
生体触媒による4−ハロ−3−ヒドロキシ酪酸エステルの光学分割法
【課題】4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物から高光学純度のS体4−ハロ−3−ヒドロキシ酪酸エステルを製造する方法、また、R体4−ハロ−3−ヒドロキシ酪酸エステルからR体3−ヒドロキシ−γ−ブチロラクトンを製造する方法、これらの製法に使用する微生物由来の酵素CHBH(4−Chloroー3−HydroxyButyrateHydrolase)、および該酵素をコードする遺伝子を提供すること。
【解決手段】(a)特定塩基配列、(b)該特定塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列であって、RhCHBH酵素活性(Rhizobium由来4−Chloroー3−HydroxyButyrateHydrolase)を有するタンパク質をコードする塩基配列を使用する。
【解決手段】(a)特定塩基配列、(b)該特定塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列であって、RhCHBH酵素活性(Rhizobium由来4−Chloroー3−HydroxyButyrateHydrolase)を有するタンパク質をコードする塩基配列を使用する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は医薬、農薬、生理活性物質および強誘電性液晶などの光学活性化合物の合成において有用な中間体となりうる光学活性S体4−ハロ−3−ヒドロキシ酪酸エステルおよびR体3−ヒドロキシ−γ−ブチロラクトンの、微生物、その処理物、微生物由来の酵素または遺伝子を用いた不斉加水分解反応による製法、およびこれらの製法に使用する微生物由来の酵素、該酵素をコードする遺伝子に関する。
【背景技術】
【0002】
光学活性化合物の製法としては、対応する光学活性出発物質から目的物へ変換する化学的合成法のほか、対応するラセミ体を光学分割剤で処理して光学活性体に分割する方法が用いられる。また近年、微生物または酵素を用いて不斉還元や不斉加水分解反応を利用する生体触媒反応により光学活性体を分離する方法が多く報告されている。
【0003】
4−ハロ−3−ヒドロキシ酪酸エステルの製法は、合成法としては、ルテニウム−光学活性ホスフィン錯体を触媒として用いる不斉還元による方法(特許文献1)が知られているが、該方法に用いる触媒の合成は容易ではなく、コスト面で問題がある。また、光学活性アミノ酸誘導体とアルコールを用いて還元する方法(特許文献2)が知られているが、生成物の光学純度が低いという欠点があり、また、−78℃の様な低温で反応させなければならないという問題がある。
【0004】
また、4−ハロ−3−ヒドロキシ酪酸エステルの生体触媒反応を用いた製法としては、微生物菌体または酵素を用いるβ−ケトエステル体の不斉還元法(特許文献3、4、5)が知られており、また遺伝子組換え技術による不斉還元法の改良も行なわれている(特許文献6)。しかし、これら微生物あるいは酵素を用いた不斉還元法によるβ−ケトエステル体からのS体4−ハロ−3−ヒドロキシ酪酸エステルの製法においては、その反応に高価なNADH(ニコチンアミドアデニンジヌクレオチド)あるいはNADPH(ニコチンアミドアデニンジヌクレオチドリン酸)などの補酵素を必要とし、さらにその酸化体から再度還元体へ変換する反応を必要とするため、グルコースオキシダーゼあるいは蟻酸デヒドロゲナーゼなどの酵素を別途必要とするうえ、その反応工程が律速反応となるなどの観点から工業的製法とは言い難い。
【0005】
また、微生物あるいは酵素を用いた4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物の不斉分解反応によるS体4−クロロ−3−ヒドロキシ酪酸エステルの製法(非特許文献1、2)が報告されているが、これらの製法では、高光学純度にて生成物を生産することはできない。またアグロバクテリウムを用いた方法(特許文献7)では、本反応に関与する酵素や遺伝子の報告がないので、生産性の向上は野性株での改良に限られてしまう。
【0006】
光学活性3−ヒドロキシ−γ−ブチロラクトンの製法としては、エチル−4−t−ブトキシ−3−オキソ酪酸をパン酵母を用いて不斉還元し、得られたR体エチル−4−t−ブトキシ−3−ヒドロキシ酪酸からフルオロ酢酸存在下においてR体3−ヒドロキシ−γ−ブチロラクトンに誘導する方法(非特許文献3)が報告されているが、この方法は化学的な合成反応を含むことから、不安定な生成物を高光学純度で得ることは困難である。また、微生物を用いた、光学活性な4−ハロ−3−ヒドロキシ酪酸エステルから対応する立体の3−ヒドロキシ−γ−ブチロラクトンの製法(特許文献8)が報告されているが、該微生物の作用の立体選択性は開示されていない。さらに、この反応に関与する酵素や遺伝子の報告がないので、生産性の向上は野性株での改良に限られてしまう。
【特許文献1】特開平1−211551号公報
【特許文献2】特開昭61−155354号公報
【特許文献3】特開昭61−146191号公報
【特許文献4】特開平6−209782号公報
【特許文献5】特開平11−187869号公報
【特許文献6】特開2000−236883号公報
【特許文献7】特開平9−322786号公報
【特許文献8】特開2002−204699号公報
【非特許文献1】Tetrahedron: Asymmetry 12 pp545-556, 2001
【非特許文献2】Tetrahedron: Asymmetry 7 pp3109-3112, 1996
【非特許文献3】Seebach, Dieter; Eberle, Martin, Synthesis (1), pp37-40, 1986
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明は、上記問題点の解決を課題とするものであり、微生物、微生物由来の酵素または酵素をコードする遺伝子により生体触媒を用いた不斉加水分解反応により4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物から高光学純度のS体4−ハロ−3−ヒドロキシ酪酸エステルを製造する方法、また、R体4−ハロ−3−ヒドロキシ酪酸エステルからR体3−ヒドロキシ−γ−ブチロラクトンを製造する方法、これらの製法に使用する微生物由来の酵素、および該酵素をコードする遺伝子を提供することを目的とする。
【0008】
本発明はまた、組換えDNAの手法を用いて立体識別性が向上し、微生物の触媒能力を従来と比較して飛躍的に増大することができ、光学活性体を大量生産することができる4−ハロ−3−ヒドロキシ酪酸エステル不斉加水分解酵素(RhCHBH(Rhizobium sp. 4-Chloro-3-HydroxyButyrate Hydrolase))およびそれをコードする遺伝子を提供することを目的とする。
本発明はまた、該遺伝子を含むベクター、形質転換体、および該酵素を用いる光学活性体の製造方法を提供することも目的とする。
【課題を解決するための手段】
【0009】
本発明者らは、S体4−ハロ−3−ヒドロキシ酪酸エステルおよびR体3−ヒドロキシ−γ−ブチロラクトンを工業的に製造する方法を見出すべく、種々研究を重ねた結果、ある種の微生物(受託番号FERM P−21129)が4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物のうち、R体に対して高度に選択的な反応性を示し、3−ヒドロキシ−γ−ブチロラクトンに変換し、S体4−クロロ−3−ヒドロキシ酪酸エステルを残存させることを見いだした。
【0010】
また、この微生物による反応において、R体4−ハロ−3−ヒドロキシ酪酸エステルを出発原料にすれば、光学純度を落とすことなくR体3−ヒドロキシ−γ−ブチロラクトンを得ることができることを見出した。
【0011】
さらに、該微生物(受託番号FERM P−21129)は反応性が低いため、該反応を触媒する酵素(以下、RhCHBHとも称する)をコードする遺伝子(以下、RhCHBH遺伝子とも称する)を単離し、組換え大腸菌により生産性を向上させることに成功した。
【0012】
したがって、本発明は、以下の(a)または(b)のいずれかの塩基配列からなる遺伝子を提供する:
(a)配列番号1の第44塩基〜第1240塩基からなる塩基配列、
(b)配列番号1の第44塩基〜第1240塩基からなる塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列であって、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用し、該混合物のR体を選択的に加水分解し、かつ、S体を選択的に残存させる酵素活性(以下、RhCHBH活性とも称する)を有するタンパク質をコードする塩基配列:
【化1】

式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【0013】
本発明はまた、
(c)配列番号1の第44塩基〜第1240塩基からなる塩基配列において、1または数個の塩基が欠失、付加または置換されている塩基配列であって、RhCHBH活性を有するタンパク質をコードする塩基配列、
(d)配列番号2に記載のアミノ酸配列をコードする塩基配列、
(e)配列番号2に記載のアミノ酸配列において1または数個のアミノ酸が欠失、置換または付加されているアミノ酸配列であって、RhCHBH活性を有するアミノ酸配列をコードする塩基配列、および、
(f)配列番号2に記載のアミノ酸配列と少なくとも75%以上の相同性を有するアミノ酸配列であって、RhCHBH活性を有するアミノ酸配列をコードする塩基配列、
のいずれかの塩基配列からなる遺伝子を包含する。
【0014】
また、本発明は、以下の(a’)または(b’)のいずれかのタンパク質を提供する:
(a’)配列番号2に記載のアミノ酸配列からなるタンパク質、
(b’)配列番号2に記載のアミノ酸配列において、1または数個のアミノ酸が欠失、置換または付加されているアミノ酸配列からなり、RhCHBH活性を有するタンパク質。
【0015】
本発明はまた、
(c’)配列番号2に記載のアミノ酸配列と少なくとも75%以上の相同性を有するアミノ酸配列からなり、RhCHBH活性を有するタンパク質も包含する。
【0016】
本発明はまた、上記遺伝子を有する細菌、好ましくは、リゾビウム属に属すると推定される細菌DS−S−51株(受託番号FERM P−21129)を提供する。
【0017】
また、本発明によれば、本発明の遺伝子を含むベクターが提供される。好ましくは、ベクターは、プラスミドpKK−RhCHBHである。ここで、プラスミドpKK−RhCHBHとは、市販のプラスミドpKK223−3(ファルマシア製)に、本発明のRhCHBH遺伝子が導入されたものをいう。
【0018】
また、本発明によれば、上記のベクターを含む形質転換体が提供される。好ましくは、宿主は、大腸菌であり、より好ましくは、大腸菌JM109株である。
【0019】
また、本発明は、RhCHBH遺伝子を発現させることを含む、RhCHBHタンパク質の製造方法を提供する。
【0020】
さらに本発明は、RhCHBHタンパク質を、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用させ、該混合物のR体を選択的に加水分解させ、かつ、S体を選択的に残存させることを含む、S体4−ハロ−3−ヒドロキシ酪酸エステルの製造方法を提供する:
【化2】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【0021】
さらに本発明は、RhCHBHタンパク質を、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのR体に作用させることを含む、下記式[2]で示されるR体3−ヒドロキシ−γ−ブチロラクトンの製造方法を提供する:
【化3】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【化4】
式[2]
【0022】
上記S体4−ハロ−3−ヒドロキシ酪酸エステルの製造方法およびR体3−ヒドロキシ−γ−ブチロラクトンの製造方法において、好ましくは式[1]のRはメチルであり、好ましくは式[1]のXは塩素原子である。
