
生体リズムの制御剤
【課題】本発明は、概日リズム周期を調節することで概日リズム障害を治療する方法、及びそのための概日リズム周期改善剤を提供することにある。
【解決手段】本発明においては、ハルミンなどのハルマラアルカロイド、又はその薬理学的に許容しうる塩を有効成分として含む、概日リズム周期改善剤を提供する。
【解決手段】本発明においては、ハルミンなどのハルマラアルカロイド、又はその薬理学的に許容しうる塩を有効成分として含む、概日リズム周期改善剤を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、概日リズムを制御することによる睡眠障害の改善剤に関する。
【背景技術】
【0002】
遺伝性の睡眠障害に加えて、社会の24時間化に伴う様々な睡眠障害が社会的問題となっている。概日リズム睡眠障害と呼ばれる一連の睡眠障害の発症には、時計遺伝子によって構成されている体内時計が関係しているものと考えられているが、その詳細なメカニズムに関しては不明である。
睡眠障害の治療法としては、特別な装置を必要とする高照度光療法や、ビタミンB12(非特許文献1)やメラトニン製剤(特許文献1)の投与が一般的である。しかし、これらの治療方法は、いずれも体内時計の位相を調節することにより生体リズムを正常化させようとするものであり、体内時計の周期異常に起因する障害の根本的な治療法とはなりえない。
また、睡眠薬としては、ベンゾジアゼピン系の薬剤(非特許文献2)が一般に用いられており、長短時間型から、短時間型、中間型、長時間型まで様々な半減期の薬剤が開発されている。しかしながら、これらの睡眠薬による睡眠障害の治療法は、対処療法的なものであり、根本的に睡眠障害を治療することは困難である。また、夢遊症状等の副作用を伴う場合も多く、その使用法には細心の注意が必要である。最近、ノシセプチン受容体の作動薬としての作用を有する4-オキソイミダゾリジン-2-スピロピペリジン誘導体に概日リズム睡眠障害治療効果が期待されている(特許文献2)が、まだ研究開発途上であり副作用も懸念される。
一方、各種飲食品の原料やそれらに含まれる天然成分を用いる睡眠改善剤の研究開発も盛んで、茶葉由来のテアニンを用いるもの(特許文献4)、内因性のメラトニン分泌効果を有する発酵ホエーなどのホエー類を添加するもの(特許文献5)の他、高麗人参エキス(非特許文献3)、イクラ油に含まれるフォスファチジルコリン(非特許文献4)などを用いた睡眠改善剤が提案されている。これらは、いずれも飲食品、嗜好品などに用いられている素材に由来しているため、安全性も高く、日常的に摂取可能であるといえるが、その詳細な睡眠改善効果や作用メカニズムの解明は十分ではなく、効果の個人差も大きい。
概日リズムは、体内時計によって制御されており、近年、時計遺伝子と呼ばれる一連の遺伝子群によって体内時計のリズムが刻まれていることが明らかとなってきた。体内時計は、概日リズムの位相調節と周期長の調節という2つの機能を有している。概日リズムの位相調節に関しては、多くの液性因子によって時計遺伝子の発現が細胞培養系において誘導されることから、生体においても、多様な調節因子が関与しているものと推測される(非特許文献5)。
【0003】
一方で、概日リズムの周期長の調節に関しても、グリコーゲンシンターゼキナーゼ3β(GSK3β)が、時計分子PERIOD2(非特許文献11)やREV-ERBα(非特許文献12)のリン酸化を介して体内時計を調節するとの報告や、概日リズムに関与する転写因子の1つであるBMAL1の抑制効果のあるc-Jun N末端キナーゼ-3(JNK3)の酵素活性調節剤を用いた睡眠改善剤も開発されてきている(特許文献3)。
睡眠障害を根本的に改善するためには、概日リズムのうちでも、とりわけ周期長を調節する睡眠改善剤の開発が強く望まれており、日常的に摂取可能な安全性の高い睡眠改善剤として、飲食品、嗜好品の素材として安全性の確立している天然成分由来の物質からの探索が望まれていた。しかしながら、これまでは詳細な概日リズム周期を検出する簡便な実験方法が確立されていなかったので、安全性の確立している天然成分由来であって概日リズムの周期長を調節する効果的な睡眠改善剤は未だ報告がない。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特表平8−502259号公報
【特許文献2】WO2003/010168
【特許文献3】WO2003/063907
【特許文献4】WO2005/097101
【特許文献5】WO2005/097101
【特許文献6】特開2009−183197号公報
【特許文献7】特表平10−505606号公報
【非特許文献】
【0005】
【非特許文献1】「サーカディアンリズム睡眠障害の臨床」千葉茂、本間研一 編、新興医学出版社、2003
【非特許文献2】「睡眠障害の対応と治療ガイドライン」、内山真 編、じほう、2002
【非特許文献3】Young Ho Rhee, et al.,Psychopharmacology,101,p.486-488(1990)
【非特許文献4】日比野英彦、Food Style21,Vol.7,No.3,p.50-53(2003)
【非特許文献5】Curr Biol 2000,10:1291-4
【非特許文献6】EMBO Reports 2007,8:366-71、Eur J Neurosci 2007,26:451-62
【非特許文献7】Science,1992,258,607-614
【非特許文献8】Cur.Biol.,2000,10,1291-1294
【非特許文献9】FEBS Lett.,1999,465,79-82
【非特許文献10】Gene to Cells,2003,8,713-720
【非特許文献11】J.Biol.Chem.,2005,280,29397-29402
【非特許文献12】Science,2006,311,1002-1005
【非特許文献13】Cell,2000,103,1009-1017
【非特許文献14】Nature,2002,418,534-539
【非特許文献15】Mol.Cell.Biol.,2008,28,3477-3488
【非特許文献16】J.Toxicol.Environ.Health A,2001,64,645-660
【非特許文献17】Pharmacology and Therapeutics,2004,102,111-129.
【非特許文献18】Neuropharmacology,1988,27,849-855
【非特許文献19】Biochemical Pharmacology,1977,26,1991-1996.
【非特許文献20】Arch.Biochem.Biophys.,1997,337,137-142
【非特許文献21】レスターグリンスプーン等著、杵渕幸子訳「サイケデリック・ドラッグ−向精神物質の科学と文化」工作舎(2000)p.36-38
【非特許文献22】Neuron.2004 Aug 19;43(4):527-37
【非特許文献23】Nuclear Receptor Signaling,2009,7,1-32
【非特許文献24】Biochim.Biophys.Acta,2006,1763,805-814
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の課題は、これまで広く摂取されてきた天然成分由来であって、かつ効果的に概日リズムの周期長を調節する概日リズム周期改善剤を提供することにある。
【課題を解決するための手段】
【0007】
最近の概日リズム関連遺伝子を用いたノックアウト実験結果によると、Bmal1遺伝子をノックアウトしたマウスでは、行動のリズムが全く見られなくなり、まるで生物リズムが全く破壊された様に振舞うのに対して、他の時計遺伝子PeriodやCryptochrome等のノックアウトマウスでは活動周期に異常が観察されるのみであり、Bmal1遺伝子は単一遺伝子の破壊において行動リズム形成を全く示さなくなる唯一つの遺伝子であることがわかった。さらにBmal1遺伝子ノックアウトマウスでは時計中枢である視交叉上核(SCN)において時計遺伝子Period1およびPeriod2の発現がほぼ完全に抑えられている。以上のことより、生物時計調節の中心となる遺伝子がBmal1遺伝子であることはほぼ間違いないと考えられている(非特許文献13)。
このBmal1遺伝子は概日リズムをもって転写調節されており、ルシフェラーゼ遺伝子を用いた培養細胞レベルでのレポーターアッセイが、哺乳類における生体内でのBmal1遺伝子を介した概日リズムを再現できることが知られていた(非特許文献14)が、従来法では、細胞内でのプラスミドの状態が不安定であり、培地中に被検物質を添加した状態で細胞培養を続けることにより、その影響を正確に測定することはできなかった。
【0008】
その後、本発明者らにより、Bmal1遺伝子の概日リズム転写をモニターするために必要最小限のプロモーター中の領域を決定することができたので(非特許文献15)、当該領域をルシフェレース遺伝子に接続しNIH3T3細胞に安定的に保持されるような細胞株を樹立した。具体的には、NIH3T3細胞にBmal1遺伝子プロモーター中の「RORE」配列及び転写開始部位を含む領域(−202〜+27位)にルシフェラーゼ遺伝子を繋いだレポータープラスミドを導入し、安定に概日リズムを刻みながらルシフェラーゼを生産する細胞株である(特願2009−175576号、特願2009−186257号)。そして、当該レポーターアッセイ法によりルシフェレース活性をモニターするだけで、転写因子BMAL1と転写因子CLOCKのヘテロ二量体によって発現が促進されたPeriod遺伝子からPERIODが合成され、核外でCRYと結合して核内移行したPERIOD-CRY複合タンパク質がBMAL1とCLOCK複合体による転写活性化を抑制するという一連の概日リズム形成サイクル自体に長期間にわたって作用する物質を、的確にしかも簡便に知ることが可能となった。
本発明者らは、当該樹立細胞株を用いて種々の物質を作用させて、ルシフェラーゼ活性をモニターしたところ、プロテインキナーゼCの活性化剤であるTPA及び同阻害剤であるG6850を作用させ、プロテインキナーゼCの活性を増減させたところ、当該活性の増減に応じて、概日リズムが延長及び短縮することが実証でき、プロテインキナーゼCがBmal1プロモーター領域の転写調節部位に作用して、概日リズム周期の調節をしている生体内物質であることを確認し、特許出願している(特願2009−175576号)。
