分類】
【出願日】平成18年11月27日(2006.11.27)
【分割の表示】特願2002−20329(P2002−20329)の分割
【原出願日】平成14年1月29日(2002.1.29)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【Fターム(参考)】
[ Back to top ]