【発明の効果】
【0023】
本発明によればリゾビウム属細菌(Rhizobium sp.)に属すると推定される微生物(DS−S−51株(受託番号FERM P−21129))を利用して4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物からR体4−ハロ−3−ヒドロキシ酪酸エステルを選択的に分解し、S体4−ハロ−3−ヒドロキシ酪酸エステルを系中に残存させることができる。また、R体4−ハロ−3−ヒドロキシ酪酸エステルを基質にすれば、光学純度を落とすことなくR体3−ヒドロキシ−γ−ブチロラクトンを得ることができる。さらに組換え大腸菌を用いれば、生産性の向上を行なうことができる。
【発明を実施するための最良の形態】
【0024】
本明細書において、「生体触媒反応」とは、微生物を含む生体、酵素、遺伝子組換え体などを利用した反応を言う。
【0025】
本明細書において、「不斉加水分解反応」とは、両エナンチオマー混合物のうちの各光学活性(異性)体に対する加水分解速度の差を利用した光学分割方法を示す。
【0026】
本明細書において、「式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステル化合物群」とは、式[1]において、Xがハロゲン原子を表し、Rが炭素原子数1〜4のアルキル基を表す、すべての化合物を含む。
【0027】
本明細書および特許請求の範囲において、「下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用し、該混合物のR体を選択的に加水分解し、かつ、S体を選択的に残存させる酵素活性」
【化5】
式[1]
とは、式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステル化合物群に属する少なくともいずれか一つの化合物のエナンチオマー混合物としてラセミ体を基質として用いた場合に、出発物質として用いたエナンチオマー混合物のうちR体基質の少なくとも95%を加水分解し、S体基質の少なくとも20%を残存させ、その結果、光学純度が90%ee(エナンチオ過剰)以上のS体エステルを生成物として生じる酵素活性をいう。該酵素は、R体基質の好ましくは少なくとも97%以上、さらに好ましくは99%以上、特に好ましくは99.5%以上加水分解する。また、該酵素は、S体基質の好ましくは40%以上、さらに好ましくは50%以上、特に好ましくは60%以上を残存させる。さらに、該酵素による反応の結果得られるS体エステルは、好ましくは95%ee以上、さらに好ましくは98%ee以上、特に好ましくは99%ee以上の光学純度である。なお、式[1]で示される化合物としては、Xが塩素原子であるものが好ましく、Rがメチルのものが好ましい。
【0028】
本明細書および特許請求の範囲において、「下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用させ、該混合物のR体を選択的に加水分解させ、かつ、S体を選択的に残存させること」とは、式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステル化合物群に属する少なくともいずれか一つの化合物のエナンチオマー混合物としてラセミ体を基質として用いた場合に、出発物質として用いたエナンチオマー混合物のうちR体基質の少なくとも95%を加水分解し、S体基質の少なくとも20%を残存させることをいう。R体基質の好ましくは少なくとも97%、さらに好ましくは99%、特に好ましくは99.5%以上を加水分解する。また、S体基質の好ましくは40%以上、さらに好ましくは50%以上、特に好ましくは60%以上を残存させる。
【0029】
本明細書および特許請求の範囲において、「光学純度を落とすことなく」とは、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステル化合物群に属する少なくともいずれか一つの化合物のR体に作用させることを含む、下記式[2]で示されるR体3−ヒドロキシ−γ−ブチロラクトンの製造方法において、基質として光学純度90%ee以上のR体の4−ハロ−3−ヒドロキシ酪酸エステルを用いた場合に、生成物として光学純度90%ee以上のR体3−ヒドロキシ−γ−ブチロラクトンを得ることをいう。基質のR体の4−ハロ−3−ヒドロキシ酪酸エステルの光学純度は、好ましくは95%ee、さらに好ましくは98%ee、特に好ましくは99%ee以上であり、得られる生成物のR体3−ヒドロキシ−γ−ブチロラクトンの光学純度は、好ましくは95%ee、さらに好ましくは98%ee、特に好ましくは99%ee以上である。
【化6】
式[1]
【化7】
式[2]
【0030】
本明細書および特許請求の範囲において、「微生物、その処理物」とは、
1)微生物の培養液、
2)遠心分離により得た微生物の菌体および/またはその菌体処理物(菌体破砕物または菌体抽出液)、あるいは
3)1)および2)を常法により固定化したものをいう。
なお、本明細書および特許請求の範囲において、「タンパク質を4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用させる」ことは、タンパク質自体を基質に作用させることのみならず、微生物、その処理物、または本発明による遺伝子を含む形質転換体を基質に作用させることも含む。
【0031】
本明細書および特許請求の範囲において、「高光学純度」とは、90%e.e.(エナンチオ過剰)以上であることを意味する。なお、高光学純度は、好ましくは95%ee、さらに好ましくは98%ee、特に好ましくは99%ee以上である。
【0032】
以下、本発明の実施の形態について詳細に説明する。
本発明によれば、下記の塩基配列:
(a)配列番号1の第44塩基〜第1240塩基からなる塩基配列、
(b)配列番号1の第44塩基〜第1240塩基からなる塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列であって、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用し、該混合物のR体を選択的に加水分解し、かつ、S体を選択的に残存させる酵素活性(以下、RhCHBH活性とも称する)を有するタンパク質をコードする塩基配列、
【化8】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
(c)配列番号1の第44塩基〜第1240塩基からなる塩基配列において、1または数個の塩基が欠失、付加または置換されている塩基配列であって、RhCHBH活性を有するタンパク質をコードする塩基配列、
(d)配列番号2に記載のアミノ酸配列をコードする塩基配列、
(e)配列番号2に記載のアミノ酸配列において1または数個のアミノ酸が欠失、置換または付加されているアミノ酸配列であって、RhCHBH活性を有するアミノ酸配列をコードする塩基配列、および、
(f)配列番号2に記載のアミノ酸配列と少なくとも75%以上の相同性を有するアミノ酸配列であって、RhCHBH活性を有するアミノ酸配列をコードする塩基配列、
のいずれかの塩基配列からなる遺伝子が提供される。
【0033】
本発明において、「1または数個のアミノ酸が欠失、置換または付加されている」とは、例えば1〜20個、好ましくは1〜15個、より好ましくは1〜10個、さらに好ましくは1〜5個の任意の数のアミノ酸が欠失、置換または付加されていることを意味する。また、「1または数個の塩基が欠失、付加または置換されている」とは、例えば1〜20個、好ましくは1〜15個、より好ましくは1〜10個、さらに好ましくは1〜5個の任意の数の塩基が欠失、置換または付加されていることを意味する。
【0034】
本発明において、「RhCHBH」とは、配列番号2のアミノ酸配列を有するタンパク質であり、分子量は、SDS−PAGEでの測定により42kDaである。
【0035】
本発明において、「ストリンジェントな条件」とは、特異的なハイブリダイゼーションのみが起こり、非特異的なハイブリダイゼーションが起きないような条件をいう。このような条件は、通常、0.2xSSC、0.1%SDS、65℃程度である。ハイブリダイゼーションにより得られるDNAは(a)の塩基配列からなるDNAと70%以上の高い相同性を有することが望ましく、さらに80%以上の相同性を有することが好ましい。ハイブリダイゼーションは、Molecular Cloning: A laboratory Mannual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY., 1989.)等に記載されている方法に準じて行うことができる。
【0036】
本発明において「相同性」とは、2つのポリペプチドあるいはポリヌクレオチド間の配列の類似の程度を意味し、比較対象のアミノ酸配列または塩基配列の領域にわたって最適な状態(配列の一致が最大となる状態)にアラインメントされた2つの配列を比較することにより決定される。相同性の数値(%)は両方の(アミノ酸または塩基)配列に存在する同一のアミノ酸または塩基を決定して、適合部位の数を決定し、次いでこの適合部位の数を比較対象の配列領域内のアミノ酸または塩基の総数で割り、得られた数値に100をかけることにより算出される。最適なアラインメントおよび相同性を得るためのアルゴリズムとしては当業者が通常利用可能な種々のアルゴリズム(例えば、BLASTアルゴリズム、FASTAアルゴリズムなど)が挙げられる。アミノ酸配列の相同性は、例えばBLASTP、FASTAなどの配列解析ソフトウェアを用いて決定される。塩基配列の相同性は、BLASTN、FASTAなどのソフトウェアを用いて決定される。
【0037】
本発明の遺伝子の取得方法は特に限定されない。本発明の遺伝子は、所望の酵素を産生する微生物の染色体からのクローニングにより、あるいはDNA合成機を用いた合成によって得ることができ、その形態は一本鎖、二本鎖のいずれでも良い。
【0038】
クローニングによって得る場合の染色体供与微生物としては、リゾビウム属に属すると推定される細菌DS−S−51株(受託番号FERM P−21129)を好適なものとして挙げることができる。
【0039】
本発明のDNAクローニングは特に限定されず、種々の方法が利用できる。好ましくは実施例に示すショットガンクローニング法を用いるとよい。
【0040】
また、本発明の遺伝子は、配列番号1に記載の塩基配列または配列番号2に記載のアミノ酸配列の情報に基づいて適当なプローブやプライマーを調製し、それらを用いて当該遺伝子が存在することが予測されるDNAライブラリーをスクリーニングすることにより単離することもできる。
【0041】
例えば、その遺伝子供与体からのRhCHBH遺伝子をもつDNA断片の取得は、精製酵素RhCHBHのポリペプチド鎖の部分アミノ酸配列をもとに行なうことができる。すなわち、精製酵素をエンドペプチダーゼにより消化し、プロテインシークエンサーにより各断片のアミノ酸配列の一部を決定し、これをもとにしてポリメラーゼ連鎖反応(PCR)用プライマーを合成する。次いでリゾビウム属に属すると推定される細菌DS−S−51株(受託番号FERM P−21129)の染色体DNAを鋳型としたPCRを行ない、RhCHBH遺伝子の一部を増幅して、その塩基配列を明らかにする。得られたRhCHBH遺伝子の部分塩基配列をプローブとして、遺伝子供与体の染色体DNAから常法により作製したDNAライブラリーよりハイブリダイゼーション法を用いて目的の遺伝子のDNA断片を得ることができる。
【0042】
取得したRhCHBH遺伝子のDNA塩基配列の決定法としては、例えばジデオキシシークエンス法が挙げられる。この方法には、PCRにより遺伝子を増幅する方法や核酸分解酵素により欠失させる方法などの、遺伝子工学分野で慣用される様々な手法が含まれる。これらの方法により、配列番号1で示される目的のDNA塩基配列中に全アミノ酸をコードするオープンリーディングフレーム(ORF)が確認できる。
【0043】