また、ヒガンバナ植物由来のアルカロイドであるリコリンならびにリコリシジノールが、濃度依存的にBmal1遺伝子の転写リズムの周期を延長させることが明らかとなったので、同日付で特許出願した(特願2009−186257号)。この本発明者らの出願以前に、リコリン、リコリシジノールを含む天然成分由来の他のアルカロイドに関しては、従来概日リズムの周期長の調節作用については報告されていない。
【0009】
本発明者らの上記樹立細胞株を用いてのさらなる探索の結果、β−カルボリン誘導体の1種であるハルマラアルカロイド(Harmala alkaloid)のハルミンが、転写リズムの周期を延長していることを観察した。この結果は、ハルミンの体内時計制御剤としての可能性を示している。ハルマラアルカロイドは、他のβ−カルボリン誘導体と同様、可逆的なMAOI(モノアミン酸化酵素阻害剤)として知られ、中枢神経系のモノアミン化合物であるセロトニンやエピネフィリンと共にメラトニン分解に対しても抑制作用があるとされている。メラトニンは概日リズム調節に影響があると期待されている物質である(特許文献1)ことから、ハルマラアルカロイドを含むβ−カルボリン誘導体一般に対してもメラトニン受容体を介した概日リズム調節への影響が期待されている(特許文献7)ものの、メラトニン自身の概日リズム調節作用に関してすら学術的にコンセンサスが得られているわけではなく、実際に概日リズム調節への影響が報告された例はない。一方、本発明者の上記知見は、ハルマラアルカロイドが直接的に時計遺伝子に作用して、概日リズム周期の延長をする化合物であることを確認したことである。さらに、ハルミン濃度を変化させて概日リズムを観察していくと、ハルミンの概日リズム延長作用に濃度依存性が存在することを確認した。加えて、ハルミンに細胞内のRORαを核内に移行させる機能があることを見出したことから、ハルミンが細胞質中のRORαを核内に移行させることでBmal1遺伝子上流のRORE配列に結合する機会を増大させ、Bmal1遺伝子発現誘導を促進した結果、概日リズムの延長を起こすという、ハルミンの概日リズム延長作用の作用機序を解明することができた。ハルミンなどのハルマラアルカロイドは、ハーブ類など食用または嗜好品用の植物成分や生体内物質であることから安全性についての懸念は低く、概日リズム周期改善用の医薬組成物としての期待はきわめて大きいものがある。
以上の知見を得て本発明を完成した。
【0010】
また、本発明により、ハルミンの概日リズム延長作用の作用機序が、直接Bmal1遺伝子に作用するのではなく、しかもRORαの全体量を増加させるのでもなく、細胞内に広く存在しているRORαのうち、細胞質中のRORαを核内に移行させることで核内のRORα量を増加させ、Bmal1遺伝子上流のRORE配列への結合量を増やし、その結果Bmal1遺伝子発現誘導を促進するという一連の工程を経て概日リズムの延長を引き起こすという作用機序であることが解明されたが、この作用機序はRORαとBmal1遺伝子との関係性においても初めての知見である。すなわち、従来からRORαがROREに結合することによりBmal1遺伝子の発現が誘導されて概日リズムの延長が引き起こされることは知られていた(非特許文献22)ものの、RORαのROREへの結合を起こすトリガーが何であるかは全く解明されていなかった。本発明における上記知見は、細胞中でのRORαの発現量が増大するのではなく、細胞中に広く分布しているRORαが何らかの物質の作用により核内への移行を促進されて核内での局在化が起こることがトリガーとなっていたことを解明したことでもある。つまり、RORαの核内局在化を観察するスクリーニング系によって、RORαの核内移行を起こさせるトリガー物質を検索することができることを示している。このような知見をもとに、スクリーニング方法についての発明も完成させた。
【0011】
すなわち、本発明は以下の発明を含むものである。
〔1〕 ハルマラアルカロイド(Harmala alkaloid)、又はその薬理学的に許容しうる塩もしくはエステルを有効成分として含む、概日リズム周期改善剤。
〔2〕 前記ハルマラアルカロイドがハルミンである、前記〔1〕に記載の概日リズム周期改善剤。
〔3〕 RORαを発現し、かつ安定して概日リズムを刻む培養細胞を用意し、当該培養細胞に対して被検物質を添加して、当該細胞の核内のRORα量を測定し、被検物質添加前の核内RORα量と比較して有意差をもって核内移行を促進させる物質を選択することを特徴とする、概日リズム周期改善物質のスクリーニング方法。
【発明の効果】
【0012】
本発明により、効果的に体内時計の周期長を調節することが可能であり、体内時計の周期異常に起因する概日リズム障害の根本的な治療のために有効でかつ安全性の高い物質を提供することができた。また、あわせて、細胞内のRORαの核内移行を観察する系を用いる概日リズム障害改善物質の優れたスクリーニング系を開発した。
【図面の簡単な説明】
【0013】
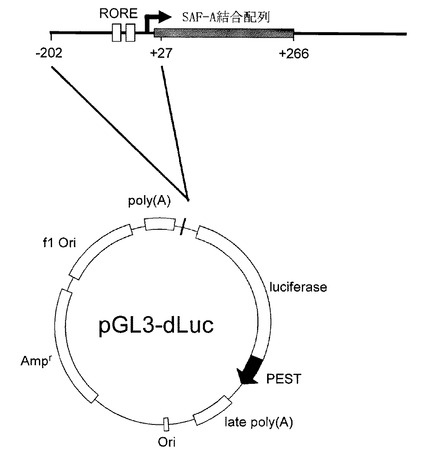
【図1】Bmal1レポータープラスミドの構築 [Bmal1プロモーター領域の遺伝子地図中でのRORE(白)ならびにSAF-A結合配列(灰)の位置を示す。ROREを含む-202〜+27の領域をプラスミドpGL3-dLucのルシフェレース遺伝子上流に挿入した。]
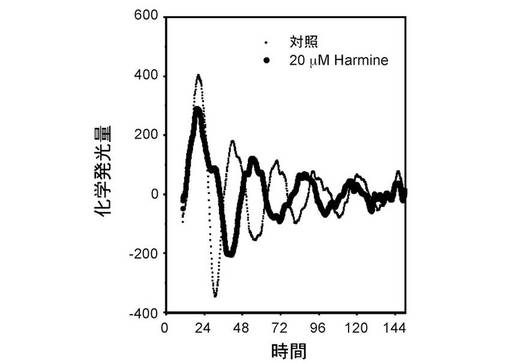
【図2】ハルミンによる概日リズム調節 [20μMのハルミン存在下(黒線)、ならびに対照(点線)の化学発光量の経時変化を示す。]
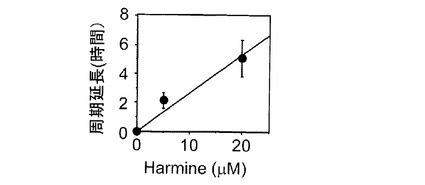
【図3】概日リズム延長作用のハルミン濃度依存性
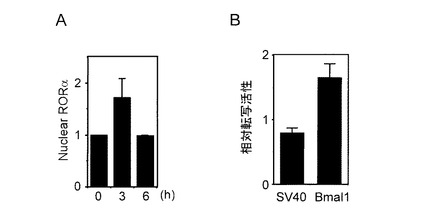
【図4】ハルミンによるRORαの核内局在化作用 [A:核内RORαタンパク量 (横軸は、20μMハルミン処理時間を表す。)] [B:ハルミン処理3時間後の相対転写活性 (図中、SV40ならびにBmal1プロモーターのハルミン処理による相対転写活性を示す。)]
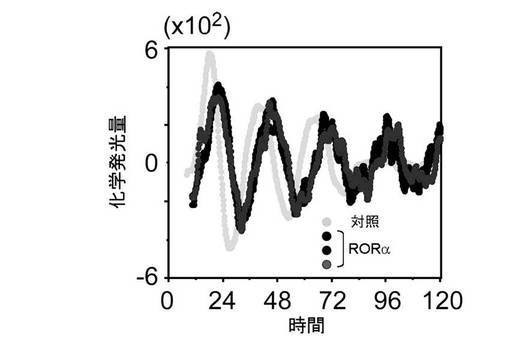
【図5】RORaによる概日リズム調節 [RORa発現(黒〜濃灰色)、ならびに対照(薄灰色)の化学発光量の経時変化を示す。]
【発明を実施するための形態】
【0014】
1.睡眠障害について
睡眠障害は、「概日リズム睡眠障害」と「睡眠呼吸障害」とに大別される。概日リズム睡眠障害には、外因性の急性症候群と内因性の慢性症候群とに区分され、睡眠相前進症候群や睡眠相後退症候群、非24時間睡眠覚醒障害、不規則型睡眠覚醒障害などは、内因性慢性症候群に含まれる。概日リズム睡眠障害には、体内時計の関与が考えられており、特に、内因性慢性症候群は、体内時計の周期異常が一因となっている。従来からの典型的な睡眠障害の治療法である光照射やメラトニン投与、あるいは様々な睡眠薬による治療法は、体内時計のリセットに作用する対処療法的なものであるのに対し、本発明が目指す睡眠障害の治療法は、体内時計の周期異常、すなわち「概日リズム障害」を改善することによる治療法である。
【0015】
2.概日リズム障害により引き起こされる疾患について
概日リズム障害に起因する疾患としては、睡眠相前進症候群(ASPS)、睡眠相後退症候群(DSPS)、非24時間睡眠相後退症候群、および季節性うつ病が典型的なものとして考えられる。その他、内因性躁鬱病、周期性緊張症、周期性高血圧症、周期性潰瘍、不規則排卵周期、およびインスリン分泌の周期性異常に起因する糖尿病などの周期性・反復性障害や、脳血管型痴呆やアルツハイマー型痴呆における夜間徘徊なども概日リズム障害が関与すると考えられている。本発明は、概日リズム障害のうちの概日リズム周期を延長することにより障害を改善しようとするものである。
【0016】
3.概日リズムとBmal1遺伝子について
体内時計の調節に係わる主要な因子としては、体内時計の刻みを促進するBmal1遺伝子とClock遺伝子、抑制因子として働くCry遺伝子とPeriod遺伝子の4種類がある。
これら遺伝子群のうちで、Bmal1遺伝子をノックアウトしたマウスでは、行動のリズムが全く見られなくなり、まるで生物リズムが全く破壊された様に振舞う。他の時計遺伝子PeriodやCry等のノックアウトマウスでは活動周期に異常が観察されるのみであり、Bmal1遺伝子は単一遺伝子の破壊において行動リズム形成を全く示さなくなる唯一つの遺伝子である。さらにBmal1遺伝子ノックアウトマウスでは時計中枢である視交叉上核(SCN)において時計遺伝子Period1およびPeriod2の発現がほぼ完全に抑えられている。以上のことより、生物時計調節の中心となるのがBmal1遺伝子であると考えられている(非特許文献13)。
最近、本発明者らは、Bmal1遺伝子プロモーター領域中の「SAF−A結合領域(-27〜+266位)」へのSAF−A結合によるBmal1遺伝子を介した概日リズムの転写調節を解明した(非特許文献15、特許文献9)。
【0017】
4.概日リズムとRORaタンパク質について
Bmal1遺伝子は、遺伝子上流のプロモータ領域中のRORE配列(6塩基のAT-rich配列とそれに続くRGGTCA配列)に結合する転写因子の作用で、正もしくは負の転写調節が行われている。最初にRORαがROREの結合によってBmal1の強い発現誘導が起こされ、当該発現の増強がREV-ERBαにより抑制されたことが観察された(非特許文献22)。ROR及びREV-ERBは、いずれも核内受容体のNR1ファミリーに属しており、RORにはα、β、γのサブタイプがあり、REV-ERBにもα、βのサブタイプが知られている。これらの転写因子はRORE配列に結合することによりBmal1遺伝子特異的に正もしくは負の転写調節を行い、発生、分化、代謝、ストレス反応等の生体調節を行っているとも考えられている(非特許文献23)。近年、RORαのリガンドがコレステロール誘導体であることが判明し、細胞膜に存在しているコレステロールと結合することにより核内のみならず、細胞膜や細胞質にも存在することが報告されている(非特許文献23)。RORαは種々の細胞によっては細胞内での局在状態が異なることが観察されてはいた(非特許文献24)ものの、局在現象の存在を示したにとどまり、RORαの核内への局在化とBmal1遺伝子発現の誘導量増大との関連性に着目して実証したのは本発明がはじめてである。
【0018】
5.概日リズムの周期を調節する物質のスクリーニング系
(1) 本発明において、概日リズムの周期を調節する物質のスクリーニングに用いるレポーターアッセイ系は、先の出願において本発明者らが確立したアッセイ系であり(特願2009−175576号、特願2009−186257号)、従来の研究でBmal1遺伝子のプロモーター領域などの特定領域に一過的に作用する物質又は遺伝子を探索するために用いられていたスクリーニング系を改変して利用するものである。
まず、プラスミドpGL3-dLuc(非特許文献14)のルシフェレース遺伝子上流に、Bmal1プロモーター領域の「RORE」を含む-202〜+27の領域を挿入して作製したレポータープラスミド(図1)を、非特許文献14の手法に従ってNIH3T3細胞に導入し、継代培養して、安定に概日リズムを刻みながらルシフェラーゼを生産する形質転換NIH3T3細胞株を樹立した。
当該樹立細胞の培地に被検物質を添加し、培養しながらルシフェラーゼの発光強度を経時的にモニターすることにより、転写因子BMAL1と転写因子CLOCKのヘテロ二量体によって発現が促進されたPeriod遺伝子からPERIODが合成され、核外でCRYと結合して核内移行したPERIOD-CRY複合タンパク質がBMAL1とCLOCK転写活性を抑制するという一連の概日リズム形成サイクル自体に長期間にわたって作用する物質、その周期を早めたり遅くしたりする物質がはじめて探索できるようになった。
【0019】
(2) 本発明により、ハルミンの概日リズム周期延長作用の作用機序が、細胞内に広く存在しているRORαのうち細胞質中の又は細胞膜に結合しているRORαに働いて核内に移行させる作用であることを解明した。つまり、RORαを核内に移行させることで核内に存在するRORα量を増加させ、Bmal1遺伝子上流のRORE配列への結合量を増やし、その結果Bmal1遺伝子発現誘導を促進するという一連の工程を経て概日リズムの延長を引き起こすという作用機序である。すなわち、このことは、細胞中に広く分布しているRORαを核内へ移行させて核内での存在量を増大させる物質を検索することで、ハルミン類と同様に、概日リズムの周期延長作用を有する物質をスクリーニングすることができることに他ならない。
細胞内でのRORαの核内移行を観察する手法としては、どのような手法でも良いが、典型的な手法を以下に2種類示す。
【0020】
(3)RORαの核内移行観察法−1
i)まず、RORαを発現し、かつ安定して概日リズムを刻む培養細胞を用意する。
その際、そのような培養細胞としては、NIH3T3細胞株などを用いることができるが、上記(1)で述べた、安定に概日リズムを刻みながらルシフェラーゼを生産する形質転換NIH3T3細胞株を用いると、概日リズムの周期延長作用の最終的な確認も同一株で行うことができるので、より好ましい。さらに、RORαを細胞質内で大量発現させるためにRORα遺伝子発現ベクターを導入することが好ましい。その場合、RORαはGFP等の融合タンパクとしておくかFLAG等の遺伝子タグにより標識しておく。
ii)RORαの当該培養細胞内での局在化状態を観察する。すなわち、細胞の核内でのRORαの存在量を測定する。核を特異的に染色しておき、標識化されたRORαの信号(発光、蛍光)量を測定することで、核内RORα量を測定することができる。通常は、下記工程iii)に先立ち工程ii)を行っておくが、同一の細胞系を用いてii)及びiii)を平行して行うこともできる。
iii)被検物質を添加後の当該細胞内の核内のRORα量を上記工程ii)と同様に測定する。
iv)工程ii)及び工程iii)の核内RORα量を比較して、被検物質添加により、有意差をもって核内のRORα量が増大している場合に、当該被検物質を、概日リズムの周期延長作用を有する物質の候補として選択する。
【0021】
(4)RORαの核内移行観察法−2
i)まず、RORαを発現し、かつ安定して概日リズムを刻むNIH3T3細胞株などの培養細胞を用意する。安定に概日リズムを刻みながらルシフェラーゼを生産する形質転換NIH3T3細胞株を用いることが好ましい点は上記手法と同様である。さらに、RORαを細胞質内で大量発現させるためにRORα遺伝子発現ベクターを導入することが好ましく、その場合、RORαはGFP等の融合タンパクとしておくかFLAG等の遺伝子タグにより標識しておくことが好ましい点も同様である。
ii)培養細胞を回収後、Dyer & Herzogの方法(BioTechniques,1995,19,192-195)に従い、遠心分離により細胞質と核タンパク画分を分離し、RORαの当該培養細胞内での局在化割合を計測する。具体的には、得られたそれぞれの画分をWestern blot解析し、抗RORa抗体(Santa Cruz Biotechnology社製)を用いてRORaタンパクを定量する。試料間の核総タンパク量の誤差を補正するために、抗ラミン抗体(Santa Cruz Biotechnology社製)を用いて核特異的マーカータンパクであるラミン量を測定し、核内RORaタンパク量を補正する。細胞質におけるRORa量は、抗アクチン抗体(Sigam社製)を用いて測定した細胞質アクチン量により補正したものを用いる。通常は、下記工程iii)に先立ち工程ii)を行っておくが、同一の細胞系を用いてii)及びiii)を平行して行うこともできる。
iii)被検物質を添加後の当該細胞内の核内のRORα量を上記工程ii)と同様に測定する。
iv)工程ii)及び工程iii)の核内RORα量を比較して、被検物質添加により、有意差をもって核内のRORα量が増大している場合に、当該被検物質を、概日リズムの周期延長作用を有する物質の候補として選択する。
【0022】
6.ハルマラアルカロイド(Harmala alkaloid)について
ハルマラアルカロイド(Harmala alkaloid)は、インドール骨格を有するβ−カルボリンアルカロイドの1種であり、MAOI(モノアミン酸化酵素阻害剤)としてインド原産のはまびし科の植物Peganum harmala(Halmala、Syrian Rue)の種子から単離された(非特許文献16)。これらハルマラアルカロイドは南米の植物の蔓の樹皮などにも含まれており、南米では古くからアヤワスカという飲料や嗅ぎタバコなどの形で摂取されていた(非特許文献17)。典型的なハルマラアルカロイドとしては、ハルミン(Harmine)、ハルマリン(Harmaline)、ハルマロール(Harmalol)があげられる(下記に化学構造を示す。)。ハルマラアルカロイドは、セロトニンやエピネフィリンというモノアミン化合物の分解を阻害する中枢神経系刺激物質であり、可逆的なMAO-A阻害活性を有し、作用が穏和で安全性が高い向精神薬として期待されている(非特許文献18〜21)。これらハルマラアルカロイドは、いずれもその作用機序も化学構造的にも非常に似通っており、特にハルミンはハルマリンの代謝産物であるとも言われている(非特許文献21)。
ハルマラアルカロイドの可逆的なMAOI(モノアミン酸化酵素阻害剤)としての作用は、メラトニン分解に対しても抑制作用があるとされおり、メラトニンは概日リズム調節に影響があると期待されている物質である(特許文献1)ため、ハルマラアルカロイドに対してもメラトニン受容体を介した概日リズム調節への影響が期待されている(特許文献7)が、メラトニン自身の概日リズム調節作用に関してすら学術的にコンセンサスが得られているわけではなく、ハルマラアルカロイドについて実際に概日リズム調節への影響が報告された例はない。一方、本発明においては、ハルマラアルカロイドのハルミンが直接体内時計遺伝子Bmal1遺伝子の転写を調節することにより概日リズム周期の延長を引き起こすことを実証したものである。
そして、本発明により、ハルミンのBmal1遺伝子の転写調節の作用機序は、直接的には細胞内に広く存在しているRORαへの作用であり、細胞質中の又は細胞膜に結合しているRORαに働いて核内に移行させることで核内に存在するRORα量を増加させ、Bmal1遺伝子上流のRORE配列への結合量を増やし、その結果Bmal1遺伝子発現誘導を促進するという一連の工程を経て概日リズムの延長を引き起こすという作用機序であることも解明した。
このことは、根本的な概日リズム改善のために必須の概日リズム周期改善剤の有効成分としてハルマラアルカロイドを用いることができることを示すものである。
そして、ハルミンなどのハルマラアルカロイドは、もともとハーブ類など食用または嗜好品用の植物成分や生体内物質であり、向精神薬として実際に投与された例もあることから、その安全性についての懸念は低い。大量に投与するとLSDのような幻覚作用を引き起こすことは知られている(非特許文献18,21)が、そもそもMAO−A阻害薬としての作用がLSDの8,000分の1であること(非特許文献21)からも、本発明における概日リズム周期改善剤としての有効量を投与する程度であれば、全く問題とはならない。概日リズム周期異常に起因する疾病を予防、治療するための優れた医薬組成物としての期待が大きい。
なお、本発明の実施態様では、典型的なハルマラアルカロイドとしてハルミンにおける、体内時計遺伝子の周期を変化させる活性について述べるが、他のハルマラアルカロイドも同様の作用を有する。
【0023】
本発明におけるハルマラアルカロイドは、以下の化学構造式で表現できる。
<式(1)>

(Rは、メトキシ基(−OCH3)または水酸基(−OH)を表す。また3位及び4位の間の点線は単結合又は2重結合であることを表す。)
典型的なハルマラアルカロイドのハルミンは、式(1)において、Rがメトキシ基であって、かつ3位と4位の間が2重結合を形成している化合物であり、以下の構造式で表される。
<式(2)>
【0024】
ハルマリン(Harmaline)は、式(1)において、Rがメトキシ基であって、かつ3位と4位の間が単結合を形成している化合物であり、以下の構造式で表される。
<式(3)>
【0025】
ハルマロール(Harmalol)は、式(1)において、Rが水酸基であって、かつ3位と4位の間が単結合を形成している化合物であり、以下の構造式で表される。
<式(4)>
【0026】
7.本発明における概日リズム周期調節の対象動物
本発明で概日リズム調節をしようとする対象動物は、ヒト等の霊長類、イヌ、ネコなどの愛玩動物、ウシ、ウマなどの家畜動物、マウス、ラットなどの実験動物を含む哺乳類及び相同性の高い時計遺伝子を持つ鳥類及び魚類である。したがって、本発明で、「Bmal1遺伝子」などというとき、ヒトを含む哺乳類、鳥類及び魚類由来の遺伝子又は遺伝子産物を指すものである。
【0027】
8.概日リズム周期改善剤について