本発明のRhCHBHをコードするDNAは、RhCHBH活性を有するアミノ酸配列が実質的に配列番号2に示されているポリペプチドをコードする塩基配列を含むことを特徴とする。ここで、RhCHBH活性を有する限り、配列番号2に示したアミノ酸配列についてアミノ酸の1個または数個の欠失、挿入、置換等があってもよい。例えば、DNAがコードするアミノ酸配列についてアミノ酸のいくつかの欠失、挿入、置換等を生じるようにDNAを改変することは、合成オリゴヌクレオチドを用いた部位特異的変異導入法などの周知の方法で適宜行うことができる。また、配列番号1に示したDNAまたは該DNAを適宜改変したDNAを鋳型にして、Mn2+イオンの存在下(通常0.5〜10mMの濃度)、または特定のヌクレオチドの濃度を低くしてPCR法を行うことによってランダムに変異が導入されたDNAを得ることができる。このようにして得られたDNAのうち、RhCHBHの活性を有するタンパク質をコードするものが、本願発明に含まれることは言うまでもない。
【0044】
また、本発明は、下記のタンパク質:
(a’)配列番号2に記載のアミノ酸配列からなるタンパク質、
(b’)配列番号2に記載のアミノ酸配列において、1または数個のアミノ酸が欠失、置換または付加されているアミノ酸配列からなり、RhCHBH活性を有するタンパク質、
(c’)配列番号2に記載のアミノ酸配列と少なくとも75%以上の相同性を有するアミノ酸配列からなり、RhCHBH活性を有するタンパク質
を提供する。
【0045】
ここで、「配列番号2に記載のアミノ酸配列と少なくとも75%以上の相同性を有するアミノ酸配列」とは、配列番号2のアミノ酸配列との相同性は75%以上であれば特に制限はなく、例えば、75%以上、好ましくは80%以上、より好ましくは85%以上、さらに好ましくは90%以上、特に好ましくは95%以上、最も好ましくは98%以上であることを意味する。
【0046】
本発明の酵素の精製方法は、特に限定されないが、通常のタンパク質の精製方法を適当に組み合わせることにより精製することができる。例えば、菌体を超音波により破砕後、硫酸アンモニウムによる塩析を行ない、沈殿の溶解物を疎水クロマトグラフィー、陰イオン交換クロマトグラフィー、ゲルろ過クロマトグラフィーを組み合わせることにより、ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動(以下SDS−PAGEと略す)で単一になるまで精製することができる。
【0047】
さらに本発明はRhCHBH活性を示すアミノ酸配列をコードする遺伝子の塩基配列を含むベクターを提供する。
【0048】
クローニング用ベクターはpUC系やpBluscript系(いずれもタカラバイオ株式会社からの市販品を利用できる)などを、特に限定されず宿主の種類に応じて用いることができる。好ましくはpUC19を用いるとよい。
【0049】
上記塩基配列(配列番号1)を有するDNA断片にコードされる構造遺伝子から、上記酵素の発現を可能とするプロモーターなどの機能領域および宿主での複製オリジンを有するベクターに組み込むことにより、組換えDNA技術を利用した本不斉加水分解反応を有する形質転換体を得るための組換えDNAを構築できる。
【0050】
発現用ベクターはpUC系やpBluscript系(タカラバイオ株式会社製等)、pET系(インビトロジェン製)など特に限定されず、宿主の種類に応じて用いることができる。発現用ベクターとして好ましいのは、宿主細胞中で自律複製可能であり、さらに組換え宿主細胞のみを選別できるような適当な選択マーカーなどが付与されたものがあげられ、適当な宿主細胞内で、本発明の遺伝子を発現できるものである。さらに、このようなベクターは公知のベクター等から公知の技術を用いて業者が容易に製造し得るようなものであってもよいし、商業的に販売されているものでも良い。特に好ましいのはpKK223−3(ファルマシア製)である。
【0051】
RhCHBH遺伝子の遺伝情報を担うDNA断片から必要な部分をベクターに組み込むことにより、宿主細胞に導入し形質転換体を得ることができる。
【0052】
用いられる宿主細胞としては、得られた組換えベクターで形質転換され、かつ本遺伝子を発現させることができるようなものであれば特に制限なく使用することができる。発現用の宿主としては、本発明の目的に沿って本遺伝子の発現を達成し得る限り、微生物、植物、動物等から限定されず用いることができる。微生物としては、例えば、大腸菌(Escherichia coli)、エンテロバクター属、サッカロミセス属、キサントモナス属、アセトバクター属、シュードモナス属、グルコノバクター属、アゾトバクター属、リゾビウム属、クレブシエラ属、サルモネラ属およびセラチア属から選ばれる微生物が挙げられるが、好ましいのは大腸菌であり、特に好ましいのは大腸菌JM109である。大腸菌JM109(pKK−RhCHBH)はFERM P−21128として独立行政法人産業技術研究所特許生物寄託センターに寄託されている。
【0053】
形質転換体は、適当な栄養培地中で培養を行なうことにより大量に得ることができる。培養に用いられる培地は微生物の生育に必要な炭素源、窒素源、無機物質等を含む通常の培地であれば何でも良い。例えば炭素源としてグルコース、フラクトース等の炭水化物、グリセロール、マンニトール、ソルビトール、プロピレングリコール等のアルコール類、酢酸、クエン酸、リンゴ酸、マレイン酸、フマル酸、グルコン酸とその塩類などの有機酸、またはそれらの混合物を用いることができる。窒素源として硫酸アンモニウム、硝酸アンモニウム、リン酸アンモニウム等の無機窒素化合物および尿素、ペプトン、カゼイン、酵母エキス、肉エキス、コーンスチープリカー等の有機窒素化合物とそれらの混合物を挙げることができる。その他、無機塩としてリン酸塩、マグネシウム塩、カリウム塩、マンガン塩、鉄塩、亜鉛塩、銅塩など、更に必要に応じてビタミン類を加えてもよい。
【0054】
また、高酵素活性を持った形質転換菌体を得るための酵素誘導添加物として、上記培地およびペプトン培地、ブイヨン培地等の栄養培地に、使用する菌株に応じて、効果的に目的のDNAを発現させるためにアンピシリン、カナマイシン、クロラムフェニコール等の抗生物質を培養液に添加してもよいし、イソプロピルβ−D(−)−チオガラクトピラノシド(IPTG)等のプロモーターの活性化誘導剤を用いることもできる。培養は好気的条件下でpH6.0〜7.5、温度25〜40℃の任意の範囲に制御して16〜24時間培養を行えばよいが、用いる形質転換体により最適な条件で培養すると更に効果的である。
【0055】
得られたRhCHBH産生形質転換体の細胞からの酵素RhCHBHの抽出は次の方法によって形質転換体を破壊して行うことができる。
1)フレンチプレスや超音波破砕による機械的(物理的)方法、
2)リゾチームなどの酵素処理方法、
3)自己溶解法、
4)浸透圧を利用した抽出法など。
【0056】
またJM109等の大腸菌(pKK−RhCHBH含有)は細胞を破砕しなくとも、培養液の状態で高活性な酵素として利用できる。
【0057】
本方法で産生される酵素は、起源のDS−S−51株(受託番FERM P−21129)におけるRhCHBHのポリペプチド配列と一致するもののみに限定されず、塩基配列変換や遺伝子変換などの遺伝子組換え手法を利用して得られるポリペプチドで、RhCHBH活性を示すものと解すべきである。すなわちペプチド配列中のアミノ酸の1個または数個が欠損したもの、あるいはそのペプチド配列中のアミノ酸の1個または数個が他のアミノ酸で置き換えられたものをも包含する。
【0058】
得られた形質転換体は、酵素RhCHBHの量産に、また形質転換体自体を用いたS体4−クロロ−3−ヒドロキシ酪酸エステルおよびR体3−ヒドロキシ−γ−ブチロラクトンの製造に好適に用い得る。
【0059】
次に、本発明のS体4−ハロ−3−ヒドロキシ酪酸エステルの製造方法およびR体3−ヒドロキシ−γ−ブチロラクトンの製造方法について説明する。
【0060】
原料の4−ハロ−3−ヒドロキシ酪酸エステル、例えば、4−クロロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物は、例えば、エピクロルヒドリンから安価に製造することができる。
【0061】
また、R体4−ハロ−3−ヒドロキシ酪酸エステルは、微生物または酵素を用いて4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物を立体選択的に加水分解させ、R体を残存させる製法(特開平9−47296号公報)により簡単に得ることができる。
【0062】
本発明で用いられるRhCHBH酵素を産生する微生物としては、上記のようなRhCHBH遺伝子を導入した形質転換体であってもよいし、天然に4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物のR体を選択的に分解し得るものであってもよく、かかる天然に4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物のR体を選択的に分解し得る微生物としては、リゾビウム属(Rhizobium sp.)に属すると推定される細菌DS−S−51株(受託番号FERM P−21129)が挙げられる。
【0063】
[分類方法詳細]
本発明で用いられる微生物の具体例であるDS−S−51株は本発明者らが土壌から分離した細菌である。本菌株の形態学的・生理学的性質に基づき委託、同定された結果、形態、生育状態、生理学的試験の結果、グラム染色陰性、カタラーゼ反応陽性、オキシダーゼ反応陽性、運動性を示す無芽胞かん菌であるなどシュードモナス(Pseudomonas )類似の性状を示した。続く生化学試験、資化性試験ではリゾビウム・ラジオバクター(Rhizobium radiobacter) に類似の性状を示した。しかしながらその性状はNAGa、GATaの試験結果でリゾビウム・ラジオバクターの典型性状とは異なるなど詳細な帰属分類群の推定には至らなかった。
【0064】
そこで、16SrRNA遺伝子の部分塩基配列約500 bpをもちいて検体の帰属分類群を委託、推定された結果、リゾビウム属に属する細菌と高い相同性を示した。16SrRNA部分塩基配列は相同性98.9%でリゾビウム・ダエジェオネンス(Rhizobium daejeonense)の16SrRNAに対し最も高い相同性を示した。さらに、分子系統樹上で系統枝を形成した。
【0065】
これらの結果および下記の菌株の諸性質により、DS−S−51株をリゾビウム属に属する細菌であると同定した。DS−S−51株の科学的性質は下記に示すとおりである。
【0066】
A.形態
細胞の形 かん菌
細胞の大きさ 0.6〜0.7 × 1.5〜2.0μm
グラム染色 陽性
胞子の有無 陰性
運動性の有無 陽性
【0067】
B.生育状態(30℃、48時間培養)
コロニーの形状 円形
コロニー表面の形状 全縁滑らか
コロニーの隆起状態 低凸状
コロニーの光沢 光沢あり
コロニーの色調 クリーム色
37℃での生育状態 反応弱
45℃の生育状態 陰性
【0068】
C.生理学的試験
カタラーゼ 陽性
オキシダーゼ 陽性
グルコース、酸産出 陰性
グルコース、ガス産出 陰性
グルコース、Oテスト 陽性
グルコース、Fテスト 陰性
【0069】
D.生化学試験
硝酸塩還元 +
インドール産出 −
ブドウ糖 酸性化 −
アルギニンジヒドロラーゼ −
ウレアーゼ +
エスクリン加水分解 +
ゼラチン加水分解 −
β‐ガラクトシダーゼ +
【0070】
E.資化性試験
ブドウ糖 +
L‐アラビノース +
D‐マンノース +
D‐マンニトール +
N‐アセチル‐D‐グルコサミン −
マルトース +
グルコン酸カリウム −
n‐カプリン酸 −
アジピン酸 −
dl‐リンゴ酸 +
クエン酸ナトリウム −
酢酸フェニル −
チトクロームオキシダーゼ +
【0071】
MacConkey寒天培地での生育試験 −
シモンズクエン酸の利用試験 −
【0072】
4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物またはR体4−ハロ−3−ヒドロキシ酪酸エステルに、RhCHBH遺伝子を含む細菌または形質転換体に含まれる酵素RhCHBHを作用させて、S体4−ハロ−3−ヒドロキシ酪酸エステルまたはR体3−ヒドロキシ−γ−ブチロラクトンを得る方法としては、酵素RhCHBHそのものを基質と接触させてもよいし、上記RhCHBH遺伝子を含む細菌または形質転換体の培養液または、それらの遠心分離により得た菌体およびその菌体処理物(菌体破砕物または菌体抽出液)あるいは、それらを常法により固定化したものを基質と接触させてもよい。
【0073】