上記ハルマラアルカロイドは、そのままで又は薬理学的に許容しうる塩もしくは摂取した生体内において容易に転換されるエステル型の前駆体の状態で、その有効量をヒトを含む動物に投与することで、概日リズム周期を延長することができるから、概日リズム周期が短い障害を有する患者に対する概日リズム周期改善剤として用いることができる。本発明に用いられる塩としては、酢酸塩、硫酸塩、塩酸塩等がある。
概日リズム周期改善剤を有効成分として含み、適当な薬理学的に許容しうる担体または賦形剤などと組み合わせることで概日リズム周期障害に起因する睡眠相前進症候群(ASPS)、睡眠相後退症候群(DSPS)、非24時間睡眠相後退症候群、および季節性うつ病の予防及び治療用の医薬組成物とすることができ、その際に他の治療用化合物または医薬と一緒に使用することができる。
また、ハルマラアルカロイドは従来から可逆性MAO−A阻害薬として、向精神作用が知られていることから、概日リズム障害に起因してうつ病を併発している場合には、特に抗うつ剤としての効果も期待される。
本発明の概日リズム周期改善剤の投与形態は、全身投与であっても局所投与であってもよく、注射、膏薬など非経口投与でも、経口投与でもよく、周知の製剤化方法が適用できる。
用量は、患者の年齢、体重等により適宜調整することができ、投与形態により異なるが、例えば、塩酸塩などとして体重1kg当たり、0.001mg〜20mg、好ましくは、0.01mg〜10mg、より好ましくは0.1〜1.0mgであり、目的に応じて投与回数を決定することが好ましい。
【0028】
また、前記5.(2)で述べた本発明の概日リズムの周期改善物質のスクリーニング系により得られた概日リズム周期改善物質も、上記ハルマラアルカロイドと同様に、適当な薬理学的に許容しうる担体または賦形剤などと組み合わせることで概日リズム周期改善剤として用いることができる。
【実施例】
【0029】
以下、実施例により本発明を具体的に説明するが、本発明は特にこれら実施例に限定されるものではない。
なお、本発明で使用されている技術的用語は、別途定義されていない限り、当業者により普通に理解されている意味を持つ。本発明の実施例で用いた遺伝子組換え技術、PCR法、その他の手法などの具体的な手順や条件は、特に断らない限り、Sambrook and Russell,Molecular Cloning:A Laboratory Manual,3rd Edition.Cold Spring Harbor Laboratory Press,Plainview,NY(2001)、Cold Spring Harbor Laboratory Press,Cold Spring Harbor,New York(1989);D.M.Glover et al.ed.,"DNA Cloning",2nd ed.,Vol.1 to 4,(The Practical Approach Series),IRL Press,Oxford University Press(1995);Ausubel,F.M.et al.,Current Protocols in Molecular Biology,John Wiley & Sons,New York,N.Y,1995;日本生化学会編、「続生化学実験講座1、遺伝子研究法II」、東京化学同人(1986);日本生化学会編、「新生化学実験講座2、核酸 III(組換えDNA技術)」、東京化学同人(1992);R.Wu ed.,"Methods in Enzymology",Vol.68(Recombinant DNA),Academic Press,New York(1980);R.Wu et al.ed.,"Methods in Enzymology",Vol.100(Recombinant DNA,PartB) & 101(Recombinant DNA,Part C),Academic Press,New York(1983);R.Wu et al.ed.,"Methods in Enzymology",Vol.153(Recombinant DNA,Part D),154(Recombinant DNA,Part E) & 155(Recombinant DNA,Part F),Academic Press,New York(1987)などに記載の方法あるいはそこで引用された文献記載の方法またはそれらと実質的に同様な方法により行うことができる。
また、本発明で引用した先行文献又は特許出願明細書の記載内容は参照して、本明細書の記載として組み入れるものとする。
【0030】
(参考例1)Bmal1プロモーターにより転写されるルシフェラーゼ遺伝子を安定的に保持しているNIH3T3細胞株の樹立
Bmal1遺伝子のリズミックな発現に必要最小限のプロモーター領域として、「RORE」を含む-202〜+27の領域を選定し、プラスミドpGL3-dLuc(非特許文献14)のルシフェラーゼ遺伝子上流に、挿入してBmal1レポータープラスミドを作製した(図1)。
当該Bmal1レポータープラスミドと共に、抗生物質(Zeocin)耐性マーカー遺伝子を持つpTracer-CMV(Invitrogen社製)をNIH3T3細胞に導入し(非特許文献14記載の手法による。)、Bmal1プロモーターにより転写調節されるルシフェラーゼ遺伝子を安定的に保持する組換えNIH3T3細胞を取得した。さらに継代培養を繰り返し、安定に概日リズムを刻みながらルシフェラーゼを生産する形質転換NIH3T3細胞株を樹立し、樹立した細胞の中でルシフェラーゼ活性の概日リズムが最も安定的に観察されるクローンを選択した。
【0031】
(実施例1)ハルミンによる概日リズムの周期延長
参考例1で樹立したNIH3T3細胞株を約5×105個を35mm培養ディッシュに播種した後、100nMデキサメサゾンにより生体リズムをリセットして用いた。
樹立したNIH3T3細胞株の発光基質ルシフェリンを含む培地に被検物質を添加し、細胞培養を続けながら、リアルタイムでレポーター遺伝子による化学発光を約150時間測定した。発光は10分毎に1分間測定した。
次いで、20μMのハルミン塩酸塩(東京化成)と、発光基質ルシフェリンを含む培地で細胞培養を続けながら、リアルタイムでレポーター遺伝子による化学発光を10分毎に1分間測定した。150時間にわたり測定したところ、概日リズムの周期は対照(点線)が25.8時間に対して20μMのハルミン存在下では30.9時間となり、20μMのハルミン存在下において概日リズム周期が約5時間延長された。(図2)
以上の結果を得たことで、ハルミンにより概日リズム周期を人為的に変化させることができることが立証できた。
【0032】
(実施例2)概日リズム延長作用のハルミン濃度依存性
参考例1で樹立したNIH3T3細胞株を約5×105個を35mm培養ディッシュに播種した後、100nMデキサメサゾンにより生体リズムをリセットして用いた。樹立したNIH3T3細胞株の発光基質ルシフェリンを含む培地に0μM(対照)、5μMおよび20μMのハルミン塩酸塩(東京化成)を加え細胞培養を続けながら、リアルタイムでレポーター遺伝子による化学発光を10分毎に1分間、150時間にわたり測定した。それぞれの概日周期時間から対象実験より得られた概日周期時間を引いた値を周期延長時間とし、添加したハルミンの濃度との関係を図3に示した。図3に示されるように、ハルミン濃度と概日リズムの延長時間との間には比例関係がみられたことから、概日リズム延長作用にハルミンの濃度依存性が存在することが示唆された。
【0033】
(実施例3)ハルミンによるRORαの核内局在化
(3−1) 参考例1の樹立NIH3T3細胞に20μMのハルミン塩酸塩を0,3,6時間作用させた後、Dyer & Herzogの方法(BioTechniques,1995,19,192-195)により細胞質と核画分を分離し、抗RORa抗体(Santa Cruz Biotechnology社)を用いたWestern blotによりRORaタンパク量を測定した。さらに同一メンブレンを抗ラミン抗体(Santa Cruz Biotechnology社)で測定したラミンタンパク(核ラミナを構成する主要タンパク質)量で補正し、核内に存在するRORaタンパク量を算出した。ハルミン作用前(時刻0)の核内RORaタンパク量を1としてその相対値を図4Aに示した。その結果、ハルミンを作用させて約3時間後に核内RORαタンパク量の増加が観察された。
【0034】
(3−2) SV40プロモーターを持つレポータープラスミドpGL3-Promoter(Promega)ならびにBmal1プロモーター(-202〜+27)を持つレポータープラスミドpGL3-Basic(Promega)を内部標準コントロールベクターpRL-CMVと共にNIH3T3細胞に導入し、24時間後20μMのハルミン塩酸塩を3時間作用させ、デュアルルシフェレースアッセイによりハルミンによる転写活性に対する影響を検討した。転写活性は内部標準ベクターにより補正した後、ハルミン無処理(対照)の転写活性を1として、その相対値で示した(図4B)。SV40プロモーター活性に対するハルミンの影響は見られなかったが、ハルミンはBmal1遺伝子プロモーター活性を増大させることが判明した。また転写活性化は、図4Aで観察された核内RORaタンパク増大量と同程度の相対値を示すことから、ハルミンによるBmal1遺伝子プロモーター活性化は核内RORaタンパク増大によることが示唆される。
【0035】
(実施例4)RORaによる概日リズムの周期延長
参考例1で樹立したNIH3T3細胞株を約5×105個を35mm培養ディッシュに播種した後、RORa発現プラスミドを導入し、24時間後に100nMデキサメサゾンにより生体リズムをリセットして用いた。樹立したNIH3T3細胞株の発光基質ルシフェリンを含む培地で細胞培養を続けながら、リアルタイムでレポーター遺伝子による化学発光を10分毎に1分間、120時間にわたり測定した。対照(薄い灰色)の概日周期が24.11時間であるのに対して、RORaを発現させた場合(3クローン:黒〜濃い灰色)は、25.16時間となり、概日リズムの周期延長が観察された。
【技術分野】
【0001】
本発明は、概日リズムを制御することによる睡眠障害の改善剤に関する。
【背景技術】
【0002】
遺伝性の睡眠障害に加えて、社会の24時間化に伴う様々な睡眠障害が社会的問題となっている。概日リズム睡眠障害と呼ばれる一連の睡眠障害の発症には、時計遺伝子によって構成されている体内時計が関係しているものと考えられているが、その詳細なメカニズムに関しては不明である。
睡眠障害の治療法としては、特別な装置を必要とする高照度光療法や、ビタミンB12(非特許文献1)やメラトニン製剤(特許文献1)の投与が一般的である。しかし、これらの治療方法は、いずれも体内時計の位相を調節することにより生体リズムを正常化させようとするものであり、体内時計の周期異常に起因する障害の根本的な治療法とはなりえない。
また、睡眠薬としては、ベンゾジアゼピン系の薬剤(非特許文献2)が一般に用いられており、長短時間型から、短時間型、中間型、長時間型まで様々な半減期の薬剤が開発されている。しかしながら、これらの睡眠薬による睡眠障害の治療法は、対処療法的なものであり、根本的に睡眠障害を治療することは困難である。また、夢遊症状等の副作用を伴う場合も多く、その使用法には細心の注意が必要である。最近、ノシセプチン受容体の作動薬としての作用を有する4-オキソイミダゾリジン-2-スピロピペリジン誘導体に概日リズム睡眠障害治療効果が期待されている(特許文献2)が、まだ研究開発途上であり副作用も懸念される。
一方、各種飲食品の原料やそれらに含まれる天然成分を用いる睡眠改善剤の研究開発も盛んで、茶葉由来のテアニンを用いるもの(特許文献4)、内因性のメラトニン分泌効果を有する発酵ホエーなどのホエー類を添加するもの(特許文献5)の他、高麗人参エキス(非特許文献3)、イクラ油に含まれるフォスファチジルコリン(非特許文献4)などを用いた睡眠改善剤が提案されている。これらは、いずれも飲食品、嗜好品などに用いられている素材に由来しているため、安全性も高く、日常的に摂取可能であるといえるが、その詳細な睡眠改善効果や作用メカニズムの解明は十分ではなく、効果の個人差も大きい。
概日リズムは、体内時計によって制御されており、近年、時計遺伝子と呼ばれる一連の遺伝子群によって体内時計のリズムが刻まれていることが明らかとなってきた。体内時計は、概日リズムの位相調節と周期長の調節という2つの機能を有している。概日リズムの位相調節に関しては、多くの液性因子によって時計遺伝子の発現が細胞培養系において誘導されることから、生体においても、多様な調節因子が関与しているものと推測される(非特許文献5)。
【0003】
一方で、概日リズムの周期長の調節に関しても、グリコーゲンシンターゼキナーゼ3β(GSK3β)が、時計分子PERIOD2(非特許文献11)やREV-ERBα(非特許文献12)のリン酸化を介して体内時計を調節するとの報告や、概日リズムに関与する転写因子の1つであるBMAL1の抑制効果のあるc-Jun N末端キナーゼ-3(JNK3)の酵素活性調節剤を用いた睡眠改善剤も開発されてきている(特許文献3)。
睡眠障害を根本的に改善するためには、概日リズムのうちでも、とりわけ周期長を調節する睡眠改善剤の開発が強く望まれており、日常的に摂取可能な安全性の高い睡眠改善剤として、飲食品、嗜好品の素材として安全性の確立している天然成分由来の物質からの探索が望まれていた。しかしながら、これまでは詳細な概日リズム周期を検出する簡便な実験方法が確立されていなかったので、安全性の確立している天然成分由来であって概日リズムの周期長を調節する効果的な睡眠改善剤は未だ報告がない。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特表平8−502259号公報
【特許文献2】WO2003/010168
【特許文献3】WO2003/063907
【特許文献4】WO2005/097101
【特許文献5】WO2005/097101
【特許文献6】特開2009−183197号公報
【特許文献7】特表平10−505606号公報
【非特許文献】
【0005】
【非特許文献1】「サーカディアンリズム睡眠障害の臨床」千葉茂、本間研一 編、新興医学出版社、2003
【非特許文献2】「睡眠障害の対応と治療ガイドライン」、内山真 編、じほう、2002
【非特許文献3】Young Ho Rhee, et al.,Psychopharmacology,101,p.486-488(1990)
【非特許文献4】日比野英彦、Food Style21,Vol.7,No.3,p.50-53(2003)
【非特許文献5】Curr Biol 2000,10:1291-4
【非特許文献6】EMBO Reports 2007,8:366-71、Eur J Neurosci 2007,26:451-62
【非特許文献7】Science,1992,258,607-614
【非特許文献8】Cur.Biol.,2000,10,1291-1294
【非特許文献9】FEBS Lett.,1999,465,79-82
【非特許文献10】Gene to Cells,2003,8,713-720
【非特許文献11】J.Biol.Chem.,2005,280,29397-29402
【非特許文献12】Science,2006,311,1002-1005
【非特許文献13】Cell,2000,103,1009-1017
【非特許文献14】Nature,2002,418,534-539
【非特許文献15】Mol.Cell.Biol.,2008,28,3477-3488
【非特許文献16】J.Toxicol.Environ.Health A,2001,64,645-660
【非特許文献17】Pharmacology and Therapeutics,2004,102,111-129.
【非特許文献18】Neuropharmacology,1988,27,849-855
【非特許文献19】Biochemical Pharmacology,1977,26,1991-1996.
【非特許文献20】Arch.Biochem.Biophys.,1997,337,137-142
【非特許文献21】レスターグリンスプーン等著、杵渕幸子訳「サイケデリック・ドラッグ−向精神物質の科学と文化」工作舎(2000)p.36-38
【非特許文献22】Neuron.2004 Aug 19;43(4):527-37
【非特許文献23】Nuclear Receptor Signaling,2009,7,1-32
【非特許文献24】Biochim.Biophys.Acta,2006,1763,805-814
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の課題は、これまで広く摂取されてきた天然成分由来であって、かつ効果的に概日リズムの周期長を調節する概日リズム周期改善剤を提供することにある。
【課題を解決するための手段】
【0007】
最近の概日リズム関連遺伝子を用いたノックアウト実験結果によると、Bmal1遺伝子をノックアウトしたマウスでは、行動のリズムが全く見られなくなり、まるで生物リズムが全く破壊された様に振舞うのに対して、他の時計遺伝子PeriodやCryptochrome等のノックアウトマウスでは活動周期に異常が観察されるのみであり、Bmal1遺伝子は単一遺伝子の破壊において行動リズム形成を全く示さなくなる唯一つの遺伝子であることがわかった。さらにBmal1遺伝子ノックアウトマウスでは時計中枢である視交叉上核(SCN)において時計遺伝子Period1およびPeriod2の発現がほぼ完全に抑えられている。以上のことより、生物時計調節の中心となる遺伝子がBmal1遺伝子であることはほぼ間違いないと考えられている(非特許文献13)。
このBmal1遺伝子は概日リズムをもって転写調節されており、ルシフェラーゼ遺伝子を用いた培養細胞レベルでのレポーターアッセイが、哺乳類における生体内でのBmal1遺伝子を介した概日リズムを再現できることが知られていた(非特許文献14)が、従来法では、細胞内でのプラスミドの状態が不安定であり、培地中に被検物質を添加した状態で細胞培養を続けることにより、その影響を正確に測定することはできなかった。
【0008】
その後、本発明者らにより、Bmal1遺伝子の概日リズム転写をモニターするために必要最小限のプロモーター中の領域を決定することができたので(非特許文献15)、当該領域をルシフェレース遺伝子に接続しNIH3T3細胞に安定的に保持されるような細胞株を樹立した。具体的には、NIH3T3細胞にBmal1遺伝子プロモーター中の「RORE」配列及び転写開始部位を含む領域(−202〜+27位)にルシフェラーゼ遺伝子を繋いだレポータープラスミドを導入し、安定に概日リズムを刻みながらルシフェラーゼを生産する細胞株である(特願2009−175576号、特願2009−186257号)。そして、当該レポーターアッセイ法によりルシフェレース活性をモニターするだけで、転写因子BMAL1と転写因子CLOCKのヘテロ二量体によって発現が促進されたPeriod遺伝子からPERIODが合成され、核外でCRYと結合して核内移行したPERIOD-CRY複合タンパク質がBMAL1とCLOCK複合体による転写活性化を抑制するという一連の概日リズム形成サイクル自体に長期間にわたって作用する物質を、的確にしかも簡便に知ることが可能となった。
本発明者らは、当該樹立細胞株を用いて種々の物質を作用させて、ルシフェラーゼ活性をモニターしたところ、プロテインキナーゼCの活性化剤であるTPA及び同阻害剤であるG6850を作用させ、プロテインキナーゼCの活性を増減させたところ、当該活性の増減に応じて、概日リズムが延長及び短縮することが実証でき、プロテインキナーゼCがBmal1プロモーター領域の転写調節部位に作用して、概日リズム周期の調節をしている生体内物質であることを確認し、特許出願している(特願2009−175576号)。
また、ヒガンバナ植物由来のアルカロイドであるリコリンならびにリコリシジノールが、濃度依存的にBmal1遺伝子の転写リズムの周期を延長させることが明らかとなったので、同日付で特許出願した(特願2009−186257号)。この本発明者らの出願以前に、リコリン、リコリシジノールを含む天然成分由来の他のアルカロイドに関しては、従来概日リズムの周期長の調節作用については報告されていない。
【0009】