基質と酵素または酵素を含む微生物との接触は、緩衝液に混合した微生物菌体混合液に基質を添加する方法などがある。反応温度は15〜50℃が好ましく、反応pHは6〜9で行なうのが好ましい。基質は初期に一括添加してもよいし、分割添加してもよい。反応は通常、攪拌あるいは振とうしながら行い、反応時間は基質濃度、微生物菌体または酵素量により異なるが、1〜120時間で終了するのがよい。好ましくはガスクロマトグラフィーなどの分析により、目的の光学活性体の濃度および光学純度を測定し終点を決定するのがよい。
【実施例】
【0074】
以下実施例をもって、本発明を詳細に説明するが、本発明はこれらに限定されるものではない。なお、実施例中の%は特に記載のない限り(w/v)で表す。
【0075】
(実施例1)DS−S−51株の培養
ペプトン、酵母エキス、グリセロールをそれぞれ1.0%含む液体栄養培地(pH7.0)5mlを入れた試験管に、凍結保存バイアルから種菌を2%(v/v)量、無菌的に植菌し、30℃で24時間培養を行なった。
【0076】
(実施例2)遺伝子のクローニング
実施例1で得られた培養液から遠心分離により菌体を回収した。菌体を490μlのTE溶液(10mMトリス−塩酸 pH8.0、1mM EDTA)に懸濁し、10%SDSを30μl、20mg/mlプロテアーゼKを50μl添加し、50℃で1時間反応させた。その後、等量のフェノール:クロロホルム:イソアミルアルコール(25:24:1)による抽出を行なった後、0.1倍量の3M酢酸ナトリウム溶液を添加し、0.6倍量の2−プロパノールを静かに重層した。その結果、界面に生じたDNAをガラス棒で糸状に巻きつけて回収した。得られたDNAを500μlのTE溶液に溶解させた。約1.1mgの染色体DNAを得た。
【0077】
制限酵素Sau3AIを用いて、上記により得られた染色体DNAの60μgを250μlの反応系で酵素0.75U用いて部分消化し、染色体DNA断片を得た。またプラスミドベクターpUC19を常法により制限酵素BamHIで消化し、BAP処理を行なった。
【0078】
上記によって消化された染色体DNA断片14μgとプラスミドベクターpUC19の800ngをT4DNAリガーゼ(タカラバイオ株式会社より購入できる)1400Uを用いて常法により16℃でライゲーション反応させ、これを用いて42℃の熱ショック法により大腸菌DH5α株(東洋紡績株式会社より購入できる)を形質転換させた。
【0079】
熱ショック後、SOC培地にて37℃で1時間培養した後、LB寒天培地(100μg/mlアンピシリン、0.5%メチル4−クロロ−3−ヒドロキシ酪酸、0.0008%BTB(ブロモチールブルー)を含む)一枚につき、上記菌体液の約100μlを塗布し、37℃で16時間インキュベートした。
【0080】
BTBにより通常pHが中性のプレートは緑色であるが、プレートに出現した合計47600個の形質転換体コロニーのうち、メチル4−クロロ−3−ヒドロキシ酪酸の脱クロルを伴った加水分解反応によりpHが低下するに伴い、周辺が黄色に変色しているコロニー1個を得た。
【0081】
得られた陽性コロニーを単離培養し、常法に従ってプラスミドを調製し、制限酵素EcoRI、HindIIIで消化し、1%アガロース電気泳動を行なったところ、約4kbのDNAがpUC19に挿入されていることがわかった。
【0082】
(実施例3)DNAの塩基配列の決定
実施例2で得られた約4kbの遺伝子断片のうち、ORF周囲1.8kbの配列決定をDNAシークエンサーにより行った。その結果、配列番号1の第44塩基〜第1240塩基に示す1197bpからなるオープンリーディングフレーム(ORF)を確認できた。これより、398残基のアミノ酸をコードしていることがわかった。アミノ酸配列を配列番号2に示す。
【0083】
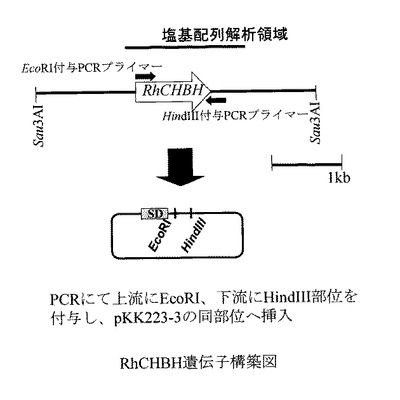
(実施例4)高発現組換え大腸菌の作製
ORFのすぐ上流にEcoRI部位、下流にHindIII部位を付与する様に配列番号3および4に示すPCRプライマー:
5'-ttgaattcatgccccataatctg-3'(配列番号3)
5'-aaaagcttgtggccgtcga-3'(配列番号4)
を設計し、増幅断片をpKK223−3のEcoRI、HindIII部位に導入し、これを用いて実施例2と同様に大腸菌JM109株を形質転換させてJM109(pKK−RhCHBH)を得た。また対照試料としてベクターpKK223−3のみを用いた形質転換により、JM109(pKK223−3)を得た。
【0084】
100μg/mlアンピシリンを含むLB培地にて作製した上記組換え大腸菌を20時間培養した。
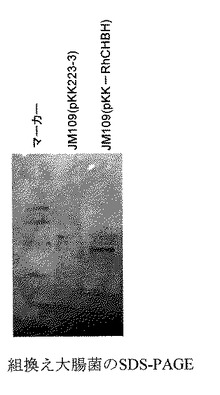
【0085】
調製した組換え大腸菌の培養液を遠心分離により集菌し、20mMリン酸カリウム緩衝液で懸濁後、超音波破砕した。5μgの抽出タンパク試料をSDS−PAGE(10%分離ゲル)に供し、クマシーブリリアントグリーンによる染色を行なった。その結果分子量は42kDaであることがわかった。用いた分子量マーカーと分子量を下記に示す。ホスホリラーゼB(94,000)、ウシ血清アルブミン(67,000)、オボアルブミン(43,000)、カルボニックアンヒドラーゼ(30,000)、トリプシンインヒビター(20,100)、α−ラクトアルブミン(14,400)。
【0086】
(実施例5)S体4−クロロ−3−ヒドロキシ酪酸エステルの生産
500ml容のバッフル付三角フラスコを用いてロータリーシェイカーで130rpmにて培養した条件以外は、実施例1または実施例4と同じ方法で培養して得たDS−S−51株の培養液100ml、および組換え大腸菌JM109(pKK−RhCHBH)の培養液100mlに、基質としてラセミ体メチル4−クロロ−3−ヒドロキシ酪酸を加え、30℃で振とうしながら反応させた。反応終了後、反応液中に残存するメチル4−クロロ−3−ヒドロキシ酪酸をガスクロマトグラフィー(カラム担体:PEG20M,60−80メッシュ)で分析した結果、その残存率(収率)はDS−S−51株については30.9%、組換え大腸菌については30.5%であった。残存率(収率)とは、添加したラセミ体量を100%とした時、反応後に残存している基質量を表す。反応液を遠心分離により菌体を除き、ついで遠心上清と等量の酢酸エチルで抽出した。この抽出物中のメチル4−クロロ−3−ヒドロキシ酪酸をアステック社(Advanced Separation Technologies Inc., NJ, USA)製のキャピラリーカラムG−TA(0.25mm×30m)を用いたガスクロマトグラフィーにより光学純度分析を行った結果、回収したメチル4−クロロ−3−ヒドロキシ酪酸は光学純度99%e.e.以上のS体であることが判明した。DS−S−51株は、1.0%(v/v)の基質を光学分割するのに40時間要した。対して、組換え大腸菌は、2.0%(v/v)の基質を4時間で光学分割した。次に実施例4で得た組換え大腸菌の培養液に基質として2.0%(v/v)になるように、エチル4−クロロ−3−ヒドロキシ酪酸、プロピル4−クロロ−3−ヒドロキシ酪酸、ブチル4−クロロ−3−ヒドロキシ酪酸のラセミ体を各々に加え、同様に4時間反応、分析した結果、各々残存率(収率)は、30.5%、30.0%、31.0%であり、各種4−クロロ−3−ヒドロキシ酪酸エステルは光学純度99%e.e.以上のS体であることが判明した。光学純度の分析条件はカラム温度:100℃、検出器温度:200℃、キャリアーガス:窒素、流速:0.5ml/min;検出器、FID;スプリット比、100/1、リテンションタイムは次に示す通りである。メチル4−クロロ−3−ヒドロキシ酪酸:R体14.4分、S体15.2分、エチル4−クロロ−3−ヒドロキシ酪酸:R体18.4分、S体19.2分、プロピル4−クロロ−3−ヒドロキシ酪酸:R体28.8分、30.2分、ブチルメチル4−クロロ−3−ヒドロキシ酪酸:R体49.2分、S体50.2分。
【0087】
(実施例6)R体3−ヒドロキシ−γ−ブチロラクトンの生産
実施例5と同様に調製したDS−S−51株の培養液100ml、および組換え大腸菌JM109(pKK−RhCHBH)の培養液100mlに、基質として光学純度99%eeのR体メチル4−クロロ−3−ヒドロキシ酪酸を加え、30℃で振とうしながら反応させた。反応終了後、生成したR体3−ヒドロキシ−γ−ブチロラクトンをガスクロマトグラフィー(カラム担体:PEG20M,60−80メッシュ)で分析した結果、その生成モル収率はDS−S−51株、組換え大腸菌とも100%であった。3−ヒドロキシ−γ−ブチロラクトンの光学純度分析は以下の様に行なった。反応液を遠心分離により菌体を除き、ついでロータリーエバポレーターにて濃縮を行なった。適当量の1,2−ジクロロエタンで抽出してシロップを得た。ジクロロメタン1mlにシロップ20μl、トリフルオロ酢酸無水物200μlを加えてよく攪拌し、30分後デシケーターで濃縮後、エタノール1mlに溶解させた後、実施例5と同じキャピラリーカラムG−TAに供した。その結果、回収した3−ヒドロキシ−γ−ブチロラクトンは光学純度99.0%e.e.のR体であることが判明した。分析条件はカラム温度:100℃、検出器温度:200℃、キャリアーガス:窒素、流速:0.5ml/min;検出器、FID;スプリット比、100/1。3−ヒドロキシ−γ−ブチロラクトンのリテンションタイムはS体29.5分、R体31.2分であった。DS−S−51株は、1.0%の基質をすべてR体に変換するのに80時間要した。対して、組換え大腸菌は、2.0%(v/v)の基質を8時間で変換した。
【図面の簡単な説明】
【0088】
【図1】図1は、RhCHBH遺伝子コンストラクトの構築を示す図である。
【図2】図2は、組換え大腸菌のSDS−PAGEを示す図である。
【技術分野】
【0001】
本発明は医薬、農薬、生理活性物質および強誘電性液晶などの光学活性化合物の合成において有用な中間体となりうる光学活性S体4−ハロ−3−ヒドロキシ酪酸エステルおよびR体3−ヒドロキシ−γ−ブチロラクトンの、微生物、その処理物、微生物由来の酵素または遺伝子を用いた不斉加水分解反応による製法、およびこれらの製法に使用する微生物由来の酵素、該酵素をコードする遺伝子に関する。
【背景技術】
【0002】
光学活性化合物の製法としては、対応する光学活性出発物質から目的物へ変換する化学的合成法のほか、対応するラセミ体を光学分割剤で処理して光学活性体に分割する方法が用いられる。また近年、微生物または酵素を用いて不斉還元や不斉加水分解反応を利用する生体触媒反応により光学活性体を分離する方法が多く報告されている。
【0003】
4−ハロ−3−ヒドロキシ酪酸エステルの製法は、合成法としては、ルテニウム−光学活性ホスフィン錯体を触媒として用いる不斉還元による方法(特許文献1)が知られているが、該方法に用いる触媒の合成は容易ではなく、コスト面で問題がある。また、光学活性アミノ酸誘導体とアルコールを用いて還元する方法(特許文献2)が知られているが、生成物の光学純度が低いという欠点があり、また、−78℃の様な低温で反応させなければならないという問題がある。
【0004】
また、4−ハロ−3−ヒドロキシ酪酸エステルの生体触媒反応を用いた製法としては、微生物菌体または酵素を用いるβ−ケトエステル体の不斉還元法(特許文献3、4、5)が知られており、また遺伝子組換え技術による不斉還元法の改良も行なわれている(特許文献6)。しかし、これら微生物あるいは酵素を用いた不斉還元法によるβ−ケトエステル体からのS体4−ハロ−3−ヒドロキシ酪酸エステルの製法においては、その反応に高価なNADH(ニコチンアミドアデニンジヌクレオチド)あるいはNADPH(ニコチンアミドアデニンジヌクレオチドリン酸)などの補酵素を必要とし、さらにその酸化体から再度還元体へ変換する反応を必要とするため、グルコースオキシダーゼあるいは蟻酸デヒドロゲナーゼなどの酵素を別途必要とするうえ、その反応工程が律速反応となるなどの観点から工業的製法とは言い難い。
【0005】
また、微生物あるいは酵素を用いた4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物の不斉分解反応によるS体4−クロロ−3−ヒドロキシ酪酸エステルの製法(非特許文献1、2)が報告されているが、これらの製法では、高光学純度にて生成物を生産することはできない。またアグロバクテリウムを用いた方法(特許文献7)では、本反応に関与する酵素や遺伝子の報告がないので、生産性の向上は野性株での改良に限られてしまう。
【0006】
光学活性3−ヒドロキシ−γ−ブチロラクトンの製法としては、エチル−4−t−ブトキシ−3−オキソ酪酸をパン酵母を用いて不斉還元し、得られたR体エチル−4−t−ブトキシ−3−ヒドロキシ酪酸からフルオロ酢酸存在下においてR体3−ヒドロキシ−γ−ブチロラクトンに誘導する方法(非特許文献3)が報告されているが、この方法は化学的な合成反応を含むことから、不安定な生成物を高光学純度で得ることは困難である。また、微生物を用いた、光学活性な4−ハロ−3−ヒドロキシ酪酸エステルから対応する立体の3−ヒドロキシ−γ−ブチロラクトンの製法(特許文献8)が報告されているが、該微生物の作用の立体選択性は開示されていない。さらに、この反応に関与する酵素や遺伝子の報告がないので、生産性の向上は野性株での改良に限られてしまう。
【特許文献1】特開平1−211551号公報
【特許文献2】特開昭61−155354号公報
【特許文献3】特開昭61−146191号公報
【特許文献4】特開平6−209782号公報
【特許文献5】特開平11−187869号公報
【特許文献6】特開2000−236883号公報
【特許文献7】特開平9−322786号公報
【特許文献8】特開2002−204699号公報
【非特許文献1】Tetrahedron: Asymmetry 12 pp545-556, 2001
【非特許文献2】Tetrahedron: Asymmetry 7 pp3109-3112, 1996
【非特許文献3】Seebach, Dieter; Eberle, Martin, Synthesis (1), pp37-40, 1986
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明は、上記問題点の解決を課題とするものであり、微生物、微生物由来の酵素または酵素をコードする遺伝子により生体触媒を用いた不斉加水分解反応により4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物から高光学純度のS体4−ハロ−3−ヒドロキシ酪酸エステルを製造する方法、また、R体4−ハロ−3−ヒドロキシ酪酸エステルからR体3−ヒドロキシ−γ−ブチロラクトンを製造する方法、これらの製法に使用する微生物由来の酵素、および該酵素をコードする遺伝子を提供することを目的とする。
【0008】
本発明はまた、組換えDNAの手法を用いて立体識別性が向上し、微生物の触媒能力を従来と比較して飛躍的に増大することができ、光学活性体を大量生産することができる4−ハロ−3−ヒドロキシ酪酸エステル不斉加水分解酵素(RhCHBH(Rhizobium sp. 4-Chloro-3-HydroxyButyrate Hydrolase))およびそれをコードする遺伝子を提供することを目的とする。
本発明はまた、該遺伝子を含むベクター、形質転換体、および該酵素を用いる光学活性体の製造方法を提供することも目的とする。
【課題を解決するための手段】
【0009】
本発明者らは、S体4−ハロ−3−ヒドロキシ酪酸エステルおよびR体3−ヒドロキシ−γ−ブチロラクトンを工業的に製造する方法を見出すべく、種々研究を重ねた結果、ある種の微生物(受託番号FERM P−21129)が4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物のうち、R体に対して高度に選択的な反応性を示し、3−ヒドロキシ−γ−ブチロラクトンに変換し、S体4−クロロ−3−ヒドロキシ酪酸エステルを残存させることを見いだした。
【0010】
また、この微生物による反応において、R体4−ハロ−3−ヒドロキシ酪酸エステルを出発原料にすれば、光学純度を落とすことなくR体3−ヒドロキシ−γ−ブチロラクトンを得ることができることを見出した。
【0011】
さらに、該微生物(受託番号FERM P−21129)は反応性が低いため、該反応を触媒する酵素(以下、RhCHBHとも称する)をコードする遺伝子(以下、RhCHBH遺伝子とも称する)を単離し、組換え大腸菌により生産性を向上させることに成功した。
【0012】
したがって、本発明は、以下の(a)または(b)のいずれかの塩基配列からなる遺伝子を提供する:
(a)配列番号1の第44塩基〜第1240塩基からなる塩基配列、
(b)配列番号1の第44塩基〜第1240塩基からなる塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列であって、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用し、該混合物のR体を選択的に加水分解し、かつ、S体を選択的に残存させる酵素活性(以下、RhCHBH活性とも称する)を有するタンパク質をコードする塩基配列:
【化1】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【0013】
本発明はまた、
(c)配列番号1の第44塩基〜第1240塩基からなる塩基配列において、1または数個の塩基が欠失、付加または置換されている塩基配列であって、RhCHBH活性を有するタンパク質をコードする塩基配列、
(d)配列番号2に記載のアミノ酸配列をコードする塩基配列、
(e)配列番号2に記載のアミノ酸配列において1または数個のアミノ酸が欠失、置換または付加されているアミノ酸配列であって、RhCHBH活性を有するアミノ酸配列をコードする塩基配列、および、
(f)配列番号2に記載のアミノ酸配列と少なくとも75%以上の相同性を有するアミノ酸配列であって、RhCHBH活性を有するアミノ酸配列をコードする塩基配列、
のいずれかの塩基配列からなる遺伝子を包含する。
【0014】
また、本発明は、以下の(a’)または(b’)のいずれかのタンパク質を提供する:
(a’)配列番号2に記載のアミノ酸配列からなるタンパク質、
(b’)配列番号2に記載のアミノ酸配列において、1または数個のアミノ酸が欠失、置換または付加されているアミノ酸配列からなり、RhCHBH活性を有するタンパク質。
【0015】
本発明はまた、
(c’)配列番号2に記載のアミノ酸配列と少なくとも75%以上の相同性を有するアミノ酸配列からなり、RhCHBH活性を有するタンパク質も包含する。
【0016】
本発明はまた、上記遺伝子を有する細菌、好ましくは、リゾビウム属に属すると推定される細菌DS−S−51株(受託番号FERM P−21129)を提供する。
【0017】
また、本発明によれば、本発明の遺伝子を含むベクターが提供される。好ましくは、ベクターは、プラスミドpKK−RhCHBHである。ここで、プラスミドpKK−RhCHBHとは、市販のプラスミドpKK223−3(ファルマシア製)に、本発明のRhCHBH遺伝子が導入されたものをいう。
【0018】
また、本発明によれば、上記のベクターを含む形質転換体が提供される。好ましくは、宿主は、大腸菌であり、より好ましくは、大腸菌JM109株である。
【0019】
また、本発明は、RhCHBH遺伝子を発現させることを含む、RhCHBHタンパク質の製造方法を提供する。
【0020】
さらに本発明は、RhCHBHタンパク質を、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用させ、該混合物のR体を選択的に加水分解させ、かつ、S体を選択的に残存させることを含む、S体4−ハロ−3−ヒドロキシ酪酸エステルの製造方法を提供する:
【化2】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【0021】
さらに本発明は、RhCHBHタンパク質を、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのR体に作用させることを含む、下記式[2]で示されるR体3−ヒドロキシ−γ−ブチロラクトンの製造方法を提供する:
【化3】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【化4】
式[2]
【0022】
上記S体4−ハロ−3−ヒドロキシ酪酸エステルの製造方法およびR体3−ヒドロキシ−γ−ブチロラクトンの製造方法において、好ましくは式[1]のRはメチルであり、好ましくは式[1]のXは塩素原子である。
【発明の効果】
【0023】
本発明によればリゾビウム属細菌(Rhizobium sp.)に属すると推定される微生物(DS−S−51株(受託番号FERM P−21129))を利用して4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物からR体4−ハロ−3−ヒドロキシ酪酸エステルを選択的に分解し、S体4−ハロ−3−ヒドロキシ酪酸エステルを系中に残存させることができる。また、R体4−ハロ−3−ヒドロキシ酪酸エステルを基質にすれば、光学純度を落とすことなくR体3−ヒドロキシ−γ−ブチロラクトンを得ることができる。さらに組換え大腸菌を用いれば、生産性の向上を行なうことができる。
【発明を実施するための最良の形態】
【0024】
本明細書において、「生体触媒反応」とは、微生物を含む生体、酵素、遺伝子組換え体などを利用した反応を言う。
【0025】
本明細書において、「不斉加水分解反応」とは、両エナンチオマー混合物のうちの各光学活性(異性)体に対する加水分解速度の差を利用した光学分割方法を示す。
【0026】
本明細書において、「式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステル化合物群」とは、式[1]において、Xがハロゲン原子を表し、Rが炭素原子数1〜4のアルキル基を表す、すべての化合物を含む。
【0027】
本明細書および特許請求の範囲において、「下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用し、該混合物のR体を選択的に加水分解し、かつ、S体を選択的に残存させる酵素活性」
【化5】
式[1]
とは、式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステル化合物群に属する少なくともいずれか一つの化合物のエナンチオマー混合物としてラセミ体を基質として用いた場合に、出発物質として用いたエナンチオマー混合物のうちR体基質の少なくとも95%を加水分解し、S体基質の少なくとも20%を残存させ、その結果、光学純度が90%ee(エナンチオ過剰)以上のS体エステルを生成物として生じる酵素活性をいう。該酵素は、R体基質の好ましくは少なくとも97%以上、さらに好ましくは99%以上、特に好ましくは99.5%以上加水分解する。また、該酵素は、S体基質の好ましくは40%以上、さらに好ましくは50%以上、特に好ましくは60%以上を残存させる。さらに、該酵素による反応の結果得られるS体エステルは、好ましくは95%ee以上、さらに好ましくは98%ee以上、特に好ましくは99%ee以上の光学純度である。なお、式[1]で示される化合物としては、Xが塩素原子であるものが好ましく、Rがメチルのものが好ましい。
【0028】
本明細書および特許請求の範囲において、「下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用させ、該混合物のR体を選択的に加水分解させ、かつ、S体を選択的に残存させること」とは、式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステル化合物群に属する少なくともいずれか一つの化合物のエナンチオマー混合物としてラセミ体を基質として用いた場合に、出発物質として用いたエナンチオマー混合物のうちR体基質の少なくとも95%を加水分解し、S体基質の少なくとも20%を残存させることをいう。R体基質の好ましくは少なくとも97%、さらに好ましくは99%、特に好ましくは99.5%以上を加水分解する。また、S体基質の好ましくは40%以上、さらに好ましくは50%以上、特に好ましくは60%以上を残存させる。
【0029】
本明細書および特許請求の範囲において、「光学純度を落とすことなく」とは、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステル化合物群に属する少なくともいずれか一つの化合物のR体に作用させることを含む、下記式[2]で示されるR体3−ヒドロキシ−γ−ブチロラクトンの製造方法において、基質として光学純度90%ee以上のR体の4−ハロ−3−ヒドロキシ酪酸エステルを用いた場合に、生成物として光学純度90%ee以上のR体3−ヒドロキシ−γ−ブチロラクトンを得ることをいう。基質のR体の4−ハロ−3−ヒドロキシ酪酸エステルの光学純度は、好ましくは95%ee、さらに好ましくは98%ee、特に好ましくは99%ee以上であり、得られる生成物のR体3−ヒドロキシ−γ−ブチロラクトンの光学純度は、好ましくは95%ee、さらに好ましくは98%ee、特に好ましくは99%ee以上である。