本発明者らの上記樹立細胞株を用いてのさらなる探索の結果、β−カルボリン誘導体の1種であるハルマラアルカロイド(Harmala alkaloid)のハルミンが、転写リズムの周期を延長していることを観察した。この結果は、ハルミンの体内時計制御剤としての可能性を示している。ハルマラアルカロイドは、他のβ−カルボリン誘導体と同様、可逆的なMAOI(モノアミン酸化酵素阻害剤)として知られ、中枢神経系のモノアミン化合物であるセロトニンやエピネフィリンと共にメラトニン分解に対しても抑制作用があるとされている。メラトニンは概日リズム調節に影響があると期待されている物質である(特許文献1)ことから、ハルマラアルカロイドを含むβ−カルボリン誘導体一般に対してもメラトニン受容体を介した概日リズム調節への影響が期待されている(特許文献7)ものの、メラトニン自身の概日リズム調節作用に関してすら学術的にコンセンサスが得られているわけではなく、実際に概日リズム調節への影響が報告された例はない。一方、本発明者の上記知見は、ハルマラアルカロイドが直接的に時計遺伝子に作用して、概日リズム周期の延長をする化合物であることを確認したことである。さらに、ハルミン濃度を変化させて概日リズムを観察していくと、ハルミンの概日リズム延長作用に濃度依存性が存在することを確認した。加えて、ハルミンに細胞内のRORαを核内に移行させる機能があることを見出したことから、ハルミンが細胞質中のRORαを核内に移行させることでBmal1遺伝子上流のRORE配列に結合する機会を増大させ、Bmal1遺伝子発現誘導を促進した結果、概日リズムの延長を起こすという、ハルミンの概日リズム延長作用の作用機序を解明することができた。ハルミンなどのハルマラアルカロイドは、ハーブ類など食用または嗜好品用の植物成分や生体内物質であることから安全性についての懸念は低く、概日リズム周期改善用の医薬組成物としての期待はきわめて大きいものがある。
以上の知見を得て本発明を完成した。
【0010】
また、本発明により、ハルミンの概日リズム延長作用の作用機序が、直接Bmal1遺伝子に作用するのではなく、しかもRORαの全体量を増加させるのでもなく、細胞内に広く存在しているRORαのうち、細胞質中のRORαを核内に移行させることで核内のRORα量を増加させ、Bmal1遺伝子上流のRORE配列への結合量を増やし、その結果Bmal1遺伝子発現誘導を促進するという一連の工程を経て概日リズムの延長を引き起こすという作用機序であることが解明されたが、この作用機序はRORαとBmal1遺伝子との関係性においても初めての知見である。すなわち、従来からRORαがROREに結合することによりBmal1遺伝子の発現が誘導されて概日リズムの延長が引き起こされることは知られていた(非特許文献22)ものの、RORαのROREへの結合を起こすトリガーが何であるかは全く解明されていなかった。本発明における上記知見は、細胞中でのRORαの発現量が増大するのではなく、細胞中に広く分布しているRORαが何らかの物質の作用により核内への移行を促進されて核内での局在化が起こることがトリガーとなっていたことを解明したことでもある。つまり、RORαの核内局在化を観察するスクリーニング系によって、RORαの核内移行を起こさせるトリガー物質を検索することができることを示している。このような知見をもとに、スクリーニング方法についての発明も完成させた。
【0011】
すなわち、本発明は以下の発明を含むものである。
〔1〕 ハルマラアルカロイド(Harmala alkaloid)、又はその薬理学的に許容しうる塩もしくはエステルを有効成分として含む、概日リズム周期改善剤。
〔2〕 前記ハルマラアルカロイドがハルミンである、前記〔1〕に記載の概日リズム周期改善剤。
〔3〕 RORαを発現し、かつ安定して概日リズムを刻む培養細胞を用意し、当該培養細胞に対して被検物質を添加して、当該細胞の核内のRORα量を測定し、被検物質添加前の核内RORα量と比較して有意差をもって核内移行を促進させる物質を選択することを特徴とする、概日リズム周期改善物質のスクリーニング方法。
【発明の効果】
【0012】
本発明により、効果的に体内時計の周期長を調節することが可能であり、体内時計の周期異常に起因する概日リズム障害の根本的な治療のために有効でかつ安全性の高い物質を提供することができた。また、あわせて、細胞内のRORαの核内移行を観察する系を用いる概日リズム障害改善物質の優れたスクリーニング系を開発した。
【図面の簡単な説明】
【0013】
【図1】Bmal1レポータープラスミドの構築 [Bmal1プロモーター領域の遺伝子地図中でのRORE(白)ならびにSAF-A結合配列(灰)の位置を示す。ROREを含む-202〜+27の領域をプラスミドpGL3-dLucのルシフェレース遺伝子上流に挿入した。]
【図2】ハルミンによる概日リズム調節 [20μMのハルミン存在下(黒線)、ならびに対照(点線)の化学発光量の経時変化を示す。]
【図3】概日リズム延長作用のハルミン濃度依存性
【図4】ハルミンによるRORαの核内局在化作用 [A:核内RORαタンパク量 (横軸は、20μMハルミン処理時間を表す。)] [B:ハルミン処理3時間後の相対転写活性 (図中、SV40ならびにBmal1プロモーターのハルミン処理による相対転写活性を示す。)]
【図5】RORaによる概日リズム調節 [RORa発現(黒〜濃灰色)、ならびに対照(薄灰色)の化学発光量の経時変化を示す。]
【発明を実施するための形態】
【0014】
1.睡眠障害について
睡眠障害は、「概日リズム睡眠障害」と「睡眠呼吸障害」とに大別される。概日リズム睡眠障害には、外因性の急性症候群と内因性の慢性症候群とに区分され、睡眠相前進症候群や睡眠相後退症候群、非24時間睡眠覚醒障害、不規則型睡眠覚醒障害などは、内因性慢性症候群に含まれる。概日リズム睡眠障害には、体内時計の関与が考えられており、特に、内因性慢性症候群は、体内時計の周期異常が一因となっている。従来からの典型的な睡眠障害の治療法である光照射やメラトニン投与、あるいは様々な睡眠薬による治療法は、体内時計のリセットに作用する対処療法的なものであるのに対し、本発明が目指す睡眠障害の治療法は、体内時計の周期異常、すなわち「概日リズム障害」を改善することによる治療法である。
【0015】
2.概日リズム障害により引き起こされる疾患について
概日リズム障害に起因する疾患としては、睡眠相前進症候群(ASPS)、睡眠相後退症候群(DSPS)、非24時間睡眠相後退症候群、および季節性うつ病が典型的なものとして考えられる。その他、内因性躁鬱病、周期性緊張症、周期性高血圧症、周期性潰瘍、不規則排卵周期、およびインスリン分泌の周期性異常に起因する糖尿病などの周期性・反復性障害や、脳血管型痴呆やアルツハイマー型痴呆における夜間徘徊なども概日リズム障害が関与すると考えられている。本発明は、概日リズム障害のうちの概日リズム周期を延長することにより障害を改善しようとするものである。
【0016】
3.概日リズムとBmal1遺伝子について
体内時計の調節に係わる主要な因子としては、体内時計の刻みを促進するBmal1遺伝子とClock遺伝子、抑制因子として働くCry遺伝子とPeriod遺伝子の4種類がある。
これら遺伝子群のうちで、Bmal1遺伝子をノックアウトしたマウスでは、行動のリズムが全く見られなくなり、まるで生物リズムが全く破壊された様に振舞う。他の時計遺伝子PeriodやCry等のノックアウトマウスでは活動周期に異常が観察されるのみであり、Bmal1遺伝子は単一遺伝子の破壊において行動リズム形成を全く示さなくなる唯一つの遺伝子である。さらにBmal1遺伝子ノックアウトマウスでは時計中枢である視交叉上核(SCN)において時計遺伝子Period1およびPeriod2の発現がほぼ完全に抑えられている。以上のことより、生物時計調節の中心となるのがBmal1遺伝子であると考えられている(非特許文献13)。
最近、本発明者らは、Bmal1遺伝子プロモーター領域中の「SAF−A結合領域(-27〜+266位)」へのSAF−A結合によるBmal1遺伝子を介した概日リズムの転写調節を解明した(非特許文献15、特許文献9)。
【0017】
4.概日リズムとRORaタンパク質について
Bmal1遺伝子は、遺伝子上流のプロモータ領域中のRORE配列(6塩基のAT-rich配列とそれに続くRGGTCA配列)に結合する転写因子の作用で、正もしくは負の転写調節が行われている。最初にRORαがROREの結合によってBmal1の強い発現誘導が起こされ、当該発現の増強がREV-ERBαにより抑制されたことが観察された(非特許文献22)。ROR及びREV-ERBは、いずれも核内受容体のNR1ファミリーに属しており、RORにはα、β、γのサブタイプがあり、REV-ERBにもα、βのサブタイプが知られている。これらの転写因子はRORE配列に結合することによりBmal1遺伝子特異的に正もしくは負の転写調節を行い、発生、分化、代謝、ストレス反応等の生体調節を行っているとも考えられている(非特許文献23)。近年、RORαのリガンドがコレステロール誘導体であることが判明し、細胞膜に存在しているコレステロールと結合することにより核内のみならず、細胞膜や細胞質にも存在することが報告されている(非特許文献23)。RORαは種々の細胞によっては細胞内での局在状態が異なることが観察されてはいた(非特許文献24)ものの、局在現象の存在を示したにとどまり、RORαの核内への局在化とBmal1遺伝子発現の誘導量増大との関連性に着目して実証したのは本発明がはじめてである。
【0018】
5.概日リズムの周期を調節する物質のスクリーニング系
(1) 本発明において、概日リズムの周期を調節する物質のスクリーニングに用いるレポーターアッセイ系は、先の出願において本発明者らが確立したアッセイ系であり(特願2009−175576号、特願2009−186257号)、従来の研究でBmal1遺伝子のプロモーター領域などの特定領域に一過的に作用する物質又は遺伝子を探索するために用いられていたスクリーニング系を改変して利用するものである。
まず、プラスミドpGL3-dLuc(非特許文献14)のルシフェレース遺伝子上流に、Bmal1プロモーター領域の「RORE」を含む-202〜+27の領域を挿入して作製したレポータープラスミド(図1)を、非特許文献14の手法に従ってNIH3T3細胞に導入し、継代培養して、安定に概日リズムを刻みながらルシフェラーゼを生産する形質転換NIH3T3細胞株を樹立した。
当該樹立細胞の培地に被検物質を添加し、培養しながらルシフェラーゼの発光強度を経時的にモニターすることにより、転写因子BMAL1と転写因子CLOCKのヘテロ二量体によって発現が促進されたPeriod遺伝子からPERIODが合成され、核外でCRYと結合して核内移行したPERIOD-CRY複合タンパク質がBMAL1とCLOCK転写活性を抑制するという一連の概日リズム形成サイクル自体に長期間にわたって作用する物質、その周期を早めたり遅くしたりする物質がはじめて探索できるようになった。
【0019】