【化6】
式[1]
【化7】
式[2]
【0030】
本明細書および特許請求の範囲において、「微生物、その処理物」とは、
1)微生物の培養液、
2)遠心分離により得た微生物の菌体および/またはその菌体処理物(菌体破砕物または菌体抽出液)、あるいは
3)1)および2)を常法により固定化したものをいう。
なお、本明細書および特許請求の範囲において、「タンパク質を4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用させる」ことは、タンパク質自体を基質に作用させることのみならず、微生物、その処理物、または本発明による遺伝子を含む形質転換体を基質に作用させることも含む。
【0031】
本明細書および特許請求の範囲において、「高光学純度」とは、90%e.e.(エナンチオ過剰)以上であることを意味する。なお、高光学純度は、好ましくは95%ee、さらに好ましくは98%ee、特に好ましくは99%ee以上である。
【0032】
以下、本発明の実施の形態について詳細に説明する。
本発明によれば、下記の塩基配列:
(a)配列番号1の第44塩基〜第1240塩基からなる塩基配列、
(b)配列番号1の第44塩基〜第1240塩基からなる塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列であって、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用し、該混合物のR体を選択的に加水分解し、かつ、S体を選択的に残存させる酵素活性(以下、RhCHBH活性とも称する)を有するタンパク質をコードする塩基配列、
【化8】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
(c)配列番号1の第44塩基〜第1240塩基からなる塩基配列において、1または数個の塩基が欠失、付加または置換されている塩基配列であって、RhCHBH活性を有するタンパク質をコードする塩基配列、
(d)配列番号2に記載のアミノ酸配列をコードする塩基配列、
(e)配列番号2に記載のアミノ酸配列において1または数個のアミノ酸が欠失、置換または付加されているアミノ酸配列であって、RhCHBH活性を有するアミノ酸配列をコードする塩基配列、および、
(f)配列番号2に記載のアミノ酸配列と少なくとも75%以上の相同性を有するアミノ酸配列であって、RhCHBH活性を有するアミノ酸配列をコードする塩基配列、
のいずれかの塩基配列からなる遺伝子が提供される。
【0033】
本発明において、「1または数個のアミノ酸が欠失、置換または付加されている」とは、例えば1〜20個、好ましくは1〜15個、より好ましくは1〜10個、さらに好ましくは1〜5個の任意の数のアミノ酸が欠失、置換または付加されていることを意味する。また、「1または数個の塩基が欠失、付加または置換されている」とは、例えば1〜20個、好ましくは1〜15個、より好ましくは1〜10個、さらに好ましくは1〜5個の任意の数の塩基が欠失、置換または付加されていることを意味する。
【0034】
本発明において、「RhCHBH」とは、配列番号2のアミノ酸配列を有するタンパク質であり、分子量は、SDS−PAGEでの測定により42kDaである。
【0035】
本発明において、「ストリンジェントな条件」とは、特異的なハイブリダイゼーションのみが起こり、非特異的なハイブリダイゼーションが起きないような条件をいう。このような条件は、通常、0.2xSSC、0.1%SDS、65℃程度である。ハイブリダイゼーションにより得られるDNAは(a)の塩基配列からなるDNAと70%以上の高い相同性を有することが望ましく、さらに80%以上の相同性を有することが好ましい。ハイブリダイゼーションは、Molecular Cloning: A laboratory Mannual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY., 1989.)等に記載されている方法に準じて行うことができる。
【0036】
本発明において「相同性」とは、2つのポリペプチドあるいはポリヌクレオチド間の配列の類似の程度を意味し、比較対象のアミノ酸配列または塩基配列の領域にわたって最適な状態(配列の一致が最大となる状態)にアラインメントされた2つの配列を比較することにより決定される。相同性の数値(%)は両方の(アミノ酸または塩基)配列に存在する同一のアミノ酸または塩基を決定して、適合部位の数を決定し、次いでこの適合部位の数を比較対象の配列領域内のアミノ酸または塩基の総数で割り、得られた数値に100をかけることにより算出される。最適なアラインメントおよび相同性を得るためのアルゴリズムとしては当業者が通常利用可能な種々のアルゴリズム(例えば、BLASTアルゴリズム、FASTAアルゴリズムなど)が挙げられる。アミノ酸配列の相同性は、例えばBLASTP、FASTAなどの配列解析ソフトウェアを用いて決定される。塩基配列の相同性は、BLASTN、FASTAなどのソフトウェアを用いて決定される。
【0037】
本発明の遺伝子の取得方法は特に限定されない。本発明の遺伝子は、所望の酵素を産生する微生物の染色体からのクローニングにより、あるいはDNA合成機を用いた合成によって得ることができ、その形態は一本鎖、二本鎖のいずれでも良い。
【0038】
クローニングによって得る場合の染色体供与微生物としては、リゾビウム属に属すると推定される細菌DS−S−51株(受託番号FERM P−21129)を好適なものとして挙げることができる。
【0039】
本発明のDNAクローニングは特に限定されず、種々の方法が利用できる。好ましくは実施例に示すショットガンクローニング法を用いるとよい。
【0040】
また、本発明の遺伝子は、配列番号1に記載の塩基配列または配列番号2に記載のアミノ酸配列の情報に基づいて適当なプローブやプライマーを調製し、それらを用いて当該遺伝子が存在することが予測されるDNAライブラリーをスクリーニングすることにより単離することもできる。
【0041】
例えば、その遺伝子供与体からのRhCHBH遺伝子をもつDNA断片の取得は、精製酵素RhCHBHのポリペプチド鎖の部分アミノ酸配列をもとに行なうことができる。すなわち、精製酵素をエンドペプチダーゼにより消化し、プロテインシークエンサーにより各断片のアミノ酸配列の一部を決定し、これをもとにしてポリメラーゼ連鎖反応(PCR)用プライマーを合成する。次いでリゾビウム属に属すると推定される細菌DS−S−51株(受託番号FERM P−21129)の染色体DNAを鋳型としたPCRを行ない、RhCHBH遺伝子の一部を増幅して、その塩基配列を明らかにする。得られたRhCHBH遺伝子の部分塩基配列をプローブとして、遺伝子供与体の染色体DNAから常法により作製したDNAライブラリーよりハイブリダイゼーション法を用いて目的の遺伝子のDNA断片を得ることができる。
【0042】
取得したRhCHBH遺伝子のDNA塩基配列の決定法としては、例えばジデオキシシークエンス法が挙げられる。この方法には、PCRにより遺伝子を増幅する方法や核酸分解酵素により欠失させる方法などの、遺伝子工学分野で慣用される様々な手法が含まれる。これらの方法により、配列番号1で示される目的のDNA塩基配列中に全アミノ酸をコードするオープンリーディングフレーム(ORF)が確認できる。
【0043】
本発明のRhCHBHをコードするDNAは、RhCHBH活性を有するアミノ酸配列が実質的に配列番号2に示されているポリペプチドをコードする塩基配列を含むことを特徴とする。ここで、RhCHBH活性を有する限り、配列番号2に示したアミノ酸配列についてアミノ酸の1個または数個の欠失、挿入、置換等があってもよい。例えば、DNAがコードするアミノ酸配列についてアミノ酸のいくつかの欠失、挿入、置換等を生じるようにDNAを改変することは、合成オリゴヌクレオチドを用いた部位特異的変異導入法などの周知の方法で適宜行うことができる。また、配列番号1に示したDNAまたは該DNAを適宜改変したDNAを鋳型にして、Mn2+イオンの存在下(通常0.5〜10mMの濃度)、または特定のヌクレオチドの濃度を低くしてPCR法を行うことによってランダムに変異が導入されたDNAを得ることができる。このようにして得られたDNAのうち、RhCHBHの活性を有するタンパク質をコードするものが、本願発明に含まれることは言うまでもない。
【0044】
また、本発明は、下記のタンパク質:
(a’)配列番号2に記載のアミノ酸配列からなるタンパク質、
(b’)配列番号2に記載のアミノ酸配列において、1または数個のアミノ酸が欠失、置換または付加されているアミノ酸配列からなり、RhCHBH活性を有するタンパク質、
(c’)配列番号2に記載のアミノ酸配列と少なくとも75%以上の相同性を有するアミノ酸配列からなり、RhCHBH活性を有するタンパク質
を提供する。
【0045】
ここで、「配列番号2に記載のアミノ酸配列と少なくとも75%以上の相同性を有するアミノ酸配列」とは、配列番号2のアミノ酸配列との相同性は75%以上であれば特に制限はなく、例えば、75%以上、好ましくは80%以上、より好ましくは85%以上、さらに好ましくは90%以上、特に好ましくは95%以上、最も好ましくは98%以上であることを意味する。
【0046】
本発明の酵素の精製方法は、特に限定されないが、通常のタンパク質の精製方法を適当に組み合わせることにより精製することができる。例えば、菌体を超音波により破砕後、硫酸アンモニウムによる塩析を行ない、沈殿の溶解物を疎水クロマトグラフィー、陰イオン交換クロマトグラフィー、ゲルろ過クロマトグラフィーを組み合わせることにより、ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動(以下SDS−PAGEと略す)で単一になるまで精製することができる。
【0047】
さらに本発明はRhCHBH活性を示すアミノ酸配列をコードする遺伝子の塩基配列を含むベクターを提供する。
【0048】
クローニング用ベクターはpUC系やpBluscript系(いずれもタカラバイオ株式会社からの市販品を利用できる)などを、特に限定されず宿主の種類に応じて用いることができる。好ましくはpUC19を用いるとよい。
【0049】
上記塩基配列(配列番号1)を有するDNA断片にコードされる構造遺伝子から、上記酵素の発現を可能とするプロモーターなどの機能領域および宿主での複製オリジンを有するベクターに組み込むことにより、組換えDNA技術を利用した本不斉加水分解反応を有する形質転換体を得るための組換えDNAを構築できる。
【0050】
発現用ベクターはpUC系やpBluscript系(タカラバイオ株式会社製等)、pET系(インビトロジェン製)など特に限定されず、宿主の種類に応じて用いることができる。発現用ベクターとして好ましいのは、宿主細胞中で自律複製可能であり、さらに組換え宿主細胞のみを選別できるような適当な選択マーカーなどが付与されたものがあげられ、適当な宿主細胞内で、本発明の遺伝子を発現できるものである。さらに、このようなベクターは公知のベクター等から公知の技術を用いて業者が容易に製造し得るようなものであってもよいし、商業的に販売されているものでも良い。特に好ましいのはpKK223−3(ファルマシア製)である。
【0051】
RhCHBH遺伝子の遺伝情報を担うDNA断片から必要な部分をベクターに組み込むことにより、宿主細胞に導入し形質転換体を得ることができる。
【0052】
用いられる宿主細胞としては、得られた組換えベクターで形質転換され、かつ本遺伝子を発現させることができるようなものであれば特に制限なく使用することができる。発現用の宿主としては、本発明の目的に沿って本遺伝子の発現を達成し得る限り、微生物、植物、動物等から限定されず用いることができる。微生物としては、例えば、大腸菌(Escherichia coli)、エンテロバクター属、サッカロミセス属、キサントモナス属、アセトバクター属、シュードモナス属、グルコノバクター属、アゾトバクター属、リゾビウム属、クレブシエラ属、サルモネラ属およびセラチア属から選ばれる微生物が挙げられるが、好ましいのは大腸菌であり、特に好ましいのは大腸菌JM109である。大腸菌JM109(pKK−RhCHBH)はFERM P−21128として独立行政法人産業技術研究所特許生物寄託センターに寄託されている。