(2) 本発明により、ハルミンの概日リズム周期延長作用の作用機序が、細胞内に広く存在しているRORαのうち細胞質中の又は細胞膜に結合しているRORαに働いて核内に移行させる作用であることを解明した。つまり、RORαを核内に移行させることで核内に存在するRORα量を増加させ、Bmal1遺伝子上流のRORE配列への結合量を増やし、その結果Bmal1遺伝子発現誘導を促進するという一連の工程を経て概日リズムの延長を引き起こすという作用機序である。すなわち、このことは、細胞中に広く分布しているRORαを核内へ移行させて核内での存在量を増大させる物質を検索することで、ハルミン類と同様に、概日リズムの周期延長作用を有する物質をスクリーニングすることができることに他ならない。
細胞内でのRORαの核内移行を観察する手法としては、どのような手法でも良いが、典型的な手法を以下に2種類示す。
【0020】
(3)RORαの核内移行観察法−1
i)まず、RORαを発現し、かつ安定して概日リズムを刻む培養細胞を用意する。
その際、そのような培養細胞としては、NIH3T3細胞株などを用いることができるが、上記(1)で述べた、安定に概日リズムを刻みながらルシフェラーゼを生産する形質転換NIH3T3細胞株を用いると、概日リズムの周期延長作用の最終的な確認も同一株で行うことができるので、より好ましい。さらに、RORαを細胞質内で大量発現させるためにRORα遺伝子発現ベクターを導入することが好ましい。その場合、RORαはGFP等の融合タンパクとしておくかFLAG等の遺伝子タグにより標識しておく。
ii)RORαの当該培養細胞内での局在化状態を観察する。すなわち、細胞の核内でのRORαの存在量を測定する。核を特異的に染色しておき、標識化されたRORαの信号(発光、蛍光)量を測定することで、核内RORα量を測定することができる。通常は、下記工程iii)に先立ち工程ii)を行っておくが、同一の細胞系を用いてii)及びiii)を平行して行うこともできる。
iii)被検物質を添加後の当該細胞内の核内のRORα量を上記工程ii)と同様に測定する。
iv)工程ii)及び工程iii)の核内RORα量を比較して、被検物質添加により、有意差をもって核内のRORα量が増大している場合に、当該被検物質を、概日リズムの周期延長作用を有する物質の候補として選択する。
【0021】
(4)RORαの核内移行観察法−2
i)まず、RORαを発現し、かつ安定して概日リズムを刻むNIH3T3細胞株などの培養細胞を用意する。安定に概日リズムを刻みながらルシフェラーゼを生産する形質転換NIH3T3細胞株を用いることが好ましい点は上記手法と同様である。さらに、RORαを細胞質内で大量発現させるためにRORα遺伝子発現ベクターを導入することが好ましく、その場合、RORαはGFP等の融合タンパクとしておくかFLAG等の遺伝子タグにより標識しておくことが好ましい点も同様である。
ii)培養細胞を回収後、Dyer & Herzogの方法(BioTechniques,1995,19,192-195)に従い、遠心分離により細胞質と核タンパク画分を分離し、RORαの当該培養細胞内での局在化割合を計測する。具体的には、得られたそれぞれの画分をWestern blot解析し、抗RORa抗体(Santa Cruz Biotechnology社製)を用いてRORaタンパクを定量する。試料間の核総タンパク量の誤差を補正するために、抗ラミン抗体(Santa Cruz Biotechnology社製)を用いて核特異的マーカータンパクであるラミン量を測定し、核内RORaタンパク量を補正する。細胞質におけるRORa量は、抗アクチン抗体(Sigam社製)を用いて測定した細胞質アクチン量により補正したものを用いる。通常は、下記工程iii)に先立ち工程ii)を行っておくが、同一の細胞系を用いてii)及びiii)を平行して行うこともできる。
iii)被検物質を添加後の当該細胞内の核内のRORα量を上記工程ii)と同様に測定する。
iv)工程ii)及び工程iii)の核内RORα量を比較して、被検物質添加により、有意差をもって核内のRORα量が増大している場合に、当該被検物質を、概日リズムの周期延長作用を有する物質の候補として選択する。
【0022】
6.ハルマラアルカロイド(Harmala alkaloid)について
ハルマラアルカロイド(Harmala alkaloid)は、インドール骨格を有するβ−カルボリンアルカロイドの1種であり、MAOI(モノアミン酸化酵素阻害剤)としてインド原産のはまびし科の植物Peganum harmala(Halmala、Syrian Rue)の種子から単離された(非特許文献16)。これらハルマラアルカロイドは南米の植物の蔓の樹皮などにも含まれており、南米では古くからアヤワスカという飲料や嗅ぎタバコなどの形で摂取されていた(非特許文献17)。典型的なハルマラアルカロイドとしては、ハルミン(Harmine)、ハルマリン(Harmaline)、ハルマロール(Harmalol)があげられる(下記に化学構造を示す。)。ハルマラアルカロイドは、セロトニンやエピネフィリンというモノアミン化合物の分解を阻害する中枢神経系刺激物質であり、可逆的なMAO-A阻害活性を有し、作用が穏和で安全性が高い向精神薬として期待されている(非特許文献18〜21)。これらハルマラアルカロイドは、いずれもその作用機序も化学構造的にも非常に似通っており、特にハルミンはハルマリンの代謝産物であるとも言われている(非特許文献21)。
ハルマラアルカロイドの可逆的なMAOI(モノアミン酸化酵素阻害剤)としての作用は、メラトニン分解に対しても抑制作用があるとされおり、メラトニンは概日リズム調節に影響があると期待されている物質である(特許文献1)ため、ハルマラアルカロイドに対してもメラトニン受容体を介した概日リズム調節への影響が期待されている(特許文献7)が、メラトニン自身の概日リズム調節作用に関してすら学術的にコンセンサスが得られているわけではなく、ハルマラアルカロイドについて実際に概日リズム調節への影響が報告された例はない。一方、本発明においては、ハルマラアルカロイドのハルミンが直接体内時計遺伝子Bmal1遺伝子の転写を調節することにより概日リズム周期の延長を引き起こすことを実証したものである。
そして、本発明により、ハルミンのBmal1遺伝子の転写調節の作用機序は、直接的には細胞内に広く存在しているRORαへの作用であり、細胞質中の又は細胞膜に結合しているRORαに働いて核内に移行させることで核内に存在するRORα量を増加させ、Bmal1遺伝子上流のRORE配列への結合量を増やし、その結果Bmal1遺伝子発現誘導を促進するという一連の工程を経て概日リズムの延長を引き起こすという作用機序であることも解明した。
このことは、根本的な概日リズム改善のために必須の概日リズム周期改善剤の有効成分としてハルマラアルカロイドを用いることができることを示すものである。
そして、ハルミンなどのハルマラアルカロイドは、もともとハーブ類など食用または嗜好品用の植物成分や生体内物質であり、向精神薬として実際に投与された例もあることから、その安全性についての懸念は低い。大量に投与するとLSDのような幻覚作用を引き起こすことは知られている(非特許文献18,21)が、そもそもMAO−A阻害薬としての作用がLSDの8,000分の1であること(非特許文献21)からも、本発明における概日リズム周期改善剤としての有効量を投与する程度であれば、全く問題とはならない。概日リズム周期異常に起因する疾病を予防、治療するための優れた医薬組成物としての期待が大きい。
なお、本発明の実施態様では、典型的なハルマラアルカロイドとしてハルミンにおける、体内時計遺伝子の周期を変化させる活性について述べるが、他のハルマラアルカロイドも同様の作用を有する。
【0023】
本発明におけるハルマラアルカロイドは、以下の化学構造式で表現できる。
<式(1)>
(Rは、メトキシ基(−OCH3)または水酸基(−OH)を表す。また3位及び4位の間の点線は単結合又は2重結合であることを表す。)
典型的なハルマラアルカロイドのハルミンは、式(1)において、Rがメトキシ基であって、かつ3位と4位の間が2重結合を形成している化合物であり、以下の構造式で表される。
<式(2)>
【0024】
ハルマリン(Harmaline)は、式(1)において、Rがメトキシ基であって、かつ3位と4位の間が単結合を形成している化合物であり、以下の構造式で表される。
<式(3)>
【0025】
ハルマロール(Harmalol)は、式(1)において、Rが水酸基であって、かつ3位と4位の間が単結合を形成している化合物であり、以下の構造式で表される。
<式(4)>
【0026】
7.本発明における概日リズム周期調節の対象動物
本発明で概日リズム調節をしようとする対象動物は、ヒト等の霊長類、イヌ、ネコなどの愛玩動物、ウシ、ウマなどの家畜動物、マウス、ラットなどの実験動物を含む哺乳類及び相同性の高い時計遺伝子を持つ鳥類及び魚類である。したがって、本発明で、「Bmal1遺伝子」などというとき、ヒトを含む哺乳類、鳥類及び魚類由来の遺伝子又は遺伝子産物を指すものである。
【0027】
8.概日リズム周期改善剤について
上記ハルマラアルカロイドは、そのままで又は薬理学的に許容しうる塩もしくは摂取した生体内において容易に転換されるエステル型の前駆体の状態で、その有効量をヒトを含む動物に投与することで、概日リズム周期を延長することができるから、概日リズム周期が短い障害を有する患者に対する概日リズム周期改善剤として用いることができる。本発明に用いられる塩としては、酢酸塩、硫酸塩、塩酸塩等がある。
概日リズム周期改善剤を有効成分として含み、適当な薬理学的に許容しうる担体または賦形剤などと組み合わせることで概日リズム周期障害に起因する睡眠相前進症候群(ASPS)、睡眠相後退症候群(DSPS)、非24時間睡眠相後退症候群、および季節性うつ病の予防及び治療用の医薬組成物とすることができ、その際に他の治療用化合物または医薬と一緒に使用することができる。
また、ハルマラアルカロイドは従来から可逆性MAO−A阻害薬として、向精神作用が知られていることから、概日リズム障害に起因してうつ病を併発している場合には、特に抗うつ剤としての効果も期待される。
本発明の概日リズム周期改善剤の投与形態は、全身投与であっても局所投与であってもよく、注射、膏薬など非経口投与でも、経口投与でもよく、周知の製剤化方法が適用できる。
用量は、患者の年齢、体重等により適宜調整することができ、投与形態により異なるが、例えば、塩酸塩などとして体重1kg当たり、0.001mg〜20mg、好ましくは、0.01mg〜10mg、より好ましくは0.1〜1.0mgであり、目的に応じて投与回数を決定することが好ましい。
【0028】