【0053】
形質転換体は、適当な栄養培地中で培養を行なうことにより大量に得ることができる。培養に用いられる培地は微生物の生育に必要な炭素源、窒素源、無機物質等を含む通常の培地であれば何でも良い。例えば炭素源としてグルコース、フラクトース等の炭水化物、グリセロール、マンニトール、ソルビトール、プロピレングリコール等のアルコール類、酢酸、クエン酸、リンゴ酸、マレイン酸、フマル酸、グルコン酸とその塩類などの有機酸、またはそれらの混合物を用いることができる。窒素源として硫酸アンモニウム、硝酸アンモニウム、リン酸アンモニウム等の無機窒素化合物および尿素、ペプトン、カゼイン、酵母エキス、肉エキス、コーンスチープリカー等の有機窒素化合物とそれらの混合物を挙げることができる。その他、無機塩としてリン酸塩、マグネシウム塩、カリウム塩、マンガン塩、鉄塩、亜鉛塩、銅塩など、更に必要に応じてビタミン類を加えてもよい。
【0054】
また、高酵素活性を持った形質転換菌体を得るための酵素誘導添加物として、上記培地およびペプトン培地、ブイヨン培地等の栄養培地に、使用する菌株に応じて、効果的に目的のDNAを発現させるためにアンピシリン、カナマイシン、クロラムフェニコール等の抗生物質を培養液に添加してもよいし、イソプロピルβ−D(−)−チオガラクトピラノシド(IPTG)等のプロモーターの活性化誘導剤を用いることもできる。培養は好気的条件下でpH6.0〜7.5、温度25〜40℃の任意の範囲に制御して16〜24時間培養を行えばよいが、用いる形質転換体により最適な条件で培養すると更に効果的である。
【0055】
得られたRhCHBH産生形質転換体の細胞からの酵素RhCHBHの抽出は次の方法によって形質転換体を破壊して行うことができる。
1)フレンチプレスや超音波破砕による機械的(物理的)方法、
2)リゾチームなどの酵素処理方法、
3)自己溶解法、
4)浸透圧を利用した抽出法など。
【0056】
またJM109等の大腸菌(pKK−RhCHBH含有)は細胞を破砕しなくとも、培養液の状態で高活性な酵素として利用できる。
【0057】
本方法で産生される酵素は、起源のDS−S−51株(受託番FERM P−21129)におけるRhCHBHのポリペプチド配列と一致するもののみに限定されず、塩基配列変換や遺伝子変換などの遺伝子組換え手法を利用して得られるポリペプチドで、RhCHBH活性を示すものと解すべきである。すなわちペプチド配列中のアミノ酸の1個または数個が欠損したもの、あるいはそのペプチド配列中のアミノ酸の1個または数個が他のアミノ酸で置き換えられたものをも包含する。
【0058】
得られた形質転換体は、酵素RhCHBHの量産に、また形質転換体自体を用いたS体4−クロロ−3−ヒドロキシ酪酸エステルおよびR体3−ヒドロキシ−γ−ブチロラクトンの製造に好適に用い得る。
【0059】
次に、本発明のS体4−ハロ−3−ヒドロキシ酪酸エステルの製造方法およびR体3−ヒドロキシ−γ−ブチロラクトンの製造方法について説明する。
【0060】
原料の4−ハロ−3−ヒドロキシ酪酸エステル、例えば、4−クロロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物は、例えば、エピクロルヒドリンから安価に製造することができる。
【0061】
また、R体4−ハロ−3−ヒドロキシ酪酸エステルは、微生物または酵素を用いて4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物を立体選択的に加水分解させ、R体を残存させる製法(特開平9−47296号公報)により簡単に得ることができる。
【0062】
本発明で用いられるRhCHBH酵素を産生する微生物としては、上記のようなRhCHBH遺伝子を導入した形質転換体であってもよいし、天然に4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物のR体を選択的に分解し得るものであってもよく、かかる天然に4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物のR体を選択的に分解し得る微生物としては、リゾビウム属(Rhizobium sp.)に属すると推定される細菌DS−S−51株(受託番号FERM P−21129)が挙げられる。
【0063】
[分類方法詳細]
本発明で用いられる微生物の具体例であるDS−S−51株は本発明者らが土壌から分離した細菌である。本菌株の形態学的・生理学的性質に基づき委託、同定された結果、形態、生育状態、生理学的試験の結果、グラム染色陰性、カタラーゼ反応陽性、オキシダーゼ反応陽性、運動性を示す無芽胞かん菌であるなどシュードモナス(Pseudomonas )類似の性状を示した。続く生化学試験、資化性試験ではリゾビウム・ラジオバクター(Rhizobium radiobacter) に類似の性状を示した。しかしながらその性状はNAGa、GATaの試験結果でリゾビウム・ラジオバクターの典型性状とは異なるなど詳細な帰属分類群の推定には至らなかった。
【0064】
そこで、16SrRNA遺伝子の部分塩基配列約500 bpをもちいて検体の帰属分類群を委託、推定された結果、リゾビウム属に属する細菌と高い相同性を示した。16SrRNA部分塩基配列は相同性98.9%でリゾビウム・ダエジェオネンス(Rhizobium daejeonense)の16SrRNAに対し最も高い相同性を示した。さらに、分子系統樹上で系統枝を形成した。
【0065】
これらの結果および下記の菌株の諸性質により、DS−S−51株をリゾビウム属に属する細菌であると同定した。DS−S−51株の科学的性質は下記に示すとおりである。
【0066】
A.形態
細胞の形 かん菌
細胞の大きさ 0.6〜0.7 × 1.5〜2.0μm
グラム染色 陽性
胞子の有無 陰性
運動性の有無 陽性
【0067】
B.生育状態(30℃、48時間培養)
コロニーの形状 円形
コロニー表面の形状 全縁滑らか
コロニーの隆起状態 低凸状
コロニーの光沢 光沢あり
コロニーの色調 クリーム色
37℃での生育状態 反応弱
45℃の生育状態 陰性
【0068】
C.生理学的試験
カタラーゼ 陽性
オキシダーゼ 陽性
グルコース、酸産出 陰性
グルコース、ガス産出 陰性
グルコース、Oテスト 陽性
グルコース、Fテスト 陰性
【0069】
D.生化学試験
硝酸塩還元 +
インドール産出 −
ブドウ糖 酸性化 −
アルギニンジヒドロラーゼ −
ウレアーゼ +
エスクリン加水分解 +
ゼラチン加水分解 −
β‐ガラクトシダーゼ +
【0070】
E.資化性試験
ブドウ糖 +
L‐アラビノース +
D‐マンノース +
D‐マンニトール +
N‐アセチル‐D‐グルコサミン −
マルトース +
グルコン酸カリウム −
n‐カプリン酸 −
アジピン酸 −
dl‐リンゴ酸 +
クエン酸ナトリウム −
酢酸フェニル −
チトクロームオキシダーゼ +
【0071】
MacConkey寒天培地での生育試験 −
シモンズクエン酸の利用試験 −
【0072】
4−ハロ−3−ヒドロキシ酪酸エステルの両エナンチオマー混合物またはR体4−ハロ−3−ヒドロキシ酪酸エステルに、RhCHBH遺伝子を含む細菌または形質転換体に含まれる酵素RhCHBHを作用させて、S体4−ハロ−3−ヒドロキシ酪酸エステルまたはR体3−ヒドロキシ−γ−ブチロラクトンを得る方法としては、酵素RhCHBHそのものを基質と接触させてもよいし、上記RhCHBH遺伝子を含む細菌または形質転換体の培養液または、それらの遠心分離により得た菌体およびその菌体処理物(菌体破砕物または菌体抽出液)あるいは、それらを常法により固定化したものを基質と接触させてもよい。
【0073】
基質と酵素または酵素を含む微生物との接触は、緩衝液に混合した微生物菌体混合液に基質を添加する方法などがある。反応温度は15〜50℃が好ましく、反応pHは6〜9で行なうのが好ましい。基質は初期に一括添加してもよいし、分割添加してもよい。反応は通常、攪拌あるいは振とうしながら行い、反応時間は基質濃度、微生物菌体または酵素量により異なるが、1〜120時間で終了するのがよい。好ましくはガスクロマトグラフィーなどの分析により、目的の光学活性体の濃度および光学純度を測定し終点を決定するのがよい。
【実施例】
【0074】
以下実施例をもって、本発明を詳細に説明するが、本発明はこれらに限定されるものではない。なお、実施例中の%は特に記載のない限り(w/v)で表す。
【0075】
(実施例1)DS−S−51株の培養
ペプトン、酵母エキス、グリセロールをそれぞれ1.0%含む液体栄養培地(pH7.0)5mlを入れた試験管に、凍結保存バイアルから種菌を2%(v/v)量、無菌的に植菌し、30℃で24時間培養を行なった。
【0076】
(実施例2)遺伝子のクローニング
実施例1で得られた培養液から遠心分離により菌体を回収した。菌体を490μlのTE溶液(10mMトリス−塩酸 pH8.0、1mM EDTA)に懸濁し、10%SDSを30μl、20mg/mlプロテアーゼKを50μl添加し、50℃で1時間反応させた。その後、等量のフェノール:クロロホルム:イソアミルアルコール(25:24:1)による抽出を行なった後、0.1倍量の3M酢酸ナトリウム溶液を添加し、0.6倍量の2−プロパノールを静かに重層した。その結果、界面に生じたDNAをガラス棒で糸状に巻きつけて回収した。得られたDNAを500μlのTE溶液に溶解させた。約1.1mgの染色体DNAを得た。
【0077】
制限酵素Sau3AIを用いて、上記により得られた染色体DNAの60μgを250μlの反応系で酵素0.75U用いて部分消化し、染色体DNA断片を得た。またプラスミドベクターpUC19を常法により制限酵素BamHIで消化し、BAP処理を行なった。
【0078】
上記によって消化された染色体DNA断片14μgとプラスミドベクターpUC19の800ngをT4DNAリガーゼ(タカラバイオ株式会社より購入できる)1400Uを用いて常法により16℃でライゲーション反応させ、これを用いて42℃の熱ショック法により大腸菌DH5α株(東洋紡績株式会社より購入できる)を形質転換させた。
【0079】
熱ショック後、SOC培地にて37℃で1時間培養した後、LB寒天培地(100μg/mlアンピシリン、0.5%メチル4−クロロ−3−ヒドロキシ酪酸、0.0008%BTB(ブロモチールブルー)を含む)一枚につき、上記菌体液の約100μlを塗布し、37℃で16時間インキュベートした。
【0080】
BTBにより通常pHが中性のプレートは緑色であるが、プレートに出現した合計47600個の形質転換体コロニーのうち、メチル4−クロロ−3−ヒドロキシ酪酸の脱クロルを伴った加水分解反応によりpHが低下するに伴い、周辺が黄色に変色しているコロニー1個を得た。
【0081】
得られた陽性コロニーを単離培養し、常法に従ってプラスミドを調製し、制限酵素EcoRI、HindIIIで消化し、1%アガロース電気泳動を行なったところ、約4kbのDNAがpUC19に挿入されていることがわかった。
【0082】
(実施例3)DNAの塩基配列の決定
実施例2で得られた約4kbの遺伝子断片のうち、ORF周囲1.8kbの配列決定をDNAシークエンサーにより行った。その結果、配列番号1の第44塩基〜第1240塩基に示す1197bpからなるオープンリーディングフレーム(ORF)を確認できた。これより、398残基のアミノ酸をコードしていることがわかった。アミノ酸配列を配列番号2に示す。
【0083】
(実施例4)高発現組換え大腸菌の作製
ORFのすぐ上流にEcoRI部位、下流にHindIII部位を付与する様に配列番号3および4に示すPCRプライマー:
5'-ttgaattcatgccccataatctg-3'(配列番号3)
5'-aaaagcttgtggccgtcga-3'(配列番号4)
を設計し、増幅断片をpKK223−3のEcoRI、HindIII部位に導入し、これを用いて実施例2と同様に大腸菌JM109株を形質転換させてJM109(pKK−RhCHBH)を得た。また対照試料としてベクターpKK223−3のみを用いた形質転換により、JM109(pKK223−3)を得た。