また、前記5.(2)で述べた本発明の概日リズムの周期改善物質のスクリーニング系により得られた概日リズム周期改善物質も、上記ハルマラアルカロイドと同様に、適当な薬理学的に許容しうる担体または賦形剤などと組み合わせることで概日リズム周期改善剤として用いることができる。
【実施例】
【0029】
以下、実施例により本発明を具体的に説明するが、本発明は特にこれら実施例に限定されるものではない。
なお、本発明で使用されている技術的用語は、別途定義されていない限り、当業者により普通に理解されている意味を持つ。本発明の実施例で用いた遺伝子組換え技術、PCR法、その他の手法などの具体的な手順や条件は、特に断らない限り、Sambrook and Russell,Molecular Cloning:A Laboratory Manual,3rd Edition.Cold Spring Harbor Laboratory Press,Plainview,NY(2001)、Cold Spring Harbor Laboratory Press,Cold Spring Harbor,New York(1989);D.M.Glover et al.ed.,"DNA Cloning",2nd ed.,Vol.1 to 4,(The Practical Approach Series),IRL Press,Oxford University Press(1995);Ausubel,F.M.et al.,Current Protocols in Molecular Biology,John Wiley & Sons,New York,N.Y,1995;日本生化学会編、「続生化学実験講座1、遺伝子研究法II」、東京化学同人(1986);日本生化学会編、「新生化学実験講座2、核酸 III(組換えDNA技術)」、東京化学同人(1992);R.Wu ed.,"Methods in Enzymology",Vol.68(Recombinant DNA),Academic Press,New York(1980);R.Wu et al.ed.,"Methods in Enzymology",Vol.100(Recombinant DNA,PartB) & 101(Recombinant DNA,Part C),Academic Press,New York(1983);R.Wu et al.ed.,"Methods in Enzymology",Vol.153(Recombinant DNA,Part D),154(Recombinant DNA,Part E) & 155(Recombinant DNA,Part F),Academic Press,New York(1987)などに記載の方法あるいはそこで引用された文献記載の方法またはそれらと実質的に同様な方法により行うことができる。
また、本発明で引用した先行文献又は特許出願明細書の記載内容は参照して、本明細書の記載として組み入れるものとする。
【0030】
(参考例1)Bmal1プロモーターにより転写されるルシフェラーゼ遺伝子を安定的に保持しているNIH3T3細胞株の樹立
Bmal1遺伝子のリズミックな発現に必要最小限のプロモーター領域として、「RORE」を含む-202〜+27の領域を選定し、プラスミドpGL3-dLuc(非特許文献14)のルシフェラーゼ遺伝子上流に、挿入してBmal1レポータープラスミドを作製した(図1)。
当該Bmal1レポータープラスミドと共に、抗生物質(Zeocin)耐性マーカー遺伝子を持つpTracer-CMV(Invitrogen社製)をNIH3T3細胞に導入し(非特許文献14記載の手法による。)、Bmal1プロモーターにより転写調節されるルシフェラーゼ遺伝子を安定的に保持する組換えNIH3T3細胞を取得した。さらに継代培養を繰り返し、安定に概日リズムを刻みながらルシフェラーゼを生産する形質転換NIH3T3細胞株を樹立し、樹立した細胞の中でルシフェラーゼ活性の概日リズムが最も安定的に観察されるクローンを選択した。
【0031】
(実施例1)ハルミンによる概日リズムの周期延長
参考例1で樹立したNIH3T3細胞株を約5×105個を35mm培養ディッシュに播種した後、100nMデキサメサゾンにより生体リズムをリセットして用いた。
樹立したNIH3T3細胞株の発光基質ルシフェリンを含む培地に被検物質を添加し、細胞培養を続けながら、リアルタイムでレポーター遺伝子による化学発光を約150時間測定した。発光は10分毎に1分間測定した。
次いで、20μMのハルミン塩酸塩(東京化成)と、発光基質ルシフェリンを含む培地で細胞培養を続けながら、リアルタイムでレポーター遺伝子による化学発光を10分毎に1分間測定した。150時間にわたり測定したところ、概日リズムの周期は対照(点線)が25.8時間に対して20μMのハルミン存在下では30.9時間となり、20μMのハルミン存在下において概日リズム周期が約5時間延長された。(図2)
以上の結果を得たことで、ハルミンにより概日リズム周期を人為的に変化させることができることが立証できた。
【0032】
(実施例2)概日リズム延長作用のハルミン濃度依存性
参考例1で樹立したNIH3T3細胞株を約5×105個を35mm培養ディッシュに播種した後、100nMデキサメサゾンにより生体リズムをリセットして用いた。樹立したNIH3T3細胞株の発光基質ルシフェリンを含む培地に0μM(対照)、5μMおよび20μMのハルミン塩酸塩(東京化成)を加え細胞培養を続けながら、リアルタイムでレポーター遺伝子による化学発光を10分毎に1分間、150時間にわたり測定した。それぞれの概日周期時間から対象実験より得られた概日周期時間を引いた値を周期延長時間とし、添加したハルミンの濃度との関係を図3に示した。図3に示されるように、ハルミン濃度と概日リズムの延長時間との間には比例関係がみられたことから、概日リズム延長作用にハルミンの濃度依存性が存在することが示唆された。
【0033】
(実施例3)ハルミンによるRORαの核内局在化
(3−1) 参考例1の樹立NIH3T3細胞に20μMのハルミン塩酸塩を0,3,6時間作用させた後、Dyer & Herzogの方法(BioTechniques,1995,19,192-195)により細胞質と核画分を分離し、抗RORa抗体(Santa Cruz Biotechnology社)を用いたWestern blotによりRORaタンパク量を測定した。さらに同一メンブレンを抗ラミン抗体(Santa Cruz Biotechnology社)で測定したラミンタンパク(核ラミナを構成する主要タンパク質)量で補正し、核内に存在するRORaタンパク量を算出した。ハルミン作用前(時刻0)の核内RORaタンパク量を1としてその相対値を図4Aに示した。その結果、ハルミンを作用させて約3時間後に核内RORαタンパク量の増加が観察された。
【0034】
(3−2) SV40プロモーターを持つレポータープラスミドpGL3-Promoter(Promega)ならびにBmal1プロモーター(-202〜+27)を持つレポータープラスミドpGL3-Basic(Promega)を内部標準コントロールベクターpRL-CMVと共にNIH3T3細胞に導入し、24時間後20μMのハルミン塩酸塩を3時間作用させ、デュアルルシフェレースアッセイによりハルミンによる転写活性に対する影響を検討した。転写活性は内部標準ベクターにより補正した後、ハルミン無処理(対照)の転写活性を1として、その相対値で示した(図4B)。SV40プロモーター活性に対するハルミンの影響は見られなかったが、ハルミンはBmal1遺伝子プロモーター活性を増大させることが判明した。また転写活性化は、図4Aで観察された核内RORaタンパク増大量と同程度の相対値を示すことから、ハルミンによるBmal1遺伝子プロモーター活性化は核内RORaタンパク増大によることが示唆される。
【0035】
(実施例4)RORaによる概日リズムの周期延長
参考例1で樹立したNIH3T3細胞株を約5×105個を35mm培養ディッシュに播種した後、RORa発現プラスミドを導入し、24時間後に100nMデキサメサゾンにより生体リズムをリセットして用いた。樹立したNIH3T3細胞株の発光基質ルシフェリンを含む培地で細胞培養を続けながら、リアルタイムでレポーター遺伝子による化学発光を10分毎に1分間、120時間にわたり測定した。対照(薄い灰色)の概日周期が24.11時間であるのに対して、RORaを発現させた場合(3クローン:黒〜濃い灰色)は、25.16時間となり、概日リズムの周期延長が観察された。
【特許請求の範囲】
【請求項1】
ハルマラアルカロイド(Harmala alkaloid)、又はその薬理学的に許容しうる塩もしくはエステルを有効成分として含む、概日リズム周期改善剤。
【請求項2】
前記ハルマラアルカロイドがハルミンである、請求項1に記載の概日リズム周期改善剤。
【請求項3】
RORαを発現し、かつ安定して概日リズムを刻む培養細胞を用意し、当該培養細胞に対して被検物質を添加して、当該細胞の核内のRORα量を測定し、被検物質添加前の核内RORα量と比較して有意差をもって核内移行を促進させる物質を選択することを特徴とする、概日リズム周期改善物質のスクリーニング方法。
【請求項1】
ハルマラアルカロイド(Harmala alkaloid)、又はその薬理学的に許容しうる塩もしくはエステルを有効成分として含む、概日リズム周期改善剤。
【請求項2】
前記ハルマラアルカロイドがハルミンである、請求項1に記載の概日リズム周期改善剤。
【請求項3】
RORαを発現し、かつ安定して概日リズムを刻む培養細胞を用意し、当該培養細胞に対して被検物質を添加して、当該細胞の核内のRORα量を測定し、被検物質添加前の核内RORα量と比較して有意差をもって核内移行を促進させる物質を選択することを特徴とする、概日リズム周期改善物質のスクリーニング方法。
【図1】

【図3】
【図4】
【図2】
【図5】
【図3】
【図4】
【図2】
【図5】
【公開番号】特開2011−195560(P2011−195560A)
【公開日】平成23年10月6日(2011.10.6)
【国際特許分類】
【出願番号】特願2010−241712(P2010−241712)
【出願日】平成22年10月28日(2010.10.28)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成23年10月6日(2011.10.6)
【国際特許分類】
【出願日】平成22年10月28日(2010.10.28)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]