【0084】
100μg/mlアンピシリンを含むLB培地にて作製した上記組換え大腸菌を20時間培養した。
【0085】
調製した組換え大腸菌の培養液を遠心分離により集菌し、20mMリン酸カリウム緩衝液で懸濁後、超音波破砕した。5μgの抽出タンパク試料をSDS−PAGE(10%分離ゲル)に供し、クマシーブリリアントグリーンによる染色を行なった。その結果分子量は42kDaであることがわかった。用いた分子量マーカーと分子量を下記に示す。ホスホリラーゼB(94,000)、ウシ血清アルブミン(67,000)、オボアルブミン(43,000)、カルボニックアンヒドラーゼ(30,000)、トリプシンインヒビター(20,100)、α−ラクトアルブミン(14,400)。
【0086】
(実施例5)S体4−クロロ−3−ヒドロキシ酪酸エステルの生産
500ml容のバッフル付三角フラスコを用いてロータリーシェイカーで130rpmにて培養した条件以外は、実施例1または実施例4と同じ方法で培養して得たDS−S−51株の培養液100ml、および組換え大腸菌JM109(pKK−RhCHBH)の培養液100mlに、基質としてラセミ体メチル4−クロロ−3−ヒドロキシ酪酸を加え、30℃で振とうしながら反応させた。反応終了後、反応液中に残存するメチル4−クロロ−3−ヒドロキシ酪酸をガスクロマトグラフィー(カラム担体:PEG20M,60−80メッシュ)で分析した結果、その残存率(収率)はDS−S−51株については30.9%、組換え大腸菌については30.5%であった。残存率(収率)とは、添加したラセミ体量を100%とした時、反応後に残存している基質量を表す。反応液を遠心分離により菌体を除き、ついで遠心上清と等量の酢酸エチルで抽出した。この抽出物中のメチル4−クロロ−3−ヒドロキシ酪酸をアステック社(Advanced Separation Technologies Inc., NJ, USA)製のキャピラリーカラムG−TA(0.25mm×30m)を用いたガスクロマトグラフィーにより光学純度分析を行った結果、回収したメチル4−クロロ−3−ヒドロキシ酪酸は光学純度99%e.e.以上のS体であることが判明した。DS−S−51株は、1.0%(v/v)の基質を光学分割するのに40時間要した。対して、組換え大腸菌は、2.0%(v/v)の基質を4時間で光学分割した。次に実施例4で得た組換え大腸菌の培養液に基質として2.0%(v/v)になるように、エチル4−クロロ−3−ヒドロキシ酪酸、プロピル4−クロロ−3−ヒドロキシ酪酸、ブチル4−クロロ−3−ヒドロキシ酪酸のラセミ体を各々に加え、同様に4時間反応、分析した結果、各々残存率(収率)は、30.5%、30.0%、31.0%であり、各種4−クロロ−3−ヒドロキシ酪酸エステルは光学純度99%e.e.以上のS体であることが判明した。光学純度の分析条件はカラム温度:100℃、検出器温度:200℃、キャリアーガス:窒素、流速:0.5ml/min;検出器、FID;スプリット比、100/1、リテンションタイムは次に示す通りである。メチル4−クロロ−3−ヒドロキシ酪酸:R体14.4分、S体15.2分、エチル4−クロロ−3−ヒドロキシ酪酸:R体18.4分、S体19.2分、プロピル4−クロロ−3−ヒドロキシ酪酸:R体28.8分、30.2分、ブチルメチル4−クロロ−3−ヒドロキシ酪酸:R体49.2分、S体50.2分。
【0087】
(実施例6)R体3−ヒドロキシ−γ−ブチロラクトンの生産
実施例5と同様に調製したDS−S−51株の培養液100ml、および組換え大腸菌JM109(pKK−RhCHBH)の培養液100mlに、基質として光学純度99%eeのR体メチル4−クロロ−3−ヒドロキシ酪酸を加え、30℃で振とうしながら反応させた。反応終了後、生成したR体3−ヒドロキシ−γ−ブチロラクトンをガスクロマトグラフィー(カラム担体:PEG20M,60−80メッシュ)で分析した結果、その生成モル収率はDS−S−51株、組換え大腸菌とも100%であった。3−ヒドロキシ−γ−ブチロラクトンの光学純度分析は以下の様に行なった。反応液を遠心分離により菌体を除き、ついでロータリーエバポレーターにて濃縮を行なった。適当量の1,2−ジクロロエタンで抽出してシロップを得た。ジクロロメタン1mlにシロップ20μl、トリフルオロ酢酸無水物200μlを加えてよく攪拌し、30分後デシケーターで濃縮後、エタノール1mlに溶解させた後、実施例5と同じキャピラリーカラムG−TAに供した。その結果、回収した3−ヒドロキシ−γ−ブチロラクトンは光学純度99.0%e.e.のR体であることが判明した。分析条件はカラム温度:100℃、検出器温度:200℃、キャリアーガス:窒素、流速:0.5ml/min;検出器、FID;スプリット比、100/1。3−ヒドロキシ−γ−ブチロラクトンのリテンションタイムはS体29.5分、R体31.2分であった。DS−S−51株は、1.0%の基質をすべてR体に変換するのに80時間要した。対して、組換え大腸菌は、2.0%(v/v)の基質を8時間で変換した。
【図面の簡単な説明】
【0088】
【図1】図1は、RhCHBH遺伝子コンストラクトの構築を示す図である。
【図2】図2は、組換え大腸菌のSDS−PAGEを示す図である。
【特許請求の範囲】
【請求項1】
以下の(a)または(b)のいずれかの塩基配列からなる遺伝子:
(a)配列番号1の第44塩基〜第1240塩基からなる塩基配列、
(b)配列番号1の第44塩基〜第1240塩基からなる塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列であって、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用し、該混合物のR体を選択的に加水分解し、かつ、S体を選択的に残存させる酵素活性を有するタンパク質をコードする塩基配列:
【化1】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【請求項2】
以下の(a’)または(b’)のいずれかのタンパク質:
(a’)配列番号2に記載のアミノ酸配列からなるタンパク質、
(b’)配列番号2に記載のアミノ酸配列において、1または数個のアミノ酸が欠失、置換または付加されているアミノ酸配列からなり、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用し、該混合物のR体を選択的に加水分解し、かつ、S体を選択的に残存させる酵素活性を有するタンパク質:
【化2】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【請求項3】
請求項1に記載の遺伝子を含むベクター。
【請求項4】
プラスミドpKK−RhCHBHである請求項3に記載のベクター。
【請求項5】
請求項1に記載の遺伝子を有する細菌。
【請求項6】
細菌DS−S−51株(受託番号FERM P−21129)である請求項5に記載の細菌。
【請求項7】
請求項1に記載の遺伝子または請求項3または4に記載のベクターを含む形質転換体。
【請求項8】
大腸菌である請求項7に記載の形質転換体。
【請求項9】
大腸菌JM109(pKK−RhCHBH)(受託番号FERM P−21128)である、請求項8に記載の形質転換体。
【請求項10】
請求項1に記載の遺伝子を発現させることを含む、請求項2に記載のタンパク質の製造方法。
【請求項11】
請求項2に記載のタンパク質を、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用させ、該混合物のR体を選択的に加水分解させ、かつ、S体を選択的に残存させることを含む、S体4−ハロ−3−ヒドロキシ酪酸エステルの製造方法:
【化3】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【請求項12】
請求項2に記載のタンパク質を、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのR体に作用させることを含む、下記式[2]で示されるR体3−ヒドロキシ−γ−ブチロラクトンの製造方法:
【化4】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【化5】
式[2]
【請求項13】
式[1]のRがメチルである化合物を用いる請求項11または12に記載の方法。
【請求項14】
Xが塩素原子である化合物を用いる請求項11〜13のいずれかに記載の方法。
【請求項1】
以下の(a)または(b)のいずれかの塩基配列からなる遺伝子:
(a)配列番号1の第44塩基〜第1240塩基からなる塩基配列、
(b)配列番号1の第44塩基〜第1240塩基からなる塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列であって、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用し、該混合物のR体を選択的に加水分解し、かつ、S体を選択的に残存させる酵素活性を有するタンパク質をコードする塩基配列:
【化1】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【請求項2】
以下の(a’)または(b’)のいずれかのタンパク質:
(a’)配列番号2に記載のアミノ酸配列からなるタンパク質、
(b’)配列番号2に記載のアミノ酸配列において、1または数個のアミノ酸が欠失、置換または付加されているアミノ酸配列からなり、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用し、該混合物のR体を選択的に加水分解し、かつ、S体を選択的に残存させる酵素活性を有するタンパク質:
【化2】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【請求項3】
請求項1に記載の遺伝子を含むベクター。
【請求項4】
プラスミドpKK−RhCHBHである請求項3に記載のベクター。
【請求項5】
請求項1に記載の遺伝子を有する細菌。
【請求項6】
細菌DS−S−51株(受託番号FERM P−21129)である請求項5に記載の細菌。
【請求項7】
請求項1に記載の遺伝子または請求項3または4に記載のベクターを含む形質転換体。
【請求項8】
大腸菌である請求項7に記載の形質転換体。
【請求項9】
大腸菌JM109(pKK−RhCHBH)(受託番号FERM P−21128)である、請求項8に記載の形質転換体。
【請求項10】
請求項1に記載の遺伝子を発現させることを含む、請求項2に記載のタンパク質の製造方法。
【請求項11】
請求項2に記載のタンパク質を、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのエナンチオマー混合物に作用させ、該混合物のR体を選択的に加水分解させ、かつ、S体を選択的に残存させることを含む、S体4−ハロ−3−ヒドロキシ酪酸エステルの製造方法:
【化3】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【請求項12】
請求項2に記載のタンパク質を、下記式[1]で示される4−ハロ−3−ヒドロキシ酪酸エステルのR体に作用させることを含む、下記式[2]で示されるR体3−ヒドロキシ−γ−ブチロラクトンの製造方法:
【化4】
式[1]
[式中、Xは、ハロゲン原子を表し、Rは炭素原子数1〜4のアルキル基を表す]。
【化5】
式[2]
【請求項13】
式[1]のRがメチルである化合物を用いる請求項11または12に記載の方法。
【請求項14】
Xが塩素原子である化合物を用いる請求項11〜13のいずれかに記載の方法。
【図1】

【図2】
【図2】
【公開番号】特開2008−194037(P2008−194037A)
【公開日】平成20年8月28日(2008.8.28)
【国際特許分類】
【出願番号】特願2008−8203(P2008−8203)
【出願日】平成20年1月17日(2008.1.17)
【出願人】(000108993)ダイソー株式会社 (229)
【Fターム(参考)】
【公開日】平成20年8月28日(2008.8.28)
【国際特許分類】
【出願日】平成20年1月17日(2008.1.17)
【出願人】(000108993)ダイソー株式会社 (229)
【Fターム(参考)】
[ Back to top ]